
花粉特異的ジンクフィンガー転写因子の遺伝子を用いて花粉稔性を低下させる方法
【課題】雄性不稔に代表される植物形質の改変に有用な、花粉に特異的な遺伝子を利用する遺伝子工学的手法の提供。
【解決手段】形質が改変された植物を作出する方法であって、特定の塩基配列のDNAを有し、小胞子および任意に葯の開裂組織に、特異的なプロモーター活性を示すDNAのいずれかを含むプロモーターと、このプロモーターに作動可能に連結された異種遺伝子とを含む、植物発現カセットを利用する方法。さらに、上記プロモーター並びに上記プロモーター及び異種遺伝子を含む植物に雄性不稔性を付与するために有用な植物発現カセット。
【解決手段】形質が改変された植物を作出する方法であって、特定の塩基配列のDNAを有し、小胞子および任意に葯の開裂組織に、特異的なプロモーター活性を示すDNAのいずれかを含むプロモーターと、このプロモーターに作動可能に連結された異種遺伝子とを含む、植物発現カセットを利用する方法。さらに、上記プロモーター並びに上記プロモーター及び異種遺伝子を含む植物に雄性不稔性を付与するために有用な植物発現カセット。
【発明の詳細な説明】
【技術分野】
【0001】
本発明は、雄しべの花粉形成組織に特異的に発現する遺伝子およびその利用に関する。本発明は、特に、小胞子に特異的に発現する、ペチュニア由来のジンクフィンガー型転写因子群(ZPT2−5、ZPT3−1およびZPT4−1)の遺伝子およびその利用に関する。
【背景技術】
【0002】
花粉の稔性は、農業・園芸における様々な局面で問題になる。たとえば、交雑育種における交配の際、自家受粉を避けるために、多くの労力がかかる除雄(雄しべの除去)の作業が行われる。種苗産業においては、交雑育種によって得られた優れた品種を商業的に保護したいという立場から、花粉稔性の無い形質が求められている。このような要請から、花粉の稔性を制御する技術(pollination control)が強く求められてきた。従来、特定の作物では、細胞質雄性不稔性の系統が交雑育種に用いられ、一定の成功を収めてきた。しかし、細胞質不稔形質は多くの場合、耐病性の低下などの好ましくない副次効果を伴い、さらに形質が不安定であること、種子の大量生産が困難なことなどの問題がある。化学薬品処理によって稔性を低下させる方法も研究されているが、安全性評価および作用機作解明が遅れており、実用化には至っていない。そこで、遺伝子工学を用いた優れた雄性不稔化技術が求められている。
【0003】
花粉は種子植物における雄性の配偶体である。支持体組織としての葯に取り囲まれた状態で進行する花粉の発達は、小胞子形成細胞(花粉母細胞)の減数分裂直後にあたる四分子期、四分子からの小胞子の放出期、花粉細胞の拡大および空胞形成に特徴付けられる一核期、有糸分裂により栄養細胞および生殖細胞への分化が生じる有糸細胞分裂期、その後の二核期という、各段階に区分される。これらの段階を経て、最終的に葯が開裂し、成熟した花粉粒が放出される。従って、小胞子は、花粉の発達を阻害し、花粉稔性を失わせるための人為的な制御に最も適した対象組織の一つといえる。
【0004】
上記のように、遺伝子工学による雄性不稔化技術には大きな期待が寄せられている。特に、小胞子のような花粉細胞の直接の前駆体において特異的に発現する遺伝子を利用できれば、植物に好ましくない形質を付与することなく、雄性不稔化を達成し得る可能性が高いと考えられる。雄しべの種々の組織に特異的なプロモーター、および、それを含む雄生不稔化のための遺伝子構築物の例は、いくつか報告されている(非特許文献1)。しかし、発現の組織特異性および時期特異性が高い、花粉稔性の制御のために有用な新たな遺伝子に対する要望が継続的に存在している。
【0005】
本発明者らは、最近、ペチュニア由来の新規な転写因子として、PEThyZPT2−5、 PEThyZPT3−1、およびPEThyZPT4−1(以下、それぞれ、ZPT2−5、ZPT3−1およびZPT4−1と略す)を含む7つのジンクフィンガー(ZF)型転写因子のcDNA配列を特定した。そして、ノーザンブロット分析から、それぞれの転写因子が葯特異的に、葯の発達の異なる段階において一過性の発現を示すことを報告した(非特許文献2)。しかし、これらの転写因子の植物における生理的な機能および作用、ならびに転写因子をコードする遺伝子の正確な発現部位およびその発現調節機構は不明であった。
【非特許文献1】ShivannaおよびSawhney編, Pollen biotechnology for crop production and improvement (Cambridge University Press)1997年、pp.237−257
【非特許文献2】Kobayashiら,Plant J.,1998年、13:571
【発明の開示】
【発明が解決しようとする課題】
【0006】
本発明の目的は、雄性不稔に代表される植物形質の改変に有用な、花粉に特異的な遺伝子を利用する遺伝子工学的手法を提供することにある。
【課題を解決するための手段】
【0007】
本発明者らは、以前にペチュニアから単離した、葯特異的な転写因子群(ZPT2−5、ZPT3−1およびZPT4−1)をコードする遺伝子をペチュニアに再導入した。その結果、花粉の正常な発達が阻害され、花粉の稔性が著しく低下するか(ZPT2−5およびZPT4−1)、または、ほぼ完全に失われる(ZPT3−1)ことを見出した。さらに本発明者らは、ZPT3−1およびZPT4−1のゲノム遺伝子からそれぞれ上流領域を単離し、そのプロモーター活性の組織特異性を調べた。その結果、一核期から二核期にかけての小胞子において、プロモーター活性が組織および時期特異的に発現されることを見出した。本発明は、これらの知見に基づいて完成された。
【0008】
本発明は、その第1の局面において、雄性不稔性の植物を作出する方法であって、以下の工程を包含する方法に関する:(i)配列番号1によって示される塩基配列の第1位から第777位までの配列を有するDNA、(ii)(i)の塩基配列を有するDNAとストリンジェントな条件下でハイブリダイズし、花粉の発達を制御する転写因子をコードするDNA、または(iii)(i)もしくは(ii)の断片であるDNAのいずれかである核酸と、上記核酸に作動可能に連結されたプロモーターとを含む、植物発現カセットを提供する工程;花粉の発達を制御する内因性の転写因子を有する植物の細胞を提供する工程であって、ここで、内因性の転写因子をコードする遺伝子は、上記核酸とストリンジェントな条件下でハイブリダイズする、工程;発現カセットを植物細胞に導入する工程;発現カセットが導入された植物細胞を植物体に再生する工程;および、再生された植物体であって、上記核酸が発現されることによって、内因性の転写因子の発現が抑制された植物体を選択する工程。
【0009】
本発明は、その第2の局面において、雄性不稔性の植物を作出する方法であって、以下の工程を包含する方法に関する:(i’)配列番号3によって示される塩基配列の第1位から第1640位までの配列を有するDNA、(ii’)(i’)の塩基配列を有するDNAとストリンジェントな条件下でハイブリダイズし、花粉の発達を制御する転写因子をコードするDNA、または(iii’)(i’)もしくは(ii’)の断片であるDNAのいずれかである核酸と、上記核酸に作動可能に連結されたプロモーターとを含む、植物発現カセットを提供する工程;花粉の発達を制御する内因性の転写因子を有する植物の細胞を提供する工程であって、ここで、内因性の転写因子をコードする遺伝子は、上記核酸とストリンジェントな条件下でハイブリダイズする、工程;発現カセットを植物細胞に導入する工程;発現カセットが導入された植物細胞を植物体に再生する工程;および、再生された植物体であって、上記核酸が発現されることによって、内因性の転写因子の発現が抑制された植物体を選択する工程。
【0010】
本発明は、その第3の局面において、雄性不稔性の植物を作出する方法であって、以下の工程を包含する方法に関する:(i”)配列番号5によって示される塩基配列の第1位から第1948位までの配列を有するDNA、(ii”)(i”)の塩基配列を有するDNAとストリンジェントな条件下でハイブリダイズし、花粉の発達を制御する転写因子をコードするDNA、または(iii”)(i”)もしくは(ii”)の断片であるDNAのいずれかである核酸と、上記核酸に作動可能に連結されたプロモーターとを含む、植物発現カセットを提供する工程;花粉の発達を制御する内因性の転写因子を有する植物の細胞を提供する工程であって、ここで、内因性の転写因子をコードする遺伝子は、上記核酸とストリンジェントな条件下でハイブリダイズする、工程;発現カセットを植物細胞に導入する工程;発現カセットが導入された植物細胞を植物体に再生する工程;および、再生された植物体であって、上記核酸が発現されることによって、内因性の転写因子の発現が抑制された植物体を選択する工程。
【0011】
ただし、上記(ii)、(ii’)および(ii”)のDNAは、それぞれ、(iv)配列番号13によって示される塩基配列の第1位から第1886位までの配列を有するDNAとストリンジェントな条件下でハイブリダイズし、花粉の発達を制御する転写因子をコードするDNAを含まない。配列番号13によって示される塩基配列は、ペチュニアから単離された別の転写因子ZPT3−2をコードするcDNA配列である(Kobayashiら、前出)。
【0012】
本発明の上記第1から第3までの局面における方法は、植物に雄性不稔性を付与する方法として、利用され得る。
【0013】
上記第1から第3までの局面における1つの実施態様において、上記核酸は上記プロモーターに対して順方向で連結され、上記植物体の細胞内でセンス方向に転写され得る。
【0014】
上記第1から第3までの局面における1つの実施態様において、上記核酸は上記プロモーターに対して逆方向で連結され、上記植物体の細胞内でアンチセンス方向に転写され得る。
【0015】
上記第1から第3までの局面における1つの実施態様において、上記植物は双子葉植物である。双子葉植物は、好ましくはナス科植物であり、より好ましくはペチュニア属植物である。
【0016】
上記第1から第3までの局面における1つの実施態様において、上記発現カセットは植物発現ベクターに組み込まれている。
【0017】
本発明の第1から第3までの局面によればまた、上記いずれかに記載の方法により作出された、雄性不稔性の植物が提供される。
【0018】
本発明は、その第4の局面において、形質が改変された植物を作出する方法であって、以下の工程を包含する方法に関する:(a’)配列番号7によって示される塩基配列の第1位から第2624位までの配列を有するDNA、または(b’)(a’)の一部の配列を有し、小胞子に特異的なプロモーター活性を示すDNAのいずれかを含むプロモーターと、上記プロモーターに作動可能に連結された異種遺伝子とを含む、植物発現カセットを提供する工程;発現カセットを植物細胞に導入する工程;および、発現カセットが導入された植物細胞を、植物体に再生する工程。
【0019】
本発明は、その第5の局面において、形質が改変された植物を作出する方法であって、以下の工程を包含する方法に関する:(a”)配列番号8によって示される塩基配列の第1位から第3631位までの配列を有するDNA、または(b”)(a”)の一部の配列を有し、小胞子に、および任意に葯の開裂組織に、特異的なプロモーター活性を示すDNAのいずれかを含むプロモーターと、上記プロモーターに作動可能に連結された異種遺伝子とを含む、植物発現カセットを提供する工程;発現カセットを植物細胞に導入する工程;および、発現カセットが導入された植物細胞を、植物体に再生する工程。
【0020】
上記第4および第5の局面における1つの実施態様において、上記形質は稔性であり、形質が改変された植物は、雄性不稔植物である。従って、本発明の上記方法は、植物に雄性不稔性を付与する方法として、利用され得る。
【0021】
上記第4および第5の局面における1つの実施態様において、上記形質は和合性であり、形質が改変された植物は自家不和合性植物である。従って、本発明の上記方法は、植物に自家不和合性を付与する方法としても、利用され得る。
【0022】
上記第4および第5の局面における1つの実施態様において、上記植物は双子葉植物である。双子葉植物は、好ましくはナス科植物であり、より好ましくはペチュニア属植物である。
【0023】
上記第4および第5の局面における1つの実施態様において、上記発現カセットは植物発現ベクターに組み込まれている。
【0024】
本発明の第4および第5の局面によればまた、上記いずれかに記載の方法により作出された、形質が改変された植物が提供される。
【0025】
本発明は、その第6の局面において、以下の(I’)または(II’)のDNAを含むプロモーターに関する:(I’)配列番号7によって示される塩基配列の第1位から第2624位までの配列を有するDNA、または(II’)(I’)の一部の配列を有し、小胞子に特異的なプロモーター活性を示すDNA。
【0026】
本発明は、その第7の局面において、以下の(I”)または(II”)のDNAを含むプロモーターに関する:(I”)配列番号8によって示される塩基配列の第1位から第3631位までの配列を有するDNA、または(II”)(I”)の一部の配列を有し、小胞子に、および任意に葯の開裂組織に特異的なプロモーター活性を示すDNA。
【0027】
本発明は、その第8の局面において上記の小胞子に特異的なプロモーターのいずれかと、上記プロモーターに作動可能に連結された異種遺伝子とを含む、植物に雄性不稔性を付与するために有用な植物発現カセットに関する。
【発明を実施するための最良の形態】
【0028】
本発明について、以下に、より詳細に説明する。
(ZPT3−2、ZPT3−1およびZPT4−1遺伝子由来の転写因子)
本発明の第1から第3までの局面である、雄性不稔性の植物を作出する方法において有用な核酸は、以下のいずれかである:
(i)配列番号1によって示される塩基配列の第1位から第777位までの配列を有するDNA、
(i’)配列番号3によって示される塩基配列の第1位から第1640位までの配列を有するDNA、
(i”)配列番号5によって示される塩基配列の第1位から第1948位までの配列を有するDNA、または、
(i)〜(i”)のいずれかの塩基配列を有するDNAとストリンジェントな条件下でハイブリダイズし、花粉の発達を制御する転写因子をコードするDNA(すなわち、(ii)、(ii’)または(ii”))、または、
上記いずれかのDNAの断片(すなわち、(iii)、(iii’)または(iii”))であるDNA。
【0029】
本発明における上記核酸は、好ましくは(i)、(i’)または(i”)のDNA、すなわち、ZPT2−5、ZPT3−1またはZPT4−1をコードするDNA、または、その断片であり、より好ましくは(i)、(i’)または(i”)のDNAである。
【0030】
本明細書において「転写因子」とは、遺伝子の調節領域のDNAに結合してmRNAの合成を制御するタンパク質をいう。ある種の転写因子は、そのDNA結合ドメインにジンクフィンガー(ZF)モチーフと呼ばれる保存性の高いアミノ酸配列を有することが知られている。ZPT2−5は、Cys2/His2型(EPFファミリー)のジンクフィンガー(ZF)タンパク質であって、176アミノ酸からなる全長のアミノ酸配列内に2つのZFモチーフを含み、さらにDLNL配列と称される疎水性領域を含む転写因子である。ZPT3−1は、同様に、EPFファミリーのZFタンパク質であって、437アミノ酸からなる全長のアミノ酸配列内に3つのZFモチーフを含み、さらにDLNL配列を含む転写因子である。ZPT4−1は、同様に、EPFファミリーのZFタンパク質であって、474アミノ酸からなる全長のアミノ酸配列内に4つのZFモチーフを含み、さらにDLNL配列を含む転写因子である。いずれも、Kobayashiら(前出)を参照のこと。ZPT2−5、ZPT3−1およびZPT4−1をコードするcDNA配列(配列番号1、3および5)を、それぞれ、対応する推定アミノ酸配列(配列番号2、4および6)と共に図1A〜1B、図2A〜2Cおよび図3A〜3Dに示す。
