
荷重感知遺伝子
【課題】高齢者の骨粗鬆症、特に麻痺等に伴う廃用性骨粗鬆症、ステロイド性骨粗鬆症または宇宙滞在時の骨量減少の予防または治療薬の開発に有用な手段の提供。
【解決手段】PDK4の発現または機能を抑制する物質を含む、骨粗鬆症の予防または治療剤、ならびに(a)骨芽細胞または骨髄由来の単球/マクロファージ系列細胞と被験物質とを接触させる工程、(b)前記被験物質を接触させた骨芽細胞または骨髄由来の単球/マクロファージ系列細胞におけるPDK4の発現量および破骨細胞形成(誘導)能を調べ、被験物質を接触させない骨芽細胞または骨髄由来の単球/マクロファージ系列細胞における発現量および破骨細胞形成(誘導)能と比較する工程、および(c)前記比較結果に基づいて、PDK4の発現を抑制する被験物質を選択する工程を含む、PDK4の発現または機能抑制を介した骨粗鬆症の予防および/または治療剤のスクリーニング方法。
【解決手段】PDK4の発現または機能を抑制する物質を含む、骨粗鬆症の予防または治療剤、ならびに(a)骨芽細胞または骨髄由来の単球/マクロファージ系列細胞と被験物質とを接触させる工程、(b)前記被験物質を接触させた骨芽細胞または骨髄由来の単球/マクロファージ系列細胞におけるPDK4の発現量および破骨細胞形成(誘導)能を調べ、被験物質を接触させない骨芽細胞または骨髄由来の単球/マクロファージ系列細胞における発現量および破骨細胞形成(誘導)能と比較する工程、および(c)前記比較結果に基づいて、PDK4の発現を抑制する被験物質を選択する工程を含む、PDK4の発現または機能抑制を介した骨粗鬆症の予防および/または治療剤のスクリーニング方法。
【発明の詳細な説明】
【技術分野】
【0001】
本発明は、荷重感知遺伝子に関する。詳しくは、重力刺激のない状態で進行する骨粗鬆症の原因の解明、その治療または予防に役立つ遺伝子およびその利用に関する。
【背景技術】
【0002】
骨量は、骨を形成する骨芽細胞の活性と骨を再吸収する破骨細胞の活性とのバランスにより決定される。現代社会における加齢に伴う主要な疾患の1つである骨粗鬆症は、これらの2つの活性のアンバランスにより生じ、食餌、身体活動(運動)、ホルモン状態、サイトカインならびに、糖尿病およびグルココルチコイド治療等の臨床状態に影響される(非特許文献1)。非荷重の姿勢、固定化(運動不足)または長期間の寝たきりにより生じる廃用性骨粗鬆症は、加齢に伴った疾患で寝たきりの患者の増加によって急速に増加している。通常、骨芽細胞と破骨細胞の活性は共役しており、骨再吸収の増加は、骨形成を促進させる。しかし、廃用性骨粗鬆症の場合、骨再吸収の増加は、骨形成を促進させない。このことは、廃用性骨粗鬆症において、骨芽細胞の活性は低下しているが破骨細胞の活性は促進していることを示している(非特許文献2)。オステオポンチン、交感神経の緊張、sclerostinおよびTRIPV4は、非荷重後の骨損失に関連していることが報告されている(非特許文献3〜6)。
【0003】
ピルビン酸デヒドロゲナーゼキナーゼのアイソザイムPDK(PDK1、PDK2、PDK3およびPDK4)は、ミトコンドリア内でピルビン酸をアセチル-CoAに変換させて解糖系をTCAサイクルのエネルギーおよび同化機能に結びつけるピルビン酸デヒドロゲナーゼ複合体(PDC)の負の調節因子である。PDCは、3つの触媒ドメイン:E1(ピルビン酸デカルボキシラーゼ)、E2(ジヒドロリポアミド アセチルトランスフェラーゼ)およびE3(ジヒドロリポアミド デヒドロゲナーゼ)からなるマルチサブユニット複合体である。PDKは、E1の特定のセリン残基をリン酸化してPDCを不活性化するが、ピルビン酸デヒドロゲナーゼホスフェートホスファターゼ(Pdp1およびPdp2)は、そのリン酸化体を脱リン酸化してPDCを活性化させる。PDKのアイソフォームはPDCに対して異なる感度を有しているが、PDK調節の短期のメカニズムには、E1の基質であるピルビン酸による阻害ならびにPDCの産物であるアセチル-CoAおよびNADHによる活性化が含まれる。哺乳動物のPDKは、組織特異的発現を示す:PDK1は、心臓、膵島および骨格筋で発現している;PDK2は、栄養供給状態ではユビキタスに発現しており、心臓、肝臓および腎臓で高発現している;PDK3は、主として精巣、腎臓および脳で発現している;ならびにPDK4は、心臓、骨格筋、肝臓、腎臓および膵島で高発現している(非特許文献7〜9)。心筋および骨格筋におけるPDK4の発現は、飢餓および糖尿病状態で迅速にアップレギュレートされ、それぞれ食餌およびインスリン投与により低下する(非特許文献10、11)。また、PDK4の発現は、グルココルチコイドによってもアップレギュレートし(非特許文献12)、インスリンによりダウンレギュレートされる(非特許文献13)。活性型PDCは、腎臓、骨格筋、横隔膜および心臓に多く、飢餓状態のPdk4-/-マウスの肝臓には多くなく、心臓におけるPDK4の過剰発現は、カルシニューリンストレス活性化経路により生じる心筋症を悪化させる(非特許文献14、15)。
【0004】
骨は、メカニカルストレスに対してその形状および強度を調節する。骨細胞は、骨における最も豊富な細胞であり、骨全体を通じて突起と骨細管を介したコミュニケーションシステムを含んでいる。骨細胞ネットワークは、理想的なメカノセンサーおよびメカノトランスダクションシステムであると考えられている(非特許文献16〜21)。
【0005】
現在、骨粗鬆症治療薬で骨吸収を抑制する薬剤として、ビスフォスフォネートとSERM(selective estrogen receptor modulator)があるが、骨吸収を常に抑制するために、骨質の低下を招き、骨強度の低下を来す場合がある。また、ビスフォスフォネートは、顎骨壊死のリスクがあり、歯の治療の際には、投与を中止しなければならない。また、無重力状態での骨量の低下を予防する薬剤は知られていない。
【先行技術文献】
【非特許文献】
【0006】
【非特許文献1】Manolagas SC, Endocr Rev 2010, 31:266-300
【非特許文献2】Bikle DD, Halloran BP, J Bone Miner Metab 1999, 17:233-244
【非特許文献3】Mizoguchi F, Mizuno A, Hayata T, Nakashima K, Heller S, Ushida T, Sokabe M, Miyasaka N, Suzuki M, Ezura Y, Noda M, J Cell Physiol 2008, 216:47-53
【非特許文献4】Ishijima M, Rittling SR, Yamashita T, Tsuji K, Kurosawa H, Nifuji A, Denhardt DT, Noda M, J Exp Med 2001, 193:399-404
【非特許文献5】Lin C, Jiang X, Dai Z, Guo X, Weng T, Wang J, Li Y, Feng G, Gao X, He L, J Bone Miner Res 2009, 24:1651-1661
【非特許文献6】Kondo H, Nifuji A, Takeda S, Ezura Y, Rittling SR, Denhardt DT, Nakashima K, Karsenty G, Noda M, J Biol Chem 2005, 280:30192-30200
【非特許文献7】Sugden, Holness MJ, Am J Physiol Endocrinol Metab 2003, 284:E855-62
【非特許文献8】Maj M, Cameron J, Robinson B, Molecular and Cellular Endocrinology 2006, 249:1-9
【非特許文献9】Roche TE, Hiromasa Y, Cellular and Molecular Life Sciences 2007, 64:830-849
【非特許文献10】Wu P, Sato J, Zhao Y, Jaskiewicz J, Popov KM, Harris RA, Biochem J 1998, 329 ( Pt 1):197-201
【非特許文献11】Wu P, Inskeep K, Bowker-Kinley MM, Popov KM, Harris RA, Diabetes 1999, 48:1593-1599
【非特許文献12】Qi D, Pulinilkunnil T, An D, Ghosh S, Abrahani A, Pospisilik JA, Brownsey R, Wambolt R, Allard M, Rodrigues B, Diabetes 2004, 53:1790-1797
【非特許文献13】Huang B, Wu P, Bowker-Kinley MM, Harris RA, Diabetes 2002, 51:276-283
【非特許文献14】Zhao, Am J Physiol Heart Circ Physiol 2008, 294:H936-43
【非特許文献15】Jeoung NH, Wu P, Joshi MA, Jaskiewicz J, Bock CB, Depaoli-Roach AA, Harris RA, Biochem J 2006, 397:417-425
【非特許文献16】Burger EH, Klein-Nulend J, FASEB J 1999, 13 Suppl:S101-112
【非特許文献17】MARTIN RB, Bone 2000, 26:71-78
【非特許文献18】Ehrlich PJ, Lanyon LE, Osteoporos Int 2002, 13:688-700
【非特許文献19】Knothe Tate ML, J Biomech 2003, 36:1409-1424
【非特許文献20】KLEIN-NULEND EHBAJ, The FASEB Journal 1999, 13:S101-11
【非特許文献21】Bonewald L, Johnson M, Bone 2008, 42:606-615
【発明の概要】
【発明が解決しようとする課題】
【0007】
本発明の目的は、高齢者の骨粗鬆症、特に麻痺等に伴う廃用性骨粗鬆症、ステロイド性骨粗鬆症または宇宙滞在時の骨量減少の予防または治療薬の開発に有用な手段を提供し、非荷重状態で進行する骨粗鬆症等の病態の解明に役立つ手段等を提供することにある。
【課題を解決するための手段】
【0008】
本発明者は、骨芽細胞におけるBCL2の過剰発現が骨細胞のアポトーシスを誘導し、骨細胞ネットワークが完全に破壊されているBCL2トランスジェニックマウスは非荷重後の骨損失に耐性であることを見出した。野生型マウスおよびBCL2トランスジェニックマウスを比較することにより、非荷重後の骨損失に応答する遺伝子を探索した。野生型マウスでは非荷重後にPDK4発現がアップレギュレートし、再荷重後にダウンレギュレートするが、これらのレギュレーションはBCL2トランスジェニックマウスでは決して観察されないことを見出し、本発明を完成するに至った。即ち、本願発明は、以下に示す通りである。
【0009】
〔1〕 PDK4の発現または機能を抑制する物質を含む、骨粗鬆症の予防または治療剤。
〔2〕 PDK4の発現または機能を抑制する物質が
(1)Pdk4遺伝子のRNAi誘導性核酸、アンチセンス核酸もしくはそれらの発現ベクター、または
(2)PDK4に対する抗体、ドミナントネガティブ変異体、アプタマーもしくはそれらの発現ベクター
である、前記〔1〕に記載の予防または治療剤。
〔3〕 骨粗鬆症が廃用性骨粗鬆症、ステロイド性骨粗鬆症および無重力時の骨量減少からなる群より選ばれる前記〔1〕または〔2〕に記載の予防または治療剤。
〔4〕 下記工程:
(a)骨芽細胞または骨髄由来の単球/マクロファージ系列細胞と被験物質とを接触させる工程、
(b)前記被験物質を接触させた骨芽細胞または骨髄由来の単球/マクロファージ系列細胞におけるPDK4の発現量を調べ、被験物質を接触させない骨芽細胞または骨髄由来の単球/マクロファージ系列細胞における発現量と比較する工程、および
(c)前記比較結果に基づいて、PDK4の発現を抑制する被験物質を選択する工程
を含む、PDK4の発現または機能抑制を介した骨粗鬆症の予防および/または治療剤のスクリーニング方法。
〔5〕 下記工程:
(a)骨芽細胞または骨髄由来の単球/マクロファージ系列細胞と被験物質とを接触させる工程、
(b)前記被験物質を接触させた骨芽細胞または骨髄由来の単球/マクロファージ系列細胞におけるPDK4の発現量を調べ、被験物質を接触させない骨芽細胞または骨髄由来の単球/マクロファージ系列細胞における発現量と比較する工程、
(c)下記工程(c1)または(c2)のいずれかにより破骨細胞への分化を誘導する工程:
(c1)前記比較工程(b)の結果に基づいて、骨芽細胞でPDK4の発現を抑制する被験物質存在下で、骨芽細胞と骨髄由来の単球/マクロファージ系列細胞とを共培養し、破骨細胞への分化を誘導する工程、さらに対照として、被験物質非存在下で、骨芽細胞と骨髄由来の単球/マクロファージ系列細胞とを共培養し、同様に破骨細胞への分化を誘導する工程、または
(c2)前記比較工程(b)の結果に基づいて、単球/マクロファージ系列細胞でPDK4の発現を抑制する被験物質存在下で、単球/マクロファージ系列細胞を、M-CSFおよびRANKL存在下で培養し、破骨細胞への分化を誘導する工程、さらに対照として、被験物質非存在下で、単球/マクロファージ系列細胞をM-CSFおよびRANKL存在下で培養し、同様に破骨細胞への分化を誘導する工程、
(d)前記工程(c)における破骨細胞への分化の程度を比較する工程、および
(e)前記比較工程(d)の結果に基づいて、骨髄由来の単球/マクロファージ系列細胞から破骨細胞への分化を抑制する被験物質を選択する工程
を含む、PDK4の発現または機能抑制を介した骨粗鬆症の予防および/または治療剤のスクリーニング方法。
〔6〕 下記工程:
(a’)被験物質存在下で、骨芽細胞と骨髄由来の単球/マクロファージ系列細胞を共培養し、破骨細胞への分化を誘導する工程、
(b’)前記工程(a’)における破骨細胞への分化の程度を比較する工程、および
(c’)前記比較工程(b’)の結果に基づいて、骨髄由来の単球/マクロファージ系列細胞から破骨細胞への分化を抑制する被験物質を選択する工程
を含む、PDK4の発現または機能抑制を介した骨粗鬆症の予防および/または治療剤のスクリーニング方法。
〔7〕 下記工程:
(a”)被験物質、M-CSFおよびRANKL存在下で、骨髄由来の単球/マクロファージ系列細胞から破骨細胞への分化を誘導する工程、
(b”)前記工程(a”)における破骨細胞への分化の程度を比較する工程、および
(c”)前記比較工程(b”)の結果に基づいて、骨髄由来の単球/マクロファージ系列細胞から破骨細胞への分化を抑制する被験物質を選択する工程
を含む、PDK4の発現または機能抑制を介した骨粗鬆症の予防および/または治療剤のスクリーニング方法。
〔8〕 Pdk4遺伝子を欠損してなる、非ヒト非荷重抵抗性骨モデル動物。
〔9〕 非荷重状態での骨量の減少の病態の解明に用いられる、前記〔8〕に記載の骨モデル動物。
〔10〕 病態が廃用性骨粗鬆症、ステロイド性骨粗鬆症および無重力時の骨量減少からなる群より選ばれる、前記〔9〕に記載の骨モデル動物。
〔11〕 前記動物がマウスである前記〔8〕〜〔10〕のいずれかに記載の骨モデル動物。
〔12〕 前記〔8〕〜〔11〕のいずれかに記載の骨モデル動物の子孫動物。
〔13〕 前記〔8〕〜〔12〕のいずれかに記載の非ヒト動物から得られる組織または細胞。
〔14〕 前記〔8〕〜〔12〕のいずれかに記載の骨モデル動物を用いることを特徴とする、骨粗鬆症の予防または治療剤のスクリーニング方法。
〔15〕 下記工程:
(a)前記〔8〕〜〔12〕のいずれかに記載の骨モデル動物の野生型対照に被験物質を投与し、当該骨モデル動物と野生型対照とを非荷重状態に供する工程、
(b)前記被験物質の投与前後の野生型対照における骨の形態、骨体積または骨密度を解析し、前記〔8〕〜〔12〕のいずれかに記載の骨モデル動物における骨の形態、骨体積または骨密度と比較する工程、および、
(c)前記(b)の比較結果に基づいて、非荷重状態における骨量の減少を抑制する被験物質を選択する工程
を含む、骨粗鬆症の予防または治療剤のスクリーニング方法。
〔16〕 下記工程:
(a)非荷重状態後に得られた前記〔13〕に記載の組織または細胞、および非荷重状態後に野生型対照から得られた組織または細胞をインビトロで生理学的条件下で維持する工程、
(b)前記野生型対照由来の組織または細胞におけるタンパク質のリン酸化の程度と、前記〔13〕由来の組織または細胞におけるタンパク質のリン酸化の程度とを比較する工程、または前記野生型対照由来の組織または細胞におけるタンパク質でPDK4に結合するタンパク質と前記〔13〕由来の組織または細胞におけるタンパク質でPDK4に結合するタンパク質を比較する工程、および
(c)前記比較結果に基づいて、リン酸化の程度が有意に変動しているタンパク質を選択する工程、または前記比較結果に基づいて、前記野生型対照由来の組織または細胞におけるタンパク質のみから得られ、前記〔13〕由来の組織または細胞におけるタンパク質からは得られないPDK4結合タンパク質を選択する工程
を含む、荷重刺激を感知し、骨粗鬆症の発症に関連するPDK4の標的タンパク質のスクリーニング方法。
【発明の効果】
【0010】
本発明は、非荷重後の骨芽細胞および骨髄細胞におけるPDK4発現のアップレギュレーションは、少なくとも一部はPDC不活性化とは別の経路を介して、非荷重後の破骨細胞形成および骨再吸収に導くという知見に基づくものである。本発明の骨粗鬆症の予防または治療剤によると、PDK4の発現または機能を抑制する物質を主成分とするものであることから、特に骨芽細胞または骨髄細胞に作用させた場合に、非荷重状態が主原因の骨粗鬆症を有効に予防または治療することができる。本発明のモデル動物によると、PDK4が欠損していることにより非荷重(無重力)に抵抗性の骨モデル動物として、無重力による骨量の減少など、無重力状態での骨組織での様々な生理学的状態を解明するモデルとして役立つ。本発明の組織または細胞によると、前記動物から得られるものであることから、in vivoのみならずin vitroで骨量減少のメカニズムの研究に利用することが可能である。本発明のスクリーニング方法によると、骨粗鬆症の予防または治療剤の開発のみならず、非荷重状態でのPDK4の基質の解明等に貢献することができる。
【図面の簡単な説明】
【0011】
【図1】PDK4発現のマイクロアレイおよびリアルタイムRT-PCR解析を示す。(A)尾懸垂実験におけるPDK4発現のマイクロアレイおよびリアルタイムRT-PCR解析。16週齢の野生型およびBCL2トランスジェニックマウスの脛骨および大腿骨を用いて、コントロール群、非荷重群(尾懸垂を8日間実施)および再荷重群(1週間の尾懸垂後、24時間尾懸垂から開放されたマウス)からmRNAを調製した。コントロール群のレベルを1に設定し、相対レベルを示す。リアルタイムRT-PCRにおけるデータは、3匹のマウスの平均±SEである。**, p<0.01 vs. コントロール。(B)16週齢の野生型およびBCL2トランスジェニックマウスのコントロール群および非荷重群由来の骨細胞または骨芽細胞画分のmRNAを用いたリアルタイムRT-PCR解析。尾懸垂を3日間実施した。野生型マウスのコントロール群の骨細胞画分のレベルを1と設定し、相対レベルを示す。データは、4匹のマウスの平均±SEである。**, p<0.01 vs. コントロール。(C)種々の組織におけるPDK4の発現。11週齢の野生型マウスの、骨格筋(SM)、心筋(CM)、腎臓(K)、肝臓(Li)、肺(Lu)、脳(B)、脾臓(Sp)、精巣(Te)、胸腺(Th)、全大腿骨(WF)、骨細胞画分(Oc)、骨芽細胞画分(Ob)および骨髄(BM)からmRNAを調製した。全大腿骨のレベルを1に設定し、相対値を示す。データは、3匹のマウスの平均±SEである。(D)骨におけるPDKアイソフォームの発現を骨格筋と比較した相対レベル。骨格筋に対する全大腿骨におけるPDKアイソフォームの相対的mRNAレベルを示す。(E)尾懸垂後のPDKアイソフォームmRNAの発現。3または7日間の尾懸垂後の12週齢の野生型マウスの骨格筋(SM)、全骨(WB)、骨細胞画分(Oc)、骨芽細胞画分(Ob)および骨髄(BM)からmRNAを調製した。コントロール群における全大腿骨のレベルを1に設定し、相対値を示す。データは、3匹のマウスの平均±SEである。*, p<0.05 vs.コントロール。
【図2】Pdk4-/-マウスの産生のための手段および産生マウスの解析結果を示す。(A)Pdk4遺伝子座、ターゲティングベクターおよび変異アレルの概略図。RI: EcoRI。(B)サザンブロット解析。DNAをEcoRIで消化し、Aに示す5’または3’プローブとハイブリダイズさせた。(C)ウエスタンブロット解析。1週間の尾懸垂後、12週齢の野生型マウスおよびPdk4-/-マウスの大腿骨からタンパク質を抽出した。(D)体重曲線。群当たりn=6。(E)マイクロ-CT解析。3、6および12月齢における野生型およびPdk4-/-マウス間の骨体積は類似していた。群当たりn=6。(F)骨芽細胞マーカー遺伝子発現のリアルタイムRT-PCR解析。5週齢の野生型およびPdk4-/-マウスの大腿骨(骨髄細胞を洗い出すことにより除去している)から試料を調製した。野生型マウスにおける値を1に設定し、相対レベルを示す。データは4匹のマウスの平均 ± SE である。(G)Col1a1およびCol1a2タンパク質の定量解析。12週齢で脛骨からタンパク質を抽出した。(H)オステオポンチンおよびオステオカルシンタンパク質のウエスタンブロット解析。12週齢で大腿骨からタンパク質を抽出した。
【図3】尾懸垂後のマイクロ-CTおよび骨組織形態計測解析を示す。11週齢の雄性野生型およびPdk4-/-マウスを1週間正常条件下(コントロール)または尾懸垂条件下(非荷重)においた。(A-E)マイクロ-CT解析。A-D、野生型(A、B)およびPdk4-/-(C, D)マウスのコントロール(A、C)および非荷重条件下(B、D)の三次元イメージ。スケールバー = 0.5mm。E、骨梁パラメータ。骨梁体積(BV/TV)、骨梁数(Tb.N)、骨梁幅(Tb.Th)、骨梁間距離(Tb.Sp)および骨梁密度(Dens)を示す。データは、8匹のマウスの平均 ± SEである。*p<0.05, **p<0.01 vs.コントロール。(F)骨組織形態計測解析。単位骨量(骨体積/組織体積、BV/TV)、類骨面(OS/BS)、類骨幅(O.Th)、骨芽細胞面(Ob.S/BS)、骨芽細胞数(N.Ob/B.Pm)、多核破骨細胞数(Mu.N.Oc/B.Pm)、破骨細胞面(Oc.S/BS)、吸収面(ES/BS)、骨石灰化速度(MAR)、石灰化面(MS/BS)および骨形成速度(BFR/BS)を示す。データは6-7匹のマウスの平均 ± SEである。**p<0.01 vs.コントロール。
【図4】非荷重時の骨形成マーカーと骨吸収マーカーの野生型マウスとPdk4-/-マウス間の比較を示す。(A)リアルタイムRT-PCR解析。コントロールまたは1週間の非荷重条件下での12週齢の野生型およびPdk4-/-マウスの大腿骨および脛骨からmRNAを調製した。データは5匹のマウスの平均 ± SEである。*p<0.05 vs.コントロール。(B)脛骨切片のTRAP染色。切片をメチルグリーンで対比染色した。スケールバー = 0.1mm。(C)TRAP5bの血清レベル。データは6-7匹のマウスの平均 ± SEである。**p<0.01 vs コントロール。
【図5】in vitroでの骨芽細胞形成を示す。(A)骨芽細胞分化中のPDK4の発現。骨芽細胞分化中の野生型初代骨芽細胞におけるPDK4発現を、コンフルエンス(0日目)ならびにコンフルエンス後3日目および5日目でリアルタイムRT-PCRで調べた。0日目のレベルを1と決定し、相対レベルを示す。データは3個のウエルの平均 ± SEである。(B)ALP活性および石灰化。野生型およびPdk4-/-マウス由来の初代骨芽細胞を、コンフルエンス後3日目および14日目に、それぞれALPおよびKossaで染色した。(C)リアルタイムRT-PCR解析。野生型およびPdk4-/-マウス由来の初代骨芽細胞をコンフルエンス後3日間培養し、mRNAを調製した。野生型骨芽細胞のレベルを1と決定し、相対レベルを示す。データは3個のウエルの平均 ± SEである。*p<0.05 vs. 野生型マウス。
【図6】in vitroでの破骨細胞形成を示す。(A)破骨細胞分化中のPDK4の発現。野生型BMMをM-CSF(20ng/ml)およびRANKL(100ng/ml)の存在下で培養した。mRNAを、M-CSFおよびRANKLの添加前(0日目)、添加後3日目および6日目に調製し、リアルタイムRT-PCRでPDK4の発現を調べた。0日目のレベルを1と決定し、相対レベルを示す。データは3個のウエルの平均 ± SEである。(B、C)BMMの破骨細胞分化。BMMを、M-CSF(20ng/ml)およびRANKL(100ng/ml)の存在下で6日間培養した(B)。スケールバー = 0.1mm。3-6の核を有するTRAP-陽性細胞の数、6を超える核を有するTRAP-陽性細胞の数およびそれらの合計を示す(C)。データは4個のウエルの平均 ± SEである。*p<0.05 vs. 野生型BMM。(D)破骨細胞分化に関連する遺伝子の発現。M-CSF(20ng/ml)およびRANKL(100ng/ml)の添加前(0日目)ならびに添加後3および6日目において、野生型およびPdk4-/- BMM由来のmRNAを用いてリアルタイムRT-PCR解析を実施した。(E-H)BMMおよび初代骨芽細胞の共培養。野生型およびPdk4-/-マウス由来のBMMを、野生型またはPdk4-/-初代骨芽細胞と共培養した。6日後にTRAP染色を実施し(E)、3-6の核を有するTRAP-陽性細胞、7-10の核を有するTRAP-陽性細胞および10を超える核を有するTRAP-陽性細胞を計数した(F)。破骨細胞の吸収活性をPitアッセイで調べ(G)、6日後に吸収pitを測定した(H)。スケールバー = 0.1mm。データは、4個のウエルの平均 ± SEである。**p<0.01 vs 野生型POBおよびBMMの共培養。
【図7】外来性PDK4によるPdk4-/-細胞における破骨細胞形成のレスキューおよびRanklプロモーターのレポーターアッセイを示す。(A-E)破骨細胞形成のレスキュー実験。A、PDK4のウエスタンブロット解析。Pdk4-/-初代骨芽細胞を、Pdk4-またはGFP-発現アデノウイルスに感染させた。B-D、BMMおよび初代骨芽細胞の共培養。Pdk4-/-マウス由来のBMMおよび初代骨芽細胞を共培養し、Pdk4-またはGFP-発現アデノウイルスに感染させた。6日後にTRAP染色を実施し(B)、3-6の核を有するTRAP-陽性細胞の数および6を超える核を有するTRAP-陽性細胞の数を計数した(C)。Pitアッセイを実施し、吸収領域を測定した(D)。スケールバー = 0.1mm。データは、4個のウエルの平均 ± SEである。*p<0.05 vs. GFP。E、リアルタイムRT-PCR解析。Pdk4-/-マウス由来の初代骨芽細胞をPdk4-またはGFP-発現アデノウイルスに感染させ、3日後にmRNAを調製した。データは、3個のウエルの平均 ± SEである。*p<0.05 vs. GFP。(F)Ranklプロモーターのレポーターアッセイ。野生型およびPdk4-/-マウス由来の初代骨芽細胞を、段階的に欠失させたRanklプロモーター領域を含むルシフェラーゼレポーターコンストラクトで形質転換した。データは、5個のウエルの平均 ± SEである。*p<0.05 vs. 野生型初代骨芽細胞。(G)PDK4によるRanklプロモーター活性の誘導。Pdk4-/-マウス由来の初代骨芽細胞に、各Ranklルシフェラーゼレポーターコンストラクトを導入、24時間後、細胞をPdk4-またはGFP-発現アデノウイルスに感染させた。データは、5個のウエルの平均 ± SEである。*p<0.05 vs. GFP。
