
自己抗原と相同性を有するエピトープに対する抗体、その調製方法およびその適用
【課題】自己抗原またはそのホモログに対するモノクローナル性自己抗体(AMAB)を得る新規の方法を提供する。
【解決手段】抗原の少なくとも1つのエピトープを生合成せず、そして抗原に特異的な抗体を産生するエピトープに対する宿主の自己寛容の欠如を利用する、遺伝的に操作された宿主動物、および、該宿主動物により産生されるAMABを提供する。さらに、細胞表面抗原に対する特異性を有するAMABの使用を含む細胞の単離方法を提供する。
【解決手段】抗原の少なくとも1つのエピトープを生合成せず、そして抗原に特異的な抗体を産生するエピトープに対する宿主の自己寛容の欠如を利用する、遺伝的に操作された宿主動物、および、該宿主動物により産生されるAMABを提供する。さらに、細胞表面抗原に対する特異性を有するAMABの使用を含む細胞の単離方法を提供する。
【発明の詳細な説明】
【技術分野】
【0001】
本発明は自己抗原またはそのホモログに対するモノクローナル性自己抗体(AMAB)を得る方法、およびこれらの抗体の細胞集団の分析における使用および細胞分離手法における使用に関する。
【背景技術】
【0002】
抗体は診断および治療の両方を含む医療用途において有用であり、そして細胞分離を包含するバイオテクノロジーの用途においても有用であることが立証されている。より一般的には、抗体を産生させ得る任意の化合物を電子顕微鏡検査および酵素免疫測定アッセイのような種々の手法を用いて、同定および位置付けをする際に抗体を使用することが、高い結合特異性を有する抗体によって容易になる。
【0003】
抗体は、鎖間のジスルフィド結合および他の分子内相互作用によって結合した重鎖および軽鎖の両ポリペプチドを含有する。個々の重鎖および軽鎖は、これらのジスルフィド結合によって対をなす。異なるクラスまたはアイソタイプの抗体のうち、3種のアイソタイプ(IgD、IgE、およびIgG)はジスルフィド結合により結合した2つの同じ重鎖/軽鎖対で構成され、そして残りの2種のアイソタイプ(IgAおよびIgM)は同じ重鎖/軽鎖対の、より複雑なポリマーで構成される。各鎖は定常部および可変部を含む。定常部はその抗体を産生する動物および抗体の特定のアイソタイプに特有であり、可変部はその抗体が結合するエピトープの構造に対応する。
【0004】
「抗原」という用語は、本明細書中では基質を意味し、これは、この基質に特異的に結合する抗体の産生を誘起し得る分子の、分子全体または分子中のドメインのいずれかである。さらに、この抗原という用語は、本明細書中では、野生型の宿主生物においては、自己認識によって抗体産生を誘起しないが、適当な遺伝学的操作を受けた宿主動物においてはこのような応答を誘起し得る基質を意味する。
【0005】
「エピトープ」という用語は、本明細書中では、Bリンパ球によって認識される、抗原上の個別の(discrete)三次元的な部位を意味する。エピトープは複雑な抗原上の免疫学的に活性な領域であり、この領域はB細胞レセプターに実際に結合し、その結果、このB細胞により産生される抗体分子によっても実際に結合される領域である。抗原は一般に少なくとも1つのエピトープを有し、そして通常1つより多いエピトープを有することが多い。タンパク質抗原のエピトープは線形または非線形であり得る。線形のエピトープとはタンパク質のアミノ酸配列中の連続したアミノ酸残基から構成されるエピトープである。線形エピトープは、立体的に折り畳まれても折り畳まれていなくてもよく、天然の三次元構造を形成し、そして免疫応答を誘起してこの抗原に対する結合特異性を有する抗体を産生する。非線形エピトープは不連続のアミノ酸残基から構成される。従って、非線形エピトープは常にある程度タンパク質が折り畳まれている必要があり、必要なアミノ酸残基を互いに近接させて、天然の三次元構造を形成し、そして免疫応答を誘起してこの抗原に対する結合特異性を有する抗体を産生する。
【0006】
「自己」という用語は、本明細書中では、通常、宿主種によって生合成される基質、または通常その宿主種が曝される基質に包含されているため、野生型宿主種のB細胞レセプターによって認識されないか、あるいはごくわずかしか認識されない抗原またはエピトープを表す。このような基質は宿主免疫系の寛容を誘導し、そしてその宿主はこの基質に対して「寛容にされた」という。
【0007】
脊椎動物の免疫系は自己抗原と外来抗原とを識別し得、外来抗原に対しては抗体を介する免疫応答を発現するが、自己抗原に対しては発現しない。抗体の応答はB細胞によって媒介される。骨髄中でB細胞が成熟する間に、整列していた配列において可変部の遺伝子再配列が起こる。このプロセスの最後には、各B細胞はイムノグロブリン重鎖をコードする単一の機能性可変部DNA配列と、イムノグロブリン軽鎖をコードする単一の機能性可変部DNA配列とを含有するようになる。このプロセスにより、各々が抗原的に単一エピトープに関与する、成熟した免疫適格性B細胞の産生に至る。まだ明らかにされていないプロセスによって、「自己」成分に対する免疫寛容は、抗原的に自己エピトープに関する可変部を有するB細胞を選択的に除去して達成される。この自己寛容は、宿主脊髄動物によって合成される抗原またはエピトープに特異的な抗体の産生を防止する。従って、宿主によって外来性と認識されるエピトープを含む抗原のみが、抗体産生のために使用され得る。
【0008】
自己エピトープと外来エピトープとが構造的に類似、あるいは「相同」である場合、宿主の免疫応答は弱くなる。従って、このようなエピトープに対して高い親和性を有する抗体を得ることは実質的に不可能である。この結果、タンパク質(例えば、NCAM、サイトカイン、およびイムノグロブリン)の高度に保存されたドメインに対する抗体を生成することは非常に困難である。なぜなら、この保存されたドメインを共有する動物は、このドメインを外来性として認識できないからである。ある種の自己免疫疾患の結果として、自己抗原に対する抗体が産生されるが、これらの抗体は非常に限られた一群の自己抗原に対して結合特異性を有するが、これらは人工的に操作し得ず、一般に結合親和性が低い。従って、自己免疫疾患の動物は、自己抗原に結合特異性を有する抗体の産生に広く有用であるわけではない。
【0009】
マウスでは、系統間のアロジェニックな差異(allogeneic difference)によ って、このようなアロジェニックな差異を生じるタンパク質に対して特異的なマウス抗マウス抗体を産生することが可能である。Kesslerら、(1979)J.Immunol.123:2772-2778; Reifお
よびAllen(1964)J. Exp. Med. 120:413-433;Marshak-Rothstein(1979)J. Immunol.122:2491-2497; ならびにOiおよびHerzenberg(1979)Molec.Immunol. 16:1005-1017を参
照のこと。最初に、T細胞表面タ ンパク質およびマウスIgD抗体に特異的なポリクロ
ーナル抗血清およびモノクローナル抗体がこの方法で得られた。しかし、これらの抗体は特定の系統のマウスの遺伝子産物を認識するに過ぎない。これらの抗体は、抗体産生宿主の自己抗原と構造的に相同ではないエピトープを認識し得るに過ぎない。さらに、これらの抗体が得られ得るエピトープは、系統間の差異およびアロタイプ系統自身の入手可能性によって制限され、従って、実際には有用性は少ない。
【0010】
細胞集団を分析および分類する多くの方法が考案されている。これらの方法には蛍光標示式細胞分取(FACS)、磁性分離(磁性ビーズに複合体化した抗体を使用する)、および特定の細胞表面「マーカー」として知られるタンパク質に対する抗体の親和性による他の方法が包含されるがこれらに限定されない。細胞の分析および分離に対するこのようなアプローチは、特に、細胞系統の決定、特定の物質を合成し得る細胞の単離、および特定の細胞タイプに伴う種々の疾患状態の治療に有用である。例えば、高度に精製した造血幹細胞は造血移植に必須であり、これにはガン患者および造血移植に関連する他の器官の移植が包含されるがこれらに限定されない。単離された細胞集団はまた、遺伝疾患、AIDS、および種々の形態のガンの治療における遺伝子治療のための重要な標的である。従って、種々の所定の細胞を実質的に純粋な形態または純粋な形態で単離する数多くの努力がなされてきた。幹細胞の単離のような場合には、このように体内で低濃度の細胞を効果的に精製するためには、幹細胞に特異的なマーカーを高い親和性で認識し、かつこのマーカーに高い親和性で結合する抗体が必要とされる。このような抗体を得ることは、ヒトとネズ
ミとで幹細胞マーカーが相同であるため、困難である。
【発明の開示】
【課題を解決するための手段】
【0011】
本発明は、自己抗原またはそのホモログに対するモノクローナル性自己抗体(AMAB)を得る新規の方法を提供する。本方法は、自己抗原内の改変形態の少なくとも1つのエピトープを生合成しないまたは合成しない遺伝的に操作された宿主動物を得ること、および少なくとも1つのエピトープに対する宿主の自己寛容の欠如を利用して抗原に特異的な抗体を産生することを包含する。本発明はまた、本方法により産生される抗体またはそのあらゆる機能性誘導体を包含する。本発明はさらに、本明細書に記載された方法により得られる、細胞表面抗原に対して特異的な抗体を利用することを含む、細胞の単離方法を包含する。
【発明を実施するための最良の形態】
【0012】
本発明は、自己抗原またはそのホモログに対して結合特異性を有するAMABを得る新規の方法を提供する。本発明は、宿主が自己として認識する抗原に対するAMABの産生の限界を克服する手段、ならびに、「標的化抗体」、すなわち公知の特定の極めて正確なエピトープに結合特異性を有するAMABを得る方法を提供する。本発明ではまた、成長および発育に不可欠と考えられる生物学的分子またはそのエピトープに対するAMABが得られる。本明細書中でさらに十分に議論されているように、標的化遺伝的置換はその分子の機能的等価物を産生させ、こうして動物が抗体を産生するに十分に成長および発育させる。
【0013】
用語「ホモログ」とは、本明細書中では、宿主動物により産生される抗原、すなわち「自己抗原」に非常に似ているため、ホモログに対する抗体の産生を排除するまたは顕著に妨げる構造を有する抗原を意味する。用語「自己抗原またはそのホモログ」とは、本明細書中では、本発明は、自己抗原に対する結合特異性を有するAMABを得ることに関するだけでなく、むしろ自己抗原に対して非常に高度な構造類似性のためあらゆる化合物に対するAMABを得ることに関すること、およびホモログは自己として十分に認識され、適切な抗体を介する免疫応答を宿主動物が生じないことを意味するために用いる。適切な抗体を介する免疫応答では、AMABが得られないか、あるいは、産生される場合、AMABが抗原に高い親和性を有さないかのいずれかのうちの1つである。
【0014】
用語「標的抗原」とは、本明細書では、宿主動物がそれに曝され、かつそれに対する免疫応答が生じるような組成物を意味する。「標的抗原エピトープ」とはAMABが結合する標的抗原上の領域である。
【0015】
用語「免疫応答」とは、本明細書中では、B細胞によるAMABの産生を意味する。これらの抗体は、病気を引き起こす因子へ免疫性を与えるような抗原変化をもたらすかどうかに関係なく特定の抗原に結合する。AMABはB細胞表面に会合し得、そしてまた自由に循環し得る。
【0016】
用語「構造的に非相同」とは、本明細書中では、遺伝的に操作した結果、宿主動物により生合成された抗原と自己抗原とを比較する記述として用いられる。一方に結合するが他方に結合しない抗体が生じ得るとき、2つの抗原は構造的に非相同である。構造的に非相同とは、2つまたはそれ以上の抗原間でいくつかの構造的な差異、おそらくわずかな差異があることを意味する。2つの抗原間の構造的に非相同な差異は、ただ1つのアミノ酸の差異、あるいはメチル基の存在または非存在のような小さなものであり得る。
【0017】
用語「機能的に等価」とは、本明細書中では、遺伝的に操作した結果、宿主動物により
生合成された標的抗原と自己抗原とを比較する記述として用いられる。多くの場合、抗体の獲得を妨害することにより、自己抗原の合成を中断させるように遺伝的に操作した宿主動物が致死的であるか、生存が低減しているか、または全身的に健康かが証明され得る。したがって、本明細書中に記載するように、遺伝的に操作することは、自己抗原の生合成を排除するために用い得、そして他の機能的に等価な抗原の産生を引き起こし得る。この機能的に等価な抗原は、AMABが得られ得る程度にまで宿主動物の生存率、全身的な健康、または免疫適格性を改良することにより、自己抗原の有害な排除を補うに十分な量の自己抗原機能をもたらす。本明細書で用いられるように、機能的に等価な抗原は、構造的に非相同であることが理解される。
【0018】
用語「生合成レパートリー」とは、本明細書中では、所定の宿主動物により生合成される化合物全体を意味する。
【0019】
用語「野生型」とは、本明細書中では、標的抗原に関しては遺伝的に操作されておらず、そしてこのような遺伝的に操作された生物系統ではない、宿主種および宿主系統の個体を意味する。
【0020】
用語「遺伝的に操作する」とは、特定の抗原または特定の一群の抗原の合成を防止または改変するように直接的に改変または排除された1つまたはそれ以上の遺伝子を有する動物を記述するのに用いられる。このような改変または削除(例えば特異的な遺伝的なノックアウトおよび置換による)は、遺伝的レベルにある。標的抗原の遺伝的操作は、一般に、本明細書中に記載されているように、抗原または一連の抗原上の少なくとも1つのエピトープに対して増強した結合特異性を有する抗体が得られ得る範囲にまで限定される。用語「遺伝的に操作された」はまた、初めから改変されている動物の子孫に適用される。
【0021】
用語「ドメイン」とは、本明細書中では、抗原のあらゆる領域または部分を意味し、これには、抗体全体、抗原全体よりも少ないあらゆる領域、または少なくとも1つのエピトープをマスクするように作用し得る追加成分によって抗原全体よりも大きくなるあらゆる領域を包含する。
【0022】
本発明は、自己抗原に対するモノクローナル性自己抗体(AMAB)を非ヒト脊椎動物宿主から得る方法に関する。
【0023】
本発明の好ましい実施態様では、以下の工程を包含する:
a)該自己抗原の少なくとも1つのエピトープを産生しないように該宿主動物のゲノムを改変する工程;
b)該自己抗原またはそのホモログを含有する組成物で該宿主動物を免疫する工程;
c)該自己抗原またはそのホモログに応答して産生される宿主細胞であって、該自己抗原またはそのホモログに対する抗体を発現する該宿主細胞を収集する工程;および
d)該収集した細胞または該細胞由来の遺伝学的材料を用いて抗体を産生する工程。
【0024】
本発明の1つの局面では、上記宿主動物は少なくとも1つのエピトープの機能的等価物を産生する。
【0025】
本発明の他の局面では、上記宿主動物は少なくとも1つのエピトープの構造的ホモログを産生する。
【0026】
本発明の1つの局面では、上記抗体は、単一の予め富化した抗原特異的B細胞および単一細胞PCRを用いて単離された、この抗体をコードする組換え遺伝子を発現することにより産生される。
【0027】
