
藻類由来のジアシルグリセロールアシルトランスフェラーゼ2遺伝子およびそれによってコードされるタンパク質
本開示は、藻類由来のジアシルグリセロールアシルトランスフェラーゼ2(DGAT2)およびDGAT2をコードする遺伝子の単離、精製、および特徴づけに関する。DGAT2は、DGAT1よりも効率的にトリアシルグリセロール内に超長鎖多価不飽和脂肪酸を取り込むことができる。本開示は、DGAT2遺伝子を用いることによって種子の油含有量、脂肪酸合成、および脂肪酸組成を調節する方法、ならびに前記遺伝子で形質転換した組織および植物に関する。本開示はまた、本開示の導入するDNA配列を含むゲノムを有する遺伝子導入植物、植物組織、および植物の種子、ならびに前記植物および植物の種子を産生する方法に関する。

【発明の詳細な説明】
【技術分野】
【0001】
優先権主張
本願は、「藻類由来のジアシルグリセロールアシルトランスフェラーゼ2遺伝子およびそれによってコードされるタンパク質」について2007年12月21日に出願された米国仮特許出願第60/008,752号の出願日の利益を主張する。
【0002】
技術分野
本開示は、概してバイオテクノロジー、とりわけ植物特性の遺伝子操作に有用な遺伝子に関する。特定の実施形態において、本開示は、単離および/または精製したポリペプチド、およびジアシルグリセロールアシルトランスフェラーゼ2(DGAT2)をコードする核酸、ならびにそれらの使用方法に関する。
【背景技術】
【0003】
背景
油料種子作物は重要な農産物である。植物の種子油は、人の食物に必須の多価不飽和脂肪酸の主要な供給源であり、かつ化学工業にとっての再生可能な原料である。発達中の種子の色素体中にある脂肪酸合成酵素複合体という酵素は、細胞質のアシルCoAプールに導かれる脂肪酸の生合成に関与して、トリアシルグリセロール蓄積を維持する。トリアシルグリセロール(TAG)の生合成は、一次基質としてのグリセロール−3−リン酸および脂肪酸アシルCoAとともに小胞体に位置する。植物の貯蔵脂質の生体集合には、3つのアシル基転移酵素、すなわちグリセロール−3−リン酸アシルトランスフェラーゼ(GPAT、EC2.3.1.15)、リゾホスファチジン酸アシルトランスフェラーゼ(LPAT、EC2.3.1.51)、およびジアシルグリセロールアシルトランスフェラーゼ(DGAT、EC2.3.1.20)がある。これら3つのアシルトランスフェラーゼは、ケネディ経路として一般的に知られる生化学過程である、TAGの形成へのDGATによるsn−1,2−ジアシルグリセロール(DAG)のアシル化という最終工程でグリセロール骨格の段階的なアシル化を触媒する。TAGを生成するためのDGATが介在するグリセロール骨格のアシル化は、植物の脂質蓄積における律速段階と提唱されている。従って、DGATは植物脂質生合成の遺伝子改変における標的である。
【発明の概要】
【0004】
発明の開示
本発明者らは、T.シュードナナ(T.pseudonana)のジアシルグリセロールアシルトランスフェラーゼ2(DGAT)と少なくとも90%の配列同一性を有するポリペプチドの属を本明細書において開示する。これらのポリペプチドを用いて、植物における多価不飽和脂肪酸の量を変えることができる。また、タラシオシラ・シュードナナ(Thalassiosira pseudonana)のDGAT2のジアシルグリセロールトランスフェラーゼ触媒ドメインを含むポリペプチド、およびDGAT2のジアシルグリセロールトランスフェラーゼ触媒ドメインと少なくとも90%の配列同一性を有するポリペプチドも開示する。さらに、T.シュードナナのDGAT2と少なくとも90%の配列同一性を有するポリペプチドをコードするポリヌクレオチド配列、およびT.シュードナナのDGAT2のジアシルグリセロールトランスフェラーゼドメインと少なくとも90%の同一性を有するポリペプチドをコードするポリヌクレオチドを記載する。
【0005】
本明細書において、本発明者らはT.シュードナナ由来の単離および精製したジアシルグリセロールアシルトランスフェラーゼ2(DGAT)遺伝子およびcDNA配列を開示する。また、T.シュードナナ由来の全長DGAT2 cDNA配列、および前記DGAT2 cDNAと少なくとも80%の配列同一性を有するcDNA配列も開示する。一部の実施形態においては、これらのcDNA配列はベクター内に含まれていてもよい。商業用植物油の産出量を増加させるため、またはそれらの組成を改変して植物および植物生成物の特定の商業的改良を達成するために、これらのポリヌクレオチドを用いて植物におけるトリアシルグリセロールの天然形成を改変することができる。
【0006】
T.シュードナナ、ならびにクラミドモナス・ラインハーディ(Chlamydomonas reinhardtii)、オストレオコッカス・ルシマリヌス(Ostreococcus lucimarinus)、オストレオコッカス・タウリ(Ostreococcus tauri)、およびファエオダクチルム・トリコルヌツム(Phaeodactylum tricornutum)を含む藻類の別の種由来のDGAT2ファミリーの別の単離および精製した遺伝子、ならびにcDNA配列も開示する。また、商業用植物油の産出量を増加させるため、またはそれらの組成を改変して植物および植物生成物の特定の商業的改良を達成するために、これらのポリヌクレオチドを用いて植物におけるトリアシルグリセロールの天然形成を改変することができる。
【0007】
核酸構築物を含む遺伝子導入植物も開示する。細胞または植物を形質転換する方法であって、細胞または植物の中に単離した、精製した、または組換えの核酸を導入することを含む方法を記載する。遺伝子的に形質転換された植物の種子を産生するプロセスには、植物の種子の中に核酸を導入することが含まれる。一部の実施形態において、これらの方法は、植物の種子油含有量を変化させるために、植物を改変するのに用いることができる。
【0008】
最も一般的に述べると、一部の例は、藻類由来のDGAT2遺伝子の単離、精製、および特徴づけ、ならびに超長鎖多価不飽和脂肪酸の生成におけるDGAT2の有用性を開示する。前述のことは、添付した図面に関連して進む、以下のいくつかの実施形態の詳細な説明からより明白になるであろう。
【図面の簡単な説明】
【0009】
【図1A】図1Aは、T.シュードナナDGAT2の全長DGAT2 cDNA配列(配列番号:2)に相当する推定アミノ酸配列(配列番号:1)を表す。
【図1B】図1Aの続き。
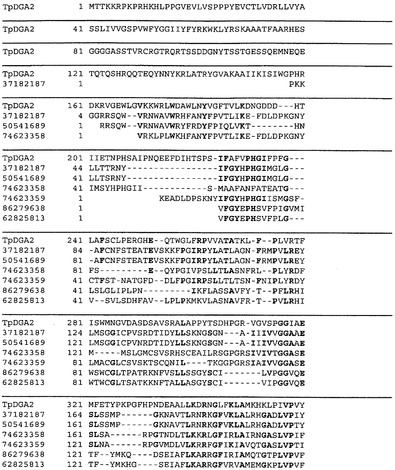
【図2A】図2Aは、配列番号:1(TpDGA2)とすべて2型ジアシルグリセロールアシルトランスフェラーゼであるgi:37182187、gi:50541689、gi:74623358、gi:74623359、gi:86279638、およびgi:62825813との間の配列アラインメントを表す。前記配列のうち、4以上に共通するアミノ酸を太字で示す。これら2型ジアシルグリセロールアシルトランスフェラーゼのジアシルグリセロールトランスフェラーゼ触媒ドメインを含むアミノ酸配列は、236−365(TpDGA2);79−208(gi:37182187);76−205(gi:50541689);76−205(gi:74623358);34−165(gi:74623359);33−165(gi:86279638);36−165(gi:62825813)残基からなる。
【図2B】図2Aの続き。
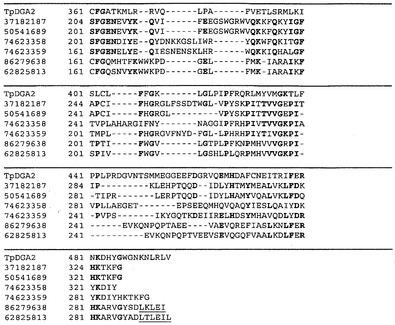
【図3】図3Aは、配列番号:1に相同なポリペプチド配列の一例(配列番号:3)を表す。図3Bは、配列番号:2に少なくとも90%相同であって、配列番号:3のポリペプチドの一部をコードするポリヌクレオチド配列(配列番号:4)の一部を表す。
【図4】図4Aは、配列番号:1に相同なポリペプチド配列の別の例(配列番号:5)を表す。図4Bは、配列番号:2に少なくとも90%相同であって、配列番号:5のポリペプチドの一部をコードするポリヌクレオチド配列(配列番号:6)の一部を表す。
【図5】図5Aは、配列番号:1に相同なポリペプチド配列の別の例(配列番号:7)を表す。図5Bは、配列番号:2に少なくとも90%相同であって、配列番号:7のポリペプチドの一部をコードするポリヌクレオチド配列(配列番号:8)の一部を表す。
【図6】図6Aは、配列番号:1に相同なポリペプチド配列の別の例(配列番号:9)を表す。図6Bは、配列番号:2に少なくとも90%相同であって、配列番号:9のポリペプチドの一部をコードするポリヌクレオチド配列(配列番号:10)の一部を表す。
【図7】図7Aは、配列番号:1に相同なポリペプチド配列の別の例(配列番号:11)を表す。図7Bは、配列番号:2に少なくとも90%相同であって、配列番号:11のポリペプチドの一部をコードするポリヌクレオチド配列(配列番号:12)の一部を表す。
【図8】図8Aは、配列番号:1に相同なポリペプチド配列の別の例(配列番号:13)を表す。図8Bは、配列番号:2に少なくとも90%相同であって、配列番号:13のポリペプチドの一部をコードするポリヌクレオチド配列(配列番号:14)の一部を表す。
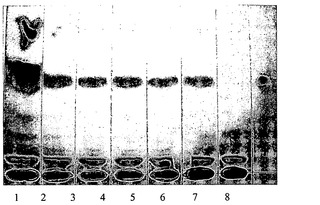
【図9】図9は、酵母変異体H1246MAT−α(DGAT、PDAT−、ASAT1−、ASAT2−、TAG形成を欠失している)でTpDGAT2およびAtDGAT1を発現させることによって生成されたTAGの薄層クロマトグラフィー(TLC)分析を表す。レーン1はAtDGAT1の発現を示し、レーン2〜6は、TpDGAT2の発現を示す。レーン2〜6ではクリアなTAG(トリアシルグリセロール)のバンドが観察された。レーン8は空ベクター(pYES2.1)コントロールを示し、このレーンにはTAG(トリアシルグリセロール)のバンドは存在しない。レーン8の右側のレーンには、TAGマーカーとして用いることができるTAG標準物質をロードした。
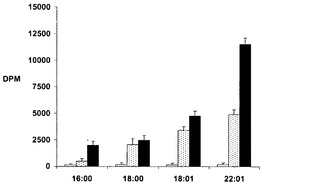
【図10】図10は、空プラスミド(pYES2.1コントロール;空のバー)、T.シュードナナDGAT2 cDNA(pYES:DGAT;点状のバー)、およびシロイヌナズナ(A.thaliana)DGATcDNA(黒無地のバー)で形質転換された酵母変異体H1246MAT−αにおけるDGAT活性を示す。誘導した酵母細胞の溶解物から調製したミクロゾーム膜画分を、アシル基供与体としての種々の14C標識されたアシルCoA、および受容体としての標識されていないsn−1,2ジオレインを用いてDGAT活性についてアッセイした。本明細書において相対的DGAT活性をDPM(TAG内に取り込まれた14C標識された基質の量)として表した。当該結果は、TpDGAT2およびAtDGAT1の基質選択性および相対的活性を示す。
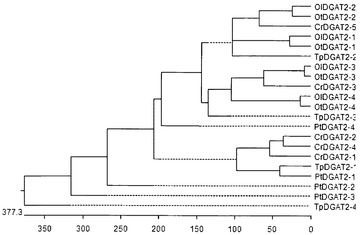
【図11】図11は、TpDGAT2のアミノ酸配列(TpDGAT2−1)とT.シュードナナ由来または他の藻類種(Cr:クラミドモナス・ラインハーディ、Ol:オストレオコッカス・ルシマリヌス、Ot:オストレオコッカス・タウリ、Pt:ファエオダクチルム・トリコルヌツム)由来のそのファミリーメンバーとのホモロジー比較を表す。TpDGAT2(TpDGAT2−1)は、そのファミリーメンバーであるTpDGAT2−2、TpDGAT2−3、およびTpDGAT2−4とそれぞれ24%、25%、および17%の配列同一性を共有している。種々の藻類種の中で、TpDGAT2(TpDGAT2−1)は、PtDGAT2−1と高い配列類似性(48%の配列同一性)を、CrDGAT2−1、CrDGAT2−2、およびCrDGAT2−4と比較的高い類似性(それぞれ20%、23%、および24%)を示す。
【発明を実施するための形態】
【0010】
I.いくつかの実施形態の概説
本明細書において、T.シュードナナの単離および精製した2型ジアシルグリセロールアシルトランスフェラーゼ(DGAT2)を開示する。他の生物および他の生物由来の細胞の中で超長鎖多価不飽和脂肪酸(VLCPUFA)の合成を改変するこのポリペプチドの驚くべき能力を用いて、所望の脂肪酸組成を有する遺伝子導入植物および植物の種子を産生するために、植物および植物の種子を形質転換する。この開示には、T.シュードナナDGAT2のものと少なくとも90%の配列同一性のアミノ酸配列を有するDGAT2活性を示すポリペプチドが含まれる。特定の実施形態において、これらのポリペプチド配列は、例えば配列番号:1、配列番号:3、配列番号:5、配列番号:7、配列番号:9、配列番号:11、または配列番号13を含む。また、T.シュードナナDGAT2のジアシルグリセロールアシルトランスフェラーゼ触媒ドメインと少なくとも90%の配列同一性を有する配列を含むポリペプチドを開示する。特定の実施形態において、これらのポリペプチド配列は、例えば配列番号15、配列番号:17、または配列番号:19を含む。図2にはT.シュードナナDGAT2のジアシルグリセロールアシルトランスフェラーゼ触媒ドメインが表されており、それはT.シュードナナDGAT2の完全な開示されているポリペプチド配列におけるアミノ酸残基236〜365からなる。
【0011】
配列番号:15のポリペプチドは、T.シュードナナDGAT2のジアシルグリセロールアシルトランスフェラーゼドメインを含む。一部の実施形態は、配列番号:15の単離または精製したポリペプチドと少なくとも約90%、91%、92%、93%、94%、95%、96%、97%、98%、99%、99.5%、99.6%、または99.7%の配列同一性を有する配列、例えば配列番号:1を含む単離または精製したポリペプチドに関する。特定の実施形態において、これらのポリペプチドはジアシルグリセロールアシルトランスフェラーゼ活性を有する。ジアシルグリセロールアシルトランスフェラーゼ活性は、例えばin vitroの酵素アッセイにより当業者によって容易に測定できる。この方法は実施例4において詳細に記載されており、このアッセイの代表的な結果は図10に示されている。当業者によって理解されるように、当該開示はまた、配列番号:15に90%以上の同一性の推定アミノ酸配列を含むタンパク質をコードする植物および藻類由来の実質的に相同なDNA配列にも関する。
【0012】
DGAT2ファミリーのメンバーである藻類由来の単離または精製した他のポリペプチドは、例えば配列番号:25(TpDGAT2−2)、配列番号:27(TpDGAT2−3)、配列番号:29(TpDGAT2−4)、配列番号:31(CrDGAT2−1)、配列番号:33(CrDGAT2−2)、配列番号:35(CrDGAT2−3)、配列番号:37(CrDGAT2−4)、配列番号:39(CrDGAT2−5)、配列番号:41(OlDGAT2−1)、配列番号:43(OlDGAT2−2)、配列番号:45(OlDGAT2−3)、配列番号:47(OlDGAT2−4)、配列番号:49(OtDGAT2−1)、配列番号:51(OtDGAT2−2)、配列番号:53(OtDGAT2−3)、配列番号:55(OtDGAT2−4)、配列番号:57(PtDGAT2−1)、配列番号:59(PtDGAT2−2)、配列番号:61(PtDGAT2−3)、または配列番号:63(PtDGAT2−4)と少なくとも90%同一であるアミノ酸配列を含む。
【0013】
一部の実施形態は、前述したポリペプチドをコードする単離または精製した核酸(ポリヌクレオチド)に関する。これらポリヌクレオチドの配列は、例えば配列番号:2、配列番号:4、配列番号:6、配列番号:8、配列番号:10、配列番号:12、配列番号:14、配列番号:16、配列番号:18、配列番号:20、配列番号:26(TpDGAT2−2)、配列番号:28(TpDGAT2−3)、配列番号:30(TpDGAT2−4)、配列番号:32(CrDGAT2−1)、配列番号:34(CrDGAT2−2)、配列番号:36(CrDGAT2−3)、配列番号:38(CrDGAT2−4)、配列番号:40(CrDGAT2−5)、配列番号:42(OlDGAT2−1)、配列番号:44(OlDGAT2−2)、配列番号:46(OlDGAT2−3)、配列番号:48(OlDGAT2−4)、配列番号:50(OtDGAT2−1)、配列番号:52(OtDGAT2−2)、配列番号:54(OtDGAT2−3)、配列番号:56(OtDGAT2−4)、配列番号:58(PtDGAT2−1)、配列番号:60(PtDGAT2−2)、配列番号:62(PtDGAT2−3)、または配列番号:64(PtDGAT2−4)を含んでもよい。一部の実施形態において、前記ポリヌクレオチド配列は、開示されているポリペプチドをコードする開示されているヌクレオチド配列の塩基と少なくとも約80%、81%、82%、83%、84%、85%、86%、87%、88%、89%、90%、91%、92%、93%、94%、95%、96%、97%、98%、99%、99.5%、99.6%、または99.7%のパーセンテージ同一性を有する。そのようなポリヌクレオチドのいくつかの例が、配列番号:21〜24である。技術を有する実践者によって理解されるように、核酸配列のわずかな変化は、コードされるポリペプチドのアミノ酸配列を必ずしも変化させない。コードされるポリペプチドのアミノ酸配列を変化させる特定の遺伝子配列におけるヌクレオチドの同一性の変化は、当該遺伝子の有効性の低下または増大をもたらし得ること、ならびに一部の応用(すなわち、アンチセンス、共抑制、またはRNAi)において、部分的配列はしばしば全長型と同じ程度に効果的に作用することは当業者によって理解されるであろう。遺伝子配列を変更または短くし得る方法は当業者に周知であり、当該改変遺伝子の有効性を調べる方法も同様である。特定の実施形態において、有効性は例えば従来のガスクロマトグラフィーによって容易に調べることができる。従って、当該遺伝子のそのようなすべての変化が本明細書の一部として含まれる。
