
調節化合物及び免疫応答を測定するためのアッセイ
【課題】TRAM(Trif関連アダプター分子)のリン酸化の測定及び定量化に用いるアッセイ方法の提供。
【解決手段】TRAMは、プロテインキナーゼCイプシロン(PKCε)によってLPS刺激により急速にリン酸化され、このリン酸化はTRAMの通常機能にとり不可欠である。TRAMのリン酸化状態の検出に適するアッセイは、TRAMを調節する活性を有する化合物を同定する際に有用性を発揮する。更に、TRAMのリン酸化を調節してToll様受容体4(TLR4)により媒介されるシグナリングを調節するのに有用性を発揮する化合物もまた開示される。
【解決手段】TRAMは、プロテインキナーゼCイプシロン(PKCε)によってLPS刺激により急速にリン酸化され、このリン酸化はTRAMの通常機能にとり不可欠である。TRAMのリン酸化状態の検出に適するアッセイは、TRAMを調節する活性を有する化合物を同定する際に有用性を発揮する。更に、TRAMのリン酸化を調節してToll様受容体4(TLR4)により媒介されるシグナリングを調節するのに有用性を発揮する化合物もまた開示される。
【発明の詳細な説明】
【技術分野】
【0001】
本発明はTRAM(Trif関連アダプター分子)のリン酸化の測定及び定量化のための新規なアッセイに関する。アッセイは特に、Toll様受容体4(TLR4)の活性化のモニタリングに用いてもよく、かかるアッセイはその活性の調節物の同定に有用性を発揮する。本発明の更なる態様は、プロテインキナーゼCイプシロンによってTRAMのリン酸化をブロック、阻害又は競争的に阻害する分子を測定するためのアッセイ方法の提供に関する。
【背景技術】
【0002】
Toll様受容体(TLR)スーパーファミリーは、病原微生物の侵入の認識、及び免疫応答の開始において中心的な役割を果たす。ヒトTLRsは現在までに10種類同定されている。各々はシグナリングカスケードの活性化につながる異なる病原関連分子パターン(PAMP)を認識し、更に次々に転写制御因子NF−κB、更には分裂促進因子活性プロテインキナーゼ(MAPKs)、p38、c−jun、N末端キナーゼ(JNK)及びp42/44(非特許文献1及び2に開示される)を活性化させる。TLR3及びTLR4も転写制御因子(IFN制御された因子3(IRF3))の活性化を介して他の経路を活性化させ、それはインターフェロン感受性反応要素(ISRE)と結合し、IFN−β(非特許文献3)を含む遺伝子サブセットを誘導する。TLRsはより大きいスーパーファミリーのメンバーであり、インターロイキン1受容体(IL−1R)/TLRスーパーファミリーとも呼ばれ、IL−1R1サブグループ及びTIR領域含有アダプターサブグループを包含する。3つの全サブグループは細胞質のToll/IL−1受容体(TIR)領域を有し、それはシグナリングにとって重要である。TLRsは細胞外のロイシンリッチリピートを有し、IL−1R1サブグループは細胞外の免疫グロビン領域を有する。アダプター分子は細胞質に存在し、細胞外領域を含まない。
【0003】
上記のように、各TLRはそれぞれ異なるPAMPを認識する。最初に同定されたTLRはTLR4であり、それはグラム陰性細菌のリポ多糖(LPS)の認識にとり重要である(非特許文献4、5)。TLR1及び6と結合したTLR2は、それぞれジアシル−及びトリアシルリポペプチドを認識する(非特許文献6)。TLR3はdsRNAに認識し(非特許文献7)、TLR5は細菌フラジェリンを認識し(非特許文献8)、TLR9は非メチル化CpGのモティーフを認識する(非特許文献9)。一度TLRがPAMPを認識すると、次のシグナリング経路を活性化させるために、TIRドメイン含有アダプターを補充する必要がある。これらのアダプターで最初に同定されたものはMyD88である。それはTLRとIL−1Rのシグナリングにおいて重要な役割を果たし(10、11、12)、それにより生じるシグナリングカスケードが詳細に研究されている(非特許文献13)。実験結果から、TLR3を除く全てのTLRsからのシグナリングに関係していることが示唆されている。MyD88欠損マウスでは、IL−1刺激への応答、又はTLR2、TLR5及びTLR9の刺激が行われない(非特許文献11)。TLR4の場合、遅い反応ではあるが、NF−κB及びMAPKの活性化が観察されている。更に、樹状細胞成熟の誘導及び転写制御因子IRF3の活性化は、MyD88欠損マウスにおいても影響を受けなかった。これは、TLR4が完全にその反応を活性化させるにはMyD88以外の因子を必要とし、この反応が2つのカテゴリー(MyD88依存性の反応と、MyD88非依存性の反応)に分類できることを示唆する。NF−κB及びTNF産生はTLR3に応答して損なわれず、それはMyD88がTLR3シグナリングとは無関係であることを示唆する。
【0004】
次に同定されたアダプターはMal(MyD88 adaptor−like)であり、それはTIRAP(TIRドメイン含有アダプタータンパク質)とも呼ばれている(非特許文献14、15)。最初は、これがTLR4へのMyD88非依存性の反応を媒介するアダプターであると考えられていたが、Mal欠損マウスによる研究の結果、これは正しくなく、またMalとMyD88は協働して作用し、MyD88依存性の経路を活性化させることが明らかとなった。MyD88欠損マウスと同様に、Mal欠損マウスではLPSに応答するNF−κB及びMAPKの遅い活性化を示し、一方樹状細胞成熟及び転写制御因子IRF3の活性化への影響は見られなかった(非特許文献15、16)。Mal欠損マウスは通常、TLR5、TLR7、TLR9、IL−1及びIL−18のリガンドに反応し、それはMyD88がこれらの受容体に必要とされる唯一のアダプターであることを裏付けるものである。TLR3シグナリングはMal欠損マウスにおいて通常観察され、それはMalもMyD88もこの経路に関係しないことを示唆するものである。興味深いことに、TLR2によって活性化するシグナリング経路はMal欠損マウスにおいて完全に喪失され、それはMal及びMyD88の両方がこの経路(16)の活性化に必要とされることを示唆するものである。
【0005】
Trif(TIRドメイン含有アダプター誘導インターフェロン−β)は第3に同定されたアダプターである(非特許文献17、18)。それはTIR含有アダプター分子−1(TICAM−1)とも呼ばれている。TrifによるNF−κB活性化(過剰発現の場合)はMal又はMyD88による場合よりもかなり小さい程度であるが、IFN−βに関しては非常に強い活性化因子として機能する(非特許文献17)。すなわち、それがMyD88非依存性の経路に関係しうることを示唆し、またTrif欠損マウスによりこの点が証明されている(非特許文献19)。LPSに応答するNF−κB活性化はこれらのマウスにおいてはほぼ正常であったが、これらの細胞でTrif及びMyD88が欠損するとき、LPSへのNF−κBの反応は完全に喪失された。Trif欠損マウスにおいてLPSに応答するIRF3の活性化が完全に喪失され、これもTrifがTLR4によって活性化するMyD88非依存性の経路に関係することを示唆するものである。TLR3によるIRF3の活性化はTrif欠損細胞においても喪失され、またNF−κBの活性化も顕著に損なわれ、それらはTrifがTLR3により用いられる唯一のアダプターであることを示唆するものである。
【0006】
またTrifが直接TLR4と結合できないことが発見され(非特許文献18)、架橋アダプターがTLR4への結合に必要であることを示唆するものである。その架橋アダプターは、現在幾つかの研究グループによって発見されており、TRAM(Trif関連アダプター分子)(非特許文献20)、TICAM−2(TIR含有アダプター分子−2)(非特許文献21)又はTIRP(TIRドメイン含有タンパク質)(非特許文献22)と呼ばれている。
【0007】
TRAMは直接TLR4と結合し、他のTLRsとは結合しない(21)。TRAMの過剰発現により、MyD88非依存的に、IRF3、IRF7及びNF−κBが穏やかに誘導された。TRAMのドミナントネガティブは、LPSによるNF−κB及びIRF3の活性化を阻害したが、TLR3リガンド、ポリ(I:C)によるこれらの転写制御因子の活性化にも影響を及ぼさなかった。TRAMの過剰発現(Trifと共に)により、IRF3の核への転座が誘導された(非特許文献20)。ドミナントネガティブ形のTrifは顕著にTRAMの活性を抑制してNF−κB及びIFN−βを活性化させ、一方ではMyD88及びMalドミナントネガティブ変異体では効果を有しなかった。
【0008】
TRAMはTrifノックダウンRAW細胞において機能することができず、すなわちTRAMがTLR4経路のTrifの上流で機能することを示唆するものである。TRAM欠損マウスの作製によりこの理論の重要性がますます増加した(非特許文献23)。これらのマウスを用いた研究により、TRAMはTLR4に応答するMyD88非依存性の経路の活性化にとって不可欠であり、他のTLR経路には関係しないことが示された。
【先行技術文献】
【非特許文献】
【0009】
【非特許文献1】Janeway,C.A.,Jr.,Medzhitov,R.,2002.Annu Rev Immunol 20,197−216.
【非特許文献2】Dunne A.,O’Neill,L.A.,2003a.Sci STKE 2003,re3.
【非特許文献3】Doyle,S.ら,(2002)Immunity 17,251−63.
【非特許文献4】Poltorak A.ら、(1998)Science 282,2085−8.
【非特許文献5】Qureshi S.T.ら、(1999)J Exp Med 189,615−25.
【非特許文献6】Takeda K.,Takeuchi O.及びAkira S.(2002)J Endotoxin Res 8,459−63.
【非特許文献7】Alexopoulou L.ら、(2001) Nature 413, 732−8.
【非特許文献8】Hayashi F.ら、(2001)Nature 410,1099−103.
【非特許文献9】Hemmi H.ら、(2000)Nature 408,740−5.
【非特許文献10】Hemmi H.ら、(2002)Nat Immunol 3,196−200.
【非特許文献11】Adachi O.ら、(1998)Immunity 9,143−50.
【非特許文献12】Takeuchi O.ら、(2000)J Immunol 164,554−7.
【非特許文献13】McGettrick A.F.及びO’Neill L.A.(2004)Mol Immunol 41,577−82.
【非特許文献14】Fitzgerald,K.A.ら、(2001)Nature 413,78−83.
【非特許文献15】Horng,T.ら、(2001)Nat Immunol 2,835−41.
【非特許文献16】Horng,T.ら、(2002)Nature 420,329−33.
【非特許文献17】Yamamoto,M.ら、(2002)J Immunol 169,6668−72.
【非特許文献18】Oshiumi,H.ら、(2003)Nat Immunol 4,161−7.
【非特許文献19】Yamamoto,M.ら、(2003) Science 301,640−3.
【非特許文献20】Fitzgerald,K.A.ら、(2003)J Exp Med 198,1043−55.
【非特許文献21】Oshiumi,H.ら、(2003)J Biol Chem 278,49751−49762.
【非特許文献22】Bin,L.H.ら、(2003)J Biol Chem 278,24526−32.
【非特許文献23】Yamamoto,M.ら、(2003) Nat Immunol 4,1144−50.
【非特許文献24】Thelen M.ら、(1991)Nature 351,320−2.
【非特許文献25】Aderem A.A.ら、(1988) Nature 332,362−4.
【非特許文献26】Wu W.C.ら、(1982)Proc Natl Acad Sci USA 79,5249−53.
【非特許文献27】Rozengurt E.ら、(1983)Proc Natl Acad Sci USA 80,7244−8.
【非特許文献28】Rozengurt E.及びSinnett−Smith J.(1983)Proc Natl Acad Sci USA 80,2936−40.
【非特許文献29】Graff J.M.ら、(1989)Science 246,503−6.
【非特許文献30】Rosen A.ら、(1990)J Exp Med 172,1211−5.
【非特許文献31】Matsubara M.ら、(2003)J Biol Chem 278,48898−902.
【非特許文献32】Takasaki A.ら、(1999)J Biol Chem 274,11848−53.
【非特許文献33】Hayashi N.,ら、(2000)Protein Sci 9,1905−13.
【発明の概要】
【発明が解決しようとする課題】
【0010】
本発明者による鋭意研究の結果、驚くべきことに、LPSのTLR4受容体(Toll様受容体4)への結合に続き、アダプター分子TRAMがプロテインキナーゼCイプシロンによって急速にリン酸化されることを見出した。すなわちTRAMがセリン16残基の部位でプロテインキナーゼCイプシロンによってリン酸化されることを確認した。TLR4の活性化を測定する際、具体的にはTLR4受容体を介するLPSシグナリングが異なる環境において適切に機能するどうかを測定する際、TRAMのリン酸化をモニターするアッセイ方法は有用なツールとなりうる。
【課題を解決するための手段】
【0011】
本発明の第1の態様は、TRAM(Trif関連アダプター分子)の活性化状態を測定する方法の提供に関し、当該方法は、TRAMを含有する細胞サンプルを提供するステップと、TRAMのリン酸化をモニターするステップを含んでなる方法であり、TRAMのリン酸化が行われない場合は、そのTRAMが活性を有しないことの指標となる。
【0012】
本願明細書における「活性化状態」の用語は、TRAMが細胞シグナリング経路に関係しているか否かを指す時に用いる。すなわち、活性TRAMはTRAMのセリン16残基のリン酸化により生じ、TRAM(活性化状態)はTLR4受容体により媒介されるシグナリングに関与する。
【0013】
アッセイの好ましい実施態様では、TRAMはリン酸化に適した条件下でキナーゼと接触する。一実施例において、当該キナーゼはプロテインキナーゼCイプシロンであってもよい。
【0014】
アッセイの更なる実施態様では、TRAMリン酸化をコントロールとの比較によりモニターし、それによりリン酸化を測定する。
【0015】
更なる態様では、TRAMリン酸化を、細胞膜領域内におけるTRAMの存在との比較により測定する。TRAMのリン酸化によって、細胞膜からのTRAMの移動が生じる。
【0016】
したがって、本発明のこの態様の更なる実施態様は、細胞膜に存在するTRAMの存在又はレベルを測定することによってTRAMのリン酸化状態を測定する方法であって、膜領域内に存在するTRAMの、膜からの移動による喪失又は減少が、リン酸化状態のTRAMの指標となる方法の提供に関する。
【0017】
一実施態様では、TRAMのリン酸化の測定方法は、細胞膜中のTRAMの存在及び/又はレベルを測定及び/又定量化する膜減少アッセイを含んでなる。
【0018】
理論に束縛されずないが、発明者は、リン酸化に続く細胞膜からのTRAMの移動が、TRAMのセリン16残基のリン酸化によって陰電荷を帯電し、膜からTRAMを移動させることとなる斥力が生じるものと予想している。
【0019】
本願明細書では、TRAM(Trif関連アダプター分子)と称する分子は、TICAM−2(TIR含有アダプター分子−2)(非特許文献21)又はTIRP(TIRドメイン含有タンパク質)(非特許文献22)と称してもよい。
【0020】
TRAMのリン酸化状態を測定するための上記のアッセイは、TRAM活性化を調節できる候補薬剤の同定のために更に変更してもよい。
【0021】
したがって、本発明の本態様の更なる実施態様は、TRAM活性の調節物質の同定方法の提供に関し、前記方法は、(i)TRAMを含有する第1及び第2の細胞サンプルを準備するステップと、(ii)TRAMの候補調節物を有する前記第1のサンプルを接触させるステップと、(iii)リン酸化に適した条件下でキナーゼと前記第1及び第2のサンプルを接触させるステップと、(iv)TRAMのリン酸化状態をモニターして、前記第1のサンプルと前記第2のサンプルとでTRAMのリン酸化を比較するステップを含んでなり、前記第1のサンプルと前記第2のサンプルとの間のTRAMリン酸化の差異を基に、TRAM活性の調節物として候補調節物質を同定することを特徴とする。
【0022】
一実施態様では、TRAMをリン酸化するキナーゼは、プロテインキナーゼCイプシロンである。
【0023】
本発明の本態様の上記のアッセイに従って同定される1つ以上の調節物質は、ペプチド分子又は非ペプチド分子(例えば化学物質又は医薬用物質)であってもよい。調節物質がペプチドである場合、抗体、抗体フラグメント又は同様の結合活性を有する分子であってもよい。更に、調節物質が抗体である場合、好ましくは単クローン抗体である。
【0024】
本発明の更なる態様は、TLR4受容体で媒介されるシグナリングを調節する薬物の調製に関する前述の態様に従って同定された調節物の使用に関する。TLR4で媒介されるシグナリングの一実施態様では、受容体は上方制御される。更なる、好ましい実施態様では、TLR4受容体で媒介されるシグナリングは下方制御される。
【0025】
更に他の本発明の態様は、TRAMのリン酸化を測定するキットの提供に関し、当該キットは参照サンプル、TRAMのリン酸化を測定するための手段、及び本発明の第1の態様の方法を使用したいずれかの本発明のアッセイを実施するための使用説明書を含んでなる。
【0026】
TRAMのリン酸化状態を評価し、それを調節するのに有用な組成物の同定のためのアッセイ方法を記載した上記の本発明の態様に加えて、本発明は更に、リガンド、特にLPSによるTLR4受容体の活性化を評価するアッセイにおいて有用性を発揮する。かかるアッセイは、TLR4受容体を介して選択的にシグナリングを上方制御又は下方制御することができる化合物の同定及び開発において有用性を発揮しうる。かかる化合物は、TLR4結合及び最も具体的にはLPS(リポ多糖)によるTLR4結合から生じるシグナリング経路の調節物質として有用性を発揮する。