【0031】
本明細書において、核酸またはDNAの「断片」とは、植物体に導入され、適切な様式で発現されたときに、当該植物の内因性の転写因子の発現を抑制し得る断片をいう。この断片は、上記(i)、(i’)、(i”)、(ii)、(ii’)または(ii”)のDNAの、ジンクフィンガーモチーフをコードする領域以外から選択され、少なくとも約40塩基対以上、好ましくは約50塩基対以上、より好ましくは約70塩基対以上、さらに好ましくは約100塩基対以上の長さを有する。
【0032】
本明細書において、ハイブリダイゼーションのための「ストリンジェントな条件」とは、特定の塩基配列(例えば、ペチュニア由来のZPT2−5、ZPT3−1またはZPT4−1をコードするDNA)と、これと相同性の高い他の塩基配列(例えば、ペチュニア以外の植物に存在する、ZPT2−5、ZPT3−1またはZPT4−1のホモログをコードするDNA)とのオリゴヌクレオチド二本鎖を形成させるのに十分な条件を意図する。本発明に適用されるストリンジェントな条件の代表的な例として、次の条件が挙げられる:1M NaCl、1%SDS、10%デキストラン硫酸、32P標識プローブDNA(1x107cpm)および50μg/mlサケ精子DNAを含む溶液中で、60℃で16時間ハイブリダイズさせた後、2xSSC/1%SDSで60℃、30分間の洗浄を2回繰り返す。
【0033】
本発明における、ZPT2−5、ZPT3−1およびZPT4−1をコードするDNA、およびこれらとストリンジェントな条件下でハイブリダイズし、花粉の発達を制御する転写因子をコードするDNAを単離するためには、公知の転写因子の遺伝子によりコードされるアミノ酸配列の保存領域に対応するデジェネレートプライマー対を用い得る。このプライマー対を用いて、植物のcDNAまたはゲノミックDNAを鋳型としてPCRを行い、その後、得られた増幅DNA断片をプローブとして用いて、同じ植物のcDNAライブラリーまたはゲノミックライブラリーをスクリーニングすることができる。そのようなプライマー対の例として、5’−CARGCNYTNGGNGGNCAY−3’(配列番号9)と、3’−RTGNCCNCCNARNGCYTG−5’(配列番号10)との組合せが挙げられる(ここで、Nはイノシンであり、RはGまたはAであり、そしてYはCまたはTである)。上記プライマー配列は、それぞれ、上記ZPT転写因子群のジンクフィンガーモチーフに含まれるアミノ酸配列QALGGHに対応する。
【0034】
従って、本発明に適用されるストリンジェントなハイブリダイゼーション条件はまた、PCRの条件としても規定され得、代表的な例においては、上記デジェネレートプライマー(配列番号9および10)が用いられ得る。この際のPCR反応条件は、94℃で5分間の変性の後、94℃で30秒間、50℃で30秒間、そして72℃で1分間を30サイクル繰り返し、最後に、72℃で7分間インキュベーションを行う条件であり得る。
【0035】
PCRは、市販のキットおよび装置の製造者の指針に基づいて行うか、当業者に周知の手法で行い得る。遺伝子ライブラリーの作製法、および遺伝子のクローニング法なども当業者に周知である。例えば、Sambrookら,Molecular Cloning:A Laboratory Manual、第2版(Cold Spring Harbor Laboratory,1989)を参照のこと。得られた遺伝子の塩基配列は、当該分野で公知のヌクレオチド配列解析法または市販されている自動シーケンサーを利用して決定し得る。
【0036】
本明細書において、転写因子が「花粉の発達を制御する」とは、代表的には、この転写因子の発現が阻害されるとき、花粉の形態または機能において有意な変化が観察されることをいう。本発明における転写因子は、代表的には、それをコードする遺伝子の発現が阻害されることにより、好ましくは約75%以上、より好ましくは約90%以上、さらに好ましくは約95%以上の花粉細胞を、成熟する前に死滅せしめる。ここで、転写因子の発現が阻害されるとは、ノーザンブロット法により測定したとき、mRNAの量が、野生型のコントロール植物と比較して約10分の1以下になることをいう。
【0037】
上記のようなスクリーニングによって単離、同定された遺伝子によってコードされる転写因子(すなわち、ZPT2−5、ZPT3−1およびZPT4−1、およびこれらのホモログ)が花粉の発達を制御することは、本明細書の開示に従って形質転換植物を作出し、その花粉の特性を観察することによって、確認し得る。
【0038】
本発明においては、花粉の発達を制御する転写因子をコードするDNAを利用して、植物細胞において、当該DNAと同一のまたは相同な塩基配列を含む内因性遺伝子の発現を抑制し得る。標的となる内因性遺伝子もまた、花粉の発達を制御する転写因子である。本発明の方法によれば、好ましくは、花粉の発達を制御する内因性転写因子以外の発現を実質的に抑制することなく、内因性転写因子の発現のみを選択的に抑制することによって、植物に雄性不稔性が付与される。
【0039】
すなわち、本発明における発現抑制技術が適用される植物細胞は、花粉の発達を制御する内因性の転写因子を有する植物の細胞であって、この内因性の転写因子をコードする遺伝子は、上記ZPT2−5、ZPT3−1またはZPT4−1、またはこれらのホモログのDNAとストリンジェントな条件下でハイブリダイズするものとして規定される。ここで「ストリンジェントな条件」の定義は、ZPT2−5、ZPT3−1およびZPT4−1のホモログの特定に関連して上述した条件と同じである。本発明の範囲を限定する意図ではないが、上記方法によって雄性不稔性を付与し得る植物は、上記ZPT遺伝子群が単離されたペチュニア、または上記ZPTホモログをコードする遺伝子が単離された植物と、進化系統学上近縁の植物であることが望ましい。ここで、進化系統学上近縁とは、代表的には同じ目に分類され、好ましくは同じ科に分類され、より好ましくは同じ属に分類され、さらに好ましくは同じ種に分類される植物である。もっとも、花粉の発達が種子植物の生殖に不可欠である事実を考慮すれば、ZPT2−5、ZPT3−1およびZPT4−1と同一または類似の機能を果たす転写因子が他の植物にも広く存在し得ることは容易に理解される。
【0040】
内因性遺伝子の発現を抑制するための手法として、代表的にはコサプレッションおよびアンチセンス技術を利用し得る。コサプレッションは、植物細胞に組み換え遺伝子を導入したとき、その遺伝子自体、およびその遺伝子の一部と相同な配列を含む内因性遺伝子の両者の発現が抑制されることをいう。コサプレッションが利用される場合、本発明における発現カセットは、転写因子をコードするDNAまたはその断片を、プロモーターに対して順方向で連結された形態で含む。転写因子をコードするDNAまたはその断片は、発現カセットとして植物細胞に導入された後、プロモーターの制御下でセンス方向に転写され得る。この導入DNAの作用により、所望とする遺伝子発現の抑制が可能となる。コサプレッションは、形質転換植物の一部の個体で認められ、他の個体では十分に起こらない場合が多い。そのため、通常、遺伝子発現が意図されたように抑制された個体が、慣用的な手順に従って選択される。
【0041】
アンチセンスは、植物細胞に組み換え遺伝子を導入したとき、導入遺伝子の転写産物(mRNA)が、内因性遺伝子の転写産物(mRNA)の相補的な配列とハイブリッドを形成して、内因性遺伝子がコードするタンパク質の翻訳が阻害されることをいう。アンチセンスが利用される場合、本発明における発現カセットは、転写因子をコードするDNAまたはその断片を、プロモーターに対して逆方向で連結された形態で含む。転写因子をコードするDNAまたはその断片は、発現カセットとして植物細胞に導入された後、プロモーターの制御下でアンチセンス方向に転写され得る。このアンチセンス転写物の作用により、所望とする遺伝子発現の抑制が可能となる。
【0042】
(ZPT3−1およびZPT4−1遺伝子由来のプロモーター)
本発明の第4および第5の局面である、形質が改変された植物を作出する方法において有用なプロモーターは、(a’)配列番号7によって示される塩基配列の第1位から第2624位までの配列を有するDNA、(a”)配列番号8によって示される塩基配列の第1位から第3631位までの配列を有するDNA、または、(a’)または(a”)の一部の配列を有し、小胞子に特異的なプロモーター活性を示すDNAのいずれかを含むプロモーターである。本発明における上記プロモーターは、好ましくは(a’)または(a”)のプロモーター、すなわち、ZPT3−1またはZPT4−1遺伝子のプロモーターである。
【0043】
ZPT3−1およびZPT4−1遺伝子のプロモーター領域の、組識特異的な発現活性に必須でない配列を除去して得られる、小胞子に特異的なプロモーター活性を有する配列は、本発明の範囲内にある。このような配列は、常法に従ってプロモーターのデリーション実験を行うことによって、得ることができる。簡潔には、ZPT3−1またはZPT4−1遺伝子のプロモーター領域の様々な欠失変異体(例えば、ZPT3−1またはZPT4−1遺伝子のプロモーター領域を、5’上流側から種々の長さに欠失させた変異体)と、適切なレポーター遺伝子(例えば、GUS遺伝子)とを融合したプラスミドを用いて、欠失変異体の組識特異的プロモーター活性を測定することによって、その活性に必須な領域を特定し得る。
【0044】
いったんプロモーター活性に必須の領域が特定されると、さらにその領域内の配列または隣接配列を改変して、プロモーターの発現活性の程度を高めることも可能である。このようにして得られる改変体もまた、小胞子に特異的なプロモーター活性を示す限り、本発明の範囲内にある。
【0045】
本発明において、「小胞子に特異的なプロモーター活性を示す」とは、プロモーターが、天然の植物中で、または任意の構造遺伝子に連結させた発現カセットとして植物に導入されたときに、DNAの転写を開始させることにより遺伝子の発現を指示する能力を、小胞子において特異的に示すことを意味する。ここで「特異的」とは、同じ植物体の花の、他のすべての組織(タペート層、花糸、花柱、花頭、花弁、萼(がく)などを含む;ただし葯の開裂組織を除く)よりも、プロモーターの発現活性が高いことをいう。上記特異的なプロモーターは、好ましくは、同じ植物体の花の他のすべての組織および花以外の部分(根、葉、茎など)よりも、プロモーターの発現活性が高く、より好ましくは、同じ植物体の花の他のすべての組織および花以外の部分において実質的に活性を示さない。「葯の開裂組織に特異的なプロモーター活性を示す」も、上記と同様に定義される。発現活性の程度は、常法に従って、小胞子におけるプロモーターの発現レベルと、花の他の組識における同じプロモータの発現レベルとを比較することによって、評価され得る。プロモーターの発現レベルは、通常、そのプロモーターの制御下で発現される遺伝子産物の産生量によって決定される。
【0046】
上記の特異的プロモーターを利用する本発明の方法は、植物の生殖に関連する形質を改変することが意図される。ここで「改変」とは、形質転換後の植物における生殖器官の少なくとも一部が、形質転換前の植物(野生種または園芸品種)に存在していた機能を失うか、形質転換前の植物に存在しなかった機能を獲得するか、または形質転換前の植物と比べて特定の機能の程度が増強もしくは低減されることをいう。このような形質の改変は、本発明におけるプロモーターに作動可能に連結された任意の異種遺伝子が、この遺伝子を導入された形質転換植物において、プロモーターの制御下で小胞子に特異的に発現される結果として生じ得る。多くの組織特異的プロモーターにおいて、その組織特異性が種間を越えて保存されていることは周知であるので、本発明のプロモーターもまた幅広い植物種に適用し得ることが容易に理解される。形質の改変の程度は、形質転換後の植物の形質を、形質転換前の植物の形質と比較することにより評価され得る。改変される好ましい形質としては、雌性不稔および自家不和合性が挙げられるが、これらに限定されない。
【0047】
本発明のプロモーターは、例えば、公知のcDNAをプローブとして用いて、植物のゲノミックライブラリーをスクリーニングし、そして対応のゲノミッククローンからコード領域の上流配列を単離することにより得ることができる。cDNAの例として、上述したペチュニア由来の転写因子であるZPT3−1およびZPT4−1のcDNAが挙げられる。
【0048】
本発明におけるプロモーターは、天然から単離されたものに限定されず、合成ポリヌクレオチドも含み得る。合成ポリヌクレオチドは、例えば、上記のようにして配列決定されたプロモーターの配列またはその活性領域を、当業者に周知の手法によって合成または改変することにより入手し得る。
【0049】
(発現カセットおよび発現ベクターの構築)
本発明における転写因子をコードするDNAは、当業者に周知の方法を用いて、適切なプロモーターに作動可能に連結された発現カセットとして、公知の遺伝子組換え技術を用いて、植物細胞に導入され得る。同様に、本発明における小胞子特異的プロモーターは、所望の異種遺伝子に作動可能に連結された発現カセットとして、植物細胞に導入され得る。
【0050】
上記転写因子に連結され得る「プロモーター」は、植物で発現する任意のプロモーターを意味し、構成的プロモーター、組織特異的プロモーター、および誘導性プロモーターのいずれをも含み得る。
【0051】
「構成的プロモーター」は、植物細胞内外の刺激と関係なく一定のレベルで構造遺伝子を発現せしめるプロモーターをいう。異種遺伝子が植物の他の組織または器官で発現しても、植物に望ましくない形質を与えない場合には、構成的プロモーターの使用が簡便であり、好ましい。構成的プロモーターの例としては、カリフラワーモザイクウイルス(CaMV)の35Sプロモーター(P35S)、およびノパリン合成酵素のプロモーター(Tnos)が挙げられるが、これらに限定されない。
【0052】
「組織特異的プロモーター」は、本発明においては、少なくとも小胞子において特異的に構造遺伝子を発現せしめるプロモーターをいう。このような組織特異的プロモーターには、本発明におけるZPT3−1およびZPT4−1遺伝子由来のプロモーターに加えて、葯特異的に発現活性を示す他の公知のプロモーターが含まれる。したがって、小胞子特異的プロモーターと転写因子をコードする配列とを含む、天然のZPT3−1およびZPT4−1遺伝子の発現カセット自体を、必要に応じて他の調節エレメントと組み合わせて、使用することも本発明の範囲に含まれる。
【0053】
「誘導性プロモーター」は、化学薬剤、物理的ストレスなどの特定の刺激を与えたときに構造遺伝子を発現せしめ、刺激の非存在下では発現活性を示さないプロモーターをいう。誘導性プロモーターの例としては、オーキシンで誘導可能な、グルタチオン S−トランスフェラーゼ(GST)プロモーター(van der Kop,D.A.ら、Plant Mol. Biol.,39:979,1999)が挙げられるが、これに限定されない。
