
転写産物の安定性と翻訳効率の向上をもたらすRNAの修飾
【課題】RNAに基づくワクチンなど、免疫療法に適用可能な、安定性および翻訳効率が向上されたRNAおよびこのようなRNAを得るための手段を提供する。
【解決手段】プロモーター、転写可能な核酸配列、及び連続するAヌクレオチド配列を含む、制限切断によって直鎖化された後、in vitro転写可能な前記配列を有する閉館分子。宿主細胞をトランスフェクトするための前記転写されたRNAの使用。前記トランスフェクトされた抗原提示細胞、特に、樹状細胞、単球またはマクロファージ。
【解決手段】プロモーター、転写可能な核酸配列、及び連続するAヌクレオチド配列を含む、制限切断によって直鎖化された後、in vitro転写可能な前記配列を有する閉館分子。宿主細胞をトランスフェクトするための前記転写されたRNAの使用。前記トランスフェクトされた抗原提示細胞、特に、樹状細胞、単球またはマクロファージ。
【発明の詳細な説明】
【技術分野】
【0001】
弱毒または不活性化された病原体を含む従来のワクチンは多くの分野で有効であるものの、いくつかの感染性病原体や腫瘍に対しては有効な防御免疫を付与しない。これには有効で、用途が広く、生産が容易かつコスト効果があり、保存が容易なワクチンが必要である。
【背景技術】
【0002】
プラスミドDNAを直接筋肉注射した後では、コードされている遺伝子の細胞表面での発現が持続することが示されており(Wolff et al., 1990)、DNAに基づくワクチンが新たな有望な免疫戦略としてみなされている。これは核酸に基づくワクチンを開発するための重要な動機となった。当初、感染性病原体に対するDNAに基づくワクチンが試験されたが(Cox et al., 1993; Davis et al., 1993; Ulmer et al., 1993; Wang et al., 1993)、すぐに、特異的抗腫瘍免疫を誘導することを目的とする対腫瘍遺伝子療法においてさらに詳細に研究されるようになった(Conry et al., 1994; Conry et al., 1995a; Spooner et al., 1995; Wang et al., 1995)。この腫瘍免疫の戦略には重要な利点がいくつかある。核酸に基づくワクチンは製造が容易で、比較的安価である。さらに、この核酸に基づくワクチンは少数の細胞から増幅させることができる。
【0003】
DNAはRNAよりも安定であるが、抗DNA抗体の誘導(Gilkeson et al., 1995)および宿主ゲノムへの導入遺伝子(transgen)の組込みなど、安全性に対する潜在的なリスクがいくつかある。これは細胞の遺伝子を不活性化し、該導入遺伝子(transgen)の制御できない長期的発現または発癌を招き、従って、例えばerb−B2(Bargmann et al., 1986)やp53(Greenblatt et al., 1994)などの発癌能を有する腫瘍関連抗原には通常適用できない。RNAの使用はこれらの潜在的リスクを回避するための魅力的な代替法となる。
【0004】
可逆的な遺伝子療法の一種としてRNAを用いることの利点には、一時的発現および非形質転換特性が含まれる。RNAはトランスジェニック的に発現させるために核に入る必要がなく、もっと言えば、宿主ゲノムに組み込まれることはなく、それにより発癌のリスクがなくなる。DNAを用いる場合 (Condon et al., 1996; Tang et al., 1992)と同様、RNAの注射もまた、in vivoで細胞性免疫応答と体液性免疫応答の双方を誘導し得る(Hoerr et al., 2000; Ying et al., 1999)。
【0005】
in vitro転写されたRNA(IVT−RNA)による免疫療法は、双方とも種々の動物モデルにおいて引き続いて試験されている2つの異なる戦略の使用に至らせる。RNAは種々の免疫経路を介して直接注入される(Hoerr et al., 2000)か、または樹状細胞(DC)がリポフェクションもしくはエレクトロポレーションにより、in vitro転写されたRNAでトランスフェクトされ、その後投与される(Heiser et al., 2000)。最近発表された研究では、RNAでトランスフェクトされたDCによる免疫化は、in vitroおよびin vivoで抗原特異的細胞傷害性Tリンパ球(CTL)を誘導ことが示されている(Su et al., 2003; Heiser et al., 2002)。T細胞介在性の免疫応答の最適な誘導の中枢的に重要な因子はとりわけ用量、すなわち、DCに対する抗原提示の密度である。導入されたIVT−RNAの長期発現を達成するためには、種々の修飾によりIVT−RNAを安定化させる、それにより、DCに対する抗原提示を高めることが試みられている。翻訳の基本要件は、その翻訳効率がポリ(A)の長さに相関する3’ポリ(A)配列が存在することである(Preiss and Hentze, 1998)。5’キャップおよび3’ポリ(A)配列は相乗作用的にin vivo翻訳を活性化する(Gallie, 1991)。グロビン遺伝子の非翻訳領域(UTR)は、RNAの安定化および翻訳効率の増強に寄与し得る他の公知のエレメントである(Malone et al., 1989)。
【0006】
いくつかのIVTベクターは文献で知られ、in vitro転写の鋳型として標準的な方法で使用され、安定化されたRNA転写産物が生成されるように遺伝的に修飾されている。これまでに文献に記載されているプロトコール(Conry et al., 1995b; Teufel et al., 2005; Strong et al., 1997; Carralot et al., 2004; Boczkowski et al., 2000)は、次の構造:RNA転写を可能とする5’RNAポリメラーゼプロモーター、それに続く、3’および/または5’のいずれかで非翻訳領域(UTR)にフランキングされた目的遺伝子、および50〜70Aヌクレオチドを含む3’ポリアデニルカセットを有するプラスミドベクターに基づく。in vitro転写前に、この環状プラスミドは、II型制限酵素(認識配列は切断部位に相当する)によりポリアデニルカセットの下流で線状化される。よって、このポリアデニルカセットは、その転写産物において後期ポリ(A)配列に相当する。この手順の結果として、いくつかのヌクレオチドが線状化後に酵素切断部位の一部として残り、その3’末端においてポリ(A)配列を延長するか、またはマスクする。この非生理学的オーバーハングがこのような構築物から細胞内で産生されるタンパク質の量に影響を及ぼすかどうかは明らかでない。
【0007】