【0027】
第4の本発明の態様は、リガンドによるTLR4活性化を検出するアッセイ方法の提供に関するものであり、当該アッセイは、TLR4発現細胞を含む細胞サンプルを準備するステップと、前記細胞をリガンドと接触させるステップと、TRAMのリン酸化を検出するステップを含んでなり、TRAMのリン酸化をTLR4へのリガンドの結合の指標とすることを特徴とする。
【0028】
好ましくは、リン酸化のレベルを、コントロール試料(例えばリガンドに曝露されない同種の細胞)のそれと比較してもよい。あるいは、試験サンプルを、周知の、予め定められた基準値をコントロールとして比較してもよい。
【0029】
リン酸化を測定することによって、TLR4受容体との相互作用を介したTRAMのリン酸化を調整する候補薬剤を同定することが可能となる。
【0030】
本発明の更なる態様は、TLR4受容体で媒介されるシグナリングを調節する薬物の調製に係る、前述の本発明の態様に従って同定されるリガンドの使用に関する。一実施態様では、TLR4受容体を介するシグナリングは上方制御される。
【0031】
本発明の第4の態様の更なる実施態様は、TLR4受容体のアゴニストを同定するためのアッセイに関し、当該アッセイは、TLR4発現細胞を含む細胞サンプルを準備するステップと、細胞を試験化合物に曝露するステップと、TRAMのリン酸化を検出するステップを含んでなり、TRAMのリン酸化レベルの上昇を、試験化合物のTLR4受容体への結合に続く活性化の指標とすることを特徴とする。
【0032】
一実施態様では、TLR4受容体のアゴニストは、TLR4受容体が媒介するシグナリングを誘導又は上方制御する。
【0033】
本発明の更なる態様は、TLR4受容体で媒介されるシグナリングを調節する薬剤の調製に係る、前述の本発明の態様に従って同定された化合物の使用に関する。一実施態様では、TLR4受容体が媒介するシグナリングは上方制御される。
【0034】
本発明のこの態様の、更に別の実施態様は、TLR4受容体のアンタゴニストを同定するアッセイ方法に関し、当該アッセイ方法は、TLR4発現細胞を含む細胞サンプルを準備するステップと、細胞を試験化合物に曝露するステップと、TRAMのリン酸化を検出するステップを含んでなり、当該方法は、試験化合物が存在する場合における、試験化合物が存在しない場合と比較したTRAMのリン酸化の減少が、当該試験化合物がアンタゴニストであることの指標となることを特徴とする。
【0035】
一実施態様では、TLR4受容体のアンタゴニストはTLR4受容体によって媒介されるシグナリングを妨害又は下方制御する。
【0036】
一実施態様では、上記アッセイ方法は更に、細胞を試験化合物に曝露する前にアゴニストに曝露する処理を含んでなる。
【0037】
本発明の更なる態様は、TLR4受容体で媒介されるシグナリングを調節する薬剤の調製に係る、前述の本発明の態様に従って同定される化合物の使用に関する。一実施態様では、TLR4受容体で媒介されるシグナリングは上方制御される。
【0038】
更に他の本発明の態様は、TLR4受容体の活性を測定するキットの提供に関し、当該キットはサンプル、及び本発明の第4の態様の方法を実施するための使用説明書を含んでなる。
【0039】
発明者は鋭意研究の結果、プロテインキナーゼCイプシロン(PKCε)が、TRAMをリン酸化するキナーゼであることを見出した。プロテインキナーゼCイプシロンによるリン酸化の阻害は、NK−κB及びIFN−βを活性化させるTRAMの能力を弱める。
【0040】
したがって、第6の本発明の態様は、プロテインキナーゼCイプシロンのインヒビターとして作用する化合物を同定するためのアッセイ方法を提供し、当該方法は、候補化合物を準備するステップと、候補化合物をプロテインキナーゼCイプシロンと接触させるステップと、プロテインキナーゼCイプシロンのTRAMをリン酸化する能力の有無を測定するステップを含んでなり、TRAMのリン酸化が行われない場合、それを候補化合物によってなされたプロテインキナーゼCイプシロンの活性のブロッキングの指標とすることを特徴とする。
【0041】
直接プロテインキナーゼCイプシロン分子を阻害することにより、TRAMのリン酸化が妨げられるだけでなく、プロテインキナーゼCイプシロンの他の細胞内における活性も阻害される。しかしながら、発明者は、プロテインキナーゼCイプシロンが結合するTRAMの特異的なドメインを同定し、これがプロテインキナーゼCイプシロンの他の細胞内での活性の阻害を引き起こすことのない、TRAMのリン酸化の選択的阻害の可能性を開拓することを見出した。
【0042】
本発明の本態様の別の実施例は、プロテインキナーゼCイプシロンによるTRAMのリン酸化を防止する化合物を同定するためのアッセイであって、当該アッセイは、候補化合物を準備するステップと、候補化合物をTRAMと接触させるステップと、TRAMを、リン酸化の開始に適する条件下でプロテインキナーゼCイプシロンに曝露するステップと、TRAMのリン酸化の有無を測定するステップを含んでなり、リン酸化が行われないことを、プロテインキナーゼCイプシロンとTRAMの間での相互作用のブロッキングの指標とすることを特徴とする。
【0043】
好ましい実施態様では、当該化合物はプロテインキナーゼCイプシロンによって選択的にTRAMのリン酸化を阻害する。
【0044】
一実施態様では、当該方法は、TRAMに存在するセリン16の残基に対応するドメイン中の部位又は領域で、化合物がTRAMと結合して、そのセリン残基がプロテインキナーゼCイプシロンによりリン酸化されるのを防止する能力を測定するステップを含んでなる。
【0045】
更なる様々な態様では、本発明はスクリーニング及びアッセイ方法、並びにそれにより同定された物質に関する。
【0046】
本発明のアッセイを用いて同定される新規な化合物は、更に別の本発明の態様を構成する。かかる化合物又は調節物質は、医薬組成物中に添加してもよい。
【0047】
調節物質、又は本発明のアッセイで同定された化合物は、ペプチド又は非ペプチド分子(例えば化学物質又は医薬物質)であってもよい。調節物質質がペプチドである場合、抗体、抗体フラグメント又は同様に結合するフラグメントであってもよい。更に、調節物質が抗体である場合、好ましくは単クローン抗体である。
【0048】
TRAMに対して特異性を有する単クローン抗体、抗体フラグメント又は同様の結合分子(プロテインキナーゼCイプシロンがTRAMのリン酸化を促進するために結合する領域におけるセリン−16残基に特に結合する、又は完全若しくは部分的にブロッキングする)は、TRAMのリン酸化の阻害に有用性を発揮し、それによりTLR4受容体へのLPSの結合に続く、その下流側への活性シグナリング伝達の促進を防止する能力を有する。
【0049】
したがって、本発明の更なる態様は、抗原結合ドメインを含む特異的な結合メンバーの提供に関し、当該抗原結合ドメインはTRAMのセリン−16残基に対する特異性を有する。
【0050】
一実施態様は、本発明はTRAMのセリン−16残基に特異的に結合するか又は結合を防ぐイムノグロブリンの提供に関する。
【0051】
更に他の実施態様は、プロテインキナーゼCイプシロンによるTRAMのセリン−16残基のリン酸化を防止するイムノグロブリンの提供に関する。
【0052】
また、本発明の本態様には、TRAMと特異的に結合してプロテインキナーゼCイプシロンによるリン酸化を防止する結合メンバーが包含される。
【0053】
内毒素は、リピドAと多糖からなるリポ多糖(LPS)複合体を含んでなる。LPSはTLR4受容体と結合し、これが適当な免疫応答を誘発する下流のシグナリングカスケードを生じさせる。
【0054】
敗血症及び敗血症性ショックなどのLPSの媒介又は内毒素の媒介による症状によりしばしば死に至る。したがって、TLR4の媒介による免疫応答経路を下方制御又は阻害する方法は、LPSの媒介による症状(例えば敗血症)の処理方法として望ましい。
【0055】
TLR4へのLPS結合に続いてTRAMがプロテインキナーゼCイプシロンによってリン酸化されるという観察から、本発明は、LPSと細胞内キナーゼとの間の相互作用を制御するためのポテンシャルルートを提供する。これにより、LPSシグナリングを無力化するための強力なメカニズムが提供される。
【0056】
したがって、本発明はLPSの媒介による症状の治療に使用できる。
【0057】
本発明のアッセイ、及び当該アッセイの使用によって解明されるTLR4シグナリング経路に対して生物学的重要性を有する化合物は、多くの医学的症状、最も具体的には内毒素及びLPSにより媒介される症状(例えば敗血症)の治療に対して特に有用性を発揮する。
【0058】
したがって、本発明の第7の態様は、内毒素により媒介される症状の治療用に適する化合物を同定するためのアッセイ方法に関し、当該アッセイは、候補化合物を準備するステップと、前記候補化合物をTRAMと接触させるステップと、候補化合物によるTRAMのリン酸化の有無を測定するステップを含んでなり、TRAMリン酸化の調節を、その化合物の有用性の指標とすることを特徴とする。
【0059】
一実施態様では、内毒素により媒介される症状は敗血症又は敗血症性ショックである。
【0060】
更に他の本発明の態様は、LPSにより媒介される症状の治療方法の提供に関し、当該方法はTRAMのリン酸化を阻害する処理を含んでなる。
【0061】
本発明の上記態様の好ましい一実施態様では、TRAMのリン酸化の阻害は、TRAMにプロテインキナーゼCイプシロンの結合を防ぐことによってなされる。
【0062】
一実施態様では、TRAMへのプロテインキナーゼCイプシロンの結合のブロッキングは、セリン16ドメインへのプロテインキナーゼCイプシロンの結合によるリン酸化を阻害する化合物によって促進される。
【0063】
更に他の実施態様では、当該化合物は、プロテインキナーゼCイプシロンがリン酸化するTRAMのセリン−16部位と直接結合する。あるいは、阻害剤はTRAM上の部位と結合し、セリン−16残基のリン酸化に必要となる、TRAM上の結合部位へのプロテインキナーゼCイプシロンのアクセスを防止する。
【0064】
TRAMの配列は、配列番号1:MGIGKSKINSCPLSLSWGとして定義される。セリン16は、配列中の最後のセリンである。
【0065】
あるいは、セリン−16残基のリン酸化に適切な部位において、TRAMへの結合に関してプロテインキナーゼCイプシロンと競合する分子の提供に関する。
【0066】
更に別の本発明の態様は、LPSにより媒介される症状(敗血症)に罹患する患者を治療する方法の提供に関し、前記患者にTRAMのリン酸化を阻害する分子を、治療上有効量で投与することを含んでなる。
【0067】
本発明の本態様の好ましい一実施態様では、当該分子はプロテインキナーゼCイプシロンによるTRAMのリン酸化を防止する。
【0068】
内毒素により媒介される症状は、特にLPSによって生じるものであり、しばしば発熱、白血球数の変化、播種性血管内凝固、低血圧、ショック及び死に至る。
【0069】
好ましい一実施態様では、内毒素により媒介される上記症状は敗血症である。
【0070】
本発明の更なる態様は、内毒素により媒介される症状の治療用薬剤の調製における、プロテインキナーゼCイプシロンの阻害剤の使用の提供に関する。
【0071】
したがって、本発明の更なる態様は抗原結合ドメインを含む特異的な結合メンバーの提供に関し、当該抗原結合ドメインはプロテインキナーゼCイプシロンに対する特異性を有する。
【0072】
一実施態様では、本発明は、プロテインキナーゼCイプシロンに特異的に結合するか又は結合をブロックするイムノグロブリンの提供に関する。
【0073】
更に他の本発明の態様は、敗血症の治療用薬剤の調製のための、プロテインキナーゼCイプシロンの阻害剤の使用に関する。
【0074】
更に別の本発明の態様は、内毒素により媒介される疾患(例えば敗血症)の治療用薬剤の調製のための、TRAMのリン酸化を防止する化合物の使用に関する。
【0075】
更に別の本発明の態様は、前述の本発明の態様のいずれか1つに従うアッセイ方法のいずれか1つによって同定される化合物を使用することを特徴とする、医学的症状の治療方法の提供に関する。
【0076】
TLR4受容体で媒介されるシグナリングは、内毒素によって媒介されるそれら以外の免疫応答と関連していてもよい。例えば、TLR4受容体を介したシグナル伝達は炎症性疾患に関係している場合もあり、それゆえ、TLR4受容体によるリガンド結合能力及びTLR4受容体の下流におけるシグナル伝達の調節は、炎症性疾患(例えば関節炎及びアテローム性動脈硬化)に影響を与えうる。若干の自己免疫性の症状では、TLR4受容体の候補リガンド又は調節物質は未だ同定されていない。しかしながら、このような場合では、炎症又は傷害を伴う組織によって産生される内因性の因子がTLR4受容体又はその関連するシグナリング経路に影響を及ぼすことが示唆される。
【0077】
したがって、更に他の本発明の態様は、炎症性又は免疫媒介性障害の治療又は予防に適する化合物を同定するためのアッセイ方法の提供に関し、当該アッセイは、候補化合物を準備するステップと、候補化合物をTRAMと接触させるステップと、候補化合物によるTRAMのリン酸化の有無を測定するステップを含んでなり、TRAMリン酸化の調節を、その化合物の有用性の指標とすることを特徴とする。
【0078】
好ましい実施態様では、当該免疫媒介性障害は関節炎又はアテローム性動脈硬化であってもよい。
【0079】
更に他の本発明の態様は、炎症性症状又は免疫媒介性障害の治療又は予防用の薬剤の調製における、プロテインキナーゼCイプシロンの阻害剤の使用に関する。
【0080】
更に別の本発明の態様は、炎症性又は免疫媒介性障害の治療又は予防用薬剤の調製における、TRAMのリン酸化を防止する化合物の使用に関する。
【0081】
更なる本発明の態様では、LPSによるTLR4受容体への結合に続くシグナリングに関連する症状の治療を必要とする患者を治療する方法であって、当該方法は本発明のアッセイのいずれか1つに従って同定される化合物の投与を含んでなる。
【0082】
他に必要としない限り、各本発明の態様の好ましい特徴は必要な変更を加えて他の態様の各々に適用してもよい。
【図面の簡単な説明】
【0083】
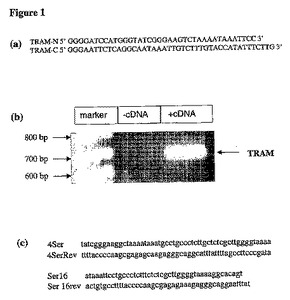
【図1】(a)TRAMのクローニングに使用した、TRAMの5’及び3’末端を標的とするプライマー。(b)得られるPCR生成物を幾つかのベクターにクローニングした。(c)特異的なプライマーを使用して、TRAMの最初の4つのセリンをアラニンに変異させ、並びに16番目のセリンのみをアラニンに変異させるため、部位特異的突然変異導入を実施した。
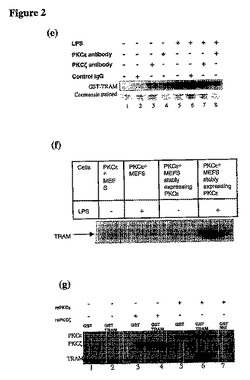
【図2a】(a)GST−TRAMは、LPSで処置されたTHP1のライゼートとの所定時間のインキュベーションによりリン酸化される。(b)細胞をポリI:Cで処理した場合、かかるTRAMのリン酸化は行われない。(c)TRAMの最初の4つのセリンをアラニンに変異させた場合、このLPS依存性のリン酸化は行われない。(d)LPSで30分間処理したTHP1細胞から調製したライゼートとのGST−TRAMプルダウンに続いて、サンプルを1時間panPKC阻害剤(ビスインドールマレイミド(ビス))の量を増加させつつインキュベートした。その結果、TRAMのリン酸化の減少が見られた。
【図2b】(e)GST−TRAMとのインキュベーションの前にPKCε抗体、PKCζ抗体又は免疫グロブリンGコントロール抗体を使用して、THP1ライゼートの免疫減少を実施した。ライゼートからPKCεの除去により、TRAMのLPS依存性のリン酸化が妨げられた。(f)PKCεを欠損するMEFsから調製したライゼートはGST−TRAMをリン酸化が行われず、一方、PKCεで再構成したMEFsではリン酸化が行われた。(g)組換えPKCε(rePKCε)又はPKCζ(rePKCζ)をGST−TRAMと15分間直接インキュベートし、その後キナーゼアッセイを実施した。rePKCεはWT−TRAMでリン酸化されたが、Malではされなかった。
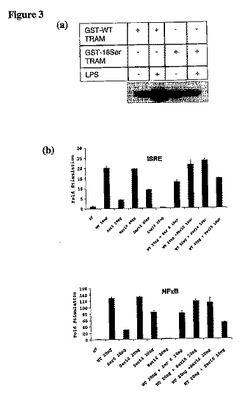
【図3】(a)セリン16の位置をアラニンに変異させたとき、GST−TRAMはTHP1ライゼートとのインキュベーションによりもはやリン酸化され得ない。(b)pcDNA3.1のみ(eV)、WT−TRAM/pcDNA3.1及びSer16−TRAM/pcDNA3.1の全てを、96穴プレートに播種したHEK293細胞にトランスフェクションした。NF−κB又はISRE−ルシフェラーゼレポーター遺伝子、並びにRenillaルシフェラーゼ内部コントロールプラスミドを同様にトランスフェクションした。24時間後、リポータ遺伝子活性を測定し、データはコントロールのレベルに対する刺激(倍数表示)の平均値として表す。グラフは、WT−TRAMはNF−κB及びISRE経路を活性化できず、一方Ser16−TRAM変異体がどちらの経路も活性化できることを示す。
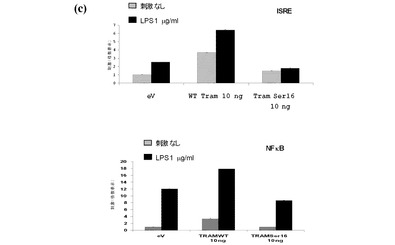
【図3c】同様の実験をHEK293−TLR4細胞において実施し、トランスフェクション後24時間において、細胞を6時間、LPS(1μg/ml)の有無においてインキュベートした。試験の結果、WT−TRAMがNF−κB及びISREを活性化させるLPSの能力を増加させ、一方Ser16−TRAM変異体がドミナントネガティブとして作用してNF−κB及びISREを刺激するLPSの能力を減少させることを示す。
【図3d】HEK293細胞を、pcDNA3.1、MyD88/pcDNA3.1又はWT−TRAM/pcDNA3.1、並びにNF−κB−ルシフェラーゼレポーター遺伝子及びRenillaルシフェラーゼ内部コントロールプラスミドでトランスフェクションする前に、1時間PKC阻害剤(ビスインドリルマレイミド(ビス))の有無によりインキュベートした。上記阻害剤はNF−κB経路を活性化させるMyD88の能力に影響を及ぼさなかったが、この経路を活性化させるTRAMの能力を阻害した。
【図4a】TRAMがセリン16残基にリン酸化されることを示す。FLAGタグ付きTRAMを過剰発現するHEK293−TLR4細胞を、30分間、1μg/mlのLPSによって刺激した。FLAGタグ付きTRAMは、抗FLAG抗体を使用して免疫沈降し、ホスホセリン抗体によりブロッティングした。
