
製麦工程におけるメチルジャスモン酸合成をモニターする方法

【課題】製麦時の発芽オオムギ種子中のメチルジャスモン酸の合成をモニターする方法を提供する。
【解決手段】製麦時のジャスモン酸メチルトランスフェラーゼ(HvJMT)の発現レベルを測定することを含む、発芽オオムギ種子中のメチルジャスモン酸の合成をモニターする方法。
【解決手段】製麦時のジャスモン酸メチルトランスフェラーゼ(HvJMT)の発現レベルを測定することを含む、発芽オオムギ種子中のメチルジャスモン酸の合成をモニターする方法。
【発明の詳細な説明】
【技術分野】
【0001】
本発明は、製麦工程において、発芽オオムギ種子中のメチルジャスモン酸の合成をモニターする方法に関する。より具体的には、本ホルモンの合成酵素をコードする遺伝子の発現レベルを測定することによって、メチルジャスモン酸の合成をモニターする方法に関する。
【背景技術】
【0002】
ホルモンの合成度合いは、植物の代謝を決定付ける重要な役割をする。ビールや発泡酒の製造の製麦工程では、原料オオムギの発芽の状態が麦芽品質に大きく影響することが知られている。製麦は、オオムギに適度の水を含ませて発芽させたのち、発芽オオムギを乾燥してその成長を止め、さらに根の部分を除去して、ビールや発泡酒の原料となる麦芽(モルト)を製造する工程をいう。
【0003】
製麦時に重要なホルモンは、ジベレリン(以下「GA」と称する)とアブシジン酸(以下「ABA」と称する)が知られており、GAとABAの合成具合、特に合成のタイミングや量が、麦芽品質を決定付ける重要なファクターであるといわれている(非特許文献1)。
【0004】
一方、製麦時の発現遺伝子と一般的な大麦発芽時の発現遺伝子を比較したところ、製麦時には、一群のジャスモン酸誘導タンパク質(以下「JIP」と称する)の遺伝子発現が顕著に高いことが解り、製麦中にはジャスモン酸(以下「JA」と称する)が合成されていることが示唆された。
【0005】
JAは、傷害によって誘導され、その防御反応をすることが知られている。つまり、製麦という非常に過密状態での発芽による接触傷害や、麦層間の温度、水分条件をなるべく均一な状態にするために、ターナーと呼ばれる攪拌装置で混合する操作が行われるが、その際の傷害により、製麦中では過剰量のJAが誘導されると考えられる。製麦時に認められる麦臭の主要原因物質であるヘキセナールは、JA合成系の中間体からの派生物であり、そのことからも製麦時にJAが合成されていることが予想される。
【0006】
JAは、種々の二次代謝産物の合成を誘導するホルモンとして知られている。その一つとして、フェニルプロパノイド合成系が知られている(非特許文献2)。ビール中のフェノール臭の原因物質として知られる4-ビニルグアイアコール(以下「4VG」と称する)の前駆体であるフェルラ酸は、前記フェニルプロパノイド合成系の中間物質である。また、JAはビール中の酸化臭原因物質の一つである、トランス-2-ノネナール合成の鍵酵素であるリポキシゲナーゼ(LOX2)の発現を誘導する(非特許文献2)ことが知られている。以上のことから、製麦時のJAの変動により麦芽品質が影響されることが推定されるが、いままでのそのような観点での研究は行われていない。
【0007】
最近になって、JAそのものよりもJAからの誘導体が植物中では活性が高いことが示された(非特許文献3)。また、JA誘導体の一つである、メチルジャスモン酸(以下「MeJA」と称する)がアラビドプシスでは生体内の活性型JAであり、JAをMeJAに変換する酵素である、ジャスモン酸カルボキシルメチルトランスフェラーゼ(以下JMTと称する)が生体内活性体JA(MeJA)合成の鍵酵素であることが解った(非特許文献4)。しかしながら、オオムギでは、活性型JA合成に関わる酵素遺伝子はクローニングされていない。
【0008】
一方、JA、およびそれらの誘導体そのものを直接測定することは不可能ではないが、非常に微量成分であり特殊な装置と経験が必要である。特に、MeJAは揮発性が高く、検出はより困難である。
【0009】
【非特許文献1】K. Yamada, Agric. Biol. Chem. 49(2): 429-434 (1985)
【非特許文献2】Wasternack C., and Pathier B (1997) Trend in plant Sci. 2, 302-307
【非特許文献3】Miersh E., et al. (1999) Phytochemistry 50, 353-361
【非特許文献4】Seo H.S. et al. (2001) PNAS 98, 4788-4793
【発明の開示】
【発明が解決しようとする課題】
【0010】
前記の事情に鑑み、製麦工程において麦芽特性を決定付けるMeJAの合成レベルを簡便な方法でモニターすることができるならば、現実の製麦工程を見直して品質の良い麦芽を得ることができるように該工程を適正に評価し管理することが可能となるだろう。このため、本発明者らは、発芽オオムギ中のMeJAの合成に関わる酵素に注目し、当該ホルモンの合成を適正に把握可能な酵素遺伝子を研究してきた。
【0011】
したがって、本発明の目的は、発芽オオムギ種子中のMeJAの合成を、当該ホルモンの合成に関わる特定の酵素遺伝子の発現を測定することによってモニターする簡便な方法を提供することである。
【課題を解決するための手段】
【0012】
本発明者らは、オオムギのMeJA合成系に関わる酵素遺伝子をクローニングし、それらの塩基配列とアミノ酸配列を決定し、この配列に基づいて、該酵素遺伝子の発現レベルと活性型JAによって誘導されることが知られているJIP遺伝子の発現の指標になる活性型JA量を比較し、これらの間に正の相関関係があること、およびビールにおけるフェノール臭原因物質として知られている4VGの前駆体であるフェルラ酸合成とも正の相関があることを見出し、本発明を完成させた。したがって、本発明は以下のものからなる。
【0013】
本発明は、第1の態様において、製麦工程における発芽オオムギ種子中のMeJAの合成をモニターする方法であって、該種子中のジャスモン酸カルボキシルメチルトランスフェラーゼ(以下「HvJMT」と称する)遺伝子の発現レベルを測定することを含む方法を提供する。
【0014】
別の実施形態において、前記HvJMT遺伝子の発現レベルは、MeJA合成量と正の相関関係にある。ここで、正の相関関係とは、発芽オオムギ種子中のHvJMT遺伝子の発現レベルが増加すると、MeJAの合成量も相対的に増加し、逆に、HvJMT遺伝子の発現レベルが減少すると、MeJAの合成量も相対的に減少することを表す。
【0015】
別の実施形態において、前記遺伝子は、配列番号2に示されるアミノ酸配列をコードする塩基配列又はその変異配列を含む。
【0016】
別の実施形態において、前記遺伝子は、配列番号1に示される塩基配列又はその変異配列を含む。
【0017】
別の実施形態において、前記遺伝子の発現レベルは、PCR法又はRT-PCR(逆転写酵素−ポリメラーゼ連鎖反応)法によって測定される。
【0018】
別の実施形態において、前記遺伝子の発現レベルは、核酸ハイブリダイゼーションによって測定される。
【発明の効果】
【0019】
本発明により、製麦工程でのMeJAの合成レベルを、当該ホルモンの合成遺伝子の発現レベルとしてモニターすることによって、非常に簡便な方法で麦芽の品質、例えばビール中の酸化臭原因物質の一つであるトランス-2-ノネナール合成の鍵酵素であるリポキシゲナーゼの発現レベルや、フェノール臭の原因物質として知られる4VGの前駆体であるフェルラ酸生成量を管理することが可能である。
【発明を実施するための最良の形態】
【0020】
本発明は、製麦工程における発芽オオムギ種子中のMeJAの合成をモニターする方法であって、該種子中のHvJMT遺伝子の発現レベルを測定することを含む方法を提供する。
【0021】
本明細書で使用する製麦とは、ビールや発泡酒の原料となる麦芽(モルト)の製造工程を指し、この工程では、オオムギに適度の水を含ませて、それを発芽させたのち、発芽オオムギを乾燥してその成長を止め、根の部分を除去して麦芽が得られる。
【0022】
本研究により、オオムギの発芽の際にMeJAの生合成に関与する酵素として、HvJMTが同定された。
【0023】
具体的には、本発明者らは、アラビドプシスJMT-cDNA(GenBank AY008434)の既知の配列に基づいて、相同性の高いオオムギEST(GenBank BI959950)を見出し、当該配列に基いてプライマーを作製し、オオムギの発芽3日目の種子からの全RNAを鋳型にして逆転写酵素-ポリメラーゼ連鎖反応(RT-PCR)を行いcDNAを得、オオムギのHvJMTの全翻訳領域を含む塩基配列と、対応するアミノ酸配列とを決定した。HvJMTの塩基配列とアミノ酸配列はそれぞれ、配列番号1及び2に示されている。決定された配列はこれまで報告がなく、したがって新規である。
【0024】
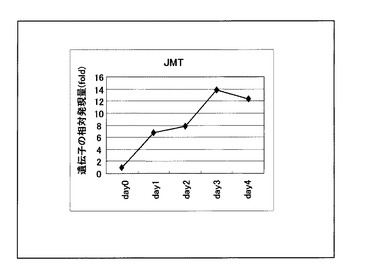
本発明におけるHvJMT遺伝子は、前記酵素JMTをコードする遺伝子を意味する。本発明者らは、製麦時のHvJMT遺伝子の発現を解析した結果、オオムギ種子の浸漬後1日目より発現の上昇が認められ、3日目にはほぼピークになった(図1)。これらの発現変動は、活性型JAによって誘導される遺伝子として知られているJIPの発現変動と非常に類似しており、このことは、製麦時の活性型JAはMeJAであり、HvJMT遺伝子の発現レベルを測定することによって、製麦時のMeJAの合成量をモニターすることが可能であることを示している。
【0025】
したがって、本発明の実施形態により、前記HvJMT遺伝子の発現レベルは、製麦時の活性型JAと想定できるMeJA合成量と正の相関関係にある。
【0026】
さらに、製麦時のHvJMTの変動は、ビールにおけるフェノール臭の原因物質である4VGの前駆体であるフェルラ酸量とも正の相関にあることが解った。
【0027】
したがって、本発明において、製麦時のMeJAの生合成レベルを、前記ホルモン合成遺伝子の発現レベルとしてモニターするならば、従来法に比べて非常に簡易な方法であるために、製麦工程における麦芽の品質等を含む工程管理が熟練者のみならず非熟練者であっても容易に行いうる。
【0028】
本発明の方法の手順について、以下にさらに詳細に説明する。
本発明によれば、前記発現レベルは、HvJMT遺伝子又は少なくとも10塩基、好ましくは少なくとも15塩基のその断片と、前記種子から得られた核酸サンプルとの核酸ハイブリダイゼーションによって測定されうる。核酸ハイブリダイゼーションは、DNA-DNAハイブリダイゼーション、DNA-RNAハイブリダイゼーション又はRNA-RNAハイブリダイゼーションのいずれでもよく、例えばノザンハイブリダイゼーション、DNAマイクロアレイ(cDNAマイクロアレイ、フィルターアレイを含む)、DNAチップ法などの手法を利用することができる。
【0029】
