
血液凝固促進剤としてのヘビ第五因子及び使用方法
【解決手段】 本発明は、血液凝固障害の治療のため、第五因子変異体を使用した組成物及び方法を提供するものである。
【発明の詳細な説明】
【技術分野】
【0001】
この出願は、35 U.S.C.§119(e)に基づいて、2008年11月21日に出願された米国仮出願第61/116,913号に対して優先権の主張を行い、その全体の開示は、完全に参照により本明細書に組み込まれる。
35 U.S.C.§202(c)に基づいて、米国政府はここに記載された発明において一定の権利を有することを認めている。本発明は、国立保健研究機構、認可番号HL−088010及びP50HD055784−01からの資金の一部により行われた。
【0002】
本発明は医学及び血液学の分野に関するものである。さらに具体的には、本発明は、ヘビ毒由来の第五因子(FV)の変異体およびそれらの誘導体を、それらを必要とする患者において凝固カスケードを調節するために使用する治療戦略について記述する。
【背景技術】
【0003】
いくつかの出版物及び特許文献は、本発明の属する技術分野について記述するため明細書に引用されている。これらの引用はそれぞれ、参照として明細書に完全に組み込まれる。
【0004】
血液凝固FVは、マルチドメインタンパク質(A1−A2−B−A3−C1−C2)であり、わずかな凝血原活性、または凝血原活性なしでプロコファクターとして循環する。補因子の活性化状態(FVa)への転換は、トロンビンによる、グリコシル化された中央Bドメインの大部分の分解除去を含む(1)。FVaは、プロトロンビナーゼの形成、プロトロンビンの生理活性のため、カルシウムイオンの存在下で負に帯電した膜表面上のセリンプロテアーゼ因子Xa(FXa)と会合する(2)。FVaがトロンビンの生成を大幅に高めるため、非活性プロコファクターとしてのFVの維持は、正常な止血の維持を発展させるうえで間違いなく重要な役割を果たす。最近では、Bドメイン内の個別の配列は、非活性プロコファクターとして持続するFVのメカニズムに寄与することが示されている(3,4)。この領域の除去は、タンパク質分解処理を必要とせずにFV活性をもたらす。
【0005】
FVは、A及びCドメイン間では高い相同性、Bドメイン間では低い相同性を脊椎動物間で広く示される(5,6)。最近のゲノムデータでは、Bドメインの長さが一般的に哺乳類の間で共通して保存されている(〜800残基)のに対し、下等脊椎動物では大きく異なる(例えば、トラフグでは〜500残基)ことを示している(5,6);これらの構造変化の機能的な意味は理解されていない。Bドメインへの主要な変化の顕著な例は、オーストラリアのヘビのグループ(タイパン(Oxyuranus microlepidotus)、コブラ・モドキ(Pseudonaja textilis)及びコースタルタイパン(Oxyuranus scutellatus))からきている。これらの種は独特であり、加えて、これらの止血システムの一部としてFVは循環しており、その毒にもFVを有している。これらの種のそれぞれからの毒由来FVは、同様のドメイン構造を有する哺乳類のFVと、〜44%の配列相同性が共通している。しかしながら、Bドメインの長さはヒトのFVと比べて飛躍的に短い(836残基に対して、〜46残基)。これらのヘビからの毒は、世界で最も有毒であるとみなされており、強力な血液凝固促進剤であり、及び、いくつかある症状のなかで特に、播種性血管内凝固障害を引き起こす(10)。これは、大量のFVと同様に、共に強力なプロトロンビン活性化因子である複合体を形成するFXa様酵素を含む。コースタルタイパン(オスキュタリンC)及びコブラ・モドキ(プスタリンC)からの複合体は部分的に精製され、及び粗毒と特徴づけられる。プロトロンビンのトロンビンへの変換もそれらの活性を強化するのもカルシウム及びリン脂質によって程度が異なるが、FVaは影響しない。さらに、プスタリンCは活性化タンパク質C(APC)に対して耐性を示し、強力な毒素としての役割に潜在的に貢献する(7)。
【発明の概要】
【課題を解決するための手段】
【0006】
本発明によって、それを必要とする患者における血液凝固障害を治療する方法が提供される。例示的な方法では、FV変異体またはそれらの誘導体の活性型の有効量を投与する工程であって、それによって前記患者の血栓形成を増進し、且つ出血性疾患の症状を改善するものである、前記投与する工程を伴う。一実施形態においては、変異第五因子は配列ID番号1の配列を有する核酸によってコードされる。他の実施形態においては、変異第五因子は、配列ID番号2の配列を有する核酸を最適化したコドンによってコードされる。
【0007】
前記方法及び第五因子凝血原は、血液凝固障害の治療に使用されることもある。血液凝固障害は、これに限定されるわけではないが、血友病AおよびB、抑制抗体に関連する血友病A及びB、凝固因子欠乏症、ビタミンKエポキシド還元塩素欠乏症、ガンマカルボキシラーゼ欠乏症、外傷に伴う出血、負傷、血栓症、血小板減少症、脳卒中、凝血障害、播種性血管内凝固障害(DIC)、過抗凝固治療障害、バーナードスーリエ症候群、グランツマンの血小板無力症、及び貯蔵プール欠乏症を含む。また、血液凝固障害は、ヘパリン、低分子量ヘパリン、五糖類、ワルファリン、低分子抗血栓及びFxa阻害剤の投与によることもある。
【0008】
本発明の変異第五因子は、患者に直接注入または局所投与することができ、または、それをコードするベクターの形で移送され得る。
【0009】
また、以下の性質の一つまたはそれ以上を示す、ヒト化及び改善型第五因子変異体が提供される。FV変異体は、活性化、補因子の状態で合成される;FV変異体は、膜表面が、セリンプロテアーゼXa因子(FXa)と高い親和性で結合するための通常の条件を回避できるよう、独自の立体構造を有する;及び、FV変異体は、その重鎖及び軽鎖を接続する固有なジスルフィド結合を含めることにより、活性化タンパク質C(APC)による不活性に対して機能的に耐性がある。
【0010】
本発明の他の実施形態においては、ヘビ第五因子をコードする配列ID番号2の単離された核酸が提供される。このような核酸は、使用のために発現ベクターに含まれ得る。
【0011】
さらに別の実施形態では、本発明は、配列ID番号1の核酸によりコードされた、単離されたヘビ第五因子タンパク質を提供する。ヘビ第五因子タンパク質は、リポソームまたはミセルに含有され得る。また、本発明は、薬学的に許容される賦形剤に含まれるヘビ第五因子タンパク質からなる薬学的組成物を提供する。
【図面の簡単な説明】
【0012】
【図1】精製タンパク質のSDS−PAGE分析 タンパク質(3μg/レーン)は、還元(パネルA、C)条件下、または、非還元(パネルB)条件下でSDS‐PAGEに曝され、及びクマシーブリリアントブルーR‐250で染色することにより可視化される。レーン1、pt−rFV、レーン2、pt−rFV−QQ、レーン3、pt−rFV プラス トロンビン、レーン4、pt−rFV‐QQ プラス トロンビン、レーン5、rFX、レーン6、rFXa、レーン7,pt−FXa。標準の見かけの分子量が左側に示されている。パネルA及びパネルBは同じマーカーを共有している。ダッシュ(−)は、収率がアミノ酸を正確に割り当てるにはあまりに低量であることを示し、及び、可能性の高いγ−カルボキシグルタミン酸の存在を示す6つの位置である。
【図2】PCPSの存在下、または非存在下におけるプロトロンビンの開裂の速度定数の測定 トロンビン生成の初期速度は、50μMのPCPS及び0.1nMのpt−FXa及び20nMのhFV−810(−◆−または−◇−)、pt−rFV(−●−または−○−)、pt−rFV−QQ(−▲−または−△−)、またはpt−rFVa(−■−または−□−)と3μMのDAPAの存在(パネルA)または非存在(パネルB)でのプロトロンビンの濃度の増加によって決定される。全てのデータセットの以下の分析は矩形双曲線に描かれ、適した速度定数は表2に示されている。データは、2つの類似した実験においての代表的なものである。
【図3】PCPSの存在下、または非存在下におけるプレトロンビン−1の開裂の速度定数の測定 トロンビン生成の初期速度は、50μMのPCPS及び0.1nMのpt−FXa及び20nMのhFV−810(−◆−または−◇−)、pt−rFV(−●−または−○−)、pt−rFV−QQ(−▲−または−△−)、またはpt−rFVa(−■−または−□−)と3μMのDAPAの存在(パネルA)または非存在(パネルB)でのプレトロンビン−1の濃度の増加によって決定される。全てのデータセットの以下の分析は矩形双曲線に描かれ、適した速度定数は表2に示されている。データは、2つの類似した実験においての代表的なものである。
【図4A】APCによる膜結合型FV(a)の不活性化 50μMのPCPS及び500nMのpt−rFV(−●−)、pt−rFVa(−(−)、hFV−810(−◆−)、またはrFVa(−(−)を含む反応混合物は10nMまたは750nM(inset)のAPCどちらかと保温培養される。選択した時間間隔において、試料は削除され、及び、「実験的手法」(パネルA)に記載された方法で、または、SDS−PAGEゲル上を走らせ、還元下(パネルB)または非還元条件下(パネルC)でクマシー ブルー R250で染色して、補因子活性をアッセイ分析した。データは二つの類似した実験においての代表的なものである。
【図4BC】APCによる膜結合型FV(a)の不活性化 50μMのPCPS及び500nMのpt−rFV(−●−)、pt−rFVa(−(−)、hFV−810(−◆−)、またはrFVa(−(−)を含む反応混合物は10nMまたは750nM(inset)のAPCどちらかと保温培養される。選択した時間間隔において、試料は削除され、及び、「実験的手法」(パネルA)に記載された方法で、または、SDS−PAGEゲル上を走らせ、還元下(パネルB)または非還元条件下(パネルC)でクマシー ブルー R250で染色して、補因子活性をアッセイ分析した。データは二つの類似した実験においての代表的なものである。
【図5A】第五因子のための核酸コーディング配列 第五因子(配列ID番号1)に誘導されたヘビ毒の核酸配列は、パネル5A及び5Bに示されている。哺乳類系での発現に最適化されたコドンを有する第五因子(配列ID番号2)に誘導されたヘビ毒の核酸配列はパネル5C及び5Dに示されている。
【図5B】第五因子のための核酸コーディング配列 第五因子(配列ID番号1)に誘導されたヘビ毒の核酸配列は、パネル5A及び5Bに示されている。哺乳類系での発現に最適化されたコドンを有する第五因子(配列ID番号2)に誘導されたヘビ毒の核酸配列はパネル5C及び5Dに示されている。
【図5C】第五因子のための核酸コーディング配列 第五因子(配列ID番号1)に誘導されたヘビ毒の核酸配列は、パネル5A及び5Bに示されている。哺乳類系での発現に最適化されたコドンを有する第五因子(配列ID番号2)に誘導されたヘビ毒の核酸配列はパネル5C及び5Dに示されている。
【図5D】第五因子のための核酸コーディング配列 第五因子(配列ID番号1)に誘導されたヘビ毒の核酸配列は、パネル5A及び5Bに示されている。哺乳類系での発現に最適化されたコドンを有する第五因子(配列ID番号2)に誘導されたヘビ毒の核酸配列はパネル5C及び5Dに示されている。
【発明を実施するための形態】
【0013】
毒ヘビは自分自身の防御のため、及び獲物を無力化するために、血液凝固促進剤などのような有毒な化合物の配列を生成する。オーストラリアのブラウンスネーク、コブラ・モドキ(Pseudonaja textilis)は、Xa因子(FXa)及び第五因子(FV)と一致する、毒由来のプロトロンビン活性化複合体を有する。前記FV組成物(pt−FV)は、ヒトFVと同様のドメイン組織を有するが、その中央ドメインはとても少ない(836アミノ酸に対して55)。ここでは、pt−FVは強力な血液凝固促進剤に変換する機能要素の固有な増加を獲得している。組み換えpt−FVを用いた測定は、それが構造的に活性補因子として合成され、ヒトFVとは異なり、活性の発現にタンパク質分解を必要としないことを明らかにした。配列の比較により、非活性プロコファクター状態を安定させるのに役立つBドメイン内の塩基性アミノ酸の保存クラスターを失っていることが明らかとなった。注目すべきことは、pt−FVは、負に帯電したリン脂質の非存在下では、高い親和性でヘビFxaを結合するように働き、及び、膜の非存在下、または存在下においては、プレトロンビン‐1と同等の運動パラメータを示すことである。さらに、pt−FVは、重鎖のタンパク質分解によってもたされた濃度であっても、活性タンパク質Cに機能的耐性がある。この安定性は、ある程度、重鎖と軽鎖を結合する固有なジスルフィド結合による可能性が高い。同時に、これらの分子の技術革新の合流点は、pt−FVを血液凝固生物兵器にしている。これらの発見は、FVがプロコファクターとしてどのように保存されるかについて重要な新しい洞察を提供し、及び、FXa‐FVa相互作用が、プロトロンビナーゼの拡張作用にどのように寄与するかよりよく理解するために広範な影響を有する。
【0014】
従って、本発明はまた、第五因子(FV)の固有な誘導体を提供する。その誘導体は、一般的なブラウンスネーク、コブラ・モドキ(Pseudonaja textilis)の毒内で、または異種発現システム(pt−FV)、または以下の固有な特徴の一つまたはそれ以上を有する同様のシステムで発現するヒトFV誘導体である。
【0015】
a)FV変異体は活性化、補因子状態で合成される
b)FV変異体は、膜表面で、そのセリンプロテアーゼXa因子(FXa)と高い親和性で結合するのに通常必要とされる条件を迂回できるよう、独自の立体構造を有する
c)FV変異体は、その重鎖及び軽鎖を接続する固有なジスルフィド結合を含めることにより、活性化タンパク質C(APC)による不活性に対して機能的に耐性がある。
【0016】
I.定義
本発明の生体分子に関連する様々な用語は、上述及び明細書及び特許請求の範囲のいたるところに用いられている。
【0017】
「変異型FV」という用語は、ヘビまたは他の異種発現システムから得られる、強力な血液凝固活性を示すFV誘導体分子に関するものである。本発明の変異型FVは配列ID番号1によってコードされ、またはその変異(例えば配列ID番号2)はヒトでの使用に適合し、望ましいFV血液凝固活性を促進する。
【0018】
「止血関連疾患」または「血液凝固障害」という用語は、血友病A及びB、抑制抗体に関連する血友病A及びB、凝固因子欠乏症、VII1 IX及びX、XI、V、XII、II因子、フォン・ヴィレブランド因子欠乏症、結合FV/FV1欠乏症、ビタミンKエポキシド還元塩素欠乏症、ガンマカルボキシラーゼ欠乏症、外傷に伴う出血、負傷、血栓症、血小板減少症、脳卒中、凝血障害、播種性血管内凝固障害(DIC)、ヘパリン、低分子量ヘパリン、五糖類、ワルファリン、低分子抗血栓(例えばFXa阻害剤)に関連する過抗凝固、及びバーナースーリエ症候群、グランツマンの血小板無力症及び貯蔵プール欠乏症のような血小板異常症などの出血障害に関連する。
【0019】
本発明の核酸を参照するにあたり、「単離された核酸」という用語が時折用いられる。この用語は、DNAに適用される際に、そこから由来する生物の天然に存在するゲノムにおいて直接に連続する(5’及び3’方向において)配列から離されているDNA分子を意味する。例えば「単離された核酸」は、プラスミドまたはウイルスベクターなどのようなベクター内に挿入された、または原核生物または真核生物のDNA内に合成されたDNAまたはcDNA分子からなる。本発明のRNA分子に関しては、「単離された核酸」という用語は、前記の単離されたDNAによりコードされたRNA分子を主に意味する。また、この用語は、「実質的に純粋な」形(用語「実質的に純粋な」に関しては、以下に定義する)で存在するように、その自然な状態(例えば、細胞または組織内)で結合されたRNA分子から十分に分離されたRNA分子に関するものである。
【0020】
タンパク質に関して、「単離されたタンパク質」または「単離及び精製されたタンパク質」という用語が、時おり本明細書において使用されている。この用語は、本発明の単離された核酸分子の発現により生成されるタンパク質を主に意味する。また、この用語は、「実質的に純粋な」形で存在するように結合された他のタンパク質から十分に分離されたタンパク質に関するものである。
【0021】
用語「プロモーター領域」は、遺伝子の転写調節領域に関するものであり、コーディング領域の5’側または3’側、またはコーディング領域内、またはイントロン内でみられることもある。
【0022】
用語「ベクター」は、そこで複製される宿主細胞内に導入するため挿入されるDNA配列内の小さな担体DNA分子に関するものである。「発現ベクター」は、宿主細胞内での発現に必要とされる調節領域と遺伝子または核酸配列を含む特殊なベクターである。用語「作動的に結合された」はコーディング配列の発現に必要な調節配列が、コーディング配列の発現がなされるようコーディング配列に適切な相対位置内のDNA分子内に配置されたことを意味する。これと同様の定義が、コーディング配列の配置及び発現ベクター内の転写制御要素(例えばプロモーター、エンハンサー、及び終止要素)に時おり適用される。この定義はまた、ハイブリッド核酸分子が生じる第一及び第二核酸分子の核酸配列の配置においても時おり適用される。
【0023】
用語「実質的に純粋な」は、少なくとも50〜60重量%の対象化合物(例えば核酸、オリゴヌクレオチド、タンパク質など)からなる標本に関するものである。標本は、より好適には少なくとも75重量%、最も好適には、90〜99重量%の対象化合物からなる。純度は対象化合物によって適切な方法(例えば、クロマトグラフィー法、アガロースまたはポリアクリルアミドゲル電気泳動法、HPLC分析など)で測定される。
【0024】
特定のヌクレオチド配列またはアミノ酸配列に関する場合の「本質的に…からなる」は、与えられた配列ID番号の性質を有する配列を意味する。例えば、アミノ酸配列を参照する場合、この用語は、配列の基本的な及び新規な特徴に作用しない配列自体及び分子の修飾を含む。
【0025】
本明細書で使用される用語「オリゴヌクレオチド」は、本発明のプライマー及びプローブに関するものであり、及び、二つまたはそれ以上、好適には三つ以上のリボ−またはデオキシリボヌクレオチドからなる核酸分子として定義される。オリゴヌクレオチドの正確な大きさは様々な要因及びオリゴヌクレオチドが使用されるそれぞれの適用に依存する。
【0026】
本明細書で使用される用語「プローブ」は制限酵素消化による精製として自然発生するものであろうと、合成であろうと、アニーリングまたはプローブと相補的な配列と核酸分子に特異的にハイブリダイズできるオリゴヌクレオチド、ポリヌクレオチドまたは核酸、RNAまたはDNAに関するものである。プローブは単一らせん構造または二重らせん構造どちらもとりうる。プローブの正確な長さは温度、プローブのソース、使用方法を含む様々な要因に依存する。例えば、診断に適用する場合では、標的配列の複雑さに依存し、典型的なオリゴヌクレオチドプローブは、15〜25またはそれ以上のヌクレオチドを含むが、それよりも少ないヌクレオチドを含むこともある。
【0027】
本明細書でのプローブは特定の標的核酸配列の異なるらせん構造と「実質的に」相補的となるよう選択される。これは、事前に定められた条件下でそれぞれの標的らせん構造と「特異的にハイブリダイズ」またはアニールをするためには、プローブが十分に相補的でなければならないということを意味する。例えば、非相補的なヌクレオチドフラグメントは、標的らせん構造と相補的であるプローブ配列の残渣によって、プローブの5’末端または3’末端に付着される。あるいは、プローブ配列が、それと共に特異的にアニールする標的核酸の配列と十分に相補的である条件下で、非相補的塩基または長い配列はプローブ内に組み入れられることができる。用語「特異的にハイブリダイズする」は、当業者において通常使用される事前に定められた条件下でハイブリダイゼーションが可能となるよう十分に相補的な(時おり「実質的に相補的な」と称される)配列の二つの一本鎖核酸分子間のつながりに関するものである。特に、この用語は、本発明の一本鎖DNAまたはRNA分子内に含まれる実質的に相補的な配列によるオリゴヌクレオチドのハイブリダイゼーションに関するものであり、非相補的配列の一本鎖核酸によるオリゴヌクレオチドのハイブリダイゼーションの実質的な除去に関するものである。
【0028】
本明細書で使用される用語「プライマー」は、生体システムからの誘導、制限酵素消化による生成であろうと、合成であろうと、適切な条件下に置かれた際に核酸合成依存テンプレートの起始因子として機能的に作用できる一本鎖または二本鎖、オリゴヌクレオチド、RNAまたはDNAに関するものである。適当な核酸テンプレート、核酸の適当なヌクレオシド三リン酸、ポリメラーゼ酵素、適当な補因子及び、適当な温度及びpHなどのような条件と提供されると、ポリメラーゼの作用またはプライマーの延長生成を増やす同様の活性により、ヌクレオチドが付加され、プライマーは31末端に延長されることもある。
【0029】
