
被子植物の四季咲き性に関与するタンパク質および遺伝子
【課題】被子植物の四季咲き性に関与する遺伝子の提供。
【解決手段】次の(a)または(b)に示されるタンパク質をコードする遺伝子:(a)植物由来の特定なアミノ酸配列を含んでなるタンパク質、(b)植物由来の特定なアミノ酸配列において、1以上のアミノ酸が置換、欠失、付加または挿入されたアミノ酸配列を含んでなり、被子植物の茎頂での花芽形成を抑制しうるタンパク質。
【解決手段】次の(a)または(b)に示されるタンパク質をコードする遺伝子:(a)植物由来の特定なアミノ酸配列を含んでなるタンパク質、(b)植物由来の特定なアミノ酸配列において、1以上のアミノ酸が置換、欠失、付加または挿入されたアミノ酸配列を含んでなり、被子植物の茎頂での花芽形成を抑制しうるタンパク質。
【発明の詳細な説明】
【0001】
【発明の背景】
発明の分野
本発明は、被子植物の四季咲き性に関与する遺伝子およびDNA断片に関する。
【0002】
背景技術
現代の花卉産業において、売上高の上位三種は、四季を通じて開花させることのできる品種である。すなわち、周年出荷できることが花卉生産にとって最も重要な因子の一つである。キクは、栽培法を工夫することによって周年生産されている。バラとカーネーションに関しては、遺伝的に四季を通じて開花する性質(四季咲き性)を有する品種が生産されている。家庭その他の庭園においても、一度植えれば四季を通じて花を楽しめることは重要な利点であるため、四季咲き性を有する品種は種苗産業においても有用である。
【0003】
バラの栽培品種の多くは四季咲き性を有し、これらは全て交雑育種によって得られたものである。ハイブリッドティー(HT)やフロリバンダ(F)等の現代の四季咲き性バラは、18世紀に始まった、中国のいくつかの四季咲き性のバラとそれまであったヨーロッパのバラとの一連の交雑によって、100年以上かけて完成したものである。
【0004】
交雑育種により作出された四季咲き性品種は、偶然の産物である。特に、バラの栽培品種の多くは4倍体以上の高次倍数体であるため、交雑による遺伝形質の伝わり方は特に複雑である。従って、特定の品種を四季咲き化することも、また四季咲き性の品種に特定の形質を導入することも、確実に行なうことは困難である。そのため、鑑賞的に価値のある品種であっても、それが四季咲き性を有さないために、実際上商品化できないものがある。また、他の多くの花卉では、適当な育種素材が無いために、四季咲き性の品種はほとんど存在しない。
【0005】
キンギョソウ(Antirrhinum)のcen(centroradialis)遺伝子及びアラビドプシス(Arabidopsis)のtfl1(terminal flower 1)遺伝子と開花との関連性について検討がなされている(特許文献1:特表平11−512289号公報)。例えば、アラビドプシスのtfl1遺伝子またはそのオルソログの作用は茎頂での花芽形成を抑制することであることが知られている。野生型の茎頂では、花芽形成が起こらず、温度、水、肥料などの栽培条件が整えば伸びつづける。一方で、人工的に誘導した、tfl1遺伝子またはそのオルソログが発現しない突然変異型では、早期に花芽形成が起こり、茎の伸長が止まり、草丈は小さくなる。
【0006】
しかし、被子植物、特にバラの四季咲き性に関しては検討されていない。
【0007】
【特許文献1】
特表平11−512289号公報
【0008】
【発明の概要】
本発明者は、被子植物の四季咲き性に関与するタンパク質およびその遺伝子、ならびに該遺伝子の発現を抑制するDNAを特定することに成功し、この遺伝子の発現が抑制されると、被子植物の茎頂での花芽形成が促進されて該植物が四季咲き性となるとの知見を得た。本発明はこの知見に基づくものである。
【0009】
従って、本発明は、四季咲き性に関与するタンパク質およびその遺伝子、ならびに該遺伝子の発現を抑制しうるDNAを提供することを目的とする。
【0010】
そして、本発明によるタンパク質は、以下の(a)または(b)に示されるものである:
(a)配列番号2で表されるアミノ酸配列を含んでなるタンパク質、
(b)配列番号2で表されるアミノ酸配列において、1以上のアミノ酸が置換、欠失、付加または挿入されたアミノ酸配列を含んでなり、被子植物の茎頂での花芽形成を抑制しうるタンパク質。
【0011】
さらに、本発明による遺伝子は、本発明によるタンパク質をコードするものである。
【0012】
さらに、本発明によるDNAは、以下の(a)または(b)に示されるDNAである:
(a)配列番号3で表されるヌクレオチド配列を含んでなるDNA、
(b)配列番号3で表されるヌクレオチド配列において、1以上のヌクレオチドが欠失、置換、付加または挿入されたヌクレオチド配列を含んでなり、被子植物におけるトランスポゾンとして機能しうるDNA。
【0013】
本発明によれば、被子植物に四季咲き性を付与することが可能となり、さらには、四季咲き性の被子植物を開発する上で有用な標的タンパク質、標的遺伝子、核酸分子および前記遺伝子の発現を抑制するDNAが提供される。
【0014】
【発明の具体的説明】
本明細書において「四季咲き性」とは、全ての季節において継続的に花を咲かせる植物の性質をいう。
【0015】
本明細書において「被子植物」とは、胚珠が子房の中に入っている植物をいい、花を観賞するための植物の殆どは、被子植物に含まれる。被子植物としては多種多様なものが挙げられ、本発明において特に限定されるものではないが、好ましくはバラ目(Rosales)に属する植物、より好ましくはバラ科(Rosaceae)に属する植物、さらに好ましくはバラ(Rosa)とされる。
【0016】
本明細書において「核酸分子」は、DNA、RNA、およびPNA(peptide nucleic acid)を含む意味で用いられる。
【0017】
本明細書において、核酸分子について用いられる「特異的」とは、その核酸分子のヌクレオチド配列が対象となる遺伝子中にのみ存在し、被子植物中の他の遺伝子中には存在しないことを意味する。
【0018】
本明細書に記載される相同性の数値は、特に明示した場合を除き、当業者に公知の相同性検索プログラムを用いて算出される数値であってよいが、好ましくはBLAST〔J. Mol. Biol., 215, 403 (1990)〕においてデフォルト(初期設定)のパラメータ(例えば、Protein-Based BLAST Search (FiHer:ON, Scoring Matrix:BLOSUM62, Word Size:3, E value:10, Gap costs:II, I, Aliguments:50))を用いて算出される数値とされる。
【0019】
本発明によるタンパク質は、配列番号2で表されるアミノ酸配列を含んでなるものであり、該タンパク質は被子植物の茎頂での花芽の形成を抑制しうるものである。また、本発明によるタンパク質は上記のものに限定されるものではなく、被子植物の茎頂での花芽の形成を抑制しうる限り、配列番号2で表されるアミノ酸配列において、1以上のアミノ酸が置換、欠失、付加または挿入されたアミノ酸配列を含んでなる変異体であってもよい。前記変異体において、置換、欠失、付加または挿入されるアミノ酸の数は特に限定されないが、好ましくは1〜9個、より好ましくは1〜5個、さらに好ましくは1〜3個とされる。また、上記変異体のアミノ酸配列は、配列番号2で表されるアミノ酸配列に対して、好ましくは少なくとも83%、より好ましくは少なくとも90%、さらに好ましくは少なくとも95%の相同性を有するものとされる。
【0020】
本発明による遺伝子は、本発明によるタンパク質をコードするものである。そのヌクレオチド配列は特に限定されないが、好ましくは配列番号1で表されるヌクレオチド配列とされる。配列番号1で表されるヌクレオチド配列は、mRNAに対するcDNAにおける翻訳開始コドンから翻訳終止コドンまでの配列であり、配列番号2で表されるアミノ酸配列をコードしている。また、本発明による遺伝子は、cDNAの配列を含んでなるものだけでなく、ゲノムDNAの配列を含んでなるものであってもよい。このようなゲノムDNAの配列を含んでなる遺伝子は、本発明のタンパク質をコードするエクソンの配列の他にイントロンの配列を含んでなるものであり、好ましくは配列番号1で表されるヌクレオチド配列中にイントロンに相当する配列が挿入されたもの、例えば、図4に示されるヌクレオチド配列(配列番号19)を含んでなるものである。さらに、本発明による遺伝子は、配列番号1で表されるヌクレオチド配列において、1以上のヌクレオチドが置換、欠失、付加または挿入されたヌクレオチド配列を含んでなるものとすることができる。ここで、置換、欠失、付加または挿入されるヌクレオチドの数は特に限定されないが、好ましくは1〜9個、より好ましくは1〜5個、さらに好ましくは1〜3個とされる。また、このようなヌクレオチド配列は、配列番号1で表されるヌクレオチド配列に対して、好ましくは少なくとも81%、より好ましくは少なくとも85%、さらに好ましくは少なくとも90%の相同性を有するものとされる。
【0021】