よって、RNAは臨床適用に特に好適であると思われる。しかしながら、遺伝子療法におけるRNAの利用は、特に細胞質ではRNAの短い半減期により著しく制限され、その結果、タンパク質発現は低くなる。
【発明の概要】
【発明が解決しようとする課題】
【0008】
本発明の目的は、安定性および翻訳効率が向上されたRNAおよびこのようなRNAを得るための手段を提供することである。該RNAを遺伝子療法アプローチに用いることで、高いグレードの発現が得られるに違いない。
【0009】
この目的は特許請求の範囲の主題により本発明に従い達成される。
【課題を解決するための手段】
【0010】
本発明は、RNA、特にmRNAの安定化およびmRNA翻訳の増強に関する。本発明は特に、転写産物の安定性および翻訳効率の向上をもたらすRNA、特にin vitro転写されたRNAの3つの修飾に関する。
【発明の効果】
【0011】
本発明によれば、オープンエンド型ポリ(A)配列を有するRNAは、末端がマスクされたポリ(A)配列を有するRNAより効率的に翻訳されることが分かった。特に約120 bpの長いポリ(A)配列は最適なRNA転写産物の安定性と翻訳効率をもたらすことが分かった。本発明はまた、RNA分子において、特にヒトβ−グロビン遺伝子の二重の3’非翻訳領域(UTR)が、個々の2つのUTRを用いた場合に予測されるトータル効果を明らかに超えるほどに翻訳効率を向上させることが実証された。本発明によれば、上記の修飾の組合せは、RNAの安定化および翻訳の増強に相乗作用を有することが分かった。
【0012】
転写産物の量およびタンパク質収量を測定するための定量的RT−PCRおよびeGFP変異体を用い、本発明は、本発明のRNA修飾が樹状細胞(DC)のトランスフェクションにおいて独立にRNAの安定性と翻訳効率増強することを実証した。よって、トランスフェクトされた細胞上での抗原特異的ペプチド/MHC複合体の密度ならびに抗原特異的CD4+およびCD8+T細胞を刺激および拡大するそれらの能力を増強することができた。よって、本発明は、本発明に記載のRNA修飾により修飾されたRNAを用いることでRNAトランスフェクトDCワクチンを至適化するための戦略に関する。
【0013】
本発明によれば、修飾、およびそれによるRNAの安定化および/または翻訳効率の向上は好ましくは、in vitroにおいてRNA転写の鋳型として好ましく働く発現ベクターを遺伝的に修飾することにより達成される。
【0014】
この種のベクターは特に、好ましくは該RNAにオープンエンドを有する、すなわち、Aヌクレオチド以外のヌクレオチドが該ポリ(A)配列の3’末端にフランキングしていないポリ(A)配列を有するRNAを転写させることを意図する。該RNAにおけるオープンエンド型ポリ(A)配列は、5’RNAポリメラーゼプロモーターの制御下でRNAを転写させ、ポリアデニルカセット(ポリ(A)配列)を含む発現ベクターにIIS型制限切断部位を導入することにより達成することができ、この認識配列はポリ(A)配列の3’に位置し、切断部位は上流、従ってポリ(A)配列内に位置する。このIIS型制限切断部位における制限切断により、プラスミドをポリ(A)配列内で線状化することができる(図2)。その後、線状化プラスミドをin vitro転写のための鋳型として用い、マスクされていないポリ(A)配列で終わる転写産物を得ることができる。
【0015】
その上、またはその代わりに、修飾、ならびにそれによるRNAの安定化および/または翻訳効率の向上は、本発明に従い、その3’末端に、好ましくはペプチドまたはタンパク質をコードする配列(オープンリーディングフレーム)とポリ(A)配列との間に2以上の3’非翻訳領域を有するRNAを転写させるように発現ベクターを遺伝的に修飾することで達成することができる。
【0016】
一態様では、本発明は、5’→3’転写方向に、(a)プロモーター;(b)転写可能な核酸配列または転写可能な核酸配列を導入するための核酸配列;(c−1)遺伝子の3’非翻訳領域に相当する、またはそれに由来する第一の核酸配列;および(c−2)遺伝子の3’非翻訳領域に相当する、またはそれに由来する第二の核酸配列を含む核酸分子に関する。
【0017】
一実施形態では、本発明の核酸分子は、(c−3)遺伝子の3’ 非翻訳領域に相当する、またはそれに由来する少なくとも1つのさらなる核酸配列をさらに含む。
【0018】
本発明の核酸分子では、プロモーター(a)の制御下の核酸配列(b)、(c−1)、(c−2)および適当であれば(c−3)は転写されて、核酸配列(c−1)および/または(c−2)および/または適当であれば(c−3)から転写された核酸配列に、好ましくは転写可能な核酸配列(b)から転写された核酸配列の翻訳効率および/または安定性を高めるような活性がある共通転写産物を生じ得ることが好ましい。
【0019】
核酸配列(c−1)、(c−2)および適当であれば(c−3)は同一であっても異なっていてもよい。
【0020】
一実施形態では、該核酸配列は、(d)プロモーター(a)の制御下で転写された際に、その転写産物に少なくとも20の連続するAヌクレオチドのヌクレオチド配列をコードする核酸配列をさらに含む。
【0021】
プロモーター(a)の制御下の核酸配列(b)、(c−1)、(c−2)、適当であれば(c−3)および(d)は転写されて、核酸配列(c−1)および/または(c−2)および/または適当であれば(c−3)および/または(d)から転写された核酸配列に、好ましくは転写可能な核酸配列(b)から転写された核酸配列の翻訳効率および/または安定性を高めるような活性がある共通転写産物を生じ得ることが好ましい。
【0022】
特定の実施形態では、核酸配列(d)は、プロモーター(a)の制御下で転写された際に、その転写産物に少なくとも40、好ましくは少なくとも80、好ましくは少なくとも100、特に約120の連続するAヌクレオチドのヌクレオチド配列をコードする。核酸配列(d)は、プロモーター(a)の制御下で転写された際に、その転写産物に500まで、好ましくは400まで、好ましくは300まで、好ましくは200まで、特に150までの連続するAヌクレオチドのヌクレオチド配列をコードするのが好ましい。
【0023】
一実施形態では、該核酸分子は、切断が5’→3’転写方向に、プロモーター(a)、核酸配列(b)、核酸配列(c−1)、(c−2)および適当であれば(c−3)、ならびに核酸配列(d)の少なくとも一部(この核酸配列(d)の少なくとも一部は、プロモーター(a)の制御下で転写された際に、その転写産物に少なくとも20の連続するAヌクレオチドのヌクレオチド配列をコードし、その転写産物において3’末端ヌクレオチドは少なくとも20の連続するAヌクレオチドの該ヌクレオチド配列のAヌクレオチドである)を含む核酸分子を生じるように、核酸配列(d)内で、好ましくは酵素的にまたは別の生化学的方法で切断され得ることを特徴とする。
