
血管増殖性疾患の治療
本発明は分子生理学の分野に関する。具体的には、本発明は、血管増殖性疾患、特に眼の血管増殖性疾患の予防および/または治療、ならびに血管増殖を示す腫瘍の治療に関する。ロイシンリッチα2糖タンパク質1(Lrg1)のレベルは、そうした疾患に罹患した患者およびそうした疾患の動物モデルにおいて増加していることが明らかになった。Lrg1のアンタゴニストを用いて、血管増殖性疾患を予防および/または治療することができる。
【発明の詳細な説明】
【技術分野】
【0001】
本発明は分子生理学の分野であって、特に眼の血管増殖性疾患の治療もしくは予防、ならびに血管増殖を示す腫瘍の治療で用いるための、ロイシンリッチα2糖タンパク質1(Lrg1)のアンタゴニストの使用に関する。
【背景技術】
【0002】
網膜血管系の異常なリモデリングは、糖尿病性網膜症、網膜静脈閉塞、未熟児網膜症、加齢黄斑変性および黄斑部毛細血管拡張症などの、失明の恐れのある疾患の顕著な特徴である。こうした血管の変化は、既存の網膜血管からの新たな毛細血管の生成(血管形成)として、ならびに既存血管の血管奇形の発達(たとえば、毛細血管拡張症)として生じる。この病原的な血管リモデリングが、視力を失う主たる要因である。
【0003】
同様の血管の病変および機能不全は、腫瘍の増殖にも付随するものであって、この場合血管新生が固形腫瘍の増大および増殖を可能にする。
【0004】
多くの資力が、(血管形成、血管新生、血管増殖、血管リモデリング、血管病変と称される)これらの血管反応を推進するメカニズムに関する知見を得ることに向けられてきた。その過程で重要な役割を果たすと確認されたさまざまな分子のうちで、血管内皮増殖因子(VEGF)およびその受容体は、非常に重要な構成要素と見なされる。抗がん剤アバスチン(または近縁のルセンティス)を用いて、眼の血管形成におけるVEGF経路を治療の標的にすると、結果としていくつかの失明の恐れのある疾患について、少なくとも短期的には、臨床的成果の改善がもたらされた。しかしながら、VEGFは神経保護活性を有し、しかも脈絡膜の開窓の維持といったハウスキーピング機能も有しているので、網膜血管の問題点を治療するために長期間、VEGFに対抗する戦略を用いることについては懸念がある。その上、VEGFは血管新生においては主要な血管新生促進因子と見なされるが、そのプロセスは多くの因子間の協調的なクロストークを必要としており、他の血管変化、たとえば拡張し蛇行した毛細血管拡張症の血管の形成、に関する生物学的根拠は不明である。
【発明の概要】
【発明が解決しようとする課題】
【0005】
したがって、代わりとなる治療標的および新規薬物を特定する必要があるが、この新規薬物は、単独で、または既存の治療法との併用で、制御されない血管増殖および/またはリモデリングが病気の一因となっているような疾患の治療のために、より有効であり、しかもオフターゲット作用はより少なくすることができる薬物である。
【課題を解決するための手段】
【0006】
本発明者らは、病気を引き起こす可能性のある血管リモデリングを調節するための、新薬の開発につながる標的として、ロイシンリッチα2糖タンパク質1(Lrg1遺伝子名:HGNC: 29480; Entrez Gene: 116844; Ensembl: ENSG00000171236; UniProtKB: P02750)を特定した。
【0007】
Lrg1は1977年に同定され(Haupt & Baudner, 1977)、その一次構造は1985年に決定された(Takahashi et al, 1985)。Lrg1はマウスとヒトの間で高度に、進化的に保存されており、ヒトLrg1に対するポリクローナル抗体が市販されているが、ある種の疾患においてトランスフォーミング増殖因子β1(TGFβ1)、TGFβ受容体II(TGFβRII)およびLrg1のレベルが同時に増加するとの報告がある(Sun et al, 1995; Li et al, 1997)。他のグループは、Lrg1を特定の疾患のバイオマーカーとして(US 2005/0064516; WO 2008/092214)ならびにシトクロムcのリガンドとして(US 2007/0184503)同定した。内皮細胞におけるTGFβシグナル伝達の機能不全は、遺伝性出血性毛細管拡張症(HHT)をもたらす。
【0008】
毛細管拡張症などの血管異常を特徴とするこの疾患群において、TGFβ内皮細胞補助受容体エンドグリンおよびTβRIコレセプターALK1の変異はそれぞれ、HHT1(McAllister et al. 1994)およびHHT2(Johnson et al. 1995)を引き起こす。TGFβは、血管リモデリングがよく見られる糖尿病性網膜症患者の網膜で増加していることも明らかになった(Spirin et al., 1999)。特定の腫瘍患者の血漿中でLrg1発現が増加することも判明しており、これは、Lrg1が有望な腫瘍バイオマーカーとして機能しうることを示唆する(Heo et al, 2007; Ferrero et al, 2009; Kakisaka et al, 2007)。しかしながら、Lrg1の生物学についてはほとんどわかっていない。
【0009】
本発明者らはこれまでに、特に眼において、ならびに血管増殖を示す腫瘍において、病気を引き起こす可能性のある血管リモデリングを調節するための、新薬につながる標的としてLrg1を特定した。
【0010】
血管の変化を伴う網膜疾患のマウスモデルを用いて、本発明者らはまず、これらの罹患網膜の血管において、他の遺伝子の中でLrg1がアップレギュレートされていることを突き止めた。次に、こうしたマウスモデルの網膜におけるLrg1の発現増加を、定量的PCRおよびウェスタンブロッティングによって検証し、in situハイブリダイゼーションおよび免疫組織化学によって、Lrg1の網膜での分布が血管であると確認した。こうしたモデルは、血管形成の標準的なモデルであり、眼以外の部位での血管形成に応用できる。
【0011】
そこで本発明者らは、Lrg1とTGFβシグナル伝達経路との関連性を調べた。
【0012】
内皮細胞において、TGFβシグナル伝達は、どこにでも存在するTGFβI型受容体アクチビン受容体様キナーゼ5(ALK5)、または内皮細胞特異的ALK1のいずれか一方と結合したTGFβ受容体IIを介して起こる可能性があり、細胞性応答は、どちらの経路が主であるかによって左右される。ALK5の場合、特定の条件下で、ECM沈着の増加および細胞休止が見られるのに対して、ALK1については、遊走および増殖の増加として現れる内皮細胞の活性化が見られる。こうした差異のあるシグナル伝達は、一つにはTGFβの濃度/バイオアベイラビリティによって制御され、また、Smadと呼ばれる下流エフェクタータンパク質ファミリーのメンバーによっても制御されるが、このSmad 2および3はALK5で活性化されSmad 1、5および8はALK1により活性化される。
【0013】
免疫沈降法により、Lrg1がTGFβRIIおよびALK1の両方に結合することが明らかになったが、このことは、Lrg1が、TGFβシグナル伝達複合体の一部として、これら2つの分子と関連して役割を有することを示唆する。
【0014】
したがって、本発明者らは、Lrg1がTGFβシグナル伝達のモジュレーターとして機能して、ALK1-およびALK5-活性化シグナル伝達カスケード間の微調整を引き起こすとの仮説を立てた。これを裏付けるものとして、siRNAを用いた内皮細胞のLrg1ノックダウンは、TGFβによって増加した細胞増殖をブロックし、Smad5リン酸化を低下させたのに対して、Lrg1の過剰発現は増殖の強化、Smad2発現のダウンレギュレーション、およびSmad5リン酸化の増加をもたらす。したがって、こうした知見は、Lrg1が血管形成を制御することができる1つの方向を明らかにする。また、Lrg1が「血管」形成に及ぼす影響を検討するMatrigel血管形成アッセイにおいて、血管形成、管形成および索状組織形成により評価される血管形成の度合いは、Lrg1過剰発現細胞から得られた馴化培地を添加したときに有意に増加し、血管新生の増加は培地中のLrg1タンパク質発現と相関していた。
【0015】
これらのデータは、TGFβRII/ALK5受容体複合体経路を介したシグナル伝達の減少と合致し、したがって、血管病変を引き起こす可能性のあるTGFβRII/ALK1シグナル伝達経路の活性化へのシフトと一致する。
【0016】
これは、TGFβシグナル伝達複合体においてLrg1をブロックすることで、TGFβを、病気を引き起こす可能性のある血管新生から逸らすことが可能であることを示唆する。この問題を検証するために、本発明者らは、TGFβによって引き起こされる内皮細胞Smad5リン酸化が、抗Lrg1抗体を用いて、もしくはALK1との結合についてLrg1と競合すると見込まれるLrg1由来ペプチド配列によって、ブロックされるかどうかを判定した。抗Lrg1抗体はリン酸化の減少を引き起こしたが、ペプチドのうち1つは特に大幅なリン酸化の減少を示した。
【0017】
したがって、Smad5のリン酸化は、Lrg1をブロックすることにより阻害することができるが、Smad5は、血管病変を生じさせるALK1で活性化されたシグナル伝達カスケードと関連しているので、これは、Lrg1のブロックが、病気を引き起こす可能性のないALK5で活性化された別のカスケードと比べて、ALK1で活性化されたカスケードをブロックする可能性があることを示している。
【0018】
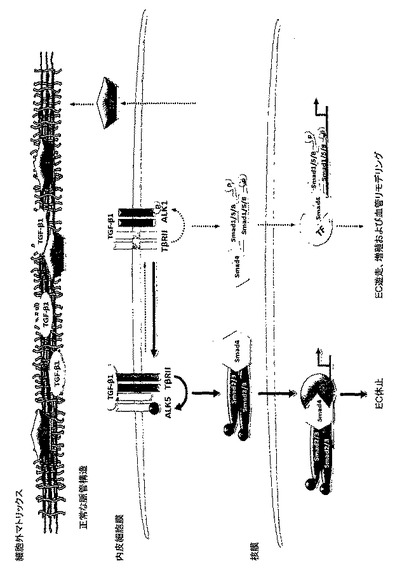
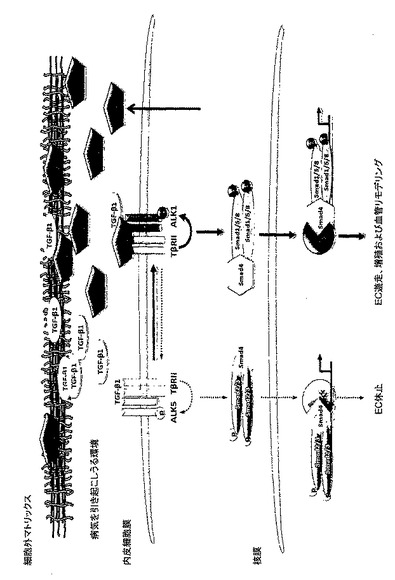
以上をまとめると、上記データは下記を示唆する:(a)Lrg1はTGFβRIIおよびALK1のどちらとも相互作用し、ALK5ではなくALK1とTGFβRIIとの相互作用を、直接、または1つもしくは複数の中間体を介して促進するものであって、その結果(b)Lrg1をブロックすることは、TGFβの活性を、血管病変を引き起こす可能性のある、ALK1で活性化されたシグナル伝達カスケードから逸らして、病気を引き起こさないALK5で活性化されたカスケードに向けることになり、その結果として(c)Lrg1は、眼および他の箇所において病気を引き起こす可能性のある血管新生を治療するための効果的な薬物標的となる。したがって、さまざまなLrg1遮断薬、特にLrg1のペプチド断片、Lrg1に対するモノクローナル抗体、およびsiRNA分子による治療を、病気を引き起こす可能性のある組織血管新生を伴う眼および他の疾患のために、構想することができる。図16は、理論に縛られることなく、本発明者らのデータによって示唆されるLrg1の役割を示すものである。
【0019】
さらに、本発明者らのデータは、VEGFとは対照的に、Lrg1が眼において病気を引き起こす可能性のある血管新生にのみ関与し、正常な発達の血管新生もしくは血管ホメオスタシスには関与しないことを示唆する。このことからLrg1は、途絶させることが望ましくないプロセスの妨害を避けるという点から見ると、VEGFよりすぐれた標的となる可能性がある。Lrg1が標的としてさらに魅力的であるのは、それが細胞外にあるので、治療のための全身経路を介してより容易にアクセスできることである。
【0020】
また、ex vivoおよびin vivoでのLrg1の役割を調べるために実験を行った。本発明者らの実験によれば、Lrg1ノックアウトマウスの大動脈輪では、対照マウスの大動脈輪と比べて血管新生の血管発芽が減少している。また、本発明者らは、網膜損傷後の脈絡膜血管新生(CNV)、および酸素誘導網膜症(OIR)後の網膜血管新生がLrg1ノックマウスでは対照マウスより減少したことも明らかにした。
【0021】
ヒトの病理においてLrg1の果たす役割の証拠として、本発明者らのデータは、増殖性糖尿病性網膜症に罹っているヒト患者においてLrg1およびTGFβの発現が増加していることを示すが、このことはマウスから得られたin vivoデータを支持する。
【0022】
本発明者らは、Lrg1に対する抗体が、Matrigel血管形成アッセイにおいて、ヒト臍帯静脈内皮細胞(HUVEC)による管形成を阻害する能力を有することも明らかにした。こうしたデータは、Lrg1に対する抗体が、血管増殖性疾患、特に眼の血管増殖性疾患、および血管増殖を示す腫瘍の治療もしくは予防に有用であることを示唆する。
【0023】
したがって、本発明は、血管増殖性疾患の治療もしくは予防に用いるロイシンリッチα2糖タンパク質1(Lrg1)のアンタゴニストを提供する。
【0024】
本発明はまた、Lrg1のアンタゴニストを同定する方法を提供するが、その方法は、候補アンタゴニストを用意すること、ならびに前記候補アンタゴニストがLrg1の機能もしくは活性をブロックするか否かを決定することを含んでおり、Lrg1の機能もしくは活性のブロックが認められれば、前記候補アンタゴニストはLrg1のアンタゴニストとして同定される。
【0025】
本発明はまた、付表2のアミノ酸配列L1-24もしくは付表3のL94-117(配列番号3)、付表2のL169-192もしくは付表3のL262-285(配列番号4)、または付表2のL227-252もしくは付表3のL320-345(配列番号5)にあるエピトープを特異的に認識して、Lrg1の活性をブロックする、モノクローナル抗体を提供する。
【0026】
本発明はまた、付表2のアミノ酸L1-24もしくは付表3のL94-117(配列番号3)、付表2のL169-192もしくは付表3のL262-285(配列番号4)、または付表2のL227-252もしくは付表3のL320-345(配列番号5)にあるエピトープを特異的に認識して、ALK1、TGFβRIIおよび/またはTGFβとLrg1との相互作用をブロックする、モノクローナル抗体を提供する。
【0027】
本発明はまた、上記抗体を作製するための方法を提供するが、その方法は、付表2のL1-24もしくは付表3のL94-117(配列番号3)、付表2のL169-192もしくは付表3のL262-285(配列番号4)、または付表2のL227-252もしくは付表3のL320-345(配列番号5)の配列内のエピトープを含んでなる免疫原でヒト以外の哺乳類を免疫化すること;ならびに前記哺乳類から抗体標品を得ること、およびその標品から、前記エピトープを特異的に認識するモノクローナル抗体を得ることを含んでなる。
【0028】
本発明はまた、Lrg1の機能もしくは活性をブロックするためにLrg1の中のどの部位を標的とすることができるかを決定するための方法を提供するが、その方法は、Lrg1タンパク質のペプチド断片を用意すること;ならびに前記のそれぞれのペプチド断片がLrg1の機能もしくは活性をブロックするか否かを判定することを含んでなる。
【0029】
本発明はまた、血管増殖の治療もしくは予防のための薬剤の製造におけるLrg1のアンタゴニストの使用を提供する。
【0030】
本発明はまた、血管増殖性疾患を治療する方法を提供するが、その方法はそれを必要とする患者に有効な量のLrg1アンタゴニストを投与することを含んでなる。
【図面の簡単な説明】
【0031】
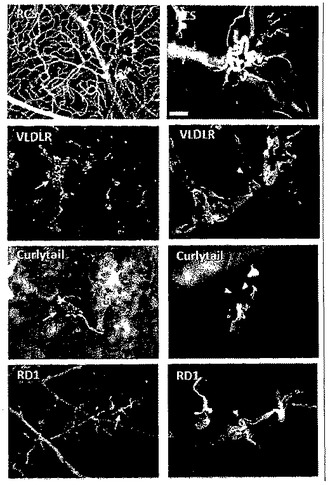
【図1】RCSラット(20週齢)、VLDLR-/-マウス(16週齢)、Curlytail-Jマウス(13週齢)およびRD1マウス(16週齢)の網膜の血管リモデリングの低倍率および高倍率画像。血管の基底膜および内皮細胞間結合をそれぞれ着色するために、網膜フラットマウントの血管を抗コラーゲンIVおよび抗クローディン5抗体で染色した。
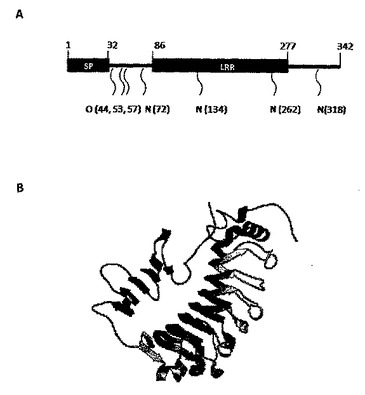
【図2】A. Lrg1タンパク質および提唱されているそのグリコシル化部位の配置図。B. ROBETTA(University of Washington, USA)により推定されたLrg1タンパク質の構造。
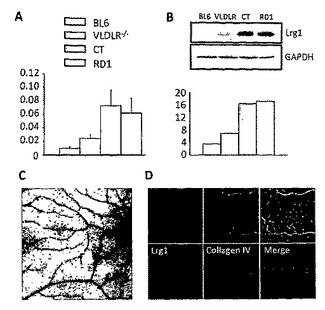
【図3】A. C57Bl6対照マウス(BL6)、VLDL受容体KOマウス(VLDLR-/-)、Curlytail-Jマウス(CT)および網膜ジストロフィー1マウス(RD1)のマウス網膜全体における、Lrg1発現の定量的RT PCR分析。B. 網膜全体のLrg1タンパク質発現のウェスタンブロット(上図)および半定量(下図)。C. Lrg1遺伝子発現を示す正常な網膜のin situハイブリダイゼーション。D. 血管の発現パターンを示す、網膜フラットマウント(上図)および網膜断面(下図)におけるLrg1の免疫組織化学染色。
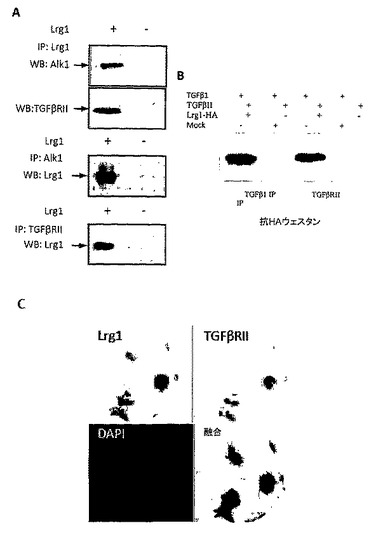
【図4A】A. Lrg1とTGFβRIIおよびALK1との、GPNT内皮細胞溶解物からの免疫共沈降。B. 組換えHA-タグ付きLrg1はTGFβおよびTGFβRIIの両者と結合する。C. GPNT内皮細胞におけるLrg1およびTGFβRII発現の共局在化。D. Lrg1ノックダウンが、TGFβによって引き起こされる増殖を弱め、Smad5リン酸化を減少させることを示す、内皮細胞増殖アッセイ(* p<0.05)。E. Lrg1の過剰発現が、TGFβによって引き起こされる増殖を強め、Smad1/5リン酸化を増加させることを示す、内皮細胞増殖アッセイ(* p<0.05)。
【図4B】A. Lrg1とTGFβRIIおよびALK1との、GPNT内皮細胞溶解物からの免疫共沈降。B. 組換えHA-タグ付きLrg1はTGFβおよびTGFβRIIの両者と結合する。C. GPNT内皮細胞におけるLrg1およびTGFβRII発現の共局在化。D. Lrg1ノックダウンが、TGFβによって引き起こされる増殖を弱め、Smad5リン酸化を減少させることを示す、内皮細胞増殖アッセイ(* p<0.05)。E. Lrg1の過剰発現が、TGFβによって引き起こされる増殖を強め、Smad1/5リン酸化を増加させることを示す、内皮細胞増殖アッセイ(* p<0.05)。
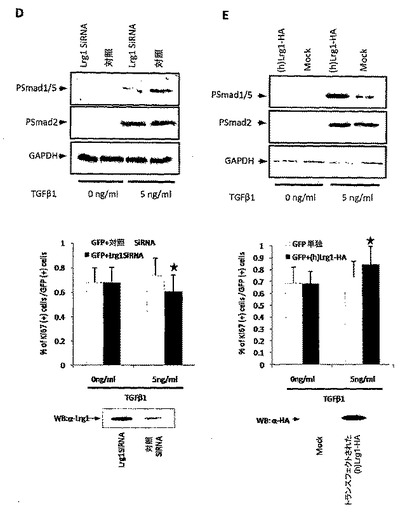
【図5】C末端に由来するLrg1ブロッキングペプチドは、GPNT内皮細胞においてLrg1/TGFβ1が誘導するSmad5リン酸化を阻害する(各条件につきn=3、p<0.0001)。右側2レーンは、C-末端ペプチドのみ、および3ペプチド、付表2のL1-24もしくは付表3のL94-117(配列番号3)、付表2のL169-192もしくは付表3のL262-285(配列番号4)、または付表2のL227-252もしくは付表3のL320-345(配列番号5)の組み合わせである。
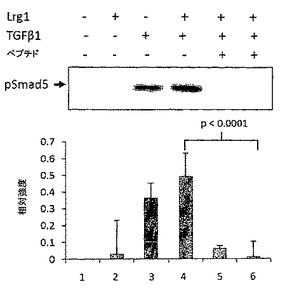
【図6】A. siRNAによるGPNT内皮細胞のLrg1ノックダウン、およびGPNT内皮細胞におけるLrg1過剰発現のウェスタンブロット。B. Lrg1ノックダウンは、TGFβ1が介在する内皮細胞増殖を低下させ(* p<0.0001)、Lrg1過剰発現は、TGFβ1が介在する内皮細胞増殖を高める(* p<0.0005)(各条件についてn=3)。
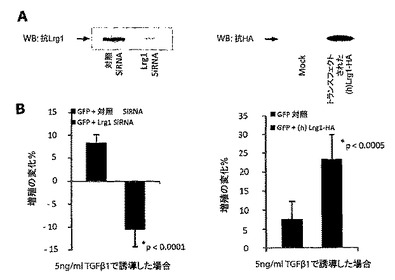
【図7】HUVECのin vivoでの「血管」形成に及ぼすLrg1の影響。A. 未処理培地、または対照の内皮細胞(EC)由来、もしくはLrg1を過剰発現するEC由来の培地をMatrigel血管形成アッセイに添加した。Lrg1馴化培地はHUVECの「索状組織」形成を高めた。B. 非馴化培地、GPNT内皮細胞馴化培地(7日)およびLrg1を過剰発現するGPNT細胞由来の馴化培地におけるLrg1のウェスタンブロット。C. さまざまな処理の後の、Matrigel内皮細胞索状組織形成複雑系の数量化(閉じた血管輪の数、および全血管面積)(各条件についてn=3)。Lrg1過剰発現細胞由来のLrg1馴化培地は、閉じた血管輪の数もしくは全血管面積で測定されるように最大の血管形成血管網を生じさせた(p<0.0001)。
【図8】TGFβおよびLrg1で処理後に、内皮細胞のSmad5リン酸化に及ぼす、Lrg1抗体(Lrg1のN-末端ドメインに対する市販のポリクローナル抗体)およびLrg1ペプチドの影響。
【図9】Lrg1ノックアウトマウス由来の大動脈輪は、野生型マウスの大動脈輪と比べて、血管新生の血管出芽の減少を示す。Lrg1 KO マウスの大動脈において血管新生の血管出芽の減少を示す、大動脈輪血管形成の代表的な画像(イソレクチンB4で緑色染色)、ならびにそれらの数量化。各群について大動脈輪n=30(** p<0.01)。
【図10】網膜疾患のCurlytail-J、RD1およびVLDLR -/-マウスモデルにおいてアップレギュレートされる遺伝子。