HvJMT遺伝子は、ゲノムDNA又はcDNA、好ましくはcDNAである。また、これらの遺伝子は、好ましくは固相に結合されているのがよい。固相として、ナイロンフィルター、シラン処理ガラス、基板、例えばポリ-L-リジンコートガラス、シリコン処理ガラス、ポリマー(メチルメタクリレートポリマー)などがある。
【0030】
ノザンハイブリダイゼーション法は、一般にRNA配列の検出、定量のために使用される。本発明では、製麦時の発芽3日目のオオムギの発芽種子から根を除去した残りの部分(胚、胚乳、皮)を液体窒素で凍結、粉砕し、既知の手法、例えばグアニジウム法、フェノール/SDS法などの手法により全RNAを得るか、あるいは、RNAをオリゴ(dT)セルロースカラムに通してpoly(A)+RNAを得て、これらのRNAをノザン法のサンプルとする(E.E. Karrerら (1991) Mol. Biol. 16: 797-805)。RNAサンプルを、アガロースゲル電気泳動にかけてサイズ分離し、その後、ナイロンもしくはニトロセルロースメンブレンにRNAを転写し、HvJMTのラベル化cDNA又はその断片をプローブとしてハイブリダイゼーションを行い、目的の遺伝子を検出、定量する(例えば、分子生物学実験プロトコルI(1997年), 西野・佐野共訳, 丸善株式会社)。
【0031】
DNAマイクロアレイ法では、ガラスやフィルターなどのアレイ(例えば、ナイロンフィルター、ポリマー、又は、ポリL-リシンコートもしくはシラン化スライドガラス)上に目的酵素をコードするcDNA又はそのセンス鎖もしくはアンチセンス鎖、あるいはそれらの断片をプローブとして固定化する。このとき市販のアレイ機(例えば、SPBIO2000(日立ソフトエンジニアリング社製)、GMS417 Arrayer(寶酒造)など)を用いて固定化を実施してもよい。一方、発芽3日目のオオムギの発芽種子を液体窒素で凍結、粉砕し、前記の既知の手法によりpoly(A)+RNAを得たのち、poly(A)+RNAから逆転写反応、(例えば32P-dNTPか又は、Cy3-dUTPもしくはCy5-dUTP(アマシャムファルマシアバイオテク社製)による)ラベルの核酸取込みを経て得られたラベル化cDNAを得る。アレイ上の固定化プローブとラベル化cDNAとのハイブリダイゼーションを行い、目的の遺伝子を検出、定量する(例えば、DNAマイクロアレイと最新PCR法(2000年)村松、那波監修、秀潤社)。
【0032】
さらに、DNAチップ法では、例えば20μmx20μmの方形の基板上に、目的酵素のゲノムDNAによってコードされるmRNAから公知の手法で作製されたcDNAの断片をプローブとして固定化し、サンプルから前記と同様に作製されたラベル化cDNAとチップ上のプローブとのハイブリダイゼーションを行うことによって、目的の遺伝子を検出、定量する(例えば、DNAマイクロアレイと最新PCR法(2000年)村松・那波監修、秀潤社)。
【0033】
前記の方法で使用されるプローブは、例えば次のようにして作製することができる。JMTをコードするcDNAは、発芽3日目のオオムギの発芽種子から全RNAを得、これを鋳型とし、アラビドプシスJMT-cDNA(GenBank AY008434)の既知の配列に基づいて、相同性の高いオオムギEST(GenBank BI959950)を見出し、当該配列に基いて設計した約10塩基〜約100塩基、好ましくは約15〜約50塩基、さらに好ましくは約20〜約30塩基のプライマーを用いてRT-PCRを行ったあと、cDNAを抽出、回収し、適当なベクターにクローニングして増幅することによって得ることができる。必要ならば、さらに5’RACE及び3’RACEを行い、cDNAの5’及び3’欠失部分を回収してもよい。決定された配列に基づいて全翻訳領域を含むプラスミド(pHvJMT-F)を作製し、これを鋳型として、例えばM13 primer M4、M13 primer RV(タカラバイオ社製)を用いてPCRを行い、PCR産物をプローブとすることができる。プローブは、cDNA又はそのセンス鎖もしくはアンチセンス鎖の全長であってもよいし、あるいはその部分長であってもよいし、あるいは5’もしくは3’非翻訳領域のみであってもよい。DNAチップでは、約10〜約50塩基長、好ましくは約15〜約30塩基長、より好ましくは約20〜約25塩基長のプローブが使用されるが、これらに限定されない。また、DNAマイクロアレイやノザンハイブリダイゼーションでは、少なくとも約10塩基長以上から全長、好ましくは約30塩基長以上から全長、より好ましくは約50塩基長以上から全長である。一般に、プローブが長いほどハイブリダイゼーション効率がよくなり、感度は上がる。逆に、プローブが短いほど感度は下がるが、特異性が上がる。目的遺伝子が遺伝子ファミリーを形成しており、それらの発現状態を区別して検出するときなどは、短いプローブを選択する。固相上のプローブは、通常0.1μg〜0.5μgの溶液を用いてスポットする。
【0034】
プローブあるいはサンプルcDNA上のラベルは、アイソトープ(例えば32P、33P、35S)又は蛍光(フルオレサミン及びその誘導体、ローダミン及びその誘導体、FITC、Cy3、Cy5)のいずれでもよい。本発明に係る遺伝子を同時に検出、定量するときには、それぞれ異なる色の蛍光でラベルすることもできる。
【0035】
ハイブリダイゼーション条件は、中程度又は高ストリンジェントな条件、好ましくは高ストリンジェントな条件、例えば、2〜6xSSC、0.1〜1%SDS溶液、50〜70℃、約1時間〜一晩の条件である。ストリンジェントな条件とは、前記サンプルcDNAと固相化プローブとのより特異的結合を可能とする条件である。一般に、塩濃度、有機溶媒(例えば、ホルムアミド)、温度条件に応じて低、中程度、高ストリンジェンシーが決まる。特に、塩濃度を減じるか、有機溶媒濃度を増加させるか、またはハイブリダイゼーション温度を上昇させるかによってストリンジェンシー(stringency)を増加させることができる。例えば、塩濃度は、塩化ナトリウム約500 mM以下およびクエン酸三ナトリウム約50 mM以下、好ましくは塩化ナトリウム約250 mM以下およびクエン酸三ナトリウム約25 mM以下である。有機溶媒濃度は、ホルムアミド約35%以上、好ましくは約50%以上である。温度は、約30℃以上、好ましくは約37℃以上、より好ましくは約42℃以上である。他の条件としては、ハイブリダイゼーション時間、洗浄剤(例えば、SDS)の濃度、およびキャリアーDNAの存否等であり、これらの条件を組み合わせることによって、様々なストリンジェンシーを設定することができる。洗浄のためのストリンジェントな温度条件は、約25℃以上、好ましくは約42℃以上、より好ましくは約60℃以上である。好ましい洗浄条件は、0.2xSSC、0.1%SDS溶液、5分の条件である。
【0036】
検出は、マイクロアレイ用の検出器、例えばScanArray 4000, 5000(General Scanning 社製、USA)、GMS418 Array Scanner(寶酒造社製)、Gene Tip Scanner(日本レーザ電子社製)などを使用することができる。定量に際しては、一定量の適当なDNA又はRNA(例えばλpolyA+RNA(タカラバイオ社製)からのラベル化cDNA、Lucidea Universal Score Card(アマシャムバイオサイエンス社製)など)、あるいは生体内で恒常的に発現している遺伝子、例えばアクチン遺伝子などをコントロールとし、バンド濃度を測定し、サンプル対コントロール比を求めることによって、サンプル中の目的の遺伝子の発現レベルを相対値として定量することができる。
【0037】
本発明の方法で特に好ましいDNAマイクロアレイの作製について、固相担体表面で直接オリゴヌクレオチドを合成する方法(オン・チップ法)及び予め調製したプローブを固相表面に固定化する方法が知られており、本発明ではそのいずれも使用できるが、後者の方法が好ましい。オン・チップ法は、光照射で選択的に除去される保護基の使用と、半導体製造に利用されるフォトリソグラフィー技術および固相合成技術とを組み合わせて、微少なマトリックスの所定の領域での選択的合成を行う方法(マスキング技術:例えば、S.P.A. Fodor(1991) Science 251:767)である。一方、予め調製したオリゴ−又はポリヌクレオチドプローブを固相表面に固定化する方法は、官能基を導入したオリゴヌクレオチドまたはポリヌクレオチドを合成し、表面処理した固相担体表面にオリゴヌクレオチドまたはポリヌクレオチドを点着し、共有結合させる(例えば、J.B. Lamtureら(1994) Nucl. Acids Res. 22:2121-2125; Z. Guoら(1994) Nucl. Acids Res. 22:5456-22:5456-5465)。オリゴ−又はポリヌクレオチドは、一般的には、表面処理した固相担体にスペーサーやクロスリンカーを介して共有結合される。ガラス表面にポリアクリルアミドゲルの微小片を整列させ、そこに合成オリゴヌクレオチドを共有結合させる方法も知られている(G. Yershovら(1996) Proc. Natl. Acad. Sci. USA 94:4913)。また、シリカマイクロアレイ上に微小電極のアレイを作製し、電極上にはストレプトアビジンを含むアガロースの浸透層を設けて反応部位とし、この部位をプラスに荷電させることでビオチン化オリゴヌクレオチドを固定し、部位の荷電を制御することで、高速で厳密なハイブリダイゼーションを可能にする方法も知られている(R.G. Sosnowskiら(1997) Proc. Natl. Acad. Sci. USA 94:1119-1123)。
【0038】
サンプルについては、前記のとおり、製麦時の発芽3日目のオオムギの発芽種子を液体窒素で凍結、粉砕し、既知の手法、例えばフェノール/クロロホルム法、グアニジウム法又はフェノール/SDS法により全RNAを得るか、又は、RNAをオリゴ(dT)セルロースカラムに通してpoly(A)+RNAを得て(E.E. Karrerら (1991) Mol. Biol. 16: 797-805)、サンプルとすることができる。
【0039】
本発明において、前記ハイブリダイゼーションは、HvJMTcDNA又はその断片と、発芽オオムギ種子の全RNA又はpoly(A)+RNAから調製されたラベル化cDNAサンプルとの間で行われる。HvJMTcDNA又はその断片の具体例は、以下のとおりである。
【0040】
HvJMTのcDNA又は断片は、配列番号2のアミノ酸配列をコードする塩基配列、その変異体、もしくはそれらの相補鎖、又は少なくとも10塩基、好ましくは少なくとも15塩基からなるそれらの断片であり、ここで該変異体は、該塩基配列において1以上、好ましくは1又は数個の塩基の置換、欠失又は付加を有するもの、あるいは該塩基配列と70%以上、80%以上、90%以上、95%以上もしくは98%以上の相同性を有するものである。さらに具体的には、HvJMTcDNA又は断片は、配列番号1の塩基配列、その変異体、もしくはそれらの相補鎖、又は少なくとも10塩基、好ましくは少なくとも15塩基からなるそれらの断片であり、ここで該変異体は、該塩基配列において1以上、好ましくは1又は数個の塩基の置換、欠失又は付加を有するもの、あるいは該塩基配列と70%以上、80%以上、90%以上、95%以上もしくは98%以上の相同性を有するものである。
