
自己免疫異常に関わるHLA−DRMHCクラスII分子に関するペプチド
HLA-DR分子と呼ばれる共有エピトープを有するMHCクラスII分子に結合する抗原性ペプチドが開示される。より具体的には、HLA-DR分子に対して増加したアフィニティーを有し、かつ慢性関節リウマチに関連する抗原性ペプチドがシトルリン化される。これらの新規なペプチドは慢性関節リウマチ診断および治療の新規な方法の基礎を提供する。
【発明の詳細な説明】
【技術分野】
【0001】
本発明は、HLA-DR分子と呼ばれる共有エピトープを有するMHCクラス II分子に結合する新規な抗原性ペプチドに関する。より詳細には、本発明は、HLA-DR分子についてのアフィニティー増加を有し、かつ特定の自己免疫障害に関連するシトルリン化抗原性ペプチドに向けられる。このようなものとして、これらの新規なペプチドは共有された抗原性(慢性関節リウマチと多発性硬化症を含むそのような自己免疫性疾患)でMHCクラスII分子を有する対象における自己免疫異常の診断および治療の新規な方法のための基礎を提供する。
【背景技術】
【0002】
本願を通して、種々の参考文献は、本発明が関係する技術水準をより完全に記載するために括弧内に引用される。これらの参考文献の開示は本明細書によって参考として本開示に組み入れられる。
【0003】
RA(慢性関節リウマチ)は、滑膜炎ならびに軟骨および骨の分解をもたらし得るパンヌス形成によって特徴付けられる一般的な自己免疫障害である。この衰弱させるような状態は人口のおよそ1%に影響する。この疾患に対する遺伝的感受性は共有エピトープ(SE)として知られるアミノ酸モチーフを含み、かつHLA-DR分子として呼ばれるMHCクラスII分子と関連している(Gregersen, P.K., J. Silver, R.J. Winchester. 1987. The shared epitope hypothesis. An approach to understanding the molecular genetics of susceptibility to rheumatoid arthritis. Arthritis Rheum. 30:1205、Zhou, Z., H.A. Menard. 2002. Autoantigenic posttranslational modifications of proteins: does it apply to rheumatoid arthritis? Curr. Opin. Rheumatol. 14:250)。アミノ酸残基Q/R、K/R、R、A, Aによって表現される、共有エピトープは、正に荷電し、かつMHCクラスII分子の主要なペプチドアンカーリングポケット(P4として知られる)の1つを形成する。
【0004】
以前の報告は、RAに関わる推定上の病原性のペプチドに関する目立った特徴は、P4(共有エピトープと相互作用する)の負に荷電した側鎖の存在であるかもしれないと示唆した(Hammer J., Gallazzi F., Bono E., Karr R.W., Guenot J., Valsasnini P., Nagy Z.A., Sinigaglia F. 1995. Peptide binding specificity of HLA-DR4 molecules: correlation with rheumatoid arthritis association. J. Exp. Med. 181:1847)。
【0005】
これは、特定のMHCクラスII分子、HLA-DR*0401、およびHLA-DR*0404(対立遺伝子多型)が、RAに関連しないHLA DR*0402分子よりも、P4ポケットにおけるアスパラギン酸およびグルタミン酸のアミノ酸残基について実質的により高いアフィニティーを有するという事実に基づいている。しかし、複数のDRB1ポケットプロフィールの分析の後で、いくつかのRA非関連対立遺伝子は、それらのP4ポケットにおける負荷電アミノ酸について、HLA-DR*0101、HLA-DR*0401、HLA-DR*0404、例えばHLA-DR*0301などでさえよりも高いアフィニティーを有することを見い出すことができる(Sturniolo T, Bono E, Ding J, Raddrizzani L, Tuereci O, Sahin U, Braxenthaler M, Gallazzi F, Protti MP, Sinigaglia F, Hammer J. 1999. Generation of tissue-specific and promiscuous HLA ligand databases using DNA microarrays and virtual HLA class II matrices. Nat. Biotechnol. 17:555)。いくつかのMHC分子は、単に非関連であるよりむしろ、疾患に対して保護的であるように見え(例えば、HLA-DR *0402)、このことは、ペプチド結合におけるこれらの対立遺伝子についての受動的な役割は存在しないかもしれないことを示唆する(Reviron, D., A. Perdriger, E. Toussirot, D. Wendling, N. Balandraud, S. Guis, G. Semana, P. Tiberghien, P. Mercier, J. Roudier. 2001. Influence of shared epitope-negative HLA-DRB1 alleles on genetic susceptibility to rheumatoid arthritis. Arthritis Rheum. 44:535)。その代わりに、保護的な対立遺伝子(MHC分子)は、ネガティブなT細胞選択を引き起こすか、または末梢寛容を確率するために十分に高いアフィニティーを有する推定の病原性ペプチドを結合することができる。
【0006】
共有エピトープを有するMHCクラスII分子は、自己反応性CD4+T細胞への提示のために関節炎性ペプチドを選択的に結合することによって疾患の病原性に関与するかもしれない。現在、RAについて原因となる自己抗原の性質は知られていない。多くの自己抗原候補がRAの関連MHCの文脈で調査されたが、CD4+T細胞およびB細胞免疫応答の共通の疾患特異的標的はとらえどころがないままで残っている。最近の研究は、RA患者がRAの診断について敏感かつ特異的(>90%)の両方であるIgG自己抗体のサブセットを有することを示した。これらの自己抗体の標的はシトルリン(翻訳後に修飾されたアルギニン(脱イミノ化アルギニン))であり、これは、特定のタンパク質/ペプチド配列の文脈の中で見い出された(van Venrooij, W. J., G. J. Pruijn. 2000. Citrullination: a small change for a protein with great consequences for rheumatoid arthritis. Arthritis Res. 2:249)。シトルリンは、抗核周囲抗体、抗ケラチン抗体、抗フィラグリン抗体、抗環状シトルリン化ペプチド抗体、および抗Sa抗体の必須の抗原性エピトープ標的である(van Venrooij, W. J., G. J. Pruijn. 2000. Citrullination: a small change for a protein with great consequences for rheumatoid arthritis. Arthritis Res. 2:249; Zhou, Z., H.A. Menard. 2002. Autoantigenic posttranslational modifications of proteins: does it apply to rheumatoid arthritis? Curr. Opin. Rheumatol. 14:250)。これらの抗体は、多くの異なったタンパク質の中でシトルリンを標的化し、結合誘導体化標的はビメンチンとフィブリン(フィブリノーゲン)であるように見える(Menard, H. A., E. Lapointe, M. D. Rochdi, Z. J. Zhou. 2000. Insights into rheumatoid arthritis derived from the Sa immune system. Arthritis Res. 2:429; Christine Masson-Bessiereら、2001. The Major Synovial Targets of the Rheumatoid arthritis-Specific Antifilaggrin Autoantibodies Are Deiminated Forms of the ( and ( Chains of Fibrin 1 The Journal of Immunology 166: 4177-4184)。抗シトルリン抗体生産が、RA患者におけるMHC共有エピトープの存在に有意に関連していることもまた、観察されている(Goldbach-Mansky R, Lee J, McCoy A, Hoxworth J, Yarboro C, Smolen JS, Steiner G, Rosen A, Zhang C, Menard HA, Zhou ZJ, Palosuo T, Van Venrooij WJ, Wilder RL, Klippel JH, Schumacher HR Jr, El-Gabalawy HS. 2000. Rheumatoid arthritis associated autoantibodies in patients with synovitis of recent onset. Arthritis Res. 2:236; Bas S, Perneger TV, Mikhnevitch E, Seitz M, Tiercy JM, Roux-Lombard P, Guerne PA. 2000. Association of rheumatoid factors and anti-filaggrin antibodies with severity of erosions in rheumatoid arthritis. Rheumatology (Oxford).39:1082)。
【特許文献1】米国特許第5,824,315号
【特許文献2】米国特許第6,184,204号
【特許文献3】米国特許第5,869,279号
【特許文献4】米国特許第5,843,456号
【特許文献5】米国特許第5,869,270号
【特許文献6】米国特許第4,609,893号
【特許文献7】米国特許第4,713,325号
【特許文献8】米国特許第4,714,681号
【特許文献9】米国特許第4,716,111号
【特許文献10】米国特許第4,716,117号
【特許文献11】米国特許第4,720,459号
【特許文献12】米国特許第5,734,02号
【特許文献13】米国特許第6,106,840号
【特許文献14】米国特許第5,635,363号
【特許文献15】米国特許第6,211,342号
【特許文献16】米国特許出願公開第20020176864号
【特許文献17】米国特許出願公開第20020122818号
【特許文献18】国際特許公開WO0129058号
【特許文献19】国際特許公開WO9932619号
【特許文献20】カナダ国特許出願第2,388,441号
【特許文献21】米国特許第5,843,445号
【特許文献22】国際特許公開WO02/14870号
【特許文献23】米国特許第6,355,617号
【非特許文献1】Gregersen, P.K., J. Silver, R.J. Winchester. 1987. The shared epitope hypothesis. An approach to understanding the molecular genetics of susceptibility to rheumatoid arthritis. Arthritis Rheum. 30:1205
【非特許文献2】Zhou, Z., H.A. Menard. 2002. Autoantigenic posttranslational modifications of proteins: does it apply to rheumatoid arthritis? Curr. Opin. Rheumatol. 14:250
【非特許文献3】Hammer J., Gallazzi F., Bono E., Karr R.W., Guenot J., Valsasnini P., Nagy Z.A., Sinigaglia F. 1995. Peptide binding specificity of HLA-DR4 molecules: correlation with rheumatoid arthritis association. J. Exp. Med. 181:1847
【非特許文献4】Sturniolo T, Bono E, Ding J, Raddrizzani L, Tuereci O, Sahin U, Braxenthaler M, Gallazzi F, Protti MP, Sinigaglia F, Hammer J. 1999. Generation of tissue-specific and promiscuous HLA ligand databases using DNA microarrays and virtual HLA class II matrices. Nat. Biotechnol. 17:555
【非特許文献5】Reviron, D., A. Perdriger, E. Toussirot, D. Wendling, N. Balandraud, S. Guis, G. Semana, P. Tiberghien, P. Mercier, J. Roudier. 2001. Influence of shared epitope-negative HLA-DRB1 alleles on genetic susceptibility to rheumatoid arthritis. Arthritis Rheum. 44:535
【非特許文献6】van Venrooij, W. J., G. J. Pruijn. 2000. Citrullination: a small change for a protein with great consequences for rheumatoid arthritis. Arthritis Res. 2:249
【非特許文献7】Zhou, Z., H.A. Menard. 2002. Autoantigenic posttranslational modifications of proteins: does it apply to rheumatoid arthritis? Curr. Opin. Rheumatol. 14:250
【非特許文献8】Menard, H. A., E. Lapointe, M. D. Rochdi, Z. J. Zhou. 2000. Insights into rheumatoid arthritis derived from the Sa immune system. Arthritis Res. 2:429
【非特許文献9】Christine Masson-Bessiereら、2001. The Major Synovial Targets of the Rheumatoid arthritis-Specific Antifilaggrin Autoantibodies Are Deiminated Forms of the ( and ( Chains of Fibrin 1 The Journal of Immunology 166: 4177-4184
【非特許文献10】Goldbach-Mansky R, Lee J, McCoy A, Hoxworth J, Yarboro C, Smolen JS, Steiner G, Rosen A, Zhang C, Menard HA, Zhou ZJ, Palosuo T, Van Venrooij WJ, Wilder RL, Klippel JH, Schumacher HR Jr, El-Gabalawy HS. 2000. Rheumatoid arthritis associated autoantibodies in patients with synovitis of recent onset. Arthritis Res. 2:236
【非特許文献11】Bas S, Perneger TV, Mikhnevitch E, Seitz M, Tiercy JM, Roux-Lombard P, Guerne PA. 2000. Association of rheumatoid factors and anti-filaggrin antibodies with severity of erosions in rheumatoid arthritis. Rheumatology (Oxford).39:1082
【非特許文献12】Dessen A, Lawrence CM, Cupo S, Zaller DM, Wiley DC. 1997. X-ray crystal structure of HLA-DR4 (DRA*0101, DRB1*0401) complexed with a peptide from human collagen II. Immunity. 7:473
【非特許文献13】Stern LJ, Brown JH, Jardetzky TS, Gorga JC, Urban RG, Strominger JL, Wiley DC.1994. Crystal structure of the human class II MHC protein HLA-DR1 complexed with an influenza virus peptide. Nature. 368:215
【非特許文献14】Hammer J, Bono E, Gallazzi F, Belunis C, Nagy Z, Sinigaglia F. 1994. Precise prediction of major histocompatibility complex class II-peptide interaction based on peptide side chain scanning. J. Exp. Med. 180:2353
【非特許文献15】Peterson DA, DiPaolo RJ, Kanagawa O, Unanue ER. 1999. Cutting edge: negative selection of immature thymocytes by a few peptide-MHC complexes: differential sensitivity of immature and mature T cells. J. Immunol. 162:3117
【非特許文献16】DiPaolo RJ, Unanue ER. 2001. The level of peptide-MHC complex determines the susceptibility to autoimmune diabetes: studies in HEL transgenic mice. Eur. J. Immunol. 31:3453
【非特許文献17】Yagi J, Janeway CA Jr. 1990. Ligand thresholds at different stages of T cell development. Int. Immunol. 2:83
【非特許文献18】Ghosh P, Amaya M, Mellins E, Wiley DC. 1995. The structure of an intermediate in class II MHC maturation: CLIP bound to HLA-DR3. Nature. 378:457
【非特許文献19】Jordan MS, Boesteanu A, Reed AJ, Petrone AL, Holenbeck AE, Lerman MA, Naji A, Caton AJ. 2001. Thymic selection of CD4+CD25+ regulatory T cells induced by an agonist self-peptide. Nat. Immunol. 2:301
【非特許文献20】Hagiwara T.ら、 Biochem & Biophy Res Comm. (2000) 290, 979-983
【非特許文献21】A. F. Spatola, in Chemistry and Biochemistry of Amino Acids, Peptides and Proteins, B. Weinstein, eds., Marcel Dekker, New York, p. 267 (1983)
【非特許文献22】Merrifield (1964) J. Am. Chem. Assoc. 65:2149
【非特許文献23】J. Amer. Chem. Soc. 85:2149 (1963)
【非特許文献24】Int. J. Peptide Protein Res. 35:161-214 (1990)
【非特許文献25】Methods of Organic Chemistry, E. Wansch (編) 第15巻, I部およびII部, Thieme, Stuttgart (1987)
【非特許文献26】Goeddel, Gene Expression Technology: Methods in Enzymology 185, Academic Press, San Diego, CA (1990)
【非特許文献27】Hugues, Sら(2002) Generation and use of alternative multimers of peptide/MHC complexes. Journal of Immunological Methods. 268:83-91
【非特許文献28】「Handbook of Pharmaceutical Additives」 (MichaelおよびIrene Ash,編 Gower Publishing Limited, Aldershot, England (1995)
【非特許文献29】「Vaccine Design-the Subunit and Adjuvant Approach」(PowellおよびNewman編, 'Plenum Press, New York, U.S.A., pp. 61-79および141-228 (1995)
【非特許文献30】BanchereauおよびSteinman, Nature 392:245-251, 1998
【非特許文献31】Zitvogelら、Nature Med. 4:594-600, 1998
【非特許文献32】Antibodies, A Laboratory Manual, Ed. Harlow および David Lane (編) Cold Spring Harbor Laboratory, N.Y. (1988)
【非特許文献33】Klein, J. Immunology: The Science of Cell-Noncell Discrimination, John Wiley & Sons, N.Y. (1982)
【非特許文献34】Dennett, R.ら Monoclonal Antibodies, Hybridoma: A New Dimension In Biological Analyses Plenum Press, N.Y. (1980)
【非特許文献35】Campbell, A.「Monoclonal Antibody Technology」Laboratory Techniques In Biochemistry And Molecular Biology, Vol. 13, Burdon.ら (編), Elsevier, Amsterdam (1984)
【非特許文献36】Baeten, D., Steenbakkers PGA., Rovers E., Veys EM., Rijnders AMW., Meijerink J., Keyser F DeおよびBoots A. Localisation of MHC Class II/HC gp-39 complexes in synovia of rheumatoid arthritis patients using complex-specific monoclonal antibodies. Abstracts of the 23rd European Workshop for Rheumatology Research, 第5巻 補遺1, 2003年2月
【非特許文献37】Baeten D., Steenbakkers P., Boots A., Veys EM., and Keyser, Fde., The presentation of the immunodominant epitope of HC gp-39 in the context of the RA-associated HLA class II molecules is specific for RA synovium. Abstracts of the 23rd European Workshop for Rheumatology Research, 第5巻 補遺1, 2003年2月)
【非特許文献38】ELISA Guidebook. Theory & Practice. Editor John R. Crowther, Author John R. Crowther, Pub. Humana Press 2000
【非特許文献39】Fire A.ら、1998 Nature 391:806-811
【非特許文献40】Fire, A. Trends Genet. 15, 358-363 (1999)
【非特許文献41】Sharp, P. A. RNA interference 2001. Genes Dev. 15, 485-490 (2001)
【非特許文献42】Hammond, S. M.ら、Nature Rev. Genet. 2, 110-1119 (2001)
【非特許文献43】Tuschl, T. Chem. Biochem. 2, 239-245 (2001)
【非特許文献44】Hamilton, A.ら、Science 286, 950-952 (1999)
【非特許文献45】Hammond, S. M.ら、Nature 404, 293-296 (2000)
【非特許文献46】Zamore, P. D.、ら、Cell 101, 25-33 (2000)
【非特許文献47】Bernstein, E.ら、Nature 409, 363-366 (2001)
【非特許文献48】Elbashir, S. M.ら、Genes Dev. 15, 188-200 (2001)
【非特許文献49】Elbashir S Mら、2001 Nature 411:494-498)
【非特許文献50】Tranquillら、2000. Enhanced T cell responsiveness to citrulline-containing myelin basic protein in multiple sclerosis patients. Mult. Scler. 6:4220-5
【非特許文献51】Nicholas A.P. 2002. Preparation of a monoclonal antibody to citrullinated epitopes: its characterization and some applications to immunohistochemistry in human brain. Glia.Mar 15;37(4):328-36
【非特許文献52】Nicholasら、2003. Immunohistochemical localization of citrullinated proteins in adult rat brain. J Comp Neurol. May 5:459(3):251-66
【非特許文献53】Vossenaar E.R.ら、(2003). PAD, a growing family of citrullinating enzymes; genes, features and involvement in disease. Bioessays. 25(11): 1106-18
【非特許文献54】Waltersら、Designing Screens: How to Make Your Hits a Hit. Nature Reviews, Drug Discovery. 第2巻, 2003年4月、259-266頁
【非特許文献55】Ito K, Bian HJ, Molina M, Han J, Magram J, Saar E, Belunis C, Bolin DR, Arceo R, Campbell R, Falcioni F, Vidovic D, Hammer J, Nagy ZA. 1996. HLA-DR4-IE chimeric class II transgenic, murine class II-deficient mice are susceptible to experimental allergic encephalomyelitis. J. Exp. Med. 183:2635
【非特許文献56】Hill, J.A., Wang, D., Jevnikar, A.M., Cairns, E., Bell, D.A. 2002. The relationship between predicted peptide-MHC class II affinity and T cell activation in a HLA-DR(1*0401 MHC class II mouse model. Arthritis Res. 5:R40
【非特許文献57】Andersson EC, Hansen BE, Jacobsen H, Madsen LS, Andersen CB, Engberg J, Rothbard JB, McDevitt GS, Malmstrom V, Holmdahl R, Svejgaard A, Fugger L. 1998. Definition of MHC and T cell receptor contacts in the HLA-DR4restricted immunodominant epitope in type II collagen and characterization of collagen-induced arthritis in HLA-DR4 and human CD4 transgenic mice. Proc. Natl. Acad. Sci. USA. 95:7574
【非特許文献58】Southwood S, Sidney J, Kondo A, del Guercio MF, Appella E, Hoffman S, Kubo RT, Chesnut RW, Grey HM, Sette A. 1998. Several common HLA-DR types share largely overlapping peptide binding repertoires. J. Immunol. 160:3363
【非特許文献59】Schllekensら、1998. J Clin Invest. Volume 101, 第1巻, 1998年1月, 273-281頁
【非特許文献60】Asaga Hら、(1998) Selective deimination of vimentin in calcium ionophore-induced apoptosis of mouse peritoneal macrophages. Biochem Biophys Res Commun. 243(3):641-6
【非特許文献61】Nakashima Kら、(1999) Molecular characterization of peptidylarginine deiminase in HL-60 cells induced by retinoic acid and 1alpha,25-dihydroxyvitamin D3. J Biol Chem. 274(39), 27786-92
【非特許文献62】Zhang Yら(2003) In vivo knockdown of gene expression in brain cancer with intravenous RNAi in adult rats. J Gene Med. 5(12):1039-45
【非特許文献63】Bestman-Smith J.ら(2000) Sterically stabilized liposomes bearing anti-HLA-DR antibodies for targeting the primary cellular reservoirs of HIV-1. Biochim Biophys Acta. 1468(1-2):161-74
【非特許文献64】Lewis D.L.ら、(2002) Efficient delivery of siRNA for inhibition of gene expression in postnatal mice. Nat Genet. 32(1): 107-8
【非特許文献65】Song E.ら、(2003) RNA interference targeting Fas protects mice from fulminant hepatitis. Nat Med. 9(3): 347-51
【非特許文献66】McCaffrey A.P.ら、(2002) RNA interference in adult mice. Nature 418(6893):38-9
【非特許文献67】Vossenaar E. R.ら、(2003) Citrullination of synovial proteins in murine models of rheumatoid arthritis. Arthritis Rheum. 48(9): 2489-500
【非特許文献68】Current Protocols in Immunology 15.5章、http://www.mrw2.interscience.wiley.com/cponline)
【発明の開示】
【発明が解決しようとする課題】
【0007】
出願人はここで、独特な相互作用がMHCクラスII分子の共有エピトープとアミノ酸シトルリンの間に存在することを実証した。この相互作用は、患者が共有エピトープを発現する自己免疫障害におけるT細胞応答およびそれに続いてこれらのシトルリン化抗原に対するB細胞応答を発生させることに関与する。さらに、出願人はここで、炎症および自己免疫障害(慢性関節リウマチならびに多発性硬化症など)の発症をもたらすT細胞応答を誘発する新規なシトルリン化抗原を同定した。
【課題を解決するための手段】
【0008】
出願人は、MHCクラスII分子の正に荷電しているP4ポケット(共有エピトープ)に対する結合を介してT細胞応答を誘発する新規なシトルリン化ペプチド抗原を同定した。出願人は、シトルリンのアミノ酸(非荷電極性アミノ酸)への正電荷を有するアミノ酸の修飾が、共有エピトープを有するMHCクラスII分子に対する抗原についてアフィニティーの増加をもたらすことを実証した。この増加した抗原アフィニティーはT細胞活性化をもたらし、これは次には、対象における慢性関節リウマチなどの自己免疫障害の発症をもたらす炎症反応を媒介する。
【0009】
このようなものとして、本発明は、T細胞活性化をもたらす共有エピトープを有するMHCクラスII分子とのシトルリン化ペプチドの結合によって特徴付けられる、自己免疫障害を診断するか、または処理するための種々の診断的および治療的なアプローチにおける本発明のシトルリン化ペプチドの使用を含む。本発明の態様において、そのような自己免疫障害は、例えば、慢性関節リウマチ(RA)および多発性硬化症(MS)を含むことができる。
【0010】
シトルリン含有ペプチド(シトルリンに対するアルギニンの変換を介して、荷電したイミノ側鎖基を非荷電カルボニルに置換するプロセス)に対するT細胞応答は、HLA-DRB1*0401トランスジェニック(DR4-IE tg)マウスで研究された。この研究は、共有エピトープと相互作用するペプチド側鎖位置のシトルリンへのアルギニンの変換が、ペプチドMHCアフィニティーを有意に増加させ、かつDR4-IE tgマウスにおいてCD4+T細胞の活性化をもたらすることを実証した。この翻訳後修飾は、DR4-IE tgマウスにおけるこれらのペプチドに対するCD4+T細胞応答を誘発するために必要であった。多くのHLA対立遺伝子についてのペプチドアフィニティーが評価され、これは、シトルリン含有ペプチドについて、共有エピトープを有するMHCクラスII分子だけがアフィニティー増加を有することを示した。
【0011】
まとめると、出願人は、RAにおける疾患関連HLA対立遺伝子の分子的基礎を説明するのを補助する、より詳細に述べると、共有エピトープを有するMHCクラスII分子がRA患者のシトルリン化自己抗原に対する自己免疫応答を開始することに関与することを明らかにする新規なペプチドMHC相互作用を同定した。シトルリン化ミエリン塩基性蛋白質(MBP)T細胞応答が以前に観察されたように、本発明はまた、MHCクラスII共有エピトープを発現する対象について、多発性硬化症(MS)についての診断的および治療的な処置を含む。
【発明を実施するための最良の形態】
【0012】
本発明は、共有エピトープを有するすべてのMHCクラスII分子を含み、その結果、シトルリンとのペプチドアフィニティーの増加が、アルギニンに対して存在している。このようなMHC分子には、HLA-DRB1*0401、HLA-DRB1*0404、HLA-DRB1*0405、HLA-DRB1*0408、HLA-DRB1*0409、HLA-DRB1*0410、HLA-DRB1*1001、HLA-DRB1*0101、HLA-DRB1*0102、HLA-DRB1*1402、HLA-DRB1*1406、HLA-DRB1*1409、及びHLA-DRB4を含むことができるがこれらに限定されない。
【0013】
シトルリン化ペプチドが、T細胞活性化および結果として生じる疾患をもたらす関節炎性抗原として作用するという知見により、T細胞活性化および自己免疫応答(例えば、RAおよびMSなどにおけるもの)をもたらす任意のMHCクラスII分子の共有エピトープに結合するシトルリン化ペプチド抗原の結合によって特徴付けられる自己免疫障害の診断のための新規かつ改善された方法(この活性化メカニズムを含む)が現在利用可能にされ、ならびにこのような自己免疫障害(RAおよびMSを含む)を発症する素因が現在利用可能にされる。
【0014】
上記に特徴付けられる自己免疫障害を処置するための新規な治療方法(RAおよびMSを含む)もまた、現在利用可能にされる。自己免疫障害の治療のためのこのような新規なアプローチは、いくつかの態様において、シトルリンへのアルギニンの変換をブロックすることを含むことができ、従って、RAまたはMSの原因となるシトルリン化ペプチド抗原の産生を阻害/減少抑制する。このようなブロッキングまたは阻害は、1つの態様において、ペプチジルアルギニンデイミナーゼ(PAD)酵素の発現、および従って、シトルリン化ペプチド抗原の生成をブロックすることを含むことができる。これはより初期の段階で疾患の進行を潜在的にブロックすることができるRAとMSのためのより費用対効果に優れた新規な治療を提供する。他の態様において、PAD酵素のブロッキングはPAD酵素の作用に対する低分子阻害の使用を含むことができる。
【0015】
本発明の態様によるものは、共有エピトープを有するMHCクラスII分子に高アフィニティーで結合する新規なシトルリン化ペプチドであり、前記ペプチドは、自己免疫障害を有する患者または自己免疫障害を発症するリスクがある患者の血液中でT細胞応答を誘発する。本発明の態様において、T細胞はCD4+T細胞である。本発明のさらなる態様において、自己免疫障害は、RAまたはMSであり得る。
【0016】
本発明の他の態様によるものは、共有エピトープを有するMHCクラスII分子に高アフィニティーで結合する新規なシトルリン化ペプチドであり、前記ペプチドは、RAを有する患者またはRAを発症するリスクがある患者の血液中でT細胞応答を誘発する。
【0017】
本発明のさらなる態様によるものは、共有エピトープを有するMHCクラスII分子に高アフィニティーで結合する新規なシトルリン化ペプチドであり、前記ペプチドは、RAを有する患者またはRAを発症するリスクがある患者の血液中でT細胞応答を誘発し、ここで、前記のT細胞はCD4+T細胞である。
【0018】
本発明のなおさらなる態様によるものは、新規なシトルリン化ペプチドであり、前記ペプチドは、内因性または外因性タンパク質に対するペプチジルアルギニンデイミナーゼの作用によって産生され、ここで前記シトルリン化ペプチドは共有エピトープを有するMHCクラスII分子に結合する。
【0019】
本発明のさらに別の側面は、共有エピトープを有するMHCクラスII分子に高アフィニティーで結合するシトルリン化ペプチドであり、前記ペプチドは、RAを有する患者またはRAを発症するリスクがある患者の血液中でT細胞応答を誘発し、前記ペプチドは、シトルリン化フィブリノーゲンペプチドおよびシトルリン化ビメンチンペプチドからなる群から選択される。いくつかの態様において、本発明におけるフィブリノーゲンペプチドおよびビメンチンペプチド、ならびにシトルリン化の標的は、これらのタンパク質の各々についての公知のヒト配列から選択される。さらに、フィブリノーゲンヒトタンパク質配列は、アルファ/アルファ鎖前駆体タンパク質、ベータ鎖前駆体タンパク質、アルファA鎖前駆体タンパク質、ガンマ鎖前駆体タンパク質またはこれらの鎖の成熟型からさらに選択できる。他の態様において、シトルリン化ペプチドは、全長フィブリノーゲンタンパク質またはビメンチンタンパク質であり得る。本発明によって含まれるタンパク質配列は、当業者によって理解されるように、Genbank (www.ncbi.nlm.nih.gov/Genbank)および SWISS-PROT (www.expasy.org)において見い出せる。
【0020】
本発明の別の態様によるものは、脱イミノ化アルギニンを含むペプチドであり、このペプチドは少なくとも約9アミノ酸長からであり、このペプチドは共有エピトープを有するMHCクラスII分子に高アフィニティーで結合し、ここでこのペプチドは、RAを有する患者またはRAを発症するリスクがある患者の血液中でT細胞応答を誘発する。
【0021】
本発明のさらに別の態様によるものは、共有エピトープを有するMHCクラスII分子に高アフィニティーで結合するシトルリン化ペプチドであり、前記ペプチドは、RAを有する患者またはRAを発症するリスクがある患者の血液中でT細胞応答を誘発し、前記ペプチドは、SAVRACitSSVPGVR (配列番号1); FSMCitIVCLV (配列番号2); VVECitHQSAC (配列番号3); FTNCitINKLK (配列番号4); LRSCitIEVLK (配列番号5); VLKCitKVIEK (配列番号6); IKICitSCRGS (配列番号7); LPSCitDRQHL (配列番号8); FRHCitHPDEA (配列番号9); FPSCitGKSSS (配列番号10); IQQCitMDGSL (配列番号11); LTQCitGSVLR (配列番号12); YHFCitVGSEA (配列番号13); YDPCitNNSPY (配列番号14); VSFCitGADYS (配列番号15); YSLCitAVRMK (配列番号16); MKICitPLVTQ (配列番号17); YRACitPAKAA (配列番号18); WQKCitQKQVK (配列番号19); IQNCitQDGSV (配列番号20); WYNCitCHAAN (配列番号21); YSMCitKMSMK (配列番号22); MKICitPFFPQ (配列番号23); LHPCitNLILY (配列番号24); VATCitDNCCI (配列番号25); LDECitFGSYC (配列番号26); LKSCitIMLEE (配列番号27); FQKCitLDGSV (配列番号28); YALCitVELED (配列番号29); WNGCitTSTA (配列番号30); WKTCitWYSMK (配列番号31); YATCitSSAVR (配列番号32); VRLCitSSVPG (配列番号33); LNDCitFANYI (配列番号34); MLQCitEEAEN (配列番号35); LNLCitETNLD (配列番号36); VETCitDGQVI (配列番号37) およびこれらの機能的類似体からなる群から選択されるアミノ酸配列の少なくとも1つを含む。
【0022】
本発明のなおさらなる態様によるものは、共有エピトープを有するMHCクラスII分子に高アフィニティーで結合するシトルリン化ペプチドであり、前記ペプチドは、RAを有する患者またはRAを発症するリスクがある患者の血液中でT細胞応答を誘発し、前記ペプチドは、SAVRACitSSVPGVR (配列番号1); FSMCitIVCLV (配列番号2); VVECitHQSAC (配列番号3); FTNCitINKLK (配列番号4); LRSCitIEVLK (配列番号5); VLKCitKVIEK (配列番号6); IKICitSCRGS (配列番号7); LPSCitDRQHL (配列番号8); FRHCitHPDEA (配列番号9); FPSCitGKSSS (配列番号10); IQQCitMDGSL (配列番号11); LTQCitGSVLR (配列番号12); YHFCitVGSEA (配列番号13); YDPCitNNSPY (配列番号14); VSFCitGADYS (配列番号15); YSLCitAVRMK (配列番号16); MKICitPLVTQ (配列番号17); YRACitPAKAA (配列番号18); WQKCitQKQVK (配列番号19); IQNCitQDGSV (配列番号20); WYNCitCHAAN (配列番号21); YSMCitKMSMK (配列番号22); MKICitPFFPQ (配列番号23); LHPCitNLILY (配列番号24); VATCitDNCCI (配列番号25); LDECitFGSYC (配列番号26); LKSCitIMLEE (配列番号27); FQKCitLDGSV (配列番号28); YALCitVELED (配列番号29); WNGCitTSTA (配列番号30); WKTCitWYSMK (配列番号31); YATCitSSAVR (配列番号32); VRLCitSSVPG (配列番号33); LNDCitFANYI (配列番号34); MLQCitEEAEN (配列番号35); LNLCitETNLD (配列番号36); VETCitDGQVI (配列番号37) およびこれらの機能的類似体からなる群から選択される。
【0023】
本発明の他の態様によると、本発明のシトルリン化ペプチドは、複数のシトルリン残基を含み、かつ抗シトルリン抗体をインビボで生成する人工的な配列であってもよい。
【0024】
1つの態様は、共有エピトープを有するMHCクラスII分子に高アフィニティーで結合するシトルリン化ペプチドであり、前記ペプチドは、哺乳動物においてインビボで抗シトルリン抗体を生成し、前記ペプチドは以下の式を有する:
Cit-Cit-X-Cit-G-Cit-Cit-Z-Cit-Cit-B-Cit-Cit(配列番号38)ここでXはY、F、W、I、L、MおよびVから選択され、ZはA、D、I、N、P、S、T、Vから選択され、そしてBはA、G、H、Q、S、TおよびVから選択される。
【0025】
本発明の態様において、このペプチドはジスルフィド結合形成によって環状化するペプチドの1つの末端または両端に対する加えられたシステインを有することができる。本発明のさらなる側面において、ペプチドはCit-Cit-Y-Cit-G-Cit-Cit-S-Cit-Cit-S-Cit-Cit (配列番号39)を含む。なおさらなる態様において、シトルリン化ペプチドは配列番号38または39の配列の始め、および/または終端に荷電したアミノ酸を含むことができる。このような荷電したアミノ酸は、アルギニン、リジン、アスパラギン、アスパラギン酸、グルタミン酸、およびグルタミン酸から選択できる。別の態様において、このペプチドは、その免疫学的アフィニティーを増加するために環状化できる。
【0026】
本発明の別の態様によるものは、共有エピトープを有するMHCクラスII分子に高アフィニティーで結合するシトルリン化ミエリン塩基性タンパク質(MBP)ペプチドであり、前記ペプチドは、MSを有する患者またはMSを発症するリスクがある患者の血液中でT細胞応答を誘発し、前記ペプチドは、FLPCitHRDTG (配列番号40), VTPCitTPPPS (配列番号41); YGGCitASKYK (配列番号42);及びLGGCitDSRSG (配列番号42) からなる群から選択される。
【0027】
本発明の別の態様によるものは、共有エピトープを有するMHCクラスII分子に高アフィニティーで結合するシトルリン化グリア線維酸性タンパク質(GFAP)ペプチドであり、前記ペプチドは、MSを有する患者またはMSを発症するリスクがある患者の血液中でT細胞応答を誘発し、前記ペプチドは、MERCitRITSA (配列番号43), LPTCitVDFSL (配列番号44), LNDCitFASYI (配列番号45), LRLCitLDQLT (配列番号46), LQICitETSLD (配列番号47),及びVEMCitDGEVI (配列番号48) からなる群から選択される。
【0028】
本発明の別の態様によるものは、好中球中でシトルリン化されるペプチドであり、こういうものとして、診断的にまたは慢性関節リウマチの治療において使用され得、前記ペプチドは、ヌクレオホスミン/B23、ヒストンH2A、ヒストンH4、およびヒストンH3からなる群から選択され、ここで前記ペプチドは1つ以上のシトルリン残基を含む。
【0029】
本発明の別の態様によるものは、LSLRTVSLG (SEQ ID NO 49); VVLRLKCGS (配列番号50); MSGRGKQGG (配列番号51); MSGRGKQGG (配列番号52); IIPRHLQLA (配列番号53); LAIRNDEEL (配列番号54); LLGRVTIAQ (配列番号55); MSGRGKGGK (配列番号56); LARRGGVKR (配列番号57); VALREIRRY (配列番号58); LLIRKLPFQ (配列番号59);及びLARRIRGER (配列番号60) からなる群から選択されるシトルリン化ペプチドであり、ここで前記「R」アルギニン残基はPADの作用によってシトルリンに変換できる。このようなペプチドは、慢性感染リウマチの徴候としてのT細胞反応性を測定するためなどの種々のアッセイにおいて使用できる。
【0030】
本発明の別の態様によるものは、対象からの試料中で抗シトルリン抗体を検出するための方法であって、前記方法は以下の工程を含む:
対象からの試料を、配列番号40から48のいずれか1つのペプチドと接触させる工程;
前記試料中で、抗シトルリン抗体への前記ペプチドの結合を検出する工程。
【0031】
本発明のなお別の態様によるものは、対象からの試料中で抗シトルリン抗体を検出するための方法であって、前記方法は以下の工程を含む:
対象からの試料を、式Cit-Cit-X-Cit-G-Cit-Cit-Z-Cit-Cit-B-Cit-Citのペプチドと接触させ、ここでXはY、F、W、I、L、MおよびVから選択され、ZはA、D、I、N、P、S、T、Vから選択され、BはA、G、H、Q、S、TおよびVから選択される工程;ならびに
前記試料中で抗シトルリン抗体への前記ペプチドの結合を検出する工程。この方法は、RAの診断において有用である。いくつかの態様において、この試料は、対象からの末梢血試料である。
【0032】
本発明のさらに別の態様によるものは、対象における自己免疫障害を診断するための方法であって、前記方法は以下の工程を含む:
前記対象から採取された試料を、式Cit-Cit-X-Cit-G-Cit-Cit-Z-Cit-Cit-B-Cit-Citのペプチドと接触させ、ここでXはY、F、W、I、L、MおよびVから選択され、ZはA、D、I、N、P、S、T、Vから選択され、BはA、G、H、Q、S、TおよびVから選択される工程;ならびに
前記試料中で抗シトルリン抗体への前記ペプチドの結合を検出し、ここで結合が自己免疫障害を示す工程。いくつかの態様において、自己免疫障害は、RAおよびMSから選択できる。
【0033】
本発明の別の態様によるものは、対象においてMSを診断するための方法であって、前記方法は以下の工程を含む:
前記対象から採取された試料を、シトルリン化MBPペプチドと接触させる工程;および
前記試料中で抗シトルリン抗体への前記ペプチドの結合を検出し、ここで結合がMSの診断を示す工程。
【0034】
本発明のペプチドのいずれか、ならびに本発明によるペプチドのマルチマー(例えば、本発明によるペプチドのダイマーまたはトリマーなど)の機能的類似体もまた、本発明の範囲内にある。本発明によるマルチマーは、複数の同じペプチドからなるホモマー、または異なるペプチドからなるヘテロマーのいずれかであり得る。本発明によるペプチドの特徴的なアミノ酸配列は、ランダムなアミノ酸配列に隣接できる。好ましいものは、ペプチドに安定化効果を有する、従って、それらの生物学的利用可能性を増加する隣接配列である。
【0035】
上記に列挙した特定のペプチドアミノ酸配列が、シトルリンに変換できる配列中でさらなるアルギニンを有することが当業者によって理解される。
【0036】
本発明の別の態様によるものは、共有エピトープを有するMHCクラスII分子に高アフィニティーで結合し、かつRAを有する患者またはRAを発症するリスクがある患者の血液中でT細胞応答を誘発することが可能であるシトルリン化ペプチド、および薬学的に許容可能な担体を含む組成物である。このシトルリン化ペプチドは、SAVRACitSSVPGVR (配列番号1); FSMCitIVCLV (配列番号2); VVECitHQSAC (配列番号3); FTNCitINKLK (配列番号4); LRSCitIEVLK (配列番号5); VLKCitKVIEK (配列番号6); IKICitSCRGS (配列番号7); LPSCitDRQHL (配列番号8); FRHCitHPDEA (配列番号9); FPSCitGKSSS (配列番号10); IQQCitMDGSL (配列番号11); LTQCitGSVLR (配列番号12); YHFCitVGSEA (配列番号13); YDPCitNNSPY (配列番号14); VSFCitGADYS (配列番号15); YSLCitAVRMK (配列番号16); MKICitPLVTQ (配列番号17); YRACitPAKAA (配列番号18); WQKCitQKQVK (配列番号19); IQNCitQDGSV (配列番号20); WYNCitCHAAN (配列番号21); YSMCitKMSMK (配列番号22); MKICitPFFPQ (配列番号23); LHPCitNLILY (配列番号24); VATCitDNCCI (配列番号25); LDECitFGSYC (配列番号26); LKSCitIMLEE (配列番号27); FQKCitLDGSV (配列番号28); YALCitVELED (配列番号29); WNGCitTSTA (配列番号30); WKTCitWYSMK (配列番号31); YATCitSSAVR (配列番号32); VRLCitSSVPG (配列番号33); LNDCitFANYI (配列番号34); MLQCitEEAEN (配列番号35); LNLCitETNLD (配列番号36); VETCitDGQVI (配列番号37); Cit-Cit-X-Cit-G-Cit-Cit-Z-Cit-Cit-B-Cit-Cit(配列番号38、ここでXはY、F、W、I、L、MおよびVから選択され、ZはA、D、I、N、P、S、T、Vから選択され、BはA、G、H、Q、S、TおよびVから選択される)およびCit-Cit-Y-Cit-G-Cit-Cit-S-Cit-Cit-S-Cit-Cit(配列番号39)からなる群から選択される。
【0037】
本発明の別の態様によるものは、共有エピトープを有するMHCクラスII分子に高アフィニティーで結合し、かつMS患者またはMSを発症するリスクがある患者の血液中でT細胞応答を誘発することが可能であるシトルリン化ペプチド、および薬学的に許容可能な担体を含む組成物である。シトルリン化ペプチドは、配列番号40から48によって表されるペプチドのいずれか1つから選択できる。
【0038】
本発明はまた、RAもしくはMSなどの自己免疫障害、またはRAもしくはMSなどの自己免疫障害に対する感受性を診断する方法における使用のための診断用手段の調製のための本発明中に開示されるシトルリン化ペプチド抗原のいずれかの使用を提供し、この方法は、個体のT細胞が、MHCクラスII共有エピトープ陽性細胞に結合したときにシトルリン化ペプチド抗原を認識するか否かを決定する工程を包含し、ここで、T細胞による認識は、個体が自己免疫障害を有するか、または自己免疫障害に対して感受性であることを示す。
【0039】
本発明のさらに別の態様によるものは、MHCクラスII共有エピトープ陽性細胞に結合するシトルリン化抗原と反応性である自己反応性T細胞の検出のための診断方法であって、前記方法は以下の工程:
患者からの末梢血単核細胞を、1種以上のシトルリン化ペプチド抗原とともにインキュベートする工程;
前記患者における活性化された自己反応性T細胞の存在を示す、T細胞の応答を検出する工程。
【0040】
いくつかの態様において、ペプチド抗原は、配列番号1から60のいずれか1つから選択できる。
【0041】
本発明の別の態様によるものは、MHCクラスII共有エピトープ陽性細胞に結合するシトルリン化抗原と反応性である活性化された自己反応性T細胞の検出のための試験キットであり、前記試験キットは、本明細書中に開示されるような1種以上のシトルリン化ペプチドを含む。いくつかの態様において、ペプチド抗原は、配列番号1から37、配列番号38から39、配列番号40から48、および配列番号49から60のいずれか1つから選択できる。
【0042】
本発明はさらに、シトルリン化ペプチド抗原を認識する前記T細胞を脱感作することによって、RAまたはMSを治療または予防する方法における使用のための、任意に担体を伴うシトルリン化ペプチド抗原を提供する。
【0043】
本発明のなおさらなる別の態様によるものは、T細胞応答をもたらす共有エピトープを有するMHCクラスII分子にシトルリン化ペプチドを結合することによって誘発される、RAに罹患している対象を治療するための方法であって、前記方法は以下の工程を含む:
薬学的に許容可能な担体とともに、SAVRACitSSVPGVR (配列番号1); FSMCitIVCLV (配列番号2); VVECitHQSAC (配列番号3); FTNCitINKLK (配列番号4); LRSCitIEVLK (配列番号5); VLKCitKVIEK (配列番号6); IKICitSCRGS (配列番号7); LPSCitDRQHL (配列番号8); FRHCitHPDEA (配列番号9); FPSCitGKSSS (配列番号10); IQQCitMDGSL (配列番号11); LTQCitGSVLR (配列番号12); YHFCitVGSEA (配列番号13); YDPCitNNSPY (配列番号14); VSFCitGADYS (配列番号15); YSLCitAVRMK (配列番号16); MKICitPLVTQ (配列番号17); YRACitPAKAA (配列番号18); WQKCitQKQVK (配列番号19); IQNCitQDGSV (配列番号20); WYNCitCHAAN (配列番号21); YSMCitKMSMK (配列番号22); MKICitPFFPQ (配列番号23); LHPCitNLILY (配列番号24); VATCitDNCCI (配列番号25); LDECitFGSYC (配列番号26); LKSCitIMLEE (配列番号27); FQKCitLDGSV (配列番号28); YALCitVELED (配列番号29); WNGCitTSTA (配列番号30); WKTCitWYSMK (配列番号31); YATCitSSAVR (配列番号32); VRLCitSSVPG (配列番号33); LNDCitFANYI (配列番号34); MLQCitEEAEN (配列番号35); LNLCitETNLD (配列番号36);及びVETCitDGQVI (配列番号37) からなる群から選択されるアミノ酸配列を含むシトルリン化ペプチドを含む組成物のT細胞寛容誘導量を投与する工程。
【0044】
本発明のなおさらなる別の態様によるものは、T細胞応答をもたらす共有エピトープを有するMHCクラスII分子にシトルリン化ペプチドを結合することによって誘発される、MSに罹患している対象を治療するための方法であって、前記方法は以下の工程を含む: FLPCitHRDTG (配列番号40), VTPCitTPPPS (配列番号41); YGGCitASKYK (配列番号42), LGGCitDSRSG (配列番号42), MERCitRITSA (配列番号43), LPTCitVDFSL (配列番号44), LNDCitFASYI (配列番号45), LRLCitLDQLT (配列番号46), LQICitETSLD (配列番号47)及びVEMCitDGEVI (配列番号48) のシトルリン化ペプチドを含む組成物のT細胞寛容誘導量を投与する工程。
【0045】
本発明のなお別の態様によるものは、シトルリン化抗原MHCクラスII細胞複合体の検出のための診断方法であって、前記方法は以下の工程を含む:
対象からの末梢血単核細胞の単離された試料を1種以上のシトルリン化ペプチド抗原とインキュベートする工程;
シトルリン化ペプチド抗原MHCクラスII細胞複合体の形成を検出し、このような検出は前記対象におけるRAまたはMSをもたらすT細胞応答を誘発する可能性を示す工程。
【0046】
本発明の態様によるものは、対象におけるシトルリン化ペプチドMHCクラスII複合体によってT細胞の活性化を予防するための方法であって、前記方法は、前記複合体に標的化される抗体を投与する工程を含む。
【0047】
本発明の別の態様によるものは、対象における潜在的に抗原性のペプチド中でのアルギニンからシトルリンへの変換、従って、シトルリン化ペプチドMHCクラスII複合体の形成を予防するための方法であって、前記方法は、ペプチジルアルギニンデイミナーゼのアンタゴニストまたは阻害剤を前記対象に投与する工程を含む。
【0048】
本発明のなお別の態様によるものは、共有エピトープを有するMHCクラスII分子へのシトルリン化ペプチドの結合をブロックし得る薬学的化合物を同定するためのスクリーニング方法であり、この方法は以下の工程を含む:
候補薬学的化合物をトランスジェニックDR4-IE tgマウスに投与する工程;および
T細胞活性を測定する工程および/またはシトルリン化ペプチド/MHCクラスII分子複合体形成を測定する工程であって、ここで、減少したT細胞活性および/または減少した複合体形成は、前記候補薬学的化合物がMHCクラスII分子への前記シトルリン化ペプチドの結合に影響を与える工程
を含む方法。
【0049】
本発明のさらなる態様によるものは、PADの阻害剤を同定するためのスクリーニング方法であって、この方法は以下の工程を含む:
(a)非シトルリン化ペプチド、PAD、および阻害剤の混合物を供給する工程;
(b)(a)に抗原提示細胞を供給する工程;ならびに
(c)ペプチド-MHC複合体に特異的なT細胞系統に(b)を適用し、ここで、PAD阻害がT細胞反応性の欠如によって特徴付けられる工程。
【0050】
本発明のさらなる態様によるものは、慢性関節リウマチの研究用の動物モデルを提供するために動物において慢性関節リウマチを誘導する方法であり、この方法は、薬学的に許容可能な担体とともに、シトルリン化ペプチドを含む組成物の慢性関節リウマチ誘導量を動物に投与する工程を含む。投与は注射によって行われ得る。
【0051】
本発明の別の態様によるものは、多発性硬化症の研究用の動物モデルを提供するために動物において多発性硬化症を誘導する方法であり、この方法は、薬学的に許容可能な担体とともに、シトルリン化ペプチドを含む組成物の多発性硬化症誘導量を動物に投与する工程を含む。投与は注射によって行われ得る。いくつかの局面において、シトルリン化ペプチドは、シトルリン化されたミエリン塩基性タンパク質(MBP)またはグリア線維酸性タンパク質(GFAP)である。
【0052】
本発明のなお別の態様によるものは、共有エピトープを有するMHCクラスII分子に結合可能であるシトルリン化ペプチド抗原を作製するための方法であって、前記方法は以下の工程を含む:
(a)アルギニンをシトルリンに変換するために十分な時間の間、少なくとも1つのアルギニンアミノ酸残基を含む内因性または外因性可溶化タンパク質にペプチジルデイミナーゼを加える工程;および
(b)(a)からタンパク質を単離する工程。
【0053】
本発明のなお別の態様によるものは、哺乳動物におけるシトルリン化ペプチド/MHCクラスII複合体の結合および形成によって特徴付けられる自己免疫障害の治療のための方法であり、この方法は、前記哺乳動物におけるペプチジルデイミナーゼ(PAD)酵素の発現をブロックする工程を包含し、ここで、このようなブロッキングは、慢性関節リウマチを誘発するシトルリン化ペプチドの生成を減少させる。
【0054】
本発明のさらなる態様によるものは、哺乳動物における慢性関節リウマチの治療のための方法であり、この方法は、ペプチジルアルギニンデイミナーゼ(PAD)遺伝子配列に標的化されるsiRNAの投与によって、哺乳動物中のPADの発現を阻害する工程を含む。
【0055】
本発明のなおさらなる態様によるものは、哺乳動物における慢性関節リウマチまたは多発性硬化症の治療のための方法であり、この方法は、PADの機能をブロックする薬剤を前記哺乳動物に投与する工程を含む。
【0056】
本発明の他の特徴および利点は、以下の詳細な説明から明らかになる。しかし、本発明の精神および範囲の中での種々の変更および改変は詳細な説明から当業者に明らかになるので、前記詳細な説明および特定の実施例は本発明の実施形態を示しているが、例示のみのために提供されることが理解されるべきである。
【0057】
好ましい実施形態の詳細な説明
出願人は、T細胞の活性化および結果として、慢性関節リウマチの発症をもたらす、共有エピトープを有するMHCクラスII分子に特異的に結合する新規なシトルリン化抗原性ペプチドを開発した。共有エピトープ(P4)と相互作用するペプチド側鎖位置のアルギニンのシトルリンへの変換は、ペプチドMHCアフィニティーを有意に増加させ、そして、CD4+T細胞の活性化をもたらす。そのようなT細胞活性化はさらに滑膜炎および慢性関節リウマチの発症をもたらす。本発明のシトルリン化ペプチド抗原は、T細胞の活性化および炎症性応答の発症を生じる、共有エピトープ複合体とのシトルリン化ペプチド/MHCクラスIIの結合および形成によって特徴付けられる、種々の自己免疫障害に関与し得ることが当業者によって理解される。それゆえに、本発明は、このような病因論に関わる自己免疫障害のためのいくつかの適用を有する。
【0058】
HLA DRB1*0401トランスジェニック(DR4-IE tg)マウスを使用して、出願人はシトルリン含有ペプチドに対する増加するT細胞応答を実証した。MHCペプチドアフィニティーを予測するのにモデルを使用して、候補T細胞エピトープは、研究について開発および選択された。これにはビメンチン、フィブリノーゲン、およびプロテオグリカンアグレカンからのエピトープが含まれていた。プロテオグリカンアグレカンの場合では、通常、このタンパク質は、P4共有エピトープと相互作用する負に荷電したアスパラギン酸(D)を含む。フィブリノーゲンペプチドおよびビメンチンペプチドは正電荷を有するP4アンカリングポケット(共有エピトープ)にアルギニンまたはシトルリンのいずれかを配置する区域におけるDRB1*0401への結合の特性に基づいて選択された。アルギニンのシトルリンへの変換(荷電しているイミノ側鎖グループを荷電されていないカルボニルに置換するプロセス)は、DRB1*0401でのペプチドのアフィニティーを劇的に増加させた。さらに、この翻訳後修飾が、DR4-IE tgマウスにおけるこれらのペプチドに対するCD4+T細胞応答を誘発するために必要であった。多くのHLA対立遺伝子についてのペプチドアフィニティーが評価され、共有エピトープを有するMHCクラスII分子のみがシトルリン含有ペプチドについてのアフィニティー増加を有することを示した。これらの研究(これは新規なペプチドMHC相互作用を同定する)は、RAにおける疾患関連HLA対立遺伝子の分子的基礎を説明することを補助し、現在、慢性関節リウマチについての新しい診断的および治療的なストラテジーを提供する。RAと関連するMHCクラスII分子の第3の超可変領域は、DRβ鎖の位置70から74にわたるアミノ酸配列Q/R、K/R、R、A、Aを含む。この共有エピトープ領域は、P4として知られている主要なペプチドアンカリングポケットの1つを形成し、抗原性ペプチドから側鎖残基に直接的に接触することが可能である71位のKまたはRに起因して正電荷を有する(Dessen A, Lawrence CM, Cupo S, Zaller DM, Wiley DC. 1997. X-ray crystal structure of HLA-DR4 (DRA*0101, DRB1*0
401) complexed with a peptide from human collagen II. Immunity. 7:473; Stern LJ, Brown JH, Jardetzky TS, Gorga JC, Urban RG, Strominger JL, Wiley DC.1994. Crystal structure of the human class II MHC protein HLA-DR1 complexed with an influenza virus peptide. Nature. 368:215)。ペプチドMHCアフィニティーに関する以前の研究は、71位のKまたはRがこのP4ポケットで相互作用し得るアミノ酸の特徴を決定付けることを示した(Hammer J., Gallazzi F., Bono E., Karr R.W., Guenot J., Valsasnini P., Nagy Z.A., Sinigaglia F. 1995. Peptide binding specificity of HLA-DR4 molecules: correlation with rheumatoid arthritis association. J. Exp. Med. 181:1847)。一般に、共有エピトープを有するMHCは、負に荷電しているか、または荷電されていない極性アミノ酸について高アフィニティーを有するのに対して、正電荷を有するアミノ酸(すなわち、アルギニン)はペプチド結合を阻害する(Hammer J, Bono E, Gallazzi F, Belunis C, Nagy Z, Sinigaglia F. 1994. Precise prediction of major histocompatibility complex class II-peptide interaction based on peptide side chain scanning. J. Exp. Med. 180:2353; Hammer J., Gallazzi F., Bono E., Karr R.W., Guenot J., Valsasnini P., Nagy Z.A., Sinigaglia F. 1995. Peptide b
inding specificity of HLA-DR4 molecules: correlation with rheumatoid arthritis association. J. Exp. Med. 181:1847)。出願人は、酵素ペプチジルアルギニンデイミナーゼ(PAD)の作用を介する脱イミノ化によって(図4)、正に荷電しているアルギニンが、極性であるが荷電していないシトルリンに変換され(翻訳後修飾)、このことが、共有エピトープP4ポケットに対するアフィニティーを増加させることを実証した。MHCアンカリングポケットにおけるアミノ酸相互作用は、残基の電荷に依存するのみならず、サイズにもまた依存するので、出願人はまた、共有エピトープによって形成されたP4ポケットがシトルリンの側鎖を収容するために十分に大きいことを確認した。これは、DRB1*0401およびDRB1*0101の結晶構造を使用する分子モデリングによって確かめられた(データ示さず)。P4共有エピトープの荷電特性およびこのポケットのサイズに基づいて、ペプチド結合シトルリンは、HLA*0401およびHLA*0101のP4アンカリングポケットで好ましく相互作用することが予測された。
【0059】
DR4-IE tgマウスからCD4+T細胞を活性化することが出願人によって実証されたペプチド配列が選択および開発された。このペプチド(軟骨プロテオグリカンアグレカンから)は、通常、P4共有エピトープと相互作用する負に荷電したアスパラギン酸(D)を含んでいる(P4D)。2つのさらなるペプチドがこの配列に基づいて合成された:1つはアルギニンによって置換されたアスパラギン酸を有した(P4R);他方は、この位置で置換されたシトルリンを有した(P4Cit)。次いで、DR4-IE tgマウスがこれらのペプチドで免疫されて、T細胞応答が10日後に評価された。ペプチドP4DはIFN-γ産生に付随した強い増殖性応答を誘導した(図1Aおよび1B)。しかし、アルギニン置換を含むペプチド(P4R)は、これらのマウスにおいてT細胞増殖またはサイトカイン産生を誘導しなかった。P4Rでの応答の非存在と対照的に、P4CitはT細胞増殖およびIFN-γ産生を誘導した。P4CitがペプチドMHC提示を通してT細胞を活性化していることを確認するために、抗DR抗体が、ペプチドMHC複合体とのTCR相互作用を阻害するために使用された(図1B)。この処理は、P4CitおよびP4Dに対する増殖性応答を阻害した。RA患者における抗シトルリン抗体のタンパク質標的からの潜在的T細胞エピトープが同定された。このタンパク質に対する自己抗体が共有エピトープを発現する患者において頻繁に見い出されるので、ビメンチンが選択された(Goldbach-Mansky R, Lee J, McCoy A, Hoxworth J, Yarboro C, Smolen JS, Steiner G, Rosen A, Zhang C, Menard HA, Zhou ZJ, Palosuo T, Van Venrooij WJ, Wilder RL, Klippel JH, Schumacher HR Jr, El-Gabalawy HS. 2000. Rheumatoid arthritis associated autoantibodies in patients with synovitis of recent onset. Arthritis Res. 2:236)。ヒトビメンチンからの候補T細胞エピトープは、ペプチドMHCアフィニティーについて予測モデルを使用して同定された(Hammer J, Bono E, Gallazzi F, Belunis C, Nagy Z, Sinigaglia F. 1994. Precise prediction of major histocompatibility complex class II-peptide interaction based on peptide side chain scanning. J. Exp. Med. 180:2353)。このペプチドは、MHCアンカリングポケットP1、P6、およびP9との好都合な相互作用を有し、かつP4共有エピトープにおいてアルギニンを有する特性に基づいて選択された。2つのペプチドが統合され、1つは修飾されていないペプチドビメンチン65-77を含み、他方はビメンチンR70Citであり、これはアルギニンがシトルリン置換されていた。次いで、これらのペプチドに対するT細胞応答は、DR4-IE tgマウスを使用して特徴付けられた。しかし、予測されるように、修飾されていないペプチドVim 65-77がT細胞活性化を誘導せず、Vim R70CitがIFN-γ産生に付随した強い増殖性応答を刺激した(図2Aおよび2C)。Vim R70Citに対するT細胞応答はまた、抗DR抗体を使用して阻害され得、MHCクラスII制限された免疫応答を確認した(図2B)。Vim R70CitによってプライムされたT細胞が未修飾ペプチドによって活性化できなかったこともまた実証され、Vim65-77がDR4結合溝と産生的に相互作用しないことをさらに支持する(図2B)。
【0060】
アルギニンのシトルリンへの変換が、共有エピトープを含んだMHCクラスII分子についてのペプチドアフィニティーを増加させることができることを確認するために、ペプチド競争アッセイが、共有エピトープ陽性(DRB1*0101、*0401、*0404)、または共有エピトープ陰性(DRB1*0301、*0701、*0802、*1101、*1302)のいずれかであった精製MHCについてVim65-77およびVim R70Citの相対的アフィニティーを決定するために行われた。Vim65-77は、すべての試験されたMHCについて低いアフィニティーから中程度のアフィニティーまでを有したのに対して、Vim R70Citは、きわだって高いアフィニティーを伴って*0101、*0401、および*0404に結合した(図3)。未修飾ペプチドと比較して、シトルリン含有ペプチドは、*0101、*0401、および*0404に、それぞれ、100倍、90倍、および20倍高いアフィニティーで結合した。最も重要なことに、アルギニンのシトルリンへの変換は、試験されたいかなる共有エピトープ陰性のMHCについてもペプチドアフィニティーを増加しなかった。
【0061】
出願人の結果は、P4におけるアルギニンまたはシトルリンのいずれかに対するDRB1対立遺伝子のディファレンシャルな結合特性が、MHCクラスII分子が疾患に関連するか、関連しないか、または保護的であるかをいかにして説明するかを示した。まず最初に、ペプチド結合アルギニンのシトルリンへの変換は、共有エピトープを有するMHCについてアフィニティーの100倍の増大を引き起こす。これは、T細胞活性化のために必要な「生化学的安全域」を超えることができる抗原提示細胞に対してより高い密度ペプチドMHC複合体を生じ得る(Peterson DA, DiPaolo RJ, Kanagawa O, Unanue ER. 1999. Cutting edge: negative selection of immature thymocytes by a few peptide-MHC complexes: differential sensitivity of immature and mature T cells. J. Immunol. 162:3117; DiPaolo RJ, Unanue ER. 2001. The level of peptide-MHC complex determines the susceptibility to autoimmune diabetes: studies in HEL transgenic mice. Eur. J. Immunol. 31:3453; Yagi J, Janeway CA Jr. 1990. Ligand thresholds at different stages of T cell development. Int. Immunol. 2:83)。第2に、関連していないMHCクラスII分子(例えば、*0301)は、アルギニンまたはシトルリンの大きな極性側鎖を生産的に収容するための適切なサイズまたは電荷を欠くP4ポケットを含み得、それゆえに修飾の状態に関わらずペプチドを結合および提示することが不可能である(Ghosh P, Amaya M, Mellins E, Wiley DC. 1995. The structure of an intermediate in class II MHC maturation: CLIP bound to HLA-DR3. Nature. 378:457)。最後に、疾患の保護的なMHCは、ネガティブ選択を引き起こすことができるペプチドMHCリガンドを生じるP4でアルギニンとシトルリンの両方と生産的に相互作用ができ、CD4+CD25+調節性T細胞の産生をもたらすか(Jordan MS, Boesteanu A, Reed AJ, Petrone AL, Holenbeck AE, Lerman MA, Naji A, Caton AJ. 2001. Thymic selection of CD4+CD25+ regulatory T cells induced by an agonist self-peptide. Nat. Immunol. 2:301)、または単に「生化学的安全域」内に留まる(Peterson DA, DiPaolo RJ, Kanagawa O, Unanue ER. 1999. Cutting edge: negative selection of immature thymocytes by a few peptide-MHC complexes: differential sensitivity of immature and mature T cells. J. Immunol. 162:3117)。出願人はまた、抗シトルリン抗体について陽性のRA患者において、増加したT細胞増殖性応答がシトルリン化フィブリノーゲンペプチドに対してなお見られたことを実証した(図5A、B、C)。
【0062】
まとめると、本発明は、ある態様において、哺乳動物のフィブリノーゲンおよびビメンチンのアミノ酸配列に基づいており、他の態様において、人工のシトルリン化ペプチド配列に基づいている一連のペプチドを実証する。用語「ペプチド」は、本明細書中で使用される場合、通常、定義された配列を有するアミノ酸残基の鎖として定義される。本明細書中で使用される場合、用語ペプチドは、用語「ペプチド」および「タンパク質」と相互に包括的である。1つの態様において、フィブリノーゲンおよびビメンチンの配列は、ヒトアミノ酸配列である。しかし、フィブリノーゲンまたはビメンチンの配列は、ヒト、ウシ、ブタ、マウス、およびラットなどであるがこれらに限定されない任意の哺乳動物種由来でであり得ることは当業者によって理解される。
【0063】
本発明はまた、シトルリンに変換される少なくとも1つのアルギニンアミノ酸を有する、全長のフィブリノーゲンとビメンチンの両方のアミノ酸配列またはそのフラグメントを含む。任意の所望の長さの任意のフラグメントであることの限界は、共有エピトープを有する任意のMHCクラスII分子に対して高いアフィニティーで結合する少なくとも1つのシトルリンを有することである。当業者は、ビメンチンならびにフィブリノーゲンのα鎖およびβ鎖中のいくつかのさらなる配列が、P4にアルギニンまたはシトルリンを位置付ける区域でHLA-DR分子を結合することが予測され、従って、本発明において具体化されることを理解する。さらに、本発明は、RA患者における抗シトルリン抗体の潜在的な標的であることが実証される任意のペプチドを含む。このことは、独特な病原性ペプチドが、不均一な特異性のアレイを有する活性化T細胞を生じることを示唆する。
【0064】
本発明の1つの局面において、ペプチジルアルギニンデイミナーゼ(PAD)による修飾に感受性であり、さらに共有エピトープを有するMHCクラスII分子を結合する特性を有する、任意の内因性または外因性のタンパク質またはペプチドが、本発明によって具体化される。このようなタンパク質またはペプチドは、1つの態様において、関節において存在することが見い出され得、これには、ビメンチンおよびフィブリノーゲンが含まれるがこれらに限定されない。
【0065】
本発明のシトルリン化ペプチドは、共有エピトープを有するMHCクラスII分子に高アフィニティーで結合する。「高アフィニティー」は、本明細書中で使用される場合、シトルリン化されていないペプチド(例えば、シトルリンではなくアルギニンアミノ酸を有するペプチドなど)と比較して、共有エピトープを有するMHCクラスII分子に、より高いかまたは増大したアフィニティーで結合するシトルリン化ペプチドの能力をいうことが当業者によって理解される。このような結合アフィニティーは、試料ペプチドが標準ペプチドに置き換わるように使用されるペプチド結合アッセイを使用して、例えばインビトロで容易に達成できることがさらに理解される(実施例の節を参照されたい)。
【0066】
1つの態様において、本発明のタンパク質は SAVRACitSSVPGVR (配列番号1); FSMCitIVCLV (配列番号2); VVECitHQSAC (配列番号3); FTNCitINKLK (配列番号4); LRSCitIEVLK (配列番号5); VLKCitKVIEK (配列番号6); IKICitSCRGS (配列番号7); LPSCitDRQHL (配列番号8); FRHCitHPDEA (配列番号9); FPSCitGKSSS (配列番号10); IQQCitMDGSL (配列番号11); LTQCitGSVLR (配列番号12); YHFCitVGSEA (配列番号13); YDPCitNNSPY (配列番号14); VSFCitGADYS (配列番号15); YSLCitAVRMK (配列番号16); MKICitPLVTQ (配列番号17); YRACitPAKAA (配列番号18); WQKCitQKQVK (配列番号19); IQNCitQDGSV (配列番号20); WYNCitCHAAN (配列番号21); YSMCitKMSMK (配列番号22); MKICitPFFPQ (配列番号23); LHPCitNLILY (配列番号24); VATCitDNCCI (配列番号25); LDECitFGSYC (配列番号26); LKSCitIMLEE (配列番号27); FQKCitLDGSV (配列番号28); YALCitVELED (配列番号29); WNGCitTSTA (配列番号30); WKTCitWYSMK (配列番号31); YATCitSSAVR (配列番号32); VRLCitSSVPG (配列番号33); LNDCitFANYI (配列番号34); MLQCitEEAEN (配列番号35); LNLCitETNLD (配列番号36);及びVETCitDGQVI (配列番号37) からなる群から選択される少なくとも1つのアミノ酸配列を含むシトルリン化ペプチドを含み得るがこれらに限定されない。
【0067】
特定のこれらのペプチド配列は、酵素ペプチジルアルギニンデイミナーゼの作用を介してシトルリンに変換できるさらなるアルギニンを含み得る。例えば、本発明のシトルリン化ペプチドは以下の式:Cit-Cit-X-Cit-G-Cit-Cit-Z-Cit-Cit-B-Cit-Cit(配列番号38)を有し、ここでXはY、F、W、I、L、MおよびVから選択され、ZはA、D、I、N、P、S、TおよびVから選択され、BはA、G、H、Q、S、TおよびVから選択される。「X」で示されるアミノ酸は、共有エピトープを有するMHCクラスII分子についてのP1アンカリングアミノ酸のために必要である。5番目の位置に存在するグリシンは、シトルリンがアミノ酸位置番号4、6、および7に配置されることを可能するために、ペプチドバックボーン上のねじれを減少することを補助する。6位のアミノ酸はP4共有エピトープポケットである。位置番号8の選択されるアミノ酸は、共有エピトープを有するMHCクラスII分子についてのP6アンカリングポケットにおいて非阻害的である。位置番号11の選択されるアミノ酸は、共有エピトープを有するMHCクラスII分子についてのP9アンカリングポケットにおいて非阻害的である。本発明の態様において、ペプチドは、Cit-Cit-Y-Cit-G-Cit-Cit-S-Cit-Cit-S-Cit-Cit(配列番号39)を含む。本発明のさらなる態様において、正に荷電したアミノ酸および/または負に荷電したアミノ酸が、ペプチド結合を増大するために、ペプチドのカルボキシ末端および/またはアミノ末端に付加できる。さらに、このペプチドは、ジスルフィド結合の形成によってペプチドを環状化するために、ペプチドの1つの末端または両方の末端にシステインを有し得る。
【0068】
式:Cit-Cit-X-Cit-G-Cit-Cit-Z-Cit-Cit-B-Cit-Cit(配列番号38)(ここでXはY、F、W、I、L、MおよびVから選択され、ZはA、D、I、N、P、S、TおよびVから選択され、BはA、G、H、Q、S、TおよびVから選択される)のペプチドは、DR4 tgマウスにおいて抗シトルリン抗体を生成することが実証された。さらに、これらのペプチドはまた、RA患者のヒト血清において自己抗体と反応することが実証された。このようなものとして、これらのペプチドは、対象におけるRAの診断のための方法およびキットにおいて使用できる。
【0069】
出願人は、シトルリン化ペプチド抗原を含む自己免疫障害がMHCクラスII分子共有エピトープを有する対象においてT細胞応答を誘発し、多発性硬化症(MS)におけるシトルリン化ペプチド誘発T細胞応答もまた、MHCクラスIIエピトープを有するMS対象のために本発明に含まれることを実証した。このようなシトルリン化ペプチドには、例えば、シトルリン化ミエリン塩基性タンパク質(MBP)およびシトルリン化グリア線維酸性タンパク質(GFAP)が含まれる。MSの診断において有用であるシトルリン化MBPペプチドおよびGFAPペプチドならびに治療方法の標的は、FLPCitHRDTG (配列番号40), VTPCitTPPPS (配列番号41); YGGCitASKYK (配列番号42), LGGCitDSRSG (配列番号42), MERCitRITSA (配列番号43), LPTCitVDFSL (配列番号44), LNDCitFASYI (配列番号45), LRLCitLDQLT (配列番号46), LQICitETSLD (配列番号47),及びVEMCitDGEVI (配列番号48) からなる群から選択されるアミノ酸配列を含み得る。
【0070】
さらに、本発明のシトルリン化ペプチドは、好中球中でシトルリン化されるペプチドをさらに含み得、このようなものとして、慢性関節リウマチの診断または治療において使用され得、前記ペプチドは、ヌクレオホスミン/B23、ヒストンH2A、ヒストンH4、およびヒストンH3からなる群から選択され、ここで、前記ペプチドは1つ以上のシトルリン残基を含む。これらのタンパク質は顆粒球中でシトルリン化されることが実証されている(Hagiwara T.ら、 Biochem & Biophy Res Comm. (2000) 290, 979-983)。好中球は、慢性関節リウマチ患者の炎症性関節に存在する。シトルリン化できるペプチドは以下の群から選択される:LSLRTVSLG (配列番号49、ヌクレオホスミン/B23);VVLRLKCGS (配列番号50、ヌクレオホスミン/B23);MSGRGKQGG、(配列番号51、ヒストンH2A);MSGRGKQGG、(配列番号52、ヒストンH2A);IIPRHLQLA、(配列番号53、ヒストンH2A);LAIRNDEEL、(配列番号54、ヒストンH2A);LLGRVTIAQ、(配列番号55、ヒストンH2A);MSGRGKGGK (配列番号56、ヒストンH4);LARRGGVKR (配列番号57、ヒストンH4);VALREIRRY (配列番号58、ヒストンH3)、LLIRKLPFQ(配列番号59、ヒストンH3)および、LARRIRGER (配列番号60、ヒストンH3)ここで前記「R」アルギニン残基は、PADの作用によってシトルリンに変換できる。このようなペプチドは、例えば、慢性関節リウマチの診断的徴候としてのT細胞反応性を測定するための種々のアッセイにおいて使用できる。
【0071】
本発明のペプチドは、少なくとも約9アミノ酸長、および約9から約55アミノ酸長であり得、当業者によって理解されるように、その中の任意の範囲の長さ(すなわち、9〜50、9〜45、9〜40、9〜35、9〜30、9〜25、9〜20、9〜15など)を含み得る。約55アミノ酸を超える長さのペプチドもまた、本発明に含まれる。ペプチドの長さは、共有エピトープを有するMHCクラスII分子へのその結合能力によってのみ制限される。本発明のペプチドはまた、当業者によって理解され、かつ例えば、米国特許第5,824,315号および米国特許第6,184,204号(これらの開示はその全体が参照として本明細書に援用される)において記載されるように、ペプチドのダイマーおよびトリマーを含み得る。本発明によるマルチマーは、複数の同じペプチドからなるホモマー、または異なるペプチドからなるヘテロマーのいずれかであり得る。言及されるように、本発明によるペプチドの特徴的なアミノ酸配列は、ランダムなアミノ酸配列に隣接できる。好ましいものは、ペプチドに安定化効果を有する、従って、それらの生物学的利用可能性を増加する隣接配列である。さらに、他のペプチド模倣物もまた、本発明のペプチドにおいて有用である。一般的な概説としては、A. F. Spatola, in Chemistry and Biochemistry of Amino Acids, Peptides and Proteins, B. Weinstein, eds., Marcel Dekker, New York, p. 267 (1983)を参照されたい。本発明のペプチドはまた、例えば、リン酸化、グリコシル化、または脂質化によって修飾されたペプチドを含む。さらに、本発明のポリペプチドはまた、ペプチドの「機能的に等価な改変体」または「類似体」を含み得る。このようなものとして、これは、部分的配列相同性を有するペプチドおよびポリペプチド、1つ以上の保存性アミノ酸変化および/または保存性でないアミノ酸変化、ならびにペプチドの生物学的特性または構造的特性(すなわち、共有エピトープを有するMHCクラスII分子に結合する能力)を変化させないペプチド結合体を含むが、これらに限定されない。
【0072】
「機能的類似体」によって、生物学的に機能的なペプチド類似体の定義における固有の性質は、分子の規定された部分内でなされ得る変化の数に限度が存在し、等価な生物学的活性(この場合は、これは共有エピトープを有するMHCクラスII分子に結合する能力を含む)の許容可能なレベルを有する分子をなお生じるという概念であることが当業者によって十分に理解される。異なる置換を有する複数の別個のペプチド/タンパク質は、本発明に従って容易に作製および使用できる。特定の残基(例えば、レセプター認識領域における残基など)が、タンパク質またはペプチドの生物学的特性または構造的特性に対して特に重要であることもまた理解され、このようなこれらの残基は、一般的に交換されなくてもよい。
【0073】
