
複数のVHおよびVκ領域を含むヒトIg遺伝子座を有するトランスジェニック哺乳動物、ならびにそれから産生される抗体
【課題】ヒト化モノクロナル抗体産生トランスジェニック動物、及びヒト化モノクローナル抗体の提供。
【解決手段】ヒト免疫グロブリン遺伝子座を含むように操作されたトランスジェニック非ヒト動物で、特に複数の可変(VHおよびVκ)遺伝子領域を含むヒトIg遺伝子座を有する。複数の可変領域遺伝子を含めることにより、その動物によって産生されるヒト抗体の特異性および多様性は高まる。さらに、このような領域を含めることにより、その動物が極めて高親和性の抗体を分泌する成熟B細胞を豊富に有するように、その動物に対してB細胞発生が増強および再構成される。
【解決手段】ヒト免疫グロブリン遺伝子座を含むように操作されたトランスジェニック非ヒト動物で、特に複数の可変(VHおよびVκ)遺伝子領域を含むヒトIg遺伝子座を有する。複数の可変領域遺伝子を含めることにより、その動物によって産生されるヒト抗体の特異性および多様性は高まる。さらに、このような領域を含めることにより、その動物が極めて高親和性の抗体を分泌する成熟B細胞を豊富に有するように、その動物に対してB細胞発生が増強および再構成される。
【発明の詳細な説明】
【技術分野】
【0001】
本発明は、ヒト免疫グロブリン遺伝子座を含むように操作されたトランスジェニック非ヒト動物に関する。特に、本発明による動物は、複数の可変(VHおよびVκ)遺伝子領域を含むヒトIg遺伝子座を有する。有利なことに、複数の可変領域遺伝子を含めることにより、その動物によって産生されるヒト抗体の特異性および多様性は高まる。さらに、このような領域を含めることにより、その動物が極めて高親和性の抗体を分泌する成熟B細胞を豊富に有するように、その動物に対してB細胞発生が増強および再構成される。
【背景技術】
【0002】
メガベースサイズのヒト遺伝子座をYAC中にクローニングおよび再構築し、それらをマウス生殖細胞系に導入しうることにより、極めて大きく、または大まかにマッピングされた遺伝子座の機能的構成要素を解明するための、ならびにヒト疾患の有用なモデルを作製するための、強力な手法が提供される。さらに、このような技術を利用してマウス遺伝子座をそれらのヒト等価物に置換することにより、発生過程におけるヒト遺伝子産物の発現および調節、それらの他の系との連絡、ならびに疾患の誘導および進行におけるそれらの関与に関して比類のない洞察が得られると考えられる。
【0003】
このような戦略の1つの重要な実用的応用は、マウス体液性免疫系の「ヒト化」である。内因性Ig遺伝子が不活性化されたマウスにヒト免疫グロブリン(Ig)遺伝子座を導入することにより、抗体のプログラム化された発現および構築の根底にある機序に加えて、B細胞の発生におけるそれらの役割をも検討する機会が提供される。さらに、このような戦略により、ヒト疾患の抗体療法という可能性の実現へ向けての重要な里程標となる、完全ヒトモノクローナル抗体(Mab)を生産するための理想的な供給源が提供されると考えられる。完全ヒト抗体は、マウスまたはマウス由来のMabに固有の免疫原性およびアレルギー性反応を最小限に抑え、それにより、投与される抗体の有効性および安全性を高めると考えられる。完全ヒト抗体の使用は、抗体の反復投与を必要とする、炎症、自己免疫および癌などの慢性的および再発性のヒト疾患の治療に、実質的な利益を提供すると考えられる。
【0004】
この目標に向けての1つの手法は、マウス抗体の産生に欠陥があってヒトIg遺伝子座の大きな断片を有するマウス系統を、組換え操作によって作製することであり、これは、このようなマウスが、マウス抗体を生じずに広範なレパートリー(repertoire)のヒト抗体を製造するであろうとの予測に立つものであった。大きなヒトIg断片は、可変遺伝子の広範な多様性に加えて、抗体の産生および発現の適切な調節も保持すると考えられる。抗体の多様化および選択のため、ならびにヒトタンパク質に対する免疫寛容を失わせるためのマウス機構を開発すれば、これらのマウス系統で再現されたヒト抗体レパートリーはヒト抗原を含む関心対象の任意の抗原に対する高親和性抗体を生じるはずである。ハイブリドーマ技術を用いることにより、望ましい特異性をもつ抗原特異的ヒトMabは容易に製造および選択しうると考えられる。
【0005】
この一般的な戦略は、本発明者らが1994年に発表した最初のXenoMouse(登録商標)系統の作出との関連で示された。グリーン(Green)ら、Nature Genetics 7:13〜21(1994)(非特許文献1)参照。XenoMouse(登録商標)系統は、可変および定常領域のコア配列を含む、ヒト重鎖およびκ軽鎖遺伝子座のそれぞれ245kbおよび190kbのサイズの生殖細胞系コンフィギュレーション断片を有するように操作された。同上。ヒトIgを含む酵母人工染色体(YAC)は、抗体の再配列および発現の双方に関してマウス系への適合性があることが実証されており、不活性化されたマウスIg遺伝子の置換も可能であった。これは、それらがB細胞発生の誘導および完全ヒト抗体の成人様ヒトレパートリーの生成、ならびに抗原特異的ヒトMabの産生の能力をもつことによって示された。また、これらの結果は、比較的多数のV遺伝子、付加的な調節要素、およびヒトIg定常領域を含むヒトIg遺伝子座のかなりの部分を導入することにより、感染および免疫化に対するヒト体液性応答の特徴である実質的に完全なレパートリーが再現される可能性があることも示唆した。
【0006】
このような手法は、1990年1月12日に提出された米国特許出願第07/466,008号(特許文献1)、1990年11月8日に提出された第07/610,515号(特許文献2)、1992年7月24日に提出された第07/919,297号(特許文献3)、1992年7月30日に提出された第07/922,649号(特許文献4)、1993年3月15日に提出された第08/031,801号(特許文献5)、1993年8月27日に提出された第08/112,848号(特許文献6)、1994年4月28日に提出された第08/234,145号(特許文献7)、1995年1月20日に提出された第08/376,279号(特許文献8)、1995年4月27日に提出された第08/430,938号(特許文献9)、1995年6月5日に提出された第08/464,584号(特許文献10)、1995年6月5日に提出された第08/464,582号(特許文献11)、1995年6月5日に提出された第08/463,191号(特許文献12)、1995年6月5日に提出された第08/462,837号(特許文献13)、1995年6月5日に提出された第08/486,853号(特許文献14)、1995年6月5日に提出された第08/486,857号(特許文献15)、1995年6月5日に提出された第08/486,859号(特許文献16)、1995年6月5日に提出された第08/462,513号(特許文献17)、および1996年10月2日に提出された第08/724,752号(特許文献18)という米国特許出願においてさらに考察および描写されている。また、1996年6月12日に認可が発行されたEP 0 463 151 B1(特許文献19)、1994年2月3日に発行された国際特許出願国際公開公報第94/02602号(特許文献20)、1996年10月31日に発行された国際特許出願国際公開公報第96/34096号(特許文献21)、および1996年4月29日に提出されたPCT/US96/05928(特許文献22)も参照されたい。上記に引用したそれぞれの特許および出願の開示は、その全体が参照として本明細書に組み入れられる。
【0007】
1つの代替的なアプローチにおいて、ジェンファームインターナショナル社(GenPharm International, Inc.)を含む他者は、「ミニ遺伝子座(minilocus)」法を用いている。ミニ遺伝子座法では、Ig遺伝子座からの複数の小片(個々の遺伝子)を含めることにより、外来性Ig遺伝子座を模倣する。このため、1つまたは複数のVH遺伝子、1つまたは複数のDH遺伝子、1つまたは複数のJH遺伝子、μ定常領域、および第2の定常領域(好ましくはγ定常領域)が、動物に挿入するための構築物(construct)中に形成される。この手法はスラニ(Surani)らに対する米国特許第5,545,807号(特許文献23)、いずれもロンバーグ(Lonberg)およびケイ(Kay)に対する米国特許第5,545,806号(特許文献24)および第5,625,825号(特許文献25)、ならびにいずれもジェンファームインターナショナル社(GenPharm International, Inc.)による米国特許出願である1990年8月29日に提出された第07/574,748号(特許文献26)、1990年8月31日に提出された第07/575,962号(特許文献27)、1991年12月17日に提出された第07/810,279号(特許文献28)、1992年3月18日に提出された第07/853,408号(特許文献29)、1992年6月23日に提出された第07/904,068号(特許文献30)、1992年12月16日に提出された第07/990,860号(特許文献31)、1993年4月26日に提出された第08/053,131号(特許文献32)、1993年7月22日に提出された第08/096,762号(特許文献33)、1993年11月18日に提出された第08/155,301号(特許文献34)、1993年12月3日に提出された第08/161,739号(特許文献35)、1993年12月10日に提出された第08/165,699号(特許文献36)、1994年3月9日に提出された第08/209,741号(特許文献37)に記載されており、それらの開示は参照として本明細書に組み入れられる。また、その開示の全体が参照として本明細書に組み入れられる1994年11月10日に発行された国際公開公報第94/25585号(特許文献38)、1993年7月24日に発行された国際公開公報第93/12227号(特許文献39)、1992年12月23日に発行された国際公開公報第92/22645号(特許文献40)、1992年3月19日に発行された国際公開公報第92/03918号(特許文献41)も参照されたい。さらに、その開示の全体が参照として本明細書に組み入れられるテイラー(Taylor)ら、1992(非特許文献2)、チェン(Chen)ら、1993(非特許文献3)、トゥアイロン(Tuaillon)ら、1993(非特許文献4)、チョイ(Choi)ら、1993(非特許文献5)、ロンバーグ(Lonberg)ら(1994)(非特許文献6)、テイラーら(1994)(非特許文献7)、およびトゥアイロンら(1995)(非特許文献8)も参照されたい。
【0008】
上記に引用し、英国医学研究会議(Medical Research Council)(「MRC」)に譲渡されたスラニ(Surani)らの発明者らは、ミニ遺伝子座法の使用により、Ig遺伝子座を有するトランスジェニックマウスを作製した。上記に引用したジェンファームインターナショナル社の業績の発明者らであるロンバーグおよびケイは、本発明者の先例にならって、内因性マウスIg遺伝子座の不活性化とスラニらの業績の実質的な重複物との組み合わせを提案した。
ミニ遺伝子座法の1つの利点は、Ig遺伝子座の一部を含む構築物の作製および動物への導入を迅速に行いうることである。しかしこれを相殺する程度に、ミニ遺伝子座法には理論的にみて、少数のV、DおよびJ遺伝子を含めることによって導入される多様性が不十分であるとの重大な欠点がある。事実、発表された研究はこの懸念を裏づけるように思われる。ミニ遺伝子座法を用いて作製された動物のB細胞発生および抗体産生は阻害されていると考えられる。このため、本発明者らは、より大きな多様性の実現を目的として、および動物の免疫レパートリーを再構築するための努力において、Ig遺伝子座の大部分を導入することを一貫して主張してきた。
したがって、ヒトIg遺伝子座のより完全な生殖細胞系配列およびコンフィギュレーションを含むトランスジェニック動物を提供することが望ましいと考えられる。内因性Igがノックアウトされたバックグラウンドに対してこのような遺伝子座を提供することはさらに望ましいと考えられる。
【先行技術文献】
【特許文献】
【0009】
【特許文献1】米国特許出願第07/466,008号
【特許文献2】米国特許出願第07/610,515号
【特許文献3】米国特許出願第07/919,297号
【特許文献4】米国特許出願第07/922,649号
【特許文献5】米国特許出願第08/031,801号
【特許文献6】米国特許出願第08/112,848号
【特許文献7】米国特許出願第08/234,145号
【特許文献8】米国特許出願第08/376,279号
【特許文献9】米国特許出願第08/430,938号
【特許文献10】米国特許出願第08/464,584号
【特許文献11】米国特許出願第08/464,582号
【特許文献12】米国特許出願第08/463,191号
【特許文献13】米国特許出願第08/462,837号
【特許文献14】米国特許出願第08/486,853号
【特許文献15】米国特許出願第08/486,857号
【特許文献16】米国特許出願第08/486,859号
【特許文献17】米国特許出願第08/462,513号
【特許文献18】米国特許出願第08/724,752号
【特許文献19】EP 0 463 151 B1
【特許文献20】国際特許出願国際公開公報第94/02602号
【特許文献21】国際特許出願国際公開公報第96/34096号
【特許文献22】PCT/US96/05928
【特許文献23】米国特許第5,545,807号
【特許文献24】米国特許第5,545,806号
【特許文献25】米国特許第5,625,825号
【特許文献26】米国特許出願第07/574,748号
【特許文献27】米国特許出願第07/575,962号
【特許文献28】米国特許出願第07/810,279号
【特許文献29】米国特許出願第07/853,408号
【特許文献30】米国特許出願第07/904,068号
【特許文献31】米国特許出願第07/990,860号
【特許文献32】米国特許出願第08/053,131号
【特許文献33】米国特許出願第08/096,762号
【特許文献34】米国特許出願第08/155,301号
【特許文献35】米国特許出願第08/161,739号
【特許文献36】米国特許出願第08/165,699号
【特許文献37】米国特許出願第08/209,741号
【特許文献38】国際公開公報第94/25585号
【特許文献39】国際公開公報第93/12227号
【特許文献40】国際公開公報第92/22645号
【特許文献41】国際公開公報第92/03918号
【非特許文献】
【0010】
【非特許文献1】グリーン(Green)ら、Nature Genetics 7:13〜21(1994)
【非特許文献2】Taylor et al., "A transgenic mouse that expresses a diversity of human sequence heavy and light chain immunoglobulins." Nucleic Acids Research 20:6287-6295 (1992).
【非特許文献3】Chen et al. "Immunoglobulin gene rearrangement in B-cell deficient mice generated by targeted deletion of the JH locus" International Immunology 5:647-656 (1993)
【非特許文献4】Tuaillon et al., "Human immunoglobulin heavy-chain minilocus recombination in transgenic mice: gene-segment use in μ and γ transcripts." Proc. Natl. Acad. Sci. USA 90:3720-3724 (1993).
【非特許文献5】Choi et al. "Transgenic mice containing a human heavy chain immunoglobulin gene fragment cloned in a yeast artificial chromosome" Nature Genetics 4:117-123 (1993)
【非特許文献6】Lonberg et al., "Antigen-specific human antibodies from mice comprising four distinct genetic modifications." Nature 368:856-859 (1994).
【非特許文献7】Taylor et al., "Human immunoglobulin transgenes undergo rearrangemet, somatic mutation and class switching in mice that lack endogenous IgM." International Immunology 6:579-591 (1994).
【非特許文献8】Tuaillon et al. "Analysis of direct and inverted DJH rearrangements in a human Ig heavy chain transgenic minilocus" J. Immunol. 154:6453-6465 (1995)
【発明の概要】
【課題を解決するための手段】
【0011】
[発明の概要]
本発明により提供されるものは、ヒト重鎖遺伝子座およびヒトκ軽鎖遺伝子座の両方を含むほぼ完全なヒトIg遺伝子座を有するトランスジェニック動物である。好ましくは、重鎖遺伝子座は、ヒト重鎖可変領域の約20%を超える、より好ましくは約40%を超える、より好ましくは約50%、さらにより好ましくは約60%を超える部分を含む。ヒトκ軽鎖に関して、遺伝子座は好ましくはヒトκ軽鎖可変領域の約20%を超える、より好ましくは約40%を超える、より好ましくは約50%、さらにより好ましくは約60%を超える部分を含む。このような比率は機能的な可変領域遺伝子の比率に関して言及することが好ましい。
【0012】
さらに好ましくは、このような動物は全DH領域、全JH領域、ヒトμ定常領域を含み、付加的なアイソタイプを作製するための他のヒト定常領域をコードする遺伝子を付加的に備えることができる。このようなアイソタイプにはγ1、γ2、γ3、α、ε、βおよび他の定常領域をコードする遺伝子を含めうる。同一の導入遺伝子上、すなわちヒトμ定常領域の下流に代替的な定常領域を含めることもでき、または別の染色体上にこのような他の定常領域を含めることもできる。このような他の定常領域が導入遺伝子をコードするヒトμ定常領域を含む染色体と同じ染色体上に含められる場合には、他の1つのアイソタイプまたは複数のアイソタイプへのシス転換(cis-switching)が達成されうることが理解されるであろう。これに対して、このような他の定常領域が導入遺伝子をコードするμ定常領域を含む染色体とは異なる染色体上に含められる場合には、他の1つのアイソタイプまたは複数のアイソタイプへのトランス転換(trans-switching)が達成されうることが理解されるであろう。このような配置は、広範な種類の抗原に対する抗体を産生させるためのマウスの設計および構築において多大な柔軟性を可能にする。
【0013】
このようなマウスは、機能的な内因性免疫グロブリンを別に産生しないことが好ましい。これは、1つの好ましい態様では、内因性の重鎖および軽鎖遺伝子座の不活性化(またはノックアウト)を通じて達成される。例えば、1つの好ましい態様では、マウス重鎖J領域ならびにマウスκ軽鎖J領域およびCκ領域が、その領域を置換または欠失させる相同的組換えベクターの利用によって不活性化される。このような技法は、本発明者らの以前の出願および刊行物に詳細に記載されている。
【0014】
予想外のことに、本発明に係るトランスジェニックマウスは、ほぼ完全に再構成された免疫系レパートリーを有すると考えられる。これは4つの別々のマウス系統を比較した際に極めて明瞭に示される:第1の系統は広範なヒト重鎖可変領域およびヒトκ軽鎖可変領域を含んでいてμアイソタイプのみをコードし、第2の系統は広範なヒト重鎖可変領域およびヒトκ軽鎖可変領域を含んでいてμおよびγ2アイソタイプをコードし、第3の系統は明らかに少ないヒト重鎖およびκ軽鎖の可変領域を含み、第4の系統は二重不活性化を受けた(double-inactivated)マウスIg遺伝子座を含む。第1および第2の系統では同一ではないが類似したB細胞発生が起こるが、第3の系統ではB細胞の発生および成熟が抑制され、第4の系統は成熟B細胞を全く含まない。さらに興味深いことに、内因性Igがノックアウトされたバックグラウンドをもつマウスではマウス抗体に比してヒト抗体の産生がかなり増強される。すなわち、ヒトIg遺伝子座および機能的に不活性化された内因性Ig遺伝子座を含むマウスは、ヒトIg遺伝子座のみを含むマウスよりもヒト抗体をほぼ100〜1000倍もの速度で効率的に産生する。
【0015】
このため、本発明の第1の局面によれば、改変を含むゲノムを有し、その改変に以下のものが含まれるトランスジェニック非ヒト哺乳動物が提供される:その哺乳動物が正常なB細胞発生を呈しないように不活性化された、内因性免疫グロブリン(Ig)遺伝子座;ヒトμ定常領域ならびにそれに対する調節およびスイッチ配列、複数のヒトJH遺伝子、複数のヒトDH遺伝子および複数のヒトVH遺伝子を含んでいて、実質的な生殖細胞系コンフィギュレーションとして挿入されたヒト重鎖Ig遺伝子座;ならびにヒトκ定常領域、複数のヒトJκ遺伝子および複数のヒトVκ遺伝子を含んでいて、実質的な生殖細胞系コンフィギュレーションとして挿入されたヒトκ軽鎖Ig遺伝子座。1つの好ましい態様において、重鎖Ig遺伝子座はヒトγ1、ヒトγ2、ヒトγ3、ヒトγ4、α、δおよびεからなる群より選択される第2の定常領域を含む。もう1つの好ましい態様において、VH遺伝子の数は約20より多い。もう1つの好ましい態様において、Vκ遺伝子の数は約15より多い。もう1つの好ましい態様では、DH遺伝子の数は約25より多く、JH遺伝子の数は約4より多く、VH遺伝子の数は約20より多く、Jκ遺伝子の数は約4より多く、Vκ遺伝子の数は約15より多い。もう1つの好ましい態様において、DH遺伝子の数、JH遺伝子の数、VH遺伝子の数、Jκ遺伝子の数およびVκ遺伝子の数は、Ig遺伝子座が約1×105種を超える、異なる機能的な抗体配列の組み合わせをコードするように選択される。1つの好ましい態様では、哺乳動物の1つの集団において、野生型と比べて平均で約50%を超えるB細胞の機能が再構成される。
【0016】
本発明の第2の局面によれば、哺乳動物で、ヒト免疫グロブリン分子を産生しうるが、機能的な内因性免疫グロブリン分子を実質的に産生しえないようにさせる改変を含むゲノムを有する、以下のように改善されたトランスジェニック非ヒト哺乳動物が提供される:哺乳動物のゲノムへの、その哺乳動物が約1×106種を超える異なる機能的なヒト免疫グロブリン配列の組み合わせをコードしうるために十分なヒトVH、DH、JH、VκおよびJκ遺伝子の挿入。
【0017】
本発明の第3の局面によれば、哺乳動物で、ヒト免疫グロブリン分子を産生しうるが、機能的な内因性免疫グロブリン分子を実質的に産生しえないようにする改変であって、その哺乳動物が機能的な内因性免疫グロブリン分子を産生する能力をもたないことに関してその哺乳動物が正常なB細胞発生を行えないようにするような改変を含むゲノムを有する、以下のように改善されたトランスジェニック非ヒト哺乳動物が提供される:哺乳動物のゲノムへの、その哺乳動物が約1×106種を超える、異なる機能的なヒト免疫グロブリン配列の組み合わせをコードしうるために十分なヒトVH、DH、JH、VκおよびJκ遺伝子、ならびに哺乳動物において正常なB細胞発生が実質的に再現されるのに十分なVHおよびVκ遺伝子の挿入。1つの好ましい態様では、哺乳動物の1つの集団において、野生型と比べて平均で約50%を超えるB細胞の機能が再構成される。
【0018】
本発明の第4の局面によれば、改変を含むゲノムを有し、その改変に以下のものが含まれるトランスジェニック非ヒト哺乳動物が提供される:不活性化された内因性重鎖免疫グロブリン(Ig)遺伝子座;不活性化された内因性κ軽鎖Ig遺伝子座;yH2のヌクレオチド配列に実質的に対応するヌクレオチド配列を含む、挿入されたヒト重鎖Ig遺伝子座;およびyK2のヌクレオチド配列に実質的に対応するヌクレオチド配列を含む、挿入されたヒトκ軽鎖Ig遺伝子座。
【0019】
本発明の第5の局面によれば、改変を含むゲノムを有し、その改変に以下のものが含まれるトランスジェニック非ヒト哺乳動物が提供される:不活性化された内因性重鎖免疫グロブリン(Ig)遺伝子座;yH2のヌクレオチド配列に実質的に対応するヌクレオチド配列を含む、挿入されたヒト重鎖Ig遺伝子座;およびyK2のヌクレオチド配列に実質的に対応するヌクレオチド配列を含む、挿入されたヒトκ軽鎖Ig遺伝子座。
【0020】
本発明の第6の局面によれば、改変を含むゲノムを有し、その改変に以下のものが含まれるトランスジェニック非ヒト哺乳動物が提供される:不活性化された内因性重鎖免疫グロブリン(Ig)遺伝子座;不活性化された内因性κ軽鎖Ig遺伝子座;ヒトγ2定常領域が存在しないyH2のヌクレオチド配列に実質的に対応するヌクレオチド配列を含む、挿入されたヒト重鎖Ig遺伝子座;およびyK2のヌクレオチド配列に実質的に対応するヌクレオチド配列を含む、挿入されたヒトκ軽鎖Ig遺伝子座。
【0021】
本発明の第7の局面によれば、改変を含むゲノムを有し、その改変に以下のものが含まれるトランスジェニック非ヒト哺乳動物が提供される:不活性化された内因性重鎖免疫グロブリン(Ig)遺伝子座;ヒトγ2定常領域が存在しないyH2のヌクレオチド配列に実質的に対応するヌクレオチド配列を含む、挿入されたヒト重鎖Ig遺伝子座;およびyK2のヌクレオチド配列に実質的に対応するヌクレオチド配列を含む、挿入されたヒトκ軽鎖Ig遺伝子座。
【0022】
本発明の第8の局面によれば、以下のものを含む、ヒト抗体を製造するための方法が提供される:上記の本発明の第1から第5の局面の哺乳動物のうち任意のものへの抗原の接種;109M-1よりも高い親和性で抗原と特異的に結合するヒト抗体を分泌する不死化細胞の集団を得るためのリンパ球細胞の回収および不死化;ならびに不死化細胞集団からの抗体の単離。
【0023】
1つの好ましい態様では、抗原はIL-8である。もう1つの好ましい態様では、抗原はEGFRである。もう1つの好ましい態様では、抗原はTNF-αである。
【0024】
本発明の第9の局面によれば、IL-8、EGFRおよびTNF-αに対する抗体を含む、本発明の第6の局面の方法によって製造される抗体が提供される。
【0025】
本発明の第10の局面によれば、複数のヒト可変領域の挿入を含む改変を含むゲノムを有するトランスジェニックマウスを製造するための改善された方法であって、改善が以下のものを含む方法が提供される:酵母人工染色体からのヒト可変領域の挿入。
【0026】
本発明の第11の局面によれば、本発明の第8の局面の改善の使用を通じて製造されたトランスジェニックマウスおよびそれらに由来するトランスジェニック子孫が提供される。
本発明の第12の局面によれば、挿入されたヒト重鎖免疫グロブリン導入遺伝子を含む改変を有するゲノムを有するトランスジェニック哺乳動物であって、改善に以下のものが含まれるトランスジェニック哺乳動物が提供される:ヒト様の接合点多様性およびヒト様の相補性決定領域3(CDR3)の長さを可能とするヒト可変領域遺伝子の選択された組(set)を含む導入遺伝子。1つの好ましい態様において、ヒト様の接合点多様性は平均7.7塩基のN付加長を含む。もう1つの好ましい態様において、ヒト様CDR3の長さは約2から約25残基の間、平均すると約14残基を含む。
本発明はまた、以下の項目を提供する:
(項目1)以下の改変を含むゲノムを有するトランスジェニック非ヒト哺乳動物:
その哺乳動物が正常なB細胞発生を呈しないように不活性化された内因性免疫グロブリン(Ig)遺伝子座;
挿入されるVHおよびVκ遺伝子の数がその哺乳動物における正常なB細胞発生が実質的に再現されるように選択される、
実質的な生殖細胞系コンフィギュレーションとして挿入されたヒト重鎖Ig遺伝子座であって、ヒトμ定常領域ならびにそれに対する調節およびスイッチ配列、複数のヒトJH遺伝子、複数のヒトDH遺伝子、ならびに複数のヒトVH遺伝子を含むヒト重鎖Ig遺伝子座;ならびに
実質的な生殖細胞系コンフィギュレーションとして挿入されたヒトκ軽鎖Ig遺伝子座であって、ヒトκ定常領域、複数のJκ遺伝子、および複数のVκ遺伝子を含むヒトκ軽鎖Ig遺伝子座。
(項目2)重鎖Ig遺伝子座がヒトγ1、ヒトγ2、ヒトγ3、ヒトγ4、α、εおよびδからなる群より選択される第2の定常領域を含む、項目1記載の哺乳動物。
(項目3)VH遺伝子の数が約20よりも多い、項目1記載の哺乳動物。
(項目4)Vκ遺伝子の数が約15よりも多い、項目1記載の哺乳動物。
(項目5)DH遺伝子の数が約25よりも多く、JH遺伝子の数が約4よりも多く、VH遺伝子の数が約20よりも多く、Jκ遺伝子の数が約4よりも多く、Vκ遺伝子の数が約15よりも多い、項目1記載の哺乳動物。
(項目6)DH遺伝子の数、JH遺伝子の数、VH遺伝子の数、Jκ遺伝子の数、およびVκ遺伝子の数が、Ig遺伝子座が約1×105種を超える異なる機能的な抗体配列の組み合わせをコードするように選択される、項目1記載の哺乳動物。
(項目7)哺乳動物の1つの集団において野生型と比べて平均で約50%を超えるB細胞の機能が再構成される、項目1記載の哺乳動物。
(項目8)哺乳動物を、ヒト免疫グロブリン分子を産生しうるが機能的な内因性抗体分子を実質的に産生しえないようにする、以下の改変を含むゲノムを有するトランスジェニック非ヒト哺乳動物:
接合点多様性または体細胞変異事象に寄与することなく、その哺乳動物が約1×106種を超える異なる機能的なヒト免疫グロブリン配列の組み合わせをコードしうるために十分なヒトVH、DH、JH、VκおよびJκ遺伝子の、哺乳動物ゲノムへの挿入。
(項目9)哺乳動物を、ヒト免疫グロブリン分子を産生しうるが機能的な内因性抗体分子を実質的に産生しえないようにする改変であって、その哺乳動物が機能的な内因性抗体分子を産生する能力をもたないことに関してその哺乳動物が正常なB細胞発生を行えないようにするような以下の改変を含むゲノムを有するトランスジェニック非ヒト哺乳動物:
その哺乳動物が約1×106種を超える異なる機能的なヒト免疫グロブリン配列の組み合わせをコードしうるために十分なヒトVH、DH、JH、VκおよびJκ遺伝子、ならびに哺乳動物において正常なB細胞発生が実質的に再現されるのに十分なVHおよびVκ遺伝子の、哺乳動物ゲノムへの挿入。
(項目10)哺乳動物の1つの集団において野生型と比べて平均で約50%を超えるB細胞の機能が再構成される、項目9記載の哺乳動物。
(項目11)以下の改変を含むゲノムを有するトランスジェニック非ヒト哺乳動物:
不活性化された内因性重鎖免疫グロブリン(Ig)遺伝子座;
不活性化された内因性κ軽鎖Ig遺伝子座;
yH2のヌクレオチド配列に実質的に対応するヌクレオチド配列を含む、挿入されたヒト重鎖Ig遺伝子座;および
yK2のヌクレオチド配列に実質的に対応するヌクレオチド配列を含む、挿入されたヒトκ軽鎖Ig遺伝子座。
(項目12)以下の改変を含むゲノムを有するトランスジェニック非ヒト哺乳動物:
不活性化された内因性重鎖免疫グロブリン(Ig)遺伝子座;
yH2のヌクレオチド配列に実質的に対応するヌクレオチド配列を含む、挿入されたヒト重鎖Ig遺伝子座;および
yK2のヌクレオチド配列に実質的に対応するヌクレオチド配列を含む、挿入されたヒトκ軽鎖Ig遺伝子座。
(項目13)以下の改変を含むゲノムを有するトランスジェニック非ヒト哺乳動物:
不活性化された内因性重鎖免疫グロブリン(Ig)遺伝子座;
不活性化された内因性κ軽鎖Ig遺伝子座;
ヒトγ2定常領域が存在しないyH2のヌクレオチド配列に実質的に対応するヌクレオチド配列を含む、挿入されたヒト重鎖Ig遺伝子座;および
yK2のヌクレオチド配列に実質的に対応するヌクレオチド配列を含む、挿入されたヒトκ軽鎖Ig遺伝子座。
(項目14)以下の改変を含むゲノムを有するトランスジェニック非ヒト哺乳動物:
不活性化された内因性重鎖免疫グロブリン(Ig)遺伝子座;
ヒトγ2定常領域が存在しないyH2のヌクレオチド配列に実質的に対応するヌクレオチド配列を含む、挿入されたヒト重鎖Ig遺伝子座;および
yK2のヌクレオチド配列に実質的に対応するヌクレオチド配列を含む、挿入されたヒトκ軽鎖Ig遺伝子座。
(項目15)以下の改変を含むゲノムを有するトランスジェニック非ヒト哺乳動物:
不活性化された内因性重鎖免疫グロブリン(Ig)遺伝子座;
ヒトγ2定常領域が存在しないyH2のヌクレオチド配列に実質的に対応するヌクレオチド配列を含む、挿入されたヒト重鎖Ig遺伝子座;および
yK2のヌクレオチド配列に実質的に対応するヌクレオチド配列を含む、挿入されたヒトκ軽鎖Ig遺伝子座。
(項目16)以下の段階を含む、ヒト抗体を製造するための方法:
項目1〜10のいずれか一項記載の哺乳動物への抗原の接種;
109M-1よりも大きい親和性で抗原と特異的に結合するヒト抗体を分泌する不死化細胞系を得るためのリンパ球細胞の回収および不死化;ならびに
不死化細胞系からの抗体の単離。
(項目17)抗原がIL-8である、項目11記載の方法。
(項目18)抗原がEGFRである、項目11記載の方法。
(項目19)抗原がTNF-αである、項目11記載の方法。
(項目20)項目11記載の方法によって製造される抗体。
(項目21)項目12記載の方法によって製造される抗IL-8抗体。
(項目22)項目13記載の方法によって製造される抗EGFR抗体。
(項目23)項目14記載の方法によって製造される抗TNF-α抗体。
(項目24)複数のヒト可変領域の挿入が含まれる以下の改変を含むゲノムを有するトランスジェニックマウスを製造するための方法:
酵母人工染色体からのヒト可変領域の挿入。
(項目25)項目19記載の改善の使用を通じて製造されたトランスジェニックマウスおよびそれらに由来するトランスジェニック子孫。
(項目26)挿入されたヒト重鎖免疫グロブリン導入遺伝子が含まれ、以下の改変を含むゲノムを有するトランスジェニック哺乳動物:
ヒト様の接合点多様性およびヒト様の相補性決定領域3(CDR3)の長さを可能とするヒト可変領域遺伝子の選択された組(set)を含む導入遺伝子。
(項目27)ヒト様の接合点多様性が平均7.7塩基のN付加長を含む、項目26記載の改変。
(項目28)ヒト様CDR3の長さが約2から約25残基の間、平均すると約14残基を含む、項目26記載の改変。
【図面の簡単な説明】
【0027】