【図8】破骨細胞形成、RanklおよびOpg発現ならびにRanklプロモーター活性におけるPDCリン酸化に対するPDK4アンタゴニストの効果を示す。(A)PDCリン酸化のウエスタンブロット解析。野生型マウス由来の初代骨芽細胞を、DCAの存在下または非存在下で0.5% FBS中、3時間培養し、抽出したタンパク質を抗-ホスホ-E1a(p-PDC)および抗-E1a(PDC)抗体と反応させた。(B)BMMの破骨細胞分化。野生型マウス由来のBMMを、M-CSF(20ng/ml)およびsRANKL(100ng/ml)の存在下、指定された濃度のDCAの存在下または非存在下で培養し、TRAP-陽性細胞を計数した。データは、3個のウエルの平均 ± SEである。(C)BMMと初代骨芽細胞との共培養における破骨細胞形成。野生型マウス由来のBMMおよび初代骨芽細胞を、指定された濃度のDCAの存在下または非存在下で共培養し、TRAP-陽性細胞を計数した。データは、3個のウエルの平均 ± SEである。(D)リアルタイムRT-PCR解析。野生型マウス由来の初代骨芽細胞を指定された濃度のDCAの存在下または非存在下で培養した。データは、3個のウエルの平均 ± SEである。(E)Ranklプロモーターのレポーターアッセイ。野生型マウス由来の初代骨芽細胞をRL(-185)で形質転換し、指定された濃度のDCAの存在下または非存在下で培養した。データは、3個のウエルの平均 ± SEである。(F、G)Ranklプロモーター活性におけるPdp1の効果。野生型マウス由来の初代骨芽細胞を0.5% FBS中で4時間培養し、Pdp1発現ベクターで形質転換した。2日後タンパク質を抽出し、抗-ホスホ-E1a(p-PDC)および抗-E1a(PDC)抗体と反応させた(F)。野生型マウス由来の初代骨芽細胞をRL(-185)と指定された量のDNAのPdp1発現ベクターで共形質転換し、36および72時間後にルシフェラーゼ活性を測定した(G)。
【図9】マウスの組織学的解析結果を示す。野生型マウス(A, C, E, G, I, K)およびPdk4-/-マウス(B, D, F, H, J, L)の4週齢(A, B)および12週齢(C, D)の脛骨切片のH-E染色、および10週齢の大腿骨の骨芽細胞(E-H)および破骨細胞(I-L)の電子顕微鏡写真。E、F、IおよびJのボックス内の領域を、それぞれG、H、KおよびLに拡大する。透過型電子顕微鏡(H-7100; Hitachi, Tokyo, Japan)を用い、既報に従って(Maruyama Z, et al., Dev Dyn 2007, 236:1876-1890)微細構造解析を実施した。スケールバー = 0.2mm(A-D)、3μm(E、F)、1μm(G、H)、5μm(I、J)、および2μm(K、L)。
【図10】血清グルコースおよびコルチゾールの測定結果を示す。11週齢の雄性野生型およびPdk4-/-マウスを、正常条件下(コントロール)または尾懸垂条件(非荷重)下で1週間飼育し、グルコースメーター(SKK, Nagoya, Japan)およびCortisol ELISAキット(Enzo, Tokyo, Japan)をそれぞれ用いて、血清グルコース(A)およびコルチゾール(B)を測定した。
【発明を実施するための形態】
【0012】
(定義)
本発明において「遺伝子」とは、2本鎖DNAのみならず、それを構成するセンス鎖およびアンチセンス鎖といった各1本鎖DNAを包含する趣旨で用いられる。またその長さによって特に制限されるものではない。本発明において遺伝子(DNA)とは、特に言及しない限り、ヒトゲノムDNAを含む2本鎖DNAおよびcDNAを含む1本鎖DNA(正鎖)並びに該正鎖と相補的な配列を有する1本鎖DNA(相補鎖)、およびこれらの断片のいずれもが含まれる。また当該「遺伝子」には、特定の塩基配列(配列番号1)で示される「遺伝子」だけでなく、これらによりコードされるタンパク質と生物学的機能が同等であるタンパク質(例えば同族体(ホモログやスプライスバリアントなど)、変異体および誘導体)をコードする「遺伝子」が包含される。かかる同族体、変異体または誘導体をコードする「遺伝子」としては、具体的には、ストリンジェントな条件下で、前記の配列番号1で示される特定塩基配列の相補配列とハイブリダイズする塩基配列を有する「遺伝子」をあげることができる。なお、ここでストリンジェントな条件は、Berger and Kimmel (1987, Guide to Molecular Cloning Techniques Methods in Enzymology, Vol. 152, Academic Press, San Diego CA) に教示されるように、核酸の融解温度(Tm)に基づいて決定することができる。例えばハイブリダイズ後の洗浄条件として、通常「1×SSC、0.1%SDS、37℃」程度の条件をあげることができる。相補鎖はかかる条件で洗浄しても対象とする正鎖とハイブリダイズ状態を維持するものであることが好ましい。特に制限されないが、より厳しいハイブリダイズ条件として「0.5×SSC、0.1%SDS、42℃」程度、さらに厳しいハイブリダイズ条件として「0.1×SSC、0.1%SDS、65℃」程度の洗浄条件をあげることができる。
【0013】
例えばヒト由来のタンパク質のホモログをコードする遺伝子としては、当該タンパク質をコードするヒト遺伝子に対応するマウスやラットなど他生物種の遺伝子が例示でき、これらの遺伝子(ホモログ)は、HomoloGene(http://www.ncbi.nlm.nih.gov/HomoloGene/)により同定することができる。具体的には、特定ヒト塩基配列をBLAST(Proc. Natl. Acad. Sci. USA 90:5873-5877, 1993、http://www.ncbi.nlm.nih.gov/BLAST/)にかけて一致する(Scoreが最も高く、E-valueが0でかつIdentityが100%を示す)配列のアクセッション番号を取得する。そのアクセッション番号をUniGene(http://www.ncbi.nlm.nih.gov/UniGene/)に入力して得られたUniGene Cluster ID(Hs.で示す番号)をHomoloGeneに入力する。結果として得られた他生物種遺伝子とヒト遺伝子との遺伝子ホモログの相関を示したリストから、特定の塩基配列で示されるヒト遺伝子に対応する遺伝子(ホモログ)としてマウスやラットなど他生物種の遺伝子を選抜することができる。
【0014】
本発明において「Pdk4遺伝子」または「Pdk4のDNA」とは、特定塩基配列(配列番号1)で示されるヒトPdk4遺伝子(DNA)ならびに、その同族体、変異体および誘導体などをコードする遺伝子(DNA)を包含する趣旨で用いられる。具体的には、配列番号1に記載のヒトPdk4遺伝子ならびに、そのマウスホモログおよびラットホモログなどが包含される。なお、遺伝子またはDNAは、機能領域の別を問うものではなく、例えば発現制御領域、コード領域、エキソン、またはイントロンを含むことができる。
【0015】
本発明において「タンパク質」または「ペプチド」には、特定のアミノ酸配列(配列番号2)で示される「タンパク質」または「ペプチド」だけでなく、これらと生物学的機能が同等であることを限度として、その同族体(ホモログやスプライスバリアント)、変異体、誘導体、成熟体およびアミノ酸修飾体などが包含される。ここでホモログとしては、ヒトのタンパク質に対応するマウスやラットなど他生物種のタンパク質が例示でき、これらはHomoloGene(http://www.ncbi.nlm.nih.gov/HomoloGene/)により同定された遺伝子の塩基配列から演繹的に同定することができる。前記変異体には、天然に存在するアレル変異体、天然に存在しない変異体、および人為的に欠失、置換、付加および挿入されることによって改変されたアミノ酸配列を有する変異体が包含される。なお、前記変異体としては、変異のないタンパク質または(ポリ)ペプチドと、少なくとも70%、好ましくは80%、より好ましくは95%、さらにより好ましくは97%相同なものをあげることができる。前記誘導体には、タンパク質のアミノ末端もしくはカルボキシ末端または側鎖を置換したものが包含される。前記アミノ酸修飾体には、天然に存在するアミノ酸修飾体、天然に存在しないアミノ酸修飾体が包含され、具体的にはアミノ酸のリン酸化体があげられる。
【0016】
本発明において「PDK4」という用語を用いる場合、配列番号で特に指定しない限り、特定アミノ酸配列(配列番号2)で示されるヒトPDK4やその同族体、変異体、誘導体、成熟体およびアミノ酸修飾体などを包含する趣旨で用いられる。
【0017】
本明細書中、PDK4は通常、温血動物(哺乳動物又は鳥類)由来のものを意味する。哺乳動物としては、例えば、マウス、ラット、ハムスター、モルモット等のげっ歯類やウサギ等の実験動物、ブタ、ウシ、ヤギ、ウマ、ヒツジ、ミンク等の家畜、イヌ、ネコ等のペット、ヒト、サル、カニクイザル、アカゲザル、マーモセット、オランウータン、チンパンジーなどの霊長類等をあげることができるが、これらに限定されるものではない。鳥類としては、ニワトリ、ウズラ、アヒル、ガチョウ、シチメンチョウ、エミュ、ダチョウ、ホロホロ鳥、ハト等を挙げることができる。PDK4は、好ましくは哺乳動物由来のものであり、より好ましくは霊長類(ヒト等)又はげっ歯類(マウス等)由来のものである。
【0018】
「PDK4が哺乳動物由来である」とは、PDK4の配列(ヌクレオチド配列又はアミノ酸配列)が哺乳動物のものであることを意味する。
【0019】
本発明においてPDK4をコードする遺伝子(以下、Pdk4遺伝子またはPdk4と省略する場合もある)は、例えば、ヒトPdk4遺伝子は、Rowlesらの文献(Rowles J. et al., 1996, J. Biol. Chem. 271 (37), pp.22376-22382)で公表されており、自体公知の方法により単離することができる。また、ヒトおよびマウスのPDK4の代表的なヌクレオチド配列及びアミノ酸配列が、NCBIに以下の通りに登録されている。
[ヒトPDK4]
ヌクレオチド配列(cDNA配列):アクセッション番号 NM_002612(バージョンNM_002612.3)(配列番号1)
アミノ酸配列:アクセッション番号 NP_002603(NP_002603.1)(配列番号2)
[マウスPDK4]
ヌクレオチド配列(cDNA配列):アクセッション番号 NM_013743(バージョンNM_013743.2)(配列番号3)
アミノ酸配列:アクセッション番号 NP_038771(バージョンNP_038771.1)(配列番号4)
【0020】
なお、本明細書においてヌクレオチド配列は、特にことわりのない限りDNAの配列として記載するが、ポリヌクレオチドがRNAである場合は、チミン(T)をウラシル(U)に適宜読み替えるものとする。
【0021】
(荷重感知遺伝子)
本明細書における「荷重感知遺伝子」とは、生体(動物)に対する荷重刺激を感知し、荷重(重力)刺激がない場合、破骨細胞の活動を活性化させる遺伝子をいう。具体的には、荷重刺激のない(非荷重)場合に骨細胞および骨細胞からのシグナルによって骨芽細胞でアップレギュレートし、再荷重後にダウンレギュレートする遺伝子であり、ピルビン酸デヒドロゲナーゼキナーゼのアイソザイムPDK4としても知られる遺伝子である。
【0022】
本発明は、Pdk4遺伝子を有効成分として含有する荷重感知遺伝子を提供する。また、本発明は、PDK4を有効成分として含有する荷重感知剤を提供する。
【0023】
(骨粗鬆症の予防または治療剤)
本発明は、PDK4の発現または活性を抑制する物質を含む、骨粗鬆症の予防または治療剤を提供する。
【0024】
本発明の予防または治療剤(本発明の医薬と称する場合もある)が対象とする疾患は、骨粗鬆症である。本発明において骨粗鬆症とは、世界保健機構の定義で、骨密度測定により若年成人の2.5標準偏差未満で診断されるものをいう。骨量の低下は種々の原因により生じるが、長期的な寝たきり状態、固定化状態、運動障害、麻痺、無重力状態での生活などによる非荷重状態が主原因である骨粗鬆症、すなわち、廃用性骨粗鬆症が具体的にあげられる。また、ステロイド性骨粗鬆症および無重力時の骨量減少もあげられる。
【0025】
PDK4の機能としては、ピルビン酸デヒドロゲナーゼキナーゼとしてこれまでに知られているキナーゼとしての機能および今後解明されるであろう様々な機能があげられるが、エンドポイントとして、荷重刺激を感知して荷重(重力)刺激がない場合、破骨細胞の活動を活性化する機能を指標とすることができる。
【0026】
PDK4の発現又は機能を抑制する物質としては、PDK4のアンタゴニストがあげられる。例えば、PDK4のPDCリン酸化に対するアンタゴニストとして知られているジクロロ酢酸およびピルビン酸デヒドロゲナーゼデホスファターゼは、破骨細胞形成、Rankl発現およびRanklプロモーター活性に影響がほとんどないことから、PDK4のアンタゴニストであって、PDCリン酸化に対するアンタゴニスト作用を有さないものが好ましく、より好ましくは低分子化合物である。このようなアンタゴニストは、本発明のスクリーニング方法により容易に選別することができる。
【0027】
PDK4の発現又は機能を抑制する物質としては、例えば、PDK4の発現を特異的に抑制し得るsiRNA、アンチセンス核酸、これらのポリヌクレオチドを発現し得る発現ベクター、低分子化合物があげられる。好ましくは、PDK4の発現を特異的に抑制し得るsiRNA、アンチセンス核酸、又はこれらのポリヌクレオチドを発現し得る発現ベクターが用いられる。
【0028】
本明細書中、「特異的な遺伝子発現の抑制」とは、標的とする遺伝子の発現を、それ以外の遺伝子の発現よりも強く抑制することを意味する。
【0029】
PDK4の発現を特異的に抑制し得るsiRNAとしては、例えば
(A)PDK4をコードするmRNA(成熟mRNA又は初期転写産物)のヌクレオチド配列又は18塩基以上のその部分配列に相補的なヌクレオチド配列を含む2本鎖のRNA、及び
(B)PDK4をコードするmRNA(成熟mRNA又は初期転写産物)と治療対象動物(好ましくはヒト)の細胞内で特異的にハイブリダイズし得る18塩基以上のヌクレオチド配列を含み、且つハイブリダイズすることによりPDK4の転写を抑制する2本鎖のRNAを挙げることができる。
【0030】
本明細書中、「特異的なハイブリダイゼーション」とは、核酸が、標的とするヌクレオチドに対して、それ以外のヌクレオチドよりも強くハイブリダイズすることを意味する。
【0031】
PDK4をコードするmRNAのヌクレオチド配列としては、例えば、配列番号1で表されるヌクレオチド配列(ヒトPDK4)、配列番号3で表されるヌクレオチド配列(マウスPDK4)をあげることができる。
【0032】
短い二本鎖RNAを細胞内に導入するとそのRNAに相補的なmRNAが分解される、いわゆるRNA干渉(RNAi)と呼ばれる現象は、以前から線虫、昆虫、植物等で知られていたが、最近、この現象が動物細胞でも起こることが確認されたことから[Nature, 411(6836): 494-498 (2001)]、リボザイムの代替技術として注目されている。
【0033】
siRNAは、代表的には、標的遺伝子のmRNAのヌクレオチド配列又はその部分配列(以下、標的ヌクレオチド配列)と相補的な配列を有するRNAとその相補鎖からなる2本鎖オリゴRNAである。また、ヘアピンループ部分を介して、標的ヌクレオチド配列に相補的な配列(第1の配列)と、その相補配列(第2の配列)とが連結された一本鎖RNAであって、ヘアピンループ型の構造をとることにより、第1の配列が第2の配列と2本鎖構造を形成するRNA(small hairpin RNA: shRNA)もsiRNAの好ましい態様の1つである。
【0034】
siRNAに含まれる、標的ヌクレオチド配列と相補的な部分の長さは、通常、約18塩基以上、好ましくは約19塩基以上、より好ましくは約21塩基以上の長さであるが、標的遺伝子の発現を特異的に抑制可能である限り、特に限定されない。siRNAが23塩基よりも長い場合には、該siRNAは細胞内で分解されて、約20塩基前後のsiRNAを生じ得るので、理論的には標的ヌクレオチド配列と相補的な部分の長さの上限は、標的遺伝子のmRNA(成熟mRNAもしくは初期転写産物)のヌクレオチド配列の全長である。しかし、インターフェロン誘導の回避、合成の容易さ、抗原性の問題等を考慮すると、該相補部分の長さは、例えば約50塩基以下、好ましくは約25塩基以下、最も好ましくは約23塩基以下である。即ち、該相補部分の長さは、通常、約18〜約50塩基、好ましくは約19〜約25塩基、より好ましくは約21〜約23塩基である。
【0035】
また、siRNAを構成する各RNA鎖の長さも、通常、約18塩基以上、好ましくは約19塩基以上、より好ましくは約21塩基以上の長さであるが、標的遺伝子の発現を特異的に抑制可能である限り、特に限定されず、理論的には各RNA鎖の長さの上限はない。しかし、インターフェロン誘導の回避、合成の容易さ、抗原性の問題等を考慮すると、siRNAの長さは、例えば約50塩基以下、好ましくは約25塩基以下、最も好ましくは約23塩基以下である。即ち、各RNA鎖の長さは、例えば通常、約18〜約50塩基、好ましくは約19〜約25塩基、より好ましくは約21〜約23塩基である。なお、shRNAの長さは、2本鎖構造をとった場合の2本鎖部分の長さとして示すものとする。
【0036】
標的ヌクレオチド配列と、siRNAに含まれるそれに相補的な配列とは、完全に相補的であることが好ましい。しかし、siRNAの中央から外れた位置についての塩基の変異(少なくとも90%以上、好ましくは95%以上の同一性の範囲内であり得る)については、完全にRNA干渉による切断活性がなくなるのではなく、部分的な活性が残存し得る。他方、siRNAの中央部の塩基の変異は影響が大きく、RNA干渉によるmRNAの切断活性が極度に低下し得る。
【0037】
siRNAは、5’及び/又は3’末端に塩基対を形成しない、付加的な塩基を有していてもよい。該付加的塩基の長さは、siRNAが標的遺伝子の発現を特異的に抑制可能である限り特に限定されないが、通常5塩基以下、例えば2〜4塩基である。該付加的塩基は、DNAでもRNAでもよいが、DNAを用いるとsiRNAの安定性を向上させることができる。このような付加的塩基の配列としては、例えばug-3’、uu-3’、tg-3’、tt-3’、ggg-3’、guuu-3’、gttt-3’、ttttt-3’、uuuuu-3’などの配列が挙げられるが、これに限定されるものではない。
【0038】
shRNAのヘアピンループのループ部分の長さは、標的遺伝子の発現を特異的に抑制可能である限り、特に限定されないが、通常、5〜25塩基程度である。該ループ部分のヌクレオチド配列は、ループを形成することができ、且つ、shRNAが標的遺伝子の発現を特異的に抑制可能である限り、特に限定されない。
【0039】
「アンチセンス核酸」とは、標的mRNA(成熟mRNA又は初期転写産物)を発現する細胞の生理的条件下で該標的mRNAと特異的にハイブリダイズし得るヌクレオチド配列を含み、且つハイブリダイズした状態で該標的mRNAにコードされるポリペプチドの翻訳を阻害し得る核酸をいう。アンチセンス核酸の種類はDNAであってもRNAであってもよいし、あるいはDNA/RNAキメラであってもよいが、好ましくはDNAである。
【0040】
PDK4の発現を特異的に抑制し得るアンチセンス核酸としては、例えば
(A)PDK4をコードするmRNA(成熟mRNA又は初期転写産物)のヌクレオチド配列又は12塩基以上のその部分配列に相補的なヌクレオチド配列を含む核酸、及び
(B)PDK4をコードするmRNA(成熟mRNA又は初期転写産物)と治療対象動物(好ましくはヒト)の細胞内で特異的にハイブリダイズし得る12塩基以上のヌクレオチド配列を含み、且つハイブリダイズした状態でPDK4ポリペプチドへの翻訳を阻害し得る核酸
等をあげることができる。
【0041】
アンチセンス核酸中の標的mRNAとハイブリダイズする部分の長さは、PDK4の発現を特異的に抑制する限り特に制限はなく、通常、約12塩基以上であり、長いものでmRNA(成熟mRNA又は初期転写産物)の全長配列と同一の長さである。ハイブリダイゼーションの特異性を考慮すると、該長さは好ましくは約15塩基以上、より好ましくは約18塩基以上である。また、合成の容易さや抗原性の問題等を考慮すると、標的mRNAとハイブリダイズする部分の長さは、通常、約200塩基以下、好ましくは約50塩基以下、より好ましくは約30塩基以下である。即ち、標的mRNAとハイブリダイズする部分の長さは、例えば約12〜約200塩基、好ましくは約15〜約50塩基、より好ましくは約18〜約30塩基である。
【0042】
アンチセンス核酸の標的ヌクレオチド配列は、PDK4の発現を特異的に抑制可能であれば特に制限はなく、PDK4のmRNA(成熟mRNA又は初期転写産物)の全長配列であっても部分配列(例えば約12塩基以上、好ましくは約15塩基以上、より好ましくは約18塩基以上)であってもよいし、あるいは初期転写産物のイントロン部分であってもよいが、好ましくは、標的配列はPDK4のmRNAの5’末端からコード領域のC末端までに位置することが望ましい。
【0043】
アンチセンス核酸中の標的mRNAとハイブリダイズする部分のヌクレオチド配列は、標的配列の塩基組成によっても異なるが、生理的条件下でPDK4のmRNAとハイブリダイズし得るために、標的配列の相補配列に対して通常約90%以上(好ましくは95%以上、最も好ましくは100%)の同一性を有するものである。
【0044】
アンチセンス核酸の大きさは、通常約12塩基以上、好ましくは約15塩基以上、より好ましくは約18塩基以上である。該大きさは、合成の容易さや抗原性の問題等から、通常約200塩基以下、好ましくは約50塩基以下、より好ましくは約30塩基以下である。
【0045】
さらに、アンチセンス核酸は、PDK4のmRNAもしくは初期転写産物とハイブリダイズして翻訳を阻害するだけでなく、二本鎖DNAであるPDLIM2遺伝子と結合して三重鎖(トリプレックス)を形成し、mRNAへの転写を阻害し得るものであってもよい。
【0046】
天然型の核酸は、細胞中に存在する核酸分解酵素によってそのリン酸ジエステル結合が容易に分解されるので、本発明において使用されるsiRNAやアンチセンス核酸は、分解酵素に安定なチオリン酸型(リン酸結合のP=OをP=Sに置換)や2’-O-メチル型等の修飾ヌクレオチドを用いて合成することもできる。siRNAやアンチセンス核酸の設計に重要な他の要素として、水溶性及び細胞膜透過性を高めること等が挙げられるが、これらはリポソームやマイクロスフェアを使用するなどの剤形の工夫によっても克服することができる。
【0047】
PDK4の発現を特異的に抑制し得るsiRNA及びアンチセンス核酸は、PDK4のmRNA配列(例えば配列番号1又は3で表されるヌクレオチド配列)や染色体DNA配列に基づいて標的配列を決定し、市販のDNA/RNA自動合成機(アプライド・バイオシステムズ社、ベックマン社等)を用いて、これに相補的なヌクレオチド配列を合成することにより調製できる。siRNAは、センス鎖及びアンチセンス鎖をDNA/RNA自動合成機でそれぞれ合成し、適当なアニーリング緩衝液中、約90〜約95℃で約1分程度変性させた後、約30〜約70℃で約1〜約8時間アニーリングさせることにより調製できる。また、相補的なオリゴヌクレオチド鎖を交互にオーバーラップするように合成して、これらをアニーリングさせた後リガーゼでライゲーションすることにより、より長い2本鎖ポリヌクレオチドを調製できる。
【0048】
本発明の医薬は、PDK4の発現を特異的に抑制するsiRNA又はアンチセンス核酸を発現し得る(コードする)発現ベクターを有効成分とすることもできる。当該発現ベクターにおいては、上述のsiRNA又はアンチセンス核酸或いはそれをコードする核酸(好ましくはDNA)が、投与対象である哺乳動物(好ましくはヒト)の細胞(例えば、骨細胞、骨芽細胞、骨髄細胞)内でプロモーター活性を発揮し得るプロモーターに機能的に連結されている。
【0049】
使用されるプロモーターは、投与対象である哺乳動物の細胞内で機能し得るものであれば特に制限はない。プロモーターとしては、polI系プロモーター、polII系プロモーター、polIII系プロモーター等を使用することができる。具体的には、SV40由来初期プロモーター、サイトメガロウイルスLTR等のウイルスプロモーター、β−アクチン遺伝子プロモーター等の哺乳動物の構成蛋白質遺伝子プロモーター、並びにtRNAプロモーター等のRNAプロモーター等が用いられる。
【0050】
本発明の医薬は、好ましくは骨芽細胞に導入されることによりその作用をより効果的に発揮するものである。したがって、有効成分としての前記発現ベクターは、発現する遺伝子が骨芽細胞特異的プロモーターの下流に作動可能に連結されているものが好ましい。
【0051】
前記骨芽細胞特異的プロモーターとは、骨芽細胞特異的に下流の遺伝子の転写開始部位を決定し、その頻度を直接的に調節する塩基配列を有するDNAをいう。前記プロモーターは、骨芽細胞が由来する動物に応じて適宜選択することができる。
【0052】
前記骨芽細胞特異的プロモーターとしては、I型コラーゲンプロモーター、オステオカルシンプロモーター等があげられるが、I型コラーゲンプロモーターが好ましい。I型コラーゲンプロモーターとしては、例えばヒト由来のプロモーターの場合、Jimenez,S.A., Varga,J., Olsen,A., Li,L., Diaz,A., Herhal,J. and Koch,J. Functional analysis of human alpha 1(I) procollagen gene promoter. Differential activity in collagen-producing and -nonproducing cells and response to transforming growth factor beta 1 J. Biol. Chem. 269 (17), 12684-12691 (1994)、マウス由来のプロモーターの場合、Ravazzolo,R., Karsenty,G. and de Crombrugghe,B. A fibroblast-specific factor binds to an upstream negative control element in the promoter of the mouse alpha 1(I) collagen gene J. Biol. Chem. 266 (12), 7382-7387 (1991)、Rossert, J., Eberspaecher, H., and de Crombrugghe, B. (1995). Separate cis-acting DNA elements of the mouse pro-a1(I) collagen promoter direct expression of reporter genes to different type I collagen-producing cells in transgenic mice. J. Cell Biol. 129, 1421-1432.などに記載されている。
【0053】
上記発現ベクターは、好ましくは上述のポリヌクレオチド又はそれをコードする核酸の下流に転写終結シグナル、すなわちターミネーター領域を含有する。さらに、形質転換細胞選択のための選択マーカー遺伝子(テトラサイクリン、アンピシリン、カナマイシン等の薬剤に対する抵抗性を付与する遺伝子、栄養要求性変異を相補する遺伝子等)をさらに含有することもできる。
【0054】
発現ベクターとしては、公知または市販のものを使用することができ、ウイルスベクターまたは非ウイルスベクターが例示される。ウイルスベクターとしては、例えば、レトロウイルス、アデノウイルス、アデノ随伴ウイルス、ヘルペスウイルス、ワクシニアウイルス、ポリオウイルス、シンドビスウイルスなどのウイルスベクターがあげられる。前記プロモーターおよび遺伝子を発現ベクターに連結する方法は、自体公知の方法により行うことができる。
【0055】
PDK4の発現または機能を抑制する物質の別の態様は、PDK4に対する抗体、ドミナントネガティブ変異体、アプタマー、もしくはそれらの発現ベクターである。
【0056】
本発明の医薬に有効成分として含有する抗体は、ヒトへの適用を考慮した場合、キメラ抗体(chimeric antibody)、ヒト化抗体(humanized antibody)、ヒト型抗体(human antibody)が好ましく、ヒト化抗体およびヒト型抗体がより好ましい。キメラ抗体は、例えば「実験医学(臨時増刊号), Vol.6, No.10, 1988」、特公平3-73280号公報等を、ヒト化抗体は、例えば特表平4-506458号公報、特開昭62-296890号公報等を、ヒト抗体は、例えば「Nature Genetics, Vol.15, p.146-156, 1997」、「Nature Genetics, Vol.7, p.13-21, 1994」、特表平4-504365号公報、国際公開第94/25585号公報、「日経サイエンス、6月号、第40〜第50頁、1995年」、「Nature, Vol.368, p.856-859, 1994」、特表平6-500233号公報等を参考にそれぞれ作製することができる。