【図11】A. WTおよびLrg1 KOマウスの網膜へのレーザー熱傷によって引き起こされた脈絡膜新血管形成の蛍光眼底血管造影図(FA)。損傷後7日で記録された血管新生増殖および漏出の大きさをそれぞれ示す、初期(B)および後期(C)FAの数値化。n=10(** p<0.01)。
【図12】(左)脳内皮細胞株GPNTはLrg1のTGFβシグナル伝達への影響を研究するために必須の成分を発現する。P = 細胞ペレット;M = 細胞培地。(右)TGFβはGPNT細胞においてLrg1遺伝子発現を誘導する。
【図13】酸素誘導網膜症(OIR)後のP17でのマウス網膜血管系の代表的な画像(イソレクチンB4で赤色染色)であって、これはLrg1 KOマウスにおいて無血管領域の増加および新生血管網の減少を示す。WTおよびLrg1 KOマウス(それぞれn=6および9)の(B)血管のない領域、および(C)新生血管網の数量化。無血管領域はLrg1ノックアウトにおいて増加し(* p<0.05)、目に見える新生血管網は野生型と比較してノックアウトにおいて少なくなっている(** p0.01)。
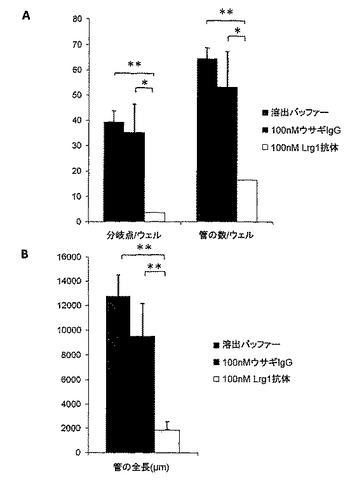
【図14】Matrigel中でのin vitroのヒト臍帯静脈内皮細胞(HUVEC)の管形成は、中和抗体である抗ヒトLrg1ポリクローナル抗体添加後に、無関係のIgGと比較して減少した。管形成は、A. 分岐点の数、管の数、およびB. 管の全長について、測定した(n=3、* p<0.05, ** p<0.01)。
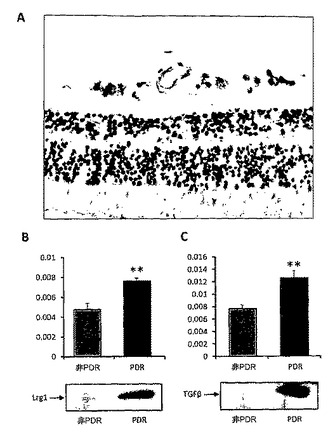
【図15】A. Lrg1について染色されたヒト網膜の断面図。B. 非糖尿病患者、および増殖性糖尿病性網膜症(PDR)患者由来の硝子体液サンプル中のLrg1(n=4)のウェスタンブロットおよび数量化。C. 非糖尿病患者およびPDR患者由来の硝子体液サンプル中のTGFβ1のウェスタンブロットおよび数量化。
【図16A】作動仮説の概略図。A. 正常な条件下で、TGFβ1シグナル伝達は主としてTGFβRII/ALK5/Smad2/3経路に向けて行われ、Lrg1は基底膜に潜んでいる。B. 病的な条件下では、Lrg1の発現増加の結果として、TGFβ1シグナル伝達は血管リモデリングの一因となるTGFβRII/ALK1/Smad1/5/8経路に向けて方向が変更される。
【図16B】作動仮説の概略図。A. 正常な条件下で、TGFβ1シグナル伝達は主としてTGFβRII/ALK5/Smad2/3経路に向けて行われ、Lrg1は基底膜に潜んでいる。B. 病的な条件下では、Lrg1の発現増加の結果として、TGFβ1シグナル伝達は血管リモデリングの一因となるTGFβRII/ALK1/Smad1/5/8経路に向けて方向が変更される。
【発明を実施するための形態】
【0032】
(発明の詳細な説明)
Lrg1のブロック
本発明のアンタゴニストはLrg1の機能をブロックする。Lrg1をブロックすることは、その活性または機能を低下させることを含んでおり、その結果として内皮細胞増殖、周皮細胞脱落、内皮細胞死、血管リモデリング、血管形成、毛細管拡張症、血管漏出などの血管増殖作用が低下するものである。
【0033】
たとえば、Lrg1のブロックは、そのLrg1とALK1、TGFβRIIおよび/またはTGFβとの相互作用のブロックを介して行うことができるが、そうしたブロックはALK1ではなくALK5とTGFβRIIとの相互作用を促進するので、TGFβの活性を、血管病変を引き起こしうるALK1活性化カスケードから逸らして、より病気の原因となりにくいALK5活性化シグナル伝達カスケードに方向転換させることが本発明者らのデータから示唆される。Lrg1のブロックは、結果としてTGFβのバイオアベイラビリティも低下させる可能性がある。
【0034】
ブロックには、Lrg1の活性もしくは機能の完全な低下および部分的な低下がいずれも含まれるが、これは、たとえばALK1-Lrg1、TGFβRII-Lrg1、および/またはTGFβ-Lrg1相互作用の完全な阻止もしくは部分的な阻止である。たとえば、本発明のブロッキングアンタゴニストは、Lrg1の活性を10から50%まで、少なくとも50%、または少なくとも70%、80%、90%、95%もしくは99%、低下させることができる。
【0035】
Lrg1活性もしくは機能のブロックは、任意の適当な手段で測定することができる。たとえば、ALK1-Lrg1、TGFβRII-Lrg1、および/またはTGFβ-Lrg1相互作用のブロックは、Smad5リン酸化への影響を測定することによって判定することができるが、これは、Smad5リン酸化がALK5活性化経路ではなくてALK1活性化経路に特有であるという根拠に基づく。
【0036】
Lrg1のブロックは、血管形成を測定するアッセイ、たとえばMatrigel中での血管増殖、大動脈輪からの血管増殖などのin vitroアッセイ、および網膜血管形成(たとえば、レーザーによって生じる脈絡膜新血管形成、酸素誘導網膜症)を測定する方法などのin vivoアッセイによって測定することもできる。
【0037】
ブロックは、たとえば使用されるアンタゴニストの性質(下記参照)に応じて、任意の適当なメカニズムで、たとえばALK1-Lrg1、TGFβRII-Lrg1、および/またはTGFβ-Lrg1の直接的もしくは間接的な相互作用における立体障害、またはLrg1発現のノックダウンによって生じるといえる。
【0038】
Lrg1のアンタゴニスト
任意の適当なアンタゴニスト、たとえばペプチドおよびペプチドミメティック、抗体、小分子阻害剤、二本鎖RNA、アプタマーおよびリボザイムを、本発明にしたがって使用することができる。好ましいアンタゴニストとしては、Lrg1のペプチド断片、二本鎖RNA、アプタマーおよび抗体がある。
【0039】
ペプチド
ペプチドアンタゴニストは、典型的には、TGFβRIIおよび/またはALK1との結合に関して全長Lrg1と競合し、したがってLrg1と拮抗する、Lrg1の断片である。こうしたペプチドは直鎖状でも環状でもよい。ペプチドアンタゴニストは典型的には、5から50、好ましくは10-40、10-30もしくは15-25アミノ酸長であって、概してLrg1内部由来の連続配列と同一であるが、ペプチドアンタゴニストがLrg1をブロックする性質を保持している限り、その同一性は100%未満でもよく、たとえば95%以上、90%以上、または80%以上であってもよい。ブロッキングペプチドは、任意の適当な方法で、たとえばLrg1配列の一部もしくはすべてに及ぶ、連続したペプチドもしくはオーバーラップしたペプチドの、系統的スクリーニングによって同定することができる。こうしたブロッキングペプチドを模倣するように、ペプチドミメティックを設計することもできる。
【0040】
二本鎖RNA
既知の技術を使用し、Lrg1の配列に関する知見を基にして、配列相同性に基づいてLrg1 RNAを標的化することにより、二本鎖RNA(dsRNA)分子を、Lrg1と拮抗するようにデザインすることができる。こうしたdsRNAは典型的には、通常ステムループ(「ヘアピン」)構造をとる低分子干渉RNA(siRNA)、またはマイクロRNAである。こうしたdsRNAの配列は、Lrg1をコードするmRNAの一部に相当する部分を含んでいる。この部分は、通常、Lrg1 mRNAの標的部分に100%相補的であるが、相補性はもっと低レベル(たとえば90%以上、または95%以上)であってもよい。
【0041】
アプタマー
アプタマーは総じて、特異的な標的分子と結合する核酸分子である。アプタマーは完全にin vitroで操作することが可能であり、化学合成によって容易に作製され、望ましい保存特性を有しており、加えて治療的応用において免疫原性をほとんど、もしくはまったく生じない。これらの特徴から、アプタマーは、薬剤および治療の実用性において特に有用である。
【0042】
本明細書で使用される「アプタマー」は、総じて、一本鎖もしくは二本鎖オリゴヌクレオチド、またはそういったオリゴヌクレオチドの混合物を表し、このオリゴヌクレオチドもしくは混合物は、標的と特異的に結合することができる。オリゴヌクレオチドアプタマーは本明細書で検討されるが、同等の結合特性を有する他のアプタマー、たとえばペプチドアプタマーなども使用できることは、当業者に当然理解されるであろう。
【0043】
概して、アプタマーには、ヌクレオチドの長さが少なくとも5、少なくとも10、または少なくとも15ヌクレオチドである、オリゴヌクレオチドが含まれるといえる。アプタマーは、ヌクレオチドが40以下、60以下、もしくは100以下、またはそれ以上の長さの配列を含有することができる。たとえば、アプタマーは5から100ヌクレオチド、10から40ヌクレオチド、または15から40ヌクレオチドまでの長さとすることができる。可能であればもっと長さの短いアプタマーが好ましいが、これは、こうしたアプタマーが他の分子もしくは材料による干渉の減少をもたらすことが多いからである。
【0044】
未修飾のアプタマーは血流から速やかに除去され、半減期は数分から数時間であるが、これは主として、ヌクレアーゼ分解および腎臓による体からのクリアランスに起因する。こうした未修飾アプタマーは、たとえば、血液凝固を刺激するような一過性疾患の治療に有用性がある。あるいはまた、アプタマーを修飾して半減期を改善することができる。いくつかのそうした修飾、たとえば2'-フッ素置換ピリミジンもしくはポリエチレングリコール(PEG)結合の付加が利用できる。
【0045】
アプタマーは、試験管内人工進化(Systematic Evolution of Ligands by Exponential enrichment)(SELEX)法のような常法によって作製することができる。SELEXは、標的分子への高度に特異的な結合による、核酸分子のin vitro進化のための方法である。それはたとえば、US 5,654,151、US 5,503,978、US 5,567,588およびWO 96/38579に記載されている。
【0046】
SELEX法は、オリゴヌクレオチド集団からの核酸アプタマーの選択、特に、望ましい標的に結合する能力を有する一本鎖核酸の選択に関わるものである。一本鎖核酸(たとえば、DNA、RNA、またはそれらのバリアント)の集団は、結合に適した条件下で標的と接触させ、混合物中で標的と結合した核酸を、結合していない核酸から分離し、その核酸-標的複合体を解離させ、標的と結合していた核酸を増幅して、望ましい結合活性を有する核酸が濃縮された集団もしくはライブラリーを生じさせるが、そこでこの一連のステップは、当該標的に特異的な結合親和性を有する核酸(アプタマー)ライブラリーをもたらすように、必要に応じて繰り返される。
【0047】
抗体
本明細書で使用される「抗体」という用語は、全抗体および任意の抗原結合フラグメント(すなわち、「抗原結合部分」)またはそれらの一本鎖を含んでいる。抗体は、ジスルフィド結合で相互に連結された、少なくとも2つの重(H)鎖および2つの軽(L)鎖を含んでなる糖タンパク質、またはその抗原結合部分を指す。それぞれの重鎖は、重鎖可変領域(本明細書ではVHと略す)および重鎖定常領域で構成される。それぞれの軽鎖は、軽鎖可変領域(本明細書ではVLと略す)および軽鎖定常領域で構成される。重鎖および軽鎖の可変領域は、抗原と相互作用する結合ドメインを含有する。VHおよびVL領域は、さらに、相補性決定領域(CDR)と呼ばれる超可変性領域に細分化することができ、フレームワーク領域(FR)と呼ばれる保存性の高い領域が散在している。
【0048】
抗体の定常領域は、免疫グロブリンと、宿主組織もしくは因子、たとえば免疫系のさまざまな細胞(例、エフェクター細胞)および古典的補体系の第1成分(C1q)など、との結合を仲介することができる。
【0049】
本発明の抗体は、モノクローナル抗体でもポリクローナル抗体でもよいが、モノクローナル抗体が好ましいといえる。本発明の抗体は、キメラ抗体、CDR-グラフト化抗体、ナノボディ、ヒトもしくはヒト化抗体、またはそれらのいずれかの抗原結合部分とすることができる。モノクローナルおよびポリクローナル抗体を作製するための実験動物は、典型的にはヒト以外の哺乳類、たとえばヤギ、ウサギ、ラットもしくはマウスであるが、他の種、たとえばラクダ科動物などで産生させることもできる。
【0050】
ポリクローナル抗体は、当該抗原を用いた適当な動物の免疫化といった常法によって作製することができる。その後、動物から血液を採取してIgG画分を精製することができる。
【0051】
本発明のモノクローナル抗体(mAb)は、従来のモノクローナル抗体の手法、たとえばKohlerおよびMilsteinの標準的な体細胞ハイブリダイゼーション法を含めて、さまざまな技術によって作製することができる。ハイブリドーマを調製するために好ましい動物系は、マウスの系である。マウスにおけるハイブリドーマ作製は、非常によく確立された手法であって、当技術分野で周知の技術を用いて達成することができる。
【0052】
本発明の抗体は、下記を含んでなる方法によって作成することができる:全長Lrg1、Lrg1のペプチド断片、Lrg1の付表2のL1-24もしくは付表3のL94-117(配列番号3)、付表2のL169-192もしくは付表3のL262-285(配列番号4)、または付表2のL227-252もしくは付表3のL320-345(配列番号5)の配列に含まれるエピトープ、またはLrg1の他の領域に含まれるエピトープなどの免疫原で、ヒト以外の哺乳類を免疫化すること;前記哺乳類から抗体標品を得ること;それらから、前記エピトープを特異的に認識するモノクローナル抗体を得ること。
【0053】
抗体の「抗原結合部分」という用語は、抗原と特異的に結合する能力を保持する、1つもしくは複数の抗体フラグメントを意味する。全長抗体のフラグメントが抗体の抗原結合機能を果たすことができることが明らかになっている。抗体の「抗原結合部分」という用語に含まれる結合フラグメントの例には、Fabフラグメント、F(ab')2 フラグメント、Fab’フラグメント、Fdフラグメント、Fvフラグメント、dAbフラグメント、および単独の相補性決定領域 (CDR)がある。scFv抗体などの一本鎖抗体も、抗体の「抗原結合部分」という用語に含まれるものとする。こうした抗体フラグメントは、当業者に知られている従来技術を用いて得ることができるが、フラグメントは完全な抗体と同様に、有用性を求めてスクリーニングすることができる。本発明の抗体は、組換え法によって調製、発現、作製または単離することができる:たとえば(a)抗体は、当該免疫グロブリン遺伝子に関するトランスジェニック動物、または染色体導入(transchromosomal)動物(たとえばマウス)から、またはそれから調製されたハイブリドーマから単離される;(b)抗体は、当該抗体を発現するよう形質転換された宿主細胞から、たとえばトランスフェクトーマから、単離される;(c)抗体は、組換えコンビナトリアル抗体ライブラリーから単離される;ならびに(d)抗体は、免疫グロブリン遺伝子配列を他のDNA配列に接合することを含む、任意の他の手法により、調製、発現、作製または単離される。
【0054】
本発明の抗体は、ヒト抗体もしくはヒト化抗体とすることができる。本発明で使用される「ヒト抗体」という用語は、フレームワーク領域およびCDR領域がともにヒト生殖細胞系列の免疫グロブリン配列に由来する、可変領域を有する抗体を含めるものとする。さらに、抗体が定常領域を含んでいる場合、その定常領域もヒト生殖細胞系列免疫グロブリン配列に由来する。本発明のヒト抗体は、ヒト生殖系列免疫グロブリン配列によってコードされないアミノ酸配列を含んでいてもよい(たとえば、in vitroのランダムもしくは部位特異的変異、またはin vitro体細胞変異によって導入される変異)。しかしながら、本明細書で使用される「ヒト抗体」という用語には、別の哺乳動物種(マウスなど)の生殖細胞系列に由来するCDR配列をヒトフレームワーク配列に接合した抗体は含まれないものとする。
【0055】
こうしたヒト抗体は、ヒトモノクローナル抗体とすることができる。このようなヒトモノクローナル抗体は、不死化細胞と融合させたB細胞などのハイブリドーマによって産生されるが、このB細胞は、ヒト重鎖導入遺伝子および軽鎖導入遺伝子を含有するゲノムを有するトランスジェニック非ヒト動物、たとえばトランスジェニックマウスから得られる。
【0056】
ヒト抗体は、ヒトリンパ球をin vitroで免疫化した後、そのリンパ球をエプスタイン-バールウイルスで形質転換することによって調製することができる。
【0057】
「ヒト抗体の誘導体」という用語は、ヒト抗体の修飾された形、たとえばヒト抗体と別の薬剤もしくは抗体とのコンジュゲートを意味する。
【0058】
「ヒト化抗体」という用語は、別の哺乳動物種、たとえばマウス、の生殖細胞系列由来のCDR配列をヒトフレームワーク配列にグラフト化した抗体を指すものとする。ヒトフレームワーク配列内に追加的なフレームワーク領域修飾を行うことができる。
【0059】
本明細書に記載のスクリーニング法を用いて、Lrg1と結合する能力を有する適当な抗体を同定することができる。したがって、本明細書に記載のスクリーニング法は、テスト化合物として、関心のある抗体を用いて実施することができる。
【0060】
本発明の抗体は、たとえば標準的なELISAもしくはウェスタンブロッティングによって、Lrg1との結合をテストすることができる。ELISAアッセイを用いて、標的タンパク質と陽性の反応性を示すハイブリドーマをスクリーニングすることもできる。抗体の結合特異性は、標的タンパク質を発現する細胞に対する抗体の結合を、たとえばフローサイトメトリーによりモニターすることによっても決定することができる。このように、本発明のスクリーニング法は、ELISAもしくはウェスタンブロッティングの実施により、またはフローサイトメトリーによって、Lrg1と結合する能力を有する抗体を同定するステップを含んでいるといえる。次に、必要とされる結合特性を有する抗体をさらにテストして、上記のLrg1の活性に及ぼす影響を明らかにすることができる。
【0061】
本発明の抗体は、上記のようにLrg1アンタゴニスト特性(Lrg1をブロックする性質)を有すると考えられる。ある実施形態において、モノクローナル抗体は、Lrg1内のエピトープを特異的に認識して、Lrg1の活性をブロックする。ある実施形態において、モノクローナル抗体は、Lrg1内のエピトープを特異的に認識して、ALK1、TGFβRIIまたはTGFβとLrg1との相互作用をブロックする。ある実施形態において、モノクローナル抗体は、付表2のアミノ酸L1-24もしくは付表3のL94-117(配列番号3)、付表2のL169-192もしくは付表3のL262-285(配列番号4)、または付表2のL227-252もしくは付表3のL320-345(配列番号5)内のエピトープを特異的に認識し、Lrg1の活性をブロックする。ある実施形態において、モノクローナル抗体は、付表2のアミノ酸L1-24もしくは付表3のL94-117(配列番号3)、付表2のL169-192もしくは付表3のL262-285(配列番号4)、または付表2のL227-252もしくは付表3のL320-345(配列番号5)内のエピトープを特異的に認識し、ALK1、TGFβRIIまたはTGFβとLrg1との相互作用をブロックする。
【0062】
本発明の抗体は、Lrg1を、すなわちLrg1内のエピトープを、特異的に認識する。抗体もしくは他の化合物が、特異的なタンパク質とは、選択的親和性もしくは高親和性をもって結合するが、他のタンパク質とは実質的に結合しないか、低い親和性でしか結合しないならば、前記の抗体もしくは他の化合物は、そのタンパク質と「特異的に結合」し、そのタンパク質を「特異的に認識」する。本発明の抗体の標的タンパク質に対する特異性は、抗体が上記の他の関連タンパク質と結合するか否か、または抗体がそれらを区別するかどうかを決定することによって、さらに検討することができる。たとえば、本発明の抗体はヒトLrg1と結合することができるが、マウスもしくは他の哺乳類のLrg1とは結合しない。
【0063】
本発明の抗体は、Lrg1と高親和性で結合することが望ましいが、好ましくはピコモルレベルで、たとえば表面プラズモン共鳴法もしくは他の任意の適当な方法で測定される親和性定数(KD)が10nM未満、1nM 未満、500pM未満または100pM 未満で結合することが望ましい。
【0064】
適当な抗体が特定され、選択されたら、その抗体のアミノ酸配列を、当技術分野で既知の方法により同定することができる。抗体をコードする遺伝子は縮重プライマーを用いてクローニングすることができる。抗体は常法により組換え技術で作製することができる。
