
自己免疫障害を治療する方法
本発明は、自己免疫疾患を治療するための方法に関する。ある具体例において、本発明は、TCCRアゴニストを投与することを含む自己免疫疾患を治療する方法に向けられる。ある具体例において、該自己免疫疾患はTh1応答によって少なくとも部分的に媒介される。ある具体例において、該自己免疫疾患はCD8+T−細胞増殖によって少なくとも部分的に媒介される。本発明は、IL−27のようなIL27R(TCCR)のアゴニストを投与することによって、多発性硬化症(MS)および慢性関節リウマチ(RA)を含めた自己免疫疾患を治療する方法を提供する。TCCRの有用なアゴニストは、IL27Rの改変体および断片、IL−27のようなIL27Rリガンド、およびその改変体および断片ならびにIL27RまたはIL27Rリガンドに結合し、IL27−媒介応答を刺激し、誘導し、または増強するアゴニスト抗体を含む。
【発明の詳細な説明】
【背景技術】
【0001】
(発明の背景)
自己免疫疾患は、複雑な相互に関連する生物学的経路の発現または結果である。正常な生理学においては、これらの生物学的経路は、傷害または負傷に応答し、傷害または負傷からの修復を開始し、外来性生物に対する先天的および獲得された防御を設置するのに臨界的である。これらの正常な生理学的経路がさらなる傷害または負傷を、応答の強度に関するものとして、異常な調節または過剰な刺激の結果として、自己に対する反応として、またはこれらの組合せとしてのいずれかで引き起こす場合、病気または病理が起こり得る。
【0002】
これらの病気の発生はしばしば多工程経路を含み、しばしば、複数の異なる生物学的系/経路に関係するが、これらの経路の1以上における臨界点での介入は軽減または治療効果を有し得る。治療的介入は有害プロセス/経路の拮抗作用または細菌プロセス/経路の組み合わせによって起こり得る。
【0003】
哺乳動物の免疫系は、細菌、ウイルス、トキシンおよび他の非−宿主物質の侵入から宿主を防御するのに協調して作用する多数のユニークな細胞よりなる。リンパ球、TおよびB細胞双方は、免疫系の特異性を大いに担っている。T細胞は胸腺において発生することから命名され、他方、B細胞は骨髄において発生する。
【0004】
Tリンパ球(T細胞)は哺乳動物免疫応答の重要な成分である。T細胞は、主要組織適合性複合体(MHC)内の遺伝子によってコードされる自己−分子に関連する抗原を認識する。抗原は抗原提示細胞、ウイルス感染細胞、癌細胞、移植片等の表面にMHC分子と共に提示され得る。T細胞系は、健康の脅威が宿主哺乳動物に曝されるこれらの改変された細胞を排除する。T細胞は、ヘルパーT細胞(CD4+)および細胞傷害性T−リンパ球(CD8+)を含む。ヘルパーT細胞(TH)は、抗原提示細胞上の抗原−MHC複合体の認識に続いてかなり増殖する。ヘルパーT細胞は、B細胞、細胞傷害性T−リンパ球、および免疫応答に参加する種々の他の細胞の活性化において中枢的な役割を演じるリンホカインのような種々のサイトカインも分泌する。細胞傷害性T−リンパ球は他の細胞の破壊を引き起こすことができる。
【0005】
液性および細胞媒介免応答双方における中枢的な事象はヘルパーT細胞の活性化およびクローン拡大である。ヘルパーT細胞活性化は、T細胞受容体(TCR)−CD3複合体の、抗原提示細胞の表面での抗原−MHCとの相互作用によって開始される。この相互作用は、休止ヘルパーT細胞が細胞周期に入るのを誘導し(G0ないしG1転移)、その結果、IL−2に対する高親和性受容体の発現をもたらす生化学的事象のカスケードを媒介する。活性化されたT細胞は、周期増殖および分化を介して記憶細胞またはエフェクター細胞を通じて進行する。
【0006】
T−ヘルパー細胞サブセット(Th1およびTh2)は免疫の2つの経路を定義する:細胞−媒介免疫性および液性免疫性。Th1およびTh2サブタイプについてのサイトカインの放出プロフィールはエフェクターメカニズムおよび細胞傷害性細胞の選択に影響する(非特許文献1;非特許文献2)。Th1細胞、CD4+細胞の機能的サブセットは、細胞−媒介免疫性を増強し、Il−2、インターフェロン−ガンマ、リンホカインべータを含めたサイトカインを生産するそれらの能力によって特徴付けられる(非特許文献1;非特許文献2)。Th1細胞によって分泌されるIl−2およびインターフェロン−ガンマはマクロファージおよび細胞傷害性細胞を活性化する。Th2細胞はCD4+細胞でもあるが、Th1細胞から区別される。Th2細胞は、抗体生産のような液性免疫性を増強するそれらの能力によって特徴付けられる。Th2細胞はIl−4、Il−5、およびIl−10を含めたサイトカインを生産する(非特許文献1;非特許文献2)。Th2細胞によって分泌されるIl−4、Il−5、およびIl−10は好酸球および肥満細胞の生産を増殖させ、ならびにIgEを含めた抗体の生産を増強し、細胞傷害性細胞の機能を減少させる(非特許文献3)。
【0007】
Th1およびTh2サイトカイン放出は、相互に抑制性のTh1およびTh2応答を変更する。例えば、IL−4はTh1細胞からのインターフェロン−ガンマの発現を阻害し、他方、インターフェロン−ガンマはTh2細胞からのIL−4の発現を阻害する(非特許文献1)。
【0008】
4つのラセン束サイトカインファミリーのメンバー(非特許文献4)は、共通の前駆体からのTヘルパー細胞のTh1およびTh2エフェクター細胞の区別される集団への増殖および最後の分化を変調する(非特許文献5)。および、IL−4はTh2細胞の発生に影響し、他方、IL−12はTh1細胞の分化に関与する(非特許文献6;非特許文献7)。
【0009】
TCCR(T−細胞サイトカイン受容体)は、IL−12 β−2受容体、G−CSFR、およびIL−6受容体に対する相同性を持つサイトカイン受容体のWS(G)XWSクラスである。これらの受容体は、細胞、特に、血液細胞の増殖および分化に関与する細胞の増殖および分化を制御できるシグナルを変換する。TCCRは、T=ヘルパー細胞応答に関与することが示唆されている。特に、TCCRおよびそのリガンドIL−27はTh1応答を促進することが仮定されている(非特許文献8;非特許文献9;非特許文献10)。
【0010】
Th1およびTh2細胞のいずれかまたは双方によって生産されるサイトカインの過剰生産は、医学的障害の宿主にインパクトを与える。例えば、Th1サイトカインの過剰生産は、多発性硬化症および慢性関節リウマチのような種々の自己免疫疾患の病因に寄与する。Th2サイトカインの過剰生産はアレルギー疾患の病因に寄与する。
【0011】
CD8+細胞傷害性T−リンパ球(CTL)は、いくつかの自己免疫疾患において組織の病原体破壊に関与する。例えば、CTLは、自己免疫I型糖尿病のクールの間に膵臓β細胞の破壊に関連する(非特許文献11)。CTLは実験的自己免疫脳脊髄炎にも関連する(非特許文献12)。CTLは移植片(GVHD)−対宿主病に関連する組織損傷を媒介する(非特許文献13)。
【0012】
多発性硬化症(MS)は、脳および脊髄に影響する中枢神経系の障害である。MSの共通の兆候および症状は、1以上の四肢において、体幹において、または顔の一方側における感覚異常;脚または手の弱化またはぎこちなさ;または(部分的な盲目および1つの目における痛みのような)目に見える乱れ、視覚のかすみ、または暗点を含む。他の共通の初期の兆候は、二重視(複視)をもたらす目の麻痺、1以上の四肢の一過的弱化、四肢のわずかな硬直または異常な脆性、わずかな歩行の乱れ、膀胱制御の困難、眩暈、および軽い情緒的乱れ(Berkow et al.(編),1999,Merck Manual of Diagnosis and Therapy:第17版(MS)についての現在の治療は、コルチコステロイド、バーターインターフェロン(Betaferon,Avonex,Rebif)、グラチラメール酢酸(Copaxone)、メトトリキセート、アザチオプリン、シクロホスファミド、クラドリビン、バクロフェン、チザニジン、アミトリプチリン、カルバマゼピン(Berkow et al.(編),1999,supra)を含む。
【0013】
慢性関節リウマチ(RA)は、典型的には小さなおよび大きな関節に影響する関節の滑膜炎によって特徴付けられる慢性自己免疫疾患であり、それらの進行性破壊に至る(Berkow et al.(編),1999,supra)。RAの兆候は硬直、圧痛、滑膜厚化、反射梗縮、内臓小節、脚潰瘍または多発性単神経炎、胸膜または心臓周囲滲出、および発熱を含むことができる(Berkow et al.(編),1999,supra)。
【0014】
RAのための現在の治療は(サリシレートを含めた)非−ステロイド、抗−炎症薬物、金化合物、メトトレキセート、ヒドロキシクロロキン、スルファサラジン、ペニシラミン、コルチコステロイド、および細胞傷害性または免疫抑制薬物(Berkow et al.(編),1999,Merck Manual of Diagnosis and Therapy:第17版)を含む。
【非特許文献1】Mosmammら、Adv.Imnunol.(1989)45:111−147
【非特許文献2】Mosmannら、Immunol.Today(1996)17:138−146
【非特許文献3】Powrieら、Immunol.Today,(1993)14:270
【非特許文献4】Bazan,PNAS,(1990)87:6934
【非特許文献5】O’Garra,A.,Immunity(1998)8:275−83
【非特許文献6】Hsiehら、Science,(1993)260:547−9
【非特許文献7】Sederら、PNAS,(1993)90:10188−92
【非特許文献8】Chenら、Nature,(2000)409:916−920
【非特許文献9】Yoshidaら、Immunity,(2001)15:569−578
【非特許文献10】Pflanzら、Immunity,(2002)16:779−790
【非特許文献11】Kagiら、J.Exp.Med.(1997)186:989−997
【非特許文献12】Husebyら、J.Exp.Med.,(2001)194(5):669−676
【非特許文献13】Graubertら、J.Clin Invest.(1997)100:904−911
【発明の開示】
【発明が解決しようとする課題】
【0015】
自己免疫疾患に対する現存の療法のいずれも、限定された効率および/または有意な毒性のため満足することが判明していない。かくして、MSおよびRAのような自己免疫疾患を治療するための新しい方法が必要である。
【課題を解決するための手段】
【0016】
(発明の要旨)
ナイーブな未分化T細胞(Th−0)は、成熟なT−ヘルパー細胞へのTh−0細胞の分化を誘導する異なるシグナルに応答する。今日、例えば、IL−27のようなTCCRのアゴニストを投与することによって、細胞受容体TCCRの活性化がT−リンパ球増殖を低下させるのに効果的であることが発見されている。T−リンパ球増殖の低下は、IL−10およびSOCS−3の増大した発現に相関した。TCCRを発現する動物は、自己免疫疾患に対して感受性が低いことが判明した。
【0017】
EAE病気モデルにおけるさらなる実験は、IL−27受容体(TCCR)−欠乏マウスは自己免疫疾患に対して過敏であることを示した。Th−細胞分化における、および免疫障害におけるIL−27の役割の研究は、IL−27が免疫抑制性であり、Th発生において複数レベルで作用するという驚くべき発見に導いた。IL−27はTh−IL17細胞の生産を抑制し、IL−6の生産を阻害し、IL−6を含めたThIL17サイトカインの生産を阻害する。IL−27はIL−10の、およびIL−4の、さらにTh−IL17細胞の阻害剤の生産を誘導し、IL12受容体の生産およびTh−1細胞の分化を刺激する。本明細書中に開示されたデータは、IL−27が、Th−1、Th−2およびTh−17細胞にわたる重要な阻害活性を含めた重要な免疫抑制機能を有することを示す。
【0018】
本発明は、IL−27のようなIL27R(TCCR)のアゴニストを投与することによって、多発性硬化症(MS)および慢性関節リウマチ(RA)を含めた自己免疫疾患を治療する方法を提供する。TCCRの有用なアゴニストは、IL27Rの改変体および断片、IL−27のようなIL27Rリガンド、およびその改変体および断片ならびにIL27RまたはIL27Rリガンドに結合し、IL27−媒介応答を刺激し、誘導し、または増強するアゴニスト抗体を含む。また、本発明は、Th−ILI7細胞を含めた、T−リンパ球および/または細胞傷害性T−リンパ球の増殖を阻害する方法を提供し、該方法はIL27/IL27R応答を刺激し、誘導し、または増強するアゴニストを投与することを含む。
【0019】
【表1】

【発明を実施するための最良の形態】
【0020】
(詳細な説明)
T−リンパ球の過剰増殖またはTh1またはTh2細胞によって生産されたサイトカインの過剰生産は、医学的障害の主に導く。例えば、Th1応答に関連するサイトカインの過剰生産、またはCD8+細胞傷害性T−リンパ球の過剰生産は同種異系移植片拒絶、(グレーヴス病および橋本甲状線炎のような)自己免疫甲状腺病、自己免疫ブドウ網膜炎、巨細胞動脈炎、(クローン病、海洋性結腸炎、局所腸炎、肉芽腫性腸炎、末端回腸炎、局所回腸炎および終末回腸炎を含めた)炎症性腸疾患、インスリン−依存性新生糖尿病、多発性硬化症、悪性貧血、乾癬、慢性関節リュウマチ、サルコイドーシス、強皮症、全身エリテマトーデスを含めた自己免疫疾患に導き得る。
【0021】
以下の実施例に詳細に説明する実験は、非−特異的T細胞刺激に応答してTCCRを発現するT細胞よりもTCCRを欠くT細胞のより大きな増殖を示す(図1参照)。以前、TCCRおよびそのリガンドIL−27がTh1応答を促進することが示唆された(Chenら、2000,Nature,407:916−920;Yoshidaら、2001,Immunity,15:569−578;Pflanzら、2002,Immunity,16:779−790)。しかしながら、驚くべきことに、TCCRを発現するマウスは、TCCRを欠くマウスよりも、実験的アレルギー性脳脊髄炎(EAE)のようなTh1応答によって部分的に特徴付けられる自己免疫疾患に対して感受性が低いことが発見された(実施例2参照)。
【0022】
以下の実施例に示すように、Tリンパ球の増殖は、細胞へのTCCRアゴニストの投与によって阻害される。また、自己免疫炎症性疾患の低下した臨床的進行およびよりひどくない兆候がTCCR(−/−)動物におけるよりもTCCR(+/+)を発現する動物に存在することが示された。
【0023】
これらのデータは、TCCRのアゴニストを用いて、T−細胞増殖を低下させることを示す。特に、データは、TCCRのアゴニストが、多発性硬化症(MS)および慢性関節リュウマチ(RA)のような自己免疫媒介障害を治療するのに有用であることを示す。
【0024】
(定義)
用語「自己免疫」とは、それを生産する生物の分子、細胞、または組織を抗体またはリンパ球のような免疫系の成分が攻撃し、または害するプロセスをいう。
【0025】
用語「自己免疫障害」は、組織損傷、または病因のような損傷が少なくとも部分的には自己免疫プロセスの結果である病気をいう。その例として、用語「自己免疫疾患」は、Th1応答またはCD8+細胞傷害性T−リンパ球によって少なくとも部分的に媒介される病気を[含む。自己免疫疾患は同種異系移植片拒絶、(グレーヴス病および橋本甲状線炎のような)自己免疫甲状腺病、自己免疫ブドウ膜網膜炎、巨細胞動脈炎、(クローン病、潰瘍性結腸炎、局所腸炎、肉芽腫腸炎、末端回腸炎、局所回腸炎、および終末回腸炎を含めた)炎症性腸疾患、インスリン−依存性真性糖尿病、多発性硬化症、悪性貧血、乾癬、慢性関節リウマチ、サルコイドーシス、強皮症、および全身エリテマトーデスを含む。
【0026】
用語「Th1応答」とは、Th1エフェクター細胞の区別される集団への前駆体からのTヘルパー細胞の分化をいい、IFN−ガンマ、IL−2、およびTNF−ベータのようなTh1細胞からのサイトカインの分泌を含む。用語「Th1バイアシング疾患」とは、Th1エフェクター細胞の区別される集団への前駆体からのTヘルパー細胞の分化に好都合な疾患をいう。
【0027】
用語「Th1サイトカイン」とは、IFN−ガンマ、IL−2、およびTNF−ベータを含めたTh1応答において発現されるサイトカインをいう(Powrieら、1993,Immunol.Today,14:270)。
【0028】
用語「Th1媒介障害」とは、Th1サイトカインの過剰生産によって圧倒的にまたは部分的に媒介される障害をいう。用語「Th1媒介障害」は、T−細胞のTh1サブタイプの分化における過剰生産または偏りに由来し得る障害を含む。そのような傷害は、自己免疫ショウガイ、例えば、RAおよびMSを含む。
【0029】
用語「Th2応答」とは、Th2エフェクター細胞の区別される集団への前駆体からのTヘルパー細胞の分化をいい、IL−4、IL−5、IL−10、およびIL−13のようなTh2細胞からのサイトカインの分泌を含む(Powrieら、1993,Immunol.Today,14:270)。用語「Th2バイアシング条件」とは、Th2エフェクター細胞の区別される集団への前駆体からのTヘルパー細胞の分化に好都合な条件をいう。
【0030】
用語「TCCRペプチド」「TCCR蛋白質」および「TCCR」は、本明細書中において用いる場合、天然配列TCCRおよびTCCRペプチド改変体を含む。TCCRペプチドはヒト組織またはもう1つの源のような種々の源から単離することができ、あるいは組換えおよび/または合成方法によって調製することができる。「天然配列TCCR」は、天然に由来するTCCRペプチドと同一のアミノ酸配列を有するペプチドである。そのような天然配列TCCRは、天然から単離することができるか、あるいは組換えおよび/または合成手段によって生産することができる。用語「天然配列TCCR」は(細胞外ドメイン配列のような)天然に生じる切形および分泌形態、(交互にスプライスされた形態のような)天然に生じる切形形態、およびTCCRの天然に生じる対立遺伝子改変体を具体的には含む。1つの具体例において、天然配列ヒトTCCRは、配列番号:1のアミノ酸1ないし636を含む成熟または全長天然配列TCCRである。同様に、天然配列マウスTCCRは、配列番号:2のアミノ酸1ないし623を含む成熟または全長天然配列TCCRである。配列番号:1および配列番号:2は、アミノ酸位置1としてここでは示されるメチオニン残基で始まることが示されるが、配列番号:1または配列番号:2のアミノ酸位置1から上流または下流いずれかに位置するもう1つのメチオニン残基は、TCCRペプチドについての出発アミノ酸残基として使用することができると考えることができ、またそれが可能である。
【0031】
「TCCRペプチド細胞外ドメイン」または「TCCR ECD」とは、膜貫通および細胞質ドメインを実質的に含まないTCCRペプチドの形態をいう。通常は、TCCRペプチドECDはそのような膜貫通および/または細胞質ドメインの約1%未満を有し、好ましくは、好ましくは、そのようなドメインの約0.5%未満を有する。本発明のTCCRペプチドについて同定されるいずれの膜貫通ドメインも、疎水性ドメインのそのタイプを同定するためにルーチン的に使用される基準に従って同定されると理解される。膜貫通ドメインの正確な境界は変化し得るが、最もありそうには、最初に同定されたドメインのいずれかの末端における約5以下のアミノ酸であり得る。それ自体、1つの具体例において、ヒトTCCRペプチドの細胞外ドメインはアミノ酸1または約33ないしX1を含ミリモル、ここに、X1は配列番号:1の残基512ないし残基522からのいずれかのアミノ酸残基である。同様に、マウスTCCRペプチドの細胞外ドメインはアミノ酸1または約25ないしX2を含ミリモル、ここに、X2は配列番号:2の残基509ないし残基519からのいずれかのアミノ酸残基である。
【0032】
用語「TCCR改変体ペプチド」は、TCCRペプチドの少なくとも1つの生物学的活性を有し、かつ:
(a1)配列番号:1のヒトTCCRペプチドの残基1または約33ないし636;
(a2)配列番号:2のマウスTCCRペプチドの残基1または約25ないし623;
(b1)配列番号:1のヒトTCCRペプチドのX3ないし636、ここに、X3は配列番号:1のいずれかのアミノ酸残基27ないし37;
(b2)配列番号:2のマウスTCCRペプチドのX4ないし623、ここに、X4は配列番号:2の20ないし30からのいずれかのアミノ酸残基であり;
(c1)1または約33ないしX1、ここに、X1は配列番号:1の残基512ないし残基522からのいずれかのアミノ酸残基であり;
(c2)1または約25ないしX2、ここに、X2は配列番号:2の残基509ないし519のいずれかのアミノ酸残基であり;
(d1)X5ないし636、ここに、X5は配列番号:1の残基533ないし543からのいずれかのアミノ酸であり;
(d2)X6ないし623、ここに、X6は配列番号:2の残基527ないし537からのいずれかのアミノ酸であり;または
(e)配列番号:1および配列番号:2のアミノ酸配列のもう1つの特異的に誘導された断片;
のアミノ酸配列に対して少なくとも約80%アミノ酸配列同一性を有するペプチドを意味する。
【0033】
そのようなTCCR改変体ペプチドは、例えば、配列番号:1および配列番号:2の配列のN−および/またはC−末端において、ならびに1以上の内部ドメイン内において1以上のアミノ酸残基が付加され、または欠失されたTCCRペプチドを含む。通常、TCCR改変体ペプチドは少なくとも80%アミノ酸配列同一性を有し、
(a1)配列番号:1のヒトTCCRペプチドの残基1または約33ないし636;
(a2)配列番号:2のマウスTCCRペプチドの残基1または約25ないし623;
(b1)配列番号:1のヒトTCCRペプチドのX3ないし636、ここに、X3は配列番号:1のいずれかのアミノ酸残基27ないし37;
(b2)配列番号:2のマウスTCCRペプチドのX4ないし623、ここに、X4は配列番号:2の20ないし30からのいずれかのアミノ酸残基であり;
(c1)1または約33ないしX1、ここに、X1は配列番号:1の残基512ないし残基522からのいずれかのアミノ酸残基であり;
(c2)1または約25ないしX2、ここに、X2は配列番号:2の残基509ないし519のいずれかのアミノ酸残基であり;
(d1)X5ないし636、ここに、X5は配列番号:1の残基533ないし543からのいずれかのアミノ酸であり;
(d2)X6ないし623、ここに、X6は配列番号:2の残基527ないし537からのいずれかのアミノ酸であり;または
(e)配列番号:1および配列番号:2のアミノ酸配列のもう1つの特異的に誘導された断片;
に対して少なくとも約81%、82%、83%、84%、85%、86%、87%、88%、89%、90%、91%、92%、93%、94%、95%、96%、97%、98%または99%アミノ酸配列同一性であり得る。
【0034】
用語「IL−27」は、本明細書中で用いる場合、天然の配列IL−27ヘテロダイマー、天然の配列IL−27成分EBI3およびp28、IL−27ヘテロダイマー改変体(本明細書中でさらに定義する、およびEBI3およびp28の改変体を含む。IL−27ヘテロダイマーおよびその成分はヒト組織タイプから、またはもう1つの源からのような種々の源から単離することができ、あるいは組換えおよび/または合成方法によって調製することができる。「天然配列IL−27」は、天然に由来するIL−27ヘテロダイマーと同一のアミノ酸配列を有するヘテロダイマーを含む。そのような天然配列IL−27ヘテロダイマーは天然から単離することができるか、あるいは組換えおよび/または合成手段によって生産することができる。用語「天然配列IL−27」は、具体的には、(細胞外ドメイン配列のような)天然に生じる切形され、かつ分泌された形態、(交互にスプライシングされた形態のような)天然に生じる切形された形態、およびIL−27ヘテロダイマーの天然に生じる対立遺伝子改変体を含む。
【0035】
用語「IL−27改変体」とは、TCCRを活性化することができる、天然配列IL−27成分EBI3およびp28を含めた、天然配列IL−27に対して相同性を有するペプチドをいう。IL−27改変体は、EBI3改変体およびp28改変体から形成されるものを含むことができる。IL−27改変体は、TCCRおよびgp130を共にかみ合わせることができるものも含むことができる。IL−27改変体は、TCCRホモダイマーを形成することができるものを含むことができる。IL−27改変体はPEG化IL−27を含む。
【0036】
用語「p28」は、本明細書中で用いる場合、天然配列p28およびp28ペプチド改変体を含む。p28はヒト組織タイプから、またはもう1つの源からのように種々の源から単離することができるか、あるいは組換えおよび/または合成方法によって調製することができる。「天然配列p28」は、天然に由来するp28ペプチドを同一のアミノ酸配列を有するペプチドを含む。そのような天然配列p28は天然から単離することができるか、あるいは組換えおよび/または合成手段によって生産することができる。用語「天然配列p28」は、具体的には、(細胞外ドメイン配列のような)天然に生じる切形され、または分泌された形態、(交互にスプライシングされた形態のような)天然に生じる切形形態、およびp28の天然に生じる対立遺伝子改変体を含む。
【0037】
用語「p28ペプチド改変体」は、天然配列ヒトp28(配列番号:3)またはマウスp28(配列番号:4)に対して少なくとも73%、75%、80%、90%、95%、または99%配列同一性を有するペプチドを含む。p28ペプチド改変体は、TCCRおよびgp130に結合することができるp28の部分を含む。p28ペプチド改変体は、TCCRを活性化することができるp28の部分を含む。p28ペプチド改変体は、TCCR上に見出されるサイトカイン受容体相同性ドメインの領域においてTCCRに結合すると考えられる、p28の第一および第三のアルファラセンからの残基、およびgp130上に見出されるIGドメインに結合すると考えられる、第一のラセンの最後において、および第四のラセンの始まりにおける残基を含む。
【0038】
用語「EBI3」は、本明細書中で用いる場合、天然配列EBI3およびEBI3ペプチド改変体を含む。EBI3ペプチドはヒト組織タイプから、またはもう1つの源からのように種々の源から単離することができるか、あるいは組換えおよび/または合成方法によって調製することができる。「天然配列EBI3」は、天然に由来するEBI3ペプチドと同一のアミノ酸配列を有するペプチドを含む。そのような天然配列EBI3は天然から単離することができるか、あるいは組換えおよび/または合成手段によって生産することができる。用語「天然配列EBI3」は、具体的には、(細胞外ドメイン配列のような)天然に生じる切形され、および分泌された形態、(交互にスプライシングされた形態のような)天然に生じる切形された形態、およびEBI3の天然に生じる対立遺伝子改変体を含む。
【0039】
用語「融合蛋白質」とは、例えば、2つの異なる蛋白質をコードする2つの遺伝子の融合から得られる発現産物をいう。該用語は、2つの異なる蛋白質の部分をコードする2つの遺伝子の部分の融合に由来する発現産物も含む。該用語は、翻訳後に起こる融合に由来する蛋白質を含む。本明細書で用いるように、該用語は、異種ペプチドに融合した、IL−27、その成分(EBI3およびp28)またはその部分を含むであろう。該用語は、異種ペプチドに融合したTCCRまたはその部分も含むであろう。該用語は、機能的な1つの鎖サイトカインを形成するためのp28に融合したEBI3も含むであろう(Pflanzら、2002,Immunity,16:779−790)。該用語は、ヒトFcタグにコンジュゲートしたIL−27を含む。
【0040】
本明細書中で用いるように、与えられたペプチドに関しては、「異種ペプチド」とは、起源に関わらず異なる配列を持つペプチドをいう。例えば、天然配列TCCRに関しては、異種ペプチドとは、天然配列TCCRのそれ以外の配列を有するペプチドをいう。天然配列IL−27に関しては、異種ペプチドとは、天然配列IL−27のそれ以外の配列を有するペプチドをいう。
【0041】
用語「アゴニスト」は、天然配列ペプチドの生物学的活性を増強する、または刺激するいずれの分子も含む。適当なアゴニストは、具体的には、アゴニストペプチド、アゴニスト抗体または抗体断片、本発明の天然ペプチドの断片またはアミノ酸配列改変体等を含む。TCCRのアゴニストを同定するための方法は、例えば、TCCRペプチドまたはTCCRペプチド−発現細胞を候補アゴニスト分子と接触させ、1以上のTCCR生物学的活性の検出可能な変化を測定することを含む。
【0042】
「TCCR生物学的活性」とは、本明細書中で用いるように、T−細胞増殖の減衰または抑制のようなTCCR媒介応答をいう。TCCR生物学的活性は、Th1応答またはTh1媒介障害の減衰または抑制を含む。TCCR生物学的活性はIL−10およびSOCS−3の発現の増加を含む。TCCR生物学的活性は、TCCRの活性化に関連するシグナリング、例えば、Stat1、Stat3、Stat4、およびStat5のようなシグナル変換および転写因子のリン酸化も含む(Lucasら、2003,PNAS,100(25):15047−52)。
【0043】
用語「抗体」および「免疫グロブリン」は最も広い意味で用いられ、具体的には、ポリクローナル抗体、(アゴニストおよびアンタゴニスト抗体を含めた)モノクローナル抗体、(二価抗体のような)多価抗体、(所望の生物学的活性を呈する二特異的抗体のような)多特異的抗体、ポリエピトープ特異性を持つ抗体組成物、アフィニティー成熟化抗体、ヒト化抗体、ヒト抗体、キメラ抗体、ならびに所望の生物学的活性を呈する(Fab、F(ab’)2、scFv、およびFvのような)抗原結合断片を含む。天然に生じる抗体は、ジスルフィド結合によって相互連結した4つのペプチド鎖、2つの同一の重(H)鎖および2つの同一の軽(L)鎖を含む。各重鎖は、重鎖可変領域ドメイン(VH)および重鎖定常領域を含む。重鎖定常領域は3つのドメインCH1、CH2およびCH3を含む。各軽鎖は、軽鎖可変領域ドメイン(VL)および軽鎖定常領域ドメインを含む。軽鎖定常領域は1つのドメインCLを含む。VHおよびVLドメインは、さらに、配列によって規定される相補性決定領域(CDR)(Kabatら、1991,Sequences of Proteins of Immunological Interest,5th Ed.Public Health Service,National Institutes of Health,Bethesda,Md.)、またはフレームワーク領域(FR)と呼ばれる、より保存された領域が散在する三次元構造によって規定される超可変ループ(HVL)(Chothiaら、1987,J.Mol.Biol.,196:901−917)に細分することができる。各VHおよびVLは典型的には、以下の順序:FR1、CDR1(HVL1)、FR2、CDR2(HVL2)、FR3、CDR3(HVL3)、FR4にてアミノ−末端ないしカルボキシ−末端に配置された3つのCDR(またはHVL)および4つのFRから構成される。
【0044】
抗体(免疫グロブリン)は、それらの重鎖の定常ドメインのアミノ酸配列に依存して異なるクラスに帰属される。免疫グロブリンの5つの主なクラス:IgA、IgD、IgE、IgG、およびIgMがあり、これらのいくつかは、IgG1、IgG2、IgA1、IgA2等のようなサブクラス(イソタイプ)にさらに分割される。免疫グロブリンの異なるクラスに対応する重鎖定常ドメインが、各々、アルファ、デルタ、イプシロン、ガンマおよびミューと呼ばれる。免疫グロブリンの異なるクラスのサブユニット構造および三次元立体配置はよく知られており、一般に、例えば、Abbasら、2000,Cellular and Mol.Immunology,4th edに記載されている。抗体は、1以上の他の蛋白質またはペプチドと抗体との共有結合または非共有結合開放によって形成されるより大きな融合分子の一部であり得る。
【0045】
用語「全長抗体」とは、少なくとも2つの重鎖および2つの軽鎖を含めたその実質的に無傷の形態の抗体をいい、以下に定義する抗体断片をいわない。該用語は、特に、Fc領域を含有する重鎖を持つ抗体をいう。全長抗体は天然配列抗体または組換え抗体であり得る。全長抗体はヒト、ヒト化、および/またはアフィニティー成熟化であり得る。
【0046】
用語「モノクローナル抗体」とは、本明細書中で用いるように、実質的に相同な抗体の集団から得られた抗体をいい、すなわち、該集団を含む個々の抗体は、該抗体の生産の間に生起し得る改変体を除いて実質的に同一である。
【0047】
本明細書中に記載されたモノクローナル抗体は、具体的には、重鎖および/または軽鎖の一部が特定の種に由来する、または特定の抗体クラスまたはサブクラスに属する抗体における対応する配列と同一である、または相同であり、他方、鎖の残りはもう1つの種に由来し、またはもう1つの抗体クラスまたはサブクラスに属する抗体における対応する配列と同一である、または相同である「キメラ」抗体、ならびに所望の生物学的活性を呈する限りそのような抗体の断片を含む(米国特許第4,816,567号;およびMorrisonら、1984,PNAS,81:6851−6855)。
【0048】
非−ヒト(例えば、マウス)抗体の「ヒト化」形態は、非−ヒト免疫グロブリンに由来する最小配列を含有する(Fv、Fab、Fab’、F(ab’)2、または抗体の他の抗原−結合サブ配列のような)キメラ免疫グロブリン、免疫グロブリン鎖、またはその断片である。ほとんどの場合、ヒト化抗体は、受容体の1以上の相補性決定領域(CDR)または超可変ループ(HVL)からの残基が、マウス、ラットまたはウサギのような非−ヒト種(ドナー抗体)の1以上のCDRまたはHVLからの残基で置き換えられ、所望の抗原特異性、アフィニティーおよび能力を有するヒト免疫グロブリン(受容体抗体)である。いくつかの場合において、ヒト免疫グロブリンの特異的Fvフレームワーク領域(FR)残基は対応する非−ヒト残基によって置き換えられている。さらに、ヒト化抗体は、受容体抗体においても、または輸入されたCDR(またはHVL)においても、あるいはフレームワーク配列においても見出されない残基を含むことができる。これらの修飾は、抗体性能をさらに改良し、最大化するように成される。一般に、ヒト化抗体は、少なくとも1つの、典型的には2つの可変ドメインの実質的に全てを含み、ここに、CDRまたはHVLの全てまたは実質的に全ては非−ヒト免疫グロブリンのそれに対応し、FRの全てまたは実質的に全てはヒト免疫グロブリン配列のそれである。
【0049】