【0057】
ドミナントネガティブ変異体は、PDK4に対する変異の導入によりその活性が低減したものである。ドミナントネガティブ変異体は、天然の翻訳産物と競合することで間接的にその機能を阻害することができる。変異としては、例えば、機能性部位における、当該部位が担う機能の低下をもたらすようなアミノ酸の変異(例、1以上のアミノ酸の欠失、置換、付加)があげられる。
【0058】
PDK4に対するアプタマーは、PDK4に特異的に結合してその機能的発現を阻害するオリゴヌクレオチドまたはペプチドである。PDK4に対するオリゴヌクレオチドアプタマーは、in vitro selection法またはSELEX法により得ることができ、例えば、以下の手順があげられる。即ち、まず、DNA/RNA自動合成機を用いてランダムにオリゴヌクレオチド(例えば、約60塩基)を合成し、オリゴヌクレオチドのプールを作製する。次に、目的の蛋白質、即ちPDK4と結合するオリゴヌクレオチドをアフィニティーカラムで分離する。分離したオリゴヌクレオチドをPCRで増幅し、前述の選抜プロセスで再度選抜する。この過程を約5回以上繰り返すことによって、PDK4に親和性の強いアプタマーを選抜することができる。PDK4に対するペプチドアプタマーは、酵母のtwo-hybrid法により得ることができる。
【0059】
本発明の医薬が発現ベクターを有効成分とする場合、当該発現ベクターは、上記タンパク質(抗体、ドミナントネガティブ変異体またはアプタマー)をコードするポリヌクレオチドが、投与対象である生体の細胞内でプロモーター活性を発揮し得るプロモーターに機能的に連結されていなければならない。ここで、発現ベクターが含み得るプロモーターは、前記RNAi誘導性核酸またはアンチセンス核酸の発現ベクターで用いられるものを限定なく利用することができる。また、発現ベクター自体も、前記RNAi誘導性核酸またはアンチセンス核酸の発現ベクターと同様のものを限定なく利用することができる。
【0060】
本発明の医薬は、PDK4の発現又は機能を抑制する物質に加え、任意の担体、例えば医薬上許容される担体を含むことができる。
【0061】
医薬上許容され得る担体としては、例えば、ショ糖、デンプン、マンニット、ソルビット、乳糖、グルコース、セルロース、タルク、リン酸カルシウム、炭酸カルシウム等の賦形剤、セルロース、メチルセルロース、ヒドロキシプロピルセルロース、ポリプロピルピロリドン、ゼラチン、アラビアゴム、ポリエチレングリコール、ショ糖、デンプン等の結合剤、デンプン、カルボキシメチルセルロース、ヒドロキシプロピルスターチ、ナトリウム−グリコール−スターチ、炭酸水素ナトリウム、リン酸カルシウム、クエン酸カルシウム等の崩壊剤、ステアリン酸マグネシウム、エアロジル、タルク、ラウリル硫酸ナトリウム等の滑剤、クエン酸、メントール、グリチルリチン・アンモニウム塩、グリシン、オレンジ粉等の芳香剤、安息香酸ナトリウム、亜硫酸水素ナトリウム、メチルパラベン、プロピルパラベン等の保存剤、クエン酸、クエン酸ナトリウム、酢酸等の安定剤、メチルセルロース、ポリビニルピロリドン、ステアリン酸アルミニウム等の懸濁剤、界面活性剤等の分散剤、水、生理食塩水、オレンジジュース等の希釈剤、カカオ脂、ポリエチレングリコール、白灯油等のベースワックスなどが挙げられるが、それらに限定されるものではない。
【0062】
PDK4の発現又は機能を抑制する物質として、PDK4の発現を特異的に抑制し得るsiRNA、アンチセンス核酸、又はこれらのポリヌクレオチドを発現し得る発現ベクターを用いる場合、核酸の細胞内への導入を促進するために、本発明の医薬はさらに核酸導入用試薬を含むことができる。
【0063】
核酸導入試薬としては、リポフェクチン、リポフェクタミン(lipofectamine)、リポフェクタミンRNAiMAX(LipofectamineRNAiMAX)、インビボフェクタミン(Invivofectamine)、DOGS(トランスフェクタム)、DOPE、DOTAP、DDAB、DHDEAB、HDEAB、ポリブレン、あるいはポリ(エチレンイミン)(PEI)等の陽イオン性脂質を用いることが出来る。また、発現ベクターとしてレトロウイルスを用いる場合には、導入試薬としてレトロネクチン、ファイブロネクチン、ポリブレン等を用いることができる。
【0064】
本発明の骨粗鬆症の予防または治療剤の投与単位形態としては、経口剤または非経口剤があげられ、非経口剤が好ましい。非経口に適した形態としては、注射剤(液剤、懸濁剤など)、坐剤、貼付剤、軟膏剤、ゼリー剤、吸入剤、クリーム剤、スプレー剤、点鼻剤、エアゾール剤などが例示される。
【0065】
医薬組成物中のPDK4の発現又は機能を抑制する物質の含有量は、特に限定されず広範囲に適宜選択されるが、例えば、医薬組成物全体の約0.001ないし100重量%である。
【0066】
本発明の医薬を所望の細胞(好ましくは骨芽細胞または骨髄細胞)に導入する方法は、例えば、in vivo法またはex vivo法があげられる。in vivo法は、本発明の医薬を、例えば注射剤などの適当な投与形態で、静脈、動脈、皮下、関節内、筋肉内などに投与することができる。ex vivo法の場合、本発明の医薬を、対象の生体内から取り出した細胞をin vitroで培養中に公知の方法によりトランスフェクション、インフェクションまたはインジェクションし、一定期間培養してPdk4遺伝子の発現抑制を確認した後、同一対象の体内に戻す方法を採用することができる。
【0067】
これら各種形態の製剤中の有効成分の投与量は、治療目的である疾患の程度、患者の年齢、体重などにより適宜調節することができる。通常、患者成人1人当たり約0.0001-100mg、好ましくは約0.001-10mgが数日ないし数カ月に1回投与される量とすればよい。レトロウイルスベクターの場合は、レトロウイルス力価として、1日患者体重1kg当たり約1×103pfuないし1×1015pfuとなる量の範囲から選ぶことができる。ex vivo法で本発明の医薬を導入した細胞の場合は、1×104細胞/bodyないし1×1015細胞/body程度を投与すればよい。
【0068】
本発明の医薬に含まれる有効成分により、生体内に投与された場合にPDK4の発現又は機能が抑制されるが、血糖値やグルココルチコイドの濃度に影響はない。これは、別のアイソザイムであるPDK1、PDK2およびPDK3が代償的に働くためであると考えられる。本発明の医薬(医薬組成物)は、哺乳動物(例えば、ラット、マウス、モルモット、ウサギ、ヒツジ、ウマ、ブタ、ウシ、サル、ヒト)に対して安全に投与されうる。
【0069】
(非荷重抵抗性骨モデル動物)
本発明は、Pdk4遺伝子を欠損してなる、非ヒト非荷重抵抗性骨モデル動物を提供する。本発明の骨モデル動物は、Pdk4遺伝子を欠損していることから、好適には、非荷重状態での骨量の減少の病態の解明に用いられる。病態としては、骨粗鬆症があげられ、好ましくは、廃用性骨粗鬆症、ステロイド性骨粗鬆症および無重力時の骨量減少からなる群より選ばれる。
【0070】
非ヒト動物としては、ヒト以外の動物であれば特に限定されるものではないが、ヒトに類似した骨細胞ネットワークを有する動物が好ましい。好適な動物としては、マウス、ラット、ハムスター、モルモット、ウサギ、イヌ、ネコ、サル等の実験動物ならびにウシ、ヒツジ、ウマ、ブタ等の家畜があげられるが、遺伝子工学的に利用が容易であるところから、マウスがより好ましい。
【0071】
本発明において、Pdk4遺伝子を欠損するとは、内因性のPdk4遺伝子の塩基配列の1以上の塩基に置換、欠失、挿入もしくは付加の変異を導入することをいい、ノックアウトとも称する。また、このような改変をした動物をノックアウト動物と称する。
【0072】
ノックアウトの場合、少なくともPdk4遺伝子の一部を欠損させる変異を導入させ、当該遺伝子の機能的欠損を有するように操作することが好ましく、具体的には、マウスPdk4遺伝子の場合、エキソンのいずれか1つを欠損させるように構築されたターゲティングベクターを用いる方法が一例としてあげられる。より具体的には、実施例3および図2に記載のターゲティングベクターを用いた方法があげられる。
【0073】
本発明の骨モデル動物には、前記遺伝子の変異が対立遺伝子の両方に導入されたホモ接合動物、前記遺伝子の変異が対立遺伝子の片方に導入されたヘテロ接合動物およびそれらの出生前の胎仔も含まれる。前記ホモ変異動物は、前記ヘテロ変異動物を交配することにより得られるものである。
【0074】
ノックアウト動物は、例えば下記の工程(a)〜(c)を含む方法により製造できる。
(a)Pdk4遺伝子の機能的欠損を含む胚性幹細胞を提供する工程;
(b)前記胚性幹細胞を胚に導入し、キメラ胚を得る工程;
(c)前記キメラ胚を動物に移植し、キメラ動物を得る工程;
(d)前記キメラ動物を交配させ、Pdk4遺伝子欠損ヘテロ接合体を得る工程。
【0075】
ノックアウト動物、胚性幹細胞、ターゲティングベクターの作製の詳細については、例えば、下記文献を参照することができる。
1.別冊 実験医学 ザ・プロトコールシリーズ 「ジーンターデティングの最新技術」(2000年、羊土社)コンディショナルターゲティング法p.115-120
2.バイオマニュアルシリーズ8 「ジーンターゲティング」−ES細胞を用いた変異マウスの作製(1995年、羊土社)p.71-77
3.Sambrookら, Molecular Cloning: A LABORATORY MANUAL, 第3版, COLD SPRING HARBOR LABORATORY PRESS, 2001年, 4.82-4.85
4.Robertson E. J. in Teratocarcinomas and embryonic stem cells-a practical approach, ed. Robertson, E. J. (IRL Press, Oxford), 1987: pp.108-112
5.Dynecki, S. M.ら, Gene Targeting -a practical approach, 2nd edition, ed. Joyner, A.L. (Oxford Univ. Press), 2000: pp.68-73
6.Dynecki, S. M. ら, Gene Targeting -a practical approach, 2nd edition, ed. Joyner, A. L. (Oxford Univ. Press), 2000: pp.75-81
【0076】
このようにして得られた本発明の骨モデル動物をさらに交配して得られる子孫動物、これら動物に由来する組織または細胞も本発明に含まれる。前記組織としては、すべての組織があげられ、長管骨、頭蓋骨、椎骨などが好ましい。また、前記細胞としては、前記組織中に含まれる細胞、組織中から単離された細胞、これら細胞から樹立した細胞株があげられ、具体的には骨芽細胞、骨細胞または骨髄細胞などが好ましい。
【0077】
本発明の非ヒト骨モデル動物、子孫動物、これら動物に由来する組織または細胞は、下記本発明のスクリーニング方法において好適に利用することができる。
【0078】
(スクリーニング方法1)
スクリーニング方法1は、細胞または組織を用いたインビトロの系で実施する。具体的には、下記工程を含む、PDK4の発現または機能抑制を介した骨粗鬆症の予防および/または治療剤のスクリーニング方法1(1)を提供する:
(a)骨芽細胞または骨髄由来の単球/マクロファージ系列細胞と被験物質とを接触させる工程、
(b)前記被験物質を接触させた骨芽細胞または骨髄由来の単球/マクロファージ系列細胞におけるPDK4の発現量を調べ、被験物質を接触させない骨芽細胞または骨髄由来の単球/マクロファージ系列細胞における発現量と比較する工程、および
(c)前記比較結果に基づいて、PDK4の発現を抑制する被験物質を選択する工程。
【0079】
前記工程(a)において、被験物質とは、いかなる公知物質および新規物質であってもよく、例えば、核酸、糖質、脂質、蛋白質、ペプチド、有機低分子化合物、コンビナトリアルケミストリー技術を用いて作製された化合物ライブラリー、固相合成やファージディスプレイ法により作製されたランダムペプチドライブラリー、あるいは微生物、動植物、海洋生物等由来の天然成分などがあげられる。また、これらの化合物の2種以上の混合物を試料として供することもできる。
【0080】
前記工程(a)において使用する骨芽細胞または骨髄由来の単球/マクロファージ系列細胞は、PDK4を発現しうる限り、いかなる動物由来の細胞であってもよい。本発明の非ヒト骨モデル動物由来の骨芽細胞または骨髄由来の単球/マクロファージ系列細胞をさらに陰性対照として用いる場合、非ヒト骨モデル動物と同種由来の細胞が望ましい。マウスの骨芽細胞および骨髄由来の単球/マクロファージ系列細胞の調製方法は、実施例7および8に記載されており、他の動物由来の細胞も実施例7および8に記載の方法に準じて調製することができる。
【0081】
工程(a)において、骨芽細胞または骨髄由来の単球/マクロファージ系列細胞をPDK4の発現が維持される条件下に置いたものを用いることができる。かかる条件下としては、骨芽細胞または骨髄由来の単球/マクロファージ系列細胞を適当な培地中に入れ、約25〜40℃のインキュベーター中で生存または培養させることが好ましい。次に、前記培地中に被験物質を添加し、インキュベートを続けることで接触がなされうる。
【0082】
前記被験物質の添加量は、有効成分の種類、培地に対する溶解性、細胞の感受性等によって適宜設定することができる。
【0083】
工程(b)において、前記被験物質を接触させた骨芽細胞または骨髄由来の単球/マクロファージ系列細胞におけるPDK4の発現量は、自体公知の方法により調べることができる。内在性のPDK4の発現量は、例えば、細胞からタンパク質を抽出してPDK4を認識する抗体を用いるウェスタンブロッテング等により測定する方法;細胞からRNAを抽出してRT−PCRまたはノーザンブロッテング等により転写されたPdk4 mRNAを測定する方法;PDK4を認識する抗体を用いて、免疫組織化学によりin situでPDK4を調べる方法等があげられる。内在性のPDK4の発現量は、Pdk4のプロモーター領域とレポーター遺伝子とを連結した発現ベクターを細胞に導入し、レポーター遺伝子の発現を測定するレポーターアッセイ等によっても推定できる。レポーターアッセイが好ましく用いられる。レポーター遺伝子は、公知の遺伝子を限定なく使用することができ、例えば、ルシフェラーゼ、GFP(緑色蛍光タンパク質)、CAT(クロラムフェニコールアセチルトランスフェラーゼ)などがあげられるが、これに限定されない。レポーター遺伝子の発現は、用いるレポーター遺伝子に応じて、自体公知の方法により測定することができる。
【0084】
前記工程(b)において、被験物質を接触させない骨芽細胞または骨髄由来の単球/マクロファージ系列細胞も同時にまたは別途調べ、接触した場合の結果と接触しない場合の結果とを比較する。また、陰性対照として、本発明の骨モデル動物由来の骨芽細胞または骨髄由来の単球/マクロファージ系列細胞も準備することが好ましい。
【0085】
前記工程(c)において、工程(b)で得られた比較結果に基づき、PDK4の発現を抑制する被験物質を選択する。選択する基準は、PDK4のタンパク質またはmRNAが低減していることを指標にすればよい。
【0086】
このようにして選択された被験物質は、骨芽細胞におけるPDK4のアンタゴニスト作用を有する剤として有用であるとともに、例えば、スクリーニング方法2のようなさらなるスクリーニングや薬理試験を実施することにより、副作用の少ない骨粗鬆症の予防または治療剤の候補薬ともなりうる。
【0087】
また、スクリーニング方法1の別の態様として、上記工程(a)および(b)に加え、下記工程を含む、PDK4の発現または機能抑制を介した骨粗鬆症の予防および/または治療剤のスクリーニング方法1(2)を提供する:
(C)下記工程(c1)または(c2)のいずれかにより破骨細胞への分化を誘導する工程:
(c1)前記比較工程(b)の結果に基づいて、骨芽細胞でPDK4の発現を抑制する被験物質存在下で、骨芽細胞と骨髄由来の単球/マクロファージ系列細胞とを共培養し、破骨細胞への分化を誘導する工程、さらに対照として、被験物質非存在下で骨芽細胞と骨髄由来の単球/マクロファージ系列細胞とを共培養し、同様に破骨細胞への分化を誘導する工程、または
(c2)前記比較工程(b)の結果に基づいて、単球/マクロファージ系列細胞でPDK4の発現を抑制する被験物質存在下で、単球/マクロファージ系列細胞を、M-CSFおよびRANKL存在下で培養し、破骨細胞への分化を誘導する工程、さらに対照として、被験物質非存在下で単球/マクロファージ系列細胞をM-CSFおよびRANKL存在下で培養し、同様に破骨細胞への分化を誘導する工程、
(d)前記工程(C)における破骨細胞への分化の程度を比較する工程、および
(e)前記比較工程(d)の結果に基づいて、骨髄由来の単球/マクロファージ系列細胞から破骨細胞への分化を抑制する被験物質を選択する工程。
【0088】
工程(C)の工程(c1)において、PDK4の発現を抑制する被験物質存在下での骨芽細胞と骨髄由来の単球/マクロファージ系列細胞との共培養は、実施例8に記載の方法に従って行うことができる。破骨細胞への分化誘導は、骨芽細胞から産生されるM-CSFおよびRANKLの存在下で培養することにより行うことができる。
【0089】
工程(C)の工程(c1)において、さらに対照として、被験物質非存在下で骨芽細胞と骨髄由来の単球/マクロファージ系列細胞とを共培養し、上記と同様の条件下で破骨細胞への分化誘導を行うことが好ましい。
【0090】
工程(C)の工程(c2)において、PDK4の発現を抑制する被験物質存在下での骨髄由来の単球/マクロファージ系列細胞の培養は、実施例8に記載の方法に従って行うことができる。破骨細胞への分化誘導は、M-CSFおよびRANKLの存在下で培養することにより行うことができる。
【0091】
工程(C)の工程(c2)において、さらに対照として、被験物質非存在下で骨髄由来の単球/マクロファージ系列細胞を培養し、上記と同様の条件下で破骨細胞への分化誘導を行うことが好ましい。
【0092】
工程(d)において、破骨細胞への分化の有無およびその程度は、実施例8に記載の方法に従って、TRAP染色し、走査型電子顕微鏡による観察によって測定することができる。
【0093】
工程(e)において、TRAP染色陽性細胞であって小核破骨細胞(3-6の核を有する)または大核破骨細胞(6を超える核を有する)の数が対照と比較して有意に減少している被験物質を選択する。
【0094】
このようにして選択された被験物質は、PDK4の発現を抑制し、かつ破骨細胞への分化を抑制しうる物質として、骨粗鬆症、特に非荷重状態における骨粗鬆症の予防または治療剤の候補薬ともなりうる。
【0095】
また、スクリーニング方法1のさらに別の態様として、下記工程を含むスクリーニング方法1(3)を実施することにより、PDK4の発現または機能抑制を介した骨粗鬆症の予防および/または治療剤の候補薬をより簡便にスクリーニングすることができる。
(a’)被験物質存在下で、骨芽細胞と骨髄由来の単球/マクロファージ系列細胞を共培養し、破骨細胞への分化を誘導する工程、
(b’)前記工程(a’)における破骨細胞への分化の程度を比較する工程、および
(c’)前記比較工程(b’)の結果に基づいて、骨髄由来の単球/マクロファージ系列細胞から破骨細胞への分化を抑制する被験物質を選択する工程。
【0096】
前記工程(a’)、(b’)および(c’)はそれぞれ、スクリーニング方法1(2)の工程(c1)、(d)および(e)と同様に実施することができる。
【0097】
また、スクリーニング方法1のさらに別の態様として、下記工程を含むスクリーニング方法1(4)を実施することにより、PDK4の発現または機能抑制を介した骨粗鬆症の予防および/または治療剤の候補薬をより簡便にスクリーニングすることができる。
(a”)被験物質、M-CSFおよびRANKL存在下で、骨髄由来の単球/マクロファージ系列細胞から破骨細胞への分化を誘導する工程、
(b”)前記工程(a”)における破骨細胞への分化の程度を比較する工程、および
(c”)前記比較工程(b”)の結果に基づいて、骨髄由来の単球/マクロファージ系列細胞から破骨細胞への分化を抑制する被験物質を選択する工程。
【0098】
前記工程(a”)、(b”)および(c”)はそれぞれ、スクリーニング方法1(2)の工程(c2)、(d)および(e)と同様に実施することができる。
【0099】
(スクリーニング方法2)
スクリーニング方法2は、本発明の骨モデル動物を用いてインビボの系で実施する。具体的には、下記工程を含む、骨粗鬆症の予防または治療剤のスクリーニング方法を提供する:
(a)本発明の骨モデル動物の野生型対照に被験物質を投与し、当該骨モデル動物と野生型対照とを非荷重状態に供する工程、
(b)前記被験物質の投与前後の野生型対照における骨の形態を解析し、本発明の骨モデル動物における骨の形態と比較する工程、および、
(c)前記(b)の比較結果に基づいて、非荷重状態における骨量の減少を抑制する被験物質を選択する工程。
【0100】
前記工程(a)において、被験物質は、スクリーニング方法1と同様のものがあげられる。被験物質を動物に投与する方法は特に限定されるものではないが、経口的または非経口的に投与され得る。非経口的投与経路としては、例えば、静脈内、動脈内、筋肉内等の全身投与、あるいは関節内、標的細胞付近への局所投与等があげられる。
【0101】
前記被験物質の投与量は、有効成分の種類、分子の大きさ、投与経路、投与対象となる動物種、投与対象の薬物受容性、体重、年齢等によって適宜設定することができる。
【0102】
前記工程(a)において、動物を非荷重状態に供する方法は、生体が重力を感知しない状態または重力の影響を受けにくい状態に維持可能であれば特に限定されるものではない。げっ歯類(マウス、ラット等)を例にとると、尾懸垂実験が好適にあげられ、実施例1に記載の方法に従って、行うことができる。尾懸垂は、通常、3〜14日間、好ましくは7日間継続して行うことが望ましい。
【0103】
前記工程(b)において、骨の形態、骨体積または骨密度を調べる方法としては、例えば、X線解析、マイクロCT(μCT)、pQCTおよび骨組織切片を用いた骨組織形態計測などがあげられ、定量的な解析方法として、μCTおよび骨組織形態計測が好ましい。μCT解析は、実施例4に記載されている。解析可能な骨梁パラメータとしては、骨梁体積、骨梁数、骨梁幅、骨梁間距離、骨梁密度、ストラクチャーモデルインデックス、骨梁連結性、異方性度があり、皮質骨パラメータとしては、皮質骨断面積、骨内膜周囲長、骨膜周囲長、皮質骨密度があげられる。骨組織形態計測の海面骨パラメータとしては、単位骨量、類骨面、類骨幅、骨芽細胞面、骨芽細胞数、多核破骨細胞数、破骨細胞面、吸収面、骨石灰化速度、石灰化面および骨形成速度があり、皮質骨パラメータとしては、骨断面積、皮質骨面積、骨髄腔面積、骨膜性骨形成率、皮質内骨形成率、皮質内骨吸収面があげられる。
【0104】
前記工程(b)において、被験物質を投与しない動物における骨の形態、骨体積または骨密度も同時にまたは別途調べ、投与動物の結果と非投与動物の結果とを比較する。また、陽性対照として、本発明の骨モデル動物を同様に解析する。
【0105】
前記工程(c)において、工程(b)で得られた比較結果に基づき、野生型動物の非荷重状態における骨量の減少を抑制する被験物質を選択する。選択する基準は、本発明の骨モデル動物を指標にすればよい。
【0106】
このようにして選択された被験物質は、非荷重状態における骨量の減少を抑制させ、骨粗鬆症、特に廃用性骨粗鬆症の候補薬となりうる。
【0107】
(スクリーニング方法3)
スクリーニング方法3は、本発明の骨モデル動物由来の組織または細胞を用いてインビトロの系で実施する。具体的には、下記工程を含む、荷重刺激を感知し骨粗鬆症の発症に関連するPDK4の標的タンパク質のスクリーニング方法を提供する:
(a)非荷重状態後に本発明の骨モデル動物から得られた組織または細胞、および非荷重状態後に野生型対照から得られた組織または細胞をインビトロで生理学的条件下で維持する工程、
(b)前記野生型対照由来の組織または細胞におけるタンパク質のリン酸化の程度と、本発明の骨モデル動物由来の組織または細胞におけるタンパク質のリン酸化の程度とを比較する工程、または前記野生型対照由来の組織または細胞におけるタンパク質でPDK4に結合するタンパク質と本発明の骨モデル動物由来の組織または細胞におけるタンパク質でPDK4に結合するタンパク質を比較する工程、および
(c)前記比較結果に基づいて、リン酸化の程度が有意に変動しているタンパク質を選択する工程、または前記比較結果に基づいて、前記野生型対照由来の組織または細胞におけるタンパク質のみから得られ、本発明の骨モデル動物由来の組織または細胞におけるタンパク質からは得られないPDK4結合タンパク質を選択する工程。
【0108】
前記工程(a)において動物を非荷重状態にするには、スクリーニング方法2と同様の条件で行えばよい。用いる組織または細胞は、動物に由来するものであれば特に限定されないが、荷重感知遺伝子PDK4の未知の基質を選択するという観点から、骨芽細胞、骨芽細胞を豊富に含む組織または骨芽細胞に分化可能な細胞もしくは組織、および破骨細胞、破骨細胞を豊富に含む組織または破骨細胞に分化可能な細胞もしくは組織が望ましい。
【0109】
前記工程(a)において、組織または細胞をインビトロで生理学的条件下で維持する方法は、野生型対照由来の組織または細胞の場合、PDK4の発現が維持される条件下に置いたものを用いることができる。かかる条件下としては、組織または細胞を適当な培地中に入れ、約25〜40℃のインキュベーター中で生存または培養させることが好ましい。本発明の骨モデル動物由来の組織または細胞も、野生型対照由来の組織または細胞と同じ条件下で維持する。
【0110】
前記工程(b)において、野生型対照由来の組織または細胞におけるタンパク質のリン酸化の程度と、本発明の骨モデル動物由来の組織または細胞におけるタンパク質のリン酸化の程度とを比較する。タンパク質のリン酸化の程度は、自体公知の方法により調べることができる。例えば、細胞抽出液の一次元あるいは二次元電気泳動により変動タンパク質を同定する方法、リン酸化抗体アフィニティカラムを用いて細胞抽出液からリン酸化蛋白を濃縮し電気泳動により比較同定する方法、細胞抽出液を酵素消化して得られたペプチドから金属キレートカラム(IMAC)を用いてリン酸化ペプチドを精製し2DLC-MS/MSによりリン酸化ペプチドを同定比較する方法、さらにこれに安定同位体標識法による定量法を組み合わせたリン酸化ペプチドのLC-MS/MSによる定量的変動解析等があげられる。さらに、PDK4と結合するタンパク質の同定も、公知の方法により調べることができる。例えば、PDK4の全部あるいは一部をベイトとした酵母を用いたツーハイブリッド法、PDK4抗体アフィニティカラムを用いて細胞抽出液からPDK4およびその結合タンパク質を濃縮し電気泳動で比較同定する方法等が挙げられる。
【0111】
前記工程(c)において、工程(b)で得られた比較結果に基づき、リン酸化の程度が有意に変動しているタンパク質を選択する。PDK4のキナーゼ活性を考慮すると、リン酸化が有意に上昇しているタンパク質を指標にすることが望ましい。また、野生型対照のみから得られ、本発明の骨モデル動物由来の組織または細胞からは得られないPDK4結合タンパク質を選択する。
【0112】
このようにして選択されたタンパク質は、荷重感知遺伝子であるPDK4の新規基質として、非荷重状態における骨低下のメカニズムの解明または特に廃用性骨粗鬆症の予防または治療剤の分子標的ともなりうる。
【実施例】
【0113】
以下、実施例により本発明をより詳細に説明するが、本発明はこれらの実施例により何ら限定されるものではない。
【0114】
[実施例1]
尾懸垂実験およびマイクロアレイ
