
赤血球上に異種補体受容体タイプ1(CR1)分子を発現する非ヒト動物及びその使用
本発明は、少なくとも部分的に、外因性補体受容体タイプ1(CR1)を赤血球上に発現するマウスに関する。更に本発明は、赤血球上で発現させる異種CR1をコードする遺伝子が発現される遺伝子コンストラクト及び非ヒト動物を特徴とする。本発明のトランスジェニック動物を用いて、対象の組織、血清及び/又は循環中の生物学的物質などの作用物質の濃度を低下させることのできる組成物を同定及び/又は評価する方法も提供される。
【発明の詳細な説明】
【背景技術】
【0001】
関連出願
本出願は、引用をもってその内容全体をここに援用することとする2003年5月9日出願の米国仮出願60/469,262号に基づく優先権を主張するものである。
【0002】
発明の背景
補体系は、外来の抗原に対する免疫系の応答で重要な数多くの様々なタンパク質から成る。補体系のタンパク質は、正常な血清中のグロブリンの約10%を成す。補体系は、その主要な成分が開裂したときに活性化し、その結果できる生成物が、単独又は他のタンパク質と一緒になって、更なる補体タンパク質を活性化することで、タンパク分解カスケードが起きる。補体系の活性化は、血管透過性の上昇、貪食細胞の化学走性、炎症性細胞の活性化、外来粒子のオプソニン化、細胞の直接の致死及び組織損傷を含め、多種の応答につながる。伝統的な経路においては、補体系の活性化は、抗原−抗体複合体により惹起される。代替的な経路では、補体系は、例えば病原性細菌の細胞壁に存在するリポ多糖などによって触媒されることがある。
【0003】
補体受容体タイプ1(CR1)は、赤血球(E)、単球/マクロファージ、顆粒球、B細胞、特定のT細胞、脾臓の瀘胞性樹状細胞、及び糸球体足細胞上に存在することが示されている膜糖タンパク質である。CR1は、活性化した補体を含有する粒子又は免疫複合体の、これらの細胞表面上への結合を媒介する。より具体的には、CR1は、補体活性化生成物C3b及びC4bに結合するため、C3b/C4b受容体とも言及されてきた。これらの相互作用の結果は、この受容体を持つ細胞種に依存する(Fearon, D. T. and Wong, W. W. (1983) Ann. Rev. Immunol. 1:243)。赤血球表面上のCR1は、肝臓への輸送に向けて免疫複合体に結合する(Cornacoff, J. B.
et al. (1983) J.
Clin. Invest. 71:236; Nedof, N. E.
et al. (1982) J.
Exp. Med.
145:1739)。好中球及び単球上のCR1は、結合した複合体を、被覆小窩を通じた吸着性エンドサイトーシスか、又は、フィブロネクチン及びラミニンなど、細胞外マトリックス中に存在するホルボールエステル、化学走性ペプチド又はタンパク質による受容体活性化後の貪食のいずれかにより、内部移行させる(Newman S. L. et al. (1980) J. Immunol.
124:2236; Wright, S. D. and Silverstein S. C. (1982) J. Exp. Med. 145:1149;
Wright S. D. et al. (1983) J. Exp. Med. 148:1338)。CR1のリン酸化は、この貪食細胞作用を獲得する際に一役を担っているかも知れない(Changelian P. S. & Fearon D. T. (1986) J. Exp. Med.
163:101)。Bリンパ球上のCR1の機能は、そこまで定義されていないが、これらの細胞を、CR1に対する抗体で処理すると、最適以下の用量のアメリカヤマゴボウマイトジェンに対するそれらの応答が亢進した(Daha, M. R. et al. (1983) Immunobiol. 164:227 (抄録))。濾胞性樹状細胞上のCR1は、抗原提示の役割を果たしているかも知れない(Klaus G. G. B. et
al. (1980) Immunol. Rev. 53.3)。
【0004】
受容体として働くことに加え、CR1はまた補体調節機能も有する。例えば伝統的及び代替的補体経路において、CR1はC3/C5コンバターゼを阻害することができ、更に、I因子によるC3b及びC4bの開裂のためのコファクタとして作用することもできる(Fearon D. T.
(1979) Proc. Natl. Acad. Sci. U. S. A. 76:5867; Liida X. & Nussenzweig V.
(1981) J. Exp. Med. 153:1138)。補体活性化の伝統的経路においては、複合体C4b、2aはC3活性化酵素、又はコンバターゼである。CR1はC4bに結合してC4b、2aの解離を促進することができる。この結合により、C4bは、不活化補体タンパク質C4c及びC4dへの開裂を通じた、I因子による不可逆的なタンパク質分解性不活化を起こしやすくなる。補体活性化の代替的経路においては、複合体C3b、4bはC3コンバターゼである。CR1はC3bに結合することができ、やはりC3b、4bの解離を促進することができる。C3b、CR1が形成されると、C3bは、I因子による不可逆的なタンパク質分解性不活化を起こして、不活化補体タンパク質C3b(iC3b)を形成しやすくなる。
【0005】
CR1は、一本のポリペプチド鎖から成る糖タンパク質である。分子量が~40,000乃至50,000ダルトンの増分ずつ異なる4つのアロタイプ型のCR1が見つかっている。2つの最もよくある形は、それぞれ250,000及び290,000ダルトンの分子量を有するF及びSアロタイプである(A及びBアロタイプとも呼ばれる)(Dykman, T. R. et al. (1983) Proc. Natl.
Acad. Sci.
U.S.A. 30:1698; Wong,
W.W. et al. (1983) J.
Clin. Invest. 72:685)。更に、210,000及び>290,000ダルトンの分子量を有する2つの珍しい形もある(Dykman, T. R. et al. (1984) J.
Exp. Med.
159:691; Dykman, R. R. et al. (1985) J. Immunol.
134:1787)。これらの違いは、明らかに、グリコシル化した状態ではなく、CR1のポリペプチド鎖の違いを表している(Wong W. W. et al. (1983) J. Clin. Invest. 72:685)。4つのCR1アロタイプの全てがC3b結合活性を有する(Dykman, T. R. et al. (1983) Proc. Natl. Acad. Sci. U.S.A. 30:1698;
Dykman, T. R. et al. (1984) J. Exp. Med. 159:691; Dykman, T. R. et al. (1985)
J. Immunol. 134:1787; Wong W. W. et al. (1983) J.
clin. Invest. 72:685)。
【0006】
CR1は多様な造血細胞上に見られるが、血中のCR1の大半は、赤血球上に特異的に存在する。このCR1は、循環免疫複合体のプロセッシング及び除去で主な役割を果たす。病原体が赤血球に付着すると、免疫複合体となった病原体は、例えば肝臓及び脾臓の固定組織マクロファージなど、アクセプタ貪食細胞に効率的に輸送される。興味深いことに、この輸送反応での病原体は、赤血球に認められる損傷を何ら及ぼさずに、赤血球から見かけ上、引き剥がされる(Lindorfer M. A. et al. (2001) Immun. Reviews 183:10-24)。この輸送反応の詳細な機序はまだ、充分には解明されていない。しかしながら、研究では、in vivoでの輸送反応は、I因子媒介性放出に依存している可能性の低いプロセスに促されていることが示されている(Lindorfer M. A. et al. (2001) Immun. Reviews 183:10-24)。
【0007】
製薬業において重要な目標は、組織又は血流中の病原体を標的とし、除去することのできる新規な治療様式の開発である。この問題に対処するためにとられてきたいくつかのアプローチは、霊長類赤血球の免疫付着機能の利点を生かすことに狙いを置いている。あるアプローチでは、二重特異的モノクローナル抗体複合体(ヘテロ重合体、HP)を、標的病原体に結合し、霊長類赤血球のCR1に固定するようにデザインする(Lindorfer M. A. et al. (2001) Immun. Reviews 183:10-24)。例えば、ヘテロ重合体を、CR1に特異的なモノクローナル抗体を、標的病原体に特異的なモノクローナル抗体に化学的に架橋したものから構成することができる。この方法で、CR1に特異的な抗体は、天然のCR1リガンドであるC3bにとってサロゲートとしての役目をする。このアプローチの利点の一つは、C3bのCR1に対する天然の親和性は低く、従って、強力な補体活性化や、複数のC3b分子を捕獲して赤血球の結合を確実にする必要があるのだが、CR1に対して大変高い親和性を持ついくつかのマウスモノクローナル抗体が作製されている点である。これらの抗体を、高親和性病原体特異的モノクローナル抗体に架橋させると、補体の非存在下でも、実質的にはあらゆる標的病原体に赤血球を結合させることができるはずである。この戦略は、数多くの細菌及びウィルスに成功裏に応用されてきた(Powers J. H. et al. (1995) Infect. Immun 63:1329-1335; Kuhn S. E. et al. (1998) J. Immunol.
160:5088-5097; Nardin A.,
et al. (1998) 211:21-31; Hahn C. S. et al. (2001) J. Immunol
166:1057-1065)。
【0008】
ある関連するアプローチでは、赤血球及び二重特異的試薬を用いた、血清及び/又は循環中の分子の標的決定を、全身性エリテマトーデス(SLE)におけるIgG抗dsDNA抗体など、自己免疫疾患に関連する数多くの自己抗体の標的決定にまで延長することができる。抗原ベースのヘテロ重合体(AHP)を用いることにより、自己抗体を赤血球媒介性除去に向けて標的決定することができる(Lindorfer
M. A. et al. (2001) Immun. Reviews 183:10-24)。抗原ベースのヘテロ重合体は、自己抗原を抗CR1モノクローナル抗体に化学的に架橋したものから成る。赤血球に結合した抗原ベースのヘテロ重合体は自己抗体を捕獲し、新たに形成されたその赤血球結合免疫複合体を、最終的な除去に向けた肝臓への輸送反応に方向付ける。
【0009】
当業においては、明らかに、組織、血清及び/又は循環から標的とされた分子を除去するために、CR1に結合するヘテロ重合体など、二重特異的組成物の能力を検査するために用いることのできる動物モデルが必要である。このような動物モデルは、二重特異的組成物や、二重特異的組成物ベースの治療法の効験及び安全性の両方を評価する上で有用であろう。残念ながら、マウスは薬物検査にとっては優れたモデルであるが、マウスはそれらの赤血球上にCR1を発現しない(Kalli and Fearon. 1994 J. Immunol.
152:2894)。
【0010】
発明の概要
本発明は、少なくとも部分的に、ヒト補体受容体タイプ1(CR1)を発現するマウスの作製と、これらの動物が赤血球上にCR1を発現するという観察とに基づくものである。従って、本発明は、異種CR1遺伝子を発現させる遺伝子コンストラクト及び非ヒト動物を特徴とする。本発明のトランスジェニック動物を用いて、対象の組織、血清及び/又は循環中の生物学的物質などの作用物質の濃度を低下させることのできる組成物を同定及び/又は評価する方法も提供される。
【0011】
ある局面では、本発明は、ヒト補体受容体1(CR1)ポリペプチドをコードするヌクレオチド配列を、プロモータに作動的に連結させて成る、非ヒト動物における発現用の遺伝子コンストラクトに関し、但し当該のコンストラクトは、赤血球上のヒトCR1の機能的発現を命令するものである。
【0012】
別の局面では、本発明は、そのゲノムが、ヒト補体受容体1(CR1)をコードするポリヌクレオチドを、プロモータに作動的に連結させて含むような非ヒト動物に関し、該ヒトCR1分子は、当該動物の赤血球上に機能的に発現するものである。好ましくは、該ヒトCR1分子が赤血球上に優先的に発現するとよい。
【0013】
好適な実施態様では、該プロモータはGATA-1プロモータである。より好ましくは、該プロモータは、GATA-1プロモータと、赤血球上の発現を活性化させる上流の制御領域とを含むとよい。
【0014】
好適な実施態様では、該非ヒト動物は、該ポリヌクレオチドについてホモ接合型又はヘテロ接合型である。好ましくは、当該の動物がトランスジェニック・マウスであるとよい。
【0015】
別の局面では、本発明は、対象の血清及び/又は循環中の作用物質の濃度を低下させることのできる二重特異的化合物をスクリーニングする方法を特徴とする。この方法は、(a)そのゲノムが、ヒト補体受容体1(CR1)をコードするポリヌクレオチドをプロモータに連結させて含むような非ヒト動物であって、前記ヒトCR1が、前記非ヒト動物の赤血球上に機能的に発現するような非ヒト動物、に、複数の検査化合物を投与するステップと、(b)前記非ヒト動物の血清及び/又は循環中の前記作用物質の濃度を低下させる上での、ある二重特異的化合物の能力を判定するステップと、(c)前記非ヒト動物の血清及び/又は循環中の前記作用物質の濃度を低下させる二重特異的化合物を選抜することで、対象の血清及び/又は循環中の作用物質の濃度を低下させることのできる二重特異的化合物を同定するステップと、を含む。
【0016】
ある関連する局面では、本発明は、対象の血清及び/又は循環中の作用物質の濃度を低下させる上での、ある二重特異的化合物の能力を評価する方法を提供するものである。この方法は、(a)そのゲノムが、ヒト補体受容体1(CR1)をコードするポリヌクレオチドをプロモータに連結させて含むような非ヒト動物であって、前記ヒトCR1が、前記非ヒト動物の赤血球上に機能的に発現するような非ヒト動物、に、当該の二重特異的化合物を投与するステップと、(b)前記非ヒト動物の血清及び/又は循環中の前記作用物質の濃度を低下させる上での、前記二重特異的化合物の能力を判定することにより、ある対象の血清及び/又は循環中の前記作用物質の濃度を低下させる上での、前記二重特異的化合物の能力を評価するステップと、を含む。
【0017】
ある実施態様では、これらの方法は、非ヒト動物に該作用物質を投与するステップを更に含む。好適な実施態様では、当該の作用物質は病原体、ウィルス、毒素、ポリヌクレオチド、細菌、又は自己免疫疾患に関連する自己抗体、である。
【0018】
好適な実施態様では、該二重特異的化合物はヒトCR1に結合するものである。好ましくは、該二重特異的化合物がヘテロ重合体であるか、又は抗原ベースのヘテロ重合体であるとよい。
【0019】
好適な実施態様では、該非ヒト動物はトランスジェニック・マウスである。
【0020】
他の好適な実施態様では、該プロモータはGATA-1プロモータか、又は、GATA-1プロモータと、赤血球上での発現を活性化させる上流の制御領域とを含む。
【0021】
更に本発明は、生物学的作用物質に曝露した対象を治療する方法を特徴とする。この方法は、対象が治療されるように、本発明により提供される方法で同定された二重特異的化合物を対象に投与するステップを含む。
【0022】
ある関連する局面では、本発明は、生物学的作用物質への曝露のリスクのある対象を治療する方法を提供する。この方法は、対象が治療されるように、本発明により提供される方法で同定された二重特異的化合物を対象に投与するステップを含む。
【0023】
更に別の関連する局面では、本発明は、感染又は自己免疫疾患のある対象を治療する方法を提供する。この方法は、対象が治療されるように、本発明により提供される方法で同定された作用物質を対象に投与するステップを含む。
【0024】
更に本発明は、ヒトCR1をその赤血球表面上に発現するトランスジェニック・マウスを作製する方法を特徴とする。この方法は、a)ヒトCR1ポリペプチドをコードするポリヌクレオチドを、マウスの卵又は胚に導入するステップであって、前記ポリヌクレオチドが、マウス赤血球上でヒトCR1を発現させるプロモータに作動的に連結されている、ステップと、b)前記CR1ポリヌクレオチドを含む卵又は胚を産期に至るまで成長させて、前記トランスジェニック・マウスを生じさせるステップと、を含む。
【0025】
ある関連する局面では、本発明は、トランスジェニック・マウスを作製する方法を提供するものである。この方法は、それぞれがヒトCR1ポリペプチドをコードするポリヌクレオチドを含む二倍体ゲノムを含む二匹のトランスジェニック・マウスを交配するステップを含み、但しこの場合、前記ポリヌクレオチドが、マウス赤血球上でヒトCR1ポリペプチドが生じるように発現することで、仔トランスジェニック・マウスが生じる。
【0026】
発明の詳細な説明
本発明は、少なくとも部分的に、そのゲノムが、ヒト補体受容体1(CR1)をコードするポリヌクレオチドをプロモータに作動的に連結させて含有するようなマウスの作製と、異種CR1分子が、該動物の赤血球上に機能的に発現するという観察とに基づく。従って、本発明は、CR1を含む遺伝子コンストラクトと、CR1が赤血球上で発現するような非ヒト動物とを特徴とする。本発明は、対象の血清及び/又は循環中の標的作用物質の濃度を低下させることのできる二重特異的化合物を同定するために前記非ヒト動物を使用する方法を提供する。更に本発明は、このような二重特異的化合物の投与が有益であろう疾患又は異常を有する対象を治療する方法を特徴とする。
【0027】
本発明がより容易に理解されるように、いくつかの用語をまず定義する。
【0028】
I. 定義
ここで用いられる場合の用語「導入遺伝子」とは、ある動物に導入された異種もしくは外来遺伝子又は組換え核酸コンストラクトを言う。該導入遺伝子は、野生型もしくは変異型遺伝子でもよく、あるいは、
それが異常なパターンで発現するように変更されたものでもよい。ここで用いる場合の用語「トランスジェニック細胞」とは、導入遺伝子を含有する細胞を言う。
【0029】
ここで用いる場合の用語「異種遺伝子」又は「外来遺伝子」には、それらが導入された先のゲノムの一部としては天然では見られなかったり、あるいは、それがゲノム中に天然で存在する一箇所又は複数の位置とは異なる位置で見られるような遺伝子が含まれる。異種DNAは、それが導入された先の細胞にとって内因性でなく、別の細胞から得られたものである。
【0030】
ここで用いる場合の用語「遺伝子コンストラクト」には、天然では同じ分子中に存在しないヌクレオチド配列が操作されて接合されているような非天然核酸分子が含まれる。
【0031】
本発明のコンストラクトは、ホスト細胞内での核酸分子の発現に適した形のものである。好ましくは、当該のコンストラクトが、発現に用いるホスト細胞に基づいて選択された一つ以上の調節配列を、発現させようとする核酸配列に作動的に連結させて含むとよい。組換え発現ベクタ内において、「作動的に連結させた」とは、(例えばin vitroの転写/翻訳系で、あるいは、ベクタをホスト細胞内に導入する場合にはホスト細胞内で)目的のヌクレオチド配列の発現が可能な態様で、該ヌクレオチド配列が調節配列に連結されていることを意味するものと、意図されている。用語「調節配列」は、プロモータ、エンハンサ及び他の発現制御配列(例えばポリアデニレーション・シグナル)を包含するものと、意図されている。このような調節配列は、例えばGoeddel; Gene Expression
Technology: Methods in Enzymology
185, Academic Press, San Diego, CA (1990) に解説されている。
【0032】
ここで用いる場合の用語「上流の制御領域」は、例えばプロモータ又は上流のエンハンサなど、発現させようとする遺伝子の5‘側にある調節配列を包含する。
【0033】
ここで用いられる場合の用語「プロモータ」は、転写を開始するためにRNAポリメラーゼの結合に関与するDNA領域を包含する。
【0034】
特定のタンパク質の発現又は特定の細胞種に関してここで用いられる用語「優先的に発現する」は、他の細胞種上よりもその細胞種で多いが、必ずしもその特定の細胞種に限定されないような発現を包含する。例えば、赤血球上で優先的に発現するタンパク質は、他の細胞種に比べ、赤血球上で少なくとも約5倍、少なくとも約10倍、少なくとも約20倍、少なくとも約30倍、少なくとも約40倍、少なくとも約50倍又はそれより高いレベルで発現するものである。
【0035】
ここで用いる場合の用語「機能的に発現する」とは、あるタンパク質の対生物活性が維持されるような該タンパク質の発現を包含する。例えば、CR1が、C3b又は抗体など、天然リガンドへの結合が可能であるときに、細胞上で機能的に発現したとする。
【0036】
ここで用いる場合の「トランスジェニック動物」とは、異種又は外来遺伝子をそれらのゲノムに導入して有するような動物を言う。用語「トランスジェニック動物」は、例えばブタ、サル、ヤギ、又はマウスなどのげっ歯類などの非ヒト哺乳動物など、当該動物の細胞のうちの一つ以上、そして好ましくは基本的に全てが導入遺伝子を含むような動物を包含する。当該の導入遺伝子は、細胞に直接、あるいは、例えばマイクロ注射、トランスフェクション、又は組換えウィルス感染などの感染などにより、当該細胞の前駆細胞への導入により間接的に、導入される。用語「遺伝子操作」は、組換えDNA分子の導入を包含する。この分子は、好ましくは、それが仔へ確実に伝えられるように染色体内に組み込まれるとよいが、あるいは、染色体外で複製するDNAであってもよい。
【0037】
好適なトランスジェニック動物は哺乳動物である。本発明によれば、哺乳動物の例には、限定はしないが、げっ歯類(例えばラット及びマウス)、イヌ、ネコ、ブタ、ヒツジ、ウシ、ヤギ、ウマ及びウサギが含まれる。好ましくは、当該の哺乳動物がヒト以外であるとよい。ここで用いる場合の用語「げっ歯類」とは、系統学的齧歯目の全てのメンバーを言う。ある好適な実施態様では、齧歯類はマウスである。
【0038】
ここで用いられる「発現」には、細胞によるタンパク質の合成が含まれる。タンパク質の発現は、例えば転写、転写後、翻訳、及び/又は翻訳後など、多様な段階で測定することができる。
【0039】
本発明によれば、トランスジェニック動物は導入遺伝子を誤発現するものである。ここで用いられる場合の用語「誤発現」には、非野生型パターンの遺伝子発現が含まれる。誤発現には、非野生型レベルでの発現、即ち、当該遺伝子が発現する時点又は段階の点で野生型とは異なる発現パターン、例えば、所定の発生期又は段階における(野生型に比較したときの)発現増加:所定の細胞種又は組織種での(野生型に比較したときの)発現増加の点で、野生型とは異なる発現パターン;スプライシング・サイズ、アミノ酸配列、翻訳後修飾、又は、発現後のポリペプチドの対生物活性の点で、野生型とは異なる発現パターン;環境刺激又は細胞外刺激が当該遺伝子の発現に及ぼす効果の点で、野生型とは異なる発現パターン、例えば、刺激強度の増減の存在下における(野生型に比較したときの)発現増加のパターン、が含まれる。誤発現には、トランスジェニック核酸からの発現が含まれる。好ましくは、本発明の導入遺伝子により、関連する内因性遺伝子が前に発現しなかったか、あるいは低レベルで発現した動物の細胞上で、外来遺伝子が発現するとよい。ある実施態様では、当該の導入遺伝子は赤血球上で発現する。ある好適な実施態様では、当該の導入遺伝子は、赤血球上で優先的に発現する。
【0040】
ここで用いられる場合の用語「マーカ配列」とは、(a)目的の遺伝子(例えばCR1遺伝子)を発現させる核酸標的決定コンストラクトの一部として用いられ、及び(b)前記標的決定コンストラクトをそれらのゲノムに導入させて有する細胞を特定するために用いられる、核酸分子を言う。例えば、マーカ配列は、細胞に検出可能な形質をもたらすタンパク質、あるいは、例えばネオマイシン耐性遺伝子などの抗生物質耐性遺伝子、又は、例えばアルカリホスファターゼ、西洋わさびペルオキシダーゼ、ルシフェラーゼ、ベータ-ガラクトシダーゼ等、当該細胞上に典型的には見られない検定可能な酵素、又は、例えば緑色蛍光タンパク質、赤色蛍光タンパク質など、当該細胞中には典型的に見られない他の便利に検出可能なタンパク質、をコードする配列であってよい。
【0041】
作用物質又は二重特異的化合物に関してここで用いられる「投与する」とは、ある組成物を動物又は細胞に分配する、送達する又は適用することを言うものと、意図されている。用語「投与する」とは、例えば非経口又は経口経路による送達、筋肉内注射、皮下/皮内注射、静脈内注射、口腔内投与、経皮送達、及び、鼻孔内もしくは呼吸管経路による投与を含め、動物の所望の位置へ当該組成物を送達するために適したいずれかの経路により、動物に当該組成物を接触させる又は分配する、送達する又は適用することを言うものと、意図されている。
【0042】
ここで用いられる場合の用語「二重特異的化合物」には、対象の血清及び/又は循環中の作用物質の濃度を潜在的に低下させることのできるあらゆる二重特異的化合物が含まれる。二重特異的化合物は、2つの異なる結合部分から成る分子である。二重特異的化合物の例には、例えば、二重特異的抗体及びヘテロ重合体がある。好適な実施態様では、当該の二重特異的化合物はヘテロ重合体か、又は抗原ベースのヘテロ重合体である。
【0043】
二重特異的抗体は、2つの特異性部分を有する単一の抗体分子である。二重特異的抗体の例は当業で公知である。ハイブリッド・ハイブリドーマ又はトリオーマにより作製される二重特異的抗体は、米国特許第4,474,893号に開示されている。二重特異的抗体は、化学的手段(Staerz et al. (1985) Nature 314:628, and Perez et
al. (1985) Nature 316:354)
及びハイブリドーマ技術(Staerz and Bevan (1986)
Proc. Natl. Acad. Sci. USA, 83:1453, 及びStaerz and Bevan (1986) Immunol. Today 7:241)により構築されてきた。二重特異的抗体は、米国特許第5,959,084号にも解説されている。二重特異的抗体のフラグメントは、米国特許第5,798,229号に解説されている。
【0044】
ここで解説されるヘテロ重合体とは、当該ヘテロ重合体の結合部分の少なくとも一つが二価であるような2つの異なる結合部分を含む分子を言う。例えば、ヘテロ重合体は、互いに接合された2つの抗体を含むことができる。本発明のある実施態様では、ヘテロ重合体は、当業で公知の数多くの架橋技術のいずれかを用いることにより架橋された2つの異なる抗体から成る。ある実施態様では、該ヘテロ重合体はヒトCR1に結合する。ある実施態様では、ヘテロ重合体は、当業で公知の数多くの架橋技術のいずれかを用いて架橋された異なる抗体から成る。
【0045】
別の実施態様では、ヘテロ重合体は、抗原ベースのヘテロ重合体である。ここで解説される、抗原ベースのヘテロ重合体とは、例えば自己免疫疾患又は異常に関連する自己抗体など、標的抗体により認識される抗原である結合部分を含有するヘテロ重合体を言い、この場合、前記結合部分は、当業で公知の数多くの架橋技術のいずれかを用いて第二の結合部分に架橋されている。好ましくは、第二の結合部分は、抗原に結合する抗体であるとよい。ある実施態様では、前記の抗原ベースのヘテロ重合体はヒトCR1に結合するものである。ここで用いられる場合の用語「作用物質」は、対象の血清及び/又は循環中に存在し、感染、異常又は疾患に関連するあらゆる物質又は生物を包含するものである。好適な作用物質は、例えば病原体、ウィルス、毒素、ポリヌクレオチド、抗体又は自己抗体などを含む生物学的物質である。ここで用いられる場合の用語「生物学的物質」は、更に、例えば炭疽菌、天然痘、ペスト、エボラ又はマーブル・ウィルスなど、生物戦争又はテロ活動で対象が曝露するかも知れない物質を包含する。
【0046】
以下の小項で本発明を更に詳述する。
【0047】
II. CR1発現コンストラクト
ある局面では、本発明は、非ヒト動物に導入されたときに、この動物の赤血球上でCR1遺伝子が機能的に発現するような核酸分子を特徴とする。好ましくは、当該CR1遺伝子はヒトCR1遺伝子であるとよいが、他の哺乳動物CR1分子を発現させることもできる。野生型ヒトCR1遺伝子のヌクレオチド配列は当業で公知であり、例えば、引用をもってその内容をここに援用することとするWilson J.G. et al, (1986) J.
Exp. Med. 164:50-59、米国特許第6,316,604号及び米国特許第5,212,071号などに解説されている。
【0048】
上記の配列のバリアントも、本発明のコンストラクトを作製する際に用いることができる。例えば、異なるアロタイプのCR1 が当業で解説されている。例えば、このようなアロタイプは、Wong et al.
1986. J. Exp. Med.
164:1531; Wong et al. 1989. J. Exp.
Med. 169:847に解説されている。他のヒトCR1配列が当業で公知であり、例えばGenBank 受託番号AF169970; J. Immunol. 151: 6214(1993) 又はJ. Biol.
Chem. 265:974-980
(1990)に見ることができる。このようなアロタイプ型のCR1をコードする核酸分子も本発明において用いることができる。
【0049】
例えばバリアント型のヒトCR1など、バリアント型のCR1も用いることができる。CR1遺伝子、改変されたCR1遺伝子、又は、ヒトCR1遺伝子と、別のヌクレオチド配列(例えば別の生物由来のCR1配列又は非CR1配列)の一部とを由来とするポリヌクレオチド配列を含むCR1遺伝子、即ちキメラCR1遺伝子、あるいは、ヒトCR1遺伝子に関連するCR1遺伝子、の部分をコードする核酸の作製及び使用も、用いることができる。このようなバリアント型のCR1を、非ヒト動物の赤血球上で機能的に発現させることができ、更に、ヒトC3b又はC4bに、及び/又は、ヒトCR1に結合する抗体にも、結合させることができる。
【0050】
ある実施態様では、CR1が相互作用する相手の分子、例えば抗体分子、C3b等への結合を高める又は低下させるように、CR1ヌクレオチド配列に変異を作ることができる。
【0051】
導入される先の非ヒト動物にとって異種のCR1遺伝子をコードする他の核酸分子の作製及び使用も提供される。例えば、非ヒト霊長類型のCR1など、ヒトCR1に対して相同性の高いCR1分子をコードする核酸配列も、このようなCR1分子又はその部分が動物の赤血球上で機能的に発現可能であり、更に、ヒトC3b及び/又はC4bに、及び/又は、ヒトCR1が結合する抗体に、結合可能であることを条件に、当該のコンストラクトに用いることができる。
【0052】
バリアント型のCR1は当業で公知の多種の方法により作製することができる。例えば、クローニングされたCR1遺伝子は、当業で公知の数多くある戦略のいずれによっても、改変することができる(Maniatis, T., 1982, Molecular Cloning, A Laboratory Manual, Cold
Spring Harbor Laboratory, Cold Spring Harbor, N.Y.)。このようなバリアントは、例えばCR1のCR1結合分子への結合を促進する又は低下させるなどのために作製することができる。CR1バリアントをコードする遺伝子の作製においては、好ましくは、改変後の遺伝子は、翻訳停止シグナルを途中に持つことなく、CR1と同じ翻訳読み枠内に留まっているとよい。別の実施態様では、CR1遺伝子を変異させて翻訳、開始、及び/又は終了配列を作製及び/又は破壊したり、あるいは、コーディング領域にばらつきを作製したり、及び/又は、新しい制限エンドヌクレアーゼ部位を形成するか、又は、既存のものを破壊して、更なるin vitro改変を容易にすることもできる。当業で公知の変異誘発のための技術を用いることができ、その中には、限定はしないが、in vitro 部位指定変異誘発法(Hutchinson, C., et al.,
1978, J.Biol. Chem. 253:6551)、TAB.RTM.リンカ(ファルマシア社)の使用等、がある。ある好適な実施態様では、CR1をコードするヌクレオチド配列の変更が、配列に対する保存的変異を含む。
【0053】
本発明の遺伝子コンストラクトの作製においては、内因性又は外因性のプロモータを用いることができる。好ましくは、当該のCR1遺伝子を、赤血球上でのCR1の発現につながる又は発現を増加させる調節配列(例えばプロモータ及び/又はエンハンサなどの転写制御配列)に作動的に連結させるとよい。CR1の発現は、必ずしも赤血球上だけでなくともよい。例えば、他の細胞に比較したときの赤血球での発現レベルを増加させるプロモータ及び/又はエンハンサを用いてもよいが、赤血球以外の細胞での発現が起きる場合もある。
【0054】
本発明での使用に適したプロモータには、限定はしないが、GATA-1遺伝子の調節配列がある。ある実施態様では、該調節配列には、DNAse I超感受性により特定され、初期赤血球系細胞での発現活性化をもたらす上流制御位置がある(例えばMcDevitt, M.A. et al., (1997) Proc.
Natl. Acad. Sci., USA, 94:7976-7981; Visvader J.E. et al., (1998) Genes & Dev. 12(4):473-479を参照されたい)。
【0055】
他の調節配列も当業で公知であり、その中には、例えばグロビン(例えばRistaldi et al.1999. Proc Natl Acad Sci U S A. 96:9654;
Ristaldi et al. 1999 Blood Cells Mol. Dis. 25:193; Tewari et al. 1996. Development. 122:3991を参照されたい)βスペクトリン(例えばSabatino et al. 1998 Mol Cell Biol. 18:6634; Gallagher et al.