機能的類似体は、保存性または非保存性のアミノ酸置換によって生成できる。アミノ酸置換は、一般的に、アミノ酸側鎖置換基の比較的な類似性、例えば、それらの疎水性、親水性、電荷、サイズなどに基づく。従って、本発明の範囲内において、保存性アミノ酸変化は、もともと存在するのと同じ型である特定の位置におけるアミノ酸変化、すなわち、疎水性アミノ酸に交換された疎水性アミノ酸、塩基性アミノ酸に交換された塩基性アミノ酸などを意味する。保存性置換の例には、非極性(疎水性)残基(例えば、イソロイシン、バリン、ロイシン、またはメチオニン)の別の残基への置換、1つの極性(親水性)残基の別の残基への置換(例えば、アルギニンとリジンの間、グルタミンとアスパラギンの間、グリシンとセリンの間)、1つの塩基性残基(例えば、リジン、アルギニン、またはヒスチジン)の別の残基への置換、または1つの酸性残基(例えば、アスパラギン酸およびグルタミン酸)の別の残基への置換、分枝鎖アミノ酸(イソロイシン、ロイシン、またはバリン)の別の残基への置換、1つの芳香族アミノ酸(例えば、フェニルアラニン、チロシン、またはトリプトファン)の別の残基への置換が含まれる。このようなアミノ酸変化は、ペプチドの全体の電荷および/または立体配置を有意に変化させないという点において、機能的類似体を生じる。このような保存性変化の例は当業者に周知であり、本発明の範囲内に含まれる。保存性置換はまた、得られるペプチドが本発明のペプチドと生物学的に機能的に等価であるという条件で、非誘導体化残基の代わりに化学的に誘導体化された残基の使用を含む。それゆえに、本発明のシトルリン化ペプチドは、配列番号1から39とは1つ以上の保存性アミノ酸置換が異なるアミノ酸配列を有するペプチドを含む。本発明のシトルリン化ペプチドはまた、配列番号1から39とは単一の変異が異なるアミノ酸配列を有するペプチドを含み、ここで、単一の変異は、単一のアミノ酸の欠失、挿入、または置換を表す。
【0074】
本発明のペプチドは、当業者の公知の方法によって、最も顕著にかつ好ましくは、合成ペプチドを生成するために、例えば、固相合成(Merrifield (1964) J. Am. Chem. Assoc. 65:2149; J. Amer. Chem. Soc. 85:2149 (1963)およびInt. J. Peptide Protein Res. 35:161-214 (1990))または均質溶液中での合成(Methods of Organic Chemistry, E. Wansch (編) 第15巻, I部およびII部, Thieme, Stuttgart (1987))などのタンパク質の化学において周知の技術を使用する化学合成によって作製できる。シトルリンは、アルギニンから正電荷を除去しかつ得られるシトルリンを事実上極性にする、酵素ペプチジルアルギニンデイミナーゼ(PAD)によって触媒される脱イミノ化のプロセスを通して作製される翻訳後修飾アルギニンである。
【0075】
シトルリン化ビメンチンおよびフィブリノーゲンについての1つの実施形態において、本発明のシトルリン化ペプチドは、公知の商業的に利用可能なビメンチンおよびフィブリノーゲンの供給源から作製できる。この態様において、凍結乾燥したビメンチンまたはフィブリノーゲンは、酵素ペプチジルアルギニンデイミナーゼが加えられる適切な緩衝液中で再構成される。この溶液は、アルギニン残基のシトルリンへの修飾を引き起こし、従って、シトルリン化されたビメンチンタンパク質またはフィブリノーゲンタンパク質を作製するために十分な時間の間、適切な温度に置かれる。次いで、シトルリン化タンパク質を、酵素を分離するための高分子量メンブレンまたは他のクロマトグラフィーの方法を使用する酵素の除去によって単離する。当業者は、インキュベーションの温度、緩衝液条件、およびインキュベーションの時間は、脱イミノ化されるタンパク質に依存して変化し得ることを理解する(Christine Masson-Bessiereら、2001. The Major Synovial Targets of the Rheumatoid arthritis-Specific Antifilaggrin Autoantibodies Are Deiminated Forms of the ( and ( Chains of Fibrin 1 The Journal of Immunology 166: 4177-4184)。本発明の1つの局面において、シトルリン化フィブリノーゲンは、プラスミノーゲン枯渇ヒトフィブリノーゲン(Calbiochem、San Diego、CA)を入手すること、およびタンパク質の脱イミノ化のために、0.86mg/mlのこのタンパク質をウサギ骨格筋PAD(7U/mgフィブリノーゲン;Sigma)とともに、0.1 M Tris-HCl(pH 7.4)、10 mM CaCl2、および5 mM DTT中で2時間、50℃でインキュベートすることによって作製された。同様のアプローチが、ミエリン塩基性タンパク質およびグリア線維酸性タンパク質を、適切な組織および細胞から作製および単離するために含まれる。
【0076】
本発明のシトルリン化タンパク質は、その配列によって明らかにされる特性に基づいて選択される方法によってさらに単離され、かつ精製できる。精製は、クロマトグラフィー方法(ゲル濾過、イオン交換、および免疫アフィニティー)などのタンパク質精製手順によって、高速液体クロマトグラフィー(HPLC、RP-HPLC、イオン交換HPLC、サイズ排除HPLC、高速等電点電気泳動、および疎水性相互作用クロマトグラフィー)によって、または沈殿(免疫沈殿)によって達成できる。ポリアクリルアミドゲル電気泳動もまた、タンパク質の分子量、電荷特性および疎水性に基づいてシトルリン化タンパク質を単離するために使用できる。精製されたシトルリン化タンパク質は、さらなる生化学的分析において使用され得、タンパク質と相互作用するため、タンパク質電荷の立体配置もしくは他のタンパク質との電荷相互作用を変化させるため、またはその機能を変化させるための医薬の設計において補助し得る、二次構造および三次構造を確立する。
【0077】
代替的には、本発明のシトルリン化ペプチドは、当業者に公知の組換えDNA技術の使用によって作製できる。シトルリンはいかなるトリヌクレオチド配列によってもコードされないので、アルギニンをコードする核酸配列が、アルギニンをシトルリンに変換するためにペプチジルアルギニンデイミナーゼ(PAD)を使用して修飾される得られるタンパク質配列とともに使用できる。シトルリンに類似するアミノ酸であるグルタミンをコードする核酸配列を使用することはさらに、本発明の範囲内にある。この様式において、アルギニンを含む天然に存在する配列が使用され得、ここで、このアルギニンは、シトルリンに類似するためにグルタミンで置換される。本発明の選択したペプチドをコードする核酸配列は、公知の様式で、適切な発現ベクター(すなわち、組換え発現ベクター)に組み込まれ得る。可能な発現ベクターには、そのベクターが使用される宿主細胞と適合性である限り、コスミド、プラスミド、または改変ウイルス(例えば、複製欠損レトロウイルス、アデノウイルス、およびアデノ関連ウイルス、レンチウイルス、ヘルペスウイルス、ポックスウイルス)が含まれる(がこれらに限定されない)。発現「ベクターが宿主細胞と適合性である」とは、発現ベクターが本発明の核酸分子(本明細書中以下に記載される)、および発現のために使用される宿主細胞に基づいて選択される付属の調節配列を含むことを意図すると定義され、前記調節配列は、核酸分子に作動可能に連結されている。「作動可能に連結される」とは、核酸の発現を可能にする様式で、核酸が調節配列に連結されることを意味する。適切な調節配列は、種々の供給源(細菌、真菌、またはウイルスの遺伝子を含む)に由来し得る。例えば、Goeddel, Gene Expression Technology: Methods in Enzymology 185, Academic Press, San Diego, CA (1990)において記載される調節配列を参照されたい。適切な調節配列の選択は、選択される宿主細胞に依存し、かつ当業者によって容易に達成できる。このような調節配列の例には以下が含まれる:転写プロモーターおよびエンハンサー、RNAポリメラーゼ結合配列、またはリボソーム結合配列(翻訳開始シグナルを含む)。選択される宿主細胞および利用される発現ベクターに依存して、他のさらなる配列(例えば、複製起点、さらなるDNA制限部位、エンハンサー、および転写の誘導性を付与する配列)が、発現ベクターに組み込まれ得る。
【0078】
本発明のペプチドはまた、当業者に公知である種々の方法(すなわち、Hugues, Sら(2002) Generation and use of alternative multimers of peptide/MHC complexes. Journal of Immunological Methods. 268:83-91、この開示はその全体が本明細書に援用される)を使用して、可溶性MHC分子と共同して組換え的に産生できる。ペプチド/MHCクラスII可溶性複合体を作製するための方法はまた、米国特許第5,869,279号(この開示はその全体が本明細書に援用される)において提供される。
【0079】
本発明が、サイトカイン、リンホカイン、および免疫調節分子からなる群からの少なくとも1つのメンバーをコードする核酸を含むベクターを含むことがさらに意図される。前記核酸配列は、本発明のシトルリン化ペプチド抗原をコードする配列と連続し得るか、または別個の核酸上にコードできる。
【0080】
本発明のペプチドは、当業者によって理解されるように、種々のアッセイにおいてそれらの検出を容易にするための標識で標識できる。このような標識には、放射活性標識および蛍光標識が含まれるがこれらに限定されない。本発明のペプチドは、例えば、ウシ血清アルブミン(BSA)またはキーホールリンペットヘモシアニンなどに結合された担体とともに提供できる。このペプチドは、固体担体(例えば、金もしくはポリスチレンのミクロスフェア、スライド、チップ)、またはマイクロタイタープレートのウェルに、共有結合的または非共有結合的に結合できる。このペプチドは、ビオチン、フルオレセイン、および西洋ワサビペルオキシダーゼなどの酵素から選択されるがこれらに限定されない標識で、直接的または間接的に標識できる。
【0081】
本発明の実施形態は、インビボでの投与のために適切である生物学的に適合可能な型である、対象への投与のための1種以上のシトルリン化ペプチドを含む薬学的組成物をさらに含む。薬学的組成物中での使用のためのシトルリン化ペプチドは、化学的にまたは組換えDNA技術によって作製できる。本発明のシトルリン化ペプチド抗原の投与は、自己応答性T細胞を有する患者における免疫系を脱感作するように(すなわち、全身性反応性低下の状態または前記ペプチドもしくは組成物に対する寛容性を誘導することによって)作用し得、従って、時間とともに炎症性応答を減少する。本発明のペプチドは、適切な薬学的組成物中で処方される、上記のようなDNA発現ベクター中で提供できる。
【0082】
「インビボでの投与のために適切である生物学的に適合可能な型」は、治療的効果が任意の毒性効果を上回る、投与される物質の型を意味する。本発明の薬学的組成物の治療的に活性な量、すなわち「有効量」は、ヒトにおける免疫応答を誘発することの所望の結果を達成するために必要である用量または時間の期間での有効な量として定義される。物質の治療的な有効量は、レシピエントの疾患状態/健康、年齢、性別、および体重などの因子、および特定のポリペプチドの固有の能力、それをコードする核酸、または所望の免疫応答を誘発するための組換えウイルスに従って変化し得る。投薬量レジメンは、最適の治療的応答を提供するために調整できる。例えば、数回の分割用量が、毎日または定期的な間隔で投与され、および/またはその用量は治療的状況の緊急度によって示されるように比例的に減少できる。投与のためのシトルリン化ペプチド抗原の量は投与の経路、投与の時間に依存し、そして個々の対象の応答に従って変動する。適切な投与経路は、筋肉内注射、皮下注射、静脈内注射、または腹腔内注射、経口投与および鼻内投与である。経口投与および鼻内投与が好ましい投与経路である。
【0083】
本明細書中に記載される組成物は、対象に投与できる薬学的に許容可能な組成物の調製のためのそれ自体公知の方法によって調製され得、その結果、活性物質(すなわち、シトルリン化ペプチド)の有効量が、薬学的に許容可能なビヒクルとの混合物中で合わせられる。適切なビヒクルは、例えば、「Handbook of Pharmaceutical Additives」 (MichaelおよびIrene Ash,編 Gower Publishing Limited, Aldershot, England (1995)において記載されている。これに基づくと、この組成物には、非限定的であるが、薬学的に許容可能なビヒクルまたは希釈物の1種以上が付随した物質の溶液が含まれ、適切なpHおよび/または生理学的液体と等張性である緩衝化溶液中に含まれ得る。この点に関して、米国特許第5,843,456号を参照できる。
【0084】
薬学的に許容可能な担体は当業者に周知であり、例えば、滅菌生理食塩水、ラクトース、スクロース、リン酸カルシウム、ゼラチン、デキストリン、寒天、ペクチン、ピーナッツ油、オリーブ油、ゴマ油、および水が含まれる。他の担体は、例えば、MHCクラスII分子であり得る。モノマー、ダイマー、トリマー、テトラマーなどを含む可溶性MHCクラスII分子、ならびにシトルリンペプチド/MHCクラスII複合体は、米国特許第5,869,270号(この開示は、参照として本明細書に援用される)において開示される方法によって作製できる。
【0085】
さらに、本発明に従う薬学的組成物は、例えば、ソルビトール、マンニトール、デンプン、スクロース、デキストリン、およびグルコースを含む炭水化物、アルブミンまたはカゼインなどのタンパク質、ならびにアルカリリン酸のような緩衝剤などの1種以上の安定剤を含み得る。
【0086】
本発明の薬学的組成物はまた、1種以上のアジュバントを含み得る。当業者に周知であるように、免疫応答を誘導/誘発する免疫原の能力は、投与形式に関わらず(すなわち、組換えウイルス、核酸、ペプチド)、免疫原がアジュバントとともに同時投与される場合に改善できる。アジュバントは、「Vaccine Design-the Subunit and Adjuvant Approach」 (PowellおよびNewman編, 'Plenum Press, New York, U.S.A., pp. 61-79および141-228 (1995)において記載および議論されている。アジュバントは、代表的には、免疫原の免疫原性を増強するが、それ自体の中でおよびそれ自体が免疫原性である必要はない。アジュバントは、免疫系の細胞への免疫因子の遅延性の、持続性の放出を容易にするデポー効果を生じるために、投与の部位に局所的に近傍に免疫原を保持することによって作用し得る。アジュバントはまた、免疫系の細胞を、免疫原デポーに引き付け得、かつこのような細胞が免疫応答を誘発することを刺激し得る。このようなものとして、本発明の実施形態は、さらにアジュバントを含む組成物を含む。
【0087】
理想的なアジュバントの望ましい特性には以下が含まれる:
1)毒性がないこと;
2)長時間続く免疫応答を刺激する能力;
3)製造の単純さおよび長期間保存における安定性;
4)必要な場合、種々の経路によって投与された抗原に対する細胞応答と体液応答の両方を誘発する能力;
5)他のアジュバントとの相乗効果;
6)抗原提示細胞(APC)の集団と選択的に相互作用する能力;
7)適切なTr、TR1、またはTH2細胞特異的免疫応答を特異的に誘発する能力;ならびに
8)抗原/免疫原に対する適切な抗体イソタイプレベル(例えば、IgA)を選択的に増加する能力。
【0088】
適切なアジュバントには、とりわけ、水酸化アルミニウム、リン酸アルミニウム、アンフィゲン、トコフェノール、モノホスフェニルリピドA、ムラミールジペプチド、およびQuill Aなどのサポニンが含まれる。好ましくは、本発明に従う寛容治療において使用されるアジュバントは、粘膜上皮に結合する、コレラ毒素Bサブユニットまたはカルボマーなどの粘膜アジュバントである。アジュバントの量は、当業者によって理解されるように、アジュバントそれ自体の性質に依存する。
【0089】
シトルリン化ペプチドが、所望の遺伝子に対してRNA干渉応答を媒介する短い二本鎖RNA(30ヌクレオチド長未満)を組み合わせて患者に投与できることが本発明においてさらに具体化される。これらの標的遺伝子は、転写因子、酵素、サイトカイン、リンホカイン、および免疫調節分子からなる群の1種以上のメンバーから選択できる。
【0090】
本発明のペプチドおよび組成物は、MHC分子の文脈において投与できる。従って、本発明のポリペプチドは、樹状細胞(DC)を含むがこれに限定されない抗原提示細胞にパルス投与できる。より具体的には、シトルリン化ペプチドが、抗原パルスされた樹状細胞(抗原提示細胞)を介して患者に投与できる。特定の実施形態において、本発明は、抗原提示細胞として樹状細胞またはその前駆細胞を使用する。これらの樹状細胞は、患者に投与する前に、転写因子、酵素、サイトカイン、リンホカイン、および免疫調節分子のそれらの産生を変化させるために、患者のPBMC(末梢血単核細胞)から増殖され、エキソビボで処理できる。樹状細胞は、高度に強力なAPCである(BanchereauおよびSteinman, Nature 392:245-251, 1998)。一般的に、樹状細胞は、それらの代表的な形状(インサイチューで星状、インビトロで見える顕著な細胞質プロセス(樹状))、高効率で抗原を取り込み、プロセスし、かつ提示するそれらの能力、および未処理のT細胞の応答を活性化するそれらの能力に基づいて同定できる。樹状細胞は、インビボまたはエキソビボで樹状細胞上に一般的に見い出されない特異的細胞表面レセプターまたはリガンドを発現するように操作され得、このような改変樹状細胞は本発明によって意図される。樹状細胞に対する代替として、分泌ベシクル抗原負荷樹状細胞(エキソソームと呼ばれる)が免疫原性組成物中で使用できる(Zitvogelら、Nature Med. 4:594-600, 1998を参照されたい)。樹状細胞および前駆細胞は、末梢血、骨髄、腫瘍浸潤細胞、腫瘍周囲組織浸潤細胞、リンパ節、脾臓、皮膚、臍帯血または他の適切な組織または体液から入手できる。例えば、樹状細胞は、末梢血から収集された単球の培養物に、GM-CSF、IL-4、IL-13および/またはTNF-αなどのサイトカインの組み合わせを加えることによって、エキソビボで分化できる。代替的には、末梢血、臍帯血、または骨髄から収集されたCD34陽性細胞が、GM-CSF、IL-3、TNFα、CD40リガンド、LPS、fit3リガンド、ならびに/または、樹状細胞の分化、成熟、および増殖を誘導する他の化合物の組み合わせを培養培地に加えることによって樹状細胞に分化できる。
【0091】
本発明のシトルリン化ペプチドは関節炎性であり、従って、RAをもたらす炎症性応答を誘発する。これは、図6AからEに実証されている。このようなものとして、これらのペプチドは、RAの治療のために有効である新規な医薬が試験および同定できる動物モデルを提供するために、非ヒト哺乳動物において臨床的関節炎を誘導するために使用できる。共有エピトープを含むMHCクラスII分子についてトランスジェニックである任意の動物が、本発明の範囲内で使用できる。本発明の1つの局面において、本発明の1種以上のシトルリン化ペプチドの投与が、関節炎の徴候関節炎、より特定には慢性関節リウマチの最終的な疾患パターンの発症をもたらすHLA-DRB1*401トランスジェニックマウス(DR4-IE tg)に提供できる。これらのDR4-IE tgトランスジェニックマウスは、ヒト抗原結合ドメイン(α1およびβ1)およびマウスCD4結合ドメイン(α2およびβ2)から構成されるキメラMHCクラスII分子を発現する。キメラDR4は、全長ヒトDR4と同一の様式でペプチドを提示し、そしてこれらのマウスは、内因性MHCクラスII分子を発現しない。さらに、これらのマウスは、野生型マウスと同じB細胞およびT細胞の補体を含み、従って、これらのトランスジェニックマウスは、インビボでT細胞に制限されるSEの免疫応答を試験するために、および従って、炎症をもたらすT細胞応答を減少または予防し得る医薬品薬剤を同定するために理想的である。DR4-IE tgマウスが使用されるが、共有エピトープを含むMHCクラスII分子についてトランスジェニックである任意のマウス系統が、本発明における薬学的化合物の試験のための動物モデルとして使用できることが理解される。
【0092】
本発明のシトルリン化されたMBPタンパク質およびGFAPタンパク質もまた、新規な医薬品がMSの治療のために試験されかつ有効であると同定できるMS動物モデルを提供するために非ヒト哺乳動物において臨床的多発性硬化症を誘導するために使用できる。このような動物モデルを作製するための方法は上記の通りである。
【0093】
トランスジェニックマウスは、1回以上、本発明の1種以上のペプチドを含むシトルリン化タンパク質(ならびに任意のアジュバントまたは他の薬学的賦形剤)を、皮下的に、皮内に、腹腔内に、または筋肉内に注射し、関節炎の徴候を発症させ得る。1つの態様において、全長シトルリン化ビメンチンまたはシトルリン化フィブリノーゲンが、関節炎の発症を誘導するためにトランスジェニックマウスに投与される。関節炎を治療するため、および関節炎の発症に対してこれらの薬学的活性の効果を研究するための炎症反応が確立されるとすぐに、潜在的な医薬品薬剤は、本発明のペプチドと同時投与されるか、または後で投与できる。好ましいマウスは、関節炎、特に慢性関節リウマチのための動物モデルとして使用される。
【0094】
本発明の実施形態において、本発明のシトルリン化ペプチドが関節炎性であり、共有エピトープを有するMHCクラスII分子への結合を介して、RAをもたらすT細胞応答を誘発するように作用するという知見は、ここで、RAおよびMSなどのシトルリン化抗原の共有エピトープ結合を含む自己免疫障害のための診断的および治療的方法を提供する。
【0095】
1つの態様において、本発明は、RAを有する患者、またはRAを有すると疑われている患者、またはRAを発症する素因がある患者からの自己反応性T細胞を検出するために使用できる。多数の方法が、シトルリン化ペプチド-MHC複合体に特異的なT細胞を検出するために、またはシトルリン化ペプチド-MHC複合体それ自体の存在のいずれかを検出するために使用できる。例えば、当業者に公知であるT細胞増殖アッセイは、対象の末梢血単核細胞(PBMC)試料中のT細胞活性化を検出するために使用できる。このような方法は、T細胞増殖の尺度として放射性同位元素(例えば、3H-チミジンなど)の取り込みによって実行される(実施例の節を参照されたい)。PBMCに存在する自己反応性T細胞活性もまた、サイトカイン特異的ELISA(すなわち、ELISPOT)を用いて、シトルリン化ペプチド-MHCクラスII複合体による活性化後のサイトカイン放出を測定することによって検出できる。別の検出方法は、例えば、II-2RのFACS分析による活性化マーカーの発現の測定である。さらに、可溶性MHCクラスII分子は、結合したシトルリン化ペプチドを伴う蛍光色素またはリポソームに連結され得、これらの複合体を認識するT細胞レセプターを有するT細胞を検出するために使用できる。
【0096】
別の実施形態において、本発明のシトルリン化ペプチドのいずれかを認識するモノクローナル抗体がまた作製され得、抗原提示細胞(APC)上の共有エピトープを有するMHCクラスII分子に提示/結合されたペプチドの存在を検出するために使用できる。これは、疾患ならびにRAを発症する素因の迅速かつ単純な診断方法を提供する。一般的に、抗体の調整のための方法は周知である。本発明のペプチドを認識するポリクローナル抗体を調製するために、ビメンチンタンパク質またはフィブリノーゲンタンパク質の規定された部分またはすべて、またはそれらの選択的転写物のいずれかが、適切なクローニングビヒクル中での対応するDNA配列の発現によって、細菌中で合成できる。次いで、タンパク質が精製され、担体タンパク質に結合されおよびフロイントアジュバントを混合され(ウサギによる抗原性応答を刺激することを補助するために)、ならびにウサギまたは他の実験用動物に注射できる。代替的には、タンパク質は、タンパク質を発現する培養細胞から単離できる。2週間間隔のブースター注射後、次いでウサギまたは他の実験用動物を、採血し、血清を単離する。血清は、直接的に使用できるか、またはアフィニティークロマトグラフィーによって精製できる。次いで、血清は、ポリアクリルアミドゲル上でタンパク質抽出物をプローブし、ビメンチンタンパク質またはフィブリノーゲンタンパク質、選択的転写物またはそれらの任意の変異体を同定するために使用できる。代替的には、合成ポリペプチド(上記で議論したようなもの)が、これらのタンパク質の抗原性部分に対して作製され得、動物に接種されるために使用できる。
【0097】
本発明のペプチド(哺乳動物または他の種のビメンチンタンパク質およびフィブリノーゲンタンパク質またはその部分)を特異的に認識するモノクローナル抗体を産生するための方法は、当業者に周知である。一般的に、活性にタンパク質を発現する細胞が、培養されるかまたは組織から単離され、そして細胞抽出物が単離される。ビメンチンタンパク質またはフィブリノーゲンタンパク質を含む、この抽出物または組換えタンパク質抽出物が、フロイントアジュバント中でマウスに注射される。3週間の期間にわたって9回注射した後、マウス脾臓が取り出され、リン酸緩衝化生理食塩水(PBS)中に再懸濁される。脾臓細胞はリンパ球の供給源として働き、それらのいくつかは適切な特異性の抗体を産生する。次いで、これらは、永久的に増殖するミエローマパートナー細胞と融合させ、融合の生成物を、HATなどの選択薬剤の存在下で多数の組織培養ウェルにプレートする。次いで、これらのウェルをELISAによってスクリーニングして、有用な抗体を作製する細胞を含むウェルを同定する。次いで、これらを新鮮にプレートする。一定の増殖期間の後、これらのウェルを再度スクリーニングして、抗体産生細胞を同定する。数回のクローニング手順が、90%より多くのウェルが抗体産生について陽性である単一クローンを含むまで、実行される。この手順から、抗体を産生する安定なクローンの系統が樹立される。次いで、モノクローナル抗体が、プロテインAまたはプロテインGセファロースを使用するアフィニティークロマトグラフィーによって精製できる。
【0098】
本発明の抗体の調製のための方法は、一般的に当該分野において公知である。例えば、Antibodies, A Laboratory Manual, Ed. Harlow および David Lane (編) Cold Spring Harbor Laboratory, N.Y. (1988) ならびにそこに引用される参考文献を参照されたい。免疫学の一般的原理を説明する標準的な参照される研究には、Klein, J. Immunology: The Science of Cell-Noncell Discrimination, John Wiley & Sons, N.Y. (1982); Dennett, R.ら Monoclonal Antibodies, Hybridoma: A New Dimension In Biological Analyses Plenum Press, N.Y. (1980) および Campbell, A. 「Monoclonal Antibody Technology」Laboratory Techniques In Biochemistry And Molecular Biology, Vol. 13, Burdon.ら (編), Elsevier, Amsterdam (1984) が含まれる。米国特許第4,609,893号、米国特許第4,713,325号、米国特許第4,714,681号、米国特許第4,716,111号、米国特許第4,716,117号、米国特許第4,720,459号(これらの開示はその全体が本明細書に援用される)もまた参照されたい。
【0099】
抗体はまた、ペプチド/MHCクラスII複合体に指向されて作製できる。ペプチド/MHCクラスII複合体に対するこのような抗体を生成する方法は公知であり、記載されている(Baeten, D., Steenbakkers PGA., Rovers E., Veys EM., Rijnders AMW., Meijerink J., Keyser F DeおよびBoots A. Localisation of MHC Class II/HC gp-39 complexes in synovia of rheumatoid arthritis patients using complex-specific monoclonal antibodies. Abstracts of the 23rd European Workshop for Rheumatology Research, 第5巻 補遺1, 2003年2月; Baeten D., Steenbakkers P., Boots A., Veys EM., and Keyser, Fde., The presentation of the immunodominant epitope of HC gp-39 in the context of the RA-associated HLA class II molecules is specific for RA synovium. Abstracts of the 23rd European Workshop for Rheumatology Research, 第5巻 補遺1, 2003年2月)。
【0100】
本発明に従う1種以上のペプチドを含む診断組成物および適切な検出剤は、従って、本発明の一部を形成する。検出の型に依存して、検出剤は、放射性同位元素、酵素、または細胞表面もしくは活性化マーカーに特異的である抗体であり得る。
【0101】
本発明に従う1種以上のペプチドを含む試験キットもまた、本発明の範囲に含まれ得る。これらの試験キットは、本発明に従う診断方法における使用のために適切である。
【0102】
式Cit-Cit-X-Cit-G-Cit-Cit-Z-Cit-Cit-B-Cit-Cit(ここでXはY、F、W、I、L、MおよびVから選択され、ZはA、D、I、N、P、S、TおよびVから選択され、BはA、G、H、Q、S、TおよびVから選択される)のペプチドは、自己免疫障害の診断のための方法において特に有用である。このような自己免疫障害には、慢性関節リウマチが含まれるがこれに限定されない。例えば、Cit-Cit-Y-Cit-G-Cit-Cit-S-Cit-Cit-S-Cit-Citなどのペプチドが、RAの診断のために患者における抗シトルリン抗体を検出するための市販の試験キット(例えば、ELISAキット)において使用できる(ELISA Guidebook. Theory & Practice. Editor John R. Crowther, Author John R. Crowther, Pub. Humana Press 2000)。手短に述べると、ペプチドは、マイクロタイタープレートのウェルに共有結合されるか、または非共有結合的に結合され、希釈された患者の血清が各ウェルに加えられ、一定の時間の間インキュベートされる。プレートの洗浄後、ペルオキシダーゼに結合体化されたIgGがウェルに加えられ、さらにインキュベートされる。さらなる洗浄の後、結合した抗体が、テトラメチルベンジジンなどの基質とともに検出される。一定の時間の間の後、反応が停止され、特定の波長(例えば450nm)でのプレートのOD読み取りが行われる。対照試料は同じ様式で同時に処理される。陽性OD読み取りは、陽性の結合応答を示す。
【0103】
本発明はまた、シトルリン化ペプチド/MHCクラスII複合体に特異的であるT細胞を標的化すること、またはこれらの複合体の形成を破壊することを含む、治療的ストラテジーを含む。これらの方法は、RAを処置するための他の公知の治療と組み合わせて使用できる。例えば、活性化T細胞は、可溶性ペプチド-MHCクラスII複合体を患者に投与することによって達成できるこれらの細胞のアポトーシスを誘導することによって除去できる(すなわち、米国特許第5,734,02号、米国特許第6,106,840号、米国特許第5,635,363号、米国特許第6,211,342号,、米国特許出願公開第20020176864号および米国特許出願公開第20020122818号(これらの開示はその全体が本明細書に援用される)。本発明はまた、対象におけるT細胞活性を調節するために、RNA干渉の方法を使用する治療ストラテジーにおいて本発明のペプチドの使用を含む。RNA干渉(RNAiまたはsiRNA)は、二本鎖RNA(dsRNA)によって引き起こされる遺伝子サイレンシングの型である、本発明の1つの態様において、このような方法を使用して、本発明のペプチドは、例えば、APC(すなわち、樹状細胞)に効果をもたらし、次には活性化された炎症性応答から調節性応答の1つに移行するようにT細胞に影響を与えるサイトカインをコードするRNA配列と合わせられる。RNA干渉(RNAi)においては、動物および植物における配列特異的、転写後遺伝子サイレンシングは、サイレンシングされる遺伝子に対して配列中の相同な二本鎖RNA(dsRNA)によって開始される。siRNA(小さな干渉RNA)は、慢性関節リウマチをもたらすタンパク質のシトルリン化に関与しているコードされたタンパク質を発現しないために、所望のmRNA(この場合はPADをコードしている)を標的化し、従ってこれを分解するように設計される。C.elegans、Drosophila、植物、および哺乳動物において遺伝子をサイレンシングするためのRNAiの使用に関連する方法は当該分野において公知である(Fire A.ら、1998 Nature 391:806-811; Fire, A. Trends Genet. 15, 358-363 (1999); Sharp, P. A. RNA interference 2001. Genes Dev. 15, 485-490 (2001); Hammond, S. M.ら、Nature Rev. Genet. 2, 110-1119 (2001); Tuschl, T. Chem. Biochem. 2, 239-245 (2001); Hamilton, A.ら、Science 286, 950-952 (1999); Hammond, S. M.ら、Nature 404, 293-296 (2000); Zamore, P. D.,ら、Cell 101, 25-33 (2000); Bernstein, E.ら、Nature 409, 363-366 (2001); Elbashir, S. M.ら、Genes Dev. 15, 188-200 (2001)、国際特許公開WO0129058号、国際特許公開WO9932619号、Elbashir S Mら、2001 Nature 411:494-498)、およびカナダ国特許出願第2,388,441号(これらの開示はその全体が本明細書に援用される))。
【0104】
本発明はまた、本明細書中で以前に記載されたような全身性免疫学的寛容性を誘導するための本発明に従う1種以上のペプチドおよび薬学的に許用可能な担体を含む治療組成物の投与による、慢性関節リウマチなどの自己免疫障害の治療の方法を含む。誘導された寛容性は、攻撃に際して組織中での局所的炎症性応答の減少をもたらす。1つの局面において、ペプチドは、例えば、米国特許第5,843,445号(この開示はその全体が参照として本明細書に援用される)において開示される方法のような全身性免疫学的寛容性を誘発するために患者の粘膜上皮を介して投与される。
【0105】
なお他の治療的ストラテジーは、細胞毒性分子または本発明のペプチドに放射核種(すなわち、125Iまたは131I)で標識した放射活性分子の結合体化を含み得る。本発明の他の実施形態は、毒素分子またはその活性フラグメントに共有結合された本発明のペプチドまたは抗体である細胞認識分子を含むイムノトキシン分子である。このようなイムノトキシン分子は、本明細書中に記載されるような慢性関節リウマチのための医薬または診断薬としての使用のために、単独で使用できるか、または組成物として使用できる。本発明のペプチドおよびMHCクラスII分子との細胞毒性分子のこのような結合体化は、CD4T細胞を標的化するために有益である。代替的には、いくつかの局面において、配列番号38または39に記載されるような特定のシトルリン化ペプチドは、B細胞を標的化する細胞毒性分子に単に標的化できる。この様式において、その表面に同じ抗体を有する抗シトルリン化抗体を産生するB細胞のみが標的化される。
【0106】
本発明の別の態様において、シトルリン化ペプチド/MHCクラスII分子複合体に特異的な抗体が複合体に結合するため、従って、T細胞による複合体認識を予防するために使用できる(すなわち、国際特許公開WO02/14870号)。代替的には、MHCクラスII分子のペプチド結合溝が、高アフィニティー非シトルリン化ペプチドと結合され得、これは次いでT細胞によって認識されず、または抗体によって結合されて、従って、シトルリン化ペプチド/MHCクラスII複合体の形成を予防する(すなわち、米国特許第6,355,617号)。
【0107】
本発明のさらなる局面は、一般的に、T細胞応答を誘発するシトルリン化ペプチド抗原を含む自己免疫障害の診断のための方法である。このような自己免疫障害には、慢性関節リウマチおよび多発性硬化症が含まれるがこれらに限定されない。多発性硬化症において、シトルリン化ミエリン塩基性タンパク質(MBP)T細胞応答が観察された(Tranquillら、2000. Enhanced T cell responsiveness to citrulline-containing myelin basic protein in multiple sclerosis patients. Mult. Scler. 6:4220-5)。それゆえに、MHCクラスII共有エピトープを発現するMS患者において、シトルリンに対する免疫応答は疾患のプロセスに寄与し得る。このようなものとして、シトルリン化MBPに対する抗シトルリン自己抗体の検出は、MSについて診断的であり得る。いくつかの態様において、T細胞反応性を阻害することは、MSについて治療的であり、この活性をアッセイすることはMSについて診断的である。さらに、慢性関節リウマチに関して本明細書中に記載される治療もまた、シトルリン化MBPが疾患をもたらすT細胞応答を誘発する、MSの治療に適用可能である。RAの治療のための、本明細書中に記載されるような、感受性のある患者(すなわち、MHCクラスII共有エピトープを発現する)において共有エピトープへのシトルリン化ペプチドの結合の阻害の方法は、MSの治療においてもまた、適用可能であり得る。MSの診断および治療方法の標的において有用なシトルリン化MBPタンパク質は、例えば、FLPCitHRDTG (配列番号40), VTPCitTPPPS (配列番号41); YGGCitASKYK (配列番号42)および LGGCitDSRSG (配列番号42)から選択できる。MSの診断において使用できるなお他のシトルリン化ペプチドは、シトルリン化グリア線維酸性タンパク質(GFAP)である。GFAPは、グリア細胞、星状細胞、アストログリアフィラメントに存在するタンパク質である(Nicholas A.P. 2002. Preparation of a monoclonal antibody to citrullinated epitopes: its characterization and some applications to immunohistochemistry in human brain. Glia.Mar 15;37(4):328-36、Nicholas ら、2003. Immunohistochemical localization of citrullinated proteins in adult rat brain. J Comp Neurol. May 5:459(3):251-66)。MSの診断の方法における使用のためのシトルリン化GFAPは、MERCitRITSA (配列番号43), LPTCitVDFSL (配列番号44), LNDCitFASYI (配列番号45), LRLCitLDQLT (配列番号46), LQICitETSLD (配列番号47)およびVEMCitDGEVI (配列番号48)から選択できる。
【0108】
本発明のシトルリン化MBPおよびGFAPは、多発性硬化症についての診断的および治療的アプローチにおける使用のためのものであることが当業者によって理解される。
【0109】
代替的には、本発明の実施形態において、アルギニンを有する特定のペプチドのシトルリンのペプチドへの修飾は、このアミノ酸反応を触媒する酵素ペプチジルアルギニンデイミナーゼ(PAD)に対するアンタゴニストおよび/または阻害剤の使用によって予防することができる。PADは同様の触媒活性を共有する酵素のファミリーのメンバーである。これらはカルシウム依存性酵素であり、これらは哺乳動物組織において4つのアイソフォームで存在する:PADIおよびIIIは毛嚢および上皮において存在する;PADIIはCNS組織、筋肉、およびある造血細胞において存在する;PADIVは造血組織、特に単球および好中球において存在する(Vossenaar E.R.ら、(2003). PAD, a growing family of citrullinating enzymes; genes, features and involvement in disease. Bioessays. 25(11): 1106-18)。PADの生理学的機能は完全には知られていないが、しかし、アルギニンの正電荷をより減少した電荷を有する極性アミノ酸シトルリンに変換するように作用する(図4)。
【0110】
PADの遺伝子は、ヒトにおいて1p36染色体、およびマウスにおいて4E1染色体に存在する。1p36染色体上の領域は、以前に慢性関節リウマチと関連すると同定されたが、この領域は、4つのPAD遺伝子に加えて4つの遺伝子を含み、慢性関節リウマチの発生をもたらす、PAD活性、ペプチドのシトルリン化の増加、または任意のHLA-DR分子への結合の間のいかなる直接的関連も存在しなかった。
【0111】
PADのインビトロおよびインビボでのシトルリン化のブロッキングは、これが、疾患プロセスにおける特異的決定的段階に影響を与えることに焦点を当てているので、慢性関節リウマチおよび多発性硬化症を治療するための新規な治療方法を提供する。本発明のこの実施形態の1つの態様において、PAD活性は、上記で手短に議論したRNA干渉(siRNA)のより特異的かつ効率的な技術を使用してブロックできる。この様式において、PAD遺伝子の機能はサイレンシングされ得、その結果、感受性のペプチド/タンパク質のシトルリン化は起こらず、慢性関節リウマチまたは多発性硬化症の発症をもたらさない。このことは、シトルリン化ペプチドの産生の結果として自己免疫障害を発症するリスクが特にあるSEおよびPADを発現する個体の治療の方法を提供する。シトルリンの産生に依存する、自己免疫障害のより初期とより後期の両方の段階が、PAD発現の阻害によって、および本発明のある態様においては、PAD IV発現の抑制によって抑制できる。
【0112】
PAD遺伝子の発現、従ってその機能をサイレンシングすることに加えて、低分子がまた、PADを阻害するために使用され得、従って、このような低分子はまた、種々の投与の経路において自己免疫障害の治療のための治療用組成物中で使用できる。組成物および組成物の投与の経路の議論は、本明細書中で以前に示した。当業者は、PADを阻害するそれらの能力について、潜在的な低分子をいかにしてスクリーニングするかの理解および知見を有する。適切なスクリーニング方法は、例えば、Waltersら (Designing Screens: How to Make Your Hits a Hit. Nature Reviews, Drug Discovery. 第2巻, 2003年4月、259-266頁;この開示はその全体が参照として本明細書に援用される)において記載されている。
【0113】
上記の開示は一般的に本発明を記載する。より完全な理解は、以下の特定の実施例を参照して得られ得る。これらの実施例は、単に例示の目的のみのために記載され、本発明の範囲を限定することを意図しない。形態の変更および等価物の置換は、状況が示唆し得または好都合にし得る場合に意図される。特定の用語が本明細書中で使用されたが、このような用語は、説明的な意味にあることが意図され、限定の目的のためではない。
【実施例】
【0114】
この実施例は、例示の目的のために記載され、本発明の範囲を限定することを意図するものではない。
【0115】
本開示および実施例において引用されるが明白に記載されない、合成化学、タンパク質およびペプチド化学、分子生物学、薬学、および免疫学の方法は、科学文献中に報告され、かつ当業者に周知である。
【0116】
(実施例1)
動物の作製
HLA-DR4-IEトランスジェニック、マウスMHCクラスII欠損マウスをこれらの実験において使用した(Ito K, Bian HJ, Molina M, Han J, Magram J, Saar E, Belunis C, Bolin DR, Arceo R, Campbell R, Falcioni F, Vidovic D, Hammer J, Nagy ZA. 1996. HLA-DR4-IE chimeric class II transgenic, murine class II-deficient mice are susceptible to experimental allergic encephalomyelitis. J. Exp. Med. 183:2635)。これらのマウスを、以前に記載されたように繁殖および維持した(Hill, J.A., Wang, D., Jevnikar, A.M., Cairns, E., Bell, D.A. 2002. The relationship between predicted peptide-MHC class II affinity and T cell activation in a HLA-DR(1*0401 MHC class II mouse model. Arthritis Res. 5:R40)。
【0117】
(実施例2)
ペプチドの生成
これらの研究において使用されるペプチドは、製造業者(Genemed Synthesis、San Francisco、CA)によって合成および精製された。ペプチドを、Hammerら (Hammer J, Bono E, Gallazzi F, Belunis C, Nagy Z, Sinigaglia F. 1994. Precise prediction of major histocompatibility complex class II-peptide interaction based on peptide side chain scanning. J. Exp. Med. 180:2353)の方法に従ってDRB1*0401についてそれらの予測されたアフィニティーに基づいて選択した。下線を付したアミノ酸は、9個のMHCクラスII分子結合ポケット(P1〜P9)と相互作用する残基を示すのに対して、太字で示すものはP4共有エピトープ位置で相互作用する。プロテオグリカンアグリカンから使用したペプチドの配列は以下の通りである:P4D = ヒトアグリカンペプチド 280-292, AGWLADRSVRYPI; P4R = 変化したヒトアグリカンペプチド280-292, AGWLARRSVRYPI; P4Cit = 変化したヒトアグリカンペプチド280-292, AGWLACitRSVRYPI。シトルリンはHammerらの予測アルゴリズムにおいて考慮されていないので、ビメンチンからの候補T細胞エピトープを同定するときには、グルタミンの値をアルギニンの代わりに置き換えた(グルタミンはシトルリンを同じ末端側鎖基を有する)。使用したビメンチンペプチドの配列は以下の通りであった:Vim 65-77 = ヒトビメンチンペプチド 65-77, SAVRARSSVPGVR, Vim R70Cit = 変化したヒトビメンチンペプチド65-77, SAVRACitSSVPGVR。
