
補助因子による受容体の活性化方法並びにリガンド活性の利用方法
【課題】 FGF受容体を介したFGF21の活性を昂進もしくは抑制する物質、FGF21活性昂進物質の昂進もしくは抑制する物質、FGF21様活性物質のスクリーニング方法及びそのためのキットの提供。
【解決手段】 FGF受容体及び/又はbetaKlothoを細胞表面に発現させた細胞系を用いることによる、betaKlotho活性の昂進又は抑制する物質、FGF21様活性物質、及びbetaKlotho様活性物質それぞれのスクリーニングの系を提供した。
【解決手段】 FGF受容体及び/又はbetaKlothoを細胞表面に発現させた細胞系を用いることによる、betaKlotho活性の昂進又は抑制する物質、FGF21様活性物質、及びbetaKlotho様活性物質それぞれのスクリーニングの系を提供した。
【発明の詳細な説明】
【技術分野】
【0001】
本発明は、受容体補助因子もしくは当該因子をコードする遺伝子又は当該因子活性を昂進もしくは抑制する物質を用いてリガンドによる受容体の活性化を促進または抑制する方法並びにリガンド活性を制御する方法(及びこれを用いた血糖値調節方法)に関する。
【背景技術】
【0002】
糖尿病で問題とされる高い血糖レベルに対しては、健康な体の中で血糖レベルが調節される仕組みを理解し、その知識を活用して、正常な値に制御することが必要であるが、未だ体内の血糖レベルの制御機構は解明できていない。現在実践的に用いられている唯一の積極的治療法は、糖尿病患者に対しインスリンを投与し、インスリンの血糖値低下作用を利用して、血糖値を正常範囲にまで低下させる方法のみである。しかし、この方法は、インスリンの分泌量が不足していることが原因の患者にとっては極めて効果的な治療法ではあるが、患者の中にはインスリンを投与しても血糖値の低下しない患者もいる。このような、所謂インスリン抵抗性の患者に対しても有効な、糖尿病薬の開発は急務であり、そのためにも体内における血糖値の制御機構を解明し、この問題を解決することが、当技術分野において大きな課題となっている。
【0003】
ところで、線維芽細胞増殖因子(FGF)とよばれるFGFファミリーの分子群は、アミノ酸配列の相同性と構造的類似性から、現在ヒト及びマウスにおいては22種類存在することが知られている。すべてのFGFファミリーメンバーの活性機能が明らかにされているわけではないが、すでに機能に関する研究が進んでいるメンバーは、いずれも線維芽細胞の増殖活性のみならず、広範な細胞に対する増殖や分化の制御活性を有することが知られ、形態形成、血管形成、神経生存維持、代謝調節などといった多様な生命現象に深く関わっている因子群である(非特許文献1,2,3,4)。このメンバーの中で比較的最近FGFファミリーメンバーに加わったFGF19、FGF21及びFGF23は、ドメイン構造からもアミノ酸配列の相同性の観点からも類似したサブグループを形成しているが、これらのFGFは、まだその作用機構の解明が十分になされているとはいえず、しかも通常のFGFファミリーで一般に用いられている細胞増殖や分化を測定するアッセイ系に供しても、ほとんど活性が検出されない。
最近、FGF21の作用として脂肪細胞のグルコーストランスポーターの発現量を昂進し、細胞内へのグルコース取り込みを上昇させ、また動物個体レベルでは、静脈内あるいは皮下投与によって血糖値の低下を引き起こすこと、血中トリグリセリド量を抑制すること、LDLコレステロールを低下、HDLコレステロールを増加させることが報告された(非特許文献1、5)。またFGF21は膵臓ランゲルハンス島に作用し、インスリンの合成量を増加させることも報告されている(非特許文献6)。このことから、FGF21は、血糖レベル調節や中性脂肪レベル調節などに重要な役割を果たしていることが考えられ、新規の作用機構を有するメタボリックシンドローム治療薬候補として浮上してきた。
しかしながら、FGF21をメタボリックシンドローム治療用医薬として用いるためには、FGF21単独では十分とはいえないことから、その作用を増強する因子の探索は必須であるが、効果的に増強できる因子はまだ発見されていない。
一般に、FGFが活性を発揮するためには、細胞膜表面のFGF受容体と結合して、これを活性化させて細胞内シグナリングを惹起することが必要であり、FGF21についても脂肪細胞に作用させた際に、細胞の表面に存在するFGFR1とFGFR2の活性化が起こることが報告されている(非特許文献1)。FGF21に対して観察された血糖値低下作用などもこれらFGF受容体との相互作用に基づくシグナル伝達の結果であると考えられ、その際のFGF21によるFGF受容体の活性化機構が解明されれば、当該機構に関連した因子を利用することで、血糖値を調節することが可能になる。しかしながら、従来の知見はFGF21のFGF受容体への結合様式の解明にも、グルコーストランスポーターや、インスリン発現などの血糖値を制御する機構の解明にも直接繋がるものではなく、このような血糖値の調節可能な因子の提供には結びつかなかった。
したがって、FGF21がどのようにしてFGF受容体を活性化するのかを早急に解明し、FGF21の血糖値等制御機能を調節する手法を確立することで、FGF21を有効成分とする効果的なメタボリックシンドローム治療薬を提供することが、当該分野の大きな課題となっていた。
【先行技術文献】
【特許文献】
【0004】
【特許文献1】特開2006-158339号公報:非特許文献7の関連特許文献
【特許文献2】米国特許第6099841号明細書
【特許文献3】米国特許第6365154号明細書
【非特許文献】
【0005】
【非特許文献1】Kharitonenkov,A.,Shiyanova,T. L.,Koester,A.,Ford,A. M.,Micanovic,R.,Galbreath,E. J.,Sandusky,G. E.,Hammond,L. J.,Moyers,J. S.,Owens,R. A.,Gromada,J.,Brozinick,J. T.,Hawkins,E. D.,Wroblewski,V. J.,Li,D. S.,Mehrbod,F.,Jaskunas,S. R.,andShanafelt,A. B. (2005) J Clin Invest 115(6),1627-1635
【非特許文献2】Itoh,N. and Ornitz,D.M. (2004) Evolution of the Fgf and Fgfr gene families. TrendsGenet,20,563-569.
【非特許文献3】Inagaki,T.,Choi,M.,Moschetta,A.,Peng,L.,Cummins,C.L.,McDonald,J.G.,Luo,G.,Jones,S.A.,Goodwin,B.,Richardson,J.A.,Gerard,R.D.,Repa,J.J.,Mangelsdorf,D.J.and Kliewer,S.A. (2005) Fibroblast growth factor 15functions as an enterohepatic signal to regulate bile acid homeostasis. CellMetab,2,217-225.
【非特許文献4】Yu,X. and White,K.E. (2005) FGF23 and disorders of phosphate homeostasis. CytokineGrowth Factor Rev,16,221-232.
【非特許文献5】Kharitonenkov,A.,Wroblewski,V. J.,Koester,A.,Chen,Y. F.,Clutinger,C. K.,Tigno,X. T.,Hansen,B. C.,Shanafelt,A. B.,andEtgen,G. J. (2007) Endocrinology 148(2),774-781
【非特許文献6】Wente,W.,Efanov,A. M.,Brenner,M.,Kharitonenkov,A.,Koster,A.,Sandusky,G. E.,Sewing,S.,Treinies,I.,Zitzer,H.,and Gromada,J.(2006) Diabetes 55(9),2470-2478
【非特許文献7】Ito,S.,Fujimori,T.,Furuya,A.,Satoh,J.,Nabeshima,Y.,andNabeshima,Y. (2005) J Clin Invest 115(8),2202-2208
【非特許文献8】Ito,S.,Kinoshita,S.,Shiraishi,N.,Nakagawa,S.,Sekine,S.,Fujimori,T. and Nabeshima,Y.I. (2000) Molecularcloning and expression analyses of mouse betaklotho,whichencodes a novel Klotho family protein. Mech Dev,98,115-119.
【非特許文献9】Nishimura,T.,Nakatake,Y.,Konishi,M. and Itoh,N. (2000) Identification of anovel FGF,FGF-21,preferentiallyexpressed in the liver. Biochim Biophys Acta,1492,203-206
【非特許文献10】LeSauteur,L.et al.(1996) J Neurosci,16,1308-1316)
【非特許文献11】Fernandez-Pol,J.A.(1985) J Biol Chem,260,5003-5011)
【発明の概要】
【発明が解決しようとする課題】
【0006】
本発明は、FGF受容体を介したFGF21活性に対する効果的な調節因子を提供することを目的とする。典型的には、FGF21によって細胞の糖取り込みが調節する際に必要とされる受容体補助因子の同定、及び当該因子を利用したFGF21の活性制御方法並びに当該活性を用いた血糖値調節方法を提供することを目的とする。
また、当該調節因子及び/又はその活性を増強若しくは抑制する物質を有効成分として含むFGF21活性調節剤を提供すると共に、血糖値の病的な上昇、低下など糖取り込み異常に関連した疾患、たとえば糖尿病の治療又は予防用医薬組成物などFGF受容体を介したFGF21活性を調節するための医薬組成物の提供も本発明の目的である。
さらに、本発明は、FGF受容体を介したFGF21活性の調節因子、当該調節因子の活性を増強若しくは抑制する物質、FGF21様活性物質などのスクリーニング系の提供も目的とする。
【課題を解決するための手段】
【0007】
上述した目的を達成するため本発明者は、鋭意検討した結果、betaKlothoこそがFGF21によって細胞の糖取り込みが調節する際に必要とされる特異的な受容体補助因子であることを見いだした。
具体的には、本発明者らは、まずFGF21がどのようにしてFGF受容体を活性化するのか、受容体を活性化するのに必要な条件は何なのかを確かめるために、本来FGF受容体を有していない細胞の表面で各FGF受容体を発現している複数の細胞を用意して、FGF21と反応させたところ、いずれのFGF受容体においてもFGF1が通常十分活性を発揮する濃度の数十倍から数百倍量のFGF21によってもFGF受容体の活性化は検出されないことを見いだした。
さらに、通常のFGFリガンドが各FGF受容体と結合する際には、受容体補助因子としてのヘパリンが必須であるため、上記の系にさらにヘパリンを添加して観察したが、ヘパリン存在下でも、非存在下と同様にFGF受容体の活性化は起こらなかった。そこで、FGF21がどのようにしてFGF受容体を活性化するのか、その必要条件は何なのかについて鋭意検討する過程で、betaKlothoが膜上に共に存在するときに、FGF21によってFGFR1cが活性化され、下流のシグナルが惹起されることを見出した。
ついで、本発明者らは、FGF受容体もbetaKlothoも内在的に発現していない細胞であるBaF3細胞に対して、FGFR1c遺伝子もしくはFGFR3c遺伝子を、又はさらにbetaKlotho遺伝子及びその構造類似体のKlotho遺伝子を導入して細胞表面に発現させた各種形質転換細胞系を用いた実験で、FGF21及びbetaKlothoがそれぞれの構造類似体に対する組み合わせを変えても互換性があるか否かを観察した結果、FGF21とbetaKlothoとの組み合わせが、FGFR1cまたはFGFR3cに対する作用において、特異的な組み合わせであることを見いだした。すなわち、このことは、betaKlothoこそがFGF21のFGF受容体に対する特異的な受容体補助因子であることを示すものである。
そして、本発明者らは、上記のFGFR1c遺伝子もしくはFGFR3c遺伝子と共にbetaKlotho遺伝子を導入して細胞表面に発現させた形質転換細胞系が、FGF受容体とbetaKlotho共存下で発揮されるFGF21によるFGF受容体の活性化の程度をきわめて鋭敏に示すものであることに着目して、当該形質転換細胞系を用いた、betaKlotho活性を昂進又は抑制する物質のスクリーニング系、FGF21様物質のスクリーニング系を提供し、また、FGFR1c遺伝子もしくはFGFR3c遺伝子のみを導入して細胞表面に発現させた形質転換細胞系を用いた、betaKlotho様の活性を示す物質のスクリーニング系を提供することができた。
以上のように、本発明者らは、betaKlothoの有無により、FGF21のFGF受容体を介した生物活性が制御できるという知見を得て、betaKlotho及びその活性の昂進又は阻害物質を用いたFGF受容体を介したFGF21活性の調節剤、医薬組成物についての本発明を完成し、同時にその際に用いた優れた形質転換細胞系を利用した各種スクリーニングに関する本発明を完成するに至った。
【0008】
すなわち、本発明は以下を包含する。
(1) betaKlotho活性を有する蛋白質を有効成分として含む、FGF受容体を介したFGF21の生物活性を増強するか又は抑制するための、FGF21活性の調節剤。
(2) betaKlotho活性を有する蛋白質が、betaKlothoの可溶性部分タンパク質である、前記(1)に記載のFGF21活性の調節剤。
(3) betaKlotho活性を有する蛋白質をコードするDNAを含む組換えDNAを有効成分として含む、FGF受容体を介したFGF21の生物活性を増強するか又は抑制するための、FGF21活性の調節剤。
(4) betaKlotho活性を有する蛋白質をコードするDNAが、betaKlothoの可溶性部分タンパク質をコードするDNAである、前記(3)に記載のFGF21活性の調節剤。
(5) betaKlotho活性の昂進物質又はbetaKlotho活性の阻害物質を有効成分として含むことを特徴とする、betaKlotho活性を有する蛋白質によるFGF受容体を介したFGF21活性の調節作用を増強するか又は抑制するためのFGF21活性の調節剤。
(6) betaKlotho活性の昂進物質がヘパリン又はヘパリン様物質である、前記(5)に記載のFGF21活性の調節剤。
(7) betaKlotho活性の昂進物質又は阻害物質が抗betaKlotho抗体である、前記(5)に記載のFGF21活性の調節剤。
(8) FGF受容体がFGFR1c又はFGFR3cである前記(1)ないし(7)のいずれかに記載のFGF21活性の調節剤。
(9) FGF受容体を介したFGF21の生物活性が、グルコース取り込み昂進、血糖値低下、グルコーストランスポーター発現昂進、インスリン発現昂進、及びメタボリズム制御のいずれかである、前記(1)ないし(8)のいずれかに記載のFGF21活性の調節剤。
(10) betaKlotho活性を有する蛋白質又は当該蛋白質をコードするDNAを含む組換えDNA、を有効成分として含む、FGF受容体を介したFGF21活性を調節するための医薬組成物。
(11) betaKlotho活性を有する蛋白質が、betaKlothoの可溶性部分タンパク質である、前記(10)に記載の医薬組成物。
(12) betaKlotho活性の昂進物質又はbetaKlotho活性の阻害物質を有効成分として含むことを特徴とする、betaKlotho活性を有する蛋白質によるFGF受容体を介したFGF21活性を調節するための医薬組成物。
(13) betaKlotho活性の昂進物質がヘパリン又はヘパリン様物質である、前記(12)に記載の医薬組成物。
(14) betaKlotho活性の昂進物質又は阻害物質が抗betaKlotho抗体である、前記(12)に記載の医薬組成物。
(15) betaKlotho活性を有する蛋白質又は当該蛋白質をコードするDNAを含む組換えDNAと共に、betaKlotho活性の昂進物質又はbetaKlotho活性の阻害物質を有効成分として含むことを特徴とする、前記(10)ないし(14)のいずれかに記載の医薬組成物。
(16) さらにFGF21を有効成分として含む、前記(10)ないし(15)のいずれかに記載の医薬組成物。
(17) FGF受容体がFGFR1c又はFGFR3cである、前記(10)ないし(16)のいずれかに記載の医薬組成物。
(18) FGF受容体を介したFGF21活性を調節するための医薬組成物が、グルコース取り込み量調節、血糖値調節、グルコーストランスポーター発現量調節、インスリン発現量調節のいずれかのための医薬組成物である、前記(10)ないし(17)のいずれかに記載の医薬組成物。
(19) 前記(1)ないし(9)のいずれかに記載のFGF21活性の調節剤を用いることを特徴とする、FGF受容体を介したFGF21の生物活性を増強するか又は抑制する方法。
(20) FGF受容体がFGFR1c又はFGFR3cである前記(19)に記載のFGF受容体を介したFGF21の生物活性を増強するか又は抑制する方法。
(21) FGF21活性の調節剤を、FGF21と共にFGF受容体に対して作用させることにより、FGF受容体の活性化を昂進又は抑制することを特徴とする、前記(19)又は(20)に記載のFGF受容体を介したFGF21の生物活性を増強するか又は抑制する方法。
(22) FGF21活性の調節剤を、FGF21及びbetaKlothoの共存下でFGF受容体に対して作用させることにより、FGF受容体の活性化を昂進又は抑制することを特徴とする、前記(19)又は(20)に記載のFGF受容体を介したFGF21の生物活性を増強するか又は抑制する方法。
(23) FGF受容体及びbetaKlothoを内在的に発現していない細胞にFGF受容体遺伝子を導入し、細胞表面にFGF受容体を発現させた形質転換細胞を用いる、FGF受容体を介したFGF21の生物活性を増強もしくは抑制する物質のスクリーニング方法であって、
細胞表面にFGF受容体を発現させた上記形質転換細胞の培養系に対して披検物質と共にFGF21を作用させる工程、又は、
FGF受容体遺伝子と共に披検物質遺伝子を上記FGF受容体及びbetaKlothoを内在的に発現していない細胞に導入して、FGF受容体と同時に披検物質も細胞表面に発現させた形質転換細胞の系に対してFGF21を作用させる工程
を含むことを特徴とする、スクリーニング方法。
(24) 前記FGF受容体及びbetaKlothoを内在的に発現していない細胞がBaF3細胞であることを特徴とする、前記(23)に記載のスクリーニング方法。
(25) 前記FGF受容体がFGFR1c又はFGFR3cである、前記(23)又は(24)に記載の、FGF21の生物活性を増強もしくは抑制する物質のスクリーニング方法。
(26) FGF受容体及びbetaKlothoを内在的に発現していない細胞にFGF受容体遺伝子を導入し、細胞表面にFGF受容体を発現させた形質転換細胞系及びFGF21を組み合わせてなる、FGF受容体を介したFGF21の生物活性を増強もしくは抑制する物質のスクリーニング用のキット。
(27) 前記FGF受容体及びbetaKlothoを内在的に発現していない細胞がBaF3細胞であることを特徴とする、前記(26)に記載のスクリーニング用のキット。
(28) 前記FGF受容体がFGFR1c又はFGFR3cである、前記(26)又は(27)に記載の、FGF21の生物活性を増強もしくは抑制する物質のスクリーニング用のキット。
(29) FGF受容体及びbetaKlothoを内在的に発現していない細胞にFGF受容体遺伝子及びbetaKlotho遺伝子を導入し、細胞表面にFGF受容体及びbetaKlothoを発現させた形質転換細胞の培養系を用いる、FGF受容体を介したFGF21活性の調節作用に係るbetaKlotho活性の昂進物質もしくは阻害物質のスクリーニング方法であって、
細胞表面にFGF受容体及びbetaKlothoを発現させた上記形質転換細胞の系に対して披検物質と共にFGF21を作用させる工程、又は、
上記FGF受容体及びbetaKlothoを内在的に発現していない細胞を宿主としてFGF受容体遺伝子及びbetaKlotho遺伝子と共に披検物質遺伝子を導入して、FGF受容体及びbetaKlothoと同時に披検物質も細胞表面に発現させた形質転換細胞の系に対してFGF21を作用させる工程
を含むことを特徴とする、スクリーニング方法。
(30) 前記FGF受容体及びbetaKlothoを内在的に発現していない細胞がBaF3細胞であることを特徴とする、前記(29)に記載のスクリーニング方法。
(31) 前記FGF受容体がFGFR1c又はFGFR3cである、前記(29)又は(30)に記載の、FGF21活性の調節作用に係るbetaKlotho活性の昂進物質もしくは阻害物質のスクリーニング方法。
(32) FGF受容体及びbetaKlothoを内在的に発現していない細胞にFGF受容体遺伝子及びbetaKlotho遺伝子を導入し、細胞表面にFGF受容体及びbetaKlothoを発現させた形質転換細胞の系、及びFGF21を組み合わせてなる、FGF受容体を介したFGF21活性の調節作用に係るbetaKlotho活性の昂進物質もしくは阻害物質のスクリーニング用のキット。
(33) 前記FGF受容体及びbetaKlothoを内在的に発現していない細胞がBaF3細胞であることを特徴とする、前記(32)に記載のスクリーニング用のキット。
(34) 前記FGF受容体がFGFR1c又はFGFR3cである、前記(32)又は(33)に記載の、FGF21活性の調節作用に係るbetaKlotho活性の昂進物質もしくは阻害物質のスクリーニング用のキット。
(35) FGF受容体及びbetaKlothoを内在的に発現していない細胞にFGF受容体遺伝子及びbetaKlotho遺伝子を導入し、細胞表面にFGF受容体及びbetaKlothoを発現させた形質転換細胞の系に対して披検物質を作用させる工程を含むことを特徴とする、FGF21様活性物質のスクリーニング方法。
(36) 前記FGF受容体及びbetaKlothoを内在的に発現していない細胞がBaF3細胞であることを特徴とする、前記(35)に記載のスクリーニング方法。
(37) 前記FGF受容体がFGFR1c又はFGFR3cである、前記(35)又は(36)に記載の、FGF21様活性物質のスクリーニング方法。
(38) FGF受容体及びbetaKlothoを内在的に発現していない細胞にFGF受容体遺伝子及びbetaKlotho遺伝子を導入し、細胞表面にFGF受容体及びbetaKlothoを発現させた形質転換細胞の系を用いることを特徴とする、FGF21様活性物質のスクリーニング用のキット。
(39) 前記FGF受容体及びbetaKlothoを内在的に発現していない細胞がBaF3細胞であることを特徴とする、前記(38)に記載のスクリーニング用のキット。
(40) 前記FGF受容体がFGFR1c又はFGFR3cである、前記(38)又は(39)に記載のFGF21様活性物質のスクリーニング用のキット。
(41) 前記(23)ないし(25)のいずれかに記載のスクリーニング方法により得られた、FGF受容体を介したFGF21の生物活性を増強もしくは抑制する物質を有効成分として含むことを特徴とする、FGF受容体とbetaKlothoの共存下で発揮される、FGF受容体を介したFGF21の生物活性を増強もしくは抑制するための、FGF21活性の調節剤。
(42) 前記(29)ないし(31)のいずれかに記載のスクリーニング方法により得られた、FGF受容体を介したFGF21活性の調節作用に係るbetaKlotho活性の昂進物質もしくは阻害物質を有効成分として含むことを特徴とする、FGF受容体とbetaKlothoの共存下で発揮される、FGF受容体を介したFGF21の生物活性を増強もしくは抑制するための、FGF21活性の調節剤。
(43) 前記(35)ないし(37)のいずれかに記載のスクリーニング方法により得られたFGF21様活性物質を有効成分とし、betaKlothoの共存下でFGF受容体に作用させることを特徴とする、FGF受容体活性化剤。
(44) 前記(23)ないし(25)のいずれかに記載のスクリーニング方法により得られた、FGF受容体を介したFGF21の生物活性を増強もしくは抑制する物質を有効成分として含むことを特徴とする、FGF受容体とbetaKlothoの共存下で発揮される、FGF受容体を介したFGF21活性を調節するための医薬組成物。
(45) 前記(29)ないし(31)のいずれかに記載のスクリーニング方法により得られた、FGF受容体を介したFGF21活性の調節作用に係るbetaKlotho活性の昂進物質もしくは阻害物質を有効成分として含むことを特徴とする、FGF受容体とbetaKlothoの共存下で発揮される、FGF受容体を介したFGF21活性を調節するための医薬組成物。
(46) 前記(35)ないし(37)のいずれかに記載のスクリーニング方法により得られたFGF21様活性物質を有効成分とし、betaKlothoの共存下でFGF受容体に作用させることを特徴とする、FGF受容体を活性化するための医薬組成物。
【発明の効果】
【0009】
本発明によれば、betaKlotho活性を有する蛋白質を用いてFGF受容体を介したFGF21活性を調節し、FGF受容体下流のシグナルを惹起もしくは抑制することでFGF21とFGF受容体との相互作用に基づく様々な反応を制御することができる。また、本発明によれば、betaKlotho活性を有する蛋白質、又はbetaKlotho活性の活性増強物質もしくは阻害物質を投与することでFGF受容体を介したFGF21活性、たとえば血糖値の低下作用を調節することができるので、betaKlotho活性を有する蛋白質、又はbetaKlotho活性の活性増強物質もしくは阻害物質をそれぞれ単独でもしくは組み合わせて、又はさらにFGF21と併用して血糖値の異常に関連した疾患、たとえば糖尿病の治療又は予防用医薬組成物を提供することができる。
また、FGF21様活性物質、betaKlothoの代替物質、及びbetaKlotho活性の活性増強物質もしくは阻害物質に対する優れたスクリーニング系も提供できる。
【図面の簡単な説明】
【0010】
【図1】FGF21の脂肪細胞特異的活性の検出の結果を示す図である。 図中、GLUT1 mRNAの量はそれぞれの細胞でFGF21未刺激時の値を1とした相対値で表した。●は分化前の繊維芽細胞を、○は分化後の脂肪細胞を示す。 FGF21が3T3-L1細胞の分化後の脂肪細胞に対して活性を発揮し、GLUT1発現を増加させることが示された。繊維芽細胞に対して活性を発揮しないことが示された。
【図2】脂肪細胞の分化に伴うFGF受容体の発現変動の検出した結果を示す特性図である。 図中、横軸は繊維芽細胞から脂肪細胞への分化誘導日数を表し、縦軸はFGF受容体mRNAの発現量を表す。 FGFR1とFGFR2が高いレベルで発現していたが、FGFR3とFGFR4は非常に発現レベルが低かった。FGFR1、FGFR2ともに、細胞における発現レベルは分化とともにやや減る傾向が認められた。