尾懸垂実験は、既報に準じて実施した(Sakai A, Nakamura T: Changes in trabecular bone turnover and bone marrow cell development in tail-suspended mice, J Musculoskelet Neuronal Interact 2001, 1:387-39222)。野生型マウスおよびBCL2トランスジェニック雄性マウス(10週齢)を、尾懸垂用の特殊なケージ(TSケージ)に置き、当該ケージに適応させるために1週間飼育した。尾懸垂は、尾懸垂群において8日間実施した。再負荷群のマウスは1週間の尾懸垂後に24時間尾懸垂から開放した。対照群は、尾懸垂を除いて同じ条件下においた。脛骨および大腿骨からRNAを抽出し、Agilent マイクロアレイを用いて、Hokkaido System Science社(Sapporo, Japan)によりマイクロアレイ解析を実施した。
【0115】
[実施例2]
リアルタイムRT-PCR
全骨から骨芽細胞、骨細胞および骨髄細胞を分離するために、大腿骨および脛骨から筋肉、結合組織および骨膜を取り除き、骨幹端で骨を切断した。大腿骨および脛骨の骨幹中の骨髄細胞をリン酸緩衝食塩水(PBS)で押し出した。次いで、マイクロ歯間ブラシ(Kobayashi Pharmaceutical Co. Ltd., Osaka, Japan)を用いて、骨芽細胞が豊富な細胞を回収した。残った骨を骨細胞が豊富な細胞として用いた。ISOGEN(Wako, Osaka, Japan)を用いて全RNAを抽出し、Light Cycler 480 SYBR Green I システム(Roche)を用いてリアルタイムRT-PCRを実施した。Gapdhの値を元にデータを正規化した。
【0116】
[実施例3]
Pdk4-/-マウスの作出
マウスPdk4遺伝子のターゲティングベクターは、BAC改変技術により作製した。BAC改変キットをGene Bridges(Heidelberg, Germany)から購入し、マウスPdk4遺伝子のすべてのエキソンを含むBACクローン、RP23-25N12をBACPAC Resource Center(Oakland, CA)から購入した。IRES-LacZおよびPGK-gb2-neoを含むカセットをPCRで増幅し、BACクローンを含むバクテリアに導入し、Pdk4遺伝子の第3エキソンと第10エキソン間でゲノムDNAを置換した。IRES-LacZおよびPGK-gb2-neoを含み、Pdk4遺伝子に相同なDNAが隣接するカセットを、PGK-tkカセットを含むプラスミドにクローニングした。最終のターゲティングコンストラクトを線状化し、ES細胞のE14ラインにエレクトロポレーションにより導入した。ターゲティングされたES細胞をC57BL/6胚盤胞にインジェクションし、キメラを作出した。既報に従って、サザンブロットによりミュータントアレルを確認した(Komori T, et al.,: Targeted disruption of Cbfa1 results in a complete lack of bone formation owing to maturational arrest of osteoblasts, Cell 1997, 89:755-764)。Pdk4+/-マウスおよびPdk4-/-マウスの全身でLacZの発現を検出することができなかったので、ゲノムDNAの欠失はmRNAの安定性またはPDK4発現それ自体に影響するよう思われた。ヘテロ接合性雄性マウスをC57BL/6J雌性マウスと6回戻し交配した。血清中の酒石酸抵抗性酸性ホスファターゼ5b(TRAP5b)レベルをELISA(IDS, Boldon, UK)により測定した。研究に先立ち、全ての実験は、長崎大学大学院医歯薬学総合研究科の動物保護と利用に関する委員会による審査と承認を受けた。
【0117】
[実施例4]
マイクロ−CT解析
定量マイクロ−CT解析は、マイクロ−CTシステム(μCT-20; Scanco Medical, Bruttisellen, Switzerland)を用いて行った。スキャンしたスライスからのデータを三次元解析に用いて、大腿骨の形態学的パラメータを計算した。骨梁のパラメータを大腿骨の遠位骨幹端で測定した。約2.4mm(成長板から0.5mm離れたところ)を頭尾方向にスキャンして、12μm増分の200枚のスライスを得た。
【0118】
[実施例5]
組織学的解析
長骨の組織学的解析のためマウスを剖検し、長骨を4%パラホルムアルデヒド/0.01M リン酸緩衝化生理食塩水中で固定し、10% EDTA(pH7.4)中で脱灰し、パラフィンに包埋した。TRAP活性について、切片(5μm厚)を染色した。骨の組織形態学的解析は、既報に従って行った(Liu W, Toyosawa S, Furuichi T, Kanatani N, Yoshida C, Liu Y, Himeno M, Narai S, Yamaguchi A, Komori T: Overexpression of Cbfa1 in osteoblasts inhibits osteoblast maturation and causes osteopenia with multiple fractures, J Cell Biol 2001, 155:157-166)。動的組織形態学的指標を評価するため、マウスを剖検する6日前と2日前に、体重10g当たり0.16mgの用量でカルセインを注射した。
【0119】
[実施例6]
ウエスタンブロット解析およびCol1a1の定量
以下の抗体を用いて、ウエスタンブロット解析を行った:ウサギ抗-オステオポンチン抗体(Santa Cruz Biotechnology, Santa Cruz, CA)、ウサギ抗-オステオカルシン抗体(IBL, Takasaki, Japan)、ウサギ抗-PDK4抗体(Abgen, San Diego, CA)、マウス抗-PDC E1a抗体(MitoSciences, Eugene, Oregon)、ウサギ抗-PDC E1a pSer293抗体(Calbiochem, Darmstadt, Germany)およびヤギ抗-アクチン抗体(Santa Cruz Biotechnology)。既報に従って、コラーゲンを抽出し、解析した(Domenicucci C, Goldberg HA, Hofmann T, Isenman D, Wasi S, Sodek J: Characterization of porcine osteonectin extracted from foetal calvariae, Biochem J 1988, 253:139-151)。
【0120】
[実施例7]
In vitroでの骨芽細胞形成
初代骨芽細胞は、既報に従って、新生マウスの頭蓋冠の小片を三次元コラーゲンゲル中で12日間培養することにより調製した(Komori T, et al., Cell 1997, 89:755-76423)。骨芽細胞の分化に関しては、50μg/ml アスコルビン酸および10mM β-グリセロールリン酸を10% FBSを補足したα-MEMに添加し、アルカリホスファターゼ(ALP)活性および石灰化に関して染色を行った。
【0121】
[実施例8]
In vitroでの破骨細胞形成
骨髄由来の単球/マクロファージ系列細胞(BMM)を、Ficoll-PaqueTM(GE Healthcare, Tokyo, Japan)を用いた密度勾配遠心分離によって12週齢のマウスの骨髄から単離し、10% FBS、20ng/ml マクロファージコロニー刺激因子(M-CSF)(Sigma, Tokyo, Japan)および100ng/mlのGST-可溶性Receptor activator of NF-κB ligand(GST-sRANKL)を含むα-MEM中で1.5x105/wellの密度で96ウエルプレートに播種し、6日間培養した。TRAP染色は、既報に従って行った(Maruyama Z, et al.: Runx2 determines bone maturity and turnover rate in postnatal bone development and is involved in bone loss in estrogen deficiency, Dev Dyn 2007, 236:1876-1890)。BMM(1×105cells/well)を、10% FBS含有α-MEM中で10-6M プロスタグランジンE2(PGE2)および10-8M 1α,25(OH)2D3の存在下、96-wellプレートにてPOB(1×104cells/well)とともに培養した。骨再吸収の評価のために、BMMおよび初代骨芽細胞を、象牙質ディスク(IDS)上で6日間共培養し、再吸収された領域を走査型電子顕微鏡(H-7100; Hitachi, Tokyo, Japan)で測定した。ジクロロ酢酸(DCA)は、Sigma-Aldrich(St. Louis, MO)から購入した。レスキュー実験に関して、Pdk4-発現アデノウイルスベクターおよびEGFP-発現アデノウイルスベクターを既報(Enomoto H, et al.,: Induction of osteoclast differentiation by Runx2 through receptor activator of nuclear factor-kappa B ligand (RANKL) and osteoprotegerin regulation and partial rescue of osteoclastogenesis in Runx2-/- mice by RANKL transgene, J Biol Chem 2003, 278:23971-23977)に従って作製し、2.5×107 plaque forming units/mlで使用した。
【0122】
[実施例9]
レポーターアッセイ
Ranklプロモーター領域のゲノムDNAを既報に従ってクローニングし(Kitazawa R, Kitazawa S, Maeda S: Promoter structure of mouse RANKL/TRANCE/OPGL/ODF gene, Biochim Biophys Acta 1999, 1445:134-141)、3.8 kbおよび1.5 kbの断片をpGL3ベクター(Promega, Madison, WI)に挿入した。ネスティッド欠損コンストラクトを、クローニングしたDNA断片のPCR増幅により作製し、シークエンスにより確認した。Pdp1 cDNAをPCRで増幅し、pSG5にライゲートし、シークエンスにより確認した。初代骨芽細胞を、Fugene 6(Roche Diagnostics, Tokyo, Japan)を用いて、それぞれRanklプロモーターコンストラクト(0.2μg)およびpRL-CMV(0.003μg)を含むDNA混合物で一過性にトランスフェクトした。72時間後、Dual Luciferase Reporter Assay System(Promega)を用いて、細胞溶解液のルシフェラーゼ活性を測定し、Renillaルシフェラーゼ活性に対して正規化した。
【0123】
結果
非荷重後の骨細胞におけるPdk4発現の誘導
本発明者は、BCL2トランスジェニックマウス(骨細胞のアポトーシスにより骨細胞ネットワークが破壊されている)および野生型マウスを用いた尾懸垂により、非荷重後にその発現がアップレギュレートされ再荷重後にダウンレギュレートする荷重感知遺伝子を探索した。脛骨および大腿骨由来のRNAを用いたマイクロアレイ解析において、野生型マウスにおいては、PDK4は非荷重後にアップレギュレートされるが再荷重によりダウンレギュレートされることを見出したが、BCL2トランスジェニックマウスでは非荷重後にPdk4はアップレギュレートされなかった(図1A)。このことは、リアルタイムRT-PCR 解析により確認され(図1A)、PDK4は野生型マウスでは破骨細胞豊富な画分および骨芽細胞豊富な画分の両方でアップレギュレートしているが、BCL2トランスジェニックマウスではアップレギュレートしなかった(図1B)。PDK4は、骨格筋および心筋で高発現し、腎臓、肝臓、肺、脾臓および骨で中等度に発現している。PDK4は、骨中では、破骨細胞豊富な画分、骨芽細胞豊富な画分および骨髄細胞において中等度に発現していた(図1C)。骨におけるPDKアイソフォームの発現レベルを比較するため、骨格筋に対して骨における発現レベルを比較した(図1D)。比率はPDK4が最高値であった。PDK4は、非荷重後に骨格筋、大腿骨全体、骨細胞豊富な画分、骨芽細胞豊富な画分および骨髄細胞でアップレギュレートしたが、PDK1、PDK2およびPDK3は、非荷重後に骨全体ならびに骨芽細胞、骨細胞および骨髄細胞の各画分においてアップレギュレートしなかった。一方、PDK2は、非荷重後の骨格筋でアップレギュレートした(図1E)。
【0124】
Pdk4-/-マウスにおける正常な骨の発達および維持
非荷重状態での骨におけるPDK4の役割を調べるため、本発明者は、Pdk4-/-マウスを作出した(図2A-C)。Pdk4-/-マウスの体重は、野生型マウスと同様であった(図2D)。μ-CT解析により、骨体積は野生型およびPdk4-/-マウス間で3、6および12月齢で同様であった(図2E)。組織学的解析において、Pdk4-/-マウスの骨は正常に発達し、骨芽細胞、骨細胞および破骨細胞の形態は野生型マウスの形態と類似していた(図9)。骨芽細胞の分化マーカー(Runx2、Osterix、ALP、Col1a1、オステオポンチン、骨シアロタンパク質(BSP)およびオステオカルシンを含む)の発現は、野生型マウスおよびPdk4-/-マウス間で類似しており、Col1a1、Col1a2、オステオポンチンおよびオステオカルシンを含む骨マトリックスタンパク質の産生は、野生型マウスおよびPdk4-/-マウス間で類似していた(図2F-H)。
【0125】
野生型マウスにおいて非荷重後の骨量が減少するが、Pdk4-/-マウスでは減少しない
尾懸垂を1週間実施し、μ-CTにより非荷重後の骨量を比較した。生理学的条件下においては、野生型マウスとPdk4-/-マウスではすべてのパラメータが類似していた(コントロール)。しかしながら、非荷重後、野生型マウスでは骨体積、脛骨の厚さおよび脛骨密度が減少したが、Pdk4-/-マウスでは減少しなかった(図3A-E)。組織形態学的解析において、すべてのパラメータ(骨体積ならびに骨形成および吸収のパラメータを含む)は、生理学的条件下では野生型マウスとPdk4-/-マウスとでは類似していた(コントロール)。非荷重後、野生型マウスでは骨体積が低下し、骨吸収パラメータ(破骨細胞数、破骨細胞面および吸収面を含む)が増加したが、Pdk4-/-マウスでは増加しなかった(図3F)。血中グルコースおよびコルチゾールのレベルは、野生型マウスとPdk4-/-マウスとでは類似しており、両マウスにおいて非荷重状態で影響がなかった(図10)。
【0126】
野生型マウスでは非荷重後に骨吸収および Ranklの発現が増加するが、Pdk4-/-マウスでは増加しない
野生型マウスおよびPdk4-/-マウスにおいて、コントロール群と非荷重群とでは骨形成速度は有意に相違しなかったが、野生型マウスおよびPdk4-/-マウスで、非荷重後にコントロール群と比較してCol1a1およびオステオカルシンの発現は同様に低下した(図3Fおよび4A)。RanklおよびCtskの発現は、野生型マウスで非荷重後に顕著に増加したが、Pdk4-/-マウスでは増加しなかった。一方、Opg、Rank、Nfatc1、DC-STAMPの発現は、野生型マウスおよびPdk4-/-マウスの両マウスにおいて非荷重後に有意に変化しなかった(図4A)。TRAP-陽性細胞およびTRAP5bの血清レベルは、野生型マウスで非荷重後に増加したが、Pdk4-/-マウスでは増加しなかった(図4B、C)。
【0127】
骨芽細胞および破骨細胞前駆体の異常から生じるPdk4-/-マウスにおける破骨細胞形成の低下
PDK4の発現は骨芽細胞の分化中に上昇するが、Pdk4-/-初代骨芽細胞は、野生型初代骨芽細胞と同様のレベルのALP活性および石灰化を示した(図5A、B)。さらに、骨芽細胞マーカー遺伝子(Runx2、Osterix、ALP、Col1a1、オステオネクチン、オステオポンチン、BSPおよびオステオカルシンがあげられる)の発現レベルは、野生型とPdk4-/-の初代骨芽細胞間で類似していた。ただし、RanklはPdk4-/-初代骨芽細胞では低下していた(図5C)。また、骨髄細胞におけるPdk4の発現は、M-CSFおよびRANKLの存在下で破骨細胞への分化中に増加し、Pdk4-/-BMMから小核破骨細胞および大核破骨細胞の形成は、野生型BMMの培養物と比較して大核破骨細胞においてより深刻な減少であった(図6A-C)。Ctskの発現は低下し、他方、Rank、Nfatc1、DC-STAMP、Sirpa、CD47およびCD9の発現には影響がなかった(図6D)。また、初代骨芽細胞とBMMとを共培養することにより、破骨細胞の分化も調べた。野性型骨芽細胞とPdk4-/-BMMとの共培養において、3-6の核を有する小核破骨細胞の数は、野性型骨芽細胞とBMMとの共培養で得られたものと同様であったが、6を超える核を含む大核破骨細胞の数は、野性型骨芽細胞とBMMとの共培養で得られたものの約半分であった。Pdk4-/-骨芽細胞と野生型BMMとの共培養も同様の結果を示した。Pdk4-/-骨芽細胞とBMMとの共培養もまた、野性型骨芽細胞とBMMとの共培養と比較して、3-6の核を有する小核破骨細胞の数は同様であったが、7-10の核を有する大核破骨細胞の数は4分の1であり、10を超える核を含む巨核破骨細胞の数は、野性型骨芽細胞とBMMとの共培養で得られたものの10分の1未満であった(図6E、F)。さらに、Pdk4-/-骨芽細胞とBMMとの共培養における吸収領域は、野性型骨芽細胞とBMMとの共培養で得られたものの10分の1未満であった(図6G、H)。
【0128】
Pdk4による破骨細胞形成とRanklプロモーターの正の調節
次に、アデノウイルストランスファーを用いて、インビトロでPDK4が破骨細胞形成を誘導するか否かを調べた(図7A-D)。Pdk4-/-骨芽細胞とBMMとの共培養物にPdk4-発現アデノウイルスを感染させると、GFP-発現アデノウイルスを感染させたときと比べて、TRAP-陽性細胞および吸収領域が増加した。Pdk4-/-初代骨芽細胞にアデノウイルスによりPdk4を導入すると、Rankl発現を強く誘導し、Opg発現を中等度に誘導した。これにより、アデノウイルスによりGFPを導入したときと比べて、Rankl/Opg比が増加した(図7E)。Rankl発現におけるPDK4の関与を調べるため、Ranklプロモーター領域のDNA断片を用いて、レポーターアッセイを行った(図7F)。3.8 kbの断片を含むレポーターベクターの活性は、野生型骨芽細胞と比較してPdk4-/-骨芽細胞で低下した。Pdk4-/-骨芽細胞でのレポーター活性の低下は、段階的に欠失させたコンストラクトを用いたアッセイでも観察された(1.5K、185、164、137および122 bp)。さらに、PDK4のPdk4-/-骨芽細胞への導入は、GFPの導入と比較して、欠失コンストラクトのレポーター活性を亢進させた(図7G)。
【0129】
PDCリン酸化に対するPdk4アンタゴニストは破骨細胞形成およびRankl誘導を阻害しない
ジクロロ酢酸(DCA)は、PDKsによるPDCのE1aサブユニットのリン酸化を阻害する(Kato M, Li J, Chuang JL, Chuang DT: Distinct Structural Mechanisms for Inhibition of Pyruvate Dehydrogenase Kinase Isoforms by AZD7545, Dichloroacetate, and Radicicol, Structure 2007, 15:992-1004)。初代骨芽細胞をDCAで処理すると、PDCのリン酸化を阻害した(図8A)。M-CSFおよびRANKLの存在下で野生型BMMを培養した場合、DCAでの処理は破骨細胞の分化に影響がなかった(図8B)。DCAでの処理は、野生型初代骨芽細胞とBMMとの共培養において破骨細胞形成にも影響がなかった(図8C)。また、RanklおよびOpgの発現ならびにRanklプロモーター活性におけるDCAの効果も調べた(図8D、E)が、DCAは影響しなかった。さらに、Pdp1の初代骨芽細胞への導入はPDCを脱リン酸化したが、Ranklプロモーター活性には影響がなかった(図8F、G)。これらの知見は、PDCのリン酸化とは別の様式で、PDK4が破骨細胞形成を調節することを示唆する。
【0130】
PDK4は、グルコースの酸化的利用を制限するため、PDCの特異的キナーゼであると考えられてきた。DCAはピルビン酸の類似体である。DCAおよびピルビン酸は、PDKのN-末端の調節ドメインの中心において共通の結合部位を共有し、DCAは、PDKの阻害およびその後のPDCの活性化につながる活性部位の変化を誘導する。DCAはPDCのリン酸化を阻害するが、破骨細胞形成、Rankl発現およびRanklプロモーター活性に影響しなかった。さらに、Pdp1によるPDCの脱リン酸化は、Ranklプロモーター活性に対して影響しなかった。これらの知見は、PDCのリン酸化とは別の様式でPDK4が破骨細胞形成を調節することを示しており、このことは、破骨細胞形成に関する骨芽細胞およびBMMにおいて、PDK4は新規な未知の基質を有していることを示唆する。破骨細胞形成に関するPDK4およびPDK4の基質は、廃用性骨粗鬆症の適切な治療標的となると考えられる。
【産業上の利用可能性】
【0131】
本発明の骨粗鬆症の予防または治療剤によると、非荷重状態が主原因の骨粗鬆症を有効に予防または治療することができる。本発明のモデル動物によると、PDK4が欠損していることにより非荷重(無重力)に抵抗性の骨モデル動物として、無重力による骨量の減少など、無重力状態での骨組織での様々な生理学的状態を解明するモデルとして役立つ。本発明の組織または細胞によると、骨量減少のメカニズムの研究に利用することが可能である。本発明のスクリーニング方法によると、骨粗鬆症の予防または治療剤の開発のみならず、非荷重状態でのPDK4の基質の解明等に貢献することができる。
【技術分野】
【0001】
本発明は、荷重感知遺伝子に関する。詳しくは、重力刺激のない状態で進行する骨粗鬆症の原因の解明、その治療または予防に役立つ遺伝子およびその利用に関する。
【背景技術】
【0002】
骨量は、骨を形成する骨芽細胞の活性と骨を再吸収する破骨細胞の活性とのバランスにより決定される。現代社会における加齢に伴う主要な疾患の1つである骨粗鬆症は、これらの2つの活性のアンバランスにより生じ、食餌、身体活動(運動)、ホルモン状態、サイトカインならびに、糖尿病およびグルココルチコイド治療等の臨床状態に影響される(非特許文献1)。非荷重の姿勢、固定化(運動不足)または長期間の寝たきりにより生じる廃用性骨粗鬆症は、加齢に伴った疾患で寝たきりの患者の増加によって急速に増加している。通常、骨芽細胞と破骨細胞の活性は共役しており、骨再吸収の増加は、骨形成を促進させる。しかし、廃用性骨粗鬆症の場合、骨再吸収の増加は、骨形成を促進させない。このことは、廃用性骨粗鬆症において、骨芽細胞の活性は低下しているが破骨細胞の活性は促進していることを示している(非特許文献2)。オステオポンチン、交感神経の緊張、sclerostinおよびTRIPV4は、非荷重後の骨損失に関連していることが報告されている(非特許文献3〜6)。
【0003】
ピルビン酸デヒドロゲナーゼキナーゼのアイソザイムPDK(PDK1、PDK2、PDK3およびPDK4)は、ミトコンドリア内でピルビン酸をアセチル-CoAに変換させて解糖系をTCAサイクルのエネルギーおよび同化機能に結びつけるピルビン酸デヒドロゲナーゼ複合体(PDC)の負の調節因子である。PDCは、3つの触媒ドメイン:E1(ピルビン酸デカルボキシラーゼ)、E2(ジヒドロリポアミド アセチルトランスフェラーゼ)およびE3(ジヒドロリポアミド デヒドロゲナーゼ)からなるマルチサブユニット複合体である。PDKは、E1の特定のセリン残基をリン酸化してPDCを不活性化するが、ピルビン酸デヒドロゲナーゼホスフェートホスファターゼ(Pdp1およびPdp2)は、そのリン酸化体を脱リン酸化してPDCを活性化させる。PDKのアイソフォームはPDCに対して異なる感度を有しているが、PDK調節の短期のメカニズムには、E1の基質であるピルビン酸による阻害ならびにPDCの産物であるアセチル-CoAおよびNADHによる活性化が含まれる。哺乳動物のPDKは、組織特異的発現を示す:PDK1は、心臓、膵島および骨格筋で発現している;PDK2は、栄養供給状態ではユビキタスに発現しており、心臓、肝臓および腎臓で高発現している;PDK3は、主として精巣、腎臓および脳で発現している;ならびにPDK4は、心臓、骨格筋、肝臓、腎臓および膵島で高発現している(非特許文献7〜9)。心筋および骨格筋におけるPDK4の発現は、飢餓および糖尿病状態で迅速にアップレギュレートされ、それぞれ食餌およびインスリン投与により低下する(非特許文献10、11)。また、PDK4の発現は、グルココルチコイドによってもアップレギュレートし(非特許文献12)、インスリンによりダウンレギュレートされる(非特許文献13)。活性型PDCは、腎臓、骨格筋、横隔膜および心臓に多く、飢餓状態のPdk4-/-マウスの肝臓には多くなく、心臓におけるPDK4の過剰発現は、カルシニューリンストレス活性化経路により生じる心筋症を悪化させる(非特許文献14、15)。
【0004】
骨は、メカニカルストレスに対してその形状および強度を調節する。骨細胞は、骨における最も豊富な細胞であり、骨全体を通じて突起と骨細管を介したコミュニケーションシステムを含んでいる。骨細胞ネットワークは、理想的なメカノセンサーおよびメカノトランスダクションシステムであると考えられている(非特許文献16〜21)。
【0005】
現在、骨粗鬆症治療薬で骨吸収を抑制する薬剤として、ビスフォスフォネートとSERM(selective estrogen receptor modulator)があるが、骨吸収を常に抑制するために、骨質の低下を招き、骨強度の低下を来す場合がある。また、ビスフォスフォネートは、顎骨壊死のリスクがあり、歯の治療の際には、投与を中止しなければならない。また、無重力状態での骨量の低下を予防する薬剤は知られていない。
【先行技術文献】
【非特許文献】
【0006】
【非特許文献1】Manolagas SC, Endocr Rev 2010, 31:266-300
【非特許文献2】Bikle DD, Halloran BP, J Bone Miner Metab 1999, 17:233-244
【非特許文献3】Mizoguchi F, Mizuno A, Hayata T, Nakashima K, Heller S, Ushida T, Sokabe M, Miyasaka N, Suzuki M, Ezura Y, Noda M, J Cell Physiol 2008, 216:47-53
【非特許文献4】Ishijima M, Rittling SR, Yamashita T, Tsuji K, Kurosawa H, Nifuji A, Denhardt DT, Noda M, J Exp Med 2001, 193:399-404
【非特許文献5】Lin C, Jiang X, Dai Z, Guo X, Weng T, Wang J, Li Y, Feng G, Gao X, He L, J Bone Miner Res 2009, 24:1651-1661
【非特許文献6】Kondo H, Nifuji A, Takeda S, Ezura Y, Rittling SR, Denhardt DT, Nakashima K, Karsenty G, Noda M, J Biol Chem 2005, 280:30192-30200
【非特許文献7】Sugden, Holness MJ, Am J Physiol Endocrinol Metab 2003, 284:E855-62
【非特許文献8】Maj M, Cameron J, Robinson B, Molecular and Cellular Endocrinology 2006, 249:1-9
【非特許文献9】Roche TE, Hiromasa Y, Cellular and Molecular Life Sciences 2007, 64:830-849
【非特許文献10】Wu P, Sato J, Zhao Y, Jaskiewicz J, Popov KM, Harris RA, Biochem J 1998, 329 ( Pt 1):197-201