【0054】
本明細書において、用語「発現カセット」または「植物発現カセット」とは、本発明における転写因子をコードするDNAと、これに作動可能に(すなわち、当該DNAの発現を制御し得るように)連結された植物発現プロモーターとを含む核酸配列、ならびに、本発明における小胞子特異的プロモーターと、これに作動可能に(すなわち、インフレームに)連結された異種遺伝子とを含む核酸配列をいう。
【0055】
上記の小胞子特異的プロモーターに連結され得る「異種遺伝子」とは、ZPT3−1およびZPT4−1遺伝子以外のペチュニアにおける内因性遺伝子、もしくはペチュニア以外の植物における内因性遺伝子、または植物に対して外来の遺伝子(例えば、動物、昆虫、細菌、および真菌に由来する遺伝子)であって、その遺伝子産物の発現が小胞子において所望される任意の遺伝子をいう。本発明における異種遺伝子の好ましい例は、コードする遺伝子産物が細胞毒性を示し、その発現によって花粉の発達を阻害する遺伝子である。このような遺伝子の具体例としてbarnase遺伝子(Beals,T.P.およびGoldberg,R.B.,Plant Cell,9:1527,1997)が挙げられるが、これに限定されない。
【0056】
「植物発現ベクター」は、発現カセットに加えて、さらに種々の調節エレメントが、宿主植物の細胞中で作動し得る状態で連結されている核酸配列をいう。好適には、ターミネーター、薬剤耐性遺伝子、およびエンハンサーを含み得る。植物発現ベクターのタイプおよび使用される調節エレメントの種類が、宿主細胞に応じて変わり得ることは、当業者に周知の事項である。本発明に用いる植物発現ベクターは、さらにT−DNA領域を有し得る。T−DNA領域は、特にアグロバクテリウムを用いて植物を形質転換する場合に遺伝子の導入の効率を高める。
【0057】
「ターミネーター」は、遺伝子のタンパク質をコードする領域の下流に位置し、DNAがmRNAに転写される際の転写の終結、およびポリA配列の付加に関与する配列である。ターミネーターは、mRNAの安定性に寄与し、そして遺伝子の発現量に影響を及ぼすことが知られている。ターミネーターの例としては、ノパリン合成酵素遺伝子のターミネーター(Tnos)、およびカリフラワーモザイクウイルス(CaMV)35Sターミネーターが挙げられるが、これらに限定されない。
【0058】
「薬剤耐性遺伝子」は、形質転換植物の選抜を容易にするものであることが望ましい。カナマイシン耐性を付与するためのネオマイシンホスホトランスフェラーゼII(NPTII)遺伝子、およびハイグロマイシン耐性を付与するためのハイグロマイシンホスホトランスフェラーゼ遺伝子などが好適に用いられ得るが、これらに限定されない。
【0059】
本発明における植物発現ベクターは、当業者に周知の遺伝子組換え技術を用いて作製され得る。植物発現ベクターの構築には、例えば、pBI系のベクターまたはpUC系のベクターが好適に用いられるが、これらに限定されない。
【0060】
(形質転換植物の作出)
上述のように構築された発現カセット、またはこれを含む発現ベクターは、公知の遺伝子組換え技術を用いて、所望の植物細胞に導入され得る。導入された発現カセットは、植物細胞中のDNAに組み込まれて存在する。なお、植物細胞中のDNAとは、染色体のみならず、植物細胞中に含まれる各種オルガネラ(例えば、ミトコンドリア、葉緑体)に含まれるDNAをも含む。
【0061】
本明細書において、用語「植物」は、単子葉植物および双子葉植物のいずれも含む。好ましい植物は、双子葉植物である。双子葉植物は、離弁花亜綱および合弁花亜綱のいずれも含む。好ましい亜綱は、合弁花亜綱である。合弁花亜綱は、リンドウ目、ナス目、シソ目、アワゴケ目、オオバコ目、キキョウ目、ゴマノハグサ目、アカネ目、マツムシソウ目、およびキク目のいずれも含む。好ましい目は、ナス目である。ナス目は、ナス科、ハゼリソウ科、ハナシノブ科、ネナシカズラ科、およびヒルガオ科のいずれも含む。好ましい科は、ナス科である。ナス科は、ペチュニア属、チョウセンアサガオ属、タバコ属、ナス属、トマト属、トウガラシ属、ホオズキ属、およびクコ属などを含む。好ましい属は、ペチュニア属、チョウセンアサガオ属、およびタバコ属であり、より好ましくは、ペチュニア属である。ペチュニア属は、P.hybrida種、P.axillaris種、P.inflata種、およびP.violacea種などを含む。好ましい種は、P.hybrida種である。「植物」は、特に他で示さない限り、花を有する植物体および植物体から得られる種子を意味する。
【0062】
「植物細胞」の例としては、花、葉、および根などの植物器官における各組織の細胞、カルスならびに懸濁培養細胞が挙げられる。
【0063】
植物細胞への植物発現ベクターの導入には、当業者に周知の方法、例えば、アグロバクテリウムを介する方法、および直接細胞に導入する方法が用いられ得る。アグロバクテリウムを介する方法としては、例えば、Nagelらの方法(FEMS Microbiol. Lett., 67:325(1990))が用いられ得る。この方法は、まず、植物発現ベクターで(例えば、エレクトロポレーションによって)アグロバクテリウムを形質転換し、次いで、形質転換されたアグロバクテリウムをリーフディスク法などの周知の方法により植物細胞に導入する方法である。植物発現ベクターを直接細胞に導入する方法としては、エレクトロポレーション法、パーティクルガン、リン酸カルシウム法、およびポリエチレングリコール法などがある。これらの方法は、当該分野において周知であり、形質転換する植物に適した方法が、当業者により適宜選択され得る。
【0064】
植物発現ベクターを導入された細胞は、例えば、カナマイシン耐性などの薬剤耐性を基準として選択される。選択された細胞は、常法により植物体に再生され得る。
【0065】
再生した植物体において、導入した植物発現ベクターが機能的であることは、当業者に周知の手法を用いて確認し得る。例えば、内因性遺伝子の発現の抑制を目的とする場合、この確認は、ノーザンブロット解析を用いた転写レベルの測定によって行い得る。このようにして、内因性の転写因子の発現が抑制された所望とする形質転換植物体を選択することができる。組織特異的プロモーターを用いた異種遺伝子の発現を目的とする場合も、異種遺伝子の発現の確認は、通常、対象組織から抽出されたRNAを試料とするノーザンブロット解析を用いて行い得る。この解析法の手順は当業者に周知である。
【0066】
本発明の方法に従って内因性の転写因子の発現が抑制され、花粉の稔性が低下したことは、例えば、転写因子をコードするDNAを含む発現ベクターで形質転換して得られた植物の花粉の形態を、必要に応じて組識化学的染色を行ったうえ、顕微鏡観察することで確認できる。
【0067】
また、本発明の方法に従ってプロモーターが小胞子に特異的に発現されたことは、例えば、プロモータとGUS遺伝子とが作動可能に連結された発現ベクターで形質転換して得られた植物における、葯を含む花の組織でのGUS活性の分布を、常法により組識化学的染色を行うことで確認できる。
【実施例】
【0068】
以下に、実施例に基づいて本発明を説明する。本発明の範囲は、実施例のみに限定されるものではない。実施例で使用される制限酵素、プラスミドなどは、商業的な供給源から入手可能である。
【0069】
(実施例1:ZPT転写因子群をコードするポリヌクレオチドを含む植物発現ベクターの構築)
以前に報告した葯特異的ZF遺伝子(Kobayashiら、前出)のうち、PEThyZPT2−5(ZPT2−5)、PEThyZPT3−1(ZPT3−1)、およびPEThyZPT4−1(ZPT4−1)のcDNAを、それぞれ、カリフラワーモザイクウイルスの35Sプロモーターの下流につないで、植物発現ベクターを作成した。具体的には、下記の通りである。
【0070】
(実施例1−1)
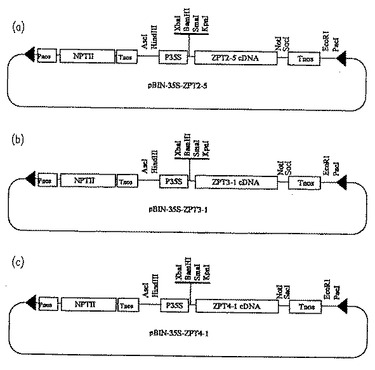
プラスミドpBI221(CLONTECH Laboratories Inc.から購入)中のカリフラワーモザイクウイルス35Sプロモーターを含むDNA断片(HindIII−XbaI断片)およびNOSターミネーターを含むDNA断片(SacI−EcoRI断片)を順次プラスミドpUCAP(van Engelen,F.A.ら、Transgenic Res.,4:288,1995)のマルチクローニングサイトに挿入し、pUCAP35Sを作製した。一方、ZPT2−5のcDNAを含むpBluescript ベクターをKpnIサイトおよびSacIサイト(いずれもベクター中のサイト)で切断し、上記pUCAP35SのKpnIおよびSacIサイトの間に挿入した。さらにこの組み替えプラスミドをEcoRIおよびHindIIIで切断し、ZPT2−5をコードするDNA断片をバイナリーベクターpBINPLUS(van Engelen, F.A.ら、前出)のEcoRIおよびHindIIIサイトに導入した。構築されたZPT2−5遺伝子は、図4(a)から明らかなように、カリフラワーモザイクウイルス(CaMV)の35Sプロモーター領域(P35S;0.9 kb)、本発明のZPT2−5をコードするポリヌクレオチド(ZPT2−5;約0.8kb)、およびノパリン合成酵素のターミネーター領域(Tnos;0.3kb)から構成されている。図4中のPnosは、ノパリン合成酵素のプロモーター領域、NPTIIはネオマイシンホスホトランスフェラーゼII遺伝子を示す。
【0071】
(実施例1−2)
ZPT3−1のcDNAを含むpBluescript ベクターをKpnIサイトおよびSacIサイト(いずれもベクター中のサイト)で切断し、pUCAP35SのKpnIおよびSacIサイトの間に挿入した。さらにこの組み換えプラスミドをEcoRIおよびHindIIIで切断し、ZPT3−1をコードするDNA断片をバイナリーベクターpBINPLUSのEcoRIおよびHindIIIサイトに導入した。構築されたZPT3−1遺伝子は、図4(b)から明らかなように、カリフラワーモザイクウイルス(CaMV)の35Sプロモーター領域(P35S;0.9kb)、本発明のZPT3−1をコードするポリヌクレオチド(ZPT3−1;約1.7kb)、およびノパリン合成酵素のターミネーター領域(Tnos;0.3kb)から構成されている。
【0072】
(実施例1−3)
ZPT4−1のcDNAを含むpBluescript ベクターをKpnIサイトおよびSacIサイト(いずれもベクター中のサイト)で切断し、上記pUCAP35SのKpnIおよびSacIサイトの間に挿入した。さらにこの組み換えプラスミドをEcoRIおよびHindIIIで切断し、ZPT4−1をコードするDNA断片をバイナリーベクターpBINPLUSのEcoRIおよびHindIIIサイトに導入した。構築されたZPT4−1遺伝子は、図4(c)から明らかなように、カリフラワーモザイクウイルス(CaMV)の35Sプロモーター領域(P35S;0.9kb)、本発明のZPT4−1をコードするポリヌクレオチド(ZPT4−1;約2.0kb),およびノパリン合成酵素のターミネーター領域(Tnos;0.3kb)から構成されている。
【0073】
(実施例2:ZPT3−1およびZPT4−1プロモーター領域の単離ならびにGUSレポーター遺伝子との連結)
ZPT3−1およびZPT4−1のcDNAをプローブにして、ペチュニアのゲノムDNAライブラリーから、対応するゲノミック・クローンを単離し、転写開始点上流のDNA断片(プロモーター領域;約2.7kbおよび約3.6kb)をサブクローニングした。それぞれのDNA断片をGUSレポーター遺伝子の上流につないでバイナリー・ベクターにクローニングした。具体的には、以下の通りである。
【0074】
(実施例2−1)
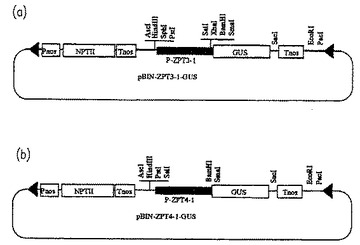
ZPT3−1のcDNAを、通常のランダムプライム法(Sambrookら、前出)を用いて[α32P]dCTPで標識して放射性標識プローブを作製し、これを用いてEMBL3ベクター(Stratagene製)中に作製したペチュニア(Petunia hybrida var. Mitchell)のゲノミックライブラリーをスクリーニングした。得られたクローンから遺伝子上流領域を含む約2.7kbのゲノムDNA断片(PstI−SacI)をpBluescriptSKベクターのPstI−SacIサイトにサブクローニングし(pBS−ZPT3−1−PS)、塩基配列を決定した(図5A〜5B)。次に、このプラスミドを鋳型とし、SalI認識配列を含むプライマー(3’−TATGGAGCTCGTCGACAG TTGATGGTTCATTTTTCTGGCTATTGTC−5’;配列番号11)および市販のM13−20プライマーを用いてPCRを行い、ZPT3−1タンパク質翻訳開始点のすぐ下流(塩基番号2661)にSalIサイトを導入した。そして、PstIおよびSalIで切断して得られたDNA断片をpUCAPGUSNTのGUSコード領域の上流に挿入した(pUCAP−ZPT3−1−GUSNT)。これによってZPT3−1遺伝子コード領域のN末端近傍の領域でin frameでGUSコード領域と接続された。さらにpUCAP−ZPT3−1−GUSNTをAscIおよびPacIで切断して得られるDNA断片(ZPT3−1プロモーター、GUSコード領域およびNOSターミネーターを含む)をpBINPLUSベクターに挿入し、pBIN−ZPT3−1−GUSを得た(図7(a))。
【0075】
(実施例2−2)
ZPT4−1についても同様にゲノムDNAを単離し、ZPT4−1遺伝子上流領域を含む約3.6kbのDNA断片(EcoRI−EcoRI)をpBluescriptSKベクターのEcoRI−EcoRIサイトにサブクローニングし(pBS−ZPT4−1−EE)、塩基配列を決定した(図6A〜6B)。このプラスミドを鋳型にし、BamHI認識配列(3’−CATGGATATAGGATCCTATATC−5’;配列番号12)を含むプライマーおよびM13−20プライマーを用いてPCRを行い、ZPT4−1タンパク質翻訳開始点のすぐ下流(塩基番号3641)にBamHIサイトを導入した。そして、EcoRIおよびBamHIで切断して得られたDNA断片をpUCAPGUSNTのGUSコード領域の上流に挿入した(pUCAP−ZPT4−1−GUSNT)。これによってZPT4−1遺伝子コード領域のN末端近傍の領域でin frameでGUSコード領域と接続された。さらに、上記と同様にしてDNA断片(AscI−PacI)をpBINPLUSベクターに挿入し、pBIN−ZPT4−1−GUSを作製した(図7(b))。
【0076】
(実施例3:各融合遺伝子のペチュニア細胞への導入)
上記の各発現ベクターを、以下の手順で、アグロバクテリウムを介して、ペチュニア(Petunia hybrida var. Mitchell)に導入した。
【0077】
(1)アグロバクテリウム・チュメファシエンスLBA4404株(CLONTECH Laboratories Inc.から購入)を250mg/mlのストレプトマイシンと50mg/mlのリファンピシンを含むL培地中、28℃で培養した。Nagelら(1990)(前出)の方法に従って、細胞懸濁液を調製し、実施例1および2で構築したプラスミドベクターをエレクトロポレーションにより、上記菌株に導入した。
【0078】
