
血管新生標的免疫複合体
【課題】癌、慢性関節リウマチ、滲出性形態の黄斑変性、及びアテローム性動脈硬化症のような血管新生に関連する疾患を治療するための免疫複合体を提供する。
【解決手段】当該免疫複合体は、典型的には、ヒンジを含めたヒトIgG1免疫グロブリンのFc領域、あるいは患者に投与したとき、標的細胞に対して細胞溶解性免疫応答又は細胞傷害作用を誘発することができる他のエフェクタードメイン(1つ又は複数)から成る。エフェクタードメインは、リシン−341がアラニンに又はセリン−344がアラニンに置換された第VII因子のような、高い親和性と特異性で組織因子に結合するが血液凝固を開始させない、第VII因子突然変異体を含む標的ドメインに複合されている。
【解決手段】当該免疫複合体は、典型的には、ヒンジを含めたヒトIgG1免疫グロブリンのFc領域、あるいは患者に投与したとき、標的細胞に対して細胞溶解性免疫応答又は細胞傷害作用を誘発することができる他のエフェクタードメイン(1つ又は複数)から成る。エフェクタードメインは、リシン−341がアラニンに又はセリン−344がアラニンに置換された第VII因子のような、高い親和性と特異性で組織因子に結合するが血液凝固を開始させない、第VII因子突然変異体を含む標的ドメインに複合されている。
【発明の詳細な説明】
【技術分野】
【0001】
関連出願の相互参照
本出願は、1999年7月1日に出願された米国特許出願番号第60/142,161に基づく優先権を主張する。
【0002】
連邦政府による資金的条を受けた研究開発の記載
本発明は、一部、アメリカ国立衛生研究所(アメリカ合衆国公衆衛生局)からの研究グランドHL-29019-17に基づく政府のサポートによって作られた。政府は、本発明に対し、一定の権利を有する。
【背景技術】
【0003】
(発明の属する技術分野)
本発明は、癌、慢性関節リウマチ、滲出性形態の黄斑変性、及びアテローム性動脈硬化症のような新しい血管の増殖(血管新生:neovascularization)に関連する疾患を有する患者を治療するための免疫複合体試薬の設計、合成及び投与に関する。本発明の免疫複合体の標的は、新生血管系の内皮細胞によって発現される膜貫通レセプタ組織因子である。組織因子はまた、多くの型の腫瘍細胞によっても発現される。それ故、本発明の治療方法は広い範囲の固体腫瘍に対する免疫療法において特に有効である。治療試薬は、標的ドメインとエフェクタードメインから成る免疫複合体である(図1)。標的ドメインは、高い親和性と特異性で組織因子に結合するが、血液凝固を開始させない、突然変異形態の第VII因子である。エフェクタードメインはIgG1免疫グロブリンのFc領域である。
【0004】
(発明の背景、従来の技術)
典型的には血管内皮細胞に対して特異的な成長因子の放出が引き金となる様々な疾患において、病的血管形成、すなわち周辺組織中の脈管からの血管増殖の誘導が認められる。病的血管形成は、固体腫瘍の成長と転移を可能にし、眼障害における視覚機能不全を引き起こし、炎症性疾患における白血球の血管外遊出を促進し、及び/又はアテローム性動脈硬化症のような心臓血管疾患の結果に影響を及ぼす、血管新生をもたらしうる。集合的に、これらは時に血管形成性疾患と称される。
【0005】
固体腫瘍の生存と成長は新生血管系の発生に決定的に依存するので、癌は主要な血管形成性疾患である(Folkman,J.(1995)N.Engl.J.Med.333,1757−1763)(非特許文献1)。多くの癌は段階的に進行し、一次腫瘍細胞の増殖から始まって、腫瘍細胞の循環系への侵入、播種性転移部位でコロニー形成、そして癌による死亡の大部分の原因である転移した腫瘍細胞の増殖へと進行する(Vogelstein,B.とKinzler,K.W.(1993)TIG9,141−143)(非特許文献2)。癌は、疾患が転移段階に達するまでしばしば検出されないままであるため、血管下部構造と転移性腫瘍細胞を根絶することができる癌療法が特に望ましい。
【0006】
血管形成はまた、慢性関節リウマチにおいても重要な役割を果たす(Szekanez,Z.ら(1998)J.Invest.Med.46,27−41)(非特許文献3)。慢性関節リウマチ(RA)は、世界中ですべての人種民族群において起こる慢性全身性炎症性疾患であり、主として滑膜性の連結及びしばしば様々な他の器官を侵す。RAの滑膜組織は広汎に血管新生される。RAでは、炎症白血球が血管の内皮層を通して滑膜内に遊出し、滑膜の炎症、そして最終的には関節の破壊をもたらす。
【0007】
血管形成は、視力の破滅的喪失をもたらす眼疾患の大半の基礎となる(Firedlander,M.ら(1996)Proc.Natl.Acad.Sci.USA 93,9764−9769)(非特許文献4)。55歳以上の個人における失明の主要な原因は滲出性(「湿性」)形態の加齢性黄斑変性(ARMD)であり、55歳未満では増殖性糖尿病性網膜症(PDR)である。ARMDとPDRはそれぞれ脈絡膜と網膜の血管新生に関する基本型の疾患であるが、他の変性又は炎症状態も、選択的にいずれかの血管系の新血管形成を引き起こしうる(同上)。
【0008】
それ故、これらの疾患状態、特に癌の治療への1つのアプローチは、主として新しい血管の増殖を阻害することによって、新生血管系の機能又は成長を抑えることであった(Chaplin,D.J.とDougherty,G.J.(1999)Br.J.Cancer,80,57−64)(非特許文献5)。血管を標的することにはいくつかの利点がある。まず第一に、血管への損傷は血流を停止させることができ、癌に適用すると、多くの依存性腫瘍細胞の死の引き金となりうる。第二に、標的細胞が血流に近接しており、薬物送達が促進される。第三に、血管内皮細胞においては治療耐性の突然変異が起こりにくい。
【0009】
薬剤、抗体及び遺伝子治療に基づくアプローチを含めて、数多くの抗血管形成療法が提案されてきた。これらは、メタロプロテイナーゼ阻害因子、ポリ硫酸ペントサン及びTNP−470、チロシンキナーゼの選択的阻害因子、そしてアンギオスタチン及びエンドスタチンのペプチド阻害因子を含む(同上、及びその中で引用される総説)。これらは典型的には、脈管形成の様々な段階、すなわち基底膜の分解、内皮細胞の移動、内皮細胞の増殖、及び管形成の段階で血管形成を妨げる。血管形成だけでなく、血管形成性疾患状態において既に形成された新生血管系も標的する、改善された治療法を開発することが望ましいであろう。
【先行技術文献】
【非特許文献】
【0010】
【非特許文献1】Folkman,J.(1995)N.Engl.J.Med.333,1757−1763
【非特許文献2】Vogelstein,B.とKinzler,K.W.(1993)TIG9,141−143
【非特許文献3】Szekanez,Z.ら(1998)J.Invest.Med.46,27−41
【非特許文献4】Firedlander,M.ら(1996)Proc.Natl.Acad.Sci.USA 93,9764−9769
【非特許文献5】Chaplin,D.J.とDougherty,G.J.(1999)Br.J.Cancer,80,57−64
【発明の概要】
【0011】
(発明が解決しようとする課題)
本発明の課題は癌、慢性関節リウマチ、滲出性形態の黄斑変性、及びアテローム性動脈硬化症を含めて、新しい血管の増殖(新生血管系)に関連する疾患のための新規治療を提供することである。より具体的な課題は、本発明はこれらの疾患状態において認められる血管形成を抑制するだけでなく、新生血管系構造も破壊する、新生血管標的療法を提供することである。
【0012】
(課題を解決するための手段)
これらやその他の目的は、組織因子に選択的に結合する標的ドメインと、標的細胞に対する細胞溶解応答又は細胞傷害性応答を動員するエフェクタードメインを含む免疫複合体を含有する組成物を提供する、本発明によって実現される。典型的な免疫療法の処置では、各々が、組織因子に結合(bind)するが血液凝固を開始させない、リシン−341がアラニンに置換された、及び/又はセリン−344がアラニンに置換された第VII因子のような、ヒト第VII因子の突然変異形態を含む標的ドメインに複合(conjugate)した免疫複合体であって、ヒンジを含めたヒトIgG1免疫グロブリンのFc領域であるエフェクタードメインを持ち、2つの同一の鎖の二量体として構築される少なくとも1つの免疫複合体を含む組成物を、血管新生に関連する疾患を有する患者に有効量で投与する。組織因子は、正常血管系ではなく腫瘍新生血管系の裏側にある内皮細胞によって、また多くの型の腫瘍細胞によっても発現されるので、本発明の第VII因子免疫複合体は広い範囲の固体腫瘍に対する免疫療法において特に有効である。新生血管系での凝固を活性化する及び/又は新生血管系に組織因子又は組織因子突然変異体を導入するというこれまでに記述されている発明(米国特許第6,001,978号においてEdgingtonとMorrisseyが述べているような)は、組織因子は新生血管系の内皮細胞によって特異的に発現されているのであるから無意味であり、本発明は2つの主要な点でこれらの発明と異なっている:組織因子は、血液凝固プロセスのイニシエーターとしてではなく、本発明の免疫複合体によって仲介される細胞溶解免疫応答を誘導するための標的として使用される、および血液凝固経路を活性化せずに新生血管系を破壊する細胞溶解免疫応答が開始される。
【0013】
従来の薬学的組成物中での精製免疫複合体蛋白質の使用、あるいは分泌形態の免疫複合体をコードするcDNAを保有するベクター系の使用を含む、当該免疫複合体の全身又は局所投与の方法も開示する。本発明に従った治療は、1つ又は複数の型の精製された免疫複合体蛋白質の有効量の、周期的又は持続的な静脈内又は腫瘍内注入、あるいは他の部位での注入を含むが、これらに限定されない。代替的な実施態様は、分泌形態の1つ又は複数の型の免疫複合体蛋白質をコードするcDNAを保有する発現ベクターの有効量の、静脈内又は腫瘍内注入、あるいは他の部位での注入によって患者を治療することを含む。後者の例は、分泌形態の本発明の免疫複合体をコードするcDNAを保有する複製欠損アデノウイルスベクター又はアデノ関連ウイルスベクターの、静脈内又は腫瘍内注入、あるいは他の部位での注入によって患者を治療することを含む。
【0014】
癌の免疫療法のような一部の実施態様では、本発明の免疫複合体を、異なる標的ドメインを持つ別の型の免疫複合体と共に投与する。これらは典型的には、腫瘍細胞、血管内皮細胞、病的状態の滑膜における侵襲細胞で発現される細胞表面分子に選択的に結合する、ヒトscFv又はVH融合ファージライブラリーから単離された一本鎖Fv又はVH分子断片である。
【図面の簡単な説明】
【0015】
【図1】図1は、本発明の免疫複合体分子の機構を例示する図表である。TD;標的ドメイン。H:2つのジスルフィド架橋を持つIgG1免疫グロブリンのヒンジ領域。CH2及びCH3:IgG1免疫グロブリンの定常領域。標的ドメインは、典型的には突然変異した活性部位を持つヒト第VII因子、あるいはヒトscFv又はVH分子から成る。エフェクタードメインは、典型的には、非修飾であるか又は放射性分子又は光活性化色素分子のような細胞傷害性物質に複合(conjugate)した、ヒトIgG1のFc領域から成る。
【図2】図2は、ヒトナチュラルキラー(NK)細胞によるヒト黒色腫細胞の免疫複合体依存性溶解を示す棒グラフである。免疫複合体の標的ドメインは黒色腫特異的ヒトscFv分子であり、エフェクタードメインはヒトIgG1免疫グロブリンのFc領域である。ヒト黒色腫細胞系統A−2058及びヒト線維芽細胞対照を蛍光色素、カルセイン−AMで標識した。NK細胞単独(棒A)又はscFv免疫複合体E26−1とNK細胞(棒B)への接触後、無傷のままである黒色腫又は線維芽細胞の分画を残留蛍光によって測定した。NKエフェクター細胞対標的細胞の比率(E/T)は3から20の範囲であった。3つの完全なセットの実験を黒色腫細胞と線維芽細胞の両方について実施した;各々の実験に関して、細胞溶解アッセイを四重に実施した。図に示す棒は、3回の実験についての細胞溶解アッセイの平均値を表わし、ほぼ10%以内で一致した。細胞溶解の割合(%)を実施例1で述べたように算定した。scFv免疫複合体G71−1に関しても同様の結果を得た。
【図3】図3は、補体によるヒト黒色腫細胞の免疫複合体依存性溶解を示す棒グラフである。手順は、NK細胞の代わりにヒト血清又は精製ウサギ補体成分を使用したことを除いて、図2の説明で述べたとおりであった。各々のアッセイについての免疫複合体と補体試薬は実施例1、表1に示されているとおりであった。
【図4A】図4は、SCIDマウスで増殖させたヒト黒色腫異種移植片における腫瘍細胞及び腫瘍血管内皮細胞への突然変異マウス第VII因子(mfVIIasm)免疫複合体の結合に関する免疫組織化学アッセイを示す。第二抗体はAPで標識した抗ヒトγ鎖であり、AP基質は青色を生じるBCIP/NBTであった;対比染色はメチルグリーンであった。パネルA:異種移植片の広い範囲に及ぶ血管形成を示す、ヘマトキシリン+エオシンで染色した対照。
【図4B】図4は、SCIDマウスで増殖させたヒト黒色腫異種移植片における腫瘍細胞及び腫瘍血管内皮細胞への突然変異マウス第VII因子(mfVIIasm)免疫複合体の結合に関する免疫組織化学アッセイを示す。第二抗体はAPで標識した抗ヒトγ鎖であり、AP基質は青色を生じるBCIP/NBTであった;対比染色はメチルグリーンであった。パネルB:血管内皮細胞と腫瘍細胞の両方の強い染色を示す、mfVIIasm免疫複合体に関する免疫組織化学。
【図4C】図4は、SCIDマウスで増殖させたヒト黒色腫異種移植片における腫瘍細胞及び腫瘍血管内皮細胞への突然変異マウス第VII因子(mfVIIasm)免疫複合体の結合に関する免疫組織化学アッセイを示す。第二抗体はAPで標識した抗ヒトγ鎖であり、AP基質は青色を生じるBCIP/NBTであった;対比染色はメチルグリーンであった。パネルC:mfVIIasm免疫複合体なしの免疫組織化学的対照。
【図5】図5は、免疫複合体をコードする複製不全アデノウイルスベクターの静脈内注入後の、SCIDマウスの血液中のG71−1 scFv及びmfVIIasm免疫複合体の濃度を示す折れ線グラフである。0日目と7日目に、G71−1免疫複合体をコードする2×1011アデノウイルス(A)又はmfVIIasm免疫複合体をコードする4×1011アデノウイルス(B)をマウスに注入した。コードされた免疫複合体の血液中の濃度をELISAによって定量した。各点は、各々の群の5匹のマウスに関する濃度の平均値である。
【図6】図6は、SCIDマウスにおけるヒト黒色腫異種移植片の成長へのG71−1及びmfVIIasm免疫複合体の阻害作用を示す折れ線グラフである。各々の曲線に関して、5匹のSCIDマウスに5×105 TF2細胞を皮下注入した。異種移植片が触知可能な大きさに成長したとき、図に示す0日目と7日目と14日目に、マウスにアデノウイルスの尾静脈注入を実施した。注入したアデノウイルスの量は、対照については4×1011、G71−1免疫複合体をコードするアデノウイルスについては2×1011、そしてmfVIIasm免疫複合体をコードするアデノウイルスについては4×1011であった。推定腫瘍容積は各群の5匹のマウスについての平均値である。
【図7】図7は、図6及び実施例2で報告した実験からの異種移植片の腫瘍重量を示す棒グラフである。20日目、すなわちアデノウイルスの最後の注入から6日後に、マウスから異種移植片を切除した。棒の高さは各群の5匹のマウスについての平均重量である。
【図8】図8は、SCIDマウスにおける大きなヒト黒色腫異種移植片の成長へのG71−1及びmfVIIasm免疫複合体の阻害作用を示す折れ線グラフである。各々のマウスに5×105の黒色腫細胞を皮下注入し、異種移植片を皮膚表面で50mm3の推定腫瘍容積まで成長させた(1日目)。1、6、12及び19日目に、G71−1免疫複合体をコードする2×1011のアデノウイルスとmfVIIasm免疫複合体をコードする7×1011のアデノウイルスの混合物を5匹のマウスの尾静脈に注入した。対照として5匹のマウスに免疫複合体をコードしていない4×1011のアデノウイルスを注入した。推定腫瘍容積は各群の5匹のマウスについての平均値である。免疫複合体をコードするアデノウイルスを注入したマウスのうちの1匹が17日目に死亡していた;その後の日にちに関する推定腫瘍容積は残りの4匹のマウスの平均値である。
【図9】図9は、低レベルの組織因子を発現するヒト黒色腫異種移植片の成長へのmfVIIasm免疫複合体の阻害作用をプロットした折れ線グラフである。マウスに5×105のLXSN細胞を皮下注入し、異種移植片が触知可能な大きさに成長したとき(0日目)、5匹のマウスにmfVIIasm免疫複合体をコードする9×1011のアデノウイルスを静脈内注入し、5匹のマウスに4×1011の対照アデノウイルスを注入した。7、13、21及び24日に追加注入を行い、25日目に形態学的及び組織化学的検査のためにマウスを解剖した。推定腫瘍容積は各群の5匹のマウスについての平均値である。
【図10A】図10は、図9及び実施例2の説明で報告した実験からのヒト黒色腫異種移植片の組織化学を示すものである。25日目に異種移植片を切除し、パラフィンに包埋して、切片をヘマトキシリン+エオシンで染色した。パネルA:免疫複合体をコードしていないアデノウイルスを注入した対照マウスからの異種移植片。
【図10B】図10は、図9及び実施例2の説明で報告した実験からのヒト黒色腫異種移植片の組織化学を示すものである。25日目に異種移植片を切除し、パラフィンに包埋して、切片をヘマトキシリン+エオシンで染色した。パネルB:mfVIIasm免疫複合体をコードするアデノウイルスを注入したマウスからの異種移植片。
【図11】図11は、SCIDマウスにおいて腫瘍内注入したアデノウイルスベクターがヒト黒色腫の成長に及ぼす用量効果を示す折れ線グラフである。最初にマウスにヒト黒色腫細胞系統TF2を皮下注入し、皮膚腫瘍が0日目に示されている大きさまで成長したとき、mfVIIasm及びG71−1免疫複合体をコードする2つのアデノウイルスベクターの混合物を腫瘍に注入した。2つのベクターの用量は次のとおりであった。○:7×108IU;▽:2×109IU;□:6×109IU。対照である◇の用量については、免疫複合体をコードしていないアデノウイルス6×109IUを注入した。
【図12】図12は、SCIDマウスにおいて腫瘍内注入した高用量のアデノウイルスベクターがヒト黒色腫の成長に及ぼす作用を示す折れ線グラフである。最初にマウスにヒト黒色腫細胞系統TF2を皮下注入し、皮膚腫瘍が0日目に示されている大きさまで成長したとき、次のアデノウイルスベクターの6×109IUを注入した:●、対照ベクター(3匹のマウス);■、mfVIIasm免疫複合体をコードするベクター(5匹のマウス);△、mfVIIasm及びG71−1免疫複合体をコードする2つのベクターの混合物(6匹のマウス)。2、5、7、9、11、13、15及び17日目に追加注入を実施した。各曲線上の点は、対応する群のすべてのマウスについての測定値の平均(±20%)である。
【図13】図13は、mfVIIasm及びG71−1免疫複合体の混合物をコードするアデノウイルスベクター又は空アデノウイルスベクター対照を腫瘍内注入したSCIDマウスからの黒色腫の写真である。実験は上記図12の説明及び実施例3の中で述べられている。最後の注入から2日後に腫瘍を切除し、写真撮影した。図の上部のものさしの原寸は1cmである。
【図14】図14は、腫瘍内注入後の腫瘍内及び肝臓におけるアデノウイルスベクターの分布を示す写真である。GFP蛋白質をコードするが免疫複合体をコードしない対照ベクターは、SCIDマウスで増殖するヒト黒色腫皮膚腫瘍の3つの部位に注入した。総ベクター用量は6×109IUであった。注入の40時間後に腫瘍と肝臓を切除し、蛍光光学系を備えた解剖顕微鏡下に無傷検査した。480nmの励起と630nmの発光でGFPシグナルが検出され、577nmの励起と630nmの発光でバックグラウンドシグナルが検出された。パネルA:腫瘍のGFP。パネルB:腫瘍のバックグラウンド。パネルC:肝臓のGFP。パネルD:肝臓のバックグラウンド。パネルAのものと同様の明るい蛍光スポットが、おそらく注入部位に対応する、他の2箇所の腫瘍部位でも検出された。写真は組織内の1つのレベルに焦点を合わせている。しかし、そのレベルの上と下に焦点を合わせると腫瘍内でもGFPスポットが検出でき、注射針が横切った経路に隣接する腫瘍細胞だけがベクターによって感染した細胞であることを示唆した。
【図15】図15は、アデノウイルスベクターを静脈内又は腫瘍内注入したSCIDマウスからの肝臓切片を示す。静脈内実験は実施例2に述べられ、腫瘍内実験は実施例3に述べられている。左側のパネルは対照ベクターを注入したものであり、右側のパネルはmfVIIasm及びG71−1免疫複合体をコードする2つのベクターの混合物を注入したものである。最後の注入から2日後に肝臓を切除し、ホルムアルデヒドで固定し、パラフィンに埋め込んだ。当該切片をヘマトキシリンとエオシンで染色して、100×の倍率で写真撮影した。
【図16】図16は、mfVIIasm免疫複合体をコードするアデノウイルスベクターの免疫コンピテントマウスへの静脈内注入がマウス皮膚黒色腫の成長に及ぼす作用を示す折れ線グラフである。C57BL/6マウスにマウス黒色腫細胞系統B16F10を皮下注入し、皮膚腫瘍が0日目に示されている大きさまで成長したとき、mfVIIasm免疫複合体をコードするベクター3×1010IPを注入した。免疫複合体のFcドメインはマウスIgG1免疫グロブリンから誘導した。5日目に再び1.5×1010IPを腫瘍に注入した。
【発明を実施するための形態】
【0016】
(発明の実施の態様)
抗血管系免疫複合体療法は、血管形成の活性状態にある増殖中の腫瘍のようなある種の疾患状態で形成される新生血管系と異なって、正常な成体哺乳類血管系は一般に静止状態(女性の生殖周期や創傷治癒のような特定のプロセスを除く)にあるという所見に基づく。それ故、静止状態にある血管内皮細胞と増殖性血管内皮細胞との分子的な相違が病的血管系についての標的なると考えられる。
【0017】
静止状態から、癌で認められるような病的血管系における血管形成状態へのスイッチは、通常、腫瘍細胞によって分泌され、血管内皮細胞上の血管内皮細胞成長因子(VEGF)レセプタに高い親和性と特異性で結合する、VEGFによって活性化される。血管内皮細胞上のレセプタへのVEGFの結合によって活性化される別の応答は、血漿第VII/VIIa因子に結合して血液凝固を開始させる、膜貫通レセプタである組織因子の発現である。結合VEGFを持つ血管内皮細胞だけが組織因子を発現するので、腫瘍血管系についての推定上の標的は、血液中を循環する第VII/VIIa因子に結合するはずである、内皮細胞上で発現される組織因子である。
【0018】
本発明は、ヒトIgG1のFcドメインに複合した標的ドメインから成る免疫複合体が、ナチュラルキラー(NK)細胞と免疫系の補体経路による標的細胞への細胞溶解応答を仲介するという所見に基づく(Wang,B.ら(1999)Proc.Natl.Acad.Sci.USA 96,1627−1632)。正常組織中に存在する安定な血管ではなく、増殖する血管の内側表面で発現される細胞表面レセプタ組織因子と、その天然のリガンドである第VII因子との間の結合は高い特異性と親和性を示すので(Hu,Z.ら(1999)Proc.Natl.Acad.Sci.USA 96,8161−8166)、本発明は、血管新生に関連する疾患の治療のための新しい免疫療法プロトコールを提供する。組織因子は多くの型の腫瘍細胞によっても発現されるので(Callander,N.S.ら(1992)Cancer,70,1194−1201)、本発明はまた、癌の治療のための高い有効性を提供する。
【0019】
しかし、組織因子への第VII因子免疫複合体の結合は血管内凝固の播種を引き起こす可能性があるので、本発明の好ましい実施態様では、組織因子への親和性に影響を及ぼさずに凝固を抑制する第VII因子突然変異体を使用する。組織因子に結合するが血液凝固を開始させないこれらの免疫複合体は、組織因子への結合に関して内因性第VII/VIIa因子と競合する。この競合は免疫複合体を強く支持する(Huら、前出)。なぜなら、免疫複合体は2つの第VII因子標的ドメイン(図1)を含み、単量体の内因性第VII因子分子が欠乏したアビディティ(結合活性)効果を与えるからである。多くの好ましい実施態様では、第VII因子が組織因子に結合したときに血液凝固プロセスを開始させる蛋白分解活性を遮断するため、ヒト第VII因子の活性部位を、Lys−341及び/又はSer−344をアラニンで置換する部位特定突然変異誘発によって突然変異させる。標的ドメインとエフェクタードメインのどちらもヒトソースから誘導できるので、ヒト患者における重要な免疫拒絶反応は最小限に抑えられる。
【0020】
