
薬剤耐性遺伝子を欠失又は不活性化させた微生物
【課題】薬剤耐性を示すロドコッカス属細菌について、より効率的且つ汎用的に薬剤耐性遺伝子が欠失又は不活性化された当該薬剤感受性株(薬剤感受性ロドコッカス属細菌)を提供する。
【解決手段】本発明は、薬剤耐性を示すロドコッカス属細菌における薬剤耐性遺伝子が欠失又は不活性化された、薬剤感受性微生物である。当該欠失又は不活性化は、例えば、ドナー微生物から薬剤耐性を有するレシピエント微生物への接合伝達を利用した、所定の工程(a)〜(d)を含む方法により行われ得る。
【解決手段】本発明は、薬剤耐性を示すロドコッカス属細菌における薬剤耐性遺伝子が欠失又は不活性化された、薬剤感受性微生物である。当該欠失又は不活性化は、例えば、ドナー微生物から薬剤耐性を有するレシピエント微生物への接合伝達を利用した、所定の工程(a)〜(d)を含む方法により行われ得る。
【発明の詳細な説明】
【技術分野】
【0001】
本発明は、ゲノム上の薬剤耐性遺伝子を欠失又は不活性化させることにより薬剤感受性にした微生物に関する。詳しくは、ロドコッカス属細菌のゲノム上の薬剤耐性遺伝子を欠失又は不活性化させた薬剤感受性微生物(薬剤感受性ロドコッカス属細菌)に関する。
【背景技術】
【0002】
ロドコッカス属細菌は、その物理的強度、有機溶媒耐性、酸化還元能、酵素などを細胞内に多量蓄積する能力などを有することから、産業的に有用な微生物触媒として知られている。例えば、ロドコッカス属細菌は、ニトリル類の酵素的水和又は加水分解によるアミドもしくは酸の生産などに利用されている(特許文献1、2)。また、難分解性化合物に対する分解能の高い微生物種としても知られており(特許文献3〜6)、その能力を活用することで、種々の有用物質生産および環境浄化への応用検討が進められている。例えば、脱硫による石油からの有用物質の生産、あるいは環境中への石油流出時におけるバイオレメディエーションなどが挙げられる。
【0003】
加えて、ロドコッカス属細菌を遺伝子組換えの方法により、さらに有用なものに改変する試みがなされている(特許文献7〜9)。例えば、ロドコッカス属細菌の遺伝子操作を効率的に推し進めるために、宿主−ベクター系の開発が進められており、新規なプラスミドの探索(特許文献10〜12)、ベクターの開発(特許文献13〜16、17〜19、非特許文献1)などが行われている。
【先行技術文献】
【特許文献】
【0004】
【特許文献1】特開平2-470号公報
【特許文献2】特開平3-251192号公報
【特許文献3】欧州特許第188316号明細書
【特許文献4】欧州特許第204555号明細書
【特許文献5】欧州特許第348901号明細書
【特許文献6】欧州特許出願第307926号明細書
【特許文献7】特開平4-211379号公報
【特許文献8】特開平6-25296号公報
【特許文献9】特開平6-303791号公報
【特許文献10】特開平4-148685号公報
【特許文献11】特開平4-330287号公報
【特許文献12】特開平7-255484号公報
【特許文献13】特開平5-64589号公報
【特許文献14】特開平8-56669号公報
【特許文献15】米国特許第4,920,054号明細書
【特許文献16】特開平8-173169号公報
【特許文献17】特開平10-248578号公報
【特許文献18】特開2006-180843号公報
【特許文献19】特開2006-50967号公報
【非特許文献】
【0005】
【非特許文献1】Journal of Bacteriology 170, 638-645 (1988)
【発明の概要】
【発明が解決しようとする課題】
【0006】
一般的に、微生物を形質転換する際、形質転換株の選抜は、導入したプラスミドの選択マーカー(例えば、ある薬剤に対する耐性遺伝子等)により行う。しかし、宿主によってはいくつかの薬剤に対して耐性を有しているものもいるため、その微生物種で適したベクター系の有する選択マーカーの機能を宿主自体も持っている場合がある。例えば、ロドコッカス属細菌を形質転換する際、カナマイシン耐性遺伝子を選択マーカーとして有するシャトルベクターがよく使用されるが、ロドコッカス属細菌はカナマイシンに耐性を示す株も多い。上記シャトルベクターを用いてそれらカナマイシン耐性ロドコッカス属細菌を形質転換する場合、高濃度のカナマイシンで形質転換株を選抜せざるを得ず、非常に効率が悪い。
【0007】
上述の場合において、プラスミド由来の選択マーカーにより、効率的な形質転換を可能にするためには、宿主微生物ゲノム上の薬剤耐性遺伝子を欠失又は不活性化し、当該薬剤に対し感受性を示すように宿主を改良すればよい。ゲノム上に存在する遺伝子を欠失又は不活性化する手法としては、相同組換え(Homologous recombination)を利用する方法が挙げられる。これは、標的遺伝子の間に他の遺伝子等を挿入して標的遺伝子を機能発現しない状態にする(不活性化)、又は、標的遺伝子の一部又は全部を含まない配列に置換することにより標的遺伝子自体を欠失させて、標的遺伝子の発現そのものを止めてしまう方法である。大腸菌や酵母などのいくつかの微生物では、効率的な相同組換え技術が確立しているため、比較的容易に標的遺伝子の欠失又は不活性化ができる。
【0008】
ロドコッカス属細菌において相同組換えを行う際の遺伝子導入法として、エレクトロポレーション法等が知られている。エレクトロポレーション法はロドコッカス属細菌における代表的な形質転換法であるが、相同組換えに適用した場合、ロドコッカス属細菌及びその類縁属細菌は非相同組換えが起こり易いという特徴を有しているため、目的の相同組換え株が極めて低確率でしか得られないという問題がある。相同組換えを行う際の他の遺伝子導入法としては、接合伝達を利用する方法(接合伝達法)が挙げられるが、当該方法はロドコッカス属細菌に対しては殆ど適用されていない。
【0009】
従って、ロドコッカス属細菌において、標的遺伝子を効率的且つ汎用的に欠失又は不活性化する方法は確立していないため、選択マーカーとして使用し得る薬剤耐性遺伝子が欠失又は不活性化したロドコッカス属細菌、すなわち、薬剤感受性のロドコッカス属細菌は十分に得られていないのが現状である。
【0010】
このような状況下において、より効率的且つ汎用的に薬剤耐性遺伝子が欠失又は不活性化された、当該薬剤感受性微生物の取得が望まれていた。
【課題を解決するための手段】
【0011】
本発明は、上記状況を考慮してなされたもので、以下に示す薬剤感受性を示すロドコッカス属細菌を提供するものである。
【0012】
すなわち、本発明は以下の通りである。
(1)ロドコッカス属細菌において、標的とする薬剤耐性遺伝子が欠失又は不活性化されたことにより当該薬剤に対して感受性を示す微生物。
【0013】
本発明の微生物としては、例えば、前記欠失又は不活性化が、ドナー微生物からレシピエント微生物への接合伝達を利用した、以下の工程(a)〜(d)を含む方法により行われるものが挙げられる。
【0014】
(a)レシピエント微生物として、ドナー微生物生育阻害マーカーが強化されたロドコッカス属細菌を作製する工程;
(b)ドナー微生物として、(i)レシピエント微生物中の標的の薬剤耐性遺伝子とその周辺の塩基配列とを含む塩基配列において当該耐性遺伝子を欠失又は不活性化させた塩基配列領域、(ii)当該ドナー微生物において機能する接合伝達開始領域、(iii)当該ドナー微生物において機能する複製開始領域、(iv) レシピエント微生物生育阻害マーカー、及び(v)レシピエント微生物に対する条件致死遺伝子を含む、遺伝子改変用プラスミドを用いて形質転換された微生物を作製する工程;
(c)前記(b)で作製されたドナー微生物から前記(a)で作製されたレシピエント微生物への接合伝達を行うことにより、当該レシピエント微生物の形質転換体を作製する工程;並びに
(d)前記(c)で作製された形質転換体を、前記条件致死遺伝子が機能し得る培養条件で培養する工程。
【0015】
本発明の微生物において、前記ロドコッカス属細菌としては、例えば、ロドコッカス・ロドクロウス(Rhodococcus rhodochrous)又はロドコッカス・エリスロポリス(Rhodococcus erythropolis)が挙げられる。
【0016】
本発明の微生物としては、例えば、前記欠失又は不活性化された薬剤耐性遺伝子がカナマイシン耐性遺伝子であるものが挙げられる。
(2)上記(1)に記載の微生物を宿主として用いて培養し、得られる培養物から目的タンパク質を採取することを特徴とする、タンパク質製造方法。
【発明の効果】
【0017】
本発明によれば、薬剤耐性を示すロドコッカス属細菌について、ロドコッカス属細菌ゲノム上の当該薬剤耐性遺伝子を欠失又は不活性化させた当該薬剤感受性微生物(当該薬剤感受性ロドコッカス細菌)を提供することができる。例えば、カナマイシン耐性遺伝子をマーカーとして有するシャトルベクターを用いて形質転換株を作製する場合であれば、本発明の薬剤感受性微生物の一態様としてのカナマイシン感受性株を使用することにより、従来のように高濃度のカナマイシンで組換え株を選抜する必要はなくなり、低濃度のカナマイシン含有培地を用いて、より効率的に形質転換株を得ることができる。また、欠失させた薬剤耐性遺伝子を導入したベクター系を構築し、当該薬剤耐性遺伝子を選択マーカーとして活用することも可能になる。よって、本発明の薬剤感受性微生物を形質転換用宿主として用いることにより、より効率的な物質生産等が行い得るようになるため、本発明は、産業的にみても極めて優れたものである。
【図面の簡単な説明】
【0018】
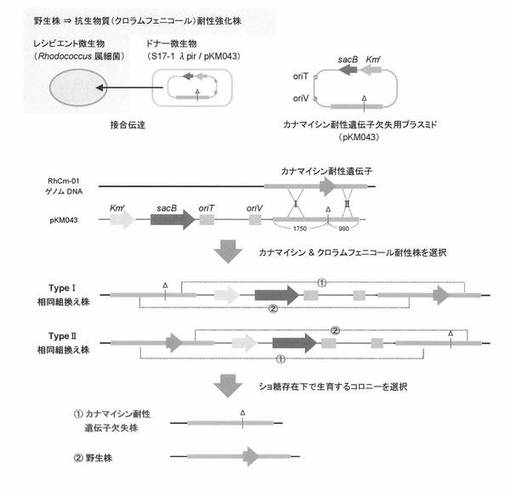
【図1】本発明によりRhodococcus属細菌のカナマイシン耐性遺伝子を欠失させる際の各工程における、当該細菌ゲノム上で起こる相同組換え等について示した概略図である。
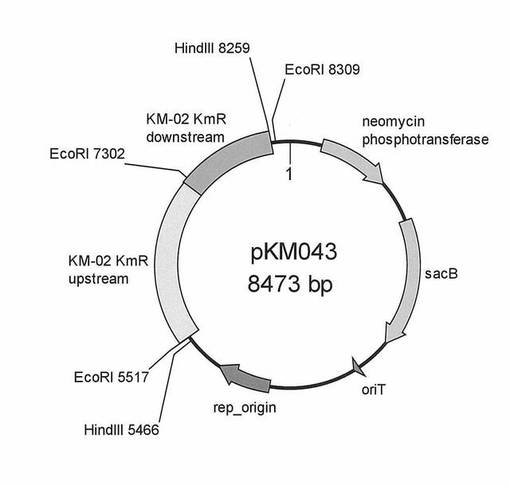
【図2】遺伝子改変用プラスミドpKM043の遺伝子地図を示す図である。
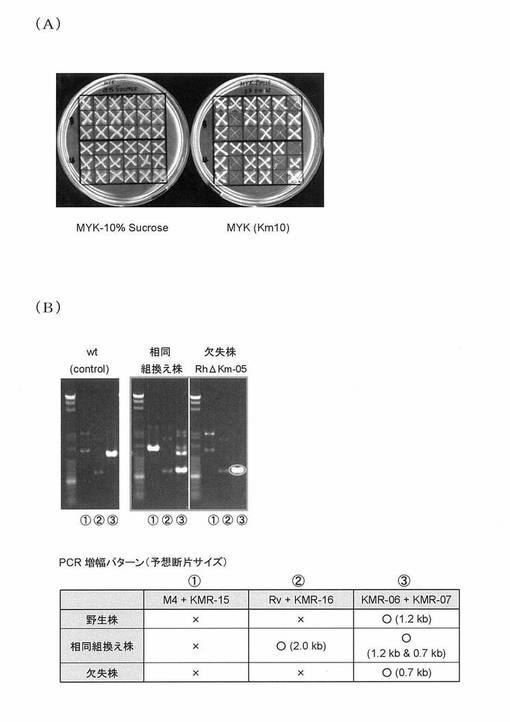
【図3】(A)カナマイシン感受性株RhΔKm-01を単離することに成功した結果を示す図である。(B)RhΔKm-01株においてカナマイシン耐性遺伝子長約0.5 kbに相当する領域がゲノムから欠失していることを確認した結果を示す図である。
【図4】(A)カナマイシン感受性株RhΔKm-10を単離することに成功した結果を示す図である。(B)RhΔKm-10のゲノムからカナマイシン耐性遺伝子が欠失していることを確認した結果を示す図である。
【発明を実施するための形態】
【0019】
以下、本発明を詳細に説明する。本発明の範囲はこれらの説明に拘束されることはなく、以下の例示以外についても、本発明の趣旨を損なわない範囲で適宜変更し実施することができる。
【0020】
なお、本明細書において引用された全ての刊行物、例えば先行技術文献、及び公開公報、特許公報その他の特許文献は、参照として本明細書に組み込まれる。
1.薬剤に対し感受性を示す微生物
本発明に係る薬剤に対し感受性を示す微生物、すなわち、薬剤に対し感受性を示すロドコッカス属細菌(以下、「本発明の微生物」又は「薬剤感受性株」ということがある)は、前述した通りある薬剤に対する耐性を有するロドコッカス属細菌において、当該薬剤に対する耐性を獲得するために必要な遺伝子(以下、「標的とする薬剤耐性遺伝子」)が欠失又は不活性化された変異株である。
【0021】
ここで、薬剤に対し感受性を示すとは、薬剤含有培地において、ロドコッカス属細菌が生育できないことをいう。また、薬剤に耐性を示すとは、薬剤含有培地においてもロドコッカス属細菌が十分に生育できることをいう。
【0022】
