
苦味を阻止する化合物を同定するための特異的T2R味覚受容体の使用
【課題】苦味を調節、望ましくは抑制する化合物を同定するためのアッセイ方法を提供する。
【解決手段】苦味化合物と特異的に結合する受容体、hT2R4、hT2R44及び/またはhT2R61の活性化に関係する化合物を同定するアッセイ方法。
【効果】このアッセイにより同定される化合物は、キニーネ、6−ニトロサッカリン、サッカリン及び/またはデナトニウムを含む苦味化合物による苦味を調節、例えば、抑制するはずであり、これらの化合物は苦味を持つ食品、飲料または医薬製品への有用な添加物である。
【解決手段】苦味化合物と特異的に結合する受容体、hT2R4、hT2R44及び/またはhT2R61の活性化に関係する化合物を同定するアッセイ方法。
【効果】このアッセイにより同定される化合物は、キニーネ、6−ニトロサッカリン、サッカリン及び/またはデナトニウムを含む苦味化合物による苦味を調節、例えば、抑制するはずであり、これらの化合物は苦味を持つ食品、飲料または医薬製品への有用な添加物である。
【発明の詳細な説明】
【技術分野】
【0001】
(関連出願)
本出願は、2001年7月10日出願の仮出願米国出願番号60/303,811及び2002年4月15日出願の仮出願米国出願番号60/372,089の優先権を主張し、その両者の全体を引用してここに取り入れる。
【0002】
(発明の分野)
本発明は、すでに報告されている味覚に関係するヒトG−タンパク共役受容体(GPCR)の一部を活性化する苦味化合物を解明することを含んでいる。特に本発明は、T2Rファミリーの特定の味覚受容体が、苦味化合物により特異的に活性化されるとの発見を含む。すなわち、ヒトT2R4味受容体が苦味アルカロイドであるキニーネによって特異的に活性化されること、ヒトT2R61味受容体がサッカリンの苦味誘導体である6−ニトロサッカリンによって特異的に活性化されること、並びにヒトT2R44受容体がデナトニウムによって特異的に活性化されるが6−ニトロサッカリンによって弱く活性化されることに関する発見を含むものである。これらの発見に基づき、そのオーソログ、スプライス変異体、一塩基多形(SNP)、及び遺伝子操作による突然変異体を含む、これらのヒト味覚受容体、フラグメントまたはそれらの変異体若しくはキメラは、キニーネ、6−ニトロサッカリンおよびデナトニウム、および構造的に関連する化合物、並びにこれらの受容体を活性化する他の化合物による苦味を阻止する化合物を同定するアッセイ、望ましくは細胞を使用するハイスループットアッセイに有用である。このアッセイを使用して同定される化合物は、食品、飲料または医薬品の添加物としてそれらの味を改善するために使用することができる。
【背景技術】
【0003】
(関連技術の説明)
ヒトが認識することができる基本的な味の種類の一つは、苦味である。苦味の生理学はごく最近までほとんど理解されていなかった。最近の研究は、味の生物学に光を注ぎ始めた(Lindemann, Nature (2001))。苦味化合物の多くは、細胞表面受容体と相互作用することにより苦味を生じることが今では認められている。これらの受容体は、細胞内Gタンパクと相互作用する7回膜貫通ドメイン受容体のファミリーに属する。T2Rと呼ばれるGPCRの新規なファミリーが、ヒト及びげっ歯類において同定されている(Adler et al., Cell 100(6): 693-702 (2000); Chandrashekar et al., Cell 100(6): 703-711 (2000); Matsunami H, Montmayeur JP, Buck LB. Nature 404(6778): 601-4 (2000))。いくつかの証拠は、T2Rが、苦味化合物に対する応答を仲介していることを示唆している。第一に、T2R遺伝子は、舌及び口蓋上皮細胞上の味覚受容体のサブセット中で特異的に発現する。第二に、ヒトT2Rの一つ(hT2R1)についての遺伝子は、ヒトにおける苦味化合物である6-n-プロピル-2-チオウラシルに対する感受性に関連する染色体遺伝子座に存在している (Adler et al., (前出)(2000))。第三に、マウスのT2Rの一つ(mT2R5)は、マウスにおける苦味化合物であるシクロヘキシミドに対する感受性に関連する染色体遺伝子座に存在している。また、mT2R5が、味覚細胞で特異的に発現し苦味刺激伝達に関連するGタンパクであるガストジューシンを、活性化できることが示された(Wong et al., Nature 381: 796-800 (1996))。mT2R5によるガストジューシンの活性化は、シクロヘキシミドに応答したときにのみ生じる(Chandrashekar et al.,(前出)(2000))。したがって、mT2Rファミリーは、マウスにおける苦味応答を仲介し、hT2Rファミリーは、ヒトにおける苦味応答を仲介すると考えられている。唯一のヒトT2Rについて、同定された苦味リガンドを持つことが示唆された。即ち、hT2R4が、デナトニウムにより活性化されることが示された(Chandrashekar et al.,(前出)2000)。しかし、その試験に使用した有効なデナトニウム濃度(1.5 mM)は、異常に高かった、すなわち、ヒトで報告されているデナトニウムについての苦味閾値よりも105倍高かった(Saroli, Naturwissenschaften 71:428-429 (1984))。したがって、hT2Rに充分に適合する特異的苦味リガンドはなかった。各hT2Rが複数の苦味リガンドに結合できるという提案もなされていた。この仮説は、hT2Rファミリーはわずか24の同定されたメンバーから構成されるが、ヒトは数百の異なる化合物を苦いと認識することができるという事実に基づいている。hT2Rの配列は既に報告されており、Zukerら(WO 01/18050 A2,(2001))及びAdlerら(WO 01/77676 A1(2001))により公開PCT出願に開示されている。その両者の全体を、引用して本明細書に取り入れる。
【0004】
T2Rの機能についての研究における困難の一つは、これらの受容体が培養哺乳動物細胞系では簡単に発現しないということである。T2R発現を改良するために、発現の良好なGPCRであるロドプシンのN−末端配列を、T2R配列に結合した(Chandrashekar et al., (前出) 2000)。このN−末端タグは、利用可能な抗体により、タンパクの発現を容易に監視することを可能とする。ロドプシンタグを組み込むことにより、哺乳動物細胞系におけるT2Rの一部の発現は改良されるが、その多くは、依然として機能研究をするほどには十分に発現しなかった。別の研究において、mT2R5は、昆虫Sf9細胞中で良好に発現し、生化学的GTPγS結合アッセイを使用する機能研究に使用された(Chandrashekar et al., (前出) 2000)。
【0005】
しかし、報告がされた内容によっても、依然、T2R遺伝子ファミリーの機能に関して確実性を欠いている。特に、どの味覚化合物が、特定のT2R受容体を活性化するかに関して理解を欠いている。個々のT2R受容体の結合特性を十分に理解することが非常に有益であり、そうすることにより所望の味覚調節特性を有する化合物を選別するためにそれを使用することが促進されるであろう。
【特許文献1】WO 01/18050 A2,(2001)
【特許文献2】WO 01/77676 A1(2001)
【非特許文献1】Lindemann, Nature (2001)
【非特許文献2】Adler et al., Cell 100(6): 693-702 (2000)
【非特許文献3】Chandrashekar et al., Cell 100(6): 703-711 (2000)
【非特許文献4】Matsunami H, Montmayeur JP, Buck LB. Nature 404(6778): 601-4 (2000)
【非特許文献5】Wong et al., Nature 381: 796-800 (1996)
【非特許文献6】Saroli, Naturwissenschaften 71:428-429 (1984)
【発明の開示】
【課題を解決するための手段】
【0006】
(発明の要約)
結論的に、本発明は、T2Rファミリーにおける幾つかの味覚受容体、特にhT2R4, hT2R61及びhT2R44は、それぞれ、苦味化合物である、キニーネ、6−ニトロサッカリン、及びデナトニウム(及び6−ニトロサッカリンには弱く)によって特異的に活性化されるという発見に関する。
【0007】
これらの発見は、特定のT2Rを発現する細胞を使用してT2Rの活性を測定する細胞によるアッセイを使用して行われた。以下に詳細に記述するように、特にその表面にhT2R4を発現するHEK-Gα15細胞系を、細胞内カルシウム濃度の変化を検出する細胞によるアッセイに使用し、他のhT2Rが同条件では活性化されないにも拘らず、同細胞系がキニーネにより特異的に活性化されることを見出した。また、キニーネは、典型的な用量‐反応相関性をもってhT2R4を活性することが示された。
【0008】
さらに、以下により詳細に記述するように、GTPγS結合アッセイを、種々の苦味化合物の存在下、バキュロウイスル発現系を用いて昆虫細胞(Sf9細胞)で発現させたヒトT2Rの活性をモニターするために使用した。このアッセイの結果は、hT2R61が、特異的に6−ニトロサッカリンにより活性化され、hT2R44が特異的にデナトニウム(及び6−ニトロサッカリンによって弱く)により活性化されることを示している。
【0009】
したがって、本発明は、キニーネ、6−ニトロサッカリン、デナトニウムまたはサッカリンのような関連化合物によるhT2R4, hT2R44及びhT2R61の受容体の活性化を阻害する化合物を同定するためのアッセイ、望ましくはハイスループットアッセイにおける、hT2R4, hT2R44及びhT2R61の使用を包含している。
【0010】
また、本発明は、これらの化合物が味覚試験において苦味を阻止することを確認することを含む方法も包含している。また、本発明は、例えば、サッカリンを含む飲料、食品または医薬品などのように、苦味を抑制するために、食品、飲料及び医薬品において同定した化合物を使用することを包含している。
【0011】
(発明の目的)
本発明の一つの目的は、キニーネまたはキニーネに構造的に関連するhT2R4を活性化する化合物によるhT2R4、またはそのフラグメント、変異体、オーソログ若しくはキメラの活性化を阻害する化合物を同定することである。
【0012】
本発明のもう一つの目的は、6−ニトロサッカリン、サッカリンまたはそれらに構造的に関連するhT2R61を活性化する化合物によるhT2R61またはそのフラグメント、変異体、オーソログ若しくはキメラの活性化を阻害する化合物を同定することである。
【0013】
本発明のそのほかの目的は、デナトニウム若しくは6−ニトロサッカリンまたはそれらに構造的に関連するhT2R44を活性化する化合物によるhT2R44の活性化を阻害する化合物を同定することである。
【0014】
本発明のさらに特別な目的は、これらの受容体により顕在化する苦味を阻止する化合物を同定する手段として、キニーネ;6−ニトロサッカリン若しくはサッカリン;またはデナトニウム若しくは6−ニトロサッカリンの夫々による上記で特定したT2R受容体の活性化を阻害する化合物を同定するための細胞を使用するアッセイにおいて、hT2R4, hT2R61若しくはhT2R44またはそれらのフラグメント、変異体、オーソログ若しくはキメラを安定してまたは一過性に発現する細胞を使用することである。
【0015】
本発明のさらに特別な目的は、キニーネまたは関連化合物によるhT2R4の活性化、6−ニトロサッカリン、サッカリンまたは関連化合物によるhT2R61の活性化、或いはデナトニウムまたは関連化合物によるhT2R44の活性化を阻害する化合物を同定するために、特に苦味化合物によるこれらのT2Rの特異的活性化を阻止する化合物を同定する手段として、望ましくはカルシウム特異的な蛍光色素を使用して細胞内カルシウムの変化を検出する細胞によるアッセイにおいて、哺乳動物または昆虫の細胞、例えば、G15タンパクを発現するHEK-293T細胞、を使用することである。
【0016】
本発明のさらに特別な目的は、特に苦味化合物によるこれらのT2Rの特異的活性化を阻止する化合物を同定する手段として、キニーネ、デナトニウム、6−ニトロサッカリンまたはサッカリンによるhT2R4, hT2R44またはhT2R61の受容体の活性化をそれぞれ阻害する化合物を同定するために、GTPγS結合アッセイにおいて、hT2R4, hT2R44またはhT2R61を発現する哺乳動物または昆虫細胞、例えば、Sf9細胞、を使用することである。
【0017】
本発明のそのほかの目的は、同定された化合物が、例えば、キニーネ、デナトニウム、6−ニトロサッカリンまたはサッカリンにより誘発される苦味を阻止することを、味覚試験において確認することである。
【0018】
本発明のそのほかの目的は、hT2R4, hT2R44またはhT2R61を特異的に活性化する化合物により誘発される苦味を阻止するための組成物中に、本明細書で記述するアッセイにおいて同定された化合物を添加物として使用することである。本発明の望ましい目的は、6−ニトロサッカリンまたはサッカリンによるhT2R61及び/またはhT2R44の活性化を阻害する化合物を、サッカリン含有組成物、特にダイエット飲料、の苦味を阻止するために使用することである。
【0019】
(発明の詳細な説明)
本発明を具体的に記述する前に、以下の定義を提示する。
【0020】
用語「T2R」ファミリーは、
(1)以下及び引用したZuker(前出)(2001)およびAdler(前出)(2001)に開示されているT2Rに対して、約25アミノ酸、最も適するのは50−100アミノ酸の枠に亘って、約30−40%のアミノ酸配列が相同、より具体的にいうと約40,50,60,70,75,80,85,90,95,96,97,98,または99%のアミノ酸配列が相同である;
(2)以下に開示するT2R配列及びその保存的に修飾された変異体からなる群から選択されるアミノ酸配列を含む免疫原に対して作成される抗体に特異的に結合する;
(3)以下に開示するT2R DNA配列及びその保存的に修飾された変異体からなる群から選択された配列にストリンジェントなハイブリッド形成条件の下に(少なくとも約100、任意に少なくとも約500-1000ヌクレオチドのサイズで)特異的にハイブリッド形成する;
(4)以下に開示するT2Rアミノ酸配列からなる群から選択されたアミノ酸配列に少なくとも約40%相同な配列を含む;または
(5)記述されたT2Rとストリンジェントなハイブリッド形成条件下に特異的にハイブリッド形成するプライマーにより増幅される
多形変異体、アレル、突然変異体、及び同属体を含む。
【0021】
すでに述べたように、T2R遺伝子は、タンパク及びDNAのレベルで実質的な配列の相違を示すが、今日までに単離されたT2Rは、全て特定の領域において一定の共通配列を含むことが認められており、それらは本明細書に引用して取り入れたAdlerら(WO 01/77676 A1(2001))及びZukerら(WO 01/18050 A2)においてすでに同定されているT2Rコンセンサス配列を持つかそれに対して少なくとも70−75%相同である。
【0022】
立体構造的にその化学感受性GPCRは、「N−末端ドメイン」、「細胞外ドメイン」、7回膜貫通領域及び対応する細胞質及び細胞外ループを含む「膜貫通ドメイン」、「細胞質領域」並びに「C−末端領域」を有する(例えば、Hoon et al., Cell, 96: 541-51 (1999); Buck & Axel, Cell, 65: 175-87 (1991)参照)。これらの領域は、例えば疎水性及び親水性ドメインを同定する配列分析プログラムのような、当業者に既知の方法を使用して構造的に同定することができる(例えば、Stryer, Biochemistry, (3rd ed. 1988)参照;多くのインターネットから入手できる配列分析プログラムのいずれか、例えば、dot.imgen.bcm.tmc.eduで見られるものも参照)。これらの領域は、キメラタンパクの作成及び本発明のインビトロアッセイ、例えば、リガンド結合アッセイに有用である。
【0023】
本発明による「T2R4」は、ヒトT2R4ポリペプチド及び対応する核酸の配列、並びに同じ機能を持つ、すなわち、キニーネまたはそのほかの苦味化合物によって特異的に活性化される、そのフラグメント、変異体、突然変異体及びオーソログを含むことが意図されている。ヒトT2R4の配列は、Zukerら(WO 01/18050) A2(2001)及びAdlerら(WO 01/77676 A1 (2001))に含まれており、下記に記述するSEQ ID NO: 1及び2(ヒトT2R4タンパク及びDNA)中に含まれる。
【0024】

【0025】
【0026】
本発明による「T2R44」は、下記のSEQ ID NO: 3及び4に含まれるタンパク及びDNA配列を持つヒトT2R44、並び同じ機能を示す、すなわち、適当なアッセイにより測定されるデナトニウムのような苦味化合物により特異的に活性化される、フラグメント、キメラ、変異体、突然変異体、オーソログを含むことが意図されている。これらフラグメント、キメラ、変異体、突然変異体、オーソログは、例えば、スプライス変異体、アレル変異体、及び一塩基多形などを含むものである。
【0027】
【0028】
【0029】
本発明による「T2R61」は、SEQ ID NO: 5 に含まれるヒトT2R61タンパク及び対応する核酸配列(SEQ ID NO: 6)、並び同じ機能を持つ、すなわち、6−ニトロサッカリン、サッカリン、または適当なアッセイを使用してスクリーニングできるような関連苦味化合物により特異的に活性化される、フラグメント、キメラ、変異体、突然変異体を含むことが意図されている。これらフラグメント、キメラ、変異体、及び突然変異体は、アレル、スプライス及び一塩基多形及びオーソログを含むものである。
【0030】
【0031】
【0032】
「細胞外ドメイン」は、したがって、細胞の膜から突き出ていて、細胞の細胞外表面に露出しているT2Rポリペプチドのドメインのことである。その領域は、細胞の細胞外表面に露出している「N−末端ドメイン」、並びに細胞の細胞外表面に露出している膜貫通ドメインの細胞外ループ、すなわち、膜貫通領域2及び3、膜貫通領域4及び5、並びに膜貫通領域6及び7の間の細胞外ループを含む。「N−末端ドメイン」は、N−末端から始まり膜貫通領域の開始点に近い領域にいたる。これらの細胞外領域は、液相及び固相におけるインビトロリガンド結合アッセイに有用である。さらに、以下に記述する膜貫通領域も、細胞外領域と組み合わせてあるいは単独でリガンド結合に関与することができるので、インビトロリガンド結合アッセイに有用である。
【0033】
7個の膜貫通「領域」を含む「膜貫通ドメイン」は、形質膜内に存在するT2Rポリペプチドのドメインのことであり、対応する細胞質(細胞内)及び細胞外ループを含むことがあり、また膜貫通「領域」とも呼ばれる。7個の膜貫通領域及び細胞外及び細胞質ループは、Kyte & Doolittle, J. Mol. Biol., 157: 105-32 (1982),またはStryer,前出に記述されているように、標準的方法を使用して同定することができる。
【0034】
「細胞質ドメイン」は、細胞の内側に面しているT2Rのドメイン、例えば、「C−末端ドメイン」及び膜貫通ドメインの細胞内ループ、例えば、膜貫通領域1及び2、膜貫通領域3及び4、及び膜貫通領域5及び6の間の細胞内ループのことである。「C−末端ドメイン」は、最後の膜貫通領域の終わりからタンパクのC−末端に達する領域のことであり、通常は細胞内に存在している。
【0035】
用語「7回膜貫通受容体」は、7回形質膜を貫通する7個の領域を持つ膜貫通タンパクのスーパーファミリーに属するポリペプチドを意味する(したがって、7個の領域は「膜貫通」ドメインまたは「TM」ドメインTM IからTM VIIと呼ばれる)。嗅覚受容体及びある種の味覚受容体は、それぞれこのスーパーファミリーに属す。7回膜貫通受容体ポリペプチドは、以下にさらに詳細に検討するように、類似する特徴的な、一次、二次、及び三次構造を有している。
【0036】
用語「リガンド結合領域」は、膜貫通ドメインIIからVII(TM IIからVII)を実質的に組み込んでいる化学感受性受容体または味覚受容体に由来する配列のことである。この領域は、リガンド、より具体的には味覚誘発化合物と結合することができる。
【0037】
用語「形質膜転移ドメイン」または単に「転移ドメイン」は、代表的な「転移ドメイン」(5'−MNGTEGPNFYVPFSNKTGVV;SEQ ID NO: 7)と機能的に同等であるポリペプチドドメインを意味する。これらのペプチドドメインは、ポリペプチドコード配列のアミノ末端に組み込まれたときに、ハイブリッド(「融合」)タンパクを、細胞形質膜へ非常に効率よく「移動」または「転移」することができる。この特殊な「転移ドメイン」は、最初、ヒトロドプシン受容体ポリペプチド、即ち7回膜貫通受容体、のアミノ末端から得られた。他の転移ドメインは、ウシロドプシン配列から得られており、同様に転移を促進するのに有用である。ロドプシン由来配列は、7回膜貫通融合タンパクを形質膜へ転移させるのに特に有効である。
【0038】
「機能的同等」とは、新規に翻訳されたタンパクを同条件において形質膜へ転移させるドメインの能力及び効果が、代表的SEQ ID NO: 7と同じであることを意味する;相対的効果は、本明細書で記述されているように、(定量的に)測定して比較される。本発明の範囲にあるドメインは、20アミノ酸鎖長の転移ドメインSEQ ID NO: 7と同じ効率で、(哺乳動物、ツメガエルなどの)細胞中において新規に合成されたポリペプチドを形質膜へ転移させるかについての所定のスクリーニングにより決定することができる。
【0039】
味覚伝導を仲介するT2Rファミリーに作用する試験化合物のアッセイにおける文脈で、語句「機能的効果」は、間接的にまたは直接的に受容体の影響下にあるパラメーター、例えば機能的、物理的及び化学的な効果、を測定することを含む。それには、インビトロ、インビボ、及びエクスビボにおけるリガンド結合、イオン流の変化、膜電位、電流、転写、Gタンパク結合、GPCRのリン酸化または脱リン酸化、信号伝達、受容体−リガンド相互作用、セカンドメッセンジャー濃度(例えば、cAMP, cGMP, IP3,または細胞内Ca2+)が含まれ、また神経伝達物質またはホルモン放出の増加または減少のようなそのほかの生理的効果も含まれる。
【0040】
「機能的効果を測定する」とは、間接的または直接的に、T2Rファミリーメンバーの影響下にあるパラメーター、例えば、機能的、物理的及び化学的効果、を増加または減少する化合物をアッセイすることを意味する。その機能的効果は、当業者に既知の方法のいずれかにより測定することができる、例えば、分光学的特徴の変化(例えば、蛍光、吸光度、屈折率)、流体力学(例えば、形状)、クロマトグラフィー特性、または溶解性、パッチクランピング、電位感受性色素、全細胞電流、放射性アイソトープ放出量、誘導マーカー、卵母細胞T2R遺伝子発現;組織培養細胞T2R発現;T2R遺伝子の転写活性化;リガンド結合アッセイ;電圧、膜電位及び伝導度の変化;イオン流アッセイ;cAMP,cGMP,及びイノシトール三燐酸(IP3)のような細胞内セカンドメッセンジャーの変化;細胞内カルシウムレベルの変化;神経伝達物質放出、などがある。
【0041】
T2Rタンパク受容体の「阻害物質」、「活性化物質」及び「活性調節物質」は、インビトロ及びインビボにおける味覚伝達のアッセイを使用して同定される阻害性分子、活性化分子、または活性調節分子、例えば、リガンド、アゴニスト、アンタゴニスト、並びにそれらの同属体及び擬似体に言及するために互換的に使用される。阻害物質は、例えば、結合して、部分的にまたは完全に刺激を阻害し、減少し、阻止し、活性化を遅らせ、不活化し、脱感作し、または味覚伝達を下方調節する化合物、例えば、アンタゴニストである。活性化物質は、例えば、結合して、刺激し、増加し、開放し、活性化し、促進し、活性化を増強し、感受性とし、または味覚伝達を上方調節する化合物、例えば、アゴニストである。活性調節物質は、例えば受容体と、活性化物質または阻害物質と結合する細胞外タンパク(例えば、エブネリン及び疎水性キャリアーファミリーのその他のメンバー);Gタンパク;キナーゼ(例えば、ロドプシンキナーゼの同属体、並びに受容体の脱活性化及び脱感作に関係するβ交感神経受容体キナーゼ);並びにやはり受容体を脱活性化及び脱感作するアレスチン、との相互作用を変化させる化合物を含む。活性調節物質は、例えば、変化した活性を持つT2Rファミリーメンバーの一般的な修飾体、並びに、天然に存在するかまたは合成の、リガンド、アンタゴニスト、アゴニスト、小型化学分子などを含む。
【0042】
阻害物質及び活性化物質のアッセイは、例えば、細胞中または細胞の膜にT2Rファミリーメンバーを発現させ、例えば、苦味化合物の活性を調節する化合物の存在下または非存在下に候補調節化合物を適用し、次いで前記のように味覚伝達における機能的効果を測定することを含む。潜在的な活性化物質、阻害物質または活性調節物質で処理したT2Rファミリーメンバーを含む検体またはアッセイは、調節の程度を調べるために、阻害物質、活性化物質または活性調節物質を含まない対照検体と比較される。対照検体(活性調節物質で処理されていない)を相対T2R活性値100%とする。対照と比較したT2R活性値が約80%、さらに50%または25−0%であればT2Rの阻害は達成される。対照と比較したT2R活性値が110%、さらに150%、さらに200−500%、または1000−3000%高いときに、T2Rの活性化は達成される。
【0043】
本明細書で使用される時、語「精製された」、「実質的に精製された」及び「単離された」は、本発明の化合物が、自然な状態では通常結合している他の異なる化合物から遊離した状態のことである。望ましくは、「精製された」、「実質的に精製された」及び「単離された」とは、その構成物が、所定の試料における総量の、少なくとも0.5重量%、1重量%、5重量%、10重量%、または20重量%であることを意味し、最も望ましくは、少なくとも50を重量%または75重量%を構成することを意味する。望ましい一態様において、これらの語は、本発明の化合物が、所定の試料における総量の少なくとも95重量%を構成することである。本明細書で使用される時、語「精製された」、「実質的に精製された」及び「単離された」は、核酸またはタンパクに関する場合、哺乳動物の体内、特にヒト、の体内に自然に存在する状態と異なる精製または濃度の状態のことである。哺乳動物の体内、特にヒトの体内に自然に存在する状態よりも高い精製または濃度とすることは、(1)他の結合構造または結合化合物からの精製、または(2)哺乳動物の体内、特にヒトの体内では通常結合しない構造または化合物との結合を含み、「単離された」の意味の範囲に入る。当業者に既知の種々の方法及び操作にしたがって、本明細書で記述する核酸若しくはタンパクまたは核酸若しくはタンパクのクラスを単離することができるし、また通常では自然に結合しない構造または化合物と結合させることができる。
【0044】
ここに使用した語「単離された」は、核酸またはポリペプチドに関する場合には、哺乳動物、特にヒト、の体内に自然に存在するのと異なる精製または濃度の状態のことである。(1)他の自然に存在する結合構造または結合化合物からの精製または(2)体内では通常結合しない構造または化合物との結合を含む、体内に自然に存在する状態よりも高い精製または濃度は、本明細書で使用される「単離された」の意味の範囲に入る。当業者に既知の種々の方法及び操作にしたがって、本明細書で記述する核酸またはポリペプチドを単離することができ、さもなければ、通常では自然に結合しない構造または化合物と結合させることができる。
【0045】
本明細書で使用される時、語句「増幅する」及び「増幅」は、下記に詳細に検討するように、組換えによりまたは自然に発現した核酸を産生または検出するために、適当な増幅方法のいずれかを使用することである。例えば、本発明は、インビボまたはインビトロで、自然発現(例えば、ゲノムまたはmRNA)または組換え(例えば、cDNA)による本発明の核酸(例えば、本発明の味覚誘発化合物結合配列)を、(例えば、ポリメラーゼ連鎖反応、PCR,により)増幅するための方法及び試薬(例えば、特異的オリゴヌクレオチドプライマー対)を提供する。
【0046】
用語「発現ベクター」とは、原核細胞、イースト、菌、植物、昆虫または哺乳動物の細胞を含むいずれかの細胞において、構成的または誘導的に、インビトロまたはインビボで、本発明の核酸配列を発現させるための組換え発現系のことである。この用語は、線状または環状発現系を含む。この用語は、エピソームに残留するまたは宿主細胞ゲノムに組み込まれた発現系を含む。この発現系は、自己複製の能力を持つものでもよく、または持たないもの、すなわち、細胞中で一過性にのみ発現するものでもよい。この用語は、組換え核酸の転写に必要な最低限の配列のみを含有する組換え発現「カセット」を含む。
【0047】
用語「ライブラリー」とは、異なる核酸またはポリペプチドの混合物である調製物を意味し、例えば、縮重プライマー対を用いて核酸を増幅することにより作成された組換え感覚受容体リガンド結合領域、特に組換え味覚受容体リガンド結合領域のライブラリー、または増幅リガンド結合領域を組み込んだ単離ベクターのコレクション、または味覚受容体をコードする少なくとも一つのベクターで無作為に遺伝子導入した各細胞の混合物がある。
【0048】
用語「核酸」または「核酸配列」とは、一本鎖または二本鎖のいずれかの形のデオキシリボヌクレオチドまたはリボヌクレオチドのオリゴヌクレオチドのことである。この用語は、天然ヌクレオチドの既知の類似体を含む、核酸、すなわち、オリゴヌクレオチドを包含する。この用語は合成骨格を持つ核酸様構造も包含する。
【0049】
特に示さない限り、特定の核酸配列は、明示された配列のみならず、その保存的に修飾された変異体(例えば、縮重コドン置換体)及び相補的配列を暗黙のうちに包含する。具体的には、縮重コドン置換は、例えば、一つ以上の選択されたコドンの三番目の位置を、混合塩基及び/またはデオキシイノシン残基で置換した配列を作成することにより行われる(Batzer et al., Nucleic Acid Res., 19: 5081 (1991); Ohtsuka et al., J. Biol. Chem., 260:2605-08 (1985); Rossolini et al., Mol. Cell. Probes, 8: 91-98 (1994))。用語核酸は、遺伝子、cDNA, mRNA, オリゴヌクレオチド、及びポリヌクレオチドと互換的に使用される。
【0050】
用語「ポリペプチド」、「ペプチド」及び「タンパク」は、アミノ酸残基の重合体を示すために本明細書において互換的に使用される。この用語は、天然アミノ酸重合体及び非天然のアミノ酸重合体のみならず、一以上のアミノ酸が天然アミノ酸の対応する人工的な化学擬似体であるアミノ酸重合体に適用される。
【0051】
本明細書で記述される「転移ドメイン」、「リガンド結合領域」またはキメラ受容体構成物は、「類似体」、または「保存的変異体」及び代表的な配列に実質的に一致する構造及び活性を持つ「擬似体」(「ペプチド擬似体」)も含む。従って、用語「保存的変異体」または「類似体」または「擬似体」は、その変化が本明細書で定義するようなポリペプチド(保存的変異体)の構造及び/または活性を実質的に変えないような修飾アミノ酸配列を持つポリペプチドのことである。これらは、アミノ酸配列の保存的修飾変異体、すなわち、タンパクの活性に重要でないアミノ酸置換、アミノ酸残基の付加若しくは欠失、或いは重要なアミノ酸の置換であっても構造及び/または活性を実質的に変えないような類似性質(例えば、酸性、塩基性、陽性または陰性荷電、極性または非極性、など)を持つ残基によるアミノ酸の置換された変異体を含む。
【0052】
より具体的には、「保存的修飾変異体」は、アミノ酸配列及び核酸配列の両者に適用する。特定の核酸配列に関しては、保存的修飾変異体とは、同じまたは本質的に同じアミノ酸配列をコードすること、或いは核酸がアミノ酸配列をコードしない場合には、本質的に同じ配列のことである。遺伝暗号の縮重のために、多数の機能的に同一の核酸が所与タンパクをコードする。
【0053】
例えば、コドンGCA, GCC, GCG及びGCUは、全てアミノ酸アラニンをコードする。したがって、コドンによってアラニンが特定されているどの位置においても、コードされているポリペプチドを変えることなく、コドンを、記述されている対応コドンのいずれかと交換することができる。
【0054】
そのような核酸変異体は「沈黙変異体」であり、保存的修飾変異体の一種である。ポリペプチドをコードする本明細書における総ての核酸配列は、該核酸の可能な沈黙変異体の全てについても言及する。当業者は、核酸の各コドン(通常はメチオニンの唯一のコドンであるAUG、及び通常トリプトファンの唯一のコドンであるTGGを除く)を修飾して機能的に同一の分子が得られることを認識している。したがって、ポリペプチドをコードする核酸の沈黙変異体は、暗黙のうちに各記述された配列の中に存在する。
【0055】
機能的に同じアミノ酸を示す保存置換表は当業者に既知である。例えば、保存的置換を選択する一つの代表的例は(元の残基、次いで代表的な置換残基を示す)、ala/glyまたはser; arg/lys; asn/glnまたはhis; asp/glu; cys/ser; gln/asn; gly/asp; gly/alaまたはpro; his/asn またはgln; ile/leu またはval;leu/ileまたはval; lys/argまたはglnまたはglu; met/leuまたはtyrまたはile; phe/met またはleuまたはtyr; ser/thr; thr/ser; trp/tyr; tyr/trpまたはphe; val/ileまたはleuである。別の代表例は下記の、相互に保存置換できるアミノ酸を含む6群を使用する:1)アラニン(A)、セリン(S)、トレオニン(T);2)アスパラギン酸(D)、グルタミン酸(E);3)アスパラギン(N)、グルタミン(Q);4)アルギニン(R)、リシン(K);5)イソロイシン(I)、ロイシン(L)、メチオニン(M)、バリン(V);及び6)フェニルアラニン(F)、チロシン(Y)、トリプトファン(W);(例えば、Creighton, Proteins, W.H.Freeman and Company (1984); Schultz and Schimer, Principles of Protein Structure, Springer-Verlag (1979)参照)。当業者は、上に示した置換だけが可能な保存的置換ではないことを理解しているであろう。例えば、ある目的のためには、陽性かまたは陰性に荷電しているアミノ酸相互の荷電アミノ酸置換をすべて保存的置換と見なすことができる。さらに、コード配列中の一つのアミノ酸または小さな割合のアミノ酸を変更、付加または削除する個別の置換、欠失または付加も「保存的修飾変異体」と考えることができる。
【0056】
用語「擬似体」及び「ペプチド擬似体」とは、ポリペプチド、例えば、転移ドメイン、リガンド結合領域、または本発明のキメラ受容体と同じ構造及び/または機能特性を実質的に有する合成化合物のことである。擬似体は、アミノ酸の合成による非天然の類似体で全体を構成することもできるし、または一部が天然のペプチドアミノ酸からなり一部が非天然のアミノ酸の類似体からなるキメラ分子としてもよい。この擬似体は、このような置換が擬似体の構造及び/または活性を変えない限り、如何なる数の天然アミノ酸保存置換を組み込むことができる。
【0057】
保存的変異体である本発明のポリペプチドと同様に、ルーチン的実験により本発明の範囲に入るか否か、すなわち、構造及び/または機能が変わっていないこと、を決定するであろう。
ポリペプチド擬似構成物は、非天然の構造成分のいずれかの組み合わせを含むことができ、それは典型的に以下の3つの構造基に由来する;a)天然アミド結合(「ペプチド結合」)以外の残基連結基;b)天然に存在するアミノ酸残基に代わる非天然残基;c)二次構造擬態を誘導する残基、すなわち、例えば、βターン、γターン、βシート、αへリックスなどの二次構造を誘導または安定化する残基。
ポリペプチドは、その残基の全てまたは一部が天然ペプチド結合以外の化学的方法により結合している場合に、擬似体として特徴付けることができる。個々のペプチド擬似残基は、ペプチド結合、そのほかの化学的結合またはカップリング手段、例えば、グルタルアルデヒド、N−ヒドロキシコハク酸イミドエステル、2価マレイミド、N,N'−ジシクロヘキシルカルボジイミド(DCC)またはN,N'−ジイソプロピルカルボジイミド(DIC)、により結合することができる。伝統的アミド結合(「ペプチド結合」)に代わりうる結合基には、例えば、ケトメチレン(例えば、-C(=O)-NH-の代わりに−C(=O)-CH2-)、アミノメチレン(CH2-NH)、エチレン、オレフィン(CH=CH)、エーテル(CH2-O)、チオエーテル(CH2-S)、テトラゾール(CN4)、チアゾール、レトロアミド、チオアミド、またはエステルがある(例えば、Spatola, Chemistry and Biochemistry of Amino Acids, Peptides and Proteins, Vol. 7, 267-357, Marcell Dekker, Peptide Backbone Modifications, NY (1983)参照)。天然に存在するアミノ酸残基に代わって、全部または一部が非天然残基を含む場合にも、ポリペプチドは、擬似体に分類される(非天然残基は、科学文献及び特許文献に詳しく記述されている)。