【0118】
(実施例3)
免疫化
DR4 tgマウスを、CFA (Difco Laboratories、Detroit, MI)中で乳化した100μlのペプチド(1μg/μl)(1:1容量比)を用いて両方の後肢の内側に皮内免疫した。10日後、マウスを屠殺し、インビトロ増殖アッセイおよびサイトカインアッセイのためにそれらの流入領域リンパ節を取り出した。
【0119】
(実施例4)
T細胞培養
細胞懸濁物を流入領域リンパ節から調製し、96ウェルプレート中で、4×105細胞/ウェルの濃度で、ペプチド抗原の存在下非存在下で、4日間培養した。抗DR抗体(BD PharMingen、Mississauga、ON)をある培養に加え(1μg/ml)、以前に記載されたようにDR制限T細胞応答を確認した(Andersson EC, Hansen BE, Jacobsen H, Madsen LS, Andersen CB, Engberg J, Rothbard JB, McDevitt GS, Malmstrom V, Holmdahl R, Svejgaard A, Fugger L. 1998. Definition of MHC and T cell receptor contacts in the HLA-DR4restricted immunodominant epitope in type II collagen and characterization of collagen-induced arthritis in HLA-DR4 and human CD4 transgenic mice. Proc. Natl. Acad. Sci. USA. 95:7574)。培養上清を78時間後に取り出し、以前に記載されたようにELISA(BD Phar Mingen, Mississauga, O.N)によってIFN-γ産生を試験した(Hill, J.A., Wang, D., Jevnikar, A.M., Cairns, E., Bell, D.A. 2002. The relationship between predicted peptide-MHC class II affinity and T cell activation in a HLA-DR(1*0401 MHC class II mouse model. Arthritis Res. 5:R40)。サイトカイン産生を2連で測定し、平均抗原特異的サイトカイン産生(対照試料中のサイトカイン産生+2SDをペプチド特異的サイトカイン産生から減算した)±SDを表す。培養終了の18時間前に、1μCiの[3H]チミジン(ICN Biomedicals、Montreal、PQ)を各ウェルに加えて、T細胞増殖を評価した。増殖実験を3連で実行し、結果をcpm±SDの平均増殖または刺激指数(実験試料のcpm/対照試料のcpm)±SEMとして表した。
【0120】
(実施例5)
ペプチド結合アッセイ
精製したHLA-DRB1*0101、*0401、*0404、*0301、*0701、*0802、*1101、および*1302に対するペプチド結合アッセイを、以前に記載されたように、放射活性標識ペプチドプローブに対して決定した(Southwood S, Sidney J, Kondo A, del Guercio MF, Appella E, Hoffman S, Kubo RT, Chesnut RW, Grey HM, Sette A. 1998. Several common HLA-DR types share largely overlapping peptide binding repertoires. J. Immunol. 160:3363)。精製したHLA-DRB1分子に対する標識ペプチドの50%阻害のために必要な未標識ビメンチンペプチドのnM濃度(IC50)を、相互作用のアフィニティー(kDa)の近似値として使用した。結果を、nMで測定したIC50値の逆数として表現する。
【0121】
(実施例6)
DR4-1E tgマウスにおける関節炎の誘導
DR4-1E tgマウスを、CFA (Difco Laboratories、Detroit, MI)中で乳化した100μlのシトルリン化ヒトフィブリノーゲン(1μg/μl)(1:1容量比)を用いて両方の後肢の内側に皮下免疫した。シトルリン化フィブリノーゲンは以前に記載されるように調製した(Christine Masson-Bessiereら、2001. The Major Synovial Targets of the Rheumatoid arthritis-Specific Antifilaggrin Autoantibodies Are Deiminated Forms of the ( and ( Chains of Fibrin 1 The Journal of Immunology 166: 4177-4184)。マウスは不完全フロイントアジュバント中の同じ抗原の第2の免疫を21日間受容し、引き続いて関節炎の徴候について観察した(図6AからF)。
【0122】
(実施例7)
DR4-IE tgマウスにおける抗シトルリン抗体の生成およびRA自己抗体の検出
DR4-IE tgマウスを、0、21、および42日めに、それぞれCFA、IFA、およびIFAで乳化した100μgのペプチドCit-Cit-Y-Cit-G-Cit-Cit-S-Cit-Cit-S-Cit-Cit(配列番号39)で免疫した。52日目にマウスを屠殺し、血清を血液から収集した。抗体反応性を、以前に記載されたように(Schllekensら、1998. J Clin Invest. Volume 101, 第1巻, 1998年1月, 273-281頁)、ELISAによって、ペプチド(配列番号39)およびcfc1ペプチドを使用して、血清試料中で検出した。ペプチド(配列番号39)またはcfc1に対する抗シトルリン抗体は、ペプチド(配列番号39)で免疫したマウスの血清中で見い出された。シトルリン化フィブリノーゲンで免疫した関節炎マウスからの抗体もまた、これらのペプチドの両方に結合した。試験した多くの対照マウスにおいて抗体は見られなかった。ペプチドCit-Cit-Y-Cit-G-Cit-Cit-S-Cit-Cit-S-Cit-Cit(配列番号39)に対するヒト抗体の反応性もまた試験され、市販のELISAキット(Quantilite、INOVA Diagnostics)と比較された。シトルリン化フィブリノーゲンに対する抗体反応性もまた実証された(図7)。
【0123】
表1は、ペプチドCit-Cit-Y-Cit-G-Cit-Cit-S-Cit-Cit-S-Cit-Cit(配列番号39)、シトルリン化フィブリノーゲンポリペプチド(トランスジェニックマウスを免疫するために以前に使用した)、および市販のCCP試薬を使用する抗シトルリン化抗体の頻度および特異性を示す。試験された患者の中でも慢性関節リウマチについてこのアッセイの感度および特異性は、Cit-Cit-Y-Cit-G-Cit-Cit-S-Cit-Cit-S-Cit-Cit(配列番号39)ペプチドおよびCCPペプチドがRAの診断のためのそれらの感度および特異性によって同様であったことを示しかつ指摘する。
【0124】
図8は、市販のCCPキットおよびCit-Cit-Y-Cit-G-Cit-Cit-S-Cit-Cit-S-Cit-Cit(配列番号39)ペプチドを使用する抗シトルリン化抗体の相関を示す。これらのアッセイを使用して試験した51例の慢性関節リウマチ患者から、0.48の相関係数が見られた。表2において見られるように、ペプチドCit-Cit-Y-Cit-G-Cit-Cit-S-Cit-Cit-S-Cit-Cit(配列番号39)は、慢性関節リウマチの診断について、98%の特異性および71%の感度を提供した。乾癬性関節炎を有する49例の患者のうちの1例のみが陽性であった。このことは、ペプチドCit-Cit-Y-Cit-G-Cit-Cit-S-Cit-Cit-S-Cit-Cit(配列番号39)および一般的には式Cit-Cit-X-Cit-G-Cit-Cit-Z-Cit-Cit-B-Cit-Cit(配列番号38)(ここでXはY、F、W、I、L、MおよびVから選択され、ZはA、D、I、N、P、S、TおよびVから選択され、BはA、G、H、Q、S、TおよびVから選択される)のペプチドが、慢性関節リウマチにおける抗シトルリン化抗体の検出のために有用であることを示す。
【0125】
(実施例8)
PAD発現についてマウスおよびヒトの細胞系統の選択のため;PAD標的化のためのsiRNAを同定するため;siRNAを使用して選択された細胞系統におけるPAD発現を阻害するため;およびPAD発現をモニターするための、siRNAを使用するPADの阻害、インビトロ研究。PAD IVに関して記載したが、当業者に理解されるように、同類のプロセスが、PAD IIまたは任意の他のPADのために使用できる。
(a)PAD(IIおよびIV)発現についてのマウスおよびヒトの細胞系統の選択
ATCCからのヒトおよびマウスの単球/マクロファージおよび顆粒球/好中球の細胞系統。これらの細胞系統は、ビタミンD3、レチノイン酸、およびデキサメタゾンなどの種々の薬剤を使用して、マクロファージおよび好中球の表現型にさらに分化できる(www.atcc.org)。好中球およびマクロファージの細胞型は、PAD IIおよびIVを発現する(Asaga Hら、(1998) Selective deimination of vimentin in calcium ionophore-induced apoptosis of mouse peritoneal macrophages. Biochem Biophys Res Commun. 243(3):641-6: Nakashima Kら、(1999) Molecular characterization of peptidylarginine deiminase in HL-60 cells induced by retinoic acid and 1alpha,25-dihydroxyvitamin D3. J Biol Chem. 274(39), 27786-92)。細胞は、RA滑膜において活性化されかつ豊富である。PAD IIおよびIVは、LPS活性化細胞系統において定量的に決定され、最高のレベルのPAD IIおよびIVを有する細胞系統はsiRNA阻害を試験するために選択される。
【0126】
(b)PAD(IIおよびIV)標的化のためのsiRNAの同定
ヒトおよびマウスのPAD IV遺伝子配列の分析は、選択判断基準を満たす全部で11個のsiRNA配列を明らかにした。11個の配列のうち、8個がヒト、3個がマウスであった(表4)。これらのsiRNAは、siRNAの安定性、有効寿命、および強度を増強する独自技術のRNA修飾(siSTABLE技術と呼ばれる)を使用する、Dharmacon Inc.から入手した(www.dharmacon.com)。
【0127】
(c)PAD(IIおよびIV)発現の阻害
siRNAは、PAD IIまたはIVを高度に発現する選択されたヒトおよびマウスの細胞系統において、これらの細胞における有効なPAD IIまたはIVの阻害のための最適条件を確立するために試験される。これは、siRNAの送達、投薬量、およびタイミングに依存する。siRNAは、当業者に周知の技術であるリポソームトランスフェクションによって細胞に送達される。トランスフェクション効率は使用される細胞型に依存して変化し得るので、いくつかのカチオン性リポソーム試薬(表5)が利用される。トランスフェクション効率および毒性は、蛍光標識したsiRNA(FL Luciferase GL2 Duplex)およびヨウ化プロピジウムをそれぞれ使用して、FACSによってモニターされる。カチオン性脂質複合体中に細胞マーカー特異的抗体を含むイムノリポソームは、PAD IVを産生するマクロファージまたは好中球のいずれかへのsiRNAの効率的な標的化を可能にするはずである(Zhang Yら(2003) In vivo knockdown of gene expression in brain cancer with intravenous RNAi in adult rats. J Gene Med. 5(12):1039-45: Bestman-Smith J.ら(2000) Sterically stabilized liposomes bearing anti-HLA-DR antibodies for targeting the primary cellular reservoirs of HIV-1. Biochim Biophys Acta. 1468(1-2):161-74)。
【0128】
(d)PAD(IIおよびIV)発現のモニタリング
未処理細胞対siRNA処理細胞でのPAD IIまたはIV mRNAおよびタンパク質発現は、RT-PCRおよびウェスタンブロッティングによってそれぞれモニターした。これらの方法の組み合わせ使用は、siRNA媒介阻害を分析するために最も良好である。PCRにおいて使用するオリゴヌクレオチドプライマーを表6および表7に列挙する。これらには、PAD IVに加えて、GAPDH、PAD I、II、およびIIIのためのプライマーが含まれる。なぜなら、ハウスキーピング遺伝子およびすべてのPADの遺伝子発現は、siRNA治療のための標的特異性を保証するからである。ヒトタンパク質とマウスタンパク質の両方を認識するポリクローナル抗PAD IV抗体は、標準として使用するために組換えPAD IVと同様に入手する。
【0129】
(実施例8)
siRNA治療の送達を実証するため;siRNA治療のための動物モデルを選択するため;およびPAD IV阻害の効果をモニターするための、siRNAを使用するPADの阻害、インビボ研究
(a)siRNA治療の送達
最も効果的かつ特異的なPAD IIおよびIVの阻害剤としてインビトロで同定されたリポソーム/イムノリポソームsiRNA複合体が静脈内注射される。このアプローチは、肝臓、肺、腎臓、脾臓、および膵臓を含む多数の器官における遺伝子発現を効果的に阻害することが以前に示された(Lewis D.L.ら、(2002) Efficient delivery of siRNA for inhibition of gene expression in postnatal mice. Nat Genet. 32(1): 107-8; Song E.ら、(2003) RNA interference targeting Fas protects mice from fulminant hepatitis. Nat Med. 9(3): 347-51)。プラスミドにコードされたsiRNAのレトロウイルス送達もまた、代替的なアプローチとして使用できる(McCaffrey A.P.ら、(2002) RNA interference in adult mice. Nature 418(6893):38-9)。
【0130】
(b)siRNA治療のための動物モデルの選択
関節炎のいくつかのマウスモデルが使用される。第1のものは、連鎖球菌細胞(SCW)誘導性関節炎であり、これは、マウスの関節内でPAD IVの随伴性のアップレギュレーションを伴う、関節炎が24時間以内に誘導される急性関節炎モデルである(Vossenaar E. R.ら、(2003) Citrullination of synovial proteins in murine models of rheumatoid arthritis. Arthritis Rheum. 48(9): 2489-500)。送達および用量を含む、siRNA治療のための最適条件の決定を表8に示す。第2のモデル、コラーゲン誘導性関節炎は、免疫の35日後に発症する持続性の関節炎を伴い、慢性的である。これは、延長した時間の期間にわたってsiRNA効果のモニタリングを可能にする(表9)。SCWおよびCIAマウスモデルは、PAD IV活性のレベルの増加およびこれらのマウスの関節におけるシトルリン化タンパク質の存在を示すが、しかし、シトルリンに標的化される自己免疫応答は存在しない。SCW関節炎がSE tgマウス中で誘導される場合、シトルリンで免疫されたときにSE tgマウスが関節炎を発症するにつれて、シトルリンに対する自己免疫応答が起こる。これは、非SE tg野生型マウスにおいてSCWで通常予測される一過性の関節炎を延長および/または悪化させる(表10)。この効果は、PAD IV阻害によって弱められる。
【0131】
(c)siRNA治療の効果のモニタリング
siRNA/リポソーム処理マウスまたは未処理マウスは、siRNA治療の効力および期間を決定するために種々の時点で屠殺される。これは、標準的なスコア付けシステムを使用して関節炎の臨床的徴候をモニタリングすることによって評価される(Current Protocols in Immunology 15.5章、http://www.mrw2.interscience.wiley.com/cponline)。疾患の病理学的徴候およびシトルリン化のプロセスもまたモニターされる。切片化された関節は、炎症およびシトルリン化タンパク質、PAD IVタンパク質およびその転写物の存在について点数付けされる。抗シトルリン抗体の産生はまた、SE tgマウスモデルにおいて評価される。
【0132】
本発明の好ましい態様を本明細書中で詳細に記載してきたが、本発明の精神または添付の特許請求の範囲から逸脱することなく本発明を変化させ得ることは当業者によって理解されよう。
【0133】
【表1】

【0134】
【表2】
【0135】
ヒト血清試料におけるJED抗原とCCP抗原に対する抗体の間の反応性の相関を見ることができる。
【0136】
【表3】
【0137】
【表4】
【0138】
すべての細胞株は本発明者らの研究室で現在使用されているか、またはATCCに注文することができる。マクロファージまたは好中球の表現型への分化は、これらの細胞型に必要である(例えば、レチノイン酸、ビタミンD3、LPS)。
【0139】
【表5】
【0140】
【表6】
【0141】
【表7】
【0142】
【表8】
【0143】
【表9】
【0144】
【表10】
【0145】
【表11】
【図面の簡単な説明】
【0146】
本発明は、本明細書中に示される詳細な説明から、および添付の図面からより完全に理解されるようになる。これらは、例示の目的のみのために示され、本明細書の意図される範囲を限定するものではない。
【図1】図1A、1B、および1Cは、共有エピトープによって形成されたP4ポケットと相互作用する位置にアルギニン(P4R)、シトルリン(P4Cit)、またはアスパラギン酸(P4D)を含有するペプチドに対する、DR4-IE tgマウスにおけるT細胞免疫応答の分析を表すグラフである。グラフ1A DR4-IE tgでは、マウスは示されたペプチド用いて免疫され、かつ10日後に流入領域リンパ節細胞に種々の濃度の同じペプチドを用いてインビトロでチャレンジした。データは、試験された各ペプチドについて8匹のマウスの平均増殖応答±SEMを表す。図1Bにおいて、リコールT細胞応答のHLA-DR制限は、抗原(コントロール)なし、10μg/mlの免疫抗原(P4Dは左のパネル、P4Citは右のパネル)とともに、または免疫抗原(10μg/ml)および抗DR抗体(DR)の存在下で、流入領域リンパ節細胞をインキュベートすることによってインビトロで決定された。結果は、各免疫抗原について4匹のマウスの平均増殖応答±SDを表す。図1Cは、10μg/mlの免疫ペプチド(P4D、P4Cit、またはP4R)を用いるインビトロチャレンジに応答したIFN-γ産生を示す。サイトカイン産生はELISAによって決定され、試験された各ペプチドについて、4匹のマウスの平均抗原特異的IFN-γ産生±SDを表す。
【図2】図2A、2B、および2Cは、修飾されていないビメンチンペプチド(Vim 65-77)またはシトルリン含有ビメンチンペプチド(Vim R70Cit)に対するDR4-IE tgマウスにおけるT細胞免疫応答の分析を示すグラフである。図2Aにおいて、DR4-IE tgマウスは、示されたペプチドで免疫され、かつ10日後に流入領域リンパ節細胞に種々の濃度の同じペプチドを用いてインビトロでチャレンジした。データは、試験された各ペプチドについて8匹のマウスの平均増殖応答±SEMを表す。図2Bは、Vim R70Cit免疫マウス(左のパネル)およびVim 65-77免疫マウス(右のパネル)からの特異性およびT細胞リコール応答のHLA-DR制限を示す。流入領域リンパ節細胞は免疫ペプチド、免疫ペプチドおよび抗DR抗体(DR)、未修飾ビメンチンペプチド(左のパネルの65-77)、またはビメンチンペプチドを含むシトルリン(右のパネルのR70Cit)でチャレンジされた。インビトロチャレンジのために使用された濃度は10μg/mlであり、結果は各免疫抗原について4匹のマウスの平均刺激指数±SEMを表す。図2Cは、種々の濃度の免疫ペプチドを用いるインビトロチャレンジに応答したIFN-γ産生を示す(Vim R70CitまたはVim 65-77)。サイトカイン産生はELISAによって決定され、試験された各ペプチドについて、4匹のマウスの平均抗原特異的IFN-γ産生±SDを表す。
【図3】図3は、精製MHCクラスII分子についてVim 65-77とVim R70Citの相対的アフィニティーを示すグラフである。共有エピトープ陽性対立遺伝子(*0101、0401、*0404)および共有エピトープ陰性対立遺伝子(*0301、*0701、*0802、*1101、*1302)に対する結合アフィニティーが、本明細書に記載される実施例において説明されるように決定された。100nM未満のIC50値を有するペプチドは高アフィニティー結合因子であると見なされる。
【図4】図4は、酵素ペプチジルアルギニンデイミナーゼ(PAD)が、いかにしてアルギニンの正電荷を有するイミノ基をシトルリンの極性の荷電されていないカルボニル基に変換するかを示す。
【図5】図5A-5Cは、(5A)抗シトルリン抗体に陰性であるRA患者;(5B)抗シトルリン抗体に陽性であることのRA患者;(5C)他のリウマチ疾患を有する非RA患者におけるシトルリン化フィブリノーゲンペプチドに対するT細胞増殖応答を示す。シトルリン化フィブリノーゲンペプチドに対するT細胞増殖は、RA患者においてのみ見られ、抗シトルリン抗体を有した患者においては、より一般的であった。末梢血単核球は、T細胞培養について実施例4で記載されているようにFicoll-Hypaque遠心沈殿によってRAドナーおよび非RAドナーから単離され、かつ培養された。これらの実験において使用されたペプチドは以下の通りである:Fib a R143Cit = VLKCitKVIEK (配列番号6); Fib a R673Cit = IQQCitMDGSL (配列番号11); Fib a R720Cit = LTQCitGSVLR (配列番号12); Fib a R847Cit = VSFCitGADYS (配列番号15); Fib b R285Cit = IQNCitQDGSV (配列番号20); Fib a R178Cit = IKICitSCRGS (配列番号7).
【図6】図6A-Fは、SE tgマウスにおいてシトルリン化フィブリノーゲンによって誘導された関節炎の臨床的および病理学的な特徴を示す写真である。図6Aは、非免疫されていないマウスからの後足の関節炎の臨床的外観を示し、6Bはヒトフィブリノーゲンで免疫された関節炎マウスであり、そして、6Cはシトルリン化ヒトフィブリノーゲンで免疫された関節炎マウスである。図6Dおよび6Eは、6Cからマウスの脛骨足根骨関節の病理学を示すH&E切片であるのに対して、6Fは6Bににおいて示されるマウスからの切片である。
【図7】図7は、DR4-IE tgマウスにおける、抗シトルリン抗体の産生を示す。
【図8】図8は、ヒト血清中で市販キット(Quantilite(商標))を使用してペプチドCit-Cit-Y-Cit-G-Cit-Cit-S-Cit-Cit-S-Cit-CitとCCPの間の相関を示す。
【技術分野】
【0001】
本発明は、HLA-DR分子と呼ばれる共有エピトープを有するMHCクラス II分子に結合する新規な抗原性ペプチドに関する。より詳細には、本発明は、HLA-DR分子についてのアフィニティー増加を有し、かつ特定の自己免疫障害に関連するシトルリン化抗原性ペプチドに向けられる。このようなものとして、これらの新規なペプチドは共有された抗原性(慢性関節リウマチと多発性硬化症を含むそのような自己免疫性疾患)でMHCクラスII分子を有する対象における自己免疫異常の診断および治療の新規な方法のための基礎を提供する。
【背景技術】
【0002】
本願を通して、種々の参考文献は、本発明が関係する技術水準をより完全に記載するために括弧内に引用される。これらの参考文献の開示は本明細書によって参考として本開示に組み入れられる。
【0003】
RA(慢性関節リウマチ)は、滑膜炎ならびに軟骨および骨の分解をもたらし得るパンヌス形成によって特徴付けられる一般的な自己免疫障害である。この衰弱させるような状態は人口のおよそ1%に影響する。この疾患に対する遺伝的感受性は共有エピトープ(SE)として知られるアミノ酸モチーフを含み、かつHLA-DR分子として呼ばれるMHCクラスII分子と関連している(Gregersen, P.K., J. Silver, R.J. Winchester. 1987. The shared epitope hypothesis. An approach to understanding the molecular genetics of susceptibility to rheumatoid arthritis. Arthritis Rheum. 30:1205、Zhou, Z., H.A. Menard. 2002. Autoantigenic posttranslational modifications of proteins: does it apply to rheumatoid arthritis? Curr. Opin. Rheumatol. 14:250)。アミノ酸残基Q/R、K/R、R、A, Aによって表現される、共有エピトープは、正に荷電し、かつMHCクラスII分子の主要なペプチドアンカーリングポケット(P4として知られる)の1つを形成する。
【0004】
以前の報告は、RAに関わる推定上の病原性のペプチドに関する目立った特徴は、P4(共有エピトープと相互作用する)の負に荷電した側鎖の存在であるかもしれないと示唆した(Hammer J., Gallazzi F., Bono E., Karr R.W., Guenot J., Valsasnini P., Nagy Z.A., Sinigaglia F. 1995. Peptide binding specificity of HLA-DR4 molecules: correlation with rheumatoid arthritis association. J. Exp. Med. 181:1847)。
【0005】
これは、特定のMHCクラスII分子、HLA-DR*0401、およびHLA-DR*0404(対立遺伝子多型)が、RAに関連しないHLA DR*0402分子よりも、P4ポケットにおけるアスパラギン酸およびグルタミン酸のアミノ酸残基について実質的により高いアフィニティーを有するという事実に基づいている。しかし、複数のDRB1ポケットプロフィールの分析の後で、いくつかのRA非関連対立遺伝子は、それらのP4ポケットにおける負荷電アミノ酸について、HLA-DR*0101、HLA-DR*0401、HLA-DR*0404、例えばHLA-DR*0301などでさえよりも高いアフィニティーを有することを見い出すことができる(Sturniolo T, Bono E, Ding J, Raddrizzani L, Tuereci O, Sahin U, Braxenthaler M, Gallazzi F, Protti MP, Sinigaglia F, Hammer J. 1999. Generation of tissue-specific and promiscuous HLA ligand databases using DNA microarrays and virtual HLA class II matrices. Nat. Biotechnol. 17:555)。いくつかのMHC分子は、単に非関連であるよりむしろ、疾患に対して保護的であるように見え(例えば、HLA-DR *0402)、このことは、ペプチド結合におけるこれらの対立遺伝子についての受動的な役割は存在しないかもしれないことを示唆する(Reviron, D., A. Perdriger, E. Toussirot, D. Wendling, N. Balandraud, S. Guis, G. Semana, P. Tiberghien, P. Mercier, J. Roudier. 2001. Influence of shared epitope-negative HLA-DRB1 alleles on genetic susceptibility to rheumatoid arthritis. Arthritis Rheum. 44:535)。その代わりに、保護的な対立遺伝子(MHC分子)は、ネガティブなT細胞選択を引き起こすか、または末梢寛容を確率するために十分に高いアフィニティーを有する推定の病原性ペプチドを結合することができる。
【0006】
共有エピトープを有するMHCクラスII分子は、自己反応性CD4+T細胞への提示のために関節炎性ペプチドを選択的に結合することによって疾患の病原性に関与するかもしれない。現在、RAについて原因となる自己抗原の性質は知られていない。多くの自己抗原候補がRAの関連MHCの文脈で調査されたが、CD4+T細胞およびB細胞免疫応答の共通の疾患特異的標的はとらえどころがないままで残っている。最近の研究は、RA患者がRAの診断について敏感かつ特異的(>90%)の両方であるIgG自己抗体のサブセットを有することを示した。これらの自己抗体の標的はシトルリン(翻訳後に修飾されたアルギニン(脱イミノ化アルギニン))であり、これは、特定のタンパク質/ペプチド配列の文脈の中で見い出された(van Venrooij, W. J., G. J. Pruijn. 2000. Citrullination: a small change for a protein with great consequences for rheumatoid arthritis. Arthritis Res. 2:249)。シトルリンは、抗核周囲抗体、抗ケラチン抗体、抗フィラグリン抗体、抗環状シトルリン化ペプチド抗体、および抗Sa抗体の必須の抗原性エピトープ標的である(van Venrooij, W. J., G. J. Pruijn. 2000. Citrullination: a small change for a protein with great consequences for rheumatoid arthritis. Arthritis Res. 2:249; Zhou, Z., H.A. Menard. 2002. Autoantigenic posttranslational modifications of proteins: does it apply to rheumatoid arthritis? Curr. Opin. Rheumatol. 14:250)。これらの抗体は、多くの異なったタンパク質の中でシトルリンを標的化し、結合誘導体化標的はビメンチンとフィブリン(フィブリノーゲン)であるように見える(Menard, H. A., E. Lapointe, M. D. Rochdi, Z. J. Zhou. 2000. Insights into rheumatoid arthritis derived from the Sa immune system. Arthritis Res. 2:429; Christine Masson-Bessiereら、2001. The Major Synovial Targets of the Rheumatoid arthritis-Specific Antifilaggrin Autoantibodies Are Deiminated Forms of the ( and ( Chains of Fibrin 1 The Journal of Immunology 166: 4177-4184)。抗シトルリン抗体生産が、RA患者におけるMHC共有エピトープの存在に有意に関連していることもまた、観察されている(Goldbach-Mansky R, Lee J, McCoy A, Hoxworth J, Yarboro C, Smolen JS, Steiner G, Rosen A, Zhang C, Menard HA, Zhou ZJ, Palosuo T, Van Venrooij WJ, Wilder RL, Klippel JH, Schumacher HR Jr, El-Gabalawy HS. 2000. Rheumatoid arthritis associated autoantibodies in patients with synovitis of recent onset. Arthritis Res. 2:236; Bas S, Perneger TV, Mikhnevitch E, Seitz M, Tiercy JM, Roux-Lombard P, Guerne PA. 2000. Association of rheumatoid factors and anti-filaggrin antibodies with severity of erosions in rheumatoid arthritis. Rheumatology (Oxford).39:1082)。
【特許文献1】米国特許第5,824,315号
【特許文献2】米国特許第6,184,204号
【特許文献3】米国特許第5,869,279号
【特許文献4】米国特許第5,843,456号
【特許文献5】米国特許第5,869,270号
【特許文献6】米国特許第4,609,893号
【特許文献7】米国特許第4,713,325号
【特許文献8】米国特許第4,714,681号
【特許文献9】米国特許第4,716,111号
【特許文献10】米国特許第4,716,117号
【特許文献11】米国特許第4,720,459号
【特許文献12】米国特許第5,734,02号
【特許文献13】米国特許第6,106,840号
【特許文献14】米国特許第5,635,363号
【特許文献15】米国特許第6,211,342号
【特許文献16】米国特許出願公開第20020176864号
【特許文献17】米国特許出願公開第20020122818号
【特許文献18】国際特許公開WO0129058号
【特許文献19】国際特許公開WO9932619号
【特許文献20】カナダ国特許出願第2,388,441号
【特許文献21】米国特許第5,843,445号
【特許文献22】国際特許公開WO02/14870号
【特許文献23】米国特許第6,355,617号
【非特許文献1】Gregersen, P.K., J. Silver, R.J. Winchester. 1987. The shared epitope hypothesis. An approach to understanding the molecular genetics of susceptibility to rheumatoid arthritis. Arthritis Rheum. 30:1205
【非特許文献2】Zhou, Z., H.A. Menard. 2002. Autoantigenic posttranslational modifications of proteins: does it apply to rheumatoid arthritis? Curr. Opin. Rheumatol. 14:250
【非特許文献3】Hammer J., Gallazzi F., Bono E., Karr R.W., Guenot J., Valsasnini P., Nagy Z.A., Sinigaglia F. 1995. Peptide binding specificity of HLA-DR4 molecules: correlation with rheumatoid arthritis association. J. Exp. Med. 181:1847
【非特許文献4】Sturniolo T, Bono E, Ding J, Raddrizzani L, Tuereci O, Sahin U, Braxenthaler M, Gallazzi F, Protti MP, Sinigaglia F, Hammer J. 1999. Generation of tissue-specific and promiscuous HLA ligand databases using DNA microarrays and virtual HLA class II matrices. Nat. Biotechnol. 17:555
【非特許文献5】Reviron, D., A. Perdriger, E. Toussirot, D. Wendling, N. Balandraud, S. Guis, G. Semana, P. Tiberghien, P. Mercier, J. Roudier. 2001. Influence of shared epitope-negative HLA-DRB1 alleles on genetic susceptibility to rheumatoid arthritis. Arthritis Rheum. 44:535
【非特許文献6】van Venrooij, W. J., G. J. Pruijn. 2000. Citrullination: a small change for a protein with great consequences for rheumatoid arthritis. Arthritis Res. 2:249
【非特許文献7】Zhou, Z., H.A. Menard. 2002. Autoantigenic posttranslational modifications of proteins: does it apply to rheumatoid arthritis? Curr. Opin. Rheumatol. 14:250
【非特許文献8】Menard, H. A., E. Lapointe, M. D. Rochdi, Z. J. Zhou. 2000. Insights into rheumatoid arthritis derived from the Sa immune system. Arthritis Res. 2:429
【非特許文献9】Christine Masson-Bessiereら、2001. The Major Synovial Targets of the Rheumatoid arthritis-Specific Antifilaggrin Autoantibodies Are Deiminated Forms of the ( and ( Chains of Fibrin 1 The Journal of Immunology 166: 4177-4184
【非特許文献10】Goldbach-Mansky R, Lee J, McCoy A, Hoxworth J, Yarboro C, Smolen JS, Steiner G, Rosen A, Zhang C, Menard HA, Zhou ZJ, Palosuo T, Van Venrooij WJ, Wilder RL, Klippel JH, Schumacher HR Jr, El-Gabalawy HS. 2000. Rheumatoid arthritis associated autoantibodies in patients with synovitis of recent onset. Arthritis Res. 2:236
【非特許文献11】Bas S, Perneger TV, Mikhnevitch E, Seitz M, Tiercy JM, Roux-Lombard P, Guerne PA. 2000. Association of rheumatoid factors and anti-filaggrin antibodies with severity of erosions in rheumatoid arthritis. Rheumatology (Oxford).39:1082
【非特許文献12】Dessen A, Lawrence CM, Cupo S, Zaller DM, Wiley DC. 1997. X-ray crystal structure of HLA-DR4 (DRA*0101, DRB1*0401) complexed with a peptide from human collagen II. Immunity. 7:473
【非特許文献13】Stern LJ, Brown JH, Jardetzky TS, Gorga JC, Urban RG, Strominger JL, Wiley DC.1994. Crystal structure of the human class II MHC protein HLA-DR1 complexed with an influenza virus peptide. Nature. 368:215
【非特許文献14】Hammer J, Bono E, Gallazzi F, Belunis C, Nagy Z, Sinigaglia F. 1994. Precise prediction of major histocompatibility complex class II-peptide interaction based on peptide side chain scanning. J. Exp. Med. 180:2353
【非特許文献15】Peterson DA, DiPaolo RJ, Kanagawa O, Unanue ER. 1999. Cutting edge: negative selection of immature thymocytes by a few peptide-MHC complexes: differential sensitivity of immature and mature T cells. J. Immunol. 162:3117
【非特許文献16】DiPaolo RJ, Unanue ER. 2001. The level of peptide-MHC complex determines the susceptibility to autoimmune diabetes: studies in HEL transgenic mice. Eur. J. Immunol. 31:3453
【非特許文献17】Yagi J, Janeway CA Jr. 1990. Ligand thresholds at different stages of T cell development. Int. Immunol. 2:83
【非特許文献18】Ghosh P, Amaya M, Mellins E, Wiley DC. 1995. The structure of an intermediate in class II MHC maturation: CLIP bound to HLA-DR3. Nature. 378:457
【非特許文献19】Jordan MS, Boesteanu A, Reed AJ, Petrone AL, Holenbeck AE, Lerman MA, Naji A, Caton AJ. 2001. Thymic selection of CD4+CD25+ regulatory T cells induced by an agonist self-peptide. Nat. Immunol. 2:301
【非特許文献20】Hagiwara T.ら、 Biochem & Biophy Res Comm. (2000) 290, 979-983
【非特許文献21】A. F. Spatola, in Chemistry and Biochemistry of Amino Acids, Peptides and Proteins, B. Weinstein, eds., Marcel Dekker, New York, p. 267 (1983)
【非特許文献22】Merrifield (1964) J. Am. Chem. Assoc. 65:2149
【非特許文献23】J. Amer. Chem. Soc. 85:2149 (1963)
【非特許文献24】Int. J. Peptide Protein Res. 35:161-214 (1990)
【非特許文献25】Methods of Organic Chemistry, E. Wansch (編) 第15巻, I部およびII部, Thieme, Stuttgart (1987)
【非特許文献26】Goeddel, Gene Expression Technology: Methods in Enzymology 185, Academic Press, San Diego, CA (1990)
【非特許文献27】Hugues, Sら(2002) Generation and use of alternative multimers of peptide/MHC complexes. Journal of Immunological Methods. 268:83-91
【非特許文献28】「Handbook of Pharmaceutical Additives」 (MichaelおよびIrene Ash,編 Gower Publishing Limited, Aldershot, England (1995)
【非特許文献29】「Vaccine Design-the Subunit and Adjuvant Approach」(PowellおよびNewman編, 'Plenum Press, New York, U.S.A., pp. 61-79および141-228 (1995)
【非特許文献30】BanchereauおよびSteinman, Nature 392:245-251, 1998
【非特許文献31】Zitvogelら、Nature Med. 4:594-600, 1998
【非特許文献32】Antibodies, A Laboratory Manual, Ed. Harlow および David Lane (編) Cold Spring Harbor Laboratory, N.Y. (1988)
【非特許文献33】Klein, J. Immunology: The Science of Cell-Noncell Discrimination, John Wiley & Sons, N.Y. (1982)
【非特許文献34】Dennett, R.ら Monoclonal Antibodies, Hybridoma: A New Dimension In Biological Analyses Plenum Press, N.Y. (1980)