本発明の一般的な実施においては、図1に示すような免疫複合体を、各々が標的ドメインに複合したエフェクタードメインを持つ、2本の鎖を含む蛋白質二量体として構築する。それらが、免疫複合体と結合したときその標的に対して細胞傷害性を示すかぎり、いくつかのタイプのエフェクタードメインが使用できる。本発明の多くの典型的な免疫複合体蛋白質は、エフェクタードメインとして、IgG1免疫グロブリンのFc領域を持つ。ここで使用するとき、これは、Fc領域と同じ生物学的機能を示す変異体及びトランケートバージョンを含む。他の実施態様では、エフェクタードメインは、放射性標識又は光活性化色素分子、例えばレーザー光線によって活性化される色素のような細胞傷害性物質でありうる。エフェクタードメインは、上述したように組織因子に結合するが血液凝固を開始させない、突然変異形態の第VII因子を含む標的ドメインに複合される。
【0021】
本発明の免疫複合体蛋白質は、癌、慢性関節リウマチ、滲出性(「湿性」)形態の黄斑変性、又はアテローム性動脈硬化症のような血管新生に関連する疾患を有する患者に投与される。投与は、治療に関わる病的状態のタイプに依存して、局所又は全身的でありうる。ここで使用するとき、「患者」の語はヒトあるいは他の種の両方を含む;本発明はそれ故、医学適用と獣医学適用の両方を持つ。獣医学的組成物及び治療においては、対応する種から誘導される標的及びエフェクタードメインを用いて免疫複合体を構築する。
【0022】
投与は、例えば、精製免疫複合体又は複製欠損アデノウイルスベクター、あるいは分泌形態の免疫複合体をコードするcDNAを保有する他のウイルスベクターの静脈内、筋肉内、腫瘍内、皮下、滑液包内、眼内、プラーク内、又は皮内注入のような、当該技術において既知の何らかの方法を通して実施しうる。他の投与経路は、液体等の非経口投与でありうる。好ましい実施態様では、1つ又は複数の免疫複合体蛋白質の静脈内又は腫瘍内注入、又は他の部位での注入によって、あるいは分泌形態の1つ又は複数の型の免疫複合体蛋白質をコードするcDNAを保有する1つ又は複数の発現ベクターの静脈内又は腫瘍内注入、又は他の部位での注入によって患者を治療する。一部の実施態様では、分泌形態の1つ又は複数の型の免疫複合体蛋白質をコードするcDNAを保有する1つ又は複数の複製欠損アデノウイルスベクター、あるいは1つ又は複数のアデノ関連ウイルスベクターの有効量の静脈内又は腫瘍内注入によって患者を治療する。複製欠損アデノウイルスベクターを用いる方法を下記で説明する。多くの典型的な実施態様は、分泌形態の免疫複合体をコードするベクターの有効量の腫瘍内及び/又は筋肉内注入を含む。癌のためにベクターを用いる場合、ベクターの腫瘍内注入は、ベクターが主として注入した腫瘍の細胞に感染するため、静脈内注入に比べて重要な安全上の恩恵を提供する。
【0023】
免疫複合体蛋白質の注入を含む投与は、免疫複合体蛋白質又は蛋白質の組合せが薬学的に許容される担体に分散又は可溶化されている組成物を用いる。一部の場合には、各々のベクターが分泌形態の免疫複合体をコードするcDNAを保有する、発現ベクターpMK33/ヒグロマイシンでトランスフェクションしたショウジョウバエ(Drosohila)S2細胞において、又は発現ベクターpcDNA3.1でトランスフェクションしたチャイニーズハムスター卵巣(CHO)細胞において、又はバキュロウイルス(Baculovirus)発現系において、免疫複合体を合成する。
【0024】
治療処置を生じさせるために必要な免疫複合体の量は、それ自体では固定されておらず、必然的に、薬学的担体と共に投与される組成物中の成分の濃度、下記で詳述する免疫系応答を増強する投与組成物中の補助化合物、そして治療する患者の年齢、体重及び臨床状態に依存する。好ましい組成物は、患者に対して許容されない毒性を生じることなく有効量の免疫複合体を供給送達する。本発明の薬学的組成物又は製剤は、他の担体、アジュバント、安定剤、防腐剤、分散剤、及び対象とする製剤の種類を考慮した当該技術において常套的な他の物質も含みうる。
【0025】
癌に適用するときには、本発明は、ヒト腫瘍細胞又は腫瘍血管系内皮細胞、又はその両方を特異的に標的する標的ドメインと、標的細胞に対する細胞溶解性免疫応答又は細胞傷害作用を活性化するエフェクタードメインを持つ免疫複合体を使用する。上述したように、血液凝固を開始させない突然変異体ヒト第VII因子に複合した、ヒンジを含めたIgG1免疫グロブリンのFc領域をエフェクタードメインとして持つ免疫複合体は、それらが腫瘍血管系と腫瘍細胞の両方で発現される組織因子を標的するので、多くの癌の治療において特に有効である。一部の実施態様では、癌患者の末梢血リンパ球から誘導されるscFv又はVH融合ファージライブラリーをこれまでに記述されている腫瘍細胞に対してパニングすることによって単離される(Cai,XとGaren,A.(1997)Proc.Natl.Acad.Sci.USA 94,9261−9266)、抗ヒト腫瘍細胞scFv又はVH抗体断片を標的ドメインとして持つ免疫複合体のような、腫瘍を選択的に標的する別のクラスの免疫複合体を患者に投与することによって治療効果をさらに高めることができる。上述したように同時に又は連続的に投与される免疫複合体の組合せは、標的細胞の細胞溶解を増強する共力作用が認められる場合に特に有益である。
【0026】
癌の治療においては、様々な癌、特に黒色腫、腎臓癌、前立腺癌、乳癌、卵巣癌、脳腫瘍、神経芽細胞腫、頭部及び頚部の癌、膵癌、膀胱癌、及び肺癌を含めた原発又は転移性固体腫瘍を治療するために抗腫瘍性免疫複合体が使用される。免疫複合体を腫瘍血管系、特に血管内皮細胞、及び/又は腫瘍細胞を標的にするために使用しうる。腫瘍血管系は、次のような、免疫療法のためのいくつかの利点を提供する。(i)組織因子を含めたいくつかの血管標的はすべての腫瘍に同じはずである。(ii)血管系を標的とする免疫複合体は、それらの標的に達するために腫瘍塊に浸潤する必要がない。(iii)各々の血管は、その生存が血管の機能的完全性に依存する数多くの腫瘍細胞に栄養分を与えるので、腫瘍血管系を標的することは治療応答の増幅を生じるはずである。(iv)血管系は免疫複合体に対する耐性を発現しにくいと考えられる。なぜなら、それには血管の内側にある内皮細胞層全体の変化を必要とするからである。新しい血管の増殖を阻害する、これまでに記述された抗血管形成法と異なって、本発明の免疫複合体は新生血管系に対する細胞溶解応答を誘発する。
【0027】
本発明の免疫複合体はまた、慢性関節リウマチ、滲出性(「湿性」)形態の黄斑変性、アテローム性動脈硬化症、及び血管新生に関連する他の疾患を治療するために有効である場合がある。IgG1免疫グロブリンのFcドメインに複合した、突然変異ヒト第VII因子によって組織因子に標的される免疫複合体を投与することは、慢性関節リウマチにおいて滑膜に侵襲し、組織因子を発現する血管内皮細胞に対して、細胞溶解性免疫応答をもたらしうる。同様に、第VII因子免疫複合体はまた、滲出性(「湿性」)形態の黄斑変性を治療するためにも有効でありうる。なぜなら、この病的状態においても広い範囲に及ぶ血管新生が認められるからである。本発明の免疫複合体はまた、プラークにおいて組織因子を発現する細胞に対して細胞溶解性免疫応答を生じさせることにより、アテローム性動脈硬化症の治療のためにも有効でありうる。
【0028】
要約すると、本発明に従った全体的な免疫療法プログラムは、血液凝固を引き起こすことなく、新生血管内皮細胞及び腫瘍細胞で発現される組織因子に高いアビディティと特異性で結合する、突然変異した活性部位を持つ第VII因子を標的ドメインとして含む免疫複合体を構築することを含む。IgG1免疫グロブリンのFc領域をエフェクタードメインとして含む免疫複合体は、NK細胞と補体によって標的細胞の溶解を仲介し、新生血管系の破壊をもたらす。本発明の重要な利点は、内因性第VII因子単量体分子と異なって、2つの第VII因子標的ドメインを含む免疫複合体が、組織因子を発現する細胞に対して外因性第VII因子よりも有意に強い結合を示し、過剰の天然リガンドの存在下で成功裡に競合するという事実である。下記の実施例2は、少なくとも約10倍モル過剰の天然リガンドの存在下で成功裡に競合した免疫複合体を例示しているが、本発明はより低い又はより高い親和性を示す他の実施態様を包含する。組織因子に対する外因性第VII因子の結合は既知の最も特異的且つ強力な反応の1つであり、ピコモル範囲のKdを示す。それにもかかわらず、本発明は、1個の免疫複合体分子中に2つの第VII因子配列を持つというアビディティ作用の結果として、組織因子への第VII因子の通常の結合を有意に改善する。このアビディティ作用は本発明の免疫複合体の標的と結合を増強するので、それらは外因性第VII因子と有効に競合し、結合がより長く持続されるため、誘発される免疫応答が増強される。新生血管系及び/又は、癌の場合には患者の型の腫瘍細胞に結合するscFv又はVH抗体断片である標的ドメインに複合したIgG1免疫グロブリンのFc領域であるエフェクタードメインを持つ別の免疫複合体を、補助療法として患者に投与することによってさらに最大化されうる。
【実施例】
【0029】
ここで示す実施例は本発明をさらに例示し、説明するものであり、いかなる意味においても限定と解釈されるべきではない。本発明の免疫複合体及び成分モノクローナル抗体を生成し、特徴づけるために使用する方法の一部は、Cai,X.とGaren,A.(1995)Proc.Natl.Acad.Sci.USA 92,6537−6541、(1996)Proc.Natl.Acad.Sci.USA 93,6280−6285、及び上記で引用した1997年の参考文献の中で、WO97/02479号として1997年1月23日に公開され、1998年4月27日に提出され、2000年3月3日に承認された特許願通し番号第08/983,607号として現在米国で審理中である、Yale UniversityとGaren及びCaiへのPCT/IB96/01032号の中で、上記で引用したWangらの中で、及び上記で引用したHu,Z.らの中で記述されている。
【0030】
実施例1
ヒトIgG1のFcエフェクタードメインに複合したヒト一本鎖Fv(scFv)標的ドメインを含む免疫複合体をこの実施例において合成し、試験する。最初に、ワクチン接種した黒色腫患者の抗体リストから誘導した、scFv融合ファージライブラリーからの黒色腫特異的クローンとしてscFv標的ドメインを単離した(上記で引用したCaiとGarenの参考文献の中で詳細に述べられている)。精製免疫複合体は融合ファージクローンと同様の結合特異性を示した:ヒト黒色腫に対しては結合が起こったが、ヒトメラノサイトあるいは他のいくつかの型の正常細胞及び腫瘍細胞に対しては結合が生じなかった。
【0031】
試験材料及び方法
細胞培養
永久ヒト黒色腫細胞系統A2058(American Type Culture Collection,Rockville,MD)及びTF2をDMEM+10%FCS中で増殖させた。ヒト微小血管内皮細胞と線維芽細胞の一次培養を新生児包皮から吸い出し、8%FBSと2%ヒト周産期血清を補足したRPMI培地中で培養した;内皮細胞には33mM 3−イソブチル−1−メチルキサンテン(IBMX)と0.5mMジブチリルcAMPをさらに補足した。新生児包皮からのヒトメラノサイトの一次培養はSkin Disease Research Center at Yale University School of Medicineによって調製された。形質転換ヒト腎細胞293−EBNA(Invitrogen)及びチャイニーズハムスター卵巣(CHO)細胞をRPMI+10%FCS中で増殖させた。(v)ショウジョウバエ細胞(Schneider S2)をEx−cell 301培地(JRH Biosciences)+10%FBS中25℃で増殖させた。静止NK細胞をロイコホレシスと免疫的選択によって健常ドナーから単離し、単離後18時間以内に使用した;細胞の大部分(>97%)がCD3−、CD56+及びCD16+であった。
【0032】
免疫複合体の調製
手順は、発現ベクターpcDNA3.1(Invitrogen)のCHO細胞へのトランスフェクション、又は発現ベクターpMK33/pMtHyのショウジョウバエ細胞へのトランスフェクションを伴った;各々のベクターは分泌された免疫複合体をコードするcDNAを保有していた(図1)。scFv標的ドメインについてのcDNAを、対応する融合ファージ(1)から次のようなSacI又はBamHI部位を含むPCRプライマーを使用して合成した:a)GTCGAGCAGAGCTCCAGGTGCAGCTGGTGCAGTCTGGGGCTGAGGTGAGGTGAAGAAGCC(配列番号:1);b)ACGTTCAGGGGATCCACCTAGGACGGTCAGCTTGGTCCC(配列番号:2)。ヒトFcエフェクタードメインは、次のようなBamHI又はSalI部位を含むPCRプライマーを使用して、ヒト末梢血リンパ球由来のcDNAライブラリーから合成した:a)ACCTTGCAGGATCCGCAAGACCCAAATCTTGTGACAAAACTCAC(配列番号:3);b)GATCACGTGTCGACTTATCATTTACCCGGAGACAGGGAGAGGCTCTTCTG(配列番号:4)。IgG1リーダーについてのcDNAを、次のようなEcoRI及びSacI末端を含む2個の相補的オリゴヌクレオチドをハイブリダイズすることによって合成した:a)AATTCATGGAGTTTGGGCTGAGCTGGCTTTTTCTTGTTGCTGCATTAAGAGGTGTCCAGTCCGAGCT(配列番号:5);b)CGGACTGGACACCTGTTAATGCAGCAACAAGAAAAGCCAGCTCAGCCCAAACTCATG(配列番号:6)。
【0033】
分泌免疫複合体をコードするこれら3つのcDNAを、最初に配列決定のためにクローニングベクターにライゲートし、次にCHO又はショウジョウバエS2細胞へのトランスフェクションのために発現ベクターpcDNA3.1及びpMK33/pMtHyにそれぞれライゲートした。CHO細胞についてのトランスフェクション手順は、細胞をRPMI+10%FCS中で増殖させ、Superfect(商標)(Qiagen)を用いて発現ベクター5μgで形質移入することを伴った。安定なトランスフェクタントをRPMI+10%FCS+G418 1mg/ml中で選択した。蛋白質の発現のために、トランスフェクションしたCHO細胞を最初にCHO血清不含培地(Sigma)中での増殖に順応させ、その後無血清培地中で懸濁液培養(2×105細胞/ml)として3日間増殖させた。ショウジョウバエS2細胞についてのトランスフェクション手順は、Ex−cell培地+10%FBS中で細胞を増殖させ、LipofectinTM(Gibco−BRL)を用いて発現ベクター10μgで形質移入することを伴った。安定なトランスフェクタントをEx−cell培地+10%FBS+ヒグロマイシン300μg/ml中で選択し、無血清Ex−cell培地中懸濁液培養としての増殖に順応させた。硫酸銅500μMを加えて、懸濁液培養においてコードされた免疫複合体の発現を誘導した。
【0034】
トランスフェクションしたCHO又はショウジョウバエS2細胞によって分泌された免疫複合体を、プロテインAマトリックス(Pierce)上でのアフィニティークロマトグラフィーによって培地から精製した。
【0035】
蛍光活性化細胞選別(FACS)によって測定した免疫複合体の結合特異性
黒色腫細胞と対照細胞を非酵素的解離培地(Sigma)において採集し、PBS/BSA+0.1%アジ化ナトリウムで洗って、免疫複合体(1μg/ml)を加えたPBS/BSA中で、又は対照として免疫複合体なしのPBS/BSA中でインキュベートした。細胞をPBS/BSAで洗い、蛍光標識抗ヒトFcγ鎖(Vector)と共に4℃で30分間インキュベートして、Becton−Dickenson FACsort装置で分析した。
【0036】
免疫複合体による黒色腫細胞抽出物の免疫沈降反応
ヒト黒色腫細胞A2058系統からの約1×107細胞のサンプルを、PBS中免疫複合体10μg/ml、1%BSA及び0.05%アジ化ナトリウムを含む溶液に懸濁し、氷上で30分間インキュベートした。細胞をPBSで2回洗い、PBS中1%NP−40、免疫複合体1μg/ml及びPMSF 0.2mMを含む溶液に20分氷上で溶解した。溶解産物をマイクロヒュージにおいて13,000RPMで5分間遠心し、上清を回収して、回転装置でひと晩プロテイン−Gビーズと共にインキュベートした。ビーズを採集し、PBS中1%NP−40を含む溶液で2回、PBSで1回洗って、ビーズを採集し、PAGEを負荷した緩衝液中で煮沸して、PAGEによって分析した。
【0037】
マトリックス援用レーザー脱着イオン化質量分析(MALDI−MS)及び液体クロマトグラフィータンデムMS(LC/MS/MS)蛋白質同定
クマシーブルーで染色した蛋白質バンドをゲルから切り出して、http://info.med.yale.edu/wmkeck/geldig3.htmに述べられているようにトリプシンで消化した。トリプシン消化物のサンプルをMicromass TofSpecSEでのMALDI−MSによって分析した。ペプチド質量検索に必要な高レベルの精度を達成するために、1060.57のプロトン化モノ同位体質量を持つブラジキニン100fmol、及び2465.2のプロトン化モノ同位体質量を持つACTHクリップを内部標準として使用した。生じたトリプシンペプチドのモノ同位体質量を、0.2ダルトンのマストレランスを用いてProFoundプログラムによりOWLデータベースに対して、及び0.015%のマストレランスを用いてPeptideSearchプログラムによりEMBL/非重複データベースに対して検索した。検索に使用した他の重要な判定基準は、140−560Kdaにわたる質量範囲、最大限1つの開裂ミス、そしてタキソノミー(分類学)に関しては制限なしとした。蛋白質化学及び質量分析試験はすべてYale UniversityのW.M.Keck Foundation & HHMI Biopolymer Laboratoryにおいて実施した。http://info.med.yale.edu/wmkekc/でさらなる情報が入手できる。
【0038】
MALDI−MS分析に使用したトリプシン消化蛋白質バンドのサンプルを、LCQイオントラップ質量分析計でも分析した。マストレランス2.0ダルトンでのタンデム質量相関アルゴリズムを用いて、MS/MSデータのSequest検索を実施し、トリプシン消化物からのペプチドと、NCBI nrデータベースでの蛋白質についての再構築した理論上のスペクトルとの間に有意の類似性が存在するかどうかを判定した。この手順についてのさらなる情報は、http://info.med.yale.edu/wmkekc/prochem.htm#ms/mspiで入手できる。
【0039】
NK細胞及び補体によるヒト黒色腫細胞の免疫複合体依存性細胞溶解に関するアッセイ
カルセイン−AM保持手順を両方のアッセイに使用した。ヒト黒色腫細胞系統又は一次ひと線維芽細胞のいずれかの標的細胞を非酵素的解離培地(Sigma)中で培養フラスコから分離し、96穴プレートに加えた(2×104細胞/穴)。付着標的細胞をPBSで1回洗い、その後免疫複合体(1μg/ml)を含む又は対照として免疫複合体を含まない無血清培地(GIBCO−BRL)中37℃で20分間インキュベートした。次に標的細胞を無血清培地中37℃で40分間、カルセイン−AM(Molecular Probes Inc.,Eugene,OR)7μMで標識した。カルセイン−AMは細胞に入り込む蛍光色素であり、そこで酵素的に変化して、細胞が溶解するまで細胞内にとどまる。NK細胞に関する細胞溶解アッセイについては、標識した標的細胞を、NK細胞対標的細胞の指示されている比率を用いてヒトNK細胞と共に37℃で3〜4時間インキュベートした。補体に関する細胞溶解アッセイについては、標識標的細胞をヒト血清又は精製ウサギ補体成分(Cedarlane laboratories,Ontario,Canada)と共に37℃で1時間インキュベートした。NK細胞又は補体とのインキュベーション後、標的細胞をPBSで2回洗い、残存する付着細胞中の蛍光(残留蛍光)をプレートリーダーで測定した。溶解緩衝液(ホウ酸ナトリウム50mM、0.1%Triton X−100、pH9.0)により37℃で3時間処理した後、標的細胞の残留蛍光を測定して、標的細胞の達成可能な最大細胞溶解を判定した。NK細胞又は補体に接触しなかった付着標的細胞の蛍光を測定して、最大残留蛍光を判定した。標的細胞の各サンプルについての%細胞溶解を次のように算定した:(サンプル標的細胞の残留蛍光−溶解標的細胞の残留蛍光)/(標的細胞の最大残留蛍光−溶解標的細胞の残留蛍光)。
【0040】
試験結果
抗黒色腫免疫複合体の合成と特性指摘
免疫複合体は、分泌についてのヒトIgG1リーダー、標的黒色腫細胞についてのヒトscFvドメイン、及びヒトIgG1−Fcエフェクタードメインをコード化した(図1)。免疫複合体分子をトランスフェクションしたCHO及びショウジョウバエS2細胞において発現させ、Fcドメインのヒンジ領域におけるジスルフィド架橋によって連結された二本鎖分子として培地から精製した(図1)。各々が融合ファージクローンE26−1又はG71−1から誘導されたscFv標的ドメインを含む、2つの免疫複合体を合成した。免疫複合体E26−1及びG71−1の結合特異性を、2つのヒト黒色腫細胞系統を用いた細胞選別(FACS)によって試験した;対照は、正常ヒトメラノサイト、線維芽細胞、微小血管及び臍血管内皮細胞、及びヒト腎細胞系統293−EBNAの一次培養であった。結果は、E26−1及びG71−1融合ファージクローンに関して得た結果と一致して、免疫複合体は黒色腫細胞系統に強く結合するが、対照には結合しないことを示した。黒色腫細胞への結合は、非酵素的解離培地(Sigma)を使用して培養フラスコから細胞を採集したときに起こったが、解離培地がトリプシンを含むとき、又は解離した細胞をその後トリプシンで処理したときには結合が起こらなかった。この所見は、免疫複合体についての同種黒色腫抗原が黒色腫細胞の表面に位置することを示唆した。同じトリプシンへの曝露が、細胞表面分子ICAM−1、MHCクラス−I及び組織因子に対する抗体の黒色腫細胞への結合には影響を及ぼさなかったことから、上記抗原はトリプシンに対して例外的に感受性であると思われる。
【0041】
免疫複合体についての同種黒色腫抗原の同定
ヒト黒色腫細胞系統A2058からの培養細胞を免疫複合体G71−1又はE26−1と平衡させ、細胞を界面活性剤で溶解した。溶解産物中の免疫複合体−抗原複合体をプロテイン−Gビーズ上に採集し、PAGEによって分析した。250kdaの見かけ分子量を持つ蛋白質バンドが黒色腫細胞において検出されたが、対照では検出されなかった。MALDI−MS法による蛋白質バンドのトリプシン消化物を分析すると、対照ゲル切片の消化物には存在しなかった75のペプチド塊が同定された。ProFoundによる蛋白質配列データベースの検索は、MCSPコア蛋白質中のペプチド塊と適合し、完全な蛋白質配列の26%に及ぶ50のペプチド塊を生じた。この同定に関するProFoundの確率スコアは1.0であり、次に最も近いスコアは2.2E−61であった。Peptide Searchによる検索では、45のペプチドがMCSPコア蛋白質に適合し、38ペプチドがそれに続く最も近い適合であった。
【0042】
250kda蛋白質のトリプシン消化物をLC/MS/MS法によっても分析した(Stone,K.L.ら(1998)Electrophoresis 19,1046−1052)。トリプシン消化物からのMS/MSデータのSequest検索は、2個又はそれ以上のペプチドについてのMS/MSスペクトルと、NCBI nrデータベースにおけるMCSPコア蛋白質からの2個又はそれ以上のペプチドについての再構築した理論上のMS/MSスペクトルの間で有意の類似性を示した。
【0043】
2つの質量分析法の結果は、免疫複合体G71−1及びE26−1によって免疫沈降した黒色腫蛋白質がMCSPコア蛋白質に適合することを示している。
【0044】
NK細胞及び補体によって仲介される黒色腫細胞の免疫複合体依存性細胞溶解
免疫系の細胞溶解経路の1つは、標的細胞に直接結合して、標的細胞の抗体依存性溶解を引き起こすことができるNK細胞を含む。NK細胞はまた、抗体のFcエフェクタードメインに結合して、抗体の標的ドメインに結合する細胞の抗体依存性溶解をもたらすことができる。この抗体依存性細胞を介する細胞溶解経路(ADCC)は同時に、Fcエフェクタードメインを含む免疫複合体の標的ドメインに結合する細胞の溶解も引き起こすはずである。免疫複合体E26−1及びG71−1に依存するADCC応答を試験するため、黒色腫細胞と線維芽細胞を蛍光色素カルセイン−AMで標識し、標識細胞をヒトNK細胞単独又はNK細胞と免疫複合体と共にインキュベートした。無傷のままで細胞中に保持された蛍光色素の量を測定することによって細胞溶解を検定した。E26−1についての結果(図2)は、免疫複合体及びNK細胞とのインキュベーション後、溶解黒色腫細胞のパーセンテージが免疫複合体なしで得られる基線値を上回って上昇し、NK細胞対黒色腫細胞の比率が20/1でほぼ100%に達した。黒色腫細胞の効率的な溶解に対し、線維芽細胞は、免疫複合体及びNK細胞とのインキュベーション後細胞溶解の有意の上昇を示さなかった。G71−1免疫複合体に関しても同様の結果が得られた。