ロドコッカス属細菌としては、ロドコッカス・ロドクロウス(Rhodococcus rhodochrous)J1、M8(SU1731814)、M33(VKM Ac-1515D)、ATCC 184、ATCC 999、ATCC 4001、ATCC 4273、ATCC 4276、ATCC 9356、ATCC 12483、ATCC 12674、ATCC 13808、ATCC 14341、ATCC 14347、ATCC 14350、ATCC 15905、ATCC 15998、ATCC 17041、ATCC 17043、ATCC 17895、ATCC 19067、ATCC 19149、ATCC 19150、ATCC 21243、ATCC 21291、ATCC 21785、ATCC 21924、ATCC 21999、ATCC 29670、ATCC 29672、ATCC 29675、ATCC 33258、ATCC 33275、ATCC 39484、IFO 3338、IFO 14894、JCM 3202、NCIMB 11215、NCIMB 11216、ロドコッカス・エリスロポリス(Rhodococcus erythropolis)PR4(NBRC 100887)、NBRC 12320、NBRC 15567、NBRC 16296、IFM 155、DSM 743、DSM 9675、DSM 43200、JCM 6821、JCM 6822、JCM 6823、JCM 6824、JCM 6827、DSM 11397、DSM 44522、JCM 2895、JCM 2893、JCM 2894、IAM 1400、IAM 1503、ロドコッカス・オパカス(Rhodococcus opacus)B4等が挙げられる。
【0023】
標的とする薬剤耐性遺伝子としては、例えば、アミノグリコシド系抗生物質耐性遺伝子、β-ラクタム系抗生物質耐性遺伝子、クロラムフェニコール系抗生物質耐性遺伝子、テトラサイクリン系抗生物質耐性遺伝子等が挙げられる。
【0024】
ここで、アミノグリコシド系抗生物質耐性遺伝子としては、カナマイシン及びネオマイシン耐性遺伝子であるアミノグリコシド3'-ホスホトランスフェラーゼ等のリン酸転移酵素遺伝子群、ストレプトマイシン耐性遺伝子であるアミノグリコシド3'-アデニリルトランスフェラーゼ等のヌクレオチジル転移酵素遺伝子群、アセチル転移酵素遺伝子群が挙げられる。
【0025】
β-ラクタム系抗生物質耐性遺伝子としては、β-ラクタマーゼ遺伝子が挙げられる。
【0026】
クロラムフェニコール系抗生物質耐性遺伝子としては、クロラムフェニコールアセチルトランスフェラーゼ遺伝子が挙げられる。
【0027】
テトラサイクリン系抗生物質耐性遺伝子としては、トランスポゾンTn10由来テトラサイクリン耐性遺伝子が挙げられる。
【0028】
また、ある薬剤に対し耐性を有するロドコッカス属細菌の中には、薬剤の修飾等による無毒化、又は細胞外への排出機能等を担う薬剤耐性遺伝子を有しているわけではなく、薬剤の標的である酵素自体が薬剤に対する耐性(薬剤抵抗性)を持つことにより、結果として薬剤耐性を示す株もある。この場合は、例えば、上記薬剤抵抗性酵素を、近縁種等の有する感受性酵素(非抵抗性酵素)に置換することにより、薬剤感受性株にすることも可能である。この場合も、薬剤耐性が発揮できなくなった、すなわち、薬剤耐性が不活性化したと考えられるため、本発明における薬剤耐性遺伝子の不活性化の範疇に入る。薬剤抵抗性を示す酵素としては、例えば、トリメトプリム耐性ジヒドロキシ葉酸レダクターゼ等が挙げられる。
2.薬剤に対し感受性を示す微生物の製造方法
標的とする薬剤耐性遺伝子を欠失又は不活性化する方法は限定されず、如何なる方法を適用してもよい。例えば、当該遺伝子又はその周辺配列と相同な配列を有するプラスミドを形質転換する方法を用いることができる。形質転換の方法としては、エレクトロポレーション法や接合伝達法が使用できる。
【0029】
上記欠失又は不活性化する方法については、本発明では以下の方法が特に好ましい。
【0030】
すなわち、ドナー微生物からレシピエント微生物への接合伝達を利用した、以下の工程(a)〜(d)を含む方法により、本発明の微生物が得られることが好ましい。なお、当該製造方法は、ドナー微生物からレシピエント微生物への接合伝達を利用した形質転換方法を用いることを含む、標的とする薬剤耐性遺伝子の欠失又は不活性化方法であって、以下の工程(a)〜(d)を含むことを特徴とする方法ということもできる。
(a)レシピエント微生物として、ドナー微生物生育阻害マーカーが強化されたロドコッカス属細菌を作製する工程;
(b)ドナー微生物として、下記(i)〜(v):
(i)レシピエント微生物中の標的とする薬剤耐性遺伝子とその周辺の塩基配列とを含む塩基配列において当該耐性遺伝子を欠失又は不活性化させた塩基配列領域、
(ii)当該ドナー微生物において機能する接合伝達開始領域、
(iii)当該ドナー微生物において機能する複製開始領域、
(iv) レシピエント微生物生育阻害マーカー、及び
(v)レシピエント微生物に対する条件致死遺伝子
を含む、遺伝子改変用プラスミドを用いて形質転換された微生物を作製する工程;
(c)工程(b)で作製されたドナー微生物から工程(a)で作製されたレシピエント微生物への接合伝達を行うことにより、当該レシピエント微生物の形質転換体を作製する工程;並びに
(d)工程(c)で作製された形質転換体を、前記条件致死遺伝子が機能し得る培養条件で培養する工程。
上記欠失又は不活性化方法においてレシピエントとして使用するロドコッカス属細菌は、限定はされないが、前述したカナマイシン耐性を有するものが例示できる。
【0031】
接合伝達を用いた遺伝子欠失又は不活性化を行う場合、ドナーとなる微生物の組換え株(ドナー微生物)とレシピエントとなる微生物(レシピエント微生物)の非組換え株の双方の生育を阻害しなければならないため、2種類の選択マーカー(ドナー微生物生育阻害マーカー及びレシピエント微生物生育阻害マーカー)が必要である。ドナー微生物生育阻害マーカーはレシピエント微生物自体が有している必要がある。レシピエント微生物生育阻害マーカーは、遺伝子改変用プラスミド中に含まれている必要がある。両マーカーの詳細については、下記の各工程において説明する。
2.1. 工程(a)
工程(a)では、ドナー微生物生育阻害マーカーが強化されたロドコッカス属細菌を作製する。
【0032】
上述の理由から、レシピエントとなるロドコッカス属細菌にはドナー微生物の生育を阻害するための薬剤耐性マーカー(ドナー微生物生育阻害マーカー)が必要である。よって、標的とする薬剤耐性遺伝子が関与する薬剤耐性以外の薬剤耐性を有していない、又は薬剤耐性の乏しいロドコッカス属細菌を使用する場合、ドナー微生物生育阻害マーカーとして機能しうる、薬剤選択可能な程度の十分な薬剤耐性を有する変異株の作製が必要となる。
【0033】
ここで、ドナー微生物生育阻害用の薬剤としては、接合伝達時にドナーとして用いる微生物(ドナー微生物)の有する薬剤耐性を考慮し、ドナー微生物が感受性を示す薬剤を選択することが好ましい。当該薬剤としては、クロラムフェニコール、アンピシリン、カナマイシン、トリメトプリム、ゲンタマイシン、ナルジクス酸、カルベニシン、チオストレプトン、テトラサイクリン、ストレプトマイシン等が好ましく、クロラムフェニコール、アンピシリンがより好ましい。
【0034】
ドナー微生物生育阻害マーカーが強化されたレシピエント微生物を作製する方法としては、特に限定はされないが、例えば、(イ)自然変異誘発法、(ロ)紫外線照射や変異誘発剤を用いる突然変異誘発法、(ハ)あらかじめ遺伝子改変用プラスミドとは別の、抗生物質耐性獲得用のプラスミドを導入する方法等が好ましく挙げられ、中でも自然変異誘発法がより好ましい。
【0035】
自然変異誘発法は、所望の薬剤を含有する培地中で対象とする微生物を継代培養等することにより、もともとは当該培地中で生育不可又は困難な微生物に自然変異を誘発させて、より高濃度の薬剤を含有する当該培地中でも生育し得る株を取得する方法である。
【0036】
レシピエント微生物となるロドコッカス属細菌の薬剤耐性をどの程度まで薬剤耐性を強化するかは、使用するロドコッカス属細菌の種類、ドナー微生物及び選択する薬剤により異なるため適宜調整することができるが、レシピエント微生物(ロドコッカス属細菌)の生育が抑制されず、且つ、ドナー微生物の生育が十分阻害される薬剤濃度を基準に強化することが好ましい。例えば、レシピエント微生物としてのロドコッカス属細菌に対して、ドナー微生物として大腸菌を、選択用薬剤としてクロラムフェニコールを用いる場合、自然突然変異により、クロラムフェニコール30〜200 mg/L、好ましくは50〜200 mg/Lを含有する培地において生育可能なロドコッカス属細菌(クロラムフェニコール耐性強化株)を得ることが望ましい。
2.2. 工程(b)
工程(b)では、接合伝達に供するドナー微生物として、所定の遺伝子改変用プラスミドを用いて形質転換された微生物を作製する。遺伝子改変用プラスミド、すなわちレシピエント微生物中の標的とする薬剤耐性遺伝子を欠失又は不活性化するためのプラスミドとしては、前述の(i)〜(v)の構成(遺伝子・塩基配列等)を含むものを用いる。
【0037】
ドナー微生物からレシピエント微生物にプラスミドを伝達するためには、oriTに加え、mob遺伝子及びtra遺伝子(群)が最小限必要である。mobはoriT特異的ニック酵素をコードする遺伝子で、この酵素がoriTに働くことによりドナー微生物からレシピエント微生物への(遺伝子改変プラスミドの)移行が開始される。traは多数の遺伝子群の総称で、性繊毛形成、接合管形成、接合制御に関与する遺伝子群で構成されている。mob遺伝子及びtra遺伝子(群)は必ずしも遺伝子改変プラスミド上になくてもよく、別のプラスミド(ヘルパープラスミド)、もしくはドナーとなる微生物ゲノム上に組み込まれていてもよい(蛋白質 核酸 酵素, vol. 38, p. 60-68 (1993))。
【0038】
本発明の製造方法において、ドナー微生物としては、上記レシピエント微生物と接合伝達可能な微生物であればよく、限定されないが、上記の接合伝達に最低限必要な構成要件を満たしている必要がある。従って、使用する遺伝子改変用プラスミド上にoriT 、mob遺伝子及びtra遺伝子(群)が一通り備わっている場合は、それぞれが機能しうる微生物であれば特に限定されないが、使用する遺伝子改変用プラスミド上にoriTのみが備わっている場合は、oriTに対応するmob遺伝子及びtra遺伝子(群)を保有する微生物を用いる必要がある。ドナーとなる微生物と遺伝子改変用プラスミドの組み合わせとしては、例えば、大腸菌S17-1とpK19mobの組み合わせが好ましい。大腸菌S17-1のゲノム上には、プラスミドRP4由来のtra遺伝子(群)が、pK19mobにはRP4由来のoriT 、mob遺伝子がそれぞれ備わっているため、ドナー微生物として使用可能である。
【0039】
ここで、前記(i)の配列は、(改変の対象とする)ロドコッカス属細菌中の標的とする薬剤耐性遺伝子と当該遺伝子の周辺の塩基配列とを含む塩基配列において、当該遺伝子を欠失又は不活性化させた塩基配列領域である。
【0040】
レシピエント微生物のゲノムから、標的とする薬剤耐性遺伝子と当該遺伝子の周辺の塩基配列とを含む塩基配列の単離(クローニング)は、遺伝子ライブラリー作製やPCR等の公知技術を用いて行うことができる。
【0041】
なお、標的とする薬剤耐性遺伝子の周辺の塩基配列としては限定されないが、当該ロドコッカス属細菌ゲノムとの相同領域(標的遺伝子周辺の塩基配列)が長いほどより効率よく相同組換えを起こすことが出来るため、クローニングに支障が出ない範囲でより長い配列を用いることが好ましい。
【0042】
例えば、当該遺伝子の上流及び下流の相同領域がそれぞれ100〜3000 bpの塩基配列を含む配列であることが好ましく、より好ましくは500〜2000 bpの塩基配列を含む配列である。単離した塩基配列を用いて、前述(i)を作製する方法は特に限定されず、PCR法や制限酵素を用いた標的遺伝子部分の切除又は置換等の公知技術を用いて行うことができる。
【0043】
欠失された遺伝子としては、ロドコッカス属細菌のゲノムにコードされる標的とする薬剤耐性遺伝子の一部又は全部が欠失されたもの、プロモーター配列の一部又は全部が欠失されたもの、当該遺伝子の発現調節に関連する遺伝子が存在する場合は、その一部又は全部が欠失されたものなどが挙げられ、これらの欠失された領域は単独でもよいし複数を組み合わせたものでもよい。