【0024】
好ましくは、切断後、該核酸分子は、少なくとも20の連続するAヌクレオチドのヌクレオチド配列の鋳型として働く鎖の末端に、その転写産物において少なくとも20の連続するAヌクレオチドのヌクレオチド配列の鋳型として働くヌクレオチド配列の一部であるTヌクレオチドを有する。
【0025】
特定の実施形態では、核酸配列(d)の少なくとも一部は、プロモーター(a)の制御下で転写された際に、その転写産物に少なくとも40、好ましくは少なくとも80、好ましくは少なくとも100、特に約120の連続するAヌクレオチドのヌクレオチド配列をコードする。核酸配列(d)は、プロモーター(a)の制御下で転写された際に、その転写産物に500まで、好ましくは400まで、好ましくは300まで、好ましくは200まで、特に150までの連続するAヌクレオチドのヌクレオチド配列をコードするのが好ましい。
【0026】
本発明の核酸分子は好ましくは、切断前には閉環分子であり、切断後には直鎖分子である。
【0027】
好ましくはIIS型制限エンドヌクレアーゼの制限切断部位である制限切断部位の助けで切断が行われるのが好ましい。
【0028】
一実施形態では、IIS型制限エンドヌクレアーゼの認識配列は核酸配列(d)の3’末端の5〜26塩基対、好ましくは24〜26塩基対下流に位置する。
【0029】
好ましい実施形態では、核酸配列(c−1)、(c−2)および適当であれば(c−3)は互いに独立にα2−グロビン、α1−グロビン、β−グロビンおよび成長ホルモン、好ましくは、ヒトβ−グロビンなどのグロビン遺伝子からなる群から選択される遺伝子に由来し、特に好ましい実施形態では、配列表の配列番号1の核酸配列またはそれに由来する核酸配列に相当する。
【0030】
さらなる態様において、本発明は、5’→3’転写方向に、(a)プロモーター;(b)転写可能な核酸配列または転写可能な核酸配列を導入するための核酸配列;および(c)プロモーター(a)の制御下で転写された際に、その転写産物に少なくとも20の連続するAヌクレオチドのヌクレオチド配列をコードする核酸配列を含む核酸分子に関する。
【0031】
プロモーター(a)の制御下の核酸配列(b)および(c)は転写されて、核酸配列(c)から転写された核酸配列に、好ましくは転写可能な核酸配列(b)から転写された核酸配列の翻訳効率および/または安定性を高めるような活性がある共通転写産物を生じ得ることが好ましい。
【0032】
特定の実施形態では、核酸配列(c)は、プロモーター(a)の制御下で転写された際に、その転写産物に少なくとも40、好ましくは少なくとも80、好ましくは少なくとも100、特に約120の連続するAヌクレオチドのヌクレオチド配列をコードする。核酸配列(c)は、プロモーター(a)の制御下で転写された際に、その転写産物に500まで、好ましくは400まで、好ましくは300まで、好ましくは200まで、特に150までの連続するAヌクレオチドのヌクレオチド配列をコードするのが好ましい。
【0033】
一実施形態では、該核酸分子は、切断が5’→3’転写方向に、プロモーター(a)、核酸配列(b)、および核酸配列(c)の少なくとも一部(この核酸配列(c)の少なくとも一部は、プロモーター(a)の制御下で転写された際に、その転写産物に少なくとも20の連続するAヌクレオチドのヌクレオチド配列をコードし、その転写産物において3’末端ヌクレオチドは少なくとも20の連続するAヌクレオチドの該ヌクレオチド配列のAヌクレオチドである)を含む核酸分子を生じるように、核酸配列(c)内で、好ましくは酵素的にまたは別の生化学的方法で切断され得る。
【0034】
好ましくは、切断後、該核酸分子は、少なくとも20の連続するAヌクレオチドのヌクレオチド配列の鋳型として働く鎖の末端に、その転写産物において少なくとも20の連続するAヌクレオチドのヌクレオチド配列の鋳型として働くヌクレオチド配列の一部であるTヌクレオチドを有する。
【0035】
特定の実施形態では、核酸配列(c)の少なくとも一部は、プロモーター(a)の制御下で転写された際に、その転写産物に少なくとも40、好ましくは少なくとも80、好ましくは少なくとも100、特に約120の連続するAヌクレオチドのヌクレオチド配列をコードする。核酸配列(c)は、プロモーター(a)の制御下で転写された際に、その転写産物に500まで、好ましくは400まで、好ましくは300まで、好ましくは200まで、特に150までの連続するAヌクレオチドのヌクレオチド配列をコードするのが好ましい。
【0036】
該核酸分子は好ましくは、切断前には閉環分子であり、切断後には直鎖分子である。
【0037】
好ましくはIIS型制限エンドヌクレアーゼの制限切断部位である制限切断部位の助けで切断が行われるのが好ましい。
【0038】
一実施形態では、IIS型制限エンドヌクレアーゼの認識配列は核酸配列(c)の3’末端の5〜26塩基対、好ましくは24〜26塩基対下流に位置する。
【0039】
本発明の1つの核酸分子では、転写可能な核酸配列は好ましくはペプチドまたはタンパク質をコードする核酸配列を含み、転写可能な核酸配列を導入するための核酸配列は好ましくは多重クローニング部位である。
【0040】
本発明の核酸分子は、(i)レポーター遺伝子;(ii)選択マーカー;および(iii)複製起点からなる群から選択される1以上のメンバーをさらに含み得る。
【0041】
一実施形態では、本発明の核酸分子は閉環コンフォメーションであり、特に線状化後に、RNA、特にmRNAのin vitro転写に好適であるのが好ましい。
【0042】
さらなる態様では、本発明は、上記の核酸分子の線状化により、好ましくは少なくとも20の連続するAヌクレオチドのヌクレオチド配列をコードする核酸配列内での切断により得られる核酸分子、ならびにプロモーター(a)の制御下、上記の核酸分子との転写、好ましくはin vitro転写により得られるRNAに関する。
【0043】
さらなる態様では、本発明は、その安定性および/または翻訳効率を高めるよう選択されたRNA分子をin vitroで転写させる方法であって、(i)遺伝子の3’非翻訳領域に相当する、またはそれに由来する第一の核酸配列(b−1)を、転写されて該RNA分子を生じ得る核酸配列(a)の3’末端に結合させること、(ii)遺伝子の3’非翻訳領域に相当する、またはそれに由来する第二の核酸配列(b−2)を、該第一の核酸配列(b−1)の3’末端に結合させること、および(iii)得られた核酸をin vitroで転写させることを含む方法に関する。
【0044】