【0041】
本明細書中で使用される「数個」とは、2〜約10個、好ましくは2〜5個である。また「変異体」は、該当の酵素の特性を実質的に変更しない程度の変異を有するものであり、限定されないが、好ましくはオオムギの異なる品種間で認められるような(突然)変異、例えば多型性に関わるような変異、スプライス変異、あるいは遺伝暗号の縮重による変異などを有するものである。さらに、本明細書中で使用される「断片」とは、前記の特定の塩基配列において少なくとも10塩基、15塩基、20塩基、25塩基、30塩基、50塩基、100塩基、もしくは150塩基から、酵素の成熟配列をコードする翻訳領域の全長未満までの翻訳領域断片、あるいは、少なくとも10塩基、15塩基、20塩基、25塩基、30塩基、50塩基、100塩基もしくは150塩基の、非翻訳領域のみ、又は翻訳領域と非翻訳領域からなる連続する塩基配列からなる断片を指す。
【0042】
本発明はさらに、配列番号1、2に示される塩基配列及びアミノ酸配列を含むMeJA合成酵素及びそれをコードする遺伝子、並びに、それらの相補鎖、それらの変異体、及びそれらの断片、好ましくは活性部位を含む断片、にも関する。ここで、変異体は、配列番号1、2に示される塩基配列又はアミノ酸配列において1以上、好ましくは1又は数個の塩基又はアミノ酸の置換、欠失又は付加を有するもの、あるいは該塩基配列又はアミノ酸配列と70%以上、80%以上、90%以上、95%以上もしくは98%以上の相同性を有するものである。また、断片は、配列番号1、2に示される塩基配列又はアミノ酸配列において少なくとも10、15、20、25、30、50、100、もしくは150塩基/アミノ酸から、酵素の翻訳領域又は成熟配列の全長未満までの断片である。
【0043】
本明細書中で使用される「相同性」は、%同一性と同義であり、公知のBLASTN、BLASTX、BLASTPなどのBLASTプログラムを用いて決定することができる(S.F. Altschulら(1990) J. Mol. Biol. 215:403-410)。
【0044】
サンプル中の対象遺伝子の発現量を定量する別の方法として、内部標準物質を用いたRT-PCR法も使用できる(例えば、PCR法最前線(1996年)関谷、藤永編、共立出版)。使用する内部標準としては細胞骨格の構成成分であるβアクチンなどの組織によって発現量に差のないと考えられるハウスキーピング遺伝子がよく用いられる。この方法では標的mRNA量が内部標準試料に対して多いか少ないかといった相対的な結果しか得られないが、mRNAの抽出効率やcDNAの合成効率を考慮しなくてもよいという利点があるため、広く用いられている。PCRによる増幅産物は、ある範囲内では指数関数的に増加するが、それを超えると頭打ちの状態になる。そのため、RT-PCRにおける発現遺伝子の定量は、増幅産物が指数関数的に増加する範囲で行わなくてはならない。また、PCRを酵素化学的に分析し、特に標準試料を用意せずに定量的PCRを行う、カイネティクス分析法も使用することができる。PCR反応は、反応がプラトーに達する前には指数関数的に生成物の量が増加する。そこで、PCRのサイクル数を横軸に、反応生成物量を対数の縦軸にプロットすると、生成物量が直線的に増加する時期があるが、この直線を延長し、y軸と交差する点を求めれば、理論的には初期鋳型量を算出することができる(バイオ実験イラストレイティット3「本当に増えるPCR」(1998年)中山広樹著、秀潤社)。実際には、一つのサンプルについてのPCR反応中に、数サイクルごとに反応液をサンプリングして、PCR産物の量を定量し、グラフにプロットしていく。そうして得られたグラフの指数増加期のポイントに対して回帰分析を行い、y切片を求める。
【0045】
本発明ではさらに、この原理を応用し、サンプリングせずにPCR反応中にリアルタイムでこの回帰分析を行うことのできる、リアルタイム定量PCRを使用することもできる。PCR生成物が特異的に蛍光標識される反応系で、蛍光強度を検出する装置の備わった温度サイクラー装置によりPCR反応を行うと、反応中の生成物の量をサンプリングの必要なくリアルタイムでモニターでき、その結果をコンピュータで回帰分析することができる。PCR生成物を標識する方法としては、蛍光標識したプローブを用いる方法と、2本鎖DNAに特異的に結合する試薬を用いる方法とがある。前者では、2種類の蛍光物質で標識し蛍光をクエンチング作用で抑えてあるプローブを、鋳型の二つのプライマーと相補的な配列の間にアニールさせる方法がよく知られている。プローブは、PCR反応が行われるとTaqポリメラーゼのもつ5’-3’エキソヌクレアーゼ活性により分解され、蛍光を発するようになる。その蛍光量は、PCR生成物の量を反映する。後者では、SYBR greenなどが用いられる。PCR反応物が検出限界に到達するときのサイクル数(CT)と初期鋳型量とは逆相関の関係にあることから、リアルタイム測定法ではCTを測定することによって初期鋳型量を定量している。数段階の既知量の鋳型を用いてCTを測定し検量線を作製すれば、未知試料の初期鋳型量の絶対値を算出することができる。
【0046】
RT-PCRで使用する逆転写酵素は、例えばM-MLV RTase、ExScript RTase(タカラバイオ社製)、AMV Reverse Transcriptase(Life Science社製)、Super Script II RT(GIBCO BRL社製)などを使用することができる。
【0047】
PCR条件は、例えばDNAの変性を94〜95℃で5秒〜5分間、プライマーのアニーリングを50〜70℃で10秒〜1分間、伸長反応を72℃で30秒〜3分間行い、これを1サイクルとして20〜40サイクルほど行い、最後に72℃で30秒〜10分間の伸長反応を行うことができる。
【0048】
プライマーは、鋳型としてのHvJMT遺伝子(配列番号1)又はその変異体(前記定義)のセンス及びアンチセンス鎖に相補的な配列である。プライマーのサイズは、通常、約15〜約30塩基長、好ましくは約17〜約25塩基長である。プライマーの例は、特に限定されないが、配列番号11及び12である。
【0049】
以下に、本発明を実施例によってさらに具体的に説明するが、本発明はこれらの実施例によって制限されないものとする。
【実施例1】
【0050】
オオムギからのRNA調製と発芽時のホルモン合成酵素をコードするcDNAの調製
(1A)オオムギからのRNAの調製
オオムギ(Hordeum vulgare cv.Harrington)を1%NaClOで10分間浸漬することにより滅菌後、水洗いし、20℃、8時間水中で浸漬した。浸漬後、水に浸した濾紙上にまき、18℃、暗所に置いた。なお、浸漬終了時点を0日目とし、以後5日間発芽を行った。
【0051】
発芽3日目の種子を液体窒素で凍結し、粉砕後、全RNAをKarrer, E.Eら((1991) Mol. Biol. 16: 797-805)の方法により抽出した。全RNAより、poly(A)+RNAをoligo-dT30<super>(Roche diagnostics)により取得した。
【0052】
温室内で育成したオオムギ幼葉を、7時間、室温、暗所で風乾させた。乾燥葉を液体窒素で粉砕後、前記した方法により、全RNA、poly(A)+RNAを抽出した。
【0053】
製麦時、浸漬直後から、1日おきに4日目までのオオムギより前記方法により全RNA、poly(A)+RNAを抽出した。
【0054】
(1B)オオムギ発芽時に発現する、JMTをコードするcDNA(HvJMT)の取得
発芽3日目全RNA1μgを鋳型として、ThermoScript RT-PCR system (Invitrogen Corp.)により、oligo(dT)20をプライマーとしてcDNAを合成した。アラビドプシスより単離された、JMT-cDNA(AY008434)の塩基配列に対して相同性の高いオオムギEST(BI959950)を見出し、BI959950に基づいて設計したTO-510:5’-ATgTTATCgCggACTTATgC-3’ (配列番号3)およびTO-511: 5’-gACCATTgggAgCCTTTC-3’ (配列番号4)を用いてRT-PCRを行った。なお、PCRは、95℃ 5分後、94℃ 30秒、60℃ 1分、72℃ 1分のセットを30サイクル、72℃ 5分の反応系で行った。
【0055】
RT-PCRによって増幅した約0.4kbpのDNAを抽出し、pCR2.1-TOPO vector (Invitrogen Corp.)にクローニング、塩基配列を確認し、pHvJMT-pを取得した。
【0056】
完全長のHvJMTcDNAを取得するために、 発芽3日目のpoly (A)+ RNA を用いて5’ 、および3’ RACE をSMART RACE cDNA amplification kit (CLONTECH) により行った。poly (A)+ RNA 0.5μgを用いて、前記SMART RACE cDNA amplification kitに基づき5’、および3’RACE用のcDNAを合成した。合成したcDNAを鋳型として、プライマーは、取得した pHvJMT-p の配列をもとに、5’RACEには TO-516: 5’-ATCTTCCggAgCCTTTgAgAg-3’ (配列番号5)、TO-517: 5’-gACCATTgggAgCCTTTCAC-3’ (配列番号6)を、3’RACEには、TO-518: 5’-gTAAATgCCATACACAgCCAATg-3’(配列番号7) 、TO-519: 5’-CATACACAgCCAATgCCTTCA-3’(配列番号8)を用いて2ステップPCRを行った。1回目のPCRは、5’RACE、および3’RACEには、それぞれTO-517、TO-518を用いて、94℃ 5秒、72℃ 3分のセットを5サイクル、94℃ 5秒、70℃ 10秒、72℃ 3分のセットを5サイクル、94℃ 5秒、68℃ 10秒、72℃ 3分のセットを20サイクルで行った。1回目の反応液を鋳型として2回目のPCRを、5’RACE、および3’RACEには、それぞれTO-516、TO-519を用いて、95℃ 5分後、94℃ 30秒、60℃ 1分、72℃ 1分のセットを30サイクル、72℃ 5分の反応系で行った。5’RACEおよび、3’RACEにより取得したそれぞれ0.5kbp、1.0kbpの断片を前記pCR2.1-TOPO vectorにクローニング、塩基配列を確認し、pHvJMT-U 、およびpHvJMT-Dを取得した。
【0057】