【0058】
「標識」または「検出分子」は、分光学的、光化学的、生化学的、免疫化学的、または化学的な方法により検出される成分である。例えば、有用な標識としては、32P,蛍光色素、電子密度試薬、(例えば、ELISAにおいて広く使用されるような)酵素、ビオチン、ジゴキシゲニン、或いは例えば、ペプチドの中に放射標識を組み込むことにより検出可能にしたハプテン及びタンパク、またはペプチドと特異的に反応する抗体を検出するために使用されるハプテン及びタンパクが含まれる。
【0059】
「標識した核酸プローブまたはオリゴヌクレオチド」は、プローブの存在がプローブに結合した標識の存在を検出することにより検出されるように、標識に、リンカーや化学結合による共有結合により、或いはイオン的な、ファンデルワールス力による、静電的な、または水素結合による非共有結合により、結合させた核酸プローブまたはオリゴヌクレオチドである。
【0060】
本明細書で使用される時、「核酸プローブまたはオリゴヌクレオチド」とは、一以上の種類の化学結合、通常は相補的塩基対による通常は水素結合形成、により、相補的配列からなる標的核酸と結合することができる核酸と定義される。本明細書で使用される時プローブは、天然塩基(すなわち、A, G, C,またはT)または修飾塩基(7−デアザグアノシン、イノシン、など)を含むことができる。さらに、プローブ中の塩基は、ハイブリッド形成を妨害しない限り、燐酸ジエステル結合以外の結合で連結することができる。したがって、例えば、プローブは燐酸ジエステル結合でなくペプチド結合により構成塩基が連結したペプチド核酸とすることができる。ハイブリッド形成条件のストリンジェンシーによってはプローブと完全な相補性を欠く標的配列とプローブが結合しうることを、当業者は理解しているであろう。プローブは任意にアイソトープ、発色団、発光団、発色源で直接標識されるか、または後にストレプトアビジン複合体が結合するビオチンのようなもので間接的に標識される。プローブの存在または不在をアッセイすることにより、選択した配列または部分配列の存在または不在を検出することができる。
【0061】
用語「異種」は、核酸の部分に関して使用された場合、天然において相互に同一の種族の中には認められない二つ以上の部分配列を核酸が含むことを示す。例えば、典型的には、新規な機能の核酸を作るために配列された無関係な遺伝子から得た二つ以上の配列、例えば、一の起源からのプロモーターと他の起源からのコード領域とを持っている核酸が、組換えにより作られる。同様に、異種タンパクとは、そのタンパクが、天然において相互に同一の種族の中には認められない二つ以上の部分配列を含むこと(例えば、融合タンパク)を示す。
【0062】
「プロモーター」は、核酸の転写を指令する核酸配列が並んだものと定義される。本明細書で使用される時、プロモーターには、例えば、ポリメラーゼII型プロモーターやTATA配列の場合における、転写開始部位の近くにある必要な核酸配列が含まれる。プロモーターは、さらに転写開始部位から数千塩基対も離れていることがある遠位エンハンサーまたはリプレッサーも含む。「構成的」プロモーターは、ほとんどの環境及び発生条件において活性であるプロモーターである。「誘導的」プロモーターは、環境または発生の制御下において活性であるプロモーターである。用語「作動的に結合している」とは、(プロモーター、または転写因子結合部位の配列のような)核酸発現調節配列と、第二の核酸配列との間での機能的連結のことであり、発現調節配列は、第二配列に相当する核酸の転写を指令する。
【0063】
本明細書で使用される時、「組換え」とは、合成されたか、インビトロで操作されたポリヌクレオチド(例えば、「組換えポリヌクレオチド」)、細胞中またはそのほかの生化学的系において遺伝子産物を生産するために組換えポリヌクレオチドを使用する方法、或いは組換えポリヌクレオチドによりコードされているポリペプチド(「組換えタンパク」)のことである。「組換え手段」は、例えば、本発明の転移ドメインと本発明のプライマーを使用して増幅された核酸配列とを含む融合タンパクの誘導的または構成的な発現のための発現カセットまたはベクターの中に、異なる起源から得た種々のコード領域またはドメインまたはプロモーター配列を持つ核酸を連結することも包含する。
【0064】
語句「選択的に(または特異的に)ハイブリッド形成する」とは、その配列が複雑な混合物(例えば、全細胞またはライブラリーDNAまたはRNA)中に存在するときに、ストリンジェントなハイブリッド形成条件の下に特別なヌクレオチド配列とのみ結合、二本鎖化または分子のハイブリッドを形成することをいう。
【0065】
語句「ストリンジェントなハイブリッド形成条件」とは、プローブが、典型的には核酸の複雑な混合物中において、標的部分配列とハイブリッドを形成し、他の配列とはハイブリッドを形成しない条件のことをいう。ストリンジェントな条件は配列に依存し、そして異なる環境では異なるであろう。長い配列は特に、高い温度でハイブリッド形成する。核酸のハイブリッド形成に関する詳細な説明は、Tijssen, Techniques in Biochemistry and Molecular Biology-Hybridisation with Nucleic Probes, “Overview of principles of hybridization and the strategy of nucleic acid assays” (1993)の中にある。一般的に、ストリンジェントな条件は、規定されているイオン強度、pHにおいて、特定の配列の融解温度(Tm)よりも約5−10℃低い温度が選択される。Tmは、(規定されたイオン強度、pH及び核酸濃度において)標的に相補的なプローブの50%が、均衡して標的配列とハイブリッド形成する温度である(標的配列が過剰に存在すると、Tmで、プローブの50%が均衡して占有される)。ストリンジェントな条件は、塩濃度が約1.0 Mナトリウムイオン未満、典型的にはpH 7.0から8.3において約0.01から1.0 Mナトリウムイオン濃度(または他の塩)であり、温度が、短いプローブ(例えば、10から50ヌクレオチド)に対して少なくとも約30℃、長いプローブ(例えば50ヌクレオチド超)に対しては少なくとも約60℃の条件であろう。ストリンジェントな条件はホルムアミドのような脱安定化試薬の添加も必要である。選択的または特異的なハイブリッド形成に対する陽性信号は、背景ハイブリッド形成の少なくとも2倍、望ましくは10倍である。代表的なストリンジェントな条件は次のようなものであろう:50%ホルムアミド、5x SSC,及び1% SDS, 42℃でインキュベート、または5x SSC, 1% SDS, 65℃でインキュベート、そして65℃において0.2x SSC及び0.1% SDS中で洗う。そのハイブリッド形成及び洗浄段階は、例えば、1,2,5,10,15,30,60分間;またはそれ以上の時間行うことができる。
【0066】
ストリンジェントな条件下に相互にハイブリッド形成しない核酸であっても、それらがコードするポリペプチドが実質的に関係あるときには、依然として実質的に関係がある。これは、例えば、核酸のコピーが遺伝子コードで許容される最大のコドン縮重を使用して作られている場合に生じる。そのような場合には、核酸は、通常、緩和なストリンジェントなハイブリッド形成条件下でハイブリッドを形成する。代表的な「緩和なストリンジェントハイブリッド形成条件」は、40%ホルムアミドの緩衝液中、1 M NaCl,1% SDS 37℃におけるハイブリッド形成、及び1x SSC 45℃における洗浄を含む。そのハイブリッド形成及び洗浄段階は、例えば、1,2,5,10,15,30,60分間またはそれ以上の時間行うことができる。ハイブリッド形成陽性は背景の少なくとも2倍である。同じストリンジェンシーにするために、別のハイブリッド形成及び洗浄条件を使用しうることを、当業者は容易に理解するであろう。
【0067】
「抗体」は、抗原に特異的に結合して認識する免疫グロブリンの遺伝子に由来する骨格領域またはそのフラグメントを含むポリペプチドのことである。認識する免疫グロブリンの遺伝子は、κ、λ、α、γ、δ、ε、及びμ定常領域、並びに無数の免疫グロブリン可変領域遺伝子を含む。軽鎖はκまたはλのいずれかに分類される。重鎖はγ、μ、α、δ、またはεに分類され、それらは順にイムノグロブリンクラス、IgG, IgM, IgA, IgD及びIgEをそれぞれ規定する。
【0068】
代表的な免疫グロブリン(抗体)構造単位は、4量体からなる。各4量体は、2対の同じポリペプチド鎖から構成される、対のそれぞれが、一つの「軽鎖」(約25 kDa)及び一つの「重鎖」(約50−70 kDa)を有している。各鎖のN−末端は、主に抗原認識の役割を担う約100から110またはそれ以上のアミノ酸からなる可変領域である。用語、可変軽鎖(VL)及び可変重鎖(VH)は、それぞれこの軽鎖及び重鎖のことである。
【0069】
「キメラ抗体」は、次のような抗体分子である:
(a)定常領域またはその一部が改変され、置換され、または交換され、その結果、抗原結合部位(可変領域)が、異なるまたは改変されたクラス、エフェクター機能及び/または種の定常領域と連結しているか、或いはキメラ抗体に新しい性質を付与する全く異なる分子、例えば、酵素、トキシン、ホルモン、成長因子、薬物などと連結している抗体分子;或いは
(b)可変領域またはその一部が、異なるまたは改変された抗原特異性を持つ可変領域により改変、置換または交換されている抗体分子。
【0070】
「抗−T2R」抗体は、T2R遺伝子、そのcDNA,または部分配列によりコードされたポリペプチドに特異的に結合する抗体または抗体フラグメントである。
【0071】
用語「免疫アッセイ」は、抗原に特異的に結合する抗体を使用するアッセイである。免疫アッセイは、抗原を単離し、標的とし、そして/または定量するために特別な抗体の特異的結合性を使用することが特徴である。
【0072】
タンパクまたはペプチドに関して、語句、抗体に「特異的に(選択的に)結合する」または「特異的に(選択的に)免疫反応する」とは、種々のタンパク及び生体成分の混合物中で、そのタンパクの存在を決定する結合反応のことである。したがって、指定された免疫アッセイ条件の下に、特定の抗体は、特定のタンパクと背景の少なくとも2倍結合し、検体中に存在するほかのタンパクとはあまり結合しない。そのような条件において抗体に特異的に結合するには、特別なタンパクに対する特異性を持つような抗体を選択する必要がある。
【0073】
例えば、ラット、マウス、またはヒトのような特定の動物種のT2Rファミリーメンバーに対して作成されたポリクロナール抗体は、T2Rポリペプチドまたはその免疫原部分と特異的に免疫反応するが、T2Rポリペプチドのオーソログまたは多形変異体及びアレル以外のほかのタンパクとは反応しないポリクロナール抗体のみを得るために選別することができる。この選別は、他の動物種のT2R分子または他のT2R分子と交差反応する抗体を取り除くことにより行われる。抗体はT2R GPCRファミリーメンバーのみを認識し、他のファミリーのGPCRを認識しないように選別することも可能である。種々の免疫アッセイ様式を、特定のタンパクと特異的に免疫反応する抗体を選別するために使用することができる。例えば、固相ELISA免疫アッセイは、タンパクと特異的に免疫反応する抗体を選別するために日常的に使用されている(例えば、Harlow & Lane, Antibodies, A Laboratory Manual, (1988)参照、免疫アッセイ様式及び特異的免疫反応性の測定に使用しうる条件を記述)。典型的には、特異的または選択的な反応は、背景信号またはノイズの少なくとも2倍であり、より典型的には背景の10から100倍以上であろう。
【0074】
語句「選択的に会合する」とは、上記に定義したように他のものと「選択的にハイブリッドを形成する」核酸の能力、または上記に定義したようにタンパクに「選択的に(または特異的に)結合する」抗体の能力のことである。
【0075】
用語「発現ベクター」とは、インビトロまたはインビボで、構成的または誘導的に、原核細胞、イースト、菌、植物、昆虫または哺乳動物細胞のいずれかの細胞中において、本発明の核酸配列を発現するための組換え発現系のことである。この用語は、線状または環状発現系を含む。この用語は、エピソームに存在するかまたは宿主細胞ゲノムに組み込まれた発現系を含む。発現系は、自己複製能力を持つものでも、持たないもの、すなわち、細胞中において一過性にのみ発現するものでもよい。この用語は、組換え核酸の転写のために最小限必要な配列のみを含む組換え発現「カセット」を含む。
【0076】
「宿主細胞」は、発現ベクターを含み、発現ベクターの複製又は発現を維持する細胞を意味する。宿主細胞は、E. coliのような原核細胞、またはイースト、昆虫、両生類のような真核細胞、またはCHO, HeLa, HEK-293のような哺乳動物細胞などとすることができ、例えば、培養細胞、外植細胞、及びインビボの細胞などがある。
【0077】
以上に基づいて、本発明は、苦味化合物によるhT2R4, hT2R44,またはhT2R61の特異的活性化を阻害する化合物を同定するアッセイを提供する。特に本発明は、キニーネまたは関連化合物によるhT2R4の活性化、6−ニトロサッカリン、サッカリン、または関連化合物によるhT2R61の活性化、またはデナトニウム、6−ニトロサッカリンまたは関連化合物によるhT2R44の活性化に影響を及ぼす(阻害する)化合物を同定するアッセイ、望ましくは細胞を使用するアッセイを提供する。上記に特定した苦味試験化合物によるこれらのhT2Rの活性化を阻害する化合物は、これらの特定のhT2Rと相互作用する他の苦味化合物により引き起こされる苦味を特異的に抑制することが期待される。
【0078】
特に、苦味アルカロイドであるキニーネは、T2R4を特異的に用量依存的に活性化することが認められた。このことは細胞内カルシウム濃度の変化を検出する、細胞によるアッセイにより発見された。(実施例で詳細に検討されるので)簡略に記述すると、大きなT細胞抗原及びG15 タンパクを安定的に発現するヒト胚腎細胞(HEK-G15)を、hT2R4発現プラスミドで(例えば、燐酸カルシウムを使用するかまたは脂質系を使用して)一過性に遺伝子導入した。さらに、他のHEK-G15細胞系を、他のヒトT2Rで一過性に遺伝子導入した。その後、蛍光を用いるアッセイを使用して一過性に遺伝子導入した細胞におけるカルシウム濃度の変化を検出した。このアッセイを使用することにより、hT2R4を発現する細胞にキニーネを加えると、PLCの活性化に至る信号伝達カスケードを誘発し、次いでカルシウム感受性蛍光色素を使用して検出される蛍光の増加を生じる細胞内カルシウム濃度の増加を生じる。これらの変化は、例えば、蛍光顕微鏡及び適当に設計されたソフトウエア(例えば、Imaging Workstation, Axon)を使用して監視される。これに対して、他のhT2Rで一過性に遺伝子導入されたHEK-G15細胞は、キニーネにより特異的に活性化しない。これらに基づくと、hT2R4は、キニーネ及び構造的に関連する化合物により特異的に活性化されるヒトT2Rであると思われる。したがって、キニーネによるhT2R4の活性化を阻害する化合物は、hT2R4により誘発される、特にhT2R4を活性化する苦味アルカロイドであるキニーネ及び関連化合物により誘発される苦味を抑制することが予期される。そのアッセイの例は後に詳細に開示する。
【0079】
また、本発明は特に、hT2R44がデナトニウム及び6−ニトロサッカリンによって特異的に活性化され、hT2R61が6−ニトロサッカリンによって特異的に活性化されるという発見を含んでいる。この発見は、図4に図解されているGTPγS結合アッセイを使用してなされた。このアッセイは、GPCR活性を監視する古典的なアッセイである(Wessling-Resnick and Johnson, J. Biol. Chem 262: 12444-12447(1987)参照)。このアッセイは、適当なリガンドの存在下、受容体がGタンパクを活性化できるという事実に基づいている。その結果、活性化されたGタンパクは、結合しているGDPをGTPに変換する。
【0080】
さらに、GTPの非加水分解性の類似体であるGTPγSを使用した場合には、ヌクレオチドは、Gタンパクにしっかりと結合したままとなり、結合していないヌクレオチドから分離することができる。したがって、このアッセイにおいて、結合したGTPγSの量は、活性化したGタンパクの量を反映し、それが受容体活性化の指標ともなる。GTPγ35S(「GTPγS」)結合アッセイは、ガストジューシン及びトランスジューシンのような種々のGタンパクを使用して行うことができる。後記の例に記述するように、Gタンパクとしてトランスジューシンを用いるこのアッセイを使用して、hT2R44 (SEQ ID NO.:3)またはhT2R61 (SEQ ID NO.:5)のいずれかを発現する昆虫細胞が、添加した苦味化合物の存在下に活性の増加を示すことが観察された。
【0081】
特に、後記の実験例に詳細に開示するように、hT2R61は、6−ニトロサッカリンの存在下に活性化され、hT2R44は、この化合物によって弱く活性化される。さらに、hT2R44は、特異的に安息香酸デナトニウムにより活性化される(安息香酸ナトリウムは、同濃度においてhT2R44を活性化しないという事実により示されるように、デナトニウム成分による活性化である)。
【0082】
これに対して、結合アッセイにより評価したときに、デナトニウムは、同じ一過性細胞発現系(Sf9バキュロウイルス発現系)を使用して発現した他のhT2Rを活性化しない。また、いくつかのデナトニウム誘導体も、hT2R44を活性化することが観察されたが、デナトニウムより程度は低かった。安息香酸デナトニウムは、ヒトに苦味を感じさせることが知られている苦味化合物の中で最も強いものの一つである。これらの結果は、hT2R44が、ヒトにおいて、デナトニウムの苦味を仲介していること、そしてhT2R44のデナトニウム活性化を阻害する化合物は、この受容体の活性化による苦味を調節(阻止)することを示唆している。
【0083】
したがって、本発明は、hT2R4活性化による苦味を阻止する化合物を同定する手段として、キニーネまたは関連化合物によるhT2R4活性化を調節(阻害)する化合物を同定するためのアッセイ、望ましくは細胞によるアッセイを提供する。
【0084】
また、本発明は、hT2R61活性化により惹起されるヒトにおける苦味を阻止する化合物を同定する手段として、6−ニトロサッカリンまたはサッカリンまたは関連化合物によるhT2R61活性化を調節(阻害)する化合物を同定するためのアッセイ、望ましくは細胞によるアッセイを提供する。
【0085】
また、本発明は、hT2R44活性化により惹起されるヒトにおける苦味を阻止する化合物を同定する手段として、デナトニウムまたは6−ニトロサッカリンまたは関連化合物によるhT2R44活性化を阻害する化合物を同定するためのアッセイ、望ましくは細胞によるアッセイを提供する。本発明に有用なアッセイ及び発現系は後に詳細に記述される。
【0086】
最も典型的に、これらのアッセイは、すでに特定したアミノ酸配列を有するhT2R4, hT2R44または hT2R61ポリペプチドをコードするDNAを発現する細胞を使用するであろう。しかし、hT2R4, hT2R44または hT2R61の機能的特性を保有するこれらの受容体ポリペプチドのフラグメント、オーソログ、変異体またはキメラもこれらのアッセイにおいて有用であることが予期される。その変異体の例としては、組換えまたは合成により調製されたか、天然に存在するスプライス変異体、一塩基多形、アレル変異体、及び突然変異体がある。本発明のアッセイにおいて使用されるT2Rの単離及び発現の方法、並びにhT2R4, hT2R44または hT2R61の活性化を抑制する化合物を同定するために、本発明において使用することが企図されているアッセイは、以下に記述される。
【0087】
T2Rの単離及び発現
本発明のT2R、またはそのフラグメント若しくは変異体の単離及び発現は、本出願に開示されたT2R核酸配列に基づいて構築されたプローブまたはプライマーを使用して、充分に確立されたクローニング方法により行うことができる。関連T2R配列は、本明細書で開示した配列及び既知のコンピューター支援探索技術、例えば、BLAST配列探索を使用して、ヒトまたは他の種のゲノムデータベースから同定することもできる。特別な態様において、本明細書で開示した偽遺伝子を、機能的アレルまたは関係遺伝子を同定するために使用することができる。
【0088】
次いで、これらの配列を機能的に発現させるための宿主細胞に感染または遺伝子導入するために、発現ベクターを使用することができる。これらの遺伝子及びベクターは、インビトロまたはインビボにおいて作製し、発現させることができる。核酸発現を変更及び調節するための望ましい表現型は、本発明のベクター中の遺伝子及び核酸(例えば、プロモーター、エンハンサーなど)の発現または活性を調節することにより得られることは、当業者はよく認識している。発現または活性を増加または減少するために、記述された既知の方法のいずれかを使用することができる。本発明は、科学および特許の文献に詳述されている当業者に既知の方法またはプロトコールのいずれとも組み合わせて実施することができる。
【0089】
そのほかに、これらの核酸は、例えば下記の既知化学合成技術によりインビトロにおいて合成することができる:Carruthers, Cold Spring Harbor Symp. Quant. Biol. 47: 411-18 (1982); Adams, Am Chem. Soc., 105: 661 (1983); Belousov, Nucleic Acids Res. 25: 3440-3444 (1997); Frenkel, Free Radic. Biol. Med. 19: 373-380 (1995); Blommers, Biochemistry 33: 7886-7896 (1994); Narang, Meth. Enzymol. 68: 90 (1979); Brown, Meth. Enzymol. 68: 109 (1979); Beaucage, Tetra. Lett. 22: 1859 (1981);米国特許番号 4,458,066。
次いで、相補鎖を合成して適当な条件下に鎖をアニーリングするか、または適当なプライマー配列とともにDNAポリメラーゼを使用して相補的鎖を加えることにより二本鎖DNAフラグメントを得ることができる。
【0090】
例えば、配列中の突然変異誘発、再クローニング、プローブの標識、配列分析、ハイブリッド形成などのような核酸の操作技術は、科学及び特許の文献に詳細に記述されており、例えば、Sambrook, ed., Molecular Cloning: A Laboratory Manual (2nd ed.), Vols. 1-3, Cold Spring Harbor Laboratory (1989); Ausubel, ed., Current Protocols in Molecular Biology, John Wiley & Sons, Inc., New York (1997); Tijssen, ed., Laboratory Techniques in Biochemistry and Molecular Biology: Hybridization With Nucleic Acid Probes, Part I, Theory and Nucleic Acid Preparation, Elsevier, N.Y. (1993)がある。
【0091】
核酸、ベクター、カプシド、ポリペプチドなどは、当業者に既知の多数の一般的な方法いずれかにより分析及び定量することができる。それらには、分析的生化学的方法、例えば、NMR,分光分析、放射線画像、電気泳動、キャピラリー電気泳動、高速液体クロマトグラフィー(HPLC)、薄層クロマトグラフィー(TLC)、及び超拡散クロマトグラフィー、種々の免疫学的方法、例えば、液相またはゲル沈降反応、免疫拡散、免疫電気泳動、放射線免疫アッセイ(RIA)、固相酵素免疫アッセイ(ELISA)、免疫蛍光アッセイ、サザン分析、ノーザン分析、ドットブロット分析、ゲル電気泳動(例えば、SDS-PAGE)、RT-PCR,定量的PCR、そのほかの核酸または標的またはシグナル増幅方法、放射線標識、シンチレーション計測、及びアフィニティークロマトグラフィーが含まれる。
【0092】
T2Rリガンド結合領域をコードする核酸を増幅するために、オリゴヌクレオチドプライマーを使用することができる。本明細書で記述する核酸は、増幅技術を使用してクローニングまたは定量的に測定することができる。増幅方法は当業者に既知であり、例えば、ポリメラーゼ連鎖反応(PCR)(Innis ed., PCR Protocols, a Guide to Methods and Applications, Academic Press, N.Y. (1990); Innis ed., PCR Strategies, Academic Press, Inc., N.Y. (1995));リガーゼ連鎖反応(LCR)(Wu, Genomics, 4: 560 (1989); Landegren, Science, 241: 1077 (1988); Barringer, Gene, 89: 117 (1990)); 転写増幅(Kwoh, PNAS, 86: 1173 (1989)); 自己維持配列複製(Guatelli, PNAS, 87: 1874(1990)); Q−βレプリカーゼ増幅(Smith, J. Clin. Microbiol., 35: 1477-91(1997)); 自動化Q−βレプリカーゼ増幅アッセイ(Burg, Mol. Cell. Probes, 10: 257-71(1996)); 及びそのほかのRNAポリメラーゼを使用する技術(例えば、NASBA,Cangene, Mississauga, Ontario)がある。Berger, Methods Enzymol., 152: 307-16(1987); Sambrook; Ausubel;米国特許番号 4,683,195及び4,683,202; Sooknanan, Biotechnology, 13: 563-64 (1995)も参照。
【0093】
核酸は、ひとたび増幅されると個別的にまたはライブラリーとして、必要に応じて種々のベクター中に通常の分子生物学的方法を使用する当業者に既知の方法に従ってクローニングすることができる。インビトロにおける増幅核酸のクローニング方法は、例えば、米国特許番号5,426,039に記述されている。増幅配列のクローニングを容易にするために、制限酵素部位をPCRプライマー対に「組み込む」こともできる。例えば、Pst I及びBsp E1部位を、本発明の代表的プライマー中に設計した。これらの特別な制限部位は、連結したときに、それらがスプライスされて得られる7回膜貫通受容体「ドナー」のコード配列に関し「フレーム単位」とする配列を有する(リガンド結合領域コード配列は、7回膜貫通ポリペプチドにとって本質的であるため、構築体が制限酵素スプライス部位の下流で翻訳されることを望むならば、フレームのずれは避けなければならない;これは挿入されたリガンド結合領域が実質的に膜貫通VII領域のほとんどを含む場合には必要ないであろう)。プライマーは、「ドナー」7回膜貫通受容体の元の配列を維持するように設計することができる。そのほかに、プライマーは、保存置換基(例えば、疎水性残基に代わる疎水性残基、前記検討を参照)または機能上害のない(例えば、形質膜挿入を妨害しない、タンパク分解酵素による切断を生じない、受容体の異常な折り畳みを生じない、など)置換基であるアミノ酸残基をコードすることができる。
【0094】
プライマー対は、T2Rタンパクのリガンド結合領域を選択的に増幅するように設計することができる。これらの領域は、異なるリガンドに対しては変わることがある;したがって、一つのリガンドに対する最小限の結合領域であるものは、第二の潜在的リガンドに対しては制限的になりすぎる。したがって、種々のドメイン構造を含む種々のサイズの結合領域を増幅するとよい;例えば、7回膜貫通T2Rの膜貫通(TM)ドメインIIからVII,IIIからVII,IIIからVIまたはIIからVI、またはそれらの変異体(例えば、個別ドメインの部分配列のみ、ドメインのオーダーの混合、など)などである。
【0095】
7回膜貫通T2Rタンパクのドメイン構造及び配列は既知であるので、当業者は、縮重プライマー対を設計するためのモデル配列として、ドメイン隣接及び内部ドメイン配列を容易に選択することができる。例えば、ドメイン領域IIからVIIをコードする核酸配列は、プライマー対を使用してPCR増幅により作製することができる。膜貫通ドメインI(TM I)配列を含む核酸を増幅するために、縮重プライマーを、前記のT2Rファミリー共通配列1のアミノ酸配列をコードする核酸から設計することができる。その縮重プライマーは、TM IからTM III,TM IからTM IV,TM IからTM V,TM IからTM VI,またはTM IからTM VIIを組み込んだ結合領域を作製するために使用することができる。他の縮重プライマーは、本明細書で提供した他のT2Rファミリー共通配列を基に設計することができる。そのような縮重プライマーは、TM IIIからTM IV,TM IIIからTM V,TM IIIからTM VIまたはTM IIIからTM VIIを組み込んだ結合領域を作製するために使用することができる。
【0096】
縮重プライマー対を設計する代表例は、当業者に良く知られている。例えば、共通配列縮重ハイブリッドオリゴヌクレオチドプライマー(CODEHOP)法コンピュータープログラムは、http://blocks.fhcrc.org/codehop.html,で接続可能であり、既知の味覚受容体リガンド結合領域として、一連の関連タンパク配列で始まるハイブリッドプライマー予測のためのBlockMaker 多重配列整列サイトと直接リンクしている(例えば、Rose, Nucleic Acids Res., 26: 1628-35 (1998); Singh, Biotechniques, 24: 318-19 (1998)参照)。
【0097】
オリゴヌクレオチドプライマー対を合成する方法は、当業者に良く知られている。「天然」塩基対または合成塩基対を使用することができる。例えば、人工核酸塩基を使用することにより、プライマー配列を操作するための多彩な方法が提供され、増幅産物のより複雑な混合物が作成される。人工核酸塩基の種々のファミリーは、内部結合回転による多重水素結合定位を予想することができ、縮重分子の認識のための手段を提供する。これらの類似体を、PCRプライマーの一つの位置に組み込むことにより、増幅産物の複雑なライブラリーを作製することができる。例えば、Hoops, Nucleic Acids Res., 25: 4866-71 (1997)参照。非極性分子も天然のDNA塩基の形状を模倣するために使用することができる。アデニンの非水素結合形状模倣体は、チミンの非極性形状模倣体に対して効率よく選択的に複製される(例えば、Morales, Nat. Struct. Biol., 5: 950-54(1998)参照)。例えば、二つの縮重塩基は、ピリミジン塩基6H,8H-3,4-ジヒドロピリミド[4,5-c][1,2]オキサジン-7-オンまたはプリン塩基N6-メトキシ-2,6-ジアミノプリンとすることができる(例えば、Hill, PNAS, 95: 4258-63 (1998)参照)。本発明の代表的縮重プライマーは、核酸塩基類似体5'-ジメトキシトリチル-N-ベンゾイル-2'-デオキシ-シチジン、3'-[(2-シアノエチル)-(N,N-ジイソプロピル)]−ホスフォルアミダイト(配列中の文字「P」、上記参照)を組み込んでいる。このピリミジン類似体は、A及びG残基を含むプリンと水素結合する。
【0098】
本明細書で開示された味覚受容体と実質的に同一である多形変異体、アレル、及び種間同属体は、上記の核酸プローブを使用して単離することができる。また、発現ライブラリーを使用して、T2R同属体を認識し、結合する、T2Rポリペプチドに対して作製された抗血清または精製抗体で、発現同属体を検出することにより、T2Rポリペプチド及びその多形変異体、アレル、及び種間同属体をクローニングすることができる。
【0099】
味覚受容体のリガンド結合領域をコードする核酸を、適当な(完全または縮重)プライマー対を使用して、適当な核酸配列を(例えば、PCRで)増幅することにより作製することができる。増幅される核酸としては、細胞若しくは組織のゲノムDNAまたは味覚受容体発現細胞に由来するmRNA若しくはcDNAがある。
【0100】
一の態様において、転移配列に融合させたT2Rをコードする核酸を含む、ハイブリッドタンパクコード配列を構築することができる。転移モチーフ及び化学的感受性受容体の他のファミリー、特に味覚受容体、の味誘発化合物結合領域を含むハイブリッドT2Rも提供される。これらの核酸配列は、転写または翻訳調節配列、例えば、転写及び翻訳開始配列、プロモーター及びエンハンサー、転写及び翻訳ターミネーター、ポリアデニル化配列、及びそのほかのDNAをRNAに転写するのに有用な配列、に作動的に連結することができる。組換え発現カセット、ベクター、及び形質転換体の構築において、目的とする細胞または組織の全てに目的とする核酸の発現を指令するためにプロモーターフラグメントを使用することができる。
【0101】
そのほかの態様において、融合タンパクは、C−末端またはN−末端転移配列を含むことができる。さらに、融合タンパクは、追加の配列、例えば、タンパク検出、精製、またはそのほかの適用のための配列を含むことができる。検出及び精製を容易にするドメインとしては、例えば、ポリヒスチジン鎖のような金属キレートペプチド、ヒスチジン−トリプトファンモジュール、または固相金属上で精製できるようなそのほかのドメイン;マルトース結合タンパク;固相免疫グロブリン上で精製できるタンパクAドメイン;またはFLAGSエクステンション/親和性精製システム(Immunex Corp,Seattle WA)に使用されるドメイン、がある。
【0102】
ファクターXa(例えば、Ottavi, Biochimie, 80: 289-93 (1998)参照)、ズブチリシンタンパク分解酵素認識モチーフ(例えば、Polyak, Protein Eng., 10: 615-19 (1997)参照);エンテロキナーゼ(Invitrogen, San Diego, CA)などの切断可能なリンカー配列を、(効率的に形質膜に発現させるための)転移配列と新規に翻訳されるポリペプチドの残りの部分との間に入れることは、精製を容易にするために有用である。例えば、一の構築体は、その後に、チオレドキシン、エンテロキナーゼ切断部位(例えば、Williams, Biochemistry, 34: 1787-97 (1995)参照)、及びC−末端転移ドメインが続く6ヒスチジン残基に連結させるポリペプチドをコードする核酸配列を含むことができる。ヒスチジン残基は、検出及び精製を容易にし、他方エンテロキナーゼ切断部位は、残余の融合タンパクから目的タンパクを精製する手段を提供する。融合タンパクをコードするベクターに関する技術及び融合タンパクの応用は、科学及び特許の文献に詳細に記述されている(例えば、Kroll, DNA Cell. Biol., 12: 441-53 (1993)参照)。
【0103】
リガンド結合領域コード配列を含む発現ベクターは、個々の発現ベクターまたは発現ベクターのライブラリーとして、科学及び特許文献に詳細に記述されている種々の通常技術により、ゲノム中または細胞質中または細胞の核中に導入し、発現させることができる。例えば、Roberts, Nature, 328: 731 (1987); Berger 前出;Schneider , Protein Expr. Purif., 6435: 10 (1995); Sambrook; Tijssen; Ausubel.参照。生化学試薬及び実験機器のメーカーの製品情報も既知の生化学方法に関する情報を提供している。ベクターは、天然資源から単離することができ、ATCCまたはGenBankライブラリーのような資源から入手することができ、或いは合成的にまたは組換えの方法により調製することができる。
【0104】
核酸は、細胞中に安定的にまたは一過性に発現する(例えば、エピソーム発現系)発現カセット、ベクターまたはウイルス中で発現させることができる。選別マーカーは、発現カセット及びベクター中に組み込まれ、形質転換された細胞及び配列に選別可能な形質を付与することができる。例えば、選別マーカーは、宿主ゲノムに組み込む必要はないように、エピソームの維持及び複製をコード化することができる。例えば、目的とするDNA配列で形質転換した細胞の選別を可能にするために、マーカーは、抗生物質耐性(例えば、クロラムフェニコール、カナマイシン、G418, ブレオマイシン、ヒグロマイシン)または除草剤耐性(例えば、クロロスルフロンまたはバスタ)をコード化することができる(例えば、Blondelet-Rouault, Gene, 190: 315-17 (1997);Aubrecht, J. Pharmacol. Exp. Ther., 281: 992-97 (1997)参照)。ネオマイシンまたはヒグロマイシンのような基質に対する耐性を付与する選別マーカーは、組織培養にのみ使用されるので、化学耐性遺伝子も、インビトロ及びインビボにおける選別マーカーとして使用される。