さらなる態様において、本発明は、その発現を増強するよう選択されたmRNA分子を転写させる方法であって、(i)遺伝子の3’非翻訳領域に相当する、またはそれに由来する第一の核酸配列(b−1)を、転写されて該RNA分子を生じ得る核酸配列(a)の3’末端に結合させること、(ii)遺伝子の3’非翻訳領域に相当する、またはそれに由来する第二の核酸配列(b−2)を、該第一の核酸配列(b−1)の3’末端に結合させること、および(iii)得られた核酸を転写させることにより得ることができるmRNAを翻訳することを含む方法に関する。転写は好ましくはin vitroで行われる。
【0045】
本発明おいて「核酸配列を核酸配列の3’末端に結合させる」とは、第一の核酸配列が第二の核酸配列の下流になり、付加的核酸配列により後者から離されてもよいような、2つの核酸配列の共有結合に関していう。
【0046】
一実施形態では、本発明の方法は、遺伝子の3’非翻訳領域に相当する、またはそれに由来する少なくとも1つのさらなる核酸配列(b−3)を第二の核酸配列(b−2)の3’末端に結合させることをさらに含む。
【0047】
核酸配列(a)、(b−1)、(b−2)および適当であれば(b−3)は転写されて、核酸配列(b−1)および/または(b−2)および/または適当であれば(b−3)から転写された核酸配列に、好ましくは核酸配列(a)により転写された核酸配列の翻訳効率および/または安定性を高めるような活性がある共通転写産物を生じ得ることが好ましい。
【0048】
さらなる実施形態では、本発明の方法は、転写された際に、少なくとも20の連続するAヌクレオチドのヌクレオチド配列をコードする核酸配列(c)を核酸配列(b−2)の、または適当であれば核酸配列(b−3)の3’末端に結合させることをさらに含む。
【0049】
核酸配列(a)、(b−1)、(b−2)および適当であれば(b−3)ならびに(c)は転写されて、核酸配列(b−1)および/または(b−2)および/または適当であれば(b−3)および/または(c)から転写された核酸配列に、好ましくは核酸配列(a)から転写された核酸配列の翻訳効率および/または安定性を高めるような活性がある共通転写産物を生じ得ることが好ましい。
【0050】
特定の実施形態では、核酸配列(c)は、その転写産物に少なくとも40、好ましくは少なくとも80、好ましくは少なくとも100、特に約120の連続するAヌクレオチドのヌクレオチド配列をコードする。核酸配列(c)は、転写された際に、その転写産物に500まで、好ましくは400まで、好ましくは300まで、好ましくは200まで、特に150までの連続するAヌクレオチドのヌクレオチド配列をコードするのが好ましい。
【0051】
特定の実施形態では、本発明の方法は、得られた核酸の転写の前に、このようにして得られた核酸の転写が、核酸配列(a)、(b−1)、(b−2)および適当であれば(b−3)ならびに少なくとも20の連続するAヌクレオチドの3’末端ヌクレオチド配列から転写された核酸配列を有する転写産物(該転写産物の3’末端ヌクレオチドは少なくとも20の連続するAヌクレオチドのヌクレオチド配列のAヌクレオチドである)を生じるように核酸配列(c)内で切断することをさらに含む。
【0052】
特定の実施形態では、該転写産物は、その3’末端に少なくとも40、好ましくは少なくとも80、好ましくは少なくとも100、特に約120の連続するAヌクレオチドのヌクレオチド配列を有する。該転写産物は、その3’末端に500まで、好ましくは400まで、好ましくは300まで、好ましくは200まで、特に150までの連続するAヌクレオチドのヌクレオチド配列を有するのが好ましい。
【0053】
好ましい実施形態では、核酸配列(b−1)、(b−2)および適当であれば(b−3)は互いに独立にα2−グロビン、α1−グロビン、β−グロビンおよび成長ホルモン、好ましくはヒトβ−グロビンなどのグロビン遺伝子からなる群から選択される遺伝子に由来し、特に好ましい実施形態では、配列表の配列番号1の核酸配列またはそれに由来する核酸配列に相当する。
【0054】
さらなる態様において、本発明は、その安定性および/または翻訳効率を高めるよう選択されたRNA分子をin vitroで転写させる方法であって、(i)転写された際に、少なくとも20の連続するAヌクレオチドのヌクレオチド配列をコードする核酸配列(b)を、転写されて前記RNA分子を生じ得る核酸配列(a)の3’末端に結合させること、および(ii)得られた核酸をin vitroで転写させることを含む方法に関する。
【0055】
さらなる態様において、本発明は、その発現を増強するよう選択されたmRNA分子を翻訳する方法であって、(i)転写された際に、少なくとも20の連続するAヌクレオチドのヌクレオチド配列をコードする核酸配列(b)を、転写されて前記mRNA分子を生じ得る核酸配列(a)の3’末端に結合させること、および(ii)得られた核酸を転写させることにより得ることができるmRNAを翻訳することを含む方法に関する。転写はin vitroで行われるのが好ましい。
【0056】
核酸配列(a)および(b)は転写されて、核酸配列(b)から転写された核酸配列に、好ましくは核酸配列(a)から転写された核酸配列の翻訳効率および/または安定性を高めるような活性がある共通転写産物を生じ得ることが好ましい。
【0057】
特定の実施形態では、核酸配列(b)は、その転写産物に少なくとも40、好ましくは少なくとも80、好ましくは少なくとも100、特に約120の連続するAヌクレオチドのヌクレオチド配列をコードする。核酸配列(b)は、転写された際に、その転写産物に500まで、好ましくは400まで、好ましくは300まで、好ましくは200まで、特に150までの連続するAヌクレオチドのヌクレオチド配列をコードするのが好ましい。
【0058】
特定の実施形態では、本発明の方法は、得られた核酸の転写の前に、このようにして得られた核酸の転写が、核酸配列(a)および少なくとも20の連続するAヌクレオチドの3’末端ヌクレオチド配列から転写された核酸配列を有する転写産物(該転写産物の3’末端ヌクレオチドは少なくとも20の連続するAヌクレオチドのヌクレオチド配列のAヌクレオチドである)を生じるように核酸配列(b)内で切断することをさらに含む。
【0059】
特定の実施形態では、該転写産物は、その3’末端に少なくとも40、好ましくは少なくとも80、好ましくは少なくとも100、特に約120の連続するAヌクレオチドを有する。該転写産物は、その3’末端に500まで、好ましくは400まで、好ましくは300まで、好ましくは200まで、特に150までの連続するAヌクレオチドを有するのが好ましい。
【0060】
本発明の方法の全ての態様において、切断は、好ましくはIIS型制限エンドヌクレアーゼの制限切断部位である制限切断部位の助けで行われるのが好ましい。