【非特許文献11】Wu P, Inskeep K, Bowker-Kinley MM, Popov KM, Harris RA, Diabetes 1999, 48:1593-1599
【非特許文献12】Qi D, Pulinilkunnil T, An D, Ghosh S, Abrahani A, Pospisilik JA, Brownsey R, Wambolt R, Allard M, Rodrigues B, Diabetes 2004, 53:1790-1797
【非特許文献13】Huang B, Wu P, Bowker-Kinley MM, Harris RA, Diabetes 2002, 51:276-283
【非特許文献14】Zhao, Am J Physiol Heart Circ Physiol 2008, 294:H936-43
【非特許文献15】Jeoung NH, Wu P, Joshi MA, Jaskiewicz J, Bock CB, Depaoli-Roach AA, Harris RA, Biochem J 2006, 397:417-425
【非特許文献16】Burger EH, Klein-Nulend J, FASEB J 1999, 13 Suppl:S101-112
【非特許文献17】MARTIN RB, Bone 2000, 26:71-78
【非特許文献18】Ehrlich PJ, Lanyon LE, Osteoporos Int 2002, 13:688-700
【非特許文献19】Knothe Tate ML, J Biomech 2003, 36:1409-1424
【非特許文献20】KLEIN-NULEND EHBAJ, The FASEB Journal 1999, 13:S101-11
【非特許文献21】Bonewald L, Johnson M, Bone 2008, 42:606-615
【発明の概要】
【発明が解決しようとする課題】
【0007】
本発明の目的は、高齢者の骨粗鬆症、特に麻痺等に伴う廃用性骨粗鬆症、ステロイド性骨粗鬆症または宇宙滞在時の骨量減少の予防または治療薬の開発に有用な手段を提供し、非荷重状態で進行する骨粗鬆症等の病態の解明に役立つ手段等を提供することにある。
【課題を解決するための手段】
【0008】
本発明者は、骨芽細胞におけるBCL2の過剰発現が骨細胞のアポトーシスを誘導し、骨細胞ネットワークが完全に破壊されているBCL2トランスジェニックマウスは非荷重後の骨損失に耐性であることを見出した。野生型マウスおよびBCL2トランスジェニックマウスを比較することにより、非荷重後の骨損失に応答する遺伝子を探索した。野生型マウスでは非荷重後にPDK4発現がアップレギュレートし、再荷重後にダウンレギュレートするが、これらのレギュレーションはBCL2トランスジェニックマウスでは決して観察されないことを見出し、本発明を完成するに至った。即ち、本願発明は、以下に示す通りである。
【0009】
〔1〕 PDK4の発現または機能を抑制する物質を含む、骨粗鬆症の予防または治療剤。
〔2〕 PDK4の発現または機能を抑制する物質が
(1)Pdk4遺伝子のRNAi誘導性核酸、アンチセンス核酸もしくはそれらの発現ベクター、または
(2)PDK4に対する抗体、ドミナントネガティブ変異体、アプタマーもしくはそれらの発現ベクター
である、前記〔1〕に記載の予防または治療剤。
〔3〕 骨粗鬆症が廃用性骨粗鬆症、ステロイド性骨粗鬆症および無重力時の骨量減少からなる群より選ばれる前記〔1〕または〔2〕に記載の予防または治療剤。
〔4〕 下記工程:
(a)骨芽細胞または骨髄由来の単球/マクロファージ系列細胞と被験物質とを接触させる工程、
(b)前記被験物質を接触させた骨芽細胞または骨髄由来の単球/マクロファージ系列細胞におけるPDK4の発現量を調べ、被験物質を接触させない骨芽細胞または骨髄由来の単球/マクロファージ系列細胞における発現量と比較する工程、および
(c)前記比較結果に基づいて、PDK4の発現を抑制する被験物質を選択する工程
を含む、PDK4の発現または機能抑制を介した骨粗鬆症の予防および/または治療剤のスクリーニング方法。
〔5〕 下記工程:
(a)骨芽細胞または骨髄由来の単球/マクロファージ系列細胞と被験物質とを接触させる工程、
(b)前記被験物質を接触させた骨芽細胞または骨髄由来の単球/マクロファージ系列細胞におけるPDK4の発現量を調べ、被験物質を接触させない骨芽細胞または骨髄由来の単球/マクロファージ系列細胞における発現量と比較する工程、
(c)下記工程(c1)または(c2)のいずれかにより破骨細胞への分化を誘導する工程:
(c1)前記比較工程(b)の結果に基づいて、骨芽細胞でPDK4の発現を抑制する被験物質存在下で、骨芽細胞と骨髄由来の単球/マクロファージ系列細胞とを共培養し、破骨細胞への分化を誘導する工程、さらに対照として、被験物質非存在下で、骨芽細胞と骨髄由来の単球/マクロファージ系列細胞とを共培養し、同様に破骨細胞への分化を誘導する工程、または
(c2)前記比較工程(b)の結果に基づいて、単球/マクロファージ系列細胞でPDK4の発現を抑制する被験物質存在下で、単球/マクロファージ系列細胞を、M-CSFおよびRANKL存在下で培養し、破骨細胞への分化を誘導する工程、さらに対照として、被験物質非存在下で、単球/マクロファージ系列細胞をM-CSFおよびRANKL存在下で培養し、同様に破骨細胞への分化を誘導する工程、
(d)前記工程(c)における破骨細胞への分化の程度を比較する工程、および
(e)前記比較工程(d)の結果に基づいて、骨髄由来の単球/マクロファージ系列細胞から破骨細胞への分化を抑制する被験物質を選択する工程
を含む、PDK4の発現または機能抑制を介した骨粗鬆症の予防および/または治療剤のスクリーニング方法。
〔6〕 下記工程:
(a’)被験物質存在下で、骨芽細胞と骨髄由来の単球/マクロファージ系列細胞を共培養し、破骨細胞への分化を誘導する工程、
(b’)前記工程(a’)における破骨細胞への分化の程度を比較する工程、および
(c’)前記比較工程(b’)の結果に基づいて、骨髄由来の単球/マクロファージ系列細胞から破骨細胞への分化を抑制する被験物質を選択する工程
を含む、PDK4の発現または機能抑制を介した骨粗鬆症の予防および/または治療剤のスクリーニング方法。
〔7〕 下記工程:
(a”)被験物質、M-CSFおよびRANKL存在下で、骨髄由来の単球/マクロファージ系列細胞から破骨細胞への分化を誘導する工程、
(b”)前記工程(a”)における破骨細胞への分化の程度を比較する工程、および
(c”)前記比較工程(b”)の結果に基づいて、骨髄由来の単球/マクロファージ系列細胞から破骨細胞への分化を抑制する被験物質を選択する工程
を含む、PDK4の発現または機能抑制を介した骨粗鬆症の予防および/または治療剤のスクリーニング方法。
〔8〕 Pdk4遺伝子を欠損してなる、非ヒト非荷重抵抗性骨モデル動物。
〔9〕 非荷重状態での骨量の減少の病態の解明に用いられる、前記〔8〕に記載の骨モデル動物。
〔10〕 病態が廃用性骨粗鬆症、ステロイド性骨粗鬆症および無重力時の骨量減少からなる群より選ばれる、前記〔9〕に記載の骨モデル動物。
〔11〕 前記動物がマウスである前記〔8〕〜〔10〕のいずれかに記載の骨モデル動物。
〔12〕 前記〔8〕〜〔11〕のいずれかに記載の骨モデル動物の子孫動物。
〔13〕 前記〔8〕〜〔12〕のいずれかに記載の非ヒト動物から得られる組織または細胞。
〔14〕 前記〔8〕〜〔12〕のいずれかに記載の骨モデル動物を用いることを特徴とする、骨粗鬆症の予防または治療剤のスクリーニング方法。
〔15〕 下記工程:
(a)前記〔8〕〜〔12〕のいずれかに記載の骨モデル動物の野生型対照に被験物質を投与し、当該骨モデル動物と野生型対照とを非荷重状態に供する工程、
(b)前記被験物質の投与前後の野生型対照における骨の形態、骨体積または骨密度を解析し、前記〔8〕〜〔12〕のいずれかに記載の骨モデル動物における骨の形態、骨体積または骨密度と比較する工程、および、
(c)前記(b)の比較結果に基づいて、非荷重状態における骨量の減少を抑制する被験物質を選択する工程
を含む、骨粗鬆症の予防または治療剤のスクリーニング方法。
〔16〕 下記工程:
(a)非荷重状態後に得られた前記〔13〕に記載の組織または細胞、および非荷重状態後に野生型対照から得られた組織または細胞をインビトロで生理学的条件下で維持する工程、
(b)前記野生型対照由来の組織または細胞におけるタンパク質のリン酸化の程度と、前記〔13〕由来の組織または細胞におけるタンパク質のリン酸化の程度とを比較する工程、または前記野生型対照由来の組織または細胞におけるタンパク質でPDK4に結合するタンパク質と前記〔13〕由来の組織または細胞におけるタンパク質でPDK4に結合するタンパク質を比較する工程、および
(c)前記比較結果に基づいて、リン酸化の程度が有意に変動しているタンパク質を選択する工程、または前記比較結果に基づいて、前記野生型対照由来の組織または細胞におけるタンパク質のみから得られ、前記〔13〕由来の組織または細胞におけるタンパク質からは得られないPDK4結合タンパク質を選択する工程
を含む、荷重刺激を感知し、骨粗鬆症の発症に関連するPDK4の標的タンパク質のスクリーニング方法。
【発明の効果】
【0010】
本発明は、非荷重後の骨芽細胞および骨髄細胞におけるPDK4発現のアップレギュレーションは、少なくとも一部はPDC不活性化とは別の経路を介して、非荷重後の破骨細胞形成および骨再吸収に導くという知見に基づくものである。本発明の骨粗鬆症の予防または治療剤によると、PDK4の発現または機能を抑制する物質を主成分とするものであることから、特に骨芽細胞または骨髄細胞に作用させた場合に、非荷重状態が主原因の骨粗鬆症を有効に予防または治療することができる。本発明のモデル動物によると、PDK4が欠損していることにより非荷重(無重力)に抵抗性の骨モデル動物として、無重力による骨量の減少など、無重力状態での骨組織での様々な生理学的状態を解明するモデルとして役立つ。本発明の組織または細胞によると、前記動物から得られるものであることから、in vivoのみならずin vitroで骨量減少のメカニズムの研究に利用することが可能である。本発明のスクリーニング方法によると、骨粗鬆症の予防または治療剤の開発のみならず、非荷重状態でのPDK4の基質の解明等に貢献することができる。
【図面の簡単な説明】
【0011】
【図1】PDK4発現のマイクロアレイおよびリアルタイムRT-PCR解析を示す。(A)尾懸垂実験におけるPDK4発現のマイクロアレイおよびリアルタイムRT-PCR解析。16週齢の野生型およびBCL2トランスジェニックマウスの脛骨および大腿骨を用いて、コントロール群、非荷重群(尾懸垂を8日間実施)および再荷重群(1週間の尾懸垂後、24時間尾懸垂から開放されたマウス)からmRNAを調製した。コントロール群のレベルを1に設定し、相対レベルを示す。リアルタイムRT-PCRにおけるデータは、3匹のマウスの平均±SEである。**, p<0.01 vs. コントロール。(B)16週齢の野生型およびBCL2トランスジェニックマウスのコントロール群および非荷重群由来の骨細胞または骨芽細胞画分のmRNAを用いたリアルタイムRT-PCR解析。尾懸垂を3日間実施した。野生型マウスのコントロール群の骨細胞画分のレベルを1と設定し、相対レベルを示す。データは、4匹のマウスの平均±SEである。**, p<0.01 vs. コントロール。(C)種々の組織におけるPDK4の発現。11週齢の野生型マウスの、骨格筋(SM)、心筋(CM)、腎臓(K)、肝臓(Li)、肺(Lu)、脳(B)、脾臓(Sp)、精巣(Te)、胸腺(Th)、全大腿骨(WF)、骨細胞画分(Oc)、骨芽細胞画分(Ob)および骨髄(BM)からmRNAを調製した。全大腿骨のレベルを1に設定し、相対値を示す。データは、3匹のマウスの平均±SEである。(D)骨におけるPDKアイソフォームの発現を骨格筋と比較した相対レベル。骨格筋に対する全大腿骨におけるPDKアイソフォームの相対的mRNAレベルを示す。(E)尾懸垂後のPDKアイソフォームmRNAの発現。3または7日間の尾懸垂後の12週齢の野生型マウスの骨格筋(SM)、全骨(WB)、骨細胞画分(Oc)、骨芽細胞画分(Ob)および骨髄(BM)からmRNAを調製した。コントロール群における全大腿骨のレベルを1に設定し、相対値を示す。データは、3匹のマウスの平均±SEである。*, p<0.05 vs.コントロール。
【図2】Pdk4-/-マウスの産生のための手段および産生マウスの解析結果を示す。(A)Pdk4遺伝子座、ターゲティングベクターおよび変異アレルの概略図。RI: EcoRI。(B)サザンブロット解析。DNAをEcoRIで消化し、Aに示す5’または3’プローブとハイブリダイズさせた。(C)ウエスタンブロット解析。1週間の尾懸垂後、12週齢の野生型マウスおよびPdk4-/-マウスの大腿骨からタンパク質を抽出した。(D)体重曲線。群当たりn=6。(E)マイクロ-CT解析。3、6および12月齢における野生型およびPdk4-/-マウス間の骨体積は類似していた。群当たりn=6。(F)骨芽細胞マーカー遺伝子発現のリアルタイムRT-PCR解析。5週齢の野生型およびPdk4-/-マウスの大腿骨(骨髄細胞を洗い出すことにより除去している)から試料を調製した。野生型マウスにおける値を1に設定し、相対レベルを示す。データは4匹のマウスの平均 ± SE である。(G)Col1a1およびCol1a2タンパク質の定量解析。12週齢で脛骨からタンパク質を抽出した。(H)オステオポンチンおよびオステオカルシンタンパク質のウエスタンブロット解析。12週齢で大腿骨からタンパク質を抽出した。
【図3】尾懸垂後のマイクロ-CTおよび骨組織形態計測解析を示す。11週齢の雄性野生型およびPdk4-/-マウスを1週間正常条件下(コントロール)または尾懸垂条件下(非荷重)においた。(A-E)マイクロ-CT解析。A-D、野生型(A、B)およびPdk4-/-(C, D)マウスのコントロール(A、C)および非荷重条件下(B、D)の三次元イメージ。スケールバー = 0.5mm。E、骨梁パラメータ。骨梁体積(BV/TV)、骨梁数(Tb.N)、骨梁幅(Tb.Th)、骨梁間距離(Tb.Sp)および骨梁密度(Dens)を示す。データは、8匹のマウスの平均 ± SEである。*p<0.05, **p<0.01 vs.コントロール。(F)骨組織形態計測解析。単位骨量(骨体積/組織体積、BV/TV)、類骨面(OS/BS)、類骨幅(O.Th)、骨芽細胞面(Ob.S/BS)、骨芽細胞数(N.Ob/B.Pm)、多核破骨細胞数(Mu.N.Oc/B.Pm)、破骨細胞面(Oc.S/BS)、吸収面(ES/BS)、骨石灰化速度(MAR)、石灰化面(MS/BS)および骨形成速度(BFR/BS)を示す。データは6-7匹のマウスの平均 ± SEである。**p<0.01 vs.コントロール。
【図4】非荷重時の骨形成マーカーと骨吸収マーカーの野生型マウスとPdk4-/-マウス間の比較を示す。(A)リアルタイムRT-PCR解析。コントロールまたは1週間の非荷重条件下での12週齢の野生型およびPdk4-/-マウスの大腿骨および脛骨からmRNAを調製した。データは5匹のマウスの平均 ± SEである。*p<0.05 vs.コントロール。(B)脛骨切片のTRAP染色。切片をメチルグリーンで対比染色した。スケールバー = 0.1mm。(C)TRAP5bの血清レベル。データは6-7匹のマウスの平均 ± SEである。**p<0.01 vs コントロール。
【図5】in vitroでの骨芽細胞形成を示す。(A)骨芽細胞分化中のPDK4の発現。骨芽細胞分化中の野生型初代骨芽細胞におけるPDK4発現を、コンフルエンス(0日目)ならびにコンフルエンス後3日目および5日目でリアルタイムRT-PCRで調べた。0日目のレベルを1と決定し、相対レベルを示す。データは3個のウエルの平均 ± SEである。(B)ALP活性および石灰化。野生型およびPdk4-/-マウス由来の初代骨芽細胞を、コンフルエンス後3日目および14日目に、それぞれALPおよびKossaで染色した。(C)リアルタイムRT-PCR解析。野生型およびPdk4-/-マウス由来の初代骨芽細胞をコンフルエンス後3日間培養し、mRNAを調製した。野生型骨芽細胞のレベルを1と決定し、相対レベルを示す。データは3個のウエルの平均 ± SEである。*p<0.05 vs. 野生型マウス。
【図6】in vitroでの破骨細胞形成を示す。(A)破骨細胞分化中のPDK4の発現。野生型BMMをM-CSF(20ng/ml)およびRANKL(100ng/ml)の存在下で培養した。mRNAを、M-CSFおよびRANKLの添加前(0日目)、添加後3日目および6日目に調製し、リアルタイムRT-PCRでPDK4の発現を調べた。0日目のレベルを1と決定し、相対レベルを示す。データは3個のウエルの平均 ± SEである。(B、C)BMMの破骨細胞分化。BMMを、M-CSF(20ng/ml)およびRANKL(100ng/ml)の存在下で6日間培養した(B)。スケールバー = 0.1mm。3-6の核を有するTRAP-陽性細胞の数、6を超える核を有するTRAP-陽性細胞の数およびそれらの合計を示す(C)。データは4個のウエルの平均 ± SEである。*p<0.05 vs. 野生型BMM。(D)破骨細胞分化に関連する遺伝子の発現。M-CSF(20ng/ml)およびRANKL(100ng/ml)の添加前(0日目)ならびに添加後3および6日目において、野生型およびPdk4-/- BMM由来のmRNAを用いてリアルタイムRT-PCR解析を実施した。(E-H)BMMおよび初代骨芽細胞の共培養。野生型およびPdk4-/-マウス由来のBMMを、野生型またはPdk4-/-初代骨芽細胞と共培養した。6日後にTRAP染色を実施し(E)、3-6の核を有するTRAP-陽性細胞、7-10の核を有するTRAP-陽性細胞および10を超える核を有するTRAP-陽性細胞を計数した(F)。破骨細胞の吸収活性をPitアッセイで調べ(G)、6日後に吸収pitを測定した(H)。スケールバー = 0.1mm。データは、4個のウエルの平均 ± SEである。**p<0.01 vs 野生型POBおよびBMMの共培養。
【図7】外来性PDK4によるPdk4-/-細胞における破骨細胞形成のレスキューおよびRanklプロモーターのレポーターアッセイを示す。(A-E)破骨細胞形成のレスキュー実験。A、PDK4のウエスタンブロット解析。Pdk4-/-初代骨芽細胞を、Pdk4-またはGFP-発現アデノウイルスに感染させた。B-D、BMMおよび初代骨芽細胞の共培養。Pdk4-/-マウス由来のBMMおよび初代骨芽細胞を共培養し、Pdk4-またはGFP-発現アデノウイルスに感染させた。6日後にTRAP染色を実施し(B)、3-6の核を有するTRAP-陽性細胞の数および6を超える核を有するTRAP-陽性細胞の数を計数した(C)。Pitアッセイを実施し、吸収領域を測定した(D)。スケールバー = 0.1mm。データは、4個のウエルの平均 ± SEである。*p<0.05 vs. GFP。E、リアルタイムRT-PCR解析。Pdk4-/-マウス由来の初代骨芽細胞をPdk4-またはGFP-発現アデノウイルスに感染させ、3日後にmRNAを調製した。データは、3個のウエルの平均 ± SEである。*p<0.05 vs. GFP。(F)Ranklプロモーターのレポーターアッセイ。野生型およびPdk4-/-マウス由来の初代骨芽細胞を、段階的に欠失させたRanklプロモーター領域を含むルシフェラーゼレポーターコンストラクトで形質転換した。データは、5個のウエルの平均 ± SEである。*p<0.05 vs. 野生型初代骨芽細胞。(G)PDK4によるRanklプロモーター活性の誘導。Pdk4-/-マウス由来の初代骨芽細胞に、各Ranklルシフェラーゼレポーターコンストラクトを導入、24時間後、細胞をPdk4-またはGFP-発現アデノウイルスに感染させた。データは、5個のウエルの平均 ± SEである。*p<0.05 vs. GFP。
【図8】破骨細胞形成、RanklおよびOpg発現ならびにRanklプロモーター活性におけるPDCリン酸化に対するPDK4アンタゴニストの効果を示す。(A)PDCリン酸化のウエスタンブロット解析。野生型マウス由来の初代骨芽細胞を、DCAの存在下または非存在下で0.5% FBS中、3時間培養し、抽出したタンパク質を抗-ホスホ-E1a(p-PDC)および抗-E1a(PDC)抗体と反応させた。(B)BMMの破骨細胞分化。野生型マウス由来のBMMを、M-CSF(20ng/ml)およびsRANKL(100ng/ml)の存在下、指定された濃度のDCAの存在下または非存在下で培養し、TRAP-陽性細胞を計数した。データは、3個のウエルの平均 ± SEである。(C)BMMと初代骨芽細胞との共培養における破骨細胞形成。野生型マウス由来のBMMおよび初代骨芽細胞を、指定された濃度のDCAの存在下または非存在下で共培養し、TRAP-陽性細胞を計数した。データは、3個のウエルの平均 ± SEである。(D)リアルタイムRT-PCR解析。野生型マウス由来の初代骨芽細胞を指定された濃度のDCAの存在下または非存在下で培養した。データは、3個のウエルの平均 ± SEである。(E)Ranklプロモーターのレポーターアッセイ。野生型マウス由来の初代骨芽細胞をRL(-185)で形質転換し、指定された濃度のDCAの存在下または非存在下で培養した。データは、3個のウエルの平均 ± SEである。(F、G)Ranklプロモーター活性におけるPdp1の効果。野生型マウス由来の初代骨芽細胞を0.5% FBS中で4時間培養し、Pdp1発現ベクターで形質転換した。2日後タンパク質を抽出し、抗-ホスホ-E1a(p-PDC)および抗-E1a(PDC)抗体と反応させた(F)。野生型マウス由来の初代骨芽細胞をRL(-185)と指定された量のDNAのPdp1発現ベクターで共形質転換し、36および72時間後にルシフェラーゼ活性を測定した(G)。
【図9】マウスの組織学的解析結果を示す。野生型マウス(A, C, E, G, I, K)およびPdk4-/-マウス(B, D, F, H, J, L)の4週齢(A, B)および12週齢(C, D)の脛骨切片のH-E染色、および10週齢の大腿骨の骨芽細胞(E-H)および破骨細胞(I-L)の電子顕微鏡写真。E、F、IおよびJのボックス内の領域を、それぞれG、H、KおよびLに拡大する。透過型電子顕微鏡(H-7100; Hitachi, Tokyo, Japan)を用い、既報に従って(Maruyama Z, et al., Dev Dyn 2007, 236:1876-1890)微細構造解析を実施した。スケールバー = 0.2mm(A-D)、3μm(E、F)、1μm(G、H)、5μm(I、J)、および2μm(K、L)。
【図10】血清グルコースおよびコルチゾールの測定結果を示す。11週齢の雄性野生型およびPdk4-/-マウスを、正常条件下(コントロール)または尾懸垂条件(非荷重)下で1週間飼育し、グルコースメーター(SKK, Nagoya, Japan)およびCortisol ELISAキット(Enzo, Tokyo, Japan)をそれぞれ用いて、血清グルコース(A)およびコルチゾール(B)を測定した。
【発明を実施するための形態】
【0012】
(定義)
本発明において「遺伝子」とは、2本鎖DNAのみならず、それを構成するセンス鎖およびアンチセンス鎖といった各1本鎖DNAを包含する趣旨で用いられる。またその長さによって特に制限されるものではない。本発明において遺伝子(DNA)とは、特に言及しない限り、ヒトゲノムDNAを含む2本鎖DNAおよびcDNAを含む1本鎖DNA(正鎖)並びに該正鎖と相補的な配列を有する1本鎖DNA(相補鎖)、およびこれらの断片のいずれもが含まれる。また当該「遺伝子」には、特定の塩基配列(配列番号1)で示される「遺伝子」だけでなく、これらによりコードされるタンパク質と生物学的機能が同等であるタンパク質(例えば同族体(ホモログやスプライスバリアントなど)、変異体および誘導体)をコードする「遺伝子」が包含される。かかる同族体、変異体または誘導体をコードする「遺伝子」としては、具体的には、ストリンジェントな条件下で、前記の配列番号1で示される特定塩基配列の相補配列とハイブリダイズする塩基配列を有する「遺伝子」をあげることができる。なお、ここでストリンジェントな条件は、Berger and Kimmel (1987, Guide to Molecular Cloning Techniques Methods in Enzymology, Vol. 152, Academic Press, San Diego CA) に教示されるように、核酸の融解温度(Tm)に基づいて決定することができる。例えばハイブリダイズ後の洗浄条件として、通常「1×SSC、0.1%SDS、37℃」程度の条件をあげることができる。相補鎖はかかる条件で洗浄しても対象とする正鎖とハイブリダイズ状態を維持するものであることが好ましい。特に制限されないが、より厳しいハイブリダイズ条件として「0.5×SSC、0.1%SDS、42℃」程度、さらに厳しいハイブリダイズ条件として「0.1×SSC、0.1%SDS、65℃」程度の洗浄条件をあげることができる。
【0013】
例えばヒト由来のタンパク質のホモログをコードする遺伝子としては、当該タンパク質をコードするヒト遺伝子に対応するマウスやラットなど他生物種の遺伝子が例示でき、これらの遺伝子(ホモログ)は、HomoloGene(http://www.ncbi.nlm.nih.gov/HomoloGene/)により同定することができる。具体的には、特定ヒト塩基配列をBLAST(Proc. Natl. Acad. Sci. USA 90:5873-5877, 1993、http://www.ncbi.nlm.nih.gov/BLAST/)にかけて一致する(Scoreが最も高く、E-valueが0でかつIdentityが100%を示す)配列のアクセッション番号を取得する。そのアクセッション番号をUniGene(http://www.ncbi.nlm.nih.gov/UniGene/)に入力して得られたUniGene Cluster ID(Hs.で示す番号)をHomoloGeneに入力する。結果として得られた他生物種遺伝子とヒト遺伝子との遺伝子ホモログの相関を示したリストから、特定の塩基配列で示されるヒト遺伝子に対応する遺伝子(ホモログ)としてマウスやラットなど他生物種の遺伝子を選抜することができる。
【0014】
本発明において「Pdk4遺伝子」または「Pdk4のDNA」とは、特定塩基配列(配列番号1)で示されるヒトPdk4遺伝子(DNA)ならびに、その同族体、変異体および誘導体などをコードする遺伝子(DNA)を包含する趣旨で用いられる。具体的には、配列番号1に記載のヒトPdk4遺伝子ならびに、そのマウスホモログおよびラットホモログなどが包含される。なお、遺伝子またはDNAは、機能領域の別を問うものではなく、例えば発現制御領域、コード領域、エキソン、またはイントロンを含むことができる。
【0015】
本発明において「タンパク質」または「ペプチド」には、特定のアミノ酸配列(配列番号2)で示される「タンパク質」または「ペプチド」だけでなく、これらと生物学的機能が同等であることを限度として、その同族体(ホモログやスプライスバリアント)、変異体、誘導体、成熟体およびアミノ酸修飾体などが包含される。ここでホモログとしては、ヒトのタンパク質に対応するマウスやラットなど他生物種のタンパク質が例示でき、これらはHomoloGene(http://www.ncbi.nlm.nih.gov/HomoloGene/)により同定された遺伝子の塩基配列から演繹的に同定することができる。前記変異体には、天然に存在するアレル変異体、天然に存在しない変異体、および人為的に欠失、置換、付加および挿入されることによって改変されたアミノ酸配列を有する変異体が包含される。なお、前記変異体としては、変異のないタンパク質または(ポリ)ペプチドと、少なくとも70%、好ましくは80%、より好ましくは95%、さらにより好ましくは97%相同なものをあげることができる。前記誘導体には、タンパク質のアミノ末端もしくはカルボキシ末端または側鎖を置換したものが包含される。前記アミノ酸修飾体には、天然に存在するアミノ酸修飾体、天然に存在しないアミノ酸修飾体が包含され、具体的にはアミノ酸のリン酸化体があげられる。
【0016】
本発明において「PDK4」という用語を用いる場合、配列番号で特に指定しない限り、特定アミノ酸配列(配列番号2)で示されるヒトPDK4やその同族体、変異体、誘導体、成熟体およびアミノ酸修飾体などを包含する趣旨で用いられる。
