
融合プロテオミクスによるNF1特異的タンパク質の同定方法、NF1特異的タンパク質発現抑制方法、NF1特異的タンパク質の腫瘍マーカー及び治療ターゲットとしての使用方法

【課題】融合プロテオミクスによるNF1(神経線維腫症1型)関連病態に関わる分子群であるNF1特異的タンパク質(TCTP)の同定方法、発現を抑制するNF1特異的タンパク質発現抑制方法、NF1の腫瘍悪性化を検査する腫瘍悪性化マーカーならびに治療ターゲットとしての使用方法の提供。
【解決手段】神経線維腫症1型(NF1)組織であって薬剤感受性組織ならびに薬剤非感受性組織から抽出したタンパク質およびmRNAをプロテオーム解析およびトランスクリプトーム解析をし、得られたデータを融合プロテオーム解析データとして一元化しGO解析するとともに、ネットワーク解析によって、NF1関連病態に関わる分子群であるNF1特異的タンパク質(TCTP)の同定、TCTPの発現を抑制することによるNF1発現の抑制、さらに、TCTPは、NF1の腫瘍悪性化に伴って発現が更新し、指標マーカーまたは治療ターゲットとして使用できる。
【解決手段】神経線維腫症1型(NF1)組織であって薬剤感受性組織ならびに薬剤非感受性組織から抽出したタンパク質およびmRNAをプロテオーム解析およびトランスクリプトーム解析をし、得られたデータを融合プロテオーム解析データとして一元化しGO解析するとともに、ネットワーク解析によって、NF1関連病態に関わる分子群であるNF1特異的タンパク質(TCTP)の同定、TCTPの発現を抑制することによるNF1発現の抑制、さらに、TCTPは、NF1の腫瘍悪性化に伴って発現が更新し、指標マーカーまたは治療ターゲットとして使用できる。
【発明の詳細な説明】
【技術分野】
【0001】
本発明は、融合プロテオミクスによるNF1特異的タンパク質の同定方法、NF1特異的タンパク質発現抑制方法、NF1特異的タンパク質の腫瘍マーカー及び治療ターゲットとしての使用方法に関するものである。更に詳細には、本発明は、融合プロテオミクスによるNF1関連病態に関わる分子群であるNF1特異的タンパク質(TCTP:Translationally Controlled Tumor Protein)の同定方法、そのNF1特異的タンパク質の発現抑制方法、およびNF1特異的タンパク質の腫瘍悪性化マーカーとして、または治療ターゲットとしての使用方法に関するものである。
【背景技術】
【0002】
神経線維腫症1型(Neurofibromatosis type 1:NF1)は、NF1原因遺伝子産物であるニューロフィブロミン(neurofibromin)というタンパク質であるニューロフィブロミンの機能が欠損してRasを介した細胞内シグナル伝達が異常をきたし、神経系細胞の増殖と分化異常を誘発し、全身の皮膚に多発性結節と色素斑、皮下の多発性神経線維腫、線維肉腫、グリオーマなどの悪性腫瘍、知能障害等の多彩な病態を伴う頻度が高い優性遺伝性疾患である。
【0003】
NF1遺伝子は、17番染色体長腕(17q11.2)上に存在する2818個のアミノ酸からなるタンパク質であるニューロフィブロミンをコードする遺伝子である。また、NF1遺伝子産物のニューロフィブロミンは、アミノ酸残基360個を中心とする領域に、Ras−GAP(Ras−GTPase-activating protein)相同領域を有し、低分子量Gタンパク質Rasと相互に作用するとともに、Ras結合GTPを加水分解してGDPに変換してRasタンパク質の機能を不活性化する、細胞内シグナル伝達の重要な調節因子と考えられている(非特許文献1)。
【0004】
近年のマウスを用いた研究により、多発性神経線維腫は、細胞−細胞間および細胞−マトリックス間の相互的なシグナルの異常が形成の誘引と考えられている。特に、これまでに、ニューロフィブロミンの発現低下に伴い活性化されたRas−MAPK 経路を介した異常な細胞増殖が大きく関与していることが知られているが、この事象のみでは多発性神経線維腫形成メカニズムの十分な解明には至っていない。また、ニューロフィブロミンの発現低下に起因すると思われる様々な所見が多々報告されていることからも、ニューロフィブロミンには、更なる未知の機能の存在が推測される。
【0005】
そこで、本発明者らは、ニューロフィブロミンの神経系細胞内機能とその欠損による細胞増殖分化異常の機構を明らかにするため、RNA 干渉法(siRNA)を用いたNF1 遺伝子発現抑制によって、神経系細胞PC12に及ぼす神経成長因子(NGF)による分化誘導への影響を解析し、生じた表現形の変化に関与する細胞内責任シグナル分子群を、融合プロテオミクス法、および細胞生物学的検証法を用いて詳細に検討して、ニューロフィブロミンの細胞内機能とNF1の病態(特に多発性神経線維腫形成)との関連性を明らかにした。
【0006】
つまり、本発明者らは、ニューロフィブロミン分子のsiRNAによる細胞内ノックダウンを行い、細胞の形態変化の詳細と融合プロテオミクスの手法によるシグナルカスケードの変化を解析した結果、そのsiRNAの作用によって細胞内actin stress fiber、focal adhesion spot、ruffling形成能が大きく変化していることを認め、細胞骨格系のダイナミックな再構築が起こっていることを見出した。これらの変化には、細胞刺激因子や接着因子からの刺激を介して、Ras-MAPKおよびPI3Kシグナル、Rho-Rock-LIM kinase-coffilinシグナル等の細胞の骨格形成に関連する分子の機能制御によって運動能を調節する機構があることを解明した(非特許文献2)。
【0007】
また、本発明者らは、NF1ノックダウンPC12細胞において、分子TCTP(Translationally controlled tumor protein)が、NF1病態の一つである神経分化異常を誘起することを明らかにして、TCTPの発現亢進がNF1関連病態に密接に関わっていることを、融合プロテオミクス手法によって確認した。
【0008】
そこで、本発明者らは、特に本発明にて新規に同定されたmTOR シグナルに関わるTCTPを中心とした分子ネットワークに注目し、TCTP 分子活性化シグナルの治療ターゲットとしての可能性、および、NF1 異常細胞の悪性化マーカーとしての可能性を検証した。
【0009】
その結果、NF1 発現抑制によりNGF 刺激PC12 内で特異的に発現が変動する活性化シグナル分子群が、総合的にNF1 病態に関わる細胞増殖分化異常に関連すると考えられた。特にmTOR 経路の新規調節因子であるTCTP の関わるネットワークがNF1 発現抑制に連動することに注目して、詳細な解析を行った。NF1 欠損細胞ではTCTP の発現が有意に上昇しており、特に、NF1 患者由来悪性腫瘍MPNST 細胞において、その発現が顕著に誘導されていることから、TCTP がNF 病態において腫瘍悪性化の指標となる可能性があることを見いだした。また、本発明者らは、TCTP の発現抑制によって、未分化細胞が正常な分化状態を形成し、細胞増殖が抑制されたことから、TCTP を中心とする活性化シグナルの治療ター
ゲットとしての可能性があることを見いだした。
【0010】
さらに、本発明者らは、NF1に関連する疾患は、NF1遺伝子産物を介したシグナルによる細胞増殖抑制と、脱落するべき細胞の生理的アポトーシスの誘導の破綻によって生成するという仮説に基づき、細胞内でこれらと相互作用してシグナルを伝達する分子の解析や、それぞれの遺伝子をターゲッティングした細胞と正常細胞の比較検討を生化学・細胞生物学的な手法を用いて行うとともに、融合プロテオミクスによってNF1特異的タンパク質(TCTP)の分子機能解析を行ったところ、TCTPの発現亢進がNF1の病態に密接に関っていることから、TCTPがNF1の治療標的になりうることを見いだした。
【0011】
さらにまた、siRNAによるNF1発現を抑制したNF1病態モデルPC12細胞と、コントロール細胞について二次元電気泳動をベースとした2D-DIGE、iTRAQを用いたLC-MALDI-ならびにLC-ESI-MS/MS解析、およびマイクロアレイを用いたトランスクリプトーム解析を同時に行い、網羅的遺伝子・タンパク質発現および翻訳後修飾情報を取得し、それらすべての同定結果をデータマイニングソフトiPEACHおよびMANGOにより統合し、統計解析マイニングソフトsubioを用いたクラスター解析によってNF1病態関連分子群を抽出した。これらを分子ネットワークソフトKeymolnetによりNF1ノックダウン細胞で異常な挙動を示すシグナル分子群の抽出を試み、TCTPがその異常共同分子群の中で重要な役割を示す事を示した。
【0012】
なお、融合プロテオミクス手法については、本発明者らによる特許文献などに記載の融合プロテオーム解析方法(特許文献1、非特許文献3)や統合プロテオミクス解析手法(特許文献2)などの解析手法を利用するのがよい。
【先行技術文献】
【特許文献】
【0013】
【特許文献1】特開2006-294014号公報
【特許文献2】特願2010-81525号公報
【非特許文献】
【0014】
【非特許文献1】Yunoue et al., J. Biol. Chem., 2003
【非特許文献2】KAKEN FY2004 Annual Research Report; Research Project Number: 16390413荒木令江他「神経線維腫症原因遺伝子群を介した腫瘍抑制機構解明と分子治療戦略の開発」
【発明の概要】
【発明が解決しようとする課題】
【0015】
そこで、本発明の目的は、融合プロテオミクスによるNF1関連病態に関わる分子群であるNF1特異的タンパク質(TCTP)の同定方法を提供することを目的としている。
【0016】
本発明はまた、TCTPの発現抑制によりNF1発現を抑制することからなるNF1特異的タンパク質発現抑制方法を提供することを別の目的としている。
【0017】
本発明はまた、siRNA、shRNAなどによるRNAiやTCTP関連遺伝子のノックダウンやノックアウトなどによりTCTPの機能の一部または全部を抑制することによりTCTP発現抑制することからなるNF1特異的タンパク質発現抑制方法を提供することをさらに別の目的としている。
【0018】
本発明はさらに、NF1特異的タンパク質をNF1腫瘍マーカーとして使用することからなるNF1特異的タンパク質の腫瘍マーカー、例えば腫瘍の悪性化を検査するマーカーとしての使用方法を提供することを別の目的としている。
【0019】
本発明はさらに、TCTP発現阻害薬、MAPK阻害剤、PI3K阻害剤およびアーテスネート(Artesunate)から選ばれるいずれか1種又は2種以上を含むことからなるNF1腫瘍の治療薬を提供することをさらに別の目的としている。
【課題を解決するための手段】
【0020】
したがって、本発明は、融合プロテオミクスによるNF1関連病態に関わる分子群であるNF1特異的タンパク質(TCTP)の同定方法、検証方法を提供する。
【0021】
また、本発明は、TCTPの発現抑制によりNF1発現を抑制することからなるNF1特異的タンパク質発現抑制方法を提供する。
【0022】
さらに、本発明は、TCTPのsiRNA、shRNAなどによるRNAiやTCTPの機能の一部または全部をノックダウンもしくはノックアウトしてTCTPの発現を抑制することからなるNF1特異的タンパク質発現抑制方法を提供する。
【0023】
さらに、本発明は、NF1特異的タンパク質をNF1腫瘍マーカーとして使用することからなるNF1特異的タンパク質の悪性化腫瘍マーカー、例えば腫瘍の悪性化を検査するマーカーとしての使用方法を提供する。
【0024】
さらにまた、本発明は、TCTP阻害剤、MAPK阻害剤、PI3K阻害剤およびアーテスネート(Artesunate)から選ばれるいずれか1種又は2種以上を含むことからなるNF1腫瘍の治療薬を提供する。
【発明の効果】
【0025】
本発明によれば、融合プロテオミクスによってNF1関連病態に関わる分子群であるNF1特異的タンパク質(TCTP)を同定することが可能であるばかりではなく、TCTPのsiRNAなどによりNF1発現を抑制可能であることから、TCTPをNF1治療標的として利用することが可能である。さらに、TCTPは、NF1患者の腫瘍マーカー、特に腫瘍の悪性化を検査するマーカーとして使用することができることから、NF1の早期発見ならびに早期治療に有用である。したがって、本発明はNF1分子標的薬の開発にとっても極めて有用である。
【0026】
さらに、本発明は、TCTP発現阻害薬、MAPK阻害剤、PI3K阻害剤およびアーテスネート(Artesunate)から選ばれるいずれか1種又は2種以上を含むことからなるNF1腫瘍の治療薬を提供することができ極めて有用である。
【図面の簡単な説明】
【0027】
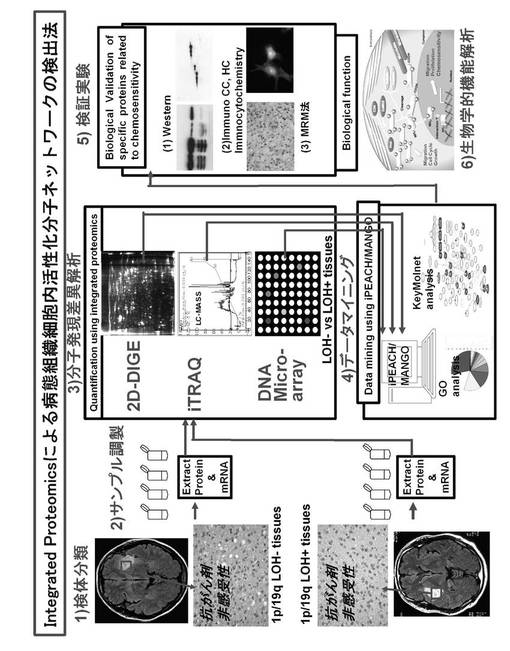
【図1】本発明の方法を示す概略図である。
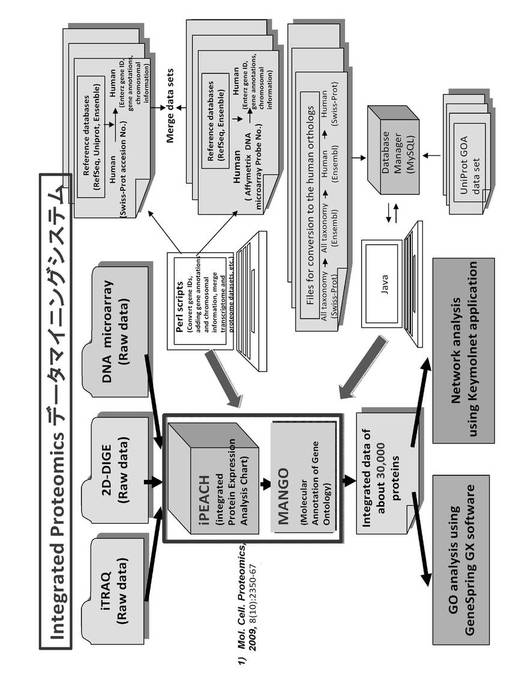
【図2】Integrated Proteomicsデータマイニングシステムの概略を示す概略図である。
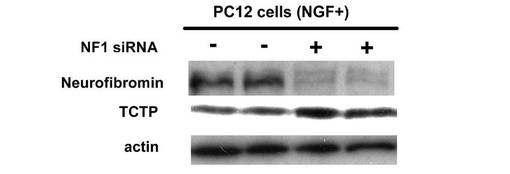
【図3】TCTP のウエスタンブロッテイングによる発現の確認を示す図である。
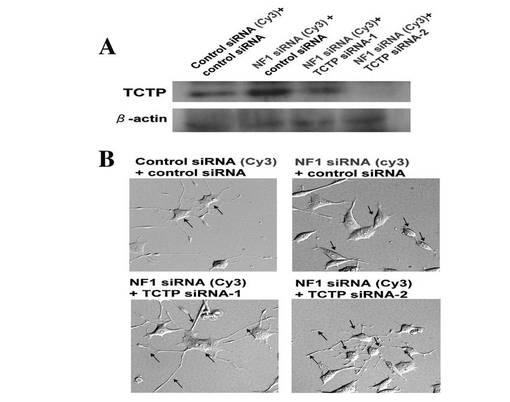
【図4】NGF NF1 KD PC12細胞内でのTCTP発現抑制による細胞形態の変化を示す図である。A: Cy3標識NF1siRNAおよびコントロールsiRNA処理PC12細胞内でのTCTP siRNA-1/-2によるTCTP の発現抑制のウエスタンブロッテイングによる発現の確認を示す図である。B: Cy3標識NF1siRNAおよびコントロールsiRNA処理PC12細胞の神経様突起発現抑制と、同細胞内でのTCTP の発現抑制による神経様突起発現と伸長を示す図である。
【図5】NF1 欠損S16 シュワン細胞内におけるTCTP の発現変動を示す図である。なお、インスリン刺激によってERK とTCTP の活性化がNF1 欠損細胞で亢進している。
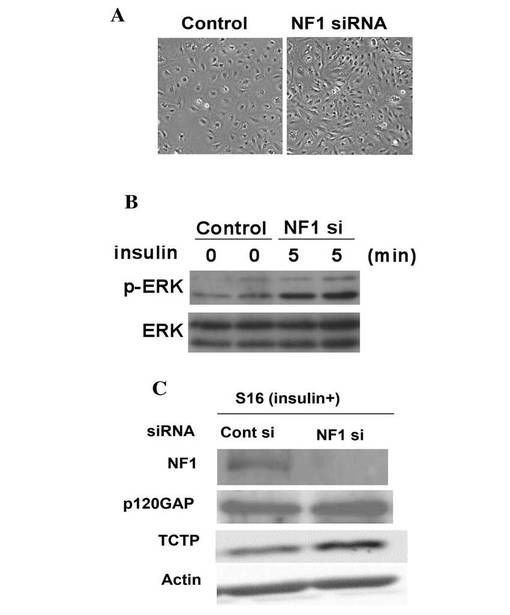
【図6】正常ならびにNF1 患者皮膚由来繊維芽細胞、およびNF1 腫瘍組織由来培養細胞(PT細胞)のニューロフィブロミンのウエスタンブロッテイングによる解析結果を示す図である。
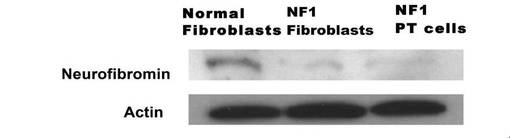
【図7】NF1患者の皮膚由来細胞(NF fibroblasts)およびニューロフィブローマ組織由来培養細胞(NF1PT細胞)における、血清刺激によるMAP kinase/ERK活性の経時的変化を示す図である。
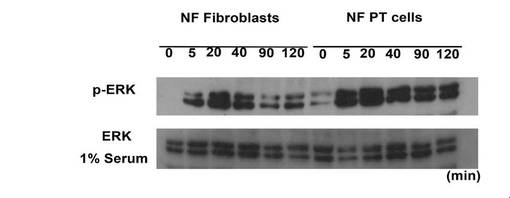
【図8】NF fibroblastsおよびNF PT細胞 のTCTP の発現をウエスタンブロッティング(A)および免疫染色(B)による解析結果を示す図である。
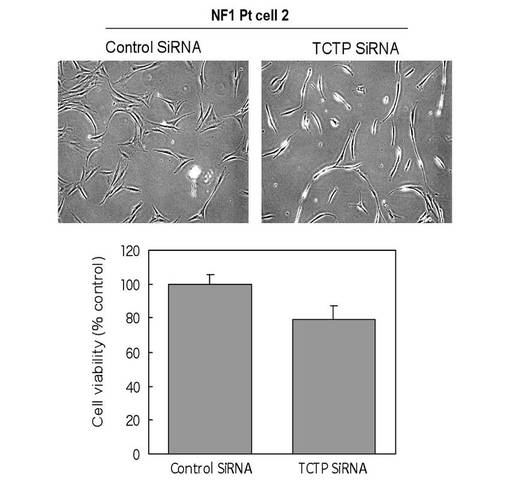
【図9】NF1PT細胞 におけるTCTP発現抑制による細胞内変化と、細胞増殖能の低下を示す図である。
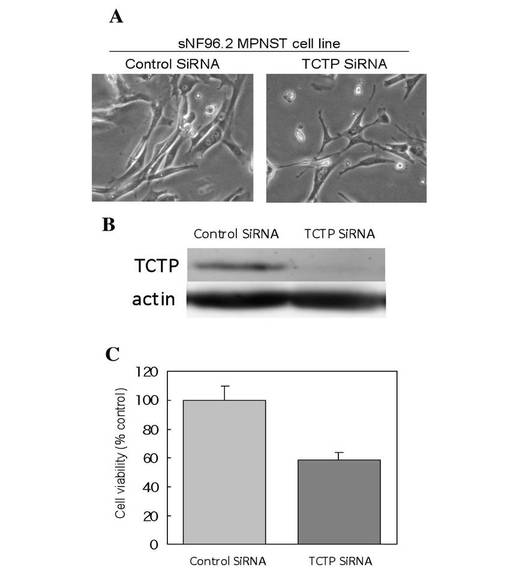
【図10】MPNST cell lineにおけるTCTP発現抑制による細胞増殖低下を示す図である。
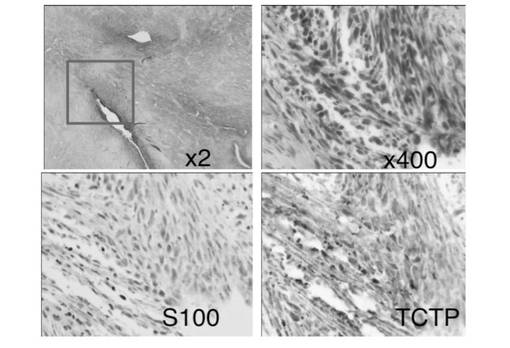
【図11】NF1 患者MPNST 組織におけるS100 およびTCTP の免疫染色の結果を示す図である。図中、MPNST 組織ではS100 の発現は低下しているが、TCTP の発現は増強しているのが観察された。
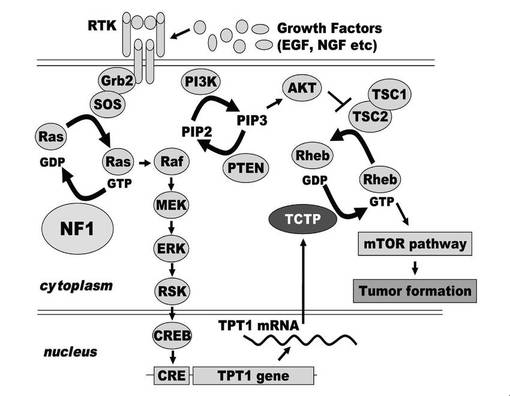
【図12】ニューロフィブロミン欠損によるTCTPを介したNF1 腫瘍形成のモデルを示す図である。
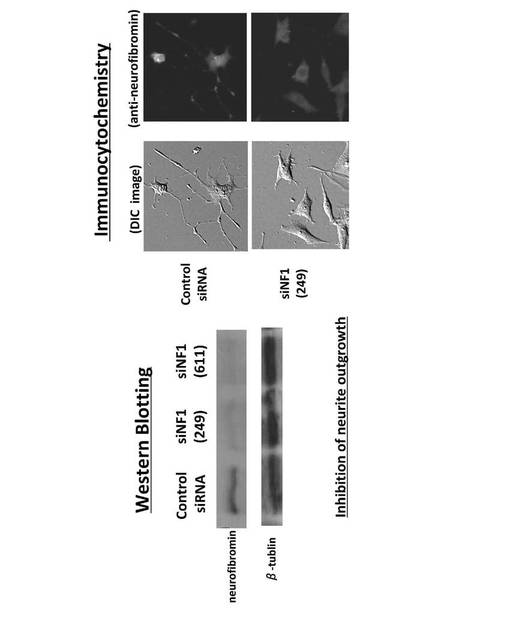
【図13】NF1−siRNA処理PC12細胞をNGFで刺激してNF1遺伝子発現抑制実験によるニューロフィブロミンの発現抑制のウエスタンブロット法および免疫細胞染色法による結果を示す図である。
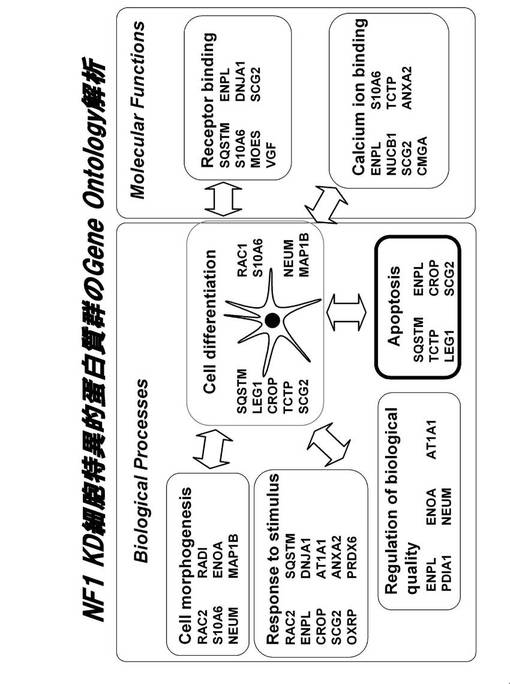
【図14】NF1 KD細胞特異的タンパク質群のGO解析結果を示す図である。
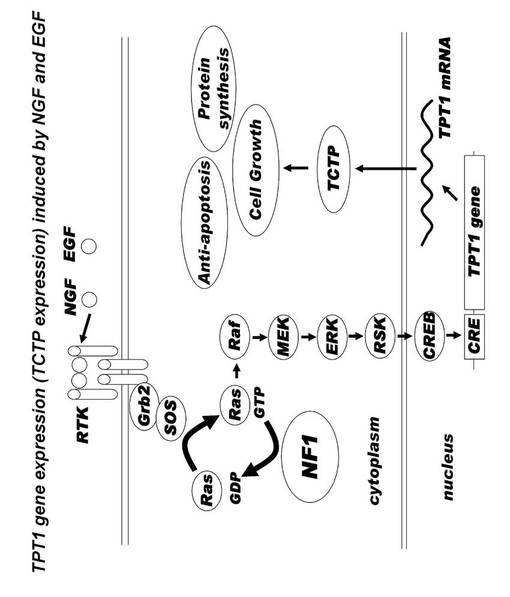
【図15】NGF およびEGFによって誘発されたTPT1遺伝子発現、つまりTCTP発現のネットワークを示す図である。
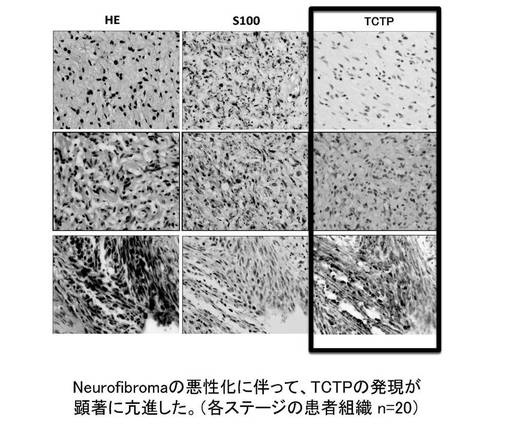
【図16】神経線維腫の悪性化に伴ってTCTP発現が更新していることを示す図である。
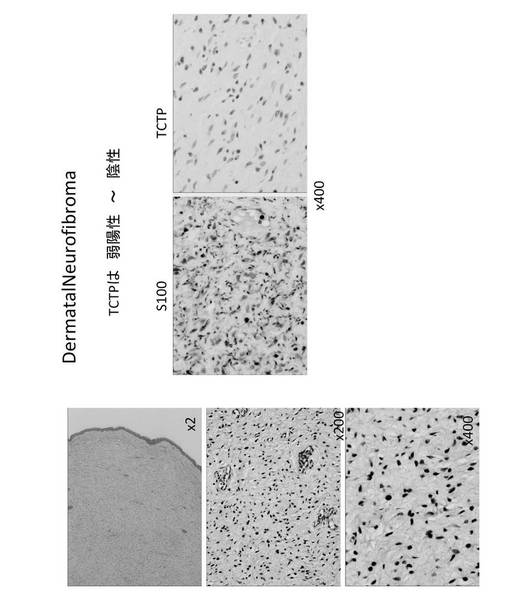
【図17】真皮性神経線維腫(dermatal neurofibroma)の細胞染色図である。
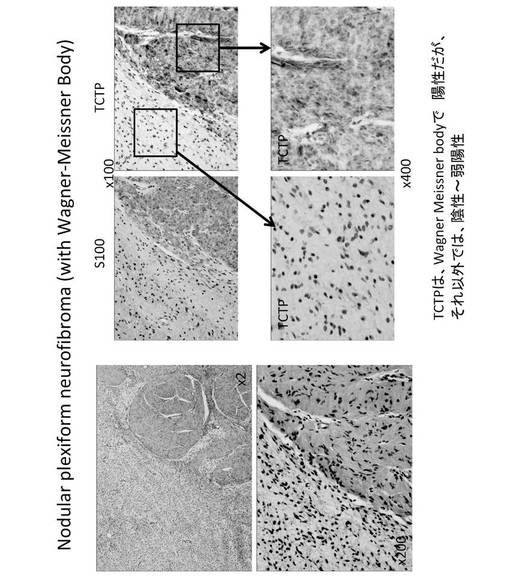