【0061】
一実施形態では、IIS型制限エンドヌクレアーゼの認識配列は、転写された際に、少なくとも20の連続するAヌクレオチドのヌクレオチド配列をコードする核酸配列の3’末端の5〜26塩基対、好ましくは24〜26塩基対下流に位置する。
【0062】
本発明はまた、選択されたRNA分子をin vitroで転写させる本発明の方法により得ることができるRNAに関する。本発明の核酸分子から、選択されたRNA分子をin vitroで転写させる本発明の方法により得ることができる、鋳型としてのRNA調製物は、そのRNAのポリ(A)配列の長さに関して好ましくは均質または本質的に均質であり、すなわち、このポリ(A)配列の長さは、10個だけ、好ましくは5個、4個、3個、2個または1個のAヌクレオチドだけ異なるRNA分子が調製物中に90%以下、好ましくは95%以下、好ましくは98%または99%以下である。
【0063】
本発明は例えば、細胞内での転写および発現において組換えタンパク質の発現を増強するために利用可能である。より具体的には、組換えタンパク質を産生する場合、本発明に記載の修飾およびその組合せを発現ベクターに導入し、細胞系において組換え核酸の転写および組換えタンパク質の発現を増強する目的でそれらを利用することができる。これには例えば、組換え抗体、ホルモン、サイトカインおよび酵素などの生産が含まれる。これによりとりわけ生産コストの低減が可能となる。
【0064】
また、遺伝子療法適用で、本発明に記載の修飾およびその組合せを利用することもできる。該修飾は遺伝子療法ベクターに導入し、それにより導入遺伝子(transgen)の発現を増強させるために利用することができる。この目的で、いずれの核酸(DNA/RNA)に基づくベクター系(例えば、プラスミド、アデノウイルス、ポックスウイルスベクター、インフルエンザウイルスベクター、およびαウイルスベクターなど)も使用可能である。例えばリンパ球または樹状細胞では、in vitroにて細胞をこれらのベクターでトランスフェクトすることができ、あるいは直接in vivoに投与することもできる。
【0065】
さらに、本発明に記載の修飾およびその組合せは、リボ核酸の安定性および/または発現を、そしてそれにより該リボ核酸によりコードされるペプチドまたはタンパク質の量を増強させることができる。コードリボ核酸は例えば遺伝子の一時的発現ために利用可能であり、可能性のある適用分野としては、in vitroにて細胞にトランスフェクトされる、または直接in vivoに投与されるRNAに基づくワクチン、例えば細胞において分化プロセスを開始させるため、またはタンパク質の機能を研究するためのin vitroにおける機能的組換えタンパク質の一時的発現、ならびにin vivoにおける、特に医薬としての、エリスロポエチン、ホルモン、血液凝固阻害剤などのような機能的組換えタンパク質の一時的発現がある。
【0066】
本発明に記載の修飾により修飾されたRNA、特にin vitroで転写されたRNAは特に抗原提示細胞をトラスフェクトするために、従って提示されるべき抗原を送達するため、また、抗原提示細胞に、該RNAから発現されたまたはそれに由来するペプチドまたはタンパク質に相当する該提示抗原を、特に切断などの細胞内プロセシングにより付加するため(すなわち、この提示抗原は例えばRNAから発現されたペプチドまたはタンパク質の断片である)のツールとして使用することができる。このような抗原提示細胞はT細胞、特にCD4+および/またはCD8+T細胞を刺激するために使用可能である。
【0067】
発明の詳細な説明
本発明によれば、組換え核酸の作製、細胞の培養および核酸、特にRNAの細胞への導入、特にエレクトロポレーションおよびリポフェクションには、標準的な方法が使用可能である。酵素反応は製造業者の使用説明書に従い、またはそれ自体公知の方法で行われる。
【0068】
本発明によれば、核酸分子または核酸配列とは、好ましくはデオキシリボ核酸(DNA)またはリボ核酸(RNA)である核酸を指す。本発明によれば、核酸はゲノムDNA、cDNA、mRNA、組換え生産された分子、および化学的に合成された分子を含む。本発明によれば、核酸は一本鎖または二本鎖、および直鎖または共有結合的に閉環した分子の形態であってよい。
【0069】
「mRNA」とは「メッセンジャーRNA」を意味し、DNAを鋳型として生産され、それ自体ペプチドまたはタンパク質をコードする「転写産物」を指す。mRNAは一般に5’非翻訳領域、タンパク質コード領域および3’非翻訳領域を含む。mRNAは、細胞内でもin vitroでも限られた半減期を有する。本発明によれば、mRNAはDNA鋳型からin vitro転写により作製することができる。このmRNAは、本発明の修飾に加え、さらなる安定化修飾およびキャッピングにより修飾してもよい。
【0070】
「核酸」とはさらにまた、ヌクレオチド塩基、糖またはリン酸基に対する核酸の化学誘導体、ならびに非天然ヌクレオチドおよびヌクレオチド類似体を含有する核酸を含む。
【0071】
本発明によれば「核酸配列に由来する核酸配列」とは、それが由来する核酸に比べて、単一または複数のヌクレオチド置換、欠失および/または付加を含み、好ましくはそれが由来する核酸と相補的である、すなわち、該核酸間にある程度の相同性があり、該核酸のヌクレオチド配列が有意な直接的または相補的様式で一致する、核酸配列を指す。本発明によれば、核酸に由来する核酸は、それが由来する核酸の機能的特性を有している。このような機能的特性としては、特に、RNAへと転写し得る核酸(転写可能な核酸配列)との機能的連結において、完全なRNA分子においてこの核酸から産生されるRNAの安定性および/または翻訳効率を高める能力が含まれる。
【0072】
本発明によれば「機能的連結」または「機能的に連結された」とは、機能的関係の範囲内の連結に関していう。核酸は、別の核酸配列と機能的に関連している場合に、「機能的に連結されている」という。例えば、プロモーターは、コード配列の転写に影響を及ぼす場合に、該コード配列に機能的に連結されているという。機能的に連結されている核酸は一般に、別の核酸と、適当であればさらなる核酸配列により離されて隣接しており、特定の実施形態では、RNAポリメラーゼにより転写されて、単一のRNA分子(共通転写産物)が得られる。
【0073】
本発明に記載される核酸は単離されていることが好ましい。「単離された核酸」とは、本発明によれば、その核酸が(i)例えばポリメラーゼ連鎖反応(PCR)によりin vitroで増幅されたものであるか、(ii)クローニングにより組換え生産されたものであるか、(iii)例えば切断およびゲル濾過分画により精製されたものであるか、または(iv)例えば化学合成により合成されたものであることを意味する。単離された核酸は組換えDNA技術による操作に利用可能な核酸である。