【0014】
一部の実施形態は、配列番号:2と少なくとも80%の相同性を有する単離または精製したポリヌクレオチド、例えば配列番号:2、配列番号:15、配列番号:16、配列番号:17、配列番号:18、配列番号:20、配列番号:21〜24を含むベクターに関する。従って、例えば配列番号:2、例えば配列番号:2、配列番号:15、配列番号:16、配列番号:17、配列番号:18、配列番号:20、または配列番号:21〜24、あるいはその一部からなる群から選択される配列を含むベクターを調製するための方法、植物細胞内へのアンチセンス方向に当該配列または部分的配列、あるいはその相補鎖を導入するための方法を提供する。
【0015】
特定の実施形態は、藻類におけるDGAT2ファミリーのメンバーと少なくとも80%の相同性を有するポリヌクレオチドを含むベクターに関する。これらのベクターは、例えば配列番号:26、配列番号:28、配列番号:30、配列番号:32、配列番号:34、配列番号:36、配列番号:38、配列番号:40、配列番号:42、配列番号:44、配列番号:46、配列番号:48、配列番号:50、配列番号:52、配列番号:54、配列番号:56、配列番号:58、配列番号:60、配列番号:62、または配列番号:64のポリヌクレオチド配列を含んでもよい。
【0016】
一部の実施形態において、DGAT2活性を備えるポリペプチドを生成する遺伝子導入植物を作り出すために、前記単離および精製したポリヌクレオチド、ならびにこれらの単離および精製したポリヌクレオチドを含むベクターを用いてもよい。従って、一実施形態は、配列番号:2と少なくとも80%の相同性を有する単離または精製したポリヌクレオチド、例えば配列番号:2、配列番号:15、配列番号:16、配列番号:17、配列番号:18、配列番号:20、または配列番号:21〜24の配列を有するデオキシリボ核酸分子を含む遺伝子導入植物および植物の種子に関する。他の実施形態は、藻類におけるDGAT2ファミリーの別のメンバーと少なくとも80%の相同性を有する単離または精製したポリヌクレオチド、例えば配列番号:26、配列番号:28、配列番号:30、配列番号:32、配列番号:34、配列番号:36、配列番号:38、配列番号:40、配列番号:42、配列番号:44、配列番号:46、配列番号:48、配列番号:50、配列番号:52、配列番号:54、配列番号:56、配列番号:58、配列番号:60、配列番号:62、または配列番号:64の配列を有するデオキシリボ核酸分子を含む遺伝子導入植物および植物の種子に関する。これら実施形態の植物は、当該核酸構築物を欠いている植物における量と比較して、種子中の多価不飽和脂肪酸の量が変化していてもよい。前記植物における脂肪酸は、約70%以上の多価不飽和脂肪酸であってよい。
【0017】
一実施形態は、そのような植物および植物の種子を産生する方法を含む。前記方法は、配列番号:1、3、5、7、9、11、13、15、17、および19からなる群から選択されるポリペプチドと少なくとも90%の配列同一性を有するポリペプチド、または藻類におけるDGAT2ファミリーのポリペプチドと少なくとも90%の配列同一性を有するポリペプチドをコードするポリヌクレオチド;例えば配列番号:25、配列番号:27、配列番号:29、配列番号:31、配列番号:33、配列番号:35、配列番号:37、配列番号:39、配列番号:41、配列番号:43、配列番号:45、配列番号:47、配列番号:49、配列番号:51、配列番号:53、配列番号:55、配列番号:57、配列番号:59、配列番号:61、または配列番号:63を含む核酸構築物を作り出すこと、ならびに植物内に当該構築物を導入することを含む。この実施形態の方法は、当業者に公知の任意の方法、非制限的な例であるアグロバクテリウム(Agrobacterium)を介して行われる形質転換の方法によって達成され得る。特定の実施形態において、前記方法は、Brusicaピルビン酸脱水素酵素キナーゼ活性を備えるポリペプチドをコードするポリヌクレオチド、ジアシルグリセロールアセチルトランスフェラーゼ活性を備えるポリペプチドをコードするポリヌクレオチド、および/またはグリセロール−3−リン酸脱水素酵素活性を備えるポリペプチドをコードするポリヌクレオチドを、当該植物内に導入することをさらに含む。この方法は、前記植物がシロイヌナズナ(Arabidopsis thaliana)、ボリジ(Borago)種、カノーラ(Canola)、リキヌス(Ricinus)種、テオブロマ(Theobroma)種、トウモロコシ(Zea)種、ゴシッピウム(Gossypium)種、ハマナ(Crambe)種、クフェア(Cuphea)種、アマ(Linum)種、レスクェレラ(Lesquerella)種、リムナンテス(Limnanthes)種、リノーラ(Linola)、ノウゼンハレン(Tropaeolum)種、マツヨイグサ(Oenothera)種、オリーブ(Olea)種、アブラヤシ(Elaeis)種、ラッカセイ(Arachis)種、ナタネ(rapeseed)、ベニバナ(Carthamus)種、ダイズ(Glycine)種、ソヤ(Soja)種、ヒマワリ(Helianthus)種、タバコ(Nicotiana)種、ベルノニア(Vernonia)種、コムギ(Triticum)種、オオムギ(Hordeum)種、オリザ(Oryza)種、カラスムギ(Avena)種、モロコシ(Sorghum)種、ライムギ(Secale)種、アブラナ(Brassicaceae)、およびイネ科(Gramineae)植物ファミリーの他のメンバーからなる群から選択されるところで実施してもよい。
【0018】
一部の実施形態において、前記方法は、導入した核酸構築物を含む植物から種子を収穫すること、および前記収穫した種子から油を抽出することをさらに含む。従って、他の実施形態は、当該方法によって産生した植物、および当該方法によって産生した植物から抽出した油を含む。
【0019】
DGAT2遺伝子またはその一部を用いることによって可能である操作および成果物のいくつかは、以下の、油含有量が増加または減少した種子、超長鎖多価不飽和脂肪酸含有量が増大した油を含む種子、および超長鎖多価不飽和脂肪酸を蓄積する増大または変化した能力を示す植物を含むが、これに制限されない。
【0020】
II.略語
CaMV カリフラワーモザイクウイルス
cDNA 相補的DNA
CERV カーネーションエッチドリングウイルス
CrDGAT2 クラミドモナス・ラインハーディ2型ジアシルグリセロールトランスフェラーゼ
DAG sn−1,2−ジアシルグリセロール
DGAT ジアシルグリセロールアシルトランスフェラーゼ
DGAT2 2型ジアシルグリセロールトランスフェラーゼ
DHA ドコサヘキサエン酸
DNA デオキシリボ核酸
EPA エイコサペンタエン酸
GPAT グリセロール3リン酸アシルトランスフェラーゼ
LPAT リゾホスファチジン酸アシルトランスフェラーゼ
OlDGAT2 オストレオコッカス・ルシマリヌス2型ジアシルグリセロールトランスフェラーゼ
OtDGAT2 オストレオコッカス・タウリ2型ジアシルグリセロールトランスフェラーゼ
PCR ポリメラーゼ連鎖反応
PtDGAT2 ファエオダクチルム・トリコルヌツ2型ジアシルグリセロールトランスフェラーゼ
RNA リボ核酸
RNAi RNA干渉
RT−PCR 逆転写PCR
T35S CaMV35Sターミネーター
TAG トリアシルグリセロール
TLC 薄層クロマトグラフィー
Tmas マンノピン合成ターミネーター
Tnos ノパリン合成ターミネーター
TpDGAT2 T.シュードナナ2型ジアシルグリセロールトランスフェラーゼ
TrbcS リブロース2リン酸カルボキシラーゼ小サブユニット終結領域
VLCPUFA 超長鎖多価不飽和脂肪酸
【0021】
III.用語
本開示の種々の実施形態の概説を容易にするために、以下の特定の用語の説明を提供する。
【0022】
相補的ヌクレオチド配列:配列の「相補的ヌクレオチド配列」とは、そのヌクレオチドが本開示の配列のものと相補的であり、その方向が反転している(逆平行配列である)任意のDNAを意味すると理解される。
【0023】
配列ホモロジーの程度またはパーセンテージ:「配列ホモロジーの程度またはパーセンテージ」という用語は、最適アラインメント後の2つの配列間の配列同一性の程度またはパーセンテージを表す。配列同一性のパーセンテージ(あるいは程度または同一性)は、比較ウィンドウ上で2つの最適に整列された配列を比較することによって決定されるものであって、前記2つの配列の最適アラインメントについて、比較ウィンドウにおけるペプチドまたはポリヌクレオチド配列の一部は、参照配列(付加または欠失を含まない)と比較して付加または欠失(すなわち、ギャップ)を含み得る。前記パーセンテージは、両配列において一致するアミノ酸残基または核酸塩基が生じている位置の数を測定してマッチした位置の数を出し、マッチした位置の数を比較ウィンドウにおける位置の総数で割り、かつ当該結果に100を掛けて配列同一性のパーセンテージを出すことによって算出される。
【0024】
単離および/または精製した相同配列:「単離および/または精製した相同配列」とは、ヌクレオチド配列の塩基またはポリペプチド配列のアミノ酸と、少なくとも約80%、81%、82%、83%、84%、85%、86%、87%、88%、89%、90%、91%、92%、93%、94%、95%、96%、97%、98%、99%、99.5%、99.6%、または99.7%のパーセンテージ同一性を有する単離および/または精製した配列を意味すると理解される。このパーセンテージは完全に統計的なものであり、無作為とそれらの長さ全体にわたる2つのヌクレオチド配列との間で差をもたらす可能性がある。例えば、シングル配列アライメントおよびマルチプル配列アライメントを行うように設計されたコンピュータープログラムによって、配列同一性を測定することができる。当業者によって理解されるように、この開示はコドン使用の縮重を包含すると理解されるであろう。さらに、ポリペプチドのアミノ酸配列において、前記ポリペプチドの構造または作用を崩壊させることなく保存的置換が行われてもよいと当業者によって理解されるであろう。保存的置換は、お互いに対して類似した疎水性、極性、およびR鎖長を有するアミノ酸を置換することによって、熟練した技術者によって達成される。また、異なる種由来の相同タンパク質の整列した配列を比較することによって、コードされたタンパク質の基本的作用を変えることなく種間で変異しているアミノ酸残基を位置づけることによって、保存的置換を特定してもよい。
【0025】
単離した:当業者によって理解されるように、「単離した」とは、それらの天然環境から「単離」されているポリペプチドを表す。
【0026】
ヌクレオチド、ポリヌクレオチド、または核酸配列:「ヌクレオチド、ポリヌクレオチド、または核酸配列」とは、単量体および二量体の(いわゆる、並行した)形態の2本鎖または1本鎖のDNA、ならびに前記DNAの転写産物の両方を意味すると理解されるであろう。
【0027】
配列同一性:2つのアミノ酸またはヌクレオチドの配列におけるアミノ酸またはヌクレオチドの残基の配列が、以下に記載するように最大の一致を目的として整列させた時に同じである場合、前記2つの配列は「同一」であると言われる。2つ(またはそれより多く)のペプチドまたはポリヌクレオチド間の配列比較は、典型的に、配列類似性について局所領域を同定および比較するために、断片または「比較ウィンドウ」にわたって最適に整列された2つの配列の配列を比較することによって行われる。比較のための配列の最適な整列は、SmithおよびWatermanの局所的ホモロジーアルゴリズム:Ad.App.Math 2:482(1981)によって、NeddlemanおよびWunschのホモロジーアラインメントアルゴリズム:J.Mol.Biol.48:443(1970)によって、PearsonおよびLipmanの類似法の探索、Proc.Natl.Acad.Sci.(U.S.A.)85:2444(1988)によって、これらアルゴリズムのコンピュータによる実施(ウィスコンシン・ジェネティクス・ソフトウェアパッケージのGAP、BESTFIT、FASTA、およびTFASTA,ジェネティクス・コンピュータ・グループ(Genetics Computer Group:GCG),575サイエンスDr.,マディソン,ウィスコンシン州)によって、または目視検査によって行われてよい。
【0028】
前記で付された配列同一性の定義は、当業者によって用いられるであろう定義である。定義それ自体はいかなるアルゴリズムの支援も必要とせず、前記アルゴリズムは、配列同一性の算出よりもむしろ、配列の最適アラインメントを達成するために唯一有用である。
【0029】
前記で付された定義から、要するに、2つの比較される配列間の配列同一性について、明確かつ唯一の数値であって、最高または最適なアラインメントに対して得られる数値に相当する数値が存在するということになる。
【0030】
http://worldwiseweb.ncbi.nlm.nih.gov/gorf/bl2.htmlのウェブサイトで入手可能であり、2つの配列間の同一性を比較および決定するために、本発明者らおよび一般的に当業者によって習慣的に用いられるソフトウェアであるBLAST NまたはBLAST P「BLAST2配列」において、比較される配列の長さに依存するギャップコストは、前記ソフトウェアによって直接選択される(すなわち、>85の長さに対する置換行列BLOSUM−62には11.2)。
【0031】
ストリンジェントなハイブリダイゼーション:ヌクレオチド配列に関するストリンジェントな条件下でのハイブリダイゼーションとは、相補的DNAの2つのフラグメント間でのハイブリダイゼーションを保持させ得るように選択された温度およびイオン強度の条件下でのハイブリダイゼーションを意味すると理解される。他の生物、例えば植物から得られる本明細書において記載されるDGAT2遺伝子のホモログは、前記ホモログを含む適切なライブラリーをスクリーニングすることによって得ることができ、前記スクリーニングは、本明細書において開示される特定のDGAT2遺伝子のヌクレオチド配列、またはその一部もしくはプローブに関して行われるものであり、あるいはBLAST、FASTA等の配列アラインメント検索プログラムを用いた配列ホモロジー検索によって同定することができる。
【0032】
III.藻類由来のDGAT2による脂肪酸量の改変
A.概説
アブラギリおよびトウゴマ由来のDGAT2に関する最近の研究は、独特な脂肪酸を含む植物において、DGAT2は独特な脂肪酸を種子貯蔵油に導くのに重要な役割を担っている可能性があることを示唆している。脂肪種子における植物脂質生合成の遺伝子改変において、DGAT2は潜在的標的になり得ると同時に、最近特徴づけされた前記酵素は、共役脂肪酸であるエレオステアリン酸(アブラギリDGAT2)およびリシノール酸(トウゴマDGAT2)の利用にそれぞれ貢献している。どちらの酵素(アブラギリDGAT2またはトウゴマDGAT2)も、商業的に所望される長鎖オメガ−3多価不飽和脂肪酸であるエイコサペンタエン酸(EPA)およびドコサヘキサエン酸(DHA)のトリアシルグリセロール(TAG)内への取り込みに関与していない。
【0033】
海の中心的な珪藻類であるT.シュードナナは、TAG内に長鎖オメガ−3多価不飽和脂肪酸であるEPAおよびDHAを生成および蓄積することができ、高レベルの超長鎖多価不飽和脂肪酸(VLCPUFA)が蓄積した油の良好な供給源である。そのため、T.シュードナナのジアシルグリセロールアシルトランスフェラーゼ2(TpDGAT2)遺伝子は、調査および特徴づけされた。驚くべきことに、アブラギリまたはトウゴマ由来のDGAT2とは異なり、TpDGAT2はTAG内に超長鎖多価不飽和脂肪酸を効率的に取り込むことができることが発見された。TpDGAT2遺伝子を用いてT.シュードナナおよび関連する藻類の種由来のポリヌクレオチド配列を調べることによって、藻類におけるDGAT2ファミリーの他のメンバーが同定された。従って、藻類DGAT2遺伝子は、遺伝子組換えツールにおいてならびに種子内のTAGの組成および蓄積の改変に対して有用であると判断された。
【0034】
B.2型ジアシルグリセロールトランスフェラーゼ活性を備えるT.シュードナナDGAT2に相同なポリペプチド
NCBIタンパク質データベース等のタンパク質データベースをBLAST等の検索エンジンを用いて検索することによって、全長T.シュードナナDGAT2に相同であるタンパク質を発見することができる。それらは合理的設計によって同定されてもよい。合理的設計の方法は、所望のポリペプチド配列の長さの範囲内で保存的アミノ酸置換を同定すること、および当該コードされるタンパク質においてそれらの置換を引き起こすことを含む。
【0035】
