
若年型糖尿病の治療
【課題】ガストリン/コレシストキニン(CCK)レセプターリガンド、特にガストリン、および表皮増殖因子(EGF)レセプターリガンド、特にトランスファー成長因子アルファ(TGFα)による組み合わせ共同作用の剌激で、膵島前駆細胞が成熟したインスリン産生細胞へと分化するのに影響を与えることによる真性糖尿病の治療を提供する。
【解決手段】ガストリン/CCKレセプターリガンドペプチドとEGFレセプターリガンドペプチドとを含むことを特徴とする。
【解決手段】ガストリン/CCKレセプターリガンドペプチドとEGFレセプターリガンドペプチドとを含むことを特徴とする。
【発明の詳細な説明】
【技術分野】
【0001】
本発明は、ガストリン/コレシストキニン(CCK)レセプターリガンド、特にガストリン、および表皮増殖因子(EGF)レセプターリガンド、特にトランスファー成長因子アルファ(TGFα)による組み合わせ共同作用の剌激で、膵島前駆細胞が成熟したインスリン産生細胞へと分化するのに影響を与えることによる真性糖尿病の治療に関連するものである。
【背景技術】
【0002】
膵島は、胎児の膵臓小管内皮にある内胚葉の幹細胞から発生するが、この膵臓小管内皮は、外分泌を行う膵臓へと発育する多能性幹細胞も含んでいる。Teitelman,G.and J.K.Lee,Developmental Biology,121:454-466(1987);Pictet,R.and W.J.Rutter,Development of the embryonic endocrine pancreas,in Endocrinology,Handbook of Physiology,ed.R.O.Greep and E.B.Astwood(1972),American Physiological Society: Washington D.C.,p.25-66.島の発生は、胎児妊娠期間中に劇的変化によって区切られる不連続な発生状態を通して進む。最初の期間は初期分化状態(protodifferentiated state)で、特徴としては、インスリンとグルカゴンの発現によって明らかな、これらの多能性幹細胞の島細胞整列への掛かり合いがみられる。これらの初期分化細胞は、掛かり合った島前駆細胞群からなり、これは島に特異的な遺伝子生成を低いレベルで発現するだけで、成熟した島細胞の細胞分化は起こらない。Pictet,R.and W.J.Rutter,上記参照。マウス妊娠期間の第16日ごろに、初期分化膵臓では急速な成長と分化の段階が始まり、特徴として、島細胞の細胞分化と島特異的遺伝子発現における数百倍の増加がみられる。組織学的には、島形成(新生)は、膵管から発達し始める島芽の増殖(膵島細胞症)で明らかになってくる。出産の直前に、島の成育速度は遅くなり、島新生と膵島細胞症は極めて少なくなる。これに伴い、島は十分に分化した状態に到達し、インスリン遺伝子発現が最高レベルとなる。このように、多くの臓器と同様、細胞の分化の完成は再生能力の低下と関連している。
【0003】
初期分化前駆体の分化は、胎児の膵臓発生の後期に起こるため、島の分化を調節する因子はこの時期に膵臓内で発現されるようである。島の発生中に発現された遺伝子のひとつが、胃腸ペプチドであるガストリンをエンコードしている。ガストリンは成人において、胃酸分泌を調整する胃ホルモンとして働くが、胎児におけるガストリン発現の主要な部位は膵島である。Brand,S.J.and P.J.Fuller,J.Biol Chem.,263:5341-5347(1988).膵島におけるガストリンの発現は一時のものである。初期分化島前駆体が分化島を形成する時期に限られている。
【0004】
膵臓ガストリンの島発生における重要性はわかっていないが、いくつかの臨床所見より、この島発生におけるガストリンの役割は次のようなものであることが示されている。例えば、ガストリン発現性島細胞腫瘍および萎縮性胃炎によって引き起こされる高ガストリン血症は、胎児の島の分化でみられるのと同様の膵島細胞症に関係している。Sacchi,T.B.,et al.,Virchows Archiv B,48:261-276(1985); and Heitz,P.U.,et al.,Diabetes,26:632-642(1977).さらに、乳児の膵島細胞症の症例で、膵臓ガストリンの異常な持続が報告されている。Hollande E.,et al.,Gastroenterology,71:255-262(1976).しかし、どちらの所見でも膵島細胞症とガストリン刺激の間の因果関係は確立されなかった。
【0005】
この中の文献引用は、これらの文献が本発明に対する先行技術と認められるとは解釈しないものとする。
【非特許文献1】Teitelman,G.and J.K.Lee,Developmental Biology,121:454-466(1987)
【非特許文献2】Pictet,R.and W.J.Rutter,Development of the embryonic endocrine pancreas,in Endocrinology,Handbook of Physiology,ed.R.O.Greep and E.B.Astwood(1972),American Physiological Society: Washington D.C.,p.25-66
【非特許文献3】Brand,S.J.and P.J.Fuller,J.Biol Chem.,263:5341-5347(1988)
【非特許文献4】Sacchi,T.B.,et al.,Virchows Archiv B,48:261-276(1985)
【非特許文献5】Heitz,P.U.,et al.,Diabetes,26:632-642(1977)
【非特許文献6】Hollande E.,et al.,Gastroenterology,71:255-262(1976)
【発明の開示】
【発明が解決しようとする課題】
【0006】
本発明は、ガストリン/CCKレセプターリガンド、例えばガストリン、およびEGFレセプターリガンド、例えばTGFαを含む組成物を、膵島前駆細胞が成熟したインスリン分泌細胞へと分化するのに影響を及ぼすのに十分な量を投与することによって、真性糖尿病を治療する方法を提供するものである。
【課題を解決するための手段】
【0007】
組成物は、全身投与するか、または発現ベクター中で核酸融合構成物を補充した細胞によって体内で発現させることができる。融合構成物には、プレプロガストリン(preprogastrin)ペプチド前駆体コード配列が含まれ、また、EGFレセプターリガンドのコード配列も含まれることがある。
【0008】
要約すると、下記に報告する研究は、完全な島細胞新生は今や、ほ乳類で、成人の膵臓小管上皮において、ガストリン/CCKレセプターリガンド、例えばガストリン、およびEGFレセプターリガンド、例えばTGFαでの剌激によって、in vivoで再活性化されたことを示すものである。これらの研究は、想定した目的を達成するには両方の成長因子が必要があり、どちらであろうと一方だけでは不十分であることを明らかにし確認する。研究は、TGFαとガストリンの膵臓におけるトランスジェニックな外来遺伝子による過剰発現について報告するもので、島発生における膵臓のガストリン発現の役割を解明し、TGFαとガストリンがそれぞれ島発生の調節に役割を果たしていることを示す。
【0009】
このように、残存する多能性膵管細胞の成熟したインスリン分泌細胞への再生的分化は、この組合せの成長因子または組成物を治療的に投与することにより、膵臓内で体内発現を実現する、真性糖尿病、特に若年発症型糖尿病の治療のための臨床上実行可能な選択肢となった。
【発明を実施するための最良の形態】
【0010】
本発明は、TGFαが誘導した化生性小管には、非常に多数のインスリン染色性細胞があることを示した研究に、部分的に基づいている。この化生性小管細胞で見られる内分泌および外分泌の低レベルの遺伝子発現は、胎児の膵臓発生早期に見られる初期分化した小管細胞での遺伝子発現と類似している。島の形成(新生)は、これらの初期分化したインスリン発現細胞が増殖・分化してできる。これは組織学的には、膵管から出る芽のように見える島として顕れる(膵細細胞症)。MT-42 TGFαトランスジェニックマウスでは、出産直後には管の化生は見られないが、わずか4週齢で化生が顕れる。このことは、胎児膵管中に見られる膵島前駆体の存続期間が延長したためではなく、TGFαが過度に発現したために、管上皮でのインスリンの発現が誘導されたことを示している。
【0011】
化生性小管には非常に多数のインスリン陽性細胞があるが、TGFαトランスジェニックマウスの島の量は、コントロール以上に増大していなかった。従ってTGFαの過発現だけでは、これらの初期分化した幹細胞を、十分に分化した島に遷移する効果は得られない。これは、島の分化には、TGFαトランスジェニックマウスの成体膵臓にはない他の因子が必要であることを意味している。初期分化した膵島前駆体の分化は胎児の発育後期に起きるので、遷移を調節する因子がこの期間に島で発現される可能性が高い。発育中の島で発現する因子には、胃腸ペプチドすなわちガストリンがある。臨床の観察でも、ガストリンの発現と膵島細胞症、つまり胎児膵管から増殖する島の出芽とに関連があるとしている。HollandeらのGastroenterology 71:255-252(1976)およびSacchi,T.B.らのVirchows Archiv B 48:261-276(1985)を参照のこと。
【0012】
ここで使用する場合、“ガストリン/CCKレセプターリガンド”という語は、同じ組織や隣接する組織または同じ個体にあるEGFレセプターも刺激したとき、インスリン産生膵島細胞の新生を誘導するような、ガストリン/CCKレセプターを刺激する化合物を含む。そのようなガストリン/CCKレセプターリガンドの例として、ガストリン34(大ガストリン)・ガストリン17(小ガストリン)・ガストリン8(ミニガストリン)といった様々な形態のガストリンや、CCK 58・CCK 33・CCK22・CCK 12・CCK 8といった様々な形態のコレシストキニンや、その他のガストリン/CCKレセプターリガンドで、EGFレセプターリガンドとの相乗作用が同じであり、カルボキシル末端のペプチドがTrp-Met-Asp-Phe-amideのもの(これはEGFレセプターリガンドと相乗的に作用するとき、成熟した膵臓中で細胞分化を誘導し、インスリン分泌性の島細胞を形成する)がある。上記化合物の活性類似体や断片およびその他の修飾物も検討した。そういったリガンドには、保存組織のある部位から、内因性ガストリンやコレシストキニンまたは同様の活性のあるペプチドの分泌を増加させる化合物も含まれている。例としては、胃酸分泌を阻害するオメプラゾールや、CCK剌激を増加する大豆トリプシン阻害物質がある。
【0013】