1999 J. Biol.
Chem. 274:6062を参照されたい)及びバンド3(例えばCox et al. 1985. J. Cell
Biol. 100:1548を参照されたい)がある。造血細胞、そして優先的には赤血球、で優先的に発現する他の遺伝子の発現を制御する調節配列も、本発明で用いてよい。
【0056】
他の適したプロモータは、例えばまず示差的発現解析を、DNAマイクロアレイ技術を用いるなどして行って、赤血球で優先的に発現する遺伝子を特定し、次に、対応する調節配列を特定するなどにより、特定することができる。例えば、肝臓又は心臓など、別の参照細胞又は組織に比べて赤血球で、例えば約2倍、5倍、又は10倍など、優先的に発現するとして特定された遺伝子を選抜し、それらのプロモータを特定し、本発明で用いることができよう。示差的発現解析を行う方法及びDNAマイクロアレイ技術は当業で公知である。
【0057】
遺伝子コンストラクトの一例は、ヒトCR1遺伝子をGATA-1プロモータの転写制御下に含むものであり、図1に示されている。造血細胞でCR1を発現させるために用いることのできる他の因子は当業で公知であり、例えば Okuno
et al. 2002. Blood. 100:4420;
Radomska et al. 2002. Blood. 100:4410; Gaines et al. 2000. J Biol Chem.275:34114; and Wilcox et al.
1999. Proc Natl
Acad Sci U S A. 96:9654に見ることができる。好ましくは、赤血球上でのCR1発現をより特異的に増加させる調節配列を当該のコンストラクトで用いるとよい。
【0058】
ある実施態様では、本発明のコンストラクトに、例えば緑色蛍光タンパク質又は抗生物質耐性遺伝子などのマーカ遺伝子を含めることができる。このようなマーカ遺伝子を用いると、当該コンストラクトを発現する細胞を特定する上で助けとなる。
【0059】
III. トランスジェニック動物の構築
ある局面では、本発明は、ヒト補体受容体1(CR1)が当該動物の赤血球上で機能的に発現するように、そのゲノムが、ヒトCR1をコードするポリヌクレオチドをプロモータに作動的に連結させて含有する非ヒト動物を提供する。更に本発明は、当該動物の赤血球上でヒトCR1を発現する非ヒトトランスジェニック動物を作製する方法も提供する。
【0060】
本発明の方法で用いられるトランスジェニック動物は、例えば哺乳動物、トリ、は虫類又は両生類などであってよい。ここで解説された用途に適した哺乳動物には、げっ歯類;反芻動物;有蹄動物;家畜哺乳動物;及び酪農動物がある。好適な動物には、げっ歯類、ヤギ、ヒツジ、ラクダ、ウシ、ブタ、ウマ、雄牛、ラマ、ニワトリ、がちょう、及び七面鳥がある。ある好適な実施態様では、該非ヒト動物はマウスである。
【0061】
トランスジェニック動物を作製する多種の方法が当業で公知である(例えばWatson, J. D., et al., "The Introduction of Foreign Genes Into
Mice," in Recombinant DNA, 2d Ed., W. H. Freeman & Co., New York
(1992), pp. 255-272; Gordon, J. W., Intl. Rev. Cytol. 115:171-229 (1989);
Jaenisch, R., Science 240: 1468-1474 (1989); Rossant, J., Neuron 2: 323-334
(1990)を参照されたい)。トランスジェニック・ブタを作製するプロトコルの一例をWhite and
Yannoutsos, Current Topics in Complement
Research: 64th Forum in Immunology, pp. 88-94; 米国特許第5,523,226号;米国特許第5,573,933号;PCT出願 WO93/25071; 及びPCT出願 WO95/04744に見ることができる。トランスジェニック・ラットを作製するためのプロトコルの一例はBader and Ganten, Clinical and
Experimental Pharmacology and Physiology, Supp. 3:S81-S87, 1996に見ることができる。トランスジェニック・ウシを作製するためのプロトコルの一例は、アカデミック・プレス社刊Transgenic Animal
Technology, A Handbook, 1994, ed. Carl A.
Pinkertに見ることができる。トランスジェニック・ヒツジを作製するためのプロトコルの一例は、アカデミック・プレス社刊Transgenic Animal Technology, A Handbook, 1994, ed., Carl A.
Pinkertに見ることができる。いくつかの例示的な方法を、以下に詳述する。
【0062】
A. 前核への注入
トランスジェニック動物は、本発明による核酸コンストラクトを卵細胞に導入することにより、作製することができる。その結果得られた卵細胞を、正常な胚発生に向けてメスの子宮に移植し、こうして発生した、導入遺伝子を持つ動物を戻し交配して、この導入遺伝子についてヘテロ接合子を作る。多様な発生上の段階での胚性標的細胞を用いて、本発明の導入遺伝子を導入する。胚性標的細胞の発生上の段階に応じて異なる方法を用いる。導入遺伝子を導入する方法の例には、限定はしないが、受精後の卵又は接合子の顕微注射(Brinster, et al., Proc. Natl. Acad. Sci.USA (1985) 82: 4438-4442)、及びウィルス組込み法(Jaenisch R., Proc. Natl. Acad. Sci. USA (1976) 73: 1260-1264;
Jahner, et al., Proc. Natl. Acad. Sci.USA (1985) 82: 6927-6931; Van der Putten,
et al., (1985) Proc. Natl. Acad. Sci. (USA) 82: 6148-6152)がある。胚の操作及び顕微注射のための手法は、例えば、Manipulating the Mouse
Embryo (引用をもってその内容をここに援用することとするCold Spring Harbor Laboratory Press, Cold Spring Harbor, NY., 1986)に解説されている。同様な方法は他のトランスジェニック動物の作製にも用いられる。
【0063】
例示的な実施態様では、トランスジェニック・マウスの作製で以下のステップを利用する。定義された近交遺伝子バックグラウンドからのオス及びメスのマウスを交配する。交配するメスのマウスは、卵胞の成長を誘導する妊娠ウマ血清PMSと、排卵を誘導するヒト絨毛性ゴナドトロピンhCGで予め処理されている。交配後、このメスをと殺し、受精卵を彼女の卵管から取り出す。この時点では、前核は融合しておらず、それらを光学顕微鏡を用いて観察することが可能である。代替的なプロトコルでは、様々な発生上の段階で胚を採集することができ、胚盤胞などを採集することができる。胚をダルベッコの改良リン酸緩衝生理食塩水(DPBS)中に回収し、10%のウシ胎児血清を添加したダルベッコの改良基本培地(DMEM)中に維持する。
【0064】
次に、外来のDNA又は組換えコンストラクト(例えばhCR1発現コンストラクト)を前核に顕微注射(卵一個当たり100-1000 分子)する。発現コンストラクトの顕微注射は、顕微鏡に取り付けられた標準的なマイクロ・マニピュレータを用いて行うことができる。例えば、胚は、典型的には、顕微注射の間中、油脂に覆われた100マイクロリットルのDPBSの液滴中に支持される。DNA溶液はオスの前核に顕微注射される。前核の膨らみにより、注射の成功を観察する。そのすぐ後に、前核(メスの前核及びオスの前核)の融合が起こり、場合によっては、外来のDNAが受精卵又は接合子の(通常は)一方の染色体内に挿入される。以下に記載するように調製された組換えES細胞を、同様の技術を用いて胚盤胞に注射することができる。
【0065】
B. 胚性幹細胞
トランスジェニック・マウスを作製する別の方法では、本発明の組換えDNA分子を、マウスの胚性幹(ES)細胞に導入することができる。次に、こうして出来た組換えES細胞をマウス胚盤胞に、前記の小項で記載したものと同様な技術を用いて顕微注射する。
【0066】
ES細胞は移植前の胚から得られ、in vitroで培養される(Evans, M J., et al., Nature 292: 154156 (1981); Bradley, M. O. et
al., Nature 309: 255-258 (1984); Gossler, et al., Proc. Natl.
Acad. Sci.
(USA)
83:9065-9069 (1986); Robertson et al., Nature 322:
445448 (1986))。標的決定コンストラクトの生殖細胞系伝播が起きるように、発生中の胚の生殖細胞に組み込ませ、その一部に成らせることができれば、いずれのES細胞株も、ここでの使用に適切である。例えば、ES細胞の作製に用いることのできるマウス株の一つが129J株である。好適なES細胞株はマウス細胞株D3(アメリカン・タイプ・カルチャー・コレクション、カタログ番号CRL 1934)である。このES細胞は、当業で公知であり、そして引用をもってその内容をここに援用することとするRobertson,
Teratocarcinomas and Embryonic Stem
Cells: A Practical Approach,
E.J. Robertson, ed. IRL Press, Washington, D.C.,
1987、Bradley et al., Current Topics in Devel. Biol., 20:357-371, 1986 及びHogan et al., Manipulating the
Mouse Embryo: A Laboratory Manual,
Cold Spring Harbor Laboratory Press, Cold Spring Harbor, NY, 1986に解説された方法を用いて培養し、DNA挿入に向けて調製することができる。
【0067】
発現コンストラクトは、例えば引用をもってここにその内容を援用することとするSambrook et al., Molecular
Cloning: A Laboratory Manual, 2nd Ed., ed., Cold Spring Harbor
laboratory Press: 1989に解説されたものなど、当業で公知の方法により、ES細胞に導入することができる。適した方法には、限定はしないが、エレクトロポレーション、顕微注射、及びリン酸カルシウム処理法がある。ES細胞に導入しようとする発現コンストラクト(例えばhCR1)は直線状であることが好ましい。直線化は、当該のDNAを、ベクタ配列内のみで切断し、当該遺伝子(例えばhCR1)又は調節配列(例えばGATA-1調節配列)内では切断しないように選択された適した制限エンドヌクレアーゼで消化することにより、達成することができる。
【0068】
発現コンストラクトの導入後に、ES細胞をコンストラクトの存在についてスクリーニングする。当該細胞は多種の方法を用いてスクリーニングすることができる。マーカ遺伝子がコンストラクトに用いられている場合、動物の細胞を、マーカ遺伝子の存在について検査することができる。例えば、マーカ遺伝子が抗生物質耐性遺伝子である場合、細胞を、さもなければ致死濃度である抗生物質(例えばneoについて選択する場合はG418)の存在下で培養することができる。生き延びる細胞は、おそらく、導入遺伝子コンストラクトを組み込ませたものであろう。マーカ遺伝子が、その活性を検出可能な酵素(例えばβ-ガラクトシダーゼ)をコードしている遺伝子である場合、この酵素の基質を適した条件下で細胞に加え、酵素活性を分析することができる。代替的又は付加的には、ES細胞ゲノムDNAを直接調べることができる。例えば、DNAをES細胞から標準的な方法を用いて抽出し、こうしてこのDNAを、導入遺伝子に特異的にハイブリダイズするようにデザインされた一つ又は複数のプローブを用いたサザン・ブロット法でプローブすることができる。更にゲノムDNAは、標的決定コンストラクトを含有する細胞のみが、適したサイズのDNA断片を生ずるように、特定の大きさと導入遺伝子の配列とを持つDNA断片を増幅するように特にデザインされたプローブを用いたPCRでも、増幅することができる。
【0069】
C. 移植
本発明の組換え核酸分子(例えばhCR1)を持つ接合子を、精管切除されたオスとの以前の交配で得られた偽妊娠のメスのマウスに移植する。一般的なプロトコルでは、レシピエントのメスを麻酔し、対腰部切開を行って輸卵管を露出させ、胚をこの輸卵管の膨大部領域に移植する。体壁を縫合し、皮膚を創傷クリップで閉じる。胚は全妊娠期間にわたって発生し、代理母は潜在的トランスジェニック・マウスを出産する。最後に、マウス新生児を、外来又は組換えDNAの存在について検査する。注射を受けた卵のうち、平均10%が正しく発生し、マウスを生じる。生まれたマウスのうち、平均で4匹のうち1匹(25%)がトランスジェニックであり、全体的な効率は2.5%となる。これらのマウスを交配すると、それらは外来の遺伝子を通常の(メンデルの)態様でマウス染色体に連鎖した状態で伝播させる。
【0070】
D. トランスジェニック・コンストラクトの存在に関するスクリーニング
トランスジェニック動物は、標準的なプロトコルにより、生後に特定することができる。尾の組織からのDNAは、例えばサザン・ブロット及び/又はPCRを用いるなどして、導入遺伝子コンストラクトの存在についてスクリーニングすることができる。次に、モザイクになっていると思われる仔を、これらが導入遺伝子を持つと考えられる場合に、ホモ接合型の動物を生じさせるために相互に交配する。仔が生殖細胞系伝播を有するかどうかが明らかでない場合、これらを親又は他の株と交配し、その仔を、ヘテロ接合型についてスクリーニングすることができる。ヘテロ接合仔を、当該DNAのサザン・ブロット及び/又はPCR増幅法により特定する。こうして、ヘテロ接合仔を互いに交配して、ホモ接合型のトランスジェニック仔を生じさせることができる。ホモ接合仔は、この交配の産物であるマウス由来や、既知のヘテロ接合仔であるマウス及び野生型マウス由来の等量のゲノムDNAのサザン・ブロット法によっても特定できよう。当該のサザン・ブロットをスクリーニングするためのプローブは、ヒトCR1遺伝子又はマーカ遺伝子あるいは両者の配列に基づき、デザインすることができる。
【0071】
トランスジェニックの仔を特定し、特徴付ける他の手段は当業で公知である。例えば、ウェスタン・ブロットを用いて、導入された遺伝子にコードされたタンパク質(例えばヒトCR1タンパク質)に対する抗体、あるいは、マーカ遺伝子が発現したときのその産物に対する抗体、によりウェスタン・ブロットをプローブすることにより、これらの仔の多様な組織に導入された遺伝子の発現レベルを評価することができる。
【0072】
細胞を固定し、抗体で標識するなどのin situ解析、及び/又は、仔の赤血球などの多様な細胞のFACS(蛍光活性化セル・ソーティング)解析は、導入遺伝子産物の存在又は非存在について調べるために適した抗体を用いて行うことができる。例えば、赤血球上のhCR1の発現を確認するためには、ヒトCR1に特異的な抗体(例えば1B4、HB8592 及び7G9 モノクローナル、米国特許出願No. 20020103343を参照されたい)を、直接結合させて用いるか、あるいは、蛍光体を結合させた、抗hCR1抗体を認識する二次抗体と併用して、フローサイトメトリを行うことができる。この解析では、ヒト赤血球をhCR1の存在に関する陽性コントロールとして用いることができ、そして正常なマウス赤血球を陰性コントロールとして用いることができる。
【0073】
E. 複数の導入遺伝子又は付加的な変異を含有するマウス
ここで解説された通りにhCR1をそれらの循環赤血球上に発現するトランスジェニック・マウスを、a)更なる導入遺伝子を持つ、又はb)他の遺伝子に変異を含有する、マウスと交配することができる。変異のそれぞれについてヘテロ接合型又はホモ接合型のマウスは、標準的な交配法を用いて作製し、維持することができる。hCR1導入遺伝子を含有するマウスと交配することのできるマウスの例には、限定はしないが、例えば マウス株NZB/W、MRL/+ 又はSJLなど、狼瘡のモデルであるマウス株など、自己免疫疾患の傾向が高いマウス株がある。
【0074】
本発明は、更に、トランスジェニック動物を由来とする細胞にも関する。突然変異又は環境による影響が原因で、後代に何らかの改変が起きる場合があるため、このような後代は、実際には親細胞と同一でないかも知れないが、それでも尚、ここで用いられるこの用語の範囲内に包含される。
【0075】
IV. 二重特異的化合物
ある局面では、本発明は、非ヒト動物の血清及び/又は循環中の作用物質の濃度を低下させる候補又は検査二重特異的化合物を同定する方法を提供するものである。該方法を用いて選択された又は最適化された化合物は、ヒトの対象など、このような化合物を投与すると有益であろう対象を治療するために用いることができる。
【0076】
本発明の方法で検査可能な候補化合物は二重特異的化合物である。ここで用いられる場合の用語「二重特異的化合物」には、2つの異なる結合特異性部分を有する化合物が包含される。二重特異的化合物の例には、例えば二重特異的抗体、ヘテロ重合体、及び抗原ベースのヘテロ重合体など、がある。
【0077】
本発明で検査可能な二重特異的分子は、好ましくは、CR1、好ましくはヒトCR1、に特異的な結合部分を、標的とされる作用物質(異なる抗体又は抗原)に特異的な第二結合部分に架橋させて含むとよい。CR1に対して特異的な結合部分の例には、限定はしないが、CR1リガンド、例えばC3b、があるが、あるいは、好適な実施態様では、CR1に対する抗体、がある。CR1に対して特異的なモノクローナル抗体の例には、限定はしないが、以下:1B4、HB8592、7G9、3D9、E-11、57F 及びYZ1のうちのいずれか一つがある。HB8592 及び1B4 はTaylor
et al., Proc. Nat. Acad. Sci.,
88:3305-3309 (1991) 及びReist et al., Eur. J. Immunol., 23:3021-3027 (1993)に開示されている。抗体3D9 及びE-11 は、Edberg
et al., (1992) Eur J Immunol. 22:
1333-9に解説されている。
【0078】
別の実施態様では、新規なCR1結合分子を、それらのCR1への結合能に基づいて同定することができる。例えば、化合物又は低分子のライブラリを無細胞結合検定法で検査することができる。例えばペプチド・ミメティック、低分子又は他の薬物などのいずれの数の検査化合物も検査に用いることができ、また、数多くのアプローチのいずれかを、生物学的ライブラリ;空間指定可能なパラレル固相又は液相ライブラリ;デコンボリューションを要する合成ライブラリ法;「ワン−ビーズ・ワン−コンパウンド」ライブラリ法;及びアフィニティ・クロマトグラフィ選抜を用いた合成ライブラリ法を含め、当業で公知のコンビナトリアル・ライブラリ法において用いて得ることができる。生物学的ライブラリのアプローチはペプチド・ライブラリに限られているが、他の四種類のアプローチは、ペプチド、非ペプチド・オリゴマ又は低分子の化合物ライブラリに応用可能である(Lam, K.S. (1997) Anticancer Drug Des. 12:145)。
【0079】
分子ライブラリの合成のための方法の例は、当業において、例えばDeWitt et al. (1993) Proc. Natl. Acad. Sci. U.S.A. 90:6909; Erb et al. (1994) Proc.
Natl. Acad. Sci. USA 91:11422;
Zuckermann et al. (1994). J. Med. Chem. 37:2678; Cho et al.
(1993) Science 261:1303; Carrell et
al. (1994) Angew. Chem. Int.
Ed. Engl.
33:2059; Carell et al. (1994) Angew. Chem.
Int. Ed.
Engl. 33:2061; and in Gallop et al. (1994) J. Med.
Chem. 37:1233に見ることができる。
【0080】
化合物のライブラリは溶液中(例えば Houghten (1992) Biotechniques 13:412-421)、又はビーズ上(Lam (1991) Nature
354:82-84)、チップ上(Fodor (1993) Nature 364:555-556)、細菌(Ladner 米国特許第5,223,409号)、胞子(Ladner 米国特許第 '409号)、プラスミド(Cull et al. (1992) Proc Natl Acad Sci USA 89:1865-1869)又はファージ上(Scott and Smith (1990) Science
249:386-390);(Devlin (1990) Science 249:404-406); (Cwirla et al. (1990) Proc. Natl. Acad. Sci. 87:6378-6382); (Felici (1991) J. Mol. Biol. 222:301-310); (上記のLadner)に提供してもよい。
【0081】
調節性物質及び天然抽出物のライブラリを検査する数多くの薬物スクリーニング・プログラムでは、所定の時間で調査される調節性物質の数を最大にするためには、高スループットの検定法が好ましい。精製済み又は半精製済みタンパク質で得られるものなど、無細胞系で行われる検定法が「一次」スクリーンとしてしばしば好ましい。なぜなら、これらは、検査対象である調節性物質により媒介される分子ターゲット中の変化を高速で進行させ、また比較的容易に検出できるように作製することが可能だからである。更に、検査対象の調節性物質の細胞毒性及び/又は生物学的利用能の効果は、in vitro系では概ね無視することができ、代わりに、当該検定の焦点を、上流又は下流の要素との結合親和性の変化に現れるであろう、当該薬物の分子ターゲットに対する効果に、主に当てることができる。
【0082】
別の実施態様では、当業で公知のファージ・ディスプレイ技術を用いて新規なCR1結合分子を同定することができる。
【0083】
ある実施態様では、本発明は、CR1又はその生物学的に活性な部分に結合する候補又は検査化合物をスクリーニングするための検定法を提供する。
【0084】
CR1に結合する分子を同定するための細胞ベースの検定法を、本発明の二重特異的化合物での使用に向けた付加的な作用物質を同定するために用いることができる。例えば、CR1を発現する細胞をスクリーニング検定法で用いることができる。例えば、CR1への結合において統計上有意な変化を生ずる化合物を同定することができる。
【0085】
本発明のある実施態様では、CR1又はその部分を、二種ハイブリッド検定法又は三種ハイブリッド検定法で「ベイト・タンパク質」として用いて(例えば米国特許第5,283,317号; Zervos et al. (1993) Cell
72:223-232; Madura et al. (1993) J. Biol.
Chem. 268:12046-12054; Bartel et al. (1993) Biotechniques
14:920-924; Iwabuchi et al. (1993) Oncogene
8:1693-1696; and Brent WO94/10300を参照されたい)、本発明のタンパク質に結合又は相互作用すると共に、それらの活性に関与するような他のタンパク質を同定することができる。前記の二種ハイブリッド系は、分離可能なDNA結合ドメイン及び活性化ドメインから成る大半の転写因子のモジュール性に基づいている。簡単に説明すると、該検定法では2つの異なるDNAコンストラクトを利用する。一方のコンストラクトでは、あるタンパク質をコードする遺伝子が、既知の転写因子(例えばGAL-4)のDNA結合ドメインをコードする遺伝子に融合されている。他方のコンストラクトでは、未知のタンパク質(「プレイ」又は「試料」)をコードする、DNA配列ライブラリ由来の一DNA配列が、該既知の転写因子の活性化ドメインをコードする遺伝子に融合されている。これら「ベイト」及び「プレイ」タンパク質がin vivoで相互作用して複合体を形成することができれば、該転写因子のDNA結合ドメイン及び活性化ドメインが近くにあることになる。この近さにより、該転写因子に対して応答性の転写調節部位に作動的に連結されたレポータ遺伝子(例えばLacZ)の転写が可能である。レポータ遺伝子の発現を検出し、機能的転写因子を含有する細胞コロニを単離し、これを用いて、本発明のタンパク質と相互作用するタンパク質をコードする、クローニングされた遺伝子を得ることができる。相互作用性タンパク質が同定されたら、本発明のタンパク質と、その相互作用性タンパク質との間の相互作用を調節する上での検査化合物の能力を、当業で公知の技術を用いて、例えばここで解説する無細胞検定法を用いて、検査することができる。
【0086】
ある実施態様では、当該の検定は、CR1結合分子が、in vitroでのCR1へのその結合能に基づいて同定される無細胞検定法である。CR1結合分子を提供することができ、CR1へのそのタンパク質の結合能 を直接的な結合を判定する当業で公知の方法を用いて検査することができる。当該タンパク質の、標的分子への結合能の判定は、例えば、リアルタイム・バイオモラキュラー・インターアクション・アナリシスBiomolecular Interaction Analysis (BIA). Sjolander, S. and Urbaniczky, C. (1991) Anal. Chem. 63:2338-2345 and Szabo et
al. (1995) Curr. Opin. Struct. Biol.
5:699-705などの技術を用いて行うことができる。ここで用いる場合の「BIA」とは、相互作用物質のいずれも標識せずに、二重特異的相互作用をリアルタイムで研究する技術である(例えばBIAcore)。表面プラズモン共鳴(SPR)の光学的現象の変化を、生物学的分子間のリアルタイムの反応の指標として用いることができる。
【0087】
本発明の無細胞検定法は、可溶性及び/又は膜結合型のタンパク質の使用の両方に適している。膜結合型のタンパク質を用いる無細胞検定法の場合、膜結合型のタンパク質が溶液中に維持されるように、可溶化剤を用いることが好ましいであろう。このような可溶化剤の例には、非イオン性の界面活性剤、例えばn-オクチルグルコシド、n-ドデシルグルコシド、n-ドデシルマルトシド、オクタノイル-N-メチルグルカミド、デカノイル-N-メチルグルカミド、Triton(R)X-100、 Triton(R)X-114、 Thesit(R)、イソトリドデシポリ(エチレングリコールエーテル)n、3-[(3-コラミドプロピル)ジメチルアミニオ]-1-プロパンスルホネート(CHAPS)、3-[(3-コラミドプロピル)ジメチルアミニオ]-2-ヒドロキシ-1-プロパンスルホネート (CHAPSO)、又はN-ドデシル=N,N-ジメチル-3-アンモニオ-1-プロパンスルホネートがある。
【0088】
タンパク質対タンパク質の相互作用を検出することができる適した検定法が当業で公知である(例えば免疫沈降法、二種ハイブリッド検定法等)。このような検定法を検査化合物の存在下及び非存在下で行うことにより、これらの検定法を用いて、本発明のタンパク質の標的分子との相互作用を調節(例えば阻害又は亢進)する化合物を同定することができる。
【0089】
当該タンパク質の、標的分子との結合能又は相互作用能の判定は、例えば直接的な結合などにより、行うことができる。直接的な結合検定法では、当該のタンパク質の標的分子への結合を、複合体中の標識済みタンパク質を検出することにより判定できるように、放射性同位元素又は酵素標識に当該タンパク質を結合させてもよいであろう。例えば、タンパク質は、125I、35S、14C、又は3Hで直接又は間接的に標識することができ、その放射性同位元素を、放射線放出を直接計数したり、あるいはシンチレーション計数により検出することができる。代替的には、分子を、例えば西洋わさびペルオキシダーゼ、アルカリホスファターゼ、又はルシフェラーゼなどで酵素標識し、その酵素標識を、適した基質の生成物への転化を判定することにより、検出することができる。
【0090】
典型的には、本発明のタンパク質又はその結合タンパク質のいずれかを固定して、複合体を形成していない形の一方又は両方のタンパク質から複合体を容易に分離できるようにし、また検定法の自動化に対応することも好ましいであろう。候補作用物質の存在下及び非存在下における、上流又は下流の結合因子への結合を、反応物を含有するのに適したいずれかの容器で行わせることもできる。例には、微量定量プレート、試験管、及びマイクロ遠心管がある。ある実施態様では、当該タンパク質をマトリックスに結合可能にするドメインを加えた融合タンパク質を提供することができる。例えば、グルタチオン-S-トランスフェラーゼ/ CR1 (GST/ CR1) 融合タンパク質をグルタチオン・セファロース・ビーズ(ミズーリ州セントルイス、シグマ・ケミカル社)又はグルタチオン誘導体化微量定量プレートに吸着させた後、これらを35Sで標識されているなどの細胞ライセートと検査対象の調節性物質とに配合し、この混合物を、例えば塩及びpHについて生理条件など、複合体形成を誘導する条件下でインキュベートすることができるが、僅かによりストリンジェントな条件が好ましいであろう。インキュベート後、ビーズを洗浄して未結合の標識を取り除き、マトリックスに固定し、放射性標識を直接(例えばビーズをシンチレーション剤中に配置する)判定するか、あるいは、複合体を解離させた後に上清中で判定する。代替的には、複合体をマトリックスから解離させ、SDS-PAGEで分離し、ビーズ画分中に見られるCR1結合タンパク質のレベルを標準的な電気泳動技術を用いてゲルから定量することができる。
【0091】
マトリックス上にタンパク質を固定する他の技術も、本検定法での使用に利用できる。例えば、ビオチン化分子をビオチン-NHS (N-ヒドロキシ-スクシンイミド)から当業で公知の技術(例えばイリノイ州ロックフォード、ピアース・ケミカルズ社のビオチン化キット)を用いて調製し、
ストレプトアビジンで被覆された96ウェル・プレート(ピアース・ケミカル社)のウェルに固定することができる。
【0092】
更に、相互作用物質のいずれも標識せずに、CR1とCR1結合分子との間の相互作用を調節する上での化合物の能力を判定することも、本発明の範囲内である。例えば、マイクロフィジオメータを用いて、本発明のタンパク質の、その標的分子との相互作用を、このタンパク質又は標的分子のいずれも標識せずに検出することができる。McConnell, H. M. et al. (1992)
Science 257:1906-1912。ここで用いられる「マイクロフィジオメータ」(例えばCytosensor)は、細胞がその環境を酸化する速度を、光指定可能な電位差センサ(LAPS)を用いて測定する分析装置である。この酸化速度の変化を、化合物と受容体との間の相互作用の指標として用いることができる。
【0093】
本発明で検査可能な抗原ベースのヘテロ重合体は、優先的には、CR1、好ましくはヒトCR1、に特異的な結合部分を、自己抗体により認識される抗原に架橋させて含む。自己抗体により認識される抗原の例には、限定はしないが、以下:VIII因子(置換組換えVIII因子による血友病治療に関連する抗体);筋アセチルコリン受容体(重症筋無力症に関連する抗体);カルジオリピン(狼瘡に関連する);血小板関連タンパク質(特発性血小板減少性紫斑病に関連する);シェーグレン症候群に関連する複数の抗原;組織移植自己免疫反応の症例への関与が示唆されている抗原;心筋に見られる抗原(自己免疫心筋炎に関連する);免疫複合体媒介性腎疾患に関連する抗原;dsDNA 及びssDNA 抗原(ループス腎炎に関連する); デズモグレイン及びデズモプラキン(天疱瘡及び類天疱瘡に関連する);又はよく特徴付けられていると共に、疾患病理に関連するいずれか他の抗原のいずれもが含まれる。
【0094】
本発明での検査の対象となるヘテロ重合体及び抗原ベースのヘテロ重合体の例や、それらを作製する方法は当業で公知である。例えば、ヘテロ重合体の例はWO 03007971A1; US 20020103343A1; US 5879679; US 5487890; US 5470570;
WO 9522977A1; WO/02075275A3、 WO/0246208A2 もしくはA3、WO/0180883A1、WO/0145669A1、WO 9205801A1、Lindorfer et al. 2001 J. Immunol. Methods. 248:125; Hahn et al. 2001. J. Immnol.
166:1057; Nardin et al. 1998. J. Immunol.
Methods. 211:21; Kuhn et
al. 1998. J. Immunol.
160:5088; Taylor et
al. 1997. Cancer
Immunol. Immunother. 45:152;
Taylor et al. 1997. J. Immunol.
159:4035; 及び Taylor et al. 1992. J. Immunol.