pHvJMT-U 、およびpHvJMT-D の塩基配列より、HvJMT-cDNAのコード領域を推定し、全コード領域を含む遺伝子の取得を、pHvJMT-U 、およびpHvJMT-Dの混合物を鋳型として、TO-543:5’- gAATTCATggCCTCCCAACCgATgg -3’ (配列番号9)およびTO-544: 5’- CCCgggATgTggTgCATggCACCTT -3’(配列番号10)を用いたPCRにより行った。なお、PCRは、95℃ 5分後、94℃ 30秒、60℃ 1分、72℃ 1分のセットを25サイクル、72℃ 5分の反応系で行った。取得した1.2kbpの断片を前記pCR2.1-TOPO vectorにクローニング、塩基配列を確認し、pHvJMT-fを取得した。取得したHvJMT-cDNAの塩基配列を配列番号1に、推定アミノ酸配列を配列番号2にそれぞれ示した。
【実施例2】
【0058】
取得したHvJMT遺伝子の翻訳産物の活性測定による確定
(2A)HvJMT-cDNAの大腸菌内発現ベクターの構築、大腸菌内発現
取得したpHvJMT-fの全コード領域を含むEcoRI-SmaI断片を、pGEX4T-1(Amersham Pharmacia Biotech.)のEcoRI-SmaI部位に導入し、pGEX-HvJMTを作成した。作成したpGEX-HvJMTは、大腸菌BLR(DE3)に形質転換した。
【0059】
pGEX-HvJMT:BLR(DE3)からGST融合したHvJMTを、定法によりIPTG誘導により大腸菌内で発現させた。大腸菌を破砕後、可溶性画分を分取し、Glutation Sepharose 4B(Amersham Pharmacia Biotech.)によりGST-HvJMTを精製した。なお、ベクターとして用いた、pGEX4T-1:BLR(DE3)より、同様の操作で調製した精製物を、コントロールとして以後の実験に用いた。
【0060】
(2B)HvJMTの活性測定による同定
GST-HvJMTを用いた、JMTの活性測定は、Seo H.S. et al. PNAS 98 p4788-4793 (2001)に記載された方法により、(±)JAを基質、S-アデノシルメチオニン(以下SAMと称する)をメチル基供与体として反応させ、GC-MSによって反応生成物のMeJAを検出することにより行った。
【0061】
10mM(±)JA、10mM SAMの共存下で、前記の操作で取得した精製物10μlを、反応バッファー(10mM 50mM Tris-HCl(pH7.5)、20mM KCl、10mMβ-mercaptoethanol)中(全液量50μl)で20℃、30分間反応させた。反応終了後、反応液を150μlのヘキサンで抽出し、ヘキサン抽出物をGC-MSで解析した。GC-MS解析は島津社製GC-MS QP-2010を使用し、カラムとしてZB-5(カラム長15m、内径0.25mm、膜厚0.25μm;Phenomenex社製)を用いた。キャリアガスにはHeガスを使用し、線速度100cm/分で一定流量流した。温度条件は気化室温度280℃、インターフェース温度280℃とし、カラム昇温プログラムは110℃(2分)→(10℃/分)→200℃→(30℃/分)→290℃(5分)とした。尚、この条件でMeJAの溶出時間はおよそ5.8分となった。MeJAの同定は、市販の質量スペクトルライブラリーデータを用いて、GC-MS分析においてm/z=70〜300のスキャン分析を行った結果をライブラリーから検索して行った。
【0062】
その結果、GST-HvJMTの反応により、MeJAが生成されることが解り、HvJMT-cDNAは、JMT活性をもつタンパク質をコードする遺伝子であることが証明された。HvJMTの基質特異性を調べるために、基質として(±)JA、ジヒドロジャスモン酸、リノレン酸、サリチル酸、ベンゼン、ケイ皮酸を用いて、[14C]methyl SAMをメチル基供与体として反応を行い、酢酸エチル抽出後の酢酸エチル層の放射線取り込み量により検討した。(±)JAを基質とした際の放射線取り込み量を100%の活性として、各基質の活性の比を表1に示した。参考として、アラビドプシス由来のJMTの基質特異性も示す。
【0063】
【表1】

【0064】
その結果、HvJMTは、アラビドプシスのJMT(AtJMT)と同様に、(±)JA、ジヒドロジャスモン酸を基質として認識することが解った。一方、AtJMT は、ジヒドロジャスモン酸を基質として認識はするものの、JAに比べてその認識は2割程度であることが報告されているが、HvJMTはジヒドロジャスモン酸に対して(±)JAと同等あるいはそれ以上に、基質として認識することが解った。
【実施例3】
【0065】
製麦時のHvJMT、ジャスモン酸誘導タンパク質遺伝子(JIP23)の発現解析
(3A)製麦時のHvJMT発現の推移
製麦時、浸漬直後(0日目)、浸漬後1、2、3、4日目のサンプルより調製した全RNA 1μgを鋳型として、ThermoScript RT-PCR system (Invitrogen Corp.)により、oligo(dT)20をプライマーとしてcDNAを合成した。合成したcDNAを鋳型として、TO-512:5’- TgCCgTAAATgCCATACACA -3’ (配列番号11)およびTO-513: 5’- CAgAACgCgTCCCTACCAg -3’ (配列番号12)を用いたRT-PCRを行った。なお、増幅断片の定量比較のための内部標準としては、前記鋳型cDNAを用いて、オオムギのアクチン遺伝子(AY14545)の配列に基づいて設計した、TO-635:5’-CCCCATgCCATTCTTCgTTT-3’ (配列番号13)およびTO-636: 5’- CTgggCCAgACTCgTCgTA-3’ (配列番号14)を用いたRT-PCR増幅産物を用いた。PCRは、95℃ 5分後、94℃ 30秒、60℃ 1分、72℃ 1分のセットを25サイクル、72℃ 5分の反応系で行った。
【0066】
製麦工程における、HvJMTの経時的な発現変動を図1に示した。図1より、HvJMTは浸漬後1日目から発現の上昇が認められ、3日目にはほぼピークとなり、4日目まで継続的に発現することが解った。
【0067】
(3B)製麦時のジャスモン酸誘導遺伝子発現の推移
ジャスモン酸誘導タンパク質として知られている、JIP23(Plant Mol Biol. 1992 May;19(2):193-204.)をコードする遺伝子の製麦時の発現を検討した。
【0068】
JIP23をコードするcDNA配列を、ガラス担体に固定化した(DNAチップ実験まるわかり(2004)、佐々木、青柳編 羊土社)。一方、製麦時、浸漬直後、浸漬後1、2、3、4日目のサンプルより調製した全RNA200ngを用いて、Agilent Low RNA Input Fluorescent Liniear Amplification Kit (Agilent Technologies)により標識、固定化したガラス担体とのハイブリダイゼーション、洗浄を行い、シグナル強度の相対値を、浸漬直後のRNAを用いた際の蛍光強度との比として求めた。
【0069】
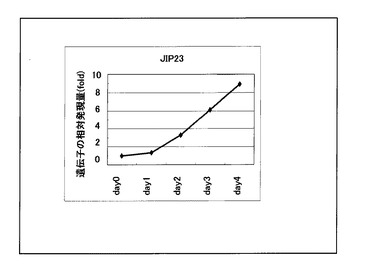
結果を図2に示した。図2より、JIP23は浸漬1日目から発現の上昇が認められ、4日目まで継続的に発現が上昇することが解り、HvJMTの発現パターンを解析することにより、大麦内部でJAによる遺伝子発現誘導の様子がモニターできることが解った。
【実施例4】
【0070】
製麦時のフェルラ酸量の推移
製麦時に大麦サンプルから熱水で抽出されるフェルラ酸量の推移を検討した。浸漬直後(0日目)、浸漬後1、2、3、4日目の大麦サンプルを凍結乾燥し、各サンプルにつき10粒を家庭用ミルで粉砕後、70度の超純水を8ml添加し1時間抽出した。抽出後、遠心により残渣を除去し、上清を回収した。回収した上清のうち1mlを分取し、1.5mlの2M塩酸を添加後、ジエチルエーテルで低極性物質を回収した。エーテル抽出物は窒素気流下で乾固した後、1mlの20% MeOH/0.1% ギ酸に溶解後、0.45μmのフィルターで濾過し、HPLC分析に供した。HPLCでの分析は、カラムとしてNucleosil C18 5μ 100Å 4.6mm×250mm (Shimadzu GLC)(カラム温度40度)、移動相として70% MeOH / 0.1% ギ酸(アイソクラティック)を用い、流速1ml/分で分離し、UV検出器を用いて310nmの吸光で検出した。定量は、フェルラ酸の標準品(和光純薬)を用いて検量線を描き、得られたピークの面積から決定した。
【0071】
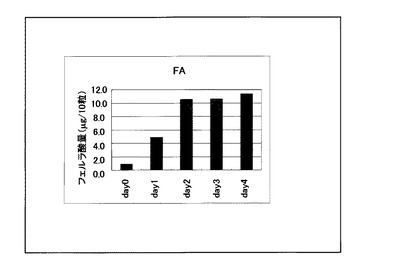
製麦工程における大麦サンプル熱水抽出物中のフェルラ酸含量の変動を図3に示した。図3より、フェルラ酸含量は、浸漬後1日目から2日目にかけて急激に上昇し、2日目以降はその含量が維持されることが解り、HvJMTの発現量と製麦時のフェルラ酸含量に、相関関係があることが解った。
【産業上の利用可能性】
【0072】
ビールや発泡酒の製造の製麦工程での麦芽品質の維持管理に利用できる。さらに、製麦工程の理論付け、麦芽品質に関与する物質合成機構の理論付け、並びに、オオムギ品種間、ロット間における製麦特性の違いの把握などに役立つ。
【図面の簡単な説明】
【0073】
【図1】図1は、製麦中のHvJMT遺伝子発現の経時的な変動を示す。
【図2】図2は、製麦中のJIP23遺伝子発現の経時的な変動を示す。
【図3】図3は、製麦中のフェルラ酸含量の経時的な変動を示す。
【配列表フリーテキスト】
【0074】
配列番号3:人工配列の説明:プライマー
配列番号4:人工配列の説明:プライマー
配列番号5:人工配列の説明:プライマー
配列番号6:人工配列の説明:プライマー
配列番号7:人工配列の説明:プライマー
配列番号8:人工配列の説明:プライマー
配列番号9:人工配列の説明:プライマー
配列番号10:人工配列の説明:プライマー
配列番号11:人工配列の説明:プライマー
配列番号12:人工配列の説明:プライマー
配列番号13:人工配列の説明:プライマー
配列番号14:人工配列の説明:プライマー
【技術分野】
【0001】
本発明は、製麦工程において、発芽オオムギ種子中のメチルジャスモン酸の合成をモニターする方法に関する。より具体的には、本ホルモンの合成酵素をコードする遺伝子の発現レベルを測定することによって、メチルジャスモン酸の合成をモニターする方法に関する。
【背景技術】
【0002】
ホルモンの合成度合いは、植物の代謝を決定付ける重要な役割をする。ビールや発泡酒の製造の製麦工程では、原料オオムギの発芽の状態が麦芽品質に大きく影響することが知られている。製麦は、オオムギに適度の水を含ませて発芽させたのち、発芽オオムギを乾燥してその成長を止め、さらに根の部分を除去して、ビールや発泡酒の原料となる麦芽(モルト)を製造する工程をいう。
【0003】
