
親水性ジオールデヒドラターゼ及び疎水性グリセロールデヒドラターゼを製造する方法
【課題】ジオールデヒドラターゼ及びグリセロールデヒドラターゼの酵素活性を維持したまま、これらの酵素の水に対する溶解性を変化する手段を提供する。
【解決手段】ジオールデヒドラターゼ又はグリセロールデヒドラターゼのβサブユニット及び/又はγサブユニットにおいてN末端アミノ酸を変異させることにより、ジオールデヒドラターゼ又はグリセロールデヒドラターゼの水に対する溶解性を変化させる方法。
【解決手段】ジオールデヒドラターゼ又はグリセロールデヒドラターゼのβサブユニット及び/又はγサブユニットにおいてN末端アミノ酸を変異させることにより、ジオールデヒドラターゼ又はグリセロールデヒドラターゼの水に対する溶解性を変化させる方法。
【発明の詳細な説明】
【技術分野】
【0001】
本発明は、ジオールデヒドラターゼ及びグリセロールデヒドラターゼの水に対する溶解性を変化させる方法、水に対する溶解性が変化したジオールデヒドラターゼ及びグリセロールデヒドラターゼ、並びにこれらの酵素により1位及び2位に水酸基を有する脂肪族ポリオールを脱水する方法に関する。
【背景技術】
【0002】
ジオールデヒドラターゼは、1,2−ジオールを脱水して対応するアルデヒドに変換する反応を、アデノシルコバラミン(補酵素B12)依存的に触媒する酵素として知られている(非特許文献1及び2)。ジオールデヒドラターゼは、Klebsiella属細菌及びCitrobacter属細菌などにおいて、特に、1,2−ジオールの存在下、好気的条件での増殖において誘導される(非特許文献3〜5)。グリセロールも基質とされうるが、1,2−ジオールに比べて基質特異性は低い。ジオールデヒドラターゼは、60kDa、30kDa及び19kDaの3つのサブユニット(それぞれαサブユニット、βサブユニット及びγサブユニット)からなる分子量約22万の酵素であり、これらのサブユニットをコードする遺伝子はタンデムに並んでいることがわかっている(非特許文献6)。
【0003】
グリセロールデヒドラターゼは、ジオールデヒドラターゼとその触媒作用及びサブユニット構造などが類似しているが、免疫学的性質や基質特異性などが異なるアイソザイムである(非特許文献7〜9)。グリセロールデヒドラターゼは、61kDa、21kDa及び16kDaの3つのサブユニット(それぞれαサブユニット、βサブユニット及びγサブユニット)からなることがわかっている。
【0004】
いずれの酵素とも既にクローニングが行われ、大腸菌による大量発現系も確立されている(非特許文献10)。しかし、ジオールデヒドラターゼは、水に対する溶解性が低い酵素であるため(非特許文献11〜12)、組換え体ジオールデヒドラターゼを大量に得ることが難しいという問題、親水性の条件下での反応に用いることができないという問題があった。ジオールデヒドラターゼを水に可溶化する方法として、界面活性剤で処理する方法が知られているが、当該方法を用いると酵素活性が失われるという問題があった。
【0005】
一方、グリセロールデヒドラターゼは、水に対する溶解性が高い酵素であるため(非特許文献7及び13)、疎水性条件下での反応には適していないという問題があった。疎水性条件下でグリセロールを基質として反応を実施したいときに、グリセロールデヒドラターゼのかわりにジオールデヒドラターゼを使用することも考えられるが、ジオールデヒドラターゼはグリセロールデヒドラターゼに比べてグリセロールに対する基質特異性が低いため、やはりグリセロールデヒドラターゼを疎水性条件下での反応に用いる手段が望まれていた。
【0006】
従って、ジオールデヒドラターゼ及びグリセロールデヒドラターゼの酵素活性を維持したまま、これらの酵素の水に対する溶解性を変化する手段が望まれていた。
【0007】
ジオールデヒドラターゼ及びグリセロールデヒドラターゼの対応するサブユニットのアミノ酸配列を比較すると、βサブユニットのN末端の32アミノ酸、及びγサブユニットのN末端の37アミノ酸を除き、高度の相同性を有することが報告されている(非特許文献14)。しかし、上記のような互いに相違するアミノ酸領域がジオールデヒドラターゼの溶解性に影響を及ぼしているかは明らかではなく、また、これらのN末端アミノ酸領域を欠失させたジオールデヒドラターゼ又は付加したグリセロールデヒドラターゼが、それぞれ親水性条件下又は疎水性条件下で触媒活性を示すかについても明らかではなかった。
【0008】
【非特許文献1】Lee, H.A. et al., Jr., and Abeles, R.H., J. Biol. Chem., 238, 2367-2373 (1963)
【非特許文献2】Toraya, T. et al., Biochem. Biophys. Res. Commun., 69, 475-480 (1976)
【非特許文献3】Toraya, T. et al., J Bacteriol., 135, 726-729 (1978)
【非特許文献4】Toraya, T. et al., J. Bacteriol., 141, 1439-1442 (1980)
【非特許文献5】Toraya, T. et al., "Chemistry and Biochemistry of B12," ed. Banerjee, R., John Wiley and Sons, New York, pp. 783-809 (1999)
【非特許文献6】Tobimatsu T, Hara T, Sakaguchi M, Kishimoto Y, Wada Y, Isoda M, Sakai T, Toraya T. J Biol Chem. 270(13), 7142-8 (1995)
【非特許文献7】Yamanishi, M. et al., J. Biochem., 269, 4484-4494 (2002)
【非特許文献8】Shibata, N. et al., Structure Fold Des., 7, 997-1008 (1999)
【非特許文献9】Toraya, T. et al., Chem. Rev., 103, 2095-2127 (2003)
【非特許文献10】Tobimatsu T, Sakai T, Hashida Y, Mizoguchi N, Miyoshi S, Toraya T. Arch Biochem Biophys. Nov 1;347(1):132-40 (1997)
【非特許文献11】McGee, D.E. et al., Biochemistry, 20, 4293-4298 (1981)
【非特許文献12】Tobimatsu, T. et al., Arch. Biochem. Biophys., 347, 132-140 (1997)
【非特許文献13】Stroinski, A. et al., Arch. Biochem. Biophys., 162, 321-330 (1974)
【非特許文献14】Tobimatsu, T. et al., J. Biol. Chem., 270, 7142-7148 (1995)
【発明の開示】
【発明が解決しようとする課題】
【0009】
本発明の課題は、ジオールデヒドラターゼ及びグリセロールデヒドラターゼの酵素活性を維持したまま、これらの酵素の水に対する溶解性を変化する手段を提供することである。
【課題を解決するための手段】
【0010】
本発明者らは、上記課題を解決すべく鋭意検討を行った結果、ジオールデヒドラターゼとグリセロールデヒドラターゼの各サブユニットのアミノ酸配列において相違するN末端アミノ酸の特定の領域がこれらの酵素の水に対する溶解性を決定づけていることを見いだし、本発明を完成するに至った。
【0011】
すなわち、本発明は以下の発明を包含する。
(1)ジオールデヒドラターゼ又はグリセロールデヒドラターゼのβサブユニット及び/又はγサブユニットにおいてN末端アミノ酸を変異させることにより、ジオールデヒドラターゼ又はグリセロールデヒドラターゼの水に対する溶解性を変化させる方法。
(2)N末端アミノ酸の変異が、
ジオールデヒドラターゼを構成するβサブユニット及び/又はγサブユニットにおけるN末端アミノ酸を欠失させること、又は
グリセロールデヒドラターゼを構成するβサブユニット及び/又はγサブユニットにおけるN末端に、ジオールデヒドラターゼの対応するサブユニットのN末端アミノ酸を付加すること、
を含む、(1)記載の方法。
(3)ジオールデヒドラターゼのβサブユニット及び/又はγサブユニットにおけるN末端アミノ酸の変異が、
ジオールデヒドラターゼのβサブユニットのN末端の9〜64個のアミノ酸を欠失させること、及び/又は
ジオールデヒドラターゼのγサブユニットのN末端の5〜42個のアミノ酸を欠失させること
を含む(2)記載の方法。
(4)グリセロールデヒドラターゼのβサブユニット及び/又はγサブユニットにおけるN末端アミノ酸の変異が、
グリセロールデヒドラターゼのβサブユニットのN末端に、ジオールデヒドラターゼのβサブユニットのN末端の9〜64個のアミノ酸を付加すること、
グリセロールデヒドラターゼのγサブユニットのN末端に、ジオールデヒドラターゼのγサブユニットのN末端の5〜42個のアミノ酸を付加すること
を含む(2)記載の方法。
(5)αサブユニット、βサブユニット及びγサブユニットを含むジオールデヒドラターゼであって、
βサブユニットのN末端の9〜64個のアミノ酸が欠失しており、及び/又はγサブユニットのN末端の5〜42個のアミノ酸が欠失しており、かつ水溶性である前記ジオールデヒドラターゼ。
(6)αサブユニット、βサブユニット及びγサブユニットを含むジオールデヒドラターゼであって、
αサブユニットが以下の(a)又は(b)のタンパク質:
(a)配列番号1のアミノ酸配列からなるタンパク質、
(b)配列番号1において1若しくは数個のアミノ酸が欠失、置換若しくは付加されたアミノ酸配列からなるタンパク質であって、ジオールデヒドラターゼのβサブユニット及びγサブユニットと会合したときにジオールデヒドラターゼ活性を有する前記タンパク質、
であり、
βサブユニットが以下の(c)〜(f)から選択されるタンパク質:
(c)配列番号3のアミノ酸配列からなるタンパク質、
(d)配列番号3において1若しくは数個のアミノ酸が欠失、置換若しくは付加されたアミノ酸配列からなるタンパク質であって、ジオールデヒドラターゼのαサブユニット及びγサブユニットと会合したときにジオールデヒドラターゼ活性を有する前記タンパク質、
(e)配列番号3においてN末端の9〜64個のアミノ酸が欠失したアミノ酸配列からなるタンパク質、
(f)(e)に記載のアミノ酸配列において1若しくは数個のアミノ酸が欠失、置換若しくは付加されたアミノ酸配列からなるタンパク質であって、ジオールデヒドラターゼのαサブユニット及びγサブユニットと会合したときにジオールデヒドラターゼ活性を有する前記タンパク質、
であり、
γサブユニットが以下の(g)〜(j)から選択されるタンパク質:
(g)配列番号5のアミノ酸配列からなるタンパク質、
(h)配列番号5において1若しくは数個のアミノ酸が欠失、置換若しくは付加されたアミノ酸配列からなるタンパク質であって、ジオールデヒドラターゼのαサブユニット及びβサブユニットと会合したときにジオールデヒドラターゼ活性を有する前記タンパク質、
(i)配列番号5においてN末端の5〜42個のアミノ酸が欠失したアミノ酸配列からなるタンパク質、
(j)(i)に記載アミノ酸配列において1若しくは数個のアミノ酸が欠失、置換若しくは付加されたアミノ酸配列からなるタンパク質であって、ジオールデヒドラターゼのαサブユニット及びβサブユニットと会合したときにジオールデヒドラターゼ活性を有する前記タンパク質
であり、
ただし、βサブユニットが(c)又は(d)のタンパク質であるときγサブユニットは(g)及び(h)のタンパク質ではない、前記ジオールデヒドラターゼ。
(7)(5)又は(6)のジオールデヒドラターゼをコードする遺伝子。
(8)(7)記載の遺伝子を含むベクター。
(9)(8)記載のベクターを含む形質転換体。
(10)αサブユニット、βサブユニット及びγサブユニットを含むグリセロールデヒドラターゼにおいて、
βサブユニットのN末端にジオールデヒドラターゼのβサブユニットのN末端の9〜64個のアミノ酸が付加しており、及び/又はγサブユニットのN末端にジオールデヒドラターゼのβサブユニットのN末端の9〜64個のアミノ酸が付加しており、かつ水に不溶性である前記グリセロールデヒドラターゼ。
(11)αサブユニット、βサブユニット及びγサブユニットを含むグリセロールデヒドラターゼにおいて、
αサブユニットが以下の(a)又は(b)のタンパク質:
(a)配列番号7のアミノ酸配列からなるタンパク質、
(b)配列番号7において1若しくは数個のアミノ酸が欠失、置換若しくは付加されたアミノ酸配列からなるタンパク質であって、グリセロールデヒドラターゼのβサブユニット及びγサブユニットと会合したときにグリセロールデヒドラターゼ活性を有する前記タンパク質、
であり、
βサブユニットが以下の(c)〜(f)から選択されるタンパク質:
(c)配列番号9のアミノ酸配列からなるタンパク質、
(d)配列番号9において1若しくは数個のアミノ酸が欠失、置換若しくは付加されたアミノ酸配列からなるタンパク質であって、グリセロールデヒドラターゼのαサブユニット及びγサブユニットと会合したときにグリセロールデヒドラターゼ活性を有する前記タンパク質、
(e)配列番号9のN末端に配列番号3のN末端の9〜64個のアミノ酸が付加してなるアミノ酸配列からなるタンパク質、
(f)(e)に記載のアミノ酸配列において1若しくは数個のアミノ酸が欠失、置換若しくは付加されたアミノ酸配列からなるタンパク質であって、グリセロールデヒドラターゼのαサブユニット及びγサブユニットと会合したときにグリセロールデヒドラターゼ活性を有する前記タンパク質、
であり、
γサブユニットが以下の(g)〜(j)から選択されるタンパク質:
(g)配列番号11のアミノ酸配列からなるタンパク質、
(h)配列番号11において1若しくは数個のアミノ酸が欠失、置換若しくは付加されたアミノ酸配列からなるタンパク質であって、グリセロールデヒドラターゼのαサブユニット及びβサブユニットと会合したときにグリセロールデヒドラターゼ活性を有する前記タンパク質、
(i)配列番号11のN末端に配列番号5のN末端の5〜42個のアミノ酸が付加してなるアミノ酸配列からなるタンパク質、
(j)(i)に記載アミノ酸配列において1若しくは数個のアミノ酸が欠失、置換若しくは付加されたアミノ酸配列からなるタンパク質であって、グリセロールデヒドラターゼのαサブユニット及びβサブユニットと会合したときにグリセロールデヒドラターゼ活性を有する前記タンパク質
であり、
ただし、βサブユニットが(c)又は(d)のタンパク質であるときγサブユニットは(g)及び(h)のタンパク質ではない、前記グリセロールデヒドラターゼ。
(12)(10)又は(11)記載のグリセロールデヒドラターゼをコードする遺伝子。
(13)(12)記載の遺伝子を含むベクター。
(14)(13)記載のベクターを含む形質転換体。
(15)以下の(a)又は(b)の遺伝子:
(a)配列番号17の塩基配列からなる遺伝子、
(b)配列番号17で表される塩基配列からなる遺伝子に対し相補的な塩基配列からなる遺伝子とストリンジェントな条件下でハイブリダイズし、かつグリセロールデヒドラターゼの活性を有するタンパク質をコードする遺伝子。
(16)(15)記載の遺伝子を含むベクター。
(17)(16)記載のベクターを含む形質転換体。
(18)αサブユニット、βサブユニット及びγサブユニットを含むグリセロールデヒドラターゼにおいて、
αサブユニットが以下の(a)又は(b)のタンパク質:
(a)配列番号18のアミノ酸配列からなるタンパク質、
(b)配列番号18において1若しくは数個のアミノ酸が欠失、置換若しくは付加されたアミノ酸配列からなるタンパク質であって、グリセロールデヒドラターゼのβサブユニット及びγサブユニットと会合したときにグリセロールデヒドラターゼ活性を有する前記タンパク質、
であり、
βサブユニットが以下の(c)〜(f)から選択されるタンパク質:
(c)配列番号9のアミノ酸配列からなるタンパク質、
(d)配列番号9において1若しくは数個のアミノ酸が欠失、置換若しくは付加されたアミノ酸配列からなるタンパク質であって、グリセロールデヒドラターゼのαサブユニット及びγサブユニットと会合したときにグリセロールデヒドラターゼ活性を有する前記タンパク質、
(e)配列番号19のアミノ酸配列からなるタンパク質、
(f)配列番号19において1若しくは数個のアミノ酸が欠失、置換若しくは付加されたアミノ酸配列からなるタンパク質であって、グリセロールデヒドラターゼのαサブユニット及びγサブユニットと会合したときにグリセロールデヒドラターゼ活性を有する前記タンパク質、
であり、
γサブユニットが以下の(g)〜(j)から選択されるタンパク質:
(g)配列番号11のアミノ酸配列からなるタンパク質、
(h)配列番号11において1若しくは数個のアミノ酸が欠失、置換若しくは付加されたアミノ酸配列からなるタンパク質であって、グリセロールデヒドラターゼのαサブユニット及びβサブユニットと会合したときにグリセロールデヒドラターゼ活性を有する前記タンパク質、
(i)配列番号20のアミノ酸配列からなるタンパク質、
(j)配列番号20において1若しくは数個のアミノ酸が欠失、置換若しくは付加されたアミノ酸配列からなるタンパク質であって、グリセロールデヒドラターゼのαサブユニット及びβサブユニットと会合したときにグリセロールデヒドラターゼ活性を有する前記タンパク質、
であり、
ただし、βサブユニットが(c)又は(d)のタンパク質であるときγサブユニットは(g)及び(h)のタンパク質ではない、前記グリセロールデヒドラターゼ。
(19)1位及び2位に水酸基を有する脂肪族ポリオールを脱水して、1位にアルデヒド基を有するアルデヒドを生成する方法あって、
1位及び2位に水酸基を有する脂肪族ポリオールと(5)若しくは(6)記載のジオールデヒドラターゼ及び/又は(10)、(11)若しくは(18)記載のグリセロールデヒドラターゼとを反応させることを含む前記方法。
(20)1位及び2位に水酸基を有する脂肪族ポリオールを脱水して、1位にアルデヒド基を有するアルデヒドを生成する方法あって、
1位及び2位に水酸基を有する脂肪族ポリオールの存在下、(7)記載のジオールデヒドラターゼをコードする遺伝子及び/又は(12)若しくは(15)記載のグリセロールデヒドラターゼをコードする遺伝子を含む形質転換体を培養することを含む前記方法。
【発明の効果】
【0012】
本発明により、ジオールデヒドラターゼ及びグリセロールデヒドラターゼの水に対する溶解性を、酵素活性を維持しつつ変化させることができる。
【発明を実施するための最良の形態】
【0013】
本発明は、グリセロールデヒドラターゼのβサブユニット及び/又はγサブユニットにおいてN末端アミノ酸を変異させることにより、ジオールデヒドラターゼ又はグリセロールデヒドラターゼの水に対する溶解性を変化させる方法に関する。
【0014】
本発明において、ジオールデヒドラターゼとは、当技術分野で通常用いられる意味を有し、すなわち、1,2−ジオールを脱水して対応するアルデヒド及び水へ変換する反応を、アデノシルコバラミン(補酵素B12)依存的に触媒する活性を有する酵素である。ジオールデヒドラターゼは、60kDa、30kDa及び19kDaの3つのサブユニット(それぞれαサブユニット、βサブユニット及びγサブユニット)を含む。
【0015】
本発明において、グリセロールデヒドラターゼは、当技術分野で通常用いられる意味を有し、すなわち、グリセロールを脱水して3−ヒドロキシプロピオンアルデヒド及び水へ変換する反応を、アデノシルコバラミン(補酵素B12)の存在下で触媒する活性を有する酵素である。グリセロールデヒドラターゼは、61kDa、21kDa及び16kDaの3つのサブユニット(それぞれαサブユニット、βサブユニット及びγサブユニット)を含む。
【0016】
ジオールデヒドラターゼ及びグリセロールデヒドラターゼにおいては、上記α、β、γサブユニットと補酵素が会合することにより、酵素活性が発揮される。
【0017】
ジオールデヒドラターゼ及びグリセロールデヒドラターゼは、上記酵素活性を有するものであれば特に限定されないが、例えば、Lactobacillus属、Citrobacter属、Clostridium属、Klebsiella属、Enterobacter属、Caloramator属、Salmonella属及びListeria属等に属する細菌に由来するものが挙げられる。
【0018】
本発明の方法では、上記ジオールデヒドラターゼ及びグリセロールデヒドラターゼのβ及び/又はγサブユニットにおいてN末端アミノ酸を変異させることにより、酵素の水に対する溶解性を変化させる。
【0019】
ジオールデヒドラターゼにおけるN末端アミノ酸の変異は、好ましくは、βサブユニット及び/又はγサブユニットにおけるN末端アミノ酸を欠失させることにより行う。βサブユニットにおいて欠失させるN末端アミノ酸は、N末端側から9〜64個、好ましくは14〜35個、より好ましくは18〜25個である。γサブユニットにおいて欠失させるN末端アミノ酸は、N末端側から5〜42個、好ましくは10〜35個、より好ましくは15〜25個である。
【0020】
グリセロールデヒドラターゼにおけるN末端アミノ酸の変異は、好ましくは、βサブユニット及び/又はγサブユニットにおけるN末端に、ジオールデヒドラターゼの対応するサブユニットのN末端アミノ酸を付加することにより実施できる。グリセロールデヒドラターゼのサブユニットに付加するジオールデヒドラターゼの対応するサブユニットのN末端アミノ酸は、どの属に属する細菌由来のものでもかまわないが、タンパク質発現を行う宿主と同じか近縁の科に属する細菌由来のもの、好ましくは同じか近縁の属に属する細菌由来のものとするのが好ましい。
【0021】
本発明の方法においては、各酵素のβサブユニット若しくはγサブユニットのいずれかにおいてのみN末端アミノ酸を変異させてもよく、又は双方のサブユニットにおいてN末端アミノ酸を変異させてもよい。いずれの場合にも本発明の効果が得られる。
【0022】
本発明はまた、上記方法によって得られた水に対する溶解性の変化した変異型ジオールデヒドラターゼ及び変異型グリセロールデヒドラターゼに関する。以下、β及び/又はγサブユニットのN末端アミノ酸が変異したジオールデヒドラターゼ及びグリセロールデヒドラターゼを、それぞれ変異型ジオールデヒドラターゼ及び変異型グリセロールデヒドラターゼと称し、N末端アミノ酸が変異していないジオールデヒドラターゼ及びグリセロールデヒドラターゼを、それぞれ野生型ジオールデヒドラターゼ及び野生型グリセロールデヒドラターゼと称する場合がある。
【0023】
すなわち本発明の一実施形態は、αサブユニット、βサブユニット及びγサブユニットを含むジオールデヒドラターゼであって、βサブユニットのN末端の9〜64個のアミノ酸が欠失しており、及び/又はγサブユニットのN末端の5〜42個のアミノ酸が欠失しており、かつ水溶性である前記ジオールデヒドラターゼに関する。ここでα又はγサブユニットにおいて欠失しているN末端アミノ酸の数については上記のとおりである。
【0024】
本発明の変異型ジオールデヒドラターゼには、βサブユニット及びγサブユニットのいずれかのN末端アミノ酸が欠失しているもの、並びにβサブユニット及びγサブユニットの双方のN末端アミノ酸が欠失しているものが包含される。
【0025】
本発明の変異型ジオールデヒドラターゼは、上記変異を含み水溶性であるとともに、ジオールデヒドラターゼ活性を有する限り特に限定されない。本発明において水溶性であるとは、変異型ジオールデヒドラターゼ遺伝子が発現したタンパク質の大部分が水に可溶であることを意味する。具体的には、酵素タンパク質を発現させた微生物菌体を適当な緩衝液に懸濁させて充分破砕した後、懸濁液を遠心分離した試料の上清部分に、酵素活性の大部分が存在することで確認できる。
【0026】
本発明の変異型ジオールデヒドラターゼとしては、例えば、Lactobacillus属、Citrobacter属、Clostridium属、Klebsiella属、Enterobacter属、Caloramator属、Salmonella属及びListeria属等に属する細菌、好ましくはKlebsiella属細菌、より好ましくはKlebsiella oxytocaに由来するジオールデヒドラターゼのβサブユニット及び/又はγサブユニットにおいて、N末端アミノ酸が欠失したジオールデヒドラターゼが挙げられる。
【0027】
野生型ジオールデヒドラターゼのα、β及びγサブユニットの公知のアミノ酸配列及び各サブユニットをコードする遺伝子の塩基配列は、公開されたデータベース、例えば、GenBank(http://www.ncbi.nlm.nih.gov)に、例えば、アクセッション番号:D45071で登録されている。
【0028】
より具体的には、野生型ジオールデヒドラターゼのαサブユニットのアミノ酸配列としては配列番号1のアミノ酸配列が挙げられ、αサブユニットをコードする遺伝子の塩基配列としては配列番号2の塩基配列が挙げられ、βサブユニットのアミノ酸配列としては配列番号3のアミノ酸配列が挙げられ、βサブユニットをコードする遺伝子の塩基配列としては配列番号4の塩基配列が挙げられ、γサブユニットのアミノ酸配列としては配列番号5のアミノ酸配列が挙げられ、γサブユニットをコードする遺伝子の塩基配列としては配列番号6の塩基配列が挙げられる。
【0029】
本発明の一実施形態において、変異型ジオールデヒドラターゼは、α、β及びγサブユニットを含み、
αサブユニットが、配列番号1のアミノ酸配列からなるタンパク質であり、
βサブユニットが、配列番号3のアミノ酸配列からなるタンパク質、又は配列番号3においてN末端の9〜64個、好ましくは14〜35個、さらに好ましくは18〜25個のアミノ酸が欠失したアミノ酸配列からなるタンパク質であり、
γサブユニットが、配列番号5のアミノ酸配列からなるタンパク質、又は配列番号5においてN末端の5〜42個、好ましくは10〜35個、さらに好ましくは15〜25個のアミノ酸が欠失したアミノ酸配列からなるタンパク質である。
【0030】
ただし、ここで、βサブユニットが配列番号3のアミノ酸配列からなるタンパク質であるときは、γサブユニットは配列番号5のアミノ酸配列からなるタンパク質ではない。
【0031】
本発明においてαサブユニット、βサブユニット及びγサブユニットには、野生型酵素における各サブユニットだけでなく、N末端アミノ酸が変異した変異型のサブユニットも包含される。
【0032】
上記のアミノ酸配列で特定される各サブユニットと機能的に同等のタンパク質を対応するサブユニットとして含む変異型ジオールデヒドラターゼもまた本発明に包含される。
【0033】
本発明において、各サブユニットと機能的に同等のタンパク質とは、当該タンパク質が各配列番号で特定されるタンパク質と同等の生物学的機能、生化学的機能を有することを指す。例えば、「配列番号1のアミノ酸配列からなるタンパク質」と機能的に同等のタンパク質としては、「配列番号1において1若しくは数個のアミノ酸が欠失、置換若しくは付加されたアミノ酸配列からなるタンパク質であって、ジオールデヒドラターゼのβサブユニット及びγサブユニットと会合したときにジオールデヒドラターゼ活性を有する前記タンパク質」が挙げられる。
【0034】
数個とは、通常2〜10個、好ましくは2〜5個、より好ましくは2〜3個をいう。また、例えば「ジオールデヒドラターゼのβサブユニット及びγサブユニットと会合したときにジオールデヒドラターゼ活性を有する前記タンパク質」とは、βサブユニット及びγサブユニット、少なくとも野生型のβサブユニット及びγサブユニットと会合して、野生型ジオールデヒドラターゼと同等の酵素活性を示すタンパク質、すなわち、さらにアデノシルコバラミン(補酵素B12)の存在下でジオールデヒドラターゼ活性を有するタンパク質を意味する。