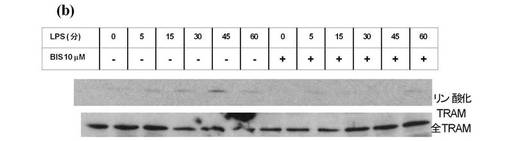
【図4b】THP1細胞を、LPSによる刺激前に1時間、PKC阻害剤ビスの有無によりインキュベートした。
【図4c】一定時間の1μg/mlのLPSによる刺激に続いて、PKCε−/−MEFs、並びにPKCεによって再構成されたPKCε−/−MEFsの結果を示す。
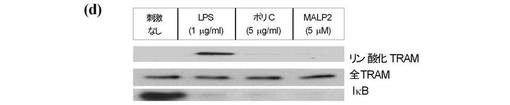
【図4d】30分間のLPS、ポリI:C若しくはMALP2による刺激に続いて、PKCε−/−MEFs、並びにPKCεによって再構成されたPKCε−/−MEFsの結果を示す。
【図5a】TRAMとPKCεの両方は、LPSシグナル伝達の遂行にとって重要であることを示す。PKCε−/−MEFs及びPKCεによって再構成されたPKCε−/−MEFs細胞(上部の4枚のパネル)、又はTRAM−/−MEFs及びWT MEFs細胞(下部の4枚のパネル)を、一定時間1μg/mlのLPS又は5μg/mlのPolyI:Cで刺激し、更にライゼートをリン酸化(チロシン180/182)及び全p38に関してイムノブロットした。
【図5b】−PKCε−/−MEFs及びPKCεによって再構成されたPKCε−/−MEFsを、1μg/mlのLPS又は5μg/mlのpolyI:Cで一定時間処理し、非還元PAGEゲル電気泳動し、IRF3に関してイムノブロットした。
【図5c】ISREルシフェラーゼレポーター遺伝子及びコントロールのRenillaルシフェラーゼレポーター遺伝子を、上記のタイプの細胞にトランスフェクションした。トランスフェクションの24時間後、細胞を1μg/mlのLPS又は5μg/mlのPolyI:Cによって6時間刺激した。データはコントロールのレベルに対するルシフェラーゼ活性の刺激(倍数表示)の平均値として表す。
【図5d】上記の細胞タイプを、LPS又はPolyI:Cの所定の濃度によって24時間刺激し、更にRANTES ELISAを実施した。示す結果は、少なくとも3つの実験の代表的なものである。
【図5e】同上。
【図6a】TRAMS16Aは、TRAM欠損MEFsの再構成がWTTRAMよりも弱かった。WTTRAM、TRAMS16A及び空のベクターを、TRAM欠損MEFsにトランスフェクションした。トランスフェクション後24時間インキュベートした。細胞を更に24時間、LPS(1μg/ml)によって刺激し、培養液上清をRANTESに関してELISA解析した。
【図6b】細胞を30分間、LPS(1μg/ml)によって刺激し、ライゼートをp38に関してウエスタンブロッティング解析した。示す結果は、少なくとも3つの実験の代表的なものである。
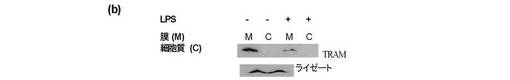
【図7】膜に存在するTRAMの量がLPS刺激により減少することを示し、TRAMが膜から消失したことを示唆するものである。
【図8a】セリン16残基がグルタミン酸に変異した場合の、膜上に存在するTRAMの量の減少を示し、この突然変異がセリン16上のTRAMのリン酸化が膜からのTRAMの消失を生じさせることを示唆するものである(レーン3と1を比較)。
【図8b】更にLPSで処理されたTHP1細胞における内因性のTRAMの減少が検出されることを示唆する。
【図8c】更にPKCε−/−MEFsFLAG−TRAMは、LPS刺激によって膜から消失しないことを示唆する。これらの結果は、PKCεによるセリン16上のTRAMのリン酸化が、膜上のTRAMの減少にとり必要であることを示唆するものである。
【発明を実施するための形態】
【0084】
アッセイ
本発明は、TRAMリン酸化の測定、更にはTRAMリン酸化の発生によるTLR4活性化のモニタリングに用いるアッセイシステム及びスクリーニング法を提供する。本発明の「アッセイシステム」の用語には、ある特定の1つ以上の現象を検出及び/又は測定するアッセイを実行して、その結果を分析するために必要な全ての構成要素が包含される。
【0085】
様々なアッセイを利用して、標的分子又はタンパク質のリン酸化状態を検出してもよい。
【0086】
一実施態様では、当該アッセイはリン酸化を受けるTRAMのドメインを認識するリン酸化特異的な抗体を使用する。最も好ましくは、この抗体は、TRAMにおけるセリン16の位置又はその近傍領域と結合する。
【0087】
TRAMのアミノ酸配列は、配列番号1:MGIGKSKINSCPLSLSWGとして定義される。
【0088】
好ましい実施態様では、本発明のアッセイではウエスタンブロッティングなどの公知の技術を使用する。当業者に公知の標準的な方法を使用して、TLR受容体に対するリガンド(例えばLPS)によって刺激した細胞から調製した抗体が、ウエスタンブロットのサンプルとして用いられる。
【0089】
更なる態様では、当該抗体を、他のアッセイのフォーマットにおいて使用してもよい。例えば、インビトロキナーゼアッセイにおいては、総タンパクの代わりにTRAM由来するペプチドフラグメントを用いてアッセイを実施できる。
【0090】
必須ではないが、好ましくは、本発明で使用するスクリーニングアッセイはハイスループット又はウルトラハイスループットアッセイであり、それによりオートメーション化された費用効果的なスクリーニング手段の提供につながる。
【発明を実施するための最良の形態】
【0091】
治療
本明細書で使用される「治療」という用語は、ヒト又は人間以外の動物に適用できるいかなる療法も意味する。治療とは、既存の症状に関するものであってもよく、又は予防的なもの(予防的処置)であってもよい。治療には、治癒的、緩和的又は予防的効果が包含される。
【0092】
医薬組成物
本発明は更に、リン酸化状態の変調又はリン酸化の防止による、TRAMのリン酸化調節用の医薬及び医薬組成物に関する。
【0093】
したがって、本発明の更なる態様は、免疫応答の調節用の医薬組成物の提供に関し、当該組成物は、主成分として、リン酸化を促進又はブロッキングすることによりTRAMのリン酸化を調節する化合物を含んでなる。
【0094】
本発明の医薬組成物、並びにその本発明での使用においては、主成分に加えて、当業者に公知の薬学的に許容できる添加剤、担体、緩衝安定剤又は他の材料を含有させてもよい。
【0095】
かかる材料は毒性を有してはならず、また主成分の有効性を妨げてはならない。担体又は他の材料の具体的な特性は投与ルートに依存する。
【0096】
投与量
組成物は好ましくは「臨床的に有効な量」(個人に利益をもたらすのに十分な量)で患者に投与される。実際の投与量、並びに投与速度及び時間は、個人の状態及び処置を受ける症状に依存する。
【0097】
至適投与量は、多くのパラメータ(例えば個人の年齢、阻害又は誘導される免疫応答の大きさ、投与される組成物の具体的な形態及び投与ルートなど)に基づいて決定できる。
【0098】
当該組成物は、血液などの特定の組織に投入されるミクロスフェア、リポソーム、他の微粒子状の輸送システム又は徐放性製剤を経て投与できる。好適な徐放性担体の例としては、例えば坐薬又はマイクロカプセルなどの一般的な剤形の半透性ポリマーマトリックスが挙げられる。
【0099】
上記の技術及びプロトコル、並びに本発明で使用できる他の技術及びプロトコルの例は、「Remington’s Pharmaceutical Sciences」,18th edition,Gennaro,A.R.,Lippincott Williams&Wilkins;第20版(December 15,2000)ISBN 0−912734−04−3、及び「Pharmaceutical Dosage Forms and Drug Delivery Systems」;Ansel,H.C.ら、第7版 ISBN 0−683305−72−7に記載され、その開示内容は本願明細書に援用される。
【0100】
抗体
本発明における「抗体」の用語は、イムノグロブリン若しくはその一部、又は抗体の結合ドメイン若しくはそのホモログであるポリペプチドを含む結合ドメインを指すものとする。
【0101】
「抗体」はイムノグロブリンであり、天然であってもよく、又は部分的又は完全に合成されたものであってもよい。当該用語はまた、抗体結合ドメイン若しくはそのホモログである結合ドメインを有するあらゆるポリペプチド、タンパク質若しくはペプチドが包含される。これらは天然給源に由来してもよく、又は部分的又は完全に合成されたものであってもよい。
【0102】
当該抗体は完全な抗体であってもよく、又はそのフラグメントであってもよい。完全な抗体に由来するフラグメントは、抗原の結合能を発揮できる。かかる結合フラグメントの例としては、(i)VL、VH、Cl及びCH1ドメインからなるFabフラグメント、(ii)VH及びCH1ドメインからなるFdフラグメント、(iii)単一の抗体のVH及びVLドメインからなるFvフラグメント、(iv)VHドメインからなるdAbフラグメント、(v)単離されたCDR領域、(vi)F(ab’)2フラグメント(2つのFabフラグメントが連結した二価のフラグメント)(vii)単鎖Fv分子(scFv)(VHドメイン及びVLドメインがペプチドリンカーで連結され、それにより2つのドメインが結合して抗原結合部位を形成する)、(viii)二重特異的な単鎖Fv二量体、及び(ix)遺伝子融合によって調製される多価性又は多重特異的フラグメント。
【0103】
抗体は多くの方法で修飾できるため、「抗体」の用語は、必要な特異性を有する結合ドメインを有するいかなる結合メンバー又は基質も包含されるものとして解釈すべきである。
【0104】
本発明の抗体は、単クローン抗体、又はそのフラグメント、誘導体、機能的均等物又はそのホモログであってもよい。抗体の定常部は、いかなる適切なイムノグロブリンサブタイプであってもよい。
【0105】
「抗体」という用語には、「ヒト化」された抗体、又はCDRグラフティングなどの技術により調製された抗体が包含される。かかる技術は、当業者にとって周知である。
【0106】
抗体作成
本発明に係る、及び本発明に用いられる特異的な結合メンバーは、いかなる適切な方法で調製してもよく、天然由来であっても人工的に合成したものであってもよい。かかる方法としては、例えば抗体ライブラリを使用した従来のハイブリドーマ技術、組換えDNA技術又はファージディスプレイ技術などが挙げられる。かかる産生技術は当業者に公知であり、またその他の抗体産生技術は、「Antibodies」:A Laboratory Manual,eds.Harlowら、Cold Spring Harbor Laboratory,1988.に記載されている。
【0107】
特に定めのない限り、本願明細書において用いられる全ての技術的及び科学的な用語は、本発明の技術分野の当業者に一般に理解されている意味を有する。
【0108】
明細書全体を通じて、特に断りのない限り、用語「含んでなる(comprise)」又は「含む(include)」又はそれらのあらゆる活用形は、特定の整数又は整数の群の包含を意味する一方で、他のいかなる整数又は整数の群を除外することを意味するものではない。
【0109】
本発明を、以下の実施例(それらは例示を目的として開示され、本発明を限定するものと解釈すべきでない)、更には図面を参照することにより例示する。
【実施例】
【0110】
材料及び方法細胞:
HEK293細胞及びTLR4(HEK293−TLR4)で安定にトランスフェクションしたHEK293細胞を、10%のウシ胎仔血清(FCS)、100単位/mlのペニシリン、100mg/mlのストレプトマイシン及び2mMのグルタミンを添加したDulbecco社の修飾Eagles培地(DMEM)で培養した。THP1細胞を、10%のFCS、100単位/mlのペニシリン、100mg/mlのストレプトマイシン及び2mMのグルタミンを添加したRPMI培地で培養した。
【0111】
逆転写酵素−ポリメラーゼ連鎖反応(RT−PCR):
鋳型として脾臓メッセンジャーRNA(BD Bioscience社)を使用してcDNAを調製した。1〜5μgRNAを、0.1μgのランダムプライマーとインキュベートし、DEPC処理水で5μLの最終液量とした。70℃で10分間、更に4℃で2分間インキュベートした。moloneyマウス白血病ウイルス逆転写酵素(MMLV−RT)を使用して逆転写反応させた。4μLの5×バッファ(250mMのトリスHCl(pH8.3)、375mM KCl及び15mM MgCl2)、2μLの100mM DTT、1μLのRNasin(40単位/ml)、1μLの10mM dNTP、1μLのMMLV−RT(200単位/μL)及び6μLのDEPC処理水を添加した。この反応液を37℃で1時間インキュベートし、酵素を不活性化するために95℃で2分間処理した。この反応液の5μLをPCR反応の鋳型として使用し、またTRAMの5’及び3’末端に特異的なプライマーを使用した。1×DNAポリメラーゼバッファ(1mMのトリスHCl(pH9.0)、5mMのKCl及び0.01%のTriton(登録商標)X−100)、各dNTP 0.2mM、2.5uのTaq DNAポリメラーゼ、特異的なオリゴヌクレオチドプライマを各0.5μM、及び0.5〜3mMのMgCl2を鋳型DNA溶液に添加し、反応混合物をPCRグレードの水を使用して50μLの最終的な液量とした。サーマルサイクラーを使用し、1分間94℃、1分間55℃及び2分間72℃の30サイクルで反応させた。PCR生成物をアガロースゲル電気泳動で分析した。
【0112】
GST発現ベクター及び哺乳類の発現ベクターへのTRAMのクローニング:
TRAMのPCR生成物を、pGEX−kgベクター(ファルマシア社製)、pcDNA3.1ベクター(インビトロゲン社)にライゲーションした。第1に、PCR生成物及びベクターを、別々の反応において切断した。5〜10単位の制限エンドヌクレアーゼ(NEB社製)、1×制限酵素バッファ、±1×BSA及び1〜10μgのDNAを混合し、滅菌水で10μLにし、2時間37℃でインキュベートした。次いで消化した生成物をPCR精製キット(Qiagen社製)を使用して精製した。1単位のT4DNAリガーゼ(1単位/μL)(プロメガ社製)、2μLのT4 10×反応バッファ、100〜150ngの切断したベクターDNA、及び200〜400ngの切断したPCR生成物を混合し、滅菌水で20μLにし、ライゲーション反応液を調製した。このライゲーション混合物を4℃で一晩静置し、更にBL21(DE3)細胞(ストラタジーン社)を形質転換した。
【0113】
TRAMの部位特異的変異導入:
Quickchange(登録商標)部位特異的変異導入キット(ストラタジーン社)を用い、TRAM遺伝子の特定の塩基を変異させた。製造業者の指示に従い、所望の突然変異を有するプライマーを使用して反応させた。
【0114】
GST−TRAMの発現及び精製:
TRAM−pGEXベクターを有するBL21(DE3)を、振盪培養器を用い、10mLのLB培地で37℃で一晩培養した。翌日、10mLを500mLのLB培地に植え継ぎ、0.6〜0.8のODとなるまで培養した。0.2mMの最終濃度でIPTGを培養液に添加し、更にこの培養液を更に30℃で4時間インキュベートした。更に培養液を、Sorvall RC5C遠心機(GSAローター使用)により、8,000回転/分で15分間遠心分離した。ペレットを25mLのNETNバッファ(20mMのトリスHCl、100mMのNaCl、1mMのEDTA、0.5%のNP40、10μg/mLのロイペプチン、10μg/mLのペプスタチン、1mMのPMSF、pH8.0)に再懸濁し、ソニケータ(Branson Sonifer 250)を使用して5分間超音波破壊し、45分間、18,000回転/分で遠心分離した。上澄みを600μLグルタチオンセファロースビーズ(アマシャム社製)に添加し、4℃で2時間インキュベートした。更にビーズを15mLのNETNバッファで5回洗浄し、600μLのNETNバッファに再懸濁した。
【0115】
ルシフェラーゼレポーター遺伝子アッセイ:
HEK293又はHEK293−TLR4細胞を、1×105細胞/mLの密度で96穴プレートに播種した。翌日、上記細胞を、Genejuice(Novagen)を用い、製造業者の指示に従い、選択したルシフェラーゼリポータプラスミド、及び選択した発現ベクターによってトランスフェクションした。NF−κB及びIRF3活性化の検出に関する実験のために、80ngのNF−κB又はIRSE−ルシフェラーゼレポーター遺伝子(ストラタジーン社)を、Renillaルシフェラーゼ内部コントロールプラスミド(ストラタジーン社)40ngと共に細胞にトランスフェクションした。24時間後に、細胞をpassive lysis buffer(プロメガ社製)に溶解し、レポーター遺伝子活性を照度計を使用して測定した。データを、コントロールのレベルに対する刺激(倍数表示)の平均値として表した。
【0116】
キナーゼアッセイ:
THP1細胞を、T175フラスコ(Sarstedt社)中に2×105細胞/mLで播種し、37℃で一晩インキュベートした。翌日、30mLの細胞懸濁液を、1時間、50mLのFalconチューブ(Sarstedt社)中でLPSの有無により処理した。細胞を遠心分離によって回収し、一度PBSで洗浄し、10分間、1mLのバッファ(10%のグリセロール(v/v)、50mMのNaF、20mMのトリスHCl(pH8.0)、2mMのEDTA、137mMのNaCl、1%のNP−40、1mMのPMSF、10μg/mLロイペプチン、1mMのNa3VO4)中で溶解させた。細胞残渣を13,000回転/分で10分間遠心分離し、上澄みをキナーゼアッセイ用に新規な試験管へ移した。グルタチオンビーズ上の精製されたGST−TRAM 50μLをエッペンドルフチューブに添加し、適当量のライゼートをチューブに添加し、4℃で2時間インキュベートした。ビーズを2,000回転数/分で5分間のスピンダウンし、更にキナーゼバッファ(20mMのHEPES(pH7.5)、10mMのMgAc、0.03%のTriton、100μg/mLのホスファチジルセリン、20mMのβ−グリセロールリン酸、0.01%(w/v)のロイペプチン及び100μMのNa3VO4)で3回洗浄した。これらのビーズを更に30μLのキナーゼバッファ(20μMの冷却したATP及び5μCi[γ32P]ATPを含有する)中に再懸濁し、37℃で30分間インキュベートした。20μLのサンプルバッファ(50mMのトリスHCl(pH6.8)、10%のグリセロール(v/v)、2%のSDS(w/v)、0.1%のブロムフェノールブルー(w/v)及び5%のβ−メルカプトエタノール)をチューブに添加し、サンプルを5分間ボイルした。サンプルを更に10%のSDS−PAGEに供し、ニトロセルロース膜へ転写した。ニトロセルロース膜はautoradカセット(コダック社製)に設置し、X線フィルムを上部に設置した。−80℃で一晩静置し、更にフィルムを感光させた。