【0065】
Lrg1内のエピトープは、当技術分野で既知の方法により同定し、特に、”PEPSCAN"法による連続ペプチドもしくはオーバーラッピングペプチドの系統的スクリーニングによって、またはLrg1をブロックすることが明らかになっているペプチド断片(上記参照)に対する抗体を作製することによって、検討することができる。こうしたペプチド内のエピトープを抗体作製のために特定することが可能であって、そうしたペプチドの例が、本明細書で検討される付表2のL1-24もしくは付表3のL94-117(配列番号3)、付表2のL169-192もしくは付表3のL262-285(配列番号4)、および付表2のL227-252もしくは付表3のL320-345(配列番号5)である。上記および他のエピトープ含有ペプチドは、抗体作製のための免疫原として使用することができる。
【0066】
治療適応症
Lrg1を介して血管増殖が生じるような、いかなる疾患も、原則的に本発明にしたがって治療、予防もしくは改善することができる。本明細書で使用される「血管増殖」「血管増殖性の」「血管増殖性疾患」および同様の用語は、血管、または血管組織もしくは細胞の、異常な、または望ましくない発達に関わる、ありとあらゆる病変を含んでいる。新血管形成および血管内皮細胞増殖を予防もしくは軽減することができるので、たとえば、(新血管の形成、たとえば既存の血管からの新たな毛細管の成長による)病気を引き起こす可能性のある血管形成も、ならびに血管奇形(たとえば、毛細血管拡張症、拡張し、蛇行した、機能不全の血管の形成、毛細血管瘤)も、予防もしくは軽減することができる。また、当技術分野で知られているように、腫瘍性成長は、増大する腫瘍に血液を供給するために新血管の形成を必要とする。したがって、Lrg1を介した血管増殖が起こる腫瘍も、本発明にしたがって治療、予防もしくは軽減することができる疾患である。
【0067】
正常な、たとえば発達上の血管新生、特に網膜における発達上の血管新生に及ぼす影響は無いか最小限であることが好ましい。眼の血管増殖性疾患の治療は好ましい実施形態である。治療可能な疾患には、糖尿病性網膜症、網膜静脈閉塞、未熟児網膜症、黄斑部毛細血管拡張症、加齢黄斑変性、または脈絡膜血管新生がある。
【0068】
腫瘍の治療、典型的には、固形腫瘍の治療は、腫瘍における血管形成の防止がその腫瘍から血液供給を奪うという点から、達成することができる。腫瘍治療の標的としては、脳、乳房、腎臓、結腸直腸、肺、前立腺、頭頸部、胃、膵臓、皮膚、子宮頸部、骨、卵巣、精巣、および肝臓の腫瘍が挙げられる。
【0069】
医薬組成物、投与量、および投与計画
本発明のアンタゴニストは、典型的には製薬上許容されるキャリアとともに製剤して、医薬組成物とする。
【0070】
本明細書で使用される「製薬上許容されるキャリア」としては、生理学的に適合する、ありとあらゆる溶媒、分散媒、コーティング、抗細菌性および抗真菌性物質、等張性および吸収遅延物質などがある。キャリアは、非経口投与、たとえば、静脈内、筋肉内、皮下、眼球内もしくは硝子体内投与(たとえば注射または点滴による)に適していることが好ましい。投与経路に応じて、モジュレーターは、酸の作用、ならびに化合物を不活化する可能性のある他の自然条件から、化合物を保護する材料に覆われていてもよい。
【0071】
本発明の医薬品は、1つもしくは複数の製薬上許容される塩を含んでいてもよい。「製薬上許容される塩」とは、親化合物の望ましい生物活性を保ち、望ましくない毒性学的影響を与えない塩を指す。こうした塩の例には、酸付加塩および塩基付加塩がある。
【0072】
好ましい製薬上許容されるキャリアには水性キャリアもしくは希釈剤が含まれる。本発明の医薬組成物中で使用することができる適当な水性キャリアの例には、水、緩衝水および生理食塩水がある。他のキャリアの例としては、エタノール、ポリオール(たとえば、グリセロール、プロピレングリコール、ポリエチレングリコールなど)、およびそれらの適当な混合物、オリーブ油などの植物油、ならびにオレイン酸エチルなどの注射可能な有機エステルが挙げられる。多くの場合、等張剤、たとえば糖類、ポリアルコール類、たとえばマンニトール、ソルビトール、または塩化ナトリウムを組成物中に含んでいることが好ましいと思われる。
【0073】
治療用組成物は典型的には無菌でなければならず、製造および保存の条件下で安定でなければならない。組成物は溶液、マイクロエマルション、リポソーム、または高い薬物濃度に適した他の規則的な構造として製造することができる。
【0074】
本発明の医薬組成物は、追加の活性成分、特に本明細書に記載のVEGFアンタゴニストを含んでいてもよい。
【0075】
本発明の範囲には、本発明のアンタゴニストおよび使用説明書を含むキットも含まれる。キットはさらに、上記の追加的な治療薬もしくは予防薬のような、1つもしくは複数の追加薬を含有することができる。
【0076】
本発明のアンタゴニストおよび組成物は、予防的および/または治療的処置のために投与することができる。
【0077】
治療的応用において、モジュレーターもしくは組成物は、以前から上記疾患に罹患している被験体に、その疾患、またはその症状のうち1つもしくは複数を治癒させ、軽減し、または部分的に進行を止めるのに十分な量が投与される。そうした治療的処置の結果、病徴の重篤度が低下し、症状のない期間の回数もしくは持続時間が増加する可能性がある。これを達成するために適切な量は、「治療に有効な量」として定義される。
【0078】
予防的応用において、製剤は、上記疾患のリスクのある被験体に、その疾患、またはその症状のうち1つもしくは複数の、その後の影響を防ぎ、または減少させるのに十分な量が投与される。これを達成するために適切な量は、「予防に有効な量」として定義される。それぞれの目的のために有効な量は、疾病もしくは傷害の重篤度、ならびに被験体の体重および全身状態によって決まってくる。本発明との関連で予防的に処置することができる疾患はたとえば滲出型AMD(加齢黄斑変性)である;一方の眼がもう一方より先に進行することがあるので、問題が認識されればその第一の眼を治療し、もう一方は予防的に処置される。
【0079】
本発明のアンタゴニストを投与するための被験体は、ヒトもしくは非ヒト動物とすることができる。「非ヒト動物」という用語には、すべての脊椎動物、たとえば哺乳類および非哺乳類、たとえば非ヒト霊長類、ヒツジ、イヌ、ネコ、ウマ、ウシ、ニワトリ、両生類、爬虫類などが含まれる。ヒトへの投与が好ましい。
【0080】
本発明のアンタゴニストは、当技術分野で知られているさまざまな方法の1つもしくはいくつかを用いて、1つもしくは複数の投与経路によって投与することができる。当業者には当然のことであるが、投与の経路および/または方法は、求められる結果に応じてさまざまである。本発明のモジュレーターに好ましい投与経路としては、たとえば、注射もしくは点滴による、静脈内、筋肉内、皮内、眼球内、腹腔内、皮下、脊髄もしくは他の非経口の投与経路が挙げられる。本明細書で使用される「非経口投与」という表現は、腸内投与および局所投与以外の、通常は注射による、投与法を意味する。あるいはまた、本発明の抗体は、非経口以外の経路、たとえば局所、表皮もしくは粘膜などの投与経路で、投与することができる。
【0081】
本発明のモジュレーターの適当な投与量は、熟練した医師ならば決定することができる。本発明の医薬組成物中の活性成分の、実際の投与量レベルは、患者に対して毒性を示すことなく、特定の患者、組成物、および投与方法に関して望ましい治療反応を達成するのに有効な活性成分の量が得られるように、さまざまに変更することができる。選択される投与量レベルは、使用される特定の本発明の組成物の活性、投与経路、投与時間、使用される特定の化合物の排泄速度、治療期間、使用される特定の組成物と併用される他の薬物、化合物および/または物質、治療すべき患者の年齢、性別、体重、疾患、全身健康状態および既往歴、ならびに医学分野で周知の類似要因などの、さまざまな薬物動態学的要因によって決まってくる。
【0082】
適当な投与量はたとえば、治療すべき患者の体重について約0.1μg/kgから約100mg/kgまでの範囲内とすることができる。たとえば、適当な投与量は、1日当たり約1μg/kgから約10mg/kg体重まで、または1日当たり約10μg/kgから約5 mg/kg体重までとすることができる。眼球内投与について、適当な投与量は、典型的には28日ごとに約1μg - 1 mgとすることができる。
【0083】
投与計画は、最適な望ましい反応(たとえば治療反応)を与えるように調整することができる。たとえば、単回投与を行ってもよく、いくつかに分割された用量を時間経過とともに投与してもよいが、治療状況の緊急性によって指示されるのに対応して用量を増減することもできる。本明細書で使用される投与単位形態は、単位投与量として治療すべき被験体に適した、物理的に個別の単位を意味する;それぞれの単位は、必要な医薬キャリアを伴って望ましい治療効果をもたらすように計算された、あらかじめ決められた量の活性化合物を含有する。
【0084】
投与は、単回投与でも複数回投与でもよい。複数回投与は、同一経路または異なる経路で、同一部位または異なる部位に投与することができる。あるいはまた、投与は、徐放性製剤によることも可能であるが、そのばあい必要な投与回数は少なくなる。投与量および回数は、患者におけるアンタゴニストの半減期、および望ましい治療期間に応じてさまざまとすることができる。
【0085】
上記のように、本発明のモジュレーターは、他の1つもしくは複数の治療薬とともに同時投与することができる。たとえば、他の薬剤は鎮痛薬、麻酔薬、免疫抑制薬もしくは抗炎症薬;またはVEGFアンタゴニストとすることができる。
【0086】
2つ以上の薬剤の併用投与は、いくつかの異なる方法で行うことができる。2つの薬剤を1つの組成物として一緒に投与してもよいが、併用療法の一環として別々の組成物として投与してもよい。たとえば、一方を、他方より先に、他方の後に、または他方と同時に投与することができる。
【0087】
併用療法
上記のように、本発明のLrg1アンタゴニストは、任意の他の適切な活性化合物と併用して投与することができる。詳細には、Lrg1およびVEGFの両者の拮抗作用は病気を引き起こす可能性のある血管新生を減少させるので、Lrg1アンタゴニスト、特に、抗VEGF抗体、たとえばアバスチンおよび/またはルセンティスな、および/または受容体に基づくVEGFトラップ、たとえばアフリベルセプトなど。
【0088】
以下の実施例は、本発明を説明するものである。
【0089】
(実施例)
【実施例1】
【0090】
1. 異常な網膜血管の遺伝子発現解析
網膜疾患についてさまざまな動物モデルがあるが、これらは遺伝的起源および細胞起源は異なるにもかかわらず、異常な血管反応を示し、この血管反応には血管形成だけでなく、他の血管変化、たとえば毛細血管拡張症(拡張し、蛇行した、機能不全の血管)も含まれる。網膜血管リモデリングの生物学的基礎について新たな知見を得るために、本発明者らはこうした4つのモデルで研究を行うことで(図1)、病気を引き起こす可能性のある網膜微小血管における遺伝子の発現差異を正常対照の微小血管と比較して検討した。血管異常が現れる段階に相当する時点で、野生型(WT)マウス、網膜ジストロフィー(RD)1マウス、Curlytail(CT)マウス、および超低比重リポタンパク質受容体(VLDLR)ノックアウトマウスから摘出し浄化した微小血管断片を調べた。3つのマウスモデルから単離されたRNAに関するマイクロアレイ遺伝子発現解析(Affymetrix)は、罹患した網膜由来の微小血管においてアップレギュレートまたはダウンレギュレートされた、全モデルに共通の63個の遺伝子を明らかにした(図10)。血管リモデリングに関する3つのマウスモデルの網膜血管系において、発現に差異のある63個の遺伝子のうち、ロイシンリッチα-2-糖タンパク質-1(Lrg1)は、もっとも重要であると評価された(FDR解析後に)。Lrg1は、タンパク質-タンパク質相互作用、シグナル伝達、ならびに細胞の接着および発生に関与する、タンパク質のロイシンリッチリピートファミリーの、分泌型糖タンパク質である(図2)。
【0091】
2. マイクロアレイデータに見られるLrg1過剰発現の検証
マウスモデルの網膜におけるLrg1の発現増加は、まず、定量PCRによって検証した。WT、RD1、CTおよびVLDLR-/-マウスの網膜全体からmRNAを抽出し、リアルタイム定量PCR(qRTPCR)に供した。マイクロアレイ解析で示されるように、対照モデルと比較すると、網膜血管病変の3モデルにおいて、Lrg1の転写産物発現が有意に(p<0.05)増加していることが、qRTPCRによって実証された(図3A)。mRNAの増加が結果としてタンパク質発現の増加につながることを確認するために、本発明者らは次に、遺伝子発現実験と同一の時点で網膜を取り出し、その組織を、ウェスタンブロットによるタンパク質分析用に調製した(図3B)。デンシトメトリー分析によるウェスタンブロットデータ(n > 3)の半定量化は、ハウスキーピングタンパク質(GAPDH)と比較して、Lrg1タンパク質発現の有意な増加を明らかにした(p<0.05)。網膜血管系におけるLrg1の分布を決定し、網膜の他の細胞がLrg1を発現するかどうか確認するために、本発明者らは、in situハイブリダイゼーションおよび免疫組織化学的検査を行って、それぞれLrg1 mRNAおよびタンパク質を検出した。正常なマウスにおいて、Lrg1 mRNA(図3C)およびタンパク質(図3D)は、大部分が血管系によって発現された。
【0092】
3. Lrg1はTGFβ受容体TGFβRIIおよびALK1と結合する
Lrg1の生物学に関しては、ほとんど何もわかっていない。多くの疾患においてTGFβ1、TGFβRIIおよびLrg1の発現レベルが同時に増加することが、いくつかの報告に記載されている(Sun et al., 1995; Li et al., 2007)。内皮細胞におけるTGFβシグナル伝達の機能不全が、遺伝性出血性毛細血管拡張症(HHT)という病気をもたらすので、上記は特に密接な関係がある。毛細血管拡張症などの血管異常を特徴とする、こうした疾病群において、TGFβ内皮補助受容体エンドグリンおよびTGFβI型受容体ALK1における変異は、HHT1(McAllister et al. 1994)およびHHT2(Johnson et al. 1995)をそれぞれ引き起こす。さらに、VLDLR-/-マウスにおいて、網膜組織ではTGFβmRNAが有意に増加しているが、RPEまたは微小血管ではそうではないという趣旨の、試験的なデータがある。さらによく当てはまることに、TGFβは、血管リモデリングがよく見られる糖尿病性網膜症患者の網膜において増加していることが明らかになった。
【0093】
内皮細胞において、TGFβシグナル伝達は、TGFβ受容体IIと、主として内皮細胞において発現されている遍在性TGFβI型受容体アクチビン受容体様キナーゼ5(ALK5)またはALK1のいずれかとの結合を介して起こり、その細胞性応答はどちらの経路が優勢であるかによって決まる。ALK5の場合、ECM沈着の増加および細胞休止が見られるのに対して、ALK1については、遊走および増殖の増加となって現れる内皮細胞の活性化が見られる。こうした差異のあるシグナル伝達は、一つにはTGFβの濃度/バイオアベイラビリティによって制御され、また、Smadと呼ばれる下流エフェクタータンパク質ファミリーのメンバーによっても制御されるが、このSmad 2および3はALK5と関連付けられ、Smad 1、5および8はALK1と関連付けられる。本発明者らは、Lrg1とTGFシグナル伝達経路の関連性を検討した。まず、ラット脳内皮細胞株(GPNT)がLrg1、TGFβRII、ならびにTGFβシグナル伝達の他成分(付表1)をいずれも発現することを確認した。GPNT細胞溶解物由来のLrg1の免疫沈降法の結果、受容体TGFβRIIおよびALK1の共沈殿が見られた(図4A)。同様に、TGFβRIIまたはALK1のいずれか一方の免疫沈降は、結果としてLrg1の共沈殿をもたらし、Lrg1が2つの受容体といずれも結合することが示された(図4A)。また、細菌由来のHA標識組換えLrg1タンパク質がTGFβRIIおよびTGFβと結合することも明らかにした(図4B)。加えて、GPNT細胞におけるLrg1およびTGFβRII発現の免疫細胞化学による可視化は、共局在化を示している(図4C)。したがって、本発明者らは、Lrg1が、TGFβRIIと、ALK1およびALK5活性化シグナル伝達カスケードとの間の微調整をもたらす、TGFβシグナル伝達のモジュレーターとして機能すると仮定する。また、TGFβがGPNT細胞においてLrg1遺伝子発現を誘導することも明らかにし、可能性のあるフィードバック機構が示唆された(図16)。
【0094】
4. Lrg1はSmadリン酸化の差異によってTGFβシグナル伝達を微調整する
Lrg1がTGFβを介した血管内皮細胞反応に影響を及ぼすかどうかを確認するために、本発明者らは次に、sirRNAを用いてGPNT細胞のLrg1をノックダウンし、それがTGFβを介した細胞増殖に及ぼす影響を測定した。対照細胞において、TGFβは、2時間にわたって内皮細胞増殖の有意な増加(p<0.05)を引き起こす(70%コンフルエント細胞)。siRNAによるGPNT内皮細胞のLrg1のノックダウンは、このTGFβを介した細胞増殖の増加をブロックする(図6AおよびB)。これは、Smad5リン酸化の減少と相関する(図4D)。反対に、Lrg1遺伝子でトランスフェクトされたGPNT細胞では、Lrg1過剰発現が、TGFβに対応した内皮細胞増殖の増強をもたらすことを示す(図6AおよびB)。この増強された反応は、Smad2発現のダウンレギュレーション、およびSmad5リン酸化の増加と相関している(図4E)。これらのデータは、TGFβRII/ALK5受容体複合経路を介したシグナル伝達の減少と符合し、したがって血管病変を引き起こしうるTGFβRII/ALK1シグナル伝達経路の活性化へのシフトと矛盾しない。
【0095】
5. Lrg1馴化培地はin vitroでの血管形成を増強する
Lrg1が内皮細胞においてTGFβシグナル伝達を微調整し、TGFβを介した細胞増殖に影響を及ぼすことが確認されたので、本発明者らは次に、Lrg1が血管形成に影響を与えるかどうかを、標準的なin vitro血管形成アッセイによって判定した。ヒト臍静脈内皮細胞(HUVEC)をMatrigel中で増殖させ、馴化されていない増殖培地、(構成的にLrg1を分泌する)GPNT細胞で馴化された培地、ならびにLrg1を過剰発現するGPNT細胞で馴化された培地に供する。対照培地がLrg1を含有しないのに対して、GPNT培地およびLrg1過剰発現GPNT培地は、それぞれ中程度、および高レベルのLrg1を含有していた(図7B)。血管形成の程度は、Lrg1過剰発現細胞から得られた馴化培地を加えたとき、最大であった(図7AおよびC)。血管新生の増加は、培地におけるLrg1タンパク質発現と相関していた。
【0096】
6. Lrg1に由来するペプチド配列、付表2のL227-252もしくは付表3のL320-345はGPNT細胞においてTGFβシグナル伝達を微調整する
本発明者らは次に、TGFβにより誘導される内皮細胞のSmad5リン酸化を、抗Lrg1抗体を用いて、またはLrg1由来ペプチド配列によって、ブロックすることができるかを確認した。Lrg1配列のロイシンリッチリピート領域に由来するペプチド(付表2のL1-24およびL169-192、もしくは付表3のL94-117およびL262-285)(タンパク質-タンパク質相互作用に関与すると考えられている)、ならびに高度に保存されたロイシンリッチC末端ドメインに由来するペプチド(付表2のL227-252もしくは付表3のL320-345)を作製した(付表2)。GPNT細胞におけるLrg1過剰発現は、結果としてSmad5リン酸化をもたらさなかった。対照細胞の5 ng/ml TGFβによる処理は、Smad5リン酸化の有意な増加をもたらす。Lrg1過剰発現細胞において、Smad5リン酸化に及ぼすTGFβの影響は有意に強められる。Lrg1過剰発現内皮細胞を抗Lrg1ポリクローナル抗体で同時に処理すると、Smad5リン酸化のレベルが減少するので、この抗体がLrg1の相互作用を妨げる能力を有することが示唆される。ペプチドとの同時処理の影響はさまざまで、付表2のペプチドL1-24(付表3のL94-117)はSmad5リン酸化に何の影響も与えず、付表2のペプチドL169-192(付表3のL262-285)は部分的な影響を与えたが、付表2のペプチドL227-252(付表3のL320-345)(図5)は劇的な阻害効果を示した。3つのペプチドをすべて組み合わせると、TGFβを介したSmad5リン酸化はほぼ完全に消失した。これらのデータは、Lrg1がTGFβを介したシグナル伝達を微調整する、ならびに、Lrg1アンタゴニストを治療薬として使用することができる、という仮説を支持する。