【図1】図1は、本発明に従って好ましいマウスに導入される、再構成されたヒト重鎖およびヒトκ軽鎖遺伝子座YACの模式図である。ヒト重鎖(1H、2H、3Hおよび4H)およびヒトκ軽鎖の近位(1K、2Kおよび3K)遺伝子座の全長にわたるYACをヒトYACライブラリーからクローニングした。ヒトIg遺伝子座に関して異なるYACの位置(CookおよびTomlinson、1995ならびにCoxら、1994から採用)、それらのサイズ、および非Ig配列を示す(一定の比率では示していない)。ヒト重鎖およびκ軽鎖YACを再構築するために、YACを2段階の手順(材料および方法の項を参照)にて酵母中に組換え導入した。ヒト重鎖を含むYACであるyH2にさらにヒトγ2遺伝子の配列によるレトロフィット(retrofit)を行った。yK2はヒトκ軽鎖を含むYACである。YACベクターの構成要素:YACベクターの腕上のテロメア:黒ぬり△、動原体●、哺乳動物(HPRT、Neo)および酵母の選択可能マーカー(TRP1、ADE2、LYS2、LEU2、URA3、HIS3)を示す。VHセグメントはオープンリーディングフレーム●、偽遺伝子□、および配列決定がなされていない領域○を有する遺伝子として分類した。Vκ部分はオープンリーディングフレーム●および偽遺伝子□を有する遺伝子として分類した。XenoMouse IIによって利用されることを本発明者らが見いだしたV遺伝子には印(*)を付した。yH2上に含まれるVH遺伝子領域は矢印で示した。
【図2】図2は、ES細胞およびXenoMouse系統に組み込まれたヒト重鎖YAC、yH2の一連のサザンブロット分析および特徴分析の結果である。図2aは、ワシントン(Washington)大学YACライブラリー供給源(Brownsteinら、1989)に由来するCGM1不死化Bリンパ芽球細胞系、yH2 YAC(0.5μgのYACを2μgの3B1 DNAに加えたもの)、非改変E14TG. 3B1(3B1)ならびにyH2含有ES細胞系:L10、J9.2、L18、L17およびJ17から調製したEcoRI(a、c)およびBamHI(b,d、e)消化DNA(2μg)の一連のサザンブロット分析の結果である。ブロッティングに用いたプローブは、ヒトVH1(a)、DH(b)[CGM1レーン中の18kb断片は第16染色体上のDセグメントである]、VH3(c)、Cμ(d)およびJH(e)である。図2bは、野生型(WT、129×B57BL/6J)、XM2A-1およびXM2A-2(2匹の独立した子孫)マウスの尾部から、または親yH2を含むES細胞系L10(他の試料と比べてややロード量が少ない)、J9.2、およびyK2を含むES細胞系J23.1から調製したEcoRI(a-b)およびBamHI(c-d)消化DNA(10μg)の一連のサザンブロット分析の結果である。用いたプローブはヒトVH1(a)、VH4(b)、ヒトγ-2(c)およびマウス3'-エンハンサー(d、5kbのバンドは内因性マウス3'-エンハンサー断片である)である。分子量マーカーの断片サイズ(kb単位)を示している。
【図3】図3は、ES細胞およびXenoMouse 2A系統に組み込まれたヒトκ軽鎖YAC、yK2の特徴を示す一連のサザンブロット分析の結果である。図3aは、CGM1細胞系(Brownsteinら、1989、前記)、yK2 YAC(0.5μgのYACを2μgの3B1 DNAに加えたもの)、非改変E14TG. 3B1(3B1)ならびにyK2含有ES細胞系:J23.1およびJ23.7から調製したEcoRI(a、c、d)およびBamHI(b、e)消化DNA(2μg)の一連のサザンブロット分析の結果である。ブロッティングに用いたプローブは、ヒトVa(a)、Kde(b)、VκII(c)、VκIII(d)およびCκ(e)である。図3bは、野生型(WT、129×B6)、XM2A-1およびXM2A-2(2匹の独立した子孫)マウスの尾部から、または親yH2を含むES細胞系L10(他の試料と比べてややロード量が少ない)、J9.2、およびyK2を含むES細胞系J23.1から調製したEcoRI消化DNA(2μg)の一連のサザンブロット分析の結果である。用いたプローブはヒトVκI(a)、VκIV(b)、VκVI(c)および3'-エンハンサー(d)である。分子量マーカーの断片サイズ(kb単位)を示している。
【図4】図4は、B細胞の再構成ならびにXenoMouse由来B細胞でのヒトμ、δおよびκ鎖の表面発現を示し、野生型マウス(WT)、二重不活性化マウス(DI)ならびにXenoMouse系統2A-1および2A-2(XM2A-1、XM2A-2)からの末梢血(図4a)および脾臓(図4b)リンパ球のフローサイトメトリー分析の結果を示す。B細胞特異的マーカーB220に対する抗体を抗ヒトμ、δ、κまたはマウスμ、δ、κもしくはλとともに用いて4色フローサイトメトリー分析を行った。4象限のそれぞれにおける陽性染色された細胞の比率を示す。細胞の単離および染色は、材料および方法の項に記載した通りに実施した。B220+μ+集団に対して指示領域でのゲーティングをまず行った後に、ヒトκ+およびマウスλ+細胞の集団を決定した。μ+およびδ+細胞の集団は、B220+集団に対してまずゲーティングをまず行った後に決定した。ある領域または4象限の内部にある陽性細胞の比率を示す。示したFACSプロフィールは、各系統に対して実施したいくつかの実験の代表例である。
【図5】図5は、XenoMouse由来のヒト抗体が、それらの特異的抗原の細胞との結合を阻止することを示す。図5aは、マウス抗ヒトIL-8抗体(R&D Systems)(□)ならびに完全ヒトMab D1.1(◆)、K2.2(●)、K4.2(黒ぬり△)およびK4.3(黒ぬり▽)による、標識[I125]IL-8のヒト好中球との結合の阻害を示す。抗体の非存在下における標識[I125]IL-8のバックグラウンド結合は2657cpmであった。図5bは、マウス抗ヒトEGFR抗体225および528(それぞれ□、▽;Calbiochem)ならびに完全ヒト抗体E1.1(●)、E2.4(黒ぬり△)、E2.5(黒ぬり▽)およびE2.11(◆)による、標識[I125]EGFのA431細胞上のその受容体との結合の阻害を示す。抗体の非存在下における[I125]EGFのバックグラウンド結合は1060cpmであった。図5cは、マウス抗ヒトTNF-α抗体(R&D Systems)(□)ならびに完全ヒトMab T22.1(◆)、T22.4(●)、T22.8(黒ぬり△)およびT22.9(黒ぬり□)による、標識[I125]TNF-αのU937細胞上のその受容体との結合の阻害を示す。抗体の非存在下における標識[I125]TNF-αのバックグラウンド結合は4010cpmであった。対照ヒトIgG2骨髄腫抗体(四角で囲まれた×印)。
【図6】図6は、XenoMouse由来の完全ヒト抗体におけるレパートリーおよび体細胞超突然変異を示す。4つの抗IL-8(図6a)および4つの抗EGFR(図6b)ヒトIgG2κ Mabの予想されるアミノ酸配列を、CDR1、CDR2およびCDR3ならびに定常領域Cγ2およびCκに分けた。各抗体のDおよびJ遺伝子を示す。生殖細胞系配列からのアミノ酸置換は太字で示している。
【図7】図7は、ヒト重鎖ゲノムおよびヒトκ軽鎖ゲノムの模式図である。
【図8】図8は、yH2(ヒト重鎖)YACの構成を示すもう1つの模式図である。
【図9】図9は、yK2(ヒトκ軽鎖)YACの構成を示すもう1つの模式図である。
【図10】図10は、yK2(ヒトκ軽鎖)YACの構成を示すもう1つの模式図である。
【図11】図11は、ES細胞内およびマウスゲノム中へのyH2(ヒト重鎖)YACの組込みが無傷でなされたことを示す一連のサザンブロット分析の結果である。詳細な説明は図2に関して提示している。
【図12】図12は、ES細胞内およびマウスゲノム中へのyK2(ヒトκ軽鎖)YACの組込みが無傷になされたことを示す一連のサザンブロット分析の結果である。詳細な説明は図3に関して提示している。
【図13】図13は、B細胞の再構成、ならびにXenoMouse由来B細胞でのヒトμ、δおよびκ鎖ならびにマウスλ鎖の表面発現を示し、末梢血のフローサイトメトリー分析を示す。詳細は図4に関して提示している。
【図14】図14は、野生型マウスによるマウス抗体産生と比較して、XenoMouse IIによるヒト抗体の産生レベルを示している。
【図15】図15は、XenoMouse II系統で発現されたヒト重鎖転写物のレパートリー解析の結果である。
【図16】図16は、XenoMouse II系統で発現されたヒトκ軽鎖転写物のレパートリー解析の結果である。
【図17】図17は、XenoMouse II系統において利用されることが観察されたヒトVHおよびVκ遺伝子が広範に利用されることに関するもう1つの描写である。
【図18】図18は、XenoMouse II系統におけるヒト抗体産生の力価を示している。
【図19】図19は、XenoMouse II系統に由来する抗IL-8抗体の遺伝子利用を描写している。
【図20】図20は、XenoMouse II系統に由来する抗IL-8抗体の重鎖アミノ酸配列を示す。
【図21】図21は、XenoMouse II系統に由来する抗IL-8抗体のκ軽鎖アミノ酸配列を示す。
【図22】図22は、XenoMouse II系統に由来するヒト抗IL-8抗体による、IL-8のヒト好中球との結合の阻止を示す。
【図23】図23は、XenoMouse II系統に由来するヒト抗IL-8抗体による、ヒト好中球上でのCD11b発現の阻害を示す。
【図24】図24は、XenoMouse II系統に由来するヒト抗IL-8抗体による、IL-8によって誘導されるカルシウム流入の阻害を示す。
【図25】図25は、XenoMouse II系統に由来するヒト抗IL-8抗体による、IL-8 RB/293走化性の阻害を示す。
【図26】図26は、ヒトIL-8誘導性皮膚炎症のウサギモデルの模式図である。
【図27】図27は、XenoMouse II系統に由来するヒト抗IL-8抗体による、図26のウサギモデルにおけるヒトIL-8誘導性皮膚炎症の阻害を示している。
【図28】図28は、XenoMouse II系統由来のヒト抗IL-8抗体による、ラット角膜ポケットモデルでの内皮細胞の血管新生の阻害を示す。
【図29】図29は、XenoMouse II系統に由来するヒト抗EGFR抗体の遺伝子利用の描写である。
【図30】図30は、XenoMouse II系統に由来するヒト抗EGFR抗体の重鎖アミノ酸配列を示している。
【図31】図31は、XenoMouse II系統に由来するヒト抗EGFR抗体による、EGFのA431細胞との結合の阻止を示している。
【図32】図32は、XenoMouse II系統に由来するヒト抗EGFR抗体による、EGFのSW948細胞との結合の阻止を示している。
【図33】図33は、XenoMouse II系統に由来するヒト抗EGFR抗体がSW948細胞のインビトロ増殖を阻害することを示す。
【図34】図34は、XenoMouse II系統に由来するヒト抗TNF-α抗体の使用による、TNF-αのU937細胞との結合の阻害を示している。
【図35】図35は、XenoMouse II系統に由来するヒト抗EGFR抗体のκ軽鎖アミノ酸配列を示している。
【発明を実施するための形態】
【0028】
[好ましい態様の詳細な説明]
本明細書において本発明者らは、実質的な生殖細胞系コンフィギュレーションにある、メガベースサイズのヒトIg遺伝子座を含むマウスの、いくつかの系統の製造および特徴分析に関して記載する。このため本発明は、YAC上での大きく複雑なヒトIg遺伝子座の再構成、および対応するマウスIg遺伝子座を機能的に置換するためのメガベースサイズのYACをマウスに首尾よく導入する方法を、初めて提供するものである。
【0029】
マウス系統
以下のマウス系統を、本明細書において記載および/または使用する。
二重不活性化(DI)系統: DI系統のマウスは、機能的な内因性マウスIgを産生しないマウスである。1つの好ましい態様において、DIマウスは不活性化されたマウスJH領域および不活性化されたマウスCκ領域を有する。この系統の構築は、別で詳細に説明されている。例えば、DI系統の作製のために用いられる技法は、1990年1月12日に提出された米国特許出願第07/466,008号、1990年11月8日に提出された第07/610,515号、1992年7月24日に提出された第07/919,297号、1993年3月15日に提出された第08/031,801号、1993年8月27日に提出された第08/112,848号、1994年4月28日に提出された第08/234,145号、1996年10月2日に提出された第08/724,752号に詳細に記載されている。1996年6月12日に認可が発行されたEP 0 463 151 B1、1994年2月3日に発行された国際特許出願国際公開公報第94/02602号、1996年10月31日に発行された国際特許出願国際公開公報第96/34096号、および1996年4月29日に提出されたPCT/US96/05928も参照されたい。上記の特許および特許出願のそれぞれの開示は、その全体が参照として本明細書に組み入れられる。DIマウスではB細胞発生が極めて未熟であることが観察および報告されている。本マウスは成熟B細胞を産生せず、プロB細胞のみを産生する。
【0030】
XenoMouse I系統: XenoMouse I系統の設計、構築および解析はグリーン(Green)ら、Nature Genetics 7:13〜21(1994)に詳細に考察されている。このようなマウスはDIバックグラウンドに対するIgMκ抗体を産生する。本マウスでは、ほとんどまたは全くB細胞発生が生じないDI系統のマウスと比べてB細胞機能の改善が認められた。XenoMouse I系統のマウスは抗原誘発刺激に対してかなり強い免疫応答を生じる能力があったが、B細胞の産生効率は低いと考えられ、異なる抗原に対する応答は制限されており、これはV遺伝子レパートリーの制限に起因するものと考えられた。
【0031】
L6系統: L6系統は、DIバックグラウンドの、内因性マウスIgに対するIgMκ抗体を産生するマウスである。L6マウスは、挿入されたヒト重鎖および挿入されたヒトκ軽鎖を含んでいる。L6系統は、二重不活性化バックグラウンドに対する重鎖挿入物を含むマウス(L6H)と、二重不活性化バックグラウンドに対するκ軽鎖挿入物を含むマウス(L6L)との交配によって作出される。重鎖挿入物は、VH6-1に始まってVH3-65に終わる約66個のVHセグメント、ならびに生殖細胞系コンフィギュレーションにある多数のD遺伝子クラスター(約32個)、JH遺伝子(6個)、イントロン性エンハンサー(Em)、CμおよびCδを越えて約25kbのところまでを含むYACに由来する、無傷の約970kbのヒトDNA挿入物を含む。軽鎖挿入物は、Vκ-B3に始まってVκ-Op11に終わる約32個のVκ遺伝子を含むYACに由来する、無傷の約800kbのヒトDNA挿入物を含む。この800kb挿入物は、Vκ-Lp-13に始まってVκ-Lp-5で終わる約100kbの欠失を含む。しかし、本DNAはVκ-Lp-13からVκ-Op-1を越えて100kbのところまで生殖細胞系コンフィギュレーションにあり、Jκ遺伝子、イントロン性の3'エンハンサー、定常Cκ遺伝子、およびKdeを含む。L6HおよびL6Lマウスは、それらのゲノム中に組み込まれた可変遺伝子の完全なスペクトルを利用しうることが示されている。L6マウスも同様に、それらのゲノム中の可変遺伝子の完全なスペクトルを利用しうると考えられる。さらにL6マウスはヒトκ軽鎖の主な発現、多数の成熟B細胞、および正常レベルのIgMκヒト抗体を呈すると考えられる。このようなマウスは多数の免疫原に対する強いヒト抗体反応を生じ、最終的にはナノモルレベル以下の親和性をもつ抗原特異的な完全ヒトMabを産生すると考えられる。
【0032】
XenoMouse IIa系統: XenoMouse IIaマウスは、マウスが機能的な内因性Igを産生しないようなDIバックグラウンドに対して生殖細胞系コンフィギュレーションでメガベースサイズのヒトIg遺伝子座を備えた、本発明者らの第2世代のXenoMouse系統(登録商標)である。本質的には本マウスの構成はL6系統と等価であるが、完全なスイッチおよび調節配列ならびにシス位にあるマウス3'エンハンサーとともにヒトγ2遺伝子をさらに含む。本マウスは、YAC上に再構成された形で約1020kbの重鎖遺伝子座および約800kbのκ軽鎖遺伝子座を含み、これには重鎖遺伝子(約66個のVH)およびκ軽鎖遺伝子(約32個のVκ)、ヒト重鎖定常領域遺伝子(μ、δおよびγ)およびκ定常領域遺伝子(Cκ)ならびに同定されたすべての主要な調節要素を含む、大半のヒト可変領域遺伝子が含まれる。これらのマウスは、それらのゲノム中に組み入れられた可変領域の完全なスペクトルを利用しうることが示されている。さらに、それらは効率的なクラス転換(class switching)および体細胞超突然変異、ヒトκ軽鎖の優位な発現、多数の成熟B細胞、ならびに正常レベルのIgMκおよびIgGκヒト抗体を呈する。このようなマウスは、ヒトIL-8、ヒトEGF受容体(EGFR)およびヒト腫瘍壊死因子-α(TNF-α)を含む多数の免疫原に対する強いヒト抗体反応を生じ、最終的にはナノモルレベルに準じる親和性をもつ抗原特異的な完全ヒトMabを産生する。この最終結果は、広範なスペクトルの抗原に対して任意の望ましい特異性をもつ高親和性の、完全ヒト治療的Mabの迅速単離のための優れた供給源としてのXenoMouse(登録商標)を決定的に示すものである。
【0033】
上記の序論から理解される通り、XenoMouse II系統は成熟B細胞の発生を経て、抗原誘発刺激に対して強力な成人様免疫応答を起こすと考えられる。L6系統も、L6LおよびL6Hマウスに関するデータから予想される通り、成熟B細胞の発生を経て、抗原誘発刺激に対して強力な成人様免疫応答を起こすと考えられる。DIマウスをXenoMouse I系統と比較し、DIおよびXenoMouse I系統をL6およびXenoMouse II系統と比較すると、顕著に異なるB細胞発生の形態が観察される。この差からみて、誘導B細胞の成熟および発生ならびに成人様免疫応答の発生には、マウスに導入される可変領域配列の量および/または質が非常に重要であると考えられる。したがって、ヒト抗体の作製における、以上の系統の明らかな用途に加えて、以上の系統は、正常免疫応答におけるヒト抗体の性質、さらには自己免疫疾患および他の障害に特徴的な異常応答を研究するための有益な手段も提供する。
【0034】
可変領域―量的多様性
抗体の特異性(すなわち、広範な抗原に対する、実際にはそれらの上にある広範な独立したエピトープに対する抗体を生成する能力)は、重鎖(VH)およびκ軽鎖(Vκ)ゲノム上の可変領域遺伝子に依存している。ヒト重鎖ゲノムは、免疫グロブリン分子のヒト重鎖の可変領域をコードする約95の機能的遺伝子を含む。さらに、ヒト軽鎖ゲノムはその近位端に、免疫グロブリン分子のヒトκ軽鎖の可変領域をコードする約40の遺伝子を含む。本発明者らは、可変軽鎖および重鎖をコードする複数の遺伝子を含めることによって抗体の特異性を高めうることを示した。
【0035】
本発明によって提供されるものは、好ましくはヒト重鎖およびヒトκ軽鎖遺伝子座を含むヒトIg遺伝子座のかなりの部分を有するトランスジェニックマウスである。このため、好ましい態様では、ヒトVHおよびVκ遺伝子の10%よりも多くが用いられる。より好ましくは、ヒトVHおよびVκ遺伝子の約20%、30%、40%、50%、60%、またはさらには70%もしくはそれよりも多くが用いられる。1つの好ましい態様では、Vκ軽鎖ゲノムの近位領域に32個の遺伝子を含む構築物が用いられ、ゲノムのVH部分にある66個の遺伝子が用いられる。理解されるであろう通り、遺伝子は連続的、すなわちヒトゲノム中で認められる順番に含まれても、異なる順序、すなわちヒトゲノム中で認められる以外の順序で含まれても、それらの組み合わせであってもよい。このため、一例として、VHもしくはVκゲノムのいずれかの完全に連続的な部分を利用することができ、またはVHもしくはVκゲノムのいずれかの種々のV遺伝子を省いて全体の連続的配置を維持することもでき、またはVHもしくはVκゲノムのいずれかの内部のV遺伝子の順序を改めることなどもできる。1つの好ましい態様において、挿入される全遺伝子座はヒトにおいて認められる通りの実質的な生殖細胞系コンフィギュレーションとして提供される。いずれの場合にも、VHおよびVκゲノムからの多様な一連の遺伝子を含めることにより、抗体特異性の増強および最終的には抗体親和性の増強に至ることが予想され、本明細書に記載される結果で示される。
【0036】
さらに、このようなマウスは、好ましくは全DH領域、全JH領域、ヒトμ定常領域を含み、さらに別のアイソタイプの抗体のコーディングおよび生成のための他のヒト定常領域を付加的に備えることができる。このようなアイソタイプには、適切なスイッチおよび調節配列を備えたγ1、γ2、γ3、γ4、α、εおよびδ、ならびに他の定常領域をコードする遺伝子が含まれうる。理解されるであろう通り、さらに以下でより詳しく考察される通り、任意の特定の定常領域の選択に関連して、種々のスイッチおよび調節配列を適切に用いることができる。
【0037】
以下の表は、厳密にV-D-J結合およびκ軽鎖とのランダムな組み合わせに基づき、N付加または体細胞変異事象を考慮しない場合に、ヒトにおいて可能な抗体の組み合わせの多様性を示している。このような検討に基づくと、ヒトでは、任意の特定のアイソタイプに380万種を超える抗体の組み合わせが存在する。
(表I)
【表1】

【0038】
本発明の1つの好ましい態様に関して、DH、JHおよびJκ遺伝子の完全な補体を備えた約66個のVH遺伝子および32個のVκ遺伝子をマウスに含めることにより、抗体産生に関して可能な多様性は、2.03×106種の異なる抗体のオーダーである。上記の通り、この種の計算にはN付加および体細胞変異事象を考慮していない。このため、L6およびXenoMouse II系統などの本発明に係るマウスは、かなりの抗体多様性をもたらすことが理解されると思われる。好ましい態様において、本マウスはN付加および体細胞変異事象を考慮に入れずとも、1×106種を超える異なる重鎖V-D-J組み合わせおよびκ軽鎖V-J組み合わせを産生する能力をもつように設計される。
【0039】
可変領域―質的多様性
量的多様性に加えて、V遺伝子の量的選択(すなわち、多くの多様な数のV遺伝子)および/またはV遺伝子の質的選択(すなわち、特定のV遺伝子の選択)も、本発明者らが本明細書で「質的多様性」と呼ぶ一定の役割を果たすように思われる。本明細書で用いる質的多様性とは、結合多様性および/または体細胞変異事象が導入されるV-D-J再配列における多様性を意味する。重鎖再配列の間には、特定の酵素(RAG-1、RAG-2、および他のものも考えられる)が、抗体遺伝子のコード領域であるDNAの切断に関与する。ターミナルデオキシヌクレオチジルトランスフェラーゼ(Tdt)活性はアップレギュレートされ、これはV-DおよびD-J遺伝子セグメントの間のヌクレオチドのN末端付加の原因となる。類似の酵素および他のもの(SCIDおよび他のDNA修復酵素)は、これらのコード区域の結合部に生じる欠失の原因となる。結合多様性に関しては、N付加事象および相補性決定領域3(CDR3)の双方がこの項目に含められる。理解されるであろう通り、CDR3はD領域にまたがって位置しており、V-DおよびD-J結合事象を含む。このため、D-J再配列およびV-D再配列の両方の過程におけるN付加および欠失がCDR3の多様性の原因となっている。
【0040】
マウスおよびヒトの結合多様性の間には、ある特定の差異があることが示されている。特に、一部の研究者は、マウスのN付加長およびCDR3長は典型的なヒトのN付加長およびCDR3長よりも一般に短いことを報告している。このようなグループは、ヒトで典型的に認められるN付加長は平均約7.7塩基であることを報告している。ヤマダ(Yamada)ら(1991)。マウス様N付加の長さは平均で約3塩基のオーダーであることが多い。フィーニー(Feeney)ら(1990)。同様に、ヒト様CDR3の長さはマウス様CDR3よりも長い。ヒトではCDR3長は2から25残基の間であり、平均14残基であることが一般的である。マウスでは平均CDR3長がこれよりも短いことをいくつかのグループが報告している。
【0041】
N付加およびCDR3付加によって生じる結合多様性は、抗体特異性を発展させるという明らかな役割を果たしている。
【0042】
本発明によれば、再配列したV-D-J遺伝子配列は、予想される成人N付加長に匹敵するN付加長を示す。さらに、CDR3配列に対応するオープンリーディングフレーム(ORF)にまたがるアミノ酸配列は、予想される成人CDR3長と同等のCDR3長を示す。このようなデータは、可変領域の量的多様性および/または可変領域の質的多様性によりヒト様の結合多様性がもたらされることを示すものである。このような結合多様性はよりヒトに類似した抗体特異性につながると考えられる。
【0043】
可変領域―親和性
本発明者らは、可変領域の含有量の増加と抗体特異性との間に、直接的な因果関係があることを決定的に示してはいないが、そうであると思われ、しかもこのような多様性が提供されることにより、マウスが広範な抗原に対する免疫応答を生じる能力が実現および増強されることが期待される。さらに、このようなマウスは、個々の抗原または免疫原上の広範なエピトープに対する免疫応答を生じる性質をさらに備えるようになると考えられる。本発明者らのデータからは、本発明に従って産生される抗体は親和性も増強されると思われる。このようなデータには、本発明によるマウスおよびXenoMouse I系統との比較のほか、ジェンファームインターナショナル(GenPharm International)社およびMRCが発表した結果の考察が含まれる。XenoMouse I系統については、上記の通り、このようなマウスではB細胞産生の効率が低く、種々の抗原に対する応答性が制限される。このような結果は、一部にはV遺伝子レパートリーの制限に起因すると思われる。同じく、ジェンファームインターナショナル社およびMRCによって発表された結果は、多様な抗原に対する応答性が制限されていることを示している。
【0044】
発明の特定の理論または作業様式に拘束されることを望むものではないが、親和性の増強は多数のV領域が供給されることによる結果であると考えられる。本発明者らのデータからは、より多くの数の供給および/またはV遺伝子配列の質の選択により、結合多様性(N付加および相補性決定領域3(「CDR3」)の多様性の形成)は増強されるが、それは成人様免疫応答に典型的であり、抗体の親和性成熟において重要な役割を果たす。このような抗体は、親和性の増強につながる体細胞変異事象においてより効果的および効率的であることも考えられる。結合多様性および体細胞変異事象のそれぞれについては、以下でさらに詳しく考察する。
【0045】
親和性に関して、複数のVHおよびVκ遺伝子の利用(すなわち、Vκ軽鎖ゲノムの近位領域にある32個の遺伝子およびゲノムのVH部分にある66個の遺伝子の使用)に由来する抗体親和性の速度および定数は、約0.50×10-6よりも大きな、好ましくは2.00×10-6よりも大きな、さらに好ましくは約4.00×10-6よりも大きな会合速度(ka、M-1S-1単位);約1.00×10-4よりも大きな、好ましくは約2.00×10-4よりも大きな、さらに好ましくは約4.00×10-4よりも大きな解離速度(kd、S-1単位);および約1.00×10-10よりも大きな、好ましくは約2.00×10-10よりも大きな、さらに好ましくは約4.00×10-10よりも大きな解離定数(M単位)をもたらす。
【0046】
このようなマウスは、その上で機能的な内因性免疫グロブリンを産生しないことが好ましい。これは、1つの好ましい態様において、内因性の重鎖および軽鎖遺伝子座の不活性化(またはノックアウト)を通じて達成される。例えば、1つの好ましい態様では、この領域を置換または欠失させる相同組換えベクターの利用により、マウス重鎖J領域ならびにマウスκ軽鎖J領域およびCκ領域が不活性化される。
【0047】
可変領域―B細胞発生
B細胞の発生はクラウス(Klaus)の、「Bリンパ球(B Lymphocytes)」(IRL Press(1990))および、「免疫グロブリン遺伝子(Immunoglobulin Genes)」(Academic Press Ltd.(1989)の1〜3章において概説されており、それらの開示は参照として本明細書に組み入れられる。一般に哺乳動物では、Bリンパ球およびTリンパ球を含む血液細胞の発生は共通の多能性幹細胞に始まる。続いてリンパ球が共通のリンパ球前駆細胞から生じる。初期の妊娠期間を経た後にB細胞の発生部位は肝臓から骨髄に移動し、哺乳動物の一生を通じてその部位に存続する。
【0048】
B細胞のライフサイクルにおいて一般に最初に認識可能な細胞は、骨髄中に認められるプロプレB細胞である。このような細胞は重鎖V-D-J再配列を開始するが、タンパク質を作ることはまだできない。続いてこの細胞は、細胞質的にはμ+細胞である、大型で急速に分裂するプレB細胞Iとなる。このプレB細胞は次いで分裂を停止し、縮小し、軽鎖V-J再配列を行って、表面IgMを発現するプレB細胞IIとなり、未熟B細胞として骨髄から遊離する。新生未熟B細胞の大部分は発生を続け、表面IgDを産生するようになるが、これは完全に成熟した免疫適格末梢B細胞としての分化および発生が完了したことを意味し、これらは主に脾臓内に存在する。しかし、δ定常領域を除去して依然として免疫適格細胞を得ることが可能である。
【0049】
B細胞の分化および発生は、表面マーカーを用いて観測および/または追跡することができる。例えば、B220抗原はプレB細胞IまたはIIと比べると成熟B細胞でより豊富に発現される。したがって、B220+であって表面IgM+(μ+)である細胞を用いて成熟B細胞の存在を判定することができる。さらに、表面IgDの発現(δ+)に関して細胞をスクリーニングすることもできる。プレB細胞IIが末梢に移行するに従って(すなわち、それらがμ+および/またはμ+、δ+になるに従って)、別の抗原である耐熱抗原がそれらによって発現される。
(表II)
【表2】
【0050】
上記のようなB細胞マーカーの使用により、B細胞の発生および分化の観測および評価が可能である。
【0051】
本発明者らは以前に、DIマウス(重鎖V-D-J再配列または軽鎖V-J再配列を起こさないマウス)が成熟B細胞を産生しないことを示した。事実、このようなマウスではプロプレB細胞の産生のところで停止しており、B細胞は骨髄から脾臓を含む末梢組織には決して移動しない。このため、B細胞の発生および抗体の産生はいずれも完全に停止する。重鎖のみが不活性化されたマウスでも同じ結果が認められており、骨髄中のB細胞の発生および分化は停止する。
【0052】
本発明者らのXenoMouse I系統は、機能的で幾分成熟したB細胞を産生した。しかし、B細胞の数は、骨髄および末梢組織のいずれにおいても野生型マウスと対比すると有意に少なかった。
【0053】
これに対して、本発明者らのXenoMouse II系統およびL6系統では予想外なことにほぼ完全なB細胞の再構築が認められた。したがって、本発明により本発明者らは、可変領域遺伝子の量的含有および質的含有によってB細胞の分化および発生をかなり再構成しうることを示した。B細胞の分化および発生の再構成は、免疫系が再構成されたことを示す。一般にB細胞の再構成は、野生型対照と比較される。この点で、本発明の好ましい態様において、挿入されたヒト可変領域を有するマウスの集団は、野生型マウスの集団と比較すると約50%を超えるB細胞機能を有する。
【0054】
さらに興味深いことに、マウス抗体からみたヒト抗体の優先的な産生は、内因性Igがノックアウトされたバックグラウンドをもつマウスではかなり増強された。すなわち、ヒトIg遺伝子座および機能的に不活性化された内因性重鎖Ig遺伝子座を含むマウスでは、ヒトIg遺伝子座のみを含んでいて内因性遺伝子座が不活性化されていないマウスと比べて、ヒト抗体の産生効率が速度にして約100倍から1000倍も効率的であった。
【0055】
アイソタイプスイッチ
本明細書において詳細に考察されるように、予想通り、XenoMouse IIマウスは、ヒト導入遺伝子にコードされるμアイソタイプから導入遺伝子にコードされるγ2アイソタイプへの効率的および効果的なアイソタイプスイッチを生じた。本発明者らは、ヒトγ4定常領域を包含およびコードするXenoMouse II系統も開発した。上記の通り、本発明に係るマウスは、ほかのアイソタイプを生成するための他のヒト定常領域をさらに備えることができる。このようなアイソタイプには、γ1、γ2、γ3、γ4、α、ε、δおよび他の定常領域をコードする遺伝子が含まれうる。代替的な定常領域は、同一の導入遺伝子上、すなわちヒトμ定常領域から下流に含めることもでき、またはこのような他の定常領域を別の染色体上に含めることもできる。このような他の定常領域がヒトμ定常領域をコードする導入遺伝子を含む染色体と同じ染色体上に含まれる場合には、他の1つのアイソタイプまたは複数のアイソタイプへのシス転換が達成されうることは理解されるであろう。これに対して、このような他の定常領域がヒトμ定常領域をコードする導入遺伝子を含む染色体とは異なる染色体上に含まれる場合には、他の1つのアイソタイプまたは複数のアイソタイプへのトランス転換が達成されうる。このような配置(arrangement)は、広範な抗原に対する抗体を生成するためのマウスの設計および作製において多大な柔軟性を可能とする。
【0056】
定常領域が、それらと関連した既知のスイッチおよび調節配列を有することは理解されると思われる。マウスおよびヒト定常領域遺伝子はすべて、1989年までに配列が決定され、発表されている。「免疫グロブリン遺伝子」(Honjoら編、Academic Press(1989))中の、その開示が参照として本明細書に組み入れられるホンジョ(Honjo)ら、「免疫グロブリン重鎖の定常領域遺伝子およびクラススイッチの分子的機序」(Constant Region Genes of the Immunoglobulin Heavy Chain and the Molecular Mechamism of Class Switching)を参照のこと。例えば、その開示が参照として本明細書に組み入れられる米国特許出願第07/574,748号では、先行技術からの既知の配列情報に基づいて、ヒトγ1定常領域のクローニングが予測されている。そこでは、再配列がなされずスイッチもなされていない遺伝子では、第1のγ1定常部エキソンの5'末端から5kbよりも近いところから始まる配列中に、全スイッチ領域が含まれることが示されている。このため、スイッチ領域は、エリソン(Ellison)ら[Nucleic Acids Res 10:4071〜4079(1982)]において開示された5'部の5.3kbのHindIII断片にも含まれる。同様に、タカハシ(Takahashi)ら[Cell 29:671〜679(1982)]も、エリソンにおいて開示された断片がスイッチ配列を含むこと、およびこの断片に7.7kbのHindIIIからBamHIまでの断片を加えたものに、重鎖アイソタイプスイッチ用の導入遺伝子の作製に必要なすべての配列が含まれるに違いないと考えられることを報告している。
【0057】
このため、過度の実験を行わずとも、選択される任意のヒト定常領域を容易に本発明に係るマウスに組み入れられることは理解されると思われる。このような定常領域は、それらの本来のスイッチ配列(すなわち、それぞれヒトγ1、2、3、4スイッチを備えたヒトγ1、2、3、4定常領域)と結合させることができ、または他のスイッチ配列(すなわち、ヒトγ2スイッチを備えたヒトγ4定常領域)と結合させることもできる。少し例を挙げるなら、マウス、ヒトまたはラットなどの種々の3'エンハンサー配列を用いることもできる。同様に他の調節配列を含めることもできる。
