
蠕虫卵、特に鞭虫卵の生物学的活性を測定する方法
本発明は、鞭虫の幼虫包蔵卵の生物学的活性を測定する方法であって、
a)ゲノムDNAのコピー数を確認するのに適したマーカー配列を使用した定量PCR分析による蠕虫卵の幼虫発育期の判定および/または確認、
b)生化学的方法および/または分子生物学的方法を用いた蠕虫の幼虫包蔵卵の代謝活性の測定、
c)蠕虫の幼虫包蔵卵における遺伝子発現の誘導能の測定、
d)高温でのプレインキュベーション後における、虫卵内に包蔵される蠕虫幼虫の運動性の、長時間観察による顕微鏡測定、および/または
e)内部標準を基準として、腸管内容物から回収した完全な形のままの幼虫包蔵卵を定量化する、実験動物における鞭虫の幼虫孵化率の測定
のうち少なくとも1種の測定を行うことを特徴とする方法に関する。
a)ゲノムDNAのコピー数を確認するのに適したマーカー配列を使用した定量PCR分析による蠕虫卵の幼虫発育期の判定および/または確認、
b)生化学的方法および/または分子生物学的方法を用いた蠕虫の幼虫包蔵卵の代謝活性の測定、
c)蠕虫の幼虫包蔵卵における遺伝子発現の誘導能の測定、
d)高温でのプレインキュベーション後における、虫卵内に包蔵される蠕虫幼虫の運動性の、長時間観察による顕微鏡測定、および/または
e)内部標準を基準として、腸管内容物から回収した完全な形のままの幼虫包蔵卵を定量化する、実験動物における鞭虫の幼虫孵化率の測定
のうち少なくとも1種の測定を行うことを特徴とする方法に関する。
【発明の詳細な説明】
【技術分野】
【0001】
本発明は、生活環の種々の発育期における、蠕虫卵、特に鞭虫卵、好ましくは豚鞭虫卵の生物学的活性を測定する方法に関する。本発明の方法により、治療用医薬品の有効成分としての蠕虫卵調製物を管理された方法で製造することができ、ヒトに対して安全かつ治療上効率的な適用を保証することができる。
【背景技術】
【0002】
寄生虫感染が、動物宿主の免疫システムの活性化に影響を及ぼすことは公知である(D M McKay, Parasitology 2006, 132: 1-12参照)。この活性化は自己免疫疾患の発症や経過にも影響する。疫学的調査によると、衛生状態が良いため寄生虫感染率が低い地域と比べて、寄生虫感染率の高い地域では自己免疫疾患の罹患率が低い。慢性炎症性腸疾患であるクローン病患者のサイトカインプロファイルにおいては、蠕虫感染によりTh2免疫細胞が刺激されうることが示されている。蠕虫に感染させることによって、Th1優位の自己免疫疾患であるクローン病を予防することができるか、またはクローン病に影響を与えることができる(Summers et al.,Am J Gastroenterol 2003, 98: 2034-2041参照)。
【0003】
1971年には既にBeer(Br. Med. J., 1971, 3: 41)により、ヒトに病原性を有するヒト鞭虫(Trichuris trichiura)による感染症などに伴って起こるような病原作用を引き起こすことなく、意図的にヒトに感染させるのに適した線虫として、豚鞭虫(Trichuris suis)が挙げられることが報告されている。より最近の研究によると、豚鞭虫のヒトへの感染は一過性のものに過ぎないと考えられ、増殖が可能となる前に豚鞭虫は排除されることが明らかとなっている。しかしながら、慢性炎症性腸疾患に対する豚鞭虫の使用に関して肯定的な臨床研究によれば、豚鞭虫をヒトに一時的に感染させることにより、免疫系を調節して治療効果をもたらすことができる(Summers et al., Am. J. Gastroenterol. 2003, 98: 2034-2041)。
【0004】
豚鞭虫の生活環は、排卵された未成熟卵(L0期)から始まり、未成熟卵は糞便とともに感染動物から排泄される。次いで、地中で3〜6ヶ月間かけて成熟し、第1幼虫期(L1)となる。L1期に達してから最初の数日間は、幼虫が虫卵内で動くのを観察することができる。その後、虫卵内の幼虫は休止状態となり、この状態を何年間も保つが、感染性を失うことはない。適切な宿主により経口摂取された後、L1幼虫は腸管腔で虫卵から孵化し、数時間以内に盲腸や結腸の粘膜に入り込む。その後、幼虫は腸粘膜内で数週間かけてさらなる幼虫発育期(L2〜L4)を経てL5成熟幼虫となり、最終的に腸管腔へと再び出て行く(Beer, Parasitol. 1973, 67: 253-262)。
【0005】
豚鞭虫卵(Trichuris suis ova=TSO)を薬学的に製造するには、in vitroまたはin situにおいて排卵された未成熟TSO(L0)を、適切な方法(WO 1999 33479、WO 2007 076868およびWO 2007 134761)を用いて、単離精製する。次いで、管理された実験室条件下で虫卵を成熟させて、生物学的に活性な(L1)TSOを形成させる。得られたTSOは、医薬品の薬学的に活性な部分を構成する。
【0006】
治療目的で提供される蠕虫卵は、生物学的医薬品として分類することができる。生物学的医薬品の製造および適用に際して評価しなければならない主要なパラメータとして、医薬品の治療有効性の最終的な指標となる生物学的活性が挙げられる。生物学的活性を分析せずには、製造工程を論理的に開発し監視することも、また、ヒトへの計画的な適用のために、治療上有効で患者にとって安全な医薬品投薬量を決定することもできない。このような理由により、蠕虫卵調製物を医薬品として安全かつ有効に使用するためには、蠕虫卵製剤の生物学的活性が十分な条件を満たしていなければならない。
【0007】
現在、豚鞭虫の分析方法として3種の方法が利用可能であるが、いずれの方法も豚鞭虫の生物学的活性を測定するには不十分である。
【0008】
1.成熟率の測定(Kringel et al., Vet. Parasitol. 2006, 139: 132-139, Paragraph 2.2(豚鞭虫);およびJohnson et al. Intl. J. Parasitol. 1998, 28, 627-633(豚回虫))
この分析は顕微鏡下で行う。形態学的基準を用いて、完全な形のままでかつ十分に成熟したL1幼虫が虫卵に含まれているかどうかを評価する。しかしながら、本出願で示される分子生物学的方法とは対照的に、形態学的評価のみでは幼虫が実際に十分に成熟しているのかどうかを確実に推定することはできない。さらに、成熟率には、幼虫を包蔵したL1−TSOの生存能力や生物学的活性に関する情報は含まれないという制限がある。さらに、この形態学的評価には観察者の経験が必要とされ、形態学的な判別が難しい事例においては主観的な分類がなされるため正確性および精度が制限される。
【0009】
2.感染率の測定
豚鞭虫に関しては、ブタを用いて分析を行う。ブタを感染させた後、特定の時点において腸粘膜に定着した幼虫数を測定し、TSOの投与量を基準にして感染率を計算する(Kringel et al., Vet. Parasitol., 2006, 139: 132-139;Summers et al., 2005, Gastroenterology, 128: 825-832;またはJohnson et al. Intl. J. Parasitol. 1998, 28, 627-633(豚回虫))。感染率を測定する際の問題点として、感染率が蠕虫卵の機能性により影響を受けるだけでなく、個々の宿主が有する要因、たとえば、実験動物の免疫システム、腸内細菌叢、腸の機能などによっても同様に影響を受けることが挙げられる。したがって、実験動物を用いた感染率の測定には大きなばらつきが本質的に伴い、試験系に生物を用いているため標準化することもできない。このため、実験動物を用いた感染率の測定は、生物学的活性試験としての適性が大幅に制限される。さらに、ブタを用いた豚鞭虫の感染率測定試験には、少なくとも3週間の長い潜伏期が伴う上、結果のばらつきが大きいため多数の動物が必要とされる。このような理由により、この試験を日常的に使用することは倫理的にも経済的にも難しい。
【0010】
さらに、Kringelらは、蠕虫卵の生物学的活性を評価する方法として、蠕虫に感染させた動物の糞便から虫卵を回収する方法を報告している。この方法では、動物に感染させてから、孵化した幼虫自らがL5期まで成熟し、交尾し、排卵するまで7〜8週間待たなければならない。また、糞便中の虫卵数から成熟幼虫の感染率または生物学的活性が示唆されるような直接的な定量的関係は存在しない。さらに、Kringelらの方法では次世代の虫卵が分析されるため、Kringelらのin vivoにおけるこの方法と本出願に記載の孵化率の測定法とは明確に区別されなければならない。
【0011】
3.WO 2007/134761には、豚鞭虫卵の生存能力を試験する方法が記載されている。この方法では、ブタの消化菅における虫卵の通過をin vitroで模擬して虫卵を孵化させる。
【0012】
しかしながら、医薬品に必要とされる精度で蠕虫卵調製物の生物学的活性を測定するには、上述した方法では不十分である。
【発明の概要】
【発明が解決しようとする課題】
【0013】
したがって、動物において生物学的活性が測定される生物学的医薬品の主な課題の一つとして、調製物の標準化が挙げられる。上述したブタへの感染のような動物モデルの使用は、非常に煩雑で、しかも相当な時間を要する。したがって、本発明の明細書では、蠕虫卵の発育やその生物学的活性に関してそれぞれ異なる側面から種々の試験を行う方法を説明する。信頼できる結果を得るためには、1試料に対し、本発明の試験のうち少なくとも1種行う必要があり、好ましくは本明細書に記載の試験のうち少なくとも3種、より好ましくは少なくとも4種、さらに好ましくは5種行い、それぞれの試験に関連するパラメータを測定する。個々の試験結果は全体として一つと見なされ、適切な蠕虫調製物は、それぞれの試験における所定の制限値を満たさなければならない。この所定の制限値を満たした場合、蠕虫卵調製物は医薬用途に適すると結論づけられる。
【0014】
したがって、産業に応用可能であって、種々の発育段階におけるTSOおよび他の蠕虫卵の生物学的活性を包括的かつ確実に分析することにより蠕虫卵の種々の生物学的機能を測定する方法に対する需要は大きい。このような方法で管理することによってのみ市場性の高い医薬品が製造できる。
【課題を解決するための手段】
【0015】
本発明の目的は、以下に詳述する方法を用いて蠕虫卵の生物学的活性を包括的に分析することである。したがって、本発明は、製造工程の厳密な管理と安全かつ治療上効率的な患者への適用とを可能にすることも目的とする。
【0016】
従来公知の方法における複雑な問題および不完全さを考慮すれば、個々の分析方法を単独で使用して、生物学的活性を確実に測定することは通常不可能であることは明らかである。
【0017】
したがって、本発明の主題は、完全に成熟した幼虫を包蔵する鞭虫卵の生物学的活性を測定する方法であって、以下の測定のうち少なくとも1種行うことを特徴とする:
【0018】
生化学的方法および/または分子生物学的方法を用いた、鞭虫卵の温度誘導性活性の測定、具体的にはATP含量の測定;
鞭虫の幼虫包蔵卵における遺伝子発現の誘導能の測定;
高温でのプレインキュベーションによる活性化後における、虫卵内に包蔵される鞭虫幼虫の運動性の長時間観察による顕微鏡測定;および/または、
内部標準を基準として、腸管内容物から回収した完全な形のままの幼虫包蔵卵を定量化する、実験動物における鞭虫幼虫の孵化率の測定。
【0019】
したがって、蠕虫の生活環の特定段階における種々の生物学的機能を分析する5種の測定法からなる試験系を開発した。個々の医薬品製造工程において1種の測定法を適用するだけでも十分であるとも考えられるが、通常、最終製品の生物学的活性を測定するためには、以下の表1に記載の測定法のうち少なくとも3種行わなければならない。
【0020】
【表1】

【0021】
薬学的に利用可能な蠕虫卵の生物学的活性を確実に測定するためには、少なくとも3種、好ましくは4種、より好ましくは5種すべての工程を組み合わせた方法を行うと好ましい。特定の生物学的活性を測定するため、または製造の副工程を監視するためであれば、本発明の方法の個々の工程を単独で実施するだけで十分な場合もある。
【0022】
5種の個々の測定法も、他の事項に関して上述した試験原理に基づく。蠕虫の幼虫包蔵卵、特に豚鞭虫の幼虫包蔵卵への適用は新規であり、これまでに報告されていない新規の一連の副工程の開発を必要とする。5種の生物学的活性を試験することにより包括的な測定が可能な本方法全体も新規であり、蠕虫卵、特に豚鞭虫卵に関してはこれまで報告されていない。本明細書に記載の5種のうち3種の方法は、休止状態の鞭虫幼虫を高温でプレインキュベートすることにより活性化することを共通の好ましい特徴とする。本発明の方法のまた別の際だった特徴としては、完全な形のままでありかつ生存可能な蠕虫卵に対して本方法を実施できることと、測定したパラメータが、宿主側の要因または本試験を実施する者の専門能力には影響を受けず、蠕虫卵の活性によってのみ影響を受けることが挙げられる。したがって、本発明の方法は、これまで用いられてきた方法に比べてより客観的である。以上のことから、薬学的利用に必要とされる標準化が可能となる。
【0023】
本発明の方法を用いることによって、蠕虫卵調製物の生物学的活性を、医薬品に必要とされる精度で確実に測定することが可能となる。本発明に記載されているように、蠕虫卵の生物学的活性の正確な測定は、適用可能な医薬品を製造するために必須の工程である。
【図面の簡単な説明】
【0024】
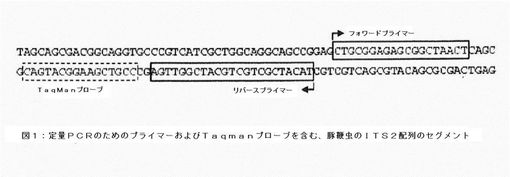
【図1】定量PCRのためのプライマーおよびTaqmanプローブを含む、豚鞭虫のITS2配列のセグメントを示す図である。
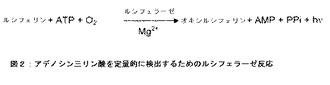
【図2】アデノシン三リン酸を定量的に検出するためのルシフェラーゼ反応の反応式を示す図である。
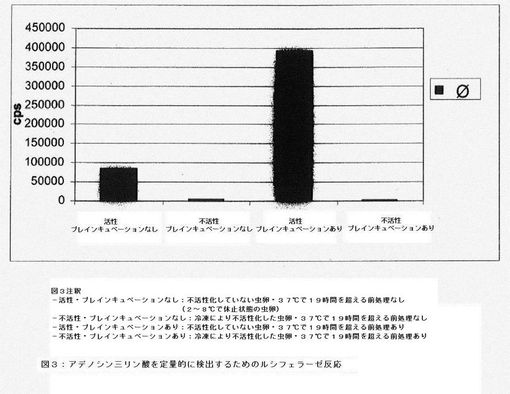
【図3】ルシフェラーゼ反応を用いたアデノシン三リン酸の定量的検出によって示される冷凍による不活性化の効果を示すグラフである。
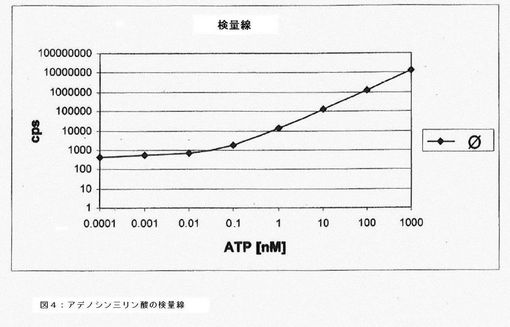
【図4】アデノシン三リン酸の検量線を示すグラフである。
【図5】豚鞭虫の蛋白質配列とセノラブディティス・エレガンス(Caenorhabhditis elegans)の相同蛋白質のアミノ酸配列とを示す図である。
【図6】in situハイブリダイゼーションに適したプローブの配列を示す図である。
【図7】運動性指数の結果を示すグラフである。
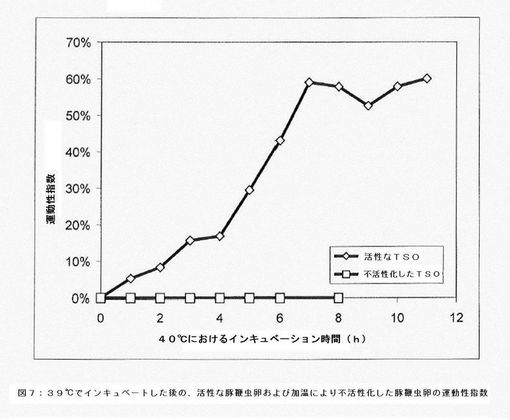
【図8】種々の観察時間における運動性指数を示すグラフである。