ここで使用する場合、“EGFレセプターリガンド”という語には、同じ組織や隣接する組織または同じ個体にあるガストリン/CCKレセプターも剌激したとき、インスリン産生膵臓島細胞の新生を誘導するような、EGFレセプターを刺激する化合物を含む。そのようなEGFレセプターの例は、EGFI-4B・EGF1-52・EGF1-49を含むEGF1-53や断片およびそれらの活性類自体などである。その他の例には、1-48や1-47などのTGFαレセプターリガンド(1-50)、アンフィレグリン(amphireglin)やポックスウイルス成長因子などのEGFレセプターリガンド、ガストリン/CCKレセプターリガンドとの相乗作用が同じであるその他のEGFレセプターリガンドである。これらには、上記物質の活性類自体や断片、およびその他の修飾物が含まれる。更に背景を知るには、CparpenterとWahl共著のPeptide Growth Factors(SpornとRoberts編:Springer Verlag、1990年)第4章を参照のこと。
【0014】
本発明の重要な態様の1つは、治療が必要な個人への糖尿病の治療法であり、膵島前駆細胞を成熟したインスリン分泌巣細胞に分化させる効果を出すために十分な量の、ガストリン/CCKレセプターリガンドとEGFレセプターリガンドを含む組成物を個人に投与するものである。この方法で分化した細胞は、膵管内に残存している潜在性の膵島前駆細胞である。本法は主に、若年型糖尿病の治療に用いる。本発明の実施態様の1つは、分化を再生する量のガストリンおよびEGFレセプターリガンドのどれか(TGFαが望ましい)を、治療が必要な個人に投与(全身性が望ましい)することから成り立っている。
【0015】
別の実施態様は、哺乳動物の外植された膵臓組織の膵島前駆細胞にガストリン/CCKレセプターリガンドとEGFレセプターリガンドを投与し、膵臓組織を再導入してその哺乳動物を剌激することから成り立っている。ここでもガストリン/CCKレセプターリガンドはガストリンが望ましく、EGFレセプターリガンドはTGFαが望ましい。
【0016】
別の実施態様では、膵島前駆細胞にキメラのインスリンプロモーターガストリン融合遺伝子構造物をトランスジェニックに膵島前駆細胞に誘導して、ガストリン/CCKレセプターリガンド刺激を引き起こす。別の実施態様では、哺乳動物にトランスジェニックに導入したEGFレセプターリガンド遺伝子が発現することによって、EGFレセプターリガンドの刺激を引き起こす。EGFレセプターリガンドはEGFαが、またEGFレセプターリガンド遺伝子はTFGα遺伝子が望ましい。
【0017】
別の実施態様では、(i)プレプロガストリンペプチド前駆体遺伝子と(ii)EGFレセプターリガンド遺伝子を哺乳動物へ安定に導入し、ガストリン/CCKレセプターリガンドとEGFレセプターリガンドの刺激を引き起こす。ここでも、EGFレセプターリガンドはTGFαが、またEGFレセプターリガンド遺伝子はTFGα遺伝子が望ましい。
【0018】
別の態様では、本発明は、ガストリン/CCKレセプターリガンド(特にガストリン)とEGFレセプターリガンド(特にTGFα)の組み合わせで膵島前駆細胞を刺激して、哺乳動物の膵島前駆細胞の分化を引き起こす方法に関する。この態様の実施態様は、哺乳動物へ安定に導入したプレプロガストリンペプチド前駆体遺伝子の発現によって、ガストリン剌激を起こすことが望ましい。この遺伝子発現は、インスリンプロモーターで制御される。例えばTGFαなどのEGFレセプターリガンド刺激は、トランスジェニックに哺乳動物に導入したEGFレセプターリガンド遺伝子の発現で起きる。上記を促進するには、哺乳動物へ安定に導入した(i)プレプロガストリンペプチド前駆体遺伝子と(ii)EGFレセプターリガンド(例えばTGFα)遺伝子の共同発現によって、ガストリンとTGFαによる刺激を起こすことが望ましい。
【0019】
本発明の別の態様は、核酸融合構造体である。この構造体は、プレプロガストリンペプチド前駆体およびインスリン転写調節配列(その配列は5'側で、プレプロガストリンペプチド前駆体をコードする配列の転写を有効にする)をコードする核酸配列を含んでいる。インスリン転写調節配列は、少なくともインスリンプロモーターを含んでいることが望ましい。プレプロガストリンペプチド前駆体をコードする配列は、ヒトのガストリン遺伝子のエクソン2および3を含む配列のポリヌクレオチド配列から成るのが望ましい具体化方法であり、任意にイントロン1および2も含む。
【0020】
本発明の別の実施態様は、(i)哺乳動物のEGFレセプターリガンド(例えばTGFα)とその転写調節配列をコードする核酸配列、および(ii)プレプロガストリンペプチド前駆体およびその転写調節配列をコードする核酸配列から成る組成物である。EGFレセプターリガンドのプロモーターは、メタロチオネン(metallothionene)プロモーターのような非組織特異性の非常に強いプロモーターが望ましい。プレプロガストリンペプチド前駆体の調節配列は、インスリンプロモーターが望ましい。この実施態様の望ましい形態は、プレプロガストリンペプチド前駆体をコードする核酸配列が、ヒトのガストリン遺伝子のイントロン1および2とエクソン2および3を含むポリヌクレオチド配列から成っているものである。
【0021】
本発明の別の態様は、プレプロガストリンペプチド前駆体をコードする配列から成る融合遺伝子構造体を含むベクターに関する。このベクターは、pGemlのようなプラスミドであったり、インスリンプロモーターを含む転写調節配列を持つファージであってもよい。
【0022】
本発明の別の態様は、(i)非組織特異性の強いプロモーター(例えばメタロチオネンプロモーター)の制御を受ける哺乳動物のEGFレセプターリガンド(例えばTGFα)をコードする配列のあるベクターと、(ii)インスリンプロモーターの制御を受けるプレプロガストリンペプチド前駆体をコードする配列のあるベクターとを含む、ベクターの混合物に関する。いずれのベクターも、プラスミドpGemlなどのプラスミドであったり、上記の性状を持つファージであってもよい。
【0023】
本発明の別の態様は、ヒト以外の哺乳動物やその細胞を含む細胞で、プレプロガストリンをコード化する組み込んだ遺伝子が安定に発現する能力があるものである。この態様の別の実施態様は、哺乳動物や哺乳動物の組織または細胞に安定に組み込まれた(i)プレプロガストリンペプチド前駆体遺伝子と、(ii)EGFレセプターリガンド(例えばTGFα)遺伝子とを発現する能力のある、ヒト以外の哺乳動物である。
【0024】
治療的投与と組成物
投与方法には、経皮・筋肉内・腹腔内・静脈内・皮下・鼻腔内・経口の経路があるが、それらに限定するものではない。化合物の投与は都合のよい経路であればどれでもよく、たとえば輸液や、ボーラス注射で上皮または粘膜皮膚内層(例えば口腔粘膜・直腸粘膜・腸粘膜など)を通る吸収によるものなどがあり、また生物学的に活性のある他の物質と併せて投与してもよい。全身性の投与が望ましい。
【0025】
本発明は製剤組成物も提供する。そのような製剤組成物は、治療的にも薬剤的にも容認できる担体または賦形剤を、治療効果が得られる分量で含んでいる。こういった担体には、食塩水、緩衝食塩水、デキストロース、水、グリセロール、エタノールやそれらの混合物があるが、それらに限定するものではない。処方は投与方法に適うものでなければならない。
【0026】
組成物には、含まれる方が良い場合には、少量の湿潤剤や乳化剤、pH緩衝剤も加えることができる。組成物は、液体溶液、懸濁液、乳化物、錠剤、丸薬、カプセル、持続放出性処方、粉末剤にできる。組成物は、従来の結合材とトリグリセリドのような担体を用いた坐薬としても処方できる。経口処方には、医薬品用のマンニトール、乳糖、澱粉、ステアリン酸マグネシウム、サッカリンナトリウム、セルロース、炭酸マグネシウムなど、標準的な担体が含まれる。
【0027】
様々な輸送方法が知られており、本発明を治療的に投与する為に使用できる。例えば、リポソーム・微少粒子・マイクロカプセルや類似の物への封入がある。
【0028】
望ましい実施態様の1つでは、ヒトの静脈内投与に適合する製剤組成物として、組成物を通常の方法に従って処方する。静脈内投与の組成物は通常、滅菌等張緩衝液の溶液である。必要に応じて、可溶化剤や注射部位での何らかの痛みを和らげるための局所麻酔剤をコンパウンドに含めてもよい。一般には、処方成分は個別にか混合するかして、1回投与量単位で供給する。例えば、凍結乾燥粉末や水分を含まない濃縮物を、アンプルやシール包(sachette)のような密閉容器に入れて、有効剤の量を表示したものなどがある。組成物を輸液で投与する場合は、医薬品用の滅菌水または滅菌食塩水が入った輸液ボトルに調剤できる。組成物を注射で投与する場合は、投与前に成分を混合できるように、注射用の滅菌水または滅菌食塩水のアンプルを供与することもできる。
【0029】
本発明の治療薬は、中性または塩の形として処方できる。薬学的に許容できる塩には、塩酸、リン酸、酢酸、シュウ酸、酒石酸などに誘導され遊離アミノ基と形成するものと、ナトリウム、カリウム、アンモニウム、カルシウム、水酸化第二鉄、イソプロピルアミン、トリエチルアミン、2-エチルアミノエタノール、ヒスチジン、プロカインなどに誘導され遊離カルボキシル基と形成するものとがある。
【0030】
本発明において、特定の疾患や状態を治療する効果があると思われる治療薬量は、疾患や状態の特性によって左右され、標準的な臨床技術で求めることができる。処方に採用する正確な用量も、投与経路と疾患や状態の重篤さに依存しており、医師の判断と各患者の状況によって決定しなければならない。しかし静脈内投与での適切な用量範囲は一般に、体重1kgあたりの活性組成物量20〜500μgである。鼻腔内投与での適切な用量範囲は一般に、体重1kgあたり0.01pg〜1mgである。有効量は、生体外または動物モデルの試験系で得た用量−反応曲線から外挿してもよい。
【0031】
坐薬は一般に活性成分を0.5〜10重量%含んでおり、経口処方では活性成分を10〜95重量%含んでいることが望ましい。
【0032】
本発明は、1つ以上の容器に、本発明の薬剤組成の1つ以上の成分を充愼した製剤パックまたはキットも提供する。そういった容器に付随するものは、薬剤製品や生物学的製剤の製造・使用および販売を取り締まる政府機関が規定した様式の注意書きなどである。その注意書きは、ヒトへの投与を目的とした製造・使用または販売の取り締まり機関による認可を反映する。
【0033】
材料と方法
下記の実施例で報告した研究には、特に注釈がない限りは、以下の材料と方法を使用した。
【0034】
実験動物: FVB系およびCD系マウスは、Taconic Farms Inc.社(ニューヨーク州Germatown)から購入した。使用したTGFαトランスジェニック系MT-42は、メタロチオニン(metallothionine)プロモーターからTGFαが高レベルで発現するが、JappanらのCell、61:1137-1146(1996)に記述がある。
【0035】
INSGASトランスジーン構造体: ラットのインスリンI遺伝子(Cordell,B.G.ら。Cell,18:533-543,1979)の-370から+38のヌクレオチドを含むPvull-Rsal断片を、pGeml(Promega Corp.社、ウイスコンシン州マディソン)に結合した。ヒトガストリン遺伝子の1.5kbのイントロン1および2とエクソン2および3を含み、プレプロガストリンペプチド前駆体をコードする4.4kbのBam Hl-EcoR1断片を分離し、pGeml(Promega社)中のラットインスリンI断片の下流のサブクローンを作成した。この断片は、Wibrog,O.がProc.Natl.Acad.Sci.USAの81: 1067-1069(1984)で、またItoらがProc.Natl.Acad.Sci.USAの81: 4662-4666(1984)で説明している。インスリンプロモーター−プレプロガストリンINSGASトランスジーン構造体は、4.8kbのXbal-EcoR1断片として切り出した。