【図18】結節性網状神経線維腫(nodular plexiform neurofibroma with Wagner-Meissner Body)の細胞染色図である。
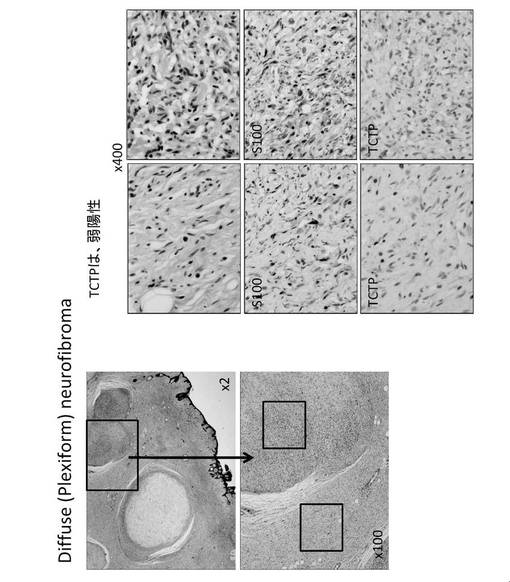
【図19】瀰漫性(網状)神経線維腫(diffuse (plexiform) neurofibroma)の細胞染色図である。
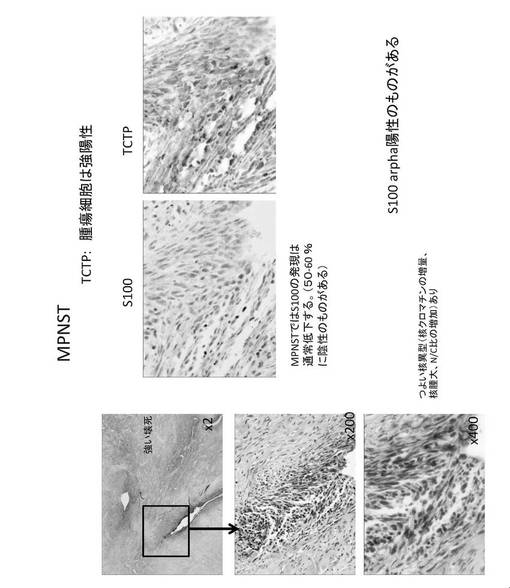
【図20】MPNST腫瘍細胞の細胞染色図である。
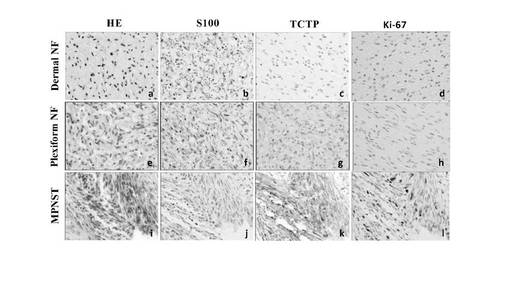
【図21】ニューロフィブローマ(NF)組織(皮膚NF、Plexiform NF、MPNST)中におけるTCTP発現のレベルとパターンを示す図。
【図22】種々のタイプのNF組織中のTCTP発現の免疫組織学的分析の結果を示す図。図中、*TCTP発現は、染色強度によってスコアした(0:染色なし;1:弱;2:中程度;3:強)。**サンプル9のTCTP発現レベルおよびKi67インデックスはPlexiform NFのdiffuse部位中の領域から取た。NDは無試験。
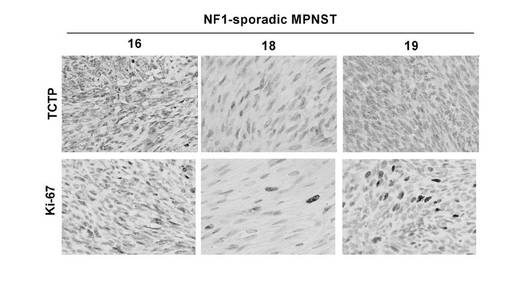
【図23】NF1散発性MPNST症例におけるTCTPの免疫組織学的分析結果を示す図。図は、Ki-67免疫染色を400倍率で示した図である。番号16,18,19は図22の組織サンプル番号に対応している。
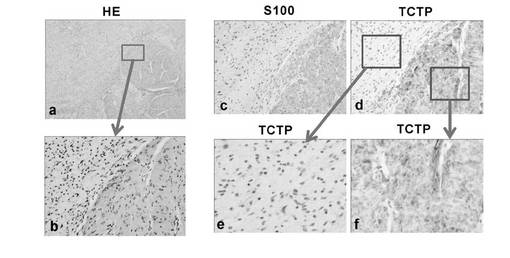
【図24】NF1-associated diffuse plexiform neurofibroma with Wagner-Meissner BodyにおけるTCTPの免疫組織学的分析結果を示す図。図中、図a(2倍率)およびb(100倍率)はH&E染色、図c(100倍率)と図d(100倍率),e(400倍率),f(400倍率)はS100と、TCTPの免疫染色を示している。なお、サンプルとしては、図22の組織サンプル番号9に相当するNFの拡散部位のサンプルを使用した。
【図25】TCTP siRNAまたはTCTP発現ベクターを用いたトランスフェクションと、sNF96.2細胞のバイアビリティーとの関連を示した図。
【図26】NF1特異的siRNAで処理した培養シュワン細胞の表現型を示す図。
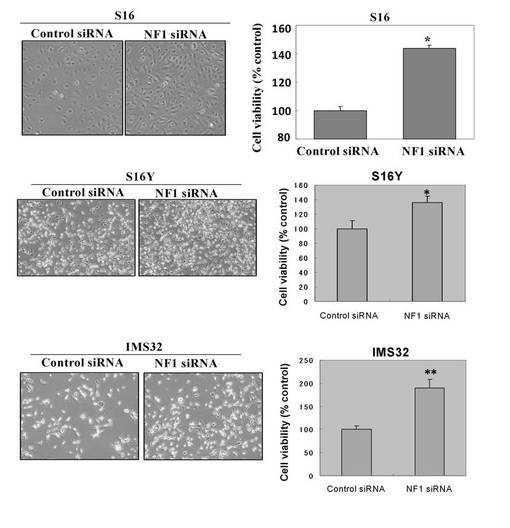
【図27】NF1欠損シュワン細胞の成長因子刺激に応答したTCTP発現上昇を示す図。
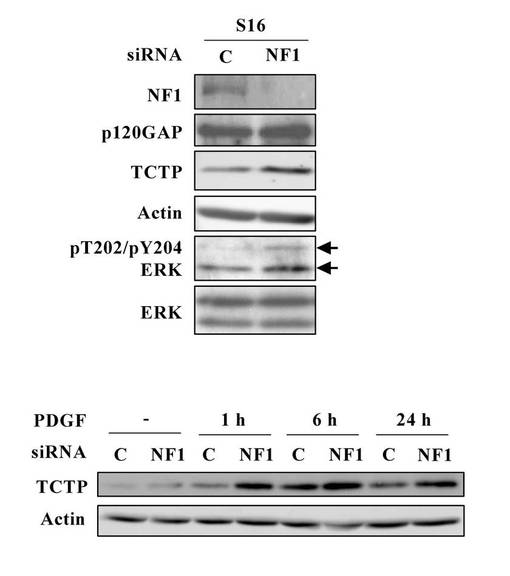
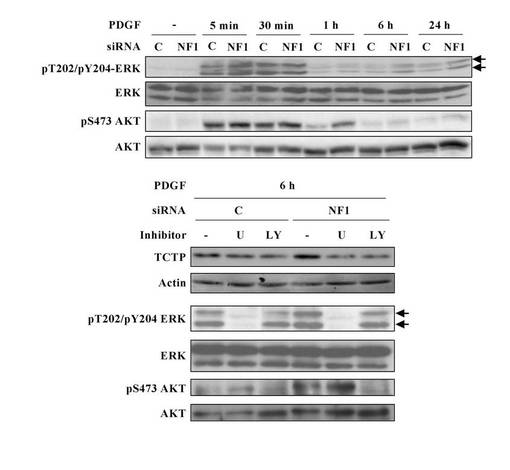
【図28】NF1欠損シュワン細胞のPDGF刺激に応答してシグナル伝達される 活性化MAPKおよびPI3K-AKT経由のTCTP発現上昇を示す図。
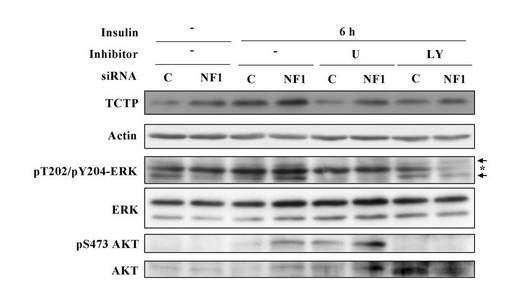
【図29】NF1欠損シュワン細胞のインスリン刺激に応答してシグナル伝達される 活性化MAPKおよびPI3K-AKT経由のTCTP発現上昇を示す図。
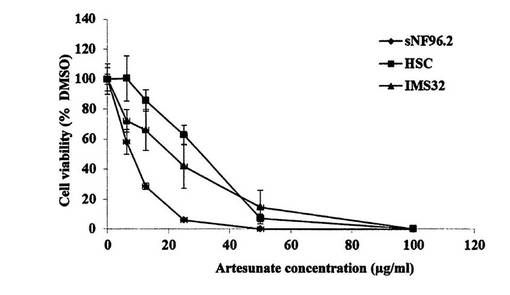
【図30】マラリアの薬として知られるアーテスネート(Artesunate)がNF1腫瘍細胞に特異的に抑制効果があることを示す図。
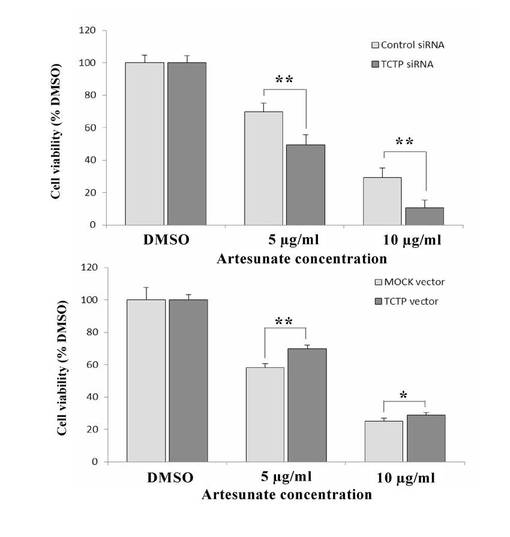
【図31】アーテスネートをTCTP siRNAまたは TCTP発現ベクターでトランスフェクトしたsNF96.2を処理した結果を示す図。
【発明を実施するための形態】
【0028】
本発明は、融合プロテオミクスによるNF1関連病態に関わる分子群であるNF1特異的タンパク質(TCTP)の同定方法、およびTCTPの発現抑制によりNF1病態発現を抑制することからなるNF1特異的タンパク質発現抑制方法に関するものである。さらに、本発明は、NF1特異的タンパク質(TCTP)を、特にNF1腫瘍の悪性化を検査する腫瘍マーカーおよび治療ターゲットとして使用する方法に関するものである。
【0029】
本発明のNF1特異的タンパク質の同定方法は次のようにして行なうことができる。まず、コントロールおよびNF1ノックダウンPC12細胞からタンパク質とmRNAを同時に抽出してサンプル調製を行う。これらの検体からタンパク質とmRNAを同時に抽出してサンプル調製を行う。得られたタンパク質サンプルについて、2種類のプロテオーム解析(2次元電気泳動法による2D-DIGE法と電気泳動を用いないiTRAQ法)、および同一のサンプルを用いたDNAマイクロアレイ法によるトランスクリプトーム解析を行う分子発現差異解析を行う。これらの解析を同時進行で行って、タンパク質レベルで差異のあった分子群を質量分析解析によって同定する。これらの得られたプロテオーム解析の全データとトランスクリプトーム解析の全データを統合してすべてのデータをデータベース化するデータマイニングを行う。次に、これらの統合データから分子クラスター解析、ネットワーク解析を含めた種々の機能解析を行い、最も重要であるTCTP分子群を抽出して検証するための検証実験を行う。検証実験としては、例えば、ウエスタンブロッテイング法、細胞免疫化学的染色法(ICC)法、組織免疫化学的染色法(IHC)法、MRM (Multiple Reaction Monitoring) 法などが使用できる。上記ステップで抽出したTCTP分子群の生物学的機能解析を行う。上記ステップで生物学的機能解説を行った後、TCTP分子群について、治療、予防、創薬開発などへの応用へと繋げていくことができる。
【0030】
まず、本発明に係る融合プロテオミクスによるNF1関連病態に関わる分子群であるTCTPの同定方法について、図1を参照しながらより詳細に説明する。ただし、以下に説明する融合プロテオミクスは、本発明の1態様に過ぎず、その他のプロテオーム解析手法も利用可能であり、本発明は下記説明に一切限定されるものではないことは留意すべきである。
【0031】
上記のようにして調製したタンパク質とmRNAとからなる解析用サンプルは、続いて、2種類のプロテオーム解析およびDNAマイクロアレイによるディファレンシャル解析に供する。つまり、タンパク質サンプルについては、2D-DIGE法ならびにiTRAQ法によるプロテオーム解析によりタンパク質変動と翻訳後修飾の解析を行った。また、mRNAサンプルについてはDNAマイクロアレイ法によるディファレンシャル解析によってmRNAの発現変動の解析を行った。本発明によれば、このように同一サンプルからタンパク質とmRNAを抽出しているので、プロテオーム解析の結果とトランスクリプトーム解析の結果を見比べることが可
能である。
【0032】
本発明における定量的プロテオミクス法としてのiTRAQ法では、RNA干渉法(siRNA)によるNF1発現抑制をそれぞれの細胞にて行い、これらのコントロール細胞と同時にタンパク質を抽出し、それぞれをトリプシン分解してペプチドを得、iTRAQ試薬で修飾した後、LC−MALDI−TOF−TOFおよびLC−ESI−QqTOFを用いてiTRAQ修飾ペプチドの同定と定量解析を行う。なお、同定したタンパク質は、ウエスタンブロッテイング法で検証し、また機能解析は、注目するべき発現変動タンパク質のsiRNAを用いて細胞生物学的および生化学的解析により行うことができる。
【0033】
次に、上記のようにして得られた2D-DIGE法ならびにiTRAQ法によるプロテオーム解析によるタンパク質変動と翻訳後修飾に関する生データと、DNAマイクロアレイ法によるディファレンシャル解析によるmRNAの発現変動に関する生データは、Perl Scriptにより、gene IDの変換、gene annotationならび染色体情報の付加、およびトランスクリプトームとプロテオーム・データセットとの併合などを行って、MANGO(Molecular Annotation of Gene Ontology)に入れてGO(Gene Ontology)アノテーション解析が行なわれる(図2)。
【0034】
MANGOによるGOアノテーション解析により得られたデータは、iPEACH(Integrated Protein Expression Analysis Chart)に入れて解析した。このiPEACHによる解析では、上記各手法で得られたデータをEntrez Gene IDに変換し、このEntrez Gene IDにタンパク質名、fold change、p値、染色体上の位置(locus)などを紐づけられる。
【0035】
このiPEACHにおける統合プロテオミクスデータマイニングシステムによる解析の結果、多数のタンパク質についての統合データか得られる。本発明においては、アットモルレベルで約8000個の細胞内タンパク質についての統合データが得られる。この統合データをさらにGeneSpring GX ソフトウエアによるGO分析および生命情報統合プラットフォーム KeyMolnetによるネットワーク分析を行なわれる。つまり、GeneSpring GX ソフトウエアによるGO分析を行った後、各クラスターの分子をKeyMolnetを用いて分子の特徴づけ(GO解析)が行われる。この統合マイニングシステムにより、複数の手法により得られるデー
タを、瞬時に一元化することが可能となり、GO解析、バスウエイ解析などに供することができるため、重要な分子群の抽出操作を容易に実施することが可能になる。
【0036】
つまり、GeneSpring GX ソフトウエアによるGO分析によりマイクロアレイデータの迅速な可視化と分析が行われ、さらに生命情報統合プラットフォームKeyMolnetによるネットワーク分析により、膨大な量の文献から生体分子、医薬、疾患、生命現象に関する情報(コンテンツ)を網羅的に収集し、分子ネットワークとして体系化することができる。
【0037】
このようにNF1 発現抑制細胞およびコントロール細胞よりタンパク質を抽出し、高感度プロテオミクス定量的発現変動解析を行うことによって、約1600 種のPC12 細胞内発現タンパク質を同定している。その内、NGF 刺激によって特異的な発現差異を示す72 種のタンパク質を同定し、さらにNF1 発現抑制により、NGF 刺激PC12 内で特異的に発現が変動する38 種の活性化シグナル分子群を同定した。これらにはRho、Rac、Cdc42、Rab、ERK、PAK、PI3K に加えて、新規の腫瘍関連ネットワーク分子群が含まれており、これらの機能解析と細胞生物学的検証を行うことによって、神経系細胞分化と細胞死抑制に関わる新規機能分子群であることが判明した(Kobayashi et al. Mol. Cell. Proteomics2009)。さらに、NF1 の発現抑制により、NGF で刺激したPC12 内で特異的に発現が変動する38 種のタンパク質を確認した。発現上昇を示したタンパク質には、mTOR 経路の調節因子であるTCTP が含まれており、TCTP の発現上昇をウエスタンブロティングにより確認した(図3)。これらは総合的にNF1 病態に関わる細胞増殖分化異常に関連すると考えられた。特にmTOR 経路の新規調節因子であるTCTPの関わるネットワークがNF1 発現抑制に連動することに注目し詳細な解析を行った。
【0038】
その結果、NF1 欠損細胞ではTCTP の発現が有意に上昇しており、特に、NF1 患者由来悪性腫瘍MPNST 細胞において、その発現が顕著に誘導されていることから、TCTP がNF 病態において腫瘍悪性化の指標となる可能性があることを見いだした。また、本発明者らは、TCTP の発現抑制によって、未分化細胞が正常な分化状態を形成し、細胞増殖が抑制されたことから、TCTP を中心とする活性化シグナルの治療ターゲットとしての可能性があることを見いだした。
【0039】
次に、NF1 siRNAで処理し、NGFで刺激したPC12細胞を、さらにTCTP siRNA で処理し、その表現形を観察した。Cy3 でラベルされたNF1 siRNA を処理したCy3陽性細胞において、神経突起伸長が阻害されていることが確認されたが、おもしろいことに、さらにその細胞にTCTP siRNA 処理すると、神経突起を特異的に伸長したNF1 欠損PC12 細胞が観察された(図4)。この結果から、NF1 のノックダウンにより阻害されていた神経突起伸長および進展維持機構が、NF1 発現抑制によるTCTP 発現亢進を抑制することによって正常化(回復)したのではないかと考えられた。
【0040】
ところで、TCTPは酵母からヒトにいたるまで、真核生物種間で構造および機能面において高度に保存されており、多彩な機能を示すタンパク質である。特にアポトーシス抑制、タンパク質合成、細胞分裂に関わる機能などの面から、TCTP は腫瘍との関連が示唆されている。しかしながら、NF1 病態の代表的な腫瘍である神経線維腫とTCTP とを関連づける報告はない。そこで神経線維腫の主体とされるシュワン細胞内におけるNF1 遺伝子機能欠損が、TCTP の発現に及ぼす影響を検証した。培養シュワン細胞S16 のsiRNA を用いたNF1 発現抑制によって、細胞増殖能の亢進およびErk の活性化を確認し、それら細胞において、インスリン刺激により有意にTCTPの発現が上昇していることを見出した(図5)。
【0041】
NF1 患者の腫瘍部位由来プライマリ培養細胞内におけるNF1 病態関連分子の発現と機能について調べるために、NF1 患者腫瘍組織より細胞の単離を行った。腫瘍部位の周囲部より繊維芽細胞様の細胞(NF fibroblasts)、腫瘍部位より肥大化している紡錘形の細胞(NF PT細胞)を単離した。ウェスタンブロットの結果、両細胞はNF1 が正常の繊維芽細胞と比べてニューロフィブロミンの発現が少なく、さらにPT細胞 においてはニューロフィブロミンが発現していないことを確認した(図6)。
【0042】
この NF Fibroblasts およびPT細胞 のNF1 活性をErk のリン酸化により評価した。無血清条件下で24 時間培養した両細胞を血清添加(1%FBS)によって刺激し、変動するリン酸化ERK をウエスタンブロッテイングにより検出したところ、無血清培養条件下において、PT細胞 はFibroblasts に比較してERK が高レベルで活性化しており、さらに血清刺激によるERK の活性化が、より長く持続していた(図7)。
【0043】
これらのことから、NF PT細胞 内では、NF1 の機能欠損によってRAS の活性化が起こり、ERK が活性化していると考えられた。これらの条件で、NF fibroblasts、NF PT細胞 両細胞のTCTP の発現をウエスタンブロッティングおよび免疫染色により検討したところ、fibroblasts に比較して、PT細胞 はTCTP が有意に高発現していることを確認した(図8)。また、TCTP siRNA を処理したPT細胞 は、顕著な形態変化を示した。この形態は正常シュワン細胞の形態に酷似していた。また、細胞増殖能の低下も同時に認められた(図9)。さらに神経線維腫の中でも最も悪性度が高い悪性末梢神経線維腫鞘MPNST 細胞sNF96.2 においても同様の実験を行った。その結果、TCTP siRNA 処理によって、PT細胞 同様に顕著な形態変化(より正常シュワン細胞に近い形態に変化)が認められると同時に、および細胞増殖能の低下が確認された(図10)。また、NF1 患者MPNST 組織におけるTCTP の発現を免疫組織化学的に観察したところ、MPNSTではS100 の発現が50%以下に低下していたが、特異的なTCTP の発現の増強が顕著に観察された(図11)。
【0044】
以上の結果よりTCTP が神経線維腫の治療ターゲットとなることのみならず、NF1 欠損による細胞の悪性化の指標となる可能性が示唆された。
【0045】
NF1 の機能欠損がTCTP の発現を上昇させる要因には、Ras-MAPK シグナル経路の活性化が考えられた。RAS-MAPK カスケードの活性化は、転写因子CREB を活性化することが知られている。CREBは、TCTP をコードする遺伝子TPT1 の転写因子であることが報告されている。すなわち、NF1欠損によるRAS-MAPK の活性化、TCTP の発現亢進が連動してmTOR経路を亢進している可能性があり、NF1 siRNA 処理したPC12 細胞、シュワン細胞およびNF1 腫瘍部位より単離されたPT細胞 内で有意にTCTP の発現が上昇し、細胞増殖を亢進しているメカニズムの一つと考えられた(図12)。
【0046】
これまでにNF1 の病態におけるmTOR経路の関与が報告されているが、どのようにしてNF1 病態においてmTOR経路が活性化する詳細な機構はよくわかっていないのが現状であった。本発明によって、NF1患者より切除された腫瘍部位より単離されたPT細胞 では、ERK の活性化が顕著に見られ、TCTPが高発現していることが明らかになったことから、TCTP の発現上昇はNF1 の病態におけるmTOR 経路の活性化に関与していることが強く示唆された。実際にラパマイシンなどのmTOR 阻害剤が、NF1の治療薬として検討されている様に、TCTP の機能やそのシグナルの上流および下流の分子を標的とした治療法の確立が期待できる。
【0047】