【0058】
インビボでのアイソタイプスイッチに対する代替および/または付加として、「キメラ」抗体の分泌に関してB細胞をスクリーニングすることもできる。例えば、L6マウスは、完全ヒトIgM抗体を産生することに加えて、種々のγ(すなわちマウスIgG1、2、3、4)などのマウス定常領域と結合した完全ヒト重鎖V、D、J領域を有する抗体を産生する。このような抗体はそれ自体で非常に有用である。例えば、当技術分野でよく知られたインビトロでのアイソタイプスイッチ法により、抗体上にヒト定常領域を含めることができる。または、および/もしくはこれに加えて、マウス定常領域をほとんどまたは全く含まないこのような抗体の断片(すなわちF(ab)およびF(ab')2断片)を調製することもできる。
【0059】
上記の通り、抗体産生に関して最も重要な因子は、望ましい抗原または抗原上のエピトープに対する特異性である。その後には、治療的必要性から抗体のクラスが重要になる。換言すれば、特定のアイソタイプまたはクラスを提供することによって、抗体の治療的指標は高まるのであろうか。この疑問の考察は補体結合などの問題を生み、それは次に特定のクラスまたはアイソタイプの抗体の選択を促す。γ定常領域は抗体の親和性の成熟を補助する。しかし、このような成熟を達成するために導入遺伝子上にヒトγ定常領域を含めることは必要でない。むしろ、この過程はμをコードする導入遺伝子上にトランス転換されるマウスγ定常領域との関連によってさらに進行すると思われる。
【0060】
材料および方法
以下の材料および方法は、本発明に係るマウスの製造および特徴に関連して用いられる。このような材料および方法は例示を意味しており、本発明の範囲を制限するものではない。
【0061】
ヒトIg由来のYACのクローニング: 以前にワシントン大学(the Washington University)(Brownsteinら、1989)およびCEPH(Abersonら、1990)のヒトYACライブラリーを、以前に記載された通りに(Mendezら、1995)ヒト重鎖およびκ軽鎖遺伝子座の配列を含むYACに関してスクリーニングした。1Hおよび1K YACのクローニングおよび特徴分析はメンデツ(Mendez)ら(1995)に記載されている。3Hおよび4H YACは、VH3プローブ(0.55kb PstI/NcoI、Bermanら、1988)を用いてワシントン大学のライブラリーから同定された。17H YACはGM1416ライブラリーから同定され、130kbの重鎖可変配列およびその3'末端に150kbのキメラ領域を含むことが明らかにされた。マツダ(Matsuda)ら、1993。2Kおよび3K YACはVκII特異的プライマーを用いてCHEFライブラリーから回収された(Albertsenら、1990)。
【0062】
YACのターゲティングおよび組換え: 酵母の増殖、交配、胞子形成および表現型の試験に関する標準的な方法を用いた(Shermanら、1986)。YACが細胞内に組み込まれた酵母内のYAC組換え体のスクリーニングを容易にするための、酵母および哺乳動物の選択可能マーカーによるYACおよびYACベクター腕のターゲティングは、酢酸リチウム形質転換法(ScheistlおよびGeitz(1989))によって行った。すべての配列の完全性を調べるために、個々のターゲティングまたは組換え段階の後にパルスフィールドゲル電気泳動法および標準的なサザンブロット法によって改変YACを分析した。
【0063】
17Hの再配向によって動原体腕および無動原体腕を相互転換させるため、ならびにその5'腕にLEU2およびURA3遺伝子の、その3'腕にHIS3遺伝子のレトロフィットを行うためにYACターゲティングベクターを用いた。図1aおよびメンデツ(Mendez)ら、1993を参照のこと。4H動原体腕には酵母ADE2遺伝子およびヒトHPRT選択可能マーカーによるレトロフィットを行った。1回目の組換え段階については、二倍体酵母株を作製し、17H、3Hおよび4Hという3種のYACがすべて存在し、無傷で安定的に維持されるものを選択した。YACが一部重複する領域の三者間相同組換え(three-way homologous recombination)を胞子形成によって誘導し、外部酵母選択可能マーカー(ADE2およびHIS3)の選択、および内部マーカーURA3の陰性選択(損失)によって望ましい組換え体を見いだした。好首尾な組換えにより、VH2-5に始まってVH3-65遺伝子の5'側に20kb伸長するIgH可変領域の80%を含む、880kbのYACが作製された。880kbのYACと1Hとの組換えには、1HをLYS2遺伝子を動原体腕に付加するpICLによるレトロフィットを行った(Hermansonら、1991)。標準的な酵母交配を用いて、1Hおよび880kbのYACの両方を含む二倍体株を選択した。胞子形成および重複相同性(overlapping homology)の使用により、YAC-酵母組換えを行った。外部酵母マーカー(ADE2およびURA3)の陽性選択および内部マーカー(TRP1、LYS2、HIS3)の損失に関するスクリーニングにより、VH6-1に始まってVH3-65で終わる約66個のVHセグメントを含む無傷の970kbのYACが見いだされた。このYACは生殖細胞系コンフィギュレーションにある主要なD遺伝子クラスター、JH遺伝子、イントロン性エンハンサー(Eμ)、Cμ、Cδを越えて25kbまでも含んでいた。続いてこの970kbのYACに、ヒトγ2遺伝子の23kbのEcoRIゲノム断片を、そのスイッチおよび調節要素、マウス重鎖3'エンハンサーの7kbのXbaI断片、メタロチオネインプロモーターの支配を受けるネオマイシン遺伝子(MMTNeo)、および酵母LYS2遺伝子を含めて、これらを含むターゲティングベクターによるレトロフィットを行った。このベクターは、3'YAC腕上にこれらの配列を運ぶ一方で、URA3遺伝子を破壊する。
【0064】
yK2 YACの作製に向けての第1段階として、標準的な酵母交配により、レトロフィットがなされた1Kおよび3K YACが両方とも存在し、無傷で安定的に維持される二倍体酵母株を選択した。IgH作製に関して記載したものと同じ過程を用いて、YAC-酵母組換えを行った。外部酵母マーカー(LYS2、TRP1)に関する陽性選択および内部マーカー(URA3、TRP1)の損失に関するスクリーニングにより、Vκ-B3に始まってVκ-Op11で終わる32個のVκを含む無傷の800kbの組換え産物を見いだした。この800kbのYACは、Vκ-Lp-13に始まってVκ-Lp-5で終わる約100kbの欠失を含む。しかし、このYACはVκ-Lp-13からVκ-Op-1を100kb越えたところまでは生殖細胞系コンフィギュレーションにある。また、このYACはJκ、イントロン性および3'エンハンサー、定常CκおよびKdeも含む。
【0065】
YACのES細胞およびマウスへの導入: YACを含む酵母スフェロプラストを、記載されている通り(Jakobovitsら、1993a;Greenら、1994)にE14.TG3B1 ES細胞と融合させた。HAT耐性コロニーを分析のために増殖させた。YACの完全性は、バーマン(Berman)ら(1988)およびメンデツ(Mendez)ら(1994)に記載された手順およびプローブ、ならびにジェミル(Gemmil)ら(1991)に記載された通りのハイブリダイゼーション条件を用いるサザンブロット分析によって評価した。キメラマウスは、C57BL/6胚盤胞へのES細胞のマイクロインジェクションによって作出した。記載されている通り(Greenら、1994)に、尾部DNAのPCR分析によってYACを含む子孫を同定した。YACの完全性は、ヒトVH3にて探索したブロットを50℃で洗うことを除き、前記のプローブおよび条件を用いるサザンブロット分析によって評価した。
【0066】
フローサイトメトリー分析: 8〜10週齢のXenoMouseおよび対照マウスから入手した末梢血および脾臓のリンパ球をLymphocyte M(Accurate)上で精製し、Fc受容体に対する非特異的な結合を阻止するために精製抗マウスCD32/CD16 Fc受容体(Pharmingen、01241D)で処理して、抗体で染色し、FACStarPLUS(Becton Dickinson、CELLQuestソフトウエア)によって分析した。用いた抗体は以下の通りである:アロフィコシアニン(APC)抗B220(Pharmingen、01129A);ビオチン抗ヒトIgM(Pharmingen、08072D);ビオチン抗マウスIgM(Pharmingen、02202D);フルオロセインイソチオシアネート(FITC)ヤギF(ab')2抗ヒトIgD(Southern Biotechnology、2032-02);FITC抗マウスIgDa(Pharmingen、05064D);FITC抗mIgDb(Pharmingen、05074D);FITC抗マウスλ(Pharmingen、02174D);PE抗ヒトκ(Pharmingen、08175A);PE抗マウスκ(Pharmingen、02155A)。ビオチン化抗体の検出にはRED613(登録商標)-ストレプトアビジン(GibcoBRL、19541-010)を用いた。
【0067】
免疫化およびハイブリドーマの作製: XenoMouse(8〜10週齢)に、一次免疫には完全フロイントアジュバント中に、2週間間隔で実施した追加免疫には不完全フロイントアジュバント中に、乳濁化した25μgの組換えヒトIL-8または5μgのTNF-α(Biosource International)の腹腔内投与による免疫化を施した。EGFR免疫化については、リン酸緩衝塩類溶液(PBS)中に再懸濁した2×107個のA431(ATCC CRL-7907)細胞の腹腔内投与による免疫化を施した。この量の投与を3回繰り返した。融合4日前に、マウスにPBS中にて抗原または細胞の最終注射を行った。免疫化マウスからの脾臓およびリンパ節のリンパ球を、非分泌型骨髄腫NSO-bcl2系(RayおよびDiamond、1994)と融合させ、以前に記載されている通り(GalfreおよびMilstein、1981)にHAT選択にかけた。
【0068】
ELISAアッセイ: マウス血清中およびハイブリドーマ上清中の抗原特異的抗体を測定するためのELISAは、抗体を捕捉するために組換えヒトIL-8およびTNF-αならびにA431細胞からアフィニティー精製されたEGFR(Sigma、E-3641)を用いて、記載されている通り(Coliganら、1994)に実施した。ヒトおよびマウスの免疫グロブリンの濃度は以下の捕捉抗体を用いて測定した:ヒトγ、κ、μIgについてはそれぞれウサギ抗ヒトIgG(Southern Biotechnology、6145-01)、ヤギ抗ヒトIgκ(Vector Laboratories、AI-3060)、マウス抗ヒトIgM(CGI/ATCC、HB-57)、ならびにマウスγ、κ、μおよびλIgの捕捉のためにはそれぞれヤギ抗マウスIgG(Caltag、M 30100)、ヤギ抗マウスIgκ(Southern Biotechnology、1050-01)、ヤギ抗マウスIgM(Southern Biotechnology、1020-01)およびヤギ抗マウスλ(Southern Biotechnology、1060-01)。ELISA実験に用いた検出用抗体は、ヤギ抗マウスIgG-HRP(Caltag、M-30107)、ヤギ抗マウスIgκ-HRT(Caltag、M 33007)、マウス抗ヒトIgG2-HRP(Southern Biotechnology、9070-05)、マウス抗ヒトIgM-HRP(Southern Biotechnology、9020-05)およびヤギ抗ヒトκ-ビオチン(Vector、BA-3060)である。ヒトおよびマウスのIgの定量化のために用いた標準は以下の通りである:ヒトIgG2(Calbiochem、400122)、ヒトIgMκ(Cappel、13000)、ヒトIgG2κ(Calbiochem、400122)、マウスIgGκ(Cappel 55939)、マウスIgMκ(Sigma、M-3795)およびマウスIgG3λ(Sigma、M-9019)。
【0069】
BIAcoreによる完全ヒトMabの親和定数の決定: 製造者によって概説された標準的な手順によりBIAcore 2000装置を用いて、精製ヒトモノクローナル抗体、Fab断片またはハイブリドーマ上清の親和性の測定をプラズモン共鳴によって実施した。
【0070】
抗体の速度論的分析は、センサー表面上に低密度に固定された抗原を用いて行った:ヒトIL-8 -81RU、A431細胞膜から精製された可溶性EGFR(Sigma、E-3641)-303RU、およびTNF-α -107RU(1,000 RUは固定されたタンパク質の約1ng/mm3に対応する)。解離速度(kd)および会合速度(ka)は、製造者によって提供されたソフトウエアであるBIAevaluation 2.1を用いて決定した。
【0071】
ラジオイムノアッセイによる親和性の測定: 0.5%BSAを含む200μlのPBS中で、125I標識ヒトIL-8(1.5×10-11Mまたは3×10-11M)を種々の濃度の精製抗IL-8ヒト抗体(5×10-13Mから4×10-9M)とともにインキュベートした。室温で15時間インキュベートした後、抗体-抗原複合体を沈降させるためにプロテインAセファロースCL-4Bを含むPBS(l/l、v/v)20μlを添加した。4℃で2時間インキュベートした後、96ウェル濾過プレート(Millipore、カタログ番号MADVN65)を用いる濾過により、プロテインAセファロースに結合した抗体-125I-IL-8複合体を遊離125I-IL-8から分離し、シンチレーションバイアルに回収して計数した。結合型および遊離型の抗体の濃度を算出し、スキャッチャード解析を用いて特異的抗原に対する抗体の結合親和性を算出した(2)。
【0072】
受容体結合アッセイ: IL-8受容体結合アッセイは、記載された通り(Lusti-Marasimhanら、1995)に、新鮮な採取血またはバフィーコートのいずれかから調製したヒト好中球を用いて実施した。0.1%ウシ血清アルブミンおよび0.02%のNaN3を含むPBS結合緩衝液による前処置を25℃で2時間行った96ウェルのMultiscreenフィルタープレート(Millipore、MADV N6550)中にて、種々の濃度の抗体を0.23nMの[125I]IL-8(Amersham、IM-249)とともに4℃で30分間インキュベートした。各ウェルに4×105個の好中球を加え、プレートを4℃で90分間インキュベートした。200μlの氷冷PBSで細胞を5回洗い、PBSを吸引によって除去した。フィルターを風乾し、シンチレーション液を添加して、シンチレーションカウンター中で計数した。抗体の存在下で検出された平均cpmを緩衝液のみの存在下で検出されたcpmで割った値として、特異的に結合した[125I]IL-8の比率を算出した。
【0073】
TNF受容体に関する結合アッセイは、上記のIL-8アッセイと同じ様式で実施した。しかし、IL-8アッセイに関して用いた好中球系の代わりにヒト単球系U937を用いた。抗体は0.25nMの[125I]TNF(Amersham、IM-206)とともにプレインキュベートした。6×105個のU937細胞を各ウェルに播いた。
【0074】
EGF受容体結合アッセイは、PBS緩衝液中の種々の濃度の抗体とともに4℃で30分間インキュベートしたA431細胞(1ウェル当り0.4×106個の細胞)を用いて実施した。0.1nMの[125I]EGF(Amersham、IM-196)を各ウェルに添加し、プレートを4℃で90分間インキュベートした。プレートを5回洗い、風乾し、シンチレーションカウンター中にて計数した。抗EGFRマウス抗体225および528(Calbiochem)を対照として用いた。
【0075】
XenoMouseにおいて発現されたヒトIg転写物およびそれらに由来するヒトMabのレパートリー解析: Fast-Trackキット(Invitrogen)を用いて、非免疫化および免疫化XenoMouseの脾臓およびリンパ節から、ポリ(A)+mRNAを単離した。ランダムプライミングによりcDNAを作製し、続いてPCRを行った。ヒトVHまたはヒトVκファミリーに特異的な可変領域プライマー(Marksら、1991)または汎用ヒトVHプライマーであるMG-30(CAGGTGCAGCTGGAGCAGTCIGG)を、以前に記載された通り(Greenら、1994)にヒトCμ(huP2)またはCκ(hκP2)定常領域に特異的なプライマーとともに、またはヒトγ2定常領域MG-40d;5'-GCTGAGGGAGTAGAGTCCTGAGGA-3'とともに用いた。TAクローニングキット(Invitrogen)を用いてPCR産物をpCRII中にクローニングし、Prismダイターミネーター配列決定キットおよびABI377配列決定装置を用いて両方のストランドの配列を決定した。ヒトMabに由来する重鎖およびκ軽鎖の転写物の配列は、上記のプライマーを用いてポリ(A+)RNAから生成されたPCR産物の直接的な配列決定によって得た。「V BASE配列ディレクトリー」(Tomlinsonら、MRC Centre for Protein Engineering、Cambridge、UK)に対するアライメントにより、MacVectorおよびGeneworksソフトウェアプログラムを用いて、すべての配列を解析した。
【0076】
抗体Fab断片の調製および精製: 抗体Fab断片は固定パパイン(Pierce)を用いることによって作製した。Fab断片を以下の2段階クロマトグラフィー方式で精製した:Fc断片およびすべての未消化抗体を捕捉するためのHiTrap(Bio-Rad)プロテインAカラム、次いで強陽イオン交換カラム(PerSeptive Biosystems)を通過させて保持されたFab断片の0.5M NaClに至る塩の直線勾配による溶出。Fab断片の特徴を、還元および非還元条件下でのSDS-PAGEおよびMALDI-TOF MSによって分析したところ、予想された〜50kDの非還元断片および〜25kDaの還元二重バンドが示された。この結果は、無傷の軽鎖および切断された重鎖を示す。還元条件下におけるMSでは、未消化抗体全体の還元によって軽鎖の質量を正確に決定しうるため、軽鎖および切断された重鎖の双方を明確に同定することが可能であった。
【実施例】
【0077】
以下の実施例は、実施した実験および得られた結果を含め、例示を目的として提供されるものであり、本発明の範囲を制限するものと考えられるべきではない。
【0078】
実施例1:YAC上でのヒト重鎖遺伝子座の再構成
本発明により、本発明者らがヒト重鎖およびヒトκ軽鎖の可変領域を再構成するために用いた戦略は、第1に大きな(メガベースサイズの)ヒトIg遺伝子座にまたがるYACに関してヒトYACライブラリーをスクリーニングし、第2にこのような領域にまたがるYACを、主として生殖細胞系コンフィギュレーションにある望ましい遺伝子座を含む単一のYAC中に組換え導入することであった。
【0079】
上記の段階的なYAC組換え方式は、酵母において減数分裂誘導性の相同組換えを高頻度にもたらし、組換えYAC上のベクター腕に存在する酵母マーカーによる望ましい組換え体の選択を可能とする(図1およびGreenら、前記を参照のこと;Silvermanら、1990およびdenDunnenら、1992も参照されたい)。
【0080】
本発明者らの戦略に関連して、本発明者らは染色体14q上にあるヒト重鎖可変領域の約1000kbのうち、約830kbに及ぶ1H(240kb)、2H(270kb)、3H(300kb)および4H(340kb)という4種のYACを同定した。このYAC 1H、2H、3Hおよび4Hを遺伝子座の再構成のために用いた(図1A参照)。パルスフィールドゲル電気泳動(PFGE)およびサザンブロット分析により、特定の非IgH配列を含むYAC 2Hの3'末端の150kbを除いて、YACが無傷であって生殖細胞系コンフィギュレーションにあることが示された(図1参照;Matsudaら、1990)。以前、本発明者らの第1世代のXenoMouse(登録商標)に導入されたYACであるYAC 1H(Greenら、前記;Mendezら、1995)は、生殖細胞系コンフィギュレーションにあるヒトCδ、Cμ、JHおよびDH領域ならびに最初の5つのVH遺伝子を含む。他の3つのYACは、VH2-5からVH3-65までのVH領域の大半をカバーし、それによってさらにほぼ61個の異なるVH遺伝子がもたらされる。組換えに先立ち、YAC 4HにはHPRT選択可能マーカーのレトロフィットを施した。YAC上に含まれる重複配列を用いることにより、段階的組換え戦略によって4種のYAC(1H、2H、3Hおよび4H)を酵母に組換え導入した(図1A参照)。このような組換え戦略によって980kbの組換えYACが作製された(図1参照)。PFGEおよびサザンブロット分析によるYACの分析から、生殖細胞系コンフィギュレーションにあるCδ領域からVH3-65遺伝子の5'側の20kbのところまでのヒト重鎖遺伝子座が存在することが確かめられた。明らかな欠失および再配列は認められなかった。
【0081】
YAC無動原体腕に対して、完全なヒトγ2定常領域、マウス3'エンハンサーおよびネオマイシン耐性遺伝子を有するベクターによるターゲティングを行い、最終的に1020kbの重鎖YACであるyH2を得た。YAC yH2は、ヒト可変領域の大半、すなわち82個のVH遺伝子のうち66個、全DH(32遺伝子)およびJH(6遺伝子)領域、ならびに3つの異なる定常領域(Cμ、CδおよびCγ)を、それらに対応する調節配列とともに含んでいた(図1A参照)。これが本発明者らのXenoMouse II系統の作製に用いた重鎖構築物である。
【0082】
実施例2:YAC上でのヒトκ軽鎖遺伝子座の再構成
ヒトκ軽鎖遺伝子座の再構成には同様の段階的組換え戦略を用いた。ヒトκ遺伝子座にまたがる3種のYACを同定した。YACは1K、2Kおよび3Kと命名した。長さ約180kbのYAC 1Kは、以前に本発明者らの第1世代のXenoMouse(登録商標)に導入された。このようなYACは、Bクラスター上にκ欠失要素(Kde)、κ3'およびイントロン性エンハンサー、Cκ、Jκならびに3種のVκ遺伝子を含む(Greenら、1994;Mendezら、1995)。YAC 2K(約480kb)および3K(約380kb)はともに、染色体2p上のκ軽鎖近位可変領域の大部分を包含する。約100kbの欠失はL13〜L5領域にまたがる(図1B;Huberら、1993)。κ遠位領域は近位領域の繰り返しであり、近位Vκ遺伝子はヒトで最も一般的に用いられるものであるため(Weicholdら、1993;Coxら、1994)、本発明者らの再構成戦略の焦点は近位領域にあった(図1B)。3種のYACの相同組換えにより、800kbの組換えYACであるyK2が回収された。この組換えYACのサイズおよび完全性を、PFGEおよびサザンブロット分析によって確認した。こうした分析から、それがLp領域に記載した通りの欠失があることを除き、生殖細胞系コンフィギュレーションにある32個のVκ遺伝子とともにヒトκ軽鎖遺伝子座の近位部をカバーすることが示された(図1B)。yK2の動原体腕および無動原体腕に、記載した通り(材料および方法)にそれぞれHPRTおよびネオマイシン選択可能マーカーを含むように改変を加えた。これが本発明者らのXenoMouse II系統の作製に用いたκ軽鎖構築物である。
【0083】
本明細書に記載されるYACであるyH2およびyK2は、主として生殖細胞系コンフィギュレーションでヒト抗体レパートリーの大半を含むように再構成された最初のメガベースサイズのヒトIg遺伝子座である。これが達成されたことは、酵母における相同組換えが、大きな複雑で不安定な遺伝子座の再構成を首尾よく行うための強力な手法であることを立証するものである。酵母内にIg遺伝子座の大部分を含む安定なYAC組換え体の選択により、本発明者らは、マウスでヒト抗体反応を再現するために必要なヒト抗体レパートリー、定常領域および調節要素をマウスに備えさせるために必要なヒトIg断片を手に入れた。
【0084】
実施例3:ES細胞へのyH2およびyK2 YACの導入
本発明者らの戦略に従い、本発明者らはYAC yH2およびyK2をマウス胚性幹(ES)細胞に導入した。YAC DNAを含むES細胞がいったん単離されれば、このようなES細胞は適切な交配を通じてマウスの作出に用いられる。
【0085】
このため、この実験では、以前に記載されている通り(Jakobovitsら、1993a;Greenら、1994)に、YACを含む酵母スフェロプラストとHPRT欠損E14.TG3B1マウスES細胞との融合を介して、YAC yH2およびyK2をES細胞に導入した。HPRT陽性のES細胞クローンは、1クローン/融合細胞15〜20×106個の割合で選択され、サザンおよびCHEFブロット分析によってYACの完全性に関する分析が行われた(図2A)。
【0086】
ES細胞とyH2を含む酵母との融合に由来する35種のES細胞クローンのうち7種(L10、J9.2、L17、L18、J17、L22、L23と命名)が、挿入物の全体にまたがるプローブによって検出される、予想されたすべてのEcoRIおよびBamHI yH2断片を含むことが明らかになった:マウス3'エンハンサー、ヒトイントロン性エンハンサー、ヒトCγ2、CδおよびCμ定常領域、DH、JHならびにすべての異なるVHファミリー:VH1、VH2、VH3、VH4、VH5およびVH6(図2Aに5つのクローンに関するデータを示した)。CHEF分析により、分析した全クローンの20%にあたるこれらのクローンが、明らかな欠失および再配列を含まない無傷のyH2 YACの全体を含むことがさらに確かめられた(非提示データ)。
【0087】
yK2を含む酵母との融合に由来するES細胞クローンについても、ヒトKde、ヒトκ3'およびイントロン性のエンハンサー、Cκ、Jκならびに異なるVκファミリーのすべて:VκI、VκII、VκIII、VκIV、VκVIに特異的なプローブを用いて、YACの完全性に関して同様に分析した。60種のクローンのうち20種は無傷であって変化のないYACを有し、これは分析した全クローンの30%であった(図3Aに2つのESクローンに関するデータを示した)。yH2およびyK2-ES細胞クローンでは種々の量の酵母ゲノム配列が検出された(非提示データ)。
【0088】
これらの結果により、主として生殖細胞系コンフィギュレーションにある再構成されたヒト遺伝子座を含むメガベースサイズの構築物が哺乳動物細胞に導入されたことが初めて示された。無傷のYACがマウスゲノムに比較的高頻度に組み込まれたことは、ES細胞-酵母スフェロプラスト融合法が、大きなヒトゲノム断片をES細胞に忠実に導入するための効果的な手法であることをさらに実証するものである。
【0089】
実施例4:XenoMouse II系統の作出
YAC DNAを含むES細胞からマウスを作出するために、胚盤胞へのマイクロインジェクションを行い、続いて交配を行った。すなわち、yH2およびyK2を保持するES細胞クローンを増殖させ、マウスC57BL/6J胚盤胞へのマイクロインジェクションを行い(Greenら、1994)、得られたキメラ雄を生殖細胞系伝達に関して評価した。伝達されたYACを有する子孫をPCR分析によって同定し、サザンブロット分析によってYACの完全性を確認した。分析したすべてのトランスジェニックマウスにおいて、YACは無傷型であることが示された(図2B、3B)。マイクロインジェクションを受けた7つのyH2-ESクローンのすべて、および8つのyK2-ESクローンのうち2つは、マウス生殖細胞系を通じて伝達された。
【0090】
内因性抗体が排除された、ヒト抗体を産生するマウスを作出するために、yH2-またはyK2-トランスジェニックマウスを二重不活性化(DI)マウス系統と交配させた。DIマウス系統は、遺伝子ターゲティングによって不活性化されたマウス重鎖およびκ鎖遺伝子座に関して同型接合的であり、このため抗体産生に欠陥がある(Jakobovitsら、1993b;Greenら、1994)。2つのyH2-トランスジェニックマウス系統であるL10およびJ9.2、ならびにyK2-トランスジェニックマウス系統の1つであるJ23.1をDIマウスと交配させ、同型接合的に不活性化されたマウス重鎖およびκ鎖バックグラウンドをもつYACを保持するマウスを作出した(yH2;DIおよびyK2;DI)。yH2;DIトランスジェニック系統のそれぞれをyK2;DIトランスジェニック系統と交配させ、同型接合的DIバックグラウンドをもつ重鎖および軽鎖YACの両方を含む2つのXenoMouse II系統である2A-1(L10;J23.1;DI)および2A-2(J9.2;J23.1;DI)をそれぞれ作出した。L10は完全に同型接合的であり、J9.2およびJ23.1は交配によって首尾よく同型接合的になる過程にある。
【0091】
XenoMouse II系統におけるヒト重鎖およびκ鎖YACの完全性は、サザンブロット分析によって確認した。図2および図3に示す通り、分析したいずれのXenoMouse系統とも、yH2およびyK2は明らかな欠失および再配列を生じずに複数世代にわたって変化せずに伝達された。
【0092】
実施例5:XenoMouse IIマウスによるB細胞発生およびヒト抗体産生
XenoMouse II系統の特徴をさらに調べるために、本発明者らはそれらのB細胞発生およびヒト抗体産生を検討した。yH2およびyK2 YACによるXenoMouse II系統におけるB細胞発生および抗体産生の再構成を、フローサイトメトリーおよびELISAによって評価した。成熟B細胞を完全に欠くDIマウスとは対照的に、XenoMouse IIは本質的に正常なB細胞発生を示し、血中の成熟B細胞を合計すると野生型マウスで認められる値の50%を上回った(図4A)。すべてのB細胞はヒトIgMおよび高レベルのB220を発現し(ヒトIgM+/B220hi)、この集団の60%はヒトIgDも発現することが示された。XenoMouseの脾臓およびリンパ節の分析でも同様の結果が得られた(提示せず)。これらの結果は野生型マウスにおける成熟B細胞の特徴とよく相関しており、XenoMouseにおけるB細胞成熟が適切であることを示す。
【0093】
XenoMouseのB細胞の大半(75〜80%)はもっぱらヒトκ軽鎖を発現し、マウスλ軽鎖を発現したものは15%に過ぎなかった(図4)。この軽鎖の分布比(hκ/mλ)は野生型マウスで認められる値と同等であり、このことは軽鎖の使用がマウス様調節を受けていることを示す。これに対して、グリーン(Green)ら、1994に記載されたXenoMouse Iで認められたhκ/mλの比は55:45であった(非提示データ)。脾臓(図4B)およびリンパ節(提示せず)由来のB細胞についても同様のことが観察され、このことはXenoMouse IIのB細胞の大部分は、もっぱら完全ヒト抗体を産生することを示す。yK2に関して同型接合的であるXenoMouse II系統におけるmλ発現B細胞のレベルは、15%から7%に減少した(非提示データ)。
【0094】
実施例6:L6系統の作出
L6系統のマウスは、XenoMouse II系統の作出に関する上記の過程と同じように作出した。しかし、L6 ES細胞系の作製に際して欠失事象があるために、ES細胞系および続いて得られるL6マウスはCδの遠位側にある配列の部分を伴わずに生じ、このため、Cγ定常領域およびその調節配列をもたない。交配の完了後に、L6マウスは欠失したCγ定常領域を除き、yK2構築物の全体およびyH2構築物の全体を含むと考えられる。
【0095】
実施例7:ヒト抗体の産生
非免疫化XenoMouseIIの血清中でヒトCμ、Cγ2およびκ軽鎖の発現が検出され、最大値はそれぞれ700、600および800μg/mlであった。これらの値がどの程度野生型に匹敵するかを明らかにするために、本発明者らは同様の無病原体条件下に保ったC57BL/6J×129マウスにおけるマウスCμ、Cγ2およびκ軽鎖の最大値を測定した。野生型マウスにおけるCμ、Cγ2およびκ軽鎖に関する値はそれぞれ400、2000および2000μg/mlであった。免疫化を施したところ、ヒトγ鎖の値は約2.5mg/mlに上昇した。マウスλの濃度は70μg/mlに過ぎず、このことからヒトκ鎖が優先的に用いられることがさらに裏づけられた。
【0096】
これらの所見は、導入されたヒトYACに適切なIg遺伝子再配列およびクラススイッチを誘導する能力、ならびに免疫化の前後ともに顕著なレベルの完全ヒトIgMおよびIgG抗体を産生する能力があることを裏づけるものである。
【0097】
実施例8:XenoMouse IIにおける多様なヒト抗体レパートリー
XenoMouse II系統における抗体レパートリーの再構成に関してさらに理解を得るために、本発明者らはマウスにいくつかの免疫原による誘発刺激を与え、このような抗体を分泌するハイブリドーマ細胞系を調製した。理解されるであろう通り、マウスでヒト抗体反応を再現させるにはyH2およびyK2 YAC上に含まれる種々のヒト可変遺伝子が多様に用いられる必要がある。XenoMouse II系統によって産生されるヒト抗体の多様性を、XenoMouseのリンパ節由来のヒト重鎖(μおよびγ)ならびにκ軽鎖転写物のクローニングおよび配列決定によって調べた。現在までの本発明者らのデータに基づくと、配列解析ではXenoMouse IIがyH2上に存在する37個の機能的VH遺伝子のうち少なくとも11個、8個の異なるDHセグメントおよび3個のJH遺伝子(JH3、JH4、JH6)を用いることが示されている(表III;本発明者らによるハイブリドーマ由来の抗体の配列決定に関してJH5も検出された)。V-D-J配列はヒトμまたはγ2定常領域と連結していた(提示せず)。
【0098】
用いられたVH遺伝子は可変領域の全体にわたって広く分布しており、7つのVHファミリーのうちの4つであった(表III)。VH3およびVH4ファミリーからのV遺伝子が主に利用された点は、ファミリーのサイズに比例する、成人におけるVH使用パターンと類似している(Yamadaら、1991;Brezinshekら、1995)。JH4が主に利用されたこともヒトB細胞で検出されるものとよく似ている(Brezinshekら、1995)。1〜12bpの範囲に及ぶ、V-DおよびD-J結合の双方での非生殖細胞系ヌクレオチドの付加(N-付加)も認められた。このようなN付加は、8個から約19個までのアミノ酸残基長をもつ相補性決定領域3(CDR3)をもたらすが、これは成人B細胞で認められるものと極めて類似している(Yamadaら、1991;Brezinshekら、1995)。XenoMouse IIで認められるこのようなCDR3長は、マウスで通常認められるCDR3長よりもはるかに長い(Feeny、1990)。
【0099】
配列決定を行った10種のκ鎖転写物においても、極めて多様なレパートリーが認められた。yK2上に存在する25個のVκ機能的オープンリーディングフレーム(ORF)のうち8個が提示されることに加えて、すべてのJκ遺伝子が検出可能であった(表IV)。用いられた異なるVκ遺伝子はyK2の全体にわたって広く分散しており、4種の主要なVκ遺伝子ファミリーのすべてが示された。すべてのVκJκ組換え産物がCκ配列と適切に連結していた。本発明者らの転写物にN付加がほとんどみられなかったことは、κ鎖再配列の段階でターミナルデオキシヌクレオチジルトランスフェラーゼ活性が大きく低下することと一致する。本発明者らがκ鎖転写物中に認めた9〜10アミノ酸の平均CDR3長は、ヒトB細胞で認められた値と同一であった(Marksら、1991)。
