
超高親和性中和抗体

【課題】呼吸合胞体ウイルス(RSV)の治療に有効な免疫グロブリンを提供する。
【解決手段】1010M−1、および1011M−1までの範囲にある結合親和性を持つ超高親和性抗体。このような抗体は新規な高親和性相補性決定領域(CDR)を持つ抗体、とりわけヒトまたはマウスから誘導された枠組み構造領域および定常部を持つ抗体を含む。このような抗体を調製し検査する方法、同様に疾病、とりわけウイルス誘導疾病を予防しおよびまたは処置するためにこれらを使用する方法。
【解決手段】1010M−1、および1011M−1までの範囲にある結合親和性を持つ超高親和性抗体。このような抗体は新規な高親和性相補性決定領域(CDR)を持つ抗体、とりわけヒトまたはマウスから誘導された枠組み構造領域および定常部を持つ抗体を含む。このような抗体を調製し検査する方法、同様に疾病、とりわけウイルス誘導疾病を予防しおよびまたは処置するためにこれらを使用する方法。
【発明の詳細な説明】
【技術分野】
【0001】
この出願は2000年1月27日に受理された合衆国暫定特許出願番号第60/178,426号の優先権を主張し、その開示はここでその全体を引用例によりここに組み込んでいる。
【0002】
本発明は新規な超高親和性中和抗体に関する。
【0003】
微生物の広がりを防止するのに妨げとなりまたはそれが困難であることにより生じる現在の感染発生数は、これらの生体を防止し、同じく既に感染した者を処置するためにより新しいアプローチの必要性を産み出した。
【背景技術】
【0004】
防止しまた処置するのにより困難な感染性外的病原因子にウイルスがある。例えば、呼吸合胞体ウイルス(RSV)は病院に収容される若い子供の急性呼吸系疾患の主な原因であり、また若い子供の下部気道感染の主な原因となる。RSVなどのような病原因子に対する有効なワクチンを産生することへの主要な障害はこれまでずっと安全性の問題であった。これに対して、このようなウイルス因子に対する免疫グロブリンの使用は何らかの価値があることを立証した。例えば高力価RSV免疫グロブリンは動物モデルでRSV感染の予防と治療の両方で有効であることが研究により示されてきた。
【0005】
別のアプローチは高い特異的中和活性を持つ抗体、とりわけモノクローナル抗体を中和する抗体の開発であった。このルートへの一つの欠点はマウスまたはラットのそれよりもむしろヒト抗体を産生し、かくしてヒト抗マウスまたはヒト抗ラット抗体応答の展開を最小にする必要性が存在したことであり、それは潜在的に更なる免疫異常を生じさせることである。
【0006】
も一つの選択的なアプローチはこれまでヒト−マウスキメラ抗体の産生であり、ここでマウス重鎖および軽鎖可変部をコードする遺伝子はキメラまたはハイブリッド抗体を産生するためにヒト重鎖および軽鎖可変部の遺伝子に結合された。
【0007】
ある場合には、マウスCDRはヒト定常部および枠組み構造領域に移植され、いくつかのマウス枠組み構造アミノ酸は「ヒト化」抗体を提供するために対応する位置のヒトアミノ酸と置換された〔クイーン、合衆国特許番号第5,693,761号および5,693,762号〕。しかしこのような抗体は無傷マウスCDR領域を含有し、混合有効性に出会い、その親和性は107乃至108M−1以下になることが多かった。
【0008】
よい親和性を持つヒト化抗RSV抗体がこれまでに調製されており、現在市販されている〔ジョンソン、合衆国特許番号第5,824,307号を参照されたい〕。
【0009】
超高親和性抗体の産生は、このような抗体の中和能力、および臨床効果の望ましい度合を達成するために抗体の産生を少なくする必要性、またそれによる産生費用の削減と同じ量での投与の臨床効果の増大というより実際の見地の両方の点から望ましいものとなろう。
【先行技術文献】
【特許文献】
【0010】
【特許文献1】米国特許番号第5,693,761号
【特許文献2】米国特許番号第5,693,762号
【特許文献3】米国特許番号第5,824,307号
【発明の概要】
【課題を解決するための手段】
【0011】
本発明は少なくとも1010M−1、また更に1011M−1の親和性定数を示す高親和性中和抗体およびその活性断片に関し、またより特異的には抗体およびまたは断片がヒト定常部を持つ抗体およびまたはその活性断片を含む中和モノクローナル免疫グロブリンに関する。
【0012】
本発明はヒト−抗マウス抗体反応(HAMA)を生じる無傷マウスCDR領域の存在なしで、また全産生の費用を削減する十分高い親和性中和活性と全産生の効率化を有する高親和性中和抗体を提供することにより前記の問題を解決する。
【0013】
本発明の一つの見地は、少なくとも1010M−1、および更には1011M−1の親和性定数を持ち、またウイルス発現タンパク質で示されるものなどのような特異的抗原決定基に向かう特異性を持つ高親和性中和抗体に関する。
【0014】
本発明の一つの目的は、抗原が哺乳類、とりわけヒトにおけるウイルス感染細胞により発現されるような場合に、ウイルスにより産生される抗原に向かう特異性を持つような高親和性中和抗体を提供することである。
【0015】
このような一つの実施例において、本発明の抗体およびまたはその活性断片、ならびにその活性断片[原文通り]は、呼吸合胞体ウイルス(RSV)により発現されまた更にRSVに感染した細胞表面(細胞表面でのその抗原の存在は細胞の合胞体への融合を生じせしめる)で発現される呼吸合胞体ウイルス、もっとも特別にはF抗原に対して特異的である。
【0016】
かくして一つの実施例において、本発明の抗体およびまたはその活性断片を含む高親和性中和免疫グロブリンは、その軽鎖可変部が(図1Aで示される)配列識別番号1の配列を持ち、その重鎖可変部が(図1Bで示される)配列識別番号2を持つ抗体と同じRSVのエピトープに結合する。
【0017】
本発明の目的は、(抗RSV抗体構造である免疫グロブリンのものと同じ特異性を持つ)図1で開示された免疫グロブリンの枠組み構造領域を実質的に持つ超高親和性中和抗体を提供することであるが、ここで本発明の抗体およびその断片を含む免疫グロブリンは1個またはそれ以上のCDR(相補性決定領域)を含有し、そのアミノ酸配列はある場合には1個のアミノ酸のみが異なり前記CDR領域の1個のみの差異に限定される場合もあり得るけれども、そのアミノ酸は所謂基準抗体のものとは独立したものである。
【0018】
本発明の望ましい実施例において、本発明の新規な免疫グロブリンは1個またはそれ以上のCDRの配列のみにおいて、図1の抗体(以下「基本抗体」または「基準抗体」もしくは「基準免疫グロブリン」と称する)とは異なり、またもっとも望ましい実施例においては、これらの差異はCDR L2、L3,H1およびH3でのみ生じる。
【0019】
本発明のとりわけ望ましい実施例は、図1で示され、従って図3,4,5,6および7で示された重鎖および軽鎖可変配列を持つ枠組み構造配列を持つ。
【0020】
一つの実施例において、本発明の高親和性中和抗体はヒト定常部を含む。
【0021】
望ましい実施例において、少なくとも1010M−1以上、また1011M−1での親和性定数(Ka)を持つ活性断片を含む本発明の高親和性RSV中和抗体は抗体またはその活性断片などのような組換え免疫グロブリンであり、それは重鎖および軽鎖のヒト定常部と枠組み構造領域を含み、ここで枠組み構造領域の少なくとも一部はヒト抗体から(またはヒト抗体枠組み構造のコンセンサスから)誘導され、前記枠組み構造領域の例は図1の抗体配列で示される。
【0022】
一つの実施例において、すべての枠組み構造領域はヒト抗体(またはヒトコンセンサス配列)から誘導される。
【0023】
も一つの高度に特異的な実施例において、少なくとも1010M−1の親和性(定数)を持つ高親和性RSV中和抗体は、非ヒト抗体から誘導されるヒト定常部、1個またはそれ以上のCDRを持つ組換え抗体であり、ここでCDRの少なくとも1個のアミノ酸は変更され、またここで枠組み構造のすべてもしくは一部はヒト抗体(あるいはヒト抗体枠組み構造のコンセンサス配列)から誘導される。
【0024】
別の実施例において、その可変鎖が図1で示され、また少なくとも1011M−1の親和性を持つ基本または基準抗体もしくは免疫グロブリンと同じエピトープに結合するヒト化中和免疫グロブリンは、少なくとも1個の下記のアミノ酸をCDRの下記の位置で、すなわちCDR H1の位置2でアラニン、CDR H3の位置で位置6でフェニルアラニン、CDR L2の位置3でフェニルアラニン、またCDR L3の位置5でフェニルアラニンを含む。他の実施例はそれぞれの位置で他の単一アミノ酸置換を含む。
【0025】
本発明のも一つの目的はここで開示された免疫グロブリンを含む組成物を提供することであり、ここで前記枠組み構造物は薬理許容希釈剤または賦形剤に懸濁される。
【0026】
本発明の更なる目的は呼吸合胞体ウイルスを予防しおよびまたは処置する方法を提供することであり、それは前記抗体またはその断片が本発明の免疫グロブリンに対しここで開示された特異性および親和性の諸性質を示すような場合に本発明の免疫グロブリンを含有する組成物の治療有効量のそれの危険にさらされ、あるいはそれに悩む患者への投与を含むものである。
【図面の簡単な説明】
【0027】
【図1】抗RSV抗体の軽鎖および重鎖可変部のアミノ酸配列を示す図。ここでCDR領域はアンダーラインされ、一方非アンダーライン残基は各鎖の可変部の枠組み構造領域を形成する。この抗体では、CDRはマウス抗RSV抗体から誘導され、一方枠組み構造領域は主としてヒト抗体から誘導された配列より成る。各CDRにとってアミノ酸置換がここで開示された高親和性CDRおよび抗体を達成するために使用された位置は太字で示されている。ここでの開示に従って、このような置換はCDR L2、L3、H1およびH3のみにあった。図1Aは、軽鎖および重鎖それぞれの軽鎖可変部を示し、図1Bは重鎖可変部を示す。定常部配列は示されていない。これらの配列は本開示を通じてIX−493と名付けられた基本クローン(表2参照)に存在する(すなわちSWSG−これは高親和性CDR H1の主要位置(表1および表3参照)にあるセリン(S)、高親和性CDR H3の主要位置にあるトリプトファン(W)、高親和性CDR L2の主要位置にあるセリン(S)、および高親和性CDR L3の主要位置にあるグリシン(G)、のSWSGを意味する)。この開示の目的のために、これは「基準抗体」である。
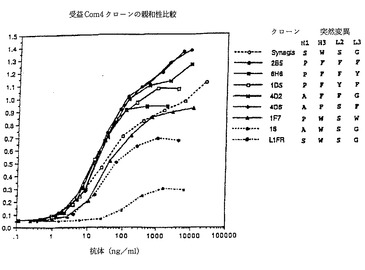
【図2】受益または高親和性クローンの特定の組の親和性の比較を示す図。クローン名称は図面の右の記号一覧の左側に記号一覧の右に示されたCDR H1、H3、L2およびL3での指示された置換と共に存在する。測定法は左軸に示された560nmでのO.D.エリザによる。クローンL1FRは図1の基準抗体の結果を表す。
【図3】ここで開示された発明のクローン1(表2)の望ましい実施例の重鎖および軽鎖可変部を示す図。CDR領域はアンダーラインされ、一方図1の抗体に対するアミノ酸の差は太字で示される。かくしてこの望ましい(すなわち高親和性の)抗体は存在するいくつかの高親和性CDR(表3)を持ち、それは基本または基準抗体より高い親和性(1010M−1以上)生じさせる。
【図4】ここで開示された発明のクローン2(表2)の望ましい実施例の重鎖および軽鎖可変部を示す図。CDR領域はアンダーラインされ、一方図1の抗体に対するアミノ酸の差は太字で示される。かくしてこの望ましい(すなわち高親和性の)抗体は存在するいくつかの高親和性CDR(表3)を持ち、それは基本または基準抗体より高い親和性(1010M−1以上)を生じさせる。
【図5】ここで開示された発明のクローン3(表2)の望ましい実施例の重鎖および軽鎖可変部を示す図。CDR領域はアンダーラインされ、一方図1の抗体に対するアミノ酸の差は太字で示される。かくしてこの望ましい(すなわち高親和性の)抗体は存在するいくつかの高親和性CDR(表3)を持ち、それは基本または基準抗体より高い親和性(1010M−1以上)を生じさせる。
【図6】ここで開示された発明のクローン22(表4)のもっとも望ましい実施例の重鎖および軽鎖可変部を示す図。CDR領域はアンダーラインされ、一方図1の抗体に対するアミノ酸の変化は太字で示される。かくしてこのもっとも望ましい(すなわち最高親和性)抗体は存在するいくつかの高親和性CDR(表3)を持ち、それは基本または基準抗体よりも高い親和性(1011M−1以上)を生じさせる。
【図7】ここで開示された発明のクローン23(表4)の望ましい実施例の重鎖および軽鎖可変部を示す図。CDR領域はアンダーラインされ、一方図1の抗体に対するアミノ酸の変化は太字で示される。かくしてこの望ましい(すなわち最高親和性)抗体は存在するいくつかの高親和性CDR(表3)を持ち、それは基本または基準抗体よりも高い親和性(1011M−1以上)を生じさせる。
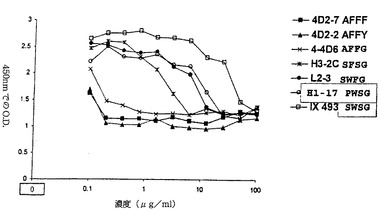
【図8】ここで開示された超高親和性抗体クローンのいくつかについての微小中和検定の結果を示す図。高親和性相補性決定領域の主要位置に存在するアミノ酸(表3参照)は(クローンが単に番号付けられるが表がその表に開示されこの図の右側の記号一覧にあるクリティカル位置で使用された実際のアミノ酸に依存してこの図と比較する表2で示されるように)H1、H3、L2およびL3の順に右側に示される。かくしてクローン4D2−7では、高親和性CDR H1の主要位置にアラニン(A)、高親和性CDR H3の主要位置にフェニルアラニン(F)、高親和性CDR L2の主要位置にフェニルアラニン(F)、また高親和性CDR L3の主要位置にフェニルアラニン(F)がある。要約すると、約25,000のHEp−2細胞がRSVおよび一定濃度の抗体と共に96ウエル平板のウエルそれぞれに加えられた(ウエル当りの抗体濃度は横座標に示される−詳細については実施例2を参照のこと)。5日成長の後、細胞は固定され、ビオチン化抗F MAbで処置され、次いでアビジン−ペルオキシダーゼ複合体に結合され、チオニトロベンゼン酸と反応する結合ペルオキシダーゼの能力が450nmのO.D.を測定することで決定された。存在するFタンパク質の量はウイルス複製の範囲の指標であったし、それによりペルオキシダーゼでの基質のより多くの反応と吸収の増加を生成した。かくしてOD450の値が低ければそれだけ、一定の抗体の指示濃度の中和能力がより大きくなる。ここでIX−493(SWSG−高親和性CDR H1の主要位置でセリン(S)、高親和性CDR H3の主要位置でトリプトファン(W)、高親和性CDR L2の主要位置でセリン(S)、および高親和性CDR L3の主要位置でグリシン(G)を意味する)は(更にL1FRおよびIX493L1FRと表示された)「基準抗体」である。
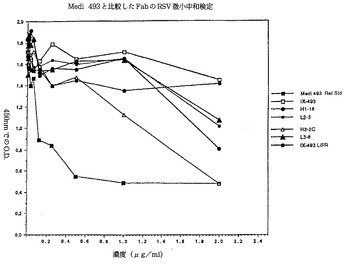
【図9】ここで開示されFab断片が抗体複製の中和化のために採用されたいくつかの抗体の微小中和検定の結果を示す図。ここでIX−493(SWSG)は基準Fab断片であり抗体Medi−493から誘導される(ジョンソン他(1997年)−引用文献23参照)。
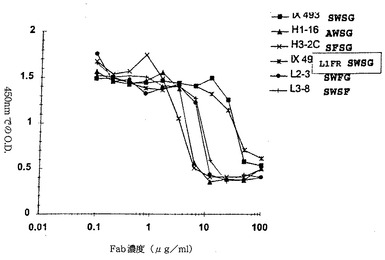
【図10】同じ抗体のFab断片への類似の実験と比較したRSVに特異的な抗体の微小中和(検定)の結果を示す図。ここでMedi−493は抗体を表し、一方IX−493L1FRは本抗体のFab断片を表す。他の系は本発明に基づき産生されたいくつかの抗体のFab断片のためのものである(内部同定のための与えられた各種文字デジタルコード表記、但し中和抗体としてのその相対的有効性には何ら関与しないもの)。
【発明を実施するための形態】
【0028】
「抗原」という用語は、抗体が親和性と特異性を持つウイルスなどのような微生物の表面に存在する構造物、しばしばポリペプチドまたはタンパク質などの構造物を引用する。
【0029】
「抗原決定基」という用語は、抗原などのような免疫グロブリンが特異性と親和性を持つ抗原構造に対する特異的な結合部位を引用する。かくしてウイルスなどの粒子は抗原を示すが、ウイルスが数多くの個別の異なる表面タンパク質を持ち、それぞれが免疫グロブリンに対する別個の潜在的結合部位を表するような場合に数多くの個別の異なる抗原部位を持つであろう。
【0030】
「免疫グロブリン」という用語は、抗原または抗原決定基に対する特異性または親和性を持つタンパク質またはポリペプチドを引用する。この用語は抗原または抗原決定基に対して特異性と親和性を持つ断片などのような一般に四量体として示される抗体ならびにその活性断片を含む。かくしてここで使用される「免疫グロブリン」は抗体およびそのすべての活性断片を含む。
【0031】
「抗体」という用語は、通常は微生物、とりわけウイルスの表面に見出されるもので抗原決定基に親和性を持つタンパク質またはポリペプチドを引用する。このような抗体は一般に4鎖よりなり、かくして四量体である。
【0032】
「中和免疫グロブリン」または「中和抗体」という用語は、生体内または細胞培養で微生物、とりわけウイルスを減少させるための本発明の抗体を含む免疫グロブリンの能力を引用する。このような能力を指示するものとしては、以下で開示される微小中和検定からのデータが存在する。このような構造は通常可変部と定常部を有し、これにより可変部は大抵抗体の特異性を決定することに責任がありそれは相補性決定領域(CDR)を含むであろう。
【0033】
「相補性決定領域」すなわち「CDR」という用語は、H(重)鎖またはL(軽)鎖のいずれかの可変部が抗原標的に特異的に結合できるアミノ酸配列を含む可変部を引用する。これらのCDR領域は特殊な抗原決定基構造のための抗体の基本的特異性を説明する。これらの領域はまた「超可変部」としても引用される。
【0034】
「活性断片」という用語は、それ自身で抗原決定基に対して高親和性を持ち、またそのような特異性の原因となる1個またはそれ以上のCDRを含む。限定されない例はFab、F(ab)’2、重−軽鎖二量体、および完全軽鎖または完全重鎖などのような一本鎖構造を含む。
【0035】
「特異性」という用語は、異なる抗原部位に対して一つの抗原部位に優先的に結合する抗体の能力を引用し、必ずしも高親和性を意味しない。
【0036】
「親和性」という用語は、その結合により形成される複合体の存在に向けて抗原と抗体の平衡状態に移行するように抗体が抗原に結合する度合を引用する。かくして抗原と抗体が相補的に同等の濃度で結合される場合には、高親和性の抗体は生成複合体の高い濃度に向けて平衡状態に移行するように利用できる抗原と結合するであろう。
【0037】
「親和性定数」という用語は、抗原に対する抗体の親和性を測定するために使用される会合定数を引用する。親和性定数が高ければそれだけ抗原または抗原決定基に対する免疫グロブリンの親和性は大きくなり、その逆も真である。親和性定数は相互モル濃度の単位で結合定数である。このような定数は抗体反応の標準動力学方法により測定されるように会合−解離反応のための速度定数から容易に計算される。
【0038】
本発明は少なくとも1010M−1の親和性定数を持つ超高親和性中和抗体およびその活性断片に向けられる。これら抗体の活性断片は、少なくとも1個の高親和性相補性決定領域(CDR)を含有する断片である。
【0039】
分子生物学と組換え技術に関する方法の到来と共に、組換え手段により抗体分子を産生し、これにより抗体のポリペプチド構造で見出される特異的アミノ酸配列をコードする遺伝子配列を生成することが今や可能となっている。このような抗体は、前記抗体のポリペプチド鎖をコード化する遺伝子配列をクローニングするか、もしくは特異的なエピトープと抗原決定基に親和性を持つ、活性四量体(H2L2)構造を形成するために合成鎖の試験管アセンブリで前記ポリペプチド鎖の直接合成により産生することができる。これは異なった種および源から中和抗体の特徴的な配列を持つ抗体の容易な産生を可能にする。
【0040】
バイオリアリターまたはプロセスのいずれの段階でも生体を何ら使用することなく直接化学合成により実験室または商業的規模での大型細胞培養を使用して試験管内または生体内でウシ、ヤギ、ヒツジなどの遺伝子導入動物を用いて免疫グロブリンの源に関係なく、またはそれらがどのように組換え技術により構築されたかまたはどのように合成されたかも関係なく、すべての免疫グロブリンは類似の全体として3次元構造を持つ。抗体の場合、この構造はH2L2として与えられ、また抗体が一般に2個の軽(L)アミノ酸鎖と2個の重(H)アミノ酸鎖を含むという事実を引用する。両方の鎖は構造的に相補性の抗原標的で相互作用できる領域を持つ。標的と相互作用する領域は「可変」または「V」部として引用され、異なる抗原特異性の抗体からアミノ酸配列での差により特徴付けられる。
【0041】