本発明による遺伝子およびタンパク質は、当業者に公知の方法に従って得ることができる。本発明による遺伝子を得る方法としては、例えば、ホスホアミダイド法等により化学的に合成する方法、ロサ・キネンシス・スポンタネア(Rosa chinensis spontanea)等の被子植物からの核酸試料を鋳型とし、目的とする遺伝子のヌクレオチド配列に基づいて設計したプライマーを用いる核酸増幅法などが挙げられる。本発明によるタンパク質を得る方法としては、例えば、被子植物の茎頂部から天然物として精製する方法、本発明による遺伝子を大腸菌や酵母などを宿主とした発現システムに組み込んでタンパク質を生産させる方法などが挙げられる。
【0022】
本発明によるタンパク質は被子植物の茎頂での花芽の形成を抑制するものであり、本発明による遺伝子は該タンパク質をコードするものである。よって、被子植物において本発明によるタンパク質または遺伝子の機能が阻害されれば、茎頂での花芽の形成抑制が解除されるため、花芽の形成が促進され、その被子植物は四季咲き性となる。従って、本発明によれば、本発明によるタンパク質または遺伝子の機能を阻害することを含んでなる、被子植物に四季咲き性を付与する方法が提供される。該方法は、当業者に公知の方法に従って行なうことができるが、好ましくは本発明による遺伝子の発現を抑制することを含んでなる。このような方法としては、例えば、アンチセンスRNA/DNA技術[バイオサイエンスとインダストリー, 50, 322 (1992)、化学, 46, 681 (1991)、Biotechnology,9, 358 (1992)、Trends in Biotechnology,10, 87 (1992) 、Trends in Biotechnology,10, 152 (1992)、細胞工学, 16, 1463 (1997)]、トリプル・ヘリックス技術[Trends in Biotechnology,10, 132 (1992)]等が挙げられる。
【0023】
本発明の好ましい実施態様によれば、本発明による遺伝子の発現の抑制は、本発明による遺伝子のアンチセンス鎖と同一のヌクレオチド配列の全部または一部を含んでなる一本鎖核酸分子を用いて行なわれる。このような方法はアンチセンス法として知られている。アンチセンス法では、発現を抑制したい遺伝子に相補的な配列をもつRNAを高レベルで発現させることにより、標的遺伝子の発現が抑制される。この方法では、本発明による遺伝子のアンチセンス鎖と同一のヌクレオチド配列の全部を含んでなる一本鎖RNAを用いることができる。また、上記の方法では、本発明による遺伝子のアンチセンス鎖と同一のヌクレオチド配列の一部を含んでなる一本鎖RNAを用いることもできる。このような部分的な一本鎖RNAは本発明による遺伝子の発現を抑制しうるものであればよく、当業者であれば適宜設計することができるが、好ましくは本発明による遺伝子に特異的なものとされ、その鎖長は、好ましくは5〜100ヌクレオチド、より好ましくは5〜50ヌクレオチド、さらに好ましくは10〜20ヌクレオチドとされる。
【0024】
本発明の他の好ましい実施態様によれば、本発明による遺伝子の発現の抑制は、本発明による遺伝子のセンス鎖と同一のヌクレオチド配列の全部または一部を含んでなる一本鎖核酸分子を用いて行なわれる。すなわち、このセンス一本鎖核酸は、上記のアンチセンス一本鎖核酸と同様に、本発明による遺伝子の発現の抑制に用いることが可能である。この方法では、本発明による遺伝子のセンス鎖と同一のヌクレオチド配列の全部を含んでなる一本鎖RNAを用いることができる。また、上記の方法では、本発明による遺伝子のセンス鎖と同一のヌクレオチド配列の一部を含んでなる一本鎖RNAを用いることもできる。このような部分的な一本鎖RNAは本発明による遺伝子の発現を抑制しうるものであればよく、当業者であれば適宜設計することができるが、好ましくは本発明による遺伝子に特異的なものとされ、その鎖長は、好ましくは5〜100ヌクレオチド、より好ましくは5〜50ヌクレオチド、さらに好ましくは10〜20ヌクレオチドとされる。
【0025】
本発明の他の好ましい実施態様によれば、本発明による遺伝子の発現の抑制は、本発明による遺伝子と同一のヌクレオチド配列の全部または一部を含んでなる二本鎖核酸分子を用いて行なわれる。例えば、この二本鎖核酸分子を用いることにより、被子植物において本発明による遺伝子のアンチセンスまたはセンス一本鎖核酸を発現させることができる。本発明による二本鎖核酸分子は、好ましくはDNAとされ、その鎖長および具体的なヌクレオチド配列は、目的とする一本鎖核酸分子の鎖長およびヌクレオチド配列に対応するものとされる。例えば、上記アンチセンス一本鎖核酸を発現させる場合には、本発明による二本鎖核酸分子は、本発明による遺伝子のアンチセンス鎖をコード鎖として含むものとされる。また、上記センス一本鎖核酸を発現させる場合には、本発明による二本鎖核酸分子は、本発明による遺伝子のセンス鎖をコード鎖として含むものとされる。
【0026】
本発明による二本鎖核酸分子は、当業者に公知の方法を用いて被子植物中で発現させることができる。例えば、プロモーター、本発明による二本鎖核酸分子、および転写ターミネーター等を含む発現用構築物を、目的とする被子植物中に導入し、得られた植物体を栽培することにより、本発明による二本鎖核酸分子を発現させることができる。植物への発現用構築物の導入は、当業者に公知の方法、例えば、アグロバクテリウム法、バイナリーベクター法、エレクトロポレーション法、PEG法、パーティクルガン法等によって行なうことができる。特に、遺伝子導入のためのバラの形質転換は、米国特許第5480789号明細書等に記載の方法によって行うことができる。
【0027】
本発明による核酸分子を用いた遺伝子発現の抑制法としては、例えば、アンチセンス鎖による形質転換法(Proc. Natl. Acad. Sci. USA 90: 6160-6164, 1993)が挙げられる。この方法では、本発明による遺伝子の全ヌクレオチド配列を有するアンチセンス二本鎖DNAが目的の植物中に導入される。これにより、本発明による遺伝子のmRNAに相補的なRNAとして、本発明によるアンチセンス一本鎖RNAが発現され、この相補的RNAにより該遺伝子の発現が極端に抑制される。
【0028】
本発明による核酸分子を用いた遺伝子発現の抑制法の他の例としては、コサプレッション法が挙げられる。この方法では、本発明による遺伝子の全ヌクレオチド配列を有するセンス二本鎖DNAが目的の植物中に導入される。これにより、本発明によるセンス一本鎖RNAが発現され、このRNAにより該遺伝子の発現が極端に抑制される(Plant Cell 9: 1357-1368, 1997)。
【0029】
本発明によるDNAは、配列番号3で表されるヌクレオチド配列を含んでなるものであり、該DNAは被子植物におけるトランスポゾンとして機能しうるものである。なお、配列番号3で表されるヌクレオチド配列において、第1〜856残基はLTRであり、第857〜6105残基はORFであり、第8069〜8925残基はLTRである。また、本発明によるDNAは上記のものに限定されるものではなく、被子植物におけるトランスポゾンとして機能しうる限り、配列番号3で表されるヌクレオチド配列において、1以上のヌクレオチドが欠失、置換、付加または挿入されたヌクレオチド配列を含んでなるものであってもよい。ここで、置換、欠失、付加または挿入されるヌクレオチドの数は特に限定されないが、好ましくは1〜9個、より好ましくは1〜5個、さらに好ましくは1〜3個とされる。また、このようなヌクレオチド配列は、配列番号3で表されるヌクレオチド配列に対して、好ましくは少なくとも70%、より好ましくは少なくとも80%、さらに好ましくは少なくとも90%の相同性を有するものとされる。
【0030】
本発明によるDNAは、当業者に公知の方法、例えば、ホスホアミダイド法等により化学的に合成する方法、オールド・ブラッシュ(Old blush)等の四季咲き性バラからの核酸試料を鋳型とし、目的とする遺伝子のヌクレオチド配列に基づいて設計したプライマーを用いる核酸増幅法などによって得ることができる。
【0031】
本発明によるDNAは、被子植物においてトランスポゾンとして機能し、本発明による遺伝子中に挿入されることにより該遺伝子の発現を抑制するものである。これにより、茎頂での花芽の形成抑制が解除されるため、花芽の形成が促進され、その被子植物は四季咲き性となる。従って、本発明によれば、本発明によるDNAを含んでなるトランスポゾンが提供され、さらには、本発明によるDNAまたはこれを含んでなるトランスポゾンを用いて本発明による遺伝子の発現を抑制することを含んでなる、被子植物に四季咲き性を付与する方法が提供される。
【0032】
この方法では、本発明によるDNAまたはトランスポゾンが被子植物に導入される。これは、本発明による遺伝子の発現が抑制されるように、当業者によって適宜行なわれるが、本発明によるDNAまたはトランスポゾンは、本発明による遺伝子中またはその近傍に導入されることが好ましく、より好ましくは本発明による遺伝子の第2イントロン中、さらに好ましくは四季咲き性バラと同じ場所に導入される。
【0033】
植物へのDNAの導入は、当業者に公知の方法、例えば、アグロバクテリウム法、バイナリーベクター法、エレクトロポレーション法、PEG法、パーティクルガン法等によって行なうことができる。特に、遺伝子導入のためのバラの形質転換は、米国特許第5480789号明細書等に記載の方法によって行うことができる。
【0034】
【実施例】
例1:四季咲き性に関与する因子の特定