【0036】
トランスジェニックマウスの世代と特性付け: 上記のようにして作成した断片を、次のようにしてマイクロインジェクション用に調整した。断片は、アガロースゲル電気泳導で分離し、塩化セシウム密度勾配で精製し、大量の注射用バッファー(NaCl 5mM、EDTA 0.1mM、Tris-HCl 5mM、pH7.4)で透析した。FVB近交系マウス(Taconic Farm,Inc.社、supra)の受精卵へ、単細胞期に標準の方法でマイクロインジェクションを行った。Hogan,B.らのManipulating the mouse embryo: A laboratory manual(マウス胚の扱い方:実験マニュアル)Cold Spring Harbor、ニューヨーク州(1986)を参照のこと。次にHoganらの方法に従って、生存した胚をCD1(Charles River Laboratoried,Inc.マサチューセッツ州ウィルミントン)里親マウスの輸卵管に移植した。遺伝子形質転換ができているマウスは、各個体の尾から分離したDNAと、32 dCTPでラベルしたヒトガストリンエクソン2プローブとをランダムプライミングして、DNAブロット法により識別した。F1マウスおよびその子孫も同様にして識別した。
【0037】
CD-1系マウス(Jappan、上記)由来のMT-TGFαトランスジーンを持つ同型接合MT-42マウスを、異型接合のINSGASマウスとかけ合わせた。離乳後、metallothionineプロモーターを誘導するために、先の記述の通りに子を50mMの酸性ZnCl2の上に置いた(Jappan、上記)。
【0038】
ノーザンブロット・ハイブリダイゼーション分析: ノーザン分析用に、CathalaらのDNA,2: 329-355(1983)の方法によって全RNAを組織から抽出した。20μgの全RNA検体を1%のアガロース変性ゲルで分離し、ニトロセルロースへ移した。RNAブロットは32PでラベルしたTGFαリボプローブか、内因性のマウスガストリンmRNAと交差ハイブリダイゼーションしないヒトガストリンのエクソン2かで、ハイブリダイゼーションした。
【0039】
ガストリンのペプチドラジオイムノアッセイ: Rehfeld,J.F.がScand.J.Clin.Lab.Investの30:361-368(1972)で述べたように、組織を摘出し、ガストリンラジオイムノアッセイにおいてC末端がアミド化した生物学的に活性のあるガストリンに特異的な抗体2604を使用して、以前の記述の通りにライジオイムノアッセイで免疫活性を分析した。どのアッセイでもチロシンをモノヨード化したヒトガストリン17トレーサーを使用し、合成ヒトガストリン17を標準として使用した。
【0040】
TGFαのペプチドラジオイムノアッセイ: TodacG.J.らがProc.Natl.Acad.Sci.USAの77:5258-5262(1980)で述べたように、組織を液体窒素で凍結し、乳鉢と乳棒で粉末にひき、酸性エタノールで抽出した。抽出物を水で調整し、タンパク濃度をクマシーブルー色素結合分析(Bio-Rad Laboratories社,カリフォルニア州 Hercules)で測定した。膵臓からとったアリコートは2つ同じものを作り、TGFαラジオイムノアッセイで試験した。アッセイでは、固相の抗ラットTGFαC末端ウサギ抗体への結合を、1251 TGFαとの競合で測定した(キットはBioTop社、ワシントン州シアトル)。
【0041】
組織学的分析: 従来法に従い、膵臓を切除し、重量を測定し、カセットに同じ向きになるように置き、ブアン液で固定し、パラフィン包埋した。
【0042】
組織の調製および免疫組織化学: 膵臓を切除してすぐに解剖し、脂肪とリンパ節を除去してブアン液で固定し、切片作成用にパラフィン包埋した。通常の方法で作成した切片を、標準法に従ってヘマトキシリン−エオシン染色した。17週齢のMT-TGFα(MT-42)トランスジェニックマウスの成体から採取した膵臓組織でインスリンの免疫染色をし、島発達に対するTGFαの過発現の効果を調べた。TGFαに誘導された化生小管にあるインスリン陽性細胞には、抗ヒトインスリン−モルモット血清(Linco社、ミズーリ州Eureka)によるイムノペルオキシダーゼ染色を用い、免疫前のモルモット血清をコントロールに使用した。免疫組織化学は5μのパラフィン切片で、ウサギ抗ガストリン単クローン抗体を使用するSternbergerのペルオキシダーゼ/抗ペルオキシダーゼ法で行った。Sternberger L.A.のImmunocytochemistry(免疫細胞化学)第2版、1979年、ニューヨーク州、Wiley、104〜170ページを参照のこと。
【0043】
ポイント計数による体型測定: 膵島・膵管・間細胞の相対的な体積を、Weibel E.R.がLab.Investig.の12:131-155(1963)で記述したポイント計数法で定量した。400倍で、切片に任意の1角から始め、25ポイントの接眼グリッドを使用して、視野1つおきに点数を付けた。対物ミクロメータの目盛りを使用して、偏りなく系統立てて視野を選択した。血管・膵管・脂肪・リンパ節や小葉間間隙が視野をさえぎっている部分は差し引いて、膵臓の部分だけにした。各ブロック毎に、最低108視野の5000ポイント(対物ミクロメーターを使用して系統立てて選択)を計数し、島組織があるポイント数を膵臓組織があるポイント数で割って相対的な島体積とした。絶対的な島の量や島数は、相対的な島体積を膵臓重量にかけて計算した。Lee,H.C.らによるEndocrinologyの124:1571-1575(1989)を参照のこと。
【0044】
統計的分析: 平均値の差について、独立データのスチューデントのt検定を用いて有意差があるか比較した。
【0045】
実施例1
TGFトランスジェニック膵臓のインスリン産生アッセイ
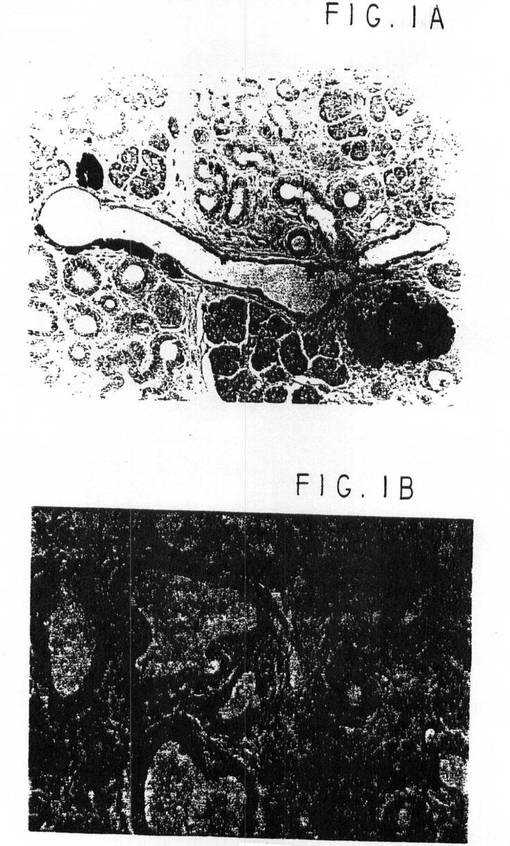
イムノペルオキシダーゼ染色では、TGFαトランスジェニック膵臓(図1A)からの化生管に多数のインスリン染色細胞が見られるが、非トランスジェニック管からはインスリン染色細胞は実質的に存在しなかった(6.1%未満)。最終倍率400×で少なくとも600個の小管細胞/動物を評価したところ、インスリン陽性細胞は、TGFαトランスジェニックマウスの化生小管中に6.0+/−0.9%(n=5)の頻度で見られた。副次的小管細胞は、隣接する島と同強度でインスリン染色されていたが、大半は染色強度が低かった(図1B)。小管細胞のインスリン染色度が低いのは、発育中の膵臓の管で報告される初期分化細胞のそれに似ている。Pictet,R.およびW.J.Rutter、「胚内分泌膵臓の発達(Development of the embryonic endocrine pancreas)」、内分泌学(Rndocrinology)、生理学ハンドブック(Handbook of Physiology)、R.O.GreepおよびE.B.Astwood編、1972、アメリカ生理学会:ワシントンD.C.25-66;およびS.Alpert他、Cell、53:295-308、1988年。
【0046】
しかし、化生管内のインスリン陽性細胞の増加に拘らず、TGFαトランスジェニックマウスの島量は増えなかった。ポイント計数体型測定により計量した島量は、非トランスジェニック同腹質量の1.93mg+/−0.46(n=5)に比べて、TGFαトランスジェニック膵臓では2.14mg+/−0.84(平均+/−se、n=6)であった。
【0047】
この結果から、TGFα過発現が初期分化前駆物質を増殖させるが、これら初期分化細胞を完全に分化した島に遷移させることはできず、分化は成人膵臓にはない他の要因に規制されると解釈できる。
【0048】
実施例2
INSGASトランスジーンからの膵臓ガストリン発現
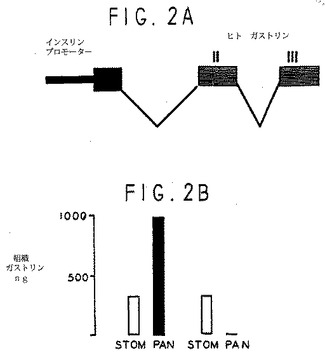
島分化規制におけるガストリンの役割を調べるため、キメラインスリンプロモータガストリン(INSGAS)トランスジーンを発現するトランスジェニックマウスを作ったが、ここではインスリンプロモータが膵臓特異的のガストリントランスジーンの発現を左右している(図2A)。ガストリン遺伝子と異なり、インスリン遺伝子表出は生後切り換えられない。そのため、INSGASトランスジーンは成人膵臓中のガストリン表出を持続させることになる。
【0049】
INSGASトランスジーンは、370bpの5’に隣接するDNAとラットインスリンI遺伝子の第1非コーティングエキソンからなる。Cordell,B.他、Cell、18:533-543、1979。これを、プレプロガストリンペプチド前駆物質をコードするヒトガストリン遺伝子の1.5kbイントロン1およびエキソン2と3を含むBam Hl-Ecorlフラグメントに結合した。Wiborg,O.他、Proc.Natl.Acad.Sci.USA、81:4662-4666、1984。4.8kbのINSGASフラグメントを分離し、同系FVB、1細胞マウス胚にマイクロインジェクションした。Hogan,B.他、「マウス胚の操作(Manipulation the mouse embryo)」:ラボラトリーマニュアル、1986、NY:コールド・スプリング・ハーバー。
【0050】
トランスジェニックマウスと非トランスジェニックマウスの膵臓および胃抽出物中のガストリン免疫反応を、ガストリンの生物活性アミド化C末端特異的な抗血清2604(Rehfeld,J.他、Scand.J.Clin.Lab.Invest.、30:361-368、1972)を使ってラジオイムノアッセイによって測定した。
【0051】
ガストリン単一クローン抗体を持つ膵臓組織の免疫染色に基づき、INSGASトランスジーンからベータ細胞特異的ガストリン発現が見られた。
【0052】
生後8週間のINSGASトランスジェニックマウスの異なる組織から分離したRNAのノザンブロットを、ヒトガストリンエキソン2プローブでハイブリダイゼーションした。膵臓には高レベルのガストリントランスジーンmRNAが見られたが他の組織にはなかった。このプローブは、ヒトガストリン遺伝子特異的で、INSGASの腔RNAにはハイブリダイゼーションは見られず、非トランスジェニックFVBマウスは、高レベルのネズミガストリンmRNAを発現する。INSGASトランスジェニックマウスの膵臓抽出物(PAN)のラジオイムノアッセイでは、内因性ネズミ遺伝子から発現した胃洞(STOM)のガストリン含有量を超える高レベルのガストリン免疫反応が見られる(図2B)。非トランスジェニック対照マウスの膵臓抽出物にはガストリンの免疫反応は検出されなかった。ガストリンのラジオイムノアッセイはカルボキシアミド化前駆物質に特異的で、ガストリンペプチド前駆物質が翻訳後に生物活性ペプチドに効率的に処理されたことを示す。ガストリン単一クローン抗体による免疫組織化学から、膵臓ベータ島細胞固有のガストリン発現がわかる(図2C)。