HまたはL鎖の可変部は抗原標的に特異的に結合できるアミノ酸配列を含有する。これらの配列内には、特性の異なる抗体と活性断片の間の極端な可変性の故で「超可変」と名付けられたより小さな配列が存在する。このような超可変部は更に「相補性決定領域」または「CDR」領域として引用される。これらのCDR領域は特定の抗原決定基構造に対する抗体の基本的特異性の原因となる。
【0042】
CDRは可変部内でのアミノ酸の隣接していない広がりを表すが、種とは拘りなく可変重鎖部および軽鎖部内の位置は可変鎖のアミノ酸配列内の類似の位置を持つものとして発現されている。すべての正統な抗体それぞれの可変重鎖および軽鎖は3CDR領域を持ち、そのそれぞれは各軽(L)鎖および重(H)鎖に対して(L1、L2、L3、H1、H2、H3)と名付けられた他のものに隣接していない。認められたCDR領域はキャバット他、ジャーナル・オブ・バイオロジカル・ケミストリー、252:6609−6616(1977)に記載されている。ナンバリングスキームは各図で示されており、ここでCDRはアンダーラインを付されたナンバーはキャバットのスキームに従っている。
【0043】
すべての哺乳類種では、抗体ペプチドは定常(すなわち高度に保存された)部と可変部を含み、また可変部内では、CDRが存在し、またCDRの外側ではあるが重鎖および軽鎖の可変部内にアミノ酸配列で構成された所謂「枠組み構造領域」が存在する。
【0044】
本発明に基づき開示された免疫グロブリンは、ウイルス表面およびウイルスで感染された細胞表面で発現されたタンパク質などのような高分子とりわけタンパク質およびもっとも特異的にはウイルスおよび他の微生物で発現されたタンパク質で見出されるエピトープ、もしくは抗原決定基に対し、(抗体およびリガンドとしての抗原の結合を示す会合定数として定義される親和性定数、すなわちKaに対し1010M−1のオーダー、更には1011M−1のオーダーで)きわめて高い親和性を提供する。本発明の高親和性抗体は中和抗体であり、かくして生体および細胞培養物でのウイルスの複製を減少し、一方逆免疫病変を予防するために受容体の抗体アミノ酸配列に対する十分な相同性を維持する。後者の特性は哺乳類および特にヒトなどのように受容生体のそれに類似する定常部の使用を通じて達成される。この特性は更に受容生体から抗体に見出されるものと同一ではないにしても類似した枠組み構造アミノ酸配列の使用を通じて達成される。後者の場合、あるアミノ酸置換は、本発明の新規なCDRと前記抗体が特異性を示す抗体との間の高親和性相互作用を促進し維持するように枠組み構造配列内で行われる。
【0045】
ここで使用されるように、「抗体」および「活性断片」または「断片」という用語は本発明の全範囲を決定する際に限定するものと見做されるべきではない。かくして「活性断片」または「免疫グロブリン」よりも抗体という用語が使用されるという事実は、特異性と親和性の必要な性質が前記構造により提示される限りにおいて、本発明またはその使用を制限するものとしてとられるものではない。
【0046】
ここで開示された発明に従って、本発明の高親和性中和抗体およびその活性断片の親和性を特徴付ける親和性定数は会合定数であり、KassocまたはKonと名付けられた複合体を形成する会合の速度定数とKdissまたはKoffと名付けられた解離定数で抗原抗体複合体系性の動態により測定された。このような定数の測定は通常の技術の範囲内にありその詳細は本発明を更に記載するためにここで与えられた実施例で引用されるように適当であれば一般的な方法論と特異的な条件を除いてここでは更に記載しないことにする。
【0047】
本発明の高親和性抗体は、前に記載の通り適切なヌクレオチド配列を配列しそれらを適切な細胞系で発現することにより適切な抗体遺伝子配列、すなわちアミノ酸配列を遺伝子操作することで達成される。何らかの望ましいヌクレオチド配列は、例えば合衆国特許番号第5,264,563号に記載されたように、突然変異誘発に基づくコドンの方法を用いて産生することができる。このような手順はオリゴヌクレオチド内で、いずれか望ましいコドン位置でのアミノ酸残基のいずれかおよびすべての頻度の産生を可能にする。これは望ましい位置で、またはこれらのいずれかの特異的なサブセット内で20アミノ酸のいずれかの完全に無作為の置換を含むことを可能にする。選択肢として、このプロセスは本発明に基づく新規なCDR配列などのように、特定のアミノ酸にアミノ酸鎖内で望ましい位置を達成するように実施することができる。要約すると、望ましいいずれかのアミノ酸配列を発現する適切なヌクレオチド配列は、本発明の新規なCDR配列を再生できる。このような手順を用いて容易に達成することができる。これはいずれか望ましいアミノ酸配列を持つ抗体などのポリペプチドを合成する能力をもたらす。例えば、選択された抗体のいずれか望ましいドメインのアミノ酸配列を決定し、また選択肢としてある範囲の置換類似体を与えるために、他の望ましいアミノ酸により置換された1個またはそれ以上のアミノ酸を持つ相同鎖を調製することを可能にする。
【0048】
このような方法を用いる際に、遺伝暗号の縮重に起因して、無作為オリゴヌクレオチド合成および部分縮重オリゴヌクレオチド合成などの方法が特定の位置で特定のアミノ酸残基を特定する遺伝暗号の冗長を取り込むであろうということは理解されねばならない。もっともこのような方法はすべてのアミノ酸配列のマスターセットを提供し、また抗体構造の最適機能または他の目的を選別するために使用することができる。このような方法はスワーラ他、全米科学アカデミー紀要、87巻、6378−6382ページ(1990年)およびデブリン他、サイエンス、249巻、404−406ページ(1990年)に記載されている。
【0049】
ここで開示された発明に従って、そのそれぞれが個別に強化結合抗体を来たす本発明に基づく1個、2個またはそれ以上の新規なCDR配列を単一ポリペプチド構造に組合わせることにより強化抗体変異体を生成することができる。このようにして同一または異なるCDRで本発明内で高親和性抗体を産生するために、いくつかの新規なアミノ酸配列を1個の抗体に結合することができる。例えば3個のこのような新規のCDR配列が採用されて、F抗原またはRSVなどのような特定の抗原構造が選別される。全体の結果はかくして順次的なやり方で各種の単一アミノ酸置換を組合せ、抗原親和性の生成結果を選別する反復過程となるであろう。このような方法は、本発明内で実施されたいくつかの抗体を調製するのに使用された。このような方法は更に本発明の抗体配列、とりわけ前記抗体のCDR領域の配列を最適化する冗長ではあっても便利な手段を表している。
【0050】
本発明の内で新規な配列と方法を用いて、このようなアプローチは最大効率で抗体を発見する仕事において、すべての可能な抗体構造の順列と組合せを生成し選別する時間と費用を回避する。これとは逆に単一10アミノ酸残基CDRの完全無作為抽出は、事実上選別の不可能な数字である10兆個以上の変異体を生成するであろう。
【0051】
この反復方法はより高親和性を持つ抗体の探究範囲を限定するために順次的過程で二種および三種のアミノ酸置換を生成するのに使用できる。
【0052】
逆に言えば、異なる抗体ドメインの配列内で必ずしもすべての位置が同じでないことは認識されねばならない。特定の位置でのいずれの種類の置換も有用であるかまたは有害である。更にある位置でのある種のアミノ酸の置換は同様に特定の抗原への親和性に影響を与えてプラスまたはマイナスのものとなる。例えば、一定の位置ですべての可能な疎水性アミノ酸を試す必要はない。いずれの疎水性アミノ酸でも同じように作用するからである。選択肢として与えられた位置での酸性または塩基性アミノ酸は測定された親和性で大きなぶれを提供する。従ってそのような置換を行う「ルール」を更に学ぶ必要があるが、しかしそのような「ルール」の決定はすべての可能な組合せと置換の研究を必要とせず−最大数の置換より少ない検討の後に傾向は明らかとなるであろう。
【0053】
前記に従って、本発明の抗体は抗RSV抗体などの超高親和性中和抗体であり、後者の場合には合衆国特許番号5,824,307号の抗体としてRSVの同じエピトープに向かう特異性を持つもっとも望ましい抗体である。
【0054】
加えて本発明の超高親和性抗体の親和性は典型的には少なくとも1010M−1である。このような高親和性はここに記載された手順による以外はた易くは測定されないが、このような値は一般に範囲の一部と見做され、また例えば1010M−1の2倍以内、あるいは1010M−1以上もしくは1010M−1に数値的に等しいこともある。この場合、親和性は親和性定数で表示され、これは相互モル濃度の単位を与えるように結合定数の状態にある。このように抗原に対する抗体の親和性はこの定数の値に比例(定数が高ければ親和性が高い)している。このような定数は抗体反応に対する標準動態方法により測定されるように、会合−解離反応への速度定数から容易に算出することができる。
【0055】
特異的実施例において、本発明の高親和性中和抗体およびその活性断片は、少なくとも1011M−1、ある場合には測定可能なもっとも高い範囲であるこの値を越えたそれぞれの抗原への親和性定数を持つ。
【0056】
本発明の高親和性中和抗体およびその活性断片は、高分子のいずれかの型に存在するエピトープとりわけタンパク質およびポリペプチド分子などの三次元構造の一部として存在するペプチドエピトープなどのようないずれかの望ましい抗原決定基に向けられる。このようなペプチドエピトープは一般に微生物および例えば癌細胞などの細胞の表面に存在し、またはその構造の一部となって存在する。このようなペプチドエピトープ発現する微生物は細菌およびウイルスを含む。ウイルスの場合には、ペプチドエピトープを発現するタンパク質およびポリペプチドはウイルス表面に発現された分子であり、またはウイルスで感染した細胞により発現される。本発明を従って開示された規則と方法の効率を評価する目的のために、1個のウイルスが本発明内で抗体を発展させる利用できる抗原源として選択された。更なる分析のために選ばれたこのウイルスは呼吸合胞体ウイルス(RSV)であった。このウイルスが選ばれたのはその複製サイクルに関連し、またその表面で見出される抗原決定基に関連してそれが十分に特徴付けられているからである。更にこれらのウイルスで感染した細胞により発現されることで知られる抗原も十分に特徴付けられている。かくしてこのウイルスはいずれもタンパク質である表面G抗原および表面F抗原の両方を発現する。G抗原はウイルスの細胞表面への結合を促進し、またFタンパク質はウイルスの細胞との融合を促進する。このように感染した細胞はその表面でF抗原を発現し、その結果ウイルスの名称となった合胞体を形成するために細胞の融合を誘導する。更にこのウイルスにより容易に感染する細胞系が十分に周知であり十分に特徴付けられているので、それを容易に試験管内で成長させ培養することができるためにこのウイルスは使いやすい対象である。更にまたこのウイルスの処置に利用できる抗体は周知であり商業的に利用可能である(例えば合衆国特許番号第5,824,307号)。これらの理由でその特許の開示が全体で引用例としてここに取り込まれている前記特許の抗体は図1で開示される「基準抗体」のアミノ酸配列を含有し、そのCDR配列は表1で要約される。かくして商業的に成功した抗体産物の利用可能性およびRSVの十分に特徴付けられた性質が、このような抗体を構築するのに使用される高親和性CDRを産生するためにここで開示された本発明と規則の抗体を試験し最適化して使用するのにこれを理想的な組合せとした。これとは別に、ここで開示された抗体、同じくこのような抗体を調製するここで開示された方法は抗原としてのRSVを限定するものではなく、抗体技術の分野で一般に容易にかつ有利に適用される。更に加えて本開示で提供され、RSVで発現される抗原決定基、およびRSVで感染した細胞に特異的に向けられる特異的実施例はまさに他のエピトープ、とりわけ関連ウイルスに存在するエピトープに高親和性を持つことができる。従って本発明に従って開示されるように、RSV、および図1で示される可変配列を持つ抗体は、本発明で教示される抗体技術の方法を展開し適用する基準点として使用される便利なモデルシステムに過ぎない。
【0057】
活性断片を含む、本発明に基づく高親和性中和抗体は、少なくとも1個の高親和性相補性決定領域(CDR)を含み、ここで前記CDRは少なくとも1010M−1の親和性定数(Ka)を持つ抗体を生じるように選択されたアミノ酸配列を持つ。望ましい実施例では、このような抗体または活性断片は少なくとも2個の高親和性CDR、または少なくとも3個の高親和性CDRもしくは更に少なくとも4個の高親和性CDRを含む。とりわけ望ましい実施例においては、このような抗体または活性断片は3乃至4個の高親和性CDRを含む。一つの望ましい実施例では、このような活性断片はFab断片である。
【0058】
本発明の高親和性抗体は一般に哺乳類、望ましくはヒト定常部および可変部を含み、前記可変部は重鎖および軽鎖枠組み構造領域と重鎖および軽鎖CDRを含み、ここで重鎖および軽鎖枠組み構造領域は哺乳類抗体、望ましくはヒト抗体から誘導され、また、ここでCDRはヒト以外の種、望ましくはマウスなどの種の抗体が誘導される。枠組み構造アミノ酸はまた非ヒトから誘導され、後者は望ましくはマウスである。
【0059】
更に本発明の高親和性抗体はCDRが誘導される抗体と同じエピトープを結合し、ここで前記超高親和性抗体の少なくとも1個のCDRはアミノ酸置換基を含有し、またここで前記置換基は非同一アミノ酸、望ましくは枠組み構造および定常ドメインに寄与するヒト抗体のCDR領域の対応配列位置のアミノ酸により、CDR領域の1個またはそれ以上のアミノ酸の置換を含む。
【0060】
高親和性CDRはそのような高親和性CDRを産生する非高親和性CDRでのアミノ酸置換基により産生することができ、または高親和性CDRはこのような高親和性CDRを形成するために直接合成することができる。かくして本発明の超高親和性中和抗体はアミノ酸置換基をCDR領域の1個のみで、望ましくは1個以上のCDR領域で、もっとも望ましくは3個乃至4個までのその領域で、多分5個または6個すべてのCDRで少なくとも1個の置換基アミノ酸を含むCDRでアミノ酸を持つであろう。
【0061】
本発明の抗体を産生するためにここで開示された方法を適用する際に、抗体を調製する方法は制限因子にはならない。かくして本発明の高親和性中和抗体は抗体をコードするポリヌクレオチド配列を生成し、前記ポリヌクレオチド配列を、そのようなペプチドを発現できるだけでなく、その時点で多分細胞により、培地に分泌された細胞からまたは細胞培養物から回収される特徴付けられた四量体抗体構造に組立てすることのできる許容細胞に挿入することにより調製することができる。このような製造手順の技術は公知のものであり、特許も与えられており、本発明を実施するために必須のものではない〔モリソン他、合衆国特許第5,807,715号〕。更に本発明の抗体のポリペプチド鎖は酵素の存在または不在の下で化学的に合成することができ、次いで通常のH2L2立体配置の四量体構造を形成するために化学結合される。かくしてここで開示された抗体を調製するいずれの方法も利用可能である。
【0062】
本発明は高親和性CDR配列を顕著に持つ結果として高親和性を持つ先に列挙された性質のある高親和性中和抗体に向けられる。本発明に従って、ここで開示された抗体のCDR配列は抗体分子に本発明の抗体に特徴的な超高親和性を与えるように最適化されている。ここで開示された枠組み構造配列、とりわけ図1、3、4、5、6および7の配列により教示されるもの、およびもっとも特別には治療用に使用された時に本発明の抗体の受容体として作用する生体の抗体に特徴的な宣常部配列と共に使用された配列と合わせてこのようなCDR配列は、そのより特異的な実施例で本発明の抗体を産生する。しかし本発明の方法はより特異的にCDRアミノ酸配列に向けられる。
【0063】
本発明、例えば高親和性中和抗体内で抗体およびまたはその活性断片などの免疫グロブリンを産生するために、本発明で教示されるルールは本発明に基づき開示される高親和性CDR配列の配列をその構造が組み込む抗体分子を産生するために有利に使用される。かくして本発明の高親和性中和抗体は、それらがいずれかのクローニングでも産生されないものであるために、本質的にはその用語が一般に使用されているような「モノクローナル」抗体では決してない。前に記載の通り、本発明の抗体の配列は直接合成されるために、いずれかの抗体配列と同一のものではなく、とりわけ現在周知のいずれのCDR配列とは同一のものではない。配列それ自身は完全に初めから新規のものであり、いずれかの生体により産生されるいずれかの抗体で完全に表すことはできない。かくしてここで開示される高親和性CDR配列はここで開示されるように最適化により見出され、または達成され、次いで一度前記高親和性CDR配列が公知であると、科学で公知のすべての手段により二量体または四量体が二官能基または単官能基であるかに拘りなく、完全に機能的抗体分子を合成するのに使用することができる。
【0064】
前記の内容を続け、またその最適化を含む本発明に基づき開示された配列をより良く記載するために、基準抗体(ここで合衆国特許番号第5,824,307号の抗RSV抗体)の軽鎖および重鎖可変部の配列が図1A(軽鎖可変部−配列識別番号1)で示され、また図1B(重鎖可変部−配列識別番号2)で示される。更に本発明に従って、新規な配列が基準抗体に比べてCDR領域のみでアミノ酸配列の差を有して産生された。この結果を達成するのに利用される一つの手段は、所謂出発または基準鎖のCDR領域に突然変異を導入し、次いで抗原(RSV Fタンパク質)親和性を得るために生成する組換えクローンを検定することであった。
【0065】
前記の内容に従って、第1の1個のCDR配列でその配列を最適化し「クリティカル」残基を決定するために変更が行われ、または前記位置よりも残基がその位置のみに限定された一連のアミノ酸置換を通じて最適化された。抗体クローンの6個のCDRそれぞれは「クリティカル」CDRが決定されるまで交代して研究された(ここで「クリティカル」CDRは表3の受益CDRまたは高親和性CDRなどのような抗体結合に十分な影響を与えるCDRを意味する)。すべてのVDRsがクリティカルとして発見されたのではなかった。この特定の研究で使用された抗体として、H1、H3、L2およびL3のみがクリティカルであるとして発見されたが、その結果は異なった抗体で異なっている。一度「高親和性」CDRが決定されると(すなわち「受益CDR」)次いで本発明の高親和性中和抗体を生じるCDR配列の組合せを最適化するためにCDRの組合せが研究された。
【0066】
非常に特異的な実施例として、ここで開示された発明は少なくとも1010M−1の親和性定数を持つ呼吸合胞体ウイルス(RSV)に対する高親和性中和抗体に関し、ここで前記親和性定数はこのような決定の変動性と抗原(ここではRSVのF抗原)に対する異なるクローン化抗体の親和性の変動性の故で、この値の少なくとも2倍以内にあることができよう。この実施例内での生成する最適化構造のいくつかは1011M−1以上のKaを有していた(例えば表4での22と23の番号を付されたクローンがそれである)。
【0067】
この高親和性中和抗体にまた、その軽鎖可変部が配列識別番号1(図1A)の配列を持ち、またその重鎖可変部が配列識別番号2(図1B)の配列を持つ抗体と同じRSVに対するエピトープに結合する抗体である。
【0068】
一般に、直前に記載した抗RSVの特異的な例に基づく本発明の抗体を同定するために使用されるアプローチは、望ましい抗体鎖を発現する遺伝子のヌクレオチド配列を生成し、次いで標準プロトコルにより大腸菌細胞を形質転換するためにそれらをベクターに挿入することであった。細胞はウエルで成長し、上澄みがサンプル抽出され捕捉リフトとエリザ技術を用いて抗原結合を測定された〔ワトキンズ他、(1997年)、分析生化学、253巻、37−45ページ、ワトキンズ他(1998年)、分析生化学、256巻169−177ページ(これらの明細は引用例としてここに組み込まれている)参照〕。これらのポリヌクレオチドはCDRでの単一アミノ酸置換を提供するように設計されていたが、それは次いで親和性増加のために選択的に結合される受益置換(増加した親和性を産出するもの)で増加した親和性を検査することができた。これらは次いでRSVのF抗体の親和性体基本または基準抗体の結合親和性を検査された。
【0069】
このプロトコルを使用して、CDR L1またはH2での単一アミノ酸置換は抗原(ここではRSVのF抗原)として使用されたエピトープへの抗体クローンの親和性に何らの増加も産生しなかったことをエリザのデータが示した。従って本発明の抗体はすべてCDR L2、L3、H1およびH3での基準抗体のみとは異なるCDR配列を包含する(ここで図1の配列を持つ基準抗体は、それに対してKaを増加することにより抗体親和性の最適化のための試験手順およびいずれか他のシステムが同じように十分使用できる単なる有用な配列であった)。