プロテオミクス手法を用いた解析により、NF1を欠損した神経系細胞に、TCTP を中心とした特異的ネットワークの活性化が関与していることを見出した。NF1 患者の神経線維腫部位より単離した培養細胞NF PT細胞では、同NF fibroblastsと比べErk の発現が有意に活性化しており、これに連動してTCTP の発現が有意に上昇していることを確認した。特に、NF1 患者由来MPNST 細胞において、TCTP の発現は顕著であり、TCTP がNF 病態において悪性化の指標となる可能性が示唆された。さらにTCTP の発現抑制することによって、NF1 由来の腫瘍性未分化細胞を正常な分化状態に誘導し、細胞増殖を抑制することを明らかにした。すなわち、NF1の機能破綻により活性化したRas-MAPK シグナルTCTP の発現上昇、およびTCTP の発現上昇が引き金となるmTOR 経路の異常な活性化が、NF1の腫瘍化を引き起こす要因の一つであることが考えられた(図12)。これらのことより、今後、NF1 の治療ターゲットとして、TCTP の機能やそのシグナルの上流および下流の分子を標的とした治療戦略が期待できる可能性が示唆された。
【0048】
さらに、本発明では、NF1の発現が抑制された腫瘍でTCTPの発現が亢進していて、その腫瘍の悪性化に伴ってTCTPの発現が有意に亢進していること、NF1由来腫瘍細胞内でTCTPを抑制すると細胞抑制効果があるが、NF1腫瘍の起源細胞である正常シュワン細胞では逆の現象が起こること、NF1を抑制した細胞で神経細胞因子によってTCTPの発現が上昇すること、このTCTPの発現上昇はMAPK阻害剤やPI3K阻害剤で抑制することができること、さらにNF1由来悪性腫瘍細胞では、TCTPを抑制することによって細胞抑制効果に加え、TCTPの過剰発現によって細胞増加効果を促進することができること、その上マラリア薬として知られるアーテスネート(Artesunate)がNF1腫瘍細胞に特異的に抑制効果があること、さらには、TCTPを抑制した上にアーテスネートを併用すると、NF1由来の腫瘍細胞の殺効果が上昇することを明らかにした。
【0049】
以下、本発明を実施例により詳細に説明するが、下記実施例は、本発明を具体的にかつ例示的に説明するだけであって、一切限定的に説明するものではないことを理解すべきである。したがって、本発明は下記実施例により一切限定されるものではない。
【実施例1】
【0050】
(NF1ノックダウンPC12細胞を用いた実験)
本実施例は、NF1の病態モデル細胞として、神経系のモデル細胞であるラット副腎褐色細胞腫の細胞株であるPC12細胞を用いた。PC12細胞は、NGF(神経成長因子)を与えると、神経突起(neurite)が伸長し、神経細胞様に分化することから神経細胞の分化モデルとして用いることができる。
【0051】
そこで、NF1−siRNAで処理したPC12細胞をNGFで刺激してNF1遺伝子発現抑制実験を行って、ニューロフィブロミンの発現抑制をウエスタンブロット法および免疫細胞染色法によって確認した。その結果、ニューロフィブロミンがタンパク質レベルで発現抑制されていることが確認された。また、NF1をノックダウンしたPC12細胞は、コントロール細胞と比べて、神経突起の伸長が抑制されるという表現型が現れた(図13)。
【0052】
(サンプル調製)
本実施例では、コントロールおよびNF1ノックダウンPC12細胞からタンパク質とmRNAを抽出してサンプルを調製した。これらのサンプルを、以後のiTRAQ法ならびに2D-DIGE法およびDNAマイクロアレイ法による解析に供した。iTRAQ(8 plex)法ならびに2D-DIGE法による解析では、コントロール細胞群のサンプルおよびコントロールサンプルのそれぞれ4検体から抽出したタンパク質を用いて、そのタンパク質変動と翻訳後修飾の解析を行った。またDNAマイクロアレイ法による解析では、それぞれ5検体から抽出したmRNAを用いて、mRNAの発現変動の解析を行った。このように同一サンプルからタンパク質とmRNAを抽出することで、プロテオミクスの結果とトランスクリプトームの結果を見比べることが可能である。
【0053】
(iTRAQ(8 plex)法による解析)
iTRAQ(8 plex)法による解析には、上記コントロール細胞群およびRNAi(RNA干渉)によるNF1ノックダウン細胞群のそれぞれ4検体よりタンパク質を抽出し、それぞれをトリプシン分解してペプチドを得た。このペプチドをiTRAQ試薬で修飾し、LC−MALDI−TOF−TOFおよび LC−ESI−QqTOFを用いて iTRAQ修飾ペプチドの同定と定量解析を行った。
【0054】
上記で調製したタンパク質サンプルをiTRAQ試薬でラベルした。各タンパク質サンプル100μgを2-D Clean-Up Kit (GE Healthcare) で処理し、得られた沈殿物を9.8 M ウレアで溶解した。iTRAQ試薬でのラベル化は、製造元のプロトコールに最小限変化させて8Plex法を用いて実施した。上記コントロール細胞群およびNF1ノックダウン細胞群からのタンパク質それぞれ4サンプルをiTRAQ試薬で処理し、各サンプルに可溶化バッファー (20 μl) と変性試薬 (1μl) を添加した。続いて、サンプルを還元剤 (2μl) で還元アルキル化後、60℃で1時間反応させた。さらに、還元されたシステイン残基をシステイン・ブロッキング剤でブロックして10分間室温で反応させた後、トリプシン水溶液を用いて37℃で16時間トリプシン分解を別個に行った。次に、それぞれのサンプルのペプチドを8種類のレポーターiTRAQ試薬(ラベル化試薬1バイアルを解凍し、エタノール80μlに再構成した)でラベルした。その後、iTRAQラベルサンプルを消化物に添加し、1時間室温で反応させた。これらのラベル化したサンプルを混合した。得られた混合サンプルをバッファー(20% v/v ACN、 10 mM リン酸カリウム、pH 3.0)で希釈し、同バッファーで平衡化したMono S column (GE Healthcare) に充填した。ペプチドは、solvent B 濃度勾配(10 mM リン酸カリウム(pH 3.0)、1 M KCl in 20% v/v ACN): 0−2 min、0 to 7% B、at 6
min、to 14% B、at 8 min、to 32% B、at 13 min、to 70% B、at 21 min、to 100% B)で溶出した。上記のようにiTRAQラベルしたサンプルからの溶出液を陽イオン交換HPLCによって40フラクションに分画した。各フラクションは真空遠心分離で乾燥し、2% ACNと0.1% トリフルオロ酢酸とを含む溶液で再水和し、ZipTip μ-C18 pipette tips (Millipore) で脱塩した。
【0055】
各フラクションの10分の1量を用いて、nano LC−MALDI−TOF−TOFおよび nano LC−ESI−QqTOFによる質量分析を行った。サンプルは、溶出フラクションをMALDIプレート上にスポットする装置を取り付けたDiNa Map (KYA Tech) を用いてC18 nano LCを行った。サンプルは、solvent A (2% ACN、 0.1 % TFA) で平衡化した C18 column (0.5 mm I.D × 1 mm length、 KYA Tech) に注入し、C18 nano-column (0.15 mm I.D × 100 mm length、KYA Tech) 中をsolvent B (70% ACN、0.1 % TFA)を300 nl/minのフロー率で90分濃度勾配 (0−10分間: 0 to 20% B; 65分まで50% B; 75分まで100% B)により分離した。
【0056】
このカラム流出液をマトリックス(2 mg/ml α−シアノ−4−ヒドロキシケイ皮酸の50% ACN、0.1% TFA)と、フロー率(1.4μl/min)で混合した。フラクションはステンレススティールMALDIターゲットプレート(192 または384ウェル/プレート(Applied Biosystems)上に30秒間隔でスポットした。ペプチドの質量スペクトルは、4700 または5800 Proteomics Analyzer(Applied Biosystems)を用いて4000 Series Explorer software (v. 3.6) で得られた。各フラクションに対する質量スペクトル(m/z 800-4000)は1,500レーザーショット放射することで得た。存在量が少ないベブチドを分析するために、各質量スペクトルにおいて、S/N 閾値が50から75まで、ならびに75から100までである全てのピークを、それぞれ5,000レーザーショットと4,000レーザーショットでのMS/MS分析のために選択した。次に、S/N 閾値が100である全てのピークを3,000レーザーショットでのMS/MS分析のために選択した。ラベル化したペプチドの断片化は、衝突ガスの環境下で圧力1 x 10-6 Torrおよび衝突エネルギー1kVの条件下で発生させた。
【0057】
さらに、上記で用いたのと同じサンプルをnano−LC−ESI MS/MSで分析した。サンプルは、5 mm RP C18プレカラム(LC Packings)に30μl/分の割合で充填し、10分間洗浄した。次に、ボアサイズ100ÅのC18 ビーズ(3μm)を充填した分離カラム(PepMap RP カラム(内径75μm、長さ150μm)(LC Packings)を用いて分離した。分離は、フロー率200 nl/分で、solvent B (85% ACN、0.1%ギ酸)の90分間濃度勾配(0-60分間は0 to 40% B;70分で100% B)を用いて行った、サンプルは一つのフラクションについて2つに分割して2回解析を行った。2回目解析は1回目の解析情報をexclusionすることによって、効率化
した。
【0058】
1つ目のフラクションについて、QSTAR Pulsar i mass spectrometer (Applied Biosystems/MDS SCIEX、CA)を用いて分析した。ソフトウエアとしては、初めに1 s MSスキャンをし、続いて最も存在量が多い3個のピークに対しては各ピークに対して5回3 s MS/MSスキャンするようにスキャンサイクルをセットしたAnalyst QS 1.1 (Applied Biosystems/MDS SCIEX)を用いて分析した。データ取得は、以前のターゲットイオンを60 s排除して行った。2つ目のフラクションは、存在量の少ないペプチドを分析するために使用した。この分析は、上記で分析したペプチド分析を除いて最初の分析と同一条件で行った。ラベルしたペプチドは、iTRAQ レポーターイオンを発生するように設計されたCID条件下で断片化された。
【0059】
MALDIまたはESIのデータ分析は、ProteinPilot(AB Sciex)を用いたデータマイニングによって行った。その結果、定量的に同定した多数のタンパク質から、コントロール細胞群で特異的に上昇しているタンパク質と、特異的に減少しているタンパク質をリストアップした。そのうちで、上昇しているもののグループで特徴的変化のあるものとしてTCTPが注目された。
【0060】
(2D-DIGEによる解析)
二次元蛍光ディファレンスゲル電気泳動(2D-DIGE)は、上記iTRAQ法で用いたサンプルと同じもの、つまり上記コントロール細胞群ならびにNF1ノックダウン細胞群をそれぞれ4検体ずつ用いて行った。各サンプル50μgの可溶化タンパク質をCy2(全検体のタンパク質を混合)、Cy3(コントロール細胞群6サンプルならびにNF1ノックダウン細胞群6サンプル)、Cy5(コントロール細胞群6サンプルならびにNF1ノックダウン細胞群6サンプル)で標識し、順列くみあわせによる3種類(Cy2、3、5) のサンプルを混合し、2次元電気泳動をそれぞれ12枚で行った後、共焦点蛍光ゲルスキャナーによる解析をして、得られた48枚の画像をDeCyderによってマイニングした。同時に、未ラベルのサンプルにて2次元電気泳動を行い、リン酸化タンパク質を特異的に染色する蛍光試薬を用いて、リン酸化タンパク質の発現の差異を同一の2次元電軌泳動ゲルで行って、リン酸化タンパク質を検出した。その結果、紫色に染色されて見えるスポットが、非感受性のサンプルにおいて、リン酸化が特異的に亢進しているタンパクスポット群、また黄色に染色して見えるスポットが、正常のサンプルにおいて、リン酸化が亢進している、つまり、非感受性のサンプルでリン酸化が減少しているタンパクスポット群であった。
【0061】
これらの方法によりタンパク質の発現変動のみならずリン酸化タンパク質のプロファイリングが可能となった。全てのタンパク質のプロファイルの定量的な統計解析結果より、タンパク質の発現に有意な変動があったスポットを電気泳動ゲルから抽出し、トリプシン分解を行ったのち、タンパクペプチドを抽出して質量分析装置により、タンパク質の同定を行った。その結果、コントロール細胞群にて特異的に上昇している全ての分子のスポットのうち、コントロール細胞群において1.2倍以上に上昇したスポットならびに1.2倍以下に減少したスポットをリストアップした。その中で等電点および分子量ともに変動しながら発現量の上昇が認められた分子として、TCTPが同定された。
【0062】
(DNAマイクロアレイによる分析)
DNAマイクロアレイによる分析は、上記と同じサンプル(各サンプルから5検体)から抽出した全RNAを用いてGene Chip3 Rat Expression assayによって常法に従って行った。まず、全RNAを単離し、リバーストランスクリプションによってcDNAを合成した。得られたcDNAをビオチンで標識した後、ビオチン標識cDNAを断片化した。このcDNAフラグメントをハイブリダイズダイズして、洗浄、スキャン、定量した。その後Gene Springs GX解析に供した。RNA の量と質は、RNA 6000 Nano Assay kitとAgilent 2100 BioAnalyzer(Agilent Technologies、CA)を用いて決定した。
【0063】
(統合プロテオミクスデータマイニングシステム)
この統合プロテオミクスデータマイニングシステムにより、iTRAQ法、2D-DIGE法およびDNAマイクロアレイ法による解析でそれぞれ得られたデータを瞬時に一元化することが可能となり、GO(Gene Ontology)アノテーション解析、バスウエイ解析等に供することができるため、重要な分子群の抽出操作が容易に可能となった。
【0064】
(MANGO によるGOアノテーション解析)
上記のiTRAQ法、2D-DIGE法およびDNAマイクロアレイ法で得られた各データをMANGO(Molecular Annotation of Gene Ontology)に入れ、GO(Gene Ontology)アノテーション解析を行った。
【0065】
(iPEACHによる解析)
GOアノテーション解析により得られたデータをiPEACH(Integrated Protein Expression Analysis Chart)に入れて解析した。このiPEACHによる解析では、上記各手法で得られたデータをEntrez Gene IDに変換し、このEntrez Gene IDにSWNo、 GeneNo、タンパク質名、発現比、fold change、p値、染色体上の位置(locus)、修飾ありなし、GO、解析方法uなどを表示し、解析方法によって異なるデータ行(同じ分子でも解析結果が異なる、2D-DIGEで複数個同定されているものは翻訳後修飾があるとしてその同定数を記載する)を含めてすべてリスト化した。このリストの表記法にフォーマットされた分子を変動率の大きいものから順に並び替えをして、統合アップリストとダウンリストを作成した。同時に、このフォーマットで書き直したプロテオミクスデータとトランスくりプトームデータを別個に抜き出す。今回の解析によるデータは定量的に解析できた分子リストとして約30000行におよび、抽出されたプロテオミクスによる解析の最も特徴的な分子がTCTPであることが分かった。
【0066】
この統合プロテオミクスデータマイニングシステムによる解析の結果、約30000個のタンパク質の統合データか得られた。この統合データをさらにGeneSpring GX ソフトウエアによるGO分析および KeyMolnetアプリケーションによるネットワーク分析を行った。
【0067】
(GeneSpring GX ソフトウエアによるGO分析)
上記のように処理したDNAマイクロアレイにより得られたデータをeneSpring GX ソフトウエアによるGO分析に供した。データを平常化して31099個のプローブを抽出し、フラグフィルタリング(P, M)で21349個のプローブに絞り込み、Fold changeを>2.0に設定して352個のプローブを抽出した。これをクラスター化した上で、GO解析をした。
【0068】
GO解析の結果、siRNAがコントロールよりアップレギュレートしている遺伝子とsiRNAがコントロールよりダウンレギュレートしている遺伝子とを合わせて352個が抽出された。
【0069】