【0044】
不活性化された遺伝子としては、ロドコッカス属細菌のゲノムにコードされる標的とする薬剤耐性遺伝子若しくは当該遺伝子の発現調節に関連する遺伝子の内部に、外来遺伝子、例えは薬剤耐性遺伝子等を導入し、本来の機能を発現しない遺伝子に改変されたもの、又は薬剤抵抗性酵素の遺伝子を薬剤非抵抗性酵素の遺伝子に置換することにより薬剤非抵抗酵素が発現するようにしたもの等が挙げられる。
【0045】
薬剤非抵抗性酵素としては、菌体内において薬剤抵抗性酵素と同様の機能を果たすが、対象の薬剤に対しては感受性を示す酵素であれば限定されないが、ロドコッカス属細菌もしくはその近縁種由来のものが好ましい。
【0046】
前記(ii)の接合伝達開始領域は、使用するドナー微生物中において接合伝達の開始点となる塩基配列を含む領域であればよく、特に限定されない。例えば、Fプラスミド由来の接合伝達開始領域を含む配列、プラスミドR6K由来のoriT、プラスミドRP4由来のoriTが好ましい。
【0047】
前記(iii)の複製開始領域は、使用するドナー微生物中において前記遺伝子改変用プラスミドの自己複製起点として機能し得る塩基配列を含む領域であればよく、特に限定はされない。例えば、プラスミドpMB1及び広宿主域プラスミドRK2由来の複製開始点並びにその派生物等を含む領域の使用が好ましい。
【0048】
前記(iv)のレシピエント微生物生育阻害マーカーは、接合伝達に供するレシピエント微生物(ロドコッカス属細菌)が感受性を示す薬剤に対する耐性遺伝子であることが好ましい。標的とする薬剤耐性遺伝子と当該生育阻害マーカーが同一の機能を有するものである場合は、標的とする薬剤耐性遺伝子のみでは耐性を獲得できない高濃度薬剤を用いることで、目的の組換え株(相同組換え株)を取得することも可能である。耐性遺伝子としては、例えば、カナマイシン耐性遺伝子、アンピシリン耐性遺伝子、ゲンタマイシン耐性遺伝子、チオストレプトン耐性遺伝子、カルベニシン耐性遺伝子等が好ましく挙げられ、中でもカナマイシン耐性遺伝子がより好ましい。
【0049】
前記(v)の条件致死遺伝子は、レシピエント微生物のゲノム上に導入され、且つある特定条件下にさらされた場合に、当該微生物を死に至らしめる作用を有し得る遺伝子であればよい。例えば、sacB遺伝子が好ましく挙げられる。sacB遺伝子は、当該遺伝子を保有し発現する微生物(例えばロドコッカス属細菌等)をスクロース含有培地で培養した場合に、スクロースを基質とし当該微生物に対して致死作用を有する有害物質を産生する酵素をコードする遺伝子である。
【0050】
接合伝達に用いる前記遺伝子改変用プラスミドは、如何なるベクターをベースとして構築されたものであってもよく、その種類は特に限定はされない。使用するベクターとしては、例えば、pBR322、pSC101、pACYC184、pACYC177、pTrc99A、pUC18、pUC19、pUC118、pUC119、pHSG298、pHSG299、pSP64、pSP65、pGEM-3、pGEM-3Z、pGEM-3Zf(-)、pGEM-4p、GEM-4Z、Bluescript M13+、Bluescript M13-、pK19mob、pK18mob、pK18mobsacB等が挙げられる。中でも、プラスミド内部に既にRP4由来のoriT及びmobを有しているpK19mob、pK18mob、pK18mobsacB(Schaefer等, Gene, vol. 45, p. 69-73(1994))を用いることがより好ましい。
前記遺伝子改変用プラスミドにおける(i)〜(v)の構成は、その配置については特に限定されず、如何なる順番で配置されていてもよい。前記(i)〜(v)の構成を含む遺伝子改変用プラスミドの構築は、公知の遺伝子組換え技術を用いて実施することができる。
【0051】
工程(b)では、上述したような各構成を有する遺伝子改変用プラスミドをドナーとして用いる微生物内に導入して形質転換された微生物、すなわち接合伝達に供するドナー微生物を作製する。その際、形質転換の方法としては、エレクトロポレーション法や塩化カルシウム法等の、微生物の形質転換方法として公知の方法を用いることができる。
2.3. 工程(c)
工程(c)では、工程(b)で得られたドナー微生物から工程(a)で得られたレシピエント微生物への接合伝達を行う。通常は、ドナー微生物及びレシピエント微生物のそれぞれの菌体懸濁液を混合し、適当なプレート培地(LB培地等)上に均一に広げて、両微生物の接合を行わせる。当該接合においては、ドナー微生物中の遺伝子改変用プラスミドがレシピエント微生物内に移動し、レシピエント微生物のゲノムと上記プラスミドとの相同配列で相同組換えが起こり、遺伝子改変用プラスミドの一部又は大部分が当該レシピエント微生物のゲノム上に導入される。この接合により、レシピエント微生物の形質転換体、すなわち相同組換え株が作製される。
【0052】
本工程(c)における所望の相同組換え株は、1重交差により標的遺伝子の上流又は下流に当該遺伝子改変用プラスミドが導入されたものである。所望の相同組換え株であるかどうかの確認は、レシピエント微生物自体の薬剤耐性、及び前記遺伝子改変用プラスミド由来の薬剤耐性を利用して行うことができる。具体的には、両薬剤を含む培地(例えば、カナマイシン及びクロラムフェニコール含有培地)において上記接合後の微生物を培養することにより、所望の相同組換え株を選択することができる。
2.4. 工程(d)
工程(d)では、工程(c)で作製されたレシピエント微生物の相同組換え株(相同組換え微生物)を、前記遺伝子改変用プラスミド由来の条件致死遺伝子が機能し得る培養条件で培養する。条件致死遺伝子が機能し得る培養条件としては、条件致死遺伝子が機能する限り、特に限定はされない。例えば、条件致死遺伝子がsacB遺伝子の場合は、スクロース含有培地を用いた培養が用いられる。
【0053】
当該培養においては、上記条件致死遺伝子を有する相同組換え微生物は生育困難であるため、当該微生物のゲノム上から2度目の相同組換えにより上記条件致死遺伝子、薬剤耐性遺伝子、接合伝達開始領域、複製開始領域を含む遺伝子改変用プラスミド由来の塩基配列領域が除かれた(脱落した)ロドコッカス属細菌を得ることができる。
【0054】
ただし、当該培養により得られた微生物の中には、レシピエント微生物中の標的の薬剤耐性遺伝子が、当初の目的通り欠失又は不活性化しているものと、そうでないもの(上記脱落の際の相同組換えにより元の薬剤耐性遺伝子の機能が復活したもの)が含まれている。よって、通常は、さらに別の培養条件でも培養し、所望の形質転換微生物を選択することができる。
【0055】
別の培養条件としては、上記薬剤遺伝子の機能を確認し得る培養条件での培養が好ましく、より詳細には、当該薬剤を含有する培地で培養したときに生育が不可又は著しく困難な株が、所望のロドコッカス属細菌、すなわち本発明の微生物(当該薬剤感受性株)として選択することができる。また、レシピエント微生物からゲノムを抽出し、当該改変領域の配列を解析することにより、所望の微生物を選択することもできる。
【0056】
上述した工程(a)〜(d)を含む方法本発明により得られた薬物感受性微生物(詳しくは、薬物感受性ロドコッカス属細菌)は、上記の通り、ゲノム上に遺伝子改変用プラスミドの痕跡が残らないため、本発明の工程(b)〜(d)を繰り返し行うことにより、同一の株を複数回改変することが可能である。
3.タンパク質の製造方法
本発明においては、前述したロドコッカス属細菌の薬剤感受性株を宿主として用いて培養し、得られる培養物から目的タンパク質を採取することを特徴とする、タンパク質の製造方法を提供することができる。宿主として用いる当該感受性株は、あるタンパク質を高発現するプラスミド、例えば、アクリルアミド製造用触媒であるニトリルヒドラターゼ等を発現し得る発現ベクターが別途導入された形質転換体として使用される。
【0057】
上記タンパク質の製造方法において、微生物を培養する方法は、通常の方法に従って行うことができる。
【0058】
微生物を培養する培地としては、微生物が資化し得る炭素源、窒素源、無機塩類等を含有し、微生物の培養を効率的に行うことができる培地であれば、天然培地、合成培地のいずれを用いてもよい。炭素源としては、グルコース、フラクトース、スクロース、デンプン等の炭水化物、酢酸、プロピオン酸等の有機酸、エタノール、プロパノール等のアルコール類、ヘキサン、デカン、ドデカン、テトラデカン等の炭化水素類が挙げられる。窒素源としては、アンモニア、塩化アンモニウム、硫酸アンモニウム、酢酸アンモニウム、リン酸アンモニウム等の無機酸若しくは有機酸のアンモニウム塩又はその他の含窒素化合物のほか、ペプトン、肉エキス、コーンスティープリカー等が挙げられる。無機物としては、リン酸第一カリウム、リン酸第二カリウム、リン酸マグネシウム、硫酸マグネシウム、塩化ナトリウム、硫酸第一鉄、硫酸マンガン、硫酸銅若しくは炭酸カルシウム等が挙げられる。
【0059】
培養条件は、限定はされないが、例えば、10℃〜45℃、好ましくは10℃〜40℃の温度下で、5〜120時間、好ましくは5〜100時間程度行うことができる。また、場合により、本培養に先立ち、少量の前培養を行うこともできる。
【0060】
例えば、ロドコッカス属細菌の形質転換体を宿主として用いて培養する場合は、振盪培養又は通気攪拌培養などの好気的条件下、4〜40℃、好ましくは20〜35℃で行うことが好ましい。培養は、無機酸又は有機酸、アルカリ溶液等を用いて適時pH調整を行うことが好ましい。
【0061】
また、培養中に当該形質転換体から発現ベクターの脱落を防ぐために、必要に応じて薬剤選択マーカーによる選択圧をかけた状態で培養してもよい。例えば、カナマイシン耐性遺伝子を含むベクターで形質転換した形質転換体を培養する場合、培養中に必要に応じてカナマイシンを培地に添加してもよい。
【0062】
上記のように、宿主となる微生物の性質に合った適当な培養条件で培養すると、高収率で異種タンパク質(目的タンパク質)を上記培養物中、すなわち、培養上清、培養菌体、又は菌体の破砕物の少なくともいずれかに蓄積することができる。これら、異種タンパク質を含有する溶液を「培養物」と称する。
【0063】
培養後、目的タンパク質が菌体内に生産される場合には、菌体のまま目的タンパク質を用いることもできるし、あるいは菌体を破砕することにより、目的タンパク質を採取することもできる。いずれの場合にも、必要であれば、遠心分離や膜ろ過などの固液分離操作により、培地除去及び洗浄を行うことができる。遠心分離は、菌体を沈降させる遠心力が供給できるものであれば特に限定されることはなく、円筒型や分離板型などを利用することができる。遠心力としては、例えば、500G〜20,000G程度で行うことができる。また、膜ろ過は、目的とする固液分離を達成できれば、精密ろ過(MF)膜、限外ろ過(UF)膜いずれでもよいが、通常、精密ろ過(MF)膜を用いることが好ましい。
【0064】
なお、菌体を破砕することにより、目的タンパク質を採取することもできる。菌体の破砕方法としては、超音波処理、フレンチプレスやホモジナイザーによる高圧処理、ビーズミルによる磨砕処理、衝撃破砕装置による衝突処理、リゾチーム、セルラーゼ、ペクチナーゼ等を用いる酵素処理、凍結融解処理、低張液処理、ファージによる溶菌誘導処理等が挙げられ、いずれかの方法を単独又は必要に応じ組み合わせて利用することができる。
【0065】
一方、目的タンパク質が菌体外に生産される場合には、培養液をそのまま使用するか、上述したような遠心分離やろ過等により菌体を除去する。その後、必要に応じて硫安沈澱による抽出等により前記培養物中から目的タンパク質を採取し、さらに必要に応じて透析、各種クロマトグラフィー(ゲルろ過、イオン交換クロマトグラフィー、アフィニティークロマトグラフィー等)を単独又は適宜組み合わせて用いることにより、精製することもできる。
以下に、実施例を挙げて本発明をより具体的に説明するが、本発明の技術的範囲はこれらに限定されるものではない。
【実施例1】
【0066】
カナマイシン耐性遺伝子欠失用プラスミドpKM043の構築(遺伝子改変用プラスミドの製造)