148:2462に教示されている。加えて、バリアント型のこれらのヘテロ重合体を作製することもできる。例えば、ある実施態様では、異なる連結化学法を用いて作製された数々の形の二重特異的分子を用いることができる。二重特異的分子の成分を架橋するために用いることのできる試薬の例には:ポリエチレングリコール、SATA、SMCC、や当業で公知の他のものがあり、例えばピアース・バイオテクノロジー社から得られる。検査することのできる二重特異的分子の形の例は、引用をもってその内容をここに援用することとする、2002年9月16日に提出されたUSSN 60/411,731に解説されている。
【0095】
別の実施態様では、様々な多量体型の二重特異的分子を作製することができる(例えば二量体、三量体、四量体、五量体、又はそれより大きな多量体型)。別の実施態様では、精製された形の二重特異的分子を、例えば引用をもってその内容をここに援用することとする、2002年5月13日に提出された USSN 60/380,211に解説されている通りに、検査することができる。
【0096】
別の実施態様では、当該のヘテロ重合体の結合部分のうちの一つが抗体である場合、異なるアイソタイプ(例えばIgA、IgD、IgE、IgG1、IgG2 (例えばIgG2a)、IgG3、IgG4、又はIgM)の抗体を用いることができる。別の実施態様では、抗体分子の部分(例えばFabフラグメント)を結合部分の一つに用いることができる。ある好適な実施態様では、結合部分のうちの少なくとも1つは、Fcドメインを含む抗体である。ある実施態様では、当該の抗体はマウス抗体である。
【0097】
別の実施態様では、抗体に対する修飾の効果を検査することができ、例えば2003年3月28日に提出されたUSSN 60/458,869に解説されたように、抗体のデイミュナイゼーションの効果を検査することができる。
【0098】
本発明の二重特異的化合物が結合することのできる標的とされる作用物質の例には、以下:ウィルス、ウィルス性粒子、毒素、細菌、ポリヌクレオチド、抗体、例えば自己免疫異常に関連する自己抗体など、のうちのいずれも含む、しかしこれらに限定はしない血液由来作用物質がある。ある実施態様では、標的とされるウィルス性作用物質の例には、限定はしないが、以下:サイトメガロウィルス、インフルエンザ、
ニューカッスル病ウィルス、水疱性口内炎ウイルス、単純疱疹ウィルス、肝炎、アデノウィルス-2、ウシウイルス性下痢ウイルス、ヒト免疫不全ウィルス (HIV)、デング熱ウィルス、マーブルグウィルス、エプスタイン-バーウィルス、のうちのいずれも含まれる。
【0099】
細菌性作用物質の例には:シュードモナス-アエルギノーサ(原語:Pseudomonas aeruginosa)、シュードモナス-フルオレセンス(原語:Pseudomonas fluorescens)、シュードモナス-アシドボランス(原語:Pseudomonas acidovorans)、シュードモナス-アルカリゲンス(原語:Pseudomonas alcaligenes)、シュードモナス-プチダ(原語:Pseudomonas putida)、ステノトロフォモナス-マルトフィリア(原語:Stenotrophomonas maltophilia)、バークホルデリア-セパシア(原語:Burkholderia cepacia)、アエロモナス-ヒドロフィリア(原語:Aeromonas hydrophilia)、エシェリヒア-コリ(原語:Escherichia coli)、シトロバクター-フロウンジ(原語:Citrobacter freundii)、サルモネラ-チフィムリウム(原語:Salmonella typhimurium)、サルモネラ-チフィ(原語:Salmonella typhi)、サルモネラ-パラチフィ(原語:Salmonella paratyphi)、サルモネラ-エンテリティディス(原語:Salmonella enteritidis)、シゲラ-ジセンテリア(原語:Shigella dysenteriae)、シゲラ-フレクスネリ(原語:Shigella flexneri)、シゲラ-ソネイ(原語:Shigella sonnei)、エンテロバクター-クロアケ(原語:Enterobacter cloacae)、エンテロバクター-アエロゲンス(原語:Enterobacter aerogenes)、クレブシエラ-ニューモニエ(原語:Klebsiella pneumoniae)、クレブシエラ-オキシトカ(原語:Klebsiella oxytoca)、セラチア-マルセセンス(原語:Serratia marcescens)、フランシセラ-ツラレンシス(原語:Francisella tularensis)、モルガネラ-モルガニイ(原語:Morganella morganii)、プロテウス-ミラビリス(原語:Proteus mirabilis)、プロテウス-ブルガリス(原語:Proteus vulgaris)、プロビデンシア-アルカリファシエンス(原語:Providencia alcalifaciens)、プロビデンシア-レットゲリ(原語:Providencia rettgeri)、プロビデンシア-スチュアーティ(原語:Providencia stuartii)、アシネトバクター-カルコアセチクス(原語:Acinetobacter calcoaceticus)、アシネトバクター-ヘモリティクス(原語:Acinetobacter haemolyticus)エルシニア-エンテコリティカ(原語:Yersinia enterocolitica)、エルシニア-ペスティス(原語:Yersinia pestis)、エルシニア-シュードツベルキュローシス(原語:Yersinia
pseudotuberculosis)、エルシニア-インテルメディア(原語:Yersinia intermedia)、ボルデテッラ-ペルツシス(原語:Bordetella pertussis)、ボルデテッラ-パラペルツシス(原語:Bordetella parapertussis)、ボルデテッラ-ブロンキセプティカ(原語:Bordetella bronchiseptica)、ヘモフィラス-インフルエンザ(原語:Haemophilus influenzae)、ヘモフィラス-パラインフルエンザ(原語:Haemophilus parainfluenzae)、ヘモフィラス-ヘモリティクス(原語:Haemophilus haemolyticus)、ヘモフィラス-パラへモリティカス(原語:Haemophilus parahaemolyticus)、ヘモフィラス-デュクレイ(原語:Haemophilus ducreyi)、パスツーレラ-マルトシダ(原語:Pasteurella multocida)、パスツーレラ-ヘモリティカ(原語:Pasteurella haemolytica)、ブランハメラ-カタラーリス(原語:Branhamella catarrhalis)、ヘリコバクター-ピロリ(原語:Helicobacter pylori)、カンピロバクター-フェツス(原語:Campylobacter fetus)、カンピロバクター-ジェジュニ(原語:Campylobacter jejuni)、カンピロバクター-コリ(原語:Campylobacter coli)、ボレリア-ブルドルフェリ(原語:Borrelia burgdorferi)、ビブリオ-コレラ(原語:Vibrio cholerae)、ビブリオ-パラヘモリティクス(原語:Vibrio parahaemolyticus)、レジオネラ-ニューモフィラ(原語:Legionella pneumophila)、リステリア-モノサイトゲンス(原語:Listeria monocytogenes)、ネイセリア-ゴノローエ(原語:Neisseria gonorrhoeae)、ネイセリア-メニンギティディス(原語:Neisseria meningitidis)、ガルドネレッラ-バギナリス(原語:Gardnerella vaginalis)、バクテロイデス-フラギリス(原語:Bacteroides fragilis)、バクテロイデス-ディスタソニス(原語:Bacteroides distasonis)、バクテロイデス3452A ホモロジー群、バクテロイデス-ブルガツス(原語:Bacteroides vulgatus)、バクテロイデス-オバルス(原語:Bacteroides ovalus)、バクテロイデス-エタイオタオミクロン(原語:Bacteroides thetaiotaomicron)、バクテロイデス-ユニフォルミス(原語:Bacteroides uniformis)、バクテロイデス-エゲルシー(原語:Bacteroides eggerthii)、バクテロイデス-スプランクニカス(原語:Bacteroides splanchnicus)、クロストリジウム-ジフィシル(原語:Clostridium difficile)、マイコバクテリウム-ツベルキュローシス(原語:Mycobacterium tuberculosis)、マイコバクテリウム-アビウム(原語:Mycobacterium avium)、マイコバクテリウム-イントラセルラーレ(原語:Mycobacterium intracellulare)、マイコバクテリウム-レプレー(原語:Mycobacterium leprae)、コリネバクテリウム-ジフテリア(原語:Corynebacterium diphtheriae)、コリネバクテリウム-ウルセランス(原語:Corynebacterium ulcerans)、スタフィロコッカス-ニューモニエ(原語:Streptococcus pneumoniae)、ストレプトコッカス-アガラクチエ(原語:Streptococcus agalactiae)、ストレプトコッカス-パイオゲンス(原語:Streptococcus pyogenes)、エンテロコッカス-フェカーリス(原語:Enterococcus faecalis)、エンテロコッカス-ファシウム(原語:Enterococcus faecium)、スタフィロコッカス-アウレウス(原語:Staphylococcus aureus)、スタフィロコッカス-エピデルミディス(原語:Staphylococcus epidermidis)、スタフィロコッカス-サプロフィティカス(原語:Staphylococcus saprophyticus)、スタフィロコッカス-インテルメディウス(原語:Staphylococcus intermedius)、スタフィロコッカス-ヒイカス亜種ヒイカス(原語:Staphylococcus hyicus subsp.
hyicus)、スタフィロコッカス-ヘモリティカス(原語:Staphylococcus
haemolyticus)、スタフィロコッカス-ホミニス(原語:Staphylococcus hominis)、スタフィロコッカス-サッカロリティクス(原語:Staphylococcus saccharolyticus)、がある。
【0100】
ある実施態様では、標的とされる作用物質は、例えば抗生物質に対して耐性であるなど、伝統的な治療法に対して耐性である。
【0101】
別の実施態様では、本発明の抗原ベースのヘテロ重合体に結合させることのできる、標的とされる作用物質の例には、限定はしないが、以下:置換組換えVII因子による血友病治療に関連する自己抗体;重症筋無力症;狼瘡、ループス腎炎、特発性血小板減少性紫斑病、シェーグレン症候群、心筋炎、又は天疱瘡及び類天疱瘡に関連する自己抗体;組織移植自己免疫反応に関連する自己抗体;免疫複合体媒介性腎疾患に関連する自己抗体;又は、よく特徴付けられていると共に、疾患病理に関連するいずれか他の自己抗体、のうちのいずれもが含まれる。
【0102】
更に他の実施態様では、本発明の二重特異的化合物に結合させることのできる生物学的作用物質の例には、限定はしないが、以下:炭疽菌、天然痘、ペスト、エボラ、及びマーブルグ・ウィルスのいずれかを含め、生物戦争に関連する可能性がある感染性作用物質及び毒素がある。
【0103】
ある実施態様では、本発明の検定法を行う際に、当該の作用物質をトランスジェニック動物に、二重特異的化合物の投与前、投与と同時又は投与後などに投与する。
【0104】
本発明の二重特異的化合物、又はそのいずれかの部分を修飾してそれらの半減期を高めてもよい。ペプチド類似体が、テンプレート・ペプチドのものと同様な性質を持つ非ペプチド薬物として、製薬業では普通に用いられている。これらの種類の非ペプチド化合物は「ペプチド・ミメティック」又は「ペプチドミメティック」と呼ばれ(引用をもってここに援用することとするFauchere, J. (1986) Adv. Drug Res. 15:29; Veber and
Freidinger (1985) TINS p.392; 及び Evans et al. (1987) J. Med. Chem
30:1229)、通常は、コンピュータ分子モデリングの助けを得て開発されている。治療上有用なペプチドに構造上似たペプチド・ミメティックを用いて、同等な治療上又は予防上の効果を生じさせることができる。概して、ペプチドミメティックは、抗原ポリペプチドなどのパラダイム・ポリペプチド(即ち、生物学的又は薬理学的活性を有するポリペプチド)に構造上似ているが、:-CH2NH-、-CH2S-、-CH2-CH2-、-CH=CH- (cis 及びtrans)、-COCH2-、-CH(OH)CH2-、及び-CH2SO-から成る群より選択される結合部に、当業で公知であり、そして更に、それぞれを引用をもってここに援用することとする以下の参考文献: Spatola, A.F. in Chemistry
and Biochemistry of Amino Acids, Peptides, and Proteins Weinstein, B., ed.,
Marcel Dekker, New York, p. 267 (1983); Spatola, A.F., Vega Data (March
1983), Vol. 1, Issue 3, “Peptide Backbone Modifications”; Morley, J.S. (1980) Trends. Pharm. Sci. pp.463-468; Hudson, D. et al.
(1979) Int. J. Pept. Prot. Res. 14:177-185 (-CH2NH-, CH2CH2-); Spatola, A.F. et
al. (1986) Life. Sci.
38:1243-1249 (-CH2-S); Hann, M.M. (1982) J. Chem.
Soc. Perkin. Trans. I 307-314
(-CH-CH-, cis 及びtrans); Almquist, R.G. et al. (1980) J. Med.
Chem.
23:1392-1398 (-COCH2-); Jennings-White, C. et
al. (1982) Tetrahedron Lett.
23:2533 (-COCH2-); Szelke, M. et al.,
European Patent Application No. EP 45665 (1982) CA: 97:39405 (-CH(OH)CH2-); Holladay, M.W. et
al. (1983) Tetrahedron. Lett.
24:4401-4404 (-C(OH)CH2-); 及びHruby, V.J. (1982) Life Sci. 31:189-199 (-CH2-S-);に記載された方法により選択的に置換された一つ以上のペプチド結合を有する。特に好適な非ペプチド結合は-CH2NH-である。このようなペプチド・ミメティックは、例えば:生産がより経済的である、化学的安定性が高い、薬理学的特性(半減期、吸収、効力、効験等)が優れている、特異性(例えば生物学的活性の範囲が幅広い)が変更されている、抗原性が減らされている、等を含め、ポリペプチドの実施態様に比較して大きな利点を有するであろう。ペプチドミメティックの標識付けは、通常、一つ以上の標識を、直接又はスペーサ(例えばアミド基)を介して、当該ペプチドミメティック上で定量的構造−活性データ及び/又は分子モデリングにより予測可能な非干渉位置に、共有結合させるステップを含む。このような非干渉位置は、一般的には治療効果を生ずるために当該ペプチドミメティックが結合する先の巨大分子との直接的な接点を形成しない位置である。ペプチドミメティックの誘導体化(例えば標識付け)は、当該ペプチドミメティックの所望の生物学的又は薬理学的活性に実質的に干渉してはならない。
【0105】
あるアミノ酸配列の一つ以上のアミノ酸を、同じ種類のD型アミノ酸(例えばL型リジンの代わりにD型リジン)に系統的に置換する方法を用いて、より安定なペプチドを作製することができる。加えて、例えば分子内でジスルフィド架橋を形成させてペプチドを環化することのできる内側のシステイン残基を加えるなどにより、当業で公知の方法により拘束されたペプチドを作製することができる(引用をもってここに援用することとするRizo and Gierasch (1992) Annu.
Rev. Biochem.
61:387)。
【0106】
このような修飾されたポリペプチドは、原核又は真核ホスト細胞内で産生させることができる。代替的には、このようなペプチドを化学法で合成することができる。異種ポリペプチドを組換えホスト内で発現させる方法、ポリペプチドの化学合成法、及びin vitro翻訳のための方法は当業で公知であり、更に、引用をもってここに援用することとするManiatis et al., Molecular Cloning: A Laboratory Manual (1989), 2nd Ed.,
Cold Spring Harbor, N.Y.; Berger and Kimmel, Methods in Enzymology, Volume 152, Guide to Molecular Cloning
Techniques (1987), Academic Press, Inc., San Diego, Calif.; Merrifield, J.
(1969) J. Am. Chem. Soc. 91:501;
Chaiken I.M. (1981) CRC Crit. Rev.
Biochem. 11:255; Kaiser et al.
(1989) Science 243:187; Merrifield,
B. (1986) Science 232:342; Kent,
S.B.H. (1988) Annu. Rev. Biochem. 57:957;
及びOfford, R.E. (1980) Semisynthetic Proteins, Wiley Publishingにも解説されている。
【0107】
ポリペプチドは典型的には直接的な化学合成で作製することができ、ヘテロ重合体の結合部分として用いることができる。 ペプチドは、非ペプチド部分を共有結合によりN末端及び/又はC末端に付着させた修飾ペプチドとして作製することができる。いくつかの好適な実施態様では、カルボキシ末端又はアミノ末端のいずれか、あるいは両者、を化学修飾する。末端のアミノ基又はカルボキシル基の最もよくある修飾法は、それぞれアセチル化及びアミド化である。アシル化(例えばアセチル化)又はアルキル化(例えばメチル化)などのアミノ末端の修飾や、アミド化などのカルボキシ末端の修飾や、環化を含む他の末端の修飾を、多様な実施態様の検査化合物に導入することができる。特定のアミノ末端及び/又はカルボキシ末端の修飾、及び/又は、コア配列へのペプチド伸長により、例えば安定性が高まる、効力及び/又は効験、血清プロテアーゼへの耐性、所望の薬物動態学的特性が高まる、等、有利な物理的、化学的、生化学的、及び薬理学的特性を提供することができる。
【0108】
V. スクリーニング検定法
本発明の方法は、非ヒト動物の血清及び/又は循環中の作用物質の濃度を低下させる候補又は検査化合物を同定するための、ここでは「スクリーニング検定法」とも呼ばれる方法を提供するものである。
【0109】
ある実施態様では、二重特異的化合物を、対象の血清、循環及び/又は組織中の作用物質の濃度を低下させる上でのそれらの能力について検査することができる。
【0110】
ある二重特異的化合物の、非ヒト動物の血清、循環及び/又は組織中の作用物質の濃度を低下させる能力は、数多くの様々な方法で評価することができる。例えば、血清、循環及び/又は組織中の作用物質の濃度は、(例えばウィルス力価を測定したり、コロニ(例えばプラーク)形成検定法を行ったり、細菌コロニの計数や、あるいは毒素等に関するELISA又は他の検出検定法を行うなどにより)組織、血液又は血清試料などの中の作用物質を計数できる当業で公知の検定法を行うなどで、直接測定することができる。
【0111】
本発明で提供される方法では、非ヒト動物の血清、循環及び/又は組織中の病原性物質などの作用物質の濃度を、少なくとも、例えば約20%、約30%、約40%、約50%、約60%、約70%、約80%、約90% 又は約100%、低下させることができる。
【0112】
別の実施態様では、対象の血清、循環及び/又は組織中の作用物質の濃度を間接的に測定することができる。例えば、血清及び/又は循環中に当該作用物質が存在することを原因とする病理を、この動物由来の組織試料を調べるなどにより、測定することができる。非ヒト動物の血清、循環及び/又は組織中の作用物質の濃度を間接的に測定するもう一つの方法は、当該作用物質が、非ヒト動物において感染を引き起こす能力を測定する方法である。例えば、感染の臨床上の兆候及び症状に対する当該二重特異的化合物の効果を測定することができる。また、例えばある臓器系から別のものへ、あるいはある個体から別の個体への感染の広がりを阻害する上での当該二重特異的化合物の能力を検査することもできる。
【0113】
別の実施態様では、当該二重特異的化合物の、非ヒト動物におけるCR1担持細胞への結合能を測定する。例えば、ある実施態様では、当該二重特異的化合物の、CR1標的分子への結合能の判定は、
リアルタイム・バイオモラキュラー・インターアクション・アナリシス (BIA) (Sjolander, S. and
Urbaniczky, C. (1991) Anal. Chem.
63:2338-2345 及びSzabo et al. (1995) Curr. Opin.
Struct. Biol. 5:699-705)などの技術を用いても行うことができる。ここで用いる場合の「BIA」とは、相互作用物質のいずれも標識せずに、二重特異的相互作用をリアルタイムで研究する技術である(例えばBIAcore)。表面プラズモン共鳴(SPR)の光学的現象の変化を、生物学的分子間のリアルタイムの反応の指標として用いることができる。
【0114】
別の実施態様では、非ヒト動物での細胞による作用物質の破壊(例えばマクロファージによる致死)を測定する。
【0115】
非ヒト動物の血清及び/又は循環中の作用物質の濃度を(当該二重特異的化合物を投与していない非ヒト動物で観察される濃度と比較して)低下させる化合物を選抜することができる。当該の検定法で検査する対象となる化合物は、検査される複数の化合物の中から選択することができる。別の実施態様では、本検定法で検査する対象の二重特異的化合物は、例えばin vitro検定法で、CR1に結合することができると既に特定されていてもよく、本検定法を用いて更に評価又は最適化されてもよい。このような場合、ある二重特異的化合物の、血清及び/又は循環中の作用物質の濃度を低下させる能力を、別の二重特異的化合物か、又は同じ化合物の最適化されていない形と比較して、 血清及び/又は循環中の当該作用物質の濃度を低下させるその能力を判定することができる。
【0116】
好適な実施態様では、本発明の二重特異的化合物を、約1μgの化合物/体重1kg乃至約100μgの化合物/体重1kgの範囲内の濃度、投与する。ここで定義するように、二重特異的化合物の治療上有効量(即ち有効な投薬量)は、約0.01 乃至5000μg/体重1kgの範囲であるが、好ましくは約0.1 乃至500μg/体重1kg、より好ましくは約2 乃至80μg/体重1kg、そして更により好ましくは約5 乃至70μg/kg、10 乃至60μg/kg、20 乃至50μg/kg、24 乃至41μg/kg、25 乃至40μg/kg、26 乃至39μg/kg、27 乃至38μg/kg、28 乃至37μg/kg、29 乃至36μg/kg、30乃至35μg/kg、31乃至34μg/kg 又は32 乃至33μg/体重1kgの範囲であるとよい。当業者であれば、疾患又は異常の重篤度、以前の治療、対象の全身の健康及び/又は年齢、及び他の既往症を含め、しかしこれらに限らず、特定の因子が、対象を有効に治療するために必要な投薬量に影響する場合があることを理解されよう。更に、治療上有効量のタンパク質、ポリペプチド、又は抗体による対象の治療には、単回の治療を含めることができるが、あるいは好ましくは一連の治療を含めることができるとよい。
【0117】
ある好適な例では、作用物質を静脈内(iv)注射した後、約1乃至500μg/体重1kgの範囲の二重特異的化合物で動物を処理する。更に、治療に用いられる二重特異的化合物の有効な投薬量を、特定の治療経過にわたって増減させてもよいことも理解されよう。投薬量の変更は、ここで解説する通りの診断検定法の結果行われてもよく、また同結果から明白となるであろう。
【0118】
検査化合物及び/又は作用物質の投与経路は、動物の循環中への静脈内(iv)注射であってよい。他の投与経路には、限定はしないが、局所、非経口、皮下、又は吸入、がある。用語「非経口」には、例えば皮下、静脈内又は筋肉内経路による注射が含まれるが、更に、例えば疾患又は傷害の部位への局所的投与も含まれる。インプラントからの化合物の持続的放出も当業で公知である。当業者であれば、適した投薬量は、治療しようとする異常の性質、患者の体重、年齢及び全身の状態や、投与経路といった因子に応じて様々であろうことは認識されよう。予備的用量は動物検査に従って決定することができ、ヒトへの投与に向けた投薬量の見積もりは当業で認められた慣例に従って行われる。
【0119】
候補化合物及び作用物質を、一定の範囲の用量にわたって動物に投与することができる。当該の作用物質も動物に投与する場合、候補化合物を、当該作用物質の投与前、投与と同時、又は投与後のいずれでも投与することができる。
【0120】
マウスなど、CR1を発現する本発明のトランスジェニック動物を用いて、例えば自己抗体、感染性物質又は毒素など、対象の血清及び/又は循環中に望ましくない作用物質が存在することに関連する、ヒトにおける異常又は疾患を治療するために有用な候補化合物をスクリーニング又は評価することができる。
【0121】
ある実施態様では、当該のスクリーニング検定法を用いて、血清及び/又は循環中の作用物質の濃度を低下させる上での二重特異的化合物の活性を評価することができる。例えば、様々な形の二重特異的化合物(例えば二量体、三量体、四量体、五量体、又はより大きな多量体型)の、血清及び/又は循環中の作用物質の濃度を低下させる上での能力を検査することができる。
【0122】
ある実施態様では、ヘテロ重合体又は抗原ヘテロ重合体の活性を従来の二重特異的抗体に比較することができる。
【0123】
ある更なる実施態様では、ヘテロ重合体又は抗原ヘテロ重合体の活性を、生理食塩水及び/又は無関係のHP(例えば本発明のある好適な例では、ФX-PA)を含む適したコントロールに比較することができる。
【0124】
別の実施態様では、様々な連結化学法を用いて調製された二重特異的化合物の能力を、血清及び/又は循環中の作用物質の濃度を低下させるそれらの能力について比較することができる。例えば、ある実施態様では、二重特異的化合物の一つの部分を、第二の部分に、SMCC/SATA 結合又はPEG/SATA 結合を用いて連結することができる。別の実施態様では、当該のトランスジェニック動物を用いて、ある特定の連結化学法が、動物において免疫応答を引き起こすかどうかを判定することができる。
【0125】
更に別の実施態様では、当該の動物及び検定法を用いて、最適化された二重特異的化合物を作製する際に用いるために好適な抗原結合部分(例えば特定の結合特異性部分を有するモノクローナル抗体)を選抜することができる。
【0126】
更に別の実施態様では、CR1に結合はするが、抗体ではない結合部分を検査することができる。
【0127】
更なる実施態様では、様々な二重特異的化合物の半減期を検査し、最適化することができる。例えば、ある実施態様では、抗体の異なる抗体アイソタイプ又はフラグメントを用いることにより、半減期を変更することができる。加えて、半減期を変更する新規な方法を判定することができる。
【0128】
別の実施態様では、多様な多量体型又は異なった態様で連結された形の二重特異的化合物のin vivoでの薬物動態を比較することができる。
【0129】
別の実施態様では、当該のトランスジェニック動物を、例えば毒素、ウィルス、細菌などに曝露した動物や、あるいは自己免疫状態を有する動物の治療における二重特異的化合物の効験に関するin vivoモデルを評価する際に用いることができる。
【0130】
別の実施態様では、当該のスクリーニング検定法を用いて、対象の治療を最適化するために、二重特異的化合物の多様な送達経路及び処方を評価することができる。
【0131】
別の実施態様では、当該の検定法を用いて、二重特異的化合物が、動物の免疫応答を亢進するかどうかを判定することができる。別の実施態様では、当該の動物及び検定法を用いて、当該動物における、二重特異的化合物のCR1担持細胞への輸送機序を評価し、そして、二重特異的化合物の活性に対して、CR1に対する抗体の親和性が及ぼす効果を評価することができる。
【0132】
更なる実施態様では、当該の動物及びスクリーニング検定法を用いて、特定の形のin vitro検査をバリデートするために、in vitro検定法とin vivoでの生物学的効果との間の相関を判定することができる。
【0133】
本発明の方法により同定された又は最適化された治療的化合物を、予防上又は治療上の用途のいずれで用いてもよく、また、単独で用いることも、あるいは、例えばワクチン、抗生物質、抗ウィルス薬、抗レトロウィルス薬(ウィルスの複製を阻害する作用薬)など、他の公知の治療法と組み合わせて用いることもできる。ある実施態様では、対象を、作用物質への曝露後に治療する。別の実施態様では、対象を、作用物質への曝露前に治療する。ある実施態様では、対象を二重特異的化合物で(必要に応じ)繰り返し、治療することができる。更に別の実施態様では、(作用物質への曝露前又は曝露後のいずれかでの)本発明の二重特異的化合物による治療の結果、当該作用物質への免疫応答が持続し、こうして更なる治療を行う必要が減るか、あるいはなくなる。
【0134】
本発明を以下の実施例で更に解説するが、以下の実施例を限定的なものと捉えられてはならない。この出願全体を通じて引用された全参考文献、特許及び公開済み特許出願の内容を、引用をもってここに援用することとする。
【0135】
実施例
以下の材料及び方法を実施例で用いた。
材料及び方法
mAb、HP、及びファージ。 以下のマウスモノクローナル抗体(mAb)を用いた: LHR-A、-B、及び-C に結合する抗CR1 mAb YZ-1 (IgG1)(Nickells et al. Clin Exp
Immunol 112, 27-33);LHR-C 及び-Dに結合する抗CR1 mAb 543(アメリカン・タイプ・カルチャー・コレクション);及びLHR-A、-B、及び-Cに結合する抗CR1 mAb 7G9 (IgG2a)(Nickells et al. Clin Exp Immunol 112, 27-33)を、ハイブリドーマから作製された腹水(Pincus et al. Clin Immunol 105,141-54)から精製した。抗ФX174 mAb 7B7 (IgG2a; Taylor et al. J Immunol 158, 842-850)及び抗炭疽菌防御抗原 mAb 14B7 (IgG2a; Little et al. Infect
Immun. 56, 1807-1813) 。バクテリオファージФX174(ФXcs70am-3 ATCC 番号49696-B1と指定されている) はアメリカン・タイプ・カルチャー・コレクション(ヴァージニア州マナサス)から得られ、ファージ・ストックは、PanVera・インビトロジェン社(カリフォルニア州カールスバッド)社により作製された。
【0136】
抗CR1 mAb 7G9 を抗ФX174 mAb 7B7(ФX −CR1 HP)にチオエーテル結合により、SMCC (スルホスクシンイミジル4−(N-マレイミドメチル) シクロヘキサン-1-カルボキシレート)、及びSATA- (N-スクシンイミジル-S-アセチルチオアセテート) (イリノイ州ロックフォード、ピアース・バイオテクノロジー社)のいずれかで活性化させた抗体を以前に解説された(Lindorfer et al. J Immunol
167, 2240-2249)ように用いて連結した。PEG HPを作製するために上記の方法の改良法を開発し、このとき一方のmAbをSMCCで活性化する代わりに、マレイミド基をNHSエステルを介して含有する生物学的に不活性なリンカ/スペーサ分子(修飾されたポリエチレングリコール〜5000MW)が用いられ、この修飾されたmAbをSATA
で活性化したmAbと反応させた。非CR1結合HP (ФX−PA HP)を、mAb 7B7 のmAb 14B7へのSMCC/SATA結合により作製した。
【0137】
トランスジェニック・マウスの作製。 ヒトCR1遺伝子の発現プラスミドを、ヒトCR1の共通の対立遺伝子CR1
(hCR1; Klickstein et al. J Exp Med 168, 1699-1717)をコードする完全cDNAを含有する9.9 kbのNot I DNA 断片を得ることにより、構築した。この断片をプラスミドpGATA-1の非反復Not I部位に挿入し、hCR1を赤血球特異的プロモータGATA1 の転写制御下に置いた(Visvader et al. Genes Dev.12,
473-479)。CR1遺伝子の方向は配列決定により確認された。GATA1-CR1 トランスジェニック・コンストラクトをSal Iで消化し、プロモータ/CR1 遺伝子断片(ほぼ22 kb)を精製し、SJLマウスの受精卵に顕微注射した後、この受精卵をC57BL/6 借り腹母に移植した。その結果得られた後代を、導入遺伝子の組込みについて、マウスの尾DNA を用いたPCRによりスクリーニングし、C57BL/6 マウスと交配することにより維持した。以下に解説するように、陽性のヘテロ接合子を、DNAのPCR分析と、FACS解析による赤血球上の発現分析により検出した。
【0138】
マウス尾からの高分子量DNAの単離を、Easy-DNA
キット(インビトロジェン社)により、このメーカのプロトコルに従って行った。ほぼ1μgのDNAに35サイクルの増幅をサーマル・サイクラで行った。トランスジェニックマウスのPCR分析に用いられた2つのトランスジェニック特異的プライマであるGATA-1正方向プライマ(5'- ACCCTTTCTGTCCTCACA -3'; 配列番号1番)及びCR1遺伝子逆方向プライマ (5'-TTTCTCCCTCCGCTTCCAGGTTG- 3'; 配列番号2番)により、653-bp のDNA 断片(図1a)が生じた。PCR産物を1%のアガロース・ゲルでの電気泳動法で分離した。
【0139】
ヒヒ。 ヒヒを、サウスウェスト・ファウンデーション・フォー・バイオメディカル・リサーチ(テキサス州サンアントニオ)で維持されたコロニから、IACUC認可との約定のもとで用いた。この研究の前に、血液を40匹の動物から得、以下に解説するようにFACSによりCR1発現レベルについて分析した。この結果を用いて、研究に含めるために、 1800-3600 のCR1/E を持つ動物を選抜した。
【0140】
赤血球。 クエン酸塩で血液凝固を阻害したマウス血を、尾の静脈から得、500×gでの遠心分離後、バフィーコート層を取り除き、赤血球を3回、カルシウム及びマグネシウムなしのハンクス液(HBSS=)(ニューヨーク州グランドアイランド、ギブコ/インビトロジェン社)で洗浄した。正常なドナーのヒト赤血球を、酸−クエン酸で血液凝固を阻害した静脈血から得、0.001 M EDTA、0.01% ゼラチンを加えた等容のHBSSで希釈した。遠心分離及びバッフィーコート層除去後、細胞を上記の緩衝液で数回、洗浄した。最後に、細胞を、カルシウム及びマグネシウム並びに0.01%ゼラチンを含有するHBSS(HBSS++/ゼラチン)に再懸濁させた。免疫複合体結合検定用の全ての赤血球は採集日に用いられ、ヘモグロブリン濃度をもとに正規化することで、細胞を水に1/30に希釈するとヘモグロビン吸光度が414nmで0.64になるようにした。
【0141】
ヒト血清。 2mlの血清(ヒト赤血球のドナーから)を5 mM EDTA を4℃に冷却して作製し、ほぼ106の重量のmEから採った洗浄した膜と60分間、混合して、ヒト抗マウスE免疫グロブリンを完全に吸収させた。次に、吸収後のヒト血清を15,000×gで遠心分離してマウス膜を取り除き、その上清をアリクォートし、免疫複合体をオプソニン化するために使用するまで−80℃で保存した。
【0142】
CR1に関する免疫ブロット法。 100μlの血液からの赤血球をHBSS=で2回、洗浄した後、ゴースティング(原語:ghosting)緩衝液(5 mM リン酸ナトリウム (pH 8.0)、1 mM EDTA、及び「プロテアーゼ阻害剤カクテル」の1/100希釈液(ミズーリ州セントルイス、シグマ社)中で一晩、溶解させた。膜をSDS-試料緩衝液(マサチューセッツ州ボストン、ボストン・バイオプロダクツ社)で可溶化させ、5分間沸騰させ、未還元のまま4-12% NuPage 勾配ゲル(カリフォルニア州カールスバッド、インビトロジェン社)で分離した。タンパク質をHybondニトロセルロース・メンブレン(イリノイ州アーリントン・ハイツ、アマーシャム・バイオスクレンセズ社)に移した。この膜を抗CR1 mAb YZ-1で免疫ブロットした後、ペルオキシダーゼ結合ヤギ抗マウス二次抗体(カリフォルニア州バーリンゲーム、ベクター・ラボラトリーズ社)と一緒にインキュベートした。ブロットをSuperSignal ウェスト・ピコ・ケムイルミネセンス基質(ピアース・バイオテクノロジー社)で可視化した。
【0143】
赤血球表面上のCR1発現のFACS分析。 抗CR1 mAb 7G9をAlexa 488TM(オレゴン州ユージーン、モラキュラー・プローブズ社)でこのメーカの指示に従って標識した。野生型mE 又はmE+CR1 又はhE をPBS/1% BSA 緩衝液 (PBS/BSA)に1%の濃度になるように懸濁させた。Alexa 488-7G9 (100μlの2μg/ml ストック)を100μlの赤血球に加え、30分間、室温でインキュベートした。PBS/BSAで2回、洗浄することにより、未結合のAlex-7G9を取り除いた。赤血球をそれらの前方散乱対側方散乱光プロファイルに基づいてフローサイトメトリで選択的に通門させ、その通門内で20,000 回の事象を、FACSCaliburTM装置及びCellQuest Pro ソフトウェア、V3.3 (カリフォルニア州サンホセ、BD バイオサイエンセズ社)を用いて分析した。
【0144】
マウス及びヒト赤血球上のCR1の分布。 ヒト又はマウス血を、HBSS=/BSA
に希釈し、赤血球を同じ緩衝液に10% (V/V)の濃度になるように再懸濁させた。抗CR1 mAb 543 を赤血球に加えた後、Alexa 488で標識されたヤギ抗マウス(モラキュラー・プローブズ社)を、抗体及び赤血球の最終濃度が、それぞれ5.0μg/ml 及び5% になるように加えた。37℃で30分間、インキュベートした後、結合しなかったmAbを遠心分離及び洗浄で取り除いた。光学及び蛍光顕微鏡検査をオリンパスBX40F4 顕微鏡(ニューヨーク州メルビル、オリンパス・アメリカ社)を1000倍の倍率で用いて行った。
【0145】
in vitroにおける免疫複合体及びHPの赤血球への結合。 BSAのV画分(シグマ社)(1mg/ml)をFITC (ウィスコンシン州ミルウォーキー、アルドリッチ・ケミカル社)で標識し、透析して、結合しなかったFITCを取り除いた。標識後のBSAの推定濃度は0.8 mg/mlだった。30μgのFITC-BSA及び100μlのウサギ抗BSA IgG (モラキュラー・プローブズ社)を最終体積250μlにして60分間、37℃で混合すると、免疫複合体を形成させることができた。30μlの免疫複合体を遠心分離管中にアリクォートした後、40μlの吸収後のヒト血清、4μlのストック・カチオン(30 mM CaCl2、100 mM MgCl2)及び150μlの赤血球懸濁液を順番に添加した。15分間、37℃でインキュベートして免疫複合体のオプソニン化と、オプソニン化後の免疫複合体の赤血球への結合の両方を起こさせた後、この混合液を3 ml の冷HBSS-0.01% ゼラチンを含有する12×75mmの試験管に移し、400×gで遠心分離し、3mlの同じ緩衝液中に再懸濁させた。細胞をFACSscanTMで分析すると、10,000回の事象がCellQuest Pro ソフトウェア V4.01を用いて記録された。
【0146】
SMCC-HPの赤血球への結合を以下の態様で判定した。親mE 又は mE+CR1又はhE をPBS/BSA 緩衝液に10%の濃度になるように懸濁させた。 HP (20 ng) を100μlの赤血球に15分間、37℃で振盪しながら加えた。PBS/BSA 緩衝液で2回、洗浄することで、結合しなかったHP を取り除いた。ヤギ抗マウスIgG-Alexa 488 (モラキュラー・プローブズ社)を10μg/ml にして試験管に加え、室温で30分間、インキュベートした後、PBS/BSA緩衝液で2回、洗浄した。次に、フローサイトメトリ検査(FACSCallibur) を試料に対して行い、赤血球を、それらの正方向散乱対側方散乱プロファイルに基づいて選択的に通門させ、20,000 回の事象をその通門内で分析した。
【0147】
in vivoのファージ除去。 ヒヒ(8-10kg)に1.5×1011PFUを静脈内注射し、20分後に0.3 mg のФX−CR1 HP (PEG 及びSMCC)、ФX-PA HP
又は等容の生理食塩水を静脈内注射した。血液試料を、大腿の静脈穿刺により、抗凝固剤としてEDTA (0.01M 最終濃度)を含有する試験管内に得、遠心分離し、血漿を−70℃で凍結させた。血漿中ファージをプラーク検定法により前に解説された(Taylor et al. J Immunol
158, 842-850)ように判定した。
【0148】
マウスに0.1 mlの体積で、3×107 PFU/動物の用量を静脈内注射した。生理食塩水のコントロール、HP又は無関係のHP(1、6、又は12μg)を0.1 ml の体積にして45分後に静脈内注射した。血液試料(50μl)を、図6bに示すようにHPの投与前及び投与後の様々な時点で、0.04MのEDTAを含有する0.5
ml Alseverの溶液中にマウスの尾から採集した。血漿を細胞から分離し、4℃で保存した。血漿中ファージをプラーク検定法により前に解説された(Taylor et al. J Immunol
158, 842-850)通りに判定した。
【0149】
実施例1: ヒト補体受容体1(CR1)ポリペプチドをコードするヌクレオチド配列をGATA-1プロモータに作動的に連結させて含有する発現コンストラクトの構築
ヒトCR1をマウス赤血球上で発現させるために、マウス赤血球で働くであろうプロモータを特定する必要があった。マウスGATA-1遺伝子の調節配列は、以前、トランスジェニック・マウスで造血系組織においていくつかの遺伝子を選択的に発現させるために用いられたことがある(McDevitt, M.A. et al., (1997) Proc. Natl. Acad. Sci., USA,
94:7976-7981; Visvader J.E. et al., (1998) Genes & Dev. 12(4):473-479)。転写因子GATA-1はいくつかの造血系及び多能前駆細胞で発現し、赤血球及び血小板の発生に必要である。赤血球系クロマチンの関係でそれらのDnase I超感受性により定義された、GATA-1遺伝子の上流にある特異的制御配列であって、トランスジェニック・マウスで導入遺伝子全体の発現を亢進させ、原始赤血球系細胞内で特異的に活性化をもたらし、ひいてはGATA-1の発生上の発現パターンを発生反復するものが特定された(McDevitt,
M.A. et al., (1997) Proc. Natl. Acad. Sci.,
USA,
94:7976-7981)。次に、この導入遺伝子カセットを用いて、マウス幹細胞白血病遺伝子(SCL/tal-1)をSCLノックアウト・マウスに再導入し、発現させた;SCL 遺伝子は通常、造血細胞、血管内皮細胞及び発生中の脳内で特異的に発現するが、該マウスGATA-1調節配列により、SCL -/- 胚の造血系の血管を救助することができた(Visvader J.E. et al., (1998) Genes & Dev. 12(4):473-479)、従って、これらのマウスGATA-1調節配列を用いて、マウス赤血球上でのヒトCR1遺伝子の発現を命令することができた。
【0150】
マウスGATA-1調節配列(近位のプロモータ;下流のイントロン配列、及び上流の超感受性部位)をウサギグロビンpoly(A)添加領域及びSV40配列と一緒に含有するコンストラクトを用いた(Visvader and Orkin.