現代の四季咲き性バラ品種では、シュートや腋芽は4〜7枚程度の葉を伸ばすと茎頂に花芽を形成する。花が咲き終わるとすぐに下の腋芽が伸び、同様にして花芽を形成する。こうして、温度、水、肥料などの栽培条件が整えば、次々にシュートを伸ばし、その先に花芽を形成し、四季を通じて開花する。これに対し、つる性のバラでは、シュートが30枚または40枚以上の葉を伸ばしても、茎頂に花芽を形成することなく伸びつづけ、一季咲きとなる。
【0035】
本発明者は、アラビドプシス(Arabidopsis)の野生型(無限花序)と突然変異型(有限花序)に関する知見、ならびに現代四季咲き性バラにおいて一季咲き性バラへの突然変異が多く見られることに基づき、以下の仮説を立て、これを証明した。
【0036】
(仮説1)一季咲き性のつる型バラではtfl1遺伝子のオルソロガス遺伝子が活性を持っているため、茎頂における花芽形成が抑制されるが、四季咲き型バラでは突然変異によって前記遺伝子の活性が低下しているため、茎頂における花芽形成が抑制されない。
【0037】
(仮説2)四季咲き性バラは、tfl1遺伝子のオルソロガス遺伝子へのトランスポゾンの挿入という突然変異によって生じ、一季咲き性のつる型バラは、該トランスポゾンの欠失という復帰突然変異によって生じる。
【0038】
(1)tfl1遺伝子のバラオルソロガス遺伝子の配列決定および該遺伝子中のトランスポゾンの存否
以下のようにして、tfl1遺伝子のバラオルソログのヌクレオチド配列を明らかにし、四季咲き型ではその遺伝子全てにトランスポゾンが挿入されており、一季咲き型では、トランスポゾンが挿入されていない遺伝子が少なくとも一つ存在することを証明した。
【0039】
tfl1遺伝子は、ホスファチジルエタノールアミン結合タンパク質(PEBP)と称する遺伝子ファミリーに属する。アラビドプシスでは6種のPEBP遺伝子が知られている。アラビドプシス、イネ、キンギョソウ、ペチュニア、トマトなどの公知のPEBP遺伝子のヌクレオチド配列に基づき、特に保存性の高い部分を選んで以下のプライマーを設計した:
フォワードプライマー(UtflF1):GATAGACCCAGATGTTCCAGGTCC(配列番号4);
リバースプライマー(UtflR1):CCTGGAATATCTGTCACCATCCT(配列番号5)。
【0040】
これらのプライマーを用いて、ロサ・キネンシス・スポンタネア(Rosa chinensis spontanea)(Rcs)のDNAを鋳型にしてPCRを行い、複数の増幅産物を得た。これらの増幅産物のヌクレオチド配列を決定し、そのヌクレオチド配列からPEBPファミリー遺伝子と考えれるもの4種を選び、これらをRPEBP1、RPEBP2、RPEBP3、およびKSNと名付けた。これらの増幅産物のうち、KSNの400bp余りの産物のヌクレオチド配列は図1に示すとおりであった。このヌクレオチド配列は、アラビドプシスtfl1遺伝子のエクソン2から3にかけてのヌクレオチド配列と高い相同性を有していた。
【0041】
上記のように決定されたKSNの部分的なヌクレオチド配列に基づき、KSN遺伝子全体のヌクレオチド配列決定を以下のようにして行なった。すなわち、RcsのDNAを鋳型として、InversePCR法により決定した。
【0042】
まず、RcsのDNA数μgをNEB社の制限酵素Cac 8Iで消化し(DNA17μl、酵素1μl、10倍バッファー2μlを37℃で4時間反応)、加熱によって酵素を不活化してから(65℃で20分間)、NEB社のT4-DNA-Ligaseを用いて環状化した(酵素消化DNA20μl、リガーゼ1μl、10倍バッファー12.5μl、および精製水91.5μlを8℃で一昼夜反応)。この環状DNAを鋳型とし、下記の二つのプライマーを用いるPCRを行なった:
KSN-IR1:CTGTAAGTTATACCAGTGCAGGTGC(配列番号6);
KSN-IF2:GGAAGTACATATTATGGCATAATC(配列番号7)。
得られた増幅産物のヌクレオチド配列を図2に示す。図2から明らかなように、Utfl-FIの上流416bpのヌクレオチド配列が決定された。
【0043】
さらに、RcsのDNA数μgをNEB社の制限酵素Sau96Iで消化し(DNA17μl、酵素1μl、10倍バッファー2μlを37℃で4時間反応)、加熱によって酵素を不活化してから(80℃で20分間)、NEB社のT4-DNA-Ligaseを用いて環状化した(酵素消化DNA20μl、リガーゼ1μl、10倍バッファー12.5μl、および精製水91.5μlを8℃で一昼夜反応)。この環状DNAを鋳型とし、上記のプライマーKSN-IR1および以下のプライマーKSN-IF1を用いるPCRを行なった:
KSN-IF1:AGGAGCTAACATTTTTGCCTTG(配列番号8)。
得られた増幅産物のヌクレオチド配列を図3に示す。図3から明らかなように、UtflR1の下流401bpのヌクレオチド配列が決定された。
【0044】
以上のように決定された配列をまとめたKSNのヌクレオチド配列を図4に示す。図4において、アラビドプシスtfl1遺伝子のヌクレオチド配列と比較してエクソンと思われる部分に囲み線を付した。さらに、KSNのコード領域のヌクレオチド配列を配列番号1に示し、これによりコードされるアミノ酸配列を配列番号2に示した。
【0045】
同様にして、RPEBP1、RPEBP2、およびRPEBP3のヌクレオチド配列も明らかにした(配列は示さない)。
【0046】
上述のように決定されたRPEBP1、RPEBP2、RPEBP3、およびKSNのヌクレオチド配列に基づき、これらの遺伝子のコーディング領域のほぼ全域を特異的に増幅するプライマーを設計した。例えば、KSN増幅用のプライマーは以下の配列とした:
KSNF1:ATTAGGAGTACAATCCTTCCTTCC(配列番号9);
KSNR2:ACCTGCCTCCTGCTAGCTGC(配列番号10)。
【0047】
四季咲き性のオールド・ブラッシュ(OBB)およびその突然変異体とされる一季咲き性のオールド・ブラッシュ・クライミング(OBCl)のDNAを鋳型とし、上記のコーディング領域増幅用プライマーを用いて、1Kbp程度の増幅物を得る条件(Promega社製のPCR Master Mixを添付プロトコルに従って使用)でのPCRを行なった。その結果、RPEBP1、RPEBP2、およびRPEBP3の増幅産物は、OBBとOBClとの間で違いが認められなかった。KSNでは予想された大きさの増幅産物が認められなかったため、PCRの条件を10Kbp程度の増幅産物が得られる条件(タカラ社製のOne Shot LAPCR Mixを添付プロトコルに従って使用)に変更して再度PCRを行った。その結果、OBBでは約10Kbpの増幅産物が、OBClでは約2Kbpの増幅産物が得られた。
【0048】
OBBおよびOBClのDNAを鋳型として得られた増幅産物は、KSNに何らかのDNAが挿入されているものと推定された。そこで、以下のようにしてその挿入物のヌクレオチド配列を明らかにした。
【0049】
まず、挿入地点と推定される前後にいくつかのプライマーを設計してPCRを行ない、増幅産物を得た。その結果、下記のプライマーが最も挿入DNAに近接したプライマーであることが明らかとなった:
KSNIF3:CATATTATGGCATAGGGTGTGGC(配列番号11);
KSNR5:CAAATGTAACATCTGTGGTGCCTG(配列番号12)。
【0050】
900bp前後と推定された短い挿入物(Si)は、上記二つのプライマーを用いてヌクレオチド配列を明らかにした。
【0051】
さらに、9Kbp前後と推定された長い挿入物(Li)のヌクレオチド配列は以下のようにして決定した。まず、PCR増幅産物を、添付のプロトコールに従って、pT7BlueT-Vector(Novagen社製)にDNA Ligation Kit Ver.2(タカラ社製)を用いて連結し、環状化した。このプラスミドを用いて、コンピテント細胞JM109株(タカラ社製)を添付プロトコールに従って形質転換した。形質転換したこの大腸菌を、添付プロトコールに従って、あらかじめ2%5−ブロモ−4−クロロ−3−インドリル−β−D−ガラクトシド(Xgal)50μlと0.1Mイソプロピル−β−D−チオガラクトピラノシド(IPTG)10μlとを塗布しておいたL−agar培地(アンピシリン50μg/mlを含む)に塗布し、生じたコロニーの中から白色のコロニーを増殖させた。
【0052】
生じたクローンのうち、クローン6が完全長のPCR増幅産物を含んでいると推定されたので、以下のようにしてそのヌクレオチド配列を明らかにした。まず、このベクターのクローニングサイトに断点を持つ制限酵素HindIIIまたはEcoRIで切断し、DNA Ligation Kit Ver.2(タカラ社製)を用いて再度連結し、環状化した。このプラスミドを用いて、コンピテント細胞JM109株(タカラ社製)を添付プロトコールに従って形質転換し、そのコロニーを得た(それぞれクローン6−12およびクローン6−16)。EcoRIを用いた大きなDNA断片を持つクローン6−16は、さらに、SpeI、Nole1、Xba1、およびPstIを用いてそれぞれに対応する箇所を切断し、同様にしてクローンを得た。得られたクローンのうち、クローン6−16−27、クローン6−16−32、クローン6−16−34、クローン6−16−40、およびクローン6−16−43の5種は互いに異なるDNA断片を含んでいると推定された。