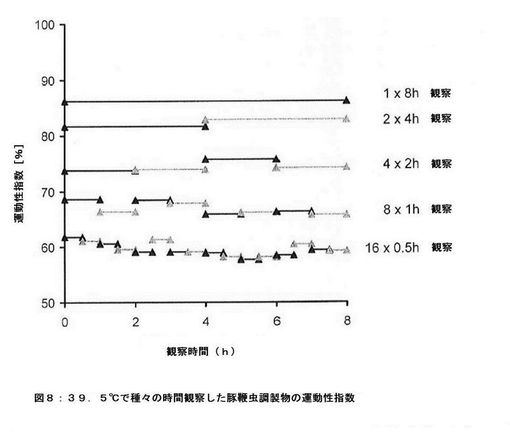
【図9】ローダミンXで標識したTSOと標識していないコントロールTSOとからなる混合物の蛍光顕微鏡写真および光学顕微鏡写真である。
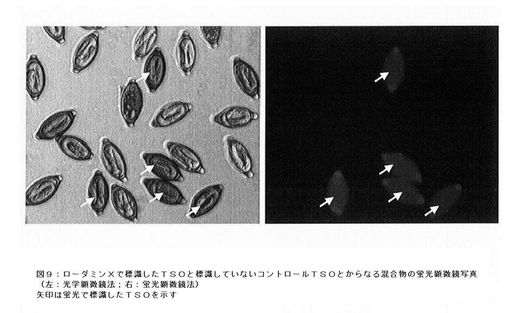
【図10】in vitro活性スコアと感染率との相関図である。
【発明を実施するための形態】
【0025】
以下、本発明の方法の一部である5種の定量法をさらに詳細に説明する。
【0026】
a)ゲノムDNAのコピー数の測定
幼虫の発育期における細胞分裂では、常に、染色体対が2倍になり、2個の娘細胞に分配される。したがって、虫卵中のゲノムDNAのコピー数は、発育途中の幼虫の体細胞数と相関すると見なすことができる。したがって、ゲノムDNAのコピー数を測定することによって、発育途中である幼虫の発育状態を表すことができる。このことは、上記の方法(成熟率の顕微鏡測定:上記参照)と比較して明らかに有利である。上記の成熟率の顕微鏡測定では、約13週間にわたる成熟の最終結果しか分析できず、幼虫が実際に完全に発育したのか(すなわち、細胞数が、完全に形成されたL1幼虫に相当するのか)に関していかなる情報も得られない。幼虫を成熟させることは蠕虫卵の医薬品製造工程の一部であり、ゲノムDNAのコピー数を測定することにより、該製造工程に伴う分析が初めて可能となる。この測定によって、蠕虫卵の医薬品製造工程の論理的な開発、および該製造工程の変更がもたらす効果の分析が可能となるだけでなく、幼虫の成熟期において製造を定期的に監視することも可能となる。
【0027】
定量PCR技術により、試料中に存在する特定の核酸配列のコピー数を測定できる。試料中に存在する特定の核酸配列中に利用可能なコピーが多ければ多いほど、より迅速に増幅プラトーに達する。コピー数は、検量線を用いることによって比較的正確に測定することができる。本明細書において、コピー数を測定するのに通常使用する方法のうちの1つは、リアルタイムPCRまたはいわゆるTaqMan PCRである。この他の核酸増幅法も当業者には知られており、同様にゲノムDNAのコピー数を測定するために使用することができる。正確なコピー数は、通常、適切な検量線を用いて求める。
【0028】
好ましくは、リボソームRNAの一部をコードするITS1−5.8S−ITS2領域が、蠕虫のゲノムDNAの好ましいマーカーとして適切である。豚鞭虫(Cutillas et al., Parasitol Res 2001, 100: 383-389)、他の鞭虫類および他の蠕虫のITS1−5.8S−ITS2領域の配列は既に報告されている。IST−1領域の定量PCRを用いて豚回虫の幼虫の発育を監視する生存能力試験も既に報告されている(Pecson et al., Appl. Envir. Microbiol., 2006, 72, 7864-7872)。
【0029】
しかしながら、この試験には蠕虫の不活性化方法を評価するという別の目的がある。
【0030】
ITS1−5.8S−ITS2領域の一部の配列が増幅される定量PCR(q−PCR)は、マーカー領域のコピー数、ひいてはゲノムDNAのコピー数を測定するのに特に適している。ITS1およびITS2の構成は種特異的であるため、この測定方法は、成熟期の分析に加えて生物を定性的に識別するのにも適している。L1期の豚鞭虫の細胞数がゲノムのコピー数を指標に約1000個であることはこれまで公知ではなかったため、このような試験は先行技術では全く報告されていない(実施例1参照)。
【0031】
図1に、ゲノムDNAのコピー数の測定に用いることができる、豚鞭虫の適切な遺伝子配列を例として示す。配列番号:1に当該遺伝子配列を示す。フォワードプライマーを配列番号:2、リバースプライマーを配列番号:3、およびTaqmanプローブの配列を配列番号:4で表す。コピー数を測定するために、豚鞭虫のゲノムの他の部分も使用できることは当業者にとって明らかである。この測定法における、これらの配列の必要条件としては、豚鞭虫において特異的に見出されるヌクレオチド配列でなければならないこと、および分析試料に混入する可能性のある他の生物が有する配列と類似性がないことが挙げられる。
【0032】
さらに、ゲノムDNAのコピー数の測定には副次的効果がある。ゲノムDNAのコピー数を測定することによって、調製物中に所望の生物が存在することを確認できるとともに、他の適切な配列を使用して他の生物の混入の有無を証明することができる。
【0033】
ゲノムDNAのコピー数を測定する際に注意すべき重要な点として、測定前に蠕虫卵を破砕しなければならないことが挙げられる。実施例に示すように、本発明においては「ポッター型ホモジナイザー」が特に適していることが判明している。好ましい実施形態においては、容量が2mlのものを使用し、間隙幅を0.01〜0.03mmに設定した。5〜15分間ホモジネートすることが好ましく、約10分間が好ましい。それぞれの分析工程において、細胞成分が分析できる状態でなければならない場合は、必ず細胞を破砕する。
【0034】
b)代謝活性の測定
細胞の生存能力を測定する方法の多くが、代謝活性の分析に基づいている。細胞代謝において、アデノシン三リン酸はエネルギー伝達物質およびエネルギー貯蔵物質として中心的な役割を果たし、細胞内代謝活性を測定するためのマーカーとして使用できる。本発明では、豚鞭虫のL1幼虫のような、分化した生物の生存能力を測定するために、このヌクレオチドの検出も利用できることが示されている。
【0035】
ルシフェラーゼ反応を用いたルミネセンス測定により、生物系におけるアデノシン三リン酸の定量が可能となる。定義によれば、ルミネセンスは「冷たい」光の放出である。ルミネセンス系は、化学的、生化学的、または電気化学的な基質の活性化に基づいており、基質が基底状態に戻る時に励起エネルギーの一部を光の形態で放出する。アデノシン三リン酸の検出には、ホタル(フォチナス・フィラリス(Photinus pyralis))から単離したルシフェラーゼを使用する。この真核生物由来の酵素は、アデノシン三リン酸、酸素、およびマグネシウムイオンの存在下で、ルシフェリンのオキシルシフェリンへの酸化を触媒して光を放出させる。反応式を図2に示す。
【0036】
豚鞭虫の生存可能な幼虫包蔵卵中に存在するアデノシン三リン酸を成功裏に測定するための必要条件としては、L1幼虫におけるアデノシン三リン酸の合成を、ルミネセンスを測定するのに十分な程度まで刺激すること;適切なホモジネート法によって虫卵を十分に破砕し、細胞内で形成されたこのヌクレオチドを放出させること;および、複雑なマトリックス由来のアデノシン三リン酸を定量的にかつ何ら妨げなく検出することが挙げられる。これまで報告されておらずかつ驚いたことに、幼虫発育が中断される前のL1幼虫におけるATP含量は低く、特定の温度で長時間にわたってインキュベートして適切に活性化させることによって初めて、ATP含量が一定の高レベルにまで増加することが判明した。さらに、生きているL1幼虫と死んでいるL1幼虫とを識別するためには、死滅させた幼虫においてアデノシン三リン酸の基準値が再現性良く測定されるような効果的な不活性化方法が必要である。
【0037】
本発明によれば、蠕虫卵群の代謝活性を分析するためにATP含量を測定することが好ましい。
【0038】
さらに、個々の蠕虫卵の代謝活性を分析する方法として、テトラゾリウム化合物を用いた染色法も適している。この染色法は、もともと、細胞培養中または組織切片中の、何ら障害なく個々の細胞が染色可能な動物細胞のために開発されたものである。テトラゾリウム化合物は、代謝が活性化している細胞において、ミトコンドリア酵素の作用により有色のホルマザンに還元され細胞内に沈殿する。この染色法を、何ら障害なく個々の細胞が染色可能な動物細胞から虫卵に包蔵され多数の細胞からなるL1蠕虫幼虫へと応用するためには、虫卵をあらかじめ処理して、虫卵内の幼虫の生存能力に影響を与えることなく基質を浸透させることが極めて重要である。豚鞭虫のL1幼虫は強固な殻に取り囲まれているため、周囲との物質移動が著しく制限され、分析に必要とされる物質の浸透が阻まれる。
【0039】
穏やかに卵殻を分解するために、次亜塩素酸で虫卵を処理すると有利であることが判明している。この方法は既に報告されている(Beer, Parasitol., 1973, 67: 263-278)。次いで、必要に応じて、キチナーゼおよびプロテアーゼを用いた酵素消化を補助的に行うこともできる。
【0040】
代謝活性の測定を可能にするためには、「ゼロ値」を測定しなければならない。本明細書において、この値は対照値であり、この値をもとにそれぞれの代謝活性を測定する。好ましい実施形態においては、「ゼロ値」は不活性な蠕虫卵を用いて測定する。
【0041】
虫卵を不活性化する方法として、冷凍による不活性化(急速冷凍後、−80℃で24時間を超えて保存)が適していることが判明した。この方法で不活性化した試料からのアデノシン三リン酸のシグナルは、バックグラウンドノイズのレベルにまで減少させることがほぼ可能であった。冷凍による虫卵試料の不活性化は、リン酸緩衝生理食塩水中で行う。言うまでもなく、冷凍による不活性化は他の測定法においても実施することができる。
【0042】
図3に、ルシフェラーゼ反応を用いたアデノシン三リン酸の定量的検出によって示される、冷凍による不活性化の効果を例として示す。プレインキュベーションの効果は、虫卵を冷凍して不活性化することによって特異的に測定できる。不活性化した(死滅させた)蠕虫卵では、プレインキュベーションを行ってもアデノシン三リン酸の増加は確認できない。これに対し、生物学的に活性な蠕虫卵は、37℃で19時間前処理後にアデノシン三リン酸が増加したことによる活性を有している。
【0043】
好ましい一実施形態では、それぞれのバッチにつき、虫卵を5500個含む550μl溶液を使用する。プレインキュベーションの後、虫卵を遠心分離(500rpmで5分間)によって分離し、溶解緩衝液225μl中に再懸濁する。インキュベーションによって形成されたアデノシン三リン酸を含む細胞内容物を放出させるため、ポッター型ホモジナイザー(容量2ml;間隙幅0.01〜0.03mm)を用いて虫卵を10分間ホモジネートする。この方法を用いるだけで虫卵が十分に破砕され、このヌクレオチドが定量的に放出される。アデノシン三リン酸を開裂する加水分解酵素を不活性化するために、虫卵の破砕は溶解緩衝液(成分:pH2のリン酸)中で行う。それぞれのバッチから50μlをピペットで取り出し、96マイクロウェルプレートに入れて、さらにリン酸緩衝液100μlで希釈する。ルシフェラーゼ反応は、市販のアデノシン三リン酸キット100μlを加えて開始する。2分間インキュベートした後、マイクロウェルプレートをルミノメーターで読み取り、ルミネセンスを測定する(図3)。リン酸緩衝液中アデノシン三リン酸の検量線(1.0μM、0.1μM、10nM、1nM、0.1nM、10pM、1.0pM、0.1pM)より、定量化を行う。代表的なATPの検量線を図4に示す。
【0044】
冷凍により不活性化した幼虫包蔵卵をホモジネートした試料に、アデノシン三リン酸を意図的に加えることにより、検量線の直線領域(1〜1000nM)において84%の回収率を達成できた。したがって、この反応に関連してくる、虫卵ホモジネート中の複雑なマトリックスとルシフェラーゼ反応およびルミネセンスとの相互作用は認められない。
【0045】
実施例において、豚鞭虫の生存可能な幼虫包蔵卵と不活性化により生存不可能になった虫卵とを識別するために、ルミネセンス測定を使用できることが明確に示されている。しかしながら、この試験を成功裏に行うには、試料の活性化、試料のホモジネート、および最適化された不活性化法による適切なコントロールの作製からなる測定パラメータをそれぞれ最適化してこれらを組み合わせなければならない。生物学的に活性な蠕虫卵中のATP含量としては、虫卵1個当たり少なくとも0.01pmolが適している。
【0046】
c)誘導性遺伝子発現の分析
この方法の原理は、遺伝子発現の誘導である。遺伝子発現は生細胞でしか誘導することができないため、これにより生きている幼虫と死んでいる幼虫とを識別できる。さらに、遺伝子発現の誘導能は、休止状態のL1幼虫が種々の活性化状態を経ることを可能にするための必要条件であると考えられ、種々の活性化状態は生活環の次の段階(孵化、粘膜への定着など)を誘導するのに必要である。
【0047】
熱ショック蛋白質は、温度を上昇させることにより容易に誘導することができ、形成されるメッセンジャーRNA(mRNA)が急速に分解せずに概ね安定していることから、熱ショック蛋白質の発現を分析することは特に有利であると考えられる。本発明の範囲内で、豚鞭虫の遺伝子配列の検出に初めて成功した。この豚鞭虫の遺伝子配列は、蠕虫であるセノラブディティス・エレガンス(Caenorhabhditis elegans)の熱ショック蛋白質hsp70と非常に相同性が高い。
【0048】
本発明の好ましい一実施形態では、豚鞭虫の熱ショック蛋白質の遺伝子発現を分析する。図5に、豚鞭虫の蛋白質配列(配列番号:5)を、セノラブディティス・エレガンスの相同蛋白質のアミノ酸配列(配列番号:6)と比較して示す。
【0049】
誘導性遺伝子発現の測定は、特定の誘導蛋白質をコードするメッセンジャーRNAの量を測定することにより行うことができる。メッセンジャーRNAのヌクレオチド配列は、この蛋白質のアミノ酸配列由来とすることができる。適切なフォワードプライマーおよびリバースプライマーを設計し、これと同時に、Taqmanプローブの配列を決定することは当業者とって問題ではない。次いで、遺伝子の誘導量をリアルタイムPCRにより測定することができる。意味のある測定を行うための必要条件としては、死滅させた虫卵を含むコントロール試料も測定することが挙げられる。さらなる必要条件としては、測定に用いる蛋白質が容易かつ確実に誘導され得るものでなければならないことが挙げられる。このような蛋白質としては熱ショック蛋白質が挙げられるが、特定の刺激によって誘導できる他の遺伝子も同様に測定に使用できる。
【0050】
個々の虫卵レベルで発現させたメッセンジャーRNAも、適切な遺伝子プローブを用いた蛍光in situハイブリダイゼーション(FISH)によって検出できる。次いで、この虫卵群に対して、フローサイトメトリーによる迅速かつ確実な分析を行うと特に有利である。
【0051】
in situハイブリダイゼーションに適したプローブの配列を図6(配列番号:7)に示す。
【0052】
d)完全な形のままの蠕虫卵内に包蔵される幼虫の運動性の測定
この方法の原理は、虫卵内のL1幼虫の機能性に関するパラメータとしての虫体の運動の検出に基づく。幼虫の運動性は、幼虫が適切な環境条件下で孵化するための必要条件である。C.エレガンスのような自由生活性の幼虫および寄生虫においては、運動性の分析は生存能力および機能性を確認するための確実な方法として有効である。