【0045】
NK細胞による溶解に対しての標的細胞の感受性は、ICAMのような付着分子の標的細胞表面での発現によって上昇し、MHCクラスI分子の発現によって低下する(Zami,L.ら(1995)Cell.Immunol.164,100−104及びStorkus,W.J.ら(1989)Proc.Natl.Acad.Sci.USA 86,2361−2364)。これらの分子の発現の差がADCCアッセイにおける黒色腫細胞及び線維芽細胞の感受性の差に寄与するかどうかを調べるため、黒色腫細胞及び線維芽細胞によるICAM−1及びMHCクラスI分子の発現をFACSによって測定した。両方の分子の発現は2つの細胞型において同様であり、NK細胞による黒色腫細胞の特異的溶解は、黒色腫細胞で発現される同種抗原への免疫複合体の結合に依存することを示唆した。
【0046】
免疫系の別の細胞溶解経路は、C1q分子が標的細胞に結合した抗体のFc領域と反応したときに活性化される補体カスケードを含む(Bruggemann M.ら(1987)J.Exp.Med.166,1351−1361)。免疫複合体E26−1及びG71−1に依存する、黒色腫細胞に対する補体仲介の細胞溶解応答を調べるため、上記で報告したNK仲介の細胞溶解応答に関して使用したのと同じアッセイ手順を、1箇所変更を加えて使用した、すなわちNK細胞の代わりに補体カスケードの成分を含むヒト血清又はウサギ血清を用いた。結果(図3及び表1)は、免疫複合体とヒト血清又はウサギ血清とのインキュベーション後、溶解した黒色腫細胞の分画が4%からほぼ100%に上昇した。黒色腫細胞の効率的な溶解に対して、線維芽細胞は免疫複合体及びヒト血清とのインキュベーション後溶解細胞分画の小さな上昇を示し、免疫複合体及びウサギ血清とのインキュベーション後には有意の上昇を示さなかった。
【0047】
【表1】

【0048】
考察
本試験については、2個のクローンからのscFv分子を、ヒトIgG1−Fcエフェクタードメインを含む免疫複合体を構築するための標的ドメインとして使用した(図1)。2つの免疫複合体によってヒト黒色腫細胞から免疫沈降した蛋白質を、黒色腫関連硫酸コンドロイチンプロテオグリカン(MCSP)のコア蛋白質として質量分析法によって同定した(Bumol,T.F.& Reisfeld,R.A.(1982)Proc.Natl.Acad.Sci.USA 79,1245−1249及びBumol,T.ら(1984)J.Biol.Chem.259,12733−12741)。MCSP分子は最初mAb 9.2.27によって認識される同種抗原として同定されたが、これはいくつかの他のmAbについての同種抗原であると思われる(Reisfeld,R.A.& Cheresh,D.A.(1987)Adv.Immunol.40,323−377;Wilson,B.S.ら(1981)Int.J.Cancer 28,293−300;Hellstrom,I.ら(1983)J.Immunol.130,1467−1472)。おそらくMCSPと同じである、黒色腫抗原HMW−MAAに結合するいくつかのscFv融合ファージクローンが、精製HMW−MAAに対するパニングによって合成ヒトscFvライブラリーから同定されている(Desai,S.A.ら(1998)Cancer Res.58,2417−2425)。MCSP分子は主として大部分のヒト黒色腫細胞の表面で発現されるが、グリア腫瘍の毛細管内皮細胞でも発現される。MCSPが、黒色腫細胞に対するパニングによって黒色腫患者のscFv融合ファージライブラリーから単離された少なくとも2つの黒色腫特異的クローンについての同種抗原であるという所見は、MCSPがin vivoでの支配的な黒色腫抗原であることを示唆する。
【0049】
2つの免疫複合体の治療上の潜在的効力の初期試験として、蛍光標識標的細胞を含むin vitroでの細胞溶解アッセイを使用して、免疫複合体がNK細胞及び補体による溶解のためにヒト黒色腫細胞を標的する能力を調べた。両方の免疫複合体が黒色腫に対するNK細胞及び補体の細胞溶解活性の明らかな上昇をもたらし、標的黒色腫細胞群の実質的に完全な溶解を生じさせたが、対照として使用した線維芽細胞に関しては、溶解のごくわずかな上昇をもたらすか又は全く上昇を生じさせなかった。NK細胞及び補体による黒色腫細胞と線維芽細胞の免疫複合体非依存性細胞溶解の有意のバックグラウンドも存在した。これは、腫瘍細胞、NK細胞及び補体を別々の個体から単離する同種異系アッセイについて予想されるものである(Ciccone,E.ら(1992)J.Exp.Med.175,709−718)。これらの成分はすべて自己由来であるので、癌患者においてはこのバックグラウンドは低下するはずである。
【0050】
in vivo細胞溶解試験の結果は、免疫複合体が、腫瘍特異的scFv又はVH標的ドメインが使用可能である黒色腫及び他の癌に関する免疫療法プロトコールにおいて潜在的な役割を持つであろうことの予備的な証拠を提供する。MCSPに結合するマウスmAb 9.2.27による黒色腫免疫療法についての限られた第I相臨床試験は、付随する毒性の証拠を伴わずに、腫瘍における抗体の特異的局在を示した(Oldham,R.K.ら(1984)J.Clin.Oncol.2,1235−1244)。より小さな分子サイズは腫瘍への侵入を改善するはずであり、また分子がヒト由来であることは免疫拒絶反応を最小限に抑えるはずであるので、MCSPに結合するscFv免疫複合体は免疫療法のためにマウスmAbよりも有効であると考えられる。G71−1及びE26−1免疫複合体は、それらのVH配列の重要な相違(配列番号:11と配列番号:12及び次の実施例参照)によって示唆されるように、おそらく異なるMCSPエピトープに結合し、それ故治療効果を高めるために一緒に投与することができるであろう。
【0051】
実施例2
この実施例は、腫瘍血管系と腫瘍細胞の両方を標的する免疫療法治療が、ヒト黒色腫の重篤な複合型免疫欠損マウス異種移植モデルにおいて有望な成績を示したことを報告する。手順は、各々がヒトIgG1のFcエフェクタードメイン領域に複合した腫瘍標的ドメインから成る2つの免疫複合体の、SCIDマウスへの全身送達を伴った。エフェクタードメインは、ナチュラルキラー(NK)細胞及び補体による標的細胞に対しての細胞溶解性免疫応答を誘導する。免疫複合体は、複製不全アデノウイルスベクターにおいて分泌分子としてコードされる。
【0052】
試験材料及び方法
細胞系統
黒色腫細胞系統LXSN、TF2及びLXSN/VEGFを、レトロウイルス仲介のトランスフェクションとクローニングによってヒト黒色腫細胞系統YU−SIT1から誘導した。LXSN系統は対照レトロウイルスでトランスフェクションされており、低レベルのTFを発現する。TF2系統はTF cDNAをコードするレトロウイルスでトランスフェクションされており、高レベルのTFを発現する。LXSN/VEGF系統はVEGF cDNAをコードするレトロウイルスでトランスフェクションされており、高レベルのVEGFを発現する。ヒト腎細胞系統293をAmerican Type Culture Collectionから購入した。
【0053】
プラスミドベクター
scFv(G71−1)免疫複合体をコードするプラスミドベクターの構築を実施例1で述べた(配列番号:11参照)。マウス第VII因子(mfVII)免疫複合体をコードするベクターの構築のために、5'プライマーACGATCTTAAGCTTCCCCACAGTCTCATCATGGTTCCA(配列番号:7)及び3'プライマーACGGTAACGGATCCCAGTAGTGGGAGTCGGAAAACCCC(配列番号:8)を使用して、マウス肝臓cDNAライブラリー(Quick−Clone cDNA,Clonetech)からのPCRによってmfVII cDNAを増幅した。停止コドンを含まないリーダー及びコード配列を含む、増幅したmfVII cDNAを、ヒトIgG1 FcドメインをコードするcDNAと共にpcDNA3.1(+)ベクター(Invitrogen)のフレーム内のHindIII及びBamHI部位にクローニングした。ベクターDNAをHB101コンピテント細胞(Life Technologies)において増幅し、配列決定した。リシン−341をアラニンコドンで置換することによってmfVII cDNAの活性部位を突然変異させた(Dickinson,C.D.ら(1996)Proc.Natl.Acad.Sci.USA 93,14379−14384)。突然変異誘発の手順は、QuickChange部位特定突然変異誘発マニュアル(Stratagene)の中で述べられているように実施した。5'プライマーはGGTACCAAGGACGCCTGCGCGGGTGACAGCGGTGGCCCA(配列番号:9)であり、3'プライマーはTGGGCCACCGCTGTCACCCGCGCAGGCGTCCTTGGTACC(配列番号:10)であった。活性部位突然変異を有するmfVII cDNAをmfVIIasmと称する。mfVIIasm cDNAを含むプラスミドをHB101コンピテント細胞に形質転換し、2×TY/カルベニシリン寒天上で形質転換コロニーを選択した。プラスミドDNAの配列は、mfVIIasm DNAにおけるlys−341コドン(AAG)のアラニンコドン(GCG)による置換を示した。
【0054】
CHO細胞における免疫複合体の合成
免疫複合体cDNAをCHO細胞に形質移入し、クローンを単離するための手順を実施例1で述べた。形質移入したCHO細胞をCHO血清不含培地(EX−CELL 301,JRH Biosciences)で培養した;mfVIIasm免疫複合体の合成のため、CHO血清不含培地に最終濃度1μg/mlまでビタミンK1(Sigma)を補足した。免疫複合体をプロテインAビーズ(Pierce)でのアフィニティークロマトグラフィーによって精製し、濃縮して、Ultrafree−15 Biomax−50フィルター(Millipore)を通しての遠心分離によって脱塩し、Tris−HCl pH8.0 10mMに調整した。Bio−Rad蛋白質アッセイ手順によって免疫複合体の濃度を測定した。
【0055】
蛍光活性化細胞選別(FACS)
黒色腫細胞を非酵素的解離溶液(Sigma)中で採集し、洗って、TBS/BSA/Ca2+(Tris−HCl pH7.4 10mM、NaCl 150mM、CaCl2 20mM、1%BSA及び0.1%NaN3)に再懸濁した。免疫複合体を加えて(最終濃度5μg/ml)、mfVIIasm免疫複合体については37℃で30分間、G71−1免疫複合体については氷上で30分間、細胞をインキュベートした;対照細胞は免疫複合体を加えずにインキュベートした。インキュベーション後、細胞をTBS/BSAで洗い、蛍光標識抗ヒトFcγ鎖(Vector Laboratories)と共に氷上で30分間インキュベートし、Becton−Dickenson FACsort装置で分析した。
【0056】
アデノウイルスベクター
アデノウイルスベクター系は、シャトルベクターpAdTrack−CMV及びpShuttle−CMVと、バックボーンベクターpAdEasy−1から成る(He,T.C.ら(1998)Proc.Natl.Acad.Sci.USA 95,2509−2514)。免疫複合体cDNAを、HindIIIで消化してpcDNA3.1プラスミドベクターから単離し、次いでクレノウ断片で消化して3'陥凹末端を埋め、その後Not1で消化してcDNA挿入物を放出させ、それをアガロースゲル電気泳動によって精製した。シャトルベクターは、最初にKpn1で消化し、次にクレノウ断片で消化して、その後Not1で消化した。免疫複合体cDNAをT4 DNAリガーゼと共に16℃でひと晩インキュベートしてシャトルベクターにライゲートし、かかるシャトルベクターを熱ショックによってHB101コンピテント細胞に形質転換した。形質転換コロニーを2×TY/カナマイシン寒天上で選択し、シャトルベクターを抽出して精製した。精製シャトルベクターとpAdTrack−CMV DNAをPmelにより37℃で2時間消化した。シャトルベクターDNA 500ngとpAdEasy−1 DNA 100ngの混合物をBJ5183コンピテント細胞に電気穿孔し、細胞を37℃で15分間振とうして、LB/カナマイシン寒天に接種した。プレートを37℃でひと晩インキュベートし、形質転換コロニーを単離した。プラスミドDNAをミニプレップから精製し、0.6%アガロースゲルでの電気泳動によって組換えアデノウイルスDNAに関してスクリーニングした。
【0057】
免疫複合体をコードする組換えアデノウイルスDNAを、CHO細胞のトランスフェクションに関して上述したプロトコールに従って、1×105の293細胞にトランスフェクションした。トランスフェクションの7日後に細胞を採集し、3回の凍結−融解サイクルによってアデノウイルスを放出させ、1つの150mm培養プレートに293細胞を感染させて増幅した。2日後、上述したようにアデノウイルスを採集し、20の培養プレートに293細胞を感染させて再び増幅した。増幅したアデノウイルスを2日後に採集し、CsCl中での遠心分離によって精製した。最終的な収量は通常、260nmでの吸光度から評価したとき約1013ウイルス粒子であった;換算は、1O.D.単位=1×1012粒子である。精製アデノウイルスをPBSで透析し、−80℃で保存した。
【0058】
SCIDマウス実験
SCIDマウスはTaconic Laboratoriesからの4週齢から5週齢の雌性であった。マウスの右ひ腹に5×105のTF2又はLXSNヒト黒色腫細胞を皮下注射した。腫瘍が、皮膚表面下で触知可能な大きさ(〜5mm3)又は皮膚表面より上のより大きなサイズ(〜50mm3)に成長したあと、尾静脈を通して免疫複合体をコードするアデノウイルスベクター、又は対照として免疫複合体をコードしないアデノウイルスベクターをマウスに注入した。血液中に分泌される免疫複合体蛋白質の濃度を、一方の眼からの血液約0.1mlをヘパリン被覆した微小毛管に採集し、血液を遠心分離にかけて細胞を取り出すことによって測定した。上清血漿を炭酸水素ナトリウム緩衝液pH9.6で希釈し、プロ結合アッセイプレート(Falcon)の穴に分配して、プレートを37℃で2時間、次いで4℃でひと晩インキュベートした。穴をPBS中5%脱脂乳で30分間遮断し、PBSで3回洗って、5%脱脂乳中1:2000に希釈したペルオキシダーゼ標識抗ヒトIgG抗体を穴に加えた。プレートを室温で1時間インキュベートし、PBS中で洗って、ペルオキシダーゼ基質OPDを加え、マイクロプレートリーダーにおいて490nmで吸光度を測定した。蛋白質標準は、プロテインAビーズでのクロマトグラフィーによって精製したヒトIgG(Sigma)であった。
【0059】
SCIDマウスの皮膚に発現する腫瘍の大きさをカリパスにより二次元で測定し、式、(幅)2(長さ)/2によって腫瘍容積を推定した。実験の最後に、マウスを解剖して腫瘍を計量した。器官を損傷の形態学的証拠に関して検討し、組織学的検査用にパラフィン切片を調製した。
【0060】
免疫組織化学
腫瘍と器官のパラフィン切片をPBS+0.3%H2O2中で30分間インキュベートし、TBS/BSA緩衝液中で30分間遮断した。TBS/BSA/Ca2+緩衝液中にmfVIIasm免疫複合体10μg/mlを含む溶液、又は対照として免疫複合体を含まない緩衝液を切片に加え、37℃で1時間インキュベートした。同じ緩衝液で3回洗った後、切片をアルカリホスファターゼで標識した抗ヒトγ鎖抗体と共に室温で1時間インキュベートして、青色を生じるBCIP/NBTで染色し、メチルグリーンで対比染色した。
【0061】
試験結果
免疫複合体の特性
scFv(G71−1)及び突然変異マウス第VII因子(mfVIIasm)免疫複合体をCHO細胞において合成し、プロテインAビーズでのアフィニティークロマトグラフィーによって培地から精製した。SDS−PAGEによる先の分析は、G71−1免疫複合体が、おそらくFcドメインのヒンジ領域間のジスルフィド架橋によって結合した、2つの同じ鎖から成ることを示した(実施例1)。mfVIIasm免疫複合体に関して同じ結果を得た。(配列番号:11及び配列番号:12参照)。mfVIIasm免疫複合体は2つの標的ドメインを持つので、単量体内因性fVII分子における1つの標的ドメインと比較して、2個のTF分子に協力的に結合し、TFを発現する細胞に対して内因性fVIIよりも強い結合をもたらすことができる。競合的FACSアッセイは、ヒトfVIIaが、ヒト黒色腫細胞上のアクセス可能部位の半数への結合に関して等モルベースでmfVIIasm免疫複合体と競合することを示した。これはおそらく、免疫複合体上の標的ドメインの1つだけがこれらの部位でTFに結合することができるからであろう。残りの部位へのmfVIIasm免疫複合体の結合は、10倍過剰のヒトfVIIaの存在下で競合することができず、免疫複合体分子の両方の標的ドメインがこれらの部位で結合することができ、強いアビディティ効果をもたらすことを示唆した。黒色腫細胞上のTF分子の約半分だけが第二TF分子に十分に近接していて、mfVIIasm免疫複合体上の両方の標的ドメインについて協力的結合部位を形成すると思われる。
【0062】
免疫療法試験のための異種移植片を、それぞれ低レベル又は高レベルのTFを発現するヒト黒色腫細胞系統LXSN及びTF2から作製した。TF2細胞によるより高いレベルのTF発現と一致して、FACSによって測定したとき、mfVIIasm免疫複合体はLXSN細胞よりもTF2細胞とより広汎に結合する。またmfVIIasm免疫複合体を、高レベルのVEGFを生成する黒色腫細胞系統LXSN/VEGFから作製したヒト黒色腫異種移植片の切片への結合に関して免疫組織化学によっても試験し、密に血管形成した異種移植片を生じた。腫瘍血管内皮細胞並びに腫瘍細胞に対して結合が起こり(図4)、異種移植片において両方の細胞型によってTFが発現されることを示唆した。正常マウスの肝臓、腎臓、肺及び脳の切片に関する免疫組織化学試験は、TFが非腫瘍性組織の血管内皮細胞によっては発現されないという他の証拠と一致して、mfVIIasm免疫複合体がこれらの組織における血管内皮細胞とは結合しないことを示した。
【0063】
免疫療法試験
SCIDマウスへの全身送達のために、各々の免疫複合体をpAdEasy−1に基づく複製不全アデノウイルスベクター系(Heら、前出)において分泌分子としてコードし、最初にヒト黒色腫細胞を皮下注入しておいたマウスの尾静脈に前記ベクターを注入した。最初の免疫療法試験は、触知可能なTF2異種移植片を発現したマウスに、各々のベクターを別個に、及び両方のベクターを一緒に注入することを伴った。合計3回の注入を週に1回の間隔で投与し、最後の注入から6日後に実験を終了した。1回目と2回目の注入後にELISAによって血液中の免疫複合体の濃度をモニターした(図5)。1回目の注入後の平均濃度は、G71−1免疫複合体については4mg/ml、mfVIIasm免疫複合体については0.04mg/mlであり、合成の速度がG71−1免疫複合体についてはmfVIIasm免疫複合体よりも約100倍高いことを示唆した。2回目の注射後各々の免疫複合体の濃度が上昇し、さらなる細胞がアデノウイルスに感染していたことを示した。皮膚表面に現われた腫瘍の大きさを二次元で測定し、腫瘍容積を評価するための測定法を用いて、異種移植片の成長をモニターした(図6)。免疫複合体をコードしないアデノウイルスを注入した対照マウスでは、腫瘍が比較的速い速度で持続的に成長し、20日後に約2,000mm3の平均容積に達した。免疫複合体をコードするアデノウイルスを注入したマウスでは、腫瘍の成長が阻害された;阻害はG71−1免疫複合体よりもmfVIIasm免疫複合体の方が強力であった。実験終了時にすべてのマウスが活動的なままであり、健康と思われ、肝臓、脾臓、肺、腎臓及び脳の組織学的検査は壊死、凝固又は出血の兆候を示さなかった。剖検後の腫瘍重量は、推定腫瘍容積と一致して、対照マウスでは免疫複合体で処置したマウスよりも低かった(図7)。腫瘍重量の最強の低下は、両方の免疫複合体で処置したマウスで起こった。
【0064】
次の2つの実験は、免疫複合体の治療効果に影響を及ぼしうる2つのパラメータ、すなわち異種移植片の初期サイズと黒色腫細胞によるTF発現のレベルを試験するためにデザインされた。(i)前記の免疫療法試験は、ヒトでの小さな腫瘍に相当する、約5mm3の推定容積に成長した触知可能な黒色腫異種移植片に関するものであった。より大きな異種移植片に対する免疫複合体の治療効果を調べるため、TF2異種移植片を推定容積約50mm3に成長させた後、2つのアデノウイルスベクターの尾静脈注入を開始した。3週間の期間中マウスに4回注入を行い、最後の注入から2日後に実験を終了した。免疫複合体をコードするアデノウイルスを注入したマウスにおける平均腫瘍容積は、実験開始時と実験終了時でほぼ同じであったのに対し、対照アデノウイルスを注入したマウスでの平均腫瘍容積は同じ期間中に約27倍上昇した(図8)。これらの結果は、腫瘍の成長が、大きな腫瘍に関しても小さな腫瘍と同じくらい有効に阻害されることを示している。免疫複合体をコードするアデノウイルスを注入した5匹のマウスのうち1匹が、3回目の注入から5日後に死亡した;マウスが検査の時点までに回収されなかったため、死亡の原因は判定できなかった。(ii)mfVIIasm免疫複合体の効果に影響を及ぼしうる1つのパラメータはTF発現のレベルであり、これは種々の腫瘍間で異なる(Callender,N.S.ら(1992)Cancer 70,1194−1201)。異種移植片での黒色腫細胞によるTFの発現が変化することの影響を調べるため、より高レベルのTFを発現する関連系統TF2から作製した異種移植片との比較のために、黒色腫細胞系統LXSNを使用して低レベルのTFを発現する異種移植片を作製した(Bromberg,M.E.ら(1999)Proc.Natl.Acad.Sci.USA 92,8205−8209)。異種移植片が触知可能な大きさに達した後、次の3週間にわたってマウスにmfVIIasm免疫複合体をコードするアデノウイルス又は対照アデノウイルスの5回の注射を実施した(図11)。対照アデノウイルスを注入した5匹のマウスでは、異種移植片は持続的に成長し、平均容積は最後の注入から2日後に1350mm3まで上昇した。同じ期間中、mfVIIasm免疫複合体を注入した5匹のマウスにおける異種移植片の平均容積は20mm3に上昇し、腫瘍の成長がLXSNとTF2異種移植片について同等であることを示した(図9と6の比較)。最後の注入の1日後に実施した剖検は、mfVIIasm免疫複合体をコードするアデノウイルスを注入した5匹のマウスのうち2匹において異種移植片が根絶されていたことを示した;他の3匹のマウスにおける平均腫瘍重量が0.11グラムであったのに対し、対照アデノウイルスを注入した5匹のマウスでは0.75グラムの平均重量であった。これら3匹のマウスから回収された小さな腫瘍は広範囲の領域の細胞壊死を示したが、対照マウスから回収したより大きな腫瘍においては壊死は生じなかった(図10)。この実験の終了時にはすべてのマウスが健康であると思われたが、解剖したマウスの形態学的検査は、mfVIIasm免疫複合体をコードするアデノウイルスを注入した5匹のマウスにおいて肝臓と脾臓への損傷を明らかにした。肝臓と脾臓の組織学的検査は、肝細胞の多くが腫脹し、脾が赤血球で広汎に浸潤されていることを示した。腫脹した肝細胞は、mfVIIasm免疫複合体をコードするアデノウイルスの3回の注入後の先の実験においても発生していたが、脾は正常であり、脾臓の欠陥が最後の2回の注入経過で発現したことを示唆した。1匹のマウスは硬膜下脳出血も有していたが、これは今回の実験からの他のマウスあるいはこれまでのいずれの実験においても起こっていなかった。この欠陥が脳血管系で発現されたTFへのmfVIIasm免疫複合体の結合によって誘発されたのかどうかは不明である。
【0065】
考察
この実施例における免疫療法処置は、各々がヒトIgG1 H鎖のFc領域に複合した腫瘍標的ドメインから成り、カメリド(Camelid)H鎖抗体に類似したホモダイマー分子を形成する、2つの免疫複合体をSCIDマウスに全身送達することを伴った。1つの型の免疫複合体については、腫瘍標的ドメインは、異種移植片において黒色腫細胞によって発現される黒色腫抗原MCSPに結合するヒトscFv分子G71−1であった。他の型の免疫複合体では、腫瘍標的ドメインは、腫瘍血管系内皮細胞によって発現されるマウスTFと異種移植片において黒色腫細胞によって発現されるヒトTFの両方の組織因子(TF)に特異的且つ強力に結合するマウス第VII因子分子であった。第VII因子免疫複合体のTFへの結合から起こりうる播種性血管内凝固(DIC)の危険性を低下させるために、活性部位の突然変異をマウスfVII標的ドメイン(mfVIIasm)に導入し、血液凝固経路を開始させるために必要な蛋白質分解活性を阻害した。
【0066】