【図3】脂肪細胞の分化に伴うbetaKlothoの発現変動の検出の結果を示す図である。 図中、横軸は繊維芽細胞から脂肪細胞への分化誘導日数を表し、縦軸はbetaKlotho、又はKlothoのmRNAの発現量を表す。 脂肪細胞分化に伴って発現するようになるbetaKlothoが脂肪細胞の機能と密接に関連することが示唆された。
【図4A】FGF21によるFGF受容体下流シグナル惹起にbetaKlothoが必要であることの解析の結果を示す図であり、図中、CL#1,#7、#10はFGFR1cとbetaKlothoの両方のcDNAを導入し形質転換したBaF3細胞クローンの名称である。下段は、それぞれの細胞から調製したライゼートを材料として、実際にタンパクとして発現しているbetaKlothoの量を、市販の抗betaKlotho抗体を用いたウェスタンブロット解析により比較した結果である。
【図4B】FGF21の量を変化させてCL#1の細胞を刺激した際のDNA合成を測定した結果であり、○はheparin存在下、●は非存在下を示す。
【図4C】FGFR1cを発現していないがbetaKlothoのみを発現しているBaF3細胞を用い、チミジン取り込み能を測定した。□はheparin存在下を、■は非存在下を示す。図4A〜Cの結果により、FGFR1cは発現していてもbetaKlothoを発現していない細胞ではFGF21に反応せず、betaKlothoを同時に発現している細胞でのみFGF21に反応する一方、FGFR1cを発現しておらずbetaKlothoのみを発現しているBaF3細胞はFGF21に対して反応しないことが示され、FGF21と反応するためには、betaKlothoとFGFR1cとの両者が必要であることが示された。
【図5】FGF21の活性発揮においてbetaKlothoとともに存在するFGF受容体の特異性の解析の結果を示す図である。 betaKlothoを発現しているBaF3細胞にFGF受容体(FGFR1b、R1c、R2b、R2c、R3b、R3c、R4)の発現ベクターを導入し、各betaKlotho/FGFR発現細胞をheparin存在下又は非存在下FGF21、FGF1で刺激してチミジンの取り込み能を測定した。 FGFR1cのほか、FGFR3cでもFGF21に反応することが示された。また、従来型のFGF(典型的にはFGF1)ではheparinの存在で急激に刺激伝達活性が高まるが、FGF21の場合はheparinの存在、非存在がそれほど活性に影響していない。
【図6】FGF21の活性発揮においてbetaKlothoとともに存在するFGF受容体の特異性をFGF19およびFGF1と比較した解析の結果を示す図である。 FGFR(FGFR1c、R2c、R3c、R4)を発現しているBaF3細胞にbetaKlothoの発現ベクターを導入し、各FGFR/betaKlotho発現細胞をheparin存在下でFGF21、FGF1及びFGF19で刺激して、チミジンの取り込み能を測定した。図中、○はFGF21、●はFGF19、▲はFGF1で刺激したことを表す。 FGF19はFGF1と同様、FGFR1c、FGFR2c、FGFR3c、FGFR4のいずれの受容体とも反応する。これに対しFGF21はFGFR1c とFGFR3cのみに反応する。高濃度のFGF21刺激によりFGFR2cに対してわずかに反応がみられる。
【図7】FGF21の刺激に伴う細胞内シグナル伝達分子の活性化の解析の結果を示す図である。 各FGFR/betaKlotho発現細胞をFGF21、FGF1及びFGF19で刺激して、FGFR下流シグナル伝達物質であるFRS2及びMARKのリン酸化を、前者では抗リン酸化FRS2抗体を用いて、後者では抗リン酸化ERK1/2抗体を用いて測定した。図中、WEHIはIL-3分泌細胞であるWEHIの培養上清を添加したことを示す。 FGF19とFGF1は、FGFR1c、FGFR2c、FGFR3c、FGFR4のいずれの受容体をbetaKlothoとともに発現する細胞のFRS2のリン酸化とMAPKのリン酸化を増加させる。これに対し、FGF21はFGFR1c/betaKlotho発現細胞とFGFR3c/betaKlotho発現細胞のみ、FRS2およびMAPKのリン酸化を増加させる。
【図8】ヒトのアミノ酸配列をもつヒト型FGFR1cとヒト型betaKlothoを共発現させた細胞に対するヒト型FGF21の活性の解析の結果を示す図である。 ヒト型FGFR1c(hR1c)発現BaF3細胞あるいはヒト型FGFR1c/ヒト型betaKlotho(hβKlotho)発現細胞を10μg/mlのheparin存在下でヒト型FGF21(hFGF21)あるいはFGF1で刺激して、チミジンの取り込み能を測定した。図中、●はhFGF21、▲はFGF1で刺激したことを表す。FGF1は両方の細胞に作用するが、ヒト型FGF21は、ヒト型FGFR1c/ヒト型betaKlotho発現細胞のみに作用して用量依存的にチミジンの取り込みを増加させた。
【図9】ヒトのアミノ酸配列をもつヒト型FGFR3cとヒト型betaKlothoを共発現させた細胞に対するヒト型FGF21の活性の解析の結果を示す図である。 ヒト型FGFR3c(hR3c)発現BaF3細胞あるいはヒト型FGFR3c/ヒト型betaKlotho(hβKlotho)発現細胞を10μg/mlのheparin存在下でヒト型FGF21(hFGF21)あるいはFGF1で刺激して、チミジンの取り込み能を測定した。図中、●はhFGF21、▲はFGF1で刺激したことを表す。FGF1は両方の細胞に作用するが、ヒト型FGF21は、ヒト型FGFR3c/ヒト型betaKlotho発現細胞のみに作用して用量依存的にチミジンの取り込みを増加させた。
【図10】FGF21の活性発揮においてFGF受容体とともに存在するKlothoの特異性の解析した結果を示す特性図である。 FGFR1cと共にbetaKlothoを発現させたBaF3細胞及びFGFR1cと共にKlothoを発現させたBaF3細胞に対して、heparinの存在下又は非存在下でFGF21及びFGF23を添加し、チミジン取り込み能を測定した。 Klothoでは、FGF21に反応しないことが示された。さらに、FGFR1cとKlothoの存在下で反応性を発揮するFGF23について、調べたところ、FGFR1cとbetaKlothoを発現していても、FGF23には反応しなかった。
【図11】BetaKlothoとFGF21あるいはFGF受容体との複合体形成の検出の結果を示す図である。 heparinの存在あるいは非存在の条件下で、可溶性betaKlothoタンパク質とFGF21をインキュベートした後、抗V5タグ抗体固定化ビーズを用いてbetaKlothoを沈降させ、FGF21がbetaKlothoに結合して共沈降するか否かを、免疫ブロッティング法にて解析した betaKlothoとFGF21はheparinの有無によらず物理的に複合体を形成する。またbetaKlothoはFGFR1c、FGFR2c、FGFR3c、FGFR4のいずれの受容体とも物理的に複合体を形成する。
【図12】BetaKlothoを強制発現させた未分化3T3-L1細胞をFGF21刺激した際のシグナル伝達の解析の結果を示す図である。 betaKlothoを発現させた3T3-L1繊維芽細胞をFGF21で刺激し、FGFRの下流のシグナル伝達分子であるFRS2とMAPKの活性化(リン酸化)をウエスタンブロッティング法で調べた。図中、PBSはダルベッコ改変リン酸緩衝生理食塩液を10μl、SU5402はFGF受容体チロシンキナーゼ阻害剤であって、系に10μM存在させることを表す。また、FGF21の添加濃度は1.1μg/ml及び4.5μg/mlの2通りで行った。 betaKlothoを発現していない未分化3T3-L1細胞はFGF21で刺激してもFRS2のリン酸化およびMAPKのリン酸化の状態は変化しない。betaKlothoを強制的に発現させた未分化3T3-L1細胞では、FGF21刺激に応じてFRS2のリン酸化およびMAPKのリン酸化が増大する。FGF受容体チロシンキナーゼ阻害剤SU5402の処理により、FGF21刺激に伴うFRS2およびMAPKのリン酸化の程度は減少する。
【図13】BetaKlothoを強制発現させた未分化3T3-L1細胞をFGF21刺激した際のGLUT1mRNAの発現解析の結果を示す図である。 betaKlothoを発現させた3T3-L1繊維芽細胞をFGF21で刺激しGLUT1のmRNA量の変化を調べた。コントロールとしてはbetaKlothoのcDNAを挿入していない空のベクターを導入した3T3-L1繊維芽細胞を用いた。 betaKlothoを発現していない未分化3T3-L1細胞はFGF21で刺激してもGLUT1mRNAの発現量は変化しない。betaKlothoを強制的に発現させた未分化3T3-L1細胞では、刺激したFGF21の濃度に応じてGLUT1mRNAの発現量が増加する。
【図14】本発明を説明する概念図。 図中、aはFGF1、FGF2などの多くのFGF、bはFGF21、cはFGF受容体、dはbetaKlotho、eはヘパリンあるいはヘパリン様糖鎖を示す。またFGF21、FGF受容体、betaKlothoの間の活性複合体の形成を、増強あるいは減弱する物質(群)をfで示す。I.従来の研究で知られていた、FGF1など多くのFGFがbetaKlothoの非存在下でFGF受容体を活性化しうる作用機構: (1)FGFリガンドが単独にFGF受容体に結合しただけでは受容体の活性化は起こらない。 (2)さらにヘパリン様の糖鎖が共存するとシグナルを惹起しうる活性複合体が形成されて、それを起点としたシグナルの伝達が行われて細胞の応答(例えば細胞の増殖など)が起きる。II.本発明において明らかにした、betaKlothoが存在することによって、FGF受容体の活性化が起きるFGF21の作用機構: (3)補助因子BetaKlothoが加わり、リガンド(FGF21)、FGF受容体、BetaKlothoの3者が存在することによって、FGF21によるFGF受容体の活性化が起きる。 (4)このシグナルを惹起しうる活性複合体形成を、増強あるいは減弱する物質は、FGF21の作用を昂進あるいは抑制する作用を有する。
【発明を実施するための形態】
【0011】
以下、本発明の実施の態様に従って、本発明を詳細に説明するが、本発明はこの実施の態様に限られない。
〔1〕betaKlothoについて
本発明は、betaKlothoの生体内での作用機能を解明し、その利用法を提供するものである。
betaKlothoは、Klothoのホモログとしてクローニングされた物質であり(非特許文献8)、マウス由来betaKlotho遺伝子の塩基配列は配列番号1、対応するアミノ酸配列は配列番号2として示され、ヒトbetaKlothoの塩基配列は配列番号7、対応するアミノ酸配列は配列番号8として示される。発達中のマウス胎児の脂肪組織などで発現していることが知られており、他生物種からの遺伝子取得方法と共に、種々の改変体、変異体などの製造方法も記載されている(非特許文献7,8、特許文献1)。
そして、上記非特許文献7および特許文献1において、遺伝子をホモで欠いたノックアウトマウスの作成とその解析の結果、betaKlotho遺伝子が「コレステロールの代謝に関与する遺伝子」の1つであり、これを用いてコレステロールの代謝を促進する物質を得るとする記述がされているものの、血糖値の調節に関連した機能にも、FGF21活性との関係についても教示する記述はない。
一方で、本発明においてはじめて、betaKlothoが、FGF21がFGF受容体と結合してFGF受容体を活性化させ、そのシグナル伝達を引き起こすために必須の物質であることが解明され、betaKlothoこそが、FGF21活性として知られていた、グルコーストランスポーターの発現量の昂進、グルコース取り込み上昇、血糖値の低下、インスリンの合成量増加などの作用において重要な調節機能を果たす因子であることが見出されたものである。
本発明においては、「betaKlotho活性」というとき、betaKlothoが有している活性である、FGF21とFGF受容体と共存して相互作用することでFGF受容体を介したシグナル伝達系を活性化させる作用、すなわち「FGF受容体を介したFGF21活性を調節する機能・活性」を表す。同様に、「betaKlotho活性を有する蛋白質」とは、betaKlotho及びその可溶性部分蛋白質のみならず、「FGF受容体を介したFGF21活性を調節する機能・活性を有する蛋白質」をさす。
betaKlothoは、betaKlotho活性にかかわる長い細胞外ドメインと短い細胞内ドメインが膜貫通ドメインでつながれた、典型的なアンカータンパク質構造をしている。細胞外ドメインのみからなる可溶性betaKlothoも「betaKlotho活性を有するタンパク質」として機能するし、さらに短い部分配列であってもbetaKlotho活性を有していれば、本発明のbetaKlotho活性を有する蛋白質として用いることができる。
【0012】
本明細書中ではbetaKlotho活性を有する蛋白質の典型例として、典型的なbetaKlothoタンパク質である、マウス全長betaKlothoタンパク質のアミノ酸配列を配列番号2として、その可溶性betaKlothoタンパク質のアミノ酸配列を配列番号3として、また、ヒトbetaKlothoアミノ酸配列を配列番号8として、可溶性ヒトbetaKlothoタンパク質を配列番号9として例示するが、これらアミノ酸配列において1個若しくは数個のアミノ酸に欠失、置換、付加等の変異が生じてもよい。また、カルボキシ末端にFlagタグ配列(配列番号4)、V5タグ配列(配列番号5)、Hisタグ配列(配列番号6)あるいはこれらの組み合わせ配列を付加してもよい。さらに、マウスbetaKlothoタンパク質とヒトbetaKlothoタンパク質のアミノ酸配列レベルでの相同性は78%であり、ヒトbetaKlothoタンパク質とマウスbetaKlothoタンパク質とで同等の機能を有しているので、進化上マウス−ヒト間程度の距離内に存在する生物種に由来するbetaKlothoも、同等のbetaKlotho活性を有しているものと期待できる。したがって、本発明におけるbetaKlothoタンパク質としては、マウス由来、及びヒト由来のbetaKlothoタンパク質だけではなく、配列番号2または配列番号8とアミノ酸の相同性が、78%以上、好ましくは85%以上、より好ましく90%以上の他の生物種のbetaKlothoタンパク質も含まれる。具体的には、「配列番号2、配列番号3、配列番号8及び配列番号9のいずれかに示されるアミノ酸配列、又はその部分断片からなるタンパク質」、「配列番号2、配列番号3、配列番号8及び配列番号9のいずれかに示されるアミノ酸配列において、1個もしくは数個のアミノ酸が欠失、置換又は付加されたアミノ酸配列からなるタンパク質」、「配列番号2、配列番号3、配列番号8及び配列番号9のいずれかに示されるアミノ酸配列と78%以上の相同性を有するアミノ酸配列からなるタンパク質」であってbetaKlotho活性を有するタンパク質も本発明における「betaKlothoタンパク質」に相当する。なお、本発明における「相同性」とは、Lipman-Pearsonの計算法(Lipman and Pearson,1985;Science 227,1435-1441)での「同一性」を示す換算数値である。
【0013】
また、典型的なbetaKlotho活性を有する蛋白質をコードするDNAとしては、マウス由来betaKlotho遺伝子(配列番号1)及びヒト由来betaKlotho遺伝子(配列番号7)があげられるが、他の生物種由来の既知のbetaKlotho遺伝子も同様に用いることができ、未知のbetaKlothoの遺伝子であっても、マウス由来またはヒト由来のbetaKlothoをコードするDNAの塩基配列(マウス由来塩基配列は配列番号1として示される。)に基づいて設計されたプローブもしくはプライマーを用いて各生物種由来のDNAライブラリーからPCR法やハイブリダイゼーション法により取得することができる。
そして、これらのbetaKlotho活性を有する蛋白質をコードするDNAに対して部位突然変異法やランダム変異導入法などで得られた突然変異DNAを用いることができる。さらに、それぞれの可溶性断片などの部分配列も「betaKlotho活性を有する蛋白質をコードするDNA」として用いることができる。
具体的には、「betaKlotho活性を有する蛋白質をコードするDNA」として、「配列番号1もしくは配列番号7に示される塩基配列からなるDNA又はその部分断片からなるDNA」、「配列番号1又は配列番号7の相補配列からなるDNAとストリンジェントな条件下でハイブリダイズするDNA」及び「配列番号2、配列番号3、配列番号8及び配列番号9のいずれかに示されるアミノ酸配列をコードするDNA」であって、「betaKlotho活性を有する蛋白質をコードするDNA」などがあげられる。
なお、上記ストリンジェントな条件とは、高い緊縮条件下であることをいい、例えば2×SSC、1×デンハルト溶液中で60℃程度の条件である。
そして、これらbetaKlothoタンパク質をコードするDNAを含む組換えDNAを用いることで、定法どおり、哺乳動物細胞宿主系などを用いた形質転換体の発現産物としてbetaKlothoタンパク質を取得することができる。また、当該「組換えDNA」は、それ自体でFGF21活性の調節剤として用いることができるが、ここで、「組換えDNA」の用語は、形質転換宿主内で発現するためのプロモーターなどと機能的に連結されていることを表し、典型的にはヒトなど哺乳動物への投与が可能なレトロウイルスなどの発現ベクターである。
また、betaKlothoの天然蛋白質の場合も、betaKlotho遺伝子を含む発現ベクターなどで形質転換した哺乳動物細胞などにより産生される組換え蛋白質の場合も、限外ろ過法、ゲルろ過法、アフィニティークロマトグラフィーなど、またはこれらを組合せた方法などの周知のタンパク質精製方法により、必要に応じて精製して用いることができる。
以下、本発明においては、各種betaKlotho活性を有する蛋白質、またはbetaKlotho活性を有するその部分断片もしくはその変異体などの同等物を含め、単に「betaKlotho」、「betaKlothoタンパク質」ともいい、これらbetaKlotho活性を有する蛋白質をコードするDNAを、単に「betaKlothoをコードするDNA」、「betaKlotho遺伝子」ともいう。
【0014】
〔2〕FGF21活性が脂肪細胞で特異的に検出されることについて
1.FGF21について
FGF21は、FGF19の配列を基にデザインしたプライマーを用いたPCRにより発見された物質である(非特許文献9)。脂肪細胞にグルコーストランスポーター(GLUT)の発現を上昇させる活性やランゲルハンス島でのインスリン合成促進活性があることも報告されている(非特許文献1、6)。そして、脂肪細胞表面のFGFR1及びFGFR2を活性化することが確認されている(非特許文献1)ことから、これら活性は、FGF受容体を介して、細胞内にシグナル伝達されて引き起こされると解される。
本発明において「FGF受容体を介したFGF21活性」又は「FGF受容体を介したFGF21の生物活性」というとき、FGF受容体、特にFGFR1cあるいはFGFR3cを介して細胞内に引き起こされる活性をいい、たとえば脂肪細胞内でグルコーストランスポーター(GLUT)の発現を上昇させる活性を指す。また、本発明におけるFGF21としては、ヒト由来のFGF21(例えば、GenBankアクセッション番号:AB021975参照)などの哺乳類由来FGF21が好ましいが、これに限られるものでなく、天然タンパク質のみならず、組換えFGF21であってもよく、さらに上記FGF21活性を有していれば、そのアミノ酸配列の一部が改変されたものであってもよい。組換えFGF21を作製するためには、大腸菌や哺乳動物細胞などの通常の形質転換宿主/ベクター系を適宜用いることができる。
【0015】
2.FGF受容体を介したFGF21活性の測定
一般に、各FGFは、細胞表面のFGF受容体に結合し、FGF受容体を活性化させ、細胞内での各種シグナル伝達機構を活性させることで、当該細胞に何らかの作用・機能を引き起こすことになるため、当該観察された作用・機能をFGF活性として測定することになる。
汎用的な測定法としては、Ornitz,D.M.,Xu,J.,Colvin,J.S.,McEwen,D.G.,MacArthur,C.A.,Coulier,F.,Gao,G.and Goldfarb,M.(1996) Receptor specificity of the fibroblast growth factor family.J Biol Chem,271,15292-15297.に記載されている細胞増殖刺激活性測定法が挙げられる。その概要は以下の通りである。表面にFGF受容体を有する培養細胞の培養液内に測定対象のFGFを加え、一定時間培養後標記チミジンを培養液中添加しさらに一定時間培養を行う。この間の標識チミジンの高分子DNAへの取り込みを測定することで、FGFによるDNA合成の活性化程度を評価する。(本発明でも、実施例4,5,6,8,9及び10においてFGF21活性を測定する際にこの手法を採用した。)
上述のように、FGF21は細胞表面のFGF受容体、特にFGFR1cを介して細胞内のグルコーストランスポーター(GLUT)発現を上昇させる活性を有しているので、本発明に係るFGF21活性の解析において、実施例1では典型的なFGF21活性の披検細胞内でのGLUTの発現の上昇能をGLUT1のmRNAの発現量として測定した。
ここで、FGF21を様々な濃度で細胞培養液に添加することにより、細胞に作用させて一定時間細胞培養を継続した後、細胞からmRNAを抽出し、GLUT1のmRNAの発現量を測定する。このmRNA発現量が増加していれば、FGF21が活性を発揮していることが示される。
GLUT1の発現上昇を解析する目的には、GLUT1抗体などを用いたウェスタンブロット法等の公知の方法によりGLUT1タンパク質発現量を測定することもできる。また、実験動物などの場合、GLUT1の転写を制御する制御領域(例えば、プロモーター領域)と、その下流に転写制御されうる形で連結されたリポーター遺伝子と組み込んだトランスジェニック動物を作製することにより、リポーター遺伝子の発現量によってFGF21活性を測定できるようにしてもよい。
なお、FGF21活性を測定するためには、これらの手法に限られることはなく、定量的な測定が可能な手法であればどのような手法でも用いることができる。たとえばFGF受容体の活性化やその下流のシグナル伝達分子の活性化をFGF受容体やシグナル伝達分子のリン酸化の増加として測定し評価する手法も用いることができる。
そして、本発明のFGF21活性のGLUT1mRNA発現量の測定の好ましい態様として、培養脂肪細胞を披検細胞として用いることができる。
培養脂肪細胞としては、ヒト、ラット初代培養細胞など各種の培養脂肪細胞を用いることができるが、本発明の実施の態様では、培養条件を操作することにより脂肪細胞に分化する能力を有する繊維芽細胞として広く当該研究領域で使用されている3T3-L1細胞を用いた。3T3-L1細胞は、分化誘導前には繊維芽細胞としての性質を有し、誘導後には脂肪細胞としての性質を有するので、FGF21を作用させた場合の効果を対比させて解析することができる。ここで「脂肪細胞の性質をもつ細胞」とは、脂肪細胞特有の細胞内代謝を行い、脂肪滴を多量に蓄積した細胞質を有する細胞を示す。3T3-L1細胞を脂肪細胞に誘導する実験系は、例えば、Kharitonenkov,A.,Shiyanova,T.L.,Koester,A.,Ford,A.M.,Micanovic,R.,Galbreath,E.J.,Sandusky,G.E.,Hammond,L.J.,Moyers,J.S.,Owens,R.A.,Gromada,J.,Brozinick,J.T.,Hawkins,E.D.,Wroblewski,V.J.,Li,D.S.,Mehrbod,F.,Jaskunas,S.R.,and Shanafelt,A.B.(2005) J Clin Invest 115(6),1627-1635に開示されている。
【0016】
3.FGF21を作用させることによるGLUT発現量の増大は脂肪細胞で特異的に検出されること
3T3-L1細胞に対して脂肪細胞に誘導分化処理を施し、誘導後の脂肪細胞の性質を有する細胞と、誘導前の繊維芽細胞としての性質を有する細胞とを用意し、FGF21の濃度を変えて作用させ、各濃度でのGLUT1のmRNAの発現量を測定し、両者を比較した。脂肪細胞に分化した細胞ではFGF21の濃度の増加に伴いGLUT1のmRNAの発現量が増加していたことから、FGF21が脂肪細胞に対して活性を発揮し、GLUT1発現を増加させることが示された。これとは対照的に、分化していない繊維芽細胞では、FGF21の濃度を変えてもGLUT1のmRNAの発現量はほとんど変化しなかった。このことから、FGF21のGLUT1発現量の増大活性は、3T3L1細胞が脂肪細胞に分化した場合に特異的に検出されることが示された。
【0017】
〔3〕脂肪細胞の分化に伴うFGF受容体の発現変動の検出
1.FGF受容体について
FGF受容体(FGFR)は細胞表面に存在する膜一回貫通型のタンパク質である。現在ではFGFR1〜5の5種類が同定されている。そのうちFGFR1〜4はチロシンキナーゼ型受容体であり、FGFが結合することにより二量体化・自己リン酸化により活性化される。活性化されたFGFRは細胞内のシグナリング分子と相互作用し、複数のシグナル伝達経路を活性化させる。FGFR1〜3には選択的スプライシングにより主に2種類のアイソフォーム(FGFR1b、FGFR1c、FGFR2b、FGFR2c、FGFR3b、FGFR3c)が存在する。
【0018】
2.脂肪細胞の分化に伴うFGF受容体の発現量の変動
上記〔1〕3.での結果は、3T3-L1細胞の脂肪細胞への分化に伴って細胞表面にFGF受容体が発現した、もしくは発現量が増加したことにより、細胞内でのシグナル伝達量が増加したことを観察している可能性もあるため、3T3-L1細胞における脂肪細胞の誘導の各段階においての、各FGF受容体発現量を測定することとした。
3T3-L1細胞の培養条件を操作して、繊維芽細胞の性質を持つ細胞から脂肪細胞の性質をもつ細胞へと徐々に誘導し、各段階において、細胞から総RNAを抽出し、そこでのFGF受容体mRNAの発現量を測定することにより、脂肪細胞の分化に伴うFGF受容体の発現変動を検出した。
その結果、脂肪細胞への分化の有無にかかわらずFGFR1とFGFR2は高レベルで発現しているのに対して、FGFR3とFGFR4の発現レベルは非常に低かった。そして、分化に伴って発現が増える受容体はなく、むしろ高レベルで発現していたFGFR1、FGFR2のいずれの受容体も、分化とともに、発現レベルがやや減る傾向が認められた。
【0019】
〔4〕脂肪細胞の分化に伴うbetaKlothoの発現変動の検出
本発明者らは、脂肪細胞へと分化する過程で脂肪細胞表面での発現量が増加する物質があれば、当該物質がFGF21と協同してFGF受容体に作用する可能性が高いと考え、発達中のマウス胎児の脂肪組織で発現することが知られていたbetaKlotho(非特許文献8、特許文献1)に着目し、3T3-L1細胞の脂肪細胞の分化に伴うbetaKlothoの発現量の変化を調べた。
具体的には、上記〔2〕3.と同様の手法により、3T3-L1細胞を脂肪細胞へ徐々に誘導し、その各段階において、細胞から総RNAを抽出し、そこでのbetaKlothoのmRNAの発現量を測定することにより、脂肪細胞の分化に伴うbetaKlothoの発現変動を検出した。
その際の対照として、betaKlothoとは別の遺伝子の発現変動を検出することができ、当該別遺伝子の方が好ましい。本発明の実施の態様としてはbetaKlothoとアミノ酸配列レベルで41%の相同性を有するKlothoの遺伝子の発現変動を対照として採用した。Klothoと比較することにより、脂肪細胞の誘導に伴うbetaKlothoの発現上昇が特徴的であることがわかる。