【0053】
本発明の範囲内で、驚くべきことに、厳密に環境温度を調節することによって、自由生活をせず虫卵内に包蔵されている蠕虫幼虫でも微細な運動を誘導できることが、豚鞭虫を例として初めて判明した。豚鞭虫の幼虫は、成熟が完了した後休止状態に陥り、適切な宿主によって経口摂取されるまで活性化および孵化しないことがこれまで報告されていた。虫卵内の幼虫の運動は非常に鈍く、本発明で開示されているように、長時間にわたり経時的に顕微鏡下で虫卵を観察しなければ検出できない。このような顕微鏡下での運動性試験は、画像分析ソフトウェアと組み合わせて自動化することができる。虫卵の生物学的活性のパラメータとしての運動性指数は、次式で計算する:
【数1】
【0054】
幼虫の運動性の測定を成功裏に行うには、温度を厳密に管理することが不可欠である。虫卵を、まず初めに、36〜42℃、好ましくは37〜41℃、特に好ましくは39.5℃に正確に設定した温度で、2〜30時間、好ましくは4〜20時間プレインキュベートする。このプレインキュベーションの後、上記の設定温度を少なくともほぼ維持できる適切な顕微鏡下に幼虫をセットする。次いで、虫卵を、36〜42℃、好ましくは38〜40℃で、2分〜4時間、好ましくは30分〜2時間にわたり経時的に記録して観察する。
【0055】
本発明に従って測定された運動性指数の結果を図7に示す。不活性化した虫卵では運動性は全く観察できないが、これとは対照的に活性な虫卵では運動性指数が時間とともに増加している。
【0056】
種々の観察時間における運動性指数を図8に示す。
【0057】
e)腸管内での孵化率の測定
この測定法の原理は、実験動物の腸管内あるいは実験動物からあらかじめ回収した腸管内容物内に含まれる蠕虫幼虫の孵化率の測定である。
【0058】
一実施形態では、実験動物から回収した腸管内容物を用いて孵化率を測定することができ、実際には、好ましくは結腸の始端または十二指腸の末端から腸管内容物を回収することが好ましい。次いで、腸管内容物を用いて孵化率を測定することができ、実験動物を用いて直接測定する必要はない。さらに、腸管を通過して糞便から回収された虫卵を分析することにより、この方法を実施することもできる。この場合、実験動物を屠殺する必要がなく、孵化率を測定するために実験動物を数回使用することができるという利点を有する。
【0059】
新規に開発されたこの測定法では、接種後、比較的短時間のうちに腸管内容物が分析され、孵化の後に残った幼虫被膜だけでなく完全な形のままの虫卵も集計される。先行技術において報告されている感染率の測定と比較した際の重大な利点としては、本発明の孵化率の測定は、個々の宿主が有する要因にはわずかにしか影響を受けない、生活環の第1段階のみを分析することが挙げられる。
【0060】
驚いたことに、適切な宿主としてブタだけでなくウサギにおいても、定量的に幼虫が孵化することを豚鞭虫について初めて示すことができた。したがって、たとえば豚鞭虫に対しては、腸の容積が小さい実験動物を利用することが可能であり、これにより顕微鏡レベルの小さな虫卵および幼虫被膜を回収することが極めて容易となる。
【0061】
驚いたことに、蛋白質を標識するために開発された市販の蛍光色素を用いて卵殻を永久的に染色できることが判明した。蛍光色素と幼虫被膜とを結合させることにより、蛍光顕微鏡法を用いて腸管内容物を分析することができるので、虫卵の回収が非常に容易となる。
【0062】
さらに重要な技術革新としては、蠕虫の未成熟卵または不活性化卵を固定の内部標準として使用することが挙げられる。このような虫卵は、生物学的に活性な幼虫包蔵卵と同一の腸内移行時間すなわち滞留時間を有するが、生物学的に活性な幼虫包蔵卵とは対照的に、腸管を通過するときも完全な形を保ったままである。内部標準として提供される虫卵を別の(異なる)蛍光色素で標識することによって、内部標準と分析対象の虫卵とを容易に識別することができる。
【0063】
本分析では、腸管内容物に含まれる、分析試料中の蛍光色素1で標識された完全な形のままの虫卵数および空の幼虫被膜数と、蛍光色素2で標識された内部標準である完全な形のままの虫卵数とを接種後の適切な時点において分析する。腸管内の孵化率は、完全な形のままの(孵化していない)虫卵を基準として、次式で計算することができる。
【数2】
【0064】
PCR反応、好ましい測定法、およびこれらより得ることができる値を下記の表2にまとめる。これらの値は、分析した調製物が、必要とされる生物学的活性を有するのかどうかを示す好ましい制限値を表す。
【0065】
【表2】
【0066】
好ましい実施形態において、本発明の新規の試験方法は、in vitroで成熟させた蠕虫卵の温度誘導性活性化能を測定することを含む。また、相補的な2種のパラメータ(運動性指数および平均ATP含量)を測定する。これらのパラメータは相俟ってin vitro活性スコアを構成する。in vitro活性スコアは、in vivoにおける生物学的活性と良い相関を示す。しかしながら、in vitro活性スコアから生物学的活性を予測することができるのは、使用する幼虫が完全に成熟している場合のみである。幼虫が完全に成熟しているかどうかは、虫卵内の豚鞭虫のゲノムのコピー数を測定する定量PCRを用いることによって分かる。さらに、並行して行われる新規のin vivoにおける孵化試験を用いることにより、分析した虫卵が生理学的条件下で実際に活性であることが保証される。以下、好ましい方法を概略的に示す:
【0067】
【0068】
本発明の重要な態様として、in vitroにおける温度誘導性活性化能を測定することが挙げられる。
【0069】
成熟後のL1−TSOは、何年にもわたって生存可能な休止状態となり、宿主による経口摂取後に活性化されて孵化するまでその状態を保つ。これまで報告されておらずかつ予想外にも、他の因子を加えなくても特定の温度範囲内で一定時間インキュベートするだけで、in vitroでも虫卵を容易に活性化できることが判明した。虫卵の活性化状態は、2種の相補的パラメータ、すなわち、完全な形のままの虫卵内の幼虫の運動性と、あらかじめ活性化した虫卵の溶解物中のATP含量とを測定することにより定量化する。文献により報告されている挙動に反しかつ予想外にも、本明細書に記載のプレインキュベーションにより虫卵内の幼虫の微細な運動が誘導される。この幼虫の運動は、幼虫が死滅することも虫卵から孵化することもなく、何時間にもおよび継続する。したがって、本明細書に記載された方法の好ましい実施形態として、活性化を引き起こす虫卵の適切なプレインキュベーションが挙げられる。活性および運動性を有する幼虫(運動性指数)を含む虫卵群の比率は、十分に多数の虫卵を顕微鏡下で経時的に観察することにより、確実に測定することができる。しかしながら、この測定では個々の幼虫の活性状態すなわち運動性/活性レベルの違いが識別されないため、分析対象の虫卵群の総活性を示す定量的な指標であるATP含量も併せて測定する。直接的であるが不連続な変数としての運動性指数と、間接的であるが連続的な変数としてのATP含量の2種の分析パラメータが互いに補い合うことによって、in vivo活性とよく相関するin vitro活性が確実に決定される。これら2種のパラメータから計算することができる、in vitro活性スコアは、虫卵のin vivo活性と良く相関する。
【0070】
本発明のin vitroにおける方法は、従来公知の方法とは明らかに異なる。虫卵内の完全に形成された幼虫が虫卵内に存在するかどうかを視覚的にしか分析しない成熟率の測定(Kringelら)では、本明細書に記載の方法とは対照的に、幼虫の生存能力または活性に関していかなる結論も得られない。また、消化菅を模擬したin vitroでの孵化率の測定は、本明細書に記載の方法と比較すると、非常に煩雑であり干渉を受けやすい。このin vitroにおける孵化試験に関しては、in vivo活性との相関性も報告されていない。
【0071】
本発明の好ましい実施形態では、幼虫の成熟状態が決定される。in vitro活性スコアは、幼虫が幼虫発育を完全に経た場合のみ、in vivoにおける状況を予測するのに役立つ意味のある結果を提供する。温度誘導性活性化能試験において、完全には発育していない幼虫でも陽性反応が示されることがあるが、これらの幼虫は機能性を持たず、宿主内で孵化することも幼虫の正常な生活環をたどることもできないと考えられる。従来公知の方法では、形態学的基準を用いて幼虫が十分に発育していることを顕微鏡下で評価する。この評価法は、幅広い経験が必要とされ、信頼性が低く、その性質上個々の虫卵ごとについてしか評価することができない。これとは対照的に、本発明による方法では、多数の虫卵を含む群のゲノムのコピー数を客観的に測定し、幼虫1匹当たりの平均体細胞数を計算することができる。
【0072】
温度誘導性活性化能試験の結果は、生理学的条件下において孵化率を容易かつ迅速に、しかも確実に測定できる新規のin vivo試験で保証することが好ましい。驚いたことに、あらかじめ不活性化した虫卵は腸管内で生存できるがそのまま変化しないのに対し、生物学的に活性な虫卵内の幼虫は腸管内において定量的に孵化することが判明した。新規に開発された方法では、調製物に含まれる活性なTSOの比率は、実験動物の腸管内を変化しないまま通過した不活性な虫卵のみを集計することよって間接的に求められる。しかしながら、この定量化を成功裏に行うには、蛍光で標識されかつ変化しないまま腸管内を通過する、不活性な虫卵からなる新規に開発された内部標準を使用しなければならない。さらに、驚いたことにかつ予想外に、生物学的に活性なTSOはウサギの腸管内でも孵化することが判明した。したがって、費用効果がより高く、飼育するのが容易で、かつ有利なことに腸の容量が極めて小さい(顕微鏡レベルの小さな虫卵を回収することが容易となる)動物モデルを、ブタの代わりに使用することができる。
【0073】
本発明によるin vivo試験と、ブタを用いた公知の感染率測定試験とは異なるものである。ブタを用いた感染率測定試験は数週間にわたって継続するのに対し、孵化率の測定に必要な日数はわずか1〜3日である。さらに、孵化率を測定する場合、宿主内における豚鞭虫の生活環うちの第一期のみが分析され、個々の宿主が有する要因にはほとんど影響を受けない。これとは対照的に、ブタを用いて感染率を測定する場合は、感染させてから3〜4週間後に測定することが好ましく、個々の宿主においてそれぞれ異なる免疫システムが形成されることを主たる原因とする、自然な生物学的変動に影響を受ける。
【0074】
本明細書に記載の分析法のさらなる利点としては、動物の糞便から虫卵を回収できることが挙げられ、これによって動物を試験のために屠殺する必要がなくなる。
【実施例】
【0075】
実施例1:
豚鞭虫のITS2配列に基づき、TaqManシステムを用いた定量PCR法を開発した。標的配列、プライマー、およびTaqmanプローブを以下に示す。遺伝子配列が公知である他種の鞭虫と豚鞭虫とを識別できるように標的配列を選択した。このようにすると、ITS2コピー数に関する情報が得られるだけでなく、分析した生物が実際に豚鞭虫であったことも定性的に証明できる。
【0076】
ポッター型ホモジナイザー(容量2ml;間隙幅0.01〜0.03mm)で虫卵(500μl溶液中1000個の虫卵)を10分間ホモジネートして、試料を調製する。驚いたことに、顕微鏡下で観察すると、遺伝子を単離する際に通例用いられる他の方法では豚鞭虫卵を破砕できないのに対して、上記の方法を用いると幼虫被膜を破損させることが可能であることが分かる。魚精子DNA1.54μgを加えた後、遺伝子単離用の市販キット(DNeasy−Blood and Tissue−Kit:キアゲン社)をメーカーの説明書に従って用い、虫卵ホモジネートと反応させる。下表に示すプロトコルに従い、単離したDNA(全量50μl)を用いてPCRを行う。
【0077】
【表3】
【0078】
PCRの増幅効率は92〜98%であり、これは虫卵1個に含まれるITS2遺伝子のコピー数を決定するのに十分な感度である。
【0079】
PCRを用いて、豚鞭虫の未成熟卵および完全に成熟した幼虫包蔵卵のITS2コピー数を分析した。結果を以下の表に示す。
【0080】
【表4】
【0081】
PCR解析の結果、幼虫を包蔵したTSOのITS2コピー数が、未成熟TSOのITS2コピー数の約1000倍であることが示された。未成熟卵(L0)は単一の細胞と見なすことができるので、この結果より、豚鞭虫のL1幼虫は約1000個の体細胞を有することが分かる。豚鞭虫のL1幼虫の体細胞数はこれまで知られていなかった。近縁種であるC.エレガンスについては、体細胞数が959であることが判明しており、この数は、本実施例において判明した豚鞭虫の体細胞数である約1000個とよく一致する。
【0082】
この方法により、幼虫発育期の相違を分析できるかどうかを確認するために、TSOを4週間成熟させ、ITS2 PCR(表5)で定期的に分析した。
【0083】
【表5】
【0084】
本実施例において、この方法を用いて、発育期がL0の幼虫と発育期がL1の幼虫とを体細胞数により識別できることが明らかに示された。したがって、この方法は、製造工程に伴う幼虫の成熟過程の分析に特に適している。蠕虫卵調製物を製造する場合、幼虫を成熟させ始めてから最初の28日間に50〜1200の相対コピー数を達成するように規定すると特に好ましいと考えられる。
【0085】
実施例2:
アデノシン三リン酸を検出するために、反応に必要なルシフェラーゼ酵素、基質であるルシフェリン、マグネシウム塩などを含む市販キットを使用することができる。幼虫包蔵卵をプレインキュベートしてアデノシン三リン酸の形成を刺激することは必須である。生存可能な幼虫包蔵卵が休止状態である場合、アデノシン三リン酸含量から、生存可能な幼虫包蔵卵と死んでいる虫卵とを十分に識別することはできない(図3)。図3の結果より、虫卵を37℃で19時間にわたりインキュベートする方法が適すると判明した。開発した分析法の重要な利点として、このインキュベーションに際して試料を調製する必要がないことが挙げられる。本発明の虫卵懸濁液は、培地の定性的および定量的組成とは無関係にそのまま使用することができる。pH価が2未満の強酸性培地でさえも試料を調製することなく使用することができ、虫卵中のアデノシン三リン酸の形成を刺激するのに何ら悪影響を与えることはない。インキュベートすることにより、生きているF1幼虫と死んでいるF1幼虫とを明確に識別することができる(図3)。
【0086】
実施例3:
セノラブディティス・エレガンスに見出される熱ショック蛋白質hsp70の配列と、ネズミ鞭虫(Trichuris muris)の相同配列と、犬鞭虫(Trichuris vulpis)の相同配列とに由来するプライマーを用いて、豚鞭虫から単離したRNAの遺伝子配列を増幅することができた。豚鞭虫由来のこの新規の配列は、蛋白質配列に翻訳すると、C.エレガンスのhsp70蛋白質の蛋白質配列と65%一致する(図5)。