【0053】
上記の8種のクローンのプラスミドに挿入されたDNAのヌクレオチド配列を、以下のプライマーを用いて前後から決定し、つなぎ合わせた:
M13R18:CAGGAAACAGCTATGACC(配列番号13);
MR13F18:TGTAAAACGACGGCCAGT(配列番号14)。
【0054】
その結果、このDNA断片(Li)は約9Kbpの大きさで、blastホモロジー検索の結果、コピアタイプのレトロトランスポゾンと相同性が高かった。このトランスポゾン(配列番号3)は、OBBのKSN遺伝子のイントロン2に挿入されていた。また、OBClの約900bpの挿入物は、このレトロトランスポゾンが消失する際にその場に残した約900bpのLTR(長い末端反復配列)であることが判明した。
【0055】
以上のように、Rcs、OBB、およびOBClのKSNとしては、挿入のない野生型(w型、約1Kbp)、レトロトランスポゾンの入った型(Li型、約10Kbp)、レトロトランスポゾンがLTRを残して出ていった型(Si型、約2Kbp)の三種が認められた。
【0056】
上記の結果から、Li型では、約9000bpのDNAの挿入によってmRNAの転写が大きく阻害されるためにKSNの発現がほとんど阻害され、Si型では、900bpという短いDNAの挿入であるためKSN発現の阻害が軽微であると考えられる。これは、OBBが四季咲きとなり、他方でOBClはつるバラのように良く伸びるが、夏から秋にかけてもぽつぽつと咲くことと一致する。
【0057】
次いで、いくつかの園芸品種を、KSN遺伝子の構造について調べた。先に設計したレトロトランスポゾンの挿入位置の前(KSNIF3)と後(KSNR5)のプライマーを用いると、w型とSi型はそれぞれ250bpと1150bpの異なる大きさの増幅産物を与える。また、KSNIF3と共に用いてレトロトランスポゾンの一部を増幅するようなプライマーKSNInsR3を、LTRよりも内側に設計した(KSNInsR3:TGTAATCTGTAGGAGATCCCATGC, 配列番号15)。KSNIF3、KSNR5およびKSNInsR3を用いて、つる性の突然変異の園芸品種が作られている6園芸品種(「フレーミング・ピース」、「ピース」、「ウエンディ・クッソンズ」、「ゴールド・バニー」、「アイス・バーグ」、および「ピンク・シホン」)のKSNを調べた。これら全ての品種において、四季咲き性の品種ではKSNIF3およびKSNR5のプライマーペアでは増幅産物がなく、KSNIF3およびKSNInsR3のプライマーペアで増幅産物が得られた。これは、四季咲き性品種ではすべてのKSNにレトロトランスポゾンが挿入されていたことを示す。一方で、つる性の突然変異品種では、KSNIF3およびKSNInsR3のプライマーペアによる増幅産物以外にKSNIF3およびKSNR5のプライマーペアによる1150bpのSi型の増幅産物が見られ、レトロトランスポゾンを含んだKSN以外に、LTRのみが残ったKSNが存在していることが明らかとなった。この様に、これら6種の園芸品種においても、四季咲き性品種ではトランスポゾンが挿入されており、つる性(一季咲き性)品種では少なくとも一つはLTRを残してトランスポゾンが転移したと思われるKSNが存在することが明らかとなった。
【0058】
(2)四季咲き性バラと一季咲き性バラとの、tfl1遺伝子バラオルソログmRNA発現レベルの比較
上記(1)で明らかとなったトランスポゾンの挿入により、四季咲き型バラのKSNのmRNA発現レベルが、一季咲き型のそれよりも著しく低いことを、以下のようにして証明した。
【0059】
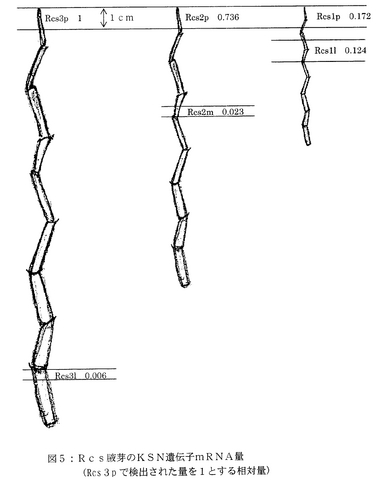
まず、野生型KSNを有するRcsの腋芽から、RNAを抽出した。Rcsの腋芽としては、15cmのもの、10cmのもの、および5cm以下のものの3種類を用い、それぞれ図5に示す位置からサンプルを採取した。同様に、OBBの腋芽の茎頂およびOBClの蕾を含む腋芽からもRNAを抽出した。RNA抽出は、Rneasy Plant Mini Kit(Qiagen社製)を用いて行なった。
次いで、KSNのmRNA量を、Quanti Tect SYBR Green RT-PCR Kit(Qiagen社製)およびLight Cycler System(Roche Molecular Biochemicals社製)を用いて比較した。また、本実験では、バラのアクチンのmRNAをコントロールとして用いた。KSNのmRNAは量が少なく、QuantiTect SYBR Green RT-PCR Kitでは測定できないことがあったので、限界希釈法によってさらに測定した。限界希釈法とは、RNA抽出液の3倍希釈系列を作り、OneStep RT-PCR Kit(Qiagen社製)を用いてKSNのmRNAが検出できなくなる希釈倍率を求め、mRNA量を推定する方法である。Rcsについての結果を下記の表1に示す。表1において、各数値は、Rcs3pについての測定値を1とした場合の相対量で示した。
【0060】
【表1】

【0061】
表1から、茎頂でのKSNのmRNA発現量は、腋芽の成長に従って増加し、茎頂から離れるにしたがって減少することが明らかとなった。さらに、OBCl茎頂におけるKSNのmRNA発現量はRcs3pの1/10000程度であり、OBB茎頂においては検出できなかった。
【0062】
(3)考察
以上の結果から、四季咲き性のバラでは、KSN遺伝子中にトランスポゾンが挿入されており、これにより該遺伝子の発現が阻害され、茎頂における花芽の形成が促進されることが明らかとなった。従って、KSN遺伝子を阻害することにより、バラに四季咲き性を付与することが可能となる。
【0063】
アラビドプシスのTFL1およびATC、キンギョソウのCEN、ならびにライ・グラス(単子葉類の牧草)のLpTFL1をアラビドプシスで発現させたときの表現形に差異は認められないため、これらの機能の違いは、タンパク質の構造の違いではなく、主に遺伝子発現の違いによると報告されている(Shepard,K.A.and Purugganon M.D., The genetics of plant morphological evolution., Current Opinion in Plant Biology 5:49-55, 2002)。さらに、KSN遺伝子の構造が真核生物全体で良く保存されていること、および単子葉植物の遺伝子が双子葉植物においても本来の機能を発揮することをも考慮すると、バラ属で発見された四季咲きのメカニズムは他の被子植物でも働くものと考えられる。
【0064】
【配列表】
【図面の簡単な説明】
【図1】図1は、部分的に決定されたKSN遺伝子のヌクレオチド配列(配列番号16)を示す図である。
【図2】図2は、部分的に決定されたKSN遺伝子のヌクレオチド配列(配列番号17)を示す図である。
【図3】図3は、部分的に決定されたKSN遺伝子のヌクレオチド配列(配列番号18)を示す図である。
【図4】図4は、KSN遺伝子のコーディング領域のヌクレオチド配列(配列番号19)を示す図である。
【図5】図5は、バラ(Rcs)腋芽の様々な部位におけるKSNのmRNA発現量を示す図である。
【0001】
【発明の背景】
発明の分野
本発明は、被子植物の四季咲き性に関与する遺伝子およびDNA断片に関する。
【0002】
背景技術
現代の花卉産業において、売上高の上位三種は、四季を通じて開花させることのできる品種である。すなわち、周年出荷できることが花卉生産にとって最も重要な因子の一つである。キクは、栽培法を工夫することによって周年生産されている。バラとカーネーションに関しては、遺伝的に四季を通じて開花する性質(四季咲き性)を有する品種が生産されている。家庭その他の庭園においても、一度植えれば四季を通じて花を楽しめることは重要な利点であるため、四季咲き性を有する品種は種苗産業においても有用である。
【0003】
バラの栽培品種の多くは四季咲き性を有し、これらは全て交雑育種によって得られたものである。ハイブリッドティー(HT)やフロリバンダ(F)等の現代の四季咲き性バラは、18世紀に始まった、中国のいくつかの四季咲き性のバラとそれまであったヨーロッパのバラとの一連の交雑によって、100年以上かけて完成したものである。
【0004】
交雑育種により作出された四季咲き性品種は、偶然の産物である。特に、バラの栽培品種の多くは4倍体以上の高次倍数体であるため、交雑による遺伝形質の伝わり方は特に複雑である。従って、特定の品種を四季咲き化することも、また四季咲き性の品種に特定の形質を導入することも、確実に行なうことは困難である。そのため、鑑賞的に価値のある品種であっても、それが四季咲き性を有さないために、実際上商品化できないものがある。また、他の多くの花卉では、適当な育種素材が無いために、四季咲き性の品種はほとんど存在しない。
【0005】
キンギョソウ(Antirrhinum)のcen(centroradialis)遺伝子及びアラビドプシス(Arabidopsis)のtfl1(terminal flower 1)遺伝子と開花との関連性について検討がなされている(特許文献1:特表平11−512289号公報)。例えば、アラビドプシスのtfl1遺伝子またはそのオルソログの作用は茎頂での花芽形成を抑制することであることが知られている。野生型の茎頂では、花芽形成が起こらず、温度、水、肥料などの栽培条件が整えば伸びつづける。一方で、人工的に誘導した、tfl1遺伝子またはそのオルソログが発現しない突然変異型では、早期に花芽形成が起こり、茎の伸長が止まり、草丈は小さくなる。
