
転写因子−E2F−5
【課題】E2F遺伝子ファミリーに属する2つの新規転写因子を開示する。
【解決手段】これらは、ヒトおよびマウスE2F−5である。それらは、DP−1およびp130と相互作用し得る。
【解決手段】これらは、ヒトおよびマウスE2F−5である。それらは、DP−1およびp130と相互作用し得る。
【発明の詳細な説明】
【技術分野】
【0001】
本発明は、新規転写因子およびその製造および用途に関する。
【背景技術】
【0002】
遺伝子発現を細胞周期の進行と同調させるためには、細胞周期中に生じる分子イベントが転写装置と統合される必要がある。
【0003】
最近、E2F(またはDRTF1)と称される転写因子が同定され、抗がん遺伝子またはがん抑制遺伝子である網膜芽細胞腫感受性遺伝子のタンパク質産物pRbに結合することが示された(例えば、非特許文献1参照)。細胞転写因子E2Fは、網膜芽細胞腫遺伝子産物(pRb)、p107、サイクリン、サイクリン依存性キナーゼなどの重要な細胞周期調節タンパク質と結合し、さらに、その転写活性は、アデノウイルスEla、SV40ラージT抗原、ヒトパピローマウイルスE7タンパク質などのある種のウイルスがんタンパク質によりモジュレーションされるため、E2Fは、細胞周期制御における鍵成分として機能していると広く考えられている。
【0004】
転写因子E2F(またはDRTF1)は、哺乳動物細胞で細胞周期が進行する間、細胞増殖の重要な調節体であることが知られている分子との一連の周期的相互作用を受けるため、細胞周期イベントと転写装置との統合において重要な役割を果たしていると考えられている。例えば、G1期からS期への進行を負に調節し腫瘍細胞中でしばしば修飾される網膜芽細胞腫がん抑制遺伝子産物(pRb)は、E2Fに結合する。同様に、pRb関連タンパク質p107は、主として、E2FとのS期複合体中に生じる。pRbおよびp107は共に、E2Fの転写活性を抑制し、一方、E2F結合部位は、c−myc、p34cdc2などの増殖に関与する種々の遺伝子の制御領域中に生じるため、E2Fの転写活性は、細胞増殖の調節に基本的に重要であると考えられている。さらに、腫瘍細胞から単離された対立遺伝子によりコードされる突然変異Rbタンパク質は、E2Fに結合しない。したがって、この突然変異Rbタンパク質は、E2F部位依存性転写活性化を阻害する能力を有さない。E2Fのもう1つの重要な特徴は、アデノウイルスEla、SV40ラージT抗原、ヒトパピローマウイルスE7などのある種のウイルスがんタンパク質が、不活性転写因子からpRbおよびp107を隔離することにより、その活性をモジュレーションすることである。この効果の発現に要求されるのは、組織培養細胞がトランスフォーメーションされ、成長制御に打ち勝つのに必要な、これらのウイルスタンパク質中の領域である。したがって、これらのがんタンパク質がE2Fを調節する能力は、該がんタンパク質が細胞成長制御の正常なメカニズムを圧倒するための手段であるかもしれないし、逆に、pRbによる転写抑制がpRb媒介性の負の成長制御の基礎であるかもしれない。
【0005】
pRbおよびp107の転写−調節特性を他の細胞周期イベントと統合するための可能性のあるメカニズムは、E2F複合体中のサイクリンAおよびcdc2関連サイクリン依存性キナーゼp33cdk2の同定から示唆された。サイクリンAはS期を経る進行(おそらく、サイクリン依存性キナーゼp33cdk2をE2FへリクルートするサイクリンAの能力により媒介されると考えられる機能)に必要である。これらのデータを総合すると、転写因子E2Fの主要な役割は、細胞周期イベントをpRb、p107、サイクリン、cdkなどの分子を介して転写装置へ中継することであると示唆され、したがってこのことは、遺伝子発現が細胞周期の進行と同調し統合されることを保証するものである。
【0006】
さらに最近になって、E2Fの性質を有する転写因子がクローニングされ配列決定された(非特許文献2及び3を参照)。
【非特許文献1】WagnerおよびGreen, Nature 352, 189-190, 1991
【非特許文献2】Helinら, Cell 70 (1992), 337-350
【非特許文献3】Kaelinら, Cell 70 (1992), 351-364
【発明の開示】
【0007】
驚くべきことに、本発明者らは、E2F遺伝子ファミリーの新規メンバーと考えられる、さらに2つの新規タンパク質を見いだし、これをE2F−5と命名した。ヒトE2F−5のcDNA配列を、このタンパク質のアミノ酸配列と共に図1Aに示す。マウスE2F−5についての対応する配列を図9Aに示す。これらの新規タンパク質は、E2F−5と称され、本明細書ではこの名称を使用することにする。
【0008】
E2F−5は、関連転写因子成分のファミリーの1つであることが見いだされた。このファミリーのメンバーは、DPタンパク質と相互作用して一連の転写因子を形成すると考えられている。DPタンパク質(またはポリペプチド)には、DP−1、DP−2およびDP−3が含まれるが、通常は、これらの最初に挙げたものが残りのものに優先すると考えられる。
【0009】
本発明は、その第1の態様において、図1Aまたは9Aに示すタンパク質、その同族体、並びに該配列および該同族体の断片であって、哺乳動物転写因子として機能しうるものを提供する。特に、本発明は、
(a)E2F−5、
(b)図1Aまたは9Aのタンパク質、
(c)(a)または(b)の突然変異体、対立遺伝子変異体または種同族体、
(d)(a)または(b)に少なくとも70%相同なタンパク質、
(e)DPタンパク質、pRb、p107および/またはp130と複合体を形成する能力を有する(a)から(d)のいずれか1つの断片、または
(f)少なくとも15のアミノ酸長の(a)から(e)のいずれかの断片、
を含んでなるポリペプチド(好ましくは実質的に単離された形態のもの)を提供する。
【0010】
以下、この定義に包含されるすべてのポリペプチドを、本発明のポリペプチドと称する。
【0011】
タンパク質pRb、p107、DPタンパク質およびp130は本発明のタンパク質と複合体を形成しうるので、本明細書では、これらを、複合体形成タンパク質(complexingproteins)または「複合体形成体(complexors)」と称する。E2F−5は、ある条件の下では、pRbと弱くしか結合できないかもしれない。
【0012】
本発明のポリペプチドが、その天然環境(例、体内)で結合しうる他のポリペプチドを含まない形態である場合、本発明のポリペプチドは、実質的に単離された形態である。本発明のポリペプチドは、該ポリペプチドの意図する目的を妨げない担体または希釈剤と混合してもよく、その場合であっても、実質的に単離されているとみなされると理解される。
【0013】
また、本発明のポリペプチドは、実質的に精製されている形態であり、この場合一般には、調製物中に含まれるポリペプチドの90%以上、例えば95%、98%または99%が本発明のポリペプチドである。
【0014】
突然変異ポリペプチドは、アミノ酸残基の付加、欠失または置換である1以上の突然変異を有する。好ましくは、該突然変異は、該ポリペプチドの構造および/または機能および/または性質に、全くあるいは実質的に影響を及ぼさない。したがって、突然変異体は、適切には、DPタンパク質、pRb、p107および/またはp130と複合体を形成しうる能力を有する。突然変異体は、天然に生じるもの(すなわち、天然源から精製または単離されている形態)であっても、合成的に得られるもの(例えば、コード化DNA上の部位特異的突然変異誘発を行うことによるもの)であってもよい。したがって、本発明のポリペプチドは、天然に生じるものであっても、組換え体(すなわち、遺伝子工学技術を用いて調製されたもの)であってもよいことは明らかである。
【0015】
対立遺伝子変異体は、ヒトまたは動物(例、マウス)において天然に生じ、図1Aまたは9Aのタンパク質と実質的に同様に遺伝子発現を調節するように機能する変異体である。
【0016】
同様に、上記タンパク質の種同族体は、別の種で天然に生じ、その種において図1Aまたは9Aのタンパク質と同等の機能を発揮する同等のタンパク質である。いずれか1つの種内に、同族体がいくつかの対立遺伝子変異体として存在していてもよく、これらはすべて上記タンパク質の同族体と考えられる。対立遺伝子変異体および種同族体は、図1Aまたは9Aのタンパク質の製造に関して本明細書に記載されている方法に従い、対立遺伝子変異体を保持する適当な細胞源(例えば、ヒトまたはげっ歯類からのもの)または別の種上でそのような方法を行うことにより得ることができる。上記タンパク質は進化的に保存されている場合があるため、対立遺伝子または種変異体をコードするクローンを得るために、本発明のポリヌクレオチドを使用して、ヒト、げっ歯類または他の細胞から作製されたライブラリーをプローブすることも可能である。上記クローンを通常の技術により操作して、本発明のポリペプチドを同定し、ついでそれ自体が公知の組換えまたは合成技術により該ポリペプチドを製造することができる。好ましい種同族体には、哺乳動物または両生類種同族体が含まれる。
【0017】
図1Aまたは9Aのタンパク質に少なくとも70%相同なタンパク質、および図1Aまたは9Aのタンパク質に少なくとも80または90%、より好ましくは少なくとも95%相同なタンパク質が本発明に包含される。これは、一般に、少なくとも20、好ましくは少なくとも30、例えば少なくとも40、60または100以上連続するアミノ酸の領域にわたる。タンパク質の相同性を測定する方法は当該技術分野で公知であるので、上述のように当業者に理解されるであろう。相同性は、通常、アミノ酸の同一性に基づいて計算される(「硬相同性(hard homology)」と称されることもある)。
【0018】
一般に、上記複合体形成体と複合体を形成する能力を有する、図1Aまたは9Aのポリペプチドまたはその対立遺伝子変異体またはその種同族体の断片は、少なくとも10、好ましくは少なくとも15、例えば少なくとも20、25、30、40、50または60のアミノ酸長である。
【0019】
断片がタンパク質の複合体と複合体を形成するか否かの判定は、複合体形成タンパク質および該断片を、それらが正常にトランス活性化転写因子を形成する条件下で用意し、そして複合体が形成されたか否かを判定することにより行うことができる。この判定は、例えば、上記複合体がE2F結合部位にin vitroで結合する能力を測定することにより、あるいは、SDS−PAGEなどの方法により推定複合体の分子量を測定することにより行ってもよい。
【0020】
好ましい断片には、DP−1または他の複合体形成体とトランス活性化複合体を形成する能力を有する断片が含まれる。本明細書中の実施例では、上記タンパク質の機能を分析するためのいくつかの方法を記載する。ポリペプチドがDP−1タンパク質とトランス活性化複合体を形成する能力を有するか否かを評価するために、上記実施例を適合させてもよい。例えば、DP−1/E2F−5複合体により活性化されるように適合されたレポーター遺伝子構築物の存在下、上記ポリペプチドを上記複合体形成体へ加えることができる(例えば、Medical Research CouncilのWO-A-94/10307の図10を参照されたし)。そのような実験により、上記ポリペプチド断片が必要な活性を有するか否かを判定することができる。
【0021】
本発明のポリペプチドは、表示的または検出可能な標識で標識してもよい。上記(表示的)標識は、上記ポリペプチドの検出を可能にする、どのような適当な標識であってもよい。適当な標識としては、放射性同位元素(例えば125I)、酵素、抗体、リンカー(例えばビオチン)などが挙げられる。サンプル中のE2F−5タンパク質の量を測定するために、イムノアッセイなどの診断方法において本発明の標識ポリペプチドを使用してもよい。
【0022】
本発明のポリペプチドまたは標識ポリペプチドを固相、例えばイムノアッセイディッシュの壁に固定してもよい。
【0023】
本発明の第2の態様は、
(a)図1Aまたは9Aに示すヌクレオチドの配列、
(b)(a)に相補的な配列、
(c)(a)または(b)のいずれかの中の配列と選択的にハイブリッド形成する能力を有する配列、
(d)第1の態様で定義されたポリペプチドをコードする配列、または、
(e)(a)から(d)の中の配列のいずれかの断片、
を含んでなるポリヌクレオチドに関する。
【0024】
したがって、本発明は、図1Aまたは9Aの配列または相補的配列と選択的にハイブリッド形成する能力を有するヌクレオチドの連続した配列を含んでなる、好ましくは実質的に単離または精製された形態のポリヌクレオチドを提供する。本発明のポリヌクレオチドは、図1Aまたは9AのDNA配列および図1Aまたは9AのDNA配列と選択的にハイブリッド形成する能力を有するその断片を含む。本発明は、そのさらに別に実施態様において、図1Aまたは9Aのタンパク質をコードするDNAまたはその断片を提供する。
【0025】
また、上記ポリヌクレオチドは、RNAを含んでなるものであってもよい。また、それは、その中に合成または修飾ヌクレオチドを含むポリヌクレオチドであってもよい。オリゴヌクレオチドに対するいくつかの異なる型の修飾が当該技術分野で公知である。これらには、メチルホスホナートおよびホスホロチオナートバックボーン、該分子の3'および/または5'末端におけるアクリジンまたはポリリシン鎖の付加が含まれる。本発明の目的のためには、本明細書に記載のオリゴヌクレオチドは、当該技術分野で利用可能ないかなる方法により修飾してもよいと理解されるべきである。治療方法で使用される本発明のオリゴヌクレオチドのin vivo活性または寿命を増加させるために、そのような修飾を行ってもよい。
【0026】
図1Aまたは9AのDNAと選択的にハイブリッド形成する能力を有するポリヌクレオチドは、一般に、少なくとも20、好ましくは少なくとも30、例えば少なくとも40、60または100以上の連続ヌクレオチドの領域内において、図1Aまたは9AのDNAに対して、少なくとも70%、好ましくは少なくとも80または90%、最適には少なくとも95%相同である。これらのポリヌクレオチドは、本発明に包含される。
【0027】
本発明のポリヌクレオチドが、その天然環境(例えば体内)で結合しうる他のポリヌクレオチドを含まない形態である場合、本発明のポリヌクレオチドは、実質的に単離された形態である。上記ポリヌクレオチドは、意図する目的を妨げない担体または希釈剤と混合してもよく、その場合であっても、該ポリヌクレオチドは実質的に単離されているとみなされると理解される。
【0028】
本発明のポリヌクレオチドは、プライマー(例えばPCRプライマー)、プローブ(例えば放射活性または非放射活性標識を用いる通常の手段により表示的または検出可能な標識で標識されているプローブ)を製造するのに使用してもよいし、該ポリヌクレオチドをベクター中にクローニングしてもよい。そのようなプライマー、プローブおよび図1Aまたは9AのDNAの他の断片は、少なくとも15、好ましくは少なくとも20、例えば少なくとも25、30または40のヌクレオチド長であり、それらも本発明に包含される。
【0029】
本発明のDNAポリヌクレオチドなどのポリヌクレオチドは、組換え的、合成的または当業者が利用可能ないかなる手段により製造してもよい。また、本明細書に開示する技術を参照して、それをクローニングしてもよい。
【0030】
本発明は、本発明のポリヌクレオチドおよびその相補体を含んでなる二本鎖ポリヌクレオチドを含む。
【0031】
本発明の第3の態様は、本発明のポリヌクレオチド、特にDNAまたはRNAポリヌクレオチドの複製および発現に適したベクター(例えば発現ベクター)に関する。上記ベクターは、例えば、複製起点、所望によりポリヌクレオチドの発現のためのプロモーターおよび所望により該プロモーターの調節体が設けられている、プラスミド、ウイルスまたはファージベクターであってもよい。上記ベクターは、1以上の選択マーカー遺伝子、例えば細菌プラスミドの場合にはアンピシリン耐性遺伝子または哺乳動物ベクターの場合にはネオマイシン耐性遺伝子を含有していてもよい。上記ベクターは、例えばRNAの製造などにin vitroで使用してもよいし、宿主細胞をトランスフェクションまたは形質転換させるために使用してもよい。また、上記ベクターは、例えば遺伝子治療方法などにin vivoで使用できるように適合させてもよい。
【0032】
この第3の態様のベクターは、好ましくは、複製可能な組換えベクターである。したがって、上記ベクターを使用して上記DNAを複製してもよい。好ましくは、上記ベクター中のDNAは、宿主細胞による上記コーディング配列の発現を付与する能力を有する制御配列に機能を発揮できるように結合している。「機能を発揮できるように結合している」という語は、記載成分が、それらが意図されるように機能できる関係で並んでいることを意味する。コーディング配列に「機能を発揮できるように結合している」制御配列は、該制御配列に適合した条件下で該コーディング配列の発現が達成されるように連結されている。本発明のポリペプチドの発現を付与するために、そのようなベクターを、適当な宿主細胞中へ形質転換またはトランスフェクションしてもよい。
【0033】
したがって、本発明のさらに別の態様は、上記第3の態様のベクターにより形質転換またはトランスフェクションされた宿主細胞に関する。これは、図1Aまたは9Aの配列またはそのオープンリーディングフレームを含む本発明のポリヌクレオチドの複製および発現を可能にする。上記細胞としては、上記ベクターに適合するものが選択され、例えば、細菌、酵母、昆虫または哺乳動物であってもよい。
【0034】
また、アンチセンスRNAを製造するために、本発明のポリヌクレオチドを上記ベクター中にアンチセンス配向で挿入してもよい。また、アンチセンスRNAまたは他のアンチセンスポリヌクレオチドを、合成的手段により製造してもよい。そのようなアンチセンスポリヌクレオチドを、細胞中のE2F−5タンパク質のレベルを制御するための方法に使用してもよい。そのような方法は、E2F−5mRNAからタンパク質への翻訳レベルを阻害または減少させるのに有効な量の上記アンチセンスポリヌクレオチドを上記細胞中に導入することを含んでいてもよい。上記細胞は、腫瘍細胞などの非制御的に増殖する細胞であってもよい。
【0035】
したがって、本発明は、第5の態様において、上記第3の態様の(発現)ベクターを用いて、本発明のポリペプチドをコードするコーディング配列の発現(該ベクターによる)を付与する条件下で形質転換またはトランスフェクションした宿主細胞を培養し、そして、発現されたポリペプチドを回収することを含んでなる、本発明のポリペプチドの製造法を提供する。
【0036】
また、本発明は、第6の態様において、本発明のポリペプチドに特異的な抗体(モノクローナルまたはポリクローナル抗体)を提供する。本発明の抗体は、該抗体の結合活性を保持するその断片および突然変異体を含む。さらに、本発明は、本発明のポリペプチドに対するモノクローナルまたはポリクローナル抗体の製造法を提供する。モノクローナル抗体は、上記タンパク質またはそのペプチド断片を免疫原として用いる通常のハイブリドーマ技術により製造してもよい。また、ポリクローナル抗体は、本発明のポリペプチドを宿主動物(例、ラット、ウサギ)に接種し、そして免疫血清を回収することを含んでなる通常の手段により製造してもよい。
【0037】
モノクローナル抗体の抗原結合活性を保持することができる該モノクローナル抗体の断片(例えば、Fv、F(ab')およびF(ab2)'断片)が、本発明のこの態様に含まれる。さらに、本発明のモノクローナル抗体を分析し(例えば、かかる抗体を発現する遺伝子のDNA配列分析による)、例えばEP-A-0239400(Winter)に開示されている方法に従い、本発明の抗体の相補性決定領域を有するヒト化抗体を作製してもよい。
【0038】
さらに本発明は、本発明の抗体またはその断片と担体または希釈剤とを含んでなる組成物を提供する。そのような組成物には医薬組成物が含まれ、この場合、上記担体または希釈剤は薬学的に許容されるものである。
【0039】
本発明のポリペプチドは、担体または希釈剤と共に組成物中に存在していてもよい。これらの組成物には医薬組成物が含まれ、この場合、上記担体または希釈剤は薬学的に許容されるものである。
【0040】
薬学的に許容される担体または希釈剤には、経口、直腸、鼻内、局所(頬、舌下など)、膣または非経口(皮下、筋内、静脈内、皮内、脊髄内、硬膜外)投与に適した製剤中で用いられるものが含まれる。上記製剤は単位投与形で提供されるのが好都合であり、薬学分野でよく知られている方法のいずれにより製造してもよい。そのような方法は、有効成分を、1以上の補助成分を構成する担体と混合する工程を含む。一般に、上記製剤は、有効成分と液体担体若しくは微小固体担体またはその両方とを均一かつ十分に混合し、ついで生成物を必要に応じて成型することにより製造する。
【0041】
例えば、非経口投与に適した製剤には、抗酸化剤、緩衝剤、静菌薬、および目的の受容者の血液と該製剤とを等張にする溶質を含有していてもよい水性または非水性の無菌注射溶液;並びに懸濁化剤、濃稠化剤、および該ポリペプチドを血液成分または1以上の器官へ標的化するように設計されているリポソームまたは他の微小粒子系を含んでいてもよい水性または非水性の無菌懸濁液が含まれる。
【0042】
本発明のポリペプチド、本発明のポリペプチドに対する抗体またはその断片および前記組成物は、ヒトなどの哺乳動物における増殖性疾患などの状態の治療、調節または診断に使用しうる。そのような状態には、DPまたはE2Fタンパク質または関連ファミリーメンバーなどの1以上の転写因子の異常な(例えば、異常に高いまたは低いレベルの)および/または異常な(例えば、遺伝子配列の突然変異によるもの)発現が含まれる。また、この状態には、図1Aまたは9Aのタンパク質により調節される遺伝子産物を与える遺伝子の異常発現により引き起こされる状態が含まれる。上記ペプチド、抗体、その断片、組成物などを用いる状態の治療または調節には、通常、そのような治療を必要とする受容者に、ポリペプチド、抗体、その断片または組成物の有効量を適宜投与することが含まれる。
【0043】
また本発明は、E2F−5−DPタンパク質複合体の形成の妨害を介して転写因子の活性化を阻害するためにこの領域に標的化されている抗体およびその断片を提供する。
【0044】
さらに本発明は、サンプル中の本発明のポリペプチドの存在または不存在を検出するためのイムノアッセイを行う方法であって、
(a)本発明の抗体を用意し、
(b)抗体−抗原複合体の形成を許容する条件下、該サンプルを該抗体と共にインキュベーションし、そして
(c)該抗体−抗原複合体が存在すれば、これを検出する、
ことを含んでなる前記方法を提供する。
【0045】
もう1つの態様において本発明は、増殖性またはウイルス性の疾患の治療のための推定化学療法剤を同定するための新規アッセイであって、DPタンパク質またはその誘導体、本発明のポリペプチドおよび推定化学療法剤を接触させ、そして、該剤がDP/E2F−5タンパク質複合体の形成を阻害する程度を測定することを含んでなる前記新規アッセイを提供する。このアッセイでは、それぞれのタンパク質が十分に用意され、上記剤の不存在下のアッセイ条件下でDP−1および/またはE2F−5タンパク質がヘテロ二量体を形成する限り、完全なDP−1および/またはE2F−5タンパク質を使用する必要はないかもしれない。
【0046】
DP−1(およびE2F1、2および3)のクローニングおよび配列決定は、当該技術分野で公知であり、DP−1に対する抗体の組換え発現および製造は、WO-A-94/10307に記載されている。
【0047】
したがって、本発明は、増殖性疾患の治療のための推定化学療法剤を同定するためのスクリーニング方法であって、
(A)(i) DPポリペプチド、
(ii) 上記第1の態様のポリペプチド、および
(iii) 推定化学療法剤、
を、成分(i)と(ii)とが(iii)の不存在下で複合体を形成する条件下で接触させ、そして
(B)成分(iii)が上記複合体を阻害しうる程度を測定する、
ことを含んでなる前記スクリーニング方法を提供する。
【0048】
このアッセイでは、アッセイの結果を測定できるように、その3つの成分のいずれか1つ以上を、例えば放射活性または比色標識により標識してもよい。推定化学療法剤には、本発明のペプチドが含まれる。
【0049】
DPタンパク質の変異体、同族体および断片は、E2F−5タンパク質の変異体、同族体および断片と対応させて定義される。
【0050】
(i)と(ii)との複合体は、例えば、それがE2F DNA結合部位にin vitroで結合する能力により測定してもよい。あるいは、上記アッセイは、レポーター遺伝子に結合したE2F結合部位を含んでなるプロモーターを上記複合体が活性化する能力を測定するin vivoアッセイであってもよい。このin vivoアッセイは、例えば、酵母、昆虫、両生類または哺乳動物細胞におけるかかるアッセイを示す実施例を参照することにより行ってもよい。
【0051】
上記アッセイで測定しうる候補治療剤には、上記第1の態様のポリペプチドだけでなく、特に、
(a)図1Aまたは9Aのタンパク質、
(b)その対立遺伝子変異体または種同族体、または
(c)(a)に少なくとも70%相同なタンパク質、
の10以上のアミノ酸の断片も含まれる。
【0052】
本発明のポリヌクレオチドまたは本発明のポリペプチドをコードする核酸を保持するベクターを、遺伝子治療方法に使用してもよい。そのような遺伝子治療は、細胞(例えば腫瘍細胞)の非制御増殖を治療するのに使用してもよい。遺伝子治療の方法は、治療が必要な患者中の細胞内で本発明のアンチセンスポリヌクレオチドを発現する能力を有するベクターの有効量を該細胞へ輸送して、E2F−5mRNAからE2F−5タンパク質またはポリペプチド(E2F−5がDPタンパク質または関連ファミリーメンバーへ結合するのを妨害するもの)への翻訳を阻害または抑制することを含む。
【0053】
上記ベクターは、好ましくは、ウイルスベクターである。このウイルスベクターは、腫瘍細胞を標的化するのに当該技術分野で利用可能ないかなる適当なベクターであってもよい。例えば、Huberら(Proc. Natl. Acad. Sci. USA (1991) 88, 8039)は、ヘパトーム、乳、結腸または皮膚細胞の形質転換のための、両種性レトロウイルスの使用を報告している。また、Culverら(Science (1992)256; 1550-1552)は、Ramら(Cancer Research (1993)53; 83-88)と同様、ウイルス特異的酵素プロドラッグ療法(virus-directed enzyme prodrug therapy)でのレトロウイルスベクターの使用を記載している。Englehardtら(Nature Genetics (1993)4;27-34)は、細胞中への嚢胞性線維症膜貫通コンダクタンス産物(CFTR)の輸送における、アデノウイルスに基づくベクターの使用を記載している。
【0054】
本発明は、いくつかのアッセイを意図する。一般には、これらは以下のように分類することができる。
【0055】
1.E2F−5トランス活性化(trans-activation)の阻害剤(すなわち、転写の活性化の阻害)を見つけるためのアッセイの実施。したがって、この阻害剤は、DNAに対するE2F−5の結合(通常はE2F結合部位で生じる)を阻害するかもしれない。適当と考えられる阻害剤はタンパク質であり、これはp107と同様または同一の効果を有していてもよい。したがって、適当な阻害分子は、上記第1の態様のタンパク質で定義したのと同様に、p107の断片、突然変異体、対立遺伝子変異体または種同族体を含んでなる。
【0056】
2.(ヘテロ)二量体形成の阻害剤のアッセイ。そのような阻害剤は、E2F−5(または上記第1の態様のポリペプチド)と複合体形成体(例えば、DP−1などのDPタンパク質)との二量体形成を妨害するかもしれない。もちろん、この阻害剤は、上記第1の態様のタンパク質で定義したのと同様のDPタンパク質の断片、突然変異体、対立遺伝子変異体または種同族体であってもよい。
【0057】
3.アッセイの第3のカテゴリーは、リン酸化の阻害剤を見つけることである。E2F−5(および上記第1の態様の他のタンパク質)はリン酸化により活性化されると考えられている。したがって、リン酸化の阻害剤は、E2F−5のトランス活性化特性を阻害すると考えられる(したがって、結局、この阻害剤は、上記の2つのアッセイのいずれかで見いだされた阻害剤と同じ効果を有するかもしれない)。リン酸化はcdkにより引き起こされ、したがって、このリン酸化の阻害剤は、そのようなアッセイにより意図されるものである。
【0058】
本発明は、いくつかの治療的用途を意図する。例えば、E2F−5に対してアンチセンスである配列の核酸を用いる遺伝子治療などである。さらに、DP−1タンパク質に結合し、それによって該DPタンパク質と不活性複合体を形成しうる分子を意図する。適当な分子には、E2F−5自体以外に上記第1の態様の分子が含まれる。そのような分子は、E2F−5の突然変異体であってもよく、該分子は、当該技術分野ではしばしば、ドミナントネガティブ分子と称される。
【0059】
本発明は、細胞の非制御増殖に基づく疾患、または非制御増殖が重要または必須の病理学的側面である疾患の治療または予防を意図する。これには、がん、ウイルス性疾患、自己増殖自体、および自己免疫疾患、例えば乾癬が含まれる。分裂細胞(例えば造血幹細胞および/または骨髄細胞)の成長を一時的に阻害したい場合があるかもしれない。これらの観点では、一般には、E2F−5の活性を予防、阻害または妨害することが求められる。
【0060】
これに対して、E2F−5発現を増加させることにより、例えば、過剰発現を促進または誘導することにより、いくつかの疾患および状態を治療することができる。これは、好ましくは、アポトーシス(プログラムされた細胞死として知られていることもある)を引き起こす。E2F−5タンパク質の過剰発現は、細胞死を引き起こすことがある。したがって、この態様は、がんの治療にも用いることができる。よって、1つの目的は、E2F−5の活性を増加させることである。E2F−1でも同様の用途が公知である(Qinら, PNAS USA 91(印刷中))。
【0061】
E2F−5遺伝子は腫瘍細胞中で突然変異しうることに留意すべきである。その場合には、突然変異遺伝子を、突然変異から生じる状態の診断に使用することができる。また、それは、突然変異遺伝子による治療に有用である。
【発明を実施するための最良の形態】
【0062】
以下の2つの実施例では、それぞれ、ヒトおよびマウス源からの本発明の新規タンパク質およびDNAの単離および特性決定について説明する。しかしながら、他の源(例えば哺乳動物源)も本発明の範囲内に包含され、上記タンパク質の他の哺乳動物同族体も同様の方法で単離されうる。本明細書に記載の実施例は例示にすぎず、本発明を何ら限定するものではない。
実施例1
概要
E2FDNA結合部位は、細胞周期中に発現が厳しく調節されている多数の遺伝子中で見いだされる。E2F転写因子の活性は、E2Fトランス活性化ドメインに結合しこれを阻害しうる特異的なリプレッサー分子との結合により調節される。E2F−1、2および3の場合、該リプレッサーは、網膜芽細胞腫遺伝子pRbの産物である。E2F−4は、pRb関連p107とは相互作用するが、pRb自体とは相互作用しない。最近、網膜芽細胞腫遺伝子ファミリーの第3のメンバーであるp130をコードするcDNAが単離された。p130も、主に細胞周期のG0期でE2FDNA結合活性と相互作用する。本発明者らは、本明細書において、E2F遺伝子ファミリーの第5のメンバーのクローニングを報告する。ヒトE2F−5cDNAは、346−アミノ酸のタンパク質(推定分子量38kDa)をコードする。E2F−5は、E2F−1(57%の類似度)よりもE2F−4(78%の類似度)と密接に関連している。E2F−5は、DP−1と協同的にコンセンサスE2F部位へ結合する点で、他のE2Fと類似している。本発明者らは、特異的なE2F−5抗血清を使用することにより、生理学的条件下でE2F−5が優先的にp130と相互作用することを示す。
緒言
E2Fは、E2F様およびDP様サブユニットよりなるヘテロ二量体転写因子のグループに与えられている名称である[27]。E2FDNA結合部位は、細胞周期中に発現が調節されるいくつかの遺伝子のプロモーター中に存在し、これらのE2F部位の存在がこれらの遺伝子の細胞周期調節発現に寄与することを示す証拠が存在する[13, 28, 38]。