【0100】
以下の表IIIおよびIVでは、XenoMouse II系統において発現されたヒト重鎖およびκ軽鎖転写物のレパートリー解析の結果を提示している。材料および方法の項で記載した通りに、ヒトμ、γおよびκに特異的なmRNAをPCRによって増幅し、クローニングして、配列決定によって解析した。表IIIには、12種のユニークなヒト重鎖クローンの一連のヌクレオチド配列を、発表された生殖細胞系配列との相同性によって同定されるVH、D、JHおよびNセグメントに分割して示している(材料および方法)。それぞれのDセグメントの指定は、少なくとも8塩基の相同性に基づいている。表IVには、8種の独立したヒトκクローンのV-J結合の一連のヌクレオチド配列を示している。配列はVκ、JκおよびNセグメントに分割され、発表されたVκおよびJκ配列との相同性に基づいて同定されている。それぞれの表におけるN付加および欠失(_として表示)は、V、DまたはJ配列との配列相同性がないことによって決定した。
(表III)
【表3】
(表IV)
【表4】
【0101】
これらの結果を、以下に記載するXenoMouse由来のハイブリドーマの配列と総合すると、V、DおよびJ遺伝子の極めて多様な成人様使用が示されるが、このことはyH2およびyK2 YAC上に存在するヒト重鎖およびκ鎖の可変領域の全体を抗体再配列のためにマウス系が利用することができ、しかも位置による偏差のない様式で利用されることを示すと考えられる。さらに重鎖およびκ鎖転写物の双方に関するN付加およびCDR3の平均長は成人B細胞で認められる値と非常に類似しており、このことは、マウスに含まれるYAC DNAがマウスにおいて成人様免疫レパートリーが生じるようにマウスの機構を指向させることを示す。
【0102】
以下の実施例に関して、本発明者らはいくつかの抗原に対する高親和性抗体を調製した。特に抗原としてはヒトIL-8およびヒトEGFRを調製した。IL-8およびEGFRを選択した根拠は以下の通りである。
【0103】
IL-8はC-X-Cケモカインファミリーのメンバーである。IL-8は、少し例を挙げればARDS、慢性関節リウマチ、炎症性腸疾患、糸球体腎炎、乾癬、アルコール性肝炎、再潅流障害を含む多くの疾患に関与する好中球に対する主な化学誘引物質として作用する。さらに、IL-8は内皮細胞に対する強力な血管新生促進因子である。図22〜28において本発明者らは、XenoMouse II系統に由来するヒト抗IL-8抗体がIL-8の作用を多くの経路において効果的に阻害することを示す。例えば、図22は、ヒト抗IL-8抗体によってIL-8のヒト好中球との結合が阻止されることを示す。図23は、ヒト抗IL-8抗体によってヒト好中球上のCD11b発現が阻害されることを示す。図24は、ヒト抗IL-8抗体によってIL-8誘導性カルシウム流入が阻害されることを示す。図25は、ヒト抗IL-8抗体によるIL-8 RB/293走化性の阻害を示す。図26は、ヒトIL-8誘導性皮膚炎症のウサギモデルの模式図を示す。図27は、図26のウサギモデルにおけるヒトIL-8誘導性皮膚炎症がヒト抗IL-8抗体によって阻害されることを示す。図28は、ラット角膜ポケットモデル上の内皮細胞の血管新生がヒト抗IL-8抗体によって阻害されることを示す。
【0104】
EGFRは、抗癌ターゲットとみなされている。例えば、種々の癌細胞上ではEGFRが最大100倍に過剰発現される。ある種の癌の開始および進行には、リガンド(EGFおよびTNF)を介した増殖刺激が極めて重要な役割を果たす。この点に関して、EGFR抗体はリガンド結合を阻害して腫瘍細胞の増殖の停止をもたらし、さらに化学療法薬と併用するとアポトーシスを誘導する。事実、複数のEGFR Mabの組み合わせがマウス異種腫瘍モデルにおける腫瘍の根絶をもたらすことが示されている。イムクローン(Imclone)社はキメラMab(C225)を用いるフェーズI臨床試験を実施しており、その安全性は実証されている。図31〜33において本発明者らは、本発明者らのヒト抗EGFR抗体に関するデータを示す。図30は、XenoMouse II系統に由来するヒト抗EGFR抗体の重鎖アミノ酸配列を示す。図31は、ヒト抗EGFR抗体によってEGFのA431細胞との結合が阻止されることを示す。図32は、ヒト抗EGFR抗体によってEGFのSW948細胞との結合が阻害されることを示す。図33は、XenoMouse II系統に由来するヒト抗EGFR抗体がインビトロでのSW948細胞の増殖を阻害することを示す。
【0105】
実施例9:XenoMouse IIによって産生される高親和性の抗原特異的ヒトMab
本発明者らは次に、XenoMouse IIにおける広範なヒトレパートリーを、多数の抗原、特に重大な臨床的関心がもたれるヒト抗原に対するヒト抗体を産生させるために利用しうるか否かを問題にした。
【0106】
このため、個々のXenoMouse IIの仔のそれぞれにヒトIL-8、ヒトEGFRおよびヒトTNF-αという3種の異なる抗原標的のうち1つによる誘発刺激を与えた。抗原はIL-8およびTNF-αの場合には可溶性タンパク質として、EGFRの場合には細胞(A431細胞)表面上に発現されたものとしていずれかの2つの異なる形式で投与された。3種の抗原のいずれについても、免疫化マウスからの血清に対して行ったELISAでは、1:3×106もの高い力価を有する強い抗原特異的ヒト抗体(IgG、Igκ)反応が示された。検出されたマウスλ反応は無視しうる程度であった。
【0107】
標準的なハイブリドーマ技術によって脾臓またはリンパ節に由来するハイブリドーマを得た上で、ELISAにより抗原特異的ヒトMabに関してスクリーニングした。
【0108】
IL-8によって免疫化した1匹のXenoMouse IIから、いずれもヒトIL-8に特異的な完全ヒト(hIgG2κ)Mabを分泌する12種の一連のハイブリドーマが得られた。これらのハイブリドーマのうち4種からの抗体であるD1.1、K2.2、K4.2およびK4.3を腹水から精製し、ヒトIL-8に対するそれらの親和性、およびIL-8のヒト好中球上の受容体との結合を阻止する効力に関して評価した。
【0109】
親和性の測定は、BIAcoreにおける表面プラズモン共鳴を用いる全抗体およびFab断片の双方の固相測定、ならびに溶液中でのラジオイムノアッセイによって行った(材料および方法)。表Vに示す通り、4種のMabに関して測定された親和性の値は1.1×109から4.8×1010M-1の範囲であった。用いた技法には若干の違いがあったものの、4種の抗体に関する親和性の値は一貫して109M-1よりも高かった。
【0110】
ELISA分析では、これらの4種の抗体がヒトIL-8に特異的であること、ならびに密接に関連したケモカインであるMIP-1α、GROα、βおよびγ、ENA-78、MCP-1またはRANTESとは交差反応しないことが確かめられた(非提示データ)。さらにBIAcore上での競合分析からは以上の抗体が少なくとも2つの異なるエピトープを認識することが示された(非提示データ)。すべての抗体がマウス抗ヒトIL-8中和抗体と同程度に効果的にIL-8のヒト好中球との結合を阻害したが、対照ヒトIgG2κ抗体はこれを阻害しなかった(図5A)。
【0111】
EGFRによって免疫化したXenoMouse IIを用いた融合実験では、いずれもEGFR特異的なヒトIgG2κMabを分泌する25種の一連のハイブリドーマが得られた。分析した13種のヒトMabのうち4種(E2.1、E2.4、E2.5,E2.11)が、マウスにおけるEGFを介した細胞増殖および腫瘍形成を阻害することが以前に示されている(Satoら、1983)EGFR特異的マウス抗体225と競合する能力に関して選択された。腹水から精製されたこれらのヒト抗体を、EGFRに対するそれらの親和性およびEGFの細胞との結合の中和に関して評価した。これらの抗体のEGFRに対する親和性は、BIAcore測定による評価では2.9×109から2.9×1010M-1の範囲であった(表V)。
【0112】
4種の抗EGFR抗体はいずれもEGFのA431細胞との結合を完全に阻止し(図5B)、それらがこれらの細胞上の高親和性および低親和性受容体(Kawamotoら、1983)の双方に対するEGFの結合を中和する能力をもつことが示された。4種の抗EGFRヒト抗体のいずれによっても、EGFのヒトSW948ヒト肺癌細胞上に発現されたEGFRとの結合が完全に阻害されることも認められた(非提示データ)。いずれの場合にも完全ヒト抗体はEGF結合の阻害の点で抗EGFRマウス抗体225と同程度に効果的であり、528抗体よりも強力であった(Gillら、1983)。いずれの細胞アッセイにおいても、対照ヒトIgG2κ抗体はEGF結合に影響を及ぼさなかった(図5Bおよび非提示データ)。
【0113】
TNF-αによって免疫化したXenoMouse IIを用いた融合実験では、12種の一連のヒトIgG2κ抗体が得られた。この12種のうち4種が、TNF-αのU937細胞上の受容体との結合を阻止する能力に関して選択された(図5C)。これらの抗体の親和性を測定したところ、1.2〜3.9×109M-1の範囲であった(表V)。
【0114】
上記のXenoMouse由来ハイブリドーマは、静置培養条件下において2〜19μg/mlの範囲の濃度で抗体を産生した。非還元条件下におけるタンパク質ゲル上での精製抗体の特徴分析では、IgG2κ抗体に関する見かけの分子量の期待値が150kDであることが明らかになった。還元条件下で検出された重鎖に関する見かけの分子量の期待値は50kDであり、軽鎖については25kDであった(非提示データ)。
【0115】
以下の表Vは、XenoMouse由来の抗原特異的完全ヒトMabの親和定数を示している。IL-8、EGFRおよびTNF-αに特異的なXenoMouse由来のヒトIgG2κ Mabの親和定数は、材料および方法の項に記載したBIAcoreまたはラジオイムノアッセイによって測定した。IL-8およびEGFRに関して示した値は精製抗体を用いて実施した独立した実験の代表値であり、TNF-αに関して示した値はハイブリドーマ上清を用いて実施した実験によるものである。
(表V)
【表5】
【0116】
実施例10:モノクローナル抗体における遺伝子使用および体細胞超突然変異
上記のIL-8およびEGFR-ヒトMabからの重鎖およびκ軽鎖転写物の配列を決定した。図6および図[[ ]]。4種のIL-8特異的抗体は、少なくとも3つの異なるVH遺伝子(VH4-34/VH4-21、VH3-30およびVH5-51)、4つの異なるDHセグメント(A1/A4、K1、ir3rcおよび21-10rc )および2つのJH(JH3およびJH4)遺伝子セグメントからなっていた。3つの異なるVκ遺伝子(012、018およびB3)がJκ3およびJκ4遺伝子と結合していた。このように多様な使用がみられることは、XenoMouse IIが多様な可変領域を備えた一連の抗IL-8中和抗体を産生しうることを示す。
【0117】
IL-8抗体転写物とは対照的に、Mab225との競合能に関して選択された抗体の配列ではVHおよびVκ遺伝子の使用が比較的制限されており、E1.1、 E2.4およびE2.5という3種の抗体は同一のVH遺伝子(4-31)を共有し、E2.11はVH4-31との相同性が高いVH4-61を含んでいた。異なるD(2、A1/A4、XP1)およびJH(JH3、JH4、JH5)セグメントが検出された。4種の抗体はいずれも同一のVκ(018)遺伝子を共有することが示された。それらのうち3つはJκ4を含み、残る1つであるE2.5はJκ2を含んでいた。
【0118】
大部分のVHおよびVκハイブリドーマ転写物では、対応する生殖細胞系セグメントからみて高度のヌクレオチド変化(7〜17箇所)が認められたが、定常領域では全く変異が検出されなかった。Vセグメントにおける変異の大部分は、予想される抗体アミノ酸配列においてアミノ酸置換をもたらし(V遺伝子1つにつき0〜12箇所)、その多くはCDR1およびCDR2領域にあった(図_)。注目すべき点は、すべての抗体で共有されていたCDR1におけるGly→Asp置換、または3つの抗体で共有されていたCDR2におけるSer→Asn置換、およびフレームワーク領域3におけるVal→Leuなどの変異が、EGFR抗体の重鎖配列で共有されていたことである。これらの結果は、XenoMouse IIにおいて抗体の成熟および選択に至る体細胞超突然変異の過程が高度に生じていることを示す。
【0119】
考察
本出願は、複雑なメガベースサイズのマウス遺伝子座を、YAC上に再構成されたサイズおよび内容の点で等価なヒトDNA断片によって、初めて機能的に置換したことを記載したものである。この手法により、マウス体液性免疫系はメガベースサイズのヒトIg遺伝子座によって「ヒト化」され、内因性抗体産生に欠陥のあるマウスにおいてヒト抗体反応が実質的に再現されるようになる。
【0120】
ヒト重鎖およびκ軽鎖遺伝子座の大部分がほぼ生殖細胞系コンフィギュレーションの形で忠実に再構成されるという本発明者らの成果は、酵母におけるYAC組換えが、Ig遺伝子座などの大きく複雑で不安定な断片を再構成するため(Mendezら、1995)、およびそれらを哺乳動物細胞内に導入するために操作するための強力な技術であることを立証するものである。さらに、2つの大きな重鎖およびκ軽鎖セグメントが無傷の形でマウス生殖細胞系に首尾よく導入されたことは、ES細胞-酵母スフェロプラスト融合がマウス生殖細胞系に異種遺伝子座を送達するための確実で効率的な手法であることを裏づける。
【0121】
XenoMouse II系統の特徴分析から、大きなIg遺伝子座には、多様性および機能性の点で野生型マウスのものに匹敵し、ヒトIgミニ遺伝子構築物(Lonbergら、1994)または小型のIg YAC(Greenら、1994)を保持するマウスで生じる体液性応答よりもはるかに優れた抗体系を再生する能力があることが示された。この差は、成熟B細胞のレベル、ヒトIg産生、クラススイッチ効率、多様性、マウスIgλ産生と対比してのヒトIgκの優位性、およびヒト抗体反応の強さ、ならびに多数の抗原に対する高親和性の抗原特異的モノクローナル抗体が産生されるとの成果として表れた。
【0122】
XenoMouse IIにおける成熟B細胞およびヒト抗体のレベルは、これまで報告されているIg-トランスジェニックマウスの中で最も高く、これまでのマウスに関して示されている値の数倍であり、野生型マウスでの値に近い。特にヒトIgGのレベルは、ヒトγ1遺伝子を備えたミニ遺伝子座Ig導入遺伝子を保持するマウスに関して報告されている値の100倍を上回る(Lonbergら、1994)。XenoMouse IIにおけるクラススイッチの効率がより高いことは、yH2上にスイッチ領域の全体を、それらのすべての調節要素ならびに付加的な調節要素とともに含めた結果である可能性が高いと考えられ、これが適切なクラススイッチの支持および維持には重要であると思われる。XenoMouse II系統における成熟B細胞のレベルの上昇は、再配列頻度がより高く、そのためにV遺伝子レパートリーが増すことによって骨髄中のB細胞発生が改善されるためである可能性が高い。B細胞の再構成は、ヒト重鎖遺伝子座に関して同型接合的であるXenoMouse II系統でははるかに顕著であると考えられる。
【0123】
循環B細胞によるヒトκとマウスλ軽鎖との発現の比は、トランスジェニックκ軽鎖遺伝子座の利用に関する有用な内部指標となる。より小さなIg YACの1対立遺伝子を含むマウスで認められるヒトκおよびマウスλの分布はほぼ等しいが、XenoMouse II系統ではヒトκの明らかな優位性が検出されている。さらに、yK2に関して同型接合的なマウスでのκ:λ比は野生型マウスと同一であった。これらの観察結果をVκ遺伝子の広範な使用と併せて考えると、XenoMouse IIにおけるヒト近位Vκ遺伝子は多様な軽鎖反応を支持するのに十分であることが強く示唆され、これはヒトで近位Vκ遺伝子がより用いられる傾向にあることとも一致する(Coxら、1994)。
【0124】
XenoMouse II系統は、組換え機構によって利用され、成熟抗体に組み入れられる遺伝子座の全長にわたるV、DおよびJ遺伝子による、極めて高度の抗体多様性を呈する。抗原結合によっていったん誘発されれば、大規模な体細胞超突然変異が起こり、抗体の親和性成熟をもたらす。
【0125】
XenoMouse IIにおけるV、D、J遺伝子の利用パターンからは、それらがヒトでの利用と類似した様式で利用可能であり、Igミニ遺伝子を保持するマウスで認められる胎児様で位置による偏差のある使用(Taylorら、1992;Taylorら、1994;Tuaillonら、1993)とは異なる、成人様ヒト抗体レパートリーが得られることも示された。マウスによって認識される多数の抗原とともに多くの機能的VHおよびVκ遺伝子が広く利用されることは、機能的な抗体反応を首尾よく再構成する上での広範なV遺伝子レパートリーの重要性を強調するものである。
【0126】
マウスにおけるヒト免疫応答の再構成の程度に関する最終的な判断材料は、マウスが抗体反応を誘発すると思われる抗原の範囲、および種々の抗原に対して抗原特異的な高親和性Mabが産生されうる容易さである。現在までに限られた数の抗原特異的ヒトMabしか得られていない、より小型のヒトIg YACまたはミニ遺伝子による操作を受けたマウスとは異なり(Lonbergら、1994;Greenら、1994;Fishwildら、1996)、XenoMouse IIは現在までに検討したすべてのヒト抗原に対するMabを産生した。XenoMouse II系統は、可溶性タンパク質または細胞表面上に発現されたもののいずれかとして提示された種々のヒト抗原に対して強いヒト抗体反応を生じた。検討した3種のヒト抗原のそれぞれによる免疫化により、10〜25種の一連の抗原特異的ヒトIgG2κ Mabが得られた。各抗原に関して、109〜1010M-1の範囲の親和性を有する抗体の一群が得られた。親和性の値が、結合活性というよりも一価結合動態を表すことを確認するために、いくつかの測定を行った:二価結合の確率を最小限に抑えるために抗原を低密度にコートしたセンサーチップを用いて無傷抗体によるBIAcore解析を行った;2種の抗体については一価Fab断片を用いて解析を再度行った;いくつかの抗体については溶液のラジオイムノアッセイによる検討も行った。これらの測定の結果から、本発明者らは、XenoMouseによって1010M-1の範囲の親和性をもつ抗体を容易に獲得しうると結論する。XenoMouse由来の抗体に関して得られた親和性の値は、遺伝子操作されたマウス(Lonbergら、Fishwildら、1996)またはコンビナトリアルライブラリー(Vaughanら、1996)から産生された、ヒト抗原に対するヒト抗体に関して報告されたものの中で最も高い。これらの高親和性と、V遺伝子の体細胞変異の結果としての高度のアミノ酸置換が組み合わされることにより、XenoMouse IIにおける親和性成熟機構が完全であり、野生型マウスにおけるものと匹敵することが裏づけられる。
【0127】
これらの結果は、抗体の多様化および選択に関するマウスの機構によってヒトIg YAC上に広範な抗体レパートリーが適切に開発されること、およびヒトタンパク質に対する免疫寛容の欠如によってヒト抗原を含む任意の関心対象の抗原に対する高親和性抗体を得ることができることを示す。これらのマウスにおけるヒト免疫グロブリン遺伝子によってヒト抗原に対する抗体が産生されうる容易性は、B細胞のレベルでの自己免疫寛容が獲得され、遺伝されないことのさらなる裏づけとなる。
【0128】
ヒト抗原に対する高親和性の完全ヒト抗体を産生しうることには、明らかな実用的意味がある。完全ヒト抗体は、マウスまたはマウス由来のMabに固有の免疫原性およびアレルギー性反応を最小限に抑え、このため投与される抗体の有効性および安全性を高めると考えられる。XenoMouse IIは、抗体の反復投与を必要とする、炎症、自己免疫および癌などの、慢性および再発性のヒト疾患の治療において実質的な利益が提供される可能性をもたらす。XenoMouse IIによって一連の完全ヒト高親和性抗体が得られる迅速性および再現性は、それがヒト抗体生産のための他の技術を上回る進歩をもたらす可能性があることを意味する。例えば、それに由来する抗体の多くの親和性を高めるために多大な努力を必要とし、1本鎖FvsまたはFabが得られるファージディスプレーとは異なり、XenoMouse IIの抗体は、さらなる操作を要せずにハイブリドーマから産生させることができる高親和性の完全に無傷な免疫グロブリンである。
【0129】
マウスにおいて、真性のヒト体液性免疫系を創出するための本明細書に記載した戦略は、マウス免疫系の他の機能を司る、T細胞受容体または主要組織適合抗原遺伝子複合体などの他の多遺伝子座のヒト化に向けても適用しうる(Jakobovits、1994)。このようなマウスは、ヒト遺伝子座の構造-機能関連および免疫系の発達におけるそれらの関与を解明する上で有用であると考えられる。
【0130】
参照としての組み入れ
特許、特許出願、論文、教科書などを含む本明細書中に引用したすべての参考文献、およびそれらの中で引用された参考文献は、それらが既知でない範囲において、その全体が参照として本明細書に組み入れられる。さらに、以下の参考文献も、このような参考文献において引用された参考文献を含め、その全体が参照として本明細書に組み入れられる。
【0131】
参考文献
【技術分野】
【0001】
本発明は、ヒト免疫グロブリン遺伝子座を含むように操作されたトランスジェニック非ヒト動物に関する。特に、本発明による動物は、複数の可変(VHおよびVκ)遺伝子領域を含むヒトIg遺伝子座を有する。有利なことに、複数の可変領域遺伝子を含めることにより、その動物によって産生されるヒト抗体の特異性および多様性は高まる。さらに、このような領域を含めることにより、その動物が極めて高親和性の抗体を分泌する成熟B細胞を豊富に有するように、その動物に対してB細胞発生が増強および再構成される。
【背景技術】
【0002】
メガベースサイズのヒト遺伝子座をYAC中にクローニングおよび再構築し、それらをマウス生殖細胞系に導入しうることにより、極めて大きく、または大まかにマッピングされた遺伝子座の機能的構成要素を解明するための、ならびにヒト疾患の有用なモデルを作製するための、強力な手法が提供される。さらに、このような技術を利用してマウス遺伝子座をそれらのヒト等価物に置換することにより、発生過程におけるヒト遺伝子産物の発現および調節、それらの他の系との連絡、ならびに疾患の誘導および進行におけるそれらの関与に関して比類のない洞察が得られると考えられる。
【0003】
このような戦略の1つの重要な実用的応用は、マウス体液性免疫系の「ヒト化」である。内因性Ig遺伝子が不活性化されたマウスにヒト免疫グロブリン(Ig)遺伝子座を導入することにより、抗体のプログラム化された発現および構築の根底にある機序に加えて、B細胞の発生におけるそれらの役割をも検討する機会が提供される。さらに、このような戦略により、ヒト疾患の抗体療法という可能性の実現へ向けての重要な里程標となる、完全ヒトモノクローナル抗体(Mab)を生産するための理想的な供給源が提供されると考えられる。完全ヒト抗体は、マウスまたはマウス由来のMabに固有の免疫原性およびアレルギー性反応を最小限に抑え、それにより、投与される抗体の有効性および安全性を高めると考えられる。完全ヒト抗体の使用は、抗体の反復投与を必要とする、炎症、自己免疫および癌などの慢性的および再発性のヒト疾患の治療に、実質的な利益を提供すると考えられる。
【0004】
この目標に向けての1つの手法は、マウス抗体の産生に欠陥があってヒトIg遺伝子座の大きな断片を有するマウス系統を、組換え操作によって作製することであり、これは、このようなマウスが、マウス抗体を生じずに広範なレパートリー(repertoire)のヒト抗体を製造するであろうとの予測に立つものであった。大きなヒトIg断片は、可変遺伝子の広範な多様性に加えて、抗体の産生および発現の適切な調節も保持すると考えられる。抗体の多様化および選択のため、ならびにヒトタンパク質に対する免疫寛容を失わせるためのマウス機構を開発すれば、これらのマウス系統で再現されたヒト抗体レパートリーはヒト抗原を含む関心対象の任意の抗原に対する高親和性抗体を生じるはずである。ハイブリドーマ技術を用いることにより、望ましい特異性をもつ抗原特異的ヒトMabは容易に製造および選択しうると考えられる。
【0005】
この一般的な戦略は、本発明者らが1994年に発表した最初のXenoMouse(登録商標)系統の作出との関連で示された。グリーン(Green)ら、Nature Genetics 7:13〜21(1994)(非特許文献1)参照。XenoMouse(登録商標)系統は、可変および定常領域のコア配列を含む、ヒト重鎖およびκ軽鎖遺伝子座のそれぞれ245kbおよび190kbのサイズの生殖細胞系コンフィギュレーション断片を有するように操作された。同上。ヒトIgを含む酵母人工染色体(YAC)は、抗体の再配列および発現の双方に関してマウス系への適合性があることが実証されており、不活性化されたマウスIg遺伝子の置換も可能であった。これは、それらがB細胞発生の誘導および完全ヒト抗体の成人様ヒトレパートリーの生成、ならびに抗原特異的ヒトMabの産生の能力をもつことによって示された。また、これらの結果は、比較的多数のV遺伝子、付加的な調節要素、およびヒトIg定常領域を含むヒトIg遺伝子座のかなりの部分を導入することにより、感染および免疫化に対するヒト体液性応答の特徴である実質的に完全なレパートリーが再現される可能性があることも示唆した。
【0006】
このような手法は、1990年1月12日に提出された米国特許出願第07/466,008号(特許文献1)、1990年11月8日に提出された第07/610,515号(特許文献2)、1992年7月24日に提出された第07/919,297号(特許文献3)、1992年7月30日に提出された第07/922,649号(特許文献4)、1993年3月15日に提出された第08/031,801号(特許文献5)、1993年8月27日に提出された第08/112,848号(特許文献6)、1994年4月28日に提出された第08/234,145号(特許文献7)、1995年1月20日に提出された第08/376,279号(特許文献8)、1995年4月27日に提出された第08/430,938号(特許文献9)、1995年6月5日に提出された第08/464,584号(特許文献10)、1995年6月5日に提出された第08/464,582号(特許文献11)、1995年6月5日に提出された第08/463,191号(特許文献12)、1995年6月5日に提出された第08/462,837号(特許文献13)、1995年6月5日に提出された第08/486,853号(特許文献14)、1995年6月5日に提出された第08/486,857号(特許文献15)、1995年6月5日に提出された第08/486,859号(特許文献16)、1995年6月5日に提出された第08/462,513号(特許文献17)、および1996年10月2日に提出された第08/724,752号(特許文献18)という米国特許出願においてさらに考察および描写されている。また、1996年6月12日に認可が発行されたEP 0 463 151 B1(特許文献19)、1994年2月3日に発行された国際特許出願国際公開公報第94/02602号(特許文献20)、1996年10月31日に発行された国際特許出願国際公開公報第96/34096号(特許文献21)、および1996年4月29日に提出されたPCT/US96/05928(特許文献22)も参照されたい。上記に引用したそれぞれの特許および出願の開示は、その全体が参照として本明細書に組み入れられる。
【0007】
1つの代替的なアプローチにおいて、ジェンファームインターナショナル社(GenPharm International, Inc.)を含む他者は、「ミニ遺伝子座(minilocus)」法を用いている。ミニ遺伝子座法では、Ig遺伝子座からの複数の小片(個々の遺伝子)を含めることにより、外来性Ig遺伝子座を模倣する。このため、1つまたは複数のVH遺伝子、1つまたは複数のDH遺伝子、1つまたは複数のJH遺伝子、μ定常領域、および第2の定常領域(好ましくはγ定常領域)が、動物に挿入するための構築物(construct)中に形成される。この手法はスラニ(Surani)らに対する米国特許第5,545,807号(特許文献23)、いずれもロンバーグ(Lonberg)およびケイ(Kay)に対する米国特許第5,545,806号(特許文献24)および第5,625,825号(特許文献25)、ならびにいずれもジェンファームインターナショナル社(GenPharm International, Inc.)による米国特許出願である1990年8月29日に提出された第07/574,748号(特許文献26)、1990年8月31日に提出された第07/575,962号(特許文献27)、1991年12月17日に提出された第07/810,279号(特許文献28)、1992年3月18日に提出された第07/853,408号(特許文献29)、1992年6月23日に提出された第07/904,068号(特許文献30)、1992年12月16日に提出された第07/990,860号(特許文献31)、1993年4月26日に提出された第08/053,131号(特許文献32)、1993年7月22日に提出された第08/096,762号(特許文献33)、1993年11月18日に提出された第08/155,301号(特許文献34)、1993年12月3日に提出された第08/161,739号(特許文献35)、1993年12月10日に提出された第08/165,699号(特許文献36)、1994年3月9日に提出された第08/209,741号(特許文献37)に記載されており、それらの開示は参照として本明細書に組み入れられる。また、その開示の全体が参照として本明細書に組み入れられる1994年11月10日に発行された国際公開公報第94/25585号(特許文献38)、1993年7月24日に発行された国際公開公報第93/12227号(特許文献39)、1992年12月23日に発行された国際公開公報第92/22645号(特許文献40)、1992年3月19日に発行された国際公開公報第92/03918号(特許文献41)も参照されたい。さらに、その開示の全体が参照として本明細書に組み入れられるテイラー(Taylor)ら、1992(非特許文献2)、チェン(Chen)ら、1993(非特許文献3)、トゥアイロン(Tuaillon)ら、1993(非特許文献4)、チョイ(Choi)ら、1993(非特許文献5)、ロンバーグ(Lonberg)ら(1994)(非特許文献6)、テイラーら(1994)(非特許文献7)、およびトゥアイロンら(1995)(非特許文献8)も参照されたい。
【0008】
上記に引用し、英国医学研究会議(Medical Research Council)(「MRC」)に譲渡されたスラニ(Surani)らの発明者らは、ミニ遺伝子座法の使用により、Ig遺伝子座を有するトランスジェニックマウスを作製した。上記に引用したジェンファームインターナショナル社の業績の発明者らであるロンバーグおよびケイは、本発明者の先例にならって、内因性マウスIg遺伝子座の不活性化とスラニらの業績の実質的な重複物との組み合わせを提案した。
ミニ遺伝子座法の1つの利点は、Ig遺伝子座の一部を含む構築物の作製および動物への導入を迅速に行いうることである。しかしこれを相殺する程度に、ミニ遺伝子座法には理論的にみて、少数のV、DおよびJ遺伝子を含めることによって導入される多様性が不十分であるとの重大な欠点がある。事実、発表された研究はこの懸念を裏づけるように思われる。ミニ遺伝子座法を用いて作製された動物のB細胞発生および抗体産生は阻害されていると考えられる。このため、本発明者らは、より大きな多様性の実現を目的として、および動物の免疫レパートリーを再構築するための努力において、Ig遺伝子座の大部分を導入することを一貫して主張してきた。
したがって、ヒトIg遺伝子座のより完全な生殖細胞系配列およびコンフィギュレーションを含むトランスジェニック動物を提供することが望ましいと考えられる。内因性Igがノックアウトされたバックグラウンドに対してこのような遺伝子座を提供することはさらに望ましいと考えられる。
【先行技術文献】
【特許文献】
【0009】
【特許文献1】米国特許出願第07/466,008号
【特許文献2】米国特許出願第07/610,515号
【特許文献3】米国特許出願第07/919,297号
【特許文献4】米国特許出願第07/922,649号
【特許文献5】米国特許出願第08/031,801号
【特許文献6】米国特許出願第08/112,848号
【特許文献7】米国特許出願第08/234,145号
【特許文献8】米国特許出願第08/376,279号
【特許文献9】米国特許出願第08/430,938号
【特許文献10】米国特許出願第08/464,584号
【特許文献11】米国特許出願第08/464,582号
【特許文献12】米国特許出願第08/463,191号
【特許文献13】米国特許出願第08/462,837号
【特許文献14】米国特許出願第08/486,853号
【特許文献15】米国特許出願第08/486,857号
【特許文献16】米国特許出願第08/486,859号
【特許文献17】米国特許出願第08/462,513号
【特許文献18】米国特許出願第08/724,752号
【特許文献19】EP 0 463 151 B1
【特許文献20】国際特許出願国際公開公報第94/02602号
【特許文献21】国際特許出願国際公開公報第96/34096号
【特許文献22】PCT/US96/05928
【特許文献23】米国特許第5,545,807号
【特許文献24】米国特許第5,545,806号
【特許文献25】米国特許第5,625,825号
【特許文献26】米国特許出願第07/574,748号
【特許文献27】米国特許出願第07/575,962号
【特許文献28】米国特許出願第07/810,279号
【特許文献29】米国特許出願第07/853,408号
【特許文献30】米国特許出願第07/904,068号
【特許文献31】米国特許出願第07/990,860号
【特許文献32】米国特許出願第08/053,131号
【特許文献33】米国特許出願第08/096,762号
【特許文献34】米国特許出願第08/155,301号
【特許文献35】米国特許出願第08/161,739号
【特許文献36】米国特許出願第08/165,699号
【特許文献37】米国特許出願第08/209,741号
【特許文献38】国際公開公報第94/25585号
【特許文献39】国際公開公報第93/12227号
【特許文献40】国際公開公報第92/22645号
【特許文献41】国際公開公報第92/03918号
【非特許文献】
【0010】
【非特許文献1】グリーン(Green)ら、Nature Genetics 7:13〜21(1994)
【非特許文献2】Taylor et al., "A transgenic mouse that expresses a diversity of human sequence heavy and light chain immunoglobulins." Nucleic Acids Research 20:6287-6295 (1992).