【0097】
7. Lrg1ノックアウトマウス由来の大動脈輪は、血管新生の血管出芽の減少を示す
次にin vivo血管形成におけるLrg1の役割を検討した。頸椎脱臼により屠殺したP14 Lrg1ノックアウトマウス、または野生型同腹仔対照から胸部大動脈を摘出し、直ちに氷冷した無血清OPTI-MEM(Invitrogen)を入れた培養皿に移した。大動脈周囲の繊維脂肪組織を細い顕微解剖用のはさみで注意深く取り除いた。長さ1 mmの大動脈輪を切開し、DMEM, pH 7.4中で調製されたラット尾部コラーゲンIゲル(1.5 mg/ml)に包埋した。大動脈輪を含有するコラーゲンゲルを96ウェルプレート中で7日間37℃に保った。各ウェルには、2.5% FCS、100 U/mlペニシリン、および100μg/mlストレプトマイシンを添加した内皮細胞基本培地が入っていた。画像はOlympus顕微鏡で撮影した。
【0098】
各大動脈輪から出芽した血管新生の血管数を数えた。Lrg1遺伝子ノックアウトに関してヘテロ欠損マウスおよびホモ欠損マウスの両者から摘出された大動脈輪は、野生型マウスの大動脈輪と比べて血管新生の血管出芽の有意な減少を示した(p<0.01)(図9)。
【0099】
8. 網膜損傷後の脈絡膜新血管形成(CNV)は、Lrg1ノックアウトマウスにおいて減少する
ブルッフ膜を、Lrg1ノックアウトマウスまたは野生型同腹仔対照のそれぞれの眼の視神経を取り囲む3カ所で、レーザーにより破断した。ブルッフ膜裂傷部位のCNV病変をレーザー処置の1週間後にin vivo蛍光眼底血管造影(FA)により測定した。蛍光は腹腔内注入により投与した。7分の間隔をおいて初期および後期の眼底血管造影図が得られた。初期の血管造影図は、注入の90秒後に得られ、脈絡膜新血管形成の大きさを示している。後期血管造影図は、脈絡膜新生血管膜からの漏出を示す。
【0100】
FAは、Lrg1ノックアウトマウスにおいて脈絡膜新血管形成が減少していることを明白に示した(図11A)。脈絡膜新血管形成の数値化は、Lrg1ノックアウトマウスにおいて、野生型マウスより血管新生増殖面積のサイズおよび漏出が有意に減少していることを明らかにした(図11BおよびC)(** p<0.01)。
【0101】
9. 酸素誘導網膜症(OIR)後の網膜新血管形成はLrg1ノックアウトマウスにおいて減少する
生後7日(P7)のLrg1ノックアウトマウスおよび野生型同腹仔対照は、哺育する親マウスとともに、酸素過剰(75%酸素)に5日間暴露したが、これは、その新生マウスにおいて、網膜血管発達の有意な阻害をもたらす。生後12日(P12)にマウスは酸素正常状態に戻され、その時、低酸素で血管のない網膜は、正常な血管再増殖および病的な新血管形成をともに引き起こすが、これは生後17日(P17)にピークとなる。網膜を摘出し、固定して、イソレクチン-B4によるホールマウント免疫染色に供した(図13A)。血管の再増殖は無血管面積を全網膜面積と比較することによって数値化した。新血管形成は、新生血管網の面積を手作業で測定することによって数値化した。
【0102】
無血管領域の大きさは、Lrg1ノックアウトマウスの網膜では有意に増加していることが判明した(* p<0.05)(図13B)。また、新生血管網の数は、Lrg1ノックアウトマウスでは、野生型マウスより減少していた(** p<0.002)(図13C)。
【0103】
10. in vitroのMatrigel中でのヒト臍静脈内皮細胞(HUVEC)の管、索状組織、および血管の形成は、ポリクローナル抗Lrg1抗体の添加によって減少する
in vitro管形成アッセイは、ヒト臍静脈内皮細胞(HUVEC)を用いてMatrigel中で行った。96ウェルプレートを、ウェル当たり60μlのMatrigelでコーティングした。各ウェルを、100 nMの(Lrg1糖タンパク質全体に対する)抗ヒトポリクローナルLrg1抗体、100 nM IgGアイソタイプ、または同量の抗体溶出バッファーの存在下で、37℃、5% CO2にて16時間、15,000 HUVECを含有する100μl EGM2培地で処理した。細胞を洗浄し、固定した。管形成は、中和抗体である抗ヒトLrg1ポリクローナル抗体の添加によって、抗体溶出バッファー添加(p<0.01)または無関係なIgG抗体の添加(p<0.05)と比べて、有意に減少した。管形成は、分岐点の数(図14A)、管の数(図14A)および管の全長(図14B)で評価した。
【0104】
11. 硝子体液におけるLrg1およびTGFβ発現は、増殖性糖尿病性網膜症(PDR)に罹っているヒト患者において増加する
ヒト網膜の免疫組織化学分析を行って、網膜血管系においてLrg1の染色を検出した(図15A)。硝子体液のサンプルは非糖尿病患者およびPDR患者から得られた。硝子体液サンプル中に存在するLrg1を、ウェスタンブロッティングによって測定し、デンシトメトリー分析で数値化した(図15B)。PDR患者の硝子体液では非糖尿病患者と比べて、Lrg1は有意に増加していた(p<0.01)。硝子体液サンプル中に存在するTGFβもウェスタンブロッティングによって測定し、Lrg1と同様に数値化した(図15C)。TGFβもPDR患者において有意に増加していた(p<0.01)。
【0105】
したがって、本発明者らは、Lrg1発現の減少が、網膜外傷に対する血管形成反応の減少と関連していることを実証した。Lrg1およびTGFβはまた、網膜の新血管形成の増加を特徴とする疾患であるPDRの患者において、アップレギュレートされていることが明らかになった。このことは、Lrg1がTGFβシグナル伝達を介して血管増殖刺激に関与するので、Lrg1アンタゴニスト、特に抗体を、望ましくない血管増殖が起こらないようにするための治療薬として使用することができるという仮説を支持する。
【0106】
(付表1)

(付表2)
ロイシンリッチリピート(赤色)および高度に保存されたC末端ドメイン(緑色)を明示するようアレンジした、マウスおよびヒトLrg1の部分配列アラインメント。
【0107】
(付表3)
ロイシンリッチα2糖タンパク質1(Lrg1)は、リモデリングされた網膜血管において最大の変化倍率を示した。ヒトおよびマウスLrg1のアラインされたアミノ酸配列。ロイシンリッチリピート領域は赤色、ブロッキングペプチドとして使用されたヒトC末端ドメイン領域は緑色で示す。
【0108】
(参考文献)
Isolation and characterization of an unknown, leucine-rich 3.1-S-alpha2-glycoprotein from human serum: Haupt H, Baudner S; Hoppe Seylers Z Physiol Chem. 1977 Jun; 358(6): 639-46. (Title translated from original German)
Periodicity of leucine and tandem repetition of a 24-amino acid segment in the primary structure of leucine-rich alpha 2-glycoprotein of human serum: Takahashi N, Takahashi Y, Putnam FW; Proc Natl Acad Sci USA. 1985 Apr;82(7):1906-10.
Differentially expressed genes in TGF-beta 1 sensitive and resistant human hepatoma cells: Sun D, Kar S, Carr BI; Cancer Lett. 1995 Feb 10; 89(1):73-9.
Expression of TGF-betas and TGF-beta type II receptor in cerebrospinal fluid of patients with idiopathic normal pressure hydrocephalus: Li X, Miyajima M, Jiang C, Arai H; Neurosci Lett. 2007 Feb 14; 413(2):141-4. Epub 2006 Dec 27.
Identification of putative serum glycoprotein biomarkers for human lung adenocarcinoma by multilectin affinity chromatography and LC-MS/MS: Heo SH, Lee SJ, Ryoo HM, Park JY, Cho JY.; Proteomics. 2007 Dec;7(23):4292-302.
Increased expression of one isoform of leucine-rich alpha-2-glycoprotein in peritoneal fluid of women with uterine leiomyomas: Ferrero S, Gillott DJ, Remorgida V, Anserini P, Ragni N, Grudzinskas JG; Arch Gynecol Obstet. 2009 Mar;279(3):365-71. Epub 2008 Jul 30.
Potential diagnostic biomarkers in serum of idiopathic pulmonary arterial hypertension: Zhang J, Zhang Y, Li N, Liu Z, Xiong C, Ni X, Pu Y, Hui R, He J, Pu J.; Respir Med. 2009 Aug 22. [Epub ahead of print]
Endoglin, a TGF-beta binding protein of endothelial cells, is the gene for hereditary haemorrhagic telangiectasia type 1. McAllister KA, Grogg KM, Johnson DW, Gallione CJ, Baldwin MA, Jackson CE, Helmbold EA, Markel DS, McKinnon WC, Murrell J, et al. Nat Genet. 1994 Dec;8(4):345-51.
A second locus for hereditary hemorrhagic telangiectasia maps to chromosome 12. Johnson DW, Berg JN, Gallione CJ, McAllister KA, Warner JP, Helmbold EA, Markel DS, Jackson CE, Porteous ME, Marchuk DA. Genome Res. 1995 Aug;5(1):21-8.
Basement membrane and growth factor gene expression in normal and diabetic human retinas. Spirin KS, Saghizadeh M, Lewin SL, Zardi L, Kenney MC, Ljubimov AV. Curr Eye Res. 1999 Jun;18(6):490-9.
Plasma proteomics of pancreatic cancer patients by multi-dimensional liquid chromatography and two-dimensional difference gel electrophoresis (2D-DIGE): up-regulation of leucine-rich alpha-2-glycoprotein in pancreatic cancer. Kakisaka T, Kondo T, Okano T, Fujii K, Honda K, Endo M, Tsuchida A, Aoki T, Itoi T, Moriyasu F, Yamada T, Kato H, Nishimura T, Todo S, Hirohashi S. J Chromatogr B Analyt Technol Biomed Life Sci. 2007 Jun 1;852(1-2):257-67. Epub 2007 Feb 1.
US 2005/0064516
WO 2008/092214
US 2007/0184503。
【0109】
配列情報
ヒトLrg1の配列
配列番号1 −ヒトLrg1のDNA配列
[配列番号2のタンパク質をコードする配列は太字で下線を付してあり、下記配列番号1内にある]
配列番号2 −ヒトLrg1のアミノ酸配列
[配列番号3-5の配列は太字で下線を付してあり、下記配列番号2 内にある]
配列番号3-5 −Lrg1内のペプチドのアミノ酸配列
配列番号3 −付表2からのヒトLrg1のアミノ酸1-24 (付表3からのL94-117)
L1-24/L94-117: LQELHLSSNGLESLSPEFLRPVPQ
配列番号4 −付表2からのヒトLrg1のアミノ酸169-192(付表3からのL262-285)
L169-192/L262-285: LDMLDLSNNSLASVPEGLWASLGQ
配列番号5 −付表2からのヒトLrg1のアミノ酸227-252(付表3からのL320-345)
L227-252/L320-345: KMFSQNDTRCAGPEAVKGQTLLAVAK
【技術分野】
【0001】
本発明は分子生理学の分野であって、特に眼の血管増殖性疾患の治療もしくは予防、ならびに血管増殖を示す腫瘍の治療で用いるための、ロイシンリッチα2糖タンパク質1(Lrg1)のアンタゴニストの使用に関する。
【背景技術】
【0002】
網膜血管系の異常なリモデリングは、糖尿病性網膜症、網膜静脈閉塞、未熟児網膜症、加齢黄斑変性および黄斑部毛細血管拡張症などの、失明の恐れのある疾患の顕著な特徴である。こうした血管の変化は、既存の網膜血管からの新たな毛細血管の生成(血管形成)として、ならびに既存血管の血管奇形の発達(たとえば、毛細血管拡張症)として生じる。この病原的な血管リモデリングが、視力を失う主たる要因である。
【0003】
同様の血管の病変および機能不全は、腫瘍の増殖にも付随するものであって、この場合血管新生が固形腫瘍の増大および増殖を可能にする。
【0004】
多くの資力が、(血管形成、血管新生、血管増殖、血管リモデリング、血管病変と称される)これらの血管反応を推進するメカニズムに関する知見を得ることに向けられてきた。その過程で重要な役割を果たすと確認されたさまざまな分子のうちで、血管内皮増殖因子(VEGF)およびその受容体は、非常に重要な構成要素と見なされる。抗がん剤アバスチン(または近縁のルセンティス)を用いて、眼の血管形成におけるVEGF経路を治療の標的にすると、結果としていくつかの失明の恐れのある疾患について、少なくとも短期的には、臨床的成果の改善がもたらされた。しかしながら、VEGFは神経保護活性を有し、しかも脈絡膜の開窓の維持といったハウスキーピング機能も有しているので、網膜血管の問題点を治療するために長期間、VEGFに対抗する戦略を用いることについては懸念がある。その上、VEGFは血管新生においては主要な血管新生促進因子と見なされるが、そのプロセスは多くの因子間の協調的なクロストークを必要としており、他の血管変化、たとえば拡張し蛇行した毛細血管拡張症の血管の形成、に関する生物学的根拠は不明である。
【発明の概要】
【発明が解決しようとする課題】
【0005】
したがって、代わりとなる治療標的および新規薬物を特定する必要があるが、この新規薬物は、単独で、または既存の治療法との併用で、制御されない血管増殖および/またはリモデリングが病気の一因となっているような疾患の治療のために、より有効であり、しかもオフターゲット作用はより少なくすることができる薬物である。
【課題を解決するための手段】
【0006】
本発明者らは、病気を引き起こす可能性のある血管リモデリングを調節するための、新薬の開発につながる標的として、ロイシンリッチα2糖タンパク質1(Lrg1遺伝子名:HGNC: 29480; Entrez Gene: 116844; Ensembl: ENSG00000171236; UniProtKB: P02750)を特定した。
【0007】
Lrg1は1977年に同定され(Haupt & Baudner, 1977)、その一次構造は1985年に決定された(Takahashi et al, 1985)。Lrg1はマウスとヒトの間で高度に、進化的に保存されており、ヒトLrg1に対するポリクローナル抗体が市販されているが、ある種の疾患においてトランスフォーミング増殖因子β1(TGFβ1)、TGFβ受容体II(TGFβRII)およびLrg1のレベルが同時に増加するとの報告がある(Sun et al, 1995; Li et al, 1997)。他のグループは、Lrg1を特定の疾患のバイオマーカーとして(US 2005/0064516; WO 2008/092214)ならびにシトクロムcのリガンドとして(US 2007/0184503)同定した。内皮細胞におけるTGFβシグナル伝達の機能不全は、遺伝性出血性毛細管拡張症(HHT)をもたらす。
【0008】
毛細管拡張症などの血管異常を特徴とするこの疾患群において、TGFβ内皮細胞補助受容体エンドグリンおよびTβRIコレセプターALK1の変異はそれぞれ、HHT1(McAllister et al. 1994)およびHHT2(Johnson et al. 1995)を引き起こす。TGFβは、血管リモデリングがよく見られる糖尿病性網膜症患者の網膜で増加していることも明らかになった(Spirin et al., 1999)。特定の腫瘍患者の血漿中でLrg1発現が増加することも判明しており、これは、Lrg1が有望な腫瘍バイオマーカーとして機能しうることを示唆する(Heo et al, 2007; Ferrero et al, 2009; Kakisaka et al, 2007)。しかしながら、Lrg1の生物学についてはほとんどわかっていない。
【0009】
本発明者らはこれまでに、特に眼において、ならびに血管増殖を示す腫瘍において、病気を引き起こす可能性のある血管リモデリングを調節するための、新薬につながる標的としてLrg1を特定した。
【0010】
血管の変化を伴う網膜疾患のマウスモデルを用いて、本発明者らはまず、これらの罹患網膜の血管において、他の遺伝子の中でLrg1がアップレギュレートされていることを突き止めた。次に、こうしたマウスモデルの網膜におけるLrg1の発現増加を、定量的PCRおよびウェスタンブロッティングによって検証し、in situハイブリダイゼーションおよび免疫組織化学によって、Lrg1の網膜での分布が血管であると確認した。こうしたモデルは、血管形成の標準的なモデルであり、眼以外の部位での血管形成に応用できる。
【0011】
そこで本発明者らは、Lrg1とTGFβシグナル伝達経路との関連性を調べた。
【0012】
内皮細胞において、TGFβシグナル伝達は、どこにでも存在するTGFβI型受容体アクチビン受容体様キナーゼ5(ALK5)、または内皮細胞特異的ALK1のいずれか一方と結合したTGFβ受容体IIを介して起こる可能性があり、細胞性応答は、どちらの経路が主であるかによって左右される。ALK5の場合、特定の条件下で、ECM沈着の増加および細胞休止が見られるのに対して、ALK1については、遊走および増殖の増加として現れる内皮細胞の活性化が見られる。こうした差異のあるシグナル伝達は、一つにはTGFβの濃度/バイオアベイラビリティによって制御され、また、Smadと呼ばれる下流エフェクタータンパク質ファミリーのメンバーによっても制御されるが、このSmad 2および3はALK5で活性化されSmad 1、5および8はALK1により活性化される。
【0013】
免疫沈降法により、Lrg1がTGFβRIIおよびALK1の両方に結合することが明らかになったが、このことは、Lrg1が、TGFβシグナル伝達複合体の一部として、これら2つの分子と関連して役割を有することを示唆する。