プライマーは、特定の条件及び適用の求めによって、様々な長さとなることがある。例えば、診断における適用では、オリゴヌクレオチドプライマーは典型的には15〜25またはそれ以上のヌクレオチド長さとなる。プライマーは、目的とする延長生成の合成を刺激する所望のテンプレートと十分に相補的である必要があり、それが、ポリメラーゼまたは同様の酵素による合成の起始因子としての使用のために、適当な近位におけるプライマーの3’ヒドロキシル部分を提供する十分な方法において、目的とするらせん構造テンプレートとアニールすることを可能としている。それは、プライマー配列が目的とするテンプレートと正確に相補的であることを示すことは必要としない。例えば、非相補的ヌクレオチド配列は、他の相補的なプライマーの5’末端に付着されることもある。あるいは、非相補的な塩基は、オリゴヌクレオチドプライマー配列内に散在され、プライマー配列は延長生成物の合成のためにテンプレート−プライマー複合体を機能的に提供するらせん構造テンプレートの配列と十分に相補的となるよう提供される。
【0030】
用語「同一比率」は、核酸またはアミノ酸配列の間での比較に関して本明細書で使用されている。核酸及びアミノ酸配列は、しばしば、核酸またはアミノ酸の配列を一列にし、それにより、二つの間の違いを明らかにするコンピュータープログラムを用いて比較される。本発明の目的のため、核酸配列の比較は、GCG Wisconsin Package version 9.1によって実行され、これは、ウィスコンシン州Madisonのthe Genetics Computer Groupによって可能となった。便宜上、プログラムによって指定された初期パラメータ(ギャップクリエーションペナルティー=12、ギャップエクステンションペナルティー=4)は、本明細書において使用するため、同一配列を比較するのに意図されている。あるいは、National Center for Biotechnology Information(ワールドワイドウェブにおいてncbi.nlm.nih.gov/blast/;Altschul et al.,1990,J MoI Biol 215:403〜410)によって提供された、初期パラメータとギャップ調整に使用するBlastn 2.0プログラムは、核酸配列及びアミノ酸配列間の同一及び類似性のレベルを測定するのに使用されることがある。
【0031】
核酸分子、ポリペプチド及びそれらの誘導体をコードするFVの変異体の調製
核酸分子
FVの変異型をコードする核酸分子または本発明のそれらの機能性誘導体は、組み換え型DNA技術の方法を用いて調製されることがある。核酸配列情報の可用性は、種々の方法によって、本発明の単離された核酸分子の調節を可能とする。例えば、FVの変異型をコードする核酸配列またはそれらの誘導体ポリペプチドは、当業者において既知である標準プロトコルを使用して適当な生体原料から分離されることがある。
【0032】
本発明の核酸は、任意の便利なクローニングベクターにおけるDNAとして維持されることがある。好適な実施形態においては、クローンは、適当なE.coli宿主細胞pBluescript(カリフォルニア州、La Jolla、Stratagene)のような、プラスミドクローニング/発現ベクター内で維持される。あるいは、核酸は、哺乳類の最奥での発現に適当なベクター内で維持されることもある。翻訳後修飾がFVの変異型またはそれらの機能性誘導体に影響を与える場合において、それは哺乳類の細胞に分子を発現するのに好適である。FV変異体または核酸分子をコードするそれらの誘導体は、cDNA、ゲノムDNA、RNA及び一本鎖または二本鎖であることがあるそれらのフラグメントを含む。従って、この発明は、本発明の核酸分子の少なくとも一つの配列とハイブリダイジングできる配列を有するオリゴヌクレオチド(センスまたはアンチセンスDNAまたはRNAのらせん構造)を提供する。このようなオリゴヌクレオチドは、FV発現の検出のためのプローブとして有用である。
【0033】
タンパク質
FVの変異体または本発明のそれらの誘導体ポリペプチドの全長は、既知の方法に従って、様々な方法で調製されることがある。タンパク質は、例えば、FVの活性型を発現する、形質転換された細菌または動物の培養された細胞または組織などの適当な材料から、免疫親和性精製によって生成されることがある。しかしながら、これは、時を選ばず与えられた細胞に存在する可能性のあるタンパク質の低分子量のために好適な方法ではない。FVの変異型またはそれらの誘導体ポリペプチドをコードする核酸分子の可用性は、当業者に既知の方法で体外での発現に使用されるFVの変異型またはそれらの誘導体の生成を可能にする。例えば、cDNAまたは遺伝子は、体外転写のためのpSP64またはpSP65のような適当な体外転写ベクター内でクローンされ、続いて、小麦胚芽またはウサギ網状赤血球溶血液のような適当な無細胞翻訳系で無細胞翻訳される。体外での転写及び翻訳システムは、例えばウィスコンシン州MadisonのPromega Biotechまたはメリーランド州RockvilleのBRLによって市販されている。
【0034】
あるいは、好適な実施形態によれば、FVの変異型またはそれらの誘導体の多くの量は、適当な原核生物または真核生物発現系においての発現により生成されることがある。例えば、FVの変異型をコードするDNA分子の一部または全部は、E.coliのような細菌性細胞または、CHOまたはHela細胞のような哺乳類の細胞内での発現に適合したプラスミドベクター内に挿入されることがある。あるいは、好適な実施形態において、FVの変異体またはそれらの誘導体からなる標識された溶融タンパク質が生成されうる。FV分子またはそれらの誘導体のような標識された溶融タンパク質は、DNA分子の一部または全部によってコードされ、E.coliのような細菌性細胞または、これに限定されるわけではないが、酵母または哺乳類の細胞のような真核細胞における発現に適合したプラスミドベクター内に挿入された目的とするポリペプチド標識の一部または全部をコードするヌクレオチド配列にリーディングフレームする正確なコドン内で連結反応される。上述されたようなベクターは、宿主細胞内にDNAの発現を可能とするような方法で配置された宿主細胞内でのDNAの発現に必要な調節要素からなる。このような調節要素は、これに限定されるわけではないが、プロモーター配列、転写起始因子配列及びエンハンサー配列を発現に必要とする。
【0035】
ヘビ毒由来FV変異体またはそれらの誘導体タンパク質は、当業者において既知の方法に従って、組換え型原核生物または真核生物系においての遺伝子発現によって生成される。好適な実施形態において、市販されている発現/分泌系が、容易に周囲媒質から精製するのに用いられることがある。市販されている発現/分泌系により、組換え型タンパク質が発現され、その後、宿主細胞から分泌される。発現/分泌ベクターが使用されない場合、代替手法は、組換え型タンパク質と特異的に結合する抗体との免疫親和性相互作用のようなアフィニティー分離による組換え型タンパク質の精製またはN末端またはC末端で6−8ヒスチジン残基により標識された組換え型タンパク質のニッケルカラムによる分離を
含む。代替標識は、FLAGエピトープ、GSTまたは赤血球凝集素エピトープからなることがある。このような方法は当業者によって通常使用されている。
【0036】
FVまたはそれらの誘導体タンパク質は、上述のような方法で調製され、標準プロトコルによって分析されることがある。例えば、このようなタンパク質は、既知の方法によってアミノ酸配列分析を受けることがある。
【0037】
上述したように、本発明によるポリペプチドを生成する便利な方法は、発現系において核酸を使用することで、それをコードする核酸を発現させることである。本発明の方法において有用である様々な発現系は、当業者にとって既知である。
【0038】
したがって、本発明は(開示された)ポリペプチドをつくる方法、ポリペプチド(一般的に核酸)をコードする核酸からの発現を含む方法も含む。これは、ポリペプチドの生成を生じるまたは可能にする適当な条件下で、ベクターを含む宿主細胞を培養することで簡便に達成されることがある。ポリペプチドは網状赤血球溶血液のような体外系において生成されることもある。
【0039】
核酸及びタンパク質またはそれらの誘導体をコードするFVの使用
FVポリペプチドまたはそれらの誘導体または血液凝固活性変化を有するのと同一のものをコードする核酸は、例えば、血液凝固カスケードを調整する治療及び/または予防薬剤として、本発明にしたがって使用されることがある。本発明はこれらの分子が凝固を増加するよう変化し得ることを明らかにした。
【0040】
本発明の好適な実施形態においては、FVまたはそれらの誘導体は、生物学的に適合した担体の点滴を介して、好適には、静脈注射を介して患者に投与されることがある。本発明のFVまたはそれらの誘導体は、任意に、リポソーム内にカプセル化され、または、分子の安定性を増加する他のリン脂質またはミセルと混合される。FVまたはそれらの誘導体は、単独で、または止血を調整するとして既知の他の薬剤と組み合わせて投与されることがある(例えば、VIIa、FIX、FVIIIまたはFX/Xa因子及びそれらの誘導体)。FVまたはそれらの誘導体を提供するのに適当な化合物は、これに限定されることはないが、患者の状態及び血行状態を含む生理学的に様々なことを考慮した上で、医師によって決定される。異なる適用及び投与経路にふさわしい様々な組成物が当業者に知られており、明細書の以下に記載されている。生成されたFVまたはそれらの誘導体を含む調製は、生理学的に許容される材料を含み、医薬品として好適に処方される。調製は、既知の先行技術の方法を実質的に使用することで定式化され得るものであり、塩化ナトリウム、塩化カルシウムのような塩とグリシン及び/またはリシンのようなアミノ酸を含むバッファーと、pH範囲6〜8で混合され得る。
【0041】
必要となるまで、第五因子またはその誘導体を含む精製された調製液は、完成した溶液または凍結乾燥した状態または冷凍状態で保管される。好適には、調製液は凍結乾燥した状態で貯蔵され、及び、適切な再構成溶液を使用して視覚的に透明な溶液に溶解する。あるいは、本発明による調製は、調製液または凍った液体として作られることも可能である。
【0042】
本発明による調製液は、特に安定的であり、例えば、適用の前に長時間溶解した形態で有効である。本発明による調製液は、一つの成分の調製液またはマルチ成分の調製液における他の要因との組み合わせの形態で、FVまたはそれらの誘導体と薬剤的調製液として手に入れられる。
【0043】
薬剤的調製液内で精製されたタンパク質を処理するのに先行して、精製されたタンパク質は従来の品質管理により管理され、及び、提示の治療形態内に形作られる。特に、精製された調製液は、組み換え製造の間、発現ベクター由来の核酸と同様に細胞核酸の非存在下でテストされ、好適には、EP0714987に記載されたような方法が用いられる。
【0044】
この発明の他の特徴は、高い安定性及び構造的及び構造的完全性を有し、及び、特に不活性因子V/Va類似中間体及びタンパク質自動分解生成物を含まず、及び、前記タイプの第五因子類似体の活性化により、及び、適当な調製液内で調剤されることによりつくられるFVまたはそれらの誘導体を含む調製液の製造を可能にすることに関するものである。薬剤的調製液は、10〜1000μg/kgの間、好適には10〜250μg/kgの間の投薬量であり、最適には10〜75μg/kgの間の投薬量と、好適には40μg/kgの変異体第五因子ポリペプチドである。患者は血液診療所で、提示があればすぐに治療を受けることができる。または、患者は1〜3時間毎にボーラス投与され、または、十分に改善がみられる場合には、一日一回のFVまたは明細書に記載されたそれらの誘導体の投与となる。
【0045】
ヘビから得られた核酸をコーディングするFVまたはそれらの誘導体は、本発明に従って様々な目的で使用されることがある。本発明の好適な実施形態において、血液凝固を調節するための核酸運搬媒体(例えば発現ベクター)は、FVまたはそれらの誘導体ポリペプチド、または本明細書に記載されたそれらの機能的フラグメントをコーディングする核酸配列からなる発現ベクターなどが提供される。発現ベクターをコードするFVまたはそれらの誘導体の患者への投与は、凝固を強化するのに役立つFVまたはそれらの誘導体ポリペプチドの発現を生じる。本発明に従って、核酸配列をコードするFVまたはそれらの誘導体は、本明細書に記載されたように、その発現が止血を調節するFVまたはそれらの誘導体をコードすることがある。
【0046】
FVまたはそれらの誘導体核酸配列からなる発現ベクターは、単独で投与されることも、または止血を調整するのに有用な他の分子と組み合わせて投与されることもある。本発明に関連して、発現ベクターまたは治療薬の組み合わせは、患者に、単独で、または薬剤的に許容な化合物で、または生物学的に親和性のある化合物で投与されることもある。
【0047】
本発明の好適な実施形態においては、FVまたはそれらの誘導体をコードする核酸配列からなる発現ベクターは、ウイルスベクターである。本発明で使用されるウイルスベクターは、これに限られるわけではないが、アデノウイルスベクター(特定プロモーター/エンハンサー組織を伴うまたは伴わない)、複数の血清型(例えば、AAV−2、AAV−5、AAV−7及びAAV−8)のアデノ随伴ウイルス(AAV)ベクター及びハイブリッドAAVベクター、レンチウイルスベクター及び擬似レンチウイルスベクター(例えば、エボラウイルス、水疱性口内炎ウイルス(VSV)、及び猫免疫不全ウイルス(FIV))、単純ヘルペスウイルスベクター、ワクシニアウイルスベクター及びレトロウイルスベクターを含む。
【0048】
本発明の最適な実施形態では、核酸配列FVまたはそれらの誘導体、またはそれらの機能的フラグメントからなるウイルスベクターを投与する方法が提供されている。本発明の方法における実用的なアデノウイルスベクターは、好適には、アデノウイルスベクターDNAの少なくとも主要な部分を含む。明細書に記載されているように、このようなアデノウイルスベクターの投与に続いて、FVまたはそれらの誘導体ポリペプチドは止血を調節するのに役立つ。
【0049】
組換え型アデノウイルスベクターは、多様な遺伝子治療適用において幅広い実用性を有する。そのような適用におけるそれらの有用性は、様々な臓器の状況で行われる体内での遺伝子転移の高効率によるところが大きい。
【0050】
アデノウイルス粒子は、適切な遺伝子運搬のための媒体として生かされることがある。このようなウイルス粒子は、二本鎖DNA非エンベロープウイルスに関連する構造的特徴及びヒト呼吸系及び消化器官系指向性のような生物学的特徴を含む、これらの適用にとって望ましい特徴を複数有する。さらに、アデノウイルスは、受容体を介した飲食作用により、生体内及び生体外の様々な細胞に影響を与えることが知られている。アデノウイルスベクターの全体的な安全性の証明、アデノウイルスの感染は、軽度のインフルエンザ様症状を含む、ヒトにおける軽度な疾患を引き起こす。
【0051】
その大きなサイズに起因して(〜36キロ塩基)、アデノウイルス遺伝子は遺伝子治療媒体として用いられるのに適している。アデノウイルスは、複製において極めて重要なアデノウイルス遺伝子及び非必須領域の除去後、外来DNAの挿入に適応するからである。このような置換は複製機能及び感染に関して正常に機能しないウイルスベクターを提供する。注目すべきは、アデノウイルスは、遺伝子治療のため及び異常遺伝子の発現のためのベクターとして用いられることである。
【0052】
遺伝子治療で用いられるアデノウイルスベクターの使用についてのさらに詳細な議論は、Berkner,1988,Biotechniques 6:616−629及びTrapnell,1993,Advanced Drug Delivery Reviews 12: 185〜199を参照のこと。
【0053】
例えば、目的とする遺伝子の複数コピー及びそれによる多量の目的遺伝子の生成物を提供できるベクターを導入することが望ましい。アデノウイルスベクターの改良及びこれらのベクターを生成する方法は、Mitani and Kubo(2002,Curr Gene Ther.2(2):135〜44);Olmsted−Davis et al.(2002,Hum Gene Ther.13(11):1337〜47);Reynolds et al.(2001,Nat Biotechnol.19(9):838〜42);米国特許番号5,998,205(多重DNAコピーからなる腫瘍特異的複製ベクターが提供されている);6,228,646(ヘルパーアデノウイルスなし、アデノウイルスベクター完全欠損が記載されている); 6,093,699(遺伝子治療のためのベクター及び方法が提供されている);6,100,242(複製欠損アデノウイルスベクターに挿入された導入遺伝子が、生体内で末梢血管疾患及び心疾患の遺伝子治療において有効に用いられることが記載されている);及び国際特許出願番号WO 94/17810及びWO 94/23744を含む参考文献、特許及び特許出願に詳しく記載されている。
【0054】
出願によっては、さらに、発現構成は、特に細胞または組織タイプでの発現を促進するのに役立つ調節要素からなる。このような調節要素は、当業者において既知であり、Sambrook et al.(1989)及びAusubel et al. (1992)において深く論じられている。本発明の発現構成における組織特定調節要素の取り込みは、FVまたはそれらの発現またはそれらの機能的フラグメント発現のための少なくとも部分的な組織指向性を提供する。例えば、サイトメガロ・ウイルス(CMV)プロモーターの制御下で、FVまたはそれらの誘導体をコードする核酸配列からなるE1欠損タイプ5アデノウイルスベクターは本発明の方法において生かされることがある。
【0055】
アデノウイルスベクターを生成するための典型的な方法
組み換え遺伝子発現のためのアデノウイルスベクターは、ヒト胚性腎細胞株293において生成される(Graham et al.,1977,J,Gen. Virol.36:59〜72)。この細胞株は、E1機能不全において、アデノウイルス2(Ad2)及びアデノウイルス5突然変異体の成長を許容する。なぜなら、この細胞株は、アデノウイルス5遺伝子の左末端からなり、従ってE1タンパク質を発現するからである。293細胞の細胞遺伝子内に導入されたE1遺伝子は、除去された遺伝子からウイルスベクターを増幅する発現系としてこれらの細胞の使用を容易にするレベルで発現される。293細胞は、独立したクローニングを助け、アデノウイルスベクターを発現するため、E1突然変異体の分離及び増殖に広範囲にわたって使用される。従って、293細胞株のような発現系は、トランスにおいて不可欠なウイルス機能を提供し、それによって、E1遺伝子に置換された外因性核酸配列において、ウイルスベクターの増殖が可能となる。Young et al.in The Adenoviruses,Ginsberg,ed.,Plenum Press,New York and London(1984),p.125〜172を参照のこと。
【0056】
アデノウイルスベクターの増殖に向いている他の発現系は、当業者において既知であり(例えば、HeLa細胞)、及び、他の場でも再検討されている。また、本発明には、FVまたはそれらの誘導体ポリペプチドをコードする核酸運搬媒体と個々の細胞を提供すること、及び、FVまたはそれらの誘導体ポリペプチドが発現する条件下で細胞を培養することからなる止血を調節する方法も含まれる。
【0057】
前述の議論より、FV変異体またはそれらの誘導体ポリペプチドまたは同一のものをコードする核酸は、異常血液凝固に関連する疾患の治療において用いられることがわかる。
【0058】
医薬組成物
本発明の発現ベクターは、生理活性タンパク質(例えば、FVまたはそれらの誘導体ポリペプチドまたは機能的フラグメントまたはそれらの誘導体)が生成できるように、対象に運搬される薬剤的化合物内に組み込まれる。本発明の特定の実施形態では、受容体がFVまたはそれらの誘導体ポリペプチドの治療的に有効な量を生成できるよう十分な遺伝子材料からなる薬学的化合物は、対象において止血作用を与えることができる。または、前述したように、変異体第五因子ポリペプチドの有効量は、それらを必要とする患者に直接的に注入される。化合物は、単独で投与、または、安定化化合物のような他の薬剤少なくとも一つと組み合わせて投与され、これに限定されるわけではないが、生理食塩水、緩衝食塩水、デキストロース及び水を含む、生体適合性のある薬剤担体に滅菌下で投与されることもある。化合物は、単独で、または、止血に作用する他の薬剤と組み合わせて患者に投与されることもある。好適な実施形態では、薬剤的化合物も薬剤的に許容な賦形剤を含む。このような賦形剤は、化合物を受容する個体に有害な免疫応答をそれ自体で導くことのない任意の薬剤を含み、及び、過度な毒性なしで投与することが可能である。薬学的に許容される賦形剤は、これに限定されるわけではないが、水、生理食塩水、グリセロール、糖類及びエタノールのような液体が含まれる。薬学的に許容される塩には、例えば、塩酸塩、臭化水素塩、リン酸塩、硫酸塩などのような鉱酸塩、及び、酢酸塩、プロピオン酸塩、マロン酸塩、安息香酸塩などのような有機酸塩が含まれる。加えて、湿潤剤、乳化剤、pH緩衝剤などのような補助剤が、このような賦形剤において存在することがある。薬学的に許容される賦形剤に関する詳細な議論は、Remington’s Pharmaceutical Sciences(Mack Pub. Co., 18th Edition,Easton,Pa.[1990]).で入手することができる。