【非特許文献35】Campbell, A.「Monoclonal Antibody Technology」Laboratory Techniques In Biochemistry And Molecular Biology, Vol. 13, Burdon.ら (編), Elsevier, Amsterdam (1984)
【非特許文献36】Baeten, D., Steenbakkers PGA., Rovers E., Veys EM., Rijnders AMW., Meijerink J., Keyser F DeおよびBoots A. Localisation of MHC Class II/HC gp-39 complexes in synovia of rheumatoid arthritis patients using complex-specific monoclonal antibodies. Abstracts of the 23rd European Workshop for Rheumatology Research, 第5巻 補遺1, 2003年2月
【非特許文献37】Baeten D., Steenbakkers P., Boots A., Veys EM., and Keyser, Fde., The presentation of the immunodominant epitope of HC gp-39 in the context of the RA-associated HLA class II molecules is specific for RA synovium. Abstracts of the 23rd European Workshop for Rheumatology Research, 第5巻 補遺1, 2003年2月)
【非特許文献38】ELISA Guidebook. Theory & Practice. Editor John R. Crowther, Author John R. Crowther, Pub. Humana Press 2000
【非特許文献39】Fire A.ら、1998 Nature 391:806-811
【非特許文献40】Fire, A. Trends Genet. 15, 358-363 (1999)
【非特許文献41】Sharp, P. A. RNA interference 2001. Genes Dev. 15, 485-490 (2001)
【非特許文献42】Hammond, S. M.ら、Nature Rev. Genet. 2, 110-1119 (2001)
【非特許文献43】Tuschl, T. Chem. Biochem. 2, 239-245 (2001)
【非特許文献44】Hamilton, A.ら、Science 286, 950-952 (1999)
【非特許文献45】Hammond, S. M.ら、Nature 404, 293-296 (2000)
【非特許文献46】Zamore, P. D.、ら、Cell 101, 25-33 (2000)
【非特許文献47】Bernstein, E.ら、Nature 409, 363-366 (2001)
【非特許文献48】Elbashir, S. M.ら、Genes Dev. 15, 188-200 (2001)
【非特許文献49】Elbashir S Mら、2001 Nature 411:494-498)
【非特許文献50】Tranquillら、2000. Enhanced T cell responsiveness to citrulline-containing myelin basic protein in multiple sclerosis patients. Mult. Scler. 6:4220-5
【非特許文献51】Nicholas A.P. 2002. Preparation of a monoclonal antibody to citrullinated epitopes: its characterization and some applications to immunohistochemistry in human brain. Glia.Mar 15;37(4):328-36
【非特許文献52】Nicholasら、2003. Immunohistochemical localization of citrullinated proteins in adult rat brain. J Comp Neurol. May 5:459(3):251-66
【非特許文献53】Vossenaar E.R.ら、(2003). PAD, a growing family of citrullinating enzymes; genes, features and involvement in disease. Bioessays. 25(11): 1106-18
【非特許文献54】Waltersら、Designing Screens: How to Make Your Hits a Hit. Nature Reviews, Drug Discovery. 第2巻, 2003年4月、259-266頁
【非特許文献55】Ito K, Bian HJ, Molina M, Han J, Magram J, Saar E, Belunis C, Bolin DR, Arceo R, Campbell R, Falcioni F, Vidovic D, Hammer J, Nagy ZA. 1996. HLA-DR4-IE chimeric class II transgenic, murine class II-deficient mice are susceptible to experimental allergic encephalomyelitis. J. Exp. Med. 183:2635
【非特許文献56】Hill, J.A., Wang, D., Jevnikar, A.M., Cairns, E., Bell, D.A. 2002. The relationship between predicted peptide-MHC class II affinity and T cell activation in a HLA-DR(1*0401 MHC class II mouse model. Arthritis Res. 5:R40
【非特許文献57】Andersson EC, Hansen BE, Jacobsen H, Madsen LS, Andersen CB, Engberg J, Rothbard JB, McDevitt GS, Malmstrom V, Holmdahl R, Svejgaard A, Fugger L. 1998. Definition of MHC and T cell receptor contacts in the HLA-DR4restricted immunodominant epitope in type II collagen and characterization of collagen-induced arthritis in HLA-DR4 and human CD4 transgenic mice. Proc. Natl. Acad. Sci. USA. 95:7574
【非特許文献58】Southwood S, Sidney J, Kondo A, del Guercio MF, Appella E, Hoffman S, Kubo RT, Chesnut RW, Grey HM, Sette A. 1998. Several common HLA-DR types share largely overlapping peptide binding repertoires. J. Immunol. 160:3363
【非特許文献59】Schllekensら、1998. J Clin Invest. Volume 101, 第1巻, 1998年1月, 273-281頁
【非特許文献60】Asaga Hら、(1998) Selective deimination of vimentin in calcium ionophore-induced apoptosis of mouse peritoneal macrophages. Biochem Biophys Res Commun. 243(3):641-6
【非特許文献61】Nakashima Kら、(1999) Molecular characterization of peptidylarginine deiminase in HL-60 cells induced by retinoic acid and 1alpha,25-dihydroxyvitamin D3. J Biol Chem. 274(39), 27786-92
【非特許文献62】Zhang Yら(2003) In vivo knockdown of gene expression in brain cancer with intravenous RNAi in adult rats. J Gene Med. 5(12):1039-45
【非特許文献63】Bestman-Smith J.ら(2000) Sterically stabilized liposomes bearing anti-HLA-DR antibodies for targeting the primary cellular reservoirs of HIV-1. Biochim Biophys Acta. 1468(1-2):161-74
【非特許文献64】Lewis D.L.ら、(2002) Efficient delivery of siRNA for inhibition of gene expression in postnatal mice. Nat Genet. 32(1): 107-8
【非特許文献65】Song E.ら、(2003) RNA interference targeting Fas protects mice from fulminant hepatitis. Nat Med. 9(3): 347-51
【非特許文献66】McCaffrey A.P.ら、(2002) RNA interference in adult mice. Nature 418(6893):38-9
【非特許文献67】Vossenaar E. R.ら、(2003) Citrullination of synovial proteins in murine models of rheumatoid arthritis. Arthritis Rheum. 48(9): 2489-500
【非特許文献68】Current Protocols in Immunology 15.5章、http://www.mrw2.interscience.wiley.com/cponline)
【発明の開示】
【発明が解決しようとする課題】
【0007】
出願人はここで、独特な相互作用がMHCクラスII分子の共有エピトープとアミノ酸シトルリンの間に存在することを実証した。この相互作用は、患者が共有エピトープを発現する自己免疫障害におけるT細胞応答およびそれに続いてこれらのシトルリン化抗原に対するB細胞応答を発生させることに関与する。さらに、出願人はここで、炎症および自己免疫障害(慢性関節リウマチならびに多発性硬化症など)の発症をもたらすT細胞応答を誘発する新規なシトルリン化抗原を同定した。
【課題を解決するための手段】
【0008】
出願人は、MHCクラスII分子の正に荷電しているP4ポケット(共有エピトープ)に対する結合を介してT細胞応答を誘発する新規なシトルリン化ペプチド抗原を同定した。出願人は、シトルリンのアミノ酸(非荷電極性アミノ酸)への正電荷を有するアミノ酸の修飾が、共有エピトープを有するMHCクラスII分子に対する抗原についてアフィニティーの増加をもたらすことを実証した。この増加した抗原アフィニティーはT細胞活性化をもたらし、これは次には、対象における慢性関節リウマチなどの自己免疫障害の発症をもたらす炎症反応を媒介する。
【0009】
このようなものとして、本発明は、T細胞活性化をもたらす共有エピトープを有するMHCクラスII分子とのシトルリン化ペプチドの結合によって特徴付けられる、自己免疫障害を診断するか、または処理するための種々の診断的および治療的なアプローチにおける本発明のシトルリン化ペプチドの使用を含む。本発明の態様において、そのような自己免疫障害は、例えば、慢性関節リウマチ(RA)および多発性硬化症(MS)を含むことができる。
【0010】
シトルリン含有ペプチド(シトルリンに対するアルギニンの変換を介して、荷電したイミノ側鎖基を非荷電カルボニルに置換するプロセス)に対するT細胞応答は、HLA-DRB1*0401トランスジェニック(DR4-IE tg)マウスで研究された。この研究は、共有エピトープと相互作用するペプチド側鎖位置のシトルリンへのアルギニンの変換が、ペプチドMHCアフィニティーを有意に増加させ、かつDR4-IE tgマウスにおいてCD4+T細胞の活性化をもたらすることを実証した。この翻訳後修飾は、DR4-IE tgマウスにおけるこれらのペプチドに対するCD4+T細胞応答を誘発するために必要であった。多くのHLA対立遺伝子についてのペプチドアフィニティーが評価され、これは、シトルリン含有ペプチドについて、共有エピトープを有するMHCクラスII分子だけがアフィニティー増加を有することを示した。
【0011】
まとめると、出願人は、RAにおける疾患関連HLA対立遺伝子の分子的基礎を説明するのを補助する、より詳細に述べると、共有エピトープを有するMHCクラスII分子がRA患者のシトルリン化自己抗原に対する自己免疫応答を開始することに関与することを明らかにする新規なペプチドMHC相互作用を同定した。シトルリン化ミエリン塩基性蛋白質(MBP)T細胞応答が以前に観察されたように、本発明はまた、MHCクラスII共有エピトープを発現する対象について、多発性硬化症(MS)についての診断的および治療的な処置を含む。
【発明を実施するための最良の形態】
【0012】
本発明は、共有エピトープを有するすべてのMHCクラスII分子を含み、その結果、シトルリンとのペプチドアフィニティーの増加が、アルギニンに対して存在している。このようなMHC分子には、HLA-DRB1*0401、HLA-DRB1*0404、HLA-DRB1*0405、HLA-DRB1*0408、HLA-DRB1*0409、HLA-DRB1*0410、HLA-DRB1*1001、HLA-DRB1*0101、HLA-DRB1*0102、HLA-DRB1*1402、HLA-DRB1*1406、HLA-DRB1*1409、及びHLA-DRB4を含むことができるがこれらに限定されない。
【0013】
シトルリン化ペプチドが、T細胞活性化および結果として生じる疾患をもたらす関節炎性抗原として作用するという知見により、T細胞活性化および自己免疫応答(例えば、RAおよびMSなどにおけるもの)をもたらす任意のMHCクラスII分子の共有エピトープに結合するシトルリン化ペプチド抗原の結合によって特徴付けられる自己免疫障害の診断のための新規かつ改善された方法(この活性化メカニズムを含む)が現在利用可能にされ、ならびにこのような自己免疫障害(RAおよびMSを含む)を発症する素因が現在利用可能にされる。
【0014】
上記に特徴付けられる自己免疫障害を処置するための新規な治療方法(RAおよびMSを含む)もまた、現在利用可能にされる。自己免疫障害の治療のためのこのような新規なアプローチは、いくつかの態様において、シトルリンへのアルギニンの変換をブロックすることを含むことができ、従って、RAまたはMSの原因となるシトルリン化ペプチド抗原の産生を阻害/減少抑制する。このようなブロッキングまたは阻害は、1つの態様において、ペプチジルアルギニンデイミナーゼ(PAD)酵素の発現、および従って、シトルリン化ペプチド抗原の生成をブロックすることを含むことができる。これはより初期の段階で疾患の進行を潜在的にブロックすることができるRAとMSのためのより費用対効果に優れた新規な治療を提供する。他の態様において、PAD酵素のブロッキングはPAD酵素の作用に対する低分子阻害の使用を含むことができる。
【0015】
本発明の態様によるものは、共有エピトープを有するMHCクラスII分子に高アフィニティーで結合する新規なシトルリン化ペプチドであり、前記ペプチドは、自己免疫障害を有する患者または自己免疫障害を発症するリスクがある患者の血液中でT細胞応答を誘発する。本発明の態様において、T細胞はCD4+T細胞である。本発明のさらなる態様において、自己免疫障害は、RAまたはMSであり得る。
【0016】
本発明の他の態様によるものは、共有エピトープを有するMHCクラスII分子に高アフィニティーで結合する新規なシトルリン化ペプチドであり、前記ペプチドは、RAを有する患者またはRAを発症するリスクがある患者の血液中でT細胞応答を誘発する。
【0017】
本発明のさらなる態様によるものは、共有エピトープを有するMHCクラスII分子に高アフィニティーで結合する新規なシトルリン化ペプチドであり、前記ペプチドは、RAを有する患者またはRAを発症するリスクがある患者の血液中でT細胞応答を誘発し、ここで、前記のT細胞はCD4+T細胞である。
【0018】
本発明のなおさらなる態様によるものは、新規なシトルリン化ペプチドであり、前記ペプチドは、内因性または外因性タンパク質に対するペプチジルアルギニンデイミナーゼの作用によって産生され、ここで前記シトルリン化ペプチドは共有エピトープを有するMHCクラスII分子に結合する。
【0019】
本発明のさらに別の側面は、共有エピトープを有するMHCクラスII分子に高アフィニティーで結合するシトルリン化ペプチドであり、前記ペプチドは、RAを有する患者またはRAを発症するリスクがある患者の血液中でT細胞応答を誘発し、前記ペプチドは、シトルリン化フィブリノーゲンペプチドおよびシトルリン化ビメンチンペプチドからなる群から選択される。いくつかの態様において、本発明におけるフィブリノーゲンペプチドおよびビメンチンペプチド、ならびにシトルリン化の標的は、これらのタンパク質の各々についての公知のヒト配列から選択される。さらに、フィブリノーゲンヒトタンパク質配列は、アルファ/アルファ鎖前駆体タンパク質、ベータ鎖前駆体タンパク質、アルファA鎖前駆体タンパク質、ガンマ鎖前駆体タンパク質またはこれらの鎖の成熟型からさらに選択できる。他の態様において、シトルリン化ペプチドは、全長フィブリノーゲンタンパク質またはビメンチンタンパク質であり得る。本発明によって含まれるタンパク質配列は、当業者によって理解されるように、Genbank (www.ncbi.nlm.nih.gov/Genbank)および SWISS-PROT (www.expasy.org)において見い出せる。
【0020】
本発明の別の態様によるものは、脱イミノ化アルギニンを含むペプチドであり、このペプチドは少なくとも約9アミノ酸長からであり、このペプチドは共有エピトープを有するMHCクラスII分子に高アフィニティーで結合し、ここでこのペプチドは、RAを有する患者またはRAを発症するリスクがある患者の血液中でT細胞応答を誘発する。
【0021】
本発明のさらに別の態様によるものは、共有エピトープを有するMHCクラスII分子に高アフィニティーで結合するシトルリン化ペプチドであり、前記ペプチドは、RAを有する患者またはRAを発症するリスクがある患者の血液中でT細胞応答を誘発し、前記ペプチドは、SAVRACitSSVPGVR (配列番号1); FSMCitIVCLV (配列番号2); VVECitHQSAC (配列番号3); FTNCitINKLK (配列番号4); LRSCitIEVLK (配列番号5); VLKCitKVIEK (配列番号6); IKICitSCRGS (配列番号7); LPSCitDRQHL (配列番号8); FRHCitHPDEA (配列番号9); FPSCitGKSSS (配列番号10); IQQCitMDGSL (配列番号11); LTQCitGSVLR (配列番号12); YHFCitVGSEA (配列番号13); YDPCitNNSPY (配列番号14); VSFCitGADYS (配列番号15); YSLCitAVRMK (配列番号16); MKICitPLVTQ (配列番号17); YRACitPAKAA (配列番号18); WQKCitQKQVK (配列番号19); IQNCitQDGSV (配列番号20); WYNCitCHAAN (配列番号21); YSMCitKMSMK (配列番号22); MKICitPFFPQ (配列番号23); LHPCitNLILY (配列番号24); VATCitDNCCI (配列番号25); LDECitFGSYC (配列番号26); LKSCitIMLEE (配列番号27); FQKCitLDGSV (配列番号28); YALCitVELED (配列番号29); WNGCitTSTA (配列番号30); WKTCitWYSMK (配列番号31); YATCitSSAVR (配列番号32); VRLCitSSVPG (配列番号33); LNDCitFANYI (配列番号34); MLQCitEEAEN (配列番号35); LNLCitETNLD (配列番号36); VETCitDGQVI (配列番号37) およびこれらの機能的類似体からなる群から選択されるアミノ酸配列の少なくとも1つを含む。
【0022】
本発明のなおさらなる態様によるものは、共有エピトープを有するMHCクラスII分子に高アフィニティーで結合するシトルリン化ペプチドであり、前記ペプチドは、RAを有する患者またはRAを発症するリスクがある患者の血液中でT細胞応答を誘発し、前記ペプチドは、SAVRACitSSVPGVR (配列番号1); FSMCitIVCLV (配列番号2); VVECitHQSAC (配列番号3); FTNCitINKLK (配列番号4); LRSCitIEVLK (配列番号5); VLKCitKVIEK (配列番号6); IKICitSCRGS (配列番号7); LPSCitDRQHL (配列番号8); FRHCitHPDEA (配列番号9); FPSCitGKSSS (配列番号10); IQQCitMDGSL (配列番号11); LTQCitGSVLR (配列番号12); YHFCitVGSEA (配列番号13); YDPCitNNSPY (配列番号14); VSFCitGADYS (配列番号15); YSLCitAVRMK (配列番号16); MKICitPLVTQ (配列番号17); YRACitPAKAA (配列番号18); WQKCitQKQVK (配列番号19); IQNCitQDGSV (配列番号20); WYNCitCHAAN (配列番号21); YSMCitKMSMK (配列番号22); MKICitPFFPQ (配列番号23); LHPCitNLILY (配列番号24); VATCitDNCCI (配列番号25); LDECitFGSYC (配列番号26); LKSCitIMLEE (配列番号27); FQKCitLDGSV (配列番号28); YALCitVELED (配列番号29); WNGCitTSTA (配列番号30); WKTCitWYSMK (配列番号31); YATCitSSAVR (配列番号32); VRLCitSSVPG (配列番号33); LNDCitFANYI (配列番号34); MLQCitEEAEN (配列番号35); LNLCitETNLD (配列番号36); VETCitDGQVI (配列番号37) およびこれらの機能的類似体からなる群から選択される。
【0023】
本発明の他の態様によると、本発明のシトルリン化ペプチドは、複数のシトルリン残基を含み、かつ抗シトルリン抗体をインビボで生成する人工的な配列であってもよい。
【0024】
1つの態様は、共有エピトープを有するMHCクラスII分子に高アフィニティーで結合するシトルリン化ペプチドであり、前記ペプチドは、哺乳動物においてインビボで抗シトルリン抗体を生成し、前記ペプチドは以下の式を有する:
Cit-Cit-X-Cit-G-Cit-Cit-Z-Cit-Cit-B-Cit-Cit(配列番号38)ここでXはY、F、W、I、L、MおよびVから選択され、ZはA、D、I、N、P、S、T、Vから選択され、そしてBはA、G、H、Q、S、TおよびVから選択される。
【0025】
本発明の態様において、このペプチドはジスルフィド結合形成によって環状化するペプチドの1つの末端または両端に対する加えられたシステインを有することができる。本発明のさらなる側面において、ペプチドはCit-Cit-Y-Cit-G-Cit-Cit-S-Cit-Cit-S-Cit-Cit (配列番号39)を含む。なおさらなる態様において、シトルリン化ペプチドは配列番号38または39の配列の始め、および/または終端に荷電したアミノ酸を含むことができる。このような荷電したアミノ酸は、アルギニン、リジン、アスパラギン、アスパラギン酸、グルタミン酸、およびグルタミン酸から選択できる。別の態様において、このペプチドは、その免疫学的アフィニティーを増加するために環状化できる。
【0026】
本発明の別の態様によるものは、共有エピトープを有するMHCクラスII分子に高アフィニティーで結合するシトルリン化ミエリン塩基性タンパク質(MBP)ペプチドであり、前記ペプチドは、MSを有する患者またはMSを発症するリスクがある患者の血液中でT細胞応答を誘発し、前記ペプチドは、FLPCitHRDTG (配列番号40), VTPCitTPPPS (配列番号41); YGGCitASKYK (配列番号42);及びLGGCitDSRSG (配列番号42) からなる群から選択される。
【0027】
本発明の別の態様によるものは、共有エピトープを有するMHCクラスII分子に高アフィニティーで結合するシトルリン化グリア線維酸性タンパク質(GFAP)ペプチドであり、前記ペプチドは、MSを有する患者またはMSを発症するリスクがある患者の血液中でT細胞応答を誘発し、前記ペプチドは、MERCitRITSA (配列番号43), LPTCitVDFSL (配列番号44), LNDCitFASYI (配列番号45), LRLCitLDQLT (配列番号46), LQICitETSLD (配列番号47),及びVEMCitDGEVI (配列番号48) からなる群から選択される。
【0028】
本発明の別の態様によるものは、好中球中でシトルリン化されるペプチドであり、こういうものとして、診断的にまたは慢性関節リウマチの治療において使用され得、前記ペプチドは、ヌクレオホスミン/B23、ヒストンH2A、ヒストンH4、およびヒストンH3からなる群から選択され、ここで前記ペプチドは1つ以上のシトルリン残基を含む。
【0029】
本発明の別の態様によるものは、LSLRTVSLG (SEQ ID NO 49); VVLRLKCGS (配列番号50); MSGRGKQGG (配列番号51); MSGRGKQGG (配列番号52); IIPRHLQLA (配列番号53); LAIRNDEEL (配列番号54); LLGRVTIAQ (配列番号55); MSGRGKGGK (配列番号56); LARRGGVKR (配列番号57); VALREIRRY (配列番号58); LLIRKLPFQ (配列番号59);及びLARRIRGER (配列番号60) からなる群から選択されるシトルリン化ペプチドであり、ここで前記「R」アルギニン残基はPADの作用によってシトルリンに変換できる。このようなペプチドは、慢性感染リウマチの徴候としてのT細胞反応性を測定するためなどの種々のアッセイにおいて使用できる。
【0030】
本発明の別の態様によるものは、対象からの試料中で抗シトルリン抗体を検出するための方法であって、前記方法は以下の工程を含む:
対象からの試料を、配列番号40から48のいずれか1つのペプチドと接触させる工程;
前記試料中で、抗シトルリン抗体への前記ペプチドの結合を検出する工程。
【0031】
本発明のなお別の態様によるものは、対象からの試料中で抗シトルリン抗体を検出するための方法であって、前記方法は以下の工程を含む:
対象からの試料を、式Cit-Cit-X-Cit-G-Cit-Cit-Z-Cit-Cit-B-Cit-Citのペプチドと接触させ、ここでXはY、F、W、I、L、MおよびVから選択され、ZはA、D、I、N、P、S、T、Vから選択され、BはA、G、H、Q、S、TおよびVから選択される工程;ならびに
前記試料中で抗シトルリン抗体への前記ペプチドの結合を検出する工程。この方法は、RAの診断において有用である。いくつかの態様において、この試料は、対象からの末梢血試料である。
【0032】
本発明のさらに別の態様によるものは、対象における自己免疫障害を診断するための方法であって、前記方法は以下の工程を含む:
前記対象から採取された試料を、式Cit-Cit-X-Cit-G-Cit-Cit-Z-Cit-Cit-B-Cit-Citのペプチドと接触させ、ここでXはY、F、W、I、L、MおよびVから選択され、ZはA、D、I、N、P、S、T、Vから選択され、BはA、G、H、Q、S、TおよびVから選択される工程;ならびに
前記試料中で抗シトルリン抗体への前記ペプチドの結合を検出し、ここで結合が自己免疫障害を示す工程。いくつかの態様において、自己免疫障害は、RAおよびMSから選択できる。
【0033】
本発明の別の態様によるものは、対象においてMSを診断するための方法であって、前記方法は以下の工程を含む:
前記対象から採取された試料を、シトルリン化MBPペプチドと接触させる工程;および
前記試料中で抗シトルリン抗体への前記ペプチドの結合を検出し、ここで結合がMSの診断を示す工程。
【0034】
本発明のペプチドのいずれか、ならびに本発明によるペプチドのマルチマー(例えば、本発明によるペプチドのダイマーまたはトリマーなど)の機能的類似体もまた、本発明の範囲内にある。本発明によるマルチマーは、複数の同じペプチドからなるホモマー、または異なるペプチドからなるヘテロマーのいずれかであり得る。本発明によるペプチドの特徴的なアミノ酸配列は、ランダムなアミノ酸配列に隣接できる。好ましいものは、ペプチドに安定化効果を有する、従って、それらの生物学的利用可能性を増加する隣接配列である。
【0035】
上記に列挙した特定のペプチドアミノ酸配列が、シトルリンに変換できる配列中でさらなるアルギニンを有することが当業者によって理解される。
【0036】
本発明の別の態様によるものは、共有エピトープを有するMHCクラスII分子に高アフィニティーで結合し、かつRAを有する患者またはRAを発症するリスクがある患者の血液中でT細胞応答を誘発することが可能であるシトルリン化ペプチド、および薬学的に許容可能な担体を含む組成物である。このシトルリン化ペプチドは、SAVRACitSSVPGVR (配列番号1); FSMCitIVCLV (配列番号2); VVECitHQSAC (配列番号3); FTNCitINKLK (配列番号4); LRSCitIEVLK (配列番号5); VLKCitKVIEK (配列番号6); IKICitSCRGS (配列番号7); LPSCitDRQHL (配列番号8); FRHCitHPDEA (配列番号9); FPSCitGKSSS (配列番号10); IQQCitMDGSL (配列番号11); LTQCitGSVLR (配列番号12); YHFCitVGSEA (配列番号13); YDPCitNNSPY (配列番号14); VSFCitGADYS (配列番号15); YSLCitAVRMK (配列番号16); MKICitPLVTQ (配列番号17); YRACitPAKAA (配列番号18); WQKCitQKQVK (配列番号19); IQNCitQDGSV (配列番号20); WYNCitCHAAN (配列番号21); YSMCitKMSMK (配列番号22); MKICitPFFPQ (配列番号23); LHPCitNLILY (配列番号24); VATCitDNCCI (配列番号25); LDECitFGSYC (配列番号26); LKSCitIMLEE (配列番号27); FQKCitLDGSV (配列番号28); YALCitVELED (配列番号29); WNGCitTSTA (配列番号30); WKTCitWYSMK (配列番号31); YATCitSSAVR (配列番号32); VRLCitSSVPG (配列番号33); LNDCitFANYI (配列番号34); MLQCitEEAEN (配列番号35); LNLCitETNLD (配列番号36); VETCitDGQVI (配列番号37); Cit-Cit-X-Cit-G-Cit-Cit-Z-Cit-Cit-B-Cit-Cit(配列番号38、ここでXはY、F、W、I、L、MおよびVから選択され、ZはA、D、I、N、P、S、T、Vから選択され、BはA、G、H、Q、S、TおよびVから選択される)およびCit-Cit-Y-Cit-G-Cit-Cit-S-Cit-Cit-S-Cit-Cit(配列番号39)からなる群から選択される。
【0037】
本発明の別の態様によるものは、共有エピトープを有するMHCクラスII分子に高アフィニティーで結合し、かつMS患者またはMSを発症するリスクがある患者の血液中でT細胞応答を誘発することが可能であるシトルリン化ペプチド、および薬学的に許容可能な担体を含む組成物である。シトルリン化ペプチドは、配列番号40から48によって表されるペプチドのいずれか1つから選択できる。
【0038】
本発明はまた、RAもしくはMSなどの自己免疫障害、またはRAもしくはMSなどの自己免疫障害に対する感受性を診断する方法における使用のための診断用手段の調製のための本発明中に開示されるシトルリン化ペプチド抗原のいずれかの使用を提供し、この方法は、個体のT細胞が、MHCクラスII共有エピトープ陽性細胞に結合したときにシトルリン化ペプチド抗原を認識するか否かを決定する工程を包含し、ここで、T細胞による認識は、個体が自己免疫障害を有するか、または自己免疫障害に対して感受性であることを示す。
【0039】
本発明のさらに別の態様によるものは、MHCクラスII共有エピトープ陽性細胞に結合するシトルリン化抗原と反応性である自己反応性T細胞の検出のための診断方法であって、前記方法は以下の工程:
患者からの末梢血単核細胞を、1種以上のシトルリン化ペプチド抗原とともにインキュベートする工程;
前記患者における活性化された自己反応性T細胞の存在を示す、T細胞の応答を検出する工程。
【0040】
いくつかの態様において、ペプチド抗原は、配列番号1から60のいずれか1つから選択できる。
【0041】
本発明の別の態様によるものは、MHCクラスII共有エピトープ陽性細胞に結合するシトルリン化抗原と反応性である活性化された自己反応性T細胞の検出のための試験キットであり、前記試験キットは、本明細書中に開示されるような1種以上のシトルリン化ペプチドを含む。いくつかの態様において、ペプチド抗原は、配列番号1から37、配列番号38から39、配列番号40から48、および配列番号49から60のいずれか1つから選択できる。
【0042】
本発明はさらに、シトルリン化ペプチド抗原を認識する前記T細胞を脱感作することによって、RAまたはMSを治療または予防する方法における使用のための、任意に担体を伴うシトルリン化ペプチド抗原を提供する。
【0043】
本発明のなおさらなる別の態様によるものは、T細胞応答をもたらす共有エピトープを有するMHCクラスII分子にシトルリン化ペプチドを結合することによって誘発される、RAに罹患している対象を治療するための方法であって、前記方法は以下の工程を含む:
薬学的に許容可能な担体とともに、SAVRACitSSVPGVR (配列番号1); FSMCitIVCLV (配列番号2); VVECitHQSAC (配列番号3); FTNCitINKLK (配列番号4); LRSCitIEVLK (配列番号5); VLKCitKVIEK (配列番号6); IKICitSCRGS (配列番号7); LPSCitDRQHL (配列番号8); FRHCitHPDEA (配列番号9); FPSCitGKSSS (配列番号10); IQQCitMDGSL (配列番号11); LTQCitGSVLR (配列番号12); YHFCitVGSEA (配列番号13); YDPCitNNSPY (配列番号14); VSFCitGADYS (配列番号15); YSLCitAVRMK (配列番号16); MKICitPLVTQ (配列番号17); YRACitPAKAA (配列番号18); WQKCitQKQVK (配列番号19); IQNCitQDGSV (配列番号20); WYNCitCHAAN (配列番号21); YSMCitKMSMK (配列番号22); MKICitPFFPQ (配列番号23); LHPCitNLILY (配列番号24); VATCitDNCCI (配列番号25); LDECitFGSYC (配列番号26); LKSCitIMLEE (配列番号27); FQKCitLDGSV (配列番号28); YALCitVELED (配列番号29); WNGCitTSTA (配列番号30); WKTCitWYSMK (配列番号31); YATCitSSAVR (配列番号32); VRLCitSSVPG (配列番号33); LNDCitFANYI (配列番号34); MLQCitEEAEN (配列番号35); LNLCitETNLD (配列番号36);及びVETCitDGQVI (配列番号37) からなる群から選択されるアミノ酸配列を含むシトルリン化ペプチドを含む組成物のT細胞寛容誘導量を投与する工程。
【0044】
本発明のなおさらなる別の態様によるものは、T細胞応答をもたらす共有エピトープを有するMHCクラスII分子にシトルリン化ペプチドを結合することによって誘発される、MSに罹患している対象を治療するための方法であって、前記方法は以下の工程を含む: FLPCitHRDTG (配列番号40), VTPCitTPPPS (配列番号41); YGGCitASKYK (配列番号42), LGGCitDSRSG (配列番号42), MERCitRITSA (配列番号43), LPTCitVDFSL (配列番号44), LNDCitFASYI (配列番号45), LRLCitLDQLT (配列番号46), LQICitETSLD (配列番号47)及びVEMCitDGEVI (配列番号48) のシトルリン化ペプチドを含む組成物のT細胞寛容誘導量を投与する工程。
【0045】
本発明のなお別の態様によるものは、シトルリン化抗原MHCクラスII細胞複合体の検出のための診断方法であって、前記方法は以下の工程を含む:
対象からの末梢血単核細胞の単離された試料を1種以上のシトルリン化ペプチド抗原とインキュベートする工程;
シトルリン化ペプチド抗原MHCクラスII細胞複合体の形成を検出し、このような検出は前記対象におけるRAまたはMSをもたらすT細胞応答を誘発する可能性を示す工程。
【0046】
本発明の態様によるものは、対象におけるシトルリン化ペプチドMHCクラスII複合体によってT細胞の活性化を予防するための方法であって、前記方法は、前記複合体に標的化される抗体を投与する工程を含む。
【0047】
本発明の別の態様によるものは、対象における潜在的に抗原性のペプチド中でのアルギニンからシトルリンへの変換、従って、シトルリン化ペプチドMHCクラスII複合体の形成を予防するための方法であって、前記方法は、ペプチジルアルギニンデイミナーゼのアンタゴニストまたは阻害剤を前記対象に投与する工程を含む。
【0048】
本発明のなお別の態様によるものは、共有エピトープを有するMHCクラスII分子へのシトルリン化ペプチドの結合をブロックし得る薬学的化合物を同定するためのスクリーニング方法であり、この方法は以下の工程を含む:
候補薬学的化合物をトランスジェニックDR4-IE tgマウスに投与する工程;および
T細胞活性を測定する工程および/またはシトルリン化ペプチド/MHCクラスII分子複合体形成を測定する工程であって、ここで、減少したT細胞活性および/または減少した複合体形成は、前記候補薬学的化合物がMHCクラスII分子への前記シトルリン化ペプチドの結合に影響を与える工程
を含む方法。
【0049】
本発明のさらなる態様によるものは、PADの阻害剤を同定するためのスクリーニング方法であって、この方法は以下の工程を含む:
(a)非シトルリン化ペプチド、PAD、および阻害剤の混合物を供給する工程;
(b)(a)に抗原提示細胞を供給する工程;ならびに
(c)ペプチド-MHC複合体に特異的なT細胞系統に(b)を適用し、ここで、PAD阻害がT細胞反応性の欠如によって特徴付けられる工程。
【0050】
本発明のさらなる態様によるものは、慢性関節リウマチの研究用の動物モデルを提供するために動物において慢性関節リウマチを誘導する方法であり、この方法は、薬学的に許容可能な担体とともに、シトルリン化ペプチドを含む組成物の慢性関節リウマチ誘導量を動物に投与する工程を含む。投与は注射によって行われ得る。
【0051】
本発明の別の態様によるものは、多発性硬化症の研究用の動物モデルを提供するために動物において多発性硬化症を誘導する方法であり、この方法は、薬学的に許容可能な担体とともに、シトルリン化ペプチドを含む組成物の多発性硬化症誘導量を動物に投与する工程を含む。投与は注射によって行われ得る。いくつかの局面において、シトルリン化ペプチドは、シトルリン化されたミエリン塩基性タンパク質(MBP)またはグリア線維酸性タンパク質(GFAP)である。
【0052】
本発明のなお別の態様によるものは、共有エピトープを有するMHCクラスII分子に結合可能であるシトルリン化ペプチド抗原を作製するための方法であって、前記方法は以下の工程を含む:
(a)アルギニンをシトルリンに変換するために十分な時間の間、少なくとも1つのアルギニンアミノ酸残基を含む内因性または外因性可溶化タンパク質にペプチジルデイミナーゼを加える工程;および
(b)(a)からタンパク質を単離する工程。
【0053】
本発明のなお別の態様によるものは、哺乳動物におけるシトルリン化ペプチド/MHCクラスII複合体の結合および形成によって特徴付けられる自己免疫障害の治療のための方法であり、この方法は、前記哺乳動物におけるペプチジルデイミナーゼ(PAD)酵素の発現をブロックする工程を包含し、ここで、このようなブロッキングは、慢性関節リウマチを誘発するシトルリン化ペプチドの生成を減少させる。
【0054】
本発明のさらなる態様によるものは、哺乳動物における慢性関節リウマチの治療のための方法であり、この方法は、ペプチジルアルギニンデイミナーゼ(PAD)遺伝子配列に標的化されるsiRNAの投与によって、哺乳動物中のPADの発現を阻害する工程を含む。
【0055】
本発明のなおさらなる態様によるものは、哺乳動物における慢性関節リウマチまたは多発性硬化症の治療のための方法であり、この方法は、PADの機能をブロックする薬剤を前記哺乳動物に投与する工程を含む。
【0056】
本発明の他の特徴および利点は、以下の詳細な説明から明らかになる。しかし、本発明の精神および範囲の中での種々の変更および改変は詳細な説明から当業者に明らかになるので、前記詳細な説明および特定の実施例は本発明の実施形態を示しているが、例示のみのために提供されることが理解されるべきである。
【0057】
好ましい実施形態の詳細な説明
出願人は、T細胞の活性化および結果として、慢性関節リウマチの発症をもたらす、共有エピトープを有するMHCクラスII分子に特異的に結合する新規なシトルリン化抗原性ペプチドを開発した。共有エピトープ(P4)と相互作用するペプチド側鎖位置のアルギニンのシトルリンへの変換は、ペプチドMHCアフィニティーを有意に増加させ、そして、CD4+T細胞の活性化をもたらす。そのようなT細胞活性化はさらに滑膜炎および慢性関節リウマチの発症をもたらす。本発明のシトルリン化ペプチド抗原は、T細胞の活性化および炎症性応答の発症を生じる、共有エピトープ複合体とのシトルリン化ペプチド/MHCクラスIIの結合および形成によって特徴付けられる、種々の自己免疫障害に関与し得ることが当業者によって理解される。それゆえに、本発明は、このような病因論に関わる自己免疫障害のためのいくつかの適用を有する。
【0058】
HLA DRB1*0401トランスジェニック(DR4-IE tg)マウスを使用して、出願人はシトルリン含有ペプチドに対する増加するT細胞応答を実証した。MHCペプチドアフィニティーを予測するのにモデルを使用して、候補T細胞エピトープは、研究について開発および選択された。これにはビメンチン、フィブリノーゲン、およびプロテオグリカンアグレカンからのエピトープが含まれていた。プロテオグリカンアグレカンの場合では、通常、このタンパク質は、P4共有エピトープと相互作用する負に荷電したアスパラギン酸(D)を含む。フィブリノーゲンペプチドおよびビメンチンペプチドは正電荷を有するP4アンカリングポケット(共有エピトープ)にアルギニンまたはシトルリンのいずれかを配置する区域におけるDRB1*0401への結合の特性に基づいて選択された。アルギニンのシトルリンへの変換(荷電しているイミノ側鎖グループを荷電されていないカルボニルに置換するプロセス)は、DRB1*0401でのペプチドのアフィニティーを劇的に増加させた。さらに、この翻訳後修飾が、DR4-IE tgマウスにおけるこれらのペプチドに対するCD4+T細胞応答を誘発するために必要であった。多くのHLA対立遺伝子についてのペプチドアフィニティーが評価され、共有エピトープを有するMHCクラスII分子のみがシトルリン含有ペプチドについてのアフィニティー増加を有することを示した。これらの研究(これは新規なペプチドMHC相互作用を同定する)は、RAにおける疾患関連HLA対立遺伝子の分子的基礎を説明することを補助し、現在、慢性関節リウマチについての新しい診断的および治療的なストラテジーを提供する。RAと関連するMHCクラスII分子の第3の超可変領域は、DRβ鎖の位置70から74にわたるアミノ酸配列Q/R、K/R、R、A、Aを含む。この共有エピトープ領域は、P4として知られている主要なペプチドアンカリングポケットの1つを形成し、抗原性ペプチドから側鎖残基に直接的に接触することが可能である71位のKまたはRに起因して正電荷を有する(Dessen A, Lawrence CM, Cupo S, Zaller DM, Wiley DC. 1997. X-ray crystal structure of HLA-DR4 (DRA*0101, DRB1*0
401) complexed with a peptide from human collagen II. Immunity. 7:473; Stern LJ, Brown JH, Jardetzky TS, Gorga JC, Urban RG, Strominger JL, Wiley DC.1994. Crystal structure of the human class II MHC protein HLA-DR1 complexed with an influenza virus peptide. Nature. 368:215)。ペプチドMHCアフィニティーに関する以前の研究は、71位のKまたはRがこのP4ポケットで相互作用し得るアミノ酸の特徴を決定付けることを示した(Hammer J., Gallazzi F., Bono E., Karr R.W., Guenot J., Valsasnini P., Nagy Z.A., Sinigaglia F. 1995. Peptide binding specificity of HLA-DR4 molecules: correlation with rheumatoid arthritis association. J. Exp. Med. 181:1847)。一般に、共有エピトープを有するMHCは、負に荷電しているか、または荷電されていない極性アミノ酸について高アフィニティーを有するのに対して、正電荷を有するアミノ酸(すなわち、アルギニン)はペプチド結合を阻害する(Hammer J, Bono E, Gallazzi F, Belunis C, Nagy Z, Sinigaglia F. 1994. Precise prediction of major histocompatibility complex class II-peptide interaction based on peptide side chain scanning. J. Exp. Med. 180:2353; Hammer J., Gallazzi F., Bono E., Karr R.W., Guenot J., Valsasnini P., Nagy Z.A., Sinigaglia F. 1995. Peptide b