【0117】
膜分画:
HEK293−TLR4細胞を1×105細胞/mLで播種し一晩インキュベートし、更に適当なプラスミドによってトランスフェクションした。トランスフェクションの24時間後、細胞を「結果」のセクションの方法に従い処理し、更に300μLの膜バッファ(20mMのトリス(pH7.5)、10mMのMgCl2、1mMのEDTA、250μMのスクロース、200μMのPMSF)中にスクラップした。細胞をdounceホモジェナイザを用いて30ストロークで溶解させ、4℃で1時間、100,000回転/分でhardwall Beckmanチューブで遠心分離した。上澄(すなわちサイトソル画分)を新規なチューブへ移し、ペレット(すなわち膜画分)を50μLのサンプルバッファ(50mMのトリスHCl(pH6.8)、10%のグリセロール(v/v)、2%のSDS(w/v)、0.1%のブロムフェノールブルー(w/v)及び5%のβ−メルカプトエタノール)中に再懸濁した。サイトソル画分を、centricon YM−10(ミリポア社製)を使用して50μLまで濃縮した。サンプルを12%のSDS−PAGEに供した。
【0118】
リン酸化特異的なTRAM抗体の産生:
Fabgennix(テキサス、米国)社に、アミノ酸16でセリンをホスホセリンで置換した、TRAM(KINSCPLSLSWGKRH)のアミノ酸7〜21に対応する合成ペプチドを使用して、セリン16でリン酸化されたTRAMを認識するリン酸化特異的な抗体の作製、精製を委託した。抗体の有効性は、リン酸化TRAMの予測されるバンドがTRAM欠損MEFs(データ示さない)から調製したサンプルでは存在しないことをもって確認した。
【0119】
RANTES ELISA
示された細胞を24穴プレート中に1×105細胞/mLで前夜に播種し、更に適当なプラスミドによってトランスフェクションした。トランスフェクションの24時間後、細胞を24時間、適当な刺激で処理した。5倍希釈した上澄みをサンプルとして使用し、R&D systems社のマウスRANTESキットを用い、製造業者の指示に従いRANTES ELISAを実施した。
【0120】
IRF3二量体化アッセイ
適当な細胞を、2×105細胞/mLで播種し、一晩インキュベートし、更に適当な刺激で処理した。細胞をPBSで洗浄し、100μL非還元サンプルバッファ(50mMのトリスHCl(pH6.8)、10%のグリセロール(v/v)、0.1%のブロムフェノールブルー(w/v)及び5%のβ−メルカプトエタノール)中にスクラップした。このうち20μLを非還元PAGEに供し、ニトロセルロース膜に転写し、IRF3に関してブロッティングした。
【0121】
結果TRAM遺伝子のクローン化及び変異体の生成:
TRAMのcDNA配列を、Genbank(Accession No.NM_021649)から検索した。TRAMの5’及び3’末端に対する特異的なプライマー(図1a)を用い、脾臓由来のmRNAを使用して、TRAMのcDNAを増幅した(図1b)。このcDNAをpGEX−kgベクターにクローニングし、細菌内でGST−TRAM融合タンパク質を発現させた。哺乳類の発現ベクターpcDNA3.1にもクローニングした。TRAMの変異体を生成するため、部位特異的変異導入を行った。位置6、10、14及び16(4Ser変異体と呼ばれる)でセリンが変異するようにプライマーを設計した(図1c)。セリン16のみを変異(Ser16変異体と呼ばれる)させるプライマーも設計した。
【0122】
LPS刺激によるTRAMのリン酸化:
TRAMをミリストイル化し、細胞を静置し、膜に移動させた(未発表データ)。幾つかのミリストイル化タンパク質を静電的なスイッチに供し、それをリン酸化することにより膜から反発される。すなわち、TRAMがリン酸化されたか否かが判定できるようにアッセイが設定されている。グルタチオンビーズ上の精製されたGST−TRAMを、THP1細胞(LPSの有無により処理)から調製したライゼートと2時間インキュベートした。更にサンプルを遠心分離し、相互作用した全てのタンパク質と共に、グルタチオンビーズ上のGST−TRAMをプルダウンした。更に[γ32P]ATPで30分間ビーズをインキュベートしてキナーゼアッセイを実施した。更にサンプルを10%のSDS−PAGEに供し、ニトロセルロース膜上に転写し、X線フィルムを使用して取り込まれた放射能を測定した。その結果、実際にTRAMがリン酸化され、すなわちこのリン酸化がLPS依存的であることを示した(図2a)。細胞が他の刺激(例えばPolyI:C)で処理されたときは、このリン酸化は起こらなかった(図2b)。
【0123】
N末端に最も近い4つのセリンを同定した。その後これらの4つのセリンを変異させた。この変異により、LPS依存性のリン酸化が見られなくなった(図2c)。
【0124】
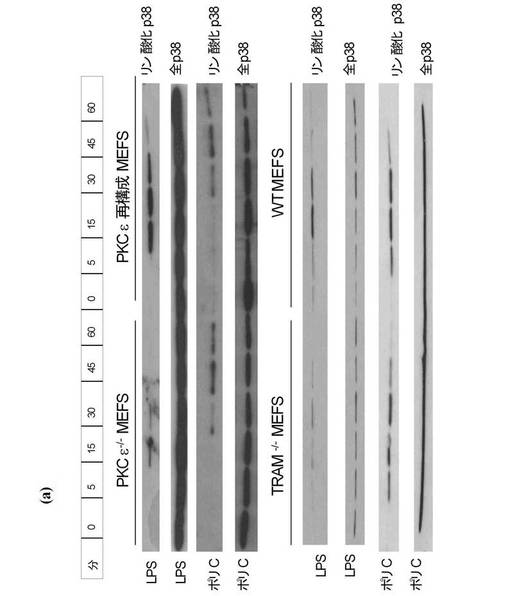
TRAMのリン酸化
プロテインキナーゼCはミリストイル化されたタンパク質(MARCKS)をリン酸化することを示したので、TRAMがPKCによってリン酸化されるか否かの試験のため、panPKC阻害剤のビスインドリルマレイミド(ビス)を使用した。上記のように、GST−TRAMをLPSによって処置したTHP1細胞から調製したライゼートとインキュベートし、グルタチオンビーズを使用してプルダウンした。ビスの量を増加させてビーズに添加し、1時間インキュベートし、更に上記のようにキナーゼアッセイを実施した。ビスによりTRAMのLPS依存性のリン酸化が阻害された(図2c)。これは、PKCキナーゼファミリーのメンバーがLPSに応答するTRAMのリン酸化の原因となることを強く示唆するものである。
【0125】
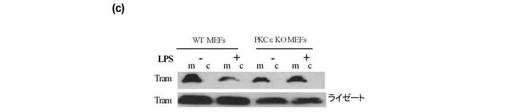
THP1ライゼートの免疫減少を、プロテインGビーズ(シグマ社)に結合させたPKCε特異抗体(サンタクルス社)とライゼートとをインキュベートすることによって実施した。特異性をチェックするコントロールとしてPKCζ抗体を用いた。抗体を遠心分離して除去し、ライゼートからPKCε又はPKCζを除去した。コントロールIgG抗体も使用した。これらのライゼートを更に上記と同様にGST−TRAMでインキュベートし、キナーゼアッセイを実施した。図2eは、ライゼートからのPKCεの除去によりTRAMのリン酸化が行われなくなることを示す。これは、PKCεがLPSに応答してTRAMをリン酸化することを示唆する。PKCζの除去はTRAMのリン酸化に影響を及ぼさず、PKCζがTRAMをリン酸化しないことを示唆する。この理論は、組換えPKCε(rePKCε)(Calbiochem社)がGST−TRAMをリン酸化したという事実(図2g)、並びにPKCε欠損MEFsから調製したライゼートではGST−TRAMをリン酸化できないという事実(図2f)によって更に裏付けられた。組換えPKCζはTRAMをリン酸化せず、重ねて特異性を示唆するものであった(図2g)。セリン16はPKCεリン酸化の候補であるため、アラニンにセリン16を変異させたところ、TRAMのLPS依存性のリン酸化が幾分損なわれることが明らかとなった(図3a)。
【0126】
位置16のセリンがマウスにおいて保存されている4つのセリンのうち唯一のものであるため、このセリンを変異させた。その結果、TRAMのLPS依存性のリン酸化が幾分損なわれた(図3a)。
【0127】
TRAMのこのリン酸化が適切な機能にとり必須か否かを試験するため、位置16のセリンをアラニンに変異させ、この変異体におけるNF−κB及びIRF3経路を機能させる能力を解析した。WT−TRAM又はSer16−TRAMを、NF−κB又はISRE−ルシフェラーゼレポーター遺伝子と共にHEK293細胞にトランスフェクションした。前述のように、TRAMはNF−κB及びISREルシフェラーゼを機能させることができる。しかしながら、セリン16が変異した場合、TRAMはいずれの経路も活性化できなくなった(図3b)。
【0128】
セリン16の変異はまた、NF−κB及びISRE経路を刺激するLPSの能力を低下させた(図3c)。この変異体は、これらの経路上でドミナントネガティブとして作用することが明らかとなった。PKCによるTRAMのリン酸化が適正な機能にとり重要であるという更なる証拠として、PKC阻害剤(ビスインドリルマレイミド)がNF−κB経路を活性化させるTRAMの能力を阻害するが、MyD88に影響を及ぼさないという事実(図3d)が挙げられる。この証拠は、セリン16が適切に機能するためには、PKCεによってTRAMがリン酸化されることが必要であることを示唆する。
【0129】
セリン16残基のリン酸化がTRAMの膜からの移動にとり十分であることを確認するため、セリン16残基をグルタミン酸に変異させた。得られる変異体(ser16Glu)は明確なグリコシル化コントロールとして機能し、すなわちグルタミン酸残基がグリコシル化された状態であるとき、セリン残基を模倣した。この変異により膜に存在するTRAMの量の重要な減少を引き起こし、セリン16上のTRAMのリン酸化が膜からのTRAMの減少を引き起こすことを示唆している(図8a、レーン3と1を比較)。
【0130】
LPSで処理したTHP1細胞の内因性のTRAMの減少も検出した(図8b)。最後に、PKCε−/−MEFsFLAG−TRAMは、LPS刺激によっても膜から消失しなかった(図8c)。この試験結果は、PKCεによるセリン16上のTRAMのリン酸化が、TRAMの膜からの消失に必要であることを示唆する。
【0131】
内因性のTRAMは、セリン16にリン酸化される
ホスホセリン抗体を使用して、過剰発現するTRAMのリン酸化の試験を行った。HEK293−TLR4細胞をFLAG−TRAMでトランスフェクションし、LPSによって刺激した。FLAG−TRAMを細胞から免疫沈降させ、ライゼートを抗ホスホセリン抗体でブロッティングした。図4aから解るように、30分間のLPSで処理した細胞において、TRAMのセリンリン酸化の増加が示された。
【0132】
内因性のTRAMがPKCεによってセリン16にリン酸化されるかどうかを立証するために、アミノ酸16でセリンをホスホセリンに置換したTRAMのアミノ酸7〜21の配列を有する合成ペプチドを用い、抗体を作製した。
【0133】
LPSで処理したTHP1細胞から調製したライゼートをイムノブロットした結果、15分後にセリン16上のTRAMリン酸化が観察され、45分でピークに達することが示された(図4b、レーン1〜6)。LPSによる刺激の1時間前に、PKC阻害剤のビスでこれらの細胞をインキュベーションした結果、TRAMのリン酸化が妨げられた(図4b、レーン7〜12)。15分間及び30分間LPSで処理した、PKCεによって再構成されたPKCε−/−MEFs細胞からのライゼートのイムノブロットの結果、TRAMの正しい分子量のバンドが確認された(図4c、上部パネル、レーン9及び10)。効果が45分(レーン11)で減衰し、60分(レーン12)に不明確となったことから、リン酸化はこの細胞タイプの場合THP1細胞よりも初期に生じることが解った。ライゼート中の全TRAMのレベルは、実験期間中一定であった(図4c、第二パネル)。重要なことに、バンドはPKCε−/−MEFsから調製したライゼートにおいて検出されず(図4c、上部パネル、レーン1〜6)、TRAM欠損細胞ではバンドは検出されなかった(示されないデータ)ことから、抗体の特異性が証明された。最後に、TLR2リガンドであるMALP2又はTLR3リガンドであるポリI:CによるPKCεを発現するMEFs細胞の30分間の処理では効果が表れなかった(図4d)。
【0134】
TRAM及びPKCε両方は、LPSシグナル伝達の遂行にとって不可欠である
更にPKCε−/−MEFsのTRAM依存性のシグナル伝達を試験した。TRAM−/−MEFs及びPKCε−/−MEFsにおける、LPSで処理した細胞のp38のリン酸化は、それらの対応する野生型MEFsと比較して著しく減少していた(図5a上部パネル、レーン9〜11とレーン3〜5を比較)。重要なことに、PKCε−/−MEFs又はTRAM−/−MEFsでは、ポリI:Cに応答したp38のリン酸化の減少が見られなかった(図5a、パネル3、右側と左側を比較)。LPSによるIRF3の活性化を、その二量体化を指標として試験した。LPSによって誘導されるIRF3の二量体化は、PKCεを発現しているMEFsと比較し、PKCε−/−MEF細胞では減少していた(図5b、レーン2及び3をレーン4及び5と比較)。ポリI:Cに応答したPKCε−/−MEFのIRF3二量体化の減少が見られなかった(図5c、レーン2及び3をレーン4及び5と比較)。Yamamotoらが示すように、TRAM欠損MEFsにおいて、この反応はLPS(ポリI:Cでない)への応答に関して損なわれていた。IRF3と連結されたレポーター遺伝子の活性化を試験した結果、LPS処理されたPKCε−/−MEFsでは損なわれたが、ポリI:Cを処置されたPKCε−/−MEFsでは正常だった(図5c、右パネル)。同様の結果がTRAM欠損MEFsにおいても得られた。更にTRAM経路の開始としてのRANTESの誘導を評価した。TRAM−/−MEFsにおいては、LPS刺激に応答して生じたRANTESのレベルが、対応する野生型MEFsと比較して激減した(図5d、左パネル)。ポリI:Cに応答するRANTES産生のレベルは、影響を受けなかった(図5d、右パネル)。重要なことに、この反応はPKCε−/−細胞においても損なわれた。図5e(左パネル)に示すように、LPSによるRANTESの誘導は、PKCε発現細胞と比較して損なわれた。両方の細胞タイプを比較したが、ポリI:Cへの反応の違いは見られなかった(図5e、右パネル)。
【0135】
RANTESの産生がTRAMのシグナリング経路と関連するマーカーであるため、PKCε−/−細胞及びTRAM−/−MEFs細胞におけるその産生の喪失は更に、TRAMがプロテインキナーゼCイプシロンによってリン酸化されなければならず、このリン酸化がその機能にとって重要であるという考察を更に支持するものである。
【0136】
TRAMS16Aは、TRAM欠損細胞のシグナル伝達を完全に再構成できない
PKCε−/−細胞のLPS処理に続くTRAM依存性の反応の顕著な喪失、及びポリI:Cで処理された細胞においては同じ反応が損なわれなかったことは、TRAMS16Aによる損なわれたシグナル伝達と組み合わせて考察すると、PKCεによるTRAMのリン酸化が、TRAMの機能において不可欠であることを強く示唆した。更なる証拠を提供するために、TRAM欠損MEFs細胞のシグナル伝達を再構成するTRAMS16Aの能力を検討した。LPSを有する野生型MEFsの処理によりRANTES産生が誘導されたが、一方ではLPSによるTRAM−/−MEFsの処理によってはRANTES産生がほとんど誘導されなかった(図6a)。TRAM欠損細胞での反応は、野生型TRAMで再構成することができた。重要なことに、TRAMS16Aはシグナリングを再構成する能力が低かったが、同様に、LPS刺激によるTRAM欠損細胞でのp38のリン酸化が、WT−TRAMの過剰発現によって再構成された。TRAMS16Aは、このシグナルを再構成することができなかった(図6b)。したがって、プロテインキナーゼCイプシロンによるセリン16のリン酸化が、LPS刺激によるTRAMの通常の機能にとり必要であると結論付けることができる。
【0137】
LPS刺激により、膜のTRAMのレベルは減少する
TRAMのミリストイル化により膜局在が促進される。特定のミリストイル化されたタンパク質がリン酸化により膜から解離することが知られている。PKCεによるTRAMのリン酸化によってTRAMの再分布が生じるか否かを更に試験した。
【0138】
FLAG−TRAM/pcDNA3.1をHEK293−TLR4細胞にトランスフェクションし、これらの細胞を更に30分間、LPSの有無により刺激した。細胞を膜画分及びサイトソル画分に分離した。膜画分に存在するTRAMの量はLPS刺激により減少し、すなわちTRAMが膜から消失することを示唆する(図7a、上部パネル、レーン1とレーン3を比較)。TRAMはサイトソル画分では検出できなかった。これは分解によるものではない。なぜなら、細胞可溶化物中のTRAMのレベルは一定だからである(図7a、第三パネル)。膜からのTRAMの減少はPKCε依存性であった。その理由は、LPS刺激の1時間前にPKC阻害剤のビスを添加することにより、LPS刺激の後でも膜へのFLAG−TRAMへの残留が見られたからである(図7a、レーン7)。TRAMS16Aの減少を観察した場合、膜からの消失は観察されなかった(図7a、第二パネル)。LPSで処理したTHP1細胞の内因性のTRAMの減少も検出した(図7b)。最後に、PKCε−/−MEFsでは、FLAG−TRAMはLPS刺激によっても膜から消失しなかった(図7c)。この実験結果は、PKCεによるセリン16上のTRAMのリン酸化が、TRAMの膜からの消失に必要であることを示唆するものである。
【0139】
(本発明の要約)
TRAMは、TLR4及びTrif間の架橋アダプタとして作用し、LPSによって活性化するシグナリングカスケードにおいて不可欠な役割を果たす。TRAMはミリストイル化され、それにより形質膜と相互作用する。TRAMはまたリン酸化される。
【0140】
LPSに応答してTRAMはリン酸化され、それはここで記載したキナーゼアッセイを使用してインヴィトロで測定されうる。このリン酸化はTRAMの通常の機能に不可欠で、静電的なスイッチングに関与し、TRAMを膜から移動させる。
【0141】
本明細書において言及される全ての文書は、参照によって本願明細書に援用される。本明細書に記載されている実施態様に対する本発明の範囲内における様々な修正や変更は、当業者にとって自明である。本発明を具体的な好ましい実施態様に関して記載したが、本発明の特許請求の範囲は、かかる具体的な実施例にのみ限定されるべきではない。本発明の実施に係る、実際に記載されている様式に対する様々な改良は当業者にとって自明であり、本発明に包含されている。
【技術分野】
【0001】
本発明はTRAM(Trif関連アダプター分子)のリン酸化の測定及び定量化のための新規なアッセイに関する。アッセイは特に、Toll様受容体4(TLR4)の活性化のモニタリングに用いてもよく、かかるアッセイはその活性の調節物の同定に有用性を発揮する。本発明の更なる態様は、プロテインキナーゼCイプシロンによってTRAMのリン酸化をブロック、阻害又は競争的に阻害する分子を測定するためのアッセイ方法の提供に関する。