【0059】
非経口投与に適した薬剤の剤形は、水溶液、好適には、ハンクス溶液、リンガー溶液または生理緩衝食塩水のような生理的に適合性のある緩衝液内で調剤される。水性懸濁注射液は、カルボキシルメチルセルロースナトリウム、ソルビトールまたはデキストランのような懸濁液の粘性を増加させるような基質を含むことがある。適当な親油性溶媒または媒体は、ゴマ油のような脂肪酸油、または、オレイン酸エチルまたはトリグリセリドまたはリポソームのような合成脂肪酸エステルを含む。随意に、懸濁液は、高濃度溶液の調製を可能にするため、化合物の溶解度を増加させる適当な安定剤または薬剤も含むことがある。局所または経鼻投与の場合、浸透されるべき特定の障壁に対して適切な浸透剤が製剤に使用されている。このような浸透剤は一般的に当業者において知られている。本発明の薬剤的化合物は当業者において既知の任意の方法(従来の混合、溶解、整粒、糖衣製造、ゲル化、乳化、カプセル化、封入、凍結乾燥処理など)で製造されている。
【0060】
薬剤的化合物は塩として提供されることがあり、また、これに限定されるわけではないが、塩酸、硫酸、酢酸、乳酸、酒石酸、リンゴ酸、コハク酸などの多くの酸と作られることがある。塩は、遊離塩基に対応する溶液よりも水または他のプロトン性溶媒に溶解しやすい傾向にある。他の場合では、好適な調製液は、4.5〜5.5のpH範囲で、1〜50μMのヒスチジン、1〜2%のスクロース、及び2〜7%のマンニトールの任意または全てを含む凍結乾燥粉末であることがある。この凍結乾燥粉末は使用前にバッファーに結合される。
【0061】
薬剤的化合物が調製された後、それらは適切な容器に入れられ、治療のために標識される。ベクターを含むFVまたはそれらの誘導体の投与のために、前述の標識には投与の量やしばしば方法が含まれることもある。
【0062】
本発明の使用に適当な医薬組成物は、活性成分が治療目的を達成するための有効量含まれている化合物を含む。治療に有効な投与量の決定は、本発明において提供される技術を使用する熟練した医療従事者の能力の範囲内で行われる。治療における投与量は、他の要因のなかでも、年齢、対象における一般的条件、異常血液凝固表現型の重症度、FVまたはそれらの誘導体ポリペプチドの発現レベルを調節する制御配列の強さに依存する。したがって、ヒトにおける治療の有効量は、ベクター基本形または、FVまたはそれらの誘導体に感染したタンパク質への個々の患者の応答に基づいて医療従事者により決定される比較的広い範囲に分類される。
【0063】
投与
FVポリペプチドの活性型変異体は、単独または他の薬剤と組み合わせて、本明細書において上述したように、適切な生理的担体において患者に直接的に注入されることもある。FVまたはそれらの誘導体、またはそれらの機能的フラグメントをコードする核酸配列からなる本発明の発現ベクターは、FVまたはそれらの誘導体ポリペプチドの活性型の予防的及び/または治療的に有効なレベルを達成する及び維持するため(以下、参照の)様々な方法により患者に投与されることもある。当業者は、特定の患者の治療のための本発明の発現ベクターをコードするFVまたはそれらの誘導体の活性型を使用するための特定の手順をすぐに見つけることができる。アデノウイルスベクターの生成及び患者への投与のための手順は、米国特許番号5,998,205、6,228,646、6,093,699、6,100,242、及び国際特許出願番号WO94/17810及びWO94/23744において記載され、これらはその全体において参照により本明細書に組み込まれている。本発明のアデノウイルスベクターをコードするFVまたはそれらの誘導体の活性型は、任意の既知の方法により投与されることがある。生体内への薬剤的化合物の直接的な運搬は、運搬を強化された対流のような運搬方法が予想されるにもかかわらず、一般的に、従来の注射器を利用する注射により達成される(例えば米国特許番号5,720,720を参照)。この方法で化合物は、皮下、表皮、皮内、髄腔内、眼窩内、粘膜内、腹腔内、静脈内、動脈内、口腔内、肝内または筋肉内に運ばれることがある。投与の他の方法には、経口投与及び経肺投与、坐薬、及び、経皮投与が含まれる。血液凝固疾患患者の治療の臨床専門医は、これに限定されるわけではないが、患者の状態及び治療の目的(例えば血液凝固の促進)を含む基準数に基づいて、活性型FVまたはそれらの誘導体核酸配列からなるアデノウイルスベクターの投与のために最適な経路を決定している。
【0064】
また、本発明の組成物は、創傷または外傷において局所的な血液凝固を促すのに使用されることもある。このアプローチでは、本発明の第五因子は局所的に適用するために製剤され、及び、戦場において、または、出血を止めなければならない、または、出血を止めることが重要とされる状況下において使用される。
【0065】
本発明は、活性型FVまたはそれらのポリペプチド誘導体をコードする核酸配列からなるAAVベクターも含む。
【0066】
活性型FVまたはそれらのポリペプチド誘導体をコードする核酸配列からなるレンチウイルスまたは疑似レンチウイルスベクターも含まれる。
【0067】
活性型FVまたはそれらのポリペプチド誘導体をエンコンディーングする核酸配列からなる裸プラスミドまたは発現ベクターも含まれる。
【0068】
以下に示す材料及び方法は、以下の実施例の実行を容易にするために提供されたものである。
【0069】
試薬
ベンズアミジン及び4−アミジノフェニルフッ化メタンスルフォニル塩酸塩(APMSF)はSigma社(St.Louis、MO)のものであり、及び、ダンシルアルギニン−N−(3−エチル−1,5−ペンタンジイル)アミド(DAPA)は、Haematologic Technologies社(バーモント州、Essex Junction)のものである。ペプチジル基質H−D−フェニルアラニル−L−ピペコリル−L−アルギニル−p―ニトロアニリド(S2238)はDiapharma社のものである。全ての組織培養試薬は、インスリン−トランスフェリン−亜セレン酸ナトリウム(ITS;インディアナ州、Indianapolis、Roche Applied Science)を除いて、Invitrogen(カリフォルニア州、Carlsbad)のものである。鶏卵L−α−ホスファチジルコリン75%(w/w)とブタ脳L−α−ホスファチジルセリン25%(w/w)からなるラメラリン脂質小胞体(PCPS)(アラバマ州、Alabaster、Avanti Polar lipids)は記載されたように調製され、及び、特徴付けられている。
【0070】
タンパク質
ヒトプロトロンビンは先述したように血漿から分離される(23)。プレトロンビン−1及びプレトロンビン2は確立されたやり方によって精製され(18)、及び、トロンビンは、血液学的技術から得られる。FV(hFV−810)、rFVa、rFX及びrFXaの構成的活性部分B−ドメインレス型が調製され、(3、24、25)に記載したように精製され、特徴付けられる。pseutarinC(pt−FXa)の精製された酵素サブユニットは、QRxPharma(オーストラリア、Sydney)から豊富に提供される。組み換え型ヒルジンは、EMD−Chemicals(カリフォルニア州、San Diego)から得られる。組み換え型ダニ抗凝固タンパク質及びヒトAPCは、Dr. Sriram Krishnaswamy(The Children’s Hospital of Philadelphia)より得られる。使用されている様々なたんぱく質の分子量及び吸光係数(E0.1%,280nm)は、プロトロンビンが72,000及び1.47(18);プレトロンビン‐1が1 49,900及び1.78(18);プレトロンビン‐2が37,500及び1.95(18);トロンビンが37,500及び1.94 (26);rFXaが46,000及び1.16(27);rFVa,が175,000及び1.78(3);rFV−810が216,000及び1.54(3),及びpt−FVが170,000,及び1.50(本研究による)である。pt−FXaにために、ヒトタンパク質のための値が使用されている。特別な記載がない限り、全ての機能的アッセイは、20mMヘペス、0.15M塩化ナトリウム、2mM塩化カルシウム、0.1%ポリエチレングリコール8000、pH7.5(アッセイバッファー)において25℃で行われる。
【0071】
分析的超遠心分離:
分子量は、光学干渉を用いた分析的超遠心分離機Beckman Optima XL−Iで測定される。沈降速度は20mMのHEPES、150mMの塩化ナトリウム、5mMの塩化カルシウム、pH7.4、AN60Tiローターによる毎分25,000回転で測定される。タンパク質濃度は、pt−rFVが0.60mg/mL(3.6μM)及びpt−FXaが0.18mg/mL(3.9μM)である。沈降係数及び分子量はg(s*)分析により測定される。減衰係数は、Babul and Stellwagen(29)に記載された手順による示差屈折率測定により測定される。
【0072】
rFV変異体の構造
毒腺由来のコブラ・モドキ(P.textilis)FV cDNA(8)は、隣接XmaI制限酵素認識部位により修飾され、それにより、pED発現プラスミド内にサブクローニングされる(3)。cDNAは、さらに、コブラ・モドキ(P.textilis)FVの信号配列及び重複によるスプライシングの技術を使用するヒトFVの信号配列をコードする配列を交換することにより修飾される(30)。最終的な構成、pED−FV−ptex、その後に成熟ベノム由来pt−FVタンパク質(残渣1から1430)が続くヒトFV信号ペプチド(残渣−28から1)のためにコードされる。
Rao et alによりレポートされるのと同一の配列を示すコブラ・モドキ(P.textilis)FV cDNA(7)は、コドン50(Lys)及び1305(Phe)を除いて、コースタルタイパン(O.scutellatus)及びタイパン(O. microlepidotus)の相応する残渣と同一である(8)。Glnにより置換されたArg742及びArg788を有するpt−FV変異体は、適当な突然変異誘発性相補的オリゴヌクレオチドを使用するQuickChange部位に誘導された突然変異生成キット(カリフォルニア州、La Jolla、Stratagene)により発生した。これらのアミノ酸は、ヒトFVにおいてアルギニン(Arg)709及びアルギニン(Arg)1545に相同であり、及び、トロンビン開裂部位を示す。突然変異生成に続いて、cDNA全体は、目的とする突然変異の存在を確認するため、及び、誤って誘導されたポリメラーゼがないことを確かめるために配列される。
【0073】
pt−rFVの発現及び精製:
FVまたはFV−QQ を含むプラスミドは、ベビーハムスター腎臓(BHK)細胞内に誘導され、及び、高生産安定クローンは、(3)に記載されたように確立される。細胞は三角フラスコ内に広がり、ITS及び2.5mMの塩化カルシウムを追加されたDMEM/F12媒体内で培養される。調製済みの媒体は、4〜6日間かけて収集され、延伸ぶりされ、及び10μM APMSF及び1mMベンズアミジンの存在下において20℃で保存される。精製のために、媒体は37℃で融解され、20mMへペス、0.15M塩化ナトリウム、5mM塩化カルシウム、pH7.4で平衡を保たれた30mLQ−セファロースカラム(ニュージャージー州、Piscataway、Amersham Biosciences)に装填される。カラムは同じバッファーで洗浄され、塩化ナトリウム0.15〜1Mの濃度勾配で溶出される。画分は、収集され、SDS−PAGE及び(3)記載のFV特定凝固アッセイにより分析される。pt−rFVを含む画分は、貯蔵され、及び20mMのMES、2mM塩化カルシウム、pH6.0に対して透析され、及び同一のバッファーで平衡を保たれたPoros HS/20カラム(10×100mm;カリフォルニア州、Foster City、Applied Biosystems)に装填される。カラムは20mMのMES、2mMの塩化カルシウム、pH6.0で洗浄され、及び、塩化ナトリウム0〜0.6M濃度勾配で溶出される。画分を含むFVは貯蔵され、及び、20mMへペス、0.15M塩化ナトリウム、2mM塩化カルシウム、pH7.4に対して透析され、及び、タンパク質は−80℃で保存される。最終収率は、調製済みの媒体1リットルあたりのpt−rFVに対して〜4mgである。
【0074】
タンパク質の特徴:
純正タンパク質は、クマシー ブリリアント ブルーR−250による染色に続くMOPSバッファーシステムを用いる非還元及び還元条件(50mMジチオスレイトール)下で、プレキャスト4〜12%勾配ゲル(Invitrogen)を用いるSDS−PAGEにより評価される。N末端配列分析は、ガルベストンのTexas Medical Branch大学のDr.Alex Kurosky及びSteve Smith、またはCenter for Disease Control(ジョージア州、Atlanta)のDr.Jan Pohlの研究室で行われる。pt−FXaにおける化学的γ−カルボキシグルタミン酸(Gla)分析は記載した研究室で行われる(24)。
【0075】
タンパク質基質開裂の反応速度論:
高分子基質開裂の定常状態における初期速度は、記載されているように、25℃で不連続に決定される(25)。プロトロンビナーゼまたはFXa−FVaに触媒作用されたプロトロンビンまたはプレトロンビン−1活性型(Km及びVmax)の速度パラメータは、高分子基質の濃度を上昇させることでトロンビン構造の初期割合を測定することにより、アッセイバッファーにおいて決定される。アッセイ混合物は、PCPS(50μM)、DAPA(3μM)、様々なpt−rFV(a)補因子種(20nM)及び様々な濃度のプロトロンビン(0〜5.0μM)またはプレトロンビン−1(0〜15μM)が含まれる。プレトロンビン−2がタンパク質基質として使用されると、それに続く反応条件は、PCPS(50μM)、DAPA(3μM)、及びプレトロンビン−2(1.4μM)が用いられ、様々なFV(a)補因子(0〜40nM)と保温培養され、及び、反応はpt−FXa(5nM)により開始される。特定の実験においては、PCPSは、反応混合物に含まれない(図の説明文を参照)。pt−rFVaが使用される実験においては、pt−rFV(600nM)は2nMトロンビンと37℃で15分間保温培養され、続いて2.4nMヒルジンが加えられる。
【0076】
pt−rFVのAPC不活性:
PCPS(50μM)存在下における様々な補因子タンパク質(500nMのpt−rFV,pt−rFVa,rhFVa,またはh−FV−810)の不活性化は、APC添加(10μMまたは750nM)により開始される。反応混合物の一定分量は指定された時間間隔で取り出され、SDS−PAGE(4〜12%ゲル)及び機能的活性の評価を通して評価される。このアッセイにおいて、残渣補因子活性は、特に(25)に記載されたようなプロトロンビン活性におけるFVaの作用を測定することにより評価される。アッセイ混合物には、1.4μMプロトロンビン、50μMのPCPS、3μMのDAPA、1nMのFVa、及び1nMのpt−FXaが含まれる。rhFVaまたはhFV−810と、ヒトrFXaは代わりに使用される。これらの条件下で、反応の初期割合は補因子の濃度と比例している。
【0077】
データ分析:
データは、マーカード アルゴリズム(Marquardt algorithm)を使用する非線形最小二乗法による回帰分析から求められる参照式によって分析される(31)。適合の質は、(32)に記載の基準により評価した。エラーの報告された推定値は、±2 S.Dを示す。pt−FXa及びpt−rFV(a)間での相互作用における解離定数(Kd)は、プレトロンビン−2活性型の初期値の1に固定された化学量論と補因子の濃度への依存から得られる(33)。プロトロンビナーゼによるプロトロンビンまたはプレトロンビン−1の開裂の初期速度測定は、Km及びVmaxの近似値を得るため、ヘンリー−ミカエリス−メンテン(Henri−Michaelis−Menten)の方程式(34)にデータを適合させることで分析される。
【0078】
以下に記載の実施例は、本発明の特定の実施形態を示すものとして提供される。これらはいかなる場合においても本発明を制限することを意図していない。
【実施例1】
【0079】
ヒトの治療における使用のためのヘビ毒由来またはヒト型誘導体の第五因子
本発明はコブラ・モドキ(P.textilis)からのヘビ毒由来FVの生物学的特性を記述する。これらの固有な性質は、以下に列記するように、凝血原特性を反復するヒトFV誘導体に設計されることもある。これらの性質とは、a)pt−FVが活性状態で存在し、補因子が自然発生の活性FV変異体の第一実施例のようであること、b)pt−FVが、セリンプロテアーゼ因子Xa(FXa)と高い親和性で結合するのを達成する膜表面のための通常要件を回避するような固有な形態を有すること、c)重鎖及び軽鎖を結合するpt−Fvの固有のジスルフィド結合により、pt−FVは、活性型タンパク質C(APC)による不活性に機能的に抵抗すること、である。これらの特徴の一つまたはそれ以上からなるヒトFV誘導体を操作することは、その凝血原特性を強化し、したがって、出血性疾患の治療において有用な治療的ツールを提供する。
【0080】
本研究においては、非常に精製されたヘビ毒由来Fxa(pt−FXa)と同様に、コブラ・モドキ(P. textilis)からの組換え型ヘビ毒由来FV(pt−rFV)による一連の機能的測定値は、FVの固有の形態の生物学的性質を調査し、ヘビ毒由来プロトロンビナーゼ複合体の強力な凝血性質における基本的メカニズムを調べるのに使用される。この働きは、pt−rFVが、哺乳類FVにおいては起こることが未知である機能的要素の複数の利益を獲得することで、有力な凝血源に適応したことを証明している。この研究結果は、FVの構造/機能関係の複数の態様に新しい光を投じ、治療的な目的のための生物学FVを容易にし、さらに、選択的優位性のためのタンパク質の既存の特性を強化する性質の驚くべき能力を強調する。
【0081】
結果
pt−rFVの発現及び物理的特徴
天然のヘビ毒の高いプロテアーゼ含有量のために、同種の単離、pt−FVの未開裂調製は得られない(12〜15)。これを回避するために、BHK細胞から精製されたpt−rFVは高い収量で示された。野生型pt−rFV及び重鎖及び軽鎖の側面に位置するトロンビン開裂部位が欠乏する変異体(pt−rFV−QQ)は、還元型SDS−PAGEにおいてシングルバンド(〜180kDa)として主に移行する(図1、パネルA)。沈降速度の研究は、pt−rFVが、170,000の分子量評価により均一であることを示した。pt−rFV−QQがトロンビンにより開裂されないのに対し、pt−rFVはpt−rFVaに処理され、特有の重鎖及び軽鎖を産出する。N−末端配列分析は、予測されたトロンビン開裂部位における的確な処理を明らかにした(図1)(7,8)。驚くべきことに、トロンビン処理の後に、pt−rFVaの大部分(>60%)は、非還元条件下においてシングルバンドとして移動し(図1、パネルB)、重鎖及び軽鎖が、固有のジスルフィド結合により結合していることを示している。この可能性は、非還元条件下における二重項としてpt−rFV及びpt−rFV−QQの移行を説明している。配列アラインメントは、重鎖に二つの遊離Cys残基(A2ドメイン、539及び641)、軽鎖に一つの遊離Cys(A3ドメイン、1032)を有するであろうことを示している(7)。
【0082】
【表1】

【0083】
コブラ・モドキ(P.textilis)またはコースタルタイパン(O.scutellatus)の天然のヘビ毒から精製されたFVは、ウシFXaの存在下において低活性である(14、15)。pt−rFVを特徴付ける初期の試みは、これらの観察と矛盾しない。しかしながら、推定値は、pt−rFVまたはpt−rFVaはヒトFVaと比較してほぼ=1/40th活性を示す。低活性は、主にpt−rFVまたはpt−rFVaが、確立された経口結合測定値を使用する膜結合ヒトFXa(Kb>400nM)との親和性を減らすことで結合されることにより起因するようである(データは示されていない)(3)。したがって、pt−rFVを特徴付けるため、pt−FXaが全てのそれに続く実験において使用された。pt−FXaのSDS−PAGE分析は、予測される結果を与えたN末端配列分析とともに図1に示される。二重項としての軽鎖の外観は、不均一なO−グリコシル化に原因があるようだ(8、16)。さらに、pt−FXaの化学的Gla分析は、配列アラインメントから予測される11モル Gla/molタンパク質から10.9±0.34モルGla/molタンパク質を産出した。
【0084】
Pt−rFVは構成的に活性型補因子である