T.シュードナナDGAT2の全長アミノ酸配列に関してNCBIタンパク質データベース(BLASTP)を検索すると、TpDGAT2に有意な配列ホモロジーを有するポリペプチドが明らかとなり、そのいくつかはTpDGAT2とともに整列して図2に示されている。保存されている2型ジアシルグリセロールトランスフェラーゼドメインは図2に整列されており、TpDGAT2におけるアミノ酸残基236〜365と示されている他のDGAT2ポリペプチド由来の相当する残基とからなる。前記保存されている2型ジアシルグリセロールトランスフェラーゼドメインは、NCBIの保存されているドメインのデータベース(http://www.ncbi.nlm.nih.gov/Structure/cdd/cdd.shtml)内に記載されている。この保存されているドメインに相同であるポリペプチド配列は、それが含まれるタンパク質にTpDGAT2の2型ジアシルグリセロール活性を付与する。
【0036】
相同配列を有するポリペプチドは、それらのホモログと同じ構造及び作用を示すように設計され得ると当業者によって理解される。熟練した技術者によって、図2の配列アラインメントにより本開示の実施例に具体的に記載されているものと相同なポリペプチドを設計することが可能である。そのような相同なポリペプチドは、本開示のポリペプチドへの保存的置換を含むもの、例えば配列番号:3、配列番号:5、配列番号:7、配列番号:9、配列番号:11、配列番号:13、配列番号:15、配列番号:17、または配列番号:19のポリペプチドであってよい。相同タンパク質がTpDGAT2と同様の2型ジアシルグリセロールトランスフェラーゼ活性を実質的に示すかどうかを判定する簡易な実験アッセイは当業者に公知である。そのようなアッセイは、過度に時間のかかるものでなく、高価でなく、または技術的に難しいものでない。TpDGAT2によって生成されたTAGを検出するために、例えば従来のガスクロマトグラフィーを用いてもよい。これらのアッセイのいくつかは、以下の詳細な実施例に記載されている。
【0037】
C.形質転換するためのDGAT2活性を有する核酸分子の使用
開示されている実施形態は、天然環境で採取されたゲノム、すなわち、T.シュードナナ、クラミドモナス・ラインハーディ、オストレオコッカス・ルシマリヌス、オストレオコッカス・タウリ、またはファエオダクチルム・トリコルヌツムの天然ゲノムのヌクレオチド配列を含まないと理解すべきである。一部の実施形態は、例えばイオン交換クロマトグラフィー、分子サイズに基づいた排除による、またはアフィニティーによる、または種々の溶媒中の溶解度に基づいた分画法等の分離方法から始めることによって、あるいは増幅、クローニング、およびそれにより当該配列がベクターによって運搬されることが可能となるサブクローニング等の遺伝子操作の方法から始めることによって、単離、精製または部分的に精製することが可能である配列に関する。
【0038】
さらに、前記に開示されている配列にハイブリダイズする核酸分子が含まれる。ハイブリダイゼーションの条件は、開示されているDGAT2分子をコードする核酸分子と少なくとも90%、95%、または97%の同一性がある場合にハイブリダイゼーションが生じるという点においてストリンジェントであってよい。ストリンジェントな条件は、公知のサザンハイブリダイゼーションに用いられているもの、例えば50%ホルムアミド、5×SSC(150mM NaCl、15mMクエン酸3ナトリウム)、50mMリン酸ナトリウム(pH7.6)、5×デンハート液、10%硫酸デキストラン、および20マイクログラム/ミリリッターの変性、せん断されたサーモンスパームDNAを有する溶液中で一晩42℃にてインキュベーション、その後0.1×SSC中で約65℃にてハイブリダイゼーション支持体の洗浄等を含んでよい。他の公知のハイブリダイゼーション条件が周知であり、Sambrook,et al.,Molecular Cloning:A Laboratory Manual,Third Edition,Cold Spring Harbor,N.Y.(2001)に記載されている。
【0039】
DNAの単離およびクローニングは、十分に確立されている。同様に、従来技術によって、単離した酵素をコードするDNAをベクター内に挿入および酵母細胞内に形質転換することができる。しかしながら、VLCPUFAを効率的に使用し得るDGAT2遺伝子がクローニングされていなかったため、DGAT2活性を調節することによる遺伝子改変の可能性に取り組むことは不可能であった。本発明者らは、DGAT2がTAG合成に関与し、かつDGATよりも効率的にVLCPUFAを利用することを確認した。
【0040】
DGAT2をコードする核酸分子、例えば配列番号:2、配列番号:16、配列番号:25、配列番号:27、配列番号:29、配列番号:31、配列番号:33、配列番号:35、配列番号:37、配列番号:39、配列番号:41、配列番号:43、配列番号:45、配列番号:47、配列番号:49、配列番号:51、配列番号:53、配列番号:55、配列番号:57、配列番号:59、または配列番号:61と少なくとも80%の同一性を有する配列を、生物、例えば植物内に形質転換してもよい。そのような相同配列は配列番号:21〜24によって例示されている。当該技術分野において公知のように、遺伝子および遺伝子構築物を生物、例えば植物内に導入し得る多数の方法があり、形質転換および組織培養の技術の組み合わせは、遺伝子組換え生物、例えば作物を作り出すための効果的戦略内に上手く取り入れられている。これらの方法は、他の場所(Potrykus,1991;Vasil,1994;WaldenおよびWingender,1995;Songstad,et al.,1995)で記載されており、当業者に周知である。例えば、減圧浸潤(Bechtold,et al.,1993)または有傷接種(Katavic,et al.,1994)によってアグロバクテリウムを介して行われるシロイヌナズナの形質転換に加えて、アグロバクテリウムTi−プラスミドを介して行われる形質転換(例えば、胚軸(DeBlock,et al.,1989)または子葉柄(Moloney,et al.,1989)の創傷感染)、粒子衝突/微粒子銃法(Sanford,et al.,1987;Nehra,et al.,1994;Becker,et al.,1994)、またはポリエチレングリコールを介したプロトプラスト形質転換法(Rhodes,et al.,1988;Shimamoto,et al.,1989)を用いることによって、他の植物および作物種を形質転換することが同様に可能であることに当業者は確実に気づくであろう。
【0041】
植物において、新規の遺伝子を伝達するまたは既存の遺伝子の発現を変えるための遺伝子操作によって達成されている植物代謝に対して成功した改変の例が多数ある。作物の性能(例えば、種子油または塊茎デンプンの含有量/組成;食物改良;除草剤、疾病、または害虫抵抗性;重金属耐性等)を改良するために、農学的に重要な多数の植物種の中に遺伝子を導入することが現在日常的に可能である(MacKenzieおよびJain,1997;Budziszewski,et al.,1996;Somerville,1993;KishoreおよびSomerville,1993)。
【0042】
当業者に明らかであり、他の場所(Meyer,1995;Dada,et al.,1997)で記載されているように、構成的プロモーター(例えば、CaMV35Sに基づくもの)を用いることによって、あるいは特定の細胞、組織(例えば、発達中の種子の子葉における導入遺伝子の発現に対するナピンプロモーター)、器官(例えば、根)に、特定の発達段階に、または特定の外部刺激(例えば、熱ショック)に応答して、遺伝子発現を標的化することができるプロモーターを用いることによって、導入遺伝子の発現についての任意に意図した上方または下方制御を管理するために、植物プロモーターを利用することができる。
【0043】
本明細書において使用するためのプロモーターは、誘導可能な、構成的な、または組織特異的な、あるいは前記特性の種々の組み合わせを有するものであってよい。有用なプロモーターは、構成的プロモーター、例えばカーネーションエッチドリングウイルス(CERV)、カリフラワーモザイクウイルス(CaMV)35Sプロモーター、またはとりわけ2つのCaMV35Sプロモーターを直列に含んでいる2倍に増強したカリフラワーモザイクウイルスプロモーター(「ダブル35S」プロモーターと称される)を含むが、これに制限されない。
【0044】
特定の状況では、構成的プロモーターの代わりに組織特異的または発生学的に調節されるプロモーターを用いるのが望ましい可能性がある。組織特異的プロモーターは、他の組織の発現に影響を及ぼすことなく、特定の組織において過剰発現を可能にする。実例として、種子組織における酵素の過剰発現に用いられるプロモーターは、1992年10月29日に公表されたPCTの国際公開公報第92/18634号に記載されているようなACPプロモーターである。
【0045】
当該プロモーターおよび終結調節領域は、宿主の植物細胞内で機能的であってもよく、前記植物細胞および遺伝子に対して異種(すなわち、天然に存在しない)または同種(前記宿主植物種に由来する)であってもよい。用いることができる適切なプロモーターは、前記に記載されている。
【0046】
前記終結調節領域は、プロモーターが得られた遺伝子または別の遺伝子の3’領域に由来してもよい。用いることができる適切な終結調節領域は当該技術分野において周知であり、アグロバクテリウム・ツメファシエンス(Agrobacterium tumefaciens)・ノパリン合成ターミネーター(Tnos)、A.ツメファシエンスマンノピン合成ターミネーター(Tmas)、およびCaMV35Sターミネーター(T35S)、エンドウリブロース2リン酸カルボキシラーゼ小サブユニット終結領域(TrbcS)、またはTnos終結領域を含んでよい。そのような遺伝子構築物は、アグロバクテリウムを介した宿主植物内への形質転換、および増加したイソプレノイド量についてのスクリーニングによって、活性について適切にスクリーニングすることができる。
【0047】
適切には、当該遺伝子に対するヌクレオチド配列は、GenBank(登録商標)(米国保健社会福祉省の登録商標)ヌクレオチドデータベースから抽出し、かつ切断しない制限酵素について検索してもよい。PCRプライマー中にこれらの制限酵素認識部位を組み込むこと等の従来の方法またはサブクローニングによって、これらの制限部位を当該遺伝子に付加してもよい。
【0048】
好ましくは、本明細書において使用するためのDNA構築物はベクター、最適には適切な宿主(植物)細胞における発現に適した発現ベクター内に包含されている。導入したDNA配列を含む植物を産生することができる任意のベクターが十分であると理解されるであろう。
【0049】
適切なベクターは当業者に周知であり、Pouwels,et al.,Cloning Vectors,A Laboratory Manual,Elsevier,Amsterdam(1986)等の一般的な技術参考文献に記載されている。特に適切なベクターは、Tiプラスミドベクターを含む。
【0050】
宿主細胞内にDNA構築物を導入するための形質転換法は当該技術分野において周知であり、マイクロインジェクション、ポリエチレングリコールの使用、エレクトロポレーション、高速弾道貫通(high velocity ballistic penetration)、またはアグロバクテリウムを介した形質転換等の方法を含む。植物細胞または植物の形質転換後、所望のDNAが組み込まれているそれらの植物細胞または植物を、抗生物質耐性、除草剤抵抗性、アミノ酸類似体に対する寛容等の方法によって、または表現型マーカーを用いることによって選択することができる。
【0051】
当該植物細胞が遺伝子発現の増加を示すかどうかを判定するために、種々のアッセイ、例えばノーザンブロッティングまたは定量的逆転写PCR(RT−PCR)を用いてもよい。従来の方法によって、形質転換された細胞から遺伝子導入植物全体を再生してもよい。同型株を作り出すために、改良されたイソプレノイド量を有するそのような遺伝子導入植物を繁殖および自家授粉させてもよい。そのような植物は、導入した特性に対する遺伝子を含む種子を産生し、選択された表現型を生じるであろう植物を産生するように成長させることができる。
【0052】
本開示に従った改変に特に好ましい植物は、シロイヌナズナ、ボリジ(ボリジ種)、カノーラ、トウゴマ(Ricinus communis)(リキヌス種)、カカオ豆(Theobroma cacao)(テオブロマ種)、トウモロコシ(Zea mays)(トウモロコシ種)、綿(ゴシッピウム種)、ハマナ種、クフェア種、亜麻(アマ種)、レスクェレラ種、およびリムナンテス種、リノーラ、ナスタチウム(ノウゼンハレン種)、マツヨイグサ種、オリーブ(オリーブ種)、ヤシ(アブラヤシ種)、ピーナッツ(ラッカセイ種)、ナタネ、ベニバナ(ベニバナ種)、大豆(ダイズ種およびソヤ種)、ヒマワリ(ヒマワリ種)、タバコ(タバコ種)、ベルノニア種、小麦(コムギ種)、大麦(オオムギ種)、米(オリザ種)、オートムギ(カラスムギ種)、モロコシ(モロコシ種)、ライ麦(ライムギ種)、またはイネ科植物ファミリーの他のメンバーを含む。
【0053】
一部の実施形態を用いて、油料種子作物から生成された油料種子の産出量または組成を改変する。油料種子作物は、食用のまたは産業的に有用な油を商業的に重要な産出量で作り出すことができる植物種であり、前記に挙げた植物種の多くを含む。そのような油料種子作物は当業者に周知である。
【0054】
一実施例において、DGAT2をコードするヌクレオチド配列で形質転換した植物を栽培する。遺伝子導入植物の種子を収穫して、前記種子の脂肪酸を抽出する。抽出した脂肪酸を、その後の組成物、例えば医薬組成物、栄養補助組成物、または食品組成物内への取り込みに用いる。
【0055】
特定の実施形態において、形質転換されるべき植物に関して、油の産出を増大または変化させる他の方法(例えば、植物における発現のために、例えばBrassicaピルビン酸脱水素酵素キナーゼ活性を有するペプチドをコードする核酸配列(例えば、Marilla,et al.に対する米国特許第7,214,859号(2007年5月8日)、Zou,et al.に対する米国特許第6,500,670号(2002年12月)、およびRandall,et al.に対する米国特許第6,256,636号(2001年7月)を参照)、ジアシルグリセロールアシルトランスフェラーゼ活性を有するペプチドをコードする核酸配列(例えば、Zou,et al.に対する米国特許第7,015,373号および米国特許第6,500,670号(2002年12月)を参照)、およびグリセロール−3−リン酸脱水素酵素活性を有するペプチドをコードする核酸配列(例えば、米国特許第7,112,724号を参照)、ならびにその組み合わせを含む群から選択される核酸配列を取り込むこと)を用いてもよい。
【0056】
実施形態は、本明細書において詳細に記載されている具体的な実施例に加えて、種々の改変および代替的形態を受け入れる余地がある。従って、実施形態は開示されている特定の形態に制限されない。むしろ、本開示の範囲は、以下の添付した特許請求の範囲に含まれるすべての改変、同等物、および代替物を包含する。
【実施例】
【0057】
実施例1:DNA操作
DNAの調製、プラスミドの増殖、および単離には、標準的方法および手順を用いた(Sambrook,et al.,1989)。配列決定は、Taq DyeDeoxy(商標)ターミネーターサイクルシークエンシングキット(Applied Biosystems社)を用いて、Applied Biosystems社のモデル373A DNAシークエンシングシステムで行った。当該ヌクレオチドおよび推定アミノ酸配列を、BLASTプログラムを用いることによって、データバングで入手可能な配列と比較した(Altschul,et al.,1990)。当該技術分野において公知であるように、NCBIのヌクレオチドおよびタンパク質データベースにおける他の脂肪酸ジアシルグリセロールアシルトランスフェラーゼ遺伝子とのホモロジーに基づいて、DGAT2クローンを同定した。
【0058】
実施例2:TpDGAT2を発現することによって、酵母形質転換体においてin vivo形成されたトリアシルグリセロール(TAG)
DGAT2遺伝子をpYES2.1(Invitrogen社)内に挿入した。当該構築物をシークエンシングによって確認し、出芽酵母サッカロマイセス・セレヴィシエ(Saccharomyces cerevisiae)の株H1246MAT−αを形質転換するために、pYES2.1/TpDGAT2を用いた。この変異株は4重の変異体である(DGAT−,PDAT−,ASAT−,ASAT2−)。プラスミドDNAを推定される形質転換体から単離し、pYES2.1/TpDGAT2の存在をサザン解析によって確認した。ベクターのみ(pYES2.1)を含むH1246MAT−α形質転換体をコントロールとして用いた。シロイヌナズナDGAT1で形質転換したH1246MAT−αはポジティブコントロールとして働いた。
【0059】
シングルコロニーを、20mLのSD培地(Ausubel,et al.,1995,Vol.2,p.13.1.3によって記載されているような、グルコースを含み、ウラシルを含まない合成デキストロース培地)中で回転式シェーカー(270rpm)上で28℃にて一晩培養した。一晩培養物からの細胞を沈殿させ、50mLの発現誘導用の培地(ガラクトースを含み、ウラシルを含まないSD培地)中に再懸濁した。細胞を270rpmで振とうさせながら、28℃にて再びインキュベーションし、4〜6時間後に回収した。GALで誘導した酵母形質転換体を、5,000rpmで5分間の遠心分離によって回収し、1mM EDTAおよび1mM DTTを含む100mM Hepes−NaOH(pH7.4)中に再懸濁した。