全てのデータをもとに、GeneSpring GX ソフトウエアによるGO分析を行った後、各クラスターの分子をKeyMolnetアプリケーションを用いて分子の特徴づけ(GO解析)を行った。その結果、Biological processにおいては、Cell morphogenesis、Response to stimulus、Regulation of biological quality、Apoptosis、ならびにCell differentiation、またMolecular functionにおいては、Receptor bindingならびにCalcium ion bindingが有意に上昇していることが分かった(図14)。
【0070】
(KeyMolnetアプリケーションによるネットワーク分析)
分子ネットワーク解析の結果、NF1 患者の神経線維腫部位より単離した培養細胞NF PT 細胞においては、NF1 の機能欠損によってRAS の活性化が起こり、ERKの発現が活性化されると、これに連動してTCTP の発現が有意に上昇することが確認された。さらに、RAS-MAPK カスケードの活性化によって、転写因子CREB が活性化することが知られている。一方、CREBはTCTP をコードする遺伝子TPT1 の転写因子であることが報告されている。したがって、このネットワーク分析は、NF1欠損によってRAS-MAPK カスケードが活性化すると、TCTP の発現が亢進され、細胞生長、抗アポトーシス効果、タンパク質合成などを促進するというネットワークを示している(図15)。
【0071】
そこで、上記で同定されたTCTPの発現のNF1に対する影響ならびに効果についてさらに調べることにする。
【0072】
TCTPのノックダウンによるNF1患者由来の腫瘍細胞(PT細胞)の表現形の変化が認められた。Controlと比べて、NF1 siRNA2で処理した細胞では、細長くなった細胞が多く観察されている。細長くなった細胞はNF1正常マウスシュワン培養細胞と形態が似ていることから、このことは、TCTPのノックダウンによりNF1の変異によって異常化していたシュワン細胞が正常シュワン細胞に戻っている可能性を示すものではないかと考察される(図15)。
【0073】
TCTP siRNAは、MPNST細胞株(sNF96.2 Cell line)においてTCTPの発現を抑制することが認められる。MPNST細胞内TCTP発現を抑制することにより、正常シュワン細胞様の表現形を示し、その細胞増殖能が抑制されている。デイッシユにMPNST細胞株を細胞数2 x 105個ずつ播種し、50 pmol のsiRNAをエレクトロポレーションによって導入し、3日後に解析した。
【0074】
またMPNST細胞をラパマイシンで処理することによりTCTPの発現が抑制されることが確認されている。ラパマイシンがTCTP発現を抑制することは、ラパマイシンの標的分子でありかつ細胞の増殖や代謝、生存における調節因子の役割を果たすセリン / スレオニン・キナーゼであるmTOR(mammalian Target Of Rapamycin)のシグナル伝達経路へのシグナル伝達を抑制することを示唆している。そこで、TCTP発現を亢進することによりmTORシグナル伝達経路を活性化することに繋がり癌や免疫膝下に対する効果的な治療方法の開発に有用と考えられる。
【0075】
なお、TCTPの発現抑制の手法としては、siRNA、shRNAなどを用いるRNAi手法の他に直接または間接的に本分子の関わる経路を阻害する薬剤など、TCTPの抑制ができる手法であればいずれも、本発明においては適用でき、例えば、TCTPをノックダウン、ノックアウトなどしてその機能の一部もしくはすべてを欠損、欠失させることも可能である。
【0076】
さらに、TCTPは、神経線維腫に対して、その悪性化に伴って TCTP 発現が顕著に亢進することが確認された(図16)。したがって、TCTPは、神経線維腫の悪性化の指標マーカーとなる可能性がある。神経線維腫は、真皮性神経線維腫(dermatal neurofibroma)である場合TCTPは弱陽性もしくは陰性であり(図17)、結節性網状神経線維腫(nodular plexiform neurofibroma with Wagner-Meissner Body)に進行した場合TCTPはWagner-Meissner Bodyでは陽性であるが、それ以外では陰性もしくは弱陽性であり(図18)、さらに進行して瀰漫性(網状)神経線維腫(diffuse (plexiform) neurofibroma)になった場合TCTPは弱陽性にある(図19)。これに対して、MPNST腫瘍細胞では、S100の発現は通常低下して、50%〜60%のものに陰性があるが、S100alphaでは陽性のものがあった。一方、TCTPは強陽性であった(図20)。
【0077】
上記融合プロテオミクス解析によって、神経幹細胞PC12を用いた神経線維腫症1型 (NF1) モデルに経時的に特異的変動をしている153種類の蛋白質を同定した。その中で、幹細胞維持およびアポトーシス調節に関わる可能性が報告されているTCTPと、その上流と下流のネットワークに注目したところ、NF1患者の腫瘍部位より単離した培養細胞NFPT細胞では、NF繊維芽細胞と比べErkの発現が有意に活性化しており、それに連動してTCTPの発現が亢進していることが確認された。特に、NF1患者由来MPNST細胞において、TCTPの発現は顕著であり、TCTPがNF1病態に関わる腫瘍の悪性化の指標となる可能性が示唆された。さらに、TCTPの発現抑制によって、未分化細胞が正常な分化状態へと誘導され、細胞増殖が抑制されることも明らかになった。さらに、TCTPの腫瘍悪性化のマーカーとしての可能性と、その分子機能やシグナルの上流および下流の分子を標的とした治療戦略が期待される。
【実施例2】
【0078】
本実施例では、NF1の発現が抑制された腫瘍でTCTPの発現が亢進していて、その腫瘍の悪性化に伴ってTCTPの発現が有意に亢進していること、を明らかにした(図21−24)。
【0079】
図21は、ニューロフイブローマ(NF)組織(皮膚NF、Plexiform NF、MPNST)中におけるTCTP発現のレベルとパターンを示している。その結果、Plexiform NFのTCTP発現レベルが、皮膚NFに比べて有意に高いこと、またNF1関連MPNST組織中においてTCTPの強い発現を観察された。
【0080】
図22は、種々のタイプのNF組織中のTCTP発現の免疫組織学的分析の結果を示している。この結果では、全ての皮膚NF患者(8名)において、僅かなまたは弱いTCTP発現が観察され、Plexiform NF患者(4名)において、弱いかまたは中程度のTCTP発現が観察され、またNF1関連MPNST患者(3名)において、中程度または強度のTCTP発現が観察された。その上、NF1散発性MPNST患者(5名)において、様々なTCTP発現レベルが観察された。また、TCTP発現は、NF1関連MPNST症例ならびにNF1散発性MPNST症例において、TCTP発現は、Ki67インデックスと関連していた。
【0081】
図23は、NF1散発性MPNST症例におけるTCTPの免疫組織学的分析結果を示す。NF1散発性MPNST症例におけるTCTPの免疫組織学的分析結果を示している。図は、400倍率で、番号16,18,19は図22の番号に対応している。図では、NF1散発性MPNST症例における種々のTCTP発現レベルが示されている。
【0082】
図24は、NF1-associated diffuse plexiform neurofibroma with Wagner-Meissner BodyにおけるTCTPの免疫組織学的分析結果を示している。この結果から、diffuse NFの特徴的症状であるWagner-Meissner body様構造中のTCTP発現レベルは、浸潤性腫瘍細胞に比べて有意に高いことが観察された。
【実施例3】
【0083】
本実施例は、NF1由来腫瘍細胞内でTCTPを抑制すると細胞抑制効果があることを確認した(図25)。
【0084】
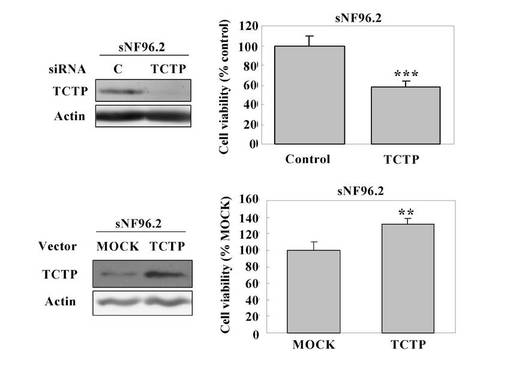
図25は、TCTP siRNAまたはTCTP発現ベクターを用いたトランスフェクションと、sNF96.2細胞の生存能との関連を示している。つまり、デイッシユにMPNST細胞株を細胞数2 x 105個ずつ播種し、50 pmol のTCTP特異的なsiRNA、あるいは1μgのTCTP発現ベクターをエレクトロポレーションによって導入し、3日後に解析した。その結果、MPNST組織由来のsNF96.2培養細胞内にTCTP特異的なsiRNA、およびTCTP発現ベクターを導入すると、それぞれ細胞生存能の減少、および上昇が確認された。
【実施例4】
【0085】
本実施例は、NF1腫瘍の起源細胞である正常シュワン細胞内のNF1遺伝子機能を抑制した場合は、NF1腫瘍にみられるシュワン細胞の異常増殖が起こることが確認された(図26)。
【0086】
図26は、NF1特異的siRNAで処理した培養正常シュワン細胞(S16, S16Y, IMS32)の表現型を示した図である。デイッシユに培養正常シュワン細胞株を細胞数2 x 105個ずつ播種し、50 pmol のNF1特異的なsiRNAをリポフェクション法によって導入し、3日後に細胞を顕微鏡で観察し、細胞の生存能を同仁堂社製のCCK-8により評価した。この結果、NF1特異的siRNA処理培養正常シュワン細胞は、コントロールsiRNAで処理した細胞に比べてより高い増殖性の表現型を示した。
【実施例5】
【0087】
本実施例は、NF1を抑制した細胞において神経系細胞の成長因子によってTCTPの発現が上昇することを示している(図27)。
【0088】
図27は、NF1欠損シュワン細胞の成長因子刺激に応答したTCTP発現上昇を示している。デイッシユに培養正常シュワン細胞株S16を細胞数2 x 105個ずつ播種し、50 pmol のNF1特異的なsiRNAをリポフェクション法によって導入し、3日後に解析した。TCTP発現および ERKリン酸化は、NF1欠損S16細胞で上昇している。デイッシユに培養正常シュワン細胞株S16を細胞数2 x 105個ずつ播種し、50 pmol のNF1特異的なsiRNAをリポフェクション法によって導入し、2日後に、無血清培地にて一晩培養して、終濃度20ng/mlになるようにPDGFで刺激後、1時間、6時間、24時間経過した細胞および刺激前の細胞より調製したサンプルのTCTPの発現変動をウエスタンブロット解析により評価した。TCTP発現は、血小板由来成長因子(PDGF)刺激に応答して、経時依存的に有意に増加した。
【実施例6】
【0089】
本実施例は、TCTPの発現上昇がMAPK阻害剤やPI3K阻害剤で抑制されることを示している(図28、29)。
【0090】
図28は、NF1欠損シュワン細胞の血小板由来成長因子(PDGF)刺激に応答してシグナル伝達される活性化MAPKおよびPI3K-AKT経由のTCTP発現上昇を示している。デイッシユに培養正常シュワン細胞株S16を細胞数2 x 105個ずつ播種し、50 pmol のNF1特異的なsiRNAをリポフェクション法によって導入し、2日後に、無血清培地にて一晩培養し、終濃度20ng/mlになるようにPDGFで刺激した。PDGF刺激後、5分、30分、1時間、6時間、24時間経過した細胞および刺激前の細胞より調製したサンプルの活性化MAPKおよびPI3K-AKTの発現変動をウエスタンブロット解析により評価した。その結果、PDGFに応答して、NF1ノックダウンS16細胞では、ERKおよび AKTリン酸化レベルが、コントロール細胞に比べて高い上昇が認められた。一方、培養正常シュワン細胞株S16を細胞数2 x 105個ずつ播種し、50 pmol のNF1特異的なsiRNAをリポフェクション法によって導入した細胞は、2日後に、無血清培地にて一晩培養し、10μMのMEK抑制剤 U0126または15μMの PI3K抑制剤LY294002で30分処理後、終濃度20ng/mlになるようにPDGFで刺激して6時間経過した細胞および刺激前の細胞より調製したサンプルのTCTPの発現変動をウエスタンブロット解析により評価した。その結果、PDGF刺激によるTCTP発現は、MEK抑制剤 U0126または PI3K抑制剤LY294002によってシグナル伝達されるMAPKまたはPI3K-AKTの抑制によって有意に抑制されることが示されている。
【0091】
図29は、NF1欠損シュワン細胞のインスリン刺激に応答してシグナル伝達される 活性化MAPKおよびPI3K-AKT経由のTCTP発現上昇を示している。つまり、デイッシユに培養正常シュワン細胞株S16を細胞数2 x 105個ずつ播種し、50 pmol のNF1特異的なsiRNAをリポフェクション法によって導入し、2日後に、無血清培地にて一晩培養し、10μMのMEK抑制剤 U0126または15μMの PI3K抑制剤LY294002で30分処理後、終濃度20ng/mlになるようにPDGFで刺激して6時間経過した細胞および刺激前の細胞より調製したサンプルのTCTPの発現変動をウエスタンブロット解析により評価した。その結果、NF1欠損シュワン細胞のインスリン刺激に応答して、TCTPの発現上昇が有意に増加し、そしてPDGF刺激によるそのTCTP発現が、MEK抑制剤 U0126または P13K抑制剤LY294002によってシグナル伝達されるMAPKまたはPI3K-AKTの抑制によって有意に抑制されることが示されている。
【実施例7】
【0092】
本実施例は、マラリア薬として知られるアーテスネート(Artesunate)がNF1腫瘍細胞に特異的に抑制効果があることを示している(図30)。
【0093】
図30は、sNF96.2 MPNST細胞、マウスIMS32細胞およびヒト原発シュワン細胞を、アーテスネート処理した。96ウェルプレートに培養シュワン細胞株を1ウェルあたり細胞数5x 103個ずつ播種し、24時間後、終濃度6.25、12.5、25、50、100μg/mlとなるようにアーテスネートを添加し、48時間培養後に細胞の生存能を同仁堂社製のCCK-8により評価した。その結果、アーテスネートは、マウスIMS32細胞およびヒト原発シュワン細胞に比べて、sNF96.2 MPNST細胞の細胞死をより高くレベルで誘発した。シュワン細胞は正常末梢神経(IC50: sNF96.2: 8.0 μg/ml;ヒト原発シュワン細胞: 30.1μg/ml;マウスIMS32: 20.2 μg/ml)。
【実施例8】
【0094】
本実施例は、TCTP siRNAまたは TCTP発現ベクターをアーテスネートと併用した場合、NF1由来の主要細胞の殺効果が上昇することを示している。つまり、50pmolのTCTP特異的な siRNAまたは1μgの TCTP発現ベクターをエレクトロポレーションにより導入したMPNST由来細胞株sNF96.2を96ウェルプレートに1ウェルあたり細胞数5x 103個ずつ播種し、24時間後、終濃度5、10μg/mlとなるようにアーテスネートを添加し、48時間培養後に細胞の生存能を同仁堂社製のCCK-8により評価した。その結果、TCTPのノックダウン、またはTCTPの過剰発現によって、sNF96.2細胞におけるアーテスネートに対する感受性の増加、または低下が確認された。
【0095】
図31は、TCTPをノックダウンしたり、またはTCTPが過剰に発現すると、sNF96.2細胞におけるアーテスネートに対する感受性が増加したり、または低下したりした。このことは、細胞毒性効果が、TCTP発現レベルと相関関係があることを示唆している。ただし、TCTPの発現上昇は、その効果を減衰させているように考えられる。
【産業上の利用可能性】
【0096】
本発明によれば、TCTPの発現は、NF1患者由来神経線維腫細胞ならびに悪性末梢神経鞘腫瘍(MPNST)の細胞質、および核に強く認められた。さらに、TCTPの発現は、NF1患者由来繊維芽細胞ならびに正常シュワン(Schwann)細胞では認められなかったが、NF1siRNAにより亢進された。さらに、TCTPのsiRNAによる発現抑制は、NF1発現抑制細胞の異常なシグナルと形態発現を正常化させた。その上、TCTPの発現は、NF1患者由来腫瘍組織においては、神経線維腫の悪性度に相関して亢進しており、悪性末梢神経鞘腫瘍(MPNST)においては最も発現が高かった。さらに、MPNST細胞は、TCTP発現を抑制することにより、正常シュワン細胞様の表現形を示し、その細胞増殖能が抑制された。さらに、NF1由来悪性腫瘍細胞に抗腫瘍薬をTCTPを抑制した状態で処理すると、その抑制効果を上昇させた。これらのことから、TCTPの発現亢進とNF1の病態とは密接に関わっていることは明白であり、またTCTPがNF1の治療標的として有用であることが確認された。
【技術分野】
【0001】
本発明は、融合プロテオミクスによるNF1特異的タンパク質の同定方法、NF1特異的タンパク質発現抑制方法、NF1特異的タンパク質の腫瘍マーカー及び治療ターゲットとしての使用方法に関するものである。更に詳細には、本発明は、融合プロテオミクスによるNF1関連病態に関わる分子群であるNF1特異的タンパク質(TCTP:Translationally Controlled Tumor Protein)の同定方法、そのNF1特異的タンパク質の発現抑制方法、およびNF1特異的タンパク質の腫瘍悪性化マーカーとして、または治療ターゲットとしての使用方法に関するものである。
【背景技術】
【0002】