【0074】
核酸は、その2つの配列が互いにハイブリダイズして安定な二重らせんを形成するこができる場合に別の核酸と「相補的」であるといい、該ハイブリダイゼーションは好ましくはポリヌクレオチド間の特異的ハイブリダイゼーションを可能とする条件(ストリンジェント条件)下で行われる。ストリンジェント条件は、例えば、Molecular Cloning: A Laboratory Manual, J. Sambrook et al., eds., 2nd edition, Cold Spring Harbor Laboratory Press, Cold Spring Harbor, New York 1989 or Current Protocols in Molecular Biology, F.M. Ausubel et al., eds., John Wiley & Sons, Inc., New Yorkに記載されており、例えば、65℃、ハイブリダイゼーションバッファー(3.5×SSC、0.02%フィコール、0.02%ポリビニルピロリドン、0.02%ウシ血清アルブミン、2.5mM NaH2PO4(pH7)、0.5%SDS、2mM EDTA)中でのハイブリダイゼーションを指す。SSCは0.15M塩化ナトリウム/0.15Mクエン酸ナトリウム、pH 7である。ハイブリダイゼーション後、DNAを写し取ったメンブランを、例えば2×SSC中、室温で洗浄した後、0.1〜0.5×SSC/0.1×SDS中、68℃までの温度で洗浄する。
【0075】
本発明によれば、相補的核酸は、少なくとも60%、少なくとも70%、少なくとも80%、少なくとも90%、好ましくは少なくとも95%、少なくとも98%または少なくとも99%同一であるヌクレオチドを有する。
【0076】
「同一性%」とは、比較する2つの配列間で最適なアライメントにおいて同一であるヌクレオチドのパーセンテージを指し、該パーセンテージは単に統計学的なものであり、この2配列間の違いは、配列の全長にわたってランダムに分布している場合もあるし、また、2配列間の最適アライメントを得るために、比較配列は参照配列に対して付加もしくは欠失を含んでいる場合もある。2配列の比較は通常、対応する配列の局部領域を特定するために、最適アライメントの後にこれらの配列をセグメントまたは「比較枠」に関して比較することにより行う。比較のための最適アライメントは手動で、またはSmith and Waterman, 1981, Ads App. Math. 2, 482によるローカルホモロジーアルゴリズム、Neddleman and Wunsch, 1970, J. Mol. Biol. 48, 443によるローカルホモロジーアルゴリズム、およびPearson and Lipman, 1988, Proc. Natl Acad. Sci. USA 85, 2444による類似性検索アルゴリズムまたは前記アルゴリズムを用いたコンピュータープログラム(GAP, BESTFIT, FASTA, BLAST P, BLAST N and TFASTA in Wisconsin Genetics Software Package, Genetics Computer Group, 575 Science Drive, Madison, Wis.)の助けで行うことができる。
【0077】
同一性パーセンテージは、比較配列が一致する位置の数を求め、この数を比較した位置の数で割り、この解に100を掛けることにより得られる。
【0078】
例えば、ウェブサイトhttp://www.ncbi.nlm.nih.gov/blast/bl2seq/wblast2.cgiで入手できるBLASTプログラム「BLAST2配列」が使用可能である。
【0079】
「核酸の3’末端」とは、本発明では、遊離ヒドロキシ基を有する末端を指す。二本鎖核酸、特にDNAの模式図では、3’末端は常に右側にある。「核酸の5’末端」とは、本発明では、遊離リン酸基を有する末端を指す。二本鎖核酸、特にDNAの模式図では、5’末端は常に左側にある。
5’末端 5’−−P−NNNNNNN−OH−3’
3’末端 3’−HO−NNNNNNN−P−−5’
【0080】
特定の実施形態では、核酸は、本発明に従い、その核酸に対して同種であっても異種であってもよい発現制御配列と機能的に連結されている。
【0081】
転写可能な核酸、特にペプチドまたはタンパク質をコードする核酸および発現制御配列は、それらが、転写可能な、特にコード核酸の転写または発現がその発現制御配列の制御下または影響下にあるように互いに共有結合されている場合に、互いに「機能的に」連結されているという。この核酸が機能的ペプチドまたはタンパク質に翻訳されるならば、そのコード配列に機能的に連結されている発現制御配列の誘導は、コード配列にフレームシフトを起こすことなく、またはコード配列が所望のペプチドまたはタンパク質の翻訳不能となることなく、コード配列の転写を起こす。
【0082】
「発現制御配列」は、本発明によれば、プロモーター、リボソーム結合配列および遺伝子の転写または誘導されるRNAの翻訳を制御する他の制御エレメントを含む。本発明の特定の実施形態では、発現制御配列は調節可能である。発現制御配列の厳密な構造は、種または細胞種によって異なるが、通常、TATAボックス、キャッピング配列およびCAAT配列など、それぞれ転写および翻訳の開始に関与する5’非転写配列ならびに5’および3’非翻訳配列を含む。より具体的には、5’非転写発現制御配列は、機能的に連結された遺伝子の転写制御のためのプロモーター配列を包含するプロモーター領域を含む。発現制御配列はまたエンハンサー配列または上流アクチベーター配列も含み得る。
【0083】
本明細書に明示される核酸、特に転写可能なコード核酸は、その核酸に対して同種であっても異種であってもよい発現制御配列、特にプロモーターのいずれと組み合わせてもよく、「同種」とは、核酸がまた発現制御配列に天然状態で機能的に連結されているということを表し、「異種」とは、核酸が発現制御配列に天然状態では機能的に連結されていないということを表す。
【0084】