【0035】
また、機能的に同等のタンパク質は、通常、アミノ酸配列レベルにおいて高い同一性を有する。高い同一性とは、アミノ酸レベルにおいて、通常、少なくとも80%以上の同一性、好ましくは90%以上の同一性、さらに好ましくは95%以上の同一性、さらに好ましくは99%以上の同一性を指す。
【0036】
より具体的には、本発明の一実施形態において、変異型ジオールデヒドラターゼは、αサブユニット、βサブユニット及びγサブユニットを含み、
αサブユニットが以下の(a)又は(b)のタンパク質:
(a)配列番号1のアミノ酸配列からなるタンパク質、
(b)配列番号1において1若しくは数個のアミノ酸が欠失、置換若しくは付加されたアミノ酸配列からなるタンパク質であって、ジオールデヒドラターゼのβサブユニット及びγサブユニットと会合したときにジオールデヒドラターゼ活性を有する前記タンパク質、
であり、
βサブユニットが以下の(c)〜(f)から選択されるタンパク質:
(c)配列番号3のアミノ酸配列からなるタンパク質、
(d)配列番号3において1若しくは数個のアミノ酸が欠失、置換若しくは付加されたアミノ酸配列からなるタンパク質であって、ジオールデヒドラターゼのαサブユニット及びγサブユニットと会合したときにジオールデヒドラターゼ活性を有する前記タンパク質、
(e)配列番号3においてN末端の9〜64個、好ましくは14〜35個、さらに好ましくは18〜25個のアミノ酸が欠失したアミノ酸配列からなるタンパク質、
(f)(e)に記載のアミノ酸配列において1若しくは数個のアミノ酸が欠失、置換若しくは付加されたアミノ酸配列からなるタンパク質であって、ジオールデヒドラターゼのαサブユニット及びγサブユニットと会合したときにジオールデヒドラターゼ活性を有する前記タンパク質、
であり、
γサブユニットが以下の(g)〜(j)から選択されるタンパク質:
(g)配列番号5のアミノ酸配列からなるタンパク質、
(h)配列番号5において1若しくは数個のアミノ酸が欠失、置換若しくは付加されたアミノ酸配列からなるタンパク質であって、ジオールデヒドラターゼのαサブユニット及びβサブユニットと会合したときにジオールデヒドラターゼ活性を有する前記タンパク質、
(i)配列番号5においてN末端の5〜42個、好ましくは10〜35個、さらに好ましくは15〜25個のアミノ酸が欠失したアミノ酸配列からなるタンパク質、
(j)(i)に記載アミノ酸配列において1若しくは数個のアミノ酸が欠失、置換若しくは付加されたアミノ酸配列からなるタンパク質であって、ジオールデヒドラターゼのαサブユニット及びβサブユニットと会合したときにジオールデヒドラターゼ活性を有する前記タンパク質
であり、
ただし、βサブユニットが(c)又は(d)のタンパク質であるときγサブユニットは(g)及び(h)のタンパク質ではない。
【0037】
本発明の変異型ジオールデヒドラターゼは、野生型ジオールデヒドラターゼと異なり水に可溶性であるが、野生型ジオールデヒドラターゼと同等の酵素活性を維持している。従って、組換え体ジオールデヒドラターゼを大量に得ることができるとともに、親水性の条件下で酵素反応を実施することができる。
【0038】
本発明の別の実施形態は、α、β及びγサブユニットを含む変異型グリセロールデヒドラターゼであって、βサブユニットのN末端にジオールデヒドラターゼのβサブユニットのN末端の9〜64個のアミノ酸が付加しており、及び/又はγサブユニットのN末端にジオールデヒドラターゼのβサブユニットのN末端の5〜42個アミノ酸が付加しており、かつ水に不溶性である前記変異型グリセロールデヒドラターゼに関する。ここでβ又はγサブユニットにおいてN末端に付加しているジオールデヒドラターゼの対応するサブユニットのN末端アミノ酸の数については上記のとおりである。
【0039】
本発明の変異型グリセロールデヒドラターゼには、βサブユニット及びγサブユニットのいずれかのN末端にアミノ酸付加を有するもの、並びにβサブユニット及びγサブユニットの双方のN末端にアミノ酸付加を有するものが包含される。いずれの場合にも本発明の効果が得られる。
【0040】
本発明の別の実施形態は、α、β及びγサブユニットを含む変異型グリセロールデヒドラターゼであって、βサブユニットのN末端の1〜50個のアミノ酸がジオールデヒドラターゼのβサブユニットのN末端の9〜70個のアミノ酸と置換されており、及び/又はγサブユニットのN末端の1〜20個のアミノ酸がジオールデヒドラターゼのβサブユニットのN末端の5〜50個のアミノ酸と置換されており、かつ水に不溶性である前記変異型グリセロールデヒドラターゼに関する。
【0041】
本発明の変異型グリセロールデヒドラターゼには、βサブユニット又はγサブユニットのいずれかのN末端のみ置換されるもの、並びにβサブユニット及びγサブユニットの双方のN末端が置換されるものが包含される。いずれの場合にも本発明の効果が得られる。
【0042】
本発明の変異型グリセロールデヒドラターゼは、上記変異を含み水に不溶性であるとともに、グリセロールデヒドラターゼ活性を有する限り特に限定されない。本発明において水に不溶性であるとは、変異型グリセロールデヒドラターゼ遺伝子が発現したタンパク質の大部分が水に不溶であることを意味する。具体的には、酵素タンパク質を発現させた微生物菌体を適当な緩衝液に懸濁させて充分破砕した後、懸濁液を遠心分離した試料の沈殿部分に、酵素活性の大部分が存在することで確認できる。
【0043】
本発明の変異型グリセロールデヒドラターゼとしては、例えば、Lactobacillus属、Citrobacter属、Clostridium属、Klebsiella属、Enterobacter属、Caloramator属、Salmonella属及びListeria属等に属する細菌、好ましくはKlebsiella属細菌、より好ましくはKlebsiella pneumoniaeに由来するグリセロールデヒドラターゼのβサブユニット及び/又はγサブユニットにおいて、N末端アミノ酸に付加又は置換を有するジオールデヒドラターゼが挙げられる。
【0044】
野生型グリセロールデヒドラターゼのα、β及びγサブユニットの公知のアミノ酸配列及び各サブユニットをコードする遺伝子の塩基配列は、公開されたデータベース、例えば、GenBank(http://www.ncbi.nlm.nih.gov)に、例えばアクセッション番号:U30903で登録されている。
【0045】
より具体的には、野生型グリセロールデヒドラターゼのαサブユニットのアミノ酸配列としては配列番号7のアミノ酸配列が挙げられ、αサブユニットをコードする遺伝子の塩基配列としては配列番号8の塩基配列が挙げられ、βサブユニットのアミノ酸配列としては配列番号9のアミノ酸配列が挙げられ、βサブユニットをコードする遺伝子の塩基配列としては配列番号10の塩基配列が挙げられ、γサブユニットのアミノ酸配列としては配列番号11のアミノ酸配列が挙げられ、γサブユニットをコードする遺伝子の塩基配列としては配列番号12の塩基配列が挙げられる。
【0046】
本発明の一実施形態において、変異型グリセロールデヒドラターゼは、α、β及びγサブユニットを含み、
αサブユニットが、配列番号7のアミノ酸配列からなるタンパク質であり、
βサブユニットが、配列番号9のアミノ酸配列からなるタンパク質、又は配列番号9のN末端に配列番号3のN末端の9〜64個、好ましくは35〜60個のアミノ酸が付加してなるアミノ酸配列からなるタンパク質、
γサブユニットが、配列番号11のアミノ酸配列からなるタンパク質、又は配列番号11のN末端に配列番号5のN末端の5〜42個、好ましくは10〜35個、さらに好ましくは15〜25個のアミノ酸が付加してなるアミノ酸配列からなるタンパク質である。
【0047】
ただし、ここで、βサブユニットが配列番号9のアミノ酸配列からなるタンパク質であるときは、γサブユニットは配列番号11のアミノ酸配列からなるタンパク質ではない。
【0048】
本発明の一実施形態において、変異型グリセロールデヒドラターゼは、α、β及びγサブユニットを含み、
αサブユニットが、配列番号7のアミノ酸配列からなるタンパク質であり、
βサブユニットが、配列番号9のアミノ酸配列からなるタンパク質、又は配列番号9のN末端の1〜50個、好ましくは20〜30個のアミノ酸が、配列番号3のN末端の9〜70個、好ましくは35〜64個、さらに好ましくは35〜62個のアミノ酸と置換されたアミノ酸配列からなるタンパク質、
γサブユニットが、配列番号11のアミノ酸配列からなるタンパク質、又は配列番号11のN末端の1〜20個、好ましくは1〜8個のアミノ酸が、配列番号5のN末端の5〜50個、好ましくは30〜45個のアミノ酸と置換されたアミノ酸配列からなるタンパク質である。
【0049】
ただし、ここで、βサブユニットが配列番号9のアミノ酸配列からなるタンパク質であるときは、γサブユニットは配列番号11のアミノ酸配列からなるタンパク質ではない。
【0050】
上記のアミノ酸配列で特定される各サブユニットと機能的に同等のタンパク質を対応するサブユニットとして含む変異型グリセロールデヒドラターゼもまた本発明に包含される。例えば、「配列番号7のアミノ酸配列からなるタンパク質」と機能的に同等のタンパク質としては、「配列番号7において1若しくは数個のアミノ酸が欠失、置換若しくは付加されたアミノ酸配列からなるタンパク質であって、グリセロールデヒドラターゼのβサブユニット及びγサブユニットと会合したときにグリセロールデヒドラターゼ活性を有する前記タンパク質」が挙げられる。上記のとおり、機能的に同等のタンパク質は、通常、アミノ酸配列レベルにおいて高い同一性を有する。
【0051】
また、例えば「グリセロールデヒドラターゼのβサブユニット及びγサブユニットと会合したときにグリセロールデヒドラターゼ活性を有する前記タンパク質」とは、βサブユニット及びγサブユニット、少なくとも野生型のβサブユニット及びγサブユニットと会合して、野生型グリセロールデヒドラターゼと同等の酵素活性を示すタンパク質、すなわち、さらにアデノシルコバラミン(補酵素B12)の存在下でグリセロールデヒドラターゼ活性を有するタンパク質を意味する。
【0052】
より具体的には、本発明の一実施形態において、変異型グリセロールデヒドラターゼは、αサブユニット、βサブユニット及びγサブユニットを含み、
αサブユニットが以下の(a)又は(b)のタンパク質:
(a)配列番号7のアミノ酸配列からなるタンパク質、
(b)配列番号7において1若しくは数個のアミノ酸が欠失、置換若しくは付加されたアミノ酸配列からなるタンパク質であって、グリセロールデヒドラターゼのβサブユニット及びγサブユニットと会合したときにグリセロールデヒドラターゼ活性を有する前記タンパク質、
であり、
βサブユニットが以下の(c)〜(f)から選択されるタンパク質:
(c)配列番号9のアミノ酸配列からなるタンパク質、
(d)配列番号9において1若しくは数個のアミノ酸が欠失、置換若しくは付加されたアミノ酸配列からなるタンパク質であって、グリセロールデヒドラターゼのαサブユニット及びγサブユニットと会合したときにグリセロールデヒドラターゼ活性を有する前記タンパク質、
(e)配列番号9のN末端に配列番号3のN末端の9〜64個、好ましくは35〜60個のアミノ酸が付加してなるアミノ酸配列からなるタンパク質、
(f)(e)に記載のアミノ酸配列において1若しくは数個のアミノ酸が欠失、置換若しくは付加されたアミノ酸配列からなるタンパク質であって、グリセロールデヒドラターゼのαサブユニット及びγサブユニットと会合したときにグリセロールデヒドラターゼ活性を有する前記タンパク質、
であり、
γサブユニットが以下の(g)〜(j)から選択されるタンパク質:
(g)配列番号11のアミノ酸配列からなるタンパク質、
(h)配列番号11において1若しくは数個のアミノ酸が欠失、置換若しくは付加されたアミノ酸配列からなるタンパク質であって、グリセロールデヒドラターゼのαサブユニット及びβサブユニットと会合したときにグリセロールデヒドラターゼ活性を有する前記タンパク質、
(i)配列番号11のN末端に配列番号5のN末端の5〜42個、好ましくは10〜35個、さらに好ましくは15〜25個のアミノ酸が付加してなるアミノ酸配列からなるタンパク質、
(j)(i)に記載アミノ酸配列において1若しくは数個のアミノ酸が欠失、置換若しくは付加されたアミノ酸配列からなるタンパク質であって、グリセロールデヒドラターゼのαサブユニット及びβサブユニットと会合したときにグリセロールデヒドラターゼ活性を有する前記タンパク質であり、
ただし、βサブユニットが(c)又は(d)のタンパク質であるときγサブユニットは(g)及び(h)のタンパク質ではない。
【0053】
一実施形態において本発明の変異型グリセロールデヒドラターゼは、αサブユニット、βサブユニット及びγサブユニットを含み、
αサブユニットが以下の(a)又は(b)のタンパク質:
(a)配列番号18のアミノ酸配列からなるタンパク質、
(b)配列番号18において1若しくは数個のアミノ酸が欠失、置換若しくは付加されたアミノ酸配列からなるタンパク質であって、グリセロールデヒドラターゼのβサブユニット及びγサブユニットと会合したときにグリセロールデヒドラターゼ活性を有する前記タンパク質、
であり、
βサブユニットが以下の(c)〜(f)から選択されるタンパク質:
(c)配列番号9のアミノ酸配列からなるタンパク質、
(d)配列番号9において1若しくは数個のアミノ酸が欠失、置換若しくは付加されたアミノ酸配列からなるタンパク質であって、グリセロールデヒドラターゼのαサブユニット及びγサブユニットと会合したときにグリセロールデヒドラターゼ活性を有する前記タンパク質、
(e)配列番号19のアミノ酸配列からなるタンパク質、
(f)配列番号19において1若しくは数個のアミノ酸が欠失、置換若しくは付加されたアミノ酸配列からなるタンパク質であって、グリセロールデヒドラターゼのαサブユニット及びγサブユニットと会合したときにグリセロールデヒドラターゼ活性を有する前記タンパク質、
であり、
γサブユニットが以下の(g)〜(j)から選択されるタンパク質:
(g)配列番号11のアミノ酸配列からなるタンパク質、
(h)配列番号11において1若しくは数個のアミノ酸が欠失、置換若しくは付加されたアミノ酸配列からなるタンパク質であって、グリセロールデヒドラターゼのαサブユニット及びβサブユニットと会合したときにグリセロールデヒドラターゼ活性を有する前記タンパク質、
(i)配列番号20のアミノ酸配列からなるタンパク質、
(j)配列番号20において1若しくは数個のアミノ酸が欠失、置換若しくは付加されたアミノ酸配列からなるタンパク質であって、グリセロールデヒドラターゼのαサブユニット及びβサブユニットと会合したときにグリセロールデヒドラターゼ活性を有する前記タンパク質、
であり、
ただし、βサブユニットが(c)又は(d)のタンパク質であるときγサブユニットは(g)及び(h)のタンパク質ではない。
【0054】
グリセロールデヒドラターゼ活性を有するタンパク質とは、野生型グリセロールデヒドラターゼと同等の酵素活性を示すタンパク質、すなわち、さらにアデノシルコバラミン(補酵素B12)の存在下でグリセロールデヒドラターゼ活性を有するタンパク質を意味する。数個とは、通常2〜10個、好ましくは2〜5個、より好ましくは2〜3個をいう。上記各サブユニットを構成するタンパク質には、各アミノ酸配列からなるタンパク質と機能的に同等のタンパク質、すなわち、アミノ酸配列レベルにおいて高い同一性を有するタンパク質であって、各サブユニットの活性を有するタンパク質も包含される。高い同一性とは、上記のとおりアミノ酸レベルにおいて、通常、少なくとも80%以上の同一性、好ましくは90%以上の同一性、さらに好ましくは95%以上の同一性、さらに好ましくは99%以上の同一性を指す。
【0055】
本発明の変異型グリセロールデヒドラターゼは、水に不溶性であるため、疎水性条件下でも酵素反応を実施することができる。従って、疎水性条件下でグリセロールを基質として反応を実施したい場合に、好適である。
【0056】
β及び/又はγサブユニットのN末端アミノ酸に変異を有するジオールデヒドラターゼ及びグリセロールデヒドラターゼは、当技術分野で公知の方法により調製できる。
【0057】
ジオールデヒドラターゼのN末端アミノ酸が欠失したサブユニットは、例えば、公知のジオールデヒドラターゼをプロテーゼで処理することにより得られる。プロテアーゼとしては、トリプシン、ペプシン、キモトリプシン、ズブチリシン、スロンビン、サーモリシン、パパイン、プラスミン等が挙げられる。例えば、ジオールデヒドラターゼをトリプシン処理し、得られた反応液の上清から水溶性のβ又はγサブユニットを得ることができる。プロテアーゼ処理の条件は、当業者であれば適宜設定することができるが、通常1.0μg/mlのトリプシンの存在下、約15℃で約10〜14時間、好ましくは約12時間反応させる。
【0058】
あるいは、本発明の変位型ジオールデヒドラターゼ及びグリセロールデヒドラターゼは、公知のジオールデヒドラターゼ又はグリセロールデヒドラターゼをコードする遺伝子、すなわち各サブユニットをコードする遺伝子の塩基配列に基づいて、当技術分野において通常用いられる方法により調製できる。すなわち、N末端アミノ酸が変異した各サブユニットをコードする遺伝子をベクターに挿入し、該ベクターで宿主細胞を形質転換し、該形質転換体を培養することにより調製できる。
【0059】
本発明の一実施形態は、本発明の変異型ジオールデヒドラターゼをコードする遺伝子、本発明の変異型グリセロールデヒドラターゼをコードする遺伝子、これら遺伝子の一方又は双方を含むベクター、該ベクターによって形質転換された、変異型ジオールデヒドラターゼをコードする遺伝子及び/又は変異型グリセロールデヒドラターゼをコードする遺伝子を含む形質転換体に関する。本発明において、「遺伝子」という用語には、DNAのみならずそのmRNAやcDNAも含むものとする。また、全長遺伝子のみならずESTも含むものとする。
【0060】
本発明の変異型ジオールデヒドラターゼをコードする遺伝子には、αサブユニットをコードする遺伝子、βサブユニットをコードする遺伝子、及びγサブユニットをコードする遺伝子を含み、N末端の9〜46個のアミノ酸が欠失しており、及び/又はγサブユニットのN末端の5〜42個のアミノ酸が欠失しており、かつ水溶性である前記ジオールデヒドラターゼをコードする遺伝子が包含される。
【0061】
一実施形態において、変異型ジオールデヒドラターゼをコードする遺伝子は、αサブユニットをコードする遺伝子、βサブユニットをコードする遺伝子、及びγサブユニットをコードする遺伝子を含み、
αサブユニットをコードする遺伝子が、配列番号2の塩基配列からなる遺伝子であり、
βサブユニットをコードする遺伝子が、配列番号4の塩基配列からなる遺伝子であるか、又は配列番号4の5’末端の27〜192個、より好ましくは42〜105個、さらに好ましくは54〜75個の塩基が欠失した塩基配列からなる遺伝子であり、
γサブユニットをコードする遺伝子が、配列番号6の塩基配列からなる遺伝子であるか、又は配列番号6の5’末端の15〜126個、より好ましくは30〜105個、さらに好ましくは45〜75個の塩基が欠失した塩基配列からなる遺伝子である。ただし、βサブユニットをコードする遺伝子が配列番号4の塩基配列からなる遺伝子である場合、γサブユニットをコードする遺伝子は配列番号6の塩基配列からなる遺伝子ではない。なお、欠失させる塩基の数は、読み枠を考慮して決定する。また、開始コドンの付加、選択した宿主内で発現しやすいようなN末端側の配列設定等の諸条件も考慮する。
【0062】
上記の各サブユニットをコードする遺伝子と機能的に同等の遺伝子を、対応するサブユニットをコードする遺伝子として含む変異型ジオールデヒドラターゼ遺伝子もまた本発明に包含される。
【0063】
あるタンパク質をコードする遺伝子と機能的に同等の遺伝子とは、当該タンパク質と機能的に同等のタンパク質をコードする遺伝子を意味し、当該遺伝子を調製する当業者によく知られた方法としては、ハイブリダイゼーション技術(Sambrook,J.et al.,Molecular Cloning 2nd ed.,9.47-9.58,Cold Spring Harbor Lab.press,1989)を利用する方法が挙げられる。
【0064】
例えば、αサブユニットをコードする遺伝子としての「配列番号2の塩基配列からなる遺伝子」と機能的に同等の遺伝子としては、「配列番号2の塩基配列の全部又は一部からなる遺伝子に対し相補的な塩基配列からなる遺伝子とストリンジェントな条件下でハイブリダイズし、かつジオールデヒドラターゼのβサブユニット及びγサブユニットと会合したときにジオールデヒドラターゼ活性を有するタンパク質をコードする遺伝子」が挙げられる。また、配列番号2の塩基配列と少なくとも65%の同一性、好ましくは少なくとも75%の同一性、より好ましくは少なくとも80%の同一性、さらに好ましくは少なくとも85%の同一性、最も好ましくは少なくとも95%の同一性を有する塩基配列からなる遺伝子も包含される。
【0065】
塩基配列の一部とは、各塩基配列の一部分の塩基配列であって、ストリンジェントな条件下でハイブリダイズさせるのに十分な塩基配列の長さを有するものであり、例えば、少なくとも15塩基、好ましくは少なくとも100塩基、より好ましくは少なくとも200塩基の配列である。好ましくは各塩基配列において連続する少なくとも15塩基、好ましくは少なくとも100塩基、より好ましくは少なくとも200塩基の配列である。ここで「連続する」とは、基準とする塩基配列のうち連続した塩基配列を含むことを意味する。
【0066】
本明細書において、ストリンジェントな条件とは、特異的なハイブリッドが形成され、非特異的なハイブリッドが形成されない条件をいい、すなわち、各塩基配列に対し高い相同性(相同性又は同一性が60%以上、好ましくは70%以上、より好ましくは80%以上、さらに好ましくは90%以上、最も好ましくは95%以上)を有するポリヌクレオチドがハイブリダイズする条件をいう。より具体的には、このような条件は、当該分野において周知慣用な手法、例えば、コロニーハイブリダイゼーション法、プラークハイブリダイゼーション法、マイクロアレイ法又はサザンブロットハイブリダイゼーション法などにおいて、具体的には、ポリヌクレオチドを固定化したメンブランを用いて、0.7〜1.0MのNaCl存在下、65℃でハイブリダイゼーションを行った後、0.1〜2倍濃度のSSC(Saline Sodium Citrate;150mM塩化ナトリウム、15mMクエン酸ナトリウム)溶液を用い、65℃でメンブランを洗浄することにより達成できる。
【0067】
また、塩基配列情報を基に合成したプライマーを用いる遺伝子増幅法、例えば、ポリメラーゼ連鎖反応(PCR)法、LAMP法などを利用して、各遺伝子と機能的に同等の遺伝子を単離することも可能である。
【0068】
より具体的には、一実施形態において、本発明の変異型ジオールデヒドラターゼをコードする遺伝子は、αサブユニットをコードする遺伝子、βサブユニットをコードする遺伝子、及びγサブユニットをコードする遺伝子を含み、
αサブユニットをコードする遺伝子が、以下の(a)又は(b)の遺伝子:
(a)配列番号2の塩基配列からなる遺伝子、
(b)配列番号2の塩基配列の全部又は一部からなる遺伝子に対し相補的な塩基配列からなる遺伝子とストリンジェントな条件下でハイブリダイズし、かつジオールデヒドラターゼのβサブユニット及びγサブユニットと会合したときにジオールデヒドラターゼ活性を有するタンパク質をコードする遺伝子
であり、
βサブユニットをコードする遺伝子が、以下の(c)〜(f)から選択される遺伝子:
(c)配列番号4の塩基配列からなる遺伝子、
(d)配列番号4の塩基配列の全部又は一部からなる遺伝子に対し相補的な塩基配列からなる遺伝子とストリンジェントな条件下でハイブリダイズし、かつジオールデヒドラターゼのαサブユニット及びγサブユニットと会合したときにジオールデヒドラターゼ活性を有するタンパク質をコードする遺伝子
(e)配列番号4の5’末端の27〜192個、より好ましくは42〜105個、さらに好ましくは54〜75個の塩基が欠失した塩基配列からなる遺伝子
(f)(e)に記載の塩基配列の全部又は一部からなる遺伝子に対し相補的な塩基配列からなる遺伝子とストリンジェントな条件下でハイブリダイズし、かつジオールデヒドラターゼのαサブユニット及びγサブユニットと会合したときにジオールデヒドラターゼ活性を有するタンパク質をコードする遺伝子
であり、
γサブユニットをコードする遺伝子が、以下の(g)〜(j)から選択される遺伝子:
(g)配列番号6の塩基配列からなる遺伝子、
(h)配列番号6の塩基配列の全部又は一部からなる遺伝子に対し相補的な塩基配列からなる遺伝子とストリンジェントな条件下でハイブリダイズし、かつジオールデヒドラターゼのαサブユニット及びβサブユニットと会合したときにジオールデヒドラターゼ活性を有するタンパク質をコードする遺伝子
(i)配列番号6の5’末端の15〜126個、より好ましくは30〜105個、さらに好ましくは45〜75個の塩基が欠失した塩基配列からなる遺伝子
(j)(i)に記載の塩基配列の全部又は一部からなる遺伝子に対し相補的な塩基配列からなる遺伝子とストリンジェントな条件下でハイブリダイズし、かつジオールデヒドラターゼのαサブユニット及びβサブユニットと会合したときにジオールデヒドラターゼ活性を有するタンパク質をコードする遺伝子
であるが、
βサブユニットをコードする遺伝子が(c)又は(d)であるときは、γサブユニットをコードする遺伝子は(g)及び(h)ではない。
【0069】
本発明の変異型グリセロールデヒドラターゼをコードする遺伝子には、αサブユニットをコードする遺伝子、βサブユニットをコードする遺伝子、及びγサブユニットをコードする遺伝子を含み、βサブユニットのN末端にジオールデヒドラターゼのβサブユニットのN末端の9〜64個のアミノ酸が付加しており、及び/又はγサブユニットのN末端にジオールデヒドラターゼのβサブユニットのN末端の5〜42個のアミノ酸が付加しており、かつ水に不溶性である前記グリセロールデヒドラターゼをコードする遺伝子が包含される。
【0070】
一実施形態において、変異型グリセロールデヒドラターゼをコードする遺伝子は、αサブユニットをコードする遺伝子、βサブユニットをコードする遺伝子、及びγサブユニットをコードする遺伝子を含み、
αサブユニットをコードする遺伝子が、配列番号8の塩基配列からなる遺伝子であり、
βサブユニットをコードする遺伝子が、配列番号10の塩基配列からなる遺伝子であるか、又は配列番号10の5’末端に、配列番号4の5’末端の27〜192個、より好ましくは105〜180個の塩基が付加した塩基配列からなる遺伝子であり、
γサブユニットをコードする遺伝子が、配列番号12の塩基配列からなる遺伝子であるか、又は配列番号12の5’末端に、配列番号6の5’末端の15〜126個、より好ましくは30〜105個、さらに好ましくは45〜75個の塩基が付加した塩基配列からなる遺伝子である。ただし、βサブユニットをコードする遺伝子が配列番号10の塩基配列からなる遺伝子である場合、γサブユニットをコードする遺伝子は配列番号12の塩基配列からなる遺伝子ではない。なお、付加する塩基の数は、読み枠を考慮して決定する。
【0071】
一実施形態において、変異型グリセロールデヒドラターゼをコードする遺伝子は、αサブユニットをコードする遺伝子、βサブユニットをコードする遺伝子、及びγサブユニットをコードする遺伝子を含み、
αサブユニットをコードする遺伝子が、配列番号8の塩基配列からなる遺伝子であり、