ヒト可変ドメイン、軽鎖および重鎖双方の選択は、ヒト化抗体を作成するにおいて用いることができる。「ベストフィット」方法に従って、げっ歯類抗体の可変ドメインの配列は、例えば、公知のヒト可変−ドメイン配列の全ライブラリーに対してスクリーニングされる。げっ歯類のそれに最も近いヒト配列は、ヒト化抗体についてのヒトフレームワーク領域(FR)として用いられる(Simsら、1993,J.Immunol.,151:2296)。別法として、受容体フレームワーク領域は軽鎖または重鎖の特定のサブグループについてのヒト抗体コンセンサス配列に由来することができる。同一フレームワークは、いくつかの異なるヒト化抗体を生産するのに用い、または修飾し、および用いることができる(Carterら、1992,Proc.Natl.Acad.Sci.USA,89:4285;Prestaら、1993,J.Immunol.,151:2623)。ヒト化抗体は、所望により、免疫グロブリン定常領域の少なくとも部分(Fc)、典型的には、ヒト免疫グロブリンのそれを含む。さらなる詳細については、例えば、Jonesら、1986,Nature,321:522−525;Reichmannら、1988,Nature,332:323−329;およびPresta,1992,Curr.Op.Struct.Biol.,2:593−596参照。ヒト化抗体はPRIMATIZED(登録商標)抗体とすることもでき、ここに、抗体の抗原−結合領域は、注目する抗原でアカゲザルを免疫化することによって生産される抗体に由来する。
【0050】
免疫化に際して、内因性免疫グロブリンの生産の不存在下でヒト抗体の十分なレパートリーを生産することができるトランスジェニック動物(例えば、マウス)を生産することができる。例えば、キメラおよび生殖系突然変異体マウスにおける抗体重鎖接合領域(JH)遺伝子のホモ接合性欠失の結果、内因性抗体生産の完全な阻害がもたらされる。そのような生殖系突然変異体マウスへのヒト生殖系免疫グロブリン遺伝子アレイの導入の結果、抗原攻撃に際してヒト抗体の生産がもたらされる。例えば、Jakobovitsら、1993,Proc.Natl.Acad.Sci.USA,90:2551;Jakobovitsら、1993,Nature,362:255−258;Bruggermannら、1993,Year in Immuno.,7:33参照。ヒト抗体は、例えば、Hoogenboomら、1991,J.Mol.Biol.,227:381;またはMarksら、1991,J.Mol.Biol.,222:581−597に記載されているようにファージ−ディスプレイライブラリーに由来することもできる。
【0051】
「ヒト抗体」は、ヒトによって生産された抗体のそれに対応するアミノ酸配列を保有し、および/または本明細書中に開示されたヒト抗体を作成するための技術のいずれかを用いて作成されたものである。
【0052】
「アフィニティー突然変異した」抗体は、それらの改変を保有しない親抗体と比較して、抗原に対する抗体のアフィニティーの改良をもたらす1以上の超可変領域において1以上の改変を有するものである。好ましいアフィニティー成熟化抗体は、標的抗原に対してナノモラーまたはピコモラーさえのアフィニティーを有するであろう。アフィニティー突然変異した抗体は公知の手法によって生産される。例えば、VHおよびVLドメインシャッフリングによるアフィニティー成熟化を記載するMarksら、1992,Bio/Technology 10:779−783参照。CDRおよび/またはフレームワーク残基のランダム突然変異誘発はBarbasら、1994,Proc.Nat.Acad.Sci.USA 91:3809−3813;Scierら、1995,Gene 169:147−155;Yeltonら、1995,J.Immunol.155:1994−2004;Jacksonら、1995,J.Immunol.154(7):3310−9;およびHawkinsら、1992,J.Mol.Biol.226:889−896に記載されている。
【0053】
「抗体断片」は、一般には、無傷抗体の抗原結合部位を含み、かつ、かくして、抗原を結合する能力を保有する無傷抗体の一部のみを含む。本定義に含まれる抗体断片の例は:
(i)重鎖および軽鎖の間に1つの鎖間ジスルフィド結合を有するVL、CL、CHおよびCH1ドメインを有するFab断片;
(ii)CH1ドメインのC−末端において1以上のシステイン残基を有するFab断片であるFab’断片;
(iii)VHおよびCH1ドメインを有するFd断片;
(iv)VHおよびCH1ドメイン、および該CH1ドメインのC−末端に1以上のシステイン残基を有するFd’断片;
(v)抗体の単一アームのVLおよびVHドメインを有するFv断片;
(vi)VHドメインよりなるdAb断片:
(vii)少なくともVL、VH、CL、CH1ドメインを含み、かつヒンジ領域を欠如する無ヒンジ抗体;
(viii)ヒンジ領域においてジスルフィドブリッジによって連結された2つのFab’断片を含む二価断片である(Fab’)2断片:
(ix)単一鎖抗体分子(例えば、単一鎖Fv’scFv);
(x)同一ペプチド鎖中の軽鎖可変ドメイン(VH)に連結された重鎖可変ドメイン(VH)を含む2つの抗原結合部位を持つ「ダイアボディー」;
(xi)単一アーム抗原結合ドメインの半減期を増大させることができるFc領域を形成するのに十分な、軽鎖、重鎖およびN−末端切形重鎖定常領域を含む単一アーム抗原結合分子;
(xii)相補性軽鎖ペプチドと一緒になって、抗原結合領域の対を形成するタンデムFdセグメント(VH−CH1−VH−CH1)の対を含む「線状抗体」;
を含む。
【0054】
本明細書中で用いるように、「治療」とは、治療すべき個体または細胞の天然コースを改変させる試みにおける臨床的介入をいい、予防のために、または臨床的病理学のコースの間に行うことができる。治療の望ましい効果は、病気の発生または再発の予防、兆候の軽減、障害のいずれかの直接的または間接的病理学的結果の減少、転移の予防、病気進行速度の減少、病気状態の軽減または緩和、および緩解または改良された予後を含む。
【0055】
TCCR
TCCR(WSX−1)は、IL−12 β−2受容体、G−CSFRおよびIL−6受容体に対する相同性を持つサイトカイン受容体のWS(G)XWSクラスのものである。最高の相同性は、IL−12 β―2受容体(26%同一性)。これらの受容体は、細胞、特に、血液細胞の増殖および分化に関与する細胞の増殖および分化を制御するシグナルを変換する。後の実施例に示されるデータは、TCCRの活性化が、直接的にまたは間接的に、CD8+T−リンパ球またはTh1応答の増殖を含めた、自己免疫プロセスの抑制を含むことを示唆する。
【0056】
自己免疫プロセスの抑制は、T−細胞を他のマイトジェン刺激に対して非−応答性とすることができるサイトカインシグナリング(SOCS)蛋白質ファミリーメンバーのサプレッサーの誘導を介して起こり得る(Alexanderら、.2004,Ann.Rev.Immunul.,22:503)。特に、SOCS−3は、ヤヌスキナーゼの活性化ループに結合し、キナーゼ活性を阻害し、それにより、サイトカインシグナリングを抑制する蛋白質である(Masuharaら、1997,Biochem.Biophys.Res.Commun.,239:439−446)。ペルオキシソームプロリファレーター−活性化受容体(PPAR)−ガンマアゴニスト(例えば、ロシグリタゾン)のようないくつかの剤の抗−炎症効果は、SOCS1およびSOCS3の転写を誘導することによって機能する(Parkら、2003,J.Biol.Chem.,278:14747−14752)。後の実施例に示したデータは、TCCR活性化がSOCS3の発現を直接的にまたは間接的に誘導することを示す。
【0057】
自己免疫抑制は、IL−10の誘導を介しても起こる。IL−10は活性化されたT細胞、B細胞、単球およびケラチノサイトによって生産されるサイトカインである。IL−10は、IL−2、IL−3、IFN−γ、GM−CSF、およびTNFを含めた多数のサイトカインの生産を阻害する。IL−10は、炎症応答を制限し、終止させるにおいて主な役割を演じる(Mooreら、2001,Ann.Rev.Immunol.,19:683)。後の実施例に示されるデータは、TCCR活性化がIL−10の発現を直接的にまたは間接的に誘導することを示す。
【0058】
ヒトTCCRのアミノ酸配列は公開されており(1996年5月23日に出願されたWO97/44455)、受託番号4759327下でGenBankから入手可能である。この配列はSprecherら、1998,Biochem.Biophys,Res.Commun.246(1):82−90にも記載されている。ヒトTCCR(hTCCR)の配列は長さが636アミノ酸であって、表1にて後に示される(配列番号:1)。単一ペプチドがアミノ酸残基1ないし32から同定されており、膜貫通ドメインが配列番号:1のアミノ酸残基517ないし538から同定されている。N−グリコシル化部位が残基51ないし54、76ないし79、302ないし305、311ないし314、374ないし377、382ないし385、467ないし470、563ないし566において同定されており、N−ミリストイル化部位が残基107ないし112、240ないし245、244ないし249、281ないし286、292ないし297、377ないし378、400ないし405、459ないし464、470ないし475、531ないし536および533ないし538において同定されている。原核生物膜リポ蛋白質脂質付着部位は残基522ないし532に存在し、成長因子およびサイトカイン受容体ファミリーシグニチャー1は残基41ないし54に存在する。また、残基183ないし191には、ヒト顆粒球−マクロファージコロニー−刺激因子(GM−CSF)に対する受容体の第二のサブユニットに対する有意な相同性の領域もある。TCCRは、配列番号:1の残基41ないし230におけるTCCR上のサイトカイン受容体相同性ドメインにてIL−27サブユニットp28に結合する。記載された全てのhTCCR残基は配列番号:1の配列に従ってナンバリングされる。
【0059】
成人において、hTCCRは胸腺で最も高度に発現されるが、発現は、末梢血液白血球(PBL)、脾臓においても観察され、および弱い発現が肺においても観察される。胎児組織は肺および腎臓において弱いTCCR発現を呈する。
【0060】
マウスTCCR(mTCCR)のアミノ酸配列は公開されており(1996年5月23日に出願されたWO97/44455)、受託番号7710109の下でGenBankから入手可能である。この配列はSprecherら、1998,Biochem.Biophys,Res.Commun.246(1):82−90にも記載されている。mTCCRに対する配列は、長さが623アミノ酸であって、表1において後に示される(配列番号:2)。単一ペプチドがアミノ酸残基1ないし24において同定され、膜貫通ドメインは配列番号:2のアミノ酸残基514ないし532において同定されている。N−グリコシル化部位は残基46ないし49、296ないし299、305ないし308、360ないし361、368ないし371、および461ないし464において同定されている。カゼインキナーゼIIリン酸化部位は残基10ないし13、93ないし96、130ないし133、172ないし175、184ないし187、235ないし238、271ないし274、272ないし275、323ないし326、606ないし609、615ないし618において同定されている。チロシンキナーゼリン酸化部位は約残基202ないし209において同定されている。N−ミリストイル化部位は約残基43ないし48、102ないし107、295ないし300、321ないし326、330ないし335、367ないし344、393ないし398、525ないし530および527ないし532において同定されており、アミド化部位は約残基240ないし243において同定されている。原核生物膜リポ蛋白質脂質付着は約残基516ないし526に存在し、成長因子およびサイトカイン受容体ファミリーシグニチャー1は約残基36ないし49に存在する。有意な相同性の領域は約残基14ないし51においてヒトエリスロポエチンと共に存在し、マウスインターロイキン−5受容体は残基211ないし219に存在する。記載された全てのmTCCR残基は配列番号:2の配列に従ってナンバリングされる。
【0061】
【化1】
【0062】
【化2】
IL−27
IL−27はTCCRに対するリガンドである(Pflanzら、2002Immunity 16(6):779−790)。IL−27はEBI3(エプスタイン−バールウィルス誘導遺伝子3)およびp28蛋白質サブユニットから構成されるヘテロダイマーサイトカインである。p28は3つの接触表面を持つ4つのラセン束サイトカインである。第一の接触表面はEBI3に結合し、第二および第四のαラセンの残基から構成される。第二の接触表面はサイトカイン受容体相同性ドメインの領域においてTCCRに結合し、第一および第三のアルファラセンの残基よりなる。第三の接触表面はgp130上で見出されるIGドメインのようなIGドメインに結合し、第一のラセンの最後および第四のラセンの始まりにおける残基を含む。
【0063】
ヒトp28のペプチド配列(配列番号:3)は243アミノ酸の長さであり、他方、マウスp28のペプチド配列(配列番号:4)は長さが234アミノ酸である(Pflanz,NCBI受託番号AAM34499)。表2にて以下に示されるこれらの配列は73%配列同一性を有する。
【0064】
【化3】
EBI3は可溶性サイトカイン受容体の構造を有し、p28上の特異的結合部位に結合する。ヒトEBI3は長さが229アミノ酸であり(Devergneら、1996,J.of Virology 70(2):1143−1153)、表3にて以下に示されるペプチド配列(配列番号:5)を有する。
【0065】
【化4】
TCCR/IL−27受容体複合体
図1はTCCR/IL−27受容体複合体の人工物を示す。IL−27に対する完全な受容体はgp130およびTCCRサブユニットを含有する。サイトカイン受容体相同性ドメインは配列番号:6の約残基126ないし323におけるgp130に存在する。gp130に存在する他の相同性ドメインは配列番号:6の約残基324ないし423、424ないし518、および519ないし614に位置する3つのフィブロネクチンIII型ドメイン、および約残基22ないし122における免疫グロブリンドメインを含む。gp130はIL−6、IL−11、CNTF、LIF、CT1、およびCLCに対する受容体の成分であることも知られている(Hibiら、1990,Cell,63(6):1149−1157)。gp130のアミノ酸配列(配列番号:6)を表4にて以下に示す。
【0066】
【化5】
【0067】
【化6】
TCCRのIL−27活性化が、主なTh1−特異的転写因子、T−betの発現を誘導する(Lucasら、2003,PNAS,100:15047−52)。TCCR活性化の効果はStats(転写のシグナルトランスジューサーおよびアクチベーター)によって媒介される。具体的には、TCCR活性化はStat1、Stat3、Stat4、およびStat5のリン酸化に導く(Lucasら、2003,supra)。後の実施例に示すデータは、TCCR活性化がCD8+T−リンパ球の増殖、またはTh1応答を含めた、自己免疫プロセスの発現を直接的または間接的に誘導することを示唆する。
TCCRおよびT−リンパ球サブタイプ
前記したように、4つのラセン束サイトカインファミリーのメンバー(Bazan,J.F.,1990,Proc Natl Acad Sci USA,87:6934−8)は、Th1およびTh2エフェクター細胞の区別される集団への共通の前駆体からのTヘルパー細胞の増殖および最後の分化において役割を演じる(O’Garra.,1998,Immunity,8:275−835)。IL−4はTh2細胞の発生に支配的に影響し、他方、IL−12はTh1細胞の分化における主な因子である。(Hsiehら、1993,Sience,260:547−9;Sederら、1993,Proc Natl Acad Sci USA,90:10188−92;Le Grosら、1990,J Exp Med,172:921−9;Swainら、1991,Immunol Rev,123:115−44)。したがって、IL−4が欠乏したマウス(Kuhnら、1991,Science,254,707−10)、IL−4受容体α鎖(Noben−Trauthら、1997,Proc Natl Acad Sci USA,94:10838−43)、またはIL−4特異的転写因子STAT6(Shimodaら、1996,Nature,380:630−3)はTh2応答が欠乏し、他方、IL−12が欠乏したマウス(Magramら、1996,Immunity4:471−81)IL−12受容体(IL−12R)β1鎖(Wuら、1997,J Immunol,159:1658−65)、またはIL−12特異的転写因子STAT4(Kaplanら、1996,Nature,382:174−7)は損なわれたTh1応答を有する。
【0068】
Th1およびTh2細胞サブタイプは共通の前駆体Th−0細胞に由来する。Th1およびTh2細胞からのサイトカイン放出プロフィールはエフェクターメカニズムおよび細胞傷害性細胞の選択に影響する。Th1細胞によって分泌されるIl−2およびインターフェロンガンマはマクロファージュおよび細胞傷害性細胞を活性化し、他方、Th2細胞によって分泌されるIl−4、Il−5、Il−6、およびIl−10は好酸球および肥満細胞の生産を増大させ、ならびにIgeを含めた抗体の生産を促進し、細胞傷害性細胞の機能を減少させる傾向がある(Powrieら、1993,Immunol.Today,14:270)。一旦樹立されれば、Th1またはTh2応答パターンは、他のサブセットの細胞によるサイトカイン生産を一般的には阻害するサイトカインの生産によって維持される。例えば、IL−4はTh1クローンからのインターフェロン−ガンマの生産を阻害し、他方、インターフェロン−ガンマはTh2クローンからのIl−4からの生産を阻害する(Mosmannら、1989,Adv.Immunol.46:111−147;Mosmannら、1989,Annu.Rev.Immunol.,7:145−173)。この負のフィードバックループは、多くの免疫応答の間における極性化サイトカインプロフィールの生産を加速する。
【0069】
細胞傷害性T−リンパ球(CD8+)は迅速に他の細胞を破壊することができる。細胞傷害性T−リンパ球は2つの主な細胞溶解経路を用いる:ペルフォリン−依存性エクソサイドーシス経路およびFasリガンド/Faskを用いる。細胞傷害性T−リンパ球はインターフェロン−ガンマのようなプロ−炎症サイトカインのプロジューサーでもある。
【0070】
Th1応答に関連するサイトカインの過剰生産またはCD8+細胞傷害性T−リンパ球の過剰増殖は、同種異系移植片拒絶、(グレーヴス病および橋本甲状腺炎のような)自己免疫甲状腺病、自己免疫ブドウ膜網膜炎、巨細胞動脈炎、(クローン病、潰瘍性結腸炎、局所腸炎、肉芽腫性腸炎、末端回腸炎、局所回腸炎、および終末回腸炎を含めた)炎症性腸疾患インスリン依存性新生糖尿病、多発性硬化症、悪性貧血、乾癬、慢性関節リウマチ、サルコイドーシス、強皮症、および全身エリテマトーデスに至りかねない。
【0071】
後の実施例に詳細に記載された実験は、非−特異的T−リンパ球刺激に応答しての、TCCRを発現するT−リンパ球と比較して、TCCRを欠如するT−リンパ球のより大きな増殖を示す(実施例1参照)。さらに、驚くべきことに、TCCRを発現するマウスは、TCCRを欠如するマウスよりも、多発性硬化症についての動物モデルにおいて、実験的アレルギー脳脊髄炎(EAE)のような自己免疫障害に対して感受性が低いことが見出された(実施例2)。
【0072】
これらのデータは、T−リンパ球増殖およびTh1媒介生物学的活性のTCCR−媒介直接的または間接的抑制を示唆する。従って、TCCRのアゴニストを用いて、T−細胞増殖を阻害し、および/または多発性硬化症および慢性関節リウマチを含めた自己免疫障害を治療することができる。
【0073】
自己免疫障害
前記したように、T−リンパ球の過剰増殖またはTh1またはTh2細胞によって生産されたサイトカインの過剰生産は医学的障害の保有者に導く。例えば、Th1応答に関連するサイトカインの過剰生産またはCD8+細胞傷害性T−リンパ球の過剰増殖は異種同系移植片拒絶、(グレーヴス病および橋本甲状腺炎のような)自己免疫甲状腺病、自己免疫ブドウ膜網膜炎、巨細胞動脈炎、(クローン病、潰瘍性結腸炎、局所腸炎、肉芽腫性炎腸炎、末端回腸炎、局所回腸炎、および終末回腸炎を含めた)炎症性腸疾患、インスリン−依存性真正糖尿病、多発性硬化症、悪性貧血、乾癬、慢性関節リウマチ、サルコイドーシス、強皮症および全身エリテマトーデスを含めた自己免疫障害に至り得る。
【0074】
多発性硬化症は、Tリンパ球依存性であると考えられる自己免疫脱髄障害である。MSは、一般には、再発−緩解コースまたは慢性進行コースを呈する。MSの病因は知られていないが、ウイルス感染、遺伝的素因、環境、および自己免疫は全て該障害に寄与しているように見える。MS患者における病巣は、圧倒的にT−リンパ球に媒介されるミクロ神経膠細胞および浸潤性マクロファージの浸潤物を含有する。CD4+Tリンパ球はこれらの病巣に存在する支配的な細胞型である。MS病巣のホールマークはプラーク、すなわち、MRIスキャンで見られる通常の白色物質から鋭く区別される脱髄の領域である。MSプラークの組織学的外観は病気の異なる段階で変化する。活性な病巣において、血液−脳関門は損傷されており、それにより、細胞外空間への血清蛋白質の血管外遊出を可能とする。炎症細胞は血管周囲カフにおいて、および白色物質全体に観察することができる。CD4+T−細胞、特にTh1はプラークのエッジにおいて毛細血管後細静脈の周りに蓄積し、白色物質にも散らばる。活性な病巣において、接着分子およびリンパ球のマーカーのアップレギュレーションおよびIL−2RおよびRおよびCD26のような単球活性化も観察された。活性病巣における脱髄には稀神経突起膠細胞の破壊は伴わない。対照的に、該病気の慢性相の間では、病巣は稀神経突起膠細胞の喪失およびしたがって、血液中のミエリン稀神経突起膠細胞糖蛋白質(MOG)抗体の存在によって特徴付けられる。
【0075】
種々のよく許容された動物モデルが、自己免疫障害について存在する。その例として、EAE(実験的アレルギー性脳脊髄炎)は、T細胞および単核細胞炎症、および中枢神経系における軸索の引き続いての脱髄によって特徴付けられるT細胞媒介自己免疫障害である。EAEは、一般的にヒトにおけるMSの関連動物モデルであると考えられている。(例えば、Bolton,C.1995,Multiple Sclerosis,143参照)。候補TCCRアゴニストのような剤は、例えば、John Wiley & Sols,Inc.によって発行され、Coligan et al.National Institutes of Healthによって編集されたCurrent Protocols in Immunology,units15.1および15.2に記載されたプロトコルを用い、免疫媒介脱髄障害に対するT細胞刺激または阻害活性について分析することができる。また、稀突起神経膠細胞またはシュワン細胞が、例えば、Duncanら、1997,Molec.Med.Today,554−561に記載されたように、中枢神経系にグラフトされたミエリン病についてのモデルも参照。
【0076】
関節炎についての動物モデルはコラーゲン−誘導関節炎である。例えば、McIndoeら、1999,Proc.Natl.Acad.Sci.USA,96:2210−2214参照。このモデルはヒト自己免疫慢性関節リウマチの臨床的、組織学的および免疫学的特徴を有し、ヒト自己免疫関節炎についての許容できるモデルである。マウスおよびラットモデルは滑膜炎、軟骨の侵食、および肋軟骨下骨によって特徴付けられる。コラーゲン−誘導関節炎は、リンパ球浸潤および滑膜肥大を含めたヒトにおける慢性関節リウマチでの多くの特徴を有する。例えば、McIndoeら、1999,Proc.Natl.Acad.Sci.USA,96:2210−2214参照。TCCRの潜在的アゴニストは、例えば、John Wiley & Sols,Inc.によって発行され、Coligan et al.National Institutes of Healthによって編集されたCurrent Protocols in Immunology,units 15.5に記載されたプロトコルを用い、これらのモデルを用いて自己免疫関節炎に対する活性について分析することができる。また、Issekutz,A.C.et al.,Immunology(1996)88:569に記載されたCD18に対するモノクローナル抗体およびVLA−4インテグリンを用いるモデルも参照されたし。
【0077】
皮膚異種同系移植片拒絶についての動物モデルは、抗−ウイルスおよび腫瘍免疫性におけるそれらの役割を示す、およびその尺度を示すイン・ビボ組織破壊を媒介するT細胞の能力をテストする手段である。ほとんどの通常かつ許容されたモデルは、マウス尾−皮膚移植片を用いる。報告された実験は、皮膚異種同系移植片拒絶はT細胞、ヘルパーT細胞およびキラー−エフェクタ−T細胞によって媒介され、抗体によっては媒介されないことを示している。例えば、Auchincloss and Sachs,1998,In:Fundamental Immunology,2nd ed.,W.E.Paul ed.,Raven Press,NY,Page889−992参照。適当な手法は、John Wiley & Sols,Inc.によって発行され、Coliganら、 1995,National Institutes of Healthによって編集されたCurrent Protocols in Immunology,unit 4.4に詳細に記載されている。他の候補TCCRアゴニストをスクリーニングするのに用いることができるトランスプラント拒絶モデルは、例えば、Tanabeら、 1994,Transplantation,58:23およびTinubuら、1994,J.Immunol.,4330−4338によって記載された同種異系心臓トランスプラントモデルを含む。
【0078】
TCCRのアゴニスト
TCCRのアゴニストは、本明細書中際書中に開示された天然配列TCCRペプチドの生物学的活性を増強し、または刺激する分子である。適当なアゴニスト分子は、具体的には、ヒト化抗体を含めたアゴニスト抗体、またはFab、Fab’、Fd、Fd’、Fv、Ab、無ヒンジ抗体、F(ab’)2断片、単一鎖抗体分子、ダイアボディー、単一アーム抗原結合分子、および線状抗体を含めたアゴニスト抗体の断片、アミノ酸配列改変体または天然ポリペプチド、ペプチド、小分子の断片などを含む。
【0079】
TCCRの適当なアゴニストは、TCCRのペプチド断片、TCCR細胞外メイン、および:
(a1)配列番号:1のヒトTCCRペプチドの残基1または約33ないし636;
(a2)配列番号:2のマウスTCCRペプチドの残基1または約25ないし623;
(b1)配列番号:1のヒトTCCRペプチドのX3ないし636、ここに、X3は配列番号:1のいずれかのアミノ酸残基27ないし37;
(b2)配列番号:2のマウスTCCRペプチドX4ないし623、ここに、X4は配列番号:2の20ないし30からのいずれかのアミノ酸残基であり;
(c1)1または約33ないしX1、ここにX1は配列番号:1の残基512ないし残基522からのいずれかのアミノ酸残基であり;
(c2)1または約25ないしX2、ここに、X2は配列番号:2の残基509ないし519からのいずれかのアミノ酸残基であり;
(d1)X5ないし636、ここに、X5は配列番号:1の残基533ないし543からのいずれかのアミノ酸であり;
(d2)X6ないし623、ここに、X6は配列番号:2の残基527ないし537からのいずれかのアミノ酸であり;または
(e)配列番号:1および配列番号:2のアミノ酸配列のもう1つの特異的に誘導された断片;
のアミノ酸配列に対して少なくとも約80%のアミノ酸配列同一性を有するTCCR改変体も含む。
【0080】
TCCRのアゴニストは、例えば、配列番号:1および配列番号:2の配列の、N−およびC−末端において、または1以上の内部ドメイン内で1以上のアミノ酸残基が付加され、または欠失されたTCCRペプチドを含む。
【0081】
TCCRのアゴニストは、天然配列IL−27、EBI3、p28、改変体、およびIL−27ヘテロダイマーに通常は関連する生物学的活性を有するその断片を含む。例えば、TCCRのアゴニストは、IL−27の天然配列成分に対して少なくとも80%、90%、95%、または99%配列同一性を有するIL−27改変体を含む。TCCRのアゴニストは、天然配列ヒトp28(配列番号:3)またはマウスp28(配列番号:4)に対して少なくとも73%、75%、80%、90%、95%、または99%配列同一性を有するp28改変体も含む。TCCRのアゴニストは、TCCRに結合することができるp28、およびgp130の部分を含む。その例として、TCCRのアゴニストは、天然配列ヒトp28(配列番号:3)またはマウスp28(配列番号:4)に対して少なくとも73%、75%、80%、90%、95%、または99%配列同一性を有し、かつTCCRおよびgp130に結合することができるp28改変体を含む。TCCRのアゴニストは、TCCR上に見出されるサイトカイン受容体相同性ドメインの領域においてTCCRに結合すると考えられる、p28の第一および第三のαラセンからの残基、およびgp130上に見出されるIGドメインに結合すると考えら得る、第一のラセンの最後および第4のラセンの始まりにおける残基を含有するp28ペプチド改変体を含む。
【0082】
TCCRのアゴニストは、例えば、TCCRに結合し、それを活性化することができる分子を含む。TCCRのアゴニストは、TCCRがホモダイマーを形成できるようにする分子、および/またはTCCRおよびgp130がヘテロダイマーを形成することができるようにする分子も含む。例えば、TCCRに対する抗体はTCCRがホモダイマーを形成できるようにすることができる。さらなる例として、TCCRおよびgp130双方に対して特異的な二価抗体はTCCRおよびgp130がヘテロダイマーを形成するようにすることができる。
【0083】
TCCRペプチドのアゴニストまたはアンタゴニストを同定するための方法は、TCCRペプチドを候補アゴニストまたはアンタゴニスト分子と接触し、次いで、TCCRに関連する1以上の生物学的活性のケン質可能な変化を測定すること含むことができる。例えば、アゴニストは、TCCRペプチドを発現する細胞を候補と接触させ、次いで、ウェスタン−ブロットまたはもう1つの適当なアッセイを用いて、Stat1、Stat3、Stat4、またはStat5のリン酸化のようなTCCRの生物学的活性について接触された細胞を分析することによって同定される。TCCRのアゴニストは、TCCRペプチドを発現するように作成されたBa/F3細胞のような細胞を候補アゴニストと接触させ、次いで、例えば、[3H]標識チミジン取込みまたはもう1つの適当なアッセイを測定することによって増殖について接触された細胞を分析することによって同定することもできる。
【0084】
モノクローナル抗体
モノクローナル抗体を生産するための多くの技術が知られている。1つの方法において、例えば、Balb/cのようなマウスは、フロイント完全アジュバントに乳化した免疫原としてのTCCRまたはその部分で免疫化し、1ないし100マイクログラムの量にて皮下または腹腔内注射される。別法として、該免疫原はMPL−TDMアジュバント(Ribi Immunochemical Research,Hamilton,MT)に乳化し、動物の後足肉趾に注射される。次いで、選択されたアジュバントに乳化されたさらなる免疫原で、免疫化されたマウスを10ないし12日後にブースター注射する。しかる後、数週間の間、マウスにさらなる免疫化注射をブースター注射することもできる。血清試料は、ELISAアッセイにおいてテストして、抗−TCCR抗体を検出するために眼窩後出血によってマウスから周期的に得ることができる。
【0085】
適当な抗体力価が検出された後、抗−TCCR抗体について「陽性」の動物に、免疫原の最終静脈内注射を注射することができる。3ないし4日後に、マウスを犠牲にし、脾臓細胞を収穫する。脾臓細胞を、次いで、(35%ポリエチレングリコールを用いて)ATCC、No.CRL 1597から入手可能な、P3X63AgU.1のような選択されたマウス骨髄腫細胞系に融合させる。該癒合によりハイブリドーマ細胞が生じ、次いで、これを、HAT(ヒポキサンチン、アミノプテリン、およびチミジン)を含有する96ウェル組織培養プレートに平板培養して、非−融合細胞、骨髄腫ハイブリッド、および脾臓細胞ハイブリッドの増殖を阻害することができる。
【0086】
選択されたハイブリドーマ細胞は、TCCRに対する反応性について、ELISA、または他の適当なアッセイでスクリーニングすることができる。陽性ハイブリドーマ細胞は、例えば、同系Balb/cマウスに腹腔内注射して、抗−TCCRモノクローナル抗体を含有する腹水液を生じさせることができる。別法としてハイブリドーマ細胞は、例えば、組織培養フラスコまたはローラーボトル中で増殖させることができる。腹水液中で生産されたモノクローナル抗体の精製は、硫酸アンモニウム沈殿、続いて、ゲル排除クロマトグラフィーまたは他の適当な方法を用いて達成することができる。別法として、抗体のプロテインAまたはプロテインGへの結合に基づくアフィニティークロマトグラフィーを使用することができる。
【0087】
TCCR−/−マウス
TCCRを発現しない「ノックアウト」マウスが構築されている(TCCR−/−)。そのようなマウスは、例えば、TCCRをコードする内因性遺伝子、および動物の胚細胞に導入された同一ペプチドをコードする改変されたゲノムDNAの間の相同組換えを介して調製することができる。
【0088】
例えば、特定のペプチドをコードするcDNAを用いて、確立された技術に従ってそのペプチドをコードするゲノムDNAをクローン化することができる。特定のペプチドをコードするゲノムDNAの部分を、欠失することができるか、あるいは取り込みをモニターするのに用いることができる選択マーカーをコードする遺伝子のようなもう1つの遺伝子を置き換えることができる。典型的には、(5’および3’末端双方における)未改変フランキングDNAの数キロベースがベクターに含まれる(相同組換えベクターの記載については、Thomasら、1987,Cell,51:503参照)。ベクターは(エレクトロポレーションによるなどして)胚性幹細胞系に導入し、導入されたDNAが内因性DNAを相同組換えされている細胞を選択する。例えば、Liら、1992,Cell,69:951参照。次いで、例えば、Braderyら、1987,In:Teratocarcinomas and Embryonic Stem Cells: A Practical Approach,E.J.Roberteon,ed.(IRL,Oxford),pp.113−152に記載されているように、選択された細胞を(マウスまたはラットのような)動物の胚盤胞に注射して、同種キメラを形成させる。次いで、キメラ胚を適当な偽妊娠雌仮腹動物に移植し、該胚を出産させて、「ノックアウト」動物を生じさせることができる。それらの生殖細胞に相同組換えDNAを保有する子孫は標準的な技術によって同定し、それを用いて、動物の全ての細胞は相同組換えDNAを含有する動物を育種することができる。