【0087】
豚鞭虫の熱ショック蛋白質と推定される蛋白質を検出するための定量RT−PCR法を開発した。この定量は、恒常的に発現される18S rRNAと比較して行った。豚鞭虫の活性な幼虫包蔵卵に熱ショック(45℃で20分間)を与え、処理していない活性な豚鞭虫と熱ショック蛋白質の発現量を比較した。
【0088】
【表6】
【0089】
相対的な発現量を分析した結果、hsp70に類似したmRNAの発現を熱ショックによって誘導できることが明らかに示された(表6)。したがって、適切な試験条件を選択することにより能動的に発現を誘導できる遺伝子は、当該活性試験に利用することができる。
【0090】
個々の虫卵における遺伝子発現の活性化は、蛍光in situハイブリダイゼーション法(FISH)を用いて検出できる。熱ショック蛋白質と推定される蛋白質を検出できるように、たとえば図6に示す、蛍光で標識したDNAプローブを作製した(配列番号:7)。
【0091】
活性な虫卵を蛍光で標識することにより、虫卵群もフローサイトメトリーを用いて分析することができる。この分析は、顕微鏡による分析よりも、迅速かつ客観的に実施できる点で有利である。
【0092】
本実施例により、本明細書に記載の方法を用いて(個々の)蠕虫卵における遺伝子発現の誘導能を分析できることが明らかに示された。したがって、休止状態のL1蠕虫幼虫の活性化能を分析できる方法が初めて利用可能となった。
【0093】
蠕虫卵調製物を製造する際に、その活性を生物学的に証明するために遺伝子発現を誘導する場合、判定基準として、蠕虫卵調製物に誘導性遺伝子発現陽性の蠕虫卵が25〜100%、好ましくは50〜100%含まれていると適している。
【0094】
実施例4:
4.1 運動性指数の分析条件に対する依存性
活性な豚鞭虫卵と、48℃で72時間インキュベートして不活性化した豚鞭虫卵とを、39℃で11時間インキュベートした。1時間毎にそれぞれの虫卵を5分間観察し、運動性を測定した。運動性指数の経時変化を図7に示す。
【0095】
本実施例により、休止状態の幼虫を活性化し、かつ一定した高い運動性指数を測定するためには、高温で比較的長時間(本実施例では39℃で7時間)のプレインキュベーションが必要であることが明らかとなった。これまで、虫卵に包蔵され完全に成熟したL1幼虫は運動しないとされていた(Parasitology 1973, 67: 253-262)ため、この結果は予想外であった。長時間におよぶインキュベーションが必要とされ、経時的に観察することにより初めて鈍い運動が見られただけだったため、虫卵内の幼虫の運動性は恐らくこれ以前に観察されたことがなかったと考えられる。
【0096】
活性な虫卵と不活性化された虫卵とを比較して運動性を分析することによって生物学的に活性な豚鞭虫卵と不活性な豚鞭虫卵とを容易に識別できることが明らかとなった。
【0097】
幼虫は同期して運動しないため、長時間にわたって観察することにより生きている虫卵内の散発的な運動を検出できる可能性が増加する。したがって、活性化段階と並んで、観察時間の長さも運動性指数の測定に影響を及ぼす。このことは、活性化段階(37℃で8時間)後に豚鞭虫卵調製物を39.5〜40℃で様々な時間観察した分析(図7)により示された。さらに、この分析により、8時間にわたる観察時間を通して運動性は安定していることも示された。
【0098】
したがって、この方法は、厳密に温度が管理された活性化段階と長時間におよぶ観察時間とを新規の特徴とする。実施例4により、虫卵内で休止状態のL1幼虫の運動を、適切な温度および観察時間からなる条件によって、誘導かつ測定できることが示された。この方法を使用すると、孵化の必要条件として、ひいては虫卵の生物学的活性のパラメータとしての運動性を容易に測定でき、活性なL1幼虫と不活性なL1幼虫とを明確に識別することができる。
【0099】
4.2 有利な試験条件
以下に挙げる試験条件が有利であると判明した。リン酸緩衝液(pH7.4)300μl中に15,000個の蠕虫卵を含む試料を、96ウェルプレート(底面積:0.31cm2)へ移す。この播種密度においては、200倍の倍率で約80〜150個の虫卵が顕微鏡の視野内に入る。37℃で8時間インキュベーションした(活性化段階)後、温度を39.5℃まで上げて、連続した4つの観察野を選択し、それぞれ2時間観察する。この観察を行うため、フィルムに毎分3画像を記録する。次いで、虫卵数と動いている幼虫数とを測定し、運動性指数を計算する。4観察野の平均値から、試料の運動性指数を得る。
【0100】
4.3 再現性および精度−感染率測定試験との比較
上記の条件下における運動性試験の再現性を調べるために、4回の実験をそれぞれ異なる日に実施し、1回の実験につき4種の豚鞭虫試料を分析した。測定結果を以下の表に示す。比較として、同一の豚鞭虫試料を用いて感染率測定試験を行った、4回の実験の結果を示す。
【0101】
【表7】
【0102】
運動性指数の4つの独立した測定実験では、最大で互いに4.2%の差異があるのに対し、感染率の4つの独立した測定実験での最大偏差は35.5%である。運動性試験での単回測定の精度は1.9〜6.3%であるのに対し、感染率測定試験での単回測定の精度は9.6〜58.4%である。したがって、運動性試験は、再現性および精度に関して感染率測定試験よりもはるかに優れている。
【0103】
4.4 運動性と生物学的活性(正確性)との相関−感染率測定試験との比較
運動性と生物学的活性との相関を分析するために、活性なTSOと加温により不活性化したTSOとからなる混合物を作製し、4.2に記載した条件に従った運動性試験により分析した。加温による不活性化は、虫卵を48℃で72時間加温することにより行った。客観的な集計結果を得るために、盲検法にて試料を測定した。
【0104】
相対生物学的活性は、測定した試料の運動性指数値とあらかじめ測定した活性なTSOの運動性指数値より計算された比率である。測定の正確性は、相対生物学的活性と試料中の活性な虫卵の実際の比率とを比較することにより求められる。
【0105】
上記と同一のTSO試料を用いた同様の測定実験を、上記の感染率測定試験についても行った。実験動物は、感染率が既知である活性なTSOを種々の量で感染させた。不活性化した虫卵との混合物は使用しなかった。結果を以下の表に示す。
【0106】
【表8】
【0107】
【表9】
【0108】
本明細書で示される運動性試験を用いることによって、TSO試料の相対生物学的活性は、0〜100%の全範囲にわたる期待値の10%未満の偏差で正確に測定することができる。この測定の精度(測定値の分散)は、相対生物学的活性が50〜100%の範囲において5%未満である。相対生物学的活性が低い場合、測定の精度は15%未満である。
【0109】
感染率測定試験の結果は、0〜100%の範囲にわたる相対生物学的活性の期待値から約25〜32%逸脱する。この測定値における分散は28〜80%である。
本実施例により、運動性指数が相対生物学的活性と直線的に相関することが明らかに示された。運動性試験は、正確性と精度に関して感染率測定試験よりはるかに優れている。
【0110】
蠕虫卵調製物の生物学的活性を測定するために運動性試験を実施する場合、運動性指数が30〜100%、好ましくは60〜100%であると蠕虫卵調製物として適している。
【0111】
実施例5:
一例として、幼虫包蔵卵と未成熟卵と不活性化した幼虫包蔵卵とからなる種々の混合物をウサギに経口投与した。8時間後、ウサギの腸管内容物を顕微鏡下で分析し、完全な形のままの虫卵数と空の幼虫被膜数とを集計した。本分析では虫卵を蛍光標識しなかったため、完全な形のままの幼虫包蔵卵と未成熟卵との比率により孵化率を計算した([IE]=幼虫包蔵卵数、[IS]=未成熟卵数として、上記の式を用いた)。分析結果と分析結果より計算した孵化率とを以下の表に示す。
【0112】
【表10】
【0113】
第1〜3群のデータを比較すると、活性な幼虫包蔵卵しかウサギの体内で孵化できなかったことが示された(完全な形のままの幼虫包蔵卵数が顕著に減少したこと、および空の幼虫被膜が多数存在したことにより確認できた)。これとは対照的に、不活性な幼虫包蔵卵は腸管内で変化しなかった(第2群)。このことにより、この方法を使用した場合、生物学的に活性な虫卵と不活性化した虫卵とは識別可能であることが明らかに証明された。さらに、未成熟卵も腸管内を通過する際に完全な形を保ったままであった(第3群)ため、分解されずに実験動物の腸管内を通過する内部標準として未成熟卵を使用することは可能である。虫卵は腸管内容物から完全には回収できず、回収した完全な形のままの虫卵の絶対数が意味を持たなくなることから、この内部標準は必要である。上記の式は、内部標準を考慮に入れて孵化率を計算するために使用する。上記の式を用いて計算すると、本実施例において分析した豚鞭虫卵は67.9%の孵化率を示したのに対し、不活性化した豚鞭虫卵は0%(数学的には−12.6%)の孵化率を示した。
【0114】
より多くの虫卵を回収し、試料である虫卵と内部標準である虫卵とをより明確に識別するために、蛍光プローブを共有結合させて豚鞭虫卵を標識した。メーカー仕様書によると蛋白質中の一級アミノ基と反応することができる、ローダミンXスクシンイミジルエステルを一例として蛍光プローブとして使用した。1000個の豚鞭虫卵を含むリン酸緩衝液(pH7.4)懸濁液200μlに、1M炭酸水素ナトリウム溶液20μlを加え、次いで、0.5%ローダミンXスクシンイミジルエステルのDMSO溶液5μlと反応させた。反応液を回転振盪しながら室温で1時間インキュベートした。次いで、遠心分離(500rpmで10分間)および上清緩衝液の交換によって、虫卵を合計8回精製した。蛍光顕微鏡法により、赤色に染色された幼虫被膜がはっきりと示された(図9)。
【0115】
蛍光標識すると、顕微鏡下で孵化率を評価するのに有利となる。さらに、蛍光活性化フローサイトメトリーを用いると、客観的かつ迅速な分析も可能となる。
【0116】
概していえば、本実施例に記載の方法を用いると孵化段階を定量的に分析することができ、この孵化段階を分析することにより生物学的活性を測定できることが本実施例において明らかに示された。蠕虫卵調製物の生物学的活性を測定するために孵化試験を実施する場合、孵化率が約25〜100%であると蠕虫卵調製物として適している。
【0117】
実施例6:
in vitroにおける温度誘導性活性化能とin vivoにおける生物学的活性との相関
温度誘導性活性化能と生物学的活性との相関性を調べるために、種々の性質を有する12種のTSO試料を分析した。まず、これらの試料の幼虫が完全に発育しているのかをPCRを用いて評価した。次いで、加温により活性化を誘導した後、運動性指数とATP含量とを測定した。これらと並行として、12種の試料それぞれに対してブタを用いた感染率測定試験を行った。感染率測定試験は、Kringelらによって報告された方法に従い、それぞれの試料に対して5匹のブタを用いて行った。運動性とATP含量の2種のin vitroパラメータを、in vitro活性スコアとしてまとめた:
【数3】
【0118】
【表11】
【0119】
in vitro活性スコアと感染率との相関を図10に示す。
【0120】
本実施例において、in vitro活性スコアはin vivoにおける生物学的活性と良く相関することが示された。個々の試料におけるわずかな偏差は、恐らく、生物学的変動(宿主が有する要因)により本質的な不安定さを抱えているという、このin vivo試験の弱点にむしろ起因すると考えられる。
【0121】
感染率測定試験を利用してプローブを測定するには、6週間かかり、5匹のブタが必要とされ、相当な人件費が伴う。本明細書に記載された5種の方法のうちの3種からなる、本実施例に記載のin vitro試験を行うのには、1〜2日間しかかからず、人件費および材料費も比較的少なくて済む。
【技術分野】
【0001】
本発明は、生活環の種々の発育期における、蠕虫卵、特に鞭虫卵、好ましくは豚鞭虫卵の生物学的活性を測定する方法に関する。本発明の方法により、治療用医薬品の有効成分としての蠕虫卵調製物を管理された方法で製造することができ、ヒトに対して安全かつ治療上効率的な適用を保証することができる。
【背景技術】
【0002】
寄生虫感染が、動物宿主の免疫システムの活性化に影響を及ぼすことは公知である(D M McKay, Parasitology 2006, 132: 1-12参照)。この活性化は自己免疫疾患の発症や経過にも影響する。疫学的調査によると、衛生状態が良いため寄生虫感染率が低い地域と比べて、寄生虫感染率の高い地域では自己免疫疾患の罹患率が低い。慢性炎症性腸疾患であるクローン病患者のサイトカインプロファイルにおいては、蠕虫感染によりTh2免疫細胞が刺激されうることが示されている。蠕虫に感染させることによって、Th1優位の自己免疫疾患であるクローン病を予防することができるか、またはクローン病に影響を与えることができる(Summers et al.,Am J Gastroenterol 2003, 98: 2034-2041参照)。
【0003】
1971年には既にBeer(Br. Med. J., 1971, 3: 41)により、ヒトに病原性を有するヒト鞭虫(Trichuris trichiura)による感染症などに伴って起こるような病原作用を引き起こすことなく、意図的にヒトに感染させるのに適した線虫として、豚鞭虫(Trichuris suis)が挙げられることが報告されている。より最近の研究によると、豚鞭虫のヒトへの感染は一過性のものに過ぎないと考えられ、増殖が可能となる前に豚鞭虫は排除されることが明らかとなっている。しかしながら、慢性炎症性腸疾患に対する豚鞭虫の使用に関して肯定的な臨床研究によれば、豚鞭虫をヒトに一時的に感染させることにより、免疫系を調節して治療効果をもたらすことができる(Summers et al., Am. J. Gastroenterol. 2003, 98: 2034-2041)。
【0004】
豚鞭虫の生活環は、排卵された未成熟卵(L0期)から始まり、未成熟卵は糞便とともに感染動物から排泄される。次いで、地中で3〜6ヶ月間かけて成熟し、第1幼虫期(L1)となる。L1期に達してから最初の数日間は、幼虫が虫卵内で動くのを観察することができる。その後、虫卵内の幼虫は休止状態となり、この状態を何年間も保つが、感染性を失うことはない。適切な宿主により経口摂取された後、L1幼虫は腸管腔で虫卵から孵化し、数時間以内に盲腸や結腸の粘膜に入り込む。その後、幼虫は腸粘膜内で数週間かけてさらなる幼虫発育期(L2〜L4)を経てL5成熟幼虫となり、最終的に腸管腔へと再び出て行く(Beer, Parasitol. 1973, 67: 253-262)。
【0005】