神経線維腫症1型(Neurofibromatosis type 1:NF1)は、NF1原因遺伝子産物であるニューロフィブロミン(neurofibromin)というタンパク質であるニューロフィブロミンの機能が欠損してRasを介した細胞内シグナル伝達が異常をきたし、神経系細胞の増殖と分化異常を誘発し、全身の皮膚に多発性結節と色素斑、皮下の多発性神経線維腫、線維肉腫、グリオーマなどの悪性腫瘍、知能障害等の多彩な病態を伴う頻度が高い優性遺伝性疾患である。
【0003】
NF1遺伝子は、17番染色体長腕(17q11.2)上に存在する2818個のアミノ酸からなるタンパク質であるニューロフィブロミンをコードする遺伝子である。また、NF1遺伝子産物のニューロフィブロミンは、アミノ酸残基360個を中心とする領域に、Ras−GAP(Ras−GTPase-activating protein)相同領域を有し、低分子量Gタンパク質Rasと相互に作用するとともに、Ras結合GTPを加水分解してGDPに変換してRasタンパク質の機能を不活性化する、細胞内シグナル伝達の重要な調節因子と考えられている(非特許文献1)。
【0004】
近年のマウスを用いた研究により、多発性神経線維腫は、細胞−細胞間および細胞−マトリックス間の相互的なシグナルの異常が形成の誘引と考えられている。特に、これまでに、ニューロフィブロミンの発現低下に伴い活性化されたRas−MAPK 経路を介した異常な細胞増殖が大きく関与していることが知られているが、この事象のみでは多発性神経線維腫形成メカニズムの十分な解明には至っていない。また、ニューロフィブロミンの発現低下に起因すると思われる様々な所見が多々報告されていることからも、ニューロフィブロミンには、更なる未知の機能の存在が推測される。
【0005】
そこで、本発明者らは、ニューロフィブロミンの神経系細胞内機能とその欠損による細胞増殖分化異常の機構を明らかにするため、RNA 干渉法(siRNA)を用いたNF1 遺伝子発現抑制によって、神経系細胞PC12に及ぼす神経成長因子(NGF)による分化誘導への影響を解析し、生じた表現形の変化に関与する細胞内責任シグナル分子群を、融合プロテオミクス法、および細胞生物学的検証法を用いて詳細に検討して、ニューロフィブロミンの細胞内機能とNF1の病態(特に多発性神経線維腫形成)との関連性を明らかにした。
【0006】
つまり、本発明者らは、ニューロフィブロミン分子のsiRNAによる細胞内ノックダウンを行い、細胞の形態変化の詳細と融合プロテオミクスの手法によるシグナルカスケードの変化を解析した結果、そのsiRNAの作用によって細胞内actin stress fiber、focal adhesion spot、ruffling形成能が大きく変化していることを認め、細胞骨格系のダイナミックな再構築が起こっていることを見出した。これらの変化には、細胞刺激因子や接着因子からの刺激を介して、Ras-MAPKおよびPI3Kシグナル、Rho-Rock-LIM kinase-coffilinシグナル等の細胞の骨格形成に関連する分子の機能制御によって運動能を調節する機構があることを解明した(非特許文献2)。
【0007】
また、本発明者らは、NF1ノックダウンPC12細胞において、分子TCTP(Translationally controlled tumor protein)が、NF1病態の一つである神経分化異常を誘起することを明らかにして、TCTPの発現亢進がNF1関連病態に密接に関わっていることを、融合プロテオミクス手法によって確認した。
【0008】
そこで、本発明者らは、特に本発明にて新規に同定されたmTOR シグナルに関わるTCTPを中心とした分子ネットワークに注目し、TCTP 分子活性化シグナルの治療ターゲットとしての可能性、および、NF1 異常細胞の悪性化マーカーとしての可能性を検証した。
【0009】
その結果、NF1 発現抑制によりNGF 刺激PC12 内で特異的に発現が変動する活性化シグナル分子群が、総合的にNF1 病態に関わる細胞増殖分化異常に関連すると考えられた。特にmTOR 経路の新規調節因子であるTCTP の関わるネットワークがNF1 発現抑制に連動することに注目して、詳細な解析を行った。NF1 欠損細胞ではTCTP の発現が有意に上昇しており、特に、NF1 患者由来悪性腫瘍MPNST 細胞において、その発現が顕著に誘導されていることから、TCTP がNF 病態において腫瘍悪性化の指標となる可能性があることを見いだした。また、本発明者らは、TCTP の発現抑制によって、未分化細胞が正常な分化状態を形成し、細胞増殖が抑制されたことから、TCTP を中心とする活性化シグナルの治療ター
ゲットとしての可能性があることを見いだした。
【0010】
さらに、本発明者らは、NF1に関連する疾患は、NF1遺伝子産物を介したシグナルによる細胞増殖抑制と、脱落するべき細胞の生理的アポトーシスの誘導の破綻によって生成するという仮説に基づき、細胞内でこれらと相互作用してシグナルを伝達する分子の解析や、それぞれの遺伝子をターゲッティングした細胞と正常細胞の比較検討を生化学・細胞生物学的な手法を用いて行うとともに、融合プロテオミクスによってNF1特異的タンパク質(TCTP)の分子機能解析を行ったところ、TCTPの発現亢進がNF1の病態に密接に関っていることから、TCTPがNF1の治療標的になりうることを見いだした。
【0011】
さらにまた、siRNAによるNF1発現を抑制したNF1病態モデルPC12細胞と、コントロール細胞について二次元電気泳動をベースとした2D-DIGE、iTRAQを用いたLC-MALDI-ならびにLC-ESI-MS/MS解析、およびマイクロアレイを用いたトランスクリプトーム解析を同時に行い、網羅的遺伝子・タンパク質発現および翻訳後修飾情報を取得し、それらすべての同定結果をデータマイニングソフトiPEACHおよびMANGOにより統合し、統計解析マイニングソフトsubioを用いたクラスター解析によってNF1病態関連分子群を抽出した。これらを分子ネットワークソフトKeymolnetによりNF1ノックダウン細胞で異常な挙動を示すシグナル分子群の抽出を試み、TCTPがその異常共同分子群の中で重要な役割を示す事を示した。
【0012】
なお、融合プロテオミクス手法については、本発明者らによる特許文献などに記載の融合プロテオーム解析方法(特許文献1、非特許文献3)や統合プロテオミクス解析手法(特許文献2)などの解析手法を利用するのがよい。
【先行技術文献】
【特許文献】
【0013】
【特許文献1】特開2006-294014号公報
【特許文献2】特願2010-81525号公報
【非特許文献】
【0014】
【非特許文献1】Yunoue et al., J. Biol. Chem., 2003
【非特許文献2】KAKEN FY2004 Annual Research Report; Research Project Number: 16390413荒木令江他「神経線維腫症原因遺伝子群を介した腫瘍抑制機構解明と分子治療戦略の開発」
【発明の概要】
【発明が解決しようとする課題】
【0015】
そこで、本発明の目的は、融合プロテオミクスによるNF1関連病態に関わる分子群であるNF1特異的タンパク質(TCTP)の同定方法を提供することを目的としている。
【0016】
本発明はまた、TCTPの発現抑制によりNF1発現を抑制することからなるNF1特異的タンパク質発現抑制方法を提供することを別の目的としている。
【0017】
本発明はまた、siRNA、shRNAなどによるRNAiやTCTP関連遺伝子のノックダウンやノックアウトなどによりTCTPの機能の一部または全部を抑制することによりTCTP発現抑制することからなるNF1特異的タンパク質発現抑制方法を提供することをさらに別の目的としている。
【0018】
本発明はさらに、NF1特異的タンパク質をNF1腫瘍マーカーとして使用することからなるNF1特異的タンパク質の腫瘍マーカー、例えば腫瘍の悪性化を検査するマーカーとしての使用方法を提供することを別の目的としている。
【0019】
本発明はさらに、TCTP発現阻害薬、MAPK阻害剤、PI3K阻害剤およびアーテスネート(Artesunate)から選ばれるいずれか1種又は2種以上を含むことからなるNF1腫瘍の治療薬を提供することをさらに別の目的としている。
【課題を解決するための手段】
【0020】
したがって、本発明は、融合プロテオミクスによるNF1関連病態に関わる分子群であるNF1特異的タンパク質(TCTP)の同定方法、検証方法を提供する。
【0021】
また、本発明は、TCTPの発現抑制によりNF1発現を抑制することからなるNF1特異的タンパク質発現抑制方法を提供する。
【0022】
さらに、本発明は、TCTPのsiRNA、shRNAなどによるRNAiやTCTPの機能の一部または全部をノックダウンもしくはノックアウトしてTCTPの発現を抑制することからなるNF1特異的タンパク質発現抑制方法を提供する。
【0023】
さらに、本発明は、NF1特異的タンパク質をNF1腫瘍マーカーとして使用することからなるNF1特異的タンパク質の悪性化腫瘍マーカー、例えば腫瘍の悪性化を検査するマーカーとしての使用方法を提供する。
【0024】
さらにまた、本発明は、TCTP阻害剤、MAPK阻害剤、PI3K阻害剤およびアーテスネート(Artesunate)から選ばれるいずれか1種又は2種以上を含むことからなるNF1腫瘍の治療薬を提供する。
【発明の効果】
【0025】
本発明によれば、融合プロテオミクスによってNF1関連病態に関わる分子群であるNF1特異的タンパク質(TCTP)を同定することが可能であるばかりではなく、TCTPのsiRNAなどによりNF1発現を抑制可能であることから、TCTPをNF1治療標的として利用することが可能である。さらに、TCTPは、NF1患者の腫瘍マーカー、特に腫瘍の悪性化を検査するマーカーとして使用することができることから、NF1の早期発見ならびに早期治療に有用である。したがって、本発明はNF1分子標的薬の開発にとっても極めて有用である。
【0026】
さらに、本発明は、TCTP発現阻害薬、MAPK阻害剤、PI3K阻害剤およびアーテスネート(Artesunate)から選ばれるいずれか1種又は2種以上を含むことからなるNF1腫瘍の治療薬を提供することができ極めて有用である。
【図面の簡単な説明】
【0027】
【図1】本発明の方法を示す概略図である。
【図2】Integrated Proteomicsデータマイニングシステムの概略を示す概略図である。
【図3】TCTP のウエスタンブロッテイングによる発現の確認を示す図である。
【図4】NGF NF1 KD PC12細胞内でのTCTP発現抑制による細胞形態の変化を示す図である。A: Cy3標識NF1siRNAおよびコントロールsiRNA処理PC12細胞内でのTCTP siRNA-1/-2によるTCTP の発現抑制のウエスタンブロッテイングによる発現の確認を示す図である。B: Cy3標識NF1siRNAおよびコントロールsiRNA処理PC12細胞の神経様突起発現抑制と、同細胞内でのTCTP の発現抑制による神経様突起発現と伸長を示す図である。
【図5】NF1 欠損S16 シュワン細胞内におけるTCTP の発現変動を示す図である。なお、インスリン刺激によってERK とTCTP の活性化がNF1 欠損細胞で亢進している。
【図6】正常ならびにNF1 患者皮膚由来繊維芽細胞、およびNF1 腫瘍組織由来培養細胞(PT細胞)のニューロフィブロミンのウエスタンブロッテイングによる解析結果を示す図である。
【図7】NF1患者の皮膚由来細胞(NF fibroblasts)およびニューロフィブローマ組織由来培養細胞(NF1PT細胞)における、血清刺激によるMAP kinase/ERK活性の経時的変化を示す図である。
【図8】NF fibroblastsおよびNF PT細胞 のTCTP の発現をウエスタンブロッティング(A)および免疫染色(B)による解析結果を示す図である。
【図9】NF1PT細胞 におけるTCTP発現抑制による細胞内変化と、細胞増殖能の低下を示す図である。
【図10】MPNST cell lineにおけるTCTP発現抑制による細胞増殖低下を示す図である。
【図11】NF1 患者MPNST 組織におけるS100 およびTCTP の免疫染色の結果を示す図である。図中、MPNST 組織ではS100 の発現は低下しているが、TCTP の発現は増強しているのが観察された。
【図12】ニューロフィブロミン欠損によるTCTPを介したNF1 腫瘍形成のモデルを示す図である。
【図13】NF1−siRNA処理PC12細胞をNGFで刺激してNF1遺伝子発現抑制実験によるニューロフィブロミンの発現抑制のウエスタンブロット法および免疫細胞染色法による結果を示す図である。
【図14】NF1 KD細胞特異的タンパク質群のGO解析結果を示す図である。
【図15】NGF およびEGFによって誘発されたTPT1遺伝子発現、つまりTCTP発現のネットワークを示す図である。
【図16】神経線維腫の悪性化に伴ってTCTP発現が更新していることを示す図である。
【図17】真皮性神経線維腫(dermatal neurofibroma)の細胞染色図である。
【図18】結節性網状神経線維腫(nodular plexiform neurofibroma with Wagner-Meissner Body)の細胞染色図である。
【図19】瀰漫性(網状)神経線維腫(diffuse (plexiform) neurofibroma)の細胞染色図である。
【図20】MPNST腫瘍細胞の細胞染色図である。
【図21】ニューロフィブローマ(NF)組織(皮膚NF、Plexiform NF、MPNST)中におけるTCTP発現のレベルとパターンを示す図。
【図22】種々のタイプのNF組織中のTCTP発現の免疫組織学的分析の結果を示す図。図中、*TCTP発現は、染色強度によってスコアした(0:染色なし;1:弱;2:中程度;3:強)。**サンプル9のTCTP発現レベルおよびKi67インデックスはPlexiform NFのdiffuse部位中の領域から取た。NDは無試験。
【図23】NF1散発性MPNST症例におけるTCTPの免疫組織学的分析結果を示す図。図は、Ki-67免疫染色を400倍率で示した図である。番号16,18,19は図22の組織サンプル番号に対応している。
【図24】NF1-associated diffuse plexiform neurofibroma with Wagner-Meissner BodyにおけるTCTPの免疫組織学的分析結果を示す図。図中、図a(2倍率)およびb(100倍率)はH&E染色、図c(100倍率)と図d(100倍率),e(400倍率),f(400倍率)はS100と、TCTPの免疫染色を示している。なお、サンプルとしては、図22の組織サンプル番号9に相当するNFの拡散部位のサンプルを使用した。
【図25】TCTP siRNAまたはTCTP発現ベクターを用いたトランスフェクションと、sNF96.2細胞のバイアビリティーとの関連を示した図。
【図26】NF1特異的siRNAで処理した培養シュワン細胞の表現型を示す図。
【図27】NF1欠損シュワン細胞の成長因子刺激に応答したTCTP発現上昇を示す図。
【図28】NF1欠損シュワン細胞のPDGF刺激に応答してシグナル伝達される 活性化MAPKおよびPI3K-AKT経由のTCTP発現上昇を示す図。
【図29】NF1欠損シュワン細胞のインスリン刺激に応答してシグナル伝達される 活性化MAPKおよびPI3K-AKT経由のTCTP発現上昇を示す図。
【図30】マラリアの薬として知られるアーテスネート(Artesunate)がNF1腫瘍細胞に特異的に抑制効果があることを示す図。
【図31】アーテスネートをTCTP siRNAまたは TCTP発現ベクターでトランスフェクトしたsNF96.2を処理した結果を示す図。
【発明を実施するための形態】
【0028】
本発明は、融合プロテオミクスによるNF1関連病態に関わる分子群であるNF1特異的タンパク質(TCTP)の同定方法、およびTCTPの発現抑制によりNF1病態発現を抑制することからなるNF1特異的タンパク質発現抑制方法に関するものである。さらに、本発明は、NF1特異的タンパク質(TCTP)を、特にNF1腫瘍の悪性化を検査する腫瘍マーカーおよび治療ターゲットとして使用する方法に関するものである。
【0029】
本発明のNF1特異的タンパク質の同定方法は次のようにして行なうことができる。まず、コントロールおよびNF1ノックダウンPC12細胞からタンパク質とmRNAを同時に抽出してサンプル調製を行う。これらの検体からタンパク質とmRNAを同時に抽出してサンプル調製を行う。得られたタンパク質サンプルについて、2種類のプロテオーム解析(2次元電気泳動法による2D-DIGE法と電気泳動を用いないiTRAQ法)、および同一のサンプルを用いたDNAマイクロアレイ法によるトランスクリプトーム解析を行う分子発現差異解析を行う。これらの解析を同時進行で行って、タンパク質レベルで差異のあった分子群を質量分析解析によって同定する。これらの得られたプロテオーム解析の全データとトランスクリプトーム解析の全データを統合してすべてのデータをデータベース化するデータマイニングを行う。次に、これらの統合データから分子クラスター解析、ネットワーク解析を含めた種々の機能解析を行い、最も重要であるTCTP分子群を抽出して検証するための検証実験を行う。検証実験としては、例えば、ウエスタンブロッテイング法、細胞免疫化学的染色法(ICC)法、組織免疫化学的染色法(IHC)法、MRM (Multiple Reaction Monitoring) 法などが使用できる。上記ステップで抽出したTCTP分子群の生物学的機能解析を行う。上記ステップで生物学的機能解説を行った後、TCTP分子群について、治療、予防、創薬開発などへの応用へと繋げていくことができる。