1998. Genes and Development.
12:473)。
【0151】
機能的ヒトCR1をコードするcDNAを含有するプラスミドpiABCDを、ロイド・クリックスタイン博士から得た(Wilson J.G. et
al, (1986) J. Exp. Med. 164:50-59; Klickstein L.B. et al. (1988) J. Exp. Med.
168:1699-1717)。CR1をコードするpiABCDの6,860 bpの断片を、マウスGATA-1調節配列(エキソン2のATG
Iから~10,000 bp上流(転写開始部位の5+上流)からこの遺伝子の3’まで延びる、>15,000bpのゲノム・マウスGATA-1遺伝子) を含有するプラスミドのNotI部位にクローニングした。形質転換後、方向の正しいクローンを選抜し、当業で公知の方法に従った配列決定により確認した。簡単に説明すると、インサート全体をベクタからSalI部位を用いて切り出した。プラスミドpSc-3Z (4.7 kb、低コピー数のプラスミド、amp)が用いられた。DNAをCsCl勾配で精製した後、SalIで消化して、GATA-1プロモータ及びCR1遺伝子をコードする15,000 bp の融合遺伝子を提供した。(図1aは、GATA1/hCR1 トランスジェニック・コンストラクトの概略的マップを示し、このとき、GATA1 の上流の7kbの配列と、エキソン3の下流の~1.5 kb の配列が、hCR1
cDNAをフランクしている。図1aで番号を付した領域は、GATA-1のエキソンを示す。エキソン2にあるGATA1-の内因性翻訳開始コドンを、Not I部位に置換した。hCAR1 のcDNAをこのNot I 部位に挿入した。シミアン・ウィルス40(pA)由来のポリアデニレーション・シグナルはこのcDNAインサートの3’側にある。導入遺伝子を検出するために用いられたPCRプライマは、GATA-1エキソン2に特異的な正方向プライマと、hCR1配列に特異的な逆方向プライマ(図1aの矢印の頭)とから成った。次に、この融合遺伝子を精製した後、トランスジェニック・マウスを作製するために用いた。
【0152】
実施例II: そのゲノムが、ヒト補体受容体1(CR1)ポリペプチドをコードするヌクレオチド配列を、GATA-1プロモータに作動的に連結させて含有するトランスジェニック・マウスの構築
GATA-1プロモータ及びCR1遺伝子をコードする15,000 bpの融合遺伝子を、B6/129 F1マウス接合子に、上に記載したように、当業で公知の方法に従って前核注射することにより、トランスジェニック・マウスを構築した。尾の一部分を取り除き、ゲノムDNAを抽出することにより、マウスを前記外来の融合遺伝子の存在についてスクリーニングした。マウス尾の生検から単離されたDNAに対するPCR分析で、創始マウスと、GATA1/hCR1コンストラクトについて生殖細胞系トランスジェニックである仔の世代が確認された。このようなマウスは、判断材料である653-bp のPCR 産物により特定された(図1b、この図においてレーン1は、テンプレートとしての蒸留水コントロールのPCR結果を示し、レーン2は、導入遺伝子マウスのゲノムDNAをテンプレートとして用いたPCR検定の結果を示す;Mは、1-kb のラダー・マーカに相当する;レーン3は、導入遺伝子を含有するプラスミドDNAのPCR結果を示す。;そしてレーン4は、既知の非トランスジェニック・マウスのゲノムDNAをテンプレートとして用いた検定結果を示す)。
【0153】
実施例III: トランスジェニックマウス赤血球上におけるヒト補体受容体CR1の発現のエビデンス
CR1抗原が、トランスジェニック・マウスの赤血球表面上で発現したことを確認するために、hE、mE 及びmE+CR1細胞をAlexa-488 結合抗CR1 mAb 7G9で染色した後にフローサイトメトリを行った(図2a)。分析では、CR1抗原がhE(右側パネル)及びトランスジェニック・マウス由来のmE(中央のパネル)の表面で発現したが、野生型(wt)マウス(左側パネル)由来のmEでは発現しなかったことが確認された。陽性コントロールとして用いられたヒトE(hE、右側パネル)は、mE+CR1上のCR1の約50%を発現した。
【0154】
mE+CR1のCR1が適したサイズであることを確認するために、免疫ブロット法を行った(図2b)。wtマウス、CR1トランスジェニック・マウス、及び正常なヒト由来の赤血球を溶解させ、可溶化し、非還元膜タンパク質をSDS-PAGEで分離した。移した後、当該ブロットを、材料及び方法の項で解説したように抗CR1
mAb YZ-1と反応させた。ヒトドナーは、200kDaのバンドを生じる共通の対立遺伝子CR1*1と、190kDaのバンドを生じるCR1*3対立遺伝子についてヘテロ接合型と推定された(図2b、レーン4;Klickstein,
L.B. & Moulds, J.M. in The Complement Facts Book (eds. B.J. Morley & M.J. Walport),
pp. 136-145にレビュー)。トランスジェニック・マウス由来の赤血球上のCR1(図2b、レーン3)は分子量においてhE CR1 CR1*1対立遺伝子と同様であり、導入遺伝子と合致した。wt mEのライセート中にCR1 抗原はYZ-1 mAb によっても何ら検出されず(図2b、レーン2);そして予備染色されたミオシン207 kDa MW マーカは図2b、レーン1に示されている。
【0155】
トランスジェニック・マウス中でGATA-1で促進されたCR1の分布を定義するために、トランスジェニック・マウス由来の組織をParagon Biotech (メリーランド州ボルチモア)で標準的手法により固定し、抗CR1 mAbで染色し、CR1の発現について調べた。CR1免疫反応性として認められる組織はなかった。
【0156】
実施例IV: 蛍光顕微鏡分析では、hE 及びmE+CR1 上のCR1のクラスタ状分布が確認された
hE一個当たりのCR1の平均レベルは大変低く(100-1,000/ E)、また一価の補体リガンドのCR1への親和性も低いため、CR1のクラスタは、補体でオプソニン化した粒子をhEに効果的にライゲートする必要があると仮定される(Klickstein et al. Immunity
7, 345-355; Edberg et al. J Immunol
139, 3739; Paccaud et al. J Immunol
141, 3889-3894; Chevalier and Kazatchkine, J
Immunol 142, 2031-2036)。このように、免疫吸着媒介性除去のモデルとしてのトランスジェニック・マウスの実用性は、CR1がmE+CR1上でクラスタになっていることに依存するかも知れない。hE (図3、左側パネル)及びmE+CR1
(図3、右側パネル)を抗CR1 mAb 543 と反応させた後、Alexa 488で標識された二次抗体と反応させて、光学(図3、下側パネル)及び蛍光顕微鏡(上側パネル)で観察した。 CR1 は、予想通り、hE上でクラスタ状に見えた(図3、左側パネル)。同じ手法をmE+CR1についても繰り返したが、CR1はやはり、クラスタ状に見えた(図3、右側パネル)が、hE上のクラスタよりも例外なく小さく、しかしより豊富だった。
【0157】
実施例V: mE+CR1の、オプソニン化したFITC標識免疫複合体及びHPへの結合能
mE+CR1上のCR1タンパク質の官能性を検査するために、これらの赤血球のオプソニン化した免疫複合体への結合能を調べた。FITC-BSA及びウサギ抗BSAから成る、FITC で標識された免疫複合体を、ヒト血清とのインキュベーションにより補体でオプソニン化し、赤血球と反応させた。次に、この試料をフローサイトメトリで分析した。免疫複合体の細胞への結合が、蛍光の正のシフトで実証されたが、これは、免疫複合体がオプソニン化され、赤血球がCR1を発現した場合、即ちhE又はトランスジェニックmE+CR1でのみ、見られた(図4a、この場合、左側のパネルは、正常なヒトE(hE)に関する結果を示し;中央のパネルは野生型マウスE (mE);右側のパネルは、ヒトCR1を発現するトランスジェニック・マウス(mE+CR1)からのものである。青色の柱状図は緩衝液で洗浄された赤血球を表す。FITCで標識された免疫複合体を作製し、その後、緩衝液(赤色の柱状図)及び正常なヒト血清(緑色の柱状図)に、赤血球の存在下で反応させた。蛍光のシフトにより、免疫複合体の赤血球への結合が示されたが、これは、免疫複合体が血清でオプソニン化され(緑色の柱状図)、CR1を発現した(hE又はmE+CR1、しかしwt mEでは見られず)場合にのみ、見られた。
【0158】
赤血球に結合したHPをAlexa 488ヤギ抗マウス抗体で染色し、フローサイトメトリで分析することにより、赤血球のHPへの結合能を調べた(図4b)。HPをhE及びmE+CR1と一緒にインキュベートした場合にはHPの結合は見られたが、wt mEへの結合は見られなかった。hE 及びmE+CR1
は本来の免疫複合体には一様に結合しなかった(図4a)が、HPはhE及びmE+CR1の総集団に結合した(図4b)ことが注目される。
【0159】
実施例VI: in vivoにおいてHPの用量及び種類がファージ除去に及ぼす効果
HP は、血流から病原体及び毒素を除去するために用いることができる。CR1トランスジェニック・マウスを、除去に向けて潜在的な病原体を赤血球CR1に結合させるHPの効験を検査するためのモデルとしてデザインした。バクテリオファージФX174が、これらの実験のモデル生物として用いられたが、なぜなら、哺乳動物細胞上にはその受容体がないため、哺乳動物にとってそれは病原性でなく(Och et al. J Clin Invest
50, 2559−2568)、またそれはプラーク検定法で容易に定量されるからである。ФX−CR1と指定されたHPは、抗ФX174 mAb(7B7)を抗CR1
mAb (7G9) に2つの異なる化学薬SMCC 又はPEGを用いて化学的に連結したものから成った(方法の項を参照されたい)。蛍光分析では、ФX−CR1 HP はhE 及びmE+hCR1 の両方に結合したが、野生型 mE (図4b)には結合しなかったことが確認された。ФX−PAと指定されたコントロールHPは、抗ФX174 mAbを炭疽菌防御抗原(PA、14B7
mAb)に対するmAbに連結することにより、作製された。hE にもmE+CR1にも結合しなかったФX−PA
HPをコントロールとして用いて、可溶性HP媒介性除去の率を判定した。この率は、液相ウィルス中和と、マウスの本来の血小板依存的免疫付着経路により誘導される(大型HP成分の)除去との和を表した。
【0160】
トランスジェニック・マウスに、バクテリオファージФX174を、3×107PFU/マウスの用量で静脈内輸注してから45分後に、PEG又はSMCC HPのいずれかを三種の異なる用量(1.0μg6μg、又は12μg/マウス)、あるいは陰性コントロールとしての生理食塩水、を静脈内輸注した。このトランスジェニック・マウスから24時間後に採血して、血漿1ml当たりのファージのPFUを判定した。1μgのHP/マウスでは、PEG及びSMCCで連結された形のФX−CR1 HP(3.8又は3.0 log)の両方で、ФX−PA HP(0.6 log)に比較したときに、血清中のファージ減少に明確な違いがあった。ФX−PA HPは、ФX-PA HP 及びФX−CR1 HPをより高い用量にしたときにファージ中和の上でより有効となった(6 及び12μgのHP /マウス、図5のグラフでは、HP注射から24時間後にマウスの尾から採取された血液試料中に存在するファージPFU/mlのlog10を表している。誤差棒は、平均の標準誤差を表す、n=3。アスタリスクは、スチューデントのt検定でp<0.01のときのФX-PA HPからの有意差を示す)。
【0161】
実施例VII: ヒヒ及びCR1トランスジェニック・マウス由来の血漿からのバクテリオファージФX174のHP媒介性除去の動態
高等霊長類はCR1を発現する赤血球を有する(参考文献30番でレビュー)ため、これらの動物は、ФX174 ファージ(Taylor et al. J Immunol
158, 842−850)を含め、様々な種類の粒子のHP媒介性除去の効験の小規模研究で用いられてきた(Taylor
et al. J Immunol 148, 2462−2468; Craig et al. Clin Immunol
92, 170-180; Nardin et al. Mol Immunol
36, 827−835; Lindorfer et al. J Immunol 167, 2240−2249)。トランスジェニック・マウスにおける除去の動態を測定する前に、本発明のHPがヒヒでどれくらい効率的に機能したかを評価することが好ましかった。なぜなら、このような研究では、トランスジェニック・マウス・モデルの予測可能性が評価されるであろうからである。ФX174(1.5×1011PFU)を t=-20分で輸注した。20分後のt=0分の時点で、SMCC又はPEGで連結されたФX−CR1 HP、又は非E結合SMCC-連結ФX−PA HP、あるいは陰性コントロールとしての生理食塩水、をヒヒに輸注した。HPを0.3 mg/動物 (それぞれ8−10 kg)という、重さベースでは1ug/マウスの用量とほぼ等しい用量、用いた。その後の100分間にわたる静脈血の試料採取(HP輸注15分及び10分前と、HP輸注後の複数の時点(5乃至90分、行われた)では、二段階の除去が明らかになった(図6aを参照されたい)。1回目は、急速な最初の段階であり、ФX−CR1 PEG HPにより3.0
log、ФX−CR1 SMCC HPにより1.6 log、そしてコントロールФX−PA
HP により0.8 logの除去が、HP輸注後5分間にあった。 HP輸注から5乃至90分後にあった、よりゆっくりとした2番目の段階では、ФX−CR1 HP (PEG)では、~1.5 log/85 分、ФX−CR1 HP (SMCC)では~1.0
log/85 分、そしてФX−PA HP では、~1.0 log/85 分の除去があった。 図6aのグラフは、治療後の様々な時点で血中に見られたファージ力価の減少を表す[可能な採血回数に限界があるため、動物は、HP投与直前の時点0では採血されていない。時点-10分における各群に見られた平均log10(PFU/ml)を減算することにより、数値を正規化した。生理食塩水群は120分間の研究全般にわたって力価に何の変化も見せないため、この時点とHP投与の時点との間の15分間の力価の変化は期待されなかった。これらの数値は、生理食塩水、ФX −CR1 HP(PEG)、ФX−CR1 HP (SMCC)及びФX −PA HP (SMCC)について、それぞれ、8.27±0.78、8.75±0.1、8.63±0.13、及び8.54±0.23、だった。プロットは平均を表し、棒は範囲を表す、n=2]
【0162】
トランスジェニック・マウスで用いるためのФX−CR1
HP(1μg)の有用な用量を定義(図5)したところで、マウスにおけるファージ除去の動態を次に定義した。CR1トランスジェニック及びwt 親C57BL/6
マウスの両者に、3×107のPFU のファージをマウス1匹当たり、注射した。ФX−CR1 HP (SMCC) 又はコントロールФX −PA HP (SMCC) を45分後に注射した。HP注射の15分前、HP注射から15分後、90分後、及び5時間後にマウスから採血した。HPの代わりに生理食塩水の注射を受けたコントロール・トランスジェニック又はwt マウスでは、循環中のファージ力価は、注射後5時間にわたって一定に留まった。CR1 トランスジェニック・マウスでは、循環ファージを減らす上で、コントロールФX −PA HPに比較したときのФX −CR1 HPの優位性が、輸注から15分以内に明白であり、輸注後5時間、続いた(図6b)。図6bのグラフは、図6aと同様に、野生型対CR-1トランスジェニック・マウスにおけるファージのHP媒介性除去が明らかとなったデータを提供する。[ФX −CR1 HP (SMCC) を、ФX−PA (SMCC) に、CR1トランスジェニック及びC57BL/6 野生型マウスにおける血流からの、注射されたバクテリオファージФX 174の除去能について比較した。ファージを−45分の時点で3×107PFU/マウスの用量、静脈内注射した。HP又は生理食塩水のコントロールを、1μg/マウスの用量、時点0で静脈内注射した。血液を、尾の採血により、HP注射から−15分、15分後、90分後、及び5時間後に採取した。グラフは、治療後の時点で血中に見られたファージ力価の減少を表す。数値は、−15分の時点で各群に見られた平均log10(PFU/ml)を除算することにより、正規化された。生理食塩水群は5時間の研究にわたって力価に何の変化も見せないため、−15分から時点0までの間の力価の変化は期待されなかった。これらの-15分の数値は、生理食塩水、CR1 マウス+ФX−CR1 HP、CR1 マウス+ФX −PA HP、wt マウス+ФX −CR1 HP、及びwt マウス+ФX −PA HP、についてそれぞれ、6.93±0.30、7.45±0.08、6.82±0.32、7.35±0.11、及び6.96±0.40だった。誤差棒は平均の標準誤差を表す、n=3。アスタリスクはステューデントのt検定によりp<0.05のときのФX−PA
HPからの有意差を示す]
【0163】
全ての時点において、ФX −CR1 HP はコントロールФX −PA HP よりも循環ファージをほぼ1.5 log、多く、減少させた。赤血球上にCR1のないwt マウスでは、ФX −CR1 HP及びコントロールФX −PA、HPは、循環ファージの力価を、全ての時点で同様なレベル減少させ、この減少は、トランスジェニック・マウスでコントロールФX−PA HPにより達成された減少レベルと同様だった。
【0164】
これらの結果は、ファージを赤血球CR1に結合させた抗体HPは、トランスジェニック・マウスにおいて血清及び/又は循環からファージを取り除く上で、可溶性ファージ結合HPよりも効率的であることを実証した。wtマウスにおけるウィルス除去では、この血管内の利点がないことは、トランスジェニック・モデルの有効性を裏付けるものである。
【0165】
均等物
当業者であれば、慣例的な実験によって、ここに解説した本発明の具体的な実施態様の均等物を数多く、認識し、又は確認できることであろう。このような均等物は以下の請求の範囲の包含するところと、意図されている。
【図面の簡単な説明】
【0166】
【図1】図1は、GATA1/hCR1 コンストラクトの概略的マップと、トランスジェニック始祖マウスを特定したPCR検定結果を示す。
【図2】図2は、トランスジェニック・マウス由来の赤血球が適した分子サイズのヒトCR1抗原を発現したことの確認を示す。
【図3】図3は、マウス及びヒト赤血球上のCR1の分布を明らかにする。
【図4】図4は、ヒト補体でオプソニン化された免疫複合体又はHPへのmE+CR1の結合能を示す。
【図5】図5は、invivoにおけるファージ除去に対するHP用量の効果を示す。
【図6】図6は、ヒヒ、CR1トランスジェニック及びwtマウスにおけるHPのファージ除去結果を示す。
【背景技術】
【0001】
関連出願
本出願は、引用をもってその内容全体をここに援用することとする2003年5月9日出願の米国仮出願60/469,262号に基づく優先権を主張するものである。
【0002】
発明の背景
補体系は、外来の抗原に対する免疫系の応答で重要な数多くの様々なタンパク質から成る。補体系のタンパク質は、正常な血清中のグロブリンの約10%を成す。補体系は、その主要な成分が開裂したときに活性化し、その結果できる生成物が、単独又は他のタンパク質と一緒になって、更なる補体タンパク質を活性化することで、タンパク分解カスケードが起きる。補体系の活性化は、血管透過性の上昇、貪食細胞の化学走性、炎症性細胞の活性化、外来粒子のオプソニン化、細胞の直接の致死及び組織損傷を含め、多種の応答につながる。伝統的な経路においては、補体系の活性化は、抗原−抗体複合体により惹起される。代替的な経路では、補体系は、例えば病原性細菌の細胞壁に存在するリポ多糖などによって触媒されることがある。
【0003】
補体受容体タイプ1(CR1)は、赤血球(E)、単球/マクロファージ、顆粒球、B細胞、特定のT細胞、脾臓の瀘胞性樹状細胞、及び糸球体足細胞上に存在することが示されている膜糖タンパク質である。CR1は、活性化した補体を含有する粒子又は免疫複合体の、これらの細胞表面上への結合を媒介する。より具体的には、CR1は、補体活性化生成物C3b及びC4bに結合するため、C3b/C4b受容体とも言及されてきた。これらの相互作用の結果は、この受容体を持つ細胞種に依存する(Fearon, D. T. and Wong, W. W. (1983) Ann. Rev. Immunol. 1:243)。赤血球表面上のCR1は、肝臓への輸送に向けて免疫複合体に結合する(Cornacoff, J. B.
et al. (1983) J.
Clin. Invest. 71:236; Nedof, N. E.
et al. (1982) J.
Exp. Med.
145:1739)。好中球及び単球上のCR1は、結合した複合体を、被覆小窩を通じた吸着性エンドサイトーシスか、又は、フィブロネクチン及びラミニンなど、細胞外マトリックス中に存在するホルボールエステル、化学走性ペプチド又はタンパク質による受容体活性化後の貪食のいずれかにより、内部移行させる(Newman S. L. et al. (1980) J. Immunol.
124:2236; Wright, S. D. and Silverstein S. C. (1982) J. Exp. Med. 145:1149;
Wright S. D. et al. (1983) J. Exp. Med. 148:1338)。CR1のリン酸化は、この貪食細胞作用を獲得する際に一役を担っているかも知れない(Changelian P. S. & Fearon D. T. (1986) J. Exp. Med.
163:101)。Bリンパ球上のCR1の機能は、そこまで定義されていないが、これらの細胞を、CR1に対する抗体で処理すると、最適以下の用量のアメリカヤマゴボウマイトジェンに対するそれらの応答が亢進した(Daha, M. R. et al. (1983) Immunobiol. 164:227 (抄録))。濾胞性樹状細胞上のCR1は、抗原提示の役割を果たしているかも知れない(Klaus G. G. B. et
al. (1980) Immunol. Rev. 53.3)。
【0004】
受容体として働くことに加え、CR1はまた補体調節機能も有する。例えば伝統的及び代替的補体経路において、CR1はC3/C5コンバターゼを阻害することができ、更に、I因子によるC3b及びC4bの開裂のためのコファクタとして作用することもできる(Fearon D. T.
(1979) Proc. Natl. Acad. Sci. U. S. A. 76:5867; Liida X. & Nussenzweig V.
(1981) J. Exp. Med. 153:1138)。補体活性化の伝統的経路においては、複合体C4b、2aはC3活性化酵素、又はコンバターゼである。CR1はC4bに結合してC4b、2aの解離を促進することができる。この結合により、C4bは、不活化補体タンパク質C4c及びC4dへの開裂を通じた、I因子による不可逆的なタンパク質分解性不活化を起こしやすくなる。補体活性化の代替的経路においては、複合体C3b、4bはC3コンバターゼである。CR1はC3bに結合することができ、やはりC3b、4bの解離を促進することができる。C3b、CR1が形成されると、C3bは、I因子による不可逆的なタンパク質分解性不活化を起こして、不活化補体タンパク質C3b(iC3b)を形成しやすくなる。
【0005】
CR1は、一本のポリペプチド鎖から成る糖タンパク質である。分子量が~40,000乃至50,000ダルトンの増分ずつ異なる4つのアロタイプ型のCR1が見つかっている。2つの最もよくある形は、それぞれ250,000及び290,000ダルトンの分子量を有するF及びSアロタイプである(A及びBアロタイプとも呼ばれる)(Dykman, T. R. et al. (1983) Proc. Natl.
Acad. Sci.
U.S.A. 30:1698; Wong,
W.W. et al. (1983) J.
Clin. Invest. 72:685)。更に、210,000及び>290,000ダルトンの分子量を有する2つの珍しい形もある(Dykman, T. R. et al. (1984) J.
Exp. Med.
159:691; Dykman, R. R. et al. (1985) J. Immunol.
134:1787)。これらの違いは、明らかに、グリコシル化した状態ではなく、CR1のポリペプチド鎖の違いを表している(Wong W. W. et al. (1983) J. Clin. Invest. 72:685)。4つのCR1アロタイプの全てがC3b結合活性を有する(Dykman, T. R. et al. (1983) Proc. Natl. Acad. Sci. U.S.A. 30:1698;
Dykman, T. R. et al. (1984) J. Exp. Med. 159:691; Dykman, T. R. et al. (1985)
J. Immunol. 134:1787; Wong W. W. et al. (1983) J.
clin. Invest. 72:685)。
【0006】
CR1は多様な造血細胞上に見られるが、血中のCR1の大半は、赤血球上に特異的に存在する。このCR1は、循環免疫複合体のプロセッシング及び除去で主な役割を果たす。病原体が赤血球に付着すると、免疫複合体となった病原体は、例えば肝臓及び脾臓の固定組織マクロファージなど、アクセプタ貪食細胞に効率的に輸送される。興味深いことに、この輸送反応での病原体は、赤血球に認められる損傷を何ら及ぼさずに、赤血球から見かけ上、引き剥がされる(Lindorfer M. A. et al. (2001) Immun. Reviews 183:10-24)。この輸送反応の詳細な機序はまだ、充分には解明されていない。しかしながら、研究では、in vivoでの輸送反応は、I因子媒介性放出に依存している可能性の低いプロセスに促されていることが示されている(Lindorfer M. A. et al. (2001) Immun. Reviews 183:10-24)。
【0007】
製薬業において重要な目標は、組織又は血流中の病原体を標的とし、除去することのできる新規な治療様式の開発である。この問題に対処するためにとられてきたいくつかのアプローチは、霊長類赤血球の免疫付着機能の利点を生かすことに狙いを置いている。あるアプローチでは、二重特異的モノクローナル抗体複合体(ヘテロ重合体、HP)を、標的病原体に結合し、霊長類赤血球のCR1に固定するようにデザインする(Lindorfer M. A. et al. (2001) Immun. Reviews 183:10-24)。例えば、ヘテロ重合体を、CR1に特異的なモノクローナル抗体を、標的病原体に特異的なモノクローナル抗体に化学的に架橋したものから構成することができる。この方法で、CR1に特異的な抗体は、天然のCR1リガンドであるC3bにとってサロゲートとしての役目をする。このアプローチの利点の一つは、C3bのCR1に対する天然の親和性は低く、従って、強力な補体活性化や、複数のC3b分子を捕獲して赤血球の結合を確実にする必要があるのだが、CR1に対して大変高い親和性を持ついくつかのマウスモノクローナル抗体が作製されている点である。これらの抗体を、高親和性病原体特異的モノクローナル抗体に架橋させると、補体の非存在下でも、実質的にはあらゆる標的病原体に赤血球を結合させることができるはずである。この戦略は、数多くの細菌及びウィルスに成功裏に応用されてきた(Powers J. H. et al. (1995) Infect. Immun 63:1329-1335; Kuhn S. E. et al. (1998) J. Immunol.