【0014】
したがって、本発明者らは、Lrg1がTGFβシグナル伝達のモジュレーターとして機能して、ALK1-およびALK5-活性化シグナル伝達カスケード間の微調整を引き起こすとの仮説を立てた。これを裏付けるものとして、siRNAを用いた内皮細胞のLrg1ノックダウンは、TGFβによって増加した細胞増殖をブロックし、Smad5リン酸化を低下させたのに対して、Lrg1の過剰発現は増殖の強化、Smad2発現のダウンレギュレーション、およびSmad5リン酸化の増加をもたらす。したがって、こうした知見は、Lrg1が血管形成を制御することができる1つの方向を明らかにする。また、Lrg1が「血管」形成に及ぼす影響を検討するMatrigel血管形成アッセイにおいて、血管形成、管形成および索状組織形成により評価される血管形成の度合いは、Lrg1過剰発現細胞から得られた馴化培地を添加したときに有意に増加し、血管新生の増加は培地中のLrg1タンパク質発現と相関していた。
【0015】
これらのデータは、TGFβRII/ALK5受容体複合体経路を介したシグナル伝達の減少と合致し、したがって、血管病変を引き起こす可能性のあるTGFβRII/ALK1シグナル伝達経路の活性化へのシフトと一致する。
【0016】
これは、TGFβシグナル伝達複合体においてLrg1をブロックすることで、TGFβを、病気を引き起こす可能性のある血管新生から逸らすことが可能であることを示唆する。この問題を検証するために、本発明者らは、TGFβによって引き起こされる内皮細胞Smad5リン酸化が、抗Lrg1抗体を用いて、もしくはALK1との結合についてLrg1と競合すると見込まれるLrg1由来ペプチド配列によって、ブロックされるかどうかを判定した。抗Lrg1抗体はリン酸化の減少を引き起こしたが、ペプチドのうち1つは特に大幅なリン酸化の減少を示した。
【0017】
したがって、Smad5のリン酸化は、Lrg1をブロックすることにより阻害することができるが、Smad5は、血管病変を生じさせるALK1で活性化されたシグナル伝達カスケードと関連しているので、これは、Lrg1のブロックが、病気を引き起こす可能性のないALK5で活性化された別のカスケードと比べて、ALK1で活性化されたカスケードをブロックする可能性があることを示している。
【0018】
以上をまとめると、上記データは下記を示唆する:(a)Lrg1はTGFβRIIおよびALK1のどちらとも相互作用し、ALK5ではなくALK1とTGFβRIIとの相互作用を、直接、または1つもしくは複数の中間体を介して促進するものであって、その結果(b)Lrg1をブロックすることは、TGFβの活性を、血管病変を引き起こす可能性のある、ALK1で活性化されたシグナル伝達カスケードから逸らして、病気を引き起こさないALK5で活性化されたカスケードに向けることになり、その結果として(c)Lrg1は、眼および他の箇所において病気を引き起こす可能性のある血管新生を治療するための効果的な薬物標的となる。したがって、さまざまなLrg1遮断薬、特にLrg1のペプチド断片、Lrg1に対するモノクローナル抗体、およびsiRNA分子による治療を、病気を引き起こす可能性のある組織血管新生を伴う眼および他の疾患のために、構想することができる。図16は、理論に縛られることなく、本発明者らのデータによって示唆されるLrg1の役割を示すものである。
【0019】
さらに、本発明者らのデータは、VEGFとは対照的に、Lrg1が眼において病気を引き起こす可能性のある血管新生にのみ関与し、正常な発達の血管新生もしくは血管ホメオスタシスには関与しないことを示唆する。このことからLrg1は、途絶させることが望ましくないプロセスの妨害を避けるという点から見ると、VEGFよりすぐれた標的となる可能性がある。Lrg1が標的としてさらに魅力的であるのは、それが細胞外にあるので、治療のための全身経路を介してより容易にアクセスできることである。
【0020】
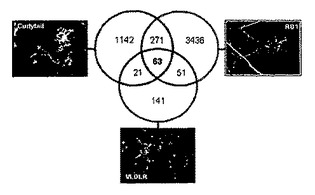
また、ex vivoおよびin vivoでのLrg1の役割を調べるために実験を行った。本発明者らの実験によれば、Lrg1ノックアウトマウスの大動脈輪では、対照マウスの大動脈輪と比べて血管新生の血管発芽が減少している。また、本発明者らは、網膜損傷後の脈絡膜血管新生(CNV)、および酸素誘導網膜症(OIR)後の網膜血管新生がLrg1ノックマウスでは対照マウスより減少したことも明らかにした。
【0021】
ヒトの病理においてLrg1の果たす役割の証拠として、本発明者らのデータは、増殖性糖尿病性網膜症に罹っているヒト患者においてLrg1およびTGFβの発現が増加していることを示すが、このことはマウスから得られたin vivoデータを支持する。
【0022】
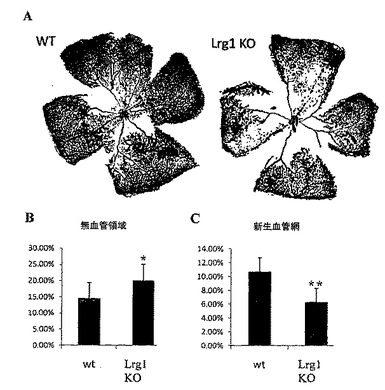
本発明者らは、Lrg1に対する抗体が、Matrigel血管形成アッセイにおいて、ヒト臍帯静脈内皮細胞(HUVEC)による管形成を阻害する能力を有することも明らかにした。こうしたデータは、Lrg1に対する抗体が、血管増殖性疾患、特に眼の血管増殖性疾患、および血管増殖を示す腫瘍の治療もしくは予防に有用であることを示唆する。
【0023】
したがって、本発明は、血管増殖性疾患の治療もしくは予防に用いるロイシンリッチα2糖タンパク質1(Lrg1)のアンタゴニストを提供する。
【0024】
本発明はまた、Lrg1のアンタゴニストを同定する方法を提供するが、その方法は、候補アンタゴニストを用意すること、ならびに前記候補アンタゴニストがLrg1の機能もしくは活性をブロックするか否かを決定することを含んでおり、Lrg1の機能もしくは活性のブロックが認められれば、前記候補アンタゴニストはLrg1のアンタゴニストとして同定される。
【0025】
本発明はまた、付表2のアミノ酸配列L1-24もしくは付表3のL94-117(配列番号3)、付表2のL169-192もしくは付表3のL262-285(配列番号4)、または付表2のL227-252もしくは付表3のL320-345(配列番号5)にあるエピトープを特異的に認識して、Lrg1の活性をブロックする、モノクローナル抗体を提供する。
【0026】
本発明はまた、付表2のアミノ酸L1-24もしくは付表3のL94-117(配列番号3)、付表2のL169-192もしくは付表3のL262-285(配列番号4)、または付表2のL227-252もしくは付表3のL320-345(配列番号5)にあるエピトープを特異的に認識して、ALK1、TGFβRIIおよび/またはTGFβとLrg1との相互作用をブロックする、モノクローナル抗体を提供する。
【0027】
本発明はまた、上記抗体を作製するための方法を提供するが、その方法は、付表2のL1-24もしくは付表3のL94-117(配列番号3)、付表2のL169-192もしくは付表3のL262-285(配列番号4)、または付表2のL227-252もしくは付表3のL320-345(配列番号5)の配列内のエピトープを含んでなる免疫原でヒト以外の哺乳類を免疫化すること;ならびに前記哺乳類から抗体標品を得ること、およびその標品から、前記エピトープを特異的に認識するモノクローナル抗体を得ることを含んでなる。
【0028】
本発明はまた、Lrg1の機能もしくは活性をブロックするためにLrg1の中のどの部位を標的とすることができるかを決定するための方法を提供するが、その方法は、Lrg1タンパク質のペプチド断片を用意すること;ならびに前記のそれぞれのペプチド断片がLrg1の機能もしくは活性をブロックするか否かを判定することを含んでなる。
【0029】
本発明はまた、血管増殖の治療もしくは予防のための薬剤の製造におけるLrg1のアンタゴニストの使用を提供する。
【0030】
本発明はまた、血管増殖性疾患を治療する方法を提供するが、その方法はそれを必要とする患者に有効な量のLrg1アンタゴニストを投与することを含んでなる。
【図面の簡単な説明】
【0031】
【図1】RCSラット(20週齢)、VLDLR-/-マウス(16週齢)、Curlytail-Jマウス(13週齢)およびRD1マウス(16週齢)の網膜の血管リモデリングの低倍率および高倍率画像。血管の基底膜および内皮細胞間結合をそれぞれ着色するために、網膜フラットマウントの血管を抗コラーゲンIVおよび抗クローディン5抗体で染色した。
【図2】A. Lrg1タンパク質および提唱されているそのグリコシル化部位の配置図。B. ROBETTA(University of Washington, USA)により推定されたLrg1タンパク質の構造。
【図3】A. C57Bl6対照マウス(BL6)、VLDL受容体KOマウス(VLDLR-/-)、Curlytail-Jマウス(CT)および網膜ジストロフィー1マウス(RD1)のマウス網膜全体における、Lrg1発現の定量的RT PCR分析。B. 網膜全体のLrg1タンパク質発現のウェスタンブロット(上図)および半定量(下図)。C. Lrg1遺伝子発現を示す正常な網膜のin situハイブリダイゼーション。D. 血管の発現パターンを示す、網膜フラットマウント(上図)および網膜断面(下図)におけるLrg1の免疫組織化学染色。
【図4A】A. Lrg1とTGFβRIIおよびALK1との、GPNT内皮細胞溶解物からの免疫共沈降。B. 組換えHA-タグ付きLrg1はTGFβおよびTGFβRIIの両者と結合する。C. GPNT内皮細胞におけるLrg1およびTGFβRII発現の共局在化。D. Lrg1ノックダウンが、TGFβによって引き起こされる増殖を弱め、Smad5リン酸化を減少させることを示す、内皮細胞増殖アッセイ(* p<0.05)。E. Lrg1の過剰発現が、TGFβによって引き起こされる増殖を強め、Smad1/5リン酸化を増加させることを示す、内皮細胞増殖アッセイ(* p<0.05)。
【図4B】A. Lrg1とTGFβRIIおよびALK1との、GPNT内皮細胞溶解物からの免疫共沈降。B. 組換えHA-タグ付きLrg1はTGFβおよびTGFβRIIの両者と結合する。C. GPNT内皮細胞におけるLrg1およびTGFβRII発現の共局在化。D. Lrg1ノックダウンが、TGFβによって引き起こされる増殖を弱め、Smad5リン酸化を減少させることを示す、内皮細胞増殖アッセイ(* p<0.05)。E. Lrg1の過剰発現が、TGFβによって引き起こされる増殖を強め、Smad1/5リン酸化を増加させることを示す、内皮細胞増殖アッセイ(* p<0.05)。
【図5】C末端に由来するLrg1ブロッキングペプチドは、GPNT内皮細胞においてLrg1/TGFβ1が誘導するSmad5リン酸化を阻害する(各条件につきn=3、p<0.0001)。右側2レーンは、C-末端ペプチドのみ、および3ペプチド、付表2のL1-24もしくは付表3のL94-117(配列番号3)、付表2のL169-192もしくは付表3のL262-285(配列番号4)、または付表2のL227-252もしくは付表3のL320-345(配列番号5)の組み合わせである。
【図6】A. siRNAによるGPNT内皮細胞のLrg1ノックダウン、およびGPNT内皮細胞におけるLrg1過剰発現のウェスタンブロット。B. Lrg1ノックダウンは、TGFβ1が介在する内皮細胞増殖を低下させ(* p<0.0001)、Lrg1過剰発現は、TGFβ1が介在する内皮細胞増殖を高める(* p<0.0005)(各条件についてn=3)。
【図7】HUVECのin vivoでの「血管」形成に及ぼすLrg1の影響。A. 未処理培地、または対照の内皮細胞(EC)由来、もしくはLrg1を過剰発現するEC由来の培地をMatrigel血管形成アッセイに添加した。Lrg1馴化培地はHUVECの「索状組織」形成を高めた。B. 非馴化培地、GPNT内皮細胞馴化培地(7日)およびLrg1を過剰発現するGPNT細胞由来の馴化培地におけるLrg1のウェスタンブロット。C. さまざまな処理の後の、Matrigel内皮細胞索状組織形成複雑系の数量化(閉じた血管輪の数、および全血管面積)(各条件についてn=3)。Lrg1過剰発現細胞由来のLrg1馴化培地は、閉じた血管輪の数もしくは全血管面積で測定されるように最大の血管形成血管網を生じさせた(p<0.0001)。
【図8】TGFβおよびLrg1で処理後に、内皮細胞のSmad5リン酸化に及ぼす、Lrg1抗体(Lrg1のN-末端ドメインに対する市販のポリクローナル抗体)およびLrg1ペプチドの影響。
【図9】Lrg1ノックアウトマウス由来の大動脈輪は、野生型マウスの大動脈輪と比べて、血管新生の血管出芽の減少を示す。Lrg1 KO マウスの大動脈において血管新生の血管出芽の減少を示す、大動脈輪血管形成の代表的な画像(イソレクチンB4で緑色染色)、ならびにそれらの数量化。各群について大動脈輪n=30(** p<0.01)。
【図10】網膜疾患のCurlytail-J、RD1およびVLDLR -/-マウスモデルにおいてアップレギュレートされる遺伝子。
【図11】A. WTおよびLrg1 KOマウスの網膜へのレーザー熱傷によって引き起こされた脈絡膜新血管形成の蛍光眼底血管造影図(FA)。損傷後7日で記録された血管新生増殖および漏出の大きさをそれぞれ示す、初期(B)および後期(C)FAの数値化。n=10(** p<0.01)。
【図12】(左)脳内皮細胞株GPNTはLrg1のTGFβシグナル伝達への影響を研究するために必須の成分を発現する。P = 細胞ペレット;M = 細胞培地。(右)TGFβはGPNT細胞においてLrg1遺伝子発現を誘導する。
【図13】酸素誘導網膜症(OIR)後のP17でのマウス網膜血管系の代表的な画像(イソレクチンB4で赤色染色)であって、これはLrg1 KOマウスにおいて無血管領域の増加および新生血管網の減少を示す。WTおよびLrg1 KOマウス(それぞれn=6および9)の(B)血管のない領域、および(C)新生血管網の数量化。無血管領域はLrg1ノックアウトにおいて増加し(* p<0.05)、目に見える新生血管網は野生型と比較してノックアウトにおいて少なくなっている(** p0.01)。
【図14】Matrigel中でのin vitroのヒト臍帯静脈内皮細胞(HUVEC)の管形成は、中和抗体である抗ヒトLrg1ポリクローナル抗体添加後に、無関係のIgGと比較して減少した。管形成は、A. 分岐点の数、管の数、およびB. 管の全長について、測定した(n=3、* p<0.05, ** p<0.01)。
【図15】A. Lrg1について染色されたヒト網膜の断面図。B. 非糖尿病患者、および増殖性糖尿病性網膜症(PDR)患者由来の硝子体液サンプル中のLrg1(n=4)のウェスタンブロットおよび数量化。C. 非糖尿病患者およびPDR患者由来の硝子体液サンプル中のTGFβ1のウェスタンブロットおよび数量化。
【図16A】作動仮説の概略図。A. 正常な条件下で、TGFβ1シグナル伝達は主としてTGFβRII/ALK5/Smad2/3経路に向けて行われ、Lrg1は基底膜に潜んでいる。B. 病的な条件下では、Lrg1の発現増加の結果として、TGFβ1シグナル伝達は血管リモデリングの一因となるTGFβRII/ALK1/Smad1/5/8経路に向けて方向が変更される。
【図16B】作動仮説の概略図。A. 正常な条件下で、TGFβ1シグナル伝達は主としてTGFβRII/ALK5/Smad2/3経路に向けて行われ、Lrg1は基底膜に潜んでいる。B. 病的な条件下では、Lrg1の発現増加の結果として、TGFβ1シグナル伝達は血管リモデリングの一因となるTGFβRII/ALK1/Smad1/5/8経路に向けて方向が変更される。
【発明を実施するための形態】
【0032】
(発明の詳細な説明)
Lrg1のブロック
本発明のアンタゴニストはLrg1の機能をブロックする。Lrg1をブロックすることは、その活性または機能を低下させることを含んでおり、その結果として内皮細胞増殖、周皮細胞脱落、内皮細胞死、血管リモデリング、血管形成、毛細管拡張症、血管漏出などの血管増殖作用が低下するものである。
【0033】
たとえば、Lrg1のブロックは、そのLrg1とALK1、TGFβRIIおよび/またはTGFβとの相互作用のブロックを介して行うことができるが、そうしたブロックはALK1ではなくALK5とTGFβRIIとの相互作用を促進するので、TGFβの活性を、血管病変を引き起こしうるALK1活性化カスケードから逸らして、より病気の原因となりにくいALK5活性化シグナル伝達カスケードに方向転換させることが本発明者らのデータから示唆される。Lrg1のブロックは、結果としてTGFβのバイオアベイラビリティも低下させる可能性がある。
【0034】
ブロックには、Lrg1の活性もしくは機能の完全な低下および部分的な低下がいずれも含まれるが、これは、たとえばALK1-Lrg1、TGFβRII-Lrg1、および/またはTGFβ-Lrg1相互作用の完全な阻止もしくは部分的な阻止である。たとえば、本発明のブロッキングアンタゴニストは、Lrg1の活性を10から50%まで、少なくとも50%、または少なくとも70%、80%、90%、95%もしくは99%、低下させることができる。
【0035】
Lrg1活性もしくは機能のブロックは、任意の適当な手段で測定することができる。たとえば、ALK1-Lrg1、TGFβRII-Lrg1、および/またはTGFβ-Lrg1相互作用のブロックは、Smad5リン酸化への影響を測定することによって判定することができるが、これは、Smad5リン酸化がALK5活性化経路ではなくてALK1活性化経路に特有であるという根拠に基づく。
【0036】
Lrg1のブロックは、血管形成を測定するアッセイ、たとえばMatrigel中での血管増殖、大動脈輪からの血管増殖などのin vitroアッセイ、および網膜血管形成(たとえば、レーザーによって生じる脈絡膜新血管形成、酸素誘導網膜症)を測定する方法などのin vivoアッセイによって測定することもできる。
【0037】
ブロックは、たとえば使用されるアンタゴニストの性質(下記参照)に応じて、任意の適当なメカニズムで、たとえばALK1-Lrg1、TGFβRII-Lrg1、および/またはTGFβ-Lrg1の直接的もしくは間接的な相互作用における立体障害、またはLrg1発現のノックダウンによって生じるといえる。
【0038】
Lrg1のアンタゴニスト
任意の適当なアンタゴニスト、たとえばペプチドおよびペプチドミメティック、抗体、小分子阻害剤、二本鎖RNA、アプタマーおよびリボザイムを、本発明にしたがって使用することができる。好ましいアンタゴニストとしては、Lrg1のペプチド断片、二本鎖RNA、アプタマーおよび抗体がある。
【0039】
ペプチド
ペプチドアンタゴニストは、典型的には、TGFβRIIおよび/またはALK1との結合に関して全長Lrg1と競合し、したがってLrg1と拮抗する、Lrg1の断片である。こうしたペプチドは直鎖状でも環状でもよい。ペプチドアンタゴニストは典型的には、5から50、好ましくは10-40、10-30もしくは15-25アミノ酸長であって、概してLrg1内部由来の連続配列と同一であるが、ペプチドアンタゴニストがLrg1をブロックする性質を保持している限り、その同一性は100%未満でもよく、たとえば95%以上、90%以上、または80%以上であってもよい。ブロッキングペプチドは、任意の適当な方法で、たとえばLrg1配列の一部もしくはすべてに及ぶ、連続したペプチドもしくはオーバーラップしたペプチドの、系統的スクリーニングによって同定することができる。こうしたブロッキングペプチドを模倣するように、ペプチドミメティックを設計することもできる。
【0040】
二本鎖RNA
既知の技術を使用し、Lrg1の配列に関する知見を基にして、配列相同性に基づいてLrg1 RNAを標的化することにより、二本鎖RNA(dsRNA)分子を、Lrg1と拮抗するようにデザインすることができる。こうしたdsRNAは典型的には、通常ステムループ(「ヘアピン」)構造をとる低分子干渉RNA(siRNA)、またはマイクロRNAである。こうしたdsRNAの配列は、Lrg1をコードするmRNAの一部に相当する部分を含んでいる。この部分は、通常、Lrg1 mRNAの標的部分に100%相補的であるが、相補性はもっと低レベル(たとえば90%以上、または95%以上)であってもよい。
【0041】
アプタマー
アプタマーは総じて、特異的な標的分子と結合する核酸分子である。アプタマーは完全にin vitroで操作することが可能であり、化学合成によって容易に作製され、望ましい保存特性を有しており、加えて治療的応用において免疫原性をほとんど、もしくはまったく生じない。これらの特徴から、アプタマーは、薬剤および治療の実用性において特に有用である。
【0042】