【0089】
後の実施例で用いるTCCR−/−マウスの作成の記載は、その内容がここに引用して援用されるWO 0129070 (de Sauvage et al.)およびChenら、2000,Nature,407:916に見出される。
【0090】
組成物および治療
自己免疫障害の治療でいうようなTCCRのアゴニストは、限定されるものではないが、免疫機能、例えば、T細胞増殖/活性化、リンホカイン放出、または免疫細胞浸潤を変調する、蛋白質、抗体、断片および改変体、小さな有機分子、ペプチド、リンペプチドなどを含む。特に、本発明中で記載されるTCCRのアゴニストは、T−リンパ球の増殖、T−リンパ球サイトカイン放出、および自己免疫障害を抑制し、減少させ、または低下させるのに有用である。
【0091】
TCCRアゴニストは、前記したスクリーニングアッセイのいずれかによって、および/またはいずれかの他の公知のスクリーニング技術によって同定することができる。
【0092】
本発明のTCCRアゴニストを公知の方法に従って処方して有用な組成物を調製し、それにより、TCCRアゴニストを許容される担体と組み合わせる。処方は、所望の程度の純度を有するTCCRアゴニストを、凍結乾燥された処方または水性溶液の形態の、任意の許容される担体、賦形剤、または安定化剤と混合することによって貯蔵のために調製される。例えば、Remington:The Science and Practice of Pharmacy 20th ed.Gennaro Ed.(2000)参照。適当な担体、賦形剤または安定化剤は使用する用量および濃度において受容者に対して非毒性であり、それは、リン酸塩、クエン酸塩、および他の有機酸のような緩衝液;アスコルビン酸を含めた抗酸化剤;低分子量(約10残基未満)ペプチド血清アルブミン、ゼラチンまたは免疫グログリンのような蛋白質;ポリビニルピヨリドンのような親水性ポリマー、グリシン、グルタミン、アスパラギン、アルギニンまたはリシンのようなアミノ酸;グルコース、マンノースまたはデキストリンを含めた単糖、二糖および他の炭水化物;EDTAのようなキレート剤;マンニトールまたはソルビトールのような糖アルコール;ナトリウムのような塩−形成対イオン;および/またはTWEEN(登録商標)PLURONICS(登録商標)PEGのようなノニオン界面活性剤を含む。
【0093】
イン・ビボ投与で用いるべき処方は滅菌されていなければならない。これは、凍結乾燥および復元に先立ってまたはそれに続いて、滅菌濾過膜を通しての濾過によって容易に達成される。
【0094】
本発明中において、組成物は、一般には、滅菌アクセスポート、例えば、皮下注射針によって刺すことができるストッパーを有する静脈内溶液バッグまたはバイアルを有する容器に入れられる。
【0095】
投与経路は、静脈内、腹腔内、脳内、筋肉内、眼内、動脈内、または病巣内経路、局所投与による、または徐放系による注射または注入のような公知の方法に従う。投与の経路は、アデノウィルスベクターのような適当なベクターでのトランスフェクションの結果としてのイン・ビボ発現を含むことができる。
【0096】
本発明の医薬組成物の量および所望の濃度は、考えられる特定のしように応じて変化させることができる。投与の適当な用量または経路の決定は、十分に、通常の技師の技量内にある。動物実験は、ヒト療法のための有効量の決定のための信頼できるガイダンスを提供する。効果的な用量の種間スケーリングは、Mordenti,J.and Chappell,W.”The use of Interspecies scaling in toxicokinetics” in Toxicokinetics and New Drug Development,Yacobi et.al.,Eds.,Pergamon Press,New York 1989,pp.42−96によって主張された原理に従って行うことができる。
【0097】
TCCRアゴニストのイン・ビボ投与を使用する場合、正常な用量は、投与経路に応じて、1日当たり約10ng/kgないし100mg/kg哺乳動物体重以上、例えば、約1mg/kg/日ないし10mg/kg/日で変化させることができる。特定の用量および送達の方法に関するガイダンスは文献中に供される;例えば、米国特許代4,657,760;第5,206,344;または第5,225,212号参照。異なる処方は異なる治療および異なる障害で効果的であり、および特異的器官または組織を治療することを意図した投与は、もう1つの器官または組織に対するものとはことなる送達を必要とするであろうと予測される。
【0098】
TCCRアゴニストの徐放投与がTCCRアゴニストの投与を必要とするいずれかの病気または障害の治療に適した放出特徴を持つ処方で望まれる場合、TCCRアゴニストのマイクロカプセル化が考えられる。徐放のための組換え蛋白質のマイクロカプセル化は、ヒト成長ホルモン(rhGH)、インターフェロン−アルファ、−ベータ、−ガンマ、インターロイキン−2、およびMN rgp120で首尾よく行われている。例えば、Johnsonら、1996,Nat.Med.2:795−199;Yasuda,1993,Biomed.Ther.,1221−1223;Horaら、1990,Bio/Technology,755−758;Cleland,1995,”Design and Production of Single Immunization Vaccines Using Polylactide Polyglycolide Microsphere Systems” in Vaccine Degsin:The Subunit and Adjuvand Approach,Powell and Newman,eds.,(Plenum Press;New York),pp.439−462;WO97/03692、WO96/40072,WO96/07399および米国特許第5,654,010参照。
【0099】
TCCRアゴニストの徐放処方は、ポリ−乳酸−コグリコール酸(PLGA)、かなりの程度の生体適合性、および広い範囲の生分解性特性を呈するポリマーを用いて開発することができる。PLGA、乳酸およびグリコール酸の分解生成物はヒト身体から迅速に除去される。さらに、このポリマーの分解性は、その分子量および組成に応じて数ヶ月ないし数年に調整することができる。さらなる情報については、Lews,”Controlled Release of Bioactive Agents from Lactide/Glycolide polymer”in Biogradable Polymers as Drug Delivery Systems,M.Chasin and R.Langdeer,editers(Marcel Dekker;New York,1990),pp.1−41参照。
【実施例】
【0100】
本発明は、以下の実施例を参照して良好に理解することができる。これらの実施例は本発明の特別な具体例の代表であることが意図され、本発明の範囲を限定することを意図しない。
【0101】
実施例1
T−細胞応答のTCCR−媒介抑制
T−細胞応答に対するTCCR活性の効果は、野生型(TCCR+/+)およびノックアウト(TCCR−/−)脾臓細胞の誘導されたT−細胞の増殖の分析によってテストした。T−細胞受容体はCD3と会合してT−細胞受容体複合体を形成する。異なる用量の抗−CD3抗体は、T−細胞受容体複合体および抗原−提示細胞(APC)のMHCクラスII分子(CD4)の相互作用に通常は関連するT−細胞の増殖を非特異的に刺激する。
【0102】
野生型(TCCR+・+)およびノックアウト(TCCR−/−)混合リンパ球、単離されたCD4+T細胞および、単離されたCD8+T細胞の増殖は、抗−CD3抗体によって刺激した(BD Pharmingen,San Digo,CA,clone 145−2c11)。細胞を湿潤化CO2インキュベータ中で3日間増殖し、増殖はアッセイの最後の8ないし16時間に測定された[3H]チミジン取り込みによって測定した。驚くべきことに、ノックアウトマウス(TCCR−/−)から得られた混合リンパ球の抗−CD3抗体誘導増殖は、表5において、および図5に示されたように、最大下用量の抗−CD3において野生型(TCCR+/+)リンパ球から得られたリンパ球のそれよりも有意に大きかった。このデータは、例えば、刺激されたT−細胞の増殖を抑制するTCCR活性の保護効果を示唆し、およびアゴニストでのTCCRの刺激は、T−細胞増殖のようなT−細胞応答を直接的にまたは間接的に抑制するのに有用であろう。
【0103】
【化7】
しかしながら、単離されたCD4+T細胞および単離されたCD8+T細胞の抗−CD3抗体誘導増殖は、各々、図6(表6)および図7(表7)に示されたように、野生型およびノックアウト細胞の間で有意には異ならなかった。このデータは、TCCRに対するリガンドであるIL−27が、CD4+T細胞およびCD8+T細胞以外のリンパ球によって生産されることを示唆する。
【0104】
【化8】
次に、抗−CD3抗体刺激に応答してのCD4+およびCD8+細胞の増殖は、CFSE(カルボキシフルオレセインジアセテート、スクシンイミジドエステル)標識アッセイで測定した。標識された細胞は、各細胞分裂後にそれらの蛍光強度の50%を喪失するので、CFSE標識は、細胞分裂の数がモニターされるのを可能とする。野生型(TCCR+/+)およびノックアウト(TCCR−/−)混合リンパ球細胞懸濁液をCFSEで標識して、細胞懸濁液中に0.5μM CFSE(Sigma,St.Louis,MO)の濃度を作り出した。次いで、細胞懸濁液を37℃で10分間インキュベートした。標識の後、FCSを5%最終濃度まで加え、細胞を直ちに遠心し、氷冷PBSで洗浄した。野生型(TCCR+/+)およびノックアウト(TCCR−/−)混合リンパ球細胞の増殖は、2.5μg/mlの濃度において抗−CD3抗体(BD Pharmingen,San Piego,Ca,clone 145−2c11)によって刺激した。
【0105】
次いで、細胞を37℃にて2日間インキュベートした。その時点において、細胞をCD4+およびCD8+に対するマーカー(CD4−CychromeまたはCD8−Cychrome)で標識し、フローサイトメトリーによって分析した。
【0106】
以下のデータは、CD4+ならびにCD8+の双方の陽性T細胞がTCCRノックアウト細胞において過剰増殖性であることを示す(表8参照)。図10および11は、インキュベーション時間の間において0、1、2、3、4または5回の細胞分裂を受けた細胞の数を示す。CD4+およびCD8+T細胞双方について、より多くの細胞が、野生型におけるよりもノックアウトにおいて3、4、5回の分裂を受けた(すなわち、ノックアウト細胞についての系はCD4+細胞(図14)およびCD8+細胞(図15)双方において右側にシフトする)。
【0107】
【化9】
実施例2
TCCRを発現するマウスはEAEに対して感受性が低い。
【0108】
実験的アレルギー性脳脊髄炎(EAE)は、多発性硬化症(MS)についての動物モデルとして働くCNSの自己免疫障害である。MSと同様に、EAEは、ミエリンに対する免疫−媒介損傷が観察可能な兆候をもたらす脱髄障害である。EAEはCD4+Th1細胞(Fifeら、2001,J.of Immun.,166:7617−7624)およびCD8+細胞傷害性T−リンパ球(CTL)(Huseby,et al.,2001,J.Exp.Med.,194(5):669−676)双方によって媒介されると考えられる。EAEに対するTCCRの効果を調べるために、EAEの臨床的進行をTCCRを発現する野生型マウス(TCCR+/+)およびTCCRを欠如するノックアウトマウス(TCCR−/−)において調べた。以下に示すように、TCCRを発現するマウスは、TCCRを欠如するマウスよりもCD4+ThおよびCD8+媒介傷害EAEに対して感受性が低い。
【0109】
ノックアウトTCCR−/−マウスはWO 0129070(de Sauvage et al.)に記載されたように創製し、C57BL/6バックグラウンドに逆−交配させ、N12創始細胞から育種した。野生型TCCR+/+対照はJackson Laboratory (Bar Harbor,ME)から購入したC57Bl/6マウスであった。
【0110】
MOG誘導EAE
MEVGWYRSPFSRVVHLYRNGKのアミノ酸配列(配列番号:7)を有するMOG35−55ペプチドは、Rainin Quartet自動ペプチド合成器(Rainin,Oaklane,CA)での9−フルオレニルメトキシカルボニル化学を用いて合成した。ペプチドを樹脂から切断し、移動相において水/アセトニトリル/0.1%TFAグラジエントを持つ分取用逆相HPLCを用いて精製した。ペプチドの同一性は、電子スプレーマススペクトロメトリーによって確認した。
【0111】
野生型TCCR+/+およびノックアウトTCCR−/―マウスを、100μlのTBSおよび100μlのCFA(フロイントの完全アジュバント)中の200μgのMOG35−55ペプチドを含有する200μlのエマルジョンで皮内免疫化して、0日にEAEを誘導した。CAFは、IFA(フロイントの不完全アジュバント)(Difco−BD Diagnostic System,Sparks,MD)を死滅し、乾燥したM,tuberculosis H37A(Difco−BD Diagnositic Systems,Sparks,MD)と8mg/ml M.tutuberculosisの濃度まで混合することによって調製した(各マウスはCFAの成分としての800μgの死滅したM.tutubersulosisを受けた)。0日に、および2日に再度、各マウスに100μlのPBS中の200ngの百日咳トキシン(List Biological Laboratories,Campbell,CA)で腹腔内注射して、血液の関門に入るのを助けた。受けた成分の用量は以下の表9にまとめる。
【0112】
【化10】
MBP誘導EAE
ASQKRPSQRHG(配列番号:8)のアミノ酸配列を有するAc 1−11ペプチドは、Rainin Quartat自動ペプチド合成器(Rainin,Oakland,CA)にて9−フルオレニルメトキシカルボニル化学を用いて合成した。該ペプチドを樹脂から切断し、移動相における水/アセトニトリル/0.1%TFAグラジエントでの分取用逆相HPLCを用いることによって精製した。ペプチドの同一性は、電子スプレーマススペクトロメトリーによって確認した。
【0113】
野生型TCCR+/+およびノックアウトTCCR−/−マウスは、100μlのCFA(フロイントの完全アジュバント)中の10μgのAc 1−11ペプチド(ASQKRPSQRHG)、ミエリン塩基性蛋白質の成分を皮内免疫化して、0日にEAEを誘導した。MOB−誘導EAEについて前記したように、CFAは、IFA(フロイントの不完全アジュバント)を、M.tuberculosis H37A(死滅しおよび乾燥した)と8mg/ml M.tuberculosisの濃度まで混合することによって調製した(各マウスは、CFAの成分として、800μgの死滅したM.tuberculosisを受けた)。2日、および3日に再度、各マウスに、100μlのPBS中の200ngの百日咳トキシンで腹腔内注射して、血液の関門に入るのを助けた。受けた成分に用量を以下の表10にまとめる。
【0114】
【化11】
全てのマウスは、1日で出発し、1週間当たり3回臨床的疾患について評価した。病気グレード4に到達したマウスを毎日評価した。グレード5におけるいずれの動物も安楽死させた。5日以内にグレード3以下まで改良されなかったものを安楽死させた。用いた臨床的グレーディングシステムを以下の表11に示す。
【0115】
【化12】
40日において、全ての残りの動物を犠牲にし、脳および脊髄を組織学的分析のために切開した。脳を切開し、(各脳についての4つのレベルの各々からの1つの切片)、H&E(ヘマトキシリンおよびエオシン染色、Sigma,St.Louis,MO)で染色して、炎症を評価した。脊髄を切片化し(各脊髄について3つの異なるレベルの各々から4つの切片)、H&Eで染色して、炎症を評価し、ルキソールファストブルー(Luxol Fast Blue)(VWR Scientific,St.Paul,MN)で染色して、脱髄を評価した。各スライドについて、最高炎症および脱髄双方についての最高スコアおよび平均スコア(各スライドでの全ての切片の平均)を報告した。用いた炎症グレーディングシステムを以下の表12に示す。
【0116】
【化13】
用いた脱髄グレーディングシステムを以下の表13に示す。
【0117】
【化14】
野生型(TCCR+/+)におけるMOG(ミエリン稀突起神経膠細胞糖蛋白質)誘導EAE(実験的アレルギー性脳脊髄炎)の臨床的進行は、ノックアウト(TCCR−/−)マウスにおける誘導されたEAEよりもひどくはなかった(図8参照)。同様に、野生型(TCCR+/+)マウスにおけるMBP(ミエリン塩基性蛋白質)誘導EAEの臨床的進行は、ノックアウト(TCCR−/−)マウスにおける誘導されたEAEよりひどくはなかった(図9参照)。
【0118】
図8および9に示すように、TCCRを欠如する動物(TCCR−/−)はEAEのよりひどい臨床的兆候を示し、他方、TCCRを発現するマウス(wt)はよりひどくない兆候および進行を示し、これは、EAEのような、自己免疫障害に対するTCCR活性の保護効果を示唆する。
【0119】
臨床データは、図10ないし13に示したように、組織学的分析によってさらに支持される。TCCR−/−マウスは、WTマウスよりも高い炎症および高い脱髄スコアを有し、TCCRを発現するマウス(wt)は、TCCRを欠如するマウス(−/−)よりもCD4+Th1およびCD8+媒介障害EAEに対して感受性が低かったことを示す。
【0120】
まとめると、データは、EAEのような、自己免疫障害に対するTCCR活性の保護的、弱める、または抑制的効果を示唆する。
【0121】
実施例3
TCCRアゴニストでの自己免疫障害の治療
実施例2について記載したように、実験的アレルギー性脳脊髄炎(EAE)は、多発性硬化症(MS)に対する動物モデルとして働くSMSのCD4+Th1またはCD+8媒介自己免疫障害である。自己免疫障害の進行およびコースに対する投与されたTCCRアゴニストの効果を調べるために、IL−27のようなTCCRアゴニストを、誘導されたEAEのようなMSの実験的モデル系において投与する。EAEは、例えば、実施例2について前記したようにマウスにおいて開始させる。EAEの臨床的進行は、TCCRを発現し(TCCR+/+)、IL−27のようなTCCRアゴニストを受けるマウスにおいて評価する。IL−27のようなTCCRアゴニストで処理したマウスは、病気の低下した臨床的兆候または進行を示し、および/または未処理TCCR+/+対照よりも自己免疫障害に対して感受性が低いと予測される。
【0122】
実施例4
TCCRアゴニストでの動物モデルにおける関節炎の治療
自己免疫障害に対するTCCRアゴニストの抑制的および/または保護的効果は、数個の利用可能な動物モデル系のうちの1つでテストすることができる。マウスにおけるコラーゲン−誘導関節炎は、自己免疫障害、慢性関節リウマチに対する1つのモデルである。このモデルは、例えば、McIndoeら、1999,PNAS USA 96:2210−2214に記載されている。マウスにおけるコラーゲン−誘導関節炎は、リンパ球浸潤および滑膜肥大を含めたヒト慢性関節リウマチでの多くの特徴を共有する。
【0123】
コラーゲン−誘導関節炎の臨床的進行は、マウス、例えば、C57BL/6マウスまたは他の適当な実験室動物において調べる。関節炎は、例えば、McIndoeら、supraで引用された方法、または他の公知の方法によってテスト動物において誘導される。一般に、モデル動物は、異なる動物種に由来するII型コラーゲンをテスト動物に注射することによって、例えば、ウシII型コラーゲンをマウスに注射することによって創生する。該コラーゲンはフロイントの完全アジュバントのようなアジュバントと組み合わせることができる。
【0124】
TCCRリガンドIL−27のようなTCCRアゴニストは、例えば、関節炎−誘導剤の投与に先立って、投与の間に、および/または投与の後に、あるいは関節炎兆候の開始に先立って、開始の間に、および/または開始後にテスト動物に投与される。投与および用量の方法は変化させることができ、例えば、1プレおよび/またはポスト用量における、2以上のプレおよび/またはポスト用量の期間にわたっての毎日の1日当たり複数用量における用量、またはTCCR受容体を発現する細胞へペプチド剤を投与するために知られた他の適当な用量における、単体中のIL−27のようなペプチドリガンドの投与を含むことができる。代替方法は、例えば、IL−27の双方のサブユニット、または連結されたIL−27サイトカインを発現する組換えアデノウイルスからのペプチドを発現させることによるIL−27のようなペプチドリガンドの送達を含む。
【0125】
臨床的疾患の進行は、例えば、McIndoeら、supraに記載されたように、テストおよび対照動物においてモニターされる。例えば、該疾患の物理的および化学的特徴をモニターし、経時的にスコア取りする。動物は、主な関節のリンパ球浸潤、滑膜肥大、滑膜液サイトカイン含有量等について分析することができる。これらのパラメータを、テスト動物および対照の間で比較する。実施例1および2に示されたTCCRの保護的/抑制的効果を保つために、TCCRアゴニストでの動物における誘導された関節炎の治療は、未処理対照と比較して、よりひどくない臨床的兆候、結果、および/または物理的または化学的特徴で証明された、抑制的および/または保護的効果を供すると予測される。
【0126】
実施例5
TCCRに対するモノクローナル抗体の調製
hTCCRに対するモノクローナル抗体は、hTCCRの細胞外ドメインを用いて調製した。免疫原は、精製目的でカルボキシ−末端に付加された8つのヒスチジン残基をタグとして加えた膜貫通部分(配列番号:1の残基517ないし538)を欠如するhTCCR(配列番号:1)であった。hTCCR(ECD)−(His)8ペプチドは、ニッケルNTAアフィニティークロマトグラフィーを介して精製した。
【0127】
hTCCR(ECD)−(His)8ペプチド(1ないし2マイクログラム)を25マイクロリットルのMPL−TDMアジュバント(Ribi Immunochemical Research,Hamilton,MT)と組み合わせ、合計12回の注射のために毎週2回、野生型balb/cマウス(Charles River Laboratories,Wilmington,MA)の肉趾に注射した。
【0128】
42日に、マウスを犠牲にし、脾臓細胞を収穫した。脾臓細胞を(35%ポリエチレングリコールを用いて)マウス骨髄腫細胞(ATCC,No.CRL1597から入手可能なP3X63AgU.1)に融合させた。融合によりハイブリドーマ細胞が創製され、これを、HAT(ヒポキサンチン、アミノプテリンおよびチミジン)培地を含有する96ウェル組織培養プレート中で平板培養して、非−融合細胞、骨髄腫ハイブリットおよび脾臓細胞ハイブリットの増殖を阻害した。
【0129】
次いで、ハイブリドーマ細胞を、TCCRへの抗体結合についてELISAアッセイにおいてスクリーニングした。TCCRに対する反応性を有する同定されたハイブリドーマ培養は培養:2685、2686、および2688を含んだ。ハイブリドーマ培養2686(抗体2686)は、2004年12月15日に、American Type Culture Collection(ATCC),Manassas,Va.に寄託し、受託番号ATCC PTA−6447を有する。
【0130】
実施例6
モノクローナルAb2686はヒトTCCRを活性化する。
【0131】
組換えTCCRを発現するBa/F3細胞を用いて、TCCRを活性化する抗−TCCR抗体の能力を分析した。Ba/F3細胞はマウスIL−3依存性細胞系である。TCCRの候補アゴニストは、該候補アゴニストに応答してのTCCRを発現するBa/F3細胞の増殖を測定することによって評価することができる。細胞増殖の結果、ポリヌクレオチドの増大した合成のため増大した[3H]―チミジンの取込がもたらされる。細胞増殖は、例えば、[3H]―チミジン接種を測定することによってモニターされる。以下に示すように、モノクローナルAB2686は、TCCRを発現するBa/F3細胞の増殖を誘導することによってTCCRアゴニスト活性を示した。
【0132】
Ba/F3細胞(Palaciosら、1985、Cell,41:727−734)は、増殖および生存双方に対してIL−3を必要とするマウス造血系因子−依存性細胞系である。Ba/F3細胞を、10%胎児子牛血清(GIBCO,Carlsbad,CA)および100pg/mLマウスIL−3(R&D Systems,Minneapolis,MN)を補足したRPMI−1640倍値(GIBCO,Carlsbad,CA)中で培養した。
【0133】
ネオマイシン耐性遺伝子を持ち、ヒトまたはマウスTCCRをコードするポリヌクレオチド配列いずれかを含有するpMSCVベクター(Clontech,Palo Alto,CA)をエレクトロポレーションによってBa/F3細胞にトランスフェクトした。安定なトランスフェクタントを1mg/mlのG418(Clontech,Palo Alto,CA)で処理して、ネオマイシン耐性に対する遺伝子を含有するベクターでトランスフェクトされている安定な真核生物細胞系を選択した。次いで、TCCRを認識するフィコ−エリスリン標識モノクローナル抗体で細胞を処理した。TCCRを発現する標識されたクローンをFACSによって選択した。
【0134】
TCCR発現細胞を、IL−3を加えていない10%胎児子牛血清を補足したRPMI−1640倍値で洗浄した。次いで、細胞を、10%胎児子牛血清を補足した100μlのRPMI−1640倍値中にてウェル当たり5×103細胞にて二連で平板培養した。精製された組換えマウスIL−3(陽性対照)または精製された抗−TCCR(ヒト)モノクローナル抗体:2685−IgG2a、2686−IgG1、2688−IgG1、対照イソタイプIgG2a(BD Pharmingen,San Diego,CA)または対照イソタイプIgG1(BD Pharmingen,San Diego,CA)を、4:1希釈シリーズとして、表14にて以下に示す濃度で加えた。
【0135】
【化15】
48時間後、1μCiの[3H]―チミジン(Amerscham−Pharmacia,Piscataway,NJ)を各ウェルに加えた。さらに6時間後、[3H]―チミジンの細胞への取り込みを、β−カウンター(Packard Topcount,PerkinElmer Life and Analytical Sciences, Boston,MA)で測定した。
【0136】
結果を図2ないし4に示す。モノクローナル抗体2685−IgG2a、2686−IgG1、2688−IgG1、および対照イソタイプIgG2aまたは対照イソタイプIgG1に応答してのヒトTCCRの発現するBa/F3細胞の増殖を図2に示す。抗体2686は、テストした他の抗体のいずれよりも[3H]―チミジンの有意により大きな取り込みを誘導し、これは、抗体2686が、Ba/F3細胞において発現されたヒトTCCRの効果的なアゴニストであることを示す。
【0137】
マウスIL−3(陽性対照)または抗体2686いずれかに応答してのヒトTCCRを発現するBa/F3細胞の増殖を図3に示す。示されたように、抗体2686は、陽性対照IL−3のそれよりも小さいに拘らず、ヒトTCCRを発現するBa/F3細胞においてTCCR応答を生じさせるのに効果的であった。
【0138】
ヒトTCCRを発現するBa/F3細胞は、図4に示すように、マウスTCCRを発現するBa/F3細胞よりも抗体2686での処理に応答しての有意により大きな量の[3H]―チミジンを取り込んだ。このデータは、抗体2686がヒトTCCRの特異的アゴニストであることを示し、マウスTCCRに対して交差反応性を示さない。
【0139】
これらの研究は、示されたアゴニスト抗体のようなTCCRのアゴニストがTCCR受容体に結合し、それを刺激して、TCCR−媒介生物学的活性、ここでは、Ba/F3細胞の増殖を誘導することができるのを示す。したがって、データは、TCCRアゴニストがイン・ビボにてTCCR−媒介活性を直接的にまたは間接的に誘導するのに有用であることを示唆する。
【0140】
実施例7
他のTCCRアゴニストの同定
TCCRアゴニスト活性を有する剤を同定し、確認するために、IL−27の断片およびTCCRの改変体を含めた推定TCCRアゴニストをTCCR結合にて分析した。TCCRの結合はイン・ビトロまたはイン・ビボ結合にて分析することができる。例えば、潜在的アゴニストを、実施例3にて前記したように、組換えTCCRを発現するように作成されたCOS細胞またはBa/F3細胞のような、TCCRを発現する細胞に投与し、潜在的アゴニストに対する細胞応答を測定する。
【0141】
受容体結合は、融合蛋白質、例えば、ヒトIgGのFcドメインを含有するイムノアドヘシンとして潜在的にペプチドアゴニストを発現させることによって分析することもできる。受容体−リンガンド結合は、例えば、該イムノアドヘシンをTCCR発現細胞と相互作用させることによって検出される。結合したイムノアドヘシンは、Fc融合ドメインを認識する蛍光試薬を用い、微視的に可視化することができる。結合は蛍光の分析によって、または他の公知の方法によって定量することができる。
【0142】
TCCRのアゴニストは、IL−10またはSOCS−3の発現のようなTCCR媒介活性を刺激する候補アゴニストの能力を分析することによってスクリーニングすることができる。例えば、TCCRを発現するT−リンパ球を候補アゴニストと接触させることができる。IL−10および/またはSOCS−3の発現は、例えば、ELISA、定量的PCR、および同様な方法によって測定することができる。対照、例えば、基礎IL−10および/またはSOCS−3レベルに対するIL−10および/またはSOCS−3の発現の増加はTCCR刺激に相関し、これは有用なTCCRアゴニストの指標となる。
【0143】
実施例8
IL−27媒介細胞増殖、およびサイトカインの誘導および抑制
野生型(TCCR+/+)およびノックアウト(TCCR−/−)CD4+細胞双方におけるサイトカイン誘導に対するIL−27の効果を中性のTh1またはTh2誘導条件下で調べた。野生型(TCCR+/+)およびノックアウト(TCCR−/−)CD4+細胞双方における細胞リコール増殖に対するIL−27の効果を、中性のTh1またはTh2誘導条件下で調べた。
【0144】
0日に、アゴニスト抗−CD3モノクローナル抗体(145−2C11,BD Pharmingen,San Diego,CA,PBS o/n中5ug/ml)で予めコートされた24ウェルプレート中で、野生型CD4+またはTCCRノックアウトCD4+細胞をウェル当たり2×105細胞にて平板培養した。次いで、個々のウェルにおける増殖を中性、Th1バイアシング、またはTh2バイアシング条件下で誘導した。中性条件はIL−2(R&D Systems,Minneapolis,MN)、抗−IL−12抗体(BD Pharmingen,San Diego、CA)、抗−IFN−γ抗体(BD Pharmingen,San Diego,CA)、抗−IL−4抗体(BD Pharmingen,San Diego,CA)およびCD−28(BD Pharmingen,San Diego,CA)の添加によって作り出した。Th1バイアシング条件は、IL−2、IL−12(R&D Systems,Minneapolis,MN)、抗−IL−4抗体、およびCD−28の添加によって作り出した。Th2バイアシング条件は、IL−2、IL−4(R&D Systems,Minneapolis,MN)、抗−IL−12抗体、抗−IFN−γ抗体、およびCD28の添加によって作り出した。これらの個々のウェルにおける処理は表15にて以下に示す。
【0145】
【化16−1】
【0146】
【化16−2】
細胞を37℃で培養した。上清の試料は24時間、48時間、および/または72時間に採取した。ELISAを、TNF−α、IL−5、IL−2、IFN−γ、IL−10、IL−6、IL−4、GM−CSFに対するプローブ(BD Pharmingen,San Diego,CAから購入したキット)にて上清試料で行った。以下の表16は、IL−27依存性誘導倍数としてのELISAデータを示す。図16AないしCは、中性(16A)、Th1バイアシング(16B)、およびTh2バイアシング条件(16C)下でのIL−2のIL−27依存性誘導を示す。図17AないしCは、中性(17A)、Th1バイアシング(17B)、およびTh2バイアシング条件(17C)下でのIL−10のIL−27依存性誘導を示す。
【0147】
データは、IL−27に応答してのTNF−α、IFN−γ、およびIL−4の誘導を示す。データはIL−27に応答してのIL−2、IL−6、およびGM−CSFの抑制も示す。データは、IL−10が、中性、Th1バイアシング、およびTh2バイアシング条件下でIL−27によって誘導されることを示す。前記したように、IL−10は、限定的かつ終了する炎症応答において主な役割を演じる。IL−10はIL−27によって誘導されるので、データは、IL−27を用いて、免疫−媒介疾患を治療できることを示唆する。
【0148】
【化17】
RNAを24時間、48時間、および/または72時間に採取した細胞試料から抽出し、次いで、以下の図17に示すように、SOCS−1、SOCS−3、PIAS−1、およびPIAS−3に対して特異的なプローブで、定量的PCR(TAQMAN(登録商標))を行った。
【0149】
【化18】
以下の表18は、IL−27依存性誘導倍数としての定量的PCRデータを示す。図18AないしCは、中性(18A)、Th1バイアシング(18B)、およびTh2バイアシング条件(18C)下でのSOCS−3のIL−27依存性誘導を示す。
【0150】
データは、SOCS−3が中性、Th1バイアシング、およびTh2バイアシング条件下でIL−27によって誘導されることを示す。前記したように、SOCS−3はサイトカインシグナリングを抑制することが知られており、いくつかの剤の抗−炎症効果のメディエーターであることが報告されている。SOCS−3はIL−27によって誘導されるので、データは、IL−27を用いて免疫−媒介疾患を治療することができるのを示唆する。
【0151】
【化19】