【0063】
E2FDNA結合活性は、網膜芽細胞腫タンパク質(pRb)およびpRb関連p107およびp130と複合していることが判明している[6, 10, 29, 37]。このグループのタンパク質は、保存モチーフである「ポケット」を共有しており、該ポケットは、細胞タンパク質およびウイルスタンパク質の両方に対する結合に関与している。このため、pRb様タンパク質のグループは、一括して、ポケットタンパク質ファミリーとして知られている。E2Fと種々のポケットタンパク質との間の複合体は、その出現が細胞周期中で異なるため、細胞周期調節において異なる機能を有すると考えられる。pRbと複合体形成しているE2Fはほとんどが、細胞周期のG1期で見いだされる[5-7, 11]。これに対して、p107とE2Fとの複合体は細胞周期中に残存するが、それらの組成は可変的である。G1では、E2Fおよびp107の他に、サイクリンEおよびcdk2が存在する。S期では、E2F/p107複合体において、サイクリンEはサイクリンAで置換される[29, 37]。p107/E2F複合体中にこれらのサイクリン/cdk複合体が存在する機能的意義は、現在のところ明らかではない。静止細胞では、E2Fとp130との複合体が、最も優勢なE2FDNA結合種である。細胞が静止状態から脱するにつれて、この複合体は消失するが、このことは、p130と相互作用するE2F活性についての細胞周期進入における役割を示唆するものである[10]。
【0064】
E2Fが転写を活性化する能力は、該ポケットタンパク質との複合体形成により調節される。E2FとpRbとの間の複合体形成は、リン酸化による調節に左右される。pRbの低リン酸化種だけがE2Fと相互作用するが、このことは、サイクリン/cdk複合体によるpRbのリン酸化が、細胞周期中のE2FとpRbとの間の相互作用を制御することを示す[5-7, 11]。
【0065】
E2FDNA結合活性の強制発現が、細胞周期のG1期からSおよびG2/M期へ細胞を進行させ[3]、E2Fが静止細胞を刺激してDNA合成を開始させうる[23]という知見により、細胞周期調節におけるE2F転写因子の非常に重要な役割が強調される。重要なことは、E2Fの過剰発現が、活性化されたras癌遺伝子と共に、初代げっ歯類繊維芽細胞の癌トランスフォーメーションを引き起こすことである[3]。
【0066】
現在までに、4つの異なるE2F様ポリペプチドが単離されている。E2F−1、2および3は、pRbとの複合体として見いだされるにすぎないが、E2F−4はp107と優先的に相互作用する[3, 15, 19, 22, 24, 30, 36]。E2Fとp107との間およびE2Fとp130との間の複合体形成がどのように調節されるかについては現在のところ知られていない。E2F/p107複合体およびE2F/p130複合体の調節の検討を始めるために、本発明者らは、E2F遺伝子ファミリーの更に別のメンバーを探索した。本発明者らは、ここで、p130と優先的に相互作用するE2F遺伝子ファミリーの第5のメンバーのクローニングを報告する。
材料および方法
酵母二ハイブリッドスクリーニング
「ベイト」プラスミドpPC97-p107を含有する酵母Y190株[17]を、酢酸リチウム法[34]を用いて、14.5日齢のCD1マウス胎児ライブラリー[8]で形質転換した。ヒスチジンが無く25mM3−アミノトリアゾールで補足されたプレート上での成長に関して200万個の形質転換体を選択し、ついで、既に記載されている[12]とおりに、β−ガラクトシダーゼ活性に関して分析した。二重陽性酵母コロニーに由来するcDNAライブラリープラスミドを、種々のGal4-DBD融合プラスミド(pPC97-p107、pPC97-bmiおよびpPC97(挿入なし))による再形質転換により標的特異性に関して試験した。部分マウスE2F−5cDNAを用いて、更に別のヒトcDNAライブラリーをスクリーニングした。本明細書に記載の完全長ヒトE2F−5cDNAを、T84結腸癌ライブラリー(Stratagene)から単離した。
【0067】
プラスミド
pPC97[8]のGal4DNA結合ドメイン(アミノ酸1〜147)とインフレーム(in frame)となっているp107のポケット領域(アミノ酸240〜816)をクローニングすることにより、pPC97-p107を得た。pGEX-2T中でアミノ酸89〜200(A)またはアミノ酸89〜346(B)をコードするヒトE2F−5cDNA断片をクローニングすることにより、pGST-E2F-5(A)および(B)を構築した。トランスフェクション実験には、以下のプラスミドを使用した。pSG-Gal4-E2F-1は、ヒトE2F−1のアミノ酸284〜437を含有している[19]。pJ3-Gal4-E2F-4およびpJ3-Gal4-E2F-5は、pJ3Ω中、Gal4のDNA結合ドメインとインフレームとなっているE2F−4またはE2F−5のヒトcDNAの断片(E2F−4の場合はアミノ酸276〜412をコードし、E2F−5の場合はアミノ酸222〜346をコードする)をクローニングすることにより得た[33]。pJ3-E2F-5は、完全長ヒトE2F−5cDNA(3'非コーディング配列の最後の184ヌクレオチドを欠く)を哺乳動物発現ベクターpJ3Ω中にクローニングすることにより構築した。E2F−5の翻訳開始コドンの後には、モノクローナル抗体12CA5により認識される10アミノ酸のエピトープ(HA)が続く。
【0068】
pCMV-DP-1、pCMV-pRb、pCMV-p107、pCMV-p107DE、PCMV-pRb△22については、既に記載されている[20, 41]。
【0069】
細胞系
U2−OSおよびCAMA細胞を、それぞれ10%または20%のウシ胎児血清で補足されたダルベッコの改変イーグル培地(Dulbecco'smodified Eagle medium)(DMEM)中で培養した。
【0070】
リン酸カルシウム沈殿法[39]を用いてトランスフェクションを行った。
【0071】
CATアッセイ
5μg (Gal4)5-CAT[25]または2μg E2F4-CAT[20]、0.2μg RSV−ルシフェラーゼおよびヘリング精子担体DNA(全量を20μg/10cmプレートとする)と共に、示されている発現ベクターでU2−OS細胞を一過性にトランスフェクションした。既に記載されているとおりに[2, 3]、CATおよびルシフェラーゼ活性に関して細胞をアッセイした。
【0072】
ノーザンブロット分析
E2F−5発現分析のために、細胞系のパネルから全細胞質RNAを調製した。20mgの全細胞RNAを、記載されているとおりに[4]、1%ホルムアルデヒドアガロースゲル中で電気泳動し、ニトロセルロースに移し、[32P]−標識部分ヒトE2F−5cDNA(nt. 666〜1038)でプローブする。ついで、各レーンにローディングされるRNAの量を制御するために、同じフィルターをラットα−チューブリンcDNAでプローブした。
【0073】
免疫試薬および免疫沈降
E2F−5に対する抗体を産生させるために、GST-E2F-5(A)および(B)(プラスミドの項を参照されたし)タンパク質を E. coli 中で産生させ、グルタチオン−セファロースビーズを用いて精製した。等量の両タンパク質をウサギに注射した。3回の免疫の後、ポリクローナル血清を得た。
【0074】
E2F−1(KH20)、E2F−4(RK13)、pRb(XZ77)およびp107(SD−4および9)に対するモノクローナル抗体については、既に記載されている[3, 20, 21, 41]。p130(C20)ウサギポリクローナル抗血清は Santa Cruz Biotechnology Inc. から入手した。既に記載されているとおりに[3]、CAMA細胞およびトランスフェクションされたU2−OS細胞を標識し、免疫沈降させた。
【0075】
ゲル遅延アッセイ
一過性にトランスフェクションされたU2−OS細胞のゲル遅延アッセイを、既に記載されている方法[20]を少し改変することにより行った。20μlの反応容量中、コンセンサスE2FDNA結合部位を特定する0.5ngの[32P]−標識オリゴヌクレオチド(Santa Cruz Biotechnology)と共に、20mMHEPES(pH7.4)、0.1M KCl、1mMMgCl2、0.1mMEDTA、7%グリセロール、1mMNaFおよび1μg 超音波処理サケ精子DNAを含有する結合緩衝液中で10μgの全細胞抽出物を使用した。室温で20分間インキュベーションする間に、DNA−タンパク質複合体を形成させる。該反応生成物を、0.25×TBE中の3.5%ポリアクリルアミドゲル上、90V、室温、2.5時間で分離した。ついで、前記ゲルを乾燥し、フィルムに露出させた。
結果
p107結合タンパク質の単離
p107と相互作用するポリペプチドをコードするcDNAを同定するために、酵母二ハイブリッドスクリーニングを行った[14]。染色体に位置する2つのGal4誘導レポーター遺伝子(HIS3およびLacZ)[12]を含有する酵母Y190株[17]を、Gal4のDNA結合ドメイン(DBD)に融合したp107のポケット領域(アミノ酸240〜816)を含有する「ベイト」プラスミド、および14.5日齢CD1マウス胎児cDNAライブラリー(ここで、それぞれのcDNAは、Gal4のトランス活性化ドメインに個々に融合されている)[8]で同時形質転換した。合計200万の形質転換体を、ヒスチジンを欠くプレート上での選択に付した。β−ガラクトシダーゼの発現に関して、87個の生存コロニーをスクリーニングした。cDNA含有プラスミドを16個の二重陽性酵母コロニーからレスキューした。他のGal4−DBD融合体をコードするプラスミドで再形質転換することにより、p107結合の特異性を確認した。16個のハイブリッドタンパク質全てが、Gal4−p107と特異的に相互作用することが判明した。該酵母からレスキューされた16個のcDNAライブラリープラスミドが10個の異なる遺伝子に由来することが、DNA配列分析により示された。3個のcDNAは、同じ遺伝子に由来しており、4個の公知E2Fに対して有意な相同性を示した。このため、本発明者らは、このcDNAによりコードされるタンパク質をE2F−5と命名した。
【0076】
ついで、該部分マウスE2F−5cDNAを使用して、ヒト結腸癌cDNAライブラリーをスクリーニングすることにより、完全長ヒトcDNAクローンを得た。最長cDNA(2.1kb)を配列決定したところ、それは、346−アミノ酸のタンパク質(推定分子量38kDa)をコードする1038bpのオープンリーディングフレームを含有していた。図1Aは、E2F−5cDNA配列および推定アミノ酸配列を示す。
【0077】
E2F−5は、E2F−1(57%の類似度)よりもE2F−4(78%の類似度)と密接に関連している。E2F−1およびE2F−4と比較すると、E2F−5中には相同な3つの領域が認められる(図1B)。該DNA結合ドメイン(E2F−5のアミノ酸43〜115)は、E2F−4DNA結合領域と93%の類似度を共有し、一方、並列するDP−1二量化ドメインは、E2F−4とE2F−5との間で81%類似する。最後に、E2F−4とE2F−5とのカルボキシル末端ポケットタンパク質相互作用ドメインは、83%類似する。E2F−4およびE2F−5は、サイクリンA結合に関与するE2F−1のアミノ末端モチーフを両タンパク質が欠いている点で、E2F−1とは異なる。E2F−5は、E2F−4のセリン反復領域を欠いている点で、E2F−4とは異なる。
【0078】
E2F−5のmRNA発現レベルを分析するために、ヒトE2F−5cDNAを使用して、いくつかのヒト細胞系から全細胞質RNAを含有するノーザンブロットをプローブした。E2F−5プローブは、ほとんどの細胞系で、低レベルの単一の2.1kbの転写産物を検出した。ヒトCAMA乳癌細胞系は、いくらか高いレベルのE2F−5を発現した(図2)。
【0079】
E2F−5は、カルボキシル末端トランス活性化ドメインを含有する
E2F−1およびE2F−4は、ポケットタンパク質結合部位と重なるカルボキシル末端トランス活性化ドメインを含有する[3, 18]。E2F−5もトランス活性化ドメインを含有するか否かを調べるために、本発明者らは、哺乳動物発現ベクターpJ3Ω中でヒトE2F−5のカルボキシル末端をGal4のDNA結合ドメインと融合させた。U2−OS骨肉腫細胞を、5つの上流Gal4部位を保持するCATレポーター遺伝子で一過性にトランスフェクションするか、あるいは、該レポーター遺伝子とGal4-E2F発現ベクターとで同時トランスフェクションした。図3に示すとおり、Gal4レポータープラスミドとGal4-E2F-5 発現ベクターとの同時トランスフェクションにより、CATレポーター遺伝子が50倍活性化された。Gal4-E2F-1またはGal4-E2F-4による同時トランスフェクションでは、CATレポーター遺伝子の活性化が2〜3倍高かった(図3)。本発明者らは、E2F−5は、強力なカルボキシル末端トランス活性化ドメインを含有すると結論する。
【0080】
E2F−5は、DNA結合のためにDP−1を必要とする
E2F−1とE2F−4とは共に、効率的なDNA結合のためには、DP−1との二量体形成を必要とする[1, 3, 20, 26]。E2F−5がコンセンサスE2FDNA結合部位に結合するか否か、およびDNAに結合するためにE2F−5がDP−1二量体形成を必要とするか否かを調べるために、本発明者らは、一過性トランスフェクション実験を行った。コアプロモーターが4つの上流E2F部位に結合しているCATレポータープラスミドで、ヒトU2−OS骨肉腫細胞をトランスフェクションした。図4に示すとおり、E2F-CATレポータープラスミドは、該骨肉腫細胞に単独でトランスフェクションされた場合には、低い活性しか有さない。DP−1またはE2F−5発現ベクターで別々にトランスフェクションしても、E2F-CATレポーターの活性化は生じなかった(図4、トラック2および6)。DP−1とE2F−5発現ベクターとの同時トランスフェクションにより、CATレポーターの強力な用量依存的相乗活性化が生じた(図4、トラック3〜5)。これらのデータが示すとおり、E2F−5はコンセンサスE2F部位に結合でき、DNA結合はDP−1依存的である。これらの結果に基づき、本発明者らは、E2F−5がE2F遺伝子ファミリーの真のメンバーであると結論する。
【0081】
E2F−5トランス活性化は、ポケットタンパク質により抑制される
E2F−1およびE2F−4のトランス活性化ドメインはポケットタンパク質相互作用表面と重なるため、これらのE2Fのトランス活性化は、ポケットタンパク質結合により抑制される。E2F−5トランス活性化に対するポケットタンパク質発現の効果を調べるために、本発明者らは、一過性トランスフェクションアッセイを用いた。E2F−1とE2F−4とは共に、それらのそれぞれのポケットタンパク質に対する効率的な結合のためにはDP−1二量体形成を必要とするため[3, 20]、本発明者らは、E2F−5およびDP−1活性化転写に対するポケットタンパク質発現の効果を測定した。E2F−5およびDP−1と共にE2F-CATレポータープラスミドでU2−OS細胞をトランスフェクションした。図5(トラック3)に示すとおり、E2F−5とDP−1との同時トランスフェクションにより、E2F-CATレポーター遺伝子が100倍以上活性化された。E2F−5刺激転写は、pRb、p107およびp130発現ベクターでの同時トランスフェクションにより、用量依存的に阻害された。無傷のポケットドメインを欠く、pRbの変異体(pRbΔ22)およびp107の変異体(p107DE)は、E2F−5トランス活性化を抑制する能力を有していなかった(図5、トラック6および9)。重要なことは、pRbおよびp107のこれらの変異形態が、成長阻害活性をも欠くことである[41]。したがって、この実験では、E2F−5の好ましい結合相手の明確な同定はできなかったが、該実験が示したところによると、ポケットタンパク質結合によりE2F−5トランス活性化が阻害され、pRbおよびp107が成長停止を起こす能力とそれらがE2F−5トランス活性化を阻害する能力との間に密接な相関が存在する。この実験で用いるU2−OS細胞がpRb−またはp107−誘導成長停止に不感受性である[41]ことを指摘しておくことは重要である。したがって、観察された、E2F−5トランス活性化に対する効果は、pRbまたはp107の非特異的細胞周期効果によるものではないらしい。
【0082】
バンドシフトアッセイにおいて、E2F−5はp130と優先的に相互作用する
E2F−5によるポケットタンパク質結合の特異性をさらに調べるために、本発明者らは、電気泳動移動度シフトアッセイ(EMSA)を行った。U2−OS細胞を、pRb、p107またはp130発現ベクターと共に又はこれらを用いることなく、DP−1およびE2F−5発現ベクターで一過性にトランスフェクションした。トランスフェクションの2日後に、全細胞抽出物を、トランスフェクションされた細胞から調製し、コンセンサスE2F部位を特定する[32P]−標識オリゴヌクレオチドと共にインキュベーションした。DNA−タンパク質複合体を、ポリアクリルアミドゲル上で分離し、ラジオグラフィーにより可視化した。図6に示すとおり、E2F−5およびDP−1発現ベクターのトランスフェクションは、模擬(mock)トランスフェクション細胞では観察されなかった新規複合体の出現につながる(図6、レーン1および2を比較されたし)。p130発現ベクターの同時トランスフェクションにより、この複合体のスーパーシフト(supershift)が生じたが、p107またはpRb発現ベクターの場合にはこれは生じなかった(図6、レーン3〜5)。これらのデータは、試験したこれらの3つのタンパク質のうちで、p130がE2F−5/DP−1ヘテロ二量体に対して最も高い親和性を有することを示唆する。
【0083】
E2F−5は、invivoでp130と優先的に相互作用する
生理学的条件下では、E2F−1はpRbに、E2F−4はp107にそれぞれ優先的に結合する[3, 15, 19, 24]。しかしながら、一過性トランスフェクション実験では、E2F−1およびE2F−4活性化遺伝子発現は、pRbとp107との両方により抑制されうる[3, 40]。この特異性の喪失は、おそらく、これらのタンパク質の一過性の過剰発現により生じるのであろう。網膜芽細胞腫タンパク質ファミリーの3つのメンバーのどれが、生理学的条件下でE2F−5と相互作用するかを検討するために、本発明者らは、ヒトE2F−5に対するポリクローナルウサギ抗血清を産生させた。in vitroで転写および翻訳されたE2F−1、E2F−4およびE2F−5を用いる最初の免疫沈降実験において、ポリクローナルE2F−5血清がE2F−5を特異的に認識することが示された(データは示していない)。ついで、E2F−5抗血清を逐次免疫沈降実験で使用した。CAMA乳癌細胞を、[32P]−オルトホスフェートで代謝的に標識し、非イオン性デタージェントライゼートを調製した。これらのライゼートを、pRb特異的抗体、p107抗体またはp130特異的抗血清を用いる免疫沈降法に付した。pRb、p107またはp130で共免疫沈降させたタンパク質を、SDS含有緩衝液中で煮沸することにより遊離させ、希釈し、E2F−5特異的抗血清で再免疫沈降させた。図6のパネルBに示すとおり、p130免疫沈降物からは、47kDaのタンパク質がE2F−5抗血清により特異的に再免疫沈降されたが、pRbまたはp107免疫沈降物からは再免疫沈降されなかった。この47kDaのタンパク質は、SDSポリアクリルアミドゲル上、一過性にトランスフェクションされたE2F−5と共移動する(データは示していない)。対照として、本発明者らは、pRbおよびp107免疫沈降物がそれらの各々のE2Fを含有するか否かを確認した。図6のパネルCに示すとおり、pRbは確かにE2F−1と共免疫沈降し、p107はE2F−4を沈降させた。総合すると、これらのデータからは、E2F−5がin vivoでp130と優先的に相互作用することが示される。
考察
本発明者らは、本明細書において、E2F遺伝子ファミリーの第5のメンバーの単離を報告する。E2F−5は、真のE2Fファミリーメンバーの特徴のすべてを有する。すなわち、それは、高度に保存されたDNA結合ドメイン、DP−1二量化ドメインおよびカルボキシル末端トランス活性化ドメインを有する。さらに、E2F−5は、DP−1と協同的にコンセンサスE2FDNA結合部位に結合し、E2F部位含有レポーター遺伝子の発現を活性化しうる。
【0084】
本発明者らは、E2F−5が3つのポケットタンパク質のどれとin vivoで優先的に相互作用するのかを検討するために、3つのタイプの実験を行った。一過性トランスフェクション実験では、E2F−5トランス活性化は、網膜芽細胞腫タンパク質ファミリーpRb、p107およびp130の3つすべてのメンバーにより抑制された。この点で、E2F−5は、E2F−1およびE2F−4と類似している。なぜなら、E2F−1およびE2F−4トランス活性化は共に、一過性トランスフェクションアッセイで、pRbおよびp107により阻害されうるからである[3, 40]。このように一過性トランスフェクションアッセイで特異性が一見ないことは、おそらく、一過性トランスフェクション細胞におけるE2Fとポケットタンパク質との両方の高い一過性発現レベルの結果であろう。一過性トランスフェクション細胞では、p107はp130と比べて10倍高いレベルで発現されることが判明しているため(データは示していない)、該一過性トランスフェクション実験(図5)におけるp130によるE2F−5トランス活性化の比較的低レベルの阻害は、低いレベルのp130発現の結果である。E2F−5のポケットタンパク質特異性を検討するために、2つの更に別の実験を行った。その第1の実験では、全3つのポケットタンパク質の発現ベクターの存在下および不存在下、E2F−5およびDP−1発現ベクターで細胞を一過性にトランスフェクションした。ついで、該トランスフェクション細胞からの抽出物と、コンセンサスE2F結合部位を特定するオリゴヌクレオチドとを用いて、バンドシフトアッセイを行った。p130の同時トランスフェクションだけが、E2F−5/DP−1複合体のスーパーシフトを起こすことができた(図6)。該バンドシフトアッセイでは、ポケットタンパク質とE2F−5との間の長時間安定な複合体だけが、E2F/ポケットタンパク質「スーパーシフト」複合体として検出される。したがって、たとえp130がp107より低レベルで発現されたとしても、E2F−5とp130との複合体は、p107/E2F−5複合体よりも安定であった(図6)。同様の実験で、本発明者らは、p107とのE2F−4DNA結合複合体を「スーパーシフト」させることはできたが、pRbとの複合体はできなかった(R.L.B.およびR.B., 未公開データ)。この結果が示唆するところによると、移動度シフト実験は、E2Fのポケットタンパク質特異性を検討するのに有用と考えられる。該移動度シフトアッセイの結果と一致して、本発明者らは、トランスフェクションされていない代謝標識CAMA乳癌細胞では、E2F−5は、p130とは共免疫沈降しうるが、p107またはpRbとは共免疫沈降しないことを見いだした(図7)。総合すると、本発明者らのデータは、生理学的条件下では、E2F−5はp130と優先的に結合することを示す。
【0085】
p130とp107とは構造的に密接に関連しており、実際、p107とp130とは共に、サイクリンAおよびEに結合する能力を有するため[16, 32, 41]、E2F−5がp130とは相互作用したが、p107とはしなかったという知見は、いくらか予想外であった。一方、p107だけが抗D型サイクリン抗体と共免疫沈降する(p130はしない)ため[32]、p107とp130とは、D型サイクリンとin vivoで相互作用する能力において異なる。重要なことは、p130/E2Fおよびp107/E2F複合体の出現が、細胞周期で異なることである[9, 10, 29, 35, 37]。このことは、p107とp130とは、細胞周期中、異なる機能を有することを示唆する。p130によるE2F−5の優先的結合は、細胞周期調節におけるp130に関するそのような異なる役割と一致する。
【0086】
E2F−5がコンセンサスE2F部位に結合しうるという本発明者らの知見は、E2F遺伝子ファミリーの残りのメンバーが結合するE2F部位とは異なるE2F部位の別個のサブセットと、E2F−5がin vivoで相互作用するという可能性を決して除外するものではない。チミジンキナーゼ遺伝子プロモーター中およびb−mybプロモーター中に存在するE2F部位がE2F/p107複合体と優先的に相互作用するという知見[28, 31]は、種々のE2Fのそのような結合部位優先性と一致する。E2Fとp130との間の複合体は、ほとんどの場合、静止細胞中で見いだされ、細胞が静止から脱した後すぐに消失するため、E2F−5応答性遺伝子は、成長因子刺激に対する静止細胞の初期応答に関与していると考えられる[10]。p130と相互作用するE2F−5が入手できれば、E2F−5応答性遺伝子の同定ができるはずである。
謝辞
マウス胎児cDNAライブラリーおよび酵母発現ベクターを贈呈していただいたP. Chevray、酵母Y1090株を贈呈していただいたS. Elledge、Gal4-bmi酵母発現ベクターを贈呈していただいたM. Alkema、p130発現ベクターを贈呈していただいたG. Hannon、ヒト細胞系RNAを贈呈していただいたA. Bes-Gennissen、およびノーザンブロットを調製していただいたY. Ramosに感謝の意を表する。
【0087】
この研究は、Netherlands Organization for Scientific Research(NWO)からの助成により支援していただいた。
実施例1の参照文献
1.Bandara,L.R., V.M.Buck, M.Zamanian, L.H.JohnstonおよびN.B. La Thangue. 1993.細胞周期調節転写因子DRTF1/E2FにおけるDP−1とE2F−1との間の機能的相乗作用(Functional synergy between DP-1 and E2F-1 in
the cell cycle-regulating transcription factor DRTF1/E2F). Embo J. 12:4317-4324.
2. Beijersbergen,R.L., E.M.Hijmans, L.Zhu およびR.Bernards1994.c−MycとpRb関連タンパク質p107との相互作用によりc−Myc媒介性トランス活性化の阻害が生じる(Interaction of c-Myc with the pRb-related protein p107 results in inhibition of c-Myc-mediated transactivation). Embo J. 13:4080-4086.
3.Beijersbergen,R.L., R.Kerkhoven, L.Zhu, L.Carlee, P.M.Voorhoeve およびR.Bernards. 1994. E2F遺伝子ファミリーの新規メンバーであるE2F−4は、発癌活性を有し、in vivoでp107と結合する(E2F-4, a new member of the E2F gene family, has oncogenic activity and associates with p107 in vivo). Genes, Dev. 8:2680-2690.
4.Bernards,R., S.K.DessainおよびR.A.Weinberg. 1986. N−myc増幅は、神経芽細胞腫においてMHCクラスI抗原発現のダウンモジュレーションを起こす(N-myc amplification causes down-modulation of MHC class I antigen expression in neuroblastoma). Cell. 47:667-674.
5.Buchkovich,K., L.A.DuffyおよびE.Harlow. 1989. 網膜芽細胞腫タンパク質は細胞周期の特定の時期にリン酸化される(The retinoblastoma protein is phosphorylated during specific phases of the cell cycle). Cell. 58:1097-1105.
6.Chellappan,S.P., S.Hiebert, M.Mudryj, J.M.HorowitzおよびJ.R.Nevins 1991.E2F転写因子はpRbタンパク質の細胞標的である(The E2F transcription factor is a cellular target for the RB protein). Cell 65:1053-1061.
7.Chen,P.L., P.Scully, J.Y.Shew, J.Y.WangおよびW.H.Lee. 1989.網膜芽細胞腫遺伝子産物のリン酸化は細胞周期および細胞分化の間にモジュレーションされる(Phosphorylation of the retinoblastoma gene product is modulated d
uring the cell cycle and cellular differentiation). Cell. 58:1193-1198.
8.Chevray,P.M.およびD.Natans. 1992. 酵母におけるタンパク質相互作用クローニング:junのロイシンジッパーと反応する哺乳動物タンパク質の同定(Proteininteraction cloning in yeast:identification of mammalian proteins that react with the leucine zipper of jun). Proc. Natl. Acad. Sci. USA. 89:5789-5793.
9.Chittenden,T., D.M.LivingstonおよびJ.A.DeCaprio.1993.一次ヒトT細胞におけるE2Fの細胞周期分析は、新規E2F複合体および遊離E2Fの生化学的に異なる形態を明らかにする(Cell cycle analysis of E2F in primary human T cells reveals novel E2F complexes and biochemically distinct forms
of free E2F). Mol Cell Biol. 13:3975-3983.
10. Cobrinik,D., P.Whyte, D.S.Peeper, T.JacksおよびR.A.Weinberg 1993.E2Fとp130E1A結合タンパク質との細胞周期特異的結合(Cell cycle-specific association of E2F with the p130 E1A-binding protein). Genes,
Dev. 7:2392-2404.
11.DeCaprio,J.A., J.W.Ludlow, D.Lynch, Y.Furukawa, J.Griffin, H.Piwnica-Worms, C.-M.HuangおよびD.M.Livingston. 1989. 網膜芽細胞腫遺伝子産物は細胞周期調節因子の特性を有する(The product of the retinoblastoma gene
has properties of a cell cycle regulatory element). Cell. 58:1085-1095.
12.Durphee.T., K.Becherer, P.-L.Chen, S.-H.Yeh, Y.Yang, A.E.Kilburn, W.-H,Lee およびE.S.J. 1993.網膜芽細胞腫タンパク質は、プロテインホスファターゼ1型触媒サブユニットと結合する(The retinoblastoma protein associates with the protein phosphatase type 1 catalytic subunit). Genes, Dev. 7:555-569.
13.Farnham,P.J., J.E.SlanskyおよびR.Kollmar. 1993.哺乳動物細胞周期におけるE2Fの役割(The role of E2F in the mammalian cell cycle). Biochim Biophys. Acta. 1155:125-131.