pDNR-1r(Clontech Laboratories社製)中のsacB遺伝子を、NspV切断サ
イトを付加したプライマーSAC-01(配列番号1)及びSAC-02(配列番号2)を使用したPCRにより増幅し、約1.9 kbのsacB遺伝子断片を得た。増幅条件は以下の通りである。
【0067】
プライマー:
SAC-01: 5'- GGTTCGAATACCTGCCGTTCACTATTATTTAGTG -3' (配列番号1)
SAC-02: 5'- GGTTCGAATCGGCATTTTCTTTTGCGTTTTTATTTG -3'(配列番号2)
反応液組成:
滅菌水 22 μl
2×PrimeSTAR(タカラバイオ社製) 25 μl
SAC-01(配列番号1) 1 μl
SAC-02(配列番号2) 1 μl
pDNR-1r(clontech社製)(100倍希釈) 1 μl
総量 50 μl
温度サイクル:
98℃ 10秒、55℃ 15秒及び72℃ 150秒の反応を30サイクル
sacB遺伝子断片を制限酵素NspV(タカラバイオ社製)で消化後、pK19mob(ナ
ショナルバイオリソースプロジェクトから分譲可能(ナショナルバイオリソースプロジェクトE. coli strain Cloning Vectorコレクション))のNspVサイトに接続し、sacB遺
伝子がカナマイシン耐性遺伝子の下流且つ同方向に導入されたプラスミドpK19mobsacB1を構築した。
【0068】
ロドコッカス・ロドクロウス(Rhodococcus rhodochrous)KM-02(産業技術総合研究所特許生物寄託センター;受託番号:FERM P-14457)を100 mlのMY培地(0.5% polypeptone、0.3% bact yeast extract、0.3% malt extract)に植菌し、30℃にて振盪培養したものを、Saito and Miura の方法(Biochim. Biophys. Acta 72, 619 (1963))によりゲノムを分離した。KM-02のゲノムを制限酵素EcoRIにより不完全
消化したものをpTrc99マルチクローニングサイト内に挿入し、プラスミドライブラリーを製造した。プラスミドライブラリーにより大腸菌JM109を形質転換し、カナマイシン硫酸塩(40μg/ml)とIPTG(イソプロピル−β−チオガラクトシド)(1mM)を含むLBプレート(1% Bact tryptone、0.5% bact yeast extract、1% NaCl、1.5%寒天)を用いてカナマイシン耐性を示す形質転換株を選択した。本形質転換株からプラスミドを抽出し、pKM031と命名した。
【0069】
pKM031を制限酵素HindIII(タカラバイオ社製)で切断し、KM-02株由来のカナマ
イシン耐性遺伝子とその周辺配列を含む約3.3 kbの断片をpUC18のHindIIIサイトに
挿入したプラスミドpKM041を構築した。
【0070】
pKM041内部のカナマイシン耐性遺伝子全長(約0.5 kb)を欠失させ、プラスミドpKM042を得た。欠失は、欠失させたい領域の両側に10数塩基ずつアニールするように設計したプライマーKMD-01(配列番号3)及びKMD-02(配列番号4)を使用したPCRにより行った。PCRの増幅条件は以下の通りである。
プライマー:
KMD-01: 5'-GACCATTGTTCCAGGTCGACTGGGACGAGT-3'(配列番号3)
KMD-02: 5'-ACTCGTCCCAGTCGACCTGGAACAATGGTC-3'(配列番号4)
反応液組成:
滅菌水 38.5 μl
10×PfuTurbo buffer 5 μl
dNTP(25 mM each) 3 μl
KMD-01(配列番号3) 1 μl
KMD-02(配列番号4) 1 μl
pKM041 1.5 μl
PfuTurbo(STRATAGENE社製) 1 μl
総量 50 μl
温度サイクル:
95℃ 60秒の反応を1サイクル、95℃ 50秒、60℃ 50秒及 び72℃ 16分の反応を30サイクル、72℃ 7分の反応を1サイクル
pKM042を制限酵素HindIIIで切断し、カナマイシン耐性遺伝子周辺配列約2.8 kb(
カナマイシン耐性遺伝子が欠失したもの)をpK19mobsacB1のHindIIIサイトに挿入したプ
ラスミドpKM043を構築した(図2参照)。
【0071】
上記プラスミドpKM043製造手順において、制限酵素により切断したDNA断片及びPCR産物の精製にはGFX PCR DNA band and Gel Band Purification kit(GE Healthcare社製)を、DNA同士の接続にはDNA Ligation Kit <Mighty Mix>(タカラバイオ社製)を、プラスミドの抽出にはQIAprep miniprep kit(QIAGEN社製)を用いた。
【実施例2】
【0072】
自然変異導入による薬剤耐性強化変異株: RhCmSR-01株の製造(レシピエント微生物の製造)
ロドコッカス・ロドクロウス(Rhodococcus rhodochrous)ATCC 12674株を、より高濃度のクロラムフェニコールを含む培地に継代培養することにより自然変異を誘発し、200 mg/Lのクロラムフェニコールに耐性を示す変異株RhCmSR-01株を単離した。継代培養の詳細は下記の通りである。
【0073】
ATCC 12674株を10 mg/Lクロラムフェニコールを含むMYKプレート(0.5% polypeptone、0.3% bact yeast extract、0.3% malt extract、0.2% KH2PO4、0.2% K2HPO4、1.5%寒天)に植菌し、30℃で3日間培養した。生育したコロニーを、10 mg/Lクロラムフェニコールを含むMYK培地(0.5% polypeptone、0.3% bact yeast extract、0.3% malt extract、0.2% KH2PO4、0.2% K2HPO4)に植菌し、30℃、200rpmで2日間培養した。次に、上記培養液を40 mg/Lクロラムフェニコールを含むMYK培地に0.1%植菌し、30℃、200rpmで2日間培養した。
【0074】
クロラムフェニコール濃度を次第に高めながら継代培養を繰り返し、最終的に培養液を200 mg/Lクロラムフェニコールを含むMYKプレートに植菌した。継代培養時、生育速度が遅い場合は、培養日数を2〜3日延長した。継代培養回数は4回である(クロラムフェニコール含有量:10、40、100、200(mg/ml)の順に継代)。出現したコロニーを200 mg/Lクロラムフェニコールを含むMYKプレートに更に2回植菌してシングルコロニー化することにより、RhCmSR-01株を単離した。
【実施例3】
【0075】
ATCC 12674株のカナマイシン感受性株: RhΔKm-01株の製造
大腸菌(Escherichia coli)S17-1λpirをpKM043により形質転換し、50 mg/Lカナマイシン硫酸塩含有LBプレートにて37℃で一晩生育させた。S17-1λpir/pKM043をLB培地に懸濁後、遠心分離(室温、4500ppm、2 min)を行い、菌体を回収した後、LBで一度洗浄し、LBに再懸濁した。
【0076】
RhCmSR-01をMYKプレート上にひき、30℃で3日間培養した。MYKプレート上のコロニーを数十個程度かきとり0.9% NaClに懸濁後、遠心分離(室温、15000ppm、2 min)を行い、菌体を回収した後、0.9% NaClで一度洗浄し、LBに再懸濁した。
【0077】
上記2種類の懸濁液を1対1の割合で穏やかに混合した後、LBプレート上に植菌し、30℃で一晩培養した(pKM043の接合伝達)。標的遺伝子と選択マーカーの一つが共にカナマイシン耐性遺伝子であるため、通常野生株が生育できないカナマイシン濃度での選抜を実施した。実際の選抜には、カナマイシン硫酸塩200 mg/L 及びクロラムフェニコール50 mg/L含有MYKプレート(0.5% polypeptone、0.3% bact yeast extract、0.3% malt extract、0.2% KH2PO4、0.2% K2HPO4、1.5%寒天)を使用した。
【0078】
プラスミドpKM043がゲノム内に相同組換えにより導入されていることをPCR法により確認した後、10%ショ糖を含むMYKプレートにて培養することにより(sacB対抗選択系(WO 01/31050)を使用)、pK19mobsacB1由来配列をATCC 12674ゲノム上から取り除いた。カナマイシン含有LBプレート上でのレプリカプレーティングにより、10 mg/L カナマイシン硫酸塩含有条件下では生育できないカナマイシン感受性株RhΔKm-01を単離することに成功した(図3A参照)。PCR法により野生株とRhΔKm-01株のゲノム分析を実施した結果、RhΔKm-01株においてカナマイシン耐性遺伝子長約0.5 kbに相当する領域がゲノムから欠失していることが確認できた(図3B参照)。
【実施例4】
【0079】
他のカナマイシン耐性ロドコッカス属細菌への適用
DB上で全ゲノム配列が公開されているロドコッカス・エリスロポリス(Rhodococcus erythropolis)PR4(製品評価技術基盤機構生物遺伝資源部門;受託番号:NBRC 100887)もATCC 12674がもつカナマイシン耐性遺伝子のホモログを有する。ホモロジー検索の結果、両株のカナマイシン耐性遺伝子が96.1%一致(塩基配列)という高い相同性を示したため、pKM043を用いた遺伝子欠失を実施した。各段階における培養時のベースとなる培地にはLB又はLBプレートを使用した。
【0080】
PR4も他のATCC 12674と同様に微弱なクロラムフェニコール耐性を持つことが確認できたため、実施例2と同様の方法と同様の方法により、120 mg/L Cmに対して耐性を示す変異株RhCmSR-09を製造し、単離した。継代培養回数は7回である(クロラムフェニコール含有量:10、20、30、60、80、100、120(mg/ml)の順に継代)。
【0081】
実施例3と同様の方法により、カナマイシン感受性株RhΔKm-10を単離した(図4A参照)。RhΔKm-10のゲノムからカナマイシン耐性遺伝子が欠失していることを確認した(図4B参照)。
【配列表フリーテキスト】
【0082】
配列番号1:合成DNA
配列番号2:合成DNA
配列番号3:合成DNA
配列番号4:合成DNA
【技術分野】
【0001】
本発明は、ゲノム上の薬剤耐性遺伝子を欠失又は不活性化させることにより薬剤感受性にした微生物に関する。詳しくは、ロドコッカス属細菌のゲノム上の薬剤耐性遺伝子を欠失又は不活性化させた薬剤感受性微生物(薬剤感受性ロドコッカス属細菌)に関する。
【背景技術】
【0002】
ロドコッカス属細菌は、その物理的強度、有機溶媒耐性、酸化還元能、酵素などを細胞内に多量蓄積する能力などを有することから、産業的に有用な微生物触媒として知られている。例えば、ロドコッカス属細菌は、ニトリル類の酵素的水和又は加水分解によるアミドもしくは酸の生産などに利用されている(特許文献1、2)。また、難分解性化合物に対する分解能の高い微生物種としても知られており(特許文献3〜6)、その能力を活用することで、種々の有用物質生産および環境浄化への応用検討が進められている。例えば、脱硫による石油からの有用物質の生産、あるいは環境中への石油流出時におけるバイオレメディエーションなどが挙げられる。
【0003】
加えて、ロドコッカス属細菌を遺伝子組換えの方法により、さらに有用なものに改変する試みがなされている(特許文献7〜9)。例えば、ロドコッカス属細菌の遺伝子操作を効率的に推し進めるために、宿主−ベクター系の開発が進められており、新規なプラスミドの探索(特許文献10〜12)、ベクターの開発(特許文献13〜16、17〜19、非特許文献1)などが行われている。
【先行技術文献】
【特許文献】
【0004】
【特許文献1】特開平2-470号公報
【特許文献2】特開平3-251192号公報
【特許文献3】欧州特許第188316号明細書
【特許文献4】欧州特許第204555号明細書