【0017】
本明細書中、PDK4は通常、温血動物(哺乳動物又は鳥類)由来のものを意味する。哺乳動物としては、例えば、マウス、ラット、ハムスター、モルモット等のげっ歯類やウサギ等の実験動物、ブタ、ウシ、ヤギ、ウマ、ヒツジ、ミンク等の家畜、イヌ、ネコ等のペット、ヒト、サル、カニクイザル、アカゲザル、マーモセット、オランウータン、チンパンジーなどの霊長類等をあげることができるが、これらに限定されるものではない。鳥類としては、ニワトリ、ウズラ、アヒル、ガチョウ、シチメンチョウ、エミュ、ダチョウ、ホロホロ鳥、ハト等を挙げることができる。PDK4は、好ましくは哺乳動物由来のものであり、より好ましくは霊長類(ヒト等)又はげっ歯類(マウス等)由来のものである。
【0018】
「PDK4が哺乳動物由来である」とは、PDK4の配列(ヌクレオチド配列又はアミノ酸配列)が哺乳動物のものであることを意味する。
【0019】
本発明においてPDK4をコードする遺伝子(以下、Pdk4遺伝子またはPdk4と省略する場合もある)は、例えば、ヒトPdk4遺伝子は、Rowlesらの文献(Rowles J. et al., 1996, J. Biol. Chem. 271 (37), pp.22376-22382)で公表されており、自体公知の方法により単離することができる。また、ヒトおよびマウスのPDK4の代表的なヌクレオチド配列及びアミノ酸配列が、NCBIに以下の通りに登録されている。
[ヒトPDK4]
ヌクレオチド配列(cDNA配列):アクセッション番号 NM_002612(バージョンNM_002612.3)(配列番号1)
アミノ酸配列:アクセッション番号 NP_002603(NP_002603.1)(配列番号2)
[マウスPDK4]
ヌクレオチド配列(cDNA配列):アクセッション番号 NM_013743(バージョンNM_013743.2)(配列番号3)
アミノ酸配列:アクセッション番号 NP_038771(バージョンNP_038771.1)(配列番号4)
【0020】
なお、本明細書においてヌクレオチド配列は、特にことわりのない限りDNAの配列として記載するが、ポリヌクレオチドがRNAである場合は、チミン(T)をウラシル(U)に適宜読み替えるものとする。
【0021】
(荷重感知遺伝子)
本明細書における「荷重感知遺伝子」とは、生体(動物)に対する荷重刺激を感知し、荷重(重力)刺激がない場合、破骨細胞の活動を活性化させる遺伝子をいう。具体的には、荷重刺激のない(非荷重)場合に骨細胞および骨細胞からのシグナルによって骨芽細胞でアップレギュレートし、再荷重後にダウンレギュレートする遺伝子であり、ピルビン酸デヒドロゲナーゼキナーゼのアイソザイムPDK4としても知られる遺伝子である。
【0022】
本発明は、Pdk4遺伝子を有効成分として含有する荷重感知遺伝子を提供する。また、本発明は、PDK4を有効成分として含有する荷重感知剤を提供する。
【0023】
(骨粗鬆症の予防または治療剤)
本発明は、PDK4の発現または活性を抑制する物質を含む、骨粗鬆症の予防または治療剤を提供する。
【0024】
本発明の予防または治療剤(本発明の医薬と称する場合もある)が対象とする疾患は、骨粗鬆症である。本発明において骨粗鬆症とは、世界保健機構の定義で、骨密度測定により若年成人の2.5標準偏差未満で診断されるものをいう。骨量の低下は種々の原因により生じるが、長期的な寝たきり状態、固定化状態、運動障害、麻痺、無重力状態での生活などによる非荷重状態が主原因である骨粗鬆症、すなわち、廃用性骨粗鬆症が具体的にあげられる。また、ステロイド性骨粗鬆症および無重力時の骨量減少もあげられる。
【0025】
PDK4の機能としては、ピルビン酸デヒドロゲナーゼキナーゼとしてこれまでに知られているキナーゼとしての機能および今後解明されるであろう様々な機能があげられるが、エンドポイントとして、荷重刺激を感知して荷重(重力)刺激がない場合、破骨細胞の活動を活性化する機能を指標とすることができる。
【0026】
PDK4の発現又は機能を抑制する物質としては、PDK4のアンタゴニストがあげられる。例えば、PDK4のPDCリン酸化に対するアンタゴニストとして知られているジクロロ酢酸およびピルビン酸デヒドロゲナーゼデホスファターゼは、破骨細胞形成、Rankl発現およびRanklプロモーター活性に影響がほとんどないことから、PDK4のアンタゴニストであって、PDCリン酸化に対するアンタゴニスト作用を有さないものが好ましく、より好ましくは低分子化合物である。このようなアンタゴニストは、本発明のスクリーニング方法により容易に選別することができる。
【0027】
PDK4の発現又は機能を抑制する物質としては、例えば、PDK4の発現を特異的に抑制し得るsiRNA、アンチセンス核酸、これらのポリヌクレオチドを発現し得る発現ベクター、低分子化合物があげられる。好ましくは、PDK4の発現を特異的に抑制し得るsiRNA、アンチセンス核酸、又はこれらのポリヌクレオチドを発現し得る発現ベクターが用いられる。
【0028】
本明細書中、「特異的な遺伝子発現の抑制」とは、標的とする遺伝子の発現を、それ以外の遺伝子の発現よりも強く抑制することを意味する。
【0029】
PDK4の発現を特異的に抑制し得るsiRNAとしては、例えば
(A)PDK4をコードするmRNA(成熟mRNA又は初期転写産物)のヌクレオチド配列又は18塩基以上のその部分配列に相補的なヌクレオチド配列を含む2本鎖のRNA、及び
(B)PDK4をコードするmRNA(成熟mRNA又は初期転写産物)と治療対象動物(好ましくはヒト)の細胞内で特異的にハイブリダイズし得る18塩基以上のヌクレオチド配列を含み、且つハイブリダイズすることによりPDK4の転写を抑制する2本鎖のRNAを挙げることができる。
【0030】
本明細書中、「特異的なハイブリダイゼーション」とは、核酸が、標的とするヌクレオチドに対して、それ以外のヌクレオチドよりも強くハイブリダイズすることを意味する。
【0031】
PDK4をコードするmRNAのヌクレオチド配列としては、例えば、配列番号1で表されるヌクレオチド配列(ヒトPDK4)、配列番号3で表されるヌクレオチド配列(マウスPDK4)をあげることができる。
【0032】
短い二本鎖RNAを細胞内に導入するとそのRNAに相補的なmRNAが分解される、いわゆるRNA干渉(RNAi)と呼ばれる現象は、以前から線虫、昆虫、植物等で知られていたが、最近、この現象が動物細胞でも起こることが確認されたことから[Nature, 411(6836): 494-498 (2001)]、リボザイムの代替技術として注目されている。
【0033】
siRNAは、代表的には、標的遺伝子のmRNAのヌクレオチド配列又はその部分配列(以下、標的ヌクレオチド配列)と相補的な配列を有するRNAとその相補鎖からなる2本鎖オリゴRNAである。また、ヘアピンループ部分を介して、標的ヌクレオチド配列に相補的な配列(第1の配列)と、その相補配列(第2の配列)とが連結された一本鎖RNAであって、ヘアピンループ型の構造をとることにより、第1の配列が第2の配列と2本鎖構造を形成するRNA(small hairpin RNA: shRNA)もsiRNAの好ましい態様の1つである。
【0034】
siRNAに含まれる、標的ヌクレオチド配列と相補的な部分の長さは、通常、約18塩基以上、好ましくは約19塩基以上、より好ましくは約21塩基以上の長さであるが、標的遺伝子の発現を特異的に抑制可能である限り、特に限定されない。siRNAが23塩基よりも長い場合には、該siRNAは細胞内で分解されて、約20塩基前後のsiRNAを生じ得るので、理論的には標的ヌクレオチド配列と相補的な部分の長さの上限は、標的遺伝子のmRNA(成熟mRNAもしくは初期転写産物)のヌクレオチド配列の全長である。しかし、インターフェロン誘導の回避、合成の容易さ、抗原性の問題等を考慮すると、該相補部分の長さは、例えば約50塩基以下、好ましくは約25塩基以下、最も好ましくは約23塩基以下である。即ち、該相補部分の長さは、通常、約18〜約50塩基、好ましくは約19〜約25塩基、より好ましくは約21〜約23塩基である。
【0035】
また、siRNAを構成する各RNA鎖の長さも、通常、約18塩基以上、好ましくは約19塩基以上、より好ましくは約21塩基以上の長さであるが、標的遺伝子の発現を特異的に抑制可能である限り、特に限定されず、理論的には各RNA鎖の長さの上限はない。しかし、インターフェロン誘導の回避、合成の容易さ、抗原性の問題等を考慮すると、siRNAの長さは、例えば約50塩基以下、好ましくは約25塩基以下、最も好ましくは約23塩基以下である。即ち、各RNA鎖の長さは、例えば通常、約18〜約50塩基、好ましくは約19〜約25塩基、より好ましくは約21〜約23塩基である。なお、shRNAの長さは、2本鎖構造をとった場合の2本鎖部分の長さとして示すものとする。
【0036】
標的ヌクレオチド配列と、siRNAに含まれるそれに相補的な配列とは、完全に相補的であることが好ましい。しかし、siRNAの中央から外れた位置についての塩基の変異(少なくとも90%以上、好ましくは95%以上の同一性の範囲内であり得る)については、完全にRNA干渉による切断活性がなくなるのではなく、部分的な活性が残存し得る。他方、siRNAの中央部の塩基の変異は影響が大きく、RNA干渉によるmRNAの切断活性が極度に低下し得る。
【0037】
siRNAは、5’及び/又は3’末端に塩基対を形成しない、付加的な塩基を有していてもよい。該付加的塩基の長さは、siRNAが標的遺伝子の発現を特異的に抑制可能である限り特に限定されないが、通常5塩基以下、例えば2〜4塩基である。該付加的塩基は、DNAでもRNAでもよいが、DNAを用いるとsiRNAの安定性を向上させることができる。このような付加的塩基の配列としては、例えばug-3’、uu-3’、tg-3’、tt-3’、ggg-3’、guuu-3’、gttt-3’、ttttt-3’、uuuuu-3’などの配列が挙げられるが、これに限定されるものではない。
【0038】
shRNAのヘアピンループのループ部分の長さは、標的遺伝子の発現を特異的に抑制可能である限り、特に限定されないが、通常、5〜25塩基程度である。該ループ部分のヌクレオチド配列は、ループを形成することができ、且つ、shRNAが標的遺伝子の発現を特異的に抑制可能である限り、特に限定されない。
【0039】
「アンチセンス核酸」とは、標的mRNA(成熟mRNA又は初期転写産物)を発現する細胞の生理的条件下で該標的mRNAと特異的にハイブリダイズし得るヌクレオチド配列を含み、且つハイブリダイズした状態で該標的mRNAにコードされるポリペプチドの翻訳を阻害し得る核酸をいう。アンチセンス核酸の種類はDNAであってもRNAであってもよいし、あるいはDNA/RNAキメラであってもよいが、好ましくはDNAである。
【0040】
PDK4の発現を特異的に抑制し得るアンチセンス核酸としては、例えば
(A)PDK4をコードするmRNA(成熟mRNA又は初期転写産物)のヌクレオチド配列又は12塩基以上のその部分配列に相補的なヌクレオチド配列を含む核酸、及び
(B)PDK4をコードするmRNA(成熟mRNA又は初期転写産物)と治療対象動物(好ましくはヒト)の細胞内で特異的にハイブリダイズし得る12塩基以上のヌクレオチド配列を含み、且つハイブリダイズした状態でPDK4ポリペプチドへの翻訳を阻害し得る核酸
等をあげることができる。
【0041】
アンチセンス核酸中の標的mRNAとハイブリダイズする部分の長さは、PDK4の発現を特異的に抑制する限り特に制限はなく、通常、約12塩基以上であり、長いものでmRNA(成熟mRNA又は初期転写産物)の全長配列と同一の長さである。ハイブリダイゼーションの特異性を考慮すると、該長さは好ましくは約15塩基以上、より好ましくは約18塩基以上である。また、合成の容易さや抗原性の問題等を考慮すると、標的mRNAとハイブリダイズする部分の長さは、通常、約200塩基以下、好ましくは約50塩基以下、より好ましくは約30塩基以下である。即ち、標的mRNAとハイブリダイズする部分の長さは、例えば約12〜約200塩基、好ましくは約15〜約50塩基、より好ましくは約18〜約30塩基である。
【0042】
アンチセンス核酸の標的ヌクレオチド配列は、PDK4の発現を特異的に抑制可能であれば特に制限はなく、PDK4のmRNA(成熟mRNA又は初期転写産物)の全長配列であっても部分配列(例えば約12塩基以上、好ましくは約15塩基以上、より好ましくは約18塩基以上)であってもよいし、あるいは初期転写産物のイントロン部分であってもよいが、好ましくは、標的配列はPDK4のmRNAの5’末端からコード領域のC末端までに位置することが望ましい。
【0043】
アンチセンス核酸中の標的mRNAとハイブリダイズする部分のヌクレオチド配列は、標的配列の塩基組成によっても異なるが、生理的条件下でPDK4のmRNAとハイブリダイズし得るために、標的配列の相補配列に対して通常約90%以上(好ましくは95%以上、最も好ましくは100%)の同一性を有するものである。
【0044】
アンチセンス核酸の大きさは、通常約12塩基以上、好ましくは約15塩基以上、より好ましくは約18塩基以上である。該大きさは、合成の容易さや抗原性の問題等から、通常約200塩基以下、好ましくは約50塩基以下、より好ましくは約30塩基以下である。
【0045】
さらに、アンチセンス核酸は、PDK4のmRNAもしくは初期転写産物とハイブリダイズして翻訳を阻害するだけでなく、二本鎖DNAであるPDLIM2遺伝子と結合して三重鎖(トリプレックス)を形成し、mRNAへの転写を阻害し得るものであってもよい。
【0046】
天然型の核酸は、細胞中に存在する核酸分解酵素によってそのリン酸ジエステル結合が容易に分解されるので、本発明において使用されるsiRNAやアンチセンス核酸は、分解酵素に安定なチオリン酸型(リン酸結合のP=OをP=Sに置換)や2’-O-メチル型等の修飾ヌクレオチドを用いて合成することもできる。siRNAやアンチセンス核酸の設計に重要な他の要素として、水溶性及び細胞膜透過性を高めること等が挙げられるが、これらはリポソームやマイクロスフェアを使用するなどの剤形の工夫によっても克服することができる。
【0047】
PDK4の発現を特異的に抑制し得るsiRNA及びアンチセンス核酸は、PDK4のmRNA配列(例えば配列番号1又は3で表されるヌクレオチド配列)や染色体DNA配列に基づいて標的配列を決定し、市販のDNA/RNA自動合成機(アプライド・バイオシステムズ社、ベックマン社等)を用いて、これに相補的なヌクレオチド配列を合成することにより調製できる。siRNAは、センス鎖及びアンチセンス鎖をDNA/RNA自動合成機でそれぞれ合成し、適当なアニーリング緩衝液中、約90〜約95℃で約1分程度変性させた後、約30〜約70℃で約1〜約8時間アニーリングさせることにより調製できる。また、相補的なオリゴヌクレオチド鎖を交互にオーバーラップするように合成して、これらをアニーリングさせた後リガーゼでライゲーションすることにより、より長い2本鎖ポリヌクレオチドを調製できる。
【0048】
本発明の医薬は、PDK4の発現を特異的に抑制するsiRNA又はアンチセンス核酸を発現し得る(コードする)発現ベクターを有効成分とすることもできる。当該発現ベクターにおいては、上述のsiRNA又はアンチセンス核酸或いはそれをコードする核酸(好ましくはDNA)が、投与対象である哺乳動物(好ましくはヒト)の細胞(例えば、骨細胞、骨芽細胞、骨髄細胞)内でプロモーター活性を発揮し得るプロモーターに機能的に連結されている。
【0049】
使用されるプロモーターは、投与対象である哺乳動物の細胞内で機能し得るものであれば特に制限はない。プロモーターとしては、polI系プロモーター、polII系プロモーター、polIII系プロモーター等を使用することができる。具体的には、SV40由来初期プロモーター、サイトメガロウイルスLTR等のウイルスプロモーター、β−アクチン遺伝子プロモーター等の哺乳動物の構成蛋白質遺伝子プロモーター、並びにtRNAプロモーター等のRNAプロモーター等が用いられる。
【0050】
本発明の医薬は、好ましくは骨芽細胞に導入されることによりその作用をより効果的に発揮するものである。したがって、有効成分としての前記発現ベクターは、発現する遺伝子が骨芽細胞特異的プロモーターの下流に作動可能に連結されているものが好ましい。
【0051】
前記骨芽細胞特異的プロモーターとは、骨芽細胞特異的に下流の遺伝子の転写開始部位を決定し、その頻度を直接的に調節する塩基配列を有するDNAをいう。前記プロモーターは、骨芽細胞が由来する動物に応じて適宜選択することができる。
【0052】
前記骨芽細胞特異的プロモーターとしては、I型コラーゲンプロモーター、オステオカルシンプロモーター等があげられるが、I型コラーゲンプロモーターが好ましい。I型コラーゲンプロモーターとしては、例えばヒト由来のプロモーターの場合、Jimenez,S.A., Varga,J., Olsen,A., Li,L., Diaz,A., Herhal,J. and Koch,J. Functional analysis of human alpha 1(I) procollagen gene promoter. Differential activity in collagen-producing and -nonproducing cells and response to transforming growth factor beta 1 J. Biol. Chem. 269 (17), 12684-12691 (1994)、マウス由来のプロモーターの場合、Ravazzolo,R., Karsenty,G. and de Crombrugghe,B. A fibroblast-specific factor binds to an upstream negative control element in the promoter of the mouse alpha 1(I) collagen gene J. Biol. Chem. 266 (12), 7382-7387 (1991)、Rossert, J., Eberspaecher, H., and de Crombrugghe, B. (1995). Separate cis-acting DNA elements of the mouse pro-a1(I) collagen promoter direct expression of reporter genes to different type I collagen-producing cells in transgenic mice. J. Cell Biol. 129, 1421-1432.などに記載されている。
【0053】
上記発現ベクターは、好ましくは上述のポリヌクレオチド又はそれをコードする核酸の下流に転写終結シグナル、すなわちターミネーター領域を含有する。さらに、形質転換細胞選択のための選択マーカー遺伝子(テトラサイクリン、アンピシリン、カナマイシン等の薬剤に対する抵抗性を付与する遺伝子、栄養要求性変異を相補する遺伝子等)をさらに含有することもできる。
【0054】
発現ベクターとしては、公知または市販のものを使用することができ、ウイルスベクターまたは非ウイルスベクターが例示される。ウイルスベクターとしては、例えば、レトロウイルス、アデノウイルス、アデノ随伴ウイルス、ヘルペスウイルス、ワクシニアウイルス、ポリオウイルス、シンドビスウイルスなどのウイルスベクターがあげられる。前記プロモーターおよび遺伝子を発現ベクターに連結する方法は、自体公知の方法により行うことができる。
【0055】
PDK4の発現または機能を抑制する物質の別の態様は、PDK4に対する抗体、ドミナントネガティブ変異体、アプタマー、もしくはそれらの発現ベクターである。
【0056】
本発明の医薬に有効成分として含有する抗体は、ヒトへの適用を考慮した場合、キメラ抗体(chimeric antibody)、ヒト化抗体(humanized antibody)、ヒト型抗体(human antibody)が好ましく、ヒト化抗体およびヒト型抗体がより好ましい。キメラ抗体は、例えば「実験医学(臨時増刊号), Vol.6, No.10, 1988」、特公平3-73280号公報等を、ヒト化抗体は、例えば特表平4-506458号公報、特開昭62-296890号公報等を、ヒト抗体は、例えば「Nature Genetics, Vol.15, p.146-156, 1997」、「Nature Genetics, Vol.7, p.13-21, 1994」、特表平4-504365号公報、国際公開第94/25585号公報、「日経サイエンス、6月号、第40〜第50頁、1995年」、「Nature, Vol.368, p.856-859, 1994」、特表平6-500233号公報等を参考にそれぞれ作製することができる。
【0057】
ドミナントネガティブ変異体は、PDK4に対する変異の導入によりその活性が低減したものである。ドミナントネガティブ変異体は、天然の翻訳産物と競合することで間接的にその機能を阻害することができる。変異としては、例えば、機能性部位における、当該部位が担う機能の低下をもたらすようなアミノ酸の変異(例、1以上のアミノ酸の欠失、置換、付加)があげられる。
【0058】
PDK4に対するアプタマーは、PDK4に特異的に結合してその機能的発現を阻害するオリゴヌクレオチドまたはペプチドである。PDK4に対するオリゴヌクレオチドアプタマーは、in vitro selection法またはSELEX法により得ることができ、例えば、以下の手順があげられる。即ち、まず、DNA/RNA自動合成機を用いてランダムにオリゴヌクレオチド(例えば、約60塩基)を合成し、オリゴヌクレオチドのプールを作製する。次に、目的の蛋白質、即ちPDK4と結合するオリゴヌクレオチドをアフィニティーカラムで分離する。分離したオリゴヌクレオチドをPCRで増幅し、前述の選抜プロセスで再度選抜する。この過程を約5回以上繰り返すことによって、PDK4に親和性の強いアプタマーを選抜することができる。PDK4に対するペプチドアプタマーは、酵母のtwo-hybrid法により得ることができる。
【0059】
本発明の医薬が発現ベクターを有効成分とする場合、当該発現ベクターは、上記タンパク質(抗体、ドミナントネガティブ変異体またはアプタマー)をコードするポリヌクレオチドが、投与対象である生体の細胞内でプロモーター活性を発揮し得るプロモーターに機能的に連結されていなければならない。ここで、発現ベクターが含み得るプロモーターは、前記RNAi誘導性核酸またはアンチセンス核酸の発現ベクターで用いられるものを限定なく利用することができる。また、発現ベクター自体も、前記RNAi誘導性核酸またはアンチセンス核酸の発現ベクターと同様のものを限定なく利用することができる。
【0060】
本発明の医薬は、PDK4の発現又は機能を抑制する物質に加え、任意の担体、例えば医薬上許容される担体を含むことができる。
【0061】
医薬上許容され得る担体としては、例えば、ショ糖、デンプン、マンニット、ソルビット、乳糖、グルコース、セルロース、タルク、リン酸カルシウム、炭酸カルシウム等の賦形剤、セルロース、メチルセルロース、ヒドロキシプロピルセルロース、ポリプロピルピロリドン、ゼラチン、アラビアゴム、ポリエチレングリコール、ショ糖、デンプン等の結合剤、デンプン、カルボキシメチルセルロース、ヒドロキシプロピルスターチ、ナトリウム−グリコール−スターチ、炭酸水素ナトリウム、リン酸カルシウム、クエン酸カルシウム等の崩壊剤、ステアリン酸マグネシウム、エアロジル、タルク、ラウリル硫酸ナトリウム等の滑剤、クエン酸、メントール、グリチルリチン・アンモニウム塩、グリシン、オレンジ粉等の芳香剤、安息香酸ナトリウム、亜硫酸水素ナトリウム、メチルパラベン、プロピルパラベン等の保存剤、クエン酸、クエン酸ナトリウム、酢酸等の安定剤、メチルセルロース、ポリビニルピロリドン、ステアリン酸アルミニウム等の懸濁剤、界面活性剤等の分散剤、水、生理食塩水、オレンジジュース等の希釈剤、カカオ脂、ポリエチレングリコール、白灯油等のベースワックスなどが挙げられるが、それらに限定されるものではない。
【0062】
PDK4の発現又は機能を抑制する物質として、PDK4の発現を特異的に抑制し得るsiRNA、アンチセンス核酸、又はこれらのポリヌクレオチドを発現し得る発現ベクターを用いる場合、核酸の細胞内への導入を促進するために、本発明の医薬はさらに核酸導入用試薬を含むことができる。
【0063】
核酸導入試薬としては、リポフェクチン、リポフェクタミン(lipofectamine)、リポフェクタミンRNAiMAX(LipofectamineRNAiMAX)、インビボフェクタミン(Invivofectamine)、DOGS(トランスフェクタム)、DOPE、DOTAP、DDAB、DHDEAB、HDEAB、ポリブレン、あるいはポリ(エチレンイミン)(PEI)等の陽イオン性脂質を用いることが出来る。また、発現ベクターとしてレトロウイルスを用いる場合には、導入試薬としてレトロネクチン、ファイブロネクチン、ポリブレン等を用いることができる。
【0064】
本発明の骨粗鬆症の予防または治療剤の投与単位形態としては、経口剤または非経口剤があげられ、非経口剤が好ましい。非経口に適した形態としては、注射剤(液剤、懸濁剤など)、坐剤、貼付剤、軟膏剤、ゼリー剤、吸入剤、クリーム剤、スプレー剤、点鼻剤、エアゾール剤などが例示される。
【0065】
医薬組成物中のPDK4の発現又は機能を抑制する物質の含有量は、特に限定されず広範囲に適宜選択されるが、例えば、医薬組成物全体の約0.001ないし100重量%である。
【0066】
本発明の医薬を所望の細胞(好ましくは骨芽細胞または骨髄細胞)に導入する方法は、例えば、in vivo法またはex vivo法があげられる。in vivo法は、本発明の医薬を、例えば注射剤などの適当な投与形態で、静脈、動脈、皮下、関節内、筋肉内などに投与することができる。ex vivo法の場合、本発明の医薬を、対象の生体内から取り出した細胞をin vitroで培養中に公知の方法によりトランスフェクション、インフェクションまたはインジェクションし、一定期間培養してPdk4遺伝子の発現抑制を確認した後、同一対象の体内に戻す方法を採用することができる。
【0067】
これら各種形態の製剤中の有効成分の投与量は、治療目的である疾患の程度、患者の年齢、体重などにより適宜調節することができる。通常、患者成人1人当たり約0.0001-100mg、好ましくは約0.001-10mgが数日ないし数カ月に1回投与される量とすればよい。レトロウイルスベクターの場合は、レトロウイルス力価として、1日患者体重1kg当たり約1×103pfuないし1×1015pfuとなる量の範囲から選ぶことができる。ex vivo法で本発明の医薬を導入した細胞の場合は、1×104細胞/bodyないし1×1015細胞/body程度を投与すればよい。
【0068】
本発明の医薬に含まれる有効成分により、生体内に投与された場合にPDK4の発現又は機能が抑制されるが、血糖値やグルココルチコイドの濃度に影響はない。これは、別のアイソザイムであるPDK1、PDK2およびPDK3が代償的に働くためであると考えられる。本発明の医薬(医薬組成物)は、哺乳動物(例えば、ラット、マウス、モルモット、ウサギ、ヒツジ、ウマ、ブタ、ウシ、サル、ヒト)に対して安全に投与されうる。
【0069】
(非荷重抵抗性骨モデル動物)
本発明は、Pdk4遺伝子を欠損してなる、非ヒト非荷重抵抗性骨モデル動物を提供する。本発明の骨モデル動物は、Pdk4遺伝子を欠損していることから、好適には、非荷重状態での骨量の減少の病態の解明に用いられる。病態としては、骨粗鬆症があげられ、好ましくは、廃用性骨粗鬆症、ステロイド性骨粗鬆症および無重力時の骨量減少からなる群より選ばれる。
【0070】
非ヒト動物としては、ヒト以外の動物であれば特に限定されるものではないが、ヒトに類似した骨細胞ネットワークを有する動物が好ましい。好適な動物としては、マウス、ラット、ハムスター、モルモット、ウサギ、イヌ、ネコ、サル等の実験動物ならびにウシ、ヒツジ、ウマ、ブタ等の家畜があげられるが、遺伝子工学的に利用が容易であるところから、マウスがより好ましい。
【0071】
本発明において、Pdk4遺伝子を欠損するとは、内因性のPdk4遺伝子の塩基配列の1以上の塩基に置換、欠失、挿入もしくは付加の変異を導入することをいい、ノックアウトとも称する。また、このような改変をした動物をノックアウト動物と称する。