pt−FV Bドメインの変わった性質のため、一連の機能的測定は、ヘビ毒由来FVのこのドメインが、プロコファクターとしてのタンパク質を維持するように機能するかどうか確定するために使用された。機能的な測定値に対する一つの懸案事項は、元の位置に生成されたトロンビンによるpt−rFVのフィードバック・タンパク質分解である。これに適応するために、反応体の濃度はタンパク質分解を最小にするように選ばれ、及び、トロンビンの可逆性抑制剤(DAPA)がアッセイに含まれる。さらに、pt−rFV−QQ、トロンビンによって媒介されるタンパク質分解に抵抗する変異体(図1)が調製された。ヒトFV(1)とは対照的に、膜結合pt−FXaを有するpt−rFvの組み立ては、プロトロンビンを急速に活性化させる(図2A)。プロトロンビン転換のための定常状態速度定数は、ヒトプロトロンビナーゼにおいて観察される定常状態速度定数と著しく類似している。(表2)(3、17)。興味深いことに、ヒトFVの誘導体で切断されたB−ドメイン、hFV−810は、pt−FV−膜によって集まり、相当する速度パラメータを有するプロトロンビンを活性化することが可能であった(表2)。pt−rFVaを生成するまたは、未開裂変異体、pt−rFV−QQを使用するトロンビンによる短いB−ドメインの除去は、相当する速度定数を与える(図2A及び表2)。これらのデータは、ヒトFVとは異なり、pt−FVは、構成的に活性型補因子であり、及び、B−ドメインのタンパク質分解除去が凝血原活性を表すことを必要としないことを示す。これは、FVの構成的活性型の自然な第一実施例を表す。
【0085】
【表2】
【0086】
アニオン性膜の非存在下でのPt−rFVの機能
血液凝固の重要な観点は、凝固タンパク質を細胞表面に(ホスファチジルセリン曝露を介して)結合する膜が正常な止血のためには重要であるということである(2)。オスキュタリン(oscutarin)及びプスタリンC(pseutarin C)によりみられる変わった観察結果は、ヘビ毒由来FXa及びFVが複合体として同時精製し、陰イオン膜非存在下において、ある程度活性を示すことである。しかしながら、オスキュタリン(oscutarin)の活性は、膜存在下において、実質的に増加する(>400−倍)ことが報告されている(12〜15)。さらにこれを調べるために、プロトロンビンを使用する溶液位相pt−FXa−pt−rFVの活性が評価された。膜非存在下において、pt−rFV、pt−rFV−QQまたはpt−rFVaを有するpt−FXaを使用するプロトロンビンの初期割合は、同じ酵素濃度で膜存在下において得られる割合と比較して7〜8倍低いものであった(図2B)。活性の減少は、プロトロンビンのKm増加(〜4倍)及びkcatの減少(〜4倍)によって反映された。対照的に、膜非存在下において、hFV−810は、膜結合補因子と比較して非常に低い活性を有した(>150倍)(図2B)。
【0087】
これらのデータの解釈の困難は、基質、プレトロンビン、トロンビンの生成の割合を高めるのに貢献する結合膜にある。さらに、pt−FXa−pt−rFVのための溶液位相解離定数に応じて、pt−FXaはpt−rFVで飽和することができず、実際の酵素複合体の濃度を低下させ、プロトロンビン活性率の減少へと至った。一点目に取り組むために、膜の存在及び非存在下でのプレトロンビン−1活性の割合が測定される。プレトロンビン−1は、γ−カルボキシグルタミン酸及びクリングル−1ドメインと結合する膜を欠失するプロトロンビンの派生物である(18)。プロトロンビンを使用している結果と整合して、pt−rFV、pt−rFV−QQまたはpt−rFVaを使用する膜結合pt−FXaを有するプレトロンビン−1の反応速度は、等しいものであり(図3A及び表2)及びヒトプロトロンビナーゼを使用する速度パラメータと同等であった(17)。しかしながら、意外にも、反応混合物からの膜の除去は、溶液位相または膜結合pt−FXa−pt−rFVにおける運動パラメータが等しいため、プレトロンビン−1活性の割合において感知できるほどの効果は有さない(図3B及び表2)。これらのデータは、pt−FXa−pt−rFVを有するプロトロンビン活性割合の減少率が、酵素機能ではなく、むしろ結合している基質膜の除去にあったことを示唆している。
【0088】
Pt−FXa及びpt−rFVが形成する高親和性複合体
膜の存在または非存在におけるプレトロンビン−1活性の割合が同等になると、データは、pt−FXaがpt−rFVと完全に飽和されることを示す。しかしながら、これを直接評価するために、いずれの膜結合または溶液位相pt−FXa−pt−rFVにおける平衡解離定数を意味する機能的な結合測定が使用された。補因子(pt−rFV、pt−rFV−QQ、pt−rFVaまたはhFV−810)の濃度の増加は、PCPSの存在下または非存在下において、pt−FXaの濃度を固定して、単一で培養された。プレトロンビン−2活性の初期割合は測定され、及び、データはpt−FXaと結合している補因子に推定された平衡解離定数を得るために分析された(表3)。PCPSの存在下において、補因子の種類に関係なく、pt−FXaと結合している解離定数はほぼ=3nMであった。この値は、膜結合ヒトFXa及びFVaを使用して得られた解離定数と類似しているものである(3)。驚くべきことに、pt−rFV、pt−FV−QQまたはpt−rFVaにおける解離定数は、膜非存在下において、わずか2〜3倍増加させた(Kdほぼ=8nM)。これは、FXa−FVaにおける溶液位相解離定数がほぼ=0.8〜3.0μMである牛におけるシステムとは対照的である(19、20)。沈降速度実験では、溶液位相pt−rFV−pt−FXaが213,000の分子量及び9.33の沈降係数(s20,w)と1:1で化学量論複合体を形成していた。これらの値はウシFXa−Fvaによって得られた結果と非常によく一致している(19)。ヘビ毒由来補因子とは対照的に、溶液位相hFV−810−pt−FXaにおける結合定数(表3)は、確立することができず、pt−FXa結合が、少なくとも溶液において、hFV−810に存在しないpt−rFVにおいて固有な性質を含むことを示唆している。これらのデータは、pt−FXa−pt−FV複合体は、プロトロンビンを急速に活性化できる1:1複合体が高い親和性を形成するため、負に荷電された膜において通常必要とされるものを回避したことを示す。
【0089】
【表3】
【0090】
Pt−rFVはAPCに機能的耐性がある
APCは止血系及びタンパク質分解の制限を介してFVa及びFVIIIaの両方を不活性化する機能における重要な抗凝血剤である(21)。ヒトFVaにとって、Arg306、Arg506及びArg679での開裂は、A2ドメインの解離において、FXa結合及び最終的には補因子機能の消失を生じる(1)。pt−FVの配列分析は、これらの部位にAPC耐性がある可能性の示唆は保存されていない(7)。プスタリンCの研究はこの論点を支持している(7);しかしながら、FXaは、データ解釈を複雑にするAPCによる不活性からFVaを保護する(1)。pt−rFVがAPCに対して耐性を示すかどうか直接調べるために、これらのタンパク質は、PCPS存在下において培養され、及び、様々な時間間隔で、SDS−PAGE及び精製されたプロトロンビンアッセイを使用する補因子活性によって、タンパク質分解を観察した。結果的にrhFVaまたはrFV−810機能を完全に失ったAPC濃度(10nM)及び反応時間(≦15分)を使用して、pt−rFvまたはpt−rFVaは完全に活性を保持し(図4A)及び、SDS−PAGE分析は、これらのタンパク質の限定された開裂のみを示した(データは示されていない)。しかしながら、長時間における高濃度APC(750nM)とのpt−rFVの培養は、結果として完全なタンパク質分解pt−rFVとなった(図4B、C)。N−末端配列分析と還元及び非還元ゲルの分析はともに、APCがA2ドメインにおいてpt−rFVを開裂することを示す。驚くべきことに、pt−rFVの完全なタンパク質分解にもかかわらず、補因子は、完全な凝血原活性を保持した(図4Aに挿入)。
第五因子変異体をコードする核酸配列は図5において提供されている。
【0091】
議論
多くの血液凝固タンパク質は、不活性な前駆体として合成され、及びその結果としてタンパク質分解活性があり、これらのタンパク質は、血管外傷(2)の場所に位置する細胞表面上に組み立てられる。本研究において、pt−FVは、これらのパラダイムを、FXa及び活性型プロトロンビンを結集させる膜表面に必要な構造的活性化補因子及びバイパスとして合成されたものと同様に回避する。さらに、Pt−FVはAPC、タンパク質分解的切断後の活性の保持に機能的耐性がある。改良された非共有結合性の相互作用と重鎖及び軽鎖を結合する独自のジスルフィド結合はおそらく機能安定性に貢献する。これらの適用の集密は、強力な凝血促進剤に形質転換されたpt−FVを有し、及び、毒物注入に続いておこる重度の播種凝固による選択的優位性のあるヘビ種をおそらく提供する。
【0092】
参考文献
1. Mann KG, Kalafatis M (2002) Factor V: A combination of Dr. Jekyll and Mr. Hyde. Blood 101:20−30.
2. Mann KG, et al. (1990) Surface dependent reactions of the vitamin K−dependent enzyme complexes. Blood 76:1−16.
3. Toso R, Camire RM (2004) Removal of B−domain sequences from factor V rather than specific proteolysis underlies the mechanism by which cofactor function is realized. J Biol Chem 279:21643−21650.
4. Zhu H, Toso R, Camire RM (2007) Inhibitory sequences within the B−domain stabilize circulating factor V in an inactive state. J Biol Chem 282:15033−15039.
5. Jiang Y, Doolittle RF (2003) The evolution of vertebrate blood coagulation as viewed from a comparison of puffer fish and sea squirt genomes. Proc Natl Acad Sci U S A 100:7527−7532.
6. Davidson CJ, et al. (2003) Molecular evolution of the vertebrate blood coagulation network. Thromb Haemost 89:420−428.
7. Rao VS, Swarup S, Kini RM (2003) The nonenzymatic subunit of pseutarin C, a prothrombin activator from eastern brown snake (Pseudonaja textilis) venom, shows structural similarity to mammalian coagulation factor V. Blood 102:1347−1354.
8. St.Pierre L, et al. (2005) Comparative analysis of prothrombin activators from the venom of Australian elapids. Mol Biol Evol 22:1853−1864.
9. Welton RE, Burnell JN (2005) Full length nucleotide sequence of a factor V−like subunit of oscutarin from Oxyuranus scutellatus scutellatus (coastal Taipan). Toxicon 46:328−336.
10. Sutherland S, Tibballs J (2001) in Australian Animal Toxins. The Creatures, Their Toxins and Care of the Poisoned Patient. (Oxford University Press, Melbourne,Australia).
11. Owen WG, Jackson CM (1973) Activation of prothrombin with Oxyuranus scutellatus scutellatus (taipan snake) venom. Thromb Res 3:705−714.
12. Walker FJ, Owen WG, Esmon CT (1980) Characterization of the prothrombin activator from the venom of Oxyuranus scutellatus scutellatus (taipan venom). Biochemistry 19:1020−1023.
13. Masci PP, Whitaker AN, de Jersey J (1988) Purification and characterization of a prothrombin activator from the venom of the Australian brown snake, Pseudonaja textilis textilis. Biochem Int 17:825−835.
14. Speijer H, Govers−Riemslag JW, Zwaal RF, Rosing J (1986) Prothrombin activation by an activator from the venom of Oxyuranus scutellatus (Taipan snake). J Biol Chem 261:13258−13267.
15. Rao VS, Kini RM (2002) Pseutarin C, a prothrombin activator from Pseudonaja textilis venom: its structural and functional similarity to mammalian coagulation factor Xa−Va complex. Thromb Haemost 88:611−619.
16. Rao VS, Swarup S, Kini RM (2004) The catalytic subunit of pseutarin C, a group C prothrombin activator from the venom of Pseudonaja textilis, is structrually similar to mammalian blood coagulation factor Xa. Thromb Haemost 92:509−521.
17. Toso R, Camire RM (2006) Role of Hirudin−like factor Va heavy chain sequences in prothrombinase function. J Biol Chem 281:8773−8779.
18. Mann KG (1976) Prothrombin. Methods Enzymol 45:123−156.
19. Pryzdial ELG, Mann KG (1991) The association of coagulation factor Xa and factor Va. J Biol Chem 266:8969−8977.
20. Boskovic DS, Giles AR, Nesheim MD (1990) Studies of the role of factor Va in the factor Xa−catalyzed activation of prothrombin, fragment 1 2−prethrombin−2, and dansyl−1−glutamyl−glycyl−l−arginine−meizothrombin in the absence of phospholipid. J Biol Chem 265:10497−10505.
21. Esmon CT (2003) The protein C pathway. Chest 124:26S−32S.
22. Higgins DL, Mann KG (1983) The interaction of bovine factor V and factor V−derived peptides with phospholipid vesicles. J Biol Chem 258:6503−6508.
23. Buddai SK, et al. (2002) Nematode anticoagulant protein c2 reveals a site on factor Xa that is important for macromolecular substrate binding to human prothrombinase. J Biol Chem 277:26689−26698.
24. Camire RM, Larson PJ, Stafford DW, High KA (2000) Enhanced g−carboxylation of recombinant factor X using a chimeric construct containing the prothrombin propeptide. Biochemistry 39:14322−14329.
25. Camire RM (2002) Prothrombinase assembly and S1 site occupation restore the catalytic activity of FXa impaired by mutation at the sodium−binding site. J Biol Chem 277:37863−37870.
26. Lundblad RL, Kingdon HS, Mann KG (1976) Thrombin Methods Enzymol 45:156−176.
27. Di Scipio RG, Hermodson MA, Yates SG, Davie EW (1977) A comparison of human prothrombin, factor IX (Christmas Factor), factor X (Stuart Factor), and protein S. Biochemistry 16:698−706.
28. Stafford WF (1992) Boundry analysis in sedimentation transport experiments: a procedure for obtaining sedimentation coefficient distributions using the time derivative of the concentration profile. Anal Biochem 203:295−301.
29. Babul J, Stellwagen E (1969) Measurement of protein concentration with interferences optics. Anal Biochem 28:216−221.
30. Horton RM, et al. (1989) Engineering hybrid genes without the use of restriction enzymes: gene splicing by overlap extension. Gene 77:61−68.
31. Bevington PR, Robinson KD (1992) in Data Reduction and Error Analysis for the Physical Sciences. (McGraw−Hill, New York).
32. Straume M, Johnson ML (1992) Analysis of residuals: criteria for determining goodness−of−fit. Methods Enzymol 210:87−105.
33. Krishnaswamy S (1990) Prothrombinase complex assembly: Contributions of protein− protein and protein−membrane interactions toward complex formation. J Biol Chem 265:3708−3718.
34. Segal IH (1975) in Enzyme Kinetics: Behavior and Analysis of Rapid Equilibrium and Steady State Enzyme Systems. (John Wiley & Sons, Inc., New York).