【0006】
しかし、被子植物、特にバラの四季咲き性に関しては検討されていない。
【0007】
【特許文献1】
特表平11−512289号公報
【0008】
【発明の概要】
本発明者は、被子植物の四季咲き性に関与するタンパク質およびその遺伝子、ならびに該遺伝子の発現を抑制するDNAを特定することに成功し、この遺伝子の発現が抑制されると、被子植物の茎頂での花芽形成が促進されて該植物が四季咲き性となるとの知見を得た。本発明はこの知見に基づくものである。
【0009】
従って、本発明は、四季咲き性に関与するタンパク質およびその遺伝子、ならびに該遺伝子の発現を抑制しうるDNAを提供することを目的とする。
【0010】
そして、本発明によるタンパク質は、以下の(a)または(b)に示されるものである:
(a)配列番号2で表されるアミノ酸配列を含んでなるタンパク質、
(b)配列番号2で表されるアミノ酸配列において、1以上のアミノ酸が置換、欠失、付加または挿入されたアミノ酸配列を含んでなり、被子植物の茎頂での花芽形成を抑制しうるタンパク質。
【0011】
さらに、本発明による遺伝子は、本発明によるタンパク質をコードするものである。
【0012】
さらに、本発明によるDNAは、以下の(a)または(b)に示されるDNAである:
(a)配列番号3で表されるヌクレオチド配列を含んでなるDNA、
(b)配列番号3で表されるヌクレオチド配列において、1以上のヌクレオチドが欠失、置換、付加または挿入されたヌクレオチド配列を含んでなり、被子植物におけるトランスポゾンとして機能しうるDNA。
【0013】
本発明によれば、被子植物に四季咲き性を付与することが可能となり、さらには、四季咲き性の被子植物を開発する上で有用な標的タンパク質、標的遺伝子、核酸分子および前記遺伝子の発現を抑制するDNAが提供される。
【0014】
【発明の具体的説明】
本明細書において「四季咲き性」とは、全ての季節において継続的に花を咲かせる植物の性質をいう。
【0015】
本明細書において「被子植物」とは、胚珠が子房の中に入っている植物をいい、花を観賞するための植物の殆どは、被子植物に含まれる。被子植物としては多種多様なものが挙げられ、本発明において特に限定されるものではないが、好ましくはバラ目(Rosales)に属する植物、より好ましくはバラ科(Rosaceae)に属する植物、さらに好ましくはバラ(Rosa)とされる。
【0016】
本明細書において「核酸分子」は、DNA、RNA、およびPNA(peptide nucleic acid)を含む意味で用いられる。
【0017】
本明細書において、核酸分子について用いられる「特異的」とは、その核酸分子のヌクレオチド配列が対象となる遺伝子中にのみ存在し、被子植物中の他の遺伝子中には存在しないことを意味する。
【0018】
本明細書に記載される相同性の数値は、特に明示した場合を除き、当業者に公知の相同性検索プログラムを用いて算出される数値であってよいが、好ましくはBLAST〔J. Mol. Biol., 215, 403 (1990)〕においてデフォルト(初期設定)のパラメータ(例えば、Protein-Based BLAST Search (FiHer:ON, Scoring Matrix:BLOSUM62, Word Size:3, E value:10, Gap costs:II, I, Aliguments:50))を用いて算出される数値とされる。
【0019】
本発明によるタンパク質は、配列番号2で表されるアミノ酸配列を含んでなるものであり、該タンパク質は被子植物の茎頂での花芽の形成を抑制しうるものである。また、本発明によるタンパク質は上記のものに限定されるものではなく、被子植物の茎頂での花芽の形成を抑制しうる限り、配列番号2で表されるアミノ酸配列において、1以上のアミノ酸が置換、欠失、付加または挿入されたアミノ酸配列を含んでなる変異体であってもよい。前記変異体において、置換、欠失、付加または挿入されるアミノ酸の数は特に限定されないが、好ましくは1〜9個、より好ましくは1〜5個、さらに好ましくは1〜3個とされる。また、上記変異体のアミノ酸配列は、配列番号2で表されるアミノ酸配列に対して、好ましくは少なくとも83%、より好ましくは少なくとも90%、さらに好ましくは少なくとも95%の相同性を有するものとされる。
【0020】
本発明による遺伝子は、本発明によるタンパク質をコードするものである。そのヌクレオチド配列は特に限定されないが、好ましくは配列番号1で表されるヌクレオチド配列とされる。配列番号1で表されるヌクレオチド配列は、mRNAに対するcDNAにおける翻訳開始コドンから翻訳終止コドンまでの配列であり、配列番号2で表されるアミノ酸配列をコードしている。また、本発明による遺伝子は、cDNAの配列を含んでなるものだけでなく、ゲノムDNAの配列を含んでなるものであってもよい。このようなゲノムDNAの配列を含んでなる遺伝子は、本発明のタンパク質をコードするエクソンの配列の他にイントロンの配列を含んでなるものであり、好ましくは配列番号1で表されるヌクレオチド配列中にイントロンに相当する配列が挿入されたもの、例えば、図4に示されるヌクレオチド配列(配列番号19)を含んでなるものである。さらに、本発明による遺伝子は、配列番号1で表されるヌクレオチド配列において、1以上のヌクレオチドが置換、欠失、付加または挿入されたヌクレオチド配列を含んでなるものとすることができる。ここで、置換、欠失、付加または挿入されるヌクレオチドの数は特に限定されないが、好ましくは1〜9個、より好ましくは1〜5個、さらに好ましくは1〜3個とされる。また、このようなヌクレオチド配列は、配列番号1で表されるヌクレオチド配列に対して、好ましくは少なくとも81%、より好ましくは少なくとも85%、さらに好ましくは少なくとも90%の相同性を有するものとされる。
【0021】
本発明による遺伝子およびタンパク質は、当業者に公知の方法に従って得ることができる。本発明による遺伝子を得る方法としては、例えば、ホスホアミダイド法等により化学的に合成する方法、ロサ・キネンシス・スポンタネア(Rosa chinensis spontanea)等の被子植物からの核酸試料を鋳型とし、目的とする遺伝子のヌクレオチド配列に基づいて設計したプライマーを用いる核酸増幅法などが挙げられる。本発明によるタンパク質を得る方法としては、例えば、被子植物の茎頂部から天然物として精製する方法、本発明による遺伝子を大腸菌や酵母などを宿主とした発現システムに組み込んでタンパク質を生産させる方法などが挙げられる。
【0022】
本発明によるタンパク質は被子植物の茎頂での花芽の形成を抑制するものであり、本発明による遺伝子は該タンパク質をコードするものである。よって、被子植物において本発明によるタンパク質または遺伝子の機能が阻害されれば、茎頂での花芽の形成抑制が解除されるため、花芽の形成が促進され、その被子植物は四季咲き性となる。従って、本発明によれば、本発明によるタンパク質または遺伝子の機能を阻害することを含んでなる、被子植物に四季咲き性を付与する方法が提供される。該方法は、当業者に公知の方法に従って行なうことができるが、好ましくは本発明による遺伝子の発現を抑制することを含んでなる。このような方法としては、例えば、アンチセンスRNA/DNA技術[バイオサイエンスとインダストリー, 50, 322 (1992)、化学, 46, 681 (1991)、Biotechnology,9, 358 (1992)、Trends in Biotechnology,10, 87 (1992) 、Trends in Biotechnology,10, 152 (1992)、細胞工学, 16, 1463 (1997)]、トリプル・ヘリックス技術[Trends in Biotechnology,10, 132 (1992)]等が挙げられる。
【0023】
本発明の好ましい実施態様によれば、本発明による遺伝子の発現の抑制は、本発明による遺伝子のアンチセンス鎖と同一のヌクレオチド配列の全部または一部を含んでなる一本鎖核酸分子を用いて行なわれる。このような方法はアンチセンス法として知られている。アンチセンス法では、発現を抑制したい遺伝子に相補的な配列をもつRNAを高レベルで発現させることにより、標的遺伝子の発現が抑制される。この方法では、本発明による遺伝子のアンチセンス鎖と同一のヌクレオチド配列の全部を含んでなる一本鎖RNAを用いることができる。また、上記の方法では、本発明による遺伝子のアンチセンス鎖と同一のヌクレオチド配列の一部を含んでなる一本鎖RNAを用いることもできる。このような部分的な一本鎖RNAは本発明による遺伝子の発現を抑制しうるものであればよく、当業者であれば適宜設計することができるが、好ましくは本発明による遺伝子に特異的なものとされ、その鎖長は、好ましくは5〜100ヌクレオチド、より好ましくは5〜50ヌクレオチド、さらに好ましくは10〜20ヌクレオチドとされる。
【0024】