【0053】
INSGASトランスジェニックマウスは、生後膵臓に高いガストリン発現があったが(図2B)、INSGASトランスジェニックマウスの膵臓組織は、対照と同一であった。ポイント計数体型測定により計量した島量(Weibel,E.R.、Lab Investig.、12:131-155、1963)は、生後5−6週間のINSGASマウス(1.78+/−0.21mg、n=1 l)や、年齢の一致する非トランスジーニックコントロール(1.74+/−0.19mg、n=1 l)と同じだった。そのため、生後膵臓のガストリン持続発現のみでは、島細胞の成長を刺激しない。
【0054】
実施例3
TGFα/INSGAS膵臓の組織学的試験
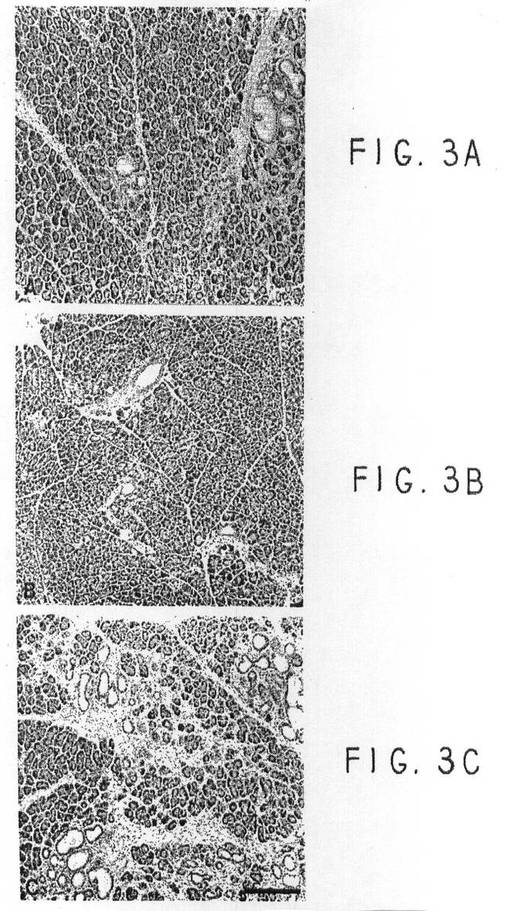
ガストリンによる島成長の刺激には、細胞の反応的群を作り出すための他の成長因子による刺激が必要と思われる。そのため、ガストリン刺激の効果を、TGFαトランスジェニックマウスで試験したが、このマウスは、初期分化した膵島前駆物質に似たインスリン発現細胞を含む化生管を持っている。ガストリンとTGFαとの間の相互作用を評価するため、同じFVB/CDI株の遺伝的背景を持つ3群のマウスを育てた。非トランスジェニック対照、TGFαシングルトランスジェニック、およびINSGAS/TGFαダブルトランスジェニックである。これら3群のマウスを生後3週間で50mM ZnCl2に入れた。生後17週間目に、マウスを犠牲にして、組織学的評価のために膵臓を取りだした。TGFαマウスとINSGAS/TGFαマウスからの膵臓には、柔らかく分散した対照用膵臓とは対照的に、弾力があって固くコンパクトであるというよく似た大きな形態的外観があった。TGFα発現は、ノザンブロット分析(データ示さず)とラジオイムノアッセイで測定したところ、TGFα群とINSGAS/TGFα群で同じだった。膵臓TGFαの免疫反応ペプチドレベルは、TGFαマウスとINSGAS/TGFαマウスで各々12.2+/−1および18.9+/−8ng/mgプロテイン(平均+/−SD)だった。
【0055】
3つのマウス群からの膵臓のヘマトキシリン染色パラフィンセクションの光顕微鏡写真を調査し、(A:INSGAS/TGFα;B:FVB/CD1対照;およびC:TGFα)を作った。INSGAS/TGFα膵臓では、一部で小管複合体が増加し、間隙細胞性がやや増加していた。図示の領域(図3A)では、5匹の動物に重大異常形態が見られ、膵臓の多くは対照と識別不可能だった(図3B)。対照的に、TGFα膵臓の領域(図3C)は典型的で、間隙細胞性と、Jappan他(上記)で述べる鮮紅色の小管化生と複合した繊維過多が見えた。
【0056】
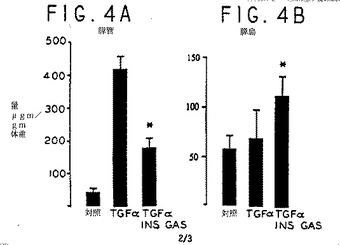
膵臓ガストリンはTGFαと相乗的に作用して島量を増やし、TGFα過発現に誘発された小管化生を抑制する。ホモ接合のMT−TGFα(MT−42)マウス(TGFα)をヘテロ接合のINSGASマウスと交配させた子孫は、ヘテロ接合のTGFαシングルトランスジェニックか、INSGASおよびTGFαトランスジーン(INSGAS/TGFα)を共に含むダブルトランスジェニックのいずれかだった。INSGASはFVB株で、TGFαはCD1株であるため、TGFαホモ接合とCD1対照(CON)を共にFVBと交配して、3つのマウス群すべてについてFVB/CD1株バックグラウンドを生成した。マウスを50mM ZnCl2で3週間処置してから、生後17週目で犠牲にした。膵臓を取り出し、計量し、同じ向きでカセットに入れ、ブワン液に固定し、パラフィンに埋め込んだ。各動物から1つのランダムセクションを使って、小管と島の相対容量をポイント計数体型測定(Weibel,E.R.、Lab Investig.、12:131-155、1963)で計量した。組織上の少なくとも200のポイントを、170×倍で50ポイントグリッドの切片として数えた。セクション全体を重なりなしにカバーした。小管または島の量は、相対容量とその動物の膵臓重量をかけて計算した。異なる平均体重を標準化するため、質量をug/g体重で表した。結果は、各群で5−6匹について平均と標準誤差である。*p<0.05(スチューデント(Student)のt不対データ)。
【0057】
INSGASトランスジーンからのガストリンの表出は、TGFα過発現による小管化生を減じた。17週目で、INSGAS/TGFαマウスの膵臓組織(図3A)は、TGFαマウス(図3C)よりも対照膵臓(図3B)のものに似ていた。
【0058】
これは、TGFαマウスとINSGAS/TGFαトランスジェニックマウスとFVB1/CD1対照の膵臓小管質量をポイント計数体型測定で計量して確認した(図4A)。INSGAS/TGFα膵臓のガストリンとTGFαの共同発現も、対照(図4B)に比べて島量を大きく増加させたが、島量は、TGFαやガストリントランスジーンのみの表出では増加しなかった。血液グルコース濃度は、3つのマウス群で大きく異ならなかった。
【0059】
本発明は、ここに述べた実施様態に限定されるものではない。前記説明および添付の図面から明かな変形は、本発明の請求の範囲内にある。
ここで様々な文献を引用したが、それらの開示内容は参考文献としてその全体が組み込まれる。
【図面の簡単な説明】
【0060】
【図1】図1Aは、イムノペルオキシダーゼ染色で、TGFαでのトランスジェニック膵臓からの化生導管における多数のインスリン染色細胞を示す写真である。図1Bは、ほとんどの小管細胞はインスリンについてあまり強く染色されていないが、たまに近接する島と同程度の強さでインスリン染色されている小管細胞があることを示す写真である。
【図2】図2Aは、インスリンプロモーター−ガストリン(INSGAS)のキメラトランスジーン(transgene)の構造を模式的に示す図である。図2Bは、INSGASトランスジェニックマウスの膵臓抽出物のラジオイムノアッセイが高レベルのガストリン免疫反応性を示し、これはネズミの内生遺伝子から発現した胃洞におけるガストリン濃度を超えていることを示している。INSGASトランスジェニックマウスは、出生直後の膵臓でガストリンの高い発現を示した。
【図3】図3Aは、実施例3の試験に用いたINSGAS/TGFαマウスの膵臓組織構造の写真である。INSGAS/TGFαの膵臓には、小管複合体が増加し、間質の細胞性がわずかに増加した領域があった。ここで示した部分は、用いた5匹の動物のうち最も異常な組織が見られたものである。図3Bは、実施例3で用いた対照マウスの膵臓組織の写真である。図3Cは、実施例3で用いたTGFαマウスの膵臓組織の写真である。実施例3の試験で報告したTGFα膵臓のこの部分は、典型的で、赤らんだ小管化生を伴う間質の細胞性と繊維形成を示した。このことは、Jhappan,et al.上記参照に記載されている。
【図4】図4Aでは、実施例3で報告した試験に基づき、第17週にはINSGAS/TGFαマウスの膵臓で、TGFαマウスのものよりも導管量が低いことを確認した、ポイント計数の体型測定データを示すヒストグラムである。図4Bでは、INSGAS/TGFα膵臓においてガストリンとTGFαを同時発現させると、対応する対照非トランスジェニックマウスと比べて、島の量が増加したことを示す、ポイント計数の体型測定データを示すヒストグラムである。さらに、TGFαの発現だけでは島の量は増加しない。これらのデータは、実施例3で示した試験に基づくものである。
【技術分野】
【0001】
本発明は、ガストリン/コレシストキニン(CCK)レセプターリガンド、特にガストリン、および表皮増殖因子(EGF)レセプターリガンド、特にトランスファー成長因子アルファ(TGFα)による組み合わせ共同作用の剌激で、膵島前駆細胞が成熟したインスリン産生細胞へと分化するのに影響を与えることによる真性糖尿病の治療に関連するものである。
【背景技術】
【0002】
膵島は、胎児の膵臓小管内皮にある内胚葉の幹細胞から発生するが、この膵臓小管内皮は、外分泌を行う膵臓へと発育する多能性幹細胞も含んでいる。Teitelman,G.and J.K.Lee,Developmental Biology,121:454-466(1987);Pictet,R.and W.J.Rutter,Development of the embryonic endocrine pancreas,in Endocrinology,Handbook of Physiology,ed.R.O.Greep and E.B.Astwood(1972),American Physiological Society: Washington D.C.,p.25-66.島の発生は、胎児妊娠期間中に劇的変化によって区切られる不連続な発生状態を通して進む。最初の期間は初期分化状態(protodifferentiated state)で、特徴としては、インスリンとグルカゴンの発現によって明らかな、これらの多能性幹細胞の島細胞整列への掛かり合いがみられる。これらの初期分化細胞は、掛かり合った島前駆細胞群からなり、これは島に特異的な遺伝子生成を低いレベルで発現するだけで、成熟した島細胞の細胞分化は起こらない。Pictet,R.and W.J.Rutter,上記参照。マウス妊娠期間の第16日ごろに、初期分化膵臓では急速な成長と分化の段階が始まり、特徴として、島細胞の細胞分化と島特異的遺伝子発現における数百倍の増加がみられる。組織学的には、島形成(新生)は、膵管から発達し始める島芽の増殖(膵島細胞症)で明らかになってくる。出産の直前に、島の成育速度は遅くなり、島新生と膵島細胞症は極めて少なくなる。これに伴い、島は十分に分化した状態に到達し、インスリン遺伝子発現が最高レベルとなる。このように、多くの臓器と同様、細胞の分化の完成は再生能力の低下と関連している。
【0003】
初期分化前駆体の分化は、胎児の膵臓発生の後期に起こるため、島の分化を調節する因子はこの時期に膵臓内で発現されるようである。島の発生中に発現された遺伝子のひとつが、胃腸ペプチドであるガストリンをエンコードしている。ガストリンは成人において、胃酸分泌を調整する胃ホルモンとして働くが、胎児におけるガストリン発現の主要な部位は膵島である。Brand,S.J.and P.J.Fuller,J.Biol Chem.,263:5341-5347(1988).膵島におけるガストリンの発現は一時のものである。初期分化島前駆体が分化島を形成する時期に限られている。
【0004】
膵臓ガストリンの島発生における重要性はわかっていないが、いくつかの臨床所見より、この島発生におけるガストリンの役割は次のようなものであることが示されている。例えば、ガストリン発現性島細胞腫瘍および萎縮性胃炎によって引き起こされる高ガストリン血症は、胎児の島の分化でみられるのと同様の膵島細胞症に関係している。Sacchi,T.B.,et al.,Virchows Archiv B,48:261-276(1985); and Heitz,P.U.,et al.,Diabetes,26:632-642(1977).さらに、乳児の膵島細胞症の症例で、膵臓ガストリンの異常な持続が報告されている。Hollande E.,et al.,Gastroenterology,71:255-262(1976).しかし、どちらの所見でも膵島細胞症とガストリン刺激の間の因果関係は確立されなかった。