【0093】
本発明の特定の好ましい実施形態が明細書において記載され、具体的に例示されているが、本発明は実施形態のように限定されるものと意図していない。以下の請求の範囲から逸脱することなく、本発明に様々な変更がなされることがある。
【技術分野】
【0001】
この出願は、35 U.S.C.§119(e)に基づいて、2008年11月21日に出願された米国仮出願第61/116,913号に対して優先権の主張を行い、その全体の開示は、完全に参照により本明細書に組み込まれる。
35 U.S.C.§202(c)に基づいて、米国政府はここに記載された発明において一定の権利を有することを認めている。本発明は、国立保健研究機構、認可番号HL−088010及びP50HD055784−01からの資金の一部により行われた。
【0002】
本発明は医学及び血液学の分野に関するものである。さらに具体的には、本発明は、ヘビ毒由来の第五因子(FV)の変異体およびそれらの誘導体を、それらを必要とする患者において凝固カスケードを調節するために使用する治療戦略について記述する。
【背景技術】
【0003】
いくつかの出版物及び特許文献は、本発明の属する技術分野について記述するため明細書に引用されている。これらの引用はそれぞれ、参照として明細書に完全に組み込まれる。
【0004】
血液凝固FVは、マルチドメインタンパク質(A1−A2−B−A3−C1−C2)であり、わずかな凝血原活性、または凝血原活性なしでプロコファクターとして循環する。補因子の活性化状態(FVa)への転換は、トロンビンによる、グリコシル化された中央Bドメインの大部分の分解除去を含む(1)。FVaは、プロトロンビナーゼの形成、プロトロンビンの生理活性のため、カルシウムイオンの存在下で負に帯電した膜表面上のセリンプロテアーゼ因子Xa(FXa)と会合する(2)。FVaがトロンビンの生成を大幅に高めるため、非活性プロコファクターとしてのFVの維持は、正常な止血の維持を発展させるうえで間違いなく重要な役割を果たす。最近では、Bドメイン内の個別の配列は、非活性プロコファクターとして持続するFVのメカニズムに寄与することが示されている(3,4)。この領域の除去は、タンパク質分解処理を必要とせずにFV活性をもたらす。
【0005】
FVは、A及びCドメイン間では高い相同性、Bドメイン間では低い相同性を脊椎動物間で広く示される(5,6)。最近のゲノムデータでは、Bドメインの長さが一般的に哺乳類の間で共通して保存されている(〜800残基)のに対し、下等脊椎動物では大きく異なる(例えば、トラフグでは〜500残基)ことを示している(5,6);これらの構造変化の機能的な意味は理解されていない。Bドメインへの主要な変化の顕著な例は、オーストラリアのヘビのグループ(タイパン(Oxyuranus microlepidotus)、コブラ・モドキ(Pseudonaja textilis)及びコースタルタイパン(Oxyuranus scutellatus))からきている。これらの種は独特であり、加えて、これらの止血システムの一部としてFVは循環しており、その毒にもFVを有している。これらの種のそれぞれからの毒由来FVは、同様のドメイン構造を有する哺乳類のFVと、〜44%の配列相同性が共通している。しかしながら、Bドメインの長さはヒトのFVと比べて飛躍的に短い(836残基に対して、〜46残基)。これらのヘビからの毒は、世界で最も有毒であるとみなされており、強力な血液凝固促進剤であり、及び、いくつかある症状のなかで特に、播種性血管内凝固障害を引き起こす(10)。これは、大量のFVと同様に、共に強力なプロトロンビン活性化因子である複合体を形成するFXa様酵素を含む。コースタルタイパン(オスキュタリンC)及びコブラ・モドキ(プスタリンC)からの複合体は部分的に精製され、及び粗毒と特徴づけられる。プロトロンビンのトロンビンへの変換もそれらの活性を強化するのもカルシウム及びリン脂質によって程度が異なるが、FVaは影響しない。さらに、プスタリンCは活性化タンパク質C(APC)に対して耐性を示し、強力な毒素としての役割に潜在的に貢献する(7)。
【発明の概要】
【課題を解決するための手段】
【0006】
本発明によって、それを必要とする患者における血液凝固障害を治療する方法が提供される。例示的な方法では、FV変異体またはそれらの誘導体の活性型の有効量を投与する工程であって、それによって前記患者の血栓形成を増進し、且つ出血性疾患の症状を改善するものである、前記投与する工程を伴う。一実施形態においては、変異第五因子は配列ID番号1の配列を有する核酸によってコードされる。他の実施形態においては、変異第五因子は、配列ID番号2の配列を有する核酸を最適化したコドンによってコードされる。
【0007】
前記方法及び第五因子凝血原は、血液凝固障害の治療に使用されることもある。血液凝固障害は、これに限定されるわけではないが、血友病AおよびB、抑制抗体に関連する血友病A及びB、凝固因子欠乏症、ビタミンKエポキシド還元塩素欠乏症、ガンマカルボキシラーゼ欠乏症、外傷に伴う出血、負傷、血栓症、血小板減少症、脳卒中、凝血障害、播種性血管内凝固障害(DIC)、過抗凝固治療障害、バーナードスーリエ症候群、グランツマンの血小板無力症、及び貯蔵プール欠乏症を含む。また、血液凝固障害は、ヘパリン、低分子量ヘパリン、五糖類、ワルファリン、低分子抗血栓及びFxa阻害剤の投与によることもある。
【0008】
本発明の変異第五因子は、患者に直接注入または局所投与することができ、または、それをコードするベクターの形で移送され得る。
【0009】
また、以下の性質の一つまたはそれ以上を示す、ヒト化及び改善型第五因子変異体が提供される。FV変異体は、活性化、補因子の状態で合成される;FV変異体は、膜表面が、セリンプロテアーゼXa因子(FXa)と高い親和性で結合するための通常の条件を回避できるよう、独自の立体構造を有する;及び、FV変異体は、その重鎖及び軽鎖を接続する固有なジスルフィド結合を含めることにより、活性化タンパク質C(APC)による不活性に対して機能的に耐性がある。
【0010】
本発明の他の実施形態においては、ヘビ第五因子をコードする配列ID番号2の単離された核酸が提供される。このような核酸は、使用のために発現ベクターに含まれ得る。
【0011】
さらに別の実施形態では、本発明は、配列ID番号1の核酸によりコードされた、単離されたヘビ第五因子タンパク質を提供する。ヘビ第五因子タンパク質は、リポソームまたはミセルに含有され得る。また、本発明は、薬学的に許容される賦形剤に含まれるヘビ第五因子タンパク質からなる薬学的組成物を提供する。
【図面の簡単な説明】
【0012】
【図1】精製タンパク質のSDS−PAGE分析 タンパク質(3μg/レーン)は、還元(パネルA、C)条件下、または、非還元(パネルB)条件下でSDS‐PAGEに曝され、及びクマシーブリリアントブルーR‐250で染色することにより可視化される。レーン1、pt−rFV、レーン2、pt−rFV−QQ、レーン3、pt−rFV プラス トロンビン、レーン4、pt−rFV‐QQ プラス トロンビン、レーン5、rFX、レーン6、rFXa、レーン7,pt−FXa。標準の見かけの分子量が左側に示されている。パネルA及びパネルBは同じマーカーを共有している。ダッシュ(−)は、収率がアミノ酸を正確に割り当てるにはあまりに低量であることを示し、及び、可能性の高いγ−カルボキシグルタミン酸の存在を示す6つの位置である。
【図2】PCPSの存在下、または非存在下におけるプロトロンビンの開裂の速度定数の測定 トロンビン生成の初期速度は、50μMのPCPS及び0.1nMのpt−FXa及び20nMのhFV−810(−◆−または−◇−)、pt−rFV(−●−または−○−)、pt−rFV−QQ(−▲−または−△−)、またはpt−rFVa(−■−または−□−)と3μMのDAPAの存在(パネルA)または非存在(パネルB)でのプロトロンビンの濃度の増加によって決定される。全てのデータセットの以下の分析は矩形双曲線に描かれ、適した速度定数は表2に示されている。データは、2つの類似した実験においての代表的なものである。
【図3】PCPSの存在下、または非存在下におけるプレトロンビン−1の開裂の速度定数の測定 トロンビン生成の初期速度は、50μMのPCPS及び0.1nMのpt−FXa及び20nMのhFV−810(−◆−または−◇−)、pt−rFV(−●−または−○−)、pt−rFV−QQ(−▲−または−△−)、またはpt−rFVa(−■−または−□−)と3μMのDAPAの存在(パネルA)または非存在(パネルB)でのプレトロンビン−1の濃度の増加によって決定される。全てのデータセットの以下の分析は矩形双曲線に描かれ、適した速度定数は表2に示されている。データは、2つの類似した実験においての代表的なものである。
【図4A】APCによる膜結合型FV(a)の不活性化 50μMのPCPS及び500nMのpt−rFV(−●−)、pt−rFVa(−(−)、hFV−810(−◆−)、またはrFVa(−(−)を含む反応混合物は10nMまたは750nM(inset)のAPCどちらかと保温培養される。選択した時間間隔において、試料は削除され、及び、「実験的手法」(パネルA)に記載された方法で、または、SDS−PAGEゲル上を走らせ、還元下(パネルB)または非還元条件下(パネルC)でクマシー ブルー R250で染色して、補因子活性をアッセイ分析した。データは二つの類似した実験においての代表的なものである。
【図4BC】APCによる膜結合型FV(a)の不活性化 50μMのPCPS及び500nMのpt−rFV(−●−)、pt−rFVa(−(−)、hFV−810(−◆−)、またはrFVa(−(−)を含む反応混合物は10nMまたは750nM(inset)のAPCどちらかと保温培養される。選択した時間間隔において、試料は削除され、及び、「実験的手法」(パネルA)に記載された方法で、または、SDS−PAGEゲル上を走らせ、還元下(パネルB)または非還元条件下(パネルC)でクマシー ブルー R250で染色して、補因子活性をアッセイ分析した。データは二つの類似した実験においての代表的なものである。
【図5A】第五因子のための核酸コーディング配列 第五因子(配列ID番号1)に誘導されたヘビ毒の核酸配列は、パネル5A及び5Bに示されている。哺乳類系での発現に最適化されたコドンを有する第五因子(配列ID番号2)に誘導されたヘビ毒の核酸配列はパネル5C及び5Dに示されている。
【図5B】第五因子のための核酸コーディング配列 第五因子(配列ID番号1)に誘導されたヘビ毒の核酸配列は、パネル5A及び5Bに示されている。哺乳類系での発現に最適化されたコドンを有する第五因子(配列ID番号2)に誘導されたヘビ毒の核酸配列はパネル5C及び5Dに示されている。
【図5C】第五因子のための核酸コーディング配列 第五因子(配列ID番号1)に誘導されたヘビ毒の核酸配列は、パネル5A及び5Bに示されている。哺乳類系での発現に最適化されたコドンを有する第五因子(配列ID番号2)に誘導されたヘビ毒の核酸配列はパネル5C及び5Dに示されている。
【図5D】第五因子のための核酸コーディング配列 第五因子(配列ID番号1)に誘導されたヘビ毒の核酸配列は、パネル5A及び5Bに示されている。哺乳類系での発現に最適化されたコドンを有する第五因子(配列ID番号2)に誘導されたヘビ毒の核酸配列はパネル5C及び5Dに示されている。
【発明を実施するための形態】
【0013】
毒ヘビは自分自身の防御のため、及び獲物を無力化するために、血液凝固促進剤などのような有毒な化合物の配列を生成する。オーストラリアのブラウンスネーク、コブラ・モドキ(Pseudonaja textilis)は、Xa因子(FXa)及び第五因子(FV)と一致する、毒由来のプロトロンビン活性化複合体を有する。前記FV組成物(pt−FV)は、ヒトFVと同様のドメイン組織を有するが、その中央ドメインはとても少ない(836アミノ酸に対して55)。ここでは、pt−FVは強力な血液凝固促進剤に変換する機能要素の固有な増加を獲得している。組み換えpt−FVを用いた測定は、それが構造的に活性補因子として合成され、ヒトFVとは異なり、活性の発現にタンパク質分解を必要としないことを明らかにした。配列の比較により、非活性プロコファクター状態を安定させるのに役立つBドメイン内の塩基性アミノ酸の保存クラスターを失っていることが明らかとなった。注目すべきことは、pt−FVは、負に帯電したリン脂質の非存在下では、高い親和性でヘビFxaを結合するように働き、及び、膜の非存在下、または存在下においては、プレトロンビン‐1と同等の運動パラメータを示すことである。さらに、pt−FVは、重鎖のタンパク質分解によってもたされた濃度であっても、活性タンパク質Cに機能的耐性がある。この安定性は、ある程度、重鎖と軽鎖を結合する固有なジスルフィド結合による可能性が高い。同時に、これらの分子の技術革新の合流点は、pt−FVを血液凝固生物兵器にしている。これらの発見は、FVがプロコファクターとしてどのように保存されるかについて重要な新しい洞察を提供し、及び、FXa‐FVa相互作用が、プロトロンビナーゼの拡張作用にどのように寄与するかよりよく理解するために広範な影響を有する。
【0014】
従って、本発明はまた、第五因子(FV)の固有な誘導体を提供する。その誘導体は、一般的なブラウンスネーク、コブラ・モドキ(Pseudonaja textilis)の毒内で、または異種発現システム(pt−FV)、または以下の固有な特徴の一つまたはそれ以上を有する同様のシステムで発現するヒトFV誘導体である。
【0015】
a)FV変異体は活性化、補因子状態で合成される
b)FV変異体は、膜表面で、そのセリンプロテアーゼXa因子(FXa)と高い親和性で結合するのに通常必要とされる条件を迂回できるよう、独自の立体構造を有する
c)FV変異体は、その重鎖及び軽鎖を接続する固有なジスルフィド結合を含めることにより、活性化タンパク質C(APC)による不活性に対して機能的に耐性がある。
【0016】
I.定義
本発明の生体分子に関連する様々な用語は、上述及び明細書及び特許請求の範囲のいたるところに用いられている。
【0017】
「変異型FV」という用語は、ヘビまたは他の異種発現システムから得られる、強力な血液凝固活性を示すFV誘導体分子に関するものである。本発明の変異型FVは配列ID番号1によってコードされ、またはその変異(例えば配列ID番号2)はヒトでの使用に適合し、望ましいFV血液凝固活性を促進する。
【0018】
「止血関連疾患」または「血液凝固障害」という用語は、血友病A及びB、抑制抗体に関連する血友病A及びB、凝固因子欠乏症、VII1 IX及びX、XI、V、XII、II因子、フォン・ヴィレブランド因子欠乏症、結合FV/FV1欠乏症、ビタミンKエポキシド還元塩素欠乏症、ガンマカルボキシラーゼ欠乏症、外傷に伴う出血、負傷、血栓症、血小板減少症、脳卒中、凝血障害、播種性血管内凝固障害(DIC)、ヘパリン、低分子量ヘパリン、五糖類、ワルファリン、低分子抗血栓(例えばFXa阻害剤)に関連する過抗凝固、及びバーナースーリエ症候群、グランツマンの血小板無力症及び貯蔵プール欠乏症のような血小板異常症などの出血障害に関連する。
【0019】
本発明の核酸を参照するにあたり、「単離された核酸」という用語が時折用いられる。この用語は、DNAに適用される際に、そこから由来する生物の天然に存在するゲノムにおいて直接に連続する(5’及び3’方向において)配列から離されているDNA分子を意味する。例えば「単離された核酸」は、プラスミドまたはウイルスベクターなどのようなベクター内に挿入された、または原核生物または真核生物のDNA内に合成されたDNAまたはcDNA分子からなる。本発明のRNA分子に関しては、「単離された核酸」という用語は、前記の単離されたDNAによりコードされたRNA分子を主に意味する。また、この用語は、「実質的に純粋な」形(用語「実質的に純粋な」に関しては、以下に定義する)で存在するように、その自然な状態(例えば、細胞または組織内)で結合されたRNA分子から十分に分離されたRNA分子に関するものである。
【0020】
タンパク質に関して、「単離されたタンパク質」または「単離及び精製されたタンパク質」という用語が、時おり本明細書において使用されている。この用語は、本発明の単離された核酸分子の発現により生成されるタンパク質を主に意味する。また、この用語は、「実質的に純粋な」形で存在するように結合された他のタンパク質から十分に分離されたタンパク質に関するものである。
【0021】
用語「プロモーター領域」は、遺伝子の転写調節領域に関するものであり、コーディング領域の5’側または3’側、またはコーディング領域内、またはイントロン内でみられることもある。
【0022】
用語「ベクター」は、そこで複製される宿主細胞内に導入するため挿入されるDNA配列内の小さな担体DNA分子に関するものである。「発現ベクター」は、宿主細胞内での発現に必要とされる調節領域と遺伝子または核酸配列を含む特殊なベクターである。用語「作動的に結合された」はコーディング配列の発現に必要な調節配列が、コーディング配列の発現がなされるようコーディング配列に適切な相対位置内のDNA分子内に配置されたことを意味する。これと同様の定義が、コーディング配列の配置及び発現ベクター内の転写制御要素(例えばプロモーター、エンハンサー、及び終止要素)に時おり適用される。この定義はまた、ハイブリッド核酸分子が生じる第一及び第二核酸分子の核酸配列の配置においても時おり適用される。
【0023】
用語「実質的に純粋な」は、少なくとも50〜60重量%の対象化合物(例えば核酸、オリゴヌクレオチド、タンパク質など)からなる標本に関するものである。標本は、より好適には少なくとも75重量%、最も好適には、90〜99重量%の対象化合物からなる。純度は対象化合物によって適切な方法(例えば、クロマトグラフィー法、アガロースまたはポリアクリルアミドゲル電気泳動法、HPLC分析など)で測定される。
【0024】
特定のヌクレオチド配列またはアミノ酸配列に関する場合の「本質的に…からなる」は、与えられた配列ID番号の性質を有する配列を意味する。例えば、アミノ酸配列を参照する場合、この用語は、配列の基本的な及び新規な特徴に作用しない配列自体及び分子の修飾を含む。
【0025】
本明細書で使用される用語「オリゴヌクレオチド」は、本発明のプライマー及びプローブに関するものであり、及び、二つまたはそれ以上、好適には三つ以上のリボ−またはデオキシリボヌクレオチドからなる核酸分子として定義される。オリゴヌクレオチドの正確な大きさは様々な要因及びオリゴヌクレオチドが使用されるそれぞれの適用に依存する。
【0026】
本明細書で使用される用語「プローブ」は制限酵素消化による精製として自然発生するものであろうと、合成であろうと、アニーリングまたはプローブと相補的な配列と核酸分子に特異的にハイブリダイズできるオリゴヌクレオチド、ポリヌクレオチドまたは核酸、RNAまたはDNAに関するものである。プローブは単一らせん構造または二重らせん構造どちらもとりうる。プローブの正確な長さは温度、プローブのソース、使用方法を含む様々な要因に依存する。例えば、診断に適用する場合では、標的配列の複雑さに依存し、典型的なオリゴヌクレオチドプローブは、15〜25またはそれ以上のヌクレオチドを含むが、それよりも少ないヌクレオチドを含むこともある。
【0027】
本明細書でのプローブは特定の標的核酸配列の異なるらせん構造と「実質的に」相補的となるよう選択される。これは、事前に定められた条件下でそれぞれの標的らせん構造と「特異的にハイブリダイズ」またはアニールをするためには、プローブが十分に相補的でなければならないということを意味する。例えば、非相補的なヌクレオチドフラグメントは、標的らせん構造と相補的であるプローブ配列の残渣によって、プローブの5’末端または3’末端に付着される。あるいは、プローブ配列が、それと共に特異的にアニールする標的核酸の配列と十分に相補的である条件下で、非相補的塩基または長い配列はプローブ内に組み入れられることができる。用語「特異的にハイブリダイズする」は、当業者において通常使用される事前に定められた条件下でハイブリダイゼーションが可能となるよう十分に相補的な(時おり「実質的に相補的な」と称される)配列の二つの一本鎖核酸分子間のつながりに関するものである。特に、この用語は、本発明の一本鎖DNAまたはRNA分子内に含まれる実質的に相補的な配列によるオリゴヌクレオチドのハイブリダイゼーションに関するものであり、非相補的配列の一本鎖核酸によるオリゴヌクレオチドのハイブリダイゼーションの実質的な除去に関するものである。
【0028】
本明細書で使用される用語「プライマー」は、生体システムからの誘導、制限酵素消化による生成であろうと、合成であろうと、適切な条件下に置かれた際に核酸合成依存テンプレートの起始因子として機能的に作用できる一本鎖または二本鎖、オリゴヌクレオチド、RNAまたはDNAに関するものである。適当な核酸テンプレート、核酸の適当なヌクレオシド三リン酸、ポリメラーゼ酵素、適当な補因子及び、適当な温度及びpHなどのような条件と提供されると、ポリメラーゼの作用またはプライマーの延長生成を増やす同様の活性により、ヌクレオチドが付加され、プライマーは31末端に延長されることもある。
【0029】
プライマーは、特定の条件及び適用の求めによって、様々な長さとなることがある。例えば、診断における適用では、オリゴヌクレオチドプライマーは典型的には15〜25またはそれ以上のヌクレオチド長さとなる。プライマーは、目的とする延長生成の合成を刺激する所望のテンプレートと十分に相補的である必要があり、それが、ポリメラーゼまたは同様の酵素による合成の起始因子としての使用のために、適当な近位におけるプライマーの3’ヒドロキシル部分を提供する十分な方法において、目的とするらせん構造テンプレートとアニールすることを可能としている。それは、プライマー配列が目的とするテンプレートと正確に相補的であることを示すことは必要としない。例えば、非相補的ヌクレオチド配列は、他の相補的なプライマーの5’末端に付着されることもある。あるいは、非相補的な塩基は、オリゴヌクレオチドプライマー配列内に散在され、プライマー配列は延長生成物の合成のためにテンプレート−プライマー複合体を機能的に提供するらせん構造テンプレートの配列と十分に相補的となるよう提供される。
【0030】
用語「同一比率」は、核酸またはアミノ酸配列の間での比較に関して本明細書で使用されている。核酸及びアミノ酸配列は、しばしば、核酸またはアミノ酸の配列を一列にし、それにより、二つの間の違いを明らかにするコンピュータープログラムを用いて比較される。本発明の目的のため、核酸配列の比較は、GCG Wisconsin Package version 9.1によって実行され、これは、ウィスコンシン州Madisonのthe Genetics Computer Groupによって可能となった。便宜上、プログラムによって指定された初期パラメータ(ギャップクリエーションペナルティー=12、ギャップエクステンションペナルティー=4)は、本明細書において使用するため、同一配列を比較するのに意図されている。あるいは、National Center for Biotechnology Information(ワールドワイドウェブにおいてncbi.nlm.nih.gov/blast/;Altschul et al.,1990,J MoI Biol 215:403〜410)によって提供された、初期パラメータとギャップ調整に使用するBlastn 2.0プログラムは、核酸配列及びアミノ酸配列間の同一及び類似性のレベルを測定するのに使用されることがある。
【0031】
核酸分子、ポリペプチド及びそれらの誘導体をコードするFVの変異体の調製
核酸分子
FVの変異型をコードする核酸分子または本発明のそれらの機能性誘導体は、組み換え型DNA技術の方法を用いて調製されることがある。核酸配列情報の可用性は、種々の方法によって、本発明の単離された核酸分子の調節を可能とする。例えば、FVの変異型をコードする核酸配列またはそれらの誘導体ポリペプチドは、当業者において既知である標準プロトコルを使用して適当な生体原料から分離されることがある。
【0032】