本発明の他の好ましい実施態様によれば、本発明による遺伝子の発現の抑制は、本発明による遺伝子のセンス鎖と同一のヌクレオチド配列の全部または一部を含んでなる一本鎖核酸分子を用いて行なわれる。すなわち、このセンス一本鎖核酸は、上記のアンチセンス一本鎖核酸と同様に、本発明による遺伝子の発現の抑制に用いることが可能である。この方法では、本発明による遺伝子のセンス鎖と同一のヌクレオチド配列の全部を含んでなる一本鎖RNAを用いることができる。また、上記の方法では、本発明による遺伝子のセンス鎖と同一のヌクレオチド配列の一部を含んでなる一本鎖RNAを用いることもできる。このような部分的な一本鎖RNAは本発明による遺伝子の発現を抑制しうるものであればよく、当業者であれば適宜設計することができるが、好ましくは本発明による遺伝子に特異的なものとされ、その鎖長は、好ましくは5〜100ヌクレオチド、より好ましくは5〜50ヌクレオチド、さらに好ましくは10〜20ヌクレオチドとされる。
【0025】
本発明の他の好ましい実施態様によれば、本発明による遺伝子の発現の抑制は、本発明による遺伝子と同一のヌクレオチド配列の全部または一部を含んでなる二本鎖核酸分子を用いて行なわれる。例えば、この二本鎖核酸分子を用いることにより、被子植物において本発明による遺伝子のアンチセンスまたはセンス一本鎖核酸を発現させることができる。本発明による二本鎖核酸分子は、好ましくはDNAとされ、その鎖長および具体的なヌクレオチド配列は、目的とする一本鎖核酸分子の鎖長およびヌクレオチド配列に対応するものとされる。例えば、上記アンチセンス一本鎖核酸を発現させる場合には、本発明による二本鎖核酸分子は、本発明による遺伝子のアンチセンス鎖をコード鎖として含むものとされる。また、上記センス一本鎖核酸を発現させる場合には、本発明による二本鎖核酸分子は、本発明による遺伝子のセンス鎖をコード鎖として含むものとされる。
【0026】
本発明による二本鎖核酸分子は、当業者に公知の方法を用いて被子植物中で発現させることができる。例えば、プロモーター、本発明による二本鎖核酸分子、および転写ターミネーター等を含む発現用構築物を、目的とする被子植物中に導入し、得られた植物体を栽培することにより、本発明による二本鎖核酸分子を発現させることができる。植物への発現用構築物の導入は、当業者に公知の方法、例えば、アグロバクテリウム法、バイナリーベクター法、エレクトロポレーション法、PEG法、パーティクルガン法等によって行なうことができる。特に、遺伝子導入のためのバラの形質転換は、米国特許第5480789号明細書等に記載の方法によって行うことができる。
【0027】
本発明による核酸分子を用いた遺伝子発現の抑制法としては、例えば、アンチセンス鎖による形質転換法(Proc. Natl. Acad. Sci. USA 90: 6160-6164, 1993)が挙げられる。この方法では、本発明による遺伝子の全ヌクレオチド配列を有するアンチセンス二本鎖DNAが目的の植物中に導入される。これにより、本発明による遺伝子のmRNAに相補的なRNAとして、本発明によるアンチセンス一本鎖RNAが発現され、この相補的RNAにより該遺伝子の発現が極端に抑制される。
【0028】
本発明による核酸分子を用いた遺伝子発現の抑制法の他の例としては、コサプレッション法が挙げられる。この方法では、本発明による遺伝子の全ヌクレオチド配列を有するセンス二本鎖DNAが目的の植物中に導入される。これにより、本発明によるセンス一本鎖RNAが発現され、このRNAにより該遺伝子の発現が極端に抑制される(Plant Cell 9: 1357-1368, 1997)。
【0029】
本発明によるDNAは、配列番号3で表されるヌクレオチド配列を含んでなるものであり、該DNAは被子植物におけるトランスポゾンとして機能しうるものである。なお、配列番号3で表されるヌクレオチド配列において、第1〜856残基はLTRであり、第857〜6105残基はORFであり、第8069〜8925残基はLTRである。また、本発明によるDNAは上記のものに限定されるものではなく、被子植物におけるトランスポゾンとして機能しうる限り、配列番号3で表されるヌクレオチド配列において、1以上のヌクレオチドが欠失、置換、付加または挿入されたヌクレオチド配列を含んでなるものであってもよい。ここで、置換、欠失、付加または挿入されるヌクレオチドの数は特に限定されないが、好ましくは1〜9個、より好ましくは1〜5個、さらに好ましくは1〜3個とされる。また、このようなヌクレオチド配列は、配列番号3で表されるヌクレオチド配列に対して、好ましくは少なくとも70%、より好ましくは少なくとも80%、さらに好ましくは少なくとも90%の相同性を有するものとされる。
【0030】
本発明によるDNAは、当業者に公知の方法、例えば、ホスホアミダイド法等により化学的に合成する方法、オールド・ブラッシュ(Old blush)等の四季咲き性バラからの核酸試料を鋳型とし、目的とする遺伝子のヌクレオチド配列に基づいて設計したプライマーを用いる核酸増幅法などによって得ることができる。
【0031】
本発明によるDNAは、被子植物においてトランスポゾンとして機能し、本発明による遺伝子中に挿入されることにより該遺伝子の発現を抑制するものである。これにより、茎頂での花芽の形成抑制が解除されるため、花芽の形成が促進され、その被子植物は四季咲き性となる。従って、本発明によれば、本発明によるDNAを含んでなるトランスポゾンが提供され、さらには、本発明によるDNAまたはこれを含んでなるトランスポゾンを用いて本発明による遺伝子の発現を抑制することを含んでなる、被子植物に四季咲き性を付与する方法が提供される。
【0032】
この方法では、本発明によるDNAまたはトランスポゾンが被子植物に導入される。これは、本発明による遺伝子の発現が抑制されるように、当業者によって適宜行なわれるが、本発明によるDNAまたはトランスポゾンは、本発明による遺伝子中またはその近傍に導入されることが好ましく、より好ましくは本発明による遺伝子の第2イントロン中、さらに好ましくは四季咲き性バラと同じ場所に導入される。
【0033】
植物へのDNAの導入は、当業者に公知の方法、例えば、アグロバクテリウム法、バイナリーベクター法、エレクトロポレーション法、PEG法、パーティクルガン法等によって行なうことができる。特に、遺伝子導入のためのバラの形質転換は、米国特許第5480789号明細書等に記載の方法によって行うことができる。
【0034】
【実施例】
例1:四季咲き性に関与する因子の特定
現代の四季咲き性バラ品種では、シュートや腋芽は4〜7枚程度の葉を伸ばすと茎頂に花芽を形成する。花が咲き終わるとすぐに下の腋芽が伸び、同様にして花芽を形成する。こうして、温度、水、肥料などの栽培条件が整えば、次々にシュートを伸ばし、その先に花芽を形成し、四季を通じて開花する。これに対し、つる性のバラでは、シュートが30枚または40枚以上の葉を伸ばしても、茎頂に花芽を形成することなく伸びつづけ、一季咲きとなる。
【0035】
本発明者は、アラビドプシス(Arabidopsis)の野生型(無限花序)と突然変異型(有限花序)に関する知見、ならびに現代四季咲き性バラにおいて一季咲き性バラへの突然変異が多く見られることに基づき、以下の仮説を立て、これを証明した。
【0036】
(仮説1)一季咲き性のつる型バラではtfl1遺伝子のオルソロガス遺伝子が活性を持っているため、茎頂における花芽形成が抑制されるが、四季咲き型バラでは突然変異によって前記遺伝子の活性が低下しているため、茎頂における花芽形成が抑制されない。
【0037】
(仮説2)四季咲き性バラは、tfl1遺伝子のオルソロガス遺伝子へのトランスポゾンの挿入という突然変異によって生じ、一季咲き性のつる型バラは、該トランスポゾンの欠失という復帰突然変異によって生じる。
【0038】
(1)tfl1遺伝子のバラオルソロガス遺伝子の配列決定および該遺伝子中のトランスポゾンの存否
以下のようにして、tfl1遺伝子のバラオルソログのヌクレオチド配列を明らかにし、四季咲き型ではその遺伝子全てにトランスポゾンが挿入されており、一季咲き型では、トランスポゾンが挿入されていない遺伝子が少なくとも一つ存在することを証明した。
【0039】
tfl1遺伝子は、ホスファチジルエタノールアミン結合タンパク質(PEBP)と称する遺伝子ファミリーに属する。アラビドプシスでは6種のPEBP遺伝子が知られている。アラビドプシス、イネ、キンギョソウ、ペチュニア、トマトなどの公知のPEBP遺伝子のヌクレオチド配列に基づき、特に保存性の高い部分を選んで以下のプライマーを設計した:
フォワードプライマー(UtflF1):GATAGACCCAGATGTTCCAGGTCC(配列番号4);
リバースプライマー(UtflR1):CCTGGAATATCTGTCACCATCCT(配列番号5)。
【0040】
これらのプライマーを用いて、ロサ・キネンシス・スポンタネア(Rosa chinensis spontanea)(Rcs)のDNAを鋳型にしてPCRを行い、複数の増幅産物を得た。これらの増幅産物のヌクレオチド配列を決定し、そのヌクレオチド配列からPEBPファミリー遺伝子と考えれるもの4種を選び、これらをRPEBP1、RPEBP2、RPEBP3、およびKSNと名付けた。これらの増幅産物のうち、KSNの400bp余りの産物のヌクレオチド配列は図1に示すとおりであった。このヌクレオチド配列は、アラビドプシスtfl1遺伝子のエクソン2から3にかけてのヌクレオチド配列と高い相同性を有していた。
【0041】