【0105】
キメラ核酸配列は、いずれかの7回膜貫通ポリペプチドの中のT2Rリガンド結合領域をコードすることができる。7回膜貫通受容体ポリペプチドは、同じ一次及び二次及び三次構造を有しているので、構造ドメイン(例えば、細胞外ドメイン、TMドメイン、細胞質ドメイン、など)は、配列分析により容易に同定することができる。例えば、同属体設計、フーリエ解析及び螺旋周期性検出により、7回膜貫通受容体配列を有する7個のドメインを同定し分析することができる。迅速フーリエ変換(FFT)アルゴリズムを、分析する配列の疎水性及び可変性を特徴付ける主要周期を評価するために使用することができる。周期性検出及びα螺旋周期性指標は、例えば、Donnelly, Protein Sci., 2: 55-70 (1993)により得ることができる。そのほかの整列及びモデルアルゴリズムは、当業者に既知である(例えば、Peitsch, Receptors Channels, 4: 161-64 (1996); Kyte & Doolittle, J. Md. Biol., 157: 105-32 (1982); Cronet, Protein Eng., 6: 59-64 (1993); http://bioinfo.weizmann.ac.il/参照)。
【0106】
本発明は、特定の核酸配列及びアミノ酸配列を有する核酸分子及びポリペプチドを含むだけでなく、そのフラグメント、特に、例えば、40,60,80,100, 150,200,または250ヌクレオチドまたはそれ以上のフラグメント、並びに例えば、10,20,30,50,70,100,または150アミノ酸、またはそれ以上のポリペプチドフラグメントも含んでいる。任意に、核酸フラグメントは、T2Rファミリーメンバーに対して作製された抗体に結合することができる抗原性ポリペプチドをコードすることができる。さらに、本発明のタンパクフラグメントは、任意にT2Rファミリーメンバーに対して作製された抗体に結合することができる抗原性フラグメントとすることができる。
【0107】
他のGPCR、望ましくは7回膜貫通スーパーファミリーのメンバーの全体または一部である追加のアミノ酸と結合した、本明細書で記述するT2Rポリペプチドの少なくとも一つの、少なくとも10,20,30,50,70,100,または150アミノ酸、またはそれ以上を含むキメラタンパクも企図される。これらのキメラは、当該受容体と他のGPCRとから作ることもでき、また、本発明の受容体の二つまたはそれ以上を組み合わせて作ることもできる。一の態様において、キメラの一部は、本発明のT2Rポリペプチドの膜貫通ドメインに相当または由来する。別の態様において、キメラの一部は、本明細書で記述するT2Rポリペプチドの膜貫通領域の一つまたはそれ以上に相当、または由来し、そして残りの部分は他のGPCRに由来し得る。キメラ受容体は、当業者に既知であり、それを作製する技術及びその中に組み込むためのGタンパク共役受容体のドメインまたはフラグメントの選別及び境界も良く知られている。したがって、当業者のこの知識をそのようなキメラ受容体を創るために容易に使用することができる。そのようなキメラ受容体を使用することにより、例えば、先行技術のアッセイ系に使用された既知の受容体のような他の受容体の信号伝達特性とともに、本明細書で具体的に開示した一の受容体の味選択特性を提供することができる。
【0108】
例えば、リガンド結合領域、細胞外ドメイン、膜貫通ドメイン、細胞質ドメイン、N−末端ドメイン、C−末端ドメインのような領域、またはそれらの組み合わせを異種タンパクに共有結合で連結することができる。例えば、T2R膜貫通領域を、異種GPCR膜貫通ドメインに連結することができ、または異種GPCR細胞外ドメインを、T2R膜貫通領域に連結することもできる。そのほかに選択される異種タンパクには、例えば、緑色蛍光タンパク、β−gal, グルタミン酸受容体、及びロドプシンN−末端が含まれる。
【0109】
本発明のT2R、フラグメント、または変異体を発現するための宿主細胞も本発明の範囲に入る。本発明のT2R、フラグメントまたは変異体をコードするcDNAのようなクローニング遺伝子または核酸の高度な発現を得るために、当業者は、通常、目的の核酸配列を、転写を指令する強力なプロモーター、転写/翻訳ターミネーター、及び、もし核酸がタンパクをコードするならば、翻訳開始のためのリボソーム結合部位、を含む発現ベクター中に再クローニングする。適当な細菌プロモーターは当業者に既知であり、例えば、Sambrookらにより記述されている。しかし、細菌発現系または真核発現系を使用することができる。
【0110】
宿主細胞中に外来核酸配列を導入する既知方法のいずれかを使用することができる。導入方法としては、リン酸カルシウムトランスフェクション、ポリブレン、プロトプラスト融合、電気穿孔、リポソーム、マイクロインジェクション、原形質ベクター、ウイルスベクター、及びそのほかのクローン化ゲノムDNA、cDNA, 合成DNAまたはそのほかの外来遺伝物質を宿主細胞に導入する既知方法がある(例えば、Sambrook et al.参照)。使用される特定の遺伝子操作方法により、本発明のT2R、フラグメント、または変異体を発現することができる宿主細胞の中に少なくとも1核酸分子を確実に導入できることだけが必要である。
【0111】
発現ベクターを細胞中に導入した後、遺伝子導入細胞を、目的の受容体、フラグメントまたは変異体の発現に適した条件の下に培養し、次いで標準的な技術を使用してその細胞を培養から回収する。そのような技術の例は、当業者に既知である。例えば、WO 00/06593が参照され、この文献は、この開示と調和するように組み込まれる。
【0112】
hT2R4, hT2R44またはhT2R61の活性を調節する化合物を検出するアッセイ
インビトロ及びインビボにおいて、試験化合物が、本発明のT2Rポリペプチドに特異的に結合するか否かを測定する方法及び成分を以下に記述する。細胞生理の多くの局面を監視して、天然に存在するかまたはキメラのT2Rに対するリガンド結合による効果を評価することができる。これらのアッセイを、T2Rポリペプチドを発現する完全な細胞、透過性細胞、または標準的方法により作製された膜フラクションで行うことができる。
【0113】
味覚受容体は、味覚誘発化合物と結合し、化学的刺激の電気信号への変換を始める。活性化された、または阻害されたGタンパクは、次には標的酵素、チャネル、及び他のエフェクタータンパクの性質を変える。いくつかの例としては、可視系におけるトランスジューシンによるcGMPホスフォジエステラーゼの活性化、刺激性Gタンパクによるアデニル酸シクラーゼの活性化、Gq及び他の同種GタンパクによるホスフォリパーゼCの活性化、並びにGi及び他のGタンパクによる種々のチャネルの調節がある。また、ホスフォリパーゼCによるジアシルグリセロールの生成及びIP3の生成、及びその次のIP3によるカルシウム移動などの下流の事象も測定することができる。
【0114】
このアッセイのT2Rタンパクまたはポリペプチドは、典型的には、SEQ ID NO.:1,3,または5に含まれる配列を持つポリペプチド、またはそのフラグメント若しくは保存的修飾変異体から選択されるであろう。
【0115】
また、このアッセイのT2Rタンパクまたはポリペプチドは、真核宿主細胞から誘導することができ、SEQ ID NO.:1,3,または5とアミノ酸相同性を持つアミノ酸配列、またはその保存的修飾変異体を含むことができる。一般的に、アミノ酸配列相同性は、少なくとも30%、望ましくは30−40%、より具体的には50−60、70%、75%、80%、85%、90%、95%、96%、97%、98%、または99%であろう。任意に、このアッセイのT2Rタンパクまたはポリペプチドは、細胞外ドメイン、膜貫通領域、細胞質ドメイン、リガンド結合ドメインなどのようなT2Rポリペプチドの領域を含むことができる。任意に、T2Rポリペプチドまたはその部分は、異種タンパクと共有結合で連結して、本明細書で記述するアッセイに使用するキメラタンパクを創ることができる。
【0116】
T2R活性の調節物質は、上記のような、組換えまたは天然に存在するT2Rタンパクまたはポリペプチドを使用して試験することができる。T2Rタンパクまたはポリペプチドは、組換え体または天然に存在するものとして、単離し、細胞中で発現させ、細胞から得た膜で発現させ、組織または動物で発現させることができる。例えば、舌のスライス、舌から分離した細胞、形質転換細胞、または膜を使用することができる。調節物質は、本明細書で記述するインビトロまたはインビボアッセイの一つを使用して試験することができる。
【0117】
調節物質の検出
インビトロ及びインビボにおいて、試験化合物が、本発明のT2R受容体に特異的に結合するか否かを調べるための成分及び方法を以下に記述する。細胞生理の多くの面をモニターして、本発明のT2Rポリペプチドへのリガンド結合による効果を評価することができる。これらのアッセイは、化学感受性受容体を発現する完全な細胞、透過性細胞、または標準的方法により若しくは新たに合成されたタンパクをインビトロで使用して作製した膜フラクションで、実施することができる。
【0118】
インビボにおいて、味覚受容体は、味調節化合物と結合し、化学的刺激を電気信号に変換することを開始する。活性化されたまたは阻害されたGタンパクは、次には標的酵素、チャネル、及びそのほかのエフェクタータンパクの性質を変える。いくつかの例としては、可視系におけるトランスジューシンによるcGMPホスフォジエステラーゼの活性化、刺激性Gタンパクによるアデニル酸シクラーゼの活性化、Gq及び他の同種GタンパクによるホスフォリパーゼCの活性化、及びGi及び他のGタンパクによる種々のチャネルの調節がある。また、ホスフォリパーゼCによるジアシルグリセロール及びIP3の生成、並びにその次のIP3によるカルシウム移動などの下流の事象も測定することができる。
【0119】
また、このアッセイのT2Rタンパクまたはポリペプチドは、真核宿主細胞から誘導することができ、本明細書で開示したT2Rポリペプチドとアミノ酸配列相同性を持つアミノ酸配列、またはそのフラグメント若しくは保存的修飾変異体を含むことができる。一般的に、アミノ酸配列相同性は、少なくとも35−50%、または任意に75%、85%、90%、95%、96%、97%、98%、若しくは99%であろう。任意に、このアッセイのT2Rタンパクまたはポリペプチドは、細胞外ドメイン、膜貫通領域、膜貫通ドメイン、細胞質ドメイン、リガンド結合ドメインなどのようなT2Rタンパクのドメインを含むことができる。さらに、T2Rポリペプチドまたはそのドメインは、異種タンパクと共有結合で連結して、本明細書で記述するアッセイに使用するキメラタンパクを創ることができる。
【0120】
T2R受容体活性の調節物質は、組換えか天然に存在する、上記のT2Rタンパクまたはポリペプチドを使用して試験される。組換え若しくは天然に存在するT2Rタンパクまたはポリペプチドは、単離し、細胞中で発現させ、細胞から得られる膜で発現させ、組織若しくは動物中で発現させることができる。例えば、舌のスライス、舌から分離した細胞、形質転換細胞、または膜を使用することができる。調節は、本明細書で記述するインビトロまたはインビボでのアッセイの一つを使用して試験することができる。
【0121】
1.インビトロ結合アッセイ
味覚変換も、本発明のT2Rポリペプチドを使用して、可溶相若しくは固相の反応で、インビトロにおいて試験することができる。特別な態様において、T2Rリガンド結合ドメインを、可溶相若しくは固相の反応において、インビトロで使用して、リガンド結合についてアッセイすることができる。
【0122】
リガンド結合ドメインは、膜貫通ドメインの細胞外ループのような細胞外ドメインの付加的部分とともに、N−末端ドメインから形成することができる。
【0123】
インビトロ結合アッセイは、代謝型グルタミン酸受容体のようなGPCRで使用されている(例えば、Han and Hampson, J. Biol. Chem. 274: 10008-10013 (1999)参照)。これらのアッセイは、放射活性または蛍光の標識リガンドで置換し、内在性の蛍光の変化またはタンパク分解感受性の変化などを測定することなどを含む。
【0124】
本発明によるT2Rポリペプチドに対するリガンド結合は、溶液中、任意に固相に付着させた二層膜、単層脂質膜、または小胞において試験することができる。調節物質の結合は、例えば、分光学的特性(例えば、蛍光、吸光度、屈折率)、流体力学的(例えば、形状)、クロマトグラフィー、または溶解特性の変化を使用して試験することができる。
【0125】
本発明の望ましい態様においては、[35S]GTPγS結合アッセイを使用する。前述のように、GPCRの活性化により、Gタンパク複合体のGαサブユニットは、結合しているGDPのGTPへの交換を促進する。Gタンパク交換活性のリガンド仲介による促進は、候補リガンドの存在下に加えられた放射活性標識[35S]GTPγSのGタンパクへの結合を測定する生化学的アッセイにおいて測定することができる。典型的には、目的の化学感受性受容体を含む膜をGタンパクと混合する。潜在的阻害物質及び/または活性化物質並びに[35S]GTPγSを、アッセイに添加し、Gタンパクへの[35S]GTPγSの結合を測定する。結合は、液体シンチレーション計測により、またはシンチレーション近接アッセイ(SPA)を含む、当業者に既知のそのほかの方法により測定することができる。他のアッセイ形態においては、蛍光標識GTPγSを利用することができる。
【0126】
2.蛍光偏光アッセイ
他の態様において、リガンド結合を検出及び監視するために蛍光偏光(「FP」)によるアッセイを使用することができる。蛍光偏光は、結合平衡、核酸ハイブリダイゼーション、及び酵素活性を測定するために広い用途で適用される技術である。蛍光偏光アッセイは、均質系であり、遠心分離、濾過、クロマトグラフィー、沈殿、または電気泳動などの分離操作を必要としない。このアッセイは、リアルタイムで、溶液中で直接行われ、固相を必要としない。偏光の測定は、迅速であり、検体を破壊しないので、偏光値は、繰り返し且つ試薬の添加後に測定することができる。一般的に、この技術は、ピコモルからマイクロモルの低レベルの蛍光の偏光値を測定するために使用することができる。本発明のT2Rポリペプチドに対するリガンドの結合を測定するための簡単にして定量的な方法において、どのように蛍光偏光を使用しうるかを、このセクションでは記述する。
【0127】
蛍光標識された分子を面偏光で励起すると、その分子の回転に反比例する程度に偏光した光を放射する。蛍光標識された大きい分子ほど、励起状態(フルオレッセインの場合に4ナノ秒)の間、静止しており、光の偏光は、相対的に励起と放射の間一定に維持される。蛍光標識された小分子は、励起状態の間、急速に回転し、偏光は、励起と放射との間で顕著に変化する。したがって、小分子は低い偏光値を示し、大分子は高い偏光値を示す。例えば、一本鎖蛍光標識オリゴヌクレオチドは、低い蛍光値を有するが、相補的鎖とハイブリッド形成すると、高い偏光値を持つ。本発明の化学感受性受容体を活性化または阻害する味覚誘発化合物の結合を検出及び監視するためにFPを使用する場合には、蛍光標識味覚誘発化合物または蛍光自発味覚誘発化合物を使用しうる。
【0128】
蛍光偏光(P)は次のように定義される:
[式中、IntΠは、励起光面に平行な放射光の強度、Int⊥は、励起光面に垂直な放射光の強度である。Pは、光の強度の比であり、無次元の数である。]。
例えば、BeaconTM及びBeacon 2000TMシステムをこのアッセイに関して使用することができる。そのシステムでは典型的に偏光をミリ偏光単位(1偏光単位=1000 mP単位)で示している。
【0129】
分子の回転とサイズの関係はPerrin式により記述されており、読者はこの式の詳細な説明を載せているJolley, M.E. (1991) Journal of Analytical Toxicology, pp.236-240,を参照されたい。要するに、Perrin式が示すところは、偏光が、直接、回転緩和時間に比例するということであり、その回転緩和時間とは、分子が約68.5度の角度を回転するのに要する時間のことである。回転緩和時間は粘度(η)、絶対温度(T)、分子容積(V)、及び気体定数(R)に関係し、次の式で示される:
【0130】
回転緩和時間は、小分子(例えば、フルオレッセイン)に対しては小さく(約1ナノ秒)、大分子(例えば、免疫グロブリン)に対しては大きい(約100ナノ秒)。粘度と温度を一定に保つ場合には、回転緩和時間、及びそれ故偏光は、分子容積に直接的に相関する。分子容積の変化は、蛍光標識分子の他の分子との相互作用、解離、重合、分解、ハイブリッド形成、または立体配座変化に起因するであろう。例えば、蛍光偏光は、プロテアーゼ、DNアーゼ、及びRNアーゼによる蛍光標識した大きな重合体の酵素分解を測定するために使用されてきた。また、タンパク/タンパク相互作用、抗体/抗原結合、及びタンパク/DNA結合の結合平衡を測定するためにも使用されてきた。
【0131】
A. 固相及び液相ハイスループットアッセイ
さらに他の態様において、本発明は、T2Rポリペプチド;またはT2Rポリペプチドを発現する細胞または組織を使用する可溶相アッセイを提供する。他の態様において、本発明は、固相を使用するハイスループット方式のインビトロアッセイを提供する。その方式において、T2RポリペプチドまたはT2Rポリペプチドを発現する細胞若しくは組織は、固相基質に付着され、味覚刺激化合物をT2R受容体と接触させ、結合を、適当なタグまたはT2Rに対して作製した抗体を使用して検出する。
【0132】
本発明のハイスループットアッセイでは、1日に異なる調節物質またはリガンドの数千個をスクリーニングすることが可能である。特に、マイクロタイタープレートの各ウエルを、選択した潜在的調節物質に対する個別のアッセイを行うために使用することができ、あるいは、濃度またはインキュベーション時間の効果を調べようとする場合には、5−10ウエル毎に1つの調節物質を試験することができる。このように、一枚の標準的マイクロタイタープレートは、約100(例えば、96)調節物質をアッセイすることができる。もし1536ウエルプレートを使用するならば、一枚のプレートで約1000から約1500の異なる化合物を容易にアッセイすることができる。各プレートウエル中において複数の化合物をアッセイすることも可能である。1日に数枚の異なるプレートをアッセイすることが可能である;本発明の集積システムを使用して約6,000−20,000の異なる化合物のスクリーニングアッセイが可能である。さらに最近、試薬操作のための微量流体法が開発された。
【0133】
目的の分子は、直接的にまたは間接的に、共有結合によりまたは非共有結合により、例えば、タグを介して固相成分に結合することができる。タグは種々の構成物とすることができる。一般的に、タグと結合する分子(タグバインダー)を固相支持層に固定し、タグをつけた目的分子(例えば、目的の味覚伝達分子)をタグ及びタグバインダーの相互作用により固相に接着する。
【0134】
文献に詳細に記述されている既知分子相互作用に基づいて、多数のタグ及びタグバインダーを使用することができる。例えば、タグが天然のバインダー、例えば、ビオチン、プロテインA、またはプロテインGを持つ場合には、適当なタグバインダー(アビジン、ストレプトアビジン、ニュートラビジン、免疫グロブリンのFc領域、など)と組み合わせて使用することができる。また、ビオチンのような天然バインダーを有する分子に対する抗体も、広く使用することができ、適当なタグバインダーである(SIGMA Immunochemicals 1998 catalogue SIGMA, St. Louis MO,参照)。
【0135】
同様に、ハプテン性または抗原性化合物は、適当な抗体と組み合わせて使用して、タグ/タグバインダー対を形成することができる。数千の特異的抗体は、市販品を入手可能であり、また多数のそのほかの抗体は文献に記述されている。例えば、よくある形態として、タグは第一抗体であり、タグバインダーは第一抗体を認識する第二抗体である。抗体−抗原相互作用に加えて、受容体−リガンド相互作用も、タグ及びタグバインダー対として適当である。例えば、細胞膜受容体のアゴニスト及びアンタゴニスト(例えば、トランスフェリン、c-kit, ウイルス受容体リガンド、サイトカイン受容体、ケモカイン受容体、インターロイキン受容体、免疫グロブリン受容体及び抗体、カドヘリンファミリー、インテグリンファミリー、セレクチンファミリーなどの細胞受容体−リガンド相互作用;例えば、Pigott & Power, The Adhesion Molecule Facts Book 1(1993)参照)。同様に、トキシン及び毒素、ウイルス抗原決定基、ホルモン(例えば、オピエート、ステロイド、など)、細胞内受容体(例えば、ステロイド、甲状腺ホルモン、レチノイド及びビタミンDを含む種々の小さなリガンドの効果を仲介するもの;ペプチド)、薬物、レクチン、糖、核酸(線状及び環状ポリマー型の両者)、オリゴ糖、タンパク、リン脂質及び抗体は全て、種々の細胞受容体と相互作用しうる。
【0136】
ポリウレタン、ポリエステル、ポリカーボネート、ポリウレア、ポリアミド、ポリエチレンイミン、ポリアリレンスルフィド、ポリシロキサン、ポリイミド、及びポリアセテートのような合成ポリマーも適当なタグまたはタグバインダーを形成することができる。この開示を通覧することにより当業者には明白であるように、そのほかの多数のタグ/タグバインダー対も本明細書で開示したアッセイシステムに有用である。
【0137】
ペプチド、ポリエーテルなどの一般的リンカーは、タグとして寄与することができ、約5から200アミノ酸の間のポリgly配列のようなポリペプチド配列を含む。そのようなフレキシブルリンカーは、当業者には既知である。例えば、ポリ(エチレングリコール)リンカーは、Shearwater Polymers, Inc., Huntsville, Alabamaから入手できる。これらのリンカーは、任意にアミド結合、スルフヒドリル結合、または異種機能の結合を有している。
【0138】
タグバインダーは、現在使用しうる種々の方法を使用して固相基質に固定される。固相基質は、通常、その基質の全てまたは部分を、タグバインダーの一部と反応する化学基を表面に固定する化学試薬に暴露することにより、誘導されまたは機能化される。例えば、長鎖部分を付着するのに適した基としては、アミン、ヒドロキシル、チオール、及びカルボキシル基がある。アミノアルキルシラン及びヒドロキシアルキルシランは、ガラス表面のような、様々な表面へと機能化するために使用することができる。そのような固相バイオポリマーを構築する方法は文献に詳細に記述されている。例えば、Merrifield, J. Am. Chem. Soc., 85: 2149-2154 (1963)(例えば、ペプチドの固相合成を記述している);Geysen et al., J. Immun. Meth., 102: 259-274 (1987)(ピン上における固相成分の合成を記述している);Frank & Doring, Tetrahedron 44: 6031-6040 (1988)(セルロース円盤上における種々のペプチド配列の合成を記述している);Fodor et al., Science, 251: 767-777 (1991); Sheldon et al., Clinical Chemistry, 39(4): 718-719(1993); 及びKozal et al., Nature Medicine, 2(7): 753-759 (1996)(全て固相に固定したバイオポリマーのアレイを記述している)参照。
基質にタグバインダーを固定する非化学的方法には、例えば、熱、UV照射による架橋結合のような他の一般的方法がある。
【0139】
3.細胞によるアッセイ
望ましい一の態様において、T2Rタンパクは、非修飾の形態において、或いは異種の、分泌経路による成熟化及び標的化を促進するシャペロン配列を持つかまたは望ましくは持たないキメラ、変異体または短縮された受容体として、真核細胞中で発現する。そのT2Rポリペプチドは、HEK-293細胞のような真核細胞で発現することができる。望ましくは、細胞は、細胞内シグナリング経路またはホスフォリパーゼCのようなシグナリングタンパクに、キメラ受容体を連結させることができる、機能的Gタンパク、例えばGα15を含む。その細胞内におけるT2R受容体の活性化は、例えば、細胞内のFURA-2による蛍光を検出することによる細胞内カルシウムの変化を検出する、標準的な方法を使用して検出することができる。そのようなアッセイが、本出願に示した実験的知見の基礎になっている。
【0140】
活性化したGPCR受容体は、受容体のC−末端(及び多分他の部位も)をリン酸化するキナーゼの基質になることがある。したがって、活性化物質は、シンチレーションカウンターでアッセイすることができる、放射標識ATPの32Pの受容体への移転を促進するであろう。C−末端のリン酸化は、アレスチン様タンパクの結合を促進し、Gタンパクの結合を妨害する。GPCR信号変換及び信号変換をアッセイする方法の一般的総説については、例えば、Methods in Enzymology, vols. 237 and 238 (1994) and volume 96 (1983); Bourne et al., Nature, 10: 349: 117-27 (1991); Bourne et al., Nature, 348: 125-32 (1990); Pitcher et al., Annu. Rev. Biochem., 67: 653-92 (1998)参照。
【0141】
T2R調節は、候補T2R調節物質で処理したT2Rポリペプチドの反応を、非処理検体または既知「陽性」対照を含む検体と比較することによりアッセイすることができる。その候補T2R調節物質は、T2Rポリペプチド活性を阻害または活性化する分子を含むことができる。一の態様において、T2Rを活性化する化合物で処理した対照検体を、相対T2R活性値100と定める。対照検体に対する相対的T2R活性値が約90%、さらに50%、さらに25−0%であるときに、T2Rポリペプチドの阻害は達成される。対照に対する相対的T2R活性値が110%、さらに150%、200−500%、または1000−2000%であるときに、T2Rポリペプチドの活性化は達成される。
【0142】
イオン流の変化は、T2Rポリペプチドを発現している細胞または膜のイオン分極(例えば、電気ポテンシャル)の変化を測定することにより検出することができる。細胞分極の変化を測定する手段の一つは、ボルテージ−クランプ及びパッチ−クランプ技術を使用して電流の変化を測定(したがって分極の変化を測定)することによるものである(例えば、「セル−アタッチド」モード、「インサイド−アウト」モード、及び「ホールセル」モード、例えば、Ackerman et al., New Engl. J Med., 336: 1575-1595 (1997)参照)。ホールセル電流は簡便に測定され、標準的に使用されている。そのほかの既知アッセイとしては、放射標識イオン流アッセイ及び電圧感受性色素を使用する蛍光アッセイが含まれる(例えば、Vestergarrd-Bogind et al., J. Membrane Biol., 88: 67-75 (1988); Gonzales & Tsien, Chem. Biol., 4: 269-277 (1997); Daniel et al., J. Pharmacol. Meth., 25: 185-193 (1991); Holevinsky et al., J. Membrane Biology, 137: 59-70 (1994)参照)。
【0143】
ポリペプチドの機能に及ぼす試験化合物の効果は、前記パラメーターのいずれかを調べることにより測定することができる。GPCR活性に影響する適当ないずれの生理学的変化も、本発明のポリペプチドに対する試験化合物の影響を評価するために使用することができる。機能的事象を完全細胞または動物を使用して測定する場合には、伝達物質放出、ホルモン放出、既知または未知遺伝子マーカーへの転写の変化(例えば、ノーザンブロット)、細胞増殖またはpHの変化のような細胞代謝の変化、及びCa2+,IP3,cGMP,または cAMPのような細胞内セカンドメッセンジャーの変化などの種々の効果を測定することができる。
【0144】
GPCRの望ましいアッセイには、受容体活性を報告させるためのイオンまたは電圧感受性色素を負荷された細胞が含まれる。その受容体の活性を測定するアッセイには、試験化合物の活性を評価するための対照として、他のGタンパク共役受容体の既知アゴニスト及びアンタゴニストも使用することができる。調節化合物(例えば、アゴニスト、アンタゴニスト)を同定するためのアッセイにおいて、細胞質中のイオンまたは膜電位のレベルの変化は、イオン感受性または膜電位蛍光指示薬をそれぞれ使用して監視されるであろう。使用することができるイオン感受性指示薬及び電圧プローブは、the Molecular Probes 1997 Catalog中に記述されている。G−タンパク共役受容体に対して、Gα15及びGα16のような相手を選ばないGタンパクを、選択アッセイにおいて使用することができる(Wilkie et al., Proc. Nat'l Acad. Sci., 88: 10049-10053 (1991))。
【0145】
受容体の活性化は、続く細胞内事象、例えば、セカンドメッセンジャーの増加、を開始する。一部のGタンパク共役受容体の活性化は、ホスフォリパーゼC仲介ホスファチジルイノシトールの加水分解によるイノシトール三リン酸(IP3)の生成を促進する(Berridge & Irvine, Nature, 312: 315-21 (1984))。IP3は、次に細胞内貯蔵カルシウムイオンの放出を促進する。このように、細胞質カルシウムイオンレベルの変化、またはIP3のようなセカンドメッセンジャーの変化をGタンパク共役受容体の機能を評価するために使用することができる。そのGタンパク共役受容体を発現する細胞は、細胞内カルシウム貯蔵からの放出及び細胞外カルシウムの形質膜イオンチャネルを経由する流入の両者による結果として細胞質カルシウムレベルの増加を示すであろう。
【0146】
望ましい態様において、T2Rポリペプチド活性は、受容体をホスフォリパーゼC信号伝達経路に連結する相手を選ばないGタンパクを持つ異種細胞内でT2R遺伝子を発現することにより測定される(Offermanns & Simon, J. Biol. Chem., 270: 15175-15180 (1995)参照)。任意に、細胞系はHEK-293(これは通常はT2R遺伝子を発現しない)であり、相手を選ばないGタンパクはGα15である(Offermanns & Simon,前出)。味覚伝達の調節は、T2Rポリペプチドに会合する分子の投与によるT2R信号伝達経路の変化に応答して変化する細胞内Ca2+レベルの変化を測定することによりアッセイされる。Ca2+レベルの変化は、任意に蛍光Ca2+指示色素及び蛍光画像を使用して測定される。
【0147】
他の態様において、本明細書に引用して取り入れた米国特許5,436,128にしたがってホスファチジルイノシトール(PI)加水分解を分析することができる。簡略に記述すると、このアッセイは、3H−ミオイノシトールで48時間またはそれ以上細胞を標識することを必要とする。標識細胞を、試験化合物で1時間処理する。処理した細胞を分解し、クロロホルム−メタノール−水で抽出し、その後イノシトールリン酸をイオン交換クロマトグラフィーにより分離し、シンチレーションカウンターで定量する。緩衝液対照の存在下におけるcpmに対するアゴニスト存在下におけるcpmの比を計算することにより活性化率が算出される。同様に、緩衝液対照(アゴニストを含んでも含まなくてもよい)存在下におけるcpmに対するアンタゴニスト存在下におけるcpmの比を計算することにより阻害率が算出される。
【0148】
他の受容体アッセイとしては、細胞内サイクリックヌクレオチド、例えば、cAMPまたは cGMPのレベルを測定するものがある。受容体の活性化によりサイクリックヌクレオチドのレベルが低下する場合には、アッセイ中の細胞に受容体活性化化合物を加える前に、細胞内サイクリックヌクレオチドのレベルを上昇させる試薬、例えば、ホルスコリン、に細胞を暴露することが望ましい。一の態様においては、免疫アッセイを使用して細胞内cAMPまたは cGMPの変化を測定することができる。Offermanns & Simon, J. Bio. Chem., 270: 15175-15180 (1995)に記述されている方法をcAMPのレベルの測定に使用することができる。また、Felley-Bosco et al., Am. J. Resp. Cell and Mol. Biol., 11: 159-164 (1994)に記述されている方法も cGMPのレベルの測定に使用することができる。さらに、cAMP及び/または cGMPを測定するアッセイキットが、本明細書に引用して取り入れた、米国特許4,115,538に記述されている。
【0149】
他の態様においては、信号変換に対する試験化合物の効果を評価するために転写レベルを測定することができる。本発明のT2Rポリペプチドを含む宿主細胞を試験化合物と相互作用をさせるために十分な時間接触させ、次いで遺伝子発現のレベルを測定する。その相互作用を行うのに十分な時間は、例えば、時間経過を追い、時間の関数として転写のレベルを測定することにより、経験的に決定される。転写量は、当業者に既知の適当な方法を使用して測定することができる。例えば、本発明のタンパクのmRNA発現は、ノーザンブロットを使用して検出することができるし、あるいはそのペプチド産物を、免疫アッセイを使用して同定することができる。また、レポーター遺伝子を使用する転写によるアッセイが、本明細書に引用して取り入れた、米国特許5,436,128に記述されているように、使用することができる。レポーター遺伝子は、例えば、クロラムフェニコールアセチル転移酵素、ルシフェラーゼ、βガラクトシダーゼ、βラクタマーゼ及びアルカリホスファターゼとすることができる。さらに、目的のタンパクを、第二のレポーター、例えば、緑色蛍光タンパクに付属させることにより間接的なレポーターとして使用することができる(例えば、Mistili & Spector, Nature Biotechnology, 15: 961-964 (1997)参照)。
【0150】
次いで転写の量を、試験化合物が存在しない同じ細胞中における転写の量と比較するか、または本発明のT2Rポリペプチドを持たない実質的に同じ細胞中における転写の量と比較する。実質的に同じ細胞は、組換え細胞が誘導された細胞と同じで、異種DNAの導入による修飾がされていない細胞から誘導することができる。転写の量に差があれば、目的のT2Rポリペプチドの活性をある程度変えたことを示している。
【0151】
4.化学感受性受容体を発現する非ヒト形質転換動物
一つまたはそれ以上の本発明の味覚受容体配列を発現する非ヒト動物も、また受容体アッセイに使用することができる。そのような発現を利用して、安定的にまたは一過性に化学感受性受容体またはそのリガンド結合領域をコードする核酸を導入した非ヒト動物を、試験化合物と接触させ、受容体ポリペプチド複合体に特異的に結合することで、試験化合物に動物が反応するか否かを測定することにより、試験化合物が哺乳動物膜貫通味覚受容体複合体に特異的に結合するか否かを測定することができる。
【0152】
本発明のベクターを導入またはそれに感染した動物は、特定のまたは一連の受容体に結び付く味覚刺激を同定及び分析するためのアッセイに特に有用である。そのようなヒト味覚受容体配列を発現するベクター感染動物は、味覚刺激、及び、例えば、細胞生理(例えば、味覚神経に対する)、中枢神経、または行動に関する味覚刺激による影響を、インビボスクリーニングするために、使用することができる。
【0153】
個別にまたはライブラリーとして、核酸及びベクターを感染/発現する方法は当業者によく知られている。個別の細胞、器官、または動物全体の種々のパラメーターは、種々の手段で測定することができる。本発明のT2R配列は、感染試薬、例えば、アデノウイルス発現ベクター、で輸送することにより動物味覚組織で発現させることができる。
【0154】
内在性味覚受容体遺伝子は、機能を保持することができ、野生(天然)型活性をそのまま存在させることができる。他の状況において、全ての味覚受容体活性が、導入した外来性ハイブリッド受容体によることを望む場合には、ノックアウト系の使用が望ましい。非ヒト形質転換動物、特に形質転換マウス、の作製方法、及び形質転換細胞を作製するための組換え構築体の選択及び調製は当業者に良く知られている。
【0155】