160:5088-5097; Nardin A.,
et al. (1998) 211:21-31; Hahn C. S. et al. (2001) J. Immunol
166:1057-1065)。
【0008】
ある関連するアプローチでは、赤血球及び二重特異的試薬を用いた、血清及び/又は循環中の分子の標的決定を、全身性エリテマトーデス(SLE)におけるIgG抗dsDNA抗体など、自己免疫疾患に関連する数多くの自己抗体の標的決定にまで延長することができる。抗原ベースのヘテロ重合体(AHP)を用いることにより、自己抗体を赤血球媒介性除去に向けて標的決定することができる(Lindorfer
M. A. et al. (2001) Immun. Reviews 183:10-24)。抗原ベースのヘテロ重合体は、自己抗原を抗CR1モノクローナル抗体に化学的に架橋したものから成る。赤血球に結合した抗原ベースのヘテロ重合体は自己抗体を捕獲し、新たに形成されたその赤血球結合免疫複合体を、最終的な除去に向けた肝臓への輸送反応に方向付ける。
【0009】
当業においては、明らかに、組織、血清及び/又は循環から標的とされた分子を除去するために、CR1に結合するヘテロ重合体など、二重特異的組成物の能力を検査するために用いることのできる動物モデルが必要である。このような動物モデルは、二重特異的組成物や、二重特異的組成物ベースの治療法の効験及び安全性の両方を評価する上で有用であろう。残念ながら、マウスは薬物検査にとっては優れたモデルであるが、マウスはそれらの赤血球上にCR1を発現しない(Kalli and Fearon. 1994 J. Immunol.
152:2894)。
【0010】
発明の概要
本発明は、少なくとも部分的に、ヒト補体受容体タイプ1(CR1)を発現するマウスの作製と、これらの動物が赤血球上にCR1を発現するという観察とに基づくものである。従って、本発明は、異種CR1遺伝子を発現させる遺伝子コンストラクト及び非ヒト動物を特徴とする。本発明のトランスジェニック動物を用いて、対象の組織、血清及び/又は循環中の生物学的物質などの作用物質の濃度を低下させることのできる組成物を同定及び/又は評価する方法も提供される。
【0011】
ある局面では、本発明は、ヒト補体受容体1(CR1)ポリペプチドをコードするヌクレオチド配列を、プロモータに作動的に連結させて成る、非ヒト動物における発現用の遺伝子コンストラクトに関し、但し当該のコンストラクトは、赤血球上のヒトCR1の機能的発現を命令するものである。
【0012】
別の局面では、本発明は、そのゲノムが、ヒト補体受容体1(CR1)をコードするポリヌクレオチドを、プロモータに作動的に連結させて含むような非ヒト動物に関し、該ヒトCR1分子は、当該動物の赤血球上に機能的に発現するものである。好ましくは、該ヒトCR1分子が赤血球上に優先的に発現するとよい。
【0013】
好適な実施態様では、該プロモータはGATA-1プロモータである。より好ましくは、該プロモータは、GATA-1プロモータと、赤血球上の発現を活性化させる上流の制御領域とを含むとよい。
【0014】
好適な実施態様では、該非ヒト動物は、該ポリヌクレオチドについてホモ接合型又はヘテロ接合型である。好ましくは、当該の動物がトランスジェニック・マウスであるとよい。
【0015】
別の局面では、本発明は、対象の血清及び/又は循環中の作用物質の濃度を低下させることのできる二重特異的化合物をスクリーニングする方法を特徴とする。この方法は、(a)そのゲノムが、ヒト補体受容体1(CR1)をコードするポリヌクレオチドをプロモータに連結させて含むような非ヒト動物であって、前記ヒトCR1が、前記非ヒト動物の赤血球上に機能的に発現するような非ヒト動物、に、複数の検査化合物を投与するステップと、(b)前記非ヒト動物の血清及び/又は循環中の前記作用物質の濃度を低下させる上での、ある二重特異的化合物の能力を判定するステップと、(c)前記非ヒト動物の血清及び/又は循環中の前記作用物質の濃度を低下させる二重特異的化合物を選抜することで、対象の血清及び/又は循環中の作用物質の濃度を低下させることのできる二重特異的化合物を同定するステップと、を含む。
【0016】
ある関連する局面では、本発明は、対象の血清及び/又は循環中の作用物質の濃度を低下させる上での、ある二重特異的化合物の能力を評価する方法を提供するものである。この方法は、(a)そのゲノムが、ヒト補体受容体1(CR1)をコードするポリヌクレオチドをプロモータに連結させて含むような非ヒト動物であって、前記ヒトCR1が、前記非ヒト動物の赤血球上に機能的に発現するような非ヒト動物、に、当該の二重特異的化合物を投与するステップと、(b)前記非ヒト動物の血清及び/又は循環中の前記作用物質の濃度を低下させる上での、前記二重特異的化合物の能力を判定することにより、ある対象の血清及び/又は循環中の前記作用物質の濃度を低下させる上での、前記二重特異的化合物の能力を評価するステップと、を含む。
【0017】
ある実施態様では、これらの方法は、非ヒト動物に該作用物質を投与するステップを更に含む。好適な実施態様では、当該の作用物質は病原体、ウィルス、毒素、ポリヌクレオチド、細菌、又は自己免疫疾患に関連する自己抗体、である。
【0018】
好適な実施態様では、該二重特異的化合物はヒトCR1に結合するものである。好ましくは、該二重特異的化合物がヘテロ重合体であるか、又は抗原ベースのヘテロ重合体であるとよい。
【0019】
好適な実施態様では、該非ヒト動物はトランスジェニック・マウスである。
【0020】
他の好適な実施態様では、該プロモータはGATA-1プロモータか、又は、GATA-1プロモータと、赤血球上での発現を活性化させる上流の制御領域とを含む。
【0021】
更に本発明は、生物学的作用物質に曝露した対象を治療する方法を特徴とする。この方法は、対象が治療されるように、本発明により提供される方法で同定された二重特異的化合物を対象に投与するステップを含む。
【0022】
ある関連する局面では、本発明は、生物学的作用物質への曝露のリスクのある対象を治療する方法を提供する。この方法は、対象が治療されるように、本発明により提供される方法で同定された二重特異的化合物を対象に投与するステップを含む。
【0023】
更に別の関連する局面では、本発明は、感染又は自己免疫疾患のある対象を治療する方法を提供する。この方法は、対象が治療されるように、本発明により提供される方法で同定された作用物質を対象に投与するステップを含む。
【0024】
更に本発明は、ヒトCR1をその赤血球表面上に発現するトランスジェニック・マウスを作製する方法を特徴とする。この方法は、a)ヒトCR1ポリペプチドをコードするポリヌクレオチドを、マウスの卵又は胚に導入するステップであって、前記ポリヌクレオチドが、マウス赤血球上でヒトCR1を発現させるプロモータに作動的に連結されている、ステップと、b)前記CR1ポリヌクレオチドを含む卵又は胚を産期に至るまで成長させて、前記トランスジェニック・マウスを生じさせるステップと、を含む。
【0025】
ある関連する局面では、本発明は、トランスジェニック・マウスを作製する方法を提供するものである。この方法は、それぞれがヒトCR1ポリペプチドをコードするポリヌクレオチドを含む二倍体ゲノムを含む二匹のトランスジェニック・マウスを交配するステップを含み、但しこの場合、前記ポリヌクレオチドが、マウス赤血球上でヒトCR1ポリペプチドが生じるように発現することで、仔トランスジェニック・マウスが生じる。
【0026】
発明の詳細な説明
本発明は、少なくとも部分的に、そのゲノムが、ヒト補体受容体1(CR1)をコードするポリヌクレオチドをプロモータに作動的に連結させて含有するようなマウスの作製と、異種CR1分子が、該動物の赤血球上に機能的に発現するという観察とに基づく。従って、本発明は、CR1を含む遺伝子コンストラクトと、CR1が赤血球上で発現するような非ヒト動物とを特徴とする。本発明は、対象の血清及び/又は循環中の標的作用物質の濃度を低下させることのできる二重特異的化合物を同定するために前記非ヒト動物を使用する方法を提供する。更に本発明は、このような二重特異的化合物の投与が有益であろう疾患又は異常を有する対象を治療する方法を特徴とする。
【0027】
本発明がより容易に理解されるように、いくつかの用語をまず定義する。
【0028】
I. 定義
ここで用いられる場合の用語「導入遺伝子」とは、ある動物に導入された異種もしくは外来遺伝子又は組換え核酸コンストラクトを言う。該導入遺伝子は、野生型もしくは変異型遺伝子でもよく、あるいは、
それが異常なパターンで発現するように変更されたものでもよい。ここで用いる場合の用語「トランスジェニック細胞」とは、導入遺伝子を含有する細胞を言う。
【0029】
ここで用いる場合の用語「異種遺伝子」又は「外来遺伝子」には、それらが導入された先のゲノムの一部としては天然では見られなかったり、あるいは、それがゲノム中に天然で存在する一箇所又は複数の位置とは異なる位置で見られるような遺伝子が含まれる。異種DNAは、それが導入された先の細胞にとって内因性でなく、別の細胞から得られたものである。
【0030】
ここで用いる場合の用語「遺伝子コンストラクト」には、天然では同じ分子中に存在しないヌクレオチド配列が操作されて接合されているような非天然核酸分子が含まれる。
【0031】
本発明のコンストラクトは、ホスト細胞内での核酸分子の発現に適した形のものである。好ましくは、当該のコンストラクトが、発現に用いるホスト細胞に基づいて選択された一つ以上の調節配列を、発現させようとする核酸配列に作動的に連結させて含むとよい。組換え発現ベクタ内において、「作動的に連結させた」とは、(例えばin vitroの転写/翻訳系で、あるいは、ベクタをホスト細胞内に導入する場合にはホスト細胞内で)目的のヌクレオチド配列の発現が可能な態様で、該ヌクレオチド配列が調節配列に連結されていることを意味するものと、意図されている。用語「調節配列」は、プロモータ、エンハンサ及び他の発現制御配列(例えばポリアデニレーション・シグナル)を包含するものと、意図されている。このような調節配列は、例えばGoeddel; Gene Expression
Technology: Methods in Enzymology
185, Academic Press, San Diego, CA (1990) に解説されている。
【0032】
ここで用いる場合の用語「上流の制御領域」は、例えばプロモータ又は上流のエンハンサなど、発現させようとする遺伝子の5‘側にある調節配列を包含する。
【0033】
ここで用いられる場合の用語「プロモータ」は、転写を開始するためにRNAポリメラーゼの結合に関与するDNA領域を包含する。
【0034】
特定のタンパク質の発現又は特定の細胞種に関してここで用いられる用語「優先的に発現する」は、他の細胞種上よりもその細胞種で多いが、必ずしもその特定の細胞種に限定されないような発現を包含する。例えば、赤血球上で優先的に発現するタンパク質は、他の細胞種に比べ、赤血球上で少なくとも約5倍、少なくとも約10倍、少なくとも約20倍、少なくとも約30倍、少なくとも約40倍、少なくとも約50倍又はそれより高いレベルで発現するものである。
【0035】
ここで用いる場合の用語「機能的に発現する」とは、あるタンパク質の対生物活性が維持されるような該タンパク質の発現を包含する。例えば、CR1が、C3b又は抗体など、天然リガンドへの結合が可能であるときに、細胞上で機能的に発現したとする。
【0036】
ここで用いる場合の「トランスジェニック動物」とは、異種又は外来遺伝子をそれらのゲノムに導入して有するような動物を言う。用語「トランスジェニック動物」は、例えばブタ、サル、ヤギ、又はマウスなどのげっ歯類などの非ヒト哺乳動物など、当該動物の細胞のうちの一つ以上、そして好ましくは基本的に全てが導入遺伝子を含むような動物を包含する。当該の導入遺伝子は、細胞に直接、あるいは、例えばマイクロ注射、トランスフェクション、又は組換えウィルス感染などの感染などにより、当該細胞の前駆細胞への導入により間接的に、導入される。用語「遺伝子操作」は、組換えDNA分子の導入を包含する。この分子は、好ましくは、それが仔へ確実に伝えられるように染色体内に組み込まれるとよいが、あるいは、染色体外で複製するDNAであってもよい。
【0037】
好適なトランスジェニック動物は哺乳動物である。本発明によれば、哺乳動物の例には、限定はしないが、げっ歯類(例えばラット及びマウス)、イヌ、ネコ、ブタ、ヒツジ、ウシ、ヤギ、ウマ及びウサギが含まれる。好ましくは、当該の哺乳動物がヒト以外であるとよい。ここで用いる場合の用語「げっ歯類」とは、系統学的齧歯目の全てのメンバーを言う。ある好適な実施態様では、齧歯類はマウスである。
【0038】
ここで用いられる「発現」には、細胞によるタンパク質の合成が含まれる。タンパク質の発現は、例えば転写、転写後、翻訳、及び/又は翻訳後など、多様な段階で測定することができる。
【0039】
本発明によれば、トランスジェニック動物は導入遺伝子を誤発現するものである。ここで用いられる場合の用語「誤発現」には、非野生型パターンの遺伝子発現が含まれる。誤発現には、非野生型レベルでの発現、即ち、当該遺伝子が発現する時点又は段階の点で野生型とは異なる発現パターン、例えば、所定の発生期又は段階における(野生型に比較したときの)発現増加:所定の細胞種又は組織種での(野生型に比較したときの)発現増加の点で、野生型とは異なる発現パターン;スプライシング・サイズ、アミノ酸配列、翻訳後修飾、又は、発現後のポリペプチドの対生物活性の点で、野生型とは異なる発現パターン;環境刺激又は細胞外刺激が当該遺伝子の発現に及ぼす効果の点で、野生型とは異なる発現パターン、例えば、刺激強度の増減の存在下における(野生型に比較したときの)発現増加のパターン、が含まれる。誤発現には、トランスジェニック核酸からの発現が含まれる。好ましくは、本発明の導入遺伝子により、関連する内因性遺伝子が前に発現しなかったか、あるいは低レベルで発現した動物の細胞上で、外来遺伝子が発現するとよい。ある実施態様では、当該の導入遺伝子は赤血球上で発現する。ある好適な実施態様では、当該の導入遺伝子は、赤血球上で優先的に発現する。
【0040】
ここで用いられる場合の用語「マーカ配列」とは、(a)目的の遺伝子(例えばCR1遺伝子)を発現させる核酸標的決定コンストラクトの一部として用いられ、及び(b)前記標的決定コンストラクトをそれらのゲノムに導入させて有する細胞を特定するために用いられる、核酸分子を言う。例えば、マーカ配列は、細胞に検出可能な形質をもたらすタンパク質、あるいは、例えばネオマイシン耐性遺伝子などの抗生物質耐性遺伝子、又は、例えばアルカリホスファターゼ、西洋わさびペルオキシダーゼ、ルシフェラーゼ、ベータ-ガラクトシダーゼ等、当該細胞上に典型的には見られない検定可能な酵素、又は、例えば緑色蛍光タンパク質、赤色蛍光タンパク質など、当該細胞中には典型的に見られない他の便利に検出可能なタンパク質、をコードする配列であってよい。
【0041】
作用物質又は二重特異的化合物に関してここで用いられる「投与する」とは、ある組成物を動物又は細胞に分配する、送達する又は適用することを言うものと、意図されている。用語「投与する」とは、例えば非経口又は経口経路による送達、筋肉内注射、皮下/皮内注射、静脈内注射、口腔内投与、経皮送達、及び、鼻孔内もしくは呼吸管経路による投与を含め、動物の所望の位置へ当該組成物を送達するために適したいずれかの経路により、動物に当該組成物を接触させる又は分配する、送達する又は適用することを言うものと、意図されている。
【0042】
ここで用いられる場合の用語「二重特異的化合物」には、対象の血清及び/又は循環中の作用物質の濃度を潜在的に低下させることのできるあらゆる二重特異的化合物が含まれる。二重特異的化合物は、2つの異なる結合部分から成る分子である。二重特異的化合物の例には、例えば、二重特異的抗体及びヘテロ重合体がある。好適な実施態様では、当該の二重特異的化合物はヘテロ重合体か、又は抗原ベースのヘテロ重合体である。
【0043】
二重特異的抗体は、2つの特異性部分を有する単一の抗体分子である。二重特異的抗体の例は当業で公知である。ハイブリッド・ハイブリドーマ又はトリオーマにより作製される二重特異的抗体は、米国特許第4,474,893号に開示されている。二重特異的抗体は、化学的手段(Staerz et al. (1985) Nature 314:628, and Perez et
al. (1985) Nature 316:354)
及びハイブリドーマ技術(Staerz and Bevan (1986)
Proc. Natl. Acad. Sci. USA, 83:1453, 及びStaerz and Bevan (1986) Immunol. Today 7:241)により構築されてきた。二重特異的抗体は、米国特許第5,959,084号にも解説されている。二重特異的抗体のフラグメントは、米国特許第5,798,229号に解説されている。
【0044】
ここで解説されるヘテロ重合体とは、当該ヘテロ重合体の結合部分の少なくとも一つが二価であるような2つの異なる結合部分を含む分子を言う。例えば、ヘテロ重合体は、互いに接合された2つの抗体を含むことができる。本発明のある実施態様では、ヘテロ重合体は、当業で公知の数多くの架橋技術のいずれかを用いることにより架橋された2つの異なる抗体から成る。ある実施態様では、該ヘテロ重合体はヒトCR1に結合する。ある実施態様では、ヘテロ重合体は、当業で公知の数多くの架橋技術のいずれかを用いて架橋された異なる抗体から成る。
【0045】
別の実施態様では、ヘテロ重合体は、抗原ベースのヘテロ重合体である。ここで解説される、抗原ベースのヘテロ重合体とは、例えば自己免疫疾患又は異常に関連する自己抗体など、標的抗体により認識される抗原である結合部分を含有するヘテロ重合体を言い、この場合、前記結合部分は、当業で公知の数多くの架橋技術のいずれかを用いて第二の結合部分に架橋されている。好ましくは、第二の結合部分は、抗原に結合する抗体であるとよい。ある実施態様では、前記の抗原ベースのヘテロ重合体はヒトCR1に結合するものである。ここで用いられる場合の用語「作用物質」は、対象の血清及び/又は循環中に存在し、感染、異常又は疾患に関連するあらゆる物質又は生物を包含するものである。好適な作用物質は、例えば病原体、ウィルス、毒素、ポリヌクレオチド、抗体又は自己抗体などを含む生物学的物質である。ここで用いられる場合の用語「生物学的物質」は、更に、例えば炭疽菌、天然痘、ペスト、エボラ又はマーブル・ウィルスなど、生物戦争又はテロ活動で対象が曝露するかも知れない物質を包含する。
【0046】
以下の小項で本発明を更に詳述する。
【0047】
II. CR1発現コンストラクト
ある局面では、本発明は、非ヒト動物に導入されたときに、この動物の赤血球上でCR1遺伝子が機能的に発現するような核酸分子を特徴とする。好ましくは、当該CR1遺伝子はヒトCR1遺伝子であるとよいが、他の哺乳動物CR1分子を発現させることもできる。野生型ヒトCR1遺伝子のヌクレオチド配列は当業で公知であり、例えば、引用をもってその内容をここに援用することとするWilson J.G. et al, (1986) J.
Exp. Med. 164:50-59、米国特許第6,316,604号及び米国特許第5,212,071号などに解説されている。
【0048】
上記の配列のバリアントも、本発明のコンストラクトを作製する際に用いることができる。例えば、異なるアロタイプのCR1 が当業で解説されている。例えば、このようなアロタイプは、Wong et al.
1986. J. Exp. Med.
164:1531; Wong et al. 1989. J. Exp.
Med. 169:847に解説されている。他のヒトCR1配列が当業で公知であり、例えばGenBank 受託番号AF169970; J. Immunol. 151: 6214(1993) 又はJ. Biol.
Chem. 265:974-980
(1990)に見ることができる。このようなアロタイプ型のCR1をコードする核酸分子も本発明において用いることができる。
【0049】
例えばバリアント型のヒトCR1など、バリアント型のCR1も用いることができる。CR1遺伝子、改変されたCR1遺伝子、又は、ヒトCR1遺伝子と、別のヌクレオチド配列(例えば別の生物由来のCR1配列又は非CR1配列)の一部とを由来とするポリヌクレオチド配列を含むCR1遺伝子、即ちキメラCR1遺伝子、あるいは、ヒトCR1遺伝子に関連するCR1遺伝子、の部分をコードする核酸の作製及び使用も、用いることができる。このようなバリアント型のCR1を、非ヒト動物の赤血球上で機能的に発現させることができ、更に、ヒトC3b又はC4bに、及び/又は、ヒトCR1に結合する抗体にも、結合させることができる。
【0050】
ある実施態様では、CR1が相互作用する相手の分子、例えば抗体分子、C3b等への結合を高める又は低下させるように、CR1ヌクレオチド配列に変異を作ることができる。
【0051】
導入される先の非ヒト動物にとって異種のCR1遺伝子をコードする他の核酸分子の作製及び使用も提供される。例えば、非ヒト霊長類型のCR1など、ヒトCR1に対して相同性の高いCR1分子をコードする核酸配列も、このようなCR1分子又はその部分が動物の赤血球上で機能的に発現可能であり、更に、ヒトC3b及び/又はC4bに、及び/又は、ヒトCR1が結合する抗体に、結合可能であることを条件に、当該のコンストラクトに用いることができる。
【0052】
バリアント型のCR1は当業で公知の多種の方法により作製することができる。例えば、クローニングされたCR1遺伝子は、当業で公知の数多くある戦略のいずれによっても、改変することができる(Maniatis, T., 1982, Molecular Cloning, A Laboratory Manual, Cold
Spring Harbor Laboratory, Cold Spring Harbor, N.Y.)。このようなバリアントは、例えばCR1のCR1結合分子への結合を促進する又は低下させるなどのために作製することができる。CR1バリアントをコードする遺伝子の作製においては、好ましくは、改変後の遺伝子は、翻訳停止シグナルを途中に持つことなく、CR1と同じ翻訳読み枠内に留まっているとよい。別の実施態様では、CR1遺伝子を変異させて翻訳、開始、及び/又は終了配列を作製及び/又は破壊したり、あるいは、コーディング領域にばらつきを作製したり、及び/又は、新しい制限エンドヌクレアーゼ部位を形成するか、又は、既存のものを破壊して、更なるin vitro改変を容易にすることもできる。当業で公知の変異誘発のための技術を用いることができ、その中には、限定はしないが、in vitro 部位指定変異誘発法(Hutchinson, C., et al.,
1978, J.Biol. Chem. 253:6551)、TAB.RTM.リンカ(ファルマシア社)の使用等、がある。ある好適な実施態様では、CR1をコードするヌクレオチド配列の変更が、配列に対する保存的変異を含む。
【0053】
本発明の遺伝子コンストラクトの作製においては、内因性又は外因性のプロモータを用いることができる。好ましくは、当該のCR1遺伝子を、赤血球上でのCR1の発現につながる又は発現を増加させる調節配列(例えばプロモータ及び/又はエンハンサなどの転写制御配列)に作動的に連結させるとよい。CR1の発現は、必ずしも赤血球上だけでなくともよい。例えば、他の細胞に比較したときの赤血球での発現レベルを増加させるプロモータ及び/又はエンハンサを用いてもよいが、赤血球以外の細胞での発現が起きる場合もある。
【0054】
本発明での使用に適したプロモータには、限定はしないが、GATA-1遺伝子の調節配列がある。ある実施態様では、該調節配列には、DNAse I超感受性により特定され、初期赤血球系細胞での発現活性化をもたらす上流制御位置がある(例えばMcDevitt, M.A. et al., (1997) Proc.
Natl. Acad. Sci., USA, 94:7976-7981; Visvader J.E. et al., (1998) Genes & Dev. 12(4):473-479を参照されたい)。
【0055】
他の調節配列も当業で公知であり、その中には、例えばグロビン(例えばRistaldi et al.1999. Proc Natl Acad Sci U S A. 96:9654;
Ristaldi et al. 1999 Blood Cells Mol. Dis. 25:193; Tewari et al. 1996. Development. 122:3991を参照されたい)βスペクトリン(例えばSabatino et al. 1998 Mol Cell Biol. 18:6634; Gallagher et al.
1999 J. Biol.
Chem. 274:6062を参照されたい)及びバンド3(例えばCox et al. 1985. J. Cell
Biol. 100:1548を参照されたい)がある。造血細胞、そして優先的には赤血球、で優先的に発現する他の遺伝子の発現を制御する調節配列も、本発明で用いてよい。
【0056】
他の適したプロモータは、例えばまず示差的発現解析を、DNAマイクロアレイ技術を用いるなどして行って、赤血球で優先的に発現する遺伝子を特定し、次に、対応する調節配列を特定するなどにより、特定することができる。例えば、肝臓又は心臓など、別の参照細胞又は組織に比べて赤血球で、例えば約2倍、5倍、又は10倍など、優先的に発現するとして特定された遺伝子を選抜し、それらのプロモータを特定し、本発明で用いることができよう。示差的発現解析を行う方法及びDNAマイクロアレイ技術は当業で公知である。
【0057】
遺伝子コンストラクトの一例は、ヒトCR1遺伝子をGATA-1プロモータの転写制御下に含むものであり、図1に示されている。造血細胞でCR1を発現させるために用いることのできる他の因子は当業で公知であり、例えば Okuno
et al. 2002. Blood. 100:4420;
Radomska et al. 2002. Blood. 100:4410; Gaines et al. 2000. J Biol Chem.275:34114; and Wilcox et al.
1999. Proc Natl
Acad Sci U S A. 96:9654に見ることができる。好ましくは、赤血球上でのCR1発現をより特異的に増加させる調節配列を当該のコンストラクトで用いるとよい。
【0058】
ある実施態様では、本発明のコンストラクトに、例えば緑色蛍光タンパク質又は抗生物質耐性遺伝子などのマーカ遺伝子を含めることができる。このようなマーカ遺伝子を用いると、当該コンストラクトを発現する細胞を特定する上で助けとなる。
【0059】
III. トランスジェニック動物の構築
ある局面では、本発明は、ヒト補体受容体1(CR1)が当該動物の赤血球上で機能的に発現するように、そのゲノムが、ヒトCR1をコードするポリヌクレオチドをプロモータに作動的に連結させて含有する非ヒト動物を提供する。更に本発明は、当該動物の赤血球上でヒトCR1を発現する非ヒトトランスジェニック動物を作製する方法も提供する。
【0060】
本発明の方法で用いられるトランスジェニック動物は、例えば哺乳動物、トリ、は虫類又は両生類などであってよい。ここで解説された用途に適した哺乳動物には、げっ歯類;反芻動物;有蹄動物;家畜哺乳動物;及び酪農動物がある。好適な動物には、げっ歯類、ヤギ、ヒツジ、ラクダ、ウシ、ブタ、ウマ、雄牛、ラマ、ニワトリ、がちょう、及び七面鳥がある。ある好適な実施態様では、該非ヒト動物はマウスである。
【0061】
トランスジェニック動物を作製する多種の方法が当業で公知である(例えばWatson, J. D., et al., "The Introduction of Foreign Genes Into
Mice," in Recombinant DNA, 2d Ed., W. H. Freeman & Co., New York
(1992), pp. 255-272; Gordon, J. W., Intl. Rev. Cytol. 115:171-229 (1989);
Jaenisch, R., Science 240: 1468-1474 (1989); Rossant, J., Neuron 2: 323-334
(1990)を参照されたい)。トランスジェニック・ブタを作製するプロトコルの一例をWhite and
Yannoutsos, Current Topics in Complement
Research: 64th Forum in Immunology, pp. 88-94; 米国特許第5,523,226号;米国特許第5,573,933号;PCT出願 WO93/25071; 及びPCT出願 WO95/04744に見ることができる。トランスジェニック・ラットを作製するためのプロトコルの一例はBader and Ganten, Clinical and
Experimental Pharmacology and Physiology, Supp. 3:S81-S87, 1996に見ることができる。トランスジェニック・ウシを作製するためのプロトコルの一例は、アカデミック・プレス社刊Transgenic Animal
Technology, A Handbook, 1994, ed. Carl A.
Pinkertに見ることができる。トランスジェニック・ヒツジを作製するためのプロトコルの一例は、アカデミック・プレス社刊Transgenic Animal Technology, A Handbook, 1994, ed., Carl A.
Pinkertに見ることができる。いくつかの例示的な方法を、以下に詳述する。
【0062】
A. 前核への注入
トランスジェニック動物は、本発明による核酸コンストラクトを卵細胞に導入することにより、作製することができる。その結果得られた卵細胞を、正常な胚発生に向けてメスの子宮に移植し、こうして発生した、導入遺伝子を持つ動物を戻し交配して、この導入遺伝子についてヘテロ接合子を作る。多様な発生上の段階での胚性標的細胞を用いて、本発明の導入遺伝子を導入する。胚性標的細胞の発生上の段階に応じて異なる方法を用いる。導入遺伝子を導入する方法の例には、限定はしないが、受精後の卵又は接合子の顕微注射(Brinster, et al., Proc. Natl. Acad. Sci.USA (1985) 82: 4438-4442)、及びウィルス組込み法(Jaenisch R., Proc. Natl. Acad. Sci. USA (1976) 73: 1260-1264;
Jahner, et al., Proc. Natl. Acad. Sci.USA (1985) 82: 6927-6931; Van der Putten,
et al., (1985) Proc. Natl. Acad. Sci. (USA) 82: 6148-6152)がある。胚の操作及び顕微注射のための手法は、例えば、Manipulating the Mouse
Embryo (引用をもってその内容をここに援用することとするCold Spring Harbor Laboratory Press, Cold Spring Harbor, NY., 1986)に解説されている。同様な方法は他のトランスジェニック動物の作製にも用いられる。
【0063】
例示的な実施態様では、トランスジェニック・マウスの作製で以下のステップを利用する。定義された近交遺伝子バックグラウンドからのオス及びメスのマウスを交配する。交配するメスのマウスは、卵胞の成長を誘導する妊娠ウマ血清PMSと、排卵を誘導するヒト絨毛性ゴナドトロピンhCGで予め処理されている。交配後、このメスをと殺し、受精卵を彼女の卵管から取り出す。この時点では、前核は融合しておらず、それらを光学顕微鏡を用いて観察することが可能である。代替的なプロトコルでは、様々な発生上の段階で胚を採集することができ、胚盤胞などを採集することができる。胚をダルベッコの改良リン酸緩衝生理食塩水(DPBS)中に回収し、10%のウシ胎児血清を添加したダルベッコの改良基本培地(DMEM)中に維持する。
【0064】
次に、外来のDNA又は組換えコンストラクト(例えばhCR1発現コンストラクト)を前核に顕微注射(卵一個当たり100-1000 分子)する。発現コンストラクトの顕微注射は、顕微鏡に取り付けられた標準的なマイクロ・マニピュレータを用いて行うことができる。例えば、胚は、典型的には、顕微注射の間中、油脂に覆われた100マイクロリットルのDPBSの液滴中に支持される。DNA溶液はオスの前核に顕微注射される。前核の膨らみにより、注射の成功を観察する。そのすぐ後に、前核(メスの前核及びオスの前核)の融合が起こり、場合によっては、外来のDNAが受精卵又は接合子の(通常は)一方の染色体内に挿入される。以下に記載するように調製された組換えES細胞を、同様の技術を用いて胚盤胞に注射することができる。
【0065】
B. 胚性幹細胞
トランスジェニック・マウスを作製する別の方法では、本発明の組換えDNA分子を、マウスの胚性幹(ES)細胞に導入することができる。次に、こうして出来た組換えES細胞をマウス胚盤胞に、前記の小項で記載したものと同様な技術を用いて顕微注射する。
【0066】
ES細胞は移植前の胚から得られ、in vitroで培養される(Evans, M J., et al., Nature 292: 154156 (1981); Bradley, M. O. et
al., Nature 309: 255-258 (1984); Gossler, et al., Proc. Natl.