中性条件下で処理した前記細胞の試料を72時間に採取し、RNAを抽出した。次いで、GENECHIP(登録商標)(Affymetrix, Santa Clara,CA)を用いて誘導された発現につき該RNAを分析した。以下の表19は、選択された遺伝子についての未処理対照に対する誘導(抑制)倍数としてのGENECHIP(登録商標)データを示す。
【0152】
ここに、データは、再度、IL−10が中性条件下でIL−27によって誘導されることを示す。前記したように、IL−10は、炎症応答を制限し、および終了させるにおいて主な役割を演じる。IL−10はIL−27によって誘導されるので、データは、IL−27を用いて、免疫−媒介疾患を治療することができることを示唆する。
【0153】
【化20】
3日に、細胞をIL−2の存在下であって、およびIL−27の存在下または不存在下で細胞を増殖させた。具体的には、IL−27に予め暴露した24ウェルプレートからの細胞を1mlの培地中に取り、次いで、2×10−4mg/ml IL−27および1×10−5mg/ml IL−2を含有する3mlの培地と共に6ウェルプレートに沈積させた。IL−27に予め暴露されていない細胞を1mlの培地中に取り、次いで、1×10−5mg/ml IL−2を含有する3mlの培地と共に6ウェルプレートに沈積させた。
【0154】
5日に、2×10−4mg/ml IL−27および1×10−5mg/ml IL−2の濃度を有する4mlの培地(IMDM(Invitrogen,Carlsbad,CA)w/10%HyClone血清)を、IL−27に予め暴露した細胞を含有するウェルに加えた。1×10−5mg/ml IL−2の濃度を有する4mlの培地を、IL−27に予め暴露していない細胞を含有するウェルに加えた。
【0155】
6日に、細胞を遠心し、次いで、培地(前記したのと同一培地)中にペレットを再度懸濁させ、カウントした。種々のウェルからの細胞カウントを表20にて以下に示し、図19に反映させる。
【0156】
データは、Th1またはTh2バイアシング条件の間に添加されたIL−27はCD4+細胞の増殖を低下させることを示し、IL−27は、多発性硬化症および慢性関節リウマチのような自己免疫疾患を含めたCD4+細胞の増殖によって特徴付けられる疾患を治療するのに有用であることを示唆する。
【0157】
【化21】
実施例9
IL−6誘導増殖のIL−27抑制
野生型(TCCR+/+)およびノックアウト(TCCR−/−)CD4+細胞のIL−6誘導増殖に対するIL−27の効果を、抗−IL−2抗体(BD Pharmingen,San Diego,CA)の存在下および不存在下で調べた。
【0158】
野生型マウスからの混合脾臓細胞(4×105)を96−ウェルプレート上のウェルに入れた。ノックアウトマウスからの混合脾臓細胞(4×105)をプレート上の別々のウェルに入れた。全てのウェルをPBS o/n中の100μlの2ug/ml 抗−CD3でコートした。ウェルを、表21にて以下に示した実験群に従って処理した。
【0159】
【化22】
細胞37℃にて培養した。48時間後、[3H]−チミジンをもう一晩の間加え、増殖を[3H]−チミジン取込みによって測定した。各群についての平均CPMを表22にて以下に示す。IL−2中和なくしての増殖を図20Aに示す。IL−2中和有りでの増殖を図20Bに示す。
【0160】
【化23】
データは、抗−IL−2抗体が存在するか否かに拘らず、IL−27が、抗−CD3によって刺激され、かつIL−6によって増強された増殖を抑制することを示す。抗−il−2抗体が存在しない場合、IL−27が、抗−CD3抗体によって刺激される増殖を抑制する。抗−IL−2抗体は、抗−CD3抗体によって刺激される増殖を低下させる。しかしながら、IL−27の添加は、この効果を部分的に和らげる。
【0161】
実施例10
IL−27受容体(TCCR)欠乏マウスはEAE超感受性である。
【0162】
IL−27は、活性化された抗原提示細胞(APC)によって生産されるリガンドである。IL−27は、特異的サブユニット、IL−27、およびIL−6Rを含めた、多数の他の受容体によって共有されるgp130よりなるヘテロダイマー受容体を通じてシグナリングを行う。本明細書中で議論するように、IL−27は種々のSTATおよびJak−1を通じてシグナルを活性化するが、支配的なシグナリング事象はSTAT−1の活性化であるように見える。STAT−1の活性化およびTH−1特異的転写因子T−betの下流誘導を介して、IL−12RB2鎖およびIFN−ガンマの発現は促進される。Il−27リガンドおよび受容体は図21にダイアグラムにより示される。
【0163】
IL−27はIL−12ファミリーのメンバーであり、サイトカインのIL−6クラスターに属する。図22参照。IL−27、EBI3およびp28の2つの成分はIL−12サブユニットに対する密接な相同性を有する。TCCRともいわれるIL−27受容体(IL−27R)の双方のサブユニットは、種々の白血球にて協調して発現される。最高の発現はT細胞およびNK細胞におけるものであるように見える。
【0164】
ナイーブな未分化T細胞(Th−0)は、ナイーブなTh−0細胞の成熟T−ヘルパー細胞への分化を誘導する異なるシグナルに応答する。一般に、2つのタイプのT−ヘルパー細胞が知られている;Th−1およびTh−2細胞。図23に図示されるように、IL−4によるTh−0細胞の刺激はTh−2細胞の発生に導き、IL−4、IL−5、IL−6、IL−10、およびIL−13を生産する。Th−2細胞およびサイトカイン産物は液性免疫および抗−蠕虫応答にインパクトを与える。IL−27および/またはIFN−ガンマによるTh−0細胞の刺激は、T−細胞におけるIL−12応答性の状態を誘導し、従って、それらはIL−12の制御下で成熟TH−1細胞に分化し、IFN−ガンマ、IL−2、およびリンホトキシン(LT)を生産することができる。Th−1細胞およびそれらのサイトカイン産物は、細胞−媒介免疫性およびマクロファージ活性化に関与する。
【0165】
Th−0細胞のTh−1およびTh−2ヘルパー細胞への分化におけるIL−27の役割のさらなる我々の理解のため、IL−27R欠乏マウス(TCCRノックアウト)を前記実施例に記載したように生産した。自己免疫疾患の間におけるIL−27についての潜在的役割を、実験的自己免疫脳炎(EAE)、多発性硬化症についてのマウスモデルを用いて調べた。EAEを持つマウスからのCD4+T細胞のみの移動がナイーブな受容体マウスにおいてEAEを引き起こしかねないので、EAEはT細胞媒介される。
【0166】
実験的EAEを誘導するために、マウスを、フロイントの完全アジュバント中のミエリン稀突起神経膠細胞糖蛋白質(MOG)35ないし55ペプチドで免疫化した。野生型(WT)およびIL−27受容体(TCCR)ノックアウトマウスをMOGで免疫化し、前記実施例に記載したようにEAEの証拠について調べた。臨床EAEスコアを処理後25日にわたって評価した。
【0167】
図24に示されたデータは、IL−27刺激を除去することによって引き起こされたEAE疾患における仮定された低下の代わりに、EAEはIL−27R欠乏マウスにおいて亢進されたことを示す。該マウスはEAE過剰感受性であるように見え、酷いEAE疾患を発生した。EAE表現型を発現する受容体欠乏マウスから採取された脊髄組織の組織学的分析を図25に示し、これは、EAEを持つIL−27受容体ノックアウトマウスにおける増強された炎症および脱髄を示す。
【0168】
この実験に加えて、種々の病原体でのIL−27受容体欠乏マウスの刺激、ならびに誘導された喘息および肝炎モデルの結果、Th−1およびTh−2媒介応答双方が亢進された。これらのデータは、IL−27が重要な免疫抑制機能を有することを示す。
【0169】
【化24】
IL−27、およびT−細胞の分化におけるその可能な役割をさらに研究するために、図27に図示した手法に従い、ナイーブなCD4+細胞をNACS−精製し、IL−27と共にまたはそれなくして抗−CD3+抗体で処理した。IL−27でのT細胞の刺激は、Th−0、Th−1またはTh−2へのT細胞分極を促進する条件下でなされた。
【0170】
簡単に述べると、24ウェル皿を5μg/ml抗−CD3(BD Pharningen)で一晩コートした。1.8×106CD4+T−細胞の容量を、IL−2(10ng/ml)および抗−CD28(1μg/ml)の存在下でウェル当たり撒いた。分化のために、以下のサイトカインおよび抗体を加えた:TH−0(各々5μg/mlの抗−IL−12、抗−IFN−ガンマ、抗−IL−4)、TH−1(3.5ng/mlのIL−12、5μg/mlの抗−IL−4)、TH−2(3.5ng/mlのIL−4、5μg/mlの抗−IFNgおよび抗−IL−12)。IL−27を200ng/mlの濃度にていくつかの培養に加えた。72時間後、上清ならびにRNAを単離し、Chip、RT−PCR、および/またはELISA分析によって特異的サイトカインの生産について分析した。得られたデータを図28に示し、これは、IL−27がT−細胞発生に対して顕著な効果を有することを示す。
【0171】
IL−27は調べたほとんどのサイトカインに対して顕著な効果を有し、この効果は、細胞が分化した条件とは一般には独立していた。IL−27は、Th−2誘導条件下でTNFαおよびIL−10、ならびにIL−4を誘導した。同時に、IL−2、IL−5、IL−6、GM−CSF、およびIL−17の生産はIL−27によってかなり抑制された。
【0172】
これらの効果のいずれかが良く知られ、かつ優れた免疫抑制サイトカインIL−10の誘導に対して二次的であるか否かを判断するために、IL−27の効果もIL−10欠乏T−細胞で調べた。図29に示すように、サイトカイン生産のIL−27誘導変調はIL−10とは独立していた。というのは、WTおよびIL−10欠乏T−細胞を比較して、IL−2またはGM−CSF生産においてほとんど差は見られなかったからである。
【0173】
イン・ビトロで見られたIL−27による免疫抑制性IL−10の強い誘導にも拘らず(図28)、EAEを持つIL−27R−/−マウスからのT−細胞においてIL−10の僅かな低下が見られたに過ぎない(図30)。しかしながら、この人工的なイン・ビトロ観察は、IL−27媒介IL−10誘導がEAEの間の重要な生物学的プロセスであるという解釈を断じて排除しない。対照的に、それは、IL−27誘導IL−10誘導をイン・ビボで研究するための我々の自由な実験技術の限界を最も反映するようである。
【0174】
実施例11
EAEはTH−17依存性である。
【0175】
最近の証拠は、ヘルパーT−細胞の新しいサブタイプ、いわゆるTH−17細胞が、EAEを含めた多くのプロ−炎症プロセスの鍵となるメディエーターであることを示唆する。これらのTh−17細胞はIL−17A、IL−17F、IL−6、TNF、およびGM−CSFを生じると報告された。図31に供されるダイアグラム参照。TH−17細胞の発生は貧弱にしか理解されていないが、それは、IL−12に対して同様性を持つもう1つのヘテロダイマーサイトカインであるIL−23に依存すると考えられる。IL−23欠乏動物はこのT−細胞表現型を効果的に発生させることができず、EAEおよびCIAに対して抵抗性である。しかしながら、IL−23は必要であるように見えるが、TH−17細胞分化でイン・ビトロでそれは十分ではない。
【0176】
前記で議論したように、IL−27R欠乏マウスはWT同腹子と比較してより酷いEAE疾患を発生させた。IL−27シグナリングに対して下流の事象を分析して、EAEの酷さに対するこの限定的効果で重要な因子を決定した。IL−27に応答しての種々のサイトカインの発現を活性化の間に調べた。
【0177】
IL−27はIFN−ガンマの生産を促進し、IFN−ガンマはIL−17を阻害することが知られている。データは、IL−27が、IFN−ガンマよりもより効果的にIL−17および他のTh−17サイトカインIL−6およびGM−CSFの生産を抑制したことを示す(図33および34)。さらに、EAEを持つTCCR−/−マウスからのリンパ節細胞は、WTよりもイン・ビトロでの再度の刺激に際してTh−17サイトカインをより多く分泌した(図37)。
【0178】
IL−17生産のIL−27媒介抑制はIFN−ガンマとは独立していた。なぜならば、INF−ガンマに対して非応答性とされたT−細胞は、以前として、IL−27での刺激に際してIL−17生産を抑制したからである。(図35)。IFN−ガンマシグナリングの不存在下において、基礎IL−17生産はより高かった。この理由は不明瞭である。なぜならば、WT培養においてさえ、FN−ガンマシグナリングはIFN−ガンマ中和抗体の添加によってブロックされるからである。かくして、IFN−ガンマR欠乏マウスにおける高いIF−17発現は発生段階の改変を反映し(すなわち、IFNgR欠乏T−細胞はIFNgRの発現よりもWT−細胞とはより多くにおいて異なり)、あるいは細胞内IFNgループを反映できるであろう。リガンドおよび受容体が共発現される細胞において、シグナリングは後期分泌経路内で起こり得るが、そのようなシグナリングは細胞内のものであり、中和抗体によってブロックされないであろう。
【0179】
Th−17細胞はIL−27R欠乏マウスにおいて調節不調であるかを判断するために、IL−27R欠乏マウスをCFAにおけるMOGで免疫化した。リンパ節の排出を14日において排除し、X・ビボにてMOGで再度刺激した。IL−27R欠乏T細胞を含有するリンパ節上清は、有意に増大したレベルのIL−17を発現した(図37および38)。
【0180】
さらに、脳および脊髄(EAEにおける炎症の現実の部位)の免疫浸潤物は、より多くの細胞がIL−27R欠乏マウスにおいて浸潤することを明らかとした。さらに、これらの細胞のより高いパーセンテージは、細胞内染色によって分析した場合にIL−17陽性であった。一緒に考え合わせると、これらの2つの観察は脊髄におけるIL−17の概略2倍発現に翻訳される(図39参照)。
【0181】
IFN−ガンマおよびIL−27双方はSTAT−1を活性化し、STAT−1ノックアウトは増大したIL−17を生じさせる。従って、IL−27は、STAT−1を活性化することによってIL−17を抑制することができる。この関係は、STAT−1ノックアウトマウスから得られた細胞におけるIL−17のIL−27媒介抑制を分析することによって調べた。STAT−1の不存在下においては、IL−27はIL−17を抑制せず、これは、該抑制がSTAT−1によって媒介されることを示す。STAT−1の不存在下においては、IL−27はIL−17のインデューサーとなる。この逆行に対するメカニズムの基礎は知られていないが、IL−27によるSTAT−3の活性化はこの効果において役割を演じると推定するのは公正である。なぜならば、他のIL−17誘導サイトカイン(顕著には、IL−23)は、STAT−1を活性化せずにSTAT−3を通じてシグナリングを行うからである(図36参照)。
【0182】
まとめると、IL−27受容体(TCCR)欠乏マウスはEAE−過剰感受性である。IL−27はイン・ビトロにてTh−17サイトカインIL−17、IL−6、およびGM−CSFを効果的に抑制した。さらに、EAEを持つIL−27受容体欠乏マウスは野生型よりも多くTh−17サイトカインを生産する。IL−27は、Th−17から免疫応答を外すことによってEAEを抑制することができる。
【0183】
実施例12
IL−6はTh−17細胞を誘導する。
【0184】
図41に図示されたように、IL−23は必要であるが、サイトカインIL−17、IL−6、GM−CSF、およびTNFを生じるTh−17細胞へのTh−0細胞の分化には十分ではない。IL−23が十分でない1つのありそうな理由は、Th−0細胞はIL−23受容体を発現せず、従って、IL−23非応答性であることである。従って、Th−0細胞においてIL−23Rを誘導することができる因子は、TH−17分化経路の絶対的成分である。
【0185】
Th−1(IFN−g)およびTH−2(IL−4)細胞のエフェクターサイトカインもまたこれらの細胞の発生に参画し、よって、フィードバックループの安定化を供するので、我々は、TH−17エフェクターサイトカインの1つは、同様に、TH−17発生に参画しなければならないと理由づけた。TH−17エフェクターサイトカインの中では、IL−6は最も有望に見える。なぜならば、その受容体はナイーブなT−細胞で発現されるからであり、および最後に分化されたT−細胞よりもIL−6の他の源(最も顕著には抗原提示細胞)があるからである。加えて、IL−6ノックアウトマウスはEAE耐性である(図42参照)。
【0186】
野生型およびIL−27受容体ノックアウトマウスは、単独で、またはIL−27およびIL−23と組み合わせて、IL−6への応答性について調べた。図43に示すように、IL−6はTh−17軸を誘導した。IL−6単独での処理はIL−23受容体を刺激し、IL−17AおよびIL−17F生産も刺激した。興味深いことには、共投与されたIL−27はIL−23受容体およびIL−17の生産のIL−6刺激増加を低下させ、または排除した(図43)。IL−23はIL−23受容体およびIl−17生産の刺激に対する僅かな効果を有し、これは、IL−6単独で示された大きな刺激に対して付加的であるように見えた。この組合せへのIL−27の添加もまた、該応答を低下させ、または排除した。再度刺激したリンパ節細胞から採取したmRNAは、野生型対照と比較してIL−23受容体ノックアウトにおけるIL−23受容体の誘導を示した(図43参照)。
【0187】
増殖アッセイにおけるIL−6の効果をさらに比較し、IL−6は野生型およびTCCRノックアウトマウス双方において精製されたT−細胞の増強された増殖を大いに刺激した。IL−27の添加は、野生型細胞においてIL−6誘導増殖を完全に中和した。しかしながら、この低下はTCCRノックアウトマウスでは見られず、これは、IL−27がIL−6の優れた増殖効果に拮抗することを示す。図44参照。従って、IL−6駆動TH−17分化を含めた、IL−27はいくつかのレベルでのIL−6アンタゴニストであるように見える。
【0188】
実施例13
Il−27の役割
図46に示すように、IL−27はT細胞の分化、特に、複数レベルにおけるTh−17細胞の発生にインパクトを与える。IL−27はIL−10、IL−4の生産、およびTh−1細胞の発生を刺激するが、それは、Th−17細胞の生産、Th−17細胞サイトカインIL−17およびGM−CSFの生産も抑制する。
【0189】
本明細書における全ての刊行物および特許出願は、本発明が関係する分野における通常の技量のレベルを示す。全ての刊行物および特許出願は、あたかも各個々の刊行物および特許出願が参照により具体的かつ個々に示されるようなのと同一程度に参照により一体化される。
【0190】
本発明を、種々の特別なおよび好ましい具体例および技術を参照して記載した。しかしながら、本発明の精神および範囲内にありつつ、多くの変形および修飾をなすことができることが理解されるべきである。
【図面の簡単な説明】
【0191】
【図1】図1は、TCCR/IL−27受容体複合体の人工物の模式図である。
【図2】図2は、モノクローナル抗体:2685−IgG2a、2686−IgG1、2688−IgG1、対照イソタイプIgG2a、および対照イソタイプIgG1に応答してヒトTCCRを発現するBa/F3細胞の増殖を示すグラフである。
【図3】図3は、マウスIL−3(陽性対照)および抗体2686に応答してのヒトTCCRを発現するBa/F3細胞の増殖を示すグラフである。
【図4】図4は、抗体2686に応答しての、ヒトTCCR、マウスTCCRを発現するBa/F3細胞、およびいずれも発現しない対照細胞の増殖を示すグラフである。
【図5】図5は、TCCRを発現しない脾臓細胞の増殖と比較した、抗―CD3刺激に応答してTCCRを発現する脾臓細胞の増殖を示すグラフである。
【図6】図6は、TCCRを発現しないCD4+T細胞の増殖と比較した、抗−CD3刺激に応答してTCCRを発現するCD4+T細胞の増殖を示すグラフである。
【図7】図7は、TCCRを発現しないCD8+T細胞の増殖と比較した、抗−CD3刺激と応答してTCCRを発現するCD8+T細胞の増殖を示すグラフである。
【図8】図8は、ノックアウト(TCCR−/−)野生型(TCCR+/+)マウスにおけるMOG誘導EAEの臨床的進行を示すグラフである。
【図9】図9は、ノックアウト(TCCR−/−)および野生型(TCCR+/+)マウスにおけるMBP誘導EAEの臨床的進行を示すグラフである。
【図10】図10は、EAEモデルにおけるノックアウト(TCCR−/−)および野生型(TCCR+/+)マウスのための脳および脊髄切片についての平均組織学的炎症スコアを示すグラフである。
【図11】図11は,EAEモデルにおけるノックアウト(TCCR−/−)および野生型(TCCR+/+)マウスについての脳および脊髄切片についての平均組織学的脱髄スコアを示すグラフである。
【図12】図12は、EAEモデルにおけるノックアウト(TCCR−/−)および野生型(TCCR+/+)マウスについての脳および脊髄切片での最大組織学的炎症スコアを示すグラフである。
【図13】図13は、EAEモデルにおけるノックアウト(TCCR−/−)および野生型(TCCR+/+)マウスについての脳および脊髄切片での最大組織学的脱髄スコアを示すグラフである。
【図14】図14は、CFSE標識アッセイにおいてTCCRを発現しないCD4+T細胞の増殖と比較した、抗−CD3刺激に応答してのTCCRを発現するCD4+T細胞の増殖を示すグラフである。
【図15】図15は、CFSE標識アッセイにおいてTCCRを発現しないCD8+T細胞の増殖と比較して、抗−CD3刺激に応答してTCCRを発現するCD8+T細胞の増殖を示すグラフである。
【図16】図16AないしCは、中性(16A)Th1バイアシング(16B)およびTh2バイアシング(16C)条件下での、種々の時点における、IL−27での処理に応答してのIL−2の誘導を示すグラフである。
【図17】図17は、中性(17A)、T51バイアシング(17B)およびTh2バイアシング(17C)条件下での、種々の時点における、IL−27での処理に応答しての、IL−10の誘導を示すグラフである。データはL−27依存性誘導の倍数として表される。
【図18】図18AないしCは、中性(18A)、Th1バイアシング(18B)、およびTh2バイアシング(18C)条件下での、種々の時点における、IL−27での処理に応答してのSOCS−3の誘導を示すグラフである。データはL−27依存性誘導の倍数として表される。
【図19】図19は、中性、Th1バイアシング、およびTh2バイアシング条件下での、IL−27処理に応答してのCD4+細胞の増殖を示すグラフである。
【図20】図20AないしBは、抗−IL−2抗体の不存在(20A)または存在(20B)におけるIL−27および/またはIL−6処理に応答しての脾臓細胞の増殖を示すグラフである。
【図21】図21は、IL−27およびその受容体IL−27R(TCCR)のダイアグラムである。
【図22】図22は、IL−12サイトカイン群内で、サイトカインのIL−27ないしIL−6クラスターの関係を示すダイアグラムである。
【図23】図23は、ヘルパーT−細胞の分化を示すダイアグラムである。
【図24】図24は、IL−27R欠乏マウスにおけるEAEに対する過敏を示すグラフである。
【図25】図25は、野生型およびIL−27RノックアウトマウスにおけるEAE表現型の組織学的分析を示す。
【図26】図26は、T−細胞におけるIL−27の関係をテストするプロトコルの模式図である。
【図27】図27は、T−細胞発生に対するIL−27の効果のテストの結果を示す。IL−27に応答してのサイトカイン低下を、以下のサイトカイン:IFN−ガンマ、IL−2、TFN、IL−4、IL−5、IL−6、IK−10、GM−CSF、およびIL−17についての野生型対照と比較する。
【図28】図28は、IL−10、IL−27、およびIL−10およびIL−27の組み合わせにおけるIL−2およびGM−CSF生産の結果をグラフで示す。
【図29】図29は、IL−27に応答してのIL−10の強力な誘導、IL−10ノックアウトマウスにおけるEAEの重症度をグラフで示す。
【図30】図30は、EAEにおけるTH−17の役割を示すグラフである。
【図31】図31は、EAE病についてのIL−23およびTH−IL−17細胞の要件をグラフで示す。
【図32】図32は、IL−27によるTH−IL−17サイトカインの抑制をグラフで示す。
【図33】図33は、IL−27によるIL−17の抑制をグラフで示す。
【図34】図34は、IL−27によって媒介されるIL−17の抑制がIFNgが非依存性であることをグラフで示す。
【図35】図35は、IL−27によるIL−17の抑制がSTAT−1によって媒介されることをグラフで示す。
【図36】図36は、IL−17−Rノックアウトマウスの再度刺激されたリンパ球からのIL−17−RノックアウトマウスからのTH−IL−17サイトカインの分泌をグラフで示す。
【図37】図37は、IL−27−R欠乏マウスにおいてMOGまたはKLHを誘導する病気に応答してのIL−17の生産を示す。
【図38】図38は、CNSに浸潤するCD4T細胞によるIL−17発現をグラフで示す。
【図39】図39は、Tヘルパー分化に対する種々のサイトカインの関係を示すグラフである。
【図40】図40は、IL−6ノックアウトマウスにおけるEAE耐性を示すグラフである。
【図41】図41は、TH−IL−17サイトカインの誘導およびIL−6による応答をグラフで示す。
【図42】図42は、IL−27によるIL−6増殖効果の拮抗作用を示すグラフである。
【図43】図43は、IL−27およびIL−6およびTH−IL−17発生の役割を示すグラフである。
【図44】図44は、IL−27の作用の複数レベルを示すグラフである。
【図45】図45は、サイトカイン発現の変化を誘導するにおけるIL−27の役割をグラフで示す。
【背景技術】
【0001】
(発明の背景)
自己免疫疾患は、複雑な相互に関連する生物学的経路の発現または結果である。正常な生理学においては、これらの生物学的経路は、傷害または負傷に応答し、傷害または負傷からの修復を開始し、外来性生物に対する先天的および獲得された防御を設置するのに臨界的である。これらの正常な生理学的経路がさらなる傷害または負傷を、応答の強度に関するものとして、異常な調節または過剰な刺激の結果として、自己に対する反応として、またはこれらの組合せとしてのいずれかで引き起こす場合、病気または病理が起こり得る。
【0002】
これらの病気の発生はしばしば多工程経路を含み、しばしば、複数の異なる生物学的系/経路に関係するが、これらの経路の1以上における臨界点での介入は軽減または治療効果を有し得る。治療的介入は有害プロセス/経路の拮抗作用または細菌プロセス/経路の組み合わせによって起こり得る。
【0003】
哺乳動物の免疫系は、細菌、ウイルス、トキシンおよび他の非−宿主物質の侵入から宿主を防御するのに協調して作用する多数のユニークな細胞よりなる。リンパ球、TおよびB細胞双方は、免疫系の特異性を大いに担っている。T細胞は胸腺において発生することから命名され、他方、B細胞は骨髄において発生する。
【0004】
Tリンパ球(T細胞)は哺乳動物免疫応答の重要な成分である。T細胞は、主要組織適合性複合体(MHC)内の遺伝子によってコードされる自己−分子に関連する抗原を認識する。抗原は抗原提示細胞、ウイルス感染細胞、癌細胞、移植片等の表面にMHC分子と共に提示され得る。T細胞系は、健康の脅威が宿主哺乳動物に曝されるこれらの改変された細胞を排除する。T細胞は、ヘルパーT細胞(CD4+)および細胞傷害性T−リンパ球(CD8+)を含む。ヘルパーT細胞(TH)は、抗原提示細胞上の抗原−MHC複合体の認識に続いてかなり増殖する。ヘルパーT細胞は、B細胞、細胞傷害性T−リンパ球、および免疫応答に参加する種々の他の細胞の活性化において中枢的な役割を演じるリンホカインのような種々のサイトカインも分泌する。細胞傷害性T−リンパ球は他の細胞の破壊を引き起こすことができる。
【0005】
液性および細胞媒介免応答双方における中枢的な事象はヘルパーT細胞の活性化およびクローン拡大である。ヘルパーT細胞活性化は、T細胞受容体(TCR)−CD3複合体の、抗原提示細胞の表面での抗原−MHCとの相互作用によって開始される。この相互作用は、休止ヘルパーT細胞が細胞周期に入るのを誘導し(G0ないしG1転移)、その結果、IL−2に対する高親和性受容体の発現をもたらす生化学的事象のカスケードを媒介する。活性化されたT細胞は、周期増殖および分化を介して記憶細胞またはエフェクター細胞を通じて進行する。
【0006】
T−ヘルパー細胞サブセット(Th1およびTh2)は免疫の2つの経路を定義する:細胞−媒介免疫性および液性免疫性。Th1およびTh2サブタイプについてのサイトカインの放出プロフィールはエフェクターメカニズムおよび細胞傷害性細胞の選択に影響する(非特許文献1;非特許文献2)。Th1細胞、CD4+細胞の機能的サブセットは、細胞−媒介免疫性を増強し、Il−2、インターフェロン−ガンマ、リンホカインべータを含めたサイトカインを生産するそれらの能力によって特徴付けられる(非特許文献1;非特許文献2)。Th1細胞によって分泌されるIl−2およびインターフェロン−ガンマはマクロファージおよび細胞傷害性細胞を活性化する。Th2細胞はCD4+細胞でもあるが、Th1細胞から区別される。Th2細胞は、抗体生産のような液性免疫性を増強するそれらの能力によって特徴付けられる。Th2細胞はIl−4、Il−5、およびIl−10を含めたサイトカインを生産する(非特許文献1;非特許文献2)。Th2細胞によって分泌されるIl−4、Il−5、およびIl−10は好酸球および肥満細胞の生産を増殖させ、ならびにIgEを含めた抗体の生産を増強し、細胞傷害性細胞の機能を減少させる(非特許文献3)。
【0007】
Th1およびTh2サイトカイン放出は、相互に抑制性のTh1およびTh2応答を変更する。例えば、IL−4はTh1細胞からのインターフェロン−ガンマの発現を阻害し、他方、インターフェロン−ガンマはTh2細胞からのIL−4の発現を阻害する(非特許文献1)。
【0008】
4つのラセン束サイトカインファミリーのメンバー(非特許文献4)は、共通の前駆体からのTヘルパー細胞のTh1およびTh2エフェクター細胞の区別される集団への増殖および最後の分化を変調する(非特許文献5)。および、IL−4はTh2細胞の発生に影響し、他方、IL−12はTh1細胞の分化に関与する(非特許文献6;非特許文献7)。
【0009】
TCCR(T−細胞サイトカイン受容体)は、IL−12 β−2受容体、G−CSFR、およびIL−6受容体に対する相同性を持つサイトカイン受容体のWS(G)XWSクラスである。これらの受容体は、細胞、特に、血液細胞の増殖および分化に関与する細胞の増殖および分化を制御できるシグナルを変換する。TCCRは、T=ヘルパー細胞応答に関与することが示唆されている。特に、TCCRおよびそのリガンドIL−27はTh1応答を促進することが仮定されている(非特許文献8;非特許文献9;非特許文献10)。
【0010】
Th1およびTh2細胞のいずれかまたは双方によって生産されるサイトカインの過剰生産は、医学的障害の宿主にインパクトを与える。例えば、Th1サイトカインの過剰生産は、多発性硬化症および慢性関節リウマチのような種々の自己免疫疾患の病因に寄与する。Th2サイトカインの過剰生産はアレルギー疾患の病因に寄与する。
【0011】
CD8+細胞傷害性T−リンパ球(CTL)は、いくつかの自己免疫疾患において組織の病原体破壊に関与する。例えば、CTLは、自己免疫I型糖尿病のクールの間に膵臓β細胞の破壊に関連する(非特許文献11)。CTLは実験的自己免疫脳脊髄炎にも関連する(非特許文献12)。CTLは移植片(GVHD)−対宿主病に関連する組織損傷を媒介する(非特許文献13)。
【0012】
多発性硬化症(MS)は、脳および脊髄に影響する中枢神経系の障害である。MSの共通の兆候および症状は、1以上の四肢において、体幹において、または顔の一方側における感覚異常;脚または手の弱化またはぎこちなさ;または(部分的な盲目および1つの目における痛みのような)目に見える乱れ、視覚のかすみ、または暗点を含む。他の共通の初期の兆候は、二重視(複視)をもたらす目の麻痺、1以上の四肢の一過的弱化、四肢のわずかな硬直または異常な脆性、わずかな歩行の乱れ、膀胱制御の困難、眩暈、および軽い情緒的乱れ(Berkow et al.(編),1999,Merck Manual of Diagnosis and Therapy:第17版(MS)についての現在の治療は、コルチコステロイド、バーターインターフェロン(Betaferon,Avonex,Rebif)、グラチラメール酢酸(Copaxone)、メトトリキセート、アザチオプリン、シクロホスファミド、クラドリビン、バクロフェン、チザニジン、アミトリプチリン、カルバマゼピン(Berkow et al.(編),1999,supra)を含む。
【0013】
慢性関節リウマチ(RA)は、典型的には小さなおよび大きな関節に影響する関節の滑膜炎によって特徴付けられる慢性自己免疫疾患であり、それらの進行性破壊に至る(Berkow et al.(編),1999,supra)。RAの兆候は硬直、圧痛、滑膜厚化、反射梗縮、内臓小節、脚潰瘍または多発性単神経炎、胸膜または心臓周囲滲出、および発熱を含むことができる(Berkow et al.(編),1999,supra)。
【0014】
RAのための現在の治療は(サリシレートを含めた)非−ステロイド、抗−炎症薬物、金化合物、メトトレキセート、ヒドロキシクロロキン、スルファサラジン、ペニシラミン、コルチコステロイド、および細胞傷害性または免疫抑制薬物(Berkow et al.(編),1999,Merck Manual of Diagnosis and Therapy:第17版)を含む。
【非特許文献1】Mosmammら、Adv.Imnunol.(1989)45:111−147
【非特許文献2】Mosmannら、Immunol.Today(1996)17:138−146
【非特許文献3】Powrieら、Immunol.Today,(1993)14:270
【非特許文献4】Bazan,PNAS,(1990)87:6934
【非特許文献5】O’Garra,A.,Immunity(1998)8:275−83
【非特許文献6】Hsiehら、Science,(1993)260:547−9
【非特許文献7】Sederら、PNAS,(1993)90:10188−92
【非特許文献8】Chenら、Nature,(2000)409:916−920
【非特許文献9】Yoshidaら、Immunity,(2001)15:569−578
【非特許文献10】Pflanzら、Immunity,(2002)16:779−790
【非特許文献11】Kagiら、J.Exp.Med.(1997)186:989−997
【非特許文献12】Husebyら、J.Exp.Med.,(2001)194(5):669−676