(2)各融合遺伝子をコードするポリヌクレオチドを、以下の方法によって、ペチュニア細胞へ導入した:上記(1)で得られたアグロバクテリウム・チュメファシエンスLBA4404株をYEB培地(DNA Cloning第2巻、78頁、Glover D.M.編、IRL Press、1985)で振とう培養(28℃、200rpm)した。得られた培養液を、滅菌水で20倍に希釈し、ペチュニア(Petunia hybrida var. Mitchell)の葉片と共存培養した。2〜3日後、抗生物質を含む培地で上記細菌を除去し、2週間ごとに選択培地で継代し、上記5種類の融合遺伝子と共に導入された、pBINPLUS由来のNPTII遺伝子発現によるカナマイシン抵抗性の有無に基づいて、形質転換されたペチュニア細胞を選抜した。選抜した細胞を、常法によりカルスを誘導した後、植物体に再分化した(Jorgensen R.A.ら,Plant Mol. Biol.,31:957,1996)。
【0079】
(実施例4:ZPT遺伝子群を導入した形質転換ペチュニアの表現型)
実施例1のベクターを導入して得られた形質転換体を用い、ZPT2−5、ZPT3−1およびZPT4−1の発現の制御に伴う花粉の形態変化を観察することによって、これらZPT遺伝子群のcDNA導入による植物への影響を検討した。具体的には、以下の通りである。
【0080】
(実施例4−1)
ZPT2−5のcDNAを35Sプロモーターの制御下に導入した形質転換体(14個体)から、ノーザンブロット解析によって、コサプレッションによる遺伝子発現の抑制が起こった個体(3個体)を選抜した。(なお、14個体のうち他の4個体では、導入したZPT2−5遺伝子の過剰発現が観察された。)ノーザンブロット解析の条件は、次の通り:7%SDS、50%ホルムアミド、5xSSC、2%ブロッキング試薬(Boehringer Mannheim製)、50mMリン酸ナトリウム・バッファー(pH7.0)、0.1%ラウリルサルコシン・ナトリウム、50μg/ml酵母tRNA、および32P標識プローブDNA(1x107cpm)を含む溶液中で、68℃で16時間ハイブリダイズさせた後、2xSSC/0.1%SDSで68℃、30分間の洗浄。
【0081】
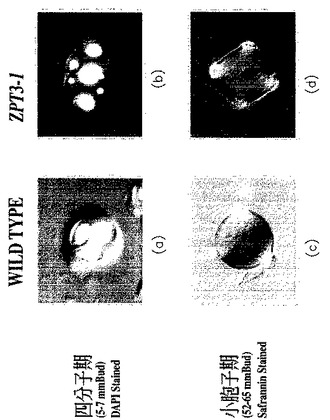
上記のコサプレッション形質転換体3個体において、以下のような表現型が観察された(図8)。
【0082】
四分子期の直前に起こる減数分裂の過程において、正常な(野生型の)ペチュニアでは、クロマチンの凝縮による細糸状構造の形成に続き(前期Iの細糸期)、相同染色体が対合する(前期Iの接合糸期)。次に、中期I(metaphaseI)に入ると、4分染色体が細胞赤道面にならび、その後、紡錘装置によって相同染色体が均等に細胞の両極に分離する。他方、ZPT2−5遺伝子のコサプレッションを起こした形質転換体では、中期Iにおいて4分染色体が細胞赤道面に正しく並ぶことのないまま、染色体の極分離が進行した。このとき、染色体の両極への分配が著しく不均等であった。
【0083】
正常な減数分裂の過程では、上記1回目の染色体分離の後、2回目の染色体分離によって、4つのハプロイドのグループが形成され、その後に細胞質分離が起こる。他方、上記のコサプレッションを起こした形質転換体では、一回目の染色体分離の直後に細胞質分離および細胞分裂が起こった。また、この不均等な細胞分裂は一回でとどまらず、少なくともさらに二回繰り返されて、最高8個の小胞子細胞が形成された。不均等な染色体分離のために、これらの小胞子細胞に含まれる染色体数は不均一であり、それに伴って、細胞の大きさも著しく不均等であった。結果として、正常なペチュニアにおいて四分子期にあたる時期に、これらの形質転換体では、正常より多い数の小胞子(最高8個)が形成されていた(図8(f);つぼみのサイズ6mmにおけるZPT2−5コサプレッション形質転換体の花粉細胞の写真。また、図9(b);四分子期における形質転換体の花粉細胞の写真も参照)。
【0084】
コサプレッション形質転換体において、小胞子の一部(10−20%)はその後も発達を続けたが、大部分の小胞子細胞は、これらを包むカロース層が分解される前に、破裂した。この段階で破裂せずに生き残った小胞子は、六面体に近い異常形態を示し、正常な小胞子の4面体形態とは明らかに異なっていた。異常形態の小胞子は、その後、一見正常な有糸分裂によって二核となって、花粉粒を形成した。しかし、これらの花粉粒は稔性をほとんど失っていた。すなわち、正常なペチュニアの雌ずいにこれらの形質転換体の花粉粒をかけると、コサプレッションを示した3系統からの花粉では、種子形成が全く無いか、または、わずかに種子が形成されるのみ(最高でも10%、つまり、ペチュニア1株あたりで産生される種子の数が、約10個の花の平均として、コントロールの1割)であった。コサプレッションを示さない形質転換体3系統からの花粉では、野生型のコントロール植物の場合と同様の、正常な種子形成が確認された。
【0085】
上記のコサプレッション形質転換体はまた、雌性配偶体形成にも異常を示し、雌性稔性が正常個体の25−35%に低下していた。すなわち、胚珠(雌性配偶体)の発達は外観上正常であったものの、野生型の花粉で受粉したとき過半数の胚珠が受精できず、受精したものも以後の発達に異常を示し、多くが死滅(abort)した。この場合も、コサプレッションを示さない形質転換体では、野生型のコントロール植物の場合と同様の、正常な雌性配偶体形成が観察された。
【0086】
(実施例4−2)
ZPT3−1のcDNAを35Sプロモーターの支配下に導入すると、ZPT2−5遺伝子を導入した場合と同様の形質変化が、15個体中、3個体に表れた(図9)。すなわち、これらの形質転換体では、減数分裂の過程でZPT2−5の場合と全く同様の異常が見られた。そして、小胞子期まで発達する細胞数は非常に少なく、生き残った小胞子は形態異常(6面体)を示した。さらに、成熟した花粉粒は稔性を失っていた。しかし、ZPT2−5の場合とは異なり、これらの個体では雌性稔性には影響がなかった。
【0087】
実施例4−1と同じ条件でのノーザンブロット法による遺伝子発現解析の結果、ZPT3−1遺伝子を導入した個体においては、ZPT3−1とZPT4−1との両方の遺伝子の発現抑制が観察された。両遺伝子は、構造的類似性が高い。すなわち、コード領域全体の塩基配列の相同性は37%であり、ZPT3−1の2番目のZF領域とZPT4−1の3番目のZF領域、およびZPT3−1の3番目のZF領域とZPT4−1の4番目のZF領域とを、相同性値が最大化されるように近傍の配列を含めて、塩基配列レベルで、それぞれ比較したときの相同性の平均値は86%である(配列比較はClustal Vプログラムによる)。従って、上記の発現抑制事象は、一つの遺伝子の導入によって二つの遺伝子の発現抑制(コサプレッション)が起こったことによる可能性が高い。このことは、これら二つの遺伝子の機能が重複していることを示唆し、両遺伝子の各々の導入によって共通した表現型の変化が見られることとも符合する。
【0088】
(実施例4−3)
ZPT4−1のcDNAを35Sプロモーターの支配下に導入すると、ZPT2−5遺伝子を導入した場合と同様の形質変化が、13個体中、2個体に表れた。すなわち、これらの形質転換体では、減数分裂の過程でZPT2−5の場合と全く同様の異常が見られた。そして、小胞子期まで発達する細胞数は非常に少なく、生き残った小胞子は形態異常(6面体)を示した。さらに、成熟した花粉粒は稔性をほとんど失っていた。しかし、ZPT3−1の場合と同様に、これらの個体では雌性稔性には影響がなかった。上述した理由から、本実施例においても、ZPT3−1とZPT4−1との両方の遺伝子の発現抑制(コサプレッション)が起こった可能性が高い。
【0089】
以上のように、ZPT2−5、ZPT3−1またはZPT4−1をコードする遺伝子の導入によって、きわめて高い効率で、花粉の発達を阻害し、稔性を喪失させることができる(ZPT3−1で99%以上、ZPT2−5およびZPT4−1で90%以上)。また、これらの遺伝子群の導入は、その効果が花粉(ZPT2−5の場合、花粉および雌性配偶体)に特異的であり、植物の他の形質に影響を与えない点で、選択的な形質改変技術として有用であり得る。
【0090】
(実施例5:ZPT3−1およびZPT4−1プロモーター活性の組織特異性)
実施例2のベクターを導入して得られた形質転換体を用い、GUS活性による組織化学的染色を行うことによって、上記DNA断片が有する組織特異的プロモーター活性を検出した。具体的には、以下の通りである。
【0091】
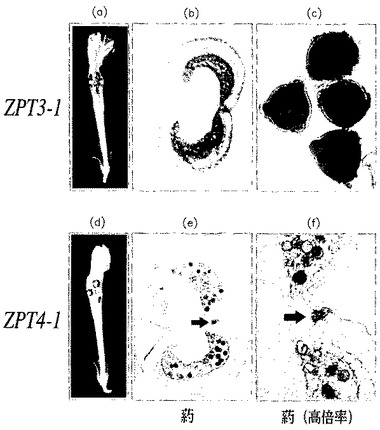
(実施例5−1)
ZPT3−1遺伝子の上流領域とGUSとの融合遺伝子を導入して得られた形質転換体の花を用い、X−GUSを基質にしてGUS活性の分布を調べた(Gallagher,S.R.編、GUS protocols:using the GUS gene as a reporter of gene expressin、Academic Press,Inc.,pp.103−114,1992)。その結果、GUS活性は1核期の小胞子において特異的に検出された(図10(a)〜(c))。
【0092】
(実施例5−2)
ZPT4−1遺伝子の上流領域とGUSとの融合遺伝子を導入して得られた形質転換体の花を用い、上記と同様にGUS活性の分布を調べた。その結果、GUS活性は、一核期から二核期にかけての小胞子および葯の開裂組織において特異的に観察された(図10(d)〜(f);葯の開裂組織を図10(e)および(f)に矢印で示す)。
【0093】
以上のように、ZPT3−1およびZPT4−1遺伝子のプロモーターは、一核期の小胞子(ZPT3−1)および一核期から二核期にかけての小胞子(ZPT4−1)にそれぞれ特異的な活性を示す。ZPT4−1遺伝子のプロモーターは、一核期から二核期にかけての葯の開裂組織にも特異的な活性を示す。
【0094】
小胞子は、のちに成熟して花粉粒を生成する前駆細胞である。従って、これらのプロモーターは、花粉の発達に関連する詳細な研究のためのツールとして有用である。さらに、これらのプロモーターまたはその活性断片を用いて、細胞毒性のある遺伝子などを小胞子に特異的に発現させ、花粉細胞を死滅させまたは機能を失わせることによって、花粉の発達を直接的かつ効率的に制御し得る。
【産業上の利用可能性】
【0095】
ZPT2−5、ZPT3−1およびZPT4−1遺伝子由来の転写因子をコードするDNA、ならびに、ZPT3−1およびZPT4−1遺伝子由来のプロモーターを利用する本件発明の方法は、遺伝子工学的に植物の形質を選択的に改変する技術、特に、雄性不稔形質を付与する技術として有用である。
【図面の簡単な説明】
【0096】
【図1A】ZPT2−5をコードする遺伝子(本明細書中、単に「ZPT2−5遺伝子」ともいう)のcDNA配列、および対応するアミノ酸配列を示す図である。2つのジンクフィンガーモチーフ、およびDLNL配列(アミノ酸145位〜155位)を下線で示す。
【図1B】図1Aの続きである。
【図2A】ZPT3−1をコードする遺伝子(本明細書中、単に「ZPT3−1遺伝子」ともいう)のcDNA配列、および対応するアミノ酸配列を示す図である。3つのジンクフィンガーモチーフ、およびDLNL配列(アミノ酸408位〜417位)を下線で示す。
【図2B】図2Aの続きである。
【図2C】図2Bの続きである。
【図3A】ZPT4−1をコードする遺伝子(本明細書中、単に「ZPT4−1遺伝子」ともいう)のcDNA配列、および対応するアミノ酸配列を示す図である。4つのジンクフィンガーモチーフ、およびDLNL配列(アミノ酸438位〜449位)を下線で示す。
【図3B】図3Aの続きである。
【図3C】図3Bの続きである。
【図3D】図3Cの続きである。
【図4】ZPT2−5、ZPT3−1およびZPT4−1の各cDNA配列を発現するための植物発現ベクター(pBIN−35S−ZPT2−5、pBIN−35S−ZPT3−1およびpBIN−35S−ZPT4−1)の構成を示す概略図である。
【図5A】ZPT3−1遺伝子のコード領域の上流配列を示す図である。転写開始点(2567位)を太矢印で、翻訳開始コドン(ATG)を下線太字で示す。
【図5B】図5Aの続きである。
【図6A】ZPT4−1遺伝子のコード領域の上流配列を示す図である。転写開始点(3503位)を太矢印で、翻訳開始コドン(ATG)を下線太字で示す。
【図6B】図6Aの続きである。
【図7】ZPT3−1およびZPT4−1遺伝子のプロモーターを解析するための植物発現ベクター(pBIN−ZPT3−1−GUSおよびpBIN−ZPT3−1−GUS)の構成を示す概略図である。
【図8】野生型のペチュニアの花粉と、pBIN−35S−ZPT2−5を導入したペチュニア(コサプレッションを起こした形質転換体)の花粉とを撮影した、生物の形態を示す写真である(倍率は400倍)。図8(a)〜(d)は野生型ペチュニア、そして図8(e)〜(h)はコサプレッション形質転換ペチュニアについての、それぞれつぼみの異なる成長段階における花粉を示す。花粉はいずれも、常法によりDAPI(4’,6−diamidino−2−phenylindole dihydrochloride n−hydrate)で染色した。
【図9】野生型のペチュニアの花粉と、pBIN−35S−ZPT3−1を導入したペチュニアの花粉とを撮影した、生物の形態を示す写真である(倍率は700倍)。図9(a)および(c)は野生型ペチュニア、図9(b)および(d)は形質転換ペチュニアについての、それぞれ四分子期および小胞子期における花粉を示す。四分子期の花粉はDAPIで、小胞子期の花粉はサフラニンで、それぞれ常法により染色した。なお、pBIN−35S−ZPT4−1を導入したペチュニアの花粉でも、図9(b)および(d)と全く同様の形態が観察された。
【図10】pBIN−ZPT3−1−GUSおよびpBIN−ZPT4−1−GUSを導入したペチュニアのGUS染色した花の器官を撮影した、生物の形態を示す写真である。いずれも、葯が一核期にある花(つぼみ)を撮影対象とした。図10(a)および(d)は、つぼみの外観を実寸大で、図10(b)および(e)は、葯の低倍率(40倍)での断面図を、図6(c)および(f)は、小胞子(図6(c);倍率は700倍)または葯の開裂組織周辺(図6(f);倍率は200倍)の高倍率での断面図を、それぞれ示す。
【技術分野】
【0001】
本発明は、雄しべの花粉形成組織に特異的に発現する遺伝子およびその利用に関する。本発明は、特に、小胞子に特異的に発現する、ペチュニア由来のジンクフィンガー型転写因子群(ZPT2−5、ZPT3−1およびZPT4−1)の遺伝子およびその利用に関する。
【背景技術】
【0002】
花粉の稔性は、農業・園芸における様々な局面で問題になる。たとえば、交雑育種における交配の際、自家受粉を避けるために、多くの労力がかかる除雄(雄しべの除去)の作業が行われる。種苗産業においては、交雑育種によって得られた優れた品種を商業的に保護したいという立場から、花粉稔性の無い形質が求められている。このような要請から、花粉の稔性を制御する技術(pollination control)が強く求められてきた。従来、特定の作物では、細胞質雄性不稔性の系統が交雑育種に用いられ、一定の成功を収めてきた。しかし、細胞質不稔形質は多くの場合、耐病性の低下などの好ましくない副次効果を伴い、さらに形質が不安定であること、種子の大量生産が困難なことなどの問題がある。化学薬品処理によって稔性を低下させる方法も研究されているが、安全性評価および作用機作解明が遅れており、実用化には至っていない。そこで、遺伝子工学を用いた優れた雄性不稔化技術が求められている。
【0003】