実施例1で報告した試験は、G71−1免疫複合体が、NK細胞と補体による培養ヒト黒色腫細胞の細胞溶解を仲介することを示した。SCIDマウスは機能的NK細胞と補体を産生する能力を保持するので、免疫複合体も、SCIDマウスにおいて成長するヒト黒色腫異種移植片の標的腫瘍細胞及び血管内皮細胞の細胞溶解を仲介することができると考えられる。免疫複合体のSCIDマウスへの全身送達を、免疫複合体をコードする複製欠損アデノウイルスベクターの尾静脈注入によって実施した。免疫複合体は各注入後少なくとも1週間血液中に分泌された。最初に低レベル又は高レベルのTFを発現するヒト黒色腫細胞系統をマウスに皮下注射し、生じた異種移植片を小(〜5mm3)又は大(〜50mm3)腫瘍に成長させた後、アデノウイルスベクターの注入を開始した。G71−1免疫複合体をコードするアデノウイルスと別々に又は一緒に投与した、mfVIIasm免疫複合体をコードするアデノウイルスの多回注入により、3〜4週間の実験期間中、すべての異種移植片のさらなる成長が妨げられた;一部のマウスでは、異種移植片は完全に退縮した。免疫複合体をコードしないアデノウイルスを注入した対照マウスでは、異種移植片の平均容積は同じ期間中に約25倍に上昇した。免疫複合体をコードするアデノウイルスベクターの5回の注入を受けたマウスでは、肝細胞の多くが腫脹し、脾が赤血球で浸潤されていた。この欠陥は、肝細胞又は脾細胞には結合しない分泌免疫複合体によって引き起こされたものではなかった。主要な原因はおそらく、主として静脈内注入されたアデノウイルスベクターに感染したマウス細胞である肝細胞による、コードされた免疫複合体の持続的な高いレベルの合成である。アデノウイルスベクターを注入したSCIDマウスの血液中の免疫複合体濃度は、mfVIIasm免疫複合体よりもG71−1免疫複合体の方が約100倍高かったが、それにもかかわらずヒト黒色腫異種移植片への阻害作用はmfVIIasm免疫複合体の方がより強力であった。mfVIIasm免疫複合体の鍵となる利点は、G71−1免疫複合体が黒色腫細胞だけに結合するのに対して、腫瘍細胞と同様に腫瘍血管内皮細胞に対しても結合が起こることである。TFが正常血管系によっては発現されないので、腫瘍血管系への結合は腫瘍特異的であるはずである。TFは、脳、肺及び腎糸球体のような他のいくつかの正常組織によって発現されるが、血管壁はより大きな血液成分を隣接細胞から分離するバリアを形成するので、これらのTF分子は内因性fVII又はfVII免疫複合体にアクセスできない。しかし、腫瘍血管は漏出性であり、腫瘍細胞によって発現されたTFへのアクセスを可能にする。それ故、ヒトfVIIasm免疫複合体は、血管内皮細胞及び腫瘍細胞上でTFを発現する広いスペクトルのヒト腫瘍にとって有効な治療薬でありうる。TF以外の腫瘍細胞標的に結合するヒトscFv免疫複合体を同時に投与することにより、ヒトfVIIasm免疫複合体の治療効果を増強することができる。
【0067】
実施例3
この実施例は、ヒト皮膚及び転移性肺黒色腫のSCIDマウスモデル、及びマウス黒色腫の免疫コンピテントマウスモデルにおいて検討した癌治療のためのプロトコールの効果と安全性の試験を報告する。プロトコールは、標的新生血管系内皮細胞及び腫瘍細胞に対する細胞溶解性免疫応答を誘導する、本発明の免疫複合体をコードする複製不全アデノウイルスベクターの腫瘍内注入を伴った。マウスモデルの実験は、単独で又は一本鎖Fv免疫複合体と共に第VII因子免疫複合体を腫瘍内送達すると、正常器官への損傷の兆候を伴わずに、注入した腫瘍、そして同時に離れた非注入転移性腫瘍の成長の阻害と退縮も生じさせることを示した。おそらく新生血管系内皮細胞上の組織因子に結合した第VII因子免疫複合体によって仲介される、腫瘍新生血管系の広い範囲に及ぶ破壊が存在した。組織因子は一般に新生血管内皮細胞と腫瘍細胞で発現されるので、第VII因子免疫複合体は広い範囲のヒト腫瘍に対する免疫療法のために使用することができる。
【0068】
試験材料及び方法
細胞系統
LXSN、TF2及びYusac2はヒト黒色腫細胞系統であり、Cakiはヒト腎腫瘍細胞系統であり、LnCapはヒト前立腺癌細胞系統であり、A204はヒト神経線維芽細胞腫系統であり、B16F10はマウス黒色腫細胞系統であり、EMT6はマウス乳腺癌細胞系統であり、BT20はヒト乳癌細胞系統であり、Colo 357はヒト膵癌細胞系統であり、MSはヒト胃癌細胞系統であり、293はアデノウイルスベクターをパッケージングするために使用されるヒト腎細胞系統(ATCC、CRL−1573)である。培地は、LnCapを除くすべての腫瘍系統についてDMEM+10%FBSであった。LnCapはRPMI 1640+10%FBSで培養した。
【0069】
アデノウイルスベクター
1)mfVIIasm及びG71−1免疫複合体をコードするアデノウイルスベクターを作製し、精製するための上記実施例2で要約した手順を、次の修正を加えて使用した。シャトルベクターDNAをPmeIで消化した後、エタノール沈殿ステップの代わりにアガロースゲルでの電気泳動によってDNAを精製し、その後QIAEX IIキット(Qiagen)を用いてDNAバンドを単離した。2)精製した試料中のベクター濃度を、次のような感染粒子(IP)及び感染単位(IU)についてのアッセイによって測定した。(i)IPアッセイは、ベクター試料を溶解緩衝液(PBS中0.1%SDS)で20倍に希釈し、260nmで吸光度を測定することを含む;IPへの換算は、1O.D.単位=1×1012IPである。(ii)IUアッセイは、293細胞培養を連続希釈ベクター試料に感染させ、感染培養を2日間インキュベートして、細胞をベクターゲノムにおけるグリーン蛍光蛋白質(Green Fluorescent Protein,GFP)遺伝子の発現に関して蛍光顕微鏡で検査することを含む。GFP遺伝子を発現する細胞数からIU力価を算定する。IP及びIU力価は±10%以内で一致した。
【0070】
腫瘍細胞におけるmfVIIasm免疫複合体の合成
腫瘍細胞を150mmの皿でほぼ集密まで増殖させ、10IU/細胞の感染多重度でmfVIIasmをコードするアデノウイルスに細胞を感染させた。感染細胞を無血清培地で4日間培養し、培地1.5mlをプロテインAビーズ(Pierce)の懸濁液10μlと混合して、4℃でひと晩回転流動した。SDS−PAGE負荷緩衝液15μl中80℃で3分間ビーズを加熱して、結合mfVIIasm免疫複合体を溶出し、溶離液をSDS−PAGEによって分別して、ニトロセルロース膜に移した。免疫複合体のバンドを、ヤギ抗ヒト又は抗マウスIgG(Fc特異的)プローブでの免疫染色によって検出した。
【0071】
免疫欠損マウスにおける免疫療法試験
4から5週齢の雌性C.B−17 SCIDマウス(Taconic Farms)を免疫欠損マウスに関するすべての実験に使用した。ヒト黒色腫細胞系統LXSN又はTFの単層培養をPBS+2mM EDTA中で解離し、洗って、PBSに再懸濁した。5×105細胞を右ひ腹に皮下注入して皮膚腫瘍を生成し、6×105TF細胞を尾静脈に静脈内注入して転移性肺腫瘍を生成した。皮膚腫瘍の大きさをカリパスにより二次元で測定し、腫瘍容積を(幅)2(長さ)/2として評価した。皮膚腫瘍が約100mm3の容積まで成長したときに、ヒトFcエフェクタードメインを含むアデノウイルスベクターの腫瘍内注入を開始した。各々の注入ステップに関して、総容量50μlのベクター試料を腫瘍の3又は4部位に注入した。実験の終了時に剖検を実施して血液サンプルを採集し、腫瘍と正常器官を形態学的及び組織学的検査用に調製した。
【0072】
免疫コンピテントマウスにおける免疫療法試験
4から5週齢の雌性C57BL/6マウス(Charles River Laboratories)を免疫コンピテントマウスに関するすべての実験に使用した。B16F10マウス黒色腫細胞の単層培養をPBS+2mM EDTAに懸濁し、洗って、PBSに再懸濁した。5×105細胞を右ひ腹に皮下注入して皮膚腫瘍を生成した。皮膚腫瘍が約140−325mm3の推定容積まで成長したときに、マウスFcエフェクタードメインを含むmfVIIasm免疫複合体をコードするアデノウイルスベクターの尾静脈注入を開始した。プロトコールの効果と安全性をモニターするための手順は、SCIDマウスについて上述したのと同じである。
【0073】
SGOTアッセイ
マウスから血清サンプルを採集し、凍結して、標準診断キットを用いてグルタミン酸オキサロ酢酸トランスアミナーゼについてアッセイした。
【0074】
試験結果
この試験に使用した2つの免疫複合体は、腫瘍標的ドメインとエフェクタードメインから成る(図1)。標的ドメインは、腫瘍血管内皮細胞及び腫瘍細胞で発現されるTFに結合するが、血液凝固を開始させない(実施例2)、突然変異マウス第VII因子分子(mfVIIasm)、若しくはヒト黒色腫細胞で選択的に発現されるその同種抗原に結合するscFv抗体G71−1のいずれかである(CaiとGaren,前出、及び実施例1)。エフェクタードメインは、標的細胞に対する細胞溶解性免疫応答を誘導するヒト又はマウスIgG1免疫グロブリンのFc領域である(実施例1)。免疫複合体を送達するためのベクターは、分泌形態の免疫複合体をコードする複製不全アデノウイルスである(実施例1)。実施例1において、免疫複合体をコードするベクターのSCIDマウスへの静脈内注入は腫瘍の成長を阻害したが、静脈内注入したベクターによる感染の主要標的である肝臓への組織学的損傷を引き起こした。一次感染が肝細胞から腫瘍細胞に再指令されうるかどうかを調べるため、ベクターをSCIDマウスにおいて増殖するヒト皮膚黒色腫に腫瘍内注入し、ベクターゲノムに挿入したグリーン蛍光蛋白質(GFP)遺伝子の発現によって腫瘍と肝臓におけるベクターの分布をマッピングした(図14)。腫瘍においては強いGFPの発現が検出されたが、肝臓では検出されず、注入した腫瘍がベクターによる感染の主要標的であることを示した。腫瘍におけるGFP発現のパターンは、注射針が横切る経路に隣接する数層の腫瘍細胞に限定されると思われた。
【0075】
mfVIIasmとG71−1免疫複合体をコードする2つのベクターの混合物を、SCIDマウスで増殖するヒト黒色腫皮膚腫瘍に注入し、その後5日間腫瘍容積を測定することによって、腫瘍成長へのベクター投与の影響を検討した。血液中に分泌されるG71−1免疫複合体の力価がより高いことを相殺するため、混合物中のmfVIIasmベクター対G71−1ベクターの比は5:1であった(実施例2)。注入に使用した2つのベクターの結合用量は、7×108から6×109感染単位(IU)の範囲であった。対照に関しては、免疫複合体をコードしていない空ベクターを6×109IUの用量で注入した。腫瘍成長の最強の阻害は、免疫複合体をコードするベクターの最高用量で得られた。このベクターの用量を下記のすべてのSCIDマウス実験に使用した。
【0076】
次の実験では、mfVIIasmとG71−1免疫複合体をコードする2つのベクターの多回腫瘍内注入での投与が腫瘍成長に及ぼす阻害作用を19日間モニターした。アデノウイルスベクターの最後の注入から2日後のマウス血液中の免疫複合体蛋白質力価は1mg/mlから2mg/mlの範囲であり、これは静脈内注入したベクターによって生じる力価の約4分の1である(実施例2)。これらの腫瘍の平均容積は、2つのベクターの1回目の注入後1日以内に約70%低下し、その後実験の残りの期間中容積は上昇しなかった。空対照ベクターを注入した腫瘍は継続的に成長し、実験終了時には、2つの免疫複合体をコードするベクターを注入した腫瘍の30mm3に対し、1900mm3の平均容積に達した。ベクターの静脈内注入に関する先の実験(実施例2)と一致して、mfVIIasm免疫複合体をコードするベクターだけを注入した腫瘍は、両方のベクターを注入した腫瘍とほぼ同じくらい強力に阻害された。マウスは実験期間を通じて活動的なままであり、最後の注入から2日後に実施した剖検では、すべての器官が形態学的及び組織学的に正常であると思われ、出血の兆候はなかった。免疫複合体をコードするベクターを静脈内注入したSCIDマウスは肝細胞への組織学的損傷を示したので(実施例2)、腫瘍内注入したSCIDマウスの肝臓を組織学的損傷、そして機能的損傷に関しても検討した。静脈内注入マウスからの肝臓切片で認められた重要な組織学的変化に対し、腫瘍内注入マウスからの肝臓切片は比較的軽度の変化を示した(図15)。剖検の際にマウスから採取した血清中のグルタミン酸オキサロ酢酸トランスアミナーゼ酵素(SGOT)のアッセイにより、肝機能をモニターした。対照マウスと免疫複合体をコードするベクターを注入したマウスのSGOTレベルは正常範囲内(210±28U/L)のままであり、肝機能がそこなわれなかったことを示した。
【0077】
mfVIIasmとG71−1免疫複合体をコードするベクターを注入したマウスから回収した腫瘍は、おそらくTFに結合したmfvVIIasm免疫複合体によって誘導される、血管内皮細胞に対する細胞溶解性免疫応答の結果として、血管がほとんど全く存在しなかった(図13)。
【0078】
腫瘍内注入プロトコールの効果は、注入した皮膚腫瘍の阻害だけでなく、注入することができない転移性腫瘍の抑制によっても決まる。転移性腫瘍への阻害作用を調べるため、ヒト黒色腫TF細胞を尾静脈に注入して、主として肺への血液運搬性転移を生じさせたSCIDマウスモデルを使用した。同時にヒト黒色腫LXSN細胞をマウスに皮下注入し、12日後に形成された皮膚腫瘍に、2つの免疫複合体をコードするベクター又は空対照ベクターを注入した。次の8週間、週に2回のスケジュールで腫瘍内注入を続け、最後の注入から2日後に剖検を行って、肺表面上の腫瘍結節の数を測定した(表2)。対照ベクターを注入した2匹のマウスはそれぞれ14個と29個の肺結節を有しており、一方2つの免疫複合体をコードするベクターを注入した5匹のマウスに関しては、3匹は全く肺結節がなく、2匹がそれぞれ1個と5個の結節を有していた。これらの結果は、2つの免疫複合体をコードするベクターの腫瘍内注入が、注入した腫瘍の成長も遠隔転移性腫瘍の成長も阻害しうることを示している。
【0079】
【表2】
【0080】
臨床環境でのプロトコールの効果に影響を及ぼしうる別のパラメータは、免疫複合体又はアデノウイルスベクターへの患者の免疫応答である。臨床用の免疫複合体の標的及びエフェクタードメインはヒト蛋白質から誘導されることになるので、患者は免疫複合体に対して有意の免疫応答を示さないはずである。しかし、アデノウイルスベクターはヒトにおいて、またマウスにおいても強い免疫原性である。マウスにおけるアデノウイルスベクターへの免疫応答の影響を調べるため、マウス黒色腫皮膚腫瘍を保有する免疫コンピテントマウスでプロトコールを試験した。マウス黒色腫細胞をベクターに感染させることはできないので、実験はベクターの腫瘍内注入ではなく静脈内注入を伴った。また、マウス黒色腫細胞はmfVIIasm免疫複合体に結合するが、G71−1免疫複合体には結合しないので、mfVIIasm免疫複合体だけしか試験することができなかった。この実験のための免疫複合体のFcエフェクタードメインは、マウスIgG1免疫グロブリンから誘導した。結果は、mfVIIasm免疫複合体をコードするアデノウイルスベクターの静脈内注入が、SCIDマウスにおいても免疫コンピテントマウスにおいても黒色腫の成長阻害をもたらすことを示している(図16)。
【0081】
TFは一般に腫瘍血管内皮細胞、及びほとんどの転移性腫瘍細胞によっても発現されるので、fVIIasm免疫複合体は、ヒト黒色腫だけでなく広範囲の他のヒト腫瘍に対しても細胞溶解性免疫応答を仲介すると考えられる。ここで報告した試験で示すように、mfVIIasm免疫複合体をコードするアデノウイルスベクターのヒト腫瘍への腫瘍内注入は、免疫複合体の高い血液力価を確立し、維持するための安全で有効なプロトコールであると思われる。しかし、注入した腫瘍の細胞はアデノウイルスによる感染に感受性であり、コードされた免疫複合体を合成し、分泌することができるはずである。ヒト黒色腫、前立腺癌、乳癌、膵癌、腎癌、胃癌及び神経芽細胞腫細胞系統、並びにマウス黒色腫及び乳癌細胞系統から成る、ヒト及びマウスの腫瘍細胞系統のパネルを、mfVIIasm免疫複合体をコードするアデノウイルスベクターの宿主として試験した。ヒト腫瘍細胞系統はすべてほぼ同じ量の免疫複合体蛋白質を産生し、分泌したことから、腫瘍内注入プロトコールが黒色腫に加えて他の種類のヒト固体腫瘍にも使用できることを示唆した。マウス腫瘍系統は、おそらくマウス細胞がアデノウイルスベクターに感染しなかったために、免疫複合体蛋白質を生成することができなかった。
【0082】
考察
この実施例で述べた癌の免疫療法プロトコールは、腫瘍血管系及び腫瘍細胞に対する細胞溶解性免疫応答を仲介する免疫複合体分子をコードする、複製不全アデノウイルスベクターの腫瘍内注入を含む(図1)。ベクターに感染した細胞はコードされた免疫複合体を合成し、それらは血液中に分泌されて、腫瘍血管系内皮細胞及び腫瘍細胞上の同種標的に結合する。ベクターは主として肝細胞ではなく注入された腫瘍の細胞に感染するので、ベクターの腫瘍内注入は静脈内注入に比べて重要な安全上の利点を提供する。マウスの肝臓も他のいずれの器官も、繰り返された腫瘍内注入から深刻な損傷を示さなかった。黒色腫以外のいくつかの型のヒト腫瘍細胞も、ベクターに感染してコードされた免疫複合体を合成し、分泌することができる。それ故、ベクターの腫瘍内注入経路はヒト固体腫瘍に一般的に適用しうる。注入のためにアクセス可能な腫瘍がない場合には、罹病性の正常組織にベクターを注入することができるであろう。
【0083】
腫瘍内注入プロトコールの治療効果をヒト皮膚黒色腫及び転移性肺黒色腫のSCIDマウスモデルで試験した。試験は、異なる標的ドメインを含む2つの免疫複合体を伴った。1)1つの標的ドメインは、腫瘍新生血管系の血管を含めた増殖する血管の内皮細胞によって、そしてまた大部分のヒト腫瘍細胞によっても発現される膜貫通レセプタ組織因子に高い親和性と特異性で結合する血液チモーゲン第VII(fVII)因子である。fVII免疫複合体は、in vivoで標的細胞上の組織因子への結合に関して内因性第VII因子と競合するはずである。2つの標的ドメインのアビディティ作用が組織因子の多数のコピーを発現する細胞への結合を増強するので、この競合はモノマー内因性分子に比べてホモダイマーである免疫複合体分子(図1)の方が圧倒的に有利である(実施例2);また、内因性第VII因子よりもベクターがコードするfVII免疫複合体の方が血液力価が高い。SCIDマウスにおいて増殖するヒト腫瘍異種移植片を含む実験には、マウスfVII標的ドメインを使用した。なぜなら、マウスfVII標的ドメインは、腫瘍細胞上のヒト組織因子及び腫瘍血管系のマウス内皮細胞上のマウス組織因子の両方に緊密に結合するからである。播種性血管内凝固を引き起こす可能性がある、fVII免疫複合体が組織因子に結合することによる血液凝固経路の開始を防ぐため、マウスfVII標的ドメインの活性部位に突然変異を導入した(mfVIIasm)。2)第二の標的ドメインは、ヒト黒色腫細胞によって選択的に発現される硫酸コンドロイチンプロテオグリカンに結合する、一本鎖Fv(scFv)抗体G71−1である(実施例1及び2)。免疫療法試験の結果は、単独で又はG71−1免疫複合体をコードするベクターと共に、mfVIIasm免疫複合体をコードするベクターを腫瘍内注入すると、注入した皮膚腫瘍、そして同時に転移性肺腫瘍の成長も阻害することを示した(図16及び表2)。免疫複合体をコードするベクターの腫瘍内注入後に存在する残留腫瘍組織にはほとんど血管が存在せず(図13)、mfVIIasm免疫複合体が、腫瘍新生血管系の広い範囲に及ぶ破壊をもたらす強力な細胞溶解性免疫応答を誘導することを示唆した。
【0084】
上記の説明は、当業者が本発明をいかにして実施するかを教示するためのものであり、説明の読後に当業者に明白となるであろう本発明の明らかな修正及びバリエーションをすべて詳述することは意図されていない。しかしながら、そのような明らかな修正及びバリエーションはすべて、下記の特許請求の範囲によって定義される本発明の範囲内に含まれることが意図されている。特許請求の範囲は、特に異なる記載がないかぎり、意図されている目的を達成するために有効な、あらゆる順序での特許請求されている成分及び段階をカバーすることが意図されている。
【0085】
本文中で引用する論文、特許及び特許出願、並びにそれらの中で引用される参考文献は、明白にその全体が参照してここに組み込まれる。
(配列表)
【技術分野】
【0001】
関連出願の相互参照
本出願は、1999年7月1日に出願された米国特許出願番号第60/142,161に基づく優先権を主張する。
【0002】
連邦政府による資金的条を受けた研究開発の記載
本発明は、一部、アメリカ国立衛生研究所(アメリカ合衆国公衆衛生局)からの研究グランドHL-29019-17に基づく政府のサポートによって作られた。政府は、本発明に対し、一定の権利を有する。
【背景技術】
【0003】
(発明の属する技術分野)
本発明は、癌、慢性関節リウマチ、滲出性形態の黄斑変性、及びアテローム性動脈硬化症のような新しい血管の増殖(血管新生:neovascularization)に関連する疾患を有する患者を治療するための免疫複合体試薬の設計、合成及び投与に関する。本発明の免疫複合体の標的は、新生血管系の内皮細胞によって発現される膜貫通レセプタ組織因子である。組織因子はまた、多くの型の腫瘍細胞によっても発現される。それ故、本発明の治療方法は広い範囲の固体腫瘍に対する免疫療法において特に有効である。治療試薬は、標的ドメインとエフェクタードメインから成る免疫複合体である(図1)。標的ドメインは、高い親和性と特異性で組織因子に結合するが、血液凝固を開始させない、突然変異形態の第VII因子である。エフェクタードメインはIgG1免疫グロブリンのFc領域である。
【0004】
(発明の背景、従来の技術)
典型的には血管内皮細胞に対して特異的な成長因子の放出が引き金となる様々な疾患において、病的血管形成、すなわち周辺組織中の脈管からの血管増殖の誘導が認められる。病的血管形成は、固体腫瘍の成長と転移を可能にし、眼障害における視覚機能不全を引き起こし、炎症性疾患における白血球の血管外遊出を促進し、及び/又はアテローム性動脈硬化症のような心臓血管疾患の結果に影響を及ぼす、血管新生をもたらしうる。集合的に、これらは時に血管形成性疾患と称される。
【0005】
固体腫瘍の生存と成長は新生血管系の発生に決定的に依存するので、癌は主要な血管形成性疾患である(Folkman,J.(1995)N.Engl.J.Med.333,1757−1763)(非特許文献1)。多くの癌は段階的に進行し、一次腫瘍細胞の増殖から始まって、腫瘍細胞の循環系への侵入、播種性転移部位でコロニー形成、そして癌による死亡の大部分の原因である転移した腫瘍細胞の増殖へと進行する(Vogelstein,B.とKinzler,K.W.(1993)TIG9,141−143)(非特許文献2)。癌は、疾患が転移段階に達するまでしばしば検出されないままであるため、血管下部構造と転移性腫瘍細胞を根絶することができる癌療法が特に望ましい。
【0006】
血管形成はまた、慢性関節リウマチにおいても重要な役割を果たす(Szekanez,Z.ら(1998)J.Invest.Med.46,27−41)(非特許文献3)。慢性関節リウマチ(RA)は、世界中ですべての人種民族群において起こる慢性全身性炎症性疾患であり、主として滑膜性の連結及びしばしば様々な他の器官を侵す。RAの滑膜組織は広汎に血管新生される。RAでは、炎症白血球が血管の内皮層を通して滑膜内に遊出し、滑膜の炎症、そして最終的には関節の破壊をもたらす。
【0007】
血管形成は、視力の破滅的喪失をもたらす眼疾患の大半の基礎となる(Firedlander,M.ら(1996)Proc.Natl.Acad.Sci.USA 93,9764−9769)(非特許文献4)。55歳以上の個人における失明の主要な原因は滲出性(「湿性」)形態の加齢性黄斑変性(ARMD)であり、55歳未満では増殖性糖尿病性網膜症(PDR)である。ARMDとPDRはそれぞれ脈絡膜と網膜の血管新生に関する基本型の疾患であるが、他の変性又は炎症状態も、選択的にいずれかの血管系の新血管形成を引き起こしうる(同上)。
【0008】
それ故、これらの疾患状態、特に癌の治療への1つのアプローチは、主として新しい血管の増殖を阻害することによって、新生血管系の機能又は成長を抑えることであった(Chaplin,D.J.とDougherty,G.J.(1999)Br.J.Cancer,80,57−64)(非特許文献5)。血管を標的することにはいくつかの利点がある。まず第一に、血管への損傷は血流を停止させることができ、癌に適用すると、多くの依存性腫瘍細胞の死の引き金となりうる。第二に、標的細胞が血流に近接しており、薬物送達が促進される。第三に、血管内皮細胞においては治療耐性の突然変異が起こりにくい。
【0009】
薬剤、抗体及び遺伝子治療に基づくアプローチを含めて、数多くの抗血管形成療法が提案されてきた。これらは、メタロプロテイナーゼ阻害因子、ポリ硫酸ペントサン及びTNP−470、チロシンキナーゼの選択的阻害因子、そしてアンギオスタチン及びエンドスタチンのペプチド阻害因子を含む(同上、及びその中で引用される総説)。これらは典型的には、脈管形成の様々な段階、すなわち基底膜の分解、内皮細胞の移動、内皮細胞の増殖、及び管形成の段階で血管形成を妨げる。血管形成だけでなく、血管形成性疾患状態において既に形成された新生血管系も標的する、改善された治療法を開発することが望ましいであろう。