製麦時に重要なホルモンは、ジベレリン(以下「GA」と称する)とアブシジン酸(以下「ABA」と称する)が知られており、GAとABAの合成具合、特に合成のタイミングや量が、麦芽品質を決定付ける重要なファクターであるといわれている(非特許文献1)。
【0004】
一方、製麦時の発現遺伝子と一般的な大麦発芽時の発現遺伝子を比較したところ、製麦時には、一群のジャスモン酸誘導タンパク質(以下「JIP」と称する)の遺伝子発現が顕著に高いことが解り、製麦中にはジャスモン酸(以下「JA」と称する)が合成されていることが示唆された。
【0005】
JAは、傷害によって誘導され、その防御反応をすることが知られている。つまり、製麦という非常に過密状態での発芽による接触傷害や、麦層間の温度、水分条件をなるべく均一な状態にするために、ターナーと呼ばれる攪拌装置で混合する操作が行われるが、その際の傷害により、製麦中では過剰量のJAが誘導されると考えられる。製麦時に認められる麦臭の主要原因物質であるヘキセナールは、JA合成系の中間体からの派生物であり、そのことからも製麦時にJAが合成されていることが予想される。
【0006】
JAは、種々の二次代謝産物の合成を誘導するホルモンとして知られている。その一つとして、フェニルプロパノイド合成系が知られている(非特許文献2)。ビール中のフェノール臭の原因物質として知られる4-ビニルグアイアコール(以下「4VG」と称する)の前駆体であるフェルラ酸は、前記フェニルプロパノイド合成系の中間物質である。また、JAはビール中の酸化臭原因物質の一つである、トランス-2-ノネナール合成の鍵酵素であるリポキシゲナーゼ(LOX2)の発現を誘導する(非特許文献2)ことが知られている。以上のことから、製麦時のJAの変動により麦芽品質が影響されることが推定されるが、いままでのそのような観点での研究は行われていない。
【0007】
最近になって、JAそのものよりもJAからの誘導体が植物中では活性が高いことが示された(非特許文献3)。また、JA誘導体の一つである、メチルジャスモン酸(以下「MeJA」と称する)がアラビドプシスでは生体内の活性型JAであり、JAをMeJAに変換する酵素である、ジャスモン酸カルボキシルメチルトランスフェラーゼ(以下JMTと称する)が生体内活性体JA(MeJA)合成の鍵酵素であることが解った(非特許文献4)。しかしながら、オオムギでは、活性型JA合成に関わる酵素遺伝子はクローニングされていない。
【0008】
一方、JA、およびそれらの誘導体そのものを直接測定することは不可能ではないが、非常に微量成分であり特殊な装置と経験が必要である。特に、MeJAは揮発性が高く、検出はより困難である。
【0009】
【非特許文献1】K. Yamada, Agric. Biol. Chem. 49(2): 429-434 (1985)
【非特許文献2】Wasternack C., and Pathier B (1997) Trend in plant Sci. 2, 302-307
【非特許文献3】Miersh E., et al. (1999) Phytochemistry 50, 353-361
【非特許文献4】Seo H.S. et al. (2001) PNAS 98, 4788-4793
【発明の開示】
【発明が解決しようとする課題】
【0010】
前記の事情に鑑み、製麦工程において麦芽特性を決定付けるMeJAの合成レベルを簡便な方法でモニターすることができるならば、現実の製麦工程を見直して品質の良い麦芽を得ることができるように該工程を適正に評価し管理することが可能となるだろう。このため、本発明者らは、発芽オオムギ中のMeJAの合成に関わる酵素に注目し、当該ホルモンの合成を適正に把握可能な酵素遺伝子を研究してきた。
【0011】
したがって、本発明の目的は、発芽オオムギ種子中のMeJAの合成を、当該ホルモンの合成に関わる特定の酵素遺伝子の発現を測定することによってモニターする簡便な方法を提供することである。
【課題を解決するための手段】
【0012】
本発明者らは、オオムギのMeJA合成系に関わる酵素遺伝子をクローニングし、それらの塩基配列とアミノ酸配列を決定し、この配列に基づいて、該酵素遺伝子の発現レベルと活性型JAによって誘導されることが知られているJIP遺伝子の発現の指標になる活性型JA量を比較し、これらの間に正の相関関係があること、およびビールにおけるフェノール臭原因物質として知られている4VGの前駆体であるフェルラ酸合成とも正の相関があることを見出し、本発明を完成させた。したがって、本発明は以下のものからなる。
【0013】
本発明は、第1の態様において、製麦工程における発芽オオムギ種子中のMeJAの合成をモニターする方法であって、該種子中のジャスモン酸カルボキシルメチルトランスフェラーゼ(以下「HvJMT」と称する)遺伝子の発現レベルを測定することを含む方法を提供する。
【0014】
別の実施形態において、前記HvJMT遺伝子の発現レベルは、MeJA合成量と正の相関関係にある。ここで、正の相関関係とは、発芽オオムギ種子中のHvJMT遺伝子の発現レベルが増加すると、MeJAの合成量も相対的に増加し、逆に、HvJMT遺伝子の発現レベルが減少すると、MeJAの合成量も相対的に減少することを表す。
【0015】
別の実施形態において、前記遺伝子は、配列番号2に示されるアミノ酸配列をコードする塩基配列又はその変異配列を含む。
【0016】
別の実施形態において、前記遺伝子は、配列番号1に示される塩基配列又はその変異配列を含む。
【0017】
別の実施形態において、前記遺伝子の発現レベルは、PCR法又はRT-PCR(逆転写酵素−ポリメラーゼ連鎖反応)法によって測定される。
【0018】
別の実施形態において、前記遺伝子の発現レベルは、核酸ハイブリダイゼーションによって測定される。
【発明の効果】
【0019】
本発明により、製麦工程でのMeJAの合成レベルを、当該ホルモンの合成遺伝子の発現レベルとしてモニターすることによって、非常に簡便な方法で麦芽の品質、例えばビール中の酸化臭原因物質の一つであるトランス-2-ノネナール合成の鍵酵素であるリポキシゲナーゼの発現レベルや、フェノール臭の原因物質として知られる4VGの前駆体であるフェルラ酸生成量を管理することが可能である。
【発明を実施するための最良の形態】
【0020】
本発明は、製麦工程における発芽オオムギ種子中のMeJAの合成をモニターする方法であって、該種子中のHvJMT遺伝子の発現レベルを測定することを含む方法を提供する。
【0021】
本明細書で使用する製麦とは、ビールや発泡酒の原料となる麦芽(モルト)の製造工程を指し、この工程では、オオムギに適度の水を含ませて、それを発芽させたのち、発芽オオムギを乾燥してその成長を止め、根の部分を除去して麦芽が得られる。
【0022】
本研究により、オオムギの発芽の際にMeJAの生合成に関与する酵素として、HvJMTが同定された。
【0023】
具体的には、本発明者らは、アラビドプシスJMT-cDNA(GenBank AY008434)の既知の配列に基づいて、相同性の高いオオムギEST(GenBank BI959950)を見出し、当該配列に基いてプライマーを作製し、オオムギの発芽3日目の種子からの全RNAを鋳型にして逆転写酵素-ポリメラーゼ連鎖反応(RT-PCR)を行いcDNAを得、オオムギのHvJMTの全翻訳領域を含む塩基配列と、対応するアミノ酸配列とを決定した。HvJMTの塩基配列とアミノ酸配列はそれぞれ、配列番号1及び2に示されている。決定された配列はこれまで報告がなく、したがって新規である。
【0024】
本発明におけるHvJMT遺伝子は、前記酵素JMTをコードする遺伝子を意味する。本発明者らは、製麦時のHvJMT遺伝子の発現を解析した結果、オオムギ種子の浸漬後1日目より発現の上昇が認められ、3日目にはほぼピークになった(図1)。これらの発現変動は、活性型JAによって誘導される遺伝子として知られているJIPの発現変動と非常に類似しており、このことは、製麦時の活性型JAはMeJAであり、HvJMT遺伝子の発現レベルを測定することによって、製麦時のMeJAの合成量をモニターすることが可能であることを示している。
【0025】
したがって、本発明の実施形態により、前記HvJMT遺伝子の発現レベルは、製麦時の活性型JAと想定できるMeJA合成量と正の相関関係にある。
【0026】
さらに、製麦時のHvJMTの変動は、ビールにおけるフェノール臭の原因物質である4VGの前駆体であるフェルラ酸量とも正の相関にあることが解った。
【0027】
したがって、本発明において、製麦時のMeJAの生合成レベルを、前記ホルモン合成遺伝子の発現レベルとしてモニターするならば、従来法に比べて非常に簡易な方法であるために、製麦工程における麦芽の品質等を含む工程管理が熟練者のみならず非熟練者であっても容易に行いうる。
【0028】
本発明の方法の手順について、以下にさらに詳細に説明する。
本発明によれば、前記発現レベルは、HvJMT遺伝子又は少なくとも10塩基、好ましくは少なくとも15塩基のその断片と、前記種子から得られた核酸サンプルとの核酸ハイブリダイゼーションによって測定されうる。核酸ハイブリダイゼーションは、DNA-DNAハイブリダイゼーション、DNA-RNAハイブリダイゼーション又はRNA-RNAハイブリダイゼーションのいずれでもよく、例えばノザンハイブリダイゼーション、DNAマイクロアレイ(cDNAマイクロアレイ、フィルターアレイを含む)、DNAチップ法などの手法を利用することができる。
【0029】
HvJMT遺伝子は、ゲノムDNA又はcDNA、好ましくはcDNAである。また、これらの遺伝子は、好ましくは固相に結合されているのがよい。固相として、ナイロンフィルター、シラン処理ガラス、基板、例えばポリ-L-リジンコートガラス、シリコン処理ガラス、ポリマー(メチルメタクリレートポリマー)などがある。
【0030】
ノザンハイブリダイゼーション法は、一般にRNA配列の検出、定量のために使用される。本発明では、製麦時の発芽3日目のオオムギの発芽種子から根を除去した残りの部分(胚、胚乳、皮)を液体窒素で凍結、粉砕し、既知の手法、例えばグアニジウム法、フェノール/SDS法などの手法により全RNAを得るか、あるいは、RNAをオリゴ(dT)セルロースカラムに通してpoly(A)+RNAを得て、これらのRNAをノザン法のサンプルとする(E.E. Karrerら (1991) Mol. Biol. 16: 797-805)。RNAサンプルを、アガロースゲル電気泳動にかけてサイズ分離し、その後、ナイロンもしくはニトロセルロースメンブレンにRNAを転写し、HvJMTのラベル化cDNA又はその断片をプローブとしてハイブリダイゼーションを行い、目的の遺伝子を検出、定量する(例えば、分子生物学実験プロトコルI(1997年), 西野・佐野共訳, 丸善株式会社)。
【0031】