「ノックアウト」細胞及び動物の作製は、抑制される遺伝子のDNA配列の一部を中断するように新規なDNA配列をゲノムの中に導入することにより哺乳動物細胞における特別な遺伝子の発現レベルを低下させるかまたは完全に停止させることができるという前提に基づいている。「遺伝子トラップ挿入」も、宿主遺伝子を破壊するために使用することができ、マウス胚幹(ES)細胞を、ノックアウト形質転換動物を作製するために使用することができる(例えば、Holzschu, Transgenic Res 6: 97-106 (1997)参照)。外来種の挿入は、典型的には相補的核酸配列の間の相同組換えによる。外来配列は、例えば、エキソン性、イントロン性若しくは転写調節配列などの修飾対象たる標的遺伝子の一部、または標的遺伝子の発現レベルに影響することができるゲノム配列;或いはそれらの組み合わせである。多能性胚性幹細胞における相同組換えによる遺伝子ターゲティングは、目的ゲノム配列を正確に修飾することを可能とする。いずれの技術もノックアウト動物を創製し、スクリーニングし、繁殖するために使用することができる、例えば、Bijvoet, Hum. Mol. Genet. 7: 53-62 (1998); Moreadith,J. Mol. Med. 75: 208-216 (1997); Tojo, Cytotechnology 19: 161-165 (1995); Mudgett, Methods Mol. Biol. 48: 167-184 (1995); Longo, Transgenic Res. 6: 321-328 (1997); 米国特許番号5,616,491; 5,464,764;5,631,153;5,487,992;5,627,059;5,272,071;WO 91/09955;WO 93/09222;WO 96/29411;WO 95/31560; WO 91/12650 参照。
【0156】
本発明の核酸は、ノックアウトヒト細胞及びその子孫を作製するための試薬として使用することもできる。同様に、本発明の核酸はマウスの「ノックイン」を作製する試薬としても使用することができる。ヒトまたはラットのT2R遺伝子配列は、マウスゲノム中のオーソログT2Rを置換することができる。このようにして、ヒトまたはラットのT2Rを発現するマウスが作製される。このマウスはヒトまたはラットのT2Rの機能を分析するために、そしてそのT2Rに対するリガンドを同定するために使用することができる。
【0157】
調節物質
T2Rファミリーメンバーの調節物質として試験される化合物は、小さな化合物、またはタンパク、糖、核酸若しくは脂質のような生化学物質のいずれでも良い。また、調節物質は、T2Rファミリーメンバーの遺伝的改変体でも良い。典型的には、試験化合物は小さな化学分子及びペプチドである。通常は、水または有機溶媒(特にDMSOを含む)に溶解しうる化合物が使用されるが、本質的に如何なる化合物も、本発明の潜在的調節物質またはリガンドとして使用することができる。アッセイは、アッセイ段階を自動化し、アッセイのために簡便な資源から化合物を供給することにより、大規模な化合物ライブラリーをスクリーニングするように設計することができ、典型的には、並行して稼動する(例えば、マイクロタイタープレート上で自動的にアッセイするマイクロタイター方式)。Sigma(St. Louis, MO), Aldrich (St. Louis, MO), Sigma-Aldrich (St. Louis, MO), Fluka Chemika-Biochemica Analytika (Buchs, Switzerland)などを含めて多数の化合物供給業者が存在することは認識されているであろう。
【0158】
一の態様において、ハイスループットスクリーニング法は、膨大な数の潜在的治療薬(潜在的調節物質またはリガンド化合物)を含むコンビナトリアル化学またはペプチドライブラリーの供給を伴う。次いで、その「コンビナトリアル化学ライブラリー」または「リガンドライブラリー」は、本明細書で記述する一つまたはそれ以上のアッセイによりスクリーニングされて、所望の特徴的活性を示すライブラリー構成員(特に化学種または亜種)を同定する。こうして同定された化合物は、よく言われる「リード化合物」とすることができ、あるいはそれ自身を潜在的なまたは実際の商品として使用することができる。
【0159】
コンビナトリアル化学ライブラリーは、化学的合成または生化学的合成のいずれかにより多数の化学的「構築ブロック」、例えば試薬、を組み合わせることにより、作製される膨大な化合物の集合である。例えば、ポリペプチドライブラリーのような線状コンビナトリアル化学ライブラリーは、与えられた化合物の長さ(すなわち、ポリペプチド化合物中のアミノ酸の数)に対して可能な方法で化学的構築ブロック(アミノ酸)のセットを組み合わせることにより形成される。化学的構築ブロックのこのようなコンビナトリアル混合により数百万の化合物を合成することができる。
【0160】
コンビナトリアル化学ライブラリーの調製及びスクリーニングは、当業者には良く知られている。そのようなコンビナトリアル化学ライブラリーには、限定はしないが、ペプチドライブラリーが含まれる(例えば、米国特許5,010,175、Furka, Int. J. Pept. Prot. Res., 37: 487-93 (1991) 及びHoughton et al., Nature, 354: 84-88 (1991) 参照)。化学的に多様なライブラリーを作製するための他の化学物質も使用しうる。そのような化学物質としては、限定はしないが:ペプトイド(例えば、WO 91/19735)、コードされたペプチド(例えば、WO 93/20242)、ランダムバイオオリゴマー(例えば、WO 92/00091)、ベンゾジアゼピン類(例えば、米国特許5,288,514)、ヒダントイン類、ベンゾジアゼピン類及びジペプチドのようなダイバーソマー(Hobbs et al., PNAS., 90: 6909-13 (1993))、ビニローグポリペプチド(Hagihara et al., J. Amer. Chem. Soc., 114: 6568 (1992))、グルコーススカホールドを持つ非ペプチド性ペプチド類似体(Hirschmann et al., J. Amer. Chem. Soc., 114: 9217-18 (1992))、小型化合物ライブラリーの類似体有機合成(Chen et al., J. Amer. Chem. Soc., 116: 2661 (1994))、オリゴカルバメート(Cho et al., Science, 261: 1303 (1993))、ペプチジルホスフォネート(Campbell et al., J. Org. Chem., 59: 658 (1994))、核酸ライブラリー(Ausubel, Berger, and Sambrook,全て前出)、ペプチド核酸ライブラリー(米国特許5,539,083)、抗体ライブラリー(Vaughn et al., Nature Biotechnology, 14(3): 309-14 (1996)及びPCT/US96/10287)、糖ライブラリー(Liang et al., Science, 274: 1520-22 (1996)及び米国特許5,593,853)、小型有機分子ライブラリー(ベンゾジアゼピン、Baum, C&EN、Jan 18, page 33 (1993);チアゾリジノン及びメタチアザノン、米国特許5,549,974;ピロリジン、米国特許5,525,735及び5,519,134;モルホリノ化合物、米国特許5,506,337;ベンゾジアゼピン、5,288,514、など)がある。
【0161】
コンビナトリアルライブラリーを調製する装置は、市販されている(例えば、357 MPS, 390 MPS(Advanced Chem Tech, Louisville KY)、Symphony(Rainin, Woburn, MA)、433A(Applied Biosystems, Foster City, CA)、9050 Plus(Millipore, Bedford, MA)参照)。さらに、多数のコンビナトリアルライブラリー自体が、市販されている(例えば、ComGenex, Princeton, NJ; Tripos, Inc., St. Louis, MO; 3D Pharmaceuticals, Exton, PA; Martek Biosciences; Columbia, MD;など参照)。
【0162】
本発明の一の態様において、T2R調節物質を、製品、組成物または成分の味を望ましい方法で調節するために、食品、菓子類、医薬組成物またはそれらの原料中に使用することができる。例えば、苦味の感覚を増強するT2R調節物質は、製品または組成物に苦味を与えるために加えることができるし、他方、苦味感覚を阻止するT2R調節物質は、製品または組成物の味を改良するために加えることができる。
【0163】
本発明により同定された化合物の使用
味覚試験においてhT2R4, hT2R44またはhT2R61の活性化により誘発される苦味を調節する、望ましくは阻止するために、本発明により同定された化合物を食品、飲料または医薬組成物に加えることができる。具体的には、キニーネまたは関連化合物によるhT2R4の活性化を阻害する化合物は、苦味を誘発する食品、飲料または医薬品への添加物として使用することができる。例えば、これらの化合物を、苦味を阻止する有効量で、キニーネまたは関連化合物を含む医薬製剤に加えるとよい。
【0164】
hT2R44またはhT2R61の6−ニトロサッカリンまたはサッカリンによる活性化を阻害する化合物を、hT2R61または hT2R44を活性化するサッカリンまたはそのほかの苦味化合物の苦味を阻止するために、食品、飲料または医薬品に加えることができる。特に、開示したアッセイを使用して同定された化合物を、サッカリンを含むダイエット飲料に入れて、サッカリンによる苦味を阻止することができることが予測される。
【0165】
最後に、デナトニウムによるhT2R44の活性化を阻害する化合物は、hT2R44受容体の活性化による苦味を阻止するための、食品、飲料または医薬品における添加物として有用である。デナトニウムは、非常に強力な苦味化合物であるので、hT2R44受容体を阻害する化合物は、種々の苦味化合物の苦味を有効に阻止するはずである。このことは、いくつかのデナトニウム誘導体によりこの受容体が活性化されるという事実により裏付けられている。
【0166】
キット
T2R遺伝子及びその同属体は、味覚受容体細胞の同定のために、法医学または父子確認のために、及び味覚伝導の試験にとって、有用な道具である。T2R核酸に特異的にハイブリッド形成するT2Rファミリーメンバー特異的試薬、例えばT2Rプローブ及びプライマー、並びにT2Rタンパクに特異的に結合するT2R特異的試薬、例えば、T2R抗体は、味覚細胞発現及び味覚伝導調節を試験するために使用される。
【0167】
T2RファミリーメンバーのDNA及びRNAの存在を調べる核酸アッセイには、当業者に既知の多数の技術が含まれ、これには、例えば、サザン分析、ノーザン分析、ドットブロット、RNアーゼプロテクション、S1分析、PCRのような増幅技術、及びin situハイブリダイゼーションがある。例えば、in situハイブリダイゼーションにあっては、後の解析及び分析のために細胞形態は保存しながら、標的核酸を、細胞内でハイブリッド形成できるように細胞環境から遊離する。以下の論文はin situハイブリダイゼーションを総説している:Singer et al., Biotechniques, 4: 230250(1986); Haase et al., Methods in Virology, vol. VII, 189-226(1984); 及びNames et al., eds., Nucleic Acid Hybridization: A Practical Approach (1987)。さらに、T2Rタンパクは、上記の種々の免疫アッセイ技術により検出することができる。試験検体は、典型的に陽性対照(例えば、組換えT2Rタンパクを発現する検体)および陰性対照と比較される。
【0168】
本発明は、T2Rファミリーメンバーの調節物質をスクリーニングするためのキットも提供する。そのキットは容易に入手できる材料及び試薬から作製することができる。例えば、そのキットは、T2R核酸またはタンパク、試験管、及びT2R活性を試験するための説明書の一つまたはそれ以上を含むことができる。任意に、キットは、機能的T2Rポリペプチドを含む。キットのユーザーの意向及びユーザーの特別なニーズに応じて、キット及び内容の幅広い種類を本発明にしたがって作製することができる。
【0169】
本発明を一般的に記述してきたが、以下の実施例を参照することにより、より容易に理解されるであろう。しかし、以下の実施例は、説明のために提供したものであり、限定を意図するものではない。本発明の精神及び範囲から離れることなく、本明細書で開示した代表例に種々の修飾及び変更を加えることができることが理解される。
【実施例】
【0170】
(実施例1)
この実施例において、キニーネ(苦味アルカロイド)が、本出願のSEQ ID NO:2に含まれるDNA配列を有するヒト苦味受容体、hT2R4を、特異的に活性化することを示す。
【0171】
hT2R4のキニーネによる活性化は、細胞内カルシウム濃度の変化を検出する細胞を使用するアッセイにおいて測定される。簡略に記述すると、大型T細胞抗原及び相手を選ばないGタンパクG15 を安定的に発現するヒト胚性腎細胞(HEK-G15)に、リン酸化カルシウムまたは脂質を使用する系により、hT2R4プラスミドを一過性に導入する。一過性に遺伝子導入した細胞を、24ウエル培養プレートに接種し、48時間機能的発現を行う。次いで、細胞を、カルシウムに特異的な蛍光色素(Fluo-4またはFura-2;Molecular Probes)とともにインキュベートし、これにより、細胞内のカルシウム濃度変化を検出するための迅速、簡便且つ信頼性のある蛍光法が提供される。キニーネを細胞に加えて、PLCの活性化及びそれに続く細胞内カルシウム濃度の増加を引き起こす、信号伝達カスケードを誘発する。このカルシウム濃度の増加は、細胞内のカルシウム色素の蛍光を変化させる。この変化を、蛍光顕微鏡及び特別に設計したソフトウエアー(Imaging Workbench, Axon)を使用して観察する。この方法により、hT2R4を発現する細胞をキニーネが特異的に活性化する(細胞内カルシウム濃度を増加する)ことが観察された。他のhT2Rを一過性に導入した親株(HEK-G15)またはHEK-G15は、キニーネに対して相当する応答を示さなかった(図1参照)。
【0172】
(実施例2)
潜在的hT2R4アンタゴニストを発見するためのハイスループットスクリーニングアッセイにも、細胞表面に高いレベルのhT2R4を安定的に発現するHEK-G15株(HEK-G15-hT2R4)を使用する。細胞表面に一過性遺伝子導入細胞よりも2-3倍多い受容体を発現する安定HEK-G15-hT2R4株を作製した(図2参照)。この場合は、アッセイの18−24時間前に細胞を96ウエルまたは384ウエル培養プレートに接種する。次いで細胞をカルシウム感受性蛍光色素(Molecular Devices)とインキュベートし、刺激し、標準的蛍光強度プレートリーダー(FLIPRまたはVIPR)で測定する。この方法を使用して、hT2R4に対するキニーネの効果をさらに特徴付けることができた。我々は、キニーネが典型的な用量反応相関でhT2R4を活性化する事を発見した(図3参照)。
【0173】
(実施例3)
この実験において、T2R受容体の活性化を監視するためにGTPγS結合アッセイを使用した(図4)。このアッセイは、GPCRの活性を監視する古典的アッセイである(Wessling-Resnick and Jonson, 1987)。これは、適当なリガンドの存在下にGPCRがGタンパクを活性化することができるという事実に基づいている。生成した活性化Gタンパクは、結合しているGDPをGTPに変換する。GTPの加水分解されない類似体、GTPγSを使用すれば、ヌクレオチドは、Gタンパクに堅く結合したままとなり、非結合ヌクレオチドから分離することができる。放射活性標識[γ-35S]GTPγSを使用することにより、結合したGTPγSの量を測定することが可能となる。結合したGTPγSの量は、活性化Gタンパクの量を反映し、それは翻って受容体の活性化を反映する。GTPγS結合アッセイにおいてGタンパクとしてトランスジューシンを使用した。トランスジューシンは、視覚系において発現し、ガストジューシンのオーソログである。トランスジューシンは、ウシ網膜から精製した(Stryer, Methods Enzymol 96: 617-627 (1983))。
【0174】
アッセイ反応は、適当なhT2Rを発現するSf9細胞膜由来の15−20μgの総タンパク、30?l緩衝液B(30 mM Tris-HCl, pH 8, 100 mM NaCl, 3 mM MgCl2, 1 mM DTT)中0.2μMトランスジューシンを含む。一部の反応は、0.05−10 mMの試験苦味化合物も含む。0.3μM [γ-35S]GTPγS (50 Ci/mmol)を加えて反応を開始し、1時間30℃で反応を行い、緩衝液B中の0.5 mM GTP液 170μlを加えて停止し、次いでニトロセルロースフィルターを通して濾過した。結合した[35S]GTPγSを液体シンチレーションカウンターで計測した。
【0175】
バキュロウイルス発現系を使用して、Sf9細胞で、全24個の既知hT2R及びmT2R5を発現した。これらのT2R構築体は、コード配列に加えたロドプシンN−末端40アミノ酸からなるN−末端タグを持っていた。Sf9細胞を、細胞106個当り増幅ウイルスストック10−20μlの比率で、適当なバキュロウイルスに感染させた。細胞を回収し、感染2日後に分析に供した。試験したT2Rの全ての発現は、抗−ロドプシンタグ抗体(B6-30)を使用して免疫ブロットにより確認した。膜の標品は、本質的に記述にしたがって、Sf9膜を5 M尿素で処理して作製した(Ryba, J. Biol Chem 270(12): 6757-6767 (1995))。次いで膜を緩衝液(20 mM Tris-HCl, pH 8, 0.5 mM EDTA, 1 mM DTT)で洗い、約2.5−3 mgの総タンパク/mlの最終濃度で再懸濁した。
【0176】
トランスジューシンを使用するGTPγS結合アッセイにおいて、10個の苦味化合物のセットを用いて全てのhT2Rを試験した。二個のhT2R、hT2R44及びhT2R61が、加えた苦味化合物の存在下で活性を増加させた。
【0177】
特に、hT2R61活性は、6−ニトロサッカリンの存在下に増加することが認められた。試験した全てのhT2Rの中では、他の受容体のたった1つ(hT2R44、下記参照)が、なお弱いながらも6−ニトロサッカリンの存在下に活性化されたが、その一方で、最も近似するhT2R、-hT2R64(hT2R61に対して89%アミノ酸配列相同)でさえも活性化されなかった(図5)。6−ニトロサッカリンは、サッカリンの苦味類似体である。それはヒトも苦味を感じる濃度(0.1−0.5 mM, 図5)で、hT2R61を活性化する。この濃度において、サッカリンは、hT2R61を明らかに活性化せず、ほとんどのヒトは苦味を感じない。生化学的データとヒトの精神物理学的データの相関は、hT2R61は、6−ニトロサッカリンの苦味を感じるヒト受容体であることを示している。
【0178】
強力な甘味剤であるサッカリン自体が、高濃度(2−10 mM)において「苦い」または「金属的」と記述される不快な味を持つことを、多くの人が認めている。低濃度においてサッカリンは、明白にhT2R61を活性化しないが、サッカリンと6−ニトロサッカリンの構造類似性から、人が感じる高濃度における苦味もhT2R61を介していることが示唆される。
【0179】
安息香酸デナトニウムの存在下に、hT2R44の活性が増加することも観察された。この化合物の存在下で、そのほかの試験hT2Rは活性上昇を示さなかった。デナトニウムによるhT2R44の活性化は、濃度に依存することが認められた(図6)。0.3 mMより高いデナトニウム濃度で、結合GTPγSの量は、このアッセイにおける、デナトニウムが有する非特異的阻害効果のために、減少する。安息香酸デナトニウムの活性成分は、デナトニウムであり、安息香酸ではない、なぜならば、100μMの安息香酸ナトリウムの存在下にhT2R44の活性は不変だからである。6−ニトロサッカリンも、かなり弱いが、hT2R44を活性化する。
【0180】
安息香酸デナトニウム(安息香酸 ベンジルジエチル[(2,6−キシリルカルバモイル)メチル]アンモニウム)は、最も強力な苦味化合物の一つとして知られており、苦味の研究に広く使用されている。そのような研究の一つとして、A. Saroli (Saroli, 前出(1984);Saroli,前出(1985))は、デナトニウム誘導体の構造とヒトにおける苦味との関係を検討した。この研究は、デナトニウム分子の選択的変化が、化合物における苦味の特異的な減少をもたらすことを実証した。hT2R44がヒトにおいてデナトニウム及びその誘導体の苦味を仲介していることを確認するために、A. Saroli(前出)(1984)及び(1985)により記述されている3個の誘導体化合物、[(カルボキシメチル)メチル]アンモニウムクロリド(DD#2)、ベンジルメチル[(2,6−キシリルカルバモイル)メチル]アンモニウムクロリド(DD#3)、及びベンジルメチルメチル[(2,6−キシリルカルバモイル)メチル]アンモニウムクロリド(DD#4)を合成した。hT2R44を活性化する能力について、これらの化合物を試験し、デナトニウムと比較した。
【0181】
図7に示すように、デナトニウム誘導体は、デナトニウムに比較してhT2R44を活性化する能力は弱い。hT2R44を活性化する化合物の能力とヒト味覚試験における相対的苦味の間に非常に良い相関がある。これらの結果は、hT2R44がヒトにおいてデナトニウム及びその誘導体の苦味を仲介していることを強く示している。すでにデナトニウムは、hT2R4を活性化することは示されていた(Chandrashekar et al., (前出)(2000))。しかし、その試験で使用されたデナトニウムの濃度(1.5 mM)は、異常に高かった。報告されているデナトニウムのヒトにおける苦味閾値より105倍高い(Saroli, (前出)(1984))。したがって、デナトニウムは、hT2R4に対して非特異的効果を有するように思われる。我々は、hT2R44も6−ニトロサッカリンにより活性化されることを示したが、6−ニトロサッカリンの存在下における活性化は、hT2R61よりもかなり低いことを示している。したがって、hT2R61は、6−ニトロサッカリンの苦味の主たるメディエーターであり、他方hT2R44は高濃度において6−ニトロサッカリンの苦味を仲介すると思われる。
【0182】
以上の詳細な記述により本発明のいくつかの態様を示したが、以上の記述は単に説明のためのものであり、開示した発明を限定するものではないことは理解されるべきである。本発明は以下の請求項によってのみ限定される。
【図面の簡単な説明】
【0183】
【図1】図1は、hT2R4を発現するHEK-G15細胞を使用する細胞によるアッセイの結果を含んでおり、その結果はキニーネがhT2R4によるカルシウム動員を誘発することを示している。このアッセイにおいて、HEK-G15細胞にhT2R4または担体DNA(pUC-18)を導入し、2日後に単細胞カルシウム画像を使用してアッセイした。パネルaからfはキニーネ(250μM最終)で刺激した後0,10,20,30,40及び50秒のhT2R4発現細胞の応答に相当する。パネルgからlはキニーネ(250μM最終)で刺激した後0,10,20,30,40及び50秒のpuc-18発現細胞の応答に相当する。
【図2】図2は、その表面にhT2R4を発現する安定なHEK-G15細胞系を使用した実験結果を含んでいる。この実験において、その細胞表面に高いレベルのhT2R4を発現するHEK-G15のクローンは、FACSを使用する2回の選別及び2回選別した集団のクローニングの後に得られた。FACS分析により選別されたクローンは、その細胞表面上に一過性に遺伝子導入された細胞のおおよそ2−3倍多いT2R受容体を発現することが明らかにされた。
【図3】図3は、96ウエルプレート上に接種したhT2R4を安定的に発現するHEK-G15のクローン(C3Cクローン)におけるhT2R4誘発カルシウム動員に対するキニーネの用量−反応相関を示す。比較の目的で、hT2R4を一過性に導入した細胞を他のプレートに接種した。キニーネの用量を増加させた効果は、FLIPRを使用して試験した。これらの結果は、hT2R4を安定的に高度に発現するクローンにおいて明らかな用量−反応相関を示す。
【図4】図4は、hT2Rを活性化する化合物を同定するために使用されるGTPγS結合アッセイの図解である。図の中で、R、はhT2Rであり、α、β、γは、Gタンパクであり、Lは、受容体を活性化するリガンドである。
【図5】図5は、6−ニトロサッカリンは、同条件下において、hT2R61を活性化するが、hT2R64を活性化しないことを示すGTPγS結合アッセイの結果を含む。hT2R61及び hT2R64の活性は、示した濃度の6−ニトロサッカリンまたはサッカリンの存在しない場合と存在する場合に、GTPγS結合アッセイを使用して測定した(この場合の活性は、加えた試験化合物が存在しない場合に対する活性のパーセントで示す)。
【図6】図6は、hT2R44がデナトニウム及び6−ニトロサッカリンによって活性化されることを示すGTPγS結合アッセイの結果を含む。活性は試験化合物を加えない場合の活性に対する相対的増加倍率として示す。
【図7】図7は、デナトニウム及びその誘導体のhT2R44活性に対する効果を評価するGTPγS結合アッセイの結果を含む。(図中の苦味閾値はA. Saroli (前出)(1984)及びA. Saroli Z.Lebensen Unters Forsch 180: 227-221 (1985))。
【技術分野】
【0001】
(関連出願)
本出願は、2001年7月10日出願の仮出願米国出願番号60/303,811及び2002年4月15日出願の仮出願米国出願番号60/372,089の優先権を主張し、その両者の全体を引用してここに取り入れる。
【0002】
(発明の分野)
本発明は、すでに報告されている味覚に関係するヒトG−タンパク共役受容体(GPCR)の一部を活性化する苦味化合物を解明することを含んでいる。特に本発明は、T2Rファミリーの特定の味覚受容体が、苦味化合物により特異的に活性化されるとの発見を含む。すなわち、ヒトT2R4味受容体が苦味アルカロイドであるキニーネによって特異的に活性化されること、ヒトT2R61味受容体がサッカリンの苦味誘導体である6−ニトロサッカリンによって特異的に活性化されること、並びにヒトT2R44受容体がデナトニウムによって特異的に活性化されるが6−ニトロサッカリンによって弱く活性化されることに関する発見を含むものである。これらの発見に基づき、そのオーソログ、スプライス変異体、一塩基多形(SNP)、及び遺伝子操作による突然変異体を含む、これらのヒト味覚受容体、フラグメントまたはそれらの変異体若しくはキメラは、キニーネ、6−ニトロサッカリンおよびデナトニウム、および構造的に関連する化合物、並びにこれらの受容体を活性化する他の化合物による苦味を阻止する化合物を同定するアッセイ、望ましくは細胞を使用するハイスループットアッセイに有用である。このアッセイを使用して同定される化合物は、食品、飲料または医薬品の添加物としてそれらの味を改善するために使用することができる。
【背景技術】
【0003】
(関連技術の説明)
ヒトが認識することができる基本的な味の種類の一つは、苦味である。苦味の生理学はごく最近までほとんど理解されていなかった。最近の研究は、味の生物学に光を注ぎ始めた(Lindemann, Nature (2001))。苦味化合物の多くは、細胞表面受容体と相互作用することにより苦味を生じることが今では認められている。これらの受容体は、細胞内Gタンパクと相互作用する7回膜貫通ドメイン受容体のファミリーに属する。T2Rと呼ばれるGPCRの新規なファミリーが、ヒト及びげっ歯類において同定されている(Adler et al., Cell 100(6): 693-702 (2000); Chandrashekar et al., Cell 100(6): 703-711 (2000); Matsunami H, Montmayeur JP, Buck LB. Nature 404(6778): 601-4 (2000))。いくつかの証拠は、T2Rが、苦味化合物に対する応答を仲介していることを示唆している。第一に、T2R遺伝子は、舌及び口蓋上皮細胞上の味覚受容体のサブセット中で特異的に発現する。第二に、ヒトT2Rの一つ(hT2R1)についての遺伝子は、ヒトにおける苦味化合物である6-n-プロピル-2-チオウラシルに対する感受性に関連する染色体遺伝子座に存在している (Adler et al., (前出)(2000))。第三に、マウスのT2Rの一つ(mT2R5)は、マウスにおける苦味化合物であるシクロヘキシミドに対する感受性に関連する染色体遺伝子座に存在している。また、mT2R5が、味覚細胞で特異的に発現し苦味刺激伝達に関連するGタンパクであるガストジューシンを、活性化できることが示された(Wong et al., Nature 381: 796-800 (1996))。mT2R5によるガストジューシンの活性化は、シクロヘキシミドに応答したときにのみ生じる(Chandrashekar et al.,(前出)(2000))。したがって、mT2Rファミリーは、マウスにおける苦味応答を仲介し、hT2Rファミリーは、ヒトにおける苦味応答を仲介すると考えられている。唯一のヒトT2Rについて、同定された苦味リガンドを持つことが示唆された。即ち、hT2R4が、デナトニウムにより活性化されることが示された(Chandrashekar et al.,(前出)2000)。しかし、その試験に使用した有効なデナトニウム濃度(1.5 mM)は、異常に高かった、すなわち、ヒトで報告されているデナトニウムについての苦味閾値よりも105倍高かった(Saroli, Naturwissenschaften 71:428-429 (1984))。したがって、hT2Rに充分に適合する特異的苦味リガンドはなかった。各hT2Rが複数の苦味リガンドに結合できるという提案もなされていた。この仮説は、hT2Rファミリーはわずか24の同定されたメンバーから構成されるが、ヒトは数百の異なる化合物を苦いと認識することができるという事実に基づいている。hT2Rの配列は既に報告されており、Zukerら(WO 01/18050 A2,(2001))及びAdlerら(WO 01/77676 A1(2001))により公開PCT出願に開示されている。その両者の全体を、引用して本明細書に取り入れる。
【0004】
T2Rの機能についての研究における困難の一つは、これらの受容体が培養哺乳動物細胞系では簡単に発現しないということである。T2R発現を改良するために、発現の良好なGPCRであるロドプシンのN−末端配列を、T2R配列に結合した(Chandrashekar et al., (前出) 2000)。このN−末端タグは、利用可能な抗体により、タンパクの発現を容易に監視することを可能とする。ロドプシンタグを組み込むことにより、哺乳動物細胞系におけるT2Rの一部の発現は改良されるが、その多くは、依然として機能研究をするほどには十分に発現しなかった。別の研究において、mT2R5は、昆虫Sf9細胞中で良好に発現し、生化学的GTPγS結合アッセイを使用する機能研究に使用された(Chandrashekar et al., (前出) 2000)。
【0005】
しかし、報告がされた内容によっても、依然、T2R遺伝子ファミリーの機能に関して確実性を欠いている。特に、どの味覚化合物が、特定のT2R受容体を活性化するかに関して理解を欠いている。個々のT2R受容体の結合特性を十分に理解することが非常に有益であり、そうすることにより所望の味覚調節特性を有する化合物を選別するためにそれを使用することが促進されるであろう。
【特許文献1】WO 01/18050 A2,(2001)
【特許文献2】WO 01/77676 A1(2001)
【非特許文献1】Lindemann, Nature (2001)
【非特許文献2】Adler et al., Cell 100(6): 693-702 (2000)
【非特許文献3】Chandrashekar et al., Cell 100(6): 703-711 (2000)
【非特許文献4】Matsunami H, Montmayeur JP, Buck LB. Nature 404(6778): 601-4 (2000)
【非特許文献5】Wong et al., Nature 381: 796-800 (1996)
【非特許文献6】Saroli, Naturwissenschaften 71:428-429 (1984)
【発明の開示】
【課題を解決するための手段】
【0006】
(発明の要約)
結論的に、本発明は、T2Rファミリーにおける幾つかの味覚受容体、特にhT2R4, hT2R61及びhT2R44は、それぞれ、苦味化合物である、キニーネ、6−ニトロサッカリン、及びデナトニウム(及び6−ニトロサッカリンには弱く)によって特異的に活性化されるという発見に関する。
【0007】
これらの発見は、特定のT2Rを発現する細胞を使用してT2Rの活性を測定する細胞によるアッセイを使用して行われた。以下に詳細に記述するように、特にその表面にhT2R4を発現するHEK-Gα15細胞系を、細胞内カルシウム濃度の変化を検出する細胞によるアッセイに使用し、他のhT2Rが同条件では活性化されないにも拘らず、同細胞系がキニーネにより特異的に活性化されることを見出した。また、キニーネは、典型的な用量‐反応相関性をもってhT2R4を活性することが示された。
【0008】
さらに、以下により詳細に記述するように、GTPγS結合アッセイを、種々の苦味化合物の存在下、バキュロウイスル発現系を用いて昆虫細胞(Sf9細胞)で発現させたヒトT2Rの活性をモニターするために使用した。このアッセイの結果は、hT2R61が、特異的に6−ニトロサッカリンにより活性化され、hT2R44が特異的にデナトニウム(及び6−ニトロサッカリンによって弱く)により活性化されることを示している。
【0009】
したがって、本発明は、キニーネ、6−ニトロサッカリン、デナトニウムまたはサッカリンのような関連化合物によるhT2R4, hT2R44及びhT2R61の受容体の活性化を阻害する化合物を同定するためのアッセイ、望ましくはハイスループットアッセイにおける、hT2R4, hT2R44及びhT2R61の使用を包含している。
【0010】
また、本発明は、これらの化合物が味覚試験において苦味を阻止することを確認することを含む方法も包含している。また、本発明は、例えば、サッカリンを含む飲料、食品または医薬品などのように、苦味を抑制するために、食品、飲料及び医薬品において同定した化合物を使用することを包含している。
【0011】
(発明の目的)
本発明の一つの目的は、キニーネまたはキニーネに構造的に関連するhT2R4を活性化する化合物によるhT2R4、またはそのフラグメント、変異体、オーソログ若しくはキメラの活性化を阻害する化合物を同定することである。
【0012】
本発明のもう一つの目的は、6−ニトロサッカリン、サッカリンまたはそれらに構造的に関連するhT2R61を活性化する化合物によるhT2R61またはそのフラグメント、変異体、オーソログ若しくはキメラの活性化を阻害する化合物を同定することである。
【0013】
本発明のそのほかの目的は、デナトニウム若しくは6−ニトロサッカリンまたはそれらに構造的に関連するhT2R44を活性化する化合物によるhT2R44の活性化を阻害する化合物を同定することである。
【0014】
本発明のさらに特別な目的は、これらの受容体により顕在化する苦味を阻止する化合物を同定する手段として、キニーネ;6−ニトロサッカリン若しくはサッカリン;またはデナトニウム若しくは6−ニトロサッカリンの夫々による上記で特定したT2R受容体の活性化を阻害する化合物を同定するための細胞を使用するアッセイにおいて、hT2R4, hT2R61若しくはhT2R44またはそれらのフラグメント、変異体、オーソログ若しくはキメラを安定してまたは一過性に発現する細胞を使用することである。
【0015】
本発明のさらに特別な目的は、キニーネまたは関連化合物によるhT2R4の活性化、6−ニトロサッカリン、サッカリンまたは関連化合物によるhT2R61の活性化、或いはデナトニウムまたは関連化合物によるhT2R44の活性化を阻害する化合物を同定するために、特に苦味化合物によるこれらのT2Rの特異的活性化を阻止する化合物を同定する手段として、望ましくはカルシウム特異的な蛍光色素を使用して細胞内カルシウムの変化を検出する細胞によるアッセイにおいて、哺乳動物または昆虫の細胞、例えば、G15タンパクを発現するHEK-293T細胞、を使用することである。
【0016】
本発明のさらに特別な目的は、特に苦味化合物によるこれらのT2Rの特異的活性化を阻止する化合物を同定する手段として、キニーネ、デナトニウム、6−ニトロサッカリンまたはサッカリンによるhT2R4, hT2R44またはhT2R61の受容体の活性化をそれぞれ阻害する化合物を同定するために、GTPγS結合アッセイにおいて、hT2R4, hT2R44またはhT2R61を発現する哺乳動物または昆虫細胞、例えば、Sf9細胞、を使用することである。
【0017】
本発明のそのほかの目的は、同定された化合物が、例えば、キニーネ、デナトニウム、6−ニトロサッカリンまたはサッカリンにより誘発される苦味を阻止することを、味覚試験において確認することである。
【0018】
本発明のそのほかの目的は、hT2R4, hT2R44またはhT2R61を特異的に活性化する化合物により誘発される苦味を阻止するための組成物中に、本明細書で記述するアッセイにおいて同定された化合物を添加物として使用することである。本発明の望ましい目的は、6−ニトロサッカリンまたはサッカリンによるhT2R61及び/またはhT2R44の活性化を阻害する化合物を、サッカリン含有組成物、特にダイエット飲料、の苦味を阻止するために使用することである。