【背景技術】
【0002】
Toll様受容体(TLR)スーパーファミリーは、病原微生物の侵入の認識、及び免疫応答の開始において中心的な役割を果たす。ヒトTLRsは現在までに10種類同定されている。各々はシグナリングカスケードの活性化につながる異なる病原関連分子パターン(PAMP)を認識し、更に次々に転写制御因子NF−κB、更には分裂促進因子活性プロテインキナーゼ(MAPKs)、p38、c−jun、N末端キナーゼ(JNK)及びp42/44(非特許文献1及び2に開示される)を活性化させる。TLR3及びTLR4も転写制御因子(IFN制御された因子3(IRF3))の活性化を介して他の経路を活性化させ、それはインターフェロン感受性反応要素(ISRE)と結合し、IFN−β(非特許文献3)を含む遺伝子サブセットを誘導する。TLRsはより大きいスーパーファミリーのメンバーであり、インターロイキン1受容体(IL−1R)/TLRスーパーファミリーとも呼ばれ、IL−1R1サブグループ及びTIR領域含有アダプターサブグループを包含する。3つの全サブグループは細胞質のToll/IL−1受容体(TIR)領域を有し、それはシグナリングにとって重要である。TLRsは細胞外のロイシンリッチリピートを有し、IL−1R1サブグループは細胞外の免疫グロビン領域を有する。アダプター分子は細胞質に存在し、細胞外領域を含まない。
【0003】
上記のように、各TLRはそれぞれ異なるPAMPを認識する。最初に同定されたTLRはTLR4であり、それはグラム陰性細菌のリポ多糖(LPS)の認識にとり重要である(非特許文献4、5)。TLR1及び6と結合したTLR2は、それぞれジアシル−及びトリアシルリポペプチドを認識する(非特許文献6)。TLR3はdsRNAに認識し(非特許文献7)、TLR5は細菌フラジェリンを認識し(非特許文献8)、TLR9は非メチル化CpGのモティーフを認識する(非特許文献9)。一度TLRがPAMPを認識すると、次のシグナリング経路を活性化させるために、TIRドメイン含有アダプターを補充する必要がある。これらのアダプターで最初に同定されたものはMyD88である。それはTLRとIL−1Rのシグナリングにおいて重要な役割を果たし(10、11、12)、それにより生じるシグナリングカスケードが詳細に研究されている(非特許文献13)。実験結果から、TLR3を除く全てのTLRsからのシグナリングに関係していることが示唆されている。MyD88欠損マウスでは、IL−1刺激への応答、又はTLR2、TLR5及びTLR9の刺激が行われない(非特許文献11)。TLR4の場合、遅い反応ではあるが、NF−κB及びMAPKの活性化が観察されている。更に、樹状細胞成熟の誘導及び転写制御因子IRF3の活性化は、MyD88欠損マウスにおいても影響を受けなかった。これは、TLR4が完全にその反応を活性化させるにはMyD88以外の因子を必要とし、この反応が2つのカテゴリー(MyD88依存性の反応と、MyD88非依存性の反応)に分類できることを示唆する。NF−κB及びTNF産生はTLR3に応答して損なわれず、それはMyD88がTLR3シグナリングとは無関係であることを示唆する。
【0004】
次に同定されたアダプターはMal(MyD88 adaptor−like)であり、それはTIRAP(TIRドメイン含有アダプタータンパク質)とも呼ばれている(非特許文献14、15)。最初は、これがTLR4へのMyD88非依存性の反応を媒介するアダプターであると考えられていたが、Mal欠損マウスによる研究の結果、これは正しくなく、またMalとMyD88は協働して作用し、MyD88依存性の経路を活性化させることが明らかとなった。MyD88欠損マウスと同様に、Mal欠損マウスではLPSに応答するNF−κB及びMAPKの遅い活性化を示し、一方樹状細胞成熟及び転写制御因子IRF3の活性化への影響は見られなかった(非特許文献15、16)。Mal欠損マウスは通常、TLR5、TLR7、TLR9、IL−1及びIL−18のリガンドに反応し、それはMyD88がこれらの受容体に必要とされる唯一のアダプターであることを裏付けるものである。TLR3シグナリングはMal欠損マウスにおいて通常観察され、それはMalもMyD88もこの経路に関係しないことを示唆するものである。興味深いことに、TLR2によって活性化するシグナリング経路はMal欠損マウスにおいて完全に喪失され、それはMal及びMyD88の両方がこの経路(16)の活性化に必要とされることを示唆するものである。
【0005】
Trif(TIRドメイン含有アダプター誘導インターフェロン−β)は第3に同定されたアダプターである(非特許文献17、18)。それはTIR含有アダプター分子−1(TICAM−1)とも呼ばれている。TrifによるNF−κB活性化(過剰発現の場合)はMal又はMyD88による場合よりもかなり小さい程度であるが、IFN−βに関しては非常に強い活性化因子として機能する(非特許文献17)。すなわち、それがMyD88非依存性の経路に関係しうることを示唆し、またTrif欠損マウスによりこの点が証明されている(非特許文献19)。LPSに応答するNF−κB活性化はこれらのマウスにおいてはほぼ正常であったが、これらの細胞でTrif及びMyD88が欠損するとき、LPSへのNF−κBの反応は完全に喪失された。Trif欠損マウスにおいてLPSに応答するIRF3の活性化が完全に喪失され、これもTrifがTLR4によって活性化するMyD88非依存性の経路に関係することを示唆するものである。TLR3によるIRF3の活性化はTrif欠損細胞においても喪失され、またNF−κBの活性化も顕著に損なわれ、それらはTrifがTLR3により用いられる唯一のアダプターであることを示唆するものである。
【0006】
またTrifが直接TLR4と結合できないことが発見され(非特許文献18)、架橋アダプターがTLR4への結合に必要であることを示唆するものである。その架橋アダプターは、現在幾つかの研究グループによって発見されており、TRAM(Trif関連アダプター分子)(非特許文献20)、TICAM−2(TIR含有アダプター分子−2)(非特許文献21)又はTIRP(TIRドメイン含有タンパク質)(非特許文献22)と呼ばれている。
【0007】
TRAMは直接TLR4と結合し、他のTLRsとは結合しない(21)。TRAMの過剰発現により、MyD88非依存的に、IRF3、IRF7及びNF−κBが穏やかに誘導された。TRAMのドミナントネガティブは、LPSによるNF−κB及びIRF3の活性化を阻害したが、TLR3リガンド、ポリ(I:C)によるこれらの転写制御因子の活性化にも影響を及ぼさなかった。TRAMの過剰発現(Trifと共に)により、IRF3の核への転座が誘導された(非特許文献20)。ドミナントネガティブ形のTrifは顕著にTRAMの活性を抑制してNF−κB及びIFN−βを活性化させ、一方ではMyD88及びMalドミナントネガティブ変異体では効果を有しなかった。
【0008】
TRAMはTrifノックダウンRAW細胞において機能することができず、すなわちTRAMがTLR4経路のTrifの上流で機能することを示唆するものである。TRAM欠損マウスの作製によりこの理論の重要性がますます増加した(非特許文献23)。これらのマウスを用いた研究により、TRAMはTLR4に応答するMyD88非依存性の経路の活性化にとって不可欠であり、他のTLR経路には関係しないことが示された。
【先行技術文献】
【非特許文献】
【0009】
【非特許文献1】Janeway,C.A.,Jr.,Medzhitov,R.,2002.Annu Rev Immunol 20,197−216.
【非特許文献2】Dunne A.,O’Neill,L.A.,2003a.Sci STKE 2003,re3.
【非特許文献3】Doyle,S.ら,(2002)Immunity 17,251−63.
【非特許文献4】Poltorak A.ら、(1998)Science 282,2085−8.
【非特許文献5】Qureshi S.T.ら、(1999)J Exp Med 189,615−25.
【非特許文献6】Takeda K.,Takeuchi O.及びAkira S.(2002)J Endotoxin Res 8,459−63.
【非特許文献7】Alexopoulou L.ら、(2001) Nature 413, 732−8.
【非特許文献8】Hayashi F.ら、(2001)Nature 410,1099−103.
【非特許文献9】Hemmi H.ら、(2000)Nature 408,740−5.
【非特許文献10】Hemmi H.ら、(2002)Nat Immunol 3,196−200.
【非特許文献11】Adachi O.ら、(1998)Immunity 9,143−50.
【非特許文献12】Takeuchi O.ら、(2000)J Immunol 164,554−7.
【非特許文献13】McGettrick A.F.及びO’Neill L.A.(2004)Mol Immunol 41,577−82.
【非特許文献14】Fitzgerald,K.A.ら、(2001)Nature 413,78−83.
【非特許文献15】Horng,T.ら、(2001)Nat Immunol 2,835−41.
【非特許文献16】Horng,T.ら、(2002)Nature 420,329−33.
【非特許文献17】Yamamoto,M.ら、(2002)J Immunol 169,6668−72.
【非特許文献18】Oshiumi,H.ら、(2003)Nat Immunol 4,161−7.
【非特許文献19】Yamamoto,M.ら、(2003) Science 301,640−3.
【非特許文献20】Fitzgerald,K.A.ら、(2003)J Exp Med 198,1043−55.
【非特許文献21】Oshiumi,H.ら、(2003)J Biol Chem 278,49751−49762.
【非特許文献22】Bin,L.H.ら、(2003)J Biol Chem 278,24526−32.
【非特許文献23】Yamamoto,M.ら、(2003) Nat Immunol 4,1144−50.
【非特許文献24】Thelen M.ら、(1991)Nature 351,320−2.
【非特許文献25】Aderem A.A.ら、(1988) Nature 332,362−4.
【非特許文献26】Wu W.C.ら、(1982)Proc Natl Acad Sci USA 79,5249−53.
【非特許文献27】Rozengurt E.ら、(1983)Proc Natl Acad Sci USA 80,7244−8.
【非特許文献28】Rozengurt E.及びSinnett−Smith J.(1983)Proc Natl Acad Sci USA 80,2936−40.
【非特許文献29】Graff J.M.ら、(1989)Science 246,503−6.
【非特許文献30】Rosen A.ら、(1990)J Exp Med 172,1211−5.
【非特許文献31】Matsubara M.ら、(2003)J Biol Chem 278,48898−902.
【非特許文献32】Takasaki A.ら、(1999)J Biol Chem 274,11848−53.
【非特許文献33】Hayashi N.,ら、(2000)Protein Sci 9,1905−13.
【発明の概要】
【発明が解決しようとする課題】
【0010】
本発明者による鋭意研究の結果、驚くべきことに、LPSのTLR4受容体(Toll様受容体4)への結合に続き、アダプター分子TRAMがプロテインキナーゼCイプシロンによって急速にリン酸化されることを見出した。すなわちTRAMがセリン16残基の部位でプロテインキナーゼCイプシロンによってリン酸化されることを確認した。TLR4の活性化を測定する際、具体的にはTLR4受容体を介するLPSシグナリングが異なる環境において適切に機能するどうかを測定する際、TRAMのリン酸化をモニターするアッセイ方法は有用なツールとなりうる。
【課題を解決するための手段】
【0011】
本発明の第1の態様は、TRAM(Trif関連アダプター分子)の活性化状態を測定する方法の提供に関し、当該方法は、TRAMを含有する細胞サンプルを提供するステップと、TRAMのリン酸化をモニターするステップを含んでなる方法であり、TRAMのリン酸化が行われない場合は、そのTRAMが活性を有しないことの指標となる。
【0012】
本願明細書における「活性化状態」の用語は、TRAMが細胞シグナリング経路に関係しているか否かを指す時に用いる。すなわち、活性TRAMはTRAMのセリン16残基のリン酸化により生じ、TRAM(活性化状態)はTLR4受容体により媒介されるシグナリングに関与する。
【0013】
アッセイの好ましい実施態様では、TRAMはリン酸化に適した条件下でキナーゼと接触する。一実施例において、当該キナーゼはプロテインキナーゼCイプシロンであってもよい。
【0014】
アッセイの更なる実施態様では、TRAMリン酸化をコントロールとの比較によりモニターし、それによりリン酸化を測定する。
【0015】
更なる態様では、TRAMリン酸化を、細胞膜領域内におけるTRAMの存在との比較により測定する。TRAMのリン酸化によって、細胞膜からのTRAMの移動が生じる。
【0016】
したがって、本発明のこの態様の更なる実施態様は、細胞膜に存在するTRAMの存在又はレベルを測定することによってTRAMのリン酸化状態を測定する方法であって、膜領域内に存在するTRAMの、膜からの移動による喪失又は減少が、リン酸化状態のTRAMの指標となる方法の提供に関する。
【0017】
一実施態様では、TRAMのリン酸化の測定方法は、細胞膜中のTRAMの存在及び/又はレベルを測定及び/又定量化する膜減少アッセイを含んでなる。
【0018】
理論に束縛されずないが、発明者は、リン酸化に続く細胞膜からのTRAMの移動が、TRAMのセリン16残基のリン酸化によって陰電荷を帯電し、膜からTRAMを移動させることとなる斥力が生じるものと予想している。
【0019】
本願明細書では、TRAM(Trif関連アダプター分子)と称する分子は、TICAM−2(TIR含有アダプター分子−2)(非特許文献21)又はTIRP(TIRドメイン含有タンパク質)(非特許文献22)と称してもよい。
【0020】
TRAMのリン酸化状態を測定するための上記のアッセイは、TRAM活性化を調節できる候補薬剤の同定のために更に変更してもよい。
【0021】
したがって、本発明の本態様の更なる実施態様は、TRAM活性の調節物質の同定方法の提供に関し、前記方法は、(i)TRAMを含有する第1及び第2の細胞サンプルを準備するステップと、(ii)TRAMの候補調節物を有する前記第1のサンプルを接触させるステップと、(iii)リン酸化に適した条件下でキナーゼと前記第1及び第2のサンプルを接触させるステップと、(iv)TRAMのリン酸化状態をモニターして、前記第1のサンプルと前記第2のサンプルとでTRAMのリン酸化を比較するステップを含んでなり、前記第1のサンプルと前記第2のサンプルとの間のTRAMリン酸化の差異を基に、TRAM活性の調節物として候補調節物質を同定することを特徴とする。
【0022】
一実施態様では、TRAMをリン酸化するキナーゼは、プロテインキナーゼCイプシロンである。
【0023】
本発明の本態様の上記のアッセイに従って同定される1つ以上の調節物質は、ペプチド分子又は非ペプチド分子(例えば化学物質又は医薬用物質)であってもよい。調節物質がペプチドである場合、抗体、抗体フラグメント又は同様の結合活性を有する分子であってもよい。更に、調節物質が抗体である場合、好ましくは単クローン抗体である。
【0024】
本発明の更なる態様は、TLR4受容体で媒介されるシグナリングを調節する薬物の調製に関する前述の態様に従って同定された調節物の使用に関する。TLR4で媒介されるシグナリングの一実施態様では、受容体は上方制御される。更なる、好ましい実施態様では、TLR4受容体で媒介されるシグナリングは下方制御される。
【0025】
更に他の本発明の態様は、TRAMのリン酸化を測定するキットの提供に関し、当該キットは参照サンプル、TRAMのリン酸化を測定するための手段、及び本発明の第1の態様の方法を使用したいずれかの本発明のアッセイを実施するための使用説明書を含んでなる。
【0026】
TRAMのリン酸化状態を評価し、それを調節するのに有用な組成物の同定のためのアッセイ方法を記載した上記の本発明の態様に加えて、本発明は更に、リガンド、特にLPSによるTLR4受容体の活性化を評価するアッセイにおいて有用性を発揮する。かかるアッセイは、TLR4受容体を介して選択的にシグナリングを上方制御又は下方制御することができる化合物の同定及び開発において有用性を発揮しうる。かかる化合物は、TLR4結合及び最も具体的にはLPS(リポ多糖)によるTLR4結合から生じるシグナリング経路の調節物質として有用性を発揮する。
【0027】
第4の本発明の態様は、リガンドによるTLR4活性化を検出するアッセイ方法の提供に関するものであり、当該アッセイは、TLR4発現細胞を含む細胞サンプルを準備するステップと、前記細胞をリガンドと接触させるステップと、TRAMのリン酸化を検出するステップを含んでなり、TRAMのリン酸化をTLR4へのリガンドの結合の指標とすることを特徴とする。
【0028】
好ましくは、リン酸化のレベルを、コントロール試料(例えばリガンドに曝露されない同種の細胞)のそれと比較してもよい。あるいは、試験サンプルを、周知の、予め定められた基準値をコントロールとして比較してもよい。
【0029】
リン酸化を測定することによって、TLR4受容体との相互作用を介したTRAMのリン酸化を調整する候補薬剤を同定することが可能となる。
【0030】
本発明の更なる態様は、TLR4受容体で媒介されるシグナリングを調節する薬物の調製に係る、前述の本発明の態様に従って同定されるリガンドの使用に関する。一実施態様では、TLR4受容体を介するシグナリングは上方制御される。
【0031】
本発明の第4の態様の更なる実施態様は、TLR4受容体のアゴニストを同定するためのアッセイに関し、当該アッセイは、TLR4発現細胞を含む細胞サンプルを準備するステップと、細胞を試験化合物に曝露するステップと、TRAMのリン酸化を検出するステップを含んでなり、TRAMのリン酸化レベルの上昇を、試験化合物のTLR4受容体への結合に続く活性化の指標とすることを特徴とする。