【0070】
このようにしてRSVに関してここで開示された抗体は、更に一般にヒト抗体から誘導され、しかしそのように誘導されない場合は望ましくはマウスから誘導される枠組み構造領域を持つ。
【0071】
基準抗体のCDRに関して、(図1の配列で与えられるように)各CDRのアミノ酸配列は表1で示される。最適化に続き本発明により教示されるアミノ酸により置換されるならば、高親和性CDR配列(高親和性中和抗体の生成)およびそれによる受益結果(親和性の増加)を産生した基本または基準抗体のCDR内のアミノ酸残基の位置は、表1で太字でアンダーラインを付して示される(基準抗体にわたり親和性増加を与えるこのような配列は「受益CDR」または「高親和性CDR」で示される)。基本または基準抗体(図1)のCDRは以下で「基本または基準CDR」としてここで引用される。かくして表1は図1で示された(すなわち最適化を監視するためにここで使用された抗RSV基準抗体に対する)CDR配列を表す。
【表1】

【0072】
ここで開示された配列と関連して、本発明の目的に適うように定義されたCDR領域はここで開示された抗体の軽鎖可変部の残基24−33(CDR L1)、49−55(CDR L2)および88−95(CDR L3)ならびに重鎖可変部の残基31−37(CDR H1)、52−67(CDR H2)および100−109(CDR H3)に一致するこれらのセグメントである。
【0073】
本発明の抗体を産生する際に、前記抗体を含むクローンを生成するまたはポリペプチド鎖をクローニングするか、それをコードするポリヌクレオチド配列を使用するかまたは使用しないポリペプチド配列の直接合成によるか、もしくはどのような方法を使用者が選択するか、何故なら前記抗体を産生する本発明の教示の限定を生じることはないために、基本抗体または基準抗体(図1で示される重鎖および軽鎖可変部(CDRプラス枠組み構造))は本発明の抗体の新規なCDR配列を生成し、また結合定数等を比較する目的のための「鋳型」として使用することができる。単一変異の6個のCDRライブラリーを特徴付け合成する標準アプローチが使用された(ウー他、全米科学アカデミー紀要、95巻:6037−6042ページ(1998年)参照)。標的CDRはヌクレオチドのアニーリングの前にライブラリーのそれぞれからまず除去された。ライブラリーの合成のために、CDRは表1にあるように定義された。本発明のCDR配列を産出するためにオリゴヌクレオチド合成用のコドンベース突然変異が(前記の通り)採用された。
【0074】
ライブラリーは最高の親和性変異体を同定するためにまず、捕捉リフトで検査された。次いでこれらのクローンは更に捕捉エリザを用いて固定化抗原への滴定で特徴付けられた。
【0075】
最高親和性変異体からのDNAは受益または高親和性置換の性質を決定するため配列化された。検査の後、CDRの4個のみに起こる8個の受益または高親和性置換が観察されたことが確定した。これらは太字でアンダーラインを付された表1の基準または基本CDRに対して差異を持つ表3のCDR配列として要約されている。かくして表3のCDR配列は、特異性がRSVのF抗原に向けられる本発明の高親和性中和抗体を産生するのに使用が可能なカセットのCDRライブラリーであると見做すことができる。
【0076】
データの分析は、選択された位置でのアミノ酸置換がエピトープ結合を大きく増加させ、とりわけ置換の性質がアミノ酸配列の挿入であり、それはフェニルアラニン、アラニン、プロリン、トリプトファンおよびチロシン、とりわけフェニルアラニンから選択され、これらすべてのそのような受益または高親和性置換が再び選択的位置にあったことを示した。
【0077】
ここに記載されRSV/抗RSVシステムを採用する最適化実施に対し、もっとも受益的な高親和性CDRは6個のCDRの3または4、また丁度4のアミノ酸位置全体で生じることが発見された。かくして本発明の高親和性中和抗体はこれまで既知の人工抗体とは違って、相補性決定領域で、またはそのような領域を共に取巻く枠組み構造領域でのみ基本または基準抗体とは異なるアミノ酸配列を含有する。かくして本発明の抗体は抗体分子に高親和性を産生するように選択された1個またはそれ以上のCDR配列を含む高親和性中和抗体である。RSVを標的として用いる特異的実施例においては、そのような差異はL2(またはCDR L2)、L3(またはCDR L3)、H1(またはCDR H1)およびH3(またはCDR H3)でのみ発見される。前に述べた通り、この抗体の最大の親和性はこれらCDRの選択されたアミノ酸位置で起こり、また本発明の実施例のいくつかでは、各CDRのT度1個のみのアミノ酸位置が高親和性を提供するために望ましいものであった。
【0078】
かくしてCDR H1にとっては、(図1Bで特別のアンダーラインを付与した特別のCDR配列のアミノ末端から数えて)CDRアミノ酸2での置換は、とりわけ基本または基準抗体のCDR H1の位置2を占めるセリンをアラニンまたはプロリンのいずれかで置換したものは、もっとも受益的であり従ってRSV抗原エピトープへの高親和性を生じることを発見した。CDR H3にとっては、CDR配列の位置6でのグリシンの置換は、とりわけフェニルアラニンまたはトリプトファンのいずれか、特にフェニルアラニンによるグリシンの置換はRSVエピトープへの親和性増加を来たしたことが発見された。CDR L2ではCDRの位置3でのセリンの置換、特にフェニルアラニンはまたチロシンのいずれかによるものはF抗原への親和性増加を来たした。CDR L3ではCDRの位置5でのグリシンの置換、とりわけフェニルアラニン、トリプトファンまたはチロシンによる置換がRSVエピトープへの親和性の増加を生じた。
【0079】
本発明に従って、このようなアミノ酸置換を組合せて1個以上が同じ交代分子内で生じるようにして、RSVのF抗体のエピトープにここで開示された抗体の親和性を大きく増加することが可能であった。
【0080】
表2は本発明に基づく数多くのクローンの(H1、H3、L2およびL3それぞれに)新規なCDR配列を用いた結果を示している。表2で示されたように、特殊な抗体が本発明の1、2または3の新規なCDR対図1Aおよび1Bで示された基本または基準抗体鎖(軽鎖(L2およびL3))および重鎖(H1およびH3)をここに取り込んでいる。新規なCDRの効果は抗原(Ag)滴定スコア(以下参照)で記載されており、ここで基本または基準抗体は表2のトップで示されそのスコアは0.1である。他のスコアはこの「基本または基準抗体」配列の0.1のスコアと比較して示されている。それぞれの位置(すなわちH1の位置2、H3の位置6、L2の位置3およびL2の位置5)での最高の親和性を与えるアミノ酸の同定は各CDRに使用される突然変異を単に示す基本または基準抗体のアミノ酸の下部に示されている。
【0081】
本実験で検討されたクローンの全数は一部重複を含めて37個であった。(括弧内の「n」値で示されたもの、例えばクローンNo.7に対する「n=4」は4個の重複クローンが検討されたことを示している)。
【0082】
一般に、親和性と受益または高親和性CDRの数との間には相関があることをデータは示しており、より高次の親和性変異体のすべては1個以上の受益または高親和性CDRを持っていた。更にすべての最良クローンは、CDR H3内の位置6にF(Phe)を有していた。また受益または高親和性CDRは疎水性、とりわけ芳香族アミノ酸が基本または基準抗体に見出される残基の位置に挿入されたものであった。
【0083】
抗体滴定検査は各測定でRSV抗原の500ngを用いて抗体の濃度を変えて使用した。表2の数多くの組合せクローンの比較データのグラフが図2で示される。
【0084】
要約すると、表2はここで記載されて手順で評価される数多くのクローンと、CDR H1、H3、L2およびL3の主要位置を(表1と表3で太字でアンダーラインされた位置)で生じるアミノ酸を示している。この表は更にこれらのクローンのCDR対基準抗体の対応するCDRの間の多くの差を要約する(表1と図1を参照)。表の右側は「Agスコア」すなわち抗原結合値を示し、それは0−4にわたる任意の性質上の値を表し、またそのそれぞれの滴定曲線に基づく異なった抗体クローンの相対的結合能力の質的測定値を表す。この値は異なる抗体の大ざっぱな質的比較のみをここで提供し結合能力の量的尺度を意図するものではない。
【0085】
表2はまた各抗体クローンの新規なCDRの数を示す。(基準抗体の対応するCDRに関連して少なくとも1個のアミノ酸の差をもつ抗体内でのCDRの数を意味する−表1参照)。新規なCDRの数は、またその抗体分子に存在する「受益」または「高親和性」CDRの数である。新規なCDRでのアミノ酸の差は基準抗体に対し表1での太字でアンダーラインされた位置で起こるであろうし、そのため表1で太字のアンダーラインされたアミノ酸は(標準単一文字アミノ酸命名法を用いて)表2のそれぞれのCDRを示すアミノ酸により置換された。
【表2】
【0086】
クローンそれぞれで表される新規なCDRはクローンを表に位置せしめ、基本または基準クローンに隣り合わせの同じCDRの対応するアミノ酸に各CDRの指示アミノ酸を適合させることで容易に決定される。便宜上アミノ酸がクローンの一つの特定CDRで異なる場合には、新しいアミノ酸は太字で示される。加えて表で示されるすべてのクローンでは、置換は新規なCDRを産出するとして前に引用したように選択された位置でのみ生じる。かくして基本または基準抗体に対するCDR H1の置換はすべてCDRの位置2にあり(図1および表1と2でアンダーラインされ太字で示されたCDR H1のN末端端部からの第2アミノ酸を意味する)、CDR H3のすべての置換は位置6にあり、CDR L2のすべての置換は位置3であり、またCDR L3のすべての置換は位置5にあり、またすべては基本または基準抗体に関連する。基本または基準抗体が選ばれたのは、それがRSVエピトープにきわめて高い親和性を持つことが既に公知であったからであるということは心に留めておかねばならない。〔ジョンソン他、(1997年)感染性疾病ジャーナル、176巻、1215−1224ページ〕。従って例えばクローン番号1では、受益または高親和性CDR H1に対し基本または基準抗体のCDR H1の位置2でセリンに代りアラニンが使用され、これによりRSVへの親和性増加を達成し、またフェニルアラニンはCDR H3の位置6でトリプトファンに代って生成し、基本または基準抗体のCDR L2の位置3でセリンが使用され、更に受益または高親和性CDR L3の位置5で発生した。
【0087】
かくして表2のクローンで試験された上澄みに産生された抗体構造の存在する本発明に基づく新規な受益CDR(すなわち対応する基本または基準CDRに代って基本または基準抗体におけるその存在が同じRSVエピトープの前記抗体の親和性を著しく増加することに役立った高親和性CDRまたはCDR配列)が表3で要約される。それぞれの場合において、太字のものは本発明のCDRが基本または基準講RSV抗体の対応するCDRとどのように新規で受益的であり、または高親和性で異なるかを示している。
【表3】
【0088】
表3のCDR配列が本発明に基づき開示された高親和性CDRの配列を表している一方、1個またはそれ以上のこれらのCDRが同じ抗体内に存在し、高親和性CDRのそれぞれに適切な配列が選択されるセットをこの表が示しているということが理解される。かくして表3で示されるように、高親和性H1 CDRがここで開示された発明の高親和性中和抗体に存在する時には、それは配列識別番号9または10の配列に一致する配列を持つ。請求された発明の中和抗体が高親和性H3 CDRを含有する時には、前記CDRは配列識別番号12を持つ。本発明の高親和性中和抗体が高親和性L2CDRを含有する時には、前記高親和性L2 CDRは配列識別番号12および13の配列より成るグループから選択されたアミノ酸配列を持つ。最後に本発明の高親和性中和抗体が高親和性L3 CDRを含有する時には、前記CDRは配列識別番号14、15,16の配列より成るグループから選択されたアミノ酸配列を持つ。
【0089】
一つの実施例において、既に述べたように高親和性中和抗体はヒト定常部を含む抗体である。
【0090】
かくして望ましい実施例において、少なくとも1010M−1、または更に少なくとも1011M−1の親和性を持つ本発明の高親和性中和抗体はヒト定常部および重鎖と軽鎖の枠組み構造を含む移植抗体であり、ここで枠組み構造の少なくとも一部はヒト抗体から(またはヒト抗体枠組み構造のコンセンサス配列から)誘導される。
【0091】
も一つの実施例において、すべての枠組み構造はヒト抗体(またはヒトコンセンサス配列)から誘導される。
【0092】
かくして少なくとも1010M−1の親和性を持つRSV中和抗体はヒト定常部、非ヒト抗体から誘導される1個またはそれ以上のCDRを持つ移植抗体であり、ここで少なくとも1個の前記CDR内の少なくとも1個のアミノ酸は変更され、またここで枠組み構造の全体または一部はヒト抗体(またはヒト抗体枠組み構造のコンセンサス配列)から誘導される。
【0093】
1個の抗体のCDR配列のも一つの抗体の非CDR領域との組合せがCDRの残りの分子への「移植」の形態から生じるために、これらのものは「CDR移植」抗体として引用されてきた。今日遺伝子工学の手法を用いることで同じ産物を実際の交代からいずれの配列を単離することなく形式することができる。望ましいCDR配列、および定常部と枠組み構造配列が公知である限りにおいて、望ましい配列をもつ遺伝子を組み立てることができ、また各種のベクターを使用して機能的四量体抗体分子の発現のために適切な細胞に挿入することができる。これを既に記載した方法を組合わせると、単一突然変異ライブラリーの組立てを可能にし、ここで抗体は対応する移植抗体と同じ配列を所有し、また従って同じ構造と結合親和性を有することになる。
【0094】
本発明の高親和性抗体は比較的純粋なまたは単離形態に、同じくウエル内でまたは平板上で成長した細胞からの上澄みに存在することができる。このような上澄みは表1のデータを生成するために使用された。本発明の抗体は画して本発明の抗体を含むひとつの組成物の形態で存在することができ、ここで前記抗体は薬理許容単体または賦形剤に懸濁される。本発明の抗体はそのような組成物に疾病、例えばRSVなどを処置する治療または薬理学的値に十分な一定の濃度で、または一定の量で存在する。前記抗体はまたより希釈された形態で組成物に存在する。
【0095】
従って本発明は更にここで記載された抗体組成物の(予防を含む)治療有効量の呼吸合胞体ウイルス感染の危険のあるまたはそれに悩む患者への投与を含むその感染症を予防しおよびまたは処置する方法を提供することに向けられる。
【0096】
本発明の高親和性抗体の一つの望ましい実施例は、その重鎖および軽鎖CDR領域が以下の配列を持つ抗体である。すなわちCDR H1は配列識別番号9の配列を持ち、CDR H3は配列識別番号11を持ち、CDR L2は配列識別番号4(図1Aの基準配列のCDR L2から変化していない)の配列を持ち、またCDR L3は配列識別番号14を持つ(表3参照)。この望ましい実施例において親和性定数は表4(クローン1)で示されるように約6.99×1010(または解離定数として約14.3pM)である。この実施例を含む抗体の重鎖および軽鎖可変部は枠組み構造配列を伴い図3で示される。
【0097】
本発明の高親和性抗体のも一つの望ましい実施例はその重鎖および軽鎖CDR領域が以下の配列を持つ抗体である。CDR H1は配列識別番号9の配列を持ち、CDR H3は配列識別番号11の配列を持ち、CDR L2は配列識別番号12の配列を持ち、またCDR L3は配列識別番号5の配列を持つ(表2、クローン2を参照のこと−図1AのCDR L3の基準配列との差はない)。この望ましい実施例において、親和性定数は表4(クローン2)で示されるように約7.30×1010(または解離定数として約13.7pM)である。この実施例を含む抗体の重鎖および軽鎖可変部は枠組み構造配列と共に図4で示される。
【0098】
本発明の高親和性抗体の追加の望ましい実施例はその重鎖および軽鎖CDR領域が以下の配列を持つ抗体である。CDR H1は配列識別番号10の配列を持ち、CDR H3は配列識別番号11の配列を持ち、CDR L2は配列識別番号12の配列を持ち、またCDR L3は配列識別番号14の配列を持ち、(CDR配列については表3を参照のこと)、そのクローンは表2で指示番号3である。この望ましい実施例において、親和性定数は表4(クローン3)で示されるように、約8.13×1010(または解離定数としては約12.3pM)である。実施例を含む抗体の重鎖および軽鎖可変部は枠組み構造配列と共に図5で示される。
【0099】
本発明の高親和性抗体のもっとも望ましい実施例はその重鎖および軽鎖CDR領域が以下の配列を持つ抗体である。CDR H1は配列識別番号9の配列を持ち、CDR H3は配列識別番号11の配列を持ち、CDR L2は配列識別番号12の配列を持ち、またCDR L3は配列識別番号14の配列を持つ(表3参照のこと)。本実施例において、親和性定数は表4(クローン22)で示されるように約3.6×1011(または解離定数として約2.8pM)である。この実施例を含む抗体の重鎖および軽鎖可変部は、枠組み構造配列と共に図6で示される。
【0100】
本発明のも一つのもっとも望ましい実施例端の重鎖および軽鎖CDR領域が以下の配列を持つ抗体である。CDR H1は配列識別番号9の配列を持ち、CDR H3は配列識別番号11の配列を持ち、CDR L2は配列識別番号12の配列を持ち、またCDR L3は配列識別番号15の配列を持つ(表3参照)。このもっとも望ましい実施例において、親和性定数は表4(クローン23)で示されるように約4×1011(解離定数として約2.5pM)である。本実施例を含む抗体の重鎖および軽鎖可変部は、枠組み構造配列と共に図7で示される。
【0101】
とりわけ望ましい実施例において、本発明の抗体は図1、3、4、5、6および7で示された枠組み構造を示す配列の枠組み構造領域を持つであろう(それぞれは同じ枠組み構造領域を含有し、CDR配列でのみ異なる)。これらのもっとも望ましい実施例は中和抗体を含みここで軽鎖可変部は配列識別番号17のアミノ酸配列を持ち、また重鎖可変部は配列識別番号18のアミノ酸配列を持つ。またもっとも望ましい実施例は中和抗体を含み、ここで軽鎖可変部は配列識別番号19のアミノ酸配列を持ち、また重鎖可変部は配列識別番号20のアミノ酸配列を持つ。(実施例はまた別の)中和抗体ではここで軽鎖可変部は配列識別番号21を持ち、重鎖可変部は配列識別番号22のアミノ酸配列を持つ。別の中和抗体では軽鎖可変部は配列識別番号23のアミノ酸配列を持ち、重鎖可変部は配列識別番号24のアミノ酸配列を持つ。別の中和抗体では軽鎖可変部は配列識別番号25のアミノ酸配列を持ち、重鎖可変部では配列識別番号26のアミノ酸配列を持つ。
【0102】
本発明の高親和性中和抗体は、共にアミノ酸セグメントをスプライスすることにより実際の中和抗体から誘導されるCDR領域および非CDR領域から組立てることができ(またこのように組立てられた抗体はここで開示された発明の範囲内にあるが)、本発明の抗体は適切な遺伝子配列をベクターに遺伝子操作することでもっとも便利に調製され、それは次いで遺伝子操作細胞による組立て抗体分子の偶発的発現のために適当な細胞系に形質移入される。実際にこのような組換え手順は、ここで開示される抗体を調製するために採用された。加えて高親和性抗体の鎖の配列は、ここでの開示から公知であるために、このような抗体は更に適切な鎖の直接合成により組立てられ、次いで四量体(H2L2)二価抗体構造に自己組立てをすることを可能にした。
【0103】
本発明の高親和性中和抗体を調製する方法は、本発明の受益CDRを含むクローン産生抗体を調製するのに使用された組合せライブラリーの創出を伴い、それは次いでRSVエピトープへの親和性を検査することができた(例えば図2)。
【実施例】
【0104】
(実施例1)
BIAcoreTMによるヒト化RSV Mabsの動態分析