上記のように決定されたKSNの部分的なヌクレオチド配列に基づき、KSN遺伝子全体のヌクレオチド配列決定を以下のようにして行なった。すなわち、RcsのDNAを鋳型として、InversePCR法により決定した。
【0042】
まず、RcsのDNA数μgをNEB社の制限酵素Cac 8Iで消化し(DNA17μl、酵素1μl、10倍バッファー2μlを37℃で4時間反応)、加熱によって酵素を不活化してから(65℃で20分間)、NEB社のT4-DNA-Ligaseを用いて環状化した(酵素消化DNA20μl、リガーゼ1μl、10倍バッファー12.5μl、および精製水91.5μlを8℃で一昼夜反応)。この環状DNAを鋳型とし、下記の二つのプライマーを用いるPCRを行なった:
KSN-IR1:CTGTAAGTTATACCAGTGCAGGTGC(配列番号6);
KSN-IF2:GGAAGTACATATTATGGCATAATC(配列番号7)。
得られた増幅産物のヌクレオチド配列を図2に示す。図2から明らかなように、Utfl-FIの上流416bpのヌクレオチド配列が決定された。
【0043】
さらに、RcsのDNA数μgをNEB社の制限酵素Sau96Iで消化し(DNA17μl、酵素1μl、10倍バッファー2μlを37℃で4時間反応)、加熱によって酵素を不活化してから(80℃で20分間)、NEB社のT4-DNA-Ligaseを用いて環状化した(酵素消化DNA20μl、リガーゼ1μl、10倍バッファー12.5μl、および精製水91.5μlを8℃で一昼夜反応)。この環状DNAを鋳型とし、上記のプライマーKSN-IR1および以下のプライマーKSN-IF1を用いるPCRを行なった:
KSN-IF1:AGGAGCTAACATTTTTGCCTTG(配列番号8)。
得られた増幅産物のヌクレオチド配列を図3に示す。図3から明らかなように、UtflR1の下流401bpのヌクレオチド配列が決定された。
【0044】
以上のように決定された配列をまとめたKSNのヌクレオチド配列を図4に示す。図4において、アラビドプシスtfl1遺伝子のヌクレオチド配列と比較してエクソンと思われる部分に囲み線を付した。さらに、KSNのコード領域のヌクレオチド配列を配列番号1に示し、これによりコードされるアミノ酸配列を配列番号2に示した。
【0045】
同様にして、RPEBP1、RPEBP2、およびRPEBP3のヌクレオチド配列も明らかにした(配列は示さない)。
【0046】
上述のように決定されたRPEBP1、RPEBP2、RPEBP3、およびKSNのヌクレオチド配列に基づき、これらの遺伝子のコーディング領域のほぼ全域を特異的に増幅するプライマーを設計した。例えば、KSN増幅用のプライマーは以下の配列とした:
KSNF1:ATTAGGAGTACAATCCTTCCTTCC(配列番号9);
KSNR2:ACCTGCCTCCTGCTAGCTGC(配列番号10)。
【0047】
四季咲き性のオールド・ブラッシュ(OBB)およびその突然変異体とされる一季咲き性のオールド・ブラッシュ・クライミング(OBCl)のDNAを鋳型とし、上記のコーディング領域増幅用プライマーを用いて、1Kbp程度の増幅物を得る条件(Promega社製のPCR Master Mixを添付プロトコルに従って使用)でのPCRを行なった。その結果、RPEBP1、RPEBP2、およびRPEBP3の増幅産物は、OBBとOBClとの間で違いが認められなかった。KSNでは予想された大きさの増幅産物が認められなかったため、PCRの条件を10Kbp程度の増幅産物が得られる条件(タカラ社製のOne Shot LAPCR Mixを添付プロトコルに従って使用)に変更して再度PCRを行った。その結果、OBBでは約10Kbpの増幅産物が、OBClでは約2Kbpの増幅産物が得られた。
【0048】
OBBおよびOBClのDNAを鋳型として得られた増幅産物は、KSNに何らかのDNAが挿入されているものと推定された。そこで、以下のようにしてその挿入物のヌクレオチド配列を明らかにした。
【0049】
まず、挿入地点と推定される前後にいくつかのプライマーを設計してPCRを行ない、増幅産物を得た。その結果、下記のプライマーが最も挿入DNAに近接したプライマーであることが明らかとなった:
KSNIF3:CATATTATGGCATAGGGTGTGGC(配列番号11);
KSNR5:CAAATGTAACATCTGTGGTGCCTG(配列番号12)。
【0050】
900bp前後と推定された短い挿入物(Si)は、上記二つのプライマーを用いてヌクレオチド配列を明らかにした。
【0051】
さらに、9Kbp前後と推定された長い挿入物(Li)のヌクレオチド配列は以下のようにして決定した。まず、PCR増幅産物を、添付のプロトコールに従って、pT7BlueT-Vector(Novagen社製)にDNA Ligation Kit Ver.2(タカラ社製)を用いて連結し、環状化した。このプラスミドを用いて、コンピテント細胞JM109株(タカラ社製)を添付プロトコールに従って形質転換した。形質転換したこの大腸菌を、添付プロトコールに従って、あらかじめ2%5−ブロモ−4−クロロ−3−インドリル−β−D−ガラクトシド(Xgal)50μlと0.1Mイソプロピル−β−D−チオガラクトピラノシド(IPTG)10μlとを塗布しておいたL−agar培地(アンピシリン50μg/mlを含む)に塗布し、生じたコロニーの中から白色のコロニーを増殖させた。
【0052】
生じたクローンのうち、クローン6が完全長のPCR増幅産物を含んでいると推定されたので、以下のようにしてそのヌクレオチド配列を明らかにした。まず、このベクターのクローニングサイトに断点を持つ制限酵素HindIIIまたはEcoRIで切断し、DNA Ligation Kit Ver.2(タカラ社製)を用いて再度連結し、環状化した。このプラスミドを用いて、コンピテント細胞JM109株(タカラ社製)を添付プロトコールに従って形質転換し、そのコロニーを得た(それぞれクローン6−12およびクローン6−16)。EcoRIを用いた大きなDNA断片を持つクローン6−16は、さらに、SpeI、Nole1、Xba1、およびPstIを用いてそれぞれに対応する箇所を切断し、同様にしてクローンを得た。得られたクローンのうち、クローン6−16−27、クローン6−16−32、クローン6−16−34、クローン6−16−40、およびクローン6−16−43の5種は互いに異なるDNA断片を含んでいると推定された。
【0053】
上記の8種のクローンのプラスミドに挿入されたDNAのヌクレオチド配列を、以下のプライマーを用いて前後から決定し、つなぎ合わせた:
M13R18:CAGGAAACAGCTATGACC(配列番号13);
MR13F18:TGTAAAACGACGGCCAGT(配列番号14)。
【0054】
その結果、このDNA断片(Li)は約9Kbpの大きさで、blastホモロジー検索の結果、コピアタイプのレトロトランスポゾンと相同性が高かった。このトランスポゾン(配列番号3)は、OBBのKSN遺伝子のイントロン2に挿入されていた。また、OBClの約900bpの挿入物は、このレトロトランスポゾンが消失する際にその場に残した約900bpのLTR(長い末端反復配列)であることが判明した。
【0055】
以上のように、Rcs、OBB、およびOBClのKSNとしては、挿入のない野生型(w型、約1Kbp)、レトロトランスポゾンの入った型(Li型、約10Kbp)、レトロトランスポゾンがLTRを残して出ていった型(Si型、約2Kbp)の三種が認められた。
【0056】
上記の結果から、Li型では、約9000bpのDNAの挿入によってmRNAの転写が大きく阻害されるためにKSNの発現がほとんど阻害され、Si型では、900bpという短いDNAの挿入であるためKSN発現の阻害が軽微であると考えられる。これは、OBBが四季咲きとなり、他方でOBClはつるバラのように良く伸びるが、夏から秋にかけてもぽつぽつと咲くことと一致する。
【0057】
次いで、いくつかの園芸品種を、KSN遺伝子の構造について調べた。先に設計したレトロトランスポゾンの挿入位置の前(KSNIF3)と後(KSNR5)のプライマーを用いると、w型とSi型はそれぞれ250bpと1150bpの異なる大きさの増幅産物を与える。また、KSNIF3と共に用いてレトロトランスポゾンの一部を増幅するようなプライマーKSNInsR3を、LTRよりも内側に設計した(KSNInsR3:TGTAATCTGTAGGAGATCCCATGC, 配列番号15)。KSNIF3、KSNR5およびKSNInsR3を用いて、つる性の突然変異の園芸品種が作られている6園芸品種(「フレーミング・ピース」、「ピース」、「ウエンディ・クッソンズ」、「ゴールド・バニー」、「アイス・バーグ」、および「ピンク・シホン」)のKSNを調べた。これら全ての品種において、四季咲き性の品種ではKSNIF3およびKSNR5のプライマーペアでは増幅産物がなく、KSNIF3およびKSNInsR3のプライマーペアで増幅産物が得られた。これは、四季咲き性品種ではすべてのKSNにレトロトランスポゾンが挿入されていたことを示す。一方で、つる性の突然変異品種では、KSNIF3およびKSNInsR3のプライマーペアによる増幅産物以外にKSNIF3およびKSNR5のプライマーペアによる1150bpのSi型の増幅産物が見られ、レトロトランスポゾンを含んだKSN以外に、LTRのみが残ったKSNが存在していることが明らかとなった。この様に、これら6種の園芸品種においても、四季咲き性品種ではトランスポゾンが挿入されており、つる性(一季咲き性)品種では少なくとも一つはLTRを残してトランスポゾンが転移したと思われるKSNが存在することが明らかとなった。