花粉は種子植物における雄性の配偶体である。支持体組織としての葯に取り囲まれた状態で進行する花粉の発達は、小胞子形成細胞(花粉母細胞)の減数分裂直後にあたる四分子期、四分子からの小胞子の放出期、花粉細胞の拡大および空胞形成に特徴付けられる一核期、有糸分裂により栄養細胞および生殖細胞への分化が生じる有糸細胞分裂期、その後の二核期という、各段階に区分される。これらの段階を経て、最終的に葯が開裂し、成熟した花粉粒が放出される。従って、小胞子は、花粉の発達を阻害し、花粉稔性を失わせるための人為的な制御に最も適した対象組織の一つといえる。
【0004】
上記のように、遺伝子工学による雄性不稔化技術には大きな期待が寄せられている。特に、小胞子のような花粉細胞の直接の前駆体において特異的に発現する遺伝子を利用できれば、植物に好ましくない形質を付与することなく、雄性不稔化を達成し得る可能性が高いと考えられる。雄しべの種々の組織に特異的なプロモーター、および、それを含む雄生不稔化のための遺伝子構築物の例は、いくつか報告されている(非特許文献1)。しかし、発現の組織特異性および時期特異性が高い、花粉稔性の制御のために有用な新たな遺伝子に対する要望が継続的に存在している。
【0005】
本発明者らは、最近、ペチュニア由来の新規な転写因子として、PEThyZPT2−5、 PEThyZPT3−1、およびPEThyZPT4−1(以下、それぞれ、ZPT2−5、ZPT3−1およびZPT4−1と略す)を含む7つのジンクフィンガー(ZF)型転写因子のcDNA配列を特定した。そして、ノーザンブロット分析から、それぞれの転写因子が葯特異的に、葯の発達の異なる段階において一過性の発現を示すことを報告した(非特許文献2)。しかし、これらの転写因子の植物における生理的な機能および作用、ならびに転写因子をコードする遺伝子の正確な発現部位およびその発現調節機構は不明であった。
【非特許文献1】ShivannaおよびSawhney編, Pollen biotechnology for crop production and improvement (Cambridge University Press)1997年、pp.237−257
【非特許文献2】Kobayashiら,Plant J.,1998年、13:571
【発明の開示】
【発明が解決しようとする課題】
【0006】
本発明の目的は、雄性不稔に代表される植物形質の改変に有用な、花粉に特異的な遺伝子を利用する遺伝子工学的手法を提供することにある。
【課題を解決するための手段】
【0007】
本発明者らは、以前にペチュニアから単離した、葯特異的な転写因子群(ZPT2−5、ZPT3−1およびZPT4−1)をコードする遺伝子をペチュニアに再導入した。その結果、花粉の正常な発達が阻害され、花粉の稔性が著しく低下するか(ZPT2−5およびZPT4−1)、または、ほぼ完全に失われる(ZPT3−1)ことを見出した。さらに本発明者らは、ZPT3−1およびZPT4−1のゲノム遺伝子からそれぞれ上流領域を単離し、そのプロモーター活性の組織特異性を調べた。その結果、一核期から二核期にかけての小胞子において、プロモーター活性が組織および時期特異的に発現されることを見出した。本発明は、これらの知見に基づいて完成された。
【0008】
本発明は、その第1の局面において、雄性不稔性の植物を作出する方法であって、以下の工程を包含する方法に関する:(i)配列番号1によって示される塩基配列の第1位から第777位までの配列を有するDNA、(ii)(i)の塩基配列を有するDNAとストリンジェントな条件下でハイブリダイズし、花粉の発達を制御する転写因子をコードするDNA、または(iii)(i)もしくは(ii)の断片であるDNAのいずれかである核酸と、上記核酸に作動可能に連結されたプロモーターとを含む、植物発現カセットを提供する工程;花粉の発達を制御する内因性の転写因子を有する植物の細胞を提供する工程であって、ここで、内因性の転写因子をコードする遺伝子は、上記核酸とストリンジェントな条件下でハイブリダイズする、工程;発現カセットを植物細胞に導入する工程;発現カセットが導入された植物細胞を植物体に再生する工程;および、再生された植物体であって、上記核酸が発現されることによって、内因性の転写因子の発現が抑制された植物体を選択する工程。
【0009】
本発明は、その第2の局面において、雄性不稔性の植物を作出する方法であって、以下の工程を包含する方法に関する:(i’)配列番号3によって示される塩基配列の第1位から第1640位までの配列を有するDNA、(ii’)(i’)の塩基配列を有するDNAとストリンジェントな条件下でハイブリダイズし、花粉の発達を制御する転写因子をコードするDNA、または(iii’)(i’)もしくは(ii’)の断片であるDNAのいずれかである核酸と、上記核酸に作動可能に連結されたプロモーターとを含む、植物発現カセットを提供する工程;花粉の発達を制御する内因性の転写因子を有する植物の細胞を提供する工程であって、ここで、内因性の転写因子をコードする遺伝子は、上記核酸とストリンジェントな条件下でハイブリダイズする、工程;発現カセットを植物細胞に導入する工程;発現カセットが導入された植物細胞を植物体に再生する工程;および、再生された植物体であって、上記核酸が発現されることによって、内因性の転写因子の発現が抑制された植物体を選択する工程。
【0010】
本発明は、その第3の局面において、雄性不稔性の植物を作出する方法であって、以下の工程を包含する方法に関する:(i”)配列番号5によって示される塩基配列の第1位から第1948位までの配列を有するDNA、(ii”)(i”)の塩基配列を有するDNAとストリンジェントな条件下でハイブリダイズし、花粉の発達を制御する転写因子をコードするDNA、または(iii”)(i”)もしくは(ii”)の断片であるDNAのいずれかである核酸と、上記核酸に作動可能に連結されたプロモーターとを含む、植物発現カセットを提供する工程;花粉の発達を制御する内因性の転写因子を有する植物の細胞を提供する工程であって、ここで、内因性の転写因子をコードする遺伝子は、上記核酸とストリンジェントな条件下でハイブリダイズする、工程;発現カセットを植物細胞に導入する工程;発現カセットが導入された植物細胞を植物体に再生する工程;および、再生された植物体であって、上記核酸が発現されることによって、内因性の転写因子の発現が抑制された植物体を選択する工程。
【0011】
ただし、上記(ii)、(ii’)および(ii”)のDNAは、それぞれ、(iv)配列番号13によって示される塩基配列の第1位から第1886位までの配列を有するDNAとストリンジェントな条件下でハイブリダイズし、花粉の発達を制御する転写因子をコードするDNAを含まない。配列番号13によって示される塩基配列は、ペチュニアから単離された別の転写因子ZPT3−2をコードするcDNA配列である(Kobayashiら、前出)。
【0012】
本発明の上記第1から第3までの局面における方法は、植物に雄性不稔性を付与する方法として、利用され得る。
【0013】
上記第1から第3までの局面における1つの実施態様において、上記核酸は上記プロモーターに対して順方向で連結され、上記植物体の細胞内でセンス方向に転写され得る。
【0014】
上記第1から第3までの局面における1つの実施態様において、上記核酸は上記プロモーターに対して逆方向で連結され、上記植物体の細胞内でアンチセンス方向に転写され得る。
【0015】
上記第1から第3までの局面における1つの実施態様において、上記植物は双子葉植物である。双子葉植物は、好ましくはナス科植物であり、より好ましくはペチュニア属植物である。
【0016】
上記第1から第3までの局面における1つの実施態様において、上記発現カセットは植物発現ベクターに組み込まれている。
【0017】
本発明の第1から第3までの局面によればまた、上記いずれかに記載の方法により作出された、雄性不稔性の植物が提供される。
【0018】
本発明は、その第4の局面において、形質が改変された植物を作出する方法であって、以下の工程を包含する方法に関する:(a’)配列番号7によって示される塩基配列の第1位から第2624位までの配列を有するDNA、または(b’)(a’)の一部の配列を有し、小胞子に特異的なプロモーター活性を示すDNAのいずれかを含むプロモーターと、上記プロモーターに作動可能に連結された異種遺伝子とを含む、植物発現カセットを提供する工程;発現カセットを植物細胞に導入する工程;および、発現カセットが導入された植物細胞を、植物体に再生する工程。
【0019】
本発明は、その第5の局面において、形質が改変された植物を作出する方法であって、以下の工程を包含する方法に関する:(a”)配列番号8によって示される塩基配列の第1位から第3631位までの配列を有するDNA、または(b”)(a”)の一部の配列を有し、小胞子に、および任意に葯の開裂組織に、特異的なプロモーター活性を示すDNAのいずれかを含むプロモーターと、上記プロモーターに作動可能に連結された異種遺伝子とを含む、植物発現カセットを提供する工程;発現カセットを植物細胞に導入する工程;および、発現カセットが導入された植物細胞を、植物体に再生する工程。
【0020】
上記第4および第5の局面における1つの実施態様において、上記形質は稔性であり、形質が改変された植物は、雄性不稔植物である。従って、本発明の上記方法は、植物に雄性不稔性を付与する方法として、利用され得る。
【0021】
上記第4および第5の局面における1つの実施態様において、上記形質は和合性であり、形質が改変された植物は自家不和合性植物である。従って、本発明の上記方法は、植物に自家不和合性を付与する方法としても、利用され得る。
【0022】
上記第4および第5の局面における1つの実施態様において、上記植物は双子葉植物である。双子葉植物は、好ましくはナス科植物であり、より好ましくはペチュニア属植物である。
【0023】
上記第4および第5の局面における1つの実施態様において、上記発現カセットは植物発現ベクターに組み込まれている。
【0024】
本発明の第4および第5の局面によればまた、上記いずれかに記載の方法により作出された、形質が改変された植物が提供される。
【0025】
本発明は、その第6の局面において、以下の(I’)または(II’)のDNAを含むプロモーターに関する:(I’)配列番号7によって示される塩基配列の第1位から第2624位までの配列を有するDNA、または(II’)(I’)の一部の配列を有し、小胞子に特異的なプロモーター活性を示すDNA。
【0026】
本発明は、その第7の局面において、以下の(I”)または(II”)のDNAを含むプロモーターに関する:(I”)配列番号8によって示される塩基配列の第1位から第3631位までの配列を有するDNA、または(II”)(I”)の一部の配列を有し、小胞子に、および任意に葯の開裂組織に特異的なプロモーター活性を示すDNA。
【0027】
本発明は、その第8の局面において上記の小胞子に特異的なプロモーターのいずれかと、上記プロモーターに作動可能に連結された異種遺伝子とを含む、植物に雄性不稔性を付与するために有用な植物発現カセットに関する。
【発明を実施するための最良の形態】
【0028】
本発明について、以下に、より詳細に説明する。
(ZPT3−2、ZPT3−1およびZPT4−1遺伝子由来の転写因子)
本発明の第1から第3までの局面である、雄性不稔性の植物を作出する方法において有用な核酸は、以下のいずれかである:
(i)配列番号1によって示される塩基配列の第1位から第777位までの配列を有するDNA、
(i’)配列番号3によって示される塩基配列の第1位から第1640位までの配列を有するDNA、
(i”)配列番号5によって示される塩基配列の第1位から第1948位までの配列を有するDNA、または、
(i)〜(i”)のいずれかの塩基配列を有するDNAとストリンジェントな条件下でハイブリダイズし、花粉の発達を制御する転写因子をコードするDNA(すなわち、(ii)、(ii’)または(ii”))、または、
上記いずれかのDNAの断片(すなわち、(iii)、(iii’)または(iii”))であるDNA。
【0029】
本発明における上記核酸は、好ましくは(i)、(i’)または(i”)のDNA、すなわち、ZPT2−5、ZPT3−1またはZPT4−1をコードするDNA、または、その断片であり、より好ましくは(i)、(i’)または(i”)のDNAである。
【0030】
本明細書において「転写因子」とは、遺伝子の調節領域のDNAに結合してmRNAの合成を制御するタンパク質をいう。ある種の転写因子は、そのDNA結合ドメインにジンクフィンガー(ZF)モチーフと呼ばれる保存性の高いアミノ酸配列を有することが知られている。ZPT2−5は、Cys2/His2型(EPFファミリー)のジンクフィンガー(ZF)タンパク質であって、176アミノ酸からなる全長のアミノ酸配列内に2つのZFモチーフを含み、さらにDLNL配列と称される疎水性領域を含む転写因子である。ZPT3−1は、同様に、EPFファミリーのZFタンパク質であって、437アミノ酸からなる全長のアミノ酸配列内に3つのZFモチーフを含み、さらにDLNL配列を含む転写因子である。ZPT4−1は、同様に、EPFファミリーのZFタンパク質であって、474アミノ酸からなる全長のアミノ酸配列内に4つのZFモチーフを含み、さらにDLNL配列を含む転写因子である。いずれも、Kobayashiら(前出)を参照のこと。ZPT2−5、ZPT3−1およびZPT4−1をコードするcDNA配列(配列番号1、3および5)を、それぞれ、対応する推定アミノ酸配列(配列番号2、4および6)と共に図1A〜1B、図2A〜2Cおよび図3A〜3Dに示す。
【0031】
本明細書において、核酸またはDNAの「断片」とは、植物体に導入され、適切な様式で発現されたときに、当該植物の内因性の転写因子の発現を抑制し得る断片をいう。この断片は、上記(i)、(i’)、(i”)、(ii)、(ii’)または(ii”)のDNAの、ジンクフィンガーモチーフをコードする領域以外から選択され、少なくとも約40塩基対以上、好ましくは約50塩基対以上、より好ましくは約70塩基対以上、さらに好ましくは約100塩基対以上の長さを有する。
【0032】
本明細書において、ハイブリダイゼーションのための「ストリンジェントな条件」とは、特定の塩基配列(例えば、ペチュニア由来のZPT2−5、ZPT3−1またはZPT4−1をコードするDNA)と、これと相同性の高い他の塩基配列(例えば、ペチュニア以外の植物に存在する、ZPT2−5、ZPT3−1またはZPT4−1のホモログをコードするDNA)とのオリゴヌクレオチド二本鎖を形成させるのに十分な条件を意図する。本発明に適用されるストリンジェントな条件の代表的な例として、次の条件が挙げられる:1M NaCl、1%SDS、10%デキストラン硫酸、32P標識プローブDNA(1x107cpm)および50μg/mlサケ精子DNAを含む溶液中で、60℃で16時間ハイブリダイズさせた後、2xSSC/1%SDSで60℃、30分間の洗浄を2回繰り返す。
【0033】