【0060】
図9を参照すると、ネガティブコントロールである空ベクターではTAGが生成されなかった(レーン8)のに対し、ポジティブコントロール(レーン1)ではTAGバンドを示した。DGAT2を含むベクターのそれぞれ(レーン2〜6)では、DGAT2がTAGを合成する能力を有することを裏付けるTAGバンドを示した。レーン8の右側のレーンには、TAGマーカーとして用いられるTAG標準物質をロードした。
【0061】
実施例3:TpDGAT2の基質選択性
Ausubel,et al.(1995)によって記載されているように、酸洗浄したガラスビーズを用いて細胞溶解物を調製した。Bradford(1976)アッセイを用いて酵母溶解物中のタンパク質を測定し、各溶解物中のタンパク質量を標準化し、アリコート(タンパク質250μg)をDGAT2活性についてアッセイした。
【0062】
30℃の水槽中で100回転/分で10分間振とうさせながら、DGATアッセイをpH7.4にて行った。アッセイ混合物(最終容量0.5mL)は、100μg溶解物タンパク質、90mM HEPES−NaOH、200μM sn−1,2ジオレイン、およびアシル基供与体としての18μM14CアシルCoA(2nCi/nmol特異的活性)を含んだ。ヘキサン:ジエチルエーテル:酢酸(70:30:1 v/v/v)中で展開させたシリカゲルGプレート上でのTLCによって14C標識されたTAGを単離し、Win−Scan2D(登録商標)ソフトウェア(Bioscan社,ワシントンDC,米国)を用いたBioscan AR−2000放射性TLCスキャナー上で、放射性標識されたTAGバンドを可視化し、当該バンドを切り出して、Taylor,et al.(1991)によって記載されているように定量化した。
【0063】
実施例4:TpDGAT2形質転換体の脂肪酸組成
S.セレヴィシエ株H1246MAT−αを、シロイヌナズナ/pYES2.1またはT.シュードナナ/pYES2.1で形質転換した。形質転換体を3日間28℃にて培養し、ガラクトースによって誘導した。前記形質転換体を、処理なし(コントロール)、50uM DHAまたは150uM DHAで処理した。AtDGAT1/pYES2.1を含む3つの形質転換体およびTpDGAT2/pYES2.1を含む3つの形質転換体についての脂肪酸プロファイルを、従来のガスクロマトグラフィー分析に基づく表1に示した。
【0064】
脂肪酸を16:0、16:1、18:0、18:1(オレイン酸)、および22:6(DHA)として同定し、それぞれの組成を総脂肪酸に対するパーセンテージで表している。DHAの発現は、コントロール株の0から150μM TpDGAT2/pYES2.1では6.01%に増加し、150μM AtDGAT/pYES2.1のものの2倍以上であった(表1)。これらの結果は、TpDGAT2がDGAT1よりもDHA脂肪酸をより効率的に利用することをさらに裏付けた。
【0065】
表1に示すように、脂肪酸組成に関して、DGAT2 cDNAを含む変異株は飽和物全体の減少および不飽和物の増加を示した。そのような変化はすべて、「より健康な」油のプロファイルに向かうものであり、カノーラ、アブラナの他の油料種子、および他の食用油料作物に直接応用して、同様の油組成の改良をもたらすことができる。
【0066】
(表1) 酵母変異体H1246MAT−αにおける、DGAT2およびDGAT1によって発現されるTAGの脂肪酸組成
【0067】
実施例5:野生型シロイヌナズナにおけるDGAT2 cDNAの過剰発現
全長DGAT2 cDNAをPCR増幅のための鋳型として用いる。フラグメントを制限エンドヌクレアーゼ消化によって切断し、ベクターの相当する部位にライゲーションする。構築物の完全性を、シークエンシングによって確認する。
【0068】
前記ベクターをA.ツメファシエンス内に導入し、野生型シロイヌナズナを形質転換するために用い、後代を分析する。
【0069】
実施例6:種子特異的発現のためのDGAT2 cDNA植物形質転換ベクターの構築
配列の各末端に新しい制限部位を提供するために、プライマーとともに、全長DGAT2 cDNAをPCR増幅のための鋳型として用いた。PCRプロファイルは以下のとおりである:94℃1分;94℃30秒,55℃30秒,72℃1分を30サイクル;および72℃5分。その後、PCR産物をPCR2.1ベクター(Invitrogen社)にライゲーションする。フラグメントを切断し、ベクターの相当する部位にライゲーションする。構築物の完全性を、シークエンシングによって確認する。
【0070】
実施例7:植物DGAT2ベクター構築物を用いたアグロバクテリウムの形質転換
エレクトロコンピテント・アグロバクテリウム細胞GV3101(pMP90)株を以下のとおりに調製する。アグロバクテリウム培養物を2YT中で24〜48時間増殖させ、600nmにおける吸光度が0.5〜0.7に到達した時点で、細胞を氷上で冷やし、遠心分離(GSAローターで5,000×g,10分,4℃)によって沈殿させる。沈殿物を1、0.5、および0.02容量の冷却した10%滅菌グリセロール中で洗浄し、0.01容量の冷却した10%グリセロールに再懸濁する。その後、当該エレクトロコンピテント細胞を液体N2中で冷凍し、−70℃で保存する。メーカーの使用説明書に従って、20〜50ngの形質転換DNAを用いたエレクトロポレーションによって、前記アグロバクテリウム細胞を形質転換し、選択培地(50μg/mLカナマイシンを含むLB)上にプレーティングし、28℃にて一晩インキュベーションする。形質転換された単一細胞を、50μg/mLカナマイシンおよび25μg/mLゲンタマイシンを含む5mL LB中で一晩増殖させる(28℃,225r.p.m)。DNAの抽出および精製を行う。植物の形質転換前に、構築物の忠実度をDNAシークエンシングによって再チェックする。
【0071】
実施例8:シロイヌナズナの形質転換
シロイヌナズナの種子を16時間・明/8時間・暗の管理の中、蛍光照明(120μE・m−2S−1)の下で22℃にて栽培する。4〜6個の植物を、湿らせたTERRA−LITE REDI−EARTH(W.R.Grace&Co.Canada社,エイジャックス,オンタリオ州,カナダ)中で10cm2植木鉢で育てる。植木鉢中の土壌混合物が接種培地中に落下するのを防ぐために、土壌をプラットホームのように積み上げて種子を頂上に植え付け、植木鉢全体をナイロン製のウィンドウスクリーンで覆い、ゴム製のバンドで固定する。最初の花が開花し始めた時点で、アグロバクテリウム懸濁液中で植物を減圧浸潤する。
【0072】
アグロバクテリウムを増殖させるために、50μg/mLカナマイシンおよび25μg/mLゲンタマイシンを含むLB培地中で5mL懸濁液を28℃にて一晩培養する。浸潤の前日に、この「種子培養物」を、50μg/mLカナマイシンおよび25μg/mLゲンタマイシンを補充した250mLのLB培地を含む4つのフラスコに分ける。これらの培養物を28℃にて一晩培養する。翌朝、600nmにおける吸光度(およそ=1.0)をチェックした後、遠心分離(GSAローターで5,000×g,10分,室温)によって細胞を回収し、浸潤培地(水中に5%スクロース,0.005% Silwet−77)に再懸濁して、600nmで0.8の吸光度を得る。
【0073】
アグロバクテリウム懸濁液をビーカーに注ぎ、花およびボルトが浸水するように鉢植えされている植物を前記ビーカーの中に逆さにする。前記ビーカーを大きなベルジャー内に置き、茎表面上で気泡が形成され、かつ溶液がわずかに泡立ち始めるまで、真空ポンプを用いて真空に吸引し、その後真空を急速に解放する。必要な時間および圧力は実験装置によって異なるが、良好な浸潤は、均一に暗くなり水浸しになる組織のように、目に見えて明らかである。植木鉢を前記ビーカーから取り出して、プラスチック製のトレイの中の側に置き、プラスチック製のドームで覆って湿度を維持する。翌日、植物から覆いを取り外して真っ直ぐに立て、Katavic,et al.(1995)によって記載されているように、連続的な光条件下、栽培箱中でおよそ4週間成長させる。長角果が成熟して乾燥した時点で、種子を収穫し、陽性の形質転換体について選択する。
【0074】
実施例9:セイヨウアブラナ(Brassica napus)の形質転換
形質転換は基本的に、Moloney,et al.,1989,Plant Cell Reports 8:238−242によって記載されているように実施する。
【0075】
A.ツメファシエンス株GV3101/pMP90(Koncz C.&Schell,J.,1986,Mol.Gen.Genet.204:383−396)を、形質転換の研究に用いる。LB培地(Difco,米国)(100mL)中の定常期の細菌培養物を遠心分離によって回収し、抗凍結剤としての1%DMSO(ジメチルスルホキシド)(Sigma社,米国)を含む10mLの新しいLB培地中に再懸濁する。細菌のアリコートを、2%スクロース、50μMアセトシリンゴン,pH5.6を含む2mLのブレインハートインフュージョン培地(Difco,米国)に添加し、28℃にて一晩インキュベーションする形質転換に用いるまで、200μLのアリコートを−20℃で保存する。細菌細胞の密度は、およそ1×109 細胞/mLである。
【0076】
Moloney,et al.(1989),Plant Cell Rep.8:238−242の方法に従って、子葉移植片を植物形質転換ベクターを含むアグロバクテリウムに暴露する。前記移植片の葉柄の切断面を細菌培養物に短時間浸す。切断面が培地に接触するように、前記移植片を共存培養培地の中に入れる。10個の移植片を、それぞれ100×15mmのペトリプレートに置く。共存培養プレートをSTRETCH’N SEAL(商標)プラスチック製の包装物で密封する。種子の発芽工程に関する前述のような温度および光周期の条件下で、プレートを3日間培養キャビネット内でインキュベーションする。その後、移植片を選択培地に移す。
【0077】
選択培地において3〜4週間後、再生している若芽(推定上の形質転換体)を切り取り、継続培養のために新しい選択培地に移す。新芽が1.5〜2.0cmの長さに到達した時点で、それらを発根培地に移す。基本的に、Jefferson,R.A.(1987),Plant Mol.Biol.Rep.5:387−405によって記載されているように、推定上の遺伝子導入された新芽をgus遺伝子の発現についてスクリーニングする。青色の染色の存在は、形質転換の証拠として見なされる。
【0078】
形質転換の確認を、カナマイシン上での選択、サザンブロッティング、PCR(ポリメラーゼ連鎖反応)、および後代分析によって立証する。
【0079】
実施例10:推定上の形質転換体(遺伝子導入植物)の選択および遺伝子導入植物の解析
各構築物に対して、種子を大量に収穫する。種子を20%漂白剤および0.01%Triton X−100を含む溶液中に種子を20分間浸すことによって表面滅菌し、その後滅菌水で3回洗浄する。滅菌した種子を、滅菌した0.1%Phytagar(500〜1,000個の種子につき約1mLのPhytagar)中に室温にて再懸濁し、および150×15mmのカナマイシン選択プレート上に2,000〜4,000個の種子を含む容量を塗布することによってプレーティングする。プレートを照明をつけずに寒い中で2日間インキュベーションし、7〜10日間制御された環境の中で(16時間・明/8時間・暗の管理の中、蛍光照明(120μE・m−2S−1)の下で22℃にて)栽培する。選択培地は、1/2MSG培地、0.8%Phytagar、3%スクロース、50μg/mLカナマイシン、および50μg/mLチメンチンを含む。ペトリプレートおよび蓋を、Micropore(商標)外科用テープ(3M カナダ,ロンドン,ON,カナダ)で密封する。7〜10日後、緑葉を有し、かつ培地中で根をしっかりと定着させている薬剤耐性植物を形質転換体として同定し、葉3〜5枚の段階で、選択された形質転換体を非常に湿らせた土壌混合物で満たした湿地の中に移植する。形質転換体を成熟させ、成熟した種子(Katavic,et al.(1994)で定義されているT2世代)を個々の植物から収穫し、さらに分析する。
【0080】
ゲノムDNAを個々のT1植物から単離する。前記T1形質転換体におけるcDNAまたは遺伝子の存在をそれぞれ確認するために、PCR増幅を行う。挿入したフラグメントのシングルコピーを含む形質転換体を選択するために、サザン分析を行う。DNAサンプルを制限酵素で消化し、1%アガロースゲル上での電気泳動によって分解し、ナイロン製のフィルター(Hybond−N+,アメルシャム社)を用いてサザンブロッティングを行う。α−[32P]dCTP(NEN/DuPont)で標識したDGAT2 cDNAフラグメントをプローブとして用いる。ハイブリダイゼーションを60℃で行う。その後、前記フィルターをKodak X−OMAT−ARフィルムに露光する。
【技術分野】
【0001】
優先権主張
本願は、「藻類由来のジアシルグリセロールアシルトランスフェラーゼ2遺伝子およびそれによってコードされるタンパク質」について2007年12月21日に出願された米国仮特許出願第60/008,752号の出願日の利益を主張する。
【0002】
技術分野
本開示は、概してバイオテクノロジー、とりわけ植物特性の遺伝子操作に有用な遺伝子に関する。特定の実施形態において、本開示は、単離および/または精製したポリペプチド、およびジアシルグリセロールアシルトランスフェラーゼ2(DGAT2)をコードする核酸、ならびにそれらの使用方法に関する。
【背景技術】
【0003】
背景
油料種子作物は重要な農産物である。植物の種子油は、人の食物に必須の多価不飽和脂肪酸の主要な供給源であり、かつ化学工業にとっての再生可能な原料である。発達中の種子の色素体中にある脂肪酸合成酵素複合体という酵素は、細胞質のアシルCoAプールに導かれる脂肪酸の生合成に関与して、トリアシルグリセロール蓄積を維持する。トリアシルグリセロール(TAG)の生合成は、一次基質としてのグリセロール−3−リン酸および脂肪酸アシルCoAとともに小胞体に位置する。植物の貯蔵脂質の生体集合には、3つのアシル基転移酵素、すなわちグリセロール−3−リン酸アシルトランスフェラーゼ(GPAT、EC2.3.1.15)、リゾホスファチジン酸アシルトランスフェラーゼ(LPAT、EC2.3.1.51)、およびジアシルグリセロールアシルトランスフェラーゼ(DGAT、EC2.3.1.20)がある。これら3つのアシルトランスフェラーゼは、ケネディ経路として一般的に知られる生化学過程である、TAGの形成へのDGATによるsn−1,2−ジアシルグリセロール(DAG)のアシル化という最終工程でグリセロール骨格の段階的なアシル化を触媒する。TAGを生成するためのDGATが介在するグリセロール骨格のアシル化は、植物の脂質蓄積における律速段階と提唱されている。従って、DGATは植物脂質生合成の遺伝子改変における標的である。
【発明の概要】
【0004】
発明の開示
本発明者らは、T.シュードナナ(T.pseudonana)のジアシルグリセロールアシルトランスフェラーゼ2(DGAT)と少なくとも90%の配列同一性を有するポリペプチドの属を本明細書において開示する。これらのポリペプチドを用いて、植物における多価不飽和脂肪酸の量を変えることができる。また、タラシオシラ・シュードナナ(Thalassiosira pseudonana)のDGAT2のジアシルグリセロールトランスフェラーゼ触媒ドメインを含むポリペプチド、およびDGAT2のジアシルグリセロールトランスフェラーゼ触媒ドメインと少なくとも90%の配列同一性を有するポリペプチドも開示する。さらに、T.シュードナナのDGAT2と少なくとも90%の配列同一性を有するポリペプチドをコードするポリヌクレオチド配列、およびT.シュードナナのDGAT2のジアシルグリセロールトランスフェラーゼドメインと少なくとも90%の同一性を有するポリペプチドをコードするポリヌクレオチドを記載する。
【0005】
本明細書において、本発明者らはT.シュードナナ由来の単離および精製したジアシルグリセロールアシルトランスフェラーゼ2(DGAT)遺伝子およびcDNA配列を開示する。また、T.シュードナナ由来の全長DGAT2 cDNA配列、および前記DGAT2 cDNAと少なくとも80%の配列同一性を有するcDNA配列も開示する。一部の実施形態においては、これらのcDNA配列はベクター内に含まれていてもよい。商業用植物油の産出量を増加させるため、またはそれらの組成を改変して植物および植物生成物の特定の商業的改良を達成するために、これらのポリヌクレオチドを用いて植物におけるトリアシルグリセロールの天然形成を改変することができる。
【0006】
T.シュードナナ、ならびにクラミドモナス・ラインハーディ(Chlamydomonas reinhardtii)、オストレオコッカス・ルシマリヌス(Ostreococcus lucimarinus)、オストレオコッカス・タウリ(Ostreococcus tauri)、およびファエオダクチルム・トリコルヌツム(Phaeodactylum tricornutum)を含む藻類の別の種由来のDGAT2ファミリーの別の単離および精製した遺伝子、ならびにcDNA配列も開示する。また、商業用植物油の産出量を増加させるため、またはそれらの組成を改変して植物および植物生成物の特定の商業的改良を達成するために、これらのポリヌクレオチドを用いて植物におけるトリアシルグリセロールの天然形成を改変することができる。