【0019】
(発明の詳細な説明)
本発明を具体的に記述する前に、以下の定義を提示する。
【0020】
用語「T2R」ファミリーは、
(1)以下及び引用したZuker(前出)(2001)およびAdler(前出)(2001)に開示されているT2Rに対して、約25アミノ酸、最も適するのは50−100アミノ酸の枠に亘って、約30−40%のアミノ酸配列が相同、より具体的にいうと約40,50,60,70,75,80,85,90,95,96,97,98,または99%のアミノ酸配列が相同である;
(2)以下に開示するT2R配列及びその保存的に修飾された変異体からなる群から選択されるアミノ酸配列を含む免疫原に対して作成される抗体に特異的に結合する;
(3)以下に開示するT2R DNA配列及びその保存的に修飾された変異体からなる群から選択された配列にストリンジェントなハイブリッド形成条件の下に(少なくとも約100、任意に少なくとも約500-1000ヌクレオチドのサイズで)特異的にハイブリッド形成する;
(4)以下に開示するT2Rアミノ酸配列からなる群から選択されたアミノ酸配列に少なくとも約40%相同な配列を含む;または
(5)記述されたT2Rとストリンジェントなハイブリッド形成条件下に特異的にハイブリッド形成するプライマーにより増幅される
多形変異体、アレル、突然変異体、及び同属体を含む。
【0021】
すでに述べたように、T2R遺伝子は、タンパク及びDNAのレベルで実質的な配列の相違を示すが、今日までに単離されたT2Rは、全て特定の領域において一定の共通配列を含むことが認められており、それらは本明細書に引用して取り入れたAdlerら(WO 01/77676 A1(2001))及びZukerら(WO 01/18050 A2)においてすでに同定されているT2Rコンセンサス配列を持つかそれに対して少なくとも70−75%相同である。
【0022】
立体構造的にその化学感受性GPCRは、「N−末端ドメイン」、「細胞外ドメイン」、7回膜貫通領域及び対応する細胞質及び細胞外ループを含む「膜貫通ドメイン」、「細胞質領域」並びに「C−末端領域」を有する(例えば、Hoon et al., Cell, 96: 541-51 (1999); Buck & Axel, Cell, 65: 175-87 (1991)参照)。これらの領域は、例えば疎水性及び親水性ドメインを同定する配列分析プログラムのような、当業者に既知の方法を使用して構造的に同定することができる(例えば、Stryer, Biochemistry, (3rd ed. 1988)参照;多くのインターネットから入手できる配列分析プログラムのいずれか、例えば、dot.imgen.bcm.tmc.eduで見られるものも参照)。これらの領域は、キメラタンパクの作成及び本発明のインビトロアッセイ、例えば、リガンド結合アッセイに有用である。
【0023】
本発明による「T2R4」は、ヒトT2R4ポリペプチド及び対応する核酸の配列、並びに同じ機能を持つ、すなわち、キニーネまたはそのほかの苦味化合物によって特異的に活性化される、そのフラグメント、変異体、突然変異体及びオーソログを含むことが意図されている。ヒトT2R4の配列は、Zukerら(WO 01/18050) A2(2001)及びAdlerら(WO 01/77676 A1 (2001))に含まれており、下記に記述するSEQ ID NO: 1及び2(ヒトT2R4タンパク及びDNA)中に含まれる。
【0024】
【0025】
【0026】
本発明による「T2R44」は、下記のSEQ ID NO: 3及び4に含まれるタンパク及びDNA配列を持つヒトT2R44、並び同じ機能を示す、すなわち、適当なアッセイにより測定されるデナトニウムのような苦味化合物により特異的に活性化される、フラグメント、キメラ、変異体、突然変異体、オーソログを含むことが意図されている。これらフラグメント、キメラ、変異体、突然変異体、オーソログは、例えば、スプライス変異体、アレル変異体、及び一塩基多形などを含むものである。
【0027】
【0028】
【0029】
本発明による「T2R61」は、SEQ ID NO: 5 に含まれるヒトT2R61タンパク及び対応する核酸配列(SEQ ID NO: 6)、並び同じ機能を持つ、すなわち、6−ニトロサッカリン、サッカリン、または適当なアッセイを使用してスクリーニングできるような関連苦味化合物により特異的に活性化される、フラグメント、キメラ、変異体、突然変異体を含むことが意図されている。これらフラグメント、キメラ、変異体、及び突然変異体は、アレル、スプライス及び一塩基多形及びオーソログを含むものである。
【0030】
【0031】
【0032】
「細胞外ドメイン」は、したがって、細胞の膜から突き出ていて、細胞の細胞外表面に露出しているT2Rポリペプチドのドメインのことである。その領域は、細胞の細胞外表面に露出している「N−末端ドメイン」、並びに細胞の細胞外表面に露出している膜貫通ドメインの細胞外ループ、すなわち、膜貫通領域2及び3、膜貫通領域4及び5、並びに膜貫通領域6及び7の間の細胞外ループを含む。「N−末端ドメイン」は、N−末端から始まり膜貫通領域の開始点に近い領域にいたる。これらの細胞外領域は、液相及び固相におけるインビトロリガンド結合アッセイに有用である。さらに、以下に記述する膜貫通領域も、細胞外領域と組み合わせてあるいは単独でリガンド結合に関与することができるので、インビトロリガンド結合アッセイに有用である。
【0033】
7個の膜貫通「領域」を含む「膜貫通ドメイン」は、形質膜内に存在するT2Rポリペプチドのドメインのことであり、対応する細胞質(細胞内)及び細胞外ループを含むことがあり、また膜貫通「領域」とも呼ばれる。7個の膜貫通領域及び細胞外及び細胞質ループは、Kyte & Doolittle, J. Mol. Biol., 157: 105-32 (1982),またはStryer,前出に記述されているように、標準的方法を使用して同定することができる。
【0034】
「細胞質ドメイン」は、細胞の内側に面しているT2Rのドメイン、例えば、「C−末端ドメイン」及び膜貫通ドメインの細胞内ループ、例えば、膜貫通領域1及び2、膜貫通領域3及び4、及び膜貫通領域5及び6の間の細胞内ループのことである。「C−末端ドメイン」は、最後の膜貫通領域の終わりからタンパクのC−末端に達する領域のことであり、通常は細胞内に存在している。
【0035】
用語「7回膜貫通受容体」は、7回形質膜を貫通する7個の領域を持つ膜貫通タンパクのスーパーファミリーに属するポリペプチドを意味する(したがって、7個の領域は「膜貫通」ドメインまたは「TM」ドメインTM IからTM VIIと呼ばれる)。嗅覚受容体及びある種の味覚受容体は、それぞれこのスーパーファミリーに属す。7回膜貫通受容体ポリペプチドは、以下にさらに詳細に検討するように、類似する特徴的な、一次、二次、及び三次構造を有している。
【0036】
用語「リガンド結合領域」は、膜貫通ドメインIIからVII(TM IIからVII)を実質的に組み込んでいる化学感受性受容体または味覚受容体に由来する配列のことである。この領域は、リガンド、より具体的には味覚誘発化合物と結合することができる。
【0037】
用語「形質膜転移ドメイン」または単に「転移ドメイン」は、代表的な「転移ドメイン」(5'−MNGTEGPNFYVPFSNKTGVV;SEQ ID NO: 7)と機能的に同等であるポリペプチドドメインを意味する。これらのペプチドドメインは、ポリペプチドコード配列のアミノ末端に組み込まれたときに、ハイブリッド(「融合」)タンパクを、細胞形質膜へ非常に効率よく「移動」または「転移」することができる。この特殊な「転移ドメイン」は、最初、ヒトロドプシン受容体ポリペプチド、即ち7回膜貫通受容体、のアミノ末端から得られた。他の転移ドメインは、ウシロドプシン配列から得られており、同様に転移を促進するのに有用である。ロドプシン由来配列は、7回膜貫通融合タンパクを形質膜へ転移させるのに特に有効である。
【0038】
「機能的同等」とは、新規に翻訳されたタンパクを同条件において形質膜へ転移させるドメインの能力及び効果が、代表的SEQ ID NO: 7と同じであることを意味する;相対的効果は、本明細書で記述されているように、(定量的に)測定して比較される。本発明の範囲にあるドメインは、20アミノ酸鎖長の転移ドメインSEQ ID NO: 7と同じ効率で、(哺乳動物、ツメガエルなどの)細胞中において新規に合成されたポリペプチドを形質膜へ転移させるかについての所定のスクリーニングにより決定することができる。
【0039】
味覚伝導を仲介するT2Rファミリーに作用する試験化合物のアッセイにおける文脈で、語句「機能的効果」は、間接的にまたは直接的に受容体の影響下にあるパラメーター、例えば機能的、物理的及び化学的な効果、を測定することを含む。それには、インビトロ、インビボ、及びエクスビボにおけるリガンド結合、イオン流の変化、膜電位、電流、転写、Gタンパク結合、GPCRのリン酸化または脱リン酸化、信号伝達、受容体−リガンド相互作用、セカンドメッセンジャー濃度(例えば、cAMP, cGMP, IP3,または細胞内Ca2+)が含まれ、また神経伝達物質またはホルモン放出の増加または減少のようなそのほかの生理的効果も含まれる。
【0040】
「機能的効果を測定する」とは、間接的または直接的に、T2Rファミリーメンバーの影響下にあるパラメーター、例えば、機能的、物理的及び化学的効果、を増加または減少する化合物をアッセイすることを意味する。その機能的効果は、当業者に既知の方法のいずれかにより測定することができる、例えば、分光学的特徴の変化(例えば、蛍光、吸光度、屈折率)、流体力学(例えば、形状)、クロマトグラフィー特性、または溶解性、パッチクランピング、電位感受性色素、全細胞電流、放射性アイソトープ放出量、誘導マーカー、卵母細胞T2R遺伝子発現;組織培養細胞T2R発現;T2R遺伝子の転写活性化;リガンド結合アッセイ;電圧、膜電位及び伝導度の変化;イオン流アッセイ;cAMP,cGMP,及びイノシトール三燐酸(IP3)のような細胞内セカンドメッセンジャーの変化;細胞内カルシウムレベルの変化;神経伝達物質放出、などがある。
【0041】
T2Rタンパク受容体の「阻害物質」、「活性化物質」及び「活性調節物質」は、インビトロ及びインビボにおける味覚伝達のアッセイを使用して同定される阻害性分子、活性化分子、または活性調節分子、例えば、リガンド、アゴニスト、アンタゴニスト、並びにそれらの同属体及び擬似体に言及するために互換的に使用される。阻害物質は、例えば、結合して、部分的にまたは完全に刺激を阻害し、減少し、阻止し、活性化を遅らせ、不活化し、脱感作し、または味覚伝達を下方調節する化合物、例えば、アンタゴニストである。活性化物質は、例えば、結合して、刺激し、増加し、開放し、活性化し、促進し、活性化を増強し、感受性とし、または味覚伝達を上方調節する化合物、例えば、アゴニストである。活性調節物質は、例えば受容体と、活性化物質または阻害物質と結合する細胞外タンパク(例えば、エブネリン及び疎水性キャリアーファミリーのその他のメンバー);Gタンパク;キナーゼ(例えば、ロドプシンキナーゼの同属体、並びに受容体の脱活性化及び脱感作に関係するβ交感神経受容体キナーゼ);並びにやはり受容体を脱活性化及び脱感作するアレスチン、との相互作用を変化させる化合物を含む。活性調節物質は、例えば、変化した活性を持つT2Rファミリーメンバーの一般的な修飾体、並びに、天然に存在するかまたは合成の、リガンド、アンタゴニスト、アゴニスト、小型化学分子などを含む。
【0042】
阻害物質及び活性化物質のアッセイは、例えば、細胞中または細胞の膜にT2Rファミリーメンバーを発現させ、例えば、苦味化合物の活性を調節する化合物の存在下または非存在下に候補調節化合物を適用し、次いで前記のように味覚伝達における機能的効果を測定することを含む。潜在的な活性化物質、阻害物質または活性調節物質で処理したT2Rファミリーメンバーを含む検体またはアッセイは、調節の程度を調べるために、阻害物質、活性化物質または活性調節物質を含まない対照検体と比較される。対照検体(活性調節物質で処理されていない)を相対T2R活性値100%とする。対照と比較したT2R活性値が約80%、さらに50%または25−0%であればT2Rの阻害は達成される。対照と比較したT2R活性値が110%、さらに150%、さらに200−500%、または1000−3000%高いときに、T2Rの活性化は達成される。
【0043】
本明細書で使用される時、語「精製された」、「実質的に精製された」及び「単離された」は、本発明の化合物が、自然な状態では通常結合している他の異なる化合物から遊離した状態のことである。望ましくは、「精製された」、「実質的に精製された」及び「単離された」とは、その構成物が、所定の試料における総量の、少なくとも0.5重量%、1重量%、5重量%、10重量%、または20重量%であることを意味し、最も望ましくは、少なくとも50を重量%または75重量%を構成することを意味する。望ましい一態様において、これらの語は、本発明の化合物が、所定の試料における総量の少なくとも95重量%を構成することである。本明細書で使用される時、語「精製された」、「実質的に精製された」及び「単離された」は、核酸またはタンパクに関する場合、哺乳動物の体内、特にヒト、の体内に自然に存在する状態と異なる精製または濃度の状態のことである。哺乳動物の体内、特にヒトの体内に自然に存在する状態よりも高い精製または濃度とすることは、(1)他の結合構造または結合化合物からの精製、または(2)哺乳動物の体内、特にヒトの体内では通常結合しない構造または化合物との結合を含み、「単離された」の意味の範囲に入る。当業者に既知の種々の方法及び操作にしたがって、本明細書で記述する核酸若しくはタンパクまたは核酸若しくはタンパクのクラスを単離することができるし、また通常では自然に結合しない構造または化合物と結合させることができる。
【0044】
ここに使用した語「単離された」は、核酸またはポリペプチドに関する場合には、哺乳動物、特にヒト、の体内に自然に存在するのと異なる精製または濃度の状態のことである。(1)他の自然に存在する結合構造または結合化合物からの精製または(2)体内では通常結合しない構造または化合物との結合を含む、体内に自然に存在する状態よりも高い精製または濃度は、本明細書で使用される「単離された」の意味の範囲に入る。当業者に既知の種々の方法及び操作にしたがって、本明細書で記述する核酸またはポリペプチドを単離することができ、さもなければ、通常では自然に結合しない構造または化合物と結合させることができる。
【0045】
本明細書で使用される時、語句「増幅する」及び「増幅」は、下記に詳細に検討するように、組換えによりまたは自然に発現した核酸を産生または検出するために、適当な増幅方法のいずれかを使用することである。例えば、本発明は、インビボまたはインビトロで、自然発現(例えば、ゲノムまたはmRNA)または組換え(例えば、cDNA)による本発明の核酸(例えば、本発明の味覚誘発化合物結合配列)を、(例えば、ポリメラーゼ連鎖反応、PCR,により)増幅するための方法及び試薬(例えば、特異的オリゴヌクレオチドプライマー対)を提供する。
【0046】
用語「発現ベクター」とは、原核細胞、イースト、菌、植物、昆虫または哺乳動物の細胞を含むいずれかの細胞において、構成的または誘導的に、インビトロまたはインビボで、本発明の核酸配列を発現させるための組換え発現系のことである。この用語は、線状または環状発現系を含む。この用語は、エピソームに残留するまたは宿主細胞ゲノムに組み込まれた発現系を含む。この発現系は、自己複製の能力を持つものでもよく、または持たないもの、すなわち、細胞中で一過性にのみ発現するものでもよい。この用語は、組換え核酸の転写に必要な最低限の配列のみを含有する組換え発現「カセット」を含む。
【0047】
用語「ライブラリー」とは、異なる核酸またはポリペプチドの混合物である調製物を意味し、例えば、縮重プライマー対を用いて核酸を増幅することにより作成された組換え感覚受容体リガンド結合領域、特に組換え味覚受容体リガンド結合領域のライブラリー、または増幅リガンド結合領域を組み込んだ単離ベクターのコレクション、または味覚受容体をコードする少なくとも一つのベクターで無作為に遺伝子導入した各細胞の混合物がある。
【0048】
用語「核酸」または「核酸配列」とは、一本鎖または二本鎖のいずれかの形のデオキシリボヌクレオチドまたはリボヌクレオチドのオリゴヌクレオチドのことである。この用語は、天然ヌクレオチドの既知の類似体を含む、核酸、すなわち、オリゴヌクレオチドを包含する。この用語は合成骨格を持つ核酸様構造も包含する。
【0049】
特に示さない限り、特定の核酸配列は、明示された配列のみならず、その保存的に修飾された変異体(例えば、縮重コドン置換体)及び相補的配列を暗黙のうちに包含する。具体的には、縮重コドン置換は、例えば、一つ以上の選択されたコドンの三番目の位置を、混合塩基及び/またはデオキシイノシン残基で置換した配列を作成することにより行われる(Batzer et al., Nucleic Acid Res., 19: 5081 (1991); Ohtsuka et al., J. Biol. Chem., 260:2605-08 (1985); Rossolini et al., Mol. Cell. Probes, 8: 91-98 (1994))。用語核酸は、遺伝子、cDNA, mRNA, オリゴヌクレオチド、及びポリヌクレオチドと互換的に使用される。
【0050】
用語「ポリペプチド」、「ペプチド」及び「タンパク」は、アミノ酸残基の重合体を示すために本明細書において互換的に使用される。この用語は、天然アミノ酸重合体及び非天然のアミノ酸重合体のみならず、一以上のアミノ酸が天然アミノ酸の対応する人工的な化学擬似体であるアミノ酸重合体に適用される。
【0051】
本明細書で記述される「転移ドメイン」、「リガンド結合領域」またはキメラ受容体構成物は、「類似体」、または「保存的変異体」及び代表的な配列に実質的に一致する構造及び活性を持つ「擬似体」(「ペプチド擬似体」)も含む。従って、用語「保存的変異体」または「類似体」または「擬似体」は、その変化が本明細書で定義するようなポリペプチド(保存的変異体)の構造及び/または活性を実質的に変えないような修飾アミノ酸配列を持つポリペプチドのことである。これらは、アミノ酸配列の保存的修飾変異体、すなわち、タンパクの活性に重要でないアミノ酸置換、アミノ酸残基の付加若しくは欠失、或いは重要なアミノ酸の置換であっても構造及び/または活性を実質的に変えないような類似性質(例えば、酸性、塩基性、陽性または陰性荷電、極性または非極性、など)を持つ残基によるアミノ酸の置換された変異体を含む。
【0052】
より具体的には、「保存的修飾変異体」は、アミノ酸配列及び核酸配列の両者に適用する。特定の核酸配列に関しては、保存的修飾変異体とは、同じまたは本質的に同じアミノ酸配列をコードすること、或いは核酸がアミノ酸配列をコードしない場合には、本質的に同じ配列のことである。遺伝暗号の縮重のために、多数の機能的に同一の核酸が所与タンパクをコードする。
【0053】
例えば、コドンGCA, GCC, GCG及びGCUは、全てアミノ酸アラニンをコードする。したがって、コドンによってアラニンが特定されているどの位置においても、コードされているポリペプチドを変えることなく、コドンを、記述されている対応コドンのいずれかと交換することができる。
【0054】
そのような核酸変異体は「沈黙変異体」であり、保存的修飾変異体の一種である。ポリペプチドをコードする本明細書における総ての核酸配列は、該核酸の可能な沈黙変異体の全てについても言及する。当業者は、核酸の各コドン(通常はメチオニンの唯一のコドンであるAUG、及び通常トリプトファンの唯一のコドンであるTGGを除く)を修飾して機能的に同一の分子が得られることを認識している。したがって、ポリペプチドをコードする核酸の沈黙変異体は、暗黙のうちに各記述された配列の中に存在する。
【0055】
機能的に同じアミノ酸を示す保存置換表は当業者に既知である。例えば、保存的置換を選択する一つの代表的例は(元の残基、次いで代表的な置換残基を示す)、ala/glyまたはser; arg/lys; asn/glnまたはhis; asp/glu; cys/ser; gln/asn; gly/asp; gly/alaまたはpro; his/asn またはgln; ile/leu またはval;leu/ileまたはval; lys/argまたはglnまたはglu; met/leuまたはtyrまたはile; phe/met またはleuまたはtyr; ser/thr; thr/ser; trp/tyr; tyr/trpまたはphe; val/ileまたはleuである。別の代表例は下記の、相互に保存置換できるアミノ酸を含む6群を使用する:1)アラニン(A)、セリン(S)、トレオニン(T);2)アスパラギン酸(D)、グルタミン酸(E);3)アスパラギン(N)、グルタミン(Q);4)アルギニン(R)、リシン(K);5)イソロイシン(I)、ロイシン(L)、メチオニン(M)、バリン(V);及び6)フェニルアラニン(F)、チロシン(Y)、トリプトファン(W);(例えば、Creighton, Proteins, W.H.Freeman and Company (1984); Schultz and Schimer, Principles of Protein Structure, Springer-Verlag (1979)参照)。当業者は、上に示した置換だけが可能な保存的置換ではないことを理解しているであろう。例えば、ある目的のためには、陽性かまたは陰性に荷電しているアミノ酸相互の荷電アミノ酸置換をすべて保存的置換と見なすことができる。さらに、コード配列中の一つのアミノ酸または小さな割合のアミノ酸を変更、付加または削除する個別の置換、欠失または付加も「保存的修飾変異体」と考えることができる。
【0056】
用語「擬似体」及び「ペプチド擬似体」とは、ポリペプチド、例えば、転移ドメイン、リガンド結合領域、または本発明のキメラ受容体と同じ構造及び/または機能特性を実質的に有する合成化合物のことである。擬似体は、アミノ酸の合成による非天然の類似体で全体を構成することもできるし、または一部が天然のペプチドアミノ酸からなり一部が非天然のアミノ酸の類似体からなるキメラ分子としてもよい。この擬似体は、このような置換が擬似体の構造及び/または活性を変えない限り、如何なる数の天然アミノ酸保存置換を組み込むことができる。
【0057】
保存的変異体である本発明のポリペプチドと同様に、ルーチン的実験により本発明の範囲に入るか否か、すなわち、構造及び/または機能が変わっていないこと、を決定するであろう。
ポリペプチド擬似構成物は、非天然の構造成分のいずれかの組み合わせを含むことができ、それは典型的に以下の3つの構造基に由来する;a)天然アミド結合(「ペプチド結合」)以外の残基連結基;b)天然に存在するアミノ酸残基に代わる非天然残基;c)二次構造擬態を誘導する残基、すなわち、例えば、βターン、γターン、βシート、αへリックスなどの二次構造を誘導または安定化する残基。
ポリペプチドは、その残基の全てまたは一部が天然ペプチド結合以外の化学的方法により結合している場合に、擬似体として特徴付けることができる。個々のペプチド擬似残基は、ペプチド結合、そのほかの化学的結合またはカップリング手段、例えば、グルタルアルデヒド、N−ヒドロキシコハク酸イミドエステル、2価マレイミド、N,N'−ジシクロヘキシルカルボジイミド(DCC)またはN,N'−ジイソプロピルカルボジイミド(DIC)、により結合することができる。伝統的アミド結合(「ペプチド結合」)に代わりうる結合基には、例えば、ケトメチレン(例えば、-C(=O)-NH-の代わりに−C(=O)-CH2-)、アミノメチレン(CH2-NH)、エチレン、オレフィン(CH=CH)、エーテル(CH2-O)、チオエーテル(CH2-S)、テトラゾール(CN4)、チアゾール、レトロアミド、チオアミド、またはエステルがある(例えば、Spatola, Chemistry and Biochemistry of Amino Acids, Peptides and Proteins, Vol. 7, 267-357, Marcell Dekker, Peptide Backbone Modifications, NY (1983)参照)。天然に存在するアミノ酸残基に代わって、全部または一部が非天然残基を含む場合にも、ポリペプチドは、擬似体に分類される(非天然残基は、科学文献及び特許文献に詳しく記述されている)。
【0058】
「標識」または「検出分子」は、分光学的、光化学的、生化学的、免疫化学的、または化学的な方法により検出される成分である。例えば、有用な標識としては、32P,蛍光色素、電子密度試薬、(例えば、ELISAにおいて広く使用されるような)酵素、ビオチン、ジゴキシゲニン、或いは例えば、ペプチドの中に放射標識を組み込むことにより検出可能にしたハプテン及びタンパク、またはペプチドと特異的に反応する抗体を検出するために使用されるハプテン及びタンパクが含まれる。
【0059】
「標識した核酸プローブまたはオリゴヌクレオチド」は、プローブの存在がプローブに結合した標識の存在を検出することにより検出されるように、標識に、リンカーや化学結合による共有結合により、或いはイオン的な、ファンデルワールス力による、静電的な、または水素結合による非共有結合により、結合させた核酸プローブまたはオリゴヌクレオチドである。
【0060】
本明細書で使用される時、「核酸プローブまたはオリゴヌクレオチド」とは、一以上の種類の化学結合、通常は相補的塩基対による通常は水素結合形成、により、相補的配列からなる標的核酸と結合することができる核酸と定義される。本明細書で使用される時プローブは、天然塩基(すなわち、A, G, C,またはT)または修飾塩基(7−デアザグアノシン、イノシン、など)を含むことができる。さらに、プローブ中の塩基は、ハイブリッド形成を妨害しない限り、燐酸ジエステル結合以外の結合で連結することができる。したがって、例えば、プローブは燐酸ジエステル結合でなくペプチド結合により構成塩基が連結したペプチド核酸とすることができる。ハイブリッド形成条件のストリンジェンシーによってはプローブと完全な相補性を欠く標的配列とプローブが結合しうることを、当業者は理解しているであろう。プローブは任意にアイソトープ、発色団、発光団、発色源で直接標識されるか、または後にストレプトアビジン複合体が結合するビオチンのようなもので間接的に標識される。プローブの存在または不在をアッセイすることにより、選択した配列または部分配列の存在または不在を検出することができる。
【0061】
用語「異種」は、核酸の部分に関して使用された場合、天然において相互に同一の種族の中には認められない二つ以上の部分配列を核酸が含むことを示す。例えば、典型的には、新規な機能の核酸を作るために配列された無関係な遺伝子から得た二つ以上の配列、例えば、一の起源からのプロモーターと他の起源からのコード領域とを持っている核酸が、組換えにより作られる。同様に、異種タンパクとは、そのタンパクが、天然において相互に同一の種族の中には認められない二つ以上の部分配列を含むこと(例えば、融合タンパク)を示す。
【0062】
「プロモーター」は、核酸の転写を指令する核酸配列が並んだものと定義される。本明細書で使用される時、プロモーターには、例えば、ポリメラーゼII型プロモーターやTATA配列の場合における、転写開始部位の近くにある必要な核酸配列が含まれる。プロモーターは、さらに転写開始部位から数千塩基対も離れていることがある遠位エンハンサーまたはリプレッサーも含む。「構成的」プロモーターは、ほとんどの環境及び発生条件において活性であるプロモーターである。「誘導的」プロモーターは、環境または発生の制御下において活性であるプロモーターである。用語「作動的に結合している」とは、(プロモーター、または転写因子結合部位の配列のような)核酸発現調節配列と、第二の核酸配列との間での機能的連結のことであり、発現調節配列は、第二配列に相当する核酸の転写を指令する。
【0063】
本明細書で使用される時、「組換え」とは、合成されたか、インビトロで操作されたポリヌクレオチド(例えば、「組換えポリヌクレオチド」)、細胞中またはそのほかの生化学的系において遺伝子産物を生産するために組換えポリヌクレオチドを使用する方法、或いは組換えポリヌクレオチドによりコードされているポリペプチド(「組換えタンパク」)のことである。「組換え手段」は、例えば、本発明の転移ドメインと本発明のプライマーを使用して増幅された核酸配列とを含む融合タンパクの誘導的または構成的な発現のための発現カセットまたはベクターの中に、異なる起源から得た種々のコード領域またはドメインまたはプロモーター配列を持つ核酸を連結することも包含する。
【0064】
語句「選択的に(または特異的に)ハイブリッド形成する」とは、その配列が複雑な混合物(例えば、全細胞またはライブラリーDNAまたはRNA)中に存在するときに、ストリンジェントなハイブリッド形成条件の下に特別なヌクレオチド配列とのみ結合、二本鎖化または分子のハイブリッドを形成することをいう。
【0065】
語句「ストリンジェントなハイブリッド形成条件」とは、プローブが、典型的には核酸の複雑な混合物中において、標的部分配列とハイブリッドを形成し、他の配列とはハイブリッドを形成しない条件のことをいう。ストリンジェントな条件は配列に依存し、そして異なる環境では異なるであろう。長い配列は特に、高い温度でハイブリッド形成する。核酸のハイブリッド形成に関する詳細な説明は、Tijssen, Techniques in Biochemistry and Molecular Biology-Hybridisation with Nucleic Probes, “Overview of principles of hybridization and the strategy of nucleic acid assays” (1993)の中にある。一般的に、ストリンジェントな条件は、規定されているイオン強度、pHにおいて、特定の配列の融解温度(Tm)よりも約5−10℃低い温度が選択される。Tmは、(規定されたイオン強度、pH及び核酸濃度において)標的に相補的なプローブの50%が、均衡して標的配列とハイブリッド形成する温度である(標的配列が過剰に存在すると、Tmで、プローブの50%が均衡して占有される)。ストリンジェントな条件は、塩濃度が約1.0 Mナトリウムイオン未満、典型的にはpH 7.0から8.3において約0.01から1.0 Mナトリウムイオン濃度(または他の塩)であり、温度が、短いプローブ(例えば、10から50ヌクレオチド)に対して少なくとも約30℃、長いプローブ(例えば50ヌクレオチド超)に対しては少なくとも約60℃の条件であろう。ストリンジェントな条件はホルムアミドのような脱安定化試薬の添加も必要である。選択的または特異的なハイブリッド形成に対する陽性信号は、背景ハイブリッド形成の少なくとも2倍、望ましくは10倍である。代表的なストリンジェントな条件は次のようなものであろう:50%ホルムアミド、5x SSC,及び1% SDS, 42℃でインキュベート、または5x SSC, 1% SDS, 65℃でインキュベート、そして65℃において0.2x SSC及び0.1% SDS中で洗う。そのハイブリッド形成及び洗浄段階は、例えば、1,2,5,10,15,30,60分間;またはそれ以上の時間行うことができる。
【0066】
ストリンジェントな条件下に相互にハイブリッド形成しない核酸であっても、それらがコードするポリペプチドが実質的に関係あるときには、依然として実質的に関係がある。これは、例えば、核酸のコピーが遺伝子コードで許容される最大のコドン縮重を使用して作られている場合に生じる。そのような場合には、核酸は、通常、緩和なストリンジェントなハイブリッド形成条件下でハイブリッドを形成する。代表的な「緩和なストリンジェントハイブリッド形成条件」は、40%ホルムアミドの緩衝液中、1 M NaCl,1% SDS 37℃におけるハイブリッド形成、及び1x SSC 45℃における洗浄を含む。そのハイブリッド形成及び洗浄段階は、例えば、1,2,5,10,15,30,60分間またはそれ以上の時間行うことができる。ハイブリッド形成陽性は背景の少なくとも2倍である。同じストリンジェンシーにするために、別のハイブリッド形成及び洗浄条件を使用しうることを、当業者は容易に理解するであろう。
【0067】
「抗体」は、抗原に特異的に結合して認識する免疫グロブリンの遺伝子に由来する骨格領域またはそのフラグメントを含むポリペプチドのことである。認識する免疫グロブリンの遺伝子は、κ、λ、α、γ、δ、ε、及びμ定常領域、並びに無数の免疫グロブリン可変領域遺伝子を含む。軽鎖はκまたはλのいずれかに分類される。重鎖はγ、μ、α、δ、またはεに分類され、それらは順にイムノグロブリンクラス、IgG, IgM, IgA, IgD及びIgEをそれぞれ規定する。
【0068】
代表的な免疫グロブリン(抗体)構造単位は、4量体からなる。各4量体は、2対の同じポリペプチド鎖から構成される、対のそれぞれが、一つの「軽鎖」(約25 kDa)及び一つの「重鎖」(約50−70 kDa)を有している。各鎖のN−末端は、主に抗原認識の役割を担う約100から110またはそれ以上のアミノ酸からなる可変領域である。用語、可変軽鎖(VL)及び可変重鎖(VH)は、それぞれこの軽鎖及び重鎖のことである。
【0069】
「キメラ抗体」は、次のような抗体分子である:
(a)定常領域またはその一部が改変され、置換され、または交換され、その結果、抗原結合部位(可変領域)が、異なるまたは改変されたクラス、エフェクター機能及び/または種の定常領域と連結しているか、或いはキメラ抗体に新しい性質を付与する全く異なる分子、例えば、酵素、トキシン、ホルモン、成長因子、薬物などと連結している抗体分子;或いは
(b)可変領域またはその一部が、異なるまたは改変された抗原特異性を持つ可変領域により改変、置換または交換されている抗体分子。
【0070】
「抗−T2R」抗体は、T2R遺伝子、そのcDNA,または部分配列によりコードされたポリペプチドに特異的に結合する抗体または抗体フラグメントである。
【0071】
用語「免疫アッセイ」は、抗原に特異的に結合する抗体を使用するアッセイである。免疫アッセイは、抗原を単離し、標的とし、そして/または定量するために特別な抗体の特異的結合性を使用することが特徴である。
【0072】
タンパクまたはペプチドに関して、語句、抗体に「特異的に(選択的に)結合する」または「特異的に(選択的に)免疫反応する」とは、種々のタンパク及び生体成分の混合物中で、そのタンパクの存在を決定する結合反応のことである。したがって、指定された免疫アッセイ条件の下に、特定の抗体は、特定のタンパクと背景の少なくとも2倍結合し、検体中に存在するほかのタンパクとはあまり結合しない。そのような条件において抗体に特異的に結合するには、特別なタンパクに対する特異性を持つような抗体を選択する必要がある。
【0073】
例えば、ラット、マウス、またはヒトのような特定の動物種のT2Rファミリーメンバーに対して作成されたポリクロナール抗体は、T2Rポリペプチドまたはその免疫原部分と特異的に免疫反応するが、T2Rポリペプチドのオーソログまたは多形変異体及びアレル以外のほかのタンパクとは反応しないポリクロナール抗体のみを得るために選別することができる。この選別は、他の動物種のT2R分子または他のT2R分子と交差反応する抗体を取り除くことにより行われる。抗体はT2R GPCRファミリーメンバーのみを認識し、他のファミリーのGPCRを認識しないように選別することも可能である。種々の免疫アッセイ様式を、特定のタンパクと特異的に免疫反応する抗体を選別するために使用することができる。例えば、固相ELISA免疫アッセイは、タンパクと特異的に免疫反応する抗体を選別するために日常的に使用されている(例えば、Harlow & Lane, Antibodies, A Laboratory Manual, (1988)参照、免疫アッセイ様式及び特異的免疫反応性の測定に使用しうる条件を記述)。典型的には、特異的または選択的な反応は、背景信号またはノイズの少なくとも2倍であり、より典型的には背景の10から100倍以上であろう。
【0074】
語句「選択的に会合する」とは、上記に定義したように他のものと「選択的にハイブリッドを形成する」核酸の能力、または上記に定義したようにタンパクに「選択的に(または特異的に)結合する」抗体の能力のことである。
【0075】
用語「発現ベクター」とは、インビトロまたはインビボで、構成的または誘導的に、原核細胞、イースト、菌、植物、昆虫または哺乳動物細胞のいずれかの細胞中において、本発明の核酸配列を発現するための組換え発現系のことである。この用語は、線状または環状発現系を含む。この用語は、エピソームに存在するかまたは宿主細胞ゲノムに組み込まれた発現系を含む。発現系は、自己複製能力を持つものでも、持たないもの、すなわち、細胞中において一過性にのみ発現するものでもよい。この用語は、組換え核酸の転写のために最小限必要な配列のみを含む組換え発現「カセット」を含む。
【0076】
「宿主細胞」は、発現ベクターを含み、発現ベクターの複製又は発現を維持する細胞を意味する。宿主細胞は、E. coliのような原核細胞、またはイースト、昆虫、両生類のような真核細胞、またはCHO, HeLa, HEK-293のような哺乳動物細胞などとすることができ、例えば、培養細胞、外植細胞、及びインビボの細胞などがある。
【0077】