本発明における、ZPT2−5、ZPT3−1およびZPT4−1をコードするDNA、およびこれらとストリンジェントな条件下でハイブリダイズし、花粉の発達を制御する転写因子をコードするDNAを単離するためには、公知の転写因子の遺伝子によりコードされるアミノ酸配列の保存領域に対応するデジェネレートプライマー対を用い得る。このプライマー対を用いて、植物のcDNAまたはゲノミックDNAを鋳型としてPCRを行い、その後、得られた増幅DNA断片をプローブとして用いて、同じ植物のcDNAライブラリーまたはゲノミックライブラリーをスクリーニングすることができる。そのようなプライマー対の例として、5’−CARGCNYTNGGNGGNCAY−3’(配列番号9)と、3’−RTGNCCNCCNARNGCYTG−5’(配列番号10)との組合せが挙げられる(ここで、Nはイノシンであり、RはGまたはAであり、そしてYはCまたはTである)。上記プライマー配列は、それぞれ、上記ZPT転写因子群のジンクフィンガーモチーフに含まれるアミノ酸配列QALGGHに対応する。
【0034】
従って、本発明に適用されるストリンジェントなハイブリダイゼーション条件はまた、PCRの条件としても規定され得、代表的な例においては、上記デジェネレートプライマー(配列番号9および10)が用いられ得る。この際のPCR反応条件は、94℃で5分間の変性の後、94℃で30秒間、50℃で30秒間、そして72℃で1分間を30サイクル繰り返し、最後に、72℃で7分間インキュベーションを行う条件であり得る。
【0035】
PCRは、市販のキットおよび装置の製造者の指針に基づいて行うか、当業者に周知の手法で行い得る。遺伝子ライブラリーの作製法、および遺伝子のクローニング法なども当業者に周知である。例えば、Sambrookら,Molecular Cloning:A Laboratory Manual、第2版(Cold Spring Harbor Laboratory,1989)を参照のこと。得られた遺伝子の塩基配列は、当該分野で公知のヌクレオチド配列解析法または市販されている自動シーケンサーを利用して決定し得る。
【0036】
本明細書において、転写因子が「花粉の発達を制御する」とは、代表的には、この転写因子の発現が阻害されるとき、花粉の形態または機能において有意な変化が観察されることをいう。本発明における転写因子は、代表的には、それをコードする遺伝子の発現が阻害されることにより、好ましくは約75%以上、より好ましくは約90%以上、さらに好ましくは約95%以上の花粉細胞を、成熟する前に死滅せしめる。ここで、転写因子の発現が阻害されるとは、ノーザンブロット法により測定したとき、mRNAの量が、野生型のコントロール植物と比較して約10分の1以下になることをいう。
【0037】
上記のようなスクリーニングによって単離、同定された遺伝子によってコードされる転写因子(すなわち、ZPT2−5、ZPT3−1およびZPT4−1、およびこれらのホモログ)が花粉の発達を制御することは、本明細書の開示に従って形質転換植物を作出し、その花粉の特性を観察することによって、確認し得る。
【0038】
本発明においては、花粉の発達を制御する転写因子をコードするDNAを利用して、植物細胞において、当該DNAと同一のまたは相同な塩基配列を含む内因性遺伝子の発現を抑制し得る。標的となる内因性遺伝子もまた、花粉の発達を制御する転写因子である。本発明の方法によれば、好ましくは、花粉の発達を制御する内因性転写因子以外の発現を実質的に抑制することなく、内因性転写因子の発現のみを選択的に抑制することによって、植物に雄性不稔性が付与される。
【0039】
すなわち、本発明における発現抑制技術が適用される植物細胞は、花粉の発達を制御する内因性の転写因子を有する植物の細胞であって、この内因性の転写因子をコードする遺伝子は、上記ZPT2−5、ZPT3−1またはZPT4−1、またはこれらのホモログのDNAとストリンジェントな条件下でハイブリダイズするものとして規定される。ここで「ストリンジェントな条件」の定義は、ZPT2−5、ZPT3−1およびZPT4−1のホモログの特定に関連して上述した条件と同じである。本発明の範囲を限定する意図ではないが、上記方法によって雄性不稔性を付与し得る植物は、上記ZPT遺伝子群が単離されたペチュニア、または上記ZPTホモログをコードする遺伝子が単離された植物と、進化系統学上近縁の植物であることが望ましい。ここで、進化系統学上近縁とは、代表的には同じ目に分類され、好ましくは同じ科に分類され、より好ましくは同じ属に分類され、さらに好ましくは同じ種に分類される植物である。もっとも、花粉の発達が種子植物の生殖に不可欠である事実を考慮すれば、ZPT2−5、ZPT3−1およびZPT4−1と同一または類似の機能を果たす転写因子が他の植物にも広く存在し得ることは容易に理解される。
【0040】
内因性遺伝子の発現を抑制するための手法として、代表的にはコサプレッションおよびアンチセンス技術を利用し得る。コサプレッションは、植物細胞に組み換え遺伝子を導入したとき、その遺伝子自体、およびその遺伝子の一部と相同な配列を含む内因性遺伝子の両者の発現が抑制されることをいう。コサプレッションが利用される場合、本発明における発現カセットは、転写因子をコードするDNAまたはその断片を、プロモーターに対して順方向で連結された形態で含む。転写因子をコードするDNAまたはその断片は、発現カセットとして植物細胞に導入された後、プロモーターの制御下でセンス方向に転写され得る。この導入DNAの作用により、所望とする遺伝子発現の抑制が可能となる。コサプレッションは、形質転換植物の一部の個体で認められ、他の個体では十分に起こらない場合が多い。そのため、通常、遺伝子発現が意図されたように抑制された個体が、慣用的な手順に従って選択される。
【0041】
アンチセンスは、植物細胞に組み換え遺伝子を導入したとき、導入遺伝子の転写産物(mRNA)が、内因性遺伝子の転写産物(mRNA)の相補的な配列とハイブリッドを形成して、内因性遺伝子がコードするタンパク質の翻訳が阻害されることをいう。アンチセンスが利用される場合、本発明における発現カセットは、転写因子をコードするDNAまたはその断片を、プロモーターに対して逆方向で連結された形態で含む。転写因子をコードするDNAまたはその断片は、発現カセットとして植物細胞に導入された後、プロモーターの制御下でアンチセンス方向に転写され得る。このアンチセンス転写物の作用により、所望とする遺伝子発現の抑制が可能となる。
【0042】
(ZPT3−1およびZPT4−1遺伝子由来のプロモーター)
本発明の第4および第5の局面である、形質が改変された植物を作出する方法において有用なプロモーターは、(a’)配列番号7によって示される塩基配列の第1位から第2624位までの配列を有するDNA、(a”)配列番号8によって示される塩基配列の第1位から第3631位までの配列を有するDNA、または、(a’)または(a”)の一部の配列を有し、小胞子に特異的なプロモーター活性を示すDNAのいずれかを含むプロモーターである。本発明における上記プロモーターは、好ましくは(a’)または(a”)のプロモーター、すなわち、ZPT3−1またはZPT4−1遺伝子のプロモーターである。
【0043】
ZPT3−1およびZPT4−1遺伝子のプロモーター領域の、組識特異的な発現活性に必須でない配列を除去して得られる、小胞子に特異的なプロモーター活性を有する配列は、本発明の範囲内にある。このような配列は、常法に従ってプロモーターのデリーション実験を行うことによって、得ることができる。簡潔には、ZPT3−1またはZPT4−1遺伝子のプロモーター領域の様々な欠失変異体(例えば、ZPT3−1またはZPT4−1遺伝子のプロモーター領域を、5’上流側から種々の長さに欠失させた変異体)と、適切なレポーター遺伝子(例えば、GUS遺伝子)とを融合したプラスミドを用いて、欠失変異体の組識特異的プロモーター活性を測定することによって、その活性に必須な領域を特定し得る。
【0044】
いったんプロモーター活性に必須の領域が特定されると、さらにその領域内の配列または隣接配列を改変して、プロモーターの発現活性の程度を高めることも可能である。このようにして得られる改変体もまた、小胞子に特異的なプロモーター活性を示す限り、本発明の範囲内にある。
【0045】
本発明において、「小胞子に特異的なプロモーター活性を示す」とは、プロモーターが、天然の植物中で、または任意の構造遺伝子に連結させた発現カセットとして植物に導入されたときに、DNAの転写を開始させることにより遺伝子の発現を指示する能力を、小胞子において特異的に示すことを意味する。ここで「特異的」とは、同じ植物体の花の、他のすべての組織(タペート層、花糸、花柱、花頭、花弁、萼(がく)などを含む;ただし葯の開裂組織を除く)よりも、プロモーターの発現活性が高いことをいう。上記特異的なプロモーターは、好ましくは、同じ植物体の花の他のすべての組織および花以外の部分(根、葉、茎など)よりも、プロモーターの発現活性が高く、より好ましくは、同じ植物体の花の他のすべての組織および花以外の部分において実質的に活性を示さない。「葯の開裂組織に特異的なプロモーター活性を示す」も、上記と同様に定義される。発現活性の程度は、常法に従って、小胞子におけるプロモーターの発現レベルと、花の他の組識における同じプロモータの発現レベルとを比較することによって、評価され得る。プロモーターの発現レベルは、通常、そのプロモーターの制御下で発現される遺伝子産物の産生量によって決定される。
【0046】
上記の特異的プロモーターを利用する本発明の方法は、植物の生殖に関連する形質を改変することが意図される。ここで「改変」とは、形質転換後の植物における生殖器官の少なくとも一部が、形質転換前の植物(野生種または園芸品種)に存在していた機能を失うか、形質転換前の植物に存在しなかった機能を獲得するか、または形質転換前の植物と比べて特定の機能の程度が増強もしくは低減されることをいう。このような形質の改変は、本発明におけるプロモーターに作動可能に連結された任意の異種遺伝子が、この遺伝子を導入された形質転換植物において、プロモーターの制御下で小胞子に特異的に発現される結果として生じ得る。多くの組織特異的プロモーターにおいて、その組織特異性が種間を越えて保存されていることは周知であるので、本発明のプロモーターもまた幅広い植物種に適用し得ることが容易に理解される。形質の改変の程度は、形質転換後の植物の形質を、形質転換前の植物の形質と比較することにより評価され得る。改変される好ましい形質としては、雌性不稔および自家不和合性が挙げられるが、これらに限定されない。
【0047】
本発明のプロモーターは、例えば、公知のcDNAをプローブとして用いて、植物のゲノミックライブラリーをスクリーニングし、そして対応のゲノミッククローンからコード領域の上流配列を単離することにより得ることができる。cDNAの例として、上述したペチュニア由来の転写因子であるZPT3−1およびZPT4−1のcDNAが挙げられる。
【0048】
本発明におけるプロモーターは、天然から単離されたものに限定されず、合成ポリヌクレオチドも含み得る。合成ポリヌクレオチドは、例えば、上記のようにして配列決定されたプロモーターの配列またはその活性領域を、当業者に周知の手法によって合成または改変することにより入手し得る。
【0049】
(発現カセットおよび発現ベクターの構築)
本発明における転写因子をコードするDNAは、当業者に周知の方法を用いて、適切なプロモーターに作動可能に連結された発現カセットとして、公知の遺伝子組換え技術を用いて、植物細胞に導入され得る。同様に、本発明における小胞子特異的プロモーターは、所望の異種遺伝子に作動可能に連結された発現カセットとして、植物細胞に導入され得る。
【0050】
上記転写因子に連結され得る「プロモーター」は、植物で発現する任意のプロモーターを意味し、構成的プロモーター、組織特異的プロモーター、および誘導性プロモーターのいずれをも含み得る。
【0051】
「構成的プロモーター」は、植物細胞内外の刺激と関係なく一定のレベルで構造遺伝子を発現せしめるプロモーターをいう。異種遺伝子が植物の他の組織または器官で発現しても、植物に望ましくない形質を与えない場合には、構成的プロモーターの使用が簡便であり、好ましい。構成的プロモーターの例としては、カリフラワーモザイクウイルス(CaMV)の35Sプロモーター(P35S)、およびノパリン合成酵素のプロモーター(Tnos)が挙げられるが、これらに限定されない。
【0052】
「組織特異的プロモーター」は、本発明においては、少なくとも小胞子において特異的に構造遺伝子を発現せしめるプロモーターをいう。このような組織特異的プロモーターには、本発明におけるZPT3−1およびZPT4−1遺伝子由来のプロモーターに加えて、葯特異的に発現活性を示す他の公知のプロモーターが含まれる。したがって、小胞子特異的プロモーターと転写因子をコードする配列とを含む、天然のZPT3−1およびZPT4−1遺伝子の発現カセット自体を、必要に応じて他の調節エレメントと組み合わせて、使用することも本発明の範囲に含まれる。
【0053】
「誘導性プロモーター」は、化学薬剤、物理的ストレスなどの特定の刺激を与えたときに構造遺伝子を発現せしめ、刺激の非存在下では発現活性を示さないプロモーターをいう。誘導性プロモーターの例としては、オーキシンで誘導可能な、グルタチオン S−トランスフェラーゼ(GST)プロモーター(van der Kop,D.A.ら、Plant Mol. Biol.,39:979,1999)が挙げられるが、これに限定されない。
【0054】
本明細書において、用語「発現カセット」または「植物発現カセット」とは、本発明における転写因子をコードするDNAと、これに作動可能に(すなわち、当該DNAの発現を制御し得るように)連結された植物発現プロモーターとを含む核酸配列、ならびに、本発明における小胞子特異的プロモーターと、これに作動可能に(すなわち、インフレームに)連結された異種遺伝子とを含む核酸配列をいう。
【0055】
上記の小胞子特異的プロモーターに連結され得る「異種遺伝子」とは、ZPT3−1およびZPT4−1遺伝子以外のペチュニアにおける内因性遺伝子、もしくはペチュニア以外の植物における内因性遺伝子、または植物に対して外来の遺伝子(例えば、動物、昆虫、細菌、および真菌に由来する遺伝子)であって、その遺伝子産物の発現が小胞子において所望される任意の遺伝子をいう。本発明における異種遺伝子の好ましい例は、コードする遺伝子産物が細胞毒性を示し、その発現によって花粉の発達を阻害する遺伝子である。このような遺伝子の具体例としてbarnase遺伝子(Beals,T.P.およびGoldberg,R.B.,Plant Cell,9:1527,1997)が挙げられるが、これに限定されない。
【0056】
「植物発現ベクター」は、発現カセットに加えて、さらに種々の調節エレメントが、宿主植物の細胞中で作動し得る状態で連結されている核酸配列をいう。好適には、ターミネーター、薬剤耐性遺伝子、およびエンハンサーを含み得る。植物発現ベクターのタイプおよび使用される調節エレメントの種類が、宿主細胞に応じて変わり得ることは、当業者に周知の事項である。本発明に用いる植物発現ベクターは、さらにT−DNA領域を有し得る。T−DNA領域は、特にアグロバクテリウムを用いて植物を形質転換する場合に遺伝子の導入の効率を高める。
【0057】
「ターミネーター」は、遺伝子のタンパク質をコードする領域の下流に位置し、DNAがmRNAに転写される際の転写の終結、およびポリA配列の付加に関与する配列である。ターミネーターは、mRNAの安定性に寄与し、そして遺伝子の発現量に影響を及ぼすことが知られている。ターミネーターの例としては、ノパリン合成酵素遺伝子のターミネーター(Tnos)、およびカリフラワーモザイクウイルス(CaMV)35Sターミネーターが挙げられるが、これらに限定されない。
【0058】