【0005】
この中の文献引用は、これらの文献が本発明に対する先行技術と認められるとは解釈しないものとする。
【非特許文献1】Teitelman,G.and J.K.Lee,Developmental Biology,121:454-466(1987)
【非特許文献2】Pictet,R.and W.J.Rutter,Development of the embryonic endocrine pancreas,in Endocrinology,Handbook of Physiology,ed.R.O.Greep and E.B.Astwood(1972),American Physiological Society: Washington D.C.,p.25-66
【非特許文献3】Brand,S.J.and P.J.Fuller,J.Biol Chem.,263:5341-5347(1988)
【非特許文献4】Sacchi,T.B.,et al.,Virchows Archiv B,48:261-276(1985)
【非特許文献5】Heitz,P.U.,et al.,Diabetes,26:632-642(1977)
【非特許文献6】Hollande E.,et al.,Gastroenterology,71:255-262(1976)
【発明の開示】
【発明が解決しようとする課題】
【0006】
本発明は、ガストリン/CCKレセプターリガンド、例えばガストリン、およびEGFレセプターリガンド、例えばTGFαを含む組成物を、膵島前駆細胞が成熟したインスリン分泌細胞へと分化するのに影響を及ぼすのに十分な量を投与することによって、真性糖尿病を治療する方法を提供するものである。
【課題を解決するための手段】
【0007】
組成物は、全身投与するか、または発現ベクター中で核酸融合構成物を補充した細胞によって体内で発現させることができる。融合構成物には、プレプロガストリン(preprogastrin)ペプチド前駆体コード配列が含まれ、また、EGFレセプターリガンドのコード配列も含まれることがある。
【0008】
要約すると、下記に報告する研究は、完全な島細胞新生は今や、ほ乳類で、成人の膵臓小管上皮において、ガストリン/CCKレセプターリガンド、例えばガストリン、およびEGFレセプターリガンド、例えばTGFαでの剌激によって、in vivoで再活性化されたことを示すものである。これらの研究は、想定した目的を達成するには両方の成長因子が必要があり、どちらであろうと一方だけでは不十分であることを明らかにし確認する。研究は、TGFαとガストリンの膵臓におけるトランスジェニックな外来遺伝子による過剰発現について報告するもので、島発生における膵臓のガストリン発現の役割を解明し、TGFαとガストリンがそれぞれ島発生の調節に役割を果たしていることを示す。
【0009】
このように、残存する多能性膵管細胞の成熟したインスリン分泌細胞への再生的分化は、この組合せの成長因子または組成物を治療的に投与することにより、膵臓内で体内発現を実現する、真性糖尿病、特に若年発症型糖尿病の治療のための臨床上実行可能な選択肢となった。
【発明を実施するための最良の形態】
【0010】
本発明は、TGFαが誘導した化生性小管には、非常に多数のインスリン染色性細胞があることを示した研究に、部分的に基づいている。この化生性小管細胞で見られる内分泌および外分泌の低レベルの遺伝子発現は、胎児の膵臓発生早期に見られる初期分化した小管細胞での遺伝子発現と類似している。島の形成(新生)は、これらの初期分化したインスリン発現細胞が増殖・分化してできる。これは組織学的には、膵管から出る芽のように見える島として顕れる(膵細細胞症)。MT-42 TGFαトランスジェニックマウスでは、出産直後には管の化生は見られないが、わずか4週齢で化生が顕れる。このことは、胎児膵管中に見られる膵島前駆体の存続期間が延長したためではなく、TGFαが過度に発現したために、管上皮でのインスリンの発現が誘導されたことを示している。
【0011】
化生性小管には非常に多数のインスリン陽性細胞があるが、TGFαトランスジェニックマウスの島の量は、コントロール以上に増大していなかった。従ってTGFαの過発現だけでは、これらの初期分化した幹細胞を、十分に分化した島に遷移する効果は得られない。これは、島の分化には、TGFαトランスジェニックマウスの成体膵臓にはない他の因子が必要であることを意味している。初期分化した膵島前駆体の分化は胎児の発育後期に起きるので、遷移を調節する因子がこの期間に島で発現される可能性が高い。発育中の島で発現する因子には、胃腸ペプチドすなわちガストリンがある。臨床の観察でも、ガストリンの発現と膵島細胞症、つまり胎児膵管から増殖する島の出芽とに関連があるとしている。HollandeらのGastroenterology 71:255-252(1976)およびSacchi,T.B.らのVirchows Archiv B 48:261-276(1985)を参照のこと。
【0012】
ここで使用する場合、“ガストリン/CCKレセプターリガンド”という語は、同じ組織や隣接する組織または同じ個体にあるEGFレセプターも刺激したとき、インスリン産生膵島細胞の新生を誘導するような、ガストリン/CCKレセプターを刺激する化合物を含む。そのようなガストリン/CCKレセプターリガンドの例として、ガストリン34(大ガストリン)・ガストリン17(小ガストリン)・ガストリン8(ミニガストリン)といった様々な形態のガストリンや、CCK 58・CCK 33・CCK22・CCK 12・CCK 8といった様々な形態のコレシストキニンや、その他のガストリン/CCKレセプターリガンドで、EGFレセプターリガンドとの相乗作用が同じであり、カルボキシル末端のペプチドがTrp-Met-Asp-Phe-amideのもの(これはEGFレセプターリガンドと相乗的に作用するとき、成熟した膵臓中で細胞分化を誘導し、インスリン分泌性の島細胞を形成する)がある。上記化合物の活性類似体や断片およびその他の修飾物も検討した。そういったリガンドには、保存組織のある部位から、内因性ガストリンやコレシストキニンまたは同様の活性のあるペプチドの分泌を増加させる化合物も含まれている。例としては、胃酸分泌を阻害するオメプラゾールや、CCK剌激を増加する大豆トリプシン阻害物質がある。
【0013】
ここで使用する場合、“EGFレセプターリガンド”という語には、同じ組織や隣接する組織または同じ個体にあるガストリン/CCKレセプターも剌激したとき、インスリン産生膵臓島細胞の新生を誘導するような、EGFレセプターを刺激する化合物を含む。そのようなEGFレセプターの例は、EGFI-4B・EGF1-52・EGF1-49を含むEGF1-53や断片およびそれらの活性類自体などである。その他の例には、1-48や1-47などのTGFαレセプターリガンド(1-50)、アンフィレグリン(amphireglin)やポックスウイルス成長因子などのEGFレセプターリガンド、ガストリン/CCKレセプターリガンドとの相乗作用が同じであるその他のEGFレセプターリガンドである。これらには、上記物質の活性類自体や断片、およびその他の修飾物が含まれる。更に背景を知るには、CparpenterとWahl共著のPeptide Growth Factors(SpornとRoberts編:Springer Verlag、1990年)第4章を参照のこと。
【0014】
本発明の重要な態様の1つは、治療が必要な個人への糖尿病の治療法であり、膵島前駆細胞を成熟したインスリン分泌巣細胞に分化させる効果を出すために十分な量の、ガストリン/CCKレセプターリガンドとEGFレセプターリガンドを含む組成物を個人に投与するものである。この方法で分化した細胞は、膵管内に残存している潜在性の膵島前駆細胞である。本法は主に、若年型糖尿病の治療に用いる。本発明の実施態様の1つは、分化を再生する量のガストリンおよびEGFレセプターリガンドのどれか(TGFαが望ましい)を、治療が必要な個人に投与(全身性が望ましい)することから成り立っている。
【0015】
別の実施態様は、哺乳動物の外植された膵臓組織の膵島前駆細胞にガストリン/CCKレセプターリガンドとEGFレセプターリガンドを投与し、膵臓組織を再導入してその哺乳動物を剌激することから成り立っている。ここでもガストリン/CCKレセプターリガンドはガストリンが望ましく、EGFレセプターリガンドはTGFαが望ましい。
【0016】
別の実施態様では、膵島前駆細胞にキメラのインスリンプロモーターガストリン融合遺伝子構造物をトランスジェニックに膵島前駆細胞に誘導して、ガストリン/CCKレセプターリガンド刺激を引き起こす。別の実施態様では、哺乳動物にトランスジェニックに導入したEGFレセプターリガンド遺伝子が発現することによって、EGFレセプターリガンドの刺激を引き起こす。EGFレセプターリガンドはEGFαが、またEGFレセプターリガンド遺伝子はTFGα遺伝子が望ましい。
【0017】
別の実施態様では、(i)プレプロガストリンペプチド前駆体遺伝子と(ii)EGFレセプターリガンド遺伝子を哺乳動物へ安定に導入し、ガストリン/CCKレセプターリガンドとEGFレセプターリガンドの刺激を引き起こす。ここでも、EGFレセプターリガンドはTGFαが、またEGFレセプターリガンド遺伝子はTFGα遺伝子が望ましい。
【0018】
別の態様では、本発明は、ガストリン/CCKレセプターリガンド(特にガストリン)とEGFレセプターリガンド(特にTGFα)の組み合わせで膵島前駆細胞を刺激して、哺乳動物の膵島前駆細胞の分化を引き起こす方法に関する。この態様の実施態様は、哺乳動物へ安定に導入したプレプロガストリンペプチド前駆体遺伝子の発現によって、ガストリン剌激を起こすことが望ましい。この遺伝子発現は、インスリンプロモーターで制御される。例えばTGFαなどのEGFレセプターリガンド刺激は、トランスジェニックに哺乳動物に導入したEGFレセプターリガンド遺伝子の発現で起きる。上記を促進するには、哺乳動物へ安定に導入した(i)プレプロガストリンペプチド前駆体遺伝子と(ii)EGFレセプターリガンド(例えばTGFα)遺伝子の共同発現によって、ガストリンとTGFαによる刺激を起こすことが望ましい。
【0019】
本発明の別の態様は、核酸融合構造体である。この構造体は、プレプロガストリンペプチド前駆体およびインスリン転写調節配列(その配列は5'側で、プレプロガストリンペプチド前駆体をコードする配列の転写を有効にする)をコードする核酸配列を含んでいる。インスリン転写調節配列は、少なくともインスリンプロモーターを含んでいることが望ましい。プレプロガストリンペプチド前駆体をコードする配列は、ヒトのガストリン遺伝子のエクソン2および3を含む配列のポリヌクレオチド配列から成るのが望ましい具体化方法であり、任意にイントロン1および2も含む。
【0020】