【0032】
一実施態様では、TLR4受容体のアゴニストは、TLR4受容体が媒介するシグナリングを誘導又は上方制御する。
【0033】
本発明の更なる態様は、TLR4受容体で媒介されるシグナリングを調節する薬剤の調製に係る、前述の本発明の態様に従って同定された化合物の使用に関する。一実施態様では、TLR4受容体が媒介するシグナリングは上方制御される。
【0034】
本発明のこの態様の、更に別の実施態様は、TLR4受容体のアンタゴニストを同定するアッセイ方法に関し、当該アッセイ方法は、TLR4発現細胞を含む細胞サンプルを準備するステップと、細胞を試験化合物に曝露するステップと、TRAMのリン酸化を検出するステップを含んでなり、当該方法は、試験化合物が存在する場合における、試験化合物が存在しない場合と比較したTRAMのリン酸化の減少が、当該試験化合物がアンタゴニストであることの指標となることを特徴とする。
【0035】
一実施態様では、TLR4受容体のアンタゴニストはTLR4受容体によって媒介されるシグナリングを妨害又は下方制御する。
【0036】
一実施態様では、上記アッセイ方法は更に、細胞を試験化合物に曝露する前にアゴニストに曝露する処理を含んでなる。
【0037】
本発明の更なる態様は、TLR4受容体で媒介されるシグナリングを調節する薬剤の調製に係る、前述の本発明の態様に従って同定される化合物の使用に関する。一実施態様では、TLR4受容体で媒介されるシグナリングは上方制御される。
【0038】
更に他の本発明の態様は、TLR4受容体の活性を測定するキットの提供に関し、当該キットはサンプル、及び本発明の第4の態様の方法を実施するための使用説明書を含んでなる。
【0039】
発明者は鋭意研究の結果、プロテインキナーゼCイプシロン(PKCε)が、TRAMをリン酸化するキナーゼであることを見出した。プロテインキナーゼCイプシロンによるリン酸化の阻害は、NK−κB及びIFN−βを活性化させるTRAMの能力を弱める。
【0040】
したがって、第6の本発明の態様は、プロテインキナーゼCイプシロンのインヒビターとして作用する化合物を同定するためのアッセイ方法を提供し、当該方法は、候補化合物を準備するステップと、候補化合物をプロテインキナーゼCイプシロンと接触させるステップと、プロテインキナーゼCイプシロンのTRAMをリン酸化する能力の有無を測定するステップを含んでなり、TRAMのリン酸化が行われない場合、それを候補化合物によってなされたプロテインキナーゼCイプシロンの活性のブロッキングの指標とすることを特徴とする。
【0041】
直接プロテインキナーゼCイプシロン分子を阻害することにより、TRAMのリン酸化が妨げられるだけでなく、プロテインキナーゼCイプシロンの他の細胞内における活性も阻害される。しかしながら、発明者は、プロテインキナーゼCイプシロンが結合するTRAMの特異的なドメインを同定し、これがプロテインキナーゼCイプシロンの他の細胞内での活性の阻害を引き起こすことのない、TRAMのリン酸化の選択的阻害の可能性を開拓することを見出した。
【0042】
本発明の本態様の別の実施例は、プロテインキナーゼCイプシロンによるTRAMのリン酸化を防止する化合物を同定するためのアッセイであって、当該アッセイは、候補化合物を準備するステップと、候補化合物をTRAMと接触させるステップと、TRAMを、リン酸化の開始に適する条件下でプロテインキナーゼCイプシロンに曝露するステップと、TRAMのリン酸化の有無を測定するステップを含んでなり、リン酸化が行われないことを、プロテインキナーゼCイプシロンとTRAMの間での相互作用のブロッキングの指標とすることを特徴とする。
【0043】
好ましい実施態様では、当該化合物はプロテインキナーゼCイプシロンによって選択的にTRAMのリン酸化を阻害する。
【0044】
一実施態様では、当該方法は、TRAMに存在するセリン16の残基に対応するドメイン中の部位又は領域で、化合物がTRAMと結合して、そのセリン残基がプロテインキナーゼCイプシロンによりリン酸化されるのを防止する能力を測定するステップを含んでなる。
【0045】
更なる様々な態様では、本発明はスクリーニング及びアッセイ方法、並びにそれにより同定された物質に関する。
【0046】
本発明のアッセイを用いて同定される新規な化合物は、更に別の本発明の態様を構成する。かかる化合物又は調節物質は、医薬組成物中に添加してもよい。
【0047】
調節物質、又は本発明のアッセイで同定された化合物は、ペプチド又は非ペプチド分子(例えば化学物質又は医薬物質)であってもよい。調節物質質がペプチドである場合、抗体、抗体フラグメント又は同様に結合するフラグメントであってもよい。更に、調節物質が抗体である場合、好ましくは単クローン抗体である。
【0048】
TRAMに対して特異性を有する単クローン抗体、抗体フラグメント又は同様の結合分子(プロテインキナーゼCイプシロンがTRAMのリン酸化を促進するために結合する領域におけるセリン−16残基に特に結合する、又は完全若しくは部分的にブロッキングする)は、TRAMのリン酸化の阻害に有用性を発揮し、それによりTLR4受容体へのLPSの結合に続く、その下流側への活性シグナリング伝達の促進を防止する能力を有する。
【0049】
したがって、本発明の更なる態様は、抗原結合ドメインを含む特異的な結合メンバーの提供に関し、当該抗原結合ドメインはTRAMのセリン−16残基に対する特異性を有する。
【0050】
一実施態様は、本発明はTRAMのセリン−16残基に特異的に結合するか又は結合を防ぐイムノグロブリンの提供に関する。
【0051】
更に他の実施態様は、プロテインキナーゼCイプシロンによるTRAMのセリン−16残基のリン酸化を防止するイムノグロブリンの提供に関する。
【0052】
また、本発明の本態様には、TRAMと特異的に結合してプロテインキナーゼCイプシロンによるリン酸化を防止する結合メンバーが包含される。
【0053】
内毒素は、リピドAと多糖からなるリポ多糖(LPS)複合体を含んでなる。LPSはTLR4受容体と結合し、これが適当な免疫応答を誘発する下流のシグナリングカスケードを生じさせる。
【0054】
敗血症及び敗血症性ショックなどのLPSの媒介又は内毒素の媒介による症状によりしばしば死に至る。したがって、TLR4の媒介による免疫応答経路を下方制御又は阻害する方法は、LPSの媒介による症状(例えば敗血症)の処理方法として望ましい。
【0055】
TLR4へのLPS結合に続いてTRAMがプロテインキナーゼCイプシロンによってリン酸化されるという観察から、本発明は、LPSと細胞内キナーゼとの間の相互作用を制御するためのポテンシャルルートを提供する。これにより、LPSシグナリングを無力化するための強力なメカニズムが提供される。
【0056】
したがって、本発明はLPSの媒介による症状の治療に使用できる。
【0057】
本発明のアッセイ、及び当該アッセイの使用によって解明されるTLR4シグナリング経路に対して生物学的重要性を有する化合物は、多くの医学的症状、最も具体的には内毒素及びLPSにより媒介される症状(例えば敗血症)の治療に対して特に有用性を発揮する。
【0058】
したがって、本発明の第7の態様は、内毒素により媒介される症状の治療用に適する化合物を同定するためのアッセイ方法に関し、当該アッセイは、候補化合物を準備するステップと、前記候補化合物をTRAMと接触させるステップと、候補化合物によるTRAMのリン酸化の有無を測定するステップを含んでなり、TRAMリン酸化の調節を、その化合物の有用性の指標とすることを特徴とする。
【0059】
一実施態様では、内毒素により媒介される症状は敗血症又は敗血症性ショックである。
【0060】
更に他の本発明の態様は、LPSにより媒介される症状の治療方法の提供に関し、当該方法はTRAMのリン酸化を阻害する処理を含んでなる。
【0061】
本発明の上記態様の好ましい一実施態様では、TRAMのリン酸化の阻害は、TRAMにプロテインキナーゼCイプシロンの結合を防ぐことによってなされる。
【0062】
一実施態様では、TRAMへのプロテインキナーゼCイプシロンの結合のブロッキングは、セリン16ドメインへのプロテインキナーゼCイプシロンの結合によるリン酸化を阻害する化合物によって促進される。
【0063】
更に他の実施態様では、当該化合物は、プロテインキナーゼCイプシロンがリン酸化するTRAMのセリン−16部位と直接結合する。あるいは、阻害剤はTRAM上の部位と結合し、セリン−16残基のリン酸化に必要となる、TRAM上の結合部位へのプロテインキナーゼCイプシロンのアクセスを防止する。
【0064】
TRAMの配列は、配列番号1:MGIGKSKINSCPLSLSWGとして定義される。セリン16は、配列中の最後のセリンである。
【0065】
あるいは、セリン−16残基のリン酸化に適切な部位において、TRAMへの結合に関してプロテインキナーゼCイプシロンと競合する分子の提供に関する。
【0066】
更に別の本発明の態様は、LPSにより媒介される症状(敗血症)に罹患する患者を治療する方法の提供に関し、前記患者にTRAMのリン酸化を阻害する分子を、治療上有効量で投与することを含んでなる。
【0067】
本発明の本態様の好ましい一実施態様では、当該分子はプロテインキナーゼCイプシロンによるTRAMのリン酸化を防止する。
【0068】
内毒素により媒介される症状は、特にLPSによって生じるものであり、しばしば発熱、白血球数の変化、播種性血管内凝固、低血圧、ショック及び死に至る。
【0069】
好ましい一実施態様では、内毒素により媒介される上記症状は敗血症である。
【0070】
本発明の更なる態様は、内毒素により媒介される症状の治療用薬剤の調製における、プロテインキナーゼCイプシロンの阻害剤の使用の提供に関する。
【0071】
したがって、本発明の更なる態様は抗原結合ドメインを含む特異的な結合メンバーの提供に関し、当該抗原結合ドメインはプロテインキナーゼCイプシロンに対する特異性を有する。
【0072】
一実施態様では、本発明は、プロテインキナーゼCイプシロンに特異的に結合するか又は結合をブロックするイムノグロブリンの提供に関する。
【0073】
更に他の本発明の態様は、敗血症の治療用薬剤の調製のための、プロテインキナーゼCイプシロンの阻害剤の使用に関する。
【0074】
更に別の本発明の態様は、内毒素により媒介される疾患(例えば敗血症)の治療用薬剤の調製のための、TRAMのリン酸化を防止する化合物の使用に関する。
【0075】
更に別の本発明の態様は、前述の本発明の態様のいずれか1つに従うアッセイ方法のいずれか1つによって同定される化合物を使用することを特徴とする、医学的症状の治療方法の提供に関する。
【0076】
TLR4受容体で媒介されるシグナリングは、内毒素によって媒介されるそれら以外の免疫応答と関連していてもよい。例えば、TLR4受容体を介したシグナル伝達は炎症性疾患に関係している場合もあり、それゆえ、TLR4受容体によるリガンド結合能力及びTLR4受容体の下流におけるシグナル伝達の調節は、炎症性疾患(例えば関節炎及びアテローム性動脈硬化)に影響を与えうる。若干の自己免疫性の症状では、TLR4受容体の候補リガンド又は調節物質は未だ同定されていない。しかしながら、このような場合では、炎症又は傷害を伴う組織によって産生される内因性の因子がTLR4受容体又はその関連するシグナリング経路に影響を及ぼすことが示唆される。
【0077】
したがって、更に他の本発明の態様は、炎症性又は免疫媒介性障害の治療又は予防に適する化合物を同定するためのアッセイ方法の提供に関し、当該アッセイは、候補化合物を準備するステップと、候補化合物をTRAMと接触させるステップと、候補化合物によるTRAMのリン酸化の有無を測定するステップを含んでなり、TRAMリン酸化の調節を、その化合物の有用性の指標とすることを特徴とする。
【0078】
好ましい実施態様では、当該免疫媒介性障害は関節炎又はアテローム性動脈硬化であってもよい。
【0079】
更に他の本発明の態様は、炎症性症状又は免疫媒介性障害の治療又は予防用の薬剤の調製における、プロテインキナーゼCイプシロンの阻害剤の使用に関する。
【0080】
更に別の本発明の態様は、炎症性又は免疫媒介性障害の治療又は予防用薬剤の調製における、TRAMのリン酸化を防止する化合物の使用に関する。
【0081】
更なる本発明の態様では、LPSによるTLR4受容体への結合に続くシグナリングに関連する症状の治療を必要とする患者を治療する方法であって、当該方法は本発明のアッセイのいずれか1つに従って同定される化合物の投与を含んでなる。
【0082】
他に必要としない限り、各本発明の態様の好ましい特徴は必要な変更を加えて他の態様の各々に適用してもよい。
【図面の簡単な説明】
【0083】
【図1】(a)TRAMのクローニングに使用した、TRAMの5’及び3’末端を標的とするプライマー。(b)得られるPCR生成物を幾つかのベクターにクローニングした。(c)特異的なプライマーを使用して、TRAMの最初の4つのセリンをアラニンに変異させ、並びに16番目のセリンのみをアラニンに変異させるため、部位特異的突然変異導入を実施した。
【図2a】(a)GST−TRAMは、LPSで処置されたTHP1のライゼートとの所定時間のインキュベーションによりリン酸化される。(b)細胞をポリI:Cで処理した場合、かかるTRAMのリン酸化は行われない。(c)TRAMの最初の4つのセリンをアラニンに変異させた場合、このLPS依存性のリン酸化は行われない。(d)LPSで30分間処理したTHP1細胞から調製したライゼートとのGST−TRAMプルダウンに続いて、サンプルを1時間panPKC阻害剤(ビスインドールマレイミド(ビス))の量を増加させつつインキュベートした。その結果、TRAMのリン酸化の減少が見られた。
【図2b】(e)GST−TRAMとのインキュベーションの前にPKCε抗体、PKCζ抗体又は免疫グロブリンGコントロール抗体を使用して、THP1ライゼートの免疫減少を実施した。ライゼートからPKCεの除去により、TRAMのLPS依存性のリン酸化が妨げられた。(f)PKCεを欠損するMEFsから調製したライゼートはGST−TRAMをリン酸化が行われず、一方、PKCεで再構成したMEFsではリン酸化が行われた。(g)組換えPKCε(rePKCε)又はPKCζ(rePKCζ)をGST−TRAMと15分間直接インキュベートし、その後キナーゼアッセイを実施した。rePKCεはWT−TRAMでリン酸化されたが、Malではされなかった。
【図3】(a)セリン16の位置をアラニンに変異させたとき、GST−TRAMはTHP1ライゼートとのインキュベーションによりもはやリン酸化され得ない。(b)pcDNA3.1のみ(eV)、WT−TRAM/pcDNA3.1及びSer16−TRAM/pcDNA3.1の全てを、96穴プレートに播種したHEK293細胞にトランスフェクションした。NF−κB又はISRE−ルシフェラーゼレポーター遺伝子、並びにRenillaルシフェラーゼ内部コントロールプラスミドを同様にトランスフェクションした。24時間後、リポータ遺伝子活性を測定し、データはコントロールのレベルに対する刺激(倍数表示)の平均値として表す。グラフは、WT−TRAMはNF−κB及びISRE経路を活性化できず、一方Ser16−TRAM変異体がどちらの経路も活性化できることを示す。
【図3c】同様の実験をHEK293−TLR4細胞において実施し、トランスフェクション後24時間において、細胞を6時間、LPS(1μg/ml)の有無においてインキュベートした。試験の結果、WT−TRAMがNF−κB及びISREを活性化させるLPSの能力を増加させ、一方Ser16−TRAM変異体がドミナントネガティブとして作用してNF−κB及びISREを刺激するLPSの能力を減少させることを示す。
【図3d】HEK293細胞を、pcDNA3.1、MyD88/pcDNA3.1又はWT−TRAM/pcDNA3.1、並びにNF−κB−ルシフェラーゼレポーター遺伝子及びRenillaルシフェラーゼ内部コントロールプラスミドでトランスフェクションする前に、1時間PKC阻害剤(ビスインドリルマレイミド(ビス))の有無によりインキュベートした。上記阻害剤はNF−κB経路を活性化させるMyD88の能力に影響を及ぼさなかったが、この経路を活性化させるTRAMの能力を阻害した。
【図4a】TRAMがセリン16残基にリン酸化されることを示す。FLAGタグ付きTRAMを過剰発現するHEK293−TLR4細胞を、30分間、1μg/mlのLPSによって刺激した。FLAGタグ付きTRAMは、抗FLAG抗体を使用して免疫沈降し、ホスホセリン抗体によりブロッティングした。
【図4b】THP1細胞を、LPSによる刺激前に1時間、PKC阻害剤ビスの有無によりインキュベートした。
【図4c】一定時間の1μg/mlのLPSによる刺激に続いて、PKCε−/−MEFs、並びにPKCεによって再構成されたPKCε−/−MEFsの結果を示す。
【図4d】30分間のLPS、ポリI:C若しくはMALP2による刺激に続いて、PKCε−/−MEFs、並びにPKCεによって再構成されたPKCε−/−MEFsの結果を示す。
【図5a】TRAMとPKCεの両方は、LPSシグナル伝達の遂行にとって重要であることを示す。PKCε−/−MEFs及びPKCεによって再構成されたPKCε−/−MEFs細胞(上部の4枚のパネル)、又はTRAM−/−MEFs及びWT MEFs細胞(下部の4枚のパネル)を、一定時間1μg/mlのLPS又は5μg/mlのPolyI:Cで刺激し、更にライゼートをリン酸化(チロシン180/182)及び全p38に関してイムノブロットした。
【図5b】−PKCε−/−MEFs及びPKCεによって再構成されたPKCε−/−MEFsを、1μg/mlのLPS又は5μg/mlのpolyI:Cで一定時間処理し、非還元PAGEゲル電気泳動し、IRF3に関してイムノブロットした。
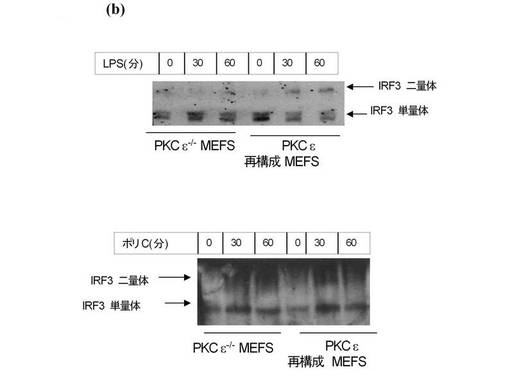
【図5c】ISREルシフェラーゼレポーター遺伝子及びコントロールのRenillaルシフェラーゼレポーター遺伝子を、上記のタイプの細胞にトランスフェクションした。トランスフェクションの24時間後、細胞を1μg/mlのLPS又は5μg/mlのPolyI:Cによって6時間刺激した。データはコントロールのレベルに対するルシフェラーゼ活性の刺激(倍数表示)の平均値として表す。
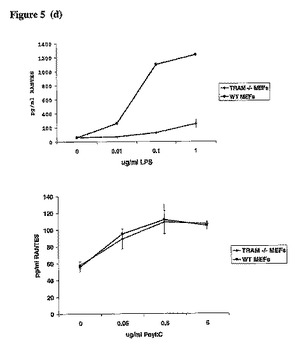