この結果から、脂肪細胞の誘導に伴うbetaKlothoの発現上昇が、脂肪細胞の機能と密接に関連することが示唆される。
【0020】
〔5〕FGF21の活性発揮におけるbetaKlothoの必要性の解析
1.FGF受容体を介したFGF21の活性発揮とbetaKlothoの関連性
以上の3T3-L1細胞及びその誘導分化した脂肪細胞を用いた実験結果から見て、FGF21の脂肪細胞特異的活性は、FGF受容体とbetaKlothoが共に存在する場合に誘導されることが示唆される。すなわち、3T3-L1細胞の表面に本来存在していたFGFR1cなどのFGF受容体と共に、脂肪細胞に分化誘導された場合に発現してくるbetaKlothoが、FGF21のFGF受容体を介した活性を飛躍的に増大させている可能性が強く示唆される。
このことを確かめるために、本発明の態様ではFGF受容体のみを発現させた細胞、FGF受容体とbetaKlothoを共に発現させた細胞、に対してFGF21が活性を有するか否かを解析する。この目的には、元来FGF受容体もbetaKlothoも発現していない細胞を用いることが必要である。
【0021】
2.FGF受容体及び/又はbetaKlothoを発現する披検細胞の作製方法
上記目的に適した、元来FGF受容体もbetaKlothoも発現していない細胞としては、マウス白血病ProB細胞であるBaF3細胞を用いることができる。しかし他の細胞でもこれらいずれの遺伝子も細胞表面に発現しておらず、かつこれら遺伝子で形質転換された場合に細胞表面に発現することができる細胞であれば、用いることができる。本来これら遺伝子を発現している細胞であっても、当該遺伝子がノックアウトされている細胞も用いることができる。
このようなFGF受容体もbetaKlothoも発現していない細胞を宿主として、FGF受容体遺伝子のみ、又はbetaKlotho遺伝子のみをBaF3細胞表面に発現させた細胞を、定法の遺伝子組換え技術を用いて用意すると共に、同様の手法で両遺伝子を用いてbetaKlothoとともにFGF受容体の種類を変えたBaF3細胞を作製した。
発現細胞の作製方法としては、特に限定されないが、例えば、リポフェクタミン試薬で発現プラスミドを細胞に導入し、プラスミド上の薬剤耐性遺伝子をマーカーとして用いて、この薬剤に耐性な細胞株を取得することで達成することができる。
【0022】
3.FGF21によるFGF受容体下流シグナル惹起にbetaKlothoが必要であることの解析
FGFR1cとbetaKlothoの両方のcDNAを用いてBaF3細胞を形質転換して複数のクローンを取得し、SDS-PAGEゲル上でbetaKlotho抗体により確認したところ、N型糖鎖の修飾に特徴的な、スメアー状態のブロードなバンドとして観察されたことから、betaKlothoはN型糖鎖の修飾を有する糖タンパクであると考えられる。
そして、FGF21を培養液に添加した場合に、FGFR1cは発現しているがbetaKlothoを発現していない細胞ではヘパリンの存在下あるいは非存在下に関わらずFGF21に反応せず、betaKlothoを共に発現している細胞でのみFGF21に反応することが示された。
また、FGF21の量を変化させてFGFR1cと共にbetaKlothoを発現しているBaF3細胞にヘパリンを作用させた実験結果からは、ヘパリンはFGF21の作用をさらに増強することがわかった。しかし、FGFR1cを発現していない場合の、betaKlothoのみを発現しているBaF3細胞は、ヘパリンの存在、非存在にかかわらずFGF21に対する反応性はない。
以上のことから、ヘパリンがFGF21に反応するためには、FGF受容体とbetaKlothoの両方が必要であることと共に、ヘパリン又はヘパリン様物質(通常のFGF受容体を介したFGF活性増強物質)には、betaKlotho活性を有する蛋白質のFGF21増強活性をさらに増強する効果があることが示された。
また、このことは、FGFR1cを発現しているBaF3細胞の系に披検物質を存在させてFGF21を作用させることで、披検物質にFGF21の活性を増強もしくは抑制する作用があるか否かを検出できることであり、FGFR1cと同時にbetaKlothoも発現しているBaF3細胞の系に披検物質を存在させてFGF21を作用させてみれば、披検物質にbetaKlothoのFGF21増強活性をさらに増強する作用があるか、もしくは抑制する作用があるかを検出できることを示している。すなわち、FGF受容体を発現しているBaF3細胞の系はFGF21の活性を増強もしくは抑制する物質のスクリーニングに、またFGF受容体と共にbetaKlothoも発現しているBaF3細胞の系は、FGF21活性調節作用にあるbetaKlotho活性を増強もしくは抑制する物質のスクリーニングに好適に用いられることが示されている。さらに、FGF受容体と共にbetaKlothoも発現しているBaF3細胞の系は、FGF21様活性物質のスクリーニングに用いることもできる。
そして、これらスクリーニングにより得られた物質は、いずれもFGF受容体とbetaKlothoの共存下で発揮されるFGF21活性の調節剤及び当該FGF21活性を調節するための医薬組成物、例えば血糖値調節用医薬組成物として用いることができる。
【0023】
〔6〕FGF21の活性発揮におけるbetaKlothoとともに存在するFGF受容体の特異性の解析
以上の実験結果から、FGF21とbetaKlothoが存在すれば、FGF受容体の一つであるFGFR1cを活性化してそれを発現している細胞の内部にシグナル伝達をすることができることが解明されたので、他のFGF受容体の場合についてのbetaKlotho共存効果を調べるために、betaKlothoとともに存在するFGF受容体の種類を変えたBaF3細胞を作製し、その細胞に対するFGF21の活性を評価した。その結果、FGFR1cのほか、FGFR3cをbetaKlothoと共に発現させた場合でもFGF21に反応することが明らかとなり、その場合もさらにヘパリンの存在によって反応性が増強される。
したがって、ヘパリンには、betaKlothoのFGF受容体を介したFGF21活性の増強効果をさらに増大する作用を有する。
また、ここで用いたFGF受容体と共にbetaKlothoも発現しているBaF3細胞の系は、当該細胞表面のFGF受容体及びbetaKlothoが共存した状態でFGF21を作用させることができるから、FGF21、betaKlotho及びFGF受容体の共存下で発揮されるFGF受容体を介したシグナル伝達系を昂進する物質又は抑制する物質のスクリーニングに用いることができる。
当該スクリーニングで得られた物質は、FGF21、betaKlotho及びFGF受容体の共存下で発揮されるFGF受容体を介したシグナル伝達系を昂進又は抑制するための医薬組成物(例えば、血糖値調節用医薬組成物)などとして用いることができる。すなわち、ヘパリンは上述のスクリーニング系により、betaKlothoのFGF受容体を介したFGF21活性の増強物質として得られた物質であるということができるから、ヘパリンを当該医薬組成物中の有効成分として用いる場合がその1つの具体例となる。
【0024】
〔7〕betaKlothoがFGF受容体との結合におけるFGF21に対する特異的な補助因子であることの確認
本発明によれば、FGF21とbetaKlothoが存在すれば、FGF受容体を活性化して細胞内部にシグナル伝達をすることができるが、そのFGF21とbetaKlothoとの関係が特異的なものか、もしくはそれぞれの構造的な類似性を有する物質であっても代替できるほどに融通性を有するものなのかを次に検討した。すなわち、具体的な実験としては、betaKlothoを、FGF21と構造的類似性の高いFGF23に作用させた場合と、betaKlothoと構造的類似性の高いKlothoを、FGF21に作用させた場合に、それぞれどの程度の互換性があるかを調べた。
その結果、FGF21の場合は、ヘパリンの存在、非存在にかかわりなく、Klothoを存在させてもFGFR1cには反応しないことが示され、反対に、FGF23の場合は、ヘパリンの存在、非存在にかかわりなく、betaKlothoを存在させてもFGFR1cには反応しないことが示された。また、いずれの場合もヘパリンが系に存在しているとFGFR1cへの反応性が高まる。
したがって、betaKlothoがFGF21に対する特異的な受容体結合補助因子であることが立証された。同時に、ヘパリンはその効果的な更なる増強因子であることも立証された。
【0025】
〔8〕betaKlotho及びbetaKlotho活性を昂進もしくは抑制する物質の利用
以上の結果を総合すれば、betaKlotho活性を有する蛋白質またはbetaKlotho活性を有する蛋白質を体内で発現できるbetaKlotho遺伝子は、FGF21活性調節剤として用いることができ、betaKlothoもしくはその遺伝子は、単独で、又はFGF21と共にそれを有効成分とする血糖値調節用医薬組成物として用いることができることが示された。
また、betaKlothoを本来発現している細胞、組織を対象とする場合、そのbetaKlotho活性を増強する物質、及び/又はbetaKlothoの発現を促進する物質を用いることも同様に有効である。betaKlothoタンパク質またはbetaKlotho遺伝子と併用することも可能である。
反対に、生体内のbetaKlotho活性を阻害する物質は、FGF受容体を介したFGF21活性の抑制剤などとして働くため、たとえばFGF21の典型的な活性である血糖値低下機能を抑え、血糖値上昇効果を呈することが期待できる。
生体内でbetaKlotho活性を昂進する物質として、実施例で当該性質が確認されたヘパリンが1例として挙げられるが、生体内におけるbetaKlothoの作用を増強する物質であれば特に限定されない。betaKlotho活性を昂進する物質は、betaKlothoとFGF受容体とを細胞表面に発現しているBaF3細胞などを用いた本発明のスクリーニング方法により、簡単に取得でき、その候補としては、例えば抗betaKlotho抗体のうちで、betaKlotho活性を模倣するような、いわゆるアゴニスト抗体があげられる。当該抗体としては、betaKlothoを認識する抗体のみならず、FGF受容体を認識する抗体、FGF21を認識する抗体、betaKlotho、FGF受容体及び/又はFGF21の複合体を認識するような抗体でありえる。
増殖因子の作用に対するアゴニスト抗体の例としては、HGF受容体のアゴニスト抗体(特許文献2)、Tie2 アゴニスト抗体(特許文献3)、EGF受容体のアゴニスト抗体(非特許文献10)、NGF受容体のアゴニスト抗体(非特許文献11)などがある。
ところで、FGF1など多くのFGFリガンドでは、それらが単独にFGF受容体に結合しただけでは受容体の活性化は起こらないが、さらにヘパリン様の糖鎖が共存すると活性複合体が形成されて、それを起点としたシグナルの伝達が行われて細胞の応答(例えば細胞の増殖など)が起きるとされている(図14下段(1)、(2))。本発明におけるFGF受容体を介したFGF21の活性発揮にあたっては、通常のFGF受容体とFGFリガンド間の(1)、(2)のような様式ではシグナル伝達は起こらない。補助因子BetaKlothoが加わり、リガンド(FGF21)、FGF受容体、BetaKlothoの3者が存在することによって、はじめてFGF21によるFGF受容体の活性化が起きる。これを概念図として表したものが図14下段(3)である。なお、概念図中の複合体形成に関わる各分子の個数はこれを限定するものではない。この活性複合体の形成を増強あるいは減弱する物質は、FGF21の作用を昂進あるいは抑制する作用を有するが、これらの物質は、本発明のbetaKlothoとFGF受容体発現細胞系を用いたスクリーニング系で簡単に取得でき、FGF21活性の調節剤あるいは調節するための医薬組成物として利用することができる(4)。
従来のアゴニスト抗体も細胞表面のリガンドと受容体との結合を促進する作用があるが、本発明におけるbetaKlotho活性を模倣した抗体も、図14(4)に示されるような、betaKlotho、FGF21、及びFGF受容体からなる複合体の形成を増強するものであって、その結果、FGF受容体を介したFGF21活性を昂進する作用があると考えられる。
生体内でbetaKlotho活性を抑制する物質の例は、生体内におけるbetaKlothoの作用を妨げる物質であれば特に限定されないが、典型的にはbetaKlotho特異的抗体があげられる。
BetaKlotho活性を模倣したアゴニスト抗体、betaKlotho活性を抑制するbetaKlotho特異的抗体のいずれも(以下、単にbetaKlotho抗体ともいう。)、ポリクローナル抗体、モノクローナル抗体のいずれであってもよく、周知の免疫学的手法により作製できる。また、該betaKlotho抗体は、抗体のフラグメント(例えば、Fab、F(ab’)2)、組換え抗体(例えば、単鎖抗体)であってもよい。さらに、betaKlotho活性を抑制する抗体をコードする核酸(プロモーター活性を有する核酸に機能可能に連結されたもの)の場合は、betaKlothoの発現を抑制する物質として好ましい。
例えば、ポリクローナル抗体は、betaKlothoあるいはそのフラグメントを、必要に応じキャリアタンパク質を用い、市販のアジュバントとともに、ラット、マウス、ウサギなどの哺乳動物を定法により免疫して得た抗血清を精製することにより取得できる。
また、モノクローナル抗体は、細胞融合法を用いて取得でき、たとえば、マウスなどをbetaKlothoあるいはそのフラグメントで免疫し、脾臓あるいはリンパ節から白血球を採取し、この白血球と骨髄腫細胞を細胞融合して抗betaKlothoモノクローナル抗体を産生するハイブリドーマを得る。
また、本発明の抗体は、キメラ抗体、ヒト化又はヒト型抗体であってもよい。キメラ抗体は、例えば「実験医学(臨時増刊号),Vol.6,No.10,1988」等を、ヒト化抗体は、例えば特表平4-506458号公報、特開昭62-296890号公報等を、ヒト抗体は、例えば「Nature Genetics,Vol.15,p.146-156,1997」、「Nature Genetics,Vol.7,p.13-21,1994」等を参考にそれぞれ作製することができる。
その他のbetaKlotho阻害物質としては、betaKlotho遺伝子のアンチセンス核酸、リボザイム、RNAi誘導性核酸、ターゲティングベクター、betaKlothoの部分的なネガティブ変異体などがあげられる。
さらに、単なるbetaKlotho阻害物質は、(特許文献1)に記載のスクリーニング方法によってもスクリーニングすることができる。
【0026】
〔9〕本発明の血糖値の調節(上昇作用、又は降下作用)にかかわる医薬組成物について
本発明の剤は、betaKlotho及びその発現又は機能を調節する物質に加え、任意の担体、例えば医薬上許容され得る担体を含むことができる。医薬上許容され得る担体としては、例えば、ショ糖、デンプン、マンニット、ソルビット、乳糖、グルコース、セルロース、タルク、リン酸カルシウム、炭酸カルシウム等の賦形剤、セルロース、メチルセルロース、ヒドロキシプロピルセルロース、ポリプロピルピロリドン、ゼラチン、アラビアゴム、ポリエチレングリコール、ショ糖、デンプン等の結合剤、デンプン、カルボキシメチルセルロース、ヒドロキシプロピルスターチ、ナトリウム−グリコール−スターチ、炭酸水素ナトリウム、リン酸カルシウム、クエン酸カルシウム等の崩壊剤、ステアリン酸マグネシウム、エアロジル、タルク、ラウリル硫酸ナトリウム等の滑剤、クエン酸、メントール、グリシルリシン・アンモニウム塩、グリシン、オレンジ粉等の芳香剤、安息香酸ナトリウム、亜硫酸水素ナトリウム、メチルパラベン、プロピルパラベン等の保存剤、クエン酸、クエン酸ナトリウム、酢酸等の安定剤、メチルセルロース、ポリビニルピロリドン、ステアリン酸アルミニウム等の懸濁剤、界面活性剤等の分散剤、水、生理食塩水、オレンジジュース等の希釈剤、カカオ脂、ポリエチレングリコール、白灯油等のベースワックスなどが挙げられるが、それらに限定されるものではない。
【0027】
経口投与に好適な製剤は、水、生理食塩水のような希釈液に有効量の物質を溶解させた液剤、有効量の物質を固体や顆粒として含んでいるカプセル剤、サッシェ剤または錠剤、適当な分散媒中に有効量の物質を懸濁させた懸濁液剤、有効量の物質を溶解させた溶液を適当な分散媒中に分散させ乳化させた乳剤等である。
非経口的な投与(例えば、静脈内注射、皮下注射、筋肉注射、局所注入など)に好適な製剤としては、水性および非水性の等張な無菌の注射液剤があり、これには抗酸化剤、緩衝液、制菌剤、等張化剤等が含まれていてもよい。また、水性および非水性の無菌の懸濁液剤が挙げられ、これには懸濁剤、可溶化剤、増粘剤、安定化剤、防腐剤等が含まれていてもよい。当該製剤は、アンプルやバイアルのように単位投与量あるいは複数回投与量ずつ容器に封入することができる。また、有効成分および医薬上許容され得る担体を凍結乾燥し、使用直前に適当な無菌のビヒクルに溶解または懸濁すればよい状態で保存することもできる。
本発明の剤の投与量は、有効成分の活性や種類、病気の重篤度、投与対象となる動物種、投与対象の薬物受容性、体重、年齢等によって異なり一概に云えないが、通常、成人1日あたり有効成分量として約0.001〜約500mg/kgである。
【0028】
また、本発明の剤において、betaKlothoをコードするDNAを含む組換えDNAを有効成分として含む場合、典型的には当該組換えDNAとしてbetaKlothoをコードするDNAを挿入した発現ベクターを有効成分として含む。当該発現ベクターは、上記の核酸分子をコードするオリゴヌクレオチドもしくはポリヌクレオチドが、投与対象である哺乳動物の細胞内でプロモーター活性を発揮し得るプロモーターに機能的に連結されていなければならない。使用されるプロモーターは、投与対象である哺乳動物で機能し得るものであれば特に制限されず、例えば、SV40由来初期プロモーターなどが挙げられ、発現ベクターとしてヒト等の哺乳動物への投与に好適なアデノウイルス、レトロウイルス、アデノ随伴ウイルス、ヘルペスウイルス等のウイルスベクターや核酸キャリア物質などと共に投与することが好ましい。
【0029】
〔10〕FGF受容体及びbetaKlothoを内在的に発現していない細胞の系を用いたスクリーニング方法及びキット
FGF受容体及びbetaKlothoを内在的に発現していない細胞としては、典型的にはマウス白血病ProB細胞であるBaF3細胞が用いられるので、以下BaF3細胞を用いて説明する。ただし、他の細胞でもこれらいずれの遺伝子も細胞表面に発現しておらず、かつこれら遺伝子で形質転換された場合に細胞表面に発現することができる細胞であれば、同様に用いることができるので、BaF3細胞には限られない。
FGF受容体を発現しているBaF3細胞の系はFGF受容体を介したFGF21の活性を増強もしくは抑制する物質のスクリーニングに用いることができる。
具体的には、まず、FGF受容体を細胞表面に発現しているBaF3細胞の培養系に対して、披検物質と共にFGF21を作用させる方法を用いることができる。その際、披検物質とFGF21の添加順序は、同時でも、何れが先でもよい。
また、FGF受容体遺伝子と共に披検物質遺伝子をBaF3細胞に導入して、FGF受容体と同時に披検物質も細胞表面に発現させたBaF3細胞の系に対してFGF21を作用させてもよい。
その際、披検物質遺伝子をFGF受容体遺伝子と同一のベクターに挿入し、又は別々のベクターに挿入してBaF3細胞に同時に導入してもよく、また、FGF受容体を細胞表面に発現しているBaF3細胞に対して、披検物質遺伝子を導入してもよい。
FGF受容体、及びその遺伝子としては、FGFR1cもしくはFGFR3c、及びそれらの遺伝子を用いることが好ましい。
したがって、FGF受容体、好ましくはFGFR1cもしくはFGFR3c、を細胞表面に発現しているBaF3細胞の系をFGF21と組み合わせることで、FGF21の生物活性を増強もしくは抑制する物質のスクリーニング用のキットとすることができる。
なお、実際のスクリーニングに際しては、BaF3細胞の増殖能を、披検物質を添加しない系などのコントロールの系と比較して評価することができる。BaF3細胞の増殖能は、標識チミジンの取り込み量の測定など常法により測定する。他のスクリーニング方法においても同様である。
また、実際のスクリーニングに際しては、BaF3細胞の増殖シグナル伝達の活性化を、披検物質を添加しない系などのコントロールの系と比較して評価することもできる。BaF3細胞の増殖シグナル伝達の活性化は、FGF受容体のチロシンリン酸化、FGF 受容体基質2(FRS2)のチロシンリン酸化、マップキナーゼ(ERK1/2)のリン酸化など常法により測定する。他のスクリーニング方法においても同様である。
【0030】
FGF受容体と共にbetaKlothoも発現しているBaF3細胞の系は、FGF21活性調節作用のあるbetaKlotho活性を増強もしくは抑制する物質のスクリーニングに用いることができる。
具体的には、まず、FGF受容体と共にbetaKlothoを細胞表面に発現しているBaF3細胞の系に対して披検物質と共にFGF21を作用させる方法を用いることができる。その際、披検物質とFGF21の添加順序は、同時でも、何れが先でもよい。
また、FGF受容体遺伝子及びbetaKlotho遺伝子と共に披検物質遺伝子をBaF3細胞に導入して、FGF受容体及びbetaKlothoと同時に披検物質も細胞表面に発現させたBaF3細胞の系に対してFGF21を作用させてもよい。
その際、披検物質遺伝子をFGF受容体遺伝子及びbetaKlotho遺伝子と同一のベクターに挿入し、又は別々のベクターに挿入してBaF3細胞に同時に導入してもよく、また、FGF受容体及びbetaKlothoを細胞表面に発現しているBaF3細胞に対して、披検物質遺伝子を導入してもよい。
FGF受容体、及びその遺伝子としては、FGFR1cもしくはFGFR3c、及びそれらの遺伝子を用いることが好ましい。
したがって、FGF受容体、好ましくはFGFR1cもしくはFGFR3c、及びbetaKlothoを細胞表面に発現しているBaF3細胞の系をFGF21と組み合わせることで、FGF21活性の調節作用に係るbetaKlotho活性の昂進物質もしくは阻害物質のスクリーニング用のキットとすることができる。
【0031】
FGF受容体と共にbetaKlothoも発現しているBaF3細胞の系は、FGF21様活性物質のスクリーニング方法に用いることができる。
具体的には、FGF受容体と共にbetaKlothoを細胞表面に発現しているBaF3細胞の系に対して披検物質を作用させ、BaF3細胞の増殖能を評価することで、FGF21様活性物質をスクリーニングすることができる。
その際、同時に、FGF受容体のみを細胞表面に発現しているBaF3細胞及び/又はbetaKlothoのみを細胞表面に発現しているBaF3細胞の系に披検物質を作用させ、その作用が、FGF21作用と同様の、FGF受容体及びbetaKlothoとの複合体を形成したことに基づく作用であることを確認することが好ましい。
FGF受容体としてはFGFR1c又はFGFR3cを用いることが好ましい。
そして、FGF受容体、好ましくはFGFR1cもしくはFGFR3c、と共にbetaKlothoを細胞表面に発現しているBaF3細胞の系を用いることで、FGF21様活性物質のスクリーニング用のキットとすることができる。
さらに、FGF受容体のみを細胞表面に発現しているBaF3細胞及び/又はbetaKlothoのみを細胞表面に発現しているBaF3細胞の系をコントロール系として組み合わせたキットがより好ましく用いることができる。
そして、これらスクリーニングにより得られた物質は、いずれもFGF受容体とbetaKlothoの共存下で発揮されるFGF21活性の調節剤及び当該FGF21活性を調節するための医薬組成物として用いることができる。
【実施例】
【0032】
以下、実施例を用いて本発明をより詳細に説明するが、本発明の技術的範囲は以下の実施例に限定されるものではない。
〔実施例1〕FGF21の脂肪細胞特異的活性の検出
FGF21は、その活性として脂肪細胞にグルコーストランスポーター1(GLUT1)の発現を上昇させることが報告されている。本実施例では、培養脂肪細胞としては、培養条件を操作することにより脂肪細胞に分化する能力を有する繊維芽細胞として3T3-L1細胞を用いた。細胞は10% ウシ胎児血清(FBS)を含有する、Dulbecco改変Eagle培地(DMEM)の培養液で培養し、繊維芽細胞としての未分化形質を維持した細胞を用意した。その細胞を試験系にて培養し、コンフルエントになるまで増殖させ、そして2日間そのままにした。その後、培養液を分化誘導培養液であるDMEM、10%FBS、0.25μMデキサメタゾン、0.5mMメチルイソブチルキサンチン、および5μg/mlのインスリンに交換して2日間培養した。その後、細胞を10%FBSおよび2.5μg/mlのインスリンを含有するDMEMにより、1日おきに培養液を交換した。このようにして分化誘導した脂肪細胞の性質をもつ細胞を得た。比較のために、分化を誘導しない繊維芽細胞も準備した。ここで「脂肪細胞の性質をもつ細胞」とは、脂肪細胞特有の細胞内代謝を行い、脂肪滴を多量に蓄積した細胞質を有する細胞を示す。
これらそれぞれの培養液にFGF21を種々の濃度で加え、細胞をさらに6時間培養後、RNAを抽出し、GLUT1のコピー数を測定した。その結果を図1に示す。
ここで、脂肪細胞に分化した細胞ではFGF21を様々な濃度で細胞培養液に添加することにより、GLUT1のmRNAの発現量が増加していたことから、FGF21が脂肪細胞に対して活性を発揮し、GLUT1発現を増加させることが示された。これとは対照的に、分化していない繊維芽細胞では、FGF21を様々な濃度で細胞培養液に添加しても、GLUT1のmRNAの発現量はほとんど変化しなかった。このことから、FGF21がGLUT1のmRNAを増加させる活性は、3T3-L1細胞では細胞が脂肪細胞に分化したあとにのみ発揮されることが示された。
【0033】
〔実施例2〕脂肪細胞の分化に伴うFGF受容体の発現変動の検出
上記実施例1で、FGF21が活性を発揮するのは3T3-L1細胞が脂肪細胞に分化した後だけであったことから、脂肪細胞に分化するとともに、細胞表面に存在するFGF受容体数が増加した結果、FGF21に対する反応性が増大した可能性が考えられた。そこで、FGFの受容体分子の発現レベルに変動があるかどうかをまず解析した。
3T3-L1細胞を、分化誘導培養液に交換すると、培養を開始してから経時的に、繊維芽細胞の性質を持つ細胞から、脂肪細胞の性質をもつ細胞に、徐々に変化する。
そこで、実施例1と同様の条件で分化誘導処理を行った後、一日おきに20日間まで3T3-L1細胞を採取し、細胞が発現するFGF受容体のmRNAの量を解析した。FGF受容体としては、膜一回貫通型チロシンキナーゼ型受容体として、4種類の遺伝子FGFR1、FGFR2、FGFR3、FGFR4が知られているので、これらのmRNA量を測定した。その結果を図2に示す。分化の有無にかかわらずFGFR1とFGFR2が高いレベルで発現しており、FGFR3とFGFR4の発現レベルは非常に低かった。そして、分化に伴って発現が増える受容体はなく、むしろ高レベルで発現していたFGFR1、FGFR2のいずれの受容体も、分化とともに、発現レベルがやや減る傾向が認められた。
【0034】
〔実施例3〕脂肪細胞の分化に伴うbetaKlothoの発現変動の検出
本発明者らは、脂肪細胞へと分化する過程で脂肪細胞表面での発現量が増加する物質があれば、当該物質がFGF21と協同してFGF受容体に作用する可能性が高いと考え、発達中のマウス胎児の脂肪組織で発現することが知られていたbetaKlothoに着目した。