本発明の別の実施態様は、(i)哺乳動物のEGFレセプターリガンド(例えばTGFα)とその転写調節配列をコードする核酸配列、および(ii)プレプロガストリンペプチド前駆体およびその転写調節配列をコードする核酸配列から成る組成物である。EGFレセプターリガンドのプロモーターは、メタロチオネン(metallothionene)プロモーターのような非組織特異性の非常に強いプロモーターが望ましい。プレプロガストリンペプチド前駆体の調節配列は、インスリンプロモーターが望ましい。この実施態様の望ましい形態は、プレプロガストリンペプチド前駆体をコードする核酸配列が、ヒトのガストリン遺伝子のイントロン1および2とエクソン2および3を含むポリヌクレオチド配列から成っているものである。
【0021】
本発明の別の態様は、プレプロガストリンペプチド前駆体をコードする配列から成る融合遺伝子構造体を含むベクターに関する。このベクターは、pGemlのようなプラスミドであったり、インスリンプロモーターを含む転写調節配列を持つファージであってもよい。
【0022】
本発明の別の態様は、(i)非組織特異性の強いプロモーター(例えばメタロチオネンプロモーター)の制御を受ける哺乳動物のEGFレセプターリガンド(例えばTGFα)をコードする配列のあるベクターと、(ii)インスリンプロモーターの制御を受けるプレプロガストリンペプチド前駆体をコードする配列のあるベクターとを含む、ベクターの混合物に関する。いずれのベクターも、プラスミドpGemlなどのプラスミドであったり、上記の性状を持つファージであってもよい。
【0023】
本発明の別の態様は、ヒト以外の哺乳動物やその細胞を含む細胞で、プレプロガストリンをコード化する組み込んだ遺伝子が安定に発現する能力があるものである。この態様の別の実施態様は、哺乳動物や哺乳動物の組織または細胞に安定に組み込まれた(i)プレプロガストリンペプチド前駆体遺伝子と、(ii)EGFレセプターリガンド(例えばTGFα)遺伝子とを発現する能力のある、ヒト以外の哺乳動物である。
【0024】
治療的投与と組成物
投与方法には、経皮・筋肉内・腹腔内・静脈内・皮下・鼻腔内・経口の経路があるが、それらに限定するものではない。化合物の投与は都合のよい経路であればどれでもよく、たとえば輸液や、ボーラス注射で上皮または粘膜皮膚内層(例えば口腔粘膜・直腸粘膜・腸粘膜など)を通る吸収によるものなどがあり、また生物学的に活性のある他の物質と併せて投与してもよい。全身性の投与が望ましい。
【0025】
本発明は製剤組成物も提供する。そのような製剤組成物は、治療的にも薬剤的にも容認できる担体または賦形剤を、治療効果が得られる分量で含んでいる。こういった担体には、食塩水、緩衝食塩水、デキストロース、水、グリセロール、エタノールやそれらの混合物があるが、それらに限定するものではない。処方は投与方法に適うものでなければならない。
【0026】
組成物には、含まれる方が良い場合には、少量の湿潤剤や乳化剤、pH緩衝剤も加えることができる。組成物は、液体溶液、懸濁液、乳化物、錠剤、丸薬、カプセル、持続放出性処方、粉末剤にできる。組成物は、従来の結合材とトリグリセリドのような担体を用いた坐薬としても処方できる。経口処方には、医薬品用のマンニトール、乳糖、澱粉、ステアリン酸マグネシウム、サッカリンナトリウム、セルロース、炭酸マグネシウムなど、標準的な担体が含まれる。
【0027】
様々な輸送方法が知られており、本発明を治療的に投与する為に使用できる。例えば、リポソーム・微少粒子・マイクロカプセルや類似の物への封入がある。
【0028】
望ましい実施態様の1つでは、ヒトの静脈内投与に適合する製剤組成物として、組成物を通常の方法に従って処方する。静脈内投与の組成物は通常、滅菌等張緩衝液の溶液である。必要に応じて、可溶化剤や注射部位での何らかの痛みを和らげるための局所麻酔剤をコンパウンドに含めてもよい。一般には、処方成分は個別にか混合するかして、1回投与量単位で供給する。例えば、凍結乾燥粉末や水分を含まない濃縮物を、アンプルやシール包(sachette)のような密閉容器に入れて、有効剤の量を表示したものなどがある。組成物を輸液で投与する場合は、医薬品用の滅菌水または滅菌食塩水が入った輸液ボトルに調剤できる。組成物を注射で投与する場合は、投与前に成分を混合できるように、注射用の滅菌水または滅菌食塩水のアンプルを供与することもできる。
【0029】
本発明の治療薬は、中性または塩の形として処方できる。薬学的に許容できる塩には、塩酸、リン酸、酢酸、シュウ酸、酒石酸などに誘導され遊離アミノ基と形成するものと、ナトリウム、カリウム、アンモニウム、カルシウム、水酸化第二鉄、イソプロピルアミン、トリエチルアミン、2-エチルアミノエタノール、ヒスチジン、プロカインなどに誘導され遊離カルボキシル基と形成するものとがある。
【0030】
本発明において、特定の疾患や状態を治療する効果があると思われる治療薬量は、疾患や状態の特性によって左右され、標準的な臨床技術で求めることができる。処方に採用する正確な用量も、投与経路と疾患や状態の重篤さに依存しており、医師の判断と各患者の状況によって決定しなければならない。しかし静脈内投与での適切な用量範囲は一般に、体重1kgあたりの活性組成物量20〜500μgである。鼻腔内投与での適切な用量範囲は一般に、体重1kgあたり0.01pg〜1mgである。有効量は、生体外または動物モデルの試験系で得た用量−反応曲線から外挿してもよい。
【0031】
坐薬は一般に活性成分を0.5〜10重量%含んでおり、経口処方では活性成分を10〜95重量%含んでいることが望ましい。
【0032】
本発明は、1つ以上の容器に、本発明の薬剤組成の1つ以上の成分を充愼した製剤パックまたはキットも提供する。そういった容器に付随するものは、薬剤製品や生物学的製剤の製造・使用および販売を取り締まる政府機関が規定した様式の注意書きなどである。その注意書きは、ヒトへの投与を目的とした製造・使用または販売の取り締まり機関による認可を反映する。
【0033】
材料と方法
下記の実施例で報告した研究には、特に注釈がない限りは、以下の材料と方法を使用した。
【0034】
実験動物: FVB系およびCD系マウスは、Taconic Farms Inc.社(ニューヨーク州Germatown)から購入した。使用したTGFαトランスジェニック系MT-42は、メタロチオニン(metallothionine)プロモーターからTGFαが高レベルで発現するが、JappanらのCell、61:1137-1146(1996)に記述がある。
【0035】
INSGASトランスジーン構造体: ラットのインスリンI遺伝子(Cordell,B.G.ら。Cell,18:533-543,1979)の-370から+38のヌクレオチドを含むPvull-Rsal断片を、pGeml(Promega Corp.社、ウイスコンシン州マディソン)に結合した。ヒトガストリン遺伝子の1.5kbのイントロン1および2とエクソン2および3を含み、プレプロガストリンペプチド前駆体をコードする4.4kbのBam Hl-EcoR1断片を分離し、pGeml(Promega社)中のラットインスリンI断片の下流のサブクローンを作成した。この断片は、Wibrog,O.がProc.Natl.Acad.Sci.USAの81: 1067-1069(1984)で、またItoらがProc.Natl.Acad.Sci.USAの81: 4662-4666(1984)で説明している。インスリンプロモーター−プレプロガストリンINSGASトランスジーン構造体は、4.8kbのXbal-EcoR1断片として切り出した。
【0036】
トランスジェニックマウスの世代と特性付け: 上記のようにして作成した断片を、次のようにしてマイクロインジェクション用に調整した。断片は、アガロースゲル電気泳導で分離し、塩化セシウム密度勾配で精製し、大量の注射用バッファー(NaCl 5mM、EDTA 0.1mM、Tris-HCl 5mM、pH7.4)で透析した。FVB近交系マウス(Taconic Farm,Inc.社、supra)の受精卵へ、単細胞期に標準の方法でマイクロインジェクションを行った。Hogan,B.らのManipulating the mouse embryo: A laboratory manual(マウス胚の扱い方:実験マニュアル)Cold Spring Harbor、ニューヨーク州(1986)を参照のこと。次にHoganらの方法に従って、生存した胚をCD1(Charles River Laboratoried,Inc.マサチューセッツ州ウィルミントン)里親マウスの輸卵管に移植した。遺伝子形質転換ができているマウスは、各個体の尾から分離したDNAと、32 dCTPでラベルしたヒトガストリンエクソン2プローブとをランダムプライミングして、DNAブロット法により識別した。F1マウスおよびその子孫も同様にして識別した。
【0037】
CD-1系マウス(Jappan、上記)由来のMT-TGFαトランスジーンを持つ同型接合MT-42マウスを、異型接合のINSGASマウスとかけ合わせた。離乳後、metallothionineプロモーターを誘導するために、先の記述の通りに子を50mMの酸性ZnCl2の上に置いた(Jappan、上記)。
【0038】
ノーザンブロット・ハイブリダイゼーション分析: ノーザン分析用に、CathalaらのDNA,2: 329-355(1983)の方法によって全RNAを組織から抽出した。20μgの全RNA検体を1%のアガロース変性ゲルで分離し、ニトロセルロースへ移した。RNAブロットは32PでラベルしたTGFαリボプローブか、内因性のマウスガストリンmRNAと交差ハイブリダイゼーションしないヒトガストリンのエクソン2かで、ハイブリダイゼーションした。
【0039】
ガストリンのペプチドラジオイムノアッセイ: Rehfeld,J.F.がScand.J.Clin.Lab.Investの30:361-368(1972)で述べたように、組織を摘出し、ガストリンラジオイムノアッセイにおいてC末端がアミド化した生物学的に活性のあるガストリンに特異的な抗体2604を使用して、以前の記述の通りにライジオイムノアッセイで免疫活性を分析した。どのアッセイでもチロシンをモノヨード化したヒトガストリン17トレーサーを使用し、合成ヒトガストリン17を標準として使用した。
【0040】
TGFαのペプチドラジオイムノアッセイ: TodacG.J.らがProc.Natl.Acad.Sci.USAの77:5258-5262(1980)で述べたように、組織を液体窒素で凍結し、乳鉢と乳棒で粉末にひき、酸性エタノールで抽出した。抽出物を水で調整し、タンパク濃度をクマシーブルー色素結合分析(Bio-Rad Laboratories社,カリフォルニア州 Hercules)で測定した。膵臓からとったアリコートは2つ同じものを作り、TGFαラジオイムノアッセイで試験した。アッセイでは、固相の抗ラットTGFαC末端ウサギ抗体への結合を、1251 TGFαとの競合で測定した(キットはBioTop社、ワシントン州シアトル)。
【0041】
組織学的分析: 従来法に従い、膵臓を切除し、重量を測定し、カセットに同じ向きになるように置き、ブアン液で固定し、パラフィン包埋した。
【0042】