「プロモーター」または「プロモーター領域」とは、RNAポリメラーゼの認識および結合部位を提供することでコード配列の発現を制御する、遺伝子のコード配列の上流(5’)DNA配列を指す。このプロモーター領域は、該遺伝子の転写の調節に関与するさらなる因子のためのさらなる認識または結合部位を含んでいてもよい。プロモーターは原核生物または真核生物の遺伝子の転写を制御し得る。プロモーターはインデューサーに応答して転写を開始する「誘導型」であるか、転写がインデューサーによって制御されなければ「構成型」である。誘導型プロモーターは、インデューサーが存在しなければ、極めて小さな程度でしか発現しないか、または全く発現しない。インデューサーの存在下では、この遺伝子は「スイッチオン」となるか、または転写レベルが高まる。これは通常、特定の転写因子の結合によって媒介される。
【0085】
本発明に従う好ましいプロモーターの例としては、SP6、T3またはT7ポリメラーゼのプロモーターがある。
【0086】
本発明によれば、「発現」とは、その最も一般的に意味で用いられ、RNAまたはRNAとタンパク質の産生を含む。また、核酸の部分的発現も含まれる。さらに、発現は一時的なものであっても安定したものであってもよい。RNAに関して「発現」または「翻訳」とは、特にペプチドまたはタンパク質の産生を指す。
【0087】
「転写されて共通転写産物を生じ得る核酸」とは、その核酸が、適当であれば該核酸を含む核酸分子の、特に、閉環核酸分子の制限酵素切断などの線状化の後、プロモーターの制御下での転写が、適当であればその間に位置する配列により離されて、互いに共有結合された該核酸の転写産物を含むRNA分子を生じるように、互いに機能的に連結されていることを意味する。
【0088】
本発明によれば、「転写」には「in vitro転写」が含まれ、この「in vitro転写」とは、RNA、特にmRNAが無細胞様式で、すなわち、好ましくは適宜調製された細胞抽出液を用いることでin vitro合成される方法に関していう。転写産物の調製には、一般に転写ベクターと呼ばれ、本発明によれば「ベクター」という用語に含まれる、クローニングベクターを用いるのが好ましい。
【0089】
「核酸配列から転写された核酸配列」とは、適当であれば後者の核酸配列の転写産物である完全RNA分子の一部としてのRNAを指す。
【0090】
「核酸配列の翻訳効率および/または安定性を高めるために活性である核酸配列」とは、第一の核酸が第二の核酸との共通転写産物において、第二の核酸の翻訳効率および/または安定性を、その翻訳効率および/または安定性が第一の核酸を用いない場合の第二の核酸の翻訳効率および/または安定性に比べて高まるように修飾することができることを意味する。これに関して、「翻訳効率」とは、特定の時間内にRNA分子がもたらす翻訳産物の量に関し、「安定性」とは、RNA分子の半減期に関していう。
【0091】
3’非翻訳領域は、タンパク質コード領域の末端コドンの下流の、遺伝子の3’末端に位置し、転写されるがアミノ酸配列へ翻訳されない領域に関していう。
【0092】
本発明によれば、第一のポリヌクレオチド領域の5’末端が第二のポリヌクレオチド領域の3’末端に最も近い第一のポリヌクレオチド領域の一部である場合に、第一のポリヌクレオチド領域は、第二のポリヌクレオチドの下流に位置するとみなされる。
【0093】
3’非翻訳領域は一般に、翻訳産物の末端コドンから、通常転写工程後に付着されるポリ(A)配列へと伸びる。哺乳類mRNAの3’非翻訳領域は一般に、AAUAAAヘキサヌクレオチド配列として知られる相同性領域を有する。この配列はおそらくポリ(A)付着シグナルであり、ポリ(A)付着部位の10〜30塩基上流に位置している場合が多い。
【0094】
3’非翻訳領域は、エキソリボヌクレアーゼに対するバリアとして働く、またはRNAの安定性を高めることが知られているタンパク質(例えば、RNA結合タンパク質)と相互作用するステム−ループ構造が得られるように折り畳み可能な1以上の逆方向反復を含み得る。
【0095】
本発明によれば5’および/または3’非翻訳領域は転写可能な、特にコード核酸と機能的に連結させることができ、これによりこれらの領域を、転写可能な核酸から転写されたRNAの安定性および/または翻訳効率が高まるように核酸と結合させる。
【0096】
免疫グロブリンmRNAの3’非翻訳領域は比較的短く(約300ヌクレオチドより短い)が、他の遺伝子の3’非翻訳領域は比較的長い。例えば、tPAの3’非翻訳領域は約800ヌクレオチドの長さであり、第VIII因子の3’非翻訳領域は約1800ヌクレオチドの長さであり、エリスロポエチンの3’非翻訳領域は約560ヌクレオチドの長さである。
【0097】
本発明によれば、3’非翻訳領域またはそれに由来する核酸配列がRNAの安定性および/または翻訳効率を高めるかどうかは、その3’非翻訳領域またはそれに由来する核酸配列を遺伝子の3’非翻訳領域に組み込み、その組み込みによりタンパク質の合成量が高まるかどうかを測定することによって判定することができる。
【0098】
上記のことは本発明において核酸が2以上の3’非翻訳領域を含む場合にも適用され、この場合には、間に好ましくは「ヘッド・トゥ・テール関係」(すなわち、これらの3’非翻訳領域は同じ配向、好ましくは核酸において天然の配向を持つ)のリンカーを用いて、または用いずに順次結合されていることが好ましい。
【0099】
本発明によれば「遺伝子」とは、1以上の細胞産物の産生および/または1以上の細胞間もしくは細胞内機能の達成を担う特定の核酸配列に関していう。より具体的には、この用語は、特定のタンパク質または機能的もしくは構造的RNA分子をコードする核酸を含むDNA部分に関する。
【0100】
「ポリアデニルカセット」または「ポリ(A)配列」は、一般にRNA分子の3’末端に位置するアデニル残基の配列を指す。本発明は、コード鎖に相補的な鎖中の反復チミジル残基に基づくDNA鋳型を介したRNA転写中に付着されるこのような配列を提供するが、このような配列は通常そのDNAにはコードされておらず、核内で転写後に鋳型依存性のRNAポリメラーゼによりRNAの空いている3’末端に付着される。本発明によれば、この種のポリ(A)配列は少なくとも20、好ましくは少なくとも40、好ましくは少なくとも80、好ましくは少なくとも100で、好ましくは500まで、好ましくは400まで、好ましくは300まで、好ましくは200まで、特に150までの連続するAヌクレオチド、特には約120の連続するAヌクレオチドのヌクレオチド配列を意味するものと理解され、「Aヌクレオチド」とはアデニル残基を指す。
【0101】
好ましい実施形態では、本発明の核酸分子はベクターである。「ベクター」とは、ここではその最も一般的な意味で用いられ、例えば核酸を原核生物および/または真核生物の宿主細胞へ導入可能とし、および適当であればゲノムへ導入可能とする核酸のいずれの中間ビヒクルも含む。このようなベクターは細胞内で複製および/または発現されることが好ましい。ベクターはプラスミド、ファージミドまたはウイルスゲノムを含む。「プラスミド」とは本明細書において一般に、染色体DNAとは独立に複製可能な染色体外遺伝物質の構築物、通常、環状DNA二重らせんに関していう。