βサブユニットをコードする遺伝子が、配列番号10の塩基配列からなる遺伝子であるか、又は配列番号10の5’末端の3〜150個の塩基、より好ましくは30〜120個の塩基、さらに好ましくは60〜90個の塩基が、特に好ましくは81個の塩基が、配列番号4の5’末端の27〜210個、より好ましくは105〜192個、さらに好ましくは105〜186の塩基、特に好ましくは180の塩基と置換された塩基配列からなる遺伝子であり、
γサブユニットをコードする遺伝子が、配列番号12の塩基配列からなる遺伝子であるか、又は配列番号12の5’末端の1〜60個の塩基、より好ましくは1〜24個の塩基、特に好ましくは3個の塩基が、配列番号6の5’末端の15〜150個、より好ましくは90〜135個、特に好ましくは99塩基と置換された塩基配列からなる遺伝子である。
【0072】
ただし、βサブユニットをコードする遺伝子が配列番号10の塩基配列からなる遺伝子である場合、γサブユニットをコードする遺伝子は配列番号12の塩基配列からなる遺伝子ではない。なお、付加又は置換する塩基の数は、読み枠を考慮して決定する。
【0073】
上記の各サブユニットをコードする遺伝子と機能的に同等の遺伝子を、対応するサブユニットをコードする遺伝子として含む変異型グリセロールデヒドラターゼ遺伝子もまた本発明に包含される。
【0074】
例えば、αサブユニットをコードする遺伝子としての「配列番号8の塩基配列からなる遺伝子」と機能的に同等の遺伝子としては、「配列番号8の塩基配列の全部又は一部からなる遺伝子に対し相補的な塩基配列からなる遺伝子とストリンジェントな条件下でハイブリダイズし、かつグリセロールデヒドラターゼのβサブユニット及びγサブユニットと会合したときにグリセロールデヒドラターゼ活性を有するタンパク質をコードする遺伝子」が挙げられる。また、各配列番号の塩基配列と少なくとも65%の同一性、好ましくは少なくとも75%の同一性、より好ましくは少なくとも80%の同一性、さらに好ましくは少なくとも85%の同一性、最も好ましくは少なくとも95%の同一性を有する塩基配列からなる遺伝子も包含される。
【0075】
より具体的には、一実施形態において、本発明の変異型グリセロールデヒドラターゼをコードする遺伝子は、αサブユニットをコードする遺伝子、βサブユニットをコードする遺伝子、及びγサブユニットをコードする遺伝子を含み、
αサブユニットをコードする遺伝子が、以下の(a)又は(b)の遺伝子:
(a)配列番号8の塩基配列からなる遺伝子、
(b)配列番号8の塩基配列の全部又は一部からなる遺伝子に対し相補的な塩基配列からなる遺伝子とストリンジェントな条件下でハイブリダイズし、かつグリセロールデヒドラターゼのβサブユニット及びγサブユニットと会合したときにグリセロールデヒドラターゼ活性を有するタンパク質をコードする遺伝子
であり、
βサブユニットをコードする遺伝子が、以下の(c)〜(f)から選択される遺伝子:
(c)配列番号10の塩基配列からなる遺伝子、
(d)配列番号10の塩基配列の全部又は一部からなる遺伝子に対し相補的な塩基配列からなる遺伝子とストリンジェントな条件下でハイブリダイズし、かつグリセロールデヒドラターゼのαサブユニット及びγサブユニットと会合したときにグリセロールデヒドラターゼ活性を有するタンパク質をコードする遺伝子
(e)配列番号10の5’末端に、配列番号4の5’末端の27〜192個、より好ましくは105〜180個の塩基が付加した塩基配列からなる遺伝子
(f)(e)に記載の塩基配列の全部又は一部からなる遺伝子に対し相補的な塩基配列からなる遺伝子とストリンジェントな条件下でハイブリダイズし、かつグリセロールデヒドラターゼのαサブユニット及びγサブユニットと会合したときにグリセロールデヒドラターゼ活性を有するタンパク質をコードする遺伝子
であり、
γサブユニットをコードする遺伝子が、以下の(g)〜(j)から選択される遺伝子:
(g)配列番号12の塩基配列からなる遺伝子、
(h)配列番号12の塩基配列の全部又は一部からなる遺伝子に対し相補的な塩基配列からなる遺伝子とストリンジェントな条件下でハイブリダイズし、かつグリセロールデヒドラターゼのαサブユニット及びβサブユニットと会合したときにグリセロールデヒドラターゼ活性を有するタンパク質をコードする遺伝子
(i)配列番号12の5’末端に、配列番号6の5’末端の15〜126個、より好ましくは30〜105個、さらに好ましくは45〜75個の塩基が付加した塩基配列からなる遺伝子
(j)(i)に記載の塩基配列の全部又は一部からなる遺伝子に対し相補的な塩基配列からなる遺伝子とストリンジェントな条件下でハイブリダイズし、かつグリセロールデヒドラターゼのαサブユニット及びβサブユニットと会合したときにグリセロールデヒドラターゼ活性を有するタンパク質をコードする遺伝子
であるが、
βサブユニットをコードする遺伝子が(c)又は(d)であるときは、γサブユニットをコードする遺伝子は(g)及び(h)ではない。
【0076】
本発明の変異型グリセロールデヒドラターゼをコードする遺伝子の具体例としては、以下の(a)又は(b)の遺伝子が挙げられる:
(a)配列番号17の塩基配列からなる遺伝子、
(b)配列番号17で表される塩基配列からなる遺伝子に対し相補的な塩基配列からなる遺伝子とストリンジェントな条件下でハイブリダイズし、かつグリセロールデヒドラターゼの活性を有するタンパク質をコードする遺伝子。
【0077】
ここでストリンジェントな条件については、上記のとおりである。従って、本発明の変異型グリセロールデヒドラターゼをコードする遺伝子には、配列番号17の塩基配列に対し高い相同性(相同性又は同一性が60%以上、好ましくは70%以上、より好ましくは80%以上、さらに好ましくは90%以上、最も好ましくは95%以上)を有する塩基配列からなる遺伝子であって、グリセロールデヒドラターゼの活性を有するタンパク質をコードする遺伝子が包含される。グリセロールデヒドラターゼの活性を有するタンパク質については、上記のとおりである。
【0078】
本発明の変異型ジオールデヒドラターゼ及びグリセロールデヒドラターゼには、他のペプチドが融合されたものも含まれる。融合に付される他のペプチドとしては、例えばFLAG(Hopp, T. P. et. al., BioTechnology 6: 1204-1210,1988)、6個のHis(ヒスチジン)残基からなる6×His、10×His、ヒトc−mycの断片、VSV−GPの断片、p18HIVの断片、T7−tag、HSV−tag、E−tag、SV40T抗原の断片、lck tag、α−tubulinの断片、B−tag、Protein Cの断片、GST(グルタチオンS−トランスフェラーゼ)、HA(インフルエンザ凝集素)、イムノグロブリン定常領域、イムノグロブリンヒンジ領域、β−ガラクトシダーゼ、MBP(マルトース結合タンパク質)等が挙げられる。これらのペプチドを融合させることにより、レセプター誘導体の精製や標識を容易に実施することができる。
【0079】
本発明の変異型ジオールデヒドラターゼ及びグリセロールデヒドラターゼの調製に使用されるベクターとしては、当技術分野において一般的に使用されるものを用いることができる。例えば、形質転換における宿主として大腸菌を用いる場合には、例えば、大腸菌内で複製させるための「ori」、及び形質転換された大腸菌を選抜するための遺伝子(例えば、アンピシリンやテトラサイクリン、カナマイシン、クロラムフェニコール等の薬剤耐性遺伝子)をベクター上に有することが望ましく、このようなベクターとして具体的には、M13系ベクター、pUC系ベクター、pBR322、pBluescript、pCR−Script、pCR2.1、pGEM−T、pDIRECT、pT7等を挙げることができる。
【0080】
特に、発現ベクターが有用である。発現ベクターとしては、例えば大腸菌での発現を目的とした場合は、lacZプロモーター(Ward et. al., Nature 341: 544-546, 1989)、araBプロモーター(Better et. al., Science 240: 1041-1043, 1988)、又はT7プロモーター等を有するベクターを例示することができる。このようなベクターとしては、上記ベクターの他にpGEX−5X−1(ファルマシア)、「QIAexpress system」(キアゲン)、pEGFP、及びpET(この場合、宿主はT7 RNAポリメラーゼを発現するBL21が好ましい)等が挙げられる。また、発現ベクターには、ポリペプチド分泌のためのシグナル配列が含まれていてもよい。
【0081】
他のベクターとしては、例えば哺乳動物由来の発現ベクター(例えばpcDNA3(インビトロゲン)や、pEGF−BOS(Nucleic Acids. Res. 18(17): 5322, 1990)、pEF、pCDM8、昆虫細胞由来の発現ベクター(例えば「Bac−to−BAC baculovairus expression system」(ギブコBRL)、pBacPAK8)、植物由来の発現ベクター(例えばpMH1、pMH2)、動物ウィルス由来の発現ベクター(例えばpHSV、pMV、pAdexLcw)、レトロウィルス由来の発現ベクター(例えばpZIPneo)、酵母由来の発現ベクター(例えば「Pichia Expression Kit」(インビトロゲン)、pNV11 、SP−Q01)、枯草菌由来の発現ベクター(例えばpPL608、pKTH50)等が挙げられる。
【0082】
CHO細胞、COS細胞、NIH3T3細胞等の動物細胞での発現を目的とした場合には、細胞内で発現させるために必要なプロモーター、例えばSV40プロモーター(Mulligan et. al., Nature 277: 108, 1979)、MMLV−LTRプロモーター、EF1プロモーター(Mizushima et. al., Nucleic Acids Res. 18: 5322, 1990)、CMVプロモーター等を持っていることが不可欠であり、細胞への形質転換を選抜するための遺伝子(例えば薬剤(ネオマイシン、G418等)耐性遺伝子)を有すればさらに好ましい。このような特性を有するベクターとしては、例えばpMAM、pDR2、pBK−RSV、pBK−CMV、pOPRSV、pOP13等が挙げられる。
【0083】
また、複製開始点としては、ポリオーマウィルス、アデノウィルス、ウシパピローマウィルス(BPV)等の由来のものを用いることもできる。さらに、宿主細胞系で遺伝子コピー数増幅のために、発現ベクターは選択マーカーとして、アミノグリコシドトランスフェラーゼ(APH)遺伝子、チミジンキナーゼ(TK)遺伝子、大腸菌キサンチングアニンホスホリボシルトランスフェラーゼ(Ecogpt)遺伝子、ジヒドロ葉酸還元酵素(dhfr)遺伝子等を含むことができる。
【0084】
ベクターが導入される宿主としては特に制限はなく、例えば大腸菌や種々の真核細胞等を用いることが可能である。真核細胞を使用する場合、例えば動物細胞、植物細胞、真菌細胞を宿主に用いることができる。動物細胞としては、哺乳類細胞、例えばCHO、COS、3T3、ミエローマ、BHK(baby hamster kidney)、HeLa、Vero、両生類細胞、例えばアフリカツメガエル卵母細胞(Valle, et. al., Nature 291: 358-340, 1981)、あるいは昆虫細胞、例えばSf9、Sf21、Tn5が知られている。CHO細胞としては、特に、DHFR遺伝子を欠損したCHO細胞であるdhfr−CHO(Proc. Natl. Acad. Sci. USA 77: 4216-4220, 1980)やCHO K−1(Proc. Natl. Acad. Sci. USA 60: 1275, 1968)を好適に使用することができる。動物細胞において、大量発現を目的とする場合には特にCHO細胞が好ましい。植物細胞としては、例えばニコチアナ・タバカム(Nicotiana tabacum)由来の細胞が挙げられる。真菌細胞としては、酵母、例えばサッカロミセス(Saccharomyces)属、例えばサッカロミセス・セレビシエ(Saccharomyces cerevisiae)、糸状菌、例えばアスペルギルス(Aspergillus)属、例えばアスペルギルス・ニガー(Aspergillus niger)が知られている。原核細胞を使用する場合、細菌細胞としては、大腸菌(E. coli)、例えばJM109、DH5α、HB101、XL1Blue、BL21等が挙げられ、その他、枯草菌が知られている。
【0085】
また、宿主として動物を使用する場合、哺乳類動物、植物、昆虫が挙げられる。哺乳類動物としては、ヤギ、ブタ、ヒツジ、マウス、ウシを用いることができる(Vicki Glaser, SPECTRUM Biotechnology Applications, 1993)。また、植物を使用する場合、例えばタバコを用いることができる。さらに、昆虫としては、例えばカイコを用いることができる。
【0086】
本発明の形質転換体を作製するためには、上記宿主に上記ベクターを導入する。そのための方法としては、大腸菌等の宿主細胞へのベクターの導入の場合、例えば塩化カルシウム法、エレクトロポレーション法(Chu, G. et. al., Nucl. Acid Res. 15: 1311-1326, 1987)を用いることができる。また、培養細胞等の宿主細胞へのベクターの導入の場合、例えばリン酸カルシウム法(Chen, C. and Okayama, H. Mol. Cell. Biol. 7: 2745-2752, 1987)、DEAEデキストラン法(Lopata, M. A. et. al., Nucl. Acids Res. 12: 5707-5717, 1984、Sussman, D. J.and Milman, G. Mol. Cell. Biol. 4: 1642-1643, 1985)、カチオニックリボソームDOTAP(ベーリンガーマンハイム)を用いた方法、リポフェクチン法(Derijard, B. Cell 7: 1025-1037, 1994、Lamb, B. T. et. al., Nature Genetics 5: 22-30, 1993、Rabindran, S. K. et. al., Science 259: 230-234, 1993)等の方法を用いることが可能である。
【0087】
さらに、動物にDNAを導入する場合、該DNAを適当なベクター(例えばアデノウイルスベクター(例えばpAdexlcw)やレトロウイルスベクター(例えばpZIPneo)等が挙げられるが、これらに制限されない)に組み込み、例えばレトロウイルス法、リポソーム法、カチオニックリポソーム法、アデノウィルス法等により生体内に導入することが可能である。
【0088】
また、昆虫にベクターを導入する場合、例えば目的のタンパク質をコードするDNAを挿入したバキュロウィルスをカイコに感染させることにより行うことができる(Susumu, M. et. al., Nature 315: 592-594, 1985)。また植物にDNAを導入する場合、例えば目的とするタンパク質をコードするDNAを植物発現用ベクター、例えばpMON 530に挿入し、このベクターをアグロバクテリウム・ツメファシエンス(Agrobacterium tumefaciens)のようなバクテリアに導入する。このバクテリアをタバコ、例えばニコチアナ・タバカムに感染させることでベクターを導入することができる(Julian K.-C. Ma et.al., Eur. J. Immunol. 24: 131-138, 1994)。
【0089】
本発明に係る変異型ジオールデヒドラターゼ及びグリセロールデヒドラターゼは、上記の形質転換体を培養することにより生産させることができる。培養は公知の方法に従って行うことができる。例えば、動物細胞の培養であれば、一般的に、培養液としては、DMEM、MEM、RPMI1640、IMDM等を使用することができる。その際、牛胎児血清(FCS)等の血清補液を併用することもできるし、無血清培養してもよい。培養時のpHは、通常、約6〜8であるのが好ましい。培養は、通常、約30〜40℃で約15〜500時間行い、必要に応じて培地の交換、通気、攪拌を加える。
【0090】
本発明の変異型ジオールデヒドラターゼ及びグリセロールデヒドラターゼは、宿主細胞内又は細胞外(培地等)から単離し、実質的に純粋で均一なタンパク質として精製することができる。タンパク質の分離、精製は、通常のタンパク質の精製で使用されている分離、精製方法を使用すればよく、何ら限定されるものではない。例えばクロマトグラフィーカラム、フィルター、限外濾過、塩析、溶媒沈殿、溶媒抽出、蒸留、免疫沈降、SDS−ポリアクリルアミドゲル電気泳動、等電点電気泳動法、透析、再結晶等を適宜選択、組み合わせればタンパク質を分離、精製することができる。又は、さらにこれらのカラムを複数組み合わせることにより精製することが可能である。
【0091】
クロマトグラフィーとしては、例えば上記のエピトープタグに対する抗体をカラムに固定したアフィニティークロマトグラフィー、イオン交換クロマトグラフィー、疎水性クロマトグラフィー、ゲル濾過、逆相クロマトグラフィー、吸着クロマトグラフィー等が挙げられる(Strategies for Protein Purification and Characterization: A Laboratory Course Manual. Ed DanielR. Marshak et al., Cold Spring Harbor Laboratory Press, 1996)。
【0092】
また、本発明の変位型ジオールデヒドラターゼ及びグリセロールデヒドラターゼをグルタチオンS−トランスフェラーゼタンパク質との融合タンパク質として、あるいはヒスチジンを複数付加させた組み換えタンパク質として宿主細胞(例えば動物細胞や大腸菌等)内で発現させた場合には、発現させた組み換えタンパク質はグルタチオンカラム又はニッケルカラムを用いて精製することができる。融合タンパク質の精製後、必要に応じて融合タンパク質のうち、目的のタンパク質以外の領域を、トロンビン又はファクターXa等により切断し、除去することも可能である。
【0093】
本発明はまた、本発明の変異型ジオールデヒドラターゼ及び/又はグリセロールデヒドラターゼにより、1位及び2位に水酸基を有する脂肪族ポリオールを脱水して、1位にアルデヒド基を有するアルデヒドを生成する方法に関する。
【0094】
上記方法は、1位及び2位に水酸基を有する脂肪族ポリオールと本発明の変異型ジオールデヒドラターゼ及び/又はグリセロールデヒドラターゼとを反応させることにより実施できる。本発明の方法は、1位及び2位に水酸基を有する脂肪族ポリオールと変異型ジオールデヒドラターゼ又はグリセロールデヒドラターゼのいずれかとを反応させてもよいし、双方の酵素と反応させてもよい。
【0095】
酵素反応条件は、当業者であれば適宜設定できるが、通常、アデノシルコバラミン(補酵素B12)の存在下、0.1M程度の1,2-プロパンジオール水溶液と酵素を接触させることにより発生するプロピオンアルデヒドを検出することで酵素活性の測定をすることが可能である。活性確定後に、基質量に応じた酵素量を設定し反応条件を決定していく。
【0096】
あるいは、1位及び2位に水酸基を有する脂肪族ポリオール及びアデノシルコバラミン(補酵素B12)の存在下で、上記変異型ジオールデヒドラターゼ及び/又はグリセロールデヒドラターゼをコードする遺伝子を含む形質転換体を培養することにより実施できる。形質転換体は、変異型ジオールデヒドラターゼをコードする遺伝子のみ又は変異型グリセロールデヒドラターゼをコードする遺伝子のみを含んでいてもよいし、双方の遺伝子を含んでいてもよい。
【0097】
培養条件は、宿主の培養に用いられる通常の方法に従って、炭素源として1位及び2位に水酸基を有する脂肪族ポリオールを用いることにより行われる。例えば、比較的リッチな培地、例えば2培地等を用いて好気培養し、菌体量を増やしてから嫌気条件にし、炭素源を与えて発酵を行う。pHは、宿主の生育を妨害せず、かつ発酵液から酸を分離するときの障害とならない試薬を用いて調整する。炭酸ナトリウム、アンモニア、ナトリウムイオン供給源、例えば塩化ナトリウムを添加してもよい。また、水酸化ナトリウム水溶液、水酸化カリウム水溶液、水酸化ナトリウム水溶液、水酸化アンモニウム水溶液、水酸化カルシウム水溶液、炭酸カリウム水溶液、炭酸ナトリウム水溶液、酢酸カリウム水溶液等の一般的なアルカリ試薬を用いてもよい。培養期間中pHは、5.0〜8.0、好ましくは5.5〜7.5に保持する。培養は、通常5%CO2存在下、30〜40℃で1〜30日間行う。
【0098】
窒素源としては、例えば、アンモニア、塩化アンモニウム、硫酸アンモニウム、リン酸アンモニウム等のアンモニウム塩の他、ペプトン、肉エキス、酵母エキス、コーンスティープリカー等が挙げられる。また、無機物としては、例えば、リン酸第一カリウム、リン酸第二カリウム、リン酸マグネシウム、硫酸マグネシウム、塩化ナトリウム等が挙げられる。
【0099】
培養中は、カナマイシン、アンピシリン、テトラサイクリン等の抗生物質を培地に添加してもよい。誘導性のプロモーターを用いた発現ベクターで形質転換した微生物を培養する場合は、インデューサーを培地に添加することもできる。例えば、イソプロピル−β−D−チオガラクトピラノシド(IPTG)、インドールアクリル酸(IAA)等を培地に添加することができる。
【0100】
あるいは、上記において得られた形質転換体の培養物から遠心分離などによって集菌を行い、適当な緩衝液に懸濁する。この菌体懸濁液を1位及び2位に水酸基を有する脂肪族ポリオールを含む緩衝液に懸濁し、反応を行ってもよい。反応の条件は、例えば、反応温度は10〜80℃、好ましくは15〜50℃、反応時間は5分〜96時間、好ましくは10分〜72時間、pHは5.0〜8.0、好ましくは5.5〜7.5である。
【0101】
また、該培養物の処理物を用いて反応を行うこともできる。該処理物としては、菌体破砕物、菌体破砕物又は培養上清から調製した粗酵素、精製酵素等が挙げられる。また、常法により担体に固定化した菌体、該処理物、酵素等を用いることができる。
【0102】
1位及び2位に水酸基を有する脂肪族ポリオールとしては、1,2−ジオール(炭素数2〜6、好ましくは2〜4の1,2−アルカンジオールおよびその誘導体、例えば、1,2−プロパンジオール)及びグリセロール、1,2-ブタンジオール、エチレングリコール、1,2,3,4−ブタンテトラオール、1,2,3-ブタントリオール、1,2,4-ブタントリオール、1−モノグリセリド等が挙げられる。1,2−ジオールを基質とする場合には、ジオールデヒドラターゼが好ましく、グリセロールを基質とする場合には、グリセロールデヒドラターゼが好ましい。
【0103】
1位及び2位に水酸基を有する脂肪族ポリオールは、本発明の変異型ジオールデヒドラターゼ又はグリセロールデヒドラターゼの触媒作用により、1位にアルデヒド基を有するアルデヒドに変換される。具体的には、本発明の方法により、1,3−プロパンジオールはプロピオンアルデヒドに変換され、グリセロールは3−ヒドロキシプロピオンアルデヒドに変換される。
【0104】
本発明の変異型ジオールデヒドラターゼは水溶性であることから、親水性の条件下では、変異型ジオールデヒドラターゼよる反応を行うのが好ましい。また、本発明の変異型グリセロールデヒドラターゼは水に不溶性であることから、疎水性の条件下では、変異型グリセロールデヒドラターゼによる反応を行うのが好ましい。変異型ジオールデヒドラターゼ及びジオールデヒドラターゼを共存させて反応を行うことにより、反応系における親水場では変異型ジオールデヒドラターゼよる触媒反応を行い、さらに反応系における疎水場では変位型グリセロールデヒドラターゼによる触媒反応を行うことができる。また、野生型ジオールデヒドラターゼと変異型ジオールデヒドラターゼを共存させて反応させたり、野生型グリセロールデヒドラターゼと野生型グリセロールデヒドラターゼを共存させて反応させることにより、反応効率を改善することができる。
【0105】
本発明の方法は、その他の反応を含む複合的反応の一部として、1位及び2位に水酸基を有する脂肪族ポリオールを脱水して、1位にアルデヒド基を有するアルデヒドを生成する場合も包含する。例えば、Klebsiella属細菌及びCitrobacter属細菌などにおいては、グリセロールは2段階の酵素触媒反応を経て、1,3−プロパンジオールに変換されるが、第1段階において、グリセロールデヒドラターゼがグリセロールを3−ヒドロキシプロピオンアルデヒド(3−HPA)及び水へ変換し(グリセロール→3−HPA+H2O)、第2段階において、3−HPAがNAD+−結合オキシドレダクターゼにより1,3−プロパンジオールに還元される(3−HPA+NADH+H+→1,3−プロパンジオール+NAD+)。本発明の方法は、上記のような複合反応における第1段階の反応において利用することもできる。
【実施例】
【0106】
実施例1 組換え型ジオールデヒドラターゼのトリプシン処理
ジオールデヒドラターゼ遺伝子を発現している組換え体大腸菌を、2% 1,2−プロパンジオールを含む50mM リン酸カリウムバッファ(pH8.0)中で、超音波処理により破砕した。沈澱を同じバッファで3回洗浄した(Tobimatsu, T. et al., Arch. Biochem. Biophys., 347, 132-140 (1997))。得られた粗製の膜画分を種々の濃度のトリプシンを含む同一のバッファ中に、タンパク質濃度が5mg/mlとなるように再懸濁した。15℃で12時間インキュベートした後、過剰のダイズトリプシン阻害剤(400μg/ml)を添加することにより反応を停止した。得られた懸濁液を遠心分離し(16,000×g、30分)、可溶性画分(上清)及び沈澱画分を分離した。その後、上清の酵素活性(白色)及び沈澱画分の酵素活性(黒色)を測定した(図1A)。1.0μg/mlのトリプシンで処理することにより得られる可溶性画分に含まれるジオールデヒドラターゼ、すなわち水溶性の変異型ジオールデヒドラターゼは、酵素活性を失うことなく可溶化されていることがわかる。
【0107】
ジオールデヒドラターゼの活性は、3−メチル−2−ベンゾチアゾリノンヒドラゾン法により、基質として1,2−プロパンジオールを用いて測定した(Toraya, T. et al., J. Biol. Chem., 252, 963-970 (1977))。ジオールデヒドラターゼの1ユニットは、1分あたり1μmolのプロピオンアルデヒドの生成を触媒する酵素活性の量として定義した。野生型ジオールデヒドラターゼの溶解性は非常に低いことから、アッセイするホモジェネート、抽出物及び酵素溶液は、2% 1,2−プロパンジオール及び0.5〜1%のBrij35を含む50mMリン酸カリウムバッファ(pH8.0)で希釈した。
【0108】
トリプシン消化した後の、上清及び沈澱画分に含まれるタンパク質を、11%ゲル上のSDS−PAGEにより分析した(図1B)。その結果、沈澱画分においては、分子量30,000のβサブユニット、分子量19,000のγサブユニットが観察されるのに対し、1.0μg/mlのトリプシンで処理することにより得られる可溶性画分においては、分子量27,000のβサブユニット、分子量17,000のγサブユニットが主に観察された。可溶性画分に含まれるβサブユニットとγサブユニットのアミノ酸配列を解析したところ、分子量27,000のβサブユニットは、βサブユニットのN末端の20アミノ酸が欠失したものであり、分子量17,000のγサブユニットは、γサブユニットのN末端の16アミノ酸が欠失したものであった。なお、αサブユニットにおいては、トリプシン処理による分子量の減少は観察されなかった。
【0109】
ジオールデヒドラターゼをトリプシンで限定的にタンパク質分解したときの、α、β及びγサブユニットのN末端領域の断片化パターンを示す(図1C)。
【0110】
実施例2 発現プラスミドの構築