【0072】
ノックアウトの場合、少なくともPdk4遺伝子の一部を欠損させる変異を導入させ、当該遺伝子の機能的欠損を有するように操作することが好ましく、具体的には、マウスPdk4遺伝子の場合、エキソンのいずれか1つを欠損させるように構築されたターゲティングベクターを用いる方法が一例としてあげられる。より具体的には、実施例3および図2に記載のターゲティングベクターを用いた方法があげられる。
【0073】
本発明の骨モデル動物には、前記遺伝子の変異が対立遺伝子の両方に導入されたホモ接合動物、前記遺伝子の変異が対立遺伝子の片方に導入されたヘテロ接合動物およびそれらの出生前の胎仔も含まれる。前記ホモ変異動物は、前記ヘテロ変異動物を交配することにより得られるものである。
【0074】
ノックアウト動物は、例えば下記の工程(a)〜(c)を含む方法により製造できる。
(a)Pdk4遺伝子の機能的欠損を含む胚性幹細胞を提供する工程;
(b)前記胚性幹細胞を胚に導入し、キメラ胚を得る工程;
(c)前記キメラ胚を動物に移植し、キメラ動物を得る工程;
(d)前記キメラ動物を交配させ、Pdk4遺伝子欠損ヘテロ接合体を得る工程。
【0075】
ノックアウト動物、胚性幹細胞、ターゲティングベクターの作製の詳細については、例えば、下記文献を参照することができる。
1.別冊 実験医学 ザ・プロトコールシリーズ 「ジーンターデティングの最新技術」(2000年、羊土社)コンディショナルターゲティング法p.115-120
2.バイオマニュアルシリーズ8 「ジーンターゲティング」−ES細胞を用いた変異マウスの作製(1995年、羊土社)p.71-77
3.Sambrookら, Molecular Cloning: A LABORATORY MANUAL, 第3版, COLD SPRING HARBOR LABORATORY PRESS, 2001年, 4.82-4.85
4.Robertson E. J. in Teratocarcinomas and embryonic stem cells-a practical approach, ed. Robertson, E. J. (IRL Press, Oxford), 1987: pp.108-112
5.Dynecki, S. M.ら, Gene Targeting -a practical approach, 2nd edition, ed. Joyner, A.L. (Oxford Univ. Press), 2000: pp.68-73
6.Dynecki, S. M. ら, Gene Targeting -a practical approach, 2nd edition, ed. Joyner, A. L. (Oxford Univ. Press), 2000: pp.75-81
【0076】
このようにして得られた本発明の骨モデル動物をさらに交配して得られる子孫動物、これら動物に由来する組織または細胞も本発明に含まれる。前記組織としては、すべての組織があげられ、長管骨、頭蓋骨、椎骨などが好ましい。また、前記細胞としては、前記組織中に含まれる細胞、組織中から単離された細胞、これら細胞から樹立した細胞株があげられ、具体的には骨芽細胞、骨細胞または骨髄細胞などが好ましい。
【0077】
本発明の非ヒト骨モデル動物、子孫動物、これら動物に由来する組織または細胞は、下記本発明のスクリーニング方法において好適に利用することができる。
【0078】
(スクリーニング方法1)
スクリーニング方法1は、細胞または組織を用いたインビトロの系で実施する。具体的には、下記工程を含む、PDK4の発現または機能抑制を介した骨粗鬆症の予防および/または治療剤のスクリーニング方法1(1)を提供する:
(a)骨芽細胞または骨髄由来の単球/マクロファージ系列細胞と被験物質とを接触させる工程、
(b)前記被験物質を接触させた骨芽細胞または骨髄由来の単球/マクロファージ系列細胞におけるPDK4の発現量を調べ、被験物質を接触させない骨芽細胞または骨髄由来の単球/マクロファージ系列細胞における発現量と比較する工程、および
(c)前記比較結果に基づいて、PDK4の発現を抑制する被験物質を選択する工程。
【0079】
前記工程(a)において、被験物質とは、いかなる公知物質および新規物質であってもよく、例えば、核酸、糖質、脂質、蛋白質、ペプチド、有機低分子化合物、コンビナトリアルケミストリー技術を用いて作製された化合物ライブラリー、固相合成やファージディスプレイ法により作製されたランダムペプチドライブラリー、あるいは微生物、動植物、海洋生物等由来の天然成分などがあげられる。また、これらの化合物の2種以上の混合物を試料として供することもできる。
【0080】
前記工程(a)において使用する骨芽細胞または骨髄由来の単球/マクロファージ系列細胞は、PDK4を発現しうる限り、いかなる動物由来の細胞であってもよい。本発明の非ヒト骨モデル動物由来の骨芽細胞または骨髄由来の単球/マクロファージ系列細胞をさらに陰性対照として用いる場合、非ヒト骨モデル動物と同種由来の細胞が望ましい。マウスの骨芽細胞および骨髄由来の単球/マクロファージ系列細胞の調製方法は、実施例7および8に記載されており、他の動物由来の細胞も実施例7および8に記載の方法に準じて調製することができる。
【0081】
工程(a)において、骨芽細胞または骨髄由来の単球/マクロファージ系列細胞をPDK4の発現が維持される条件下に置いたものを用いることができる。かかる条件下としては、骨芽細胞または骨髄由来の単球/マクロファージ系列細胞を適当な培地中に入れ、約25〜40℃のインキュベーター中で生存または培養させることが好ましい。次に、前記培地中に被験物質を添加し、インキュベートを続けることで接触がなされうる。
【0082】
前記被験物質の添加量は、有効成分の種類、培地に対する溶解性、細胞の感受性等によって適宜設定することができる。
【0083】
工程(b)において、前記被験物質を接触させた骨芽細胞または骨髄由来の単球/マクロファージ系列細胞におけるPDK4の発現量は、自体公知の方法により調べることができる。内在性のPDK4の発現量は、例えば、細胞からタンパク質を抽出してPDK4を認識する抗体を用いるウェスタンブロッテング等により測定する方法;細胞からRNAを抽出してRT−PCRまたはノーザンブロッテング等により転写されたPdk4 mRNAを測定する方法;PDK4を認識する抗体を用いて、免疫組織化学によりin situでPDK4を調べる方法等があげられる。内在性のPDK4の発現量は、Pdk4のプロモーター領域とレポーター遺伝子とを連結した発現ベクターを細胞に導入し、レポーター遺伝子の発現を測定するレポーターアッセイ等によっても推定できる。レポーターアッセイが好ましく用いられる。レポーター遺伝子は、公知の遺伝子を限定なく使用することができ、例えば、ルシフェラーゼ、GFP(緑色蛍光タンパク質)、CAT(クロラムフェニコールアセチルトランスフェラーゼ)などがあげられるが、これに限定されない。レポーター遺伝子の発現は、用いるレポーター遺伝子に応じて、自体公知の方法により測定することができる。
【0084】
前記工程(b)において、被験物質を接触させない骨芽細胞または骨髄由来の単球/マクロファージ系列細胞も同時にまたは別途調べ、接触した場合の結果と接触しない場合の結果とを比較する。また、陰性対照として、本発明の骨モデル動物由来の骨芽細胞または骨髄由来の単球/マクロファージ系列細胞も準備することが好ましい。
【0085】
前記工程(c)において、工程(b)で得られた比較結果に基づき、PDK4の発現を抑制する被験物質を選択する。選択する基準は、PDK4のタンパク質またはmRNAが低減していることを指標にすればよい。
【0086】
このようにして選択された被験物質は、骨芽細胞におけるPDK4のアンタゴニスト作用を有する剤として有用であるとともに、例えば、スクリーニング方法2のようなさらなるスクリーニングや薬理試験を実施することにより、副作用の少ない骨粗鬆症の予防または治療剤の候補薬ともなりうる。
【0087】
また、スクリーニング方法1の別の態様として、上記工程(a)および(b)に加え、下記工程を含む、PDK4の発現または機能抑制を介した骨粗鬆症の予防および/または治療剤のスクリーニング方法1(2)を提供する:
(C)下記工程(c1)または(c2)のいずれかにより破骨細胞への分化を誘導する工程:
(c1)前記比較工程(b)の結果に基づいて、骨芽細胞でPDK4の発現を抑制する被験物質存在下で、骨芽細胞と骨髄由来の単球/マクロファージ系列細胞とを共培養し、破骨細胞への分化を誘導する工程、さらに対照として、被験物質非存在下で骨芽細胞と骨髄由来の単球/マクロファージ系列細胞とを共培養し、同様に破骨細胞への分化を誘導する工程、または
(c2)前記比較工程(b)の結果に基づいて、単球/マクロファージ系列細胞でPDK4の発現を抑制する被験物質存在下で、単球/マクロファージ系列細胞を、M-CSFおよびRANKL存在下で培養し、破骨細胞への分化を誘導する工程、さらに対照として、被験物質非存在下で単球/マクロファージ系列細胞をM-CSFおよびRANKL存在下で培養し、同様に破骨細胞への分化を誘導する工程、
(d)前記工程(C)における破骨細胞への分化の程度を比較する工程、および
(e)前記比較工程(d)の結果に基づいて、骨髄由来の単球/マクロファージ系列細胞から破骨細胞への分化を抑制する被験物質を選択する工程。
【0088】
工程(C)の工程(c1)において、PDK4の発現を抑制する被験物質存在下での骨芽細胞と骨髄由来の単球/マクロファージ系列細胞との共培養は、実施例8に記載の方法に従って行うことができる。破骨細胞への分化誘導は、骨芽細胞から産生されるM-CSFおよびRANKLの存在下で培養することにより行うことができる。
【0089】
工程(C)の工程(c1)において、さらに対照として、被験物質非存在下で骨芽細胞と骨髄由来の単球/マクロファージ系列細胞とを共培養し、上記と同様の条件下で破骨細胞への分化誘導を行うことが好ましい。
【0090】
工程(C)の工程(c2)において、PDK4の発現を抑制する被験物質存在下での骨髄由来の単球/マクロファージ系列細胞の培養は、実施例8に記載の方法に従って行うことができる。破骨細胞への分化誘導は、M-CSFおよびRANKLの存在下で培養することにより行うことができる。
【0091】
工程(C)の工程(c2)において、さらに対照として、被験物質非存在下で骨髄由来の単球/マクロファージ系列細胞を培養し、上記と同様の条件下で破骨細胞への分化誘導を行うことが好ましい。
【0092】
工程(d)において、破骨細胞への分化の有無およびその程度は、実施例8に記載の方法に従って、TRAP染色し、走査型電子顕微鏡による観察によって測定することができる。
【0093】
工程(e)において、TRAP染色陽性細胞であって小核破骨細胞(3-6の核を有する)または大核破骨細胞(6を超える核を有する)の数が対照と比較して有意に減少している被験物質を選択する。
【0094】
このようにして選択された被験物質は、PDK4の発現を抑制し、かつ破骨細胞への分化を抑制しうる物質として、骨粗鬆症、特に非荷重状態における骨粗鬆症の予防または治療剤の候補薬ともなりうる。
【0095】
また、スクリーニング方法1のさらに別の態様として、下記工程を含むスクリーニング方法1(3)を実施することにより、PDK4の発現または機能抑制を介した骨粗鬆症の予防および/または治療剤の候補薬をより簡便にスクリーニングすることができる。
(a’)被験物質存在下で、骨芽細胞と骨髄由来の単球/マクロファージ系列細胞を共培養し、破骨細胞への分化を誘導する工程、
(b’)前記工程(a’)における破骨細胞への分化の程度を比較する工程、および
(c’)前記比較工程(b’)の結果に基づいて、骨髄由来の単球/マクロファージ系列細胞から破骨細胞への分化を抑制する被験物質を選択する工程。
【0096】
前記工程(a’)、(b’)および(c’)はそれぞれ、スクリーニング方法1(2)の工程(c1)、(d)および(e)と同様に実施することができる。
【0097】
また、スクリーニング方法1のさらに別の態様として、下記工程を含むスクリーニング方法1(4)を実施することにより、PDK4の発現または機能抑制を介した骨粗鬆症の予防および/または治療剤の候補薬をより簡便にスクリーニングすることができる。
(a”)被験物質、M-CSFおよびRANKL存在下で、骨髄由来の単球/マクロファージ系列細胞から破骨細胞への分化を誘導する工程、
(b”)前記工程(a”)における破骨細胞への分化の程度を比較する工程、および
(c”)前記比較工程(b”)の結果に基づいて、骨髄由来の単球/マクロファージ系列細胞から破骨細胞への分化を抑制する被験物質を選択する工程。
【0098】
前記工程(a”)、(b”)および(c”)はそれぞれ、スクリーニング方法1(2)の工程(c2)、(d)および(e)と同様に実施することができる。
【0099】
(スクリーニング方法2)
スクリーニング方法2は、本発明の骨モデル動物を用いてインビボの系で実施する。具体的には、下記工程を含む、骨粗鬆症の予防または治療剤のスクリーニング方法を提供する:
(a)本発明の骨モデル動物の野生型対照に被験物質を投与し、当該骨モデル動物と野生型対照とを非荷重状態に供する工程、
(b)前記被験物質の投与前後の野生型対照における骨の形態を解析し、本発明の骨モデル動物における骨の形態と比較する工程、および、
(c)前記(b)の比較結果に基づいて、非荷重状態における骨量の減少を抑制する被験物質を選択する工程。
【0100】
前記工程(a)において、被験物質は、スクリーニング方法1と同様のものがあげられる。被験物質を動物に投与する方法は特に限定されるものではないが、経口的または非経口的に投与され得る。非経口的投与経路としては、例えば、静脈内、動脈内、筋肉内等の全身投与、あるいは関節内、標的細胞付近への局所投与等があげられる。
【0101】
前記被験物質の投与量は、有効成分の種類、分子の大きさ、投与経路、投与対象となる動物種、投与対象の薬物受容性、体重、年齢等によって適宜設定することができる。
【0102】
前記工程(a)において、動物を非荷重状態に供する方法は、生体が重力を感知しない状態または重力の影響を受けにくい状態に維持可能であれば特に限定されるものではない。げっ歯類(マウス、ラット等)を例にとると、尾懸垂実験が好適にあげられ、実施例1に記載の方法に従って、行うことができる。尾懸垂は、通常、3〜14日間、好ましくは7日間継続して行うことが望ましい。
【0103】
前記工程(b)において、骨の形態、骨体積または骨密度を調べる方法としては、例えば、X線解析、マイクロCT(μCT)、pQCTおよび骨組織切片を用いた骨組織形態計測などがあげられ、定量的な解析方法として、μCTおよび骨組織形態計測が好ましい。μCT解析は、実施例4に記載されている。解析可能な骨梁パラメータとしては、骨梁体積、骨梁数、骨梁幅、骨梁間距離、骨梁密度、ストラクチャーモデルインデックス、骨梁連結性、異方性度があり、皮質骨パラメータとしては、皮質骨断面積、骨内膜周囲長、骨膜周囲長、皮質骨密度があげられる。骨組織形態計測の海面骨パラメータとしては、単位骨量、類骨面、類骨幅、骨芽細胞面、骨芽細胞数、多核破骨細胞数、破骨細胞面、吸収面、骨石灰化速度、石灰化面および骨形成速度があり、皮質骨パラメータとしては、骨断面積、皮質骨面積、骨髄腔面積、骨膜性骨形成率、皮質内骨形成率、皮質内骨吸収面があげられる。
【0104】
前記工程(b)において、被験物質を投与しない動物における骨の形態、骨体積または骨密度も同時にまたは別途調べ、投与動物の結果と非投与動物の結果とを比較する。また、陽性対照として、本発明の骨モデル動物を同様に解析する。
【0105】
前記工程(c)において、工程(b)で得られた比較結果に基づき、野生型動物の非荷重状態における骨量の減少を抑制する被験物質を選択する。選択する基準は、本発明の骨モデル動物を指標にすればよい。
【0106】
このようにして選択された被験物質は、非荷重状態における骨量の減少を抑制させ、骨粗鬆症、特に廃用性骨粗鬆症の候補薬となりうる。
【0107】
(スクリーニング方法3)
スクリーニング方法3は、本発明の骨モデル動物由来の組織または細胞を用いてインビトロの系で実施する。具体的には、下記工程を含む、荷重刺激を感知し骨粗鬆症の発症に関連するPDK4の標的タンパク質のスクリーニング方法を提供する:
(a)非荷重状態後に本発明の骨モデル動物から得られた組織または細胞、および非荷重状態後に野生型対照から得られた組織または細胞をインビトロで生理学的条件下で維持する工程、
(b)前記野生型対照由来の組織または細胞におけるタンパク質のリン酸化の程度と、本発明の骨モデル動物由来の組織または細胞におけるタンパク質のリン酸化の程度とを比較する工程、または前記野生型対照由来の組織または細胞におけるタンパク質でPDK4に結合するタンパク質と本発明の骨モデル動物由来の組織または細胞におけるタンパク質でPDK4に結合するタンパク質を比較する工程、および
(c)前記比較結果に基づいて、リン酸化の程度が有意に変動しているタンパク質を選択する工程、または前記比較結果に基づいて、前記野生型対照由来の組織または細胞におけるタンパク質のみから得られ、本発明の骨モデル動物由来の組織または細胞におけるタンパク質からは得られないPDK4結合タンパク質を選択する工程。
【0108】
前記工程(a)において動物を非荷重状態にするには、スクリーニング方法2と同様の条件で行えばよい。用いる組織または細胞は、動物に由来するものであれば特に限定されないが、荷重感知遺伝子PDK4の未知の基質を選択するという観点から、骨芽細胞、骨芽細胞を豊富に含む組織または骨芽細胞に分化可能な細胞もしくは組織、および破骨細胞、破骨細胞を豊富に含む組織または破骨細胞に分化可能な細胞もしくは組織が望ましい。
【0109】
前記工程(a)において、組織または細胞をインビトロで生理学的条件下で維持する方法は、野生型対照由来の組織または細胞の場合、PDK4の発現が維持される条件下に置いたものを用いることができる。かかる条件下としては、組織または細胞を適当な培地中に入れ、約25〜40℃のインキュベーター中で生存または培養させることが好ましい。本発明の骨モデル動物由来の組織または細胞も、野生型対照由来の組織または細胞と同じ条件下で維持する。
【0110】
前記工程(b)において、野生型対照由来の組織または細胞におけるタンパク質のリン酸化の程度と、本発明の骨モデル動物由来の組織または細胞におけるタンパク質のリン酸化の程度とを比較する。タンパク質のリン酸化の程度は、自体公知の方法により調べることができる。例えば、細胞抽出液の一次元あるいは二次元電気泳動により変動タンパク質を同定する方法、リン酸化抗体アフィニティカラムを用いて細胞抽出液からリン酸化蛋白を濃縮し電気泳動により比較同定する方法、細胞抽出液を酵素消化して得られたペプチドから金属キレートカラム(IMAC)を用いてリン酸化ペプチドを精製し2DLC-MS/MSによりリン酸化ペプチドを同定比較する方法、さらにこれに安定同位体標識法による定量法を組み合わせたリン酸化ペプチドのLC-MS/MSによる定量的変動解析等があげられる。さらに、PDK4と結合するタンパク質の同定も、公知の方法により調べることができる。例えば、PDK4の全部あるいは一部をベイトとした酵母を用いたツーハイブリッド法、PDK4抗体アフィニティカラムを用いて細胞抽出液からPDK4およびその結合タンパク質を濃縮し電気泳動で比較同定する方法等が挙げられる。
【0111】
前記工程(c)において、工程(b)で得られた比較結果に基づき、リン酸化の程度が有意に変動しているタンパク質を選択する。PDK4のキナーゼ活性を考慮すると、リン酸化が有意に上昇しているタンパク質を指標にすることが望ましい。また、野生型対照のみから得られ、本発明の骨モデル動物由来の組織または細胞からは得られないPDK4結合タンパク質を選択する。
【0112】
このようにして選択されたタンパク質は、荷重感知遺伝子であるPDK4の新規基質として、非荷重状態における骨低下のメカニズムの解明または特に廃用性骨粗鬆症の予防または治療剤の分子標的ともなりうる。
【実施例】
【0113】
以下、実施例により本発明をより詳細に説明するが、本発明はこれらの実施例により何ら限定されるものではない。
【0114】
[実施例1]
尾懸垂実験およびマイクロアレイ
尾懸垂実験は、既報に準じて実施した(Sakai A, Nakamura T: Changes in trabecular bone turnover and bone marrow cell development in tail-suspended mice, J Musculoskelet Neuronal Interact 2001, 1:387-39222)。野生型マウスおよびBCL2トランスジェニック雄性マウス(10週齢)を、尾懸垂用の特殊なケージ(TSケージ)に置き、当該ケージに適応させるために1週間飼育した。尾懸垂は、尾懸垂群において8日間実施した。再負荷群のマウスは1週間の尾懸垂後に24時間尾懸垂から開放した。対照群は、尾懸垂を除いて同じ条件下においた。脛骨および大腿骨からRNAを抽出し、Agilent マイクロアレイを用いて、Hokkaido System Science社(Sapporo, Japan)によりマイクロアレイ解析を実施した。
【0115】
[実施例2]
リアルタイムRT-PCR
全骨から骨芽細胞、骨細胞および骨髄細胞を分離するために、大腿骨および脛骨から筋肉、結合組織および骨膜を取り除き、骨幹端で骨を切断した。大腿骨および脛骨の骨幹中の骨髄細胞をリン酸緩衝食塩水(PBS)で押し出した。次いで、マイクロ歯間ブラシ(Kobayashi Pharmaceutical Co. Ltd., Osaka, Japan)を用いて、骨芽細胞が豊富な細胞を回収した。残った骨を骨細胞が豊富な細胞として用いた。ISOGEN(Wako, Osaka, Japan)を用いて全RNAを抽出し、Light Cycler 480 SYBR Green I システム(Roche)を用いてリアルタイムRT-PCRを実施した。Gapdhの値を元にデータを正規化した。
【0116】
[実施例3]
Pdk4-/-マウスの作出
マウスPdk4遺伝子のターゲティングベクターは、BAC改変技術により作製した。BAC改変キットをGene Bridges(Heidelberg, Germany)から購入し、マウスPdk4遺伝子のすべてのエキソンを含むBACクローン、RP23-25N12をBACPAC Resource Center(Oakland, CA)から購入した。IRES-LacZおよびPGK-gb2-neoを含むカセットをPCRで増幅し、BACクローンを含むバクテリアに導入し、Pdk4遺伝子の第3エキソンと第10エキソン間でゲノムDNAを置換した。IRES-LacZおよびPGK-gb2-neoを含み、Pdk4遺伝子に相同なDNAが隣接するカセットを、PGK-tkカセットを含むプラスミドにクローニングした。最終のターゲティングコンストラクトを線状化し、ES細胞のE14ラインにエレクトロポレーションにより導入した。ターゲティングされたES細胞をC57BL/6胚盤胞にインジェクションし、キメラを作出した。既報に従って、サザンブロットによりミュータントアレルを確認した(Komori T, et al.,: Targeted disruption of Cbfa1 results in a complete lack of bone formation owing to maturational arrest of osteoblasts, Cell 1997, 89:755-764)。Pdk4+/-マウスおよびPdk4-/-マウスの全身でLacZの発現を検出することができなかったので、ゲノムDNAの欠失はmRNAの安定性またはPDK4発現それ自体に影響するよう思われた。ヘテロ接合性雄性マウスをC57BL/6J雌性マウスと6回戻し交配した。血清中の酒石酸抵抗性酸性ホスファターゼ5b(TRAP5b)レベルをELISA(IDS, Boldon, UK)により測定した。研究に先立ち、全ての実験は、長崎大学大学院医歯薬学総合研究科の動物保護と利用に関する委員会による審査と承認を受けた。
【0117】
[実施例4]
マイクロ−CT解析
定量マイクロ−CT解析は、マイクロ−CTシステム(μCT-20; Scanco Medical, Bruttisellen, Switzerland)を用いて行った。スキャンしたスライスからのデータを三次元解析に用いて、大腿骨の形態学的パラメータを計算した。骨梁のパラメータを大腿骨の遠位骨幹端で測定した。約2.4mm(成長板から0.5mm離れたところ)を頭尾方向にスキャンして、12μm増分の200枚のスライスを得た。
【0118】
[実施例5]
組織学的解析
長骨の組織学的解析のためマウスを剖検し、長骨を4%パラホルムアルデヒド/0.01M リン酸緩衝化生理食塩水中で固定し、10% EDTA(pH7.4)中で脱灰し、パラフィンに包埋した。TRAP活性について、切片(5μm厚)を染色した。骨の組織形態学的解析は、既報に従って行った(Liu W, Toyosawa S, Furuichi T, Kanatani N, Yoshida C, Liu Y, Himeno M, Narai S, Yamaguchi A, Komori T: Overexpression of Cbfa1 in osteoblasts inhibits osteoblast maturation and causes osteopenia with multiple fractures, J Cell Biol 2001, 155:157-166)。動的組織形態学的指標を評価するため、マウスを剖検する6日前と2日前に、体重10g当たり0.16mgの用量でカルセインを注射した。
【0119】
[実施例6]
ウエスタンブロット解析およびCol1a1の定量
以下の抗体を用いて、ウエスタンブロット解析を行った:ウサギ抗-オステオポンチン抗体(Santa Cruz Biotechnology, Santa Cruz, CA)、ウサギ抗-オステオカルシン抗体(IBL, Takasaki, Japan)、ウサギ抗-PDK4抗体(Abgen, San Diego, CA)、マウス抗-PDC E1a抗体(MitoSciences, Eugene, Oregon)、ウサギ抗-PDC E1a pSer293抗体(Calbiochem, Darmstadt, Germany)およびヤギ抗-アクチン抗体(Santa Cruz Biotechnology)。既報に従って、コラーゲンを抽出し、解析した(Domenicucci C, Goldberg HA, Hofmann T, Isenman D, Wasi S, Sodek J: Characterization of porcine osteonectin extracted from foetal calvariae, Biochem J 1988, 253:139-151)。
【0120】
[実施例7]
In vitroでの骨芽細胞形成
初代骨芽細胞は、既報に従って、新生マウスの頭蓋冠の小片を三次元コラーゲンゲル中で12日間培養することにより調製した(Komori T, et al., Cell 1997, 89:755-76423)。骨芽細胞の分化に関しては、50μg/ml アスコルビン酸および10mM β-グリセロールリン酸を10% FBSを補足したα-MEMに添加し、アルカリホスファターゼ(ALP)活性および石灰化に関して染色を行った。
【0121】
[実施例8]
In vitroでの破骨細胞形成