高親和性抗RSV MabsとRSV Fタンパク質の間の相互作用の動態がファルマシアのBIAcoreTMバイオセンサーを用いる表面プラスモン(細胞質遺伝子)共鳴により研究された。C末端切形Fタンパク質を発現する組換えバキュロウイルスは動態研究のための抗原の豊富な源を提供した。分泌Fタンパク質を含有する上澄みはコンカナバリンA(conA)とQセファロースカラムでの連続クロマトグラフィーにより約20倍富化された。プールされた小部分はクエン酸ナトリウム10mM(pH5.5)に対して透析され、約0.1mg/mlに濃縮された。典型的な実験では、Fタンパク質のアリコート(100ml)が、BIAcoreセンサーチップにアミン結合された。固定された量はマウスモノクローナル抗体H1129またはH1308Fで飽和された時約2000応答単位(Rmax)のシグナルを提供した(抗体H1129あるいはH1308は合衆国特許番号第5,824307号で提供され、その開示はここで引用例として取り込まれている)。これは、結合手順に次いでFタンパク質に同じ数の「A」および「C」抗原部位が存在したことを示していた。2個の無関連Mabs(RVFV 4D4とCMV H758)は固定化Fタンパク質と相互作用を示さなかった。典型的な動態研究は異なる濃度(25−300nM)でのMab35mlの0.05%トウィーン−20(PBS/トウィーン)0.05%を含むPBS緩衝液への注入を含んでいた。流速は5ml/分に維持され、7分の結合フェーズを与えられた。Mabの注入に続き、流動物は解離速度を決定するために30分間PBS/トウィーン緩衝液で変換された。センサーチップは塩酸10mMの2分パルスのサイクル間で再生された。再生段階は固定Fタンパク質の結合能力を僅かではあるが最小ロスとして生じた(サイクルごと4%のロス)。この小さな減少は結合および解離の速度定数(それぞれKonおよびKoffと称されるもの)の計算値を変えることはなかった。
【0105】
より特異的には、Kassoc(またはKon)の測定のために、Fタンパク質がEDC/NHS活性化センサーチップ全体に注入されるEDC/NHS法(EDC=N−エチル−N′−〔3−ジエチルアミノ−プロピル〕カルボジイミド;NHS=N−ヒドロキシスクシンイミド)により直接固定された。要約すると、NaOAc、10mM、pH4.0でFタンパク質4μg/mlが調製され、約30μmlの注入は前記引用条件の下で約500RU(応答ユニット)の固定Fタンパク質を提供した。未加工流動細胞(VnR固定化−CMデキストラン表面)が動態分析のために取り出された。カラムは塩酸100mMを用いて(完全再生に必要とされる72秒の接触時間で)再生された。この処置は固定抗原を何ら損傷することなく結合Fabを完全に除去し40回もの再生に使用することができた。Kon測定では、Fab濃度は12.5nM、25nM、50nM、100nM、200nMおよび400nMであった。解離フェーズは(解離フェーズの開始30秒後から)230秒乃至900秒まで分析された。動態は1:1のラングミュアフィッティング(グローバルフィッティング)で解析された。測定はHBS−EP緩衝液(ヘペス10mM、pH7.4、塩化ナトリウム150mM、EDTA3mM、0.005%(容量/容量)界面活性剤P20)内で行われた。
【0106】
ここで開示されたように組合せクローン内測定に関しては、KonおよびKoffが個別に測定された。Konは単一変異クローンに対するのと同じ条件で測定され、同様に分析された。
【0107】
Kdissoc(またはKoff)の測定については、下記の条件が採用された。要約すると、Fタンパク質4100RUが未加工として使用されるCM−デキストランと共に(前の通り)固定化された。ここで3000RUのFabは(機械の変動を相殺するためにきわめて十分な解離Fabと)結合した。HBSとFタンパク質5nM(KdissocまたはKd−解離平衡定数よりも約350−2000倍高いもの)が緩衝液として使用された。解離フェーズは5μl/分の流速で6−15時間であった。ここで使用された条件の下で解離Fabの再結合は最小であった。その詳細については、バイオセンサーのマニュアルを参照されたい。
【0108】
ここで開示されたFタンパク質またはRSVの他のエピトープ部位への高親和性抗RSV抗体の結合は、結合または会合の第2オーダー比率定数に対する解離の第1オーダー比率定数(Kd=Kassoc)の割合から算出された。Kassocの値は下記の等式に基づき計算される。
dR/dt=Kassoc〔Mab〕Rmax−(Kassoc〔Mab〕+Kdiss)R
ここでRとRmaxはそれぞれ時間tと無限での応答単位である。Rの関数としてのdr/dtのプロットは(Kassoc〔Mab〕+Kdiss)の勾配を与える。これらの勾配は〔Mab〕に線型に関連しているため、Kassocは〔Mab〕に対する勾配の再プロットから誘導することができる。新しいラインの勾配はKassocに等しい。Kdissの値はY線分から外挿することができるけれども、より正確な値はKdissの直線測定により決定された。Mabの注入フェーズに続き、PBS/トウィーン緩衝液が、センサーチップを横断流動する。この点から〔Mab〕=0である。dR/dtのための前述の等式は従って
dr/dt=k または dR/R=Kdissdt
となり、この等式を統合すると
In(Ro/Rt)=Kdisst
となり、ここでRo/Rtはそれぞれ時間0(解離フェーズ開始)および時間tでの応答ユニットである。最後にtの関数としてのプロッティングIn(Ro/Rt)はKdissの勾配を提供する。
【0109】
ここでの望ましい実施例において、このような抗体変異体からの数値は以下の通りである。
【表4】
【0110】
ここでCDRは基準抗体として表1で(太字とアンダーラインで示されたCDRの主要位置(クリティカル位置)で基準アミノ酸を置換するアミノ酸を表す。かくして例えば、クローン22は表1(配列識別番号6)のその位置で示されるセリンに代ってCDR H1(重鎖可変部−配列識別番号24の残基32)の位置2でアラニンを持つ。表1(配列識別番号8)のその位置で示されるトリプトファンに代ってCDR H3(重鎖可変部−配列識別番号24の残基105)の位置6でフェニルアラニン、表1(配列識別番号4)のその位置で示されるセリンに代ってCDR L2(軽鎖可変部−配列識別番号23の残基51)の位置3でのフェニルアラニン、および表1(配列識別番号5)のその位置で示されたグリシンに代ってCDR L3(軽鎖可変部−配列識別番号23)の位置5でのフェニルアラニンなどを持つ。
【0111】
もちろんこれらのクローンを形成する際には、表3は本発明の抗体の高親和性CDRが取り出される潜在的CDRのプールを表す。例えば、表4のクローン23はCDR L3の例外を除きクローン22と同じCDRを使用し、それは表1(配列識別番号5)のその位置で示されるグリシンに代えてCDR L3(表3−配列識別番号15)の位置5でチロシンを持つ。対応するクリティカル位置での置換は同じように表2で示される。
【0112】
(実施例2)
微小中和検定
本発明の抗体の中和は微小中和検定で測定された。この微小中和検定はアンダースン他(1985年)に記載された手順の修飾である。呼吸合胞体ウイルス(RSV−ロング株)の10個のTCID50が96ウエル平板のウエルで2時間37℃で試験される抗体(またはFabs)の連続希釈液で保温された。RSV感染性HEp−2細胞(2.5×104)は次いで各ウエルに加えられ、5日間37℃で5%炭酸ガスの下で培養された。5日後に培地は吸引され細胞は洗浄されメタノール80%PBS20%で平板に固定された。RSV複製は次いでFタンパク質発現で測定された。固定細胞はビオチン接合抗Fタンパク質モノクローナル抗体(パンFタンパク質、C部位特異的MAb 133−1H)で保温され、洗浄され西洋ワサビペルオキシダーゼ接合アビジンがウエルに加えられた。ウエルは再洗浄され基質TMB(チオニトロベンゼン酸)の出来高が450nmで測定された。中和力価はウイルスのみの対照細胞から450nm(OD450)の吸光度で少なくとも50%の減少を起こす抗体濃度で表現された。本発明のいくつかの抗体の結果は図8のグラフで示されており、一方Fab断片を使用した結果は図9のグラフで描かれている。
(背景および引用文献)
【0113】
1.C.B.ホール,R.G.ダグラス,J.M.ガイマン他、ニューイングランド・ジャーナル・オブ・メディスン293巻:1343ページ、1975年。
【0114】
2.C.B.ホール,J.T.マクブライド,E.E.ウォルシュ他、ニューイングランド・ジャーナル・オブ・メディスン308巻:1443ページ、1983年。
【0115】
3.C.B.ホール,J.T.マクブライド,C.L.ギャラ他、JAMA254巻:3047ページ、1985年。
【0116】
4.E.R.ウォルド他、小児科ジャーナル112巻:154ページ、1988年。
【0117】
5.A.Z.キャピキャン,R.H.ミッチェル,R.M.チャノック他、疫学アメリカン・ジャーナル89巻:405ページ、1969年。
【0118】
6.G.A.プリンス,V.G.へミング,R.L.ホースウッド他、ウイルス研究3巻:193ページ、1985年。
【0119】
7.V.G.へミング,G.A.プリンス,R.L.ホースウッド他、感染疾病ジャーナル152巻:1083ページ、1985年。
【0120】
8.P.F.ライト,R.B.ベルシュ他、感染と免疫、37巻:397ページ、1982年。
【0121】
9.D.A.コンラッド,J.C.クリステンソン他、小児科感染疾病ジャーナル、6巻:152ページ、1987年。
【0122】
10.A.F.ロビューリョ,R.L.ホイーラー,J.トラン他、全米科学アカデミー紀要、86巻:4220ページ、1989年。
【0123】
11.Z.ステップレフスキー,L.K.サン,C.W.シアマン他、全米科学アカデミー紀要、85巻:4852ページ、1988年。
【0124】
12.G.L.ブーリアンヌ,N.穂積,M.J.シュルマン、ネイチャー、312巻:643ページ、1984年。
【0125】
13.L.K.サン,P.カーチス,E.ラコヴィッツ−シュルチンスカ他、全米科学アカデミー紀要、84巻:214ページ、1987年。
【0126】
14.A.Y.リュ,P.W.マック,C.I.チャンピオン,R.P.ロビンソン、遺伝子、54巻:33ページ、1987年。
【0127】
15.S.L.モリソン,M.J.ジョンソン,L.A.ハーセンバー,V.T.オイ、全米科学アカデミー紀要、81巻:6851ページ、1984年。
【0128】
16.S.L.モリソン、サイエンス、229巻:1202ページ、1985年。
【0129】
17.B.G.サヘーガン,H.ドーライ,J.ザルツガーバー−ミュラー他、免疫学ジャーナル、137巻:1066ページ、1986年。
【0130】
18.S.武田,T.内藤,K.浜,T.野間,T.本城、ネイチャー、314巻:452ページ、1985年。
【0131】
19.D.A.カーソン,B.D.フライマーク、先端免疫学、38巻:275ページ、1986年。
【0132】
20.T.A.ビーラー他、ウイルス学ジャーナル、63巻:2941−2950ページ、1989年。
【0133】
21.コーリン他、ウイルス学、143巻:569−582ページ、1989年。
【0134】
22.アンダースン他、酵素免疫検定に基づく呼吸合胞体ウイルスの微小中和検定、臨床微生物学ジャーナル(1985年)22巻:1050−1052ページ。
【0135】
23.ジョンソン他、呼吸合胞体ウイルスに対し有力な試験管内生体内活性を持つヒト化モノクローナル抗体(MEDI−493)の開発、感染性疾病ジャーナル(1997年)、176巻:1215−1224ページ。
【技術分野】
【0001】
この出願は2000年1月27日に受理された合衆国暫定特許出願番号第60/178,426号の優先権を主張し、その開示はここでその全体を引用例によりここに組み込んでいる。
【0002】
本発明は新規な超高親和性中和抗体に関する。
【0003】
微生物の広がりを防止するのに妨げとなりまたはそれが困難であることにより生じる現在の感染発生数は、これらの生体を防止し、同じく既に感染した者を処置するためにより新しいアプローチの必要性を産み出した。
【背景技術】
【0004】
防止しまた処置するのにより困難な感染性外的病原因子にウイルスがある。例えば、呼吸合胞体ウイルス(RSV)は病院に収容される若い子供の急性呼吸系疾患の主な原因であり、また若い子供の下部気道感染の主な原因となる。RSVなどのような病原因子に対する有効なワクチンを産生することへの主要な障害はこれまでずっと安全性の問題であった。これに対して、このようなウイルス因子に対する免疫グロブリンの使用は何らかの価値があることを立証した。例えば高力価RSV免疫グロブリンは動物モデルでRSV感染の予防と治療の両方で有効であることが研究により示されてきた。
【0005】
別のアプローチは高い特異的中和活性を持つ抗体、とりわけモノクローナル抗体を中和する抗体の開発であった。このルートへの一つの欠点はマウスまたはラットのそれよりもむしろヒト抗体を産生し、かくしてヒト抗マウスまたはヒト抗ラット抗体応答の展開を最小にする必要性が存在したことであり、それは潜在的に更なる免疫異常を生じさせることである。
【0006】
も一つの選択的なアプローチはこれまでヒト−マウスキメラ抗体の産生であり、ここでマウス重鎖および軽鎖可変部をコードする遺伝子はキメラまたはハイブリッド抗体を産生するためにヒト重鎖および軽鎖可変部の遺伝子に結合された。
【0007】
ある場合には、マウスCDRはヒト定常部および枠組み構造領域に移植され、いくつかのマウス枠組み構造アミノ酸は「ヒト化」抗体を提供するために対応する位置のヒトアミノ酸と置換された〔クイーン、合衆国特許番号第5,693,761号および5,693,762号〕。しかしこのような抗体は無傷マウスCDR領域を含有し、混合有効性に出会い、その親和性は107乃至108M−1以下になることが多かった。
【0008】
よい親和性を持つヒト化抗RSV抗体がこれまでに調製されており、現在市販されている〔ジョンソン、合衆国特許番号第5,824,307号を参照されたい〕。
【0009】
超高親和性抗体の産生は、このような抗体の中和能力、および臨床効果の望ましい度合を達成するために抗体の産生を少なくする必要性、またそれによる産生費用の削減と同じ量での投与の臨床効果の増大というより実際の見地の両方の点から望ましいものとなろう。
【先行技術文献】
【特許文献】
【0010】
【特許文献1】米国特許番号第5,693,761号
【特許文献2】米国特許番号第5,693,762号
【特許文献3】米国特許番号第5,824,307号
【発明の概要】
【課題を解決するための手段】
【0011】
本発明は少なくとも1010M−1、また更に1011M−1の親和性定数を示す高親和性中和抗体およびその活性断片に関し、またより特異的には抗体およびまたは断片がヒト定常部を持つ抗体およびまたはその活性断片を含む中和モノクローナル免疫グロブリンに関する。
【0012】
本発明はヒト−抗マウス抗体反応(HAMA)を生じる無傷マウスCDR領域の存在なしで、また全産生の費用を削減する十分高い親和性中和活性と全産生の効率化を有する高親和性中和抗体を提供することにより前記の問題を解決する。
【0013】
本発明の一つの見地は、少なくとも1010M−1、および更には1011M−1の親和性定数を持ち、またウイルス発現タンパク質で示されるものなどのような特異的抗原決定基に向かう特異性を持つ高親和性中和抗体に関する。
【0014】
本発明の一つの目的は、抗原が哺乳類、とりわけヒトにおけるウイルス感染細胞により発現されるような場合に、ウイルスにより産生される抗原に向かう特異性を持つような高親和性中和抗体を提供することである。
【0015】
このような一つの実施例において、本発明の抗体およびまたはその活性断片、ならびにその活性断片[原文通り]は、呼吸合胞体ウイルス(RSV)により発現されまた更にRSVに感染した細胞表面(細胞表面でのその抗原の存在は細胞の合胞体への融合を生じせしめる)で発現される呼吸合胞体ウイルス、もっとも特別にはF抗原に対して特異的である。
【0016】
かくして一つの実施例において、本発明の抗体およびまたはその活性断片を含む高親和性中和免疫グロブリンは、その軽鎖可変部が(図1Aで示される)配列識別番号1の配列を持ち、その重鎖可変部が(図1Bで示される)配列識別番号2を持つ抗体と同じRSVのエピトープに結合する。
【0017】
本発明の目的は、(抗RSV抗体構造である免疫グロブリンのものと同じ特異性を持つ)図1で開示された免疫グロブリンの枠組み構造領域を実質的に持つ超高親和性中和抗体を提供することであるが、ここで本発明の抗体およびその断片を含む免疫グロブリンは1個またはそれ以上のCDR(相補性決定領域)を含有し、そのアミノ酸配列はある場合には1個のアミノ酸のみが異なり前記CDR領域の1個のみの差異に限定される場合もあり得るけれども、そのアミノ酸は所謂基準抗体のものとは独立したものである。
【0018】
本発明の望ましい実施例において、本発明の新規な免疫グロブリンは1個またはそれ以上のCDRの配列のみにおいて、図1の抗体(以下「基本抗体」または「基準抗体」もしくは「基準免疫グロブリン」と称する)とは異なり、またもっとも望ましい実施例においては、これらの差異はCDR L2、L3,H1およびH3でのみ生じる。
【0019】
本発明のとりわけ望ましい実施例は、図1で示され、従って図3,4,5,6および7で示された重鎖および軽鎖可変配列を持つ枠組み構造配列を持つ。
【0020】
一つの実施例において、本発明の高親和性中和抗体はヒト定常部を含む。
【0021】
望ましい実施例において、少なくとも1010M−1以上、また1011M−1での親和性定数(Ka)を持つ活性断片を含む本発明の高親和性RSV中和抗体は抗体またはその活性断片などのような組換え免疫グロブリンであり、それは重鎖および軽鎖のヒト定常部と枠組み構造領域を含み、ここで枠組み構造領域の少なくとも一部はヒト抗体から(またはヒト抗体枠組み構造のコンセンサスから)誘導され、前記枠組み構造領域の例は図1の抗体配列で示される。
【0022】
一つの実施例において、すべての枠組み構造領域はヒト抗体(またはヒトコンセンサス配列)から誘導される。
【0023】
も一つの高度に特異的な実施例において、少なくとも1010M−1の親和性(定数)を持つ高親和性RSV中和抗体は、非ヒト抗体から誘導されるヒト定常部、1個またはそれ以上のCDRを持つ組換え抗体であり、ここでCDRの少なくとも1個のアミノ酸は変更され、またここで枠組み構造のすべてもしくは一部はヒト抗体(あるいはヒト抗体枠組み構造のコンセンサス配列)から誘導される。
【0024】
別の実施例において、その可変鎖が図1で示され、また少なくとも1011M−1の親和性を持つ基本または基準抗体もしくは免疫グロブリンと同じエピトープに結合するヒト化中和免疫グロブリンは、少なくとも1個の下記のアミノ酸をCDRの下記の位置で、すなわちCDR H1の位置2でアラニン、CDR H3の位置で位置6でフェニルアラニン、CDR L2の位置3でフェニルアラニン、またCDR L3の位置5でフェニルアラニンを含む。他の実施例はそれぞれの位置で他の単一アミノ酸置換を含む。
【0025】
本発明のも一つの目的はここで開示された免疫グロブリンを含む組成物を提供することであり、ここで前記枠組み構造物は薬理許容希釈剤または賦形剤に懸濁される。
【0026】
本発明の更なる目的は呼吸合胞体ウイルスを予防しおよびまたは処置する方法を提供することであり、それは前記抗体またはその断片が本発明の免疫グロブリンに対しここで開示された特異性および親和性の諸性質を示すような場合に本発明の免疫グロブリンを含有する組成物の治療有効量のそれの危険にさらされ、あるいはそれに悩む患者への投与を含むものである。
【図面の簡単な説明】
【0027】
【図1】抗RSV抗体の軽鎖および重鎖可変部のアミノ酸配列を示す図。ここでCDR領域はアンダーラインされ、一方非アンダーライン残基は各鎖の可変部の枠組み構造領域を形成する。この抗体では、CDRはマウス抗RSV抗体から誘導され、一方枠組み構造領域は主としてヒト抗体から誘導された配列より成る。各CDRにとってアミノ酸置換がここで開示された高親和性CDRおよび抗体を達成するために使用された位置は太字で示されている。ここでの開示に従って、このような置換はCDR L2、L3、H1およびH3のみにあった。図1Aは、軽鎖および重鎖それぞれの軽鎖可変部を示し、図1Bは重鎖可変部を示す。定常部配列は示されていない。これらの配列は本開示を通じてIX−493と名付けられた基本クローン(表2参照)に存在する(すなわちSWSG−これは高親和性CDR H1の主要位置(表1および表3参照)にあるセリン(S)、高親和性CDR H3の主要位置にあるトリプトファン(W)、高親和性CDR L2の主要位置にあるセリン(S)、および高親和性CDR L3の主要位置にあるグリシン(G)、のSWSGを意味する)。この開示の目的のために、これは「基準抗体」である。
【図2】受益または高親和性クローンの特定の組の親和性の比較を示す図。クローン名称は図面の右の記号一覧の左側に記号一覧の右に示されたCDR H1、H3、L2およびL3での指示された置換と共に存在する。測定法は左軸に示された560nmでのO.D.エリザによる。クローンL1FRは図1の基準抗体の結果を表す。
【図3】ここで開示された発明のクローン1(表2)の望ましい実施例の重鎖および軽鎖可変部を示す図。CDR領域はアンダーラインされ、一方図1の抗体に対するアミノ酸の差は太字で示される。かくしてこの望ましい(すなわち高親和性の)抗体は存在するいくつかの高親和性CDR(表3)を持ち、それは基本または基準抗体より高い親和性(1010M−1以上)生じさせる。