14.Fields,S.およびO.Song. 1989. タンパク質−タンパク質相互作用を検出するための新規遺伝系(A novel genetic system to detect protein-protein
interactions). Nature. 340:245-246.
15. Ginsberg,D., G.Vairo, T.Chittenden, X.Zhi-Xiong, G.Xu, W.K.L., J.A.DeCaprio, L.J.B.およびD.M.Livingston. 1994. E2F転写因子ファミリーの新規メンバーであるE2F−4はp107と相互作用する(E2F-4, a new member of the E2F transcription factor family, interacts with p107). Genes,
Dev. 8:2665-2679.
16.Hannon,G.J., D.DemetrickおよびD.Beach. 1993. CDK2およびサイクリンとの相互作用によるRb関連p130の単離(Isolation of the Rb-related p130 through its interaction with CDK2 and cyclins). Genes, Dev. 7:2378-2391.
17. Harper,J.W., G.R.Adami, N.Wei, K.KeyomarsiおよびS.J.Elledge. 1993.p21Cdk相互作用タンパク質Cip1は、G1サイクリン依存性キナーゼの強力な阻害剤である(The p21 Cdk-interacting protein Cip1 is a potent inhibitor of G1 cyclin-dependent kinases). Cell. 75:805-816.
18.Helin,K., E.HarlowおよびA.Fattaey 1993. 網膜芽細胞腫タンパク質の直接結合によるE2F−1トランス活性化の阻害(Inhibition of E2F-1 transactivation by direct binding of the retinoblastoma protein). Mol Cell Biol. 13:6501-6508.
19. Helin,K., J.A.Lees, M.Vidal, N.Dyson, E.HarlowおよびA.Fattey. 1992. 転写因子E2Fの性質を有するpRb結合タンパク質をコードするcDNA(A cDNA encoding a pRB-binding protein with properties of the transcription factor E2F). Cell. 70:337-350.
20. Helin,K., C.-L.Wu, A.R.Fattaey, J.A.Lees, B.D.Dynlacht, C.NgwuおよびE.Harlow. 1993. 転写因子E2F−1とDP−1とのヘテロ二量体形成は協同的トランス活性化につながる(Heterodimerization of the transcription factor E2F-1 and DP-1 leads to cooperative trans-activation ). Genes, Dev.
7:1850-1861.
21. Hu,Q., C.Bautista, G.Edwards, D.Defeo-Jones, R.JonesおよびE.Harlow, 1991. ヒト網膜芽細胞腫タンパク質に特異的な抗体は関連ポリペプチドのファミリーを同定する(Antibodies specific for the human retinoblastoma protein identify a family of related polypeptides). Mol. Cell. Biol. 11:5792-5799.
22. Ivey-Hoyle,M., R.Conroy, H.E.Huber, P.J.Goodhart, A.OliffおよびD.C.Heimbrook. 1993. E2F−2のクローニングおよび特徴づけ:転写因子E2Fの生化学的性質を有する新規タンパク質(Cloningand Characterization of E2F-2, a novel protein with the biochemical properties of transcription factor E2F). Mol Cell Biol. 13:7802-7812.
23. Johnson,D.G., J.K.Schwarz, W.D.CressおよびJ.R.Nevins. 1993. 転写因子E2F1の発現は静止細胞がS期に進入するのを誘導する(Expression of transcription factor E2F1 induces quiescent cells to enter S phase). Nature. 365:349-352.
24. Kaelin,W.G., W.Krek, W.R.Sellers, J.A.DeCaprio, F.Ajchenbaum, C.S.Fuchs, T.Chittenden, Y.Li.P.Farnham, M.A.Blanar, D.M.LivingstonおよびE.K.Flemington. 1992. E2F様性質を有する網膜芽細胞腫結合タンパク質をコードするcDNAの発現クローニング(Expression cloning of a cDNA encoding a retinoblastoma-binding protein with E2F-like properties). Cell. 70:351-364.
25.Kato,G.J., J.Barrett, G.M.VillaおよびC.V.Dang.1990. 腫瘍性トランスフォーメーションに要するアミノ末端c−mycドメインは転写を活性化する(An amino-terminal c-myc domain required for neoplastic transformation activates transcription). Mol.Cell. Biol. 10:5914-5920.
26. Krek,W., D.M.LivingstonおよびS.Shirodkar. 1993. 種々のE2Fファミリーメンバーの複合体形成により促進されるDNAおよび網膜芽細胞腫遺伝子産物に対する結合(Bindingto DNA and the retinoblastoma gene product promoted by complex formation of different E2F family members). Science. 262: 1557-1560.
27. La Thangue, N.B. 1994.DPおよびE2Fタンパク質:細胞周期制御に関与するヘテロ二量体転写因子の成分(DP and E2F proteins: components of a
heterodimeric transcription factor implicated in cell cycle control). Curr. Opin. Cell Biol. 6:443-450.
28.Lam,E.W.およびR.J.Watson. 1993. E2F結合部位はマウスβ−myb転写の細胞周期調節抑制を媒介する(An E2F-binding site mediates cell-cycle regulated repression of mouse B-myb transcription). Embo J. 12:2705-2713.
29. Lees,E., B.Faha, V.Dulic, S.I.ReedおよびE.Harlow. 1992. サイクリンE/cdk2およびサイクリンA/cdk2キナーゼはp107およびE2Fに、一時的に異なる態様で結合する(Cyclin E/cdk2 and cyclin A/cdk2 kinases associate with p107 and E2F in a temporally distinct manner). Genes Dev. 6:1874-1885.
30. Lees,J.A., M.Saito, M.Vidal, M.Valentine, T.Look, E.Harlow, N.DysonおよびK.Helin. 1993.網膜芽細胞腫タンパク質はE2F転写因子のファミリーに結合する(The retinoblastoma protein binds to a family of E2F transcription factors). Mol Cell Biol. 13:7813-7825.
31.Li,L.J., G.S.NaeveおよびA.S.Lee.1993.G1−S期転写調節に重要なヒトチミジンキナーゼプロモーター因子に結合するサイクリンA−p107およびp33cdk2複合体の一時的調節(Temporal regulation of cyclin A-p107
and p33cdk2 complexes binding to a human thymidine kinase promoter element important for G1-S phase transcriptional regulation). Proc Natl Acad
Sci USA. 90:3554-3558.
32.Li,Y., C.Graham, S.Lacy, A.M.V.DuncanおよびP.Whyte. 1993.アデノウイルスE1A関連130kDタンパク質は網膜芽細胞腫遺伝子ファミリーのメンバーによりコードされ、サイクリンAおよびEと物理的に相互作用する(The adenovirus E1A-associated 130-kD protein is encoded by a member of the retinoblastoma gene family and physically interacts with cyclins A and E). Genes, Dev. 7:2366-2377.
33.Morgenstern,J.およびH.Land. 1990. 一連の哺乳動物発現ベクターおよび安定におよび一過性にトランスフェクションされた細胞におけるレポーター遺伝子の該ベクターによる発現の特徴づけ(A series of mammalian expression vectors and characterisation of their expression of a reporter gene in stably and transiently transfected cells). Nucleic Acid Res. 18:1068.
34. Schiestl,R.HおよびR.D.Gietz. 1989.キャリアーとして一本鎖核酸を用いる無傷酵母細胞の高効率形質転換(High efficiency transformation of intact yeast cell using single stranded nucleic acid as a carrier). Curr. Genet. 16:339-346.
35. Schwarz,J.K., S.H.Devoto, E.J.Smith, S.P.Chellappan, L.JakoiおよびJ.R.Nevins. 1993. 細胞増殖応答期のp107およびRbタンパク質とE2Fとの相互作用(Interactions of the p107 and Rb proteins with E2F during the cell proliferation response). Embo J. 12:1013-1020.
36. Shan,B., X.Zhu, P.L.Chen, T.Durfee, Y.Yang, D.SharpおよびW.H.Lee. 1992.網膜芽細胞腫関連タンパク質をコードする細胞遺伝子の分子クローニング:転写因子E2Fの性質を有する遺伝子の同定(Molecular cloning of cellular genes encoding retinoblastoma-associated proteins:identification of a gene with properties of the transcription factor E2F). Mol Cell Biol. 12:5620-5631.
37. Shirodkar,S., M.Ewen, J.A.DeCaprio, J.Morgan, D.M.LivingstonおよびT.Chittenden. 1992. 転写因子E2Fは網膜芽細胞腫産物およびp107サイクリンA複合体と細胞周期調節的に相互作用する(The transcription factor E2F interacts with the retinoblastoma product and a p107-cyclin A complex
in a cell cycle-regulated manner). Cell. 68:157-166.
38. Slansky,J.E., Y.Li, W.G.KaelinおよびP.J.Farnham. 1993.E2F1mRNAのタンパク質合成依存的増加は、ジヒドロ葉酸レダクターゼプロモーターの成長調節と相関する(A protein synthesis-dependent increase in E2F1 mRNA correlates with growth regulation of the dihydrofolate reductase promoter). Mol Cell Biol. 13:1610-1618.
39. Van der Eb, A.J.およびF.L.Graham. 1980. 腫瘍ウイルスDNAのトランスフォーメーション活性のアッセイ(Assay of transforming activity of tumor virus DNA). Meth. Enzymol. 65:826-839.
40. Zamanian,M.およびN.B.La Thangue. 1993. Rb関連タンパク質p107による転写抑制(Transcriptional repression by the Rb-related protein p107). Mol Biol of Cell. 4:389-396.
41. Zhu,L., S. van den Heuvel, K.Helin, A Fattaey, M.Ewen, D.Livingston, N.DysonおよびE.Harlow. 1993. 網膜芽細胞腫タンパク質の同族体であるp107による細胞増殖の阻害(Inhibition of cell proliferation by p107, a relative of the retinoblastoma protein). Genes Dev. 7:1111-1125.
実施例2
概要
転写因子DRTF1/E2Fは、それが細胞周期進行のキーとなるレギュレーター(例、網膜芽細胞腫抑制遺伝子産物および関連ポケットタンパク質、サイクリンおよびサイクリン依存性キナーゼ)と相互作用することにより、細胞増殖の制御に関与している。2つの異なるタンパク質ファミリー(DPおよびE2F)のメンバーがDP/E2Fヘテロ二量体として相互作用する場合に、DRTF1/E2FDNA結合活性が生じる。ここでは、本発明者らは、E2F−5と称される、E2Fタンパク質ファミリーの新規メンバーの単離および特徴づけを報告する。E2F−5は、マウスDP−1に対する結合能を有する分子に関して14.5d.p.c.マウス胎児ライブラリーをスクリーニングする酵母二ハイブリッドアッセイにより単離されたが、DPタンパク質ファミリーのすべての公知メンバーとも相互作用する。哺乳動物細胞抽出物に存在する一般的なDRTF1/E2FDNA結合活性(E2F部位を介して協同的DNA結合活性および転写活性化を起こす相互作用)により、E2F−5はDP−1との生理学的ヘテロ二量体として存在する。強力な転写活性化ドメインは、酵母および哺乳動物細胞の両方で機能し、E2F−5のC末端領域に存在する。pRbよりむしろpRb関連タンパク質p107の発現が、E2F−5の転写活性を不活性化する。E2F−5と該ファミリーの他のメンバーとの配列の比較から、E2F−5は、E2F−1、−2および−3よりも、E2F−4に対する類似度が大きいことが示される。E2F−5とE2F−4との構造的および機能的類似性は、E2Fタンパク質のサブファミリーを定義づけるものである。
緒言
細胞転写因子DRTF1/E2Fは、細胞周期進行と同調した転写に関与していることを示す証拠がかなり存在する。例えば、DRTF1/E2Fは、主要な標的の1つであり、それを介して、網膜芽細胞腫癌抑制遺伝子産物(pRb)が、細胞増殖に対するその負の効果を奏すると考えられる(La Thangue, 1994)。したがって、DRTF1/E2Fの転写活性および標的遺伝子[その多くは、細胞周期進行に必要なタンパク質をコードしている(Nevins, 1992)]の活性を調節することにより、pRbは、初期細胞周期を経る進行に影響を及ぼしうる。ヒト腫瘍細胞中に生じる、Rb中の天然の突然変異は、DRTF1/E2Fに結合しないタンパク質をコードする(Bandaraら, 1992; Heibertら, 1992; ZamanianおよびLa Thangue, 1992)が、このことは、調節解除するDRTF1/E2Fと異常細胞成長との間の相関性を強調するものである。さらに、アデノウイルスE1a、ヒトパピローマウイルスE7、SV40ラージT抗原などのウイルス性癌タンパク質の形質転換活性は、それらがpRbおよび関連タンパク質の隔離によりDRTF1/E2Fを調節解除する能力と相関しており(Nevins, 1992)、このことは、この見解をさらに支持するものである。
【0088】
細胞周期において中心的な役割を果たす他の分子も、DRTF1/E2Fと相互作用する。サイクリンAおよびEは触媒サブユニットcdk2と共に、該転写因子中のDNA結合成分と接触することにより直接的に(Krekら, 1994)、あるいは、pRb関連ポケットタンパク質p107またはp130のスペーサー領域中で生じる接触を介して間接的に(Leesら, 1992; Cobrinikら, 1993)、DRTF1/E2Fに結合する。p107およびp130と結合するサイクリン−cdk複合体の役割については未だ解明されていないが、サイクリンA/cdk2キナーゼ複合体とDRTF1/E2Fとの間の直接的相互作用は、そのDNA結合活性に影響を及ぼすことが示されている(Krekら, 1994)。
【0089】
DRTF1/E2Fの分子組成が明らかにされはじめている。始原型分子(prototypemolecules)がDP−1(Girlingら, 1993)およびE2F−1(Helinら, 1992; Kaelinら, 1992; Shanら, 1992)である2つの異なるタンパク質ファミリーのメンバーがDP/E2Fヘテロ二量体として相互作用する場合に(La Thangue, 1994)、一般的なDNA結合活性DRTF1/E2Fが生じることが現在明らかとなっている。両タンパク質の間で類似する小さな領域が、現在単離されているすべてのDPおよびE2Fファミリーメンバーで保存されており(Girlingら, 1994)、この領域は、それらのタンパク質がヘテロ二量体として相互作用するのを可能にし(Bandaraら, 1993; Helinら, 1993; Krekら, 1993)、それにより、多様な組み合わせの相互作用の発生が可能となる。
【0090】
それぞれ、DRTF1/E2Fとの相互作用の各々の特徴的なプロフィールを有するpRb、p107およびp130の結合が、細胞周期進行中に一時的調節的に生じる(Shirodkarら, 1992; Schwarzら, 1993; Cobrinikら, 1993)。現在までに単離されているE2Fファミリーメンバーのうち、E2F−1、−2および−3はpRbを(Ivey-Hoyleら, 1993; Leesら, 1993)、E2F−4はp107タンパク質を認識するが(Beijersbergenら, 1994; Ginsbergら, 1994)、その考えられる説明は、ポケットタンパク質の一時的な相互作用が、E2F/DPヘテロ二量体中のE2Fファミリーメンバーの組成および/または利用性の調節を反映しているというものである。
【0091】
多数の細胞型において、DP−1は、細胞周期進行中に生じる種々の形態で存在するDRTF1/E2Fの通常のDNA結合成分であるが(Bandaraら,1994)、DP−2などの他のDPファミリーメンバーは、組織限定的に発現される(Girlingら, 1994)。したがって、DRTF1/E2Fの分子組成が、細胞周期進行および細胞表現型により影響を受ける可能性があると考えられる。
【0092】
E2Fタンパク質ファミリーの複雑性については、未だ解明されていない。この問題を検討するために、本発明者らは、酵母に基づく二ハイブリッドスクリーニングを行って、該ファミリーの新規メンバーを定義づけした。ここで、本発明者らは、E2Fファミリーの新規メンバーであるマウスE2F−5の単離および特徴づけを報告する。E2F−5は、DPタンパク質ファミリーの公知メンバーのすべてと相互作用する。哺乳動物細胞抽出物中で、E2F−5はDP−1との生理学的ヘテロ二量体として存在し、その相互作用は、E2F部位を介する協同的DNA結合および転写活性化を起こす。E2F−5は、ポケット−タンパク質結合に際して特異的に不活性化される強力なトランス活性化ドメインを有する。E2F−5のタンパク質配列および分子構成は、E2F−4との関連性が、該ファミリーの他のメンバーとの関連性より密接であり、これは、機能的および構造的に関連しているE2Fタンパク質のサブファミリーを最初に定義づけるものである。
結果
E2F−5の単離
E2Fタンパク質ファミリーの多様性を調べるために、本発明者らは、酵母二ハイブリッドに基づく方法(FieldsおよびSong, 1989)により、新規メンバーを同定した(図8)。DP−1はE2Fファミリーメンバーの通常の生理学的反応の相手であるため(Bandaraら, 1993; Bandaraら, 1994)、本発明者らは、ベイトとしてDP−1を使用することを選択した。14.5d.p.c.マウス胎児(ChevrayおよびNathans, 1992)から調製された活性化ドメインタグ付きcDNAライブラリーを、LexA−DP−1と相互作用する能力を有するハイブリッドタンパク質に関してスクリーニングした。同定されたクローンの1つは、いくつかの基準によりLexA−DP−1と特異的に相互作用するハイブリッドタンパク質をコードしていた。該cDNA配列の部分分析は、E2Fファミリーメンバーとの広範な類似性を示し、したがって、完全タンパク質配列をコードするcDNAクローンを、F9ECcDNAライブラリーからさらに単離した。E2Fファミリーの他のメンバーとのタンパク質配列の比較により、該cDNAクローンが新規メンバーをコードしていることが示された。E2F−1、−2、−3および−4として既に単離されているE2Fタンパク質に採用されている名称に従い、本発明者らは、このクローンをE2F−5と称する。
【0093】
マウスE2F−5の予想サイズは、335残基である(図9a)。この予想は、考えられる最初の開始メチオニンの位置、および同じメチオニンから翻訳が開始されるヒトE2F−5配列に対して存在する広範な相同性に基づく(Hijmansら,投稿中)。E2F−5は、他のE2Fファミリーメンバーの間で保存されているドメインとの広範な配列類似性を含有する(Ivey-Hoyleら, 1993; Leesら,1993; Beijersbegenら, 1994; Ginsbergら, 1994)。例えば、該DNA結合ドメインは、E2F−1に対して48%、E2F−4に対して87%の同一性を示す(図9bおよびc)。この領域内で、C末端サブドメインは、DPファミリーのメンバーと類似する唯一の領域を含有する(図9bおよびcに示す)。DEFボックスとして知られる(Girlingら, 1994; LamおよびLa Thangue, 1994)この領域は、DP/E2Fヘテロ二量体の形成に密接に関与している(Bandaraら, 1993; BandaraおよびLa Thangue, 投稿予定)。また、DPとE2Fとのタンパク質間でDEFボックス内で保存されている残基は、E2F−5内で完全に保存されており(図9c)、このことは、DP/E2Fヘテロ二量体の形成におけるこのサブドメインの潜在的重要性を強調するものである。
【0094】
E2F−5と残りのファミリーメンバーとの間では、いくつかの更に別の領域が保存されている。マークトボックス(marked box)(Leesら, 1993)およびポケット−タンパク質結合ドメインは、E2F−1中のこれらの領域と58%および50%の同一性を、E2F−4中の同じ領域と75%および72%の同一性を示す(図9bおよび9c)。また、該ロイシンジップ領域中の疎水性残基の位置は、他のE2Fファミリーメンバーに対して保存されている(図9c)。E2F−5中の疎水性残基は、2つのさらなるC末端位置(L144およびV151;図9cを参照されたし)で7個の反復と重なるため、実際に、E2F−5は、E2F−1、−2および−3より長いジップを形成しうる。
【0095】
E2F−5の特徴は、E2F−1、−2および−3よりもE2F−4との密接な関係を示唆する。該タンパク質は、該DNA結合ドメインのN末端ほどは伸長せず、両タンパク質は、残りのE2F中に生じるN末端サイクリンA結合ドメインを欠く点で(図9b)、E2F−5の構成はE2F−4のものと類似する。さらに、該タンパク質配列の比較により、E2F−5とE2F−4との関係は、その各々と該ファミリーの残りのメンバーとの関係よりも密接であることが示される。これは、E2F−5とE2F−4との間では多数の残基が共通であるがE2F−1、−2および−3間ではそうでない該保存ドメインから特に明らかである(図9c)。全体的には、E2F−5は、E2F−4とは70%、E2F−1とは38%のアミノ酸残基が同一である。したがって、この類似性に基づくと、E2F−5および−4は、E2Fタンパク質ファミリーの1つのサブファミリーを代表し、E2F−1、−2および−3は、それらの密接な類似性のため、別のサブファミリーを代表する。
【0096】
DPファミリーメンバーとの結合および転写協同作用
DPファミリーメンバーがE2Fファミリーメンバーと相互作用する場合に、一般的なDRTF1/E2FDNA結合活性が生じる(La Thangue, 1994)。DP−1およびE2F−1では、該相互作用により協同的転写活性化、DNA結合、およびpRbとの相互作用が生じる(Bandaraら, 1993; Helinら, 1993; Krekら, 1993)。したがって、本発明者らは、E2F−5がDPファミリーメンバーと協同作用しうるか否かの判定に関心があった。
【0097】
これらの問題に答えるために、本発明者らはまず、LexA融合タンパク質として該ハイブリッドベイト中に提示されている種々のDP分子を使用する酵母二ハイブリッドアッセイを用いた(図8)。E2F−5またはE2F−1のいずれかを、活性化ドメイン(GAD)タグ付きハイブリッドタンパク質として発現させ、LexAレポーター構築物の転写活性化の程度を、β−ガラクトシダーゼ活性を測定することにより評価した。E2F−5およびE2F−1は共に、DPタンパク質ファミリーのすべての公知メンバー(すなわち、DP−1、−2、−3およびショウジョウバエDP)と同等に相互作用する能力を有する(データは示していない)。
【0098】
ついで、本発明者らは、E2F−5がDP−1と協同して、E2F結合部位を介して転写を活性化しうるか否かを評価した。このために、本発明者らは、E2F−5およびDP−1を一緒にまたは単独で発現させ、E2F部位レポーター構築物p4xWT CYC1の転写活性を測定する酵母アッセイを用いた(図10a)。以前の研究では、このアッセイは、E2F−1とDP−1との間の協同作用を測定するために使用されている(Bandaraら, 1993)。該DP−1ハイブリッドタンパク質の発現の後、該レポーターの転写活性は有意な影響を受けず、E2F−5ハイブリッドによりわずかに影響されたにすぎなかった(図10b)。しかしながら、両方を一緒に発現させた場合には、レポーター活性の刺激は6倍を越えた(図10b)。突然変異E2F結合部位を担持する、p4xWT CYC1の誘導体であるp4xMT CYC1(図10a; Bandaraら, 1993)の活性は、同じ条件で影響を受けなかった(データは示していない)。したがって、本発明者らは、E2F−5がDP−1と協同作用すると結論する。
【0099】
E2F−5の転写活性化およびポケットタンパク質調節
酵母および哺乳動物細胞の両方で、E2F−5の転写活性化能を評価した。酵母で転写活性をアッセイするために、E2F−5のC末端領域(残基198〜335)をLexAに融合し、LexA結合部位により駆動されるレポーター構築物の活性を評価した(図11a)。pLEX.E2F-5の存在下では、該ベクター単独で発現させた場合よりも、該レポーターの活性がかなり大きかったので(図11b)、E2F−5タンパク質は強力なトランス活性化ドメインを含有する。同様に、LexA E2F-1は、転写活性化能を有していた(図11b)。したがって、E2F−5は、酵母において転写を効率的に活性化する。
【0100】
これらの結果を哺乳動物細胞で確認し、ポケット−タンパク質とE2F−5との相互作用の機能的影響を評価するために、本発明者らは、E2F−5の同じ領域をGal4 DNA結合ドメインに融合し、一過性トランスフェクションアッセイを用いて、Gal4結合部位により駆動されるレポーター構築物(pGAL-CAT)の転写活性を調べた(図12a)。pGAL-E2F-5の存在下でpGAL-CATの転写活性はpG4と比べて15倍大きかったので、3T3細胞では、E2F−5は、Gal4DNA結合ドメイン単独の活性と比べて、転写を効率的に活性化した(図12b)。種々の他の細胞型でも同様の結果が得られたが(データは示していない)、このことは、哺乳動物細胞中で機能するトランス活性化ドメインをE2F−5が含有することを示すものである。
【0101】
ついで、本発明者らは、E2F−5の転写活性を用いて、pRbまたはp107のいずれかとの相互作用の機能的影響を評価した。野生型pRbおよびp107についての対照として、本発明者らは、DRTF1/E2Fとは相互作用しないRbの天然に存在する突然変異対立遺伝子によりコードされるタンパク質であるRbΔ22の活性(ZamanianおよびLa Thangue, 1992)、およびアンチセンスp107RNAの活性(ZamanianおよびLa Thangue, 1993)を調べた。pCMVHRbまたはpCMVHRbΔ22の存在下でpGAL-CATの活性は影響を受けなかったため(図12b)、野生型pRbまたはRbΔ22はいずれも、E2F−5の活性に有意な影響を及ぼさなかった。これに対して、E2F−5と共発現するp107(pCMV107から)はE2F−5の転写活性を減少させたが、アンチセンスp107(pCMV107ASから)は影響を及ぼさなかったため、この効果は特異的であった(図12b)。本発明者らは、哺乳動物細胞において、p107はE2F−5の転写活性を不活性化すると結論する。同様の実験方法を用いて、p130タンパク質がE2F−5の転写活性を不活性化することが示された(データは示していない)。
【0102】
E2F−5は、DRTF1/E2Fの生理学的DNA結合成分である
E2F−5が、哺乳動物細胞から調製された抽出物中に限定される一般的なDRTF1/E2FDNA結合活性の生理学的DNA結合成分であるか否かを判定するために、異なるペプチド配列に対する2つの異なる抗E2F−5ペプチド血清(共に、GST−E2F−5融合タンパク質と特異的に反応する抗血清である)を調製した(データは示していない)。
【0103】
マウスF9胎児性癌(F9EC)細胞からの抽出物を用いて行うゲル遅延アッセイにより、DRTF1/E2FDNA結合活性に対するこれらの抗血清の効果を調べた。DP−1が、F9EC細胞抽出物中のDRTF1/E2FDNA結合活性の通常の、おそらく普遍的な成分であることが、以前の研究で示されており(Girlingら,1993; Bandaraら, 1993)、その一例を、図13(トラック1〜4)に示す。抗E2F−5血清は共にDRTF1/E2FDNA結合活性を阻害したが、この効果は、相同性ペプチドの存在下では明らかでないため(図13、トラック5〜12)、特異的なものである。抗E2F−5はDNA結合活性の有意な減少を引き起こすが、この効果は、抗DP−1により生じるものほど顕著でないことは明らかである(図13、トラック1〜4と5〜12とを比較されたし)。これは、F9EC細胞抽出物中のDP−1/E2Fヘテロ二量体の亜集団中にE2F−5が存在するためと考えられ、DP−1で観察されるものと対照的である。
【0104】
E2F−5のDNA結合特性
E2F−5のDNA結合特性を調べるために、本発明者らは、E2F−5をGST融合タンパク質として発現させ精製した。E2F−1は、単独でアッセイした場合には、有意なDNA結合活性を有していたが(図14、トラック1〜4を比較されたし)、既に得られている結果(Bandaraら,1993)と一致して、GST−DP−1は、E2F部位に対する結合においてGST−E2F−1と協同作用した。これに対して、E2F−5は単独では、DNA結合活性はほとんど検出できなかった(図14、トラック5〜7)。しかしながら、E2F−5とDP−1との間の協同作用は、E2F−1とDP−1との間のものよりも相当大きかった(図13、トラック8〜10)。したがって、E2F−5とDP−1とはDNA結合活性について協同作用する。
【0105】
E2F−5RNAのレベル
本発明者らは、種々の細胞系でE2F−5RNAのレベルを測定し、さらに、E2F−5レベルとE2Fファミリーの他のメンバーとを比較することに関心があった。このために、種々の白血病細胞系およびF9EC細胞の非同時培養物からRNAを調製し、ノーザンブロッティングによりE2F−5RNAのレベルを評価した。E2F−5RNAの量は、細胞系によってかなり異なっていた。F9EC細胞およびいくつかの白血病細胞系、例えばDAUDIおよびRAGIは、高レベルで発現した(図15、トラック1、7および8)。これに対して、HL60およびTF1は、低レベルのE2F−5RNAを含有していた(図15、トラック4および6)。E2F−5RNAレベルのこのプロフィールは、E2F−1のレベルとかなり異なっていた。例えば、K562、HL60およびTF1細胞には、E2F−5と比べて、有意なレベルのE2F−1RNAが存在していた(図15、トラック3、4および6)。EL4細胞では、明らかにこの逆であり、E2F−5レベルが高く、E2F−1が低かった(図15、トラック10)。E2F−5RNAレベルは細胞型に影響され、E2F−5とE2F−1との間のRNAレベルには相関性がほとんどないと、本発明者らは結論する。