実施例1の結果から、βサブユニットのN末端から20アミノ酸を欠失させることにより、及び/又はγサブユニットのN末端から16アミノ酸を欠失させることにより、ジオールデヒドラターゼを水溶性に変化させることができるかどうかを試験するため、βサブユニット及び/又はγサブユニットにおいてN末端アミノ酸配列が欠失した変異型ジオールデヒドラターゼを産生するための発現プラスミドを以下のように構築した。
【0111】
βサブユニットのN末端20アミノ酸のみが欠失したジオールデヒドラターゼに対する発現プラスミドをpUSI2E(αDβ’DγD)とし、γサブユニットのN末端16アミノ酸のみが欠失したジオールデヒドラターゼに対する発現プラスミドをpUSI2E(αDβDγD)と記載し、β及びγサブユニットの双方のN末端アミノ酸が欠失したジオールデヒドラターゼに対する発現プラスミドをpUSI2E(αDβ’Dγ’D)とする。また、N末端20アミノ酸が欠失したβサブユニットをβ’サブユニットと記載し、N末端16アミノ酸が欠失したγサブユニットをγ’サブユニットと記載する場合もある。
【0112】
pUSI2E(Tobimatsu, T.et al., J. Biol. Chem., 270, 7142-7148 (1995))をNdeIを用いて部分的に消化し、4種のdNTPの存在下、DNAポリメラーゼIのKlenowフラグメントで処理した。そしてセルフライゲーションにより、プラスミドpUSI2ENdを作成した。pUSI2E(DD)からの3.0kbのBamHI−BglII DNAフラグメントを、pUSI2ENdのBamHI−BglII領域に挿入し、pUSI2ENd(DD)を作成した。ジオールデヒドラターゼの各サブユニットにおいて、N末端20アミノ酸が欠失したβ’サブユニットをコードするDNAセグメント及びN末端16アミノ酸が欠失したγ’サブユニットをコードするDNAセグメントを、TaqDNAポリメラーゼ(インビトロジェン)及び以下のプライマー対を用いたPCRにより増幅した。
【0113】
β’サブユニット:
5’−GCGAGCATATGGGCAGCGATAAACCG−3’(配列番号13)
5’−TCAGATCTTAAAGCGCCACGCGCAG−3’(配列番号14)
γ’サブユニット:
5’−TTGAGCCATATGAACAGCCTGC−3’(配列番号15)
5’−AGAGATCTTAATCGTCGCCTTTGAG−3’(配列番号16)
【0114】
増幅した640bp及び500bpのDNAフラグメントを、TAクローニングキット(インビトロジェン)を用いてpCR2.1ベクターにクローニングし、NdeI及びBglIIで消化し、pUSI2ENd(DD)からの5.0kbのNdeI−BglIIフラグメントに連結し、プラスミドpUSI2ENd(β’D)及びpUSI2ENd(γ’D)をそれぞれ作成した。プラスミドpUSI2E(αD)(Tobimatsu, T. et al., Arch. Biochem. Biophys., 347, 132-140 (1997))を、BglIIで消化し、pUSI2ENd(β’D)から得られた0.7kbのBamHI−BglIIフラグメントに連結し、プラスミドpUSI2E(αDβ’D)を得た。プラスミドpUSI2E(αDβ’D)をBglIIで消化し、pUSI2ENd(γ’D)及びpUSI2ENd(γD)から得られた0.5kbのBamHI−BglIIフラグメントに連結し、プラスミドpUSI2E(αDβ’Dγ’D)及びpUSI2E(αDβ’DγD)をそれぞれ作成した。プラスミドpUSI2E(αDβD)をBglIIで消化し、pUSI2ENd(γ’D)から得られた0.5kbのBamHI−BglIIフラグメントに連結し、プラスミドpUSI2E(αDβDγ’D)を得た。配列決定により、PCR増幅中に望ましくない変異が起こっていないことを確認した。
【0115】
発現プラスミドによる大腸菌の形質転換及び形質転換体の培養は、Tobimatsu, T.et al., J. Biol. Chem., 270, 7142-7148 (1995)に記載された方法と同様に実施した。
【0116】
各変異型酵素を発現している組換え体大腸菌の細胞ホモジェネート(homo)、上清(sup)及び沈澱画分(ppt)に含まれるタンパク質を、11%ゲル上のSDS−PAGEに付し、タンパク質染色を行った。分子量マーカーとしてSDS−7(Sigma)を用いた。α、β、β’、γ及びγ’サブユニットの位置を、右側の矢印によって示す(図2)。
【0117】
沈澱画分(ppt)の結果を見ると、ジオールデヒドラターゼをコードする遺伝子を含む組換え体大腸菌(以下、変異株と称する場合もある)においてβ’及びγ’サブユニットのバンドは、野生株のβ及びγサブユニットのバンドほど濃いバンドではないことがわかる。一方、上清(sup)の結果を見ると、変異株におけるβ’及びγ’サブユニットのバンドは、野生型ジオールデヒドラターゼをコードする遺伝子を含む組換え体大腸菌(以下、野生株と称する場合もある)のβ及びγサブユニットのバンドより濃いことがわかる。以上からβ及び/又はγサブユニットのN末端が欠失したジオールデヒドラターゼをコードする遺伝子を含む変異株において、水溶性のジオールデヒドラターゼが産生されたことがわかる。
【0118】
実施例3 上清及び沈澱におけるジオールデヒドラターゼ活性
実施例2で得られた各発現プラスミドを有する組換え体大腸菌BL21(DE3)(9.2〜9.8×1010)を洗浄し、2% 1,2−プロパンジオールを含む50mMリン酸カリウムバッファ(pH8.0)2.6mlに再懸濁し、超音波処理により破砕した。得られた細胞ホモジェネートを遠心分離し(17,500×g、30分)、上清及び沈澱画分を分離した。得られた沈澱を2.6mlの同じバッファで洗浄し、2% 1,2−プロパンジオール及び1% Brij35を含む10mM リン酸カリウムバッファ(pH8.0)に再懸濁し同じ体積とした。ジオールデヒドラターゼ活性は、適切に希釈した後で実施例1と同様に測定した。結果を以下の表1に示す。
【0119】
【表1】

【0120】
表1の結果から、可溶性画分に回収されたジオールデヒドラターゼの割合は、いずれの変異株においても96%を超えていたが、野生株においては、2%に過ぎなかった。以上から本発明の変異型ジオールデヒドラターゼが高度に水溶性であることがわかる。また、野生型ジオールデヒドラターゼにおいては、β及びγサブユニットのN末端アミノ酸領域が、この酵素の沈澱や凝集に必須であることがわかる。また、変異型ジオールデヒドラターゼが酵素活性を有することから、水溶性の変異型ジオールデヒドラターゼが酵素活性を有することもわかる。
【0121】
実施例4 変異型ジオールデヒドラターゼと野生型ジオールデヒドラターゼの比較
αサブユニット、β’サブユニット及びγ’サブユニットを含む変異型ジオールデヒドラターゼ、野生型ジオールデヒドラターゼ、並びに1.0μg/mlのトリプシンで、15℃で12時間処理したジオールデヒドラターゼを比較するため、プラスミドpUSI2E(αDβ’Dγ’D)を有する大腸菌の無細胞抽出物をPAGEに付した。無細胞抽出物のPAGEは、0.1Mの1,2−プロパンジオールの存在下、Davis, B.J., Ann. N. Y. Acad. Sci., 121, 404-427 (1964))に記載の非変性条件下で、又はLaemmli, U.K., Nature, 227, 680-685 (1970)に記載の変性条件下で実施した。ウェスタンブロット分析は、Tobimatsu, T. et al., J. Biol. Chem., 270, 7142-7148 (1995)に記載の方法に従って実施した(図3)。
【0122】
非変性条件下の5%ゲル上のPAGEの後、タンパク質染色(図3A)又は活性染色(図3B)を実施した。野生型及びトリプシン消化ジオールデヒドラターゼ及びグリセロールデヒドラターゼの位置を、右側の矢印により示す。酵素活性を示すタンパク質バンドを切り出し、11%ゲル上のSDS−PAGEによって分析し、タンパク質染色した(図3C)。タンパク質染色は、クマシーブリリアントブルー R−250を用いて実施した。サブユニットの位置を、右側の矢印で示す。分子量マーカーとしてSDS−7(Sigma)を用いた。
【0123】
図3A及びBに示されるように、ジオールデヒドラターゼ活性を示すバンドは、変異型ジオールデヒドラターゼを含む無細胞抽出物にのみ観察された。この活性なバンドをゲルから切り出してSDS−PAGEに付したところ、観察された3種のペプチドのバンドは、ジオールデヒドラターゼのトリプシン処理によって得られる分子量60,000、23,000及び17,000のバンドと一致した(図3C)。
【0124】
以上からプラスミドpUSI2E(αDβ’Dγ’D)を有する大腸菌によって産生された組換え体変異型ジオールデヒドラターゼが触媒活性を有すること、並びに、分子量60、000、23,000及び17,000のサブユニットを含むことが確認された。
【0125】
実施例5 変異型ジオールデヒドラターゼの精製及び分析
pUSI2E(αDβ’Dγ’D)を有する大腸菌3.3gから、超音波処理により無細胞抽出物を調製した。該無細胞抽出物の硫安分画(40〜50%)により得られた沈澱を2% 1,2−プロパンジオールを含むバッファに溶解し、Sepharose CL6Bカラム(350mL)にロードした。4ユニット/mgを超える比活性を有する酵素を含む画分を回収した。該酵素溶液をCentricon(Millipore Corp.)で濃縮し、4volの2% 1,2−プロパンジオールと混合した。該溶液を、予め2% 1,2−プロパンジオールを含む10mM KPB(pH8)で平衡化しておいたDEAEセルロース(Serva)カラム(1mL)にロードした。カラムを同じバッファの5volで洗浄し、50mM KClを含む同じバッファ、続いて100mM KClを含む同じバッファの5volで展開した。35ユニット/mgを超える比活性を有する精製酵素を、酵素の特性決定に使用した。タンパク質は、Lowry, O.H. et al., J. Biol. Chem., 193, 265-275 (1951)に記載の方法により、結晶化ウシ血清アルブミンを標準として用いて、アッセイした。比活性はユニット/タンパク質(mg)として記載した。
【0126】
得られた変異型ジオールデヒドラターゼ(αβ’γ’)調製物の比活性は47ユニット/mgであった。デンシトメトリー分析により、得られた変異型ジオールデヒドラターゼの純度は55%であったことから、この変異型ジオールデヒドラターゼの比活性は85ユニット/mgであると考えられる。この比活性の値は、K. oxytocaから水溶性の形態で精製されたジオールデヒドラターゼの比活性(91.5ユニット/mg)及び野生型のジオールデヒドラターゼの比活性(106ユニット/mg)に匹敵するものであった。
【0127】
また、基質としてのDL−1,2−プロパンジオールに対するKm値は、変異型ジオールデヒドラターゼで0.11mMであり、野生型ジオールデヒドラターゼで0.10mMであった。以上から、本発明の変異型ジオールデヒドラターゼが、野生型のジオールデヒドラターゼと比べ、触媒活性において同等であることがわかる。
【0128】
実施例6 疎水化グリセロールデヒドラターゼの調製
グリセロールデヒドラターゼ(GD)とジオールデヒドラターゼ(DD)のサブユニットは高い相同性を有しているが、βとγサブユニットのN末端領域は、GDが短く、更にβサブユニットはそれに続く30数アミノ酸残基がなくなっている(図4)。DDで余分に付いているN末端領域だけでなく、βサブユニットの相同性が低い領域も酵素の低溶解性に関与する可能性がある。
【0129】
変異型酵素として、βおよびγサブユニットがそれぞれ下向き矢印#2と#3までDDの配列を持ちそれ以降をGDの配列を持つ変異体2(αGNβD2G2γDG)を作成した。すなわち、変異体2は、グリセロールデヒドラターゼ(GD)のβサブユニットのN末端の1〜27番目のアミノ酸をジオールデヒドラターゼ(DD)のN末端の1〜60番目のアミノ酸に置き換え、さらにγサブユニットのN末端の1番目のアミノ酸をジオールデヒドラターゼ(DD)のN末端の1〜33番目のアミノ酸に置き換えたグリセロールデヒドラターゼ変異体である。上記変異体の遺伝子構造の概略を図5に示す。また得られる変異体2の塩基配列を配列番号17に示す。変異体2においてαサブユニットのアミノ酸配列を配列番号18に、βサブユニットのアミノ酸配列を配列番号19に、γサブユニットのアミノ酸配列を配列番号20に示す。
【0130】
上記変異型グリセロールデヒドラターゼ(変異体2)を産生するための発現プラスミド:pNEX(αGNβD2G2γDG)を実施例2と同様に構築した。
【0131】
pNEXベクターの構造を図6に示す。またpNEX(αG)の作成手順を図7に示す。pUSI2E(αG)も同様に作成した。挿入DNA断片(図8)は、TaqDNAポリメラーゼ(インビトロジェン)及び図9に示すプライマー対を用いたPCRにより増幅した。得られるPCR断片を図10に示す。PCRはKOD Dashにて、以下の条件のstep down PCR法で行った。
【0132】
【表2】
【0133】
増幅断片をpNEX(αG)またはpUSI2E(αG)に連結することにより、発現プラスミドを作成した。
【0134】
発現プラスミドによる大腸菌の形質転換及び形質転換体の培養は、Tobimatsu, T.et al., J. Biol. Chem., 270, 7142-7148 (1995)に記載された方法と同様に実施した。
【0135】
各変異型酵素を発現している組換え体大腸菌をトルエン処理し(図11)、細胞破壊と分離操作を行った(図12)。組換え体大腸菌の細胞ホモジェネート(homo)、上清(sup)、沈澱画分(ppt)及び洗液(wash)に含まれるタンパク質を、11%ゲル上のSDS−PAGEに付し、タンパク質染色を行った(図13)。
【0136】
またそれぞれの画分についてグリセロールデヒドラターゼ活性を測定した。コントロールとして野生型(WT)のジオールデヒドラターゼを用いた。結果を以下の表に示す。
【0137】
【表3】
【0138】
大腸菌で組み換え体として発現させた野生型のグリセロールデヒドラターゼ(GD)の場合は、酵素活性の大部分が上清画分にみられ可溶型で存在することが報告されているが(Yamanishi M, et al., Eur. J. Biochem. 269,4484-494 (2002); Tobimatsu T, et al., J. Biol. Chem., 271(37), 22352-22357(1996))、表3より、変異体2では、酵素活性の大半が沈澱画分にみられたことから、GDが低溶解性化していることが明らかである。すなわち、本発明により、変異型グリセロールデヒドラターゼの水に対する溶解性が変化し、疎水性のグリセロールデヒドラターゼが得られたことが示された。
【図面の簡単な説明】
【0139】
【図1】トリプシン消化がジオールデヒドラターゼの活性と溶解性に及ぼす影響を示す。
【図2】N末端アミノ酸が欠失したβ及び/又はγサブユニットを有する変異型ジオールデヒドラターゼの大腸菌における発現及び分布を示す。
【図3】αβ’γ’変異型ジオールデヒドラターゼとトリプシン消化ジオールデヒドラターゼとの比較を示す。
【図4】Klebsiella pneumoniae由来のグリセロールデヒドラターゼ(GD)とKlebsiella oxytoca由来のジオールデヒドラターゼ(DD)の各サブユニットのアミノ酸配列の相同性を示す。
【図5】変異体2の遺伝子構造の概略図を示す。
【図6】pNEXベクターの構造を示す。
【図7】pNEX(αG)の作成手順を示す。
【図8】挿入DNA断片(NβDGγDG)を示す。
【図9】実施例6で発現プラスミドの作成においてPCRに用いたプライマー対及びプライマー領域の相補性を示す。↓のところがジオールデヒドラターゼとグリセロールデヒドラターゼとがつながる場所を示す。
【図10】図9に示したプライマー対を用いて得られるPCR断片を示す。
【図11】組換え体大腸菌をトルエン処理した手順を示す。
【図12】組換え体大腸菌をトルエン処理した後、細胞破壊と分離操作を行った手順を示す。
【図13】変異型グリセロールデヒドラターゼをSDS−PAGEに付し、タンパク質染色を行った結果を示す。
【技術分野】
【0001】
本発明は、ジオールデヒドラターゼ及びグリセロールデヒドラターゼの水に対する溶解性を変化させる方法、水に対する溶解性が変化したジオールデヒドラターゼ及びグリセロールデヒドラターゼ、並びにこれらの酵素により1位及び2位に水酸基を有する脂肪族ポリオールを脱水する方法に関する。
【背景技術】
【0002】
ジオールデヒドラターゼは、1,2−ジオールを脱水して対応するアルデヒドに変換する反応を、アデノシルコバラミン(補酵素B12)依存的に触媒する酵素として知られている(非特許文献1及び2)。ジオールデヒドラターゼは、Klebsiella属細菌及びCitrobacter属細菌などにおいて、特に、1,2−ジオールの存在下、好気的条件での増殖において誘導される(非特許文献3〜5)。グリセロールも基質とされうるが、1,2−ジオールに比べて基質特異性は低い。ジオールデヒドラターゼは、60kDa、30kDa及び19kDaの3つのサブユニット(それぞれαサブユニット、βサブユニット及びγサブユニット)からなる分子量約22万の酵素であり、これらのサブユニットをコードする遺伝子はタンデムに並んでいることがわかっている(非特許文献6)。
【0003】
グリセロールデヒドラターゼは、ジオールデヒドラターゼとその触媒作用及びサブユニット構造などが類似しているが、免疫学的性質や基質特異性などが異なるアイソザイムである(非特許文献7〜9)。グリセロールデヒドラターゼは、61kDa、21kDa及び16kDaの3つのサブユニット(それぞれαサブユニット、βサブユニット及びγサブユニット)からなることがわかっている。
【0004】
いずれの酵素とも既にクローニングが行われ、大腸菌による大量発現系も確立されている(非特許文献10)。しかし、ジオールデヒドラターゼは、水に対する溶解性が低い酵素であるため(非特許文献11〜12)、組換え体ジオールデヒドラターゼを大量に得ることが難しいという問題、親水性の条件下での反応に用いることができないという問題があった。ジオールデヒドラターゼを水に可溶化する方法として、界面活性剤で処理する方法が知られているが、当該方法を用いると酵素活性が失われるという問題があった。
【0005】
一方、グリセロールデヒドラターゼは、水に対する溶解性が高い酵素であるため(非特許文献7及び13)、疎水性条件下での反応には適していないという問題があった。疎水性条件下でグリセロールを基質として反応を実施したいときに、グリセロールデヒドラターゼのかわりにジオールデヒドラターゼを使用することも考えられるが、ジオールデヒドラターゼはグリセロールデヒドラターゼに比べてグリセロールに対する基質特異性が低いため、やはりグリセロールデヒドラターゼを疎水性条件下での反応に用いる手段が望まれていた。
【0006】
従って、ジオールデヒドラターゼ及びグリセロールデヒドラターゼの酵素活性を維持したまま、これらの酵素の水に対する溶解性を変化する手段が望まれていた。
【0007】
ジオールデヒドラターゼ及びグリセロールデヒドラターゼの対応するサブユニットのアミノ酸配列を比較すると、βサブユニットのN末端の32アミノ酸、及びγサブユニットのN末端の37アミノ酸を除き、高度の相同性を有することが報告されている(非特許文献14)。しかし、上記のような互いに相違するアミノ酸領域がジオールデヒドラターゼの溶解性に影響を及ぼしているかは明らかではなく、また、これらのN末端アミノ酸領域を欠失させたジオールデヒドラターゼ又は付加したグリセロールデヒドラターゼが、それぞれ親水性条件下又は疎水性条件下で触媒活性を示すかについても明らかではなかった。
【0008】
【非特許文献1】Lee, H.A. et al., Jr., and Abeles, R.H., J. Biol. Chem., 238, 2367-2373 (1963)
【非特許文献2】Toraya, T. et al., Biochem. Biophys. Res. Commun., 69, 475-480 (1976)
【非特許文献3】Toraya, T. et al., J Bacteriol., 135, 726-729 (1978)
【非特許文献4】Toraya, T. et al., J. Bacteriol., 141, 1439-1442 (1980)
【非特許文献5】Toraya, T. et al., "Chemistry and Biochemistry of B12," ed. Banerjee, R., John Wiley and Sons, New York, pp. 783-809 (1999)
【非特許文献6】Tobimatsu T, Hara T, Sakaguchi M, Kishimoto Y, Wada Y, Isoda M, Sakai T, Toraya T. J Biol Chem. 270(13), 7142-8 (1995)
【非特許文献7】Yamanishi, M. et al., J. Biochem., 269, 4484-4494 (2002)
【非特許文献8】Shibata, N. et al., Structure Fold Des., 7, 997-1008 (1999)
【非特許文献9】Toraya, T. et al., Chem. Rev., 103, 2095-2127 (2003)
【非特許文献10】Tobimatsu T, Sakai T, Hashida Y, Mizoguchi N, Miyoshi S, Toraya T. Arch Biochem Biophys. Nov 1;347(1):132-40 (1997)
【非特許文献11】McGee, D.E. et al., Biochemistry, 20, 4293-4298 (1981)
【非特許文献12】Tobimatsu, T. et al., Arch. Biochem. Biophys., 347, 132-140 (1997)
【非特許文献13】Stroinski, A. et al., Arch. Biochem. Biophys., 162, 321-330 (1974)
【非特許文献14】Tobimatsu, T. et al., J. Biol. Chem., 270, 7142-7148 (1995)
【発明の開示】
【発明が解決しようとする課題】
【0009】
本発明の課題は、ジオールデヒドラターゼ及びグリセロールデヒドラターゼの酵素活性を維持したまま、これらの酵素の水に対する溶解性を変化する手段を提供することである。
【課題を解決するための手段】
【0010】
本発明者らは、上記課題を解決すべく鋭意検討を行った結果、ジオールデヒドラターゼとグリセロールデヒドラターゼの各サブユニットのアミノ酸配列において相違するN末端アミノ酸の特定の領域がこれらの酵素の水に対する溶解性を決定づけていることを見いだし、本発明を完成するに至った。
【0011】
すなわち、本発明は以下の発明を包含する。
(1)ジオールデヒドラターゼ又はグリセロールデヒドラターゼのβサブユニット及び/又はγサブユニットにおいてN末端アミノ酸を変異させることにより、ジオールデヒドラターゼ又はグリセロールデヒドラターゼの水に対する溶解性を変化させる方法。
(2)N末端アミノ酸の変異が、
ジオールデヒドラターゼを構成するβサブユニット及び/又はγサブユニットにおけるN末端アミノ酸を欠失させること、又は
グリセロールデヒドラターゼを構成するβサブユニット及び/又はγサブユニットにおけるN末端に、ジオールデヒドラターゼの対応するサブユニットのN末端アミノ酸を付加すること、
を含む、(1)記載の方法。
(3)ジオールデヒドラターゼのβサブユニット及び/又はγサブユニットにおけるN末端アミノ酸の変異が、
ジオールデヒドラターゼのβサブユニットのN末端の9〜64個のアミノ酸を欠失させること、及び/又は
ジオールデヒドラターゼのγサブユニットのN末端の5〜42個のアミノ酸を欠失させること
を含む(2)記載の方法。
(4)グリセロールデヒドラターゼのβサブユニット及び/又はγサブユニットにおけるN末端アミノ酸の変異が、
グリセロールデヒドラターゼのβサブユニットのN末端に、ジオールデヒドラターゼのβサブユニットのN末端の9〜64個のアミノ酸を付加すること、
グリセロールデヒドラターゼのγサブユニットのN末端に、ジオールデヒドラターゼのγサブユニットのN末端の5〜42個のアミノ酸を付加すること
を含む(2)記載の方法。
(5)αサブユニット、βサブユニット及びγサブユニットを含むジオールデヒドラターゼであって、
βサブユニットのN末端の9〜64個のアミノ酸が欠失しており、及び/又はγサブユニットのN末端の5〜42個のアミノ酸が欠失しており、かつ水溶性である前記ジオールデヒドラターゼ。
(6)αサブユニット、βサブユニット及びγサブユニットを含むジオールデヒドラターゼであって、
αサブユニットが以下の(a)又は(b)のタンパク質:
(a)配列番号1のアミノ酸配列からなるタンパク質、
(b)配列番号1において1若しくは数個のアミノ酸が欠失、置換若しくは付加されたアミノ酸配列からなるタンパク質であって、ジオールデヒドラターゼのβサブユニット及びγサブユニットと会合したときにジオールデヒドラターゼ活性を有する前記タンパク質、
であり、
βサブユニットが以下の(c)〜(f)から選択されるタンパク質:
(c)配列番号3のアミノ酸配列からなるタンパク質、
(d)配列番号3において1若しくは数個のアミノ酸が欠失、置換若しくは付加されたアミノ酸配列からなるタンパク質であって、ジオールデヒドラターゼのαサブユニット及びγサブユニットと会合したときにジオールデヒドラターゼ活性を有する前記タンパク質、
(e)配列番号3においてN末端の9〜64個のアミノ酸が欠失したアミノ酸配列からなるタンパク質、
(f)(e)に記載のアミノ酸配列において1若しくは数個のアミノ酸が欠失、置換若しくは付加されたアミノ酸配列からなるタンパク質であって、ジオールデヒドラターゼのαサブユニット及びγサブユニットと会合したときにジオールデヒドラターゼ活性を有する前記タンパク質、
であり、
γサブユニットが以下の(g)〜(j)から選択されるタンパク質:
(g)配列番号5のアミノ酸配列からなるタンパク質、
(h)配列番号5において1若しくは数個のアミノ酸が欠失、置換若しくは付加されたアミノ酸配列からなるタンパク質であって、ジオールデヒドラターゼのαサブユニット及びβサブユニットと会合したときにジオールデヒドラターゼ活性を有する前記タンパク質、
(i)配列番号5においてN末端の5〜42個のアミノ酸が欠失したアミノ酸配列からなるタンパク質、
(j)(i)に記載アミノ酸配列において1若しくは数個のアミノ酸が欠失、置換若しくは付加されたアミノ酸配列からなるタンパク質であって、ジオールデヒドラターゼのαサブユニット及びβサブユニットと会合したときにジオールデヒドラターゼ活性を有する前記タンパク質
であり、
ただし、βサブユニットが(c)又は(d)のタンパク質であるときγサブユニットは(g)及び(h)のタンパク質ではない、前記ジオールデヒドラターゼ。
(7)(5)又は(6)のジオールデヒドラターゼをコードする遺伝子。
(8)(7)記載の遺伝子を含むベクター。
(9)(8)記載のベクターを含む形質転換体。
(10)αサブユニット、βサブユニット及びγサブユニットを含むグリセロールデヒドラターゼにおいて、
βサブユニットのN末端にジオールデヒドラターゼのβサブユニットのN末端の9〜64個のアミノ酸が付加しており、及び/又はγサブユニットのN末端にジオールデヒドラターゼのβサブユニットのN末端の9〜64個のアミノ酸が付加しており、かつ水に不溶性である前記グリセロールデヒドラターゼ。