【先行技術文献】
【非特許文献】
【0010】
【非特許文献1】Folkman,J.(1995)N.Engl.J.Med.333,1757−1763
【非特許文献2】Vogelstein,B.とKinzler,K.W.(1993)TIG9,141−143
【非特許文献3】Szekanez,Z.ら(1998)J.Invest.Med.46,27−41
【非特許文献4】Firedlander,M.ら(1996)Proc.Natl.Acad.Sci.USA 93,9764−9769
【非特許文献5】Chaplin,D.J.とDougherty,G.J.(1999)Br.J.Cancer,80,57−64
【発明の概要】
【0011】
(発明が解決しようとする課題)
本発明の課題は癌、慢性関節リウマチ、滲出性形態の黄斑変性、及びアテローム性動脈硬化症を含めて、新しい血管の増殖(新生血管系)に関連する疾患のための新規治療を提供することである。より具体的な課題は、本発明はこれらの疾患状態において認められる血管形成を抑制するだけでなく、新生血管系構造も破壊する、新生血管標的療法を提供することである。
【0012】
(課題を解決するための手段)
これらやその他の目的は、組織因子に選択的に結合する標的ドメインと、標的細胞に対する細胞溶解応答又は細胞傷害性応答を動員するエフェクタードメインを含む免疫複合体を含有する組成物を提供する、本発明によって実現される。典型的な免疫療法の処置では、各々が、組織因子に結合(bind)するが血液凝固を開始させない、リシン−341がアラニンに置換された、及び/又はセリン−344がアラニンに置換された第VII因子のような、ヒト第VII因子の突然変異形態を含む標的ドメインに複合(conjugate)した免疫複合体であって、ヒンジを含めたヒトIgG1免疫グロブリンのFc領域であるエフェクタードメインを持ち、2つの同一の鎖の二量体として構築される少なくとも1つの免疫複合体を含む組成物を、血管新生に関連する疾患を有する患者に有効量で投与する。組織因子は、正常血管系ではなく腫瘍新生血管系の裏側にある内皮細胞によって、また多くの型の腫瘍細胞によっても発現されるので、本発明の第VII因子免疫複合体は広い範囲の固体腫瘍に対する免疫療法において特に有効である。新生血管系での凝固を活性化する及び/又は新生血管系に組織因子又は組織因子突然変異体を導入するというこれまでに記述されている発明(米国特許第6,001,978号においてEdgingtonとMorrisseyが述べているような)は、組織因子は新生血管系の内皮細胞によって特異的に発現されているのであるから無意味であり、本発明は2つの主要な点でこれらの発明と異なっている:組織因子は、血液凝固プロセスのイニシエーターとしてではなく、本発明の免疫複合体によって仲介される細胞溶解免疫応答を誘導するための標的として使用される、および血液凝固経路を活性化せずに新生血管系を破壊する細胞溶解免疫応答が開始される。
【0013】
従来の薬学的組成物中での精製免疫複合体蛋白質の使用、あるいは分泌形態の免疫複合体をコードするcDNAを保有するベクター系の使用を含む、当該免疫複合体の全身又は局所投与の方法も開示する。本発明に従った治療は、1つ又は複数の型の精製された免疫複合体蛋白質の有効量の、周期的又は持続的な静脈内又は腫瘍内注入、あるいは他の部位での注入を含むが、これらに限定されない。代替的な実施態様は、分泌形態の1つ又は複数の型の免疫複合体蛋白質をコードするcDNAを保有する発現ベクターの有効量の、静脈内又は腫瘍内注入、あるいは他の部位での注入によって患者を治療することを含む。後者の例は、分泌形態の本発明の免疫複合体をコードするcDNAを保有する複製欠損アデノウイルスベクター又はアデノ関連ウイルスベクターの、静脈内又は腫瘍内注入、あるいは他の部位での注入によって患者を治療することを含む。
【0014】
癌の免疫療法のような一部の実施態様では、本発明の免疫複合体を、異なる標的ドメインを持つ別の型の免疫複合体と共に投与する。これらは典型的には、腫瘍細胞、血管内皮細胞、病的状態の滑膜における侵襲細胞で発現される細胞表面分子に選択的に結合する、ヒトscFv又はVH融合ファージライブラリーから単離された一本鎖Fv又はVH分子断片である。
【図面の簡単な説明】
【0015】
【図1】図1は、本発明の免疫複合体分子の機構を例示する図表である。TD;標的ドメイン。H:2つのジスルフィド架橋を持つIgG1免疫グロブリンのヒンジ領域。CH2及びCH3:IgG1免疫グロブリンの定常領域。標的ドメインは、典型的には突然変異した活性部位を持つヒト第VII因子、あるいはヒトscFv又はVH分子から成る。エフェクタードメインは、典型的には、非修飾であるか又は放射性分子又は光活性化色素分子のような細胞傷害性物質に複合(conjugate)した、ヒトIgG1のFc領域から成る。
【図2】図2は、ヒトナチュラルキラー(NK)細胞によるヒト黒色腫細胞の免疫複合体依存性溶解を示す棒グラフである。免疫複合体の標的ドメインは黒色腫特異的ヒトscFv分子であり、エフェクタードメインはヒトIgG1免疫グロブリンのFc領域である。ヒト黒色腫細胞系統A−2058及びヒト線維芽細胞対照を蛍光色素、カルセイン−AMで標識した。NK細胞単独(棒A)又はscFv免疫複合体E26−1とNK細胞(棒B)への接触後、無傷のままである黒色腫又は線維芽細胞の分画を残留蛍光によって測定した。NKエフェクター細胞対標的細胞の比率(E/T)は3から20の範囲であった。3つの完全なセットの実験を黒色腫細胞と線維芽細胞の両方について実施した;各々の実験に関して、細胞溶解アッセイを四重に実施した。図に示す棒は、3回の実験についての細胞溶解アッセイの平均値を表わし、ほぼ10%以内で一致した。細胞溶解の割合(%)を実施例1で述べたように算定した。scFv免疫複合体G71−1に関しても同様の結果を得た。
【図3】図3は、補体によるヒト黒色腫細胞の免疫複合体依存性溶解を示す棒グラフである。手順は、NK細胞の代わりにヒト血清又は精製ウサギ補体成分を使用したことを除いて、図2の説明で述べたとおりであった。各々のアッセイについての免疫複合体と補体試薬は実施例1、表1に示されているとおりであった。
【図4A】図4は、SCIDマウスで増殖させたヒト黒色腫異種移植片における腫瘍細胞及び腫瘍血管内皮細胞への突然変異マウス第VII因子(mfVIIasm)免疫複合体の結合に関する免疫組織化学アッセイを示す。第二抗体はAPで標識した抗ヒトγ鎖であり、AP基質は青色を生じるBCIP/NBTであった;対比染色はメチルグリーンであった。パネルA:異種移植片の広い範囲に及ぶ血管形成を示す、ヘマトキシリン+エオシンで染色した対照。
【図4B】図4は、SCIDマウスで増殖させたヒト黒色腫異種移植片における腫瘍細胞及び腫瘍血管内皮細胞への突然変異マウス第VII因子(mfVIIasm)免疫複合体の結合に関する免疫組織化学アッセイを示す。第二抗体はAPで標識した抗ヒトγ鎖であり、AP基質は青色を生じるBCIP/NBTであった;対比染色はメチルグリーンであった。パネルB:血管内皮細胞と腫瘍細胞の両方の強い染色を示す、mfVIIasm免疫複合体に関する免疫組織化学。
【図4C】図4は、SCIDマウスで増殖させたヒト黒色腫異種移植片における腫瘍細胞及び腫瘍血管内皮細胞への突然変異マウス第VII因子(mfVIIasm)免疫複合体の結合に関する免疫組織化学アッセイを示す。第二抗体はAPで標識した抗ヒトγ鎖であり、AP基質は青色を生じるBCIP/NBTであった;対比染色はメチルグリーンであった。パネルC:mfVIIasm免疫複合体なしの免疫組織化学的対照。
【図5】図5は、免疫複合体をコードする複製不全アデノウイルスベクターの静脈内注入後の、SCIDマウスの血液中のG71−1 scFv及びmfVIIasm免疫複合体の濃度を示す折れ線グラフである。0日目と7日目に、G71−1免疫複合体をコードする2×1011アデノウイルス(A)又はmfVIIasm免疫複合体をコードする4×1011アデノウイルス(B)をマウスに注入した。コードされた免疫複合体の血液中の濃度をELISAによって定量した。各点は、各々の群の5匹のマウスに関する濃度の平均値である。
【図6】図6は、SCIDマウスにおけるヒト黒色腫異種移植片の成長へのG71−1及びmfVIIasm免疫複合体の阻害作用を示す折れ線グラフである。各々の曲線に関して、5匹のSCIDマウスに5×105 TF2細胞を皮下注入した。異種移植片が触知可能な大きさに成長したとき、図に示す0日目と7日目と14日目に、マウスにアデノウイルスの尾静脈注入を実施した。注入したアデノウイルスの量は、対照については4×1011、G71−1免疫複合体をコードするアデノウイルスについては2×1011、そしてmfVIIasm免疫複合体をコードするアデノウイルスについては4×1011であった。推定腫瘍容積は各群の5匹のマウスについての平均値である。
【図7】図7は、図6及び実施例2で報告した実験からの異種移植片の腫瘍重量を示す棒グラフである。20日目、すなわちアデノウイルスの最後の注入から6日後に、マウスから異種移植片を切除した。棒の高さは各群の5匹のマウスについての平均重量である。
【図8】図8は、SCIDマウスにおける大きなヒト黒色腫異種移植片の成長へのG71−1及びmfVIIasm免疫複合体の阻害作用を示す折れ線グラフである。各々のマウスに5×105の黒色腫細胞を皮下注入し、異種移植片を皮膚表面で50mm3の推定腫瘍容積まで成長させた(1日目)。1、6、12及び19日目に、G71−1免疫複合体をコードする2×1011のアデノウイルスとmfVIIasm免疫複合体をコードする7×1011のアデノウイルスの混合物を5匹のマウスの尾静脈に注入した。対照として5匹のマウスに免疫複合体をコードしていない4×1011のアデノウイルスを注入した。推定腫瘍容積は各群の5匹のマウスについての平均値である。免疫複合体をコードするアデノウイルスを注入したマウスのうちの1匹が17日目に死亡していた;その後の日にちに関する推定腫瘍容積は残りの4匹のマウスの平均値である。
【図9】図9は、低レベルの組織因子を発現するヒト黒色腫異種移植片の成長へのmfVIIasm免疫複合体の阻害作用をプロットした折れ線グラフである。マウスに5×105のLXSN細胞を皮下注入し、異種移植片が触知可能な大きさに成長したとき(0日目)、5匹のマウスにmfVIIasm免疫複合体をコードする9×1011のアデノウイルスを静脈内注入し、5匹のマウスに4×1011の対照アデノウイルスを注入した。7、13、21及び24日に追加注入を行い、25日目に形態学的及び組織化学的検査のためにマウスを解剖した。推定腫瘍容積は各群の5匹のマウスについての平均値である。
【図10A】図10は、図9及び実施例2の説明で報告した実験からのヒト黒色腫異種移植片の組織化学を示すものである。25日目に異種移植片を切除し、パラフィンに包埋して、切片をヘマトキシリン+エオシンで染色した。パネルA:免疫複合体をコードしていないアデノウイルスを注入した対照マウスからの異種移植片。
【図10B】図10は、図9及び実施例2の説明で報告した実験からのヒト黒色腫異種移植片の組織化学を示すものである。25日目に異種移植片を切除し、パラフィンに包埋して、切片をヘマトキシリン+エオシンで染色した。パネルB:mfVIIasm免疫複合体をコードするアデノウイルスを注入したマウスからの異種移植片。
【図11】図11は、SCIDマウスにおいて腫瘍内注入したアデノウイルスベクターがヒト黒色腫の成長に及ぼす用量効果を示す折れ線グラフである。最初にマウスにヒト黒色腫細胞系統TF2を皮下注入し、皮膚腫瘍が0日目に示されている大きさまで成長したとき、mfVIIasm及びG71−1免疫複合体をコードする2つのアデノウイルスベクターの混合物を腫瘍に注入した。2つのベクターの用量は次のとおりであった。○:7×108IU;▽:2×109IU;□:6×109IU。対照である◇の用量については、免疫複合体をコードしていないアデノウイルス6×109IUを注入した。
【図12】図12は、SCIDマウスにおいて腫瘍内注入した高用量のアデノウイルスベクターがヒト黒色腫の成長に及ぼす作用を示す折れ線グラフである。最初にマウスにヒト黒色腫細胞系統TF2を皮下注入し、皮膚腫瘍が0日目に示されている大きさまで成長したとき、次のアデノウイルスベクターの6×109IUを注入した:●、対照ベクター(3匹のマウス);■、mfVIIasm免疫複合体をコードするベクター(5匹のマウス);△、mfVIIasm及びG71−1免疫複合体をコードする2つのベクターの混合物(6匹のマウス)。2、5、7、9、11、13、15及び17日目に追加注入を実施した。各曲線上の点は、対応する群のすべてのマウスについての測定値の平均(±20%)である。
【図13】図13は、mfVIIasm及びG71−1免疫複合体の混合物をコードするアデノウイルスベクター又は空アデノウイルスベクター対照を腫瘍内注入したSCIDマウスからの黒色腫の写真である。実験は上記図12の説明及び実施例3の中で述べられている。最後の注入から2日後に腫瘍を切除し、写真撮影した。図の上部のものさしの原寸は1cmである。
【図14】図14は、腫瘍内注入後の腫瘍内及び肝臓におけるアデノウイルスベクターの分布を示す写真である。GFP蛋白質をコードするが免疫複合体をコードしない対照ベクターは、SCIDマウスで増殖するヒト黒色腫皮膚腫瘍の3つの部位に注入した。総ベクター用量は6×109IUであった。注入の40時間後に腫瘍と肝臓を切除し、蛍光光学系を備えた解剖顕微鏡下に無傷検査した。480nmの励起と630nmの発光でGFPシグナルが検出され、577nmの励起と630nmの発光でバックグラウンドシグナルが検出された。パネルA:腫瘍のGFP。パネルB:腫瘍のバックグラウンド。パネルC:肝臓のGFP。パネルD:肝臓のバックグラウンド。パネルAのものと同様の明るい蛍光スポットが、おそらく注入部位に対応する、他の2箇所の腫瘍部位でも検出された。写真は組織内の1つのレベルに焦点を合わせている。しかし、そのレベルの上と下に焦点を合わせると腫瘍内でもGFPスポットが検出でき、注射針が横切った経路に隣接する腫瘍細胞だけがベクターによって感染した細胞であることを示唆した。
【図15】図15は、アデノウイルスベクターを静脈内又は腫瘍内注入したSCIDマウスからの肝臓切片を示す。静脈内実験は実施例2に述べられ、腫瘍内実験は実施例3に述べられている。左側のパネルは対照ベクターを注入したものであり、右側のパネルはmfVIIasm及びG71−1免疫複合体をコードする2つのベクターの混合物を注入したものである。最後の注入から2日後に肝臓を切除し、ホルムアルデヒドで固定し、パラフィンに埋め込んだ。当該切片をヘマトキシリンとエオシンで染色して、100×の倍率で写真撮影した。
【図16】図16は、mfVIIasm免疫複合体をコードするアデノウイルスベクターの免疫コンピテントマウスへの静脈内注入がマウス皮膚黒色腫の成長に及ぼす作用を示す折れ線グラフである。C57BL/6マウスにマウス黒色腫細胞系統B16F10を皮下注入し、皮膚腫瘍が0日目に示されている大きさまで成長したとき、mfVIIasm免疫複合体をコードするベクター3×1010IPを注入した。免疫複合体のFcドメインはマウスIgG1免疫グロブリンから誘導した。5日目に再び1.5×1010IPを腫瘍に注入した。
【発明を実施するための形態】
【0016】
(発明の実施の態様)
抗血管系免疫複合体療法は、血管形成の活性状態にある増殖中の腫瘍のようなある種の疾患状態で形成される新生血管系と異なって、正常な成体哺乳類血管系は一般に静止状態(女性の生殖周期や創傷治癒のような特定のプロセスを除く)にあるという所見に基づく。それ故、静止状態にある血管内皮細胞と増殖性血管内皮細胞との分子的な相違が病的血管系についての標的なると考えられる。
【0017】
静止状態から、癌で認められるような病的血管系における血管形成状態へのスイッチは、通常、腫瘍細胞によって分泌され、血管内皮細胞上の血管内皮細胞成長因子(VEGF)レセプタに高い親和性と特異性で結合する、VEGFによって活性化される。血管内皮細胞上のレセプタへのVEGFの結合によって活性化される別の応答は、血漿第VII/VIIa因子に結合して血液凝固を開始させる、膜貫通レセプタである組織因子の発現である。結合VEGFを持つ血管内皮細胞だけが組織因子を発現するので、腫瘍血管系についての推定上の標的は、血液中を循環する第VII/VIIa因子に結合するはずである、内皮細胞上で発現される組織因子である。
【0018】
本発明は、ヒトIgG1のFcドメインに複合した標的ドメインから成る免疫複合体が、ナチュラルキラー(NK)細胞と免疫系の補体経路による標的細胞への細胞溶解応答を仲介するという所見に基づく(Wang,B.ら(1999)Proc.Natl.Acad.Sci.USA 96,1627−1632)。正常組織中に存在する安定な血管ではなく、増殖する血管の内側表面で発現される細胞表面レセプタ組織因子と、その天然のリガンドである第VII因子との間の結合は高い特異性と親和性を示すので(Hu,Z.ら(1999)Proc.Natl.Acad.Sci.USA 96,8161−8166)、本発明は、血管新生に関連する疾患の治療のための新しい免疫療法プロトコールを提供する。組織因子は多くの型の腫瘍細胞によっても発現されるので(Callander,N.S.ら(1992)Cancer,70,1194−1201)、本発明はまた、癌の治療のための高い有効性を提供する。
【0019】
しかし、組織因子への第VII因子免疫複合体の結合は血管内凝固の播種を引き起こす可能性があるので、本発明の好ましい実施態様では、組織因子への親和性に影響を及ぼさずに凝固を抑制する第VII因子突然変異体を使用する。組織因子に結合するが血液凝固を開始させないこれらの免疫複合体は、組織因子への結合に関して内因性第VII/VIIa因子と競合する。この競合は免疫複合体を強く支持する(Huら、前出)。なぜなら、免疫複合体は2つの第VII因子標的ドメイン(図1)を含み、単量体の内因性第VII因子分子が欠乏したアビディティ(結合活性)効果を与えるからである。多くの好ましい実施態様では、第VII因子が組織因子に結合したときに血液凝固プロセスを開始させる蛋白分解活性を遮断するため、ヒト第VII因子の活性部位を、Lys−341及び/又はSer−344をアラニンで置換する部位特定突然変異誘発によって突然変異させる。標的ドメインとエフェクタードメインのどちらもヒトソースから誘導できるので、ヒト患者における重要な免疫拒絶反応は最小限に抑えられる。
【0020】
本発明の一般的な実施においては、図1に示すような免疫複合体を、各々が標的ドメインに複合したエフェクタードメインを持つ、2本の鎖を含む蛋白質二量体として構築する。それらが、免疫複合体と結合したときその標的に対して細胞傷害性を示すかぎり、いくつかのタイプのエフェクタードメインが使用できる。本発明の多くの典型的な免疫複合体蛋白質は、エフェクタードメインとして、IgG1免疫グロブリンのFc領域を持つ。ここで使用するとき、これは、Fc領域と同じ生物学的機能を示す変異体及びトランケートバージョンを含む。他の実施態様では、エフェクタードメインは、放射性標識又は光活性化色素分子、例えばレーザー光線によって活性化される色素のような細胞傷害性物質でありうる。エフェクタードメインは、上述したように組織因子に結合するが血液凝固を開始させない、突然変異形態の第VII因子を含む標的ドメインに複合される。
【0021】
本発明の免疫複合体蛋白質は、癌、慢性関節リウマチ、滲出性(「湿性」)形態の黄斑変性、又はアテローム性動脈硬化症のような血管新生に関連する疾患を有する患者に投与される。投与は、治療に関わる病的状態のタイプに依存して、局所又は全身的でありうる。ここで使用するとき、「患者」の語はヒトあるいは他の種の両方を含む;本発明はそれ故、医学適用と獣医学適用の両方を持つ。獣医学的組成物及び治療においては、対応する種から誘導される標的及びエフェクタードメインを用いて免疫複合体を構築する。
【0022】
投与は、例えば、精製免疫複合体又は複製欠損アデノウイルスベクター、あるいは分泌形態の免疫複合体をコードするcDNAを保有する他のウイルスベクターの静脈内、筋肉内、腫瘍内、皮下、滑液包内、眼内、プラーク内、又は皮内注入のような、当該技術において既知の何らかの方法を通して実施しうる。他の投与経路は、液体等の非経口投与でありうる。好ましい実施態様では、1つ又は複数の免疫複合体蛋白質の静脈内又は腫瘍内注入、又は他の部位での注入によって、あるいは分泌形態の1つ又は複数の型の免疫複合体蛋白質をコードするcDNAを保有する1つ又は複数の発現ベクターの静脈内又は腫瘍内注入、又は他の部位での注入によって患者を治療する。一部の実施態様では、分泌形態の1つ又は複数の型の免疫複合体蛋白質をコードするcDNAを保有する1つ又は複数の複製欠損アデノウイルスベクター、あるいは1つ又は複数のアデノ関連ウイルスベクターの有効量の静脈内又は腫瘍内注入によって患者を治療する。複製欠損アデノウイルスベクターを用いる方法を下記で説明する。多くの典型的な実施態様は、分泌形態の免疫複合体をコードするベクターの有効量の腫瘍内及び/又は筋肉内注入を含む。癌のためにベクターを用いる場合、ベクターの腫瘍内注入は、ベクターが主として注入した腫瘍の細胞に感染するため、静脈内注入に比べて重要な安全上の恩恵を提供する。
【0023】
免疫複合体蛋白質の注入を含む投与は、免疫複合体蛋白質又は蛋白質の組合せが薬学的に許容される担体に分散又は可溶化されている組成物を用いる。一部の場合には、各々のベクターが分泌形態の免疫複合体をコードするcDNAを保有する、発現ベクターpMK33/ヒグロマイシンでトランスフェクションしたショウジョウバエ(Drosohila)S2細胞において、又は発現ベクターpcDNA3.1でトランスフェクションしたチャイニーズハムスター卵巣(CHO)細胞において、又はバキュロウイルス(Baculovirus)発現系において、免疫複合体を合成する。
【0024】
治療処置を生じさせるために必要な免疫複合体の量は、それ自体では固定されておらず、必然的に、薬学的担体と共に投与される組成物中の成分の濃度、下記で詳述する免疫系応答を増強する投与組成物中の補助化合物、そして治療する患者の年齢、体重及び臨床状態に依存する。好ましい組成物は、患者に対して許容されない毒性を生じることなく有効量の免疫複合体を供給送達する。本発明の薬学的組成物又は製剤は、他の担体、アジュバント、安定剤、防腐剤、分散剤、及び対象とする製剤の種類を考慮した当該技術において常套的な他の物質も含みうる。
【0025】
癌に適用するときには、本発明は、ヒト腫瘍細胞又は腫瘍血管系内皮細胞、又はその両方を特異的に標的する標的ドメインと、標的細胞に対する細胞溶解性免疫応答又は細胞傷害作用を活性化するエフェクタードメインを持つ免疫複合体を使用する。上述したように、血液凝固を開始させない突然変異体ヒト第VII因子に複合した、ヒンジを含めたIgG1免疫グロブリンのFc領域をエフェクタードメインとして持つ免疫複合体は、それらが腫瘍血管系と腫瘍細胞の両方で発現される組織因子を標的するので、多くの癌の治療において特に有効である。一部の実施態様では、癌患者の末梢血リンパ球から誘導されるscFv又はVH融合ファージライブラリーをこれまでに記述されている腫瘍細胞に対してパニングすることによって単離される(Cai,XとGaren,A.(1997)Proc.Natl.Acad.Sci.USA 94,9261−9266)、抗ヒト腫瘍細胞scFv又はVH抗体断片を標的ドメインとして持つ免疫複合体のような、腫瘍を選択的に標的する別のクラスの免疫複合体を患者に投与することによって治療効果をさらに高めることができる。上述したように同時に又は連続的に投与される免疫複合体の組合せは、標的細胞の細胞溶解を増強する共力作用が認められる場合に特に有益である。
【0026】
癌の治療においては、様々な癌、特に黒色腫、腎臓癌、前立腺癌、乳癌、卵巣癌、脳腫瘍、神経芽細胞腫、頭部及び頚部の癌、膵癌、膀胱癌、及び肺癌を含めた原発又は転移性固体腫瘍を治療するために抗腫瘍性免疫複合体が使用される。免疫複合体を腫瘍血管系、特に血管内皮細胞、及び/又は腫瘍細胞を標的にするために使用しうる。腫瘍血管系は、次のような、免疫療法のためのいくつかの利点を提供する。(i)組織因子を含めたいくつかの血管標的はすべての腫瘍に同じはずである。(ii)血管系を標的とする免疫複合体は、それらの標的に達するために腫瘍塊に浸潤する必要がない。(iii)各々の血管は、その生存が血管の機能的完全性に依存する数多くの腫瘍細胞に栄養分を与えるので、腫瘍血管系を標的することは治療応答の増幅を生じるはずである。(iv)血管系は免疫複合体に対する耐性を発現しにくいと考えられる。なぜなら、それには血管の内側にある内皮細胞層全体の変化を必要とするからである。新しい血管の増殖を阻害する、これまでに記述された抗血管形成法と異なって、本発明の免疫複合体は新生血管系に対する細胞溶解応答を誘発する。
【0027】