本発明の1つの実施態様では、宿主動物は特定の自己抗原を合成しないように遺伝的に操作されている。遺伝的に改変された宿主が自己抗原またはそのホモログで免疫される場合、宿主の免疫系は、自己抗原を自己として認識せず、したがってAMABが得られ得る抗体媒介免疫応答を生じ得る。このような「ノックアウト」変異体を得る方法は当該技術分野で公知であり、例えば、Mansourら、Nature336:348-352(1986)に記載されている。本明細書中で用いられるほとんどの遺伝的に操作された宿主動物では、ホモ接合変異体
を得るために、または何回も遺伝的に操作した宿主動物を得るために繁殖が用いられ得る。したがって、宿主に対する遺伝的改変は最終的に生殖系列伝達を介して得られ得る。
【0028】
遺伝的に操作されたマウスの子孫もまた本発明に包含される。トランスジェニック動物を産生する当該技術分野で公知のあらゆる他の方法も、本明細書中での使用に適切である。適切な方法には、Kitamuraら、(1991)Nature350:423-426;Shinkaiら、(1993)Science259:822-825;およびKomoriら、(1993)Science261:1171-1175の記載が包含される
がこれらに限定されない。簡単に述べると 、免疫系の自己抗原の場合では、遺伝子改変
がホモ接合の胚幹(ES)細胞を、RAG−欠失マウスのような免疫不全マウスの未分化胚芽細胞に移植し得る。多くの場合、再形成した動物は免疫欠失の表現型を欠き、そのリンパ球に遺伝子改変を含む。
【0029】
本発明の他の実施態様では、宿主動物を特定の自己抗原の合成が機能的に等価な抗原の合成により置換されるように遺伝的に操作する。従って、排除されることによって致死的になり、劇的に生存が低減することが証明される抗原に対して抗体が得られ得るか、さもなくば抗体を獲得する努力を妨げ得る。
【0030】
宿主の生合成レパートリーから除かれる自己抗原は、あらゆる化合物、ドメイン、またはそれらのエピトープであり得、通常は野生型宿主種により合成される。本発明により得られる抗体は、タンパク質、ペプチド、炭水化物、脂質、核酸、酵素補因子、あるいはあらゆる天然に生じる凝集物またはそれらの共有結合複合物、あるいはそれらのあらゆるリン酸化種または硫酸化種を含むがこれらに限定されないあらゆる抗原に対する。本発明のこれらの実施態様では、宿主は抗原の生合成が排除または改変されるようなあらゆる方法で遺伝的に操作され得る。例えば、抗原の生合成に包含される特定の酵素の発現をノックアウトすることで、抗原の非産生または機能的に等価な抗原の産生が生じ得る。例えば、自己抗原またはそのホモログの生合成は、この抗原またはそのホモログを産生する代謝経路に影響を与える酵素をコードする遺伝子の変異により排除される。このような酵素には重合化に含まれる酵素が含まれるがそれらに限定されず、糖タンパク質の合成におけるような炭水化物、リン酸基、脂質、硫黄含有基のタンパク質への付加が排除される結果、タンパク質に共有結合する化合物の構造を除去または変化させる。従って、特定の自己糖タンパク質または糖タンパク質上の炭水化物構造に結合特性を有する抗体が、本発明の方法により得られ得る。また、このような酵素には、自己抗原またはそのホモログを産生する代謝経路に影響を与える酵素が包含される。
【0031】
抗原の産生を排除または変化させるより全体的な方法が利用され得る。これらの方法には、特定の遺伝子または遺伝子ファミリーの合成を排除すること、および特定の細胞タイプを除去することが包含されるがこれらに限定されない。適切な遺伝子または遺伝子ファミリーの例には、ステロイドまたはサイトカインのような特定の因子の存在により調節されるものあるいは細胞タイプ特異的に発現される遺伝子が含まれる。これらの細胞タイプの1例は、褐色脂肪細胞である。
【0032】
本発明の好ましい実施態様では、抗原はタンパク質である。タンパク質についてのノックアウト変異には、完全なタンパク質の合成を妨げること(完全な遺伝子を除去すること
によって、あるいは転写制御因子または翻訳制御因子を改変することによって)、または唯一の抗原性ドメインの合成を排除することが包含されるが、残りのタンパク質部分は正常に合成される。当該技術分野で公知のあらゆる形態の置換が本明細書中での使用に適切であるが、タンパク質抗原の場合では、自己抗原と機能的に等価な抗原との置換は少なくとも3つの異なるタイプの置換を伴う。
【0033】
第1に、自己抗原をコードする遺伝子が、自己抗原をコードする遺伝子に由来する組換え遺伝子により置換され得る。タンパク質をコードする遺伝子は、抗原領域が欠失し、改変アミノ酸配列により置換されるように改変されるか、または新規アミノ酸配列の追加によりマスクされるように改変される。従って、自己タンパク質抗原の改変における遺伝的に操作された抗原の除去は、単一のアミノ酸の追加、排除、または置換のような小さなことであり得る。重要なことは、自己タンパク質抗原の構造におけるこのような小さな変化により、エピトープに対して非常に正確なAMABが自己抗原の機能を大きく妨害することなく得られ得る。適切なドメインにおける単一のアミノ酸の変化の結果、その単一のアミノ酸を含むエピトープに対するAMABを生じる。
【0034】
第2に、自己抗原をコードする遺伝子が、コードするタンパク質が機能的に等価だが少なくとも一部は抗原的に非等価となるように、宿主種に関連のある生物から得られる相同なタンパク質をコードする遺伝子により置換され得る。非常に関連のある種は、通常同じタンパク質に高度な配列相同性(典型的には90%より大きい)を有する。従ってこの方法は、抗原の発現が生物の健康に重要である場合に本発明を実施するには理想的であり、そして少数のエピトープに結合特異性を有するAMABが望ましい。この方法はまた、自己抗原をコードする遺伝子の1つの領域のみを、関連する種に由来する同じ抗原をコードする遺伝子の対応する領域に置換することにより実施され得る。
【0035】
第3に、自己抗原をコードする遺伝子が、同じかまたは類似の機能を有するが構造的に非相同であることが知られているタンパク質をコードする遺伝子により置換され得る。この方法は、抗原の発現が生物の健康に重要であり、特定のエピトープに結合特異性を有するAMABが必要ではない場合に特に有用である。
【0036】
本発明により得られるAMABが結合特異性を有し得るタンパク質抗原の例には、細胞表面抗原が包含されるが、これに限定されない。細胞表面抗原には、細胞接着分子、MHCクラスI分子およびクラスII分子、インテグリン、サイトカイン、セレクチン、サイトカインレセプター、およびイムノグロブリン(例えばIgD抗体)が包含されるがこれらに限定されない。
【0037】
本発明は、魚類、は虫類、両生類、鳥類、および哺乳類を包含する任意の、非ヒト脊椎動物宿主を用いて実施され得る。しかし、宿主はほとんどの場合、哺乳類であり、通常、齧歯類、兎類、霊長類、食肉類、奇蹄類、および偶蹄類が包含されるがこれらに限定されない種類に属する。好ましくは、宿主はネズミであり、より好ましくはマウスである。
【0038】
AMABを作製する方法は当該技術分野で公知であり、本明細書中では詳細に述べない。モノクローナル抗体産生の分野で公知のあらゆる方法を、ここで使用し得る。このような方法には、所望の特異性を有する細胞表面抗体によりB細胞を分離すること、軽鎖および重鎖の可変部を発現するDNAをクローニングすること、および適切な宿主細胞中で組換え遺伝子を発現させることが包含されるがこれらに限定されない。標準モノクローナル抗体産生手法を使用し得、ここで、AMABは不死化抗体産生ハイブリドーマ細胞から得られる。これらのハイブリドーマは、Bリンパ球、好ましくは免疫された宿主の脾臓から単離されたBリンパ球と、それと適合する不死化細胞、好ましくはB細胞ミエローマとを融合することにより産生され得る。
【0039】
本発明はさらに、本明細書中に記載された方法によって得られるAMABを含有する物質の組成物をも包含する。本明細書中で用いるように、「AMAB」または「抗体」という用語は、抗体全体、およびその機能性部分を含む抗体フラグメントを包含する。「AMAB」という用語は、抗体全体が結合特異性を有しているエピトープへ結合するに充分な、軽鎖可変部および/または重鎖可変部の部分から構成される、あらゆる単一特異性化合物を包含する。上記フラグメントは少なくとも1つの重鎖イムノグロブリンポリペプチドまたは軽鎖イムノグロブリンポリペプチドの可変部を含み得、そしてFabフラグメント、Fab2フラグメント、およびFvフラグメントを包含するがこれらに限定されない。
【0040】
さらに、単一特異性ドメインは、当該技術分野で公知のあらゆる方法により、他の適切な分子に付着し得る。この付着は、例えば、化学的あるいは遺伝学的操作によって行われ得る。AMABは、当該技術分野で公知のあらゆる組換え手段によって産生され得る。このような組換えAMABには、細菌において産生されたフラグメント、そしてその定常部の大部分がヒト抗体の定常部によって置換されたAMABが包含されるがこれらに限定されない。さらに、このような「ヒト化」AMABは、本明細書中に記載の遺伝的に操作された宿主脊椎動物を、野生型Ig遺伝子座の代わりに機能性ヒトIg遺伝子座を発現する適合するトランスジェニック動物で作り出すことによって得られ得る。ヒトIg遺伝子座を発現するトランスジェニック動物に関しては、WIPO公開公報WO91/10741号、およびRajewskyらのDEP4228162.8を参照のこと。うまく交配すると、特別に自己抗原を発現しない
が、ヒト化Igタンパク質を発現する宿主動物が得られ得る。このような動物が免疫されると、この動物は特定の自己抗原に対するヒト化AMABまたは部分ヒト化AMABを産生する。このようなヒト化AMABは、治療指示薬(therapeutic indication)に使用するのに好ましい。
【0041】
AMABは他の化合物と複合体化し得る。この化合物には、酵素、リポーター酵素、リポソーム、磁性ビーズ、コロイド磁性ビーズ、固体キャリア、毒素、ハプテン、蛍光色素、蛍光粒子、金属化合物、放射性の化合物、薬剤、またはハプテンが包含されるがこれらに限定されない。AMABに複合体化し得る酵素には、アルカリホスファターゼ、ペルオキシダーゼ、ウレアーゼ、およびβ−ガラクトシダーゼが包含されるがこれらに限定されない。AMABに複合体化し得る蛍光色素には、フルオレセインイソチオシアネート、テトラメチルローダミンイソチオシアネート、フィコエリトリン、アロフィコシアニン類(allophycocyanins)、およびテキサスレッドが包含されるがこれらに限定されない。抗体に複合体化し得るその他の蛍光色素に関しては、Haugland,R.P. Molecular Probes: Handbook of Fluorescent Probes and Research Chemicals(1992-1994)を参照のこと。AMABに複合体化し得る金属化合物には、フェリチン、金コロイド、および特にコロイド超常磁性粒子(ビーズ)が包含されるが、これらに限定されない。AMABに複合体化し得るハプテンには、ビオチン、ジゴキシゲニン、オキサゾロン(oxazalone)、およびニト
ロフェノールが包含されるが、これらに限定されない。AMABに複合体化し得るかまたは導入し得る放射性化合物は当該技術分野で公知であり、放射性同位元素のテクネチウム99m(99TC)、125I、および任意の放射性核(14C、3Hおよび35Sが包含されるが、これらに限定されない)を含有するアミノ酸が包含されるが、これらに限定されない
。当該技術分野で公知の任意の方法によってタンパク質に複合体化し得る当該技術分野で公知のあらゆる薬剤が、本発明における使用に適している。このような薬剤は、この薬剤を標的分子へ高度に特異的に送達するために、AMABに複合体化され得る。
【0042】
本発明はさらにAMABを使用する方法を提供し、この方法にはイムノアッセイおよび細胞分離が包含されるがこれらに限定されない。適切なイムノアッセイは当該技術分野で公知であり、本明細書中で詳細に記述する必要はない。イムノアッセイには、ELISAおよびRIAが包含されるがこれらに限定されない。細胞の分離方法には、分子の分泌に
基づく分離方法および細胞表面分子に基づく分離方法が包含されるがこれらに限定されない。分子の分泌に基づく細胞の分離方法は、米国特許出願第07/965,934号および国際出願番号PCT/US93/10126に記載されている。特異的な細胞表面マーカーに基づいて細胞を分離する方法は一般に、本明細書中に記載される方法によってAMABを得る工程、AMABを異種細胞集団に接触させる工程、および細胞表面抗原に対するAMABの親和性に基づく細胞分離手法を行う工程を包含する。まず特定の細胞表面抗原を有する細胞を除去するか、あるいは特定の細胞系統を除去することによる、当該技術分野で公知のあらゆる方法が、細胞の分離または単離のために用いられ得る。
【0043】
AMABは、特定の細胞系統および/または特定の分化段階に関連するマーカーを同定するために特に有用である。AMABは固体支持体に直接または間接に付着し分離を可能とする。使用する分離手法は、採集する画分の生存率を最大限に保持するような方法であるべきである。分離するために、異なる効果を有する種々の手法が用いられ得る。このような分離では、マーカーを持たないで存在する細胞全体の、10%まで、通常約5%以下、好ましくは約1%以下が、保持される細胞集団と共に残存し得る。用いられる所定の手法は、分離効率、方法の細胞毒性、実行の容易さおよび速さ、そして精巧な器具および/または技術の必要性に依存する。
【0044】
細胞分離の手順には、コロイド磁性粒子に結合した抗体を用いる磁性分離、アフィニティークロマトグラフィー、およびモノクローナル抗体に結合した細胞毒性試薬、または当該技術分野で公知の任意の抗体依存性分離手法と共に用いられる細胞毒性試薬が包含されるがこれらに限定されない。さらに、細胞は固体マトリックス(例えばプレート)に付着した抗体を用いる「パンニング」によって分離し得る。蛍光標示式細胞分取器(FACS)もまた使用し得る。これは、異なる程度の精巧さを有し得、複数のカラーチャネル、鋭角(lowangle)および鈍 角(obtuse)光散乱検出チャネル、およびインピーダンスチャネルが包含されるがこれらに限定されない。当該技術分野で公知のあらゆる抗体依存性分離手法を、抗体の親和性よりもむしろ細胞の物理的性質に依存するポジティブおよびネガティブ両方の分離手法と共に使用し得る。このような細胞の物理的性質に依存する分離手法には、溶離および密度勾配遠心分離法が包含されるがこれらに限定されない。
【0045】
細胞の分離方法は、Dynal(Oslo、Norway)、Cellpro(Seattle)、またはAdvanced Magnetics(Boston)から市販されている。例えば、モノクローナル性自己抗体は、DynalM
450のような磁性ポリスチレン粒子または同様の磁性粒子に 直接結合し得、そして例え
ば、細胞分離に使用され得る。あるいは、抗体はビオチン化され得、あるいはジゴキシゲニンと結合し得、そしてアビジンコーティングまたは抗ジゴキシゲニンコーティングされた、SEPARATELC(Cellpro)のようなアフィニティーカラムと共に使用し得る。しかし、
好ましい実施態様では、モノクローナル性自己抗体は、例えば多糖類による有機コーティングを有するコロイド超常磁性粒子と組み合わせて使用される。Miltenyiら、(1990)Cytometry11:231-238を参照のこと。これらの粒子の大きさは10nm〜200nm、好ま