【非特許文献13】Graubertら、J.Clin Invest.(1997)100:904−911
【発明の開示】
【発明が解決しようとする課題】
【0015】
自己免疫疾患に対する現存の療法のいずれも、限定された効率および/または有意な毒性のため満足することが判明していない。かくして、MSおよびRAのような自己免疫疾患を治療するための新しい方法が必要である。
【課題を解決するための手段】
【0016】
(発明の要旨)
ナイーブな未分化T細胞(Th−0)は、成熟なT−ヘルパー細胞へのTh−0細胞の分化を誘導する異なるシグナルに応答する。今日、例えば、IL−27のようなTCCRのアゴニストを投与することによって、細胞受容体TCCRの活性化がT−リンパ球増殖を低下させるのに効果的であることが発見されている。T−リンパ球増殖の低下は、IL−10およびSOCS−3の増大した発現に相関した。TCCRを発現する動物は、自己免疫疾患に対して感受性が低いことが判明した。
【0017】
EAE病気モデルにおけるさらなる実験は、IL−27受容体(TCCR)−欠乏マウスは自己免疫疾患に対して過敏であることを示した。Th−細胞分化における、および免疫障害におけるIL−27の役割の研究は、IL−27が免疫抑制性であり、Th発生において複数レベルで作用するという驚くべき発見に導いた。IL−27はTh−IL17細胞の生産を抑制し、IL−6の生産を阻害し、IL−6を含めたThIL17サイトカインの生産を阻害する。IL−27はIL−10の、およびIL−4の、さらにTh−IL17細胞の阻害剤の生産を誘導し、IL12受容体の生産およびTh−1細胞の分化を刺激する。本明細書中に開示されたデータは、IL−27が、Th−1、Th−2およびTh−17細胞にわたる重要な阻害活性を含めた重要な免疫抑制機能を有することを示す。
【0018】
本発明は、IL−27のようなIL27R(TCCR)のアゴニストを投与することによって、多発性硬化症(MS)および慢性関節リウマチ(RA)を含めた自己免疫疾患を治療する方法を提供する。TCCRの有用なアゴニストは、IL27Rの改変体および断片、IL−27のようなIL27Rリガンド、およびその改変体および断片ならびにIL27RまたはIL27Rリガンドに結合し、IL27−媒介応答を刺激し、誘導し、または増強するアゴニスト抗体を含む。また、本発明は、Th−ILI7細胞を含めた、T−リンパ球および/または細胞傷害性T−リンパ球の増殖を阻害する方法を提供し、該方法はIL27/IL27R応答を刺激し、誘導し、または増強するアゴニストを投与することを含む。
【0019】
【表1】
【発明を実施するための最良の形態】
【0020】
(詳細な説明)
T−リンパ球の過剰増殖またはTh1またはTh2細胞によって生産されたサイトカインの過剰生産は、医学的障害の主に導く。例えば、Th1応答に関連するサイトカインの過剰生産、またはCD8+細胞傷害性T−リンパ球の過剰生産は同種異系移植片拒絶、(グレーヴス病および橋本甲状線炎のような)自己免疫甲状腺病、自己免疫ブドウ網膜炎、巨細胞動脈炎、(クローン病、海洋性結腸炎、局所腸炎、肉芽腫性腸炎、末端回腸炎、局所回腸炎および終末回腸炎を含めた)炎症性腸疾患、インスリン−依存性新生糖尿病、多発性硬化症、悪性貧血、乾癬、慢性関節リュウマチ、サルコイドーシス、強皮症、全身エリテマトーデスを含めた自己免疫疾患に導き得る。
【0021】
以下の実施例に詳細に説明する実験は、非−特異的T細胞刺激に応答してTCCRを発現するT細胞よりもTCCRを欠くT細胞のより大きな増殖を示す(図1参照)。以前、TCCRおよびそのリガンドIL−27がTh1応答を促進することが示唆された(Chenら、2000,Nature,407:916−920;Yoshidaら、2001,Immunity,15:569−578;Pflanzら、2002,Immunity,16:779−790)。しかしながら、驚くべきことに、TCCRを発現するマウスは、TCCRを欠くマウスよりも、実験的アレルギー性脳脊髄炎(EAE)のようなTh1応答によって部分的に特徴付けられる自己免疫疾患に対して感受性が低いことが発見された(実施例2参照)。
【0022】
以下の実施例に示すように、Tリンパ球の増殖は、細胞へのTCCRアゴニストの投与によって阻害される。また、自己免疫炎症性疾患の低下した臨床的進行およびよりひどくない兆候がTCCR(−/−)動物におけるよりもTCCR(+/+)を発現する動物に存在することが示された。
【0023】
これらのデータは、TCCRのアゴニストを用いて、T−細胞増殖を低下させることを示す。特に、データは、TCCRのアゴニストが、多発性硬化症(MS)および慢性関節リュウマチ(RA)のような自己免疫媒介障害を治療するのに有用であることを示す。
【0024】
(定義)
用語「自己免疫」とは、それを生産する生物の分子、細胞、または組織を抗体またはリンパ球のような免疫系の成分が攻撃し、または害するプロセスをいう。
【0025】
用語「自己免疫障害」は、組織損傷、または病因のような損傷が少なくとも部分的には自己免疫プロセスの結果である病気をいう。その例として、用語「自己免疫疾患」は、Th1応答またはCD8+細胞傷害性T−リンパ球によって少なくとも部分的に媒介される病気を[含む。自己免疫疾患は同種異系移植片拒絶、(グレーヴス病および橋本甲状線炎のような)自己免疫甲状腺病、自己免疫ブドウ膜網膜炎、巨細胞動脈炎、(クローン病、潰瘍性結腸炎、局所腸炎、肉芽腫腸炎、末端回腸炎、局所回腸炎、および終末回腸炎を含めた)炎症性腸疾患、インスリン−依存性真性糖尿病、多発性硬化症、悪性貧血、乾癬、慢性関節リウマチ、サルコイドーシス、強皮症、および全身エリテマトーデスを含む。
【0026】
用語「Th1応答」とは、Th1エフェクター細胞の区別される集団への前駆体からのTヘルパー細胞の分化をいい、IFN−ガンマ、IL−2、およびTNF−ベータのようなTh1細胞からのサイトカインの分泌を含む。用語「Th1バイアシング疾患」とは、Th1エフェクター細胞の区別される集団への前駆体からのTヘルパー細胞の分化に好都合な疾患をいう。
【0027】
用語「Th1サイトカイン」とは、IFN−ガンマ、IL−2、およびTNF−ベータを含めたTh1応答において発現されるサイトカインをいう(Powrieら、1993,Immunol.Today,14:270)。
【0028】
用語「Th1媒介障害」とは、Th1サイトカインの過剰生産によって圧倒的にまたは部分的に媒介される障害をいう。用語「Th1媒介障害」は、T−細胞のTh1サブタイプの分化における過剰生産または偏りに由来し得る障害を含む。そのような傷害は、自己免疫ショウガイ、例えば、RAおよびMSを含む。
【0029】
用語「Th2応答」とは、Th2エフェクター細胞の区別される集団への前駆体からのTヘルパー細胞の分化をいい、IL−4、IL−5、IL−10、およびIL−13のようなTh2細胞からのサイトカインの分泌を含む(Powrieら、1993,Immunol.Today,14:270)。用語「Th2バイアシング条件」とは、Th2エフェクター細胞の区別される集団への前駆体からのTヘルパー細胞の分化に好都合な条件をいう。
【0030】
用語「TCCRペプチド」「TCCR蛋白質」および「TCCR」は、本明細書中において用いる場合、天然配列TCCRおよびTCCRペプチド改変体を含む。TCCRペプチドはヒト組織またはもう1つの源のような種々の源から単離することができ、あるいは組換えおよび/または合成方法によって調製することができる。「天然配列TCCR」は、天然に由来するTCCRペプチドと同一のアミノ酸配列を有するペプチドである。そのような天然配列TCCRは、天然から単離することができるか、あるいは組換えおよび/または合成手段によって生産することができる。用語「天然配列TCCR」は(細胞外ドメイン配列のような)天然に生じる切形および分泌形態、(交互にスプライスされた形態のような)天然に生じる切形形態、およびTCCRの天然に生じる対立遺伝子改変体を具体的には含む。1つの具体例において、天然配列ヒトTCCRは、配列番号:1のアミノ酸1ないし636を含む成熟または全長天然配列TCCRである。同様に、天然配列マウスTCCRは、配列番号:2のアミノ酸1ないし623を含む成熟または全長天然配列TCCRである。配列番号:1および配列番号:2は、アミノ酸位置1としてここでは示されるメチオニン残基で始まることが示されるが、配列番号:1または配列番号:2のアミノ酸位置1から上流または下流いずれかに位置するもう1つのメチオニン残基は、TCCRペプチドについての出発アミノ酸残基として使用することができると考えることができ、またそれが可能である。
【0031】
「TCCRペプチド細胞外ドメイン」または「TCCR ECD」とは、膜貫通および細胞質ドメインを実質的に含まないTCCRペプチドの形態をいう。通常は、TCCRペプチドECDはそのような膜貫通および/または細胞質ドメインの約1%未満を有し、好ましくは、好ましくは、そのようなドメインの約0.5%未満を有する。本発明のTCCRペプチドについて同定されるいずれの膜貫通ドメインも、疎水性ドメインのそのタイプを同定するためにルーチン的に使用される基準に従って同定されると理解される。膜貫通ドメインの正確な境界は変化し得るが、最もありそうには、最初に同定されたドメインのいずれかの末端における約5以下のアミノ酸であり得る。それ自体、1つの具体例において、ヒトTCCRペプチドの細胞外ドメインはアミノ酸1または約33ないしX1を含ミリモル、ここに、X1は配列番号:1の残基512ないし残基522からのいずれかのアミノ酸残基である。同様に、マウスTCCRペプチドの細胞外ドメインはアミノ酸1または約25ないしX2を含ミリモル、ここに、X2は配列番号:2の残基509ないし残基519からのいずれかのアミノ酸残基である。
【0032】
用語「TCCR改変体ペプチド」は、TCCRペプチドの少なくとも1つの生物学的活性を有し、かつ:
(a1)配列番号:1のヒトTCCRペプチドの残基1または約33ないし636;
(a2)配列番号:2のマウスTCCRペプチドの残基1または約25ないし623;
(b1)配列番号:1のヒトTCCRペプチドのX3ないし636、ここに、X3は配列番号:1のいずれかのアミノ酸残基27ないし37;
(b2)配列番号:2のマウスTCCRペプチドのX4ないし623、ここに、X4は配列番号:2の20ないし30からのいずれかのアミノ酸残基であり;
(c1)1または約33ないしX1、ここに、X1は配列番号:1の残基512ないし残基522からのいずれかのアミノ酸残基であり;
(c2)1または約25ないしX2、ここに、X2は配列番号:2の残基509ないし519のいずれかのアミノ酸残基であり;
(d1)X5ないし636、ここに、X5は配列番号:1の残基533ないし543からのいずれかのアミノ酸であり;
(d2)X6ないし623、ここに、X6は配列番号:2の残基527ないし537からのいずれかのアミノ酸であり;または
(e)配列番号:1および配列番号:2のアミノ酸配列のもう1つの特異的に誘導された断片;
のアミノ酸配列に対して少なくとも約80%アミノ酸配列同一性を有するペプチドを意味する。
【0033】
そのようなTCCR改変体ペプチドは、例えば、配列番号:1および配列番号:2の配列のN−および/またはC−末端において、ならびに1以上の内部ドメイン内において1以上のアミノ酸残基が付加され、または欠失されたTCCRペプチドを含む。通常、TCCR改変体ペプチドは少なくとも80%アミノ酸配列同一性を有し、
(a1)配列番号:1のヒトTCCRペプチドの残基1または約33ないし636;
(a2)配列番号:2のマウスTCCRペプチドの残基1または約25ないし623;
(b1)配列番号:1のヒトTCCRペプチドのX3ないし636、ここに、X3は配列番号:1のいずれかのアミノ酸残基27ないし37;
(b2)配列番号:2のマウスTCCRペプチドのX4ないし623、ここに、X4は配列番号:2の20ないし30からのいずれかのアミノ酸残基であり;
(c1)1または約33ないしX1、ここに、X1は配列番号:1の残基512ないし残基522からのいずれかのアミノ酸残基であり;
(c2)1または約25ないしX2、ここに、X2は配列番号:2の残基509ないし519のいずれかのアミノ酸残基であり;
(d1)X5ないし636、ここに、X5は配列番号:1の残基533ないし543からのいずれかのアミノ酸であり;
(d2)X6ないし623、ここに、X6は配列番号:2の残基527ないし537からのいずれかのアミノ酸であり;または
(e)配列番号:1および配列番号:2のアミノ酸配列のもう1つの特異的に誘導された断片;
に対して少なくとも約81%、82%、83%、84%、85%、86%、87%、88%、89%、90%、91%、92%、93%、94%、95%、96%、97%、98%または99%アミノ酸配列同一性であり得る。
【0034】
用語「IL−27」は、本明細書中で用いる場合、天然の配列IL−27ヘテロダイマー、天然の配列IL−27成分EBI3およびp28、IL−27ヘテロダイマー改変体(本明細書中でさらに定義する、およびEBI3およびp28の改変体を含む。IL−27ヘテロダイマーおよびその成分はヒト組織タイプから、またはもう1つの源からのような種々の源から単離することができ、あるいは組換えおよび/または合成方法によって調製することができる。「天然配列IL−27」は、天然に由来するIL−27ヘテロダイマーと同一のアミノ酸配列を有するヘテロダイマーを含む。そのような天然配列IL−27ヘテロダイマーは天然から単離することができるか、あるいは組換えおよび/または合成手段によって生産することができる。用語「天然配列IL−27」は、具体的には、(細胞外ドメイン配列のような)天然に生じる切形され、かつ分泌された形態、(交互にスプライシングされた形態のような)天然に生じる切形された形態、およびIL−27ヘテロダイマーの天然に生じる対立遺伝子改変体を含む。
【0035】
用語「IL−27改変体」とは、TCCRを活性化することができる、天然配列IL−27成分EBI3およびp28を含めた、天然配列IL−27に対して相同性を有するペプチドをいう。IL−27改変体は、EBI3改変体およびp28改変体から形成されるものを含むことができる。IL−27改変体は、TCCRおよびgp130を共にかみ合わせることができるものも含むことができる。IL−27改変体は、TCCRホモダイマーを形成することができるものを含むことができる。IL−27改変体はPEG化IL−27を含む。
【0036】
用語「p28」は、本明細書中で用いる場合、天然配列p28およびp28ペプチド改変体を含む。p28はヒト組織タイプから、またはもう1つの源からのように種々の源から単離することができるか、あるいは組換えおよび/または合成方法によって調製することができる。「天然配列p28」は、天然に由来するp28ペプチドを同一のアミノ酸配列を有するペプチドを含む。そのような天然配列p28は天然から単離することができるか、あるいは組換えおよび/または合成手段によって生産することができる。用語「天然配列p28」は、具体的には、(細胞外ドメイン配列のような)天然に生じる切形され、または分泌された形態、(交互にスプライシングされた形態のような)天然に生じる切形形態、およびp28の天然に生じる対立遺伝子改変体を含む。
【0037】
用語「p28ペプチド改変体」は、天然配列ヒトp28(配列番号:3)またはマウスp28(配列番号:4)に対して少なくとも73%、75%、80%、90%、95%、または99%配列同一性を有するペプチドを含む。p28ペプチド改変体は、TCCRおよびgp130に結合することができるp28の部分を含む。p28ペプチド改変体は、TCCRを活性化することができるp28の部分を含む。p28ペプチド改変体は、TCCR上に見出されるサイトカイン受容体相同性ドメインの領域においてTCCRに結合すると考えられる、p28の第一および第三のアルファラセンからの残基、およびgp130上に見出されるIGドメインに結合すると考えられる、第一のラセンの最後において、および第四のラセンの始まりにおける残基を含む。
【0038】
用語「EBI3」は、本明細書中で用いる場合、天然配列EBI3およびEBI3ペプチド改変体を含む。EBI3ペプチドはヒト組織タイプから、またはもう1つの源からのように種々の源から単離することができるか、あるいは組換えおよび/または合成方法によって調製することができる。「天然配列EBI3」は、天然に由来するEBI3ペプチドと同一のアミノ酸配列を有するペプチドを含む。そのような天然配列EBI3は天然から単離することができるか、あるいは組換えおよび/または合成手段によって生産することができる。用語「天然配列EBI3」は、具体的には、(細胞外ドメイン配列のような)天然に生じる切形され、および分泌された形態、(交互にスプライシングされた形態のような)天然に生じる切形された形態、およびEBI3の天然に生じる対立遺伝子改変体を含む。
【0039】
用語「融合蛋白質」とは、例えば、2つの異なる蛋白質をコードする2つの遺伝子の融合から得られる発現産物をいう。該用語は、2つの異なる蛋白質の部分をコードする2つの遺伝子の部分の融合に由来する発現産物も含む。該用語は、翻訳後に起こる融合に由来する蛋白質を含む。本明細書で用いるように、該用語は、異種ペプチドに融合した、IL−27、その成分(EBI3およびp28)またはその部分を含むであろう。該用語は、異種ペプチドに融合したTCCRまたはその部分も含むであろう。該用語は、機能的な1つの鎖サイトカインを形成するためのp28に融合したEBI3も含むであろう(Pflanzら、2002,Immunity,16:779−790)。該用語は、ヒトFcタグにコンジュゲートしたIL−27を含む。
【0040】
本明細書中で用いるように、与えられたペプチドに関しては、「異種ペプチド」とは、起源に関わらず異なる配列を持つペプチドをいう。例えば、天然配列TCCRに関しては、異種ペプチドとは、天然配列TCCRのそれ以外の配列を有するペプチドをいう。天然配列IL−27に関しては、異種ペプチドとは、天然配列IL−27のそれ以外の配列を有するペプチドをいう。
【0041】
用語「アゴニスト」は、天然配列ペプチドの生物学的活性を増強する、または刺激するいずれの分子も含む。適当なアゴニストは、具体的には、アゴニストペプチド、アゴニスト抗体または抗体断片、本発明の天然ペプチドの断片またはアミノ酸配列改変体等を含む。TCCRのアゴニストを同定するための方法は、例えば、TCCRペプチドまたはTCCRペプチド−発現細胞を候補アゴニスト分子と接触させ、1以上のTCCR生物学的活性の検出可能な変化を測定することを含む。
【0042】
「TCCR生物学的活性」とは、本明細書中で用いるように、T−細胞増殖の減衰または抑制のようなTCCR媒介応答をいう。TCCR生物学的活性は、Th1応答またはTh1媒介障害の減衰または抑制を含む。TCCR生物学的活性はIL−10およびSOCS−3の発現の増加を含む。TCCR生物学的活性は、TCCRの活性化に関連するシグナリング、例えば、Stat1、Stat3、Stat4、およびStat5のようなシグナル変換および転写因子のリン酸化も含む(Lucasら、2003,PNAS,100(25):15047−52)。
【0043】
用語「抗体」および「免疫グロブリン」は最も広い意味で用いられ、具体的には、ポリクローナル抗体、(アゴニストおよびアンタゴニスト抗体を含めた)モノクローナル抗体、(二価抗体のような)多価抗体、(所望の生物学的活性を呈する二特異的抗体のような)多特異的抗体、ポリエピトープ特異性を持つ抗体組成物、アフィニティー成熟化抗体、ヒト化抗体、ヒト抗体、キメラ抗体、ならびに所望の生物学的活性を呈する(Fab、F(ab’)2、scFv、およびFvのような)抗原結合断片を含む。天然に生じる抗体は、ジスルフィド結合によって相互連結した4つのペプチド鎖、2つの同一の重(H)鎖および2つの同一の軽(L)鎖を含む。各重鎖は、重鎖可変領域ドメイン(VH)および重鎖定常領域を含む。重鎖定常領域は3つのドメインCH1、CH2およびCH3を含む。各軽鎖は、軽鎖可変領域ドメイン(VL)および軽鎖定常領域ドメインを含む。軽鎖定常領域は1つのドメインCLを含む。VHおよびVLドメインは、さらに、配列によって規定される相補性決定領域(CDR)(Kabatら、1991,Sequences of Proteins of Immunological Interest,5th Ed.Public Health Service,National Institutes of Health,Bethesda,Md.)、またはフレームワーク領域(FR)と呼ばれる、より保存された領域が散在する三次元構造によって規定される超可変ループ(HVL)(Chothiaら、1987,J.Mol.Biol.,196:901−917)に細分することができる。各VHおよびVLは典型的には、以下の順序:FR1、CDR1(HVL1)、FR2、CDR2(HVL2)、FR3、CDR3(HVL3)、FR4にてアミノ−末端ないしカルボキシ−末端に配置された3つのCDR(またはHVL)および4つのFRから構成される。
【0044】
抗体(免疫グロブリン)は、それらの重鎖の定常ドメインのアミノ酸配列に依存して異なるクラスに帰属される。免疫グロブリンの5つの主なクラス:IgA、IgD、IgE、IgG、およびIgMがあり、これらのいくつかは、IgG1、IgG2、IgA1、IgA2等のようなサブクラス(イソタイプ)にさらに分割される。免疫グロブリンの異なるクラスに対応する重鎖定常ドメインが、各々、アルファ、デルタ、イプシロン、ガンマおよびミューと呼ばれる。免疫グロブリンの異なるクラスのサブユニット構造および三次元立体配置はよく知られており、一般に、例えば、Abbasら、2000,Cellular and Mol.Immunology,4th edに記載されている。抗体は、1以上の他の蛋白質またはペプチドと抗体との共有結合または非共有結合開放によって形成されるより大きな融合分子の一部であり得る。
【0045】
用語「全長抗体」とは、少なくとも2つの重鎖および2つの軽鎖を含めたその実質的に無傷の形態の抗体をいい、以下に定義する抗体断片をいわない。該用語は、特に、Fc領域を含有する重鎖を持つ抗体をいう。全長抗体は天然配列抗体または組換え抗体であり得る。全長抗体はヒト、ヒト化、および/またはアフィニティー成熟化であり得る。
【0046】
用語「モノクローナル抗体」とは、本明細書中で用いるように、実質的に相同な抗体の集団から得られた抗体をいい、すなわち、該集団を含む個々の抗体は、該抗体の生産の間に生起し得る改変体を除いて実質的に同一である。
【0047】
本明細書中に記載されたモノクローナル抗体は、具体的には、重鎖および/または軽鎖の一部が特定の種に由来する、または特定の抗体クラスまたはサブクラスに属する抗体における対応する配列と同一である、または相同であり、他方、鎖の残りはもう1つの種に由来し、またはもう1つの抗体クラスまたはサブクラスに属する抗体における対応する配列と同一である、または相同である「キメラ」抗体、ならびに所望の生物学的活性を呈する限りそのような抗体の断片を含む(米国特許第4,816,567号;およびMorrisonら、1984,PNAS,81:6851−6855)。
【0048】
非−ヒト(例えば、マウス)抗体の「ヒト化」形態は、非−ヒト免疫グロブリンに由来する最小配列を含有する(Fv、Fab、Fab’、F(ab’)2、または抗体の他の抗原−結合サブ配列のような)キメラ免疫グロブリン、免疫グロブリン鎖、またはその断片である。ほとんどの場合、ヒト化抗体は、受容体の1以上の相補性決定領域(CDR)または超可変ループ(HVL)からの残基が、マウス、ラットまたはウサギのような非−ヒト種(ドナー抗体)の1以上のCDRまたはHVLからの残基で置き換えられ、所望の抗原特異性、アフィニティーおよび能力を有するヒト免疫グロブリン(受容体抗体)である。いくつかの場合において、ヒト免疫グロブリンの特異的Fvフレームワーク領域(FR)残基は対応する非−ヒト残基によって置き換えられている。さらに、ヒト化抗体は、受容体抗体においても、または輸入されたCDR(またはHVL)においても、あるいはフレームワーク配列においても見出されない残基を含むことができる。これらの修飾は、抗体性能をさらに改良し、最大化するように成される。一般に、ヒト化抗体は、少なくとも1つの、典型的には2つの可変ドメインの実質的に全てを含み、ここに、CDRまたはHVLの全てまたは実質的に全ては非−ヒト免疫グロブリンのそれに対応し、FRの全てまたは実質的に全てはヒト免疫グロブリン配列のそれである。
【0049】
ヒト可変ドメイン、軽鎖および重鎖双方の選択は、ヒト化抗体を作成するにおいて用いることができる。「ベストフィット」方法に従って、げっ歯類抗体の可変ドメインの配列は、例えば、公知のヒト可変−ドメイン配列の全ライブラリーに対してスクリーニングされる。げっ歯類のそれに最も近いヒト配列は、ヒト化抗体についてのヒトフレームワーク領域(FR)として用いられる(Simsら、1993,J.Immunol.,151:2296)。別法として、受容体フレームワーク領域は軽鎖または重鎖の特定のサブグループについてのヒト抗体コンセンサス配列に由来することができる。同一フレームワークは、いくつかの異なるヒト化抗体を生産するのに用い、または修飾し、および用いることができる(Carterら、1992,Proc.Natl.Acad.Sci.USA,89:4285;Prestaら、1993,J.Immunol.,151:2623)。ヒト化抗体は、所望により、免疫グロブリン定常領域の少なくとも部分(Fc)、典型的には、ヒト免疫グロブリンのそれを含む。さらなる詳細については、例えば、Jonesら、1986,Nature,321:522−525;Reichmannら、1988,Nature,332:323−329;およびPresta,1992,Curr.Op.Struct.Biol.,2:593−596参照。ヒト化抗体はPRIMATIZED(登録商標)抗体とすることもでき、ここに、抗体の抗原−結合領域は、注目する抗原でアカゲザルを免疫化することによって生産される抗体に由来する。
【0050】
免疫化に際して、内因性免疫グロブリンの生産の不存在下でヒト抗体の十分なレパートリーを生産することができるトランスジェニック動物(例えば、マウス)を生産することができる。例えば、キメラおよび生殖系突然変異体マウスにおける抗体重鎖接合領域(JH)遺伝子のホモ接合性欠失の結果、内因性抗体生産の完全な阻害がもたらされる。そのような生殖系突然変異体マウスへのヒト生殖系免疫グロブリン遺伝子アレイの導入の結果、抗原攻撃に際してヒト抗体の生産がもたらされる。例えば、Jakobovitsら、1993,Proc.Natl.Acad.Sci.USA,90:2551;Jakobovitsら、1993,Nature,362:255−258;Bruggermannら、1993,Year in Immuno.,7:33参照。ヒト抗体は、例えば、Hoogenboomら、1991,J.Mol.Biol.,227:381;またはMarksら、1991,J.Mol.Biol.,222:581−597に記載されているようにファージ−ディスプレイライブラリーに由来することもできる。
【0051】
「ヒト抗体」は、ヒトによって生産された抗体のそれに対応するアミノ酸配列を保有し、および/または本明細書中に開示されたヒト抗体を作成するための技術のいずれかを用いて作成されたものである。
【0052】
「アフィニティー突然変異した」抗体は、それらの改変を保有しない親抗体と比較して、抗原に対する抗体のアフィニティーの改良をもたらす1以上の超可変領域において1以上の改変を有するものである。好ましいアフィニティー成熟化抗体は、標的抗原に対してナノモラーまたはピコモラーさえのアフィニティーを有するであろう。アフィニティー突然変異した抗体は公知の手法によって生産される。例えば、VHおよびVLドメインシャッフリングによるアフィニティー成熟化を記載するMarksら、1992,Bio/Technology 10:779−783参照。CDRおよび/またはフレームワーク残基のランダム突然変異誘発はBarbasら、1994,Proc.Nat.Acad.Sci.USA 91:3809−3813;Scierら、1995,Gene 169:147−155;Yeltonら、1995,J.Immunol.155:1994−2004;Jacksonら、1995,J.Immunol.154(7):3310−9;およびHawkinsら、1992,J.Mol.Biol.226:889−896に記載されている。
【0053】
「抗体断片」は、一般には、無傷抗体の抗原結合部位を含み、かつ、かくして、抗原を結合する能力を保有する無傷抗体の一部のみを含む。本定義に含まれる抗体断片の例は:
(i)重鎖および軽鎖の間に1つの鎖間ジスルフィド結合を有するVL、CL、CHおよびCH1ドメインを有するFab断片;
(ii)CH1ドメインのC−末端において1以上のシステイン残基を有するFab断片であるFab’断片;
(iii)VHおよびCH1ドメインを有するFd断片;
(iv)VHおよびCH1ドメイン、および該CH1ドメインのC−末端に1以上のシステイン残基を有するFd’断片;
(v)抗体の単一アームのVLおよびVHドメインを有するFv断片;
(vi)VHドメインよりなるdAb断片:
(vii)少なくともVL、VH、CL、CH1ドメインを含み、かつヒンジ領域を欠如する無ヒンジ抗体;
(viii)ヒンジ領域においてジスルフィドブリッジによって連結された2つのFab’断片を含む二価断片である(Fab’)2断片:
(ix)単一鎖抗体分子(例えば、単一鎖Fv’scFv);
(x)同一ペプチド鎖中の軽鎖可変ドメイン(VH)に連結された重鎖可変ドメイン(VH)を含む2つの抗原結合部位を持つ「ダイアボディー」;
(xi)単一アーム抗原結合ドメインの半減期を増大させることができるFc領域を形成するのに十分な、軽鎖、重鎖およびN−末端切形重鎖定常領域を含む単一アーム抗原結合分子;
(xii)相補性軽鎖ペプチドと一緒になって、抗原結合領域の対を形成するタンデムFdセグメント(VH−CH1−VH−CH1)の対を含む「線状抗体」;
を含む。
【0054】
本明細書中で用いるように、「治療」とは、治療すべき個体または細胞の天然コースを改変させる試みにおける臨床的介入をいい、予防のために、または臨床的病理学のコースの間に行うことができる。治療の望ましい効果は、病気の発生または再発の予防、兆候の軽減、障害のいずれかの直接的または間接的病理学的結果の減少、転移の予防、病気進行速度の減少、病気状態の軽減または緩和、および緩解または改良された予後を含む。
【0055】
TCCR
TCCR(WSX−1)は、IL−12 β−2受容体、G−CSFRおよびIL−6受容体に対する相同性を持つサイトカイン受容体のWS(G)XWSクラスのものである。最高の相同性は、IL−12 β―2受容体(26%同一性)。これらの受容体は、細胞、特に、血液細胞の増殖および分化に関与する細胞の増殖および分化を制御するシグナルを変換する。後の実施例に示されるデータは、TCCRの活性化が、直接的にまたは間接的に、CD8+T−リンパ球またはTh1応答の増殖を含めた、自己免疫プロセスの抑制を含むことを示唆する。
【0056】
自己免疫プロセスの抑制は、T−細胞を他のマイトジェン刺激に対して非−応答性とすることができるサイトカインシグナリング(SOCS)蛋白質ファミリーメンバーのサプレッサーの誘導を介して起こり得る(Alexanderら、.2004,Ann.Rev.Immunul.,22:503)。特に、SOCS−3は、ヤヌスキナーゼの活性化ループに結合し、キナーゼ活性を阻害し、それにより、サイトカインシグナリングを抑制する蛋白質である(Masuharaら、1997,Biochem.Biophys.Res.Commun.,239:439−446)。ペルオキシソームプロリファレーター−活性化受容体(PPAR)−ガンマアゴニスト(例えば、ロシグリタゾン)のようないくつかの剤の抗−炎症効果は、SOCS1およびSOCS3の転写を誘導することによって機能する(Parkら、2003,J.Biol.Chem.,278:14747−14752)。後の実施例に示したデータは、TCCR活性化がSOCS3の発現を直接的にまたは間接的に誘導することを示す。
【0057】
自己免疫抑制は、IL−10の誘導を介しても起こる。IL−10は活性化されたT細胞、B細胞、単球およびケラチノサイトによって生産されるサイトカインである。IL−10は、IL−2、IL−3、IFN−γ、GM−CSF、およびTNFを含めた多数のサイトカインの生産を阻害する。IL−10は、炎症応答を制限し、終止させるにおいて主な役割を演じる(Mooreら、2001,Ann.Rev.Immunol.,19:683)。後の実施例に示されるデータは、TCCR活性化がIL−10の発現を直接的にまたは間接的に誘導することを示す。
【0058】
ヒトTCCRのアミノ酸配列は公開されており(1996年5月23日に出願されたWO97/44455)、受託番号4759327下でGenBankから入手可能である。この配列はSprecherら、1998,Biochem.Biophys,Res.Commun.246(1):82−90にも記載されている。ヒトTCCR(hTCCR)の配列は長さが636アミノ酸であって、表1にて後に示される(配列番号:1)。単一ペプチドがアミノ酸残基1ないし32から同定されており、膜貫通ドメインが配列番号:1のアミノ酸残基517ないし538から同定されている。N−グリコシル化部位が残基51ないし54、76ないし79、302ないし305、311ないし314、374ないし377、382ないし385、467ないし470、563ないし566において同定されており、N−ミリストイル化部位が残基107ないし112、240ないし245、244ないし249、281ないし286、292ないし297、377ないし378、400ないし405、459ないし464、470ないし475、531ないし536および533ないし538において同定されている。原核生物膜リポ蛋白質脂質付着部位は残基522ないし532に存在し、成長因子およびサイトカイン受容体ファミリーシグニチャー1は残基41ないし54に存在する。また、残基183ないし191には、ヒト顆粒球−マクロファージコロニー−刺激因子(GM−CSF)に対する受容体の第二のサブユニットに対する有意な相同性の領域もある。TCCRは、配列番号:1の残基41ないし230におけるTCCR上のサイトカイン受容体相同性ドメインにてIL−27サブユニットp28に結合する。記載された全てのhTCCR残基は配列番号:1の配列に従ってナンバリングされる。
【0059】
成人において、hTCCRは胸腺で最も高度に発現されるが、発現は、末梢血液白血球(PBL)、脾臓においても観察され、および弱い発現が肺においても観察される。胎児組織は肺および腎臓において弱いTCCR発現を呈する。
【0060】