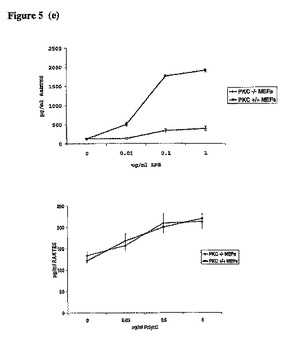
【図5d】上記の細胞タイプを、LPS又はPolyI:Cの所定の濃度によって24時間刺激し、更にRANTES ELISAを実施した。示す結果は、少なくとも3つの実験の代表的なものである。
【図5e】同上。
【図6a】TRAMS16Aは、TRAM欠損MEFsの再構成がWTTRAMよりも弱かった。WTTRAM、TRAMS16A及び空のベクターを、TRAM欠損MEFsにトランスフェクションした。トランスフェクション後24時間インキュベートした。細胞を更に24時間、LPS(1μg/ml)によって刺激し、培養液上清をRANTESに関してELISA解析した。
【図6b】細胞を30分間、LPS(1μg/ml)によって刺激し、ライゼートをp38に関してウエスタンブロッティング解析した。示す結果は、少なくとも3つの実験の代表的なものである。
【図7】膜に存在するTRAMの量がLPS刺激により減少することを示し、TRAMが膜から消失したことを示唆するものである。
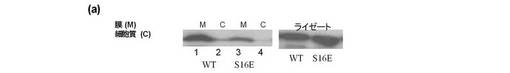
【図8a】セリン16残基がグルタミン酸に変異した場合の、膜上に存在するTRAMの量の減少を示し、この突然変異がセリン16上のTRAMのリン酸化が膜からのTRAMの消失を生じさせることを示唆するものである(レーン3と1を比較)。
【図8b】更にLPSで処理されたTHP1細胞における内因性のTRAMの減少が検出されることを示唆する。
【図8c】更にPKCε−/−MEFsFLAG−TRAMは、LPS刺激によって膜から消失しないことを示唆する。これらの結果は、PKCεによるセリン16上のTRAMのリン酸化が、膜上のTRAMの減少にとり必要であることを示唆するものである。
【発明を実施するための形態】
【0084】
アッセイ
本発明は、TRAMリン酸化の測定、更にはTRAMリン酸化の発生によるTLR4活性化のモニタリングに用いるアッセイシステム及びスクリーニング法を提供する。本発明の「アッセイシステム」の用語には、ある特定の1つ以上の現象を検出及び/又は測定するアッセイを実行して、その結果を分析するために必要な全ての構成要素が包含される。
【0085】
様々なアッセイを利用して、標的分子又はタンパク質のリン酸化状態を検出してもよい。
【0086】
一実施態様では、当該アッセイはリン酸化を受けるTRAMのドメインを認識するリン酸化特異的な抗体を使用する。最も好ましくは、この抗体は、TRAMにおけるセリン16の位置又はその近傍領域と結合する。
【0087】
TRAMのアミノ酸配列は、配列番号1:MGIGKSKINSCPLSLSWGとして定義される。
【0088】
好ましい実施態様では、本発明のアッセイではウエスタンブロッティングなどの公知の技術を使用する。当業者に公知の標準的な方法を使用して、TLR受容体に対するリガンド(例えばLPS)によって刺激した細胞から調製した抗体が、ウエスタンブロットのサンプルとして用いられる。
【0089】
更なる態様では、当該抗体を、他のアッセイのフォーマットにおいて使用してもよい。例えば、インビトロキナーゼアッセイにおいては、総タンパクの代わりにTRAM由来するペプチドフラグメントを用いてアッセイを実施できる。
【0090】
必須ではないが、好ましくは、本発明で使用するスクリーニングアッセイはハイスループット又はウルトラハイスループットアッセイであり、それによりオートメーション化された費用効果的なスクリーニング手段の提供につながる。
【発明を実施するための最良の形態】
【0091】
治療
本明細書で使用される「治療」という用語は、ヒト又は人間以外の動物に適用できるいかなる療法も意味する。治療とは、既存の症状に関するものであってもよく、又は予防的なもの(予防的処置)であってもよい。治療には、治癒的、緩和的又は予防的効果が包含される。
【0092】
医薬組成物
本発明は更に、リン酸化状態の変調又はリン酸化の防止による、TRAMのリン酸化調節用の医薬及び医薬組成物に関する。
【0093】
したがって、本発明の更なる態様は、免疫応答の調節用の医薬組成物の提供に関し、当該組成物は、主成分として、リン酸化を促進又はブロッキングすることによりTRAMのリン酸化を調節する化合物を含んでなる。
【0094】
本発明の医薬組成物、並びにその本発明での使用においては、主成分に加えて、当業者に公知の薬学的に許容できる添加剤、担体、緩衝安定剤又は他の材料を含有させてもよい。
【0095】
かかる材料は毒性を有してはならず、また主成分の有効性を妨げてはならない。担体又は他の材料の具体的な特性は投与ルートに依存する。
【0096】
投与量
組成物は好ましくは「臨床的に有効な量」(個人に利益をもたらすのに十分な量)で患者に投与される。実際の投与量、並びに投与速度及び時間は、個人の状態及び処置を受ける症状に依存する。
【0097】
至適投与量は、多くのパラメータ(例えば個人の年齢、阻害又は誘導される免疫応答の大きさ、投与される組成物の具体的な形態及び投与ルートなど)に基づいて決定できる。
【0098】
当該組成物は、血液などの特定の組織に投入されるミクロスフェア、リポソーム、他の微粒子状の輸送システム又は徐放性製剤を経て投与できる。好適な徐放性担体の例としては、例えば坐薬又はマイクロカプセルなどの一般的な剤形の半透性ポリマーマトリックスが挙げられる。
【0099】
上記の技術及びプロトコル、並びに本発明で使用できる他の技術及びプロトコルの例は、「Remington’s Pharmaceutical Sciences」,18th edition,Gennaro,A.R.,Lippincott Williams&Wilkins;第20版(December 15,2000)ISBN 0−912734−04−3、及び「Pharmaceutical Dosage Forms and Drug Delivery Systems」;Ansel,H.C.ら、第7版 ISBN 0−683305−72−7に記載され、その開示内容は本願明細書に援用される。
【0100】
抗体
本発明における「抗体」の用語は、イムノグロブリン若しくはその一部、又は抗体の結合ドメイン若しくはそのホモログであるポリペプチドを含む結合ドメインを指すものとする。
【0101】
「抗体」はイムノグロブリンであり、天然であってもよく、又は部分的又は完全に合成されたものであってもよい。当該用語はまた、抗体結合ドメイン若しくはそのホモログである結合ドメインを有するあらゆるポリペプチド、タンパク質若しくはペプチドが包含される。これらは天然給源に由来してもよく、又は部分的又は完全に合成されたものであってもよい。
【0102】
当該抗体は完全な抗体であってもよく、又はそのフラグメントであってもよい。完全な抗体に由来するフラグメントは、抗原の結合能を発揮できる。かかる結合フラグメントの例としては、(i)VL、VH、Cl及びCH1ドメインからなるFabフラグメント、(ii)VH及びCH1ドメインからなるFdフラグメント、(iii)単一の抗体のVH及びVLドメインからなるFvフラグメント、(iv)VHドメインからなるdAbフラグメント、(v)単離されたCDR領域、(vi)F(ab’)2フラグメント(2つのFabフラグメントが連結した二価のフラグメント)(vii)単鎖Fv分子(scFv)(VHドメイン及びVLドメインがペプチドリンカーで連結され、それにより2つのドメインが結合して抗原結合部位を形成する)、(viii)二重特異的な単鎖Fv二量体、及び(ix)遺伝子融合によって調製される多価性又は多重特異的フラグメント。
【0103】
抗体は多くの方法で修飾できるため、「抗体」の用語は、必要な特異性を有する結合ドメインを有するいかなる結合メンバー又は基質も包含されるものとして解釈すべきである。
【0104】
本発明の抗体は、単クローン抗体、又はそのフラグメント、誘導体、機能的均等物又はそのホモログであってもよい。抗体の定常部は、いかなる適切なイムノグロブリンサブタイプであってもよい。
【0105】
「抗体」という用語には、「ヒト化」された抗体、又はCDRグラフティングなどの技術により調製された抗体が包含される。かかる技術は、当業者にとって周知である。
【0106】
抗体作成
本発明に係る、及び本発明に用いられる特異的な結合メンバーは、いかなる適切な方法で調製してもよく、天然由来であっても人工的に合成したものであってもよい。かかる方法としては、例えば抗体ライブラリを使用した従来のハイブリドーマ技術、組換えDNA技術又はファージディスプレイ技術などが挙げられる。かかる産生技術は当業者に公知であり、またその他の抗体産生技術は、「Antibodies」:A Laboratory Manual,eds.Harlowら、Cold Spring Harbor Laboratory,1988.に記載されている。
【0107】
特に定めのない限り、本願明細書において用いられる全ての技術的及び科学的な用語は、本発明の技術分野の当業者に一般に理解されている意味を有する。
【0108】
明細書全体を通じて、特に断りのない限り、用語「含んでなる(comprise)」又は「含む(include)」又はそれらのあらゆる活用形は、特定の整数又は整数の群の包含を意味する一方で、他のいかなる整数又は整数の群を除外することを意味するものではない。
【0109】
本発明を、以下の実施例(それらは例示を目的として開示され、本発明を限定するものと解釈すべきでない)、更には図面を参照することにより例示する。
【実施例】
【0110】
材料及び方法細胞:
HEK293細胞及びTLR4(HEK293−TLR4)で安定にトランスフェクションしたHEK293細胞を、10%のウシ胎仔血清(FCS)、100単位/mlのペニシリン、100mg/mlのストレプトマイシン及び2mMのグルタミンを添加したDulbecco社の修飾Eagles培地(DMEM)で培養した。THP1細胞を、10%のFCS、100単位/mlのペニシリン、100mg/mlのストレプトマイシン及び2mMのグルタミンを添加したRPMI培地で培養した。
【0111】
逆転写酵素−ポリメラーゼ連鎖反応(RT−PCR):
鋳型として脾臓メッセンジャーRNA(BD Bioscience社)を使用してcDNAを調製した。1〜5μgRNAを、0.1μgのランダムプライマーとインキュベートし、DEPC処理水で5μLの最終液量とした。70℃で10分間、更に4℃で2分間インキュベートした。moloneyマウス白血病ウイルス逆転写酵素(MMLV−RT)を使用して逆転写反応させた。4μLの5×バッファ(250mMのトリスHCl(pH8.3)、375mM KCl及び15mM MgCl2)、2μLの100mM DTT、1μLのRNasin(40単位/ml)、1μLの10mM dNTP、1μLのMMLV−RT(200単位/μL)及び6μLのDEPC処理水を添加した。この反応液を37℃で1時間インキュベートし、酵素を不活性化するために95℃で2分間処理した。この反応液の5μLをPCR反応の鋳型として使用し、またTRAMの5’及び3’末端に特異的なプライマーを使用した。1×DNAポリメラーゼバッファ(1mMのトリスHCl(pH9.0)、5mMのKCl及び0.01%のTriton(登録商標)X−100)、各dNTP 0.2mM、2.5uのTaq DNAポリメラーゼ、特異的なオリゴヌクレオチドプライマを各0.5μM、及び0.5〜3mMのMgCl2を鋳型DNA溶液に添加し、反応混合物をPCRグレードの水を使用して50μLの最終的な液量とした。サーマルサイクラーを使用し、1分間94℃、1分間55℃及び2分間72℃の30サイクルで反応させた。PCR生成物をアガロースゲル電気泳動で分析した。
【0112】
GST発現ベクター及び哺乳類の発現ベクターへのTRAMのクローニング:
TRAMのPCR生成物を、pGEX−kgベクター(ファルマシア社製)、pcDNA3.1ベクター(インビトロゲン社)にライゲーションした。第1に、PCR生成物及びベクターを、別々の反応において切断した。5〜10単位の制限エンドヌクレアーゼ(NEB社製)、1×制限酵素バッファ、±1×BSA及び1〜10μgのDNAを混合し、滅菌水で10μLにし、2時間37℃でインキュベートした。次いで消化した生成物をPCR精製キット(Qiagen社製)を使用して精製した。1単位のT4DNAリガーゼ(1単位/μL)(プロメガ社製)、2μLのT4 10×反応バッファ、100〜150ngの切断したベクターDNA、及び200〜400ngの切断したPCR生成物を混合し、滅菌水で20μLにし、ライゲーション反応液を調製した。このライゲーション混合物を4℃で一晩静置し、更にBL21(DE3)細胞(ストラタジーン社)を形質転換した。
【0113】
TRAMの部位特異的変異導入:
Quickchange(登録商標)部位特異的変異導入キット(ストラタジーン社)を用い、TRAM遺伝子の特定の塩基を変異させた。製造業者の指示に従い、所望の突然変異を有するプライマーを使用して反応させた。
【0114】
GST−TRAMの発現及び精製:
TRAM−pGEXベクターを有するBL21(DE3)を、振盪培養器を用い、10mLのLB培地で37℃で一晩培養した。翌日、10mLを500mLのLB培地に植え継ぎ、0.6〜0.8のODとなるまで培養した。0.2mMの最終濃度でIPTGを培養液に添加し、更にこの培養液を更に30℃で4時間インキュベートした。更に培養液を、Sorvall RC5C遠心機(GSAローター使用)により、8,000回転/分で15分間遠心分離した。ペレットを25mLのNETNバッファ(20mMのトリスHCl、100mMのNaCl、1mMのEDTA、0.5%のNP40、10μg/mLのロイペプチン、10μg/mLのペプスタチン、1mMのPMSF、pH8.0)に再懸濁し、ソニケータ(Branson Sonifer 250)を使用して5分間超音波破壊し、45分間、18,000回転/分で遠心分離した。上澄みを600μLグルタチオンセファロースビーズ(アマシャム社製)に添加し、4℃で2時間インキュベートした。更にビーズを15mLのNETNバッファで5回洗浄し、600μLのNETNバッファに再懸濁した。
【0115】
ルシフェラーゼレポーター遺伝子アッセイ:
HEK293又はHEK293−TLR4細胞を、1×105細胞/mLの密度で96穴プレートに播種した。翌日、上記細胞を、Genejuice(Novagen)を用い、製造業者の指示に従い、選択したルシフェラーゼリポータプラスミド、及び選択した発現ベクターによってトランスフェクションした。NF−κB及びIRF3活性化の検出に関する実験のために、80ngのNF−κB又はIRSE−ルシフェラーゼレポーター遺伝子(ストラタジーン社)を、Renillaルシフェラーゼ内部コントロールプラスミド(ストラタジーン社)40ngと共に細胞にトランスフェクションした。24時間後に、細胞をpassive lysis buffer(プロメガ社製)に溶解し、レポーター遺伝子活性を照度計を使用して測定した。データを、コントロールのレベルに対する刺激(倍数表示)の平均値として表した。
【0116】
キナーゼアッセイ:
THP1細胞を、T175フラスコ(Sarstedt社)中に2×105細胞/mLで播種し、37℃で一晩インキュベートした。翌日、30mLの細胞懸濁液を、1時間、50mLのFalconチューブ(Sarstedt社)中でLPSの有無により処理した。細胞を遠心分離によって回収し、一度PBSで洗浄し、10分間、1mLのバッファ(10%のグリセロール(v/v)、50mMのNaF、20mMのトリスHCl(pH8.0)、2mMのEDTA、137mMのNaCl、1%のNP−40、1mMのPMSF、10μg/mLロイペプチン、1mMのNa3VO4)中で溶解させた。細胞残渣を13,000回転/分で10分間遠心分離し、上澄みをキナーゼアッセイ用に新規な試験管へ移した。グルタチオンビーズ上の精製されたGST−TRAM 50μLをエッペンドルフチューブに添加し、適当量のライゼートをチューブに添加し、4℃で2時間インキュベートした。ビーズを2,000回転数/分で5分間のスピンダウンし、更にキナーゼバッファ(20mMのHEPES(pH7.5)、10mMのMgAc、0.03%のTriton、100μg/mLのホスファチジルセリン、20mMのβ−グリセロールリン酸、0.01%(w/v)のロイペプチン及び100μMのNa3VO4)で3回洗浄した。これらのビーズを更に30μLのキナーゼバッファ(20μMの冷却したATP及び5μCi[γ32P]ATPを含有する)中に再懸濁し、37℃で30分間インキュベートした。20μLのサンプルバッファ(50mMのトリスHCl(pH6.8)、10%のグリセロール(v/v)、2%のSDS(w/v)、0.1%のブロムフェノールブルー(w/v)及び5%のβ−メルカプトエタノール)をチューブに添加し、サンプルを5分間ボイルした。サンプルを更に10%のSDS−PAGEに供し、ニトロセルロース膜へ転写した。ニトロセルロース膜はautoradカセット(コダック社製)に設置し、X線フィルムを上部に設置した。−80℃で一晩静置し、更にフィルムを感光させた。
【0117】
膜分画:
HEK293−TLR4細胞を1×105細胞/mLで播種し一晩インキュベートし、更に適当なプラスミドによってトランスフェクションした。トランスフェクションの24時間後、細胞を「結果」のセクションの方法に従い処理し、更に300μLの膜バッファ(20mMのトリス(pH7.5)、10mMのMgCl2、1mMのEDTA、250μMのスクロース、200μMのPMSF)中にスクラップした。細胞をdounceホモジェナイザを用いて30ストロークで溶解させ、4℃で1時間、100,000回転/分でhardwall Beckmanチューブで遠心分離した。上澄(すなわちサイトソル画分)を新規なチューブへ移し、ペレット(すなわち膜画分)を50μLのサンプルバッファ(50mMのトリスHCl(pH6.8)、10%のグリセロール(v/v)、2%のSDS(w/v)、0.1%のブロムフェノールブルー(w/v)及び5%のβ−メルカプトエタノール)中に再懸濁した。サイトソル画分を、centricon YM−10(ミリポア社製)を使用して50μLまで濃縮した。サンプルを12%のSDS−PAGEに供した。
【0118】
リン酸化特異的なTRAM抗体の産生:
Fabgennix(テキサス、米国)社に、アミノ酸16でセリンをホスホセリンで置換した、TRAM(KINSCPLSLSWGKRH)のアミノ酸7〜21に対応する合成ペプチドを使用して、セリン16でリン酸化されたTRAMを認識するリン酸化特異的な抗体の作製、精製を委託した。抗体の有効性は、リン酸化TRAMの予測されるバンドがTRAM欠損MEFs(データ示さない)から調製したサンプルでは存在しないことをもって確認した。