豚鞭虫卵(Trichuris suis ova=TSO)を薬学的に製造するには、in vitroまたはin situにおいて排卵された未成熟TSO(L0)を、適切な方法(WO 1999 33479、WO 2007 076868およびWO 2007 134761)を用いて、単離精製する。次いで、管理された実験室条件下で虫卵を成熟させて、生物学的に活性な(L1)TSOを形成させる。得られたTSOは、医薬品の薬学的に活性な部分を構成する。
【0006】
治療目的で提供される蠕虫卵は、生物学的医薬品として分類することができる。生物学的医薬品の製造および適用に際して評価しなければならない主要なパラメータとして、医薬品の治療有効性の最終的な指標となる生物学的活性が挙げられる。生物学的活性を分析せずには、製造工程を論理的に開発し監視することも、また、ヒトへの計画的な適用のために、治療上有効で患者にとって安全な医薬品投薬量を決定することもできない。このような理由により、蠕虫卵調製物を医薬品として安全かつ有効に使用するためには、蠕虫卵製剤の生物学的活性が十分な条件を満たしていなければならない。
【0007】
現在、豚鞭虫の分析方法として3種の方法が利用可能であるが、いずれの方法も豚鞭虫の生物学的活性を測定するには不十分である。
【0008】
1.成熟率の測定(Kringel et al., Vet. Parasitol. 2006, 139: 132-139, Paragraph 2.2(豚鞭虫);およびJohnson et al. Intl. J. Parasitol. 1998, 28, 627-633(豚回虫))
この分析は顕微鏡下で行う。形態学的基準を用いて、完全な形のままでかつ十分に成熟したL1幼虫が虫卵に含まれているかどうかを評価する。しかしながら、本出願で示される分子生物学的方法とは対照的に、形態学的評価のみでは幼虫が実際に十分に成熟しているのかどうかを確実に推定することはできない。さらに、成熟率には、幼虫を包蔵したL1−TSOの生存能力や生物学的活性に関する情報は含まれないという制限がある。さらに、この形態学的評価には観察者の経験が必要とされ、形態学的な判別が難しい事例においては主観的な分類がなされるため正確性および精度が制限される。
【0009】
2.感染率の測定
豚鞭虫に関しては、ブタを用いて分析を行う。ブタを感染させた後、特定の時点において腸粘膜に定着した幼虫数を測定し、TSOの投与量を基準にして感染率を計算する(Kringel et al., Vet. Parasitol., 2006, 139: 132-139;Summers et al., 2005, Gastroenterology, 128: 825-832;またはJohnson et al. Intl. J. Parasitol. 1998, 28, 627-633(豚回虫))。感染率を測定する際の問題点として、感染率が蠕虫卵の機能性により影響を受けるだけでなく、個々の宿主が有する要因、たとえば、実験動物の免疫システム、腸内細菌叢、腸の機能などによっても同様に影響を受けることが挙げられる。したがって、実験動物を用いた感染率の測定には大きなばらつきが本質的に伴い、試験系に生物を用いているため標準化することもできない。このため、実験動物を用いた感染率の測定は、生物学的活性試験としての適性が大幅に制限される。さらに、ブタを用いた豚鞭虫の感染率測定試験には、少なくとも3週間の長い潜伏期が伴う上、結果のばらつきが大きいため多数の動物が必要とされる。このような理由により、この試験を日常的に使用することは倫理的にも経済的にも難しい。
【0010】
さらに、Kringelらは、蠕虫卵の生物学的活性を評価する方法として、蠕虫に感染させた動物の糞便から虫卵を回収する方法を報告している。この方法では、動物に感染させてから、孵化した幼虫自らがL5期まで成熟し、交尾し、排卵するまで7〜8週間待たなければならない。また、糞便中の虫卵数から成熟幼虫の感染率または生物学的活性が示唆されるような直接的な定量的関係は存在しない。さらに、Kringelらの方法では次世代の虫卵が分析されるため、Kringelらのin vivoにおけるこの方法と本出願に記載の孵化率の測定法とは明確に区別されなければならない。
【0011】
3.WO 2007/134761には、豚鞭虫卵の生存能力を試験する方法が記載されている。この方法では、ブタの消化菅における虫卵の通過をin vitroで模擬して虫卵を孵化させる。
【0012】
しかしながら、医薬品に必要とされる精度で蠕虫卵調製物の生物学的活性を測定するには、上述した方法では不十分である。
【発明の概要】
【発明が解決しようとする課題】
【0013】
したがって、動物において生物学的活性が測定される生物学的医薬品の主な課題の一つとして、調製物の標準化が挙げられる。上述したブタへの感染のような動物モデルの使用は、非常に煩雑で、しかも相当な時間を要する。したがって、本発明の明細書では、蠕虫卵の発育やその生物学的活性に関してそれぞれ異なる側面から種々の試験を行う方法を説明する。信頼できる結果を得るためには、1試料に対し、本発明の試験のうち少なくとも1種行う必要があり、好ましくは本明細書に記載の試験のうち少なくとも3種、より好ましくは少なくとも4種、さらに好ましくは5種行い、それぞれの試験に関連するパラメータを測定する。個々の試験結果は全体として一つと見なされ、適切な蠕虫調製物は、それぞれの試験における所定の制限値を満たさなければならない。この所定の制限値を満たした場合、蠕虫卵調製物は医薬用途に適すると結論づけられる。
【0014】
したがって、産業に応用可能であって、種々の発育段階におけるTSOおよび他の蠕虫卵の生物学的活性を包括的かつ確実に分析することにより蠕虫卵の種々の生物学的機能を測定する方法に対する需要は大きい。このような方法で管理することによってのみ市場性の高い医薬品が製造できる。
【課題を解決するための手段】
【0015】
本発明の目的は、以下に詳述する方法を用いて蠕虫卵の生物学的活性を包括的に分析することである。したがって、本発明は、製造工程の厳密な管理と安全かつ治療上効率的な患者への適用とを可能にすることも目的とする。
【0016】
従来公知の方法における複雑な問題および不完全さを考慮すれば、個々の分析方法を単独で使用して、生物学的活性を確実に測定することは通常不可能であることは明らかである。
【0017】
したがって、本発明の主題は、完全に成熟した幼虫を包蔵する鞭虫卵の生物学的活性を測定する方法であって、以下の測定のうち少なくとも1種行うことを特徴とする:
【0018】
生化学的方法および/または分子生物学的方法を用いた、鞭虫卵の温度誘導性活性の測定、具体的にはATP含量の測定;
鞭虫の幼虫包蔵卵における遺伝子発現の誘導能の測定;
高温でのプレインキュベーションによる活性化後における、虫卵内に包蔵される鞭虫幼虫の運動性の長時間観察による顕微鏡測定;および/または、
内部標準を基準として、腸管内容物から回収した完全な形のままの幼虫包蔵卵を定量化する、実験動物における鞭虫幼虫の孵化率の測定。
【0019】
したがって、蠕虫の生活環の特定段階における種々の生物学的機能を分析する5種の測定法からなる試験系を開発した。個々の医薬品製造工程において1種の測定法を適用するだけでも十分であるとも考えられるが、通常、最終製品の生物学的活性を測定するためには、以下の表1に記載の測定法のうち少なくとも3種行わなければならない。
【0020】
【表1】
【0021】
薬学的に利用可能な蠕虫卵の生物学的活性を確実に測定するためには、少なくとも3種、好ましくは4種、より好ましくは5種すべての工程を組み合わせた方法を行うと好ましい。特定の生物学的活性を測定するため、または製造の副工程を監視するためであれば、本発明の方法の個々の工程を単独で実施するだけで十分な場合もある。
【0022】
5種の個々の測定法も、他の事項に関して上述した試験原理に基づく。蠕虫の幼虫包蔵卵、特に豚鞭虫の幼虫包蔵卵への適用は新規であり、これまでに報告されていない新規の一連の副工程の開発を必要とする。5種の生物学的活性を試験することにより包括的な測定が可能な本方法全体も新規であり、蠕虫卵、特に豚鞭虫卵に関してはこれまで報告されていない。本明細書に記載の5種のうち3種の方法は、休止状態の鞭虫幼虫を高温でプレインキュベートすることにより活性化することを共通の好ましい特徴とする。本発明の方法のまた別の際だった特徴としては、完全な形のままでありかつ生存可能な蠕虫卵に対して本方法を実施できることと、測定したパラメータが、宿主側の要因または本試験を実施する者の専門能力には影響を受けず、蠕虫卵の活性によってのみ影響を受けることが挙げられる。したがって、本発明の方法は、これまで用いられてきた方法に比べてより客観的である。以上のことから、薬学的利用に必要とされる標準化が可能となる。
【0023】
本発明の方法を用いることによって、蠕虫卵調製物の生物学的活性を、医薬品に必要とされる精度で確実に測定することが可能となる。本発明に記載されているように、蠕虫卵の生物学的活性の正確な測定は、適用可能な医薬品を製造するために必須の工程である。
【図面の簡単な説明】
【0024】
【図1】定量PCRのためのプライマーおよびTaqmanプローブを含む、豚鞭虫のITS2配列のセグメントを示す図である。
【図2】アデノシン三リン酸を定量的に検出するためのルシフェラーゼ反応の反応式を示す図である。
【図3】ルシフェラーゼ反応を用いたアデノシン三リン酸の定量的検出によって示される冷凍による不活性化の効果を示すグラフである。
【図4】アデノシン三リン酸の検量線を示すグラフである。
【図5】豚鞭虫の蛋白質配列とセノラブディティス・エレガンス(Caenorhabhditis elegans)の相同蛋白質のアミノ酸配列とを示す図である。
【図6】in situハイブリダイゼーションに適したプローブの配列を示す図である。
【図7】運動性指数の結果を示すグラフである。
【図8】種々の観察時間における運動性指数を示すグラフである。
【図9】ローダミンXで標識したTSOと標識していないコントロールTSOとからなる混合物の蛍光顕微鏡写真および光学顕微鏡写真である。
【図10】in vitro活性スコアと感染率との相関図である。
【発明を実施するための形態】
【0025】
以下、本発明の方法の一部である5種の定量法をさらに詳細に説明する。
【0026】
a)ゲノムDNAのコピー数の測定
幼虫の発育期における細胞分裂では、常に、染色体対が2倍になり、2個の娘細胞に分配される。したがって、虫卵中のゲノムDNAのコピー数は、発育途中の幼虫の体細胞数と相関すると見なすことができる。したがって、ゲノムDNAのコピー数を測定することによって、発育途中である幼虫の発育状態を表すことができる。このことは、上記の方法(成熟率の顕微鏡測定:上記参照)と比較して明らかに有利である。上記の成熟率の顕微鏡測定では、約13週間にわたる成熟の最終結果しか分析できず、幼虫が実際に完全に発育したのか(すなわち、細胞数が、完全に形成されたL1幼虫に相当するのか)に関していかなる情報も得られない。幼虫を成熟させることは蠕虫卵の医薬品製造工程の一部であり、ゲノムDNAのコピー数を測定することにより、該製造工程に伴う分析が初めて可能となる。この測定によって、蠕虫卵の医薬品製造工程の論理的な開発、および該製造工程の変更がもたらす効果の分析が可能となるだけでなく、幼虫の成熟期において製造を定期的に監視することも可能となる。
【0027】
定量PCR技術により、試料中に存在する特定の核酸配列のコピー数を測定できる。試料中に存在する特定の核酸配列中に利用可能なコピーが多ければ多いほど、より迅速に増幅プラトーに達する。コピー数は、検量線を用いることによって比較的正確に測定することができる。本明細書において、コピー数を測定するのに通常使用する方法のうちの1つは、リアルタイムPCRまたはいわゆるTaqMan PCRである。この他の核酸増幅法も当業者には知られており、同様にゲノムDNAのコピー数を測定するために使用することができる。正確なコピー数は、通常、適切な検量線を用いて求める。
【0028】
好ましくは、リボソームRNAの一部をコードするITS1−5.8S−ITS2領域が、蠕虫のゲノムDNAの好ましいマーカーとして適切である。豚鞭虫(Cutillas et al., Parasitol Res 2001, 100: 383-389)、他の鞭虫類および他の蠕虫のITS1−5.8S−ITS2領域の配列は既に報告されている。IST−1領域の定量PCRを用いて豚回虫の幼虫の発育を監視する生存能力試験も既に報告されている(Pecson et al., Appl. Envir. Microbiol., 2006, 72, 7864-7872)。
【0029】
しかしながら、この試験には蠕虫の不活性化方法を評価するという別の目的がある。
【0030】
ITS1−5.8S−ITS2領域の一部の配列が増幅される定量PCR(q−PCR)は、マーカー領域のコピー数、ひいてはゲノムDNAのコピー数を測定するのに特に適している。ITS1およびITS2の構成は種特異的であるため、この測定方法は、成熟期の分析に加えて生物を定性的に識別するのにも適している。L1期の豚鞭虫の細胞数がゲノムのコピー数を指標に約1000個であることはこれまで公知ではなかったため、このような試験は先行技術では全く報告されていない(実施例1参照)。
【0031】
図1に、ゲノムDNAのコピー数の測定に用いることができる、豚鞭虫の適切な遺伝子配列を例として示す。配列番号:1に当該遺伝子配列を示す。フォワードプライマーを配列番号:2、リバースプライマーを配列番号:3、およびTaqmanプローブの配列を配列番号:4で表す。コピー数を測定するために、豚鞭虫のゲノムの他の部分も使用できることは当業者にとって明らかである。この測定法における、これらの配列の必要条件としては、豚鞭虫において特異的に見出されるヌクレオチド配列でなければならないこと、および分析試料に混入する可能性のある他の生物が有する配列と類似性がないことが挙げられる。
【0032】
さらに、ゲノムDNAのコピー数の測定には副次的効果がある。ゲノムDNAのコピー数を測定することによって、調製物中に所望の生物が存在することを確認できるとともに、他の適切な配列を使用して他の生物の混入の有無を証明することができる。
【0033】
ゲノムDNAのコピー数を測定する際に注意すべき重要な点として、測定前に蠕虫卵を破砕しなければならないことが挙げられる。実施例に示すように、本発明においては「ポッター型ホモジナイザー」が特に適していることが判明している。好ましい実施形態においては、容量が2mlのものを使用し、間隙幅を0.01〜0.03mmに設定した。5〜15分間ホモジネートすることが好ましく、約10分間が好ましい。それぞれの分析工程において、細胞成分が分析できる状態でなければならない場合は、必ず細胞を破砕する。
【0034】
b)代謝活性の測定