本明細書で使用される「アプタマー」は、総じて、一本鎖もしくは二本鎖オリゴヌクレオチド、またはそういったオリゴヌクレオチドの混合物を表し、このオリゴヌクレオチドもしくは混合物は、標的と特異的に結合することができる。オリゴヌクレオチドアプタマーは本明細書で検討されるが、同等の結合特性を有する他のアプタマー、たとえばペプチドアプタマーなども使用できることは、当業者に当然理解されるであろう。
【0043】
概して、アプタマーには、ヌクレオチドの長さが少なくとも5、少なくとも10、または少なくとも15ヌクレオチドである、オリゴヌクレオチドが含まれるといえる。アプタマーは、ヌクレオチドが40以下、60以下、もしくは100以下、またはそれ以上の長さの配列を含有することができる。たとえば、アプタマーは5から100ヌクレオチド、10から40ヌクレオチド、または15から40ヌクレオチドまでの長さとすることができる。可能であればもっと長さの短いアプタマーが好ましいが、これは、こうしたアプタマーが他の分子もしくは材料による干渉の減少をもたらすことが多いからである。
【0044】
未修飾のアプタマーは血流から速やかに除去され、半減期は数分から数時間であるが、これは主として、ヌクレアーゼ分解および腎臓による体からのクリアランスに起因する。こうした未修飾アプタマーは、たとえば、血液凝固を刺激するような一過性疾患の治療に有用性がある。あるいはまた、アプタマーを修飾して半減期を改善することができる。いくつかのそうした修飾、たとえば2'-フッ素置換ピリミジンもしくはポリエチレングリコール(PEG)結合の付加が利用できる。
【0045】
アプタマーは、試験管内人工進化(Systematic Evolution of Ligands by Exponential enrichment)(SELEX)法のような常法によって作製することができる。SELEXは、標的分子への高度に特異的な結合による、核酸分子のin vitro進化のための方法である。それはたとえば、US 5,654,151、US 5,503,978、US 5,567,588およびWO 96/38579に記載されている。
【0046】
SELEX法は、オリゴヌクレオチド集団からの核酸アプタマーの選択、特に、望ましい標的に結合する能力を有する一本鎖核酸の選択に関わるものである。一本鎖核酸(たとえば、DNA、RNA、またはそれらのバリアント)の集団は、結合に適した条件下で標的と接触させ、混合物中で標的と結合した核酸を、結合していない核酸から分離し、その核酸-標的複合体を解離させ、標的と結合していた核酸を増幅して、望ましい結合活性を有する核酸が濃縮された集団もしくはライブラリーを生じさせるが、そこでこの一連のステップは、当該標的に特異的な結合親和性を有する核酸(アプタマー)ライブラリーをもたらすように、必要に応じて繰り返される。
【0047】
抗体
本明細書で使用される「抗体」という用語は、全抗体および任意の抗原結合フラグメント(すなわち、「抗原結合部分」)またはそれらの一本鎖を含んでいる。抗体は、ジスルフィド結合で相互に連結された、少なくとも2つの重(H)鎖および2つの軽(L)鎖を含んでなる糖タンパク質、またはその抗原結合部分を指す。それぞれの重鎖は、重鎖可変領域(本明細書ではVHと略す)および重鎖定常領域で構成される。それぞれの軽鎖は、軽鎖可変領域(本明細書ではVLと略す)および軽鎖定常領域で構成される。重鎖および軽鎖の可変領域は、抗原と相互作用する結合ドメインを含有する。VHおよびVL領域は、さらに、相補性決定領域(CDR)と呼ばれる超可変性領域に細分化することができ、フレームワーク領域(FR)と呼ばれる保存性の高い領域が散在している。
【0048】
抗体の定常領域は、免疫グロブリンと、宿主組織もしくは因子、たとえば免疫系のさまざまな細胞(例、エフェクター細胞)および古典的補体系の第1成分(C1q)など、との結合を仲介することができる。
【0049】
本発明の抗体は、モノクローナル抗体でもポリクローナル抗体でもよいが、モノクローナル抗体が好ましいといえる。本発明の抗体は、キメラ抗体、CDR-グラフト化抗体、ナノボディ、ヒトもしくはヒト化抗体、またはそれらのいずれかの抗原結合部分とすることができる。モノクローナルおよびポリクローナル抗体を作製するための実験動物は、典型的にはヒト以外の哺乳類、たとえばヤギ、ウサギ、ラットもしくはマウスであるが、他の種、たとえばラクダ科動物などで産生させることもできる。
【0050】
ポリクローナル抗体は、当該抗原を用いた適当な動物の免疫化といった常法によって作製することができる。その後、動物から血液を採取してIgG画分を精製することができる。
【0051】
本発明のモノクローナル抗体(mAb)は、従来のモノクローナル抗体の手法、たとえばKohlerおよびMilsteinの標準的な体細胞ハイブリダイゼーション法を含めて、さまざまな技術によって作製することができる。ハイブリドーマを調製するために好ましい動物系は、マウスの系である。マウスにおけるハイブリドーマ作製は、非常によく確立された手法であって、当技術分野で周知の技術を用いて達成することができる。
【0052】
本発明の抗体は、下記を含んでなる方法によって作成することができる:全長Lrg1、Lrg1のペプチド断片、Lrg1の付表2のL1-24もしくは付表3のL94-117(配列番号3)、付表2のL169-192もしくは付表3のL262-285(配列番号4)、または付表2のL227-252もしくは付表3のL320-345(配列番号5)の配列に含まれるエピトープ、またはLrg1の他の領域に含まれるエピトープなどの免疫原で、ヒト以外の哺乳類を免疫化すること;前記哺乳類から抗体標品を得ること;それらから、前記エピトープを特異的に認識するモノクローナル抗体を得ること。
【0053】
抗体の「抗原結合部分」という用語は、抗原と特異的に結合する能力を保持する、1つもしくは複数の抗体フラグメントを意味する。全長抗体のフラグメントが抗体の抗原結合機能を果たすことができることが明らかになっている。抗体の「抗原結合部分」という用語に含まれる結合フラグメントの例には、Fabフラグメント、F(ab')2 フラグメント、Fab’フラグメント、Fdフラグメント、Fvフラグメント、dAbフラグメント、および単独の相補性決定領域 (CDR)がある。scFv抗体などの一本鎖抗体も、抗体の「抗原結合部分」という用語に含まれるものとする。こうした抗体フラグメントは、当業者に知られている従来技術を用いて得ることができるが、フラグメントは完全な抗体と同様に、有用性を求めてスクリーニングすることができる。本発明の抗体は、組換え法によって調製、発現、作製または単離することができる:たとえば(a)抗体は、当該免疫グロブリン遺伝子に関するトランスジェニック動物、または染色体導入(transchromosomal)動物(たとえばマウス)から、またはそれから調製されたハイブリドーマから単離される;(b)抗体は、当該抗体を発現するよう形質転換された宿主細胞から、たとえばトランスフェクトーマから、単離される;(c)抗体は、組換えコンビナトリアル抗体ライブラリーから単離される;ならびに(d)抗体は、免疫グロブリン遺伝子配列を他のDNA配列に接合することを含む、任意の他の手法により、調製、発現、作製または単離される。
【0054】
本発明の抗体は、ヒト抗体もしくはヒト化抗体とすることができる。本発明で使用される「ヒト抗体」という用語は、フレームワーク領域およびCDR領域がともにヒト生殖細胞系列の免疫グロブリン配列に由来する、可変領域を有する抗体を含めるものとする。さらに、抗体が定常領域を含んでいる場合、その定常領域もヒト生殖細胞系列免疫グロブリン配列に由来する。本発明のヒト抗体は、ヒト生殖系列免疫グロブリン配列によってコードされないアミノ酸配列を含んでいてもよい(たとえば、in vitroのランダムもしくは部位特異的変異、またはin vitro体細胞変異によって導入される変異)。しかしながら、本明細書で使用される「ヒト抗体」という用語には、別の哺乳動物種(マウスなど)の生殖細胞系列に由来するCDR配列をヒトフレームワーク配列に接合した抗体は含まれないものとする。
【0055】
こうしたヒト抗体は、ヒトモノクローナル抗体とすることができる。このようなヒトモノクローナル抗体は、不死化細胞と融合させたB細胞などのハイブリドーマによって産生されるが、このB細胞は、ヒト重鎖導入遺伝子および軽鎖導入遺伝子を含有するゲノムを有するトランスジェニック非ヒト動物、たとえばトランスジェニックマウスから得られる。
【0056】
ヒト抗体は、ヒトリンパ球をin vitroで免疫化した後、そのリンパ球をエプスタイン-バールウイルスで形質転換することによって調製することができる。
【0057】
「ヒト抗体の誘導体」という用語は、ヒト抗体の修飾された形、たとえばヒト抗体と別の薬剤もしくは抗体とのコンジュゲートを意味する。
【0058】
「ヒト化抗体」という用語は、別の哺乳動物種、たとえばマウス、の生殖細胞系列由来のCDR配列をヒトフレームワーク配列にグラフト化した抗体を指すものとする。ヒトフレームワーク配列内に追加的なフレームワーク領域修飾を行うことができる。
【0059】
本明細書に記載のスクリーニング法を用いて、Lrg1と結合する能力を有する適当な抗体を同定することができる。したがって、本明細書に記載のスクリーニング法は、テスト化合物として、関心のある抗体を用いて実施することができる。
【0060】
本発明の抗体は、たとえば標準的なELISAもしくはウェスタンブロッティングによって、Lrg1との結合をテストすることができる。ELISAアッセイを用いて、標的タンパク質と陽性の反応性を示すハイブリドーマをスクリーニングすることもできる。抗体の結合特異性は、標的タンパク質を発現する細胞に対する抗体の結合を、たとえばフローサイトメトリーによりモニターすることによっても決定することができる。このように、本発明のスクリーニング法は、ELISAもしくはウェスタンブロッティングの実施により、またはフローサイトメトリーによって、Lrg1と結合する能力を有する抗体を同定するステップを含んでいるといえる。次に、必要とされる結合特性を有する抗体をさらにテストして、上記のLrg1の活性に及ぼす影響を明らかにすることができる。
【0061】
本発明の抗体は、上記のようにLrg1アンタゴニスト特性(Lrg1をブロックする性質)を有すると考えられる。ある実施形態において、モノクローナル抗体は、Lrg1内のエピトープを特異的に認識して、Lrg1の活性をブロックする。ある実施形態において、モノクローナル抗体は、Lrg1内のエピトープを特異的に認識して、ALK1、TGFβRIIまたはTGFβとLrg1との相互作用をブロックする。ある実施形態において、モノクローナル抗体は、付表2のアミノ酸L1-24もしくは付表3のL94-117(配列番号3)、付表2のL169-192もしくは付表3のL262-285(配列番号4)、または付表2のL227-252もしくは付表3のL320-345(配列番号5)内のエピトープを特異的に認識し、Lrg1の活性をブロックする。ある実施形態において、モノクローナル抗体は、付表2のアミノ酸L1-24もしくは付表3のL94-117(配列番号3)、付表2のL169-192もしくは付表3のL262-285(配列番号4)、または付表2のL227-252もしくは付表3のL320-345(配列番号5)内のエピトープを特異的に認識し、ALK1、TGFβRIIまたはTGFβとLrg1との相互作用をブロックする。
【0062】
本発明の抗体は、Lrg1を、すなわちLrg1内のエピトープを、特異的に認識する。抗体もしくは他の化合物が、特異的なタンパク質とは、選択的親和性もしくは高親和性をもって結合するが、他のタンパク質とは実質的に結合しないか、低い親和性でしか結合しないならば、前記の抗体もしくは他の化合物は、そのタンパク質と「特異的に結合」し、そのタンパク質を「特異的に認識」する。本発明の抗体の標的タンパク質に対する特異性は、抗体が上記の他の関連タンパク質と結合するか否か、または抗体がそれらを区別するかどうかを決定することによって、さらに検討することができる。たとえば、本発明の抗体はヒトLrg1と結合することができるが、マウスもしくは他の哺乳類のLrg1とは結合しない。
【0063】
本発明の抗体は、Lrg1と高親和性で結合することが望ましいが、好ましくはピコモルレベルで、たとえば表面プラズモン共鳴法もしくは他の任意の適当な方法で測定される親和性定数(KD)が10nM未満、1nM 未満、500pM未満または100pM 未満で結合することが望ましい。
【0064】
適当な抗体が特定され、選択されたら、その抗体のアミノ酸配列を、当技術分野で既知の方法により同定することができる。抗体をコードする遺伝子は縮重プライマーを用いてクローニングすることができる。抗体は常法により組換え技術で作製することができる。
【0065】
Lrg1内のエピトープは、当技術分野で既知の方法により同定し、特に、”PEPSCAN"法による連続ペプチドもしくはオーバーラッピングペプチドの系統的スクリーニングによって、またはLrg1をブロックすることが明らかになっているペプチド断片(上記参照)に対する抗体を作製することによって、検討することができる。こうしたペプチド内のエピトープを抗体作製のために特定することが可能であって、そうしたペプチドの例が、本明細書で検討される付表2のL1-24もしくは付表3のL94-117(配列番号3)、付表2のL169-192もしくは付表3のL262-285(配列番号4)、および付表2のL227-252もしくは付表3のL320-345(配列番号5)である。上記および他のエピトープ含有ペプチドは、抗体作製のための免疫原として使用することができる。
【0066】
治療適応症
Lrg1を介して血管増殖が生じるような、いかなる疾患も、原則的に本発明にしたがって治療、予防もしくは改善することができる。本明細書で使用される「血管増殖」「血管増殖性の」「血管増殖性疾患」および同様の用語は、血管、または血管組織もしくは細胞の、異常な、または望ましくない発達に関わる、ありとあらゆる病変を含んでいる。新血管形成および血管内皮細胞増殖を予防もしくは軽減することができるので、たとえば、(新血管の形成、たとえば既存の血管からの新たな毛細管の成長による)病気を引き起こす可能性のある血管形成も、ならびに血管奇形(たとえば、毛細血管拡張症、拡張し、蛇行した、機能不全の血管の形成、毛細血管瘤)も、予防もしくは軽減することができる。また、当技術分野で知られているように、腫瘍性成長は、増大する腫瘍に血液を供給するために新血管の形成を必要とする。したがって、Lrg1を介した血管増殖が起こる腫瘍も、本発明にしたがって治療、予防もしくは軽減することができる疾患である。
【0067】
正常な、たとえば発達上の血管新生、特に網膜における発達上の血管新生に及ぼす影響は無いか最小限であることが好ましい。眼の血管増殖性疾患の治療は好ましい実施形態である。治療可能な疾患には、糖尿病性網膜症、網膜静脈閉塞、未熟児網膜症、黄斑部毛細血管拡張症、加齢黄斑変性、または脈絡膜血管新生がある。
【0068】
腫瘍の治療、典型的には、固形腫瘍の治療は、腫瘍における血管形成の防止がその腫瘍から血液供給を奪うという点から、達成することができる。腫瘍治療の標的としては、脳、乳房、腎臓、結腸直腸、肺、前立腺、頭頸部、胃、膵臓、皮膚、子宮頸部、骨、卵巣、精巣、および肝臓の腫瘍が挙げられる。
【0069】
医薬組成物、投与量、および投与計画
本発明のアンタゴニストは、典型的には製薬上許容されるキャリアとともに製剤して、医薬組成物とする。
【0070】
本明細書で使用される「製薬上許容されるキャリア」としては、生理学的に適合する、ありとあらゆる溶媒、分散媒、コーティング、抗細菌性および抗真菌性物質、等張性および吸収遅延物質などがある。キャリアは、非経口投与、たとえば、静脈内、筋肉内、皮下、眼球内もしくは硝子体内投与(たとえば注射または点滴による)に適していることが好ましい。投与経路に応じて、モジュレーターは、酸の作用、ならびに化合物を不活化する可能性のある他の自然条件から、化合物を保護する材料に覆われていてもよい。
【0071】
本発明の医薬品は、1つもしくは複数の製薬上許容される塩を含んでいてもよい。「製薬上許容される塩」とは、親化合物の望ましい生物活性を保ち、望ましくない毒性学的影響を与えない塩を指す。こうした塩の例には、酸付加塩および塩基付加塩がある。
【0072】
好ましい製薬上許容されるキャリアには水性キャリアもしくは希釈剤が含まれる。本発明の医薬組成物中で使用することができる適当な水性キャリアの例には、水、緩衝水および生理食塩水がある。他のキャリアの例としては、エタノール、ポリオール(たとえば、グリセロール、プロピレングリコール、ポリエチレングリコールなど)、およびそれらの適当な混合物、オリーブ油などの植物油、ならびにオレイン酸エチルなどの注射可能な有機エステルが挙げられる。多くの場合、等張剤、たとえば糖類、ポリアルコール類、たとえばマンニトール、ソルビトール、または塩化ナトリウムを組成物中に含んでいることが好ましいと思われる。
【0073】
治療用組成物は典型的には無菌でなければならず、製造および保存の条件下で安定でなければならない。組成物は溶液、マイクロエマルション、リポソーム、または高い薬物濃度に適した他の規則的な構造として製造することができる。
【0074】
本発明の医薬組成物は、追加の活性成分、特に本明細書に記載のVEGFアンタゴニストを含んでいてもよい。
【0075】
本発明の範囲には、本発明のアンタゴニストおよび使用説明書を含むキットも含まれる。キットはさらに、上記の追加的な治療薬もしくは予防薬のような、1つもしくは複数の追加薬を含有することができる。
【0076】
本発明のアンタゴニストおよび組成物は、予防的および/または治療的処置のために投与することができる。
【0077】
治療的応用において、モジュレーターもしくは組成物は、以前から上記疾患に罹患している被験体に、その疾患、またはその症状のうち1つもしくは複数を治癒させ、軽減し、または部分的に進行を止めるのに十分な量が投与される。そうした治療的処置の結果、病徴の重篤度が低下し、症状のない期間の回数もしくは持続時間が増加する可能性がある。これを達成するために適切な量は、「治療に有効な量」として定義される。
【0078】
予防的応用において、製剤は、上記疾患のリスクのある被験体に、その疾患、またはその症状のうち1つもしくは複数の、その後の影響を防ぎ、または減少させるのに十分な量が投与される。これを達成するために適切な量は、「予防に有効な量」として定義される。それぞれの目的のために有効な量は、疾病もしくは傷害の重篤度、ならびに被験体の体重および全身状態によって決まってくる。本発明との関連で予防的に処置することができる疾患はたとえば滲出型AMD(加齢黄斑変性)である;一方の眼がもう一方より先に進行することがあるので、問題が認識されればその第一の眼を治療し、もう一方は予防的に処置される。
【0079】
本発明のアンタゴニストを投与するための被験体は、ヒトもしくは非ヒト動物とすることができる。「非ヒト動物」という用語には、すべての脊椎動物、たとえば哺乳類および非哺乳類、たとえば非ヒト霊長類、ヒツジ、イヌ、ネコ、ウマ、ウシ、ニワトリ、両生類、爬虫類などが含まれる。ヒトへの投与が好ましい。
【0080】
本発明のアンタゴニストは、当技術分野で知られているさまざまな方法の1つもしくはいくつかを用いて、1つもしくは複数の投与経路によって投与することができる。当業者には当然のことであるが、投与の経路および/または方法は、求められる結果に応じてさまざまである。本発明のモジュレーターに好ましい投与経路としては、たとえば、注射もしくは点滴による、静脈内、筋肉内、皮内、眼球内、腹腔内、皮下、脊髄もしくは他の非経口の投与経路が挙げられる。本明細書で使用される「非経口投与」という表現は、腸内投与および局所投与以外の、通常は注射による、投与法を意味する。あるいはまた、本発明の抗体は、非経口以外の経路、たとえば局所、表皮もしくは粘膜などの投与経路で、投与することができる。
【0081】
本発明のモジュレーターの適当な投与量は、熟練した医師ならば決定することができる。本発明の医薬組成物中の活性成分の、実際の投与量レベルは、患者に対して毒性を示すことなく、特定の患者、組成物、および投与方法に関して望ましい治療反応を達成するのに有効な活性成分の量が得られるように、さまざまに変更することができる。選択される投与量レベルは、使用される特定の本発明の組成物の活性、投与経路、投与時間、使用される特定の化合物の排泄速度、治療期間、使用される特定の組成物と併用される他の薬物、化合物および/または物質、治療すべき患者の年齢、性別、体重、疾患、全身健康状態および既往歴、ならびに医学分野で周知の類似要因などの、さまざまな薬物動態学的要因によって決まってくる。