【図4】ここで開示された発明のクローン2(表2)の望ましい実施例の重鎖および軽鎖可変部を示す図。CDR領域はアンダーラインされ、一方図1の抗体に対するアミノ酸の差は太字で示される。かくしてこの望ましい(すなわち高親和性の)抗体は存在するいくつかの高親和性CDR(表3)を持ち、それは基本または基準抗体より高い親和性(1010M−1以上)を生じさせる。
【図5】ここで開示された発明のクローン3(表2)の望ましい実施例の重鎖および軽鎖可変部を示す図。CDR領域はアンダーラインされ、一方図1の抗体に対するアミノ酸の差は太字で示される。かくしてこの望ましい(すなわち高親和性の)抗体は存在するいくつかの高親和性CDR(表3)を持ち、それは基本または基準抗体より高い親和性(1010M−1以上)を生じさせる。
【図6】ここで開示された発明のクローン22(表4)のもっとも望ましい実施例の重鎖および軽鎖可変部を示す図。CDR領域はアンダーラインされ、一方図1の抗体に対するアミノ酸の変化は太字で示される。かくしてこのもっとも望ましい(すなわち最高親和性)抗体は存在するいくつかの高親和性CDR(表3)を持ち、それは基本または基準抗体よりも高い親和性(1011M−1以上)を生じさせる。
【図7】ここで開示された発明のクローン23(表4)の望ましい実施例の重鎖および軽鎖可変部を示す図。CDR領域はアンダーラインされ、一方図1の抗体に対するアミノ酸の変化は太字で示される。かくしてこの望ましい(すなわち最高親和性)抗体は存在するいくつかの高親和性CDR(表3)を持ち、それは基本または基準抗体よりも高い親和性(1011M−1以上)を生じさせる。
【図8】ここで開示された超高親和性抗体クローンのいくつかについての微小中和検定の結果を示す図。高親和性相補性決定領域の主要位置に存在するアミノ酸(表3参照)は(クローンが単に番号付けられるが表がその表に開示されこの図の右側の記号一覧にあるクリティカル位置で使用された実際のアミノ酸に依存してこの図と比較する表2で示されるように)H1、H3、L2およびL3の順に右側に示される。かくしてクローン4D2−7では、高親和性CDR H1の主要位置にアラニン(A)、高親和性CDR H3の主要位置にフェニルアラニン(F)、高親和性CDR L2の主要位置にフェニルアラニン(F)、また高親和性CDR L3の主要位置にフェニルアラニン(F)がある。要約すると、約25,000のHEp−2細胞がRSVおよび一定濃度の抗体と共に96ウエル平板のウエルそれぞれに加えられた(ウエル当りの抗体濃度は横座標に示される−詳細については実施例2を参照のこと)。5日成長の後、細胞は固定され、ビオチン化抗F MAbで処置され、次いでアビジン−ペルオキシダーゼ複合体に結合され、チオニトロベンゼン酸と反応する結合ペルオキシダーゼの能力が450nmのO.D.を測定することで決定された。存在するFタンパク質の量はウイルス複製の範囲の指標であったし、それによりペルオキシダーゼでの基質のより多くの反応と吸収の増加を生成した。かくしてOD450の値が低ければそれだけ、一定の抗体の指示濃度の中和能力がより大きくなる。ここでIX−493(SWSG−高親和性CDR H1の主要位置でセリン(S)、高親和性CDR H3の主要位置でトリプトファン(W)、高親和性CDR L2の主要位置でセリン(S)、および高親和性CDR L3の主要位置でグリシン(G)を意味する)は(更にL1FRおよびIX493L1FRと表示された)「基準抗体」である。
【図9】ここで開示されFab断片が抗体複製の中和化のために採用されたいくつかの抗体の微小中和検定の結果を示す図。ここでIX−493(SWSG)は基準Fab断片であり抗体Medi−493から誘導される(ジョンソン他(1997年)−引用文献23参照)。
【図10】同じ抗体のFab断片への類似の実験と比較したRSVに特異的な抗体の微小中和(検定)の結果を示す図。ここでMedi−493は抗体を表し、一方IX−493L1FRは本抗体のFab断片を表す。他の系は本発明に基づき産生されたいくつかの抗体のFab断片のためのものである(内部同定のための与えられた各種文字デジタルコード表記、但し中和抗体としてのその相対的有効性には何ら関与しないもの)。
【発明を実施するための形態】
【0028】
「抗原」という用語は、抗体が親和性と特異性を持つウイルスなどのような微生物の表面に存在する構造物、しばしばポリペプチドまたはタンパク質などの構造物を引用する。
【0029】
「抗原決定基」という用語は、抗原などのような免疫グロブリンが特異性と親和性を持つ抗原構造に対する特異的な結合部位を引用する。かくしてウイルスなどの粒子は抗原を示すが、ウイルスが数多くの個別の異なる表面タンパク質を持ち、それぞれが免疫グロブリンに対する別個の潜在的結合部位を表するような場合に数多くの個別の異なる抗原部位を持つであろう。
【0030】
「免疫グロブリン」という用語は、抗原または抗原決定基に対する特異性または親和性を持つタンパク質またはポリペプチドを引用する。この用語は抗原または抗原決定基に対して特異性と親和性を持つ断片などのような一般に四量体として示される抗体ならびにその活性断片を含む。かくしてここで使用される「免疫グロブリン」は抗体およびそのすべての活性断片を含む。
【0031】
「抗体」という用語は、通常は微生物、とりわけウイルスの表面に見出されるもので抗原決定基に親和性を持つタンパク質またはポリペプチドを引用する。このような抗体は一般に4鎖よりなり、かくして四量体である。
【0032】
「中和免疫グロブリン」または「中和抗体」という用語は、生体内または細胞培養で微生物、とりわけウイルスを減少させるための本発明の抗体を含む免疫グロブリンの能力を引用する。このような能力を指示するものとしては、以下で開示される微小中和検定からのデータが存在する。このような構造は通常可変部と定常部を有し、これにより可変部は大抵抗体の特異性を決定することに責任がありそれは相補性決定領域(CDR)を含むであろう。
【0033】
「相補性決定領域」すなわち「CDR」という用語は、H(重)鎖またはL(軽)鎖のいずれかの可変部が抗原標的に特異的に結合できるアミノ酸配列を含む可変部を引用する。これらのCDR領域は特殊な抗原決定基構造のための抗体の基本的特異性を説明する。これらの領域はまた「超可変部」としても引用される。
【0034】
「活性断片」という用語は、それ自身で抗原決定基に対して高親和性を持ち、またそのような特異性の原因となる1個またはそれ以上のCDRを含む。限定されない例はFab、F(ab)’2、重−軽鎖二量体、および完全軽鎖または完全重鎖などのような一本鎖構造を含む。
【0035】
「特異性」という用語は、異なる抗原部位に対して一つの抗原部位に優先的に結合する抗体の能力を引用し、必ずしも高親和性を意味しない。
【0036】
「親和性」という用語は、その結合により形成される複合体の存在に向けて抗原と抗体の平衡状態に移行するように抗体が抗原に結合する度合を引用する。かくして抗原と抗体が相補的に同等の濃度で結合される場合には、高親和性の抗体は生成複合体の高い濃度に向けて平衡状態に移行するように利用できる抗原と結合するであろう。
【0037】
「親和性定数」という用語は、抗原に対する抗体の親和性を測定するために使用される会合定数を引用する。親和性定数が高ければそれだけ抗原または抗原決定基に対する免疫グロブリンの親和性は大きくなり、その逆も真である。親和性定数は相互モル濃度の単位で結合定数である。このような定数は抗体反応の標準動力学方法により測定されるように会合−解離反応のための速度定数から容易に計算される。
【0038】
本発明は少なくとも1010M−1の親和性定数を持つ超高親和性中和抗体およびその活性断片に向けられる。これら抗体の活性断片は、少なくとも1個の高親和性相補性決定領域(CDR)を含有する断片である。
【0039】
分子生物学と組換え技術に関する方法の到来と共に、組換え手段により抗体分子を産生し、これにより抗体のポリペプチド構造で見出される特異的アミノ酸配列をコードする遺伝子配列を生成することが今や可能となっている。このような抗体は、前記抗体のポリペプチド鎖をコード化する遺伝子配列をクローニングするか、もしくは特異的なエピトープと抗原決定基に親和性を持つ、活性四量体(H2L2)構造を形成するために合成鎖の試験管アセンブリで前記ポリペプチド鎖の直接合成により産生することができる。これは異なった種および源から中和抗体の特徴的な配列を持つ抗体の容易な産生を可能にする。
【0040】
バイオリアリターまたはプロセスのいずれの段階でも生体を何ら使用することなく直接化学合成により実験室または商業的規模での大型細胞培養を使用して試験管内または生体内でウシ、ヤギ、ヒツジなどの遺伝子導入動物を用いて免疫グロブリンの源に関係なく、またはそれらがどのように組換え技術により構築されたかまたはどのように合成されたかも関係なく、すべての免疫グロブリンは類似の全体として3次元構造を持つ。抗体の場合、この構造はH2L2として与えられ、また抗体が一般に2個の軽(L)アミノ酸鎖と2個の重(H)アミノ酸鎖を含むという事実を引用する。両方の鎖は構造的に相補性の抗原標的で相互作用できる領域を持つ。標的と相互作用する領域は「可変」または「V」部として引用され、異なる抗原特異性の抗体からアミノ酸配列での差により特徴付けられる。
【0041】
HまたはL鎖の可変部は抗原標的に特異的に結合できるアミノ酸配列を含有する。これらの配列内には、特性の異なる抗体と活性断片の間の極端な可変性の故で「超可変」と名付けられたより小さな配列が存在する。このような超可変部は更に「相補性決定領域」または「CDR」領域として引用される。これらのCDR領域は特定の抗原決定基構造に対する抗体の基本的特異性の原因となる。
【0042】
CDRは可変部内でのアミノ酸の隣接していない広がりを表すが、種とは拘りなく可変重鎖部および軽鎖部内の位置は可変鎖のアミノ酸配列内の類似の位置を持つものとして発現されている。すべての正統な抗体それぞれの可変重鎖および軽鎖は3CDR領域を持ち、そのそれぞれは各軽(L)鎖および重(H)鎖に対して(L1、L2、L3、H1、H2、H3)と名付けられた他のものに隣接していない。認められたCDR領域はキャバット他、ジャーナル・オブ・バイオロジカル・ケミストリー、252:6609−6616(1977)に記載されている。ナンバリングスキームは各図で示されており、ここでCDRはアンダーラインを付されたナンバーはキャバットのスキームに従っている。
【0043】
すべての哺乳類種では、抗体ペプチドは定常(すなわち高度に保存された)部と可変部を含み、また可変部内では、CDRが存在し、またCDRの外側ではあるが重鎖および軽鎖の可変部内にアミノ酸配列で構成された所謂「枠組み構造領域」が存在する。
【0044】
本発明に基づき開示された免疫グロブリンは、ウイルス表面およびウイルスで感染された細胞表面で発現されたタンパク質などのような高分子とりわけタンパク質およびもっとも特異的にはウイルスおよび他の微生物で発現されたタンパク質で見出されるエピトープ、もしくは抗原決定基に対し、(抗体およびリガンドとしての抗原の結合を示す会合定数として定義される親和性定数、すなわちKaに対し1010M−1のオーダー、更には1011M−1のオーダーで)きわめて高い親和性を提供する。本発明の高親和性抗体は中和抗体であり、かくして生体および細胞培養物でのウイルスの複製を減少し、一方逆免疫病変を予防するために受容体の抗体アミノ酸配列に対する十分な相同性を維持する。後者の特性は哺乳類および特にヒトなどのように受容生体のそれに類似する定常部の使用を通じて達成される。この特性は更に受容生体から抗体に見出されるものと同一ではないにしても類似した枠組み構造アミノ酸配列の使用を通じて達成される。後者の場合、あるアミノ酸置換は、本発明の新規なCDRと前記抗体が特異性を示す抗体との間の高親和性相互作用を促進し維持するように枠組み構造配列内で行われる。
【0045】
ここで使用されるように、「抗体」および「活性断片」または「断片」という用語は本発明の全範囲を決定する際に限定するものと見做されるべきではない。かくして「活性断片」または「免疫グロブリン」よりも抗体という用語が使用されるという事実は、特異性と親和性の必要な性質が前記構造により提示される限りにおいて、本発明またはその使用を制限するものとしてとられるものではない。
【0046】
ここで開示された発明に従って、本発明の高親和性中和抗体およびその活性断片の親和性を特徴付ける親和性定数は会合定数であり、KassocまたはKonと名付けられた複合体を形成する会合の速度定数とKdissまたはKoffと名付けられた解離定数で抗原抗体複合体系性の動態により測定された。このような定数の測定は通常の技術の範囲内にありその詳細は本発明を更に記載するためにここで与えられた実施例で引用されるように適当であれば一般的な方法論と特異的な条件を除いてここでは更に記載しないことにする。
【0047】
本発明の高親和性抗体は、前に記載の通り適切なヌクレオチド配列を配列しそれらを適切な細胞系で発現することにより適切な抗体遺伝子配列、すなわちアミノ酸配列を遺伝子操作することで達成される。何らかの望ましいヌクレオチド配列は、例えば合衆国特許番号第5,264,563号に記載されたように、突然変異誘発に基づくコドンの方法を用いて産生することができる。このような手順はオリゴヌクレオチド内で、いずれか望ましいコドン位置でのアミノ酸残基のいずれかおよびすべての頻度の産生を可能にする。これは望ましい位置で、またはこれらのいずれかの特異的なサブセット内で20アミノ酸のいずれかの完全に無作為の置換を含むことを可能にする。選択肢として、このプロセスは本発明に基づく新規なCDR配列などのように、特定のアミノ酸にアミノ酸鎖内で望ましい位置を達成するように実施することができる。要約すると、望ましいいずれかのアミノ酸配列を発現する適切なヌクレオチド配列は、本発明の新規なCDR配列を再生できる。このような手順を用いて容易に達成することができる。これはいずれか望ましいアミノ酸配列を持つ抗体などのポリペプチドを合成する能力をもたらす。例えば、選択された抗体のいずれか望ましいドメインのアミノ酸配列を決定し、また選択肢としてある範囲の置換類似体を与えるために、他の望ましいアミノ酸により置換された1個またはそれ以上のアミノ酸を持つ相同鎖を調製することを可能にする。
【0048】
このような方法を用いる際に、遺伝暗号の縮重に起因して、無作為オリゴヌクレオチド合成および部分縮重オリゴヌクレオチド合成などの方法が特定の位置で特定のアミノ酸残基を特定する遺伝暗号の冗長を取り込むであろうということは理解されねばならない。もっともこのような方法はすべてのアミノ酸配列のマスターセットを提供し、また抗体構造の最適機能または他の目的を選別するために使用することができる。このような方法はスワーラ他、全米科学アカデミー紀要、87巻、6378−6382ページ(1990年)およびデブリン他、サイエンス、249巻、404−406ページ(1990年)に記載されている。
【0049】
ここで開示された発明に従って、そのそれぞれが個別に強化結合抗体を来たす本発明に基づく1個、2個またはそれ以上の新規なCDR配列を単一ポリペプチド構造に組合わせることにより強化抗体変異体を生成することができる。このようにして同一または異なるCDRで本発明内で高親和性抗体を産生するために、いくつかの新規なアミノ酸配列を1個の抗体に結合することができる。例えば3個のこのような新規のCDR配列が採用されて、F抗原またはRSVなどのような特定の抗原構造が選別される。全体の結果はかくして順次的なやり方で各種の単一アミノ酸置換を組合せ、抗原親和性の生成結果を選別する反復過程となるであろう。このような方法は、本発明内で実施されたいくつかの抗体を調製するのに使用された。このような方法は更に本発明の抗体配列、とりわけ前記抗体のCDR領域の配列を最適化する冗長ではあっても便利な手段を表している。
【0050】
本発明の内で新規な配列と方法を用いて、このようなアプローチは最大効率で抗体を発見する仕事において、すべての可能な抗体構造の順列と組合せを生成し選別する時間と費用を回避する。これとは逆に単一10アミノ酸残基CDRの完全無作為抽出は、事実上選別の不可能な数字である10兆個以上の変異体を生成するであろう。
【0051】
この反復方法はより高親和性を持つ抗体の探究範囲を限定するために順次的過程で二種および三種のアミノ酸置換を生成するのに使用できる。
【0052】
逆に言えば、異なる抗体ドメインの配列内で必ずしもすべての位置が同じでないことは認識されねばならない。特定の位置でのいずれの種類の置換も有用であるかまたは有害である。更にある位置でのある種のアミノ酸の置換は同様に特定の抗原への親和性に影響を与えてプラスまたはマイナスのものとなる。例えば、一定の位置ですべての可能な疎水性アミノ酸を試す必要はない。いずれの疎水性アミノ酸でも同じように作用するからである。選択肢として与えられた位置での酸性または塩基性アミノ酸は測定された親和性で大きなぶれを提供する。従ってそのような置換を行う「ルール」を更に学ぶ必要があるが、しかしそのような「ルール」の決定はすべての可能な組合せと置換の研究を必要とせず−最大数の置換より少ない検討の後に傾向は明らかとなるであろう。
【0053】
前記に従って、本発明の抗体は抗RSV抗体などの超高親和性中和抗体であり、後者の場合には合衆国特許番号5,824,307号の抗体としてRSVの同じエピトープに向かう特異性を持つもっとも望ましい抗体である。
【0054】
加えて本発明の超高親和性抗体の親和性は典型的には少なくとも1010M−1である。このような高親和性はここに記載された手順による以外はた易くは測定されないが、このような値は一般に範囲の一部と見做され、また例えば1010M−1の2倍以内、あるいは1010M−1以上もしくは1010M−1に数値的に等しいこともある。この場合、親和性は親和性定数で表示され、これは相互モル濃度の単位を与えるように結合定数の状態にある。このように抗原に対する抗体の親和性はこの定数の値に比例(定数が高ければ親和性が高い)している。このような定数は抗体反応に対する標準動態方法により測定されるように、会合−解離反応への速度定数から容易に算出することができる。
【0055】
特異的実施例において、本発明の高親和性中和抗体およびその活性断片は、少なくとも1011M−1、ある場合には測定可能なもっとも高い範囲であるこの値を越えたそれぞれの抗原への親和性定数を持つ。
【0056】
本発明の高親和性中和抗体およびその活性断片は、高分子のいずれかの型に存在するエピトープとりわけタンパク質およびポリペプチド分子などの三次元構造の一部として存在するペプチドエピトープなどのようないずれかの望ましい抗原決定基に向けられる。このようなペプチドエピトープは一般に微生物および例えば癌細胞などの細胞の表面に存在し、またはその構造の一部となって存在する。このようなペプチドエピトープ発現する微生物は細菌およびウイルスを含む。ウイルスの場合には、ペプチドエピトープを発現するタンパク質およびポリペプチドはウイルス表面に発現された分子であり、またはウイルスで感染した細胞により発現される。本発明を従って開示された規則と方法の効率を評価する目的のために、1個のウイルスが本発明内で抗体を発展させる利用できる抗原源として選択された。更なる分析のために選ばれたこのウイルスは呼吸合胞体ウイルス(RSV)であった。このウイルスが選ばれたのはその複製サイクルに関連し、またその表面で見出される抗原決定基に関連してそれが十分に特徴付けられているからである。更にこれらのウイルスで感染した細胞により発現されることで知られる抗原も十分に特徴付けられている。かくしてこのウイルスはいずれもタンパク質である表面G抗原および表面F抗原の両方を発現する。G抗原はウイルスの細胞表面への結合を促進し、またFタンパク質はウイルスの細胞との融合を促進する。このように感染した細胞はその表面でF抗原を発現し、その結果ウイルスの名称となった合胞体を形成するために細胞の融合を誘導する。更にこのウイルスにより容易に感染する細胞系が十分に周知であり十分に特徴付けられているので、それを容易に試験管内で成長させ培養することができるためにこのウイルスは使いやすい対象である。更にまたこのウイルスの処置に利用できる抗体は周知であり商業的に利用可能である(例えば合衆国特許番号第5,824,307号)。これらの理由でその特許の開示が全体で引用例としてここに取り込まれている前記特許の抗体は図1で開示される「基準抗体」のアミノ酸配列を含有し、そのCDR配列は表1で要約される。かくして商業的に成功した抗体産物の利用可能性およびRSVの十分に特徴付けられた性質が、このような抗体を構築するのに使用される高親和性CDRを産生するためにここで開示された本発明と規則の抗体を試験し最適化して使用するのにこれを理想的な組合せとした。これとは別に、ここで開示された抗体、同じくこのような抗体を調製するここで開示された方法は抗原としてのRSVを限定するものではなく、抗体技術の分野で一般に容易にかつ有利に適用される。更に加えて本開示で提供され、RSVで発現される抗原決定基、およびRSVで感染した細胞に特異的に向けられる特異的実施例はまさに他のエピトープ、とりわけ関連ウイルスに存在するエピトープに高親和性を持つことができる。従って本発明に従って開示されるように、RSV、および図1で示される可変配列を持つ抗体は、本発明で教示される抗体技術の方法を展開し適用する基準点として使用される便利なモデルシステムに過ぎない。