【特許文献5】欧州特許第348901号明細書
【特許文献6】欧州特許出願第307926号明細書
【特許文献7】特開平4-211379号公報
【特許文献8】特開平6-25296号公報
【特許文献9】特開平6-303791号公報
【特許文献10】特開平4-148685号公報
【特許文献11】特開平4-330287号公報
【特許文献12】特開平7-255484号公報
【特許文献13】特開平5-64589号公報
【特許文献14】特開平8-56669号公報
【特許文献15】米国特許第4,920,054号明細書
【特許文献16】特開平8-173169号公報
【特許文献17】特開平10-248578号公報
【特許文献18】特開2006-180843号公報
【特許文献19】特開2006-50967号公報
【非特許文献】
【0005】
【非特許文献1】Journal of Bacteriology 170, 638-645 (1988)
【発明の概要】
【発明が解決しようとする課題】
【0006】
一般的に、微生物を形質転換する際、形質転換株の選抜は、導入したプラスミドの選択マーカー(例えば、ある薬剤に対する耐性遺伝子等)により行う。しかし、宿主によってはいくつかの薬剤に対して耐性を有しているものもいるため、その微生物種で適したベクター系の有する選択マーカーの機能を宿主自体も持っている場合がある。例えば、ロドコッカス属細菌を形質転換する際、カナマイシン耐性遺伝子を選択マーカーとして有するシャトルベクターがよく使用されるが、ロドコッカス属細菌はカナマイシンに耐性を示す株も多い。上記シャトルベクターを用いてそれらカナマイシン耐性ロドコッカス属細菌を形質転換する場合、高濃度のカナマイシンで形質転換株を選抜せざるを得ず、非常に効率が悪い。
【0007】
上述の場合において、プラスミド由来の選択マーカーにより、効率的な形質転換を可能にするためには、宿主微生物ゲノム上の薬剤耐性遺伝子を欠失又は不活性化し、当該薬剤に対し感受性を示すように宿主を改良すればよい。ゲノム上に存在する遺伝子を欠失又は不活性化する手法としては、相同組換え(Homologous recombination)を利用する方法が挙げられる。これは、標的遺伝子の間に他の遺伝子等を挿入して標的遺伝子を機能発現しない状態にする(不活性化)、又は、標的遺伝子の一部又は全部を含まない配列に置換することにより標的遺伝子自体を欠失させて、標的遺伝子の発現そのものを止めてしまう方法である。大腸菌や酵母などのいくつかの微生物では、効率的な相同組換え技術が確立しているため、比較的容易に標的遺伝子の欠失又は不活性化ができる。
【0008】
ロドコッカス属細菌において相同組換えを行う際の遺伝子導入法として、エレクトロポレーション法等が知られている。エレクトロポレーション法はロドコッカス属細菌における代表的な形質転換法であるが、相同組換えに適用した場合、ロドコッカス属細菌及びその類縁属細菌は非相同組換えが起こり易いという特徴を有しているため、目的の相同組換え株が極めて低確率でしか得られないという問題がある。相同組換えを行う際の他の遺伝子導入法としては、接合伝達を利用する方法(接合伝達法)が挙げられるが、当該方法はロドコッカス属細菌に対しては殆ど適用されていない。
【0009】
従って、ロドコッカス属細菌において、標的遺伝子を効率的且つ汎用的に欠失又は不活性化する方法は確立していないため、選択マーカーとして使用し得る薬剤耐性遺伝子が欠失又は不活性化したロドコッカス属細菌、すなわち、薬剤感受性のロドコッカス属細菌は十分に得られていないのが現状である。
【0010】
このような状況下において、より効率的且つ汎用的に薬剤耐性遺伝子が欠失又は不活性化された、当該薬剤感受性微生物の取得が望まれていた。
【課題を解決するための手段】
【0011】
本発明は、上記状況を考慮してなされたもので、以下に示す薬剤感受性を示すロドコッカス属細菌を提供するものである。
【0012】
すなわち、本発明は以下の通りである。
(1)ロドコッカス属細菌において、標的とする薬剤耐性遺伝子が欠失又は不活性化されたことにより当該薬剤に対して感受性を示す微生物。
【0013】
本発明の微生物としては、例えば、前記欠失又は不活性化が、ドナー微生物からレシピエント微生物への接合伝達を利用した、以下の工程(a)〜(d)を含む方法により行われるものが挙げられる。
【0014】
(a)レシピエント微生物として、ドナー微生物生育阻害マーカーが強化されたロドコッカス属細菌を作製する工程;
(b)ドナー微生物として、(i)レシピエント微生物中の標的の薬剤耐性遺伝子とその周辺の塩基配列とを含む塩基配列において当該耐性遺伝子を欠失又は不活性化させた塩基配列領域、(ii)当該ドナー微生物において機能する接合伝達開始領域、(iii)当該ドナー微生物において機能する複製開始領域、(iv) レシピエント微生物生育阻害マーカー、及び(v)レシピエント微生物に対する条件致死遺伝子を含む、遺伝子改変用プラスミドを用いて形質転換された微生物を作製する工程;
(c)前記(b)で作製されたドナー微生物から前記(a)で作製されたレシピエント微生物への接合伝達を行うことにより、当該レシピエント微生物の形質転換体を作製する工程;並びに
(d)前記(c)で作製された形質転換体を、前記条件致死遺伝子が機能し得る培養条件で培養する工程。
【0015】
本発明の微生物において、前記ロドコッカス属細菌としては、例えば、ロドコッカス・ロドクロウス(Rhodococcus rhodochrous)又はロドコッカス・エリスロポリス(Rhodococcus erythropolis)が挙げられる。
【0016】
本発明の微生物としては、例えば、前記欠失又は不活性化された薬剤耐性遺伝子がカナマイシン耐性遺伝子であるものが挙げられる。
(2)上記(1)に記載の微生物を宿主として用いて培養し、得られる培養物から目的タンパク質を採取することを特徴とする、タンパク質製造方法。
【発明の効果】
【0017】
本発明によれば、薬剤耐性を示すロドコッカス属細菌について、ロドコッカス属細菌ゲノム上の当該薬剤耐性遺伝子を欠失又は不活性化させた当該薬剤感受性微生物(当該薬剤感受性ロドコッカス細菌)を提供することができる。例えば、カナマイシン耐性遺伝子をマーカーとして有するシャトルベクターを用いて形質転換株を作製する場合であれば、本発明の薬剤感受性微生物の一態様としてのカナマイシン感受性株を使用することにより、従来のように高濃度のカナマイシンで組換え株を選抜する必要はなくなり、低濃度のカナマイシン含有培地を用いて、より効率的に形質転換株を得ることができる。また、欠失させた薬剤耐性遺伝子を導入したベクター系を構築し、当該薬剤耐性遺伝子を選択マーカーとして活用することも可能になる。よって、本発明の薬剤感受性微生物を形質転換用宿主として用いることにより、より効率的な物質生産等が行い得るようになるため、本発明は、産業的にみても極めて優れたものである。
【図面の簡単な説明】
【0018】
【図1】本発明によりRhodococcus属細菌のカナマイシン耐性遺伝子を欠失させる際の各工程における、当該細菌ゲノム上で起こる相同組換え等について示した概略図である。
【図2】遺伝子改変用プラスミドpKM043の遺伝子地図を示す図である。
【図3】(A)カナマイシン感受性株RhΔKm-01を単離することに成功した結果を示す図である。(B)RhΔKm-01株においてカナマイシン耐性遺伝子長約0.5 kbに相当する領域がゲノムから欠失していることを確認した結果を示す図である。
【図4】(A)カナマイシン感受性株RhΔKm-10を単離することに成功した結果を示す図である。(B)RhΔKm-10のゲノムからカナマイシン耐性遺伝子が欠失していることを確認した結果を示す図である。
【発明を実施するための形態】
【0019】
以下、本発明を詳細に説明する。本発明の範囲はこれらの説明に拘束されることはなく、以下の例示以外についても、本発明の趣旨を損なわない範囲で適宜変更し実施することができる。
【0020】
なお、本明細書において引用された全ての刊行物、例えば先行技術文献、及び公開公報、特許公報その他の特許文献は、参照として本明細書に組み込まれる。
1.薬剤に対し感受性を示す微生物
本発明に係る薬剤に対し感受性を示す微生物、すなわち、薬剤に対し感受性を示すロドコッカス属細菌(以下、「本発明の微生物」又は「薬剤感受性株」ということがある)は、前述した通りある薬剤に対する耐性を有するロドコッカス属細菌において、当該薬剤に対する耐性を獲得するために必要な遺伝子(以下、「標的とする薬剤耐性遺伝子」)が欠失又は不活性化された変異株である。
【0021】
ここで、薬剤に対し感受性を示すとは、薬剤含有培地において、ロドコッカス属細菌が生育できないことをいう。また、薬剤に耐性を示すとは、薬剤含有培地においてもロドコッカス属細菌が十分に生育できることをいう。
【0022】
ロドコッカス属細菌としては、ロドコッカス・ロドクロウス(Rhodococcus rhodochrous)J1、M8(SU1731814)、M33(VKM Ac-1515D)、ATCC 184、ATCC 999、ATCC 4001、ATCC 4273、ATCC 4276、ATCC 9356、ATCC 12483、ATCC 12674、ATCC 13808、ATCC 14341、ATCC 14347、ATCC 14350、ATCC 15905、ATCC 15998、ATCC 17041、ATCC 17043、ATCC 17895、ATCC 19067、ATCC 19149、ATCC 19150、ATCC 21243、ATCC 21291、ATCC 21785、ATCC 21924、ATCC 21999、ATCC 29670、ATCC 29672、ATCC 29675、ATCC 33258、ATCC 33275、ATCC 39484、IFO 3338、IFO 14894、JCM 3202、NCIMB 11215、NCIMB 11216、ロドコッカス・エリスロポリス(Rhodococcus erythropolis)PR4(NBRC 100887)、NBRC 12320、NBRC 15567、NBRC 16296、IFM 155、DSM 743、DSM 9675、DSM 43200、JCM 6821、JCM 6822、JCM 6823、JCM 6824、JCM 6827、DSM 11397、DSM 44522、JCM 2895、JCM 2893、JCM 2894、IAM 1400、IAM 1503、ロドコッカス・オパカス(Rhodococcus opacus)B4等が挙げられる。
【0023】
標的とする薬剤耐性遺伝子としては、例えば、アミノグリコシド系抗生物質耐性遺伝子、β-ラクタム系抗生物質耐性遺伝子、クロラムフェニコール系抗生物質耐性遺伝子、テトラサイクリン系抗生物質耐性遺伝子等が挙げられる。
【0024】
ここで、アミノグリコシド系抗生物質耐性遺伝子としては、カナマイシン及びネオマイシン耐性遺伝子であるアミノグリコシド3'-ホスホトランスフェラーゼ等のリン酸転移酵素遺伝子群、ストレプトマイシン耐性遺伝子であるアミノグリコシド3'-アデニリルトランスフェラーゼ等のヌクレオチジル転移酵素遺伝子群、アセチル転移酵素遺伝子群が挙げられる。
【0025】
β-ラクタム系抗生物質耐性遺伝子としては、β-ラクタマーゼ遺伝子が挙げられる。
【0026】
クロラムフェニコール系抗生物質耐性遺伝子としては、クロラムフェニコールアセチルトランスフェラーゼ遺伝子が挙げられる。
【0027】
テトラサイクリン系抗生物質耐性遺伝子としては、トランスポゾンTn10由来テトラサイクリン耐性遺伝子が挙げられる。
【0028】
また、ある薬剤に対し耐性を有するロドコッカス属細菌の中には、薬剤の修飾等による無毒化、又は細胞外への排出機能等を担う薬剤耐性遺伝子を有しているわけではなく、薬剤の標的である酵素自体が薬剤に対する耐性(薬剤抵抗性)を持つことにより、結果として薬剤耐性を示す株もある。この場合は、例えば、上記薬剤抵抗性酵素を、近縁種等の有する感受性酵素(非抵抗性酵素)に置換することにより、薬剤感受性株にすることも可能である。この場合も、薬剤耐性が発揮できなくなった、すなわち、薬剤耐性が不活性化したと考えられるため、本発明における薬剤耐性遺伝子の不活性化の範疇に入る。薬剤抵抗性を示す酵素としては、例えば、トリメトプリム耐性ジヒドロキシ葉酸レダクターゼ等が挙げられる。