しくは40nmと100nmとの間であり得、そして自己抗体に直接結合していてもよく、または抗イムノグロブリン、アビジン、または抗ハプテン特異的マイクロビーズと組み合わせて使用されてもよい。例えば、抗体をハプテンにより修飾し、そしてコロイド超常磁性粒子がハプテン修飾物を認識する。多糖類コーティングされた超常磁性粒子はMiltenyiBiotec GmbH、Germanyから市販されてい る。また、抗体結合コロイド超常磁性粒子は、目的の細胞に結合し、この細胞が高勾配磁性分離を用いて単離される。また、細胞の亜群が上記抗体に結合し、さらにこの抗体は、抗イムノグロブリン特異的コロイド超常磁性粒子により認識される。
【0046】
用い得る手順の1つは、まず細胞を低温(通常約4℃)で短時間、所定の細胞表面抗原に特異的な飽和レベルのAMABと共にインキュベートし、次いでその細胞をPBSおよ
びウシ胎児血清(FCS)緩衝剤(cushion)で洗浄する方法である。次いで、細胞を上
記のような緩衝液に懸濁し、そして、AMABまたはAMAB抗原複合体に特異的な種々のタンパク質を用いて、特定の抗原決定基に対するAMABに基づいて細胞を分離する。
【0047】
AMABは、特定の細胞タイプの分離を容易にするマーカーと複合体化され得る。このマーカーには、磁性ビーズ(これは直接分離される)、ビオチン(これは支持体に結合したアビジンまたはストレプトアビジンで除去し得る)、ジゴキシゲニン(これは抗ジゴキシゲニン抗体で検出される)、蛍光色素(これはFACSに使用され得る)などが包含されるがこれらに限定されない。残留する細胞の生存率を過度に損なわない限り、いかなる手法も使用し得る。
【0048】
細胞表面抗原を含む細胞(例えば抗原特異的B細胞)の実質的な富化(一般に少なくとも約50%、好ましくは少なくとも約70%)後、細胞は、FACSまたは当該技術分野で公知の他の方法によって分離され得る。種々の方法による多色分析を使用し得、これらにはFACSおよび蛍光顕微鏡が包含されるがこれらに限定されない。この細胞は、特定の抗原に対する染色の程度に基づいて分離され得る。
【0049】
本発明はさらに、本明細書中に記載された方法によって、レセプターである自己抗原に対するAMABを得ること、およびこのようなAMABを薬剤として使用することを包含し、ここでAMABはレセプターアゴニストまたはアンタゴニストとしての有効性を示す。
【0050】
以下の実施例により本発明を説明するが、限定はしない。
【実施例】
【0051】
(実施例1)
(遺伝子の標的化)
Cδ遺伝子の標的化を、RoesおよびRajewsky、J. Exp. Med. 177:45-55(1993);な
らびにRoesおよびRajewsky、Int.Immunol. 3:1367(1991)に記載さ れるように行った。簡単に述べると、全部で108個のE14−1ES細胞を、Cδ1エキソンの大部分を置換するように、そしてこのエキソンに存在する制限部位を充たすことによって、Cδ3中にフレームシフト変異を挿入するように設計された標的化ベクターを用いてトランスフェクトした。生殖系列へ変異を導入した結果、両方のδ鎖Igドメインエキソンを機能的に不活性化した。これは、L鎖のμと競合し得る、かつ分泌され得る短縮型δH鎖の発現の可能性を除くのに重要であると考えられた。マウス生殖系列におけるフレームシフトの存在が、Cδ3中のHindIII部位を充たした結果生じるNheI制限部位により示さ
れた。Cδ2エキソンは、非機能性スプライシングアクセプターに帰因する偽エキソンである。
【0052】
選択で生き残ったコロニーをPCRで分析し、そして陽性クローンをさらにサザンブロッティングにより分析して、標的化された遺伝子座の構造を確認した。相同組換え体は、二重耐性クローンの1/17またはG418耐性クローンの1/103の頻度で得られた。野生型Cδ遺伝子座および変異型Cδ遺伝子座の制限マップ、ならびに5つの別個の組換え体候補およびコントロール細胞に由来するHindIII消化したゲノムDNAの制限
分析が示された。相同組換えの結果、HindIII部位の欠損を生じるCδ3中のフレー
ムシフト変異の存在または 欠如に依って、3.8kb生殖系列バンドに加えて、4.4
kbまたは6.0kbのバンドを生じた。10個の相同組換え体クローンのうちの9個が6.0kbのフラグメントを示し、組換えの開始点がCδ3エキソンの3’に位置することが示された。このように、Cδ1エキソンおよびCδ3エキソンは、これらのクローン中で機能しなかった。1つのクローンは、Cδ3エキソン中にHindIII制限部位を保
持し、4.4kbの診断用制限フラグメントを生じた。標的化された遺伝子座の構造を、種々の他のプローブおよび制限酵素を用いて確認した。サザンブロット分析を、Sambrook、Fritsch、およびManiatis、Molecular Cloning, A Laboratory Manual、第2版、Cold Spring Harbor Laboratory、NY(1989)に記載のように行った。
【0053】
(実施例2)
(IgD欠損マウスの作製)
IgD欠損マウスの作製を、RoesおよびRajewsky(1993)、J. Exp. Med. 177:45-55
に記載のように行った。簡単に述べると、Cδ遺伝子不活性化の方法および陽性クローンのスクリーニング手順は、実施例1に記載のようにした。標的化ES細胞クローンを、C57BL/6マウスから単離した胚盤胞に注入し、そして(C57BL/6×BALB/c)の里親(foster)に移した。雄のキメラの子を、δT変異の生殖系列伝達のためにC57BL/6の雌とつがいにした。ES細胞由来の子を毛色により識別し、そしてδTと呼ばれる変異の存在についてサザンブロッティングによるかまたは表現型をフローサイトメトリーにより分析した。ホモ接合変異マウス(δT/δT)を、ヘテロ接合の子の異種交配により得た。
【0054】
(実施例3)
(推定(δT/δT)変異マウスのノーザンブロット分析)
推定(δT/δT)変異マウスのノーザンブロット分析を、RoesおよびRajewsky(1993)による記載のように行った。簡単に述べると、δT変異の結果、H鎖のIgドメインをコードする両方のエキソンが機能的に不活性化される。しかし、トランスメンブレンエキソンおよびヒンジ部エキソンは、完全なままであり、かつ潜在的に機能性を維持する。CμおよびCδ遺伝子の両方を含有する前駆体RNAの異常なスプライシングの結果、Cμ遺伝子の細胞外ドメインならびにCδのトランスメンブレン部分および細胞質部分をコードするかなりの量のキメラIg転写物が産生される可能性を除くために、ホモ接合変異(δT/δT)マウスおよび野生型マウスの脾臓から単離したポリ(A)+RNAを、ノー
ザンブロッティングで分析した。CδトランスメンブレンエキソンにスプライシングされたCμエキソンを含むmRNAは、2.4kb(μs)または2.7kb(μm)の正常なCμ転写物よりも大きい。なぜなら、δメッセージの3’非翻訳領域は、μメッセージのものよりも600bp長いからである。
【0055】
Cδトランスメンブレン特異的プローブを用いるホモ接合変異マウスの脾臓ポリ(A)+RNAのハイブリダイゼーションでは、4.8kb、4.0kb、3.8kb、および
3.0kbのバンドが示された。しかし、これらのバンドはいずれも、Cμ特異的プローブとハイブリダイズしなかった。2つのプローブの検出限界は、標準プラスミドDNAへのハイブリダイゼーションから得られたシグナルにより判断されるように、2つの因子(Cμでは1.2×106コピーおよびδmプローブでは0.6×106コピー)の限りでは類似していた。Cμプローブは、mRNAおよび標準プラスミドに完全に適合する1k
bのcDNAフラグメントであるが、δmプローブはδm1/m2イントロンを含むため、標準プラスミドにおける700bp以外は480bpの長さを越えるmRNAとハイブリダイズするにすぎない。従って、δmエキソンにスプライシングされたCμエキソンを示すmRNAは、δmプローブで検出される場合は、Cμプローブでも表れるべきである。なぜなら、3.0kb、3.8kb、4.0kb、および4.8kbのバンドは明らかにδmプローブの検出限界を上回るため、それらがCμ配列を含む場合は、さらにCμプローブでも検出されるべきだからである。しかし、これはその場合ではない。
【0056】
さらに、Cδ3エキソンに対して特異的なneo+遺伝子およびneo+プロー ブを用いた同じブロッティングの一連のハイブリダイゼーションにより、3.8kb、4.0kb、および4.8kbのバンドも、neo+遺伝子由来の配列を含むことが示され、それら
が異常なスプライシング産物を表すことが示された。3.0kbのバンドは、実施例1に記載のフレームシフト変異のために、標的化対立遺伝子中で機能しないCδ3エキソンに対して特異的なプローブとハイブリダイズした。さらに、neo+プローブを用いて、2
.4kbのmRNA(これはδmプローブとハイブリダイズしなかった)もまた検出された。Cδm特異的プローブとハイブリダイズする1.6kbおよび2.0kbの少量のmRNAが、正常マウスおよび変異マウスの両方に由来する10μgのポリ(A)+RNA
中で検出可能である。しかし、これらのmRNAは、正常のCμメッセージよりも小さく、従って、機能性Ig分子をコードしそうにない。
【0057】
潜在的な機能性キメラμ/δ分子を表すmRNA種は、10μg程度のδT/δTマウスの脾臓ポリ(A)+RNAを用いるノーザンブロットで検出し得なかった。δH鎖をコ
ードするmRNAは、Cδm特異的プローブを用いて、300ng程度の少量の正常マウスの脾臓ポリ(A)+RNAで検出され得るので、μの細胞外ドメインおよびδのトラン
スメンブレン部分をコードする推定mRNAは、それが少しでも存在するなら、正常マウスのδH鎖メッセージよりも少なくとも30倍より少ない。従って、IgD様分子をコードし得る可能性のあるmRNAは、ノーザンブロット分析によりホモ接合変異マウス(δT/δT)中で検出不可能である。
【0058】
(実施例4)
(マウス−抗マウス−IgDモノクローナル抗体の産生)
IgD欠損マウスを、実施例1および2に記載のように遺伝子の標的化により作製した。こうして得られた1匹の動物を、ミョウバン中に沈降したIgDクラス(267.7δ「a」アロタイプ)のマウスモノクローナル抗体で、腹腔内(i.p.)に免疫した。
【0059】
6週間後、この動物に、「b」アロタイプの可溶性B1−8で、腹腔内に追加免疫を行い、両アロタイプを認識するモノクローナル抗体を得た。
【0060】
3日後、免疫したマウスの脾臓細胞を、標準PEG融合プロトコルを用いて、X63 Ag8.6.5.3と融合した。ハイブリッドを、8枚の96ウェルプレートに、直接クローニングし、そしてHAT選択培地を用いて選択した。得られたハイブリドーマを、Blー8(IgD)、BSA(ウシ血清アルブミン)、267.7(IgD)、R33−24−12(抗IgM)でコートしたプレートを用いて、ELISAによる抗IgD抗体の産生についてスクリーニングを行い、そして、R33−18−10.1−ビオチン抗体(ラット抗マウスκ)およびR33−60−ビオチン抗体(抗IgM)を用いて発色させた。17個のクローンがB1−8(IgDb)への反応性を示し、これらのうち13個のク
ローンが267.7(IgDa)への反応性を示し、これらのうちの1個がIgMクラス のものであった。他の2つのクローンは、IgMへの反応性を示したが、IgDには反応性を示さなかった。さらに、ハイブリドーマを、相対結合親和性およびアイソタイプについて特徴付けた。試験した21個のクローンのうち15個は、IgGlアイソタイプを示した。さらに、このクローンを、ビオチン化抗マウスIgGlとストレプトアビジン−フィコエリトリンとを用いてトリプシン処理およびトリプシン未処理のマウス脾臓細胞への結合を特徴付けた。トリプシンは表面のIgDをカットし、そしてIgD分子上のエピトープを制限し得る。
【0061】
最も親和性が高くかつIgGlアイソタイプの3個のクローン(δ1.2、δ1.3、およびδ3.5)を、さらなる実験のために用いた。これらのクローンにより産生された数mgの精製AMABを、標準の方法によりローラーボトルを用いて産生した。16個のクローンを、培養上清でマウス脾臓細胞をインキュベートすることによる染色アッセイで試験した。結合した抗IgDAMABを、ビオチン化抗マウスIgGlを用いて検出し、そしてストレプトアビジン−PEにより発色させた。
【0062】
(実施例5)
(B細胞富化領域でのマウス脾臓組織のIgDAMAB染色)
C57BL/6マウス由来の凍結脾臓切片を、実施例4に記載のように調製したビオチン化マウス抗マウスIgD AMAB δ1.3を用いて染色し、そして供給者の指示に従ってPierceNHSビオチンを用いてビオチン化した。染色パターンを、製造者の指示に従って、SigmaのAEC染色キットを用い、ストレプトア ビジンに結合したペルオキシダーゼを用いて発色させた。得られた結果を図1に示す。染色パターンは、ポリクローナルヤギ抗マウスIgD抗血清について見られるパターンと一致し、本発明により得られたIgDAMABが所望の自己抗原に対して結合特異性を有することを示す。
【0063】
(実施例6)
(フルオレセインに対する抗体の複合体化および種々の濃度下での脾臓細胞の染色)
実施例4からそれぞれ得られた0.5mgの精製AMAB(δ1.3およびδ3.5)を、1.5mlの0.1M NaHCO3中に再緩衝化させ、そしてDMSO中に溶解させた15μlのカルボキシルフルオレセイン−ヒドロキシ−スクシンイミドエステル(Boehringer Mannheim)(1mg/ml)と1時間反応さ せた。非結合フルオレセインを、ゲル濾過により除去した。F/P比は、両方の結合について約3〜3.5であることが示された。100万個のマウス脾臓細胞を、図2に示す複合体化AMABの種々の希釈液と15分間インキュベートし、そしてフローサイトメトリーにより分析した。δ1.3について得られた結果を図2(a)に示し、そしてδ3.5について得られた結果を、図2(b)に示す。データは、IgD陽性細胞の検出には1μg/mlほどの低濃度で充分であること、およびAMABが高い親和性を有することを示す。
【0064】
(実施例7)
(マウス脾臓細胞の二重染色)
B6×129マウス由来の脾臓細胞を、フルオレセイン複合体化AMABδ 1.3お
よびフィコエリトリン複合体化抗マウスIgM抗体(R33−24)を用いて染色し、洗浄し、そしてリンパ球についてゲート(gate)し、そしてヨウ化プロピジウム染色により死細胞を除外するFACScanフローサイトメーターで分析した。
【0065】
得られた結果を図3に示す。この結果により、ほとんどのIgM産生細胞はIgDを共発現するが、わずかなIgM+ IgD-細胞が得られたことが示される。この染色は、I
gDの予想された染色パターンによく一致する。またこれにより、AMABが正常マウスに通常存在する分子に結合し、従って、モノクローナル性自己抗体であることが示される。
【0066】
(実施例8)
(コロイド磁性粒子複合体化モノクローナル性自己抗体を用いた細胞の磁性分離)
抗体の磁性粒子への複合体化:
精製AMAB δ1.3およびδ3.5を、供給者の指示に従って、SPDP (Pierce)カップリング化学物質を用いてMACSアミノマイクロビーズ(MiltenyiBiotec GmbH、Bergisch Gladbach、Germany)に複合体化させた。約200μgの活性化した抗体を、1mlのSPDP修飾MACSアミノマイクロビーズ(OD450=10)に複合体化させた。結合したビーズを、MACSA1カラムを用いて2回精製し、そしてPBSでOD450=1の濃度に再緩衝化させた(Miltenyiら,1990, Cytometry, 11:231-238を参照のこと)。