考察
E2F−5およびE2F−4は、E2Fタンパク質のサブファミリーである
本発明者らは、E2Fタンパク質ファミリーの第5のメンバーであるE2F−5の単離および特徴づけを報告する。潜在的ロイシンジップ、マークトボックス(marked box)およびポケットタンパク質結合領域などの、E2F−5中のドメインの多くは、E2Fファミリーの他のメンバーと保存されているが、DNA結合ドメインの外のN末端配列が欠けていることは、E2F−4と同様の構造的構成を示すものである(Beijersbergenら, 1994; Ginsbergら, 1994)。E2Fファミリーの残りのメンバー、すなわちE2F−1、−2および−3は、伸長したN−末端を有し、その内部に、サイクリンAと相互作用する能力を有するドメインが存在する(Krekら, 1994)。このドメインの役割は、サイクリンA/cdk2キナーゼをDP−1/E2Fヘテロ二量体へリクルートすることであり、ついでそれは、DP−1のリン酸化を引き起こし、その機能的影響は、DP−1/E2Fヘテロ二量体のDNA結合活性の減少であると示唆されている(Krekら, 1994)。そのような機構は、細胞周期進行のより後期におけるE2F部位依存性遺伝子の転写活性の調節に重要かもしれない。E2F−5(およびE2F−4)中にサイクリンA結合ドメインが存在しないことは、E2F−5/DP−1ヘテロ二量体のDNA結合活性が、他の機構によりダウンレギュレーションされうることを示唆する。確かに、これが達成される可能なシナリオは、p107および/またはp130タンパク質によるものであろう。なぜなら、これらのタンパク質のスペーサー領域は、サイクリンA/cdk2複合体またはサイクリンE/cdk2複合体のいずれかに結合しうるからである(Leesら, 1993; Cobrinikら, 1993; Hannonら, 1993; Liら, 1993)。これらのポケットタンパク質が、サイクリンA結合E2Fファミリーメンバーの役割を代替し、サイクリン/cdk複合体をDP/E2Fヘテロ二量体へリクルートしている可能性がある。
【0106】
E2F−5のタンパク質配列をE2Fファミリーの他のメンバーのものと比較すると、E2F−1、−2および−3よりもE2F−4に対する密接な関係が示された。興味深いことに、E2F−4は、生理学的条件下でp107と相互作用する能力を有する該ファミリーの唯一の公知メンバーである(Beijersbergenら, 1994; Ginsbergら, 1994)。本発明者らは、E2F−5の転写活性が、pRbよりもむしろp107またはp130により不活性化されうることを示した。しかしながら、これは、p107が、pRbよりもp130に対して密接な関係を有することを反映しているのかもしれず(Ewenら, 1991; Cobrinikら, 1993; Liら, 1993)、したがって、生理学的相互作用を完全には反映していないのかもしれない。ヒトE2F−5が生理学的条件下でp130と相互作用することが示されている結果(Hijmansら, 投稿中)は、この考え方と一致する。
【0107】
E2F−5とDP−1との間の協同作用
DNA結合および転写活性において、E2F−5はDP−1と協同作用する。この点で、E2F−5は、E2Fファミリーの他のメンバーと同様の性質を有する。しかしながら、注目すべき興味深いことは、E2F−5とDP−1との間の協同作用が、例えば同等の実験条件でE2F−1とDP−1との間で観察される協同作用よりも、かなり大きいことである(例えば、図14を参照されたし)。このアッセイが細胞内の機能的特性を反映するのであれば、過剰のDP−1が存在する細胞内環境では、E2F−5およびE2F−1が同等に増加した場合、E2F−5/DP−1DNA結合活性レベルの方が比較的大きくなると考えられる。さらに、特定のE2F/DPヘテロ二量体にとって好ましい標的遺伝子があれば、DNA結合活性のこれらの相違は、転写活性の相違を反映するかもしれない。
【0108】
それらの種々のE2Fタンパク質の細胞周期制御における正確な役割は、未だ確定されていない。しかしながら、それらは、細胞周期進行の異なる時期で標的遺伝子の転写活性を調節すると考えられる。例えば、E2F−1、−2および−3がpRbと相互作用することは(Helinら, 1992; Kaelinら, 1992; Ivey-Hoyle ら, 1993; Leesら, 1993)、それらが、G1を経る細胞周期進行を調節することを示唆する。これに対して、G1の終わりに向かってp107はDRTF1/E2Fと結合し、そしてS期でピークを迎えるが(Shirodkarら, 1992)、p130は、細胞周期進行の初期に、ほとんどはG0の間に優先的に結合する(Cobrinikら, 1993)。しかしながら、種々の細胞型中のE2F−1およびE2F−5のレベルは、E2Fファミリーメンバーの発現が、細胞周期の時期だけでなく、表現型によっても影響されることを示唆する。細胞は、その細胞周期要件に最も適した、E2Fタンパク質の好ましいサブセットを利用するという可能性がある。
【0109】
ここで報告しているE2F−5の分子的および機能的特徴、およびDPファミリーメンバーとのその相互作用から示されることは、E2FとDPファミリーメンバーとの間の種々のヘテロ二量体が理論的に可能なことである。将来の研究の大きな目標は、細胞周期制御におけるこれらの転写因子のそれぞれの生理学的役割を理解することである。
材料および方法
酵母株、培地および方法
この研究で使用したサッカロミセス・セレビシエ(Saccharomyces cerevisiae)株は、以下のとおりであった:W3031a(Mata ade2-100trp1-1 leu2-3, 112 his3-11 ura3); CTY10-5d(Matα ade2 trp1-901 leu2-3, 112 his3-200 gal4 gal80 URA3::lexAop-lacZ)およびPCY2(Matαgal14 gal80 URA3::GAL1-lacZ lys2-801 his3-200 trp1-63 leu2 ade2-101 )およびJZ1(Joossら, 1994; Matα lys2-801 ade2-10 leu2Δ1 trpΔ63 his3Δ200 URA3::lexAop-CYC1-lacZ)。酵母株をYPDまたはYNB培地中で増殖させ、酢酸リチウム法の改変法により形質転換した。コロニー着色β−ガラクトシダーゼ活性アッセイ(colony colourβ-galactosidase activity assay)を通常の方法で行った。少なくとも3つの別個の形質転換体について、中期対数期培養物中、個々の形質転換体のβ−ガラクトシダーゼ活性を定量した。
【0110】
ライブラリーDNA、プラスミドおよびオリゴヌクレオチド
pPC67は、GAL4タンパク質のトランス活性化ドメインをコードする酵母配列の下流と融合した14.5d.p.c.のCD−1マウス胎児オリゴdTプライミングcDNAライブラリーである(GAD; ChevrayおよびNathans, 1992)。特異的にクローニングしたポリdTプライミングcDNAのλZapII F9ECライブラリーから、完全なcDNAクローンを単離した(Scholerら, 1990)。
【0111】
プラスミドpTR27は、ポリリンカー配列が伸長しているpBTM116(Bandaraら, 1993)の誘導体であり、pLEX.DP-1、pGAD.E2F-1、p4xWT CYC1およびp4xMT CYC1については、既に記載されている(Bandaraら, 1993)。pLEX(HIS).DP-1は、プラスミドpLEX(HIS)(これは、TRP1がHIS3で置換されているpBTM116の誘導体)中のDP−1(アミノ酸残基59〜410)と完全細菌LexAタンパク質との融合体をコードする。pGAD.E2F-5は、プラスミドpACTII(Durfeeら, 1993)中の酵母GAL4タンパク質(768〜881)の活性化ドメインを有するハイブリッドタンパク質として発現されるE2F−5の完全コーディング配列を含有する。pLEX.E2F-5は、プラスミドpTR27中のLexAタンパク質の完全コーディング配列の下流に、ハイブリッドタンパク質として発現されるアミノ酸残基198〜335のE2F−5コーディング配列を含有する。pLEX.E2F-1は、pTR27中に完全長E2F−1(1〜437)を担持する。プラスミドpG4(以前はpG4mポリIIと呼ばれていた; Websterら, 1989)は、SV40初期プロモーターの制御下、GAL4DNA結合ドメイン(1〜148)をコードする。プラスミドpGAL4.E2F-5は、pG4中のGAL4配列の下流で融合した残基198〜335のE2F−5コーディング配列を含有する。プラスミドpCMVHRb、pCMVHRb△22、pCMV107およびpCMV107ASについては、既に記載されている(ZamanianおよびLa Thangue, 1992; 1993)。
【0112】
ライブラリースクリーニング
40μg pPC67ライブラリーDNAを、40μg pLEX(HIS).DP-1と共にCTY10-5d中に同時形質転換した。選択寒天プレート上で成長している約400,000の形質転換体を、in situ濾紙β−ガラクトシダーゼアッセイによりスクリーニングした。該ライブラリープラスミドをレスキュー(rescue)するために、青色のコロニーを単離し、ヒスチジンの存在下で選択液体培地中で飽和するまで成長させ、pLEX(HIS).DP-1をキュアリングした。選択最少寒天上にレプリカプレーティングした後、β−ガラクトシダーゼについてアッセイした場合に青色を与えないTrp+His-コロニーからのプラスミドDNAを、E. coli HW87中にエレクトロポレーションした。プラスミドを回収し、pLEX(HIS).DP-1または対照プラスミド(pLEX(HIS))のいずれかでCTY10-5d中に再度形質転換した。pLEX(HIS).DP-1が存在する場合にのみ青色コロニーを与えるTrp+表現型を付与するプラスミドを、さらなる分析のために選択した。完全長cDNAを得るために、該挿入物を切り出し、放射能標識し、これを用いて、λZapII F9ECライブラリーからの約105のプラークをスクリーニングし、それから完全長E2F−5cDNAを単離し、pBluescript中にレスキューした。
【0113】
3T3細胞の一過性トランスフェクション
既に記載されている通常のリン酸カルシウム沈殿法により(ZamanianおよびLa Thangue, 1992)、トランスフェクションおよびアッセイを行った。内部対照としてのpCMV-βgalに由来するβ−ガラクトシダーゼ活性を、既に記載されているとおりに測定した(ZamanianおよびLa Thangue, 1992)。
【0114】
抗血清およびゲル遅延分析
E2F−5に由来する2つの異なるペプチド配列に対して産生させたウサギ抗血清(抗−E2F−5(1)、抗−E2F−5(2)と称する)を調製し、F9EC細胞抽出物中のDRTF1/E2FDNA結合活性に対する効果を評価した[既に記載されているとおりに行った(Girlingら,1993)]。E2F結合部位は、アデノウイルスE2aプロモーターから取った(La Thangueら, 1990)。相同性(+)または無関係な(−)ペプチドのいずれかを該DNA結合アッセイへ加えて、既に記載されているとおりに(Girlingら, 1993)特異性を評価した。DP−1のC末端配列由来のペプチドに対して、抗DP−1(24)抗血清を産生させた。GST融合タンパク質を用いて行うDNA結合アッセイは、既に記載されているとおりである(Bandaraら, 1993; 1994)。通常の方法により、GST−DP−1、−E2F−1(Bandaraら, 1993)および−E2F−5(アミノ酸残基2〜335)を発現させ精製した。
【0115】
ノーザン分析
通常の方法により、示されている細胞系から調製したRNA上で、RNAレベルのノーザン分析を行った。E2F−5プローブは、3'非翻訳領域中に伸長する840のヌクレオチドを含有していた。E2F−1プローブは、PCRにより調製された遺伝子の完全コーディング配列を含有していた。GAPDH用のプローブは、内部対照として使用した。
実施例2の参照文献
Bandara,L.R., Adamczewski,J.P., Hunt,T.およびLa Thangue,N.B. (1991). Nature 352, 249-251.
Bandara,L.R., Adamczewski,J.P., Poon,R.Y., Zamanian,M., Hunt,T.およびLa Thangue,N.B. (1992). J. Cell Sci. (Suppl.) 16, 77-85.
Bandara,L.R., Buck,V.M., Zamanian,M., Johnston,L.H.およびLa Thangue,N.B. (1993). EMBO J. 12, 4317-4324.
Bandara,L.R., Lam,E.W.-F., Sφrensen,T.S., Zamanian,M., Girling,R.およびLa Thangue,N.B. (1994). EMBO J. 13, 3104-3114.
Beijersbergen,R.L., Kerkhoven,R.M., Zhu,L., Carlee,L., Voorhoeve,P.M.およびBernards,R. (1994). Genes. Dev. 8:2680-2690.
Chevray,P.およびD.Nathans. (1992). Proc. Natl. Acad. Sci. USA 89:5789-5793.
Cobrinik,D., Whyte,P., Peeper,D.S., Jacks T.およびWeinberg,R.A. (1993). Genes Dev. 7, 2392-2404
Durfee,T., Becherer,K., Chen,P.-L., Yeh,S.-H., Yang,Y., Kilburn, A.E., Lee, W.-H.およびElledge,S.J.(1993). Genes Dev. 7, 555-569.
Ewen,M.E., Xing,Y., Lawrence,J.B.およびLivingston,D.M. (1991). Cell 66:1155-1164.
Fields,S.およびSong,O. (1989). Nature 340:245-246.
Ginsberg,D., Vairo,G., Chittenden,T., Xiao,Z.-X., Xu.G., Wydner,K.L.,
DeCaprio,J.A., Lawrence,J.B.およびLivingston,D.M. (1994). Genes. Dev. 8:2665-2679.
Girling,R., Partridge,J.F., Bandara,L.R., Burden,N., Totty,N.F., Hsuan, J.J.およびLa Thangue,N.B. (1993). Nature 362, 83-87.
Girling,R., Bandara,L.R., Ormondroyd,E., Lam,E.W.-F., Kotecha.S., Mohun,T.およびLa Thangue,N.B. (1994). Mol. Biol. Cell. 5, 1081-1092.
Hannon,G.J., Demetrick,D.およびBeach,D. (1993). Genes Dev. 7:2378-2391
Heibert,S.W., Chellappan,S.P., Horowitz,J.M.およびNevins,J.R. (1992).
Genes Dev. 6, 177-185.
Helin,K., Lees,J.A., Vidal,M., Dyson,N., Harlow,E.およびFattaey,A. (1992). Cell 70, 337-350.
Helin,K., Wu,C.-L., Fattaey,A.R., Lees,J.A., Dynlacht,B.D., Ngwu,C.およびHarlow,E.(1993). Genes Dev. 7, 1850-1861.
Ivey-Hoyle,M., Conroy,R., Huber,H.E., Goodhart,P.J., Oliff,A.およびHeimbrook,D.C. (1993). Mol. Cell. Biol. 13:7802-7812.
Jooss,K.U., Funk,M.およびMuller,R. (1994). EMBO J. 13, 1467-1475.
Kaelin,W.G., Krek,W., Sellers,W.R., DeCaprio,J.A., Ajchenbaum,F., Fuchs,C.S., Chittenden,T., Li,Y., Farnham,P,J., Blanar,M.A., Livingston,D.M.およびFlemington,E.K. (1992). Cell 70, 351-364.
Krek,W., Livingston,D.M.およびShirodkar,S. (1993). Science 262, 1557-1560.
Krek,W., Ewen,M.E., Shirodkar,S., Arany,Z., Kaelin,W.G.およびLivingston,D.M. (1994). Cell 78, 161-172.
La Thangue,N.B. (1994). Trends Biochem. Sci. 19, 108-114.
La Thangue,N., Thimmappaya,B.およびRigby,P.W.J. (1990). Nucl. Acids. Res. 18, 2929-2938.
Lam,E.W.-F.およびLa Thangue,N.B. (1994). Curr. Op. Cell Biol. 6, 859-866.
Lees,E., Fahar,B., Dulic,V., Reed,S.I.およびHarlow,E. (1992). Genes Dev. 6, 1874-1885.
Lees,J.A., Saito,M., Vidal,M., Valentine,M., Look,T., Harlow,E., Dyson,N.およびHelin,K.(1993). Mol. Cell. Biol. 13:7813-7825.
Li.Y., Graham,C., Lacy,S., Duncan,A.M.V.およびWhyte,P. (1993). Genes Dev. 7:2366-2377.
Nevins,J.R. (1992). Science 258, 424-429.
Schwarz,J.K., Devoto,S.H., Smith,E.J., Chellappan,S.P., Jakoi,L.およびNevins,J.R. (1993). EMBO J. 12, 1013-1020.
Scholer,H.R., Ruppert,S., Suzuki,N., Chowdhury,K.およびGruss,P. (1990). Nature 344, 435-439.
Shan,B., Zhu,X, Chen,P.L., Durfee,T., Yang,Y., Sharp,D.およびLee,W.H. (1992). Mol.Cell.Biol. 12, 5620-5631.
Shirodkar,S., Ewen,M., DeCaprio,J.A., Morgan,J., Livingston,D.M.およびChittenden,T. (1992). Cell 68, 157-166.
Webster,N.J.G., Green,S., Tasset,D., Ponglikitmongkol,M.およびChambon,P. (1989). EMBO.J., 8, 1441-1446.
Zamanian,M,およびLa Thangue,N.B. (1992). EMBO J. 11:2603-2610.
Zamanian,M.およびLa Thangue,N.B. (1993). Mol. Biol. Cell. 4:389-396.
【図面の簡単な説明】
【0116】
【図1A−1】図1A-1はヒトE2F−5の構造を示す図であり、ヒトE2F−5cDNAのヌクレオチド配列および推定アミノ酸配列を示す図である。
【図1A−2】図1A-2はヒトE2F−5の構造を示す図であり、ヒトE2F−5cDNAのヌクレオチド配列および推定アミノ酸配列を示す図である。
【図1B】図1Bは、E2F−1およびE2F−4と比較してE2F−5を示す図であり、該保存ドメインの境界をアミノ酸番号で示す。E2F−4中のSSは、セリンに富むモチーフを示す。
【図2】図2は、ヒト細胞系でのE2F−5の発現パターンを示す図である。 (A)示されているヒト細胞系からの全細胞質RNAを含有するノーザンブロットを、ヒトE2F−5cDNAプローブとハイブリッド形成させた。以下のヒト細胞系からのRNAを使用した:CAMA、ヒト乳がん;A549、肺がん;MDA、MDA−MD157乳がん;OVCAR、卵巣がん;HS578T、乳がん;LUCY、卵巣がん;HT29、結腸がん;CEM、T−細胞白血病;K562、赤白血病。(B)ラットα−チューブリンプローブとハイブリッド形成させた同じフィルター。
【図3】図3は、E2F−5は、カルボキシル末端トランス活性化ドメインを有することを示す図である。Gal-4-E2F発現ベクター(1μg)および0.2μg pRSV-ルシフェラーゼ(内部対照)の存在下または不存在下、上流のGal4部位を保持するCATレポータープラスミド(5μg)で、U−2OS細胞をトランスフェクションした。それぞれのサンプルのCAT活性を、ルシフェラーゼ活性に対して標準化した。トランスフェクションの2日後に、CAT活性をアッセイした。CATレポーター遺伝子単独に対するGal4-E2Fの活性化倍率を表す。データは、少なくとも3つの独立した実験を2回行った結果を表す。
【図4】図4は、E2F−5およびDP−1はトランス活性化で協同することを示す図である。示されているとおり、増加する量のpJ3-E2F-5発現ベクター(1、2または5μg)および100ngのpCMV-DP-1で、U2−OS骨肉腫細胞をトランスフェクションした。それぞれのトランスフェクションでは、2μgのレポーター構築物(E2F4-CAT)および0.2μgのpRSV-ルシフェラーゼを加えた。それぞれのサンプルのCAT活性を、ルシフェラーゼ活性に対して標準化した。データは、少なくとも3つの独立した実験を2回行った結果を表す。
【図5】図5は、ポケットタンパク質によるE2F−5トランス活性化の阻害を示す図である。5μgのpJ3-E2F-5および100ngのpCMV-DP-1と、pCMV-Rb(50および100ng)、pCMV-RbΔ22(100ng)、pCMV-p107(50および100ng)、pCMVp107DE(100ng)またはpCMV-HA-p130(50、100および500ng)とを組み合わせて、US−OS細胞をトランスフェクションした。発現プラスミドと共に2μgのE2F4-CATおよび0.2μgのpRSV-ルシフェラーゼで、上記細胞をトランスフェクションした。ルシフェラーゼ内部対照に対して、CAT活性を標準化した。E2F4-CATの基準レベル(これを1.0とする)に対する活性化倍率を計算した。データは、少なくとも3つの独立した実験を2回行った結果を表す。
【図6】図6は、一過性にトランスフェクションされたU2−OS細胞中のE2F含有複合体を示す図である。示されているとおり、pRb-p107またはp130発現ベクターの存在下または不存在下、E2F−5およびDP−1発現ベクターで、U2−OS骨肉腫細胞を一過性にトランスフェクションした。2日後、全細胞抽出物を調製し、コンセンサスE2FDNA結合部位を含有する[32P]−標識オリゴヌクレオチドと共にインキュベーションし、ゲル電気泳動に供した。遊離プローブ、E2F−5/DP−1DNA複合体およびE2F−5/ポケットタンパク質複合体の位置を示す。
【図7】図7は、E2F−5は、in vivoでp130と優先的に相互作用することを示す図である。ヒトCAMA乳がん細胞を[32P]−オルトホスファートで標識し、非イオン性デタージェントライゼートを逐次免疫沈降法に供した。第1の免疫沈降法では、ライゼートを、示されているとおり、pRb、p107またはp130抗体と共にインキュベーションした。パネルAは、CAMA細胞からのpRb−p107およびp130特異的抗体による第1の免疫沈降法を示す。pRb、p107およびp130結合タンパク質を、SDS含有緩衝液中で煮沸することにより、その免疫沈降したポケットタンパク質から遊離させ、E2F−5特異的抗血清で再度免疫沈降させた(パネルB)。対照として、pRbおよびp107免疫沈降物から遊離したタンパク質を、抗E2F−1(KH20)またはE2F−4(RK13)抗体で再度免疫沈降させた(パネルC)。免疫沈降したタンパク質を7.5%SDS−ポリアクリルアミドゲル上で分離し、該ゲルを乾燥し、タンパク質をオートラジオグラフィーで検出した。
【図8】図8は、マウスE2F−5を単離するための戦略を示す図である。「餌」となるLEXA DP-1を使用して、14.5d.p.c.のマウス胎児活性化ドメイン標識(tagged)cDNAライブラリーをスクリーニングした。
【図9A−1】図9A-1は、マウスE2F−5の配列およびE2Fファミリーの他のメンバーとの比較を示す図であり、E2F−5のヌクレオチド配列および推定アミノ酸残基配列を示す図である。
【図9A−2】図9A-2は、マウスE2F−5の配列およびE2Fファミリーの他のメンバーとの比較を示す図であり、E2F−5のヌクレオチド配列および推定アミノ酸残基配列を示す図である。
【図9B】図9Bは、E2F−5(中段)を、E2F−1(上段)およびE2F−4(下段)と比較して示す図であり、E2F−5とE2F−1との間およびE2F−5とE2F−4との間のタンパク質配列レベルの同一性の割合(%)を示す。E2Fファミリーメンバーの間で共有されているドメインを示す。(Leesら, 1993)
【図9C】図9Cは、E2Fファミリーメンバー内での保存ドメイン中のアミノ酸残基配列の比較を示す図である。ハイライトが付されている残基はすべてのファミリーメンバーに共通であり、枠で囲まれている残基はE2F−4とE2F−5とで共通である。ロイシンジップ領域中の太字の残基は、7個からなる反復中の疎水性残基を示す。E2FファミリーメンバーとDP−1との間で共有されているDEFボックス領域中の残基を、線により示す。
【図10】図10は、酵母におけるE2F−5およびDP−1によるE2F部位依存的転写の活性化を示す図である。 (a)レポーターおよびエフェクター構築物の概要 (b)示されているE2F−5およびDP−1発現ベクターを、単独で(レーン2および3)または一緒に(レーン4)、酵母中に形質転換し、4つの野生型E2F部位を保持するp4xWT CYC1の活性を評価した。平行実験において、p4xMTCYC1の活性は、いずれの条件においても影響を受けなかった。示しているデータは、3回の読み取りから得たものである。
【図11】図11は、酵母におけるE2F−5による転写活性化を示す図である。 (a)レポ−ターおよびエフェクター構築物の概要 (b)pLexA-CYC1-lacZの活性をアッセイすることにより、pLEX.E2F-1(トラック2)またはpLEX.E2F-5(トラック3)のいずれかの転写活性を酵母で評価した。示しているデータは、3回の読み取りから得たものである。
【図12】図12は、ポケットタンパク質調節E2F−5を示す図である。 (a)レポーターおよびエフェクター構築物の概要。 (b)野生型pRb(トラック3)、pRbΔ22(トラック4)、p107(トラック5)およびp107AS(トラック6)の存在下、Gal-E2F-5(トラック2)の転写活性を評価した。比較のために、pG4(トラック7〜10)で同様の処理を行った。示しているデータは、3回の読み取りから得たものである。
【図13】図13は、E2F−5は、F9EC細胞中のDRTF1/E2Fの生理学的DNA結合成分であることを示す図である。E2F−5内の異なる領域からの別個のペプチドに対して産生させた2つのタイプの抗血清がF9EC細胞DRTF1/E2F DNA結合活性に及ぼす効果について評価した。抗E2F−5血清、抗ペプチド1(トラック5〜8)および抗ペプチド2(トラック9〜12)を、相同性(+;トラック5、7、9および11)または無関係な(−;トラック6、8、10および12)ペプチドのいずれかの存在下で評価した。比較のために、相同性(トラック1および3)または無関係なペプチド(トラック2および4)のいずれかの存在下、抗DP−1(24)(トラック1〜4)の効果を評価した。それぞれの対またはトラック(+および−)は、異なる抗血清調製物での処理を表す。DRTF1/E2Fb/cDNA結合複合体(La Thangueら, 1990)について示す。
【図14】図14は、E2F−5のDNA結合特性を示す図である。E2F−5、E2F−1およびDP−1をGST融合タンパク質として発現させ、精製し、それらのDNA結合活性をゲル遅延法により評価した。E2F−1またはE2F−5を単独で(それぞれ、トラック1および2、並びに5、6および7)または一定濃度のDP−1(トラック3および4、並びに8、9および10)と共にアッセイした。トラック11は、プローブ単独の場合を示す。E2F−1として加えたタンパク質は、全体で、約50ng(トラック1および3)または100ng(トラック2および4)、E2F−5としては25ng(トラック5および8)、50ng(トラック6および9)または100ng(トラック7および10)、およびDP−1としては50ngであった。
【図15】図15は、E2F−5RNAのレベルを示す図である。示されている細胞系で、E2F−5RNAのレベルをE2F−1と比較した。内部対照として、GAPDH RNAのレベルを評価した。
【技術分野】
【0001】
本発明は、新規転写因子およびその製造および用途に関する。
【背景技術】
【0002】
遺伝子発現を細胞周期の進行と同調させるためには、細胞周期中に生じる分子イベントが転写装置と統合される必要がある。
【0003】
最近、E2F(またはDRTF1)と称される転写因子が同定され、抗がん遺伝子またはがん抑制遺伝子である網膜芽細胞腫感受性遺伝子のタンパク質産物pRbに結合することが示された(例えば、非特許文献1参照)。細胞転写因子E2Fは、網膜芽細胞腫遺伝子産物(pRb)、p107、サイクリン、サイクリン依存性キナーゼなどの重要な細胞周期調節タンパク質と結合し、さらに、その転写活性は、アデノウイルスEla、SV40ラージT抗原、ヒトパピローマウイルスE7タンパク質などのある種のウイルスがんタンパク質によりモジュレーションされるため、E2Fは、細胞周期制御における鍵成分として機能していると広く考えられている。
【0004】
転写因子E2F(またはDRTF1)は、哺乳動物細胞で細胞周期が進行する間、細胞増殖の重要な調節体であることが知られている分子との一連の周期的相互作用を受けるため、細胞周期イベントと転写装置との統合において重要な役割を果たしていると考えられている。例えば、G1期からS期への進行を負に調節し腫瘍細胞中でしばしば修飾される網膜芽細胞腫がん抑制遺伝子産物(pRb)は、E2Fに結合する。同様に、pRb関連タンパク質p107は、主として、E2FとのS期複合体中に生じる。pRbおよびp107は共に、E2Fの転写活性を抑制し、一方、E2F結合部位は、c−myc、p34cdc2などの増殖に関与する種々の遺伝子の制御領域中に生じるため、E2Fの転写活性は、細胞増殖の調節に基本的に重要であると考えられている。さらに、腫瘍細胞から単離された対立遺伝子によりコードされる突然変異Rbタンパク質は、E2Fに結合しない。したがって、この突然変異Rbタンパク質は、E2F部位依存性転写活性化を阻害する能力を有さない。E2Fのもう1つの重要な特徴は、アデノウイルスEla、SV40ラージT抗原、ヒトパピローマウイルスE7などのある種のウイルスがんタンパク質が、不活性転写因子からpRbおよびp107を隔離することにより、その活性をモジュレーションすることである。この効果の発現に要求されるのは、組織培養細胞がトランスフォーメーションされ、成長制御に打ち勝つのに必要な、これらのウイルスタンパク質中の領域である。したがって、これらのがんタンパク質がE2Fを調節する能力は、該がんタンパク質が細胞成長制御の正常なメカニズムを圧倒するための手段であるかもしれないし、逆に、pRbによる転写抑制がpRb媒介性の負の成長制御の基礎であるかもしれない。
【0005】
pRbおよびp107の転写−調節特性を他の細胞周期イベントと統合するための可能性のあるメカニズムは、E2F複合体中のサイクリンAおよびcdc2関連サイクリン依存性キナーゼp33cdk2の同定から示唆された。サイクリンAはS期を経る進行(おそらく、サイクリン依存性キナーゼp33cdk2をE2FへリクルートするサイクリンAの能力により媒介されると考えられる機能)に必要である。これらのデータを総合すると、転写因子E2Fの主要な役割は、細胞周期イベントをpRb、p107、サイクリン、cdkなどの分子を介して転写装置へ中継することであると示唆され、したがってこのことは、遺伝子発現が細胞周期の進行と同調し統合されることを保証するものである。
【0006】
さらに最近になって、E2Fの性質を有する転写因子がクローニングされ配列決定された(非特許文献2及び3を参照)。
【非特許文献1】WagnerおよびGreen, Nature 352, 189-190, 1991
【非特許文献2】Helinら, Cell 70 (1992), 337-350
【非特許文献3】Kaelinら, Cell 70 (1992), 351-364
【発明の開示】
【0007】
驚くべきことに、本発明者らは、E2F遺伝子ファミリーの新規メンバーと考えられる、さらに2つの新規タンパク質を見いだし、これをE2F−5と命名した。ヒトE2F−5のcDNA配列を、このタンパク質のアミノ酸配列と共に図1Aに示す。マウスE2F−5についての対応する配列を図9Aに示す。これらの新規タンパク質は、E2F−5と称され、本明細書ではこの名称を使用することにする。