2.薬剤に対し感受性を示す微生物の製造方法
標的とする薬剤耐性遺伝子を欠失又は不活性化する方法は限定されず、如何なる方法を適用してもよい。例えば、当該遺伝子又はその周辺配列と相同な配列を有するプラスミドを形質転換する方法を用いることができる。形質転換の方法としては、エレクトロポレーション法や接合伝達法が使用できる。
【0029】
上記欠失又は不活性化する方法については、本発明では以下の方法が特に好ましい。
【0030】
すなわち、ドナー微生物からレシピエント微生物への接合伝達を利用した、以下の工程(a)〜(d)を含む方法により、本発明の微生物が得られることが好ましい。なお、当該製造方法は、ドナー微生物からレシピエント微生物への接合伝達を利用した形質転換方法を用いることを含む、標的とする薬剤耐性遺伝子の欠失又は不活性化方法であって、以下の工程(a)〜(d)を含むことを特徴とする方法ということもできる。
(a)レシピエント微生物として、ドナー微生物生育阻害マーカーが強化されたロドコッカス属細菌を作製する工程;
(b)ドナー微生物として、下記(i)〜(v):
(i)レシピエント微生物中の標的とする薬剤耐性遺伝子とその周辺の塩基配列とを含む塩基配列において当該耐性遺伝子を欠失又は不活性化させた塩基配列領域、
(ii)当該ドナー微生物において機能する接合伝達開始領域、
(iii)当該ドナー微生物において機能する複製開始領域、
(iv) レシピエント微生物生育阻害マーカー、及び
(v)レシピエント微生物に対する条件致死遺伝子
を含む、遺伝子改変用プラスミドを用いて形質転換された微生物を作製する工程;
(c)工程(b)で作製されたドナー微生物から工程(a)で作製されたレシピエント微生物への接合伝達を行うことにより、当該レシピエント微生物の形質転換体を作製する工程;並びに
(d)工程(c)で作製された形質転換体を、前記条件致死遺伝子が機能し得る培養条件で培養する工程。
上記欠失又は不活性化方法においてレシピエントとして使用するロドコッカス属細菌は、限定はされないが、前述したカナマイシン耐性を有するものが例示できる。
【0031】
接合伝達を用いた遺伝子欠失又は不活性化を行う場合、ドナーとなる微生物の組換え株(ドナー微生物)とレシピエントとなる微生物(レシピエント微生物)の非組換え株の双方の生育を阻害しなければならないため、2種類の選択マーカー(ドナー微生物生育阻害マーカー及びレシピエント微生物生育阻害マーカー)が必要である。ドナー微生物生育阻害マーカーはレシピエント微生物自体が有している必要がある。レシピエント微生物生育阻害マーカーは、遺伝子改変用プラスミド中に含まれている必要がある。両マーカーの詳細については、下記の各工程において説明する。
2.1. 工程(a)
工程(a)では、ドナー微生物生育阻害マーカーが強化されたロドコッカス属細菌を作製する。
【0032】
上述の理由から、レシピエントとなるロドコッカス属細菌にはドナー微生物の生育を阻害するための薬剤耐性マーカー(ドナー微生物生育阻害マーカー)が必要である。よって、標的とする薬剤耐性遺伝子が関与する薬剤耐性以外の薬剤耐性を有していない、又は薬剤耐性の乏しいロドコッカス属細菌を使用する場合、ドナー微生物生育阻害マーカーとして機能しうる、薬剤選択可能な程度の十分な薬剤耐性を有する変異株の作製が必要となる。
【0033】
ここで、ドナー微生物生育阻害用の薬剤としては、接合伝達時にドナーとして用いる微生物(ドナー微生物)の有する薬剤耐性を考慮し、ドナー微生物が感受性を示す薬剤を選択することが好ましい。当該薬剤としては、クロラムフェニコール、アンピシリン、カナマイシン、トリメトプリム、ゲンタマイシン、ナルジクス酸、カルベニシン、チオストレプトン、テトラサイクリン、ストレプトマイシン等が好ましく、クロラムフェニコール、アンピシリンがより好ましい。
【0034】
ドナー微生物生育阻害マーカーが強化されたレシピエント微生物を作製する方法としては、特に限定はされないが、例えば、(イ)自然変異誘発法、(ロ)紫外線照射や変異誘発剤を用いる突然変異誘発法、(ハ)あらかじめ遺伝子改変用プラスミドとは別の、抗生物質耐性獲得用のプラスミドを導入する方法等が好ましく挙げられ、中でも自然変異誘発法がより好ましい。
【0035】
自然変異誘発法は、所望の薬剤を含有する培地中で対象とする微生物を継代培養等することにより、もともとは当該培地中で生育不可又は困難な微生物に自然変異を誘発させて、より高濃度の薬剤を含有する当該培地中でも生育し得る株を取得する方法である。
【0036】
レシピエント微生物となるロドコッカス属細菌の薬剤耐性をどの程度まで薬剤耐性を強化するかは、使用するロドコッカス属細菌の種類、ドナー微生物及び選択する薬剤により異なるため適宜調整することができるが、レシピエント微生物(ロドコッカス属細菌)の生育が抑制されず、且つ、ドナー微生物の生育が十分阻害される薬剤濃度を基準に強化することが好ましい。例えば、レシピエント微生物としてのロドコッカス属細菌に対して、ドナー微生物として大腸菌を、選択用薬剤としてクロラムフェニコールを用いる場合、自然突然変異により、クロラムフェニコール30〜200 mg/L、好ましくは50〜200 mg/Lを含有する培地において生育可能なロドコッカス属細菌(クロラムフェニコール耐性強化株)を得ることが望ましい。
2.2. 工程(b)
工程(b)では、接合伝達に供するドナー微生物として、所定の遺伝子改変用プラスミドを用いて形質転換された微生物を作製する。遺伝子改変用プラスミド、すなわちレシピエント微生物中の標的とする薬剤耐性遺伝子を欠失又は不活性化するためのプラスミドとしては、前述の(i)〜(v)の構成(遺伝子・塩基配列等)を含むものを用いる。
【0037】
ドナー微生物からレシピエント微生物にプラスミドを伝達するためには、oriTに加え、mob遺伝子及びtra遺伝子(群)が最小限必要である。mobはoriT特異的ニック酵素をコードする遺伝子で、この酵素がoriTに働くことによりドナー微生物からレシピエント微生物への(遺伝子改変プラスミドの)移行が開始される。traは多数の遺伝子群の総称で、性繊毛形成、接合管形成、接合制御に関与する遺伝子群で構成されている。mob遺伝子及びtra遺伝子(群)は必ずしも遺伝子改変プラスミド上になくてもよく、別のプラスミド(ヘルパープラスミド)、もしくはドナーとなる微生物ゲノム上に組み込まれていてもよい(蛋白質 核酸 酵素, vol. 38, p. 60-68 (1993))。
【0038】
本発明の製造方法において、ドナー微生物としては、上記レシピエント微生物と接合伝達可能な微生物であればよく、限定されないが、上記の接合伝達に最低限必要な構成要件を満たしている必要がある。従って、使用する遺伝子改変用プラスミド上にoriT 、mob遺伝子及びtra遺伝子(群)が一通り備わっている場合は、それぞれが機能しうる微生物であれば特に限定されないが、使用する遺伝子改変用プラスミド上にoriTのみが備わっている場合は、oriTに対応するmob遺伝子及びtra遺伝子(群)を保有する微生物を用いる必要がある。ドナーとなる微生物と遺伝子改変用プラスミドの組み合わせとしては、例えば、大腸菌S17-1とpK19mobの組み合わせが好ましい。大腸菌S17-1のゲノム上には、プラスミドRP4由来のtra遺伝子(群)が、pK19mobにはRP4由来のoriT 、mob遺伝子がそれぞれ備わっているため、ドナー微生物として使用可能である。
【0039】
ここで、前記(i)の配列は、(改変の対象とする)ロドコッカス属細菌中の標的とする薬剤耐性遺伝子と当該遺伝子の周辺の塩基配列とを含む塩基配列において、当該遺伝子を欠失又は不活性化させた塩基配列領域である。
【0040】
レシピエント微生物のゲノムから、標的とする薬剤耐性遺伝子と当該遺伝子の周辺の塩基配列とを含む塩基配列の単離(クローニング)は、遺伝子ライブラリー作製やPCR等の公知技術を用いて行うことができる。
【0041】
なお、標的とする薬剤耐性遺伝子の周辺の塩基配列としては限定されないが、当該ロドコッカス属細菌ゲノムとの相同領域(標的遺伝子周辺の塩基配列)が長いほどより効率よく相同組換えを起こすことが出来るため、クローニングに支障が出ない範囲でより長い配列を用いることが好ましい。
【0042】
例えば、当該遺伝子の上流及び下流の相同領域がそれぞれ100〜3000 bpの塩基配列を含む配列であることが好ましく、より好ましくは500〜2000 bpの塩基配列を含む配列である。単離した塩基配列を用いて、前述(i)を作製する方法は特に限定されず、PCR法や制限酵素を用いた標的遺伝子部分の切除又は置換等の公知技術を用いて行うことができる。
【0043】
欠失された遺伝子としては、ロドコッカス属細菌のゲノムにコードされる標的とする薬剤耐性遺伝子の一部又は全部が欠失されたもの、プロモーター配列の一部又は全部が欠失されたもの、当該遺伝子の発現調節に関連する遺伝子が存在する場合は、その一部又は全部が欠失されたものなどが挙げられ、これらの欠失された領域は単独でもよいし複数を組み合わせたものでもよい。
【0044】
不活性化された遺伝子としては、ロドコッカス属細菌のゲノムにコードされる標的とする薬剤耐性遺伝子若しくは当該遺伝子の発現調節に関連する遺伝子の内部に、外来遺伝子、例えは薬剤耐性遺伝子等を導入し、本来の機能を発現しない遺伝子に改変されたもの、又は薬剤抵抗性酵素の遺伝子を薬剤非抵抗性酵素の遺伝子に置換することにより薬剤非抵抗酵素が発現するようにしたもの等が挙げられる。
【0045】
薬剤非抵抗性酵素としては、菌体内において薬剤抵抗性酵素と同様の機能を果たすが、対象の薬剤に対しては感受性を示す酵素であれば限定されないが、ロドコッカス属細菌もしくはその近縁種由来のものが好ましい。
【0046】
前記(ii)の接合伝達開始領域は、使用するドナー微生物中において接合伝達の開始点となる塩基配列を含む領域であればよく、特に限定されない。例えば、Fプラスミド由来の接合伝達開始領域を含む配列、プラスミドR6K由来のoriT、プラスミドRP4由来のoriTが好ましい。
【0047】
前記(iii)の複製開始領域は、使用するドナー微生物中において前記遺伝子改変用プラスミドの自己複製起点として機能し得る塩基配列を含む領域であればよく、特に限定はされない。例えば、プラスミドpMB1及び広宿主域プラスミドRK2由来の複製開始点並びにその派生物等を含む領域の使用が好ましい。
【0048】
前記(iv)のレシピエント微生物生育阻害マーカーは、接合伝達に供するレシピエント微生物(ロドコッカス属細菌)が感受性を示す薬剤に対する耐性遺伝子であることが好ましい。標的とする薬剤耐性遺伝子と当該生育阻害マーカーが同一の機能を有するものである場合は、標的とする薬剤耐性遺伝子のみでは耐性を獲得できない高濃度薬剤を用いることで、目的の組換え株(相同組換え株)を取得することも可能である。耐性遺伝子としては、例えば、カナマイシン耐性遺伝子、アンピシリン耐性遺伝子、ゲンタマイシン耐性遺伝子、チオストレプトン耐性遺伝子、カルベニシン耐性遺伝子等が好ましく挙げられ、中でもカナマイシン耐性遺伝子がより好ましい。
【0049】
前記(v)の条件致死遺伝子は、レシピエント微生物のゲノム上に導入され、且つある特定条件下にさらされた場合に、当該微生物を死に至らしめる作用を有し得る遺伝子であればよい。例えば、sacB遺伝子が好ましく挙げられる。sacB遺伝子は、当該遺伝子を保有し発現する微生物(例えばロドコッカス属細菌等)をスクロース含有培地で培養した場合に、スクロースを基質とし当該微生物に対して致死作用を有する有害物質を産生する酵素をコードする遺伝子である。
【0050】