DNAマイクロアレイ法では、ガラスやフィルターなどのアレイ(例えば、ナイロンフィルター、ポリマー、又は、ポリL-リシンコートもしくはシラン化スライドガラス)上に目的酵素をコードするcDNA又はそのセンス鎖もしくはアンチセンス鎖、あるいはそれらの断片をプローブとして固定化する。このとき市販のアレイ機(例えば、SPBIO2000(日立ソフトエンジニアリング社製)、GMS417 Arrayer(寶酒造)など)を用いて固定化を実施してもよい。一方、発芽3日目のオオムギの発芽種子を液体窒素で凍結、粉砕し、前記の既知の手法によりpoly(A)+RNAを得たのち、poly(A)+RNAから逆転写反応、(例えば32P-dNTPか又は、Cy3-dUTPもしくはCy5-dUTP(アマシャムファルマシアバイオテク社製)による)ラベルの核酸取込みを経て得られたラベル化cDNAを得る。アレイ上の固定化プローブとラベル化cDNAとのハイブリダイゼーションを行い、目的の遺伝子を検出、定量する(例えば、DNAマイクロアレイと最新PCR法(2000年)村松、那波監修、秀潤社)。
【0032】
さらに、DNAチップ法では、例えば20μmx20μmの方形の基板上に、目的酵素のゲノムDNAによってコードされるmRNAから公知の手法で作製されたcDNAの断片をプローブとして固定化し、サンプルから前記と同様に作製されたラベル化cDNAとチップ上のプローブとのハイブリダイゼーションを行うことによって、目的の遺伝子を検出、定量する(例えば、DNAマイクロアレイと最新PCR法(2000年)村松・那波監修、秀潤社)。
【0033】
前記の方法で使用されるプローブは、例えば次のようにして作製することができる。JMTをコードするcDNAは、発芽3日目のオオムギの発芽種子から全RNAを得、これを鋳型とし、アラビドプシスJMT-cDNA(GenBank AY008434)の既知の配列に基づいて、相同性の高いオオムギEST(GenBank BI959950)を見出し、当該配列に基いて設計した約10塩基〜約100塩基、好ましくは約15〜約50塩基、さらに好ましくは約20〜約30塩基のプライマーを用いてRT-PCRを行ったあと、cDNAを抽出、回収し、適当なベクターにクローニングして増幅することによって得ることができる。必要ならば、さらに5’RACE及び3’RACEを行い、cDNAの5’及び3’欠失部分を回収してもよい。決定された配列に基づいて全翻訳領域を含むプラスミド(pHvJMT-F)を作製し、これを鋳型として、例えばM13 primer M4、M13 primer RV(タカラバイオ社製)を用いてPCRを行い、PCR産物をプローブとすることができる。プローブは、cDNA又はそのセンス鎖もしくはアンチセンス鎖の全長であってもよいし、あるいはその部分長であってもよいし、あるいは5’もしくは3’非翻訳領域のみであってもよい。DNAチップでは、約10〜約50塩基長、好ましくは約15〜約30塩基長、より好ましくは約20〜約25塩基長のプローブが使用されるが、これらに限定されない。また、DNAマイクロアレイやノザンハイブリダイゼーションでは、少なくとも約10塩基長以上から全長、好ましくは約30塩基長以上から全長、より好ましくは約50塩基長以上から全長である。一般に、プローブが長いほどハイブリダイゼーション効率がよくなり、感度は上がる。逆に、プローブが短いほど感度は下がるが、特異性が上がる。目的遺伝子が遺伝子ファミリーを形成しており、それらの発現状態を区別して検出するときなどは、短いプローブを選択する。固相上のプローブは、通常0.1μg〜0.5μgの溶液を用いてスポットする。
【0034】
プローブあるいはサンプルcDNA上のラベルは、アイソトープ(例えば32P、33P、35S)又は蛍光(フルオレサミン及びその誘導体、ローダミン及びその誘導体、FITC、Cy3、Cy5)のいずれでもよい。本発明に係る遺伝子を同時に検出、定量するときには、それぞれ異なる色の蛍光でラベルすることもできる。
【0035】
ハイブリダイゼーション条件は、中程度又は高ストリンジェントな条件、好ましくは高ストリンジェントな条件、例えば、2〜6xSSC、0.1〜1%SDS溶液、50〜70℃、約1時間〜一晩の条件である。ストリンジェントな条件とは、前記サンプルcDNAと固相化プローブとのより特異的結合を可能とする条件である。一般に、塩濃度、有機溶媒(例えば、ホルムアミド)、温度条件に応じて低、中程度、高ストリンジェンシーが決まる。特に、塩濃度を減じるか、有機溶媒濃度を増加させるか、またはハイブリダイゼーション温度を上昇させるかによってストリンジェンシー(stringency)を増加させることができる。例えば、塩濃度は、塩化ナトリウム約500 mM以下およびクエン酸三ナトリウム約50 mM以下、好ましくは塩化ナトリウム約250 mM以下およびクエン酸三ナトリウム約25 mM以下である。有機溶媒濃度は、ホルムアミド約35%以上、好ましくは約50%以上である。温度は、約30℃以上、好ましくは約37℃以上、より好ましくは約42℃以上である。他の条件としては、ハイブリダイゼーション時間、洗浄剤(例えば、SDS)の濃度、およびキャリアーDNAの存否等であり、これらの条件を組み合わせることによって、様々なストリンジェンシーを設定することができる。洗浄のためのストリンジェントな温度条件は、約25℃以上、好ましくは約42℃以上、より好ましくは約60℃以上である。好ましい洗浄条件は、0.2xSSC、0.1%SDS溶液、5分の条件である。
【0036】
検出は、マイクロアレイ用の検出器、例えばScanArray 4000, 5000(General Scanning 社製、USA)、GMS418 Array Scanner(寶酒造社製)、Gene Tip Scanner(日本レーザ電子社製)などを使用することができる。定量に際しては、一定量の適当なDNA又はRNA(例えばλpolyA+RNA(タカラバイオ社製)からのラベル化cDNA、Lucidea Universal Score Card(アマシャムバイオサイエンス社製)など)、あるいは生体内で恒常的に発現している遺伝子、例えばアクチン遺伝子などをコントロールとし、バンド濃度を測定し、サンプル対コントロール比を求めることによって、サンプル中の目的の遺伝子の発現レベルを相対値として定量することができる。
【0037】
本発明の方法で特に好ましいDNAマイクロアレイの作製について、固相担体表面で直接オリゴヌクレオチドを合成する方法(オン・チップ法)及び予め調製したプローブを固相表面に固定化する方法が知られており、本発明ではそのいずれも使用できるが、後者の方法が好ましい。オン・チップ法は、光照射で選択的に除去される保護基の使用と、半導体製造に利用されるフォトリソグラフィー技術および固相合成技術とを組み合わせて、微少なマトリックスの所定の領域での選択的合成を行う方法(マスキング技術:例えば、S.P.A. Fodor(1991) Science 251:767)である。一方、予め調製したオリゴ−又はポリヌクレオチドプローブを固相表面に固定化する方法は、官能基を導入したオリゴヌクレオチドまたはポリヌクレオチドを合成し、表面処理した固相担体表面にオリゴヌクレオチドまたはポリヌクレオチドを点着し、共有結合させる(例えば、J.B. Lamtureら(1994) Nucl. Acids Res. 22:2121-2125; Z. Guoら(1994) Nucl. Acids Res. 22:5456-22:5456-5465)。オリゴ−又はポリヌクレオチドは、一般的には、表面処理した固相担体にスペーサーやクロスリンカーを介して共有結合される。ガラス表面にポリアクリルアミドゲルの微小片を整列させ、そこに合成オリゴヌクレオチドを共有結合させる方法も知られている(G. Yershovら(1996) Proc. Natl. Acad. Sci. USA 94:4913)。また、シリカマイクロアレイ上に微小電極のアレイを作製し、電極上にはストレプトアビジンを含むアガロースの浸透層を設けて反応部位とし、この部位をプラスに荷電させることでビオチン化オリゴヌクレオチドを固定し、部位の荷電を制御することで、高速で厳密なハイブリダイゼーションを可能にする方法も知られている(R.G. Sosnowskiら(1997) Proc. Natl. Acad. Sci. USA 94:1119-1123)。
【0038】
サンプルについては、前記のとおり、製麦時の発芽3日目のオオムギの発芽種子を液体窒素で凍結、粉砕し、既知の手法、例えばフェノール/クロロホルム法、グアニジウム法又はフェノール/SDS法により全RNAを得るか、又は、RNAをオリゴ(dT)セルロースカラムに通してpoly(A)+RNAを得て(E.E. Karrerら (1991) Mol. Biol. 16: 797-805)、サンプルとすることができる。
【0039】
本発明において、前記ハイブリダイゼーションは、HvJMTcDNA又はその断片と、発芽オオムギ種子の全RNA又はpoly(A)+RNAから調製されたラベル化cDNAサンプルとの間で行われる。HvJMTcDNA又はその断片の具体例は、以下のとおりである。
【0040】
HvJMTのcDNA又は断片は、配列番号2のアミノ酸配列をコードする塩基配列、その変異体、もしくはそれらの相補鎖、又は少なくとも10塩基、好ましくは少なくとも15塩基からなるそれらの断片であり、ここで該変異体は、該塩基配列において1以上、好ましくは1又は数個の塩基の置換、欠失又は付加を有するもの、あるいは該塩基配列と70%以上、80%以上、90%以上、95%以上もしくは98%以上の相同性を有するものである。さらに具体的には、HvJMTcDNA又は断片は、配列番号1の塩基配列、その変異体、もしくはそれらの相補鎖、又は少なくとも10塩基、好ましくは少なくとも15塩基からなるそれらの断片であり、ここで該変異体は、該塩基配列において1以上、好ましくは1又は数個の塩基の置換、欠失又は付加を有するもの、あるいは該塩基配列と70%以上、80%以上、90%以上、95%以上もしくは98%以上の相同性を有するものである。
【0041】
本明細書中で使用される「数個」とは、2〜約10個、好ましくは2〜5個である。また「変異体」は、該当の酵素の特性を実質的に変更しない程度の変異を有するものであり、限定されないが、好ましくはオオムギの異なる品種間で認められるような(突然)変異、例えば多型性に関わるような変異、スプライス変異、あるいは遺伝暗号の縮重による変異などを有するものである。さらに、本明細書中で使用される「断片」とは、前記の特定の塩基配列において少なくとも10塩基、15塩基、20塩基、25塩基、30塩基、50塩基、100塩基、もしくは150塩基から、酵素の成熟配列をコードする翻訳領域の全長未満までの翻訳領域断片、あるいは、少なくとも10塩基、15塩基、20塩基、25塩基、30塩基、50塩基、100塩基もしくは150塩基の、非翻訳領域のみ、又は翻訳領域と非翻訳領域からなる連続する塩基配列からなる断片を指す。
【0042】