【0058】
(2)四季咲き性バラと一季咲き性バラとの、tfl1遺伝子バラオルソログmRNA発現レベルの比較
上記(1)で明らかとなったトランスポゾンの挿入により、四季咲き型バラのKSNのmRNA発現レベルが、一季咲き型のそれよりも著しく低いことを、以下のようにして証明した。
【0059】
まず、野生型KSNを有するRcsの腋芽から、RNAを抽出した。Rcsの腋芽としては、15cmのもの、10cmのもの、および5cm以下のものの3種類を用い、それぞれ図5に示す位置からサンプルを採取した。同様に、OBBの腋芽の茎頂およびOBClの蕾を含む腋芽からもRNAを抽出した。RNA抽出は、Rneasy Plant Mini Kit(Qiagen社製)を用いて行なった。
次いで、KSNのmRNA量を、Quanti Tect SYBR Green RT-PCR Kit(Qiagen社製)およびLight Cycler System(Roche Molecular Biochemicals社製)を用いて比較した。また、本実験では、バラのアクチンのmRNAをコントロールとして用いた。KSNのmRNAは量が少なく、QuantiTect SYBR Green RT-PCR Kitでは測定できないことがあったので、限界希釈法によってさらに測定した。限界希釈法とは、RNA抽出液の3倍希釈系列を作り、OneStep RT-PCR Kit(Qiagen社製)を用いてKSNのmRNAが検出できなくなる希釈倍率を求め、mRNA量を推定する方法である。Rcsについての結果を下記の表1に示す。表1において、各数値は、Rcs3pについての測定値を1とした場合の相対量で示した。
【0060】
【表1】
【0061】
表1から、茎頂でのKSNのmRNA発現量は、腋芽の成長に従って増加し、茎頂から離れるにしたがって減少することが明らかとなった。さらに、OBCl茎頂におけるKSNのmRNA発現量はRcs3pの1/10000程度であり、OBB茎頂においては検出できなかった。
【0062】
(3)考察
以上の結果から、四季咲き性のバラでは、KSN遺伝子中にトランスポゾンが挿入されており、これにより該遺伝子の発現が阻害され、茎頂における花芽の形成が促進されることが明らかとなった。従って、KSN遺伝子を阻害することにより、バラに四季咲き性を付与することが可能となる。
【0063】
アラビドプシスのTFL1およびATC、キンギョソウのCEN、ならびにライ・グラス(単子葉類の牧草)のLpTFL1をアラビドプシスで発現させたときの表現形に差異は認められないため、これらの機能の違いは、タンパク質の構造の違いではなく、主に遺伝子発現の違いによると報告されている(Shepard,K.A.and Purugganon M.D., The genetics of plant morphological evolution., Current Opinion in Plant Biology 5:49-55, 2002)。さらに、KSN遺伝子の構造が真核生物全体で良く保存されていること、および単子葉植物の遺伝子が双子葉植物においても本来の機能を発揮することをも考慮すると、バラ属で発見された四季咲きのメカニズムは他の被子植物でも働くものと考えられる。
【0064】
【配列表】
【図面の簡単な説明】
【図1】図1は、部分的に決定されたKSN遺伝子のヌクレオチド配列(配列番号16)を示す図である。
【図2】図2は、部分的に決定されたKSN遺伝子のヌクレオチド配列(配列番号17)を示す図である。
【図3】図3は、部分的に決定されたKSN遺伝子のヌクレオチド配列(配列番号18)を示す図である。
【図4】図4は、KSN遺伝子のコーディング領域のヌクレオチド配列(配列番号19)を示す図である。
【図5】図5は、バラ(Rcs)腋芽の様々な部位におけるKSNのmRNA発現量を示す図である。
【特許請求の範囲】
【請求項1】
以下の(a)または(b)に示されるタンパク質:
(a)配列番号2で表されるアミノ酸配列を含んでなるタンパク質、
(b)配列番号2で表されるアミノ酸配列において、1以上のアミノ酸が置換、欠失、付加または挿入されたアミノ酸配列を含んでなり、被子植物の茎頂での花芽形成を抑制しうるタンパク質。
【請求項2】
前記被子植物がバラ科に属するものである、請求項1に記載のタンパク質。
【請求項3】
請求項1または2に記載のタンパク質をコードする遺伝子。
【請求項4】
配列番号1で表されるヌクレオチド配列を含んでなる、請求項3に記載の遺伝子。
【請求項5】
請求項3または4に記載の遺伝子のセンス鎖と同一のヌクレオチド配列の全部または一部を含んでなる、一本鎖核酸分子。
【請求項6】
請求項3または4に記載の遺伝子のアンチセンス鎖と同一のヌクレオチド配列の全部または一部を含んでなる、一本鎖核酸分子。
【請求項7】
請求項3または4に記載の遺伝子と同一のヌクレオチド配列の全部または一部を含んでなる、二本鎖核酸分子。
【請求項8】
請求項5もしくは6に記載の一本鎖核酸分子または請求項7に記載の二本鎖核酸分子を用いて、請求項3または4に記載の遺伝子の発現を抑制することを含んでなる、被子植物に四季咲き性を付与する方法。
【請求項9】
前記被子植物がバラ科に属するものである、請求項8に記載の方法。
【請求項10】
以下の(a)または(b)に示されるDNA:
(a)配列番号3で表されるヌクレオチド配列を含んでなるDNA、
(b)配列番号3で表されるヌクレオチド配列において、1以上のヌクレオチドが欠失、置換、付加または挿入されたヌクレオチド配列を含んでなり、被子植物におけるトランスポゾンとして機能しうるDNA。
【請求項11】
前記被子植物がバラ科に属するものである、請求項10に記載のDNA。
【請求項12】
請求項10または11に記載のDNAを含んでなるトランスポゾン。
【請求項13】
請求項10もしくは11に記載のDNAまたは請求項12に記載のトランスポゾンを用いて、請求項3または4に記載の遺伝子の発現を抑制することを含んでなる、被子植物に四季咲き性を付与する方法。
【請求項14】
前記被子植物がバラ科に属するものである、請求項13に記載の方法。
【請求項1】
以下の(a)または(b)に示されるタンパク質:
(a)配列番号2で表されるアミノ酸配列を含んでなるタンパク質、
(b)配列番号2で表されるアミノ酸配列において、1以上のアミノ酸が置換、欠失、付加または挿入されたアミノ酸配列を含んでなり、被子植物の茎頂での花芽形成を抑制しうるタンパク質。
【請求項2】
前記被子植物がバラ科に属するものである、請求項1に記載のタンパク質。
【請求項3】
請求項1または2に記載のタンパク質をコードする遺伝子。
【請求項4】
配列番号1で表されるヌクレオチド配列を含んでなる、請求項3に記載の遺伝子。
【請求項5】
請求項3または4に記載の遺伝子のセンス鎖と同一のヌクレオチド配列の全部または一部を含んでなる、一本鎖核酸分子。
【請求項6】
請求項3または4に記載の遺伝子のアンチセンス鎖と同一のヌクレオチド配列の全部または一部を含んでなる、一本鎖核酸分子。
【請求項7】
請求項3または4に記載の遺伝子と同一のヌクレオチド配列の全部または一部を含んでなる、二本鎖核酸分子。
【請求項8】
請求項5もしくは6に記載の一本鎖核酸分子または請求項7に記載の二本鎖核酸分子を用いて、請求項3または4に記載の遺伝子の発現を抑制することを含んでなる、被子植物に四季咲き性を付与する方法。
【請求項9】
前記被子植物がバラ科に属するものである、請求項8に記載の方法。
【請求項10】
以下の(a)または(b)に示されるDNA:
(a)配列番号3で表されるヌクレオチド配列を含んでなるDNA、
(b)配列番号3で表されるヌクレオチド配列において、1以上のヌクレオチドが欠失、置換、付加または挿入されたヌクレオチド配列を含んでなり、被子植物におけるトランスポゾンとして機能しうるDNA。
【請求項11】
前記被子植物がバラ科に属するものである、請求項10に記載のDNA。
【請求項12】
請求項10または11に記載のDNAを含んでなるトランスポゾン。
【請求項13】
請求項10もしくは11に記載のDNAまたは請求項12に記載のトランスポゾンを用いて、請求項3または4に記載の遺伝子の発現を抑制することを含んでなる、被子植物に四季咲き性を付与する方法。
【請求項14】
前記被子植物がバラ科に属するものである、請求項13に記載の方法。
【図1】


【図2】

【図3】

【図4】

【図5】
【図2】
【図3】
【図4】
【図5】
【公開番号】特開2006−149202(P2006−149202A)
【公開日】平成18年6月15日(2006.6.15)
【国際特許分類】
【出願番号】特願2003−27372(P2003−27372)
【出願日】平成15年2月4日(2003.2.4)
【出願人】(000250100)湧永製薬株式会社 (51)
【Fターム(参考)】
【公開日】平成18年6月15日(2006.6.15)
【国際特許分類】
【出願日】平成15年2月4日(2003.2.4)
【出願人】(000250100)湧永製薬株式会社 (51)
【Fターム(参考)】
[ Back to top ]