【0008】
E2F−5は、関連転写因子成分のファミリーの1つであることが見いだされた。このファミリーのメンバーは、DPタンパク質と相互作用して一連の転写因子を形成すると考えられている。DPタンパク質(またはポリペプチド)には、DP−1、DP−2およびDP−3が含まれるが、通常は、これらの最初に挙げたものが残りのものに優先すると考えられる。
【0009】
本発明は、その第1の態様において、図1Aまたは9Aに示すタンパク質、その同族体、並びに該配列および該同族体の断片であって、哺乳動物転写因子として機能しうるものを提供する。特に、本発明は、
(a)E2F−5、
(b)図1Aまたは9Aのタンパク質、
(c)(a)または(b)の突然変異体、対立遺伝子変異体または種同族体、
(d)(a)または(b)に少なくとも70%相同なタンパク質、
(e)DPタンパク質、pRb、p107および/またはp130と複合体を形成する能力を有する(a)から(d)のいずれか1つの断片、または
(f)少なくとも15のアミノ酸長の(a)から(e)のいずれかの断片、
を含んでなるポリペプチド(好ましくは実質的に単離された形態のもの)を提供する。
【0010】
以下、この定義に包含されるすべてのポリペプチドを、本発明のポリペプチドと称する。
【0011】
タンパク質pRb、p107、DPタンパク質およびp130は本発明のタンパク質と複合体を形成しうるので、本明細書では、これらを、複合体形成タンパク質(complexingproteins)または「複合体形成体(complexors)」と称する。E2F−5は、ある条件の下では、pRbと弱くしか結合できないかもしれない。
【0012】
本発明のポリペプチドが、その天然環境(例、体内)で結合しうる他のポリペプチドを含まない形態である場合、本発明のポリペプチドは、実質的に単離された形態である。本発明のポリペプチドは、該ポリペプチドの意図する目的を妨げない担体または希釈剤と混合してもよく、その場合であっても、実質的に単離されているとみなされると理解される。
【0013】
また、本発明のポリペプチドは、実質的に精製されている形態であり、この場合一般には、調製物中に含まれるポリペプチドの90%以上、例えば95%、98%または99%が本発明のポリペプチドである。
【0014】
突然変異ポリペプチドは、アミノ酸残基の付加、欠失または置換である1以上の突然変異を有する。好ましくは、該突然変異は、該ポリペプチドの構造および/または機能および/または性質に、全くあるいは実質的に影響を及ぼさない。したがって、突然変異体は、適切には、DPタンパク質、pRb、p107および/またはp130と複合体を形成しうる能力を有する。突然変異体は、天然に生じるもの(すなわち、天然源から精製または単離されている形態)であっても、合成的に得られるもの(例えば、コード化DNA上の部位特異的突然変異誘発を行うことによるもの)であってもよい。したがって、本発明のポリペプチドは、天然に生じるものであっても、組換え体(すなわち、遺伝子工学技術を用いて調製されたもの)であってもよいことは明らかである。
【0015】
対立遺伝子変異体は、ヒトまたは動物(例、マウス)において天然に生じ、図1Aまたは9Aのタンパク質と実質的に同様に遺伝子発現を調節するように機能する変異体である。
【0016】
同様に、上記タンパク質の種同族体は、別の種で天然に生じ、その種において図1Aまたは9Aのタンパク質と同等の機能を発揮する同等のタンパク質である。いずれか1つの種内に、同族体がいくつかの対立遺伝子変異体として存在していてもよく、これらはすべて上記タンパク質の同族体と考えられる。対立遺伝子変異体および種同族体は、図1Aまたは9Aのタンパク質の製造に関して本明細書に記載されている方法に従い、対立遺伝子変異体を保持する適当な細胞源(例えば、ヒトまたはげっ歯類からのもの)または別の種上でそのような方法を行うことにより得ることができる。上記タンパク質は進化的に保存されている場合があるため、対立遺伝子または種変異体をコードするクローンを得るために、本発明のポリヌクレオチドを使用して、ヒト、げっ歯類または他の細胞から作製されたライブラリーをプローブすることも可能である。上記クローンを通常の技術により操作して、本発明のポリペプチドを同定し、ついでそれ自体が公知の組換えまたは合成技術により該ポリペプチドを製造することができる。好ましい種同族体には、哺乳動物または両生類種同族体が含まれる。
【0017】
図1Aまたは9Aのタンパク質に少なくとも70%相同なタンパク質、および図1Aまたは9Aのタンパク質に少なくとも80または90%、より好ましくは少なくとも95%相同なタンパク質が本発明に包含される。これは、一般に、少なくとも20、好ましくは少なくとも30、例えば少なくとも40、60または100以上連続するアミノ酸の領域にわたる。タンパク質の相同性を測定する方法は当該技術分野で公知であるので、上述のように当業者に理解されるであろう。相同性は、通常、アミノ酸の同一性に基づいて計算される(「硬相同性(hard homology)」と称されることもある)。
【0018】
一般に、上記複合体形成体と複合体を形成する能力を有する、図1Aまたは9Aのポリペプチドまたはその対立遺伝子変異体またはその種同族体の断片は、少なくとも10、好ましくは少なくとも15、例えば少なくとも20、25、30、40、50または60のアミノ酸長である。
【0019】
断片がタンパク質の複合体と複合体を形成するか否かの判定は、複合体形成タンパク質および該断片を、それらが正常にトランス活性化転写因子を形成する条件下で用意し、そして複合体が形成されたか否かを判定することにより行うことができる。この判定は、例えば、上記複合体がE2F結合部位にin vitroで結合する能力を測定することにより、あるいは、SDS−PAGEなどの方法により推定複合体の分子量を測定することにより行ってもよい。
【0020】
好ましい断片には、DP−1または他の複合体形成体とトランス活性化複合体を形成する能力を有する断片が含まれる。本明細書中の実施例では、上記タンパク質の機能を分析するためのいくつかの方法を記載する。ポリペプチドがDP−1タンパク質とトランス活性化複合体を形成する能力を有するか否かを評価するために、上記実施例を適合させてもよい。例えば、DP−1/E2F−5複合体により活性化されるように適合されたレポーター遺伝子構築物の存在下、上記ポリペプチドを上記複合体形成体へ加えることができる(例えば、Medical Research CouncilのWO-A-94/10307の図10を参照されたし)。そのような実験により、上記ポリペプチド断片が必要な活性を有するか否かを判定することができる。
【0021】
本発明のポリペプチドは、表示的または検出可能な標識で標識してもよい。上記(表示的)標識は、上記ポリペプチドの検出を可能にする、どのような適当な標識であってもよい。適当な標識としては、放射性同位元素(例えば125I)、酵素、抗体、リンカー(例えばビオチン)などが挙げられる。サンプル中のE2F−5タンパク質の量を測定するために、イムノアッセイなどの診断方法において本発明の標識ポリペプチドを使用してもよい。
【0022】
本発明のポリペプチドまたは標識ポリペプチドを固相、例えばイムノアッセイディッシュの壁に固定してもよい。
【0023】
本発明の第2の態様は、
(a)図1Aまたは9Aに示すヌクレオチドの配列、
(b)(a)に相補的な配列、
(c)(a)または(b)のいずれかの中の配列と選択的にハイブリッド形成する能力を有する配列、
(d)第1の態様で定義されたポリペプチドをコードする配列、または、
(e)(a)から(d)の中の配列のいずれかの断片、
を含んでなるポリヌクレオチドに関する。
【0024】
したがって、本発明は、図1Aまたは9Aの配列または相補的配列と選択的にハイブリッド形成する能力を有するヌクレオチドの連続した配列を含んでなる、好ましくは実質的に単離または精製された形態のポリヌクレオチドを提供する。本発明のポリヌクレオチドは、図1Aまたは9AのDNA配列および図1Aまたは9AのDNA配列と選択的にハイブリッド形成する能力を有するその断片を含む。本発明は、そのさらに別に実施態様において、図1Aまたは9Aのタンパク質をコードするDNAまたはその断片を提供する。
【0025】
また、上記ポリヌクレオチドは、RNAを含んでなるものであってもよい。また、それは、その中に合成または修飾ヌクレオチドを含むポリヌクレオチドであってもよい。オリゴヌクレオチドに対するいくつかの異なる型の修飾が当該技術分野で公知である。これらには、メチルホスホナートおよびホスホロチオナートバックボーン、該分子の3'および/または5'末端におけるアクリジンまたはポリリシン鎖の付加が含まれる。本発明の目的のためには、本明細書に記載のオリゴヌクレオチドは、当該技術分野で利用可能ないかなる方法により修飾してもよいと理解されるべきである。治療方法で使用される本発明のオリゴヌクレオチドのin vivo活性または寿命を増加させるために、そのような修飾を行ってもよい。
【0026】
図1Aまたは9AのDNAと選択的にハイブリッド形成する能力を有するポリヌクレオチドは、一般に、少なくとも20、好ましくは少なくとも30、例えば少なくとも40、60または100以上の連続ヌクレオチドの領域内において、図1Aまたは9AのDNAに対して、少なくとも70%、好ましくは少なくとも80または90%、最適には少なくとも95%相同である。これらのポリヌクレオチドは、本発明に包含される。
【0027】
本発明のポリヌクレオチドが、その天然環境(例えば体内)で結合しうる他のポリヌクレオチドを含まない形態である場合、本発明のポリヌクレオチドは、実質的に単離された形態である。上記ポリヌクレオチドは、意図する目的を妨げない担体または希釈剤と混合してもよく、その場合であっても、該ポリヌクレオチドは実質的に単離されているとみなされると理解される。
【0028】
本発明のポリヌクレオチドは、プライマー(例えばPCRプライマー)、プローブ(例えば放射活性または非放射活性標識を用いる通常の手段により表示的または検出可能な標識で標識されているプローブ)を製造するのに使用してもよいし、該ポリヌクレオチドをベクター中にクローニングしてもよい。そのようなプライマー、プローブおよび図1Aまたは9AのDNAの他の断片は、少なくとも15、好ましくは少なくとも20、例えば少なくとも25、30または40のヌクレオチド長であり、それらも本発明に包含される。
【0029】
本発明のDNAポリヌクレオチドなどのポリヌクレオチドは、組換え的、合成的または当業者が利用可能ないかなる手段により製造してもよい。また、本明細書に開示する技術を参照して、それをクローニングしてもよい。
【0030】
本発明は、本発明のポリヌクレオチドおよびその相補体を含んでなる二本鎖ポリヌクレオチドを含む。
【0031】
本発明の第3の態様は、本発明のポリヌクレオチド、特にDNAまたはRNAポリヌクレオチドの複製および発現に適したベクター(例えば発現ベクター)に関する。上記ベクターは、例えば、複製起点、所望によりポリヌクレオチドの発現のためのプロモーターおよび所望により該プロモーターの調節体が設けられている、プラスミド、ウイルスまたはファージベクターであってもよい。上記ベクターは、1以上の選択マーカー遺伝子、例えば細菌プラスミドの場合にはアンピシリン耐性遺伝子または哺乳動物ベクターの場合にはネオマイシン耐性遺伝子を含有していてもよい。上記ベクターは、例えばRNAの製造などにin vitroで使用してもよいし、宿主細胞をトランスフェクションまたは形質転換させるために使用してもよい。また、上記ベクターは、例えば遺伝子治療方法などにin vivoで使用できるように適合させてもよい。
【0032】
この第3の態様のベクターは、好ましくは、複製可能な組換えベクターである。したがって、上記ベクターを使用して上記DNAを複製してもよい。好ましくは、上記ベクター中のDNAは、宿主細胞による上記コーディング配列の発現を付与する能力を有する制御配列に機能を発揮できるように結合している。「機能を発揮できるように結合している」という語は、記載成分が、それらが意図されるように機能できる関係で並んでいることを意味する。コーディング配列に「機能を発揮できるように結合している」制御配列は、該制御配列に適合した条件下で該コーディング配列の発現が達成されるように連結されている。本発明のポリペプチドの発現を付与するために、そのようなベクターを、適当な宿主細胞中へ形質転換またはトランスフェクションしてもよい。
【0033】
したがって、本発明のさらに別の態様は、上記第3の態様のベクターにより形質転換またはトランスフェクションされた宿主細胞に関する。これは、図1Aまたは9Aの配列またはそのオープンリーディングフレームを含む本発明のポリヌクレオチドの複製および発現を可能にする。上記細胞としては、上記ベクターに適合するものが選択され、例えば、細菌、酵母、昆虫または哺乳動物であってもよい。
【0034】
また、アンチセンスRNAを製造するために、本発明のポリヌクレオチドを上記ベクター中にアンチセンス配向で挿入してもよい。また、アンチセンスRNAまたは他のアンチセンスポリヌクレオチドを、合成的手段により製造してもよい。そのようなアンチセンスポリヌクレオチドを、細胞中のE2F−5タンパク質のレベルを制御するための方法に使用してもよい。そのような方法は、E2F−5mRNAからタンパク質への翻訳レベルを阻害または減少させるのに有効な量の上記アンチセンスポリヌクレオチドを上記細胞中に導入することを含んでいてもよい。上記細胞は、腫瘍細胞などの非制御的に増殖する細胞であってもよい。
【0035】
したがって、本発明は、第5の態様において、上記第3の態様の(発現)ベクターを用いて、本発明のポリペプチドをコードするコーディング配列の発現(該ベクターによる)を付与する条件下で形質転換またはトランスフェクションした宿主細胞を培養し、そして、発現されたポリペプチドを回収することを含んでなる、本発明のポリペプチドの製造法を提供する。
【0036】
また、本発明は、第6の態様において、本発明のポリペプチドに特異的な抗体(モノクローナルまたはポリクローナル抗体)を提供する。本発明の抗体は、該抗体の結合活性を保持するその断片および突然変異体を含む。さらに、本発明は、本発明のポリペプチドに対するモノクローナルまたはポリクローナル抗体の製造法を提供する。モノクローナル抗体は、上記タンパク質またはそのペプチド断片を免疫原として用いる通常のハイブリドーマ技術により製造してもよい。また、ポリクローナル抗体は、本発明のポリペプチドを宿主動物(例、ラット、ウサギ)に接種し、そして免疫血清を回収することを含んでなる通常の手段により製造してもよい。
【0037】
モノクローナル抗体の抗原結合活性を保持することができる該モノクローナル抗体の断片(例えば、Fv、F(ab')およびF(ab2)'断片)が、本発明のこの態様に含まれる。さらに、本発明のモノクローナル抗体を分析し(例えば、かかる抗体を発現する遺伝子のDNA配列分析による)、例えばEP-A-0239400(Winter)に開示されている方法に従い、本発明の抗体の相補性決定領域を有するヒト化抗体を作製してもよい。
【0038】
さらに本発明は、本発明の抗体またはその断片と担体または希釈剤とを含んでなる組成物を提供する。そのような組成物には医薬組成物が含まれ、この場合、上記担体または希釈剤は薬学的に許容されるものである。
【0039】
本発明のポリペプチドは、担体または希釈剤と共に組成物中に存在していてもよい。これらの組成物には医薬組成物が含まれ、この場合、上記担体または希釈剤は薬学的に許容されるものである。
【0040】
薬学的に許容される担体または希釈剤には、経口、直腸、鼻内、局所(頬、舌下など)、膣または非経口(皮下、筋内、静脈内、皮内、脊髄内、硬膜外)投与に適した製剤中で用いられるものが含まれる。上記製剤は単位投与形で提供されるのが好都合であり、薬学分野でよく知られている方法のいずれにより製造してもよい。そのような方法は、有効成分を、1以上の補助成分を構成する担体と混合する工程を含む。一般に、上記製剤は、有効成分と液体担体若しくは微小固体担体またはその両方とを均一かつ十分に混合し、ついで生成物を必要に応じて成型することにより製造する。
【0041】
例えば、非経口投与に適した製剤には、抗酸化剤、緩衝剤、静菌薬、および目的の受容者の血液と該製剤とを等張にする溶質を含有していてもよい水性または非水性の無菌注射溶液;並びに懸濁化剤、濃稠化剤、および該ポリペプチドを血液成分または1以上の器官へ標的化するように設計されているリポソームまたは他の微小粒子系を含んでいてもよい水性または非水性の無菌懸濁液が含まれる。
【0042】
本発明のポリペプチド、本発明のポリペプチドに対する抗体またはその断片および前記組成物は、ヒトなどの哺乳動物における増殖性疾患などの状態の治療、調節または診断に使用しうる。そのような状態には、DPまたはE2Fタンパク質または関連ファミリーメンバーなどの1以上の転写因子の異常な(例えば、異常に高いまたは低いレベルの)および/または異常な(例えば、遺伝子配列の突然変異によるもの)発現が含まれる。また、この状態には、図1Aまたは9Aのタンパク質により調節される遺伝子産物を与える遺伝子の異常発現により引き起こされる状態が含まれる。上記ペプチド、抗体、その断片、組成物などを用いる状態の治療または調節には、通常、そのような治療を必要とする受容者に、ポリペプチド、抗体、その断片または組成物の有効量を適宜投与することが含まれる。
【0043】
また本発明は、E2F−5−DPタンパク質複合体の形成の妨害を介して転写因子の活性化を阻害するためにこの領域に標的化されている抗体およびその断片を提供する。
【0044】
さらに本発明は、サンプル中の本発明のポリペプチドの存在または不存在を検出するためのイムノアッセイを行う方法であって、
(a)本発明の抗体を用意し、
(b)抗体−抗原複合体の形成を許容する条件下、該サンプルを該抗体と共にインキュベーションし、そして
(c)該抗体−抗原複合体が存在すれば、これを検出する、
ことを含んでなる前記方法を提供する。
【0045】
もう1つの態様において本発明は、増殖性またはウイルス性の疾患の治療のための推定化学療法剤を同定するための新規アッセイであって、DPタンパク質またはその誘導体、本発明のポリペプチドおよび推定化学療法剤を接触させ、そして、該剤がDP/E2F−5タンパク質複合体の形成を阻害する程度を測定することを含んでなる前記新規アッセイを提供する。このアッセイでは、それぞれのタンパク質が十分に用意され、上記剤の不存在下のアッセイ条件下でDP−1および/またはE2F−5タンパク質がヘテロ二量体を形成する限り、完全なDP−1および/またはE2F−5タンパク質を使用する必要はないかもしれない。
【0046】
DP−1(およびE2F1、2および3)のクローニングおよび配列決定は、当該技術分野で公知であり、DP−1に対する抗体の組換え発現および製造は、WO-A-94/10307に記載されている。
【0047】
したがって、本発明は、増殖性疾患の治療のための推定化学療法剤を同定するためのスクリーニング方法であって、
(A)(i) DPポリペプチド、
(ii) 上記第1の態様のポリペプチド、および
(iii) 推定化学療法剤、
を、成分(i)と(ii)とが(iii)の不存在下で複合体を形成する条件下で接触させ、そして
(B)成分(iii)が上記複合体を阻害しうる程度を測定する、
ことを含んでなる前記スクリーニング方法を提供する。
【0048】
このアッセイでは、アッセイの結果を測定できるように、その3つの成分のいずれか1つ以上を、例えば放射活性または比色標識により標識してもよい。推定化学療法剤には、本発明のペプチドが含まれる。
【0049】
DPタンパク質の変異体、同族体および断片は、E2F−5タンパク質の変異体、同族体および断片と対応させて定義される。
【0050】
(i)と(ii)との複合体は、例えば、それがE2F DNA結合部位にin vitroで結合する能力により測定してもよい。あるいは、上記アッセイは、レポーター遺伝子に結合したE2F結合部位を含んでなるプロモーターを上記複合体が活性化する能力を測定するin vivoアッセイであってもよい。このin vivoアッセイは、例えば、酵母、昆虫、両生類または哺乳動物細胞におけるかかるアッセイを示す実施例を参照することにより行ってもよい。
【0051】
上記アッセイで測定しうる候補治療剤には、上記第1の態様のポリペプチドだけでなく、特に、
(a)図1Aまたは9Aのタンパク質、
(b)その対立遺伝子変異体または種同族体、または
(c)(a)に少なくとも70%相同なタンパク質、
の10以上のアミノ酸の断片も含まれる。
【0052】
本発明のポリヌクレオチドまたは本発明のポリペプチドをコードする核酸を保持するベクターを、遺伝子治療方法に使用してもよい。そのような遺伝子治療は、細胞(例えば腫瘍細胞)の非制御増殖を治療するのに使用してもよい。遺伝子治療の方法は、治療が必要な患者中の細胞内で本発明のアンチセンスポリヌクレオチドを発現する能力を有するベクターの有効量を該細胞へ輸送して、E2F−5mRNAからE2F−5タンパク質またはポリペプチド(E2F−5がDPタンパク質または関連ファミリーメンバーへ結合するのを妨害するもの)への翻訳を阻害または抑制することを含む。
【0053】
上記ベクターは、好ましくは、ウイルスベクターである。このウイルスベクターは、腫瘍細胞を標的化するのに当該技術分野で利用可能ないかなる適当なベクターであってもよい。例えば、Huberら(Proc. Natl. Acad. Sci. USA (1991) 88, 8039)は、ヘパトーム、乳、結腸または皮膚細胞の形質転換のための、両種性レトロウイルスの使用を報告している。また、Culverら(Science (1992)256; 1550-1552)は、Ramら(Cancer Research (1993)53; 83-88)と同様、ウイルス特異的酵素プロドラッグ療法(virus-directed enzyme prodrug therapy)でのレトロウイルスベクターの使用を記載している。Englehardtら(Nature Genetics (1993)4;27-34)は、細胞中への嚢胞性線維症膜貫通コンダクタンス産物(CFTR)の輸送における、アデノウイルスに基づくベクターの使用を記載している。
【0054】
本発明は、いくつかのアッセイを意図する。一般には、これらは以下のように分類することができる。
【0055】
1.E2F−5トランス活性化(trans-activation)の阻害剤(すなわち、転写の活性化の阻害)を見つけるためのアッセイの実施。したがって、この阻害剤は、DNAに対するE2F−5の結合(通常はE2F結合部位で生じる)を阻害するかもしれない。適当と考えられる阻害剤はタンパク質であり、これはp107と同様または同一の効果を有していてもよい。したがって、適当な阻害分子は、上記第1の態様のタンパク質で定義したのと同様に、p107の断片、突然変異体、対立遺伝子変異体または種同族体を含んでなる。
【0056】
2.(ヘテロ)二量体形成の阻害剤のアッセイ。そのような阻害剤は、E2F−5(または上記第1の態様のポリペプチド)と複合体形成体(例えば、DP−1などのDPタンパク質)との二量体形成を妨害するかもしれない。もちろん、この阻害剤は、上記第1の態様のタンパク質で定義したのと同様のDPタンパク質の断片、突然変異体、対立遺伝子変異体または種同族体であってもよい。
【0057】
3.アッセイの第3のカテゴリーは、リン酸化の阻害剤を見つけることである。E2F−5(および上記第1の態様の他のタンパク質)はリン酸化により活性化されると考えられている。したがって、リン酸化の阻害剤は、E2F−5のトランス活性化特性を阻害すると考えられる(したがって、結局、この阻害剤は、上記の2つのアッセイのいずれかで見いだされた阻害剤と同じ効果を有するかもしれない)。リン酸化はcdkにより引き起こされ、したがって、このリン酸化の阻害剤は、そのようなアッセイにより意図されるものである。
【0058】
本発明は、いくつかの治療的用途を意図する。例えば、E2F−5に対してアンチセンスである配列の核酸を用いる遺伝子治療などである。さらに、DP−1タンパク質に結合し、それによって該DPタンパク質と不活性複合体を形成しうる分子を意図する。適当な分子には、E2F−5自体以外に上記第1の態様の分子が含まれる。そのような分子は、E2F−5の突然変異体であってもよく、該分子は、当該技術分野ではしばしば、ドミナントネガティブ分子と称される。
【0059】
本発明は、細胞の非制御増殖に基づく疾患、または非制御増殖が重要または必須の病理学的側面である疾患の治療または予防を意図する。これには、がん、ウイルス性疾患、自己増殖自体、および自己免疫疾患、例えば乾癬が含まれる。分裂細胞(例えば造血幹細胞および/または骨髄細胞)の成長を一時的に阻害したい場合があるかもしれない。これらの観点では、一般には、E2F−5の活性を予防、阻害または妨害することが求められる。
【0060】
これに対して、E2F−5発現を増加させることにより、例えば、過剰発現を促進または誘導することにより、いくつかの疾患および状態を治療することができる。これは、好ましくは、アポトーシス(プログラムされた細胞死として知られていることもある)を引き起こす。E2F−5タンパク質の過剰発現は、細胞死を引き起こすことがある。したがって、この態様は、がんの治療にも用いることができる。よって、1つの目的は、E2F−5の活性を増加させることである。E2F−1でも同様の用途が公知である(Qinら, PNAS USA 91(印刷中))。
【0061】
E2F−5遺伝子は腫瘍細胞中で突然変異しうることに留意すべきである。その場合には、突然変異遺伝子を、突然変異から生じる状態の診断に使用することができる。また、それは、突然変異遺伝子による治療に有用である。
【発明を実施するための最良の形態】
【0062】
以下の2つの実施例では、それぞれ、ヒトおよびマウス源からの本発明の新規タンパク質およびDNAの単離および特性決定について説明する。しかしながら、他の源(例えば哺乳動物源)も本発明の範囲内に包含され、上記タンパク質の他の哺乳動物同族体も同様の方法で単離されうる。本明細書に記載の実施例は例示にすぎず、本発明を何ら限定するものではない。
実施例1
概要
E2FDNA結合部位は、細胞周期中に発現が厳しく調節されている多数の遺伝子中で見いだされる。E2F転写因子の活性は、E2Fトランス活性化ドメインに結合しこれを阻害しうる特異的なリプレッサー分子との結合により調節される。E2F−1、2および3の場合、該リプレッサーは、網膜芽細胞腫遺伝子pRbの産物である。E2F−4は、pRb関連p107とは相互作用するが、pRb自体とは相互作用しない。最近、網膜芽細胞腫遺伝子ファミリーの第3のメンバーであるp130をコードするcDNAが単離された。p130も、主に細胞周期のG0期でE2FDNA結合活性と相互作用する。本発明者らは、本明細書において、E2F遺伝子ファミリーの第5のメンバーのクローニングを報告する。ヒトE2F−5cDNAは、346−アミノ酸のタンパク質(推定分子量38kDa)をコードする。E2F−5は、E2F−1(57%の類似度)よりもE2F−4(78%の類似度)と密接に関連している。E2F−5は、DP−1と協同的にコンセンサスE2F部位へ結合する点で、他のE2Fと類似している。本発明者らは、特異的なE2F−5抗血清を使用することにより、生理学的条件下でE2F−5が優先的にp130と相互作用することを示す。
緒言
E2Fは、E2F様およびDP様サブユニットよりなるヘテロ二量体転写因子のグループに与えられている名称である[27]。E2FDNA結合部位は、細胞周期中に発現が調節されるいくつかの遺伝子のプロモーター中に存在し、これらのE2F部位の存在がこれらの遺伝子の細胞周期調節発現に寄与することを示す証拠が存在する[13, 28, 38]。
【0063】
E2FDNA結合活性は、網膜芽細胞腫タンパク質(pRb)およびpRb関連p107およびp130と複合していることが判明している[6, 10, 29, 37]。このグループのタンパク質は、保存モチーフである「ポケット」を共有しており、該ポケットは、細胞タンパク質およびウイルスタンパク質の両方に対する結合に関与している。このため、pRb様タンパク質のグループは、一括して、ポケットタンパク質ファミリーとして知られている。E2Fと種々のポケットタンパク質との間の複合体は、その出現が細胞周期中で異なるため、細胞周期調節において異なる機能を有すると考えられる。pRbと複合体形成しているE2Fはほとんどが、細胞周期のG1期で見いだされる[5-7, 11]。これに対して、p107とE2Fとの複合体は細胞周期中に残存するが、それらの組成は可変的である。G1では、E2Fおよびp107の他に、サイクリンEおよびcdk2が存在する。S期では、E2F/p107複合体において、サイクリンEはサイクリンAで置換される[29, 37]。p107/E2F複合体中にこれらのサイクリン/cdk複合体が存在する機能的意義は、現在のところ明らかではない。静止細胞では、E2Fとp130との複合体が、最も優勢なE2FDNA結合種である。細胞が静止状態から脱するにつれて、この複合体は消失するが、このことは、p130と相互作用するE2F活性についての細胞周期進入における役割を示唆するものである[10]。
【0064】
E2Fが転写を活性化する能力は、該ポケットタンパク質との複合体形成により調節される。E2FとpRbとの間の複合体形成は、リン酸化による調節に左右される。pRbの低リン酸化種だけがE2Fと相互作用するが、このことは、サイクリン/cdk複合体によるpRbのリン酸化が、細胞周期中のE2FとpRbとの間の相互作用を制御することを示す[5-7, 11]。
【0065】
E2FDNA結合活性の強制発現が、細胞周期のG1期からSおよびG2/M期へ細胞を進行させ[3]、E2Fが静止細胞を刺激してDNA合成を開始させうる[23]という知見により、細胞周期調節におけるE2F転写因子の非常に重要な役割が強調される。重要なことは、E2Fの過剰発現が、活性化されたras癌遺伝子と共に、初代げっ歯類繊維芽細胞の癌トランスフォーメーションを引き起こすことである[3]。
【0066】
現在までに、4つの異なるE2F様ポリペプチドが単離されている。E2F−1、2および3は、pRbとの複合体として見いだされるにすぎないが、E2F−4はp107と優先的に相互作用する[3, 15, 19, 22, 24, 30, 36]。E2Fとp107との間およびE2Fとp130との間の複合体形成がどのように調節されるかについては現在のところ知られていない。E2F/p107複合体およびE2F/p130複合体の調節の検討を始めるために、本発明者らは、E2F遺伝子ファミリーの更に別のメンバーを探索した。本発明者らは、ここで、p130と優先的に相互作用するE2F遺伝子ファミリーの第5のメンバーのクローニングを報告する。
材料および方法
酵母二ハイブリッドスクリーニング
「ベイト」プラスミドpPC97-p107を含有する酵母Y190株[17]を、酢酸リチウム法[34]を用いて、14.5日齢のCD1マウス胎児ライブラリー[8]で形質転換した。ヒスチジンが無く25mM3−アミノトリアゾールで補足されたプレート上での成長に関して200万個の形質転換体を選択し、ついで、既に記載されている[12]とおりに、β−ガラクトシダーゼ活性に関して分析した。二重陽性酵母コロニーに由来するcDNAライブラリープラスミドを、種々のGal4-DBD融合プラスミド(pPC97-p107、pPC97-bmiおよびpPC97(挿入なし))による再形質転換により標的特異性に関して試験した。部分マウスE2F−5cDNAを用いて、更に別のヒトcDNAライブラリーをスクリーニングした。本明細書に記載の完全長ヒトE2F−5cDNAを、T84結腸癌ライブラリー(Stratagene)から単離した。
【0067】
プラスミド
pPC97[8]のGal4DNA結合ドメイン(アミノ酸1〜147)とインフレーム(in frame)となっているp107のポケット領域(アミノ酸240〜816)をクローニングすることにより、pPC97-p107を得た。pGEX-2T中でアミノ酸89〜200(A)またはアミノ酸89〜346(B)をコードするヒトE2F−5cDNA断片をクローニングすることにより、pGST-E2F-5(A)および(B)を構築した。トランスフェクション実験には、以下のプラスミドを使用した。pSG-Gal4-E2F-1は、ヒトE2F−1のアミノ酸284〜437を含有している[19]。pJ3-Gal4-E2F-4およびpJ3-Gal4-E2F-5は、pJ3Ω中、Gal4のDNA結合ドメインとインフレームとなっているE2F−4またはE2F−5のヒトcDNAの断片(E2F−4の場合はアミノ酸276〜412をコードし、E2F−5の場合はアミノ酸222〜346をコードする)をクローニングすることにより得た[33]。pJ3-E2F-5は、完全長ヒトE2F−5cDNA(3'非コーディング配列の最後の184ヌクレオチドを欠く)を哺乳動物発現ベクターpJ3Ω中にクローニングすることにより構築した。E2F−5の翻訳開始コドンの後には、モノクローナル抗体12CA5により認識される10アミノ酸のエピトープ(HA)が続く。
【0068】
pCMV-DP-1、pCMV-pRb、pCMV-p107、pCMV-p107DE、PCMV-pRb△22については、既に記載されている[20, 41]。
【0069】
細胞系
U2−OSおよびCAMA細胞を、それぞれ10%または20%のウシ胎児血清で補足されたダルベッコの改変イーグル培地(Dulbecco'smodified Eagle medium)(DMEM)中で培養した。
【0070】
リン酸カルシウム沈殿法[39]を用いてトランスフェクションを行った。
【0071】
CATアッセイ
5μg (Gal4)5-CAT[25]または2μg E2F4-CAT[20]、0.2μg RSV−ルシフェラーゼおよびヘリング精子担体DNA(全量を20μg/10cmプレートとする)と共に、示されている発現ベクターでU2−OS細胞を一過性にトランスフェクションした。既に記載されているとおりに[2, 3]、CATおよびルシフェラーゼ活性に関して細胞をアッセイした。
【0072】
ノーザンブロット分析