接合伝達に用いる前記遺伝子改変用プラスミドは、如何なるベクターをベースとして構築されたものであってもよく、その種類は特に限定はされない。使用するベクターとしては、例えば、pBR322、pSC101、pACYC184、pACYC177、pTrc99A、pUC18、pUC19、pUC118、pUC119、pHSG298、pHSG299、pSP64、pSP65、pGEM-3、pGEM-3Z、pGEM-3Zf(-)、pGEM-4p、GEM-4Z、Bluescript M13+、Bluescript M13-、pK19mob、pK18mob、pK18mobsacB等が挙げられる。中でも、プラスミド内部に既にRP4由来のoriT及びmobを有しているpK19mob、pK18mob、pK18mobsacB(Schaefer等, Gene, vol. 45, p. 69-73(1994))を用いることがより好ましい。
前記遺伝子改変用プラスミドにおける(i)〜(v)の構成は、その配置については特に限定されず、如何なる順番で配置されていてもよい。前記(i)〜(v)の構成を含む遺伝子改変用プラスミドの構築は、公知の遺伝子組換え技術を用いて実施することができる。
【0051】
工程(b)では、上述したような各構成を有する遺伝子改変用プラスミドをドナーとして用いる微生物内に導入して形質転換された微生物、すなわち接合伝達に供するドナー微生物を作製する。その際、形質転換の方法としては、エレクトロポレーション法や塩化カルシウム法等の、微生物の形質転換方法として公知の方法を用いることができる。
2.3. 工程(c)
工程(c)では、工程(b)で得られたドナー微生物から工程(a)で得られたレシピエント微生物への接合伝達を行う。通常は、ドナー微生物及びレシピエント微生物のそれぞれの菌体懸濁液を混合し、適当なプレート培地(LB培地等)上に均一に広げて、両微生物の接合を行わせる。当該接合においては、ドナー微生物中の遺伝子改変用プラスミドがレシピエント微生物内に移動し、レシピエント微生物のゲノムと上記プラスミドとの相同配列で相同組換えが起こり、遺伝子改変用プラスミドの一部又は大部分が当該レシピエント微生物のゲノム上に導入される。この接合により、レシピエント微生物の形質転換体、すなわち相同組換え株が作製される。
【0052】
本工程(c)における所望の相同組換え株は、1重交差により標的遺伝子の上流又は下流に当該遺伝子改変用プラスミドが導入されたものである。所望の相同組換え株であるかどうかの確認は、レシピエント微生物自体の薬剤耐性、及び前記遺伝子改変用プラスミド由来の薬剤耐性を利用して行うことができる。具体的には、両薬剤を含む培地(例えば、カナマイシン及びクロラムフェニコール含有培地)において上記接合後の微生物を培養することにより、所望の相同組換え株を選択することができる。
2.4. 工程(d)
工程(d)では、工程(c)で作製されたレシピエント微生物の相同組換え株(相同組換え微生物)を、前記遺伝子改変用プラスミド由来の条件致死遺伝子が機能し得る培養条件で培養する。条件致死遺伝子が機能し得る培養条件としては、条件致死遺伝子が機能する限り、特に限定はされない。例えば、条件致死遺伝子がsacB遺伝子の場合は、スクロース含有培地を用いた培養が用いられる。
【0053】
当該培養においては、上記条件致死遺伝子を有する相同組換え微生物は生育困難であるため、当該微生物のゲノム上から2度目の相同組換えにより上記条件致死遺伝子、薬剤耐性遺伝子、接合伝達開始領域、複製開始領域を含む遺伝子改変用プラスミド由来の塩基配列領域が除かれた(脱落した)ロドコッカス属細菌を得ることができる。
【0054】
ただし、当該培養により得られた微生物の中には、レシピエント微生物中の標的の薬剤耐性遺伝子が、当初の目的通り欠失又は不活性化しているものと、そうでないもの(上記脱落の際の相同組換えにより元の薬剤耐性遺伝子の機能が復活したもの)が含まれている。よって、通常は、さらに別の培養条件でも培養し、所望の形質転換微生物を選択することができる。
【0055】
別の培養条件としては、上記薬剤遺伝子の機能を確認し得る培養条件での培養が好ましく、より詳細には、当該薬剤を含有する培地で培養したときに生育が不可又は著しく困難な株が、所望のロドコッカス属細菌、すなわち本発明の微生物(当該薬剤感受性株)として選択することができる。また、レシピエント微生物からゲノムを抽出し、当該改変領域の配列を解析することにより、所望の微生物を選択することもできる。
【0056】
上述した工程(a)〜(d)を含む方法本発明により得られた薬物感受性微生物(詳しくは、薬物感受性ロドコッカス属細菌)は、上記の通り、ゲノム上に遺伝子改変用プラスミドの痕跡が残らないため、本発明の工程(b)〜(d)を繰り返し行うことにより、同一の株を複数回改変することが可能である。
3.タンパク質の製造方法
本発明においては、前述したロドコッカス属細菌の薬剤感受性株を宿主として用いて培養し、得られる培養物から目的タンパク質を採取することを特徴とする、タンパク質の製造方法を提供することができる。宿主として用いる当該感受性株は、あるタンパク質を高発現するプラスミド、例えば、アクリルアミド製造用触媒であるニトリルヒドラターゼ等を発現し得る発現ベクターが別途導入された形質転換体として使用される。
【0057】
上記タンパク質の製造方法において、微生物を培養する方法は、通常の方法に従って行うことができる。
【0058】
微生物を培養する培地としては、微生物が資化し得る炭素源、窒素源、無機塩類等を含有し、微生物の培養を効率的に行うことができる培地であれば、天然培地、合成培地のいずれを用いてもよい。炭素源としては、グルコース、フラクトース、スクロース、デンプン等の炭水化物、酢酸、プロピオン酸等の有機酸、エタノール、プロパノール等のアルコール類、ヘキサン、デカン、ドデカン、テトラデカン等の炭化水素類が挙げられる。窒素源としては、アンモニア、塩化アンモニウム、硫酸アンモニウム、酢酸アンモニウム、リン酸アンモニウム等の無機酸若しくは有機酸のアンモニウム塩又はその他の含窒素化合物のほか、ペプトン、肉エキス、コーンスティープリカー等が挙げられる。無機物としては、リン酸第一カリウム、リン酸第二カリウム、リン酸マグネシウム、硫酸マグネシウム、塩化ナトリウム、硫酸第一鉄、硫酸マンガン、硫酸銅若しくは炭酸カルシウム等が挙げられる。
【0059】
培養条件は、限定はされないが、例えば、10℃〜45℃、好ましくは10℃〜40℃の温度下で、5〜120時間、好ましくは5〜100時間程度行うことができる。また、場合により、本培養に先立ち、少量の前培養を行うこともできる。
【0060】
例えば、ロドコッカス属細菌の形質転換体を宿主として用いて培養する場合は、振盪培養又は通気攪拌培養などの好気的条件下、4〜40℃、好ましくは20〜35℃で行うことが好ましい。培養は、無機酸又は有機酸、アルカリ溶液等を用いて適時pH調整を行うことが好ましい。
【0061】
また、培養中に当該形質転換体から発現ベクターの脱落を防ぐために、必要に応じて薬剤選択マーカーによる選択圧をかけた状態で培養してもよい。例えば、カナマイシン耐性遺伝子を含むベクターで形質転換した形質転換体を培養する場合、培養中に必要に応じてカナマイシンを培地に添加してもよい。
【0062】
上記のように、宿主となる微生物の性質に合った適当な培養条件で培養すると、高収率で異種タンパク質(目的タンパク質)を上記培養物中、すなわち、培養上清、培養菌体、又は菌体の破砕物の少なくともいずれかに蓄積することができる。これら、異種タンパク質を含有する溶液を「培養物」と称する。
【0063】
培養後、目的タンパク質が菌体内に生産される場合には、菌体のまま目的タンパク質を用いることもできるし、あるいは菌体を破砕することにより、目的タンパク質を採取することもできる。いずれの場合にも、必要であれば、遠心分離や膜ろ過などの固液分離操作により、培地除去及び洗浄を行うことができる。遠心分離は、菌体を沈降させる遠心力が供給できるものであれば特に限定されることはなく、円筒型や分離板型などを利用することができる。遠心力としては、例えば、500G〜20,000G程度で行うことができる。また、膜ろ過は、目的とする固液分離を達成できれば、精密ろ過(MF)膜、限外ろ過(UF)膜いずれでもよいが、通常、精密ろ過(MF)膜を用いることが好ましい。
【0064】
なお、菌体を破砕することにより、目的タンパク質を採取することもできる。菌体の破砕方法としては、超音波処理、フレンチプレスやホモジナイザーによる高圧処理、ビーズミルによる磨砕処理、衝撃破砕装置による衝突処理、リゾチーム、セルラーゼ、ペクチナーゼ等を用いる酵素処理、凍結融解処理、低張液処理、ファージによる溶菌誘導処理等が挙げられ、いずれかの方法を単独又は必要に応じ組み合わせて利用することができる。
【0065】
一方、目的タンパク質が菌体外に生産される場合には、培養液をそのまま使用するか、上述したような遠心分離やろ過等により菌体を除去する。その後、必要に応じて硫安沈澱による抽出等により前記培養物中から目的タンパク質を採取し、さらに必要に応じて透析、各種クロマトグラフィー(ゲルろ過、イオン交換クロマトグラフィー、アフィニティークロマトグラフィー等)を単独又は適宜組み合わせて用いることにより、精製することもできる。
以下に、実施例を挙げて本発明をより具体的に説明するが、本発明の技術的範囲はこれらに限定されるものではない。
【実施例1】
【0066】
カナマイシン耐性遺伝子欠失用プラスミドpKM043の構築(遺伝子改変用プラスミドの製造)
pDNR-1r(Clontech Laboratories社製)中のsacB遺伝子を、NspV切断サ
イトを付加したプライマーSAC-01(配列番号1)及びSAC-02(配列番号2)を使用したPCRにより増幅し、約1.9 kbのsacB遺伝子断片を得た。増幅条件は以下の通りである。
【0067】
プライマー:
SAC-01: 5'- GGTTCGAATACCTGCCGTTCACTATTATTTAGTG -3' (配列番号1)
SAC-02: 5'- GGTTCGAATCGGCATTTTCTTTTGCGTTTTTATTTG -3'(配列番号2)
反応液組成:
滅菌水 22 μl
2×PrimeSTAR(タカラバイオ社製) 25 μl
SAC-01(配列番号1) 1 μl
SAC-02(配列番号2) 1 μl
pDNR-1r(clontech社製)(100倍希釈) 1 μl
総量 50 μl
温度サイクル:
98℃ 10秒、55℃ 15秒及び72℃ 150秒の反応を30サイクル
sacB遺伝子断片を制限酵素NspV(タカラバイオ社製)で消化後、pK19mob(ナ
ショナルバイオリソースプロジェクトから分譲可能(ナショナルバイオリソースプロジェクトE. coli strain Cloning Vectorコレクション))のNspVサイトに接続し、sacB遺
伝子がカナマイシン耐性遺伝子の下流且つ同方向に導入されたプラスミドpK19mobsacB1を構築した。
【0068】
ロドコッカス・ロドクロウス(Rhodococcus rhodochrous)KM-02(産業技術総合研究所特許生物寄託センター;受託番号:FERM P-14457)を100 mlのMY培地(0.5% polypeptone、0.3% bact yeast extract、0.3% malt extract)に植菌し、30℃にて振盪培養したものを、Saito and Miura の方法(Biochim. Biophys. Acta 72, 619 (1963))によりゲノムを分離した。KM-02のゲノムを制限酵素EcoRIにより不完全
消化したものをpTrc99マルチクローニングサイト内に挿入し、プラスミドライブラリーを製造した。プラスミドライブラリーにより大腸菌JM109を形質転換し、カナマイシン硫酸塩(40μg/ml)とIPTG(イソプロピル−β−チオガラクトシド)(1mM)を含むLBプレート(1% Bact tryptone、0.5% bact yeast extract、1% NaCl、1.5%寒天)を用いてカナマイシン耐性を示す形質転換株を選択した。本形質転換株からプラスミドを抽出し、pKM031と命名した。
【0069】
pKM031を制限酵素HindIII(タカラバイオ社製)で切断し、KM-02株由来のカナマ
イシン耐性遺伝子とその周辺配列を含む約3.3 kbの断片をpUC18のHindIIIサイトに
挿入したプラスミドpKM041を構築した。