【非特許文献3】Chen et al. "Immunoglobulin gene rearrangement in B-cell deficient mice generated by targeted deletion of the JH locus" International Immunology 5:647-656 (1993)
【非特許文献4】Tuaillon et al., "Human immunoglobulin heavy-chain minilocus recombination in transgenic mice: gene-segment use in μ and γ transcripts." Proc. Natl. Acad. Sci. USA 90:3720-3724 (1993).
【非特許文献5】Choi et al. "Transgenic mice containing a human heavy chain immunoglobulin gene fragment cloned in a yeast artificial chromosome" Nature Genetics 4:117-123 (1993)
【非特許文献6】Lonberg et al., "Antigen-specific human antibodies from mice comprising four distinct genetic modifications." Nature 368:856-859 (1994).
【非特許文献7】Taylor et al., "Human immunoglobulin transgenes undergo rearrangemet, somatic mutation and class switching in mice that lack endogenous IgM." International Immunology 6:579-591 (1994).
【非特許文献8】Tuaillon et al. "Analysis of direct and inverted DJH rearrangements in a human Ig heavy chain transgenic minilocus" J. Immunol. 154:6453-6465 (1995)
【発明の概要】
【課題を解決するための手段】
【0011】
[発明の概要]
本発明により提供されるものは、ヒト重鎖遺伝子座およびヒトκ軽鎖遺伝子座の両方を含むほぼ完全なヒトIg遺伝子座を有するトランスジェニック動物である。好ましくは、重鎖遺伝子座は、ヒト重鎖可変領域の約20%を超える、より好ましくは約40%を超える、より好ましくは約50%、さらにより好ましくは約60%を超える部分を含む。ヒトκ軽鎖に関して、遺伝子座は好ましくはヒトκ軽鎖可変領域の約20%を超える、より好ましくは約40%を超える、より好ましくは約50%、さらにより好ましくは約60%を超える部分を含む。このような比率は機能的な可変領域遺伝子の比率に関して言及することが好ましい。
【0012】
さらに好ましくは、このような動物は全DH領域、全JH領域、ヒトμ定常領域を含み、付加的なアイソタイプを作製するための他のヒト定常領域をコードする遺伝子を付加的に備えることができる。このようなアイソタイプにはγ1、γ2、γ3、α、ε、βおよび他の定常領域をコードする遺伝子を含めうる。同一の導入遺伝子上、すなわちヒトμ定常領域の下流に代替的な定常領域を含めることもでき、または別の染色体上にこのような他の定常領域を含めることもできる。このような他の定常領域が導入遺伝子をコードするヒトμ定常領域を含む染色体と同じ染色体上に含められる場合には、他の1つのアイソタイプまたは複数のアイソタイプへのシス転換(cis-switching)が達成されうることが理解されるであろう。これに対して、このような他の定常領域が導入遺伝子をコードするμ定常領域を含む染色体とは異なる染色体上に含められる場合には、他の1つのアイソタイプまたは複数のアイソタイプへのトランス転換(trans-switching)が達成されうることが理解されるであろう。このような配置は、広範な種類の抗原に対する抗体を産生させるためのマウスの設計および構築において多大な柔軟性を可能にする。
【0013】
このようなマウスは、機能的な内因性免疫グロブリンを別に産生しないことが好ましい。これは、1つの好ましい態様では、内因性の重鎖および軽鎖遺伝子座の不活性化(またはノックアウト)を通じて達成される。例えば、1つの好ましい態様では、マウス重鎖J領域ならびにマウスκ軽鎖J領域およびCκ領域が、その領域を置換または欠失させる相同的組換えベクターの利用によって不活性化される。このような技法は、本発明者らの以前の出願および刊行物に詳細に記載されている。
【0014】
予想外のことに、本発明に係るトランスジェニックマウスは、ほぼ完全に再構成された免疫系レパートリーを有すると考えられる。これは4つの別々のマウス系統を比較した際に極めて明瞭に示される:第1の系統は広範なヒト重鎖可変領域およびヒトκ軽鎖可変領域を含んでいてμアイソタイプのみをコードし、第2の系統は広範なヒト重鎖可変領域およびヒトκ軽鎖可変領域を含んでいてμおよびγ2アイソタイプをコードし、第3の系統は明らかに少ないヒト重鎖およびκ軽鎖の可変領域を含み、第4の系統は二重不活性化を受けた(double-inactivated)マウスIg遺伝子座を含む。第1および第2の系統では同一ではないが類似したB細胞発生が起こるが、第3の系統ではB細胞の発生および成熟が抑制され、第4の系統は成熟B細胞を全く含まない。さらに興味深いことに、内因性Igがノックアウトされたバックグラウンドをもつマウスではマウス抗体に比してヒト抗体の産生がかなり増強される。すなわち、ヒトIg遺伝子座および機能的に不活性化された内因性Ig遺伝子座を含むマウスは、ヒトIg遺伝子座のみを含むマウスよりもヒト抗体をほぼ100〜1000倍もの速度で効率的に産生する。
【0015】
このため、本発明の第1の局面によれば、改変を含むゲノムを有し、その改変に以下のものが含まれるトランスジェニック非ヒト哺乳動物が提供される:その哺乳動物が正常なB細胞発生を呈しないように不活性化された、内因性免疫グロブリン(Ig)遺伝子座;ヒトμ定常領域ならびにそれに対する調節およびスイッチ配列、複数のヒトJH遺伝子、複数のヒトDH遺伝子および複数のヒトVH遺伝子を含んでいて、実質的な生殖細胞系コンフィギュレーションとして挿入されたヒト重鎖Ig遺伝子座;ならびにヒトκ定常領域、複数のヒトJκ遺伝子および複数のヒトVκ遺伝子を含んでいて、実質的な生殖細胞系コンフィギュレーションとして挿入されたヒトκ軽鎖Ig遺伝子座。1つの好ましい態様において、重鎖Ig遺伝子座はヒトγ1、ヒトγ2、ヒトγ3、ヒトγ4、α、δおよびεからなる群より選択される第2の定常領域を含む。もう1つの好ましい態様において、VH遺伝子の数は約20より多い。もう1つの好ましい態様において、Vκ遺伝子の数は約15より多い。もう1つの好ましい態様では、DH遺伝子の数は約25より多く、JH遺伝子の数は約4より多く、VH遺伝子の数は約20より多く、Jκ遺伝子の数は約4より多く、Vκ遺伝子の数は約15より多い。もう1つの好ましい態様において、DH遺伝子の数、JH遺伝子の数、VH遺伝子の数、Jκ遺伝子の数およびVκ遺伝子の数は、Ig遺伝子座が約1×105種を超える、異なる機能的な抗体配列の組み合わせをコードするように選択される。1つの好ましい態様では、哺乳動物の1つの集団において、野生型と比べて平均で約50%を超えるB細胞の機能が再構成される。
【0016】
本発明の第2の局面によれば、哺乳動物で、ヒト免疫グロブリン分子を産生しうるが、機能的な内因性免疫グロブリン分子を実質的に産生しえないようにさせる改変を含むゲノムを有する、以下のように改善されたトランスジェニック非ヒト哺乳動物が提供される:哺乳動物のゲノムへの、その哺乳動物が約1×106種を超える異なる機能的なヒト免疫グロブリン配列の組み合わせをコードしうるために十分なヒトVH、DH、JH、VκおよびJκ遺伝子の挿入。
【0017】
本発明の第3の局面によれば、哺乳動物で、ヒト免疫グロブリン分子を産生しうるが、機能的な内因性免疫グロブリン分子を実質的に産生しえないようにする改変であって、その哺乳動物が機能的な内因性免疫グロブリン分子を産生する能力をもたないことに関してその哺乳動物が正常なB細胞発生を行えないようにするような改変を含むゲノムを有する、以下のように改善されたトランスジェニック非ヒト哺乳動物が提供される:哺乳動物のゲノムへの、その哺乳動物が約1×106種を超える、異なる機能的なヒト免疫グロブリン配列の組み合わせをコードしうるために十分なヒトVH、DH、JH、VκおよびJκ遺伝子、ならびに哺乳動物において正常なB細胞発生が実質的に再現されるのに十分なVHおよびVκ遺伝子の挿入。1つの好ましい態様では、哺乳動物の1つの集団において、野生型と比べて平均で約50%を超えるB細胞の機能が再構成される。
【0018】
本発明の第4の局面によれば、改変を含むゲノムを有し、その改変に以下のものが含まれるトランスジェニック非ヒト哺乳動物が提供される:不活性化された内因性重鎖免疫グロブリン(Ig)遺伝子座;不活性化された内因性κ軽鎖Ig遺伝子座;yH2のヌクレオチド配列に実質的に対応するヌクレオチド配列を含む、挿入されたヒト重鎖Ig遺伝子座;およびyK2のヌクレオチド配列に実質的に対応するヌクレオチド配列を含む、挿入されたヒトκ軽鎖Ig遺伝子座。
【0019】
本発明の第5の局面によれば、改変を含むゲノムを有し、その改変に以下のものが含まれるトランスジェニック非ヒト哺乳動物が提供される:不活性化された内因性重鎖免疫グロブリン(Ig)遺伝子座;yH2のヌクレオチド配列に実質的に対応するヌクレオチド配列を含む、挿入されたヒト重鎖Ig遺伝子座;およびyK2のヌクレオチド配列に実質的に対応するヌクレオチド配列を含む、挿入されたヒトκ軽鎖Ig遺伝子座。
【0020】
本発明の第6の局面によれば、改変を含むゲノムを有し、その改変に以下のものが含まれるトランスジェニック非ヒト哺乳動物が提供される:不活性化された内因性重鎖免疫グロブリン(Ig)遺伝子座;不活性化された内因性κ軽鎖Ig遺伝子座;ヒトγ2定常領域が存在しないyH2のヌクレオチド配列に実質的に対応するヌクレオチド配列を含む、挿入されたヒト重鎖Ig遺伝子座;およびyK2のヌクレオチド配列に実質的に対応するヌクレオチド配列を含む、挿入されたヒトκ軽鎖Ig遺伝子座。
【0021】
本発明の第7の局面によれば、改変を含むゲノムを有し、その改変に以下のものが含まれるトランスジェニック非ヒト哺乳動物が提供される:不活性化された内因性重鎖免疫グロブリン(Ig)遺伝子座;ヒトγ2定常領域が存在しないyH2のヌクレオチド配列に実質的に対応するヌクレオチド配列を含む、挿入されたヒト重鎖Ig遺伝子座;およびyK2のヌクレオチド配列に実質的に対応するヌクレオチド配列を含む、挿入されたヒトκ軽鎖Ig遺伝子座。
【0022】
本発明の第8の局面によれば、以下のものを含む、ヒト抗体を製造するための方法が提供される:上記の本発明の第1から第5の局面の哺乳動物のうち任意のものへの抗原の接種;109M-1よりも高い親和性で抗原と特異的に結合するヒト抗体を分泌する不死化細胞の集団を得るためのリンパ球細胞の回収および不死化;ならびに不死化細胞集団からの抗体の単離。
【0023】
1つの好ましい態様では、抗原はIL-8である。もう1つの好ましい態様では、抗原はEGFRである。もう1つの好ましい態様では、抗原はTNF-αである。
【0024】
本発明の第9の局面によれば、IL-8、EGFRおよびTNF-αに対する抗体を含む、本発明の第6の局面の方法によって製造される抗体が提供される。
【0025】
本発明の第10の局面によれば、複数のヒト可変領域の挿入を含む改変を含むゲノムを有するトランスジェニックマウスを製造するための改善された方法であって、改善が以下のものを含む方法が提供される:酵母人工染色体からのヒト可変領域の挿入。
【0026】
本発明の第11の局面によれば、本発明の第8の局面の改善の使用を通じて製造されたトランスジェニックマウスおよびそれらに由来するトランスジェニック子孫が提供される。
本発明の第12の局面によれば、挿入されたヒト重鎖免疫グロブリン導入遺伝子を含む改変を有するゲノムを有するトランスジェニック哺乳動物であって、改善に以下のものが含まれるトランスジェニック哺乳動物が提供される:ヒト様の接合点多様性およびヒト様の相補性決定領域3(CDR3)の長さを可能とするヒト可変領域遺伝子の選択された組(set)を含む導入遺伝子。1つの好ましい態様において、ヒト様の接合点多様性は平均7.7塩基のN付加長を含む。もう1つの好ましい態様において、ヒト様CDR3の長さは約2から約25残基の間、平均すると約14残基を含む。
本発明はまた、以下の項目を提供する:
(項目1)以下の改変を含むゲノムを有するトランスジェニック非ヒト哺乳動物:
その哺乳動物が正常なB細胞発生を呈しないように不活性化された内因性免疫グロブリン(Ig)遺伝子座;
挿入されるVHおよびVκ遺伝子の数がその哺乳動物における正常なB細胞発生が実質的に再現されるように選択される、
実質的な生殖細胞系コンフィギュレーションとして挿入されたヒト重鎖Ig遺伝子座であって、ヒトμ定常領域ならびにそれに対する調節およびスイッチ配列、複数のヒトJH遺伝子、複数のヒトDH遺伝子、ならびに複数のヒトVH遺伝子を含むヒト重鎖Ig遺伝子座;ならびに
実質的な生殖細胞系コンフィギュレーションとして挿入されたヒトκ軽鎖Ig遺伝子座であって、ヒトκ定常領域、複数のJκ遺伝子、および複数のVκ遺伝子を含むヒトκ軽鎖Ig遺伝子座。
(項目2)重鎖Ig遺伝子座がヒトγ1、ヒトγ2、ヒトγ3、ヒトγ4、α、εおよびδからなる群より選択される第2の定常領域を含む、項目1記載の哺乳動物。
(項目3)VH遺伝子の数が約20よりも多い、項目1記載の哺乳動物。
(項目4)Vκ遺伝子の数が約15よりも多い、項目1記載の哺乳動物。
(項目5)DH遺伝子の数が約25よりも多く、JH遺伝子の数が約4よりも多く、VH遺伝子の数が約20よりも多く、Jκ遺伝子の数が約4よりも多く、Vκ遺伝子の数が約15よりも多い、項目1記載の哺乳動物。
(項目6)DH遺伝子の数、JH遺伝子の数、VH遺伝子の数、Jκ遺伝子の数、およびVκ遺伝子の数が、Ig遺伝子座が約1×105種を超える異なる機能的な抗体配列の組み合わせをコードするように選択される、項目1記載の哺乳動物。
(項目7)哺乳動物の1つの集団において野生型と比べて平均で約50%を超えるB細胞の機能が再構成される、項目1記載の哺乳動物。
(項目8)哺乳動物を、ヒト免疫グロブリン分子を産生しうるが機能的な内因性抗体分子を実質的に産生しえないようにする、以下の改変を含むゲノムを有するトランスジェニック非ヒト哺乳動物:
接合点多様性または体細胞変異事象に寄与することなく、その哺乳動物が約1×106種を超える異なる機能的なヒト免疫グロブリン配列の組み合わせをコードしうるために十分なヒトVH、DH、JH、VκおよびJκ遺伝子の、哺乳動物ゲノムへの挿入。
(項目9)哺乳動物を、ヒト免疫グロブリン分子を産生しうるが機能的な内因性抗体分子を実質的に産生しえないようにする改変であって、その哺乳動物が機能的な内因性抗体分子を産生する能力をもたないことに関してその哺乳動物が正常なB細胞発生を行えないようにするような以下の改変を含むゲノムを有するトランスジェニック非ヒト哺乳動物:
その哺乳動物が約1×106種を超える異なる機能的なヒト免疫グロブリン配列の組み合わせをコードしうるために十分なヒトVH、DH、JH、VκおよびJκ遺伝子、ならびに哺乳動物において正常なB細胞発生が実質的に再現されるのに十分なVHおよびVκ遺伝子の、哺乳動物ゲノムへの挿入。
(項目10)哺乳動物の1つの集団において野生型と比べて平均で約50%を超えるB細胞の機能が再構成される、項目9記載の哺乳動物。
(項目11)以下の改変を含むゲノムを有するトランスジェニック非ヒト哺乳動物:
不活性化された内因性重鎖免疫グロブリン(Ig)遺伝子座;
不活性化された内因性κ軽鎖Ig遺伝子座;
yH2のヌクレオチド配列に実質的に対応するヌクレオチド配列を含む、挿入されたヒト重鎖Ig遺伝子座;および
yK2のヌクレオチド配列に実質的に対応するヌクレオチド配列を含む、挿入されたヒトκ軽鎖Ig遺伝子座。
(項目12)以下の改変を含むゲノムを有するトランスジェニック非ヒト哺乳動物:
不活性化された内因性重鎖免疫グロブリン(Ig)遺伝子座;
yH2のヌクレオチド配列に実質的に対応するヌクレオチド配列を含む、挿入されたヒト重鎖Ig遺伝子座;および
yK2のヌクレオチド配列に実質的に対応するヌクレオチド配列を含む、挿入されたヒトκ軽鎖Ig遺伝子座。
(項目13)以下の改変を含むゲノムを有するトランスジェニック非ヒト哺乳動物:
不活性化された内因性重鎖免疫グロブリン(Ig)遺伝子座;
不活性化された内因性κ軽鎖Ig遺伝子座;
ヒトγ2定常領域が存在しないyH2のヌクレオチド配列に実質的に対応するヌクレオチド配列を含む、挿入されたヒト重鎖Ig遺伝子座;および
yK2のヌクレオチド配列に実質的に対応するヌクレオチド配列を含む、挿入されたヒトκ軽鎖Ig遺伝子座。
(項目14)以下の改変を含むゲノムを有するトランスジェニック非ヒト哺乳動物:
不活性化された内因性重鎖免疫グロブリン(Ig)遺伝子座;
ヒトγ2定常領域が存在しないyH2のヌクレオチド配列に実質的に対応するヌクレオチド配列を含む、挿入されたヒト重鎖Ig遺伝子座;および
yK2のヌクレオチド配列に実質的に対応するヌクレオチド配列を含む、挿入されたヒトκ軽鎖Ig遺伝子座。
(項目15)以下の改変を含むゲノムを有するトランスジェニック非ヒト哺乳動物:
不活性化された内因性重鎖免疫グロブリン(Ig)遺伝子座;
ヒトγ2定常領域が存在しないyH2のヌクレオチド配列に実質的に対応するヌクレオチド配列を含む、挿入されたヒト重鎖Ig遺伝子座;および
yK2のヌクレオチド配列に実質的に対応するヌクレオチド配列を含む、挿入されたヒトκ軽鎖Ig遺伝子座。
(項目16)以下の段階を含む、ヒト抗体を製造するための方法:
項目1〜10のいずれか一項記載の哺乳動物への抗原の接種;
109M-1よりも大きい親和性で抗原と特異的に結合するヒト抗体を分泌する不死化細胞系を得るためのリンパ球細胞の回収および不死化;ならびに
不死化細胞系からの抗体の単離。
(項目17)抗原がIL-8である、項目11記載の方法。
(項目18)抗原がEGFRである、項目11記載の方法。
(項目19)抗原がTNF-αである、項目11記載の方法。
(項目20)項目11記載の方法によって製造される抗体。
(項目21)項目12記載の方法によって製造される抗IL-8抗体。
(項目22)項目13記載の方法によって製造される抗EGFR抗体。
(項目23)項目14記載の方法によって製造される抗TNF-α抗体。
(項目24)複数のヒト可変領域の挿入が含まれる以下の改変を含むゲノムを有するトランスジェニックマウスを製造するための方法:
酵母人工染色体からのヒト可変領域の挿入。
(項目25)項目19記載の改善の使用を通じて製造されたトランスジェニックマウスおよびそれらに由来するトランスジェニック子孫。
(項目26)挿入されたヒト重鎖免疫グロブリン導入遺伝子が含まれ、以下の改変を含むゲノムを有するトランスジェニック哺乳動物:
ヒト様の接合点多様性およびヒト様の相補性決定領域3(CDR3)の長さを可能とするヒト可変領域遺伝子の選択された組(set)を含む導入遺伝子。
(項目27)ヒト様の接合点多様性が平均7.7塩基のN付加長を含む、項目26記載の改変。
(項目28)ヒト様CDR3の長さが約2から約25残基の間、平均すると約14残基を含む、項目26記載の改変。
【図面の簡単な説明】
【0027】
【図1】図1は、本発明に従って好ましいマウスに導入される、再構成されたヒト重鎖およびヒトκ軽鎖遺伝子座YACの模式図である。ヒト重鎖(1H、2H、3Hおよび4H)およびヒトκ軽鎖の近位(1K、2Kおよび3K)遺伝子座の全長にわたるYACをヒトYACライブラリーからクローニングした。ヒトIg遺伝子座に関して異なるYACの位置(CookおよびTomlinson、1995ならびにCoxら、1994から採用)、それらのサイズ、および非Ig配列を示す(一定の比率では示していない)。ヒト重鎖およびκ軽鎖YACを再構築するために、YACを2段階の手順(材料および方法の項を参照)にて酵母中に組換え導入した。ヒト重鎖を含むYACであるyH2にさらにヒトγ2遺伝子の配列によるレトロフィット(retrofit)を行った。yK2はヒトκ軽鎖を含むYACである。YACベクターの構成要素:YACベクターの腕上のテロメア:黒ぬり△、動原体●、哺乳動物(HPRT、Neo)および酵母の選択可能マーカー(TRP1、ADE2、LYS2、LEU2、URA3、HIS3)を示す。VHセグメントはオープンリーディングフレーム●、偽遺伝子□、および配列決定がなされていない領域○を有する遺伝子として分類した。Vκ部分はオープンリーディングフレーム●および偽遺伝子□を有する遺伝子として分類した。XenoMouse IIによって利用されることを本発明者らが見いだしたV遺伝子には印(*)を付した。yH2上に含まれるVH遺伝子領域は矢印で示した。
【図2】図2は、ES細胞およびXenoMouse系統に組み込まれたヒト重鎖YAC、yH2の一連のサザンブロット分析および特徴分析の結果である。図2aは、ワシントン(Washington)大学YACライブラリー供給源(Brownsteinら、1989)に由来するCGM1不死化Bリンパ芽球細胞系、yH2 YAC(0.5μgのYACを2μgの3B1 DNAに加えたもの)、非改変E14TG. 3B1(3B1)ならびにyH2含有ES細胞系:L10、J9.2、L18、L17およびJ17から調製したEcoRI(a、c)およびBamHI(b,d、e)消化DNA(2μg)の一連のサザンブロット分析の結果である。ブロッティングに用いたプローブは、ヒトVH1(a)、DH(b)[CGM1レーン中の18kb断片は第16染色体上のDセグメントである]、VH3(c)、Cμ(d)およびJH(e)である。図2bは、野生型(WT、129×B57BL/6J)、XM2A-1およびXM2A-2(2匹の独立した子孫)マウスの尾部から、または親yH2を含むES細胞系L10(他の試料と比べてややロード量が少ない)、J9.2、およびyK2を含むES細胞系J23.1から調製したEcoRI(a-b)およびBamHI(c-d)消化DNA(10μg)の一連のサザンブロット分析の結果である。用いたプローブはヒトVH1(a)、VH4(b)、ヒトγ-2(c)およびマウス3'-エンハンサー(d、5kbのバンドは内因性マウス3'-エンハンサー断片である)である。分子量マーカーの断片サイズ(kb単位)を示している。
【図3】図3は、ES細胞およびXenoMouse 2A系統に組み込まれたヒトκ軽鎖YAC、yK2の特徴を示す一連のサザンブロット分析の結果である。図3aは、CGM1細胞系(Brownsteinら、1989、前記)、yK2 YAC(0.5μgのYACを2μgの3B1 DNAに加えたもの)、非改変E14TG. 3B1(3B1)ならびにyK2含有ES細胞系:J23.1およびJ23.7から調製したEcoRI(a、c、d)およびBamHI(b、e)消化DNA(2μg)の一連のサザンブロット分析の結果である。ブロッティングに用いたプローブは、ヒトVa(a)、Kde(b)、VκII(c)、VκIII(d)およびCκ(e)である。図3bは、野生型(WT、129×B6)、XM2A-1およびXM2A-2(2匹の独立した子孫)マウスの尾部から、または親yH2を含むES細胞系L10(他の試料と比べてややロード量が少ない)、J9.2、およびyK2を含むES細胞系J23.1から調製したEcoRI消化DNA(2μg)の一連のサザンブロット分析の結果である。用いたプローブはヒトVκI(a)、VκIV(b)、VκVI(c)および3'-エンハンサー(d)である。分子量マーカーの断片サイズ(kb単位)を示している。
【図4】図4は、B細胞の再構成ならびにXenoMouse由来B細胞でのヒトμ、δおよびκ鎖の表面発現を示し、野生型マウス(WT)、二重不活性化マウス(DI)ならびにXenoMouse系統2A-1および2A-2(XM2A-1、XM2A-2)からの末梢血(図4a)および脾臓(図4b)リンパ球のフローサイトメトリー分析の結果を示す。B細胞特異的マーカーB220に対する抗体を抗ヒトμ、δ、κまたはマウスμ、δ、κもしくはλとともに用いて4色フローサイトメトリー分析を行った。4象限のそれぞれにおける陽性染色された細胞の比率を示す。細胞の単離および染色は、材料および方法の項に記載した通りに実施した。B220+μ+集団に対して指示領域でのゲーティングをまず行った後に、ヒトκ+およびマウスλ+細胞の集団を決定した。μ+およびδ+細胞の集団は、B220+集団に対してまずゲーティングをまず行った後に決定した。ある領域または4象限の内部にある陽性細胞の比率を示す。示したFACSプロフィールは、各系統に対して実施したいくつかの実験の代表例である。
【図5】図5は、XenoMouse由来のヒト抗体が、それらの特異的抗原の細胞との結合を阻止することを示す。図5aは、マウス抗ヒトIL-8抗体(R&D Systems)(□)ならびに完全ヒトMab D1.1(◆)、K2.2(●)、K4.2(黒ぬり△)およびK4.3(黒ぬり▽)による、標識[I125]IL-8のヒト好中球との結合の阻害を示す。抗体の非存在下における標識[I125]IL-8のバックグラウンド結合は2657cpmであった。図5bは、マウス抗ヒトEGFR抗体225および528(それぞれ□、▽;Calbiochem)ならびに完全ヒト抗体E1.1(●)、E2.4(黒ぬり△)、E2.5(黒ぬり▽)およびE2.11(◆)による、標識[I125]EGFのA431細胞上のその受容体との結合の阻害を示す。抗体の非存在下における[I125]EGFのバックグラウンド結合は1060cpmであった。図5cは、マウス抗ヒトTNF-α抗体(R&D Systems)(□)ならびに完全ヒトMab T22.1(◆)、T22.4(●)、T22.8(黒ぬり△)およびT22.9(黒ぬり□)による、標識[I125]TNF-αのU937細胞上のその受容体との結合の阻害を示す。抗体の非存在下における標識[I125]TNF-αのバックグラウンド結合は4010cpmであった。対照ヒトIgG2骨髄腫抗体(四角で囲まれた×印)。
【図6】図6は、XenoMouse由来の完全ヒト抗体におけるレパートリーおよび体細胞超突然変異を示す。4つの抗IL-8(図6a)および4つの抗EGFR(図6b)ヒトIgG2κ Mabの予想されるアミノ酸配列を、CDR1、CDR2およびCDR3ならびに定常領域Cγ2およびCκに分けた。各抗体のDおよびJ遺伝子を示す。生殖細胞系配列からのアミノ酸置換は太字で示している。
【図7】図7は、ヒト重鎖ゲノムおよびヒトκ軽鎖ゲノムの模式図である。
【図8】図8は、yH2(ヒト重鎖)YACの構成を示すもう1つの模式図である。
【図9】図9は、yK2(ヒトκ軽鎖)YACの構成を示すもう1つの模式図である。
【図10】図10は、yK2(ヒトκ軽鎖)YACの構成を示すもう1つの模式図である。
【図11】図11は、ES細胞内およびマウスゲノム中へのyH2(ヒト重鎖)YACの組込みが無傷でなされたことを示す一連のサザンブロット分析の結果である。詳細な説明は図2に関して提示している。
【図12】図12は、ES細胞内およびマウスゲノム中へのyK2(ヒトκ軽鎖)YACの組込みが無傷になされたことを示す一連のサザンブロット分析の結果である。詳細な説明は図3に関して提示している。
【図13】図13は、B細胞の再構成、ならびにXenoMouse由来B細胞でのヒトμ、δおよびκ鎖ならびにマウスλ鎖の表面発現を示し、末梢血のフローサイトメトリー分析を示す。詳細は図4に関して提示している。
【図14】図14は、野生型マウスによるマウス抗体産生と比較して、XenoMouse IIによるヒト抗体の産生レベルを示している。
【図15】図15は、XenoMouse II系統で発現されたヒト重鎖転写物のレパートリー解析の結果である。
【図16】図16は、XenoMouse II系統で発現されたヒトκ軽鎖転写物のレパートリー解析の結果である。
【図17】図17は、XenoMouse II系統において利用されることが観察されたヒトVHおよびVκ遺伝子が広範に利用されることに関するもう1つの描写である。
【図18】図18は、XenoMouse II系統におけるヒト抗体産生の力価を示している。
【図19】図19は、XenoMouse II系統に由来する抗IL-8抗体の遺伝子利用を描写している。
【図20】図20は、XenoMouse II系統に由来する抗IL-8抗体の重鎖アミノ酸配列を示す。
【図21】図21は、XenoMouse II系統に由来する抗IL-8抗体のκ軽鎖アミノ酸配列を示す。
【図22】図22は、XenoMouse II系統に由来するヒト抗IL-8抗体による、IL-8のヒト好中球との結合の阻止を示す。
【図23】図23は、XenoMouse II系統に由来するヒト抗IL-8抗体による、ヒト好中球上でのCD11b発現の阻害を示す。