E2F−5発現分析のために、細胞系のパネルから全細胞質RNAを調製した。20mgの全細胞RNAを、記載されているとおりに[4]、1%ホルムアルデヒドアガロースゲル中で電気泳動し、ニトロセルロースに移し、[32P]−標識部分ヒトE2F−5cDNA(nt. 666〜1038)でプローブする。ついで、各レーンにローディングされるRNAの量を制御するために、同じフィルターをラットα−チューブリンcDNAでプローブした。
【0073】
免疫試薬および免疫沈降
E2F−5に対する抗体を産生させるために、GST-E2F-5(A)および(B)(プラスミドの項を参照されたし)タンパク質を E. coli 中で産生させ、グルタチオン−セファロースビーズを用いて精製した。等量の両タンパク質をウサギに注射した。3回の免疫の後、ポリクローナル血清を得た。
【0074】
E2F−1(KH20)、E2F−4(RK13)、pRb(XZ77)およびp107(SD−4および9)に対するモノクローナル抗体については、既に記載されている[3, 20, 21, 41]。p130(C20)ウサギポリクローナル抗血清は Santa Cruz Biotechnology Inc. から入手した。既に記載されているとおりに[3]、CAMA細胞およびトランスフェクションされたU2−OS細胞を標識し、免疫沈降させた。
【0075】
ゲル遅延アッセイ
一過性にトランスフェクションされたU2−OS細胞のゲル遅延アッセイを、既に記載されている方法[20]を少し改変することにより行った。20μlの反応容量中、コンセンサスE2FDNA結合部位を特定する0.5ngの[32P]−標識オリゴヌクレオチド(Santa Cruz Biotechnology)と共に、20mMHEPES(pH7.4)、0.1M KCl、1mMMgCl2、0.1mMEDTA、7%グリセロール、1mMNaFおよび1μg 超音波処理サケ精子DNAを含有する結合緩衝液中で10μgの全細胞抽出物を使用した。室温で20分間インキュベーションする間に、DNA−タンパク質複合体を形成させる。該反応生成物を、0.25×TBE中の3.5%ポリアクリルアミドゲル上、90V、室温、2.5時間で分離した。ついで、前記ゲルを乾燥し、フィルムに露出させた。
結果
p107結合タンパク質の単離
p107と相互作用するポリペプチドをコードするcDNAを同定するために、酵母二ハイブリッドスクリーニングを行った[14]。染色体に位置する2つのGal4誘導レポーター遺伝子(HIS3およびLacZ)[12]を含有する酵母Y190株[17]を、Gal4のDNA結合ドメイン(DBD)に融合したp107のポケット領域(アミノ酸240〜816)を含有する「ベイト」プラスミド、および14.5日齢CD1マウス胎児cDNAライブラリー(ここで、それぞれのcDNAは、Gal4のトランス活性化ドメインに個々に融合されている)[8]で同時形質転換した。合計200万の形質転換体を、ヒスチジンを欠くプレート上での選択に付した。β−ガラクトシダーゼの発現に関して、87個の生存コロニーをスクリーニングした。cDNA含有プラスミドを16個の二重陽性酵母コロニーからレスキューした。他のGal4−DBD融合体をコードするプラスミドで再形質転換することにより、p107結合の特異性を確認した。16個のハイブリッドタンパク質全てが、Gal4−p107と特異的に相互作用することが判明した。該酵母からレスキューされた16個のcDNAライブラリープラスミドが10個の異なる遺伝子に由来することが、DNA配列分析により示された。3個のcDNAは、同じ遺伝子に由来しており、4個の公知E2Fに対して有意な相同性を示した。このため、本発明者らは、このcDNAによりコードされるタンパク質をE2F−5と命名した。
【0076】
ついで、該部分マウスE2F−5cDNAを使用して、ヒト結腸癌cDNAライブラリーをスクリーニングすることにより、完全長ヒトcDNAクローンを得た。最長cDNA(2.1kb)を配列決定したところ、それは、346−アミノ酸のタンパク質(推定分子量38kDa)をコードする1038bpのオープンリーディングフレームを含有していた。図1Aは、E2F−5cDNA配列および推定アミノ酸配列を示す。
【0077】
E2F−5は、E2F−1(57%の類似度)よりもE2F−4(78%の類似度)と密接に関連している。E2F−1およびE2F−4と比較すると、E2F−5中には相同な3つの領域が認められる(図1B)。該DNA結合ドメイン(E2F−5のアミノ酸43〜115)は、E2F−4DNA結合領域と93%の類似度を共有し、一方、並列するDP−1二量化ドメインは、E2F−4とE2F−5との間で81%類似する。最後に、E2F−4とE2F−5とのカルボキシル末端ポケットタンパク質相互作用ドメインは、83%類似する。E2F−4およびE2F−5は、サイクリンA結合に関与するE2F−1のアミノ末端モチーフを両タンパク質が欠いている点で、E2F−1とは異なる。E2F−5は、E2F−4のセリン反復領域を欠いている点で、E2F−4とは異なる。
【0078】
E2F−5のmRNA発現レベルを分析するために、ヒトE2F−5cDNAを使用して、いくつかのヒト細胞系から全細胞質RNAを含有するノーザンブロットをプローブした。E2F−5プローブは、ほとんどの細胞系で、低レベルの単一の2.1kbの転写産物を検出した。ヒトCAMA乳癌細胞系は、いくらか高いレベルのE2F−5を発現した(図2)。
【0079】
E2F−5は、カルボキシル末端トランス活性化ドメインを含有する
E2F−1およびE2F−4は、ポケットタンパク質結合部位と重なるカルボキシル末端トランス活性化ドメインを含有する[3, 18]。E2F−5もトランス活性化ドメインを含有するか否かを調べるために、本発明者らは、哺乳動物発現ベクターpJ3Ω中でヒトE2F−5のカルボキシル末端をGal4のDNA結合ドメインと融合させた。U2−OS骨肉腫細胞を、5つの上流Gal4部位を保持するCATレポーター遺伝子で一過性にトランスフェクションするか、あるいは、該レポーター遺伝子とGal4-E2F発現ベクターとで同時トランスフェクションした。図3に示すとおり、Gal4レポータープラスミドとGal4-E2F-5 発現ベクターとの同時トランスフェクションにより、CATレポーター遺伝子が50倍活性化された。Gal4-E2F-1またはGal4-E2F-4による同時トランスフェクションでは、CATレポーター遺伝子の活性化が2〜3倍高かった(図3)。本発明者らは、E2F−5は、強力なカルボキシル末端トランス活性化ドメインを含有すると結論する。
【0080】
E2F−5は、DNA結合のためにDP−1を必要とする
E2F−1とE2F−4とは共に、効率的なDNA結合のためには、DP−1との二量体形成を必要とする[1, 3, 20, 26]。E2F−5がコンセンサスE2FDNA結合部位に結合するか否か、およびDNAに結合するためにE2F−5がDP−1二量体形成を必要とするか否かを調べるために、本発明者らは、一過性トランスフェクション実験を行った。コアプロモーターが4つの上流E2F部位に結合しているCATレポータープラスミドで、ヒトU2−OS骨肉腫細胞をトランスフェクションした。図4に示すとおり、E2F-CATレポータープラスミドは、該骨肉腫細胞に単独でトランスフェクションされた場合には、低い活性しか有さない。DP−1またはE2F−5発現ベクターで別々にトランスフェクションしても、E2F-CATレポーターの活性化は生じなかった(図4、トラック2および6)。DP−1とE2F−5発現ベクターとの同時トランスフェクションにより、CATレポーターの強力な用量依存的相乗活性化が生じた(図4、トラック3〜5)。これらのデータが示すとおり、E2F−5はコンセンサスE2F部位に結合でき、DNA結合はDP−1依存的である。これらの結果に基づき、本発明者らは、E2F−5がE2F遺伝子ファミリーの真のメンバーであると結論する。
【0081】
E2F−5トランス活性化は、ポケットタンパク質により抑制される
E2F−1およびE2F−4のトランス活性化ドメインはポケットタンパク質相互作用表面と重なるため、これらのE2Fのトランス活性化は、ポケットタンパク質結合により抑制される。E2F−5トランス活性化に対するポケットタンパク質発現の効果を調べるために、本発明者らは、一過性トランスフェクションアッセイを用いた。E2F−1とE2F−4とは共に、それらのそれぞれのポケットタンパク質に対する効率的な結合のためにはDP−1二量体形成を必要とするため[3, 20]、本発明者らは、E2F−5およびDP−1活性化転写に対するポケットタンパク質発現の効果を測定した。E2F−5およびDP−1と共にE2F-CATレポータープラスミドでU2−OS細胞をトランスフェクションした。図5(トラック3)に示すとおり、E2F−5とDP−1との同時トランスフェクションにより、E2F-CATレポーター遺伝子が100倍以上活性化された。E2F−5刺激転写は、pRb、p107およびp130発現ベクターでの同時トランスフェクションにより、用量依存的に阻害された。無傷のポケットドメインを欠く、pRbの変異体(pRbΔ22)およびp107の変異体(p107DE)は、E2F−5トランス活性化を抑制する能力を有していなかった(図5、トラック6および9)。重要なことは、pRbおよびp107のこれらの変異形態が、成長阻害活性をも欠くことである[41]。したがって、この実験では、E2F−5の好ましい結合相手の明確な同定はできなかったが、該実験が示したところによると、ポケットタンパク質結合によりE2F−5トランス活性化が阻害され、pRbおよびp107が成長停止を起こす能力とそれらがE2F−5トランス活性化を阻害する能力との間に密接な相関が存在する。この実験で用いるU2−OS細胞がpRb−またはp107−誘導成長停止に不感受性である[41]ことを指摘しておくことは重要である。したがって、観察された、E2F−5トランス活性化に対する効果は、pRbまたはp107の非特異的細胞周期効果によるものではないらしい。
【0082】
バンドシフトアッセイにおいて、E2F−5はp130と優先的に相互作用する
E2F−5によるポケットタンパク質結合の特異性をさらに調べるために、本発明者らは、電気泳動移動度シフトアッセイ(EMSA)を行った。U2−OS細胞を、pRb、p107またはp130発現ベクターと共に又はこれらを用いることなく、DP−1およびE2F−5発現ベクターで一過性にトランスフェクションした。トランスフェクションの2日後に、全細胞抽出物を、トランスフェクションされた細胞から調製し、コンセンサスE2F部位を特定する[32P]−標識オリゴヌクレオチドと共にインキュベーションした。DNA−タンパク質複合体を、ポリアクリルアミドゲル上で分離し、ラジオグラフィーにより可視化した。図6に示すとおり、E2F−5およびDP−1発現ベクターのトランスフェクションは、模擬(mock)トランスフェクション細胞では観察されなかった新規複合体の出現につながる(図6、レーン1および2を比較されたし)。p130発現ベクターの同時トランスフェクションにより、この複合体のスーパーシフト(supershift)が生じたが、p107またはpRb発現ベクターの場合にはこれは生じなかった(図6、レーン3〜5)。これらのデータは、試験したこれらの3つのタンパク質のうちで、p130がE2F−5/DP−1ヘテロ二量体に対して最も高い親和性を有することを示唆する。
【0083】
E2F−5は、invivoでp130と優先的に相互作用する
生理学的条件下では、E2F−1はpRbに、E2F−4はp107にそれぞれ優先的に結合する[3, 15, 19, 24]。しかしながら、一過性トランスフェクション実験では、E2F−1およびE2F−4活性化遺伝子発現は、pRbとp107との両方により抑制されうる[3, 40]。この特異性の喪失は、おそらく、これらのタンパク質の一過性の過剰発現により生じるのであろう。網膜芽細胞腫タンパク質ファミリーの3つのメンバーのどれが、生理学的条件下でE2F−5と相互作用するかを検討するために、本発明者らは、ヒトE2F−5に対するポリクローナルウサギ抗血清を産生させた。in vitroで転写および翻訳されたE2F−1、E2F−4およびE2F−5を用いる最初の免疫沈降実験において、ポリクローナルE2F−5血清がE2F−5を特異的に認識することが示された(データは示していない)。ついで、E2F−5抗血清を逐次免疫沈降実験で使用した。CAMA乳癌細胞を、[32P]−オルトホスフェートで代謝的に標識し、非イオン性デタージェントライゼートを調製した。これらのライゼートを、pRb特異的抗体、p107抗体またはp130特異的抗血清を用いる免疫沈降法に付した。pRb、p107またはp130で共免疫沈降させたタンパク質を、SDS含有緩衝液中で煮沸することにより遊離させ、希釈し、E2F−5特異的抗血清で再免疫沈降させた。図6のパネルBに示すとおり、p130免疫沈降物からは、47kDaのタンパク質がE2F−5抗血清により特異的に再免疫沈降されたが、pRbまたはp107免疫沈降物からは再免疫沈降されなかった。この47kDaのタンパク質は、SDSポリアクリルアミドゲル上、一過性にトランスフェクションされたE2F−5と共移動する(データは示していない)。対照として、本発明者らは、pRbおよびp107免疫沈降物がそれらの各々のE2Fを含有するか否かを確認した。図6のパネルCに示すとおり、pRbは確かにE2F−1と共免疫沈降し、p107はE2F−4を沈降させた。総合すると、これらのデータからは、E2F−5がin vivoでp130と優先的に相互作用することが示される。
考察
本発明者らは、本明細書において、E2F遺伝子ファミリーの第5のメンバーの単離を報告する。E2F−5は、真のE2Fファミリーメンバーの特徴のすべてを有する。すなわち、それは、高度に保存されたDNA結合ドメイン、DP−1二量化ドメインおよびカルボキシル末端トランス活性化ドメインを有する。さらに、E2F−5は、DP−1と協同的にコンセンサスE2FDNA結合部位に結合し、E2F部位含有レポーター遺伝子の発現を活性化しうる。
【0084】
本発明者らは、E2F−5が3つのポケットタンパク質のどれとin vivoで優先的に相互作用するのかを検討するために、3つのタイプの実験を行った。一過性トランスフェクション実験では、E2F−5トランス活性化は、網膜芽細胞腫タンパク質ファミリーpRb、p107およびp130の3つすべてのメンバーにより抑制された。この点で、E2F−5は、E2F−1およびE2F−4と類似している。なぜなら、E2F−1およびE2F−4トランス活性化は共に、一過性トランスフェクションアッセイで、pRbおよびp107により阻害されうるからである[3, 40]。このように一過性トランスフェクションアッセイで特異性が一見ないことは、おそらく、一過性トランスフェクション細胞におけるE2Fとポケットタンパク質との両方の高い一過性発現レベルの結果であろう。一過性トランスフェクション細胞では、p107はp130と比べて10倍高いレベルで発現されることが判明しているため(データは示していない)、該一過性トランスフェクション実験(図5)におけるp130によるE2F−5トランス活性化の比較的低レベルの阻害は、低いレベルのp130発現の結果である。E2F−5のポケットタンパク質特異性を検討するために、2つの更に別の実験を行った。その第1の実験では、全3つのポケットタンパク質の発現ベクターの存在下および不存在下、E2F−5およびDP−1発現ベクターで細胞を一過性にトランスフェクションした。ついで、該トランスフェクション細胞からの抽出物と、コンセンサスE2F結合部位を特定するオリゴヌクレオチドとを用いて、バンドシフトアッセイを行った。p130の同時トランスフェクションだけが、E2F−5/DP−1複合体のスーパーシフトを起こすことができた(図6)。該バンドシフトアッセイでは、ポケットタンパク質とE2F−5との間の長時間安定な複合体だけが、E2F/ポケットタンパク質「スーパーシフト」複合体として検出される。したがって、たとえp130がp107より低レベルで発現されたとしても、E2F−5とp130との複合体は、p107/E2F−5複合体よりも安定であった(図6)。同様の実験で、本発明者らは、p107とのE2F−4DNA結合複合体を「スーパーシフト」させることはできたが、pRbとの複合体はできなかった(R.L.B.およびR.B., 未公開データ)。この結果が示唆するところによると、移動度シフト実験は、E2Fのポケットタンパク質特異性を検討するのに有用と考えられる。該移動度シフトアッセイの結果と一致して、本発明者らは、トランスフェクションされていない代謝標識CAMA乳癌細胞では、E2F−5は、p130とは共免疫沈降しうるが、p107またはpRbとは共免疫沈降しないことを見いだした(図7)。総合すると、本発明者らのデータは、生理学的条件下では、E2F−5はp130と優先的に結合することを示す。
【0085】
p130とp107とは構造的に密接に関連しており、実際、p107とp130とは共に、サイクリンAおよびEに結合する能力を有するため[16, 32, 41]、E2F−5がp130とは相互作用したが、p107とはしなかったという知見は、いくらか予想外であった。一方、p107だけが抗D型サイクリン抗体と共免疫沈降する(p130はしない)ため[32]、p107とp130とは、D型サイクリンとin vivoで相互作用する能力において異なる。重要なことは、p130/E2Fおよびp107/E2F複合体の出現が、細胞周期で異なることである[9, 10, 29, 35, 37]。このことは、p107とp130とは、細胞周期中、異なる機能を有することを示唆する。p130によるE2F−5の優先的結合は、細胞周期調節におけるp130に関するそのような異なる役割と一致する。
【0086】
E2F−5がコンセンサスE2F部位に結合しうるという本発明者らの知見は、E2F遺伝子ファミリーの残りのメンバーが結合するE2F部位とは異なるE2F部位の別個のサブセットと、E2F−5がin vivoで相互作用するという可能性を決して除外するものではない。チミジンキナーゼ遺伝子プロモーター中およびb−mybプロモーター中に存在するE2F部位がE2F/p107複合体と優先的に相互作用するという知見[28, 31]は、種々のE2Fのそのような結合部位優先性と一致する。E2Fとp130との間の複合体は、ほとんどの場合、静止細胞中で見いだされ、細胞が静止から脱した後すぐに消失するため、E2F−5応答性遺伝子は、成長因子刺激に対する静止細胞の初期応答に関与していると考えられる[10]。p130と相互作用するE2F−5が入手できれば、E2F−5応答性遺伝子の同定ができるはずである。
謝辞
マウス胎児cDNAライブラリーおよび酵母発現ベクターを贈呈していただいたP. Chevray、酵母Y1090株を贈呈していただいたS. Elledge、Gal4-bmi酵母発現ベクターを贈呈していただいたM. Alkema、p130発現ベクターを贈呈していただいたG. Hannon、ヒト細胞系RNAを贈呈していただいたA. Bes-Gennissen、およびノーザンブロットを調製していただいたY. Ramosに感謝の意を表する。
【0087】
この研究は、Netherlands Organization for Scientific Research(NWO)からの助成により支援していただいた。
実施例1の参照文献
1.Bandara,L.R., V.M.Buck, M.Zamanian, L.H.JohnstonおよびN.B. La Thangue. 1993.細胞周期調節転写因子DRTF1/E2FにおけるDP−1とE2F−1との間の機能的相乗作用(Functional synergy between DP-1 and E2F-1 in
the cell cycle-regulating transcription factor DRTF1/E2F). Embo J. 12:4317-4324.
2. Beijersbergen,R.L., E.M.Hijmans, L.Zhu およびR.Bernards1994.c−MycとpRb関連タンパク質p107との相互作用によりc−Myc媒介性トランス活性化の阻害が生じる(Interaction of c-Myc with the pRb-related protein p107 results in inhibition of c-Myc-mediated transactivation). Embo J. 13:4080-4086.
3.Beijersbergen,R.L., R.Kerkhoven, L.Zhu, L.Carlee, P.M.Voorhoeve およびR.Bernards. 1994. E2F遺伝子ファミリーの新規メンバーであるE2F−4は、発癌活性を有し、in vivoでp107と結合する(E2F-4, a new member of the E2F gene family, has oncogenic activity and associates with p107 in vivo). Genes, Dev. 8:2680-2690.
4.Bernards,R., S.K.DessainおよびR.A.Weinberg. 1986. N−myc増幅は、神経芽細胞腫においてMHCクラスI抗原発現のダウンモジュレーションを起こす(N-myc amplification causes down-modulation of MHC class I antigen expression in neuroblastoma). Cell. 47:667-674.
5.Buchkovich,K., L.A.DuffyおよびE.Harlow. 1989. 網膜芽細胞腫タンパク質は細胞周期の特定の時期にリン酸化される(The retinoblastoma protein is phosphorylated during specific phases of the cell cycle). Cell. 58:1097-1105.
6.Chellappan,S.P., S.Hiebert, M.Mudryj, J.M.HorowitzおよびJ.R.Nevins 1991.E2F転写因子はpRbタンパク質の細胞標的である(The E2F transcription factor is a cellular target for the RB protein). Cell 65:1053-1061.
7.Chen,P.L., P.Scully, J.Y.Shew, J.Y.WangおよびW.H.Lee. 1989.網膜芽細胞腫遺伝子産物のリン酸化は細胞周期および細胞分化の間にモジュレーションされる(Phosphorylation of the retinoblastoma gene product is modulated d
uring the cell cycle and cellular differentiation). Cell. 58:1193-1198.
8.Chevray,P.M.およびD.Natans. 1992. 酵母におけるタンパク質相互作用クローニング:junのロイシンジッパーと反応する哺乳動物タンパク質の同定(Proteininteraction cloning in yeast:identification of mammalian proteins that react with the leucine zipper of jun). Proc. Natl. Acad. Sci. USA. 89:5789-5793.
9.Chittenden,T., D.M.LivingstonおよびJ.A.DeCaprio.1993.一次ヒトT細胞におけるE2Fの細胞周期分析は、新規E2F複合体および遊離E2Fの生化学的に異なる形態を明らかにする(Cell cycle analysis of E2F in primary human T cells reveals novel E2F complexes and biochemically distinct forms
of free E2F). Mol Cell Biol. 13:3975-3983.
10. Cobrinik,D., P.Whyte, D.S.Peeper, T.JacksおよびR.A.Weinberg 1993.E2Fとp130E1A結合タンパク質との細胞周期特異的結合(Cell cycle-specific association of E2F with the p130 E1A-binding protein). Genes,
Dev. 7:2392-2404.
11.DeCaprio,J.A., J.W.Ludlow, D.Lynch, Y.Furukawa, J.Griffin, H.Piwnica-Worms, C.-M.HuangおよびD.M.Livingston. 1989. 網膜芽細胞腫遺伝子産物は細胞周期調節因子の特性を有する(The product of the retinoblastoma gene
has properties of a cell cycle regulatory element). Cell. 58:1085-1095.
12.Durphee.T., K.Becherer, P.-L.Chen, S.-H.Yeh, Y.Yang, A.E.Kilburn, W.-H,Lee およびE.S.J. 1993.網膜芽細胞腫タンパク質は、プロテインホスファターゼ1型触媒サブユニットと結合する(The retinoblastoma protein associates with the protein phosphatase type 1 catalytic subunit). Genes, Dev. 7:555-569.
13.Farnham,P.J., J.E.SlanskyおよびR.Kollmar. 1993.哺乳動物細胞周期におけるE2Fの役割(The role of E2F in the mammalian cell cycle). Biochim Biophys. Acta. 1155:125-131.
14.Fields,S.およびO.Song. 1989. タンパク質−タンパク質相互作用を検出するための新規遺伝系(A novel genetic system to detect protein-protein
interactions). Nature. 340:245-246.
15. Ginsberg,D., G.Vairo, T.Chittenden, X.Zhi-Xiong, G.Xu, W.K.L., J.A.DeCaprio, L.J.B.およびD.M.Livingston. 1994. E2F転写因子ファミリーの新規メンバーであるE2F−4はp107と相互作用する(E2F-4, a new member of the E2F transcription factor family, interacts with p107). Genes,
Dev. 8:2665-2679.
16.Hannon,G.J., D.DemetrickおよびD.Beach. 1993. CDK2およびサイクリンとの相互作用によるRb関連p130の単離(Isolation of the Rb-related p130 through its interaction with CDK2 and cyclins). Genes, Dev. 7:2378-2391.
17. Harper,J.W., G.R.Adami, N.Wei, K.KeyomarsiおよびS.J.Elledge. 1993.p21Cdk相互作用タンパク質Cip1は、G1サイクリン依存性キナーゼの強力な阻害剤である(The p21 Cdk-interacting protein Cip1 is a potent inhibitor of G1 cyclin-dependent kinases). Cell. 75:805-816.
18.Helin,K., E.HarlowおよびA.Fattaey 1993. 網膜芽細胞腫タンパク質の直接結合によるE2F−1トランス活性化の阻害(Inhibition of E2F-1 transactivation by direct binding of the retinoblastoma protein). Mol Cell Biol. 13:6501-6508.
19. Helin,K., J.A.Lees, M.Vidal, N.Dyson, E.HarlowおよびA.Fattey. 1992. 転写因子E2Fの性質を有するpRb結合タンパク質をコードするcDNA(A cDNA encoding a pRB-binding protein with properties of the transcription factor E2F). Cell. 70:337-350.
20. Helin,K., C.-L.Wu, A.R.Fattaey, J.A.Lees, B.D.Dynlacht, C.NgwuおよびE.Harlow. 1993. 転写因子E2F−1とDP−1とのヘテロ二量体形成は協同的トランス活性化につながる(Heterodimerization of the transcription factor E2F-1 and DP-1 leads to cooperative trans-activation ). Genes, Dev.
7:1850-1861.
21. Hu,Q., C.Bautista, G.Edwards, D.Defeo-Jones, R.JonesおよびE.Harlow, 1991. ヒト網膜芽細胞腫タンパク質に特異的な抗体は関連ポリペプチドのファミリーを同定する(Antibodies specific for the human retinoblastoma protein identify a family of related polypeptides). Mol. Cell. Biol. 11:5792-5799.
22. Ivey-Hoyle,M., R.Conroy, H.E.Huber, P.J.Goodhart, A.OliffおよびD.C.Heimbrook. 1993. E2F−2のクローニングおよび特徴づけ:転写因子E2Fの生化学的性質を有する新規タンパク質(Cloningand Characterization of E2F-2, a novel protein with the biochemical properties of transcription factor E2F). Mol Cell Biol. 13:7802-7812.
23. Johnson,D.G., J.K.Schwarz, W.D.CressおよびJ.R.Nevins. 1993. 転写因子E2F1の発現は静止細胞がS期に進入するのを誘導する(Expression of transcription factor E2F1 induces quiescent cells to enter S phase). Nature. 365:349-352.
24. Kaelin,W.G., W.Krek, W.R.Sellers, J.A.DeCaprio, F.Ajchenbaum, C.S.Fuchs, T.Chittenden, Y.Li.P.Farnham, M.A.Blanar, D.M.LivingstonおよびE.K.Flemington. 1992. E2F様性質を有する網膜芽細胞腫結合タンパク質をコードするcDNAの発現クローニング(Expression cloning of a cDNA encoding a retinoblastoma-binding protein with E2F-like properties). Cell. 70:351-364.
25.Kato,G.J., J.Barrett, G.M.VillaおよびC.V.Dang.1990. 腫瘍性トランスフォーメーションに要するアミノ末端c−mycドメインは転写を活性化する(An amino-terminal c-myc domain required for neoplastic transformation activates transcription). Mol.Cell. Biol. 10:5914-5920.
26. Krek,W., D.M.LivingstonおよびS.Shirodkar. 1993. 種々のE2Fファミリーメンバーの複合体形成により促進されるDNAおよび網膜芽細胞腫遺伝子産物に対する結合(Bindingto DNA and the retinoblastoma gene product promoted by complex formation of different E2F family members). Science. 262: 1557-1560.
27. La Thangue, N.B. 1994.DPおよびE2Fタンパク質:細胞周期制御に関与するヘテロ二量体転写因子の成分(DP and E2F proteins: components of a
heterodimeric transcription factor implicated in cell cycle control). Curr. Opin. Cell Biol. 6:443-450.
28.Lam,E.W.およびR.J.Watson. 1993. E2F結合部位はマウスβ−myb転写の細胞周期調節抑制を媒介する(An E2F-binding site mediates cell-cycle regulated repression of mouse B-myb transcription). Embo J. 12:2705-2713.
29. Lees,E., B.Faha, V.Dulic, S.I.ReedおよびE.Harlow. 1992. サイクリンE/cdk2およびサイクリンA/cdk2キナーゼはp107およびE2Fに、一時的に異なる態様で結合する(Cyclin E/cdk2 and cyclin A/cdk2 kinases associate with p107 and E2F in a temporally distinct manner). Genes Dev. 6:1874-1885.
30. Lees,J.A., M.Saito, M.Vidal, M.Valentine, T.Look, E.Harlow, N.DysonおよびK.Helin. 1993.網膜芽細胞腫タンパク質はE2F転写因子のファミリーに結合する(The retinoblastoma protein binds to a family of E2F transcription factors). Mol Cell Biol. 13:7813-7825.