【0070】
pKM041内部のカナマイシン耐性遺伝子全長(約0.5 kb)を欠失させ、プラスミドpKM042を得た。欠失は、欠失させたい領域の両側に10数塩基ずつアニールするように設計したプライマーKMD-01(配列番号3)及びKMD-02(配列番号4)を使用したPCRにより行った。PCRの増幅条件は以下の通りである。
プライマー:
KMD-01: 5'-GACCATTGTTCCAGGTCGACTGGGACGAGT-3'(配列番号3)
KMD-02: 5'-ACTCGTCCCAGTCGACCTGGAACAATGGTC-3'(配列番号4)
反応液組成:
滅菌水 38.5 μl
10×PfuTurbo buffer 5 μl
dNTP(25 mM each) 3 μl
KMD-01(配列番号3) 1 μl
KMD-02(配列番号4) 1 μl
pKM041 1.5 μl
PfuTurbo(STRATAGENE社製) 1 μl
総量 50 μl
温度サイクル:
95℃ 60秒の反応を1サイクル、95℃ 50秒、60℃ 50秒及 び72℃ 16分の反応を30サイクル、72℃ 7分の反応を1サイクル
pKM042を制限酵素HindIIIで切断し、カナマイシン耐性遺伝子周辺配列約2.8 kb(
カナマイシン耐性遺伝子が欠失したもの)をpK19mobsacB1のHindIIIサイトに挿入したプ
ラスミドpKM043を構築した(図2参照)。
【0071】
上記プラスミドpKM043製造手順において、制限酵素により切断したDNA断片及びPCR産物の精製にはGFX PCR DNA band and Gel Band Purification kit(GE Healthcare社製)を、DNA同士の接続にはDNA Ligation Kit <Mighty Mix>(タカラバイオ社製)を、プラスミドの抽出にはQIAprep miniprep kit(QIAGEN社製)を用いた。
【実施例2】
【0072】
自然変異導入による薬剤耐性強化変異株: RhCmSR-01株の製造(レシピエント微生物の製造)
ロドコッカス・ロドクロウス(Rhodococcus rhodochrous)ATCC 12674株を、より高濃度のクロラムフェニコールを含む培地に継代培養することにより自然変異を誘発し、200 mg/Lのクロラムフェニコールに耐性を示す変異株RhCmSR-01株を単離した。継代培養の詳細は下記の通りである。
【0073】
ATCC 12674株を10 mg/Lクロラムフェニコールを含むMYKプレート(0.5% polypeptone、0.3% bact yeast extract、0.3% malt extract、0.2% KH2PO4、0.2% K2HPO4、1.5%寒天)に植菌し、30℃で3日間培養した。生育したコロニーを、10 mg/Lクロラムフェニコールを含むMYK培地(0.5% polypeptone、0.3% bact yeast extract、0.3% malt extract、0.2% KH2PO4、0.2% K2HPO4)に植菌し、30℃、200rpmで2日間培養した。次に、上記培養液を40 mg/Lクロラムフェニコールを含むMYK培地に0.1%植菌し、30℃、200rpmで2日間培養した。
【0074】
クロラムフェニコール濃度を次第に高めながら継代培養を繰り返し、最終的に培養液を200 mg/Lクロラムフェニコールを含むMYKプレートに植菌した。継代培養時、生育速度が遅い場合は、培養日数を2〜3日延長した。継代培養回数は4回である(クロラムフェニコール含有量:10、40、100、200(mg/ml)の順に継代)。出現したコロニーを200 mg/Lクロラムフェニコールを含むMYKプレートに更に2回植菌してシングルコロニー化することにより、RhCmSR-01株を単離した。
【実施例3】
【0075】
ATCC 12674株のカナマイシン感受性株: RhΔKm-01株の製造
大腸菌(Escherichia coli)S17-1λpirをpKM043により形質転換し、50 mg/Lカナマイシン硫酸塩含有LBプレートにて37℃で一晩生育させた。S17-1λpir/pKM043をLB培地に懸濁後、遠心分離(室温、4500ppm、2 min)を行い、菌体を回収した後、LBで一度洗浄し、LBに再懸濁した。
【0076】
RhCmSR-01をMYKプレート上にひき、30℃で3日間培養した。MYKプレート上のコロニーを数十個程度かきとり0.9% NaClに懸濁後、遠心分離(室温、15000ppm、2 min)を行い、菌体を回収した後、0.9% NaClで一度洗浄し、LBに再懸濁した。
【0077】
上記2種類の懸濁液を1対1の割合で穏やかに混合した後、LBプレート上に植菌し、30℃で一晩培養した(pKM043の接合伝達)。標的遺伝子と選択マーカーの一つが共にカナマイシン耐性遺伝子であるため、通常野生株が生育できないカナマイシン濃度での選抜を実施した。実際の選抜には、カナマイシン硫酸塩200 mg/L 及びクロラムフェニコール50 mg/L含有MYKプレート(0.5% polypeptone、0.3% bact yeast extract、0.3% malt extract、0.2% KH2PO4、0.2% K2HPO4、1.5%寒天)を使用した。
【0078】
プラスミドpKM043がゲノム内に相同組換えにより導入されていることをPCR法により確認した後、10%ショ糖を含むMYKプレートにて培養することにより(sacB対抗選択系(WO 01/31050)を使用)、pK19mobsacB1由来配列をATCC 12674ゲノム上から取り除いた。カナマイシン含有LBプレート上でのレプリカプレーティングにより、10 mg/L カナマイシン硫酸塩含有条件下では生育できないカナマイシン感受性株RhΔKm-01を単離することに成功した(図3A参照)。PCR法により野生株とRhΔKm-01株のゲノム分析を実施した結果、RhΔKm-01株においてカナマイシン耐性遺伝子長約0.5 kbに相当する領域がゲノムから欠失していることが確認できた(図3B参照)。
【実施例4】
【0079】
他のカナマイシン耐性ロドコッカス属細菌への適用
DB上で全ゲノム配列が公開されているロドコッカス・エリスロポリス(Rhodococcus erythropolis)PR4(製品評価技術基盤機構生物遺伝資源部門;受託番号:NBRC 100887)もATCC 12674がもつカナマイシン耐性遺伝子のホモログを有する。ホモロジー検索の結果、両株のカナマイシン耐性遺伝子が96.1%一致(塩基配列)という高い相同性を示したため、pKM043を用いた遺伝子欠失を実施した。各段階における培養時のベースとなる培地にはLB又はLBプレートを使用した。
【0080】
PR4も他のATCC 12674と同様に微弱なクロラムフェニコール耐性を持つことが確認できたため、実施例2と同様の方法と同様の方法により、120 mg/L Cmに対して耐性を示す変異株RhCmSR-09を製造し、単離した。継代培養回数は7回である(クロラムフェニコール含有量:10、20、30、60、80、100、120(mg/ml)の順に継代)。
【0081】
実施例3と同様の方法により、カナマイシン感受性株RhΔKm-10を単離した(図4A参照)。RhΔKm-10のゲノムからカナマイシン耐性遺伝子が欠失していることを確認した(図4B参照)。
【配列表フリーテキスト】
【0082】
配列番号1:合成DNA
配列番号2:合成DNA
配列番号3:合成DNA
配列番号4:合成DNA
【特許請求の範囲】
【請求項1】
ロドコッカス属細菌において、標的とする薬剤耐性遺伝子が欠失又は不活性化されたことにより当該薬剤に対して感受性を示す微生物。
【請求項2】
前記欠失又は不活性化が、ドナー微生物からレシピエント微生物への接合伝達を利用した、以下の工程(a)〜(d)を含む方法により行われるものである、請求項1記載の微生物。
(a)レシピエント微生物として、ドナー微生物生育阻害マーカーが強化されたロドコッカス属細菌を作製する工程;
(b)ドナー微生物として、(i)レシピエント微生物中の標的の薬剤耐性遺伝子とその周辺の塩基配列とを含む塩基配列において当該耐性遺伝子を欠失又は不活性化させた塩基配列領域、(ii)当該ドナー微生物において機能する接合伝達開始領域、(iii)当該ドナー微生物において機能する複製開始領域、(iv) レシピエント微生物生育阻害マーカー、及び(v)レシピエント微生物に対する条件致死遺伝子を含む、遺伝子改変用プラスミドを用いて形質転換された微生物を作製する工程;
(c)前記(b)で作製されたドナー微生物から前記(a)で作製されたレシピエント微生物への接合伝達を行うことにより、当該レシピエント微生物の形質転換体を作製する工程;並びに
(d)前記(c)で作製された形質転換体を、前記条件致死遺伝子が機能し得る培養条件で培養する工程。
【請求項3】
ロドコッカス属細菌が、ロドコッカス・ロドクロウス(Rhodococcus rhodochrous)又はロドコッカス・エリスロポリス(Rhodococcus erythropolis)である、請求項1又は2記載の微生物。
【請求項4】
欠失又は不活性化された薬剤耐性遺伝子が、カナマイシン耐性遺伝子である、請求項1〜3のいずれか1項に記載の微生物
【請求項5】
請求項1〜4のいずれか1項に記載の微生物を宿主として用いて培養し、得られる培養物から目的タンパク質を採取することを特徴とする、タンパク質製造方法。
【請求項1】
ロドコッカス属細菌において、標的とする薬剤耐性遺伝子が欠失又は不活性化されたことにより当該薬剤に対して感受性を示す微生物。
【請求項2】
前記欠失又は不活性化が、ドナー微生物からレシピエント微生物への接合伝達を利用した、以下の工程(a)〜(d)を含む方法により行われるものである、請求項1記載の微生物。
(a)レシピエント微生物として、ドナー微生物生育阻害マーカーが強化されたロドコッカス属細菌を作製する工程;
(b)ドナー微生物として、(i)レシピエント微生物中の標的の薬剤耐性遺伝子とその周辺の塩基配列とを含む塩基配列において当該耐性遺伝子を欠失又は不活性化させた塩基配列領域、(ii)当該ドナー微生物において機能する接合伝達開始領域、(iii)当該ドナー微生物において機能する複製開始領域、(iv) レシピエント微生物生育阻害マーカー、及び(v)レシピエント微生物に対する条件致死遺伝子を含む、遺伝子改変用プラスミドを用いて形質転換された微生物を作製する工程;
(c)前記(b)で作製されたドナー微生物から前記(a)で作製されたレシピエント微生物への接合伝達を行うことにより、当該レシピエント微生物の形質転換体を作製する工程;並びに
(d)前記(c)で作製された形質転換体を、前記条件致死遺伝子が機能し得る培養条件で培養する工程。
【請求項3】
ロドコッカス属細菌が、ロドコッカス・ロドクロウス(Rhodococcus rhodochrous)又はロドコッカス・エリスロポリス(Rhodococcus erythropolis)である、請求項1又は2記載の微生物。
【請求項4】
欠失又は不活性化された薬剤耐性遺伝子が、カナマイシン耐性遺伝子である、請求項1〜3のいずれか1項に記載の微生物
【請求項5】
請求項1〜4のいずれか1項に記載の微生物を宿主として用いて培養し、得られる培養物から目的タンパク質を採取することを特徴とする、タンパク質製造方法。
【図1】


【図2】
【図3】

【図4】
【図2】
【図3】
【図4】
【公開番号】特開2011−200134(P2011−200134A)
【公開日】平成23年10月13日(2011.10.13)
【国際特許分類】
【出願番号】特願2010−68406(P2010−68406)
【出願日】平成22年3月24日(2010.3.24)
【出願人】(000006035)三菱レイヨン株式会社 (2,875)
【Fターム(参考)】
【公開日】平成23年10月13日(2011.10.13)
【国際特許分類】
【出願日】平成22年3月24日(2010.3.24)
【出願人】(000006035)三菱レイヨン株式会社 (2,875)
【Fターム(参考)】
[ Back to top ]