本発明の免疫複合体はまた、慢性関節リウマチ、滲出性(「湿性」)形態の黄斑変性、アテローム性動脈硬化症、及び血管新生に関連する他の疾患を治療するために有効である場合がある。IgG1免疫グロブリンのFcドメインに複合した、突然変異ヒト第VII因子によって組織因子に標的される免疫複合体を投与することは、慢性関節リウマチにおいて滑膜に侵襲し、組織因子を発現する血管内皮細胞に対して、細胞溶解性免疫応答をもたらしうる。同様に、第VII因子免疫複合体はまた、滲出性(「湿性」)形態の黄斑変性を治療するためにも有効でありうる。なぜなら、この病的状態においても広い範囲に及ぶ血管新生が認められるからである。本発明の免疫複合体はまた、プラークにおいて組織因子を発現する細胞に対して細胞溶解性免疫応答を生じさせることにより、アテローム性動脈硬化症の治療のためにも有効でありうる。
【0028】
要約すると、本発明に従った全体的な免疫療法プログラムは、血液凝固を引き起こすことなく、新生血管内皮細胞及び腫瘍細胞で発現される組織因子に高いアビディティと特異性で結合する、突然変異した活性部位を持つ第VII因子を標的ドメインとして含む免疫複合体を構築することを含む。IgG1免疫グロブリンのFc領域をエフェクタードメインとして含む免疫複合体は、NK細胞と補体によって標的細胞の溶解を仲介し、新生血管系の破壊をもたらす。本発明の重要な利点は、内因性第VII因子単量体分子と異なって、2つの第VII因子標的ドメインを含む免疫複合体が、組織因子を発現する細胞に対して外因性第VII因子よりも有意に強い結合を示し、過剰の天然リガンドの存在下で成功裡に競合するという事実である。下記の実施例2は、少なくとも約10倍モル過剰の天然リガンドの存在下で成功裡に競合した免疫複合体を例示しているが、本発明はより低い又はより高い親和性を示す他の実施態様を包含する。組織因子に対する外因性第VII因子の結合は既知の最も特異的且つ強力な反応の1つであり、ピコモル範囲のKdを示す。それにもかかわらず、本発明は、1個の免疫複合体分子中に2つの第VII因子配列を持つというアビディティ作用の結果として、組織因子への第VII因子の通常の結合を有意に改善する。このアビディティ作用は本発明の免疫複合体の標的と結合を増強するので、それらは外因性第VII因子と有効に競合し、結合がより長く持続されるため、誘発される免疫応答が増強される。新生血管系及び/又は、癌の場合には患者の型の腫瘍細胞に結合するscFv又はVH抗体断片である標的ドメインに複合したIgG1免疫グロブリンのFc領域であるエフェクタードメインを持つ別の免疫複合体を、補助療法として患者に投与することによってさらに最大化されうる。
【実施例】
【0029】
ここで示す実施例は本発明をさらに例示し、説明するものであり、いかなる意味においても限定と解釈されるべきではない。本発明の免疫複合体及び成分モノクローナル抗体を生成し、特徴づけるために使用する方法の一部は、Cai,X.とGaren,A.(1995)Proc.Natl.Acad.Sci.USA 92,6537−6541、(1996)Proc.Natl.Acad.Sci.USA 93,6280−6285、及び上記で引用した1997年の参考文献の中で、WO97/02479号として1997年1月23日に公開され、1998年4月27日に提出され、2000年3月3日に承認された特許願通し番号第08/983,607号として現在米国で審理中である、Yale UniversityとGaren及びCaiへのPCT/IB96/01032号の中で、上記で引用したWangらの中で、及び上記で引用したHu,Z.らの中で記述されている。
【0030】
実施例1
ヒトIgG1のFcエフェクタードメインに複合したヒト一本鎖Fv(scFv)標的ドメインを含む免疫複合体をこの実施例において合成し、試験する。最初に、ワクチン接種した黒色腫患者の抗体リストから誘導した、scFv融合ファージライブラリーからの黒色腫特異的クローンとしてscFv標的ドメインを単離した(上記で引用したCaiとGarenの参考文献の中で詳細に述べられている)。精製免疫複合体は融合ファージクローンと同様の結合特異性を示した:ヒト黒色腫に対しては結合が起こったが、ヒトメラノサイトあるいは他のいくつかの型の正常細胞及び腫瘍細胞に対しては結合が生じなかった。
【0031】
試験材料及び方法
細胞培養
永久ヒト黒色腫細胞系統A2058(American Type Culture Collection,Rockville,MD)及びTF2をDMEM+10%FCS中で増殖させた。ヒト微小血管内皮細胞と線維芽細胞の一次培養を新生児包皮から吸い出し、8%FBSと2%ヒト周産期血清を補足したRPMI培地中で培養した;内皮細胞には33mM 3−イソブチル−1−メチルキサンテン(IBMX)と0.5mMジブチリルcAMPをさらに補足した。新生児包皮からのヒトメラノサイトの一次培養はSkin Disease Research Center at Yale University School of Medicineによって調製された。形質転換ヒト腎細胞293−EBNA(Invitrogen)及びチャイニーズハムスター卵巣(CHO)細胞をRPMI+10%FCS中で増殖させた。(v)ショウジョウバエ細胞(Schneider S2)をEx−cell 301培地(JRH Biosciences)+10%FBS中25℃で増殖させた。静止NK細胞をロイコホレシスと免疫的選択によって健常ドナーから単離し、単離後18時間以内に使用した;細胞の大部分(>97%)がCD3−、CD56+及びCD16+であった。
【0032】
免疫複合体の調製
手順は、発現ベクターpcDNA3.1(Invitrogen)のCHO細胞へのトランスフェクション、又は発現ベクターpMK33/pMtHyのショウジョウバエ細胞へのトランスフェクションを伴った;各々のベクターは分泌された免疫複合体をコードするcDNAを保有していた(図1)。scFv標的ドメインについてのcDNAを、対応する融合ファージ(1)から次のようなSacI又はBamHI部位を含むPCRプライマーを使用して合成した:a)GTCGAGCAGAGCTCCAGGTGCAGCTGGTGCAGTCTGGGGCTGAGGTGAGGTGAAGAAGCC(配列番号:1);b)ACGTTCAGGGGATCCACCTAGGACGGTCAGCTTGGTCCC(配列番号:2)。ヒトFcエフェクタードメインは、次のようなBamHI又はSalI部位を含むPCRプライマーを使用して、ヒト末梢血リンパ球由来のcDNAライブラリーから合成した:a)ACCTTGCAGGATCCGCAAGACCCAAATCTTGTGACAAAACTCAC(配列番号:3);b)GATCACGTGTCGACTTATCATTTACCCGGAGACAGGGAGAGGCTCTTCTG(配列番号:4)。IgG1リーダーについてのcDNAを、次のようなEcoRI及びSacI末端を含む2個の相補的オリゴヌクレオチドをハイブリダイズすることによって合成した:a)AATTCATGGAGTTTGGGCTGAGCTGGCTTTTTCTTGTTGCTGCATTAAGAGGTGTCCAGTCCGAGCT(配列番号:5);b)CGGACTGGACACCTGTTAATGCAGCAACAAGAAAAGCCAGCTCAGCCCAAACTCATG(配列番号:6)。
【0033】
分泌免疫複合体をコードするこれら3つのcDNAを、最初に配列決定のためにクローニングベクターにライゲートし、次にCHO又はショウジョウバエS2細胞へのトランスフェクションのために発現ベクターpcDNA3.1及びpMK33/pMtHyにそれぞれライゲートした。CHO細胞についてのトランスフェクション手順は、細胞をRPMI+10%FCS中で増殖させ、Superfect(商標)(Qiagen)を用いて発現ベクター5μgで形質移入することを伴った。安定なトランスフェクタントをRPMI+10%FCS+G418 1mg/ml中で選択した。蛋白質の発現のために、トランスフェクションしたCHO細胞を最初にCHO血清不含培地(Sigma)中での増殖に順応させ、その後無血清培地中で懸濁液培養(2×105細胞/ml)として3日間増殖させた。ショウジョウバエS2細胞についてのトランスフェクション手順は、Ex−cell培地+10%FBS中で細胞を増殖させ、LipofectinTM(Gibco−BRL)を用いて発現ベクター10μgで形質移入することを伴った。安定なトランスフェクタントをEx−cell培地+10%FBS+ヒグロマイシン300μg/ml中で選択し、無血清Ex−cell培地中懸濁液培養としての増殖に順応させた。硫酸銅500μMを加えて、懸濁液培養においてコードされた免疫複合体の発現を誘導した。
【0034】
トランスフェクションしたCHO又はショウジョウバエS2細胞によって分泌された免疫複合体を、プロテインAマトリックス(Pierce)上でのアフィニティークロマトグラフィーによって培地から精製した。
【0035】
蛍光活性化細胞選別(FACS)によって測定した免疫複合体の結合特異性
黒色腫細胞と対照細胞を非酵素的解離培地(Sigma)において採集し、PBS/BSA+0.1%アジ化ナトリウムで洗って、免疫複合体(1μg/ml)を加えたPBS/BSA中で、又は対照として免疫複合体なしのPBS/BSA中でインキュベートした。細胞をPBS/BSAで洗い、蛍光標識抗ヒトFcγ鎖(Vector)と共に4℃で30分間インキュベートして、Becton−Dickenson FACsort装置で分析した。
【0036】
免疫複合体による黒色腫細胞抽出物の免疫沈降反応
ヒト黒色腫細胞A2058系統からの約1×107細胞のサンプルを、PBS中免疫複合体10μg/ml、1%BSA及び0.05%アジ化ナトリウムを含む溶液に懸濁し、氷上で30分間インキュベートした。細胞をPBSで2回洗い、PBS中1%NP−40、免疫複合体1μg/ml及びPMSF 0.2mMを含む溶液に20分氷上で溶解した。溶解産物をマイクロヒュージにおいて13,000RPMで5分間遠心し、上清を回収して、回転装置でひと晩プロテイン−Gビーズと共にインキュベートした。ビーズを採集し、PBS中1%NP−40を含む溶液で2回、PBSで1回洗って、ビーズを採集し、PAGEを負荷した緩衝液中で煮沸して、PAGEによって分析した。
【0037】
マトリックス援用レーザー脱着イオン化質量分析(MALDI−MS)及び液体クロマトグラフィータンデムMS(LC/MS/MS)蛋白質同定
クマシーブルーで染色した蛋白質バンドをゲルから切り出して、http://info.med.yale.edu/wmkeck/geldig3.htmに述べられているようにトリプシンで消化した。トリプシン消化物のサンプルをMicromass TofSpecSEでのMALDI−MSによって分析した。ペプチド質量検索に必要な高レベルの精度を達成するために、1060.57のプロトン化モノ同位体質量を持つブラジキニン100fmol、及び2465.2のプロトン化モノ同位体質量を持つACTHクリップを内部標準として使用した。生じたトリプシンペプチドのモノ同位体質量を、0.2ダルトンのマストレランスを用いてProFoundプログラムによりOWLデータベースに対して、及び0.015%のマストレランスを用いてPeptideSearchプログラムによりEMBL/非重複データベースに対して検索した。検索に使用した他の重要な判定基準は、140−560Kdaにわたる質量範囲、最大限1つの開裂ミス、そしてタキソノミー(分類学)に関しては制限なしとした。蛋白質化学及び質量分析試験はすべてYale UniversityのW.M.Keck Foundation & HHMI Biopolymer Laboratoryにおいて実施した。http://info.med.yale.edu/wmkekc/でさらなる情報が入手できる。
【0038】
MALDI−MS分析に使用したトリプシン消化蛋白質バンドのサンプルを、LCQイオントラップ質量分析計でも分析した。マストレランス2.0ダルトンでのタンデム質量相関アルゴリズムを用いて、MS/MSデータのSequest検索を実施し、トリプシン消化物からのペプチドと、NCBI nrデータベースでの蛋白質についての再構築した理論上のスペクトルとの間に有意の類似性が存在するかどうかを判定した。この手順についてのさらなる情報は、http://info.med.yale.edu/wmkekc/prochem.htm#ms/mspiで入手できる。
【0039】
NK細胞及び補体によるヒト黒色腫細胞の免疫複合体依存性細胞溶解に関するアッセイ
カルセイン−AM保持手順を両方のアッセイに使用した。ヒト黒色腫細胞系統又は一次ひと線維芽細胞のいずれかの標的細胞を非酵素的解離培地(Sigma)中で培養フラスコから分離し、96穴プレートに加えた(2×104細胞/穴)。付着標的細胞をPBSで1回洗い、その後免疫複合体(1μg/ml)を含む又は対照として免疫複合体を含まない無血清培地(GIBCO−BRL)中37℃で20分間インキュベートした。次に標的細胞を無血清培地中37℃で40分間、カルセイン−AM(Molecular Probes Inc.,Eugene,OR)7μMで標識した。カルセイン−AMは細胞に入り込む蛍光色素であり、そこで酵素的に変化して、細胞が溶解するまで細胞内にとどまる。NK細胞に関する細胞溶解アッセイについては、標識した標的細胞を、NK細胞対標的細胞の指示されている比率を用いてヒトNK細胞と共に37℃で3〜4時間インキュベートした。補体に関する細胞溶解アッセイについては、標識標的細胞をヒト血清又は精製ウサギ補体成分(Cedarlane laboratories,Ontario,Canada)と共に37℃で1時間インキュベートした。NK細胞又は補体とのインキュベーション後、標的細胞をPBSで2回洗い、残存する付着細胞中の蛍光(残留蛍光)をプレートリーダーで測定した。溶解緩衝液(ホウ酸ナトリウム50mM、0.1%Triton X−100、pH9.0)により37℃で3時間処理した後、標的細胞の残留蛍光を測定して、標的細胞の達成可能な最大細胞溶解を判定した。NK細胞又は補体に接触しなかった付着標的細胞の蛍光を測定して、最大残留蛍光を判定した。標的細胞の各サンプルについての%細胞溶解を次のように算定した:(サンプル標的細胞の残留蛍光−溶解標的細胞の残留蛍光)/(標的細胞の最大残留蛍光−溶解標的細胞の残留蛍光)。
【0040】
試験結果
抗黒色腫免疫複合体の合成と特性指摘
免疫複合体は、分泌についてのヒトIgG1リーダー、標的黒色腫細胞についてのヒトscFvドメイン、及びヒトIgG1−Fcエフェクタードメインをコード化した(図1)。免疫複合体分子をトランスフェクションしたCHO及びショウジョウバエS2細胞において発現させ、Fcドメインのヒンジ領域におけるジスルフィド架橋によって連結された二本鎖分子として培地から精製した(図1)。各々が融合ファージクローンE26−1又はG71−1から誘導されたscFv標的ドメインを含む、2つの免疫複合体を合成した。免疫複合体E26−1及びG71−1の結合特異性を、2つのヒト黒色腫細胞系統を用いた細胞選別(FACS)によって試験した;対照は、正常ヒトメラノサイト、線維芽細胞、微小血管及び臍血管内皮細胞、及びヒト腎細胞系統293−EBNAの一次培養であった。結果は、E26−1及びG71−1融合ファージクローンに関して得た結果と一致して、免疫複合体は黒色腫細胞系統に強く結合するが、対照には結合しないことを示した。黒色腫細胞への結合は、非酵素的解離培地(Sigma)を使用して培養フラスコから細胞を採集したときに起こったが、解離培地がトリプシンを含むとき、又は解離した細胞をその後トリプシンで処理したときには結合が起こらなかった。この所見は、免疫複合体についての同種黒色腫抗原が黒色腫細胞の表面に位置することを示唆した。同じトリプシンへの曝露が、細胞表面分子ICAM−1、MHCクラス−I及び組織因子に対する抗体の黒色腫細胞への結合には影響を及ぼさなかったことから、上記抗原はトリプシンに対して例外的に感受性であると思われる。
【0041】
免疫複合体についての同種黒色腫抗原の同定
ヒト黒色腫細胞系統A2058からの培養細胞を免疫複合体G71−1又はE26−1と平衡させ、細胞を界面活性剤で溶解した。溶解産物中の免疫複合体−抗原複合体をプロテイン−Gビーズ上に採集し、PAGEによって分析した。250kdaの見かけ分子量を持つ蛋白質バンドが黒色腫細胞において検出されたが、対照では検出されなかった。MALDI−MS法による蛋白質バンドのトリプシン消化物を分析すると、対照ゲル切片の消化物には存在しなかった75のペプチド塊が同定された。ProFoundによる蛋白質配列データベースの検索は、MCSPコア蛋白質中のペプチド塊と適合し、完全な蛋白質配列の26%に及ぶ50のペプチド塊を生じた。この同定に関するProFoundの確率スコアは1.0であり、次に最も近いスコアは2.2E−61であった。Peptide Searchによる検索では、45のペプチドがMCSPコア蛋白質に適合し、38ペプチドがそれに続く最も近い適合であった。
【0042】
250kda蛋白質のトリプシン消化物をLC/MS/MS法によっても分析した(Stone,K.L.ら(1998)Electrophoresis 19,1046−1052)。トリプシン消化物からのMS/MSデータのSequest検索は、2個又はそれ以上のペプチドについてのMS/MSスペクトルと、NCBI nrデータベースにおけるMCSPコア蛋白質からの2個又はそれ以上のペプチドについての再構築した理論上のMS/MSスペクトルの間で有意の類似性を示した。
【0043】
2つの質量分析法の結果は、免疫複合体G71−1及びE26−1によって免疫沈降した黒色腫蛋白質がMCSPコア蛋白質に適合することを示している。
【0044】
NK細胞及び補体によって仲介される黒色腫細胞の免疫複合体依存性細胞溶解
免疫系の細胞溶解経路の1つは、標的細胞に直接結合して、標的細胞の抗体依存性溶解を引き起こすことができるNK細胞を含む。NK細胞はまた、抗体のFcエフェクタードメインに結合して、抗体の標的ドメインに結合する細胞の抗体依存性溶解をもたらすことができる。この抗体依存性細胞を介する細胞溶解経路(ADCC)は同時に、Fcエフェクタードメインを含む免疫複合体の標的ドメインに結合する細胞の溶解も引き起こすはずである。免疫複合体E26−1及びG71−1に依存するADCC応答を試験するため、黒色腫細胞と線維芽細胞を蛍光色素カルセイン−AMで標識し、標識細胞をヒトNK細胞単独又はNK細胞と免疫複合体と共にインキュベートした。無傷のままで細胞中に保持された蛍光色素の量を測定することによって細胞溶解を検定した。E26−1についての結果(図2)は、免疫複合体及びNK細胞とのインキュベーション後、溶解黒色腫細胞のパーセンテージが免疫複合体なしで得られる基線値を上回って上昇し、NK細胞対黒色腫細胞の比率が20/1でほぼ100%に達した。黒色腫細胞の効率的な溶解に対し、線維芽細胞は、免疫複合体及びNK細胞とのインキュベーション後細胞溶解の有意の上昇を示さなかった。G71−1免疫複合体に関しても同様の結果が得られた。
【0045】
NK細胞による溶解に対しての標的細胞の感受性は、ICAMのような付着分子の標的細胞表面での発現によって上昇し、MHCクラスI分子の発現によって低下する(Zami,L.ら(1995)Cell.Immunol.164,100−104及びStorkus,W.J.ら(1989)Proc.Natl.Acad.Sci.USA 86,2361−2364)。これらの分子の発現の差がADCCアッセイにおける黒色腫細胞及び線維芽細胞の感受性の差に寄与するかどうかを調べるため、黒色腫細胞及び線維芽細胞によるICAM−1及びMHCクラスI分子の発現をFACSによって測定した。両方の分子の発現は2つの細胞型において同様であり、NK細胞による黒色腫細胞の特異的溶解は、黒色腫細胞で発現される同種抗原への免疫複合体の結合に依存することを示唆した。
【0046】
免疫系の別の細胞溶解経路は、C1q分子が標的細胞に結合した抗体のFc領域と反応したときに活性化される補体カスケードを含む(Bruggemann M.ら(1987)J.Exp.Med.166,1351−1361)。免疫複合体E26−1及びG71−1に依存する、黒色腫細胞に対する補体仲介の細胞溶解応答を調べるため、上記で報告したNK仲介の細胞溶解応答に関して使用したのと同じアッセイ手順を、1箇所変更を加えて使用した、すなわちNK細胞の代わりに補体カスケードの成分を含むヒト血清又はウサギ血清を用いた。結果(図3及び表1)は、免疫複合体とヒト血清又はウサギ血清とのインキュベーション後、溶解した黒色腫細胞の分画が4%からほぼ100%に上昇した。黒色腫細胞の効率的な溶解に対して、線維芽細胞は免疫複合体及びヒト血清とのインキュベーション後溶解細胞分画の小さな上昇を示し、免疫複合体及びウサギ血清とのインキュベーション後には有意の上昇を示さなかった。
【0047】
【表1】
【0048】
考察
本試験については、2個のクローンからのscFv分子を、ヒトIgG1−Fcエフェクタードメインを含む免疫複合体を構築するための標的ドメインとして使用した(図1)。2つの免疫複合体によってヒト黒色腫細胞から免疫沈降した蛋白質を、黒色腫関連硫酸コンドロイチンプロテオグリカン(MCSP)のコア蛋白質として質量分析法によって同定した(Bumol,T.F.& Reisfeld,R.A.(1982)Proc.Natl.Acad.Sci.USA 79,1245−1249及びBumol,T.ら(1984)J.Biol.Chem.259,12733−12741)。MCSP分子は最初mAb 9.2.27によって認識される同種抗原として同定されたが、これはいくつかの他のmAbについての同種抗原であると思われる(Reisfeld,R.A.& Cheresh,D.A.(1987)Adv.Immunol.40,323−377;Wilson,B.S.ら(1981)Int.J.Cancer 28,293−300;Hellstrom,I.ら(1983)J.Immunol.130,1467−1472)。おそらくMCSPと同じである、黒色腫抗原HMW−MAAに結合するいくつかのscFv融合ファージクローンが、精製HMW−MAAに対するパニングによって合成ヒトscFvライブラリーから同定されている(Desai,S.A.ら(1998)Cancer Res.58,2417−2425)。MCSP分子は主として大部分のヒト黒色腫細胞の表面で発現されるが、グリア腫瘍の毛細管内皮細胞でも発現される。MCSPが、黒色腫細胞に対するパニングによって黒色腫患者のscFv融合ファージライブラリーから単離された少なくとも2つの黒色腫特異的クローンについての同種抗原であるという所見は、MCSPがin vivoでの支配的な黒色腫抗原であることを示唆する。
【0049】
2つの免疫複合体の治療上の潜在的効力の初期試験として、蛍光標識標的細胞を含むin vitroでの細胞溶解アッセイを使用して、免疫複合体がNK細胞及び補体による溶解のためにヒト黒色腫細胞を標的する能力を調べた。両方の免疫複合体が黒色腫に対するNK細胞及び補体の細胞溶解活性の明らかな上昇をもたらし、標的黒色腫細胞群の実質的に完全な溶解を生じさせたが、対照として使用した線維芽細胞に関しては、溶解のごくわずかな上昇をもたらすか又は全く上昇を生じさせなかった。NK細胞及び補体による黒色腫細胞と線維芽細胞の免疫複合体非依存性細胞溶解の有意のバックグラウンドも存在した。これは、腫瘍細胞、NK細胞及び補体を別々の個体から単離する同種異系アッセイについて予想されるものである(Ciccone,E.ら(1992)J.Exp.Med.175,709−718)。これらの成分はすべて自己由来であるので、癌患者においてはこのバックグラウンドは低下するはずである。
【0050】
in vivo細胞溶解試験の結果は、免疫複合体が、腫瘍特異的scFv又はVH標的ドメインが使用可能である黒色腫及び他の癌に関する免疫療法プロトコールにおいて潜在的な役割を持つであろうことの予備的な証拠を提供する。MCSPに結合するマウスmAb 9.2.27による黒色腫免疫療法についての限られた第I相臨床試験は、付随する毒性の証拠を伴わずに、腫瘍における抗体の特異的局在を示した(Oldham,R.K.ら(1984)J.Clin.Oncol.2,1235−1244)。より小さな分子サイズは腫瘍への侵入を改善するはずであり、また分子がヒト由来であることは免疫拒絶反応を最小限に抑えるはずであるので、MCSPに結合するscFv免疫複合体は免疫療法のためにマウスmAbよりも有効であると考えられる。G71−1及びE26−1免疫複合体は、それらのVH配列の重要な相違(配列番号:11と配列番号:12及び次の実施例参照)によって示唆されるように、おそらく異なるMCSPエピトープに結合し、それ故治療効果を高めるために一緒に投与することができるであろう。
【0051】
実施例2