inding specificity of HLA-DR4 molecules: correlation with rheumatoid arthritis association. J. Exp. Med. 181:1847)。出願人は、酵素ペプチジルアルギニンデイミナーゼ(PAD)の作用を介する脱イミノ化によって(図4)、正に荷電しているアルギニンが、極性であるが荷電していないシトルリンに変換され(翻訳後修飾)、このことが、共有エピトープP4ポケットに対するアフィニティーを増加させることを実証した。MHCアンカリングポケットにおけるアミノ酸相互作用は、残基の電荷に依存するのみならず、サイズにもまた依存するので、出願人はまた、共有エピトープによって形成されたP4ポケットがシトルリンの側鎖を収容するために十分に大きいことを確認した。これは、DRB1*0401およびDRB1*0101の結晶構造を使用する分子モデリングによって確かめられた(データ示さず)。P4共有エピトープの荷電特性およびこのポケットのサイズに基づいて、ペプチド結合シトルリンは、HLA*0401およびHLA*0101のP4アンカリングポケットで好ましく相互作用することが予測された。
【0059】
DR4-IE tgマウスからCD4+T細胞を活性化することが出願人によって実証されたペプチド配列が選択および開発された。このペプチド(軟骨プロテオグリカンアグレカンから)は、通常、P4共有エピトープと相互作用する負に荷電したアスパラギン酸(D)を含んでいる(P4D)。2つのさらなるペプチドがこの配列に基づいて合成された:1つはアルギニンによって置換されたアスパラギン酸を有した(P4R);他方は、この位置で置換されたシトルリンを有した(P4Cit)。次いで、DR4-IE tgマウスがこれらのペプチドで免疫されて、T細胞応答が10日後に評価された。ペプチドP4DはIFN-γ産生に付随した強い増殖性応答を誘導した(図1Aおよび1B)。しかし、アルギニン置換を含むペプチド(P4R)は、これらのマウスにおいてT細胞増殖またはサイトカイン産生を誘導しなかった。P4Rでの応答の非存在と対照的に、P4CitはT細胞増殖およびIFN-γ産生を誘導した。P4CitがペプチドMHC提示を通してT細胞を活性化していることを確認するために、抗DR抗体が、ペプチドMHC複合体とのTCR相互作用を阻害するために使用された(図1B)。この処理は、P4CitおよびP4Dに対する増殖性応答を阻害した。RA患者における抗シトルリン抗体のタンパク質標的からの潜在的T細胞エピトープが同定された。このタンパク質に対する自己抗体が共有エピトープを発現する患者において頻繁に見い出されるので、ビメンチンが選択された(Goldbach-Mansky R, Lee J, McCoy A, Hoxworth J, Yarboro C, Smolen JS, Steiner G, Rosen A, Zhang C, Menard HA, Zhou ZJ, Palosuo T, Van Venrooij WJ, Wilder RL, Klippel JH, Schumacher HR Jr, El-Gabalawy HS. 2000. Rheumatoid arthritis associated autoantibodies in patients with synovitis of recent onset. Arthritis Res. 2:236)。ヒトビメンチンからの候補T細胞エピトープは、ペプチドMHCアフィニティーについて予測モデルを使用して同定された(Hammer J, Bono E, Gallazzi F, Belunis C, Nagy Z, Sinigaglia F. 1994. Precise prediction of major histocompatibility complex class II-peptide interaction based on peptide side chain scanning. J. Exp. Med. 180:2353)。このペプチドは、MHCアンカリングポケットP1、P6、およびP9との好都合な相互作用を有し、かつP4共有エピトープにおいてアルギニンを有する特性に基づいて選択された。2つのペプチドが統合され、1つは修飾されていないペプチドビメンチン65-77を含み、他方はビメンチンR70Citであり、これはアルギニンがシトルリン置換されていた。次いで、これらのペプチドに対するT細胞応答は、DR4-IE tgマウスを使用して特徴付けられた。しかし、予測されるように、修飾されていないペプチドVim 65-77がT細胞活性化を誘導せず、Vim R70CitがIFN-γ産生に付随した強い増殖性応答を刺激した(図2Aおよび2C)。Vim R70Citに対するT細胞応答はまた、抗DR抗体を使用して阻害され得、MHCクラスII制限された免疫応答を確認した(図2B)。Vim R70CitによってプライムされたT細胞が未修飾ペプチドによって活性化できなかったこともまた実証され、Vim65-77がDR4結合溝と産生的に相互作用しないことをさらに支持する(図2B)。
【0060】
アルギニンのシトルリンへの変換が、共有エピトープを含んだMHCクラスII分子についてのペプチドアフィニティーを増加させることができることを確認するために、ペプチド競争アッセイが、共有エピトープ陽性(DRB1*0101、*0401、*0404)、または共有エピトープ陰性(DRB1*0301、*0701、*0802、*1101、*1302)のいずれかであった精製MHCについてVim65-77およびVim R70Citの相対的アフィニティーを決定するために行われた。Vim65-77は、すべての試験されたMHCについて低いアフィニティーから中程度のアフィニティーまでを有したのに対して、Vim R70Citは、きわだって高いアフィニティーを伴って*0101、*0401、および*0404に結合した(図3)。未修飾ペプチドと比較して、シトルリン含有ペプチドは、*0101、*0401、および*0404に、それぞれ、100倍、90倍、および20倍高いアフィニティーで結合した。最も重要なことに、アルギニンのシトルリンへの変換は、試験されたいかなる共有エピトープ陰性のMHCについてもペプチドアフィニティーを増加しなかった。
【0061】
出願人の結果は、P4におけるアルギニンまたはシトルリンのいずれかに対するDRB1対立遺伝子のディファレンシャルな結合特性が、MHCクラスII分子が疾患に関連するか、関連しないか、または保護的であるかをいかにして説明するかを示した。まず最初に、ペプチド結合アルギニンのシトルリンへの変換は、共有エピトープを有するMHCについてアフィニティーの100倍の増大を引き起こす。これは、T細胞活性化のために必要な「生化学的安全域」を超えることができる抗原提示細胞に対してより高い密度ペプチドMHC複合体を生じ得る(Peterson DA, DiPaolo RJ, Kanagawa O, Unanue ER. 1999. Cutting edge: negative selection of immature thymocytes by a few peptide-MHC complexes: differential sensitivity of immature and mature T cells. J. Immunol. 162:3117; DiPaolo RJ, Unanue ER. 2001. The level of peptide-MHC complex determines the susceptibility to autoimmune diabetes: studies in HEL transgenic mice. Eur. J. Immunol. 31:3453; Yagi J, Janeway CA Jr. 1990. Ligand thresholds at different stages of T cell development. Int. Immunol. 2:83)。第2に、関連していないMHCクラスII分子(例えば、*0301)は、アルギニンまたはシトルリンの大きな極性側鎖を生産的に収容するための適切なサイズまたは電荷を欠くP4ポケットを含み得、それゆえに修飾の状態に関わらずペプチドを結合および提示することが不可能である(Ghosh P, Amaya M, Mellins E, Wiley DC. 1995. The structure of an intermediate in class II MHC maturation: CLIP bound to HLA-DR3. Nature. 378:457)。最後に、疾患の保護的なMHCは、ネガティブ選択を引き起こすことができるペプチドMHCリガンドを生じるP4でアルギニンとシトルリンの両方と生産的に相互作用ができ、CD4+CD25+調節性T細胞の産生をもたらすか(Jordan MS, Boesteanu A, Reed AJ, Petrone AL, Holenbeck AE, Lerman MA, Naji A, Caton AJ. 2001. Thymic selection of CD4+CD25+ regulatory T cells induced by an agonist self-peptide. Nat. Immunol. 2:301)、または単に「生化学的安全域」内に留まる(Peterson DA, DiPaolo RJ, Kanagawa O, Unanue ER. 1999. Cutting edge: negative selection of immature thymocytes by a few peptide-MHC complexes: differential sensitivity of immature and mature T cells. J. Immunol. 162:3117)。出願人はまた、抗シトルリン抗体について陽性のRA患者において、増加したT細胞増殖性応答がシトルリン化フィブリノーゲンペプチドに対してなお見られたことを実証した(図5A、B、C)。
【0062】
まとめると、本発明は、ある態様において、哺乳動物のフィブリノーゲンおよびビメンチンのアミノ酸配列に基づいており、他の態様において、人工のシトルリン化ペプチド配列に基づいている一連のペプチドを実証する。用語「ペプチド」は、本明細書中で使用される場合、通常、定義された配列を有するアミノ酸残基の鎖として定義される。本明細書中で使用される場合、用語ペプチドは、用語「ペプチド」および「タンパク質」と相互に包括的である。1つの態様において、フィブリノーゲンおよびビメンチンの配列は、ヒトアミノ酸配列である。しかし、フィブリノーゲンまたはビメンチンの配列は、ヒト、ウシ、ブタ、マウス、およびラットなどであるがこれらに限定されない任意の哺乳動物種由来でであり得ることは当業者によって理解される。
【0063】
本発明はまた、シトルリンに変換される少なくとも1つのアルギニンアミノ酸を有する、全長のフィブリノーゲンとビメンチンの両方のアミノ酸配列またはそのフラグメントを含む。任意の所望の長さの任意のフラグメントであることの限界は、共有エピトープを有する任意のMHCクラスII分子に対して高いアフィニティーで結合する少なくとも1つのシトルリンを有することである。当業者は、ビメンチンならびにフィブリノーゲンのα鎖およびβ鎖中のいくつかのさらなる配列が、P4にアルギニンまたはシトルリンを位置付ける区域でHLA-DR分子を結合することが予測され、従って、本発明において具体化されることを理解する。さらに、本発明は、RA患者における抗シトルリン抗体の潜在的な標的であることが実証される任意のペプチドを含む。このことは、独特な病原性ペプチドが、不均一な特異性のアレイを有する活性化T細胞を生じることを示唆する。
【0064】
本発明の1つの局面において、ペプチジルアルギニンデイミナーゼ(PAD)による修飾に感受性であり、さらに共有エピトープを有するMHCクラスII分子を結合する特性を有する、任意の内因性または外因性のタンパク質またはペプチドが、本発明によって具体化される。このようなタンパク質またはペプチドは、1つの態様において、関節において存在することが見い出され得、これには、ビメンチンおよびフィブリノーゲンが含まれるがこれらに限定されない。
【0065】
本発明のシトルリン化ペプチドは、共有エピトープを有するMHCクラスII分子に高アフィニティーで結合する。「高アフィニティー」は、本明細書中で使用される場合、シトルリン化されていないペプチド(例えば、シトルリンではなくアルギニンアミノ酸を有するペプチドなど)と比較して、共有エピトープを有するMHCクラスII分子に、より高いかまたは増大したアフィニティーで結合するシトルリン化ペプチドの能力をいうことが当業者によって理解される。このような結合アフィニティーは、試料ペプチドが標準ペプチドに置き換わるように使用されるペプチド結合アッセイを使用して、例えばインビトロで容易に達成できることがさらに理解される(実施例の節を参照されたい)。
【0066】
1つの態様において、本発明のタンパク質は SAVRACitSSVPGVR (配列番号1); FSMCitIVCLV (配列番号2); VVECitHQSAC (配列番号3); FTNCitINKLK (配列番号4); LRSCitIEVLK (配列番号5); VLKCitKVIEK (配列番号6); IKICitSCRGS (配列番号7); LPSCitDRQHL (配列番号8); FRHCitHPDEA (配列番号9); FPSCitGKSSS (配列番号10); IQQCitMDGSL (配列番号11); LTQCitGSVLR (配列番号12); YHFCitVGSEA (配列番号13); YDPCitNNSPY (配列番号14); VSFCitGADYS (配列番号15); YSLCitAVRMK (配列番号16); MKICitPLVTQ (配列番号17); YRACitPAKAA (配列番号18); WQKCitQKQVK (配列番号19); IQNCitQDGSV (配列番号20); WYNCitCHAAN (配列番号21); YSMCitKMSMK (配列番号22); MKICitPFFPQ (配列番号23); LHPCitNLILY (配列番号24); VATCitDNCCI (配列番号25); LDECitFGSYC (配列番号26); LKSCitIMLEE (配列番号27); FQKCitLDGSV (配列番号28); YALCitVELED (配列番号29); WNGCitTSTA (配列番号30); WKTCitWYSMK (配列番号31); YATCitSSAVR (配列番号32); VRLCitSSVPG (配列番号33); LNDCitFANYI (配列番号34); MLQCitEEAEN (配列番号35); LNLCitETNLD (配列番号36);及びVETCitDGQVI (配列番号37) からなる群から選択される少なくとも1つのアミノ酸配列を含むシトルリン化ペプチドを含み得るがこれらに限定されない。
【0067】
特定のこれらのペプチド配列は、酵素ペプチジルアルギニンデイミナーゼの作用を介してシトルリンに変換できるさらなるアルギニンを含み得る。例えば、本発明のシトルリン化ペプチドは以下の式:Cit-Cit-X-Cit-G-Cit-Cit-Z-Cit-Cit-B-Cit-Cit(配列番号38)を有し、ここでXはY、F、W、I、L、MおよびVから選択され、ZはA、D、I、N、P、S、TおよびVから選択され、BはA、G、H、Q、S、TおよびVから選択される。「X」で示されるアミノ酸は、共有エピトープを有するMHCクラスII分子についてのP1アンカリングアミノ酸のために必要である。5番目の位置に存在するグリシンは、シトルリンがアミノ酸位置番号4、6、および7に配置されることを可能するために、ペプチドバックボーン上のねじれを減少することを補助する。6位のアミノ酸はP4共有エピトープポケットである。位置番号8の選択されるアミノ酸は、共有エピトープを有するMHCクラスII分子についてのP6アンカリングポケットにおいて非阻害的である。位置番号11の選択されるアミノ酸は、共有エピトープを有するMHCクラスII分子についてのP9アンカリングポケットにおいて非阻害的である。本発明の態様において、ペプチドは、Cit-Cit-Y-Cit-G-Cit-Cit-S-Cit-Cit-S-Cit-Cit(配列番号39)を含む。本発明のさらなる態様において、正に荷電したアミノ酸および/または負に荷電したアミノ酸が、ペプチド結合を増大するために、ペプチドのカルボキシ末端および/またはアミノ末端に付加できる。さらに、このペプチドは、ジスルフィド結合の形成によってペプチドを環状化するために、ペプチドの1つの末端または両方の末端にシステインを有し得る。
【0068】
式:Cit-Cit-X-Cit-G-Cit-Cit-Z-Cit-Cit-B-Cit-Cit(配列番号38)(ここでXはY、F、W、I、L、MおよびVから選択され、ZはA、D、I、N、P、S、TおよびVから選択され、BはA、G、H、Q、S、TおよびVから選択される)のペプチドは、DR4 tgマウスにおいて抗シトルリン抗体を生成することが実証された。さらに、これらのペプチドはまた、RA患者のヒト血清において自己抗体と反応することが実証された。このようなものとして、これらのペプチドは、対象におけるRAの診断のための方法およびキットにおいて使用できる。
【0069】
出願人は、シトルリン化ペプチド抗原を含む自己免疫障害がMHCクラスII分子共有エピトープを有する対象においてT細胞応答を誘発し、多発性硬化症(MS)におけるシトルリン化ペプチド誘発T細胞応答もまた、MHCクラスIIエピトープを有するMS対象のために本発明に含まれることを実証した。このようなシトルリン化ペプチドには、例えば、シトルリン化ミエリン塩基性タンパク質(MBP)およびシトルリン化グリア線維酸性タンパク質(GFAP)が含まれる。MSの診断において有用であるシトルリン化MBPペプチドおよびGFAPペプチドならびに治療方法の標的は、FLPCitHRDTG (配列番号40), VTPCitTPPPS (配列番号41); YGGCitASKYK (配列番号42), LGGCitDSRSG (配列番号42), MERCitRITSA (配列番号43), LPTCitVDFSL (配列番号44), LNDCitFASYI (配列番号45), LRLCitLDQLT (配列番号46), LQICitETSLD (配列番号47),及びVEMCitDGEVI (配列番号48) からなる群から選択されるアミノ酸配列を含み得る。
【0070】
さらに、本発明のシトルリン化ペプチドは、好中球中でシトルリン化されるペプチドをさらに含み得、このようなものとして、慢性関節リウマチの診断または治療において使用され得、前記ペプチドは、ヌクレオホスミン/B23、ヒストンH2A、ヒストンH4、およびヒストンH3からなる群から選択され、ここで、前記ペプチドは1つ以上のシトルリン残基を含む。これらのタンパク質は顆粒球中でシトルリン化されることが実証されている(Hagiwara T.ら、 Biochem & Biophy Res Comm. (2000) 290, 979-983)。好中球は、慢性関節リウマチ患者の炎症性関節に存在する。シトルリン化できるペプチドは以下の群から選択される:LSLRTVSLG (配列番号49、ヌクレオホスミン/B23);VVLRLKCGS (配列番号50、ヌクレオホスミン/B23);MSGRGKQGG、(配列番号51、ヒストンH2A);MSGRGKQGG、(配列番号52、ヒストンH2A);IIPRHLQLA、(配列番号53、ヒストンH2A);LAIRNDEEL、(配列番号54、ヒストンH2A);LLGRVTIAQ、(配列番号55、ヒストンH2A);MSGRGKGGK (配列番号56、ヒストンH4);LARRGGVKR (配列番号57、ヒストンH4);VALREIRRY (配列番号58、ヒストンH3)、LLIRKLPFQ(配列番号59、ヒストンH3)および、LARRIRGER (配列番号60、ヒストンH3)ここで前記「R」アルギニン残基は、PADの作用によってシトルリンに変換できる。このようなペプチドは、例えば、慢性関節リウマチの診断的徴候としてのT細胞反応性を測定するための種々のアッセイにおいて使用できる。
【0071】
本発明のペプチドは、少なくとも約9アミノ酸長、および約9から約55アミノ酸長であり得、当業者によって理解されるように、その中の任意の範囲の長さ(すなわち、9〜50、9〜45、9〜40、9〜35、9〜30、9〜25、9〜20、9〜15など)を含み得る。約55アミノ酸を超える長さのペプチドもまた、本発明に含まれる。ペプチドの長さは、共有エピトープを有するMHCクラスII分子へのその結合能力によってのみ制限される。本発明のペプチドはまた、当業者によって理解され、かつ例えば、米国特許第5,824,315号および米国特許第6,184,204号(これらの開示はその全体が参照として本明細書に援用される)において記載されるように、ペプチドのダイマーおよびトリマーを含み得る。本発明によるマルチマーは、複数の同じペプチドからなるホモマー、または異なるペプチドからなるヘテロマーのいずれかであり得る。言及されるように、本発明によるペプチドの特徴的なアミノ酸配列は、ランダムなアミノ酸配列に隣接できる。好ましいものは、ペプチドに安定化効果を有する、従って、それらの生物学的利用可能性を増加する隣接配列である。さらに、他のペプチド模倣物もまた、本発明のペプチドにおいて有用である。一般的な概説としては、A. F. Spatola, in Chemistry and Biochemistry of Amino Acids, Peptides and Proteins, B. Weinstein, eds., Marcel Dekker, New York, p. 267 (1983)を参照されたい。本発明のペプチドはまた、例えば、リン酸化、グリコシル化、または脂質化によって修飾されたペプチドを含む。さらに、本発明のポリペプチドはまた、ペプチドの「機能的に等価な改変体」または「類似体」を含み得る。このようなものとして、これは、部分的配列相同性を有するペプチドおよびポリペプチド、1つ以上の保存性アミノ酸変化および/または保存性でないアミノ酸変化、ならびにペプチドの生物学的特性または構造的特性(すなわち、共有エピトープを有するMHCクラスII分子に結合する能力)を変化させないペプチド結合体を含むが、これらに限定されない。
【0072】
「機能的類似体」によって、生物学的に機能的なペプチド類似体の定義における固有の性質は、分子の規定された部分内でなされ得る変化の数に限度が存在し、等価な生物学的活性(この場合は、これは共有エピトープを有するMHCクラスII分子に結合する能力を含む)の許容可能なレベルを有する分子をなお生じるという概念であることが当業者によって十分に理解される。異なる置換を有する複数の別個のペプチド/タンパク質は、本発明に従って容易に作製および使用できる。特定の残基(例えば、レセプター認識領域における残基など)が、タンパク質またはペプチドの生物学的特性または構造的特性に対して特に重要であることもまた理解され、このようなこれらの残基は、一般的に交換されなくてもよい。
【0073】
機能的類似体は、保存性または非保存性のアミノ酸置換によって生成できる。アミノ酸置換は、一般的に、アミノ酸側鎖置換基の比較的な類似性、例えば、それらの疎水性、親水性、電荷、サイズなどに基づく。従って、本発明の範囲内において、保存性アミノ酸変化は、もともと存在するのと同じ型である特定の位置におけるアミノ酸変化、すなわち、疎水性アミノ酸に交換された疎水性アミノ酸、塩基性アミノ酸に交換された塩基性アミノ酸などを意味する。保存性置換の例には、非極性(疎水性)残基(例えば、イソロイシン、バリン、ロイシン、またはメチオニン)の別の残基への置換、1つの極性(親水性)残基の別の残基への置換(例えば、アルギニンとリジンの間、グルタミンとアスパラギンの間、グリシンとセリンの間)、1つの塩基性残基(例えば、リジン、アルギニン、またはヒスチジン)の別の残基への置換、または1つの酸性残基(例えば、アスパラギン酸およびグルタミン酸)の別の残基への置換、分枝鎖アミノ酸(イソロイシン、ロイシン、またはバリン)の別の残基への置換、1つの芳香族アミノ酸(例えば、フェニルアラニン、チロシン、またはトリプトファン)の別の残基への置換が含まれる。このようなアミノ酸変化は、ペプチドの全体の電荷および/または立体配置を有意に変化させないという点において、機能的類似体を生じる。このような保存性変化の例は当業者に周知であり、本発明の範囲内に含まれる。保存性置換はまた、得られるペプチドが本発明のペプチドと生物学的に機能的に等価であるという条件で、非誘導体化残基の代わりに化学的に誘導体化された残基の使用を含む。それゆえに、本発明のシトルリン化ペプチドは、配列番号1から39とは1つ以上の保存性アミノ酸置換が異なるアミノ酸配列を有するペプチドを含む。本発明のシトルリン化ペプチドはまた、配列番号1から39とは単一の変異が異なるアミノ酸配列を有するペプチドを含み、ここで、単一の変異は、単一のアミノ酸の欠失、挿入、または置換を表す。
【0074】
本発明のペプチドは、当業者の公知の方法によって、最も顕著にかつ好ましくは、合成ペプチドを生成するために、例えば、固相合成(Merrifield (1964) J. Am. Chem. Assoc. 65:2149; J. Amer. Chem. Soc. 85:2149 (1963)およびInt. J. Peptide Protein Res. 35:161-214 (1990))または均質溶液中での合成(Methods of Organic Chemistry, E. Wansch (編) 第15巻, I部およびII部, Thieme, Stuttgart (1987))などのタンパク質の化学において周知の技術を使用する化学合成によって作製できる。シトルリンは、アルギニンから正電荷を除去しかつ得られるシトルリンを事実上極性にする、酵素ペプチジルアルギニンデイミナーゼ(PAD)によって触媒される脱イミノ化のプロセスを通して作製される翻訳後修飾アルギニンである。
【0075】
シトルリン化ビメンチンおよびフィブリノーゲンについての1つの実施形態において、本発明のシトルリン化ペプチドは、公知の商業的に利用可能なビメンチンおよびフィブリノーゲンの供給源から作製できる。この態様において、凍結乾燥したビメンチンまたはフィブリノーゲンは、酵素ペプチジルアルギニンデイミナーゼが加えられる適切な緩衝液中で再構成される。この溶液は、アルギニン残基のシトルリンへの修飾を引き起こし、従って、シトルリン化されたビメンチンタンパク質またはフィブリノーゲンタンパク質を作製するために十分な時間の間、適切な温度に置かれる。次いで、シトルリン化タンパク質を、酵素を分離するための高分子量メンブレンまたは他のクロマトグラフィーの方法を使用する酵素の除去によって単離する。当業者は、インキュベーションの温度、緩衝液条件、およびインキュベーションの時間は、脱イミノ化されるタンパク質に依存して変化し得ることを理解する(Christine Masson-Bessiereら、2001. The Major Synovial Targets of the Rheumatoid arthritis-Specific Antifilaggrin Autoantibodies Are Deiminated Forms of the ( and ( Chains of Fibrin 1 The Journal of Immunology 166: 4177-4184)。本発明の1つの局面において、シトルリン化フィブリノーゲンは、プラスミノーゲン枯渇ヒトフィブリノーゲン(Calbiochem、San Diego、CA)を入手すること、およびタンパク質の脱イミノ化のために、0.86mg/mlのこのタンパク質をウサギ骨格筋PAD(7U/mgフィブリノーゲン;Sigma)とともに、0.1 M Tris-HCl(pH 7.4)、10 mM CaCl2、および5 mM DTT中で2時間、50℃でインキュベートすることによって作製された。同様のアプローチが、ミエリン塩基性タンパク質およびグリア線維酸性タンパク質を、適切な組織および細胞から作製および単離するために含まれる。
【0076】
本発明のシトルリン化タンパク質は、その配列によって明らかにされる特性に基づいて選択される方法によってさらに単離され、かつ精製できる。精製は、クロマトグラフィー方法(ゲル濾過、イオン交換、および免疫アフィニティー)などのタンパク質精製手順によって、高速液体クロマトグラフィー(HPLC、RP-HPLC、イオン交換HPLC、サイズ排除HPLC、高速等電点電気泳動、および疎水性相互作用クロマトグラフィー)によって、または沈殿(免疫沈殿)によって達成できる。ポリアクリルアミドゲル電気泳動もまた、タンパク質の分子量、電荷特性および疎水性に基づいてシトルリン化タンパク質を単離するために使用できる。精製されたシトルリン化タンパク質は、さらなる生化学的分析において使用され得、タンパク質と相互作用するため、タンパク質電荷の立体配置もしくは他のタンパク質との電荷相互作用を変化させるため、またはその機能を変化させるための医薬の設計において補助し得る、二次構造および三次構造を確立する。
【0077】
代替的には、本発明のシトルリン化ペプチドは、当業者に公知の組換えDNA技術の使用によって作製できる。シトルリンはいかなるトリヌクレオチド配列によってもコードされないので、アルギニンをコードする核酸配列が、アルギニンをシトルリンに変換するためにペプチジルアルギニンデイミナーゼ(PAD)を使用して修飾される得られるタンパク質配列とともに使用できる。シトルリンに類似するアミノ酸であるグルタミンをコードする核酸配列を使用することはさらに、本発明の範囲内にある。この様式において、アルギニンを含む天然に存在する配列が使用され得、ここで、このアルギニンは、シトルリンに類似するためにグルタミンで置換される。本発明の選択したペプチドをコードする核酸配列は、公知の様式で、適切な発現ベクター(すなわち、組換え発現ベクター)に組み込まれ得る。可能な発現ベクターには、そのベクターが使用される宿主細胞と適合性である限り、コスミド、プラスミド、または改変ウイルス(例えば、複製欠損レトロウイルス、アデノウイルス、およびアデノ関連ウイルス、レンチウイルス、ヘルペスウイルス、ポックスウイルス)が含まれる(がこれらに限定されない)。発現「ベクターが宿主細胞と適合性である」とは、発現ベクターが本発明の核酸分子(本明細書中以下に記載される)、および発現のために使用される宿主細胞に基づいて選択される付属の調節配列を含むことを意図すると定義され、前記調節配列は、核酸分子に作動可能に連結されている。「作動可能に連結される」とは、核酸の発現を可能にする様式で、核酸が調節配列に連結されることを意味する。適切な調節配列は、種々の供給源(細菌、真菌、またはウイルスの遺伝子を含む)に由来し得る。例えば、Goeddel, Gene Expression Technology: Methods in Enzymology 185, Academic Press, San Diego, CA (1990)において記載される調節配列を参照されたい。適切な調節配列の選択は、選択される宿主細胞に依存し、かつ当業者によって容易に達成できる。このような調節配列の例には以下が含まれる:転写プロモーターおよびエンハンサー、RNAポリメラーゼ結合配列、またはリボソーム結合配列(翻訳開始シグナルを含む)。選択される宿主細胞および利用される発現ベクターに依存して、他のさらなる配列(例えば、複製起点、さらなるDNA制限部位、エンハンサー、および転写の誘導性を付与する配列)が、発現ベクターに組み込まれ得る。
【0078】
本発明のペプチドはまた、当業者に公知である種々の方法(すなわち、Hugues, Sら(2002) Generation and use of alternative multimers of peptide/MHC complexes. Journal of Immunological Methods. 268:83-91、この開示はその全体が本明細書に援用される)を使用して、可溶性MHC分子と共同して組換え的に産生できる。ペプチド/MHCクラスII可溶性複合体を作製するための方法はまた、米国特許第5,869,279号(この開示はその全体が本明細書に援用される)において提供される。
【0079】
本発明が、サイトカイン、リンホカイン、および免疫調節分子からなる群からの少なくとも1つのメンバーをコードする核酸を含むベクターを含むことがさらに意図される。前記核酸配列は、本発明のシトルリン化ペプチド抗原をコードする配列と連続し得るか、または別個の核酸上にコードできる。
【0080】
本発明のペプチドは、当業者によって理解されるように、種々のアッセイにおいてそれらの検出を容易にするための標識で標識できる。このような標識には、放射活性標識および蛍光標識が含まれるがこれらに限定されない。本発明のペプチドは、例えば、ウシ血清アルブミン(BSA)またはキーホールリンペットヘモシアニンなどに結合された担体とともに提供できる。このペプチドは、固体担体(例えば、金もしくはポリスチレンのミクロスフェア、スライド、チップ)、またはマイクロタイタープレートのウェルに、共有結合的または非共有結合的に結合できる。このペプチドは、ビオチン、フルオレセイン、および西洋ワサビペルオキシダーゼなどの酵素から選択されるがこれらに限定されない標識で、直接的または間接的に標識できる。
【0081】
本発明の実施形態は、インビボでの投与のために適切である生物学的に適合可能な型である、対象への投与のための1種以上のシトルリン化ペプチドを含む薬学的組成物をさらに含む。薬学的組成物中での使用のためのシトルリン化ペプチドは、化学的にまたは組換えDNA技術によって作製できる。本発明のシトルリン化ペプチド抗原の投与は、自己応答性T細胞を有する患者における免疫系を脱感作するように(すなわち、全身性反応性低下の状態または前記ペプチドもしくは組成物に対する寛容性を誘導することによって)作用し得、従って、時間とともに炎症性応答を減少する。本発明のペプチドは、適切な薬学的組成物中で処方される、上記のようなDNA発現ベクター中で提供できる。
【0082】
「インビボでの投与のために適切である生物学的に適合可能な型」は、治療的効果が任意の毒性効果を上回る、投与される物質の型を意味する。本発明の薬学的組成物の治療的に活性な量、すなわち「有効量」は、ヒトにおける免疫応答を誘発することの所望の結果を達成するために必要である用量または時間の期間での有効な量として定義される。物質の治療的な有効量は、レシピエントの疾患状態/健康、年齢、性別、および体重などの因子、および特定のポリペプチドの固有の能力、それをコードする核酸、または所望の免疫応答を誘発するための組換えウイルスに従って変化し得る。投薬量レジメンは、最適の治療的応答を提供するために調整できる。例えば、数回の分割用量が、毎日または定期的な間隔で投与され、および/またはその用量は治療的状況の緊急度によって示されるように比例的に減少できる。投与のためのシトルリン化ペプチド抗原の量は投与の経路、投与の時間に依存し、そして個々の対象の応答に従って変動する。適切な投与経路は、筋肉内注射、皮下注射、静脈内注射、または腹腔内注射、経口投与および鼻内投与である。経口投与および鼻内投与が好ましい投与経路である。
【0083】
本明細書中に記載される組成物は、対象に投与できる薬学的に許容可能な組成物の調製のためのそれ自体公知の方法によって調製され得、その結果、活性物質(すなわち、シトルリン化ペプチド)の有効量が、薬学的に許容可能なビヒクルとの混合物中で合わせられる。適切なビヒクルは、例えば、「Handbook of Pharmaceutical Additives」 (MichaelおよびIrene Ash,編 Gower Publishing Limited, Aldershot, England (1995)において記載されている。これに基づくと、この組成物には、非限定的であるが、薬学的に許容可能なビヒクルまたは希釈物の1種以上が付随した物質の溶液が含まれ、適切なpHおよび/または生理学的液体と等張性である緩衝化溶液中に含まれ得る。この点に関して、米国特許第5,843,456号を参照できる。
【0084】
薬学的に許容可能な担体は当業者に周知であり、例えば、滅菌生理食塩水、ラクトース、スクロース、リン酸カルシウム、ゼラチン、デキストリン、寒天、ペクチン、ピーナッツ油、オリーブ油、ゴマ油、および水が含まれる。他の担体は、例えば、MHCクラスII分子であり得る。モノマー、ダイマー、トリマー、テトラマーなどを含む可溶性MHCクラスII分子、ならびにシトルリンペプチド/MHCクラスII複合体は、米国特許第5,869,270号(この開示は、参照として本明細書に援用される)において開示される方法によって作製できる。
【0085】
さらに、本発明に従う薬学的組成物は、例えば、ソルビトール、マンニトール、デンプン、スクロース、デキストリン、およびグルコースを含む炭水化物、アルブミンまたはカゼインなどのタンパク質、ならびにアルカリリン酸のような緩衝剤などの1種以上の安定剤を含み得る。
【0086】
本発明の薬学的組成物はまた、1種以上のアジュバントを含み得る。当業者に周知であるように、免疫応答を誘導/誘発する免疫原の能力は、投与形式に関わらず(すなわち、組換えウイルス、核酸、ペプチド)、免疫原がアジュバントとともに同時投与される場合に改善できる。アジュバントは、「Vaccine Design-the Subunit and Adjuvant Approach」 (PowellおよびNewman編, 'Plenum Press, New York, U.S.A., pp. 61-79および141-228 (1995)において記載および議論されている。アジュバントは、代表的には、免疫原の免疫原性を増強するが、それ自体の中でおよびそれ自体が免疫原性である必要はない。アジュバントは、免疫系の細胞への免疫因子の遅延性の、持続性の放出を容易にするデポー効果を生じるために、投与の部位に局所的に近傍に免疫原を保持することによって作用し得る。アジュバントはまた、免疫系の細胞を、免疫原デポーに引き付け得、かつこのような細胞が免疫応答を誘発することを刺激し得る。このようなものとして、本発明の実施形態は、さらにアジュバントを含む組成物を含む。
【0087】
理想的なアジュバントの望ましい特性には以下が含まれる:
1)毒性がないこと;
2)長時間続く免疫応答を刺激する能力;
3)製造の単純さおよび長期間保存における安定性;
4)必要な場合、種々の経路によって投与された抗原に対する細胞応答と体液応答の両方を誘発する能力;
5)他のアジュバントとの相乗効果;
6)抗原提示細胞(APC)の集団と選択的に相互作用する能力;
7)適切なTr、TR1、またはTH2細胞特異的免疫応答を特異的に誘発する能力;ならびに
8)抗原/免疫原に対する適切な抗体イソタイプレベル(例えば、IgA)を選択的に増加する能力。
【0088】
適切なアジュバントには、とりわけ、水酸化アルミニウム、リン酸アルミニウム、アンフィゲン、トコフェノール、モノホスフェニルリピドA、ムラミールジペプチド、およびQuill Aなどのサポニンが含まれる。好ましくは、本発明に従う寛容治療において使用されるアジュバントは、粘膜上皮に結合する、コレラ毒素Bサブユニットまたはカルボマーなどの粘膜アジュバントである。アジュバントの量は、当業者によって理解されるように、アジュバントそれ自体の性質に依存する。
【0089】
シトルリン化ペプチドが、所望の遺伝子に対してRNA干渉応答を媒介する短い二本鎖RNA(30ヌクレオチド長未満)を組み合わせて患者に投与できることが本発明においてさらに具体化される。これらの標的遺伝子は、転写因子、酵素、サイトカイン、リンホカイン、および免疫調節分子からなる群の1種以上のメンバーから選択できる。
【0090】
本発明のペプチドおよび組成物は、MHC分子の文脈において投与できる。従って、本発明のポリペプチドは、樹状細胞(DC)を含むがこれに限定されない抗原提示細胞にパルス投与できる。より具体的には、シトルリン化ペプチドが、抗原パルスされた樹状細胞(抗原提示細胞)を介して患者に投与できる。特定の実施形態において、本発明は、抗原提示細胞として樹状細胞またはその前駆細胞を使用する。これらの樹状細胞は、患者に投与する前に、転写因子、酵素、サイトカイン、リンホカイン、および免疫調節分子のそれらの産生を変化させるために、患者のPBMC(末梢血単核細胞)から増殖され、エキソビボで処理できる。樹状細胞は、高度に強力なAPCである(BanchereauおよびSteinman, Nature 392:245-251, 1998)。一般的に、樹状細胞は、それらの代表的な形状(インサイチューで星状、インビトロで見える顕著な細胞質プロセス(樹状))、高効率で抗原を取り込み、プロセスし、かつ提示するそれらの能力、および未処理のT細胞の応答を活性化するそれらの能力に基づいて同定できる。樹状細胞は、インビボまたはエキソビボで樹状細胞上に一般的に見い出されない特異的細胞表面レセプターまたはリガンドを発現するように操作され得、このような改変樹状細胞は本発明によって意図される。樹状細胞に対する代替として、分泌ベシクル抗原負荷樹状細胞(エキソソームと呼ばれる)が免疫原性組成物中で使用できる(Zitvogelら、Nature Med. 4:594-600, 1998を参照されたい)。樹状細胞および前駆細胞は、末梢血、骨髄、腫瘍浸潤細胞、腫瘍周囲組織浸潤細胞、リンパ節、脾臓、皮膚、臍帯血または他の適切な組織または体液から入手できる。例えば、樹状細胞は、末梢血から収集された単球の培養物に、GM-CSF、IL-4、IL-13および/またはTNF-αなどのサイトカインの組み合わせを加えることによって、エキソビボで分化できる。代替的には、末梢血、臍帯血、または骨髄から収集されたCD34陽性細胞が、GM-CSF、IL-3、TNFα、CD40リガンド、LPS、fit3リガンド、ならびに/または、樹状細胞の分化、成熟、および増殖を誘導する他の化合物の組み合わせを培養培地に加えることによって樹状細胞に分化できる。
【0091】
本発明のシトルリン化ペプチドは関節炎性であり、従って、RAをもたらす炎症性応答を誘発する。これは、図6AからEに実証されている。このようなものとして、これらのペプチドは、RAの治療のために有効である新規な医薬が試験および同定できる動物モデルを提供するために、非ヒト哺乳動物において臨床的関節炎を誘導するために使用できる。共有エピトープを含むMHCクラスII分子についてトランスジェニックである任意の動物が、本発明の範囲内で使用できる。本発明の1つの局面において、本発明の1種以上のシトルリン化ペプチドの投与が、関節炎の徴候関節炎、より特定には慢性関節リウマチの最終的な疾患パターンの発症をもたらすHLA-DRB1*401トランスジェニックマウス(DR4-IE tg)に提供できる。これらのDR4-IE tgトランスジェニックマウスは、ヒト抗原結合ドメイン(α1およびβ1)およびマウスCD4結合ドメイン(α2およびβ2)から構成されるキメラMHCクラスII分子を発現する。キメラDR4は、全長ヒトDR4と同一の様式でペプチドを提示し、そしてこれらのマウスは、内因性MHCクラスII分子を発現しない。さらに、これらのマウスは、野生型マウスと同じB細胞およびT細胞の補体を含み、従って、これらのトランスジェニックマウスは、インビボでT細胞に制限されるSEの免疫応答を試験するために、および従って、炎症をもたらすT細胞応答を減少または予防し得る医薬品薬剤を同定するために理想的である。DR4-IE tgマウスが使用されるが、共有エピトープを含むMHCクラスII分子についてトランスジェニックである任意のマウス系統が、本発明における薬学的化合物の試験のための動物モデルとして使用できることが理解される。
【0092】
本発明のシトルリン化されたMBPタンパク質およびGFAPタンパク質もまた、新規な医薬品がMSの治療のために試験されかつ有効であると同定できるMS動物モデルを提供するために非ヒト哺乳動物において臨床的多発性硬化症を誘導するために使用できる。このような動物モデルを作製するための方法は上記の通りである。
【0093】
トランスジェニックマウスは、1回以上、本発明の1種以上のペプチドを含むシトルリン化タンパク質(ならびに任意のアジュバントまたは他の薬学的賦形剤)を、皮下的に、皮内に、腹腔内に、または筋肉内に注射し、関節炎の徴候を発症させ得る。1つの態様において、全長シトルリン化ビメンチンまたはシトルリン化フィブリノーゲンが、関節炎の発症を誘導するためにトランスジェニックマウスに投与される。関節炎を治療するため、および関節炎の発症に対してこれらの薬学的活性の効果を研究するための炎症反応が確立されるとすぐに、潜在的な医薬品薬剤は、本発明のペプチドと同時投与されるか、または後で投与できる。好ましいマウスは、関節炎、特に慢性関節リウマチのための動物モデルとして使用される。
【0094】
本発明の実施形態において、本発明のシトルリン化ペプチドが関節炎性であり、共有エピトープを有するMHCクラスII分子への結合を介して、RAをもたらすT細胞応答を誘発するように作用するという知見は、ここで、RAおよびMSなどのシトルリン化抗原の共有エピトープ結合を含む自己免疫障害のための診断的および治療的方法を提供する。
【0095】
1つの態様において、本発明は、RAを有する患者、またはRAを有すると疑われている患者、またはRAを発症する素因がある患者からの自己反応性T細胞を検出するために使用できる。多数の方法が、シトルリン化ペプチド-MHC複合体に特異的なT細胞を検出するために、またはシトルリン化ペプチド-MHC複合体それ自体の存在のいずれかを検出するために使用できる。例えば、当業者に公知であるT細胞増殖アッセイは、対象の末梢血単核細胞(PBMC)試料中のT細胞活性化を検出するために使用できる。このような方法は、T細胞増殖の尺度として放射性同位元素(例えば、3H-チミジンなど)の取り込みによって実行される(実施例の節を参照されたい)。PBMCに存在する自己反応性T細胞活性もまた、サイトカイン特異的ELISA(すなわち、ELISPOT)を用いて、シトルリン化ペプチド-MHCクラスII複合体による活性化後のサイトカイン放出を測定することによって検出できる。別の検出方法は、例えば、II-2RのFACS分析による活性化マーカーの発現の測定である。さらに、可溶性MHCクラスII分子は、結合したシトルリン化ペプチドを伴う蛍光色素またはリポソームに連結され得、これらの複合体を認識するT細胞レセプターを有するT細胞を検出するために使用できる。
【0096】