【0067】
細胞の磁性標識:
80μl中1千万個の脾臓細胞を、δ1.3およびδ3.5複合体化磁性粒子の1〜5の希釈した液と4℃で15分間インキュベートした。次いで、同じフルオレセイン複合体
化AMABを添加して最終濃度を8μg/mlにした。細胞を抗体と5分間反応させ、次いで、PBSで1回洗浄した。磁性標識およびフルオレセイン標識された細胞を、供給者の指示に従って、MACS磁性細胞分取器(MiltenyiBiotec GmbH)を用いて分離した。細胞を、25Gフローリストリクター(flow ristrictor)で動作する予め充填されたA
2 MACSカラムに付与し、カラムを4mlの緩衝液で洗浄し、そしてカラムを通過す
る細胞を非磁性画分として収集した。カラムを、23Gフローレジスター(flow resistor)を用いて 、1回の逆流手順を用いて洗浄し、そして残った細胞を溶離した。すべての画分を、フローサイトメトリー(FACScan)により分析した。死細胞をヨウ化プロピジウム
染色により同定し、分析から除外した。
【0068】
図4(a)〜(f)は、分離前または分離後のFACS分析のヒストグラムを示す。図4(a)および(b)は分離前の細胞を示し、図4(c)および(d)は非磁性画分を示し、そして図4(e)および(f)は陽性細胞を示す。図(a)、(c)、および(e)は、δ1.3の結果を示し、図(b)、(d)、および(f)が、δ3.5の結果を示す。データにより、少なくとも95%のIgD発現細胞がカラムによって保持され、保持されてカラムから溶離されるIgD細胞は少なくとも92%の純度を有することが示される。この分離および分析におけるバックグラウンド染色は、主に非特異的に抗体を取り込むマクロファージにより生じる。
【0069】
(実施例9)
(NCAMノックアウトマウス)
神経細胞接着分子(NCAM)は、同種親和性および異種親和性の細胞−細胞相互作用を媒介するイムノグロブリンスーパーファミリーのメンバーである。NCAMは、異なるスプライシングにより発生した種々のイソ体である。Hemperlyら、(1986)Proc.Natl.Acad. Sci. USA 83:3037-3041;Barthelsら、(1987)EMBOJ. 6:907-914;およびBarthelsら、(1992)Eur.J.Neurosci. 4:327-337を参照のこと。胚発生の間は、NCAMは
3つの全胚葉(germ layer)の派生部で発現するが、成熟した動物では、主に神経組織中に存在する。神経胚形成、軸索成長(outgrowth)、網膜の組織形成、および嗅覚系の発達の
ようなプロセスが、NCAMの制御発現と関連付けられる。Crossinら、(1990)Exp. Neurol.,109:5-15;Tosneyら、(1986)Dev.Biol., 114:437-452;Thieryら、(1977)J.Biol. Chem., 252:6841-6845;Keyら、(1990)J.CellBiol., 110:172
9-1743;およびChungら、(1991)J. Comp. Neurol., 314:290-305を参照のこと。遺伝子標的化により作製されたホモ接合NCAM陰性マウスは、健康で、繁殖性である。成熟した変異体は、全脳重量の10%の減少、および36%の嗅球の大きさの減衰を示す。NCAMの欠失は、神経発生および神経形成性と関連すると考えられる炭水化物構造である、タンパク質結合α−(2,8)−連結ポリシアル酸のほとんどすべての欠損を伴う。Theodosisら、(1991)Proc.Natl.Acad. Sci. USA 88:5494-5498を参照のこと。Morris、(1981)Learn. Motiv. 12:239-260に記載されるようなMorris水迷路中での動物試験に
より、空間学習の欠落が示されたが、変異マウスの活動および運動能は正常であった。
【0070】
ホモ接合変異マウスの遺伝子標的化および作製を、標準プロトコルを用いて行い、そしてサザンブロッティングおよび対立遺伝子特異的PCRにより確認した。ヌクレアーゼSI保護アッセイ、ノーザンブロッティング、およびウエスタンブロッティングにより、標的化遺伝子座が無効対立遺伝子として確認された。モノクローナル抗体およびポリクローナル血清を用いた、NCAMについての脳切片の免疫細胞化学分析により、野生型動物およびヘテロ接合動物中の嗅球の糸球体および顆粒細胞層に最も強い染色が示された。すべての染色は、成熟した脳におけるNCAM発現についての報告とよく一致した。Chungら
、(1991)を参照のこと。ホモ接合変異マウスでは、予想通りNCAM免疫反応性が全欠損していた。
【0071】
2種の系統のヘテロ接合動物をつがいにし、そして78匹の子を得た。これらのうち交配した対立遺伝子について、38匹の動物(49%)がヘテロ接合型であり。22匹(28%)が野生型であり、そして18匹(23%)がホモ接合型であり、ほとんど完全なメンデル分配を示した。ホモ接合変異動物は繁殖性であり、そして、これらの動物は野生型およびヘテロ接合体の同腹子よりも約10重量%小さいが、生後4カ月まで健康である。変異体およびヘテロ接合動物から分離した脳は、明らかに解剖学的差異を有した:嗅球は、+/+動物および+/−動物と比較して、変異体では大きさが減少し;そして脳重量は体重の補正後約10%まで減少した。
【0072】
NCAMに対して高親和性のAMABを生成させるために、上記のマウスに適切なアジュバントに懸濁した抗原量のNCAMを接種した。追加免疫注射を必要に応じて与え、そして抗体力価を定期的に測定する。一旦、力価が充分であると、AMAB生成のための適切な方法が続けられる。
【0073】
(発明の要旨)
本発明は具体的には以下を提供する。
(1) 以下の工程を包含する、自己抗原に対するモノクローナル性自己抗体を非ヒト脊椎動物宿主から得る方法:a)上記自己抗原の少なくとも1つのエピトープを産生しないように上記宿主動物のゲノムを改変する工程;
b)上記自己抗原またはそのホモログを含有する組成物で上記宿主動物を免疫する工程;
c)上記自己抗原またはそのホモログに応答して産生される宿主細胞であって、上記自己抗原またはそのホモログに対する抗体を発現する上記宿主細胞を収集する工程;およびd)上記収集した細胞または上記細胞由来の遺伝学的材料を用いて抗体を産生する工程。
(2) 上記宿主動物が少なくとも1つのエピトープの機能的等価物を産生する、項目1に記載の方法。
(3) 上記宿主動物が少なくとも1つのエピトープの構造的ホモログを産生する、項目1に記載の方法。
(4) 上記自己抗原が、タンパク質、ペプチド、炭水化物、脂質、酵素補因子、および天然に生じる凝集物またはそれらの共有結合複合物からなる群より選択される、項目1に記載の方法。
(5) 上記自己抗原が細胞表面抗原である、項目4に記載の方法。
(6) 上記自己抗原がサイトカインである、項目4に記載の方法。
(7) 上記自己抗原が細胞接着分子およびイムノグロブリンからなる群より選択される、項目5に記載の方法。
(8) 上記自己抗原がIgD抗体である、項目7に記載の方法。
(9) 上記自己抗原が細胞接着分子である、項目7に記載の方法。
(10) 上記細胞の収集が、抗原特異的B細胞を富化または単離する工程をさらに包含する、項目1に記載の方法。
(11) 上記抗体が、さらに上記抗体をコードする組換え遺伝子を発現することにより産生される、項目1に記載の方法。
(12) 上記組換え遺伝子が、単一の予め富化した抗原特異的B細胞および単一細胞PCRを用いて単離される、項目11に記載の方法。
(13) 上記抗体が少なくとも一部でヒト化されている、項目1に記載の方法。
(14) 上記宿主動物が哺乳類であり、そして、齧歯類、兎類、霊長類、食肉類、奇蹄類、および偶蹄類からなる群より選択される、項目1に記載の方法。
(15) 上記宿主動物が齧歯類またはネズミである、項目14に記載の方法。
(16) 上記宿主動物がマウスである、項目15に記載の方法。
(17) 遺伝的改変が生殖系列伝達を介して得られるように、上記工程b)の前に、遺伝的に操作された宿主動物を繁殖させる工程をさらに包含する、項目1に記載の方法。
(18) 上記抗原またはそのホモログの生合成が、上記抗原またはそのホモログを産生する代謝経路に影響を与える酵素をコードする遺伝子の変異により排除される、項目1に記載の方法。
(19) 項目1に記載の方法により得られる抗体を含有する組成物。
(20) ELISAに使用するための、項目19に記載の組成物。
(21) 項目1に記載の方法により得られる抗体フラグメントを含有する組成物であって、上記フラグメントが上記自己抗原に結合するに十分である、組成物。
(22) 上記フラグメントがFabフラグメントである、項目21に記載の組成物。
(23) 上記抗体が他の化合物と複合体化される、項目20に記載の組成物。
(24) 上記化合物が、リポーター酵素、蛍光色素、金属化合物、毒素、ハプテン、磁性粒子、固体キャリア、リポソーム、および蛍光粒子からなる群より選択される、項目23に記載の組成物。
(25) 上記磁性粒子がコロイド超常磁性粒子である、項目24に記載の組成物。
(26) 上記化合物が、アルカリホスファターゼ、ペルオキシダーゼ、およびウレアーゼからなる群より選択される酵素である、項目24に記載の組成物。
(27) 上記化合物が、フルオレセインイソチオシアネート、テトラメチルローダミンイソチオシアネート、フィコエリトリン、アロフィコシアニン、およびテキサスレッドからなる群より選択される蛍光色素である、項目24に記載の組成物。
(28) 上記化合物が、フェリチン、金コロイド、および磁性ビーズからなる群より選択される金属化合物である、項目24に記載の組成物。
(29) 上記化合物が放射性同位元素を含有する、項目23に記載の組成物。
(30) 上記化合物がビオチンである、項目23に記載の組成物。
(31) 以下の工程を包含する、細胞表面抗原の存在に基づいて細胞を分離する方法:a)項目1に記載の方法により細胞表面抗原に対する結合特異性を有するモノクローナル性自己抗体を得る工程;
b)上記抗体を異種細胞集団と接触させる工程であって、ここで、上記細胞の亜群が、上記抗体が結合特異性を有する上記細胞表面抗原を発現する、工程;およびc)上記抗体により認識される細胞を、上記抗体により認識されない細胞から分離する工程。
(32) 上記抗体がコロイド超常磁性粒子とともに用いられ、そして上記細胞が高勾配磁性分離を用いて単離される、項目31に記載の方法。
(33) 上記抗体がコロイド超常磁性粒子に結合される、項目32に記載の方法。
(34) 上記細胞の亜群が上記抗体に結合され、そして上記抗体が抗イムノグロブリン特異的コロイド超常磁性粒子により認識される、項目32に記載の方法。
(35) 上記抗体がハプテンにより改変され、そして上記コロイド超常磁性粒子が上記ハプテン改変物を認識する、項目32に記載の方法。
【0074】
本明細書中に引用したすべての刊行物および特許出願は、個々の刊行物または特許出願が具体的かつ個別に参考として援用されているのと同じ程度まで、参考として本明細書中に援用されている。
【0075】
上記発明は、明瞭にそして理解されることを目的とする例示および実施例により詳細に記載されているが、所定の改変が実施され得ることは当業者に明らかである。従って、説明および実施例は本発明の範囲を限定するように解釈されるべきではなく、本発明の範囲は添付の請求の範囲により記載される。
【図面の簡単な説明】
【0076】
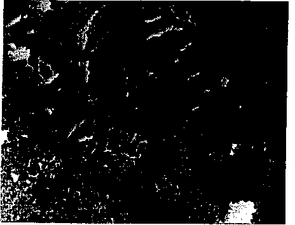
【図1】B細胞富化領域中のマウス脾臓組織の抗IgD抗体染色を表す。
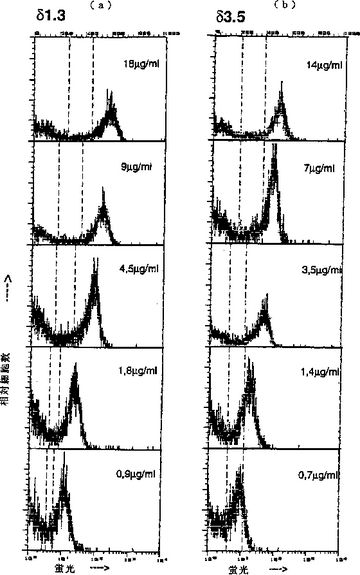
【図2】(a)は、マウス脾臓組織におけるδ1.3 AMABの滴定の結果を表し、(b)は、マウス脾臓細胞におけるδ3.5 AMABの滴定の結果を表す。
【図3】抗IgM抗体および抗IgD抗体で二重染色したマウス脾臓細胞のFACS分析の結果を表す。
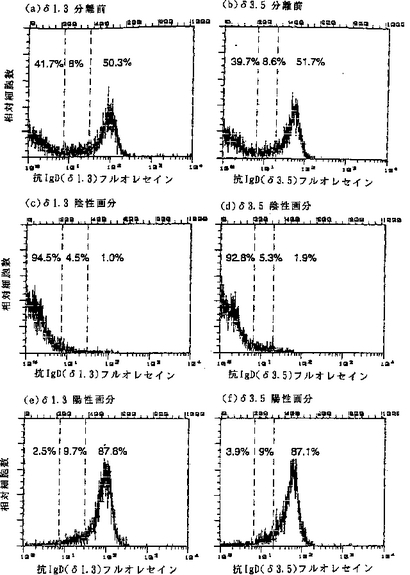
【図4】(a)はδ1.3におけるコロイド超常磁性粒子に結合した抗IgD抗体を用いたマウス脾臓細胞の分離前の結果を表し、(b)はδ3.5における分離前の結果を表し、(c)はδ1.3における分離後の陰性画分の結果を表し、(d)はδ3.5における分離後の陰性画分の結果を表し、(e)はδ1.3における分離後の陽性画分の結果を表し、そして(f)はδ3.5における分離後の陽性画分の結果を表す。
【技術分野】
【0001】
本発明は自己抗原またはそのホモログに対するモノクローナル性自己抗体(AMAB)を得る方法、およびこれらの抗体の細胞集団の分析における使用および細胞分離手法における使用に関する。
【背景技術】
【0002】
抗体は診断および治療の両方を含む医療用途において有用であり、そして細胞分離を包含するバイオテクノロジーの用途においても有用であることが立証されている。より一般的には、抗体を産生させ得る任意の化合物を電子顕微鏡検査および酵素免疫測定アッセイのような種々の手法を用いて、同定および位置付けをする際に抗体を使用することが、高い結合特異性を有する抗体によって容易になる。
【0003】
抗体は、鎖間のジスルフィド結合および他の分子内相互作用によって結合した重鎖および軽鎖の両ポリペプチドを含有する。個々の重鎖および軽鎖は、これらのジスルフィド結合によって対をなす。異なるクラスまたはアイソタイプの抗体のうち、3種のアイソタイプ(IgD、IgE、およびIgG)はジスルフィド結合により結合した2つの同じ重鎖/軽鎖対で構成され、そして残りの2種のアイソタイプ(IgAおよびIgM)は同じ重鎖/軽鎖対の、より複雑なポリマーで構成される。各鎖は定常部および可変部を含む。定常部はその抗体を産生する動物および抗体の特定のアイソタイプに特有であり、可変部はその抗体が結合するエピトープの構造に対応する。
【0004】
「抗原」という用語は、本明細書中では基質を意味し、これは、この基質に特異的に結合する抗体の産生を誘起し得る分子の、分子全体または分子中のドメインのいずれかである。さらに、この抗原という用語は、本明細書中では、野生型の宿主生物においては、自己認識によって抗体産生を誘起しないが、適当な遺伝学的操作を受けた宿主動物においてはこのような応答を誘起し得る基質を意味する。
【0005】
「エピトープ」という用語は、本明細書中では、Bリンパ球によって認識される、抗原上の個別の(discrete)三次元的な部位を意味する。エピトープは複雑な抗原上の免疫学的に活性な領域であり、この領域はB細胞レセプターに実際に結合し、その結果、このB細胞により産生される抗体分子によっても実際に結合される領域である。抗原は一般に少なくとも1つのエピトープを有し、そして通常1つより多いエピトープを有することが多い。タンパク質抗原のエピトープは線形または非線形であり得る。線形のエピトープとはタンパク質のアミノ酸配列中の連続したアミノ酸残基から構成されるエピトープである。線形エピトープは、立体的に折り畳まれても折り畳まれていなくてもよく、天然の三次元構造を形成し、そして免疫応答を誘起してこの抗原に対する結合特異性を有する抗体を産生する。非線形エピトープは不連続のアミノ酸残基から構成される。従って、非線形エピトープは常にある程度タンパク質が折り畳まれている必要があり、必要なアミノ酸残基を互いに近接させて、天然の三次元構造を形成し、そして免疫応答を誘起してこの抗原に対する結合特異性を有する抗体を産生する。