【0030】
まず、本発明に係る融合プロテオミクスによるNF1関連病態に関わる分子群であるTCTPの同定方法について、図1を参照しながらより詳細に説明する。ただし、以下に説明する融合プロテオミクスは、本発明の1態様に過ぎず、その他のプロテオーム解析手法も利用可能であり、本発明は下記説明に一切限定されるものではないことは留意すべきである。
【0031】
上記のようにして調製したタンパク質とmRNAとからなる解析用サンプルは、続いて、2種類のプロテオーム解析およびDNAマイクロアレイによるディファレンシャル解析に供する。つまり、タンパク質サンプルについては、2D-DIGE法ならびにiTRAQ法によるプロテオーム解析によりタンパク質変動と翻訳後修飾の解析を行った。また、mRNAサンプルについてはDNAマイクロアレイ法によるディファレンシャル解析によってmRNAの発現変動の解析を行った。本発明によれば、このように同一サンプルからタンパク質とmRNAを抽出しているので、プロテオーム解析の結果とトランスクリプトーム解析の結果を見比べることが可
能である。
【0032】
本発明における定量的プロテオミクス法としてのiTRAQ法では、RNA干渉法(siRNA)によるNF1発現抑制をそれぞれの細胞にて行い、これらのコントロール細胞と同時にタンパク質を抽出し、それぞれをトリプシン分解してペプチドを得、iTRAQ試薬で修飾した後、LC−MALDI−TOF−TOFおよびLC−ESI−QqTOFを用いてiTRAQ修飾ペプチドの同定と定量解析を行う。なお、同定したタンパク質は、ウエスタンブロッテイング法で検証し、また機能解析は、注目するべき発現変動タンパク質のsiRNAを用いて細胞生物学的および生化学的解析により行うことができる。
【0033】
次に、上記のようにして得られた2D-DIGE法ならびにiTRAQ法によるプロテオーム解析によるタンパク質変動と翻訳後修飾に関する生データと、DNAマイクロアレイ法によるディファレンシャル解析によるmRNAの発現変動に関する生データは、Perl Scriptにより、gene IDの変換、gene annotationならび染色体情報の付加、およびトランスクリプトームとプロテオーム・データセットとの併合などを行って、MANGO(Molecular Annotation of Gene Ontology)に入れてGO(Gene Ontology)アノテーション解析が行なわれる(図2)。
【0034】
MANGOによるGOアノテーション解析により得られたデータは、iPEACH(Integrated Protein Expression Analysis Chart)に入れて解析した。このiPEACHによる解析では、上記各手法で得られたデータをEntrez Gene IDに変換し、このEntrez Gene IDにタンパク質名、fold change、p値、染色体上の位置(locus)などを紐づけられる。
【0035】
このiPEACHにおける統合プロテオミクスデータマイニングシステムによる解析の結果、多数のタンパク質についての統合データか得られる。本発明においては、アットモルレベルで約8000個の細胞内タンパク質についての統合データが得られる。この統合データをさらにGeneSpring GX ソフトウエアによるGO分析および生命情報統合プラットフォーム KeyMolnetによるネットワーク分析を行なわれる。つまり、GeneSpring GX ソフトウエアによるGO分析を行った後、各クラスターの分子をKeyMolnetを用いて分子の特徴づけ(GO解析)が行われる。この統合マイニングシステムにより、複数の手法により得られるデー
タを、瞬時に一元化することが可能となり、GO解析、バスウエイ解析などに供することができるため、重要な分子群の抽出操作を容易に実施することが可能になる。
【0036】
つまり、GeneSpring GX ソフトウエアによるGO分析によりマイクロアレイデータの迅速な可視化と分析が行われ、さらに生命情報統合プラットフォームKeyMolnetによるネットワーク分析により、膨大な量の文献から生体分子、医薬、疾患、生命現象に関する情報(コンテンツ)を網羅的に収集し、分子ネットワークとして体系化することができる。
【0037】
このようにNF1 発現抑制細胞およびコントロール細胞よりタンパク質を抽出し、高感度プロテオミクス定量的発現変動解析を行うことによって、約1600 種のPC12 細胞内発現タンパク質を同定している。その内、NGF 刺激によって特異的な発現差異を示す72 種のタンパク質を同定し、さらにNF1 発現抑制により、NGF 刺激PC12 内で特異的に発現が変動する38 種の活性化シグナル分子群を同定した。これらにはRho、Rac、Cdc42、Rab、ERK、PAK、PI3K に加えて、新規の腫瘍関連ネットワーク分子群が含まれており、これらの機能解析と細胞生物学的検証を行うことによって、神経系細胞分化と細胞死抑制に関わる新規機能分子群であることが判明した(Kobayashi et al. Mol. Cell. Proteomics2009)。さらに、NF1 の発現抑制により、NGF で刺激したPC12 内で特異的に発現が変動する38 種のタンパク質を確認した。発現上昇を示したタンパク質には、mTOR 経路の調節因子であるTCTP が含まれており、TCTP の発現上昇をウエスタンブロティングにより確認した(図3)。これらは総合的にNF1 病態に関わる細胞増殖分化異常に関連すると考えられた。特にmTOR 経路の新規調節因子であるTCTPの関わるネットワークがNF1 発現抑制に連動することに注目し詳細な解析を行った。
【0038】
その結果、NF1 欠損細胞ではTCTP の発現が有意に上昇しており、特に、NF1 患者由来悪性腫瘍MPNST 細胞において、その発現が顕著に誘導されていることから、TCTP がNF 病態において腫瘍悪性化の指標となる可能性があることを見いだした。また、本発明者らは、TCTP の発現抑制によって、未分化細胞が正常な分化状態を形成し、細胞増殖が抑制されたことから、TCTP を中心とする活性化シグナルの治療ターゲットとしての可能性があることを見いだした。
【0039】
次に、NF1 siRNAで処理し、NGFで刺激したPC12細胞を、さらにTCTP siRNA で処理し、その表現形を観察した。Cy3 でラベルされたNF1 siRNA を処理したCy3陽性細胞において、神経突起伸長が阻害されていることが確認されたが、おもしろいことに、さらにその細胞にTCTP siRNA 処理すると、神経突起を特異的に伸長したNF1 欠損PC12 細胞が観察された(図4)。この結果から、NF1 のノックダウンにより阻害されていた神経突起伸長および進展維持機構が、NF1 発現抑制によるTCTP 発現亢進を抑制することによって正常化(回復)したのではないかと考えられた。
【0040】
ところで、TCTPは酵母からヒトにいたるまで、真核生物種間で構造および機能面において高度に保存されており、多彩な機能を示すタンパク質である。特にアポトーシス抑制、タンパク質合成、細胞分裂に関わる機能などの面から、TCTP は腫瘍との関連が示唆されている。しかしながら、NF1 病態の代表的な腫瘍である神経線維腫とTCTP とを関連づける報告はない。そこで神経線維腫の主体とされるシュワン細胞内におけるNF1 遺伝子機能欠損が、TCTP の発現に及ぼす影響を検証した。培養シュワン細胞S16 のsiRNA を用いたNF1 発現抑制によって、細胞増殖能の亢進およびErk の活性化を確認し、それら細胞において、インスリン刺激により有意にTCTPの発現が上昇していることを見出した(図5)。
【0041】
NF1 患者の腫瘍部位由来プライマリ培養細胞内におけるNF1 病態関連分子の発現と機能について調べるために、NF1 患者腫瘍組織より細胞の単離を行った。腫瘍部位の周囲部より繊維芽細胞様の細胞(NF fibroblasts)、腫瘍部位より肥大化している紡錘形の細胞(NF PT細胞)を単離した。ウェスタンブロットの結果、両細胞はNF1 が正常の繊維芽細胞と比べてニューロフィブロミンの発現が少なく、さらにPT細胞 においてはニューロフィブロミンが発現していないことを確認した(図6)。
【0042】
この NF Fibroblasts およびPT細胞 のNF1 活性をErk のリン酸化により評価した。無血清条件下で24 時間培養した両細胞を血清添加(1%FBS)によって刺激し、変動するリン酸化ERK をウエスタンブロッテイングにより検出したところ、無血清培養条件下において、PT細胞 はFibroblasts に比較してERK が高レベルで活性化しており、さらに血清刺激によるERK の活性化が、より長く持続していた(図7)。
【0043】
これらのことから、NF PT細胞 内では、NF1 の機能欠損によってRAS の活性化が起こり、ERK が活性化していると考えられた。これらの条件で、NF fibroblasts、NF PT細胞 両細胞のTCTP の発現をウエスタンブロッティングおよび免疫染色により検討したところ、fibroblasts に比較して、PT細胞 はTCTP が有意に高発現していることを確認した(図8)。また、TCTP siRNA を処理したPT細胞 は、顕著な形態変化を示した。この形態は正常シュワン細胞の形態に酷似していた。また、細胞増殖能の低下も同時に認められた(図9)。さらに神経線維腫の中でも最も悪性度が高い悪性末梢神経線維腫鞘MPNST 細胞sNF96.2 においても同様の実験を行った。その結果、TCTP siRNA 処理によって、PT細胞 同様に顕著な形態変化(より正常シュワン細胞に近い形態に変化)が認められると同時に、および細胞増殖能の低下が確認された(図10)。また、NF1 患者MPNST 組織におけるTCTP の発現を免疫組織化学的に観察したところ、MPNSTではS100 の発現が50%以下に低下していたが、特異的なTCTP の発現の増強が顕著に観察された(図11)。
【0044】
以上の結果よりTCTP が神経線維腫の治療ターゲットとなることのみならず、NF1 欠損による細胞の悪性化の指標となる可能性が示唆された。
【0045】
NF1 の機能欠損がTCTP の発現を上昇させる要因には、Ras-MAPK シグナル経路の活性化が考えられた。RAS-MAPK カスケードの活性化は、転写因子CREB を活性化することが知られている。CREBは、TCTP をコードする遺伝子TPT1 の転写因子であることが報告されている。すなわち、NF1欠損によるRAS-MAPK の活性化、TCTP の発現亢進が連動してmTOR経路を亢進している可能性があり、NF1 siRNA 処理したPC12 細胞、シュワン細胞およびNF1 腫瘍部位より単離されたPT細胞 内で有意にTCTP の発現が上昇し、細胞増殖を亢進しているメカニズムの一つと考えられた(図12)。
【0046】
これまでにNF1 の病態におけるmTOR経路の関与が報告されているが、どのようにしてNF1 病態においてmTOR経路が活性化する詳細な機構はよくわかっていないのが現状であった。本発明によって、NF1患者より切除された腫瘍部位より単離されたPT細胞 では、ERK の活性化が顕著に見られ、TCTPが高発現していることが明らかになったことから、TCTP の発現上昇はNF1 の病態におけるmTOR 経路の活性化に関与していることが強く示唆された。実際にラパマイシンなどのmTOR 阻害剤が、NF1の治療薬として検討されている様に、TCTP の機能やそのシグナルの上流および下流の分子を標的とした治療法の確立が期待できる。
【0047】
プロテオミクス手法を用いた解析により、NF1を欠損した神経系細胞に、TCTP を中心とした特異的ネットワークの活性化が関与していることを見出した。NF1 患者の神経線維腫部位より単離した培養細胞NF PT細胞では、同NF fibroblastsと比べErk の発現が有意に活性化しており、これに連動してTCTP の発現が有意に上昇していることを確認した。特に、NF1 患者由来MPNST 細胞において、TCTP の発現は顕著であり、TCTP がNF 病態において悪性化の指標となる可能性が示唆された。さらにTCTP の発現抑制することによって、NF1 由来の腫瘍性未分化細胞を正常な分化状態に誘導し、細胞増殖を抑制することを明らかにした。すなわち、NF1の機能破綻により活性化したRas-MAPK シグナルTCTP の発現上昇、およびTCTP の発現上昇が引き金となるmTOR 経路の異常な活性化が、NF1の腫瘍化を引き起こす要因の一つであることが考えられた(図12)。これらのことより、今後、NF1 の治療ターゲットとして、TCTP の機能やそのシグナルの上流および下流の分子を標的とした治療戦略が期待できる可能性が示唆された。
【0048】
さらに、本発明では、NF1の発現が抑制された腫瘍でTCTPの発現が亢進していて、その腫瘍の悪性化に伴ってTCTPの発現が有意に亢進していること、NF1由来腫瘍細胞内でTCTPを抑制すると細胞抑制効果があるが、NF1腫瘍の起源細胞である正常シュワン細胞では逆の現象が起こること、NF1を抑制した細胞で神経細胞因子によってTCTPの発現が上昇すること、このTCTPの発現上昇はMAPK阻害剤やPI3K阻害剤で抑制することができること、さらにNF1由来悪性腫瘍細胞では、TCTPを抑制することによって細胞抑制効果に加え、TCTPの過剰発現によって細胞増加効果を促進することができること、その上マラリア薬として知られるアーテスネート(Artesunate)がNF1腫瘍細胞に特異的に抑制効果があること、さらには、TCTPを抑制した上にアーテスネートを併用すると、NF1由来の腫瘍細胞の殺効果が上昇することを明らかにした。
【0049】
以下、本発明を実施例により詳細に説明するが、下記実施例は、本発明を具体的にかつ例示的に説明するだけであって、一切限定的に説明するものではないことを理解すべきである。したがって、本発明は下記実施例により一切限定されるものではない。
【実施例1】
【0050】
(NF1ノックダウンPC12細胞を用いた実験)
本実施例は、NF1の病態モデル細胞として、神経系のモデル細胞であるラット副腎褐色細胞腫の細胞株であるPC12細胞を用いた。PC12細胞は、NGF(神経成長因子)を与えると、神経突起(neurite)が伸長し、神経細胞様に分化することから神経細胞の分化モデルとして用いることができる。
【0051】
そこで、NF1−siRNAで処理したPC12細胞をNGFで刺激してNF1遺伝子発現抑制実験を行って、ニューロフィブロミンの発現抑制をウエスタンブロット法および免疫細胞染色法によって確認した。その結果、ニューロフィブロミンがタンパク質レベルで発現抑制されていることが確認された。また、NF1をノックダウンしたPC12細胞は、コントロール細胞と比べて、神経突起の伸長が抑制されるという表現型が現れた(図13)。
【0052】
(サンプル調製)
本実施例では、コントロールおよびNF1ノックダウンPC12細胞からタンパク質とmRNAを抽出してサンプルを調製した。これらのサンプルを、以後のiTRAQ法ならびに2D-DIGE法およびDNAマイクロアレイ法による解析に供した。iTRAQ(8 plex)法ならびに2D-DIGE法による解析では、コントロール細胞群のサンプルおよびコントロールサンプルのそれぞれ4検体から抽出したタンパク質を用いて、そのタンパク質変動と翻訳後修飾の解析を行った。またDNAマイクロアレイ法による解析では、それぞれ5検体から抽出したmRNAを用いて、mRNAの発現変動の解析を行った。このように同一サンプルからタンパク質とmRNAを抽出することで、プロテオミクスの結果とトランスクリプトームの結果を見比べることが可能である。
【0053】
(iTRAQ(8 plex)法による解析)
iTRAQ(8 plex)法による解析には、上記コントロール細胞群およびRNAi(RNA干渉)によるNF1ノックダウン細胞群のそれぞれ4検体よりタンパク質を抽出し、それぞれをトリプシン分解してペプチドを得た。このペプチドをiTRAQ試薬で修飾し、LC−MALDI−TOF−TOFおよび LC−ESI−QqTOFを用いて iTRAQ修飾ペプチドの同定と定量解析を行った。
【0054】