本発明の核酸は、任意の便利なクローニングベクターにおけるDNAとして維持されることがある。好適な実施形態においては、クローンは、適当なE.coli宿主細胞pBluescript(カリフォルニア州、La Jolla、Stratagene)のような、プラスミドクローニング/発現ベクター内で維持される。あるいは、核酸は、哺乳類の最奥での発現に適当なベクター内で維持されることもある。翻訳後修飾がFVの変異型またはそれらの機能性誘導体に影響を与える場合において、それは哺乳類の細胞に分子を発現するのに好適である。FV変異体または核酸分子をコードするそれらの誘導体は、cDNA、ゲノムDNA、RNA及び一本鎖または二本鎖であることがあるそれらのフラグメントを含む。従って、この発明は、本発明の核酸分子の少なくとも一つの配列とハイブリダイジングできる配列を有するオリゴヌクレオチド(センスまたはアンチセンスDNAまたはRNAのらせん構造)を提供する。このようなオリゴヌクレオチドは、FV発現の検出のためのプローブとして有用である。
【0033】
タンパク質
FVの変異体または本発明のそれらの誘導体ポリペプチドの全長は、既知の方法に従って、様々な方法で調製されることがある。タンパク質は、例えば、FVの活性型を発現する、形質転換された細菌または動物の培養された細胞または組織などの適当な材料から、免疫親和性精製によって生成されることがある。しかしながら、これは、時を選ばず与えられた細胞に存在する可能性のあるタンパク質の低分子量のために好適な方法ではない。FVの変異型またはそれらの誘導体ポリペプチドをコードする核酸分子の可用性は、当業者に既知の方法で体外での発現に使用されるFVの変異型またはそれらの誘導体の生成を可能にする。例えば、cDNAまたは遺伝子は、体外転写のためのpSP64またはpSP65のような適当な体外転写ベクター内でクローンされ、続いて、小麦胚芽またはウサギ網状赤血球溶血液のような適当な無細胞翻訳系で無細胞翻訳される。体外での転写及び翻訳システムは、例えばウィスコンシン州MadisonのPromega Biotechまたはメリーランド州RockvilleのBRLによって市販されている。
【0034】
あるいは、好適な実施形態によれば、FVの変異型またはそれらの誘導体の多くの量は、適当な原核生物または真核生物発現系においての発現により生成されることがある。例えば、FVの変異型をコードするDNA分子の一部または全部は、E.coliのような細菌性細胞または、CHOまたはHela細胞のような哺乳類の細胞内での発現に適合したプラスミドベクター内に挿入されることがある。あるいは、好適な実施形態において、FVの変異体またはそれらの誘導体からなる標識された溶融タンパク質が生成されうる。FV分子またはそれらの誘導体のような標識された溶融タンパク質は、DNA分子の一部または全部によってコードされ、E.coliのような細菌性細胞または、これに限定されるわけではないが、酵母または哺乳類の細胞のような真核細胞における発現に適合したプラスミドベクター内に挿入された目的とするポリペプチド標識の一部または全部をコードするヌクレオチド配列にリーディングフレームする正確なコドン内で連結反応される。上述されたようなベクターは、宿主細胞内にDNAの発現を可能とするような方法で配置された宿主細胞内でのDNAの発現に必要な調節要素からなる。このような調節要素は、これに限定されるわけではないが、プロモーター配列、転写起始因子配列及びエンハンサー配列を発現に必要とする。
【0035】
ヘビ毒由来FV変異体またはそれらの誘導体タンパク質は、当業者において既知の方法に従って、組換え型原核生物または真核生物系においての遺伝子発現によって生成される。好適な実施形態において、市販されている発現/分泌系が、容易に周囲媒質から精製するのに用いられることがある。市販されている発現/分泌系により、組換え型タンパク質が発現され、その後、宿主細胞から分泌される。発現/分泌ベクターが使用されない場合、代替手法は、組換え型タンパク質と特異的に結合する抗体との免疫親和性相互作用のようなアフィニティー分離による組換え型タンパク質の精製またはN末端またはC末端で6−8ヒスチジン残基により標識された組換え型タンパク質のニッケルカラムによる分離を
含む。代替標識は、FLAGエピトープ、GSTまたは赤血球凝集素エピトープからなることがある。このような方法は当業者によって通常使用されている。
【0036】
FVまたはそれらの誘導体タンパク質は、上述のような方法で調製され、標準プロトコルによって分析されることがある。例えば、このようなタンパク質は、既知の方法によってアミノ酸配列分析を受けることがある。
【0037】
上述したように、本発明によるポリペプチドを生成する便利な方法は、発現系において核酸を使用することで、それをコードする核酸を発現させることである。本発明の方法において有用である様々な発現系は、当業者にとって既知である。
【0038】
したがって、本発明は(開示された)ポリペプチドをつくる方法、ポリペプチド(一般的に核酸)をコードする核酸からの発現を含む方法も含む。これは、ポリペプチドの生成を生じるまたは可能にする適当な条件下で、ベクターを含む宿主細胞を培養することで簡便に達成されることがある。ポリペプチドは網状赤血球溶血液のような体外系において生成されることもある。
【0039】
核酸及びタンパク質またはそれらの誘導体をコードするFVの使用
FVポリペプチドまたはそれらの誘導体または血液凝固活性変化を有するのと同一のものをコードする核酸は、例えば、血液凝固カスケードを調整する治療及び/または予防薬剤として、本発明にしたがって使用されることがある。本発明はこれらの分子が凝固を増加するよう変化し得ることを明らかにした。
【0040】
本発明の好適な実施形態においては、FVまたはそれらの誘導体は、生物学的に適合した担体の点滴を介して、好適には、静脈注射を介して患者に投与されることがある。本発明のFVまたはそれらの誘導体は、任意に、リポソーム内にカプセル化され、または、分子の安定性を増加する他のリン脂質またはミセルと混合される。FVまたはそれらの誘導体は、単独で、または止血を調整するとして既知の他の薬剤と組み合わせて投与されることがある(例えば、VIIa、FIX、FVIIIまたはFX/Xa因子及びそれらの誘導体)。FVまたはそれらの誘導体を提供するのに適当な化合物は、これに限定されることはないが、患者の状態及び血行状態を含む生理学的に様々なことを考慮した上で、医師によって決定される。異なる適用及び投与経路にふさわしい様々な組成物が当業者に知られており、明細書の以下に記載されている。生成されたFVまたはそれらの誘導体を含む調製は、生理学的に許容される材料を含み、医薬品として好適に処方される。調製は、既知の先行技術の方法を実質的に使用することで定式化され得るものであり、塩化ナトリウム、塩化カルシウムのような塩とグリシン及び/またはリシンのようなアミノ酸を含むバッファーと、pH範囲6〜8で混合され得る。
【0041】
必要となるまで、第五因子またはその誘導体を含む精製された調製液は、完成した溶液または凍結乾燥した状態または冷凍状態で保管される。好適には、調製液は凍結乾燥した状態で貯蔵され、及び、適切な再構成溶液を使用して視覚的に透明な溶液に溶解する。あるいは、本発明による調製は、調製液または凍った液体として作られることも可能である。
【0042】
本発明による調製液は、特に安定的であり、例えば、適用の前に長時間溶解した形態で有効である。本発明による調製液は、一つの成分の調製液またはマルチ成分の調製液における他の要因との組み合わせの形態で、FVまたはそれらの誘導体と薬剤的調製液として手に入れられる。
【0043】
薬剤的調製液内で精製されたタンパク質を処理するのに先行して、精製されたタンパク質は従来の品質管理により管理され、及び、提示の治療形態内に形作られる。特に、精製された調製液は、組み換え製造の間、発現ベクター由来の核酸と同様に細胞核酸の非存在下でテストされ、好適には、EP0714987に記載されたような方法が用いられる。
【0044】
この発明の他の特徴は、高い安定性及び構造的及び構造的完全性を有し、及び、特に不活性因子V/Va類似中間体及びタンパク質自動分解生成物を含まず、及び、前記タイプの第五因子類似体の活性化により、及び、適当な調製液内で調剤されることによりつくられるFVまたはそれらの誘導体を含む調製液の製造を可能にすることに関するものである。薬剤的調製液は、10〜1000μg/kgの間、好適には10〜250μg/kgの間の投薬量であり、最適には10〜75μg/kgの間の投薬量と、好適には40μg/kgの変異体第五因子ポリペプチドである。患者は血液診療所で、提示があればすぐに治療を受けることができる。または、患者は1〜3時間毎にボーラス投与され、または、十分に改善がみられる場合には、一日一回のFVまたは明細書に記載されたそれらの誘導体の投与となる。
【0045】
ヘビから得られた核酸をコーディングするFVまたはそれらの誘導体は、本発明に従って様々な目的で使用されることがある。本発明の好適な実施形態において、血液凝固を調節するための核酸運搬媒体(例えば発現ベクター)は、FVまたはそれらの誘導体ポリペプチド、または本明細書に記載されたそれらの機能的フラグメントをコーディングする核酸配列からなる発現ベクターなどが提供される。発現ベクターをコードするFVまたはそれらの誘導体の患者への投与は、凝固を強化するのに役立つFVまたはそれらの誘導体ポリペプチドの発現を生じる。本発明に従って、核酸配列をコードするFVまたはそれらの誘導体は、本明細書に記載されたように、その発現が止血を調節するFVまたはそれらの誘導体をコードすることがある。
【0046】
FVまたはそれらの誘導体核酸配列からなる発現ベクターは、単独で投与されることも、または止血を調整するのに有用な他の分子と組み合わせて投与されることもある。本発明に関連して、発現ベクターまたは治療薬の組み合わせは、患者に、単独で、または薬剤的に許容な化合物で、または生物学的に親和性のある化合物で投与されることもある。
【0047】
本発明の好適な実施形態においては、FVまたはそれらの誘導体をコードする核酸配列からなる発現ベクターは、ウイルスベクターである。本発明で使用されるウイルスベクターは、これに限られるわけではないが、アデノウイルスベクター(特定プロモーター/エンハンサー組織を伴うまたは伴わない)、複数の血清型(例えば、AAV−2、AAV−5、AAV−7及びAAV−8)のアデノ随伴ウイルス(AAV)ベクター及びハイブリッドAAVベクター、レンチウイルスベクター及び擬似レンチウイルスベクター(例えば、エボラウイルス、水疱性口内炎ウイルス(VSV)、及び猫免疫不全ウイルス(FIV))、単純ヘルペスウイルスベクター、ワクシニアウイルスベクター及びレトロウイルスベクターを含む。
【0048】
本発明の最適な実施形態では、核酸配列FVまたはそれらの誘導体、またはそれらの機能的フラグメントからなるウイルスベクターを投与する方法が提供されている。本発明の方法における実用的なアデノウイルスベクターは、好適には、アデノウイルスベクターDNAの少なくとも主要な部分を含む。明細書に記載されているように、このようなアデノウイルスベクターの投与に続いて、FVまたはそれらの誘導体ポリペプチドは止血を調節するのに役立つ。
【0049】
組換え型アデノウイルスベクターは、多様な遺伝子治療適用において幅広い実用性を有する。そのような適用におけるそれらの有用性は、様々な臓器の状況で行われる体内での遺伝子転移の高効率によるところが大きい。
【0050】
アデノウイルス粒子は、適切な遺伝子運搬のための媒体として生かされることがある。このようなウイルス粒子は、二本鎖DNA非エンベロープウイルスに関連する構造的特徴及びヒト呼吸系及び消化器官系指向性のような生物学的特徴を含む、これらの適用にとって望ましい特徴を複数有する。さらに、アデノウイルスは、受容体を介した飲食作用により、生体内及び生体外の様々な細胞に影響を与えることが知られている。アデノウイルスベクターの全体的な安全性の証明、アデノウイルスの感染は、軽度のインフルエンザ様症状を含む、ヒトにおける軽度な疾患を引き起こす。
【0051】
その大きなサイズに起因して(〜36キロ塩基)、アデノウイルス遺伝子は遺伝子治療媒体として用いられるのに適している。アデノウイルスは、複製において極めて重要なアデノウイルス遺伝子及び非必須領域の除去後、外来DNAの挿入に適応するからである。このような置換は複製機能及び感染に関して正常に機能しないウイルスベクターを提供する。注目すべきは、アデノウイルスは、遺伝子治療のため及び異常遺伝子の発現のためのベクターとして用いられることである。
【0052】
遺伝子治療で用いられるアデノウイルスベクターの使用についてのさらに詳細な議論は、Berkner,1988,Biotechniques 6:616−629及びTrapnell,1993,Advanced Drug Delivery Reviews 12: 185〜199を参照のこと。
【0053】
例えば、目的とする遺伝子の複数コピー及びそれによる多量の目的遺伝子の生成物を提供できるベクターを導入することが望ましい。アデノウイルスベクターの改良及びこれらのベクターを生成する方法は、Mitani and Kubo(2002,Curr Gene Ther.2(2):135〜44);Olmsted−Davis et al.(2002,Hum Gene Ther.13(11):1337〜47);Reynolds et al.(2001,Nat Biotechnol.19(9):838〜42);米国特許番号5,998,205(多重DNAコピーからなる腫瘍特異的複製ベクターが提供されている);6,228,646(ヘルパーアデノウイルスなし、アデノウイルスベクター完全欠損が記載されている); 6,093,699(遺伝子治療のためのベクター及び方法が提供されている);6,100,242(複製欠損アデノウイルスベクターに挿入された導入遺伝子が、生体内で末梢血管疾患及び心疾患の遺伝子治療において有効に用いられることが記載されている);及び国際特許出願番号WO 94/17810及びWO 94/23744を含む参考文献、特許及び特許出願に詳しく記載されている。
【0054】
出願によっては、さらに、発現構成は、特に細胞または組織タイプでの発現を促進するのに役立つ調節要素からなる。このような調節要素は、当業者において既知であり、Sambrook et al.(1989)及びAusubel et al. (1992)において深く論じられている。本発明の発現構成における組織特定調節要素の取り込みは、FVまたはそれらの発現またはそれらの機能的フラグメント発現のための少なくとも部分的な組織指向性を提供する。例えば、サイトメガロ・ウイルス(CMV)プロモーターの制御下で、FVまたはそれらの誘導体をコードする核酸配列からなるE1欠損タイプ5アデノウイルスベクターは本発明の方法において生かされることがある。
【0055】
アデノウイルスベクターを生成するための典型的な方法
組み換え遺伝子発現のためのアデノウイルスベクターは、ヒト胚性腎細胞株293において生成される(Graham et al.,1977,J,Gen. Virol.36:59〜72)。この細胞株は、E1機能不全において、アデノウイルス2(Ad2)及びアデノウイルス5突然変異体の成長を許容する。なぜなら、この細胞株は、アデノウイルス5遺伝子の左末端からなり、従ってE1タンパク質を発現するからである。293細胞の細胞遺伝子内に導入されたE1遺伝子は、除去された遺伝子からウイルスベクターを増幅する発現系としてこれらの細胞の使用を容易にするレベルで発現される。293細胞は、独立したクローニングを助け、アデノウイルスベクターを発現するため、E1突然変異体の分離及び増殖に広範囲にわたって使用される。従って、293細胞株のような発現系は、トランスにおいて不可欠なウイルス機能を提供し、それによって、E1遺伝子に置換された外因性核酸配列において、ウイルスベクターの増殖が可能となる。Young et al.in The Adenoviruses,Ginsberg,ed.,Plenum Press,New York and London(1984),p.125〜172を参照のこと。
【0056】
アデノウイルスベクターの増殖に向いている他の発現系は、当業者において既知であり(例えば、HeLa細胞)、及び、他の場でも再検討されている。また、本発明には、FVまたはそれらの誘導体ポリペプチドをコードする核酸運搬媒体と個々の細胞を提供すること、及び、FVまたはそれらの誘導体ポリペプチドが発現する条件下で細胞を培養することからなる止血を調節する方法も含まれる。
【0057】
前述の議論より、FV変異体またはそれらの誘導体ポリペプチドまたは同一のものをコードする核酸は、異常血液凝固に関連する疾患の治療において用いられることがわかる。
【0058】
医薬組成物
本発明の発現ベクターは、生理活性タンパク質(例えば、FVまたはそれらの誘導体ポリペプチドまたは機能的フラグメントまたはそれらの誘導体)が生成できるように、対象に運搬される薬剤的化合物内に組み込まれる。本発明の特定の実施形態では、受容体がFVまたはそれらの誘導体ポリペプチドの治療的に有効な量を生成できるよう十分な遺伝子材料からなる薬学的化合物は、対象において止血作用を与えることができる。または、前述したように、変異体第五因子ポリペプチドの有効量は、それらを必要とする患者に直接的に注入される。化合物は、単独で投与、または、安定化化合物のような他の薬剤少なくとも一つと組み合わせて投与され、これに限定されるわけではないが、生理食塩水、緩衝食塩水、デキストロース及び水を含む、生体適合性のある薬剤担体に滅菌下で投与されることもある。化合物は、単独で、または、止血に作用する他の薬剤と組み合わせて患者に投与されることもある。好適な実施形態では、薬剤的化合物も薬剤的に許容な賦形剤を含む。このような賦形剤は、化合物を受容する個体に有害な免疫応答をそれ自体で導くことのない任意の薬剤を含み、及び、過度な毒性なしで投与することが可能である。薬学的に許容される賦形剤は、これに限定されるわけではないが、水、生理食塩水、グリセロール、糖類及びエタノールのような液体が含まれる。薬学的に許容される塩には、例えば、塩酸塩、臭化水素塩、リン酸塩、硫酸塩などのような鉱酸塩、及び、酢酸塩、プロピオン酸塩、マロン酸塩、安息香酸塩などのような有機酸塩が含まれる。加えて、湿潤剤、乳化剤、pH緩衝剤などのような補助剤が、このような賦形剤において存在することがある。薬学的に許容される賦形剤に関する詳細な議論は、Remington’s Pharmaceutical Sciences(Mack Pub. Co., 18th Edition,Easton,Pa.[1990]).で入手することができる。
【0059】
非経口投与に適した薬剤の剤形は、水溶液、好適には、ハンクス溶液、リンガー溶液または生理緩衝食塩水のような生理的に適合性のある緩衝液内で調剤される。水性懸濁注射液は、カルボキシルメチルセルロースナトリウム、ソルビトールまたはデキストランのような懸濁液の粘性を増加させるような基質を含むことがある。適当な親油性溶媒または媒体は、ゴマ油のような脂肪酸油、または、オレイン酸エチルまたはトリグリセリドまたはリポソームのような合成脂肪酸エステルを含む。随意に、懸濁液は、高濃度溶液の調製を可能にするため、化合物の溶解度を増加させる適当な安定剤または薬剤も含むことがある。局所または経鼻投与の場合、浸透されるべき特定の障壁に対して適切な浸透剤が製剤に使用されている。このような浸透剤は一般的に当業者において知られている。本発明の薬剤的化合物は当業者において既知の任意の方法(従来の混合、溶解、整粒、糖衣製造、ゲル化、乳化、カプセル化、封入、凍結乾燥処理など)で製造されている。
【0060】
薬剤的化合物は塩として提供されることがあり、また、これに限定されるわけではないが、塩酸、硫酸、酢酸、乳酸、酒石酸、リンゴ酸、コハク酸などの多くの酸と作られることがある。塩は、遊離塩基に対応する溶液よりも水または他のプロトン性溶媒に溶解しやすい傾向にある。他の場合では、好適な調製液は、4.5〜5.5のpH範囲で、1〜50μMのヒスチジン、1〜2%のスクロース、及び2〜7%のマンニトールの任意または全てを含む凍結乾燥粉末であることがある。この凍結乾燥粉末は使用前にバッファーに結合される。
【0061】
薬剤的化合物が調製された後、それらは適切な容器に入れられ、治療のために標識される。ベクターを含むFVまたはそれらの誘導体の投与のために、前述の標識には投与の量やしばしば方法が含まれることもある。
【0062】
本発明の使用に適当な医薬組成物は、活性成分が治療目的を達成するための有効量含まれている化合物を含む。治療に有効な投与量の決定は、本発明において提供される技術を使用する熟練した医療従事者の能力の範囲内で行われる。治療における投与量は、他の要因のなかでも、年齢、対象における一般的条件、異常血液凝固表現型の重症度、FVまたはそれらの誘導体ポリペプチドの発現レベルを調節する制御配列の強さに依存する。したがって、ヒトにおける治療の有効量は、ベクター基本形または、FVまたはそれらの誘導体に感染したタンパク質への個々の患者の応答に基づいて医療従事者により決定される比較的広い範囲に分類される。
【0063】
投与
FVポリペプチドの活性型変異体は、単独または他の薬剤と組み合わせて、本明細書において上述したように、適切な生理的担体において患者に直接的に注入されることもある。FVまたはそれらの誘導体、またはそれらの機能的フラグメントをコードする核酸配列からなる本発明の発現ベクターは、FVまたはそれらの誘導体ポリペプチドの活性型の予防的及び/または治療的に有効なレベルを達成する及び維持するため(以下、参照の)様々な方法により患者に投与されることもある。当業者は、特定の患者の治療のための本発明の発現ベクターをコードするFVまたはそれらの誘導体の活性型を使用するための特定の手順をすぐに見つけることができる。アデノウイルスベクターの生成及び患者への投与のための手順は、米国特許番号5,998,205、6,228,646、6,093,699、6,100,242、及び国際特許出願番号WO94/17810及びWO94/23744において記載され、これらはその全体において参照により本明細書に組み込まれている。本発明のアデノウイルスベクターをコードするFVまたはそれらの誘導体の活性型は、任意の既知の方法により投与されることがある。生体内への薬剤的化合物の直接的な運搬は、運搬を強化された対流のような運搬方法が予想されるにもかかわらず、一般的に、従来の注射器を利用する注射により達成される(例えば米国特許番号5,720,720を参照)。この方法で化合物は、皮下、表皮、皮内、髄腔内、眼窩内、粘膜内、腹腔内、静脈内、動脈内、口腔内、肝内または筋肉内に運ばれることがある。投与の他の方法には、経口投与及び経肺投与、坐薬、及び、経皮投与が含まれる。血液凝固疾患患者の治療の臨床専門医は、これに限定されるわけではないが、患者の状態及び治療の目的(例えば血液凝固の促進)を含む基準数に基づいて、活性型FVまたはそれらの誘導体核酸配列からなるアデノウイルスベクターの投与のために最適な経路を決定している。
【0064】
また、本発明の組成物は、創傷または外傷において局所的な血液凝固を促すのに使用されることもある。このアプローチでは、本発明の第五因子は局所的に適用するために製剤され、及び、戦場において、または、出血を止めなければならない、または、出血を止めることが重要とされる状況下において使用される。
【0065】
本発明は、活性型FVまたはそれらのポリペプチド誘導体をコードする核酸配列からなるAAVベクターも含む。
【0066】
活性型FVまたはそれらのポリペプチド誘導体をコードする核酸配列からなるレンチウイルスまたは疑似レンチウイルスベクターも含まれる。
【0067】
活性型FVまたはそれらのポリペプチド誘導体をエンコンディーングする核酸配列からなる裸プラスミドまたは発現ベクターも含まれる。
【0068】
以下に示す材料及び方法は、以下の実施例の実行を容易にするために提供されたものである。
【0069】
試薬