骨髄由来の単球/マクロファージ系列細胞(BMM)を、Ficoll-PaqueTM(GE Healthcare, Tokyo, Japan)を用いた密度勾配遠心分離によって12週齢のマウスの骨髄から単離し、10% FBS、20ng/ml マクロファージコロニー刺激因子(M-CSF)(Sigma, Tokyo, Japan)および100ng/mlのGST-可溶性Receptor activator of NF-κB ligand(GST-sRANKL)を含むα-MEM中で1.5x105/wellの密度で96ウエルプレートに播種し、6日間培養した。TRAP染色は、既報に従って行った(Maruyama Z, et al.: Runx2 determines bone maturity and turnover rate in postnatal bone development and is involved in bone loss in estrogen deficiency, Dev Dyn 2007, 236:1876-1890)。BMM(1×105cells/well)を、10% FBS含有α-MEM中で10-6M プロスタグランジンE2(PGE2)および10-8M 1α,25(OH)2D3の存在下、96-wellプレートにてPOB(1×104cells/well)とともに培養した。骨再吸収の評価のために、BMMおよび初代骨芽細胞を、象牙質ディスク(IDS)上で6日間共培養し、再吸収された領域を走査型電子顕微鏡(H-7100; Hitachi, Tokyo, Japan)で測定した。ジクロロ酢酸(DCA)は、Sigma-Aldrich(St. Louis, MO)から購入した。レスキュー実験に関して、Pdk4-発現アデノウイルスベクターおよびEGFP-発現アデノウイルスベクターを既報(Enomoto H, et al.,: Induction of osteoclast differentiation by Runx2 through receptor activator of nuclear factor-kappa B ligand (RANKL) and osteoprotegerin regulation and partial rescue of osteoclastogenesis in Runx2-/- mice by RANKL transgene, J Biol Chem 2003, 278:23971-23977)に従って作製し、2.5×107 plaque forming units/mlで使用した。
【0122】
[実施例9]
レポーターアッセイ
Ranklプロモーター領域のゲノムDNAを既報に従ってクローニングし(Kitazawa R, Kitazawa S, Maeda S: Promoter structure of mouse RANKL/TRANCE/OPGL/ODF gene, Biochim Biophys Acta 1999, 1445:134-141)、3.8 kbおよび1.5 kbの断片をpGL3ベクター(Promega, Madison, WI)に挿入した。ネスティッド欠損コンストラクトを、クローニングしたDNA断片のPCR増幅により作製し、シークエンスにより確認した。Pdp1 cDNAをPCRで増幅し、pSG5にライゲートし、シークエンスにより確認した。初代骨芽細胞を、Fugene 6(Roche Diagnostics, Tokyo, Japan)を用いて、それぞれRanklプロモーターコンストラクト(0.2μg)およびpRL-CMV(0.003μg)を含むDNA混合物で一過性にトランスフェクトした。72時間後、Dual Luciferase Reporter Assay System(Promega)を用いて、細胞溶解液のルシフェラーゼ活性を測定し、Renillaルシフェラーゼ活性に対して正規化した。
【0123】
結果
非荷重後の骨細胞におけるPdk4発現の誘導
本発明者は、BCL2トランスジェニックマウス(骨細胞のアポトーシスにより骨細胞ネットワークが破壊されている)および野生型マウスを用いた尾懸垂により、非荷重後にその発現がアップレギュレートされ再荷重後にダウンレギュレートする荷重感知遺伝子を探索した。脛骨および大腿骨由来のRNAを用いたマイクロアレイ解析において、野生型マウスにおいては、PDK4は非荷重後にアップレギュレートされるが再荷重によりダウンレギュレートされることを見出したが、BCL2トランスジェニックマウスでは非荷重後にPdk4はアップレギュレートされなかった(図1A)。このことは、リアルタイムRT-PCR 解析により確認され(図1A)、PDK4は野生型マウスでは破骨細胞豊富な画分および骨芽細胞豊富な画分の両方でアップレギュレートしているが、BCL2トランスジェニックマウスではアップレギュレートしなかった(図1B)。PDK4は、骨格筋および心筋で高発現し、腎臓、肝臓、肺、脾臓および骨で中等度に発現している。PDK4は、骨中では、破骨細胞豊富な画分、骨芽細胞豊富な画分および骨髄細胞において中等度に発現していた(図1C)。骨におけるPDKアイソフォームの発現レベルを比較するため、骨格筋に対して骨における発現レベルを比較した(図1D)。比率はPDK4が最高値であった。PDK4は、非荷重後に骨格筋、大腿骨全体、骨細胞豊富な画分、骨芽細胞豊富な画分および骨髄細胞でアップレギュレートしたが、PDK1、PDK2およびPDK3は、非荷重後に骨全体ならびに骨芽細胞、骨細胞および骨髄細胞の各画分においてアップレギュレートしなかった。一方、PDK2は、非荷重後の骨格筋でアップレギュレートした(図1E)。
【0124】
Pdk4-/-マウスにおける正常な骨の発達および維持
非荷重状態での骨におけるPDK4の役割を調べるため、本発明者は、Pdk4-/-マウスを作出した(図2A-C)。Pdk4-/-マウスの体重は、野生型マウスと同様であった(図2D)。μ-CT解析により、骨体積は野生型およびPdk4-/-マウス間で3、6および12月齢で同様であった(図2E)。組織学的解析において、Pdk4-/-マウスの骨は正常に発達し、骨芽細胞、骨細胞および破骨細胞の形態は野生型マウスの形態と類似していた(図9)。骨芽細胞の分化マーカー(Runx2、Osterix、ALP、Col1a1、オステオポンチン、骨シアロタンパク質(BSP)およびオステオカルシンを含む)の発現は、野生型マウスおよびPdk4-/-マウス間で類似しており、Col1a1、Col1a2、オステオポンチンおよびオステオカルシンを含む骨マトリックスタンパク質の産生は、野生型マウスおよびPdk4-/-マウス間で類似していた(図2F-H)。
【0125】
野生型マウスにおいて非荷重後の骨量が減少するが、Pdk4-/-マウスでは減少しない
尾懸垂を1週間実施し、μ-CTにより非荷重後の骨量を比較した。生理学的条件下においては、野生型マウスとPdk4-/-マウスではすべてのパラメータが類似していた(コントロール)。しかしながら、非荷重後、野生型マウスでは骨体積、脛骨の厚さおよび脛骨密度が減少したが、Pdk4-/-マウスでは減少しなかった(図3A-E)。組織形態学的解析において、すべてのパラメータ(骨体積ならびに骨形成および吸収のパラメータを含む)は、生理学的条件下では野生型マウスとPdk4-/-マウスとでは類似していた(コントロール)。非荷重後、野生型マウスでは骨体積が低下し、骨吸収パラメータ(破骨細胞数、破骨細胞面および吸収面を含む)が増加したが、Pdk4-/-マウスでは増加しなかった(図3F)。血中グルコースおよびコルチゾールのレベルは、野生型マウスとPdk4-/-マウスとでは類似しており、両マウスにおいて非荷重状態で影響がなかった(図10)。
【0126】
野生型マウスでは非荷重後に骨吸収および Ranklの発現が増加するが、Pdk4-/-マウスでは増加しない
野生型マウスおよびPdk4-/-マウスにおいて、コントロール群と非荷重群とでは骨形成速度は有意に相違しなかったが、野生型マウスおよびPdk4-/-マウスで、非荷重後にコントロール群と比較してCol1a1およびオステオカルシンの発現は同様に低下した(図3Fおよび4A)。RanklおよびCtskの発現は、野生型マウスで非荷重後に顕著に増加したが、Pdk4-/-マウスでは増加しなかった。一方、Opg、Rank、Nfatc1、DC-STAMPの発現は、野生型マウスおよびPdk4-/-マウスの両マウスにおいて非荷重後に有意に変化しなかった(図4A)。TRAP-陽性細胞およびTRAP5bの血清レベルは、野生型マウスで非荷重後に増加したが、Pdk4-/-マウスでは増加しなかった(図4B、C)。
【0127】
骨芽細胞および破骨細胞前駆体の異常から生じるPdk4-/-マウスにおける破骨細胞形成の低下
PDK4の発現は骨芽細胞の分化中に上昇するが、Pdk4-/-初代骨芽細胞は、野生型初代骨芽細胞と同様のレベルのALP活性および石灰化を示した(図5A、B)。さらに、骨芽細胞マーカー遺伝子(Runx2、Osterix、ALP、Col1a1、オステオネクチン、オステオポンチン、BSPおよびオステオカルシンがあげられる)の発現レベルは、野生型とPdk4-/-の初代骨芽細胞間で類似していた。ただし、RanklはPdk4-/-初代骨芽細胞では低下していた(図5C)。また、骨髄細胞におけるPdk4の発現は、M-CSFおよびRANKLの存在下で破骨細胞への分化中に増加し、Pdk4-/-BMMから小核破骨細胞および大核破骨細胞の形成は、野生型BMMの培養物と比較して大核破骨細胞においてより深刻な減少であった(図6A-C)。Ctskの発現は低下し、他方、Rank、Nfatc1、DC-STAMP、Sirpa、CD47およびCD9の発現には影響がなかった(図6D)。また、初代骨芽細胞とBMMとを共培養することにより、破骨細胞の分化も調べた。野性型骨芽細胞とPdk4-/-BMMとの共培養において、3-6の核を有する小核破骨細胞の数は、野性型骨芽細胞とBMMとの共培養で得られたものと同様であったが、6を超える核を含む大核破骨細胞の数は、野性型骨芽細胞とBMMとの共培養で得られたものの約半分であった。Pdk4-/-骨芽細胞と野生型BMMとの共培養も同様の結果を示した。Pdk4-/-骨芽細胞とBMMとの共培養もまた、野性型骨芽細胞とBMMとの共培養と比較して、3-6の核を有する小核破骨細胞の数は同様であったが、7-10の核を有する大核破骨細胞の数は4分の1であり、10を超える核を含む巨核破骨細胞の数は、野性型骨芽細胞とBMMとの共培養で得られたものの10分の1未満であった(図6E、F)。さらに、Pdk4-/-骨芽細胞とBMMとの共培養における吸収領域は、野性型骨芽細胞とBMMとの共培養で得られたものの10分の1未満であった(図6G、H)。
【0128】
Pdk4による破骨細胞形成とRanklプロモーターの正の調節
次に、アデノウイルストランスファーを用いて、インビトロでPDK4が破骨細胞形成を誘導するか否かを調べた(図7A-D)。Pdk4-/-骨芽細胞とBMMとの共培養物にPdk4-発現アデノウイルスを感染させると、GFP-発現アデノウイルスを感染させたときと比べて、TRAP-陽性細胞および吸収領域が増加した。Pdk4-/-初代骨芽細胞にアデノウイルスによりPdk4を導入すると、Rankl発現を強く誘導し、Opg発現を中等度に誘導した。これにより、アデノウイルスによりGFPを導入したときと比べて、Rankl/Opg比が増加した(図7E)。Rankl発現におけるPDK4の関与を調べるため、Ranklプロモーター領域のDNA断片を用いて、レポーターアッセイを行った(図7F)。3.8 kbの断片を含むレポーターベクターの活性は、野生型骨芽細胞と比較してPdk4-/-骨芽細胞で低下した。Pdk4-/-骨芽細胞でのレポーター活性の低下は、段階的に欠失させたコンストラクトを用いたアッセイでも観察された(1.5K、185、164、137および122 bp)。さらに、PDK4のPdk4-/-骨芽細胞への導入は、GFPの導入と比較して、欠失コンストラクトのレポーター活性を亢進させた(図7G)。
【0129】
PDCリン酸化に対するPdk4アンタゴニストは破骨細胞形成およびRankl誘導を阻害しない
ジクロロ酢酸(DCA)は、PDKsによるPDCのE1aサブユニットのリン酸化を阻害する(Kato M, Li J, Chuang JL, Chuang DT: Distinct Structural Mechanisms for Inhibition of Pyruvate Dehydrogenase Kinase Isoforms by AZD7545, Dichloroacetate, and Radicicol, Structure 2007, 15:992-1004)。初代骨芽細胞をDCAで処理すると、PDCのリン酸化を阻害した(図8A)。M-CSFおよびRANKLの存在下で野生型BMMを培養した場合、DCAでの処理は破骨細胞の分化に影響がなかった(図8B)。DCAでの処理は、野生型初代骨芽細胞とBMMとの共培養において破骨細胞形成にも影響がなかった(図8C)。また、RanklおよびOpgの発現ならびにRanklプロモーター活性におけるDCAの効果も調べた(図8D、E)が、DCAは影響しなかった。さらに、Pdp1の初代骨芽細胞への導入はPDCを脱リン酸化したが、Ranklプロモーター活性には影響がなかった(図8F、G)。これらの知見は、PDCのリン酸化とは別の様式で、PDK4が破骨細胞形成を調節することを示唆する。
【0130】
PDK4は、グルコースの酸化的利用を制限するため、PDCの特異的キナーゼであると考えられてきた。DCAはピルビン酸の類似体である。DCAおよびピルビン酸は、PDKのN-末端の調節ドメインの中心において共通の結合部位を共有し、DCAは、PDKの阻害およびその後のPDCの活性化につながる活性部位の変化を誘導する。DCAはPDCのリン酸化を阻害するが、破骨細胞形成、Rankl発現およびRanklプロモーター活性に影響しなかった。さらに、Pdp1によるPDCの脱リン酸化は、Ranklプロモーター活性に対して影響しなかった。これらの知見は、PDCのリン酸化とは別の様式でPDK4が破骨細胞形成を調節することを示しており、このことは、破骨細胞形成に関する骨芽細胞およびBMMにおいて、PDK4は新規な未知の基質を有していることを示唆する。破骨細胞形成に関するPDK4およびPDK4の基質は、廃用性骨粗鬆症の適切な治療標的となると考えられる。
【産業上の利用可能性】
【0131】
本発明の骨粗鬆症の予防または治療剤によると、非荷重状態が主原因の骨粗鬆症を有効に予防または治療することができる。本発明のモデル動物によると、PDK4が欠損していることにより非荷重(無重力)に抵抗性の骨モデル動物として、無重力による骨量の減少など、無重力状態での骨組織での様々な生理学的状態を解明するモデルとして役立つ。本発明の組織または細胞によると、骨量減少のメカニズムの研究に利用することが可能である。本発明のスクリーニング方法によると、骨粗鬆症の予防または治療剤の開発のみならず、非荷重状態でのPDK4の基質の解明等に貢献することができる。
【特許請求の範囲】
【請求項1】
PDK4の発現または機能を抑制する物質を含む、骨粗鬆症の予防または治療剤。
【請求項2】
PDK4の発現または機能を抑制する物質が
(1)Pdk4遺伝子のRNAi誘導性核酸、アンチセンス核酸もしくはそれらの発現ベクター、または
(2)PDK4に対する抗体、ドミナントネガティブ変異体、アプタマーもしくはそれらの発現ベクター
である、請求項1に記載の予防または治療剤。
【請求項3】
骨粗鬆症が廃用性骨粗鬆症、ステロイド性骨粗鬆症および無重力時の骨量減少からなる群より選ばれる請求項1または2に記載の予防または治療剤。
【請求項4】
下記工程:
(a)骨芽細胞または骨髄由来の単球/マクロファージ系列細胞と被験物質とを接触させる工程、
(b)前記被験物質を接触させた骨芽細胞または骨髄由来の単球/マクロファージ系列細胞におけるPDK4の発現量を調べ、被験物質を接触させない骨芽細胞または骨髄由来の単球/マクロファージ系列細胞における発現量と比較する工程、および
(c)前記比較結果に基づいて、PDK4の発現を抑制する被験物質を選択する工程
を含む、PDK4の発現または機能抑制を介した骨粗鬆症の予防および/または治療剤のスクリーニング方法。
【請求項5】
下記工程:
(a)骨芽細胞または骨髄由来の単球/マクロファージ系列細胞と被験物質とを接触させる工程、
(b)前記被験物質を接触させた骨芽細胞または骨髄由来の単球/マクロファージ系列細胞におけるPDK4の発現量を調べ、被験物質を接触させない骨芽細胞または骨髄由来の単球/マクロファージ系列細胞における発現量と比較する工程、
(c)下記工程(c1)または(c2)のいずれかにより破骨細胞への分化を誘導する工程:
(c1)前記比較工程(b)の結果に基づいて、骨芽細胞でPDK4の発現を抑制する被験物質存在下で、骨芽細胞と骨髄由来の単球/マクロファージ系列細胞とを共培養し、破骨細胞への分化を誘導する工程、さらに対照として、被験物質非存在下で、骨芽細胞と骨髄由来の単球/マクロファージ系列細胞とを共培養し、同様に破骨細胞への分化を誘導する工程、または
(c2)前記比較工程(b)の結果に基づいて、単球/マクロファージ系列細胞でPDK4の発現を抑制する被験物質存在下で、単球/マクロファージ系列細胞を、M-CSFおよびRANKL存在下で培養し、破骨細胞への分化を誘導する工程、さらに対照として、被験物質非存在下で単球/マクロファージ系列細胞をM-CSFおよびRANKL存在下で培養し、同様に破骨細胞への分化を誘導する工程、
(d)前記工程(c)における破骨細胞への分化の程度を比較する工程、および
(e)前記比較工程(d)の結果に基づいて、骨髄由来の単球/マクロファージ系列細胞から破骨細胞への分化を抑制する被験物質を選択する工程
を含む、PDK4の発現または機能抑制を介した骨粗鬆症の予防および/または治療剤のスクリーニング方法。
【請求項6】
下記工程:
(a’)被験物質存在下で、骨芽細胞と骨髄由来の単球/マクロファージ系列細胞を共培養し、破骨細胞への分化を誘導する工程、
(b’)前記工程(a’)における破骨細胞への分化の程度を比較する工程、および
(c’)前記比較工程(b’)の結果に基づいて、骨髄由来の単球/マクロファージ系列細胞から破骨細胞への分化を抑制する被験物質を選択する工程
を含む、PDK4の発現または機能抑制を介した骨粗鬆症の予防および/または治療剤のスクリーニング方法。
【請求項7】
下記工程:
(a”)被験物質、M-CSFおよびRANKL存在下で、骨髄由来の単球/マクロファージ系列細胞から破骨細胞への分化を誘導する工程、
(b”)前記工程(a”)における破骨細胞への分化の程度を比較する工程、および
(c”)前記比較工程(b”)の結果に基づいて、骨髄由来の単球/マクロファージ系列細胞から破骨細胞への分化を抑制する被験物質を選択する工程
を含む、PDK4の発現または機能抑制を介した骨粗鬆症の予防および/または治療剤のスクリーニング方法。
【請求項8】
Pdk4遺伝子を欠損してなる、非ヒト非荷重抵抗性骨モデル動物。
【請求項9】
非荷重状態での骨量の減少の病態の解明に用いられる、請求項8に記載の骨モデル動物。
【請求項10】
病態が廃用性骨粗鬆症、ステロイド性骨粗鬆症および無重力時の骨量減少からなる群より選ばれる、請求項9に記載の骨モデル動物。
【請求項11】
前記動物がマウスである請求項8〜10のいずれか1項に記載の骨モデル動物。
【請求項12】
請求項8〜11のいずれか1項に記載の骨モデル動物の子孫動物。
【請求項13】
請求項8〜12のいずれか1項に記載の非ヒト動物から得られる組織または細胞。
【請求項14】
請求項8〜12のいずれか1項に記載の骨モデル動物を用いることを特徴とする、骨粗鬆症の予防または治療剤のスクリーニング方法。
【請求項15】
下記工程:
(a)請求項8〜12のいずれか1項に記載の骨モデル動物の野生型対照に被験物質を投与し、当該骨モデル動物と野生型対照とを非荷重状態に供する工程、
(b)前記被験物質の投与前後の野生型対照における骨の形態、骨体積または骨密度を解析し、請求項8〜12のいずれか1項に記載の骨モデル動物における骨の形態、骨体積または骨密度と比較する工程、および、
(c)前記(b)の比較結果に基づいて、非荷重状態における骨量の減少を抑制する被験物質を選択する工程
を含む、骨粗鬆症の予防または治療剤のスクリーニング方法。
【請求項16】
下記工程:
(a)非荷重状態後に得られた請求項13に記載の組織または細胞、および非荷重状態後に野生型対照から得られた組織または細胞をインビトロで生理学的条件下で維持する工程、
(b)前記野生型対照由来の組織または細胞におけるタンパク質のリン酸化の程度と、請求項13由来の組織または細胞におけるタンパク質のリン酸化の程度とを比較する工程、または前記野生型対照由来の組織または細胞におけるタンパク質でPDK4に結合するタンパク質と請求項13由来の組織または細胞におけるタンパク質でPDK4に結合するタンパク質を比較する工程、および
(c)前記比較結果に基づいて、リン酸化の程度が有意に変動しているタンパク質を選択する工程、または前記比較結果に基づいて、前記野生型対照由来の組織または細胞におけるタンパク質のみから得られ、請求項13由来の組織または細胞におけるタンパク質からは得られないPDK4結合タンパク質を選択する工程
を含む、荷重刺激を感知し、骨粗鬆症の発症に関連するPDK4の標的タンパク質のスクリーニング方法。
【請求項1】
PDK4の発現または機能を抑制する物質を含む、骨粗鬆症の予防または治療剤。
【請求項2】
PDK4の発現または機能を抑制する物質が
(1)Pdk4遺伝子のRNAi誘導性核酸、アンチセンス核酸もしくはそれらの発現ベクター、または
(2)PDK4に対する抗体、ドミナントネガティブ変異体、アプタマーもしくはそれらの発現ベクター
である、請求項1に記載の予防または治療剤。
【請求項3】
骨粗鬆症が廃用性骨粗鬆症、ステロイド性骨粗鬆症および無重力時の骨量減少からなる群より選ばれる請求項1または2に記載の予防または治療剤。
【請求項4】
下記工程:
(a)骨芽細胞または骨髄由来の単球/マクロファージ系列細胞と被験物質とを接触させる工程、
(b)前記被験物質を接触させた骨芽細胞または骨髄由来の単球/マクロファージ系列細胞におけるPDK4の発現量を調べ、被験物質を接触させない骨芽細胞または骨髄由来の単球/マクロファージ系列細胞における発現量と比較する工程、および
(c)前記比較結果に基づいて、PDK4の発現を抑制する被験物質を選択する工程
を含む、PDK4の発現または機能抑制を介した骨粗鬆症の予防および/または治療剤のスクリーニング方法。
【請求項5】
下記工程:
(a)骨芽細胞または骨髄由来の単球/マクロファージ系列細胞と被験物質とを接触させる工程、
(b)前記被験物質を接触させた骨芽細胞または骨髄由来の単球/マクロファージ系列細胞におけるPDK4の発現量を調べ、被験物質を接触させない骨芽細胞または骨髄由来の単球/マクロファージ系列細胞における発現量と比較する工程、
(c)下記工程(c1)または(c2)のいずれかにより破骨細胞への分化を誘導する工程:
(c1)前記比較工程(b)の結果に基づいて、骨芽細胞でPDK4の発現を抑制する被験物質存在下で、骨芽細胞と骨髄由来の単球/マクロファージ系列細胞とを共培養し、破骨細胞への分化を誘導する工程、さらに対照として、被験物質非存在下で、骨芽細胞と骨髄由来の単球/マクロファージ系列細胞とを共培養し、同様に破骨細胞への分化を誘導する工程、または
(c2)前記比較工程(b)の結果に基づいて、単球/マクロファージ系列細胞でPDK4の発現を抑制する被験物質存在下で、単球/マクロファージ系列細胞を、M-CSFおよびRANKL存在下で培養し、破骨細胞への分化を誘導する工程、さらに対照として、被験物質非存在下で単球/マクロファージ系列細胞をM-CSFおよびRANKL存在下で培養し、同様に破骨細胞への分化を誘導する工程、
(d)前記工程(c)における破骨細胞への分化の程度を比較する工程、および
(e)前記比較工程(d)の結果に基づいて、骨髄由来の単球/マクロファージ系列細胞から破骨細胞への分化を抑制する被験物質を選択する工程
を含む、PDK4の発現または機能抑制を介した骨粗鬆症の予防および/または治療剤のスクリーニング方法。
【請求項6】
下記工程:
(a’)被験物質存在下で、骨芽細胞と骨髄由来の単球/マクロファージ系列細胞を共培養し、破骨細胞への分化を誘導する工程、
(b’)前記工程(a’)における破骨細胞への分化の程度を比較する工程、および
(c’)前記比較工程(b’)の結果に基づいて、骨髄由来の単球/マクロファージ系列細胞から破骨細胞への分化を抑制する被験物質を選択する工程
を含む、PDK4の発現または機能抑制を介した骨粗鬆症の予防および/または治療剤のスクリーニング方法。
【請求項7】
下記工程:
(a”)被験物質、M-CSFおよびRANKL存在下で、骨髄由来の単球/マクロファージ系列細胞から破骨細胞への分化を誘導する工程、
(b”)前記工程(a”)における破骨細胞への分化の程度を比較する工程、および
(c”)前記比較工程(b”)の結果に基づいて、骨髄由来の単球/マクロファージ系列細胞から破骨細胞への分化を抑制する被験物質を選択する工程
を含む、PDK4の発現または機能抑制を介した骨粗鬆症の予防および/または治療剤のスクリーニング方法。
【請求項8】
Pdk4遺伝子を欠損してなる、非ヒト非荷重抵抗性骨モデル動物。
【請求項9】
非荷重状態での骨量の減少の病態の解明に用いられる、請求項8に記載の骨モデル動物。
【請求項10】
病態が廃用性骨粗鬆症、ステロイド性骨粗鬆症および無重力時の骨量減少からなる群より選ばれる、請求項9に記載の骨モデル動物。
【請求項11】
前記動物がマウスである請求項8〜10のいずれか1項に記載の骨モデル動物。
【請求項12】
請求項8〜11のいずれか1項に記載の骨モデル動物の子孫動物。
【請求項13】
請求項8〜12のいずれか1項に記載の非ヒト動物から得られる組織または細胞。
【請求項14】
請求項8〜12のいずれか1項に記載の骨モデル動物を用いることを特徴とする、骨粗鬆症の予防または治療剤のスクリーニング方法。
【請求項15】
下記工程:
(a)請求項8〜12のいずれか1項に記載の骨モデル動物の野生型対照に被験物質を投与し、当該骨モデル動物と野生型対照とを非荷重状態に供する工程、
(b)前記被験物質の投与前後の野生型対照における骨の形態、骨体積または骨密度を解析し、請求項8〜12のいずれか1項に記載の骨モデル動物における骨の形態、骨体積または骨密度と比較する工程、および、
(c)前記(b)の比較結果に基づいて、非荷重状態における骨量の減少を抑制する被験物質を選択する工程
を含む、骨粗鬆症の予防または治療剤のスクリーニング方法。
【請求項16】
下記工程:
(a)非荷重状態後に得られた請求項13に記載の組織または細胞、および非荷重状態後に野生型対照から得られた組織または細胞をインビトロで生理学的条件下で維持する工程、
(b)前記野生型対照由来の組織または細胞におけるタンパク質のリン酸化の程度と、請求項13由来の組織または細胞におけるタンパク質のリン酸化の程度とを比較する工程、または前記野生型対照由来の組織または細胞におけるタンパク質でPDK4に結合するタンパク質と請求項13由来の組織または細胞におけるタンパク質でPDK4に結合するタンパク質を比較する工程、および
(c)前記比較結果に基づいて、リン酸化の程度が有意に変動しているタンパク質を選択する工程、または前記比較結果に基づいて、前記野生型対照由来の組織または細胞におけるタンパク質のみから得られ、請求項13由来の組織または細胞におけるタンパク質からは得られないPDK4結合タンパク質を選択する工程
を含む、荷重刺激を感知し、骨粗鬆症の発症に関連するPDK4の標的タンパク質のスクリーニング方法。
【図1】


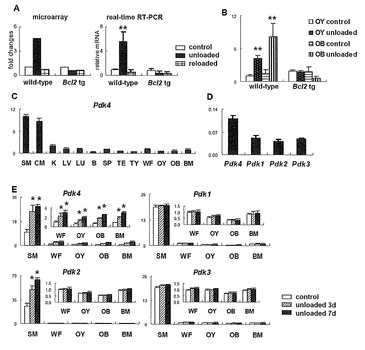
【図10】
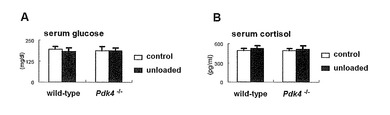
【図2】
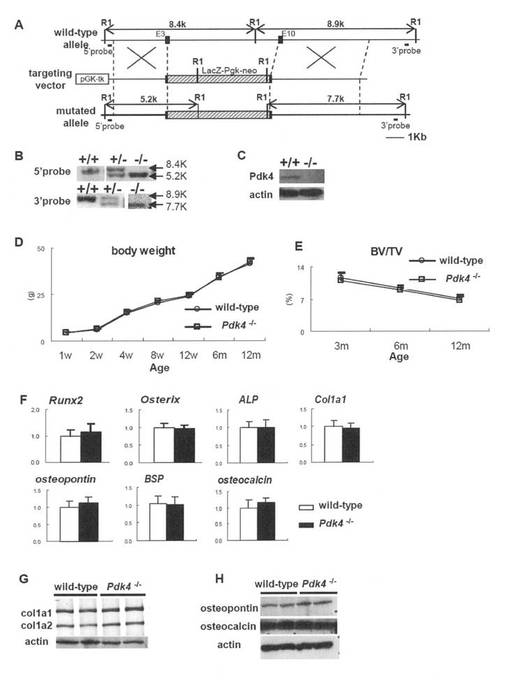
【図3】
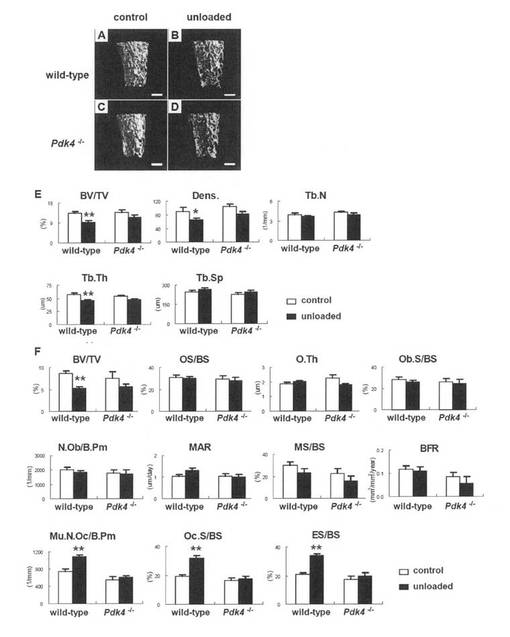
【図4】
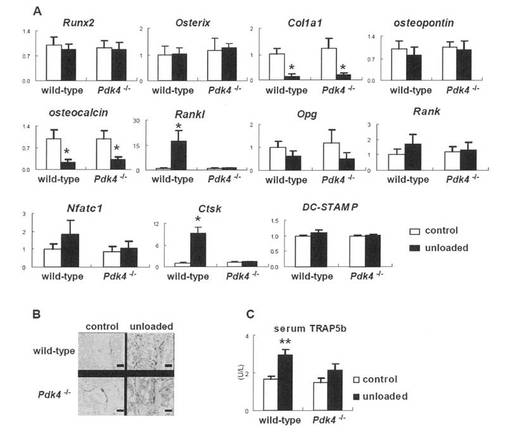
【図5】
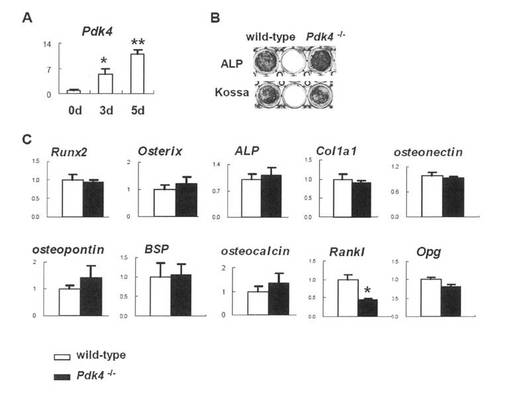
【図6】
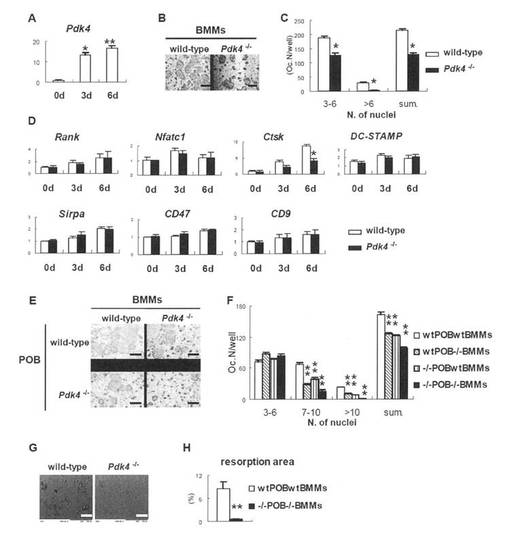
【図7】
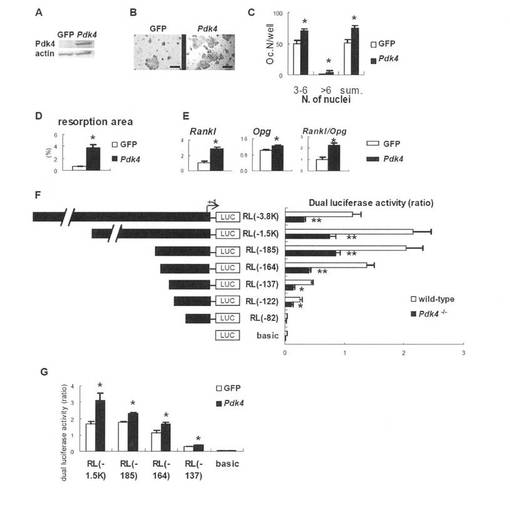
【図8】
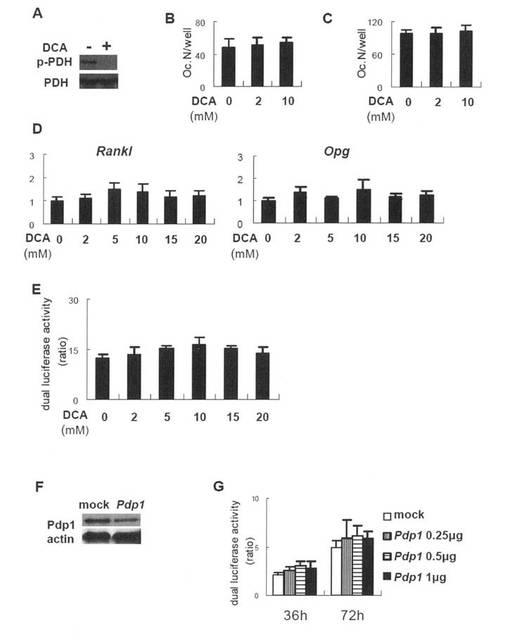
【図9】
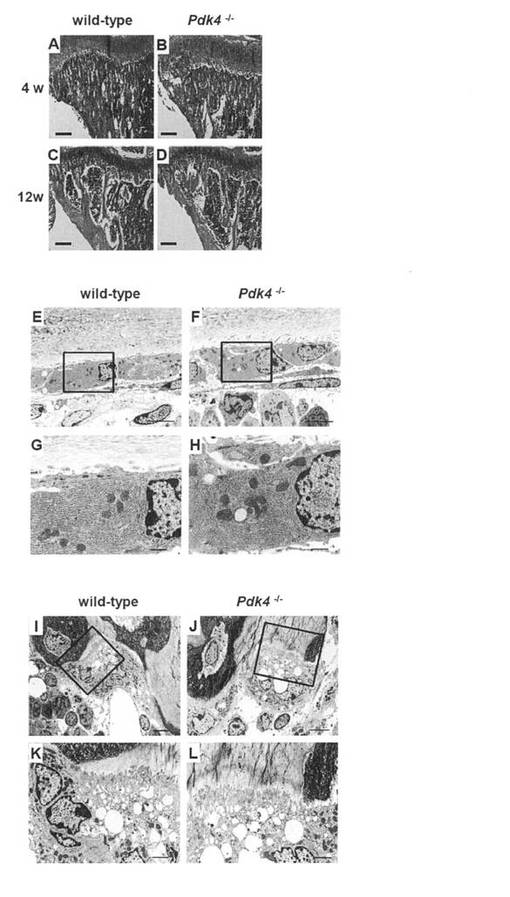
【図10】
【図2】
【図3】
【図4】
【図5】
【図6】
【図7】
【図8】
【図9】
【公開番号】特開2013−6783(P2013−6783A)
【公開日】平成25年1月10日(2013.1.10)
【国際特許分類】
【出願番号】特願2011−138935(P2011−138935)
【出願日】平成23年6月22日(2011.6.22)
【出願人】(504205521)国立大学法人 長崎大学 (226)
【Fターム(参考)】
【公開日】平成25年1月10日(2013.1.10)
【国際特許分類】
【出願日】平成23年6月22日(2011.6.22)
【出願人】(504205521)国立大学法人 長崎大学 (226)
【Fターム(参考)】
[ Back to top ]