【0006】
「自己」という用語は、本明細書中では、通常、宿主種によって生合成される基質、または通常その宿主種が曝される基質に包含されているため、野生型宿主種のB細胞レセプターによって認識されないか、あるいはごくわずかしか認識されない抗原またはエピトープを表す。このような基質は宿主免疫系の寛容を誘導し、そしてその宿主はこの基質に対して「寛容にされた」という。
【0007】
脊椎動物の免疫系は自己抗原と外来抗原とを識別し得、外来抗原に対しては抗体を介する免疫応答を発現するが、自己抗原に対しては発現しない。抗体の応答はB細胞によって媒介される。骨髄中でB細胞が成熟する間に、整列していた配列において可変部の遺伝子再配列が起こる。このプロセスの最後には、各B細胞はイムノグロブリン重鎖をコードする単一の機能性可変部DNA配列と、イムノグロブリン軽鎖をコードする単一の機能性可変部DNA配列とを含有するようになる。このプロセスにより、各々が抗原的に単一エピトープに関与する、成熟した免疫適格性B細胞の産生に至る。まだ明らかにされていないプロセスによって、「自己」成分に対する免疫寛容は、抗原的に自己エピトープに関する可変部を有するB細胞を選択的に除去して達成される。この自己寛容は、宿主脊髄動物によって合成される抗原またはエピトープに特異的な抗体の産生を防止する。従って、宿主によって外来性と認識されるエピトープを含む抗原のみが、抗体産生のために使用され得る。
【0008】
自己エピトープと外来エピトープとが構造的に類似、あるいは「相同」である場合、宿主の免疫応答は弱くなる。従って、このようなエピトープに対して高い親和性を有する抗体を得ることは実質的に不可能である。この結果、タンパク質(例えば、NCAM、サイトカイン、およびイムノグロブリン)の高度に保存されたドメインに対する抗体を生成することは非常に困難である。なぜなら、この保存されたドメインを共有する動物は、このドメインを外来性として認識できないからである。ある種の自己免疫疾患の結果として、自己抗原に対する抗体が産生されるが、これらの抗体は非常に限られた一群の自己抗原に対して結合特異性を有するが、これらは人工的に操作し得ず、一般に結合親和性が低い。従って、自己免疫疾患の動物は、自己抗原に結合特異性を有する抗体の産生に広く有用であるわけではない。
【0009】
マウスでは、系統間のアロジェニックな差異(allogeneic difference)によ って、このようなアロジェニックな差異を生じるタンパク質に対して特異的なマウス抗マウス抗体を産生することが可能である。Kesslerら、(1979)J.Immunol.123:2772-2778; Reifお
よびAllen(1964)J. Exp. Med. 120:413-433;Marshak-Rothstein(1979)J. Immunol.122:2491-2497; ならびにOiおよびHerzenberg(1979)Molec.Immunol. 16:1005-1017を参
照のこと。最初に、T細胞表面タ ンパク質およびマウスIgD抗体に特異的なポリクロ
ーナル抗血清およびモノクローナル抗体がこの方法で得られた。しかし、これらの抗体は特定の系統のマウスの遺伝子産物を認識するに過ぎない。これらの抗体は、抗体産生宿主の自己抗原と構造的に相同ではないエピトープを認識し得るに過ぎない。さらに、これらの抗体が得られ得るエピトープは、系統間の差異およびアロタイプ系統自身の入手可能性によって制限され、従って、実際には有用性は少ない。
【0010】
細胞集団を分析および分類する多くの方法が考案されている。これらの方法には蛍光標示式細胞分取(FACS)、磁性分離(磁性ビーズに複合体化した抗体を使用する)、および特定の細胞表面「マーカー」として知られるタンパク質に対する抗体の親和性による他の方法が包含されるがこれらに限定されない。細胞の分析および分離に対するこのようなアプローチは、特に、細胞系統の決定、特定の物質を合成し得る細胞の単離、および特定の細胞タイプに伴う種々の疾患状態の治療に有用である。例えば、高度に精製した造血幹細胞は造血移植に必須であり、これにはガン患者および造血移植に関連する他の器官の移植が包含されるがこれらに限定されない。単離された細胞集団はまた、遺伝疾患、AIDS、および種々の形態のガンの治療における遺伝子治療のための重要な標的である。従って、種々の所定の細胞を実質的に純粋な形態または純粋な形態で単離する数多くの努力がなされてきた。幹細胞の単離のような場合には、このように体内で低濃度の細胞を効果的に精製するためには、幹細胞に特異的なマーカーを高い親和性で認識し、かつこのマーカーに高い親和性で結合する抗体が必要とされる。このような抗体を得ることは、ヒトとネズ
ミとで幹細胞マーカーが相同であるため、困難である。
【発明の開示】
【課題を解決するための手段】
【0011】
本発明は、自己抗原またはそのホモログに対するモノクローナル性自己抗体(AMAB)を得る新規の方法を提供する。本方法は、自己抗原内の改変形態の少なくとも1つのエピトープを生合成しないまたは合成しない遺伝的に操作された宿主動物を得ること、および少なくとも1つのエピトープに対する宿主の自己寛容の欠如を利用して抗原に特異的な抗体を産生することを包含する。本発明はまた、本方法により産生される抗体またはそのあらゆる機能性誘導体を包含する。本発明はさらに、本明細書に記載された方法により得られる、細胞表面抗原に対して特異的な抗体を利用することを含む、細胞の単離方法を包含する。
【発明を実施するための最良の形態】
【0012】
本発明は、自己抗原またはそのホモログに対して結合特異性を有するAMABを得る新規の方法を提供する。本発明は、宿主が自己として認識する抗原に対するAMABの産生の限界を克服する手段、ならびに、「標的化抗体」、すなわち公知の特定の極めて正確なエピトープに結合特異性を有するAMABを得る方法を提供する。本発明ではまた、成長および発育に不可欠と考えられる生物学的分子またはそのエピトープに対するAMABが得られる。本明細書中でさらに十分に議論されているように、標的化遺伝的置換はその分子の機能的等価物を産生させ、こうして動物が抗体を産生するに十分に成長および発育させる。
【0013】
用語「ホモログ」とは、本明細書中では、宿主動物により産生される抗原、すなわち「自己抗原」に非常に似ているため、ホモログに対する抗体の産生を排除するまたは顕著に妨げる構造を有する抗原を意味する。用語「自己抗原またはそのホモログ」とは、本明細書中では、本発明は、自己抗原に対する結合特異性を有するAMABを得ることに関するだけでなく、むしろ自己抗原に対して非常に高度な構造類似性のためあらゆる化合物に対するAMABを得ることに関すること、およびホモログは自己として十分に認識され、適切な抗体を介する免疫応答を宿主動物が生じないことを意味するために用いる。適切な抗体を介する免疫応答では、AMABが得られないか、あるいは、産生される場合、AMABが抗原に高い親和性を有さないかのいずれかのうちの1つである。
【0014】
用語「標的抗原」とは、本明細書では、宿主動物がそれに曝され、かつそれに対する免疫応答が生じるような組成物を意味する。「標的抗原エピトープ」とはAMABが結合する標的抗原上の領域である。
【0015】
用語「免疫応答」とは、本明細書中では、B細胞によるAMABの産生を意味する。これらの抗体は、病気を引き起こす因子へ免疫性を与えるような抗原変化をもたらすかどうかに関係なく特定の抗原に結合する。AMABはB細胞表面に会合し得、そしてまた自由に循環し得る。
【0016】
用語「構造的に非相同」とは、本明細書中では、遺伝的に操作した結果、宿主動物により生合成された抗原と自己抗原とを比較する記述として用いられる。一方に結合するが他方に結合しない抗体が生じ得るとき、2つの抗原は構造的に非相同である。構造的に非相同とは、2つまたはそれ以上の抗原間でいくつかの構造的な差異、おそらくわずかな差異があることを意味する。2つの抗原間の構造的に非相同な差異は、ただ1つのアミノ酸の差異、あるいはメチル基の存在または非存在のような小さなものであり得る。
【0017】
用語「機能的に等価」とは、本明細書中では、遺伝的に操作した結果、宿主動物により
生合成された標的抗原と自己抗原とを比較する記述として用いられる。多くの場合、抗体の獲得を妨害することにより、自己抗原の合成を中断させるように遺伝的に操作した宿主動物が致死的であるか、生存が低減しているか、または全身的に健康かが証明され得る。したがって、本明細書中に記載するように、遺伝的に操作することは、自己抗原の生合成を排除するために用い得、そして他の機能的に等価な抗原の産生を引き起こし得る。この機能的に等価な抗原は、AMABが得られ得る程度にまで宿主動物の生存率、全身的な健康、または免疫適格性を改良することにより、自己抗原の有害な排除を補うに十分な量の自己抗原機能をもたらす。本明細書で用いられるように、機能的に等価な抗原は、構造的に非相同であることが理解される。
【0018】
用語「生合成レパートリー」とは、本明細書中では、所定の宿主動物により生合成される化合物全体を意味する。
【0019】
用語「野生型」とは、本明細書中では、標的抗原に関しては遺伝的に操作されておらず、そしてこのような遺伝的に操作された生物系統ではない、宿主種および宿主系統の個体を意味する。
【0020】
用語「遺伝的に操作する」とは、特定の抗原または特定の一群の抗原の合成を防止または改変するように直接的に改変または排除された1つまたはそれ以上の遺伝子を有する動物を記述するのに用いられる。このような改変または削除(例えば特異的な遺伝的なノックアウトおよび置換による)は、遺伝的レベルにある。標的抗原の遺伝的操作は、一般に、本明細書中に記載されているように、抗原または一連の抗原上の少なくとも1つのエピトープに対して増強した結合特異性を有する抗体が得られ得る範囲にまで限定される。用語「遺伝的に操作された」はまた、初めから改変されている動物の子孫に適用される。
【0021】
用語「ドメイン」とは、本明細書中では、抗原のあらゆる領域または部分を意味し、これには、抗体全体、抗原全体よりも少ないあらゆる領域、または少なくとも1つのエピトープをマスクするように作用し得る追加成分によって抗原全体よりも大きくなるあらゆる領域を包含する。
【0022】
本発明は、自己抗原に対するモノクローナル性自己抗体(AMAB)を非ヒト脊椎動物宿主から得る方法に関する。
【0023】
本発明の好ましい実施態様では、以下の工程を包含する:
a)該自己抗原の少なくとも1つのエピトープを産生しないように該宿主動物のゲノムを改変する工程;
b)該自己抗原またはそのホモログを含有する組成物で該宿主動物を免疫する工程;
c)該自己抗原またはそのホモログに応答して産生される宿主細胞であって、該自己抗原またはそのホモログに対する抗体を発現する該宿主細胞を収集する工程;および
d)該収集した細胞または該細胞由来の遺伝学的材料を用いて抗体を産生する工程。
【0024】
本発明の1つの局面では、上記宿主動物は少なくとも1つのエピトープの機能的等価物を産生する。
【0025】
本発明の他の局面では、上記宿主動物は少なくとも1つのエピトープの構造的ホモログを産生する。
【0026】
本発明の1つの局面では、上記抗体は、単一の予め富化した抗原特異的B細胞および単一細胞PCRを用いて単離された、この抗体をコードする組換え遺伝子を発現することにより産生される。
【0027】
本発明の1つの実施態様では、宿主動物は特定の自己抗原を合成しないように遺伝的に操作されている。遺伝的に改変された宿主が自己抗原またはそのホモログで免疫される場合、宿主の免疫系は、自己抗原を自己として認識せず、したがってAMABが得られ得る抗体媒介免疫応答を生じ得る。このような「ノックアウト」変異体を得る方法は当該技術分野で公知であり、例えば、Mansourら、Nature336:348-352(1986)に記載されている。本明細書中で用いられるほとんどの遺伝的に操作された宿主動物では、ホモ接合変異体
を得るために、または何回も遺伝的に操作した宿主動物を得るために繁殖が用いられ得る。したがって、宿主に対する遺伝的改変は最終的に生殖系列伝達を介して得られ得る。
【0028】
遺伝的に操作されたマウスの子孫もまた本発明に包含される。トランスジェニック動物を産生する当該技術分野で公知のあらゆる他の方法も、本明細書中での使用に適切である。適切な方法には、Kitamuraら、(1991)Nature350:423-426;Shinkaiら、(1993)Science259:822-825;およびKomoriら、(1993)Science261:1171-1175の記載が包含される
がこれらに限定されない。簡単に述べると 、免疫系の自己抗原の場合では、遺伝子改変
がホモ接合の胚幹(ES)細胞を、RAG−欠失マウスのような免疫不全マウスの未分化胚芽細胞に移植し得る。多くの場合、再形成した動物は免疫欠失の表現型を欠き、そのリンパ球に遺伝子改変を含む。
【0029】
本発明の他の実施態様では、宿主動物を特定の自己抗原の合成が機能的に等価な抗原の合成により置換されるように遺伝的に操作する。従って、排除されることによって致死的になり、劇的に生存が低減することが証明される抗原に対して抗体が得られ得るか、さもなくば抗体を獲得する努力を妨げ得る。
【0030】
宿主の生合成レパートリーから除かれる自己抗原は、あらゆる化合物、ドメイン、またはそれらのエピトープであり得、通常は野生型宿主種により合成される。本発明により得られる抗体は、タンパク質、ペプチド、炭水化物、脂質、核酸、酵素補因子、あるいはあらゆる天然に生じる凝集物またはそれらの共有結合複合物、あるいはそれらのあらゆるリン酸化種または硫酸化種を含むがこれらに限定されないあらゆる抗原に対する。本発明のこれらの実施態様では、宿主は抗原の生合成が排除または改変されるようなあらゆる方法で遺伝的に操作され得る。例えば、抗原の生合成に包含される特定の酵素の発現をノックアウトすることで、抗原の非産生または機能的に等価な抗原の産生が生じ得る。例えば、自己抗原またはそのホモログの生合成は、この抗原またはそのホモログを産生する代謝経路に影響を与える酵素をコードする遺伝子の変異により排除される。このような酵素には重合化に含まれる酵素が含まれるがそれらに限定されず、糖タンパク質の合成におけるような炭水化物、リン酸基、脂質、硫黄含有基のタンパク質への付加が排除される結果、タンパク質に共有結合する化合物の構造を除去または変化させる。従って、特定の自己糖タンパク質または糖タンパク質上の炭水化物構造に結合特性を有する抗体が、本発明の方法により得られ得る。また、このような酵素には、自己抗原またはそのホモログを産生する代謝経路に影響を与える酵素が包含される。
【0031】
抗原の産生を排除または変化させるより全体的な方法が利用され得る。これらの方法には、特定の遺伝子または遺伝子ファミリーの合成を排除すること、および特定の細胞タイプを除去することが包含されるがこれらに限定されない。適切な遺伝子または遺伝子ファミリーの例には、ステロイドまたはサイトカインのような特定の因子の存在により調節されるものあるいは細胞タイプ特異的に発現される遺伝子が含まれる。これらの細胞タイプの1例は、褐色脂肪細胞である。
【0032】
本発明の好ましい実施態様では、抗原はタンパク質である。タンパク質についてのノックアウト変異には、完全なタンパク質の合成を妨げること(完全な遺伝子を除去すること