【0057】
活性断片を含む、本発明に基づく高親和性中和抗体は、少なくとも1個の高親和性相補性決定領域(CDR)を含み、ここで前記CDRは少なくとも1010M−1の親和性定数(Ka)を持つ抗体を生じるように選択されたアミノ酸配列を持つ。望ましい実施例では、このような抗体または活性断片は少なくとも2個の高親和性CDR、または少なくとも3個の高親和性CDRもしくは更に少なくとも4個の高親和性CDRを含む。とりわけ望ましい実施例においては、このような抗体または活性断片は3乃至4個の高親和性CDRを含む。一つの望ましい実施例では、このような活性断片はFab断片である。
【0058】
本発明の高親和性抗体は一般に哺乳類、望ましくはヒト定常部および可変部を含み、前記可変部は重鎖および軽鎖枠組み構造領域と重鎖および軽鎖CDRを含み、ここで重鎖および軽鎖枠組み構造領域は哺乳類抗体、望ましくはヒト抗体から誘導され、また、ここでCDRはヒト以外の種、望ましくはマウスなどの種の抗体が誘導される。枠組み構造アミノ酸はまた非ヒトから誘導され、後者は望ましくはマウスである。
【0059】
更に本発明の高親和性抗体はCDRが誘導される抗体と同じエピトープを結合し、ここで前記超高親和性抗体の少なくとも1個のCDRはアミノ酸置換基を含有し、またここで前記置換基は非同一アミノ酸、望ましくは枠組み構造および定常ドメインに寄与するヒト抗体のCDR領域の対応配列位置のアミノ酸により、CDR領域の1個またはそれ以上のアミノ酸の置換を含む。
【0060】
高親和性CDRはそのような高親和性CDRを産生する非高親和性CDRでのアミノ酸置換基により産生することができ、または高親和性CDRはこのような高親和性CDRを形成するために直接合成することができる。かくして本発明の超高親和性中和抗体はアミノ酸置換基をCDR領域の1個のみで、望ましくは1個以上のCDR領域で、もっとも望ましくは3個乃至4個までのその領域で、多分5個または6個すべてのCDRで少なくとも1個の置換基アミノ酸を含むCDRでアミノ酸を持つであろう。
【0061】
本発明の抗体を産生するためにここで開示された方法を適用する際に、抗体を調製する方法は制限因子にはならない。かくして本発明の高親和性中和抗体は抗体をコードするポリヌクレオチド配列を生成し、前記ポリヌクレオチド配列を、そのようなペプチドを発現できるだけでなく、その時点で多分細胞により、培地に分泌された細胞からまたは細胞培養物から回収される特徴付けられた四量体抗体構造に組立てすることのできる許容細胞に挿入することにより調製することができる。このような製造手順の技術は公知のものであり、特許も与えられており、本発明を実施するために必須のものではない〔モリソン他、合衆国特許第5,807,715号〕。更に本発明の抗体のポリペプチド鎖は酵素の存在または不在の下で化学的に合成することができ、次いで通常のH2L2立体配置の四量体構造を形成するために化学結合される。かくしてここで開示された抗体を調製するいずれの方法も利用可能である。
【0062】
本発明は高親和性CDR配列を顕著に持つ結果として高親和性を持つ先に列挙された性質のある高親和性中和抗体に向けられる。本発明に従って、ここで開示された抗体のCDR配列は抗体分子に本発明の抗体に特徴的な超高親和性を与えるように最適化されている。ここで開示された枠組み構造配列、とりわけ図1、3、4、5、6および7の配列により教示されるもの、およびもっとも特別には治療用に使用された時に本発明の抗体の受容体として作用する生体の抗体に特徴的な宣常部配列と共に使用された配列と合わせてこのようなCDR配列は、そのより特異的な実施例で本発明の抗体を産生する。しかし本発明の方法はより特異的にCDRアミノ酸配列に向けられる。
【0063】
本発明、例えば高親和性中和抗体内で抗体およびまたはその活性断片などの免疫グロブリンを産生するために、本発明で教示されるルールは本発明に基づき開示される高親和性CDR配列の配列をその構造が組み込む抗体分子を産生するために有利に使用される。かくして本発明の高親和性中和抗体は、それらがいずれかのクローニングでも産生されないものであるために、本質的にはその用語が一般に使用されているような「モノクローナル」抗体では決してない。前に記載の通り、本発明の抗体の配列は直接合成されるために、いずれかの抗体配列と同一のものではなく、とりわけ現在周知のいずれのCDR配列とは同一のものではない。配列それ自身は完全に初めから新規のものであり、いずれかの生体により産生されるいずれかの抗体で完全に表すことはできない。かくしてここで開示される高親和性CDR配列はここで開示されるように最適化により見出され、または達成され、次いで一度前記高親和性CDR配列が公知であると、科学で公知のすべての手段により二量体または四量体が二官能基または単官能基であるかに拘りなく、完全に機能的抗体分子を合成するのに使用することができる。
【0064】
前記の内容を続け、またその最適化を含む本発明に基づき開示された配列をより良く記載するために、基準抗体(ここで合衆国特許番号第5,824,307号の抗RSV抗体)の軽鎖および重鎖可変部の配列が図1A(軽鎖可変部−配列識別番号1)で示され、また図1B(重鎖可変部−配列識別番号2)で示される。更に本発明に従って、新規な配列が基準抗体に比べてCDR領域のみでアミノ酸配列の差を有して産生された。この結果を達成するのに利用される一つの手段は、所謂出発または基準鎖のCDR領域に突然変異を導入し、次いで抗原(RSV Fタンパク質)親和性を得るために生成する組換えクローンを検定することであった。
【0065】
前記の内容に従って、第1の1個のCDR配列でその配列を最適化し「クリティカル」残基を決定するために変更が行われ、または前記位置よりも残基がその位置のみに限定された一連のアミノ酸置換を通じて最適化された。抗体クローンの6個のCDRそれぞれは「クリティカル」CDRが決定されるまで交代して研究された(ここで「クリティカル」CDRは表3の受益CDRまたは高親和性CDRなどのような抗体結合に十分な影響を与えるCDRを意味する)。すべてのVDRsがクリティカルとして発見されたのではなかった。この特定の研究で使用された抗体として、H1、H3、L2およびL3のみがクリティカルであるとして発見されたが、その結果は異なった抗体で異なっている。一度「高親和性」CDRが決定されると(すなわち「受益CDR」)次いで本発明の高親和性中和抗体を生じるCDR配列の組合せを最適化するためにCDRの組合せが研究された。
【0066】
非常に特異的な実施例として、ここで開示された発明は少なくとも1010M−1の親和性定数を持つ呼吸合胞体ウイルス(RSV)に対する高親和性中和抗体に関し、ここで前記親和性定数はこのような決定の変動性と抗原(ここではRSVのF抗原)に対する異なるクローン化抗体の親和性の変動性の故で、この値の少なくとも2倍以内にあることができよう。この実施例内での生成する最適化構造のいくつかは1011M−1以上のKaを有していた(例えば表4での22と23の番号を付されたクローンがそれである)。
【0067】
この高親和性中和抗体にまた、その軽鎖可変部が配列識別番号1(図1A)の配列を持ち、またその重鎖可変部が配列識別番号2(図1B)の配列を持つ抗体と同じRSVに対するエピトープに結合する抗体である。
【0068】
一般に、直前に記載した抗RSVの特異的な例に基づく本発明の抗体を同定するために使用されるアプローチは、望ましい抗体鎖を発現する遺伝子のヌクレオチド配列を生成し、次いで標準プロトコルにより大腸菌細胞を形質転換するためにそれらをベクターに挿入することであった。細胞はウエルで成長し、上澄みがサンプル抽出され捕捉リフトとエリザ技術を用いて抗原結合を測定された〔ワトキンズ他、(1997年)、分析生化学、253巻、37−45ページ、ワトキンズ他(1998年)、分析生化学、256巻169−177ページ(これらの明細は引用例としてここに組み込まれている)参照〕。これらのポリヌクレオチドはCDRでの単一アミノ酸置換を提供するように設計されていたが、それは次いで親和性増加のために選択的に結合される受益置換(増加した親和性を産出するもの)で増加した親和性を検査することができた。これらは次いでRSVのF抗体の親和性体基本または基準抗体の結合親和性を検査された。
【0069】
このプロトコルを使用して、CDR L1またはH2での単一アミノ酸置換は抗原(ここではRSVのF抗原)として使用されたエピトープへの抗体クローンの親和性に何らの増加も産生しなかったことをエリザのデータが示した。従って本発明の抗体はすべてCDR L2、L3、H1およびH3での基準抗体のみとは異なるCDR配列を包含する(ここで図1の配列を持つ基準抗体は、それに対してKaを増加することにより抗体親和性の最適化のための試験手順およびいずれか他のシステムが同じように十分使用できる単なる有用な配列であった)。
【0070】
このようにしてRSVに関してここで開示された抗体は、更に一般にヒト抗体から誘導され、しかしそのように誘導されない場合は望ましくはマウスから誘導される枠組み構造領域を持つ。
【0071】
基準抗体のCDRに関して、(図1の配列で与えられるように)各CDRのアミノ酸配列は表1で示される。最適化に続き本発明により教示されるアミノ酸により置換されるならば、高親和性CDR配列(高親和性中和抗体の生成)およびそれによる受益結果(親和性の増加)を産生した基本または基準抗体のCDR内のアミノ酸残基の位置は、表1で太字でアンダーラインを付して示される(基準抗体にわたり親和性増加を与えるこのような配列は「受益CDR」または「高親和性CDR」で示される)。基本または基準抗体(図1)のCDRは以下で「基本または基準CDR」としてここで引用される。かくして表1は図1で示された(すなわち最適化を監視するためにここで使用された抗RSV基準抗体に対する)CDR配列を表す。
【表1】
【0072】
ここで開示された配列と関連して、本発明の目的に適うように定義されたCDR領域はここで開示された抗体の軽鎖可変部の残基24−33(CDR L1)、49−55(CDR L2)および88−95(CDR L3)ならびに重鎖可変部の残基31−37(CDR H1)、52−67(CDR H2)および100−109(CDR H3)に一致するこれらのセグメントである。
【0073】
本発明の抗体を産生する際に、前記抗体を含むクローンを生成するまたはポリペプチド鎖をクローニングするか、それをコードするポリヌクレオチド配列を使用するかまたは使用しないポリペプチド配列の直接合成によるか、もしくはどのような方法を使用者が選択するか、何故なら前記抗体を産生する本発明の教示の限定を生じることはないために、基本抗体または基準抗体(図1で示される重鎖および軽鎖可変部(CDRプラス枠組み構造))は本発明の抗体の新規なCDR配列を生成し、また結合定数等を比較する目的のための「鋳型」として使用することができる。単一変異の6個のCDRライブラリーを特徴付け合成する標準アプローチが使用された(ウー他、全米科学アカデミー紀要、95巻:6037−6042ページ(1998年)参照)。標的CDRはヌクレオチドのアニーリングの前にライブラリーのそれぞれからまず除去された。ライブラリーの合成のために、CDRは表1にあるように定義された。本発明のCDR配列を産出するためにオリゴヌクレオチド合成用のコドンベース突然変異が(前記の通り)採用された。
【0074】
ライブラリーは最高の親和性変異体を同定するためにまず、捕捉リフトで検査された。次いでこれらのクローンは更に捕捉エリザを用いて固定化抗原への滴定で特徴付けられた。
【0075】
最高親和性変異体からのDNAは受益または高親和性置換の性質を決定するため配列化された。検査の後、CDRの4個のみに起こる8個の受益または高親和性置換が観察されたことが確定した。これらは太字でアンダーラインを付された表1の基準または基本CDRに対して差異を持つ表3のCDR配列として要約されている。かくして表3のCDR配列は、特異性がRSVのF抗原に向けられる本発明の高親和性中和抗体を産生するのに使用が可能なカセットのCDRライブラリーであると見做すことができる。
【0076】
データの分析は、選択された位置でのアミノ酸置換がエピトープ結合を大きく増加させ、とりわけ置換の性質がアミノ酸配列の挿入であり、それはフェニルアラニン、アラニン、プロリン、トリプトファンおよびチロシン、とりわけフェニルアラニンから選択され、これらすべてのそのような受益または高親和性置換が再び選択的位置にあったことを示した。
【0077】
ここに記載されRSV/抗RSVシステムを採用する最適化実施に対し、もっとも受益的な高親和性CDRは6個のCDRの3または4、また丁度4のアミノ酸位置全体で生じることが発見された。かくして本発明の高親和性中和抗体はこれまで既知の人工抗体とは違って、相補性決定領域で、またはそのような領域を共に取巻く枠組み構造領域でのみ基本または基準抗体とは異なるアミノ酸配列を含有する。かくして本発明の抗体は抗体分子に高親和性を産生するように選択された1個またはそれ以上のCDR配列を含む高親和性中和抗体である。RSVを標的として用いる特異的実施例においては、そのような差異はL2(またはCDR L2)、L3(またはCDR L3)、H1(またはCDR H1)およびH3(またはCDR H3)でのみ発見される。前に述べた通り、この抗体の最大の親和性はこれらCDRの選択されたアミノ酸位置で起こり、また本発明の実施例のいくつかでは、各CDRのT度1個のみのアミノ酸位置が高親和性を提供するために望ましいものであった。
【0078】
かくしてCDR H1にとっては、(図1Bで特別のアンダーラインを付与した特別のCDR配列のアミノ末端から数えて)CDRアミノ酸2での置換は、とりわけ基本または基準抗体のCDR H1の位置2を占めるセリンをアラニンまたはプロリンのいずれかで置換したものは、もっとも受益的であり従ってRSV抗原エピトープへの高親和性を生じることを発見した。CDR H3にとっては、CDR配列の位置6でのグリシンの置換は、とりわけフェニルアラニンまたはトリプトファンのいずれか、特にフェニルアラニンによるグリシンの置換はRSVエピトープへの親和性増加を来たしたことが発見された。CDR L2ではCDRの位置3でのセリンの置換、特にフェニルアラニンはまたチロシンのいずれかによるものはF抗原への親和性増加を来たした。CDR L3ではCDRの位置5でのグリシンの置換、とりわけフェニルアラニン、トリプトファンまたはチロシンによる置換がRSVエピトープへの親和性の増加を生じた。
【0079】
本発明に従って、このようなアミノ酸置換を組合せて1個以上が同じ交代分子内で生じるようにして、RSVのF抗体のエピトープにここで開示された抗体の親和性を大きく増加することが可能であった。
【0080】
表2は本発明に基づく数多くのクローンの(H1、H3、L2およびL3それぞれに)新規なCDR配列を用いた結果を示している。表2で示されたように、特殊な抗体が本発明の1、2または3の新規なCDR対図1Aおよび1Bで示された基本または基準抗体鎖(軽鎖(L2およびL3))および重鎖(H1およびH3)をここに取り込んでいる。新規なCDRの効果は抗原(Ag)滴定スコア(以下参照)で記載されており、ここで基本または基準抗体は表2のトップで示されそのスコアは0.1である。他のスコアはこの「基本または基準抗体」配列の0.1のスコアと比較して示されている。それぞれの位置(すなわちH1の位置2、H3の位置6、L2の位置3およびL2の位置5)での最高の親和性を与えるアミノ酸の同定は各CDRに使用される突然変異を単に示す基本または基準抗体のアミノ酸の下部に示されている。
【0081】
本実験で検討されたクローンの全数は一部重複を含めて37個であった。(括弧内の「n」値で示されたもの、例えばクローンNo.7に対する「n=4」は4個の重複クローンが検討されたことを示している)。
【0082】
一般に、親和性と受益または高親和性CDRの数との間には相関があることをデータは示しており、より高次の親和性変異体のすべては1個以上の受益または高親和性CDRを持っていた。更にすべての最良クローンは、CDR H3内の位置6にF(Phe)を有していた。また受益または高親和性CDRは疎水性、とりわけ芳香族アミノ酸が基本または基準抗体に見出される残基の位置に挿入されたものであった。
【0083】
抗体滴定検査は各測定でRSV抗原の500ngを用いて抗体の濃度を変えて使用した。表2の数多くの組合せクローンの比較データのグラフが図2で示される。
【0084】
要約すると、表2はここで記載されて手順で評価される数多くのクローンと、CDR H1、H3、L2およびL3の主要位置を(表1と表3で太字でアンダーラインされた位置)で生じるアミノ酸を示している。この表は更にこれらのクローンのCDR対基準抗体の対応するCDRの間の多くの差を要約する(表1と図1を参照)。表の右側は「Agスコア」すなわち抗原結合値を示し、それは0−4にわたる任意の性質上の値を表し、またそのそれぞれの滴定曲線に基づく異なった抗体クローンの相対的結合能力の質的測定値を表す。この値は異なる抗体の大ざっぱな質的比較のみをここで提供し結合能力の量的尺度を意図するものではない。
【0085】
表2はまた各抗体クローンの新規なCDRの数を示す。(基準抗体の対応するCDRに関連して少なくとも1個のアミノ酸の差をもつ抗体内でのCDRの数を意味する−表1参照)。新規なCDRの数は、またその抗体分子に存在する「受益」または「高親和性」CDRの数である。新規なCDRでのアミノ酸の差は基準抗体に対し表1での太字でアンダーラインされた位置で起こるであろうし、そのため表1で太字のアンダーラインされたアミノ酸は(標準単一文字アミノ酸命名法を用いて)表2のそれぞれのCDRを示すアミノ酸により置換された。
【表2】
【0086】
クローンそれぞれで表される新規なCDRはクローンを表に位置せしめ、基本または基準クローンに隣り合わせの同じCDRの対応するアミノ酸に各CDRの指示アミノ酸を適合させることで容易に決定される。便宜上アミノ酸がクローンの一つの特定CDRで異なる場合には、新しいアミノ酸は太字で示される。加えて表で示されるすべてのクローンでは、置換は新規なCDRを産出するとして前に引用したように選択された位置でのみ生じる。かくして基本または基準抗体に対するCDR H1の置換はすべてCDRの位置2にあり(図1および表1と2でアンダーラインされ太字で示されたCDR H1のN末端端部からの第2アミノ酸を意味する)、CDR H3のすべての置換は位置6にあり、CDR L2のすべての置換は位置3であり、またCDR L3のすべての置換は位置5にあり、またすべては基本または基準抗体に関連する。基本または基準抗体が選ばれたのは、それがRSVエピトープにきわめて高い親和性を持つことが既に公知であったからであるということは心に留めておかねばならない。〔ジョンソン他、(1997年)感染性疾病ジャーナル、176巻、1215−1224ページ〕。従って例えばクローン番号1では、受益または高親和性CDR H1に対し基本または基準抗体のCDR H1の位置2でセリンに代りアラニンが使用され、これによりRSVへの親和性増加を達成し、またフェニルアラニンはCDR H3の位置6でトリプトファンに代って生成し、基本または基準抗体のCDR L2の位置3でセリンが使用され、更に受益または高親和性CDR L3の位置5で発生した。
【0087】
かくして表2のクローンで試験された上澄みに産生された抗体構造の存在する本発明に基づく新規な受益CDR(すなわち対応する基本または基準CDRに代って基本または基準抗体におけるその存在が同じRSVエピトープの前記抗体の親和性を著しく増加することに役立った高親和性CDRまたはCDR配列)が表3で要約される。それぞれの場合において、太字のものは本発明のCDRが基本または基準講RSV抗体の対応するCDRとどのように新規で受益的であり、または高親和性で異なるかを示している。
【表3】
【0088】