上記実施例2と同様にして、3T3-L1細胞が脂肪細胞に分化する過程でのbetaKlothoの発現レベルを経時的に解析した。
さらに、比較のために、最近FGF23に対する受容体補助因子として働くことが報告されたKlothoの発現レベルも同様にして解析した。KlothoはbetaKlothoと構造的に類似性があるが、アミノ酸配列レベルで41%の相同性(塩基配列レベルで55%)であり、両者の性質はまったく異なるもので、明らかな別遺伝子にコードされた分子であると位置づけられる。
その解析結果を、図3に示す。betaKlothoは脂肪細胞に分化した細胞でのみ高レベルで発現していた。これに対して、Klothoは分化前も分化後もほとんど発現していなかった。このことから、脂肪細胞分化に伴って発現するようになるbetaKlothoが脂肪細胞の機能と密接に関連することが示唆されたため、FGF21に対するFGF受容体補助因子である可能性を探るため解析する計画をした。
【0035】
〔実施例4〕FGF21によるFGF受容体下流シグナル惹起にbetaKlothoが必要であることの解析
上記実施例2と3から、実施例1で示されるようなFGF21の脂肪細胞特異的活性は、FGF受容体とbetaKlothoが共に存在する場合に誘導されることが示唆される。そこで本実施例において、元来はFGF受容体を持たない細胞BaF3にFGFR1cを強制発現させた細胞に対し、さらにbetaKlothoを強制発現させたときとさせないときで、FGF21に対する反応性がどのように変わるかを解析した。
FGFR1c発現BaF3細胞、FGFR1c及びbetaKlotho発現BaF3細胞は以下のように調製した。FGFR1c発現BaF3細胞にマウスFGFR1cの全長cDNAがクローニングされた動物細胞用発現ベクター(Ornitz,D.M.,Xu,J.,Colvin,J.S.,McEwen,D.G.,MacArthur,C.A.,Coulier,F.,Gao,G.and Goldfarb,M.(1996) Receptor specificity of the fibroblast growth factor family.J Biol Chem,271,15292-15297.に記載されている)をエレクトロポレーション法によりBaF3細胞に導入した。G418存在下で約2週間培養を行い、薬剤耐性クローンを得た後、FGF1刺激により増殖させることができるクローンとして取得した。FGFR1cとbetaKlothoを共に発現するBaF3細胞は、動物細胞用発現ベクターにマウスbetaKlothoの全長cDNAをクローニングし、これをFGFR1c発現BaF3細胞に導入した後、Puromycin及びG418存在下で約2週間培養を行い、耐性クローンとして取得した。betaKlothoを単独で発現するBaF3細胞はbetaKlotho発現ベクターをBaF3細胞に導入した後、Puromycin耐性クローンとして取得した。これらの遺伝子導入したBaF3細胞は10%FBSとインターロイキン3(IL3、WEHI-3細胞のコンディションメディウムを5%加えることにより供給)を含むRPMI1640培地で維持した。増殖刺激実験時は細胞をRPMI1640培地で洗浄後10%FBSを含むRPMI1640培地に懸濁し培養プレートに播種して用いた。
解析結果を、図4に示す。
A)で、CL#1,#7、#10はFGFR1cとbetaKlothoの両方のcDNAをBaF3細胞に導入し取得したクローンの名称である。A)の下段は、それぞれの細胞から調製したライゼートを材料として、実際にタンパクとして発現しているbetaKlothoの量を、市販の抗betaKlotho抗体を用いたウエスタンプロット解析により比較した結果である。CL#10ではbetaKlothoは検出レベル以下であり、CL#1、CL#7では同程度のレベルのbetaKlothoの発現がみられた。BetaKlothoは主に2本のブロードなバンドとして観察され、N型糖鎖の修飾が起こっている可能性が考えられた。
それぞれのクローンおよびFGFR1cのみを発現しているBaF3細胞は播種直後、ヘパリン存在下でFGF21(白色バー)あるいはコントロールとしてPBS(黒白色バー)の添加により刺激し、42時間培養した。その後トリチウムラベルされたチミジンを培養液中に添加し、更に6時間培養を継続し、細胞に取り込まれたカウントを測定することによりDNA合成を評価した。それぞれFGF21未刺激のDNA合成値を1とした場合のFGF21によるDNA合成の割合を倍率として表した。その結果FGFR1cは発現しているがbetaKlothoを発現していない細胞ではFGF21に反応せず、betaKlothoを共に発現している細胞でのみFGF21に反応することが示された。尚、図には示していないが、FGFR1cのみを発現しているBaF細胞はヘパリンが存在している条件でもFGF21に反応しない。
B)はFGF21の量を変化させてCL#1の細胞を刺激した際のDNA合成を測定した結果である。ヘパリン存在下(白丸)、非存在下(黒丸)ともに、FGF21による用量依存的なDNA合成刺激活性がみられた。またヘパリンはFGF21の作用を増強することがわかった。C)はFGFR1cを発現していないがbetaKlothoのみを発現しているBaF3細胞のFGF21に対する反応性を調べた結果で、ヘパリン存在下(白色バー)、非存在下(黒色バー)の両条件下ともに、FGF21には反応しなかった。BaF3の維持に用いているインターロイキン3(IL3)刺激では顕著な増殖が観られた。
以上のことから、FGF21に反応するためには、FGF受容体とbetaKlothoの両方が必要であることが示された。
【0036】
〔実施例5〕FGF21の活性発揮においてbetaKlothoとともに存在するFGF受容体の特異性の解析(1)
上記実施例4において、FGF21に反応するためには、FGF受容体であるFGFR1cとbetaKlothoの両方が必要であることが示されたが、FGF受容体として、ほかの受容体分子ではどうであるかを調べた。betaKlotho発現BaF3細胞に各FGFR(FGFR1b、R1c、R2b、R2c、R3b、R3c、R4)の発現ベクター(Ornitz,D.M.,Xu,J.,Colvin,J.S.,McEwen,D.G.,MacArthur,C.A.,Coulier,F.,Gao,G.and Goldfarb,M.(1996) Receptor specificity of the fibroblast growth factor family.J Biol Chem,271,15292-15297.に記載されている)を導入して、Puromycin及びG418耐性クローンを取得し、FGFR/betaKlotho発現細胞を得た。それぞれをFGF21で刺激しDNA合成を測定した。その結果図5に示すように、FGFR1cのほか、FGFR3cをbetaKlothoと共に発現させた場合でもFGF21に反応することが明らかとなった。さらにヘパリンの存在により反応性は増強されることが示された。
【0037】
〔実施例6〕FGF21の活性発揮においてbetaKlothoとともに存在するFGF受容体の特異性の解析(2)
betaKlothoとともに各FGFR(FGFR1c、R2c、R3c、R4)を発現しているBaF3細胞に対するFGF21のDNA合成刺激活性を、進化的にFGF21に近縁のFGF19の活性と比較した。実施例5と逆の手順で、すなわち各FGFRを発現しているBaF3細胞にbetaKlothoの発現ベクターを導入してPuromycin及びG418耐性クローンを取得し、FGFR/betaKlotho発現細胞を得た。細胞をヘパリン存在下で、FGF21、FGF19、FGF1の量を図のように変化させて刺激し、DNA合成を測定した。図に示されるように、FGF19はいずれの細胞に対しても強い活性を示すのに対して、FGF21はR1c/betaKlotho発現細胞とR3c/betaKlotho発現細胞のみを活性化することが明らかとなった。(図6)
【0038】
〔実施例7〕FGF21の刺激に伴う細胞内シグナル伝達分子の活性化の解析
実施例6で取得した各FGFR/betaKlotho発現細胞をFGFで刺激した際の、FGFRの下流のシグナル伝達分子FRS2(fibroblast growth factor receptor substrate 2、FGF受容体基質2)とMAPK(mitogen activated protein kinase、マイトージェン活性化プロテインキナーゼ、別名ERK(extracellular signal-regulated kinaseで通常分子量44kDaのERK1と42kDaのERK2の2種類を指す)の活性化状態(リン酸化状態)を解析した。それぞれの細胞をヘパリン存在下、各FGFで10分間刺激した後、細胞ライゼートを調製し、市販の抗リン酸化FRS2抗体あるいは抗リン酸化MAPK抗体を用いてリン酸化の程度をウエスタンブロッティング法で測定することにより評価した。
その結果を図7に示す。図においてMAPKのカラムは抗ERK1/2抗体を用いたウエスタンブロッティングによる細胞ライゼート中の全MAPKの量を示すが、刺激あるいはコントロールの無刺激の細胞においてMAPK(ERK1/2)の総量自体の変化はない。FGF1とFGF19はいずれの細胞においてもFRS2とMAPKのリン酸化を顕著に増加させるのに対し、FGF21は、R1c/betaKlotho発現細胞とR3c/betaKlotho発現細胞においてFRS2とMAPKのリン酸化を増大させることが示された。この結果は実施例6の結果をよく反映している。
【0039】
〔実施例8〕ヒトのアミノ酸配列をもつヒト型FGFR1cとヒト型betaKlothoを共発現させた細胞に対するヒト型FGF21の活性の解析
実験例4、5、6、7において、FGF21はbetaKlotho存在下にFGFR1cに反応することを示したが、実際の実験系で用いたFGFR1c、betaKlotho、FGF21などはすべて哺乳類の典型的なモデルとしてマウス由来のものを用いている。これら各分子のヒト、マウス間でのアミノ酸配列の相同性はFGF21が80%、betaKlothoが78%、FGFR1が98%と極めて高いことからも当然にヒト、マウス間の互換性に問題なく、ヒト型の分子でもマウス型と同様の結論が得られることは明らかである。本実施例では、念のために、その点を確認するためのヒト型分子を用いた実験系を構築して実証を行った。具体的には、FGFR1c、betaKlothoのヒト型のcDNAは市販のヒトRNA試料(タカラバイオ社)からPCRにより調製し、FGFR1cはSRalphaプロモーターを有する発現ベクター(pSRalpha)に、betaKlothoはCMVプロモーターを有する発現ベクター(pcDNA3.1;インビトロージェン社)に導入した。実験例4と同様にエレクトロポレーション法によりFGFR1c発現ベクターをBaF3細胞に導入し、G418耐性クローンとしてFGFR1c発現BaF3細胞を取得した。FGFR1cとbetaKlothoの共発現細胞はFGFR1c発現BaF3細胞にbetaKlotho発現ベクターを導入し、Puromycin耐性クローンとして取得した。ヒト型FGF21タンパク質は以下のように調製した。ヒト型FGF21のcDNAを市販のヒトRNA試料(タカラバイオ社)からPCRにより調製し、大腸菌用の発現ベクターに導入した。このベクターを用いて非特許文献1に記載の手法に準拠して、タンパク質を調製した。ヘパリン存在下で種々の濃度でヒト型FGF21を FGFR1c発現細胞あるいはFGFR1cとbetaKlothoの共発現細胞に添加した際のDNA合成を測定した結果を図8に示す。コントロールとして用いたFGF1はbetaKlothoの発現の有無にかかわらず、細胞のDNA合成を促進したが、FGF21はbetaKlothoを共発現している細胞のみを活性化することが示された。
このことは、ヒト由来分子を用いた場合にも、マウス由来分子を用いた各実施例の実験結果と同様の結果が導かれるであろうという当初の予想を、あらためて裏付けるものであった。
【0040】
〔実施例9〕ヒトのアミノ酸配列をもつヒト型FGFR3cとヒト型betaKlothoを共発現させた細胞に対するヒト型FGF21の活性の解析
FGFR3のヒト、マウス間でのアミノ酸配列の相同性は93%である。ヒト型FGFR3cの場合においても、マウス型と同様の結論が得られることを実証する実験を行った。FGFR3cのヒト型のcDNAは市販のヒトRNA試料(タカラバイオ社)からPCRにより調製し、細胞内部分の配列をヒト型FGFR1の配列と置換してSRalphaプロモーターを有する発現ベクター(pSRalpha)に導入した。エレクトロポレーション法によりFGFR3c発現ベクターをBaF3細胞に導入し、G418耐性クローンとしてFGFR3c発現BaF3細胞を取得した。FGFR3cとbetaKlothoの共発現細胞はFGFR3c発現BaF3細胞にbetaKlotho発現ベクターを導入し、Puromycin耐性クローンとして取得した。ヘパリン存在下で種々の濃度でヒト型FGF21を FGFR3c発現細胞あるいはFGFR3cとbetaKlothoの共発現細胞に添加した際のDNA合成を測定した結果を図9に示す。コントロールとして用いたFGF1はbetaKlothoの発現の有無にかかわらず、細胞のDNA合成を促進したが、FGF21はbetaKlothoを共発現している細胞のみを活性化することが示された。この実験結果からも、ヒト型の分子でもマウス型と同様の結論が得られることが実証された。
【0041】
〔実施例10〕FGF21の活性発揮においてFGF受容体とともに存在するbetaKlothoの特異性の解析
betaKlothoがFGF受容体との結合におけるFGF21に対する特異的な補助因子であることを確認するために、betaKlothoを、FGF21と構造的類似性の高いFGF23に作用させた場合と、betaKlothoと構造的類似性の高いKlothoを、FGF21に作用させた場合に、それぞれどの程度の互換性があるかを調べた。
具体的には、FGFR1cと共にbetaKlothoを発現させたBaF3細胞及びFGFR1cと共にKlothoを発現させたBaF3細胞を、実施例4と同様の形質転換方法及び同一の培養条件を用いて調製した。そして、各形質転換細胞に対して、ヘパリンの存在下、及び非存在下でFGF21及びFGF23を添加し、DNA合成量を測定した。
その結果を図10に示す。FGF21の場合は、ヘパリンの存在、非存在にかかわりなく、klothoを存在させてもFGFR1cには反応しないことが示され、反対に、FGF23の場合は、ヘパリンの存在、非存在にかかわりなく、betaKlothoを存在させてもFGFR1cには反応しないことが示された。
このように、FGF21とbetaKlotho、FGF23とKlothoそれぞれの組合せでのみFGFR1c発現細胞に活性を発揮できることが示されたことからも、betaKlothoがFGF21に対する特異的な受容体結合補助因子であることが立証された。
【0042】
〔実施例11〕BetaKlothoとFGF21あるいはFGF受容体との複合体形成の検出
実施例4から実施例10においては、FGF21のFGF受容体を介した作用発現のためにbetaKlothoが必要であることを細胞生物学的に示した。
本実施例においては、betaKlothoとFGF受容体あるいはFGF21の間の複合体形成を生化学的に調べた。具体的には、betaKlothoの細胞外ドメインのC末端にV5タグとHisタグをタンデムに付加した可溶性タンパク質を調製し、試験管内においてヘパリンの存在あるいは非存在の条件下で、可溶性betaKlothoタンパク質とFGF21をインキュベートした後、抗V5タグ抗体固定化ビーズを用いてbetaKlothoを沈降させ、FGF21がbetaKlothoに結合して共沈降するか否かを、免疫ブロッティング法にて解析した。
その結果を図11A)に示す。ヘパリンの有無によらず、betaKlothoの存在下でFGF21が沈降してくることから、betaKlothoはFGF21との結合が、ヘパリンの有無によらず形成されることが確認された。
また、可溶性betaKlothoタンパク質とFGF受容体の細胞外ドメインに免疫グロブリンのFcドメインを付加した融合タンパク(FGFR-Fc:R&D Systems社製)を、インキュベートした後、プロテインAビーズを用いてFGFR-Fcを沈降させ、betaKlothoがFGFR-Fcに結合して共沈降するか否かを解析した。
その結果を図11B)に示す。FGFR-Fcの存在に依存してbetaKlothoが沈降してくることが示され、betaKlothoはFGF受容体サブタイプのR1c、R2c、R3c、R4のいずれとも結合することが明らかになった。
【0043】
〔実施例12〕BetaKlothoを強制発現させた未分化3T3-L1細胞をFGF21刺激した際のシグナル伝達の解析
実施例1において、FGF21は脂肪細胞に作用してGLUT1のmRNA量を増大させる活性を有するが、分化前の細胞に対してはこの活性を発揮しないことを示した。実施例2では、betaKlothoが脂肪細胞の分化に伴って発現が増大することを示した。実施例3から10では、BaF3細胞を用いてbetaKlothoがFGF21に対するFGF受容体補助因子であることを示した。
そこで本実施例では、脂肪細胞へ分化する前の3T3-L1細胞においてもbetaKlothoがFGF21に対するFGF受容体補助因子として機能するか否かを調べた。実施例1に記載した通りに培養した3T3-L1繊維芽細胞に、実施例4に記載した方法で作成したbetaKlothoの発現ベクター、及びコントロールとしてbetaKlothoのcDNAを挿入していない空のベクターを、リポフェクション法により導入し、Puromycin耐性クローンを得た。それぞれの細胞をFGF21で刺激し、10分後に細胞ライゼートを調製し、FGFRの下流のシグナル伝達分子であるFRS2とMAPKの活性化をウエスタンブロッティング法で調べた。
その結果を図12に示す。図のようにbetaKlothoの発現ベクターを導入した細胞においてのみFGF21の刺激に反応してFRS2とMAPKのリン酸化が増加することが示された。また、FGFRのチロシンキナーゼ阻害剤であるSU5402(メルク社製)で3時間前処理した細胞をFGF21で刺激した場合には、リン酸化の程度が減少することから、FGF21の活性はFGFRを介することにより伝達されていることが示された。
【0044】
〔実施例13〕BetaKlothoを強制発現させた未分化3T3-L1細胞をFGF21刺激した際のGLUT1mRNAの発現解析
実施例12で作出したbetaKlothoを強制発現させた3T3-L1繊維芽細胞を用い、刺激するFGF21量に応じたGLUT1(グルコーストランスポーター1)のmRNA量の変化を調べた。
betaKlothoの発現ベクターあるいはコントロールとしてbetaKlothoのcDNAを挿入していない空のベクターを導入した3T3-L1繊維芽細胞の培養液にFGF21を種々の濃度で加え6時間培養した後にRNAを抽出し、GLUT1のmRNA量を測定した。
図13に示すように、betaKlothoを強制発現させた3T3-L1繊維芽細胞では培養液に添加するFGF21の量に従って、GLUT1mRNAが増加することが有意にみとめられた。(図の*はスチューデントのt検定によって5%未満の危険率で、**は1%未満の危険率で有意差があることを示す。)一方、コントロールの3T3-L1繊維芽細胞ではFGF21の添加によってGLUT1mRNA量に変化は見られなかった。この結果から、脂肪細胞に分化する前の3T3-L1細胞でもbetaKlothoを強制発現させることによりFGF21刺激を受容してGLUT1の発現量を増加することが明らかになった。
【技術分野】
【0001】
本発明は、受容体補助因子もしくは当該因子をコードする遺伝子又は当該因子活性を昂進もしくは抑制する物質を用いてリガンドによる受容体の活性化を促進または抑制する方法並びにリガンド活性を制御する方法(及びこれを用いた血糖値調節方法)に関する。
【背景技術】
【0002】
糖尿病で問題とされる高い血糖レベルに対しては、健康な体の中で血糖レベルが調節される仕組みを理解し、その知識を活用して、正常な値に制御することが必要であるが、未だ体内の血糖レベルの制御機構は解明できていない。現在実践的に用いられている唯一の積極的治療法は、糖尿病患者に対しインスリンを投与し、インスリンの血糖値低下作用を利用して、血糖値を正常範囲にまで低下させる方法のみである。しかし、この方法は、インスリンの分泌量が不足していることが原因の患者にとっては極めて効果的な治療法ではあるが、患者の中にはインスリンを投与しても血糖値の低下しない患者もいる。このような、所謂インスリン抵抗性の患者に対しても有効な、糖尿病薬の開発は急務であり、そのためにも体内における血糖値の制御機構を解明し、この問題を解決することが、当技術分野において大きな課題となっている。
【0003】
ところで、線維芽細胞増殖因子(FGF)とよばれるFGFファミリーの分子群は、アミノ酸配列の相同性と構造的類似性から、現在ヒト及びマウスにおいては22種類存在することが知られている。すべてのFGFファミリーメンバーの活性機能が明らかにされているわけではないが、すでに機能に関する研究が進んでいるメンバーは、いずれも線維芽細胞の増殖活性のみならず、広範な細胞に対する増殖や分化の制御活性を有することが知られ、形態形成、血管形成、神経生存維持、代謝調節などといった多様な生命現象に深く関わっている因子群である(非特許文献1,2,3,4)。このメンバーの中で比較的最近FGFファミリーメンバーに加わったFGF19、FGF21及びFGF23は、ドメイン構造からもアミノ酸配列の相同性の観点からも類似したサブグループを形成しているが、これらのFGFは、まだその作用機構の解明が十分になされているとはいえず、しかも通常のFGFファミリーで一般に用いられている細胞増殖や分化を測定するアッセイ系に供しても、ほとんど活性が検出されない。
最近、FGF21の作用として脂肪細胞のグルコーストランスポーターの発現量を昂進し、細胞内へのグルコース取り込みを上昇させ、また動物個体レベルでは、静脈内あるいは皮下投与によって血糖値の低下を引き起こすこと、血中トリグリセリド量を抑制すること、LDLコレステロールを低下、HDLコレステロールを増加させることが報告された(非特許文献1、5)。またFGF21は膵臓ランゲルハンス島に作用し、インスリンの合成量を増加させることも報告されている(非特許文献6)。このことから、FGF21は、血糖レベル調節や中性脂肪レベル調節などに重要な役割を果たしていることが考えられ、新規の作用機構を有するメタボリックシンドローム治療薬候補として浮上してきた。
しかしながら、FGF21をメタボリックシンドローム治療用医薬として用いるためには、FGF21単独では十分とはいえないことから、その作用を増強する因子の探索は必須であるが、効果的に増強できる因子はまだ発見されていない。
一般に、FGFが活性を発揮するためには、細胞膜表面のFGF受容体と結合して、これを活性化させて細胞内シグナリングを惹起することが必要であり、FGF21についても脂肪細胞に作用させた際に、細胞の表面に存在するFGFR1とFGFR2の活性化が起こることが報告されている(非特許文献1)。FGF21に対して観察された血糖値低下作用などもこれらFGF受容体との相互作用に基づくシグナル伝達の結果であると考えられ、その際のFGF21によるFGF受容体の活性化機構が解明されれば、当該機構に関連した因子を利用することで、血糖値を調節することが可能になる。しかしながら、従来の知見はFGF21のFGF受容体への結合様式の解明にも、グルコーストランスポーターや、インスリン発現などの血糖値を制御する機構の解明にも直接繋がるものではなく、このような血糖値の調節可能な因子の提供には結びつかなかった。
したがって、FGF21がどのようにしてFGF受容体を活性化するのかを早急に解明し、FGF21の血糖値等制御機能を調節する手法を確立することで、FGF21を有効成分とする効果的なメタボリックシンドローム治療薬を提供することが、当該分野の大きな課題となっていた。
【先行技術文献】
【特許文献】
【0004】
【特許文献1】特開2006-158339号公報:非特許文献7の関連特許文献
【特許文献2】米国特許第6099841号明細書
【特許文献3】米国特許第6365154号明細書
【非特許文献】
【0005】
【非特許文献1】Kharitonenkov,A.,Shiyanova,T. L.,Koester,A.,Ford,A. M.,Micanovic,R.,Galbreath,E. J.,Sandusky,G. E.,Hammond,L. J.,Moyers,J. S.,Owens,R. A.,Gromada,J.,Brozinick,J. T.,Hawkins,E. D.,Wroblewski,V. J.,Li,D. S.,Mehrbod,F.,Jaskunas,S. R.,andShanafelt,A. B. (2005) J Clin Invest 115(6),1627-1635
【非特許文献2】Itoh,N. and Ornitz,D.M. (2004) Evolution of the Fgf and Fgfr gene families. TrendsGenet,20,563-569.
【非特許文献3】Inagaki,T.,Choi,M.,Moschetta,A.,Peng,L.,Cummins,C.L.,McDonald,J.G.,Luo,G.,Jones,S.A.,Goodwin,B.,Richardson,J.A.,Gerard,R.D.,Repa,J.J.,Mangelsdorf,D.J.and Kliewer,S.A. (2005) Fibroblast growth factor 15functions as an enterohepatic signal to regulate bile acid homeostasis. CellMetab,2,217-225.
【非特許文献4】Yu,X. and White,K.E. (2005) FGF23 and disorders of phosphate homeostasis. CytokineGrowth Factor Rev,16,221-232.
【非特許文献5】Kharitonenkov,A.,Wroblewski,V. J.,Koester,A.,Chen,Y. F.,Clutinger,C. K.,Tigno,X. T.,Hansen,B. C.,Shanafelt,A. B.,andEtgen,G. J. (2007) Endocrinology 148(2),774-781
【非特許文献6】Wente,W.,Efanov,A. M.,Brenner,M.,Kharitonenkov,A.,Koster,A.,Sandusky,G. E.,Sewing,S.,Treinies,I.,Zitzer,H.,and Gromada,J.(2006) Diabetes 55(9),2470-2478
【非特許文献7】Ito,S.,Fujimori,T.,Furuya,A.,Satoh,J.,Nabeshima,Y.,andNabeshima,Y. (2005) J Clin Invest 115(8),2202-2208
【非特許文献8】Ito,S.,Kinoshita,S.,Shiraishi,N.,Nakagawa,S.,Sekine,S.,Fujimori,T. and Nabeshima,Y.I. (2000) Molecularcloning and expression analyses of mouse betaklotho,whichencodes a novel Klotho family protein. Mech Dev,98,115-119.