【0007】
核酸構築物を含む遺伝子導入植物も開示する。細胞または植物を形質転換する方法であって、細胞または植物の中に単離した、精製した、または組換えの核酸を導入することを含む方法を記載する。遺伝子的に形質転換された植物の種子を産生するプロセスには、植物の種子の中に核酸を導入することが含まれる。一部の実施形態において、これらの方法は、植物の種子油含有量を変化させるために、植物を改変するのに用いることができる。
【0008】
最も一般的に述べると、一部の例は、藻類由来のDGAT2遺伝子の単離、精製、および特徴づけ、ならびに超長鎖多価不飽和脂肪酸の生成におけるDGAT2の有用性を開示する。前述のことは、添付した図面に関連して進む、以下のいくつかの実施形態の詳細な説明からより明白になるであろう。
【図面の簡単な説明】
【0009】
【図1A】図1Aは、T.シュードナナDGAT2の全長DGAT2 cDNA配列(配列番号:2)に相当する推定アミノ酸配列(配列番号:1)を表す。
【図1B】図1Aの続き。
【図2A】図2Aは、配列番号:1(TpDGA2)とすべて2型ジアシルグリセロールアシルトランスフェラーゼであるgi:37182187、gi:50541689、gi:74623358、gi:74623359、gi:86279638、およびgi:62825813との間の配列アラインメントを表す。前記配列のうち、4以上に共通するアミノ酸を太字で示す。これら2型ジアシルグリセロールアシルトランスフェラーゼのジアシルグリセロールトランスフェラーゼ触媒ドメインを含むアミノ酸配列は、236−365(TpDGA2);79−208(gi:37182187);76−205(gi:50541689);76−205(gi:74623358);34−165(gi:74623359);33−165(gi:86279638);36−165(gi:62825813)残基からなる。
【図2B】図2Aの続き。
【図3】図3Aは、配列番号:1に相同なポリペプチド配列の一例(配列番号:3)を表す。図3Bは、配列番号:2に少なくとも90%相同であって、配列番号:3のポリペプチドの一部をコードするポリヌクレオチド配列(配列番号:4)の一部を表す。
【図4】図4Aは、配列番号:1に相同なポリペプチド配列の別の例(配列番号:5)を表す。図4Bは、配列番号:2に少なくとも90%相同であって、配列番号:5のポリペプチドの一部をコードするポリヌクレオチド配列(配列番号:6)の一部を表す。
【図5】図5Aは、配列番号:1に相同なポリペプチド配列の別の例(配列番号:7)を表す。図5Bは、配列番号:2に少なくとも90%相同であって、配列番号:7のポリペプチドの一部をコードするポリヌクレオチド配列(配列番号:8)の一部を表す。
【図6】図6Aは、配列番号:1に相同なポリペプチド配列の別の例(配列番号:9)を表す。図6Bは、配列番号:2に少なくとも90%相同であって、配列番号:9のポリペプチドの一部をコードするポリヌクレオチド配列(配列番号:10)の一部を表す。
【図7】図7Aは、配列番号:1に相同なポリペプチド配列の別の例(配列番号:11)を表す。図7Bは、配列番号:2に少なくとも90%相同であって、配列番号:11のポリペプチドの一部をコードするポリヌクレオチド配列(配列番号:12)の一部を表す。
【図8】図8Aは、配列番号:1に相同なポリペプチド配列の別の例(配列番号:13)を表す。図8Bは、配列番号:2に少なくとも90%相同であって、配列番号:13のポリペプチドの一部をコードするポリヌクレオチド配列(配列番号:14)の一部を表す。
【図9】図9は、酵母変異体H1246MAT−α(DGAT、PDAT−、ASAT1−、ASAT2−、TAG形成を欠失している)でTpDGAT2およびAtDGAT1を発現させることによって生成されたTAGの薄層クロマトグラフィー(TLC)分析を表す。レーン1はAtDGAT1の発現を示し、レーン2〜6は、TpDGAT2の発現を示す。レーン2〜6ではクリアなTAG(トリアシルグリセロール)のバンドが観察された。レーン8は空ベクター(pYES2.1)コントロールを示し、このレーンにはTAG(トリアシルグリセロール)のバンドは存在しない。レーン8の右側のレーンには、TAGマーカーとして用いることができるTAG標準物質をロードした。
【図10】図10は、空プラスミド(pYES2.1コントロール;空のバー)、T.シュードナナDGAT2 cDNA(pYES:DGAT;点状のバー)、およびシロイヌナズナ(A.thaliana)DGATcDNA(黒無地のバー)で形質転換された酵母変異体H1246MAT−αにおけるDGAT活性を示す。誘導した酵母細胞の溶解物から調製したミクロゾーム膜画分を、アシル基供与体としての種々の14C標識されたアシルCoA、および受容体としての標識されていないsn−1,2ジオレインを用いてDGAT活性についてアッセイした。本明細書において相対的DGAT活性をDPM(TAG内に取り込まれた14C標識された基質の量)として表した。当該結果は、TpDGAT2およびAtDGAT1の基質選択性および相対的活性を示す。
【図11】図11は、TpDGAT2のアミノ酸配列(TpDGAT2−1)とT.シュードナナ由来または他の藻類種(Cr:クラミドモナス・ラインハーディ、Ol:オストレオコッカス・ルシマリヌス、Ot:オストレオコッカス・タウリ、Pt:ファエオダクチルム・トリコルヌツム)由来のそのファミリーメンバーとのホモロジー比較を表す。TpDGAT2(TpDGAT2−1)は、そのファミリーメンバーであるTpDGAT2−2、TpDGAT2−3、およびTpDGAT2−4とそれぞれ24%、25%、および17%の配列同一性を共有している。種々の藻類種の中で、TpDGAT2(TpDGAT2−1)は、PtDGAT2−1と高い配列類似性(48%の配列同一性)を、CrDGAT2−1、CrDGAT2−2、およびCrDGAT2−4と比較的高い類似性(それぞれ20%、23%、および24%)を示す。
【発明を実施するための形態】
【0010】
I.いくつかの実施形態の概説
本明細書において、T.シュードナナの単離および精製した2型ジアシルグリセロールアシルトランスフェラーゼ(DGAT2)を開示する。他の生物および他の生物由来の細胞の中で超長鎖多価不飽和脂肪酸(VLCPUFA)の合成を改変するこのポリペプチドの驚くべき能力を用いて、所望の脂肪酸組成を有する遺伝子導入植物および植物の種子を産生するために、植物および植物の種子を形質転換する。この開示には、T.シュードナナDGAT2のものと少なくとも90%の配列同一性のアミノ酸配列を有するDGAT2活性を示すポリペプチドが含まれる。特定の実施形態において、これらのポリペプチド配列は、例えば配列番号:1、配列番号:3、配列番号:5、配列番号:7、配列番号:9、配列番号:11、または配列番号13を含む。また、T.シュードナナDGAT2のジアシルグリセロールアシルトランスフェラーゼ触媒ドメインと少なくとも90%の配列同一性を有する配列を含むポリペプチドを開示する。特定の実施形態において、これらのポリペプチド配列は、例えば配列番号15、配列番号:17、または配列番号:19を含む。図2にはT.シュードナナDGAT2のジアシルグリセロールアシルトランスフェラーゼ触媒ドメインが表されており、それはT.シュードナナDGAT2の完全な開示されているポリペプチド配列におけるアミノ酸残基236〜365からなる。
【0011】
配列番号:15のポリペプチドは、T.シュードナナDGAT2のジアシルグリセロールアシルトランスフェラーゼドメインを含む。一部の実施形態は、配列番号:15の単離または精製したポリペプチドと少なくとも約90%、91%、92%、93%、94%、95%、96%、97%、98%、99%、99.5%、99.6%、または99.7%の配列同一性を有する配列、例えば配列番号:1を含む単離または精製したポリペプチドに関する。特定の実施形態において、これらのポリペプチドはジアシルグリセロールアシルトランスフェラーゼ活性を有する。ジアシルグリセロールアシルトランスフェラーゼ活性は、例えばin vitroの酵素アッセイにより当業者によって容易に測定できる。この方法は実施例4において詳細に記載されており、このアッセイの代表的な結果は図10に示されている。当業者によって理解されるように、当該開示はまた、配列番号:15に90%以上の同一性の推定アミノ酸配列を含むタンパク質をコードする植物および藻類由来の実質的に相同なDNA配列にも関する。
【0012】
DGAT2ファミリーのメンバーである藻類由来の単離または精製した他のポリペプチドは、例えば配列番号:25(TpDGAT2−2)、配列番号:27(TpDGAT2−3)、配列番号:29(TpDGAT2−4)、配列番号:31(CrDGAT2−1)、配列番号:33(CrDGAT2−2)、配列番号:35(CrDGAT2−3)、配列番号:37(CrDGAT2−4)、配列番号:39(CrDGAT2−5)、配列番号:41(OlDGAT2−1)、配列番号:43(OlDGAT2−2)、配列番号:45(OlDGAT2−3)、配列番号:47(OlDGAT2−4)、配列番号:49(OtDGAT2−1)、配列番号:51(OtDGAT2−2)、配列番号:53(OtDGAT2−3)、配列番号:55(OtDGAT2−4)、配列番号:57(PtDGAT2−1)、配列番号:59(PtDGAT2−2)、配列番号:61(PtDGAT2−3)、または配列番号:63(PtDGAT2−4)と少なくとも90%同一であるアミノ酸配列を含む。
【0013】
一部の実施形態は、前述したポリペプチドをコードする単離または精製した核酸(ポリヌクレオチド)に関する。これらポリヌクレオチドの配列は、例えば配列番号:2、配列番号:4、配列番号:6、配列番号:8、配列番号:10、配列番号:12、配列番号:14、配列番号:16、配列番号:18、配列番号:20、配列番号:26(TpDGAT2−2)、配列番号:28(TpDGAT2−3)、配列番号:30(TpDGAT2−4)、配列番号:32(CrDGAT2−1)、配列番号:34(CrDGAT2−2)、配列番号:36(CrDGAT2−3)、配列番号:38(CrDGAT2−4)、配列番号:40(CrDGAT2−5)、配列番号:42(OlDGAT2−1)、配列番号:44(OlDGAT2−2)、配列番号:46(OlDGAT2−3)、配列番号:48(OlDGAT2−4)、配列番号:50(OtDGAT2−1)、配列番号:52(OtDGAT2−2)、配列番号:54(OtDGAT2−3)、配列番号:56(OtDGAT2−4)、配列番号:58(PtDGAT2−1)、配列番号:60(PtDGAT2−2)、配列番号:62(PtDGAT2−3)、または配列番号:64(PtDGAT2−4)を含んでもよい。一部の実施形態において、前記ポリヌクレオチド配列は、開示されているポリペプチドをコードする開示されているヌクレオチド配列の塩基と少なくとも約80%、81%、82%、83%、84%、85%、86%、87%、88%、89%、90%、91%、92%、93%、94%、95%、96%、97%、98%、99%、99.5%、99.6%、または99.7%のパーセンテージ同一性を有する。そのようなポリヌクレオチドのいくつかの例が、配列番号:21〜24である。技術を有する実践者によって理解されるように、核酸配列のわずかな変化は、コードされるポリペプチドのアミノ酸配列を必ずしも変化させない。コードされるポリペプチドのアミノ酸配列を変化させる特定の遺伝子配列におけるヌクレオチドの同一性の変化は、当該遺伝子の有効性の低下または増大をもたらし得ること、ならびに一部の応用(すなわち、アンチセンス、共抑制、またはRNAi)において、部分的配列はしばしば全長型と同じ程度に効果的に作用することは当業者によって理解されるであろう。遺伝子配列を変更または短くし得る方法は当業者に周知であり、当該改変遺伝子の有効性を調べる方法も同様である。特定の実施形態において、有効性は例えば従来のガスクロマトグラフィーによって容易に調べることができる。従って、当該遺伝子のそのようなすべての変化が本明細書の一部として含まれる。
【0014】
一部の実施形態は、配列番号:2と少なくとも80%の相同性を有する単離または精製したポリヌクレオチド、例えば配列番号:2、配列番号:15、配列番号:16、配列番号:17、配列番号:18、配列番号:20、配列番号:21〜24を含むベクターに関する。従って、例えば配列番号:2、例えば配列番号:2、配列番号:15、配列番号:16、配列番号:17、配列番号:18、配列番号:20、または配列番号:21〜24、あるいはその一部からなる群から選択される配列を含むベクターを調製するための方法、植物細胞内へのアンチセンス方向に当該配列または部分的配列、あるいはその相補鎖を導入するための方法を提供する。
【0015】
特定の実施形態は、藻類におけるDGAT2ファミリーのメンバーと少なくとも80%の相同性を有するポリヌクレオチドを含むベクターに関する。これらのベクターは、例えば配列番号:26、配列番号:28、配列番号:30、配列番号:32、配列番号:34、配列番号:36、配列番号:38、配列番号:40、配列番号:42、配列番号:44、配列番号:46、配列番号:48、配列番号:50、配列番号:52、配列番号:54、配列番号:56、配列番号:58、配列番号:60、配列番号:62、または配列番号:64のポリヌクレオチド配列を含んでもよい。
【0016】
一部の実施形態において、DGAT2活性を備えるポリペプチドを生成する遺伝子導入植物を作り出すために、前記単離および精製したポリヌクレオチド、ならびにこれらの単離および精製したポリヌクレオチドを含むベクターを用いてもよい。従って、一実施形態は、配列番号:2と少なくとも80%の相同性を有する単離または精製したポリヌクレオチド、例えば配列番号:2、配列番号:15、配列番号:16、配列番号:17、配列番号:18、配列番号:20、または配列番号:21〜24の配列を有するデオキシリボ核酸分子を含む遺伝子導入植物および植物の種子に関する。他の実施形態は、藻類におけるDGAT2ファミリーの別のメンバーと少なくとも80%の相同性を有する単離または精製したポリヌクレオチド、例えば配列番号:26、配列番号:28、配列番号:30、配列番号:32、配列番号:34、配列番号:36、配列番号:38、配列番号:40、配列番号:42、配列番号:44、配列番号:46、配列番号:48、配列番号:50、配列番号:52、配列番号:54、配列番号:56、配列番号:58、配列番号:60、配列番号:62、または配列番号:64の配列を有するデオキシリボ核酸分子を含む遺伝子導入植物および植物の種子に関する。これら実施形態の植物は、当該核酸構築物を欠いている植物における量と比較して、種子中の多価不飽和脂肪酸の量が変化していてもよい。前記植物における脂肪酸は、約70%以上の多価不飽和脂肪酸であってよい。
【0017】
一実施形態は、そのような植物および植物の種子を産生する方法を含む。前記方法は、配列番号:1、3、5、7、9、11、13、15、17、および19からなる群から選択されるポリペプチドと少なくとも90%の配列同一性を有するポリペプチド、または藻類におけるDGAT2ファミリーのポリペプチドと少なくとも90%の配列同一性を有するポリペプチドをコードするポリヌクレオチド;例えば配列番号:25、配列番号:27、配列番号:29、配列番号:31、配列番号:33、配列番号:35、配列番号:37、配列番号:39、配列番号:41、配列番号:43、配列番号:45、配列番号:47、配列番号:49、配列番号:51、配列番号:53、配列番号:55、配列番号:57、配列番号:59、配列番号:61、または配列番号:63を含む核酸構築物を作り出すこと、ならびに植物内に当該構築物を導入することを含む。この実施形態の方法は、当業者に公知の任意の方法、非制限的な例であるアグロバクテリウム(Agrobacterium)を介して行われる形質転換の方法によって達成され得る。特定の実施形態において、前記方法は、Brusicaピルビン酸脱水素酵素キナーゼ活性を備えるポリペプチドをコードするポリヌクレオチド、ジアシルグリセロールアセチルトランスフェラーゼ活性を備えるポリペプチドをコードするポリヌクレオチド、および/またはグリセロール−3−リン酸脱水素酵素活性を備えるポリペプチドをコードするポリヌクレオチドを、当該植物内に導入することをさらに含む。この方法は、前記植物がシロイヌナズナ(Arabidopsis thaliana)、ボリジ(Borago)種、カノーラ(Canola)、リキヌス(Ricinus)種、テオブロマ(Theobroma)種、トウモロコシ(Zea)種、ゴシッピウム(Gossypium)種、ハマナ(Crambe)種、クフェア(Cuphea)種、アマ(Linum)種、レスクェレラ(Lesquerella)種、リムナンテス(Limnanthes)種、リノーラ(Linola)、ノウゼンハレン(Tropaeolum)種、マツヨイグサ(Oenothera)種、オリーブ(Olea)種、アブラヤシ(Elaeis)種、ラッカセイ(Arachis)種、ナタネ(rapeseed)、ベニバナ(Carthamus)種、ダイズ(Glycine)種、ソヤ(Soja)種、ヒマワリ(Helianthus)種、タバコ(Nicotiana)種、ベルノニア(Vernonia)種、コムギ(Triticum)種、オオムギ(Hordeum)種、オリザ(Oryza)種、カラスムギ(Avena)種、モロコシ(Sorghum)種、ライムギ(Secale)種、アブラナ(Brassicaceae)、およびイネ科(Gramineae)植物ファミリーの他のメンバーからなる群から選択されるところで実施してもよい。