本発明はさらに、配列番号1、2に示される塩基配列及びアミノ酸配列を含むMeJA合成酵素及びそれをコードする遺伝子、並びに、それらの相補鎖、それらの変異体、及びそれらの断片、好ましくは活性部位を含む断片、にも関する。ここで、変異体は、配列番号1、2に示される塩基配列又はアミノ酸配列において1以上、好ましくは1又は数個の塩基又はアミノ酸の置換、欠失又は付加を有するもの、あるいは該塩基配列又はアミノ酸配列と70%以上、80%以上、90%以上、95%以上もしくは98%以上の相同性を有するものである。また、断片は、配列番号1、2に示される塩基配列又はアミノ酸配列において少なくとも10、15、20、25、30、50、100、もしくは150塩基/アミノ酸から、酵素の翻訳領域又は成熟配列の全長未満までの断片である。
【0043】
本明細書中で使用される「相同性」は、%同一性と同義であり、公知のBLASTN、BLASTX、BLASTPなどのBLASTプログラムを用いて決定することができる(S.F. Altschulら(1990) J. Mol. Biol. 215:403-410)。
【0044】
サンプル中の対象遺伝子の発現量を定量する別の方法として、内部標準物質を用いたRT-PCR法も使用できる(例えば、PCR法最前線(1996年)関谷、藤永編、共立出版)。使用する内部標準としては細胞骨格の構成成分であるβアクチンなどの組織によって発現量に差のないと考えられるハウスキーピング遺伝子がよく用いられる。この方法では標的mRNA量が内部標準試料に対して多いか少ないかといった相対的な結果しか得られないが、mRNAの抽出効率やcDNAの合成効率を考慮しなくてもよいという利点があるため、広く用いられている。PCRによる増幅産物は、ある範囲内では指数関数的に増加するが、それを超えると頭打ちの状態になる。そのため、RT-PCRにおける発現遺伝子の定量は、増幅産物が指数関数的に増加する範囲で行わなくてはならない。また、PCRを酵素化学的に分析し、特に標準試料を用意せずに定量的PCRを行う、カイネティクス分析法も使用することができる。PCR反応は、反応がプラトーに達する前には指数関数的に生成物の量が増加する。そこで、PCRのサイクル数を横軸に、反応生成物量を対数の縦軸にプロットすると、生成物量が直線的に増加する時期があるが、この直線を延長し、y軸と交差する点を求めれば、理論的には初期鋳型量を算出することができる(バイオ実験イラストレイティット3「本当に増えるPCR」(1998年)中山広樹著、秀潤社)。実際には、一つのサンプルについてのPCR反応中に、数サイクルごとに反応液をサンプリングして、PCR産物の量を定量し、グラフにプロットしていく。そうして得られたグラフの指数増加期のポイントに対して回帰分析を行い、y切片を求める。
【0045】
本発明ではさらに、この原理を応用し、サンプリングせずにPCR反応中にリアルタイムでこの回帰分析を行うことのできる、リアルタイム定量PCRを使用することもできる。PCR生成物が特異的に蛍光標識される反応系で、蛍光強度を検出する装置の備わった温度サイクラー装置によりPCR反応を行うと、反応中の生成物の量をサンプリングの必要なくリアルタイムでモニターでき、その結果をコンピュータで回帰分析することができる。PCR生成物を標識する方法としては、蛍光標識したプローブを用いる方法と、2本鎖DNAに特異的に結合する試薬を用いる方法とがある。前者では、2種類の蛍光物質で標識し蛍光をクエンチング作用で抑えてあるプローブを、鋳型の二つのプライマーと相補的な配列の間にアニールさせる方法がよく知られている。プローブは、PCR反応が行われるとTaqポリメラーゼのもつ5’-3’エキソヌクレアーゼ活性により分解され、蛍光を発するようになる。その蛍光量は、PCR生成物の量を反映する。後者では、SYBR greenなどが用いられる。PCR反応物が検出限界に到達するときのサイクル数(CT)と初期鋳型量とは逆相関の関係にあることから、リアルタイム測定法ではCTを測定することによって初期鋳型量を定量している。数段階の既知量の鋳型を用いてCTを測定し検量線を作製すれば、未知試料の初期鋳型量の絶対値を算出することができる。
【0046】
RT-PCRで使用する逆転写酵素は、例えばM-MLV RTase、ExScript RTase(タカラバイオ社製)、AMV Reverse Transcriptase(Life Science社製)、Super Script II RT(GIBCO BRL社製)などを使用することができる。
【0047】
PCR条件は、例えばDNAの変性を94〜95℃で5秒〜5分間、プライマーのアニーリングを50〜70℃で10秒〜1分間、伸長反応を72℃で30秒〜3分間行い、これを1サイクルとして20〜40サイクルほど行い、最後に72℃で30秒〜10分間の伸長反応を行うことができる。
【0048】
プライマーは、鋳型としてのHvJMT遺伝子(配列番号1)又はその変異体(前記定義)のセンス及びアンチセンス鎖に相補的な配列である。プライマーのサイズは、通常、約15〜約30塩基長、好ましくは約17〜約25塩基長である。プライマーの例は、特に限定されないが、配列番号11及び12である。
【0049】
以下に、本発明を実施例によってさらに具体的に説明するが、本発明はこれらの実施例によって制限されないものとする。
【実施例1】
【0050】
オオムギからのRNA調製と発芽時のホルモン合成酵素をコードするcDNAの調製
(1A)オオムギからのRNAの調製
オオムギ(Hordeum vulgare cv.Harrington)を1%NaClOで10分間浸漬することにより滅菌後、水洗いし、20℃、8時間水中で浸漬した。浸漬後、水に浸した濾紙上にまき、18℃、暗所に置いた。なお、浸漬終了時点を0日目とし、以後5日間発芽を行った。
【0051】
発芽3日目の種子を液体窒素で凍結し、粉砕後、全RNAをKarrer, E.Eら((1991) Mol. Biol. 16: 797-805)の方法により抽出した。全RNAより、poly(A)+RNAをoligo-dT30<super>(Roche diagnostics)により取得した。
【0052】
温室内で育成したオオムギ幼葉を、7時間、室温、暗所で風乾させた。乾燥葉を液体窒素で粉砕後、前記した方法により、全RNA、poly(A)+RNAを抽出した。
【0053】
製麦時、浸漬直後から、1日おきに4日目までのオオムギより前記方法により全RNA、poly(A)+RNAを抽出した。
【0054】
(1B)オオムギ発芽時に発現する、JMTをコードするcDNA(HvJMT)の取得
発芽3日目全RNA1μgを鋳型として、ThermoScript RT-PCR system (Invitrogen Corp.)により、oligo(dT)20をプライマーとしてcDNAを合成した。アラビドプシスより単離された、JMT-cDNA(AY008434)の塩基配列に対して相同性の高いオオムギEST(BI959950)を見出し、BI959950に基づいて設計したTO-510:5’-ATgTTATCgCggACTTATgC-3’ (配列番号3)およびTO-511: 5’-gACCATTgggAgCCTTTC-3’ (配列番号4)を用いてRT-PCRを行った。なお、PCRは、95℃ 5分後、94℃ 30秒、60℃ 1分、72℃ 1分のセットを30サイクル、72℃ 5分の反応系で行った。
【0055】
RT-PCRによって増幅した約0.4kbpのDNAを抽出し、pCR2.1-TOPO vector (Invitrogen Corp.)にクローニング、塩基配列を確認し、pHvJMT-pを取得した。
【0056】
完全長のHvJMTcDNAを取得するために、 発芽3日目のpoly (A)+ RNA を用いて5’ 、および3’ RACE をSMART RACE cDNA amplification kit (CLONTECH) により行った。poly (A)+ RNA 0.5μgを用いて、前記SMART RACE cDNA amplification kitに基づき5’、および3’RACE用のcDNAを合成した。合成したcDNAを鋳型として、プライマーは、取得した pHvJMT-p の配列をもとに、5’RACEには TO-516: 5’-ATCTTCCggAgCCTTTgAgAg-3’ (配列番号5)、TO-517: 5’-gACCATTgggAgCCTTTCAC-3’ (配列番号6)を、3’RACEには、TO-518: 5’-gTAAATgCCATACACAgCCAATg-3’(配列番号7) 、TO-519: 5’-CATACACAgCCAATgCCTTCA-3’(配列番号8)を用いて2ステップPCRを行った。1回目のPCRは、5’RACE、および3’RACEには、それぞれTO-517、TO-518を用いて、94℃ 5秒、72℃ 3分のセットを5サイクル、94℃ 5秒、70℃ 10秒、72℃ 3分のセットを5サイクル、94℃ 5秒、68℃ 10秒、72℃ 3分のセットを20サイクルで行った。1回目の反応液を鋳型として2回目のPCRを、5’RACE、および3’RACEには、それぞれTO-516、TO-519を用いて、95℃ 5分後、94℃ 30秒、60℃ 1分、72℃ 1分のセットを30サイクル、72℃ 5分の反応系で行った。5’RACEおよび、3’RACEにより取得したそれぞれ0.5kbp、1.0kbpの断片を前記pCR2.1-TOPO vectorにクローニング、塩基配列を確認し、pHvJMT-U 、およびpHvJMT-Dを取得した。
【0057】
pHvJMT-U 、およびpHvJMT-D の塩基配列より、HvJMT-cDNAのコード領域を推定し、全コード領域を含む遺伝子の取得を、pHvJMT-U 、およびpHvJMT-Dの混合物を鋳型として、TO-543:5’- gAATTCATggCCTCCCAACCgATgg -3’ (配列番号9)およびTO-544: 5’- CCCgggATgTggTgCATggCACCTT -3’(配列番号10)を用いたPCRにより行った。なお、PCRは、95℃ 5分後、94℃ 30秒、60℃ 1分、72℃ 1分のセットを25サイクル、72℃ 5分の反応系で行った。取得した1.2kbpの断片を前記pCR2.1-TOPO vectorにクローニング、塩基配列を確認し、pHvJMT-fを取得した。取得したHvJMT-cDNAの塩基配列を配列番号1に、推定アミノ酸配列を配列番号2にそれぞれ示した。
【実施例2】
【0058】
取得したHvJMT遺伝子の翻訳産物の活性測定による確定
(2A)HvJMT-cDNAの大腸菌内発現ベクターの構築、大腸菌内発現
取得したpHvJMT-fの全コード領域を含むEcoRI-SmaI断片を、pGEX4T-1(Amersham Pharmacia Biotech.)のEcoRI-SmaI部位に導入し、pGEX-HvJMTを作成した。作成したpGEX-HvJMTは、大腸菌BLR(DE3)に形質転換した。
【0059】