【非特許文献9】Nishimura,T.,Nakatake,Y.,Konishi,M. and Itoh,N. (2000) Identification of anovel FGF,FGF-21,preferentiallyexpressed in the liver. Biochim Biophys Acta,1492,203-206
【非特許文献10】LeSauteur,L.et al.(1996) J Neurosci,16,1308-1316)
【非特許文献11】Fernandez-Pol,J.A.(1985) J Biol Chem,260,5003-5011)
【発明の概要】
【発明が解決しようとする課題】
【0006】
本発明は、FGF受容体を介したFGF21活性に対する効果的な調節因子を提供することを目的とする。典型的には、FGF21によって細胞の糖取り込みが調節する際に必要とされる受容体補助因子の同定、及び当該因子を利用したFGF21の活性制御方法並びに当該活性を用いた血糖値調節方法を提供することを目的とする。
また、当該調節因子及び/又はその活性を増強若しくは抑制する物質を有効成分として含むFGF21活性調節剤を提供すると共に、血糖値の病的な上昇、低下など糖取り込み異常に関連した疾患、たとえば糖尿病の治療又は予防用医薬組成物などFGF受容体を介したFGF21活性を調節するための医薬組成物の提供も本発明の目的である。
さらに、本発明は、FGF受容体を介したFGF21活性の調節因子、当該調節因子の活性を増強若しくは抑制する物質、FGF21様活性物質などのスクリーニング系の提供も目的とする。
【課題を解決するための手段】
【0007】
上述した目的を達成するため本発明者は、鋭意検討した結果、betaKlothoこそがFGF21によって細胞の糖取り込みが調節する際に必要とされる特異的な受容体補助因子であることを見いだした。
具体的には、本発明者らは、まずFGF21がどのようにしてFGF受容体を活性化するのか、受容体を活性化するのに必要な条件は何なのかを確かめるために、本来FGF受容体を有していない細胞の表面で各FGF受容体を発現している複数の細胞を用意して、FGF21と反応させたところ、いずれのFGF受容体においてもFGF1が通常十分活性を発揮する濃度の数十倍から数百倍量のFGF21によってもFGF受容体の活性化は検出されないことを見いだした。
さらに、通常のFGFリガンドが各FGF受容体と結合する際には、受容体補助因子としてのヘパリンが必須であるため、上記の系にさらにヘパリンを添加して観察したが、ヘパリン存在下でも、非存在下と同様にFGF受容体の活性化は起こらなかった。そこで、FGF21がどのようにしてFGF受容体を活性化するのか、その必要条件は何なのかについて鋭意検討する過程で、betaKlothoが膜上に共に存在するときに、FGF21によってFGFR1cが活性化され、下流のシグナルが惹起されることを見出した。
ついで、本発明者らは、FGF受容体もbetaKlothoも内在的に発現していない細胞であるBaF3細胞に対して、FGFR1c遺伝子もしくはFGFR3c遺伝子を、又はさらにbetaKlotho遺伝子及びその構造類似体のKlotho遺伝子を導入して細胞表面に発現させた各種形質転換細胞系を用いた実験で、FGF21及びbetaKlothoがそれぞれの構造類似体に対する組み合わせを変えても互換性があるか否かを観察した結果、FGF21とbetaKlothoとの組み合わせが、FGFR1cまたはFGFR3cに対する作用において、特異的な組み合わせであることを見いだした。すなわち、このことは、betaKlothoこそがFGF21のFGF受容体に対する特異的な受容体補助因子であることを示すものである。
そして、本発明者らは、上記のFGFR1c遺伝子もしくはFGFR3c遺伝子と共にbetaKlotho遺伝子を導入して細胞表面に発現させた形質転換細胞系が、FGF受容体とbetaKlotho共存下で発揮されるFGF21によるFGF受容体の活性化の程度をきわめて鋭敏に示すものであることに着目して、当該形質転換細胞系を用いた、betaKlotho活性を昂進又は抑制する物質のスクリーニング系、FGF21様物質のスクリーニング系を提供し、また、FGFR1c遺伝子もしくはFGFR3c遺伝子のみを導入して細胞表面に発現させた形質転換細胞系を用いた、betaKlotho様の活性を示す物質のスクリーニング系を提供することができた。
以上のように、本発明者らは、betaKlothoの有無により、FGF21のFGF受容体を介した生物活性が制御できるという知見を得て、betaKlotho及びその活性の昂進又は阻害物質を用いたFGF受容体を介したFGF21活性の調節剤、医薬組成物についての本発明を完成し、同時にその際に用いた優れた形質転換細胞系を利用した各種スクリーニングに関する本発明を完成するに至った。
【0008】
すなわち、本発明は以下を包含する。
(1) betaKlotho活性を有する蛋白質を有効成分として含む、FGF受容体を介したFGF21の生物活性を増強するか又は抑制するための、FGF21活性の調節剤。
(2) betaKlotho活性を有する蛋白質が、betaKlothoの可溶性部分タンパク質である、前記(1)に記載のFGF21活性の調節剤。
(3) betaKlotho活性を有する蛋白質をコードするDNAを含む組換えDNAを有効成分として含む、FGF受容体を介したFGF21の生物活性を増強するか又は抑制するための、FGF21活性の調節剤。
(4) betaKlotho活性を有する蛋白質をコードするDNAが、betaKlothoの可溶性部分タンパク質をコードするDNAである、前記(3)に記載のFGF21活性の調節剤。
(5) betaKlotho活性の昂進物質又はbetaKlotho活性の阻害物質を有効成分として含むことを特徴とする、betaKlotho活性を有する蛋白質によるFGF受容体を介したFGF21活性の調節作用を増強するか又は抑制するためのFGF21活性の調節剤。
(6) betaKlotho活性の昂進物質がヘパリン又はヘパリン様物質である、前記(5)に記載のFGF21活性の調節剤。
(7) betaKlotho活性の昂進物質又は阻害物質が抗betaKlotho抗体である、前記(5)に記載のFGF21活性の調節剤。
(8) FGF受容体がFGFR1c又はFGFR3cである前記(1)ないし(7)のいずれかに記載のFGF21活性の調節剤。
(9) FGF受容体を介したFGF21の生物活性が、グルコース取り込み昂進、血糖値低下、グルコーストランスポーター発現昂進、インスリン発現昂進、及びメタボリズム制御のいずれかである、前記(1)ないし(8)のいずれかに記載のFGF21活性の調節剤。
(10) betaKlotho活性を有する蛋白質又は当該蛋白質をコードするDNAを含む組換えDNA、を有効成分として含む、FGF受容体を介したFGF21活性を調節するための医薬組成物。
(11) betaKlotho活性を有する蛋白質が、betaKlothoの可溶性部分タンパク質である、前記(10)に記載の医薬組成物。
(12) betaKlotho活性の昂進物質又はbetaKlotho活性の阻害物質を有効成分として含むことを特徴とする、betaKlotho活性を有する蛋白質によるFGF受容体を介したFGF21活性を調節するための医薬組成物。
(13) betaKlotho活性の昂進物質がヘパリン又はヘパリン様物質である、前記(12)に記載の医薬組成物。
(14) betaKlotho活性の昂進物質又は阻害物質が抗betaKlotho抗体である、前記(12)に記載の医薬組成物。
(15) betaKlotho活性を有する蛋白質又は当該蛋白質をコードするDNAを含む組換えDNAと共に、betaKlotho活性の昂進物質又はbetaKlotho活性の阻害物質を有効成分として含むことを特徴とする、前記(10)ないし(14)のいずれかに記載の医薬組成物。
(16) さらにFGF21を有効成分として含む、前記(10)ないし(15)のいずれかに記載の医薬組成物。
(17) FGF受容体がFGFR1c又はFGFR3cである、前記(10)ないし(16)のいずれかに記載の医薬組成物。
(18) FGF受容体を介したFGF21活性を調節するための医薬組成物が、グルコース取り込み量調節、血糖値調節、グルコーストランスポーター発現量調節、インスリン発現量調節のいずれかのための医薬組成物である、前記(10)ないし(17)のいずれかに記載の医薬組成物。
(19) 前記(1)ないし(9)のいずれかに記載のFGF21活性の調節剤を用いることを特徴とする、FGF受容体を介したFGF21の生物活性を増強するか又は抑制する方法。
(20) FGF受容体がFGFR1c又はFGFR3cである前記(19)に記載のFGF受容体を介したFGF21の生物活性を増強するか又は抑制する方法。
(21) FGF21活性の調節剤を、FGF21と共にFGF受容体に対して作用させることにより、FGF受容体の活性化を昂進又は抑制することを特徴とする、前記(19)又は(20)に記載のFGF受容体を介したFGF21の生物活性を増強するか又は抑制する方法。
(22) FGF21活性の調節剤を、FGF21及びbetaKlothoの共存下でFGF受容体に対して作用させることにより、FGF受容体の活性化を昂進又は抑制することを特徴とする、前記(19)又は(20)に記載のFGF受容体を介したFGF21の生物活性を増強するか又は抑制する方法。
(23) FGF受容体及びbetaKlothoを内在的に発現していない細胞にFGF受容体遺伝子を導入し、細胞表面にFGF受容体を発現させた形質転換細胞を用いる、FGF受容体を介したFGF21の生物活性を増強もしくは抑制する物質のスクリーニング方法であって、
細胞表面にFGF受容体を発現させた上記形質転換細胞の培養系に対して披検物質と共にFGF21を作用させる工程、又は、
FGF受容体遺伝子と共に披検物質遺伝子を上記FGF受容体及びbetaKlothoを内在的に発現していない細胞に導入して、FGF受容体と同時に披検物質も細胞表面に発現させた形質転換細胞の系に対してFGF21を作用させる工程
を含むことを特徴とする、スクリーニング方法。
(24) 前記FGF受容体及びbetaKlothoを内在的に発現していない細胞がBaF3細胞であることを特徴とする、前記(23)に記載のスクリーニング方法。
(25) 前記FGF受容体がFGFR1c又はFGFR3cである、前記(23)又は(24)に記載の、FGF21の生物活性を増強もしくは抑制する物質のスクリーニング方法。
(26) FGF受容体及びbetaKlothoを内在的に発現していない細胞にFGF受容体遺伝子を導入し、細胞表面にFGF受容体を発現させた形質転換細胞系及びFGF21を組み合わせてなる、FGF受容体を介したFGF21の生物活性を増強もしくは抑制する物質のスクリーニング用のキット。
(27) 前記FGF受容体及びbetaKlothoを内在的に発現していない細胞がBaF3細胞であることを特徴とする、前記(26)に記載のスクリーニング用のキット。
(28) 前記FGF受容体がFGFR1c又はFGFR3cである、前記(26)又は(27)に記載の、FGF21の生物活性を増強もしくは抑制する物質のスクリーニング用のキット。
(29) FGF受容体及びbetaKlothoを内在的に発現していない細胞にFGF受容体遺伝子及びbetaKlotho遺伝子を導入し、細胞表面にFGF受容体及びbetaKlothoを発現させた形質転換細胞の培養系を用いる、FGF受容体を介したFGF21活性の調節作用に係るbetaKlotho活性の昂進物質もしくは阻害物質のスクリーニング方法であって、
細胞表面にFGF受容体及びbetaKlothoを発現させた上記形質転換細胞の系に対して披検物質と共にFGF21を作用させる工程、又は、
上記FGF受容体及びbetaKlothoを内在的に発現していない細胞を宿主としてFGF受容体遺伝子及びbetaKlotho遺伝子と共に披検物質遺伝子を導入して、FGF受容体及びbetaKlothoと同時に披検物質も細胞表面に発現させた形質転換細胞の系に対してFGF21を作用させる工程
を含むことを特徴とする、スクリーニング方法。
(30) 前記FGF受容体及びbetaKlothoを内在的に発現していない細胞がBaF3細胞であることを特徴とする、前記(29)に記載のスクリーニング方法。
(31) 前記FGF受容体がFGFR1c又はFGFR3cである、前記(29)又は(30)に記載の、FGF21活性の調節作用に係るbetaKlotho活性の昂進物質もしくは阻害物質のスクリーニング方法。
(32) FGF受容体及びbetaKlothoを内在的に発現していない細胞にFGF受容体遺伝子及びbetaKlotho遺伝子を導入し、細胞表面にFGF受容体及びbetaKlothoを発現させた形質転換細胞の系、及びFGF21を組み合わせてなる、FGF受容体を介したFGF21活性の調節作用に係るbetaKlotho活性の昂進物質もしくは阻害物質のスクリーニング用のキット。
(33) 前記FGF受容体及びbetaKlothoを内在的に発現していない細胞がBaF3細胞であることを特徴とする、前記(32)に記載のスクリーニング用のキット。
(34) 前記FGF受容体がFGFR1c又はFGFR3cである、前記(32)又は(33)に記載の、FGF21活性の調節作用に係るbetaKlotho活性の昂進物質もしくは阻害物質のスクリーニング用のキット。
(35) FGF受容体及びbetaKlothoを内在的に発現していない細胞にFGF受容体遺伝子及びbetaKlotho遺伝子を導入し、細胞表面にFGF受容体及びbetaKlothoを発現させた形質転換細胞の系に対して披検物質を作用させる工程を含むことを特徴とする、FGF21様活性物質のスクリーニング方法。
(36) 前記FGF受容体及びbetaKlothoを内在的に発現していない細胞がBaF3細胞であることを特徴とする、前記(35)に記載のスクリーニング方法。
(37) 前記FGF受容体がFGFR1c又はFGFR3cである、前記(35)又は(36)に記載の、FGF21様活性物質のスクリーニング方法。
(38) FGF受容体及びbetaKlothoを内在的に発現していない細胞にFGF受容体遺伝子及びbetaKlotho遺伝子を導入し、細胞表面にFGF受容体及びbetaKlothoを発現させた形質転換細胞の系を用いることを特徴とする、FGF21様活性物質のスクリーニング用のキット。
(39) 前記FGF受容体及びbetaKlothoを内在的に発現していない細胞がBaF3細胞であることを特徴とする、前記(38)に記載のスクリーニング用のキット。
(40) 前記FGF受容体がFGFR1c又はFGFR3cである、前記(38)又は(39)に記載のFGF21様活性物質のスクリーニング用のキット。
(41) 前記(23)ないし(25)のいずれかに記載のスクリーニング方法により得られた、FGF受容体を介したFGF21の生物活性を増強もしくは抑制する物質を有効成分として含むことを特徴とする、FGF受容体とbetaKlothoの共存下で発揮される、FGF受容体を介したFGF21の生物活性を増強もしくは抑制するための、FGF21活性の調節剤。
(42) 前記(29)ないし(31)のいずれかに記載のスクリーニング方法により得られた、FGF受容体を介したFGF21活性の調節作用に係るbetaKlotho活性の昂進物質もしくは阻害物質を有効成分として含むことを特徴とする、FGF受容体とbetaKlothoの共存下で発揮される、FGF受容体を介したFGF21の生物活性を増強もしくは抑制するための、FGF21活性の調節剤。
(43) 前記(35)ないし(37)のいずれかに記載のスクリーニング方法により得られたFGF21様活性物質を有効成分とし、betaKlothoの共存下でFGF受容体に作用させることを特徴とする、FGF受容体活性化剤。
(44) 前記(23)ないし(25)のいずれかに記載のスクリーニング方法により得られた、FGF受容体を介したFGF21の生物活性を増強もしくは抑制する物質を有効成分として含むことを特徴とする、FGF受容体とbetaKlothoの共存下で発揮される、FGF受容体を介したFGF21活性を調節するための医薬組成物。
(45) 前記(29)ないし(31)のいずれかに記載のスクリーニング方法により得られた、FGF受容体を介したFGF21活性の調節作用に係るbetaKlotho活性の昂進物質もしくは阻害物質を有効成分として含むことを特徴とする、FGF受容体とbetaKlothoの共存下で発揮される、FGF受容体を介したFGF21活性を調節するための医薬組成物。
(46) 前記(35)ないし(37)のいずれかに記載のスクリーニング方法により得られたFGF21様活性物質を有効成分とし、betaKlothoの共存下でFGF受容体に作用させることを特徴とする、FGF受容体を活性化するための医薬組成物。
【発明の効果】
【0009】
本発明によれば、betaKlotho活性を有する蛋白質を用いてFGF受容体を介したFGF21活性を調節し、FGF受容体下流のシグナルを惹起もしくは抑制することでFGF21とFGF受容体との相互作用に基づく様々な反応を制御することができる。また、本発明によれば、betaKlotho活性を有する蛋白質、又はbetaKlotho活性の活性増強物質もしくは阻害物質を投与することでFGF受容体を介したFGF21活性、たとえば血糖値の低下作用を調節することができるので、betaKlotho活性を有する蛋白質、又はbetaKlotho活性の活性増強物質もしくは阻害物質をそれぞれ単独でもしくは組み合わせて、又はさらにFGF21と併用して血糖値の異常に関連した疾患、たとえば糖尿病の治療又は予防用医薬組成物を提供することができる。
また、FGF21様活性物質、betaKlothoの代替物質、及びbetaKlotho活性の活性増強物質もしくは阻害物質に対する優れたスクリーニング系も提供できる。
【図面の簡単な説明】
【0010】
【図1】FGF21の脂肪細胞特異的活性の検出の結果を示す図である。 図中、GLUT1 mRNAの量はそれぞれの細胞でFGF21未刺激時の値を1とした相対値で表した。●は分化前の繊維芽細胞を、○は分化後の脂肪細胞を示す。 FGF21が3T3-L1細胞の分化後の脂肪細胞に対して活性を発揮し、GLUT1発現を増加させることが示された。繊維芽細胞に対して活性を発揮しないことが示された。
【図2】脂肪細胞の分化に伴うFGF受容体の発現変動の検出した結果を示す特性図である。 図中、横軸は繊維芽細胞から脂肪細胞への分化誘導日数を表し、縦軸はFGF受容体mRNAの発現量を表す。 FGFR1とFGFR2が高いレベルで発現していたが、FGFR3とFGFR4は非常に発現レベルが低かった。FGFR1、FGFR2ともに、細胞における発現レベルは分化とともにやや減る傾向が認められた。
【図3】脂肪細胞の分化に伴うbetaKlothoの発現変動の検出の結果を示す図である。 図中、横軸は繊維芽細胞から脂肪細胞への分化誘導日数を表し、縦軸はbetaKlotho、又はKlothoのmRNAの発現量を表す。 脂肪細胞分化に伴って発現するようになるbetaKlothoが脂肪細胞の機能と密接に関連することが示唆された。
【図4A】FGF21によるFGF受容体下流シグナル惹起にbetaKlothoが必要であることの解析の結果を示す図であり、図中、CL#1,#7、#10はFGFR1cとbetaKlothoの両方のcDNAを導入し形質転換したBaF3細胞クローンの名称である。下段は、それぞれの細胞から調製したライゼートを材料として、実際にタンパクとして発現しているbetaKlothoの量を、市販の抗betaKlotho抗体を用いたウェスタンブロット解析により比較した結果である。
【図4B】FGF21の量を変化させてCL#1の細胞を刺激した際のDNA合成を測定した結果であり、○はheparin存在下、●は非存在下を示す。
【図4C】FGFR1cを発現していないがbetaKlothoのみを発現しているBaF3細胞を用い、チミジン取り込み能を測定した。□はheparin存在下を、■は非存在下を示す。図4A〜Cの結果により、FGFR1cは発現していてもbetaKlothoを発現していない細胞ではFGF21に反応せず、betaKlothoを同時に発現している細胞でのみFGF21に反応する一方、FGFR1cを発現しておらずbetaKlothoのみを発現しているBaF3細胞はFGF21に対して反応しないことが示され、FGF21と反応するためには、betaKlothoとFGFR1cとの両者が必要であることが示された。
【図5】FGF21の活性発揮においてbetaKlothoとともに存在するFGF受容体の特異性の解析の結果を示す図である。 betaKlothoを発現しているBaF3細胞にFGF受容体(FGFR1b、R1c、R2b、R2c、R3b、R3c、R4)の発現ベクターを導入し、各betaKlotho/FGFR発現細胞をheparin存在下又は非存在下FGF21、FGF1で刺激してチミジンの取り込み能を測定した。 FGFR1cのほか、FGFR3cでもFGF21に反応することが示された。また、従来型のFGF(典型的にはFGF1)ではheparinの存在で急激に刺激伝達活性が高まるが、FGF21の場合はheparinの存在、非存在がそれほど活性に影響していない。
【図6】FGF21の活性発揮においてbetaKlothoとともに存在するFGF受容体の特異性をFGF19およびFGF1と比較した解析の結果を示す図である。 FGFR(FGFR1c、R2c、R3c、R4)を発現しているBaF3細胞にbetaKlothoの発現ベクターを導入し、各FGFR/betaKlotho発現細胞をheparin存在下でFGF21、FGF1及びFGF19で刺激して、チミジンの取り込み能を測定した。図中、○はFGF21、●はFGF19、▲はFGF1で刺激したことを表す。 FGF19はFGF1と同様、FGFR1c、FGFR2c、FGFR3c、FGFR4のいずれの受容体とも反応する。これに対しFGF21はFGFR1c とFGFR3cのみに反応する。高濃度のFGF21刺激によりFGFR2cに対してわずかに反応がみられる。
【図7】FGF21の刺激に伴う細胞内シグナル伝達分子の活性化の解析の結果を示す図である。 各FGFR/betaKlotho発現細胞をFGF21、FGF1及びFGF19で刺激して、FGFR下流シグナル伝達物質であるFRS2及びMARKのリン酸化を、前者では抗リン酸化FRS2抗体を用いて、後者では抗リン酸化ERK1/2抗体を用いて測定した。図中、WEHIはIL-3分泌細胞であるWEHIの培養上清を添加したことを示す。 FGF19とFGF1は、FGFR1c、FGFR2c、FGFR3c、FGFR4のいずれの受容体をbetaKlothoとともに発現する細胞のFRS2のリン酸化とMAPKのリン酸化を増加させる。これに対し、FGF21はFGFR1c/betaKlotho発現細胞とFGFR3c/betaKlotho発現細胞のみ、FRS2およびMAPKのリン酸化を増加させる。
【図8】ヒトのアミノ酸配列をもつヒト型FGFR1cとヒト型betaKlothoを共発現させた細胞に対するヒト型FGF21の活性の解析の結果を示す図である。 ヒト型FGFR1c(hR1c)発現BaF3細胞あるいはヒト型FGFR1c/ヒト型betaKlotho(hβKlotho)発現細胞を10μg/mlのheparin存在下でヒト型FGF21(hFGF21)あるいはFGF1で刺激して、チミジンの取り込み能を測定した。図中、●はhFGF21、▲はFGF1で刺激したことを表す。FGF1は両方の細胞に作用するが、ヒト型FGF21は、ヒト型FGFR1c/ヒト型betaKlotho発現細胞のみに作用して用量依存的にチミジンの取り込みを増加させた。
【図9】ヒトのアミノ酸配列をもつヒト型FGFR3cとヒト型betaKlothoを共発現させた細胞に対するヒト型FGF21の活性の解析の結果を示す図である。 ヒト型FGFR3c(hR3c)発現BaF3細胞あるいはヒト型FGFR3c/ヒト型betaKlotho(hβKlotho)発現細胞を10μg/mlのheparin存在下でヒト型FGF21(hFGF21)あるいはFGF1で刺激して、チミジンの取り込み能を測定した。図中、●はhFGF21、▲はFGF1で刺激したことを表す。FGF1は両方の細胞に作用するが、ヒト型FGF21は、ヒト型FGFR3c/ヒト型betaKlotho発現細胞のみに作用して用量依存的にチミジンの取り込みを増加させた。
【図10】FGF21の活性発揮においてFGF受容体とともに存在するKlothoの特異性の解析した結果を示す特性図である。 FGFR1cと共にbetaKlothoを発現させたBaF3細胞及びFGFR1cと共にKlothoを発現させたBaF3細胞に対して、heparinの存在下又は非存在下でFGF21及びFGF23を添加し、チミジン取り込み能を測定した。 Klothoでは、FGF21に反応しないことが示された。さらに、FGFR1cとKlothoの存在下で反応性を発揮するFGF23について、調べたところ、FGFR1cとbetaKlothoを発現していても、FGF23には反応しなかった。
【図11】BetaKlothoとFGF21あるいはFGF受容体との複合体形成の検出の結果を示す図である。 heparinの存在あるいは非存在の条件下で、可溶性betaKlothoタンパク質とFGF21をインキュベートした後、抗V5タグ抗体固定化ビーズを用いてbetaKlothoを沈降させ、FGF21がbetaKlothoに結合して共沈降するか否かを、免疫ブロッティング法にて解析した betaKlothoとFGF21はheparinの有無によらず物理的に複合体を形成する。またbetaKlothoはFGFR1c、FGFR2c、FGFR3c、FGFR4のいずれの受容体とも物理的に複合体を形成する。
【図12】BetaKlothoを強制発現させた未分化3T3-L1細胞をFGF21刺激した際のシグナル伝達の解析の結果を示す図である。 betaKlothoを発現させた3T3-L1繊維芽細胞をFGF21で刺激し、FGFRの下流のシグナル伝達分子であるFRS2とMAPKの活性化(リン酸化)をウエスタンブロッティング法で調べた。図中、PBSはダルベッコ改変リン酸緩衝生理食塩液を10μl、SU5402はFGF受容体チロシンキナーゼ阻害剤であって、系に10μM存在させることを表す。また、FGF21の添加濃度は1.1μg/ml及び4.5μg/mlの2通りで行った。 betaKlothoを発現していない未分化3T3-L1細胞はFGF21で刺激してもFRS2のリン酸化およびMAPKのリン酸化の状態は変化しない。betaKlothoを強制的に発現させた未分化3T3-L1細胞では、FGF21刺激に応じてFRS2のリン酸化およびMAPKのリン酸化が増大する。FGF受容体チロシンキナーゼ阻害剤SU5402の処理により、FGF21刺激に伴うFRS2およびMAPKのリン酸化の程度は減少する。