【0119】
RANTES ELISA
示された細胞を24穴プレート中に1×105細胞/mLで前夜に播種し、更に適当なプラスミドによってトランスフェクションした。トランスフェクションの24時間後、細胞を24時間、適当な刺激で処理した。5倍希釈した上澄みをサンプルとして使用し、R&D systems社のマウスRANTESキットを用い、製造業者の指示に従いRANTES ELISAを実施した。
【0120】
IRF3二量体化アッセイ
適当な細胞を、2×105細胞/mLで播種し、一晩インキュベートし、更に適当な刺激で処理した。細胞をPBSで洗浄し、100μL非還元サンプルバッファ(50mMのトリスHCl(pH6.8)、10%のグリセロール(v/v)、0.1%のブロムフェノールブルー(w/v)及び5%のβ−メルカプトエタノール)中にスクラップした。このうち20μLを非還元PAGEに供し、ニトロセルロース膜に転写し、IRF3に関してブロッティングした。
【0121】
結果TRAM遺伝子のクローン化及び変異体の生成:
TRAMのcDNA配列を、Genbank(Accession No.NM_021649)から検索した。TRAMの5’及び3’末端に対する特異的なプライマー(図1a)を用い、脾臓由来のmRNAを使用して、TRAMのcDNAを増幅した(図1b)。このcDNAをpGEX−kgベクターにクローニングし、細菌内でGST−TRAM融合タンパク質を発現させた。哺乳類の発現ベクターpcDNA3.1にもクローニングした。TRAMの変異体を生成するため、部位特異的変異導入を行った。位置6、10、14及び16(4Ser変異体と呼ばれる)でセリンが変異するようにプライマーを設計した(図1c)。セリン16のみを変異(Ser16変異体と呼ばれる)させるプライマーも設計した。
【0122】
LPS刺激によるTRAMのリン酸化:
TRAMをミリストイル化し、細胞を静置し、膜に移動させた(未発表データ)。幾つかのミリストイル化タンパク質を静電的なスイッチに供し、それをリン酸化することにより膜から反発される。すなわち、TRAMがリン酸化されたか否かが判定できるようにアッセイが設定されている。グルタチオンビーズ上の精製されたGST−TRAMを、THP1細胞(LPSの有無により処理)から調製したライゼートと2時間インキュベートした。更にサンプルを遠心分離し、相互作用した全てのタンパク質と共に、グルタチオンビーズ上のGST−TRAMをプルダウンした。更に[γ32P]ATPで30分間ビーズをインキュベートしてキナーゼアッセイを実施した。更にサンプルを10%のSDS−PAGEに供し、ニトロセルロース膜上に転写し、X線フィルムを使用して取り込まれた放射能を測定した。その結果、実際にTRAMがリン酸化され、すなわちこのリン酸化がLPS依存的であることを示した(図2a)。細胞が他の刺激(例えばPolyI:C)で処理されたときは、このリン酸化は起こらなかった(図2b)。
【0123】
N末端に最も近い4つのセリンを同定した。その後これらの4つのセリンを変異させた。この変異により、LPS依存性のリン酸化が見られなくなった(図2c)。
【0124】
TRAMのリン酸化
プロテインキナーゼCはミリストイル化されたタンパク質(MARCKS)をリン酸化することを示したので、TRAMがPKCによってリン酸化されるか否かの試験のため、panPKC阻害剤のビスインドリルマレイミド(ビス)を使用した。上記のように、GST−TRAMをLPSによって処置したTHP1細胞から調製したライゼートとインキュベートし、グルタチオンビーズを使用してプルダウンした。ビスの量を増加させてビーズに添加し、1時間インキュベートし、更に上記のようにキナーゼアッセイを実施した。ビスによりTRAMのLPS依存性のリン酸化が阻害された(図2c)。これは、PKCキナーゼファミリーのメンバーがLPSに応答するTRAMのリン酸化の原因となることを強く示唆するものである。
【0125】
THP1ライゼートの免疫減少を、プロテインGビーズ(シグマ社)に結合させたPKCε特異抗体(サンタクルス社)とライゼートとをインキュベートすることによって実施した。特異性をチェックするコントロールとしてPKCζ抗体を用いた。抗体を遠心分離して除去し、ライゼートからPKCε又はPKCζを除去した。コントロールIgG抗体も使用した。これらのライゼートを更に上記と同様にGST−TRAMでインキュベートし、キナーゼアッセイを実施した。図2eは、ライゼートからのPKCεの除去によりTRAMのリン酸化が行われなくなることを示す。これは、PKCεがLPSに応答してTRAMをリン酸化することを示唆する。PKCζの除去はTRAMのリン酸化に影響を及ぼさず、PKCζがTRAMをリン酸化しないことを示唆する。この理論は、組換えPKCε(rePKCε)(Calbiochem社)がGST−TRAMをリン酸化したという事実(図2g)、並びにPKCε欠損MEFsから調製したライゼートではGST−TRAMをリン酸化できないという事実(図2f)によって更に裏付けられた。組換えPKCζはTRAMをリン酸化せず、重ねて特異性を示唆するものであった(図2g)。セリン16はPKCεリン酸化の候補であるため、アラニンにセリン16を変異させたところ、TRAMのLPS依存性のリン酸化が幾分損なわれることが明らかとなった(図3a)。
【0126】
位置16のセリンがマウスにおいて保存されている4つのセリンのうち唯一のものであるため、このセリンを変異させた。その結果、TRAMのLPS依存性のリン酸化が幾分損なわれた(図3a)。
【0127】
TRAMのこのリン酸化が適切な機能にとり必須か否かを試験するため、位置16のセリンをアラニンに変異させ、この変異体におけるNF−κB及びIRF3経路を機能させる能力を解析した。WT−TRAM又はSer16−TRAMを、NF−κB又はISRE−ルシフェラーゼレポーター遺伝子と共にHEK293細胞にトランスフェクションした。前述のように、TRAMはNF−κB及びISREルシフェラーゼを機能させることができる。しかしながら、セリン16が変異した場合、TRAMはいずれの経路も活性化できなくなった(図3b)。
【0128】
セリン16の変異はまた、NF−κB及びISRE経路を刺激するLPSの能力を低下させた(図3c)。この変異体は、これらの経路上でドミナントネガティブとして作用することが明らかとなった。PKCによるTRAMのリン酸化が適正な機能にとり重要であるという更なる証拠として、PKC阻害剤(ビスインドリルマレイミド)がNF−κB経路を活性化させるTRAMの能力を阻害するが、MyD88に影響を及ぼさないという事実(図3d)が挙げられる。この証拠は、セリン16が適切に機能するためには、PKCεによってTRAMがリン酸化されることが必要であることを示唆する。
【0129】
セリン16残基のリン酸化がTRAMの膜からの移動にとり十分であることを確認するため、セリン16残基をグルタミン酸に変異させた。得られる変異体(ser16Glu)は明確なグリコシル化コントロールとして機能し、すなわちグルタミン酸残基がグリコシル化された状態であるとき、セリン残基を模倣した。この変異により膜に存在するTRAMの量の重要な減少を引き起こし、セリン16上のTRAMのリン酸化が膜からのTRAMの減少を引き起こすことを示唆している(図8a、レーン3と1を比較)。
【0130】
LPSで処理したTHP1細胞の内因性のTRAMの減少も検出した(図8b)。最後に、PKCε−/−MEFsFLAG−TRAMは、LPS刺激によっても膜から消失しなかった(図8c)。この試験結果は、PKCεによるセリン16上のTRAMのリン酸化が、TRAMの膜からの消失に必要であることを示唆する。
【0131】
内因性のTRAMは、セリン16にリン酸化される
ホスホセリン抗体を使用して、過剰発現するTRAMのリン酸化の試験を行った。HEK293−TLR4細胞をFLAG−TRAMでトランスフェクションし、LPSによって刺激した。FLAG−TRAMを細胞から免疫沈降させ、ライゼートを抗ホスホセリン抗体でブロッティングした。図4aから解るように、30分間のLPSで処理した細胞において、TRAMのセリンリン酸化の増加が示された。
【0132】
内因性のTRAMがPKCεによってセリン16にリン酸化されるかどうかを立証するために、アミノ酸16でセリンをホスホセリンに置換したTRAMのアミノ酸7〜21の配列を有する合成ペプチドを用い、抗体を作製した。
【0133】
LPSで処理したTHP1細胞から調製したライゼートをイムノブロットした結果、15分後にセリン16上のTRAMリン酸化が観察され、45分でピークに達することが示された(図4b、レーン1〜6)。LPSによる刺激の1時間前に、PKC阻害剤のビスでこれらの細胞をインキュベーションした結果、TRAMのリン酸化が妨げられた(図4b、レーン7〜12)。15分間及び30分間LPSで処理した、PKCεによって再構成されたPKCε−/−MEFs細胞からのライゼートのイムノブロットの結果、TRAMの正しい分子量のバンドが確認された(図4c、上部パネル、レーン9及び10)。効果が45分(レーン11)で減衰し、60分(レーン12)に不明確となったことから、リン酸化はこの細胞タイプの場合THP1細胞よりも初期に生じることが解った。ライゼート中の全TRAMのレベルは、実験期間中一定であった(図4c、第二パネル)。重要なことに、バンドはPKCε−/−MEFsから調製したライゼートにおいて検出されず(図4c、上部パネル、レーン1〜6)、TRAM欠損細胞ではバンドは検出されなかった(示されないデータ)ことから、抗体の特異性が証明された。最後に、TLR2リガンドであるMALP2又はTLR3リガンドであるポリI:CによるPKCεを発現するMEFs細胞の30分間の処理では効果が表れなかった(図4d)。
【0134】
TRAM及びPKCε両方は、LPSシグナル伝達の遂行にとって不可欠である
更にPKCε−/−MEFsのTRAM依存性のシグナル伝達を試験した。TRAM−/−MEFs及びPKCε−/−MEFsにおける、LPSで処理した細胞のp38のリン酸化は、それらの対応する野生型MEFsと比較して著しく減少していた(図5a上部パネル、レーン9〜11とレーン3〜5を比較)。重要なことに、PKCε−/−MEFs又はTRAM−/−MEFsでは、ポリI:Cに応答したp38のリン酸化の減少が見られなかった(図5a、パネル3、右側と左側を比較)。LPSによるIRF3の活性化を、その二量体化を指標として試験した。LPSによって誘導されるIRF3の二量体化は、PKCεを発現しているMEFsと比較し、PKCε−/−MEF細胞では減少していた(図5b、レーン2及び3をレーン4及び5と比較)。ポリI:Cに応答したPKCε−/−MEFのIRF3二量体化の減少が見られなかった(図5c、レーン2及び3をレーン4及び5と比較)。Yamamotoらが示すように、TRAM欠損MEFsにおいて、この反応はLPS(ポリI:Cでない)への応答に関して損なわれていた。IRF3と連結されたレポーター遺伝子の活性化を試験した結果、LPS処理されたPKCε−/−MEFsでは損なわれたが、ポリI:Cを処置されたPKCε−/−MEFsでは正常だった(図5c、右パネル)。同様の結果がTRAM欠損MEFsにおいても得られた。更にTRAM経路の開始としてのRANTESの誘導を評価した。TRAM−/−MEFsにおいては、LPS刺激に応答して生じたRANTESのレベルが、対応する野生型MEFsと比較して激減した(図5d、左パネル)。ポリI:Cに応答するRANTES産生のレベルは、影響を受けなかった(図5d、右パネル)。重要なことに、この反応はPKCε−/−細胞においても損なわれた。図5e(左パネル)に示すように、LPSによるRANTESの誘導は、PKCε発現細胞と比較して損なわれた。両方の細胞タイプを比較したが、ポリI:Cへの反応の違いは見られなかった(図5e、右パネル)。
【0135】
RANTESの産生がTRAMのシグナリング経路と関連するマーカーであるため、PKCε−/−細胞及びTRAM−/−MEFs細胞におけるその産生の喪失は更に、TRAMがプロテインキナーゼCイプシロンによってリン酸化されなければならず、このリン酸化がその機能にとって重要であるという考察を更に支持するものである。
【0136】
TRAMS16Aは、TRAM欠損細胞のシグナル伝達を完全に再構成できない
PKCε−/−細胞のLPS処理に続くTRAM依存性の反応の顕著な喪失、及びポリI:Cで処理された細胞においては同じ反応が損なわれなかったことは、TRAMS16Aによる損なわれたシグナル伝達と組み合わせて考察すると、PKCεによるTRAMのリン酸化が、TRAMの機能において不可欠であることを強く示唆した。更なる証拠を提供するために、TRAM欠損MEFs細胞のシグナル伝達を再構成するTRAMS16Aの能力を検討した。LPSを有する野生型MEFsの処理によりRANTES産生が誘導されたが、一方ではLPSによるTRAM−/−MEFsの処理によってはRANTES産生がほとんど誘導されなかった(図6a)。TRAM欠損細胞での反応は、野生型TRAMで再構成することができた。重要なことに、TRAMS16Aはシグナリングを再構成する能力が低かったが、同様に、LPS刺激によるTRAM欠損細胞でのp38のリン酸化が、WT−TRAMの過剰発現によって再構成された。TRAMS16Aは、このシグナルを再構成することができなかった(図6b)。したがって、プロテインキナーゼCイプシロンによるセリン16のリン酸化が、LPS刺激によるTRAMの通常の機能にとり必要であると結論付けることができる。
【0137】
LPS刺激により、膜のTRAMのレベルは減少する
TRAMのミリストイル化により膜局在が促進される。特定のミリストイル化されたタンパク質がリン酸化により膜から解離することが知られている。PKCεによるTRAMのリン酸化によってTRAMの再分布が生じるか否かを更に試験した。
【0138】
FLAG−TRAM/pcDNA3.1をHEK293−TLR4細胞にトランスフェクションし、これらの細胞を更に30分間、LPSの有無により刺激した。細胞を膜画分及びサイトソル画分に分離した。膜画分に存在するTRAMの量はLPS刺激により減少し、すなわちTRAMが膜から消失することを示唆する(図7a、上部パネル、レーン1とレーン3を比較)。TRAMはサイトソル画分では検出できなかった。これは分解によるものではない。なぜなら、細胞可溶化物中のTRAMのレベルは一定だからである(図7a、第三パネル)。膜からのTRAMの減少はPKCε依存性であった。その理由は、LPS刺激の1時間前にPKC阻害剤のビスを添加することにより、LPS刺激の後でも膜へのFLAG−TRAMへの残留が見られたからである(図7a、レーン7)。TRAMS16Aの減少を観察した場合、膜からの消失は観察されなかった(図7a、第二パネル)。LPSで処理したTHP1細胞の内因性のTRAMの減少も検出した(図7b)。最後に、PKCε−/−MEFsでは、FLAG−TRAMはLPS刺激によっても膜から消失しなかった(図7c)。この実験結果は、PKCεによるセリン16上のTRAMのリン酸化が、TRAMの膜からの消失に必要であることを示唆するものである。
【0139】
(本発明の要約)
TRAMは、TLR4及びTrif間の架橋アダプタとして作用し、LPSによって活性化するシグナリングカスケードにおいて不可欠な役割を果たす。TRAMはミリストイル化され、それにより形質膜と相互作用する。TRAMはまたリン酸化される。
【0140】
LPSに応答してTRAMはリン酸化され、それはここで記載したキナーゼアッセイを使用してインヴィトロで測定されうる。このリン酸化はTRAMの通常の機能に不可欠で、静電的なスイッチングに関与し、TRAMを膜から移動させる。
【0141】
本明細書において言及される全ての文書は、参照によって本願明細書に援用される。本明細書に記載されている実施態様に対する本発明の範囲内における様々な修正や変更は、当業者にとって自明である。本発明を具体的な好ましい実施態様に関して記載したが、本発明の特許請求の範囲は、かかる具体的な実施例にのみ限定されるべきではない。本発明の実施に係る、実際に記載されている様式に対する様々な改良は当業者にとって自明であり、本発明に包含されている。
【特許請求の範囲】
【請求項1】
TRAMの活性化状態を測定するアッセイ方法であって、前記方法が、細胞サンプルを含むTRAMを準備するステップと、TRAMのリン酸化をモニタリングするステップを含んでなり、TRAMがリン酸化されないことを、そのTRAMが不活性であることの指標とする方法。
【請求項1】
TRAMの活性化状態を測定するアッセイ方法であって、前記方法が、細胞サンプルを含むTRAMを準備するステップと、TRAMのリン酸化をモニタリングするステップを含んでなり、TRAMがリン酸化されないことを、そのTRAMが不活性であることの指標とする方法。
【図1】


【図2a】

【図2b】
【図3】
【図3c】
【図3d】
【図5d】
【図5e】
【図4a】

【図4b】
【図4c】

【図4d】
【図5a】
【図5b】
【図5c】

【図6a】

【図6b】

【図7】

【図8a】
【図8b】
【図8c】
【図2a】
【図2b】
【図3】
【図3c】
【図3d】
【図5d】
【図5e】
【図4a】
【図4b】
【図4c】
【図4d】
【図5a】
【図5b】
【図5c】
【図6a】
【図6b】
【図7】
【図8a】
【図8b】
【図8c】
【公開番号】特開2012−157358(P2012−157358A)
【公開日】平成24年8月23日(2012.8.23)
【国際特許分類】
【出願番号】特願2012−71368(P2012−71368)
【出願日】平成24年3月27日(2012.3.27)
【分割の表示】特願2008−503674(P2008−503674)の分割
【原出願日】平成18年4月3日(2006.4.3)
【出願人】(510001962)オプソナ セラピューティクス リミテッド (5)
【Fターム(参考)】
【公開日】平成24年8月23日(2012.8.23)
【国際特許分類】
【出願日】平成24年3月27日(2012.3.27)
【分割の表示】特願2008−503674(P2008−503674)の分割
【原出願日】平成18年4月3日(2006.4.3)
【出願人】(510001962)オプソナ セラピューティクス リミテッド (5)
【Fターム(参考)】
[ Back to top ]