【0018】
一部の実施形態において、前記方法は、導入した核酸構築物を含む植物から種子を収穫すること、および前記収穫した種子から油を抽出することをさらに含む。従って、他の実施形態は、当該方法によって産生した植物、および当該方法によって産生した植物から抽出した油を含む。
【0019】
DGAT2遺伝子またはその一部を用いることによって可能である操作および成果物のいくつかは、以下の、油含有量が増加または減少した種子、超長鎖多価不飽和脂肪酸含有量が増大した油を含む種子、および超長鎖多価不飽和脂肪酸を蓄積する増大または変化した能力を示す植物を含むが、これに制限されない。
【0020】
II.略語
CaMV カリフラワーモザイクウイルス
cDNA 相補的DNA
CERV カーネーションエッチドリングウイルス
CrDGAT2 クラミドモナス・ラインハーディ2型ジアシルグリセロールトランスフェラーゼ
DAG sn−1,2−ジアシルグリセロール
DGAT ジアシルグリセロールアシルトランスフェラーゼ
DGAT2 2型ジアシルグリセロールトランスフェラーゼ
DHA ドコサヘキサエン酸
DNA デオキシリボ核酸
EPA エイコサペンタエン酸
GPAT グリセロール3リン酸アシルトランスフェラーゼ
LPAT リゾホスファチジン酸アシルトランスフェラーゼ
OlDGAT2 オストレオコッカス・ルシマリヌス2型ジアシルグリセロールトランスフェラーゼ
OtDGAT2 オストレオコッカス・タウリ2型ジアシルグリセロールトランスフェラーゼ
PCR ポリメラーゼ連鎖反応
PtDGAT2 ファエオダクチルム・トリコルヌツ2型ジアシルグリセロールトランスフェラーゼ
RNA リボ核酸
RNAi RNA干渉
RT−PCR 逆転写PCR
T35S CaMV35Sターミネーター
TAG トリアシルグリセロール
TLC 薄層クロマトグラフィー
Tmas マンノピン合成ターミネーター
Tnos ノパリン合成ターミネーター
TpDGAT2 T.シュードナナ2型ジアシルグリセロールトランスフェラーゼ
TrbcS リブロース2リン酸カルボキシラーゼ小サブユニット終結領域
VLCPUFA 超長鎖多価不飽和脂肪酸
【0021】
III.用語
本開示の種々の実施形態の概説を容易にするために、以下の特定の用語の説明を提供する。
【0022】
相補的ヌクレオチド配列:配列の「相補的ヌクレオチド配列」とは、そのヌクレオチドが本開示の配列のものと相補的であり、その方向が反転している(逆平行配列である)任意のDNAを意味すると理解される。
【0023】
配列ホモロジーの程度またはパーセンテージ:「配列ホモロジーの程度またはパーセンテージ」という用語は、最適アラインメント後の2つの配列間の配列同一性の程度またはパーセンテージを表す。配列同一性のパーセンテージ(あるいは程度または同一性)は、比較ウィンドウ上で2つの最適に整列された配列を比較することによって決定されるものであって、前記2つの配列の最適アラインメントについて、比較ウィンドウにおけるペプチドまたはポリヌクレオチド配列の一部は、参照配列(付加または欠失を含まない)と比較して付加または欠失(すなわち、ギャップ)を含み得る。前記パーセンテージは、両配列において一致するアミノ酸残基または核酸塩基が生じている位置の数を測定してマッチした位置の数を出し、マッチした位置の数を比較ウィンドウにおける位置の総数で割り、かつ当該結果に100を掛けて配列同一性のパーセンテージを出すことによって算出される。
【0024】
単離および/または精製した相同配列:「単離および/または精製した相同配列」とは、ヌクレオチド配列の塩基またはポリペプチド配列のアミノ酸と、少なくとも約80%、81%、82%、83%、84%、85%、86%、87%、88%、89%、90%、91%、92%、93%、94%、95%、96%、97%、98%、99%、99.5%、99.6%、または99.7%のパーセンテージ同一性を有する単離および/または精製した配列を意味すると理解される。このパーセンテージは完全に統計的なものであり、無作為とそれらの長さ全体にわたる2つのヌクレオチド配列との間で差をもたらす可能性がある。例えば、シングル配列アライメントおよびマルチプル配列アライメントを行うように設計されたコンピュータープログラムによって、配列同一性を測定することができる。当業者によって理解されるように、この開示はコドン使用の縮重を包含すると理解されるであろう。さらに、ポリペプチドのアミノ酸配列において、前記ポリペプチドの構造または作用を崩壊させることなく保存的置換が行われてもよいと当業者によって理解されるであろう。保存的置換は、お互いに対して類似した疎水性、極性、およびR鎖長を有するアミノ酸を置換することによって、熟練した技術者によって達成される。また、異なる種由来の相同タンパク質の整列した配列を比較することによって、コードされたタンパク質の基本的作用を変えることなく種間で変異しているアミノ酸残基を位置づけることによって、保存的置換を特定してもよい。
【0025】
単離した:当業者によって理解されるように、「単離した」とは、それらの天然環境から「単離」されているポリペプチドを表す。
【0026】
ヌクレオチド、ポリヌクレオチド、または核酸配列:「ヌクレオチド、ポリヌクレオチド、または核酸配列」とは、単量体および二量体の(いわゆる、並行した)形態の2本鎖または1本鎖のDNA、ならびに前記DNAの転写産物の両方を意味すると理解されるであろう。
【0027】
配列同一性:2つのアミノ酸またはヌクレオチドの配列におけるアミノ酸またはヌクレオチドの残基の配列が、以下に記載するように最大の一致を目的として整列させた時に同じである場合、前記2つの配列は「同一」であると言われる。2つ(またはそれより多く)のペプチドまたはポリヌクレオチド間の配列比較は、典型的に、配列類似性について局所領域を同定および比較するために、断片または「比較ウィンドウ」にわたって最適に整列された2つの配列の配列を比較することによって行われる。比較のための配列の最適な整列は、SmithおよびWatermanの局所的ホモロジーアルゴリズム:Ad.App.Math 2:482(1981)によって、NeddlemanおよびWunschのホモロジーアラインメントアルゴリズム:J.Mol.Biol.48:443(1970)によって、PearsonおよびLipmanの類似法の探索、Proc.Natl.Acad.Sci.(U.S.A.)85:2444(1988)によって、これらアルゴリズムのコンピュータによる実施(ウィスコンシン・ジェネティクス・ソフトウェアパッケージのGAP、BESTFIT、FASTA、およびTFASTA,ジェネティクス・コンピュータ・グループ(Genetics Computer Group:GCG),575サイエンスDr.,マディソン,ウィスコンシン州)によって、または目視検査によって行われてよい。
【0028】
前記で付された配列同一性の定義は、当業者によって用いられるであろう定義である。定義それ自体はいかなるアルゴリズムの支援も必要とせず、前記アルゴリズムは、配列同一性の算出よりもむしろ、配列の最適アラインメントを達成するために唯一有用である。
【0029】
前記で付された定義から、要するに、2つの比較される配列間の配列同一性について、明確かつ唯一の数値であって、最高または最適なアラインメントに対して得られる数値に相当する数値が存在するということになる。
【0030】
http://worldwiseweb.ncbi.nlm.nih.gov/gorf/bl2.htmlのウェブサイトで入手可能であり、2つの配列間の同一性を比較および決定するために、本発明者らおよび一般的に当業者によって習慣的に用いられるソフトウェアであるBLAST NまたはBLAST P「BLAST2配列」において、比較される配列の長さに依存するギャップコストは、前記ソフトウェアによって直接選択される(すなわち、>85の長さに対する置換行列BLOSUM−62には11.2)。
【0031】
ストリンジェントなハイブリダイゼーション:ヌクレオチド配列に関するストリンジェントな条件下でのハイブリダイゼーションとは、相補的DNAの2つのフラグメント間でのハイブリダイゼーションを保持させ得るように選択された温度およびイオン強度の条件下でのハイブリダイゼーションを意味すると理解される。他の生物、例えば植物から得られる本明細書において記載されるDGAT2遺伝子のホモログは、前記ホモログを含む適切なライブラリーをスクリーニングすることによって得ることができ、前記スクリーニングは、本明細書において開示される特定のDGAT2遺伝子のヌクレオチド配列、またはその一部もしくはプローブに関して行われるものであり、あるいはBLAST、FASTA等の配列アラインメント検索プログラムを用いた配列ホモロジー検索によって同定することができる。
【0032】
III.藻類由来のDGAT2による脂肪酸量の改変
A.概説
アブラギリおよびトウゴマ由来のDGAT2に関する最近の研究は、独特な脂肪酸を含む植物において、DGAT2は独特な脂肪酸を種子貯蔵油に導くのに重要な役割を担っている可能性があることを示唆している。脂肪種子における植物脂質生合成の遺伝子改変において、DGAT2は潜在的標的になり得ると同時に、最近特徴づけされた前記酵素は、共役脂肪酸であるエレオステアリン酸(アブラギリDGAT2)およびリシノール酸(トウゴマDGAT2)の利用にそれぞれ貢献している。どちらの酵素(アブラギリDGAT2またはトウゴマDGAT2)も、商業的に所望される長鎖オメガ−3多価不飽和脂肪酸であるエイコサペンタエン酸(EPA)およびドコサヘキサエン酸(DHA)のトリアシルグリセロール(TAG)内への取り込みに関与していない。
【0033】
海の中心的な珪藻類であるT.シュードナナは、TAG内に長鎖オメガ−3多価不飽和脂肪酸であるEPAおよびDHAを生成および蓄積することができ、高レベルの超長鎖多価不飽和脂肪酸(VLCPUFA)が蓄積した油の良好な供給源である。そのため、T.シュードナナのジアシルグリセロールアシルトランスフェラーゼ2(TpDGAT2)遺伝子は、調査および特徴づけされた。驚くべきことに、アブラギリまたはトウゴマ由来のDGAT2とは異なり、TpDGAT2はTAG内に超長鎖多価不飽和脂肪酸を効率的に取り込むことができることが発見された。TpDGAT2遺伝子を用いてT.シュードナナおよび関連する藻類の種由来のポリヌクレオチド配列を調べることによって、藻類におけるDGAT2ファミリーの他のメンバーが同定された。従って、藻類DGAT2遺伝子は、遺伝子組換えツールにおいてならびに種子内のTAGの組成および蓄積の改変に対して有用であると判断された。
【0034】
B.2型ジアシルグリセロールトランスフェラーゼ活性を備えるT.シュードナナDGAT2に相同なポリペプチド
NCBIタンパク質データベース等のタンパク質データベースをBLAST等の検索エンジンを用いて検索することによって、全長T.シュードナナDGAT2に相同であるタンパク質を発見することができる。それらは合理的設計によって同定されてもよい。合理的設計の方法は、所望のポリペプチド配列の長さの範囲内で保存的アミノ酸置換を同定すること、および当該コードされるタンパク質においてそれらの置換を引き起こすことを含む。
【0035】
T.シュードナナDGAT2の全長アミノ酸配列に関してNCBIタンパク質データベース(BLASTP)を検索すると、TpDGAT2に有意な配列ホモロジーを有するポリペプチドが明らかとなり、そのいくつかはTpDGAT2とともに整列して図2に示されている。保存されている2型ジアシルグリセロールトランスフェラーゼドメインは図2に整列されており、TpDGAT2におけるアミノ酸残基236〜365と示されている他のDGAT2ポリペプチド由来の相当する残基とからなる。前記保存されている2型ジアシルグリセロールトランスフェラーゼドメインは、NCBIの保存されているドメインのデータベース(http://www.ncbi.nlm.nih.gov/Structure/cdd/cdd.shtml)内に記載されている。この保存されているドメインに相同であるポリペプチド配列は、それが含まれるタンパク質にTpDGAT2の2型ジアシルグリセロール活性を付与する。
【0036】
相同配列を有するポリペプチドは、それらのホモログと同じ構造及び作用を示すように設計され得ると当業者によって理解される。熟練した技術者によって、図2の配列アラインメントにより本開示の実施例に具体的に記載されているものと相同なポリペプチドを設計することが可能である。そのような相同なポリペプチドは、本開示のポリペプチドへの保存的置換を含むもの、例えば配列番号:3、配列番号:5、配列番号:7、配列番号:9、配列番号:11、配列番号:13、配列番号:15、配列番号:17、または配列番号:19のポリペプチドであってよい。相同タンパク質がTpDGAT2と同様の2型ジアシルグリセロールトランスフェラーゼ活性を実質的に示すかどうかを判定する簡易な実験アッセイは当業者に公知である。そのようなアッセイは、過度に時間のかかるものでなく、高価でなく、または技術的に難しいものでない。TpDGAT2によって生成されたTAGを検出するために、例えば従来のガスクロマトグラフィーを用いてもよい。これらのアッセイのいくつかは、以下の詳細な実施例に記載されている。
【0037】
C.形質転換するためのDGAT2活性を有する核酸分子の使用
開示されている実施形態は、天然環境で採取されたゲノム、すなわち、T.シュードナナ、クラミドモナス・ラインハーディ、オストレオコッカス・ルシマリヌス、オストレオコッカス・タウリ、またはファエオダクチルム・トリコルヌツムの天然ゲノムのヌクレオチド配列を含まないと理解すべきである。一部の実施形態は、例えばイオン交換クロマトグラフィー、分子サイズに基づいた排除による、またはアフィニティーによる、または種々の溶媒中の溶解度に基づいた分画法等の分離方法から始めることによって、あるいは増幅、クローニング、およびそれにより当該配列がベクターによって運搬されることが可能となるサブクローニング等の遺伝子操作の方法から始めることによって、単離、精製または部分的に精製することが可能である配列に関する。
【0038】
さらに、前記に開示されている配列にハイブリダイズする核酸分子が含まれる。ハイブリダイゼーションの条件は、開示されているDGAT2分子をコードする核酸分子と少なくとも90%、95%、または97%の同一性がある場合にハイブリダイゼーションが生じるという点においてストリンジェントであってよい。ストリンジェントな条件は、公知のサザンハイブリダイゼーションに用いられているもの、例えば50%ホルムアミド、5×SSC(150mM NaCl、15mMクエン酸3ナトリウム)、50mMリン酸ナトリウム(pH7.6)、5×デンハート液、10%硫酸デキストラン、および20マイクログラム/ミリリッターの変性、せん断されたサーモンスパームDNAを有する溶液中で一晩42℃にてインキュベーション、その後0.1×SSC中で約65℃にてハイブリダイゼーション支持体の洗浄等を含んでよい。他の公知のハイブリダイゼーション条件が周知であり、Sambrook,et al.,Molecular Cloning:A Laboratory Manual,Third Edition,Cold Spring Harbor,N.Y.(2001)に記載されている。
【0039】
DNAの単離およびクローニングは、十分に確立されている。同様に、従来技術によって、単離した酵素をコードするDNAをベクター内に挿入および酵母細胞内に形質転換することができる。しかしながら、VLCPUFAを効率的に使用し得るDGAT2遺伝子がクローニングされていなかったため、DGAT2活性を調節することによる遺伝子改変の可能性に取り組むことは不可能であった。本発明者らは、DGAT2がTAG合成に関与し、かつDGATよりも効率的にVLCPUFAを利用することを確認した。
【0040】
DGAT2をコードする核酸分子、例えば配列番号:2、配列番号:16、配列番号:25、配列番号:27、配列番号:29、配列番号:31、配列番号:33、配列番号:35、配列番号:37、配列番号:39、配列番号:41、配列番号:43、配列番号:45、配列番号:47、配列番号:49、配列番号:51、配列番号:53、配列番号:55、配列番号:57、配列番号:59、または配列番号:61と少なくとも80%の同一性を有する配列を、生物、例えば植物内に形質転換してもよい。そのような相同配列は配列番号:21〜24によって例示されている。当該技術分野において公知のように、遺伝子および遺伝子構築物を生物、例えば植物内に導入し得る多数の方法があり、形質転換および組織培養の技術の組み合わせは、遺伝子組換え生物、例えば作物を作り出すための効果的戦略内に上手く取り入れられている。これらの方法は、他の場所(Potrykus,1991;Vasil,1994;WaldenおよびWingender,1995;Songstad,et al.,1995)で記載されており、当業者に周知である。例えば、減圧浸潤(Bechtold,et al.,1993)または有傷接種(Katavic,et al.,1994)によってアグロバクテリウムを介して行われるシロイヌナズナの形質転換に加えて、アグロバクテリウムTi−プラスミドを介して行われる形質転換(例えば、胚軸(DeBlock,et al.,1989)または子葉柄(Moloney,et al.,1989)の創傷感染)、粒子衝突/微粒子銃法(Sanford,et al.,1987;Nehra,et al.,1994;Becker,et al.,1994)、またはポリエチレングリコールを介したプロトプラスト形質転換法(Rhodes,et al.,1988;Shimamoto,et al.,1989)を用いることによって、他の植物および作物種を形質転換することが同様に可能であることに当業者は確実に気づくであろう。