【図13】BetaKlothoを強制発現させた未分化3T3-L1細胞をFGF21刺激した際のGLUT1mRNAの発現解析の結果を示す図である。 betaKlothoを発現させた3T3-L1繊維芽細胞をFGF21で刺激しGLUT1のmRNA量の変化を調べた。コントロールとしてはbetaKlothoのcDNAを挿入していない空のベクターを導入した3T3-L1繊維芽細胞を用いた。 betaKlothoを発現していない未分化3T3-L1細胞はFGF21で刺激してもGLUT1mRNAの発現量は変化しない。betaKlothoを強制的に発現させた未分化3T3-L1細胞では、刺激したFGF21の濃度に応じてGLUT1mRNAの発現量が増加する。
【図14】本発明を説明する概念図。 図中、aはFGF1、FGF2などの多くのFGF、bはFGF21、cはFGF受容体、dはbetaKlotho、eはヘパリンあるいはヘパリン様糖鎖を示す。またFGF21、FGF受容体、betaKlothoの間の活性複合体の形成を、増強あるいは減弱する物質(群)をfで示す。I.従来の研究で知られていた、FGF1など多くのFGFがbetaKlothoの非存在下でFGF受容体を活性化しうる作用機構: (1)FGFリガンドが単独にFGF受容体に結合しただけでは受容体の活性化は起こらない。 (2)さらにヘパリン様の糖鎖が共存するとシグナルを惹起しうる活性複合体が形成されて、それを起点としたシグナルの伝達が行われて細胞の応答(例えば細胞の増殖など)が起きる。II.本発明において明らかにした、betaKlothoが存在することによって、FGF受容体の活性化が起きるFGF21の作用機構: (3)補助因子BetaKlothoが加わり、リガンド(FGF21)、FGF受容体、BetaKlothoの3者が存在することによって、FGF21によるFGF受容体の活性化が起きる。 (4)このシグナルを惹起しうる活性複合体形成を、増強あるいは減弱する物質は、FGF21の作用を昂進あるいは抑制する作用を有する。
【発明を実施するための形態】
【0011】
以下、本発明の実施の態様に従って、本発明を詳細に説明するが、本発明はこの実施の態様に限られない。
〔1〕betaKlothoについて
本発明は、betaKlothoの生体内での作用機能を解明し、その利用法を提供するものである。
betaKlothoは、Klothoのホモログとしてクローニングされた物質であり(非特許文献8)、マウス由来betaKlotho遺伝子の塩基配列は配列番号1、対応するアミノ酸配列は配列番号2として示され、ヒトbetaKlothoの塩基配列は配列番号7、対応するアミノ酸配列は配列番号8として示される。発達中のマウス胎児の脂肪組織などで発現していることが知られており、他生物種からの遺伝子取得方法と共に、種々の改変体、変異体などの製造方法も記載されている(非特許文献7,8、特許文献1)。
そして、上記非特許文献7および特許文献1において、遺伝子をホモで欠いたノックアウトマウスの作成とその解析の結果、betaKlotho遺伝子が「コレステロールの代謝に関与する遺伝子」の1つであり、これを用いてコレステロールの代謝を促進する物質を得るとする記述がされているものの、血糖値の調節に関連した機能にも、FGF21活性との関係についても教示する記述はない。
一方で、本発明においてはじめて、betaKlothoが、FGF21がFGF受容体と結合してFGF受容体を活性化させ、そのシグナル伝達を引き起こすために必須の物質であることが解明され、betaKlothoこそが、FGF21活性として知られていた、グルコーストランスポーターの発現量の昂進、グルコース取り込み上昇、血糖値の低下、インスリンの合成量増加などの作用において重要な調節機能を果たす因子であることが見出されたものである。
本発明においては、「betaKlotho活性」というとき、betaKlothoが有している活性である、FGF21とFGF受容体と共存して相互作用することでFGF受容体を介したシグナル伝達系を活性化させる作用、すなわち「FGF受容体を介したFGF21活性を調節する機能・活性」を表す。同様に、「betaKlotho活性を有する蛋白質」とは、betaKlotho及びその可溶性部分蛋白質のみならず、「FGF受容体を介したFGF21活性を調節する機能・活性を有する蛋白質」をさす。
betaKlothoは、betaKlotho活性にかかわる長い細胞外ドメインと短い細胞内ドメインが膜貫通ドメインでつながれた、典型的なアンカータンパク質構造をしている。細胞外ドメインのみからなる可溶性betaKlothoも「betaKlotho活性を有するタンパク質」として機能するし、さらに短い部分配列であってもbetaKlotho活性を有していれば、本発明のbetaKlotho活性を有する蛋白質として用いることができる。
【0012】
本明細書中ではbetaKlotho活性を有する蛋白質の典型例として、典型的なbetaKlothoタンパク質である、マウス全長betaKlothoタンパク質のアミノ酸配列を配列番号2として、その可溶性betaKlothoタンパク質のアミノ酸配列を配列番号3として、また、ヒトbetaKlothoアミノ酸配列を配列番号8として、可溶性ヒトbetaKlothoタンパク質を配列番号9として例示するが、これらアミノ酸配列において1個若しくは数個のアミノ酸に欠失、置換、付加等の変異が生じてもよい。また、カルボキシ末端にFlagタグ配列(配列番号4)、V5タグ配列(配列番号5)、Hisタグ配列(配列番号6)あるいはこれらの組み合わせ配列を付加してもよい。さらに、マウスbetaKlothoタンパク質とヒトbetaKlothoタンパク質のアミノ酸配列レベルでの相同性は78%であり、ヒトbetaKlothoタンパク質とマウスbetaKlothoタンパク質とで同等の機能を有しているので、進化上マウス−ヒト間程度の距離内に存在する生物種に由来するbetaKlothoも、同等のbetaKlotho活性を有しているものと期待できる。したがって、本発明におけるbetaKlothoタンパク質としては、マウス由来、及びヒト由来のbetaKlothoタンパク質だけではなく、配列番号2または配列番号8とアミノ酸の相同性が、78%以上、好ましくは85%以上、より好ましく90%以上の他の生物種のbetaKlothoタンパク質も含まれる。具体的には、「配列番号2、配列番号3、配列番号8及び配列番号9のいずれかに示されるアミノ酸配列、又はその部分断片からなるタンパク質」、「配列番号2、配列番号3、配列番号8及び配列番号9のいずれかに示されるアミノ酸配列において、1個もしくは数個のアミノ酸が欠失、置換又は付加されたアミノ酸配列からなるタンパク質」、「配列番号2、配列番号3、配列番号8及び配列番号9のいずれかに示されるアミノ酸配列と78%以上の相同性を有するアミノ酸配列からなるタンパク質」であってbetaKlotho活性を有するタンパク質も本発明における「betaKlothoタンパク質」に相当する。なお、本発明における「相同性」とは、Lipman-Pearsonの計算法(Lipman and Pearson,1985;Science 227,1435-1441)での「同一性」を示す換算数値である。
【0013】
また、典型的なbetaKlotho活性を有する蛋白質をコードするDNAとしては、マウス由来betaKlotho遺伝子(配列番号1)及びヒト由来betaKlotho遺伝子(配列番号7)があげられるが、他の生物種由来の既知のbetaKlotho遺伝子も同様に用いることができ、未知のbetaKlothoの遺伝子であっても、マウス由来またはヒト由来のbetaKlothoをコードするDNAの塩基配列(マウス由来塩基配列は配列番号1として示される。)に基づいて設計されたプローブもしくはプライマーを用いて各生物種由来のDNAライブラリーからPCR法やハイブリダイゼーション法により取得することができる。
そして、これらのbetaKlotho活性を有する蛋白質をコードするDNAに対して部位突然変異法やランダム変異導入法などで得られた突然変異DNAを用いることができる。さらに、それぞれの可溶性断片などの部分配列も「betaKlotho活性を有する蛋白質をコードするDNA」として用いることができる。
具体的には、「betaKlotho活性を有する蛋白質をコードするDNA」として、「配列番号1もしくは配列番号7に示される塩基配列からなるDNA又はその部分断片からなるDNA」、「配列番号1又は配列番号7の相補配列からなるDNAとストリンジェントな条件下でハイブリダイズするDNA」及び「配列番号2、配列番号3、配列番号8及び配列番号9のいずれかに示されるアミノ酸配列をコードするDNA」であって、「betaKlotho活性を有する蛋白質をコードするDNA」などがあげられる。
なお、上記ストリンジェントな条件とは、高い緊縮条件下であることをいい、例えば2×SSC、1×デンハルト溶液中で60℃程度の条件である。
そして、これらbetaKlothoタンパク質をコードするDNAを含む組換えDNAを用いることで、定法どおり、哺乳動物細胞宿主系などを用いた形質転換体の発現産物としてbetaKlothoタンパク質を取得することができる。また、当該「組換えDNA」は、それ自体でFGF21活性の調節剤として用いることができるが、ここで、「組換えDNA」の用語は、形質転換宿主内で発現するためのプロモーターなどと機能的に連結されていることを表し、典型的にはヒトなど哺乳動物への投与が可能なレトロウイルスなどの発現ベクターである。
また、betaKlothoの天然蛋白質の場合も、betaKlotho遺伝子を含む発現ベクターなどで形質転換した哺乳動物細胞などにより産生される組換え蛋白質の場合も、限外ろ過法、ゲルろ過法、アフィニティークロマトグラフィーなど、またはこれらを組合せた方法などの周知のタンパク質精製方法により、必要に応じて精製して用いることができる。
以下、本発明においては、各種betaKlotho活性を有する蛋白質、またはbetaKlotho活性を有するその部分断片もしくはその変異体などの同等物を含め、単に「betaKlotho」、「betaKlothoタンパク質」ともいい、これらbetaKlotho活性を有する蛋白質をコードするDNAを、単に「betaKlothoをコードするDNA」、「betaKlotho遺伝子」ともいう。
【0014】
〔2〕FGF21活性が脂肪細胞で特異的に検出されることについて
1.FGF21について
FGF21は、FGF19の配列を基にデザインしたプライマーを用いたPCRにより発見された物質である(非特許文献9)。脂肪細胞にグルコーストランスポーター(GLUT)の発現を上昇させる活性やランゲルハンス島でのインスリン合成促進活性があることも報告されている(非特許文献1、6)。そして、脂肪細胞表面のFGFR1及びFGFR2を活性化することが確認されている(非特許文献1)ことから、これら活性は、FGF受容体を介して、細胞内にシグナル伝達されて引き起こされると解される。
本発明において「FGF受容体を介したFGF21活性」又は「FGF受容体を介したFGF21の生物活性」というとき、FGF受容体、特にFGFR1cあるいはFGFR3cを介して細胞内に引き起こされる活性をいい、たとえば脂肪細胞内でグルコーストランスポーター(GLUT)の発現を上昇させる活性を指す。また、本発明におけるFGF21としては、ヒト由来のFGF21(例えば、GenBankアクセッション番号:AB021975参照)などの哺乳類由来FGF21が好ましいが、これに限られるものでなく、天然タンパク質のみならず、組換えFGF21であってもよく、さらに上記FGF21活性を有していれば、そのアミノ酸配列の一部が改変されたものであってもよい。組換えFGF21を作製するためには、大腸菌や哺乳動物細胞などの通常の形質転換宿主/ベクター系を適宜用いることができる。
【0015】
2.FGF受容体を介したFGF21活性の測定
一般に、各FGFは、細胞表面のFGF受容体に結合し、FGF受容体を活性化させ、細胞内での各種シグナル伝達機構を活性させることで、当該細胞に何らかの作用・機能を引き起こすことになるため、当該観察された作用・機能をFGF活性として測定することになる。
汎用的な測定法としては、Ornitz,D.M.,Xu,J.,Colvin,J.S.,McEwen,D.G.,MacArthur,C.A.,Coulier,F.,Gao,G.and Goldfarb,M.(1996) Receptor specificity of the fibroblast growth factor family.J Biol Chem,271,15292-15297.に記載されている細胞増殖刺激活性測定法が挙げられる。その概要は以下の通りである。表面にFGF受容体を有する培養細胞の培養液内に測定対象のFGFを加え、一定時間培養後標記チミジンを培養液中添加しさらに一定時間培養を行う。この間の標識チミジンの高分子DNAへの取り込みを測定することで、FGFによるDNA合成の活性化程度を評価する。(本発明でも、実施例4,5,6,8,9及び10においてFGF21活性を測定する際にこの手法を採用した。)
上述のように、FGF21は細胞表面のFGF受容体、特にFGFR1cを介して細胞内のグルコーストランスポーター(GLUT)発現を上昇させる活性を有しているので、本発明に係るFGF21活性の解析において、実施例1では典型的なFGF21活性の披検細胞内でのGLUTの発現の上昇能をGLUT1のmRNAの発現量として測定した。
ここで、FGF21を様々な濃度で細胞培養液に添加することにより、細胞に作用させて一定時間細胞培養を継続した後、細胞からmRNAを抽出し、GLUT1のmRNAの発現量を測定する。このmRNA発現量が増加していれば、FGF21が活性を発揮していることが示される。
GLUT1の発現上昇を解析する目的には、GLUT1抗体などを用いたウェスタンブロット法等の公知の方法によりGLUT1タンパク質発現量を測定することもできる。また、実験動物などの場合、GLUT1の転写を制御する制御領域(例えば、プロモーター領域)と、その下流に転写制御されうる形で連結されたリポーター遺伝子と組み込んだトランスジェニック動物を作製することにより、リポーター遺伝子の発現量によってFGF21活性を測定できるようにしてもよい。
なお、FGF21活性を測定するためには、これらの手法に限られることはなく、定量的な測定が可能な手法であればどのような手法でも用いることができる。たとえばFGF受容体の活性化やその下流のシグナル伝達分子の活性化をFGF受容体やシグナル伝達分子のリン酸化の増加として測定し評価する手法も用いることができる。
そして、本発明のFGF21活性のGLUT1mRNA発現量の測定の好ましい態様として、培養脂肪細胞を披検細胞として用いることができる。
培養脂肪細胞としては、ヒト、ラット初代培養細胞など各種の培養脂肪細胞を用いることができるが、本発明の実施の態様では、培養条件を操作することにより脂肪細胞に分化する能力を有する繊維芽細胞として広く当該研究領域で使用されている3T3-L1細胞を用いた。3T3-L1細胞は、分化誘導前には繊維芽細胞としての性質を有し、誘導後には脂肪細胞としての性質を有するので、FGF21を作用させた場合の効果を対比させて解析することができる。ここで「脂肪細胞の性質をもつ細胞」とは、脂肪細胞特有の細胞内代謝を行い、脂肪滴を多量に蓄積した細胞質を有する細胞を示す。3T3-L1細胞を脂肪細胞に誘導する実験系は、例えば、Kharitonenkov,A.,Shiyanova,T.L.,Koester,A.,Ford,A.M.,Micanovic,R.,Galbreath,E.J.,Sandusky,G.E.,Hammond,L.J.,Moyers,J.S.,Owens,R.A.,Gromada,J.,Brozinick,J.T.,Hawkins,E.D.,Wroblewski,V.J.,Li,D.S.,Mehrbod,F.,Jaskunas,S.R.,and Shanafelt,A.B.(2005) J Clin Invest 115(6),1627-1635に開示されている。
【0016】
3.FGF21を作用させることによるGLUT発現量の増大は脂肪細胞で特異的に検出されること
3T3-L1細胞に対して脂肪細胞に誘導分化処理を施し、誘導後の脂肪細胞の性質を有する細胞と、誘導前の繊維芽細胞としての性質を有する細胞とを用意し、FGF21の濃度を変えて作用させ、各濃度でのGLUT1のmRNAの発現量を測定し、両者を比較した。脂肪細胞に分化した細胞ではFGF21の濃度の増加に伴いGLUT1のmRNAの発現量が増加していたことから、FGF21が脂肪細胞に対して活性を発揮し、GLUT1発現を増加させることが示された。これとは対照的に、分化していない繊維芽細胞では、FGF21の濃度を変えてもGLUT1のmRNAの発現量はほとんど変化しなかった。このことから、FGF21のGLUT1発現量の増大活性は、3T3L1細胞が脂肪細胞に分化した場合に特異的に検出されることが示された。
【0017】
〔3〕脂肪細胞の分化に伴うFGF受容体の発現変動の検出
1.FGF受容体について
FGF受容体(FGFR)は細胞表面に存在する膜一回貫通型のタンパク質である。現在ではFGFR1〜5の5種類が同定されている。そのうちFGFR1〜4はチロシンキナーゼ型受容体であり、FGFが結合することにより二量体化・自己リン酸化により活性化される。活性化されたFGFRは細胞内のシグナリング分子と相互作用し、複数のシグナル伝達経路を活性化させる。FGFR1〜3には選択的スプライシングにより主に2種類のアイソフォーム(FGFR1b、FGFR1c、FGFR2b、FGFR2c、FGFR3b、FGFR3c)が存在する。
【0018】
2.脂肪細胞の分化に伴うFGF受容体の発現量の変動
上記〔1〕3.での結果は、3T3-L1細胞の脂肪細胞への分化に伴って細胞表面にFGF受容体が発現した、もしくは発現量が増加したことにより、細胞内でのシグナル伝達量が増加したことを観察している可能性もあるため、3T3-L1細胞における脂肪細胞の誘導の各段階においての、各FGF受容体発現量を測定することとした。
3T3-L1細胞の培養条件を操作して、繊維芽細胞の性質を持つ細胞から脂肪細胞の性質をもつ細胞へと徐々に誘導し、各段階において、細胞から総RNAを抽出し、そこでのFGF受容体mRNAの発現量を測定することにより、脂肪細胞の分化に伴うFGF受容体の発現変動を検出した。
その結果、脂肪細胞への分化の有無にかかわらずFGFR1とFGFR2は高レベルで発現しているのに対して、FGFR3とFGFR4の発現レベルは非常に低かった。そして、分化に伴って発現が増える受容体はなく、むしろ高レベルで発現していたFGFR1、FGFR2のいずれの受容体も、分化とともに、発現レベルがやや減る傾向が認められた。
【0019】
〔4〕脂肪細胞の分化に伴うbetaKlothoの発現変動の検出
本発明者らは、脂肪細胞へと分化する過程で脂肪細胞表面での発現量が増加する物質があれば、当該物質がFGF21と協同してFGF受容体に作用する可能性が高いと考え、発達中のマウス胎児の脂肪組織で発現することが知られていたbetaKlotho(非特許文献8、特許文献1)に着目し、3T3-L1細胞の脂肪細胞の分化に伴うbetaKlothoの発現量の変化を調べた。
具体的には、上記〔2〕3.と同様の手法により、3T3-L1細胞を脂肪細胞へ徐々に誘導し、その各段階において、細胞から総RNAを抽出し、そこでのbetaKlothoのmRNAの発現量を測定することにより、脂肪細胞の分化に伴うbetaKlothoの発現変動を検出した。
その際の対照として、betaKlothoとは別の遺伝子の発現変動を検出することができ、当該別遺伝子の方が好ましい。本発明の実施の態様としてはbetaKlothoとアミノ酸配列レベルで41%の相同性を有するKlothoの遺伝子の発現変動を対照として採用した。Klothoと比較することにより、脂肪細胞の誘導に伴うbetaKlothoの発現上昇が特徴的であることがわかる。
この結果から、脂肪細胞の誘導に伴うbetaKlothoの発現上昇が、脂肪細胞の機能と密接に関連することが示唆される。
【0020】
〔5〕FGF21の活性発揮におけるbetaKlothoの必要性の解析
1.FGF受容体を介したFGF21の活性発揮とbetaKlothoの関連性
以上の3T3-L1細胞及びその誘導分化した脂肪細胞を用いた実験結果から見て、FGF21の脂肪細胞特異的活性は、FGF受容体とbetaKlothoが共に存在する場合に誘導されることが示唆される。すなわち、3T3-L1細胞の表面に本来存在していたFGFR1cなどのFGF受容体と共に、脂肪細胞に分化誘導された場合に発現してくるbetaKlothoが、FGF21のFGF受容体を介した活性を飛躍的に増大させている可能性が強く示唆される。
このことを確かめるために、本発明の態様ではFGF受容体のみを発現させた細胞、FGF受容体とbetaKlothoを共に発現させた細胞、に対してFGF21が活性を有するか否かを解析する。この目的には、元来FGF受容体もbetaKlothoも発現していない細胞を用いることが必要である。
【0021】
2.FGF受容体及び/又はbetaKlothoを発現する披検細胞の作製方法
上記目的に適した、元来FGF受容体もbetaKlothoも発現していない細胞としては、マウス白血病ProB細胞であるBaF3細胞を用いることができる。しかし他の細胞でもこれらいずれの遺伝子も細胞表面に発現しておらず、かつこれら遺伝子で形質転換された場合に細胞表面に発現することができる細胞であれば、用いることができる。本来これら遺伝子を発現している細胞であっても、当該遺伝子がノックアウトされている細胞も用いることができる。
このようなFGF受容体もbetaKlothoも発現していない細胞を宿主として、FGF受容体遺伝子のみ、又はbetaKlotho遺伝子のみをBaF3細胞表面に発現させた細胞を、定法の遺伝子組換え技術を用いて用意すると共に、同様の手法で両遺伝子を用いてbetaKlothoとともにFGF受容体の種類を変えたBaF3細胞を作製した。
発現細胞の作製方法としては、特に限定されないが、例えば、リポフェクタミン試薬で発現プラスミドを細胞に導入し、プラスミド上の薬剤耐性遺伝子をマーカーとして用いて、この薬剤に耐性な細胞株を取得することで達成することができる。
【0022】
3.FGF21によるFGF受容体下流シグナル惹起にbetaKlothoが必要であることの解析
FGFR1cとbetaKlothoの両方のcDNAを用いてBaF3細胞を形質転換して複数のクローンを取得し、SDS-PAGEゲル上でbetaKlotho抗体により確認したところ、N型糖鎖の修飾に特徴的な、スメアー状態のブロードなバンドとして観察されたことから、betaKlothoはN型糖鎖の修飾を有する糖タンパクであると考えられる。
そして、FGF21を培養液に添加した場合に、FGFR1cは発現しているがbetaKlothoを発現していない細胞ではヘパリンの存在下あるいは非存在下に関わらずFGF21に反応せず、betaKlothoを共に発現している細胞でのみFGF21に反応することが示された。
また、FGF21の量を変化させてFGFR1cと共にbetaKlothoを発現しているBaF3細胞にヘパリンを作用させた実験結果からは、ヘパリンはFGF21の作用をさらに増強することがわかった。しかし、FGFR1cを発現していない場合の、betaKlothoのみを発現しているBaF3細胞は、ヘパリンの存在、非存在にかかわらずFGF21に対する反応性はない。
以上のことから、ヘパリンがFGF21に反応するためには、FGF受容体とbetaKlothoの両方が必要であることと共に、ヘパリン又はヘパリン様物質(通常のFGF受容体を介したFGF活性増強物質)には、betaKlotho活性を有する蛋白質のFGF21増強活性をさらに増強する効果があることが示された。
また、このことは、FGFR1cを発現しているBaF3細胞の系に披検物質を存在させてFGF21を作用させることで、披検物質にFGF21の活性を増強もしくは抑制する作用があるか否かを検出できることであり、FGFR1cと同時にbetaKlothoも発現しているBaF3細胞の系に披検物質を存在させてFGF21を作用させてみれば、披検物質にbetaKlothoのFGF21増強活性をさらに増強する作用があるか、もしくは抑制する作用があるかを検出できることを示している。すなわち、FGF受容体を発現しているBaF3細胞の系はFGF21の活性を増強もしくは抑制する物質のスクリーニングに、またFGF受容体と共にbetaKlothoも発現しているBaF3細胞の系は、FGF21活性調節作用にあるbetaKlotho活性を増強もしくは抑制する物質のスクリーニングに好適に用いられることが示されている。さらに、FGF受容体と共にbetaKlothoも発現しているBaF3細胞の系は、FGF21様活性物質のスクリーニングに用いることもできる。
そして、これらスクリーニングにより得られた物質は、いずれもFGF受容体とbetaKlothoの共存下で発揮されるFGF21活性の調節剤及び当該FGF21活性を調節するための医薬組成物、例えば血糖値調節用医薬組成物として用いることができる。
【0023】
〔6〕FGF21の活性発揮におけるbetaKlothoとともに存在するFGF受容体の特異性の解析
以上の実験結果から、FGF21とbetaKlothoが存在すれば、FGF受容体の一つであるFGFR1cを活性化してそれを発現している細胞の内部にシグナル伝達をすることができることが解明されたので、他のFGF受容体の場合についてのbetaKlotho共存効果を調べるために、betaKlothoとともに存在するFGF受容体の種類を変えたBaF3細胞を作製し、その細胞に対するFGF21の活性を評価した。その結果、FGFR1cのほか、FGFR3cをbetaKlothoと共に発現させた場合でもFGF21に反応することが明らかとなり、その場合もさらにヘパリンの存在によって反応性が増強される。