以上に基づいて、本発明は、苦味化合物によるhT2R4, hT2R44,またはhT2R61の特異的活性化を阻害する化合物を同定するアッセイを提供する。特に本発明は、キニーネまたは関連化合物によるhT2R4の活性化、6−ニトロサッカリン、サッカリン、または関連化合物によるhT2R61の活性化、またはデナトニウム、6−ニトロサッカリンまたは関連化合物によるhT2R44の活性化に影響を及ぼす(阻害する)化合物を同定するアッセイ、望ましくは細胞を使用するアッセイを提供する。上記に特定した苦味試験化合物によるこれらのhT2Rの活性化を阻害する化合物は、これらの特定のhT2Rと相互作用する他の苦味化合物により引き起こされる苦味を特異的に抑制することが期待される。
【0078】
特に、苦味アルカロイドであるキニーネは、T2R4を特異的に用量依存的に活性化することが認められた。このことは細胞内カルシウム濃度の変化を検出する、細胞によるアッセイにより発見された。(実施例で詳細に検討されるので)簡略に記述すると、大きなT細胞抗原及びG15 タンパクを安定的に発現するヒト胚腎細胞(HEK-G15)を、hT2R4発現プラスミドで(例えば、燐酸カルシウムを使用するかまたは脂質系を使用して)一過性に遺伝子導入した。さらに、他のHEK-G15細胞系を、他のヒトT2Rで一過性に遺伝子導入した。その後、蛍光を用いるアッセイを使用して一過性に遺伝子導入した細胞におけるカルシウム濃度の変化を検出した。このアッセイを使用することにより、hT2R4を発現する細胞にキニーネを加えると、PLCの活性化に至る信号伝達カスケードを誘発し、次いでカルシウム感受性蛍光色素を使用して検出される蛍光の増加を生じる細胞内カルシウム濃度の増加を生じる。これらの変化は、例えば、蛍光顕微鏡及び適当に設計されたソフトウエア(例えば、Imaging Workstation, Axon)を使用して監視される。これに対して、他のhT2Rで一過性に遺伝子導入されたHEK-G15細胞は、キニーネにより特異的に活性化しない。これらに基づくと、hT2R4は、キニーネ及び構造的に関連する化合物により特異的に活性化されるヒトT2Rであると思われる。したがって、キニーネによるhT2R4の活性化を阻害する化合物は、hT2R4により誘発される、特にhT2R4を活性化する苦味アルカロイドであるキニーネ及び関連化合物により誘発される苦味を抑制することが予期される。そのアッセイの例は後に詳細に開示する。
【0079】
また、本発明は特に、hT2R44がデナトニウム及び6−ニトロサッカリンによって特異的に活性化され、hT2R61が6−ニトロサッカリンによって特異的に活性化されるという発見を含んでいる。この発見は、図4に図解されているGTPγS結合アッセイを使用してなされた。このアッセイは、GPCR活性を監視する古典的なアッセイである(Wessling-Resnick and Johnson, J. Biol. Chem 262: 12444-12447(1987)参照)。このアッセイは、適当なリガンドの存在下、受容体がGタンパクを活性化できるという事実に基づいている。その結果、活性化されたGタンパクは、結合しているGDPをGTPに変換する。
【0080】
さらに、GTPの非加水分解性の類似体であるGTPγSを使用した場合には、ヌクレオチドは、Gタンパクにしっかりと結合したままとなり、結合していないヌクレオチドから分離することができる。したがって、このアッセイにおいて、結合したGTPγSの量は、活性化したGタンパクの量を反映し、それが受容体活性化の指標ともなる。GTPγ35S(「GTPγS」)結合アッセイは、ガストジューシン及びトランスジューシンのような種々のGタンパクを使用して行うことができる。後記の例に記述するように、Gタンパクとしてトランスジューシンを用いるこのアッセイを使用して、hT2R44 (SEQ ID NO.:3)またはhT2R61 (SEQ ID NO.:5)のいずれかを発現する昆虫細胞が、添加した苦味化合物の存在下に活性の増加を示すことが観察された。
【0081】
特に、後記の実験例に詳細に開示するように、hT2R61は、6−ニトロサッカリンの存在下に活性化され、hT2R44は、この化合物によって弱く活性化される。さらに、hT2R44は、特異的に安息香酸デナトニウムにより活性化される(安息香酸ナトリウムは、同濃度においてhT2R44を活性化しないという事実により示されるように、デナトニウム成分による活性化である)。
【0082】
これに対して、結合アッセイにより評価したときに、デナトニウムは、同じ一過性細胞発現系(Sf9バキュロウイルス発現系)を使用して発現した他のhT2Rを活性化しない。また、いくつかのデナトニウム誘導体も、hT2R44を活性化することが観察されたが、デナトニウムより程度は低かった。安息香酸デナトニウムは、ヒトに苦味を感じさせることが知られている苦味化合物の中で最も強いものの一つである。これらの結果は、hT2R44が、ヒトにおいて、デナトニウムの苦味を仲介していること、そしてhT2R44のデナトニウム活性化を阻害する化合物は、この受容体の活性化による苦味を調節(阻止)することを示唆している。
【0083】
したがって、本発明は、hT2R4活性化による苦味を阻止する化合物を同定する手段として、キニーネまたは関連化合物によるhT2R4活性化を調節(阻害)する化合物を同定するためのアッセイ、望ましくは細胞によるアッセイを提供する。
【0084】
また、本発明は、hT2R61活性化により惹起されるヒトにおける苦味を阻止する化合物を同定する手段として、6−ニトロサッカリンまたはサッカリンまたは関連化合物によるhT2R61活性化を調節(阻害)する化合物を同定するためのアッセイ、望ましくは細胞によるアッセイを提供する。
【0085】
また、本発明は、hT2R44活性化により惹起されるヒトにおける苦味を阻止する化合物を同定する手段として、デナトニウムまたは6−ニトロサッカリンまたは関連化合物によるhT2R44活性化を阻害する化合物を同定するためのアッセイ、望ましくは細胞によるアッセイを提供する。本発明に有用なアッセイ及び発現系は後に詳細に記述される。
【0086】
最も典型的に、これらのアッセイは、すでに特定したアミノ酸配列を有するhT2R4, hT2R44または hT2R61ポリペプチドをコードするDNAを発現する細胞を使用するであろう。しかし、hT2R4, hT2R44または hT2R61の機能的特性を保有するこれらの受容体ポリペプチドのフラグメント、オーソログ、変異体またはキメラもこれらのアッセイにおいて有用であることが予期される。その変異体の例としては、組換えまたは合成により調製されたか、天然に存在するスプライス変異体、一塩基多形、アレル変異体、及び突然変異体がある。本発明のアッセイにおいて使用されるT2Rの単離及び発現の方法、並びにhT2R4, hT2R44または hT2R61の活性化を抑制する化合物を同定するために、本発明において使用することが企図されているアッセイは、以下に記述される。
【0087】
T2Rの単離及び発現
本発明のT2R、またはそのフラグメント若しくは変異体の単離及び発現は、本出願に開示されたT2R核酸配列に基づいて構築されたプローブまたはプライマーを使用して、充分に確立されたクローニング方法により行うことができる。関連T2R配列は、本明細書で開示した配列及び既知のコンピューター支援探索技術、例えば、BLAST配列探索を使用して、ヒトまたは他の種のゲノムデータベースから同定することもできる。特別な態様において、本明細書で開示した偽遺伝子を、機能的アレルまたは関係遺伝子を同定するために使用することができる。
【0088】
次いで、これらの配列を機能的に発現させるための宿主細胞に感染または遺伝子導入するために、発現ベクターを使用することができる。これらの遺伝子及びベクターは、インビトロまたはインビボにおいて作製し、発現させることができる。核酸発現を変更及び調節するための望ましい表現型は、本発明のベクター中の遺伝子及び核酸(例えば、プロモーター、エンハンサーなど)の発現または活性を調節することにより得られることは、当業者はよく認識している。発現または活性を増加または減少するために、記述された既知の方法のいずれかを使用することができる。本発明は、科学および特許の文献に詳述されている当業者に既知の方法またはプロトコールのいずれとも組み合わせて実施することができる。
【0089】
そのほかに、これらの核酸は、例えば下記の既知化学合成技術によりインビトロにおいて合成することができる:Carruthers, Cold Spring Harbor Symp. Quant. Biol. 47: 411-18 (1982); Adams, Am Chem. Soc., 105: 661 (1983); Belousov, Nucleic Acids Res. 25: 3440-3444 (1997); Frenkel, Free Radic. Biol. Med. 19: 373-380 (1995); Blommers, Biochemistry 33: 7886-7896 (1994); Narang, Meth. Enzymol. 68: 90 (1979); Brown, Meth. Enzymol. 68: 109 (1979); Beaucage, Tetra. Lett. 22: 1859 (1981);米国特許番号 4,458,066。
次いで、相補鎖を合成して適当な条件下に鎖をアニーリングするか、または適当なプライマー配列とともにDNAポリメラーゼを使用して相補的鎖を加えることにより二本鎖DNAフラグメントを得ることができる。
【0090】
例えば、配列中の突然変異誘発、再クローニング、プローブの標識、配列分析、ハイブリッド形成などのような核酸の操作技術は、科学及び特許の文献に詳細に記述されており、例えば、Sambrook, ed., Molecular Cloning: A Laboratory Manual (2nd ed.), Vols. 1-3, Cold Spring Harbor Laboratory (1989); Ausubel, ed., Current Protocols in Molecular Biology, John Wiley & Sons, Inc., New York (1997); Tijssen, ed., Laboratory Techniques in Biochemistry and Molecular Biology: Hybridization With Nucleic Acid Probes, Part I, Theory and Nucleic Acid Preparation, Elsevier, N.Y. (1993)がある。
【0091】
核酸、ベクター、カプシド、ポリペプチドなどは、当業者に既知の多数の一般的な方法いずれかにより分析及び定量することができる。それらには、分析的生化学的方法、例えば、NMR,分光分析、放射線画像、電気泳動、キャピラリー電気泳動、高速液体クロマトグラフィー(HPLC)、薄層クロマトグラフィー(TLC)、及び超拡散クロマトグラフィー、種々の免疫学的方法、例えば、液相またはゲル沈降反応、免疫拡散、免疫電気泳動、放射線免疫アッセイ(RIA)、固相酵素免疫アッセイ(ELISA)、免疫蛍光アッセイ、サザン分析、ノーザン分析、ドットブロット分析、ゲル電気泳動(例えば、SDS-PAGE)、RT-PCR,定量的PCR、そのほかの核酸または標的またはシグナル増幅方法、放射線標識、シンチレーション計測、及びアフィニティークロマトグラフィーが含まれる。
【0092】
T2Rリガンド結合領域をコードする核酸を増幅するために、オリゴヌクレオチドプライマーを使用することができる。本明細書で記述する核酸は、増幅技術を使用してクローニングまたは定量的に測定することができる。増幅方法は当業者に既知であり、例えば、ポリメラーゼ連鎖反応(PCR)(Innis ed., PCR Protocols, a Guide to Methods and Applications, Academic Press, N.Y. (1990); Innis ed., PCR Strategies, Academic Press, Inc., N.Y. (1995));リガーゼ連鎖反応(LCR)(Wu, Genomics, 4: 560 (1989); Landegren, Science, 241: 1077 (1988); Barringer, Gene, 89: 117 (1990)); 転写増幅(Kwoh, PNAS, 86: 1173 (1989)); 自己維持配列複製(Guatelli, PNAS, 87: 1874(1990)); Q−βレプリカーゼ増幅(Smith, J. Clin. Microbiol., 35: 1477-91(1997)); 自動化Q−βレプリカーゼ増幅アッセイ(Burg, Mol. Cell. Probes, 10: 257-71(1996)); 及びそのほかのRNAポリメラーゼを使用する技術(例えば、NASBA,Cangene, Mississauga, Ontario)がある。Berger, Methods Enzymol., 152: 307-16(1987); Sambrook; Ausubel;米国特許番号 4,683,195及び4,683,202; Sooknanan, Biotechnology, 13: 563-64 (1995)も参照。
【0093】
核酸は、ひとたび増幅されると個別的にまたはライブラリーとして、必要に応じて種々のベクター中に通常の分子生物学的方法を使用する当業者に既知の方法に従ってクローニングすることができる。インビトロにおける増幅核酸のクローニング方法は、例えば、米国特許番号5,426,039に記述されている。増幅配列のクローニングを容易にするために、制限酵素部位をPCRプライマー対に「組み込む」こともできる。例えば、Pst I及びBsp E1部位を、本発明の代表的プライマー中に設計した。これらの特別な制限部位は、連結したときに、それらがスプライスされて得られる7回膜貫通受容体「ドナー」のコード配列に関し「フレーム単位」とする配列を有する(リガンド結合領域コード配列は、7回膜貫通ポリペプチドにとって本質的であるため、構築体が制限酵素スプライス部位の下流で翻訳されることを望むならば、フレームのずれは避けなければならない;これは挿入されたリガンド結合領域が実質的に膜貫通VII領域のほとんどを含む場合には必要ないであろう)。プライマーは、「ドナー」7回膜貫通受容体の元の配列を維持するように設計することができる。そのほかに、プライマーは、保存置換基(例えば、疎水性残基に代わる疎水性残基、前記検討を参照)または機能上害のない(例えば、形質膜挿入を妨害しない、タンパク分解酵素による切断を生じない、受容体の異常な折り畳みを生じない、など)置換基であるアミノ酸残基をコードすることができる。
【0094】
プライマー対は、T2Rタンパクのリガンド結合領域を選択的に増幅するように設計することができる。これらの領域は、異なるリガンドに対しては変わることがある;したがって、一つのリガンドに対する最小限の結合領域であるものは、第二の潜在的リガンドに対しては制限的になりすぎる。したがって、種々のドメイン構造を含む種々のサイズの結合領域を増幅するとよい;例えば、7回膜貫通T2Rの膜貫通(TM)ドメインIIからVII,IIIからVII,IIIからVIまたはIIからVI、またはそれらの変異体(例えば、個別ドメインの部分配列のみ、ドメインのオーダーの混合、など)などである。
【0095】
7回膜貫通T2Rタンパクのドメイン構造及び配列は既知であるので、当業者は、縮重プライマー対を設計するためのモデル配列として、ドメイン隣接及び内部ドメイン配列を容易に選択することができる。例えば、ドメイン領域IIからVIIをコードする核酸配列は、プライマー対を使用してPCR増幅により作製することができる。膜貫通ドメインI(TM I)配列を含む核酸を増幅するために、縮重プライマーを、前記のT2Rファミリー共通配列1のアミノ酸配列をコードする核酸から設計することができる。その縮重プライマーは、TM IからTM III,TM IからTM IV,TM IからTM V,TM IからTM VI,またはTM IからTM VIIを組み込んだ結合領域を作製するために使用することができる。他の縮重プライマーは、本明細書で提供した他のT2Rファミリー共通配列を基に設計することができる。そのような縮重プライマーは、TM IIIからTM IV,TM IIIからTM V,TM IIIからTM VIまたはTM IIIからTM VIIを組み込んだ結合領域を作製するために使用することができる。
【0096】
縮重プライマー対を設計する代表例は、当業者に良く知られている。例えば、共通配列縮重ハイブリッドオリゴヌクレオチドプライマー(CODEHOP)法コンピュータープログラムは、http://blocks.fhcrc.org/codehop.html,で接続可能であり、既知の味覚受容体リガンド結合領域として、一連の関連タンパク配列で始まるハイブリッドプライマー予測のためのBlockMaker 多重配列整列サイトと直接リンクしている(例えば、Rose, Nucleic Acids Res., 26: 1628-35 (1998); Singh, Biotechniques, 24: 318-19 (1998)参照)。
【0097】
オリゴヌクレオチドプライマー対を合成する方法は、当業者に良く知られている。「天然」塩基対または合成塩基対を使用することができる。例えば、人工核酸塩基を使用することにより、プライマー配列を操作するための多彩な方法が提供され、増幅産物のより複雑な混合物が作成される。人工核酸塩基の種々のファミリーは、内部結合回転による多重水素結合定位を予想することができ、縮重分子の認識のための手段を提供する。これらの類似体を、PCRプライマーの一つの位置に組み込むことにより、増幅産物の複雑なライブラリーを作製することができる。例えば、Hoops, Nucleic Acids Res., 25: 4866-71 (1997)参照。非極性分子も天然のDNA塩基の形状を模倣するために使用することができる。アデニンの非水素結合形状模倣体は、チミンの非極性形状模倣体に対して効率よく選択的に複製される(例えば、Morales, Nat. Struct. Biol., 5: 950-54(1998)参照)。例えば、二つの縮重塩基は、ピリミジン塩基6H,8H-3,4-ジヒドロピリミド[4,5-c][1,2]オキサジン-7-オンまたはプリン塩基N6-メトキシ-2,6-ジアミノプリンとすることができる(例えば、Hill, PNAS, 95: 4258-63 (1998)参照)。本発明の代表的縮重プライマーは、核酸塩基類似体5'-ジメトキシトリチル-N-ベンゾイル-2'-デオキシ-シチジン、3'-[(2-シアノエチル)-(N,N-ジイソプロピル)]−ホスフォルアミダイト(配列中の文字「P」、上記参照)を組み込んでいる。このピリミジン類似体は、A及びG残基を含むプリンと水素結合する。
【0098】
本明細書で開示された味覚受容体と実質的に同一である多形変異体、アレル、及び種間同属体は、上記の核酸プローブを使用して単離することができる。また、発現ライブラリーを使用して、T2R同属体を認識し、結合する、T2Rポリペプチドに対して作製された抗血清または精製抗体で、発現同属体を検出することにより、T2Rポリペプチド及びその多形変異体、アレル、及び種間同属体をクローニングすることができる。
【0099】
味覚受容体のリガンド結合領域をコードする核酸を、適当な(完全または縮重)プライマー対を使用して、適当な核酸配列を(例えば、PCRで)増幅することにより作製することができる。増幅される核酸としては、細胞若しくは組織のゲノムDNAまたは味覚受容体発現細胞に由来するmRNA若しくはcDNAがある。
【0100】
一の態様において、転移配列に融合させたT2Rをコードする核酸を含む、ハイブリッドタンパクコード配列を構築することができる。転移モチーフ及び化学的感受性受容体の他のファミリー、特に味覚受容体、の味誘発化合物結合領域を含むハイブリッドT2Rも提供される。これらの核酸配列は、転写または翻訳調節配列、例えば、転写及び翻訳開始配列、プロモーター及びエンハンサー、転写及び翻訳ターミネーター、ポリアデニル化配列、及びそのほかのDNAをRNAに転写するのに有用な配列、に作動的に連結することができる。組換え発現カセット、ベクター、及び形質転換体の構築において、目的とする細胞または組織の全てに目的とする核酸の発現を指令するためにプロモーターフラグメントを使用することができる。
【0101】
そのほかの態様において、融合タンパクは、C−末端またはN−末端転移配列を含むことができる。さらに、融合タンパクは、追加の配列、例えば、タンパク検出、精製、またはそのほかの適用のための配列を含むことができる。検出及び精製を容易にするドメインとしては、例えば、ポリヒスチジン鎖のような金属キレートペプチド、ヒスチジン−トリプトファンモジュール、または固相金属上で精製できるようなそのほかのドメイン;マルトース結合タンパク;固相免疫グロブリン上で精製できるタンパクAドメイン;またはFLAGSエクステンション/親和性精製システム(Immunex Corp,Seattle WA)に使用されるドメイン、がある。
【0102】
ファクターXa(例えば、Ottavi, Biochimie, 80: 289-93 (1998)参照)、ズブチリシンタンパク分解酵素認識モチーフ(例えば、Polyak, Protein Eng., 10: 615-19 (1997)参照);エンテロキナーゼ(Invitrogen, San Diego, CA)などの切断可能なリンカー配列を、(効率的に形質膜に発現させるための)転移配列と新規に翻訳されるポリペプチドの残りの部分との間に入れることは、精製を容易にするために有用である。例えば、一の構築体は、その後に、チオレドキシン、エンテロキナーゼ切断部位(例えば、Williams, Biochemistry, 34: 1787-97 (1995)参照)、及びC−末端転移ドメインが続く6ヒスチジン残基に連結させるポリペプチドをコードする核酸配列を含むことができる。ヒスチジン残基は、検出及び精製を容易にし、他方エンテロキナーゼ切断部位は、残余の融合タンパクから目的タンパクを精製する手段を提供する。融合タンパクをコードするベクターに関する技術及び融合タンパクの応用は、科学及び特許の文献に詳細に記述されている(例えば、Kroll, DNA Cell. Biol., 12: 441-53 (1993)参照)。
【0103】
リガンド結合領域コード配列を含む発現ベクターは、個々の発現ベクターまたは発現ベクターのライブラリーとして、科学及び特許文献に詳細に記述されている種々の通常技術により、ゲノム中または細胞質中または細胞の核中に導入し、発現させることができる。例えば、Roberts, Nature, 328: 731 (1987); Berger 前出;Schneider , Protein Expr. Purif., 6435: 10 (1995); Sambrook; Tijssen; Ausubel.参照。生化学試薬及び実験機器のメーカーの製品情報も既知の生化学方法に関する情報を提供している。ベクターは、天然資源から単離することができ、ATCCまたはGenBankライブラリーのような資源から入手することができ、或いは合成的にまたは組換えの方法により調製することができる。
【0104】
核酸は、細胞中に安定的にまたは一過性に発現する(例えば、エピソーム発現系)発現カセット、ベクターまたはウイルス中で発現させることができる。選別マーカーは、発現カセット及びベクター中に組み込まれ、形質転換された細胞及び配列に選別可能な形質を付与することができる。例えば、選別マーカーは、宿主ゲノムに組み込む必要はないように、エピソームの維持及び複製をコード化することができる。例えば、目的とするDNA配列で形質転換した細胞の選別を可能にするために、マーカーは、抗生物質耐性(例えば、クロラムフェニコール、カナマイシン、G418, ブレオマイシン、ヒグロマイシン)または除草剤耐性(例えば、クロロスルフロンまたはバスタ)をコード化することができる(例えば、Blondelet-Rouault, Gene, 190: 315-17 (1997);Aubrecht, J. Pharmacol. Exp. Ther., 281: 992-97 (1997)参照)。ネオマイシンまたはヒグロマイシンのような基質に対する耐性を付与する選別マーカーは、組織培養にのみ使用されるので、化学耐性遺伝子も、インビトロ及びインビボにおける選別マーカーとして使用される。
【0105】
キメラ核酸配列は、いずれかの7回膜貫通ポリペプチドの中のT2Rリガンド結合領域をコードすることができる。7回膜貫通受容体ポリペプチドは、同じ一次及び二次及び三次構造を有しているので、構造ドメイン(例えば、細胞外ドメイン、TMドメイン、細胞質ドメイン、など)は、配列分析により容易に同定することができる。例えば、同属体設計、フーリエ解析及び螺旋周期性検出により、7回膜貫通受容体配列を有する7個のドメインを同定し分析することができる。迅速フーリエ変換(FFT)アルゴリズムを、分析する配列の疎水性及び可変性を特徴付ける主要周期を評価するために使用することができる。周期性検出及びα螺旋周期性指標は、例えば、Donnelly, Protein Sci., 2: 55-70 (1993)により得ることができる。そのほかの整列及びモデルアルゴリズムは、当業者に既知である(例えば、Peitsch, Receptors Channels, 4: 161-64 (1996); Kyte & Doolittle, J. Md. Biol., 157: 105-32 (1982); Cronet, Protein Eng., 6: 59-64 (1993); http://bioinfo.weizmann.ac.il/参照)。
【0106】
本発明は、特定の核酸配列及びアミノ酸配列を有する核酸分子及びポリペプチドを含むだけでなく、そのフラグメント、特に、例えば、40,60,80,100, 150,200,または250ヌクレオチドまたはそれ以上のフラグメント、並びに例えば、10,20,30,50,70,100,または150アミノ酸、またはそれ以上のポリペプチドフラグメントも含んでいる。任意に、核酸フラグメントは、T2Rファミリーメンバーに対して作製された抗体に結合することができる抗原性ポリペプチドをコードすることができる。さらに、本発明のタンパクフラグメントは、任意にT2Rファミリーメンバーに対して作製された抗体に結合することができる抗原性フラグメントとすることができる。
【0107】
他のGPCR、望ましくは7回膜貫通スーパーファミリーのメンバーの全体または一部である追加のアミノ酸と結合した、本明細書で記述するT2Rポリペプチドの少なくとも一つの、少なくとも10,20,30,50,70,100,または150アミノ酸、またはそれ以上を含むキメラタンパクも企図される。これらのキメラは、当該受容体と他のGPCRとから作ることもでき、また、本発明の受容体の二つまたはそれ以上を組み合わせて作ることもできる。一の態様において、キメラの一部は、本発明のT2Rポリペプチドの膜貫通ドメインに相当または由来する。別の態様において、キメラの一部は、本明細書で記述するT2Rポリペプチドの膜貫通領域の一つまたはそれ以上に相当、または由来し、そして残りの部分は他のGPCRに由来し得る。キメラ受容体は、当業者に既知であり、それを作製する技術及びその中に組み込むためのGタンパク共役受容体のドメインまたはフラグメントの選別及び境界も良く知られている。したがって、当業者のこの知識をそのようなキメラ受容体を創るために容易に使用することができる。そのようなキメラ受容体を使用することにより、例えば、先行技術のアッセイ系に使用された既知の受容体のような他の受容体の信号伝達特性とともに、本明細書で具体的に開示した一の受容体の味選択特性を提供することができる。
【0108】
例えば、リガンド結合領域、細胞外ドメイン、膜貫通ドメイン、細胞質ドメイン、N−末端ドメイン、C−末端ドメインのような領域、またはそれらの組み合わせを異種タンパクに共有結合で連結することができる。例えば、T2R膜貫通領域を、異種GPCR膜貫通ドメインに連結することができ、または異種GPCR細胞外ドメインを、T2R膜貫通領域に連結することもできる。そのほかに選択される異種タンパクには、例えば、緑色蛍光タンパク、β−gal, グルタミン酸受容体、及びロドプシンN−末端が含まれる。
【0109】
本発明のT2R、フラグメント、または変異体を発現するための宿主細胞も本発明の範囲に入る。本発明のT2R、フラグメントまたは変異体をコードするcDNAのようなクローニング遺伝子または核酸の高度な発現を得るために、当業者は、通常、目的の核酸配列を、転写を指令する強力なプロモーター、転写/翻訳ターミネーター、及び、もし核酸がタンパクをコードするならば、翻訳開始のためのリボソーム結合部位、を含む発現ベクター中に再クローニングする。適当な細菌プロモーターは当業者に既知であり、例えば、Sambrookらにより記述されている。しかし、細菌発現系または真核発現系を使用することができる。
【0110】
宿主細胞中に外来核酸配列を導入する既知方法のいずれかを使用することができる。導入方法としては、リン酸カルシウムトランスフェクション、ポリブレン、プロトプラスト融合、電気穿孔、リポソーム、マイクロインジェクション、原形質ベクター、ウイルスベクター、及びそのほかのクローン化ゲノムDNA、cDNA, 合成DNAまたはそのほかの外来遺伝物質を宿主細胞に導入する既知方法がある(例えば、Sambrook et al.参照)。使用される特定の遺伝子操作方法により、本発明のT2R、フラグメント、または変異体を発現することができる宿主細胞の中に少なくとも1核酸分子を確実に導入できることだけが必要である。
【0111】
発現ベクターを細胞中に導入した後、遺伝子導入細胞を、目的の受容体、フラグメントまたは変異体の発現に適した条件の下に培養し、次いで標準的な技術を使用してその細胞を培養から回収する。そのような技術の例は、当業者に既知である。例えば、WO 00/06593が参照され、この文献は、この開示と調和するように組み込まれる。
【0112】
hT2R4, hT2R44またはhT2R61の活性を調節する化合物を検出するアッセイ
インビトロ及びインビボにおいて、試験化合物が、本発明のT2Rポリペプチドに特異的に結合するか否かを測定する方法及び成分を以下に記述する。細胞生理の多くの局面を監視して、天然に存在するかまたはキメラのT2Rに対するリガンド結合による効果を評価することができる。これらのアッセイを、T2Rポリペプチドを発現する完全な細胞、透過性細胞、または標準的方法により作製された膜フラクションで行うことができる。
【0113】
味覚受容体は、味覚誘発化合物と結合し、化学的刺激の電気信号への変換を始める。活性化された、または阻害されたGタンパクは、次には標的酵素、チャネル、及び他のエフェクタータンパクの性質を変える。いくつかの例としては、可視系におけるトランスジューシンによるcGMPホスフォジエステラーゼの活性化、刺激性Gタンパクによるアデニル酸シクラーゼの活性化、Gq及び他の同種GタンパクによるホスフォリパーゼCの活性化、並びにGi及び他のGタンパクによる種々のチャネルの調節がある。また、ホスフォリパーゼCによるジアシルグリセロールの生成及びIP3の生成、及びその次のIP3によるカルシウム移動などの下流の事象も測定することができる。
【0114】
このアッセイのT2Rタンパクまたはポリペプチドは、典型的には、SEQ ID NO.:1,3,または5に含まれる配列を持つポリペプチド、またはそのフラグメント若しくは保存的修飾変異体から選択されるであろう。
【0115】
また、このアッセイのT2Rタンパクまたはポリペプチドは、真核宿主細胞から誘導することができ、SEQ ID NO.:1,3,または5とアミノ酸相同性を持つアミノ酸配列、またはその保存的修飾変異体を含むことができる。一般的に、アミノ酸配列相同性は、少なくとも30%、望ましくは30−40%、より具体的には50−60、70%、75%、80%、85%、90%、95%、96%、97%、98%、または99%であろう。任意に、このアッセイのT2Rタンパクまたはポリペプチドは、細胞外ドメイン、膜貫通領域、細胞質ドメイン、リガンド結合ドメインなどのようなT2Rポリペプチドの領域を含むことができる。任意に、T2Rポリペプチドまたはその部分は、異種タンパクと共有結合で連結して、本明細書で記述するアッセイに使用するキメラタンパクを創ることができる。
【0116】
T2R活性の調節物質は、上記のような、組換えまたは天然に存在するT2Rタンパクまたはポリペプチドを使用して試験することができる。T2Rタンパクまたはポリペプチドは、組換え体または天然に存在するものとして、単離し、細胞中で発現させ、細胞から得た膜で発現させ、組織または動物で発現させることができる。例えば、舌のスライス、舌から分離した細胞、形質転換細胞、または膜を使用することができる。調節物質は、本明細書で記述するインビトロまたはインビボアッセイの一つを使用して試験することができる。
【0117】
調節物質の検出
インビトロ及びインビボにおいて、試験化合物が、本発明のT2R受容体に特異的に結合するか否かを調べるための成分及び方法を以下に記述する。細胞生理の多くの面をモニターして、本発明のT2Rポリペプチドへのリガンド結合による効果を評価することができる。これらのアッセイは、化学感受性受容体を発現する完全な細胞、透過性細胞、または標準的方法により若しくは新たに合成されたタンパクをインビトロで使用して作製した膜フラクションで、実施することができる。
【0118】
インビボにおいて、味覚受容体は、味調節化合物と結合し、化学的刺激を電気信号に変換することを開始する。活性化されたまたは阻害されたGタンパクは、次には標的酵素、チャネル、及びそのほかのエフェクタータンパクの性質を変える。いくつかの例としては、可視系におけるトランスジューシンによるcGMPホスフォジエステラーゼの活性化、刺激性Gタンパクによるアデニル酸シクラーゼの活性化、Gq及び他の同種GタンパクによるホスフォリパーゼCの活性化、及びGi及び他のGタンパクによる種々のチャネルの調節がある。また、ホスフォリパーゼCによるジアシルグリセロール及びIP3の生成、並びにその次のIP3によるカルシウム移動などの下流の事象も測定することができる。
【0119】
また、このアッセイのT2Rタンパクまたはポリペプチドは、真核宿主細胞から誘導することができ、本明細書で開示したT2Rポリペプチドとアミノ酸配列相同性を持つアミノ酸配列、またはそのフラグメント若しくは保存的修飾変異体を含むことができる。一般的に、アミノ酸配列相同性は、少なくとも35−50%、または任意に75%、85%、90%、95%、96%、97%、98%、若しくは99%であろう。任意に、このアッセイのT2Rタンパクまたはポリペプチドは、細胞外ドメイン、膜貫通領域、膜貫通ドメイン、細胞質ドメイン、リガンド結合ドメインなどのようなT2Rタンパクのドメインを含むことができる。さらに、T2Rポリペプチドまたはそのドメインは、異種タンパクと共有結合で連結して、本明細書で記述するアッセイに使用するキメラタンパクを創ることができる。
【0120】
T2R受容体活性の調節物質は、組換えか天然に存在する、上記のT2Rタンパクまたはポリペプチドを使用して試験される。組換え若しくは天然に存在するT2Rタンパクまたはポリペプチドは、単離し、細胞中で発現させ、細胞から得られる膜で発現させ、組織若しくは動物中で発現させることができる。例えば、舌のスライス、舌から分離した細胞、形質転換細胞、または膜を使用することができる。調節は、本明細書で記述するインビトロまたはインビボでのアッセイの一つを使用して試験することができる。
【0121】
1.インビトロ結合アッセイ
味覚変換も、本発明のT2Rポリペプチドを使用して、可溶相若しくは固相の反応で、インビトロにおいて試験することができる。特別な態様において、T2Rリガンド結合ドメインを、可溶相若しくは固相の反応において、インビトロで使用して、リガンド結合についてアッセイすることができる。
【0122】
リガンド結合ドメインは、膜貫通ドメインの細胞外ループのような細胞外ドメインの付加的部分とともに、N−末端ドメインから形成することができる。
【0123】
インビトロ結合アッセイは、代謝型グルタミン酸受容体のようなGPCRで使用されている(例えば、Han and Hampson, J. Biol. Chem. 274: 10008-10013 (1999)参照)。これらのアッセイは、放射活性または蛍光の標識リガンドで置換し、内在性の蛍光の変化またはタンパク分解感受性の変化などを測定することなどを含む。
【0124】
本発明によるT2Rポリペプチドに対するリガンド結合は、溶液中、任意に固相に付着させた二層膜、単層脂質膜、または小胞において試験することができる。調節物質の結合は、例えば、分光学的特性(例えば、蛍光、吸光度、屈折率)、流体力学的(例えば、形状)、クロマトグラフィー、または溶解特性の変化を使用して試験することができる。
【0125】
本発明の望ましい態様においては、[35S]GTPγS結合アッセイを使用する。前述のように、GPCRの活性化により、Gタンパク複合体のGαサブユニットは、結合しているGDPのGTPへの交換を促進する。Gタンパク交換活性のリガンド仲介による促進は、候補リガンドの存在下に加えられた放射活性標識[35S]GTPγSのGタンパクへの結合を測定する生化学的アッセイにおいて測定することができる。典型的には、目的の化学感受性受容体を含む膜をGタンパクと混合する。潜在的阻害物質及び/または活性化物質並びに[35S]GTPγSを、アッセイに添加し、Gタンパクへの[35S]GTPγSの結合を測定する。結合は、液体シンチレーション計測により、またはシンチレーション近接アッセイ(SPA)を含む、当業者に既知のそのほかの方法により測定することができる。他のアッセイ形態においては、蛍光標識GTPγSを利用することができる。