細胞の生存能力を測定する方法の多くが、代謝活性の分析に基づいている。細胞代謝において、アデノシン三リン酸はエネルギー伝達物質およびエネルギー貯蔵物質として中心的な役割を果たし、細胞内代謝活性を測定するためのマーカーとして使用できる。本発明では、豚鞭虫のL1幼虫のような、分化した生物の生存能力を測定するために、このヌクレオチドの検出も利用できることが示されている。
【0035】
ルシフェラーゼ反応を用いたルミネセンス測定により、生物系におけるアデノシン三リン酸の定量が可能となる。定義によれば、ルミネセンスは「冷たい」光の放出である。ルミネセンス系は、化学的、生化学的、または電気化学的な基質の活性化に基づいており、基質が基底状態に戻る時に励起エネルギーの一部を光の形態で放出する。アデノシン三リン酸の検出には、ホタル(フォチナス・フィラリス(Photinus pyralis))から単離したルシフェラーゼを使用する。この真核生物由来の酵素は、アデノシン三リン酸、酸素、およびマグネシウムイオンの存在下で、ルシフェリンのオキシルシフェリンへの酸化を触媒して光を放出させる。反応式を図2に示す。
【0036】
豚鞭虫の生存可能な幼虫包蔵卵中に存在するアデノシン三リン酸を成功裏に測定するための必要条件としては、L1幼虫におけるアデノシン三リン酸の合成を、ルミネセンスを測定するのに十分な程度まで刺激すること;適切なホモジネート法によって虫卵を十分に破砕し、細胞内で形成されたこのヌクレオチドを放出させること;および、複雑なマトリックス由来のアデノシン三リン酸を定量的にかつ何ら妨げなく検出することが挙げられる。これまで報告されておらずかつ驚いたことに、幼虫発育が中断される前のL1幼虫におけるATP含量は低く、特定の温度で長時間にわたってインキュベートして適切に活性化させることによって初めて、ATP含量が一定の高レベルにまで増加することが判明した。さらに、生きているL1幼虫と死んでいるL1幼虫とを識別するためには、死滅させた幼虫においてアデノシン三リン酸の基準値が再現性良く測定されるような効果的な不活性化方法が必要である。
【0037】
本発明によれば、蠕虫卵群の代謝活性を分析するためにATP含量を測定することが好ましい。
【0038】
さらに、個々の蠕虫卵の代謝活性を分析する方法として、テトラゾリウム化合物を用いた染色法も適している。この染色法は、もともと、細胞培養中または組織切片中の、何ら障害なく個々の細胞が染色可能な動物細胞のために開発されたものである。テトラゾリウム化合物は、代謝が活性化している細胞において、ミトコンドリア酵素の作用により有色のホルマザンに還元され細胞内に沈殿する。この染色法を、何ら障害なく個々の細胞が染色可能な動物細胞から虫卵に包蔵され多数の細胞からなるL1蠕虫幼虫へと応用するためには、虫卵をあらかじめ処理して、虫卵内の幼虫の生存能力に影響を与えることなく基質を浸透させることが極めて重要である。豚鞭虫のL1幼虫は強固な殻に取り囲まれているため、周囲との物質移動が著しく制限され、分析に必要とされる物質の浸透が阻まれる。
【0039】
穏やかに卵殻を分解するために、次亜塩素酸で虫卵を処理すると有利であることが判明している。この方法は既に報告されている(Beer, Parasitol., 1973, 67: 263-278)。次いで、必要に応じて、キチナーゼおよびプロテアーゼを用いた酵素消化を補助的に行うこともできる。
【0040】
代謝活性の測定を可能にするためには、「ゼロ値」を測定しなければならない。本明細書において、この値は対照値であり、この値をもとにそれぞれの代謝活性を測定する。好ましい実施形態においては、「ゼロ値」は不活性な蠕虫卵を用いて測定する。
【0041】
虫卵を不活性化する方法として、冷凍による不活性化(急速冷凍後、−80℃で24時間を超えて保存)が適していることが判明した。この方法で不活性化した試料からのアデノシン三リン酸のシグナルは、バックグラウンドノイズのレベルにまで減少させることがほぼ可能であった。冷凍による虫卵試料の不活性化は、リン酸緩衝生理食塩水中で行う。言うまでもなく、冷凍による不活性化は他の測定法においても実施することができる。
【0042】
図3に、ルシフェラーゼ反応を用いたアデノシン三リン酸の定量的検出によって示される、冷凍による不活性化の効果を例として示す。プレインキュベーションの効果は、虫卵を冷凍して不活性化することによって特異的に測定できる。不活性化した(死滅させた)蠕虫卵では、プレインキュベーションを行ってもアデノシン三リン酸の増加は確認できない。これに対し、生物学的に活性な蠕虫卵は、37℃で19時間前処理後にアデノシン三リン酸が増加したことによる活性を有している。
【0043】
好ましい一実施形態では、それぞれのバッチにつき、虫卵を5500個含む550μl溶液を使用する。プレインキュベーションの後、虫卵を遠心分離(500rpmで5分間)によって分離し、溶解緩衝液225μl中に再懸濁する。インキュベーションによって形成されたアデノシン三リン酸を含む細胞内容物を放出させるため、ポッター型ホモジナイザー(容量2ml;間隙幅0.01〜0.03mm)を用いて虫卵を10分間ホモジネートする。この方法を用いるだけで虫卵が十分に破砕され、このヌクレオチドが定量的に放出される。アデノシン三リン酸を開裂する加水分解酵素を不活性化するために、虫卵の破砕は溶解緩衝液(成分:pH2のリン酸)中で行う。それぞれのバッチから50μlをピペットで取り出し、96マイクロウェルプレートに入れて、さらにリン酸緩衝液100μlで希釈する。ルシフェラーゼ反応は、市販のアデノシン三リン酸キット100μlを加えて開始する。2分間インキュベートした後、マイクロウェルプレートをルミノメーターで読み取り、ルミネセンスを測定する(図3)。リン酸緩衝液中アデノシン三リン酸の検量線(1.0μM、0.1μM、10nM、1nM、0.1nM、10pM、1.0pM、0.1pM)より、定量化を行う。代表的なATPの検量線を図4に示す。
【0044】
冷凍により不活性化した幼虫包蔵卵をホモジネートした試料に、アデノシン三リン酸を意図的に加えることにより、検量線の直線領域(1〜1000nM)において84%の回収率を達成できた。したがって、この反応に関連してくる、虫卵ホモジネート中の複雑なマトリックスとルシフェラーゼ反応およびルミネセンスとの相互作用は認められない。
【0045】
実施例において、豚鞭虫の生存可能な幼虫包蔵卵と不活性化により生存不可能になった虫卵とを識別するために、ルミネセンス測定を使用できることが明確に示されている。しかしながら、この試験を成功裏に行うには、試料の活性化、試料のホモジネート、および最適化された不活性化法による適切なコントロールの作製からなる測定パラメータをそれぞれ最適化してこれらを組み合わせなければならない。生物学的に活性な蠕虫卵中のATP含量としては、虫卵1個当たり少なくとも0.01pmolが適している。
【0046】
c)誘導性遺伝子発現の分析
この方法の原理は、遺伝子発現の誘導である。遺伝子発現は生細胞でしか誘導することができないため、これにより生きている幼虫と死んでいる幼虫とを識別できる。さらに、遺伝子発現の誘導能は、休止状態のL1幼虫が種々の活性化状態を経ることを可能にするための必要条件であると考えられ、種々の活性化状態は生活環の次の段階(孵化、粘膜への定着など)を誘導するのに必要である。
【0047】
熱ショック蛋白質は、温度を上昇させることにより容易に誘導することができ、形成されるメッセンジャーRNA(mRNA)が急速に分解せずに概ね安定していることから、熱ショック蛋白質の発現を分析することは特に有利であると考えられる。本発明の範囲内で、豚鞭虫の遺伝子配列の検出に初めて成功した。この豚鞭虫の遺伝子配列は、蠕虫であるセノラブディティス・エレガンス(Caenorhabhditis elegans)の熱ショック蛋白質hsp70と非常に相同性が高い。
【0048】
本発明の好ましい一実施形態では、豚鞭虫の熱ショック蛋白質の遺伝子発現を分析する。図5に、豚鞭虫の蛋白質配列(配列番号:5)を、セノラブディティス・エレガンスの相同蛋白質のアミノ酸配列(配列番号:6)と比較して示す。
【0049】
誘導性遺伝子発現の測定は、特定の誘導蛋白質をコードするメッセンジャーRNAの量を測定することにより行うことができる。メッセンジャーRNAのヌクレオチド配列は、この蛋白質のアミノ酸配列由来とすることができる。適切なフォワードプライマーおよびリバースプライマーを設計し、これと同時に、Taqmanプローブの配列を決定することは当業者とって問題ではない。次いで、遺伝子の誘導量をリアルタイムPCRにより測定することができる。意味のある測定を行うための必要条件としては、死滅させた虫卵を含むコントロール試料も測定することが挙げられる。さらなる必要条件としては、測定に用いる蛋白質が容易かつ確実に誘導され得るものでなければならないことが挙げられる。このような蛋白質としては熱ショック蛋白質が挙げられるが、特定の刺激によって誘導できる他の遺伝子も同様に測定に使用できる。
【0050】
個々の虫卵レベルで発現させたメッセンジャーRNAも、適切な遺伝子プローブを用いた蛍光in situハイブリダイゼーション(FISH)によって検出できる。次いで、この虫卵群に対して、フローサイトメトリーによる迅速かつ確実な分析を行うと特に有利である。
【0051】
in situハイブリダイゼーションに適したプローブの配列を図6(配列番号:7)に示す。
【0052】
d)完全な形のままの蠕虫卵内に包蔵される幼虫の運動性の測定
この方法の原理は、虫卵内のL1幼虫の機能性に関するパラメータとしての虫体の運動の検出に基づく。幼虫の運動性は、幼虫が適切な環境条件下で孵化するための必要条件である。C.エレガンスのような自由生活性の幼虫および寄生虫においては、運動性の分析は生存能力および機能性を確認するための確実な方法として有効である。
【0053】
本発明の範囲内で、驚くべきことに、厳密に環境温度を調節することによって、自由生活をせず虫卵内に包蔵されている蠕虫幼虫でも微細な運動を誘導できることが、豚鞭虫を例として初めて判明した。豚鞭虫の幼虫は、成熟が完了した後休止状態に陥り、適切な宿主によって経口摂取されるまで活性化および孵化しないことがこれまで報告されていた。虫卵内の幼虫の運動は非常に鈍く、本発明で開示されているように、長時間にわたり経時的に顕微鏡下で虫卵を観察しなければ検出できない。このような顕微鏡下での運動性試験は、画像分析ソフトウェアと組み合わせて自動化することができる。虫卵の生物学的活性のパラメータとしての運動性指数は、次式で計算する:
【数1】
【0054】
幼虫の運動性の測定を成功裏に行うには、温度を厳密に管理することが不可欠である。虫卵を、まず初めに、36〜42℃、好ましくは37〜41℃、特に好ましくは39.5℃に正確に設定した温度で、2〜30時間、好ましくは4〜20時間プレインキュベートする。このプレインキュベーションの後、上記の設定温度を少なくともほぼ維持できる適切な顕微鏡下に幼虫をセットする。次いで、虫卵を、36〜42℃、好ましくは38〜40℃で、2分〜4時間、好ましくは30分〜2時間にわたり経時的に記録して観察する。
【0055】
本発明に従って測定された運動性指数の結果を図7に示す。不活性化した虫卵では運動性は全く観察できないが、これとは対照的に活性な虫卵では運動性指数が時間とともに増加している。
【0056】
種々の観察時間における運動性指数を図8に示す。
【0057】
e)腸管内での孵化率の測定
この測定法の原理は、実験動物の腸管内あるいは実験動物からあらかじめ回収した腸管内容物内に含まれる蠕虫幼虫の孵化率の測定である。
【0058】
一実施形態では、実験動物から回収した腸管内容物を用いて孵化率を測定することができ、実際には、好ましくは結腸の始端または十二指腸の末端から腸管内容物を回収することが好ましい。次いで、腸管内容物を用いて孵化率を測定することができ、実験動物を用いて直接測定する必要はない。さらに、腸管を通過して糞便から回収された虫卵を分析することにより、この方法を実施することもできる。この場合、実験動物を屠殺する必要がなく、孵化率を測定するために実験動物を数回使用することができるという利点を有する。
【0059】
新規に開発されたこの測定法では、接種後、比較的短時間のうちに腸管内容物が分析され、孵化の後に残った幼虫被膜だけでなく完全な形のままの虫卵も集計される。先行技術において報告されている感染率の測定と比較した際の重大な利点としては、本発明の孵化率の測定は、個々の宿主が有する要因にはわずかにしか影響を受けない、生活環の第1段階のみを分析することが挙げられる。
【0060】
驚いたことに、適切な宿主としてブタだけでなくウサギにおいても、定量的に幼虫が孵化することを豚鞭虫について初めて示すことができた。したがって、たとえば豚鞭虫に対しては、腸の容積が小さい実験動物を利用することが可能であり、これにより顕微鏡レベルの小さな虫卵および幼虫被膜を回収することが極めて容易となる。
【0061】
驚いたことに、蛋白質を標識するために開発された市販の蛍光色素を用いて卵殻を永久的に染色できることが判明した。蛍光色素と幼虫被膜とを結合させることにより、蛍光顕微鏡法を用いて腸管内容物を分析することができるので、虫卵の回収が非常に容易となる。
【0062】
さらに重要な技術革新としては、蠕虫の未成熟卵または不活性化卵を固定の内部標準として使用することが挙げられる。このような虫卵は、生物学的に活性な幼虫包蔵卵と同一の腸内移行時間すなわち滞留時間を有するが、生物学的に活性な幼虫包蔵卵とは対照的に、腸管を通過するときも完全な形を保ったままである。内部標準として提供される虫卵を別の(異なる)蛍光色素で標識することによって、内部標準と分析対象の虫卵とを容易に識別することができる。
【0063】
本分析では、腸管内容物に含まれる、分析試料中の蛍光色素1で標識された完全な形のままの虫卵数および空の幼虫被膜数と、蛍光色素2で標識された内部標準である完全な形のままの虫卵数とを接種後の適切な時点において分析する。腸管内の孵化率は、完全な形のままの(孵化していない)虫卵を基準として、次式で計算することができる。
【数2】
【0064】
PCR反応、好ましい測定法、およびこれらより得ることができる値を下記の表2にまとめる。これらの値は、分析した調製物が、必要とされる生物学的活性を有するのかどうかを示す好ましい制限値を表す。
【0065】
【表2】
【0066】
好ましい実施形態において、本発明の新規の試験方法は、in vitroで成熟させた蠕虫卵の温度誘導性活性化能を測定することを含む。また、相補的な2種のパラメータ(運動性指数および平均ATP含量)を測定する。これらのパラメータは相俟ってin vitro活性スコアを構成する。in vitro活性スコアは、in vivoにおける生物学的活性と良い相関を示す。しかしながら、in vitro活性スコアから生物学的活性を予測することができるのは、使用する幼虫が完全に成熟している場合のみである。幼虫が完全に成熟しているかどうかは、虫卵内の豚鞭虫のゲノムのコピー数を測定する定量PCRを用いることによって分かる。さらに、並行して行われる新規のin vivoにおける孵化試験を用いることにより、分析した虫卵が生理学的条件下で実際に活性であることが保証される。以下、好ましい方法を概略的に示す:
【0067】
【0068】
本発明の重要な態様として、in vitroにおける温度誘導性活性化能を測定することが挙げられる。
【0069】
成熟後のL1−TSOは、何年にもわたって生存可能な休止状態となり、宿主による経口摂取後に活性化されて孵化するまでその状態を保つ。これまで報告されておらずかつ予想外にも、他の因子を加えなくても特定の温度範囲内で一定時間インキュベートするだけで、in vitroでも虫卵を容易に活性化できることが判明した。虫卵の活性化状態は、2種の相補的パラメータ、すなわち、完全な形のままの虫卵内の幼虫の運動性と、あらかじめ活性化した虫卵の溶解物中のATP含量とを測定することにより定量化する。文献により報告されている挙動に反しかつ予想外にも、本明細書に記載のプレインキュベーションにより虫卵内の幼虫の微細な運動が誘導される。この幼虫の運動は、幼虫が死滅することも虫卵から孵化することもなく、何時間にもおよび継続する。したがって、本明細書に記載された方法の好ましい実施形態として、活性化を引き起こす虫卵の適切なプレインキュベーションが挙げられる。活性および運動性を有する幼虫(運動性指数)を含む虫卵群の比率は、十分に多数の虫卵を顕微鏡下で経時的に観察することにより、確実に測定することができる。しかしながら、この測定では個々の幼虫の活性状態すなわち運動性/活性レベルの違いが識別されないため、分析対象の虫卵群の総活性を示す定量的な指標であるATP含量も併せて測定する。直接的であるが不連続な変数としての運動性指数と、間接的であるが連続的な変数としてのATP含量の2種の分析パラメータが互いに補い合うことによって、in vivo活性とよく相関するin vitro活性が確実に決定される。これら2種のパラメータから計算することができる、in vitro活性スコアは、虫卵のin vivo活性と良く相関する。
【0070】
本発明のin vitroにおける方法は、従来公知の方法とは明らかに異なる。虫卵内の完全に形成された幼虫が虫卵内に存在するかどうかを視覚的にしか分析しない成熟率の測定(Kringelら)では、本明細書に記載の方法とは対照的に、幼虫の生存能力または活性に関していかなる結論も得られない。また、消化菅を模擬したin vitroでの孵化率の測定は、本明細書に記載の方法と比較すると、非常に煩雑であり干渉を受けやすい。このin vitroにおける孵化試験に関しては、in vivo活性との相関性も報告されていない。
【0071】
本発明の好ましい実施形態では、幼虫の成熟状態が決定される。in vitro活性スコアは、幼虫が幼虫発育を完全に経た場合のみ、in vivoにおける状況を予測するのに役立つ意味のある結果を提供する。温度誘導性活性化能試験において、完全には発育していない幼虫でも陽性反応が示されることがあるが、これらの幼虫は機能性を持たず、宿主内で孵化することも幼虫の正常な生活環をたどることもできないと考えられる。従来公知の方法では、形態学的基準を用いて幼虫が十分に発育していることを顕微鏡下で評価する。この評価法は、幅広い経験が必要とされ、信頼性が低く、その性質上個々の虫卵ごとについてしか評価することができない。これとは対照的に、本発明による方法では、多数の虫卵を含む群のゲノムのコピー数を客観的に測定し、幼虫1匹当たりの平均体細胞数を計算することができる。
【0072】
温度誘導性活性化能試験の結果は、生理学的条件下において孵化率を容易かつ迅速に、しかも確実に測定できる新規のin vivo試験で保証することが好ましい。驚いたことに、あらかじめ不活性化した虫卵は腸管内で生存できるがそのまま変化しないのに対し、生物学的に活性な虫卵内の幼虫は腸管内において定量的に孵化することが判明した。新規に開発された方法では、調製物に含まれる活性なTSOの比率は、実験動物の腸管内を変化しないまま通過した不活性な虫卵のみを集計することよって間接的に求められる。しかしながら、この定量化を成功裏に行うには、蛍光で標識されかつ変化しないまま腸管内を通過する、不活性な虫卵からなる新規に開発された内部標準を使用しなければならない。さらに、驚いたことにかつ予想外に、生物学的に活性なTSOはウサギの腸管内でも孵化することが判明した。したがって、費用効果がより高く、飼育するのが容易で、かつ有利なことに腸の容量が極めて小さい(顕微鏡レベルの小さな虫卵を回収することが容易となる)動物モデルを、ブタの代わりに使用することができる。
【0073】
本発明によるin vivo試験と、ブタを用いた公知の感染率測定試験とは異なるものである。ブタを用いた感染率測定試験は数週間にわたって継続するのに対し、孵化率の測定に必要な日数はわずか1〜3日である。さらに、孵化率を測定する場合、宿主内における豚鞭虫の生活環うちの第一期のみが分析され、個々の宿主が有する要因にはほとんど影響を受けない。これとは対照的に、ブタを用いて感染率を測定する場合は、感染させてから3〜4週間後に測定することが好ましく、個々の宿主においてそれぞれ異なる免疫システムが形成されることを主たる原因とする、自然な生物学的変動に影響を受ける。
【0074】
本明細書に記載の分析法のさらなる利点としては、動物の糞便から虫卵を回収できることが挙げられ、これによって動物を試験のために屠殺する必要がなくなる。
【実施例】
【0075】
実施例1:
豚鞭虫のITS2配列に基づき、TaqManシステムを用いた定量PCR法を開発した。標的配列、プライマー、およびTaqmanプローブを以下に示す。遺伝子配列が公知である他種の鞭虫と豚鞭虫とを識別できるように標的配列を選択した。このようにすると、ITS2コピー数に関する情報が得られるだけでなく、分析した生物が実際に豚鞭虫であったことも定性的に証明できる。
【0076】
ポッター型ホモジナイザー(容量2ml;間隙幅0.01〜0.03mm)で虫卵(500μl溶液中1000個の虫卵)を10分間ホモジネートして、試料を調製する。驚いたことに、顕微鏡下で観察すると、遺伝子を単離する際に通例用いられる他の方法では豚鞭虫卵を破砕できないのに対して、上記の方法を用いると幼虫被膜を破損させることが可能であることが分かる。魚精子DNA1.54μgを加えた後、遺伝子単離用の市販キット(DNeasy−Blood and Tissue−Kit:キアゲン社)をメーカーの説明書に従って用い、虫卵ホモジネートと反応させる。下表に示すプロトコルに従い、単離したDNA(全量50μl)を用いてPCRを行う。
【0077】
【表3】
【0078】
PCRの増幅効率は92〜98%であり、これは虫卵1個に含まれるITS2遺伝子のコピー数を決定するのに十分な感度である。
【0079】
PCRを用いて、豚鞭虫の未成熟卵および完全に成熟した幼虫包蔵卵のITS2コピー数を分析した。結果を以下の表に示す。
【0080】
【表4】
【0081】
PCR解析の結果、幼虫を包蔵したTSOのITS2コピー数が、未成熟TSOのITS2コピー数の約1000倍であることが示された。未成熟卵(L0)は単一の細胞と見なすことができるので、この結果より、豚鞭虫のL1幼虫は約1000個の体細胞を有することが分かる。豚鞭虫のL1幼虫の体細胞数はこれまで知られていなかった。近縁種であるC.エレガンスについては、体細胞数が959であることが判明しており、この数は、本実施例において判明した豚鞭虫の体細胞数である約1000個とよく一致する。
【0082】
この方法により、幼虫発育期の相違を分析できるかどうかを確認するために、TSOを4週間成熟させ、ITS2 PCR(表5)で定期的に分析した。
【0083】
【表5】
【0084】
本実施例において、この方法を用いて、発育期がL0の幼虫と発育期がL1の幼虫とを体細胞数により識別できることが明らかに示された。したがって、この方法は、製造工程に伴う幼虫の成熟過程の分析に特に適している。蠕虫卵調製物を製造する場合、幼虫を成熟させ始めてから最初の28日間に50〜1200の相対コピー数を達成するように規定すると特に好ましいと考えられる。
【0085】
実施例2:
アデノシン三リン酸を検出するために、反応に必要なルシフェラーゼ酵素、基質であるルシフェリン、マグネシウム塩などを含む市販キットを使用することができる。幼虫包蔵卵をプレインキュベートしてアデノシン三リン酸の形成を刺激することは必須である。生存可能な幼虫包蔵卵が休止状態である場合、アデノシン三リン酸含量から、生存可能な幼虫包蔵卵と死んでいる虫卵とを十分に識別することはできない(図3)。図3の結果より、虫卵を37℃で19時間にわたりインキュベートする方法が適すると判明した。開発した分析法の重要な利点として、このインキュベーションに際して試料を調製する必要がないことが挙げられる。本発明の虫卵懸濁液は、培地の定性的および定量的組成とは無関係にそのまま使用することができる。pH価が2未満の強酸性培地でさえも試料を調製することなく使用することができ、虫卵中のアデノシン三リン酸の形成を刺激するのに何ら悪影響を与えることはない。インキュベートすることにより、生きているF1幼虫と死んでいるF1幼虫とを明確に識別することができる(図3)。
【0086】
実施例3:
セノラブディティス・エレガンスに見出される熱ショック蛋白質hsp70の配列と、ネズミ鞭虫(Trichuris muris)の相同配列と、犬鞭虫(Trichuris vulpis)の相同配列とに由来するプライマーを用いて、豚鞭虫から単離したRNAの遺伝子配列を増幅することができた。豚鞭虫由来のこの新規の配列は、蛋白質配列に翻訳すると、C.エレガンスのhsp70蛋白質の蛋白質配列と65%一致する(図5)。
【0087】
豚鞭虫の熱ショック蛋白質と推定される蛋白質を検出するための定量RT−PCR法を開発した。この定量は、恒常的に発現される18S rRNAと比較して行った。豚鞭虫の活性な幼虫包蔵卵に熱ショック(45℃で20分間)を与え、処理していない活性な豚鞭虫と熱ショック蛋白質の発現量を比較した。
【0088】
【表6】
【0089】
相対的な発現量を分析した結果、hsp70に類似したmRNAの発現を熱ショックによって誘導できることが明らかに示された(表6)。したがって、適切な試験条件を選択することにより能動的に発現を誘導できる遺伝子は、当該活性試験に利用することができる。
【0090】
個々の虫卵における遺伝子発現の活性化は、蛍光in situハイブリダイゼーション法(FISH)を用いて検出できる。熱ショック蛋白質と推定される蛋白質を検出できるように、たとえば図6に示す、蛍光で標識したDNAプローブを作製した(配列番号:7)。
【0091】
活性な虫卵を蛍光で標識することにより、虫卵群もフローサイトメトリーを用いて分析することができる。この分析は、顕微鏡による分析よりも、迅速かつ客観的に実施できる点で有利である。
【0092】
本実施例により、本明細書に記載の方法を用いて(個々の)蠕虫卵における遺伝子発現の誘導能を分析できることが明らかに示された。したがって、休止状態のL1蠕虫幼虫の活性化能を分析できる方法が初めて利用可能となった。
【0093】
蠕虫卵調製物を製造する際に、その活性を生物学的に証明するために遺伝子発現を誘導する場合、判定基準として、蠕虫卵調製物に誘導性遺伝子発現陽性の蠕虫卵が25〜100%、好ましくは50〜100%含まれていると適している。
【0094】
実施例4:
4.1 運動性指数の分析条件に対する依存性
活性な豚鞭虫卵と、48℃で72時間インキュベートして不活性化した豚鞭虫卵とを、39℃で11時間インキュベートした。1時間毎にそれぞれの虫卵を5分間観察し、運動性を測定した。運動性指数の経時変化を図7に示す。
【0095】
本実施例により、休止状態の幼虫を活性化し、かつ一定した高い運動性指数を測定するためには、高温で比較的長時間(本実施例では39℃で7時間)のプレインキュベーションが必要であることが明らかとなった。これまで、虫卵に包蔵され完全に成熟したL1幼虫は運動しないとされていた(Parasitology 1973, 67: 253-262)ため、この結果は予想外であった。長時間におよぶインキュベーションが必要とされ、経時的に観察することにより初めて鈍い運動が見られただけだったため、虫卵内の幼虫の運動性は恐らくこれ以前に観察されたことがなかったと考えられる。
【0096】
活性な虫卵と不活性化された虫卵とを比較して運動性を分析することによって生物学的に活性な豚鞭虫卵と不活性な豚鞭虫卵とを容易に識別できることが明らかとなった。
【0097】
幼虫は同期して運動しないため、長時間にわたって観察することにより生きている虫卵内の散発的な運動を検出できる可能性が増加する。したがって、活性化段階と並んで、観察時間の長さも運動性指数の測定に影響を及ぼす。このことは、活性化段階(37℃で8時間)後に豚鞭虫卵調製物を39.5〜40℃で様々な時間観察した分析(図7)により示された。さらに、この分析により、8時間にわたる観察時間を通して運動性は安定していることも示された。
【0098】
したがって、この方法は、厳密に温度が管理された活性化段階と長時間におよぶ観察時間とを新規の特徴とする。実施例4により、虫卵内で休止状態のL1幼虫の運動を、適切な温度および観察時間からなる条件によって、誘導かつ測定できることが示された。この方法を使用すると、孵化の必要条件として、ひいては虫卵の生物学的活性のパラメータとしての運動性を容易に測定でき、活性なL1幼虫と不活性なL1幼虫とを明確に識別することができる。
【0099】
4.2 有利な試験条件
以下に挙げる試験条件が有利であると判明した。リン酸緩衝液(pH7.4)300μl中に15,000個の蠕虫卵を含む試料を、96ウェルプレート(底面積:0.31cm2)へ移す。この播種密度においては、200倍の倍率で約80〜150個の虫卵が顕微鏡の視野内に入る。37℃で8時間インキュベーションした(活性化段階)後、温度を39.5℃まで上げて、連続した4つの観察野を選択し、それぞれ2時間観察する。この観察を行うため、フィルムに毎分3画像を記録する。次いで、虫卵数と動いている幼虫数とを測定し、運動性指数を計算する。4観察野の平均値から、試料の運動性指数を得る。
【0100】
4.3 再現性および精度−感染率測定試験との比較
上記の条件下における運動性試験の再現性を調べるために、4回の実験をそれぞれ異なる日に実施し、1回の実験につき4種の豚鞭虫試料を分析した。測定結果を以下の表に示す。比較として、同一の豚鞭虫試料を用いて感染率測定試験を行った、4回の実験の結果を示す。
【0101】
【表7】
【0102】