表3のCDR配列が本発明に基づき開示された高親和性CDRの配列を表している一方、1個またはそれ以上のこれらのCDRが同じ抗体内に存在し、高親和性CDRのそれぞれに適切な配列が選択されるセットをこの表が示しているということが理解される。かくして表3で示されるように、高親和性H1 CDRがここで開示された発明の高親和性中和抗体に存在する時には、それは配列識別番号9または10の配列に一致する配列を持つ。請求された発明の中和抗体が高親和性H3 CDRを含有する時には、前記CDRは配列識別番号12を持つ。本発明の高親和性中和抗体が高親和性L2CDRを含有する時には、前記高親和性L2 CDRは配列識別番号12および13の配列より成るグループから選択されたアミノ酸配列を持つ。最後に本発明の高親和性中和抗体が高親和性L3 CDRを含有する時には、前記CDRは配列識別番号14、15,16の配列より成るグループから選択されたアミノ酸配列を持つ。
【0089】
一つの実施例において、既に述べたように高親和性中和抗体はヒト定常部を含む抗体である。
【0090】
かくして望ましい実施例において、少なくとも1010M−1、または更に少なくとも1011M−1の親和性を持つ本発明の高親和性中和抗体はヒト定常部および重鎖と軽鎖の枠組み構造を含む移植抗体であり、ここで枠組み構造の少なくとも一部はヒト抗体から(またはヒト抗体枠組み構造のコンセンサス配列から)誘導される。
【0091】
も一つの実施例において、すべての枠組み構造はヒト抗体(またはヒトコンセンサス配列)から誘導される。
【0092】
かくして少なくとも1010M−1の親和性を持つRSV中和抗体はヒト定常部、非ヒト抗体から誘導される1個またはそれ以上のCDRを持つ移植抗体であり、ここで少なくとも1個の前記CDR内の少なくとも1個のアミノ酸は変更され、またここで枠組み構造の全体または一部はヒト抗体(またはヒト抗体枠組み構造のコンセンサス配列)から誘導される。
【0093】
1個の抗体のCDR配列のも一つの抗体の非CDR領域との組合せがCDRの残りの分子への「移植」の形態から生じるために、これらのものは「CDR移植」抗体として引用されてきた。今日遺伝子工学の手法を用いることで同じ産物を実際の交代からいずれの配列を単離することなく形式することができる。望ましいCDR配列、および定常部と枠組み構造配列が公知である限りにおいて、望ましい配列をもつ遺伝子を組み立てることができ、また各種のベクターを使用して機能的四量体抗体分子の発現のために適切な細胞に挿入することができる。これを既に記載した方法を組合わせると、単一突然変異ライブラリーの組立てを可能にし、ここで抗体は対応する移植抗体と同じ配列を所有し、また従って同じ構造と結合親和性を有することになる。
【0094】
本発明の高親和性抗体は比較的純粋なまたは単離形態に、同じくウエル内でまたは平板上で成長した細胞からの上澄みに存在することができる。このような上澄みは表1のデータを生成するために使用された。本発明の抗体は画して本発明の抗体を含むひとつの組成物の形態で存在することができ、ここで前記抗体は薬理許容単体または賦形剤に懸濁される。本発明の抗体はそのような組成物に疾病、例えばRSVなどを処置する治療または薬理学的値に十分な一定の濃度で、または一定の量で存在する。前記抗体はまたより希釈された形態で組成物に存在する。
【0095】
従って本発明は更にここで記載された抗体組成物の(予防を含む)治療有効量の呼吸合胞体ウイルス感染の危険のあるまたはそれに悩む患者への投与を含むその感染症を予防しおよびまたは処置する方法を提供することに向けられる。
【0096】
本発明の高親和性抗体の一つの望ましい実施例は、その重鎖および軽鎖CDR領域が以下の配列を持つ抗体である。すなわちCDR H1は配列識別番号9の配列を持ち、CDR H3は配列識別番号11を持ち、CDR L2は配列識別番号4(図1Aの基準配列のCDR L2から変化していない)の配列を持ち、またCDR L3は配列識別番号14を持つ(表3参照)。この望ましい実施例において親和性定数は表4(クローン1)で示されるように約6.99×1010(または解離定数として約14.3pM)である。この実施例を含む抗体の重鎖および軽鎖可変部は枠組み構造配列を伴い図3で示される。
【0097】
本発明の高親和性抗体のも一つの望ましい実施例はその重鎖および軽鎖CDR領域が以下の配列を持つ抗体である。CDR H1は配列識別番号9の配列を持ち、CDR H3は配列識別番号11の配列を持ち、CDR L2は配列識別番号12の配列を持ち、またCDR L3は配列識別番号5の配列を持つ(表2、クローン2を参照のこと−図1AのCDR L3の基準配列との差はない)。この望ましい実施例において、親和性定数は表4(クローン2)で示されるように約7.30×1010(または解離定数として約13.7pM)である。この実施例を含む抗体の重鎖および軽鎖可変部は枠組み構造配列と共に図4で示される。
【0098】
本発明の高親和性抗体の追加の望ましい実施例はその重鎖および軽鎖CDR領域が以下の配列を持つ抗体である。CDR H1は配列識別番号10の配列を持ち、CDR H3は配列識別番号11の配列を持ち、CDR L2は配列識別番号12の配列を持ち、またCDR L3は配列識別番号14の配列を持ち、(CDR配列については表3を参照のこと)、そのクローンは表2で指示番号3である。この望ましい実施例において、親和性定数は表4(クローン3)で示されるように、約8.13×1010(または解離定数としては約12.3pM)である。実施例を含む抗体の重鎖および軽鎖可変部は枠組み構造配列と共に図5で示される。
【0099】
本発明の高親和性抗体のもっとも望ましい実施例はその重鎖および軽鎖CDR領域が以下の配列を持つ抗体である。CDR H1は配列識別番号9の配列を持ち、CDR H3は配列識別番号11の配列を持ち、CDR L2は配列識別番号12の配列を持ち、またCDR L3は配列識別番号14の配列を持つ(表3参照のこと)。本実施例において、親和性定数は表4(クローン22)で示されるように約3.6×1011(または解離定数として約2.8pM)である。この実施例を含む抗体の重鎖および軽鎖可変部は、枠組み構造配列と共に図6で示される。
【0100】
本発明のも一つのもっとも望ましい実施例端の重鎖および軽鎖CDR領域が以下の配列を持つ抗体である。CDR H1は配列識別番号9の配列を持ち、CDR H3は配列識別番号11の配列を持ち、CDR L2は配列識別番号12の配列を持ち、またCDR L3は配列識別番号15の配列を持つ(表3参照)。このもっとも望ましい実施例において、親和性定数は表4(クローン23)で示されるように約4×1011(解離定数として約2.5pM)である。本実施例を含む抗体の重鎖および軽鎖可変部は、枠組み構造配列と共に図7で示される。
【0101】
とりわけ望ましい実施例において、本発明の抗体は図1、3、4、5、6および7で示された枠組み構造を示す配列の枠組み構造領域を持つであろう(それぞれは同じ枠組み構造領域を含有し、CDR配列でのみ異なる)。これらのもっとも望ましい実施例は中和抗体を含みここで軽鎖可変部は配列識別番号17のアミノ酸配列を持ち、また重鎖可変部は配列識別番号18のアミノ酸配列を持つ。またもっとも望ましい実施例は中和抗体を含み、ここで軽鎖可変部は配列識別番号19のアミノ酸配列を持ち、また重鎖可変部は配列識別番号20のアミノ酸配列を持つ。(実施例はまた別の)中和抗体ではここで軽鎖可変部は配列識別番号21を持ち、重鎖可変部は配列識別番号22のアミノ酸配列を持つ。別の中和抗体では軽鎖可変部は配列識別番号23のアミノ酸配列を持ち、重鎖可変部は配列識別番号24のアミノ酸配列を持つ。別の中和抗体では軽鎖可変部は配列識別番号25のアミノ酸配列を持ち、重鎖可変部では配列識別番号26のアミノ酸配列を持つ。
【0102】
本発明の高親和性中和抗体は、共にアミノ酸セグメントをスプライスすることにより実際の中和抗体から誘導されるCDR領域および非CDR領域から組立てることができ(またこのように組立てられた抗体はここで開示された発明の範囲内にあるが)、本発明の抗体は適切な遺伝子配列をベクターに遺伝子操作することでもっとも便利に調製され、それは次いで遺伝子操作細胞による組立て抗体分子の偶発的発現のために適当な細胞系に形質移入される。実際にこのような組換え手順は、ここで開示される抗体を調製するために採用された。加えて高親和性抗体の鎖の配列は、ここでの開示から公知であるために、このような抗体は更に適切な鎖の直接合成により組立てられ、次いで四量体(H2L2)二価抗体構造に自己組立てをすることを可能にした。
【0103】
本発明の高親和性中和抗体を調製する方法は、本発明の受益CDRを含むクローン産生抗体を調製するのに使用された組合せライブラリーの創出を伴い、それは次いでRSVエピトープへの親和性を検査することができた(例えば図2)。
【実施例】
【0104】
(実施例1)
BIAcoreTMによるヒト化RSV Mabsの動態分析
高親和性抗RSV MabsとRSV Fタンパク質の間の相互作用の動態がファルマシアのBIAcoreTMバイオセンサーを用いる表面プラスモン(細胞質遺伝子)共鳴により研究された。C末端切形Fタンパク質を発現する組換えバキュロウイルスは動態研究のための抗原の豊富な源を提供した。分泌Fタンパク質を含有する上澄みはコンカナバリンA(conA)とQセファロースカラムでの連続クロマトグラフィーにより約20倍富化された。プールされた小部分はクエン酸ナトリウム10mM(pH5.5)に対して透析され、約0.1mg/mlに濃縮された。典型的な実験では、Fタンパク質のアリコート(100ml)が、BIAcoreセンサーチップにアミン結合された。固定された量はマウスモノクローナル抗体H1129またはH1308Fで飽和された時約2000応答単位(Rmax)のシグナルを提供した(抗体H1129あるいはH1308は合衆国特許番号第5,824307号で提供され、その開示はここで引用例として取り込まれている)。これは、結合手順に次いでFタンパク質に同じ数の「A」および「C」抗原部位が存在したことを示していた。2個の無関連Mabs(RVFV 4D4とCMV H758)は固定化Fタンパク質と相互作用を示さなかった。典型的な動態研究は異なる濃度(25−300nM)でのMab35mlの0.05%トウィーン−20(PBS/トウィーン)0.05%を含むPBS緩衝液への注入を含んでいた。流速は5ml/分に維持され、7分の結合フェーズを与えられた。Mabの注入に続き、流動物は解離速度を決定するために30分間PBS/トウィーン緩衝液で変換された。センサーチップは塩酸10mMの2分パルスのサイクル間で再生された。再生段階は固定Fタンパク質の結合能力を僅かではあるが最小ロスとして生じた(サイクルごと4%のロス)。この小さな減少は結合および解離の速度定数(それぞれKonおよびKoffと称されるもの)の計算値を変えることはなかった。
【0105】
より特異的には、Kassoc(またはKon)の測定のために、Fタンパク質がEDC/NHS活性化センサーチップ全体に注入されるEDC/NHS法(EDC=N−エチル−N′−〔3−ジエチルアミノ−プロピル〕カルボジイミド;NHS=N−ヒドロキシスクシンイミド)により直接固定された。要約すると、NaOAc、10mM、pH4.0でFタンパク質4μg/mlが調製され、約30μmlの注入は前記引用条件の下で約500RU(応答ユニット)の固定Fタンパク質を提供した。未加工流動細胞(VnR固定化−CMデキストラン表面)が動態分析のために取り出された。カラムは塩酸100mMを用いて(完全再生に必要とされる72秒の接触時間で)再生された。この処置は固定抗原を何ら損傷することなく結合Fabを完全に除去し40回もの再生に使用することができた。Kon測定では、Fab濃度は12.5nM、25nM、50nM、100nM、200nMおよび400nMであった。解離フェーズは(解離フェーズの開始30秒後から)230秒乃至900秒まで分析された。動態は1:1のラングミュアフィッティング(グローバルフィッティング)で解析された。測定はHBS−EP緩衝液(ヘペス10mM、pH7.4、塩化ナトリウム150mM、EDTA3mM、0.005%(容量/容量)界面活性剤P20)内で行われた。
【0106】
ここで開示されたように組合せクローン内測定に関しては、KonおよびKoffが個別に測定された。Konは単一変異クローンに対するのと同じ条件で測定され、同様に分析された。
【0107】
Kdissoc(またはKoff)の測定については、下記の条件が採用された。要約すると、Fタンパク質4100RUが未加工として使用されるCM−デキストランと共に(前の通り)固定化された。ここで3000RUのFabは(機械の変動を相殺するためにきわめて十分な解離Fabと)結合した。HBSとFタンパク質5nM(KdissocまたはKd−解離平衡定数よりも約350−2000倍高いもの)が緩衝液として使用された。解離フェーズは5μl/分の流速で6−15時間であった。ここで使用された条件の下で解離Fabの再結合は最小であった。その詳細については、バイオセンサーのマニュアルを参照されたい。
【0108】
ここで開示されたFタンパク質またはRSVの他のエピトープ部位への高親和性抗RSV抗体の結合は、結合または会合の第2オーダー比率定数に対する解離の第1オーダー比率定数(Kd=Kassoc)の割合から算出された。Kassocの値は下記の等式に基づき計算される。
dR/dt=Kassoc〔Mab〕Rmax−(Kassoc〔Mab〕+Kdiss)R
ここでRとRmaxはそれぞれ時間tと無限での応答単位である。Rの関数としてのdr/dtのプロットは(Kassoc〔Mab〕+Kdiss)の勾配を与える。これらの勾配は〔Mab〕に線型に関連しているため、Kassocは〔Mab〕に対する勾配の再プロットから誘導することができる。新しいラインの勾配はKassocに等しい。Kdissの値はY線分から外挿することができるけれども、より正確な値はKdissの直線測定により決定された。Mabの注入フェーズに続き、PBS/トウィーン緩衝液が、センサーチップを横断流動する。この点から〔Mab〕=0である。dR/dtのための前述の等式は従って
dr/dt=k または dR/R=Kdissdt
となり、この等式を統合すると
In(Ro/Rt)=Kdisst
となり、ここでRo/Rtはそれぞれ時間0(解離フェーズ開始)および時間tでの応答ユニットである。最後にtの関数としてのプロッティングIn(Ro/Rt)はKdissの勾配を提供する。
【0109】
ここでの望ましい実施例において、このような抗体変異体からの数値は以下の通りである。
【表4】
【0110】
ここでCDRは基準抗体として表1で(太字とアンダーラインで示されたCDRの主要位置(クリティカル位置)で基準アミノ酸を置換するアミノ酸を表す。かくして例えば、クローン22は表1(配列識別番号6)のその位置で示されるセリンに代ってCDR H1(重鎖可変部−配列識別番号24の残基32)の位置2でアラニンを持つ。表1(配列識別番号8)のその位置で示されるトリプトファンに代ってCDR H3(重鎖可変部−配列識別番号24の残基105)の位置6でフェニルアラニン、表1(配列識別番号4)のその位置で示されるセリンに代ってCDR L2(軽鎖可変部−配列識別番号23の残基51)の位置3でのフェニルアラニン、および表1(配列識別番号5)のその位置で示されたグリシンに代ってCDR L3(軽鎖可変部−配列識別番号23)の位置5でのフェニルアラニンなどを持つ。
【0111】
もちろんこれらのクローンを形成する際には、表3は本発明の抗体の高親和性CDRが取り出される潜在的CDRのプールを表す。例えば、表4のクローン23はCDR L3の例外を除きクローン22と同じCDRを使用し、それは表1(配列識別番号5)のその位置で示されるグリシンに代えてCDR L3(表3−配列識別番号15)の位置5でチロシンを持つ。対応するクリティカル位置での置換は同じように表2で示される。
【0112】
(実施例2)
微小中和検定
本発明の抗体の中和は微小中和検定で測定された。この微小中和検定はアンダースン他(1985年)に記載された手順の修飾である。呼吸合胞体ウイルス(RSV−ロング株)の10個のTCID50が96ウエル平板のウエルで2時間37℃で試験される抗体(またはFabs)の連続希釈液で保温された。RSV感染性HEp−2細胞(2.5×104)は次いで各ウエルに加えられ、5日間37℃で5%炭酸ガスの下で培養された。5日後に培地は吸引され細胞は洗浄されメタノール80%PBS20%で平板に固定された。RSV複製は次いでFタンパク質発現で測定された。固定細胞はビオチン接合抗Fタンパク質モノクローナル抗体(パンFタンパク質、C部位特異的MAb 133−1H)で保温され、洗浄され西洋ワサビペルオキシダーゼ接合アビジンがウエルに加えられた。ウエルは再洗浄され基質TMB(チオニトロベンゼン酸)の出来高が450nmで測定された。中和力価はウイルスのみの対照細胞から450nm(OD450)の吸光度で少なくとも50%の減少を起こす抗体濃度で表現された。本発明のいくつかの抗体の結果は図8のグラフで示されており、一方Fab断片を使用した結果は図9のグラフで描かれている。
(背景および引用文献)
【0113】
1.C.B.ホール,R.G.ダグラス,J.M.ガイマン他、ニューイングランド・ジャーナル・オブ・メディスン293巻:1343ページ、1975年。
【0114】
2.C.B.ホール,J.T.マクブライド,E.E.ウォルシュ他、ニューイングランド・ジャーナル・オブ・メディスン308巻:1443ページ、1983年。
【0115】
3.C.B.ホール,J.T.マクブライド,C.L.ギャラ他、JAMA254巻:3047ページ、1985年。
【0116】
4.E.R.ウォルド他、小児科ジャーナル112巻:154ページ、1988年。
【0117】
5.A.Z.キャピキャン,R.H.ミッチェル,R.M.チャノック他、疫学アメリカン・ジャーナル89巻:405ページ、1969年。
【0118】
6.G.A.プリンス,V.G.へミング,R.L.ホースウッド他、ウイルス研究3巻:193ページ、1985年。
【0119】
7.V.G.へミング,G.A.プリンス,R.L.ホースウッド他、感染疾病ジャーナル152巻:1083ページ、1985年。
【0120】
8.P.F.ライト,R.B.ベルシュ他、感染と免疫、37巻:397ページ、1982年。
【0121】
9.D.A.コンラッド,J.C.クリステンソン他、小児科感染疾病ジャーナル、6巻:152ページ、1987年。
【0122】
10.A.F.ロビューリョ,R.L.ホイーラー,J.トラン他、全米科学アカデミー紀要、86巻:4220ページ、1989年。
【0123】
11.Z.ステップレフスキー,L.K.サン,C.W.シアマン他、全米科学アカデミー紀要、85巻:4852ページ、1988年。
【0124】
12.G.L.ブーリアンヌ,N.穂積,M.J.シュルマン、ネイチャー、312巻:643ページ、1984年。
【0125】
13.L.K.サン,P.カーチス,E.ラコヴィッツ−シュルチンスカ他、全米科学アカデミー紀要、84巻:214ページ、1987年。
【0126】
14.A.Y.リュ,P.W.マック,C.I.チャンピオン,R.P.ロビンソン、遺伝子、54巻:33ページ、1987年。
【0127】
15.S.L.モリソン,M.J.ジョンソン,L.A.ハーセンバー,V.T.オイ、全米科学アカデミー紀要、81巻:6851ページ、1984年。
【0128】
16.S.L.モリソン、サイエンス、229巻:1202ページ、1985年。
【0129】
17.B.G.サヘーガン,H.ドーライ,J.ザルツガーバー−ミュラー他、免疫学ジャーナル、137巻:1066ページ、1986年。
【0130】
18.S.武田,T.内藤,K.浜,T.野間,T.本城、ネイチャー、314巻:452ページ、1985年。
【0131】
19.D.A.カーソン,B.D.フライマーク、先端免疫学、38巻:275ページ、1986年。
【0132】
20.T.A.ビーラー他、ウイルス学ジャーナル、63巻:2941−2950ページ、1989年。
【0133】
21.コーリン他、ウイルス学、143巻:569−582ページ、1989年。
【0134】
22.アンダースン他、酵素免疫検定に基づく呼吸合胞体ウイルスの微小中和検定、臨床微生物学ジャーナル(1985年)22巻:1050−1052ページ。
【0135】
23.ジョンソン他、呼吸合胞体ウイルスに対し有力な試験管内生体内活性を持つヒト化モノクローナル抗体(MEDI−493)の開発、感染性疾病ジャーナル(1997年)、176巻:1215−1224ページ。
【特許請求の範囲】
【請求項1】
少なくとも3個の高親和性相補性決定領域(CDR)を含む一つの高親和性中和免疫グロブリンであって、ここで前記各高親和性CDRが少なくとも1個の抗原決定基に向かう特異性を持ち、また前記抗原決定基に対し少なくとも1010M−1の親和性定数(Ka)を有する免疫グロブリンを生じるように選択されたアミノ酸配列を持つことを特徴とする高親和性中和免疫グロブリン。
【請求項2】