【図24】図24は、XenoMouse II系統に由来するヒト抗IL-8抗体による、IL-8によって誘導されるカルシウム流入の阻害を示す。
【図25】図25は、XenoMouse II系統に由来するヒト抗IL-8抗体による、IL-8 RB/293走化性の阻害を示す。
【図26】図26は、ヒトIL-8誘導性皮膚炎症のウサギモデルの模式図である。
【図27】図27は、XenoMouse II系統に由来するヒト抗IL-8抗体による、図26のウサギモデルにおけるヒトIL-8誘導性皮膚炎症の阻害を示している。
【図28】図28は、XenoMouse II系統由来のヒト抗IL-8抗体による、ラット角膜ポケットモデルでの内皮細胞の血管新生の阻害を示す。
【図29】図29は、XenoMouse II系統に由来するヒト抗EGFR抗体の遺伝子利用の描写である。
【図30】図30は、XenoMouse II系統に由来するヒト抗EGFR抗体の重鎖アミノ酸配列を示している。
【図31】図31は、XenoMouse II系統に由来するヒト抗EGFR抗体による、EGFのA431細胞との結合の阻止を示している。
【図32】図32は、XenoMouse II系統に由来するヒト抗EGFR抗体による、EGFのSW948細胞との結合の阻止を示している。
【図33】図33は、XenoMouse II系統に由来するヒト抗EGFR抗体がSW948細胞のインビトロ増殖を阻害することを示す。
【図34】図34は、XenoMouse II系統に由来するヒト抗TNF-α抗体の使用による、TNF-αのU937細胞との結合の阻害を示している。
【図35】図35は、XenoMouse II系統に由来するヒト抗EGFR抗体のκ軽鎖アミノ酸配列を示している。
【発明を実施するための形態】
【0028】
[好ましい態様の詳細な説明]
本明細書において本発明者らは、実質的な生殖細胞系コンフィギュレーションにある、メガベースサイズのヒトIg遺伝子座を含むマウスの、いくつかの系統の製造および特徴分析に関して記載する。このため本発明は、YAC上での大きく複雑なヒトIg遺伝子座の再構成、および対応するマウスIg遺伝子座を機能的に置換するためのメガベースサイズのYACをマウスに首尾よく導入する方法を、初めて提供するものである。
【0029】
マウス系統
以下のマウス系統を、本明細書において記載および/または使用する。
二重不活性化(DI)系統: DI系統のマウスは、機能的な内因性マウスIgを産生しないマウスである。1つの好ましい態様において、DIマウスは不活性化されたマウスJH領域および不活性化されたマウスCκ領域を有する。この系統の構築は、別で詳細に説明されている。例えば、DI系統の作製のために用いられる技法は、1990年1月12日に提出された米国特許出願第07/466,008号、1990年11月8日に提出された第07/610,515号、1992年7月24日に提出された第07/919,297号、1993年3月15日に提出された第08/031,801号、1993年8月27日に提出された第08/112,848号、1994年4月28日に提出された第08/234,145号、1996年10月2日に提出された第08/724,752号に詳細に記載されている。1996年6月12日に認可が発行されたEP 0 463 151 B1、1994年2月3日に発行された国際特許出願国際公開公報第94/02602号、1996年10月31日に発行された国際特許出願国際公開公報第96/34096号、および1996年4月29日に提出されたPCT/US96/05928も参照されたい。上記の特許および特許出願のそれぞれの開示は、その全体が参照として本明細書に組み入れられる。DIマウスではB細胞発生が極めて未熟であることが観察および報告されている。本マウスは成熟B細胞を産生せず、プロB細胞のみを産生する。
【0030】
XenoMouse I系統: XenoMouse I系統の設計、構築および解析はグリーン(Green)ら、Nature Genetics 7:13〜21(1994)に詳細に考察されている。このようなマウスはDIバックグラウンドに対するIgMκ抗体を産生する。本マウスでは、ほとんどまたは全くB細胞発生が生じないDI系統のマウスと比べてB細胞機能の改善が認められた。XenoMouse I系統のマウスは抗原誘発刺激に対してかなり強い免疫応答を生じる能力があったが、B細胞の産生効率は低いと考えられ、異なる抗原に対する応答は制限されており、これはV遺伝子レパートリーの制限に起因するものと考えられた。
【0031】
L6系統: L6系統は、DIバックグラウンドの、内因性マウスIgに対するIgMκ抗体を産生するマウスである。L6マウスは、挿入されたヒト重鎖および挿入されたヒトκ軽鎖を含んでいる。L6系統は、二重不活性化バックグラウンドに対する重鎖挿入物を含むマウス(L6H)と、二重不活性化バックグラウンドに対するκ軽鎖挿入物を含むマウス(L6L)との交配によって作出される。重鎖挿入物は、VH6-1に始まってVH3-65に終わる約66個のVHセグメント、ならびに生殖細胞系コンフィギュレーションにある多数のD遺伝子クラスター(約32個)、JH遺伝子(6個)、イントロン性エンハンサー(Em)、CμおよびCδを越えて約25kbのところまでを含むYACに由来する、無傷の約970kbのヒトDNA挿入物を含む。軽鎖挿入物は、Vκ-B3に始まってVκ-Op11に終わる約32個のVκ遺伝子を含むYACに由来する、無傷の約800kbのヒトDNA挿入物を含む。この800kb挿入物は、Vκ-Lp-13に始まってVκ-Lp-5で終わる約100kbの欠失を含む。しかし、本DNAはVκ-Lp-13からVκ-Op-1を越えて100kbのところまで生殖細胞系コンフィギュレーションにあり、Jκ遺伝子、イントロン性の3'エンハンサー、定常Cκ遺伝子、およびKdeを含む。L6HおよびL6Lマウスは、それらのゲノム中に組み込まれた可変遺伝子の完全なスペクトルを利用しうることが示されている。L6マウスも同様に、それらのゲノム中の可変遺伝子の完全なスペクトルを利用しうると考えられる。さらにL6マウスはヒトκ軽鎖の主な発現、多数の成熟B細胞、および正常レベルのIgMκヒト抗体を呈すると考えられる。このようなマウスは多数の免疫原に対する強いヒト抗体反応を生じ、最終的にはナノモルレベル以下の親和性をもつ抗原特異的な完全ヒトMabを産生すると考えられる。
【0032】
XenoMouse IIa系統: XenoMouse IIaマウスは、マウスが機能的な内因性Igを産生しないようなDIバックグラウンドに対して生殖細胞系コンフィギュレーションでメガベースサイズのヒトIg遺伝子座を備えた、本発明者らの第2世代のXenoMouse系統(登録商標)である。本質的には本マウスの構成はL6系統と等価であるが、完全なスイッチおよび調節配列ならびにシス位にあるマウス3'エンハンサーとともにヒトγ2遺伝子をさらに含む。本マウスは、YAC上に再構成された形で約1020kbの重鎖遺伝子座および約800kbのκ軽鎖遺伝子座を含み、これには重鎖遺伝子(約66個のVH)およびκ軽鎖遺伝子(約32個のVκ)、ヒト重鎖定常領域遺伝子(μ、δおよびγ)およびκ定常領域遺伝子(Cκ)ならびに同定されたすべての主要な調節要素を含む、大半のヒト可変領域遺伝子が含まれる。これらのマウスは、それらのゲノム中に組み入れられた可変領域の完全なスペクトルを利用しうることが示されている。さらに、それらは効率的なクラス転換(class switching)および体細胞超突然変異、ヒトκ軽鎖の優位な発現、多数の成熟B細胞、ならびに正常レベルのIgMκおよびIgGκヒト抗体を呈する。このようなマウスは、ヒトIL-8、ヒトEGF受容体(EGFR)およびヒト腫瘍壊死因子-α(TNF-α)を含む多数の免疫原に対する強いヒト抗体反応を生じ、最終的にはナノモルレベルに準じる親和性をもつ抗原特異的な完全ヒトMabを産生する。この最終結果は、広範なスペクトルの抗原に対して任意の望ましい特異性をもつ高親和性の、完全ヒト治療的Mabの迅速単離のための優れた供給源としてのXenoMouse(登録商標)を決定的に示すものである。
【0033】
上記の序論から理解される通り、XenoMouse II系統は成熟B細胞の発生を経て、抗原誘発刺激に対して強力な成人様免疫応答を起こすと考えられる。L6系統も、L6LおよびL6Hマウスに関するデータから予想される通り、成熟B細胞の発生を経て、抗原誘発刺激に対して強力な成人様免疫応答を起こすと考えられる。DIマウスをXenoMouse I系統と比較し、DIおよびXenoMouse I系統をL6およびXenoMouse II系統と比較すると、顕著に異なるB細胞発生の形態が観察される。この差からみて、誘導B細胞の成熟および発生ならびに成人様免疫応答の発生には、マウスに導入される可変領域配列の量および/または質が非常に重要であると考えられる。したがって、ヒト抗体の作製における、以上の系統の明らかな用途に加えて、以上の系統は、正常免疫応答におけるヒト抗体の性質、さらには自己免疫疾患および他の障害に特徴的な異常応答を研究するための有益な手段も提供する。
【0034】
可変領域―量的多様性
抗体の特異性(すなわち、広範な抗原に対する、実際にはそれらの上にある広範な独立したエピトープに対する抗体を生成する能力)は、重鎖(VH)およびκ軽鎖(Vκ)ゲノム上の可変領域遺伝子に依存している。ヒト重鎖ゲノムは、免疫グロブリン分子のヒト重鎖の可変領域をコードする約95の機能的遺伝子を含む。さらに、ヒト軽鎖ゲノムはその近位端に、免疫グロブリン分子のヒトκ軽鎖の可変領域をコードする約40の遺伝子を含む。本発明者らは、可変軽鎖および重鎖をコードする複数の遺伝子を含めることによって抗体の特異性を高めうることを示した。
【0035】
本発明によって提供されるものは、好ましくはヒト重鎖およびヒトκ軽鎖遺伝子座を含むヒトIg遺伝子座のかなりの部分を有するトランスジェニックマウスである。このため、好ましい態様では、ヒトVHおよびVκ遺伝子の10%よりも多くが用いられる。より好ましくは、ヒトVHおよびVκ遺伝子の約20%、30%、40%、50%、60%、またはさらには70%もしくはそれよりも多くが用いられる。1つの好ましい態様では、Vκ軽鎖ゲノムの近位領域に32個の遺伝子を含む構築物が用いられ、ゲノムのVH部分にある66個の遺伝子が用いられる。理解されるであろう通り、遺伝子は連続的、すなわちヒトゲノム中で認められる順番に含まれても、異なる順序、すなわちヒトゲノム中で認められる以外の順序で含まれても、それらの組み合わせであってもよい。このため、一例として、VHもしくはVκゲノムのいずれかの完全に連続的な部分を利用することができ、またはVHもしくはVκゲノムのいずれかの種々のV遺伝子を省いて全体の連続的配置を維持することもでき、またはVHもしくはVκゲノムのいずれかの内部のV遺伝子の順序を改めることなどもできる。1つの好ましい態様において、挿入される全遺伝子座はヒトにおいて認められる通りの実質的な生殖細胞系コンフィギュレーションとして提供される。いずれの場合にも、VHおよびVκゲノムからの多様な一連の遺伝子を含めることにより、抗体特異性の増強および最終的には抗体親和性の増強に至ることが予想され、本明細書に記載される結果で示される。
【0036】
さらに、このようなマウスは、好ましくは全DH領域、全JH領域、ヒトμ定常領域を含み、さらに別のアイソタイプの抗体のコーディングおよび生成のための他のヒト定常領域を付加的に備えることができる。このようなアイソタイプには、適切なスイッチおよび調節配列を備えたγ1、γ2、γ3、γ4、α、εおよびδ、ならびに他の定常領域をコードする遺伝子が含まれうる。理解されるであろう通り、さらに以下でより詳しく考察される通り、任意の特定の定常領域の選択に関連して、種々のスイッチおよび調節配列を適切に用いることができる。
【0037】
以下の表は、厳密にV-D-J結合およびκ軽鎖とのランダムな組み合わせに基づき、N付加または体細胞変異事象を考慮しない場合に、ヒトにおいて可能な抗体の組み合わせの多様性を示している。このような検討に基づくと、ヒトでは、任意の特定のアイソタイプに380万種を超える抗体の組み合わせが存在する。
(表I)
【表1】
【0038】
本発明の1つの好ましい態様に関して、DH、JHおよびJκ遺伝子の完全な補体を備えた約66個のVH遺伝子および32個のVκ遺伝子をマウスに含めることにより、抗体産生に関して可能な多様性は、2.03×106種の異なる抗体のオーダーである。上記の通り、この種の計算にはN付加および体細胞変異事象を考慮していない。このため、L6およびXenoMouse II系統などの本発明に係るマウスは、かなりの抗体多様性をもたらすことが理解されると思われる。好ましい態様において、本マウスはN付加および体細胞変異事象を考慮に入れずとも、1×106種を超える異なる重鎖V-D-J組み合わせおよびκ軽鎖V-J組み合わせを産生する能力をもつように設計される。
【0039】
可変領域―質的多様性
量的多様性に加えて、V遺伝子の量的選択(すなわち、多くの多様な数のV遺伝子)および/またはV遺伝子の質的選択(すなわち、特定のV遺伝子の選択)も、本発明者らが本明細書で「質的多様性」と呼ぶ一定の役割を果たすように思われる。本明細書で用いる質的多様性とは、結合多様性および/または体細胞変異事象が導入されるV-D-J再配列における多様性を意味する。重鎖再配列の間には、特定の酵素(RAG-1、RAG-2、および他のものも考えられる)が、抗体遺伝子のコード領域であるDNAの切断に関与する。ターミナルデオキシヌクレオチジルトランスフェラーゼ(Tdt)活性はアップレギュレートされ、これはV-DおよびD-J遺伝子セグメントの間のヌクレオチドのN末端付加の原因となる。類似の酵素および他のもの(SCIDおよび他のDNA修復酵素)は、これらのコード区域の結合部に生じる欠失の原因となる。結合多様性に関しては、N付加事象および相補性決定領域3(CDR3)の双方がこの項目に含められる。理解されるであろう通り、CDR3はD領域にまたがって位置しており、V-DおよびD-J結合事象を含む。このため、D-J再配列およびV-D再配列の両方の過程におけるN付加および欠失がCDR3の多様性の原因となっている。
【0040】
マウスおよびヒトの結合多様性の間には、ある特定の差異があることが示されている。特に、一部の研究者は、マウスのN付加長およびCDR3長は典型的なヒトのN付加長およびCDR3長よりも一般に短いことを報告している。このようなグループは、ヒトで典型的に認められるN付加長は平均約7.7塩基であることを報告している。ヤマダ(Yamada)ら(1991)。マウス様N付加の長さは平均で約3塩基のオーダーであることが多い。フィーニー(Feeney)ら(1990)。同様に、ヒト様CDR3の長さはマウス様CDR3よりも長い。ヒトではCDR3長は2から25残基の間であり、平均14残基であることが一般的である。マウスでは平均CDR3長がこれよりも短いことをいくつかのグループが報告している。
【0041】
N付加およびCDR3付加によって生じる結合多様性は、抗体特異性を発展させるという明らかな役割を果たしている。
【0042】
本発明によれば、再配列したV-D-J遺伝子配列は、予想される成人N付加長に匹敵するN付加長を示す。さらに、CDR3配列に対応するオープンリーディングフレーム(ORF)にまたがるアミノ酸配列は、予想される成人CDR3長と同等のCDR3長を示す。このようなデータは、可変領域の量的多様性および/または可変領域の質的多様性によりヒト様の結合多様性がもたらされることを示すものである。このような結合多様性はよりヒトに類似した抗体特異性につながると考えられる。
【0043】
可変領域―親和性
本発明者らは、可変領域の含有量の増加と抗体特異性との間に、直接的な因果関係があることを決定的に示してはいないが、そうであると思われ、しかもこのような多様性が提供されることにより、マウスが広範な抗原に対する免疫応答を生じる能力が実現および増強されることが期待される。さらに、このようなマウスは、個々の抗原または免疫原上の広範なエピトープに対する免疫応答を生じる性質をさらに備えるようになると考えられる。本発明者らのデータからは、本発明に従って産生される抗体は親和性も増強されると思われる。このようなデータには、本発明によるマウスおよびXenoMouse I系統との比較のほか、ジェンファームインターナショナル(GenPharm International)社およびMRCが発表した結果の考察が含まれる。XenoMouse I系統については、上記の通り、このようなマウスではB細胞産生の効率が低く、種々の抗原に対する応答性が制限される。このような結果は、一部にはV遺伝子レパートリーの制限に起因すると思われる。同じく、ジェンファームインターナショナル社およびMRCによって発表された結果は、多様な抗原に対する応答性が制限されていることを示している。
【0044】
発明の特定の理論または作業様式に拘束されることを望むものではないが、親和性の増強は多数のV領域が供給されることによる結果であると考えられる。本発明者らのデータからは、より多くの数の供給および/またはV遺伝子配列の質の選択により、結合多様性(N付加および相補性決定領域3(「CDR3」)の多様性の形成)は増強されるが、それは成人様免疫応答に典型的であり、抗体の親和性成熟において重要な役割を果たす。このような抗体は、親和性の増強につながる体細胞変異事象においてより効果的および効率的であることも考えられる。結合多様性および体細胞変異事象のそれぞれについては、以下でさらに詳しく考察する。
【0045】
親和性に関して、複数のVHおよびVκ遺伝子の利用(すなわち、Vκ軽鎖ゲノムの近位領域にある32個の遺伝子およびゲノムのVH部分にある66個の遺伝子の使用)に由来する抗体親和性の速度および定数は、約0.50×10-6よりも大きな、好ましくは2.00×10-6よりも大きな、さらに好ましくは約4.00×10-6よりも大きな会合速度(ka、M-1S-1単位);約1.00×10-4よりも大きな、好ましくは約2.00×10-4よりも大きな、さらに好ましくは約4.00×10-4よりも大きな解離速度(kd、S-1単位);および約1.00×10-10よりも大きな、好ましくは約2.00×10-10よりも大きな、さらに好ましくは約4.00×10-10よりも大きな解離定数(M単位)をもたらす。
【0046】
このようなマウスは、その上で機能的な内因性免疫グロブリンを産生しないことが好ましい。これは、1つの好ましい態様において、内因性の重鎖および軽鎖遺伝子座の不活性化(またはノックアウト)を通じて達成される。例えば、1つの好ましい態様では、この領域を置換または欠失させる相同組換えベクターの利用により、マウス重鎖J領域ならびにマウスκ軽鎖J領域およびCκ領域が不活性化される。
【0047】
可変領域―B細胞発生
B細胞の発生はクラウス(Klaus)の、「Bリンパ球(B Lymphocytes)」(IRL Press(1990))および、「免疫グロブリン遺伝子(Immunoglobulin Genes)」(Academic Press Ltd.(1989)の1〜3章において概説されており、それらの開示は参照として本明細書に組み入れられる。一般に哺乳動物では、Bリンパ球およびTリンパ球を含む血液細胞の発生は共通の多能性幹細胞に始まる。続いてリンパ球が共通のリンパ球前駆細胞から生じる。初期の妊娠期間を経た後にB細胞の発生部位は肝臓から骨髄に移動し、哺乳動物の一生を通じてその部位に存続する。
【0048】
B細胞のライフサイクルにおいて一般に最初に認識可能な細胞は、骨髄中に認められるプロプレB細胞である。このような細胞は重鎖V-D-J再配列を開始するが、タンパク質を作ることはまだできない。続いてこの細胞は、細胞質的にはμ+細胞である、大型で急速に分裂するプレB細胞Iとなる。このプレB細胞は次いで分裂を停止し、縮小し、軽鎖V-J再配列を行って、表面IgMを発現するプレB細胞IIとなり、未熟B細胞として骨髄から遊離する。新生未熟B細胞の大部分は発生を続け、表面IgDを産生するようになるが、これは完全に成熟した免疫適格末梢B細胞としての分化および発生が完了したことを意味し、これらは主に脾臓内に存在する。しかし、δ定常領域を除去して依然として免疫適格細胞を得ることが可能である。
【0049】
B細胞の分化および発生は、表面マーカーを用いて観測および/または追跡することができる。例えば、B220抗原はプレB細胞IまたはIIと比べると成熟B細胞でより豊富に発現される。したがって、B220+であって表面IgM+(μ+)である細胞を用いて成熟B細胞の存在を判定することができる。さらに、表面IgDの発現(δ+)に関して細胞をスクリーニングすることもできる。プレB細胞IIが末梢に移行するに従って(すなわち、それらがμ+および/またはμ+、δ+になるに従って)、別の抗原である耐熱抗原がそれらによって発現される。
(表II)
【表2】
【0050】
上記のようなB細胞マーカーの使用により、B細胞の発生および分化の観測および評価が可能である。
【0051】
本発明者らは以前に、DIマウス(重鎖V-D-J再配列または軽鎖V-J再配列を起こさないマウス)が成熟B細胞を産生しないことを示した。事実、このようなマウスではプロプレB細胞の産生のところで停止しており、B細胞は骨髄から脾臓を含む末梢組織には決して移動しない。このため、B細胞の発生および抗体の産生はいずれも完全に停止する。重鎖のみが不活性化されたマウスでも同じ結果が認められており、骨髄中のB細胞の発生および分化は停止する。
【0052】
本発明者らのXenoMouse I系統は、機能的で幾分成熟したB細胞を産生した。しかし、B細胞の数は、骨髄および末梢組織のいずれにおいても野生型マウスと対比すると有意に少なかった。
【0053】
これに対して、本発明者らのXenoMouse II系統およびL6系統では予想外なことにほぼ完全なB細胞の再構築が認められた。したがって、本発明により本発明者らは、可変領域遺伝子の量的含有および質的含有によってB細胞の分化および発生をかなり再構成しうることを示した。B細胞の分化および発生の再構成は、免疫系が再構成されたことを示す。一般にB細胞の再構成は、野生型対照と比較される。この点で、本発明の好ましい態様において、挿入されたヒト可変領域を有するマウスの集団は、野生型マウスの集団と比較すると約50%を超えるB細胞機能を有する。
【0054】
さらに興味深いことに、マウス抗体からみたヒト抗体の優先的な産生は、内因性Igがノックアウトされたバックグラウンドをもつマウスではかなり増強された。すなわち、ヒトIg遺伝子座および機能的に不活性化された内因性重鎖Ig遺伝子座を含むマウスでは、ヒトIg遺伝子座のみを含んでいて内因性遺伝子座が不活性化されていないマウスと比べて、ヒト抗体の産生効率が速度にして約100倍から1000倍も効率的であった。
【0055】
アイソタイプスイッチ
本明細書において詳細に考察されるように、予想通り、XenoMouse IIマウスは、ヒト導入遺伝子にコードされるμアイソタイプから導入遺伝子にコードされるγ2アイソタイプへの効率的および効果的なアイソタイプスイッチを生じた。本発明者らは、ヒトγ4定常領域を包含およびコードするXenoMouse II系統も開発した。上記の通り、本発明に係るマウスは、ほかのアイソタイプを生成するための他のヒト定常領域をさらに備えることができる。このようなアイソタイプには、γ1、γ2、γ3、γ4、α、ε、δおよび他の定常領域をコードする遺伝子が含まれうる。代替的な定常領域は、同一の導入遺伝子上、すなわちヒトμ定常領域から下流に含めることもでき、またはこのような他の定常領域を別の染色体上に含めることもできる。このような他の定常領域がヒトμ定常領域をコードする導入遺伝子を含む染色体と同じ染色体上に含まれる場合には、他の1つのアイソタイプまたは複数のアイソタイプへのシス転換が達成されうることは理解されるであろう。これに対して、このような他の定常領域がヒトμ定常領域をコードする導入遺伝子を含む染色体とは異なる染色体上に含まれる場合には、他の1つのアイソタイプまたは複数のアイソタイプへのトランス転換が達成されうる。このような配置(arrangement)は、広範な抗原に対する抗体を生成するためのマウスの設計および作製において多大な柔軟性を可能とする。
【0056】
定常領域が、それらと関連した既知のスイッチおよび調節配列を有することは理解されると思われる。マウスおよびヒト定常領域遺伝子はすべて、1989年までに配列が決定され、発表されている。「免疫グロブリン遺伝子」(Honjoら編、Academic Press(1989))中の、その開示が参照として本明細書に組み入れられるホンジョ(Honjo)ら、「免疫グロブリン重鎖の定常領域遺伝子およびクラススイッチの分子的機序」(Constant Region Genes of the Immunoglobulin Heavy Chain and the Molecular Mechamism of Class Switching)を参照のこと。例えば、その開示が参照として本明細書に組み入れられる米国特許出願第07/574,748号では、先行技術からの既知の配列情報に基づいて、ヒトγ1定常領域のクローニングが予測されている。そこでは、再配列がなされずスイッチもなされていない遺伝子では、第1のγ1定常部エキソンの5'末端から5kbよりも近いところから始まる配列中に、全スイッチ領域が含まれることが示されている。このため、スイッチ領域は、エリソン(Ellison)ら[Nucleic Acids Res 10:4071〜4079(1982)]において開示された5'部の5.3kbのHindIII断片にも含まれる。同様に、タカハシ(Takahashi)ら[Cell 29:671〜679(1982)]も、エリソンにおいて開示された断片がスイッチ配列を含むこと、およびこの断片に7.7kbのHindIIIからBamHIまでの断片を加えたものに、重鎖アイソタイプスイッチ用の導入遺伝子の作製に必要なすべての配列が含まれるに違いないと考えられることを報告している。
【0057】
このため、過度の実験を行わずとも、選択される任意のヒト定常領域を容易に本発明に係るマウスに組み入れられることは理解されると思われる。このような定常領域は、それらの本来のスイッチ配列(すなわち、それぞれヒトγ1、2、3、4スイッチを備えたヒトγ1、2、3、4定常領域)と結合させることができ、または他のスイッチ配列(すなわち、ヒトγ2スイッチを備えたヒトγ4定常領域)と結合させることもできる。少し例を挙げるなら、マウス、ヒトまたはラットなどの種々の3'エンハンサー配列を用いることもできる。同様に他の調節配列を含めることもできる。
【0058】
インビボでのアイソタイプスイッチに対する代替および/または付加として、「キメラ」抗体の分泌に関してB細胞をスクリーニングすることもできる。例えば、L6マウスは、完全ヒトIgM抗体を産生することに加えて、種々のγ(すなわちマウスIgG1、2、3、4)などのマウス定常領域と結合した完全ヒト重鎖V、D、J領域を有する抗体を産生する。このような抗体はそれ自体で非常に有用である。例えば、当技術分野でよく知られたインビトロでのアイソタイプスイッチ法により、抗体上にヒト定常領域を含めることができる。または、および/もしくはこれに加えて、マウス定常領域をほとんどまたは全く含まないこのような抗体の断片(すなわちF(ab)およびF(ab')2断片)を調製することもできる。
【0059】
上記の通り、抗体産生に関して最も重要な因子は、望ましい抗原または抗原上のエピトープに対する特異性である。その後には、治療的必要性から抗体のクラスが重要になる。換言すれば、特定のアイソタイプまたはクラスを提供することによって、抗体の治療的指標は高まるのであろうか。この疑問の考察は補体結合などの問題を生み、それは次に特定のクラスまたはアイソタイプの抗体の選択を促す。γ定常領域は抗体の親和性の成熟を補助する。しかし、このような成熟を達成するために導入遺伝子上にヒトγ定常領域を含めることは必要でない。むしろ、この過程はμをコードする導入遺伝子上にトランス転換されるマウスγ定常領域との関連によってさらに進行すると思われる。
【0060】
材料および方法
以下の材料および方法は、本発明に係るマウスの製造および特徴に関連して用いられる。このような材料および方法は例示を意味しており、本発明の範囲を制限するものではない。
【0061】
ヒトIg由来のYACのクローニング: 以前にワシントン大学(the Washington University)(Brownsteinら、1989)およびCEPH(Abersonら、1990)のヒトYACライブラリーを、以前に記載された通りに(Mendezら、1995)ヒト重鎖およびκ軽鎖遺伝子座の配列を含むYACに関してスクリーニングした。1Hおよび1K YACのクローニングおよび特徴分析はメンデツ(Mendez)ら(1995)に記載されている。3Hおよび4H YACは、VH3プローブ(0.55kb PstI/NcoI、Bermanら、1988)を用いてワシントン大学のライブラリーから同定された。17H YACはGM1416ライブラリーから同定され、130kbの重鎖可変配列およびその3'末端に150kbのキメラ領域を含むことが明らかにされた。マツダ(Matsuda)ら、1993。2Kおよび3K YACはVκII特異的プライマーを用いてCHEFライブラリーから回収された(Albertsenら、1990)。