運動性指数の4つの独立した測定実験では、最大で互いに4.2%の差異があるのに対し、感染率の4つの独立した測定実験での最大偏差は35.5%である。運動性試験での単回測定の精度は1.9〜6.3%であるのに対し、感染率測定試験での単回測定の精度は9.6〜58.4%である。したがって、運動性試験は、再現性および精度に関して感染率測定試験よりもはるかに優れている。
【0103】
4.4 運動性と生物学的活性(正確性)との相関−感染率測定試験との比較
運動性と生物学的活性との相関を分析するために、活性なTSOと加温により不活性化したTSOとからなる混合物を作製し、4.2に記載した条件に従った運動性試験により分析した。加温による不活性化は、虫卵を48℃で72時間加温することにより行った。客観的な集計結果を得るために、盲検法にて試料を測定した。
【0104】
相対生物学的活性は、測定した試料の運動性指数値とあらかじめ測定した活性なTSOの運動性指数値より計算された比率である。測定の正確性は、相対生物学的活性と試料中の活性な虫卵の実際の比率とを比較することにより求められる。
【0105】
上記と同一のTSO試料を用いた同様の測定実験を、上記の感染率測定試験についても行った。実験動物は、感染率が既知である活性なTSOを種々の量で感染させた。不活性化した虫卵との混合物は使用しなかった。結果を以下の表に示す。
【0106】
【表8】
【0107】
【表9】
【0108】
本明細書で示される運動性試験を用いることによって、TSO試料の相対生物学的活性は、0〜100%の全範囲にわたる期待値の10%未満の偏差で正確に測定することができる。この測定の精度(測定値の分散)は、相対生物学的活性が50〜100%の範囲において5%未満である。相対生物学的活性が低い場合、測定の精度は15%未満である。
【0109】
感染率測定試験の結果は、0〜100%の範囲にわたる相対生物学的活性の期待値から約25〜32%逸脱する。この測定値における分散は28〜80%である。
本実施例により、運動性指数が相対生物学的活性と直線的に相関することが明らかに示された。運動性試験は、正確性と精度に関して感染率測定試験よりはるかに優れている。
【0110】
蠕虫卵調製物の生物学的活性を測定するために運動性試験を実施する場合、運動性指数が30〜100%、好ましくは60〜100%であると蠕虫卵調製物として適している。
【0111】
実施例5:
一例として、幼虫包蔵卵と未成熟卵と不活性化した幼虫包蔵卵とからなる種々の混合物をウサギに経口投与した。8時間後、ウサギの腸管内容物を顕微鏡下で分析し、完全な形のままの虫卵数と空の幼虫被膜数とを集計した。本分析では虫卵を蛍光標識しなかったため、完全な形のままの幼虫包蔵卵と未成熟卵との比率により孵化率を計算した([IE]=幼虫包蔵卵数、[IS]=未成熟卵数として、上記の式を用いた)。分析結果と分析結果より計算した孵化率とを以下の表に示す。
【0112】
【表10】
【0113】
第1〜3群のデータを比較すると、活性な幼虫包蔵卵しかウサギの体内で孵化できなかったことが示された(完全な形のままの幼虫包蔵卵数が顕著に減少したこと、および空の幼虫被膜が多数存在したことにより確認できた)。これとは対照的に、不活性な幼虫包蔵卵は腸管内で変化しなかった(第2群)。このことにより、この方法を使用した場合、生物学的に活性な虫卵と不活性化した虫卵とは識別可能であることが明らかに証明された。さらに、未成熟卵も腸管内を通過する際に完全な形を保ったままであった(第3群)ため、分解されずに実験動物の腸管内を通過する内部標準として未成熟卵を使用することは可能である。虫卵は腸管内容物から完全には回収できず、回収した完全な形のままの虫卵の絶対数が意味を持たなくなることから、この内部標準は必要である。上記の式は、内部標準を考慮に入れて孵化率を計算するために使用する。上記の式を用いて計算すると、本実施例において分析した豚鞭虫卵は67.9%の孵化率を示したのに対し、不活性化した豚鞭虫卵は0%(数学的には−12.6%)の孵化率を示した。
【0114】
より多くの虫卵を回収し、試料である虫卵と内部標準である虫卵とをより明確に識別するために、蛍光プローブを共有結合させて豚鞭虫卵を標識した。メーカー仕様書によると蛋白質中の一級アミノ基と反応することができる、ローダミンXスクシンイミジルエステルを一例として蛍光プローブとして使用した。1000個の豚鞭虫卵を含むリン酸緩衝液(pH7.4)懸濁液200μlに、1M炭酸水素ナトリウム溶液20μlを加え、次いで、0.5%ローダミンXスクシンイミジルエステルのDMSO溶液5μlと反応させた。反応液を回転振盪しながら室温で1時間インキュベートした。次いで、遠心分離(500rpmで10分間)および上清緩衝液の交換によって、虫卵を合計8回精製した。蛍光顕微鏡法により、赤色に染色された幼虫被膜がはっきりと示された(図9)。
【0115】
蛍光標識すると、顕微鏡下で孵化率を評価するのに有利となる。さらに、蛍光活性化フローサイトメトリーを用いると、客観的かつ迅速な分析も可能となる。
【0116】
概していえば、本実施例に記載の方法を用いると孵化段階を定量的に分析することができ、この孵化段階を分析することにより生物学的活性を測定できることが本実施例において明らかに示された。蠕虫卵調製物の生物学的活性を測定するために孵化試験を実施する場合、孵化率が約25〜100%であると蠕虫卵調製物として適している。
【0117】
実施例6:
in vitroにおける温度誘導性活性化能とin vivoにおける生物学的活性との相関
温度誘導性活性化能と生物学的活性との相関性を調べるために、種々の性質を有する12種のTSO試料を分析した。まず、これらの試料の幼虫が完全に発育しているのかをPCRを用いて評価した。次いで、加温により活性化を誘導した後、運動性指数とATP含量とを測定した。これらと並行として、12種の試料それぞれに対してブタを用いた感染率測定試験を行った。感染率測定試験は、Kringelらによって報告された方法に従い、それぞれの試料に対して5匹のブタを用いて行った。運動性とATP含量の2種のin vitroパラメータを、in vitro活性スコアとしてまとめた:
【数3】
【0118】
【表11】
【0119】
in vitro活性スコアと感染率との相関を図10に示す。
【0120】
本実施例において、in vitro活性スコアはin vivoにおける生物学的活性と良く相関することが示された。個々の試料におけるわずかな偏差は、恐らく、生物学的変動(宿主が有する要因)により本質的な不安定さを抱えているという、このin vivo試験の弱点にむしろ起因すると考えられる。
【0121】
感染率測定試験を利用してプローブを測定するには、6週間かかり、5匹のブタが必要とされ、相当な人件費が伴う。本明細書に記載された5種の方法のうちの3種からなる、本実施例に記載のin vitro試験を行うのには、1〜2日間しかかからず、人件費および材料費も比較的少なくて済む。
【特許請求の範囲】
【請求項1】
鞭虫の幼虫包蔵卵の生物学的活性を測定する方法であって、
a)ゲノムDNAのコピー数を確認するのに適したマーカー配列を使用した定量PCR分析による蠕虫卵の幼虫発育期の判定および/または確認、
b)生化学的方法および/または分子生物学的方法を用いた蠕虫の幼虫包蔵卵の代謝活性の測定、
c)蠕虫の幼虫包蔵卵における遺伝子発現の誘導能の測定、
d)高温でのプレインキュベーション後における、虫卵内に包蔵される蠕虫幼虫の運動性の、長時間観察による顕微鏡測定、および/または
e)内部標準を基準として、腸管内容物から回収した完全な形のままの幼虫包蔵卵を定量化する、実験動物における鞭虫の幼虫孵化率の測定
のうち少なくとも1種の測定を行うことを特徴とする方法。
【請求項2】
豚鞭虫に適した特異的配列を使用した定量PCR分析によりゲノムDNAのコピー数を測定することを特徴とする、請求項1a)に記載の方法。
【請求項3】
豚鞭虫の幼虫包蔵卵の代謝活性を測定するためにATP含量を測定することを特徴とする、請求項1b)に記載の方法。
【請求項4】
ルミネセンス測定の前に、
aa)36〜42℃
bb)2〜30時間
cc)pH0.1〜3の懸濁培地中
の条件下で鞭虫卵をプレインキュベートすることを特徴とする、請求項3に記載の方法。
【請求項5】
鞭虫卵をまず、次亜塩素酸、キチナーゼ、および/またはプロテアーゼから選択される前処理剤で処理し、次いでテトラゾリウム塩で染色することを特徴とする、請求項1b)に記載の方法。
【請求項6】
熱ショック蛋白質の誘導能を測定することを特徴とする、請求項1c)に記載の方法。
【請求項7】
前記発現を、蛍光標識されたヌクレオチドプローブを用いたハイブリダイゼーションにより検出することを特徴とする、請求項1c)に記載の方法。
【請求項8】
前記ハイブリダイゼーションをフローサイトメトリーにより検出することを特徴とする、請求項1c)に記載の方法。
【請求項9】
2分〜8時間にわたって経時的に記録することにより、虫卵内の蠕虫幼虫の運動性を顕微鏡下で測定することを特徴とする、請求項1d)に記載の方法。
【請求項10】
試験対象の蠕虫卵を蛍光プローブで標識し、内部標準を異なる色の蛍光プローブで標識することを特徴とする、請求項1e)に記載の方法。
【請求項11】
ウサギおよび/またはブタの腸管内容物を試験系として使用することを特徴とする、請求項1e)に記載の方法。
【請求項12】
前記孵化率を確認するために、鞭虫の未成熟卵または不活性化卵を内部標準として使用することを特徴とする、請求項1e)に記載の方法。
【請求項13】
1a)、1b)、1c)、1d)、または1e)から選択される少なくとも3種の測定を行うことを特徴とする、先行する請求項のいずれか1項に記載の方法。
【請求項14】
測定を行う前に、厳密に標準化された条件下で少なくとも30分〜24時間にわたって鞭虫卵をプレインキュベートし、このプレインキュベーションにおいて、必要であれば、必須パラメータ、特に温度を変更できることを特徴とする、先行する請求項のいずれか1項に記載の方法。
【請求項15】
請求項1〜14のいずれか1項に記載の方法により測定された鞭虫卵のうち少なくとも50%が生物学的活性を有することを特徴とする鞭虫卵。
【請求項16】
自己免疫疾患治療用医薬品を製造するための請求項15に記載の鞭虫卵の使用。
【請求項1】
鞭虫の幼虫包蔵卵の生物学的活性を測定する方法であって、
a)ゲノムDNAのコピー数を確認するのに適したマーカー配列を使用した定量PCR分析による蠕虫卵の幼虫発育期の判定および/または確認、
b)生化学的方法および/または分子生物学的方法を用いた蠕虫の幼虫包蔵卵の代謝活性の測定、
c)蠕虫の幼虫包蔵卵における遺伝子発現の誘導能の測定、
d)高温でのプレインキュベーション後における、虫卵内に包蔵される蠕虫幼虫の運動性の、長時間観察による顕微鏡測定、および/または
e)内部標準を基準として、腸管内容物から回収した完全な形のままの幼虫包蔵卵を定量化する、実験動物における鞭虫の幼虫孵化率の測定
のうち少なくとも1種の測定を行うことを特徴とする方法。
【請求項2】
豚鞭虫に適した特異的配列を使用した定量PCR分析によりゲノムDNAのコピー数を測定することを特徴とする、請求項1a)に記載の方法。
【請求項3】
豚鞭虫の幼虫包蔵卵の代謝活性を測定するためにATP含量を測定することを特徴とする、請求項1b)に記載の方法。
【請求項4】
ルミネセンス測定の前に、
aa)36〜42℃
bb)2〜30時間
cc)pH0.1〜3の懸濁培地中
の条件下で鞭虫卵をプレインキュベートすることを特徴とする、請求項3に記載の方法。
【請求項5】
鞭虫卵をまず、次亜塩素酸、キチナーゼ、および/またはプロテアーゼから選択される前処理剤で処理し、次いでテトラゾリウム塩で染色することを特徴とする、請求項1b)に記載の方法。
【請求項6】
熱ショック蛋白質の誘導能を測定することを特徴とする、請求項1c)に記載の方法。
【請求項7】
前記発現を、蛍光標識されたヌクレオチドプローブを用いたハイブリダイゼーションにより検出することを特徴とする、請求項1c)に記載の方法。
【請求項8】
前記ハイブリダイゼーションをフローサイトメトリーにより検出することを特徴とする、請求項1c)に記載の方法。
【請求項9】
2分〜8時間にわたって経時的に記録することにより、虫卵内の蠕虫幼虫の運動性を顕微鏡下で測定することを特徴とする、請求項1d)に記載の方法。
【請求項10】
試験対象の蠕虫卵を蛍光プローブで標識し、内部標準を異なる色の蛍光プローブで標識することを特徴とする、請求項1e)に記載の方法。
【請求項11】
ウサギおよび/またはブタの腸管内容物を試験系として使用することを特徴とする、請求項1e)に記載の方法。
【請求項12】
前記孵化率を確認するために、鞭虫の未成熟卵または不活性化卵を内部標準として使用することを特徴とする、請求項1e)に記載の方法。
【請求項13】
1a)、1b)、1c)、1d)、または1e)から選択される少なくとも3種の測定を行うことを特徴とする、先行する請求項のいずれか1項に記載の方法。
【請求項14】
測定を行う前に、厳密に標準化された条件下で少なくとも30分〜24時間にわたって鞭虫卵をプレインキュベートし、このプレインキュベーションにおいて、必要であれば、必須パラメータ、特に温度を変更できることを特徴とする、先行する請求項のいずれか1項に記載の方法。
【請求項15】
請求項1〜14のいずれか1項に記載の方法により測定された鞭虫卵のうち少なくとも50%が生物学的活性を有することを特徴とする鞭虫卵。
【請求項16】
自己免疫疾患治療用医薬品を製造するための請求項15に記載の鞭虫卵の使用。
【図2】

【図5】

【図6】

【図1】
【図3】
【図4】
【図7】
【図8】
【図9】
【図10】
【図5】
【図6】
【図1】
【図3】
【図4】
【図7】
【図8】
【図9】
【図10】
【公表番号】特表2011−523847(P2011−523847A)
【公表日】平成23年8月25日(2011.8.25)
【国際特許分類】
【出願番号】特願2011−509973(P2011−509973)
【出願日】平成21年5月20日(2009.5.20)
【国際出願番号】PCT/EP2009/056106
【国際公開番号】WO2009/156232
【国際公開日】平成21年12月30日(2009.12.30)
【出願人】(509274005)ドクトル ファルク ファルマ ゲーエムベーハー (4)
【Fターム(参考)】
【公表日】平成23年8月25日(2011.8.25)
【国際特許分類】
【出願日】平成21年5月20日(2009.5.20)
【国際出願番号】PCT/EP2009/056106
【国際公開番号】WO2009/156232
【国際公開日】平成21年12月30日(2009.12.30)
【出願人】(509274005)ドクトル ファルク ファルマ ゲーエムベーハー (4)
【Fターム(参考)】
[ Back to top ]