31.Li,L.J., G.S.NaeveおよびA.S.Lee.1993.G1−S期転写調節に重要なヒトチミジンキナーゼプロモーター因子に結合するサイクリンA−p107およびp33cdk2複合体の一時的調節(Temporal regulation of cyclin A-p107
and p33cdk2 complexes binding to a human thymidine kinase promoter element important for G1-S phase transcriptional regulation). Proc Natl Acad
Sci USA. 90:3554-3558.
32.Li,Y., C.Graham, S.Lacy, A.M.V.DuncanおよびP.Whyte. 1993.アデノウイルスE1A関連130kDタンパク質は網膜芽細胞腫遺伝子ファミリーのメンバーによりコードされ、サイクリンAおよびEと物理的に相互作用する(The adenovirus E1A-associated 130-kD protein is encoded by a member of the retinoblastoma gene family and physically interacts with cyclins A and E). Genes, Dev. 7:2366-2377.
33.Morgenstern,J.およびH.Land. 1990. 一連の哺乳動物発現ベクターおよび安定におよび一過性にトランスフェクションされた細胞におけるレポーター遺伝子の該ベクターによる発現の特徴づけ(A series of mammalian expression vectors and characterisation of their expression of a reporter gene in stably and transiently transfected cells). Nucleic Acid Res. 18:1068.
34. Schiestl,R.HおよびR.D.Gietz. 1989.キャリアーとして一本鎖核酸を用いる無傷酵母細胞の高効率形質転換(High efficiency transformation of intact yeast cell using single stranded nucleic acid as a carrier). Curr. Genet. 16:339-346.
35. Schwarz,J.K., S.H.Devoto, E.J.Smith, S.P.Chellappan, L.JakoiおよびJ.R.Nevins. 1993. 細胞増殖応答期のp107およびRbタンパク質とE2Fとの相互作用(Interactions of the p107 and Rb proteins with E2F during the cell proliferation response). Embo J. 12:1013-1020.
36. Shan,B., X.Zhu, P.L.Chen, T.Durfee, Y.Yang, D.SharpおよびW.H.Lee. 1992.網膜芽細胞腫関連タンパク質をコードする細胞遺伝子の分子クローニング:転写因子E2Fの性質を有する遺伝子の同定(Molecular cloning of cellular genes encoding retinoblastoma-associated proteins:identification of a gene with properties of the transcription factor E2F). Mol Cell Biol. 12:5620-5631.
37. Shirodkar,S., M.Ewen, J.A.DeCaprio, J.Morgan, D.M.LivingstonおよびT.Chittenden. 1992. 転写因子E2Fは網膜芽細胞腫産物およびp107サイクリンA複合体と細胞周期調節的に相互作用する(The transcription factor E2F interacts with the retinoblastoma product and a p107-cyclin A complex
in a cell cycle-regulated manner). Cell. 68:157-166.
38. Slansky,J.E., Y.Li, W.G.KaelinおよびP.J.Farnham. 1993.E2F1mRNAのタンパク質合成依存的増加は、ジヒドロ葉酸レダクターゼプロモーターの成長調節と相関する(A protein synthesis-dependent increase in E2F1 mRNA correlates with growth regulation of the dihydrofolate reductase promoter). Mol Cell Biol. 13:1610-1618.
39. Van der Eb, A.J.およびF.L.Graham. 1980. 腫瘍ウイルスDNAのトランスフォーメーション活性のアッセイ(Assay of transforming activity of tumor virus DNA). Meth. Enzymol. 65:826-839.
40. Zamanian,M.およびN.B.La Thangue. 1993. Rb関連タンパク質p107による転写抑制(Transcriptional repression by the Rb-related protein p107). Mol Biol of Cell. 4:389-396.
41. Zhu,L., S. van den Heuvel, K.Helin, A Fattaey, M.Ewen, D.Livingston, N.DysonおよびE.Harlow. 1993. 網膜芽細胞腫タンパク質の同族体であるp107による細胞増殖の阻害(Inhibition of cell proliferation by p107, a relative of the retinoblastoma protein). Genes Dev. 7:1111-1125.
実施例2
概要
転写因子DRTF1/E2Fは、それが細胞周期進行のキーとなるレギュレーター(例、網膜芽細胞腫抑制遺伝子産物および関連ポケットタンパク質、サイクリンおよびサイクリン依存性キナーゼ)と相互作用することにより、細胞増殖の制御に関与している。2つの異なるタンパク質ファミリー(DPおよびE2F)のメンバーがDP/E2Fヘテロ二量体として相互作用する場合に、DRTF1/E2FDNA結合活性が生じる。ここでは、本発明者らは、E2F−5と称される、E2Fタンパク質ファミリーの新規メンバーの単離および特徴づけを報告する。E2F−5は、マウスDP−1に対する結合能を有する分子に関して14.5d.p.c.マウス胎児ライブラリーをスクリーニングする酵母二ハイブリッドアッセイにより単離されたが、DPタンパク質ファミリーのすべての公知メンバーとも相互作用する。哺乳動物細胞抽出物に存在する一般的なDRTF1/E2FDNA結合活性(E2F部位を介して協同的DNA結合活性および転写活性化を起こす相互作用)により、E2F−5はDP−1との生理学的ヘテロ二量体として存在する。強力な転写活性化ドメインは、酵母および哺乳動物細胞の両方で機能し、E2F−5のC末端領域に存在する。pRbよりむしろpRb関連タンパク質p107の発現が、E2F−5の転写活性を不活性化する。E2F−5と該ファミリーの他のメンバーとの配列の比較から、E2F−5は、E2F−1、−2および−3よりも、E2F−4に対する類似度が大きいことが示される。E2F−5とE2F−4との構造的および機能的類似性は、E2Fタンパク質のサブファミリーを定義づけるものである。
緒言
細胞転写因子DRTF1/E2Fは、細胞周期進行と同調した転写に関与していることを示す証拠がかなり存在する。例えば、DRTF1/E2Fは、主要な標的の1つであり、それを介して、網膜芽細胞腫癌抑制遺伝子産物(pRb)が、細胞増殖に対するその負の効果を奏すると考えられる(La Thangue, 1994)。したがって、DRTF1/E2Fの転写活性および標的遺伝子[その多くは、細胞周期進行に必要なタンパク質をコードしている(Nevins, 1992)]の活性を調節することにより、pRbは、初期細胞周期を経る進行に影響を及ぼしうる。ヒト腫瘍細胞中に生じる、Rb中の天然の突然変異は、DRTF1/E2Fに結合しないタンパク質をコードする(Bandaraら, 1992; Heibertら, 1992; ZamanianおよびLa Thangue, 1992)が、このことは、調節解除するDRTF1/E2Fと異常細胞成長との間の相関性を強調するものである。さらに、アデノウイルスE1a、ヒトパピローマウイルスE7、SV40ラージT抗原などのウイルス性癌タンパク質の形質転換活性は、それらがpRbおよび関連タンパク質の隔離によりDRTF1/E2Fを調節解除する能力と相関しており(Nevins, 1992)、このことは、この見解をさらに支持するものである。
【0088】
細胞周期において中心的な役割を果たす他の分子も、DRTF1/E2Fと相互作用する。サイクリンAおよびEは触媒サブユニットcdk2と共に、該転写因子中のDNA結合成分と接触することにより直接的に(Krekら, 1994)、あるいは、pRb関連ポケットタンパク質p107またはp130のスペーサー領域中で生じる接触を介して間接的に(Leesら, 1992; Cobrinikら, 1993)、DRTF1/E2Fに結合する。p107およびp130と結合するサイクリン−cdk複合体の役割については未だ解明されていないが、サイクリンA/cdk2キナーゼ複合体とDRTF1/E2Fとの間の直接的相互作用は、そのDNA結合活性に影響を及ぼすことが示されている(Krekら, 1994)。
【0089】
DRTF1/E2Fの分子組成が明らかにされはじめている。始原型分子(prototypemolecules)がDP−1(Girlingら, 1993)およびE2F−1(Helinら, 1992; Kaelinら, 1992; Shanら, 1992)である2つの異なるタンパク質ファミリーのメンバーがDP/E2Fヘテロ二量体として相互作用する場合に(La Thangue, 1994)、一般的なDNA結合活性DRTF1/E2Fが生じることが現在明らかとなっている。両タンパク質の間で類似する小さな領域が、現在単離されているすべてのDPおよびE2Fファミリーメンバーで保存されており(Girlingら, 1994)、この領域は、それらのタンパク質がヘテロ二量体として相互作用するのを可能にし(Bandaraら, 1993; Helinら, 1993; Krekら, 1993)、それにより、多様な組み合わせの相互作用の発生が可能となる。
【0090】
それぞれ、DRTF1/E2Fとの相互作用の各々の特徴的なプロフィールを有するpRb、p107およびp130の結合が、細胞周期進行中に一時的調節的に生じる(Shirodkarら, 1992; Schwarzら, 1993; Cobrinikら, 1993)。現在までに単離されているE2Fファミリーメンバーのうち、E2F−1、−2および−3はpRbを(Ivey-Hoyleら, 1993; Leesら, 1993)、E2F−4はp107タンパク質を認識するが(Beijersbergenら, 1994; Ginsbergら, 1994)、その考えられる説明は、ポケットタンパク質の一時的な相互作用が、E2F/DPヘテロ二量体中のE2Fファミリーメンバーの組成および/または利用性の調節を反映しているというものである。
【0091】
多数の細胞型において、DP−1は、細胞周期進行中に生じる種々の形態で存在するDRTF1/E2Fの通常のDNA結合成分であるが(Bandaraら,1994)、DP−2などの他のDPファミリーメンバーは、組織限定的に発現される(Girlingら, 1994)。したがって、DRTF1/E2Fの分子組成が、細胞周期進行および細胞表現型により影響を受ける可能性があると考えられる。
【0092】
E2Fタンパク質ファミリーの複雑性については、未だ解明されていない。この問題を検討するために、本発明者らは、酵母に基づく二ハイブリッドスクリーニングを行って、該ファミリーの新規メンバーを定義づけした。ここで、本発明者らは、E2Fファミリーの新規メンバーであるマウスE2F−5の単離および特徴づけを報告する。E2F−5は、DPタンパク質ファミリーの公知メンバーのすべてと相互作用する。哺乳動物細胞抽出物中で、E2F−5はDP−1との生理学的ヘテロ二量体として存在し、その相互作用は、E2F部位を介する協同的DNA結合および転写活性化を起こす。E2F−5は、ポケット−タンパク質結合に際して特異的に不活性化される強力なトランス活性化ドメインを有する。E2F−5のタンパク質配列および分子構成は、E2F−4との関連性が、該ファミリーの他のメンバーとの関連性より密接であり、これは、機能的および構造的に関連しているE2Fタンパク質のサブファミリーを最初に定義づけるものである。
結果
E2F−5の単離
E2Fタンパク質ファミリーの多様性を調べるために、本発明者らは、酵母二ハイブリッドに基づく方法(FieldsおよびSong, 1989)により、新規メンバーを同定した(図8)。DP−1はE2Fファミリーメンバーの通常の生理学的反応の相手であるため(Bandaraら, 1993; Bandaraら, 1994)、本発明者らは、ベイトとしてDP−1を使用することを選択した。14.5d.p.c.マウス胎児(ChevrayおよびNathans, 1992)から調製された活性化ドメインタグ付きcDNAライブラリーを、LexA−DP−1と相互作用する能力を有するハイブリッドタンパク質に関してスクリーニングした。同定されたクローンの1つは、いくつかの基準によりLexA−DP−1と特異的に相互作用するハイブリッドタンパク質をコードしていた。該cDNA配列の部分分析は、E2Fファミリーメンバーとの広範な類似性を示し、したがって、完全タンパク質配列をコードするcDNAクローンを、F9ECcDNAライブラリーからさらに単離した。E2Fファミリーの他のメンバーとのタンパク質配列の比較により、該cDNAクローンが新規メンバーをコードしていることが示された。E2F−1、−2、−3および−4として既に単離されているE2Fタンパク質に採用されている名称に従い、本発明者らは、このクローンをE2F−5と称する。
【0093】
マウスE2F−5の予想サイズは、335残基である(図9a)。この予想は、考えられる最初の開始メチオニンの位置、および同じメチオニンから翻訳が開始されるヒトE2F−5配列に対して存在する広範な相同性に基づく(Hijmansら,投稿中)。E2F−5は、他のE2Fファミリーメンバーの間で保存されているドメインとの広範な配列類似性を含有する(Ivey-Hoyleら, 1993; Leesら,1993; Beijersbegenら, 1994; Ginsbergら, 1994)。例えば、該DNA結合ドメインは、E2F−1に対して48%、E2F−4に対して87%の同一性を示す(図9bおよびc)。この領域内で、C末端サブドメインは、DPファミリーのメンバーと類似する唯一の領域を含有する(図9bおよびcに示す)。DEFボックスとして知られる(Girlingら, 1994; LamおよびLa Thangue, 1994)この領域は、DP/E2Fヘテロ二量体の形成に密接に関与している(Bandaraら, 1993; BandaraおよびLa Thangue, 投稿予定)。また、DPとE2Fとのタンパク質間でDEFボックス内で保存されている残基は、E2F−5内で完全に保存されており(図9c)、このことは、DP/E2Fヘテロ二量体の形成におけるこのサブドメインの潜在的重要性を強調するものである。
【0094】
E2F−5と残りのファミリーメンバーとの間では、いくつかの更に別の領域が保存されている。マークトボックス(marked box)(Leesら, 1993)およびポケット−タンパク質結合ドメインは、E2F−1中のこれらの領域と58%および50%の同一性を、E2F−4中の同じ領域と75%および72%の同一性を示す(図9bおよび9c)。また、該ロイシンジップ領域中の疎水性残基の位置は、他のE2Fファミリーメンバーに対して保存されている(図9c)。E2F−5中の疎水性残基は、2つのさらなるC末端位置(L144およびV151;図9cを参照されたし)で7個の反復と重なるため、実際に、E2F−5は、E2F−1、−2および−3より長いジップを形成しうる。
【0095】
E2F−5の特徴は、E2F−1、−2および−3よりもE2F−4との密接な関係を示唆する。該タンパク質は、該DNA結合ドメインのN末端ほどは伸長せず、両タンパク質は、残りのE2F中に生じるN末端サイクリンA結合ドメインを欠く点で(図9b)、E2F−5の構成はE2F−4のものと類似する。さらに、該タンパク質配列の比較により、E2F−5とE2F−4との関係は、その各々と該ファミリーの残りのメンバーとの関係よりも密接であることが示される。これは、E2F−5とE2F−4との間では多数の残基が共通であるがE2F−1、−2および−3間ではそうでない該保存ドメインから特に明らかである(図9c)。全体的には、E2F−5は、E2F−4とは70%、E2F−1とは38%のアミノ酸残基が同一である。したがって、この類似性に基づくと、E2F−5および−4は、E2Fタンパク質ファミリーの1つのサブファミリーを代表し、E2F−1、−2および−3は、それらの密接な類似性のため、別のサブファミリーを代表する。
【0096】
DPファミリーメンバーとの結合および転写協同作用
DPファミリーメンバーがE2Fファミリーメンバーと相互作用する場合に、一般的なDRTF1/E2FDNA結合活性が生じる(La Thangue, 1994)。DP−1およびE2F−1では、該相互作用により協同的転写活性化、DNA結合、およびpRbとの相互作用が生じる(Bandaraら, 1993; Helinら, 1993; Krekら, 1993)。したがって、本発明者らは、E2F−5がDPファミリーメンバーと協同作用しうるか否かの判定に関心があった。
【0097】
これらの問題に答えるために、本発明者らはまず、LexA融合タンパク質として該ハイブリッドベイト中に提示されている種々のDP分子を使用する酵母二ハイブリッドアッセイを用いた(図8)。E2F−5またはE2F−1のいずれかを、活性化ドメイン(GAD)タグ付きハイブリッドタンパク質として発現させ、LexAレポーター構築物の転写活性化の程度を、β−ガラクトシダーゼ活性を測定することにより評価した。E2F−5およびE2F−1は共に、DPタンパク質ファミリーのすべての公知メンバー(すなわち、DP−1、−2、−3およびショウジョウバエDP)と同等に相互作用する能力を有する(データは示していない)。
【0098】
ついで、本発明者らは、E2F−5がDP−1と協同して、E2F結合部位を介して転写を活性化しうるか否かを評価した。このために、本発明者らは、E2F−5およびDP−1を一緒にまたは単独で発現させ、E2F部位レポーター構築物p4xWT CYC1の転写活性を測定する酵母アッセイを用いた(図10a)。以前の研究では、このアッセイは、E2F−1とDP−1との間の協同作用を測定するために使用されている(Bandaraら, 1993)。該DP−1ハイブリッドタンパク質の発現の後、該レポーターの転写活性は有意な影響を受けず、E2F−5ハイブリッドによりわずかに影響されたにすぎなかった(図10b)。しかしながら、両方を一緒に発現させた場合には、レポーター活性の刺激は6倍を越えた(図10b)。突然変異E2F結合部位を担持する、p4xWT CYC1の誘導体であるp4xMT CYC1(図10a; Bandaraら, 1993)の活性は、同じ条件で影響を受けなかった(データは示していない)。したがって、本発明者らは、E2F−5がDP−1と協同作用すると結論する。
【0099】
E2F−5の転写活性化およびポケットタンパク質調節
酵母および哺乳動物細胞の両方で、E2F−5の転写活性化能を評価した。酵母で転写活性をアッセイするために、E2F−5のC末端領域(残基198〜335)をLexAに融合し、LexA結合部位により駆動されるレポーター構築物の活性を評価した(図11a)。pLEX.E2F-5の存在下では、該ベクター単独で発現させた場合よりも、該レポーターの活性がかなり大きかったので(図11b)、E2F−5タンパク質は強力なトランス活性化ドメインを含有する。同様に、LexA E2F-1は、転写活性化能を有していた(図11b)。したがって、E2F−5は、酵母において転写を効率的に活性化する。
【0100】
これらの結果を哺乳動物細胞で確認し、ポケット−タンパク質とE2F−5との相互作用の機能的影響を評価するために、本発明者らは、E2F−5の同じ領域をGal4 DNA結合ドメインに融合し、一過性トランスフェクションアッセイを用いて、Gal4結合部位により駆動されるレポーター構築物(pGAL-CAT)の転写活性を調べた(図12a)。pGAL-E2F-5の存在下でpGAL-CATの転写活性はpG4と比べて15倍大きかったので、3T3細胞では、E2F−5は、Gal4DNA結合ドメイン単独の活性と比べて、転写を効率的に活性化した(図12b)。種々の他の細胞型でも同様の結果が得られたが(データは示していない)、このことは、哺乳動物細胞中で機能するトランス活性化ドメインをE2F−5が含有することを示すものである。
【0101】
ついで、本発明者らは、E2F−5の転写活性を用いて、pRbまたはp107のいずれかとの相互作用の機能的影響を評価した。野生型pRbおよびp107についての対照として、本発明者らは、DRTF1/E2Fとは相互作用しないRbの天然に存在する突然変異対立遺伝子によりコードされるタンパク質であるRbΔ22の活性(ZamanianおよびLa Thangue, 1992)、およびアンチセンスp107RNAの活性(ZamanianおよびLa Thangue, 1993)を調べた。pCMVHRbまたはpCMVHRbΔ22の存在下でpGAL-CATの活性は影響を受けなかったため(図12b)、野生型pRbまたはRbΔ22はいずれも、E2F−5の活性に有意な影響を及ぼさなかった。これに対して、E2F−5と共発現するp107(pCMV107から)はE2F−5の転写活性を減少させたが、アンチセンスp107(pCMV107ASから)は影響を及ぼさなかったため、この効果は特異的であった(図12b)。本発明者らは、哺乳動物細胞において、p107はE2F−5の転写活性を不活性化すると結論する。同様の実験方法を用いて、p130タンパク質がE2F−5の転写活性を不活性化することが示された(データは示していない)。
【0102】
E2F−5は、DRTF1/E2Fの生理学的DNA結合成分である
E2F−5が、哺乳動物細胞から調製された抽出物中に限定される一般的なDRTF1/E2FDNA結合活性の生理学的DNA結合成分であるか否かを判定するために、異なるペプチド配列に対する2つの異なる抗E2F−5ペプチド血清(共に、GST−E2F−5融合タンパク質と特異的に反応する抗血清である)を調製した(データは示していない)。
【0103】
マウスF9胎児性癌(F9EC)細胞からの抽出物を用いて行うゲル遅延アッセイにより、DRTF1/E2FDNA結合活性に対するこれらの抗血清の効果を調べた。DP−1が、F9EC細胞抽出物中のDRTF1/E2FDNA結合活性の通常の、おそらく普遍的な成分であることが、以前の研究で示されており(Girlingら,1993; Bandaraら, 1993)、その一例を、図13(トラック1〜4)に示す。抗E2F−5血清は共にDRTF1/E2FDNA結合活性を阻害したが、この効果は、相同性ペプチドの存在下では明らかでないため(図13、トラック5〜12)、特異的なものである。抗E2F−5はDNA結合活性の有意な減少を引き起こすが、この効果は、抗DP−1により生じるものほど顕著でないことは明らかである(図13、トラック1〜4と5〜12とを比較されたし)。これは、F9EC細胞抽出物中のDP−1/E2Fヘテロ二量体の亜集団中にE2F−5が存在するためと考えられ、DP−1で観察されるものと対照的である。
【0104】
E2F−5のDNA結合特性
E2F−5のDNA結合特性を調べるために、本発明者らは、E2F−5をGST融合タンパク質として発現させ精製した。E2F−1は、単独でアッセイした場合には、有意なDNA結合活性を有していたが(図14、トラック1〜4を比較されたし)、既に得られている結果(Bandaraら,1993)と一致して、GST−DP−1は、E2F部位に対する結合においてGST−E2F−1と協同作用した。これに対して、E2F−5は単独では、DNA結合活性はほとんど検出できなかった(図14、トラック5〜7)。しかしながら、E2F−5とDP−1との間の協同作用は、E2F−1とDP−1との間のものよりも相当大きかった(図13、トラック8〜10)。したがって、E2F−5とDP−1とはDNA結合活性について協同作用する。
【0105】
E2F−5RNAのレベル
本発明者らは、種々の細胞系でE2F−5RNAのレベルを測定し、さらに、E2F−5レベルとE2Fファミリーの他のメンバーとを比較することに関心があった。このために、種々の白血病細胞系およびF9EC細胞の非同時培養物からRNAを調製し、ノーザンブロッティングによりE2F−5RNAのレベルを評価した。E2F−5RNAの量は、細胞系によってかなり異なっていた。F9EC細胞およびいくつかの白血病細胞系、例えばDAUDIおよびRAGIは、高レベルで発現した(図15、トラック1、7および8)。これに対して、HL60およびTF1は、低レベルのE2F−5RNAを含有していた(図15、トラック4および6)。E2F−5RNAレベルのこのプロフィールは、E2F−1のレベルとかなり異なっていた。例えば、K562、HL60およびTF1細胞には、E2F−5と比べて、有意なレベルのE2F−1RNAが存在していた(図15、トラック3、4および6)。EL4細胞では、明らかにこの逆であり、E2F−5レベルが高く、E2F−1が低かった(図15、トラック10)。E2F−5RNAレベルは細胞型に影響され、E2F−5とE2F−1との間のRNAレベルには相関性がほとんどないと、本発明者らは結論する。
考察
E2F−5およびE2F−4は、E2Fタンパク質のサブファミリーである
本発明者らは、E2Fタンパク質ファミリーの第5のメンバーであるE2F−5の単離および特徴づけを報告する。潜在的ロイシンジップ、マークトボックス(marked box)およびポケットタンパク質結合領域などの、E2F−5中のドメインの多くは、E2Fファミリーの他のメンバーと保存されているが、DNA結合ドメインの外のN末端配列が欠けていることは、E2F−4と同様の構造的構成を示すものである(Beijersbergenら, 1994; Ginsbergら, 1994)。E2Fファミリーの残りのメンバー、すなわちE2F−1、−2および−3は、伸長したN−末端を有し、その内部に、サイクリンAと相互作用する能力を有するドメインが存在する(Krekら, 1994)。このドメインの役割は、サイクリンA/cdk2キナーゼをDP−1/E2Fヘテロ二量体へリクルートすることであり、ついでそれは、DP−1のリン酸化を引き起こし、その機能的影響は、DP−1/E2Fヘテロ二量体のDNA結合活性の減少であると示唆されている(Krekら, 1994)。そのような機構は、細胞周期進行のより後期におけるE2F部位依存性遺伝子の転写活性の調節に重要かもしれない。E2F−5(およびE2F−4)中にサイクリンA結合ドメインが存在しないことは、E2F−5/DP−1ヘテロ二量体のDNA結合活性が、他の機構によりダウンレギュレーションされうることを示唆する。確かに、これが達成される可能なシナリオは、p107および/またはp130タンパク質によるものであろう。なぜなら、これらのタンパク質のスペーサー領域は、サイクリンA/cdk2複合体またはサイクリンE/cdk2複合体のいずれかに結合しうるからである(Leesら, 1993; Cobrinikら, 1993; Hannonら, 1993; Liら, 1993)。これらのポケットタンパク質が、サイクリンA結合E2Fファミリーメンバーの役割を代替し、サイクリン/cdk複合体をDP/E2Fヘテロ二量体へリクルートしている可能性がある。
【0106】
E2F−5のタンパク質配列をE2Fファミリーの他のメンバーのものと比較すると、E2F−1、−2および−3よりもE2F−4に対する密接な関係が示された。興味深いことに、E2F−4は、生理学的条件下でp107と相互作用する能力を有する該ファミリーの唯一の公知メンバーである(Beijersbergenら, 1994; Ginsbergら, 1994)。本発明者らは、E2F−5の転写活性が、pRbよりもむしろp107またはp130により不活性化されうることを示した。しかしながら、これは、p107が、pRbよりもp130に対して密接な関係を有することを反映しているのかもしれず(Ewenら, 1991; Cobrinikら, 1993; Liら, 1993)、したがって、生理学的相互作用を完全には反映していないのかもしれない。ヒトE2F−5が生理学的条件下でp130と相互作用することが示されている結果(Hijmansら, 投稿中)は、この考え方と一致する。
【0107】
E2F−5とDP−1との間の協同作用
DNA結合および転写活性において、E2F−5はDP−1と協同作用する。この点で、E2F−5は、E2Fファミリーの他のメンバーと同様の性質を有する。しかしながら、注目すべき興味深いことは、E2F−5とDP−1との間の協同作用が、例えば同等の実験条件でE2F−1とDP−1との間で観察される協同作用よりも、かなり大きいことである(例えば、図14を参照されたし)。このアッセイが細胞内の機能的特性を反映するのであれば、過剰のDP−1が存在する細胞内環境では、E2F−5およびE2F−1が同等に増加した場合、E2F−5/DP−1DNA結合活性レベルの方が比較的大きくなると考えられる。さらに、特定のE2F/DPヘテロ二量体にとって好ましい標的遺伝子があれば、DNA結合活性のこれらの相違は、転写活性の相違を反映するかもしれない。
【0108】
それらの種々のE2Fタンパク質の細胞周期制御における正確な役割は、未だ確定されていない。しかしながら、それらは、細胞周期進行の異なる時期で標的遺伝子の転写活性を調節すると考えられる。例えば、E2F−1、−2および−3がpRbと相互作用することは(Helinら, 1992; Kaelinら, 1992; Ivey-Hoyle ら, 1993; Leesら, 1993)、それらが、G1を経る細胞周期進行を調節することを示唆する。これに対して、G1の終わりに向かってp107はDRTF1/E2Fと結合し、そしてS期でピークを迎えるが(Shirodkarら, 1992)、p130は、細胞周期進行の初期に、ほとんどはG0の間に優先的に結合する(Cobrinikら, 1993)。しかしながら、種々の細胞型中のE2F−1およびE2F−5のレベルは、E2Fファミリーメンバーの発現が、細胞周期の時期だけでなく、表現型によっても影響されることを示唆する。細胞は、その細胞周期要件に最も適した、E2Fタンパク質の好ましいサブセットを利用するという可能性がある。
【0109】
ここで報告しているE2F−5の分子的および機能的特徴、およびDPファミリーメンバーとのその相互作用から示されることは、E2FとDPファミリーメンバーとの間の種々のヘテロ二量体が理論的に可能なことである。将来の研究の大きな目標は、細胞周期制御におけるこれらの転写因子のそれぞれの生理学的役割を理解することである。
材料および方法
酵母株、培地および方法
この研究で使用したサッカロミセス・セレビシエ(Saccharomyces cerevisiae)株は、以下のとおりであった:W3031a(Mata ade2-100trp1-1 leu2-3, 112 his3-11 ura3); CTY10-5d(Matα ade2 trp1-901 leu2-3, 112 his3-200 gal4 gal80 URA3::lexAop-lacZ)およびPCY2(Matαgal14 gal80 URA3::GAL1-lacZ lys2-801 his3-200 trp1-63 leu2 ade2-101 )およびJZ1(Joossら, 1994; Matα lys2-801 ade2-10 leu2Δ1 trpΔ63 his3Δ200 URA3::lexAop-CYC1-lacZ)。酵母株をYPDまたはYNB培地中で増殖させ、酢酸リチウム法の改変法により形質転換した。コロニー着色β−ガラクトシダーゼ活性アッセイ(colony colourβ-galactosidase activity assay)を通常の方法で行った。少なくとも3つの別個の形質転換体について、中期対数期培養物中、個々の形質転換体のβ−ガラクトシダーゼ活性を定量した。
【0110】
ライブラリーDNA、プラスミドおよびオリゴヌクレオチド
pPC67は、GAL4タンパク質のトランス活性化ドメインをコードする酵母配列の下流と融合した14.5d.p.c.のCD−1マウス胎児オリゴdTプライミングcDNAライブラリーである(GAD; ChevrayおよびNathans, 1992)。特異的にクローニングしたポリdTプライミングcDNAのλZapII F9ECライブラリーから、完全なcDNAクローンを単離した(Scholerら, 1990)。
【0111】
プラスミドpTR27は、ポリリンカー配列が伸長しているpBTM116(Bandaraら, 1993)の誘導体であり、pLEX.DP-1、pGAD.E2F-1、p4xWT CYC1およびp4xMT CYC1については、既に記載されている(Bandaraら, 1993)。pLEX(HIS).DP-1は、プラスミドpLEX(HIS)(これは、TRP1がHIS3で置換されているpBTM116の誘導体)中のDP−1(アミノ酸残基59〜410)と完全細菌LexAタンパク質との融合体をコードする。pGAD.E2F-5は、プラスミドpACTII(Durfeeら, 1993)中の酵母GAL4タンパク質(768〜881)の活性化ドメインを有するハイブリッドタンパク質として発現されるE2F−5の完全コーディング配列を含有する。pLEX.E2F-5は、プラスミドpTR27中のLexAタンパク質の完全コーディング配列の下流に、ハイブリッドタンパク質として発現されるアミノ酸残基198〜335のE2F−5コーディング配列を含有する。pLEX.E2F-1は、pTR27中に完全長E2F−1(1〜437)を担持する。プラスミドpG4(以前はpG4mポリIIと呼ばれていた; Websterら, 1989)は、SV40初期プロモーターの制御下、GAL4DNA結合ドメイン(1〜148)をコードする。プラスミドpGAL4.E2F-5は、pG4中のGAL4配列の下流で融合した残基198〜335のE2F−5コーディング配列を含有する。プラスミドpCMVHRb、pCMVHRb△22、pCMV107およびpCMV107ASについては、既に記載されている(ZamanianおよびLa Thangue, 1992; 1993)。
【0112】
ライブラリースクリーニング
40μg pPC67ライブラリーDNAを、40μg pLEX(HIS).DP-1と共にCTY10-5d中に同時形質転換した。選択寒天プレート上で成長している約400,000の形質転換体を、in situ濾紙β−ガラクトシダーゼアッセイによりスクリーニングした。該ライブラリープラスミドをレスキュー(rescue)するために、青色のコロニーを単離し、ヒスチジンの存在下で選択液体培地中で飽和するまで成長させ、pLEX(HIS).DP-1をキュアリングした。選択最少寒天上にレプリカプレーティングした後、β−ガラクトシダーゼについてアッセイした場合に青色を与えないTrp+His-コロニーからのプラスミドDNAを、E. coli HW87中にエレクトロポレーションした。プラスミドを回収し、pLEX(HIS).DP-1または対照プラスミド(pLEX(HIS))のいずれかでCTY10-5d中に再度形質転換した。pLEX(HIS).DP-1が存在する場合にのみ青色コロニーを与えるTrp+表現型を付与するプラスミドを、さらなる分析のために選択した。完全長cDNAを得るために、該挿入物を切り出し、放射能標識し、これを用いて、λZapII F9ECライブラリーからの約105のプラークをスクリーニングし、それから完全長E2F−5cDNAを単離し、pBluescript中にレスキューした。
【0113】
3T3細胞の一過性トランスフェクション
既に記載されている通常のリン酸カルシウム沈殿法により(ZamanianおよびLa Thangue, 1992)、トランスフェクションおよびアッセイを行った。内部対照としてのpCMV-βgalに由来するβ−ガラクトシダーゼ活性を、既に記載されているとおりに測定した(ZamanianおよびLa Thangue, 1992)。
【0114】
抗血清およびゲル遅延分析
E2F−5に由来する2つの異なるペプチド配列に対して産生させたウサギ抗血清(抗−E2F−5(1)、抗−E2F−5(2)と称する)を調製し、F9EC細胞抽出物中のDRTF1/E2FDNA結合活性に対する効果を評価した[既に記載されているとおりに行った(Girlingら,1993)]。E2F結合部位は、アデノウイルスE2aプロモーターから取った(La Thangueら, 1990)。相同性(+)または無関係な(−)ペプチドのいずれかを該DNA結合アッセイへ加えて、既に記載されているとおりに(Girlingら, 1993)特異性を評価した。DP−1のC末端配列由来のペプチドに対して、抗DP−1(24)抗血清を産生させた。GST融合タンパク質を用いて行うDNA結合アッセイは、既に記載されているとおりである(Bandaraら, 1993; 1994)。通常の方法により、GST−DP−1、−E2F−1(Bandaraら, 1993)および−E2F−5(アミノ酸残基2〜335)を発現させ精製した。
【0115】
ノーザン分析
通常の方法により、示されている細胞系から調製したRNA上で、RNAレベルのノーザン分析を行った。E2F−5プローブは、3'非翻訳領域中に伸長する840のヌクレオチドを含有していた。E2F−1プローブは、PCRにより調製された遺伝子の完全コーディング配列を含有していた。GAPDH用のプローブは、内部対照として使用した。
実施例2の参照文献
Bandara,L.R., Adamczewski,J.P., Hunt,T.およびLa Thangue,N.B. (1991). Nature 352, 249-251.