Acad. Sci.
(USA)
83:9065-9069 (1986); Robertson et al., Nature 322:
445448 (1986))。標的決定コンストラクトの生殖細胞系伝播が起きるように、発生中の胚の生殖細胞に組み込ませ、その一部に成らせることができれば、いずれのES細胞株も、ここでの使用に適切である。例えば、ES細胞の作製に用いることのできるマウス株の一つが129J株である。好適なES細胞株はマウス細胞株D3(アメリカン・タイプ・カルチャー・コレクション、カタログ番号CRL 1934)である。このES細胞は、当業で公知であり、そして引用をもってその内容をここに援用することとするRobertson,
Teratocarcinomas and Embryonic Stem
Cells: A Practical Approach,
E.J. Robertson, ed. IRL Press, Washington, D.C.,
1987、Bradley et al., Current Topics in Devel. Biol., 20:357-371, 1986 及びHogan et al., Manipulating the
Mouse Embryo: A Laboratory Manual,
Cold Spring Harbor Laboratory Press, Cold Spring Harbor, NY, 1986に解説された方法を用いて培養し、DNA挿入に向けて調製することができる。
【0067】
発現コンストラクトは、例えば引用をもってここにその内容を援用することとするSambrook et al., Molecular
Cloning: A Laboratory Manual, 2nd Ed., ed., Cold Spring Harbor
laboratory Press: 1989に解説されたものなど、当業で公知の方法により、ES細胞に導入することができる。適した方法には、限定はしないが、エレクトロポレーション、顕微注射、及びリン酸カルシウム処理法がある。ES細胞に導入しようとする発現コンストラクト(例えばhCR1)は直線状であることが好ましい。直線化は、当該のDNAを、ベクタ配列内のみで切断し、当該遺伝子(例えばhCR1)又は調節配列(例えばGATA-1調節配列)内では切断しないように選択された適した制限エンドヌクレアーゼで消化することにより、達成することができる。
【0068】
発現コンストラクトの導入後に、ES細胞をコンストラクトの存在についてスクリーニングする。当該細胞は多種の方法を用いてスクリーニングすることができる。マーカ遺伝子がコンストラクトに用いられている場合、動物の細胞を、マーカ遺伝子の存在について検査することができる。例えば、マーカ遺伝子が抗生物質耐性遺伝子である場合、細胞を、さもなければ致死濃度である抗生物質(例えばneoについて選択する場合はG418)の存在下で培養することができる。生き延びる細胞は、おそらく、導入遺伝子コンストラクトを組み込ませたものであろう。マーカ遺伝子が、その活性を検出可能な酵素(例えばβ-ガラクトシダーゼ)をコードしている遺伝子である場合、この酵素の基質を適した条件下で細胞に加え、酵素活性を分析することができる。代替的又は付加的には、ES細胞ゲノムDNAを直接調べることができる。例えば、DNAをES細胞から標準的な方法を用いて抽出し、こうしてこのDNAを、導入遺伝子に特異的にハイブリダイズするようにデザインされた一つ又は複数のプローブを用いたサザン・ブロット法でプローブすることができる。更にゲノムDNAは、標的決定コンストラクトを含有する細胞のみが、適したサイズのDNA断片を生ずるように、特定の大きさと導入遺伝子の配列とを持つDNA断片を増幅するように特にデザインされたプローブを用いたPCRでも、増幅することができる。
【0069】
C. 移植
本発明の組換え核酸分子(例えばhCR1)を持つ接合子を、精管切除されたオスとの以前の交配で得られた偽妊娠のメスのマウスに移植する。一般的なプロトコルでは、レシピエントのメスを麻酔し、対腰部切開を行って輸卵管を露出させ、胚をこの輸卵管の膨大部領域に移植する。体壁を縫合し、皮膚を創傷クリップで閉じる。胚は全妊娠期間にわたって発生し、代理母は潜在的トランスジェニック・マウスを出産する。最後に、マウス新生児を、外来又は組換えDNAの存在について検査する。注射を受けた卵のうち、平均10%が正しく発生し、マウスを生じる。生まれたマウスのうち、平均で4匹のうち1匹(25%)がトランスジェニックであり、全体的な効率は2.5%となる。これらのマウスを交配すると、それらは外来の遺伝子を通常の(メンデルの)態様でマウス染色体に連鎖した状態で伝播させる。
【0070】
D. トランスジェニック・コンストラクトの存在に関するスクリーニング
トランスジェニック動物は、標準的なプロトコルにより、生後に特定することができる。尾の組織からのDNAは、例えばサザン・ブロット及び/又はPCRを用いるなどして、導入遺伝子コンストラクトの存在についてスクリーニングすることができる。次に、モザイクになっていると思われる仔を、これらが導入遺伝子を持つと考えられる場合に、ホモ接合型の動物を生じさせるために相互に交配する。仔が生殖細胞系伝播を有するかどうかが明らかでない場合、これらを親又は他の株と交配し、その仔を、ヘテロ接合型についてスクリーニングすることができる。ヘテロ接合仔を、当該DNAのサザン・ブロット及び/又はPCR増幅法により特定する。こうして、ヘテロ接合仔を互いに交配して、ホモ接合型のトランスジェニック仔を生じさせることができる。ホモ接合仔は、この交配の産物であるマウス由来や、既知のヘテロ接合仔であるマウス及び野生型マウス由来の等量のゲノムDNAのサザン・ブロット法によっても特定できよう。当該のサザン・ブロットをスクリーニングするためのプローブは、ヒトCR1遺伝子又はマーカ遺伝子あるいは両者の配列に基づき、デザインすることができる。
【0071】
トランスジェニックの仔を特定し、特徴付ける他の手段は当業で公知である。例えば、ウェスタン・ブロットを用いて、導入された遺伝子にコードされたタンパク質(例えばヒトCR1タンパク質)に対する抗体、あるいは、マーカ遺伝子が発現したときのその産物に対する抗体、によりウェスタン・ブロットをプローブすることにより、これらの仔の多様な組織に導入された遺伝子の発現レベルを評価することができる。
【0072】
細胞を固定し、抗体で標識するなどのin situ解析、及び/又は、仔の赤血球などの多様な細胞のFACS(蛍光活性化セル・ソーティング)解析は、導入遺伝子産物の存在又は非存在について調べるために適した抗体を用いて行うことができる。例えば、赤血球上のhCR1の発現を確認するためには、ヒトCR1に特異的な抗体(例えば1B4、HB8592 及び7G9 モノクローナル、米国特許出願No. 20020103343を参照されたい)を、直接結合させて用いるか、あるいは、蛍光体を結合させた、抗hCR1抗体を認識する二次抗体と併用して、フローサイトメトリを行うことができる。この解析では、ヒト赤血球をhCR1の存在に関する陽性コントロールとして用いることができ、そして正常なマウス赤血球を陰性コントロールとして用いることができる。
【0073】
E. 複数の導入遺伝子又は付加的な変異を含有するマウス
ここで解説された通りにhCR1をそれらの循環赤血球上に発現するトランスジェニック・マウスを、a)更なる導入遺伝子を持つ、又はb)他の遺伝子に変異を含有する、マウスと交配することができる。変異のそれぞれについてヘテロ接合型又はホモ接合型のマウスは、標準的な交配法を用いて作製し、維持することができる。hCR1導入遺伝子を含有するマウスと交配することのできるマウスの例には、限定はしないが、例えば マウス株NZB/W、MRL/+ 又はSJLなど、狼瘡のモデルであるマウス株など、自己免疫疾患の傾向が高いマウス株がある。
【0074】
本発明は、更に、トランスジェニック動物を由来とする細胞にも関する。突然変異又は環境による影響が原因で、後代に何らかの改変が起きる場合があるため、このような後代は、実際には親細胞と同一でないかも知れないが、それでも尚、ここで用いられるこの用語の範囲内に包含される。
【0075】
IV. 二重特異的化合物
ある局面では、本発明は、非ヒト動物の血清及び/又は循環中の作用物質の濃度を低下させる候補又は検査二重特異的化合物を同定する方法を提供するものである。該方法を用いて選択された又は最適化された化合物は、ヒトの対象など、このような化合物を投与すると有益であろう対象を治療するために用いることができる。
【0076】
本発明の方法で検査可能な候補化合物は二重特異的化合物である。ここで用いられる場合の用語「二重特異的化合物」には、2つの異なる結合特異性部分を有する化合物が包含される。二重特異的化合物の例には、例えば二重特異的抗体、ヘテロ重合体、及び抗原ベースのヘテロ重合体など、がある。
【0077】
本発明で検査可能な二重特異的分子は、好ましくは、CR1、好ましくはヒトCR1、に特異的な結合部分を、標的とされる作用物質(異なる抗体又は抗原)に特異的な第二結合部分に架橋させて含むとよい。CR1に対して特異的な結合部分の例には、限定はしないが、CR1リガンド、例えばC3b、があるが、あるいは、好適な実施態様では、CR1に対する抗体、がある。CR1に対して特異的なモノクローナル抗体の例には、限定はしないが、以下:1B4、HB8592、7G9、3D9、E-11、57F 及びYZ1のうちのいずれか一つがある。HB8592 及び1B4 はTaylor
et al., Proc. Nat. Acad. Sci.,
88:3305-3309 (1991) 及びReist et al., Eur. J. Immunol., 23:3021-3027 (1993)に開示されている。抗体3D9 及びE-11 は、Edberg
et al., (1992) Eur J Immunol. 22:
1333-9に解説されている。
【0078】
別の実施態様では、新規なCR1結合分子を、それらのCR1への結合能に基づいて同定することができる。例えば、化合物又は低分子のライブラリを無細胞結合検定法で検査することができる。例えばペプチド・ミメティック、低分子又は他の薬物などのいずれの数の検査化合物も検査に用いることができ、また、数多くのアプローチのいずれかを、生物学的ライブラリ;空間指定可能なパラレル固相又は液相ライブラリ;デコンボリューションを要する合成ライブラリ法;「ワン−ビーズ・ワン−コンパウンド」ライブラリ法;及びアフィニティ・クロマトグラフィ選抜を用いた合成ライブラリ法を含め、当業で公知のコンビナトリアル・ライブラリ法において用いて得ることができる。生物学的ライブラリのアプローチはペプチド・ライブラリに限られているが、他の四種類のアプローチは、ペプチド、非ペプチド・オリゴマ又は低分子の化合物ライブラリに応用可能である(Lam, K.S. (1997) Anticancer Drug Des. 12:145)。
【0079】
分子ライブラリの合成のための方法の例は、当業において、例えばDeWitt et al. (1993) Proc. Natl. Acad. Sci. U.S.A. 90:6909; Erb et al. (1994) Proc.
Natl. Acad. Sci. USA 91:11422;
Zuckermann et al. (1994). J. Med. Chem. 37:2678; Cho et al.
(1993) Science 261:1303; Carrell et
al. (1994) Angew. Chem. Int.
Ed. Engl.
33:2059; Carell et al. (1994) Angew. Chem.
Int. Ed.
Engl. 33:2061; and in Gallop et al. (1994) J. Med.
Chem. 37:1233に見ることができる。
【0080】
化合物のライブラリは溶液中(例えば Houghten (1992) Biotechniques 13:412-421)、又はビーズ上(Lam (1991) Nature
354:82-84)、チップ上(Fodor (1993) Nature 364:555-556)、細菌(Ladner 米国特許第5,223,409号)、胞子(Ladner 米国特許第 '409号)、プラスミド(Cull et al. (1992) Proc Natl Acad Sci USA 89:1865-1869)又はファージ上(Scott and Smith (1990) Science
249:386-390);(Devlin (1990) Science 249:404-406); (Cwirla et al. (1990) Proc. Natl. Acad. Sci. 87:6378-6382); (Felici (1991) J. Mol. Biol. 222:301-310); (上記のLadner)に提供してもよい。
【0081】
調節性物質及び天然抽出物のライブラリを検査する数多くの薬物スクリーニング・プログラムでは、所定の時間で調査される調節性物質の数を最大にするためには、高スループットの検定法が好ましい。精製済み又は半精製済みタンパク質で得られるものなど、無細胞系で行われる検定法が「一次」スクリーンとしてしばしば好ましい。なぜなら、これらは、検査対象である調節性物質により媒介される分子ターゲット中の変化を高速で進行させ、また比較的容易に検出できるように作製することが可能だからである。更に、検査対象の調節性物質の細胞毒性及び/又は生物学的利用能の効果は、in vitro系では概ね無視することができ、代わりに、当該検定の焦点を、上流又は下流の要素との結合親和性の変化に現れるであろう、当該薬物の分子ターゲットに対する効果に、主に当てることができる。
【0082】
別の実施態様では、当業で公知のファージ・ディスプレイ技術を用いて新規なCR1結合分子を同定することができる。
【0083】
ある実施態様では、本発明は、CR1又はその生物学的に活性な部分に結合する候補又は検査化合物をスクリーニングするための検定法を提供する。
【0084】
CR1に結合する分子を同定するための細胞ベースの検定法を、本発明の二重特異的化合物での使用に向けた付加的な作用物質を同定するために用いることができる。例えば、CR1を発現する細胞をスクリーニング検定法で用いることができる。例えば、CR1への結合において統計上有意な変化を生ずる化合物を同定することができる。
【0085】
本発明のある実施態様では、CR1又はその部分を、二種ハイブリッド検定法又は三種ハイブリッド検定法で「ベイト・タンパク質」として用いて(例えば米国特許第5,283,317号; Zervos et al. (1993) Cell
72:223-232; Madura et al. (1993) J. Biol.
Chem. 268:12046-12054; Bartel et al. (1993) Biotechniques
14:920-924; Iwabuchi et al. (1993) Oncogene
8:1693-1696; and Brent WO94/10300を参照されたい)、本発明のタンパク質に結合又は相互作用すると共に、それらの活性に関与するような他のタンパク質を同定することができる。前記の二種ハイブリッド系は、分離可能なDNA結合ドメイン及び活性化ドメインから成る大半の転写因子のモジュール性に基づいている。簡単に説明すると、該検定法では2つの異なるDNAコンストラクトを利用する。一方のコンストラクトでは、あるタンパク質をコードする遺伝子が、既知の転写因子(例えばGAL-4)のDNA結合ドメインをコードする遺伝子に融合されている。他方のコンストラクトでは、未知のタンパク質(「プレイ」又は「試料」)をコードする、DNA配列ライブラリ由来の一DNA配列が、該既知の転写因子の活性化ドメインをコードする遺伝子に融合されている。これら「ベイト」及び「プレイ」タンパク質がin vivoで相互作用して複合体を形成することができれば、該転写因子のDNA結合ドメイン及び活性化ドメインが近くにあることになる。この近さにより、該転写因子に対して応答性の転写調節部位に作動的に連結されたレポータ遺伝子(例えばLacZ)の転写が可能である。レポータ遺伝子の発現を検出し、機能的転写因子を含有する細胞コロニを単離し、これを用いて、本発明のタンパク質と相互作用するタンパク質をコードする、クローニングされた遺伝子を得ることができる。相互作用性タンパク質が同定されたら、本発明のタンパク質と、その相互作用性タンパク質との間の相互作用を調節する上での検査化合物の能力を、当業で公知の技術を用いて、例えばここで解説する無細胞検定法を用いて、検査することができる。
【0086】
ある実施態様では、当該の検定は、CR1結合分子が、in vitroでのCR1へのその結合能に基づいて同定される無細胞検定法である。CR1結合分子を提供することができ、CR1へのそのタンパク質の結合能 を直接的な結合を判定する当業で公知の方法を用いて検査することができる。当該タンパク質の、標的分子への結合能の判定は、例えば、リアルタイム・バイオモラキュラー・インターアクション・アナリシスBiomolecular Interaction Analysis (BIA). Sjolander, S. and Urbaniczky, C. (1991) Anal. Chem. 63:2338-2345 and Szabo et
al. (1995) Curr. Opin. Struct. Biol.
5:699-705などの技術を用いて行うことができる。ここで用いる場合の「BIA」とは、相互作用物質のいずれも標識せずに、二重特異的相互作用をリアルタイムで研究する技術である(例えばBIAcore)。表面プラズモン共鳴(SPR)の光学的現象の変化を、生物学的分子間のリアルタイムの反応の指標として用いることができる。
【0087】
本発明の無細胞検定法は、可溶性及び/又は膜結合型のタンパク質の使用の両方に適している。膜結合型のタンパク質を用いる無細胞検定法の場合、膜結合型のタンパク質が溶液中に維持されるように、可溶化剤を用いることが好ましいであろう。このような可溶化剤の例には、非イオン性の界面活性剤、例えばn-オクチルグルコシド、n-ドデシルグルコシド、n-ドデシルマルトシド、オクタノイル-N-メチルグルカミド、デカノイル-N-メチルグルカミド、Triton(R)X-100、 Triton(R)X-114、 Thesit(R)、イソトリドデシポリ(エチレングリコールエーテル)n、3-[(3-コラミドプロピル)ジメチルアミニオ]-1-プロパンスルホネート(CHAPS)、3-[(3-コラミドプロピル)ジメチルアミニオ]-2-ヒドロキシ-1-プロパンスルホネート (CHAPSO)、又はN-ドデシル=N,N-ジメチル-3-アンモニオ-1-プロパンスルホネートがある。
【0088】
タンパク質対タンパク質の相互作用を検出することができる適した検定法が当業で公知である(例えば免疫沈降法、二種ハイブリッド検定法等)。このような検定法を検査化合物の存在下及び非存在下で行うことにより、これらの検定法を用いて、本発明のタンパク質の標的分子との相互作用を調節(例えば阻害又は亢進)する化合物を同定することができる。
【0089】
当該タンパク質の、標的分子との結合能又は相互作用能の判定は、例えば直接的な結合などにより、行うことができる。直接的な結合検定法では、当該のタンパク質の標的分子への結合を、複合体中の標識済みタンパク質を検出することにより判定できるように、放射性同位元素又は酵素標識に当該タンパク質を結合させてもよいであろう。例えば、タンパク質は、125I、35S、14C、又は3Hで直接又は間接的に標識することができ、その放射性同位元素を、放射線放出を直接計数したり、あるいはシンチレーション計数により検出することができる。代替的には、分子を、例えば西洋わさびペルオキシダーゼ、アルカリホスファターゼ、又はルシフェラーゼなどで酵素標識し、その酵素標識を、適した基質の生成物への転化を判定することにより、検出することができる。
【0090】
典型的には、本発明のタンパク質又はその結合タンパク質のいずれかを固定して、複合体を形成していない形の一方又は両方のタンパク質から複合体を容易に分離できるようにし、また検定法の自動化に対応することも好ましいであろう。候補作用物質の存在下及び非存在下における、上流又は下流の結合因子への結合を、反応物を含有するのに適したいずれかの容器で行わせることもできる。例には、微量定量プレート、試験管、及びマイクロ遠心管がある。ある実施態様では、当該タンパク質をマトリックスに結合可能にするドメインを加えた融合タンパク質を提供することができる。例えば、グルタチオン-S-トランスフェラーゼ/ CR1 (GST/ CR1) 融合タンパク質をグルタチオン・セファロース・ビーズ(ミズーリ州セントルイス、シグマ・ケミカル社)又はグルタチオン誘導体化微量定量プレートに吸着させた後、これらを35Sで標識されているなどの細胞ライセートと検査対象の調節性物質とに配合し、この混合物を、例えば塩及びpHについて生理条件など、複合体形成を誘導する条件下でインキュベートすることができるが、僅かによりストリンジェントな条件が好ましいであろう。インキュベート後、ビーズを洗浄して未結合の標識を取り除き、マトリックスに固定し、放射性標識を直接(例えばビーズをシンチレーション剤中に配置する)判定するか、あるいは、複合体を解離させた後に上清中で判定する。代替的には、複合体をマトリックスから解離させ、SDS-PAGEで分離し、ビーズ画分中に見られるCR1結合タンパク質のレベルを標準的な電気泳動技術を用いてゲルから定量することができる。
【0091】
マトリックス上にタンパク質を固定する他の技術も、本検定法での使用に利用できる。例えば、ビオチン化分子をビオチン-NHS (N-ヒドロキシ-スクシンイミド)から当業で公知の技術(例えばイリノイ州ロックフォード、ピアース・ケミカルズ社のビオチン化キット)を用いて調製し、
ストレプトアビジンで被覆された96ウェル・プレート(ピアース・ケミカル社)のウェルに固定することができる。
【0092】
更に、相互作用物質のいずれも標識せずに、CR1とCR1結合分子との間の相互作用を調節する上での化合物の能力を判定することも、本発明の範囲内である。例えば、マイクロフィジオメータを用いて、本発明のタンパク質の、その標的分子との相互作用を、このタンパク質又は標的分子のいずれも標識せずに検出することができる。McConnell, H. M. et al. (1992)
Science 257:1906-1912。ここで用いられる「マイクロフィジオメータ」(例えばCytosensor)は、細胞がその環境を酸化する速度を、光指定可能な電位差センサ(LAPS)を用いて測定する分析装置である。この酸化速度の変化を、化合物と受容体との間の相互作用の指標として用いることができる。
【0093】
本発明で検査可能な抗原ベースのヘテロ重合体は、優先的には、CR1、好ましくはヒトCR1、に特異的な結合部分を、自己抗体により認識される抗原に架橋させて含む。自己抗体により認識される抗原の例には、限定はしないが、以下:VIII因子(置換組換えVIII因子による血友病治療に関連する抗体);筋アセチルコリン受容体(重症筋無力症に関連する抗体);カルジオリピン(狼瘡に関連する);血小板関連タンパク質(特発性血小板減少性紫斑病に関連する);シェーグレン症候群に関連する複数の抗原;組織移植自己免疫反応の症例への関与が示唆されている抗原;心筋に見られる抗原(自己免疫心筋炎に関連する);免疫複合体媒介性腎疾患に関連する抗原;dsDNA 及びssDNA 抗原(ループス腎炎に関連する); デズモグレイン及びデズモプラキン(天疱瘡及び類天疱瘡に関連する);又はよく特徴付けられていると共に、疾患病理に関連するいずれか他の抗原のいずれもが含まれる。
【0094】
本発明での検査の対象となるヘテロ重合体及び抗原ベースのヘテロ重合体の例や、それらを作製する方法は当業で公知である。例えば、ヘテロ重合体の例はWO 03007971A1; US 20020103343A1; US 5879679; US 5487890; US 5470570;
WO 9522977A1; WO/02075275A3、 WO/0246208A2 もしくはA3、WO/0180883A1、WO/0145669A1、WO 9205801A1、Lindorfer et al. 2001 J. Immunol. Methods. 248:125; Hahn et al. 2001. J. Immnol.
166:1057; Nardin et al. 1998. J. Immunol.
Methods. 211:21; Kuhn et
al. 1998. J. Immunol.
160:5088; Taylor et
al. 1997. Cancer
Immunol. Immunother. 45:152;
Taylor et al. 1997. J. Immunol.
159:4035; 及び Taylor et al. 1992. J. Immunol.
148:2462に教示されている。加えて、バリアント型のこれらのヘテロ重合体を作製することもできる。例えば、ある実施態様では、異なる連結化学法を用いて作製された数々の形の二重特異的分子を用いることができる。二重特異的分子の成分を架橋するために用いることのできる試薬の例には:ポリエチレングリコール、SATA、SMCC、や当業で公知の他のものがあり、例えばピアース・バイオテクノロジー社から得られる。検査することのできる二重特異的分子の形の例は、引用をもってその内容をここに援用することとする、2002年9月16日に提出されたUSSN 60/411,731に解説されている。
【0095】
別の実施態様では、様々な多量体型の二重特異的分子を作製することができる(例えば二量体、三量体、四量体、五量体、又はそれより大きな多量体型)。別の実施態様では、精製された形の二重特異的分子を、例えば引用をもってその内容をここに援用することとする、2002年5月13日に提出された USSN 60/380,211に解説されている通りに、検査することができる。
【0096】
別の実施態様では、当該のヘテロ重合体の結合部分のうちの一つが抗体である場合、異なるアイソタイプ(例えばIgA、IgD、IgE、IgG1、IgG2 (例えばIgG2a)、IgG3、IgG4、又はIgM)の抗体を用いることができる。別の実施態様では、抗体分子の部分(例えばFabフラグメント)を結合部分の一つに用いることができる。ある好適な実施態様では、結合部分のうちの少なくとも1つは、Fcドメインを含む抗体である。ある実施態様では、当該の抗体はマウス抗体である。
【0097】
別の実施態様では、抗体に対する修飾の効果を検査することができ、例えば2003年3月28日に提出されたUSSN 60/458,869に解説されたように、抗体のデイミュナイゼーションの効果を検査することができる。
【0098】
本発明の二重特異的化合物が結合することのできる標的とされる作用物質の例には、以下:ウィルス、ウィルス性粒子、毒素、細菌、ポリヌクレオチド、抗体、例えば自己免疫異常に関連する自己抗体など、のうちのいずれも含む、しかしこれらに限定はしない血液由来作用物質がある。ある実施態様では、標的とされるウィルス性作用物質の例には、限定はしないが、以下:サイトメガロウィルス、インフルエンザ、
ニューカッスル病ウィルス、水疱性口内炎ウイルス、単純疱疹ウィルス、肝炎、アデノウィルス-2、ウシウイルス性下痢ウイルス、ヒト免疫不全ウィルス (HIV)、デング熱ウィルス、マーブルグウィルス、エプスタイン-バーウィルス、のうちのいずれも含まれる。
【0099】
細菌性作用物質の例には:シュードモナス-アエルギノーサ(原語:Pseudomonas aeruginosa)、シュードモナス-フルオレセンス(原語:Pseudomonas fluorescens)、シュードモナス-アシドボランス(原語:Pseudomonas acidovorans)、シュードモナス-アルカリゲンス(原語:Pseudomonas alcaligenes)、シュードモナス-プチダ(原語:Pseudomonas putida)、ステノトロフォモナス-マルトフィリア(原語:Stenotrophomonas maltophilia)、バークホルデリア-セパシア(原語:Burkholderia cepacia)、アエロモナス-ヒドロフィリア(原語:Aeromonas hydrophilia)、エシェリヒア-コリ(原語:Escherichia coli)、シトロバクター-フロウンジ(原語:Citrobacter freundii)、サルモネラ-チフィムリウム(原語:Salmonella typhimurium)、サルモネラ-チフィ(原語:Salmonella typhi)、サルモネラ-パラチフィ(原語:Salmonella paratyphi)、サルモネラ-エンテリティディス(原語:Salmonella enteritidis)、シゲラ-ジセンテリア(原語:Shigella dysenteriae)、シゲラ-フレクスネリ(原語:Shigella flexneri)、シゲラ-ソネイ(原語:Shigella sonnei)、エンテロバクター-クロアケ(原語:Enterobacter cloacae)、エンテロバクター-アエロゲンス(原語:Enterobacter aerogenes)、クレブシエラ-ニューモニエ(原語:Klebsiella pneumoniae)、クレブシエラ-オキシトカ(原語:Klebsiella oxytoca)、セラチア-マルセセンス(原語:Serratia marcescens)、フランシセラ-ツラレンシス(原語:Francisella tularensis)、モルガネラ-モルガニイ(原語:Morganella morganii)、プロテウス-ミラビリス(原語:Proteus mirabilis)、プロテウス-ブルガリス(原語:Proteus vulgaris)、プロビデンシア-アルカリファシエンス(原語:Providencia alcalifaciens)、プロビデンシア-レットゲリ(原語:Providencia rettgeri)、プロビデンシア-スチュアーティ(原語:Providencia stuartii)、アシネトバクター-カルコアセチクス(原語:Acinetobacter calcoaceticus)、アシネトバクター-ヘモリティクス(原語:Acinetobacter haemolyticus)エルシニア-エンテコリティカ(原語:Yersinia enterocolitica)、エルシニア-ペスティス(原語:Yersinia pestis)、エルシニア-シュードツベルキュローシス(原語:Yersinia
pseudotuberculosis)、エルシニア-インテルメディア(原語:Yersinia intermedia)、ボルデテッラ-ペルツシス(原語:Bordetella pertussis)、ボルデテッラ-パラペルツシス(原語:Bordetella parapertussis)、ボルデテッラ-ブロンキセプティカ(原語:Bordetella bronchiseptica)、ヘモフィラス-インフルエンザ(原語:Haemophilus influenzae)、ヘモフィラス-パラインフルエンザ(原語:Haemophilus parainfluenzae)、ヘモフィラス-ヘモリティクス(原語:Haemophilus haemolyticus)、ヘモフィラス-パラへモリティカス(原語:Haemophilus parahaemolyticus)、ヘモフィラス-デュクレイ(原語:Haemophilus ducreyi)、パスツーレラ-マルトシダ(原語:Pasteurella multocida)、パスツーレラ-ヘモリティカ(原語:Pasteurella haemolytica)、ブランハメラ-カタラーリス(原語:Branhamella catarrhalis)、ヘリコバクター-ピロリ(原語:Helicobacter pylori)、カンピロバクター-フェツス(原語:Campylobacter fetus)、カンピロバクター-ジェジュニ(原語:Campylobacter jejuni)、カンピロバクター-コリ(原語:Campylobacter coli)、ボレリア-ブルドルフェリ(原語:Borrelia burgdorferi)、ビブリオ-コレラ(原語:Vibrio cholerae)、ビブリオ-パラヘモリティクス(原語:Vibrio parahaemolyticus)、レジオネラ-ニューモフィラ(原語:Legionella pneumophila)、リステリア-モノサイトゲンス(原語:Listeria monocytogenes)、ネイセリア-ゴノローエ(原語:Neisseria gonorrhoeae)、ネイセリア-メニンギティディス(原語:Neisseria meningitidis)、ガルドネレッラ-バギナリス(原語:Gardnerella vaginalis)、バクテロイデス-フラギリス(原語:Bacteroides fragilis)、バクテロイデス-ディスタソニス(原語:Bacteroides distasonis)、バクテロイデス3452A ホモロジー群、バクテロイデス-ブルガツス(原語:Bacteroides vulgatus)、バクテロイデス-オバルス(原語:Bacteroides ovalus)、バクテロイデス-エタイオタオミクロン(原語:Bacteroides thetaiotaomicron)、バクテロイデス-ユニフォルミス(原語:Bacteroides uniformis)、バクテロイデス-エゲルシー(原語:Bacteroides eggerthii)、バクテロイデス-スプランクニカス(原語:Bacteroides splanchnicus)、クロストリジウム-ジフィシル(原語:Clostridium difficile)、マイコバクテリウム-ツベルキュローシス(原語:Mycobacterium tuberculosis)、マイコバクテリウム-アビウム(原語:Mycobacterium avium)、マイコバクテリウム-イントラセルラーレ(原語:Mycobacterium intracellulare)、マイコバクテリウム-レプレー(原語:Mycobacterium leprae)、コリネバクテリウム-ジフテリア(原語:Corynebacterium diphtheriae)、コリネバクテリウム-ウルセランス(原語:Corynebacterium ulcerans)、スタフィロコッカス-ニューモニエ(原語:Streptococcus pneumoniae)、ストレプトコッカス-アガラクチエ(原語:Streptococcus agalactiae)、ストレプトコッカス-パイオゲンス(原語:Streptococcus pyogenes)、エンテロコッカス-フェカーリス(原語:Enterococcus faecalis)、エンテロコッカス-ファシウム(原語:Enterococcus faecium)、スタフィロコッカス-アウレウス(原語:Staphylococcus aureus)、スタフィロコッカス-エピデルミディス(原語:Staphylococcus epidermidis)、スタフィロコッカス-サプロフィティカス(原語:Staphylococcus saprophyticus)、スタフィロコッカス-インテルメディウス(原語:Staphylococcus intermedius)、スタフィロコッカス-ヒイカス亜種ヒイカス(原語:Staphylococcus hyicus subsp.
hyicus)、スタフィロコッカス-ヘモリティカス(原語:Staphylococcus
haemolyticus)、スタフィロコッカス-ホミニス(原語:Staphylococcus hominis)、スタフィロコッカス-サッカロリティクス(原語:Staphylococcus saccharolyticus)、がある。
【0100】
ある実施態様では、標的とされる作用物質は、例えば抗生物質に対して耐性であるなど、伝統的な治療法に対して耐性である。
【0101】
別の実施態様では、本発明の抗原ベースのヘテロ重合体に結合させることのできる、標的とされる作用物質の例には、限定はしないが、以下:置換組換えVII因子による血友病治療に関連する自己抗体;重症筋無力症;狼瘡、ループス腎炎、特発性血小板減少性紫斑病、シェーグレン症候群、心筋炎、又は天疱瘡及び類天疱瘡に関連する自己抗体;組織移植自己免疫反応に関連する自己抗体;免疫複合体媒介性腎疾患に関連する自己抗体;又は、よく特徴付けられていると共に、疾患病理に関連するいずれか他の自己抗体、のうちのいずれもが含まれる。
【0102】
更に他の実施態様では、本発明の二重特異的化合物に結合させることのできる生物学的作用物質の例には、限定はしないが、以下:炭疽菌、天然痘、ペスト、エボラ、及びマーブルグ・ウィルスのいずれかを含め、生物戦争に関連する可能性がある感染性作用物質及び毒素がある。
【0103】
ある実施態様では、本発明の検定法を行う際に、当該の作用物質をトランスジェニック動物に、二重特異的化合物の投与前、投与と同時又は投与後などに投与する。
【0104】
本発明の二重特異的化合物、又はそのいずれかの部分を修飾してそれらの半減期を高めてもよい。ペプチド類似体が、テンプレート・ペプチドのものと同様な性質を持つ非ペプチド薬物として、製薬業では普通に用いられている。これらの種類の非ペプチド化合物は「ペプチド・ミメティック」又は「ペプチドミメティック」と呼ばれ(引用をもってここに援用することとするFauchere, J. (1986) Adv. Drug Res. 15:29; Veber and
Freidinger (1985) TINS p.392; 及び Evans et al. (1987) J. Med. Chem
30:1229)、通常は、コンピュータ分子モデリングの助けを得て開発されている。治療上有用なペプチドに構造上似たペプチド・ミメティックを用いて、同等な治療上又は予防上の効果を生じさせることができる。概して、ペプチドミメティックは、抗原ポリペプチドなどのパラダイム・ポリペプチド(即ち、生物学的又は薬理学的活性を有するポリペプチド)に構造上似ているが、:-CH2NH-、-CH2S-、-CH2-CH2-、-CH=CH- (cis 及びtrans)、-COCH2-、-CH(OH)CH2-、及び-CH2SO-から成る群より選択される結合部に、当業で公知であり、そして更に、それぞれを引用をもってここに援用することとする以下の参考文献: Spatola, A.F. in Chemistry
and Biochemistry of Amino Acids, Peptides, and Proteins Weinstein, B., ed.,
Marcel Dekker, New York, p. 267 (1983); Spatola, A.F., Vega Data (March
1983), Vol. 1, Issue 3, “Peptide Backbone Modifications”; Morley, J.S. (1980) Trends. Pharm. Sci. pp.463-468; Hudson, D. et al.