したがって、ヘパリンには、betaKlothoのFGF受容体を介したFGF21活性の増強効果をさらに増大する作用を有する。
また、ここで用いたFGF受容体と共にbetaKlothoも発現しているBaF3細胞の系は、当該細胞表面のFGF受容体及びbetaKlothoが共存した状態でFGF21を作用させることができるから、FGF21、betaKlotho及びFGF受容体の共存下で発揮されるFGF受容体を介したシグナル伝達系を昂進する物質又は抑制する物質のスクリーニングに用いることができる。
当該スクリーニングで得られた物質は、FGF21、betaKlotho及びFGF受容体の共存下で発揮されるFGF受容体を介したシグナル伝達系を昂進又は抑制するための医薬組成物(例えば、血糖値調節用医薬組成物)などとして用いることができる。すなわち、ヘパリンは上述のスクリーニング系により、betaKlothoのFGF受容体を介したFGF21活性の増強物質として得られた物質であるということができるから、ヘパリンを当該医薬組成物中の有効成分として用いる場合がその1つの具体例となる。
【0024】
〔7〕betaKlothoがFGF受容体との結合におけるFGF21に対する特異的な補助因子であることの確認
本発明によれば、FGF21とbetaKlothoが存在すれば、FGF受容体を活性化して細胞内部にシグナル伝達をすることができるが、そのFGF21とbetaKlothoとの関係が特異的なものか、もしくはそれぞれの構造的な類似性を有する物質であっても代替できるほどに融通性を有するものなのかを次に検討した。すなわち、具体的な実験としては、betaKlothoを、FGF21と構造的類似性の高いFGF23に作用させた場合と、betaKlothoと構造的類似性の高いKlothoを、FGF21に作用させた場合に、それぞれどの程度の互換性があるかを調べた。
その結果、FGF21の場合は、ヘパリンの存在、非存在にかかわりなく、Klothoを存在させてもFGFR1cには反応しないことが示され、反対に、FGF23の場合は、ヘパリンの存在、非存在にかかわりなく、betaKlothoを存在させてもFGFR1cには反応しないことが示された。また、いずれの場合もヘパリンが系に存在しているとFGFR1cへの反応性が高まる。
したがって、betaKlothoがFGF21に対する特異的な受容体結合補助因子であることが立証された。同時に、ヘパリンはその効果的な更なる増強因子であることも立証された。
【0025】
〔8〕betaKlotho及びbetaKlotho活性を昂進もしくは抑制する物質の利用
以上の結果を総合すれば、betaKlotho活性を有する蛋白質またはbetaKlotho活性を有する蛋白質を体内で発現できるbetaKlotho遺伝子は、FGF21活性調節剤として用いることができ、betaKlothoもしくはその遺伝子は、単独で、又はFGF21と共にそれを有効成分とする血糖値調節用医薬組成物として用いることができることが示された。
また、betaKlothoを本来発現している細胞、組織を対象とする場合、そのbetaKlotho活性を増強する物質、及び/又はbetaKlothoの発現を促進する物質を用いることも同様に有効である。betaKlothoタンパク質またはbetaKlotho遺伝子と併用することも可能である。
反対に、生体内のbetaKlotho活性を阻害する物質は、FGF受容体を介したFGF21活性の抑制剤などとして働くため、たとえばFGF21の典型的な活性である血糖値低下機能を抑え、血糖値上昇効果を呈することが期待できる。
生体内でbetaKlotho活性を昂進する物質として、実施例で当該性質が確認されたヘパリンが1例として挙げられるが、生体内におけるbetaKlothoの作用を増強する物質であれば特に限定されない。betaKlotho活性を昂進する物質は、betaKlothoとFGF受容体とを細胞表面に発現しているBaF3細胞などを用いた本発明のスクリーニング方法により、簡単に取得でき、その候補としては、例えば抗betaKlotho抗体のうちで、betaKlotho活性を模倣するような、いわゆるアゴニスト抗体があげられる。当該抗体としては、betaKlothoを認識する抗体のみならず、FGF受容体を認識する抗体、FGF21を認識する抗体、betaKlotho、FGF受容体及び/又はFGF21の複合体を認識するような抗体でありえる。
増殖因子の作用に対するアゴニスト抗体の例としては、HGF受容体のアゴニスト抗体(特許文献2)、Tie2 アゴニスト抗体(特許文献3)、EGF受容体のアゴニスト抗体(非特許文献10)、NGF受容体のアゴニスト抗体(非特許文献11)などがある。
ところで、FGF1など多くのFGFリガンドでは、それらが単独にFGF受容体に結合しただけでは受容体の活性化は起こらないが、さらにヘパリン様の糖鎖が共存すると活性複合体が形成されて、それを起点としたシグナルの伝達が行われて細胞の応答(例えば細胞の増殖など)が起きるとされている(図14下段(1)、(2))。本発明におけるFGF受容体を介したFGF21の活性発揮にあたっては、通常のFGF受容体とFGFリガンド間の(1)、(2)のような様式ではシグナル伝達は起こらない。補助因子BetaKlothoが加わり、リガンド(FGF21)、FGF受容体、BetaKlothoの3者が存在することによって、はじめてFGF21によるFGF受容体の活性化が起きる。これを概念図として表したものが図14下段(3)である。なお、概念図中の複合体形成に関わる各分子の個数はこれを限定するものではない。この活性複合体の形成を増強あるいは減弱する物質は、FGF21の作用を昂進あるいは抑制する作用を有するが、これらの物質は、本発明のbetaKlothoとFGF受容体発現細胞系を用いたスクリーニング系で簡単に取得でき、FGF21活性の調節剤あるいは調節するための医薬組成物として利用することができる(4)。
従来のアゴニスト抗体も細胞表面のリガンドと受容体との結合を促進する作用があるが、本発明におけるbetaKlotho活性を模倣した抗体も、図14(4)に示されるような、betaKlotho、FGF21、及びFGF受容体からなる複合体の形成を増強するものであって、その結果、FGF受容体を介したFGF21活性を昂進する作用があると考えられる。
生体内でbetaKlotho活性を抑制する物質の例は、生体内におけるbetaKlothoの作用を妨げる物質であれば特に限定されないが、典型的にはbetaKlotho特異的抗体があげられる。
BetaKlotho活性を模倣したアゴニスト抗体、betaKlotho活性を抑制するbetaKlotho特異的抗体のいずれも(以下、単にbetaKlotho抗体ともいう。)、ポリクローナル抗体、モノクローナル抗体のいずれであってもよく、周知の免疫学的手法により作製できる。また、該betaKlotho抗体は、抗体のフラグメント(例えば、Fab、F(ab’)2)、組換え抗体(例えば、単鎖抗体)であってもよい。さらに、betaKlotho活性を抑制する抗体をコードする核酸(プロモーター活性を有する核酸に機能可能に連結されたもの)の場合は、betaKlothoの発現を抑制する物質として好ましい。
例えば、ポリクローナル抗体は、betaKlothoあるいはそのフラグメントを、必要に応じキャリアタンパク質を用い、市販のアジュバントとともに、ラット、マウス、ウサギなどの哺乳動物を定法により免疫して得た抗血清を精製することにより取得できる。
また、モノクローナル抗体は、細胞融合法を用いて取得でき、たとえば、マウスなどをbetaKlothoあるいはそのフラグメントで免疫し、脾臓あるいはリンパ節から白血球を採取し、この白血球と骨髄腫細胞を細胞融合して抗betaKlothoモノクローナル抗体を産生するハイブリドーマを得る。
また、本発明の抗体は、キメラ抗体、ヒト化又はヒト型抗体であってもよい。キメラ抗体は、例えば「実験医学(臨時増刊号),Vol.6,No.10,1988」等を、ヒト化抗体は、例えば特表平4-506458号公報、特開昭62-296890号公報等を、ヒト抗体は、例えば「Nature Genetics,Vol.15,p.146-156,1997」、「Nature Genetics,Vol.7,p.13-21,1994」等を参考にそれぞれ作製することができる。
その他のbetaKlotho阻害物質としては、betaKlotho遺伝子のアンチセンス核酸、リボザイム、RNAi誘導性核酸、ターゲティングベクター、betaKlothoの部分的なネガティブ変異体などがあげられる。
さらに、単なるbetaKlotho阻害物質は、(特許文献1)に記載のスクリーニング方法によってもスクリーニングすることができる。
【0026】
〔9〕本発明の血糖値の調節(上昇作用、又は降下作用)にかかわる医薬組成物について
本発明の剤は、betaKlotho及びその発現又は機能を調節する物質に加え、任意の担体、例えば医薬上許容され得る担体を含むことができる。医薬上許容され得る担体としては、例えば、ショ糖、デンプン、マンニット、ソルビット、乳糖、グルコース、セルロース、タルク、リン酸カルシウム、炭酸カルシウム等の賦形剤、セルロース、メチルセルロース、ヒドロキシプロピルセルロース、ポリプロピルピロリドン、ゼラチン、アラビアゴム、ポリエチレングリコール、ショ糖、デンプン等の結合剤、デンプン、カルボキシメチルセルロース、ヒドロキシプロピルスターチ、ナトリウム−グリコール−スターチ、炭酸水素ナトリウム、リン酸カルシウム、クエン酸カルシウム等の崩壊剤、ステアリン酸マグネシウム、エアロジル、タルク、ラウリル硫酸ナトリウム等の滑剤、クエン酸、メントール、グリシルリシン・アンモニウム塩、グリシン、オレンジ粉等の芳香剤、安息香酸ナトリウム、亜硫酸水素ナトリウム、メチルパラベン、プロピルパラベン等の保存剤、クエン酸、クエン酸ナトリウム、酢酸等の安定剤、メチルセルロース、ポリビニルピロリドン、ステアリン酸アルミニウム等の懸濁剤、界面活性剤等の分散剤、水、生理食塩水、オレンジジュース等の希釈剤、カカオ脂、ポリエチレングリコール、白灯油等のベースワックスなどが挙げられるが、それらに限定されるものではない。
【0027】
経口投与に好適な製剤は、水、生理食塩水のような希釈液に有効量の物質を溶解させた液剤、有効量の物質を固体や顆粒として含んでいるカプセル剤、サッシェ剤または錠剤、適当な分散媒中に有効量の物質を懸濁させた懸濁液剤、有効量の物質を溶解させた溶液を適当な分散媒中に分散させ乳化させた乳剤等である。
非経口的な投与(例えば、静脈内注射、皮下注射、筋肉注射、局所注入など)に好適な製剤としては、水性および非水性の等張な無菌の注射液剤があり、これには抗酸化剤、緩衝液、制菌剤、等張化剤等が含まれていてもよい。また、水性および非水性の無菌の懸濁液剤が挙げられ、これには懸濁剤、可溶化剤、増粘剤、安定化剤、防腐剤等が含まれていてもよい。当該製剤は、アンプルやバイアルのように単位投与量あるいは複数回投与量ずつ容器に封入することができる。また、有効成分および医薬上許容され得る担体を凍結乾燥し、使用直前に適当な無菌のビヒクルに溶解または懸濁すればよい状態で保存することもできる。
本発明の剤の投与量は、有効成分の活性や種類、病気の重篤度、投与対象となる動物種、投与対象の薬物受容性、体重、年齢等によって異なり一概に云えないが、通常、成人1日あたり有効成分量として約0.001〜約500mg/kgである。
【0028】
また、本発明の剤において、betaKlothoをコードするDNAを含む組換えDNAを有効成分として含む場合、典型的には当該組換えDNAとしてbetaKlothoをコードするDNAを挿入した発現ベクターを有効成分として含む。当該発現ベクターは、上記の核酸分子をコードするオリゴヌクレオチドもしくはポリヌクレオチドが、投与対象である哺乳動物の細胞内でプロモーター活性を発揮し得るプロモーターに機能的に連結されていなければならない。使用されるプロモーターは、投与対象である哺乳動物で機能し得るものであれば特に制限されず、例えば、SV40由来初期プロモーターなどが挙げられ、発現ベクターとしてヒト等の哺乳動物への投与に好適なアデノウイルス、レトロウイルス、アデノ随伴ウイルス、ヘルペスウイルス等のウイルスベクターや核酸キャリア物質などと共に投与することが好ましい。
【0029】
〔10〕FGF受容体及びbetaKlothoを内在的に発現していない細胞の系を用いたスクリーニング方法及びキット
FGF受容体及びbetaKlothoを内在的に発現していない細胞としては、典型的にはマウス白血病ProB細胞であるBaF3細胞が用いられるので、以下BaF3細胞を用いて説明する。ただし、他の細胞でもこれらいずれの遺伝子も細胞表面に発現しておらず、かつこれら遺伝子で形質転換された場合に細胞表面に発現することができる細胞であれば、同様に用いることができるので、BaF3細胞には限られない。
FGF受容体を発現しているBaF3細胞の系はFGF受容体を介したFGF21の活性を増強もしくは抑制する物質のスクリーニングに用いることができる。
具体的には、まず、FGF受容体を細胞表面に発現しているBaF3細胞の培養系に対して、披検物質と共にFGF21を作用させる方法を用いることができる。その際、披検物質とFGF21の添加順序は、同時でも、何れが先でもよい。
また、FGF受容体遺伝子と共に披検物質遺伝子をBaF3細胞に導入して、FGF受容体と同時に披検物質も細胞表面に発現させたBaF3細胞の系に対してFGF21を作用させてもよい。
その際、披検物質遺伝子をFGF受容体遺伝子と同一のベクターに挿入し、又は別々のベクターに挿入してBaF3細胞に同時に導入してもよく、また、FGF受容体を細胞表面に発現しているBaF3細胞に対して、披検物質遺伝子を導入してもよい。
FGF受容体、及びその遺伝子としては、FGFR1cもしくはFGFR3c、及びそれらの遺伝子を用いることが好ましい。
したがって、FGF受容体、好ましくはFGFR1cもしくはFGFR3c、を細胞表面に発現しているBaF3細胞の系をFGF21と組み合わせることで、FGF21の生物活性を増強もしくは抑制する物質のスクリーニング用のキットとすることができる。
なお、実際のスクリーニングに際しては、BaF3細胞の増殖能を、披検物質を添加しない系などのコントロールの系と比較して評価することができる。BaF3細胞の増殖能は、標識チミジンの取り込み量の測定など常法により測定する。他のスクリーニング方法においても同様である。
また、実際のスクリーニングに際しては、BaF3細胞の増殖シグナル伝達の活性化を、披検物質を添加しない系などのコントロールの系と比較して評価することもできる。BaF3細胞の増殖シグナル伝達の活性化は、FGF受容体のチロシンリン酸化、FGF 受容体基質2(FRS2)のチロシンリン酸化、マップキナーゼ(ERK1/2)のリン酸化など常法により測定する。他のスクリーニング方法においても同様である。
【0030】
FGF受容体と共にbetaKlothoも発現しているBaF3細胞の系は、FGF21活性調節作用のあるbetaKlotho活性を増強もしくは抑制する物質のスクリーニングに用いることができる。
具体的には、まず、FGF受容体と共にbetaKlothoを細胞表面に発現しているBaF3細胞の系に対して披検物質と共にFGF21を作用させる方法を用いることができる。その際、披検物質とFGF21の添加順序は、同時でも、何れが先でもよい。
また、FGF受容体遺伝子及びbetaKlotho遺伝子と共に披検物質遺伝子をBaF3細胞に導入して、FGF受容体及びbetaKlothoと同時に披検物質も細胞表面に発現させたBaF3細胞の系に対してFGF21を作用させてもよい。
その際、披検物質遺伝子をFGF受容体遺伝子及びbetaKlotho遺伝子と同一のベクターに挿入し、又は別々のベクターに挿入してBaF3細胞に同時に導入してもよく、また、FGF受容体及びbetaKlothoを細胞表面に発現しているBaF3細胞に対して、披検物質遺伝子を導入してもよい。
FGF受容体、及びその遺伝子としては、FGFR1cもしくはFGFR3c、及びそれらの遺伝子を用いることが好ましい。
したがって、FGF受容体、好ましくはFGFR1cもしくはFGFR3c、及びbetaKlothoを細胞表面に発現しているBaF3細胞の系をFGF21と組み合わせることで、FGF21活性の調節作用に係るbetaKlotho活性の昂進物質もしくは阻害物質のスクリーニング用のキットとすることができる。
【0031】
FGF受容体と共にbetaKlothoも発現しているBaF3細胞の系は、FGF21様活性物質のスクリーニング方法に用いることができる。
具体的には、FGF受容体と共にbetaKlothoを細胞表面に発現しているBaF3細胞の系に対して披検物質を作用させ、BaF3細胞の増殖能を評価することで、FGF21様活性物質をスクリーニングすることができる。
その際、同時に、FGF受容体のみを細胞表面に発現しているBaF3細胞及び/又はbetaKlothoのみを細胞表面に発現しているBaF3細胞の系に披検物質を作用させ、その作用が、FGF21作用と同様の、FGF受容体及びbetaKlothoとの複合体を形成したことに基づく作用であることを確認することが好ましい。
FGF受容体としてはFGFR1c又はFGFR3cを用いることが好ましい。
そして、FGF受容体、好ましくはFGFR1cもしくはFGFR3c、と共にbetaKlothoを細胞表面に発現しているBaF3細胞の系を用いることで、FGF21様活性物質のスクリーニング用のキットとすることができる。
さらに、FGF受容体のみを細胞表面に発現しているBaF3細胞及び/又はbetaKlothoのみを細胞表面に発現しているBaF3細胞の系をコントロール系として組み合わせたキットがより好ましく用いることができる。
そして、これらスクリーニングにより得られた物質は、いずれもFGF受容体とbetaKlothoの共存下で発揮されるFGF21活性の調節剤及び当該FGF21活性を調節するための医薬組成物として用いることができる。
【実施例】
【0032】
以下、実施例を用いて本発明をより詳細に説明するが、本発明の技術的範囲は以下の実施例に限定されるものではない。
〔実施例1〕FGF21の脂肪細胞特異的活性の検出
FGF21は、その活性として脂肪細胞にグルコーストランスポーター1(GLUT1)の発現を上昇させることが報告されている。本実施例では、培養脂肪細胞としては、培養条件を操作することにより脂肪細胞に分化する能力を有する繊維芽細胞として3T3-L1細胞を用いた。細胞は10% ウシ胎児血清(FBS)を含有する、Dulbecco改変Eagle培地(DMEM)の培養液で培養し、繊維芽細胞としての未分化形質を維持した細胞を用意した。その細胞を試験系にて培養し、コンフルエントになるまで増殖させ、そして2日間そのままにした。その後、培養液を分化誘導培養液であるDMEM、10%FBS、0.25μMデキサメタゾン、0.5mMメチルイソブチルキサンチン、および5μg/mlのインスリンに交換して2日間培養した。その後、細胞を10%FBSおよび2.5μg/mlのインスリンを含有するDMEMにより、1日おきに培養液を交換した。このようにして分化誘導した脂肪細胞の性質をもつ細胞を得た。比較のために、分化を誘導しない繊維芽細胞も準備した。ここで「脂肪細胞の性質をもつ細胞」とは、脂肪細胞特有の細胞内代謝を行い、脂肪滴を多量に蓄積した細胞質を有する細胞を示す。
これらそれぞれの培養液にFGF21を種々の濃度で加え、細胞をさらに6時間培養後、RNAを抽出し、GLUT1のコピー数を測定した。その結果を図1に示す。
ここで、脂肪細胞に分化した細胞ではFGF21を様々な濃度で細胞培養液に添加することにより、GLUT1のmRNAの発現量が増加していたことから、FGF21が脂肪細胞に対して活性を発揮し、GLUT1発現を増加させることが示された。これとは対照的に、分化していない繊維芽細胞では、FGF21を様々な濃度で細胞培養液に添加しても、GLUT1のmRNAの発現量はほとんど変化しなかった。このことから、FGF21がGLUT1のmRNAを増加させる活性は、3T3-L1細胞では細胞が脂肪細胞に分化したあとにのみ発揮されることが示された。
【0033】
〔実施例2〕脂肪細胞の分化に伴うFGF受容体の発現変動の検出
上記実施例1で、FGF21が活性を発揮するのは3T3-L1細胞が脂肪細胞に分化した後だけであったことから、脂肪細胞に分化するとともに、細胞表面に存在するFGF受容体数が増加した結果、FGF21に対する反応性が増大した可能性が考えられた。そこで、FGFの受容体分子の発現レベルに変動があるかどうかをまず解析した。
3T3-L1細胞を、分化誘導培養液に交換すると、培養を開始してから経時的に、繊維芽細胞の性質を持つ細胞から、脂肪細胞の性質をもつ細胞に、徐々に変化する。
そこで、実施例1と同様の条件で分化誘導処理を行った後、一日おきに20日間まで3T3-L1細胞を採取し、細胞が発現するFGF受容体のmRNAの量を解析した。FGF受容体としては、膜一回貫通型チロシンキナーゼ型受容体として、4種類の遺伝子FGFR1、FGFR2、FGFR3、FGFR4が知られているので、これらのmRNA量を測定した。その結果を図2に示す。分化の有無にかかわらずFGFR1とFGFR2が高いレベルで発現しており、FGFR3とFGFR4の発現レベルは非常に低かった。そして、分化に伴って発現が増える受容体はなく、むしろ高レベルで発現していたFGFR1、FGFR2のいずれの受容体も、分化とともに、発現レベルがやや減る傾向が認められた。
【0034】
〔実施例3〕脂肪細胞の分化に伴うbetaKlothoの発現変動の検出
本発明者らは、脂肪細胞へと分化する過程で脂肪細胞表面での発現量が増加する物質があれば、当該物質がFGF21と協同してFGF受容体に作用する可能性が高いと考え、発達中のマウス胎児の脂肪組織で発現することが知られていたbetaKlothoに着目した。
上記実施例2と同様にして、3T3-L1細胞が脂肪細胞に分化する過程でのbetaKlothoの発現レベルを経時的に解析した。
さらに、比較のために、最近FGF23に対する受容体補助因子として働くことが報告されたKlothoの発現レベルも同様にして解析した。KlothoはbetaKlothoと構造的に類似性があるが、アミノ酸配列レベルで41%の相同性(塩基配列レベルで55%)であり、両者の性質はまったく異なるもので、明らかな別遺伝子にコードされた分子であると位置づけられる。
その解析結果を、図3に示す。betaKlothoは脂肪細胞に分化した細胞でのみ高レベルで発現していた。これに対して、Klothoは分化前も分化後もほとんど発現していなかった。このことから、脂肪細胞分化に伴って発現するようになるbetaKlothoが脂肪細胞の機能と密接に関連することが示唆されたため、FGF21に対するFGF受容体補助因子である可能性を探るため解析する計画をした。
【0035】
〔実施例4〕FGF21によるFGF受容体下流シグナル惹起にbetaKlothoが必要であることの解析
上記実施例2と3から、実施例1で示されるようなFGF21の脂肪細胞特異的活性は、FGF受容体とbetaKlothoが共に存在する場合に誘導されることが示唆される。そこで本実施例において、元来はFGF受容体を持たない細胞BaF3にFGFR1cを強制発現させた細胞に対し、さらにbetaKlothoを強制発現させたときとさせないときで、FGF21に対する反応性がどのように変わるかを解析した。
FGFR1c発現BaF3細胞、FGFR1c及びbetaKlotho発現BaF3細胞は以下のように調製した。FGFR1c発現BaF3細胞にマウスFGFR1cの全長cDNAがクローニングされた動物細胞用発現ベクター(Ornitz,D.M.,Xu,J.,Colvin,J.S.,McEwen,D.G.,MacArthur,C.A.,Coulier,F.,Gao,G.and Goldfarb,M.(1996) Receptor specificity of the fibroblast growth factor family.J Biol Chem,271,15292-15297.に記載されている)をエレクトロポレーション法によりBaF3細胞に導入した。G418存在下で約2週間培養を行い、薬剤耐性クローンを得た後、FGF1刺激により増殖させることができるクローンとして取得した。FGFR1cとbetaKlothoを共に発現するBaF3細胞は、動物細胞用発現ベクターにマウスbetaKlothoの全長cDNAをクローニングし、これをFGFR1c発現BaF3細胞に導入した後、Puromycin及びG418存在下で約2週間培養を行い、耐性クローンとして取得した。betaKlothoを単独で発現するBaF3細胞はbetaKlotho発現ベクターをBaF3細胞に導入した後、Puromycin耐性クローンとして取得した。これらの遺伝子導入したBaF3細胞は10%FBSとインターロイキン3(IL3、WEHI-3細胞のコンディションメディウムを5%加えることにより供給)を含むRPMI1640培地で維持した。増殖刺激実験時は細胞をRPMI1640培地で洗浄後10%FBSを含むRPMI1640培地に懸濁し培養プレートに播種して用いた。
解析結果を、図4に示す。
A)で、CL#1,#7、#10はFGFR1cとbetaKlothoの両方のcDNAをBaF3細胞に導入し取得したクローンの名称である。A)の下段は、それぞれの細胞から調製したライゼートを材料として、実際にタンパクとして発現しているbetaKlothoの量を、市販の抗betaKlotho抗体を用いたウエスタンプロット解析により比較した結果である。CL#10ではbetaKlothoは検出レベル以下であり、CL#1、CL#7では同程度のレベルのbetaKlothoの発現がみられた。BetaKlothoは主に2本のブロードなバンドとして観察され、N型糖鎖の修飾が起こっている可能性が考えられた。
それぞれのクローンおよびFGFR1cのみを発現しているBaF3細胞は播種直後、ヘパリン存在下でFGF21(白色バー)あるいはコントロールとしてPBS(黒白色バー)の添加により刺激し、42時間培養した。その後トリチウムラベルされたチミジンを培養液中に添加し、更に6時間培養を継続し、細胞に取り込まれたカウントを測定することによりDNA合成を評価した。それぞれFGF21未刺激のDNA合成値を1とした場合のFGF21によるDNA合成の割合を倍率として表した。その結果FGFR1cは発現しているがbetaKlothoを発現していない細胞ではFGF21に反応せず、betaKlothoを共に発現している細胞でのみFGF21に反応することが示された。尚、図には示していないが、FGFR1cのみを発現しているBaF細胞はヘパリンが存在している条件でもFGF21に反応しない。