pGEX-HvJMT:BLR(DE3)からGST融合したHvJMTを、定法によりIPTG誘導により大腸菌内で発現させた。大腸菌を破砕後、可溶性画分を分取し、Glutation Sepharose 4B(Amersham Pharmacia Biotech.)によりGST-HvJMTを精製した。なお、ベクターとして用いた、pGEX4T-1:BLR(DE3)より、同様の操作で調製した精製物を、コントロールとして以後の実験に用いた。
【0060】
(2B)HvJMTの活性測定による同定
GST-HvJMTを用いた、JMTの活性測定は、Seo H.S. et al. PNAS 98 p4788-4793 (2001)に記載された方法により、(±)JAを基質、S-アデノシルメチオニン(以下SAMと称する)をメチル基供与体として反応させ、GC-MSによって反応生成物のMeJAを検出することにより行った。
【0061】
10mM(±)JA、10mM SAMの共存下で、前記の操作で取得した精製物10μlを、反応バッファー(10mM 50mM Tris-HCl(pH7.5)、20mM KCl、10mMβ-mercaptoethanol)中(全液量50μl)で20℃、30分間反応させた。反応終了後、反応液を150μlのヘキサンで抽出し、ヘキサン抽出物をGC-MSで解析した。GC-MS解析は島津社製GC-MS QP-2010を使用し、カラムとしてZB-5(カラム長15m、内径0.25mm、膜厚0.25μm;Phenomenex社製)を用いた。キャリアガスにはHeガスを使用し、線速度100cm/分で一定流量流した。温度条件は気化室温度280℃、インターフェース温度280℃とし、カラム昇温プログラムは110℃(2分)→(10℃/分)→200℃→(30℃/分)→290℃(5分)とした。尚、この条件でMeJAの溶出時間はおよそ5.8分となった。MeJAの同定は、市販の質量スペクトルライブラリーデータを用いて、GC-MS分析においてm/z=70〜300のスキャン分析を行った結果をライブラリーから検索して行った。
【0062】
その結果、GST-HvJMTの反応により、MeJAが生成されることが解り、HvJMT-cDNAは、JMT活性をもつタンパク質をコードする遺伝子であることが証明された。HvJMTの基質特異性を調べるために、基質として(±)JA、ジヒドロジャスモン酸、リノレン酸、サリチル酸、ベンゼン、ケイ皮酸を用いて、[14C]methyl SAMをメチル基供与体として反応を行い、酢酸エチル抽出後の酢酸エチル層の放射線取り込み量により検討した。(±)JAを基質とした際の放射線取り込み量を100%の活性として、各基質の活性の比を表1に示した。参考として、アラビドプシス由来のJMTの基質特異性も示す。
【0063】
【表1】
【0064】
その結果、HvJMTは、アラビドプシスのJMT(AtJMT)と同様に、(±)JA、ジヒドロジャスモン酸を基質として認識することが解った。一方、AtJMT は、ジヒドロジャスモン酸を基質として認識はするものの、JAに比べてその認識は2割程度であることが報告されているが、HvJMTはジヒドロジャスモン酸に対して(±)JAと同等あるいはそれ以上に、基質として認識することが解った。
【実施例3】
【0065】
製麦時のHvJMT、ジャスモン酸誘導タンパク質遺伝子(JIP23)の発現解析
(3A)製麦時のHvJMT発現の推移
製麦時、浸漬直後(0日目)、浸漬後1、2、3、4日目のサンプルより調製した全RNA 1μgを鋳型として、ThermoScript RT-PCR system (Invitrogen Corp.)により、oligo(dT)20をプライマーとしてcDNAを合成した。合成したcDNAを鋳型として、TO-512:5’- TgCCgTAAATgCCATACACA -3’ (配列番号11)およびTO-513: 5’- CAgAACgCgTCCCTACCAg -3’ (配列番号12)を用いたRT-PCRを行った。なお、増幅断片の定量比較のための内部標準としては、前記鋳型cDNAを用いて、オオムギのアクチン遺伝子(AY14545)の配列に基づいて設計した、TO-635:5’-CCCCATgCCATTCTTCgTTT-3’ (配列番号13)およびTO-636: 5’- CTgggCCAgACTCgTCgTA-3’ (配列番号14)を用いたRT-PCR増幅産物を用いた。PCRは、95℃ 5分後、94℃ 30秒、60℃ 1分、72℃ 1分のセットを25サイクル、72℃ 5分の反応系で行った。
【0066】
製麦工程における、HvJMTの経時的な発現変動を図1に示した。図1より、HvJMTは浸漬後1日目から発現の上昇が認められ、3日目にはほぼピークとなり、4日目まで継続的に発現することが解った。
【0067】
(3B)製麦時のジャスモン酸誘導遺伝子発現の推移
ジャスモン酸誘導タンパク質として知られている、JIP23(Plant Mol Biol. 1992 May;19(2):193-204.)をコードする遺伝子の製麦時の発現を検討した。
【0068】
JIP23をコードするcDNA配列を、ガラス担体に固定化した(DNAチップ実験まるわかり(2004)、佐々木、青柳編 羊土社)。一方、製麦時、浸漬直後、浸漬後1、2、3、4日目のサンプルより調製した全RNA200ngを用いて、Agilent Low RNA Input Fluorescent Liniear Amplification Kit (Agilent Technologies)により標識、固定化したガラス担体とのハイブリダイゼーション、洗浄を行い、シグナル強度の相対値を、浸漬直後のRNAを用いた際の蛍光強度との比として求めた。
【0069】
結果を図2に示した。図2より、JIP23は浸漬1日目から発現の上昇が認められ、4日目まで継続的に発現が上昇することが解り、HvJMTの発現パターンを解析することにより、大麦内部でJAによる遺伝子発現誘導の様子がモニターできることが解った。
【実施例4】
【0070】
製麦時のフェルラ酸量の推移
製麦時に大麦サンプルから熱水で抽出されるフェルラ酸量の推移を検討した。浸漬直後(0日目)、浸漬後1、2、3、4日目の大麦サンプルを凍結乾燥し、各サンプルにつき10粒を家庭用ミルで粉砕後、70度の超純水を8ml添加し1時間抽出した。抽出後、遠心により残渣を除去し、上清を回収した。回収した上清のうち1mlを分取し、1.5mlの2M塩酸を添加後、ジエチルエーテルで低極性物質を回収した。エーテル抽出物は窒素気流下で乾固した後、1mlの20% MeOH/0.1% ギ酸に溶解後、0.45μmのフィルターで濾過し、HPLC分析に供した。HPLCでの分析は、カラムとしてNucleosil C18 5μ 100Å 4.6mm×250mm (Shimadzu GLC)(カラム温度40度)、移動相として70% MeOH / 0.1% ギ酸(アイソクラティック)を用い、流速1ml/分で分離し、UV検出器を用いて310nmの吸光で検出した。定量は、フェルラ酸の標準品(和光純薬)を用いて検量線を描き、得られたピークの面積から決定した。
【0071】
製麦工程における大麦サンプル熱水抽出物中のフェルラ酸含量の変動を図3に示した。図3より、フェルラ酸含量は、浸漬後1日目から2日目にかけて急激に上昇し、2日目以降はその含量が維持されることが解り、HvJMTの発現量と製麦時のフェルラ酸含量に、相関関係があることが解った。
【産業上の利用可能性】
【0072】
ビールや発泡酒の製造の製麦工程での麦芽品質の維持管理に利用できる。さらに、製麦工程の理論付け、麦芽品質に関与する物質合成機構の理論付け、並びに、オオムギ品種間、ロット間における製麦特性の違いの把握などに役立つ。
【図面の簡単な説明】
【0073】
【図1】図1は、製麦中のHvJMT遺伝子発現の経時的な変動を示す。
【図2】図2は、製麦中のJIP23遺伝子発現の経時的な変動を示す。
【図3】図3は、製麦中のフェルラ酸含量の経時的な変動を示す。
【配列表フリーテキスト】
【0074】
配列番号3:人工配列の説明:プライマー
配列番号4:人工配列の説明:プライマー
配列番号5:人工配列の説明:プライマー
配列番号6:人工配列の説明:プライマー
配列番号7:人工配列の説明:プライマー
配列番号8:人工配列の説明:プライマー
配列番号9:人工配列の説明:プライマー
配列番号10:人工配列の説明:プライマー
配列番号11:人工配列の説明:プライマー
配列番号12:人工配列の説明:プライマー
配列番号13:人工配列の説明:プライマー
配列番号14:人工配列の説明:プライマー
【特許請求の範囲】
【請求項1】
製麦工程における発芽オオムギ種子中のメチルジャスモン酸の合成をモニターする方法であって、該種子中のジャスモン酸カルボキシルメチルトランスフェラーゼ(HvJMT)遺伝子の発現レベルを測定することを含む、前記方法。
【請求項2】
前記遺伝子の発現レベルが、メチルジャスモン酸合成量と正の相関関係にある、請求項1に記載の方法。
【請求項3】
前記遺伝子が、配列番号2に示されるアミノ酸配列をコードする塩基配列又はその変異配列を含む、請求項1又は2に記載の方法。
【請求項4】
前記遺伝子が、配列番号1に示される塩基配列又はその変異配列を含む、請求項3に記載の方法。
【請求項5】
前記遺伝子の発現レベルが、PCR法又はRT-PCR法によって測定される、請求項1〜4のいずれか1項に記載の方法。
【請求項6】
前記遺伝子の発現レベルが、核酸ハイブリダイゼーションによって測定される、請求項1〜4のいずれか1項に記載の方法。
【請求項1】
製麦工程における発芽オオムギ種子中のメチルジャスモン酸の合成をモニターする方法であって、該種子中のジャスモン酸カルボキシルメチルトランスフェラーゼ(HvJMT)遺伝子の発現レベルを測定することを含む、前記方法。
【請求項2】
前記遺伝子の発現レベルが、メチルジャスモン酸合成量と正の相関関係にある、請求項1に記載の方法。
【請求項3】
前記遺伝子が、配列番号2に示されるアミノ酸配列をコードする塩基配列又はその変異配列を含む、請求項1又は2に記載の方法。
【請求項4】
前記遺伝子が、配列番号1に示される塩基配列又はその変異配列を含む、請求項3に記載の方法。
【請求項5】
前記遺伝子の発現レベルが、PCR法又はRT-PCR法によって測定される、請求項1〜4のいずれか1項に記載の方法。
【請求項6】
前記遺伝子の発現レベルが、核酸ハイブリダイゼーションによって測定される、請求項1〜4のいずれか1項に記載の方法。
【図1】

【図2】
【図3】
【図2】
【図3】
【公開番号】特開2007−174956(P2007−174956A)
【公開日】平成19年7月12日(2007.7.12)
【国際特許分類】
【出願番号】特願2005−376291(P2005−376291)
【出願日】平成17年12月27日(2005.12.27)
【出願人】(000253503)麒麟麦酒株式会社 (247)
【Fターム(参考)】
【公開日】平成19年7月12日(2007.7.12)
【国際特許分類】
【出願日】平成17年12月27日(2005.12.27)
【出願人】(000253503)麒麟麦酒株式会社 (247)
【Fターム(参考)】
[ Back to top ]