(11)αサブユニット、βサブユニット及びγサブユニットを含むグリセロールデヒドラターゼにおいて、
αサブユニットが以下の(a)又は(b)のタンパク質:
(a)配列番号7のアミノ酸配列からなるタンパク質、
(b)配列番号7において1若しくは数個のアミノ酸が欠失、置換若しくは付加されたアミノ酸配列からなるタンパク質であって、グリセロールデヒドラターゼのβサブユニット及びγサブユニットと会合したときにグリセロールデヒドラターゼ活性を有する前記タンパク質、
であり、
βサブユニットが以下の(c)〜(f)から選択されるタンパク質:
(c)配列番号9のアミノ酸配列からなるタンパク質、
(d)配列番号9において1若しくは数個のアミノ酸が欠失、置換若しくは付加されたアミノ酸配列からなるタンパク質であって、グリセロールデヒドラターゼのαサブユニット及びγサブユニットと会合したときにグリセロールデヒドラターゼ活性を有する前記タンパク質、
(e)配列番号9のN末端に配列番号3のN末端の9〜64個のアミノ酸が付加してなるアミノ酸配列からなるタンパク質、
(f)(e)に記載のアミノ酸配列において1若しくは数個のアミノ酸が欠失、置換若しくは付加されたアミノ酸配列からなるタンパク質であって、グリセロールデヒドラターゼのαサブユニット及びγサブユニットと会合したときにグリセロールデヒドラターゼ活性を有する前記タンパク質、
であり、
γサブユニットが以下の(g)〜(j)から選択されるタンパク質:
(g)配列番号11のアミノ酸配列からなるタンパク質、
(h)配列番号11において1若しくは数個のアミノ酸が欠失、置換若しくは付加されたアミノ酸配列からなるタンパク質であって、グリセロールデヒドラターゼのαサブユニット及びβサブユニットと会合したときにグリセロールデヒドラターゼ活性を有する前記タンパク質、
(i)配列番号11のN末端に配列番号5のN末端の5〜42個のアミノ酸が付加してなるアミノ酸配列からなるタンパク質、
(j)(i)に記載アミノ酸配列において1若しくは数個のアミノ酸が欠失、置換若しくは付加されたアミノ酸配列からなるタンパク質であって、グリセロールデヒドラターゼのαサブユニット及びβサブユニットと会合したときにグリセロールデヒドラターゼ活性を有する前記タンパク質
であり、
ただし、βサブユニットが(c)又は(d)のタンパク質であるときγサブユニットは(g)及び(h)のタンパク質ではない、前記グリセロールデヒドラターゼ。
(12)(10)又は(11)記載のグリセロールデヒドラターゼをコードする遺伝子。
(13)(12)記載の遺伝子を含むベクター。
(14)(13)記載のベクターを含む形質転換体。
(15)以下の(a)又は(b)の遺伝子:
(a)配列番号17の塩基配列からなる遺伝子、
(b)配列番号17で表される塩基配列からなる遺伝子に対し相補的な塩基配列からなる遺伝子とストリンジェントな条件下でハイブリダイズし、かつグリセロールデヒドラターゼの活性を有するタンパク質をコードする遺伝子。
(16)(15)記載の遺伝子を含むベクター。
(17)(16)記載のベクターを含む形質転換体。
(18)αサブユニット、βサブユニット及びγサブユニットを含むグリセロールデヒドラターゼにおいて、
αサブユニットが以下の(a)又は(b)のタンパク質:
(a)配列番号18のアミノ酸配列からなるタンパク質、
(b)配列番号18において1若しくは数個のアミノ酸が欠失、置換若しくは付加されたアミノ酸配列からなるタンパク質であって、グリセロールデヒドラターゼのβサブユニット及びγサブユニットと会合したときにグリセロールデヒドラターゼ活性を有する前記タンパク質、
であり、
βサブユニットが以下の(c)〜(f)から選択されるタンパク質:
(c)配列番号9のアミノ酸配列からなるタンパク質、
(d)配列番号9において1若しくは数個のアミノ酸が欠失、置換若しくは付加されたアミノ酸配列からなるタンパク質であって、グリセロールデヒドラターゼのαサブユニット及びγサブユニットと会合したときにグリセロールデヒドラターゼ活性を有する前記タンパク質、
(e)配列番号19のアミノ酸配列からなるタンパク質、
(f)配列番号19において1若しくは数個のアミノ酸が欠失、置換若しくは付加されたアミノ酸配列からなるタンパク質であって、グリセロールデヒドラターゼのαサブユニット及びγサブユニットと会合したときにグリセロールデヒドラターゼ活性を有する前記タンパク質、
であり、
γサブユニットが以下の(g)〜(j)から選択されるタンパク質:
(g)配列番号11のアミノ酸配列からなるタンパク質、
(h)配列番号11において1若しくは数個のアミノ酸が欠失、置換若しくは付加されたアミノ酸配列からなるタンパク質であって、グリセロールデヒドラターゼのαサブユニット及びβサブユニットと会合したときにグリセロールデヒドラターゼ活性を有する前記タンパク質、
(i)配列番号20のアミノ酸配列からなるタンパク質、
(j)配列番号20において1若しくは数個のアミノ酸が欠失、置換若しくは付加されたアミノ酸配列からなるタンパク質であって、グリセロールデヒドラターゼのαサブユニット及びβサブユニットと会合したときにグリセロールデヒドラターゼ活性を有する前記タンパク質、
であり、
ただし、βサブユニットが(c)又は(d)のタンパク質であるときγサブユニットは(g)及び(h)のタンパク質ではない、前記グリセロールデヒドラターゼ。
(19)1位及び2位に水酸基を有する脂肪族ポリオールを脱水して、1位にアルデヒド基を有するアルデヒドを生成する方法あって、
1位及び2位に水酸基を有する脂肪族ポリオールと(5)若しくは(6)記載のジオールデヒドラターゼ及び/又は(10)、(11)若しくは(18)記載のグリセロールデヒドラターゼとを反応させることを含む前記方法。
(20)1位及び2位に水酸基を有する脂肪族ポリオールを脱水して、1位にアルデヒド基を有するアルデヒドを生成する方法あって、
1位及び2位に水酸基を有する脂肪族ポリオールの存在下、(7)記載のジオールデヒドラターゼをコードする遺伝子及び/又は(12)若しくは(15)記載のグリセロールデヒドラターゼをコードする遺伝子を含む形質転換体を培養することを含む前記方法。
【発明の効果】
【0012】
本発明により、ジオールデヒドラターゼ及びグリセロールデヒドラターゼの水に対する溶解性を、酵素活性を維持しつつ変化させることができる。
【発明を実施するための最良の形態】
【0013】
本発明は、グリセロールデヒドラターゼのβサブユニット及び/又はγサブユニットにおいてN末端アミノ酸を変異させることにより、ジオールデヒドラターゼ又はグリセロールデヒドラターゼの水に対する溶解性を変化させる方法に関する。
【0014】
本発明において、ジオールデヒドラターゼとは、当技術分野で通常用いられる意味を有し、すなわち、1,2−ジオールを脱水して対応するアルデヒド及び水へ変換する反応を、アデノシルコバラミン(補酵素B12)依存的に触媒する活性を有する酵素である。ジオールデヒドラターゼは、60kDa、30kDa及び19kDaの3つのサブユニット(それぞれαサブユニット、βサブユニット及びγサブユニット)を含む。
【0015】
本発明において、グリセロールデヒドラターゼは、当技術分野で通常用いられる意味を有し、すなわち、グリセロールを脱水して3−ヒドロキシプロピオンアルデヒド及び水へ変換する反応を、アデノシルコバラミン(補酵素B12)の存在下で触媒する活性を有する酵素である。グリセロールデヒドラターゼは、61kDa、21kDa及び16kDaの3つのサブユニット(それぞれαサブユニット、βサブユニット及びγサブユニット)を含む。
【0016】
ジオールデヒドラターゼ及びグリセロールデヒドラターゼにおいては、上記α、β、γサブユニットと補酵素が会合することにより、酵素活性が発揮される。
【0017】
ジオールデヒドラターゼ及びグリセロールデヒドラターゼは、上記酵素活性を有するものであれば特に限定されないが、例えば、Lactobacillus属、Citrobacter属、Clostridium属、Klebsiella属、Enterobacter属、Caloramator属、Salmonella属及びListeria属等に属する細菌に由来するものが挙げられる。
【0018】
本発明の方法では、上記ジオールデヒドラターゼ及びグリセロールデヒドラターゼのβ及び/又はγサブユニットにおいてN末端アミノ酸を変異させることにより、酵素の水に対する溶解性を変化させる。
【0019】
ジオールデヒドラターゼにおけるN末端アミノ酸の変異は、好ましくは、βサブユニット及び/又はγサブユニットにおけるN末端アミノ酸を欠失させることにより行う。βサブユニットにおいて欠失させるN末端アミノ酸は、N末端側から9〜64個、好ましくは14〜35個、より好ましくは18〜25個である。γサブユニットにおいて欠失させるN末端アミノ酸は、N末端側から5〜42個、好ましくは10〜35個、より好ましくは15〜25個である。
【0020】
グリセロールデヒドラターゼにおけるN末端アミノ酸の変異は、好ましくは、βサブユニット及び/又はγサブユニットにおけるN末端に、ジオールデヒドラターゼの対応するサブユニットのN末端アミノ酸を付加することにより実施できる。グリセロールデヒドラターゼのサブユニットに付加するジオールデヒドラターゼの対応するサブユニットのN末端アミノ酸は、どの属に属する細菌由来のものでもかまわないが、タンパク質発現を行う宿主と同じか近縁の科に属する細菌由来のもの、好ましくは同じか近縁の属に属する細菌由来のものとするのが好ましい。
【0021】
本発明の方法においては、各酵素のβサブユニット若しくはγサブユニットのいずれかにおいてのみN末端アミノ酸を変異させてもよく、又は双方のサブユニットにおいてN末端アミノ酸を変異させてもよい。いずれの場合にも本発明の効果が得られる。
【0022】
本発明はまた、上記方法によって得られた水に対する溶解性の変化した変異型ジオールデヒドラターゼ及び変異型グリセロールデヒドラターゼに関する。以下、β及び/又はγサブユニットのN末端アミノ酸が変異したジオールデヒドラターゼ及びグリセロールデヒドラターゼを、それぞれ変異型ジオールデヒドラターゼ及び変異型グリセロールデヒドラターゼと称し、N末端アミノ酸が変異していないジオールデヒドラターゼ及びグリセロールデヒドラターゼを、それぞれ野生型ジオールデヒドラターゼ及び野生型グリセロールデヒドラターゼと称する場合がある。
【0023】
すなわち本発明の一実施形態は、αサブユニット、βサブユニット及びγサブユニットを含むジオールデヒドラターゼであって、βサブユニットのN末端の9〜64個のアミノ酸が欠失しており、及び/又はγサブユニットのN末端の5〜42個のアミノ酸が欠失しており、かつ水溶性である前記ジオールデヒドラターゼに関する。ここでα又はγサブユニットにおいて欠失しているN末端アミノ酸の数については上記のとおりである。
【0024】
本発明の変異型ジオールデヒドラターゼには、βサブユニット及びγサブユニットのいずれかのN末端アミノ酸が欠失しているもの、並びにβサブユニット及びγサブユニットの双方のN末端アミノ酸が欠失しているものが包含される。
【0025】
本発明の変異型ジオールデヒドラターゼは、上記変異を含み水溶性であるとともに、ジオールデヒドラターゼ活性を有する限り特に限定されない。本発明において水溶性であるとは、変異型ジオールデヒドラターゼ遺伝子が発現したタンパク質の大部分が水に可溶であることを意味する。具体的には、酵素タンパク質を発現させた微生物菌体を適当な緩衝液に懸濁させて充分破砕した後、懸濁液を遠心分離した試料の上清部分に、酵素活性の大部分が存在することで確認できる。
【0026】
本発明の変異型ジオールデヒドラターゼとしては、例えば、Lactobacillus属、Citrobacter属、Clostridium属、Klebsiella属、Enterobacter属、Caloramator属、Salmonella属及びListeria属等に属する細菌、好ましくはKlebsiella属細菌、より好ましくはKlebsiella oxytocaに由来するジオールデヒドラターゼのβサブユニット及び/又はγサブユニットにおいて、N末端アミノ酸が欠失したジオールデヒドラターゼが挙げられる。
【0027】
野生型ジオールデヒドラターゼのα、β及びγサブユニットの公知のアミノ酸配列及び各サブユニットをコードする遺伝子の塩基配列は、公開されたデータベース、例えば、GenBank(http://www.ncbi.nlm.nih.gov)に、例えば、アクセッション番号:D45071で登録されている。
【0028】
より具体的には、野生型ジオールデヒドラターゼのαサブユニットのアミノ酸配列としては配列番号1のアミノ酸配列が挙げられ、αサブユニットをコードする遺伝子の塩基配列としては配列番号2の塩基配列が挙げられ、βサブユニットのアミノ酸配列としては配列番号3のアミノ酸配列が挙げられ、βサブユニットをコードする遺伝子の塩基配列としては配列番号4の塩基配列が挙げられ、γサブユニットのアミノ酸配列としては配列番号5のアミノ酸配列が挙げられ、γサブユニットをコードする遺伝子の塩基配列としては配列番号6の塩基配列が挙げられる。
【0029】
本発明の一実施形態において、変異型ジオールデヒドラターゼは、α、β及びγサブユニットを含み、
αサブユニットが、配列番号1のアミノ酸配列からなるタンパク質であり、
βサブユニットが、配列番号3のアミノ酸配列からなるタンパク質、又は配列番号3においてN末端の9〜64個、好ましくは14〜35個、さらに好ましくは18〜25個のアミノ酸が欠失したアミノ酸配列からなるタンパク質であり、
γサブユニットが、配列番号5のアミノ酸配列からなるタンパク質、又は配列番号5においてN末端の5〜42個、好ましくは10〜35個、さらに好ましくは15〜25個のアミノ酸が欠失したアミノ酸配列からなるタンパク質である。
【0030】
ただし、ここで、βサブユニットが配列番号3のアミノ酸配列からなるタンパク質であるときは、γサブユニットは配列番号5のアミノ酸配列からなるタンパク質ではない。
【0031】
本発明においてαサブユニット、βサブユニット及びγサブユニットには、野生型酵素における各サブユニットだけでなく、N末端アミノ酸が変異した変異型のサブユニットも包含される。
【0032】
上記のアミノ酸配列で特定される各サブユニットと機能的に同等のタンパク質を対応するサブユニットとして含む変異型ジオールデヒドラターゼもまた本発明に包含される。
【0033】
本発明において、各サブユニットと機能的に同等のタンパク質とは、当該タンパク質が各配列番号で特定されるタンパク質と同等の生物学的機能、生化学的機能を有することを指す。例えば、「配列番号1のアミノ酸配列からなるタンパク質」と機能的に同等のタンパク質としては、「配列番号1において1若しくは数個のアミノ酸が欠失、置換若しくは付加されたアミノ酸配列からなるタンパク質であって、ジオールデヒドラターゼのβサブユニット及びγサブユニットと会合したときにジオールデヒドラターゼ活性を有する前記タンパク質」が挙げられる。
【0034】
数個とは、通常2〜10個、好ましくは2〜5個、より好ましくは2〜3個をいう。また、例えば「ジオールデヒドラターゼのβサブユニット及びγサブユニットと会合したときにジオールデヒドラターゼ活性を有する前記タンパク質」とは、βサブユニット及びγサブユニット、少なくとも野生型のβサブユニット及びγサブユニットと会合して、野生型ジオールデヒドラターゼと同等の酵素活性を示すタンパク質、すなわち、さらにアデノシルコバラミン(補酵素B12)の存在下でジオールデヒドラターゼ活性を有するタンパク質を意味する。
【0035】
また、機能的に同等のタンパク質は、通常、アミノ酸配列レベルにおいて高い同一性を有する。高い同一性とは、アミノ酸レベルにおいて、通常、少なくとも80%以上の同一性、好ましくは90%以上の同一性、さらに好ましくは95%以上の同一性、さらに好ましくは99%以上の同一性を指す。
【0036】
より具体的には、本発明の一実施形態において、変異型ジオールデヒドラターゼは、αサブユニット、βサブユニット及びγサブユニットを含み、
αサブユニットが以下の(a)又は(b)のタンパク質:
(a)配列番号1のアミノ酸配列からなるタンパク質、
(b)配列番号1において1若しくは数個のアミノ酸が欠失、置換若しくは付加されたアミノ酸配列からなるタンパク質であって、ジオールデヒドラターゼのβサブユニット及びγサブユニットと会合したときにジオールデヒドラターゼ活性を有する前記タンパク質、
であり、
βサブユニットが以下の(c)〜(f)から選択されるタンパク質:
(c)配列番号3のアミノ酸配列からなるタンパク質、
(d)配列番号3において1若しくは数個のアミノ酸が欠失、置換若しくは付加されたアミノ酸配列からなるタンパク質であって、ジオールデヒドラターゼのαサブユニット及びγサブユニットと会合したときにジオールデヒドラターゼ活性を有する前記タンパク質、
(e)配列番号3においてN末端の9〜64個、好ましくは14〜35個、さらに好ましくは18〜25個のアミノ酸が欠失したアミノ酸配列からなるタンパク質、
(f)(e)に記載のアミノ酸配列において1若しくは数個のアミノ酸が欠失、置換若しくは付加されたアミノ酸配列からなるタンパク質であって、ジオールデヒドラターゼのαサブユニット及びγサブユニットと会合したときにジオールデヒドラターゼ活性を有する前記タンパク質、
であり、
γサブユニットが以下の(g)〜(j)から選択されるタンパク質:
(g)配列番号5のアミノ酸配列からなるタンパク質、
(h)配列番号5において1若しくは数個のアミノ酸が欠失、置換若しくは付加されたアミノ酸配列からなるタンパク質であって、ジオールデヒドラターゼのαサブユニット及びβサブユニットと会合したときにジオールデヒドラターゼ活性を有する前記タンパク質、
(i)配列番号5においてN末端の5〜42個、好ましくは10〜35個、さらに好ましくは15〜25個のアミノ酸が欠失したアミノ酸配列からなるタンパク質、
(j)(i)に記載アミノ酸配列において1若しくは数個のアミノ酸が欠失、置換若しくは付加されたアミノ酸配列からなるタンパク質であって、ジオールデヒドラターゼのαサブユニット及びβサブユニットと会合したときにジオールデヒドラターゼ活性を有する前記タンパク質
であり、
ただし、βサブユニットが(c)又は(d)のタンパク質であるときγサブユニットは(g)及び(h)のタンパク質ではない。
【0037】
本発明の変異型ジオールデヒドラターゼは、野生型ジオールデヒドラターゼと異なり水に可溶性であるが、野生型ジオールデヒドラターゼと同等の酵素活性を維持している。従って、組換え体ジオールデヒドラターゼを大量に得ることができるとともに、親水性の条件下で酵素反応を実施することができる。
【0038】
本発明の別の実施形態は、α、β及びγサブユニットを含む変異型グリセロールデヒドラターゼであって、βサブユニットのN末端にジオールデヒドラターゼのβサブユニットのN末端の9〜64個のアミノ酸が付加しており、及び/又はγサブユニットのN末端にジオールデヒドラターゼのβサブユニットのN末端の5〜42個アミノ酸が付加しており、かつ水に不溶性である前記変異型グリセロールデヒドラターゼに関する。ここでβ又はγサブユニットにおいてN末端に付加しているジオールデヒドラターゼの対応するサブユニットのN末端アミノ酸の数については上記のとおりである。
【0039】
本発明の変異型グリセロールデヒドラターゼには、βサブユニット及びγサブユニットのいずれかのN末端にアミノ酸付加を有するもの、並びにβサブユニット及びγサブユニットの双方のN末端にアミノ酸付加を有するものが包含される。いずれの場合にも本発明の効果が得られる。
【0040】
本発明の別の実施形態は、α、β及びγサブユニットを含む変異型グリセロールデヒドラターゼであって、βサブユニットのN末端の1〜50個のアミノ酸がジオールデヒドラターゼのβサブユニットのN末端の9〜70個のアミノ酸と置換されており、及び/又はγサブユニットのN末端の1〜20個のアミノ酸がジオールデヒドラターゼのβサブユニットのN末端の5〜50個のアミノ酸と置換されており、かつ水に不溶性である前記変異型グリセロールデヒドラターゼに関する。
【0041】
本発明の変異型グリセロールデヒドラターゼには、βサブユニット又はγサブユニットのいずれかのN末端のみ置換されるもの、並びにβサブユニット及びγサブユニットの双方のN末端が置換されるものが包含される。いずれの場合にも本発明の効果が得られる。
【0042】
本発明の変異型グリセロールデヒドラターゼは、上記変異を含み水に不溶性であるとともに、グリセロールデヒドラターゼ活性を有する限り特に限定されない。本発明において水に不溶性であるとは、変異型グリセロールデヒドラターゼ遺伝子が発現したタンパク質の大部分が水に不溶であることを意味する。具体的には、酵素タンパク質を発現させた微生物菌体を適当な緩衝液に懸濁させて充分破砕した後、懸濁液を遠心分離した試料の沈殿部分に、酵素活性の大部分が存在することで確認できる。
【0043】
本発明の変異型グリセロールデヒドラターゼとしては、例えば、Lactobacillus属、Citrobacter属、Clostridium属、Klebsiella属、Enterobacter属、Caloramator属、Salmonella属及びListeria属等に属する細菌、好ましくはKlebsiella属細菌、より好ましくはKlebsiella pneumoniaeに由来するグリセロールデヒドラターゼのβサブユニット及び/又はγサブユニットにおいて、N末端アミノ酸に付加又は置換を有するジオールデヒドラターゼが挙げられる。
【0044】
野生型グリセロールデヒドラターゼのα、β及びγサブユニットの公知のアミノ酸配列及び各サブユニットをコードする遺伝子の塩基配列は、公開されたデータベース、例えば、GenBank(http://www.ncbi.nlm.nih.gov)に、例えばアクセッション番号:U30903で登録されている。
【0045】
より具体的には、野生型グリセロールデヒドラターゼのαサブユニットのアミノ酸配列としては配列番号7のアミノ酸配列が挙げられ、αサブユニットをコードする遺伝子の塩基配列としては配列番号8の塩基配列が挙げられ、βサブユニットのアミノ酸配列としては配列番号9のアミノ酸配列が挙げられ、βサブユニットをコードする遺伝子の塩基配列としては配列番号10の塩基配列が挙げられ、γサブユニットのアミノ酸配列としては配列番号11のアミノ酸配列が挙げられ、γサブユニットをコードする遺伝子の塩基配列としては配列番号12の塩基配列が挙げられる。
【0046】
本発明の一実施形態において、変異型グリセロールデヒドラターゼは、α、β及びγサブユニットを含み、
αサブユニットが、配列番号7のアミノ酸配列からなるタンパク質であり、
βサブユニットが、配列番号9のアミノ酸配列からなるタンパク質、又は配列番号9のN末端に配列番号3のN末端の9〜64個、好ましくは35〜60個のアミノ酸が付加してなるアミノ酸配列からなるタンパク質、
γサブユニットが、配列番号11のアミノ酸配列からなるタンパク質、又は配列番号11のN末端に配列番号5のN末端の5〜42個、好ましくは10〜35個、さらに好ましくは15〜25個のアミノ酸が付加してなるアミノ酸配列からなるタンパク質である。
【0047】
ただし、ここで、βサブユニットが配列番号9のアミノ酸配列からなるタンパク質であるときは、γサブユニットは配列番号11のアミノ酸配列からなるタンパク質ではない。
【0048】
本発明の一実施形態において、変異型グリセロールデヒドラターゼは、α、β及びγサブユニットを含み、
αサブユニットが、配列番号7のアミノ酸配列からなるタンパク質であり、
βサブユニットが、配列番号9のアミノ酸配列からなるタンパク質、又は配列番号9のN末端の1〜50個、好ましくは20〜30個のアミノ酸が、配列番号3のN末端の9〜70個、好ましくは35〜64個、さらに好ましくは35〜62個のアミノ酸と置換されたアミノ酸配列からなるタンパク質、
γサブユニットが、配列番号11のアミノ酸配列からなるタンパク質、又は配列番号11のN末端の1〜20個、好ましくは1〜8個のアミノ酸が、配列番号5のN末端の5〜50個、好ましくは30〜45個のアミノ酸と置換されたアミノ酸配列からなるタンパク質である。
【0049】
ただし、ここで、βサブユニットが配列番号9のアミノ酸配列からなるタンパク質であるときは、γサブユニットは配列番号11のアミノ酸配列からなるタンパク質ではない。
【0050】
上記のアミノ酸配列で特定される各サブユニットと機能的に同等のタンパク質を対応するサブユニットとして含む変異型グリセロールデヒドラターゼもまた本発明に包含される。例えば、「配列番号7のアミノ酸配列からなるタンパク質」と機能的に同等のタンパク質としては、「配列番号7において1若しくは数個のアミノ酸が欠失、置換若しくは付加されたアミノ酸配列からなるタンパク質であって、グリセロールデヒドラターゼのβサブユニット及びγサブユニットと会合したときにグリセロールデヒドラターゼ活性を有する前記タンパク質」が挙げられる。上記のとおり、機能的に同等のタンパク質は、通常、アミノ酸配列レベルにおいて高い同一性を有する。
【0051】
また、例えば「グリセロールデヒドラターゼのβサブユニット及びγサブユニットと会合したときにグリセロールデヒドラターゼ活性を有する前記タンパク質」とは、βサブユニット及びγサブユニット、少なくとも野生型のβサブユニット及びγサブユニットと会合して、野生型グリセロールデヒドラターゼと同等の酵素活性を示すタンパク質、すなわち、さらにアデノシルコバラミン(補酵素B12)の存在下でグリセロールデヒドラターゼ活性を有するタンパク質を意味する。
【0052】
より具体的には、本発明の一実施形態において、変異型グリセロールデヒドラターゼは、αサブユニット、βサブユニット及びγサブユニットを含み、
αサブユニットが以下の(a)又は(b)のタンパク質:
(a)配列番号7のアミノ酸配列からなるタンパク質、
(b)配列番号7において1若しくは数個のアミノ酸が欠失、置換若しくは付加されたアミノ酸配列からなるタンパク質であって、グリセロールデヒドラターゼのβサブユニット及びγサブユニットと会合したときにグリセロールデヒドラターゼ活性を有する前記タンパク質、
であり、
βサブユニットが以下の(c)〜(f)から選択されるタンパク質:
(c)配列番号9のアミノ酸配列からなるタンパク質、
(d)配列番号9において1若しくは数個のアミノ酸が欠失、置換若しくは付加されたアミノ酸配列からなるタンパク質であって、グリセロールデヒドラターゼのαサブユニット及びγサブユニットと会合したときにグリセロールデヒドラターゼ活性を有する前記タンパク質、
(e)配列番号9のN末端に配列番号3のN末端の9〜64個、好ましくは35〜60個のアミノ酸が付加してなるアミノ酸配列からなるタンパク質、
(f)(e)に記載のアミノ酸配列において1若しくは数個のアミノ酸が欠失、置換若しくは付加されたアミノ酸配列からなるタンパク質であって、グリセロールデヒドラターゼのαサブユニット及びγサブユニットと会合したときにグリセロールデヒドラターゼ活性を有する前記タンパク質、
であり、
γサブユニットが以下の(g)〜(j)から選択されるタンパク質:
(g)配列番号11のアミノ酸配列からなるタンパク質、
(h)配列番号11において1若しくは数個のアミノ酸が欠失、置換若しくは付加されたアミノ酸配列からなるタンパク質であって、グリセロールデヒドラターゼのαサブユニット及びβサブユニットと会合したときにグリセロールデヒドラターゼ活性を有する前記タンパク質、
(i)配列番号11のN末端に配列番号5のN末端の5〜42個、好ましくは10〜35個、さらに好ましくは15〜25個のアミノ酸が付加してなるアミノ酸配列からなるタンパク質、