上記で調製したタンパク質サンプルをiTRAQ試薬でラベルした。各タンパク質サンプル100μgを2-D Clean-Up Kit (GE Healthcare) で処理し、得られた沈殿物を9.8 M ウレアで溶解した。iTRAQ試薬でのラベル化は、製造元のプロトコールに最小限変化させて8Plex法を用いて実施した。上記コントロール細胞群およびNF1ノックダウン細胞群からのタンパク質それぞれ4サンプルをiTRAQ試薬で処理し、各サンプルに可溶化バッファー (20 μl) と変性試薬 (1μl) を添加した。続いて、サンプルを還元剤 (2μl) で還元アルキル化後、60℃で1時間反応させた。さらに、還元されたシステイン残基をシステイン・ブロッキング剤でブロックして10分間室温で反応させた後、トリプシン水溶液を用いて37℃で16時間トリプシン分解を別個に行った。次に、それぞれのサンプルのペプチドを8種類のレポーターiTRAQ試薬(ラベル化試薬1バイアルを解凍し、エタノール80μlに再構成した)でラベルした。その後、iTRAQラベルサンプルを消化物に添加し、1時間室温で反応させた。これらのラベル化したサンプルを混合した。得られた混合サンプルをバッファー(20% v/v ACN、 10 mM リン酸カリウム、pH 3.0)で希釈し、同バッファーで平衡化したMono S column (GE Healthcare) に充填した。ペプチドは、solvent B 濃度勾配(10 mM リン酸カリウム(pH 3.0)、1 M KCl in 20% v/v ACN): 0−2 min、0 to 7% B、at 6
min、to 14% B、at 8 min、to 32% B、at 13 min、to 70% B、at 21 min、to 100% B)で溶出した。上記のようにiTRAQラベルしたサンプルからの溶出液を陽イオン交換HPLCによって40フラクションに分画した。各フラクションは真空遠心分離で乾燥し、2% ACNと0.1% トリフルオロ酢酸とを含む溶液で再水和し、ZipTip μ-C18 pipette tips (Millipore) で脱塩した。
【0055】
各フラクションの10分の1量を用いて、nano LC−MALDI−TOF−TOFおよび nano LC−ESI−QqTOFによる質量分析を行った。サンプルは、溶出フラクションをMALDIプレート上にスポットする装置を取り付けたDiNa Map (KYA Tech) を用いてC18 nano LCを行った。サンプルは、solvent A (2% ACN、 0.1 % TFA) で平衡化した C18 column (0.5 mm I.D × 1 mm length、 KYA Tech) に注入し、C18 nano-column (0.15 mm I.D × 100 mm length、KYA Tech) 中をsolvent B (70% ACN、0.1 % TFA)を300 nl/minのフロー率で90分濃度勾配 (0−10分間: 0 to 20% B; 65分まで50% B; 75分まで100% B)により分離した。
【0056】
このカラム流出液をマトリックス(2 mg/ml α−シアノ−4−ヒドロキシケイ皮酸の50% ACN、0.1% TFA)と、フロー率(1.4μl/min)で混合した。フラクションはステンレススティールMALDIターゲットプレート(192 または384ウェル/プレート(Applied Biosystems)上に30秒間隔でスポットした。ペプチドの質量スペクトルは、4700 または5800 Proteomics Analyzer(Applied Biosystems)を用いて4000 Series Explorer software (v. 3.6) で得られた。各フラクションに対する質量スペクトル(m/z 800-4000)は1,500レーザーショット放射することで得た。存在量が少ないベブチドを分析するために、各質量スペクトルにおいて、S/N 閾値が50から75まで、ならびに75から100までである全てのピークを、それぞれ5,000レーザーショットと4,000レーザーショットでのMS/MS分析のために選択した。次に、S/N 閾値が100である全てのピークを3,000レーザーショットでのMS/MS分析のために選択した。ラベル化したペプチドの断片化は、衝突ガスの環境下で圧力1 x 10-6 Torrおよび衝突エネルギー1kVの条件下で発生させた。
【0057】
さらに、上記で用いたのと同じサンプルをnano−LC−ESI MS/MSで分析した。サンプルは、5 mm RP C18プレカラム(LC Packings)に30μl/分の割合で充填し、10分間洗浄した。次に、ボアサイズ100ÅのC18 ビーズ(3μm)を充填した分離カラム(PepMap RP カラム(内径75μm、長さ150μm)(LC Packings)を用いて分離した。分離は、フロー率200 nl/分で、solvent B (85% ACN、0.1%ギ酸)の90分間濃度勾配(0-60分間は0 to 40% B;70分で100% B)を用いて行った、サンプルは一つのフラクションについて2つに分割して2回解析を行った。2回目解析は1回目の解析情報をexclusionすることによって、効率化
した。
【0058】
1つ目のフラクションについて、QSTAR Pulsar i mass spectrometer (Applied Biosystems/MDS SCIEX、CA)を用いて分析した。ソフトウエアとしては、初めに1 s MSスキャンをし、続いて最も存在量が多い3個のピークに対しては各ピークに対して5回3 s MS/MSスキャンするようにスキャンサイクルをセットしたAnalyst QS 1.1 (Applied Biosystems/MDS SCIEX)を用いて分析した。データ取得は、以前のターゲットイオンを60 s排除して行った。2つ目のフラクションは、存在量の少ないペプチドを分析するために使用した。この分析は、上記で分析したペプチド分析を除いて最初の分析と同一条件で行った。ラベルしたペプチドは、iTRAQ レポーターイオンを発生するように設計されたCID条件下で断片化された。
【0059】
MALDIまたはESIのデータ分析は、ProteinPilot(AB Sciex)を用いたデータマイニングによって行った。その結果、定量的に同定した多数のタンパク質から、コントロール細胞群で特異的に上昇しているタンパク質と、特異的に減少しているタンパク質をリストアップした。そのうちで、上昇しているもののグループで特徴的変化のあるものとしてTCTPが注目された。
【0060】
(2D-DIGEによる解析)
二次元蛍光ディファレンスゲル電気泳動(2D-DIGE)は、上記iTRAQ法で用いたサンプルと同じもの、つまり上記コントロール細胞群ならびにNF1ノックダウン細胞群をそれぞれ4検体ずつ用いて行った。各サンプル50μgの可溶化タンパク質をCy2(全検体のタンパク質を混合)、Cy3(コントロール細胞群6サンプルならびにNF1ノックダウン細胞群6サンプル)、Cy5(コントロール細胞群6サンプルならびにNF1ノックダウン細胞群6サンプル)で標識し、順列くみあわせによる3種類(Cy2、3、5) のサンプルを混合し、2次元電気泳動をそれぞれ12枚で行った後、共焦点蛍光ゲルスキャナーによる解析をして、得られた48枚の画像をDeCyderによってマイニングした。同時に、未ラベルのサンプルにて2次元電気泳動を行い、リン酸化タンパク質を特異的に染色する蛍光試薬を用いて、リン酸化タンパク質の発現の差異を同一の2次元電軌泳動ゲルで行って、リン酸化タンパク質を検出した。その結果、紫色に染色されて見えるスポットが、非感受性のサンプルにおいて、リン酸化が特異的に亢進しているタンパクスポット群、また黄色に染色して見えるスポットが、正常のサンプルにおいて、リン酸化が亢進している、つまり、非感受性のサンプルでリン酸化が減少しているタンパクスポット群であった。
【0061】
これらの方法によりタンパク質の発現変動のみならずリン酸化タンパク質のプロファイリングが可能となった。全てのタンパク質のプロファイルの定量的な統計解析結果より、タンパク質の発現に有意な変動があったスポットを電気泳動ゲルから抽出し、トリプシン分解を行ったのち、タンパクペプチドを抽出して質量分析装置により、タンパク質の同定を行った。その結果、コントロール細胞群にて特異的に上昇している全ての分子のスポットのうち、コントロール細胞群において1.2倍以上に上昇したスポットならびに1.2倍以下に減少したスポットをリストアップした。その中で等電点および分子量ともに変動しながら発現量の上昇が認められた分子として、TCTPが同定された。
【0062】
(DNAマイクロアレイによる分析)
DNAマイクロアレイによる分析は、上記と同じサンプル(各サンプルから5検体)から抽出した全RNAを用いてGene Chip3 Rat Expression assayによって常法に従って行った。まず、全RNAを単離し、リバーストランスクリプションによってcDNAを合成した。得られたcDNAをビオチンで標識した後、ビオチン標識cDNAを断片化した。このcDNAフラグメントをハイブリダイズダイズして、洗浄、スキャン、定量した。その後Gene Springs GX解析に供した。RNA の量と質は、RNA 6000 Nano Assay kitとAgilent 2100 BioAnalyzer(Agilent Technologies、CA)を用いて決定した。
【0063】
(統合プロテオミクスデータマイニングシステム)
この統合プロテオミクスデータマイニングシステムにより、iTRAQ法、2D-DIGE法およびDNAマイクロアレイ法による解析でそれぞれ得られたデータを瞬時に一元化することが可能となり、GO(Gene Ontology)アノテーション解析、バスウエイ解析等に供することができるため、重要な分子群の抽出操作が容易に可能となった。
【0064】
(MANGO によるGOアノテーション解析)
上記のiTRAQ法、2D-DIGE法およびDNAマイクロアレイ法で得られた各データをMANGO(Molecular Annotation of Gene Ontology)に入れ、GO(Gene Ontology)アノテーション解析を行った。
【0065】
(iPEACHによる解析)
GOアノテーション解析により得られたデータをiPEACH(Integrated Protein Expression Analysis Chart)に入れて解析した。このiPEACHによる解析では、上記各手法で得られたデータをEntrez Gene IDに変換し、このEntrez Gene IDにSWNo、 GeneNo、タンパク質名、発現比、fold change、p値、染色体上の位置(locus)、修飾ありなし、GO、解析方法uなどを表示し、解析方法によって異なるデータ行(同じ分子でも解析結果が異なる、2D-DIGEで複数個同定されているものは翻訳後修飾があるとしてその同定数を記載する)を含めてすべてリスト化した。このリストの表記法にフォーマットされた分子を変動率の大きいものから順に並び替えをして、統合アップリストとダウンリストを作成した。同時に、このフォーマットで書き直したプロテオミクスデータとトランスくりプトームデータを別個に抜き出す。今回の解析によるデータは定量的に解析できた分子リストとして約30000行におよび、抽出されたプロテオミクスによる解析の最も特徴的な分子がTCTPであることが分かった。
【0066】
この統合プロテオミクスデータマイニングシステムによる解析の結果、約30000個のタンパク質の統合データか得られた。この統合データをさらにGeneSpring GX ソフトウエアによるGO分析および KeyMolnetアプリケーションによるネットワーク分析を行った。
【0067】
(GeneSpring GX ソフトウエアによるGO分析)
上記のように処理したDNAマイクロアレイにより得られたデータをeneSpring GX ソフトウエアによるGO分析に供した。データを平常化して31099個のプローブを抽出し、フラグフィルタリング(P, M)で21349個のプローブに絞り込み、Fold changeを>2.0に設定して352個のプローブを抽出した。これをクラスター化した上で、GO解析をした。
【0068】
GO解析の結果、siRNAがコントロールよりアップレギュレートしている遺伝子とsiRNAがコントロールよりダウンレギュレートしている遺伝子とを合わせて352個が抽出された。
【0069】
全てのデータをもとに、GeneSpring GX ソフトウエアによるGO分析を行った後、各クラスターの分子をKeyMolnetアプリケーションを用いて分子の特徴づけ(GO解析)を行った。その結果、Biological processにおいては、Cell morphogenesis、Response to stimulus、Regulation of biological quality、Apoptosis、ならびにCell differentiation、またMolecular functionにおいては、Receptor bindingならびにCalcium ion bindingが有意に上昇していることが分かった(図14)。
【0070】
(KeyMolnetアプリケーションによるネットワーク分析)
分子ネットワーク解析の結果、NF1 患者の神経線維腫部位より単離した培養細胞NF PT 細胞においては、NF1 の機能欠損によってRAS の活性化が起こり、ERKの発現が活性化されると、これに連動してTCTP の発現が有意に上昇することが確認された。さらに、RAS-MAPK カスケードの活性化によって、転写因子CREB が活性化することが知られている。一方、CREBはTCTP をコードする遺伝子TPT1 の転写因子であることが報告されている。したがって、このネットワーク分析は、NF1欠損によってRAS-MAPK カスケードが活性化すると、TCTP の発現が亢進され、細胞生長、抗アポトーシス効果、タンパク質合成などを促進するというネットワークを示している(図15)。
【0071】
そこで、上記で同定されたTCTPの発現のNF1に対する影響ならびに効果についてさらに調べることにする。
【0072】
TCTPのノックダウンによるNF1患者由来の腫瘍細胞(PT細胞)の表現形の変化が認められた。Controlと比べて、NF1 siRNA2で処理した細胞では、細長くなった細胞が多く観察されている。細長くなった細胞はNF1正常マウスシュワン培養細胞と形態が似ていることから、このことは、TCTPのノックダウンによりNF1の変異によって異常化していたシュワン細胞が正常シュワン細胞に戻っている可能性を示すものではないかと考察される(図15)。
【0073】
TCTP siRNAは、MPNST細胞株(sNF96.2 Cell line)においてTCTPの発現を抑制することが認められる。MPNST細胞内TCTP発現を抑制することにより、正常シュワン細胞様の表現形を示し、その細胞増殖能が抑制されている。デイッシユにMPNST細胞株を細胞数2 x 105個ずつ播種し、50 pmol のsiRNAをエレクトロポレーションによって導入し、3日後に解析した。
【0074】
またMPNST細胞をラパマイシンで処理することによりTCTPの発現が抑制されることが確認されている。ラパマイシンがTCTP発現を抑制することは、ラパマイシンの標的分子でありかつ細胞の増殖や代謝、生存における調節因子の役割を果たすセリン / スレオニン・キナーゼであるmTOR(mammalian Target Of Rapamycin)のシグナル伝達経路へのシグナル伝達を抑制することを示唆している。そこで、TCTP発現を亢進することによりmTORシグナル伝達経路を活性化することに繋がり癌や免疫膝下に対する効果的な治療方法の開発に有用と考えられる。
【0075】
なお、TCTPの発現抑制の手法としては、siRNA、shRNAなどを用いるRNAi手法の他に直接または間接的に本分子の関わる経路を阻害する薬剤など、TCTPの抑制ができる手法であればいずれも、本発明においては適用でき、例えば、TCTPをノックダウン、ノックアウトなどしてその機能の一部もしくはすべてを欠損、欠失させることも可能である。
【0076】
さらに、TCTPは、神経線維腫に対して、その悪性化に伴って TCTP 発現が顕著に亢進することが確認された(図16)。したがって、TCTPは、神経線維腫の悪性化の指標マーカーとなる可能性がある。神経線維腫は、真皮性神経線維腫(dermatal neurofibroma)である場合TCTPは弱陽性もしくは陰性であり(図17)、結節性網状神経線維腫(nodular plexiform neurofibroma with Wagner-Meissner Body)に進行した場合TCTPはWagner-Meissner Bodyでは陽性であるが、それ以外では陰性もしくは弱陽性であり(図18)、さらに進行して瀰漫性(網状)神経線維腫(diffuse (plexiform) neurofibroma)になった場合TCTPは弱陽性にある(図19)。これに対して、MPNST腫瘍細胞では、S100の発現は通常低下して、50%〜60%のものに陰性があるが、S100alphaでは陽性のものがあった。一方、TCTPは強陽性であった(図20)。
【0077】