によって、あるいは転写制御因子または翻訳制御因子を改変することによって)、または唯一の抗原性ドメインの合成を排除することが包含されるが、残りのタンパク質部分は正常に合成される。当該技術分野で公知のあらゆる形態の置換が本明細書中での使用に適切であるが、タンパク質抗原の場合では、自己抗原と機能的に等価な抗原との置換は少なくとも3つの異なるタイプの置換を伴う。
【0033】
第1に、自己抗原をコードする遺伝子が、自己抗原をコードする遺伝子に由来する組換え遺伝子により置換され得る。タンパク質をコードする遺伝子は、抗原領域が欠失し、改変アミノ酸配列により置換されるように改変されるか、または新規アミノ酸配列の追加によりマスクされるように改変される。従って、自己タンパク質抗原の改変における遺伝的に操作された抗原の除去は、単一のアミノ酸の追加、排除、または置換のような小さなことであり得る。重要なことは、自己タンパク質抗原の構造におけるこのような小さな変化により、エピトープに対して非常に正確なAMABが自己抗原の機能を大きく妨害することなく得られ得る。適切なドメインにおける単一のアミノ酸の変化の結果、その単一のアミノ酸を含むエピトープに対するAMABを生じる。
【0034】
第2に、自己抗原をコードする遺伝子が、コードするタンパク質が機能的に等価だが少なくとも一部は抗原的に非等価となるように、宿主種に関連のある生物から得られる相同なタンパク質をコードする遺伝子により置換され得る。非常に関連のある種は、通常同じタンパク質に高度な配列相同性(典型的には90%より大きい)を有する。従ってこの方法は、抗原の発現が生物の健康に重要である場合に本発明を実施するには理想的であり、そして少数のエピトープに結合特異性を有するAMABが望ましい。この方法はまた、自己抗原をコードする遺伝子の1つの領域のみを、関連する種に由来する同じ抗原をコードする遺伝子の対応する領域に置換することにより実施され得る。
【0035】
第3に、自己抗原をコードする遺伝子が、同じかまたは類似の機能を有するが構造的に非相同であることが知られているタンパク質をコードする遺伝子により置換され得る。この方法は、抗原の発現が生物の健康に重要であり、特定のエピトープに結合特異性を有するAMABが必要ではない場合に特に有用である。
【0036】
本発明により得られるAMABが結合特異性を有し得るタンパク質抗原の例には、細胞表面抗原が包含されるが、これに限定されない。細胞表面抗原には、細胞接着分子、MHCクラスI分子およびクラスII分子、インテグリン、サイトカイン、セレクチン、サイトカインレセプター、およびイムノグロブリン(例えばIgD抗体)が包含されるがこれらに限定されない。
【0037】
本発明は、魚類、は虫類、両生類、鳥類、および哺乳類を包含する任意の、非ヒト脊椎動物宿主を用いて実施され得る。しかし、宿主はほとんどの場合、哺乳類であり、通常、齧歯類、兎類、霊長類、食肉類、奇蹄類、および偶蹄類が包含されるがこれらに限定されない種類に属する。好ましくは、宿主はネズミであり、より好ましくはマウスである。
【0038】
AMABを作製する方法は当該技術分野で公知であり、本明細書中では詳細に述べない。モノクローナル抗体産生の分野で公知のあらゆる方法を、ここで使用し得る。このような方法には、所望の特異性を有する細胞表面抗体によりB細胞を分離すること、軽鎖および重鎖の可変部を発現するDNAをクローニングすること、および適切な宿主細胞中で組換え遺伝子を発現させることが包含されるがこれらに限定されない。標準モノクローナル抗体産生手法を使用し得、ここで、AMABは不死化抗体産生ハイブリドーマ細胞から得られる。これらのハイブリドーマは、Bリンパ球、好ましくは免疫された宿主の脾臓から単離されたBリンパ球と、それと適合する不死化細胞、好ましくはB細胞ミエローマとを融合することにより産生され得る。
【0039】
本発明はさらに、本明細書中に記載された方法によって得られるAMABを含有する物質の組成物をも包含する。本明細書中で用いるように、「AMAB」または「抗体」という用語は、抗体全体、およびその機能性部分を含む抗体フラグメントを包含する。「AMAB」という用語は、抗体全体が結合特異性を有しているエピトープへ結合するに充分な、軽鎖可変部および/または重鎖可変部の部分から構成される、あらゆる単一特異性化合物を包含する。上記フラグメントは少なくとも1つの重鎖イムノグロブリンポリペプチドまたは軽鎖イムノグロブリンポリペプチドの可変部を含み得、そしてFabフラグメント、Fab2フラグメント、およびFvフラグメントを包含するがこれらに限定されない。
【0040】
さらに、単一特異性ドメインは、当該技術分野で公知のあらゆる方法により、他の適切な分子に付着し得る。この付着は、例えば、化学的あるいは遺伝学的操作によって行われ得る。AMABは、当該技術分野で公知のあらゆる組換え手段によって産生され得る。このような組換えAMABには、細菌において産生されたフラグメント、そしてその定常部の大部分がヒト抗体の定常部によって置換されたAMABが包含されるがこれらに限定されない。さらに、このような「ヒト化」AMABは、本明細書中に記載の遺伝的に操作された宿主脊椎動物を、野生型Ig遺伝子座の代わりに機能性ヒトIg遺伝子座を発現する適合するトランスジェニック動物で作り出すことによって得られ得る。ヒトIg遺伝子座を発現するトランスジェニック動物に関しては、WIPO公開公報WO91/10741号、およびRajewskyらのDEP4228162.8を参照のこと。うまく交配すると、特別に自己抗原を発現しない
が、ヒト化Igタンパク質を発現する宿主動物が得られ得る。このような動物が免疫されると、この動物は特定の自己抗原に対するヒト化AMABまたは部分ヒト化AMABを産生する。このようなヒト化AMABは、治療指示薬(therapeutic indication)に使用するのに好ましい。
【0041】
AMABは他の化合物と複合体化し得る。この化合物には、酵素、リポーター酵素、リポソーム、磁性ビーズ、コロイド磁性ビーズ、固体キャリア、毒素、ハプテン、蛍光色素、蛍光粒子、金属化合物、放射性の化合物、薬剤、またはハプテンが包含されるがこれらに限定されない。AMABに複合体化し得る酵素には、アルカリホスファターゼ、ペルオキシダーゼ、ウレアーゼ、およびβ−ガラクトシダーゼが包含されるがこれらに限定されない。AMABに複合体化し得る蛍光色素には、フルオレセインイソチオシアネート、テトラメチルローダミンイソチオシアネート、フィコエリトリン、アロフィコシアニン類(allophycocyanins)、およびテキサスレッドが包含されるがこれらに限定されない。抗体に複合体化し得るその他の蛍光色素に関しては、Haugland,R.P. Molecular Probes: Handbook of Fluorescent Probes and Research Chemicals(1992-1994)を参照のこと。AMABに複合体化し得る金属化合物には、フェリチン、金コロイド、および特にコロイド超常磁性粒子(ビーズ)が包含されるが、これらに限定されない。AMABに複合体化し得るハプテンには、ビオチン、ジゴキシゲニン、オキサゾロン(oxazalone)、およびニト
ロフェノールが包含されるが、これらに限定されない。AMABに複合体化し得るかまたは導入し得る放射性化合物は当該技術分野で公知であり、放射性同位元素のテクネチウム99m(99TC)、125I、および任意の放射性核(14C、3Hおよび35Sが包含されるが、これらに限定されない)を含有するアミノ酸が包含されるが、これらに限定されない
。当該技術分野で公知の任意の方法によってタンパク質に複合体化し得る当該技術分野で公知のあらゆる薬剤が、本発明における使用に適している。このような薬剤は、この薬剤を標的分子へ高度に特異的に送達するために、AMABに複合体化され得る。
【0042】
本発明はさらにAMABを使用する方法を提供し、この方法にはイムノアッセイおよび細胞分離が包含されるがこれらに限定されない。適切なイムノアッセイは当該技術分野で公知であり、本明細書中で詳細に記述する必要はない。イムノアッセイには、ELISAおよびRIAが包含されるがこれらに限定されない。細胞の分離方法には、分子の分泌に
基づく分離方法および細胞表面分子に基づく分離方法が包含されるがこれらに限定されない。分子の分泌に基づく細胞の分離方法は、米国特許出願第07/965,934号および国際出願番号PCT/US93/10126に記載されている。特異的な細胞表面マーカーに基づいて細胞を分離する方法は一般に、本明細書中に記載される方法によってAMABを得る工程、AMABを異種細胞集団に接触させる工程、および細胞表面抗原に対するAMABの親和性に基づく細胞分離手法を行う工程を包含する。まず特定の細胞表面抗原を有する細胞を除去するか、あるいは特定の細胞系統を除去することによる、当該技術分野で公知のあらゆる方法が、細胞の分離または単離のために用いられ得る。
【0043】
AMABは、特定の細胞系統および/または特定の分化段階に関連するマーカーを同定するために特に有用である。AMABは固体支持体に直接または間接に付着し分離を可能とする。使用する分離手法は、採集する画分の生存率を最大限に保持するような方法であるべきである。分離するために、異なる効果を有する種々の手法が用いられ得る。このような分離では、マーカーを持たないで存在する細胞全体の、10%まで、通常約5%以下、好ましくは約1%以下が、保持される細胞集団と共に残存し得る。用いられる所定の手法は、分離効率、方法の細胞毒性、実行の容易さおよび速さ、そして精巧な器具および/または技術の必要性に依存する。
【0044】
細胞分離の手順には、コロイド磁性粒子に結合した抗体を用いる磁性分離、アフィニティークロマトグラフィー、およびモノクローナル抗体に結合した細胞毒性試薬、または当該技術分野で公知の任意の抗体依存性分離手法と共に用いられる細胞毒性試薬が包含されるがこれらに限定されない。さらに、細胞は固体マトリックス(例えばプレート)に付着した抗体を用いる「パンニング」によって分離し得る。蛍光標示式細胞分取器(FACS)もまた使用し得る。これは、異なる程度の精巧さを有し得、複数のカラーチャネル、鋭角(lowangle)および鈍 角(obtuse)光散乱検出チャネル、およびインピーダンスチャネルが包含されるがこれらに限定されない。当該技術分野で公知のあらゆる抗体依存性分離手法を、抗体の親和性よりもむしろ細胞の物理的性質に依存するポジティブおよびネガティブ両方の分離手法と共に使用し得る。このような細胞の物理的性質に依存する分離手法には、溶離および密度勾配遠心分離法が包含されるがこれらに限定されない。
【0045】
細胞の分離方法は、Dynal(Oslo、Norway)、Cellpro(Seattle)、またはAdvanced Magnetics(Boston)から市販されている。例えば、モノクローナル性自己抗体は、DynalM
450のような磁性ポリスチレン粒子または同様の磁性粒子に 直接結合し得、そして例え
ば、細胞分離に使用され得る。あるいは、抗体はビオチン化され得、あるいはジゴキシゲニンと結合し得、そしてアビジンコーティングまたは抗ジゴキシゲニンコーティングされた、SEPARATELC(Cellpro)のようなアフィニティーカラムと共に使用し得る。しかし、
好ましい実施態様では、モノクローナル性自己抗体は、例えば多糖類による有機コーティングを有するコロイド超常磁性粒子と組み合わせて使用される。Miltenyiら、(1990)Cytometry11:231-238を参照のこと。これらの粒子の大きさは10nm〜200nm、好ま
しくは40nmと100nmとの間であり得、そして自己抗体に直接結合していてもよく、または抗イムノグロブリン、アビジン、または抗ハプテン特異的マイクロビーズと組み合わせて使用されてもよい。例えば、抗体をハプテンにより修飾し、そしてコロイド超常磁性粒子がハプテン修飾物を認識する。多糖類コーティングされた超常磁性粒子はMiltenyiBiotec GmbH、Germanyから市販されてい る。また、抗体結合コロイド超常磁性粒子は、目的の細胞に結合し、この細胞が高勾配磁性分離を用いて単離される。また、細胞の亜群が上記抗体に結合し、さらにこの抗体は、抗イムノグロブリン特異的コロイド超常磁性粒子により認識される。
【0046】
用い得る手順の1つは、まず細胞を低温(通常約4℃)で短時間、所定の細胞表面抗原に特異的な飽和レベルのAMABと共にインキュベートし、次いでその細胞をPBSおよ
びウシ胎児血清(FCS)緩衝剤(cushion)で洗浄する方法である。次いで、細胞を上
記のような緩衝液に懸濁し、そして、AMABまたはAMAB抗原複合体に特異的な種々のタンパク質を用いて、特定の抗原決定基に対するAMABに基づいて細胞を分離する。
【0047】
AMABは、特定の細胞タイプの分離を容易にするマーカーと複合体化され得る。このマーカーには、磁性ビーズ(これは直接分離される)、ビオチン(これは支持体に結合したアビジンまたはストレプトアビジンで除去し得る)、ジゴキシゲニン(これは抗ジゴキシゲニン抗体で検出される)、蛍光色素(これはFACSに使用され得る)などが包含されるがこれらに限定されない。残留する細胞の生存率を過度に損なわない限り、いかなる手法も使用し得る。
【0048】
細胞表面抗原を含む細胞(例えば抗原特異的B細胞)の実質的な富化(一般に少なくとも約50%、好ましくは少なくとも約70%)後、細胞は、FACSまたは当該技術分野で公知の他の方法によって分離され得る。種々の方法による多色分析を使用し得、これらにはFACSおよび蛍光顕微鏡が包含されるがこれらに限定されない。この細胞は、特定の抗原に対する染色の程度に基づいて分離され得る。
【0049】
本発明はさらに、本明細書中に記載された方法によって、レセプターである自己抗原に対するAMABを得ること、およびこのようなAMABを薬剤として使用することを包含し、ここでAMABはレセプターアゴニストまたはアンタゴニストとしての有効性を示す。
【0050】
以下の実施例により本発明を説明するが、限定はしない。
【実施例】
【0051】
(実施例1)
(遺伝子の標的化)
Cδ遺伝子の標的化を、RoesおよびRajewsky、J. Exp. Med. 177:45-55(1993);な
らびにRoesおよびRajewsky、Int.Immunol. 3:1367(1991)に記載さ れるように行った。簡単に述べると、全部で108個のE14−1ES細胞を、Cδ1エキソンの大部分を置換するように、そしてこのエキソンに存在する制限部位を充たすことによって、Cδ3中にフレームシフト変異を挿入するように設計された標的化ベクターを用いてトランスフェクトした。生殖系列へ変異を導入した結果、両方のδ鎖Igドメインエキソンを機能的に不活性化した。これは、L鎖のμと競合し得る、かつ分泌され得る短縮型δH鎖の発現の可能性を除くのに重要であると考えられた。マウス生殖系列におけるフレームシフトの存在が、Cδ3中のHindIII部位を充たした結果生じるNheI制限部位により示さ
れた。Cδ2エキソンは、非機能性スプライシングアクセプターに帰因する偽エキソンである。
【0052】
選択で生き残ったコロニーをPCRで分析し、そして陽性クローンをさらにサザンブロッティングにより分析して、標的化された遺伝子座の構造を確認した。相同組換え体は、二重耐性クローンの1/17またはG418耐性クローンの1/103の頻度で得られた。野生型Cδ遺伝子座および変異型Cδ遺伝子座の制限マップ、ならびに5つの別個の組換え体候補およびコントロール細胞に由来するHindIII消化したゲノムDNAの制限