(j)(i)に記載アミノ酸配列において1若しくは数個のアミノ酸が欠失、置換若しくは付加されたアミノ酸配列からなるタンパク質であって、グリセロールデヒドラターゼのαサブユニット及びβサブユニットと会合したときにグリセロールデヒドラターゼ活性を有する前記タンパク質であり、
ただし、βサブユニットが(c)又は(d)のタンパク質であるときγサブユニットは(g)及び(h)のタンパク質ではない。
【0053】
一実施形態において本発明の変異型グリセロールデヒドラターゼは、αサブユニット、βサブユニット及びγサブユニットを含み、
αサブユニットが以下の(a)又は(b)のタンパク質:
(a)配列番号18のアミノ酸配列からなるタンパク質、
(b)配列番号18において1若しくは数個のアミノ酸が欠失、置換若しくは付加されたアミノ酸配列からなるタンパク質であって、グリセロールデヒドラターゼのβサブユニット及びγサブユニットと会合したときにグリセロールデヒドラターゼ活性を有する前記タンパク質、
であり、
βサブユニットが以下の(c)〜(f)から選択されるタンパク質:
(c)配列番号9のアミノ酸配列からなるタンパク質、
(d)配列番号9において1若しくは数個のアミノ酸が欠失、置換若しくは付加されたアミノ酸配列からなるタンパク質であって、グリセロールデヒドラターゼのαサブユニット及びγサブユニットと会合したときにグリセロールデヒドラターゼ活性を有する前記タンパク質、
(e)配列番号19のアミノ酸配列からなるタンパク質、
(f)配列番号19において1若しくは数個のアミノ酸が欠失、置換若しくは付加されたアミノ酸配列からなるタンパク質であって、グリセロールデヒドラターゼのαサブユニット及びγサブユニットと会合したときにグリセロールデヒドラターゼ活性を有する前記タンパク質、
であり、
γサブユニットが以下の(g)〜(j)から選択されるタンパク質:
(g)配列番号11のアミノ酸配列からなるタンパク質、
(h)配列番号11において1若しくは数個のアミノ酸が欠失、置換若しくは付加されたアミノ酸配列からなるタンパク質であって、グリセロールデヒドラターゼのαサブユニット及びβサブユニットと会合したときにグリセロールデヒドラターゼ活性を有する前記タンパク質、
(i)配列番号20のアミノ酸配列からなるタンパク質、
(j)配列番号20において1若しくは数個のアミノ酸が欠失、置換若しくは付加されたアミノ酸配列からなるタンパク質であって、グリセロールデヒドラターゼのαサブユニット及びβサブユニットと会合したときにグリセロールデヒドラターゼ活性を有する前記タンパク質、
であり、
ただし、βサブユニットが(c)又は(d)のタンパク質であるときγサブユニットは(g)及び(h)のタンパク質ではない。
【0054】
グリセロールデヒドラターゼ活性を有するタンパク質とは、野生型グリセロールデヒドラターゼと同等の酵素活性を示すタンパク質、すなわち、さらにアデノシルコバラミン(補酵素B12)の存在下でグリセロールデヒドラターゼ活性を有するタンパク質を意味する。数個とは、通常2〜10個、好ましくは2〜5個、より好ましくは2〜3個をいう。上記各サブユニットを構成するタンパク質には、各アミノ酸配列からなるタンパク質と機能的に同等のタンパク質、すなわち、アミノ酸配列レベルにおいて高い同一性を有するタンパク質であって、各サブユニットの活性を有するタンパク質も包含される。高い同一性とは、上記のとおりアミノ酸レベルにおいて、通常、少なくとも80%以上の同一性、好ましくは90%以上の同一性、さらに好ましくは95%以上の同一性、さらに好ましくは99%以上の同一性を指す。
【0055】
本発明の変異型グリセロールデヒドラターゼは、水に不溶性であるため、疎水性条件下でも酵素反応を実施することができる。従って、疎水性条件下でグリセロールを基質として反応を実施したい場合に、好適である。
【0056】
β及び/又はγサブユニットのN末端アミノ酸に変異を有するジオールデヒドラターゼ及びグリセロールデヒドラターゼは、当技術分野で公知の方法により調製できる。
【0057】
ジオールデヒドラターゼのN末端アミノ酸が欠失したサブユニットは、例えば、公知のジオールデヒドラターゼをプロテーゼで処理することにより得られる。プロテアーゼとしては、トリプシン、ペプシン、キモトリプシン、ズブチリシン、スロンビン、サーモリシン、パパイン、プラスミン等が挙げられる。例えば、ジオールデヒドラターゼをトリプシン処理し、得られた反応液の上清から水溶性のβ又はγサブユニットを得ることができる。プロテアーゼ処理の条件は、当業者であれば適宜設定することができるが、通常1.0μg/mlのトリプシンの存在下、約15℃で約10〜14時間、好ましくは約12時間反応させる。
【0058】
あるいは、本発明の変位型ジオールデヒドラターゼ及びグリセロールデヒドラターゼは、公知のジオールデヒドラターゼ又はグリセロールデヒドラターゼをコードする遺伝子、すなわち各サブユニットをコードする遺伝子の塩基配列に基づいて、当技術分野において通常用いられる方法により調製できる。すなわち、N末端アミノ酸が変異した各サブユニットをコードする遺伝子をベクターに挿入し、該ベクターで宿主細胞を形質転換し、該形質転換体を培養することにより調製できる。
【0059】
本発明の一実施形態は、本発明の変異型ジオールデヒドラターゼをコードする遺伝子、本発明の変異型グリセロールデヒドラターゼをコードする遺伝子、これら遺伝子の一方又は双方を含むベクター、該ベクターによって形質転換された、変異型ジオールデヒドラターゼをコードする遺伝子及び/又は変異型グリセロールデヒドラターゼをコードする遺伝子を含む形質転換体に関する。本発明において、「遺伝子」という用語には、DNAのみならずそのmRNAやcDNAも含むものとする。また、全長遺伝子のみならずESTも含むものとする。
【0060】
本発明の変異型ジオールデヒドラターゼをコードする遺伝子には、αサブユニットをコードする遺伝子、βサブユニットをコードする遺伝子、及びγサブユニットをコードする遺伝子を含み、N末端の9〜46個のアミノ酸が欠失しており、及び/又はγサブユニットのN末端の5〜42個のアミノ酸が欠失しており、かつ水溶性である前記ジオールデヒドラターゼをコードする遺伝子が包含される。
【0061】
一実施形態において、変異型ジオールデヒドラターゼをコードする遺伝子は、αサブユニットをコードする遺伝子、βサブユニットをコードする遺伝子、及びγサブユニットをコードする遺伝子を含み、
αサブユニットをコードする遺伝子が、配列番号2の塩基配列からなる遺伝子であり、
βサブユニットをコードする遺伝子が、配列番号4の塩基配列からなる遺伝子であるか、又は配列番号4の5’末端の27〜192個、より好ましくは42〜105個、さらに好ましくは54〜75個の塩基が欠失した塩基配列からなる遺伝子であり、
γサブユニットをコードする遺伝子が、配列番号6の塩基配列からなる遺伝子であるか、又は配列番号6の5’末端の15〜126個、より好ましくは30〜105個、さらに好ましくは45〜75個の塩基が欠失した塩基配列からなる遺伝子である。ただし、βサブユニットをコードする遺伝子が配列番号4の塩基配列からなる遺伝子である場合、γサブユニットをコードする遺伝子は配列番号6の塩基配列からなる遺伝子ではない。なお、欠失させる塩基の数は、読み枠を考慮して決定する。また、開始コドンの付加、選択した宿主内で発現しやすいようなN末端側の配列設定等の諸条件も考慮する。
【0062】
上記の各サブユニットをコードする遺伝子と機能的に同等の遺伝子を、対応するサブユニットをコードする遺伝子として含む変異型ジオールデヒドラターゼ遺伝子もまた本発明に包含される。
【0063】
あるタンパク質をコードする遺伝子と機能的に同等の遺伝子とは、当該タンパク質と機能的に同等のタンパク質をコードする遺伝子を意味し、当該遺伝子を調製する当業者によく知られた方法としては、ハイブリダイゼーション技術(Sambrook,J.et al.,Molecular Cloning 2nd ed.,9.47-9.58,Cold Spring Harbor Lab.press,1989)を利用する方法が挙げられる。
【0064】
例えば、αサブユニットをコードする遺伝子としての「配列番号2の塩基配列からなる遺伝子」と機能的に同等の遺伝子としては、「配列番号2の塩基配列の全部又は一部からなる遺伝子に対し相補的な塩基配列からなる遺伝子とストリンジェントな条件下でハイブリダイズし、かつジオールデヒドラターゼのβサブユニット及びγサブユニットと会合したときにジオールデヒドラターゼ活性を有するタンパク質をコードする遺伝子」が挙げられる。また、配列番号2の塩基配列と少なくとも65%の同一性、好ましくは少なくとも75%の同一性、より好ましくは少なくとも80%の同一性、さらに好ましくは少なくとも85%の同一性、最も好ましくは少なくとも95%の同一性を有する塩基配列からなる遺伝子も包含される。
【0065】
塩基配列の一部とは、各塩基配列の一部分の塩基配列であって、ストリンジェントな条件下でハイブリダイズさせるのに十分な塩基配列の長さを有するものであり、例えば、少なくとも15塩基、好ましくは少なくとも100塩基、より好ましくは少なくとも200塩基の配列である。好ましくは各塩基配列において連続する少なくとも15塩基、好ましくは少なくとも100塩基、より好ましくは少なくとも200塩基の配列である。ここで「連続する」とは、基準とする塩基配列のうち連続した塩基配列を含むことを意味する。
【0066】
本明細書において、ストリンジェントな条件とは、特異的なハイブリッドが形成され、非特異的なハイブリッドが形成されない条件をいい、すなわち、各塩基配列に対し高い相同性(相同性又は同一性が60%以上、好ましくは70%以上、より好ましくは80%以上、さらに好ましくは90%以上、最も好ましくは95%以上)を有するポリヌクレオチドがハイブリダイズする条件をいう。より具体的には、このような条件は、当該分野において周知慣用な手法、例えば、コロニーハイブリダイゼーション法、プラークハイブリダイゼーション法、マイクロアレイ法又はサザンブロットハイブリダイゼーション法などにおいて、具体的には、ポリヌクレオチドを固定化したメンブランを用いて、0.7〜1.0MのNaCl存在下、65℃でハイブリダイゼーションを行った後、0.1〜2倍濃度のSSC(Saline Sodium Citrate;150mM塩化ナトリウム、15mMクエン酸ナトリウム)溶液を用い、65℃でメンブランを洗浄することにより達成できる。
【0067】
また、塩基配列情報を基に合成したプライマーを用いる遺伝子増幅法、例えば、ポリメラーゼ連鎖反応(PCR)法、LAMP法などを利用して、各遺伝子と機能的に同等の遺伝子を単離することも可能である。
【0068】
より具体的には、一実施形態において、本発明の変異型ジオールデヒドラターゼをコードする遺伝子は、αサブユニットをコードする遺伝子、βサブユニットをコードする遺伝子、及びγサブユニットをコードする遺伝子を含み、
αサブユニットをコードする遺伝子が、以下の(a)又は(b)の遺伝子:
(a)配列番号2の塩基配列からなる遺伝子、
(b)配列番号2の塩基配列の全部又は一部からなる遺伝子に対し相補的な塩基配列からなる遺伝子とストリンジェントな条件下でハイブリダイズし、かつジオールデヒドラターゼのβサブユニット及びγサブユニットと会合したときにジオールデヒドラターゼ活性を有するタンパク質をコードする遺伝子
であり、
βサブユニットをコードする遺伝子が、以下の(c)〜(f)から選択される遺伝子:
(c)配列番号4の塩基配列からなる遺伝子、
(d)配列番号4の塩基配列の全部又は一部からなる遺伝子に対し相補的な塩基配列からなる遺伝子とストリンジェントな条件下でハイブリダイズし、かつジオールデヒドラターゼのαサブユニット及びγサブユニットと会合したときにジオールデヒドラターゼ活性を有するタンパク質をコードする遺伝子
(e)配列番号4の5’末端の27〜192個、より好ましくは42〜105個、さらに好ましくは54〜75個の塩基が欠失した塩基配列からなる遺伝子
(f)(e)に記載の塩基配列の全部又は一部からなる遺伝子に対し相補的な塩基配列からなる遺伝子とストリンジェントな条件下でハイブリダイズし、かつジオールデヒドラターゼのαサブユニット及びγサブユニットと会合したときにジオールデヒドラターゼ活性を有するタンパク質をコードする遺伝子
であり、
γサブユニットをコードする遺伝子が、以下の(g)〜(j)から選択される遺伝子:
(g)配列番号6の塩基配列からなる遺伝子、
(h)配列番号6の塩基配列の全部又は一部からなる遺伝子に対し相補的な塩基配列からなる遺伝子とストリンジェントな条件下でハイブリダイズし、かつジオールデヒドラターゼのαサブユニット及びβサブユニットと会合したときにジオールデヒドラターゼ活性を有するタンパク質をコードする遺伝子
(i)配列番号6の5’末端の15〜126個、より好ましくは30〜105個、さらに好ましくは45〜75個の塩基が欠失した塩基配列からなる遺伝子
(j)(i)に記載の塩基配列の全部又は一部からなる遺伝子に対し相補的な塩基配列からなる遺伝子とストリンジェントな条件下でハイブリダイズし、かつジオールデヒドラターゼのαサブユニット及びβサブユニットと会合したときにジオールデヒドラターゼ活性を有するタンパク質をコードする遺伝子
であるが、
βサブユニットをコードする遺伝子が(c)又は(d)であるときは、γサブユニットをコードする遺伝子は(g)及び(h)ではない。
【0069】
本発明の変異型グリセロールデヒドラターゼをコードする遺伝子には、αサブユニットをコードする遺伝子、βサブユニットをコードする遺伝子、及びγサブユニットをコードする遺伝子を含み、βサブユニットのN末端にジオールデヒドラターゼのβサブユニットのN末端の9〜64個のアミノ酸が付加しており、及び/又はγサブユニットのN末端にジオールデヒドラターゼのβサブユニットのN末端の5〜42個のアミノ酸が付加しており、かつ水に不溶性である前記グリセロールデヒドラターゼをコードする遺伝子が包含される。
【0070】
一実施形態において、変異型グリセロールデヒドラターゼをコードする遺伝子は、αサブユニットをコードする遺伝子、βサブユニットをコードする遺伝子、及びγサブユニットをコードする遺伝子を含み、
αサブユニットをコードする遺伝子が、配列番号8の塩基配列からなる遺伝子であり、
βサブユニットをコードする遺伝子が、配列番号10の塩基配列からなる遺伝子であるか、又は配列番号10の5’末端に、配列番号4の5’末端の27〜192個、より好ましくは105〜180個の塩基が付加した塩基配列からなる遺伝子であり、
γサブユニットをコードする遺伝子が、配列番号12の塩基配列からなる遺伝子であるか、又は配列番号12の5’末端に、配列番号6の5’末端の15〜126個、より好ましくは30〜105個、さらに好ましくは45〜75個の塩基が付加した塩基配列からなる遺伝子である。ただし、βサブユニットをコードする遺伝子が配列番号10の塩基配列からなる遺伝子である場合、γサブユニットをコードする遺伝子は配列番号12の塩基配列からなる遺伝子ではない。なお、付加する塩基の数は、読み枠を考慮して決定する。
【0071】
一実施形態において、変異型グリセロールデヒドラターゼをコードする遺伝子は、αサブユニットをコードする遺伝子、βサブユニットをコードする遺伝子、及びγサブユニットをコードする遺伝子を含み、
αサブユニットをコードする遺伝子が、配列番号8の塩基配列からなる遺伝子であり、
βサブユニットをコードする遺伝子が、配列番号10の塩基配列からなる遺伝子であるか、又は配列番号10の5’末端の3〜150個の塩基、より好ましくは30〜120個の塩基、さらに好ましくは60〜90個の塩基が、特に好ましくは81個の塩基が、配列番号4の5’末端の27〜210個、より好ましくは105〜192個、さらに好ましくは105〜186の塩基、特に好ましくは180の塩基と置換された塩基配列からなる遺伝子であり、
γサブユニットをコードする遺伝子が、配列番号12の塩基配列からなる遺伝子であるか、又は配列番号12の5’末端の1〜60個の塩基、より好ましくは1〜24個の塩基、特に好ましくは3個の塩基が、配列番号6の5’末端の15〜150個、より好ましくは90〜135個、特に好ましくは99塩基と置換された塩基配列からなる遺伝子である。
【0072】
ただし、βサブユニットをコードする遺伝子が配列番号10の塩基配列からなる遺伝子である場合、γサブユニットをコードする遺伝子は配列番号12の塩基配列からなる遺伝子ではない。なお、付加又は置換する塩基の数は、読み枠を考慮して決定する。
【0073】
上記の各サブユニットをコードする遺伝子と機能的に同等の遺伝子を、対応するサブユニットをコードする遺伝子として含む変異型グリセロールデヒドラターゼ遺伝子もまた本発明に包含される。
【0074】
例えば、αサブユニットをコードする遺伝子としての「配列番号8の塩基配列からなる遺伝子」と機能的に同等の遺伝子としては、「配列番号8の塩基配列の全部又は一部からなる遺伝子に対し相補的な塩基配列からなる遺伝子とストリンジェントな条件下でハイブリダイズし、かつグリセロールデヒドラターゼのβサブユニット及びγサブユニットと会合したときにグリセロールデヒドラターゼ活性を有するタンパク質をコードする遺伝子」が挙げられる。また、各配列番号の塩基配列と少なくとも65%の同一性、好ましくは少なくとも75%の同一性、より好ましくは少なくとも80%の同一性、さらに好ましくは少なくとも85%の同一性、最も好ましくは少なくとも95%の同一性を有する塩基配列からなる遺伝子も包含される。
【0075】
より具体的には、一実施形態において、本発明の変異型グリセロールデヒドラターゼをコードする遺伝子は、αサブユニットをコードする遺伝子、βサブユニットをコードする遺伝子、及びγサブユニットをコードする遺伝子を含み、
αサブユニットをコードする遺伝子が、以下の(a)又は(b)の遺伝子:
(a)配列番号8の塩基配列からなる遺伝子、
(b)配列番号8の塩基配列の全部又は一部からなる遺伝子に対し相補的な塩基配列からなる遺伝子とストリンジェントな条件下でハイブリダイズし、かつグリセロールデヒドラターゼのβサブユニット及びγサブユニットと会合したときにグリセロールデヒドラターゼ活性を有するタンパク質をコードする遺伝子
であり、
βサブユニットをコードする遺伝子が、以下の(c)〜(f)から選択される遺伝子:
(c)配列番号10の塩基配列からなる遺伝子、
(d)配列番号10の塩基配列の全部又は一部からなる遺伝子に対し相補的な塩基配列からなる遺伝子とストリンジェントな条件下でハイブリダイズし、かつグリセロールデヒドラターゼのαサブユニット及びγサブユニットと会合したときにグリセロールデヒドラターゼ活性を有するタンパク質をコードする遺伝子
(e)配列番号10の5’末端に、配列番号4の5’末端の27〜192個、より好ましくは105〜180個の塩基が付加した塩基配列からなる遺伝子
(f)(e)に記載の塩基配列の全部又は一部からなる遺伝子に対し相補的な塩基配列からなる遺伝子とストリンジェントな条件下でハイブリダイズし、かつグリセロールデヒドラターゼのαサブユニット及びγサブユニットと会合したときにグリセロールデヒドラターゼ活性を有するタンパク質をコードする遺伝子
であり、
γサブユニットをコードする遺伝子が、以下の(g)〜(j)から選択される遺伝子:
(g)配列番号12の塩基配列からなる遺伝子、
(h)配列番号12の塩基配列の全部又は一部からなる遺伝子に対し相補的な塩基配列からなる遺伝子とストリンジェントな条件下でハイブリダイズし、かつグリセロールデヒドラターゼのαサブユニット及びβサブユニットと会合したときにグリセロールデヒドラターゼ活性を有するタンパク質をコードする遺伝子
(i)配列番号12の5’末端に、配列番号6の5’末端の15〜126個、より好ましくは30〜105個、さらに好ましくは45〜75個の塩基が付加した塩基配列からなる遺伝子
(j)(i)に記載の塩基配列の全部又は一部からなる遺伝子に対し相補的な塩基配列からなる遺伝子とストリンジェントな条件下でハイブリダイズし、かつグリセロールデヒドラターゼのαサブユニット及びβサブユニットと会合したときにグリセロールデヒドラターゼ活性を有するタンパク質をコードする遺伝子
であるが、
βサブユニットをコードする遺伝子が(c)又は(d)であるときは、γサブユニットをコードする遺伝子は(g)及び(h)ではない。
【0076】
本発明の変異型グリセロールデヒドラターゼをコードする遺伝子の具体例としては、以下の(a)又は(b)の遺伝子が挙げられる:
(a)配列番号17の塩基配列からなる遺伝子、
(b)配列番号17で表される塩基配列からなる遺伝子に対し相補的な塩基配列からなる遺伝子とストリンジェントな条件下でハイブリダイズし、かつグリセロールデヒドラターゼの活性を有するタンパク質をコードする遺伝子。
【0077】
ここでストリンジェントな条件については、上記のとおりである。従って、本発明の変異型グリセロールデヒドラターゼをコードする遺伝子には、配列番号17の塩基配列に対し高い相同性(相同性又は同一性が60%以上、好ましくは70%以上、より好ましくは80%以上、さらに好ましくは90%以上、最も好ましくは95%以上)を有する塩基配列からなる遺伝子であって、グリセロールデヒドラターゼの活性を有するタンパク質をコードする遺伝子が包含される。グリセロールデヒドラターゼの活性を有するタンパク質については、上記のとおりである。
【0078】
本発明の変異型ジオールデヒドラターゼ及びグリセロールデヒドラターゼには、他のペプチドが融合されたものも含まれる。融合に付される他のペプチドとしては、例えばFLAG(Hopp, T. P. et. al., BioTechnology 6: 1204-1210,1988)、6個のHis(ヒスチジン)残基からなる6×His、10×His、ヒトc−mycの断片、VSV−GPの断片、p18HIVの断片、T7−tag、HSV−tag、E−tag、SV40T抗原の断片、lck tag、α−tubulinの断片、B−tag、Protein Cの断片、GST(グルタチオンS−トランスフェラーゼ)、HA(インフルエンザ凝集素)、イムノグロブリン定常領域、イムノグロブリンヒンジ領域、β−ガラクトシダーゼ、MBP(マルトース結合タンパク質)等が挙げられる。これらのペプチドを融合させることにより、レセプター誘導体の精製や標識を容易に実施することができる。
【0079】
本発明の変異型ジオールデヒドラターゼ及びグリセロールデヒドラターゼの調製に使用されるベクターとしては、当技術分野において一般的に使用されるものを用いることができる。例えば、形質転換における宿主として大腸菌を用いる場合には、例えば、大腸菌内で複製させるための「ori」、及び形質転換された大腸菌を選抜するための遺伝子(例えば、アンピシリンやテトラサイクリン、カナマイシン、クロラムフェニコール等の薬剤耐性遺伝子)をベクター上に有することが望ましく、このようなベクターとして具体的には、M13系ベクター、pUC系ベクター、pBR322、pBluescript、pCR−Script、pCR2.1、pGEM−T、pDIRECT、pT7等を挙げることができる。
【0080】
特に、発現ベクターが有用である。発現ベクターとしては、例えば大腸菌での発現を目的とした場合は、lacZプロモーター(Ward et. al., Nature 341: 544-546, 1989)、araBプロモーター(Better et. al., Science 240: 1041-1043, 1988)、又はT7プロモーター等を有するベクターを例示することができる。このようなベクターとしては、上記ベクターの他にpGEX−5X−1(ファルマシア)、「QIAexpress system」(キアゲン)、pEGFP、及びpET(この場合、宿主はT7 RNAポリメラーゼを発現するBL21が好ましい)等が挙げられる。また、発現ベクターには、ポリペプチド分泌のためのシグナル配列が含まれていてもよい。
【0081】
他のベクターとしては、例えば哺乳動物由来の発現ベクター(例えばpcDNA3(インビトロゲン)や、pEGF−BOS(Nucleic Acids. Res. 18(17): 5322, 1990)、pEF、pCDM8、昆虫細胞由来の発現ベクター(例えば「Bac−to−BAC baculovairus expression system」(ギブコBRL)、pBacPAK8)、植物由来の発現ベクター(例えばpMH1、pMH2)、動物ウィルス由来の発現ベクター(例えばpHSV、pMV、pAdexLcw)、レトロウィルス由来の発現ベクター(例えばpZIPneo)、酵母由来の発現ベクター(例えば「Pichia Expression Kit」(インビトロゲン)、pNV11 、SP−Q01)、枯草菌由来の発現ベクター(例えばpPL608、pKTH50)等が挙げられる。
【0082】
CHO細胞、COS細胞、NIH3T3細胞等の動物細胞での発現を目的とした場合には、細胞内で発現させるために必要なプロモーター、例えばSV40プロモーター(Mulligan et. al., Nature 277: 108, 1979)、MMLV−LTRプロモーター、EF1プロモーター(Mizushima et. al., Nucleic Acids Res. 18: 5322, 1990)、CMVプロモーター等を持っていることが不可欠であり、細胞への形質転換を選抜するための遺伝子(例えば薬剤(ネオマイシン、G418等)耐性遺伝子)を有すればさらに好ましい。このような特性を有するベクターとしては、例えばpMAM、pDR2、pBK−RSV、pBK−CMV、pOPRSV、pOP13等が挙げられる。
【0083】
また、複製開始点としては、ポリオーマウィルス、アデノウィルス、ウシパピローマウィルス(BPV)等の由来のものを用いることもできる。さらに、宿主細胞系で遺伝子コピー数増幅のために、発現ベクターは選択マーカーとして、アミノグリコシドトランスフェラーゼ(APH)遺伝子、チミジンキナーゼ(TK)遺伝子、大腸菌キサンチングアニンホスホリボシルトランスフェラーゼ(Ecogpt)遺伝子、ジヒドロ葉酸還元酵素(dhfr)遺伝子等を含むことができる。
【0084】
ベクターが導入される宿主としては特に制限はなく、例えば大腸菌や種々の真核細胞等を用いることが可能である。真核細胞を使用する場合、例えば動物細胞、植物細胞、真菌細胞を宿主に用いることができる。動物細胞としては、哺乳類細胞、例えばCHO、COS、3T3、ミエローマ、BHK(baby hamster kidney)、HeLa、Vero、両生類細胞、例えばアフリカツメガエル卵母細胞(Valle, et. al., Nature 291: 358-340, 1981)、あるいは昆虫細胞、例えばSf9、Sf21、Tn5が知られている。CHO細胞としては、特に、DHFR遺伝子を欠損したCHO細胞であるdhfr−CHO(Proc. Natl. Acad. Sci. USA 77: 4216-4220, 1980)やCHO K−1(Proc. Natl. Acad. Sci. USA 60: 1275, 1968)を好適に使用することができる。動物細胞において、大量発現を目的とする場合には特にCHO細胞が好ましい。植物細胞としては、例えばニコチアナ・タバカム(Nicotiana tabacum)由来の細胞が挙げられる。真菌細胞としては、酵母、例えばサッカロミセス(Saccharomyces)属、例えばサッカロミセス・セレビシエ(Saccharomyces cerevisiae)、糸状菌、例えばアスペルギルス(Aspergillus)属、例えばアスペルギルス・ニガー(Aspergillus niger)が知られている。原核細胞を使用する場合、細菌細胞としては、大腸菌(E. coli)、例えばJM109、DH5α、HB101、XL1Blue、BL21等が挙げられ、その他、枯草菌が知られている。
【0085】