別の実施形態において、本発明のシトルリン化ペプチドのいずれかを認識するモノクローナル抗体がまた作製され得、抗原提示細胞(APC)上の共有エピトープを有するMHCクラスII分子に提示/結合されたペプチドの存在を検出するために使用できる。これは、疾患ならびにRAを発症する素因の迅速かつ単純な診断方法を提供する。一般的に、抗体の調整のための方法は周知である。本発明のペプチドを認識するポリクローナル抗体を調製するために、ビメンチンタンパク質またはフィブリノーゲンタンパク質の規定された部分またはすべて、またはそれらの選択的転写物のいずれかが、適切なクローニングビヒクル中での対応するDNA配列の発現によって、細菌中で合成できる。次いで、タンパク質が精製され、担体タンパク質に結合されおよびフロイントアジュバントを混合され(ウサギによる抗原性応答を刺激することを補助するために)、ならびにウサギまたは他の実験用動物に注射できる。代替的には、タンパク質は、タンパク質を発現する培養細胞から単離できる。2週間間隔のブースター注射後、次いでウサギまたは他の実験用動物を、採血し、血清を単離する。血清は、直接的に使用できるか、またはアフィニティークロマトグラフィーによって精製できる。次いで、血清は、ポリアクリルアミドゲル上でタンパク質抽出物をプローブし、ビメンチンタンパク質またはフィブリノーゲンタンパク質、選択的転写物またはそれらの任意の変異体を同定するために使用できる。代替的には、合成ポリペプチド(上記で議論したようなもの)が、これらのタンパク質の抗原性部分に対して作製され得、動物に接種されるために使用できる。
【0097】
本発明のペプチド(哺乳動物または他の種のビメンチンタンパク質およびフィブリノーゲンタンパク質またはその部分)を特異的に認識するモノクローナル抗体を産生するための方法は、当業者に周知である。一般的に、活性にタンパク質を発現する細胞が、培養されるかまたは組織から単離され、そして細胞抽出物が単離される。ビメンチンタンパク質またはフィブリノーゲンタンパク質を含む、この抽出物または組換えタンパク質抽出物が、フロイントアジュバント中でマウスに注射される。3週間の期間にわたって9回注射した後、マウス脾臓が取り出され、リン酸緩衝化生理食塩水(PBS)中に再懸濁される。脾臓細胞はリンパ球の供給源として働き、それらのいくつかは適切な特異性の抗体を産生する。次いで、これらは、永久的に増殖するミエローマパートナー細胞と融合させ、融合の生成物を、HATなどの選択薬剤の存在下で多数の組織培養ウェルにプレートする。次いで、これらのウェルをELISAによってスクリーニングして、有用な抗体を作製する細胞を含むウェルを同定する。次いで、これらを新鮮にプレートする。一定の増殖期間の後、これらのウェルを再度スクリーニングして、抗体産生細胞を同定する。数回のクローニング手順が、90%より多くのウェルが抗体産生について陽性である単一クローンを含むまで、実行される。この手順から、抗体を産生する安定なクローンの系統が樹立される。次いで、モノクローナル抗体が、プロテインAまたはプロテインGセファロースを使用するアフィニティークロマトグラフィーによって精製できる。
【0098】
本発明の抗体の調製のための方法は、一般的に当該分野において公知である。例えば、Antibodies, A Laboratory Manual, Ed. Harlow および David Lane (編) Cold Spring Harbor Laboratory, N.Y. (1988) ならびにそこに引用される参考文献を参照されたい。免疫学の一般的原理を説明する標準的な参照される研究には、Klein, J. Immunology: The Science of Cell-Noncell Discrimination, John Wiley & Sons, N.Y. (1982); Dennett, R.ら Monoclonal Antibodies, Hybridoma: A New Dimension In Biological Analyses Plenum Press, N.Y. (1980) および Campbell, A. 「Monoclonal Antibody Technology」Laboratory Techniques In Biochemistry And Molecular Biology, Vol. 13, Burdon.ら (編), Elsevier, Amsterdam (1984) が含まれる。米国特許第4,609,893号、米国特許第4,713,325号、米国特許第4,714,681号、米国特許第4,716,111号、米国特許第4,716,117号、米国特許第4,720,459号(これらの開示はその全体が本明細書に援用される)もまた参照されたい。
【0099】
抗体はまた、ペプチド/MHCクラスII複合体に指向されて作製できる。ペプチド/MHCクラスII複合体に対するこのような抗体を生成する方法は公知であり、記載されている(Baeten, D., Steenbakkers PGA., Rovers E., Veys EM., Rijnders AMW., Meijerink J., Keyser F DeおよびBoots A. Localisation of MHC Class II/HC gp-39 complexes in synovia of rheumatoid arthritis patients using complex-specific monoclonal antibodies. Abstracts of the 23rd European Workshop for Rheumatology Research, 第5巻 補遺1, 2003年2月; Baeten D., Steenbakkers P., Boots A., Veys EM., and Keyser, Fde., The presentation of the immunodominant epitope of HC gp-39 in the context of the RA-associated HLA class II molecules is specific for RA synovium. Abstracts of the 23rd European Workshop for Rheumatology Research, 第5巻 補遺1, 2003年2月)。
【0100】
本発明に従う1種以上のペプチドを含む診断組成物および適切な検出剤は、従って、本発明の一部を形成する。検出の型に依存して、検出剤は、放射性同位元素、酵素、または細胞表面もしくは活性化マーカーに特異的である抗体であり得る。
【0101】
本発明に従う1種以上のペプチドを含む試験キットもまた、本発明の範囲に含まれ得る。これらの試験キットは、本発明に従う診断方法における使用のために適切である。
【0102】
式Cit-Cit-X-Cit-G-Cit-Cit-Z-Cit-Cit-B-Cit-Cit(ここでXはY、F、W、I、L、MおよびVから選択され、ZはA、D、I、N、P、S、TおよびVから選択され、BはA、G、H、Q、S、TおよびVから選択される)のペプチドは、自己免疫障害の診断のための方法において特に有用である。このような自己免疫障害には、慢性関節リウマチが含まれるがこれに限定されない。例えば、Cit-Cit-Y-Cit-G-Cit-Cit-S-Cit-Cit-S-Cit-Citなどのペプチドが、RAの診断のために患者における抗シトルリン抗体を検出するための市販の試験キット(例えば、ELISAキット)において使用できる(ELISA Guidebook. Theory & Practice. Editor John R. Crowther, Author John R. Crowther, Pub. Humana Press 2000)。手短に述べると、ペプチドは、マイクロタイタープレートのウェルに共有結合されるか、または非共有結合的に結合され、希釈された患者の血清が各ウェルに加えられ、一定の時間の間インキュベートされる。プレートの洗浄後、ペルオキシダーゼに結合体化されたIgGがウェルに加えられ、さらにインキュベートされる。さらなる洗浄の後、結合した抗体が、テトラメチルベンジジンなどの基質とともに検出される。一定の時間の間の後、反応が停止され、特定の波長(例えば450nm)でのプレートのOD読み取りが行われる。対照試料は同じ様式で同時に処理される。陽性OD読み取りは、陽性の結合応答を示す。
【0103】
本発明はまた、シトルリン化ペプチド/MHCクラスII複合体に特異的であるT細胞を標的化すること、またはこれらの複合体の形成を破壊することを含む、治療的ストラテジーを含む。これらの方法は、RAを処置するための他の公知の治療と組み合わせて使用できる。例えば、活性化T細胞は、可溶性ペプチド-MHCクラスII複合体を患者に投与することによって達成できるこれらの細胞のアポトーシスを誘導することによって除去できる(すなわち、米国特許第5,734,02号、米国特許第6,106,840号、米国特許第5,635,363号、米国特許第6,211,342号,、米国特許出願公開第20020176864号および米国特許出願公開第20020122818号(これらの開示はその全体が本明細書に援用される)。本発明はまた、対象におけるT細胞活性を調節するために、RNA干渉の方法を使用する治療ストラテジーにおいて本発明のペプチドの使用を含む。RNA干渉(RNAiまたはsiRNA)は、二本鎖RNA(dsRNA)によって引き起こされる遺伝子サイレンシングの型である、本発明の1つの態様において、このような方法を使用して、本発明のペプチドは、例えば、APC(すなわち、樹状細胞)に効果をもたらし、次には活性化された炎症性応答から調節性応答の1つに移行するようにT細胞に影響を与えるサイトカインをコードするRNA配列と合わせられる。RNA干渉(RNAi)においては、動物および植物における配列特異的、転写後遺伝子サイレンシングは、サイレンシングされる遺伝子に対して配列中の相同な二本鎖RNA(dsRNA)によって開始される。siRNA(小さな干渉RNA)は、慢性関節リウマチをもたらすタンパク質のシトルリン化に関与しているコードされたタンパク質を発現しないために、所望のmRNA(この場合はPADをコードしている)を標的化し、従ってこれを分解するように設計される。C.elegans、Drosophila、植物、および哺乳動物において遺伝子をサイレンシングするためのRNAiの使用に関連する方法は当該分野において公知である(Fire A.ら、1998 Nature 391:806-811; Fire, A. Trends Genet. 15, 358-363 (1999); Sharp, P. A. RNA interference 2001. Genes Dev. 15, 485-490 (2001); Hammond, S. M.ら、Nature Rev. Genet. 2, 110-1119 (2001); Tuschl, T. Chem. Biochem. 2, 239-245 (2001); Hamilton, A.ら、Science 286, 950-952 (1999); Hammond, S. M.ら、Nature 404, 293-296 (2000); Zamore, P. D.,ら、Cell 101, 25-33 (2000); Bernstein, E.ら、Nature 409, 363-366 (2001); Elbashir, S. M.ら、Genes Dev. 15, 188-200 (2001)、国際特許公開WO0129058号、国際特許公開WO9932619号、Elbashir S Mら、2001 Nature 411:494-498)、およびカナダ国特許出願第2,388,441号(これらの開示はその全体が本明細書に援用される))。
【0104】
本発明はまた、本明細書中で以前に記載されたような全身性免疫学的寛容性を誘導するための本発明に従う1種以上のペプチドおよび薬学的に許用可能な担体を含む治療組成物の投与による、慢性関節リウマチなどの自己免疫障害の治療の方法を含む。誘導された寛容性は、攻撃に際して組織中での局所的炎症性応答の減少をもたらす。1つの局面において、ペプチドは、例えば、米国特許第5,843,445号(この開示はその全体が参照として本明細書に援用される)において開示される方法のような全身性免疫学的寛容性を誘発するために患者の粘膜上皮を介して投与される。
【0105】
なお他の治療的ストラテジーは、細胞毒性分子または本発明のペプチドに放射核種(すなわち、125Iまたは131I)で標識した放射活性分子の結合体化を含み得る。本発明の他の実施形態は、毒素分子またはその活性フラグメントに共有結合された本発明のペプチドまたは抗体である細胞認識分子を含むイムノトキシン分子である。このようなイムノトキシン分子は、本明細書中に記載されるような慢性関節リウマチのための医薬または診断薬としての使用のために、単独で使用できるか、または組成物として使用できる。本発明のペプチドおよびMHCクラスII分子との細胞毒性分子のこのような結合体化は、CD4T細胞を標的化するために有益である。代替的には、いくつかの局面において、配列番号38または39に記載されるような特定のシトルリン化ペプチドは、B細胞を標的化する細胞毒性分子に単に標的化できる。この様式において、その表面に同じ抗体を有する抗シトルリン化抗体を産生するB細胞のみが標的化される。
【0106】
本発明の別の態様において、シトルリン化ペプチド/MHCクラスII分子複合体に特異的な抗体が複合体に結合するため、従って、T細胞による複合体認識を予防するために使用できる(すなわち、国際特許公開WO02/14870号)。代替的には、MHCクラスII分子のペプチド結合溝が、高アフィニティー非シトルリン化ペプチドと結合され得、これは次いでT細胞によって認識されず、または抗体によって結合されて、従って、シトルリン化ペプチド/MHCクラスII複合体の形成を予防する(すなわち、米国特許第6,355,617号)。
【0107】
本発明のさらなる局面は、一般的に、T細胞応答を誘発するシトルリン化ペプチド抗原を含む自己免疫障害の診断のための方法である。このような自己免疫障害には、慢性関節リウマチおよび多発性硬化症が含まれるがこれらに限定されない。多発性硬化症において、シトルリン化ミエリン塩基性タンパク質(MBP)T細胞応答が観察された(Tranquillら、2000. Enhanced T cell responsiveness to citrulline-containing myelin basic protein in multiple sclerosis patients. Mult. Scler. 6:4220-5)。それゆえに、MHCクラスII共有エピトープを発現するMS患者において、シトルリンに対する免疫応答は疾患のプロセスに寄与し得る。このようなものとして、シトルリン化MBPに対する抗シトルリン自己抗体の検出は、MSについて診断的であり得る。いくつかの態様において、T細胞反応性を阻害することは、MSについて治療的であり、この活性をアッセイすることはMSについて診断的である。さらに、慢性関節リウマチに関して本明細書中に記載される治療もまた、シトルリン化MBPが疾患をもたらすT細胞応答を誘発する、MSの治療に適用可能である。RAの治療のための、本明細書中に記載されるような、感受性のある患者(すなわち、MHCクラスII共有エピトープを発現する)において共有エピトープへのシトルリン化ペプチドの結合の阻害の方法は、MSの治療においてもまた、適用可能であり得る。MSの診断および治療方法の標的において有用なシトルリン化MBPタンパク質は、例えば、FLPCitHRDTG (配列番号40), VTPCitTPPPS (配列番号41); YGGCitASKYK (配列番号42)および LGGCitDSRSG (配列番号42)から選択できる。MSの診断において使用できるなお他のシトルリン化ペプチドは、シトルリン化グリア線維酸性タンパク質(GFAP)である。GFAPは、グリア細胞、星状細胞、アストログリアフィラメントに存在するタンパク質である(Nicholas A.P. 2002. Preparation of a monoclonal antibody to citrullinated epitopes: its characterization and some applications to immunohistochemistry in human brain. Glia.Mar 15;37(4):328-36、Nicholas ら、2003. Immunohistochemical localization of citrullinated proteins in adult rat brain. J Comp Neurol. May 5:459(3):251-66)。MSの診断の方法における使用のためのシトルリン化GFAPは、MERCitRITSA (配列番号43), LPTCitVDFSL (配列番号44), LNDCitFASYI (配列番号45), LRLCitLDQLT (配列番号46), LQICitETSLD (配列番号47)およびVEMCitDGEVI (配列番号48)から選択できる。
【0108】
本発明のシトルリン化MBPおよびGFAPは、多発性硬化症についての診断的および治療的アプローチにおける使用のためのものであることが当業者によって理解される。
【0109】
代替的には、本発明の実施形態において、アルギニンを有する特定のペプチドのシトルリンのペプチドへの修飾は、このアミノ酸反応を触媒する酵素ペプチジルアルギニンデイミナーゼ(PAD)に対するアンタゴニストおよび/または阻害剤の使用によって予防することができる。PADは同様の触媒活性を共有する酵素のファミリーのメンバーである。これらはカルシウム依存性酵素であり、これらは哺乳動物組織において4つのアイソフォームで存在する:PADIおよびIIIは毛嚢および上皮において存在する;PADIIはCNS組織、筋肉、およびある造血細胞において存在する;PADIVは造血組織、特に単球および好中球において存在する(Vossenaar E.R.ら、(2003). PAD, a growing family of citrullinating enzymes; genes, features and involvement in disease. Bioessays. 25(11): 1106-18)。PADの生理学的機能は完全には知られていないが、しかし、アルギニンの正電荷をより減少した電荷を有する極性アミノ酸シトルリンに変換するように作用する(図4)。
【0110】
PADの遺伝子は、ヒトにおいて1p36染色体、およびマウスにおいて4E1染色体に存在する。1p36染色体上の領域は、以前に慢性関節リウマチと関連すると同定されたが、この領域は、4つのPAD遺伝子に加えて4つの遺伝子を含み、慢性関節リウマチの発生をもたらす、PAD活性、ペプチドのシトルリン化の増加、または任意のHLA-DR分子への結合の間のいかなる直接的関連も存在しなかった。
【0111】
PADのインビトロおよびインビボでのシトルリン化のブロッキングは、これが、疾患プロセスにおける特異的決定的段階に影響を与えることに焦点を当てているので、慢性関節リウマチおよび多発性硬化症を治療するための新規な治療方法を提供する。本発明のこの実施形態の1つの態様において、PAD活性は、上記で手短に議論したRNA干渉(siRNA)のより特異的かつ効率的な技術を使用してブロックできる。この様式において、PAD遺伝子の機能はサイレンシングされ得、その結果、感受性のペプチド/タンパク質のシトルリン化は起こらず、慢性関節リウマチまたは多発性硬化症の発症をもたらさない。このことは、シトルリン化ペプチドの産生の結果として自己免疫障害を発症するリスクが特にあるSEおよびPADを発現する個体の治療の方法を提供する。シトルリンの産生に依存する、自己免疫障害のより初期とより後期の両方の段階が、PAD発現の阻害によって、および本発明のある態様においては、PAD IV発現の抑制によって抑制できる。
【0112】
PAD遺伝子の発現、従ってその機能をサイレンシングすることに加えて、低分子がまた、PADを阻害するために使用され得、従って、このような低分子はまた、種々の投与の経路において自己免疫障害の治療のための治療用組成物中で使用できる。組成物および組成物の投与の経路の議論は、本明細書中で以前に示した。当業者は、PADを阻害するそれらの能力について、潜在的な低分子をいかにしてスクリーニングするかの理解および知見を有する。適切なスクリーニング方法は、例えば、Waltersら (Designing Screens: How to Make Your Hits a Hit. Nature Reviews, Drug Discovery. 第2巻, 2003年4月、259-266頁;この開示はその全体が参照として本明細書に援用される)において記載されている。
【0113】
上記の開示は一般的に本発明を記載する。より完全な理解は、以下の特定の実施例を参照して得られ得る。これらの実施例は、単に例示の目的のみのために記載され、本発明の範囲を限定することを意図しない。形態の変更および等価物の置換は、状況が示唆し得または好都合にし得る場合に意図される。特定の用語が本明細書中で使用されたが、このような用語は、説明的な意味にあることが意図され、限定の目的のためではない。
【実施例】
【0114】
この実施例は、例示の目的のために記載され、本発明の範囲を限定することを意図するものではない。
【0115】
本開示および実施例において引用されるが明白に記載されない、合成化学、タンパク質およびペプチド化学、分子生物学、薬学、および免疫学の方法は、科学文献中に報告され、かつ当業者に周知である。
【0116】
(実施例1)
動物の作製
HLA-DR4-IEトランスジェニック、マウスMHCクラスII欠損マウスをこれらの実験において使用した(Ito K, Bian HJ, Molina M, Han J, Magram J, Saar E, Belunis C, Bolin DR, Arceo R, Campbell R, Falcioni F, Vidovic D, Hammer J, Nagy ZA. 1996. HLA-DR4-IE chimeric class II transgenic, murine class II-deficient mice are susceptible to experimental allergic encephalomyelitis. J. Exp. Med. 183:2635)。これらのマウスを、以前に記載されたように繁殖および維持した(Hill, J.A., Wang, D., Jevnikar, A.M., Cairns, E., Bell, D.A. 2002. The relationship between predicted peptide-MHC class II affinity and T cell activation in a HLA-DR(1*0401 MHC class II mouse model. Arthritis Res. 5:R40)。
【0117】
(実施例2)
ペプチドの生成
これらの研究において使用されるペプチドは、製造業者(Genemed Synthesis、San Francisco、CA)によって合成および精製された。ペプチドを、Hammerら (Hammer J, Bono E, Gallazzi F, Belunis C, Nagy Z, Sinigaglia F. 1994. Precise prediction of major histocompatibility complex class II-peptide interaction based on peptide side chain scanning. J. Exp. Med. 180:2353)の方法に従ってDRB1*0401についてそれらの予測されたアフィニティーに基づいて選択した。下線を付したアミノ酸は、9個のMHCクラスII分子結合ポケット(P1〜P9)と相互作用する残基を示すのに対して、太字で示すものはP4共有エピトープ位置で相互作用する。プロテオグリカンアグリカンから使用したペプチドの配列は以下の通りである:P4D = ヒトアグリカンペプチド 280-292, AGWLADRSVRYPI; P4R = 変化したヒトアグリカンペプチド280-292, AGWLARRSVRYPI; P4Cit = 変化したヒトアグリカンペプチド280-292, AGWLACitRSVRYPI。シトルリンはHammerらの予測アルゴリズムにおいて考慮されていないので、ビメンチンからの候補T細胞エピトープを同定するときには、グルタミンの値をアルギニンの代わりに置き換えた(グルタミンはシトルリンを同じ末端側鎖基を有する)。使用したビメンチンペプチドの配列は以下の通りであった:Vim 65-77 = ヒトビメンチンペプチド 65-77, SAVRARSSVPGVR, Vim R70Cit = 変化したヒトビメンチンペプチド65-77, SAVRACitSSVPGVR。
【0118】
(実施例3)
免疫化
DR4 tgマウスを、CFA (Difco Laboratories、Detroit, MI)中で乳化した100μlのペプチド(1μg/μl)(1:1容量比)を用いて両方の後肢の内側に皮内免疫した。10日後、マウスを屠殺し、インビトロ増殖アッセイおよびサイトカインアッセイのためにそれらの流入領域リンパ節を取り出した。
【0119】
(実施例4)
T細胞培養
細胞懸濁物を流入領域リンパ節から調製し、96ウェルプレート中で、4×105細胞/ウェルの濃度で、ペプチド抗原の存在下非存在下で、4日間培養した。抗DR抗体(BD PharMingen、Mississauga、ON)をある培養に加え(1μg/ml)、以前に記載されたようにDR制限T細胞応答を確認した(Andersson EC, Hansen BE, Jacobsen H, Madsen LS, Andersen CB, Engberg J, Rothbard JB, McDevitt GS, Malmstrom V, Holmdahl R, Svejgaard A, Fugger L. 1998. Definition of MHC and T cell receptor contacts in the HLA-DR4restricted immunodominant epitope in type II collagen and characterization of collagen-induced arthritis in HLA-DR4 and human CD4 transgenic mice. Proc. Natl. Acad. Sci. USA. 95:7574)。培養上清を78時間後に取り出し、以前に記載されたようにELISA(BD Phar Mingen, Mississauga, O.N)によってIFN-γ産生を試験した(Hill, J.A., Wang, D., Jevnikar, A.M., Cairns, E., Bell, D.A. 2002. The relationship between predicted peptide-MHC class II affinity and T cell activation in a HLA-DR(1*0401 MHC class II mouse model. Arthritis Res. 5:R40)。サイトカイン産生を2連で測定し、平均抗原特異的サイトカイン産生(対照試料中のサイトカイン産生+2SDをペプチド特異的サイトカイン産生から減算した)±SDを表す。培養終了の18時間前に、1μCiの[3H]チミジン(ICN Biomedicals、Montreal、PQ)を各ウェルに加えて、T細胞増殖を評価した。増殖実験を3連で実行し、結果をcpm±SDの平均増殖または刺激指数(実験試料のcpm/対照試料のcpm)±SEMとして表した。
【0120】
(実施例5)
ペプチド結合アッセイ
精製したHLA-DRB1*0101、*0401、*0404、*0301、*0701、*0802、*1101、および*1302に対するペプチド結合アッセイを、以前に記載されたように、放射活性標識ペプチドプローブに対して決定した(Southwood S, Sidney J, Kondo A, del Guercio MF, Appella E, Hoffman S, Kubo RT, Chesnut RW, Grey HM, Sette A. 1998. Several common HLA-DR types share largely overlapping peptide binding repertoires. J. Immunol. 160:3363)。精製したHLA-DRB1分子に対する標識ペプチドの50%阻害のために必要な未標識ビメンチンペプチドのnM濃度(IC50)を、相互作用のアフィニティー(kDa)の近似値として使用した。結果を、nMで測定したIC50値の逆数として表現する。
【0121】
(実施例6)
DR4-1E tgマウスにおける関節炎の誘導
DR4-1E tgマウスを、CFA (Difco Laboratories、Detroit, MI)中で乳化した100μlのシトルリン化ヒトフィブリノーゲン(1μg/μl)(1:1容量比)を用いて両方の後肢の内側に皮下免疫した。シトルリン化フィブリノーゲンは以前に記載されるように調製した(Christine Masson-Bessiereら、2001. The Major Synovial Targets of the Rheumatoid arthritis-Specific Antifilaggrin Autoantibodies Are Deiminated Forms of the ( and ( Chains of Fibrin 1 The Journal of Immunology 166: 4177-4184)。マウスは不完全フロイントアジュバント中の同じ抗原の第2の免疫を21日間受容し、引き続いて関節炎の徴候について観察した(図6AからF)。
【0122】
(実施例7)
DR4-IE tgマウスにおける抗シトルリン抗体の生成およびRA自己抗体の検出
DR4-IE tgマウスを、0、21、および42日めに、それぞれCFA、IFA、およびIFAで乳化した100μgのペプチドCit-Cit-Y-Cit-G-Cit-Cit-S-Cit-Cit-S-Cit-Cit(配列番号39)で免疫した。52日目にマウスを屠殺し、血清を血液から収集した。抗体反応性を、以前に記載されたように(Schllekensら、1998. J Clin Invest. Volume 101, 第1巻, 1998年1月, 273-281頁)、ELISAによって、ペプチド(配列番号39)およびcfc1ペプチドを使用して、血清試料中で検出した。ペプチド(配列番号39)またはcfc1に対する抗シトルリン抗体は、ペプチド(配列番号39)で免疫したマウスの血清中で見い出された。シトルリン化フィブリノーゲンで免疫した関節炎マウスからの抗体もまた、これらのペプチドの両方に結合した。試験した多くの対照マウスにおいて抗体は見られなかった。ペプチドCit-Cit-Y-Cit-G-Cit-Cit-S-Cit-Cit-S-Cit-Cit(配列番号39)に対するヒト抗体の反応性もまた試験され、市販のELISAキット(Quantilite、INOVA Diagnostics)と比較された。シトルリン化フィブリノーゲンに対する抗体反応性もまた実証された(図7)。
【0123】
表1は、ペプチドCit-Cit-Y-Cit-G-Cit-Cit-S-Cit-Cit-S-Cit-Cit(配列番号39)、シトルリン化フィブリノーゲンポリペプチド(トランスジェニックマウスを免疫するために以前に使用した)、および市販のCCP試薬を使用する抗シトルリン化抗体の頻度および特異性を示す。試験された患者の中でも慢性関節リウマチについてこのアッセイの感度および特異性は、Cit-Cit-Y-Cit-G-Cit-Cit-S-Cit-Cit-S-Cit-Cit(配列番号39)ペプチドおよびCCPペプチドがRAの診断のためのそれらの感度および特異性によって同様であったことを示しかつ指摘する。
【0124】
図8は、市販のCCPキットおよびCit-Cit-Y-Cit-G-Cit-Cit-S-Cit-Cit-S-Cit-Cit(配列番号39)ペプチドを使用する抗シトルリン化抗体の相関を示す。これらのアッセイを使用して試験した51例の慢性関節リウマチ患者から、0.48の相関係数が見られた。表2において見られるように、ペプチドCit-Cit-Y-Cit-G-Cit-Cit-S-Cit-Cit-S-Cit-Cit(配列番号39)は、慢性関節リウマチの診断について、98%の特異性および71%の感度を提供した。乾癬性関節炎を有する49例の患者のうちの1例のみが陽性であった。このことは、ペプチドCit-Cit-Y-Cit-G-Cit-Cit-S-Cit-Cit-S-Cit-Cit(配列番号39)および一般的には式Cit-Cit-X-Cit-G-Cit-Cit-Z-Cit-Cit-B-Cit-Cit(配列番号38)(ここでXはY、F、W、I、L、MおよびVから選択され、ZはA、D、I、N、P、S、TおよびVから選択され、BはA、G、H、Q、S、TおよびVから選択される)のペプチドが、慢性関節リウマチにおける抗シトルリン化抗体の検出のために有用であることを示す。
【0125】
(実施例8)
PAD発現についてマウスおよびヒトの細胞系統の選択のため;PAD標的化のためのsiRNAを同定するため;siRNAを使用して選択された細胞系統におけるPAD発現を阻害するため;およびPAD発現をモニターするための、siRNAを使用するPADの阻害、インビトロ研究。PAD IVに関して記載したが、当業者に理解されるように、同類のプロセスが、PAD IIまたは任意の他のPADのために使用できる。
(a)PAD(IIおよびIV)発現についてのマウスおよびヒトの細胞系統の選択
ATCCからのヒトおよびマウスの単球/マクロファージおよび顆粒球/好中球の細胞系統。これらの細胞系統は、ビタミンD3、レチノイン酸、およびデキサメタゾンなどの種々の薬剤を使用して、マクロファージおよび好中球の表現型にさらに分化できる(www.atcc.org)。好中球およびマクロファージの細胞型は、PAD IIおよびIVを発現する(Asaga Hら、(1998) Selective deimination of vimentin in calcium ionophore-induced apoptosis of mouse peritoneal macrophages. Biochem Biophys Res Commun. 243(3):641-6: Nakashima Kら、(1999) Molecular characterization of peptidylarginine deiminase in HL-60 cells induced by retinoic acid and 1alpha,25-dihydroxyvitamin D3. J Biol Chem. 274(39), 27786-92)。細胞は、RA滑膜において活性化されかつ豊富である。PAD IIおよびIVは、LPS活性化細胞系統において定量的に決定され、最高のレベルのPAD IIおよびIVを有する細胞系統はsiRNA阻害を試験するために選択される。
【0126】
(b)PAD(IIおよびIV)標的化のためのsiRNAの同定
ヒトおよびマウスのPAD IV遺伝子配列の分析は、選択判断基準を満たす全部で11個のsiRNA配列を明らかにした。11個の配列のうち、8個がヒト、3個がマウスであった(表4)。これらのsiRNAは、siRNAの安定性、有効寿命、および強度を増強する独自技術のRNA修飾(siSTABLE技術と呼ばれる)を使用する、Dharmacon Inc.から入手した(www.dharmacon.com)。
【0127】
(c)PAD(IIおよびIV)発現の阻害
siRNAは、PAD IIまたはIVを高度に発現する選択されたヒトおよびマウスの細胞系統において、これらの細胞における有効なPAD IIまたはIVの阻害のための最適条件を確立するために試験される。これは、siRNAの送達、投薬量、およびタイミングに依存する。siRNAは、当業者に周知の技術であるリポソームトランスフェクションによって細胞に送達される。トランスフェクション効率は使用される細胞型に依存して変化し得るので、いくつかのカチオン性リポソーム試薬(表5)が利用される。トランスフェクション効率および毒性は、蛍光標識したsiRNA(FL Luciferase GL2 Duplex)およびヨウ化プロピジウムをそれぞれ使用して、FACSによってモニターされる。カチオン性脂質複合体中に細胞マーカー特異的抗体を含むイムノリポソームは、PAD IVを産生するマクロファージまたは好中球のいずれかへのsiRNAの効率的な標的化を可能にするはずである(Zhang Yら(2003) In vivo knockdown of gene expression in brain cancer with intravenous RNAi in adult rats. J Gene Med. 5(12):1039-45: Bestman-Smith J.ら(2000) Sterically stabilized liposomes bearing anti-HLA-DR antibodies for targeting the primary cellular reservoirs of HIV-1. Biochim Biophys Acta. 1468(1-2):161-74)。
【0128】
(d)PAD(IIおよびIV)発現のモニタリング
未処理細胞対siRNA処理細胞でのPAD IIまたはIV mRNAおよびタンパク質発現は、RT-PCRおよびウェスタンブロッティングによってそれぞれモニターした。これらの方法の組み合わせ使用は、siRNA媒介阻害を分析するために最も良好である。PCRにおいて使用するオリゴヌクレオチドプライマーを表6および表7に列挙する。これらには、PAD IVに加えて、GAPDH、PAD I、II、およびIIIのためのプライマーが含まれる。なぜなら、ハウスキーピング遺伝子およびすべてのPADの遺伝子発現は、siRNA治療のための標的特異性を保証するからである。ヒトタンパク質とマウスタンパク質の両方を認識するポリクローナル抗PAD IV抗体は、標準として使用するために組換えPAD IVと同様に入手する。
【0129】
(実施例8)
siRNA治療の送達を実証するため;siRNA治療のための動物モデルを選択するため;およびPAD IV阻害の効果をモニターするための、siRNAを使用するPADの阻害、インビボ研究
(a)siRNA治療の送達
最も効果的かつ特異的なPAD IIおよびIVの阻害剤としてインビトロで同定されたリポソーム/イムノリポソームsiRNA複合体が静脈内注射される。このアプローチは、肝臓、肺、腎臓、脾臓、および膵臓を含む多数の器官における遺伝子発現を効果的に阻害することが以前に示された(Lewis D.L.ら、(2002) Efficient delivery of siRNA for inhibition of gene expression in postnatal mice. Nat Genet. 32(1): 107-8; Song E.ら、(2003) RNA interference targeting Fas protects mice from fulminant hepatitis. Nat Med. 9(3): 347-51)。プラスミドにコードされたsiRNAのレトロウイルス送達もまた、代替的なアプローチとして使用できる(McCaffrey A.P.ら、(2002) RNA interference in adult mice. Nature 418(6893):38-9)。
【0130】
(b)siRNA治療のための動物モデルの選択
関節炎のいくつかのマウスモデルが使用される。第1のものは、連鎖球菌細胞(SCW)誘導性関節炎であり、これは、マウスの関節内でPAD IVの随伴性のアップレギュレーションを伴う、関節炎が24時間以内に誘導される急性関節炎モデルである(Vossenaar E. R.ら、(2003) Citrullination of synovial proteins in murine models of rheumatoid arthritis. Arthritis Rheum. 48(9): 2489-500)。送達および用量を含む、siRNA治療のための最適条件の決定を表8に示す。第2のモデル、コラーゲン誘導性関節炎は、免疫の35日後に発症する持続性の関節炎を伴い、慢性的である。これは、延長した時間の期間にわたってsiRNA効果のモニタリングを可能にする(表9)。SCWおよびCIAマウスモデルは、PAD IV活性のレベルの増加およびこれらのマウスの関節におけるシトルリン化タンパク質の存在を示すが、しかし、シトルリンに標的化される自己免疫応答は存在しない。SCW関節炎がSE tgマウス中で誘導される場合、シトルリンで免疫されたときにSE tgマウスが関節炎を発症するにつれて、シトルリンに対する自己免疫応答が起こる。これは、非SE tg野生型マウスにおいてSCWで通常予測される一過性の関節炎を延長および/または悪化させる(表10)。この効果は、PAD IV阻害によって弱められる。
【0131】
(c)siRNA治療の効果のモニタリング
siRNA/リポソーム処理マウスまたは未処理マウスは、siRNA治療の効力および期間を決定するために種々の時点で屠殺される。これは、標準的なスコア付けシステムを使用して関節炎の臨床的徴候をモニタリングすることによって評価される(Current Protocols in Immunology 15.5章、http://www.mrw2.interscience.wiley.com/cponline)。疾患の病理学的徴候およびシトルリン化のプロセスもまたモニターされる。切片化された関節は、炎症およびシトルリン化タンパク質、PAD IVタンパク質およびその転写物の存在について点数付けされる。抗シトルリン抗体の産生はまた、SE tgマウスモデルにおいて評価される。
【0132】
本発明の好ましい態様を本明細書中で詳細に記載してきたが、本発明の精神または添付の特許請求の範囲から逸脱することなく本発明を変化させ得ることは当業者によって理解されよう。
【0133】
【表1】
【0134】
【表2】
【0135】
ヒト血清試料におけるJED抗原とCCP抗原に対する抗体の間の反応性の相関を見ることができる。
【0136】
【表3】
【0137】
【表4】
【0138】
すべての細胞株は本発明者らの研究室で現在使用されているか、またはATCCに注文することができる。マクロファージまたは好中球の表現型への分化は、これらの細胞型に必要である(例えば、レチノイン酸、ビタミンD3、LPS)。
【0139】
【表5】
【0140】
【表6】
【0141】
【表7】
【0142】
【表8】
【0143】
【表9】
【0144】
【表10】
【0145】
【表11】
【図面の簡単な説明】
【0146】
本発明は、本明細書中に示される詳細な説明から、および添付の図面からより完全に理解されるようになる。これらは、例示の目的のみのために示され、本明細書の意図される範囲を限定するものではない。
【図1】図1A、1B、および1Cは、共有エピトープによって形成されたP4ポケットと相互作用する位置にアルギニン(P4R)、シトルリン(P4Cit)、またはアスパラギン酸(P4D)を含有するペプチドに対する、DR4-IE tgマウスにおけるT細胞免疫応答の分析を表すグラフである。グラフ1A DR4-IE tgでは、マウスは示されたペプチド用いて免疫され、かつ10日後に流入領域リンパ節細胞に種々の濃度の同じペプチドを用いてインビトロでチャレンジした。データは、試験された各ペプチドについて8匹のマウスの平均増殖応答±SEMを表す。図1Bにおいて、リコールT細胞応答のHLA-DR制限は、抗原(コントロール)なし、10μg/mlの免疫抗原(P4Dは左のパネル、P4Citは右のパネル)とともに、または免疫抗原(10μg/ml)および抗DR抗体(DR)の存在下で、流入領域リンパ節細胞をインキュベートすることによってインビトロで決定された。結果は、各免疫抗原について4匹のマウスの平均増殖応答±SDを表す。図1Cは、10μg/mlの免疫ペプチド(P4D、P4Cit、またはP4R)を用いるインビトロチャレンジに応答したIFN-γ産生を示す。サイトカイン産生はELISAによって決定され、試験された各ペプチドについて、4匹のマウスの平均抗原特異的IFN-γ産生±SDを表す。
【図2】図2A、2B、および2Cは、修飾されていないビメンチンペプチド(Vim 65-77)またはシトルリン含有ビメンチンペプチド(Vim R70Cit)に対するDR4-IE tgマウスにおけるT細胞免疫応答の分析を示すグラフである。図2Aにおいて、DR4-IE tgマウスは、示されたペプチドで免疫され、かつ10日後に流入領域リンパ節細胞に種々の濃度の同じペプチドを用いてインビトロでチャレンジした。データは、試験された各ペプチドについて8匹のマウスの平均増殖応答±SEMを表す。図2Bは、Vim R70Cit免疫マウス(左のパネル)およびVim 65-77免疫マウス(右のパネル)からの特異性およびT細胞リコール応答のHLA-DR制限を示す。流入領域リンパ節細胞は免疫ペプチド、免疫ペプチドおよび抗DR抗体(DR)、未修飾ビメンチンペプチド(左のパネルの65-77)、またはビメンチンペプチドを含むシトルリン(右のパネルのR70Cit)でチャレンジされた。インビトロチャレンジのために使用された濃度は10μg/mlであり、結果は各免疫抗原について4匹のマウスの平均刺激指数±SEMを表す。図2Cは、種々の濃度の免疫ペプチドを用いるインビトロチャレンジに応答したIFN-γ産生を示す(Vim R70CitまたはVim 65-77)。サイトカイン産生はELISAによって決定され、試験された各ペプチドについて、4匹のマウスの平均抗原特異的IFN-γ産生±SDを表す。
【図3】図3は、精製MHCクラスII分子についてVim 65-77とVim R70Citの相対的アフィニティーを示すグラフである。共有エピトープ陽性対立遺伝子(*0101、0401、*0404)および共有エピトープ陰性対立遺伝子(*0301、*0701、*0802、*1101、*1302)に対する結合アフィニティーが、本明細書に記載される実施例において説明されるように決定された。100nM未満のIC50値を有するペプチドは高アフィニティー結合因子であると見なされる。
【図4】図4は、酵素ペプチジルアルギニンデイミナーゼ(PAD)が、いかにしてアルギニンの正電荷を有するイミノ基をシトルリンの極性の荷電されていないカルボニル基に変換するかを示す。
【図5】図5A-5Cは、(5A)抗シトルリン抗体に陰性であるRA患者;(5B)抗シトルリン抗体に陽性であることのRA患者;(5C)他のリウマチ疾患を有する非RA患者におけるシトルリン化フィブリノーゲンペプチドに対するT細胞増殖応答を示す。シトルリン化フィブリノーゲンペプチドに対するT細胞増殖は、RA患者においてのみ見られ、抗シトルリン抗体を有した患者においては、より一般的であった。末梢血単核球は、T細胞培養について実施例4で記載されているようにFicoll-Hypaque遠心沈殿によってRAドナーおよび非RAドナーから単離され、かつ培養された。これらの実験において使用されたペプチドは以下の通りである:Fib a R143Cit = VLKCitKVIEK (配列番号6); Fib a R673Cit = IQQCitMDGSL (配列番号11); Fib a R720Cit = LTQCitGSVLR (配列番号12); Fib a R847Cit = VSFCitGADYS (配列番号15); Fib b R285Cit = IQNCitQDGSV (配列番号20); Fib a R178Cit = IKICitSCRGS (配列番号7).