この実施例は、腫瘍血管系と腫瘍細胞の両方を標的する免疫療法治療が、ヒト黒色腫の重篤な複合型免疫欠損マウス異種移植モデルにおいて有望な成績を示したことを報告する。手順は、各々がヒトIgG1のFcエフェクタードメイン領域に複合した腫瘍標的ドメインから成る2つの免疫複合体の、SCIDマウスへの全身送達を伴った。エフェクタードメインは、ナチュラルキラー(NK)細胞及び補体による標的細胞に対しての細胞溶解性免疫応答を誘導する。免疫複合体は、複製不全アデノウイルスベクターにおいて分泌分子としてコードされる。
【0052】
試験材料及び方法
細胞系統
黒色腫細胞系統LXSN、TF2及びLXSN/VEGFを、レトロウイルス仲介のトランスフェクションとクローニングによってヒト黒色腫細胞系統YU−SIT1から誘導した。LXSN系統は対照レトロウイルスでトランスフェクションされており、低レベルのTFを発現する。TF2系統はTF cDNAをコードするレトロウイルスでトランスフェクションされており、高レベルのTFを発現する。LXSN/VEGF系統はVEGF cDNAをコードするレトロウイルスでトランスフェクションされており、高レベルのVEGFを発現する。ヒト腎細胞系統293をAmerican Type Culture Collectionから購入した。
【0053】
プラスミドベクター
scFv(G71−1)免疫複合体をコードするプラスミドベクターの構築を実施例1で述べた(配列番号:11参照)。マウス第VII因子(mfVII)免疫複合体をコードするベクターの構築のために、5'プライマーACGATCTTAAGCTTCCCCACAGTCTCATCATGGTTCCA(配列番号:7)及び3'プライマーACGGTAACGGATCCCAGTAGTGGGAGTCGGAAAACCCC(配列番号:8)を使用して、マウス肝臓cDNAライブラリー(Quick−Clone cDNA,Clonetech)からのPCRによってmfVII cDNAを増幅した。停止コドンを含まないリーダー及びコード配列を含む、増幅したmfVII cDNAを、ヒトIgG1 FcドメインをコードするcDNAと共にpcDNA3.1(+)ベクター(Invitrogen)のフレーム内のHindIII及びBamHI部位にクローニングした。ベクターDNAをHB101コンピテント細胞(Life Technologies)において増幅し、配列決定した。リシン−341をアラニンコドンで置換することによってmfVII cDNAの活性部位を突然変異させた(Dickinson,C.D.ら(1996)Proc.Natl.Acad.Sci.USA 93,14379−14384)。突然変異誘発の手順は、QuickChange部位特定突然変異誘発マニュアル(Stratagene)の中で述べられているように実施した。5'プライマーはGGTACCAAGGACGCCTGCGCGGGTGACAGCGGTGGCCCA(配列番号:9)であり、3'プライマーはTGGGCCACCGCTGTCACCCGCGCAGGCGTCCTTGGTACC(配列番号:10)であった。活性部位突然変異を有するmfVII cDNAをmfVIIasmと称する。mfVIIasm cDNAを含むプラスミドをHB101コンピテント細胞に形質転換し、2×TY/カルベニシリン寒天上で形質転換コロニーを選択した。プラスミドDNAの配列は、mfVIIasm DNAにおけるlys−341コドン(AAG)のアラニンコドン(GCG)による置換を示した。
【0054】
CHO細胞における免疫複合体の合成
免疫複合体cDNAをCHO細胞に形質移入し、クローンを単離するための手順を実施例1で述べた。形質移入したCHO細胞をCHO血清不含培地(EX−CELL 301,JRH Biosciences)で培養した;mfVIIasm免疫複合体の合成のため、CHO血清不含培地に最終濃度1μg/mlまでビタミンK1(Sigma)を補足した。免疫複合体をプロテインAビーズ(Pierce)でのアフィニティークロマトグラフィーによって精製し、濃縮して、Ultrafree−15 Biomax−50フィルター(Millipore)を通しての遠心分離によって脱塩し、Tris−HCl pH8.0 10mMに調整した。Bio−Rad蛋白質アッセイ手順によって免疫複合体の濃度を測定した。
【0055】
蛍光活性化細胞選別(FACS)
黒色腫細胞を非酵素的解離溶液(Sigma)中で採集し、洗って、TBS/BSA/Ca2+(Tris−HCl pH7.4 10mM、NaCl 150mM、CaCl2 20mM、1%BSA及び0.1%NaN3)に再懸濁した。免疫複合体を加えて(最終濃度5μg/ml)、mfVIIasm免疫複合体については37℃で30分間、G71−1免疫複合体については氷上で30分間、細胞をインキュベートした;対照細胞は免疫複合体を加えずにインキュベートした。インキュベーション後、細胞をTBS/BSAで洗い、蛍光標識抗ヒトFcγ鎖(Vector Laboratories)と共に氷上で30分間インキュベートし、Becton−Dickenson FACsort装置で分析した。
【0056】
アデノウイルスベクター
アデノウイルスベクター系は、シャトルベクターpAdTrack−CMV及びpShuttle−CMVと、バックボーンベクターpAdEasy−1から成る(He,T.C.ら(1998)Proc.Natl.Acad.Sci.USA 95,2509−2514)。免疫複合体cDNAを、HindIIIで消化してpcDNA3.1プラスミドベクターから単離し、次いでクレノウ断片で消化して3'陥凹末端を埋め、その後Not1で消化してcDNA挿入物を放出させ、それをアガロースゲル電気泳動によって精製した。シャトルベクターは、最初にKpn1で消化し、次にクレノウ断片で消化して、その後Not1で消化した。免疫複合体cDNAをT4 DNAリガーゼと共に16℃でひと晩インキュベートしてシャトルベクターにライゲートし、かかるシャトルベクターを熱ショックによってHB101コンピテント細胞に形質転換した。形質転換コロニーを2×TY/カナマイシン寒天上で選択し、シャトルベクターを抽出して精製した。精製シャトルベクターとpAdTrack−CMV DNAをPmelにより37℃で2時間消化した。シャトルベクターDNA 500ngとpAdEasy−1 DNA 100ngの混合物をBJ5183コンピテント細胞に電気穿孔し、細胞を37℃で15分間振とうして、LB/カナマイシン寒天に接種した。プレートを37℃でひと晩インキュベートし、形質転換コロニーを単離した。プラスミドDNAをミニプレップから精製し、0.6%アガロースゲルでの電気泳動によって組換えアデノウイルスDNAに関してスクリーニングした。
【0057】
免疫複合体をコードする組換えアデノウイルスDNAを、CHO細胞のトランスフェクションに関して上述したプロトコールに従って、1×105の293細胞にトランスフェクションした。トランスフェクションの7日後に細胞を採集し、3回の凍結−融解サイクルによってアデノウイルスを放出させ、1つの150mm培養プレートに293細胞を感染させて増幅した。2日後、上述したようにアデノウイルスを採集し、20の培養プレートに293細胞を感染させて再び増幅した。増幅したアデノウイルスを2日後に採集し、CsCl中での遠心分離によって精製した。最終的な収量は通常、260nmでの吸光度から評価したとき約1013ウイルス粒子であった;換算は、1O.D.単位=1×1012粒子である。精製アデノウイルスをPBSで透析し、−80℃で保存した。
【0058】
SCIDマウス実験
SCIDマウスはTaconic Laboratoriesからの4週齢から5週齢の雌性であった。マウスの右ひ腹に5×105のTF2又はLXSNヒト黒色腫細胞を皮下注射した。腫瘍が、皮膚表面下で触知可能な大きさ(〜5mm3)又は皮膚表面より上のより大きなサイズ(〜50mm3)に成長したあと、尾静脈を通して免疫複合体をコードするアデノウイルスベクター、又は対照として免疫複合体をコードしないアデノウイルスベクターをマウスに注入した。血液中に分泌される免疫複合体蛋白質の濃度を、一方の眼からの血液約0.1mlをヘパリン被覆した微小毛管に採集し、血液を遠心分離にかけて細胞を取り出すことによって測定した。上清血漿を炭酸水素ナトリウム緩衝液pH9.6で希釈し、プロ結合アッセイプレート(Falcon)の穴に分配して、プレートを37℃で2時間、次いで4℃でひと晩インキュベートした。穴をPBS中5%脱脂乳で30分間遮断し、PBSで3回洗って、5%脱脂乳中1:2000に希釈したペルオキシダーゼ標識抗ヒトIgG抗体を穴に加えた。プレートを室温で1時間インキュベートし、PBS中で洗って、ペルオキシダーゼ基質OPDを加え、マイクロプレートリーダーにおいて490nmで吸光度を測定した。蛋白質標準は、プロテインAビーズでのクロマトグラフィーによって精製したヒトIgG(Sigma)であった。
【0059】
SCIDマウスの皮膚に発現する腫瘍の大きさをカリパスにより二次元で測定し、式、(幅)2(長さ)/2によって腫瘍容積を推定した。実験の最後に、マウスを解剖して腫瘍を計量した。器官を損傷の形態学的証拠に関して検討し、組織学的検査用にパラフィン切片を調製した。
【0060】
免疫組織化学
腫瘍と器官のパラフィン切片をPBS+0.3%H2O2中で30分間インキュベートし、TBS/BSA緩衝液中で30分間遮断した。TBS/BSA/Ca2+緩衝液中にmfVIIasm免疫複合体10μg/mlを含む溶液、又は対照として免疫複合体を含まない緩衝液を切片に加え、37℃で1時間インキュベートした。同じ緩衝液で3回洗った後、切片をアルカリホスファターゼで標識した抗ヒトγ鎖抗体と共に室温で1時間インキュベートして、青色を生じるBCIP/NBTで染色し、メチルグリーンで対比染色した。
【0061】
試験結果
免疫複合体の特性
scFv(G71−1)及び突然変異マウス第VII因子(mfVIIasm)免疫複合体をCHO細胞において合成し、プロテインAビーズでのアフィニティークロマトグラフィーによって培地から精製した。SDS−PAGEによる先の分析は、G71−1免疫複合体が、おそらくFcドメインのヒンジ領域間のジスルフィド架橋によって結合した、2つの同じ鎖から成ることを示した(実施例1)。mfVIIasm免疫複合体に関して同じ結果を得た。(配列番号:11及び配列番号:12参照)。mfVIIasm免疫複合体は2つの標的ドメインを持つので、単量体内因性fVII分子における1つの標的ドメインと比較して、2個のTF分子に協力的に結合し、TFを発現する細胞に対して内因性fVIIよりも強い結合をもたらすことができる。競合的FACSアッセイは、ヒトfVIIaが、ヒト黒色腫細胞上のアクセス可能部位の半数への結合に関して等モルベースでmfVIIasm免疫複合体と競合することを示した。これはおそらく、免疫複合体上の標的ドメインの1つだけがこれらの部位でTFに結合することができるからであろう。残りの部位へのmfVIIasm免疫複合体の結合は、10倍過剰のヒトfVIIaの存在下で競合することができず、免疫複合体分子の両方の標的ドメインがこれらの部位で結合することができ、強いアビディティ効果をもたらすことを示唆した。黒色腫細胞上のTF分子の約半分だけが第二TF分子に十分に近接していて、mfVIIasm免疫複合体上の両方の標的ドメインについて協力的結合部位を形成すると思われる。
【0062】
免疫療法試験のための異種移植片を、それぞれ低レベル又は高レベルのTFを発現するヒト黒色腫細胞系統LXSN及びTF2から作製した。TF2細胞によるより高いレベルのTF発現と一致して、FACSによって測定したとき、mfVIIasm免疫複合体はLXSN細胞よりもTF2細胞とより広汎に結合する。またmfVIIasm免疫複合体を、高レベルのVEGFを生成する黒色腫細胞系統LXSN/VEGFから作製したヒト黒色腫異種移植片の切片への結合に関して免疫組織化学によっても試験し、密に血管形成した異種移植片を生じた。腫瘍血管内皮細胞並びに腫瘍細胞に対して結合が起こり(図4)、異種移植片において両方の細胞型によってTFが発現されることを示唆した。正常マウスの肝臓、腎臓、肺及び脳の切片に関する免疫組織化学試験は、TFが非腫瘍性組織の血管内皮細胞によっては発現されないという他の証拠と一致して、mfVIIasm免疫複合体がこれらの組織における血管内皮細胞とは結合しないことを示した。
【0063】
免疫療法試験
SCIDマウスへの全身送達のために、各々の免疫複合体をpAdEasy−1に基づく複製不全アデノウイルスベクター系(Heら、前出)において分泌分子としてコードし、最初にヒト黒色腫細胞を皮下注入しておいたマウスの尾静脈に前記ベクターを注入した。最初の免疫療法試験は、触知可能なTF2異種移植片を発現したマウスに、各々のベクターを別個に、及び両方のベクターを一緒に注入することを伴った。合計3回の注入を週に1回の間隔で投与し、最後の注入から6日後に実験を終了した。1回目と2回目の注入後にELISAによって血液中の免疫複合体の濃度をモニターした(図5)。1回目の注入後の平均濃度は、G71−1免疫複合体については4mg/ml、mfVIIasm免疫複合体については0.04mg/mlであり、合成の速度がG71−1免疫複合体についてはmfVIIasm免疫複合体よりも約100倍高いことを示唆した。2回目の注射後各々の免疫複合体の濃度が上昇し、さらなる細胞がアデノウイルスに感染していたことを示した。皮膚表面に現われた腫瘍の大きさを二次元で測定し、腫瘍容積を評価するための測定法を用いて、異種移植片の成長をモニターした(図6)。免疫複合体をコードしないアデノウイルスを注入した対照マウスでは、腫瘍が比較的速い速度で持続的に成長し、20日後に約2,000mm3の平均容積に達した。免疫複合体をコードするアデノウイルスを注入したマウスでは、腫瘍の成長が阻害された;阻害はG71−1免疫複合体よりもmfVIIasm免疫複合体の方が強力であった。実験終了時にすべてのマウスが活動的なままであり、健康と思われ、肝臓、脾臓、肺、腎臓及び脳の組織学的検査は壊死、凝固又は出血の兆候を示さなかった。剖検後の腫瘍重量は、推定腫瘍容積と一致して、対照マウスでは免疫複合体で処置したマウスよりも低かった(図7)。腫瘍重量の最強の低下は、両方の免疫複合体で処置したマウスで起こった。
【0064】
次の2つの実験は、免疫複合体の治療効果に影響を及ぼしうる2つのパラメータ、すなわち異種移植片の初期サイズと黒色腫細胞によるTF発現のレベルを試験するためにデザインされた。(i)前記の免疫療法試験は、ヒトでの小さな腫瘍に相当する、約5mm3の推定容積に成長した触知可能な黒色腫異種移植片に関するものであった。より大きな異種移植片に対する免疫複合体の治療効果を調べるため、TF2異種移植片を推定容積約50mm3に成長させた後、2つのアデノウイルスベクターの尾静脈注入を開始した。3週間の期間中マウスに4回注入を行い、最後の注入から2日後に実験を終了した。免疫複合体をコードするアデノウイルスを注入したマウスにおける平均腫瘍容積は、実験開始時と実験終了時でほぼ同じであったのに対し、対照アデノウイルスを注入したマウスでの平均腫瘍容積は同じ期間中に約27倍上昇した(図8)。これらの結果は、腫瘍の成長が、大きな腫瘍に関しても小さな腫瘍と同じくらい有効に阻害されることを示している。免疫複合体をコードするアデノウイルスを注入した5匹のマウスのうち1匹が、3回目の注入から5日後に死亡した;マウスが検査の時点までに回収されなかったため、死亡の原因は判定できなかった。(ii)mfVIIasm免疫複合体の効果に影響を及ぼしうる1つのパラメータはTF発現のレベルであり、これは種々の腫瘍間で異なる(Callender,N.S.ら(1992)Cancer 70,1194−1201)。異種移植片での黒色腫細胞によるTFの発現が変化することの影響を調べるため、より高レベルのTFを発現する関連系統TF2から作製した異種移植片との比較のために、黒色腫細胞系統LXSNを使用して低レベルのTFを発現する異種移植片を作製した(Bromberg,M.E.ら(1999)Proc.Natl.Acad.Sci.USA 92,8205−8209)。異種移植片が触知可能な大きさに達した後、次の3週間にわたってマウスにmfVIIasm免疫複合体をコードするアデノウイルス又は対照アデノウイルスの5回の注射を実施した(図11)。対照アデノウイルスを注入した5匹のマウスでは、異種移植片は持続的に成長し、平均容積は最後の注入から2日後に1350mm3まで上昇した。同じ期間中、mfVIIasm免疫複合体を注入した5匹のマウスにおける異種移植片の平均容積は20mm3に上昇し、腫瘍の成長がLXSNとTF2異種移植片について同等であることを示した(図9と6の比較)。最後の注入の1日後に実施した剖検は、mfVIIasm免疫複合体をコードするアデノウイルスを注入した5匹のマウスのうち2匹において異種移植片が根絶されていたことを示した;他の3匹のマウスにおける平均腫瘍重量が0.11グラムであったのに対し、対照アデノウイルスを注入した5匹のマウスでは0.75グラムの平均重量であった。これら3匹のマウスから回収された小さな腫瘍は広範囲の領域の細胞壊死を示したが、対照マウスから回収したより大きな腫瘍においては壊死は生じなかった(図10)。この実験の終了時にはすべてのマウスが健康であると思われたが、解剖したマウスの形態学的検査は、mfVIIasm免疫複合体をコードするアデノウイルスを注入した5匹のマウスにおいて肝臓と脾臓への損傷を明らかにした。肝臓と脾臓の組織学的検査は、肝細胞の多くが腫脹し、脾が赤血球で広汎に浸潤されていることを示した。腫脹した肝細胞は、mfVIIasm免疫複合体をコードするアデノウイルスの3回の注入後の先の実験においても発生していたが、脾は正常であり、脾臓の欠陥が最後の2回の注入経過で発現したことを示唆した。1匹のマウスは硬膜下脳出血も有していたが、これは今回の実験からの他のマウスあるいはこれまでのいずれの実験においても起こっていなかった。この欠陥が脳血管系で発現されたTFへのmfVIIasm免疫複合体の結合によって誘発されたのかどうかは不明である。
【0065】
考察
この実施例における免疫療法処置は、各々がヒトIgG1 H鎖のFc領域に複合した腫瘍標的ドメインから成り、カメリド(Camelid)H鎖抗体に類似したホモダイマー分子を形成する、2つの免疫複合体をSCIDマウスに全身送達することを伴った。1つの型の免疫複合体については、腫瘍標的ドメインは、異種移植片において黒色腫細胞によって発現される黒色腫抗原MCSPに結合するヒトscFv分子G71−1であった。他の型の免疫複合体では、腫瘍標的ドメインは、腫瘍血管系内皮細胞によって発現されるマウスTFと異種移植片において黒色腫細胞によって発現されるヒトTFの両方の組織因子(TF)に特異的且つ強力に結合するマウス第VII因子分子であった。第VII因子免疫複合体のTFへの結合から起こりうる播種性血管内凝固(DIC)の危険性を低下させるために、活性部位の突然変異をマウスfVII標的ドメイン(mfVIIasm)に導入し、血液凝固経路を開始させるために必要な蛋白質分解活性を阻害した。
【0066】
実施例1で報告した試験は、G71−1免疫複合体が、NK細胞と補体による培養ヒト黒色腫細胞の細胞溶解を仲介することを示した。SCIDマウスは機能的NK細胞と補体を産生する能力を保持するので、免疫複合体も、SCIDマウスにおいて成長するヒト黒色腫異種移植片の標的腫瘍細胞及び血管内皮細胞の細胞溶解を仲介することができると考えられる。免疫複合体のSCIDマウスへの全身送達を、免疫複合体をコードする複製欠損アデノウイルスベクターの尾静脈注入によって実施した。免疫複合体は各注入後少なくとも1週間血液中に分泌された。最初に低レベル又は高レベルのTFを発現するヒト黒色腫細胞系統をマウスに皮下注射し、生じた異種移植片を小(〜5mm3)又は大(〜50mm3)腫瘍に成長させた後、アデノウイルスベクターの注入を開始した。G71−1免疫複合体をコードするアデノウイルスと別々に又は一緒に投与した、mfVIIasm免疫複合体をコードするアデノウイルスの多回注入により、3〜4週間の実験期間中、すべての異種移植片のさらなる成長が妨げられた;一部のマウスでは、異種移植片は完全に退縮した。免疫複合体をコードしないアデノウイルスを注入した対照マウスでは、異種移植片の平均容積は同じ期間中に約25倍に上昇した。免疫複合体をコードするアデノウイルスベクターの5回の注入を受けたマウスでは、肝細胞の多くが腫脹し、脾が赤血球で浸潤されていた。この欠陥は、肝細胞又は脾細胞には結合しない分泌免疫複合体によって引き起こされたものではなかった。主要な原因はおそらく、主として静脈内注入されたアデノウイルスベクターに感染したマウス細胞である肝細胞による、コードされた免疫複合体の持続的な高いレベルの合成である。アデノウイルスベクターを注入したSCIDマウスの血液中の免疫複合体濃度は、mfVIIasm免疫複合体よりもG71−1免疫複合体の方が約100倍高かったが、それにもかかわらずヒト黒色腫異種移植片への阻害作用はmfVIIasm免疫複合体の方がより強力であった。mfVIIasm免疫複合体の鍵となる利点は、G71−1免疫複合体が黒色腫細胞だけに結合するのに対して、腫瘍細胞と同様に腫瘍血管内皮細胞に対しても結合が起こることである。TFが正常血管系によっては発現されないので、腫瘍血管系への結合は腫瘍特異的であるはずである。TFは、脳、肺及び腎糸球体のような他のいくつかの正常組織によって発現されるが、血管壁はより大きな血液成分を隣接細胞から分離するバリアを形成するので、これらのTF分子は内因性fVII又はfVII免疫複合体にアクセスできない。しかし、腫瘍血管は漏出性であり、腫瘍細胞によって発現されたTFへのアクセスを可能にする。それ故、ヒトfVIIasm免疫複合体は、血管内皮細胞及び腫瘍細胞上でTFを発現する広いスペクトルのヒト腫瘍にとって有効な治療薬でありうる。TF以外の腫瘍細胞標的に結合するヒトscFv免疫複合体を同時に投与することにより、ヒトfVIIasm免疫複合体の治療効果を増強することができる。
【0067】
実施例3
この実施例は、ヒト皮膚及び転移性肺黒色腫のSCIDマウスモデル、及びマウス黒色腫の免疫コンピテントマウスモデルにおいて検討した癌治療のためのプロトコールの効果と安全性の試験を報告する。プロトコールは、標的新生血管系内皮細胞及び腫瘍細胞に対する細胞溶解性免疫応答を誘導する、本発明の免疫複合体をコードする複製不全アデノウイルスベクターの腫瘍内注入を伴った。マウスモデルの実験は、単独で又は一本鎖Fv免疫複合体と共に第VII因子免疫複合体を腫瘍内送達すると、正常器官への損傷の兆候を伴わずに、注入した腫瘍、そして同時に離れた非注入転移性腫瘍の成長の阻害と退縮も生じさせることを示した。おそらく新生血管系内皮細胞上の組織因子に結合した第VII因子免疫複合体によって仲介される、腫瘍新生血管系の広い範囲に及ぶ破壊が存在した。組織因子は一般に新生血管内皮細胞と腫瘍細胞で発現されるので、第VII因子免疫複合体は広い範囲のヒト腫瘍に対する免疫療法のために使用することができる。
【0068】
試験材料及び方法
細胞系統
LXSN、TF2及びYusac2はヒト黒色腫細胞系統であり、Cakiはヒト腎腫瘍細胞系統であり、LnCapはヒト前立腺癌細胞系統であり、A204はヒト神経線維芽細胞腫系統であり、B16F10はマウス黒色腫細胞系統であり、EMT6はマウス乳腺癌細胞系統であり、BT20はヒト乳癌細胞系統であり、Colo 357はヒト膵癌細胞系統であり、MSはヒト胃癌細胞系統であり、293はアデノウイルスベクターをパッケージングするために使用されるヒト腎細胞系統(ATCC、CRL−1573)である。培地は、LnCapを除くすべての腫瘍系統についてDMEM+10%FBSであった。LnCapはRPMI 1640+10%FBSで培養した。
【0069】
アデノウイルスベクター
1)mfVIIasm及びG71−1免疫複合体をコードするアデノウイルスベクターを作製し、精製するための上記実施例2で要約した手順を、次の修正を加えて使用した。シャトルベクターDNAをPmeIで消化した後、エタノール沈殿ステップの代わりにアガロースゲルでの電気泳動によってDNAを精製し、その後QIAEX IIキット(Qiagen)を用いてDNAバンドを単離した。2)精製した試料中のベクター濃度を、次のような感染粒子(IP)及び感染単位(IU)についてのアッセイによって測定した。(i)IPアッセイは、ベクター試料を溶解緩衝液(PBS中0.1%SDS)で20倍に希釈し、260nmで吸光度を測定することを含む;IPへの換算は、1O.D.単位=1×1012IPである。(ii)IUアッセイは、293細胞培養を連続希釈ベクター試料に感染させ、感染培養を2日間インキュベートして、細胞をベクターゲノムにおけるグリーン蛍光蛋白質(Green Fluorescent Protein,GFP)遺伝子の発現に関して蛍光顕微鏡で検査することを含む。GFP遺伝子を発現する細胞数からIU力価を算定する。IP及びIU力価は±10%以内で一致した。
【0070】
腫瘍細胞におけるmfVIIasm免疫複合体の合成
腫瘍細胞を150mmの皿でほぼ集密まで増殖させ、10IU/細胞の感染多重度でmfVIIasmをコードするアデノウイルスに細胞を感染させた。感染細胞を無血清培地で4日間培養し、培地1.5mlをプロテインAビーズ(Pierce)の懸濁液10μlと混合して、4℃でひと晩回転流動した。SDS−PAGE負荷緩衝液15μl中80℃で3分間ビーズを加熱して、結合mfVIIasm免疫複合体を溶出し、溶離液をSDS−PAGEによって分別して、ニトロセルロース膜に移した。免疫複合体のバンドを、ヤギ抗ヒト又は抗マウスIgG(Fc特異的)プローブでの免疫染色によって検出した。