組織の調製および免疫組織化学: 膵臓を切除してすぐに解剖し、脂肪とリンパ節を除去してブアン液で固定し、切片作成用にパラフィン包埋した。通常の方法で作成した切片を、標準法に従ってヘマトキシリン−エオシン染色した。17週齢のMT-TGFα(MT-42)トランスジェニックマウスの成体から採取した膵臓組織でインスリンの免疫染色をし、島発達に対するTGFαの過発現の効果を調べた。TGFαに誘導された化生小管にあるインスリン陽性細胞には、抗ヒトインスリン−モルモット血清(Linco社、ミズーリ州Eureka)によるイムノペルオキシダーゼ染色を用い、免疫前のモルモット血清をコントロールに使用した。免疫組織化学は5μのパラフィン切片で、ウサギ抗ガストリン単クローン抗体を使用するSternbergerのペルオキシダーゼ/抗ペルオキシダーゼ法で行った。Sternberger L.A.のImmunocytochemistry(免疫細胞化学)第2版、1979年、ニューヨーク州、Wiley、104〜170ページを参照のこと。
【0043】
ポイント計数による体型測定: 膵島・膵管・間細胞の相対的な体積を、Weibel E.R.がLab.Investig.の12:131-155(1963)で記述したポイント計数法で定量した。400倍で、切片に任意の1角から始め、25ポイントの接眼グリッドを使用して、視野1つおきに点数を付けた。対物ミクロメータの目盛りを使用して、偏りなく系統立てて視野を選択した。血管・膵管・脂肪・リンパ節や小葉間間隙が視野をさえぎっている部分は差し引いて、膵臓の部分だけにした。各ブロック毎に、最低108視野の5000ポイント(対物ミクロメーターを使用して系統立てて選択)を計数し、島組織があるポイント数を膵臓組織があるポイント数で割って相対的な島体積とした。絶対的な島の量や島数は、相対的な島体積を膵臓重量にかけて計算した。Lee,H.C.らによるEndocrinologyの124:1571-1575(1989)を参照のこと。
【0044】
統計的分析: 平均値の差について、独立データのスチューデントのt検定を用いて有意差があるか比較した。
【0045】
実施例1
TGFトランスジェニック膵臓のインスリン産生アッセイ
イムノペルオキシダーゼ染色では、TGFαトランスジェニック膵臓(図1A)からの化生管に多数のインスリン染色細胞が見られるが、非トランスジェニック管からはインスリン染色細胞は実質的に存在しなかった(6.1%未満)。最終倍率400×で少なくとも600個の小管細胞/動物を評価したところ、インスリン陽性細胞は、TGFαトランスジェニックマウスの化生小管中に6.0+/−0.9%(n=5)の頻度で見られた。副次的小管細胞は、隣接する島と同強度でインスリン染色されていたが、大半は染色強度が低かった(図1B)。小管細胞のインスリン染色度が低いのは、発育中の膵臓の管で報告される初期分化細胞のそれに似ている。Pictet,R.およびW.J.Rutter、「胚内分泌膵臓の発達(Development of the embryonic endocrine pancreas)」、内分泌学(Rndocrinology)、生理学ハンドブック(Handbook of Physiology)、R.O.GreepおよびE.B.Astwood編、1972、アメリカ生理学会:ワシントンD.C.25-66;およびS.Alpert他、Cell、53:295-308、1988年。
【0046】
しかし、化生管内のインスリン陽性細胞の増加に拘らず、TGFαトランスジェニックマウスの島量は増えなかった。ポイント計数体型測定により計量した島量は、非トランスジェニック同腹質量の1.93mg+/−0.46(n=5)に比べて、TGFαトランスジェニック膵臓では2.14mg+/−0.84(平均+/−se、n=6)であった。
【0047】
この結果から、TGFα過発現が初期分化前駆物質を増殖させるが、これら初期分化細胞を完全に分化した島に遷移させることはできず、分化は成人膵臓にはない他の要因に規制されると解釈できる。
【0048】
実施例2
INSGASトランスジーンからの膵臓ガストリン発現
島分化規制におけるガストリンの役割を調べるため、キメラインスリンプロモータガストリン(INSGAS)トランスジーンを発現するトランスジェニックマウスを作ったが、ここではインスリンプロモータが膵臓特異的のガストリントランスジーンの発現を左右している(図2A)。ガストリン遺伝子と異なり、インスリン遺伝子表出は生後切り換えられない。そのため、INSGASトランスジーンは成人膵臓中のガストリン表出を持続させることになる。
【0049】
INSGASトランスジーンは、370bpの5’に隣接するDNAとラットインスリンI遺伝子の第1非コーティングエキソンからなる。Cordell,B.他、Cell、18:533-543、1979。これを、プレプロガストリンペプチド前駆物質をコードするヒトガストリン遺伝子の1.5kbイントロン1およびエキソン2と3を含むBam Hl-Ecorlフラグメントに結合した。Wiborg,O.他、Proc.Natl.Acad.Sci.USA、81:4662-4666、1984。4.8kbのINSGASフラグメントを分離し、同系FVB、1細胞マウス胚にマイクロインジェクションした。Hogan,B.他、「マウス胚の操作(Manipulation the mouse embryo)」:ラボラトリーマニュアル、1986、NY:コールド・スプリング・ハーバー。
【0050】
トランスジェニックマウスと非トランスジェニックマウスの膵臓および胃抽出物中のガストリン免疫反応を、ガストリンの生物活性アミド化C末端特異的な抗血清2604(Rehfeld,J.他、Scand.J.Clin.Lab.Invest.、30:361-368、1972)を使ってラジオイムノアッセイによって測定した。
【0051】
ガストリン単一クローン抗体を持つ膵臓組織の免疫染色に基づき、INSGASトランスジーンからベータ細胞特異的ガストリン発現が見られた。
【0052】
生後8週間のINSGASトランスジェニックマウスの異なる組織から分離したRNAのノザンブロットを、ヒトガストリンエキソン2プローブでハイブリダイゼーションした。膵臓には高レベルのガストリントランスジーンmRNAが見られたが他の組織にはなかった。このプローブは、ヒトガストリン遺伝子特異的で、INSGASの腔RNAにはハイブリダイゼーションは見られず、非トランスジェニックFVBマウスは、高レベルのネズミガストリンmRNAを発現する。INSGASトランスジェニックマウスの膵臓抽出物(PAN)のラジオイムノアッセイでは、内因性ネズミ遺伝子から発現した胃洞(STOM)のガストリン含有量を超える高レベルのガストリン免疫反応が見られる(図2B)。非トランスジェニック対照マウスの膵臓抽出物にはガストリンの免疫反応は検出されなかった。ガストリンのラジオイムノアッセイはカルボキシアミド化前駆物質に特異的で、ガストリンペプチド前駆物質が翻訳後に生物活性ペプチドに効率的に処理されたことを示す。ガストリン単一クローン抗体による免疫組織化学から、膵臓ベータ島細胞固有のガストリン発現がわかる(図2C)。
【0053】
INSGASトランスジェニックマウスは、生後膵臓に高いガストリン発現があったが(図2B)、INSGASトランスジェニックマウスの膵臓組織は、対照と同一であった。ポイント計数体型測定により計量した島量(Weibel,E.R.、Lab Investig.、12:131-155、1963)は、生後5−6週間のINSGASマウス(1.78+/−0.21mg、n=1 l)や、年齢の一致する非トランスジーニックコントロール(1.74+/−0.19mg、n=1 l)と同じだった。そのため、生後膵臓のガストリン持続発現のみでは、島細胞の成長を刺激しない。
【0054】
実施例3
TGFα/INSGAS膵臓の組織学的試験
ガストリンによる島成長の刺激には、細胞の反応的群を作り出すための他の成長因子による刺激が必要と思われる。そのため、ガストリン刺激の効果を、TGFαトランスジェニックマウスで試験したが、このマウスは、初期分化した膵島前駆物質に似たインスリン発現細胞を含む化生管を持っている。ガストリンとTGFαとの間の相互作用を評価するため、同じFVB/CDI株の遺伝的背景を持つ3群のマウスを育てた。非トランスジェニック対照、TGFαシングルトランスジェニック、およびINSGAS/TGFαダブルトランスジェニックである。これら3群のマウスを生後3週間で50mM ZnCl2に入れた。生後17週間目に、マウスを犠牲にして、組織学的評価のために膵臓を取りだした。TGFαマウスとINSGAS/TGFαマウスからの膵臓には、柔らかく分散した対照用膵臓とは対照的に、弾力があって固くコンパクトであるというよく似た大きな形態的外観があった。TGFα発現は、ノザンブロット分析(データ示さず)とラジオイムノアッセイで測定したところ、TGFα群とINSGAS/TGFα群で同じだった。膵臓TGFαの免疫反応ペプチドレベルは、TGFαマウスとINSGAS/TGFαマウスで各々12.2+/−1および18.9+/−8ng/mgプロテイン(平均+/−SD)だった。
【0055】
3つのマウス群からの膵臓のヘマトキシリン染色パラフィンセクションの光顕微鏡写真を調査し、(A:INSGAS/TGFα;B:FVB/CD1対照;およびC:TGFα)を作った。INSGAS/TGFα膵臓では、一部で小管複合体が増加し、間隙細胞性がやや増加していた。図示の領域(図3A)では、5匹の動物に重大異常形態が見られ、膵臓の多くは対照と識別不可能だった(図3B)。対照的に、TGFα膵臓の領域(図3C)は典型的で、間隙細胞性と、Jappan他(上記)で述べる鮮紅色の小管化生と複合した繊維過多が見えた。
【0056】
膵臓ガストリンはTGFαと相乗的に作用して島量を増やし、TGFα過発現に誘発された小管化生を抑制する。ホモ接合のMT−TGFα(MT−42)マウス(TGFα)をヘテロ接合のINSGASマウスと交配させた子孫は、ヘテロ接合のTGFαシングルトランスジェニックか、INSGASおよびTGFαトランスジーン(INSGAS/TGFα)を共に含むダブルトランスジェニックのいずれかだった。INSGASはFVB株で、TGFαはCD1株であるため、TGFαホモ接合とCD1対照(CON)を共にFVBと交配して、3つのマウス群すべてについてFVB/CD1株バックグラウンドを生成した。マウスを50mM ZnCl2で3週間処置してから、生後17週目で犠牲にした。膵臓を取り出し、計量し、同じ向きでカセットに入れ、ブワン液に固定し、パラフィンに埋め込んだ。各動物から1つのランダムセクションを使って、小管と島の相対容量をポイント計数体型測定(Weibel,E.R.、Lab Investig.、12:131-155、1963)で計量した。組織上の少なくとも200のポイントを、170×倍で50ポイントグリッドの切片として数えた。セクション全体を重なりなしにカバーした。小管または島の量は、相対容量とその動物の膵臓重量をかけて計算した。異なる平均体重を標準化するため、質量をug/g体重で表した。結果は、各群で5−6匹について平均と標準誤差である。*p<0.05(スチューデント(Student)のt不対データ)。
【0057】