マウスTCCR(mTCCR)のアミノ酸配列は公開されており(1996年5月23日に出願されたWO97/44455)、受託番号7710109の下でGenBankから入手可能である。この配列はSprecherら、1998,Biochem.Biophys,Res.Commun.246(1):82−90にも記載されている。mTCCRに対する配列は、長さが623アミノ酸であって、表1において後に示される(配列番号:2)。単一ペプチドがアミノ酸残基1ないし24において同定され、膜貫通ドメインは配列番号:2のアミノ酸残基514ないし532において同定されている。N−グリコシル化部位は残基46ないし49、296ないし299、305ないし308、360ないし361、368ないし371、および461ないし464において同定されている。カゼインキナーゼIIリン酸化部位は残基10ないし13、93ないし96、130ないし133、172ないし175、184ないし187、235ないし238、271ないし274、272ないし275、323ないし326、606ないし609、615ないし618において同定されている。チロシンキナーゼリン酸化部位は約残基202ないし209において同定されている。N−ミリストイル化部位は約残基43ないし48、102ないし107、295ないし300、321ないし326、330ないし335、367ないし344、393ないし398、525ないし530および527ないし532において同定されており、アミド化部位は約残基240ないし243において同定されている。原核生物膜リポ蛋白質脂質付着は約残基516ないし526に存在し、成長因子およびサイトカイン受容体ファミリーシグニチャー1は約残基36ないし49に存在する。有意な相同性の領域は約残基14ないし51においてヒトエリスロポエチンと共に存在し、マウスインターロイキン−5受容体は残基211ないし219に存在する。記載された全てのmTCCR残基は配列番号:2の配列に従ってナンバリングされる。
【0061】
【化1】
【0062】
【化2】
IL−27
IL−27はTCCRに対するリガンドである(Pflanzら、2002Immunity 16(6):779−790)。IL−27はEBI3(エプスタイン−バールウィルス誘導遺伝子3)およびp28蛋白質サブユニットから構成されるヘテロダイマーサイトカインである。p28は3つの接触表面を持つ4つのラセン束サイトカインである。第一の接触表面はEBI3に結合し、第二および第四のαラセンの残基から構成される。第二の接触表面はサイトカイン受容体相同性ドメインの領域においてTCCRに結合し、第一および第三のアルファラセンの残基よりなる。第三の接触表面はgp130上で見出されるIGドメインのようなIGドメインに結合し、第一のラセンの最後および第四のラセンの始まりにおける残基を含む。
【0063】
ヒトp28のペプチド配列(配列番号:3)は243アミノ酸の長さであり、他方、マウスp28のペプチド配列(配列番号:4)は長さが234アミノ酸である(Pflanz,NCBI受託番号AAM34499)。表2にて以下に示されるこれらの配列は73%配列同一性を有する。
【0064】
【化3】
EBI3は可溶性サイトカイン受容体の構造を有し、p28上の特異的結合部位に結合する。ヒトEBI3は長さが229アミノ酸であり(Devergneら、1996,J.of Virology 70(2):1143−1153)、表3にて以下に示されるペプチド配列(配列番号:5)を有する。
【0065】
【化4】
TCCR/IL−27受容体複合体
図1はTCCR/IL−27受容体複合体の人工物を示す。IL−27に対する完全な受容体はgp130およびTCCRサブユニットを含有する。サイトカイン受容体相同性ドメインは配列番号:6の約残基126ないし323におけるgp130に存在する。gp130に存在する他の相同性ドメインは配列番号:6の約残基324ないし423、424ないし518、および519ないし614に位置する3つのフィブロネクチンIII型ドメイン、および約残基22ないし122における免疫グロブリンドメインを含む。gp130はIL−6、IL−11、CNTF、LIF、CT1、およびCLCに対する受容体の成分であることも知られている(Hibiら、1990,Cell,63(6):1149−1157)。gp130のアミノ酸配列(配列番号:6)を表4にて以下に示す。
【0066】
【化5】
【0067】
【化6】
TCCRのIL−27活性化が、主なTh1−特異的転写因子、T−betの発現を誘導する(Lucasら、2003,PNAS,100:15047−52)。TCCR活性化の効果はStats(転写のシグナルトランスジューサーおよびアクチベーター)によって媒介される。具体的には、TCCR活性化はStat1、Stat3、Stat4、およびStat5のリン酸化に導く(Lucasら、2003,supra)。後の実施例に示すデータは、TCCR活性化がCD8+T−リンパ球の増殖、またはTh1応答を含めた、自己免疫プロセスの発現を直接的または間接的に誘導することを示唆する。
TCCRおよびT−リンパ球サブタイプ
前記したように、4つのラセン束サイトカインファミリーのメンバー(Bazan,J.F.,1990,Proc Natl Acad Sci USA,87:6934−8)は、Th1およびTh2エフェクター細胞の区別される集団への共通の前駆体からのTヘルパー細胞の増殖および最後の分化において役割を演じる(O’Garra.,1998,Immunity,8:275−835)。IL−4はTh2細胞の発生に支配的に影響し、他方、IL−12はTh1細胞の分化における主な因子である。(Hsiehら、1993,Sience,260:547−9;Sederら、1993,Proc Natl Acad Sci USA,90:10188−92;Le Grosら、1990,J Exp Med,172:921−9;Swainら、1991,Immunol Rev,123:115−44)。したがって、IL−4が欠乏したマウス(Kuhnら、1991,Science,254,707−10)、IL−4受容体α鎖(Noben−Trauthら、1997,Proc Natl Acad Sci USA,94:10838−43)、またはIL−4特異的転写因子STAT6(Shimodaら、1996,Nature,380:630−3)はTh2応答が欠乏し、他方、IL−12が欠乏したマウス(Magramら、1996,Immunity4:471−81)IL−12受容体(IL−12R)β1鎖(Wuら、1997,J Immunol,159:1658−65)、またはIL−12特異的転写因子STAT4(Kaplanら、1996,Nature,382:174−7)は損なわれたTh1応答を有する。
【0068】
Th1およびTh2細胞サブタイプは共通の前駆体Th−0細胞に由来する。Th1およびTh2細胞からのサイトカイン放出プロフィールはエフェクターメカニズムおよび細胞傷害性細胞の選択に影響する。Th1細胞によって分泌されるIl−2およびインターフェロンガンマはマクロファージュおよび細胞傷害性細胞を活性化し、他方、Th2細胞によって分泌されるIl−4、Il−5、Il−6、およびIl−10は好酸球および肥満細胞の生産を増大させ、ならびにIgeを含めた抗体の生産を促進し、細胞傷害性細胞の機能を減少させる傾向がある(Powrieら、1993,Immunol.Today,14:270)。一旦樹立されれば、Th1またはTh2応答パターンは、他のサブセットの細胞によるサイトカイン生産を一般的には阻害するサイトカインの生産によって維持される。例えば、IL−4はTh1クローンからのインターフェロン−ガンマの生産を阻害し、他方、インターフェロン−ガンマはTh2クローンからのIl−4からの生産を阻害する(Mosmannら、1989,Adv.Immunol.46:111−147;Mosmannら、1989,Annu.Rev.Immunol.,7:145−173)。この負のフィードバックループは、多くの免疫応答の間における極性化サイトカインプロフィールの生産を加速する。
【0069】
細胞傷害性T−リンパ球(CD8+)は迅速に他の細胞を破壊することができる。細胞傷害性T−リンパ球は2つの主な細胞溶解経路を用いる:ペルフォリン−依存性エクソサイドーシス経路およびFasリガンド/Faskを用いる。細胞傷害性T−リンパ球はインターフェロン−ガンマのようなプロ−炎症サイトカインのプロジューサーでもある。
【0070】
Th1応答に関連するサイトカインの過剰生産またはCD8+細胞傷害性T−リンパ球の過剰増殖は、同種異系移植片拒絶、(グレーヴス病および橋本甲状腺炎のような)自己免疫甲状腺病、自己免疫ブドウ膜網膜炎、巨細胞動脈炎、(クローン病、潰瘍性結腸炎、局所腸炎、肉芽腫性腸炎、末端回腸炎、局所回腸炎、および終末回腸炎を含めた)炎症性腸疾患インスリン依存性新生糖尿病、多発性硬化症、悪性貧血、乾癬、慢性関節リウマチ、サルコイドーシス、強皮症、および全身エリテマトーデスに至りかねない。
【0071】
後の実施例に詳細に記載された実験は、非−特異的T−リンパ球刺激に応答しての、TCCRを発現するT−リンパ球と比較して、TCCRを欠如するT−リンパ球のより大きな増殖を示す(実施例1参照)。さらに、驚くべきことに、TCCRを発現するマウスは、TCCRを欠如するマウスよりも、多発性硬化症についての動物モデルにおいて、実験的アレルギー脳脊髄炎(EAE)のような自己免疫障害に対して感受性が低いことが見出された(実施例2)。
【0072】
これらのデータは、T−リンパ球増殖およびTh1媒介生物学的活性のTCCR−媒介直接的または間接的抑制を示唆する。従って、TCCRのアゴニストを用いて、T−細胞増殖を阻害し、および/または多発性硬化症および慢性関節リウマチを含めた自己免疫障害を治療することができる。
【0073】
自己免疫障害
前記したように、T−リンパ球の過剰増殖またはTh1またはTh2細胞によって生産されたサイトカインの過剰生産は医学的障害の保有者に導く。例えば、Th1応答に関連するサイトカインの過剰生産またはCD8+細胞傷害性T−リンパ球の過剰増殖は異種同系移植片拒絶、(グレーヴス病および橋本甲状腺炎のような)自己免疫甲状腺病、自己免疫ブドウ膜網膜炎、巨細胞動脈炎、(クローン病、潰瘍性結腸炎、局所腸炎、肉芽腫性炎腸炎、末端回腸炎、局所回腸炎、および終末回腸炎を含めた)炎症性腸疾患、インスリン−依存性真正糖尿病、多発性硬化症、悪性貧血、乾癬、慢性関節リウマチ、サルコイドーシス、強皮症および全身エリテマトーデスを含めた自己免疫障害に至り得る。
【0074】
多発性硬化症は、Tリンパ球依存性であると考えられる自己免疫脱髄障害である。MSは、一般には、再発−緩解コースまたは慢性進行コースを呈する。MSの病因は知られていないが、ウイルス感染、遺伝的素因、環境、および自己免疫は全て該障害に寄与しているように見える。MS患者における病巣は、圧倒的にT−リンパ球に媒介されるミクロ神経膠細胞および浸潤性マクロファージの浸潤物を含有する。CD4+Tリンパ球はこれらの病巣に存在する支配的な細胞型である。MS病巣のホールマークはプラーク、すなわち、MRIスキャンで見られる通常の白色物質から鋭く区別される脱髄の領域である。MSプラークの組織学的外観は病気の異なる段階で変化する。活性な病巣において、血液−脳関門は損傷されており、それにより、細胞外空間への血清蛋白質の血管外遊出を可能とする。炎症細胞は血管周囲カフにおいて、および白色物質全体に観察することができる。CD4+T−細胞、特にTh1はプラークのエッジにおいて毛細血管後細静脈の周りに蓄積し、白色物質にも散らばる。活性な病巣において、接着分子およびリンパ球のマーカーのアップレギュレーションおよびIL−2RおよびRおよびCD26のような単球活性化も観察された。活性病巣における脱髄には稀神経突起膠細胞の破壊は伴わない。対照的に、該病気の慢性相の間では、病巣は稀神経突起膠細胞の喪失およびしたがって、血液中のミエリン稀神経突起膠細胞糖蛋白質(MOG)抗体の存在によって特徴付けられる。
【0075】
種々のよく許容された動物モデルが、自己免疫障害について存在する。その例として、EAE(実験的アレルギー性脳脊髄炎)は、T細胞および単核細胞炎症、および中枢神経系における軸索の引き続いての脱髄によって特徴付けられるT細胞媒介自己免疫障害である。EAEは、一般的にヒトにおけるMSの関連動物モデルであると考えられている。(例えば、Bolton,C.1995,Multiple Sclerosis,143参照)。候補TCCRアゴニストのような剤は、例えば、John Wiley & Sols,Inc.によって発行され、Coligan et al.National Institutes of Healthによって編集されたCurrent Protocols in Immunology,units15.1および15.2に記載されたプロトコルを用い、免疫媒介脱髄障害に対するT細胞刺激または阻害活性について分析することができる。また、稀突起神経膠細胞またはシュワン細胞が、例えば、Duncanら、1997,Molec.Med.Today,554−561に記載されたように、中枢神経系にグラフトされたミエリン病についてのモデルも参照。
【0076】
関節炎についての動物モデルはコラーゲン−誘導関節炎である。例えば、McIndoeら、1999,Proc.Natl.Acad.Sci.USA,96:2210−2214参照。このモデルはヒト自己免疫慢性関節リウマチの臨床的、組織学的および免疫学的特徴を有し、ヒト自己免疫関節炎についての許容できるモデルである。マウスおよびラットモデルは滑膜炎、軟骨の侵食、および肋軟骨下骨によって特徴付けられる。コラーゲン−誘導関節炎は、リンパ球浸潤および滑膜肥大を含めたヒトにおける慢性関節リウマチでの多くの特徴を有する。例えば、McIndoeら、1999,Proc.Natl.Acad.Sci.USA,96:2210−2214参照。TCCRの潜在的アゴニストは、例えば、John Wiley & Sols,Inc.によって発行され、Coligan et al.National Institutes of Healthによって編集されたCurrent Protocols in Immunology,units 15.5に記載されたプロトコルを用い、これらのモデルを用いて自己免疫関節炎に対する活性について分析することができる。また、Issekutz,A.C.et al.,Immunology(1996)88:569に記載されたCD18に対するモノクローナル抗体およびVLA−4インテグリンを用いるモデルも参照されたし。
【0077】
皮膚異種同系移植片拒絶についての動物モデルは、抗−ウイルスおよび腫瘍免疫性におけるそれらの役割を示す、およびその尺度を示すイン・ビボ組織破壊を媒介するT細胞の能力をテストする手段である。ほとんどの通常かつ許容されたモデルは、マウス尾−皮膚移植片を用いる。報告された実験は、皮膚異種同系移植片拒絶はT細胞、ヘルパーT細胞およびキラー−エフェクタ−T細胞によって媒介され、抗体によっては媒介されないことを示している。例えば、Auchincloss and Sachs,1998,In:Fundamental Immunology,2nd ed.,W.E.Paul ed.,Raven Press,NY,Page889−992参照。適当な手法は、John Wiley & Sols,Inc.によって発行され、Coliganら、 1995,National Institutes of Healthによって編集されたCurrent Protocols in Immunology,unit 4.4に詳細に記載されている。他の候補TCCRアゴニストをスクリーニングするのに用いることができるトランスプラント拒絶モデルは、例えば、Tanabeら、 1994,Transplantation,58:23およびTinubuら、1994,J.Immunol.,4330−4338によって記載された同種異系心臓トランスプラントモデルを含む。
【0078】
TCCRのアゴニスト
TCCRのアゴニストは、本明細書中際書中に開示された天然配列TCCRペプチドの生物学的活性を増強し、または刺激する分子である。適当なアゴニスト分子は、具体的には、ヒト化抗体を含めたアゴニスト抗体、またはFab、Fab’、Fd、Fd’、Fv、Ab、無ヒンジ抗体、F(ab’)2断片、単一鎖抗体分子、ダイアボディー、単一アーム抗原結合分子、および線状抗体を含めたアゴニスト抗体の断片、アミノ酸配列改変体または天然ポリペプチド、ペプチド、小分子の断片などを含む。
【0079】
TCCRの適当なアゴニストは、TCCRのペプチド断片、TCCR細胞外メイン、および:
(a1)配列番号:1のヒトTCCRペプチドの残基1または約33ないし636;
(a2)配列番号:2のマウスTCCRペプチドの残基1または約25ないし623;
(b1)配列番号:1のヒトTCCRペプチドのX3ないし636、ここに、X3は配列番号:1のいずれかのアミノ酸残基27ないし37;
(b2)配列番号:2のマウスTCCRペプチドX4ないし623、ここに、X4は配列番号:2の20ないし30からのいずれかのアミノ酸残基であり;
(c1)1または約33ないしX1、ここにX1は配列番号:1の残基512ないし残基522からのいずれかのアミノ酸残基であり;
(c2)1または約25ないしX2、ここに、X2は配列番号:2の残基509ないし519からのいずれかのアミノ酸残基であり;
(d1)X5ないし636、ここに、X5は配列番号:1の残基533ないし543からのいずれかのアミノ酸であり;
(d2)X6ないし623、ここに、X6は配列番号:2の残基527ないし537からのいずれかのアミノ酸であり;または
(e)配列番号:1および配列番号:2のアミノ酸配列のもう1つの特異的に誘導された断片;
のアミノ酸配列に対して少なくとも約80%のアミノ酸配列同一性を有するTCCR改変体も含む。
【0080】
TCCRのアゴニストは、例えば、配列番号:1および配列番号:2の配列の、N−およびC−末端において、または1以上の内部ドメイン内で1以上のアミノ酸残基が付加され、または欠失されたTCCRペプチドを含む。
【0081】
TCCRのアゴニストは、天然配列IL−27、EBI3、p28、改変体、およびIL−27ヘテロダイマーに通常は関連する生物学的活性を有するその断片を含む。例えば、TCCRのアゴニストは、IL−27の天然配列成分に対して少なくとも80%、90%、95%、または99%配列同一性を有するIL−27改変体を含む。TCCRのアゴニストは、天然配列ヒトp28(配列番号:3)またはマウスp28(配列番号:4)に対して少なくとも73%、75%、80%、90%、95%、または99%配列同一性を有するp28改変体も含む。TCCRのアゴニストは、TCCRに結合することができるp28、およびgp130の部分を含む。その例として、TCCRのアゴニストは、天然配列ヒトp28(配列番号:3)またはマウスp28(配列番号:4)に対して少なくとも73%、75%、80%、90%、95%、または99%配列同一性を有し、かつTCCRおよびgp130に結合することができるp28改変体を含む。TCCRのアゴニストは、TCCR上に見出されるサイトカイン受容体相同性ドメインの領域においてTCCRに結合すると考えられる、p28の第一および第三のαラセンからの残基、およびgp130上に見出されるIGドメインに結合すると考えら得る、第一のラセンの最後および第4のラセンの始まりにおける残基を含有するp28ペプチド改変体を含む。
【0082】
TCCRのアゴニストは、例えば、TCCRに結合し、それを活性化することができる分子を含む。TCCRのアゴニストは、TCCRがホモダイマーを形成できるようにする分子、および/またはTCCRおよびgp130がヘテロダイマーを形成することができるようにする分子も含む。例えば、TCCRに対する抗体はTCCRがホモダイマーを形成できるようにすることができる。さらなる例として、TCCRおよびgp130双方に対して特異的な二価抗体はTCCRおよびgp130がヘテロダイマーを形成するようにすることができる。
【0083】
TCCRペプチドのアゴニストまたはアンタゴニストを同定するための方法は、TCCRペプチドを候補アゴニストまたはアンタゴニスト分子と接触し、次いで、TCCRに関連する1以上の生物学的活性のケン質可能な変化を測定すること含むことができる。例えば、アゴニストは、TCCRペプチドを発現する細胞を候補と接触させ、次いで、ウェスタン−ブロットまたはもう1つの適当なアッセイを用いて、Stat1、Stat3、Stat4、またはStat5のリン酸化のようなTCCRの生物学的活性について接触された細胞を分析することによって同定される。TCCRのアゴニストは、TCCRペプチドを発現するように作成されたBa/F3細胞のような細胞を候補アゴニストと接触させ、次いで、例えば、[3H]標識チミジン取込みまたはもう1つの適当なアッセイを測定することによって増殖について接触された細胞を分析することによって同定することもできる。
【0084】
モノクローナル抗体
モノクローナル抗体を生産するための多くの技術が知られている。1つの方法において、例えば、Balb/cのようなマウスは、フロイント完全アジュバントに乳化した免疫原としてのTCCRまたはその部分で免疫化し、1ないし100マイクログラムの量にて皮下または腹腔内注射される。別法として、該免疫原はMPL−TDMアジュバント(Ribi Immunochemical Research,Hamilton,MT)に乳化し、動物の後足肉趾に注射される。次いで、選択されたアジュバントに乳化されたさらなる免疫原で、免疫化されたマウスを10ないし12日後にブースター注射する。しかる後、数週間の間、マウスにさらなる免疫化注射をブースター注射することもできる。血清試料は、ELISAアッセイにおいてテストして、抗−TCCR抗体を検出するために眼窩後出血によってマウスから周期的に得ることができる。
【0085】
適当な抗体力価が検出された後、抗−TCCR抗体について「陽性」の動物に、免疫原の最終静脈内注射を注射することができる。3ないし4日後に、マウスを犠牲にし、脾臓細胞を収穫する。脾臓細胞を、次いで、(35%ポリエチレングリコールを用いて)ATCC、No.CRL 1597から入手可能な、P3X63AgU.1のような選択されたマウス骨髄腫細胞系に融合させる。該癒合によりハイブリドーマ細胞が生じ、次いで、これを、HAT(ヒポキサンチン、アミノプテリン、およびチミジン)を含有する96ウェル組織培養プレートに平板培養して、非−融合細胞、骨髄腫ハイブリッド、および脾臓細胞ハイブリッドの増殖を阻害することができる。
【0086】
選択されたハイブリドーマ細胞は、TCCRに対する反応性について、ELISA、または他の適当なアッセイでスクリーニングすることができる。陽性ハイブリドーマ細胞は、例えば、同系Balb/cマウスに腹腔内注射して、抗−TCCRモノクローナル抗体を含有する腹水液を生じさせることができる。別法としてハイブリドーマ細胞は、例えば、組織培養フラスコまたはローラーボトル中で増殖させることができる。腹水液中で生産されたモノクローナル抗体の精製は、硫酸アンモニウム沈殿、続いて、ゲル排除クロマトグラフィーまたは他の適当な方法を用いて達成することができる。別法として、抗体のプロテインAまたはプロテインGへの結合に基づくアフィニティークロマトグラフィーを使用することができる。
【0087】
TCCR−/−マウス
TCCRを発現しない「ノックアウト」マウスが構築されている(TCCR−/−)。そのようなマウスは、例えば、TCCRをコードする内因性遺伝子、および動物の胚細胞に導入された同一ペプチドをコードする改変されたゲノムDNAの間の相同組換えを介して調製することができる。
【0088】
例えば、特定のペプチドをコードするcDNAを用いて、確立された技術に従ってそのペプチドをコードするゲノムDNAをクローン化することができる。特定のペプチドをコードするゲノムDNAの部分を、欠失することができるか、あるいは取り込みをモニターするのに用いることができる選択マーカーをコードする遺伝子のようなもう1つの遺伝子を置き換えることができる。典型的には、(5’および3’末端双方における)未改変フランキングDNAの数キロベースがベクターに含まれる(相同組換えベクターの記載については、Thomasら、1987,Cell,51:503参照)。ベクターは(エレクトロポレーションによるなどして)胚性幹細胞系に導入し、導入されたDNAが内因性DNAを相同組換えされている細胞を選択する。例えば、Liら、1992,Cell,69:951参照。次いで、例えば、Braderyら、1987,In:Teratocarcinomas and Embryonic Stem Cells: A Practical Approach,E.J.Roberteon,ed.(IRL,Oxford),pp.113−152に記載されているように、選択された細胞を(マウスまたはラットのような)動物の胚盤胞に注射して、同種キメラを形成させる。次いで、キメラ胚を適当な偽妊娠雌仮腹動物に移植し、該胚を出産させて、「ノックアウト」動物を生じさせることができる。それらの生殖細胞に相同組換えDNAを保有する子孫は標準的な技術によって同定し、それを用いて、動物の全ての細胞は相同組換えDNAを含有する動物を育種することができる。
【0089】
後の実施例で用いるTCCR−/−マウスの作成の記載は、その内容がここに引用して援用されるWO 0129070 (de Sauvage et al.)およびChenら、2000,Nature,407:916に見出される。
【0090】
組成物および治療
自己免疫障害の治療でいうようなTCCRのアゴニストは、限定されるものではないが、免疫機能、例えば、T細胞増殖/活性化、リンホカイン放出、または免疫細胞浸潤を変調する、蛋白質、抗体、断片および改変体、小さな有機分子、ペプチド、リンペプチドなどを含む。特に、本発明中で記載されるTCCRのアゴニストは、T−リンパ球の増殖、T−リンパ球サイトカイン放出、および自己免疫障害を抑制し、減少させ、または低下させるのに有用である。
【0091】
TCCRアゴニストは、前記したスクリーニングアッセイのいずれかによって、および/またはいずれかの他の公知のスクリーニング技術によって同定することができる。
【0092】
本発明のTCCRアゴニストを公知の方法に従って処方して有用な組成物を調製し、それにより、TCCRアゴニストを許容される担体と組み合わせる。処方は、所望の程度の純度を有するTCCRアゴニストを、凍結乾燥された処方または水性溶液の形態の、任意の許容される担体、賦形剤、または安定化剤と混合することによって貯蔵のために調製される。例えば、Remington:The Science and Practice of Pharmacy 20th ed.Gennaro Ed.(2000)参照。適当な担体、賦形剤または安定化剤は使用する用量および濃度において受容者に対して非毒性であり、それは、リン酸塩、クエン酸塩、および他の有機酸のような緩衝液;アスコルビン酸を含めた抗酸化剤;低分子量(約10残基未満)ペプチド血清アルブミン、ゼラチンまたは免疫グログリンのような蛋白質;ポリビニルピヨリドンのような親水性ポリマー、グリシン、グルタミン、アスパラギン、アルギニンまたはリシンのようなアミノ酸;グルコース、マンノースまたはデキストリンを含めた単糖、二糖および他の炭水化物;EDTAのようなキレート剤;マンニトールまたはソルビトールのような糖アルコール;ナトリウムのような塩−形成対イオン;および/またはTWEEN(登録商標)PLURONICS(登録商標)PEGのようなノニオン界面活性剤を含む。
【0093】
イン・ビボ投与で用いるべき処方は滅菌されていなければならない。これは、凍結乾燥および復元に先立ってまたはそれに続いて、滅菌濾過膜を通しての濾過によって容易に達成される。
【0094】
本発明中において、組成物は、一般には、滅菌アクセスポート、例えば、皮下注射針によって刺すことができるストッパーを有する静脈内溶液バッグまたはバイアルを有する容器に入れられる。
【0095】
投与経路は、静脈内、腹腔内、脳内、筋肉内、眼内、動脈内、または病巣内経路、局所投与による、または徐放系による注射または注入のような公知の方法に従う。投与の経路は、アデノウィルスベクターのような適当なベクターでのトランスフェクションの結果としてのイン・ビボ発現を含むことができる。
【0096】
本発明の医薬組成物の量および所望の濃度は、考えられる特定のしように応じて変化させることができる。投与の適当な用量または経路の決定は、十分に、通常の技師の技量内にある。動物実験は、ヒト療法のための有効量の決定のための信頼できるガイダンスを提供する。効果的な用量の種間スケーリングは、Mordenti,J.and Chappell,W.”The use of Interspecies scaling in toxicokinetics” in Toxicokinetics and New Drug Development,Yacobi et.al.,Eds.,Pergamon Press,New York 1989,pp.42−96によって主張された原理に従って行うことができる。
【0097】
TCCRアゴニストのイン・ビボ投与を使用する場合、正常な用量は、投与経路に応じて、1日当たり約10ng/kgないし100mg/kg哺乳動物体重以上、例えば、約1mg/kg/日ないし10mg/kg/日で変化させることができる。特定の用量および送達の方法に関するガイダンスは文献中に供される;例えば、米国特許代4,657,760;第5,206,344;または第5,225,212号参照。異なる処方は異なる治療および異なる障害で効果的であり、および特異的器官または組織を治療することを意図した投与は、もう1つの器官または組織に対するものとはことなる送達を必要とするであろうと予測される。
【0098】
TCCRアゴニストの徐放投与がTCCRアゴニストの投与を必要とするいずれかの病気または障害の治療に適した放出特徴を持つ処方で望まれる場合、TCCRアゴニストのマイクロカプセル化が考えられる。徐放のための組換え蛋白質のマイクロカプセル化は、ヒト成長ホルモン(rhGH)、インターフェロン−アルファ、−ベータ、−ガンマ、インターロイキン−2、およびMN rgp120で首尾よく行われている。例えば、Johnsonら、1996,Nat.Med.2:795−199;Yasuda,1993,Biomed.Ther.,1221−1223;Horaら、1990,Bio/Technology,755−758;Cleland,1995,”Design and Production of Single Immunization Vaccines Using Polylactide Polyglycolide Microsphere Systems” in Vaccine Degsin:The Subunit and Adjuvand Approach,Powell and Newman,eds.,(Plenum Press;New York),pp.439−462;WO97/03692、WO96/40072,WO96/07399および米国特許第5,654,010参照。
【0099】
TCCRアゴニストの徐放処方は、ポリ−乳酸−コグリコール酸(PLGA)、かなりの程度の生体適合性、および広い範囲の生分解性特性を呈するポリマーを用いて開発することができる。PLGA、乳酸およびグリコール酸の分解生成物はヒト身体から迅速に除去される。さらに、このポリマーの分解性は、その分子量および組成に応じて数ヶ月ないし数年に調整することができる。さらなる情報については、Lews,”Controlled Release of Bioactive Agents from Lactide/Glycolide polymer”in Biogradable Polymers as Drug Delivery Systems,M.Chasin and R.Langdeer,editers(Marcel Dekker;New York,1990),pp.1−41参照。
【実施例】
【0100】
本発明は、以下の実施例を参照して良好に理解することができる。これらの実施例は本発明の特別な具体例の代表であることが意図され、本発明の範囲を限定することを意図しない。
【0101】
実施例1
T−細胞応答のTCCR−媒介抑制
T−細胞応答に対するTCCR活性の効果は、野生型(TCCR+/+)およびノックアウト(TCCR−/−)脾臓細胞の誘導されたT−細胞の増殖の分析によってテストした。T−細胞受容体はCD3と会合してT−細胞受容体複合体を形成する。異なる用量の抗−CD3抗体は、T−細胞受容体複合体および抗原−提示細胞(APC)のMHCクラスII分子(CD4)の相互作用に通常は関連するT−細胞の増殖を非特異的に刺激する。
【0102】
野生型(TCCR+・+)およびノックアウト(TCCR−/−)混合リンパ球、単離されたCD4+T細胞および、単離されたCD8+T細胞の増殖は、抗−CD3抗体によって刺激した(BD Pharmingen,San Digo,CA,clone 145−2c11)。細胞を湿潤化CO2インキュベータ中で3日間増殖し、増殖はアッセイの最後の8ないし16時間に測定された[3H]チミジン取り込みによって測定した。驚くべきことに、ノックアウトマウス(TCCR−/−)から得られた混合リンパ球の抗−CD3抗体誘導増殖は、表5において、および図5に示されたように、最大下用量の抗−CD3において野生型(TCCR+/+)リンパ球から得られたリンパ球のそれよりも有意に大きかった。このデータは、例えば、刺激されたT−細胞の増殖を抑制するTCCR活性の保護効果を示唆し、およびアゴニストでのTCCRの刺激は、T−細胞増殖のようなT−細胞応答を直接的にまたは間接的に抑制するのに有用であろう。
【0103】
【化7】
しかしながら、単離されたCD4+T細胞および単離されたCD8+T細胞の抗−CD3抗体誘導増殖は、各々、図6(表6)および図7(表7)に示されたように、野生型およびノックアウト細胞の間で有意には異ならなかった。このデータは、TCCRに対するリガンドであるIL−27が、CD4+T細胞およびCD8+T細胞以外のリンパ球によって生産されることを示唆する。
【0104】
【化8】
次に、抗−CD3抗体刺激に応答してのCD4+およびCD8+細胞の増殖は、CFSE(カルボキシフルオレセインジアセテート、スクシンイミジドエステル)標識アッセイで測定した。標識された細胞は、各細胞分裂後にそれらの蛍光強度の50%を喪失するので、CFSE標識は、細胞分裂の数がモニターされるのを可能とする。野生型(TCCR+/+)およびノックアウト(TCCR−/−)混合リンパ球細胞懸濁液をCFSEで標識して、細胞懸濁液中に0.5μM CFSE(Sigma,St.Louis,MO)の濃度を作り出した。次いで、細胞懸濁液を37℃で10分間インキュベートした。標識の後、FCSを5%最終濃度まで加え、細胞を直ちに遠心し、氷冷PBSで洗浄した。野生型(TCCR+/+)およびノックアウト(TCCR−/−)混合リンパ球細胞の増殖は、2.5μg/mlの濃度において抗−CD3抗体(BD Pharmingen,San Piego,Ca,clone 145−2c11)によって刺激した。