【0126】
2.蛍光偏光アッセイ
他の態様において、リガンド結合を検出及び監視するために蛍光偏光(「FP」)によるアッセイを使用することができる。蛍光偏光は、結合平衡、核酸ハイブリダイゼーション、及び酵素活性を測定するために広い用途で適用される技術である。蛍光偏光アッセイは、均質系であり、遠心分離、濾過、クロマトグラフィー、沈殿、または電気泳動などの分離操作を必要としない。このアッセイは、リアルタイムで、溶液中で直接行われ、固相を必要としない。偏光の測定は、迅速であり、検体を破壊しないので、偏光値は、繰り返し且つ試薬の添加後に測定することができる。一般的に、この技術は、ピコモルからマイクロモルの低レベルの蛍光の偏光値を測定するために使用することができる。本発明のT2Rポリペプチドに対するリガンドの結合を測定するための簡単にして定量的な方法において、どのように蛍光偏光を使用しうるかを、このセクションでは記述する。
【0127】
蛍光標識された分子を面偏光で励起すると、その分子の回転に反比例する程度に偏光した光を放射する。蛍光標識された大きい分子ほど、励起状態(フルオレッセインの場合に4ナノ秒)の間、静止しており、光の偏光は、相対的に励起と放射の間一定に維持される。蛍光標識された小分子は、励起状態の間、急速に回転し、偏光は、励起と放射との間で顕著に変化する。したがって、小分子は低い偏光値を示し、大分子は高い偏光値を示す。例えば、一本鎖蛍光標識オリゴヌクレオチドは、低い蛍光値を有するが、相補的鎖とハイブリッド形成すると、高い偏光値を持つ。本発明の化学感受性受容体を活性化または阻害する味覚誘発化合物の結合を検出及び監視するためにFPを使用する場合には、蛍光標識味覚誘発化合物または蛍光自発味覚誘発化合物を使用しうる。
【0128】
蛍光偏光(P)は次のように定義される:
[式中、IntΠは、励起光面に平行な放射光の強度、Int⊥は、励起光面に垂直な放射光の強度である。Pは、光の強度の比であり、無次元の数である。]。
例えば、BeaconTM及びBeacon 2000TMシステムをこのアッセイに関して使用することができる。そのシステムでは典型的に偏光をミリ偏光単位(1偏光単位=1000 mP単位)で示している。
【0129】
分子の回転とサイズの関係はPerrin式により記述されており、読者はこの式の詳細な説明を載せているJolley, M.E. (1991) Journal of Analytical Toxicology, pp.236-240,を参照されたい。要するに、Perrin式が示すところは、偏光が、直接、回転緩和時間に比例するということであり、その回転緩和時間とは、分子が約68.5度の角度を回転するのに要する時間のことである。回転緩和時間は粘度(η)、絶対温度(T)、分子容積(V)、及び気体定数(R)に関係し、次の式で示される:
【0130】
回転緩和時間は、小分子(例えば、フルオレッセイン)に対しては小さく(約1ナノ秒)、大分子(例えば、免疫グロブリン)に対しては大きい(約100ナノ秒)。粘度と温度を一定に保つ場合には、回転緩和時間、及びそれ故偏光は、分子容積に直接的に相関する。分子容積の変化は、蛍光標識分子の他の分子との相互作用、解離、重合、分解、ハイブリッド形成、または立体配座変化に起因するであろう。例えば、蛍光偏光は、プロテアーゼ、DNアーゼ、及びRNアーゼによる蛍光標識した大きな重合体の酵素分解を測定するために使用されてきた。また、タンパク/タンパク相互作用、抗体/抗原結合、及びタンパク/DNA結合の結合平衡を測定するためにも使用されてきた。
【0131】
A. 固相及び液相ハイスループットアッセイ
さらに他の態様において、本発明は、T2Rポリペプチド;またはT2Rポリペプチドを発現する細胞または組織を使用する可溶相アッセイを提供する。他の態様において、本発明は、固相を使用するハイスループット方式のインビトロアッセイを提供する。その方式において、T2RポリペプチドまたはT2Rポリペプチドを発現する細胞若しくは組織は、固相基質に付着され、味覚刺激化合物をT2R受容体と接触させ、結合を、適当なタグまたはT2Rに対して作製した抗体を使用して検出する。
【0132】
本発明のハイスループットアッセイでは、1日に異なる調節物質またはリガンドの数千個をスクリーニングすることが可能である。特に、マイクロタイタープレートの各ウエルを、選択した潜在的調節物質に対する個別のアッセイを行うために使用することができ、あるいは、濃度またはインキュベーション時間の効果を調べようとする場合には、5−10ウエル毎に1つの調節物質を試験することができる。このように、一枚の標準的マイクロタイタープレートは、約100(例えば、96)調節物質をアッセイすることができる。もし1536ウエルプレートを使用するならば、一枚のプレートで約1000から約1500の異なる化合物を容易にアッセイすることができる。各プレートウエル中において複数の化合物をアッセイすることも可能である。1日に数枚の異なるプレートをアッセイすることが可能である;本発明の集積システムを使用して約6,000−20,000の異なる化合物のスクリーニングアッセイが可能である。さらに最近、試薬操作のための微量流体法が開発された。
【0133】
目的の分子は、直接的にまたは間接的に、共有結合によりまたは非共有結合により、例えば、タグを介して固相成分に結合することができる。タグは種々の構成物とすることができる。一般的に、タグと結合する分子(タグバインダー)を固相支持層に固定し、タグをつけた目的分子(例えば、目的の味覚伝達分子)をタグ及びタグバインダーの相互作用により固相に接着する。
【0134】
文献に詳細に記述されている既知分子相互作用に基づいて、多数のタグ及びタグバインダーを使用することができる。例えば、タグが天然のバインダー、例えば、ビオチン、プロテインA、またはプロテインGを持つ場合には、適当なタグバインダー(アビジン、ストレプトアビジン、ニュートラビジン、免疫グロブリンのFc領域、など)と組み合わせて使用することができる。また、ビオチンのような天然バインダーを有する分子に対する抗体も、広く使用することができ、適当なタグバインダーである(SIGMA Immunochemicals 1998 catalogue SIGMA, St. Louis MO,参照)。
【0135】
同様に、ハプテン性または抗原性化合物は、適当な抗体と組み合わせて使用して、タグ/タグバインダー対を形成することができる。数千の特異的抗体は、市販品を入手可能であり、また多数のそのほかの抗体は文献に記述されている。例えば、よくある形態として、タグは第一抗体であり、タグバインダーは第一抗体を認識する第二抗体である。抗体−抗原相互作用に加えて、受容体−リガンド相互作用も、タグ及びタグバインダー対として適当である。例えば、細胞膜受容体のアゴニスト及びアンタゴニスト(例えば、トランスフェリン、c-kit, ウイルス受容体リガンド、サイトカイン受容体、ケモカイン受容体、インターロイキン受容体、免疫グロブリン受容体及び抗体、カドヘリンファミリー、インテグリンファミリー、セレクチンファミリーなどの細胞受容体−リガンド相互作用;例えば、Pigott & Power, The Adhesion Molecule Facts Book 1(1993)参照)。同様に、トキシン及び毒素、ウイルス抗原決定基、ホルモン(例えば、オピエート、ステロイド、など)、細胞内受容体(例えば、ステロイド、甲状腺ホルモン、レチノイド及びビタミンDを含む種々の小さなリガンドの効果を仲介するもの;ペプチド)、薬物、レクチン、糖、核酸(線状及び環状ポリマー型の両者)、オリゴ糖、タンパク、リン脂質及び抗体は全て、種々の細胞受容体と相互作用しうる。
【0136】
ポリウレタン、ポリエステル、ポリカーボネート、ポリウレア、ポリアミド、ポリエチレンイミン、ポリアリレンスルフィド、ポリシロキサン、ポリイミド、及びポリアセテートのような合成ポリマーも適当なタグまたはタグバインダーを形成することができる。この開示を通覧することにより当業者には明白であるように、そのほかの多数のタグ/タグバインダー対も本明細書で開示したアッセイシステムに有用である。
【0137】
ペプチド、ポリエーテルなどの一般的リンカーは、タグとして寄与することができ、約5から200アミノ酸の間のポリgly配列のようなポリペプチド配列を含む。そのようなフレキシブルリンカーは、当業者には既知である。例えば、ポリ(エチレングリコール)リンカーは、Shearwater Polymers, Inc., Huntsville, Alabamaから入手できる。これらのリンカーは、任意にアミド結合、スルフヒドリル結合、または異種機能の結合を有している。
【0138】
タグバインダーは、現在使用しうる種々の方法を使用して固相基質に固定される。固相基質は、通常、その基質の全てまたは部分を、タグバインダーの一部と反応する化学基を表面に固定する化学試薬に暴露することにより、誘導されまたは機能化される。例えば、長鎖部分を付着するのに適した基としては、アミン、ヒドロキシル、チオール、及びカルボキシル基がある。アミノアルキルシラン及びヒドロキシアルキルシランは、ガラス表面のような、様々な表面へと機能化するために使用することができる。そのような固相バイオポリマーを構築する方法は文献に詳細に記述されている。例えば、Merrifield, J. Am. Chem. Soc., 85: 2149-2154 (1963)(例えば、ペプチドの固相合成を記述している);Geysen et al., J. Immun. Meth., 102: 259-274 (1987)(ピン上における固相成分の合成を記述している);Frank & Doring, Tetrahedron 44: 6031-6040 (1988)(セルロース円盤上における種々のペプチド配列の合成を記述している);Fodor et al., Science, 251: 767-777 (1991); Sheldon et al., Clinical Chemistry, 39(4): 718-719(1993); 及びKozal et al., Nature Medicine, 2(7): 753-759 (1996)(全て固相に固定したバイオポリマーのアレイを記述している)参照。
基質にタグバインダーを固定する非化学的方法には、例えば、熱、UV照射による架橋結合のような他の一般的方法がある。
【0139】
3.細胞によるアッセイ
望ましい一の態様において、T2Rタンパクは、非修飾の形態において、或いは異種の、分泌経路による成熟化及び標的化を促進するシャペロン配列を持つかまたは望ましくは持たないキメラ、変異体または短縮された受容体として、真核細胞中で発現する。そのT2Rポリペプチドは、HEK-293細胞のような真核細胞で発現することができる。望ましくは、細胞は、細胞内シグナリング経路またはホスフォリパーゼCのようなシグナリングタンパクに、キメラ受容体を連結させることができる、機能的Gタンパク、例えばGα15を含む。その細胞内におけるT2R受容体の活性化は、例えば、細胞内のFURA-2による蛍光を検出することによる細胞内カルシウムの変化を検出する、標準的な方法を使用して検出することができる。そのようなアッセイが、本出願に示した実験的知見の基礎になっている。
【0140】
活性化したGPCR受容体は、受容体のC−末端(及び多分他の部位も)をリン酸化するキナーゼの基質になることがある。したがって、活性化物質は、シンチレーションカウンターでアッセイすることができる、放射標識ATPの32Pの受容体への移転を促進するであろう。C−末端のリン酸化は、アレスチン様タンパクの結合を促進し、Gタンパクの結合を妨害する。GPCR信号変換及び信号変換をアッセイする方法の一般的総説については、例えば、Methods in Enzymology, vols. 237 and 238 (1994) and volume 96 (1983); Bourne et al., Nature, 10: 349: 117-27 (1991); Bourne et al., Nature, 348: 125-32 (1990); Pitcher et al., Annu. Rev. Biochem., 67: 653-92 (1998)参照。
【0141】
T2R調節は、候補T2R調節物質で処理したT2Rポリペプチドの反応を、非処理検体または既知「陽性」対照を含む検体と比較することによりアッセイすることができる。その候補T2R調節物質は、T2Rポリペプチド活性を阻害または活性化する分子を含むことができる。一の態様において、T2Rを活性化する化合物で処理した対照検体を、相対T2R活性値100と定める。対照検体に対する相対的T2R活性値が約90%、さらに50%、さらに25−0%であるときに、T2Rポリペプチドの阻害は達成される。対照に対する相対的T2R活性値が110%、さらに150%、200−500%、または1000−2000%であるときに、T2Rポリペプチドの活性化は達成される。
【0142】
イオン流の変化は、T2Rポリペプチドを発現している細胞または膜のイオン分極(例えば、電気ポテンシャル)の変化を測定することにより検出することができる。細胞分極の変化を測定する手段の一つは、ボルテージ−クランプ及びパッチ−クランプ技術を使用して電流の変化を測定(したがって分極の変化を測定)することによるものである(例えば、「セル−アタッチド」モード、「インサイド−アウト」モード、及び「ホールセル」モード、例えば、Ackerman et al., New Engl. J Med., 336: 1575-1595 (1997)参照)。ホールセル電流は簡便に測定され、標準的に使用されている。そのほかの既知アッセイとしては、放射標識イオン流アッセイ及び電圧感受性色素を使用する蛍光アッセイが含まれる(例えば、Vestergarrd-Bogind et al., J. Membrane Biol., 88: 67-75 (1988); Gonzales & Tsien, Chem. Biol., 4: 269-277 (1997); Daniel et al., J. Pharmacol. Meth., 25: 185-193 (1991); Holevinsky et al., J. Membrane Biology, 137: 59-70 (1994)参照)。
【0143】
ポリペプチドの機能に及ぼす試験化合物の効果は、前記パラメーターのいずれかを調べることにより測定することができる。GPCR活性に影響する適当ないずれの生理学的変化も、本発明のポリペプチドに対する試験化合物の影響を評価するために使用することができる。機能的事象を完全細胞または動物を使用して測定する場合には、伝達物質放出、ホルモン放出、既知または未知遺伝子マーカーへの転写の変化(例えば、ノーザンブロット)、細胞増殖またはpHの変化のような細胞代謝の変化、及びCa2+,IP3,cGMP,または cAMPのような細胞内セカンドメッセンジャーの変化などの種々の効果を測定することができる。
【0144】
GPCRの望ましいアッセイには、受容体活性を報告させるためのイオンまたは電圧感受性色素を負荷された細胞が含まれる。その受容体の活性を測定するアッセイには、試験化合物の活性を評価するための対照として、他のGタンパク共役受容体の既知アゴニスト及びアンタゴニストも使用することができる。調節化合物(例えば、アゴニスト、アンタゴニスト)を同定するためのアッセイにおいて、細胞質中のイオンまたは膜電位のレベルの変化は、イオン感受性または膜電位蛍光指示薬をそれぞれ使用して監視されるであろう。使用することができるイオン感受性指示薬及び電圧プローブは、the Molecular Probes 1997 Catalog中に記述されている。G−タンパク共役受容体に対して、Gα15及びGα16のような相手を選ばないGタンパクを、選択アッセイにおいて使用することができる(Wilkie et al., Proc. Nat'l Acad. Sci., 88: 10049-10053 (1991))。
【0145】
受容体の活性化は、続く細胞内事象、例えば、セカンドメッセンジャーの増加、を開始する。一部のGタンパク共役受容体の活性化は、ホスフォリパーゼC仲介ホスファチジルイノシトールの加水分解によるイノシトール三リン酸(IP3)の生成を促進する(Berridge & Irvine, Nature, 312: 315-21 (1984))。IP3は、次に細胞内貯蔵カルシウムイオンの放出を促進する。このように、細胞質カルシウムイオンレベルの変化、またはIP3のようなセカンドメッセンジャーの変化をGタンパク共役受容体の機能を評価するために使用することができる。そのGタンパク共役受容体を発現する細胞は、細胞内カルシウム貯蔵からの放出及び細胞外カルシウムの形質膜イオンチャネルを経由する流入の両者による結果として細胞質カルシウムレベルの増加を示すであろう。
【0146】
望ましい態様において、T2Rポリペプチド活性は、受容体をホスフォリパーゼC信号伝達経路に連結する相手を選ばないGタンパクを持つ異種細胞内でT2R遺伝子を発現することにより測定される(Offermanns & Simon, J. Biol. Chem., 270: 15175-15180 (1995)参照)。任意に、細胞系はHEK-293(これは通常はT2R遺伝子を発現しない)であり、相手を選ばないGタンパクはGα15である(Offermanns & Simon,前出)。味覚伝達の調節は、T2Rポリペプチドに会合する分子の投与によるT2R信号伝達経路の変化に応答して変化する細胞内Ca2+レベルの変化を測定することによりアッセイされる。Ca2+レベルの変化は、任意に蛍光Ca2+指示色素及び蛍光画像を使用して測定される。
【0147】
他の態様において、本明細書に引用して取り入れた米国特許5,436,128にしたがってホスファチジルイノシトール(PI)加水分解を分析することができる。簡略に記述すると、このアッセイは、3H−ミオイノシトールで48時間またはそれ以上細胞を標識することを必要とする。標識細胞を、試験化合物で1時間処理する。処理した細胞を分解し、クロロホルム−メタノール−水で抽出し、その後イノシトールリン酸をイオン交換クロマトグラフィーにより分離し、シンチレーションカウンターで定量する。緩衝液対照の存在下におけるcpmに対するアゴニスト存在下におけるcpmの比を計算することにより活性化率が算出される。同様に、緩衝液対照(アゴニストを含んでも含まなくてもよい)存在下におけるcpmに対するアンタゴニスト存在下におけるcpmの比を計算することにより阻害率が算出される。
【0148】
他の受容体アッセイとしては、細胞内サイクリックヌクレオチド、例えば、cAMPまたは cGMPのレベルを測定するものがある。受容体の活性化によりサイクリックヌクレオチドのレベルが低下する場合には、アッセイ中の細胞に受容体活性化化合物を加える前に、細胞内サイクリックヌクレオチドのレベルを上昇させる試薬、例えば、ホルスコリン、に細胞を暴露することが望ましい。一の態様においては、免疫アッセイを使用して細胞内cAMPまたは cGMPの変化を測定することができる。Offermanns & Simon, J. Bio. Chem., 270: 15175-15180 (1995)に記述されている方法をcAMPのレベルの測定に使用することができる。また、Felley-Bosco et al., Am. J. Resp. Cell and Mol. Biol., 11: 159-164 (1994)に記述されている方法も cGMPのレベルの測定に使用することができる。さらに、cAMP及び/または cGMPを測定するアッセイキットが、本明細書に引用して取り入れた、米国特許4,115,538に記述されている。
【0149】
他の態様においては、信号変換に対する試験化合物の効果を評価するために転写レベルを測定することができる。本発明のT2Rポリペプチドを含む宿主細胞を試験化合物と相互作用をさせるために十分な時間接触させ、次いで遺伝子発現のレベルを測定する。その相互作用を行うのに十分な時間は、例えば、時間経過を追い、時間の関数として転写のレベルを測定することにより、経験的に決定される。転写量は、当業者に既知の適当な方法を使用して測定することができる。例えば、本発明のタンパクのmRNA発現は、ノーザンブロットを使用して検出することができるし、あるいはそのペプチド産物を、免疫アッセイを使用して同定することができる。また、レポーター遺伝子を使用する転写によるアッセイが、本明細書に引用して取り入れた、米国特許5,436,128に記述されているように、使用することができる。レポーター遺伝子は、例えば、クロラムフェニコールアセチル転移酵素、ルシフェラーゼ、βガラクトシダーゼ、βラクタマーゼ及びアルカリホスファターゼとすることができる。さらに、目的のタンパクを、第二のレポーター、例えば、緑色蛍光タンパクに付属させることにより間接的なレポーターとして使用することができる(例えば、Mistili & Spector, Nature Biotechnology, 15: 961-964 (1997)参照)。
【0150】
次いで転写の量を、試験化合物が存在しない同じ細胞中における転写の量と比較するか、または本発明のT2Rポリペプチドを持たない実質的に同じ細胞中における転写の量と比較する。実質的に同じ細胞は、組換え細胞が誘導された細胞と同じで、異種DNAの導入による修飾がされていない細胞から誘導することができる。転写の量に差があれば、目的のT2Rポリペプチドの活性をある程度変えたことを示している。
【0151】
4.化学感受性受容体を発現する非ヒト形質転換動物
一つまたはそれ以上の本発明の味覚受容体配列を発現する非ヒト動物も、また受容体アッセイに使用することができる。そのような発現を利用して、安定的にまたは一過性に化学感受性受容体またはそのリガンド結合領域をコードする核酸を導入した非ヒト動物を、試験化合物と接触させ、受容体ポリペプチド複合体に特異的に結合することで、試験化合物に動物が反応するか否かを測定することにより、試験化合物が哺乳動物膜貫通味覚受容体複合体に特異的に結合するか否かを測定することができる。
【0152】
本発明のベクターを導入またはそれに感染した動物は、特定のまたは一連の受容体に結び付く味覚刺激を同定及び分析するためのアッセイに特に有用である。そのようなヒト味覚受容体配列を発現するベクター感染動物は、味覚刺激、及び、例えば、細胞生理(例えば、味覚神経に対する)、中枢神経、または行動に関する味覚刺激による影響を、インビボスクリーニングするために、使用することができる。
【0153】
個別にまたはライブラリーとして、核酸及びベクターを感染/発現する方法は当業者によく知られている。個別の細胞、器官、または動物全体の種々のパラメーターは、種々の手段で測定することができる。本発明のT2R配列は、感染試薬、例えば、アデノウイルス発現ベクター、で輸送することにより動物味覚組織で発現させることができる。
【0154】
内在性味覚受容体遺伝子は、機能を保持することができ、野生(天然)型活性をそのまま存在させることができる。他の状況において、全ての味覚受容体活性が、導入した外来性ハイブリッド受容体によることを望む場合には、ノックアウト系の使用が望ましい。非ヒト形質転換動物、特に形質転換マウス、の作製方法、及び形質転換細胞を作製するための組換え構築体の選択及び調製は当業者に良く知られている。
【0155】
「ノックアウト」細胞及び動物の作製は、抑制される遺伝子のDNA配列の一部を中断するように新規なDNA配列をゲノムの中に導入することにより哺乳動物細胞における特別な遺伝子の発現レベルを低下させるかまたは完全に停止させることができるという前提に基づいている。「遺伝子トラップ挿入」も、宿主遺伝子を破壊するために使用することができ、マウス胚幹(ES)細胞を、ノックアウト形質転換動物を作製するために使用することができる(例えば、Holzschu, Transgenic Res 6: 97-106 (1997)参照)。外来種の挿入は、典型的には相補的核酸配列の間の相同組換えによる。外来配列は、例えば、エキソン性、イントロン性若しくは転写調節配列などの修飾対象たる標的遺伝子の一部、または標的遺伝子の発現レベルに影響することができるゲノム配列;或いはそれらの組み合わせである。多能性胚性幹細胞における相同組換えによる遺伝子ターゲティングは、目的ゲノム配列を正確に修飾することを可能とする。いずれの技術もノックアウト動物を創製し、スクリーニングし、繁殖するために使用することができる、例えば、Bijvoet, Hum. Mol. Genet. 7: 53-62 (1998); Moreadith,J. Mol. Med. 75: 208-216 (1997); Tojo, Cytotechnology 19: 161-165 (1995); Mudgett, Methods Mol. Biol. 48: 167-184 (1995); Longo, Transgenic Res. 6: 321-328 (1997); 米国特許番号5,616,491; 5,464,764;5,631,153;5,487,992;5,627,059;5,272,071;WO 91/09955;WO 93/09222;WO 96/29411;WO 95/31560; WO 91/12650 参照。
【0156】
本発明の核酸は、ノックアウトヒト細胞及びその子孫を作製するための試薬として使用することもできる。同様に、本発明の核酸はマウスの「ノックイン」を作製する試薬としても使用することができる。ヒトまたはラットのT2R遺伝子配列は、マウスゲノム中のオーソログT2Rを置換することができる。このようにして、ヒトまたはラットのT2Rを発現するマウスが作製される。このマウスはヒトまたはラットのT2Rの機能を分析するために、そしてそのT2Rに対するリガンドを同定するために使用することができる。
【0157】
調節物質
T2Rファミリーメンバーの調節物質として試験される化合物は、小さな化合物、またはタンパク、糖、核酸若しくは脂質のような生化学物質のいずれでも良い。また、調節物質は、T2Rファミリーメンバーの遺伝的改変体でも良い。典型的には、試験化合物は小さな化学分子及びペプチドである。通常は、水または有機溶媒(特にDMSOを含む)に溶解しうる化合物が使用されるが、本質的に如何なる化合物も、本発明の潜在的調節物質またはリガンドとして使用することができる。アッセイは、アッセイ段階を自動化し、アッセイのために簡便な資源から化合物を供給することにより、大規模な化合物ライブラリーをスクリーニングするように設計することができ、典型的には、並行して稼動する(例えば、マイクロタイタープレート上で自動的にアッセイするマイクロタイター方式)。Sigma(St. Louis, MO), Aldrich (St. Louis, MO), Sigma-Aldrich (St. Louis, MO), Fluka Chemika-Biochemica Analytika (Buchs, Switzerland)などを含めて多数の化合物供給業者が存在することは認識されているであろう。
【0158】
一の態様において、ハイスループットスクリーニング法は、膨大な数の潜在的治療薬(潜在的調節物質またはリガンド化合物)を含むコンビナトリアル化学またはペプチドライブラリーの供給を伴う。次いで、その「コンビナトリアル化学ライブラリー」または「リガンドライブラリー」は、本明細書で記述する一つまたはそれ以上のアッセイによりスクリーニングされて、所望の特徴的活性を示すライブラリー構成員(特に化学種または亜種)を同定する。こうして同定された化合物は、よく言われる「リード化合物」とすることができ、あるいはそれ自身を潜在的なまたは実際の商品として使用することができる。
【0159】
コンビナトリアル化学ライブラリーは、化学的合成または生化学的合成のいずれかにより多数の化学的「構築ブロック」、例えば試薬、を組み合わせることにより、作製される膨大な化合物の集合である。例えば、ポリペプチドライブラリーのような線状コンビナトリアル化学ライブラリーは、与えられた化合物の長さ(すなわち、ポリペプチド化合物中のアミノ酸の数)に対して可能な方法で化学的構築ブロック(アミノ酸)のセットを組み合わせることにより形成される。化学的構築ブロックのこのようなコンビナトリアル混合により数百万の化合物を合成することができる。
【0160】
コンビナトリアル化学ライブラリーの調製及びスクリーニングは、当業者には良く知られている。そのようなコンビナトリアル化学ライブラリーには、限定はしないが、ペプチドライブラリーが含まれる(例えば、米国特許5,010,175、Furka, Int. J. Pept. Prot. Res., 37: 487-93 (1991) 及びHoughton et al., Nature, 354: 84-88 (1991) 参照)。化学的に多様なライブラリーを作製するための他の化学物質も使用しうる。そのような化学物質としては、限定はしないが:ペプトイド(例えば、WO 91/19735)、コードされたペプチド(例えば、WO 93/20242)、ランダムバイオオリゴマー(例えば、WO 92/00091)、ベンゾジアゼピン類(例えば、米国特許5,288,514)、ヒダントイン類、ベンゾジアゼピン類及びジペプチドのようなダイバーソマー(Hobbs et al., PNAS., 90: 6909-13 (1993))、ビニローグポリペプチド(Hagihara et al., J. Amer. Chem. Soc., 114: 6568 (1992))、グルコーススカホールドを持つ非ペプチド性ペプチド類似体(Hirschmann et al., J. Amer. Chem. Soc., 114: 9217-18 (1992))、小型化合物ライブラリーの類似体有機合成(Chen et al., J. Amer. Chem. Soc., 116: 2661 (1994))、オリゴカルバメート(Cho et al., Science, 261: 1303 (1993))、ペプチジルホスフォネート(Campbell et al., J. Org. Chem., 59: 658 (1994))、核酸ライブラリー(Ausubel, Berger, and Sambrook,全て前出)、ペプチド核酸ライブラリー(米国特許5,539,083)、抗体ライブラリー(Vaughn et al., Nature Biotechnology, 14(3): 309-14 (1996)及びPCT/US96/10287)、糖ライブラリー(Liang et al., Science, 274: 1520-22 (1996)及び米国特許5,593,853)、小型有機分子ライブラリー(ベンゾジアゼピン、Baum, C&EN、Jan 18, page 33 (1993);チアゾリジノン及びメタチアザノン、米国特許5,549,974;ピロリジン、米国特許5,525,735及び5,519,134;モルホリノ化合物、米国特許5,506,337;ベンゾジアゼピン、5,288,514、など)がある。
【0161】
コンビナトリアルライブラリーを調製する装置は、市販されている(例えば、357 MPS, 390 MPS(Advanced Chem Tech, Louisville KY)、Symphony(Rainin, Woburn, MA)、433A(Applied Biosystems, Foster City, CA)、9050 Plus(Millipore, Bedford, MA)参照)。さらに、多数のコンビナトリアルライブラリー自体が、市販されている(例えば、ComGenex, Princeton, NJ; Tripos, Inc., St. Louis, MO; 3D Pharmaceuticals, Exton, PA; Martek Biosciences; Columbia, MD;など参照)。
【0162】
本発明の一の態様において、T2R調節物質を、製品、組成物または成分の味を望ましい方法で調節するために、食品、菓子類、医薬組成物またはそれらの原料中に使用することができる。例えば、苦味の感覚を増強するT2R調節物質は、製品または組成物に苦味を与えるために加えることができるし、他方、苦味感覚を阻止するT2R調節物質は、製品または組成物の味を改良するために加えることができる。
【0163】
本発明により同定された化合物の使用
味覚試験においてhT2R4, hT2R44またはhT2R61の活性化により誘発される苦味を調節する、望ましくは阻止するために、本発明により同定された化合物を食品、飲料または医薬組成物に加えることができる。具体的には、キニーネまたは関連化合物によるhT2R4の活性化を阻害する化合物は、苦味を誘発する食品、飲料または医薬品への添加物として使用することができる。例えば、これらの化合物を、苦味を阻止する有効量で、キニーネまたは関連化合物を含む医薬製剤に加えるとよい。
【0164】
hT2R44またはhT2R61の6−ニトロサッカリンまたはサッカリンによる活性化を阻害する化合物を、hT2R61または hT2R44を活性化するサッカリンまたはそのほかの苦味化合物の苦味を阻止するために、食品、飲料または医薬品に加えることができる。特に、開示したアッセイを使用して同定された化合物を、サッカリンを含むダイエット飲料に入れて、サッカリンによる苦味を阻止することができることが予測される。
【0165】
最後に、デナトニウムによるhT2R44の活性化を阻害する化合物は、hT2R44受容体の活性化による苦味を阻止するための、食品、飲料または医薬品における添加物として有用である。デナトニウムは、非常に強力な苦味化合物であるので、hT2R44受容体を阻害する化合物は、種々の苦味化合物の苦味を有効に阻止するはずである。このことは、いくつかのデナトニウム誘導体によりこの受容体が活性化されるという事実により裏付けられている。
【0166】
キット
T2R遺伝子及びその同属体は、味覚受容体細胞の同定のために、法医学または父子確認のために、及び味覚伝導の試験にとって、有用な道具である。T2R核酸に特異的にハイブリッド形成するT2Rファミリーメンバー特異的試薬、例えばT2Rプローブ及びプライマー、並びにT2Rタンパクに特異的に結合するT2R特異的試薬、例えば、T2R抗体は、味覚細胞発現及び味覚伝導調節を試験するために使用される。
【0167】
T2RファミリーメンバーのDNA及びRNAの存在を調べる核酸アッセイには、当業者に既知の多数の技術が含まれ、これには、例えば、サザン分析、ノーザン分析、ドットブロット、RNアーゼプロテクション、S1分析、PCRのような増幅技術、及びin situハイブリダイゼーションがある。例えば、in situハイブリダイゼーションにあっては、後の解析及び分析のために細胞形態は保存しながら、標的核酸を、細胞内でハイブリッド形成できるように細胞環境から遊離する。以下の論文はin situハイブリダイゼーションを総説している:Singer et al., Biotechniques, 4: 230250(1986); Haase et al., Methods in Virology, vol. VII, 189-226(1984); 及びNames et al., eds., Nucleic Acid Hybridization: A Practical Approach (1987)。さらに、T2Rタンパクは、上記の種々の免疫アッセイ技術により検出することができる。試験検体は、典型的に陽性対照(例えば、組換えT2Rタンパクを発現する検体)および陰性対照と比較される。
【0168】
本発明は、T2Rファミリーメンバーの調節物質をスクリーニングするためのキットも提供する。そのキットは容易に入手できる材料及び試薬から作製することができる。例えば、そのキットは、T2R核酸またはタンパク、試験管、及びT2R活性を試験するための説明書の一つまたはそれ以上を含むことができる。任意に、キットは、機能的T2Rポリペプチドを含む。キットのユーザーの意向及びユーザーの特別なニーズに応じて、キット及び内容の幅広い種類を本発明にしたがって作製することができる。
【0169】
本発明を一般的に記述してきたが、以下の実施例を参照することにより、より容易に理解されるであろう。しかし、以下の実施例は、説明のために提供したものであり、限定を意図するものではない。本発明の精神及び範囲から離れることなく、本明細書で開示した代表例に種々の修飾及び変更を加えることができることが理解される。
【実施例】
【0170】
(実施例1)
この実施例において、キニーネ(苦味アルカロイド)が、本出願のSEQ ID NO:2に含まれるDNA配列を有するヒト苦味受容体、hT2R4を、特異的に活性化することを示す。
【0171】
hT2R4のキニーネによる活性化は、細胞内カルシウム濃度の変化を検出する細胞を使用するアッセイにおいて測定される。簡略に記述すると、大型T細胞抗原及び相手を選ばないGタンパクG15 を安定的に発現するヒト胚性腎細胞(HEK-G15)に、リン酸化カルシウムまたは脂質を使用する系により、hT2R4プラスミドを一過性に導入する。一過性に遺伝子導入した細胞を、24ウエル培養プレートに接種し、48時間機能的発現を行う。次いで、細胞を、カルシウムに特異的な蛍光色素(Fluo-4またはFura-2;Molecular Probes)とともにインキュベートし、これにより、細胞内のカルシウム濃度変化を検出するための迅速、簡便且つ信頼性のある蛍光法が提供される。キニーネを細胞に加えて、PLCの活性化及びそれに続く細胞内カルシウム濃度の増加を引き起こす、信号伝達カスケードを誘発する。このカルシウム濃度の増加は、細胞内のカルシウム色素の蛍光を変化させる。この変化を、蛍光顕微鏡及び特別に設計したソフトウエアー(Imaging Workbench, Axon)を使用して観察する。この方法により、hT2R4を発現する細胞をキニーネが特異的に活性化する(細胞内カルシウム濃度を増加する)ことが観察された。他のhT2Rを一過性に導入した親株(HEK-G15)またはHEK-G15は、キニーネに対して相当する応答を示さなかった(図1参照)。