(1979) Int. J. Pept. Prot. Res. 14:177-185 (-CH2NH-, CH2CH2-); Spatola, A.F. et
al. (1986) Life. Sci.
38:1243-1249 (-CH2-S); Hann, M.M. (1982) J. Chem.
Soc. Perkin. Trans. I 307-314
(-CH-CH-, cis 及びtrans); Almquist, R.G. et al. (1980) J. Med.
Chem.
23:1392-1398 (-COCH2-); Jennings-White, C. et
al. (1982) Tetrahedron Lett.
23:2533 (-COCH2-); Szelke, M. et al.,
European Patent Application No. EP 45665 (1982) CA: 97:39405 (-CH(OH)CH2-); Holladay, M.W. et
al. (1983) Tetrahedron. Lett.
24:4401-4404 (-C(OH)CH2-); 及びHruby, V.J. (1982) Life Sci. 31:189-199 (-CH2-S-);に記載された方法により選択的に置換された一つ以上のペプチド結合を有する。特に好適な非ペプチド結合は-CH2NH-である。このようなペプチド・ミメティックは、例えば:生産がより経済的である、化学的安定性が高い、薬理学的特性(半減期、吸収、効力、効験等)が優れている、特異性(例えば生物学的活性の範囲が幅広い)が変更されている、抗原性が減らされている、等を含め、ポリペプチドの実施態様に比較して大きな利点を有するであろう。ペプチドミメティックの標識付けは、通常、一つ以上の標識を、直接又はスペーサ(例えばアミド基)を介して、当該ペプチドミメティック上で定量的構造−活性データ及び/又は分子モデリングにより予測可能な非干渉位置に、共有結合させるステップを含む。このような非干渉位置は、一般的には治療効果を生ずるために当該ペプチドミメティックが結合する先の巨大分子との直接的な接点を形成しない位置である。ペプチドミメティックの誘導体化(例えば標識付け)は、当該ペプチドミメティックの所望の生物学的又は薬理学的活性に実質的に干渉してはならない。
【0105】
あるアミノ酸配列の一つ以上のアミノ酸を、同じ種類のD型アミノ酸(例えばL型リジンの代わりにD型リジン)に系統的に置換する方法を用いて、より安定なペプチドを作製することができる。加えて、例えば分子内でジスルフィド架橋を形成させてペプチドを環化することのできる内側のシステイン残基を加えるなどにより、当業で公知の方法により拘束されたペプチドを作製することができる(引用をもってここに援用することとするRizo and Gierasch (1992) Annu.
Rev. Biochem.
61:387)。
【0106】
このような修飾されたポリペプチドは、原核又は真核ホスト細胞内で産生させることができる。代替的には、このようなペプチドを化学法で合成することができる。異種ポリペプチドを組換えホスト内で発現させる方法、ポリペプチドの化学合成法、及びin vitro翻訳のための方法は当業で公知であり、更に、引用をもってここに援用することとするManiatis et al., Molecular Cloning: A Laboratory Manual (1989), 2nd Ed.,
Cold Spring Harbor, N.Y.; Berger and Kimmel, Methods in Enzymology, Volume 152, Guide to Molecular Cloning
Techniques (1987), Academic Press, Inc., San Diego, Calif.; Merrifield, J.
(1969) J. Am. Chem. Soc. 91:501;
Chaiken I.M. (1981) CRC Crit. Rev.
Biochem. 11:255; Kaiser et al.
(1989) Science 243:187; Merrifield,
B. (1986) Science 232:342; Kent,
S.B.H. (1988) Annu. Rev. Biochem. 57:957;
及びOfford, R.E. (1980) Semisynthetic Proteins, Wiley Publishingにも解説されている。
【0107】
ポリペプチドは典型的には直接的な化学合成で作製することができ、ヘテロ重合体の結合部分として用いることができる。 ペプチドは、非ペプチド部分を共有結合によりN末端及び/又はC末端に付着させた修飾ペプチドとして作製することができる。いくつかの好適な実施態様では、カルボキシ末端又はアミノ末端のいずれか、あるいは両者、を化学修飾する。末端のアミノ基又はカルボキシル基の最もよくある修飾法は、それぞれアセチル化及びアミド化である。アシル化(例えばアセチル化)又はアルキル化(例えばメチル化)などのアミノ末端の修飾や、アミド化などのカルボキシ末端の修飾や、環化を含む他の末端の修飾を、多様な実施態様の検査化合物に導入することができる。特定のアミノ末端及び/又はカルボキシ末端の修飾、及び/又は、コア配列へのペプチド伸長により、例えば安定性が高まる、効力及び/又は効験、血清プロテアーゼへの耐性、所望の薬物動態学的特性が高まる、等、有利な物理的、化学的、生化学的、及び薬理学的特性を提供することができる。
【0108】
V. スクリーニング検定法
本発明の方法は、非ヒト動物の血清及び/又は循環中の作用物質の濃度を低下させる候補又は検査化合物を同定するための、ここでは「スクリーニング検定法」とも呼ばれる方法を提供するものである。
【0109】
ある実施態様では、二重特異的化合物を、対象の血清、循環及び/又は組織中の作用物質の濃度を低下させる上でのそれらの能力について検査することができる。
【0110】
ある二重特異的化合物の、非ヒト動物の血清、循環及び/又は組織中の作用物質の濃度を低下させる能力は、数多くの様々な方法で評価することができる。例えば、血清、循環及び/又は組織中の作用物質の濃度は、(例えばウィルス力価を測定したり、コロニ(例えばプラーク)形成検定法を行ったり、細菌コロニの計数や、あるいは毒素等に関するELISA又は他の検出検定法を行うなどにより)組織、血液又は血清試料などの中の作用物質を計数できる当業で公知の検定法を行うなどで、直接測定することができる。
【0111】
本発明で提供される方法では、非ヒト動物の血清、循環及び/又は組織中の病原性物質などの作用物質の濃度を、少なくとも、例えば約20%、約30%、約40%、約50%、約60%、約70%、約80%、約90% 又は約100%、低下させることができる。
【0112】
別の実施態様では、対象の血清、循環及び/又は組織中の作用物質の濃度を間接的に測定することができる。例えば、血清及び/又は循環中に当該作用物質が存在することを原因とする病理を、この動物由来の組織試料を調べるなどにより、測定することができる。非ヒト動物の血清、循環及び/又は組織中の作用物質の濃度を間接的に測定するもう一つの方法は、当該作用物質が、非ヒト動物において感染を引き起こす能力を測定する方法である。例えば、感染の臨床上の兆候及び症状に対する当該二重特異的化合物の効果を測定することができる。また、例えばある臓器系から別のものへ、あるいはある個体から別の個体への感染の広がりを阻害する上での当該二重特異的化合物の能力を検査することもできる。
【0113】
別の実施態様では、当該二重特異的化合物の、非ヒト動物におけるCR1担持細胞への結合能を測定する。例えば、ある実施態様では、当該二重特異的化合物の、CR1標的分子への結合能の判定は、
リアルタイム・バイオモラキュラー・インターアクション・アナリシス (BIA) (Sjolander, S. and
Urbaniczky, C. (1991) Anal. Chem.
63:2338-2345 及びSzabo et al. (1995) Curr. Opin.
Struct. Biol. 5:699-705)などの技術を用いても行うことができる。ここで用いる場合の「BIA」とは、相互作用物質のいずれも標識せずに、二重特異的相互作用をリアルタイムで研究する技術である(例えばBIAcore)。表面プラズモン共鳴(SPR)の光学的現象の変化を、生物学的分子間のリアルタイムの反応の指標として用いることができる。
【0114】
別の実施態様では、非ヒト動物での細胞による作用物質の破壊(例えばマクロファージによる致死)を測定する。
【0115】
非ヒト動物の血清及び/又は循環中の作用物質の濃度を(当該二重特異的化合物を投与していない非ヒト動物で観察される濃度と比較して)低下させる化合物を選抜することができる。当該の検定法で検査する対象となる化合物は、検査される複数の化合物の中から選択することができる。別の実施態様では、本検定法で検査する対象の二重特異的化合物は、例えばin vitro検定法で、CR1に結合することができると既に特定されていてもよく、本検定法を用いて更に評価又は最適化されてもよい。このような場合、ある二重特異的化合物の、血清及び/又は循環中の作用物質の濃度を低下させる能力を、別の二重特異的化合物か、又は同じ化合物の最適化されていない形と比較して、 血清及び/又は循環中の当該作用物質の濃度を低下させるその能力を判定することができる。
【0116】
好適な実施態様では、本発明の二重特異的化合物を、約1μgの化合物/体重1kg乃至約100μgの化合物/体重1kgの範囲内の濃度、投与する。ここで定義するように、二重特異的化合物の治療上有効量(即ち有効な投薬量)は、約0.01 乃至5000μg/体重1kgの範囲であるが、好ましくは約0.1 乃至500μg/体重1kg、より好ましくは約2 乃至80μg/体重1kg、そして更により好ましくは約5 乃至70μg/kg、10 乃至60μg/kg、20 乃至50μg/kg、24 乃至41μg/kg、25 乃至40μg/kg、26 乃至39μg/kg、27 乃至38μg/kg、28 乃至37μg/kg、29 乃至36μg/kg、30乃至35μg/kg、31乃至34μg/kg 又は32 乃至33μg/体重1kgの範囲であるとよい。当業者であれば、疾患又は異常の重篤度、以前の治療、対象の全身の健康及び/又は年齢、及び他の既往症を含め、しかしこれらに限らず、特定の因子が、対象を有効に治療するために必要な投薬量に影響する場合があることを理解されよう。更に、治療上有効量のタンパク質、ポリペプチド、又は抗体による対象の治療には、単回の治療を含めることができるが、あるいは好ましくは一連の治療を含めることができるとよい。
【0117】
ある好適な例では、作用物質を静脈内(iv)注射した後、約1乃至500μg/体重1kgの範囲の二重特異的化合物で動物を処理する。更に、治療に用いられる二重特異的化合物の有効な投薬量を、特定の治療経過にわたって増減させてもよいことも理解されよう。投薬量の変更は、ここで解説する通りの診断検定法の結果行われてもよく、また同結果から明白となるであろう。
【0118】
検査化合物及び/又は作用物質の投与経路は、動物の循環中への静脈内(iv)注射であってよい。他の投与経路には、限定はしないが、局所、非経口、皮下、又は吸入、がある。用語「非経口」には、例えば皮下、静脈内又は筋肉内経路による注射が含まれるが、更に、例えば疾患又は傷害の部位への局所的投与も含まれる。インプラントからの化合物の持続的放出も当業で公知である。当業者であれば、適した投薬量は、治療しようとする異常の性質、患者の体重、年齢及び全身の状態や、投与経路といった因子に応じて様々であろうことは認識されよう。予備的用量は動物検査に従って決定することができ、ヒトへの投与に向けた投薬量の見積もりは当業で認められた慣例に従って行われる。
【0119】
候補化合物及び作用物質を、一定の範囲の用量にわたって動物に投与することができる。当該の作用物質も動物に投与する場合、候補化合物を、当該作用物質の投与前、投与と同時、又は投与後のいずれでも投与することができる。
【0120】
マウスなど、CR1を発現する本発明のトランスジェニック動物を用いて、例えば自己抗体、感染性物質又は毒素など、対象の血清及び/又は循環中に望ましくない作用物質が存在することに関連する、ヒトにおける異常又は疾患を治療するために有用な候補化合物をスクリーニング又は評価することができる。
【0121】
ある実施態様では、当該のスクリーニング検定法を用いて、血清及び/又は循環中の作用物質の濃度を低下させる上での二重特異的化合物の活性を評価することができる。例えば、様々な形の二重特異的化合物(例えば二量体、三量体、四量体、五量体、又はより大きな多量体型)の、血清及び/又は循環中の作用物質の濃度を低下させる上での能力を検査することができる。
【0122】
ある実施態様では、ヘテロ重合体又は抗原ヘテロ重合体の活性を従来の二重特異的抗体に比較することができる。
【0123】
ある更なる実施態様では、ヘテロ重合体又は抗原ヘテロ重合体の活性を、生理食塩水及び/又は無関係のHP(例えば本発明のある好適な例では、ФX-PA)を含む適したコントロールに比較することができる。
【0124】
別の実施態様では、様々な連結化学法を用いて調製された二重特異的化合物の能力を、血清及び/又は循環中の作用物質の濃度を低下させるそれらの能力について比較することができる。例えば、ある実施態様では、二重特異的化合物の一つの部分を、第二の部分に、SMCC/SATA 結合又はPEG/SATA 結合を用いて連結することができる。別の実施態様では、当該のトランスジェニック動物を用いて、ある特定の連結化学法が、動物において免疫応答を引き起こすかどうかを判定することができる。
【0125】
更に別の実施態様では、当該の動物及び検定法を用いて、最適化された二重特異的化合物を作製する際に用いるために好適な抗原結合部分(例えば特定の結合特異性部分を有するモノクローナル抗体)を選抜することができる。
【0126】
更に別の実施態様では、CR1に結合はするが、抗体ではない結合部分を検査することができる。
【0127】
更なる実施態様では、様々な二重特異的化合物の半減期を検査し、最適化することができる。例えば、ある実施態様では、抗体の異なる抗体アイソタイプ又はフラグメントを用いることにより、半減期を変更することができる。加えて、半減期を変更する新規な方法を判定することができる。
【0128】
別の実施態様では、多様な多量体型又は異なった態様で連結された形の二重特異的化合物のin vivoでの薬物動態を比較することができる。
【0129】
別の実施態様では、当該のトランスジェニック動物を、例えば毒素、ウィルス、細菌などに曝露した動物や、あるいは自己免疫状態を有する動物の治療における二重特異的化合物の効験に関するin vivoモデルを評価する際に用いることができる。
【0130】
別の実施態様では、当該のスクリーニング検定法を用いて、対象の治療を最適化するために、二重特異的化合物の多様な送達経路及び処方を評価することができる。
【0131】
別の実施態様では、当該の検定法を用いて、二重特異的化合物が、動物の免疫応答を亢進するかどうかを判定することができる。別の実施態様では、当該の動物及び検定法を用いて、当該動物における、二重特異的化合物のCR1担持細胞への輸送機序を評価し、そして、二重特異的化合物の活性に対して、CR1に対する抗体の親和性が及ぼす効果を評価することができる。
【0132】
更なる実施態様では、当該の動物及びスクリーニング検定法を用いて、特定の形のin vitro検査をバリデートするために、in vitro検定法とin vivoでの生物学的効果との間の相関を判定することができる。
【0133】
本発明の方法により同定された又は最適化された治療的化合物を、予防上又は治療上の用途のいずれで用いてもよく、また、単独で用いることも、あるいは、例えばワクチン、抗生物質、抗ウィルス薬、抗レトロウィルス薬(ウィルスの複製を阻害する作用薬)など、他の公知の治療法と組み合わせて用いることもできる。ある実施態様では、対象を、作用物質への曝露後に治療する。別の実施態様では、対象を、作用物質への曝露前に治療する。ある実施態様では、対象を二重特異的化合物で(必要に応じ)繰り返し、治療することができる。更に別の実施態様では、(作用物質への曝露前又は曝露後のいずれかでの)本発明の二重特異的化合物による治療の結果、当該作用物質への免疫応答が持続し、こうして更なる治療を行う必要が減るか、あるいはなくなる。
【0134】
本発明を以下の実施例で更に解説するが、以下の実施例を限定的なものと捉えられてはならない。この出願全体を通じて引用された全参考文献、特許及び公開済み特許出願の内容を、引用をもってここに援用することとする。
【0135】
実施例
以下の材料及び方法を実施例で用いた。
材料及び方法
mAb、HP、及びファージ。 以下のマウスモノクローナル抗体(mAb)を用いた: LHR-A、-B、及び-C に結合する抗CR1 mAb YZ-1 (IgG1)(Nickells et al. Clin Exp
Immunol 112, 27-33);LHR-C 及び-Dに結合する抗CR1 mAb 543(アメリカン・タイプ・カルチャー・コレクション);及びLHR-A、-B、及び-Cに結合する抗CR1 mAb 7G9 (IgG2a)(Nickells et al. Clin Exp Immunol 112, 27-33)を、ハイブリドーマから作製された腹水(Pincus et al. Clin Immunol 105,141-54)から精製した。抗ФX174 mAb 7B7 (IgG2a; Taylor et al. J Immunol 158, 842-850)及び抗炭疽菌防御抗原 mAb 14B7 (IgG2a; Little et al. Infect
Immun. 56, 1807-1813) 。バクテリオファージФX174(ФXcs70am-3 ATCC 番号49696-B1と指定されている) はアメリカン・タイプ・カルチャー・コレクション(ヴァージニア州マナサス)から得られ、ファージ・ストックは、PanVera・インビトロジェン社(カリフォルニア州カールスバッド)社により作製された。
【0136】
抗CR1 mAb 7G9 を抗ФX174 mAb 7B7(ФX −CR1 HP)にチオエーテル結合により、SMCC (スルホスクシンイミジル4−(N-マレイミドメチル) シクロヘキサン-1-カルボキシレート)、及びSATA- (N-スクシンイミジル-S-アセチルチオアセテート) (イリノイ州ロックフォード、ピアース・バイオテクノロジー社)のいずれかで活性化させた抗体を以前に解説された(Lindorfer et al. J Immunol
167, 2240-2249)ように用いて連結した。PEG HPを作製するために上記の方法の改良法を開発し、このとき一方のmAbをSMCCで活性化する代わりに、マレイミド基をNHSエステルを介して含有する生物学的に不活性なリンカ/スペーサ分子(修飾されたポリエチレングリコール〜5000MW)が用いられ、この修飾されたmAbをSATA
で活性化したmAbと反応させた。非CR1結合HP (ФX−PA HP)を、mAb 7B7 のmAb 14B7へのSMCC/SATA結合により作製した。
【0137】
トランスジェニック・マウスの作製。 ヒトCR1遺伝子の発現プラスミドを、ヒトCR1の共通の対立遺伝子CR1
(hCR1; Klickstein et al. J Exp Med 168, 1699-1717)をコードする完全cDNAを含有する9.9 kbのNot I DNA 断片を得ることにより、構築した。この断片をプラスミドpGATA-1の非反復Not I部位に挿入し、hCR1を赤血球特異的プロモータGATA1 の転写制御下に置いた(Visvader et al. Genes Dev.12,
473-479)。CR1遺伝子の方向は配列決定により確認された。GATA1-CR1 トランスジェニック・コンストラクトをSal Iで消化し、プロモータ/CR1 遺伝子断片(ほぼ22 kb)を精製し、SJLマウスの受精卵に顕微注射した後、この受精卵をC57BL/6 借り腹母に移植した。その結果得られた後代を、導入遺伝子の組込みについて、マウスの尾DNA を用いたPCRによりスクリーニングし、C57BL/6 マウスと交配することにより維持した。以下に解説するように、陽性のヘテロ接合子を、DNAのPCR分析と、FACS解析による赤血球上の発現分析により検出した。
【0138】
マウス尾からの高分子量DNAの単離を、Easy-DNA
キット(インビトロジェン社)により、このメーカのプロトコルに従って行った。ほぼ1μgのDNAに35サイクルの増幅をサーマル・サイクラで行った。トランスジェニックマウスのPCR分析に用いられた2つのトランスジェニック特異的プライマであるGATA-1正方向プライマ(5'- ACCCTTTCTGTCCTCACA -3'; 配列番号1番)及びCR1遺伝子逆方向プライマ (5'-TTTCTCCCTCCGCTTCCAGGTTG- 3'; 配列番号2番)により、653-bp のDNA 断片(図1a)が生じた。PCR産物を1%のアガロース・ゲルでの電気泳動法で分離した。
【0139】
ヒヒ。 ヒヒを、サウスウェスト・ファウンデーション・フォー・バイオメディカル・リサーチ(テキサス州サンアントニオ)で維持されたコロニから、IACUC認可との約定のもとで用いた。この研究の前に、血液を40匹の動物から得、以下に解説するようにFACSによりCR1発現レベルについて分析した。この結果を用いて、研究に含めるために、 1800-3600 のCR1/E を持つ動物を選抜した。
【0140】
赤血球。 クエン酸塩で血液凝固を阻害したマウス血を、尾の静脈から得、500×gでの遠心分離後、バフィーコート層を取り除き、赤血球を3回、カルシウム及びマグネシウムなしのハンクス液(HBSS=)(ニューヨーク州グランドアイランド、ギブコ/インビトロジェン社)で洗浄した。正常なドナーのヒト赤血球を、酸−クエン酸で血液凝固を阻害した静脈血から得、0.001 M EDTA、0.01% ゼラチンを加えた等容のHBSSで希釈した。遠心分離及びバッフィーコート層除去後、細胞を上記の緩衝液で数回、洗浄した。最後に、細胞を、カルシウム及びマグネシウム並びに0.01%ゼラチンを含有するHBSS(HBSS++/ゼラチン)に再懸濁させた。免疫複合体結合検定用の全ての赤血球は採集日に用いられ、ヘモグロブリン濃度をもとに正規化することで、細胞を水に1/30に希釈するとヘモグロビン吸光度が414nmで0.64になるようにした。
【0141】
ヒト血清。 2mlの血清(ヒト赤血球のドナーから)を5 mM EDTA を4℃に冷却して作製し、ほぼ106の重量のmEから採った洗浄した膜と60分間、混合して、ヒト抗マウスE免疫グロブリンを完全に吸収させた。次に、吸収後のヒト血清を15,000×gで遠心分離してマウス膜を取り除き、その上清をアリクォートし、免疫複合体をオプソニン化するために使用するまで−80℃で保存した。
【0142】
CR1に関する免疫ブロット法。 100μlの血液からの赤血球をHBSS=で2回、洗浄した後、ゴースティング(原語:ghosting)緩衝液(5 mM リン酸ナトリウム (pH 8.0)、1 mM EDTA、及び「プロテアーゼ阻害剤カクテル」の1/100希釈液(ミズーリ州セントルイス、シグマ社)中で一晩、溶解させた。膜をSDS-試料緩衝液(マサチューセッツ州ボストン、ボストン・バイオプロダクツ社)で可溶化させ、5分間沸騰させ、未還元のまま4-12% NuPage 勾配ゲル(カリフォルニア州カールスバッド、インビトロジェン社)で分離した。タンパク質をHybondニトロセルロース・メンブレン(イリノイ州アーリントン・ハイツ、アマーシャム・バイオスクレンセズ社)に移した。この膜を抗CR1 mAb YZ-1で免疫ブロットした後、ペルオキシダーゼ結合ヤギ抗マウス二次抗体(カリフォルニア州バーリンゲーム、ベクター・ラボラトリーズ社)と一緒にインキュベートした。ブロットをSuperSignal ウェスト・ピコ・ケムイルミネセンス基質(ピアース・バイオテクノロジー社)で可視化した。
【0143】
赤血球表面上のCR1発現のFACS分析。 抗CR1 mAb 7G9をAlexa 488TM(オレゴン州ユージーン、モラキュラー・プローブズ社)でこのメーカの指示に従って標識した。野生型mE 又はmE+CR1 又はhE をPBS/1% BSA 緩衝液 (PBS/BSA)に1%の濃度になるように懸濁させた。Alexa 488-7G9 (100μlの2μg/ml ストック)を100μlの赤血球に加え、30分間、室温でインキュベートした。PBS/BSAで2回、洗浄することにより、未結合のAlex-7G9を取り除いた。赤血球をそれらの前方散乱対側方散乱光プロファイルに基づいてフローサイトメトリで選択的に通門させ、その通門内で20,000 回の事象を、FACSCaliburTM装置及びCellQuest Pro ソフトウェア、V3.3 (カリフォルニア州サンホセ、BD バイオサイエンセズ社)を用いて分析した。
【0144】
マウス及びヒト赤血球上のCR1の分布。 ヒト又はマウス血を、HBSS=/BSA
に希釈し、赤血球を同じ緩衝液に10% (V/V)の濃度になるように再懸濁させた。抗CR1 mAb 543 を赤血球に加えた後、Alexa 488で標識されたヤギ抗マウス(モラキュラー・プローブズ社)を、抗体及び赤血球の最終濃度が、それぞれ5.0μg/ml 及び5% になるように加えた。37℃で30分間、インキュベートした後、結合しなかったmAbを遠心分離及び洗浄で取り除いた。光学及び蛍光顕微鏡検査をオリンパスBX40F4 顕微鏡(ニューヨーク州メルビル、オリンパス・アメリカ社)を1000倍の倍率で用いて行った。
【0145】
in vitroにおける免疫複合体及びHPの赤血球への結合。 BSAのV画分(シグマ社)(1mg/ml)をFITC (ウィスコンシン州ミルウォーキー、アルドリッチ・ケミカル社)で標識し、透析して、結合しなかったFITCを取り除いた。標識後のBSAの推定濃度は0.8 mg/mlだった。30μgのFITC-BSA及び100μlのウサギ抗BSA IgG (モラキュラー・プローブズ社)を最終体積250μlにして60分間、37℃で混合すると、免疫複合体を形成させることができた。30μlの免疫複合体を遠心分離管中にアリクォートした後、40μlの吸収後のヒト血清、4μlのストック・カチオン(30 mM CaCl2、100 mM MgCl2)及び150μlの赤血球懸濁液を順番に添加した。15分間、37℃でインキュベートして免疫複合体のオプソニン化と、オプソニン化後の免疫複合体の赤血球への結合の両方を起こさせた後、この混合液を3 ml の冷HBSS-0.01% ゼラチンを含有する12×75mmの試験管に移し、400×gで遠心分離し、3mlの同じ緩衝液中に再懸濁させた。細胞をFACSscanTMで分析すると、10,000回の事象がCellQuest Pro ソフトウェア V4.01を用いて記録された。
【0146】
SMCC-HPの赤血球への結合を以下の態様で判定した。親mE 又は mE+CR1又はhE をPBS/BSA 緩衝液に10%の濃度になるように懸濁させた。 HP (20 ng) を100μlの赤血球に15分間、37℃で振盪しながら加えた。PBS/BSA 緩衝液で2回、洗浄することで、結合しなかったHP を取り除いた。ヤギ抗マウスIgG-Alexa 488 (モラキュラー・プローブズ社)を10μg/ml にして試験管に加え、室温で30分間、インキュベートした後、PBS/BSA緩衝液で2回、洗浄した。次に、フローサイトメトリ検査(FACSCallibur) を試料に対して行い、赤血球を、それらの正方向散乱対側方散乱プロファイルに基づいて選択的に通門させ、20,000 回の事象をその通門内で分析した。
【0147】
in vivoのファージ除去。 ヒヒ(8-10kg)に1.5×1011PFUを静脈内注射し、20分後に0.3 mg のФX−CR1 HP (PEG 及びSMCC)、ФX-PA HP
又は等容の生理食塩水を静脈内注射した。血液試料を、大腿の静脈穿刺により、抗凝固剤としてEDTA (0.01M 最終濃度)を含有する試験管内に得、遠心分離し、血漿を−70℃で凍結させた。血漿中ファージをプラーク検定法により前に解説された(Taylor et al. J Immunol
158, 842-850)ように判定した。
【0148】
マウスに0.1 mlの体積で、3×107 PFU/動物の用量を静脈内注射した。生理食塩水のコントロール、HP又は無関係のHP(1、6、又は12μg)を0.1 ml の体積にして45分後に静脈内注射した。血液試料(50μl)を、図6bに示すようにHPの投与前及び投与後の様々な時点で、0.04MのEDTAを含有する0.5
ml Alseverの溶液中にマウスの尾から採集した。血漿を細胞から分離し、4℃で保存した。血漿中ファージをプラーク検定法により前に解説された(Taylor et al. J Immunol
158, 842-850)通りに判定した。
【0149】
実施例1: ヒト補体受容体1(CR1)ポリペプチドをコードするヌクレオチド配列をGATA-1プロモータに作動的に連結させて含有する発現コンストラクトの構築
ヒトCR1をマウス赤血球上で発現させるために、マウス赤血球で働くであろうプロモータを特定する必要があった。マウスGATA-1遺伝子の調節配列は、以前、トランスジェニック・マウスで造血系組織においていくつかの遺伝子を選択的に発現させるために用いられたことがある(McDevitt, M.A. et al., (1997) Proc. Natl. Acad. Sci., USA,
94:7976-7981; Visvader J.E. et al., (1998) Genes & Dev. 12(4):473-479)。転写因子GATA-1はいくつかの造血系及び多能前駆細胞で発現し、赤血球及び血小板の発生に必要である。赤血球系クロマチンの関係でそれらのDnase I超感受性により定義された、GATA-1遺伝子の上流にある特異的制御配列であって、トランスジェニック・マウスで導入遺伝子全体の発現を亢進させ、原始赤血球系細胞内で特異的に活性化をもたらし、ひいてはGATA-1の発生上の発現パターンを発生反復するものが特定された(McDevitt,
M.A. et al., (1997) Proc. Natl. Acad. Sci.,
USA,
94:7976-7981)。次に、この導入遺伝子カセットを用いて、マウス幹細胞白血病遺伝子(SCL/tal-1)をSCLノックアウト・マウスに再導入し、発現させた;SCL 遺伝子は通常、造血細胞、血管内皮細胞及び発生中の脳内で特異的に発現するが、該マウスGATA-1調節配列により、SCL -/- 胚の造血系の血管を救助することができた(Visvader J.E. et al., (1998) Genes & Dev. 12(4):473-479)、従って、これらのマウスGATA-1調節配列を用いて、マウス赤血球上でのヒトCR1遺伝子の発現を命令することができた。
【0150】
マウスGATA-1調節配列(近位のプロモータ;下流のイントロン配列、及び上流の超感受性部位)をウサギグロビンpoly(A)添加領域及びSV40配列と一緒に含有するコンストラクトを用いた(Visvader and Orkin.
1998. Genes and Development.
12:473)。
【0151】
機能的ヒトCR1をコードするcDNAを含有するプラスミドpiABCDを、ロイド・クリックスタイン博士から得た(Wilson J.G. et
al, (1986) J. Exp. Med. 164:50-59; Klickstein L.B. et al. (1988) J. Exp. Med.