「薬剤耐性遺伝子」は、形質転換植物の選抜を容易にするものであることが望ましい。カナマイシン耐性を付与するためのネオマイシンホスホトランスフェラーゼII(NPTII)遺伝子、およびハイグロマイシン耐性を付与するためのハイグロマイシンホスホトランスフェラーゼ遺伝子などが好適に用いられ得るが、これらに限定されない。
【0059】
本発明における植物発現ベクターは、当業者に周知の遺伝子組換え技術を用いて作製され得る。植物発現ベクターの構築には、例えば、pBI系のベクターまたはpUC系のベクターが好適に用いられるが、これらに限定されない。
【0060】
(形質転換植物の作出)
上述のように構築された発現カセット、またはこれを含む発現ベクターは、公知の遺伝子組換え技術を用いて、所望の植物細胞に導入され得る。導入された発現カセットは、植物細胞中のDNAに組み込まれて存在する。なお、植物細胞中のDNAとは、染色体のみならず、植物細胞中に含まれる各種オルガネラ(例えば、ミトコンドリア、葉緑体)に含まれるDNAをも含む。
【0061】
本明細書において、用語「植物」は、単子葉植物および双子葉植物のいずれも含む。好ましい植物は、双子葉植物である。双子葉植物は、離弁花亜綱および合弁花亜綱のいずれも含む。好ましい亜綱は、合弁花亜綱である。合弁花亜綱は、リンドウ目、ナス目、シソ目、アワゴケ目、オオバコ目、キキョウ目、ゴマノハグサ目、アカネ目、マツムシソウ目、およびキク目のいずれも含む。好ましい目は、ナス目である。ナス目は、ナス科、ハゼリソウ科、ハナシノブ科、ネナシカズラ科、およびヒルガオ科のいずれも含む。好ましい科は、ナス科である。ナス科は、ペチュニア属、チョウセンアサガオ属、タバコ属、ナス属、トマト属、トウガラシ属、ホオズキ属、およびクコ属などを含む。好ましい属は、ペチュニア属、チョウセンアサガオ属、およびタバコ属であり、より好ましくは、ペチュニア属である。ペチュニア属は、P.hybrida種、P.axillaris種、P.inflata種、およびP.violacea種などを含む。好ましい種は、P.hybrida種である。「植物」は、特に他で示さない限り、花を有する植物体および植物体から得られる種子を意味する。
【0062】
「植物細胞」の例としては、花、葉、および根などの植物器官における各組織の細胞、カルスならびに懸濁培養細胞が挙げられる。
【0063】
植物細胞への植物発現ベクターの導入には、当業者に周知の方法、例えば、アグロバクテリウムを介する方法、および直接細胞に導入する方法が用いられ得る。アグロバクテリウムを介する方法としては、例えば、Nagelらの方法(FEMS Microbiol. Lett., 67:325(1990))が用いられ得る。この方法は、まず、植物発現ベクターで(例えば、エレクトロポレーションによって)アグロバクテリウムを形質転換し、次いで、形質転換されたアグロバクテリウムをリーフディスク法などの周知の方法により植物細胞に導入する方法である。植物発現ベクターを直接細胞に導入する方法としては、エレクトロポレーション法、パーティクルガン、リン酸カルシウム法、およびポリエチレングリコール法などがある。これらの方法は、当該分野において周知であり、形質転換する植物に適した方法が、当業者により適宜選択され得る。
【0064】
植物発現ベクターを導入された細胞は、例えば、カナマイシン耐性などの薬剤耐性を基準として選択される。選択された細胞は、常法により植物体に再生され得る。
【0065】
再生した植物体において、導入した植物発現ベクターが機能的であることは、当業者に周知の手法を用いて確認し得る。例えば、内因性遺伝子の発現の抑制を目的とする場合、この確認は、ノーザンブロット解析を用いた転写レベルの測定によって行い得る。このようにして、内因性の転写因子の発現が抑制された所望とする形質転換植物体を選択することができる。組織特異的プロモーターを用いた異種遺伝子の発現を目的とする場合も、異種遺伝子の発現の確認は、通常、対象組織から抽出されたRNAを試料とするノーザンブロット解析を用いて行い得る。この解析法の手順は当業者に周知である。
【0066】
本発明の方法に従って内因性の転写因子の発現が抑制され、花粉の稔性が低下したことは、例えば、転写因子をコードするDNAを含む発現ベクターで形質転換して得られた植物の花粉の形態を、必要に応じて組識化学的染色を行ったうえ、顕微鏡観察することで確認できる。
【0067】
また、本発明の方法に従ってプロモーターが小胞子に特異的に発現されたことは、例えば、プロモータとGUS遺伝子とが作動可能に連結された発現ベクターで形質転換して得られた植物における、葯を含む花の組織でのGUS活性の分布を、常法により組識化学的染色を行うことで確認できる。
【実施例】
【0068】
以下に、実施例に基づいて本発明を説明する。本発明の範囲は、実施例のみに限定されるものではない。実施例で使用される制限酵素、プラスミドなどは、商業的な供給源から入手可能である。
【0069】
(実施例1:ZPT転写因子群をコードするポリヌクレオチドを含む植物発現ベクターの構築)
以前に報告した葯特異的ZF遺伝子(Kobayashiら、前出)のうち、PEThyZPT2−5(ZPT2−5)、PEThyZPT3−1(ZPT3−1)、およびPEThyZPT4−1(ZPT4−1)のcDNAを、それぞれ、カリフラワーモザイクウイルスの35Sプロモーターの下流につないで、植物発現ベクターを作成した。具体的には、下記の通りである。
【0070】
(実施例1−1)
プラスミドpBI221(CLONTECH Laboratories Inc.から購入)中のカリフラワーモザイクウイルス35Sプロモーターを含むDNA断片(HindIII−XbaI断片)およびNOSターミネーターを含むDNA断片(SacI−EcoRI断片)を順次プラスミドpUCAP(van Engelen,F.A.ら、Transgenic Res.,4:288,1995)のマルチクローニングサイトに挿入し、pUCAP35Sを作製した。一方、ZPT2−5のcDNAを含むpBluescript ベクターをKpnIサイトおよびSacIサイト(いずれもベクター中のサイト)で切断し、上記pUCAP35SのKpnIおよびSacIサイトの間に挿入した。さらにこの組み替えプラスミドをEcoRIおよびHindIIIで切断し、ZPT2−5をコードするDNA断片をバイナリーベクターpBINPLUS(van Engelen, F.A.ら、前出)のEcoRIおよびHindIIIサイトに導入した。構築されたZPT2−5遺伝子は、図4(a)から明らかなように、カリフラワーモザイクウイルス(CaMV)の35Sプロモーター領域(P35S;0.9 kb)、本発明のZPT2−5をコードするポリヌクレオチド(ZPT2−5;約0.8kb)、およびノパリン合成酵素のターミネーター領域(Tnos;0.3kb)から構成されている。図4中のPnosは、ノパリン合成酵素のプロモーター領域、NPTIIはネオマイシンホスホトランスフェラーゼII遺伝子を示す。
【0071】
(実施例1−2)
ZPT3−1のcDNAを含むpBluescript ベクターをKpnIサイトおよびSacIサイト(いずれもベクター中のサイト)で切断し、pUCAP35SのKpnIおよびSacIサイトの間に挿入した。さらにこの組み換えプラスミドをEcoRIおよびHindIIIで切断し、ZPT3−1をコードするDNA断片をバイナリーベクターpBINPLUSのEcoRIおよびHindIIIサイトに導入した。構築されたZPT3−1遺伝子は、図4(b)から明らかなように、カリフラワーモザイクウイルス(CaMV)の35Sプロモーター領域(P35S;0.9kb)、本発明のZPT3−1をコードするポリヌクレオチド(ZPT3−1;約1.7kb)、およびノパリン合成酵素のターミネーター領域(Tnos;0.3kb)から構成されている。
【0072】
(実施例1−3)
ZPT4−1のcDNAを含むpBluescript ベクターをKpnIサイトおよびSacIサイト(いずれもベクター中のサイト)で切断し、上記pUCAP35SのKpnIおよびSacIサイトの間に挿入した。さらにこの組み換えプラスミドをEcoRIおよびHindIIIで切断し、ZPT4−1をコードするDNA断片をバイナリーベクターpBINPLUSのEcoRIおよびHindIIIサイトに導入した。構築されたZPT4−1遺伝子は、図4(c)から明らかなように、カリフラワーモザイクウイルス(CaMV)の35Sプロモーター領域(P35S;0.9kb)、本発明のZPT4−1をコードするポリヌクレオチド(ZPT4−1;約2.0kb),およびノパリン合成酵素のターミネーター領域(Tnos;0.3kb)から構成されている。
【0073】
(実施例2:ZPT3−1およびZPT4−1プロモーター領域の単離ならびにGUSレポーター遺伝子との連結)
ZPT3−1およびZPT4−1のcDNAをプローブにして、ペチュニアのゲノムDNAライブラリーから、対応するゲノミック・クローンを単離し、転写開始点上流のDNA断片(プロモーター領域;約2.7kbおよび約3.6kb)をサブクローニングした。それぞれのDNA断片をGUSレポーター遺伝子の上流につないでバイナリー・ベクターにクローニングした。具体的には、以下の通りである。
【0074】
(実施例2−1)
ZPT3−1のcDNAを、通常のランダムプライム法(Sambrookら、前出)を用いて[α32P]dCTPで標識して放射性標識プローブを作製し、これを用いてEMBL3ベクター(Stratagene製)中に作製したペチュニア(Petunia hybrida var. Mitchell)のゲノミックライブラリーをスクリーニングした。得られたクローンから遺伝子上流領域を含む約2.7kbのゲノムDNA断片(PstI−SacI)をpBluescriptSKベクターのPstI−SacIサイトにサブクローニングし(pBS−ZPT3−1−PS)、塩基配列を決定した(図5A〜5B)。次に、このプラスミドを鋳型とし、SalI認識配列を含むプライマー(3’−TATGGAGCTCGTCGACAG TTGATGGTTCATTTTTCTGGCTATTGTC−5’;配列番号11)および市販のM13−20プライマーを用いてPCRを行い、ZPT3−1タンパク質翻訳開始点のすぐ下流(塩基番号2661)にSalIサイトを導入した。そして、PstIおよびSalIで切断して得られたDNA断片をpUCAPGUSNTのGUSコード領域の上流に挿入した(pUCAP−ZPT3−1−GUSNT)。これによってZPT3−1遺伝子コード領域のN末端近傍の領域でin frameでGUSコード領域と接続された。さらにpUCAP−ZPT3−1−GUSNTをAscIおよびPacIで切断して得られるDNA断片(ZPT3−1プロモーター、GUSコード領域およびNOSターミネーターを含む)をpBINPLUSベクターに挿入し、pBIN−ZPT3−1−GUSを得た(図7(a))。
【0075】
(実施例2−2)
ZPT4−1についても同様にゲノムDNAを単離し、ZPT4−1遺伝子上流領域を含む約3.6kbのDNA断片(EcoRI−EcoRI)をpBluescriptSKベクターのEcoRI−EcoRIサイトにサブクローニングし(pBS−ZPT4−1−EE)、塩基配列を決定した(図6A〜6B)。このプラスミドを鋳型にし、BamHI認識配列(3’−CATGGATATAGGATCCTATATC−5’;配列番号12)を含むプライマーおよびM13−20プライマーを用いてPCRを行い、ZPT4−1タンパク質翻訳開始点のすぐ下流(塩基番号3641)にBamHIサイトを導入した。そして、EcoRIおよびBamHIで切断して得られたDNA断片をpUCAPGUSNTのGUSコード領域の上流に挿入した(pUCAP−ZPT4−1−GUSNT)。これによってZPT4−1遺伝子コード領域のN末端近傍の領域でin frameでGUSコード領域と接続された。さらに、上記と同様にしてDNA断片(AscI−PacI)をpBINPLUSベクターに挿入し、pBIN−ZPT4−1−GUSを作製した(図7(b))。
【0076】
(実施例3:各融合遺伝子のペチュニア細胞への導入)
上記の各発現ベクターを、以下の手順で、アグロバクテリウムを介して、ペチュニア(Petunia hybrida var. Mitchell)に導入した。
【0077】
(1)アグロバクテリウム・チュメファシエンスLBA4404株(CLONTECH Laboratories Inc.から購入)を250mg/mlのストレプトマイシンと50mg/mlのリファンピシンを含むL培地中、28℃で培養した。Nagelら(1990)(前出)の方法に従って、細胞懸濁液を調製し、実施例1および2で構築したプラスミドベクターをエレクトロポレーションにより、上記菌株に導入した。
【0078】
(2)各融合遺伝子をコードするポリヌクレオチドを、以下の方法によって、ペチュニア細胞へ導入した:上記(1)で得られたアグロバクテリウム・チュメファシエンスLBA4404株をYEB培地(DNA Cloning第2巻、78頁、Glover D.M.編、IRL Press、1985)で振とう培養(28℃、200rpm)した。得られた培養液を、滅菌水で20倍に希釈し、ペチュニア(Petunia hybrida var. Mitchell)の葉片と共存培養した。2〜3日後、抗生物質を含む培地で上記細菌を除去し、2週間ごとに選択培地で継代し、上記5種類の融合遺伝子と共に導入された、pBINPLUS由来のNPTII遺伝子発現によるカナマイシン抵抗性の有無に基づいて、形質転換されたペチュニア細胞を選抜した。選抜した細胞を、常法によりカルスを誘導した後、植物体に再分化した(Jorgensen R.A.ら,Plant Mol. Biol.,31:957,1996)。
【0079】
(実施例4:ZPT遺伝子群を導入した形質転換ペチュニアの表現型)
実施例1のベクターを導入して得られた形質転換体を用い、ZPT2−5、ZPT3−1およびZPT4−1の発現の制御に伴う花粉の形態変化を観察することによって、これらZPT遺伝子群のcDNA導入による植物への影響を検討した。具体的には、以下の通りである。
【0080】
(実施例4−1)
ZPT2−5のcDNAを35Sプロモーターの制御下に導入した形質転換体(14個体)から、ノーザンブロット解析によって、コサプレッションによる遺伝子発現の抑制が起こった個体(3個体)を選抜した。(なお、14個体のうち他の4個体では、導入したZPT2−5遺伝子の過剰発現が観察された。)ノーザンブロット解析の条件は、次の通り:7%SDS、50%ホルムアミド、5xSSC、2%ブロッキング試薬(Boehringer Mannheim製)、50mMリン酸ナトリウム・バッファー(pH7.0)、0.1%ラウリルサルコシン・ナトリウム、50μg/ml酵母tRNA、および32P標識プローブDNA(1x107cpm)を含む溶液中で、68℃で16時間ハイブリダイズさせた後、2xSSC/0.1%SDSで68℃、30分間の洗浄。
【0081】
上記のコサプレッション形質転換体3個体において、以下のような表現型が観察された(図8)。
【0082】
四分子期の直前に起こる減数分裂の過程において、正常な(野生型の)ペチュニアでは、クロマチンの凝縮による細糸状構造の形成に続き(前期Iの細糸期)、相同染色体が対合する(前期Iの接合糸期)。次に、中期I(metaphaseI)に入ると、4分染色体が細胞赤道面にならび、その後、紡錘装置によって相同染色体が均等に細胞の両極に分離する。他方、ZPT2−5遺伝子のコサプレッションを起こした形質転換体では、中期Iにおいて4分染色体が細胞赤道面に正しく並ぶことのないまま、染色体の極分離が進行した。このとき、染色体の両極への分配が著しく不均等であった。