【0062】
YACのターゲティングおよび組換え: 酵母の増殖、交配、胞子形成および表現型の試験に関する標準的な方法を用いた(Shermanら、1986)。YACが細胞内に組み込まれた酵母内のYAC組換え体のスクリーニングを容易にするための、酵母および哺乳動物の選択可能マーカーによるYACおよびYACベクター腕のターゲティングは、酢酸リチウム形質転換法(ScheistlおよびGeitz(1989))によって行った。すべての配列の完全性を調べるために、個々のターゲティングまたは組換え段階の後にパルスフィールドゲル電気泳動法および標準的なサザンブロット法によって改変YACを分析した。
【0063】
17Hの再配向によって動原体腕および無動原体腕を相互転換させるため、ならびにその5'腕にLEU2およびURA3遺伝子の、その3'腕にHIS3遺伝子のレトロフィットを行うためにYACターゲティングベクターを用いた。図1aおよびメンデツ(Mendez)ら、1993を参照のこと。4H動原体腕には酵母ADE2遺伝子およびヒトHPRT選択可能マーカーによるレトロフィットを行った。1回目の組換え段階については、二倍体酵母株を作製し、17H、3Hおよび4Hという3種のYACがすべて存在し、無傷で安定的に維持されるものを選択した。YACが一部重複する領域の三者間相同組換え(three-way homologous recombination)を胞子形成によって誘導し、外部酵母選択可能マーカー(ADE2およびHIS3)の選択、および内部マーカーURA3の陰性選択(損失)によって望ましい組換え体を見いだした。好首尾な組換えにより、VH2-5に始まってVH3-65遺伝子の5'側に20kb伸長するIgH可変領域の80%を含む、880kbのYACが作製された。880kbのYACと1Hとの組換えには、1HをLYS2遺伝子を動原体腕に付加するpICLによるレトロフィットを行った(Hermansonら、1991)。標準的な酵母交配を用いて、1Hおよび880kbのYACの両方を含む二倍体株を選択した。胞子形成および重複相同性(overlapping homology)の使用により、YAC-酵母組換えを行った。外部酵母マーカー(ADE2およびURA3)の陽性選択および内部マーカー(TRP1、LYS2、HIS3)の損失に関するスクリーニングにより、VH6-1に始まってVH3-65で終わる約66個のVHセグメントを含む無傷の970kbのYACが見いだされた。このYACは生殖細胞系コンフィギュレーションにある主要なD遺伝子クラスター、JH遺伝子、イントロン性エンハンサー(Eμ)、Cμ、Cδを越えて25kbまでも含んでいた。続いてこの970kbのYACに、ヒトγ2遺伝子の23kbのEcoRIゲノム断片を、そのスイッチおよび調節要素、マウス重鎖3'エンハンサーの7kbのXbaI断片、メタロチオネインプロモーターの支配を受けるネオマイシン遺伝子(MMTNeo)、および酵母LYS2遺伝子を含めて、これらを含むターゲティングベクターによるレトロフィットを行った。このベクターは、3'YAC腕上にこれらの配列を運ぶ一方で、URA3遺伝子を破壊する。
【0064】
yK2 YACの作製に向けての第1段階として、標準的な酵母交配により、レトロフィットがなされた1Kおよび3K YACが両方とも存在し、無傷で安定的に維持される二倍体酵母株を選択した。IgH作製に関して記載したものと同じ過程を用いて、YAC-酵母組換えを行った。外部酵母マーカー(LYS2、TRP1)に関する陽性選択および内部マーカー(URA3、TRP1)の損失に関するスクリーニングにより、Vκ-B3に始まってVκ-Op11で終わる32個のVκを含む無傷の800kbの組換え産物を見いだした。この800kbのYACは、Vκ-Lp-13に始まってVκ-Lp-5で終わる約100kbの欠失を含む。しかし、このYACはVκ-Lp-13からVκ-Op-1を100kb越えたところまでは生殖細胞系コンフィギュレーションにある。また、このYACはJκ、イントロン性および3'エンハンサー、定常CκおよびKdeも含む。
【0065】
YACのES細胞およびマウスへの導入: YACを含む酵母スフェロプラストを、記載されている通り(Jakobovitsら、1993a;Greenら、1994)にE14.TG3B1 ES細胞と融合させた。HAT耐性コロニーを分析のために増殖させた。YACの完全性は、バーマン(Berman)ら(1988)およびメンデツ(Mendez)ら(1994)に記載された手順およびプローブ、ならびにジェミル(Gemmil)ら(1991)に記載された通りのハイブリダイゼーション条件を用いるサザンブロット分析によって評価した。キメラマウスは、C57BL/6胚盤胞へのES細胞のマイクロインジェクションによって作出した。記載されている通り(Greenら、1994)に、尾部DNAのPCR分析によってYACを含む子孫を同定した。YACの完全性は、ヒトVH3にて探索したブロットを50℃で洗うことを除き、前記のプローブおよび条件を用いるサザンブロット分析によって評価した。
【0066】
フローサイトメトリー分析: 8〜10週齢のXenoMouseおよび対照マウスから入手した末梢血および脾臓のリンパ球をLymphocyte M(Accurate)上で精製し、Fc受容体に対する非特異的な結合を阻止するために精製抗マウスCD32/CD16 Fc受容体(Pharmingen、01241D)で処理して、抗体で染色し、FACStarPLUS(Becton Dickinson、CELLQuestソフトウエア)によって分析した。用いた抗体は以下の通りである:アロフィコシアニン(APC)抗B220(Pharmingen、01129A);ビオチン抗ヒトIgM(Pharmingen、08072D);ビオチン抗マウスIgM(Pharmingen、02202D);フルオロセインイソチオシアネート(FITC)ヤギF(ab')2抗ヒトIgD(Southern Biotechnology、2032-02);FITC抗マウスIgDa(Pharmingen、05064D);FITC抗mIgDb(Pharmingen、05074D);FITC抗マウスλ(Pharmingen、02174D);PE抗ヒトκ(Pharmingen、08175A);PE抗マウスκ(Pharmingen、02155A)。ビオチン化抗体の検出にはRED613(登録商標)-ストレプトアビジン(GibcoBRL、19541-010)を用いた。
【0067】
免疫化およびハイブリドーマの作製: XenoMouse(8〜10週齢)に、一次免疫には完全フロイントアジュバント中に、2週間間隔で実施した追加免疫には不完全フロイントアジュバント中に、乳濁化した25μgの組換えヒトIL-8または5μgのTNF-α(Biosource International)の腹腔内投与による免疫化を施した。EGFR免疫化については、リン酸緩衝塩類溶液(PBS)中に再懸濁した2×107個のA431(ATCC CRL-7907)細胞の腹腔内投与による免疫化を施した。この量の投与を3回繰り返した。融合4日前に、マウスにPBS中にて抗原または細胞の最終注射を行った。免疫化マウスからの脾臓およびリンパ節のリンパ球を、非分泌型骨髄腫NSO-bcl2系(RayおよびDiamond、1994)と融合させ、以前に記載されている通り(GalfreおよびMilstein、1981)にHAT選択にかけた。
【0068】
ELISAアッセイ: マウス血清中およびハイブリドーマ上清中の抗原特異的抗体を測定するためのELISAは、抗体を捕捉するために組換えヒトIL-8およびTNF-αならびにA431細胞からアフィニティー精製されたEGFR(Sigma、E-3641)を用いて、記載されている通り(Coliganら、1994)に実施した。ヒトおよびマウスの免疫グロブリンの濃度は以下の捕捉抗体を用いて測定した:ヒトγ、κ、μIgについてはそれぞれウサギ抗ヒトIgG(Southern Biotechnology、6145-01)、ヤギ抗ヒトIgκ(Vector Laboratories、AI-3060)、マウス抗ヒトIgM(CGI/ATCC、HB-57)、ならびにマウスγ、κ、μおよびλIgの捕捉のためにはそれぞれヤギ抗マウスIgG(Caltag、M 30100)、ヤギ抗マウスIgκ(Southern Biotechnology、1050-01)、ヤギ抗マウスIgM(Southern Biotechnology、1020-01)およびヤギ抗マウスλ(Southern Biotechnology、1060-01)。ELISA実験に用いた検出用抗体は、ヤギ抗マウスIgG-HRP(Caltag、M-30107)、ヤギ抗マウスIgκ-HRT(Caltag、M 33007)、マウス抗ヒトIgG2-HRP(Southern Biotechnology、9070-05)、マウス抗ヒトIgM-HRP(Southern Biotechnology、9020-05)およびヤギ抗ヒトκ-ビオチン(Vector、BA-3060)である。ヒトおよびマウスのIgの定量化のために用いた標準は以下の通りである:ヒトIgG2(Calbiochem、400122)、ヒトIgMκ(Cappel、13000)、ヒトIgG2κ(Calbiochem、400122)、マウスIgGκ(Cappel 55939)、マウスIgMκ(Sigma、M-3795)およびマウスIgG3λ(Sigma、M-9019)。
【0069】
BIAcoreによる完全ヒトMabの親和定数の決定: 製造者によって概説された標準的な手順によりBIAcore 2000装置を用いて、精製ヒトモノクローナル抗体、Fab断片またはハイブリドーマ上清の親和性の測定をプラズモン共鳴によって実施した。
【0070】
抗体の速度論的分析は、センサー表面上に低密度に固定された抗原を用いて行った:ヒトIL-8 -81RU、A431細胞膜から精製された可溶性EGFR(Sigma、E-3641)-303RU、およびTNF-α -107RU(1,000 RUは固定されたタンパク質の約1ng/mm3に対応する)。解離速度(kd)および会合速度(ka)は、製造者によって提供されたソフトウエアであるBIAevaluation 2.1を用いて決定した。
【0071】
ラジオイムノアッセイによる親和性の測定: 0.5%BSAを含む200μlのPBS中で、125I標識ヒトIL-8(1.5×10-11Mまたは3×10-11M)を種々の濃度の精製抗IL-8ヒト抗体(5×10-13Mから4×10-9M)とともにインキュベートした。室温で15時間インキュベートした後、抗体-抗原複合体を沈降させるためにプロテインAセファロースCL-4Bを含むPBS(l/l、v/v)20μlを添加した。4℃で2時間インキュベートした後、96ウェル濾過プレート(Millipore、カタログ番号MADVN65)を用いる濾過により、プロテインAセファロースに結合した抗体-125I-IL-8複合体を遊離125I-IL-8から分離し、シンチレーションバイアルに回収して計数した。結合型および遊離型の抗体の濃度を算出し、スキャッチャード解析を用いて特異的抗原に対する抗体の結合親和性を算出した(2)。
【0072】
受容体結合アッセイ: IL-8受容体結合アッセイは、記載された通り(Lusti-Marasimhanら、1995)に、新鮮な採取血またはバフィーコートのいずれかから調製したヒト好中球を用いて実施した。0.1%ウシ血清アルブミンおよび0.02%のNaN3を含むPBS結合緩衝液による前処置を25℃で2時間行った96ウェルのMultiscreenフィルタープレート(Millipore、MADV N6550)中にて、種々の濃度の抗体を0.23nMの[125I]IL-8(Amersham、IM-249)とともに4℃で30分間インキュベートした。各ウェルに4×105個の好中球を加え、プレートを4℃で90分間インキュベートした。200μlの氷冷PBSで細胞を5回洗い、PBSを吸引によって除去した。フィルターを風乾し、シンチレーション液を添加して、シンチレーションカウンター中で計数した。抗体の存在下で検出された平均cpmを緩衝液のみの存在下で検出されたcpmで割った値として、特異的に結合した[125I]IL-8の比率を算出した。
【0073】
TNF受容体に関する結合アッセイは、上記のIL-8アッセイと同じ様式で実施した。しかし、IL-8アッセイに関して用いた好中球系の代わりにヒト単球系U937を用いた。抗体は0.25nMの[125I]TNF(Amersham、IM-206)とともにプレインキュベートした。6×105個のU937細胞を各ウェルに播いた。
【0074】
EGF受容体結合アッセイは、PBS緩衝液中の種々の濃度の抗体とともに4℃で30分間インキュベートしたA431細胞(1ウェル当り0.4×106個の細胞)を用いて実施した。0.1nMの[125I]EGF(Amersham、IM-196)を各ウェルに添加し、プレートを4℃で90分間インキュベートした。プレートを5回洗い、風乾し、シンチレーションカウンター中にて計数した。抗EGFRマウス抗体225および528(Calbiochem)を対照として用いた。
【0075】
XenoMouseにおいて発現されたヒトIg転写物およびそれらに由来するヒトMabのレパートリー解析: Fast-Trackキット(Invitrogen)を用いて、非免疫化および免疫化XenoMouseの脾臓およびリンパ節から、ポリ(A)+mRNAを単離した。ランダムプライミングによりcDNAを作製し、続いてPCRを行った。ヒトVHまたはヒトVκファミリーに特異的な可変領域プライマー(Marksら、1991)または汎用ヒトVHプライマーであるMG-30(CAGGTGCAGCTGGAGCAGTCIGG)を、以前に記載された通り(Greenら、1994)にヒトCμ(huP2)またはCκ(hκP2)定常領域に特異的なプライマーとともに、またはヒトγ2定常領域MG-40d;5'-GCTGAGGGAGTAGAGTCCTGAGGA-3'とともに用いた。TAクローニングキット(Invitrogen)を用いてPCR産物をpCRII中にクローニングし、Prismダイターミネーター配列決定キットおよびABI377配列決定装置を用いて両方のストランドの配列を決定した。ヒトMabに由来する重鎖およびκ軽鎖の転写物の配列は、上記のプライマーを用いてポリ(A+)RNAから生成されたPCR産物の直接的な配列決定によって得た。「V BASE配列ディレクトリー」(Tomlinsonら、MRC Centre for Protein Engineering、Cambridge、UK)に対するアライメントにより、MacVectorおよびGeneworksソフトウェアプログラムを用いて、すべての配列を解析した。
【0076】
抗体Fab断片の調製および精製: 抗体Fab断片は固定パパイン(Pierce)を用いることによって作製した。Fab断片を以下の2段階クロマトグラフィー方式で精製した:Fc断片およびすべての未消化抗体を捕捉するためのHiTrap(Bio-Rad)プロテインAカラム、次いで強陽イオン交換カラム(PerSeptive Biosystems)を通過させて保持されたFab断片の0.5M NaClに至る塩の直線勾配による溶出。Fab断片の特徴を、還元および非還元条件下でのSDS-PAGEおよびMALDI-TOF MSによって分析したところ、予想された〜50kDの非還元断片および〜25kDaの還元二重バンドが示された。この結果は、無傷の軽鎖および切断された重鎖を示す。還元条件下におけるMSでは、未消化抗体全体の還元によって軽鎖の質量を正確に決定しうるため、軽鎖および切断された重鎖の双方を明確に同定することが可能であった。
【実施例】
【0077】
以下の実施例は、実施した実験および得られた結果を含め、例示を目的として提供されるものであり、本発明の範囲を制限するものと考えられるべきではない。
【0078】
実施例1:YAC上でのヒト重鎖遺伝子座の再構成
本発明により、本発明者らがヒト重鎖およびヒトκ軽鎖の可変領域を再構成するために用いた戦略は、第1に大きな(メガベースサイズの)ヒトIg遺伝子座にまたがるYACに関してヒトYACライブラリーをスクリーニングし、第2にこのような領域にまたがるYACを、主として生殖細胞系コンフィギュレーションにある望ましい遺伝子座を含む単一のYAC中に組換え導入することであった。
【0079】
上記の段階的なYAC組換え方式は、酵母において減数分裂誘導性の相同組換えを高頻度にもたらし、組換えYAC上のベクター腕に存在する酵母マーカーによる望ましい組換え体の選択を可能とする(図1およびGreenら、前記を参照のこと;Silvermanら、1990およびdenDunnenら、1992も参照されたい)。
【0080】
本発明者らの戦略に関連して、本発明者らは染色体14q上にあるヒト重鎖可変領域の約1000kbのうち、約830kbに及ぶ1H(240kb)、2H(270kb)、3H(300kb)および4H(340kb)という4種のYACを同定した。このYAC 1H、2H、3Hおよび4Hを遺伝子座の再構成のために用いた(図1A参照)。パルスフィールドゲル電気泳動(PFGE)およびサザンブロット分析により、特定の非IgH配列を含むYAC 2Hの3'末端の150kbを除いて、YACが無傷であって生殖細胞系コンフィギュレーションにあることが示された(図1参照;Matsudaら、1990)。以前、本発明者らの第1世代のXenoMouse(登録商標)に導入されたYACであるYAC 1H(Greenら、前記;Mendezら、1995)は、生殖細胞系コンフィギュレーションにあるヒトCδ、Cμ、JHおよびDH領域ならびに最初の5つのVH遺伝子を含む。他の3つのYACは、VH2-5からVH3-65までのVH領域の大半をカバーし、それによってさらにほぼ61個の異なるVH遺伝子がもたらされる。組換えに先立ち、YAC 4HにはHPRT選択可能マーカーのレトロフィットを施した。YAC上に含まれる重複配列を用いることにより、段階的組換え戦略によって4種のYAC(1H、2H、3Hおよび4H)を酵母に組換え導入した(図1A参照)。このような組換え戦略によって980kbの組換えYACが作製された(図1参照)。PFGEおよびサザンブロット分析によるYACの分析から、生殖細胞系コンフィギュレーションにあるCδ領域からVH3-65遺伝子の5'側の20kbのところまでのヒト重鎖遺伝子座が存在することが確かめられた。明らかな欠失および再配列は認められなかった。
【0081】
YAC無動原体腕に対して、完全なヒトγ2定常領域、マウス3'エンハンサーおよびネオマイシン耐性遺伝子を有するベクターによるターゲティングを行い、最終的に1020kbの重鎖YACであるyH2を得た。YAC yH2は、ヒト可変領域の大半、すなわち82個のVH遺伝子のうち66個、全DH(32遺伝子)およびJH(6遺伝子)領域、ならびに3つの異なる定常領域(Cμ、CδおよびCγ)を、それらに対応する調節配列とともに含んでいた(図1A参照)。これが本発明者らのXenoMouse II系統の作製に用いた重鎖構築物である。
【0082】
実施例2:YAC上でのヒトκ軽鎖遺伝子座の再構成
ヒトκ軽鎖遺伝子座の再構成には同様の段階的組換え戦略を用いた。ヒトκ遺伝子座にまたがる3種のYACを同定した。YACは1K、2Kおよび3Kと命名した。長さ約180kbのYAC 1Kは、以前に本発明者らの第1世代のXenoMouse(登録商標)に導入された。このようなYACは、Bクラスター上にκ欠失要素(Kde)、κ3'およびイントロン性エンハンサー、Cκ、Jκならびに3種のVκ遺伝子を含む(Greenら、1994;Mendezら、1995)。YAC 2K(約480kb)および3K(約380kb)はともに、染色体2p上のκ軽鎖近位可変領域の大部分を包含する。約100kbの欠失はL13〜L5領域にまたがる(図1B;Huberら、1993)。κ遠位領域は近位領域の繰り返しであり、近位Vκ遺伝子はヒトで最も一般的に用いられるものであるため(Weicholdら、1993;Coxら、1994)、本発明者らの再構成戦略の焦点は近位領域にあった(図1B)。3種のYACの相同組換えにより、800kbの組換えYACであるyK2が回収された。この組換えYACのサイズおよび完全性を、PFGEおよびサザンブロット分析によって確認した。こうした分析から、それがLp領域に記載した通りの欠失があることを除き、生殖細胞系コンフィギュレーションにある32個のVκ遺伝子とともにヒトκ軽鎖遺伝子座の近位部をカバーすることが示された(図1B)。yK2の動原体腕および無動原体腕に、記載した通り(材料および方法)にそれぞれHPRTおよびネオマイシン選択可能マーカーを含むように改変を加えた。これが本発明者らのXenoMouse II系統の作製に用いたκ軽鎖構築物である。
【0083】
本明細書に記載されるYACであるyH2およびyK2は、主として生殖細胞系コンフィギュレーションでヒト抗体レパートリーの大半を含むように再構成された最初のメガベースサイズのヒトIg遺伝子座である。これが達成されたことは、酵母における相同組換えが、大きな複雑で不安定な遺伝子座の再構成を首尾よく行うための強力な手法であることを立証するものである。酵母内にIg遺伝子座の大部分を含む安定なYAC組換え体の選択により、本発明者らは、マウスでヒト抗体反応を再現するために必要なヒト抗体レパートリー、定常領域および調節要素をマウスに備えさせるために必要なヒトIg断片を手に入れた。
【0084】
実施例3:ES細胞へのyH2およびyK2 YACの導入
本発明者らの戦略に従い、本発明者らはYAC yH2およびyK2をマウス胚性幹(ES)細胞に導入した。YAC DNAを含むES細胞がいったん単離されれば、このようなES細胞は適切な交配を通じてマウスの作出に用いられる。
【0085】
このため、この実験では、以前に記載されている通り(Jakobovitsら、1993a;Greenら、1994)に、YACを含む酵母スフェロプラストとHPRT欠損E14.TG3B1マウスES細胞との融合を介して、YAC yH2およびyK2をES細胞に導入した。HPRT陽性のES細胞クローンは、1クローン/融合細胞15〜20×106個の割合で選択され、サザンおよびCHEFブロット分析によってYACの完全性に関する分析が行われた(図2A)。
【0086】
ES細胞とyH2を含む酵母との融合に由来する35種のES細胞クローンのうち7種(L10、J9.2、L17、L18、J17、L22、L23と命名)が、挿入物の全体にまたがるプローブによって検出される、予想されたすべてのEcoRIおよびBamHI yH2断片を含むことが明らかになった:マウス3'エンハンサー、ヒトイントロン性エンハンサー、ヒトCγ2、CδおよびCμ定常領域、DH、JHならびにすべての異なるVHファミリー:VH1、VH2、VH3、VH4、VH5およびVH6(図2Aに5つのクローンに関するデータを示した)。CHEF分析により、分析した全クローンの20%にあたるこれらのクローンが、明らかな欠失および再配列を含まない無傷のyH2 YACの全体を含むことがさらに確かめられた(非提示データ)。
【0087】
yK2を含む酵母との融合に由来するES細胞クローンについても、ヒトKde、ヒトκ3'およびイントロン性のエンハンサー、Cκ、Jκならびに異なるVκファミリーのすべて:VκI、VκII、VκIII、VκIV、VκVIに特異的なプローブを用いて、YACの完全性に関して同様に分析した。60種のクローンのうち20種は無傷であって変化のないYACを有し、これは分析した全クローンの30%であった(図3Aに2つのESクローンに関するデータを示した)。yH2およびyK2-ES細胞クローンでは種々の量の酵母ゲノム配列が検出された(非提示データ)。
【0088】
これらの結果により、主として生殖細胞系コンフィギュレーションにある再構成されたヒト遺伝子座を含むメガベースサイズの構築物が哺乳動物細胞に導入されたことが初めて示された。無傷のYACがマウスゲノムに比較的高頻度に組み込まれたことは、ES細胞-酵母スフェロプラスト融合法が、大きなヒトゲノム断片をES細胞に忠実に導入するための効果的な手法であることをさらに実証するものである。
【0089】
実施例4:XenoMouse II系統の作出
YAC DNAを含むES細胞からマウスを作出するために、胚盤胞へのマイクロインジェクションを行い、続いて交配を行った。すなわち、yH2およびyK2を保持するES細胞クローンを増殖させ、マウスC57BL/6J胚盤胞へのマイクロインジェクションを行い(Greenら、1994)、得られたキメラ雄を生殖細胞系伝達に関して評価した。伝達されたYACを有する子孫をPCR分析によって同定し、サザンブロット分析によってYACの完全性を確認した。分析したすべてのトランスジェニックマウスにおいて、YACは無傷型であることが示された(図2B、3B)。マイクロインジェクションを受けた7つのyH2-ESクローンのすべて、および8つのyK2-ESクローンのうち2つは、マウス生殖細胞系を通じて伝達された。
【0090】
内因性抗体が排除された、ヒト抗体を産生するマウスを作出するために、yH2-またはyK2-トランスジェニックマウスを二重不活性化(DI)マウス系統と交配させた。DIマウス系統は、遺伝子ターゲティングによって不活性化されたマウス重鎖およびκ鎖遺伝子座に関して同型接合的であり、このため抗体産生に欠陥がある(Jakobovitsら、1993b;Greenら、1994)。2つのyH2-トランスジェニックマウス系統であるL10およびJ9.2、ならびにyK2-トランスジェニックマウス系統の1つであるJ23.1をDIマウスと交配させ、同型接合的に不活性化されたマウス重鎖およびκ鎖バックグラウンドをもつYACを保持するマウスを作出した(yH2;DIおよびyK2;DI)。yH2;DIトランスジェニック系統のそれぞれをyK2;DIトランスジェニック系統と交配させ、同型接合的DIバックグラウンドをもつ重鎖および軽鎖YACの両方を含む2つのXenoMouse II系統である2A-1(L10;J23.1;DI)および2A-2(J9.2;J23.1;DI)をそれぞれ作出した。L10は完全に同型接合的であり、J9.2およびJ23.1は交配によって首尾よく同型接合的になる過程にある。
【0091】
XenoMouse II系統におけるヒト重鎖およびκ鎖YACの完全性は、サザンブロット分析によって確認した。図2および図3に示す通り、分析したいずれのXenoMouse系統とも、yH2およびyK2は明らかな欠失および再配列を生じずに複数世代にわたって変化せずに伝達された。
【0092】
実施例5:XenoMouse IIマウスによるB細胞発生およびヒト抗体産生
XenoMouse II系統の特徴をさらに調べるために、本発明者らはそれらのB細胞発生およびヒト抗体産生を検討した。yH2およびyK2 YACによるXenoMouse II系統におけるB細胞発生および抗体産生の再構成を、フローサイトメトリーおよびELISAによって評価した。成熟B細胞を完全に欠くDIマウスとは対照的に、XenoMouse IIは本質的に正常なB細胞発生を示し、血中の成熟B細胞を合計すると野生型マウスで認められる値の50%を上回った(図4A)。すべてのB細胞はヒトIgMおよび高レベルのB220を発現し(ヒトIgM+/B220hi)、この集団の60%はヒトIgDも発現することが示された。XenoMouseの脾臓およびリンパ節の分析でも同様の結果が得られた(提示せず)。これらの結果は野生型マウスにおける成熟B細胞の特徴とよく相関しており、XenoMouseにおけるB細胞成熟が適切であることを示す。
【0093】
XenoMouseのB細胞の大半(75〜80%)はもっぱらヒトκ軽鎖を発現し、マウスλ軽鎖を発現したものは15%に過ぎなかった(図4)。この軽鎖の分布比(hκ/mλ)は野生型マウスで認められる値と同等であり、このことは軽鎖の使用がマウス様調節を受けていることを示す。これに対して、グリーン(Green)ら、1994に記載されたXenoMouse Iで認められたhκ/mλの比は55:45であった(非提示データ)。脾臓(図4B)およびリンパ節(提示せず)由来のB細胞についても同様のことが観察され、このことはXenoMouse IIのB細胞の大部分は、もっぱら完全ヒト抗体を産生することを示す。yK2に関して同型接合的であるXenoMouse II系統におけるmλ発現B細胞のレベルは、15%から7%に減少した(非提示データ)。
【0094】
実施例6:L6系統の作出
L6系統のマウスは、XenoMouse II系統の作出に関する上記の過程と同じように作出した。しかし、L6 ES細胞系の作製に際して欠失事象があるために、ES細胞系および続いて得られるL6マウスはCδの遠位側にある配列の部分を伴わずに生じ、このため、Cγ定常領域およびその調節配列をもたない。交配の完了後に、L6マウスは欠失したCγ定常領域を除き、yK2構築物の全体およびyH2構築物の全体を含むと考えられる。
【0095】
実施例7:ヒト抗体の産生