Bandara,L.R., Adamczewski,J.P., Poon,R.Y., Zamanian,M., Hunt,T.およびLa Thangue,N.B. (1992). J. Cell Sci. (Suppl.) 16, 77-85.
Bandara,L.R., Buck,V.M., Zamanian,M., Johnston,L.H.およびLa Thangue,N.B. (1993). EMBO J. 12, 4317-4324.
Bandara,L.R., Lam,E.W.-F., Sφrensen,T.S., Zamanian,M., Girling,R.およびLa Thangue,N.B. (1994). EMBO J. 13, 3104-3114.
Beijersbergen,R.L., Kerkhoven,R.M., Zhu,L., Carlee,L., Voorhoeve,P.M.およびBernards,R. (1994). Genes. Dev. 8:2680-2690.
Chevray,P.およびD.Nathans. (1992). Proc. Natl. Acad. Sci. USA 89:5789-5793.
Cobrinik,D., Whyte,P., Peeper,D.S., Jacks T.およびWeinberg,R.A. (1993). Genes Dev. 7, 2392-2404
Durfee,T., Becherer,K., Chen,P.-L., Yeh,S.-H., Yang,Y., Kilburn, A.E., Lee, W.-H.およびElledge,S.J.(1993). Genes Dev. 7, 555-569.
Ewen,M.E., Xing,Y., Lawrence,J.B.およびLivingston,D.M. (1991). Cell 66:1155-1164.
Fields,S.およびSong,O. (1989). Nature 340:245-246.
Ginsberg,D., Vairo,G., Chittenden,T., Xiao,Z.-X., Xu.G., Wydner,K.L.,
DeCaprio,J.A., Lawrence,J.B.およびLivingston,D.M. (1994). Genes. Dev. 8:2665-2679.
Girling,R., Partridge,J.F., Bandara,L.R., Burden,N., Totty,N.F., Hsuan, J.J.およびLa Thangue,N.B. (1993). Nature 362, 83-87.
Girling,R., Bandara,L.R., Ormondroyd,E., Lam,E.W.-F., Kotecha.S., Mohun,T.およびLa Thangue,N.B. (1994). Mol. Biol. Cell. 5, 1081-1092.
Hannon,G.J., Demetrick,D.およびBeach,D. (1993). Genes Dev. 7:2378-2391
Heibert,S.W., Chellappan,S.P., Horowitz,J.M.およびNevins,J.R. (1992).
Genes Dev. 6, 177-185.
Helin,K., Lees,J.A., Vidal,M., Dyson,N., Harlow,E.およびFattaey,A. (1992). Cell 70, 337-350.
Helin,K., Wu,C.-L., Fattaey,A.R., Lees,J.A., Dynlacht,B.D., Ngwu,C.およびHarlow,E.(1993). Genes Dev. 7, 1850-1861.
Ivey-Hoyle,M., Conroy,R., Huber,H.E., Goodhart,P.J., Oliff,A.およびHeimbrook,D.C. (1993). Mol. Cell. Biol. 13:7802-7812.
Jooss,K.U., Funk,M.およびMuller,R. (1994). EMBO J. 13, 1467-1475.
Kaelin,W.G., Krek,W., Sellers,W.R., DeCaprio,J.A., Ajchenbaum,F., Fuchs,C.S., Chittenden,T., Li,Y., Farnham,P,J., Blanar,M.A., Livingston,D.M.およびFlemington,E.K. (1992). Cell 70, 351-364.
Krek,W., Livingston,D.M.およびShirodkar,S. (1993). Science 262, 1557-1560.
Krek,W., Ewen,M.E., Shirodkar,S., Arany,Z., Kaelin,W.G.およびLivingston,D.M. (1994). Cell 78, 161-172.
La Thangue,N.B. (1994). Trends Biochem. Sci. 19, 108-114.
La Thangue,N., Thimmappaya,B.およびRigby,P.W.J. (1990). Nucl. Acids. Res. 18, 2929-2938.
Lam,E.W.-F.およびLa Thangue,N.B. (1994). Curr. Op. Cell Biol. 6, 859-866.
Lees,E., Fahar,B., Dulic,V., Reed,S.I.およびHarlow,E. (1992). Genes Dev. 6, 1874-1885.
Lees,J.A., Saito,M., Vidal,M., Valentine,M., Look,T., Harlow,E., Dyson,N.およびHelin,K.(1993). Mol. Cell. Biol. 13:7813-7825.
Li.Y., Graham,C., Lacy,S., Duncan,A.M.V.およびWhyte,P. (1993). Genes Dev. 7:2366-2377.
Nevins,J.R. (1992). Science 258, 424-429.
Schwarz,J.K., Devoto,S.H., Smith,E.J., Chellappan,S.P., Jakoi,L.およびNevins,J.R. (1993). EMBO J. 12, 1013-1020.
Scholer,H.R., Ruppert,S., Suzuki,N., Chowdhury,K.およびGruss,P. (1990). Nature 344, 435-439.
Shan,B., Zhu,X, Chen,P.L., Durfee,T., Yang,Y., Sharp,D.およびLee,W.H. (1992). Mol.Cell.Biol. 12, 5620-5631.
Shirodkar,S., Ewen,M., DeCaprio,J.A., Morgan,J., Livingston,D.M.およびChittenden,T. (1992). Cell 68, 157-166.
Webster,N.J.G., Green,S., Tasset,D., Ponglikitmongkol,M.およびChambon,P. (1989). EMBO.J., 8, 1441-1446.
Zamanian,M,およびLa Thangue,N.B. (1992). EMBO J. 11:2603-2610.
Zamanian,M.およびLa Thangue,N.B. (1993). Mol. Biol. Cell. 4:389-396.
【図面の簡単な説明】
【0116】
【図1A−1】図1A-1はヒトE2F−5の構造を示す図であり、ヒトE2F−5cDNAのヌクレオチド配列および推定アミノ酸配列を示す図である。
【図1A−2】図1A-2はヒトE2F−5の構造を示す図であり、ヒトE2F−5cDNAのヌクレオチド配列および推定アミノ酸配列を示す図である。
【図1B】図1Bは、E2F−1およびE2F−4と比較してE2F−5を示す図であり、該保存ドメインの境界をアミノ酸番号で示す。E2F−4中のSSは、セリンに富むモチーフを示す。
【図2】図2は、ヒト細胞系でのE2F−5の発現パターンを示す図である。 (A)示されているヒト細胞系からの全細胞質RNAを含有するノーザンブロットを、ヒトE2F−5cDNAプローブとハイブリッド形成させた。以下のヒト細胞系からのRNAを使用した:CAMA、ヒト乳がん;A549、肺がん;MDA、MDA−MD157乳がん;OVCAR、卵巣がん;HS578T、乳がん;LUCY、卵巣がん;HT29、結腸がん;CEM、T−細胞白血病;K562、赤白血病。(B)ラットα−チューブリンプローブとハイブリッド形成させた同じフィルター。
【図3】図3は、E2F−5は、カルボキシル末端トランス活性化ドメインを有することを示す図である。Gal-4-E2F発現ベクター(1μg)および0.2μg pRSV-ルシフェラーゼ(内部対照)の存在下または不存在下、上流のGal4部位を保持するCATレポータープラスミド(5μg)で、U−2OS細胞をトランスフェクションした。それぞれのサンプルのCAT活性を、ルシフェラーゼ活性に対して標準化した。トランスフェクションの2日後に、CAT活性をアッセイした。CATレポーター遺伝子単独に対するGal4-E2Fの活性化倍率を表す。データは、少なくとも3つの独立した実験を2回行った結果を表す。
【図4】図4は、E2F−5およびDP−1はトランス活性化で協同することを示す図である。示されているとおり、増加する量のpJ3-E2F-5発現ベクター(1、2または5μg)および100ngのpCMV-DP-1で、U2−OS骨肉腫細胞をトランスフェクションした。それぞれのトランスフェクションでは、2μgのレポーター構築物(E2F4-CAT)および0.2μgのpRSV-ルシフェラーゼを加えた。それぞれのサンプルのCAT活性を、ルシフェラーゼ活性に対して標準化した。データは、少なくとも3つの独立した実験を2回行った結果を表す。
【図5】図5は、ポケットタンパク質によるE2F−5トランス活性化の阻害を示す図である。5μgのpJ3-E2F-5および100ngのpCMV-DP-1と、pCMV-Rb(50および100ng)、pCMV-RbΔ22(100ng)、pCMV-p107(50および100ng)、pCMVp107DE(100ng)またはpCMV-HA-p130(50、100および500ng)とを組み合わせて、US−OS細胞をトランスフェクションした。発現プラスミドと共に2μgのE2F4-CATおよび0.2μgのpRSV-ルシフェラーゼで、上記細胞をトランスフェクションした。ルシフェラーゼ内部対照に対して、CAT活性を標準化した。E2F4-CATの基準レベル(これを1.0とする)に対する活性化倍率を計算した。データは、少なくとも3つの独立した実験を2回行った結果を表す。
【図6】図6は、一過性にトランスフェクションされたU2−OS細胞中のE2F含有複合体を示す図である。示されているとおり、pRb-p107またはp130発現ベクターの存在下または不存在下、E2F−5およびDP−1発現ベクターで、U2−OS骨肉腫細胞を一過性にトランスフェクションした。2日後、全細胞抽出物を調製し、コンセンサスE2FDNA結合部位を含有する[32P]−標識オリゴヌクレオチドと共にインキュベーションし、ゲル電気泳動に供した。遊離プローブ、E2F−5/DP−1DNA複合体およびE2F−5/ポケットタンパク質複合体の位置を示す。
【図7】図7は、E2F−5は、in vivoでp130と優先的に相互作用することを示す図である。ヒトCAMA乳がん細胞を[32P]−オルトホスファートで標識し、非イオン性デタージェントライゼートを逐次免疫沈降法に供した。第1の免疫沈降法では、ライゼートを、示されているとおり、pRb、p107またはp130抗体と共にインキュベーションした。パネルAは、CAMA細胞からのpRb−p107およびp130特異的抗体による第1の免疫沈降法を示す。pRb、p107およびp130結合タンパク質を、SDS含有緩衝液中で煮沸することにより、その免疫沈降したポケットタンパク質から遊離させ、E2F−5特異的抗血清で再度免疫沈降させた(パネルB)。対照として、pRbおよびp107免疫沈降物から遊離したタンパク質を、抗E2F−1(KH20)またはE2F−4(RK13)抗体で再度免疫沈降させた(パネルC)。免疫沈降したタンパク質を7.5%SDS−ポリアクリルアミドゲル上で分離し、該ゲルを乾燥し、タンパク質をオートラジオグラフィーで検出した。
【図8】図8は、マウスE2F−5を単離するための戦略を示す図である。「餌」となるLEXA DP-1を使用して、14.5d.p.c.のマウス胎児活性化ドメイン標識(tagged)cDNAライブラリーをスクリーニングした。
【図9A−1】図9A-1は、マウスE2F−5の配列およびE2Fファミリーの他のメンバーとの比較を示す図であり、E2F−5のヌクレオチド配列および推定アミノ酸残基配列を示す図である。
【図9A−2】図9A-2は、マウスE2F−5の配列およびE2Fファミリーの他のメンバーとの比較を示す図であり、E2F−5のヌクレオチド配列および推定アミノ酸残基配列を示す図である。
【図9B】図9Bは、E2F−5(中段)を、E2F−1(上段)およびE2F−4(下段)と比較して示す図であり、E2F−5とE2F−1との間およびE2F−5とE2F−4との間のタンパク質配列レベルの同一性の割合(%)を示す。E2Fファミリーメンバーの間で共有されているドメインを示す。(Leesら, 1993)
【図9C】図9Cは、E2Fファミリーメンバー内での保存ドメイン中のアミノ酸残基配列の比較を示す図である。ハイライトが付されている残基はすべてのファミリーメンバーに共通であり、枠で囲まれている残基はE2F−4とE2F−5とで共通である。ロイシンジップ領域中の太字の残基は、7個からなる反復中の疎水性残基を示す。E2FファミリーメンバーとDP−1との間で共有されているDEFボックス領域中の残基を、線により示す。
【図10】図10は、酵母におけるE2F−5およびDP−1によるE2F部位依存的転写の活性化を示す図である。 (a)レポーターおよびエフェクター構築物の概要 (b)示されているE2F−5およびDP−1発現ベクターを、単独で(レーン2および3)または一緒に(レーン4)、酵母中に形質転換し、4つの野生型E2F部位を保持するp4xWT CYC1の活性を評価した。平行実験において、p4xMTCYC1の活性は、いずれの条件においても影響を受けなかった。示しているデータは、3回の読み取りから得たものである。
【図11】図11は、酵母におけるE2F−5による転写活性化を示す図である。 (a)レポ−ターおよびエフェクター構築物の概要 (b)pLexA-CYC1-lacZの活性をアッセイすることにより、pLEX.E2F-1(トラック2)またはpLEX.E2F-5(トラック3)のいずれかの転写活性を酵母で評価した。示しているデータは、3回の読み取りから得たものである。
【図12】図12は、ポケットタンパク質調節E2F−5を示す図である。 (a)レポーターおよびエフェクター構築物の概要。 (b)野生型pRb(トラック3)、pRbΔ22(トラック4)、p107(トラック5)およびp107AS(トラック6)の存在下、Gal-E2F-5(トラック2)の転写活性を評価した。比較のために、pG4(トラック7〜10)で同様の処理を行った。示しているデータは、3回の読み取りから得たものである。
【図13】図13は、E2F−5は、F9EC細胞中のDRTF1/E2Fの生理学的DNA結合成分であることを示す図である。E2F−5内の異なる領域からの別個のペプチドに対して産生させた2つのタイプの抗血清がF9EC細胞DRTF1/E2F DNA結合活性に及ぼす効果について評価した。抗E2F−5血清、抗ペプチド1(トラック5〜8)および抗ペプチド2(トラック9〜12)を、相同性(+;トラック5、7、9および11)または無関係な(−;トラック6、8、10および12)ペプチドのいずれかの存在下で評価した。比較のために、相同性(トラック1および3)または無関係なペプチド(トラック2および4)のいずれかの存在下、抗DP−1(24)(トラック1〜4)の効果を評価した。それぞれの対またはトラック(+および−)は、異なる抗血清調製物での処理を表す。DRTF1/E2Fb/cDNA結合複合体(La Thangueら, 1990)について示す。
【図14】図14は、E2F−5のDNA結合特性を示す図である。E2F−5、E2F−1およびDP−1をGST融合タンパク質として発現させ、精製し、それらのDNA結合活性をゲル遅延法により評価した。E2F−1またはE2F−5を単独で(それぞれ、トラック1および2、並びに5、6および7)または一定濃度のDP−1(トラック3および4、並びに8、9および10)と共にアッセイした。トラック11は、プローブ単独の場合を示す。E2F−1として加えたタンパク質は、全体で、約50ng(トラック1および3)または100ng(トラック2および4)、E2F−5としては25ng(トラック5および8)、50ng(トラック6および9)または100ng(トラック7および10)、およびDP−1としては50ngであった。
【図15】図15は、E2F−5RNAのレベルを示す図である。示されている細胞系で、E2F−5RNAのレベルをE2F−1と比較した。内部対照として、GAPDH RNAのレベルを評価した。
【特許請求の範囲】
【請求項1】
(a) 配列番号2または配列番号4で表される配列からなるタンパク質、
(b)配列番号2または配列番号4で表される配列において、1個のアミノ酸が付加、欠失または置換された(a)に記載のタンパク質の突然変異体、
(c)(a)のタンパク質に対して少なくとも90%のアミノ酸同一性を有し、哺乳類転写因子として機能し得るタンパク質、または
(d)DPタンパク質、p107またはp130と複合体を形成する能力を有する、少なくとも30アミノ酸の(a)から(c)のいずれか1つの断片、
を含んでなるポリペプチド。
【請求項2】
(a)配列番号1または配列番号3で表されるヌクレオチドの配列、
(b)(a)に相補的な配列、
(c)請求項1に記載のポリペプチドをコードする配列、または
(d)少なくとも25ヌクレオチド長の(a)から(c)のいずれかの配列の断片、
を含んでなるポリヌクレオチド。
【請求項3】
請求項2に記載のポリヌクレオチドおよびその相補的配列を含んでなる二本鎖ポリヌクレオチド。
【請求項4】
検出可能な標識を有する、請求項1に記載のポリペプチドまたは請求項2または3に記載のポリヌクレオチド。
【請求項5】
請求項3または4に記載のポリヌクレオチドを含んでなるベクター。
【請求項6】
請求項1に記載のポリペプチドをコードするコーディング配列を含んでなる複製可能な組換えベクターである、請求項5に記載のベクター。
【請求項7】
請求項5に記載のベクターを含んでなる宿主細胞または請求項6に記載の組換えベクターにより形質転換またはトランスフェクションされた宿主細胞。
【請求項8】
請求項5に記載の組換えベクターにより形質転換された宿主細胞であって、コーディング配列が、該宿主細胞による該コーディング配列の発現を可能にする能力を有する制御配列に機能を発揮できるように結合していることを特徴とする前記宿主細胞。
【請求項9】
組換えベクターのコーディング配列の発現を可能にする条件下で請求項7または8に記載の宿主細胞を培養し、発現されたポリペプチドを回収することを含んでなる、請求項1に記載のポリペプチドの製造法。
【請求項1】
(a) 配列番号2または配列番号4で表される配列からなるタンパク質、
(b)配列番号2または配列番号4で表される配列において、1個のアミノ酸が付加、欠失または置換された(a)に記載のタンパク質の突然変異体、
(c)(a)のタンパク質に対して少なくとも90%のアミノ酸同一性を有し、哺乳類転写因子として機能し得るタンパク質、または
(d)DPタンパク質、p107またはp130と複合体を形成する能力を有する、少なくとも30アミノ酸の(a)から(c)のいずれか1つの断片、
を含んでなるポリペプチド。
【請求項2】
(a)配列番号1または配列番号3で表されるヌクレオチドの配列、
(b)(a)に相補的な配列、
(c)請求項1に記載のポリペプチドをコードする配列、または
(d)少なくとも25ヌクレオチド長の(a)から(c)のいずれかの配列の断片、
を含んでなるポリヌクレオチド。
【請求項3】
請求項2に記載のポリヌクレオチドおよびその相補的配列を含んでなる二本鎖ポリヌクレオチド。
【請求項4】
検出可能な標識を有する、請求項1に記載のポリペプチドまたは請求項2または3に記載のポリヌクレオチド。
【請求項5】
請求項3または4に記載のポリヌクレオチドを含んでなるベクター。
【請求項6】
請求項1に記載のポリペプチドをコードするコーディング配列を含んでなる複製可能な組換えベクターである、請求項5に記載のベクター。
【請求項7】
請求項5に記載のベクターを含んでなる宿主細胞または請求項6に記載の組換えベクターにより形質転換またはトランスフェクションされた宿主細胞。
【請求項8】
請求項5に記載の組換えベクターにより形質転換された宿主細胞であって、コーディング配列が、該宿主細胞による該コーディング配列の発現を可能にする能力を有する制御配列に機能を発揮できるように結合していることを特徴とする前記宿主細胞。
【請求項9】
組換えベクターのコーディング配列の発現を可能にする条件下で請求項7または8に記載の宿主細胞を培養し、発現されたポリペプチドを回収することを含んでなる、請求項1に記載のポリペプチドの製造法。
【図1A−1】



【図1A−2】

【図1B】
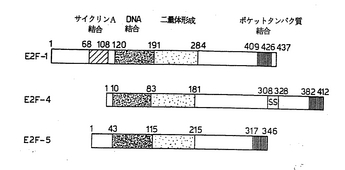
【図2】
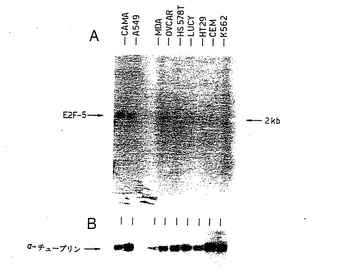
【図3】
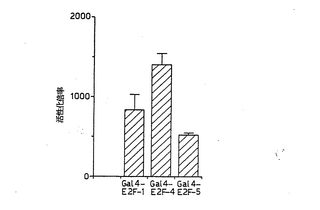
【図4】
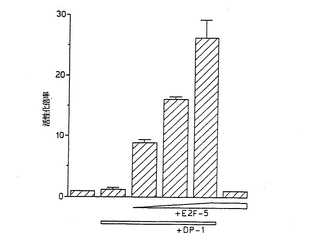
【図5】
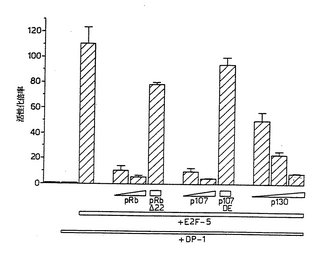
【図6】
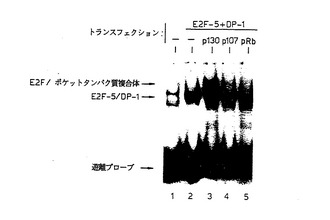
【図7】
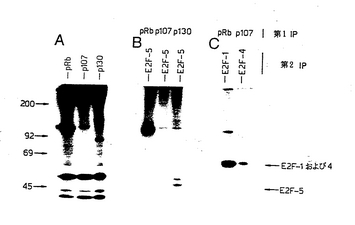
【図8】
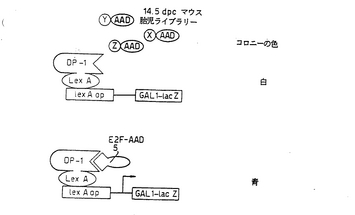
【図9A−1】

【図9A−2】

【図9B】
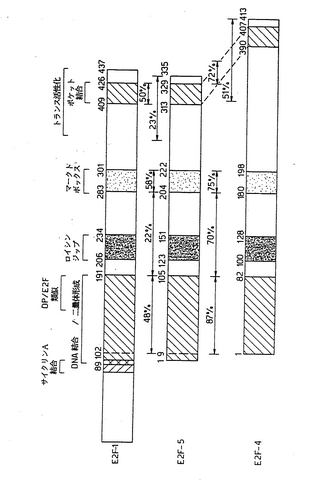
【図9C】

【図10】
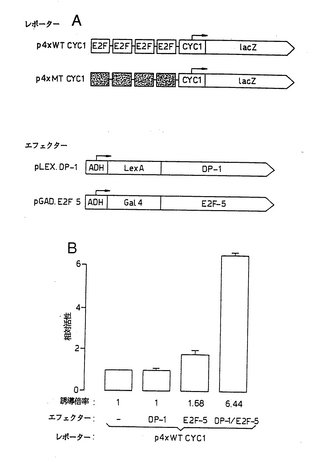
【図11】
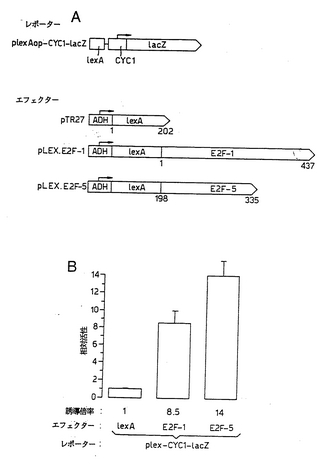
【図12】
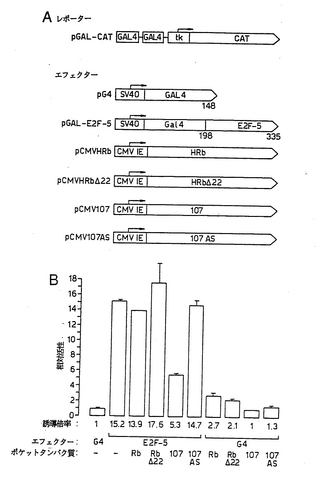
【図13】
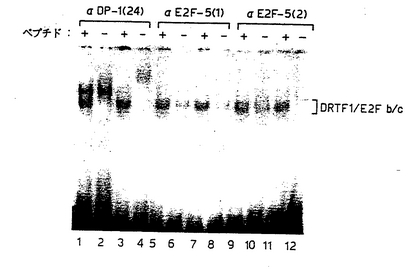
【図14】
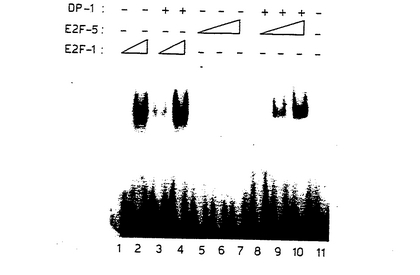
【図15】
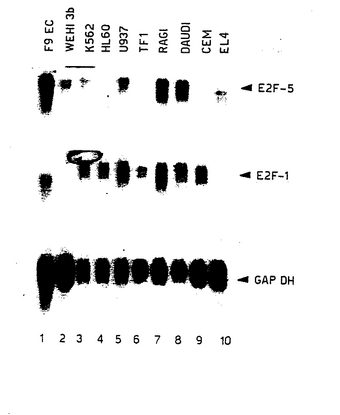
【図1A−2】
【図1B】
【図2】
【図3】
【図4】
【図5】
【図6】
【図7】
【図8】
【図9A−1】
【図9A−2】
【図9B】
【図9C】
【図10】
【図11】
【図12】
【図13】
【図14】
【図15】
【公開番号】特開2006−340730(P2006−340730A)
【公開日】平成18年12月21日(2006.12.21)
【国際特許分類】
【出願番号】特願2006−222739(P2006−222739)
【出願日】平成18年8月17日(2006.8.17)
【分割の表示】特願平8−524730の分割
【原出願日】平成7年4月18日(1995.4.18)
【出願人】(504367645)トポターゲット ユーケー リミテッド (12)
【Fターム(参考)】
【公開日】平成18年12月21日(2006.12.21)
【国際特許分類】
【出願日】平成18年8月17日(2006.8.17)
【分割の表示】特願平8−524730の分割
【原出願日】平成7年4月18日(1995.4.18)
【出願人】(504367645)トポターゲット ユーケー リミテッド (12)
【Fターム(参考)】
[ Back to top ]