ベンズアミジン及び4−アミジノフェニルフッ化メタンスルフォニル塩酸塩(APMSF)はSigma社(St.Louis、MO)のものであり、及び、ダンシルアルギニン−N−(3−エチル−1,5−ペンタンジイル)アミド(DAPA)は、Haematologic Technologies社(バーモント州、Essex Junction)のものである。ペプチジル基質H−D−フェニルアラニル−L−ピペコリル−L−アルギニル−p―ニトロアニリド(S2238)はDiapharma社のものである。全ての組織培養試薬は、インスリン−トランスフェリン−亜セレン酸ナトリウム(ITS;インディアナ州、Indianapolis、Roche Applied Science)を除いて、Invitrogen(カリフォルニア州、Carlsbad)のものである。鶏卵L−α−ホスファチジルコリン75%(w/w)とブタ脳L−α−ホスファチジルセリン25%(w/w)からなるラメラリン脂質小胞体(PCPS)(アラバマ州、Alabaster、Avanti Polar lipids)は記載されたように調製され、及び、特徴付けられている。
【0070】
タンパク質
ヒトプロトロンビンは先述したように血漿から分離される(23)。プレトロンビン−1及びプレトロンビン2は確立されたやり方によって精製され(18)、及び、トロンビンは、血液学的技術から得られる。FV(hFV−810)、rFVa、rFX及びrFXaの構成的活性部分B−ドメインレス型が調製され、(3、24、25)に記載したように精製され、特徴付けられる。pseutarinC(pt−FXa)の精製された酵素サブユニットは、QRxPharma(オーストラリア、Sydney)から豊富に提供される。組み換え型ヒルジンは、EMD−Chemicals(カリフォルニア州、San Diego)から得られる。組み換え型ダニ抗凝固タンパク質及びヒトAPCは、Dr. Sriram Krishnaswamy(The Children’s Hospital of Philadelphia)より得られる。使用されている様々なたんぱく質の分子量及び吸光係数(E0.1%,280nm)は、プロトロンビンが72,000及び1.47(18);プレトロンビン‐1が1 49,900及び1.78(18);プレトロンビン‐2が37,500及び1.95(18);トロンビンが37,500及び1.94 (26);rFXaが46,000及び1.16(27);rFVa,が175,000及び1.78(3);rFV−810が216,000及び1.54(3),及びpt−FVが170,000,及び1.50(本研究による)である。pt−FXaにために、ヒトタンパク質のための値が使用されている。特別な記載がない限り、全ての機能的アッセイは、20mMヘペス、0.15M塩化ナトリウム、2mM塩化カルシウム、0.1%ポリエチレングリコール8000、pH7.5(アッセイバッファー)において25℃で行われる。
【0071】
分析的超遠心分離:
分子量は、光学干渉を用いた分析的超遠心分離機Beckman Optima XL−Iで測定される。沈降速度は20mMのHEPES、150mMの塩化ナトリウム、5mMの塩化カルシウム、pH7.4、AN60Tiローターによる毎分25,000回転で測定される。タンパク質濃度は、pt−rFVが0.60mg/mL(3.6μM)及びpt−FXaが0.18mg/mL(3.9μM)である。沈降係数及び分子量はg(s*)分析により測定される。減衰係数は、Babul and Stellwagen(29)に記載された手順による示差屈折率測定により測定される。
【0072】
rFV変異体の構造
毒腺由来のコブラ・モドキ(P.textilis)FV cDNA(8)は、隣接XmaI制限酵素認識部位により修飾され、それにより、pED発現プラスミド内にサブクローニングされる(3)。cDNAは、さらに、コブラ・モドキ(P.textilis)FVの信号配列及び重複によるスプライシングの技術を使用するヒトFVの信号配列をコードする配列を交換することにより修飾される(30)。最終的な構成、pED−FV−ptex、その後に成熟ベノム由来pt−FVタンパク質(残渣1から1430)が続くヒトFV信号ペプチド(残渣−28から1)のためにコードされる。
Rao et alによりレポートされるのと同一の配列を示すコブラ・モドキ(P.textilis)FV cDNA(7)は、コドン50(Lys)及び1305(Phe)を除いて、コースタルタイパン(O.scutellatus)及びタイパン(O. microlepidotus)の相応する残渣と同一である(8)。Glnにより置換されたArg742及びArg788を有するpt−FV変異体は、適当な突然変異誘発性相補的オリゴヌクレオチドを使用するQuickChange部位に誘導された突然変異生成キット(カリフォルニア州、La Jolla、Stratagene)により発生した。これらのアミノ酸は、ヒトFVにおいてアルギニン(Arg)709及びアルギニン(Arg)1545に相同であり、及び、トロンビン開裂部位を示す。突然変異生成に続いて、cDNA全体は、目的とする突然変異の存在を確認するため、及び、誤って誘導されたポリメラーゼがないことを確かめるために配列される。
【0073】
pt−rFVの発現及び精製:
FVまたはFV−QQ を含むプラスミドは、ベビーハムスター腎臓(BHK)細胞内に誘導され、及び、高生産安定クローンは、(3)に記載されたように確立される。細胞は三角フラスコ内に広がり、ITS及び2.5mMの塩化カルシウムを追加されたDMEM/F12媒体内で培養される。調製済みの媒体は、4〜6日間かけて収集され、延伸ぶりされ、及び10μM APMSF及び1mMベンズアミジンの存在下において20℃で保存される。精製のために、媒体は37℃で融解され、20mMへペス、0.15M塩化ナトリウム、5mM塩化カルシウム、pH7.4で平衡を保たれた30mLQ−セファロースカラム(ニュージャージー州、Piscataway、Amersham Biosciences)に装填される。カラムは同じバッファーで洗浄され、塩化ナトリウム0.15〜1Mの濃度勾配で溶出される。画分は、収集され、SDS−PAGE及び(3)記載のFV特定凝固アッセイにより分析される。pt−rFVを含む画分は、貯蔵され、及び20mMのMES、2mM塩化カルシウム、pH6.0に対して透析され、及び同一のバッファーで平衡を保たれたPoros HS/20カラム(10×100mm;カリフォルニア州、Foster City、Applied Biosystems)に装填される。カラムは20mMのMES、2mMの塩化カルシウム、pH6.0で洗浄され、及び、塩化ナトリウム0〜0.6M濃度勾配で溶出される。画分を含むFVは貯蔵され、及び、20mMへペス、0.15M塩化ナトリウム、2mM塩化カルシウム、pH7.4に対して透析され、及び、タンパク質は−80℃で保存される。最終収率は、調製済みの媒体1リットルあたりのpt−rFVに対して〜4mgである。
【0074】
タンパク質の特徴:
純正タンパク質は、クマシー ブリリアント ブルーR−250による染色に続くMOPSバッファーシステムを用いる非還元及び還元条件(50mMジチオスレイトール)下で、プレキャスト4〜12%勾配ゲル(Invitrogen)を用いるSDS−PAGEにより評価される。N末端配列分析は、ガルベストンのTexas Medical Branch大学のDr.Alex Kurosky及びSteve Smith、またはCenter for Disease Control(ジョージア州、Atlanta)のDr.Jan Pohlの研究室で行われる。pt−FXaにおける化学的γ−カルボキシグルタミン酸(Gla)分析は記載した研究室で行われる(24)。
【0075】
タンパク質基質開裂の反応速度論:
高分子基質開裂の定常状態における初期速度は、記載されているように、25℃で不連続に決定される(25)。プロトロンビナーゼまたはFXa−FVaに触媒作用されたプロトロンビンまたはプレトロンビン−1活性型(Km及びVmax)の速度パラメータは、高分子基質の濃度を上昇させることでトロンビン構造の初期割合を測定することにより、アッセイバッファーにおいて決定される。アッセイ混合物は、PCPS(50μM)、DAPA(3μM)、様々なpt−rFV(a)補因子種(20nM)及び様々な濃度のプロトロンビン(0〜5.0μM)またはプレトロンビン−1(0〜15μM)が含まれる。プレトロンビン−2がタンパク質基質として使用されると、それに続く反応条件は、PCPS(50μM)、DAPA(3μM)、及びプレトロンビン−2(1.4μM)が用いられ、様々なFV(a)補因子(0〜40nM)と保温培養され、及び、反応はpt−FXa(5nM)により開始される。特定の実験においては、PCPSは、反応混合物に含まれない(図の説明文を参照)。pt−rFVaが使用される実験においては、pt−rFV(600nM)は2nMトロンビンと37℃で15分間保温培養され、続いて2.4nMヒルジンが加えられる。
【0076】
pt−rFVのAPC不活性:
PCPS(50μM)存在下における様々な補因子タンパク質(500nMのpt−rFV,pt−rFVa,rhFVa,またはh−FV−810)の不活性化は、APC添加(10μMまたは750nM)により開始される。反応混合物の一定分量は指定された時間間隔で取り出され、SDS−PAGE(4〜12%ゲル)及び機能的活性の評価を通して評価される。このアッセイにおいて、残渣補因子活性は、特に(25)に記載されたようなプロトロンビン活性におけるFVaの作用を測定することにより評価される。アッセイ混合物には、1.4μMプロトロンビン、50μMのPCPS、3μMのDAPA、1nMのFVa、及び1nMのpt−FXaが含まれる。rhFVaまたはhFV−810と、ヒトrFXaは代わりに使用される。これらの条件下で、反応の初期割合は補因子の濃度と比例している。
【0077】
データ分析:
データは、マーカード アルゴリズム(Marquardt algorithm)を使用する非線形最小二乗法による回帰分析から求められる参照式によって分析される(31)。適合の質は、(32)に記載の基準により評価した。エラーの報告された推定値は、±2 S.Dを示す。pt−FXa及びpt−rFV(a)間での相互作用における解離定数(Kd)は、プレトロンビン−2活性型の初期値の1に固定された化学量論と補因子の濃度への依存から得られる(33)。プロトロンビナーゼによるプロトロンビンまたはプレトロンビン−1の開裂の初期速度測定は、Km及びVmaxの近似値を得るため、ヘンリー−ミカエリス−メンテン(Henri−Michaelis−Menten)の方程式(34)にデータを適合させることで分析される。
【0078】
以下に記載の実施例は、本発明の特定の実施形態を示すものとして提供される。これらはいかなる場合においても本発明を制限することを意図していない。
【実施例1】
【0079】
ヒトの治療における使用のためのヘビ毒由来またはヒト型誘導体の第五因子
本発明はコブラ・モドキ(P.textilis)からのヘビ毒由来FVの生物学的特性を記述する。これらの固有な性質は、以下に列記するように、凝血原特性を反復するヒトFV誘導体に設計されることもある。これらの性質とは、a)pt−FVが活性状態で存在し、補因子が自然発生の活性FV変異体の第一実施例のようであること、b)pt−FVが、セリンプロテアーゼ因子Xa(FXa)と高い親和性で結合するのを達成する膜表面のための通常要件を回避するような固有な形態を有すること、c)重鎖及び軽鎖を結合するpt−Fvの固有のジスルフィド結合により、pt−FVは、活性型タンパク質C(APC)による不活性に機能的に抵抗すること、である。これらの特徴の一つまたはそれ以上からなるヒトFV誘導体を操作することは、その凝血原特性を強化し、したがって、出血性疾患の治療において有用な治療的ツールを提供する。
【0080】
本研究においては、非常に精製されたヘビ毒由来Fxa(pt−FXa)と同様に、コブラ・モドキ(P. textilis)からの組換え型ヘビ毒由来FV(pt−rFV)による一連の機能的測定値は、FVの固有の形態の生物学的性質を調査し、ヘビ毒由来プロトロンビナーゼ複合体の強力な凝血性質における基本的メカニズムを調べるのに使用される。この働きは、pt−rFVが、哺乳類FVにおいては起こることが未知である機能的要素の複数の利益を獲得することで、有力な凝血源に適応したことを証明している。この研究結果は、FVの構造/機能関係の複数の態様に新しい光を投じ、治療的な目的のための生物学FVを容易にし、さらに、選択的優位性のためのタンパク質の既存の特性を強化する性質の驚くべき能力を強調する。
【0081】
結果
pt−rFVの発現及び物理的特徴
天然のヘビ毒の高いプロテアーゼ含有量のために、同種の単離、pt−FVの未開裂調製は得られない(12〜15)。これを回避するために、BHK細胞から精製されたpt−rFVは高い収量で示された。野生型pt−rFV及び重鎖及び軽鎖の側面に位置するトロンビン開裂部位が欠乏する変異体(pt−rFV−QQ)は、還元型SDS−PAGEにおいてシングルバンド(〜180kDa)として主に移行する(図1、パネルA)。沈降速度の研究は、pt−rFVが、170,000の分子量評価により均一であることを示した。pt−rFV−QQがトロンビンにより開裂されないのに対し、pt−rFVはpt−rFVaに処理され、特有の重鎖及び軽鎖を産出する。N−末端配列分析は、予測されたトロンビン開裂部位における的確な処理を明らかにした(図1)(7,8)。驚くべきことに、トロンビン処理の後に、pt−rFVaの大部分(>60%)は、非還元条件下においてシングルバンドとして移動し(図1、パネルB)、重鎖及び軽鎖が、固有のジスルフィド結合により結合していることを示している。この可能性は、非還元条件下における二重項としてpt−rFV及びpt−rFV−QQの移行を説明している。配列アラインメントは、重鎖に二つの遊離Cys残基(A2ドメイン、539及び641)、軽鎖に一つの遊離Cys(A3ドメイン、1032)を有するであろうことを示している(7)。
【0082】
【表1】
【0083】
コブラ・モドキ(P.textilis)またはコースタルタイパン(O.scutellatus)の天然のヘビ毒から精製されたFVは、ウシFXaの存在下において低活性である(14、15)。pt−rFVを特徴付ける初期の試みは、これらの観察と矛盾しない。しかしながら、推定値は、pt−rFVまたはpt−rFVaはヒトFVaと比較してほぼ=1/40th活性を示す。低活性は、主にpt−rFVまたはpt−rFVaが、確立された経口結合測定値を使用する膜結合ヒトFXa(Kb>400nM)との親和性を減らすことで結合されることにより起因するようである(データは示されていない)(3)。したがって、pt−rFVを特徴付けるため、pt−FXaが全てのそれに続く実験において使用された。pt−FXaのSDS−PAGE分析は、予測される結果を与えたN末端配列分析とともに図1に示される。二重項としての軽鎖の外観は、不均一なO−グリコシル化に原因があるようだ(8、16)。さらに、pt−FXaの化学的Gla分析は、配列アラインメントから予測される11モル Gla/molタンパク質から10.9±0.34モルGla/molタンパク質を産出した。
【0084】
Pt−rFVは構成的に活性型補因子である
pt−FV Bドメインの変わった性質のため、一連の機能的測定は、ヘビ毒由来FVのこのドメインが、プロコファクターとしてのタンパク質を維持するように機能するかどうか確定するために使用された。機能的な測定値に対する一つの懸案事項は、元の位置に生成されたトロンビンによるpt−rFVのフィードバック・タンパク質分解である。これに適応するために、反応体の濃度はタンパク質分解を最小にするように選ばれ、及び、トロンビンの可逆性抑制剤(DAPA)がアッセイに含まれる。さらに、pt−rFV−QQ、トロンビンによって媒介されるタンパク質分解に抵抗する変異体(図1)が調製された。ヒトFV(1)とは対照的に、膜結合pt−FXaを有するpt−rFvの組み立ては、プロトロンビンを急速に活性化させる(図2A)。プロトロンビン転換のための定常状態速度定数は、ヒトプロトロンビナーゼにおいて観察される定常状態速度定数と著しく類似している。(表2)(3、17)。興味深いことに、ヒトFVの誘導体で切断されたB−ドメイン、hFV−810は、pt−FV−膜によって集まり、相当する速度パラメータを有するプロトロンビンを活性化することが可能であった(表2)。pt−rFVaを生成するまたは、未開裂変異体、pt−rFV−QQを使用するトロンビンによる短いB−ドメインの除去は、相当する速度定数を与える(図2A及び表2)。これらのデータは、ヒトFVとは異なり、pt−FVは、構成的に活性型補因子であり、及び、B−ドメインのタンパク質分解除去が凝血原活性を表すことを必要としないことを示す。これは、FVの構成的活性型の自然な第一実施例を表す。
【0085】
【表2】
【0086】
アニオン性膜の非存在下でのPt−rFVの機能
血液凝固の重要な観点は、凝固タンパク質を細胞表面に(ホスファチジルセリン曝露を介して)結合する膜が正常な止血のためには重要であるということである(2)。オスキュタリン(oscutarin)及びプスタリンC(pseutarin C)によりみられる変わった観察結果は、ヘビ毒由来FXa及びFVが複合体として同時精製し、陰イオン膜非存在下において、ある程度活性を示すことである。しかしながら、オスキュタリン(oscutarin)の活性は、膜存在下において、実質的に増加する(>400−倍)ことが報告されている(12〜15)。さらにこれを調べるために、プロトロンビンを使用する溶液位相pt−FXa−pt−rFVの活性が評価された。膜非存在下において、pt−rFV、pt−rFV−QQまたはpt−rFVaを有するpt−FXaを使用するプロトロンビンの初期割合は、同じ酵素濃度で膜存在下において得られる割合と比較して7〜8倍低いものであった(図2B)。活性の減少は、プロトロンビンのKm増加(〜4倍)及びkcatの減少(〜4倍)によって反映された。対照的に、膜非存在下において、hFV−810は、膜結合補因子と比較して非常に低い活性を有した(>150倍)(図2B)。
【0087】
これらのデータの解釈の困難は、基質、プレトロンビン、トロンビンの生成の割合を高めるのに貢献する結合膜にある。さらに、pt−FXa−pt−rFVのための溶液位相解離定数に応じて、pt−FXaはpt−rFVで飽和することができず、実際の酵素複合体の濃度を低下させ、プロトロンビン活性率の減少へと至った。一点目に取り組むために、膜の存在及び非存在下でのプレトロンビン−1活性の割合が測定される。プレトロンビン−1は、γ−カルボキシグルタミン酸及びクリングル−1ドメインと結合する膜を欠失するプロトロンビンの派生物である(18)。プロトロンビンを使用している結果と整合して、pt−rFV、pt−rFV−QQまたはpt−rFVaを使用する膜結合pt−FXaを有するプレトロンビン−1の反応速度は、等しいものであり(図3A及び表2)及びヒトプロトロンビナーゼを使用する速度パラメータと同等であった(17)。しかしながら、意外にも、反応混合物からの膜の除去は、溶液位相または膜結合pt−FXa−pt−rFVにおける運動パラメータが等しいため、プレトロンビン−1活性の割合において感知できるほどの効果は有さない(図3B及び表2)。これらのデータは、pt−FXa−pt−rFVを有するプロトロンビン活性割合の減少率が、酵素機能ではなく、むしろ結合している基質膜の除去にあったことを示唆している。
【0088】
Pt−FXa及びpt−rFVが形成する高親和性複合体
膜の存在または非存在におけるプレトロンビン−1活性の割合が同等になると、データは、pt−FXaがpt−rFVと完全に飽和されることを示す。しかしながら、これを直接評価するために、いずれの膜結合または溶液位相pt−FXa−pt−rFVにおける平衡解離定数を意味する機能的な結合測定が使用された。補因子(pt−rFV、pt−rFV−QQ、pt−rFVaまたはhFV−810)の濃度の増加は、PCPSの存在下または非存在下において、pt−FXaの濃度を固定して、単一で培養された。プレトロンビン−2活性の初期割合は測定され、及び、データはpt−FXaと結合している補因子に推定された平衡解離定数を得るために分析された(表3)。PCPSの存在下において、補因子の種類に関係なく、pt−FXaと結合している解離定数はほぼ=3nMであった。この値は、膜結合ヒトFXa及びFVaを使用して得られた解離定数と類似しているものである(3)。驚くべきことに、pt−rFV、pt−FV−QQまたはpt−rFVaにおける解離定数は、膜非存在下において、わずか2〜3倍増加させた(Kdほぼ=8nM)。これは、FXa−FVaにおける溶液位相解離定数がほぼ=0.8〜3.0μMである牛におけるシステムとは対照的である(19、20)。沈降速度実験では、溶液位相pt−rFV−pt−FXaが213,000の分子量及び9.33の沈降係数(s20,w)と1:1で化学量論複合体を形成していた。これらの値はウシFXa−Fvaによって得られた結果と非常によく一致している(19)。ヘビ毒由来補因子とは対照的に、溶液位相hFV−810−pt−FXaにおける結合定数(表3)は、確立することができず、pt−FXa結合が、少なくとも溶液において、hFV−810に存在しないpt−rFVにおいて固有な性質を含むことを示唆している。これらのデータは、pt−FXa−pt−FV複合体は、プロトロンビンを急速に活性化できる1:1複合体が高い親和性を形成するため、負に荷電された膜において通常必要とされるものを回避したことを示す。
【0089】
【表3】
【0090】
Pt−rFVはAPCに機能的耐性がある
APCは止血系及びタンパク質分解の制限を介してFVa及びFVIIIaの両方を不活性化する機能における重要な抗凝血剤である(21)。ヒトFVaにとって、Arg306、Arg506及びArg679での開裂は、A2ドメインの解離において、FXa結合及び最終的には補因子機能の消失を生じる(1)。pt−FVの配列分析は、これらの部位にAPC耐性がある可能性の示唆は保存されていない(7)。プスタリンCの研究はこの論点を支持している(7);しかしながら、FXaは、データ解釈を複雑にするAPCによる不活性からFVaを保護する(1)。pt−rFVがAPCに対して耐性を示すかどうか直接調べるために、これらのタンパク質は、PCPS存在下において培養され、及び、様々な時間間隔で、SDS−PAGE及び精製されたプロトロンビンアッセイを使用する補因子活性によって、タンパク質分解を観察した。結果的にrhFVaまたはrFV−810機能を完全に失ったAPC濃度(10nM)及び反応時間(≦15分)を使用して、pt−rFvまたはpt−rFVaは完全に活性を保持し(図4A)及び、SDS−PAGE分析は、これらのタンパク質の限定された開裂のみを示した(データは示されていない)。しかしながら、長時間における高濃度APC(750nM)とのpt−rFVの培養は、結果として完全なタンパク質分解pt−rFVとなった(図4B、C)。N−末端配列分析と還元及び非還元ゲルの分析はともに、APCがA2ドメインにおいてpt−rFVを開裂することを示す。驚くべきことに、pt−rFVの完全なタンパク質分解にもかかわらず、補因子は、完全な凝血原活性を保持した(図4Aに挿入)。
第五因子変異体をコードする核酸配列は図5において提供されている。
【0091】
議論
多くの血液凝固タンパク質は、不活性な前駆体として合成され、及びその結果としてタンパク質分解活性があり、これらのタンパク質は、血管外傷(2)の場所に位置する細胞表面上に組み立てられる。本研究において、pt−FVは、これらのパラダイムを、FXa及び活性型プロトロンビンを結集させる膜表面に必要な構造的活性化補因子及びバイパスとして合成されたものと同様に回避する。さらに、Pt−FVはAPC、タンパク質分解的切断後の活性の保持に機能的耐性がある。改良された非共有結合性の相互作用と重鎖及び軽鎖を結合する独自のジスルフィド結合はおそらく機能安定性に貢献する。これらの適用の集密は、強力な凝血促進剤に形質転換されたpt−FVを有し、及び、毒物注入に続いておこる重度の播種凝固による選択的優位性のあるヘビ種をおそらく提供する。