上記融合プロテオミクス解析によって、神経幹細胞PC12を用いた神経線維腫症1型 (NF1) モデルに経時的に特異的変動をしている153種類の蛋白質を同定した。その中で、幹細胞維持およびアポトーシス調節に関わる可能性が報告されているTCTPと、その上流と下流のネットワークに注目したところ、NF1患者の腫瘍部位より単離した培養細胞NFPT細胞では、NF繊維芽細胞と比べErkの発現が有意に活性化しており、それに連動してTCTPの発現が亢進していることが確認された。特に、NF1患者由来MPNST細胞において、TCTPの発現は顕著であり、TCTPがNF1病態に関わる腫瘍の悪性化の指標となる可能性が示唆された。さらに、TCTPの発現抑制によって、未分化細胞が正常な分化状態へと誘導され、細胞増殖が抑制されることも明らかになった。さらに、TCTPの腫瘍悪性化のマーカーとしての可能性と、その分子機能やシグナルの上流および下流の分子を標的とした治療戦略が期待される。
【実施例2】
【0078】
本実施例では、NF1の発現が抑制された腫瘍でTCTPの発現が亢進していて、その腫瘍の悪性化に伴ってTCTPの発現が有意に亢進していること、を明らかにした(図21−24)。
【0079】
図21は、ニューロフイブローマ(NF)組織(皮膚NF、Plexiform NF、MPNST)中におけるTCTP発現のレベルとパターンを示している。その結果、Plexiform NFのTCTP発現レベルが、皮膚NFに比べて有意に高いこと、またNF1関連MPNST組織中においてTCTPの強い発現を観察された。
【0080】
図22は、種々のタイプのNF組織中のTCTP発現の免疫組織学的分析の結果を示している。この結果では、全ての皮膚NF患者(8名)において、僅かなまたは弱いTCTP発現が観察され、Plexiform NF患者(4名)において、弱いかまたは中程度のTCTP発現が観察され、またNF1関連MPNST患者(3名)において、中程度または強度のTCTP発現が観察された。その上、NF1散発性MPNST患者(5名)において、様々なTCTP発現レベルが観察された。また、TCTP発現は、NF1関連MPNST症例ならびにNF1散発性MPNST症例において、TCTP発現は、Ki67インデックスと関連していた。
【0081】
図23は、NF1散発性MPNST症例におけるTCTPの免疫組織学的分析結果を示す。NF1散発性MPNST症例におけるTCTPの免疫組織学的分析結果を示している。図は、400倍率で、番号16,18,19は図22の番号に対応している。図では、NF1散発性MPNST症例における種々のTCTP発現レベルが示されている。
【0082】
図24は、NF1-associated diffuse plexiform neurofibroma with Wagner-Meissner BodyにおけるTCTPの免疫組織学的分析結果を示している。この結果から、diffuse NFの特徴的症状であるWagner-Meissner body様構造中のTCTP発現レベルは、浸潤性腫瘍細胞に比べて有意に高いことが観察された。
【実施例3】
【0083】
本実施例は、NF1由来腫瘍細胞内でTCTPを抑制すると細胞抑制効果があることを確認した(図25)。
【0084】
図25は、TCTP siRNAまたはTCTP発現ベクターを用いたトランスフェクションと、sNF96.2細胞の生存能との関連を示している。つまり、デイッシユにMPNST細胞株を細胞数2 x 105個ずつ播種し、50 pmol のTCTP特異的なsiRNA、あるいは1μgのTCTP発現ベクターをエレクトロポレーションによって導入し、3日後に解析した。その結果、MPNST組織由来のsNF96.2培養細胞内にTCTP特異的なsiRNA、およびTCTP発現ベクターを導入すると、それぞれ細胞生存能の減少、および上昇が確認された。
【実施例4】
【0085】
本実施例は、NF1腫瘍の起源細胞である正常シュワン細胞内のNF1遺伝子機能を抑制した場合は、NF1腫瘍にみられるシュワン細胞の異常増殖が起こることが確認された(図26)。
【0086】
図26は、NF1特異的siRNAで処理した培養正常シュワン細胞(S16, S16Y, IMS32)の表現型を示した図である。デイッシユに培養正常シュワン細胞株を細胞数2 x 105個ずつ播種し、50 pmol のNF1特異的なsiRNAをリポフェクション法によって導入し、3日後に細胞を顕微鏡で観察し、細胞の生存能を同仁堂社製のCCK-8により評価した。この結果、NF1特異的siRNA処理培養正常シュワン細胞は、コントロールsiRNAで処理した細胞に比べてより高い増殖性の表現型を示した。
【実施例5】
【0087】
本実施例は、NF1を抑制した細胞において神経系細胞の成長因子によってTCTPの発現が上昇することを示している(図27)。
【0088】
図27は、NF1欠損シュワン細胞の成長因子刺激に応答したTCTP発現上昇を示している。デイッシユに培養正常シュワン細胞株S16を細胞数2 x 105個ずつ播種し、50 pmol のNF1特異的なsiRNAをリポフェクション法によって導入し、3日後に解析した。TCTP発現および ERKリン酸化は、NF1欠損S16細胞で上昇している。デイッシユに培養正常シュワン細胞株S16を細胞数2 x 105個ずつ播種し、50 pmol のNF1特異的なsiRNAをリポフェクション法によって導入し、2日後に、無血清培地にて一晩培養して、終濃度20ng/mlになるようにPDGFで刺激後、1時間、6時間、24時間経過した細胞および刺激前の細胞より調製したサンプルのTCTPの発現変動をウエスタンブロット解析により評価した。TCTP発現は、血小板由来成長因子(PDGF)刺激に応答して、経時依存的に有意に増加した。
【実施例6】
【0089】
本実施例は、TCTPの発現上昇がMAPK阻害剤やPI3K阻害剤で抑制されることを示している(図28、29)。
【0090】
図28は、NF1欠損シュワン細胞の血小板由来成長因子(PDGF)刺激に応答してシグナル伝達される活性化MAPKおよびPI3K-AKT経由のTCTP発現上昇を示している。デイッシユに培養正常シュワン細胞株S16を細胞数2 x 105個ずつ播種し、50 pmol のNF1特異的なsiRNAをリポフェクション法によって導入し、2日後に、無血清培地にて一晩培養し、終濃度20ng/mlになるようにPDGFで刺激した。PDGF刺激後、5分、30分、1時間、6時間、24時間経過した細胞および刺激前の細胞より調製したサンプルの活性化MAPKおよびPI3K-AKTの発現変動をウエスタンブロット解析により評価した。その結果、PDGFに応答して、NF1ノックダウンS16細胞では、ERKおよび AKTリン酸化レベルが、コントロール細胞に比べて高い上昇が認められた。一方、培養正常シュワン細胞株S16を細胞数2 x 105個ずつ播種し、50 pmol のNF1特異的なsiRNAをリポフェクション法によって導入した細胞は、2日後に、無血清培地にて一晩培養し、10μMのMEK抑制剤 U0126または15μMの PI3K抑制剤LY294002で30分処理後、終濃度20ng/mlになるようにPDGFで刺激して6時間経過した細胞および刺激前の細胞より調製したサンプルのTCTPの発現変動をウエスタンブロット解析により評価した。その結果、PDGF刺激によるTCTP発現は、MEK抑制剤 U0126または PI3K抑制剤LY294002によってシグナル伝達されるMAPKまたはPI3K-AKTの抑制によって有意に抑制されることが示されている。
【0091】
図29は、NF1欠損シュワン細胞のインスリン刺激に応答してシグナル伝達される 活性化MAPKおよびPI3K-AKT経由のTCTP発現上昇を示している。つまり、デイッシユに培養正常シュワン細胞株S16を細胞数2 x 105個ずつ播種し、50 pmol のNF1特異的なsiRNAをリポフェクション法によって導入し、2日後に、無血清培地にて一晩培養し、10μMのMEK抑制剤 U0126または15μMの PI3K抑制剤LY294002で30分処理後、終濃度20ng/mlになるようにPDGFで刺激して6時間経過した細胞および刺激前の細胞より調製したサンプルのTCTPの発現変動をウエスタンブロット解析により評価した。その結果、NF1欠損シュワン細胞のインスリン刺激に応答して、TCTPの発現上昇が有意に増加し、そしてPDGF刺激によるそのTCTP発現が、MEK抑制剤 U0126または P13K抑制剤LY294002によってシグナル伝達されるMAPKまたはPI3K-AKTの抑制によって有意に抑制されることが示されている。
【実施例7】
【0092】
本実施例は、マラリア薬として知られるアーテスネート(Artesunate)がNF1腫瘍細胞に特異的に抑制効果があることを示している(図30)。
【0093】
図30は、sNF96.2 MPNST細胞、マウスIMS32細胞およびヒト原発シュワン細胞を、アーテスネート処理した。96ウェルプレートに培養シュワン細胞株を1ウェルあたり細胞数5x 103個ずつ播種し、24時間後、終濃度6.25、12.5、25、50、100μg/mlとなるようにアーテスネートを添加し、48時間培養後に細胞の生存能を同仁堂社製のCCK-8により評価した。その結果、アーテスネートは、マウスIMS32細胞およびヒト原発シュワン細胞に比べて、sNF96.2 MPNST細胞の細胞死をより高くレベルで誘発した。シュワン細胞は正常末梢神経(IC50: sNF96.2: 8.0 μg/ml;ヒト原発シュワン細胞: 30.1μg/ml;マウスIMS32: 20.2 μg/ml)。
【実施例8】
【0094】
本実施例は、TCTP siRNAまたは TCTP発現ベクターをアーテスネートと併用した場合、NF1由来の主要細胞の殺効果が上昇することを示している。つまり、50pmolのTCTP特異的な siRNAまたは1μgの TCTP発現ベクターをエレクトロポレーションにより導入したMPNST由来細胞株sNF96.2を96ウェルプレートに1ウェルあたり細胞数5x 103個ずつ播種し、24時間後、終濃度5、10μg/mlとなるようにアーテスネートを添加し、48時間培養後に細胞の生存能を同仁堂社製のCCK-8により評価した。その結果、TCTPのノックダウン、またはTCTPの過剰発現によって、sNF96.2細胞におけるアーテスネートに対する感受性の増加、または低下が確認された。
【0095】
図31は、TCTPをノックダウンしたり、またはTCTPが過剰に発現すると、sNF96.2細胞におけるアーテスネートに対する感受性が増加したり、または低下したりした。このことは、細胞毒性効果が、TCTP発現レベルと相関関係があることを示唆している。ただし、TCTPの発現上昇は、その効果を減衰させているように考えられる。
【産業上の利用可能性】
【0096】
本発明によれば、TCTPの発現は、NF1患者由来神経線維腫細胞ならびに悪性末梢神経鞘腫瘍(MPNST)の細胞質、および核に強く認められた。さらに、TCTPの発現は、NF1患者由来繊維芽細胞ならびに正常シュワン(Schwann)細胞では認められなかったが、NF1siRNAにより亢進された。さらに、TCTPのsiRNAによる発現抑制は、NF1発現抑制細胞の異常なシグナルと形態発現を正常化させた。その上、TCTPの発現は、NF1患者由来腫瘍組織においては、神経線維腫の悪性度に相関して亢進しており、悪性末梢神経鞘腫瘍(MPNST)においては最も発現が高かった。さらに、MPNST細胞は、TCTP発現を抑制することにより、正常シュワン細胞様の表現形を示し、その細胞増殖能が抑制された。さらに、NF1由来悪性腫瘍細胞に抗腫瘍薬をTCTPを抑制した状態で処理すると、その抑制効果を上昇させた。これらのことから、TCTPの発現亢進とNF1の病態とは密接に関わっていることは明白であり、またTCTPがNF1の治療標的として有用であることが確認された。
【特許請求の範囲】
【請求項1】
神経線維腫症1型(NF1)組織であって薬剤感受性組織ならびに薬剤非感受性組織から抽出したタンパク質およびmRNAをプロテオーム解析およびトランスクリプトーム解析をしてそれぞれ得られたデータを融合プロテオーム解析データとして一元化し、得られたデータをGO解析するとともに、ネットワーク解析することによって、NF1関連病態に関わる分子群であるNF1特異的タンパク質を同定することを特徴とするNF1特異的タンパク質の同定方法。
【請求項2】
請求項1に記載のNF1特異的タンパク質の同定方法であって、該NF1特異的タンパク質がTCTPであることを特徴とするNF1特異的タンパク質の同定方法。
【請求項3】
NF1特異的タンパク質の発現を抑制することによりNF1病態発現を抑制することを特徴とするNF1特異的タンパク質発現抑制方法。
【請求項4】
請求項1に記載のNF1特異的タンパク質のNF1特異的タンパク質発現抑制方法であって、該NF1特異的タンパク質がTCTPであることを特徴とするNF1特異的タンパク質発現抑制方法。
【請求項5】
請求項3または4に記載のNF1特異的タンパク質発現抑制方法であって、NF1病態発現抑制がTCTPのRNAiまたはTCTPのノックダウンもしくはノックアウトによって行うことを特徴とするNF1特異的タンパク質発現抑制方法。
【請求項6】
請求項2ないし5項のいずれか1項に記載のNF1特異的タンパク質発現抑制方法であって、該RNAiがTCTPのsiRNA またはshRNAを用いて行うことを特徴とするNF1特異的タンパク質発現抑制方法。
【請求項7】
NF1特異的タンパク質をNF1の腫瘍悪性化を検査する指標マーカーとして使用することを特徴とするNF1特異的タンパク質の使用方法。
【請求項8】
NF1特異的タンパク質をNF1の腫瘍悪性化を指標マーカーとして使用してNF1の腫瘍悪性化を検査することを特徴とするNF1特異的タンパク質によるNF1の腫瘍悪性化検査方法。
【請求項9】
TCTP阻害薬(siRNA)、MAPK阻害剤、PI3K阻害剤、Artesunateから選ばれるいずれか1種又は2種以上を有効成分として含むことを特徴とするNF1腫瘍の治療薬。
【請求項1】
神経線維腫症1型(NF1)組織であって薬剤感受性組織ならびに薬剤非感受性組織から抽出したタンパク質およびmRNAをプロテオーム解析およびトランスクリプトーム解析をしてそれぞれ得られたデータを融合プロテオーム解析データとして一元化し、得られたデータをGO解析するとともに、ネットワーク解析することによって、NF1関連病態に関わる分子群であるNF1特異的タンパク質を同定することを特徴とするNF1特異的タンパク質の同定方法。
【請求項2】
請求項1に記載のNF1特異的タンパク質の同定方法であって、該NF1特異的タンパク質がTCTPであることを特徴とするNF1特異的タンパク質の同定方法。
【請求項3】
NF1特異的タンパク質の発現を抑制することによりNF1病態発現を抑制することを特徴とするNF1特異的タンパク質発現抑制方法。
【請求項4】
請求項1に記載のNF1特異的タンパク質のNF1特異的タンパク質発現抑制方法であって、該NF1特異的タンパク質がTCTPであることを特徴とするNF1特異的タンパク質発現抑制方法。
【請求項5】
請求項3または4に記載のNF1特異的タンパク質発現抑制方法であって、NF1病態発現抑制がTCTPのRNAiまたはTCTPのノックダウンもしくはノックアウトによって行うことを特徴とするNF1特異的タンパク質発現抑制方法。
【請求項6】
請求項2ないし5項のいずれか1項に記載のNF1特異的タンパク質発現抑制方法であって、該RNAiがTCTPのsiRNA またはshRNAを用いて行うことを特徴とするNF1特異的タンパク質発現抑制方法。
【請求項7】
NF1特異的タンパク質をNF1の腫瘍悪性化を検査する指標マーカーとして使用することを特徴とするNF1特異的タンパク質の使用方法。
【請求項8】
NF1特異的タンパク質をNF1の腫瘍悪性化を指標マーカーとして使用してNF1の腫瘍悪性化を検査することを特徴とするNF1特異的タンパク質によるNF1の腫瘍悪性化検査方法。
【請求項9】
TCTP阻害薬(siRNA)、MAPK阻害剤、PI3K阻害剤、Artesunateから選ばれるいずれか1種又は2種以上を有効成分として含むことを特徴とするNF1腫瘍の治療薬。
【図1】


【図2】
【図3】
【図4】
【図5】
【図6】
【図7】
【図8】
【図9】
【図10】
【図11】
【図12】
【図13】
【図14】
【図15】
【図16】
【図17】
【図18】
【図19】
【図20】
【図21】
【図22】

【図23】
【図24】
【図25】
【図26】
【図27】
【図28】
【図29】
【図30】
【図31】
【図2】
【図3】
【図4】
【図5】
【図6】
【図7】
【図8】
【図9】
【図10】
【図11】
【図12】
【図13】
【図14】
【図15】
【図16】
【図17】
【図18】
【図19】
【図20】
【図21】
【図22】
【図23】
【図24】
【図25】
【図26】
【図27】
【図28】
【図29】
【図30】
【図31】
【公開番号】特開2012−213391(P2012−213391A)
【公開日】平成24年11月8日(2012.11.8)
【国際特許分類】
【出願番号】特願2012−75242(P2012−75242)
【出願日】平成24年3月28日(2012.3.28)
【出願人】(504159235)国立大学法人 熊本大学 (314)
【Fターム(参考)】
【公開日】平成24年11月8日(2012.11.8)
【国際特許分類】
【出願日】平成24年3月28日(2012.3.28)
【出願人】(504159235)国立大学法人 熊本大学 (314)
【Fターム(参考)】
[ Back to top ]