【0041】
植物において、新規の遺伝子を伝達するまたは既存の遺伝子の発現を変えるための遺伝子操作によって達成されている植物代謝に対して成功した改変の例が多数ある。作物の性能(例えば、種子油または塊茎デンプンの含有量/組成;食物改良;除草剤、疾病、または害虫抵抗性;重金属耐性等)を改良するために、農学的に重要な多数の植物種の中に遺伝子を導入することが現在日常的に可能である(MacKenzieおよびJain,1997;Budziszewski,et al.,1996;Somerville,1993;KishoreおよびSomerville,1993)。
【0042】
当業者に明らかであり、他の場所(Meyer,1995;Dada,et al.,1997)で記載されているように、構成的プロモーター(例えば、CaMV35Sに基づくもの)を用いることによって、あるいは特定の細胞、組織(例えば、発達中の種子の子葉における導入遺伝子の発現に対するナピンプロモーター)、器官(例えば、根)に、特定の発達段階に、または特定の外部刺激(例えば、熱ショック)に応答して、遺伝子発現を標的化することができるプロモーターを用いることによって、導入遺伝子の発現についての任意に意図した上方または下方制御を管理するために、植物プロモーターを利用することができる。
【0043】
本明細書において使用するためのプロモーターは、誘導可能な、構成的な、または組織特異的な、あるいは前記特性の種々の組み合わせを有するものであってよい。有用なプロモーターは、構成的プロモーター、例えばカーネーションエッチドリングウイルス(CERV)、カリフラワーモザイクウイルス(CaMV)35Sプロモーター、またはとりわけ2つのCaMV35Sプロモーターを直列に含んでいる2倍に増強したカリフラワーモザイクウイルスプロモーター(「ダブル35S」プロモーターと称される)を含むが、これに制限されない。
【0044】
特定の状況では、構成的プロモーターの代わりに組織特異的または発生学的に調節されるプロモーターを用いるのが望ましい可能性がある。組織特異的プロモーターは、他の組織の発現に影響を及ぼすことなく、特定の組織において過剰発現を可能にする。実例として、種子組織における酵素の過剰発現に用いられるプロモーターは、1992年10月29日に公表されたPCTの国際公開公報第92/18634号に記載されているようなACPプロモーターである。
【0045】
当該プロモーターおよび終結調節領域は、宿主の植物細胞内で機能的であってもよく、前記植物細胞および遺伝子に対して異種(すなわち、天然に存在しない)または同種(前記宿主植物種に由来する)であってもよい。用いることができる適切なプロモーターは、前記に記載されている。
【0046】
前記終結調節領域は、プロモーターが得られた遺伝子または別の遺伝子の3’領域に由来してもよい。用いることができる適切な終結調節領域は当該技術分野において周知であり、アグロバクテリウム・ツメファシエンス(Agrobacterium tumefaciens)・ノパリン合成ターミネーター(Tnos)、A.ツメファシエンスマンノピン合成ターミネーター(Tmas)、およびCaMV35Sターミネーター(T35S)、エンドウリブロース2リン酸カルボキシラーゼ小サブユニット終結領域(TrbcS)、またはTnos終結領域を含んでよい。そのような遺伝子構築物は、アグロバクテリウムを介した宿主植物内への形質転換、および増加したイソプレノイド量についてのスクリーニングによって、活性について適切にスクリーニングすることができる。
【0047】
適切には、当該遺伝子に対するヌクレオチド配列は、GenBank(登録商標)(米国保健社会福祉省の登録商標)ヌクレオチドデータベースから抽出し、かつ切断しない制限酵素について検索してもよい。PCRプライマー中にこれらの制限酵素認識部位を組み込むこと等の従来の方法またはサブクローニングによって、これらの制限部位を当該遺伝子に付加してもよい。
【0048】
好ましくは、本明細書において使用するためのDNA構築物はベクター、最適には適切な宿主(植物)細胞における発現に適した発現ベクター内に包含されている。導入したDNA配列を含む植物を産生することができる任意のベクターが十分であると理解されるであろう。
【0049】
適切なベクターは当業者に周知であり、Pouwels,et al.,Cloning Vectors,A Laboratory Manual,Elsevier,Amsterdam(1986)等の一般的な技術参考文献に記載されている。特に適切なベクターは、Tiプラスミドベクターを含む。
【0050】
宿主細胞内にDNA構築物を導入するための形質転換法は当該技術分野において周知であり、マイクロインジェクション、ポリエチレングリコールの使用、エレクトロポレーション、高速弾道貫通(high velocity ballistic penetration)、またはアグロバクテリウムを介した形質転換等の方法を含む。植物細胞または植物の形質転換後、所望のDNAが組み込まれているそれらの植物細胞または植物を、抗生物質耐性、除草剤抵抗性、アミノ酸類似体に対する寛容等の方法によって、または表現型マーカーを用いることによって選択することができる。
【0051】
当該植物細胞が遺伝子発現の増加を示すかどうかを判定するために、種々のアッセイ、例えばノーザンブロッティングまたは定量的逆転写PCR(RT−PCR)を用いてもよい。従来の方法によって、形質転換された細胞から遺伝子導入植物全体を再生してもよい。同型株を作り出すために、改良されたイソプレノイド量を有するそのような遺伝子導入植物を繁殖および自家授粉させてもよい。そのような植物は、導入した特性に対する遺伝子を含む種子を産生し、選択された表現型を生じるであろう植物を産生するように成長させることができる。
【0052】
本開示に従った改変に特に好ましい植物は、シロイヌナズナ、ボリジ(ボリジ種)、カノーラ、トウゴマ(Ricinus communis)(リキヌス種)、カカオ豆(Theobroma cacao)(テオブロマ種)、トウモロコシ(Zea mays)(トウモロコシ種)、綿(ゴシッピウム種)、ハマナ種、クフェア種、亜麻(アマ種)、レスクェレラ種、およびリムナンテス種、リノーラ、ナスタチウム(ノウゼンハレン種)、マツヨイグサ種、オリーブ(オリーブ種)、ヤシ(アブラヤシ種)、ピーナッツ(ラッカセイ種)、ナタネ、ベニバナ(ベニバナ種)、大豆(ダイズ種およびソヤ種)、ヒマワリ(ヒマワリ種)、タバコ(タバコ種)、ベルノニア種、小麦(コムギ種)、大麦(オオムギ種)、米(オリザ種)、オートムギ(カラスムギ種)、モロコシ(モロコシ種)、ライ麦(ライムギ種)、またはイネ科植物ファミリーの他のメンバーを含む。
【0053】
一部の実施形態を用いて、油料種子作物から生成された油料種子の産出量または組成を改変する。油料種子作物は、食用のまたは産業的に有用な油を商業的に重要な産出量で作り出すことができる植物種であり、前記に挙げた植物種の多くを含む。そのような油料種子作物は当業者に周知である。
【0054】
一実施例において、DGAT2をコードするヌクレオチド配列で形質転換した植物を栽培する。遺伝子導入植物の種子を収穫して、前記種子の脂肪酸を抽出する。抽出した脂肪酸を、その後の組成物、例えば医薬組成物、栄養補助組成物、または食品組成物内への取り込みに用いる。
【0055】
特定の実施形態において、形質転換されるべき植物に関して、油の産出を増大または変化させる他の方法(例えば、植物における発現のために、例えばBrassicaピルビン酸脱水素酵素キナーゼ活性を有するペプチドをコードする核酸配列(例えば、Marilla,et al.に対する米国特許第7,214,859号(2007年5月8日)、Zou,et al.に対する米国特許第6,500,670号(2002年12月)、およびRandall,et al.に対する米国特許第6,256,636号(2001年7月)を参照)、ジアシルグリセロールアシルトランスフェラーゼ活性を有するペプチドをコードする核酸配列(例えば、Zou,et al.に対する米国特許第7,015,373号および米国特許第6,500,670号(2002年12月)を参照)、およびグリセロール−3−リン酸脱水素酵素活性を有するペプチドをコードする核酸配列(例えば、米国特許第7,112,724号を参照)、ならびにその組み合わせを含む群から選択される核酸配列を取り込むこと)を用いてもよい。
【0056】
実施形態は、本明細書において詳細に記載されている具体的な実施例に加えて、種々の改変および代替的形態を受け入れる余地がある。従って、実施形態は開示されている特定の形態に制限されない。むしろ、本開示の範囲は、以下の添付した特許請求の範囲に含まれるすべての改変、同等物、および代替物を包含する。
【実施例】
【0057】
実施例1:DNA操作
DNAの調製、プラスミドの増殖、および単離には、標準的方法および手順を用いた(Sambrook,et al.,1989)。配列決定は、Taq DyeDeoxy(商標)ターミネーターサイクルシークエンシングキット(Applied Biosystems社)を用いて、Applied Biosystems社のモデル373A DNAシークエンシングシステムで行った。当該ヌクレオチドおよび推定アミノ酸配列を、BLASTプログラムを用いることによって、データバングで入手可能な配列と比較した(Altschul,et al.,1990)。当該技術分野において公知であるように、NCBIのヌクレオチドおよびタンパク質データベースにおける他の脂肪酸ジアシルグリセロールアシルトランスフェラーゼ遺伝子とのホモロジーに基づいて、DGAT2クローンを同定した。
【0058】
実施例2:TpDGAT2を発現することによって、酵母形質転換体においてin vivo形成されたトリアシルグリセロール(TAG)
DGAT2遺伝子をpYES2.1(Invitrogen社)内に挿入した。当該構築物をシークエンシングによって確認し、出芽酵母サッカロマイセス・セレヴィシエ(Saccharomyces cerevisiae)の株H1246MAT−αを形質転換するために、pYES2.1/TpDGAT2を用いた。この変異株は4重の変異体である(DGAT−,PDAT−,ASAT−,ASAT2−)。プラスミドDNAを推定される形質転換体から単離し、pYES2.1/TpDGAT2の存在をサザン解析によって確認した。ベクターのみ(pYES2.1)を含むH1246MAT−α形質転換体をコントロールとして用いた。シロイヌナズナDGAT1で形質転換したH1246MAT−αはポジティブコントロールとして働いた。
【0059】
シングルコロニーを、20mLのSD培地(Ausubel,et al.,1995,Vol.2,p.13.1.3によって記載されているような、グルコースを含み、ウラシルを含まない合成デキストロース培地)中で回転式シェーカー(270rpm)上で28℃にて一晩培養した。一晩培養物からの細胞を沈殿させ、50mLの発現誘導用の培地(ガラクトースを含み、ウラシルを含まないSD培地)中に再懸濁した。細胞を270rpmで振とうさせながら、28℃にて再びインキュベーションし、4〜6時間後に回収した。GALで誘導した酵母形質転換体を、5,000rpmで5分間の遠心分離によって回収し、1mM EDTAおよび1mM DTTを含む100mM Hepes−NaOH(pH7.4)中に再懸濁した。
【0060】
図9を参照すると、ネガティブコントロールである空ベクターではTAGが生成されなかった(レーン8)のに対し、ポジティブコントロール(レーン1)ではTAGバンドを示した。DGAT2を含むベクターのそれぞれ(レーン2〜6)では、DGAT2がTAGを合成する能力を有することを裏付けるTAGバンドを示した。レーン8の右側のレーンには、TAGマーカーとして用いられるTAG標準物質をロードした。
【0061】
実施例3:TpDGAT2の基質選択性
Ausubel,et al.(1995)によって記載されているように、酸洗浄したガラスビーズを用いて細胞溶解物を調製した。Bradford(1976)アッセイを用いて酵母溶解物中のタンパク質を測定し、各溶解物中のタンパク質量を標準化し、アリコート(タンパク質250μg)をDGAT2活性についてアッセイした。
【0062】
30℃の水槽中で100回転/分で10分間振とうさせながら、DGATアッセイをpH7.4にて行った。アッセイ混合物(最終容量0.5mL)は、100μg溶解物タンパク質、90mM HEPES−NaOH、200μM sn−1,2ジオレイン、およびアシル基供与体としての18μM14CアシルCoA(2nCi/nmol特異的活性)を含んだ。ヘキサン:ジエチルエーテル:酢酸(70:30:1 v/v/v)中で展開させたシリカゲルGプレート上でのTLCによって14C標識されたTAGを単離し、Win−Scan2D(登録商標)ソフトウェア(Bioscan社,ワシントンDC,米国)を用いたBioscan AR−2000放射性TLCスキャナー上で、放射性標識されたTAGバンドを可視化し、当該バンドを切り出して、Taylor,et al.(1991)によって記載されているように定量化した。
【0063】
実施例4:TpDGAT2形質転換体の脂肪酸組成
S.セレヴィシエ株H1246MAT−αを、シロイヌナズナ/pYES2.1またはT.シュードナナ/pYES2.1で形質転換した。形質転換体を3日間28℃にて培養し、ガラクトースによって誘導した。前記形質転換体を、処理なし(コントロール)、50uM DHAまたは150uM DHAで処理した。AtDGAT1/pYES2.1を含む3つの形質転換体およびTpDGAT2/pYES2.1を含む3つの形質転換体についての脂肪酸プロファイルを、従来のガスクロマトグラフィー分析に基づく表1に示した。
【0064】
脂肪酸を16:0、16:1、18:0、18:1(オレイン酸)、および22:6(DHA)として同定し、それぞれの組成を総脂肪酸に対するパーセンテージで表している。DHAの発現は、コントロール株の0から150μM TpDGAT2/pYES2.1では6.01%に増加し、150μM AtDGAT/pYES2.1のものの2倍以上であった(表1)。これらの結果は、TpDGAT2がDGAT1よりもDHA脂肪酸をより効率的に利用することをさらに裏付けた。
【0065】
表1に示すように、脂肪酸組成に関して、DGAT2 cDNAを含む変異株は飽和物全体の減少および不飽和物の増加を示した。そのような変化はすべて、「より健康な」油のプロファイルに向かうものであり、カノーラ、アブラナの他の油料種子、および他の食用油料作物に直接応用して、同様の油組成の改良をもたらすことができる。
【0066】
(表1) 酵母変異体H1246MAT−αにおける、DGAT2およびDGAT1によって発現されるTAGの脂肪酸組成
【0067】
実施例5:野生型シロイヌナズナにおけるDGAT2 cDNAの過剰発現
全長DGAT2 cDNAをPCR増幅のための鋳型として用いる。フラグメントを制限エンドヌクレアーゼ消化によって切断し、ベクターの相当する部位にライゲーションする。構築物の完全性を、シークエンシングによって確認する。
【0068】
前記ベクターをA.ツメファシエンス内に導入し、野生型シロイヌナズナを形質転換するために用い、後代を分析する。
【0069】
実施例6:種子特異的発現のためのDGAT2 cDNA植物形質転換ベクターの構築
配列の各末端に新しい制限部位を提供するために、プライマーとともに、全長DGAT2 cDNAをPCR増幅のための鋳型として用いた。PCRプロファイルは以下のとおりである:94℃1分;94℃30秒,55℃30秒,72℃1分を30サイクル;および72℃5分。その後、PCR産物をPCR2.1ベクター(Invitrogen社)にライゲーションする。フラグメントを切断し、ベクターの相当する部位にライゲーションする。構築物の完全性を、シークエンシングによって確認する。
【0070】
実施例7:植物DGAT2ベクター構築物を用いたアグロバクテリウムの形質転換
エレクトロコンピテント・アグロバクテリウム細胞GV3101(pMP90)株を以下のとおりに調製する。アグロバクテリウム培養物を2YT中で24〜48時間増殖させ、600nmにおける吸光度が0.5〜0.7に到達した時点で、細胞を氷上で冷やし、遠心分離(GSAローターで5,000×g,10分,4℃)によって沈殿させる。沈殿物を1、0.5、および0.02容量の冷却した10%滅菌グリセロール中で洗浄し、0.01容量の冷却した10%グリセロールに再懸濁する。その後、当該エレクトロコンピテント細胞を液体N2中で冷凍し、−70℃で保存する。メーカーの使用説明書に従って、20〜50ngの形質転換DNAを用いたエレクトロポレーションによって、前記アグロバクテリウム細胞を形質転換し、選択培地(50μg/mLカナマイシンを含むLB)上にプレーティングし、28℃にて一晩インキュベーションする。形質転換された単一細胞を、50μg/mLカナマイシンおよび25μg/mLゲンタマイシンを含む5mL LB中で一晩増殖させる(28℃,225r.p.m)。DNAの抽出および精製を行う。植物の形質転換前に、構築物の忠実度をDNAシークエンシングによって再チェックする。