分析が示された。相同組換えの結果、HindIII部位の欠損を生じるCδ3中のフレー
ムシフト変異の存在または 欠如に依って、3.8kb生殖系列バンドに加えて、4.4
kbまたは6.0kbのバンドを生じた。10個の相同組換え体クローンのうちの9個が6.0kbのフラグメントを示し、組換えの開始点がCδ3エキソンの3’に位置することが示された。このように、Cδ1エキソンおよびCδ3エキソンは、これらのクローン中で機能しなかった。1つのクローンは、Cδ3エキソン中にHindIII制限部位を保
持し、4.4kbの診断用制限フラグメントを生じた。標的化された遺伝子座の構造を、種々の他のプローブおよび制限酵素を用いて確認した。サザンブロット分析を、Sambrook、Fritsch、およびManiatis、Molecular Cloning, A Laboratory Manual、第2版、Cold Spring Harbor Laboratory、NY(1989)に記載のように行った。
【0053】
(実施例2)
(IgD欠損マウスの作製)
IgD欠損マウスの作製を、RoesおよびRajewsky(1993)、J. Exp. Med. 177:45-55
に記載のように行った。簡単に述べると、Cδ遺伝子不活性化の方法および陽性クローンのスクリーニング手順は、実施例1に記載のようにした。標的化ES細胞クローンを、C57BL/6マウスから単離した胚盤胞に注入し、そして(C57BL/6×BALB/c)の里親(foster)に移した。雄のキメラの子を、δT変異の生殖系列伝達のためにC57BL/6の雌とつがいにした。ES細胞由来の子を毛色により識別し、そしてδTと呼ばれる変異の存在についてサザンブロッティングによるかまたは表現型をフローサイトメトリーにより分析した。ホモ接合変異マウス(δT/δT)を、ヘテロ接合の子の異種交配により得た。
【0054】
(実施例3)
(推定(δT/δT)変異マウスのノーザンブロット分析)
推定(δT/δT)変異マウスのノーザンブロット分析を、RoesおよびRajewsky(1993)による記載のように行った。簡単に述べると、δT変異の結果、H鎖のIgドメインをコードする両方のエキソンが機能的に不活性化される。しかし、トランスメンブレンエキソンおよびヒンジ部エキソンは、完全なままであり、かつ潜在的に機能性を維持する。CμおよびCδ遺伝子の両方を含有する前駆体RNAの異常なスプライシングの結果、Cμ遺伝子の細胞外ドメインならびにCδのトランスメンブレン部分および細胞質部分をコードするかなりの量のキメラIg転写物が産生される可能性を除くために、ホモ接合変異(δT/δT)マウスおよび野生型マウスの脾臓から単離したポリ(A)+RNAを、ノー
ザンブロッティングで分析した。CδトランスメンブレンエキソンにスプライシングされたCμエキソンを含むmRNAは、2.4kb(μs)または2.7kb(μm)の正常なCμ転写物よりも大きい。なぜなら、δメッセージの3’非翻訳領域は、μメッセージのものよりも600bp長いからである。
【0055】
Cδトランスメンブレン特異的プローブを用いるホモ接合変異マウスの脾臓ポリ(A)+RNAのハイブリダイゼーションでは、4.8kb、4.0kb、3.8kb、および
3.0kbのバンドが示された。しかし、これらのバンドはいずれも、Cμ特異的プローブとハイブリダイズしなかった。2つのプローブの検出限界は、標準プラスミドDNAへのハイブリダイゼーションから得られたシグナルにより判断されるように、2つの因子(Cμでは1.2×106コピーおよびδmプローブでは0.6×106コピー)の限りでは類似していた。Cμプローブは、mRNAおよび標準プラスミドに完全に適合する1k
bのcDNAフラグメントであるが、δmプローブはδm1/m2イントロンを含むため、標準プラスミドにおける700bp以外は480bpの長さを越えるmRNAとハイブリダイズするにすぎない。従って、δmエキソンにスプライシングされたCμエキソンを示すmRNAは、δmプローブで検出される場合は、Cμプローブでも表れるべきである。なぜなら、3.0kb、3.8kb、4.0kb、および4.8kbのバンドは明らかにδmプローブの検出限界を上回るため、それらがCμ配列を含む場合は、さらにCμプローブでも検出されるべきだからである。しかし、これはその場合ではない。
【0056】
さらに、Cδ3エキソンに対して特異的なneo+遺伝子およびneo+プロー ブを用いた同じブロッティングの一連のハイブリダイゼーションにより、3.8kb、4.0kb、および4.8kbのバンドも、neo+遺伝子由来の配列を含むことが示され、それら
が異常なスプライシング産物を表すことが示された。3.0kbのバンドは、実施例1に記載のフレームシフト変異のために、標的化対立遺伝子中で機能しないCδ3エキソンに対して特異的なプローブとハイブリダイズした。さらに、neo+プローブを用いて、2
.4kbのmRNA(これはδmプローブとハイブリダイズしなかった)もまた検出された。Cδm特異的プローブとハイブリダイズする1.6kbおよび2.0kbの少量のmRNAが、正常マウスおよび変異マウスの両方に由来する10μgのポリ(A)+RNA
中で検出可能である。しかし、これらのmRNAは、正常のCμメッセージよりも小さく、従って、機能性Ig分子をコードしそうにない。
【0057】
潜在的な機能性キメラμ/δ分子を表すmRNA種は、10μg程度のδT/δTマウスの脾臓ポリ(A)+RNAを用いるノーザンブロットで検出し得なかった。δH鎖をコ
ードするmRNAは、Cδm特異的プローブを用いて、300ng程度の少量の正常マウスの脾臓ポリ(A)+RNAで検出され得るので、μの細胞外ドメインおよびδのトラン
スメンブレン部分をコードする推定mRNAは、それが少しでも存在するなら、正常マウスのδH鎖メッセージよりも少なくとも30倍より少ない。従って、IgD様分子をコードし得る可能性のあるmRNAは、ノーザンブロット分析によりホモ接合変異マウス(δT/δT)中で検出不可能である。
【0058】
(実施例4)
(マウス−抗マウス−IgDモノクローナル抗体の産生)
IgD欠損マウスを、実施例1および2に記載のように遺伝子の標的化により作製した。こうして得られた1匹の動物を、ミョウバン中に沈降したIgDクラス(267.7δ「a」アロタイプ)のマウスモノクローナル抗体で、腹腔内(i.p.)に免疫した。
【0059】
6週間後、この動物に、「b」アロタイプの可溶性B1−8で、腹腔内に追加免疫を行い、両アロタイプを認識するモノクローナル抗体を得た。
【0060】
3日後、免疫したマウスの脾臓細胞を、標準PEG融合プロトコルを用いて、X63 Ag8.6.5.3と融合した。ハイブリッドを、8枚の96ウェルプレートに、直接クローニングし、そしてHAT選択培地を用いて選択した。得られたハイブリドーマを、Blー8(IgD)、BSA(ウシ血清アルブミン)、267.7(IgD)、R33−24−12(抗IgM)でコートしたプレートを用いて、ELISAによる抗IgD抗体の産生についてスクリーニングを行い、そして、R33−18−10.1−ビオチン抗体(ラット抗マウスκ)およびR33−60−ビオチン抗体(抗IgM)を用いて発色させた。17個のクローンがB1−8(IgDb)への反応性を示し、これらのうち13個のク
ローンが267.7(IgDa)への反応性を示し、これらのうちの1個がIgMクラス のものであった。他の2つのクローンは、IgMへの反応性を示したが、IgDには反応性を示さなかった。さらに、ハイブリドーマを、相対結合親和性およびアイソタイプについて特徴付けた。試験した21個のクローンのうち15個は、IgGlアイソタイプを示した。さらに、このクローンを、ビオチン化抗マウスIgGlとストレプトアビジン−フィコエリトリンとを用いてトリプシン処理およびトリプシン未処理のマウス脾臓細胞への結合を特徴付けた。トリプシンは表面のIgDをカットし、そしてIgD分子上のエピトープを制限し得る。
【0061】
最も親和性が高くかつIgGlアイソタイプの3個のクローン(δ1.2、δ1.3、およびδ3.5)を、さらなる実験のために用いた。これらのクローンにより産生された数mgの精製AMABを、標準の方法によりローラーボトルを用いて産生した。16個のクローンを、培養上清でマウス脾臓細胞をインキュベートすることによる染色アッセイで試験した。結合した抗IgDAMABを、ビオチン化抗マウスIgGlを用いて検出し、そしてストレプトアビジン−PEにより発色させた。
【0062】
(実施例5)
(B細胞富化領域でのマウス脾臓組織のIgDAMAB染色)
C57BL/6マウス由来の凍結脾臓切片を、実施例4に記載のように調製したビオチン化マウス抗マウスIgD AMAB δ1.3を用いて染色し、そして供給者の指示に従ってPierceNHSビオチンを用いてビオチン化した。染色パターンを、製造者の指示に従って、SigmaのAEC染色キットを用い、ストレプトア ビジンに結合したペルオキシダーゼを用いて発色させた。得られた結果を図1に示す。染色パターンは、ポリクローナルヤギ抗マウスIgD抗血清について見られるパターンと一致し、本発明により得られたIgDAMABが所望の自己抗原に対して結合特異性を有することを示す。
【0063】
(実施例6)
(フルオレセインに対する抗体の複合体化および種々の濃度下での脾臓細胞の染色)
実施例4からそれぞれ得られた0.5mgの精製AMAB(δ1.3およびδ3.5)を、1.5mlの0.1M NaHCO3中に再緩衝化させ、そしてDMSO中に溶解させた15μlのカルボキシルフルオレセイン−ヒドロキシ−スクシンイミドエステル(Boehringer Mannheim)(1mg/ml)と1時間反応さ せた。非結合フルオレセインを、ゲル濾過により除去した。F/P比は、両方の結合について約3〜3.5であることが示された。100万個のマウス脾臓細胞を、図2に示す複合体化AMABの種々の希釈液と15分間インキュベートし、そしてフローサイトメトリーにより分析した。δ1.3について得られた結果を図2(a)に示し、そしてδ3.5について得られた結果を、図2(b)に示す。データは、IgD陽性細胞の検出には1μg/mlほどの低濃度で充分であること、およびAMABが高い親和性を有することを示す。
【0064】
(実施例7)
(マウス脾臓細胞の二重染色)
B6×129マウス由来の脾臓細胞を、フルオレセイン複合体化AMABδ 1.3お
よびフィコエリトリン複合体化抗マウスIgM抗体(R33−24)を用いて染色し、洗浄し、そしてリンパ球についてゲート(gate)し、そしてヨウ化プロピジウム染色により死細胞を除外するFACScanフローサイトメーターで分析した。
【0065】
得られた結果を図3に示す。この結果により、ほとんどのIgM産生細胞はIgDを共発現するが、わずかなIgM+ IgD-細胞が得られたことが示される。この染色は、I
gDの予想された染色パターンによく一致する。またこれにより、AMABが正常マウスに通常存在する分子に結合し、従って、モノクローナル性自己抗体であることが示される。
【0066】
(実施例8)
(コロイド磁性粒子複合体化モノクローナル性自己抗体を用いた細胞の磁性分離)
抗体の磁性粒子への複合体化:
精製AMAB δ1.3およびδ3.5を、供給者の指示に従って、SPDP (Pierce)カップリング化学物質を用いてMACSアミノマイクロビーズ(MiltenyiBiotec GmbH、Bergisch Gladbach、Germany)に複合体化させた。約200μgの活性化した抗体を、1mlのSPDP修飾MACSアミノマイクロビーズ(OD450=10)に複合体化させた。結合したビーズを、MACSA1カラムを用いて2回精製し、そしてPBSでOD450=1の濃度に再緩衝化させた(Miltenyiら,1990, Cytometry, 11:231-238を参照のこと)。
【0067】
細胞の磁性標識:
80μl中1千万個の脾臓細胞を、δ1.3およびδ3.5複合体化磁性粒子の1〜5の希釈した液と4℃で15分間インキュベートした。次いで、同じフルオレセイン複合体
化AMABを添加して最終濃度を8μg/mlにした。細胞を抗体と5分間反応させ、次いで、PBSで1回洗浄した。磁性標識およびフルオレセイン標識された細胞を、供給者の指示に従って、MACS磁性細胞分取器(MiltenyiBiotec GmbH)を用いて分離した。細胞を、25Gフローリストリクター(flow ristrictor)で動作する予め充填されたA
2 MACSカラムに付与し、カラムを4mlの緩衝液で洗浄し、そしてカラムを通過す
る細胞を非磁性画分として収集した。カラムを、23Gフローレジスター(flow resistor)を用いて 、1回の逆流手順を用いて洗浄し、そして残った細胞を溶離した。すべての画分を、フローサイトメトリー(FACScan)により分析した。死細胞をヨウ化プロピジウム
染色により同定し、分析から除外した。
【0068】
図4(a)〜(f)は、分離前または分離後のFACS分析のヒストグラムを示す。図4(a)および(b)は分離前の細胞を示し、図4(c)および(d)は非磁性画分を示し、そして図4(e)および(f)は陽性細胞を示す。図(a)、(c)、および(e)は、δ1.3の結果を示し、図(b)、(d)、および(f)が、δ3.5の結果を示す。データにより、少なくとも95%のIgD発現細胞がカラムによって保持され、保持されてカラムから溶離されるIgD細胞は少なくとも92%の純度を有することが示される。この分離および分析におけるバックグラウンド染色は、主に非特異的に抗体を取り込むマクロファージにより生じる。
【0069】
(実施例9)
(NCAMノックアウトマウス)
神経細胞接着分子(NCAM)は、同種親和性および異種親和性の細胞−細胞相互作用を媒介するイムノグロブリンスーパーファミリーのメンバーである。NCAMは、異なるスプライシングにより発生した種々のイソ体である。Hemperlyら、(1986)Proc.Natl.Acad. Sci. USA 83:3037-3041;Barthelsら、(1987)EMBOJ. 6:907-914;およびBarthelsら、(1992)Eur.J.Neurosci. 4:327-337を参照のこと。胚発生の間は、NCAMは
3つの全胚葉(germ layer)の派生部で発現するが、成熟した動物では、主に神経組織中に存在する。神経胚形成、軸索成長(outgrowth)、網膜の組織形成、および嗅覚系の発達の
ようなプロセスが、NCAMの制御発現と関連付けられる。Crossinら、(1990)Exp. Neurol.,109:5-15;Tosneyら、(1986)Dev.Biol., 114:437-452;Thieryら、(1977)J.Biol. Chem., 252:6841-6845;Keyら、(1990)J.CellBiol., 110:172
9-1743;およびChungら、(1991)J. Comp. Neurol., 314:290-305を参照のこと。遺伝子標的化により作製されたホモ接合NCAM陰性マウスは、健康で、繁殖性である。成熟した変異体は、全脳重量の10%の減少、および36%の嗅球の大きさの減衰を示す。NCAMの欠失は、神経発生および神経形成性と関連すると考えられる炭水化物構造である、タンパク質結合α−(2,8)−連結ポリシアル酸のほとんどすべての欠損を伴う。Theodosisら、(1991)Proc.Natl.Acad. Sci. USA 88:5494-5498を参照のこと。Morris、(1981)Learn. Motiv. 12:239-260に記載されるようなMorris水迷路中での動物試験に