【0092】
参考文献
1. Mann KG, Kalafatis M (2002) Factor V: A combination of Dr. Jekyll and Mr. Hyde. Blood 101:20−30.
2. Mann KG, et al. (1990) Surface dependent reactions of the vitamin K−dependent enzyme complexes. Blood 76:1−16.
3. Toso R, Camire RM (2004) Removal of B−domain sequences from factor V rather than specific proteolysis underlies the mechanism by which cofactor function is realized. J Biol Chem 279:21643−21650.
4. Zhu H, Toso R, Camire RM (2007) Inhibitory sequences within the B−domain stabilize circulating factor V in an inactive state. J Biol Chem 282:15033−15039.
5. Jiang Y, Doolittle RF (2003) The evolution of vertebrate blood coagulation as viewed from a comparison of puffer fish and sea squirt genomes. Proc Natl Acad Sci U S A 100:7527−7532.
6. Davidson CJ, et al. (2003) Molecular evolution of the vertebrate blood coagulation network. Thromb Haemost 89:420−428.
7. Rao VS, Swarup S, Kini RM (2003) The nonenzymatic subunit of pseutarin C, a prothrombin activator from eastern brown snake (Pseudonaja textilis) venom, shows structural similarity to mammalian coagulation factor V. Blood 102:1347−1354.
8. St.Pierre L, et al. (2005) Comparative analysis of prothrombin activators from the venom of Australian elapids. Mol Biol Evol 22:1853−1864.
9. Welton RE, Burnell JN (2005) Full length nucleotide sequence of a factor V−like subunit of oscutarin from Oxyuranus scutellatus scutellatus (coastal Taipan). Toxicon 46:328−336.
10. Sutherland S, Tibballs J (2001) in Australian Animal Toxins. The Creatures, Their Toxins and Care of the Poisoned Patient. (Oxford University Press, Melbourne,Australia).
11. Owen WG, Jackson CM (1973) Activation of prothrombin with Oxyuranus scutellatus scutellatus (taipan snake) venom. Thromb Res 3:705−714.
12. Walker FJ, Owen WG, Esmon CT (1980) Characterization of the prothrombin activator from the venom of Oxyuranus scutellatus scutellatus (taipan venom). Biochemistry 19:1020−1023.
13. Masci PP, Whitaker AN, de Jersey J (1988) Purification and characterization of a prothrombin activator from the venom of the Australian brown snake, Pseudonaja textilis textilis. Biochem Int 17:825−835.
14. Speijer H, Govers−Riemslag JW, Zwaal RF, Rosing J (1986) Prothrombin activation by an activator from the venom of Oxyuranus scutellatus (Taipan snake). J Biol Chem 261:13258−13267.
15. Rao VS, Kini RM (2002) Pseutarin C, a prothrombin activator from Pseudonaja textilis venom: its structural and functional similarity to mammalian coagulation factor Xa−Va complex. Thromb Haemost 88:611−619.
16. Rao VS, Swarup S, Kini RM (2004) The catalytic subunit of pseutarin C, a group C prothrombin activator from the venom of Pseudonaja textilis, is structrually similar to mammalian blood coagulation factor Xa. Thromb Haemost 92:509−521.
17. Toso R, Camire RM (2006) Role of Hirudin−like factor Va heavy chain sequences in prothrombinase function. J Biol Chem 281:8773−8779.
18. Mann KG (1976) Prothrombin. Methods Enzymol 45:123−156.
19. Pryzdial ELG, Mann KG (1991) The association of coagulation factor Xa and factor Va. J Biol Chem 266:8969−8977.
20. Boskovic DS, Giles AR, Nesheim MD (1990) Studies of the role of factor Va in the factor Xa−catalyzed activation of prothrombin, fragment 1 2−prethrombin−2, and dansyl−1−glutamyl−glycyl−l−arginine−meizothrombin in the absence of phospholipid. J Biol Chem 265:10497−10505.
21. Esmon CT (2003) The protein C pathway. Chest 124:26S−32S.
22. Higgins DL, Mann KG (1983) The interaction of bovine factor V and factor V−derived peptides with phospholipid vesicles. J Biol Chem 258:6503−6508.
23. Buddai SK, et al. (2002) Nematode anticoagulant protein c2 reveals a site on factor Xa that is important for macromolecular substrate binding to human prothrombinase. J Biol Chem 277:26689−26698.
24. Camire RM, Larson PJ, Stafford DW, High KA (2000) Enhanced g−carboxylation of recombinant factor X using a chimeric construct containing the prothrombin propeptide. Biochemistry 39:14322−14329.
25. Camire RM (2002) Prothrombinase assembly and S1 site occupation restore the catalytic activity of FXa impaired by mutation at the sodium−binding site. J Biol Chem 277:37863−37870.
26. Lundblad RL, Kingdon HS, Mann KG (1976) Thrombin Methods Enzymol 45:156−176.
27. Di Scipio RG, Hermodson MA, Yates SG, Davie EW (1977) A comparison of human prothrombin, factor IX (Christmas Factor), factor X (Stuart Factor), and protein S. Biochemistry 16:698−706.
28. Stafford WF (1992) Boundry analysis in sedimentation transport experiments: a procedure for obtaining sedimentation coefficient distributions using the time derivative of the concentration profile. Anal Biochem 203:295−301.
29. Babul J, Stellwagen E (1969) Measurement of protein concentration with interferences optics. Anal Biochem 28:216−221.
30. Horton RM, et al. (1989) Engineering hybrid genes without the use of restriction enzymes: gene splicing by overlap extension. Gene 77:61−68.
31. Bevington PR, Robinson KD (1992) in Data Reduction and Error Analysis for the Physical Sciences. (McGraw−Hill, New York).
32. Straume M, Johnson ML (1992) Analysis of residuals: criteria for determining goodness−of−fit. Methods Enzymol 210:87−105.
33. Krishnaswamy S (1990) Prothrombinase complex assembly: Contributions of protein− protein and protein−membrane interactions toward complex formation. J Biol Chem 265:3708−3718.
34. Segal IH (1975) in Enzyme Kinetics: Behavior and Analysis of Rapid Equilibrium and Steady State Enzyme Systems. (John Wiley & Sons, Inc., New York).
【0093】
本発明の特定の好ましい実施形態が明細書において記載され、具体的に例示されているが、本発明は実施形態のように限定されるものと意図していない。以下の請求の範囲から逸脱することなく、本発明に様々な変更がなされることがある。
【特許請求の範囲】
【請求項1】
それを必要とする患者における血液凝固障害を治療する方法であって、FV変異体またはその誘導体の活性型の有効量を投与する工程であって、これにより前記患者の血栓形成を増進し、且つ前記血液凝固障害の症状を改善するものである、前記投与する工程を有する、方法。
【請求項2】
請求項1記載の方法において、前記活性変異体は、配列ID番号1及び配列ID番号2からなる群から選択される核酸によってコードされたものである、方法。
【請求項3】
血友病Aを治療するための請求項1記載の方法。
【請求項4】
血友病Bを治療するための請求項1記載の方法。
【請求項5】
請求項1記載の方法において、前記FVまたはその誘導体の活性型は静脈内にまたは局所的に移送されるものである、方法。
【請求項6】
請求項1記載の方法において、前記FVまたはその誘導体の活性型をコードするベクターは、前記患者に投与されるものである、方法。
【請求項7】
請求項1記載の方法において、前記変異体は血液凝固促進剤であり、及び、前記疾患は血友病A及びB、抑制抗体が関連する血友病A及びB、血液凝固因子欠乏症、ビタミンKエポキシド還元塩素欠乏症、ガンマカルボキシラーゼ欠乏症、外傷に伴う出血、負傷、血栓症、血小板減少症、脳卒中、凝血障害、播種性血管内凝固障害(DIC)、過抗凝固治療障害、バーナードスーリエ症候群、グランツマンの血小板無力症、及び貯蔵プール欠乏症からなる群から選択されるものである、方法。
【請求項8】
請求項7記載の方法において、前記過抗凝固治療障害は、ヘパリン、低分子量ヘパリン、五糖類、ワルファリン、低分子抗血栓及びFXa阻害剤の投与によるものである、方法。
【請求項9】
請求項1記載の方法において、前記変異体は、一日に少なくとも一回、約10〜500μg/kgの用量で静脈内投与されるものである、方法。
【請求項10】
ヘビ第五因子をコードする配列ID番号2の単離された核酸。
【請求項11】
発現ベクターに含有される請求項10記載の核酸。
【請求項12】
請求項11記載の核酸を有する単離された宿主細胞。
【請求項13】
配列ID番号1の核酸によってコードされた、単離されたヘビ第五因子タンパク質。
【請求項14】
リポソームまたはミセルに含有される請求項13記載のタンパク質。
【請求項15】
請求項13記載のタンパク質を有する薬学的組成物であって、薬学的に許容される賦形剤に含まれるものである、組成物。
【請求項1】
それを必要とする患者における血液凝固障害を治療する方法であって、FV変異体またはその誘導体の活性型の有効量を投与する工程であって、これにより前記患者の血栓形成を増進し、且つ前記血液凝固障害の症状を改善するものである、前記投与する工程を有する、方法。
【請求項2】
請求項1記載の方法において、前記活性変異体は、配列ID番号1及び配列ID番号2からなる群から選択される核酸によってコードされたものである、方法。
【請求項3】
血友病Aを治療するための請求項1記載の方法。
【請求項4】
血友病Bを治療するための請求項1記載の方法。
【請求項5】
請求項1記載の方法において、前記FVまたはその誘導体の活性型は静脈内にまたは局所的に移送されるものである、方法。
【請求項6】
請求項1記載の方法において、前記FVまたはその誘導体の活性型をコードするベクターは、前記患者に投与されるものである、方法。
【請求項7】
請求項1記載の方法において、前記変異体は血液凝固促進剤であり、及び、前記疾患は血友病A及びB、抑制抗体が関連する血友病A及びB、血液凝固因子欠乏症、ビタミンKエポキシド還元塩素欠乏症、ガンマカルボキシラーゼ欠乏症、外傷に伴う出血、負傷、血栓症、血小板減少症、脳卒中、凝血障害、播種性血管内凝固障害(DIC)、過抗凝固治療障害、バーナードスーリエ症候群、グランツマンの血小板無力症、及び貯蔵プール欠乏症からなる群から選択されるものである、方法。
【請求項8】
請求項7記載の方法において、前記過抗凝固治療障害は、ヘパリン、低分子量ヘパリン、五糖類、ワルファリン、低分子抗血栓及びFXa阻害剤の投与によるものである、方法。
【請求項9】
請求項1記載の方法において、前記変異体は、一日に少なくとも一回、約10〜500μg/kgの用量で静脈内投与されるものである、方法。
【請求項10】
ヘビ第五因子をコードする配列ID番号2の単離された核酸。
【請求項11】
発現ベクターに含有される請求項10記載の核酸。
【請求項12】
請求項11記載の核酸を有する単離された宿主細胞。
【請求項13】
配列ID番号1の核酸によってコードされた、単離されたヘビ第五因子タンパク質。
【請求項14】
リポソームまたはミセルに含有される請求項13記載のタンパク質。
【請求項15】
請求項13記載のタンパク質を有する薬学的組成物であって、薬学的に許容される賦形剤に含まれるものである、組成物。
【図1】

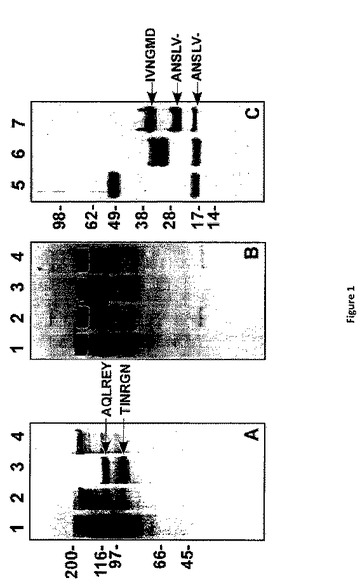
【図2】
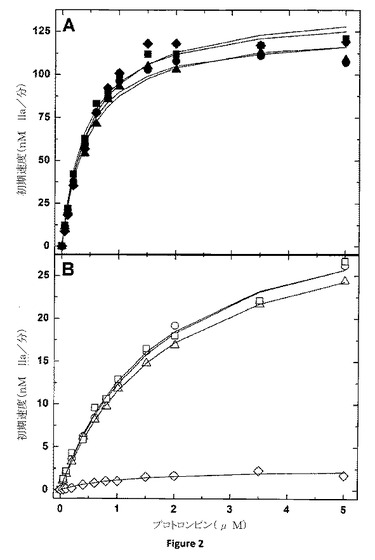
【図3】
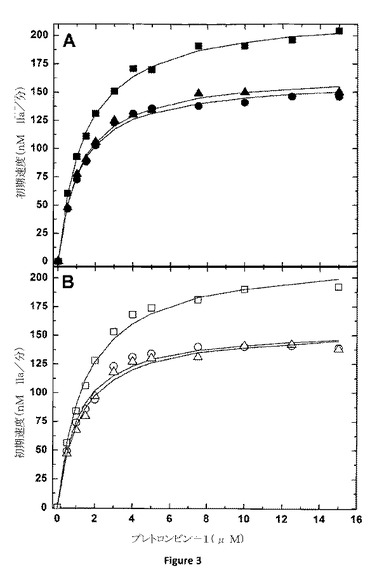
【図4A】
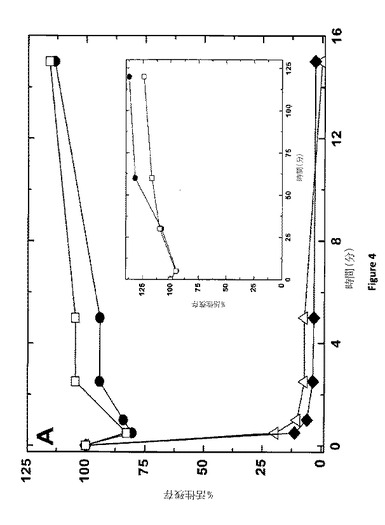
【図4BC】
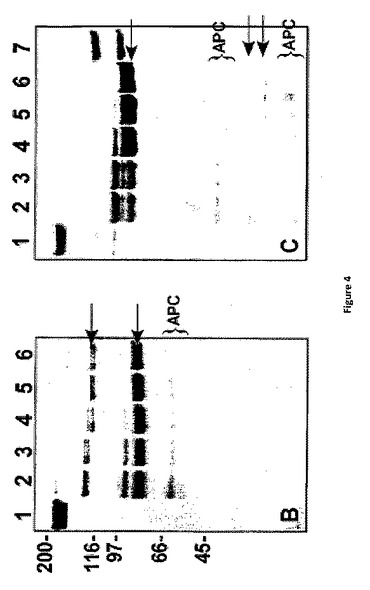
【図5A】

【図5B】

【図5C】

【図5D】

【図2】
【図3】
【図4A】
【図4BC】
【図5A】
【図5B】
【図5C】
【図5D】
【公表番号】特表2012−509670(P2012−509670A)
【公表日】平成24年4月26日(2012.4.26)
【国際特許分類】
【出願番号】特願2011−537682(P2011−537682)
【出願日】平成21年11月23日(2009.11.23)
【国際出願番号】PCT/US2009/065515
【国際公開番号】WO2010/060035
【国際公開日】平成22年5月27日(2010.5.27)
【出願人】(510226358)
【Fターム(参考)】
【公表日】平成24年4月26日(2012.4.26)
【国際特許分類】
【出願日】平成21年11月23日(2009.11.23)
【国際出願番号】PCT/US2009/065515
【国際公開番号】WO2010/060035
【国際公開日】平成22年5月27日(2010.5.27)
【出願人】(510226358)
【Fターム(参考)】
[ Back to top ]