また、宿主として動物を使用する場合、哺乳類動物、植物、昆虫が挙げられる。哺乳類動物としては、ヤギ、ブタ、ヒツジ、マウス、ウシを用いることができる(Vicki Glaser, SPECTRUM Biotechnology Applications, 1993)。また、植物を使用する場合、例えばタバコを用いることができる。さらに、昆虫としては、例えばカイコを用いることができる。
【0086】
本発明の形質転換体を作製するためには、上記宿主に上記ベクターを導入する。そのための方法としては、大腸菌等の宿主細胞へのベクターの導入の場合、例えば塩化カルシウム法、エレクトロポレーション法(Chu, G. et. al., Nucl. Acid Res. 15: 1311-1326, 1987)を用いることができる。また、培養細胞等の宿主細胞へのベクターの導入の場合、例えばリン酸カルシウム法(Chen, C. and Okayama, H. Mol. Cell. Biol. 7: 2745-2752, 1987)、DEAEデキストラン法(Lopata, M. A. et. al., Nucl. Acids Res. 12: 5707-5717, 1984、Sussman, D. J.and Milman, G. Mol. Cell. Biol. 4: 1642-1643, 1985)、カチオニックリボソームDOTAP(ベーリンガーマンハイム)を用いた方法、リポフェクチン法(Derijard, B. Cell 7: 1025-1037, 1994、Lamb, B. T. et. al., Nature Genetics 5: 22-30, 1993、Rabindran, S. K. et. al., Science 259: 230-234, 1993)等の方法を用いることが可能である。
【0087】
さらに、動物にDNAを導入する場合、該DNAを適当なベクター(例えばアデノウイルスベクター(例えばpAdexlcw)やレトロウイルスベクター(例えばpZIPneo)等が挙げられるが、これらに制限されない)に組み込み、例えばレトロウイルス法、リポソーム法、カチオニックリポソーム法、アデノウィルス法等により生体内に導入することが可能である。
【0088】
また、昆虫にベクターを導入する場合、例えば目的のタンパク質をコードするDNAを挿入したバキュロウィルスをカイコに感染させることにより行うことができる(Susumu, M. et. al., Nature 315: 592-594, 1985)。また植物にDNAを導入する場合、例えば目的とするタンパク質をコードするDNAを植物発現用ベクター、例えばpMON 530に挿入し、このベクターをアグロバクテリウム・ツメファシエンス(Agrobacterium tumefaciens)のようなバクテリアに導入する。このバクテリアをタバコ、例えばニコチアナ・タバカムに感染させることでベクターを導入することができる(Julian K.-C. Ma et.al., Eur. J. Immunol. 24: 131-138, 1994)。
【0089】
本発明に係る変異型ジオールデヒドラターゼ及びグリセロールデヒドラターゼは、上記の形質転換体を培養することにより生産させることができる。培養は公知の方法に従って行うことができる。例えば、動物細胞の培養であれば、一般的に、培養液としては、DMEM、MEM、RPMI1640、IMDM等を使用することができる。その際、牛胎児血清(FCS)等の血清補液を併用することもできるし、無血清培養してもよい。培養時のpHは、通常、約6〜8であるのが好ましい。培養は、通常、約30〜40℃で約15〜500時間行い、必要に応じて培地の交換、通気、攪拌を加える。
【0090】
本発明の変異型ジオールデヒドラターゼ及びグリセロールデヒドラターゼは、宿主細胞内又は細胞外(培地等)から単離し、実質的に純粋で均一なタンパク質として精製することができる。タンパク質の分離、精製は、通常のタンパク質の精製で使用されている分離、精製方法を使用すればよく、何ら限定されるものではない。例えばクロマトグラフィーカラム、フィルター、限外濾過、塩析、溶媒沈殿、溶媒抽出、蒸留、免疫沈降、SDS−ポリアクリルアミドゲル電気泳動、等電点電気泳動法、透析、再結晶等を適宜選択、組み合わせればタンパク質を分離、精製することができる。又は、さらにこれらのカラムを複数組み合わせることにより精製することが可能である。
【0091】
クロマトグラフィーとしては、例えば上記のエピトープタグに対する抗体をカラムに固定したアフィニティークロマトグラフィー、イオン交換クロマトグラフィー、疎水性クロマトグラフィー、ゲル濾過、逆相クロマトグラフィー、吸着クロマトグラフィー等が挙げられる(Strategies for Protein Purification and Characterization: A Laboratory Course Manual. Ed DanielR. Marshak et al., Cold Spring Harbor Laboratory Press, 1996)。
【0092】
また、本発明の変位型ジオールデヒドラターゼ及びグリセロールデヒドラターゼをグルタチオンS−トランスフェラーゼタンパク質との融合タンパク質として、あるいはヒスチジンを複数付加させた組み換えタンパク質として宿主細胞(例えば動物細胞や大腸菌等)内で発現させた場合には、発現させた組み換えタンパク質はグルタチオンカラム又はニッケルカラムを用いて精製することができる。融合タンパク質の精製後、必要に応じて融合タンパク質のうち、目的のタンパク質以外の領域を、トロンビン又はファクターXa等により切断し、除去することも可能である。
【0093】
本発明はまた、本発明の変異型ジオールデヒドラターゼ及び/又はグリセロールデヒドラターゼにより、1位及び2位に水酸基を有する脂肪族ポリオールを脱水して、1位にアルデヒド基を有するアルデヒドを生成する方法に関する。
【0094】
上記方法は、1位及び2位に水酸基を有する脂肪族ポリオールと本発明の変異型ジオールデヒドラターゼ及び/又はグリセロールデヒドラターゼとを反応させることにより実施できる。本発明の方法は、1位及び2位に水酸基を有する脂肪族ポリオールと変異型ジオールデヒドラターゼ又はグリセロールデヒドラターゼのいずれかとを反応させてもよいし、双方の酵素と反応させてもよい。
【0095】
酵素反応条件は、当業者であれば適宜設定できるが、通常、アデノシルコバラミン(補酵素B12)の存在下、0.1M程度の1,2-プロパンジオール水溶液と酵素を接触させることにより発生するプロピオンアルデヒドを検出することで酵素活性の測定をすることが可能である。活性確定後に、基質量に応じた酵素量を設定し反応条件を決定していく。
【0096】
あるいは、1位及び2位に水酸基を有する脂肪族ポリオール及びアデノシルコバラミン(補酵素B12)の存在下で、上記変異型ジオールデヒドラターゼ及び/又はグリセロールデヒドラターゼをコードする遺伝子を含む形質転換体を培養することにより実施できる。形質転換体は、変異型ジオールデヒドラターゼをコードする遺伝子のみ又は変異型グリセロールデヒドラターゼをコードする遺伝子のみを含んでいてもよいし、双方の遺伝子を含んでいてもよい。
【0097】
培養条件は、宿主の培養に用いられる通常の方法に従って、炭素源として1位及び2位に水酸基を有する脂肪族ポリオールを用いることにより行われる。例えば、比較的リッチな培地、例えば2培地等を用いて好気培養し、菌体量を増やしてから嫌気条件にし、炭素源を与えて発酵を行う。pHは、宿主の生育を妨害せず、かつ発酵液から酸を分離するときの障害とならない試薬を用いて調整する。炭酸ナトリウム、アンモニア、ナトリウムイオン供給源、例えば塩化ナトリウムを添加してもよい。また、水酸化ナトリウム水溶液、水酸化カリウム水溶液、水酸化ナトリウム水溶液、水酸化アンモニウム水溶液、水酸化カルシウム水溶液、炭酸カリウム水溶液、炭酸ナトリウム水溶液、酢酸カリウム水溶液等の一般的なアルカリ試薬を用いてもよい。培養期間中pHは、5.0〜8.0、好ましくは5.5〜7.5に保持する。培養は、通常5%CO2存在下、30〜40℃で1〜30日間行う。
【0098】
窒素源としては、例えば、アンモニア、塩化アンモニウム、硫酸アンモニウム、リン酸アンモニウム等のアンモニウム塩の他、ペプトン、肉エキス、酵母エキス、コーンスティープリカー等が挙げられる。また、無機物としては、例えば、リン酸第一カリウム、リン酸第二カリウム、リン酸マグネシウム、硫酸マグネシウム、塩化ナトリウム等が挙げられる。
【0099】
培養中は、カナマイシン、アンピシリン、テトラサイクリン等の抗生物質を培地に添加してもよい。誘導性のプロモーターを用いた発現ベクターで形質転換した微生物を培養する場合は、インデューサーを培地に添加することもできる。例えば、イソプロピル−β−D−チオガラクトピラノシド(IPTG)、インドールアクリル酸(IAA)等を培地に添加することができる。
【0100】
あるいは、上記において得られた形質転換体の培養物から遠心分離などによって集菌を行い、適当な緩衝液に懸濁する。この菌体懸濁液を1位及び2位に水酸基を有する脂肪族ポリオールを含む緩衝液に懸濁し、反応を行ってもよい。反応の条件は、例えば、反応温度は10〜80℃、好ましくは15〜50℃、反応時間は5分〜96時間、好ましくは10分〜72時間、pHは5.0〜8.0、好ましくは5.5〜7.5である。
【0101】
また、該培養物の処理物を用いて反応を行うこともできる。該処理物としては、菌体破砕物、菌体破砕物又は培養上清から調製した粗酵素、精製酵素等が挙げられる。また、常法により担体に固定化した菌体、該処理物、酵素等を用いることができる。
【0102】
1位及び2位に水酸基を有する脂肪族ポリオールとしては、1,2−ジオール(炭素数2〜6、好ましくは2〜4の1,2−アルカンジオールおよびその誘導体、例えば、1,2−プロパンジオール)及びグリセロール、1,2-ブタンジオール、エチレングリコール、1,2,3,4−ブタンテトラオール、1,2,3-ブタントリオール、1,2,4-ブタントリオール、1−モノグリセリド等が挙げられる。1,2−ジオールを基質とする場合には、ジオールデヒドラターゼが好ましく、グリセロールを基質とする場合には、グリセロールデヒドラターゼが好ましい。
【0103】
1位及び2位に水酸基を有する脂肪族ポリオールは、本発明の変異型ジオールデヒドラターゼ又はグリセロールデヒドラターゼの触媒作用により、1位にアルデヒド基を有するアルデヒドに変換される。具体的には、本発明の方法により、1,3−プロパンジオールはプロピオンアルデヒドに変換され、グリセロールは3−ヒドロキシプロピオンアルデヒドに変換される。
【0104】
本発明の変異型ジオールデヒドラターゼは水溶性であることから、親水性の条件下では、変異型ジオールデヒドラターゼよる反応を行うのが好ましい。また、本発明の変異型グリセロールデヒドラターゼは水に不溶性であることから、疎水性の条件下では、変異型グリセロールデヒドラターゼによる反応を行うのが好ましい。変異型ジオールデヒドラターゼ及びジオールデヒドラターゼを共存させて反応を行うことにより、反応系における親水場では変異型ジオールデヒドラターゼよる触媒反応を行い、さらに反応系における疎水場では変位型グリセロールデヒドラターゼによる触媒反応を行うことができる。また、野生型ジオールデヒドラターゼと変異型ジオールデヒドラターゼを共存させて反応させたり、野生型グリセロールデヒドラターゼと野生型グリセロールデヒドラターゼを共存させて反応させることにより、反応効率を改善することができる。
【0105】
本発明の方法は、その他の反応を含む複合的反応の一部として、1位及び2位に水酸基を有する脂肪族ポリオールを脱水して、1位にアルデヒド基を有するアルデヒドを生成する場合も包含する。例えば、Klebsiella属細菌及びCitrobacter属細菌などにおいては、グリセロールは2段階の酵素触媒反応を経て、1,3−プロパンジオールに変換されるが、第1段階において、グリセロールデヒドラターゼがグリセロールを3−ヒドロキシプロピオンアルデヒド(3−HPA)及び水へ変換し(グリセロール→3−HPA+H2O)、第2段階において、3−HPAがNAD+−結合オキシドレダクターゼにより1,3−プロパンジオールに還元される(3−HPA+NADH+H+→1,3−プロパンジオール+NAD+)。本発明の方法は、上記のような複合反応における第1段階の反応において利用することもできる。
【実施例】
【0106】
実施例1 組換え型ジオールデヒドラターゼのトリプシン処理
ジオールデヒドラターゼ遺伝子を発現している組換え体大腸菌を、2% 1,2−プロパンジオールを含む50mM リン酸カリウムバッファ(pH8.0)中で、超音波処理により破砕した。沈澱を同じバッファで3回洗浄した(Tobimatsu, T. et al., Arch. Biochem. Biophys., 347, 132-140 (1997))。得られた粗製の膜画分を種々の濃度のトリプシンを含む同一のバッファ中に、タンパク質濃度が5mg/mlとなるように再懸濁した。15℃で12時間インキュベートした後、過剰のダイズトリプシン阻害剤(400μg/ml)を添加することにより反応を停止した。得られた懸濁液を遠心分離し(16,000×g、30分)、可溶性画分(上清)及び沈澱画分を分離した。その後、上清の酵素活性(白色)及び沈澱画分の酵素活性(黒色)を測定した(図1A)。1.0μg/mlのトリプシンで処理することにより得られる可溶性画分に含まれるジオールデヒドラターゼ、すなわち水溶性の変異型ジオールデヒドラターゼは、酵素活性を失うことなく可溶化されていることがわかる。
【0107】
ジオールデヒドラターゼの活性は、3−メチル−2−ベンゾチアゾリノンヒドラゾン法により、基質として1,2−プロパンジオールを用いて測定した(Toraya, T. et al., J. Biol. Chem., 252, 963-970 (1977))。ジオールデヒドラターゼの1ユニットは、1分あたり1μmolのプロピオンアルデヒドの生成を触媒する酵素活性の量として定義した。野生型ジオールデヒドラターゼの溶解性は非常に低いことから、アッセイするホモジェネート、抽出物及び酵素溶液は、2% 1,2−プロパンジオール及び0.5〜1%のBrij35を含む50mMリン酸カリウムバッファ(pH8.0)で希釈した。
【0108】
トリプシン消化した後の、上清及び沈澱画分に含まれるタンパク質を、11%ゲル上のSDS−PAGEにより分析した(図1B)。その結果、沈澱画分においては、分子量30,000のβサブユニット、分子量19,000のγサブユニットが観察されるのに対し、1.0μg/mlのトリプシンで処理することにより得られる可溶性画分においては、分子量27,000のβサブユニット、分子量17,000のγサブユニットが主に観察された。可溶性画分に含まれるβサブユニットとγサブユニットのアミノ酸配列を解析したところ、分子量27,000のβサブユニットは、βサブユニットのN末端の20アミノ酸が欠失したものであり、分子量17,000のγサブユニットは、γサブユニットのN末端の16アミノ酸が欠失したものであった。なお、αサブユニットにおいては、トリプシン処理による分子量の減少は観察されなかった。
【0109】
ジオールデヒドラターゼをトリプシンで限定的にタンパク質分解したときの、α、β及びγサブユニットのN末端領域の断片化パターンを示す(図1C)。
【0110】
実施例2 発現プラスミドの構築
実施例1の結果から、βサブユニットのN末端から20アミノ酸を欠失させることにより、及び/又はγサブユニットのN末端から16アミノ酸を欠失させることにより、ジオールデヒドラターゼを水溶性に変化させることができるかどうかを試験するため、βサブユニット及び/又はγサブユニットにおいてN末端アミノ酸配列が欠失した変異型ジオールデヒドラターゼを産生するための発現プラスミドを以下のように構築した。
【0111】
βサブユニットのN末端20アミノ酸のみが欠失したジオールデヒドラターゼに対する発現プラスミドをpUSI2E(αDβ’DγD)とし、γサブユニットのN末端16アミノ酸のみが欠失したジオールデヒドラターゼに対する発現プラスミドをpUSI2E(αDβDγD)と記載し、β及びγサブユニットの双方のN末端アミノ酸が欠失したジオールデヒドラターゼに対する発現プラスミドをpUSI2E(αDβ’Dγ’D)とする。また、N末端20アミノ酸が欠失したβサブユニットをβ’サブユニットと記載し、N末端16アミノ酸が欠失したγサブユニットをγ’サブユニットと記載する場合もある。
【0112】
pUSI2E(Tobimatsu, T.et al., J. Biol. Chem., 270, 7142-7148 (1995))をNdeIを用いて部分的に消化し、4種のdNTPの存在下、DNAポリメラーゼIのKlenowフラグメントで処理した。そしてセルフライゲーションにより、プラスミドpUSI2ENdを作成した。pUSI2E(DD)からの3.0kbのBamHI−BglII DNAフラグメントを、pUSI2ENdのBamHI−BglII領域に挿入し、pUSI2ENd(DD)を作成した。ジオールデヒドラターゼの各サブユニットにおいて、N末端20アミノ酸が欠失したβ’サブユニットをコードするDNAセグメント及びN末端16アミノ酸が欠失したγ’サブユニットをコードするDNAセグメントを、TaqDNAポリメラーゼ(インビトロジェン)及び以下のプライマー対を用いたPCRにより増幅した。
【0113】
β’サブユニット:
5’−GCGAGCATATGGGCAGCGATAAACCG−3’(配列番号13)
5’−TCAGATCTTAAAGCGCCACGCGCAG−3’(配列番号14)
γ’サブユニット:
5’−TTGAGCCATATGAACAGCCTGC−3’(配列番号15)
5’−AGAGATCTTAATCGTCGCCTTTGAG−3’(配列番号16)
【0114】
増幅した640bp及び500bpのDNAフラグメントを、TAクローニングキット(インビトロジェン)を用いてpCR2.1ベクターにクローニングし、NdeI及びBglIIで消化し、pUSI2ENd(DD)からの5.0kbのNdeI−BglIIフラグメントに連結し、プラスミドpUSI2ENd(β’D)及びpUSI2ENd(γ’D)をそれぞれ作成した。プラスミドpUSI2E(αD)(Tobimatsu, T. et al., Arch. Biochem. Biophys., 347, 132-140 (1997))を、BglIIで消化し、pUSI2ENd(β’D)から得られた0.7kbのBamHI−BglIIフラグメントに連結し、プラスミドpUSI2E(αDβ’D)を得た。プラスミドpUSI2E(αDβ’D)をBglIIで消化し、pUSI2ENd(γ’D)及びpUSI2ENd(γD)から得られた0.5kbのBamHI−BglIIフラグメントに連結し、プラスミドpUSI2E(αDβ’Dγ’D)及びpUSI2E(αDβ’DγD)をそれぞれ作成した。プラスミドpUSI2E(αDβD)をBglIIで消化し、pUSI2ENd(γ’D)から得られた0.5kbのBamHI−BglIIフラグメントに連結し、プラスミドpUSI2E(αDβDγ’D)を得た。配列決定により、PCR増幅中に望ましくない変異が起こっていないことを確認した。
【0115】
発現プラスミドによる大腸菌の形質転換及び形質転換体の培養は、Tobimatsu, T.et al., J. Biol. Chem., 270, 7142-7148 (1995)に記載された方法と同様に実施した。
【0116】
各変異型酵素を発現している組換え体大腸菌の細胞ホモジェネート(homo)、上清(sup)及び沈澱画分(ppt)に含まれるタンパク質を、11%ゲル上のSDS−PAGEに付し、タンパク質染色を行った。分子量マーカーとしてSDS−7(Sigma)を用いた。α、β、β’、γ及びγ’サブユニットの位置を、右側の矢印によって示す(図2)。
【0117】
沈澱画分(ppt)の結果を見ると、ジオールデヒドラターゼをコードする遺伝子を含む組換え体大腸菌(以下、変異株と称する場合もある)においてβ’及びγ’サブユニットのバンドは、野生株のβ及びγサブユニットのバンドほど濃いバンドではないことがわかる。一方、上清(sup)の結果を見ると、変異株におけるβ’及びγ’サブユニットのバンドは、野生型ジオールデヒドラターゼをコードする遺伝子を含む組換え体大腸菌(以下、野生株と称する場合もある)のβ及びγサブユニットのバンドより濃いことがわかる。以上からβ及び/又はγサブユニットのN末端が欠失したジオールデヒドラターゼをコードする遺伝子を含む変異株において、水溶性のジオールデヒドラターゼが産生されたことがわかる。
【0118】
実施例3 上清及び沈澱におけるジオールデヒドラターゼ活性
実施例2で得られた各発現プラスミドを有する組換え体大腸菌BL21(DE3)(9.2〜9.8×1010)を洗浄し、2% 1,2−プロパンジオールを含む50mMリン酸カリウムバッファ(pH8.0)2.6mlに再懸濁し、超音波処理により破砕した。得られた細胞ホモジェネートを遠心分離し(17,500×g、30分)、上清及び沈澱画分を分離した。得られた沈澱を2.6mlの同じバッファで洗浄し、2% 1,2−プロパンジオール及び1% Brij35を含む10mM リン酸カリウムバッファ(pH8.0)に再懸濁し同じ体積とした。ジオールデヒドラターゼ活性は、適切に希釈した後で実施例1と同様に測定した。結果を以下の表1に示す。
【0119】
【表1】
【0120】
表1の結果から、可溶性画分に回収されたジオールデヒドラターゼの割合は、いずれの変異株においても96%を超えていたが、野生株においては、2%に過ぎなかった。以上から本発明の変異型ジオールデヒドラターゼが高度に水溶性であることがわかる。また、野生型ジオールデヒドラターゼにおいては、β及びγサブユニットのN末端アミノ酸領域が、この酵素の沈澱や凝集に必須であることがわかる。また、変異型ジオールデヒドラターゼが酵素活性を有することから、水溶性の変異型ジオールデヒドラターゼが酵素活性を有することもわかる。
【0121】
実施例4 変異型ジオールデヒドラターゼと野生型ジオールデヒドラターゼの比較
αサブユニット、β’サブユニット及びγ’サブユニットを含む変異型ジオールデヒドラターゼ、野生型ジオールデヒドラターゼ、並びに1.0μg/mlのトリプシンで、15℃で12時間処理したジオールデヒドラターゼを比較するため、プラスミドpUSI2E(αDβ’Dγ’D)を有する大腸菌の無細胞抽出物をPAGEに付した。無細胞抽出物のPAGEは、0.1Mの1,2−プロパンジオールの存在下、Davis, B.J., Ann. N. Y. Acad. Sci., 121, 404-427 (1964))に記載の非変性条件下で、又はLaemmli, U.K., Nature, 227, 680-685 (1970)に記載の変性条件下で実施した。ウェスタンブロット分析は、Tobimatsu, T. et al., J. Biol. Chem., 270, 7142-7148 (1995)に記載の方法に従って実施した(図3)。
【0122】
非変性条件下の5%ゲル上のPAGEの後、タンパク質染色(図3A)又は活性染色(図3B)を実施した。野生型及びトリプシン消化ジオールデヒドラターゼ及びグリセロールデヒドラターゼの位置を、右側の矢印により示す。酵素活性を示すタンパク質バンドを切り出し、11%ゲル上のSDS−PAGEによって分析し、タンパク質染色した(図3C)。タンパク質染色は、クマシーブリリアントブルー R−250を用いて実施した。サブユニットの位置を、右側の矢印で示す。分子量マーカーとしてSDS−7(Sigma)を用いた。
【0123】
図3A及びBに示されるように、ジオールデヒドラターゼ活性を示すバンドは、変異型ジオールデヒドラターゼを含む無細胞抽出物にのみ観察された。この活性なバンドをゲルから切り出してSDS−PAGEに付したところ、観察された3種のペプチドのバンドは、ジオールデヒドラターゼのトリプシン処理によって得られる分子量60,000、23,000及び17,000のバンドと一致した(図3C)。
【0124】
以上からプラスミドpUSI2E(αDβ’Dγ’D)を有する大腸菌によって産生された組換え体変異型ジオールデヒドラターゼが触媒活性を有すること、並びに、分子量60、000、23,000及び17,000のサブユニットを含むことが確認された。
【0125】
実施例5 変異型ジオールデヒドラターゼの精製及び分析
pUSI2E(αDβ’Dγ’D)を有する大腸菌3.3gから、超音波処理により無細胞抽出物を調製した。該無細胞抽出物の硫安分画(40〜50%)により得られた沈澱を2% 1,2−プロパンジオールを含むバッファに溶解し、Sepharose CL6Bカラム(350mL)にロードした。4ユニット/mgを超える比活性を有する酵素を含む画分を回収した。該酵素溶液をCentricon(Millipore Corp.)で濃縮し、4volの2% 1,2−プロパンジオールと混合した。該溶液を、予め2% 1,2−プロパンジオールを含む10mM KPB(pH8)で平衡化しておいたDEAEセルロース(Serva)カラム(1mL)にロードした。カラムを同じバッファの5volで洗浄し、50mM KClを含む同じバッファ、続いて100mM KClを含む同じバッファの5volで展開した。35ユニット/mgを超える比活性を有する精製酵素を、酵素の特性決定に使用した。タンパク質は、Lowry, O.H. et al., J. Biol. Chem., 193, 265-275 (1951)に記載の方法により、結晶化ウシ血清アルブミンを標準として用いて、アッセイした。比活性はユニット/タンパク質(mg)として記載した。
【0126】
得られた変異型ジオールデヒドラターゼ(αβ’γ’)調製物の比活性は47ユニット/mgであった。デンシトメトリー分析により、得られた変異型ジオールデヒドラターゼの純度は55%であったことから、この変異型ジオールデヒドラターゼの比活性は85ユニット/mgであると考えられる。この比活性の値は、K. oxytocaから水溶性の形態で精製されたジオールデヒドラターゼの比活性(91.5ユニット/mg)及び野生型のジオールデヒドラターゼの比活性(106ユニット/mg)に匹敵するものであった。
【0127】
また、基質としてのDL−1,2−プロパンジオールに対するKm値は、変異型ジオールデヒドラターゼで0.11mMであり、野生型ジオールデヒドラターゼで0.10mMであった。以上から、本発明の変異型ジオールデヒドラターゼが、野生型のジオールデヒドラターゼと比べ、触媒活性において同等であることがわかる。
【0128】
実施例6 疎水化グリセロールデヒドラターゼの調製
グリセロールデヒドラターゼ(GD)とジオールデヒドラターゼ(DD)のサブユニットは高い相同性を有しているが、βとγサブユニットのN末端領域は、GDが短く、更にβサブユニットはそれに続く30数アミノ酸残基がなくなっている(図4)。DDで余分に付いているN末端領域だけでなく、βサブユニットの相同性が低い領域も酵素の低溶解性に関与する可能性がある。
【0129】
変異型酵素として、βおよびγサブユニットがそれぞれ下向き矢印#2と#3までDDの配列を持ちそれ以降をGDの配列を持つ変異体2(αGNβD2G2γDG)を作成した。すなわち、変異体2は、グリセロールデヒドラターゼ(GD)のβサブユニットのN末端の1〜27番目のアミノ酸をジオールデヒドラターゼ(DD)のN末端の1〜60番目のアミノ酸に置き換え、さらにγサブユニットのN末端の1番目のアミノ酸をジオールデヒドラターゼ(DD)のN末端の1〜33番目のアミノ酸に置き換えたグリセロールデヒドラターゼ変異体である。上記変異体の遺伝子構造の概略を図5に示す。また得られる変異体2の塩基配列を配列番号17に示す。変異体2においてαサブユニットのアミノ酸配列を配列番号18に、βサブユニットのアミノ酸配列を配列番号19に、γサブユニットのアミノ酸配列を配列番号20に示す。