【0105】
次いで、細胞を37℃にて2日間インキュベートした。その時点において、細胞をCD4+およびCD8+に対するマーカー(CD4−CychromeまたはCD8−Cychrome)で標識し、フローサイトメトリーによって分析した。
【0106】
以下のデータは、CD4+ならびにCD8+の双方の陽性T細胞がTCCRノックアウト細胞において過剰増殖性であることを示す(表8参照)。図10および11は、インキュベーション時間の間において0、1、2、3、4または5回の細胞分裂を受けた細胞の数を示す。CD4+およびCD8+T細胞双方について、より多くの細胞が、野生型におけるよりもノックアウトにおいて3、4、5回の分裂を受けた(すなわち、ノックアウト細胞についての系はCD4+細胞(図14)およびCD8+細胞(図15)双方において右側にシフトする)。
【0107】
【化9】
実施例2
TCCRを発現するマウスはEAEに対して感受性が低い。
【0108】
実験的アレルギー性脳脊髄炎(EAE)は、多発性硬化症(MS)についての動物モデルとして働くCNSの自己免疫障害である。MSと同様に、EAEは、ミエリンに対する免疫−媒介損傷が観察可能な兆候をもたらす脱髄障害である。EAEはCD4+Th1細胞(Fifeら、2001,J.of Immun.,166:7617−7624)およびCD8+細胞傷害性T−リンパ球(CTL)(Huseby,et al.,2001,J.Exp.Med.,194(5):669−676)双方によって媒介されると考えられる。EAEに対するTCCRの効果を調べるために、EAEの臨床的進行をTCCRを発現する野生型マウス(TCCR+/+)およびTCCRを欠如するノックアウトマウス(TCCR−/−)において調べた。以下に示すように、TCCRを発現するマウスは、TCCRを欠如するマウスよりもCD4+ThおよびCD8+媒介傷害EAEに対して感受性が低い。
【0109】
ノックアウトTCCR−/−マウスはWO 0129070(de Sauvage et al.)に記載されたように創製し、C57BL/6バックグラウンドに逆−交配させ、N12創始細胞から育種した。野生型TCCR+/+対照はJackson Laboratory (Bar Harbor,ME)から購入したC57Bl/6マウスであった。
【0110】
MOG誘導EAE
MEVGWYRSPFSRVVHLYRNGKのアミノ酸配列(配列番号:7)を有するMOG35−55ペプチドは、Rainin Quartet自動ペプチド合成器(Rainin,Oaklane,CA)での9−フルオレニルメトキシカルボニル化学を用いて合成した。ペプチドを樹脂から切断し、移動相において水/アセトニトリル/0.1%TFAグラジエントを持つ分取用逆相HPLCを用いて精製した。ペプチドの同一性は、電子スプレーマススペクトロメトリーによって確認した。
【0111】
野生型TCCR+/+およびノックアウトTCCR−/―マウスを、100μlのTBSおよび100μlのCFA(フロイントの完全アジュバント)中の200μgのMOG35−55ペプチドを含有する200μlのエマルジョンで皮内免疫化して、0日にEAEを誘導した。CAFは、IFA(フロイントの不完全アジュバント)(Difco−BD Diagnostic System,Sparks,MD)を死滅し、乾燥したM,tuberculosis H37A(Difco−BD Diagnositic Systems,Sparks,MD)と8mg/ml M.tutuberculosisの濃度まで混合することによって調製した(各マウスはCFAの成分としての800μgの死滅したM.tutubersulosisを受けた)。0日に、および2日に再度、各マウスに100μlのPBS中の200ngの百日咳トキシン(List Biological Laboratories,Campbell,CA)で腹腔内注射して、血液の関門に入るのを助けた。受けた成分の用量は以下の表9にまとめる。
【0112】
【化10】
MBP誘導EAE
ASQKRPSQRHG(配列番号:8)のアミノ酸配列を有するAc 1−11ペプチドは、Rainin Quartat自動ペプチド合成器(Rainin,Oakland,CA)にて9−フルオレニルメトキシカルボニル化学を用いて合成した。該ペプチドを樹脂から切断し、移動相における水/アセトニトリル/0.1%TFAグラジエントでの分取用逆相HPLCを用いることによって精製した。ペプチドの同一性は、電子スプレーマススペクトロメトリーによって確認した。
【0113】
野生型TCCR+/+およびノックアウトTCCR−/−マウスは、100μlのCFA(フロイントの完全アジュバント)中の10μgのAc 1−11ペプチド(ASQKRPSQRHG)、ミエリン塩基性蛋白質の成分を皮内免疫化して、0日にEAEを誘導した。MOB−誘導EAEについて前記したように、CFAは、IFA(フロイントの不完全アジュバント)を、M.tuberculosis H37A(死滅しおよび乾燥した)と8mg/ml M.tuberculosisの濃度まで混合することによって調製した(各マウスは、CFAの成分として、800μgの死滅したM.tuberculosisを受けた)。2日、および3日に再度、各マウスに、100μlのPBS中の200ngの百日咳トキシンで腹腔内注射して、血液の関門に入るのを助けた。受けた成分に用量を以下の表10にまとめる。
【0114】
【化11】
全てのマウスは、1日で出発し、1週間当たり3回臨床的疾患について評価した。病気グレード4に到達したマウスを毎日評価した。グレード5におけるいずれの動物も安楽死させた。5日以内にグレード3以下まで改良されなかったものを安楽死させた。用いた臨床的グレーディングシステムを以下の表11に示す。
【0115】
【化12】
40日において、全ての残りの動物を犠牲にし、脳および脊髄を組織学的分析のために切開した。脳を切開し、(各脳についての4つのレベルの各々からの1つの切片)、H&E(ヘマトキシリンおよびエオシン染色、Sigma,St.Louis,MO)で染色して、炎症を評価した。脊髄を切片化し(各脊髄について3つの異なるレベルの各々から4つの切片)、H&Eで染色して、炎症を評価し、ルキソールファストブルー(Luxol Fast Blue)(VWR Scientific,St.Paul,MN)で染色して、脱髄を評価した。各スライドについて、最高炎症および脱髄双方についての最高スコアおよび平均スコア(各スライドでの全ての切片の平均)を報告した。用いた炎症グレーディングシステムを以下の表12に示す。
【0116】
【化13】
用いた脱髄グレーディングシステムを以下の表13に示す。
【0117】
【化14】
野生型(TCCR+/+)におけるMOG(ミエリン稀突起神経膠細胞糖蛋白質)誘導EAE(実験的アレルギー性脳脊髄炎)の臨床的進行は、ノックアウト(TCCR−/−)マウスにおける誘導されたEAEよりもひどくはなかった(図8参照)。同様に、野生型(TCCR+/+)マウスにおけるMBP(ミエリン塩基性蛋白質)誘導EAEの臨床的進行は、ノックアウト(TCCR−/−)マウスにおける誘導されたEAEよりひどくはなかった(図9参照)。
【0118】
図8および9に示すように、TCCRを欠如する動物(TCCR−/−)はEAEのよりひどい臨床的兆候を示し、他方、TCCRを発現するマウス(wt)はよりひどくない兆候および進行を示し、これは、EAEのような、自己免疫障害に対するTCCR活性の保護効果を示唆する。
【0119】
臨床データは、図10ないし13に示したように、組織学的分析によってさらに支持される。TCCR−/−マウスは、WTマウスよりも高い炎症および高い脱髄スコアを有し、TCCRを発現するマウス(wt)は、TCCRを欠如するマウス(−/−)よりもCD4+Th1およびCD8+媒介障害EAEに対して感受性が低かったことを示す。
【0120】
まとめると、データは、EAEのような、自己免疫障害に対するTCCR活性の保護的、弱める、または抑制的効果を示唆する。
【0121】
実施例3
TCCRアゴニストでの自己免疫障害の治療
実施例2について記載したように、実験的アレルギー性脳脊髄炎(EAE)は、多発性硬化症(MS)に対する動物モデルとして働くSMSのCD4+Th1またはCD+8媒介自己免疫障害である。自己免疫障害の進行およびコースに対する投与されたTCCRアゴニストの効果を調べるために、IL−27のようなTCCRアゴニストを、誘導されたEAEのようなMSの実験的モデル系において投与する。EAEは、例えば、実施例2について前記したようにマウスにおいて開始させる。EAEの臨床的進行は、TCCRを発現し(TCCR+/+)、IL−27のようなTCCRアゴニストを受けるマウスにおいて評価する。IL−27のようなTCCRアゴニストで処理したマウスは、病気の低下した臨床的兆候または進行を示し、および/または未処理TCCR+/+対照よりも自己免疫障害に対して感受性が低いと予測される。
【0122】
実施例4
TCCRアゴニストでの動物モデルにおける関節炎の治療
自己免疫障害に対するTCCRアゴニストの抑制的および/または保護的効果は、数個の利用可能な動物モデル系のうちの1つでテストすることができる。マウスにおけるコラーゲン−誘導関節炎は、自己免疫障害、慢性関節リウマチに対する1つのモデルである。このモデルは、例えば、McIndoeら、1999,PNAS USA 96:2210−2214に記載されている。マウスにおけるコラーゲン−誘導関節炎は、リンパ球浸潤および滑膜肥大を含めたヒト慢性関節リウマチでの多くの特徴を共有する。
【0123】
コラーゲン−誘導関節炎の臨床的進行は、マウス、例えば、C57BL/6マウスまたは他の適当な実験室動物において調べる。関節炎は、例えば、McIndoeら、supraで引用された方法、または他の公知の方法によってテスト動物において誘導される。一般に、モデル動物は、異なる動物種に由来するII型コラーゲンをテスト動物に注射することによって、例えば、ウシII型コラーゲンをマウスに注射することによって創生する。該コラーゲンはフロイントの完全アジュバントのようなアジュバントと組み合わせることができる。
【0124】
TCCRリガンドIL−27のようなTCCRアゴニストは、例えば、関節炎−誘導剤の投与に先立って、投与の間に、および/または投与の後に、あるいは関節炎兆候の開始に先立って、開始の間に、および/または開始後にテスト動物に投与される。投与および用量の方法は変化させることができ、例えば、1プレおよび/またはポスト用量における、2以上のプレおよび/またはポスト用量の期間にわたっての毎日の1日当たり複数用量における用量、またはTCCR受容体を発現する細胞へペプチド剤を投与するために知られた他の適当な用量における、単体中のIL−27のようなペプチドリガンドの投与を含むことができる。代替方法は、例えば、IL−27の双方のサブユニット、または連結されたIL−27サイトカインを発現する組換えアデノウイルスからのペプチドを発現させることによるIL−27のようなペプチドリガンドの送達を含む。
【0125】
臨床的疾患の進行は、例えば、McIndoeら、supraに記載されたように、テストおよび対照動物においてモニターされる。例えば、該疾患の物理的および化学的特徴をモニターし、経時的にスコア取りする。動物は、主な関節のリンパ球浸潤、滑膜肥大、滑膜液サイトカイン含有量等について分析することができる。これらのパラメータを、テスト動物および対照の間で比較する。実施例1および2に示されたTCCRの保護的/抑制的効果を保つために、TCCRアゴニストでの動物における誘導された関節炎の治療は、未処理対照と比較して、よりひどくない臨床的兆候、結果、および/または物理的または化学的特徴で証明された、抑制的および/または保護的効果を供すると予測される。
【0126】
実施例5
TCCRに対するモノクローナル抗体の調製
hTCCRに対するモノクローナル抗体は、hTCCRの細胞外ドメインを用いて調製した。免疫原は、精製目的でカルボキシ−末端に付加された8つのヒスチジン残基をタグとして加えた膜貫通部分(配列番号:1の残基517ないし538)を欠如するhTCCR(配列番号:1)であった。hTCCR(ECD)−(His)8ペプチドは、ニッケルNTAアフィニティークロマトグラフィーを介して精製した。
【0127】
hTCCR(ECD)−(His)8ペプチド(1ないし2マイクログラム)を25マイクロリットルのMPL−TDMアジュバント(Ribi Immunochemical Research,Hamilton,MT)と組み合わせ、合計12回の注射のために毎週2回、野生型balb/cマウス(Charles River Laboratories,Wilmington,MA)の肉趾に注射した。
【0128】
42日に、マウスを犠牲にし、脾臓細胞を収穫した。脾臓細胞を(35%ポリエチレングリコールを用いて)マウス骨髄腫細胞(ATCC,No.CRL1597から入手可能なP3X63AgU.1)に融合させた。融合によりハイブリドーマ細胞が創製され、これを、HAT(ヒポキサンチン、アミノプテリンおよびチミジン)培地を含有する96ウェル組織培養プレート中で平板培養して、非−融合細胞、骨髄腫ハイブリットおよび脾臓細胞ハイブリットの増殖を阻害した。
【0129】
次いで、ハイブリドーマ細胞を、TCCRへの抗体結合についてELISAアッセイにおいてスクリーニングした。TCCRに対する反応性を有する同定されたハイブリドーマ培養は培養:2685、2686、および2688を含んだ。ハイブリドーマ培養2686(抗体2686)は、2004年12月15日に、American Type Culture Collection(ATCC),Manassas,Va.に寄託し、受託番号ATCC PTA−6447を有する。
【0130】
実施例6
モノクローナルAb2686はヒトTCCRを活性化する。
【0131】
組換えTCCRを発現するBa/F3細胞を用いて、TCCRを活性化する抗−TCCR抗体の能力を分析した。Ba/F3細胞はマウスIL−3依存性細胞系である。TCCRの候補アゴニストは、該候補アゴニストに応答してのTCCRを発現するBa/F3細胞の増殖を測定することによって評価することができる。細胞増殖の結果、ポリヌクレオチドの増大した合成のため増大した[3H]―チミジンの取込がもたらされる。細胞増殖は、例えば、[3H]―チミジン接種を測定することによってモニターされる。以下に示すように、モノクローナルAB2686は、TCCRを発現するBa/F3細胞の増殖を誘導することによってTCCRアゴニスト活性を示した。
【0132】
Ba/F3細胞(Palaciosら、1985、Cell,41:727−734)は、増殖および生存双方に対してIL−3を必要とするマウス造血系因子−依存性細胞系である。Ba/F3細胞を、10%胎児子牛血清(GIBCO,Carlsbad,CA)および100pg/mLマウスIL−3(R&D Systems,Minneapolis,MN)を補足したRPMI−1640倍値(GIBCO,Carlsbad,CA)中で培養した。
【0133】
ネオマイシン耐性遺伝子を持ち、ヒトまたはマウスTCCRをコードするポリヌクレオチド配列いずれかを含有するpMSCVベクター(Clontech,Palo Alto,CA)をエレクトロポレーションによってBa/F3細胞にトランスフェクトした。安定なトランスフェクタントを1mg/mlのG418(Clontech,Palo Alto,CA)で処理して、ネオマイシン耐性に対する遺伝子を含有するベクターでトランスフェクトされている安定な真核生物細胞系を選択した。次いで、TCCRを認識するフィコ−エリスリン標識モノクローナル抗体で細胞を処理した。TCCRを発現する標識されたクローンをFACSによって選択した。
【0134】
TCCR発現細胞を、IL−3を加えていない10%胎児子牛血清を補足したRPMI−1640倍値で洗浄した。次いで、細胞を、10%胎児子牛血清を補足した100μlのRPMI−1640倍値中にてウェル当たり5×103細胞にて二連で平板培養した。精製された組換えマウスIL−3(陽性対照)または精製された抗−TCCR(ヒト)モノクローナル抗体:2685−IgG2a、2686−IgG1、2688−IgG1、対照イソタイプIgG2a(BD Pharmingen,San Diego,CA)または対照イソタイプIgG1(BD Pharmingen,San Diego,CA)を、4:1希釈シリーズとして、表14にて以下に示す濃度で加えた。
【0135】
【化15】
48時間後、1μCiの[3H]―チミジン(Amerscham−Pharmacia,Piscataway,NJ)を各ウェルに加えた。さらに6時間後、[3H]―チミジンの細胞への取り込みを、β−カウンター(Packard Topcount,PerkinElmer Life and Analytical Sciences, Boston,MA)で測定した。
【0136】
結果を図2ないし4に示す。モノクローナル抗体2685−IgG2a、2686−IgG1、2688−IgG1、および対照イソタイプIgG2aまたは対照イソタイプIgG1に応答してのヒトTCCRの発現するBa/F3細胞の増殖を図2に示す。抗体2686は、テストした他の抗体のいずれよりも[3H]―チミジンの有意により大きな取り込みを誘導し、これは、抗体2686が、Ba/F3細胞において発現されたヒトTCCRの効果的なアゴニストであることを示す。
【0137】
マウスIL−3(陽性対照)または抗体2686いずれかに応答してのヒトTCCRを発現するBa/F3細胞の増殖を図3に示す。示されたように、抗体2686は、陽性対照IL−3のそれよりも小さいに拘らず、ヒトTCCRを発現するBa/F3細胞においてTCCR応答を生じさせるのに効果的であった。
【0138】
ヒトTCCRを発現するBa/F3細胞は、図4に示すように、マウスTCCRを発現するBa/F3細胞よりも抗体2686での処理に応答しての有意により大きな量の[3H]―チミジンを取り込んだ。このデータは、抗体2686がヒトTCCRの特異的アゴニストであることを示し、マウスTCCRに対して交差反応性を示さない。
【0139】
これらの研究は、示されたアゴニスト抗体のようなTCCRのアゴニストがTCCR受容体に結合し、それを刺激して、TCCR−媒介生物学的活性、ここでは、Ba/F3細胞の増殖を誘導することができるのを示す。したがって、データは、TCCRアゴニストがイン・ビボにてTCCR−媒介活性を直接的にまたは間接的に誘導するのに有用であることを示唆する。
【0140】
実施例7
他のTCCRアゴニストの同定
TCCRアゴニスト活性を有する剤を同定し、確認するために、IL−27の断片およびTCCRの改変体を含めた推定TCCRアゴニストをTCCR結合にて分析した。TCCRの結合はイン・ビトロまたはイン・ビボ結合にて分析することができる。例えば、潜在的アゴニストを、実施例3にて前記したように、組換えTCCRを発現するように作成されたCOS細胞またはBa/F3細胞のような、TCCRを発現する細胞に投与し、潜在的アゴニストに対する細胞応答を測定する。
【0141】
受容体結合は、融合蛋白質、例えば、ヒトIgGのFcドメインを含有するイムノアドヘシンとして潜在的にペプチドアゴニストを発現させることによって分析することもできる。受容体−リンガンド結合は、例えば、該イムノアドヘシンをTCCR発現細胞と相互作用させることによって検出される。結合したイムノアドヘシンは、Fc融合ドメインを認識する蛍光試薬を用い、微視的に可視化することができる。結合は蛍光の分析によって、または他の公知の方法によって定量することができる。
【0142】
TCCRのアゴニストは、IL−10またはSOCS−3の発現のようなTCCR媒介活性を刺激する候補アゴニストの能力を分析することによってスクリーニングすることができる。例えば、TCCRを発現するT−リンパ球を候補アゴニストと接触させることができる。IL−10および/またはSOCS−3の発現は、例えば、ELISA、定量的PCR、および同様な方法によって測定することができる。対照、例えば、基礎IL−10および/またはSOCS−3レベルに対するIL−10および/またはSOCS−3の発現の増加はTCCR刺激に相関し、これは有用なTCCRアゴニストの指標となる。
【0143】
実施例8
IL−27媒介細胞増殖、およびサイトカインの誘導および抑制
野生型(TCCR+/+)およびノックアウト(TCCR−/−)CD4+細胞双方におけるサイトカイン誘導に対するIL−27の効果を中性のTh1またはTh2誘導条件下で調べた。野生型(TCCR+/+)およびノックアウト(TCCR−/−)CD4+細胞双方における細胞リコール増殖に対するIL−27の効果を、中性のTh1またはTh2誘導条件下で調べた。
【0144】
0日に、アゴニスト抗−CD3モノクローナル抗体(145−2C11,BD Pharmingen,San Diego,CA,PBS o/n中5ug/ml)で予めコートされた24ウェルプレート中で、野生型CD4+またはTCCRノックアウトCD4+細胞をウェル当たり2×105細胞にて平板培養した。次いで、個々のウェルにおける増殖を中性、Th1バイアシング、またはTh2バイアシング条件下で誘導した。中性条件はIL−2(R&D Systems,Minneapolis,MN)、抗−IL−12抗体(BD Pharmingen,San Diego、CA)、抗−IFN−γ抗体(BD Pharmingen,San Diego,CA)、抗−IL−4抗体(BD Pharmingen,San Diego,CA)およびCD−28(BD Pharmingen,San Diego,CA)の添加によって作り出した。Th1バイアシング条件は、IL−2、IL−12(R&D Systems,Minneapolis,MN)、抗−IL−4抗体、およびCD−28の添加によって作り出した。Th2バイアシング条件は、IL−2、IL−4(R&D Systems,Minneapolis,MN)、抗−IL−12抗体、抗−IFN−γ抗体、およびCD28の添加によって作り出した。これらの個々のウェルにおける処理は表15にて以下に示す。
【0145】
【化16−1】
【0146】
【化16−2】
細胞を37℃で培養した。上清の試料は24時間、48時間、および/または72時間に採取した。ELISAを、TNF−α、IL−5、IL−2、IFN−γ、IL−10、IL−6、IL−4、GM−CSFに対するプローブ(BD Pharmingen,San Diego,CAから購入したキット)にて上清試料で行った。以下の表16は、IL−27依存性誘導倍数としてのELISAデータを示す。図16AないしCは、中性(16A)、Th1バイアシング(16B)、およびTh2バイアシング条件(16C)下でのIL−2のIL−27依存性誘導を示す。図17AないしCは、中性(17A)、Th1バイアシング(17B)、およびTh2バイアシング条件(17C)下でのIL−10のIL−27依存性誘導を示す。
【0147】
データは、IL−27に応答してのTNF−α、IFN−γ、およびIL−4の誘導を示す。データはIL−27に応答してのIL−2、IL−6、およびGM−CSFの抑制も示す。データは、IL−10が、中性、Th1バイアシング、およびTh2バイアシング条件下でIL−27によって誘導されることを示す。前記したように、IL−10は、限定的かつ終了する炎症応答において主な役割を演じる。IL−10はIL−27によって誘導されるので、データは、IL−27を用いて、免疫−媒介疾患を治療できることを示唆する。
【0148】
【化17】
RNAを24時間、48時間、および/または72時間に採取した細胞試料から抽出し、次いで、以下の図17に示すように、SOCS−1、SOCS−3、PIAS−1、およびPIAS−3に対して特異的なプローブで、定量的PCR(TAQMAN(登録商標))を行った。
【0149】
【化18】
以下の表18は、IL−27依存性誘導倍数としての定量的PCRデータを示す。図18AないしCは、中性(18A)、Th1バイアシング(18B)、およびTh2バイアシング条件(18C)下でのSOCS−3のIL−27依存性誘導を示す。
【0150】
データは、SOCS−3が中性、Th1バイアシング、およびTh2バイアシング条件下でIL−27によって誘導されることを示す。前記したように、SOCS−3はサイトカインシグナリングを抑制することが知られており、いくつかの剤の抗−炎症効果のメディエーターであることが報告されている。SOCS−3はIL−27によって誘導されるので、データは、IL−27を用いて免疫−媒介疾患を治療することができるのを示唆する。
【0151】
【化19】
中性条件下で処理した前記細胞の試料を72時間に採取し、RNAを抽出した。次いで、GENECHIP(登録商標)(Affymetrix, Santa Clara,CA)を用いて誘導された発現につき該RNAを分析した。以下の表19は、選択された遺伝子についての未処理対照に対する誘導(抑制)倍数としてのGENECHIP(登録商標)データを示す。
【0152】
ここに、データは、再度、IL−10が中性条件下でIL−27によって誘導されることを示す。前記したように、IL−10は、炎症応答を制限し、および終了させるにおいて主な役割を演じる。IL−10はIL−27によって誘導されるので、データは、IL−27を用いて、免疫−媒介疾患を治療することができることを示唆する。
【0153】
【化20】
3日に、細胞をIL−2の存在下であって、およびIL−27の存在下または不存在下で細胞を増殖させた。具体的には、IL−27に予め暴露した24ウェルプレートからの細胞を1mlの培地中に取り、次いで、2×10−4mg/ml IL−27および1×10−5mg/ml IL−2を含有する3mlの培地と共に6ウェルプレートに沈積させた。IL−27に予め暴露されていない細胞を1mlの培地中に取り、次いで、1×10−5mg/ml IL−2を含有する3mlの培地と共に6ウェルプレートに沈積させた。
【0154】
5日に、2×10−4mg/ml IL−27および1×10−5mg/ml IL−2の濃度を有する4mlの培地(IMDM(Invitrogen,Carlsbad,CA)w/10%HyClone血清)を、IL−27に予め暴露した細胞を含有するウェルに加えた。1×10−5mg/ml IL−2の濃度を有する4mlの培地を、IL−27に予め暴露していない細胞を含有するウェルに加えた。
【0155】
6日に、細胞を遠心し、次いで、培地(前記したのと同一培地)中にペレットを再度懸濁させ、カウントした。種々のウェルからの細胞カウントを表20にて以下に示し、図19に反映させる。
【0156】
データは、Th1またはTh2バイアシング条件の間に添加されたIL−27はCD4+細胞の増殖を低下させることを示し、IL−27は、多発性硬化症および慢性関節リウマチのような自己免疫疾患を含めたCD4+細胞の増殖によって特徴付けられる疾患を治療するのに有用であることを示唆する。
【0157】
【化21】
実施例9
IL−6誘導増殖のIL−27抑制
野生型(TCCR+/+)およびノックアウト(TCCR−/−)CD4+細胞のIL−6誘導増殖に対するIL−27の効果を、抗−IL−2抗体(BD Pharmingen,San Diego,CA)の存在下および不存在下で調べた。
【0158】
野生型マウスからの混合脾臓細胞(4×105)を96−ウェルプレート上のウェルに入れた。ノックアウトマウスからの混合脾臓細胞(4×105)をプレート上の別々のウェルに入れた。全てのウェルをPBS o/n中の100μlの2ug/ml 抗−CD3でコートした。ウェルを、表21にて以下に示した実験群に従って処理した。
【0159】
【化22】
細胞37℃にて培養した。48時間後、[3H]−チミジンをもう一晩の間加え、増殖を[3H]−チミジン取込みによって測定した。各群についての平均CPMを表22にて以下に示す。IL−2中和なくしての増殖を図20Aに示す。IL−2中和有りでの増殖を図20Bに示す。
【0160】
【化23】
データは、抗−IL−2抗体が存在するか否かに拘らず、IL−27が、抗−CD3によって刺激され、かつIL−6によって増強された増殖を抑制することを示す。抗−il−2抗体が存在しない場合、IL−27が、抗−CD3抗体によって刺激される増殖を抑制する。抗−IL−2抗体は、抗−CD3抗体によって刺激される増殖を低下させる。しかしながら、IL−27の添加は、この効果を部分的に和らげる。
【0161】
実施例10
IL−27受容体(TCCR)欠乏マウスはEAE超感受性である。
【0162】
IL−27は、活性化された抗原提示細胞(APC)によって生産されるリガンドである。IL−27は、特異的サブユニット、IL−27、およびIL−6Rを含めた、多数の他の受容体によって共有されるgp130よりなるヘテロダイマー受容体を通じてシグナリングを行う。本明細書中で議論するように、IL−27は種々のSTATおよびJak−1を通じてシグナルを活性化するが、支配的なシグナリング事象はSTAT−1の活性化であるように見える。STAT−1の活性化およびTH−1特異的転写因子T−betの下流誘導を介して、IL−12RB2鎖およびIFN−ガンマの発現は促進される。Il−27リガンドおよび受容体は図21にダイアグラムにより示される。
【0163】
IL−27はIL−12ファミリーのメンバーであり、サイトカインのIL−6クラスターに属する。図22参照。IL−27、EBI3およびp28の2つの成分はIL−12サブユニットに対する密接な相同性を有する。TCCRともいわれるIL−27受容体(IL−27R)の双方のサブユニットは、種々の白血球にて協調して発現される。最高の発現はT細胞およびNK細胞におけるものであるように見える。
【0164】
ナイーブな未分化T細胞(Th−0)は、ナイーブなTh−0細胞の成熟T−ヘルパー細胞への分化を誘導する異なるシグナルに応答する。一般に、2つのタイプのT−ヘルパー細胞が知られている;Th−1およびTh−2細胞。図23に図示されるように、IL−4によるTh−0細胞の刺激はTh−2細胞の発生に導き、IL−4、IL−5、IL−6、IL−10、およびIL−13を生産する。Th−2細胞およびサイトカイン産物は液性免疫および抗−蠕虫応答にインパクトを与える。IL−27および/またはIFN−ガンマによるTh−0細胞の刺激は、T−細胞におけるIL−12応答性の状態を誘導し、従って、それらはIL−12の制御下で成熟TH−1細胞に分化し、IFN−ガンマ、IL−2、およびリンホトキシン(LT)を生産することができる。Th−1細胞およびそれらのサイトカイン産物は、細胞−媒介免疫性およびマクロファージ活性化に関与する。
【0165】
Th−0細胞のTh−1およびTh−2ヘルパー細胞への分化におけるIL−27の役割のさらなる我々の理解のため、IL−27R欠乏マウス(TCCRノックアウト)を前記実施例に記載したように生産した。自己免疫疾患の間におけるIL−27についての潜在的役割を、実験的自己免疫脳炎(EAE)、多発性硬化症についてのマウスモデルを用いて調べた。EAEを持つマウスからのCD4+T細胞のみの移動がナイーブな受容体マウスにおいてEAEを引き起こしかねないので、EAEはT細胞媒介される。
【0166】
実験的EAEを誘導するために、マウスを、フロイントの完全アジュバント中のミエリン稀突起神経膠細胞糖蛋白質(MOG)35ないし55ペプチドで免疫化した。野生型(WT)およびIL−27受容体(TCCR)ノックアウトマウスをMOGで免疫化し、前記実施例に記載したようにEAEの証拠について調べた。臨床EAEスコアを処理後25日にわたって評価した。
【0167】
図24に示されたデータは、IL−27刺激を除去することによって引き起こされたEAE疾患における仮定された低下の代わりに、EAEはIL−27R欠乏マウスにおいて亢進されたことを示す。該マウスはEAE過剰感受性であるように見え、酷いEAE疾患を発生した。EAE表現型を発現する受容体欠乏マウスから採取された脊髄組織の組織学的分析を図25に示し、これは、EAEを持つIL−27受容体ノックアウトマウスにおける増強された炎症および脱髄を示す。
【0168】
この実験に加えて、種々の病原体でのIL−27受容体欠乏マウスの刺激、ならびに誘導された喘息および肝炎モデルの結果、Th−1およびTh−2媒介応答双方が亢進された。これらのデータは、IL−27が重要な免疫抑制機能を有することを示す。
【0169】
【化24】
IL−27、およびT−細胞の分化におけるその可能な役割をさらに研究するために、図27に図示した手法に従い、ナイーブなCD4+細胞をNACS−精製し、IL−27と共にまたはそれなくして抗−CD3+抗体で処理した。IL−27でのT細胞の刺激は、Th−0、Th−1またはTh−2へのT細胞分極を促進する条件下でなされた。
【0170】
簡単に述べると、24ウェル皿を5μg/ml抗−CD3(BD Pharningen)で一晩コートした。1.8×106CD4+T−細胞の容量を、IL−2(10ng/ml)および抗−CD28(1μg/ml)の存在下でウェル当たり撒いた。分化のために、以下のサイトカインおよび抗体を加えた:TH−0(各々5μg/mlの抗−IL−12、抗−IFN−ガンマ、抗−IL−4)、TH−1(3.5ng/mlのIL−12、5μg/mlの抗−IL−4)、TH−2(3.5ng/mlのIL−4、5μg/mlの抗−IFNgおよび抗−IL−12)。IL−27を200ng/mlの濃度にていくつかの培養に加えた。72時間後、上清ならびにRNAを単離し、Chip、RT−PCR、および/またはELISA分析によって特異的サイトカインの生産について分析した。得られたデータを図28に示し、これは、IL−27がT−細胞発生に対して顕著な効果を有することを示す。
【0171】
IL−27は調べたほとんどのサイトカインに対して顕著な効果を有し、この効果は、細胞が分化した条件とは一般には独立していた。IL−27は、Th−2誘導条件下でTNFαおよびIL−10、ならびにIL−4を誘導した。同時に、IL−2、IL−5、IL−6、GM−CSF、およびIL−17の生産はIL−27によってかなり抑制された。
【0172】
これらの効果のいずれかが良く知られ、かつ優れた免疫抑制サイトカインIL−10の誘導に対して二次的であるか否かを判断するために、IL−27の効果もIL−10欠乏T−細胞で調べた。図29に示すように、サイトカイン生産のIL−27誘導変調はIL−10とは独立していた。というのは、WTおよびIL−10欠乏T−細胞を比較して、IL−2またはGM−CSF生産においてほとんど差は見られなかったからである。
【0173】
イン・ビトロで見られたIL−27による免疫抑制性IL−10の強い誘導にも拘らず(図28)、EAEを持つIL−27R−/−マウスからのT−細胞においてIL−10の僅かな低下が見られたに過ぎない(図30)。しかしながら、この人工的なイン・ビトロ観察は、IL−27媒介IL−10誘導がEAEの間の重要な生物学的プロセスであるという解釈を断じて排除しない。対照的に、それは、IL−27誘導IL−10誘導をイン・ビボで研究するための我々の自由な実験技術の限界を最も反映するようである。