より、空間学習の欠落が示されたが、変異マウスの活動および運動能は正常であった。
【0070】
ホモ接合変異マウスの遺伝子標的化および作製を、標準プロトコルを用いて行い、そしてサザンブロッティングおよび対立遺伝子特異的PCRにより確認した。ヌクレアーゼSI保護アッセイ、ノーザンブロッティング、およびウエスタンブロッティングにより、標的化遺伝子座が無効対立遺伝子として確認された。モノクローナル抗体およびポリクローナル血清を用いた、NCAMについての脳切片の免疫細胞化学分析により、野生型動物およびヘテロ接合動物中の嗅球の糸球体および顆粒細胞層に最も強い染色が示された。すべての染色は、成熟した脳におけるNCAM発現についての報告とよく一致した。Chungら
、(1991)を参照のこと。ホモ接合変異マウスでは、予想通りNCAM免疫反応性が全欠損していた。
【0071】
2種の系統のヘテロ接合動物をつがいにし、そして78匹の子を得た。これらのうち交配した対立遺伝子について、38匹の動物(49%)がヘテロ接合型であり。22匹(28%)が野生型であり、そして18匹(23%)がホモ接合型であり、ほとんど完全なメンデル分配を示した。ホモ接合変異動物は繁殖性であり、そして、これらの動物は野生型およびヘテロ接合体の同腹子よりも約10重量%小さいが、生後4カ月まで健康である。変異体およびヘテロ接合動物から分離した脳は、明らかに解剖学的差異を有した:嗅球は、+/+動物および+/−動物と比較して、変異体では大きさが減少し;そして脳重量は体重の補正後約10%まで減少した。
【0072】
NCAMに対して高親和性のAMABを生成させるために、上記のマウスに適切なアジュバントに懸濁した抗原量のNCAMを接種した。追加免疫注射を必要に応じて与え、そして抗体力価を定期的に測定する。一旦、力価が充分であると、AMAB生成のための適切な方法が続けられる。
【0073】
(発明の要旨)
本発明は具体的には以下を提供する。
(1) 以下の工程を包含する、自己抗原に対するモノクローナル性自己抗体を非ヒト脊椎動物宿主から得る方法:a)上記自己抗原の少なくとも1つのエピトープを産生しないように上記宿主動物のゲノムを改変する工程;
b)上記自己抗原またはそのホモログを含有する組成物で上記宿主動物を免疫する工程;
c)上記自己抗原またはそのホモログに応答して産生される宿主細胞であって、上記自己抗原またはそのホモログに対する抗体を発現する上記宿主細胞を収集する工程;およびd)上記収集した細胞または上記細胞由来の遺伝学的材料を用いて抗体を産生する工程。
(2) 上記宿主動物が少なくとも1つのエピトープの機能的等価物を産生する、項目1に記載の方法。
(3) 上記宿主動物が少なくとも1つのエピトープの構造的ホモログを産生する、項目1に記載の方法。
(4) 上記自己抗原が、タンパク質、ペプチド、炭水化物、脂質、酵素補因子、および天然に生じる凝集物またはそれらの共有結合複合物からなる群より選択される、項目1に記載の方法。
(5) 上記自己抗原が細胞表面抗原である、項目4に記載の方法。
(6) 上記自己抗原がサイトカインである、項目4に記載の方法。
(7) 上記自己抗原が細胞接着分子およびイムノグロブリンからなる群より選択される、項目5に記載の方法。
(8) 上記自己抗原がIgD抗体である、項目7に記載の方法。
(9) 上記自己抗原が細胞接着分子である、項目7に記載の方法。
(10) 上記細胞の収集が、抗原特異的B細胞を富化または単離する工程をさらに包含する、項目1に記載の方法。
(11) 上記抗体が、さらに上記抗体をコードする組換え遺伝子を発現することにより産生される、項目1に記載の方法。
(12) 上記組換え遺伝子が、単一の予め富化した抗原特異的B細胞および単一細胞PCRを用いて単離される、項目11に記載の方法。
(13) 上記抗体が少なくとも一部でヒト化されている、項目1に記載の方法。
(14) 上記宿主動物が哺乳類であり、そして、齧歯類、兎類、霊長類、食肉類、奇蹄類、および偶蹄類からなる群より選択される、項目1に記載の方法。
(15) 上記宿主動物が齧歯類またはネズミである、項目14に記載の方法。
(16) 上記宿主動物がマウスである、項目15に記載の方法。
(17) 遺伝的改変が生殖系列伝達を介して得られるように、上記工程b)の前に、遺伝的に操作された宿主動物を繁殖させる工程をさらに包含する、項目1に記載の方法。
(18) 上記抗原またはそのホモログの生合成が、上記抗原またはそのホモログを産生する代謝経路に影響を与える酵素をコードする遺伝子の変異により排除される、項目1に記載の方法。
(19) 項目1に記載の方法により得られる抗体を含有する組成物。
(20) ELISAに使用するための、項目19に記載の組成物。
(21) 項目1に記載の方法により得られる抗体フラグメントを含有する組成物であって、上記フラグメントが上記自己抗原に結合するに十分である、組成物。
(22) 上記フラグメントがFabフラグメントである、項目21に記載の組成物。
(23) 上記抗体が他の化合物と複合体化される、項目20に記載の組成物。
(24) 上記化合物が、リポーター酵素、蛍光色素、金属化合物、毒素、ハプテン、磁性粒子、固体キャリア、リポソーム、および蛍光粒子からなる群より選択される、項目23に記載の組成物。
(25) 上記磁性粒子がコロイド超常磁性粒子である、項目24に記載の組成物。
(26) 上記化合物が、アルカリホスファターゼ、ペルオキシダーゼ、およびウレアーゼからなる群より選択される酵素である、項目24に記載の組成物。
(27) 上記化合物が、フルオレセインイソチオシアネート、テトラメチルローダミンイソチオシアネート、フィコエリトリン、アロフィコシアニン、およびテキサスレッドからなる群より選択される蛍光色素である、項目24に記載の組成物。
(28) 上記化合物が、フェリチン、金コロイド、および磁性ビーズからなる群より選択される金属化合物である、項目24に記載の組成物。
(29) 上記化合物が放射性同位元素を含有する、項目23に記載の組成物。
(30) 上記化合物がビオチンである、項目23に記載の組成物。
(31) 以下の工程を包含する、細胞表面抗原の存在に基づいて細胞を分離する方法:a)項目1に記載の方法により細胞表面抗原に対する結合特異性を有するモノクローナル性自己抗体を得る工程;
b)上記抗体を異種細胞集団と接触させる工程であって、ここで、上記細胞の亜群が、上記抗体が結合特異性を有する上記細胞表面抗原を発現する、工程;およびc)上記抗体により認識される細胞を、上記抗体により認識されない細胞から分離する工程。
(32) 上記抗体がコロイド超常磁性粒子とともに用いられ、そして上記細胞が高勾配磁性分離を用いて単離される、項目31に記載の方法。
(33) 上記抗体がコロイド超常磁性粒子に結合される、項目32に記載の方法。
(34) 上記細胞の亜群が上記抗体に結合され、そして上記抗体が抗イムノグロブリン特異的コロイド超常磁性粒子により認識される、項目32に記載の方法。
(35) 上記抗体がハプテンにより改変され、そして上記コロイド超常磁性粒子が上記ハプテン改変物を認識する、項目32に記載の方法。
【0074】
本明細書中に引用したすべての刊行物および特許出願は、個々の刊行物または特許出願が具体的かつ個別に参考として援用されているのと同じ程度まで、参考として本明細書中に援用されている。
【0075】
上記発明は、明瞭にそして理解されることを目的とする例示および実施例により詳細に記載されているが、所定の改変が実施され得ることは当業者に明らかである。従って、説明および実施例は本発明の範囲を限定するように解釈されるべきではなく、本発明の範囲は添付の請求の範囲により記載される。
【図面の簡単な説明】
【0076】
【図1】B細胞富化領域中のマウス脾臓組織の抗IgD抗体染色を表す。
【図2】(a)は、マウス脾臓組織におけるδ1.3 AMABの滴定の結果を表し、(b)は、マウス脾臓細胞におけるδ3.5 AMABの滴定の結果を表す。
【図3】抗IgM抗体および抗IgD抗体で二重染色したマウス脾臓細胞のFACS分析の結果を表す。
【図4】(a)はδ1.3におけるコロイド超常磁性粒子に結合した抗IgD抗体を用いたマウス脾臓細胞の分離前の結果を表し、(b)はδ3.5における分離前の結果を表し、(c)はδ1.3における分離後の陰性画分の結果を表し、(d)はδ3.5における分離後の陰性画分の結果を表し、(e)はδ1.3における分離後の陽性画分の結果を表し、そして(f)はδ3.5における分離後の陽性画分の結果を表す。
【特許請求の範囲】
【請求項1】
以下の工程:
a)該自己抗原の少なくとも1つのエピトープを産生しないようにゲノムが改変された動物を得る工程;
b)該動物を免疫して該エピトープに対する抗体応答を惹起させる工程;
c)該エピトープに応答して産生される細胞であって、該エピトープに対する抗体を発現する該宿主細胞を収集する工程;および
d)該抗体産生細胞を不死化して該エピトープに対するハイブリドーマを作製する工程
e)該収集した細胞または該細胞由来の遺伝学的材料を用いて抗体を産生する工程
を包含する、自己抗原のエピトープに対するモノクローナル性自己抗体を非ヒト脊椎動物宿主から得る方法により得られる自己抗原のエピトープに対する抗体を含有する組成物。
【請求項2】
ELISAに使用するための、請求項1に記載の組成物。
【請求項3】
以下の工程:
a)該自己抗原の少なくとも1つのエピトープを産生しないようにゲノムが改変された動物を得る工程;
b)該動物を免疫して該エピトープに対する抗体応答を惹起させる工程;
c)該エピトープに応答して産生される細胞であって、該エピトープに対する抗体を発現する該宿主細胞を収集する工程;および
d)該抗体産生細胞を不死化して該エピトープに対するハイブリドーマを作製する工程
e)該収集した細胞または該細胞由来の遺伝学的材料を用いて抗体を産生する工程
を包含する、自己抗原のエピトープに対するモノクローナル性自己抗体を非ヒト脊椎動物宿主から得る方法により得られる自己抗原のエピトープに対する抗体のフラグメントを含有する組成物であって、該フラグメントが前記自己抗原に結合するに十分である、組成物。
【請求項4】
前記フラグメントがFabフラグメントである、請求項3に記載の組成物。
【請求項5】
前記抗体が他の化合物と複合体化される、請求項2に記載の組成物。
【請求項6】
前記化合物が、リポーター酵素、蛍光色素、金属化合物、毒素、ハプテン、磁性粒子、固体キャリア、リポソーム、および蛍光粒子からなる群より選択される、請求項5に記載の組成物。
【請求項7】
前記磁性粒子がコロイド超常磁性粒子である、請求項6に記載の組成物。
【請求項8】
前記化合物が、アルカリホスファターゼ、ペルオキシダーゼ、およびウレアーゼからなる群より選択される酵素である、請求項7に記載の組成物。
【請求項9】
前記化合物が、フルオレセインイソチオシアネート、テトラメチルローダミンイソチオシアネート、フィコエリトリン、アロフィコシアニン、およびテキサスレッドからなる群より選択される蛍光色素である、請求項6に記載の組成物。
【請求項10】
前記化合物が、フェリチン、金コロイド、および磁性ビーズからなる群より選択される金属化合物である、請求項6に記載の組成物。
【請求項11】
前記化合物が放射性同位元素を含有する、請求項5に記載の組成物。
【請求項12】
前記化合物がビオチンである、請求項5に記載の組成物。
【請求項1】
以下の工程:
a)該自己抗原の少なくとも1つのエピトープを産生しないようにゲノムが改変された動物を得る工程;
b)該動物を免疫して該エピトープに対する抗体応答を惹起させる工程;
c)該エピトープに応答して産生される細胞であって、該エピトープに対する抗体を発現する該宿主細胞を収集する工程;および
d)該抗体産生細胞を不死化して該エピトープに対するハイブリドーマを作製する工程
e)該収集した細胞または該細胞由来の遺伝学的材料を用いて抗体を産生する工程
を包含する、自己抗原のエピトープに対するモノクローナル性自己抗体を非ヒト脊椎動物宿主から得る方法により得られる自己抗原のエピトープに対する抗体を含有する組成物。
【請求項2】
ELISAに使用するための、請求項1に記載の組成物。
【請求項3】
以下の工程:
a)該自己抗原の少なくとも1つのエピトープを産生しないようにゲノムが改変された動物を得る工程;
b)該動物を免疫して該エピトープに対する抗体応答を惹起させる工程;
c)該エピトープに応答して産生される細胞であって、該エピトープに対する抗体を発現する該宿主細胞を収集する工程;および
d)該抗体産生細胞を不死化して該エピトープに対するハイブリドーマを作製する工程
e)該収集した細胞または該細胞由来の遺伝学的材料を用いて抗体を産生する工程
を包含する、自己抗原のエピトープに対するモノクローナル性自己抗体を非ヒト脊椎動物宿主から得る方法により得られる自己抗原のエピトープに対する抗体のフラグメントを含有する組成物であって、該フラグメントが前記自己抗原に結合するに十分である、組成物。
【請求項4】
前記フラグメントがFabフラグメントである、請求項3に記載の組成物。
【請求項5】
前記抗体が他の化合物と複合体化される、請求項2に記載の組成物。
【請求項6】
前記化合物が、リポーター酵素、蛍光色素、金属化合物、毒素、ハプテン、磁性粒子、固体キャリア、リポソーム、および蛍光粒子からなる群より選択される、請求項5に記載の組成物。
【請求項7】
前記磁性粒子がコロイド超常磁性粒子である、請求項6に記載の組成物。
【請求項8】
前記化合物が、アルカリホスファターゼ、ペルオキシダーゼ、およびウレアーゼからなる群より選択される酵素である、請求項7に記載の組成物。
【請求項9】
前記化合物が、フルオレセインイソチオシアネート、テトラメチルローダミンイソチオシアネート、フィコエリトリン、アロフィコシアニン、およびテキサスレッドからなる群より選択される蛍光色素である、請求項6に記載の組成物。
【請求項10】
前記化合物が、フェリチン、金コロイド、および磁性ビーズからなる群より選択される金属化合物である、請求項6に記載の組成物。
【請求項11】
前記化合物が放射性同位元素を含有する、請求項5に記載の組成物。
【請求項12】
前記化合物がビオチンである、請求項5に記載の組成物。
【図1】


【図2】
【図3】

【図4】
【図2】
【図3】
【図4】
【公開番号】特開2006−151991(P2006−151991A)
【公開日】平成18年6月15日(2006.6.15)
【国際特許分類】
【出願番号】特願2006−4229(P2006−4229)
【出願日】平成18年1月11日(2006.1.11)
【分割の表示】特願平7−87269の分割
【原出願日】平成7年4月12日(1995.4.12)
【出願人】(595053892)
【氏名又は名称原語表記】STEFAN MILTENYI
【住所又は居所原語表記】Friedrich−Ebert−Straβe 68 51429 Bergisch Gladbach Germany
【Fターム(参考)】
【公開日】平成18年6月15日(2006.6.15)
【国際特許分類】
【出願日】平成18年1月11日(2006.1.11)
【分割の表示】特願平7−87269の分割
【原出願日】平成7年4月12日(1995.4.12)
【出願人】(595053892)
【氏名又は名称原語表記】STEFAN MILTENYI
【住所又は居所原語表記】Friedrich−Ebert−Straβe 68 51429 Bergisch Gladbach Germany
【Fターム(参考)】
[ Back to top ]