前記免疫グロブリンが少なくとも4個の高親和性CDR含むことを特徴とする請求項1記載の高親和性中和免疫グロブリン。
【請求項3】
前記免疫グロブリンが3個の高親和性CDRを持つことを特徴とする請求項1記載の高親和性中和免疫グロブリン。
【請求項4】
前記免疫グロブリンが4個の高親和性CDRを持つことを特徴とする請求項2記載の高親和性中和免疫グロブリン。
【請求項5】
前記親和性定数(Ka)が少なくとも1011であることを特徴とする請求項2記載の高親和性中和免疫グロブリン。
【請求項6】
前記免疫グロブリンがウイルスにより発現される少なくとも1個のタンパク質に特異的であることを特徴とする請求項1記載の高親和性中和免疫グロブリン。
【請求項7】
前記ウイルスが呼吸合胞体ウイルス(RSV)であることを特徴とする請求項6記載の高親和性中和免疫グロブリン。
【請求項8】
前記タンパク質がRSVのFタンパク質であることを特徴とする請求項7記載の高親和性中和免疫グロブリン。
【請求項9】
前記免疫グロブリンがヒト定常部を持つことを特徴とする請求項1記載の高親和性中和免疫グロブリン。
【請求項10】
前記免疫グロブリンが図1の基準免疫グロブリンと同じRSVに対するエピトープと結合することを特徴とする請求項1記載の高親和性中和免疫グロブリン。
【請求項11】
前記免疫グロブリンがヒト免疫グロブリンのみから誘導される枠組み構造を含むことを特徴とする高親和性中和免疫グロブリン。
【請求項12】
枠組み構造の少なくとも一部がマウス免疫グロブリンから誘導されることを特徴とする請求項1記載の高親和性中和免疫グロブリン。
【請求項13】
少なくとも1個の高親和性CDRが、図1の基本抗体または基準抗体の対応するCDRで非フェニルアラニン残基を生じる位置でフェニルアラニン残基を含有することを特徴とする請求項1記載の高親和性中和免疫グロブリン。
【請求項14】
前記免疫グロブリンが高親和性H1 CDRを含み、また前記CDRが配列識別番号9および10より成るグループから選択されるアミノ酸配列を持つことを特徴とする請求項1記載の高親和性中和免疫グロブリン。
【請求項15】
前記免疫グロブリンが配列識別番号11のアミノ酸配列を持つ高親和性H3 CDRを含むことを特徴とする請求項1記載の高親和性中和免疫グロブリン。
【請求項16】
前記免疫グロブリンが高親和性L2 CDRを含み、また前記CDRが配列識別番号12および13より成るグループから選択されるアミノ酸配列を持つことを特徴とする請求項1記載の高親和性中和免疫グロブリン。
【請求項17】
前記免疫グロブリンが配列識別番号14,15および16より成るグループから選択されるアミノ酸配列を有する高親和性L3 CDRを持つことを特徴とする請求項1記載の高親和性中和免疫グロブリン。
【請求項18】
H1 CDRが配列識別番号9のアミノ酸配列を持ち、H3 CDRが配列識別番号11のアミノ酸配列を持ち、L2 CDRが配列識別番号4のアミノ酸配列を持ち、またL3 CDRが配列識別番号14のアミノ酸配列を持つことを特徴とする請求項1記載の高親和性中和免疫グロブリン。
【請求項19】
H1 CDRが配列識別番号9のアミノ酸配列を持ち、H3 CDRが配列識別番号11のアミノ酸配列を持ち、L2 CDRが配列識別番号12のアミノ酸配列を持ち、またL3 CDRが配列識別番号5のアミノ酸配列を持つことを特徴とする請求項1記載の高親和性中和免疫グロブリン。
【請求項20】
H1 CDRが配列識別番号10のアミノ酸配列を持ち、H3 CDRが配列識別番号11のアミノ酸配列を持ち、L2 CDRが配列識別番号12のアミノ酸配列を持ち、またL3 CDRが配列識別番号14のアミノ酸配列を持つことを特徴とする請求項2記載の高親和性中和免疫グロブリン。
【請求項21】
H1 CDRが配列識別番号9のアミノ酸配列を持ち、H3 CDRが配列識別番号11のアミノ酸配列を持ち、L2 CDRが配列識別番号12のアミノ酸配列を持ち、またL3 CDRが配列識別番号14のアミノ酸配列を持つことを特徴とする請求項2記載の高親和性中和免疫グロブリン。
【請求項22】
H1 CDRが配列識別番号9のアミノ酸配列を持ち、H3 CDRが配列識別番号11のアミノ酸配列を持ち、L2 CDRが配列識別番号12のアミノ酸配列を持ち、またL3 CDRが配列識別番号15のアミノ酸配列を持つことを特徴とする請求項2記載の高親和性中和免疫グロブリン。
【請求項23】
重鎖可変部が配列識別番号17を持ち、また重鎖可変部が配列識別番号18を持つことを特徴とする請求項1記載の高親和性中和免疫グロブリン。
【請求項24】
重鎖可変部が配列識別番号19を持ち、また重鎖可変部が配列識別番号20を持つことを特徴とする請求項1記載の高親和性中和免疫グロブリン。
【請求項25】
重鎖可変部が配列識別番号21を持ち、また重鎖可変部が配列識別番号22を持つことを特徴とする請求項2記載の高親和性中和免疫グロブリン。
【請求項26】
重鎖可変部が配列識別番号23を持ち、また重鎖可変部が配列識別番号24を持つことを特徴とする請求項2記載の高親和性中和免疫グロブリン。
【請求項27】
重鎖可変部が配列識別番号25を持ち、また重鎖可変部が配列識別番号26を持つことを特徴とする請求項2記載の高親和性中和免疫グロブリン。
【請求項28】
少なくとも1010M−1の親和性定数を持つ一つの組換え高親和性中和免疫グロブリンであって、ここで前記免疫グロブリンが、少なくともその部分がヒト抗体から誘導されるヒト定常部とヒト重鎖および軽鎖枠組み構造領域を含むことを特徴とする組換え高親和性中和免疫グロブリン。
【請求項29】
重鎖および軽鎖枠組み構造領域がヒト抗体のコンセンサス配列から誘導されることを特徴とする請求項28記載の組換え高親和性中和免疫グロブリン。
【請求項30】
親和性定数が少なくとも1011M−1であることを特徴とする請求項29記載の組換え高親和性中和免疫グロブリン。
【請求項31】
重鎖および軽鎖枠組み構造領域がヒト抗体のコンセンサス配列から誘導されることを特徴とする請求項30記載の組換え高親和性中和免疫グロブリン。
【請求項32】
請求項1記載の免疫グロブリンが薬理許容担体に懸濁されていることを特徴とする前記免疫グロブリンを含む一つの組成物。
【請求項33】
請求項6記載の免疫グロブリンが薬理許容担体に懸濁されていることを特徴とする前記免疫グロブリンを含む一つの組成物。
【請求項34】
疾病を予防しおよびまたは処置する一つの方法であって、請求項32記載の免疫グロブリン組成物の治療有効量をその疾病の危険にさらされまたはそれに悩む患者に投与することを含むことを特徴とする方法。
【請求項35】
ウイルス誘導疾病を予防しおよびまたは処置する一つの方法であって、請求項33記載の免疫グロブリン組成物の治療有効量をその疾病の危険にさらされまたはそれに悩む患者に投与することを含むことを特徴とする方法。
【請求項36】
呼吸合胞体ウイルスを予防しおよびまたは処置する一つの方法であって、薬理許容担体に懸濁された請求項7記載の免疫グロブリンの治療有効量をそのウイルスの危険にさらされまたはそれに悩む患者に投与することを含むことを特徴とする方法。
【請求項37】
請求項3記載の高親和性中和免疫グロブリンであって、ここで前記免疫グロブリンがFab、F(ab)’2、重鎖−軽鎖二量体、重鎖および軽鎖より成るグループから選択されることを特徴とする高親和性中和免疫グロブリン。
【請求項38】
請求項4記載の高親和性中和免疫グロブリンであって、ここで前記免疫グロブリンがFab、F(ab)’2、重鎖−軽鎖二量体、重鎖および軽鎖より成るグループから選択されることを特徴とする高親和性中和免疫グロブリン。
【請求項39】
請求項1記載の高親和性中和免疫グロブリンであって、ここで前記免疫グロブリンが抗体であることを特徴とする高親和性中和免疫グロブリン。
【請求項40】
請求項2記載の高親和性中和免疫グロブリンであって、ここで前記免疫グロブリンが抗体であることを特徴とする高親和性中和免疫グロブリン。
【請求項41】
請求項6記載の高親和性中和免疫グロブリンであって、ここで前記免疫グロブリンがFab、F(ab)’2、重鎖−軽鎖二量体、重鎖および軽鎖より成るグループから選択されることを特徴とする高親和性中和免疫グロブリン。
【請求項42】
請求項7記載の高親和性中和免疫グロブリンであって、ここで前記免疫グロブリンがFab、F(ab)’2、重鎖−軽鎖二量体、重鎖および軽鎖より成るグループから選択されることを特徴とする高親和性中和免疫グロブリン。
【請求項43】
請求項8記載の高親和性中和免疫グロブリンであって、ここで前記免疫グロブリンがFab、F(ab)’2、重鎖−軽鎖二量体、重鎖および軽鎖より成るグループから選択されることを特徴とする高親和性中和免疫グロブリン。
【請求項44】
疾病を予防およびまたは処置する一つの方法であって、薬理許容担体に懸濁された請求項37または38記載の免疫グロブリンの治療有効量をその疾病の危険にさらされまたはそれに悩む患者に投与することを含むことを特徴とする方法。
【請求項45】
ウイルス誘導疾病を予防およびまたは処置する一つの方法であって、薬理許容担体に懸濁された請求項41記載の免疫グロブリンの治療有効量をその疾病の危険にさらされまたはそれに悩む患者に投与することを含むことを特徴とする方法。
【請求項46】
呼吸合胞体ウイルスを予防およびまたは処置する一つの方法であって、薬理許容担体に懸濁された請求項42または43記載の免疫グロブリンの治療有効量をその疾病の危険にさらされまたはそれに悩む患者に投与することを含むことを特徴とする方法。
【請求項1】
少なくとも3個の高親和性相補性決定領域(CDR)を含む一つの高親和性中和免疫グロブリンであって、ここで前記各高親和性CDRが少なくとも1個の抗原決定基に向かう特異性を持ち、また前記抗原決定基に対し少なくとも1010M−1の親和性定数(Ka)を有する免疫グロブリンを生じるように選択されたアミノ酸配列を持つことを特徴とする高親和性中和免疫グロブリン。
【請求項2】
前記免疫グロブリンが少なくとも4個の高親和性CDR含むことを特徴とする請求項1記載の高親和性中和免疫グロブリン。
【請求項3】
前記免疫グロブリンが3個の高親和性CDRを持つことを特徴とする請求項1記載の高親和性中和免疫グロブリン。
【請求項4】
前記免疫グロブリンが4個の高親和性CDRを持つことを特徴とする請求項2記載の高親和性中和免疫グロブリン。
【請求項5】
前記親和性定数(Ka)が少なくとも1011であることを特徴とする請求項2記載の高親和性中和免疫グロブリン。
【請求項6】
前記免疫グロブリンがウイルスにより発現される少なくとも1個のタンパク質に特異的であることを特徴とする請求項1記載の高親和性中和免疫グロブリン。
【請求項7】
前記ウイルスが呼吸合胞体ウイルス(RSV)であることを特徴とする請求項6記載の高親和性中和免疫グロブリン。
【請求項8】
前記タンパク質がRSVのFタンパク質であることを特徴とする請求項7記載の高親和性中和免疫グロブリン。
【請求項9】
前記免疫グロブリンがヒト定常部を持つことを特徴とする請求項1記載の高親和性中和免疫グロブリン。
【請求項10】
前記免疫グロブリンが図1の基準免疫グロブリンと同じRSVに対するエピトープと結合することを特徴とする請求項1記載の高親和性中和免疫グロブリン。
【請求項11】
前記免疫グロブリンがヒト免疫グロブリンのみから誘導される枠組み構造を含むことを特徴とする高親和性中和免疫グロブリン。
【請求項12】
枠組み構造の少なくとも一部がマウス免疫グロブリンから誘導されることを特徴とする請求項1記載の高親和性中和免疫グロブリン。
【請求項13】
少なくとも1個の高親和性CDRが、図1の基本抗体または基準抗体の対応するCDRで非フェニルアラニン残基を生じる位置でフェニルアラニン残基を含有することを特徴とする請求項1記載の高親和性中和免疫グロブリン。
【請求項14】
前記免疫グロブリンが高親和性H1 CDRを含み、また前記CDRが配列識別番号9および10より成るグループから選択されるアミノ酸配列を持つことを特徴とする請求項1記載の高親和性中和免疫グロブリン。
【請求項15】
前記免疫グロブリンが配列識別番号11のアミノ酸配列を持つ高親和性H3 CDRを含むことを特徴とする請求項1記載の高親和性中和免疫グロブリン。
【請求項16】
前記免疫グロブリンが高親和性L2 CDRを含み、また前記CDRが配列識別番号12および13より成るグループから選択されるアミノ酸配列を持つことを特徴とする請求項1記載の高親和性中和免疫グロブリン。
【請求項17】
前記免疫グロブリンが配列識別番号14,15および16より成るグループから選択されるアミノ酸配列を有する高親和性L3 CDRを持つことを特徴とする請求項1記載の高親和性中和免疫グロブリン。
【請求項18】
H1 CDRが配列識別番号9のアミノ酸配列を持ち、H3 CDRが配列識別番号11のアミノ酸配列を持ち、L2 CDRが配列識別番号4のアミノ酸配列を持ち、またL3 CDRが配列識別番号14のアミノ酸配列を持つことを特徴とする請求項1記載の高親和性中和免疫グロブリン。
【請求項19】
H1 CDRが配列識別番号9のアミノ酸配列を持ち、H3 CDRが配列識別番号11のアミノ酸配列を持ち、L2 CDRが配列識別番号12のアミノ酸配列を持ち、またL3 CDRが配列識別番号5のアミノ酸配列を持つことを特徴とする請求項1記載の高親和性中和免疫グロブリン。
【請求項20】
H1 CDRが配列識別番号10のアミノ酸配列を持ち、H3 CDRが配列識別番号11のアミノ酸配列を持ち、L2 CDRが配列識別番号12のアミノ酸配列を持ち、またL3 CDRが配列識別番号14のアミノ酸配列を持つことを特徴とする請求項2記載の高親和性中和免疫グロブリン。
【請求項21】
H1 CDRが配列識別番号9のアミノ酸配列を持ち、H3 CDRが配列識別番号11のアミノ酸配列を持ち、L2 CDRが配列識別番号12のアミノ酸配列を持ち、またL3 CDRが配列識別番号14のアミノ酸配列を持つことを特徴とする請求項2記載の高親和性中和免疫グロブリン。
【請求項22】
H1 CDRが配列識別番号9のアミノ酸配列を持ち、H3 CDRが配列識別番号11のアミノ酸配列を持ち、L2 CDRが配列識別番号12のアミノ酸配列を持ち、またL3 CDRが配列識別番号15のアミノ酸配列を持つことを特徴とする請求項2記載の高親和性中和免疫グロブリン。
【請求項23】
重鎖可変部が配列識別番号17を持ち、また重鎖可変部が配列識別番号18を持つことを特徴とする請求項1記載の高親和性中和免疫グロブリン。
【請求項24】
重鎖可変部が配列識別番号19を持ち、また重鎖可変部が配列識別番号20を持つことを特徴とする請求項1記載の高親和性中和免疫グロブリン。
【請求項25】
重鎖可変部が配列識別番号21を持ち、また重鎖可変部が配列識別番号22を持つことを特徴とする請求項2記載の高親和性中和免疫グロブリン。
【請求項26】
重鎖可変部が配列識別番号23を持ち、また重鎖可変部が配列識別番号24を持つことを特徴とする請求項2記載の高親和性中和免疫グロブリン。
【請求項27】
重鎖可変部が配列識別番号25を持ち、また重鎖可変部が配列識別番号26を持つことを特徴とする請求項2記載の高親和性中和免疫グロブリン。
【請求項28】
少なくとも1010M−1の親和性定数を持つ一つの組換え高親和性中和免疫グロブリンであって、ここで前記免疫グロブリンが、少なくともその部分がヒト抗体から誘導されるヒト定常部とヒト重鎖および軽鎖枠組み構造領域を含むことを特徴とする組換え高親和性中和免疫グロブリン。
【請求項29】
重鎖および軽鎖枠組み構造領域がヒト抗体のコンセンサス配列から誘導されることを特徴とする請求項28記載の組換え高親和性中和免疫グロブリン。
【請求項30】
親和性定数が少なくとも1011M−1であることを特徴とする請求項29記載の組換え高親和性中和免疫グロブリン。
【請求項31】
重鎖および軽鎖枠組み構造領域がヒト抗体のコンセンサス配列から誘導されることを特徴とする請求項30記載の組換え高親和性中和免疫グロブリン。
【請求項32】
請求項1記載の免疫グロブリンが薬理許容担体に懸濁されていることを特徴とする前記免疫グロブリンを含む一つの組成物。
【請求項33】
請求項6記載の免疫グロブリンが薬理許容担体に懸濁されていることを特徴とする前記免疫グロブリンを含む一つの組成物。
【請求項34】
疾病を予防しおよびまたは処置する一つの方法であって、請求項32記載の免疫グロブリン組成物の治療有効量をその疾病の危険にさらされまたはそれに悩む患者に投与することを含むことを特徴とする方法。
【請求項35】
ウイルス誘導疾病を予防しおよびまたは処置する一つの方法であって、請求項33記載の免疫グロブリン組成物の治療有効量をその疾病の危険にさらされまたはそれに悩む患者に投与することを含むことを特徴とする方法。
【請求項36】
呼吸合胞体ウイルスを予防しおよびまたは処置する一つの方法であって、薬理許容担体に懸濁された請求項7記載の免疫グロブリンの治療有効量をそのウイルスの危険にさらされまたはそれに悩む患者に投与することを含むことを特徴とする方法。
【請求項37】
請求項3記載の高親和性中和免疫グロブリンであって、ここで前記免疫グロブリンがFab、F(ab)’2、重鎖−軽鎖二量体、重鎖および軽鎖より成るグループから選択されることを特徴とする高親和性中和免疫グロブリン。
【請求項38】
請求項4記載の高親和性中和免疫グロブリンであって、ここで前記免疫グロブリンがFab、F(ab)’2、重鎖−軽鎖二量体、重鎖および軽鎖より成るグループから選択されることを特徴とする高親和性中和免疫グロブリン。
【請求項39】
請求項1記載の高親和性中和免疫グロブリンであって、ここで前記免疫グロブリンが抗体であることを特徴とする高親和性中和免疫グロブリン。
【請求項40】
請求項2記載の高親和性中和免疫グロブリンであって、ここで前記免疫グロブリンが抗体であることを特徴とする高親和性中和免疫グロブリン。
【請求項41】
請求項6記載の高親和性中和免疫グロブリンであって、ここで前記免疫グロブリンがFab、F(ab)’2、重鎖−軽鎖二量体、重鎖および軽鎖より成るグループから選択されることを特徴とする高親和性中和免疫グロブリン。
【請求項42】
請求項7記載の高親和性中和免疫グロブリンであって、ここで前記免疫グロブリンがFab、F(ab)’2、重鎖−軽鎖二量体、重鎖および軽鎖より成るグループから選択されることを特徴とする高親和性中和免疫グロブリン。
【請求項43】
請求項8記載の高親和性中和免疫グロブリンであって、ここで前記免疫グロブリンがFab、F(ab)’2、重鎖−軽鎖二量体、重鎖および軽鎖より成るグループから選択されることを特徴とする高親和性中和免疫グロブリン。
【請求項44】
疾病を予防およびまたは処置する一つの方法であって、薬理許容担体に懸濁された請求項37または38記載の免疫グロブリンの治療有効量をその疾病の危険にさらされまたはそれに悩む患者に投与することを含むことを特徴とする方法。
【請求項45】
ウイルス誘導疾病を予防およびまたは処置する一つの方法であって、薬理許容担体に懸濁された請求項41記載の免疫グロブリンの治療有効量をその疾病の危険にさらされまたはそれに悩む患者に投与することを含むことを特徴とする方法。
【請求項46】
呼吸合胞体ウイルスを予防およびまたは処置する一つの方法であって、薬理許容担体に懸濁された請求項42または43記載の免疫グロブリンの治療有効量をその疾病の危険にさらされまたはそれに悩む患者に投与することを含むことを特徴とする方法。
【図1】


【図2】
【図3】

【図4】

【図5】

【図6】

【図7】

【図8】
【図9】
【図10】
【図2】
【図3】
【図4】
【図5】
【図6】
【図7】
【図8】
【図9】
【図10】
【公開番号】特開2010−180207(P2010−180207A)
【公開日】平成22年8月19日(2010.8.19)
【国際特許分類】
【出願番号】特願2010−26299(P2010−26299)
【出願日】平成22年2月9日(2010.2.9)
【分割の表示】特願2001−561064(P2001−561064)の分割
【原出願日】平成13年1月26日(2001.1.26)
【出願人】(504333972)メディミューン,エルエルシー (108)
【出願人】(502274174)アプライド モレキュラー エボルーション,インコーポレイテッド (2)
【Fターム(参考)】
【公開日】平成22年8月19日(2010.8.19)
【国際特許分類】
【出願日】平成22年2月9日(2010.2.9)
【分割の表示】特願2001−561064(P2001−561064)の分割
【原出願日】平成13年1月26日(2001.1.26)
【出願人】(504333972)メディミューン,エルエルシー (108)
【出願人】(502274174)アプライド モレキュラー エボルーション,インコーポレイテッド (2)
【Fターム(参考)】
[ Back to top ]