【0071】
実施例8:シロイヌナズナの形質転換
シロイヌナズナの種子を16時間・明/8時間・暗の管理の中、蛍光照明(120μE・m−2S−1)の下で22℃にて栽培する。4〜6個の植物を、湿らせたTERRA−LITE REDI−EARTH(W.R.Grace&Co.Canada社,エイジャックス,オンタリオ州,カナダ)中で10cm2植木鉢で育てる。植木鉢中の土壌混合物が接種培地中に落下するのを防ぐために、土壌をプラットホームのように積み上げて種子を頂上に植え付け、植木鉢全体をナイロン製のウィンドウスクリーンで覆い、ゴム製のバンドで固定する。最初の花が開花し始めた時点で、アグロバクテリウム懸濁液中で植物を減圧浸潤する。
【0072】
アグロバクテリウムを増殖させるために、50μg/mLカナマイシンおよび25μg/mLゲンタマイシンを含むLB培地中で5mL懸濁液を28℃にて一晩培養する。浸潤の前日に、この「種子培養物」を、50μg/mLカナマイシンおよび25μg/mLゲンタマイシンを補充した250mLのLB培地を含む4つのフラスコに分ける。これらの培養物を28℃にて一晩培養する。翌朝、600nmにおける吸光度(およそ=1.0)をチェックした後、遠心分離(GSAローターで5,000×g,10分,室温)によって細胞を回収し、浸潤培地(水中に5%スクロース,0.005% Silwet−77)に再懸濁して、600nmで0.8の吸光度を得る。
【0073】
アグロバクテリウム懸濁液をビーカーに注ぎ、花およびボルトが浸水するように鉢植えされている植物を前記ビーカーの中に逆さにする。前記ビーカーを大きなベルジャー内に置き、茎表面上で気泡が形成され、かつ溶液がわずかに泡立ち始めるまで、真空ポンプを用いて真空に吸引し、その後真空を急速に解放する。必要な時間および圧力は実験装置によって異なるが、良好な浸潤は、均一に暗くなり水浸しになる組織のように、目に見えて明らかである。植木鉢を前記ビーカーから取り出して、プラスチック製のトレイの中の側に置き、プラスチック製のドームで覆って湿度を維持する。翌日、植物から覆いを取り外して真っ直ぐに立て、Katavic,et al.(1995)によって記載されているように、連続的な光条件下、栽培箱中でおよそ4週間成長させる。長角果が成熟して乾燥した時点で、種子を収穫し、陽性の形質転換体について選択する。
【0074】
実施例9:セイヨウアブラナ(Brassica napus)の形質転換
形質転換は基本的に、Moloney,et al.,1989,Plant Cell Reports 8:238−242によって記載されているように実施する。
【0075】
A.ツメファシエンス株GV3101/pMP90(Koncz C.&Schell,J.,1986,Mol.Gen.Genet.204:383−396)を、形質転換の研究に用いる。LB培地(Difco,米国)(100mL)中の定常期の細菌培養物を遠心分離によって回収し、抗凍結剤としての1%DMSO(ジメチルスルホキシド)(Sigma社,米国)を含む10mLの新しいLB培地中に再懸濁する。細菌のアリコートを、2%スクロース、50μMアセトシリンゴン,pH5.6を含む2mLのブレインハートインフュージョン培地(Difco,米国)に添加し、28℃にて一晩インキュベーションする形質転換に用いるまで、200μLのアリコートを−20℃で保存する。細菌細胞の密度は、およそ1×109 細胞/mLである。
【0076】
Moloney,et al.(1989),Plant Cell Rep.8:238−242の方法に従って、子葉移植片を植物形質転換ベクターを含むアグロバクテリウムに暴露する。前記移植片の葉柄の切断面を細菌培養物に短時間浸す。切断面が培地に接触するように、前記移植片を共存培養培地の中に入れる。10個の移植片を、それぞれ100×15mmのペトリプレートに置く。共存培養プレートをSTRETCH’N SEAL(商標)プラスチック製の包装物で密封する。種子の発芽工程に関する前述のような温度および光周期の条件下で、プレートを3日間培養キャビネット内でインキュベーションする。その後、移植片を選択培地に移す。
【0077】
選択培地において3〜4週間後、再生している若芽(推定上の形質転換体)を切り取り、継続培養のために新しい選択培地に移す。新芽が1.5〜2.0cmの長さに到達した時点で、それらを発根培地に移す。基本的に、Jefferson,R.A.(1987),Plant Mol.Biol.Rep.5:387−405によって記載されているように、推定上の遺伝子導入された新芽をgus遺伝子の発現についてスクリーニングする。青色の染色の存在は、形質転換の証拠として見なされる。
【0078】
形質転換の確認を、カナマイシン上での選択、サザンブロッティング、PCR(ポリメラーゼ連鎖反応)、および後代分析によって立証する。
【0079】
実施例10:推定上の形質転換体(遺伝子導入植物)の選択および遺伝子導入植物の解析
各構築物に対して、種子を大量に収穫する。種子を20%漂白剤および0.01%Triton X−100を含む溶液中に種子を20分間浸すことによって表面滅菌し、その後滅菌水で3回洗浄する。滅菌した種子を、滅菌した0.1%Phytagar(500〜1,000個の種子につき約1mLのPhytagar)中に室温にて再懸濁し、および150×15mmのカナマイシン選択プレート上に2,000〜4,000個の種子を含む容量を塗布することによってプレーティングする。プレートを照明をつけずに寒い中で2日間インキュベーションし、7〜10日間制御された環境の中で(16時間・明/8時間・暗の管理の中、蛍光照明(120μE・m−2S−1)の下で22℃にて)栽培する。選択培地は、1/2MSG培地、0.8%Phytagar、3%スクロース、50μg/mLカナマイシン、および50μg/mLチメンチンを含む。ペトリプレートおよび蓋を、Micropore(商標)外科用テープ(3M カナダ,ロンドン,ON,カナダ)で密封する。7〜10日後、緑葉を有し、かつ培地中で根をしっかりと定着させている薬剤耐性植物を形質転換体として同定し、葉3〜5枚の段階で、選択された形質転換体を非常に湿らせた土壌混合物で満たした湿地の中に移植する。形質転換体を成熟させ、成熟した種子(Katavic,et al.(1994)で定義されているT2世代)を個々の植物から収穫し、さらに分析する。
【0080】
ゲノムDNAを個々のT1植物から単離する。前記T1形質転換体におけるcDNAまたは遺伝子の存在をそれぞれ確認するために、PCR増幅を行う。挿入したフラグメントのシングルコピーを含む形質転換体を選択するために、サザン分析を行う。DNAサンプルを制限酵素で消化し、1%アガロースゲル上での電気泳動によって分解し、ナイロン製のフィルター(Hybond−N+,アメルシャム社)を用いてサザンブロッティングを行う。α−[32P]dCTP(NEN/DuPont)で標識したDGAT2 cDNAフラグメントをプローブとして用いる。ハイブリダイゼーションを60℃で行う。その後、前記フィルターをKodak X−OMAT−ARフィルムに露光する。
【特許請求の範囲】
【請求項1】
配列番号:15と少なくとも90%の配列同一性を有する配列を含む単離または精製したポリペプチド。
【請求項2】
配列番号:1と少なくとも90%の配列同一性を有する、請求項1に記載の単離または精製したポリペプチド。
【請求項3】
ジアシルグリセロールアシルトランスフェラーゼ活性を有する、請求項1に記載の単離または精製したポリペプチド。
【請求項4】
請求項1に記載の単離または精製したポリペプチドをコードする、単離または精製した核酸配列。
【請求項5】
ベクター内に存在する、請求項4に記載の核酸配列。
【請求項6】
請求項4に記載の核酸配列を含む遺伝子導入植物。
【請求項7】
前記核酸配列を欠いている植物における量と比較して、種子中の多価不飽和脂肪酸の量が変化している、請求項4に記載の核酸配列を含む遺伝子導入植物。
【請求項8】
前記植物中の脂肪酸が約70%以上の多価不飽和脂肪酸である、請求項4に記載の核酸配列を含む遺伝子導入植物。
【請求項9】
請求項4に記載の核酸配列で形質転換した酵母細胞。
【請求項10】
配列番号:1、15、25、27、28、31、33、35、37、39、41、43、45、47、49、51、53、55、57、59、および61からなる群から選択されるポリペプチドと少なくとも90%の配列同一性を有するポリペプチドをコードするポリヌクレオチドを含む核酸を植物内に導入する段階を含む、植物における超長鎖多価不飽和脂肪酸の量を変化させる方法。
【請求項11】
アグロバクテリウム(Agrobacterium)を介した形質転換によって前記植物内に核酸構築物を導入する、請求項10に記載の方法。
【請求項12】
Brusicaピルビン酸脱水素酵素キナーゼ活性を備えるポリペプチドをコードするポリヌクレオチドを前記植物内に導入する段階をさらに含む、請求項10に記載の方法。
【請求項13】
ジアシルグリセロールアシルトランスフェラーゼ活性を備えるポリペプチドをコードするポリヌクレオチドを導入する段階をさらに含む、請求項10に記載の方法。
【請求項14】
グリセロール−3−リン酸脱水素酵素活性を備えるポリペプチドをコードするポリヌクレオチドを導入する段階をさらに含む、請求項10に記載の方法。
【請求項15】
前記植物が、シロイヌナズナ(Arabidopsis thaliana)、ボリジ種(Borago spp.)、カノーラ(Canola)、リキヌス種(Ricinus spp.)、テオブロマ種(Theobroma spp.)、トウモロコシ種(Zea spp.)、ゴシッピウム種(Gossypium spp.)、ハマナ種(Crambe spp.)、クフェア種(Cuphea spp.)、アマ種(Linum spp.)、レスクェレラ種(Lesquerella spp.)、リムナンテス種(Limnanthes spp.)、リノーラ(Linola)、ノウゼンハレン種(Tropaeolum spp.)、マツヨイグサ種(Oenothera spp.)、オリーブ種(Olea spp.)、アブラヤシ種(Elaeis spp.)、ラッカセイ種(Arachis spp.)、ナタネ(rapeseed)、ベニバナ種(Carthamus spp.)、ダイズ種(Glycine spp.)、ソヤ種(SoJa spp.)、ヒマワリ種(Helianthus spp.)、タバコ種(Nicotiana spp.)、ベルノニア種(Vernonia spp.)、コムギ種(Triticum spp.)、オオムギ種(Hordeum spp.)、オリザ種(Oryza spp.)、カラスムギ種(Avena spp.)、モロコシ種(Sorghum spp.)、ライムギ種(Secale spp.)、アブラナ(Brassicaceae)、およびイネ科(Gramineae)植物ファミリーの他のメンバーからなる群から選択される、請求項10に記載の方法。
【請求項16】
請求項10に記載の方法によって産生した植物。
【請求項17】
請求項10に記載の方法によって産生した前記植物から収穫した種子。
【請求項18】
請求項10に記載の方法によって産生した前記植物から抽出した油。
【請求項19】
前記核酸構築物を含む植物から種子を収穫する段階、および前記収穫した種子から油を抽出する段階をさらに含む、請求項10に記載の方法。
【請求項1】
配列番号:15と少なくとも90%の配列同一性を有する配列を含む単離または精製したポリペプチド。
【請求項2】
配列番号:1と少なくとも90%の配列同一性を有する、請求項1に記載の単離または精製したポリペプチド。
【請求項3】
ジアシルグリセロールアシルトランスフェラーゼ活性を有する、請求項1に記載の単離または精製したポリペプチド。
【請求項4】
請求項1に記載の単離または精製したポリペプチドをコードする、単離または精製した核酸配列。
【請求項5】
ベクター内に存在する、請求項4に記載の核酸配列。
【請求項6】
請求項4に記載の核酸配列を含む遺伝子導入植物。
【請求項7】
前記核酸配列を欠いている植物における量と比較して、種子中の多価不飽和脂肪酸の量が変化している、請求項4に記載の核酸配列を含む遺伝子導入植物。
【請求項8】
前記植物中の脂肪酸が約70%以上の多価不飽和脂肪酸である、請求項4に記載の核酸配列を含む遺伝子導入植物。
【請求項9】
請求項4に記載の核酸配列で形質転換した酵母細胞。
【請求項10】
配列番号:1、15、25、27、28、31、33、35、37、39、41、43、45、47、49、51、53、55、57、59、および61からなる群から選択されるポリペプチドと少なくとも90%の配列同一性を有するポリペプチドをコードするポリヌクレオチドを含む核酸を植物内に導入する段階を含む、植物における超長鎖多価不飽和脂肪酸の量を変化させる方法。
【請求項11】
アグロバクテリウム(Agrobacterium)を介した形質転換によって前記植物内に核酸構築物を導入する、請求項10に記載の方法。
【請求項12】
Brusicaピルビン酸脱水素酵素キナーゼ活性を備えるポリペプチドをコードするポリヌクレオチドを前記植物内に導入する段階をさらに含む、請求項10に記載の方法。
【請求項13】
ジアシルグリセロールアシルトランスフェラーゼ活性を備えるポリペプチドをコードするポリヌクレオチドを導入する段階をさらに含む、請求項10に記載の方法。
【請求項14】
グリセロール−3−リン酸脱水素酵素活性を備えるポリペプチドをコードするポリヌクレオチドを導入する段階をさらに含む、請求項10に記載の方法。
【請求項15】
前記植物が、シロイヌナズナ(Arabidopsis thaliana)、ボリジ種(Borago spp.)、カノーラ(Canola)、リキヌス種(Ricinus spp.)、テオブロマ種(Theobroma spp.)、トウモロコシ種(Zea spp.)、ゴシッピウム種(Gossypium spp.)、ハマナ種(Crambe spp.)、クフェア種(Cuphea spp.)、アマ種(Linum spp.)、レスクェレラ種(Lesquerella spp.)、リムナンテス種(Limnanthes spp.)、リノーラ(Linola)、ノウゼンハレン種(Tropaeolum spp.)、マツヨイグサ種(Oenothera spp.)、オリーブ種(Olea spp.)、アブラヤシ種(Elaeis spp.)、ラッカセイ種(Arachis spp.)、ナタネ(rapeseed)、ベニバナ種(Carthamus spp.)、ダイズ種(Glycine spp.)、ソヤ種(SoJa spp.)、ヒマワリ種(Helianthus spp.)、タバコ種(Nicotiana spp.)、ベルノニア種(Vernonia spp.)、コムギ種(Triticum spp.)、オオムギ種(Hordeum spp.)、オリザ種(Oryza spp.)、カラスムギ種(Avena spp.)、モロコシ種(Sorghum spp.)、ライムギ種(Secale spp.)、アブラナ(Brassicaceae)、およびイネ科(Gramineae)植物ファミリーの他のメンバーからなる群から選択される、請求項10に記載の方法。
【請求項16】
請求項10に記載の方法によって産生した植物。
【請求項17】
請求項10に記載の方法によって産生した前記植物から収穫した種子。
【請求項18】
請求項10に記載の方法によって産生した前記植物から抽出した油。
【請求項19】
前記核酸構築物を含む植物から種子を収穫する段階、および前記収穫した種子から油を抽出する段階をさらに含む、請求項10に記載の方法。
【図1A】


【図1B】

【図2A】
【図2B】
【図3】

【図4】

【図5】

【図6】

【図7】

【図8】

【図9】
【図10】
【図11】
【図1B】
【図2A】
【図2B】
【図3】
【図4】
【図5】
【図6】
【図7】
【図8】
【図9】
【図10】
【図11】
【公表番号】特表2011−507513(P2011−507513A)
【公表日】平成23年3月10日(2011.3.10)
【国際特許分類】
【出願番号】特願2010−539472(P2010−539472)
【出願日】平成20年12月16日(2008.12.16)
【国際出願番号】PCT/US2008/013811
【国際公開番号】WO2009/085169
【国際公開日】平成21年7月9日(2009.7.9)
【出願人】(510170693)ナショナル リサーチ カウンシル オブ カナダ (1)
【出願人】(505412443)ダウ アグロサイエンシズ リミテッド ライアビリティー カンパニー (10)
【Fターム(参考)】
【公表日】平成23年3月10日(2011.3.10)
【国際特許分類】
【出願日】平成20年12月16日(2008.12.16)
【国際出願番号】PCT/US2008/013811
【国際公開番号】WO2009/085169
【国際公開日】平成21年7月9日(2009.7.9)
【出願人】(510170693)ナショナル リサーチ カウンシル オブ カナダ (1)
【出願人】(505412443)ダウ アグロサイエンシズ リミテッド ライアビリティー カンパニー (10)
【Fターム(参考)】
[ Back to top ]