【0174】
実施例11
EAEはTH−17依存性である。
【0175】
最近の証拠は、ヘルパーT−細胞の新しいサブタイプ、いわゆるTH−17細胞が、EAEを含めた多くのプロ−炎症プロセスの鍵となるメディエーターであることを示唆する。これらのTh−17細胞はIL−17A、IL−17F、IL−6、TNF、およびGM−CSFを生じると報告された。図31に供されるダイアグラム参照。TH−17細胞の発生は貧弱にしか理解されていないが、それは、IL−12に対して同様性を持つもう1つのヘテロダイマーサイトカインであるIL−23に依存すると考えられる。IL−23欠乏動物はこのT−細胞表現型を効果的に発生させることができず、EAEおよびCIAに対して抵抗性である。しかしながら、IL−23は必要であるように見えるが、TH−17細胞分化でイン・ビトロでそれは十分ではない。
【0176】
前記で議論したように、IL−27R欠乏マウスはWT同腹子と比較してより酷いEAE疾患を発生させた。IL−27シグナリングに対して下流の事象を分析して、EAEの酷さに対するこの限定的効果で重要な因子を決定した。IL−27に応答しての種々のサイトカインの発現を活性化の間に調べた。
【0177】
IL−27はIFN−ガンマの生産を促進し、IFN−ガンマはIL−17を阻害することが知られている。データは、IL−27が、IFN−ガンマよりもより効果的にIL−17および他のTh−17サイトカインIL−6およびGM−CSFの生産を抑制したことを示す(図33および34)。さらに、EAEを持つTCCR−/−マウスからのリンパ節細胞は、WTよりもイン・ビトロでの再度の刺激に際してTh−17サイトカインをより多く分泌した(図37)。
【0178】
IL−17生産のIL−27媒介抑制はIFN−ガンマとは独立していた。なぜならば、INF−ガンマに対して非応答性とされたT−細胞は、以前として、IL−27での刺激に際してIL−17生産を抑制したからである。(図35)。IFN−ガンマシグナリングの不存在下において、基礎IL−17生産はより高かった。この理由は不明瞭である。なぜならば、WT培養においてさえ、FN−ガンマシグナリングはIFN−ガンマ中和抗体の添加によってブロックされるからである。かくして、IFN−ガンマR欠乏マウスにおける高いIF−17発現は発生段階の改変を反映し(すなわち、IFNgR欠乏T−細胞はIFNgRの発現よりもWT−細胞とはより多くにおいて異なり)、あるいは細胞内IFNgループを反映できるであろう。リガンドおよび受容体が共発現される細胞において、シグナリングは後期分泌経路内で起こり得るが、そのようなシグナリングは細胞内のものであり、中和抗体によってブロックされないであろう。
【0179】
Th−17細胞はIL−27R欠乏マウスにおいて調節不調であるかを判断するために、IL−27R欠乏マウスをCFAにおけるMOGで免疫化した。リンパ節の排出を14日において排除し、X・ビボにてMOGで再度刺激した。IL−27R欠乏T細胞を含有するリンパ節上清は、有意に増大したレベルのIL−17を発現した(図37および38)。
【0180】
さらに、脳および脊髄(EAEにおける炎症の現実の部位)の免疫浸潤物は、より多くの細胞がIL−27R欠乏マウスにおいて浸潤することを明らかとした。さらに、これらの細胞のより高いパーセンテージは、細胞内染色によって分析した場合にIL−17陽性であった。一緒に考え合わせると、これらの2つの観察は脊髄におけるIL−17の概略2倍発現に翻訳される(図39参照)。
【0181】
IFN−ガンマおよびIL−27双方はSTAT−1を活性化し、STAT−1ノックアウトは増大したIL−17を生じさせる。従って、IL−27は、STAT−1を活性化することによってIL−17を抑制することができる。この関係は、STAT−1ノックアウトマウスから得られた細胞におけるIL−17のIL−27媒介抑制を分析することによって調べた。STAT−1の不存在下においては、IL−27はIL−17を抑制せず、これは、該抑制がSTAT−1によって媒介されることを示す。STAT−1の不存在下においては、IL−27はIL−17のインデューサーとなる。この逆行に対するメカニズムの基礎は知られていないが、IL−27によるSTAT−3の活性化はこの効果において役割を演じると推定するのは公正である。なぜならば、他のIL−17誘導サイトカイン(顕著には、IL−23)は、STAT−1を活性化せずにSTAT−3を通じてシグナリングを行うからである(図36参照)。
【0182】
まとめると、IL−27受容体(TCCR)欠乏マウスはEAE−過剰感受性である。IL−27はイン・ビトロにてTh−17サイトカインIL−17、IL−6、およびGM−CSFを効果的に抑制した。さらに、EAEを持つIL−27受容体欠乏マウスは野生型よりも多くTh−17サイトカインを生産する。IL−27は、Th−17から免疫応答を外すことによってEAEを抑制することができる。
【0183】
実施例12
IL−6はTh−17細胞を誘導する。
【0184】
図41に図示されたように、IL−23は必要であるが、サイトカインIL−17、IL−6、GM−CSF、およびTNFを生じるTh−17細胞へのTh−0細胞の分化には十分ではない。IL−23が十分でない1つのありそうな理由は、Th−0細胞はIL−23受容体を発現せず、従って、IL−23非応答性であることである。従って、Th−0細胞においてIL−23Rを誘導することができる因子は、TH−17分化経路の絶対的成分である。
【0185】
Th−1(IFN−g)およびTH−2(IL−4)細胞のエフェクターサイトカインもまたこれらの細胞の発生に参画し、よって、フィードバックループの安定化を供するので、我々は、TH−17エフェクターサイトカインの1つは、同様に、TH−17発生に参画しなければならないと理由づけた。TH−17エフェクターサイトカインの中では、IL−6は最も有望に見える。なぜならば、その受容体はナイーブなT−細胞で発現されるからであり、および最後に分化されたT−細胞よりもIL−6の他の源(最も顕著には抗原提示細胞)があるからである。加えて、IL−6ノックアウトマウスはEAE耐性である(図42参照)。
【0186】
野生型およびIL−27受容体ノックアウトマウスは、単独で、またはIL−27およびIL−23と組み合わせて、IL−6への応答性について調べた。図43に示すように、IL−6はTh−17軸を誘導した。IL−6単独での処理はIL−23受容体を刺激し、IL−17AおよびIL−17F生産も刺激した。興味深いことには、共投与されたIL−27はIL−23受容体およびIL−17の生産のIL−6刺激増加を低下させ、または排除した(図43)。IL−23はIL−23受容体およびIl−17生産の刺激に対する僅かな効果を有し、これは、IL−6単独で示された大きな刺激に対して付加的であるように見えた。この組合せへのIL−27の添加もまた、該応答を低下させ、または排除した。再度刺激したリンパ節細胞から採取したmRNAは、野生型対照と比較してIL−23受容体ノックアウトにおけるIL−23受容体の誘導を示した(図43参照)。
【0187】
増殖アッセイにおけるIL−6の効果をさらに比較し、IL−6は野生型およびTCCRノックアウトマウス双方において精製されたT−細胞の増強された増殖を大いに刺激した。IL−27の添加は、野生型細胞においてIL−6誘導増殖を完全に中和した。しかしながら、この低下はTCCRノックアウトマウスでは見られず、これは、IL−27がIL−6の優れた増殖効果に拮抗することを示す。図44参照。従って、IL−6駆動TH−17分化を含めた、IL−27はいくつかのレベルでのIL−6アンタゴニストであるように見える。
【0188】
実施例13
Il−27の役割
図46に示すように、IL−27はT細胞の分化、特に、複数レベルにおけるTh−17細胞の発生にインパクトを与える。IL−27はIL−10、IL−4の生産、およびTh−1細胞の発生を刺激するが、それは、Th−17細胞の生産、Th−17細胞サイトカインIL−17およびGM−CSFの生産も抑制する。
【0189】
本明細書における全ての刊行物および特許出願は、本発明が関係する分野における通常の技量のレベルを示す。全ての刊行物および特許出願は、あたかも各個々の刊行物および特許出願が参照により具体的かつ個々に示されるようなのと同一程度に参照により一体化される。
【0190】
本発明を、種々の特別なおよび好ましい具体例および技術を参照して記載した。しかしながら、本発明の精神および範囲内にありつつ、多くの変形および修飾をなすことができることが理解されるべきである。
【図面の簡単な説明】
【0191】
【図1】図1は、TCCR/IL−27受容体複合体の人工物の模式図である。
【図2】図2は、モノクローナル抗体:2685−IgG2a、2686−IgG1、2688−IgG1、対照イソタイプIgG2a、および対照イソタイプIgG1に応答してヒトTCCRを発現するBa/F3細胞の増殖を示すグラフである。
【図3】図3は、マウスIL−3(陽性対照)および抗体2686に応答してのヒトTCCRを発現するBa/F3細胞の増殖を示すグラフである。
【図4】図4は、抗体2686に応答しての、ヒトTCCR、マウスTCCRを発現するBa/F3細胞、およびいずれも発現しない対照細胞の増殖を示すグラフである。
【図5】図5は、TCCRを発現しない脾臓細胞の増殖と比較した、抗―CD3刺激に応答してTCCRを発現する脾臓細胞の増殖を示すグラフである。
【図6】図6は、TCCRを発現しないCD4+T細胞の増殖と比較した、抗−CD3刺激に応答してTCCRを発現するCD4+T細胞の増殖を示すグラフである。
【図7】図7は、TCCRを発現しないCD8+T細胞の増殖と比較した、抗−CD3刺激と応答してTCCRを発現するCD8+T細胞の増殖を示すグラフである。
【図8】図8は、ノックアウト(TCCR−/−)野生型(TCCR+/+)マウスにおけるMOG誘導EAEの臨床的進行を示すグラフである。
【図9】図9は、ノックアウト(TCCR−/−)および野生型(TCCR+/+)マウスにおけるMBP誘導EAEの臨床的進行を示すグラフである。
【図10】図10は、EAEモデルにおけるノックアウト(TCCR−/−)および野生型(TCCR+/+)マウスのための脳および脊髄切片についての平均組織学的炎症スコアを示すグラフである。
【図11】図11は,EAEモデルにおけるノックアウト(TCCR−/−)および野生型(TCCR+/+)マウスについての脳および脊髄切片についての平均組織学的脱髄スコアを示すグラフである。
【図12】図12は、EAEモデルにおけるノックアウト(TCCR−/−)および野生型(TCCR+/+)マウスについての脳および脊髄切片での最大組織学的炎症スコアを示すグラフである。
【図13】図13は、EAEモデルにおけるノックアウト(TCCR−/−)および野生型(TCCR+/+)マウスについての脳および脊髄切片での最大組織学的脱髄スコアを示すグラフである。
【図14】図14は、CFSE標識アッセイにおいてTCCRを発現しないCD4+T細胞の増殖と比較した、抗−CD3刺激に応答してのTCCRを発現するCD4+T細胞の増殖を示すグラフである。
【図15】図15は、CFSE標識アッセイにおいてTCCRを発現しないCD8+T細胞の増殖と比較して、抗−CD3刺激に応答してTCCRを発現するCD8+T細胞の増殖を示すグラフである。
【図16】図16AないしCは、中性(16A)Th1バイアシング(16B)およびTh2バイアシング(16C)条件下での、種々の時点における、IL−27での処理に応答してのIL−2の誘導を示すグラフである。
【図17】図17は、中性(17A)、T51バイアシング(17B)およびTh2バイアシング(17C)条件下での、種々の時点における、IL−27での処理に応答しての、IL−10の誘導を示すグラフである。データはL−27依存性誘導の倍数として表される。
【図18】図18AないしCは、中性(18A)、Th1バイアシング(18B)、およびTh2バイアシング(18C)条件下での、種々の時点における、IL−27での処理に応答してのSOCS−3の誘導を示すグラフである。データはL−27依存性誘導の倍数として表される。
【図19】図19は、中性、Th1バイアシング、およびTh2バイアシング条件下での、IL−27処理に応答してのCD4+細胞の増殖を示すグラフである。
【図20】図20AないしBは、抗−IL−2抗体の不存在(20A)または存在(20B)におけるIL−27および/またはIL−6処理に応答しての脾臓細胞の増殖を示すグラフである。
【図21】図21は、IL−27およびその受容体IL−27R(TCCR)のダイアグラムである。
【図22】図22は、IL−12サイトカイン群内で、サイトカインのIL−27ないしIL−6クラスターの関係を示すダイアグラムである。
【図23】図23は、ヘルパーT−細胞の分化を示すダイアグラムである。
【図24】図24は、IL−27R欠乏マウスにおけるEAEに対する過敏を示すグラフである。
【図25】図25は、野生型およびIL−27RノックアウトマウスにおけるEAE表現型の組織学的分析を示す。
【図26】図26は、T−細胞におけるIL−27の関係をテストするプロトコルの模式図である。
【図27】図27は、T−細胞発生に対するIL−27の効果のテストの結果を示す。IL−27に応答してのサイトカイン低下を、以下のサイトカイン:IFN−ガンマ、IL−2、TFN、IL−4、IL−5、IL−6、IK−10、GM−CSF、およびIL−17についての野生型対照と比較する。
【図28】図28は、IL−10、IL−27、およびIL−10およびIL−27の組み合わせにおけるIL−2およびGM−CSF生産の結果をグラフで示す。
【図29】図29は、IL−27に応答してのIL−10の強力な誘導、IL−10ノックアウトマウスにおけるEAEの重症度をグラフで示す。
【図30】図30は、EAEにおけるTH−17の役割を示すグラフである。
【図31】図31は、EAE病についてのIL−23およびTH−IL−17細胞の要件をグラフで示す。
【図32】図32は、IL−27によるTH−IL−17サイトカインの抑制をグラフで示す。
【図33】図33は、IL−27によるIL−17の抑制をグラフで示す。
【図34】図34は、IL−27によって媒介されるIL−17の抑制がIFNgが非依存性であることをグラフで示す。
【図35】図35は、IL−27によるIL−17の抑制がSTAT−1によって媒介されることをグラフで示す。
【図36】図36は、IL−17−Rノックアウトマウスの再度刺激されたリンパ球からのIL−17−RノックアウトマウスからのTH−IL−17サイトカインの分泌をグラフで示す。
【図37】図37は、IL−27−R欠乏マウスにおいてMOGまたはKLHを誘導する病気に応答してのIL−17の生産を示す。
【図38】図38は、CNSに浸潤するCD4T細胞によるIL−17発現をグラフで示す。
【図39】図39は、Tヘルパー分化に対する種々のサイトカインの関係を示すグラフである。
【図40】図40は、IL−6ノックアウトマウスにおけるEAE耐性を示すグラフである。
【図41】図41は、TH−IL−17サイトカインの誘導およびIL−6による応答をグラフで示す。
【図42】図42は、IL−27によるIL−6増殖効果の拮抗作用を示すグラフである。
【図43】図43は、IL−27およびIL−6およびTH−IL−17発生の役割を示すグラフである。
【図44】図44は、IL−27の作用の複数レベルを示すグラフである。
【図45】図45は、サイトカイン発現の変化を誘導するにおけるIL−27の役割をグラフで示す。
【特許請求の範囲】
【請求項1】
TCCRアゴニストを投与することを含む自己免疫障害を治療する方法。
【請求項2】
自己免疫障害の治療用の医薬の製造のためのTCCRアゴニストの使用。
【請求項3】
TCCRアゴニストをリンパ球に投与することを含むリンパ球においてIL−10発現を増加させる方法。
【請求項4】
TCCRアゴニストをリンパ球に投与することを含むリンパ球においてSOCS3発現を増加させる方法。
【請求項5】
前記TCCRアゴニストが抗体である請求項1ないし4いずれか一項に記載の方法または使用。
【請求項6】
前記TCCRアゴニストがモノクローナル抗体である請求項1ないし4いずれか一項に記載の方法または使用。
【請求項7】
前記モノクローナル抗体が、American Type Culture Collection受託番号ATCC PTA−6447下で寄託されたハイブリドーマ細胞株によって生産される請求項6記載の方法または使用。
【請求項8】
前記TCCRアゴニストがヒト化抗体である請求項1ないし4いずれか一項に記載の方法または使用。
【請求項9】
前記TCCRアゴニストが、TH1応答を減少させ、または抑制することができるTCCR改変体である請求項1ないし4いずれか一項に記載の方法または使用。
【請求項10】
前記TCCRアゴニストが抗体断片または単一鎖抗体である請求項1ないし4いずれか一項に記載の方法または使用。
【請求項11】
前記アゴニストがTCCR細胞外ドメインである請求項1ないし4いずれか一項に記載の方法または使用。
【請求項12】
前記アゴニストがIL−27またはその部分を含む請求項1ないし4いずれの1記載の方法または使用。
【請求項13】
前記アゴニストがIL−27改変体を含む、請求項1ないし4いずれか一項に記載の方法または使用。
【請求項14】
前記アゴニストが、TCCRおよびgp130に結合することができるp28の部分を含むIL−27改変体を含む、請求項1ないし4いずれか一項に記載の方法または使用。
【請求項15】
前記アゴニストが、TH1応答を減少または抑制することができるIL−27の部分、および異種ペプチドを含む融合蛋白質である請求項1ないし4いずれか一項に記載の方法または使用。
【請求項16】
前記異種ペプチドが抗体のFc部分を含む請求項15記載の方法または使用。
【請求項17】
前記自己免疫障害が同種異系移植片拒絶である請求項1ないし2いずれか一項に記載の方法または使用。
【請求項18】
前記自己免疫障害が自己免疫甲状腺疾患である請求項1ないし2いずれか一項に記載の方法または使用。
【請求項19】
前記自己免疫障害が自己免疫ブドウ膜網膜炎である請求項1ないし2いずれか一項に記載の方法または使用。
【請求項20】
前記自己免疫障害が巨細胞動脈炎である請求項1ないし2いずれか一項に記載の方法または使用。
【請求項21】
前記自己免疫障害が炎症性腸疾患である請求項1ないし2いずれか一項に記載の方法または使用。
【請求項22】
前記自己免疫障害がインスリン−依存性真性糖尿病である請求項1ないし2いずれか一項に記載の方法または使用。
【請求項23】
前記自己免疫疾患が多発性硬化症である請求項1ないし2いずれか一項に記載の方法または使用。
【請求項24】
前記自己免疫障害が悪性貧血である請求項1ないし2いずれか一項に記載の方法または使用。
【請求項25】
前記自己免疫障害が乾癬である請求項1ないし2いずれか一項に記載の方法または使用。
【請求項26】
前記自己免疫障害が慢性関節リウマチである請求項1ないし2いずれか一項に記載の方法または使用。
【請求項27】
前記自己免疫障害がサルコイドーシスである請求項1ないし2いずれか一項に記載の方法または使用。
【請求項28】
前記自己免疫障害が強皮症である請求項1ないし2いずれか一項に記載の方法または使用。
【請求項29】
前記自己免疫障害が全身エリテマトーデスである請求項1ないし2いずれか一項に記載の方法または使用。
【請求項30】
前記自己免疫障害がTh1応答によって少なくとも部分的に媒介される請求項1ないし2いずれか一項に記載の方法または使用。
【請求項31】
前記自己免疫障害がCD8+T−細胞増殖によって少なくとも部分的に媒介される請求項1ないし2いずれか一項に記載の方法または使用。
【請求項32】
TCCRアゴニストをスクリーニングする方法であって、
TCCRを発現する細胞を候補TCCRアゴニストと接触させる工程;
該候補TCCRアゴニストに応答してTCCR−活性化遺伝子の発現を分析する工程;そして、
該TCCR−活性化遺伝子の発現の増加を該候補TCCRアゴニストの活性と相関させる工程;
を含む、方法。
【請求項33】
前記TCCR−活性化遺伝子がIL−10、SOCS−3、または双方を含む請求項32記載の方法。
【請求項34】
前記TCCR−活性化遺伝子がSOCS−3を含む請求項32記載の方法。
【請求項35】
前記細胞がT−リンパ球である請求項32記載の方法。
【請求項36】
前記分析が定量的PCR分析を含む請求項32記載の方法。
【請求項37】
前記分析がイムノアッセイ分析を含む請求項32記載の方法。
【請求項38】
American Type Culture Collection受託番号ATCC PTA−6447の下で寄託されたハイブリドーマ細胞株によって生産されるモノクローナル抗体。
【請求項39】
American Type Culture Collection受託番号ATCC PTA−6447の下で寄託されたハイブリドーマ細胞株。
【請求項40】
IL−27またはそのアゴニストを投与することを含む免疫応答を治療または抑制する方法。
【請求項41】
前記免疫応答がTh−17細胞によって媒介される請求項1記載の方法。
【請求項42】
前記免疫応答がTh−1またはTh2媒介応答である請求項1記載の方法。
【請求項43】
前記免疫応答が過剰炎症応答である請求項1記載の方法。
【請求項44】
前記免疫応答が自己免疫応答である請求項1記載の方法。
【請求項45】
前記IL−27がIL−17生産を抑制する請求項1記載の方法。
【請求項46】
IL−27またはそのアゴニストを投与することを含むIL−17、IL−6、またはGM−CSF生産を阻害する方法。
【請求項47】
IL−6のアンタゴニストまたはその受容体を投与することを含む免疫応答を治療または抑制する方法。
【請求項48】
前記アンタゴニストが抗体、アプトマー、またはその受容体でIL−6の活性化をブロックする小分子アンタゴニストである請求項47記載の方法。
【請求項1】
TCCRアゴニストを投与することを含む自己免疫障害を治療する方法。
【請求項2】
自己免疫障害の治療用の医薬の製造のためのTCCRアゴニストの使用。
【請求項3】
TCCRアゴニストをリンパ球に投与することを含むリンパ球においてIL−10発現を増加させる方法。
【請求項4】
TCCRアゴニストをリンパ球に投与することを含むリンパ球においてSOCS3発現を増加させる方法。
【請求項5】
前記TCCRアゴニストが抗体である請求項1ないし4いずれか一項に記載の方法または使用。
【請求項6】
前記TCCRアゴニストがモノクローナル抗体である請求項1ないし4いずれか一項に記載の方法または使用。
【請求項7】
前記モノクローナル抗体が、American Type Culture Collection受託番号ATCC PTA−6447下で寄託されたハイブリドーマ細胞株によって生産される請求項6記載の方法または使用。
【請求項8】
前記TCCRアゴニストがヒト化抗体である請求項1ないし4いずれか一項に記載の方法または使用。
【請求項9】
前記TCCRアゴニストが、TH1応答を減少させ、または抑制することができるTCCR改変体である請求項1ないし4いずれか一項に記載の方法または使用。
【請求項10】
前記TCCRアゴニストが抗体断片または単一鎖抗体である請求項1ないし4いずれか一項に記載の方法または使用。
【請求項11】
前記アゴニストがTCCR細胞外ドメインである請求項1ないし4いずれか一項に記載の方法または使用。
【請求項12】
前記アゴニストがIL−27またはその部分を含む請求項1ないし4いずれの1記載の方法または使用。
【請求項13】
前記アゴニストがIL−27改変体を含む、請求項1ないし4いずれか一項に記載の方法または使用。
【請求項14】
前記アゴニストが、TCCRおよびgp130に結合することができるp28の部分を含むIL−27改変体を含む、請求項1ないし4いずれか一項に記載の方法または使用。
【請求項15】
前記アゴニストが、TH1応答を減少または抑制することができるIL−27の部分、および異種ペプチドを含む融合蛋白質である請求項1ないし4いずれか一項に記載の方法または使用。
【請求項16】
前記異種ペプチドが抗体のFc部分を含む請求項15記載の方法または使用。
【請求項17】
前記自己免疫障害が同種異系移植片拒絶である請求項1ないし2いずれか一項に記載の方法または使用。
【請求項18】
前記自己免疫障害が自己免疫甲状腺疾患である請求項1ないし2いずれか一項に記載の方法または使用。
【請求項19】
前記自己免疫障害が自己免疫ブドウ膜網膜炎である請求項1ないし2いずれか一項に記載の方法または使用。
【請求項20】
前記自己免疫障害が巨細胞動脈炎である請求項1ないし2いずれか一項に記載の方法または使用。
【請求項21】
前記自己免疫障害が炎症性腸疾患である請求項1ないし2いずれか一項に記載の方法または使用。
【請求項22】
前記自己免疫障害がインスリン−依存性真性糖尿病である請求項1ないし2いずれか一項に記載の方法または使用。
【請求項23】
前記自己免疫疾患が多発性硬化症である請求項1ないし2いずれか一項に記載の方法または使用。
【請求項24】
前記自己免疫障害が悪性貧血である請求項1ないし2いずれか一項に記載の方法または使用。
【請求項25】
前記自己免疫障害が乾癬である請求項1ないし2いずれか一項に記載の方法または使用。
【請求項26】
前記自己免疫障害が慢性関節リウマチである請求項1ないし2いずれか一項に記載の方法または使用。
【請求項27】
前記自己免疫障害がサルコイドーシスである請求項1ないし2いずれか一項に記載の方法または使用。
【請求項28】
前記自己免疫障害が強皮症である請求項1ないし2いずれか一項に記載の方法または使用。
【請求項29】
前記自己免疫障害が全身エリテマトーデスである請求項1ないし2いずれか一項に記載の方法または使用。
【請求項30】
前記自己免疫障害がTh1応答によって少なくとも部分的に媒介される請求項1ないし2いずれか一項に記載の方法または使用。
【請求項31】
前記自己免疫障害がCD8+T−細胞増殖によって少なくとも部分的に媒介される請求項1ないし2いずれか一項に記載の方法または使用。
【請求項32】
TCCRアゴニストをスクリーニングする方法であって、
TCCRを発現する細胞を候補TCCRアゴニストと接触させる工程;
該候補TCCRアゴニストに応答してTCCR−活性化遺伝子の発現を分析する工程;そして、
該TCCR−活性化遺伝子の発現の増加を該候補TCCRアゴニストの活性と相関させる工程;
を含む、方法。
【請求項33】
前記TCCR−活性化遺伝子がIL−10、SOCS−3、または双方を含む請求項32記載の方法。
【請求項34】
前記TCCR−活性化遺伝子がSOCS−3を含む請求項32記載の方法。
【請求項35】
前記細胞がT−リンパ球である請求項32記載の方法。
【請求項36】
前記分析が定量的PCR分析を含む請求項32記載の方法。
【請求項37】
前記分析がイムノアッセイ分析を含む請求項32記載の方法。
【請求項38】
American Type Culture Collection受託番号ATCC PTA−6447の下で寄託されたハイブリドーマ細胞株によって生産されるモノクローナル抗体。
【請求項39】
American Type Culture Collection受託番号ATCC PTA−6447の下で寄託されたハイブリドーマ細胞株。
【請求項40】
IL−27またはそのアゴニストを投与することを含む免疫応答を治療または抑制する方法。
【請求項41】
前記免疫応答がTh−17細胞によって媒介される請求項1記載の方法。
【請求項42】
前記免疫応答がTh−1またはTh2媒介応答である請求項1記載の方法。
【請求項43】
前記免疫応答が過剰炎症応答である請求項1記載の方法。
【請求項44】
前記免疫応答が自己免疫応答である請求項1記載の方法。
【請求項45】
前記IL−27がIL−17生産を抑制する請求項1記載の方法。
【請求項46】
IL−27またはそのアゴニストを投与することを含むIL−17、IL−6、またはGM−CSF生産を阻害する方法。
【請求項47】
IL−6のアンタゴニストまたはその受容体を投与することを含む免疫応答を治療または抑制する方法。
【請求項48】
前記アンタゴニストが抗体、アプトマー、またはその受容体でIL−6の活性化をブロックする小分子アンタゴニストである請求項47記載の方法。
【図1】

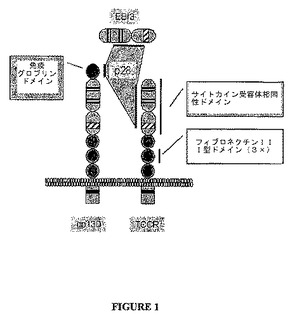
【図2】
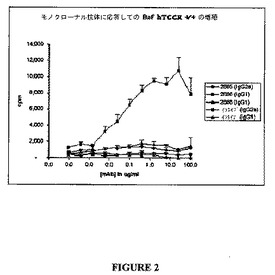
【図3】
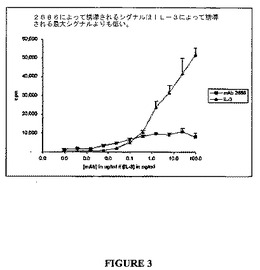
【図4】
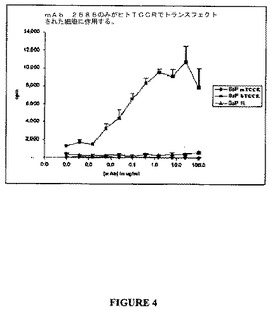
【図5】
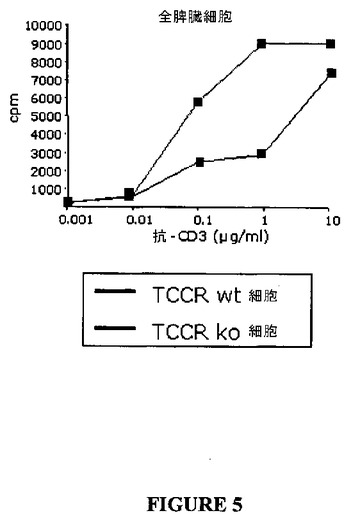
【図6】
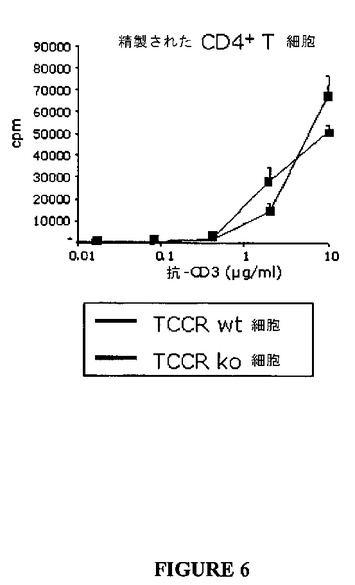
【図7】
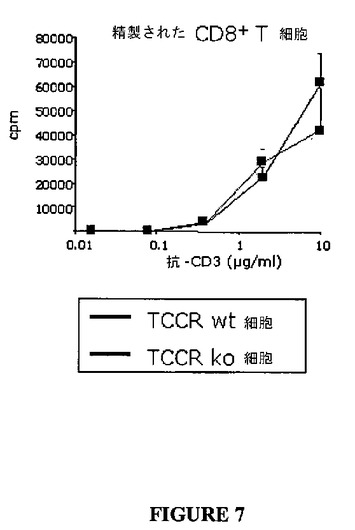
【図8】
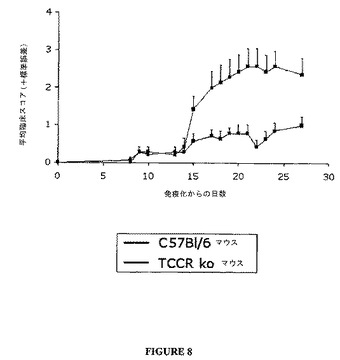
【図9】
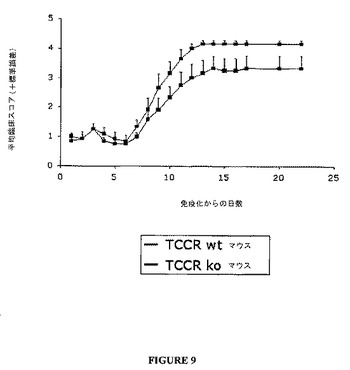
【図10】
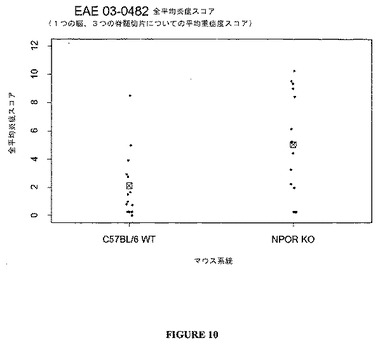
【図11】
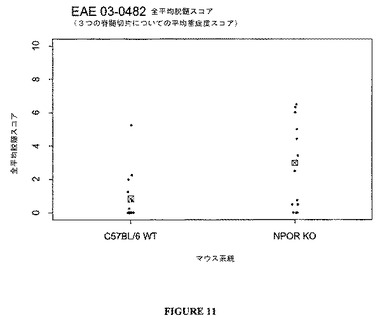
【図12】
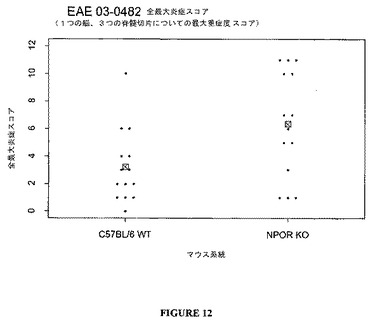
【図13】
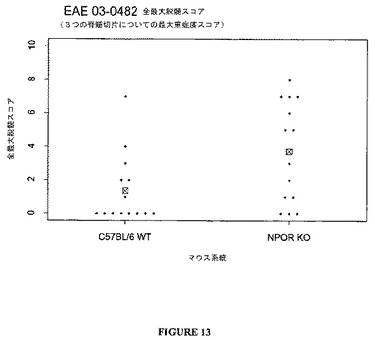
【図14】
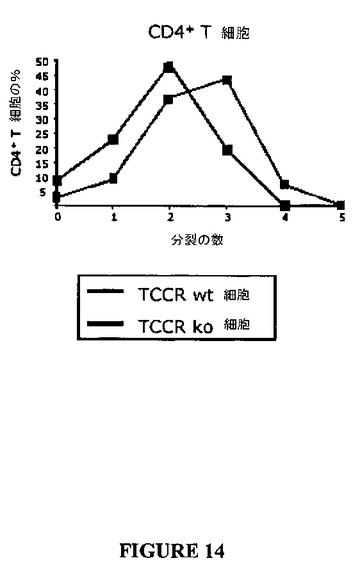
【図15】
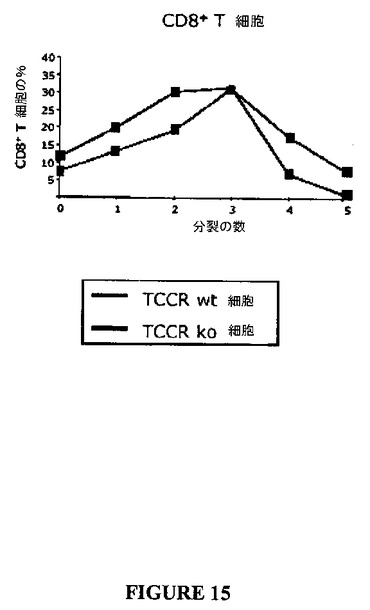
【図16】
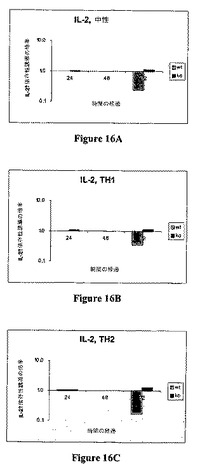
【図17】
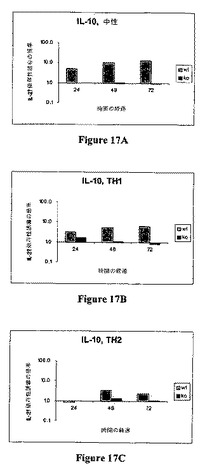
【図18】
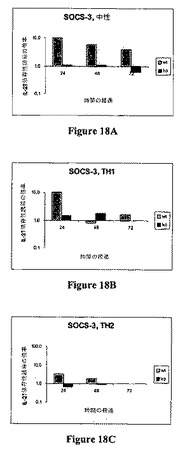
【図19】
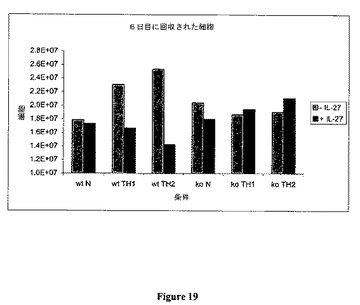
【図20】
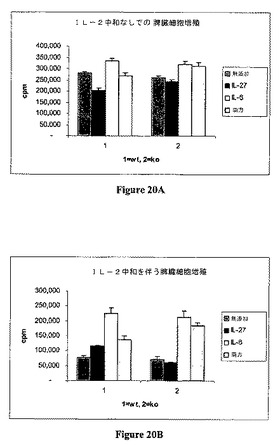
【図21】
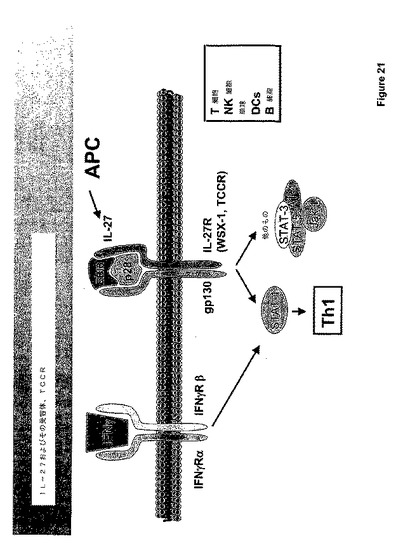
【図22】
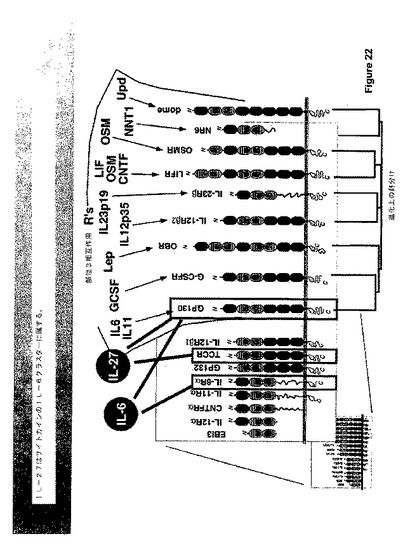
【図23】
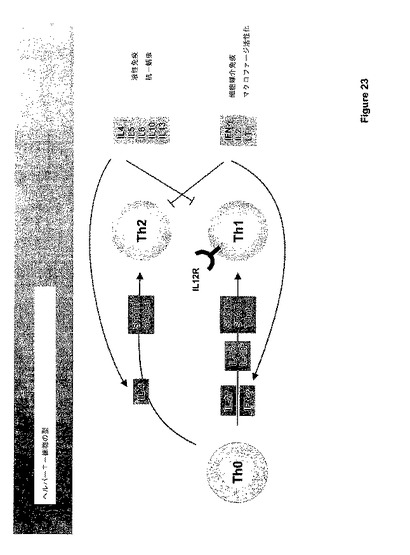
【図24】
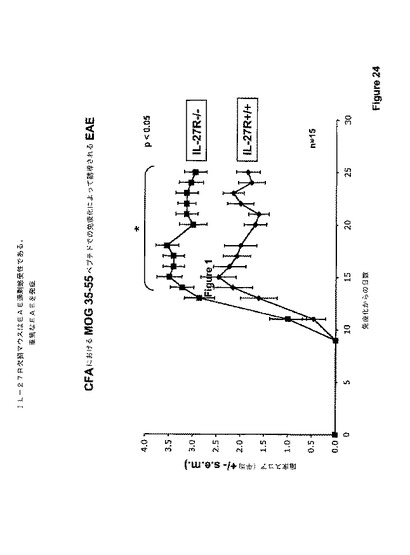
【図25】
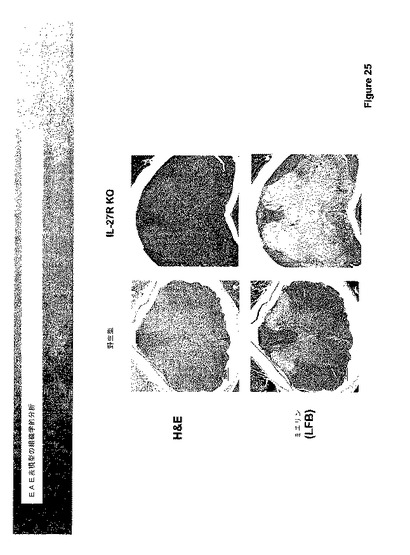
【図26】
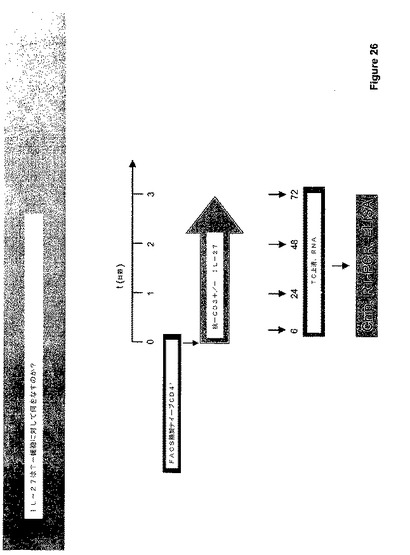
【図27】
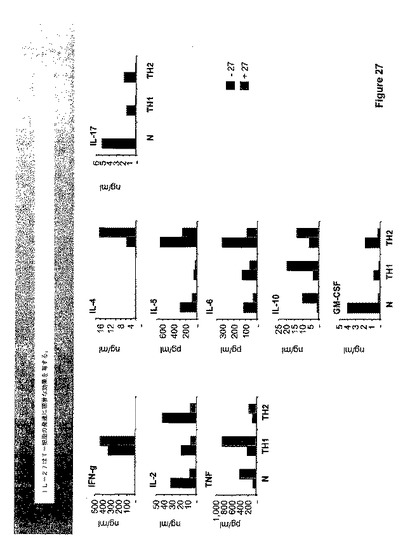
【図28】
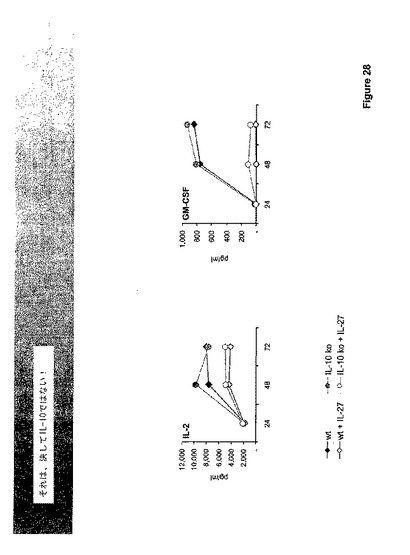
【図29】
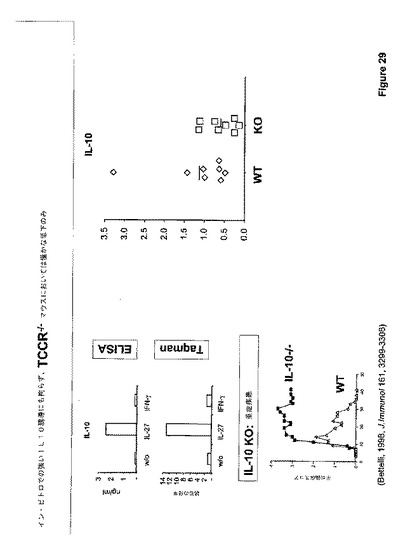
【図30】
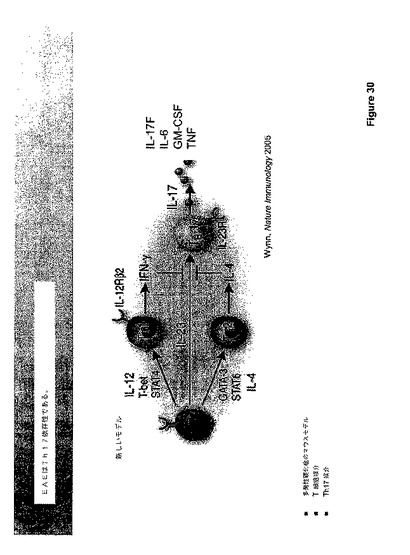
【図31】
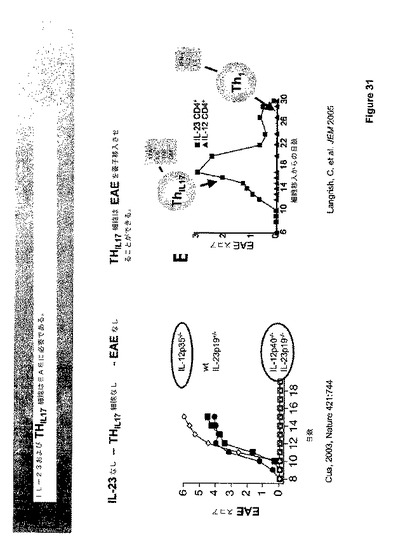
【図32】
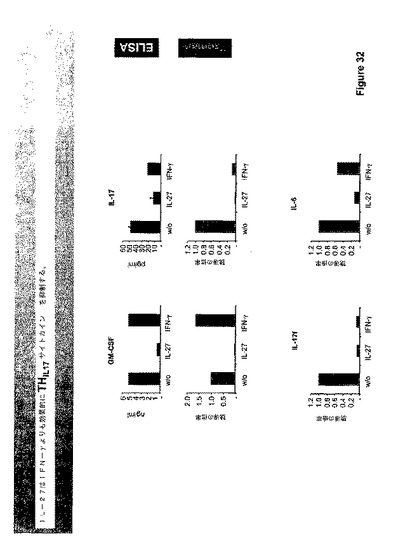
【図33】
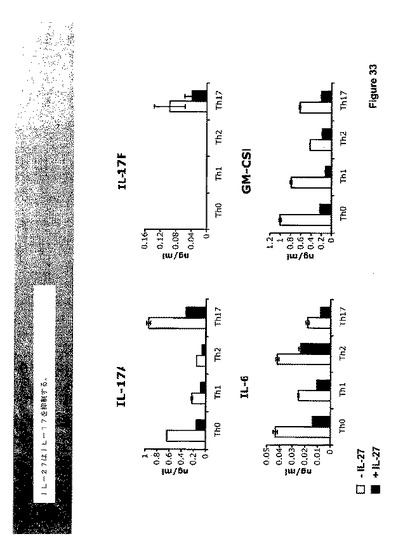
【図34】
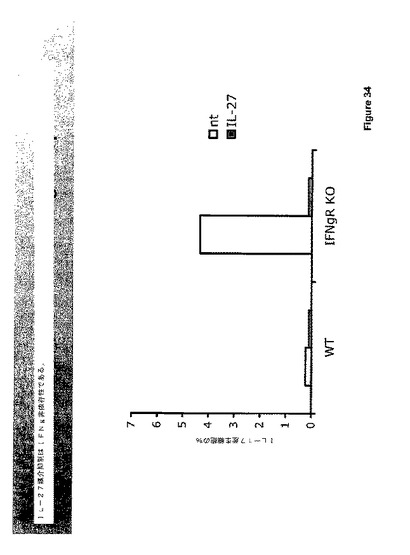
【図35】
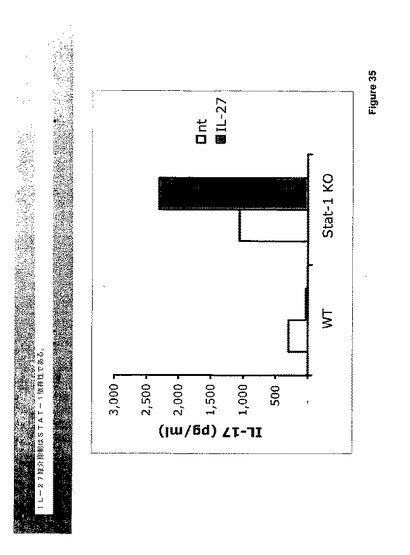
【図36】

【図37】
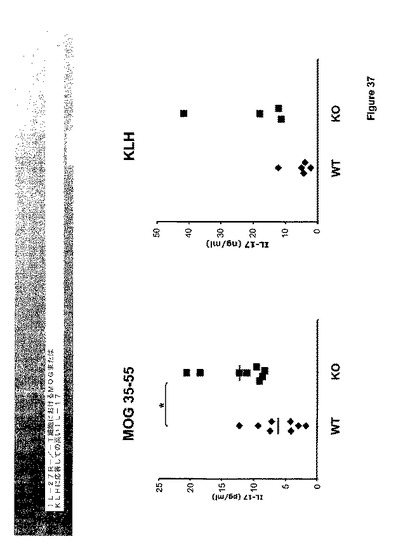
【図38】
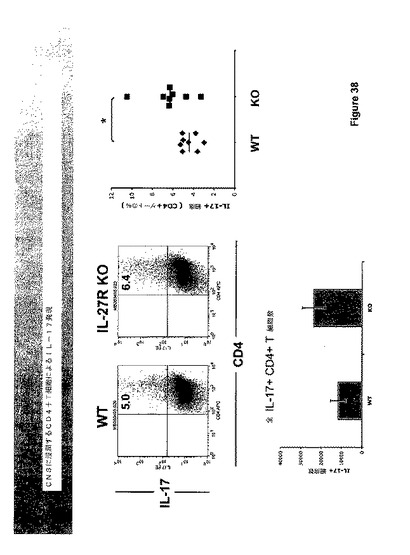
【図39】
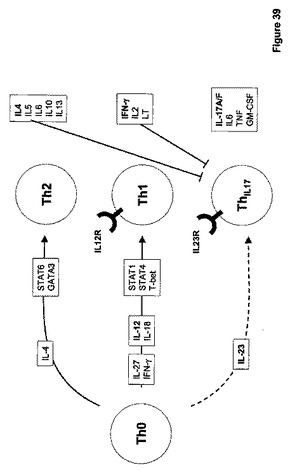
【図40】
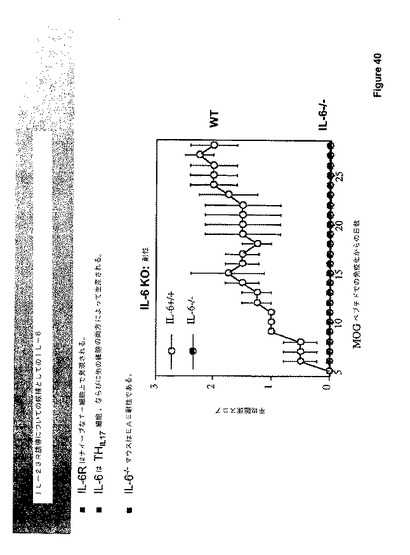
【図41】
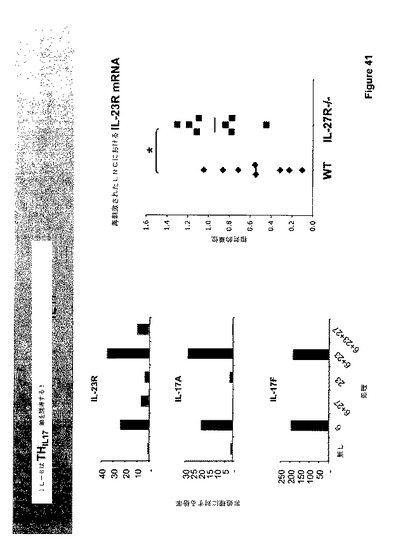
【図42】

【図43】
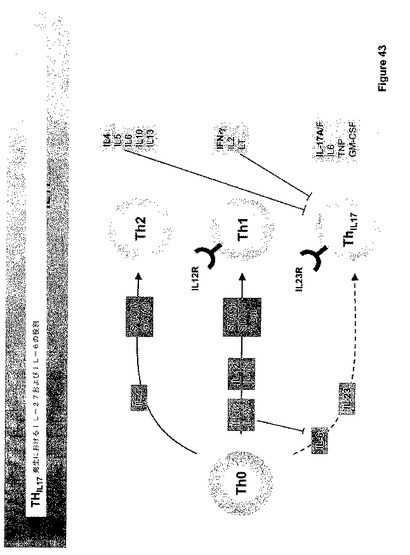
【図44】
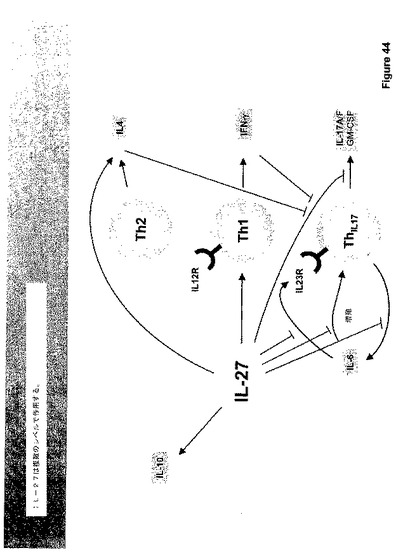
【図45】
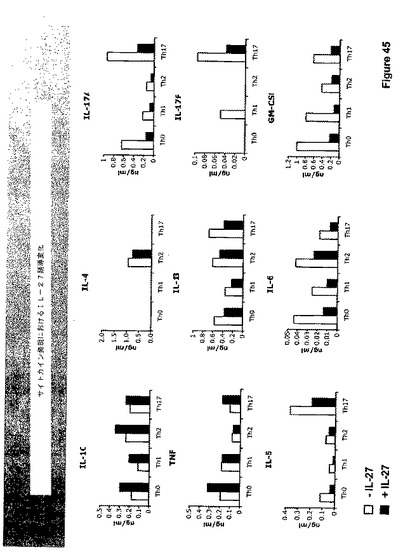
【図2】
【図3】
【図4】
【図5】
【図6】
【図7】
【図8】
【図9】
【図10】
【図11】
【図12】
【図13】
【図14】
【図15】
【図16】
【図17】
【図18】
【図19】
【図20】
【図21】
【図22】
【図23】
【図24】
【図25】
【図26】
【図27】
【図28】
【図29】
【図30】
【図31】
【図32】
【図33】
【図34】
【図35】
【図36】
【図37】
【図38】
【図39】
【図40】
【図41】
【図42】
【図43】
【図44】
【図45】
【公表番号】特表2008−524242(P2008−524242A)
【公表日】平成20年7月10日(2008.7.10)
【国際特許分類】
【出願番号】特願2007−546935(P2007−546935)
【出願日】平成17年12月16日(2005.12.16)
【国際出願番号】PCT/US2005/045603
【国際公開番号】WO2006/066088
【国際公開日】平成18年6月22日(2006.6.22)
【出願人】(596168317)ジェネンテック・インコーポレーテッド (372)
【氏名又は名称原語表記】GENENTECH,INC.
【Fターム(参考)】
【公表日】平成20年7月10日(2008.7.10)
【国際特許分類】
【出願日】平成17年12月16日(2005.12.16)
【国際出願番号】PCT/US2005/045603
【国際公開番号】WO2006/066088
【国際公開日】平成18年6月22日(2006.6.22)
【出願人】(596168317)ジェネンテック・インコーポレーテッド (372)
【氏名又は名称原語表記】GENENTECH,INC.
【Fターム(参考)】
[ Back to top ]