INSGASトランスジーンからのガストリンの表出は、TGFα過発現による小管化生を減じた。17週目で、INSGAS/TGFαマウスの膵臓組織(図3A)は、TGFαマウス(図3C)よりも対照膵臓(図3B)のものに似ていた。
【0058】
これは、TGFαマウスとINSGAS/TGFαトランスジェニックマウスとFVB1/CD1対照の膵臓小管質量をポイント計数体型測定で計量して確認した(図4A)。INSGAS/TGFα膵臓のガストリンとTGFαの共同発現も、対照(図4B)に比べて島量を大きく増加させたが、島量は、TGFαやガストリントランスジーンのみの表出では増加しなかった。血液グルコース濃度は、3つのマウス群で大きく異ならなかった。
【0059】
本発明は、ここに述べた実施様態に限定されるものではない。前記説明および添付の図面から明かな変形は、本発明の請求の範囲内にある。
ここで様々な文献を引用したが、それらの開示内容は参考文献としてその全体が組み込まれる。
【図面の簡単な説明】
【0060】
【図1】図1Aは、イムノペルオキシダーゼ染色で、TGFαでのトランスジェニック膵臓からの化生導管における多数のインスリン染色細胞を示す写真である。図1Bは、ほとんどの小管細胞はインスリンについてあまり強く染色されていないが、たまに近接する島と同程度の強さでインスリン染色されている小管細胞があることを示す写真である。
【図2】図2Aは、インスリンプロモーター−ガストリン(INSGAS)のキメラトランスジーン(transgene)の構造を模式的に示す図である。図2Bは、INSGASトランスジェニックマウスの膵臓抽出物のラジオイムノアッセイが高レベルのガストリン免疫反応性を示し、これはネズミの内生遺伝子から発現した胃洞におけるガストリン濃度を超えていることを示している。INSGASトランスジェニックマウスは、出生直後の膵臓でガストリンの高い発現を示した。
【図3】図3Aは、実施例3の試験に用いたINSGAS/TGFαマウスの膵臓組織構造の写真である。INSGAS/TGFαの膵臓には、小管複合体が増加し、間質の細胞性がわずかに増加した領域があった。ここで示した部分は、用いた5匹の動物のうち最も異常な組織が見られたものである。図3Bは、実施例3で用いた対照マウスの膵臓組織の写真である。図3Cは、実施例3で用いたTGFαマウスの膵臓組織の写真である。実施例3の試験で報告したTGFα膵臓のこの部分は、典型的で、赤らんだ小管化生を伴う間質の細胞性と繊維形成を示した。このことは、Jhappan,et al.上記参照に記載されている。
【図4】図4Aでは、実施例3で報告した試験に基づき、第17週にはINSGAS/TGFαマウスの膵臓で、TGFαマウスのものよりも導管量が低いことを確認した、ポイント計数の体型測定データを示すヒストグラムである。図4Bでは、INSGAS/TGFα膵臓においてガストリンとTGFαを同時発現させると、対応する対照非トランスジェニックマウスと比べて、島の量が増加したことを示す、ポイント計数の体型測定データを示すヒストグラムである。さらに、TGFαの発現だけでは島の量は増加しない。これらのデータは、実施例3で示した試験に基づくものである。
【特許請求の範囲】
【請求項1】
ガストリン/CCKレセプターリガンドペプチドとEGFレセプターリガンドペプチドとの組み合わせを用いて、ex vivoで膵島前駆細胞を刺激することを含む、哺乳動物の膵島前駆細胞を成熟インスリン分泌細胞へと分化させるためのex vivoでの方法。
【請求項2】
ガストリンとEGFレセプターリガンドペプチドとを用いて、単離した膵島前駆細胞を処理する、請求項1記載のex vivoでの方法。
【請求項3】
ガストリンがガストリン8、17または34である、請求項2記載のex vivoでの方法。
【請求項4】
EGFレセプターリガンドペプチドがEGF類似体である、請求項1、2または3記載のex vivoでの方法。
【請求項5】
EGFレセプターリガンドペプチドがTGFαである、請求項1、2または3記載のex vivoでの方法。
【請求項6】
EGFレセプターリガンドペプチドがアンフィレグリンまたはポックスウイルス成長因子である、請求項1、2または3記載のex vivoでの方法。
【請求項7】
哺乳動物が糖尿病を患っている、請求項1ないし6のいずれか一項に記載のex vivoでの方法。
【請求項8】
哺乳動物がヒトである、請求項7記載のex vivoでの方法。
【請求項9】
膵臓前駆細胞を、分化を引き起こす量のガストリンおよびEGFレセプターリガンドペプチドを用いて処理する、請求項2ないし8のいずれか一項に記載のex vivoでの方法。
【請求項10】
膵島前駆細胞が哺乳動物の体外移植された膵臓組織に由来する、請求項1ないし9のいずれか一項に記載のex vivoでの方法。
【請求項11】
哺乳動物に、刺激した膵島前駆細胞を導入することをさらに含む、請求項1ないし10のいずれか一項に記載のex vivoでの方法。
【請求項12】
哺乳動物において膵島前駆細胞を成熟インスリン分泌膵島細胞へと分化させる全身投与用医薬の調製における、全身的に有効量のガストリン/CCKレセプターリガンドペプチドとEGFレセプターリガンドペプチドの使用。
【請求項13】
膵島前駆細胞が体外移植された膵臓組織に由来し、分化後に哺乳動物に導入される、請求項12記載の使用。
【請求項14】
糖尿病治療用の全身投与用医薬の調製における、全身的に有効量のガストリン/CCKレセプターリガンドペプチドとEGFレセプターリガンドペプチドの使用。
【請求項15】
単離された膵島前駆細胞がガストリンとEGFレセプターリガンドペプチドを用いて処理される、請求項12、13または14記載の使用。
【請求項16】
ガストリンがガストリン8、17または34である、請求項15記載の使用。
【請求項17】
EGFレセプターリガンドペプチドがEGF類似体である、請求項12ないし16のいずれか一項に記載の使用。
【請求項18】
EGFレセプターリガンドペプチドがTGFαである、請求項12ないし16のいずれか一項に記載の使用。
【請求項19】
EGFレセプターリガンドペプチドがアンフィレグリンまたはポックスウイルス成長因子である、請求項12ないし18のいずれか一項に記載の使用。
【請求項20】
ヒトにおいて膵島前駆細胞から成熟インスリン分泌膵島細胞への分化を刺激する全身投与に有効な量でガストリンとEGFレセプターリガンドペプチドとを含む、新生糖尿病を治療するための組成物。
【請求項21】
約20から500マイクログラムのガストリンとEGFレセプターリガンドペプチドとを含む、静脈内投与用の請求項20記載の組成物。
【請求項1】
ガストリン/CCKレセプターリガンドペプチドとEGFレセプターリガンドペプチドとの組み合わせを用いて、ex vivoで膵島前駆細胞を刺激することを含む、哺乳動物の膵島前駆細胞を成熟インスリン分泌細胞へと分化させるためのex vivoでの方法。
【請求項2】
ガストリンとEGFレセプターリガンドペプチドとを用いて、単離した膵島前駆細胞を処理する、請求項1記載のex vivoでの方法。
【請求項3】
ガストリンがガストリン8、17または34である、請求項2記載のex vivoでの方法。
【請求項4】
EGFレセプターリガンドペプチドがEGF類似体である、請求項1、2または3記載のex vivoでの方法。
【請求項5】
EGFレセプターリガンドペプチドがTGFαである、請求項1、2または3記載のex vivoでの方法。
【請求項6】
EGFレセプターリガンドペプチドがアンフィレグリンまたはポックスウイルス成長因子である、請求項1、2または3記載のex vivoでの方法。
【請求項7】
哺乳動物が糖尿病を患っている、請求項1ないし6のいずれか一項に記載のex vivoでの方法。
【請求項8】
哺乳動物がヒトである、請求項7記載のex vivoでの方法。
【請求項9】
膵臓前駆細胞を、分化を引き起こす量のガストリンおよびEGFレセプターリガンドペプチドを用いて処理する、請求項2ないし8のいずれか一項に記載のex vivoでの方法。
【請求項10】
膵島前駆細胞が哺乳動物の体外移植された膵臓組織に由来する、請求項1ないし9のいずれか一項に記載のex vivoでの方法。
【請求項11】
哺乳動物に、刺激した膵島前駆細胞を導入することをさらに含む、請求項1ないし10のいずれか一項に記載のex vivoでの方法。
【請求項12】
哺乳動物において膵島前駆細胞を成熟インスリン分泌膵島細胞へと分化させる全身投与用医薬の調製における、全身的に有効量のガストリン/CCKレセプターリガンドペプチドとEGFレセプターリガンドペプチドの使用。
【請求項13】
膵島前駆細胞が体外移植された膵臓組織に由来し、分化後に哺乳動物に導入される、請求項12記載の使用。
【請求項14】
糖尿病治療用の全身投与用医薬の調製における、全身的に有効量のガストリン/CCKレセプターリガンドペプチドとEGFレセプターリガンドペプチドの使用。
【請求項15】
単離された膵島前駆細胞がガストリンとEGFレセプターリガンドペプチドを用いて処理される、請求項12、13または14記載の使用。
【請求項16】
ガストリンがガストリン8、17または34である、請求項15記載の使用。
【請求項17】
EGFレセプターリガンドペプチドがEGF類似体である、請求項12ないし16のいずれか一項に記載の使用。
【請求項18】
EGFレセプターリガンドペプチドがTGFαである、請求項12ないし16のいずれか一項に記載の使用。
【請求項19】
EGFレセプターリガンドペプチドがアンフィレグリンまたはポックスウイルス成長因子である、請求項12ないし18のいずれか一項に記載の使用。
【請求項20】
ヒトにおいて膵島前駆細胞から成熟インスリン分泌膵島細胞への分化を刺激する全身投与に有効な量でガストリンとEGFレセプターリガンドペプチドとを含む、新生糖尿病を治療するための組成物。
【請求項21】
約20から500マイクログラムのガストリンとEGFレセプターリガンドペプチドとを含む、静脈内投与用の請求項20記載の組成物。
【図2】


【図4】
【図1】
【図3】
【図4】
【図1】
【図3】
【公開番号】特開2008−307053(P2008−307053A)
【公開日】平成20年12月25日(2008.12.25)
【国際特許分類】
【出願番号】特願2008−177330(P2008−177330)
【出願日】平成20年7月7日(2008.7.7)
【分割の表示】特願2004−234827(P2004−234827)の分割
【原出願日】平成6年1月24日(1994.1.24)
【出願人】(500373873)アールティーピー・ファーマ・コーポレーション (1)
【出願人】(592017633)ザ ジェネラル ホスピタル コーポレイション (177)
【Fターム(参考)】
【公開日】平成20年12月25日(2008.12.25)
【国際特許分類】
【出願日】平成20年7月7日(2008.7.7)
【分割の表示】特願2004−234827(P2004−234827)の分割
【原出願日】平成6年1月24日(1994.1.24)
【出願人】(500373873)アールティーピー・ファーマ・コーポレーション (1)
【出願人】(592017633)ザ ジェネラル ホスピタル コーポレイション (177)
【Fターム(参考)】
[ Back to top ]