【0071】
免疫欠損マウスにおける免疫療法試験
4から5週齢の雌性C.B−17 SCIDマウス(Taconic Farms)を免疫欠損マウスに関するすべての実験に使用した。ヒト黒色腫細胞系統LXSN又はTFの単層培養をPBS+2mM EDTA中で解離し、洗って、PBSに再懸濁した。5×105細胞を右ひ腹に皮下注入して皮膚腫瘍を生成し、6×105TF細胞を尾静脈に静脈内注入して転移性肺腫瘍を生成した。皮膚腫瘍の大きさをカリパスにより二次元で測定し、腫瘍容積を(幅)2(長さ)/2として評価した。皮膚腫瘍が約100mm3の容積まで成長したときに、ヒトFcエフェクタードメインを含むアデノウイルスベクターの腫瘍内注入を開始した。各々の注入ステップに関して、総容量50μlのベクター試料を腫瘍の3又は4部位に注入した。実験の終了時に剖検を実施して血液サンプルを採集し、腫瘍と正常器官を形態学的及び組織学的検査用に調製した。
【0072】
免疫コンピテントマウスにおける免疫療法試験
4から5週齢の雌性C57BL/6マウス(Charles River Laboratories)を免疫コンピテントマウスに関するすべての実験に使用した。B16F10マウス黒色腫細胞の単層培養をPBS+2mM EDTAに懸濁し、洗って、PBSに再懸濁した。5×105細胞を右ひ腹に皮下注入して皮膚腫瘍を生成した。皮膚腫瘍が約140−325mm3の推定容積まで成長したときに、マウスFcエフェクタードメインを含むmfVIIasm免疫複合体をコードするアデノウイルスベクターの尾静脈注入を開始した。プロトコールの効果と安全性をモニターするための手順は、SCIDマウスについて上述したのと同じである。
【0073】
SGOTアッセイ
マウスから血清サンプルを採集し、凍結して、標準診断キットを用いてグルタミン酸オキサロ酢酸トランスアミナーゼについてアッセイした。
【0074】
試験結果
この試験に使用した2つの免疫複合体は、腫瘍標的ドメインとエフェクタードメインから成る(図1)。標的ドメインは、腫瘍血管内皮細胞及び腫瘍細胞で発現されるTFに結合するが、血液凝固を開始させない(実施例2)、突然変異マウス第VII因子分子(mfVIIasm)、若しくはヒト黒色腫細胞で選択的に発現されるその同種抗原に結合するscFv抗体G71−1のいずれかである(CaiとGaren,前出、及び実施例1)。エフェクタードメインは、標的細胞に対する細胞溶解性免疫応答を誘導するヒト又はマウスIgG1免疫グロブリンのFc領域である(実施例1)。免疫複合体を送達するためのベクターは、分泌形態の免疫複合体をコードする複製不全アデノウイルスである(実施例1)。実施例1において、免疫複合体をコードするベクターのSCIDマウスへの静脈内注入は腫瘍の成長を阻害したが、静脈内注入したベクターによる感染の主要標的である肝臓への組織学的損傷を引き起こした。一次感染が肝細胞から腫瘍細胞に再指令されうるかどうかを調べるため、ベクターをSCIDマウスにおいて増殖するヒト皮膚黒色腫に腫瘍内注入し、ベクターゲノムに挿入したグリーン蛍光蛋白質(GFP)遺伝子の発現によって腫瘍と肝臓におけるベクターの分布をマッピングした(図14)。腫瘍においては強いGFPの発現が検出されたが、肝臓では検出されず、注入した腫瘍がベクターによる感染の主要標的であることを示した。腫瘍におけるGFP発現のパターンは、注射針が横切る経路に隣接する数層の腫瘍細胞に限定されると思われた。
【0075】
mfVIIasmとG71−1免疫複合体をコードする2つのベクターの混合物を、SCIDマウスで増殖するヒト黒色腫皮膚腫瘍に注入し、その後5日間腫瘍容積を測定することによって、腫瘍成長へのベクター投与の影響を検討した。血液中に分泌されるG71−1免疫複合体の力価がより高いことを相殺するため、混合物中のmfVIIasmベクター対G71−1ベクターの比は5:1であった(実施例2)。注入に使用した2つのベクターの結合用量は、7×108から6×109感染単位(IU)の範囲であった。対照に関しては、免疫複合体をコードしていない空ベクターを6×109IUの用量で注入した。腫瘍成長の最強の阻害は、免疫複合体をコードするベクターの最高用量で得られた。このベクターの用量を下記のすべてのSCIDマウス実験に使用した。
【0076】
次の実験では、mfVIIasmとG71−1免疫複合体をコードする2つのベクターの多回腫瘍内注入での投与が腫瘍成長に及ぼす阻害作用を19日間モニターした。アデノウイルスベクターの最後の注入から2日後のマウス血液中の免疫複合体蛋白質力価は1mg/mlから2mg/mlの範囲であり、これは静脈内注入したベクターによって生じる力価の約4分の1である(実施例2)。これらの腫瘍の平均容積は、2つのベクターの1回目の注入後1日以内に約70%低下し、その後実験の残りの期間中容積は上昇しなかった。空対照ベクターを注入した腫瘍は継続的に成長し、実験終了時には、2つの免疫複合体をコードするベクターを注入した腫瘍の30mm3に対し、1900mm3の平均容積に達した。ベクターの静脈内注入に関する先の実験(実施例2)と一致して、mfVIIasm免疫複合体をコードするベクターだけを注入した腫瘍は、両方のベクターを注入した腫瘍とほぼ同じくらい強力に阻害された。マウスは実験期間を通じて活動的なままであり、最後の注入から2日後に実施した剖検では、すべての器官が形態学的及び組織学的に正常であると思われ、出血の兆候はなかった。免疫複合体をコードするベクターを静脈内注入したSCIDマウスは肝細胞への組織学的損傷を示したので(実施例2)、腫瘍内注入したSCIDマウスの肝臓を組織学的損傷、そして機能的損傷に関しても検討した。静脈内注入マウスからの肝臓切片で認められた重要な組織学的変化に対し、腫瘍内注入マウスからの肝臓切片は比較的軽度の変化を示した(図15)。剖検の際にマウスから採取した血清中のグルタミン酸オキサロ酢酸トランスアミナーゼ酵素(SGOT)のアッセイにより、肝機能をモニターした。対照マウスと免疫複合体をコードするベクターを注入したマウスのSGOTレベルは正常範囲内(210±28U/L)のままであり、肝機能がそこなわれなかったことを示した。
【0077】
mfVIIasmとG71−1免疫複合体をコードするベクターを注入したマウスから回収した腫瘍は、おそらくTFに結合したmfvVIIasm免疫複合体によって誘導される、血管内皮細胞に対する細胞溶解性免疫応答の結果として、血管がほとんど全く存在しなかった(図13)。
【0078】
腫瘍内注入プロトコールの効果は、注入した皮膚腫瘍の阻害だけでなく、注入することができない転移性腫瘍の抑制によっても決まる。転移性腫瘍への阻害作用を調べるため、ヒト黒色腫TF細胞を尾静脈に注入して、主として肺への血液運搬性転移を生じさせたSCIDマウスモデルを使用した。同時にヒト黒色腫LXSN細胞をマウスに皮下注入し、12日後に形成された皮膚腫瘍に、2つの免疫複合体をコードするベクター又は空対照ベクターを注入した。次の8週間、週に2回のスケジュールで腫瘍内注入を続け、最後の注入から2日後に剖検を行って、肺表面上の腫瘍結節の数を測定した(表2)。対照ベクターを注入した2匹のマウスはそれぞれ14個と29個の肺結節を有しており、一方2つの免疫複合体をコードするベクターを注入した5匹のマウスに関しては、3匹は全く肺結節がなく、2匹がそれぞれ1個と5個の結節を有していた。これらの結果は、2つの免疫複合体をコードするベクターの腫瘍内注入が、注入した腫瘍の成長も遠隔転移性腫瘍の成長も阻害しうることを示している。
【0079】
【表2】
【0080】
臨床環境でのプロトコールの効果に影響を及ぼしうる別のパラメータは、免疫複合体又はアデノウイルスベクターへの患者の免疫応答である。臨床用の免疫複合体の標的及びエフェクタードメインはヒト蛋白質から誘導されることになるので、患者は免疫複合体に対して有意の免疫応答を示さないはずである。しかし、アデノウイルスベクターはヒトにおいて、またマウスにおいても強い免疫原性である。マウスにおけるアデノウイルスベクターへの免疫応答の影響を調べるため、マウス黒色腫皮膚腫瘍を保有する免疫コンピテントマウスでプロトコールを試験した。マウス黒色腫細胞をベクターに感染させることはできないので、実験はベクターの腫瘍内注入ではなく静脈内注入を伴った。また、マウス黒色腫細胞はmfVIIasm免疫複合体に結合するが、G71−1免疫複合体には結合しないので、mfVIIasm免疫複合体だけしか試験することができなかった。この実験のための免疫複合体のFcエフェクタードメインは、マウスIgG1免疫グロブリンから誘導した。結果は、mfVIIasm免疫複合体をコードするアデノウイルスベクターの静脈内注入が、SCIDマウスにおいても免疫コンピテントマウスにおいても黒色腫の成長阻害をもたらすことを示している(図16)。
【0081】
TFは一般に腫瘍血管内皮細胞、及びほとんどの転移性腫瘍細胞によっても発現されるので、fVIIasm免疫複合体は、ヒト黒色腫だけでなく広範囲の他のヒト腫瘍に対しても細胞溶解性免疫応答を仲介すると考えられる。ここで報告した試験で示すように、mfVIIasm免疫複合体をコードするアデノウイルスベクターのヒト腫瘍への腫瘍内注入は、免疫複合体の高い血液力価を確立し、維持するための安全で有効なプロトコールであると思われる。しかし、注入した腫瘍の細胞はアデノウイルスによる感染に感受性であり、コードされた免疫複合体を合成し、分泌することができるはずである。ヒト黒色腫、前立腺癌、乳癌、膵癌、腎癌、胃癌及び神経芽細胞腫細胞系統、並びにマウス黒色腫及び乳癌細胞系統から成る、ヒト及びマウスの腫瘍細胞系統のパネルを、mfVIIasm免疫複合体をコードするアデノウイルスベクターの宿主として試験した。ヒト腫瘍細胞系統はすべてほぼ同じ量の免疫複合体蛋白質を産生し、分泌したことから、腫瘍内注入プロトコールが黒色腫に加えて他の種類のヒト固体腫瘍にも使用できることを示唆した。マウス腫瘍系統は、おそらくマウス細胞がアデノウイルスベクターに感染しなかったために、免疫複合体蛋白質を生成することができなかった。
【0082】
考察
この実施例で述べた癌の免疫療法プロトコールは、腫瘍血管系及び腫瘍細胞に対する細胞溶解性免疫応答を仲介する免疫複合体分子をコードする、複製不全アデノウイルスベクターの腫瘍内注入を含む(図1)。ベクターに感染した細胞はコードされた免疫複合体を合成し、それらは血液中に分泌されて、腫瘍血管系内皮細胞及び腫瘍細胞上の同種標的に結合する。ベクターは主として肝細胞ではなく注入された腫瘍の細胞に感染するので、ベクターの腫瘍内注入は静脈内注入に比べて重要な安全上の利点を提供する。マウスの肝臓も他のいずれの器官も、繰り返された腫瘍内注入から深刻な損傷を示さなかった。黒色腫以外のいくつかの型のヒト腫瘍細胞も、ベクターに感染してコードされた免疫複合体を合成し、分泌することができる。それ故、ベクターの腫瘍内注入経路はヒト固体腫瘍に一般的に適用しうる。注入のためにアクセス可能な腫瘍がない場合には、罹病性の正常組織にベクターを注入することができるであろう。
【0083】
腫瘍内注入プロトコールの治療効果をヒト皮膚黒色腫及び転移性肺黒色腫のSCIDマウスモデルで試験した。試験は、異なる標的ドメインを含む2つの免疫複合体を伴った。1)1つの標的ドメインは、腫瘍新生血管系の血管を含めた増殖する血管の内皮細胞によって、そしてまた大部分のヒト腫瘍細胞によっても発現される膜貫通レセプタ組織因子に高い親和性と特異性で結合する血液チモーゲン第VII(fVII)因子である。fVII免疫複合体は、in vivoで標的細胞上の組織因子への結合に関して内因性第VII因子と競合するはずである。2つの標的ドメインのアビディティ作用が組織因子の多数のコピーを発現する細胞への結合を増強するので、この競合はモノマー内因性分子に比べてホモダイマーである免疫複合体分子(図1)の方が圧倒的に有利である(実施例2);また、内因性第VII因子よりもベクターがコードするfVII免疫複合体の方が血液力価が高い。SCIDマウスにおいて増殖するヒト腫瘍異種移植片を含む実験には、マウスfVII標的ドメインを使用した。なぜなら、マウスfVII標的ドメインは、腫瘍細胞上のヒト組織因子及び腫瘍血管系のマウス内皮細胞上のマウス組織因子の両方に緊密に結合するからである。播種性血管内凝固を引き起こす可能性がある、fVII免疫複合体が組織因子に結合することによる血液凝固経路の開始を防ぐため、マウスfVII標的ドメインの活性部位に突然変異を導入した(mfVIIasm)。2)第二の標的ドメインは、ヒト黒色腫細胞によって選択的に発現される硫酸コンドロイチンプロテオグリカンに結合する、一本鎖Fv(scFv)抗体G71−1である(実施例1及び2)。免疫療法試験の結果は、単独で又はG71−1免疫複合体をコードするベクターと共に、mfVIIasm免疫複合体をコードするベクターを腫瘍内注入すると、注入した皮膚腫瘍、そして同時に転移性肺腫瘍の成長も阻害することを示した(図16及び表2)。免疫複合体をコードするベクターの腫瘍内注入後に存在する残留腫瘍組織にはほとんど血管が存在せず(図13)、mfVIIasm免疫複合体が、腫瘍新生血管系の広い範囲に及ぶ破壊をもたらす強力な細胞溶解性免疫応答を誘導することを示唆した。
【0084】
上記の説明は、当業者が本発明をいかにして実施するかを教示するためのものであり、説明の読後に当業者に明白となるであろう本発明の明らかな修正及びバリエーションをすべて詳述することは意図されていない。しかしながら、そのような明らかな修正及びバリエーションはすべて、下記の特許請求の範囲によって定義される本発明の範囲内に含まれることが意図されている。特許請求の範囲は、特に異なる記載がないかぎり、意図されている目的を達成するために有効な、あらゆる順序での特許請求されている成分及び段階をカバーすることが意図されている。
【0085】
本文中で引用する論文、特許及び特許出願、並びにそれらの中で引用される参考文献は、明白にその全体が参照してここに組み込まれる。
(配列表)
【特許請求の範囲】
【請求項1】
各々が、組織因子に結合するが血液凝固を開始させない、第VII因子の突然変異形態である標的ドメインに複合したIgG1免疫グロブリンのFc領域であるエフェクタードメインを持つ、2つの同一の鎖の二量体として構築される免疫複合体蛋白質を含む血管新生を特徴とする、病的状態の治療において有用な組成物。
【請求項2】
免疫複合体蛋白質の標的ドメインが、リシン−341がアラニンに置換されたヒト第VII因子を含む、請求項1記載の組成物。
【請求項3】
免疫複合体蛋白質の標的ドメインが、セリン−344がアラニンに置換されたヒト第VII因子を含む、請求項1記載の組成物。
【請求項4】
各々が、新生血管系に結合するヒトscFv又はVH抗体断片である標的ドメインに複合したIgG1免疫グロブリンのFc領域であるエフェクタードメインを持つ、2つの同一の鎖の二量体として構築される第二免疫複合体蛋白質をさらに含む、請求項1記載の組成物。
【請求項5】
各々が、特定の型の腫瘍細胞に結合するヒトscFv又はVH抗体断片である標的ドメインに複合したIgG1免疫グロブリンのFc領域であるエフェクタードメインを持つ、2つの同一の鎖の二量体として構築される第二免疫複合体蛋白質をさらに含む、請求項1記載の組成物。
【請求項6】
免疫複合体蛋白質が発現ベクターにおける分泌分子としてコードされる、請求項1から5のいずれか1項記載の組成物。
【請求項7】
発現ベクターが複製欠損アデノウイルスベクターである、請求項6記載の組成物。
【請求項8】
発現ベクターがアデノ関連ウイルス発現ベクターである、請求項6記載の組成物。
【請求項9】
組織因子に結合するが血液凝固を開始させない、第VII因子の突然変異形態を含む標的ドメインに複合したIgG1免疫グロブリンのFc領域を含む、少なくとも1つの型の免疫複合体蛋白質の有効量を、当該疾患を有する患者に投与することを含む、血管新生に関連する疾患を治療するための方法。
【請求項10】
免疫複合体蛋白質の標的ドメインが、リシン−341がアラニンに置換されたヒト第VII因子を含む、請求項9記載の方法。
【請求項11】
免疫複合体蛋白質の標的ドメインが、セリン−344がアラニンに置換されたヒト第VII因子を含む、請求項9記載の方法。
【請求項12】
新生血管系又は腫瘍細胞に結合するヒトscFv又はVH抗体断片である標的ドメインに複合したIgG1免疫グロブリンのFc領域であるエフェクタードメインを持つ第二免疫複合体蛋白質を、補助療法として患者に投与する、請求項9記載の方法。
【請求項13】
血管新生化腫瘍を含む癌、慢性関節リウマチ、及び滲出性形態の黄斑変性から成る群から選択される疾患のための治療である、請求項9記載の方法。
【請求項14】
薬学的に許容される担体中の免疫複合体蛋白質の投与によって患者を治療する、請求項9〜13のいずれか1項記載の方法。
【請求項15】
1つ又は複数の型の免疫複合体蛋白質の分泌形態をコードするcDNAを保有する複製欠損アデノウイルスベクター又はアデノ関連ウイルスベクターの投与によって患者を治療する、請求項9〜13のいずれか1項記載の方法。
【請求項16】
複製欠損アデノウイルスベクターを使用する、請求項15記載の方法。
【請求項17】
リシン−341がアラニンに置換された天然第VII因子、セリン−344がアラニンに置換された天然第VII因子、及びそれらの混合物から成る群から選択されるヒト第VII因子の突然変異形態を含む標的ドメインに複合したヒトIgG1免疫グロブリンのFc領域を含む、少なくとも1つの型の免疫複合体蛋白質の有効量を当該患者に投与することを含む、患者における癌を治療するための方法。
【請求項18】
患者の型の腫瘍細胞に結合するヒトscFv又はVH抗体断片である標的ドメインに複合したヒトIgG1免疫グロブリンのFc領域であるエフェクタードメインを持つ第二免疫複合体蛋白質を、補助療法として患者に投与する、請求項17記載の方法。
【請求項19】
薬学的に許容される担体中の免疫複合体を投与することによって患者を治療する、請求項17又は18記載の方法。
【請求項20】
1つ又は複数の型の免疫複合体蛋白質の分泌形態をコードするcDNAを運ぶ複製欠損アデノウイルスベクターを投与することによって患者を治療する、請求項17又は18記載の方法。
【請求項1】
各々が、組織因子に結合するが血液凝固を開始させない、第VII因子の突然変異形態である標的ドメインに複合したIgG1免疫グロブリンのFc領域であるエフェクタードメインを持つ、2つの同一の鎖の二量体として構築される免疫複合体蛋白質を含む血管新生を特徴とする、病的状態の治療において有用な組成物。
【請求項2】
免疫複合体蛋白質の標的ドメインが、リシン−341がアラニンに置換されたヒト第VII因子を含む、請求項1記載の組成物。
【請求項3】
免疫複合体蛋白質の標的ドメインが、セリン−344がアラニンに置換されたヒト第VII因子を含む、請求項1記載の組成物。
【請求項4】
各々が、新生血管系に結合するヒトscFv又はVH抗体断片である標的ドメインに複合したIgG1免疫グロブリンのFc領域であるエフェクタードメインを持つ、2つの同一の鎖の二量体として構築される第二免疫複合体蛋白質をさらに含む、請求項1記載の組成物。
【請求項5】
各々が、特定の型の腫瘍細胞に結合するヒトscFv又はVH抗体断片である標的ドメインに複合したIgG1免疫グロブリンのFc領域であるエフェクタードメインを持つ、2つの同一の鎖の二量体として構築される第二免疫複合体蛋白質をさらに含む、請求項1記載の組成物。
【請求項6】
免疫複合体蛋白質が発現ベクターにおける分泌分子としてコードされる、請求項1から5のいずれか1項記載の組成物。
【請求項7】
発現ベクターが複製欠損アデノウイルスベクターである、請求項6記載の組成物。
【請求項8】
発現ベクターがアデノ関連ウイルス発現ベクターである、請求項6記載の組成物。
【請求項9】
組織因子に結合するが血液凝固を開始させない、第VII因子の突然変異形態を含む標的ドメインに複合したIgG1免疫グロブリンのFc領域を含む、少なくとも1つの型の免疫複合体蛋白質の有効量を、当該疾患を有する患者に投与することを含む、血管新生に関連する疾患を治療するための方法。
【請求項10】
免疫複合体蛋白質の標的ドメインが、リシン−341がアラニンに置換されたヒト第VII因子を含む、請求項9記載の方法。
【請求項11】
免疫複合体蛋白質の標的ドメインが、セリン−344がアラニンに置換されたヒト第VII因子を含む、請求項9記載の方法。
【請求項12】
新生血管系又は腫瘍細胞に結合するヒトscFv又はVH抗体断片である標的ドメインに複合したIgG1免疫グロブリンのFc領域であるエフェクタードメインを持つ第二免疫複合体蛋白質を、補助療法として患者に投与する、請求項9記載の方法。
【請求項13】
血管新生化腫瘍を含む癌、慢性関節リウマチ、及び滲出性形態の黄斑変性から成る群から選択される疾患のための治療である、請求項9記載の方法。
【請求項14】
薬学的に許容される担体中の免疫複合体蛋白質の投与によって患者を治療する、請求項9〜13のいずれか1項記載の方法。
【請求項15】
1つ又は複数の型の免疫複合体蛋白質の分泌形態をコードするcDNAを保有する複製欠損アデノウイルスベクター又はアデノ関連ウイルスベクターの投与によって患者を治療する、請求項9〜13のいずれか1項記載の方法。
【請求項16】
複製欠損アデノウイルスベクターを使用する、請求項15記載の方法。
【請求項17】
リシン−341がアラニンに置換された天然第VII因子、セリン−344がアラニンに置換された天然第VII因子、及びそれらの混合物から成る群から選択されるヒト第VII因子の突然変異形態を含む標的ドメインに複合したヒトIgG1免疫グロブリンのFc領域を含む、少なくとも1つの型の免疫複合体蛋白質の有効量を当該患者に投与することを含む、患者における癌を治療するための方法。
【請求項18】
患者の型の腫瘍細胞に結合するヒトscFv又はVH抗体断片である標的ドメインに複合したヒトIgG1免疫グロブリンのFc領域であるエフェクタードメインを持つ第二免疫複合体蛋白質を、補助療法として患者に投与する、請求項17記載の方法。
【請求項19】
薬学的に許容される担体中の免疫複合体を投与することによって患者を治療する、請求項17又は18記載の方法。
【請求項20】
1つ又は複数の型の免疫複合体蛋白質の分泌形態をコードするcDNAを運ぶ複製欠損アデノウイルスベクターを投与することによって患者を治療する、請求項17又は18記載の方法。
【図1】

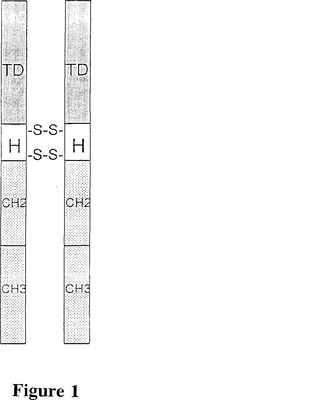
【図2】
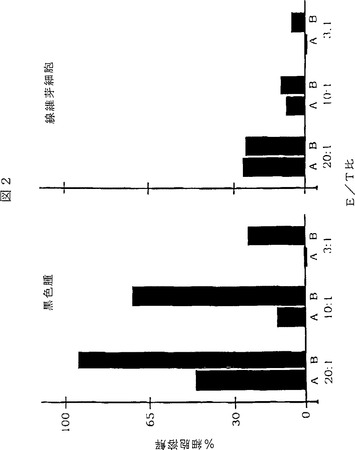
【図3】
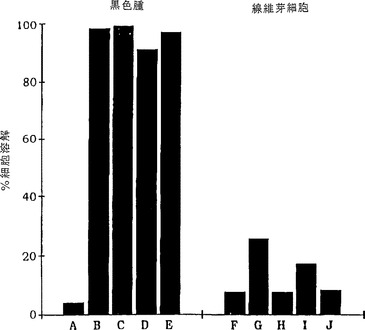
【図4A】

【図4B】

【図4C】

【図5】

【図6】
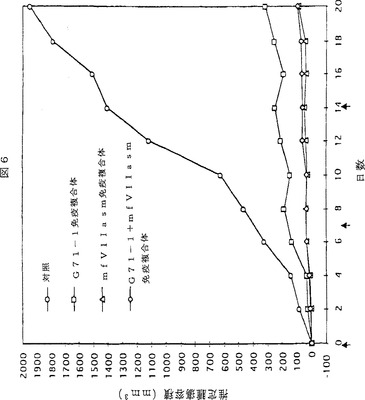
【図7】
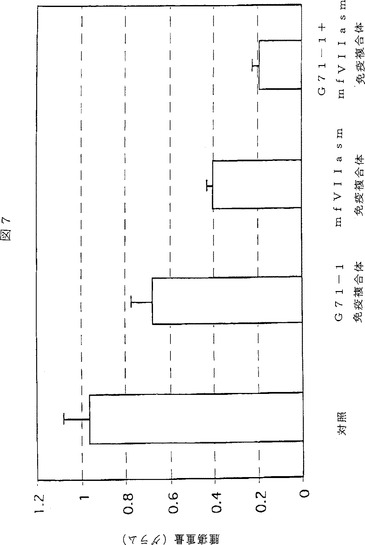
【図8】
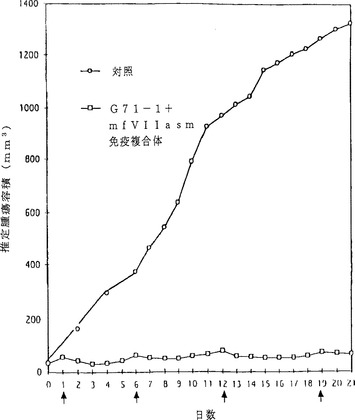
【図9】
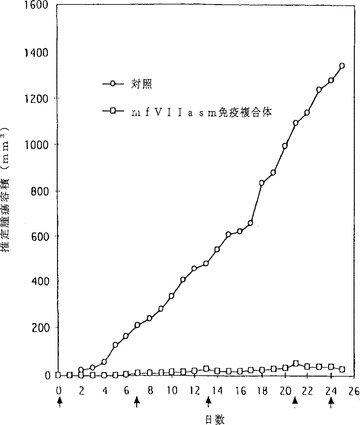
【図10A】

【図10B】

【図11】
【図12】
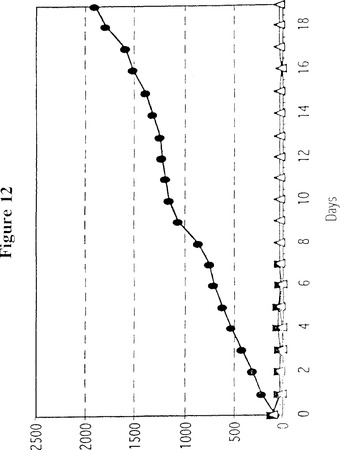
【図13】
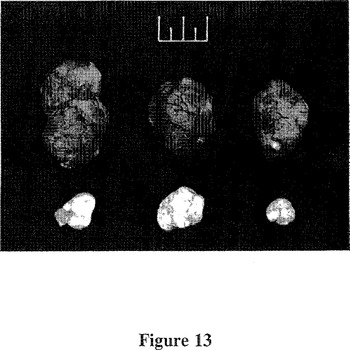
【図14】
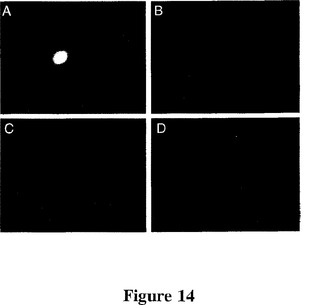
【図15】

【図16】
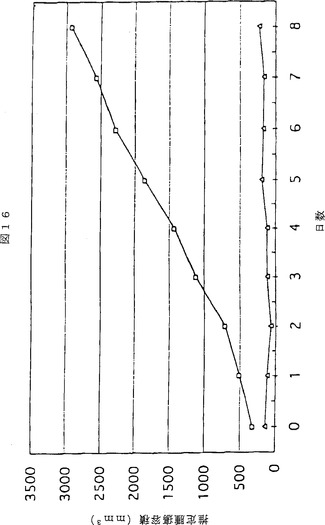
【図2】
【図3】
【図4A】
【図4B】
【図4C】
【図5】
【図6】
【図7】
【図8】
【図9】
【図10A】
【図10B】
【図11】
【図12】
【図13】
【図14】
【図15】
【図16】
【公開番号】特開2012−255032(P2012−255032A)
【公開日】平成24年12月27日(2012.12.27)
【国際特許分類】
【外国語出願】
【出願番号】特願2012−213338(P2012−213338)
【出願日】平成24年9月27日(2012.9.27)
【分割の表示】特願2001−508226(P2001−508226)の分割
【原出願日】平成12年6月14日(2000.6.14)
【出願人】(500121300)エール ユニバーシティ (2)
【Fターム(参考)】
【公開日】平成24年12月27日(2012.12.27)
【国際特許分類】
【出願番号】特願2012−213338(P2012−213338)
【出願日】平成24年9月27日(2012.9.27)
【分割の表示】特願2001−508226(P2001−508226)の分割
【原出願日】平成12年6月14日(2000.6.14)
【出願人】(500121300)エール ユニバーシティ (2)
【Fターム(参考)】
[ Back to top ]