【0082】
適当な投与量はたとえば、治療すべき患者の体重について約0.1μg/kgから約100mg/kgまでの範囲内とすることができる。たとえば、適当な投与量は、1日当たり約1μg/kgから約10mg/kg体重まで、または1日当たり約10μg/kgから約5 mg/kg体重までとすることができる。眼球内投与について、適当な投与量は、典型的には28日ごとに約1μg - 1 mgとすることができる。
【0083】
投与計画は、最適な望ましい反応(たとえば治療反応)を与えるように調整することができる。たとえば、単回投与を行ってもよく、いくつかに分割された用量を時間経過とともに投与してもよいが、治療状況の緊急性によって指示されるのに対応して用量を増減することもできる。本明細書で使用される投与単位形態は、単位投与量として治療すべき被験体に適した、物理的に個別の単位を意味する;それぞれの単位は、必要な医薬キャリアを伴って望ましい治療効果をもたらすように計算された、あらかじめ決められた量の活性化合物を含有する。
【0084】
投与は、単回投与でも複数回投与でもよい。複数回投与は、同一経路または異なる経路で、同一部位または異なる部位に投与することができる。あるいはまた、投与は、徐放性製剤によることも可能であるが、そのばあい必要な投与回数は少なくなる。投与量および回数は、患者におけるアンタゴニストの半減期、および望ましい治療期間に応じてさまざまとすることができる。
【0085】
上記のように、本発明のモジュレーターは、他の1つもしくは複数の治療薬とともに同時投与することができる。たとえば、他の薬剤は鎮痛薬、麻酔薬、免疫抑制薬もしくは抗炎症薬;またはVEGFアンタゴニストとすることができる。
【0086】
2つ以上の薬剤の併用投与は、いくつかの異なる方法で行うことができる。2つの薬剤を1つの組成物として一緒に投与してもよいが、併用療法の一環として別々の組成物として投与してもよい。たとえば、一方を、他方より先に、他方の後に、または他方と同時に投与することができる。
【0087】
併用療法
上記のように、本発明のLrg1アンタゴニストは、任意の他の適切な活性化合物と併用して投与することができる。詳細には、Lrg1およびVEGFの両者の拮抗作用は病気を引き起こす可能性のある血管新生を減少させるので、Lrg1アンタゴニスト、特に、抗VEGF抗体、たとえばアバスチンおよび/またはルセンティスな、および/または受容体に基づくVEGFトラップ、たとえばアフリベルセプトなど。
【0088】
以下の実施例は、本発明を説明するものである。
【0089】
(実施例)
【実施例1】
【0090】
1. 異常な網膜血管の遺伝子発現解析
網膜疾患についてさまざまな動物モデルがあるが、これらは遺伝的起源および細胞起源は異なるにもかかわらず、異常な血管反応を示し、この血管反応には血管形成だけでなく、他の血管変化、たとえば毛細血管拡張症(拡張し、蛇行した、機能不全の血管)も含まれる。網膜血管リモデリングの生物学的基礎について新たな知見を得るために、本発明者らはこうした4つのモデルで研究を行うことで(図1)、病気を引き起こす可能性のある網膜微小血管における遺伝子の発現差異を正常対照の微小血管と比較して検討した。血管異常が現れる段階に相当する時点で、野生型(WT)マウス、網膜ジストロフィー(RD)1マウス、Curlytail(CT)マウス、および超低比重リポタンパク質受容体(VLDLR)ノックアウトマウスから摘出し浄化した微小血管断片を調べた。3つのマウスモデルから単離されたRNAに関するマイクロアレイ遺伝子発現解析(Affymetrix)は、罹患した網膜由来の微小血管においてアップレギュレートまたはダウンレギュレートされた、全モデルに共通の63個の遺伝子を明らかにした(図10)。血管リモデリングに関する3つのマウスモデルの網膜血管系において、発現に差異のある63個の遺伝子のうち、ロイシンリッチα-2-糖タンパク質-1(Lrg1)は、もっとも重要であると評価された(FDR解析後に)。Lrg1は、タンパク質-タンパク質相互作用、シグナル伝達、ならびに細胞の接着および発生に関与する、タンパク質のロイシンリッチリピートファミリーの、分泌型糖タンパク質である(図2)。
【0091】
2. マイクロアレイデータに見られるLrg1過剰発現の検証
マウスモデルの網膜におけるLrg1の発現増加は、まず、定量PCRによって検証した。WT、RD1、CTおよびVLDLR-/-マウスの網膜全体からmRNAを抽出し、リアルタイム定量PCR(qRTPCR)に供した。マイクロアレイ解析で示されるように、対照モデルと比較すると、網膜血管病変の3モデルにおいて、Lrg1の転写産物発現が有意に(p<0.05)増加していることが、qRTPCRによって実証された(図3A)。mRNAの増加が結果としてタンパク質発現の増加につながることを確認するために、本発明者らは次に、遺伝子発現実験と同一の時点で網膜を取り出し、その組織を、ウェスタンブロットによるタンパク質分析用に調製した(図3B)。デンシトメトリー分析によるウェスタンブロットデータ(n > 3)の半定量化は、ハウスキーピングタンパク質(GAPDH)と比較して、Lrg1タンパク質発現の有意な増加を明らかにした(p<0.05)。網膜血管系におけるLrg1の分布を決定し、網膜の他の細胞がLrg1を発現するかどうか確認するために、本発明者らは、in situハイブリダイゼーションおよび免疫組織化学的検査を行って、それぞれLrg1 mRNAおよびタンパク質を検出した。正常なマウスにおいて、Lrg1 mRNA(図3C)およびタンパク質(図3D)は、大部分が血管系によって発現された。
【0092】
3. Lrg1はTGFβ受容体TGFβRIIおよびALK1と結合する
Lrg1の生物学に関しては、ほとんど何もわかっていない。多くの疾患においてTGFβ1、TGFβRIIおよびLrg1の発現レベルが同時に増加することが、いくつかの報告に記載されている(Sun et al., 1995; Li et al., 2007)。内皮細胞におけるTGFβシグナル伝達の機能不全が、遺伝性出血性毛細血管拡張症(HHT)という病気をもたらすので、上記は特に密接な関係がある。毛細血管拡張症などの血管異常を特徴とする、こうした疾病群において、TGFβ内皮補助受容体エンドグリンおよびTGFβI型受容体ALK1における変異は、HHT1(McAllister et al. 1994)およびHHT2(Johnson et al. 1995)をそれぞれ引き起こす。さらに、VLDLR-/-マウスにおいて、網膜組織ではTGFβmRNAが有意に増加しているが、RPEまたは微小血管ではそうではないという趣旨の、試験的なデータがある。さらによく当てはまることに、TGFβは、血管リモデリングがよく見られる糖尿病性網膜症患者の網膜において増加していることが明らかになった。
【0093】
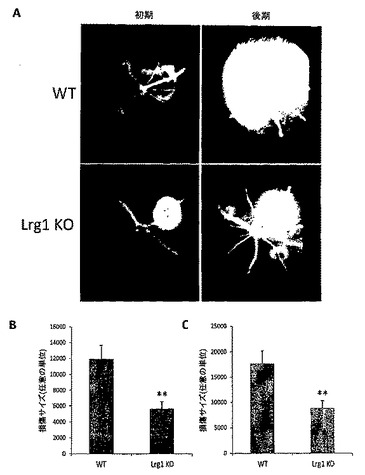
内皮細胞において、TGFβシグナル伝達は、TGFβ受容体IIと、主として内皮細胞において発現されている遍在性TGFβI型受容体アクチビン受容体様キナーゼ5(ALK5)またはALK1のいずれかとの結合を介して起こり、その細胞性応答はどちらの経路が優勢であるかによって決まる。ALK5の場合、ECM沈着の増加および細胞休止が見られるのに対して、ALK1については、遊走および増殖の増加となって現れる内皮細胞の活性化が見られる。こうした差異のあるシグナル伝達は、一つにはTGFβの濃度/バイオアベイラビリティによって制御され、また、Smadと呼ばれる下流エフェクタータンパク質ファミリーのメンバーによっても制御されるが、このSmad 2および3はALK5と関連付けられ、Smad 1、5および8はALK1と関連付けられる。本発明者らは、Lrg1とTGFシグナル伝達経路の関連性を検討した。まず、ラット脳内皮細胞株(GPNT)がLrg1、TGFβRII、ならびにTGFβシグナル伝達の他成分(付表1)をいずれも発現することを確認した。GPNT細胞溶解物由来のLrg1の免疫沈降法の結果、受容体TGFβRIIおよびALK1の共沈殿が見られた(図4A)。同様に、TGFβRIIまたはALK1のいずれか一方の免疫沈降は、結果としてLrg1の共沈殿をもたらし、Lrg1が2つの受容体といずれも結合することが示された(図4A)。また、細菌由来のHA標識組換えLrg1タンパク質がTGFβRIIおよびTGFβと結合することも明らかにした(図4B)。加えて、GPNT細胞におけるLrg1およびTGFβRII発現の免疫細胞化学による可視化は、共局在化を示している(図4C)。したがって、本発明者らは、Lrg1が、TGFβRIIと、ALK1およびALK5活性化シグナル伝達カスケードとの間の微調整をもたらす、TGFβシグナル伝達のモジュレーターとして機能すると仮定する。また、TGFβがGPNT細胞においてLrg1遺伝子発現を誘導することも明らかにし、可能性のあるフィードバック機構が示唆された(図16)。
【0094】
4. Lrg1はSmadリン酸化の差異によってTGFβシグナル伝達を微調整する
Lrg1がTGFβを介した血管内皮細胞反応に影響を及ぼすかどうかを確認するために、本発明者らは次に、sirRNAを用いてGPNT細胞のLrg1をノックダウンし、それがTGFβを介した細胞増殖に及ぼす影響を測定した。対照細胞において、TGFβは、2時間にわたって内皮細胞増殖の有意な増加(p<0.05)を引き起こす(70%コンフルエント細胞)。siRNAによるGPNT内皮細胞のLrg1のノックダウンは、このTGFβを介した細胞増殖の増加をブロックする(図6AおよびB)。これは、Smad5リン酸化の減少と相関する(図4D)。反対に、Lrg1遺伝子でトランスフェクトされたGPNT細胞では、Lrg1過剰発現が、TGFβに対応した内皮細胞増殖の増強をもたらすことを示す(図6AおよびB)。この増強された反応は、Smad2発現のダウンレギュレーション、およびSmad5リン酸化の増加と相関している(図4E)。これらのデータは、TGFβRII/ALK5受容体複合経路を介したシグナル伝達の減少と符合し、したがって血管病変を引き起こしうるTGFβRII/ALK1シグナル伝達経路の活性化へのシフトと矛盾しない。
【0095】
5. Lrg1馴化培地はin vitroでの血管形成を増強する
Lrg1が内皮細胞においてTGFβシグナル伝達を微調整し、TGFβを介した細胞増殖に影響を及ぼすことが確認されたので、本発明者らは次に、Lrg1が血管形成に影響を与えるかどうかを、標準的なin vitro血管形成アッセイによって判定した。ヒト臍静脈内皮細胞(HUVEC)をMatrigel中で増殖させ、馴化されていない増殖培地、(構成的にLrg1を分泌する)GPNT細胞で馴化された培地、ならびにLrg1を過剰発現するGPNT細胞で馴化された培地に供する。対照培地がLrg1を含有しないのに対して、GPNT培地およびLrg1過剰発現GPNT培地は、それぞれ中程度、および高レベルのLrg1を含有していた(図7B)。血管形成の程度は、Lrg1過剰発現細胞から得られた馴化培地を加えたとき、最大であった(図7AおよびC)。血管新生の増加は、培地におけるLrg1タンパク質発現と相関していた。
【0096】
6. Lrg1に由来するペプチド配列、付表2のL227-252もしくは付表3のL320-345はGPNT細胞においてTGFβシグナル伝達を微調整する
本発明者らは次に、TGFβにより誘導される内皮細胞のSmad5リン酸化を、抗Lrg1抗体を用いて、またはLrg1由来ペプチド配列によって、ブロックすることができるかを確認した。Lrg1配列のロイシンリッチリピート領域に由来するペプチド(付表2のL1-24およびL169-192、もしくは付表3のL94-117およびL262-285)(タンパク質-タンパク質相互作用に関与すると考えられている)、ならびに高度に保存されたロイシンリッチC末端ドメインに由来するペプチド(付表2のL227-252もしくは付表3のL320-345)を作製した(付表2)。GPNT細胞におけるLrg1過剰発現は、結果としてSmad5リン酸化をもたらさなかった。対照細胞の5 ng/ml TGFβによる処理は、Smad5リン酸化の有意な増加をもたらす。Lrg1過剰発現細胞において、Smad5リン酸化に及ぼすTGFβの影響は有意に強められる。Lrg1過剰発現内皮細胞を抗Lrg1ポリクローナル抗体で同時に処理すると、Smad5リン酸化のレベルが減少するので、この抗体がLrg1の相互作用を妨げる能力を有することが示唆される。ペプチドとの同時処理の影響はさまざまで、付表2のペプチドL1-24(付表3のL94-117)はSmad5リン酸化に何の影響も与えず、付表2のペプチドL169-192(付表3のL262-285)は部分的な影響を与えたが、付表2のペプチドL227-252(付表3のL320-345)(図5)は劇的な阻害効果を示した。3つのペプチドをすべて組み合わせると、TGFβを介したSmad5リン酸化はほぼ完全に消失した。これらのデータは、Lrg1がTGFβを介したシグナル伝達を微調整する、ならびに、Lrg1アンタゴニストを治療薬として使用することができる、という仮説を支持する。
【0097】
7. Lrg1ノックアウトマウス由来の大動脈輪は、血管新生の血管出芽の減少を示す
次にin vivo血管形成におけるLrg1の役割を検討した。頸椎脱臼により屠殺したP14 Lrg1ノックアウトマウス、または野生型同腹仔対照から胸部大動脈を摘出し、直ちに氷冷した無血清OPTI-MEM(Invitrogen)を入れた培養皿に移した。大動脈周囲の繊維脂肪組織を細い顕微解剖用のはさみで注意深く取り除いた。長さ1 mmの大動脈輪を切開し、DMEM, pH 7.4中で調製されたラット尾部コラーゲンIゲル(1.5 mg/ml)に包埋した。大動脈輪を含有するコラーゲンゲルを96ウェルプレート中で7日間37℃に保った。各ウェルには、2.5% FCS、100 U/mlペニシリン、および100μg/mlストレプトマイシンを添加した内皮細胞基本培地が入っていた。画像はOlympus顕微鏡で撮影した。
【0098】
各大動脈輪から出芽した血管新生の血管数を数えた。Lrg1遺伝子ノックアウトに関してヘテロ欠損マウスおよびホモ欠損マウスの両者から摘出された大動脈輪は、野生型マウスの大動脈輪と比べて血管新生の血管出芽の有意な減少を示した(p<0.01)(図9)。
【0099】
8. 網膜損傷後の脈絡膜新血管形成(CNV)は、Lrg1ノックアウトマウスにおいて減少する
ブルッフ膜を、Lrg1ノックアウトマウスまたは野生型同腹仔対照のそれぞれの眼の視神経を取り囲む3カ所で、レーザーにより破断した。ブルッフ膜裂傷部位のCNV病変をレーザー処置の1週間後にin vivo蛍光眼底血管造影(FA)により測定した。蛍光は腹腔内注入により投与した。7分の間隔をおいて初期および後期の眼底血管造影図が得られた。初期の血管造影図は、注入の90秒後に得られ、脈絡膜新血管形成の大きさを示している。後期血管造影図は、脈絡膜新生血管膜からの漏出を示す。
【0100】
FAは、Lrg1ノックアウトマウスにおいて脈絡膜新血管形成が減少していることを明白に示した(図11A)。脈絡膜新血管形成の数値化は、Lrg1ノックアウトマウスにおいて、野生型マウスより血管新生増殖面積のサイズおよび漏出が有意に減少していることを明らかにした(図11BおよびC)(** p<0.01)。
【0101】
9. 酸素誘導網膜症(OIR)後の網膜新血管形成はLrg1ノックアウトマウスにおいて減少する
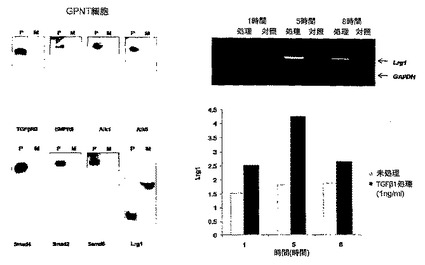
生後7日(P7)のLrg1ノックアウトマウスおよび野生型同腹仔対照は、哺育する親マウスとともに、酸素過剰(75%酸素)に5日間暴露したが、これは、その新生マウスにおいて、網膜血管発達の有意な阻害をもたらす。生後12日(P12)にマウスは酸素正常状態に戻され、その時、低酸素で血管のない網膜は、正常な血管再増殖および病的な新血管形成をともに引き起こすが、これは生後17日(P17)にピークとなる。網膜を摘出し、固定して、イソレクチン-B4によるホールマウント免疫染色に供した(図13A)。血管の再増殖は無血管面積を全網膜面積と比較することによって数値化した。新血管形成は、新生血管網の面積を手作業で測定することによって数値化した。
【0102】
無血管領域の大きさは、Lrg1ノックアウトマウスの網膜では有意に増加していることが判明した(* p<0.05)(図13B)。また、新生血管網の数は、Lrg1ノックアウトマウスでは、野生型マウスより減少していた(** p<0.002)(図13C)。
【0103】
10. in vitroのMatrigel中でのヒト臍静脈内皮細胞(HUVEC)の管、索状組織、および血管の形成は、ポリクローナル抗Lrg1抗体の添加によって減少する
in vitro管形成アッセイは、ヒト臍静脈内皮細胞(HUVEC)を用いてMatrigel中で行った。96ウェルプレートを、ウェル当たり60μlのMatrigelでコーティングした。各ウェルを、100 nMの(Lrg1糖タンパク質全体に対する)抗ヒトポリクローナルLrg1抗体、100 nM IgGアイソタイプ、または同量の抗体溶出バッファーの存在下で、37℃、5% CO2にて16時間、15,000 HUVECを含有する100μl EGM2培地で処理した。細胞を洗浄し、固定した。管形成は、中和抗体である抗ヒトLrg1ポリクローナル抗体の添加によって、抗体溶出バッファー添加(p<0.01)または無関係なIgG抗体の添加(p<0.05)と比べて、有意に減少した。管形成は、分岐点の数(図14A)、管の数(図14A)および管の全長(図14B)で評価した。
【0104】
11. 硝子体液におけるLrg1およびTGFβ発現は、増殖性糖尿病性網膜症(PDR)に罹っているヒト患者において増加する
ヒト網膜の免疫組織化学分析を行って、網膜血管系においてLrg1の染色を検出した(図15A)。硝子体液のサンプルは非糖尿病患者およびPDR患者から得られた。硝子体液サンプル中に存在するLrg1を、ウェスタンブロッティングによって測定し、デンシトメトリー分析で数値化した(図15B)。PDR患者の硝子体液では非糖尿病患者と比べて、Lrg1は有意に増加していた(p<0.01)。硝子体液サンプル中に存在するTGFβもウェスタンブロッティングによって測定し、Lrg1と同様に数値化した(図15C)。TGFβもPDR患者において有意に増加していた(p<0.01)。
【0105】
したがって、本発明者らは、Lrg1発現の減少が、網膜外傷に対する血管形成反応の減少と関連していることを実証した。Lrg1およびTGFβはまた、網膜の新血管形成の増加を特徴とする疾患であるPDRの患者において、アップレギュレートされていることが明らかになった。このことは、Lrg1がTGFβシグナル伝達を介して血管増殖刺激に関与するので、Lrg1アンタゴニスト、特に抗体を、望ましくない血管増殖が起こらないようにするための治療薬として使用することができるという仮説を支持する。
【0106】
(付表1)
(付表2)
ロイシンリッチリピート(赤色)および高度に保存されたC末端ドメイン(緑色)を明示するようアレンジした、マウスおよびヒトLrg1の部分配列アラインメント。
【0107】
(付表3)
ロイシンリッチα2糖タンパク質1(Lrg1)は、リモデリングされた網膜血管において最大の変化倍率を示した。ヒトおよびマウスLrg1のアラインされたアミノ酸配列。ロイシンリッチリピート領域は赤色、ブロッキングペプチドとして使用されたヒトC末端ドメイン領域は緑色で示す。
【0108】
(参考文献)
Isolation and characterization of an unknown, leucine-rich 3.1-S-alpha2-glycoprotein from human serum: Haupt H, Baudner S; Hoppe Seylers Z Physiol Chem. 1977 Jun; 358(6): 639-46. (Title translated from original German)
Periodicity of leucine and tandem repetition of a 24-amino acid segment in the primary structure of leucine-rich alpha 2-glycoprotein of human serum: Takahashi N, Takahashi Y, Putnam FW; Proc Natl Acad Sci USA. 1985 Apr;82(7):1906-10.