【0083】
正常な減数分裂の過程では、上記1回目の染色体分離の後、2回目の染色体分離によって、4つのハプロイドのグループが形成され、その後に細胞質分離が起こる。他方、上記のコサプレッションを起こした形質転換体では、一回目の染色体分離の直後に細胞質分離および細胞分裂が起こった。また、この不均等な細胞分裂は一回でとどまらず、少なくともさらに二回繰り返されて、最高8個の小胞子細胞が形成された。不均等な染色体分離のために、これらの小胞子細胞に含まれる染色体数は不均一であり、それに伴って、細胞の大きさも著しく不均等であった。結果として、正常なペチュニアにおいて四分子期にあたる時期に、これらの形質転換体では、正常より多い数の小胞子(最高8個)が形成されていた(図8(f);つぼみのサイズ6mmにおけるZPT2−5コサプレッション形質転換体の花粉細胞の写真。また、図9(b);四分子期における形質転換体の花粉細胞の写真も参照)。
【0084】
コサプレッション形質転換体において、小胞子の一部(10−20%)はその後も発達を続けたが、大部分の小胞子細胞は、これらを包むカロース層が分解される前に、破裂した。この段階で破裂せずに生き残った小胞子は、六面体に近い異常形態を示し、正常な小胞子の4面体形態とは明らかに異なっていた。異常形態の小胞子は、その後、一見正常な有糸分裂によって二核となって、花粉粒を形成した。しかし、これらの花粉粒は稔性をほとんど失っていた。すなわち、正常なペチュニアの雌ずいにこれらの形質転換体の花粉粒をかけると、コサプレッションを示した3系統からの花粉では、種子形成が全く無いか、または、わずかに種子が形成されるのみ(最高でも10%、つまり、ペチュニア1株あたりで産生される種子の数が、約10個の花の平均として、コントロールの1割)であった。コサプレッションを示さない形質転換体3系統からの花粉では、野生型のコントロール植物の場合と同様の、正常な種子形成が確認された。
【0085】
上記のコサプレッション形質転換体はまた、雌性配偶体形成にも異常を示し、雌性稔性が正常個体の25−35%に低下していた。すなわち、胚珠(雌性配偶体)の発達は外観上正常であったものの、野生型の花粉で受粉したとき過半数の胚珠が受精できず、受精したものも以後の発達に異常を示し、多くが死滅(abort)した。この場合も、コサプレッションを示さない形質転換体では、野生型のコントロール植物の場合と同様の、正常な雌性配偶体形成が観察された。
【0086】
(実施例4−2)
ZPT3−1のcDNAを35Sプロモーターの支配下に導入すると、ZPT2−5遺伝子を導入した場合と同様の形質変化が、15個体中、3個体に表れた(図9)。すなわち、これらの形質転換体では、減数分裂の過程でZPT2−5の場合と全く同様の異常が見られた。そして、小胞子期まで発達する細胞数は非常に少なく、生き残った小胞子は形態異常(6面体)を示した。さらに、成熟した花粉粒は稔性を失っていた。しかし、ZPT2−5の場合とは異なり、これらの個体では雌性稔性には影響がなかった。
【0087】
実施例4−1と同じ条件でのノーザンブロット法による遺伝子発現解析の結果、ZPT3−1遺伝子を導入した個体においては、ZPT3−1とZPT4−1との両方の遺伝子の発現抑制が観察された。両遺伝子は、構造的類似性が高い。すなわち、コード領域全体の塩基配列の相同性は37%であり、ZPT3−1の2番目のZF領域とZPT4−1の3番目のZF領域、およびZPT3−1の3番目のZF領域とZPT4−1の4番目のZF領域とを、相同性値が最大化されるように近傍の配列を含めて、塩基配列レベルで、それぞれ比較したときの相同性の平均値は86%である(配列比較はClustal Vプログラムによる)。従って、上記の発現抑制事象は、一つの遺伝子の導入によって二つの遺伝子の発現抑制(コサプレッション)が起こったことによる可能性が高い。このことは、これら二つの遺伝子の機能が重複していることを示唆し、両遺伝子の各々の導入によって共通した表現型の変化が見られることとも符合する。
【0088】
(実施例4−3)
ZPT4−1のcDNAを35Sプロモーターの支配下に導入すると、ZPT2−5遺伝子を導入した場合と同様の形質変化が、13個体中、2個体に表れた。すなわち、これらの形質転換体では、減数分裂の過程でZPT2−5の場合と全く同様の異常が見られた。そして、小胞子期まで発達する細胞数は非常に少なく、生き残った小胞子は形態異常(6面体)を示した。さらに、成熟した花粉粒は稔性をほとんど失っていた。しかし、ZPT3−1の場合と同様に、これらの個体では雌性稔性には影響がなかった。上述した理由から、本実施例においても、ZPT3−1とZPT4−1との両方の遺伝子の発現抑制(コサプレッション)が起こった可能性が高い。
【0089】
以上のように、ZPT2−5、ZPT3−1またはZPT4−1をコードする遺伝子の導入によって、きわめて高い効率で、花粉の発達を阻害し、稔性を喪失させることができる(ZPT3−1で99%以上、ZPT2−5およびZPT4−1で90%以上)。また、これらの遺伝子群の導入は、その効果が花粉(ZPT2−5の場合、花粉および雌性配偶体)に特異的であり、植物の他の形質に影響を与えない点で、選択的な形質改変技術として有用であり得る。
【0090】
(実施例5:ZPT3−1およびZPT4−1プロモーター活性の組織特異性)
実施例2のベクターを導入して得られた形質転換体を用い、GUS活性による組織化学的染色を行うことによって、上記DNA断片が有する組織特異的プロモーター活性を検出した。具体的には、以下の通りである。
【0091】
(実施例5−1)
ZPT3−1遺伝子の上流領域とGUSとの融合遺伝子を導入して得られた形質転換体の花を用い、X−GUSを基質にしてGUS活性の分布を調べた(Gallagher,S.R.編、GUS protocols:using the GUS gene as a reporter of gene expressin、Academic Press,Inc.,pp.103−114,1992)。その結果、GUS活性は1核期の小胞子において特異的に検出された(図10(a)〜(c))。
【0092】
(実施例5−2)
ZPT4−1遺伝子の上流領域とGUSとの融合遺伝子を導入して得られた形質転換体の花を用い、上記と同様にGUS活性の分布を調べた。その結果、GUS活性は、一核期から二核期にかけての小胞子および葯の開裂組織において特異的に観察された(図10(d)〜(f);葯の開裂組織を図10(e)および(f)に矢印で示す)。
【0093】
以上のように、ZPT3−1およびZPT4−1遺伝子のプロモーターは、一核期の小胞子(ZPT3−1)および一核期から二核期にかけての小胞子(ZPT4−1)にそれぞれ特異的な活性を示す。ZPT4−1遺伝子のプロモーターは、一核期から二核期にかけての葯の開裂組織にも特異的な活性を示す。
【0094】
小胞子は、のちに成熟して花粉粒を生成する前駆細胞である。従って、これらのプロモーターは、花粉の発達に関連する詳細な研究のためのツールとして有用である。さらに、これらのプロモーターまたはその活性断片を用いて、細胞毒性のある遺伝子などを小胞子に特異的に発現させ、花粉細胞を死滅させまたは機能を失わせることによって、花粉の発達を直接的かつ効率的に制御し得る。
【産業上の利用可能性】
【0095】
ZPT2−5、ZPT3−1およびZPT4−1遺伝子由来の転写因子をコードするDNA、ならびに、ZPT3−1およびZPT4−1遺伝子由来のプロモーターを利用する本件発明の方法は、遺伝子工学的に植物の形質を選択的に改変する技術、特に、雄性不稔形質を付与する技術として有用である。
【図面の簡単な説明】
【0096】
【図1A】ZPT2−5をコードする遺伝子(本明細書中、単に「ZPT2−5遺伝子」ともいう)のcDNA配列、および対応するアミノ酸配列を示す図である。2つのジンクフィンガーモチーフ、およびDLNL配列(アミノ酸145位〜155位)を下線で示す。
【図1B】図1Aの続きである。
【図2A】ZPT3−1をコードする遺伝子(本明細書中、単に「ZPT3−1遺伝子」ともいう)のcDNA配列、および対応するアミノ酸配列を示す図である。3つのジンクフィンガーモチーフ、およびDLNL配列(アミノ酸408位〜417位)を下線で示す。
【図2B】図2Aの続きである。
【図2C】図2Bの続きである。
【図3A】ZPT4−1をコードする遺伝子(本明細書中、単に「ZPT4−1遺伝子」ともいう)のcDNA配列、および対応するアミノ酸配列を示す図である。4つのジンクフィンガーモチーフ、およびDLNL配列(アミノ酸438位〜449位)を下線で示す。
【図3B】図3Aの続きである。
【図3C】図3Bの続きである。
【図3D】図3Cの続きである。
【図4】ZPT2−5、ZPT3−1およびZPT4−1の各cDNA配列を発現するための植物発現ベクター(pBIN−35S−ZPT2−5、pBIN−35S−ZPT3−1およびpBIN−35S−ZPT4−1)の構成を示す概略図である。
【図5A】ZPT3−1遺伝子のコード領域の上流配列を示す図である。転写開始点(2567位)を太矢印で、翻訳開始コドン(ATG)を下線太字で示す。
【図5B】図5Aの続きである。
【図6A】ZPT4−1遺伝子のコード領域の上流配列を示す図である。転写開始点(3503位)を太矢印で、翻訳開始コドン(ATG)を下線太字で示す。
【図6B】図6Aの続きである。
【図7】ZPT3−1およびZPT4−1遺伝子のプロモーターを解析するための植物発現ベクター(pBIN−ZPT3−1−GUSおよびpBIN−ZPT3−1−GUS)の構成を示す概略図である。
【図8】野生型のペチュニアの花粉と、pBIN−35S−ZPT2−5を導入したペチュニア(コサプレッションを起こした形質転換体)の花粉とを撮影した、生物の形態を示す写真である(倍率は400倍)。図8(a)〜(d)は野生型ペチュニア、そして図8(e)〜(h)はコサプレッション形質転換ペチュニアについての、それぞれつぼみの異なる成長段階における花粉を示す。花粉はいずれも、常法によりDAPI(4’,6−diamidino−2−phenylindole dihydrochloride n−hydrate)で染色した。
【図9】野生型のペチュニアの花粉と、pBIN−35S−ZPT3−1を導入したペチュニアの花粉とを撮影した、生物の形態を示す写真である(倍率は700倍)。図9(a)および(c)は野生型ペチュニア、図9(b)および(d)は形質転換ペチュニアについての、それぞれ四分子期および小胞子期における花粉を示す。四分子期の花粉はDAPIで、小胞子期の花粉はサフラニンで、それぞれ常法により染色した。なお、pBIN−35S−ZPT4−1を導入したペチュニアの花粉でも、図9(b)および(d)と全く同様の形態が観察された。
【図10】pBIN−ZPT3−1−GUSおよびpBIN−ZPT4−1−GUSを導入したペチュニアのGUS染色した花の器官を撮影した、生物の形態を示す写真である。いずれも、葯が一核期にある花(つぼみ)を撮影対象とした。図10(a)および(d)は、つぼみの外観を実寸大で、図10(b)および(e)は、葯の低倍率(40倍)での断面図を、図6(c)および(f)は、小胞子(図6(c);倍率は700倍)または葯の開裂組織周辺(図6(f);倍率は200倍)の高倍率での断面図を、それぞれ示す。
【特許請求の範囲】
【請求項1】
雄性不稔性の植物を作出する方法であって:
(i)配列番号1によって示される塩基配列の第1位から第777位までの配列からなるDNA、(ii)(i)の塩基配列からなるDNAとストリンジェントな条件下でハイブリダイズし、花粉の発達を制御する転写因子をコードするDNA、または(iii)(i)もしくは(ii)の断片であるDNAのいずれかである核酸と、該核酸に作動可能に連結されたプロモーターとを含む、植物発現カセットを提供する工程、
花粉の発達を制御する内因性の転写因子を有する植物の細胞を提供する工程であって、ここで、該内因性の転写因子をコードする遺伝子は、該核酸とストリンジェントな条件下でハイブリダイズする、工程、
該発現カセットを該植物細胞に導入する工程、
該発現カセットが導入された植物細胞を植物体に再生する工程、および
該再生された植物体であって、該核酸が発現されることによって、該内因性の転写因子の発現が抑制された植物体を選択する工程、
を包含し、
ただし、上記(ii)のDNAは、(iv)配列番号13によって示される塩基配列の第1位から第1886位までの配列からなるDNAとストリンジェントな条件下でハイブリダイズし、花粉の発達を制御する転写因子をコードするDNAを含まない、
方法。
【請求項1】
雄性不稔性の植物を作出する方法であって:
(i)配列番号1によって示される塩基配列の第1位から第777位までの配列からなるDNA、(ii)(i)の塩基配列からなるDNAとストリンジェントな条件下でハイブリダイズし、花粉の発達を制御する転写因子をコードするDNA、または(iii)(i)もしくは(ii)の断片であるDNAのいずれかである核酸と、該核酸に作動可能に連結されたプロモーターとを含む、植物発現カセットを提供する工程、
花粉の発達を制御する内因性の転写因子を有する植物の細胞を提供する工程であって、ここで、該内因性の転写因子をコードする遺伝子は、該核酸とストリンジェントな条件下でハイブリダイズする、工程、
該発現カセットを該植物細胞に導入する工程、
該発現カセットが導入された植物細胞を植物体に再生する工程、および
該再生された植物体であって、該核酸が発現されることによって、該内因性の転写因子の発現が抑制された植物体を選択する工程、
を包含し、
ただし、上記(ii)のDNAは、(iv)配列番号13によって示される塩基配列の第1位から第1886位までの配列からなるDNAとストリンジェントな条件下でハイブリダイズし、花粉の発達を制御する転写因子をコードするDNAを含まない、
方法。
【図1A】



【図1B】

【図2A】

【図2B】

【図2C】

【図3A】

【図3B】

【図3C】

【図3D】

【図4】
【図5A】

【図5B】

【図6A】

【図6B】

【図7】
【図8】

【図9】
【図10】
【図1B】
【図2A】
【図2B】
【図2C】
【図3A】
【図3B】
【図3C】
【図3D】
【図4】
【図5A】
【図5B】
【図6A】
【図6B】
【図7】
【図8】
【図9】
【図10】
【公開番号】特開2007−202561(P2007−202561A)
【公開日】平成19年8月16日(2007.8.16)
【国際特許分類】
【出願番号】特願2007−44747(P2007−44747)
【出願日】平成19年2月23日(2007.2.23)
【分割の表示】特願2003−110912(P2003−110912)の分割
【原出願日】平成11年11月19日(1999.11.19)
【出願人】(501167644)独立行政法人農業生物資源研究所 (200)
【出願人】(501203344)独立行政法人農業・食品産業技術総合研究機構 (827)
【Fターム(参考)】
【公開日】平成19年8月16日(2007.8.16)
【国際特許分類】
【出願日】平成19年2月23日(2007.2.23)
【分割の表示】特願2003−110912(P2003−110912)の分割
【原出願日】平成11年11月19日(1999.11.19)
【出願人】(501167644)独立行政法人農業生物資源研究所 (200)
【出願人】(501203344)独立行政法人農業・食品産業技術総合研究機構 (827)
【Fターム(参考)】
[ Back to top ]