168:1699-1717)。CR1をコードするpiABCDの6,860 bpの断片を、マウスGATA-1調節配列(エキソン2のATG
Iから~10,000 bp上流(転写開始部位の5+上流)からこの遺伝子の3’まで延びる、>15,000bpのゲノム・マウスGATA-1遺伝子) を含有するプラスミドのNotI部位にクローニングした。形質転換後、方向の正しいクローンを選抜し、当業で公知の方法に従った配列決定により確認した。簡単に説明すると、インサート全体をベクタからSalI部位を用いて切り出した。プラスミドpSc-3Z (4.7 kb、低コピー数のプラスミド、amp)が用いられた。DNAをCsCl勾配で精製した後、SalIで消化して、GATA-1プロモータ及びCR1遺伝子をコードする15,000 bp の融合遺伝子を提供した。(図1aは、GATA1/hCR1 トランスジェニック・コンストラクトの概略的マップを示し、このとき、GATA1 の上流の7kbの配列と、エキソン3の下流の~1.5 kb の配列が、hCR1
cDNAをフランクしている。図1aで番号を付した領域は、GATA-1のエキソンを示す。エキソン2にあるGATA1-の内因性翻訳開始コドンを、Not I部位に置換した。hCAR1 のcDNAをこのNot I 部位に挿入した。シミアン・ウィルス40(pA)由来のポリアデニレーション・シグナルはこのcDNAインサートの3’側にある。導入遺伝子を検出するために用いられたPCRプライマは、GATA-1エキソン2に特異的な正方向プライマと、hCR1配列に特異的な逆方向プライマ(図1aの矢印の頭)とから成った。次に、この融合遺伝子を精製した後、トランスジェニック・マウスを作製するために用いた。
【0152】
実施例II: そのゲノムが、ヒト補体受容体1(CR1)ポリペプチドをコードするヌクレオチド配列を、GATA-1プロモータに作動的に連結させて含有するトランスジェニック・マウスの構築
GATA-1プロモータ及びCR1遺伝子をコードする15,000 bpの融合遺伝子を、B6/129 F1マウス接合子に、上に記載したように、当業で公知の方法に従って前核注射することにより、トランスジェニック・マウスを構築した。尾の一部分を取り除き、ゲノムDNAを抽出することにより、マウスを前記外来の融合遺伝子の存在についてスクリーニングした。マウス尾の生検から単離されたDNAに対するPCR分析で、創始マウスと、GATA1/hCR1コンストラクトについて生殖細胞系トランスジェニックである仔の世代が確認された。このようなマウスは、判断材料である653-bp のPCR 産物により特定された(図1b、この図においてレーン1は、テンプレートとしての蒸留水コントロールのPCR結果を示し、レーン2は、導入遺伝子マウスのゲノムDNAをテンプレートとして用いたPCR検定の結果を示す;Mは、1-kb のラダー・マーカに相当する;レーン3は、導入遺伝子を含有するプラスミドDNAのPCR結果を示す。;そしてレーン4は、既知の非トランスジェニック・マウスのゲノムDNAをテンプレートとして用いた検定結果を示す)。
【0153】
実施例III: トランスジェニックマウス赤血球上におけるヒト補体受容体CR1の発現のエビデンス
CR1抗原が、トランスジェニック・マウスの赤血球表面上で発現したことを確認するために、hE、mE 及びmE+CR1細胞をAlexa-488 結合抗CR1 mAb 7G9で染色した後にフローサイトメトリを行った(図2a)。分析では、CR1抗原がhE(右側パネル)及びトランスジェニック・マウス由来のmE(中央のパネル)の表面で発現したが、野生型(wt)マウス(左側パネル)由来のmEでは発現しなかったことが確認された。陽性コントロールとして用いられたヒトE(hE、右側パネル)は、mE+CR1上のCR1の約50%を発現した。
【0154】
mE+CR1のCR1が適したサイズであることを確認するために、免疫ブロット法を行った(図2b)。wtマウス、CR1トランスジェニック・マウス、及び正常なヒト由来の赤血球を溶解させ、可溶化し、非還元膜タンパク質をSDS-PAGEで分離した。移した後、当該ブロットを、材料及び方法の項で解説したように抗CR1
mAb YZ-1と反応させた。ヒトドナーは、200kDaのバンドを生じる共通の対立遺伝子CR1*1と、190kDaのバンドを生じるCR1*3対立遺伝子についてヘテロ接合型と推定された(図2b、レーン4;Klickstein,
L.B. & Moulds, J.M. in The Complement Facts Book (eds. B.J. Morley & M.J. Walport),
pp. 136-145にレビュー)。トランスジェニック・マウス由来の赤血球上のCR1(図2b、レーン3)は分子量においてhE CR1 CR1*1対立遺伝子と同様であり、導入遺伝子と合致した。wt mEのライセート中にCR1 抗原はYZ-1 mAb によっても何ら検出されず(図2b、レーン2);そして予備染色されたミオシン207 kDa MW マーカは図2b、レーン1に示されている。
【0155】
トランスジェニック・マウス中でGATA-1で促進されたCR1の分布を定義するために、トランスジェニック・マウス由来の組織をParagon Biotech (メリーランド州ボルチモア)で標準的手法により固定し、抗CR1 mAbで染色し、CR1の発現について調べた。CR1免疫反応性として認められる組織はなかった。
【0156】
実施例IV: 蛍光顕微鏡分析では、hE 及びmE+CR1 上のCR1のクラスタ状分布が確認された
hE一個当たりのCR1の平均レベルは大変低く(100-1,000/ E)、また一価の補体リガンドのCR1への親和性も低いため、CR1のクラスタは、補体でオプソニン化した粒子をhEに効果的にライゲートする必要があると仮定される(Klickstein et al. Immunity
7, 345-355; Edberg et al. J Immunol
139, 3739; Paccaud et al. J Immunol
141, 3889-3894; Chevalier and Kazatchkine, J
Immunol 142, 2031-2036)。このように、免疫吸着媒介性除去のモデルとしてのトランスジェニック・マウスの実用性は、CR1がmE+CR1上でクラスタになっていることに依存するかも知れない。hE (図3、左側パネル)及びmE+CR1
(図3、右側パネル)を抗CR1 mAb 543 と反応させた後、Alexa 488で標識された二次抗体と反応させて、光学(図3、下側パネル)及び蛍光顕微鏡(上側パネル)で観察した。 CR1 は、予想通り、hE上でクラスタ状に見えた(図3、左側パネル)。同じ手法をmE+CR1についても繰り返したが、CR1はやはり、クラスタ状に見えた(図3、右側パネル)が、hE上のクラスタよりも例外なく小さく、しかしより豊富だった。
【0157】
実施例V: mE+CR1の、オプソニン化したFITC標識免疫複合体及びHPへの結合能
mE+CR1上のCR1タンパク質の官能性を検査するために、これらの赤血球のオプソニン化した免疫複合体への結合能を調べた。FITC-BSA及びウサギ抗BSAから成る、FITC で標識された免疫複合体を、ヒト血清とのインキュベーションにより補体でオプソニン化し、赤血球と反応させた。次に、この試料をフローサイトメトリで分析した。免疫複合体の細胞への結合が、蛍光の正のシフトで実証されたが、これは、免疫複合体がオプソニン化され、赤血球がCR1を発現した場合、即ちhE又はトランスジェニックmE+CR1でのみ、見られた(図4a、この場合、左側のパネルは、正常なヒトE(hE)に関する結果を示し;中央のパネルは野生型マウスE (mE);右側のパネルは、ヒトCR1を発現するトランスジェニック・マウス(mE+CR1)からのものである。青色の柱状図は緩衝液で洗浄された赤血球を表す。FITCで標識された免疫複合体を作製し、その後、緩衝液(赤色の柱状図)及び正常なヒト血清(緑色の柱状図)に、赤血球の存在下で反応させた。蛍光のシフトにより、免疫複合体の赤血球への結合が示されたが、これは、免疫複合体が血清でオプソニン化され(緑色の柱状図)、CR1を発現した(hE又はmE+CR1、しかしwt mEでは見られず)場合にのみ、見られた。
【0158】
赤血球に結合したHPをAlexa 488ヤギ抗マウス抗体で染色し、フローサイトメトリで分析することにより、赤血球のHPへの結合能を調べた(図4b)。HPをhE及びmE+CR1と一緒にインキュベートした場合にはHPの結合は見られたが、wt mEへの結合は見られなかった。hE 及びmE+CR1
は本来の免疫複合体には一様に結合しなかった(図4a)が、HPはhE及びmE+CR1の総集団に結合した(図4b)ことが注目される。
【0159】
実施例VI: in vivoにおいてHPの用量及び種類がファージ除去に及ぼす効果
HP は、血流から病原体及び毒素を除去するために用いることができる。CR1トランスジェニック・マウスを、除去に向けて潜在的な病原体を赤血球CR1に結合させるHPの効験を検査するためのモデルとしてデザインした。バクテリオファージФX174が、これらの実験のモデル生物として用いられたが、なぜなら、哺乳動物細胞上にはその受容体がないため、哺乳動物にとってそれは病原性でなく(Och et al. J Clin Invest
50, 2559−2568)、またそれはプラーク検定法で容易に定量されるからである。ФX−CR1と指定されたHPは、抗ФX174 mAb(7B7)を抗CR1
mAb (7G9) に2つの異なる化学薬SMCC 又はPEGを用いて化学的に連結したものから成った(方法の項を参照されたい)。蛍光分析では、ФX−CR1 HP はhE 及びmE+hCR1 の両方に結合したが、野生型 mE (図4b)には結合しなかったことが確認された。ФX−PAと指定されたコントロールHPは、抗ФX174 mAbを炭疽菌防御抗原(PA、14B7
mAb)に対するmAbに連結することにより、作製された。hE にもmE+CR1にも結合しなかったФX−PA
HPをコントロールとして用いて、可溶性HP媒介性除去の率を判定した。この率は、液相ウィルス中和と、マウスの本来の血小板依存的免疫付着経路により誘導される(大型HP成分の)除去との和を表した。
【0160】
トランスジェニック・マウスに、バクテリオファージФX174を、3×107PFU/マウスの用量で静脈内輸注してから45分後に、PEG又はSMCC HPのいずれかを三種の異なる用量(1.0μg6μg、又は12μg/マウス)、あるいは陰性コントロールとしての生理食塩水、を静脈内輸注した。このトランスジェニック・マウスから24時間後に採血して、血漿1ml当たりのファージのPFUを判定した。1μgのHP/マウスでは、PEG及びSMCCで連結された形のФX−CR1 HP(3.8又は3.0 log)の両方で、ФX−PA HP(0.6 log)に比較したときに、血清中のファージ減少に明確な違いがあった。ФX−PA HPは、ФX-PA HP 及びФX−CR1 HPをより高い用量にしたときにファージ中和の上でより有効となった(6 及び12μgのHP /マウス、図5のグラフでは、HP注射から24時間後にマウスの尾から採取された血液試料中に存在するファージPFU/mlのlog10を表している。誤差棒は、平均の標準誤差を表す、n=3。アスタリスクは、スチューデントのt検定でp<0.01のときのФX-PA HPからの有意差を示す)。
【0161】
実施例VII: ヒヒ及びCR1トランスジェニック・マウス由来の血漿からのバクテリオファージФX174のHP媒介性除去の動態
高等霊長類はCR1を発現する赤血球を有する(参考文献30番でレビュー)ため、これらの動物は、ФX174 ファージ(Taylor et al. J Immunol
158, 842−850)を含め、様々な種類の粒子のHP媒介性除去の効験の小規模研究で用いられてきた(Taylor
et al. J Immunol 148, 2462−2468; Craig et al. Clin Immunol
92, 170-180; Nardin et al. Mol Immunol
36, 827−835; Lindorfer et al. J Immunol 167, 2240−2249)。トランスジェニック・マウスにおける除去の動態を測定する前に、本発明のHPがヒヒでどれくらい効率的に機能したかを評価することが好ましかった。なぜなら、このような研究では、トランスジェニック・マウス・モデルの予測可能性が評価されるであろうからである。ФX174(1.5×1011PFU)を t=-20分で輸注した。20分後のt=0分の時点で、SMCC又はPEGで連結されたФX−CR1 HP、又は非E結合SMCC-連結ФX−PA HP、あるいは陰性コントロールとしての生理食塩水、をヒヒに輸注した。HPを0.3 mg/動物 (それぞれ8−10 kg)という、重さベースでは1ug/マウスの用量とほぼ等しい用量、用いた。その後の100分間にわたる静脈血の試料採取(HP輸注15分及び10分前と、HP輸注後の複数の時点(5乃至90分、行われた)では、二段階の除去が明らかになった(図6aを参照されたい)。1回目は、急速な最初の段階であり、ФX−CR1 PEG HPにより3.0
log、ФX−CR1 SMCC HPにより1.6 log、そしてコントロールФX−PA
HP により0.8 logの除去が、HP輸注後5分間にあった。 HP輸注から5乃至90分後にあった、よりゆっくりとした2番目の段階では、ФX−CR1 HP (PEG)では、~1.5 log/85 分、ФX−CR1 HP (SMCC)では~1.0
log/85 分、そしてФX−PA HP では、~1.0 log/85 分の除去があった。 図6aのグラフは、治療後の様々な時点で血中に見られたファージ力価の減少を表す[可能な採血回数に限界があるため、動物は、HP投与直前の時点0では採血されていない。時点-10分における各群に見られた平均log10(PFU/ml)を減算することにより、数値を正規化した。生理食塩水群は120分間の研究全般にわたって力価に何の変化も見せないため、この時点とHP投与の時点との間の15分間の力価の変化は期待されなかった。これらの数値は、生理食塩水、ФX −CR1 HP(PEG)、ФX−CR1 HP (SMCC)及びФX −PA HP (SMCC)について、それぞれ、8.27±0.78、8.75±0.1、8.63±0.13、及び8.54±0.23、だった。プロットは平均を表し、棒は範囲を表す、n=2]
【0162】
トランスジェニック・マウスで用いるためのФX−CR1
HP(1μg)の有用な用量を定義(図5)したところで、マウスにおけるファージ除去の動態を次に定義した。CR1トランスジェニック及びwt 親C57BL/6
マウスの両者に、3×107のPFU のファージをマウス1匹当たり、注射した。ФX−CR1 HP (SMCC) 又はコントロールФX −PA HP (SMCC) を45分後に注射した。HP注射の15分前、HP注射から15分後、90分後、及び5時間後にマウスから採血した。HPの代わりに生理食塩水の注射を受けたコントロール・トランスジェニック又はwt マウスでは、循環中のファージ力価は、注射後5時間にわたって一定に留まった。CR1 トランスジェニック・マウスでは、循環ファージを減らす上で、コントロールФX −PA HPに比較したときのФX −CR1 HPの優位性が、輸注から15分以内に明白であり、輸注後5時間、続いた(図6b)。図6bのグラフは、図6aと同様に、野生型対CR-1トランスジェニック・マウスにおけるファージのHP媒介性除去が明らかとなったデータを提供する。[ФX −CR1 HP (SMCC) を、ФX−PA (SMCC) に、CR1トランスジェニック及びC57BL/6 野生型マウスにおける血流からの、注射されたバクテリオファージФX 174の除去能について比較した。ファージを−45分の時点で3×107PFU/マウスの用量、静脈内注射した。HP又は生理食塩水のコントロールを、1μg/マウスの用量、時点0で静脈内注射した。血液を、尾の採血により、HP注射から−15分、15分後、90分後、及び5時間後に採取した。グラフは、治療後の時点で血中に見られたファージ力価の減少を表す。数値は、−15分の時点で各群に見られた平均log10(PFU/ml)を除算することにより、正規化された。生理食塩水群は5時間の研究にわたって力価に何の変化も見せないため、−15分から時点0までの間の力価の変化は期待されなかった。これらの-15分の数値は、生理食塩水、CR1 マウス+ФX−CR1 HP、CR1 マウス+ФX −PA HP、wt マウス+ФX −CR1 HP、及びwt マウス+ФX −PA HP、についてそれぞれ、6.93±0.30、7.45±0.08、6.82±0.32、7.35±0.11、及び6.96±0.40だった。誤差棒は平均の標準誤差を表す、n=3。アスタリスクはステューデントのt検定によりp<0.05のときのФX−PA
HPからの有意差を示す]
【0163】
全ての時点において、ФX −CR1 HP はコントロールФX −PA HP よりも循環ファージをほぼ1.5 log、多く、減少させた。赤血球上にCR1のないwt マウスでは、ФX −CR1 HP及びコントロールФX −PA、HPは、循環ファージの力価を、全ての時点で同様なレベル減少させ、この減少は、トランスジェニック・マウスでコントロールФX−PA HPにより達成された減少レベルと同様だった。
【0164】
これらの結果は、ファージを赤血球CR1に結合させた抗体HPは、トランスジェニック・マウスにおいて血清及び/又は循環からファージを取り除く上で、可溶性ファージ結合HPよりも効率的であることを実証した。wtマウスにおけるウィルス除去では、この血管内の利点がないことは、トランスジェニック・モデルの有効性を裏付けるものである。
【0165】
均等物
当業者であれば、慣例的な実験によって、ここに解説した本発明の具体的な実施態様の均等物を数多く、認識し、又は確認できることであろう。このような均等物は以下の請求の範囲の包含するところと、意図されている。
【図面の簡単な説明】
【0166】
【図1】図1は、GATA1/hCR1 コンストラクトの概略的マップと、トランスジェニック始祖マウスを特定したPCR検定結果を示す。
【図2】図2は、トランスジェニック・マウス由来の赤血球が適した分子サイズのヒトCR1抗原を発現したことの確認を示す。
【図3】図3は、マウス及びヒト赤血球上のCR1の分布を明らかにする。
【図4】図4は、ヒト補体でオプソニン化された免疫複合体又はHPへのmE+CR1の結合能を示す。
【図5】図5は、invivoにおけるファージ除去に対するHP用量の効果を示す。
【図6】図6は、ヒヒ、CR1トランスジェニック及びwtマウスにおけるHPのファージ除去結果を示す。
【特許請求の範囲】
【請求項1】
ヒト補体受容体1(CR1)ポリペプチドをコードするヌクレオチド配列を、プロモータに作動的に連結させて含む、非ヒト動物で発現させるための遺伝子コンストラクトであって、赤血球上でのヒトCR1の機能的発現を命令する遺伝子コンストラクト。
【請求項2】
前記プロモータがGATA-1プロモータである、請求項1に記載のコンストラクト。
【請求項3】
前記プロモータが、GATA-1プロモータと、赤血球上での発現を活性化させる上流の制御領域とを含む、請求項1に記載のコンストラクト。
【請求項4】
そのゲノムが、ヒト補体受容体1(CR1)をコードするポリヌクレオチドをプロモータに作動的に連結させて含む非ヒト動物であって、前記ヒトCR1分子が、前記動物の赤血球上で機能的に発現する、非ヒト動物。
【請求項5】
前記ヒトCR1分子が赤血球上で優先的に発現する、請求項4に記載の非ヒト動物。
【請求項6】
前記非ヒト動物がトランスジェニック・マウスである、請求項4に記載の非ヒト動物。
【請求項7】
前記プロモータがGATA-1プロモータである、請求項4に記載の非ヒト動物。
【請求項8】
前記プロモータが、GATA-1プロモータと、赤血球上での発現を活性化させる上流の制御領域とを含む、請求項4に記載の非ヒト動物。
【請求項9】
前記動物が前記ポリヌクレオチドについてホモ接合型である、請求項4に記載の非ヒト動物。
【請求項10】
前記動物が前記ポリヌクレオチドについてヘテロ接合型である、請求項4に記載の非ヒト動物。
【請求項11】
対象の血清及び/又は循環中の作用物質の濃度を低下させることのできる二重特異的化合物をスクリーニングするための方法であって:
そのゲノムが、ヒト補体受容体1(CR1)をコードするポリヌクレオチドをプロモータに作動的に連結させて含む非ヒト動物に、複数の検査二重特異的化合物を投与するステップであって、前記ヒトCR1が前記非ヒト動物の赤血球上で機能的に発現する、ステップと;
前記非ヒト動物の血清及び/又は循環中の前記作用物質の濃度を低下させる上での二重特異的化合物の能力を判定するステップと;
前記非ヒト動物の血清及び/又は循環中の前記作用物質の濃度を低下させる二重特異的化合物を選抜することで、対象の血清及び/又は循環中の作用物質の濃度を低下させることのできる二重特異的化合物を同定するステップと
を含む、方法。
【請求項12】
対象の血清及び/又は循環中の作用物質の濃度を低下させる上での二重特異的化合物の能力を評価する方法であって:
そのゲノムが、ヒト補体受容体1(CR1)をコードするポリヌクレオチドをプロモータに作動的に連結させて含む非ヒト動物に、前記二重特異的化合物を投与するステップであって、前記ヒトCR1が前記非ヒト動物の赤血球上で機能的に発現する、ステップと;
前記非ヒト動物の血清及び/又は循環中の前記作用物質の濃度を低下させる上での前記二重特異的化合物の能力を判定することで、前記非ヒト動物の血清及び/又は循環中の前記作用物質の濃度を低下させる上での前記二重特異的化合物の能力を評価するステップと
を含む、方法。
【請求項13】
前記作用物質を前記非ヒト動物に投与するステップを更に含む、請求項11又は12に記載の方法。
【請求項14】
前記作用物質が病原体である、請求項13に記載の方法。
【請求項15】
前記作用物質がウィルスである、請求項13に記載の方法。
【請求項16】
前記作用物質が毒素である、請求項13に記載の方法。
【請求項17】
前記作用物質がポリヌクレオチドである、請求項11又は12に記載の方法。
【請求項18】
前記作用物質が細菌である、請求項13に記載の方法。
【請求項19】
前記作用物質が自己免疫疾患に関連する自己抗体である、請求項13に記載の方法。
【請求項20】
前記二重特異的化合物が、ヒトCR1に結合する二重特異的化合物である、請求項11又は12に記載の方法。
【請求項21】
前記二重特異的化合物がヘテロ重合体である、請求項20に記載の方法。
【請求項22】
前記二重特異的化合物が抗原ベースのヘテロ重合体である、請求項20に記載の方法。
【請求項23】
前記非ヒト動物がトランスジェニック・マウスである、請求項11又は12に記載の方法。
【請求項24】
前記プロモータがGATA-1プロモータである、請求項11又は12に記載の方法。
【請求項25】
前記プロモータが、GATA-1プロモータと、赤血球上での発現を活性化させる上流の制御領域とを含む、請求項11又は12に記載の方法。
【請求項26】
対象が治療されるように、請求項11又は12で同定された二重特異的化合物を対象に投与するステップを含む、生物学的作用物質に曝露した対象を治療する方法。
【請求項27】
対象が治療されるように、請求項11又は12で同定された二重特異的化合物を対象に投与するステップを含む、生物学的作用物質への曝露のリスクのある対象を治療する方法。
【請求項28】
対象が治療されるように、請求項11又は12で同定された作用物質を対象に投与するステップを含む、感染又は自己免疫疾患を持つ対象を治療する方法。
【請求項29】
ヒトCR1をその赤血球表面上に発現するトランスジェニック・マウスを作製する方法であって:
a)ヒトCR1ポリペプチドをコードするポリヌクレオチドを、マウスの卵又は胚に導入するステップであって、前記ポリヌクレオチドが、ヒトCR1をマウス赤血球上で発現させるプロモータに作動的に連結されている、ステップと;
b)前記CR1ポリヌクレオチドを含む卵又は胚を産期まで成長させて、トランスジェニック・マウスを生じさせるステップと
を含む、方法。
【請求項30】
それぞれがヒトCR1ポリペプチドをコードするポリヌクレオチドを含む二倍体ゲノムを含む二匹のトランスジェニック・マウスを交配するステップを含み、前記ポリヌクレオチドが、マウス赤血球上でヒトCR1ポリペプチドを生ずるように発現することで、仔トランスジェニック・マウスが生ずる、トランスジェニック・マウスを作製する方法。
【請求項1】
ヒト補体受容体1(CR1)ポリペプチドをコードするヌクレオチド配列を、プロモータに作動的に連結させて含む、非ヒト動物で発現させるための遺伝子コンストラクトであって、赤血球上でのヒトCR1の機能的発現を命令する遺伝子コンストラクト。
【請求項2】
前記プロモータがGATA-1プロモータである、請求項1に記載のコンストラクト。
【請求項3】
前記プロモータが、GATA-1プロモータと、赤血球上での発現を活性化させる上流の制御領域とを含む、請求項1に記載のコンストラクト。
【請求項4】
そのゲノムが、ヒト補体受容体1(CR1)をコードするポリヌクレオチドをプロモータに作動的に連結させて含む非ヒト動物であって、前記ヒトCR1分子が、前記動物の赤血球上で機能的に発現する、非ヒト動物。
【請求項5】
前記ヒトCR1分子が赤血球上で優先的に発現する、請求項4に記載の非ヒト動物。
【請求項6】
前記非ヒト動物がトランスジェニック・マウスである、請求項4に記載の非ヒト動物。
【請求項7】
前記プロモータがGATA-1プロモータである、請求項4に記載の非ヒト動物。
【請求項8】
前記プロモータが、GATA-1プロモータと、赤血球上での発現を活性化させる上流の制御領域とを含む、請求項4に記載の非ヒト動物。
【請求項9】
前記動物が前記ポリヌクレオチドについてホモ接合型である、請求項4に記載の非ヒト動物。
【請求項10】
前記動物が前記ポリヌクレオチドについてヘテロ接合型である、請求項4に記載の非ヒト動物。
【請求項11】
対象の血清及び/又は循環中の作用物質の濃度を低下させることのできる二重特異的化合物をスクリーニングするための方法であって:
そのゲノムが、ヒト補体受容体1(CR1)をコードするポリヌクレオチドをプロモータに作動的に連結させて含む非ヒト動物に、複数の検査二重特異的化合物を投与するステップであって、前記ヒトCR1が前記非ヒト動物の赤血球上で機能的に発現する、ステップと;
前記非ヒト動物の血清及び/又は循環中の前記作用物質の濃度を低下させる上での二重特異的化合物の能力を判定するステップと;
前記非ヒト動物の血清及び/又は循環中の前記作用物質の濃度を低下させる二重特異的化合物を選抜することで、対象の血清及び/又は循環中の作用物質の濃度を低下させることのできる二重特異的化合物を同定するステップと
を含む、方法。
【請求項12】
対象の血清及び/又は循環中の作用物質の濃度を低下させる上での二重特異的化合物の能力を評価する方法であって:
そのゲノムが、ヒト補体受容体1(CR1)をコードするポリヌクレオチドをプロモータに作動的に連結させて含む非ヒト動物に、前記二重特異的化合物を投与するステップであって、前記ヒトCR1が前記非ヒト動物の赤血球上で機能的に発現する、ステップと;
前記非ヒト動物の血清及び/又は循環中の前記作用物質の濃度を低下させる上での前記二重特異的化合物の能力を判定することで、前記非ヒト動物の血清及び/又は循環中の前記作用物質の濃度を低下させる上での前記二重特異的化合物の能力を評価するステップと
を含む、方法。
【請求項13】
前記作用物質を前記非ヒト動物に投与するステップを更に含む、請求項11又は12に記載の方法。
【請求項14】
前記作用物質が病原体である、請求項13に記載の方法。
【請求項15】
前記作用物質がウィルスである、請求項13に記載の方法。
【請求項16】
前記作用物質が毒素である、請求項13に記載の方法。
【請求項17】
前記作用物質がポリヌクレオチドである、請求項11又は12に記載の方法。
【請求項18】
前記作用物質が細菌である、請求項13に記載の方法。
【請求項19】
前記作用物質が自己免疫疾患に関連する自己抗体である、請求項13に記載の方法。
【請求項20】
前記二重特異的化合物が、ヒトCR1に結合する二重特異的化合物である、請求項11又は12に記載の方法。
【請求項21】
前記二重特異的化合物がヘテロ重合体である、請求項20に記載の方法。
【請求項22】
前記二重特異的化合物が抗原ベースのヘテロ重合体である、請求項20に記載の方法。
【請求項23】
前記非ヒト動物がトランスジェニック・マウスである、請求項11又は12に記載の方法。
【請求項24】
前記プロモータがGATA-1プロモータである、請求項11又は12に記載の方法。
【請求項25】
前記プロモータが、GATA-1プロモータと、赤血球上での発現を活性化させる上流の制御領域とを含む、請求項11又は12に記載の方法。
【請求項26】
対象が治療されるように、請求項11又は12で同定された二重特異的化合物を対象に投与するステップを含む、生物学的作用物質に曝露した対象を治療する方法。
【請求項27】
対象が治療されるように、請求項11又は12で同定された二重特異的化合物を対象に投与するステップを含む、生物学的作用物質への曝露のリスクのある対象を治療する方法。
【請求項28】
対象が治療されるように、請求項11又は12で同定された作用物質を対象に投与するステップを含む、感染又は自己免疫疾患を持つ対象を治療する方法。
【請求項29】
ヒトCR1をその赤血球表面上に発現するトランスジェニック・マウスを作製する方法であって:
a)ヒトCR1ポリペプチドをコードするポリヌクレオチドを、マウスの卵又は胚に導入するステップであって、前記ポリヌクレオチドが、ヒトCR1をマウス赤血球上で発現させるプロモータに作動的に連結されている、ステップと;
b)前記CR1ポリヌクレオチドを含む卵又は胚を産期まで成長させて、トランスジェニック・マウスを生じさせるステップと
を含む、方法。
【請求項30】
それぞれがヒトCR1ポリペプチドをコードするポリヌクレオチドを含む二倍体ゲノムを含む二匹のトランスジェニック・マウスを交配するステップを含み、前記ポリヌクレオチドが、マウス赤血球上でヒトCR1ポリペプチドを生ずるように発現することで、仔トランスジェニック・マウスが生ずる、トランスジェニック・マウスを作製する方法。
【図1】


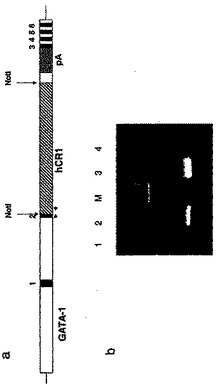
【図2】
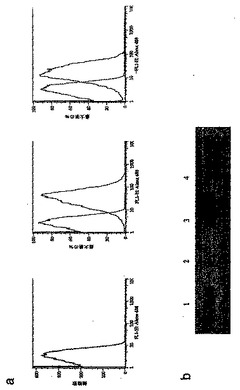
【図3】

【図4】
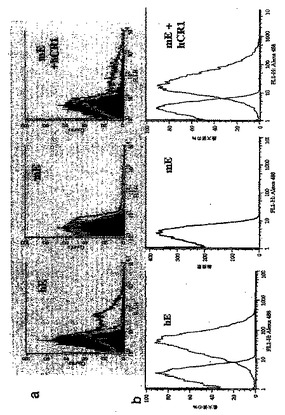
【図5】
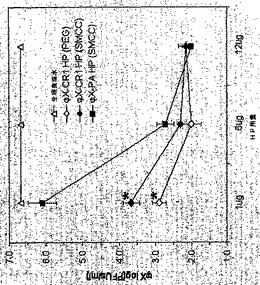
【図6】
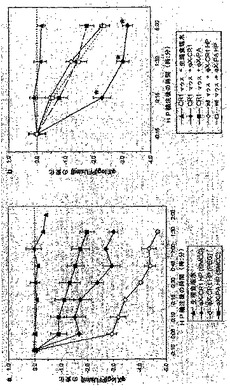
【図2】
【図3】
【図4】
【図5】
【図6】
【公表番号】特表2008−500011(P2008−500011A)
【公表日】平成20年1月10日(2008.1.10)
【国際特許分類】
【出願番号】特願2006−532968(P2006−532968)
【出願日】平成16年5月10日(2004.5.10)
【国際出願番号】PCT/US2004/014781
【国際公開番号】WO2005/005479
【国際公開日】平成17年1月20日(2005.1.20)
【出願人】(505220170)ユニバーシティー オブ マサチューセッツ (6)
【氏名又は名称原語表記】University of Massachusetts
【住所又は居所原語表記】1 Beacon Street, 26th Floor,Boston, MA 02108 (US).
【Fターム(参考)】
【公表日】平成20年1月10日(2008.1.10)
【国際特許分類】
【出願日】平成16年5月10日(2004.5.10)
【国際出願番号】PCT/US2004/014781
【国際公開番号】WO2005/005479
【国際公開日】平成17年1月20日(2005.1.20)
【出願人】(505220170)ユニバーシティー オブ マサチューセッツ (6)
【氏名又は名称原語表記】University of Massachusetts
【住所又は居所原語表記】1 Beacon Street, 26th Floor,Boston, MA 02108 (US).
【Fターム(参考)】
[ Back to top ]