非免疫化XenoMouseIIの血清中でヒトCμ、Cγ2およびκ軽鎖の発現が検出され、最大値はそれぞれ700、600および800μg/mlであった。これらの値がどの程度野生型に匹敵するかを明らかにするために、本発明者らは同様の無病原体条件下に保ったC57BL/6J×129マウスにおけるマウスCμ、Cγ2およびκ軽鎖の最大値を測定した。野生型マウスにおけるCμ、Cγ2およびκ軽鎖に関する値はそれぞれ400、2000および2000μg/mlであった。免疫化を施したところ、ヒトγ鎖の値は約2.5mg/mlに上昇した。マウスλの濃度は70μg/mlに過ぎず、このことからヒトκ鎖が優先的に用いられることがさらに裏づけられた。
【0096】
これらの所見は、導入されたヒトYACに適切なIg遺伝子再配列およびクラススイッチを誘導する能力、ならびに免疫化の前後ともに顕著なレベルの完全ヒトIgMおよびIgG抗体を産生する能力があることを裏づけるものである。
【0097】
実施例8:XenoMouse IIにおける多様なヒト抗体レパートリー
XenoMouse II系統における抗体レパートリーの再構成に関してさらに理解を得るために、本発明者らはマウスにいくつかの免疫原による誘発刺激を与え、このような抗体を分泌するハイブリドーマ細胞系を調製した。理解されるであろう通り、マウスでヒト抗体反応を再現させるにはyH2およびyK2 YAC上に含まれる種々のヒト可変遺伝子が多様に用いられる必要がある。XenoMouse II系統によって産生されるヒト抗体の多様性を、XenoMouseのリンパ節由来のヒト重鎖(μおよびγ)ならびにκ軽鎖転写物のクローニングおよび配列決定によって調べた。現在までの本発明者らのデータに基づくと、配列解析ではXenoMouse IIがyH2上に存在する37個の機能的VH遺伝子のうち少なくとも11個、8個の異なるDHセグメントおよび3個のJH遺伝子(JH3、JH4、JH6)を用いることが示されている(表III;本発明者らによるハイブリドーマ由来の抗体の配列決定に関してJH5も検出された)。V-D-J配列はヒトμまたはγ2定常領域と連結していた(提示せず)。
【0098】
用いられたVH遺伝子は可変領域の全体にわたって広く分布しており、7つのVHファミリーのうちの4つであった(表III)。VH3およびVH4ファミリーからのV遺伝子が主に利用された点は、ファミリーのサイズに比例する、成人におけるVH使用パターンと類似している(Yamadaら、1991;Brezinshekら、1995)。JH4が主に利用されたこともヒトB細胞で検出されるものとよく似ている(Brezinshekら、1995)。1〜12bpの範囲に及ぶ、V-DおよびD-J結合の双方での非生殖細胞系ヌクレオチドの付加(N-付加)も認められた。このようなN付加は、8個から約19個までのアミノ酸残基長をもつ相補性決定領域3(CDR3)をもたらすが、これは成人B細胞で認められるものと極めて類似している(Yamadaら、1991;Brezinshekら、1995)。XenoMouse IIで認められるこのようなCDR3長は、マウスで通常認められるCDR3長よりもはるかに長い(Feeny、1990)。
【0099】
配列決定を行った10種のκ鎖転写物においても、極めて多様なレパートリーが認められた。yK2上に存在する25個のVκ機能的オープンリーディングフレーム(ORF)のうち8個が提示されることに加えて、すべてのJκ遺伝子が検出可能であった(表IV)。用いられた異なるVκ遺伝子はyK2の全体にわたって広く分散しており、4種の主要なVκ遺伝子ファミリーのすべてが示された。すべてのVκJκ組換え産物がCκ配列と適切に連結していた。本発明者らの転写物にN付加がほとんどみられなかったことは、κ鎖再配列の段階でターミナルデオキシヌクレオチジルトランスフェラーゼ活性が大きく低下することと一致する。本発明者らがκ鎖転写物中に認めた9〜10アミノ酸の平均CDR3長は、ヒトB細胞で認められた値と同一であった(Marksら、1991)。
【0100】
以下の表IIIおよびIVでは、XenoMouse II系統において発現されたヒト重鎖およびκ軽鎖転写物のレパートリー解析の結果を提示している。材料および方法の項で記載した通りに、ヒトμ、γおよびκに特異的なmRNAをPCRによって増幅し、クローニングして、配列決定によって解析した。表IIIには、12種のユニークなヒト重鎖クローンの一連のヌクレオチド配列を、発表された生殖細胞系配列との相同性によって同定されるVH、D、JHおよびNセグメントに分割して示している(材料および方法)。それぞれのDセグメントの指定は、少なくとも8塩基の相同性に基づいている。表IVには、8種の独立したヒトκクローンのV-J結合の一連のヌクレオチド配列を示している。配列はVκ、JκおよびNセグメントに分割され、発表されたVκおよびJκ配列との相同性に基づいて同定されている。それぞれの表におけるN付加および欠失(_として表示)は、V、DまたはJ配列との配列相同性がないことによって決定した。
(表III)
【表3】
(表IV)
【表4】
【0101】
これらの結果を、以下に記載するXenoMouse由来のハイブリドーマの配列と総合すると、V、DおよびJ遺伝子の極めて多様な成人様使用が示されるが、このことはyH2およびyK2 YAC上に存在するヒト重鎖およびκ鎖の可変領域の全体を抗体再配列のためにマウス系が利用することができ、しかも位置による偏差のない様式で利用されることを示すと考えられる。さらに重鎖およびκ鎖転写物の双方に関するN付加およびCDR3の平均長は成人B細胞で認められる値と非常に類似しており、このことは、マウスに含まれるYAC DNAがマウスにおいて成人様免疫レパートリーが生じるようにマウスの機構を指向させることを示す。
【0102】
以下の実施例に関して、本発明者らはいくつかの抗原に対する高親和性抗体を調製した。特に抗原としてはヒトIL-8およびヒトEGFRを調製した。IL-8およびEGFRを選択した根拠は以下の通りである。
【0103】
IL-8はC-X-Cケモカインファミリーのメンバーである。IL-8は、少し例を挙げればARDS、慢性関節リウマチ、炎症性腸疾患、糸球体腎炎、乾癬、アルコール性肝炎、再潅流障害を含む多くの疾患に関与する好中球に対する主な化学誘引物質として作用する。さらに、IL-8は内皮細胞に対する強力な血管新生促進因子である。図22〜28において本発明者らは、XenoMouse II系統に由来するヒト抗IL-8抗体がIL-8の作用を多くの経路において効果的に阻害することを示す。例えば、図22は、ヒト抗IL-8抗体によってIL-8のヒト好中球との結合が阻止されることを示す。図23は、ヒト抗IL-8抗体によってヒト好中球上のCD11b発現が阻害されることを示す。図24は、ヒト抗IL-8抗体によってIL-8誘導性カルシウム流入が阻害されることを示す。図25は、ヒト抗IL-8抗体によるIL-8 RB/293走化性の阻害を示す。図26は、ヒトIL-8誘導性皮膚炎症のウサギモデルの模式図を示す。図27は、図26のウサギモデルにおけるヒトIL-8誘導性皮膚炎症がヒト抗IL-8抗体によって阻害されることを示す。図28は、ラット角膜ポケットモデル上の内皮細胞の血管新生がヒト抗IL-8抗体によって阻害されることを示す。
【0104】
EGFRは、抗癌ターゲットとみなされている。例えば、種々の癌細胞上ではEGFRが最大100倍に過剰発現される。ある種の癌の開始および進行には、リガンド(EGFおよびTNF)を介した増殖刺激が極めて重要な役割を果たす。この点に関して、EGFR抗体はリガンド結合を阻害して腫瘍細胞の増殖の停止をもたらし、さらに化学療法薬と併用するとアポトーシスを誘導する。事実、複数のEGFR Mabの組み合わせがマウス異種腫瘍モデルにおける腫瘍の根絶をもたらすことが示されている。イムクローン(Imclone)社はキメラMab(C225)を用いるフェーズI臨床試験を実施しており、その安全性は実証されている。図31〜33において本発明者らは、本発明者らのヒト抗EGFR抗体に関するデータを示す。図30は、XenoMouse II系統に由来するヒト抗EGFR抗体の重鎖アミノ酸配列を示す。図31は、ヒト抗EGFR抗体によってEGFのA431細胞との結合が阻止されることを示す。図32は、ヒト抗EGFR抗体によってEGFのSW948細胞との結合が阻害されることを示す。図33は、XenoMouse II系統に由来するヒト抗EGFR抗体がインビトロでのSW948細胞の増殖を阻害することを示す。
【0105】
実施例9:XenoMouse IIによって産生される高親和性の抗原特異的ヒトMab
本発明者らは次に、XenoMouse IIにおける広範なヒトレパートリーを、多数の抗原、特に重大な臨床的関心がもたれるヒト抗原に対するヒト抗体を産生させるために利用しうるか否かを問題にした。
【0106】
このため、個々のXenoMouse IIの仔のそれぞれにヒトIL-8、ヒトEGFRおよびヒトTNF-αという3種の異なる抗原標的のうち1つによる誘発刺激を与えた。抗原はIL-8およびTNF-αの場合には可溶性タンパク質として、EGFRの場合には細胞(A431細胞)表面上に発現されたものとしていずれかの2つの異なる形式で投与された。3種の抗原のいずれについても、免疫化マウスからの血清に対して行ったELISAでは、1:3×106もの高い力価を有する強い抗原特異的ヒト抗体(IgG、Igκ)反応が示された。検出されたマウスλ反応は無視しうる程度であった。
【0107】
標準的なハイブリドーマ技術によって脾臓またはリンパ節に由来するハイブリドーマを得た上で、ELISAにより抗原特異的ヒトMabに関してスクリーニングした。
【0108】
IL-8によって免疫化した1匹のXenoMouse IIから、いずれもヒトIL-8に特異的な完全ヒト(hIgG2κ)Mabを分泌する12種の一連のハイブリドーマが得られた。これらのハイブリドーマのうち4種からの抗体であるD1.1、K2.2、K4.2およびK4.3を腹水から精製し、ヒトIL-8に対するそれらの親和性、およびIL-8のヒト好中球上の受容体との結合を阻止する効力に関して評価した。
【0109】
親和性の測定は、BIAcoreにおける表面プラズモン共鳴を用いる全抗体およびFab断片の双方の固相測定、ならびに溶液中でのラジオイムノアッセイによって行った(材料および方法)。表Vに示す通り、4種のMabに関して測定された親和性の値は1.1×109から4.8×1010M-1の範囲であった。用いた技法には若干の違いがあったものの、4種の抗体に関する親和性の値は一貫して109M-1よりも高かった。
【0110】
ELISA分析では、これらの4種の抗体がヒトIL-8に特異的であること、ならびに密接に関連したケモカインであるMIP-1α、GROα、βおよびγ、ENA-78、MCP-1またはRANTESとは交差反応しないことが確かめられた(非提示データ)。さらにBIAcore上での競合分析からは以上の抗体が少なくとも2つの異なるエピトープを認識することが示された(非提示データ)。すべての抗体がマウス抗ヒトIL-8中和抗体と同程度に効果的にIL-8のヒト好中球との結合を阻害したが、対照ヒトIgG2κ抗体はこれを阻害しなかった(図5A)。
【0111】
EGFRによって免疫化したXenoMouse IIを用いた融合実験では、いずれもEGFR特異的なヒトIgG2κMabを分泌する25種の一連のハイブリドーマが得られた。分析した13種のヒトMabのうち4種(E2.1、E2.4、E2.5,E2.11)が、マウスにおけるEGFを介した細胞増殖および腫瘍形成を阻害することが以前に示されている(Satoら、1983)EGFR特異的マウス抗体225と競合する能力に関して選択された。腹水から精製されたこれらのヒト抗体を、EGFRに対するそれらの親和性およびEGFの細胞との結合の中和に関して評価した。これらの抗体のEGFRに対する親和性は、BIAcore測定による評価では2.9×109から2.9×1010M-1の範囲であった(表V)。
【0112】
4種の抗EGFR抗体はいずれもEGFのA431細胞との結合を完全に阻止し(図5B)、それらがこれらの細胞上の高親和性および低親和性受容体(Kawamotoら、1983)の双方に対するEGFの結合を中和する能力をもつことが示された。4種の抗EGFRヒト抗体のいずれによっても、EGFのヒトSW948ヒト肺癌細胞上に発現されたEGFRとの結合が完全に阻害されることも認められた(非提示データ)。いずれの場合にも完全ヒト抗体はEGF結合の阻害の点で抗EGFRマウス抗体225と同程度に効果的であり、528抗体よりも強力であった(Gillら、1983)。いずれの細胞アッセイにおいても、対照ヒトIgG2κ抗体はEGF結合に影響を及ぼさなかった(図5Bおよび非提示データ)。
【0113】
TNF-αによって免疫化したXenoMouse IIを用いた融合実験では、12種の一連のヒトIgG2κ抗体が得られた。この12種のうち4種が、TNF-αのU937細胞上の受容体との結合を阻止する能力に関して選択された(図5C)。これらの抗体の親和性を測定したところ、1.2〜3.9×109M-1の範囲であった(表V)。
【0114】
上記のXenoMouse由来ハイブリドーマは、静置培養条件下において2〜19μg/mlの範囲の濃度で抗体を産生した。非還元条件下におけるタンパク質ゲル上での精製抗体の特徴分析では、IgG2κ抗体に関する見かけの分子量の期待値が150kDであることが明らかになった。還元条件下で検出された重鎖に関する見かけの分子量の期待値は50kDであり、軽鎖については25kDであった(非提示データ)。
【0115】
以下の表Vは、XenoMouse由来の抗原特異的完全ヒトMabの親和定数を示している。IL-8、EGFRおよびTNF-αに特異的なXenoMouse由来のヒトIgG2κ Mabの親和定数は、材料および方法の項に記載したBIAcoreまたはラジオイムノアッセイによって測定した。IL-8およびEGFRに関して示した値は精製抗体を用いて実施した独立した実験の代表値であり、TNF-αに関して示した値はハイブリドーマ上清を用いて実施した実験によるものである。
(表V)
【表5】
【0116】
実施例10:モノクローナル抗体における遺伝子使用および体細胞超突然変異
上記のIL-8およびEGFR-ヒトMabからの重鎖およびκ軽鎖転写物の配列を決定した。図6および図[[ ]]。4種のIL-8特異的抗体は、少なくとも3つの異なるVH遺伝子(VH4-34/VH4-21、VH3-30およびVH5-51)、4つの異なるDHセグメント(A1/A4、K1、ir3rcおよび21-10rc )および2つのJH(JH3およびJH4)遺伝子セグメントからなっていた。3つの異なるVκ遺伝子(012、018およびB3)がJκ3およびJκ4遺伝子と結合していた。このように多様な使用がみられることは、XenoMouse IIが多様な可変領域を備えた一連の抗IL-8中和抗体を産生しうることを示す。
【0117】
IL-8抗体転写物とは対照的に、Mab225との競合能に関して選択された抗体の配列ではVHおよびVκ遺伝子の使用が比較的制限されており、E1.1、 E2.4およびE2.5という3種の抗体は同一のVH遺伝子(4-31)を共有し、E2.11はVH4-31との相同性が高いVH4-61を含んでいた。異なるD(2、A1/A4、XP1)およびJH(JH3、JH4、JH5)セグメントが検出された。4種の抗体はいずれも同一のVκ(018)遺伝子を共有することが示された。それらのうち3つはJκ4を含み、残る1つであるE2.5はJκ2を含んでいた。
【0118】
大部分のVHおよびVκハイブリドーマ転写物では、対応する生殖細胞系セグメントからみて高度のヌクレオチド変化(7〜17箇所)が認められたが、定常領域では全く変異が検出されなかった。Vセグメントにおける変異の大部分は、予想される抗体アミノ酸配列においてアミノ酸置換をもたらし(V遺伝子1つにつき0〜12箇所)、その多くはCDR1およびCDR2領域にあった(図_)。注目すべき点は、すべての抗体で共有されていたCDR1におけるGly→Asp置換、または3つの抗体で共有されていたCDR2におけるSer→Asn置換、およびフレームワーク領域3におけるVal→Leuなどの変異が、EGFR抗体の重鎖配列で共有されていたことである。これらの結果は、XenoMouse IIにおいて抗体の成熟および選択に至る体細胞超突然変異の過程が高度に生じていることを示す。
【0119】
考察
本出願は、複雑なメガベースサイズのマウス遺伝子座を、YAC上に再構成されたサイズおよび内容の点で等価なヒトDNA断片によって、初めて機能的に置換したことを記載したものである。この手法により、マウス体液性免疫系はメガベースサイズのヒトIg遺伝子座によって「ヒト化」され、内因性抗体産生に欠陥のあるマウスにおいてヒト抗体反応が実質的に再現されるようになる。
【0120】
ヒト重鎖およびκ軽鎖遺伝子座の大部分がほぼ生殖細胞系コンフィギュレーションの形で忠実に再構成されるという本発明者らの成果は、酵母におけるYAC組換えが、Ig遺伝子座などの大きく複雑で不安定な断片を再構成するため(Mendezら、1995)、およびそれらを哺乳動物細胞内に導入するために操作するための強力な技術であることを立証するものである。さらに、2つの大きな重鎖およびκ軽鎖セグメントが無傷の形でマウス生殖細胞系に首尾よく導入されたことは、ES細胞-酵母スフェロプラスト融合がマウス生殖細胞系に異種遺伝子座を送達するための確実で効率的な手法であることを裏づける。
【0121】
XenoMouse II系統の特徴分析から、大きなIg遺伝子座には、多様性および機能性の点で野生型マウスのものに匹敵し、ヒトIgミニ遺伝子構築物(Lonbergら、1994)または小型のIg YAC(Greenら、1994)を保持するマウスで生じる体液性応答よりもはるかに優れた抗体系を再生する能力があることが示された。この差は、成熟B細胞のレベル、ヒトIg産生、クラススイッチ効率、多様性、マウスIgλ産生と対比してのヒトIgκの優位性、およびヒト抗体反応の強さ、ならびに多数の抗原に対する高親和性の抗原特異的モノクローナル抗体が産生されるとの成果として表れた。
【0122】
XenoMouse IIにおける成熟B細胞およびヒト抗体のレベルは、これまで報告されているIg-トランスジェニックマウスの中で最も高く、これまでのマウスに関して示されている値の数倍であり、野生型マウスでの値に近い。特にヒトIgGのレベルは、ヒトγ1遺伝子を備えたミニ遺伝子座Ig導入遺伝子を保持するマウスに関して報告されている値の100倍を上回る(Lonbergら、1994)。XenoMouse IIにおけるクラススイッチの効率がより高いことは、yH2上にスイッチ領域の全体を、それらのすべての調節要素ならびに付加的な調節要素とともに含めた結果である可能性が高いと考えられ、これが適切なクラススイッチの支持および維持には重要であると思われる。XenoMouse II系統における成熟B細胞のレベルの上昇は、再配列頻度がより高く、そのためにV遺伝子レパートリーが増すことによって骨髄中のB細胞発生が改善されるためである可能性が高い。B細胞の再構成は、ヒト重鎖遺伝子座に関して同型接合的であるXenoMouse II系統でははるかに顕著であると考えられる。
【0123】
循環B細胞によるヒトκとマウスλ軽鎖との発現の比は、トランスジェニックκ軽鎖遺伝子座の利用に関する有用な内部指標となる。より小さなIg YACの1対立遺伝子を含むマウスで認められるヒトκおよびマウスλの分布はほぼ等しいが、XenoMouse II系統ではヒトκの明らかな優位性が検出されている。さらに、yK2に関して同型接合的なマウスでのκ:λ比は野生型マウスと同一であった。これらの観察結果をVκ遺伝子の広範な使用と併せて考えると、XenoMouse IIにおけるヒト近位Vκ遺伝子は多様な軽鎖反応を支持するのに十分であることが強く示唆され、これはヒトで近位Vκ遺伝子がより用いられる傾向にあることとも一致する(Coxら、1994)。
【0124】
XenoMouse II系統は、組換え機構によって利用され、成熟抗体に組み入れられる遺伝子座の全長にわたるV、DおよびJ遺伝子による、極めて高度の抗体多様性を呈する。抗原結合によっていったん誘発されれば、大規模な体細胞超突然変異が起こり、抗体の親和性成熟をもたらす。
【0125】
XenoMouse IIにおけるV、D、J遺伝子の利用パターンからは、それらがヒトでの利用と類似した様式で利用可能であり、Igミニ遺伝子を保持するマウスで認められる胎児様で位置による偏差のある使用(Taylorら、1992;Taylorら、1994;Tuaillonら、1993)とは異なる、成人様ヒト抗体レパートリーが得られることも示された。マウスによって認識される多数の抗原とともに多くの機能的VHおよびVκ遺伝子が広く利用されることは、機能的な抗体反応を首尾よく再構成する上での広範なV遺伝子レパートリーの重要性を強調するものである。
【0126】
マウスにおけるヒト免疫応答の再構成の程度に関する最終的な判断材料は、マウスが抗体反応を誘発すると思われる抗原の範囲、および種々の抗原に対して抗原特異的な高親和性Mabが産生されうる容易さである。現在までに限られた数の抗原特異的ヒトMabしか得られていない、より小型のヒトIg YACまたはミニ遺伝子による操作を受けたマウスとは異なり(Lonbergら、1994;Greenら、1994;Fishwildら、1996)、XenoMouse IIは現在までに検討したすべてのヒト抗原に対するMabを産生した。XenoMouse II系統は、可溶性タンパク質または細胞表面上に発現されたもののいずれかとして提示された種々のヒト抗原に対して強いヒト抗体反応を生じた。検討した3種のヒト抗原のそれぞれによる免疫化により、10〜25種の一連の抗原特異的ヒトIgG2κ Mabが得られた。各抗原に関して、109〜1010M-1の範囲の親和性を有する抗体の一群が得られた。親和性の値が、結合活性というよりも一価結合動態を表すことを確認するために、いくつかの測定を行った:二価結合の確率を最小限に抑えるために抗原を低密度にコートしたセンサーチップを用いて無傷抗体によるBIAcore解析を行った;2種の抗体については一価Fab断片を用いて解析を再度行った;いくつかの抗体については溶液のラジオイムノアッセイによる検討も行った。これらの測定の結果から、本発明者らは、XenoMouseによって1010M-1の範囲の親和性をもつ抗体を容易に獲得しうると結論する。XenoMouse由来の抗体に関して得られた親和性の値は、遺伝子操作されたマウス(Lonbergら、Fishwildら、1996)またはコンビナトリアルライブラリー(Vaughanら、1996)から産生された、ヒト抗原に対するヒト抗体に関して報告されたものの中で最も高い。これらの高親和性と、V遺伝子の体細胞変異の結果としての高度のアミノ酸置換が組み合わされることにより、XenoMouse IIにおける親和性成熟機構が完全であり、野生型マウスにおけるものと匹敵することが裏づけられる。
【0127】
これらの結果は、抗体の多様化および選択に関するマウスの機構によってヒトIg YAC上に広範な抗体レパートリーが適切に開発されること、およびヒトタンパク質に対する免疫寛容の欠如によってヒト抗原を含む任意の関心対象の抗原に対する高親和性抗体を得ることができることを示す。これらのマウスにおけるヒト免疫グロブリン遺伝子によってヒト抗原に対する抗体が産生されうる容易性は、B細胞のレベルでの自己免疫寛容が獲得され、遺伝されないことのさらなる裏づけとなる。
【0128】
ヒト抗原に対する高親和性の完全ヒト抗体を産生しうることには、明らかな実用的意味がある。完全ヒト抗体は、マウスまたはマウス由来のMabに固有の免疫原性およびアレルギー性反応を最小限に抑え、このため投与される抗体の有効性および安全性を高めると考えられる。XenoMouse IIは、抗体の反復投与を必要とする、炎症、自己免疫および癌などの、慢性および再発性のヒト疾患の治療において実質的な利益が提供される可能性をもたらす。XenoMouse IIによって一連の完全ヒト高親和性抗体が得られる迅速性および再現性は、それがヒト抗体生産のための他の技術を上回る進歩をもたらす可能性があることを意味する。例えば、それに由来する抗体の多くの親和性を高めるために多大な努力を必要とし、1本鎖FvsまたはFabが得られるファージディスプレーとは異なり、XenoMouse IIの抗体は、さらなる操作を要せずにハイブリドーマから産生させることができる高親和性の完全に無傷な免疫グロブリンである。
【0129】
マウスにおいて、真性のヒト体液性免疫系を創出するための本明細書に記載した戦略は、マウス免疫系の他の機能を司る、T細胞受容体または主要組織適合抗原遺伝子複合体などの他の多遺伝子座のヒト化に向けても適用しうる(Jakobovits、1994)。このようなマウスは、ヒト遺伝子座の構造-機能関連および免疫系の発達におけるそれらの関与を解明する上で有用であると考えられる。
【0130】
参照としての組み入れ
特許、特許出願、論文、教科書などを含む本明細書中に引用したすべての参考文献、およびそれらの中で引用された参考文献は、それらが既知でない範囲において、その全体が参照として本明細書に組み入れられる。さらに、以下の参考文献も、このような参考文献において引用された参考文献を含め、その全体が参照として本明細書に組み入れられる。
【0131】
参考文献
【特許請求の範囲】
【請求項1】
明細書中に記載の発明。
【請求項1】
明細書中に記載の発明。
【図1】


【図2】
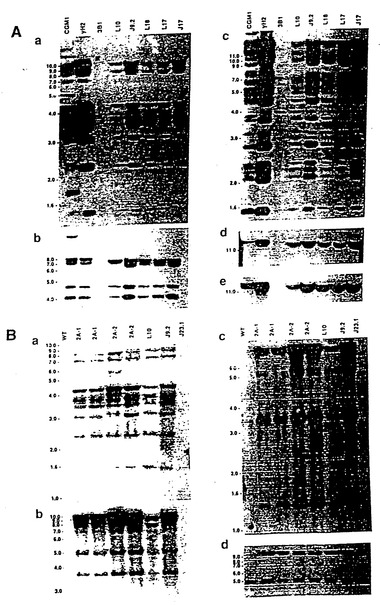
【図3】
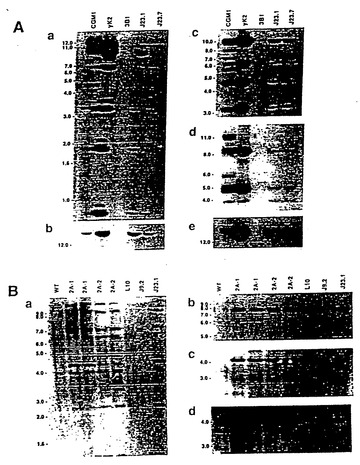
【図4】
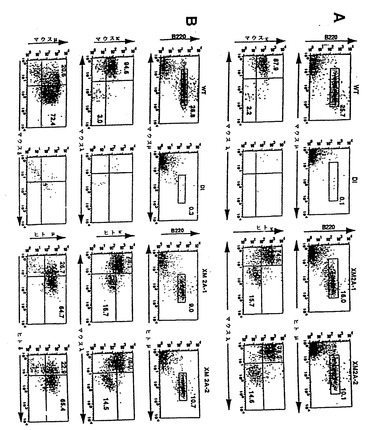
【図5】
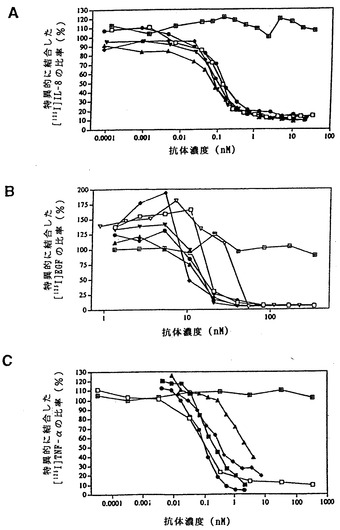
【図7】

【図8】

【図9】

【図10】
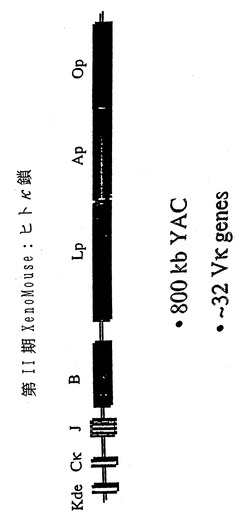
【図11】
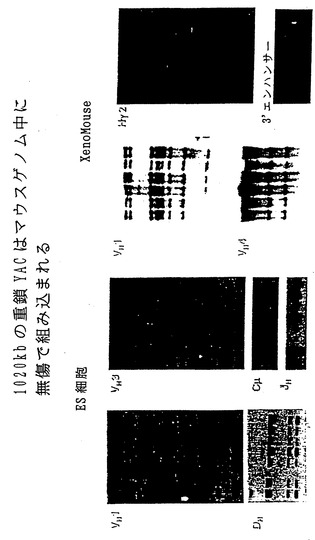
【図12】
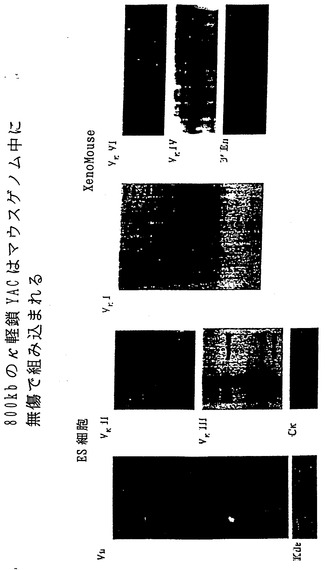
【図13】
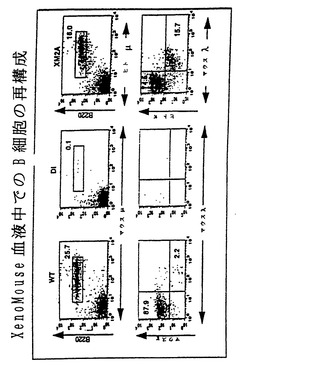
【図14】

【図15】

【図16】

【図17】
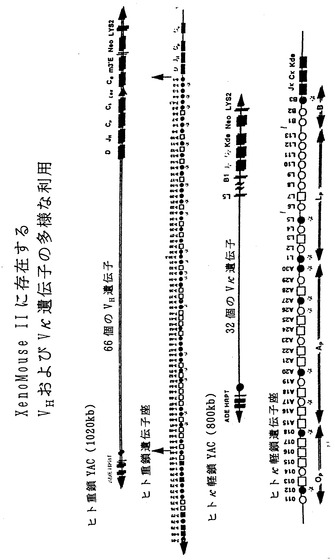
【図18】
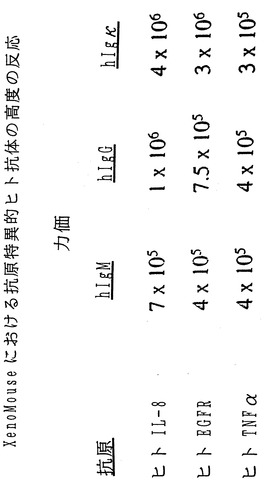
【図19】

【図20】

【図21】
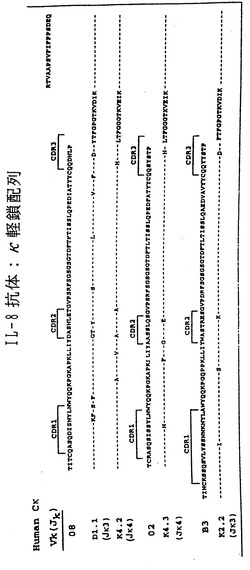
【図22】
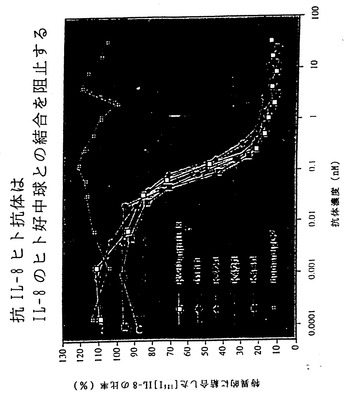
【図23】
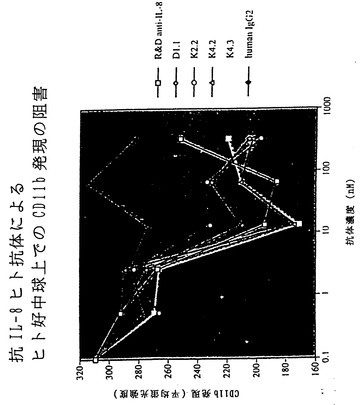
【図24】
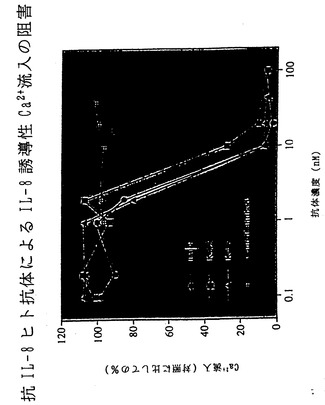
【図25】
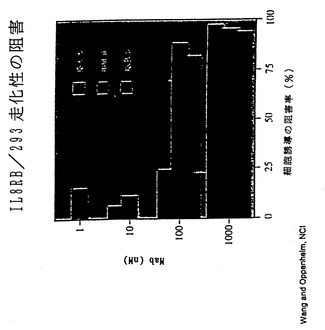
【図26】
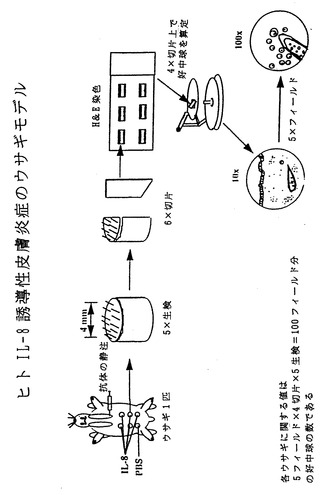
【図27】
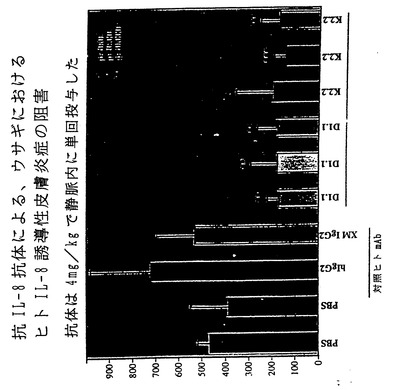
【図28】

【図29】
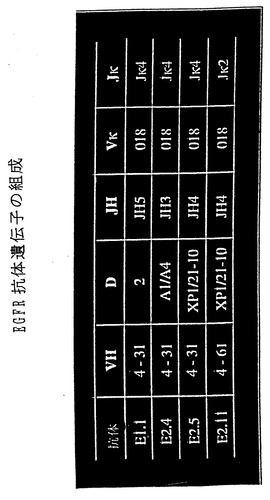
【図31】
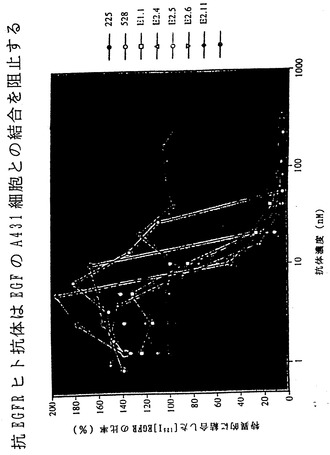
【図32】
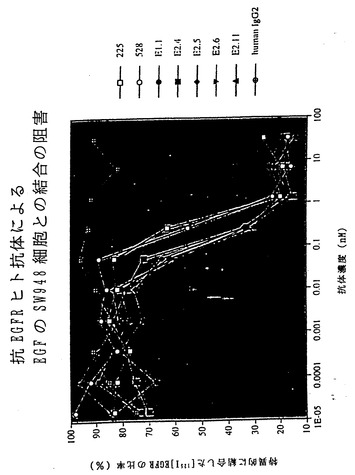
【図33】
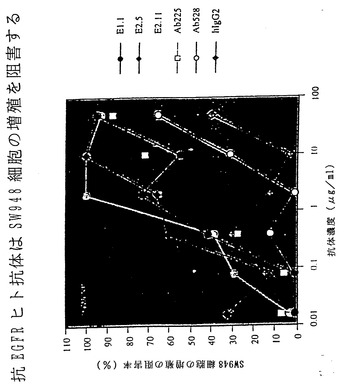
【図34】
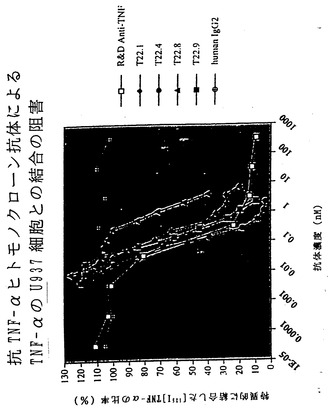
【図35】

【図6】
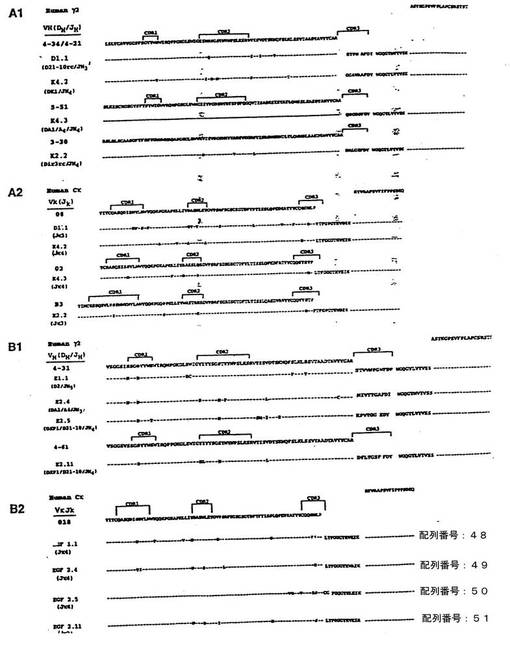
【図30】

【図2】
【図3】
【図4】
【図5】
【図7】
【図8】
【図9】
【図10】
【図11】
【図12】
【図13】
【図14】
【図15】
【図16】
【図17】
【図18】
【図19】
【図20】
【図21】
【図22】
【図23】
【図24】
【図25】
【図26】
【図27】
【図28】
【図29】
【図31】
【図32】
【図33】
【図34】
【図35】
【図6】
【図30】
【公開番号】特開2011−212019(P2011−212019A)
【公開日】平成23年10月27日(2011.10.27)
【国際特許分類】
【出願番号】特願2011−146571(P2011−146571)
【出願日】平成23年6月30日(2011.6.30)
【分割の表示】特願2008−124406(P2008−124406)の分割
【原出願日】平成9年12月3日(1997.12.3)
【出願人】(398005777)アムジェン フレモント インク. (40)
【Fターム(参考)】
【公開日】平成23年10月27日(2011.10.27)
【国際特許分類】
【出願日】平成23年6月30日(2011.6.30)
【分割の表示】特願2008−124406(P2008−124406)の分割
【原出願日】平成9年12月3日(1997.12.3)
【出願人】(398005777)アムジェン フレモント インク. (40)
【Fターム(参考)】
[ Back to top ]