【図6】図6A-Fは、SE tgマウスにおいてシトルリン化フィブリノーゲンによって誘導された関節炎の臨床的および病理学的な特徴を示す写真である。図6Aは、非免疫されていないマウスからの後足の関節炎の臨床的外観を示し、6Bはヒトフィブリノーゲンで免疫された関節炎マウスであり、そして、6Cはシトルリン化ヒトフィブリノーゲンで免疫された関節炎マウスである。図6Dおよび6Eは、6Cからマウスの脛骨足根骨関節の病理学を示すH&E切片であるのに対して、6Fは6Bににおいて示されるマウスからの切片である。
【図7】図7は、DR4-IE tgマウスにおける、抗シトルリン抗体の産生を示す。
【図8】図8は、ヒト血清中で市販キット(Quantilite(商標))を使用してペプチドCit-Cit-Y-Cit-G-Cit-Cit-S-Cit-Cit-S-Cit-CitとCCPの間の相関を示す。
【特許請求の範囲】
【請求項1】
共有エピトープを有するMHCクラスII分子に増加したアフィニティーで結合するシトルリン化ペプチドであって、SAVRACitSSVPGVR (配列番号1); FSMCitIVCLV (配列番号2); VVECitHQSAC (配列番号3); FTNCitINKLK (配列番号4); LRSCitIEVLK (配列番号5); VLKCitKVIEK (配列番号6); IKICitSCRGS (配列番号7); LPSCitDRQHL (配列番号8); FRHCitHPDEA (配列番号9); FPSCitGKSSS (配列番号10); IQQCitMDGSL (配列番号11); LTQCitGSVLR (配列番号12); YHFCitVGSEA (配列番号13); YDPCitNNSPY (配列番号14); VSFCitGADYS (配列番号15); YSLCitAVRMK (配列番号16); MKICitPLVTQ (配列番号17); YRACitPAKAA (配列番号18); WQKCitQKQVK (配列番号19); IQNCitQDGSV (配列番号20); WYNCitCHAAN (配列番号21); YSMCitKMSMK (配列番号22); MKICitPFFPQ (配列番号23); LHPCitNLILY (配列番号24); VATCitDNCCI (配列番号25); LDECitFGSYC (配列番号26); LKSCitIMLEE (配列番号27); FQKCitLDGSV (配列番号28); YALCitVELED (配列番号29); WNGCitTSTA (配列番号30); WKTCitWYSMK (配列番号31); YATCitSSAVR (配列番号32); VRLCitSSVPG (配列番号33); LNDCitFANYI (配列番号34); MLQCitEEAEN (配列番号35); LNLCitETNLD (配列番号36);及びVETCitDGQVI (配列番号37) からなる群から選択される少なくとも1つのアミノ酸配列を含むペプチド。
【請求項2】
前記ペプチドがさらなるシトルリン残基を含む、請求項1に記載のペプチド。
【請求項3】
請求項1または2に記載のペプチドの機能的類似体。
【請求項4】
請求項1、2、または3に記載のペプチドに特異的な抗体。
【請求項5】
共有エピトープを有するMHCクラスII分子に増加したアフィニティーで結合するペプチドであって、前記ペプチドは式Cit-Cit-X-Cit-G-Cit-Cit-Z-Cit-Cit-B-Cit-Citを有し、ここでXはY、F、W、I、L、MおよびVから選択され、ZはA、D、I、N、P、S、TおよびVから選択され、BはA、G、H、Q、S、TおよびVから選択されるペプチド。
【請求項6】
前記ペプチドが配列Cit-Cit-Y-Cit-G-Cit-Cit-S-Cit-Cit-S-Cit-Citを含む、請求項5に記載のペプチド。
【請求項7】
前記ペプチドがさらなるシトルリン残基を含む、請求項5または6に記載のペプチド。
【請求項8】
前記ペプチドが、前記ペプチドの先頭または終端にさらなる荷電したアミノ酸を含み、前記荷電したアミノ酸がアルギニン、リジン、アルパラギン、およびグルタミン酸からなる群から選択される、請求項5、6、または7に記載のペプチド。
【請求項9】
請求項5、6、7または8に記載のペプチドの機能的類似体。
【請求項10】
請求項5から9のいずれか一項に記載のペプチドに特異的な抗体。
【請求項11】
前記ペプチドが環状化されている請求項1から3および5から9のいずれか一項に記載のペプチド。
【請求項12】
前記ペプチドが検出可能な標識で標識されている、請求項1、5、または11に記載のペプチド。
【請求項13】
前記ペプチドが細胞毒性分子または放射活性分子に結合体化されている、請求項1から12のいずれか一項に記載のペプチド。
【請求項14】
請求項1から13のいずれか一項に記載のペプチドの1種以上の有効量および薬学的に許容可能な担体を含む薬学的組成物。
【請求項15】
前記組成物がアジュバントをさらに含む、請求項14に記載の薬学的組成物。
【請求項16】
慢性関節リウマチを診断する方法であって、
(a)対象からの血液試料をシトルリン化ペプチドと接触させる工程;
(b)前記試料中のT細胞が活性化されているか否か、および/または前記シトルリン化ペプチドと、共有エピトープを有する前記MHCクラスII分子との間に複合体が形成されるか否かを決定する工程
を含む方法。
【請求項17】
前記シトルリン化ペプチドが、
(a)配列番号1から37のペプチド;
(b)配列番号38から39のペプチド;および
(c)配列番号49から60のペプチド
からなる群から選択され、ここでアルギニンがシトルリンで置換されている請求項16に記載の方法。
【請求項18】
対象における慢性関節リウマチ、または慢性関節リウマチに対する感受性を診断する方法であって、
前記対象の抗原提示細胞上に共有エピトープを有するMHCクラスII分子に結合した請求項1から3、5から9、または11から13のいずれか一項に記載のシトルリン化ペプチド抗原への、前記対象のT細胞による認識を決定する工程を含み、ここで、前記T細胞による認識が、前記対象が慢性関節リウマチを有するか、または慢性関節リウマチに感受性であることを示す
方法。
【請求項19】
共有エピトープを有するMHCクラスII分子に結合したシトルリン化ペプチド抗原と反応性である自己反応性T細胞の検出のための診断方法であって、
末梢血単核細胞の単離した試料を、請求項1から3、5から9、または11から13のいずれか一項に記載の1つの以上のシトルリン化ペプチドとインキュベートする工程;
前記対象における活性化自己反応性T細胞の存在を示す、T細胞の応答を検出する工程
を含む方法。
【請求項20】
共有エピトープを有するMHCクラスII分子に結合したシトルリン化ペプチド抗原と反応性である自己反応性T細胞の検出のための診断方法であって、
末梢血単核細胞の単離した試料を、配列番号49から60のいずれか1つの1つ以上のシトルリン化ペプチドとインキュベートする工程;
前記対象における活性化自己反応性T細胞の存在を示す、T細胞の応答を検出する工程
を含む方法。
【請求項21】
前記T細胞がCD4+である、請求項19または20に記載の方法。
【請求項22】
活性化T細胞が、前記対象が自己免疫障害を有するか、または自己免疫障害に感受性であることを示す、請求項21に記載の方法。
【請求項23】
前記自己免疫障害が慢性関節リウマチである、請求項22に記載の方法。
【請求項24】
対象からの試料中で抗シトルリン自己抗体を検出するための方法であって、
試料を式Cit-Cit-X-Cit-G-Cit-Cit-Z-Cit-Cit-B-Cit-Citのペプチドと接触させ、ここでXはY、F、W、I、L、MおよびVから選択され、ZはA、D、I、N、P、S、T、Vから選択され、BはA、G、H、Q、S、TおよびVから選択される工程;ならびに
前記試料中で抗シトルリン抗体への前記ペプチドの結合を検出する工程
を含む方法。
【請求項25】
前記ペプチドが配列Cit-Cit-Y-Cit-G-Cit-Cit-S-Cit-Cit-S-Cit-Citを含む、請求項24に記載の方法。
【請求項26】
前記ペプチドが環状化されている、請求項25に記載の方法。
【請求項27】
抗シトルリン抗体への前記ペプチドの結合の検出が自己免疫障害の診断用である、請求項24、25、または26に記載の方法。
【請求項28】
前記自己免疫障害が慢性関節リウマチである、請求項27に記載の方法。
【請求項29】
前記自己免疫障害が多発性硬化症である、請求項27に記載の方法。
【請求項30】
慢性関節リウマチを有する対象を治療するための方法であって、前記対象における共有エピトープを有する前記MHCクラスII分子への、請求項1から5に記載のペプチドの結合を妨害する工程を含む方法。
【請求項31】
前記対象におけるペプチジルアルギニンデイミナーゼの発現を阻害する工程を含む、請求項30に記載の方法。
【請求項32】
前記阻害が、前記ペプチジルアルギニンデイミナーゼの遺伝子配列に標的化されるsiRNAの前記対象への投与を含む、請求項31に記載の方法。
【請求項33】
前記対象中でのペプチジルアルギニンデイミナーゼの機能を阻害する工程を含む、請求項31に記載の方法。
【請求項34】
前記阻害が、ペプチジルアルギニンデイミナーゼの機能をブロックする、前記対象への薬剤の投与を含む、請求項33に記載の方法。
【請求項35】
T細胞活性化および炎症性応答をもたらす、共有エピトープを有するMHCクラスII分子へのシトルリン化ペプチドの結合によって誘発される慢性関節リウマチに罹患している対象を治療するための方法であって、
薬学的に許容可能な担体とともに、請求項1または5に記載のペプチドを含む組成物のT細胞寛容誘導量を前記対象に投与する工程
を含む方法。
【請求項36】
前記薬学的に許容可能な担体が可溶性MHCクラスII分子である、請求項35に記載の方法。
【請求項37】
共有エピトープ細胞とのシトルリン化抗原MHCクラスII複合体の形成を含む、自己免疫障害の診断方法であって、
対象からの末梢血単核細胞の単離された試料を1種以上のシトルリン化ペプチド抗原とインキュベートする工程;
共有エピトープ細胞複合体とのシトルリン化抗原MHCクラスIIの形成を検出する工程であって、このような検出が前記対象において自己免疫障害をもたらすT細胞応答を誘発する可能性を示す
方法。
【請求項38】
前記シトルリン化ペプチドが、
(a)配列番号1から37のペプチド;
(b)配列番号38から39のペプチド;
(c)配列番号40から48のペプチド;および
(d)配列番号49から60のペプチド
から選択され、ここでアルギニンがシトルリンで置換されている請求項37に記載の方法。
【請求項39】
自己免疫障害が慢性関節リウマチである、請求項37または38に記載の方法。
【請求項40】
前記シトルリン化ペプチドがシトルリン化BMPペプチドである、請求項37に記載の方法。
【請求項41】
前記シトルリン化BMPペプチドが配列番号40から48からなる群から選択される、請求項40に記載の方法。
【請求項42】
前記自己免疫障害が多発性硬化症である、請求項37または38に記載の方法。
【請求項43】
対象における共有エピトープ複合体を有するシトルリン化ペプチドMHCクラスIIによってT細胞の活性化を最小化または予防するための方法であって、前記T細胞の活性化を阻害するために、前記複合体を標的化し、これに結合する抗体を前記対象に投与する工程を含む方法。
【請求項44】
対象における潜在的に抗原性のペプチド中でアルギニンのシトルリンへの変換を予防するための方法であって、前記対象にペプチジルアルギニンデイミナーゼのアンタゴニストまたは阻害剤の有効量を投与する工程を含む方法。
【請求項45】
共有エピトープを有するMHCクラスII分子へのシトルリン化ペプチドの結合を阻害する薬学的化合物を同定するためのスクリーニング方法であって、
候補薬学的化合物をトランスジェニックDR4-IE tgマウスに投与する工程;および
T細胞活性を測定する工程および/またはシトルリン化ペプチド/共有エピトープを有するMHCクラスII分子の複合体形成を測定する工程
を含む方法。
【請求項46】
慢性関節リウマチの研究のための動物モデルを提供するために動物において慢性関節リウマチを誘導する方法であって、薬学的に許容可能な担体とともに、請求項1または請求項5に記載の1種以上のシトルリン化ペプチドを含む組成物の慢性関節リウマチ誘導量を前記動物に投与する工程を含む方法。
【請求項47】
投与が注射による、請求項46に記載の方法。
【請求項48】
多発性硬化症の研究のための動物モデルを提供するために動物において多発性硬化症を誘導する方法であって、配列番号40から48から選択される1種以上のシトルリン化ペプチドを含む組成物の多発性硬化症誘導量を前記動物に投与する工程を含む方法。
【請求項49】
多発性硬化症の診断のための方法における、配列番号40から48のペプチドのいずれか1つの使用。
【請求項50】
MSの診断において有用であるシトルリン化MBPペプチドおよび/またはGFAPペプチドであって、FLPCitHRDTG (配列番号40), VTPCitTPPPS (配列番号41); YGGCitASKYK (配列番号42), LGGCitDSRSG (配列番号42), MERCitRITSA (配列番号43), LPTCitVDFSL (配列番号44), LNDCitFASYI (配列番号45), LRLCitLDQLT (配列番号46), LQICitETSLD (配列番号47),及びVEMCitDGEVI (配列番号48) およびその類似体からなる群から選択されるペプチド。
【請求項51】
請求項50のペプチド、およびそのための薬学的に許容可能な担体を含む組成物。
【請求項52】
共有エピトープを有するMHCクラスII分子に結合したシトルリン化ペプチド抗原と反応性である自己反応性T細胞の検出のための診断方法であって、
対象からの末梢血単核細胞の単離された試料を、配列番号40から48のいずれか1つに記載の1つ以上のシトルリン化ペプチドとインキュベートする工程;
前記対象における活性化自己反応性T細胞の存在を示す、T細胞の応答を検出する工程
を含む方法。
【請求項1】
共有エピトープを有するMHCクラスII分子に増加したアフィニティーで結合するシトルリン化ペプチドであって、SAVRACitSSVPGVR (配列番号1); FSMCitIVCLV (配列番号2); VVECitHQSAC (配列番号3); FTNCitINKLK (配列番号4); LRSCitIEVLK (配列番号5); VLKCitKVIEK (配列番号6); IKICitSCRGS (配列番号7); LPSCitDRQHL (配列番号8); FRHCitHPDEA (配列番号9); FPSCitGKSSS (配列番号10); IQQCitMDGSL (配列番号11); LTQCitGSVLR (配列番号12); YHFCitVGSEA (配列番号13); YDPCitNNSPY (配列番号14); VSFCitGADYS (配列番号15); YSLCitAVRMK (配列番号16); MKICitPLVTQ (配列番号17); YRACitPAKAA (配列番号18); WQKCitQKQVK (配列番号19); IQNCitQDGSV (配列番号20); WYNCitCHAAN (配列番号21); YSMCitKMSMK (配列番号22); MKICitPFFPQ (配列番号23); LHPCitNLILY (配列番号24); VATCitDNCCI (配列番号25); LDECitFGSYC (配列番号26); LKSCitIMLEE (配列番号27); FQKCitLDGSV (配列番号28); YALCitVELED (配列番号29); WNGCitTSTA (配列番号30); WKTCitWYSMK (配列番号31); YATCitSSAVR (配列番号32); VRLCitSSVPG (配列番号33); LNDCitFANYI (配列番号34); MLQCitEEAEN (配列番号35); LNLCitETNLD (配列番号36);及びVETCitDGQVI (配列番号37) からなる群から選択される少なくとも1つのアミノ酸配列を含むペプチド。
【請求項2】
前記ペプチドがさらなるシトルリン残基を含む、請求項1に記載のペプチド。
【請求項3】
請求項1または2に記載のペプチドの機能的類似体。
【請求項4】
請求項1、2、または3に記載のペプチドに特異的な抗体。
【請求項5】
共有エピトープを有するMHCクラスII分子に増加したアフィニティーで結合するペプチドであって、前記ペプチドは式Cit-Cit-X-Cit-G-Cit-Cit-Z-Cit-Cit-B-Cit-Citを有し、ここでXはY、F、W、I、L、MおよびVから選択され、ZはA、D、I、N、P、S、TおよびVから選択され、BはA、G、H、Q、S、TおよびVから選択されるペプチド。
【請求項6】
前記ペプチドが配列Cit-Cit-Y-Cit-G-Cit-Cit-S-Cit-Cit-S-Cit-Citを含む、請求項5に記載のペプチド。
【請求項7】
前記ペプチドがさらなるシトルリン残基を含む、請求項5または6に記載のペプチド。
【請求項8】
前記ペプチドが、前記ペプチドの先頭または終端にさらなる荷電したアミノ酸を含み、前記荷電したアミノ酸がアルギニン、リジン、アルパラギン、およびグルタミン酸からなる群から選択される、請求項5、6、または7に記載のペプチド。
【請求項9】
請求項5、6、7または8に記載のペプチドの機能的類似体。
【請求項10】
請求項5から9のいずれか一項に記載のペプチドに特異的な抗体。
【請求項11】
前記ペプチドが環状化されている請求項1から3および5から9のいずれか一項に記載のペプチド。
【請求項12】
前記ペプチドが検出可能な標識で標識されている、請求項1、5、または11に記載のペプチド。
【請求項13】
前記ペプチドが細胞毒性分子または放射活性分子に結合体化されている、請求項1から12のいずれか一項に記載のペプチド。
【請求項14】
請求項1から13のいずれか一項に記載のペプチドの1種以上の有効量および薬学的に許容可能な担体を含む薬学的組成物。
【請求項15】
前記組成物がアジュバントをさらに含む、請求項14に記載の薬学的組成物。
【請求項16】
慢性関節リウマチを診断する方法であって、
(a)対象からの血液試料をシトルリン化ペプチドと接触させる工程;
(b)前記試料中のT細胞が活性化されているか否か、および/または前記シトルリン化ペプチドと、共有エピトープを有する前記MHCクラスII分子との間に複合体が形成されるか否かを決定する工程
を含む方法。
【請求項17】
前記シトルリン化ペプチドが、
(a)配列番号1から37のペプチド;
(b)配列番号38から39のペプチド;および
(c)配列番号49から60のペプチド
からなる群から選択され、ここでアルギニンがシトルリンで置換されている請求項16に記載の方法。
【請求項18】
対象における慢性関節リウマチ、または慢性関節リウマチに対する感受性を診断する方法であって、
前記対象の抗原提示細胞上に共有エピトープを有するMHCクラスII分子に結合した請求項1から3、5から9、または11から13のいずれか一項に記載のシトルリン化ペプチド抗原への、前記対象のT細胞による認識を決定する工程を含み、ここで、前記T細胞による認識が、前記対象が慢性関節リウマチを有するか、または慢性関節リウマチに感受性であることを示す
方法。
【請求項19】
共有エピトープを有するMHCクラスII分子に結合したシトルリン化ペプチド抗原と反応性である自己反応性T細胞の検出のための診断方法であって、
末梢血単核細胞の単離した試料を、請求項1から3、5から9、または11から13のいずれか一項に記載の1つの以上のシトルリン化ペプチドとインキュベートする工程;
前記対象における活性化自己反応性T細胞の存在を示す、T細胞の応答を検出する工程
を含む方法。
【請求項20】
共有エピトープを有するMHCクラスII分子に結合したシトルリン化ペプチド抗原と反応性である自己反応性T細胞の検出のための診断方法であって、
末梢血単核細胞の単離した試料を、配列番号49から60のいずれか1つの1つ以上のシトルリン化ペプチドとインキュベートする工程;
前記対象における活性化自己反応性T細胞の存在を示す、T細胞の応答を検出する工程
を含む方法。
【請求項21】
前記T細胞がCD4+である、請求項19または20に記載の方法。
【請求項22】
活性化T細胞が、前記対象が自己免疫障害を有するか、または自己免疫障害に感受性であることを示す、請求項21に記載の方法。
【請求項23】
前記自己免疫障害が慢性関節リウマチである、請求項22に記載の方法。
【請求項24】
対象からの試料中で抗シトルリン自己抗体を検出するための方法であって、
試料を式Cit-Cit-X-Cit-G-Cit-Cit-Z-Cit-Cit-B-Cit-Citのペプチドと接触させ、ここでXはY、F、W、I、L、MおよびVから選択され、ZはA、D、I、N、P、S、T、Vから選択され、BはA、G、H、Q、S、TおよびVから選択される工程;ならびに
前記試料中で抗シトルリン抗体への前記ペプチドの結合を検出する工程
を含む方法。
【請求項25】
前記ペプチドが配列Cit-Cit-Y-Cit-G-Cit-Cit-S-Cit-Cit-S-Cit-Citを含む、請求項24に記載の方法。
【請求項26】
前記ペプチドが環状化されている、請求項25に記載の方法。
【請求項27】
抗シトルリン抗体への前記ペプチドの結合の検出が自己免疫障害の診断用である、請求項24、25、または26に記載の方法。
【請求項28】
前記自己免疫障害が慢性関節リウマチである、請求項27に記載の方法。
【請求項29】
前記自己免疫障害が多発性硬化症である、請求項27に記載の方法。
【請求項30】
慢性関節リウマチを有する対象を治療するための方法であって、前記対象における共有エピトープを有する前記MHCクラスII分子への、請求項1から5に記載のペプチドの結合を妨害する工程を含む方法。
【請求項31】
前記対象におけるペプチジルアルギニンデイミナーゼの発現を阻害する工程を含む、請求項30に記載の方法。
【請求項32】
前記阻害が、前記ペプチジルアルギニンデイミナーゼの遺伝子配列に標的化されるsiRNAの前記対象への投与を含む、請求項31に記載の方法。
【請求項33】
前記対象中でのペプチジルアルギニンデイミナーゼの機能を阻害する工程を含む、請求項31に記載の方法。
【請求項34】
前記阻害が、ペプチジルアルギニンデイミナーゼの機能をブロックする、前記対象への薬剤の投与を含む、請求項33に記載の方法。
【請求項35】
T細胞活性化および炎症性応答をもたらす、共有エピトープを有するMHCクラスII分子へのシトルリン化ペプチドの結合によって誘発される慢性関節リウマチに罹患している対象を治療するための方法であって、
薬学的に許容可能な担体とともに、請求項1または5に記載のペプチドを含む組成物のT細胞寛容誘導量を前記対象に投与する工程
を含む方法。
【請求項36】
前記薬学的に許容可能な担体が可溶性MHCクラスII分子である、請求項35に記載の方法。
【請求項37】
共有エピトープ細胞とのシトルリン化抗原MHCクラスII複合体の形成を含む、自己免疫障害の診断方法であって、
対象からの末梢血単核細胞の単離された試料を1種以上のシトルリン化ペプチド抗原とインキュベートする工程;
共有エピトープ細胞複合体とのシトルリン化抗原MHCクラスIIの形成を検出する工程であって、このような検出が前記対象において自己免疫障害をもたらすT細胞応答を誘発する可能性を示す
方法。
【請求項38】
前記シトルリン化ペプチドが、
(a)配列番号1から37のペプチド;
(b)配列番号38から39のペプチド;
(c)配列番号40から48のペプチド;および
(d)配列番号49から60のペプチド
から選択され、ここでアルギニンがシトルリンで置換されている請求項37に記載の方法。
【請求項39】
自己免疫障害が慢性関節リウマチである、請求項37または38に記載の方法。
【請求項40】
前記シトルリン化ペプチドがシトルリン化BMPペプチドである、請求項37に記載の方法。
【請求項41】
前記シトルリン化BMPペプチドが配列番号40から48からなる群から選択される、請求項40に記載の方法。
【請求項42】
前記自己免疫障害が多発性硬化症である、請求項37または38に記載の方法。
【請求項43】
対象における共有エピトープ複合体を有するシトルリン化ペプチドMHCクラスIIによってT細胞の活性化を最小化または予防するための方法であって、前記T細胞の活性化を阻害するために、前記複合体を標的化し、これに結合する抗体を前記対象に投与する工程を含む方法。
【請求項44】
対象における潜在的に抗原性のペプチド中でアルギニンのシトルリンへの変換を予防するための方法であって、前記対象にペプチジルアルギニンデイミナーゼのアンタゴニストまたは阻害剤の有効量を投与する工程を含む方法。
【請求項45】
共有エピトープを有するMHCクラスII分子へのシトルリン化ペプチドの結合を阻害する薬学的化合物を同定するためのスクリーニング方法であって、
候補薬学的化合物をトランスジェニックDR4-IE tgマウスに投与する工程;および
T細胞活性を測定する工程および/またはシトルリン化ペプチド/共有エピトープを有するMHCクラスII分子の複合体形成を測定する工程
を含む方法。
【請求項46】
慢性関節リウマチの研究のための動物モデルを提供するために動物において慢性関節リウマチを誘導する方法であって、薬学的に許容可能な担体とともに、請求項1または請求項5に記載の1種以上のシトルリン化ペプチドを含む組成物の慢性関節リウマチ誘導量を前記動物に投与する工程を含む方法。
【請求項47】
投与が注射による、請求項46に記載の方法。
【請求項48】
多発性硬化症の研究のための動物モデルを提供するために動物において多発性硬化症を誘導する方法であって、配列番号40から48から選択される1種以上のシトルリン化ペプチドを含む組成物の多発性硬化症誘導量を前記動物に投与する工程を含む方法。
【請求項49】
多発性硬化症の診断のための方法における、配列番号40から48のペプチドのいずれか1つの使用。
【請求項50】
MSの診断において有用であるシトルリン化MBPペプチドおよび/またはGFAPペプチドであって、FLPCitHRDTG (配列番号40), VTPCitTPPPS (配列番号41); YGGCitASKYK (配列番号42), LGGCitDSRSG (配列番号42), MERCitRITSA (配列番号43), LPTCitVDFSL (配列番号44), LNDCitFASYI (配列番号45), LRLCitLDQLT (配列番号46), LQICitETSLD (配列番号47),及びVEMCitDGEVI (配列番号48) およびその類似体からなる群から選択されるペプチド。
【請求項51】
請求項50のペプチド、およびそのための薬学的に許容可能な担体を含む組成物。
【請求項52】
共有エピトープを有するMHCクラスII分子に結合したシトルリン化ペプチド抗原と反応性である自己反応性T細胞の検出のための診断方法であって、
対象からの末梢血単核細胞の単離された試料を、配列番号40から48のいずれか1つに記載の1つ以上のシトルリン化ペプチドとインキュベートする工程;
前記対象における活性化自己反応性T細胞の存在を示す、T細胞の応答を検出する工程
を含む方法。
【図3】

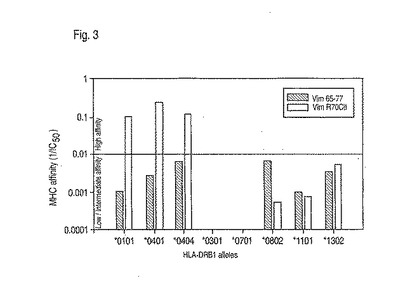
【図4】
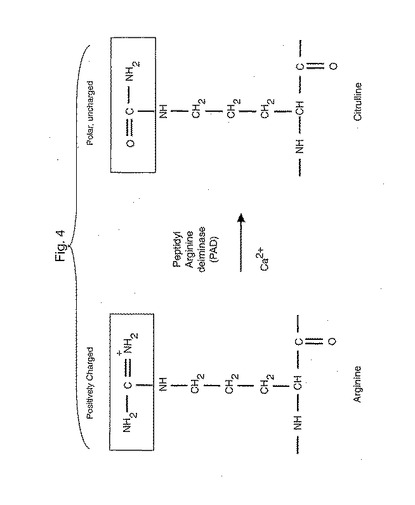
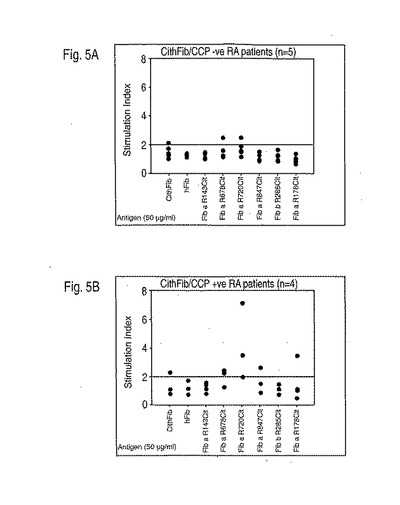
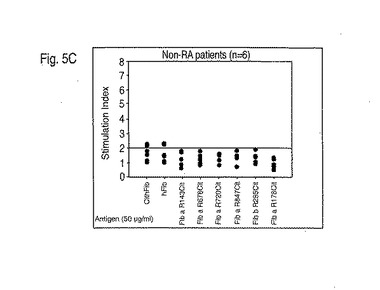
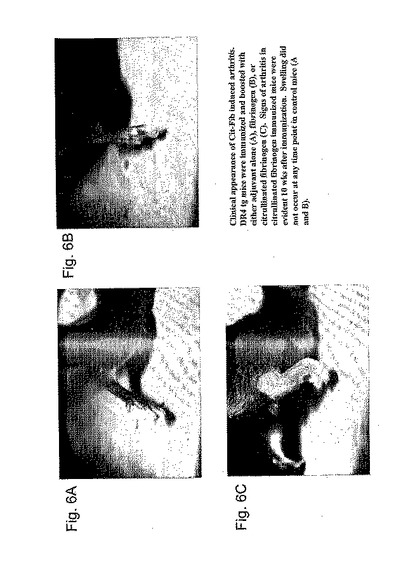
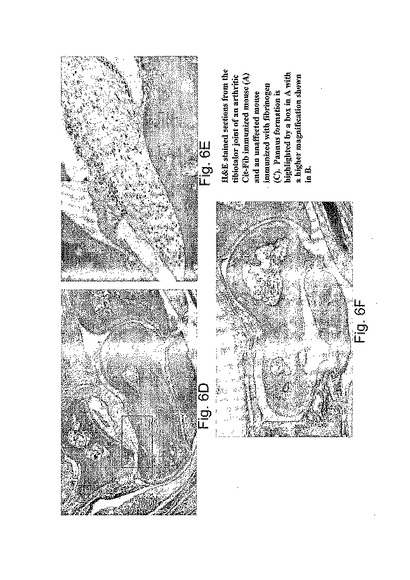
【図7】
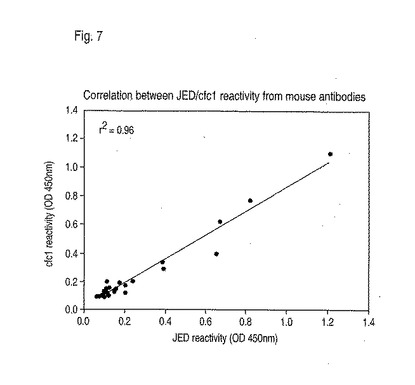
【図8】
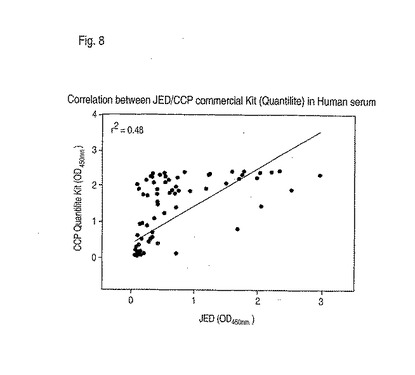
【図4】
【図7】
【図8】
【公表番号】特表2007−524583(P2007−524583A)
【公表日】平成19年8月30日(2007.8.30)
【国際特許分類】
【出願番号】特願2006−504070(P2006−504070)
【出願日】平成16年3月5日(2004.3.5)
【国際出願番号】PCT/CA2004/000337
【国際公開番号】WO2004/078098
【国際公開日】平成16年9月16日(2004.9.16)
【出願人】(505335980)ロンドン・ヘルス・サイエンシズ・センター・リサーチ・インコーポレーテッド (2)
【Fターム(参考)】
【公表日】平成19年8月30日(2007.8.30)
【国際特許分類】
【出願日】平成16年3月5日(2004.3.5)
【国際出願番号】PCT/CA2004/000337
【国際公開番号】WO2004/078098
【国際公開日】平成16年9月16日(2004.9.16)
【出願人】(505335980)ロンドン・ヘルス・サイエンシズ・センター・リサーチ・インコーポレーテッド (2)
【Fターム(参考)】
[ Back to top ]