B)はFGF21の量を変化させてCL#1の細胞を刺激した際のDNA合成を測定した結果である。ヘパリン存在下(白丸)、非存在下(黒丸)ともに、FGF21による用量依存的なDNA合成刺激活性がみられた。またヘパリンはFGF21の作用を増強することがわかった。C)はFGFR1cを発現していないがbetaKlothoのみを発現しているBaF3細胞のFGF21に対する反応性を調べた結果で、ヘパリン存在下(白色バー)、非存在下(黒色バー)の両条件下ともに、FGF21には反応しなかった。BaF3の維持に用いているインターロイキン3(IL3)刺激では顕著な増殖が観られた。
以上のことから、FGF21に反応するためには、FGF受容体とbetaKlothoの両方が必要であることが示された。
【0036】
〔実施例5〕FGF21の活性発揮においてbetaKlothoとともに存在するFGF受容体の特異性の解析(1)
上記実施例4において、FGF21に反応するためには、FGF受容体であるFGFR1cとbetaKlothoの両方が必要であることが示されたが、FGF受容体として、ほかの受容体分子ではどうであるかを調べた。betaKlotho発現BaF3細胞に各FGFR(FGFR1b、R1c、R2b、R2c、R3b、R3c、R4)の発現ベクター(Ornitz,D.M.,Xu,J.,Colvin,J.S.,McEwen,D.G.,MacArthur,C.A.,Coulier,F.,Gao,G.and Goldfarb,M.(1996) Receptor specificity of the fibroblast growth factor family.J Biol Chem,271,15292-15297.に記載されている)を導入して、Puromycin及びG418耐性クローンを取得し、FGFR/betaKlotho発現細胞を得た。それぞれをFGF21で刺激しDNA合成を測定した。その結果図5に示すように、FGFR1cのほか、FGFR3cをbetaKlothoと共に発現させた場合でもFGF21に反応することが明らかとなった。さらにヘパリンの存在により反応性は増強されることが示された。
【0037】
〔実施例6〕FGF21の活性発揮においてbetaKlothoとともに存在するFGF受容体の特異性の解析(2)
betaKlothoとともに各FGFR(FGFR1c、R2c、R3c、R4)を発現しているBaF3細胞に対するFGF21のDNA合成刺激活性を、進化的にFGF21に近縁のFGF19の活性と比較した。実施例5と逆の手順で、すなわち各FGFRを発現しているBaF3細胞にbetaKlothoの発現ベクターを導入してPuromycin及びG418耐性クローンを取得し、FGFR/betaKlotho発現細胞を得た。細胞をヘパリン存在下で、FGF21、FGF19、FGF1の量を図のように変化させて刺激し、DNA合成を測定した。図に示されるように、FGF19はいずれの細胞に対しても強い活性を示すのに対して、FGF21はR1c/betaKlotho発現細胞とR3c/betaKlotho発現細胞のみを活性化することが明らかとなった。(図6)
【0038】
〔実施例7〕FGF21の刺激に伴う細胞内シグナル伝達分子の活性化の解析
実施例6で取得した各FGFR/betaKlotho発現細胞をFGFで刺激した際の、FGFRの下流のシグナル伝達分子FRS2(fibroblast growth factor receptor substrate 2、FGF受容体基質2)とMAPK(mitogen activated protein kinase、マイトージェン活性化プロテインキナーゼ、別名ERK(extracellular signal-regulated kinaseで通常分子量44kDaのERK1と42kDaのERK2の2種類を指す)の活性化状態(リン酸化状態)を解析した。それぞれの細胞をヘパリン存在下、各FGFで10分間刺激した後、細胞ライゼートを調製し、市販の抗リン酸化FRS2抗体あるいは抗リン酸化MAPK抗体を用いてリン酸化の程度をウエスタンブロッティング法で測定することにより評価した。
その結果を図7に示す。図においてMAPKのカラムは抗ERK1/2抗体を用いたウエスタンブロッティングによる細胞ライゼート中の全MAPKの量を示すが、刺激あるいはコントロールの無刺激の細胞においてMAPK(ERK1/2)の総量自体の変化はない。FGF1とFGF19はいずれの細胞においてもFRS2とMAPKのリン酸化を顕著に増加させるのに対し、FGF21は、R1c/betaKlotho発現細胞とR3c/betaKlotho発現細胞においてFRS2とMAPKのリン酸化を増大させることが示された。この結果は実施例6の結果をよく反映している。
【0039】
〔実施例8〕ヒトのアミノ酸配列をもつヒト型FGFR1cとヒト型betaKlothoを共発現させた細胞に対するヒト型FGF21の活性の解析
実験例4、5、6、7において、FGF21はbetaKlotho存在下にFGFR1cに反応することを示したが、実際の実験系で用いたFGFR1c、betaKlotho、FGF21などはすべて哺乳類の典型的なモデルとしてマウス由来のものを用いている。これら各分子のヒト、マウス間でのアミノ酸配列の相同性はFGF21が80%、betaKlothoが78%、FGFR1が98%と極めて高いことからも当然にヒト、マウス間の互換性に問題なく、ヒト型の分子でもマウス型と同様の結論が得られることは明らかである。本実施例では、念のために、その点を確認するためのヒト型分子を用いた実験系を構築して実証を行った。具体的には、FGFR1c、betaKlothoのヒト型のcDNAは市販のヒトRNA試料(タカラバイオ社)からPCRにより調製し、FGFR1cはSRalphaプロモーターを有する発現ベクター(pSRalpha)に、betaKlothoはCMVプロモーターを有する発現ベクター(pcDNA3.1;インビトロージェン社)に導入した。実験例4と同様にエレクトロポレーション法によりFGFR1c発現ベクターをBaF3細胞に導入し、G418耐性クローンとしてFGFR1c発現BaF3細胞を取得した。FGFR1cとbetaKlothoの共発現細胞はFGFR1c発現BaF3細胞にbetaKlotho発現ベクターを導入し、Puromycin耐性クローンとして取得した。ヒト型FGF21タンパク質は以下のように調製した。ヒト型FGF21のcDNAを市販のヒトRNA試料(タカラバイオ社)からPCRにより調製し、大腸菌用の発現ベクターに導入した。このベクターを用いて非特許文献1に記載の手法に準拠して、タンパク質を調製した。ヘパリン存在下で種々の濃度でヒト型FGF21を FGFR1c発現細胞あるいはFGFR1cとbetaKlothoの共発現細胞に添加した際のDNA合成を測定した結果を図8に示す。コントロールとして用いたFGF1はbetaKlothoの発現の有無にかかわらず、細胞のDNA合成を促進したが、FGF21はbetaKlothoを共発現している細胞のみを活性化することが示された。
このことは、ヒト由来分子を用いた場合にも、マウス由来分子を用いた各実施例の実験結果と同様の結果が導かれるであろうという当初の予想を、あらためて裏付けるものであった。
【0040】
〔実施例9〕ヒトのアミノ酸配列をもつヒト型FGFR3cとヒト型betaKlothoを共発現させた細胞に対するヒト型FGF21の活性の解析
FGFR3のヒト、マウス間でのアミノ酸配列の相同性は93%である。ヒト型FGFR3cの場合においても、マウス型と同様の結論が得られることを実証する実験を行った。FGFR3cのヒト型のcDNAは市販のヒトRNA試料(タカラバイオ社)からPCRにより調製し、細胞内部分の配列をヒト型FGFR1の配列と置換してSRalphaプロモーターを有する発現ベクター(pSRalpha)に導入した。エレクトロポレーション法によりFGFR3c発現ベクターをBaF3細胞に導入し、G418耐性クローンとしてFGFR3c発現BaF3細胞を取得した。FGFR3cとbetaKlothoの共発現細胞はFGFR3c発現BaF3細胞にbetaKlotho発現ベクターを導入し、Puromycin耐性クローンとして取得した。ヘパリン存在下で種々の濃度でヒト型FGF21を FGFR3c発現細胞あるいはFGFR3cとbetaKlothoの共発現細胞に添加した際のDNA合成を測定した結果を図9に示す。コントロールとして用いたFGF1はbetaKlothoの発現の有無にかかわらず、細胞のDNA合成を促進したが、FGF21はbetaKlothoを共発現している細胞のみを活性化することが示された。この実験結果からも、ヒト型の分子でもマウス型と同様の結論が得られることが実証された。
【0041】
〔実施例10〕FGF21の活性発揮においてFGF受容体とともに存在するbetaKlothoの特異性の解析
betaKlothoがFGF受容体との結合におけるFGF21に対する特異的な補助因子であることを確認するために、betaKlothoを、FGF21と構造的類似性の高いFGF23に作用させた場合と、betaKlothoと構造的類似性の高いKlothoを、FGF21に作用させた場合に、それぞれどの程度の互換性があるかを調べた。
具体的には、FGFR1cと共にbetaKlothoを発現させたBaF3細胞及びFGFR1cと共にKlothoを発現させたBaF3細胞を、実施例4と同様の形質転換方法及び同一の培養条件を用いて調製した。そして、各形質転換細胞に対して、ヘパリンの存在下、及び非存在下でFGF21及びFGF23を添加し、DNA合成量を測定した。
その結果を図10に示す。FGF21の場合は、ヘパリンの存在、非存在にかかわりなく、klothoを存在させてもFGFR1cには反応しないことが示され、反対に、FGF23の場合は、ヘパリンの存在、非存在にかかわりなく、betaKlothoを存在させてもFGFR1cには反応しないことが示された。
このように、FGF21とbetaKlotho、FGF23とKlothoそれぞれの組合せでのみFGFR1c発現細胞に活性を発揮できることが示されたことからも、betaKlothoがFGF21に対する特異的な受容体結合補助因子であることが立証された。
【0042】
〔実施例11〕BetaKlothoとFGF21あるいはFGF受容体との複合体形成の検出
実施例4から実施例10においては、FGF21のFGF受容体を介した作用発現のためにbetaKlothoが必要であることを細胞生物学的に示した。
本実施例においては、betaKlothoとFGF受容体あるいはFGF21の間の複合体形成を生化学的に調べた。具体的には、betaKlothoの細胞外ドメインのC末端にV5タグとHisタグをタンデムに付加した可溶性タンパク質を調製し、試験管内においてヘパリンの存在あるいは非存在の条件下で、可溶性betaKlothoタンパク質とFGF21をインキュベートした後、抗V5タグ抗体固定化ビーズを用いてbetaKlothoを沈降させ、FGF21がbetaKlothoに結合して共沈降するか否かを、免疫ブロッティング法にて解析した。
その結果を図11A)に示す。ヘパリンの有無によらず、betaKlothoの存在下でFGF21が沈降してくることから、betaKlothoはFGF21との結合が、ヘパリンの有無によらず形成されることが確認された。
また、可溶性betaKlothoタンパク質とFGF受容体の細胞外ドメインに免疫グロブリンのFcドメインを付加した融合タンパク(FGFR-Fc:R&D Systems社製)を、インキュベートした後、プロテインAビーズを用いてFGFR-Fcを沈降させ、betaKlothoがFGFR-Fcに結合して共沈降するか否かを解析した。
その結果を図11B)に示す。FGFR-Fcの存在に依存してbetaKlothoが沈降してくることが示され、betaKlothoはFGF受容体サブタイプのR1c、R2c、R3c、R4のいずれとも結合することが明らかになった。
【0043】
〔実施例12〕BetaKlothoを強制発現させた未分化3T3-L1細胞をFGF21刺激した際のシグナル伝達の解析
実施例1において、FGF21は脂肪細胞に作用してGLUT1のmRNA量を増大させる活性を有するが、分化前の細胞に対してはこの活性を発揮しないことを示した。実施例2では、betaKlothoが脂肪細胞の分化に伴って発現が増大することを示した。実施例3から10では、BaF3細胞を用いてbetaKlothoがFGF21に対するFGF受容体補助因子であることを示した。
そこで本実施例では、脂肪細胞へ分化する前の3T3-L1細胞においてもbetaKlothoがFGF21に対するFGF受容体補助因子として機能するか否かを調べた。実施例1に記載した通りに培養した3T3-L1繊維芽細胞に、実施例4に記載した方法で作成したbetaKlothoの発現ベクター、及びコントロールとしてbetaKlothoのcDNAを挿入していない空のベクターを、リポフェクション法により導入し、Puromycin耐性クローンを得た。それぞれの細胞をFGF21で刺激し、10分後に細胞ライゼートを調製し、FGFRの下流のシグナル伝達分子であるFRS2とMAPKの活性化をウエスタンブロッティング法で調べた。
その結果を図12に示す。図のようにbetaKlothoの発現ベクターを導入した細胞においてのみFGF21の刺激に反応してFRS2とMAPKのリン酸化が増加することが示された。また、FGFRのチロシンキナーゼ阻害剤であるSU5402(メルク社製)で3時間前処理した細胞をFGF21で刺激した場合には、リン酸化の程度が減少することから、FGF21の活性はFGFRを介することにより伝達されていることが示された。
【0044】
〔実施例13〕BetaKlothoを強制発現させた未分化3T3-L1細胞をFGF21刺激した際のGLUT1mRNAの発現解析
実施例12で作出したbetaKlothoを強制発現させた3T3-L1繊維芽細胞を用い、刺激するFGF21量に応じたGLUT1(グルコーストランスポーター1)のmRNA量の変化を調べた。
betaKlothoの発現ベクターあるいはコントロールとしてbetaKlothoのcDNAを挿入していない空のベクターを導入した3T3-L1繊維芽細胞の培養液にFGF21を種々の濃度で加え6時間培養した後にRNAを抽出し、GLUT1のmRNA量を測定した。
図13に示すように、betaKlothoを強制発現させた3T3-L1繊維芽細胞では培養液に添加するFGF21の量に従って、GLUT1mRNAが増加することが有意にみとめられた。(図の*はスチューデントのt検定によって5%未満の危険率で、**は1%未満の危険率で有意差があることを示す。)一方、コントロールの3T3-L1繊維芽細胞ではFGF21の添加によってGLUT1mRNA量に変化は見られなかった。この結果から、脂肪細胞に分化する前の3T3-L1細胞でもbetaKlothoを強制発現させることによりFGF21刺激を受容してGLUT1の発現量を増加することが明らかになった。
【特許請求の範囲】
【請求項1】
FGF受容体及びbetaKlothoを内在的に発現していない細胞にFGF受容体遺伝子を導入し、細胞表面にFGF受容体を発現させた形質転換細胞を用いる、FGF受容体を介したFGF21の生物活性を増強又は抑制する物質のスクリーニング方法であって、
細胞表面にFGF受容体を発現させた上記形質転換細胞の培養系に対して披検物質と共にFGF21を作用させる工程、又は、
FGF受容体遺伝子と共に披検物質遺伝子を上記FGF受容体及びbetaKlothoを内在的に発現していない細胞に導入して、FGF受容体と同時に披検物質も細胞表面に発現させた形質転換細胞の系に対してFGF21を作用させる工程、
を含むことを特徴とする、スクリーニング方法。
【請求項2】
FGF受容体及びbetaKlothoを内在的に発現していない細胞にFGF受容体遺伝子を導入し、細胞表面にFGF受容体を発現させた形質転換細胞系及びFGF21を組み合わせてなる、FGF受容体を介したFGF21の生物活性を調節する物質のスクリーニング用のキット。
【請求項3】
FGF受容体及びbetaKlothoを内在的に発現していない細胞にFGF受容体遺伝子及びbetaKlotho遺伝子を導入し、細胞表面にFGF受容体及びbetaKlothoを発現させた形質転換細胞の培養系を用いる、FGF受容体を介したFGF21活性の調節作用に係るbetaKlotho活性の昂進物質もしくは阻害物質のスクリーニング方法であって、
細胞表面にFGF受容体及びbetaKlothoを発現させた上記形質転換細胞の系に対して披検物質と共にFGF21を作用させる工程、又は、
上記FGF受容体及びbetaKlothoを内在的に発現していない細胞を宿主としてFGF受容体遺伝子及びbetaKlotho遺伝子と共に披検物質遺伝子を導入して、FGF受容体及びbetaKlothoと同時に披検物質も細胞表面に発現させた形質転換細胞の系に対してFGF21を作用させる工程、
を含むことを特徴とする、スクリーニング方法。
【請求項4】
FGF受容体及びbetaKlothoを内在的に発現していない細胞にFGF受容体遺伝子及びbetaKlotho遺伝子を導入し、細胞表面にFGF受容体及びbetaKlothoを発現させた形質転換細胞の系、及びFGF21を組み合わせてなる、FGF受容体を介したFGF21の生物活性の調節作用に係るbetaKlotho活性の昂進物質もしくは阻害物質のスクリーニング用のキット。
【請求項5】
FGF受容体及びbetaKlothoを内在的に発現していない細胞にFGF受容体遺伝子及びbetaKlotho遺伝子を導入し、細胞表面にFGF受容体及びbetaKlothoを発現させた形質転換細胞の系に対して披検物質を作用させる工程を含むことを特徴とする、FGF21様活性物質のスクリーニング方法。
【請求項6】
FGF受容体及びbetaKlothoを内在的に発現していない細胞にFGF受容体遺伝子及びbetaKlotho遺伝子を導入し、細胞表面にFGF受容体及びbetaKlothoを発現させた形質転換細胞の系を用いることを特徴とする、FGF21様活性物質のスクリーニング用のキット。
【請求項1】
FGF受容体及びbetaKlothoを内在的に発現していない細胞にFGF受容体遺伝子を導入し、細胞表面にFGF受容体を発現させた形質転換細胞を用いる、FGF受容体を介したFGF21の生物活性を増強又は抑制する物質のスクリーニング方法であって、
細胞表面にFGF受容体を発現させた上記形質転換細胞の培養系に対して披検物質と共にFGF21を作用させる工程、又は、
FGF受容体遺伝子と共に披検物質遺伝子を上記FGF受容体及びbetaKlothoを内在的に発現していない細胞に導入して、FGF受容体と同時に披検物質も細胞表面に発現させた形質転換細胞の系に対してFGF21を作用させる工程、
を含むことを特徴とする、スクリーニング方法。
【請求項2】
FGF受容体及びbetaKlothoを内在的に発現していない細胞にFGF受容体遺伝子を導入し、細胞表面にFGF受容体を発現させた形質転換細胞系及びFGF21を組み合わせてなる、FGF受容体を介したFGF21の生物活性を調節する物質のスクリーニング用のキット。
【請求項3】
FGF受容体及びbetaKlothoを内在的に発現していない細胞にFGF受容体遺伝子及びbetaKlotho遺伝子を導入し、細胞表面にFGF受容体及びbetaKlothoを発現させた形質転換細胞の培養系を用いる、FGF受容体を介したFGF21活性の調節作用に係るbetaKlotho活性の昂進物質もしくは阻害物質のスクリーニング方法であって、
細胞表面にFGF受容体及びbetaKlothoを発現させた上記形質転換細胞の系に対して披検物質と共にFGF21を作用させる工程、又は、
上記FGF受容体及びbetaKlothoを内在的に発現していない細胞を宿主としてFGF受容体遺伝子及びbetaKlotho遺伝子と共に披検物質遺伝子を導入して、FGF受容体及びbetaKlothoと同時に披検物質も細胞表面に発現させた形質転換細胞の系に対してFGF21を作用させる工程、
を含むことを特徴とする、スクリーニング方法。
【請求項4】
FGF受容体及びbetaKlothoを内在的に発現していない細胞にFGF受容体遺伝子及びbetaKlotho遺伝子を導入し、細胞表面にFGF受容体及びbetaKlothoを発現させた形質転換細胞の系、及びFGF21を組み合わせてなる、FGF受容体を介したFGF21の生物活性の調節作用に係るbetaKlotho活性の昂進物質もしくは阻害物質のスクリーニング用のキット。
【請求項5】
FGF受容体及びbetaKlothoを内在的に発現していない細胞にFGF受容体遺伝子及びbetaKlotho遺伝子を導入し、細胞表面にFGF受容体及びbetaKlothoを発現させた形質転換細胞の系に対して披検物質を作用させる工程を含むことを特徴とする、FGF21様活性物質のスクリーニング方法。
【請求項6】
FGF受容体及びbetaKlothoを内在的に発現していない細胞にFGF受容体遺伝子及びbetaKlotho遺伝子を導入し、細胞表面にFGF受容体及びbetaKlothoを発現させた形質転換細胞の系を用いることを特徴とする、FGF21様活性物質のスクリーニング用のキット。
【図1】


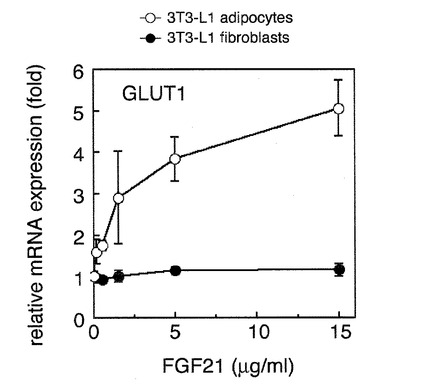
【図2】
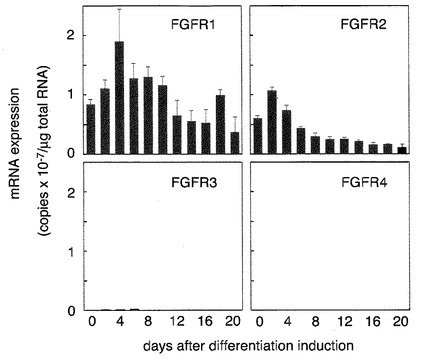
【図3】
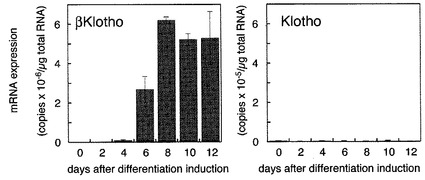
【図4A】
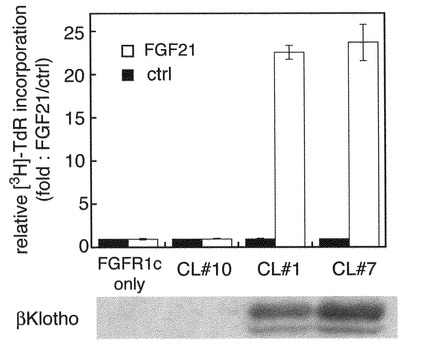
【図4B】
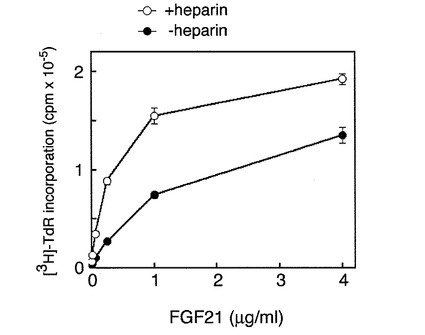
【図4C】
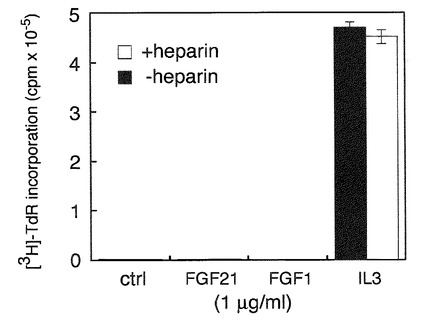
【図5】
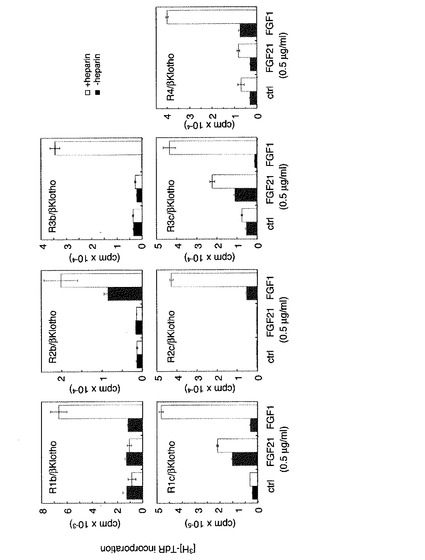
【図6】
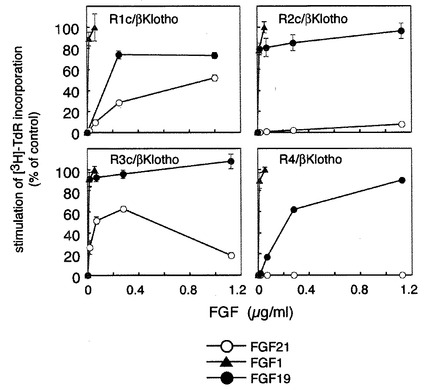
【図7】
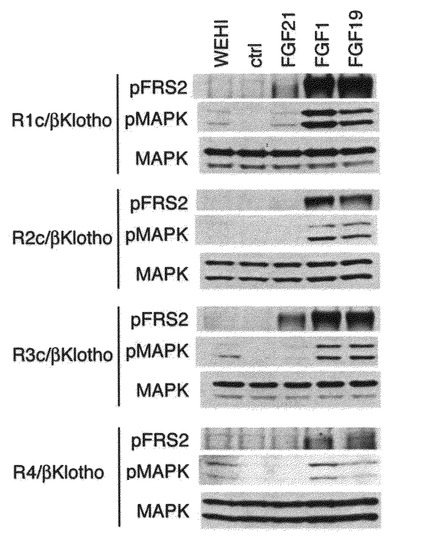
【図8】
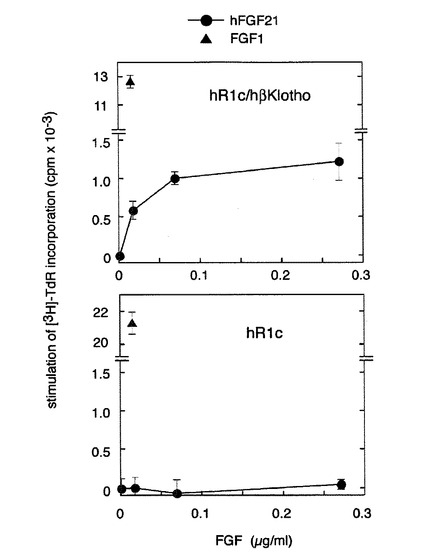
【図9】
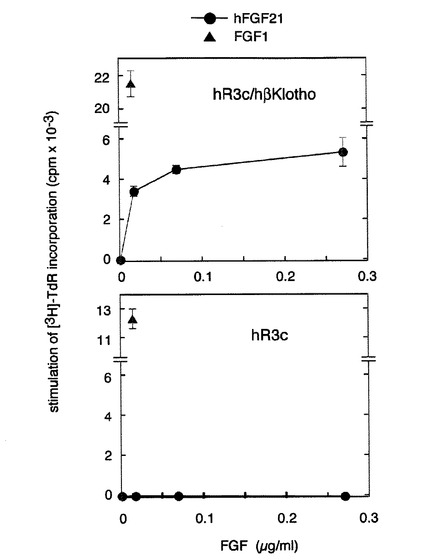
【図10】
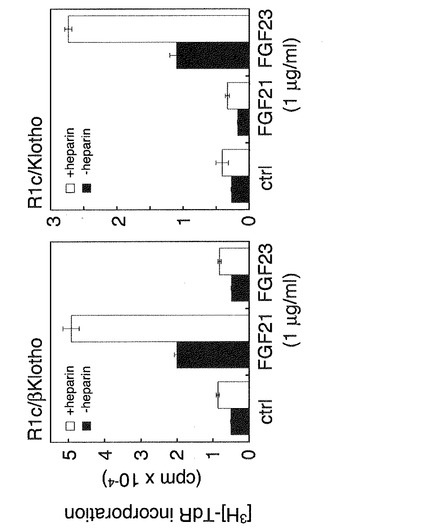
【図11】
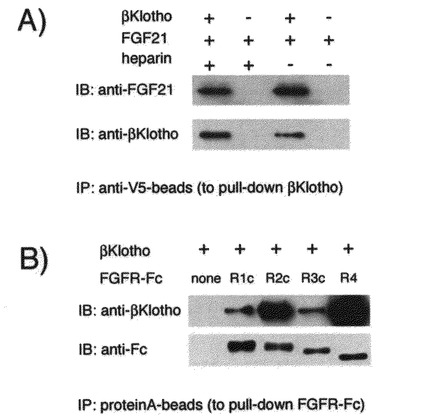
【図12】
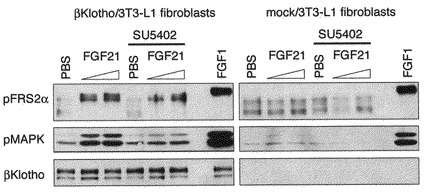
【図13】
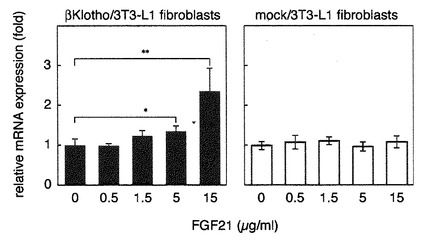
【図14】
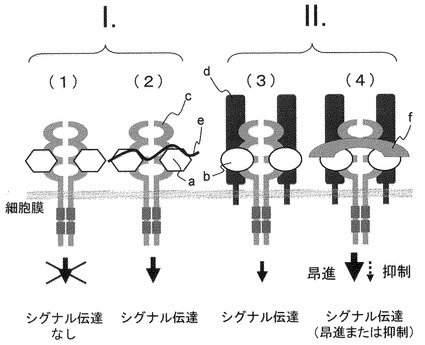
【図2】
【図3】
【図4A】
【図4B】
【図4C】
【図5】
【図6】
【図7】
【図8】
【図9】
【図10】
【図11】
【図12】
【図13】
【図14】
【公開番号】特開2012−230113(P2012−230113A)
【公開日】平成24年11月22日(2012.11.22)
【国際特許分類】
【出願番号】特願2012−142533(P2012−142533)
【出願日】平成24年6月25日(2012.6.25)
【分割の表示】特願2008−99837(P2008−99837)の分割
【原出願日】平成20年4月7日(2008.4.7)
【新規性喪失の例外の表示】特許法第30条第1項適用申請有り 2007年11月25日 BMB2007発行の「第30回日本分子生物学会年会 第80回日本生化学会大会 合同大会 BMB2007 講演要旨集」に発表
【新規性喪失の例外の表示】特許法第30条第1項適用申請有り 2008年1月10日 インターネットアドレス「http://mend.endojournals.org/cgi/rapidpdf/me.2007−0313vl」に発表
【新規性喪失の例外の表示】特許法第30条第1項適用申請有り 2008年1月22日 独立行政法人 産業技術総合研究所、インド生物工学庁主催の「産総研(日本)−生物工学庁(インド)ワークショップ」に文書をもって発表
【新規性喪失の例外の表示】特許法第30条第1項適用申請有り 2008年1月31日〜2月1日 独立行政法人 産業技術総合研究所 ライフサイエンス分野融合会議、産業技術連携推進会議 ライフサイエンス部会・バイオテクノロジー分科会主催の「ライフサイエンス分野融合会議 ライフサイエンス部会・バイオテクノロジー分科会 合同研究発表会」に文書をもって発表
【出願人】(301021533)独立行政法人産業技術総合研究所 (6,529)
【Fターム(参考)】
【公開日】平成24年11月22日(2012.11.22)
【国際特許分類】
【出願日】平成24年6月25日(2012.6.25)
【分割の表示】特願2008−99837(P2008−99837)の分割
【原出願日】平成20年4月7日(2008.4.7)
【新規性喪失の例外の表示】特許法第30条第1項適用申請有り 2007年11月25日 BMB2007発行の「第30回日本分子生物学会年会 第80回日本生化学会大会 合同大会 BMB2007 講演要旨集」に発表
【新規性喪失の例外の表示】特許法第30条第1項適用申請有り 2008年1月10日 インターネットアドレス「http://mend.endojournals.org/cgi/rapidpdf/me.2007−0313vl」に発表
【新規性喪失の例外の表示】特許法第30条第1項適用申請有り 2008年1月22日 独立行政法人 産業技術総合研究所、インド生物工学庁主催の「産総研(日本)−生物工学庁(インド)ワークショップ」に文書をもって発表
【新規性喪失の例外の表示】特許法第30条第1項適用申請有り 2008年1月31日〜2月1日 独立行政法人 産業技術総合研究所 ライフサイエンス分野融合会議、産業技術連携推進会議 ライフサイエンス部会・バイオテクノロジー分科会主催の「ライフサイエンス分野融合会議 ライフサイエンス部会・バイオテクノロジー分科会 合同研究発表会」に文書をもって発表
【出願人】(301021533)独立行政法人産業技術総合研究所 (6,529)
【Fターム(参考)】
[ Back to top ]