【0130】
上記変異型グリセロールデヒドラターゼ(変異体2)を産生するための発現プラスミド:pNEX(αGNβD2G2γDG)を実施例2と同様に構築した。
【0131】
pNEXベクターの構造を図6に示す。またpNEX(αG)の作成手順を図7に示す。pUSI2E(αG)も同様に作成した。挿入DNA断片(図8)は、TaqDNAポリメラーゼ(インビトロジェン)及び図9に示すプライマー対を用いたPCRにより増幅した。得られるPCR断片を図10に示す。PCRはKOD Dashにて、以下の条件のstep down PCR法で行った。
【0132】
【表2】
【0133】
増幅断片をpNEX(αG)またはpUSI2E(αG)に連結することにより、発現プラスミドを作成した。
【0134】
発現プラスミドによる大腸菌の形質転換及び形質転換体の培養は、Tobimatsu, T.et al., J. Biol. Chem., 270, 7142-7148 (1995)に記載された方法と同様に実施した。
【0135】
各変異型酵素を発現している組換え体大腸菌をトルエン処理し(図11)、細胞破壊と分離操作を行った(図12)。組換え体大腸菌の細胞ホモジェネート(homo)、上清(sup)、沈澱画分(ppt)及び洗液(wash)に含まれるタンパク質を、11%ゲル上のSDS−PAGEに付し、タンパク質染色を行った(図13)。
【0136】
またそれぞれの画分についてグリセロールデヒドラターゼ活性を測定した。コントロールとして野生型(WT)のジオールデヒドラターゼを用いた。結果を以下の表に示す。
【0137】
【表3】
【0138】
大腸菌で組み換え体として発現させた野生型のグリセロールデヒドラターゼ(GD)の場合は、酵素活性の大部分が上清画分にみられ可溶型で存在することが報告されているが(Yamanishi M, et al., Eur. J. Biochem. 269,4484-494 (2002); Tobimatsu T, et al., J. Biol. Chem., 271(37), 22352-22357(1996))、表3より、変異体2では、酵素活性の大半が沈澱画分にみられたことから、GDが低溶解性化していることが明らかである。すなわち、本発明により、変異型グリセロールデヒドラターゼの水に対する溶解性が変化し、疎水性のグリセロールデヒドラターゼが得られたことが示された。
【図面の簡単な説明】
【0139】
【図1】トリプシン消化がジオールデヒドラターゼの活性と溶解性に及ぼす影響を示す。
【図2】N末端アミノ酸が欠失したβ及び/又はγサブユニットを有する変異型ジオールデヒドラターゼの大腸菌における発現及び分布を示す。
【図3】αβ’γ’変異型ジオールデヒドラターゼとトリプシン消化ジオールデヒドラターゼとの比較を示す。
【図4】Klebsiella pneumoniae由来のグリセロールデヒドラターゼ(GD)とKlebsiella oxytoca由来のジオールデヒドラターゼ(DD)の各サブユニットのアミノ酸配列の相同性を示す。
【図5】変異体2の遺伝子構造の概略図を示す。
【図6】pNEXベクターの構造を示す。
【図7】pNEX(αG)の作成手順を示す。
【図8】挿入DNA断片(NβDGγDG)を示す。
【図9】実施例6で発現プラスミドの作成においてPCRに用いたプライマー対及びプライマー領域の相補性を示す。↓のところがジオールデヒドラターゼとグリセロールデヒドラターゼとがつながる場所を示す。
【図10】図9に示したプライマー対を用いて得られるPCR断片を示す。
【図11】組換え体大腸菌をトルエン処理した手順を示す。
【図12】組換え体大腸菌をトルエン処理した後、細胞破壊と分離操作を行った手順を示す。
【図13】変異型グリセロールデヒドラターゼをSDS−PAGEに付し、タンパク質染色を行った結果を示す。
【特許請求の範囲】
【請求項1】
ジオールデヒドラターゼ又はグリセロールデヒドラターゼのβサブユニット及び/又はγサブユニットにおいてN末端アミノ酸を変異させることにより、ジオールデヒドラターゼ又はグリセロールデヒドラターゼの水に対する溶解性を変化させる方法。
【請求項2】
N末端アミノ酸の変異が、
ジオールデヒドラターゼを構成するβサブユニット及び/又はγサブユニットにおけるN末端アミノ酸を欠失させること、又は
グリセロールデヒドラターゼを構成するβサブユニット及び/又はγサブユニットにおけるN末端に、ジオールデヒドラターゼの対応するサブユニットのN末端アミノ酸を付加すること、
を含む、請求項1記載の方法。
【請求項3】
ジオールデヒドラターゼのβサブユニット及び/又はγサブユニットにおけるN末端アミノ酸の変異が、
ジオールデヒドラターゼのβサブユニットのN末端の9〜64個のアミノ酸を欠失させること、及び/又は
ジオールデヒドラターゼのγサブユニットのN末端の5〜42個のアミノ酸を欠失させること
を含む請求項2記載の方法。
【請求項4】
グリセロールデヒドラターゼのβサブユニット及び/又はγサブユニットにおけるN末端アミノ酸の変異が、
グリセロールデヒドラターゼのβサブユニットのN末端に、ジオールデヒドラターゼのβサブユニットのN末端の9〜64個のアミノ酸を付加すること、
グリセロールデヒドラターゼのγサブユニットのN末端に、ジオールデヒドラターゼのγサブユニットのN末端の5〜42個のアミノ酸を付加すること
を含む請求項2記載の方法。
【請求項5】
αサブユニット、βサブユニット及びγサブユニットを含むジオールデヒドラターゼであって、
βサブユニットのN末端の9〜64個のアミノ酸が欠失しており、及び/又はγサブユニットのN末端の5〜42個のアミノ酸が欠失しており、かつ水溶性である前記ジオールデヒドラターゼ。
【請求項6】
αサブユニット、βサブユニット及びγサブユニットを含むジオールデヒドラターゼであって、
αサブユニットが以下の(a)又は(b)のタンパク質:
(a)配列番号1のアミノ酸配列からなるタンパク質、
(b)配列番号1において1若しくは数個のアミノ酸が欠失、置換若しくは付加されたアミノ酸配列からなるタンパク質であって、ジオールデヒドラターゼのβサブユニット及びγサブユニットと会合したときにジオールデヒドラターゼ活性を有する前記タンパク質、
であり、
βサブユニットが以下の(c)〜(f)から選択されるタンパク質:
(c)配列番号3のアミノ酸配列からなるタンパク質、
(d)配列番号3において1若しくは数個のアミノ酸が欠失、置換若しくは付加されたアミノ酸配列からなるタンパク質であって、ジオールデヒドラターゼのαサブユニット及びγサブユニットと会合したときにジオールデヒドラターゼ活性を有する前記タンパク質、
(e)配列番号3においてN末端の9〜64個のアミノ酸が欠失したアミノ酸配列からなるタンパク質、
(f)(e)に記載のアミノ酸配列において1若しくは数個のアミノ酸が欠失、置換若しくは付加されたアミノ酸配列からなるタンパク質であって、ジオールデヒドラターゼのαサブユニット及びγサブユニットと会合したときにジオールデヒドラターゼ活性を有する前記タンパク質、
であり、
γサブユニットが以下の(g)〜(j)から選択されるタンパク質:
(g)配列番号5のアミノ酸配列からなるタンパク質、
(h)配列番号5において1若しくは数個のアミノ酸が欠失、置換若しくは付加されたアミノ酸配列からなるタンパク質であって、ジオールデヒドラターゼのαサブユニット及びβサブユニットと会合したときにジオールデヒドラターゼ活性を有する前記タンパク質、
(i)配列番号5においてN末端の5〜42個のアミノ酸が欠失したアミノ酸配列からなるタンパク質、
(j)(i)に記載アミノ酸配列において1若しくは数個のアミノ酸が欠失、置換若しくは付加されたアミノ酸配列からなるタンパク質であって、ジオールデヒドラターゼのαサブユニット及びβサブユニットと会合したときにジオールデヒドラターゼ活性を有する前記タンパク質
であり、
ただし、βサブユニットが(c)又は(d)のタンパク質であるときγサブユニットは(g)及び(h)のタンパク質ではない、前記ジオールデヒドラターゼ。
【請求項7】
請求項5又は6記載のジオールデヒドラターゼをコードする遺伝子。
【請求項8】
請求項7記載の遺伝子を含むベクター。
【請求項9】
請求項8記載のベクターを含む形質転換体。
【請求項10】
αサブユニット、βサブユニット及びγサブユニットを含むグリセロールデヒドラターゼにおいて、
βサブユニットのN末端にジオールデヒドラターゼのβサブユニットのN末端の9〜64個のアミノ酸が付加しており、及び/又はγサブユニットのN末端にジオールデヒドラターゼのβサブユニットのN末端の5〜42個のアミノ酸が付加しており、かつ水に不溶性である前記グリセロールデヒドラターゼ。
【請求項11】
αサブユニット、βサブユニット及びγサブユニットを含むグリセロールデヒドラターゼにおいて、
αサブユニットが以下の(a)又は(b)のタンパク質:
(a)配列番号7のアミノ酸配列からなるタンパク質、
(b)配列番号7において1若しくは数個のアミノ酸が欠失、置換若しくは付加されたアミノ酸配列からなるタンパク質であって、グリセロールデヒドラターゼのβサブユニット及びγサブユニットと会合したときにグリセロールデヒドラターゼ活性を有する前記タンパク質、
であり、
βサブユニットが以下の(c)〜(f)から選択されるタンパク質:
(c)配列番号9のアミノ酸配列からなるタンパク質、
(d)配列番号9において1若しくは数個のアミノ酸が欠失、置換若しくは付加されたアミノ酸配列からなるタンパク質であって、グリセロールデヒドラターゼのαサブユニット及びγサブユニットと会合したときにグリセロールデヒドラターゼ活性を有する前記タンパク質、
(e)配列番号9のN末端に配列番号3のN末端の9〜64個のアミノ酸が付加してなるアミノ酸配列からなるタンパク質、
(f)(e)に記載のアミノ酸配列において1若しくは数個のアミノ酸が欠失、置換若しくは付加されたアミノ酸配列からなるタンパク質であって、グリセロールデヒドラターゼのαサブユニット及びγサブユニットと会合したときにグリセロールデヒドラターゼ活性を有する前記タンパク質、
であり、
γサブユニットが以下の(g)〜(j)から選択されるタンパク質:
(g)配列番号11のアミノ酸配列からなるタンパク質、
(h)配列番号11において1若しくは数個のアミノ酸が欠失、置換若しくは付加されたアミノ酸配列からなるタンパク質であって、グリセロールデヒドラターゼのαサブユニット及びβサブユニットと会合したときにグリセロールデヒドラターゼ活性を有する前記タンパク質、
(i)配列番号11のN末端に配列番号5のN末端の5〜42個のアミノ酸が付加してなるアミノ酸配列からなるタンパク質、
(j)(i)に記載アミノ酸配列において1若しくは数個のアミノ酸が欠失、置換若しくは付加されたアミノ酸配列からなるタンパク質であって、グリセロールデヒドラターゼのαサブユニット及びβサブユニットと会合したときにグリセロールデヒドラターゼ活性を有する前記タンパク質
であり、
ただし、βサブユニットが(c)又は(d)のタンパク質であるときγサブユニットは(g)及び(h)のタンパク質ではない、前記グリセロールデヒドラターゼ。
【請求項12】
請求項10又は11記載のグリセロールデヒドラターゼをコードする遺伝子。
【請求項13】
請求項12記載の遺伝子を含むベクター。
【請求項14】
請求項13記載のベクターを含む形質転換体。
【請求項15】
以下の(a)又は(b)の遺伝子:
(a)配列番号17の塩基配列からなる遺伝子、
(b)配列番号17で表される塩基配列からなる遺伝子に対し相補的な塩基配列からなる遺伝子とストリンジェントな条件下でハイブリダイズし、かつグリセロールデヒドラターゼの活性を有するタンパク質をコードする遺伝子。
【請求項16】
請求項15記載の遺伝子を含むベクター。
【請求項17】
請求項16記載のベクターを含む形質転換体。
【請求項18】
αサブユニット、βサブユニット及びγサブユニットを含むグリセロールデヒドラターゼにおいて、
αサブユニットが以下の(a)又は(b)のタンパク質:
(a)配列番号18のアミノ酸配列からなるタンパク質、
(b)配列番号18において1若しくは数個のアミノ酸が欠失、置換若しくは付加されたアミノ酸配列からなるタンパク質であって、グリセロールデヒドラターゼのβサブユニット及びγサブユニットと会合したときにグリセロールデヒドラターゼ活性を有する前記タンパク質、
であり、
βサブユニットが以下の(c)〜(f)から選択されるタンパク質:
(c)配列番号9のアミノ酸配列からなるタンパク質、
(d)配列番号9において1若しくは数個のアミノ酸が欠失、置換若しくは付加されたアミノ酸配列からなるタンパク質であって、グリセロールデヒドラターゼのαサブユニット及びγサブユニットと会合したときにグリセロールデヒドラターゼ活性を有する前記タンパク質、
(e)配列番号19のアミノ酸配列からなるタンパク質、
(f)配列番号19において1若しくは数個のアミノ酸が欠失、置換若しくは付加されたアミノ酸配列からなるタンパク質であって、グリセロールデヒドラターゼのαサブユニット及びγサブユニットと会合したときにグリセロールデヒドラターゼ活性を有する前記タンパク質、
であり、
γサブユニットが以下の(g)〜(j)から選択されるタンパク質:
(g)配列番号11のアミノ酸配列からなるタンパク質、
(h)配列番号11において1若しくは数個のアミノ酸が欠失、置換若しくは付加されたアミノ酸配列からなるタンパク質であって、グリセロールデヒドラターゼのαサブユニット及びβサブユニットと会合したときにグリセロールデヒドラターゼ活性を有する前記タンパク質、
(i)配列番号20のアミノ酸配列からなるタンパク質、
(j)配列番号20において1若しくは数個のアミノ酸が欠失、置換若しくは付加されたアミノ酸配列からなるタンパク質であって、グリセロールデヒドラターゼのαサブユニット及びβサブユニットと会合したときにグリセロールデヒドラターゼ活性を有する前記タンパク質、
であり、
ただし、βサブユニットが(c)又は(d)のタンパク質であるときγサブユニットは(g)及び(h)のタンパク質ではない、前記グリセロールデヒドラターゼ。
【請求項19】
1位及び2位に水酸基を有する脂肪族ポリオールを脱水して、1位にアルデヒド基を有するアルデヒドを生成する方法あって、
1位及び2位に水酸基を有する脂肪族ポリオールと請求項5若しくは6記載のジオールデヒドラターゼ及び/又は請求項10、11若しくは18記載のグリセロールデヒドラターゼとを反応させることを含む前記方法。
【請求項20】
1位及び2位に水酸基を有する脂肪族ポリオールを脱水して、1位にアルデヒド基を有するアルデヒドを生成する方法あって、
1位及び2位に水酸基を有する脂肪族ポリオールの存在下、請求項7記載のジオールデヒドラターゼをコードする遺伝子及び/又は請求項12若しくは15記載のグリセロールデヒドラターゼをコードする遺伝子を含む形質転換体を培養することを含む前記方法。
【請求項1】
ジオールデヒドラターゼ又はグリセロールデヒドラターゼのβサブユニット及び/又はγサブユニットにおいてN末端アミノ酸を変異させることにより、ジオールデヒドラターゼ又はグリセロールデヒドラターゼの水に対する溶解性を変化させる方法。
【請求項2】
N末端アミノ酸の変異が、
ジオールデヒドラターゼを構成するβサブユニット及び/又はγサブユニットにおけるN末端アミノ酸を欠失させること、又は
グリセロールデヒドラターゼを構成するβサブユニット及び/又はγサブユニットにおけるN末端に、ジオールデヒドラターゼの対応するサブユニットのN末端アミノ酸を付加すること、
を含む、請求項1記載の方法。
【請求項3】
ジオールデヒドラターゼのβサブユニット及び/又はγサブユニットにおけるN末端アミノ酸の変異が、
ジオールデヒドラターゼのβサブユニットのN末端の9〜64個のアミノ酸を欠失させること、及び/又は
ジオールデヒドラターゼのγサブユニットのN末端の5〜42個のアミノ酸を欠失させること
を含む請求項2記載の方法。
【請求項4】
グリセロールデヒドラターゼのβサブユニット及び/又はγサブユニットにおけるN末端アミノ酸の変異が、
グリセロールデヒドラターゼのβサブユニットのN末端に、ジオールデヒドラターゼのβサブユニットのN末端の9〜64個のアミノ酸を付加すること、
グリセロールデヒドラターゼのγサブユニットのN末端に、ジオールデヒドラターゼのγサブユニットのN末端の5〜42個のアミノ酸を付加すること
を含む請求項2記載の方法。
【請求項5】
αサブユニット、βサブユニット及びγサブユニットを含むジオールデヒドラターゼであって、
βサブユニットのN末端の9〜64個のアミノ酸が欠失しており、及び/又はγサブユニットのN末端の5〜42個のアミノ酸が欠失しており、かつ水溶性である前記ジオールデヒドラターゼ。
【請求項6】
αサブユニット、βサブユニット及びγサブユニットを含むジオールデヒドラターゼであって、
αサブユニットが以下の(a)又は(b)のタンパク質:
(a)配列番号1のアミノ酸配列からなるタンパク質、
(b)配列番号1において1若しくは数個のアミノ酸が欠失、置換若しくは付加されたアミノ酸配列からなるタンパク質であって、ジオールデヒドラターゼのβサブユニット及びγサブユニットと会合したときにジオールデヒドラターゼ活性を有する前記タンパク質、
であり、
βサブユニットが以下の(c)〜(f)から選択されるタンパク質:
(c)配列番号3のアミノ酸配列からなるタンパク質、
(d)配列番号3において1若しくは数個のアミノ酸が欠失、置換若しくは付加されたアミノ酸配列からなるタンパク質であって、ジオールデヒドラターゼのαサブユニット及びγサブユニットと会合したときにジオールデヒドラターゼ活性を有する前記タンパク質、
(e)配列番号3においてN末端の9〜64個のアミノ酸が欠失したアミノ酸配列からなるタンパク質、
(f)(e)に記載のアミノ酸配列において1若しくは数個のアミノ酸が欠失、置換若しくは付加されたアミノ酸配列からなるタンパク質であって、ジオールデヒドラターゼのαサブユニット及びγサブユニットと会合したときにジオールデヒドラターゼ活性を有する前記タンパク質、
であり、
γサブユニットが以下の(g)〜(j)から選択されるタンパク質:
(g)配列番号5のアミノ酸配列からなるタンパク質、
(h)配列番号5において1若しくは数個のアミノ酸が欠失、置換若しくは付加されたアミノ酸配列からなるタンパク質であって、ジオールデヒドラターゼのαサブユニット及びβサブユニットと会合したときにジオールデヒドラターゼ活性を有する前記タンパク質、
(i)配列番号5においてN末端の5〜42個のアミノ酸が欠失したアミノ酸配列からなるタンパク質、
(j)(i)に記載アミノ酸配列において1若しくは数個のアミノ酸が欠失、置換若しくは付加されたアミノ酸配列からなるタンパク質であって、ジオールデヒドラターゼのαサブユニット及びβサブユニットと会合したときにジオールデヒドラターゼ活性を有する前記タンパク質
であり、
ただし、βサブユニットが(c)又は(d)のタンパク質であるときγサブユニットは(g)及び(h)のタンパク質ではない、前記ジオールデヒドラターゼ。
【請求項7】
請求項5又は6記載のジオールデヒドラターゼをコードする遺伝子。
【請求項8】
請求項7記載の遺伝子を含むベクター。
【請求項9】
請求項8記載のベクターを含む形質転換体。
【請求項10】
αサブユニット、βサブユニット及びγサブユニットを含むグリセロールデヒドラターゼにおいて、
βサブユニットのN末端にジオールデヒドラターゼのβサブユニットのN末端の9〜64個のアミノ酸が付加しており、及び/又はγサブユニットのN末端にジオールデヒドラターゼのβサブユニットのN末端の5〜42個のアミノ酸が付加しており、かつ水に不溶性である前記グリセロールデヒドラターゼ。
【請求項11】
αサブユニット、βサブユニット及びγサブユニットを含むグリセロールデヒドラターゼにおいて、
αサブユニットが以下の(a)又は(b)のタンパク質:
(a)配列番号7のアミノ酸配列からなるタンパク質、
(b)配列番号7において1若しくは数個のアミノ酸が欠失、置換若しくは付加されたアミノ酸配列からなるタンパク質であって、グリセロールデヒドラターゼのβサブユニット及びγサブユニットと会合したときにグリセロールデヒドラターゼ活性を有する前記タンパク質、
であり、
βサブユニットが以下の(c)〜(f)から選択されるタンパク質:
(c)配列番号9のアミノ酸配列からなるタンパク質、
(d)配列番号9において1若しくは数個のアミノ酸が欠失、置換若しくは付加されたアミノ酸配列からなるタンパク質であって、グリセロールデヒドラターゼのαサブユニット及びγサブユニットと会合したときにグリセロールデヒドラターゼ活性を有する前記タンパク質、
(e)配列番号9のN末端に配列番号3のN末端の9〜64個のアミノ酸が付加してなるアミノ酸配列からなるタンパク質、
(f)(e)に記載のアミノ酸配列において1若しくは数個のアミノ酸が欠失、置換若しくは付加されたアミノ酸配列からなるタンパク質であって、グリセロールデヒドラターゼのαサブユニット及びγサブユニットと会合したときにグリセロールデヒドラターゼ活性を有する前記タンパク質、
であり、
γサブユニットが以下の(g)〜(j)から選択されるタンパク質:
(g)配列番号11のアミノ酸配列からなるタンパク質、
(h)配列番号11において1若しくは数個のアミノ酸が欠失、置換若しくは付加されたアミノ酸配列からなるタンパク質であって、グリセロールデヒドラターゼのαサブユニット及びβサブユニットと会合したときにグリセロールデヒドラターゼ活性を有する前記タンパク質、
(i)配列番号11のN末端に配列番号5のN末端の5〜42個のアミノ酸が付加してなるアミノ酸配列からなるタンパク質、
(j)(i)に記載アミノ酸配列において1若しくは数個のアミノ酸が欠失、置換若しくは付加されたアミノ酸配列からなるタンパク質であって、グリセロールデヒドラターゼのαサブユニット及びβサブユニットと会合したときにグリセロールデヒドラターゼ活性を有する前記タンパク質
であり、
ただし、βサブユニットが(c)又は(d)のタンパク質であるときγサブユニットは(g)及び(h)のタンパク質ではない、前記グリセロールデヒドラターゼ。
【請求項12】
請求項10又は11記載のグリセロールデヒドラターゼをコードする遺伝子。
【請求項13】
請求項12記載の遺伝子を含むベクター。
【請求項14】
請求項13記載のベクターを含む形質転換体。
【請求項15】
以下の(a)又は(b)の遺伝子:
(a)配列番号17の塩基配列からなる遺伝子、
(b)配列番号17で表される塩基配列からなる遺伝子に対し相補的な塩基配列からなる遺伝子とストリンジェントな条件下でハイブリダイズし、かつグリセロールデヒドラターゼの活性を有するタンパク質をコードする遺伝子。
【請求項16】
請求項15記載の遺伝子を含むベクター。
【請求項17】
請求項16記載のベクターを含む形質転換体。
【請求項18】
αサブユニット、βサブユニット及びγサブユニットを含むグリセロールデヒドラターゼにおいて、
αサブユニットが以下の(a)又は(b)のタンパク質:
(a)配列番号18のアミノ酸配列からなるタンパク質、
(b)配列番号18において1若しくは数個のアミノ酸が欠失、置換若しくは付加されたアミノ酸配列からなるタンパク質であって、グリセロールデヒドラターゼのβサブユニット及びγサブユニットと会合したときにグリセロールデヒドラターゼ活性を有する前記タンパク質、
であり、
βサブユニットが以下の(c)〜(f)から選択されるタンパク質:
(c)配列番号9のアミノ酸配列からなるタンパク質、
(d)配列番号9において1若しくは数個のアミノ酸が欠失、置換若しくは付加されたアミノ酸配列からなるタンパク質であって、グリセロールデヒドラターゼのαサブユニット及びγサブユニットと会合したときにグリセロールデヒドラターゼ活性を有する前記タンパク質、
(e)配列番号19のアミノ酸配列からなるタンパク質、
(f)配列番号19において1若しくは数個のアミノ酸が欠失、置換若しくは付加されたアミノ酸配列からなるタンパク質であって、グリセロールデヒドラターゼのαサブユニット及びγサブユニットと会合したときにグリセロールデヒドラターゼ活性を有する前記タンパク質、
であり、
γサブユニットが以下の(g)〜(j)から選択されるタンパク質:
(g)配列番号11のアミノ酸配列からなるタンパク質、
(h)配列番号11において1若しくは数個のアミノ酸が欠失、置換若しくは付加されたアミノ酸配列からなるタンパク質であって、グリセロールデヒドラターゼのαサブユニット及びβサブユニットと会合したときにグリセロールデヒドラターゼ活性を有する前記タンパク質、
(i)配列番号20のアミノ酸配列からなるタンパク質、
(j)配列番号20において1若しくは数個のアミノ酸が欠失、置換若しくは付加されたアミノ酸配列からなるタンパク質であって、グリセロールデヒドラターゼのαサブユニット及びβサブユニットと会合したときにグリセロールデヒドラターゼ活性を有する前記タンパク質、
であり、
ただし、βサブユニットが(c)又は(d)のタンパク質であるときγサブユニットは(g)及び(h)のタンパク質ではない、前記グリセロールデヒドラターゼ。
【請求項19】
1位及び2位に水酸基を有する脂肪族ポリオールを脱水して、1位にアルデヒド基を有するアルデヒドを生成する方法あって、
1位及び2位に水酸基を有する脂肪族ポリオールと請求項5若しくは6記載のジオールデヒドラターゼ及び/又は請求項10、11若しくは18記載のグリセロールデヒドラターゼとを反応させることを含む前記方法。
【請求項20】
1位及び2位に水酸基を有する脂肪族ポリオールを脱水して、1位にアルデヒド基を有するアルデヒドを生成する方法あって、
1位及び2位に水酸基を有する脂肪族ポリオールの存在下、請求項7記載のジオールデヒドラターゼをコードする遺伝子及び/又は請求項12若しくは15記載のグリセロールデヒドラターゼをコードする遺伝子を含む形質転換体を培養することを含む前記方法。
【図1】

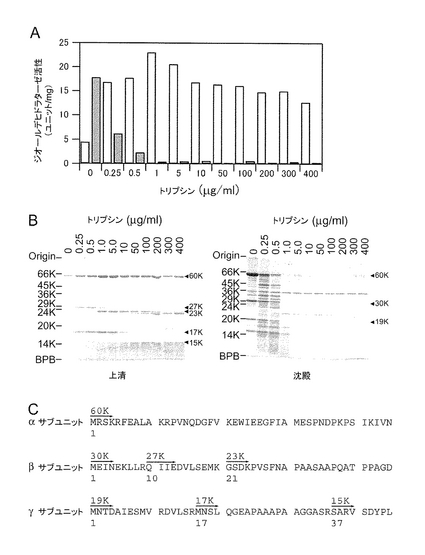
【図2】
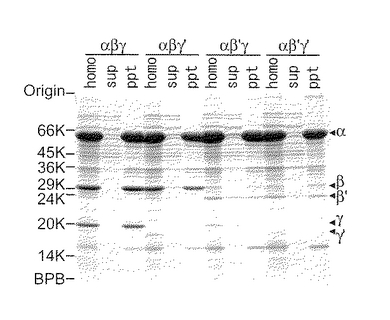
【図3】
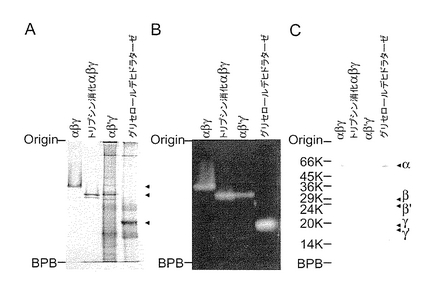
【図4】

【図5】
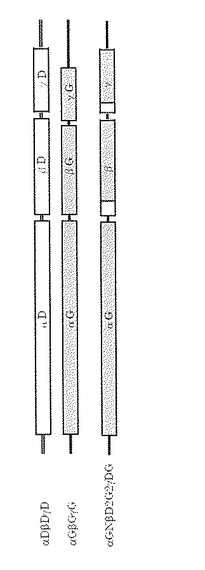
【図6】
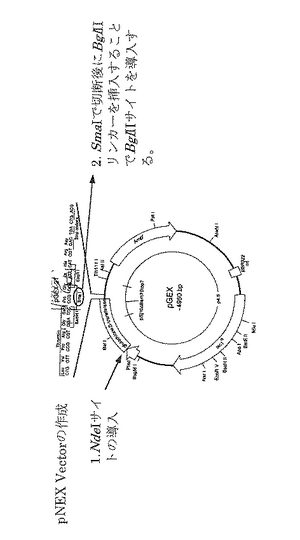
【図7】
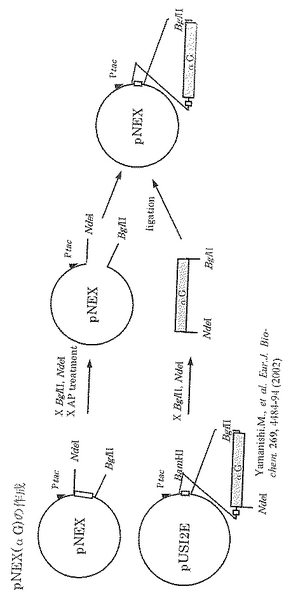
【図8】
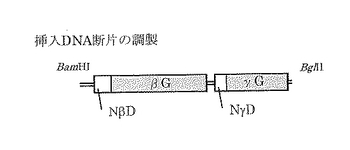
【図9】

【図10】
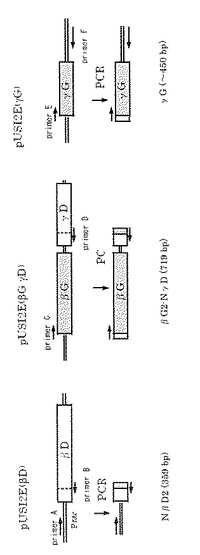
【図11】

【図12】
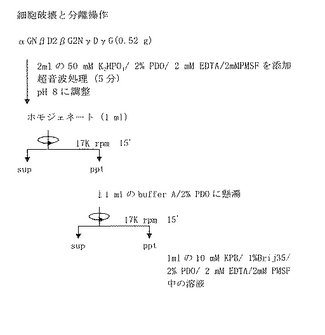
【図13】

【図2】
【図3】
【図4】
【図5】
【図6】
【図7】
【図8】
【図9】
【図10】
【図11】
【図12】
【図13】
【公開番号】特開2006−296420(P2006−296420A)
【公開日】平成18年11月2日(2006.11.2)
【国際特許分類】
【出願番号】特願2006−78916(P2006−78916)
【出願日】平成18年3月22日(2006.3.22)
【出願人】(000004628)株式会社日本触媒 (2,292)
【出願人】(504147243)国立大学法人 岡山大学 (444)
【Fターム(参考)】
【公開日】平成18年11月2日(2006.11.2)
【国際特許分類】
【出願日】平成18年3月22日(2006.3.22)
【出願人】(000004628)株式会社日本触媒 (2,292)
【出願人】(504147243)国立大学法人 岡山大学 (444)
【Fターム(参考)】
[ Back to top ]