Differentially expressed genes in TGF-beta 1 sensitive and resistant human hepatoma cells: Sun D, Kar S, Carr BI; Cancer Lett. 1995 Feb 10; 89(1):73-9.
Expression of TGF-betas and TGF-beta type II receptor in cerebrospinal fluid of patients with idiopathic normal pressure hydrocephalus: Li X, Miyajima M, Jiang C, Arai H; Neurosci Lett. 2007 Feb 14; 413(2):141-4. Epub 2006 Dec 27.
Identification of putative serum glycoprotein biomarkers for human lung adenocarcinoma by multilectin affinity chromatography and LC-MS/MS: Heo SH, Lee SJ, Ryoo HM, Park JY, Cho JY.; Proteomics. 2007 Dec;7(23):4292-302.
Increased expression of one isoform of leucine-rich alpha-2-glycoprotein in peritoneal fluid of women with uterine leiomyomas: Ferrero S, Gillott DJ, Remorgida V, Anserini P, Ragni N, Grudzinskas JG; Arch Gynecol Obstet. 2009 Mar;279(3):365-71. Epub 2008 Jul 30.
Potential diagnostic biomarkers in serum of idiopathic pulmonary arterial hypertension: Zhang J, Zhang Y, Li N, Liu Z, Xiong C, Ni X, Pu Y, Hui R, He J, Pu J.; Respir Med. 2009 Aug 22. [Epub ahead of print]
Endoglin, a TGF-beta binding protein of endothelial cells, is the gene for hereditary haemorrhagic telangiectasia type 1. McAllister KA, Grogg KM, Johnson DW, Gallione CJ, Baldwin MA, Jackson CE, Helmbold EA, Markel DS, McKinnon WC, Murrell J, et al. Nat Genet. 1994 Dec;8(4):345-51.
A second locus for hereditary hemorrhagic telangiectasia maps to chromosome 12. Johnson DW, Berg JN, Gallione CJ, McAllister KA, Warner JP, Helmbold EA, Markel DS, Jackson CE, Porteous ME, Marchuk DA. Genome Res. 1995 Aug;5(1):21-8.
Basement membrane and growth factor gene expression in normal and diabetic human retinas. Spirin KS, Saghizadeh M, Lewin SL, Zardi L, Kenney MC, Ljubimov AV. Curr Eye Res. 1999 Jun;18(6):490-9.
Plasma proteomics of pancreatic cancer patients by multi-dimensional liquid chromatography and two-dimensional difference gel electrophoresis (2D-DIGE): up-regulation of leucine-rich alpha-2-glycoprotein in pancreatic cancer. Kakisaka T, Kondo T, Okano T, Fujii K, Honda K, Endo M, Tsuchida A, Aoki T, Itoi T, Moriyasu F, Yamada T, Kato H, Nishimura T, Todo S, Hirohashi S. J Chromatogr B Analyt Technol Biomed Life Sci. 2007 Jun 1;852(1-2):257-67. Epub 2007 Feb 1.
US 2005/0064516
WO 2008/092214
US 2007/0184503。
【0109】
配列情報
ヒトLrg1の配列
配列番号1 −ヒトLrg1のDNA配列
[配列番号2のタンパク質をコードする配列は太字で下線を付してあり、下記配列番号1内にある]
配列番号2 −ヒトLrg1のアミノ酸配列
[配列番号3-5の配列は太字で下線を付してあり、下記配列番号2 内にある]
配列番号3-5 −Lrg1内のペプチドのアミノ酸配列
配列番号3 −付表2からのヒトLrg1のアミノ酸1-24 (付表3からのL94-117)
L1-24/L94-117: LQELHLSSNGLESLSPEFLRPVPQ
配列番号4 −付表2からのヒトLrg1のアミノ酸169-192(付表3からのL262-285)
L169-192/L262-285: LDMLDLSNNSLASVPEGLWASLGQ
配列番号5 −付表2からのヒトLrg1のアミノ酸227-252(付表3からのL320-345)
L227-252/L320-345: KMFSQNDTRCAGPEAVKGQTLLAVAK
【特許請求の範囲】
【請求項1】
血管増殖性疾患の治療または予防に使用するためのロイシンリッチα2糖タンパク質1(Lrg1)のアンタゴニスト。
【請求項2】
前記アンタゴニストが、TGFβシグナル伝達複合体において、
(a) アクチビン受容体様キナーゼ1(ALK1)とLrg1との相互作用;
(b) Lrg1とTGFβ受容体II(TGFβRII)との相互作用;および/または
(c) Lrg1とTGFβとの相互作用
をブロックする、請求項1に記載のLrg1アンタゴニスト。
【請求項3】
前記アンタゴニストによる前記のブロックがALKとLrg1との相互作用を減少させ、それによってALK1とTGFβ受容体II(TGFβRII)との相互作用が低下し、TGFβRIIとアクチビン受容体様キナーゼ5(ALK5)との相互作用を促進し、それによりALK1活性化シグナル伝達カスケードにおけるTGFβの効果が、ALK5活性化シグナル伝達カスケードにおけるTGFβの効果と比べて低下する、請求項2に記載のLrg1アンタゴニスト。
【請求項4】
Lrg1機能をブロックする抗体、二本鎖RNA、アプタマー、またはペプチドもしくはペプチドミメティックを含む、請求項1〜3のいずれか1つに記載のLrg1アンタゴニスト。
【請求項5】
Lrg1の断片である、請求項4に記載のアンタゴニストペプチド。
【請求項6】
配列L1-24(配列番号3)、L169-192(配列番号4)、およびL227-252(配列番号5)の1つもしくは複数を含んでなる、請求項5に記載のアンタゴニストペプチド断片。
【請求項7】
Lrg1のアミノ酸227-252を含んでなる、またはそのアミノ酸からなる、請求項6に記載のアンタゴニストペプチド断片。
【請求項8】
モノクローナル抗体である、請求項4に記載のアンタゴニスト抗体。
【請求項9】
Lrg1のL1-24(配列番号3)、L169-192(配列番号4)、またはL227-252(配列番号5)の配列内のエピトープを特異的に認識する、請求項8に記載のアンタゴニストモノクローナル抗体。
【請求項10】
Lrg1のL227-252(配列番号5)内のエピトープを特異的に認識する、請求項9に記載のアンタゴニストモノクローナル抗体。
【請求項11】
低分子干渉RNA(siRNA)またはマイクロRNA(miRNA)である、請求項4に記載のアンタゴニスト二本鎖RNA。
【請求項12】
疾患の治療に使用するための、請求項1〜11のいずれか1つに記載のLrg1アンタゴニストであって、その疾患において前記血管増殖性疾患が、新血管形成、血管内皮細胞増殖、血管新生、毛細血管拡張症、または毛細血管瘤を含む、前記アンタゴニスト。
【請求項13】
眼の血管増殖性疾患の治療に使用するための、請求項1〜12のいずれか1つに記載のLrg1アンタゴニスト。
【請求項14】
眼の疾患の治療に使用するための、請求項13に記載のLrg1アンタゴニストであって、その疾患が糖尿病性網膜症、網膜静脈閉塞、未熟児網膜症、黄斑部毛細血管拡張症、加齢黄斑変性、または脈絡膜血管新生から選択される、前記アンタゴニスト。
【請求項15】
血管増殖を示す腫瘍の治療に使用するための、請求項1〜12のいずれか1つに記載のLrg1アンタゴニスト。
【請求項16】
腫瘍の治療に使用するための、請求項15に記載のLrg1アンタゴニストであって、その腫瘍が脳腫瘍、乳房腫瘍、腎臓腫瘍、大腸腫瘍、肺腫瘍、前立腺腫瘍、頭頸部腫瘍、胃腫瘍、膵臓腫瘍、皮膚腫瘍、頸部腫瘤、骨腫瘍、卵巣腫瘍、精巣腫瘍、および肝臓腫瘍から選択される、前記アンタゴニスト。
【請求項17】
抗血管形成化合物と併用して使用するための、請求項1〜16のいずれか1つに記載のLrg1アンタゴニスト。
【請求項18】
前記抗血管形成化合物が血管内皮細胞増殖因子(VEGF)のアンタゴニストである、請求項17に記載のLrg1アンタゴニスト。
【請求項19】
前記VEGFアンタゴニストが抗VEGF抗体である、請求項18に記載のLrg1アンタゴニスト。
【請求項20】
Lrg1のアンタゴニストを同定する方法であって、その方法は、
(a) アンタゴニスト候補を用意すること、ならびに
(b) 前記アンタゴニスト候補がLrg1の機能もしくは活性をブロックするか否かを判定すること;
を含んでなり、Lrg1の機能もしくは活性をブロックすることが観察されれば、前記アンタゴニスト候補がLrg1のアンタゴニストとして同定される、前記方法。
【請求項21】
Lrg1アンタゴニストが、
(a) アクチビン受容体様キナーゼ1(ALK1)とLrg1との相互作用;
(b) Lrg1とTGFβ受容体II(TGFβRII)との相互作用;および/または
(c) Lrg1とTGFβとの相互作用
をブロックする、請求項20に記載の方法。
【請求項22】
アミノ酸L1-24(配列番号3)、L169-192(配列番号4)、またはL227-252(配列番号5)内のエピトープを特異的に認識し、Lrg1活性をブロックする、モノクローナル抗体。
【請求項23】
アミノ酸L1-24(配列番号3)、L169-192(配列番号4)、またはL227-252(配列番号5)内のエピトープを特異的に認識し、
(a) ALK1とLrg1との相互作用;
(b) Lrg1とTGFβ受容体II(TGFβRII)との相互作用;および/または
(c) Lrg1とTGFβとの相互作用
をブロックする、モノクローナル抗体。
【請求項24】
請求項22または23に記載の抗体を作製するための方法であって、
(a) 非ヒト哺乳類を、Lrg1のL1-24(配列番号3)、L169-192(配列番号4)、またはL227-252(配列番号5)配列内のエピトープを含有する免疫原で免疫化すること;ならびに
(b) 前記哺乳類から抗体標品を得ること、および、その標品から前記エピトープを特異的に認識するモノクローナル抗体を得ること、
を含んでなる、前記方法。
【請求項25】
Lrg1内のどの部位がLrg1の機能もしくは活性をブロックする標的となり得るかを決定するための方法であって、
(a) Lrg1タンパク質のペプチド断片を用意すること;ならびに
(b) 前記ペプチド断片のそれぞれがLrg1の機能もしくは活性をブロックするかどうかを判定すること
を含んでなる、前記方法。
【請求項26】
前記ステップ(b)でLrg1の機能もしくは活性をブロックすると判明したペプチド断片を特異的に認識する、抗体またはアプタマーを得ることをさらに含んでなり;さらに必要に応じて、前記抗体もしくはアプタマーがLrg1の機能もしくは活性をブロックするかどうか判定することを含んでなる、請求項25に記載の方法。
【請求項27】
血管増殖性疾患の治療もしくは予防のための、薬剤の製造におけるLrg1アンタゴニストの使用。
【請求項28】
必要な患者に有効量のLrg1アンタゴニストを投与することを含んでなる、血管増殖性疾患を治療する方法。
【請求項1】
血管増殖性疾患の治療または予防に使用するためのロイシンリッチα2糖タンパク質1(Lrg1)のアンタゴニスト。
【請求項2】
前記アンタゴニストが、TGFβシグナル伝達複合体において、
(a) アクチビン受容体様キナーゼ1(ALK1)とLrg1との相互作用;
(b) Lrg1とTGFβ受容体II(TGFβRII)との相互作用;および/または
(c) Lrg1とTGFβとの相互作用
をブロックする、請求項1に記載のLrg1アンタゴニスト。
【請求項3】
前記アンタゴニストによる前記のブロックがALKとLrg1との相互作用を減少させ、それによってALK1とTGFβ受容体II(TGFβRII)との相互作用が低下し、TGFβRIIとアクチビン受容体様キナーゼ5(ALK5)との相互作用を促進し、それによりALK1活性化シグナル伝達カスケードにおけるTGFβの効果が、ALK5活性化シグナル伝達カスケードにおけるTGFβの効果と比べて低下する、請求項2に記載のLrg1アンタゴニスト。
【請求項4】
Lrg1機能をブロックする抗体、二本鎖RNA、アプタマー、またはペプチドもしくはペプチドミメティックを含む、請求項1〜3のいずれか1つに記載のLrg1アンタゴニスト。
【請求項5】
Lrg1の断片である、請求項4に記載のアンタゴニストペプチド。
【請求項6】
配列L1-24(配列番号3)、L169-192(配列番号4)、およびL227-252(配列番号5)の1つもしくは複数を含んでなる、請求項5に記載のアンタゴニストペプチド断片。
【請求項7】
Lrg1のアミノ酸227-252を含んでなる、またはそのアミノ酸からなる、請求項6に記載のアンタゴニストペプチド断片。
【請求項8】
モノクローナル抗体である、請求項4に記載のアンタゴニスト抗体。
【請求項9】
Lrg1のL1-24(配列番号3)、L169-192(配列番号4)、またはL227-252(配列番号5)の配列内のエピトープを特異的に認識する、請求項8に記載のアンタゴニストモノクローナル抗体。
【請求項10】
Lrg1のL227-252(配列番号5)内のエピトープを特異的に認識する、請求項9に記載のアンタゴニストモノクローナル抗体。
【請求項11】
低分子干渉RNA(siRNA)またはマイクロRNA(miRNA)である、請求項4に記載のアンタゴニスト二本鎖RNA。
【請求項12】
疾患の治療に使用するための、請求項1〜11のいずれか1つに記載のLrg1アンタゴニストであって、その疾患において前記血管増殖性疾患が、新血管形成、血管内皮細胞増殖、血管新生、毛細血管拡張症、または毛細血管瘤を含む、前記アンタゴニスト。
【請求項13】
眼の血管増殖性疾患の治療に使用するための、請求項1〜12のいずれか1つに記載のLrg1アンタゴニスト。
【請求項14】
眼の疾患の治療に使用するための、請求項13に記載のLrg1アンタゴニストであって、その疾患が糖尿病性網膜症、網膜静脈閉塞、未熟児網膜症、黄斑部毛細血管拡張症、加齢黄斑変性、または脈絡膜血管新生から選択される、前記アンタゴニスト。
【請求項15】
血管増殖を示す腫瘍の治療に使用するための、請求項1〜12のいずれか1つに記載のLrg1アンタゴニスト。
【請求項16】
腫瘍の治療に使用するための、請求項15に記載のLrg1アンタゴニストであって、その腫瘍が脳腫瘍、乳房腫瘍、腎臓腫瘍、大腸腫瘍、肺腫瘍、前立腺腫瘍、頭頸部腫瘍、胃腫瘍、膵臓腫瘍、皮膚腫瘍、頸部腫瘤、骨腫瘍、卵巣腫瘍、精巣腫瘍、および肝臓腫瘍から選択される、前記アンタゴニスト。
【請求項17】
抗血管形成化合物と併用して使用するための、請求項1〜16のいずれか1つに記載のLrg1アンタゴニスト。
【請求項18】
前記抗血管形成化合物が血管内皮細胞増殖因子(VEGF)のアンタゴニストである、請求項17に記載のLrg1アンタゴニスト。
【請求項19】
前記VEGFアンタゴニストが抗VEGF抗体である、請求項18に記載のLrg1アンタゴニスト。
【請求項20】
Lrg1のアンタゴニストを同定する方法であって、その方法は、
(a) アンタゴニスト候補を用意すること、ならびに
(b) 前記アンタゴニスト候補がLrg1の機能もしくは活性をブロックするか否かを判定すること;
を含んでなり、Lrg1の機能もしくは活性をブロックすることが観察されれば、前記アンタゴニスト候補がLrg1のアンタゴニストとして同定される、前記方法。
【請求項21】
Lrg1アンタゴニストが、
(a) アクチビン受容体様キナーゼ1(ALK1)とLrg1との相互作用;
(b) Lrg1とTGFβ受容体II(TGFβRII)との相互作用;および/または
(c) Lrg1とTGFβとの相互作用
をブロックする、請求項20に記載の方法。
【請求項22】
アミノ酸L1-24(配列番号3)、L169-192(配列番号4)、またはL227-252(配列番号5)内のエピトープを特異的に認識し、Lrg1活性をブロックする、モノクローナル抗体。
【請求項23】
アミノ酸L1-24(配列番号3)、L169-192(配列番号4)、またはL227-252(配列番号5)内のエピトープを特異的に認識し、
(a) ALK1とLrg1との相互作用;
(b) Lrg1とTGFβ受容体II(TGFβRII)との相互作用;および/または
(c) Lrg1とTGFβとの相互作用
をブロックする、モノクローナル抗体。
【請求項24】
請求項22または23に記載の抗体を作製するための方法であって、
(a) 非ヒト哺乳類を、Lrg1のL1-24(配列番号3)、L169-192(配列番号4)、またはL227-252(配列番号5)配列内のエピトープを含有する免疫原で免疫化すること;ならびに
(b) 前記哺乳類から抗体標品を得ること、および、その標品から前記エピトープを特異的に認識するモノクローナル抗体を得ること、
を含んでなる、前記方法。
【請求項25】
Lrg1内のどの部位がLrg1の機能もしくは活性をブロックする標的となり得るかを決定するための方法であって、
(a) Lrg1タンパク質のペプチド断片を用意すること;ならびに
(b) 前記ペプチド断片のそれぞれがLrg1の機能もしくは活性をブロックするかどうかを判定すること
を含んでなる、前記方法。
【請求項26】
前記ステップ(b)でLrg1の機能もしくは活性をブロックすると判明したペプチド断片を特異的に認識する、抗体またはアプタマーを得ることをさらに含んでなり;さらに必要に応じて、前記抗体もしくはアプタマーがLrg1の機能もしくは活性をブロックするかどうか判定することを含んでなる、請求項25に記載の方法。
【請求項27】
血管増殖性疾患の治療もしくは予防のための、薬剤の製造におけるLrg1アンタゴニストの使用。
【請求項28】
必要な患者に有効量のLrg1アンタゴニストを投与することを含んでなる、血管増殖性疾患を治療する方法。
【図1】

【図2】
【図3】
【図4A】
【図4B】
【図5】
【図6】
【図7】
【図8】

【図9】

【図10】
【図11】
【図12】
【図13】
【図14】
【図15】
【図16A】
【図16B】
【図2】
【図3】
【図4A】
【図4B】
【図5】
【図6】
【図7】
【図8】
【図9】
【図10】
【図11】
【図12】
【図13】
【図14】
【図15】
【図16A】
【図16B】
【公表番号】特表2013−503621(P2013−503621A)
【公表日】平成25年2月4日(2013.2.4)
【国際特許分類】
【出願番号】特願2012−527383(P2012−527383)
【出願日】平成22年9月6日(2010.9.6)
【国際出願番号】PCT/GB2010/001681
【国際公開番号】WO2011/027129
【国際公開日】平成23年3月10日(2011.3.10)
【出願人】(505367464)ユーシーエル ビジネス ピーエルシー (20)
【Fターム(参考)】
【公表日】平成25年2月4日(2013.2.4)
【国際特許分類】
【出願日】平成22年9月6日(2010.9.6)
【国際出願番号】PCT/GB2010/001681
【国際公開番号】WO2011/027129
【国際公開日】平成23年3月10日(2011.3.10)
【出願人】(505367464)ユーシーエル ビジネス ピーエルシー (20)
【Fターム(参考)】
[ Back to top ]