【0172】
(実施例2)
潜在的hT2R4アンタゴニストを発見するためのハイスループットスクリーニングアッセイにも、細胞表面に高いレベルのhT2R4を安定的に発現するHEK-G15株(HEK-G15-hT2R4)を使用する。細胞表面に一過性遺伝子導入細胞よりも2-3倍多い受容体を発現する安定HEK-G15-hT2R4株を作製した(図2参照)。この場合は、アッセイの18−24時間前に細胞を96ウエルまたは384ウエル培養プレートに接種する。次いで細胞をカルシウム感受性蛍光色素(Molecular Devices)とインキュベートし、刺激し、標準的蛍光強度プレートリーダー(FLIPRまたはVIPR)で測定する。この方法を使用して、hT2R4に対するキニーネの効果をさらに特徴付けることができた。我々は、キニーネが典型的な用量反応相関でhT2R4を活性化する事を発見した(図3参照)。
【0173】
(実施例3)
この実験において、T2R受容体の活性化を監視するためにGTPγS結合アッセイを使用した(図4)。このアッセイは、GPCRの活性を監視する古典的アッセイである(Wessling-Resnick and Jonson, 1987)。これは、適当なリガンドの存在下にGPCRがGタンパクを活性化することができるという事実に基づいている。生成した活性化Gタンパクは、結合しているGDPをGTPに変換する。GTPの加水分解されない類似体、GTPγSを使用すれば、ヌクレオチドは、Gタンパクに堅く結合したままとなり、非結合ヌクレオチドから分離することができる。放射活性標識[γ-35S]GTPγSを使用することにより、結合したGTPγSの量を測定することが可能となる。結合したGTPγSの量は、活性化Gタンパクの量を反映し、それは翻って受容体の活性化を反映する。GTPγS結合アッセイにおいてGタンパクとしてトランスジューシンを使用した。トランスジューシンは、視覚系において発現し、ガストジューシンのオーソログである。トランスジューシンは、ウシ網膜から精製した(Stryer, Methods Enzymol 96: 617-627 (1983))。
【0174】
アッセイ反応は、適当なhT2Rを発現するSf9細胞膜由来の15−20μgの総タンパク、30?l緩衝液B(30 mM Tris-HCl, pH 8, 100 mM NaCl, 3 mM MgCl2, 1 mM DTT)中0.2μMトランスジューシンを含む。一部の反応は、0.05−10 mMの試験苦味化合物も含む。0.3μM [γ-35S]GTPγS (50 Ci/mmol)を加えて反応を開始し、1時間30℃で反応を行い、緩衝液B中の0.5 mM GTP液 170μlを加えて停止し、次いでニトロセルロースフィルターを通して濾過した。結合した[35S]GTPγSを液体シンチレーションカウンターで計測した。
【0175】
バキュロウイルス発現系を使用して、Sf9細胞で、全24個の既知hT2R及びmT2R5を発現した。これらのT2R構築体は、コード配列に加えたロドプシンN−末端40アミノ酸からなるN−末端タグを持っていた。Sf9細胞を、細胞106個当り増幅ウイルスストック10−20μlの比率で、適当なバキュロウイルスに感染させた。細胞を回収し、感染2日後に分析に供した。試験したT2Rの全ての発現は、抗−ロドプシンタグ抗体(B6-30)を使用して免疫ブロットにより確認した。膜の標品は、本質的に記述にしたがって、Sf9膜を5 M尿素で処理して作製した(Ryba, J. Biol Chem 270(12): 6757-6767 (1995))。次いで膜を緩衝液(20 mM Tris-HCl, pH 8, 0.5 mM EDTA, 1 mM DTT)で洗い、約2.5−3 mgの総タンパク/mlの最終濃度で再懸濁した。
【0176】
トランスジューシンを使用するGTPγS結合アッセイにおいて、10個の苦味化合物のセットを用いて全てのhT2Rを試験した。二個のhT2R、hT2R44及びhT2R61が、加えた苦味化合物の存在下で活性を増加させた。
【0177】
特に、hT2R61活性は、6−ニトロサッカリンの存在下に増加することが認められた。試験した全てのhT2Rの中では、他の受容体のたった1つ(hT2R44、下記参照)が、なお弱いながらも6−ニトロサッカリンの存在下に活性化されたが、その一方で、最も近似するhT2R、-hT2R64(hT2R61に対して89%アミノ酸配列相同)でさえも活性化されなかった(図5)。6−ニトロサッカリンは、サッカリンの苦味類似体である。それはヒトも苦味を感じる濃度(0.1−0.5 mM, 図5)で、hT2R61を活性化する。この濃度において、サッカリンは、hT2R61を明らかに活性化せず、ほとんどのヒトは苦味を感じない。生化学的データとヒトの精神物理学的データの相関は、hT2R61は、6−ニトロサッカリンの苦味を感じるヒト受容体であることを示している。
【0178】
強力な甘味剤であるサッカリン自体が、高濃度(2−10 mM)において「苦い」または「金属的」と記述される不快な味を持つことを、多くの人が認めている。低濃度においてサッカリンは、明白にhT2R61を活性化しないが、サッカリンと6−ニトロサッカリンの構造類似性から、人が感じる高濃度における苦味もhT2R61を介していることが示唆される。
【0179】
安息香酸デナトニウムの存在下に、hT2R44の活性が増加することも観察された。この化合物の存在下で、そのほかの試験hT2Rは活性上昇を示さなかった。デナトニウムによるhT2R44の活性化は、濃度に依存することが認められた(図6)。0.3 mMより高いデナトニウム濃度で、結合GTPγSの量は、このアッセイにおける、デナトニウムが有する非特異的阻害効果のために、減少する。安息香酸デナトニウムの活性成分は、デナトニウムであり、安息香酸ではない、なぜならば、100μMの安息香酸ナトリウムの存在下にhT2R44の活性は不変だからである。6−ニトロサッカリンも、かなり弱いが、hT2R44を活性化する。
【0180】
安息香酸デナトニウム(安息香酸 ベンジルジエチル[(2,6−キシリルカルバモイル)メチル]アンモニウム)は、最も強力な苦味化合物の一つとして知られており、苦味の研究に広く使用されている。そのような研究の一つとして、A. Saroli (Saroli, 前出(1984);Saroli,前出(1985))は、デナトニウム誘導体の構造とヒトにおける苦味との関係を検討した。この研究は、デナトニウム分子の選択的変化が、化合物における苦味の特異的な減少をもたらすことを実証した。hT2R44がヒトにおいてデナトニウム及びその誘導体の苦味を仲介していることを確認するために、A. Saroli(前出)(1984)及び(1985)により記述されている3個の誘導体化合物、[(カルボキシメチル)メチル]アンモニウムクロリド(DD#2)、ベンジルメチル[(2,6−キシリルカルバモイル)メチル]アンモニウムクロリド(DD#3)、及びベンジルメチルメチル[(2,6−キシリルカルバモイル)メチル]アンモニウムクロリド(DD#4)を合成した。hT2R44を活性化する能力について、これらの化合物を試験し、デナトニウムと比較した。
【0181】
図7に示すように、デナトニウム誘導体は、デナトニウムに比較してhT2R44を活性化する能力は弱い。hT2R44を活性化する化合物の能力とヒト味覚試験における相対的苦味の間に非常に良い相関がある。これらの結果は、hT2R44がヒトにおいてデナトニウム及びその誘導体の苦味を仲介していることを強く示している。すでにデナトニウムは、hT2R4を活性化することは示されていた(Chandrashekar et al., (前出)(2000))。しかし、その試験で使用されたデナトニウムの濃度(1.5 mM)は、異常に高かった。報告されているデナトニウムのヒトにおける苦味閾値より105倍高い(Saroli, (前出)(1984))。したがって、デナトニウムは、hT2R4に対して非特異的効果を有するように思われる。我々は、hT2R44も6−ニトロサッカリンにより活性化されることを示したが、6−ニトロサッカリンの存在下における活性化は、hT2R61よりもかなり低いことを示している。したがって、hT2R61は、6−ニトロサッカリンの苦味の主たるメディエーターであり、他方hT2R44は高濃度において6−ニトロサッカリンの苦味を仲介すると思われる。
【0182】
以上の詳細な記述により本発明のいくつかの態様を示したが、以上の記述は単に説明のためのものであり、開示した発明を限定するものではないことは理解されるべきである。本発明は以下の請求項によってのみ限定される。
【図面の簡単な説明】
【0183】
【図1】図1は、hT2R4を発現するHEK-G15細胞を使用する細胞によるアッセイの結果を含んでおり、その結果はキニーネがhT2R4によるカルシウム動員を誘発することを示している。このアッセイにおいて、HEK-G15細胞にhT2R4または担体DNA(pUC-18)を導入し、2日後に単細胞カルシウム画像を使用してアッセイした。パネルaからfはキニーネ(250μM最終)で刺激した後0,10,20,30,40及び50秒のhT2R4発現細胞の応答に相当する。パネルgからlはキニーネ(250μM最終)で刺激した後0,10,20,30,40及び50秒のpuc-18発現細胞の応答に相当する。
【図2】図2は、その表面にhT2R4を発現する安定なHEK-G15細胞系を使用した実験結果を含んでいる。この実験において、その細胞表面に高いレベルのhT2R4を発現するHEK-G15のクローンは、FACSを使用する2回の選別及び2回選別した集団のクローニングの後に得られた。FACS分析により選別されたクローンは、その細胞表面上に一過性に遺伝子導入された細胞のおおよそ2−3倍多いT2R受容体を発現することが明らかにされた。
【図3】図3は、96ウエルプレート上に接種したhT2R4を安定的に発現するHEK-G15のクローン(C3Cクローン)におけるhT2R4誘発カルシウム動員に対するキニーネの用量−反応相関を示す。比較の目的で、hT2R4を一過性に導入した細胞を他のプレートに接種した。キニーネの用量を増加させた効果は、FLIPRを使用して試験した。これらの結果は、hT2R4を安定的に高度に発現するクローンにおいて明らかな用量−反応相関を示す。
【図4】図4は、hT2Rを活性化する化合物を同定するために使用されるGTPγS結合アッセイの図解である。図の中で、R、はhT2Rであり、α、β、γは、Gタンパクであり、Lは、受容体を活性化するリガンドである。
【図5】図5は、6−ニトロサッカリンは、同条件下において、hT2R61を活性化するが、hT2R64を活性化しないことを示すGTPγS結合アッセイの結果を含む。hT2R61及び hT2R64の活性は、示した濃度の6−ニトロサッカリンまたはサッカリンの存在しない場合と存在する場合に、GTPγS結合アッセイを使用して測定した(この場合の活性は、加えた試験化合物が存在しない場合に対する活性のパーセントで示す)。
【図6】図6は、hT2R44がデナトニウム及び6−ニトロサッカリンによって活性化されることを示すGTPγS結合アッセイの結果を含む。活性は試験化合物を加えない場合の活性に対する相対的増加倍率として示す。
【図7】図7は、デナトニウム及びその誘導体のhT2R44活性に対する効果を評価するGTPγS結合アッセイの結果を含む。(図中の苦味閾値はA. Saroli (前出)(1984)及びA. Saroli Z.Lebensen Unters Forsch 180: 227-221 (1985))。
【特許請求の範囲】
【請求項1】
hT2R61による苦味を特に調節する化合物を同定するアッセイ方法であって、
(i) hT2R61、または6−ニトロサッカリン、またはサッカリンと特異的に結合する、該受容体の変異体、フラグメント、オーソログ、突然変異体若しくはキメラの活性化に対する影響について、該化合物をスクリーニングし;
(ii)6−ニトロサッカリン、またはサッカリンによる該受容体の活性化に対する影響に基づいて、hT2R61による苦味を該化合物が調節するか否かを測定する
ことを含む、アッセイ方法。
【請求項2】
該hT2R61が、SEQ ID NO.:5に含まれるアミノ酸配列を有する、請求項1に記載のアッセイ。
【請求項3】
該hT2R61が、細胞膜に発現している、請求項1に記載のアッセイ。
【請求項4】
該細胞膜が単離されている、請求項1に記載のアッセイ。
【請求項5】
該細胞膜が、昆虫細胞上に存在する、請求項1に記載のアッセイ。
【請求項6】
該細胞が真核細胞である、請求項1に記載のアッセイ。
【請求項7】
該細胞が、哺乳動物細胞または昆虫細胞である、請求項1に記載のアッセイ。
【請求項8】
該哺乳動物細胞が、HEK細胞、BHK細胞、COS細胞及びCHO細胞から選択される、請求項1に記載のアッセイ。
【請求項9】
該化合物の調節効果が、さらに味覚試験により確認される、請求項1に記載のアッセイ。
【請求項10】
hT2R61による苦味を調節する化合物を同定する方法であって、
(i) hT2R61タンパク、または苦味に関与するその変異体、オーソログ、キメラ若しくはフラグメントを発現し、任意に内在性または外来性Gタンパクを含む細胞系または細胞膜を得;
(ii)該細胞系を、hT2R61を活性化する6−ニトロサッカリンに接触させ;
(iii) 該細胞系を、苦味に影響する可能性についてスクリーニングする少なくとも一つの化合物と接触させ;
(iv) 該化合物が苦味を調節するか否かを、6−ニトロサッカリンによる該hT2R61タンパク、又はその変異体若しくはフラグメントの活性化に対する影響に基づいて同定する
ことを含む、同定方法。
【請求項11】
該化合物に対する該影響が、細胞内イオン濃度の変化に基づいて選択される、請求項10に記載の方法。
【請求項12】
該イオンが、カルシウムである請求項11に記載の方法。
【請求項13】
該イオンが、ナトリウムである請求項11に記載の方法。
【請求項14】
該化合物が、細胞膜電位に関する効果に基づいて選択される、請求項10に記載の方法。
【請求項15】
該化合物が、該T2Rの転写に関する効果に基づいて選択される、請求項10に記載の方法。
【請求項16】
該化合物が、該T2Rの6−ニトロサッカリンとの相互作用を阻害する能力に基づいて選択される、請求項10に記載の方法。
【請求項17】
該hT2R61が、形質膜輸送を促進する配列に作動的に結合している、請求項10に記載の方法。
【請求項18】
該細胞系が、HEK-293細胞系である請求項10に記載の方法。
【請求項19】
該hT2R61ポリペプチドに対する該化合物の影響を、細胞内cAMP, cGMPまたはIP3の変化に基づいてアッセイする、請求項10に記載の方法。
【請求項20】
該hT2R61フラグメントが、細胞外ドメインまたは膜貫通領域である、請求項43に記載の方法。
【請求項21】
レポーター分子を使用して、カルシウムイオン濃度の相違をモニターする、請求項10に記載の方法。
【請求項22】
該レポーター分子が、カルシウムに特異的な蛍光色素である、請求項10に記載の方法。
【請求項23】
該色素が、Fluo-3, Fluo-4またはFura-2である、請求項22に記載の方法。
【請求項24】
該hT2R61、変異体またはフラグメントが、レポーター分子に付属している、請求項10に記載の方法。
【請求項25】
該レポーターが、緑色蛍光タンパクまたはその変異体である、請求項24に記載の方法。
【請求項26】
該hT2R61ポリペプチド、変異体またはフラグメントが、溶液中に含まれる、請求項10に記載の方法。
【請求項27】
該hT2R61ポリペプチド、変異体またはフラグメントが、固相に付着した二層膜中に、脂質単層中に、または小胞中に存在する、請求項10に記載の方法。
【請求項28】
該化合物の結合が、分光学的特性、流体力学的特性、クロマトグラフィーによる特性または溶解性の変化に基づいて評価される、請求項1に記載の方法。
【請求項29】
該化合物が、該hT2R61ポリペプチド、変異体またはフラグメントとGタンパクとの複合体形成に対する影響に基づいて選択される、請求項1に記載の方法。
【請求項30】
該化合物が、該hT2R61ポリペプチド、変異体またはフラグメントとガストジューシンまたはトランスジューシンとの相互作用に対する影響に基づいて選択される、請求項10に記載の方法。
【請求項31】
該化合物が、蛍光偏光アッセイを使用して選択される、請求項1に記載の方法。
【請求項32】
該hT2R61ポリペプチドが、固相基質に付着している、請求項1に記載の方法。
【請求項33】
ハイスループットアッセイを含む、請求項1に記載の方法。
【請求項34】
該細胞系が、該受容体を安定的に発現する、請求項10に記載の方法。
【請求項35】
該細胞系が、該受容体を一過性に発現する、請求項10に記載の方法。
【請求項36】
該細胞系が、哺乳動物または昆虫の細胞である、請求項34または35に記載の方法。
【請求項37】
該昆虫細胞がSf9細胞である、請求項36に記載の方法。
【請求項38】
該アッセイが、GTPγS結合アッセイである、請求項10に記載の方法。
【請求項39】
該アッセイが、Gタンパクとしてトランスジューシンまたはガストジューシンを使用する、請求項38に記載の方法。
【請求項40】
該受容体が、バキュロウイルス発現系を使用して発現される、請求項39に記載の方法。
【請求項41】
hT2R61による苦味を阻害する該化合物の能力を、さらに味覚試験により確認する、請求項10に記載の方法。
【請求項42】
味覚試験が、サッカリンを含む組成物を使用する、請求項41に記載の方法。
【請求項43】
hT2R44による苦味を特に調節する化合物を同定するアッセイ方法であって、
(i) デナトニウム及び/または6−ニトロサッカリンによるhT2R44、または該受容体の変異体、フラグメント、キメラ、オーソログ若しくは突然変異体の活性化に対する影響について、該化合物をスクリーニングし;
(ii) 該化合物がhT2R44による苦味を調節するか否かを、デナトニウム及び/または6−ニトロサッカリンによる該受容体の活性化を調節する能力に基づいて測定する:
ことを含む、アッセイ方法。
【請求項44】
該hT2R44が、SEQ ID NO.:3に含まれるアミノ酸配列を有する、請求項43に記載のアッセイ。
【請求項45】
該hT2R44が、細胞膜に発現している請求項43に記載のアッセイ。
【請求項46】
該細胞膜が、単離されている請求項45に記載のアッセイ。
【請求項47】
該細胞膜が、昆虫細胞に由来する請求項45に記載のアッセイ。
【請求項48】
該細胞が、真核細胞である請求項45に記載のアッセイ。
【請求項49】
該細胞が、哺乳動物細胞または昆虫細胞である請求項48に記載のアッセイ。
【請求項50】
該哺乳動物細胞が、HEK細胞、BHK細胞、COS細胞及びCHO細胞から選択される、請求項49に記載のアッセイ。
【請求項51】
該化合物の調節効果が、さらに味覚試験によって確認される、請求項43に記載の方法。
【請求項52】
hT2R44による苦味を調節する化合物を同定する方法であって、
(i) hT2R44タンパク、又は苦味に関与するその変異体、オーソログ、キメラ若しくはフラグメントを発現し、任意に内在性または外来性Gタンパクを含む、細胞系または細胞膜を得;
(ii) 該細胞系を、hT2R44を活性化するデナトニウムに接触させ;
(iii) 該細胞系を、苦味に影響する可能性についてスクリーニングされる少なくとも一つの化合物と接触させ;
(iv) 該化合物が苦味を調節するか否かを、デナトニウムによる該T2Rタンパク、又はその変異体若しくはフラグメントの活性化に対する影響に基づいて同定する
ことを含む、同定方法。
【請求項53】
該化合物に対する該影響が、細胞内イオン濃度の変化に基づいて選択される請求項52に記載の方法。
【請求項54】
該イオンが、カルシウムである、請求項53に記載の方法。
【請求項55】
該イオンが、ナトリウムである、請求項53に記載の方法。
【請求項56】
該化合物が、細胞膜電位に関する効果に基づいて選択される、請求項52に記載の方法。
【請求項57】
該化合物が、該T2Rの転写に関する効果に基づいて選択される、請求項52に記載の方法。
【請求項58】
該化合物が、該T2Rとデナトニウム及び/または6−ニトロサッカリンとの相互作用を阻害する能力に基づいて選択される請求項52に記載の方法。
【請求項59】
該hT2R44が、形質膜輸送を促進する配列に作動的に結合している、請求項52に記載の方法。
【請求項60】
該細胞系が、HEK-293細胞系である、請求項52に記載の方法。
【請求項61】
該hT2R44に対する該化合物の影響が、細胞内cAMP, cGMPまたはIP3の変化に基づいてアッセイされる、請求項52に記載の方法。
【請求項62】
該hT2R44フラグメントが、細胞外ドメインまたは膜貫通領域である、請求項52に記載の方法。
【請求項63】
レポーター分子を使用して、カルシウムイオン濃度の相違をモニターする、請求項52に記載の方法。
【請求項64】
該レポーター分子が、カルシウムに特異的な蛍光色素である、請求項52に記載の方法。
【請求項65】
該色素が、Fluor-3, Fluo-4またはFura-2である、請求項64に記載の方法。
【請求項66】
該hT2R44ポリペプチド、又はその変異体若しくはフラグメントが、レポーター分子に付属している、請求項52に記載の方法。
【請求項67】
該レポーターが、緑色蛍光タンパクまたはその変異体である、請求項66に記載の方法。
【請求項68】
該hT2R44ポリペプチド、又はその変異体若しくはフラグメントが、溶液中に含まれる請求項43に記載の方法。
【請求項69】
該hT2R44ポリペプチド、又はその変異体若しくはフラグメントが、固相に付着された二層膜中、脂質単層膜中、または小胞中に存在する、請求項43に記載の方法。
【請求項70】
該化合物の結合が、分光学的特性、流体力学的特性、クロマトグラフィーによる特性または溶解性の変化に基づいて評価される、請求項43に記載の方法。
【請求項71】
該化合物が、該hT2R44ポリペプチド、又はその変異体若しくはフラグメントとGタンパクとの複合体形成に対する影響に基づいて選択される、請求項43に記載の方法。
【請求項72】
該化合物が、該hT2R44ポリペプチド、又はその変異体若しくはフラグメントとガストジューシンまたはトランスジューシンとの相互作用に対する影響に基づいて選択される、請求項52に記載の方法。
【請求項73】
該化合物が、蛍光偏光アッセイを使用して選択される、請求項43に記載の方法。
【請求項74】
該hT2R44ポリペプチドが、固相基質に付着している、請求項43に記載の方法。
【請求項75】
ハイスループットアッセイを含む、請求項43に記載の方法。
【請求項76】
該細胞系が、該受容体を安定的に発現する、請求項52に記載の方法。
【請求項77】
該細胞系が、該受容体を一過性に発現する、請求項52に記載の方法。
【請求項78】
該細胞系が、哺乳動物または昆虫の細胞である、請求項76または77に記載の方法。
【請求項79】
該昆虫細胞が、Sf9細胞である、請求項78に記載の方法。
【請求項80】
該アッセイが、GTPγS結合アッセイである、請求項76に記載の方法。
【請求項81】
該アッセイが、Gタンパクとしてトランスジューシンまたはガストジューシンを使用する、請求項80に記載の方法。
【請求項82】
該受容体が、バキュロウイルス発現系を使用して発現される、請求項79に記載の方法。
【請求項83】
hT2R44による苦味を阻害する該化合物の能力を、さらに味覚試験により確認する、請求項52に記載の方法。
【請求項1】
hT2R61による苦味を特に調節する化合物を同定するアッセイ方法であって、
(i) hT2R61、または6−ニトロサッカリン、またはサッカリンと特異的に結合する、該受容体の変異体、フラグメント、オーソログ、突然変異体若しくはキメラの活性化に対する影響について、該化合物をスクリーニングし;
(ii)6−ニトロサッカリン、またはサッカリンによる該受容体の活性化に対する影響に基づいて、hT2R61による苦味を該化合物が調節するか否かを測定する
ことを含む、アッセイ方法。
【請求項2】
該hT2R61が、SEQ ID NO.:5に含まれるアミノ酸配列を有する、請求項1に記載のアッセイ。
【請求項3】
該hT2R61が、細胞膜に発現している、請求項1に記載のアッセイ。
【請求項4】
該細胞膜が単離されている、請求項1に記載のアッセイ。
【請求項5】
該細胞膜が、昆虫細胞上に存在する、請求項1に記載のアッセイ。
【請求項6】
該細胞が真核細胞である、請求項1に記載のアッセイ。
【請求項7】
該細胞が、哺乳動物細胞または昆虫細胞である、請求項1に記載のアッセイ。
【請求項8】
該哺乳動物細胞が、HEK細胞、BHK細胞、COS細胞及びCHO細胞から選択される、請求項1に記載のアッセイ。
【請求項9】
該化合物の調節効果が、さらに味覚試験により確認される、請求項1に記載のアッセイ。
【請求項10】
hT2R61による苦味を調節する化合物を同定する方法であって、
(i) hT2R61タンパク、または苦味に関与するその変異体、オーソログ、キメラ若しくはフラグメントを発現し、任意に内在性または外来性Gタンパクを含む細胞系または細胞膜を得;
(ii)該細胞系を、hT2R61を活性化する6−ニトロサッカリンに接触させ;
(iii) 該細胞系を、苦味に影響する可能性についてスクリーニングする少なくとも一つの化合物と接触させ;
(iv) 該化合物が苦味を調節するか否かを、6−ニトロサッカリンによる該hT2R61タンパク、又はその変異体若しくはフラグメントの活性化に対する影響に基づいて同定する
ことを含む、同定方法。
【請求項11】
該化合物に対する該影響が、細胞内イオン濃度の変化に基づいて選択される、請求項10に記載の方法。
【請求項12】
該イオンが、カルシウムである請求項11に記載の方法。
【請求項13】
該イオンが、ナトリウムである請求項11に記載の方法。
【請求項14】
該化合物が、細胞膜電位に関する効果に基づいて選択される、請求項10に記載の方法。
【請求項15】
該化合物が、該T2Rの転写に関する効果に基づいて選択される、請求項10に記載の方法。
【請求項16】
該化合物が、該T2Rの6−ニトロサッカリンとの相互作用を阻害する能力に基づいて選択される、請求項10に記載の方法。
【請求項17】
該hT2R61が、形質膜輸送を促進する配列に作動的に結合している、請求項10に記載の方法。
【請求項18】
該細胞系が、HEK-293細胞系である請求項10に記載の方法。
【請求項19】
該hT2R61ポリペプチドに対する該化合物の影響を、細胞内cAMP, cGMPまたはIP3の変化に基づいてアッセイする、請求項10に記載の方法。
【請求項20】
該hT2R61フラグメントが、細胞外ドメインまたは膜貫通領域である、請求項43に記載の方法。
【請求項21】
レポーター分子を使用して、カルシウムイオン濃度の相違をモニターする、請求項10に記載の方法。
【請求項22】
該レポーター分子が、カルシウムに特異的な蛍光色素である、請求項10に記載の方法。
【請求項23】
該色素が、Fluo-3, Fluo-4またはFura-2である、請求項22に記載の方法。
【請求項24】
該hT2R61、変異体またはフラグメントが、レポーター分子に付属している、請求項10に記載の方法。
【請求項25】
該レポーターが、緑色蛍光タンパクまたはその変異体である、請求項24に記載の方法。
【請求項26】
該hT2R61ポリペプチド、変異体またはフラグメントが、溶液中に含まれる、請求項10に記載の方法。
【請求項27】
該hT2R61ポリペプチド、変異体またはフラグメントが、固相に付着した二層膜中に、脂質単層中に、または小胞中に存在する、請求項10に記載の方法。
【請求項28】
該化合物の結合が、分光学的特性、流体力学的特性、クロマトグラフィーによる特性または溶解性の変化に基づいて評価される、請求項1に記載の方法。
【請求項29】
該化合物が、該hT2R61ポリペプチド、変異体またはフラグメントとGタンパクとの複合体形成に対する影響に基づいて選択される、請求項1に記載の方法。
【請求項30】
該化合物が、該hT2R61ポリペプチド、変異体またはフラグメントとガストジューシンまたはトランスジューシンとの相互作用に対する影響に基づいて選択される、請求項10に記載の方法。
【請求項31】
該化合物が、蛍光偏光アッセイを使用して選択される、請求項1に記載の方法。
【請求項32】
該hT2R61ポリペプチドが、固相基質に付着している、請求項1に記載の方法。
【請求項33】
ハイスループットアッセイを含む、請求項1に記載の方法。
【請求項34】
該細胞系が、該受容体を安定的に発現する、請求項10に記載の方法。
【請求項35】
該細胞系が、該受容体を一過性に発現する、請求項10に記載の方法。
【請求項36】
該細胞系が、哺乳動物または昆虫の細胞である、請求項34または35に記載の方法。
【請求項37】
該昆虫細胞がSf9細胞である、請求項36に記載の方法。
【請求項38】
該アッセイが、GTPγS結合アッセイである、請求項10に記載の方法。
【請求項39】
該アッセイが、Gタンパクとしてトランスジューシンまたはガストジューシンを使用する、請求項38に記載の方法。
【請求項40】
該受容体が、バキュロウイルス発現系を使用して発現される、請求項39に記載の方法。
【請求項41】
hT2R61による苦味を阻害する該化合物の能力を、さらに味覚試験により確認する、請求項10に記載の方法。
【請求項42】
味覚試験が、サッカリンを含む組成物を使用する、請求項41に記載の方法。
【請求項43】
hT2R44による苦味を特に調節する化合物を同定するアッセイ方法であって、
(i) デナトニウム及び/または6−ニトロサッカリンによるhT2R44、または該受容体の変異体、フラグメント、キメラ、オーソログ若しくは突然変異体の活性化に対する影響について、該化合物をスクリーニングし;
(ii) 該化合物がhT2R44による苦味を調節するか否かを、デナトニウム及び/または6−ニトロサッカリンによる該受容体の活性化を調節する能力に基づいて測定する:
ことを含む、アッセイ方法。
【請求項44】
該hT2R44が、SEQ ID NO.:3に含まれるアミノ酸配列を有する、請求項43に記載のアッセイ。
【請求項45】
該hT2R44が、細胞膜に発現している請求項43に記載のアッセイ。
【請求項46】
該細胞膜が、単離されている請求項45に記載のアッセイ。
【請求項47】
該細胞膜が、昆虫細胞に由来する請求項45に記載のアッセイ。
【請求項48】
該細胞が、真核細胞である請求項45に記載のアッセイ。
【請求項49】
該細胞が、哺乳動物細胞または昆虫細胞である請求項48に記載のアッセイ。
【請求項50】
該哺乳動物細胞が、HEK細胞、BHK細胞、COS細胞及びCHO細胞から選択される、請求項49に記載のアッセイ。
【請求項51】
該化合物の調節効果が、さらに味覚試験によって確認される、請求項43に記載の方法。
【請求項52】
hT2R44による苦味を調節する化合物を同定する方法であって、
(i) hT2R44タンパク、又は苦味に関与するその変異体、オーソログ、キメラ若しくはフラグメントを発現し、任意に内在性または外来性Gタンパクを含む、細胞系または細胞膜を得;
(ii) 該細胞系を、hT2R44を活性化するデナトニウムに接触させ;
(iii) 該細胞系を、苦味に影響する可能性についてスクリーニングされる少なくとも一つの化合物と接触させ;
(iv) 該化合物が苦味を調節するか否かを、デナトニウムによる該T2Rタンパク、又はその変異体若しくはフラグメントの活性化に対する影響に基づいて同定する
ことを含む、同定方法。
【請求項53】
該化合物に対する該影響が、細胞内イオン濃度の変化に基づいて選択される請求項52に記載の方法。
【請求項54】
該イオンが、カルシウムである、請求項53に記載の方法。
【請求項55】
該イオンが、ナトリウムである、請求項53に記載の方法。
【請求項56】
該化合物が、細胞膜電位に関する効果に基づいて選択される、請求項52に記載の方法。
【請求項57】
該化合物が、該T2Rの転写に関する効果に基づいて選択される、請求項52に記載の方法。
【請求項58】
該化合物が、該T2Rとデナトニウム及び/または6−ニトロサッカリンとの相互作用を阻害する能力に基づいて選択される請求項52に記載の方法。
【請求項59】
該hT2R44が、形質膜輸送を促進する配列に作動的に結合している、請求項52に記載の方法。
【請求項60】
該細胞系が、HEK-293細胞系である、請求項52に記載の方法。
【請求項61】
該hT2R44に対する該化合物の影響が、細胞内cAMP, cGMPまたはIP3の変化に基づいてアッセイされる、請求項52に記載の方法。
【請求項62】
該hT2R44フラグメントが、細胞外ドメインまたは膜貫通領域である、請求項52に記載の方法。
【請求項63】
レポーター分子を使用して、カルシウムイオン濃度の相違をモニターする、請求項52に記載の方法。
【請求項64】
該レポーター分子が、カルシウムに特異的な蛍光色素である、請求項52に記載の方法。
【請求項65】
該色素が、Fluor-3, Fluo-4またはFura-2である、請求項64に記載の方法。
【請求項66】
該hT2R44ポリペプチド、又はその変異体若しくはフラグメントが、レポーター分子に付属している、請求項52に記載の方法。
【請求項67】
該レポーターが、緑色蛍光タンパクまたはその変異体である、請求項66に記載の方法。
【請求項68】
該hT2R44ポリペプチド、又はその変異体若しくはフラグメントが、溶液中に含まれる請求項43に記載の方法。
【請求項69】
該hT2R44ポリペプチド、又はその変異体若しくはフラグメントが、固相に付着された二層膜中、脂質単層膜中、または小胞中に存在する、請求項43に記載の方法。
【請求項70】
該化合物の結合が、分光学的特性、流体力学的特性、クロマトグラフィーによる特性または溶解性の変化に基づいて評価される、請求項43に記載の方法。
【請求項71】
該化合物が、該hT2R44ポリペプチド、又はその変異体若しくはフラグメントとGタンパクとの複合体形成に対する影響に基づいて選択される、請求項43に記載の方法。
【請求項72】
該化合物が、該hT2R44ポリペプチド、又はその変異体若しくはフラグメントとガストジューシンまたはトランスジューシンとの相互作用に対する影響に基づいて選択される、請求項52に記載の方法。
【請求項73】
該化合物が、蛍光偏光アッセイを使用して選択される、請求項43に記載の方法。
【請求項74】
該hT2R44ポリペプチドが、固相基質に付着している、請求項43に記載の方法。
【請求項75】
ハイスループットアッセイを含む、請求項43に記載の方法。
【請求項76】
該細胞系が、該受容体を安定的に発現する、請求項52に記載の方法。
【請求項77】
該細胞系が、該受容体を一過性に発現する、請求項52に記載の方法。
【請求項78】
該細胞系が、哺乳動物または昆虫の細胞である、請求項76または77に記載の方法。
【請求項79】
該昆虫細胞が、Sf9細胞である、請求項78に記載の方法。
【請求項80】
該アッセイが、GTPγS結合アッセイである、請求項76に記載の方法。
【請求項81】
該アッセイが、Gタンパクとしてトランスジューシンまたはガストジューシンを使用する、請求項80に記載の方法。
【請求項82】
該受容体が、バキュロウイルス発現系を使用して発現される、請求項79に記載の方法。
【請求項83】
hT2R44による苦味を阻害する該化合物の能力を、さらに味覚試験により確認する、請求項52に記載の方法。
【図1】

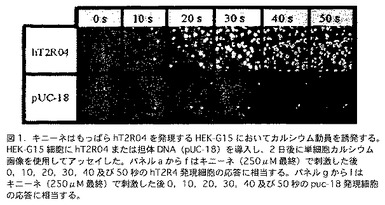
【図2】
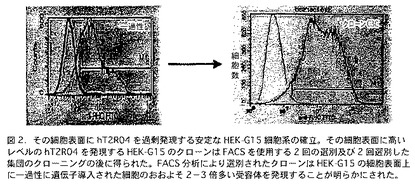
【図3】
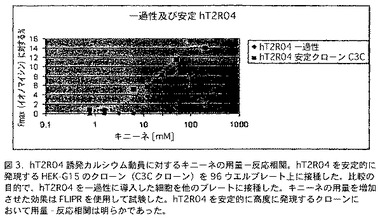
【図4】
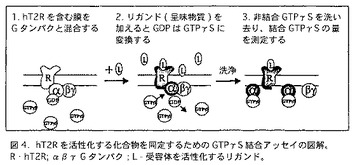
【図5】
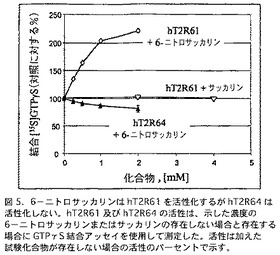
【図6】
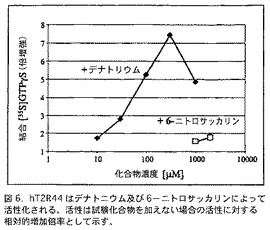
【図7】
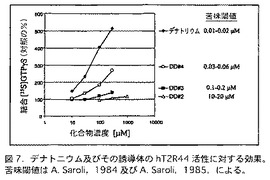
【図2】
【図3】
【図4】
【図5】
【図6】
【図7】
【公開番号】特開2009−109(P2009−109A)
【公開日】平成21年1月8日(2009.1.8)
【国際特許分類】
【出願番号】特願2008−150862(P2008−150862)
【出願日】平成20年6月9日(2008.6.9)
【分割の表示】特願2003−512252(P2003−512252)の分割
【原出願日】平成14年7月10日(2002.7.10)
【出願人】(502362806)セノミックス、インコーポレイテッド (9)
【Fターム(参考)】
【公開日】平成21年1月8日(2009.1.8)
【国際特許分類】
【出願日】平成20年6月9日(2008.6.9)
【分割の表示】特願2003−512252(P2003−512252)の分割
【原出願日】平成14年7月10日(2002.7.10)
【出願人】(502362806)セノミックス、インコーポレイテッド (9)
【Fターム(参考)】
[ Back to top ]