【0102】
本発明によれば、「宿主細胞」とは、外来核酸で形質転換またはトランスフェクション可能ないずれの細胞も指す。「宿主細胞」とは、本発明によれば、原核生物(例えば、大腸菌(E. coli))または真核細胞(例えば、酵母細胞および昆虫細胞)を含む。ヒト、マウス、ハムスター、ブタ、ヤギ、霊長類などの哺乳類細胞が特に好ましい。これらの細胞は多数の細胞種に由来し、一次細胞と細胞系統を含む。特定の例としては、ケラチノサイト、末梢血白血球、骨髄幹細胞および胚幹細胞が挙げられる。他の実施形態では、宿主細胞は抗原提示細胞、特に樹状細胞、単球またはマクロファージである。核酸は一コピーまたは数個ピーで宿主細胞に存在してもよく、一実施形態では宿主細胞内で発現される。
【0103】
本発明によれば、核酸によりコードされるペプチドまたはタンパク質は、細胞質、核、膜、オルガネラに存在するペプチドまたはタンパク質であってもよいし、あるいは分泌されるものでもよい。これらのものとして構造タンパク質、調節タンパク質、ホルモン、神経伝達物質、成長調節因子、分化因子、遺伝子発現調節因子、DNA会合タンパク質、酵素、血清タンパク質、受容体、薬剤、免疫調節剤、癌遺伝子、毒素、腫瘍抗原または抗原が挙げられる。これらのペプチドまたはタンパク質は天然配列を持っていてもよいし、あるいはそれらの生物活性を増強、阻害、調節または排除するために変異された配列を持っていてもよい。
【0104】
「ペプチド」とは、ペプチド結合を介して互いに連結された2以上、好ましくは3以上、好ましくは4以上、好ましくは6以上、好ましくは8以上、好ましくは10以上、好ましくは13以上、好ましくは16以上、好ましくは20以上で、好ましくは50まで、好ましくは100までまたは好ましくは150までの連続するアミノ酸を含む物質を指す。「タンパク質」とは、大きなペプチド、好ましくは少なくとも151のアミノ酸を有するペプチドを指すが、本明細書では「ペプチド」と「タンパク質」は通常同義で用いる。本発明において「ペプチド」および「タンパク質」は、アミノ酸成分だけでなく、糖およびリン酸構造などの非アミノ酸成分も含む物質を含み、またエステル、チオエーテルまたはジスルフィド結合などの結合を含む物質も含む。
【0105】
本発明は、患者に投与する核酸、特にRNAを提供する。一実施形態では、核酸はex vivo法、すなわち、患者から細胞を取り出し、それらの細胞を遺伝的に修飾し、修飾された細胞を患者に再び導入することにより投与される。トランスフェクションおよび形質導入法は当業者に公知である。本発明はまた、in vivo投与される核酸も提供する。
【0106】
本発明によれば、「トランスフェクション」とは、1以上の核酸を生物体または宿主細胞に導入することを指す。本発明に従いin vitroまたはin vivoで核酸を細胞に導入するためには、様々な方法が使用できる。このような方法としては、核酸 CaPO4沈降物のトランスフェクション、DEAEと会合した核酸のトランスフェクション、目的の核酸を運ぶウイルスによるトランスフェクションまたは感染、およびリポソームに媒介されるトランスフェクションなどが挙げられる。特定の実施形態では、核酸を特定の細胞に向けることが好ましい。このような実施形態では、核酸を細胞へ投与するために用いる担体(例えば、レトロウイルスまたはリポソーム)にターゲティング分子を結合させてもよい。例えば、標的細胞上の表面膜タンパク質に特異的な抗体または標的細胞上の受容体に対するリガンドなどの分子を、核酸担体に組み込むか結合させればよい。リポソームによる核酸の投与が望ましければ、ターゲティングおよび/または吸収を可能とするためにエンドサイトーシスに関連する表面膜タンパク質に結合するタンパク質をリポソーム製剤に組み込めばよい。このようなタンパク質としては、特定の細胞種に特異的なキャプシドタンパク質またはその断片、インターナライズされるタンパク質に対する抗体、および細胞内部位をターゲティングするタンパク質などが挙げられる。
【0107】
「レポーター」とは、レポーター遺伝子によりコードされ、レポーターアッセイで測定される分子、一般にペプチドまたはタンパク質に関していう。慣例の系としては、通常、酵素レポーターを用い、そのレポーターの活性を測定する。
【0108】
「多重クローニング部位」とは、そのいずれか1つが例えばベクターの切断および核酸の挿入に使用できる制限酵素部位を含む核酸領域に関していう。
【0109】
本発明によれば、ヌクレオチドまたはアミノ酸などの2つのエレメントは、互いに直接隣接していれば連続のものである。例えば、x個の連続するヌクレオチドNの配列は、配列(N)xと表す。
【0110】
「制限エンドヌクレアーゼ」または「制限酵素」とは、DNA分子の両鎖のホスホジエステル結合を特定の塩基配列内で切断する酵素種を指す。これらの酵素は、二本鎖DNA分子上の、認識配列と呼ばれる特定の結合部位を認識する。DNA中のホスホジエステル結合がこのような酵素により切断される部位は切断部位と呼ばれる。IIS型酵素の場合、この切断部位はDNA結合部位から所定の距離離れたところに存在する。本発明によれば、「制限エンドヌクレアーゼ」には、例えば、酵素SapI、EciI、BpiI、AarI、AloI、BaeI、BbvCI、PpiIおよびPsrI、BsrD1、BtsI、EarI、BmrI、BsaI、BsmBI、FauI、BbsI、BciVI、BfuAI、BspMI、BseRI、EciI、BtgZI、BpuEI、BsgI、MmeI、CspCI、BaeI、BsaMI、Mva1269I、PctI、Bse3DI、BseMI、Bst6I、Eam1104I、Ksp632I、BfiI、Bso31I、BspTNI、Eco31I、Esp3I、BfuI、Acc36I、AarI、Eco57I、Eco57MI、GsuI、AloI、Hin4I、PpiI、およびPsrIが含まれる。
【0111】
「半減期」とは、分子の活性、量または数の半分が消失するのにかかる時間を指す。
【0112】
以下、本発明を図面および実施例により詳しく説明するが、これらは単に例示であって、限定を示すものではない。これらの説明および実施例に基づいて当業者ならばさらなる実施例を利用することができるが、それらも同様に本発明の範囲内にある。
【図面の簡単な説明】
【0113】
【図1】図1:さらなるクローニングのため本発明に従って用いられる基本ベクター これらのベクターはRNAポリメラーゼ5’プロモーターの制御下でRNA転写が可能であり、ポリアデニルカセットを含む。
【0114】
【図2】図2:II型制限酵素(例えば、SpeI)とIIS型制限酵素(例えば、SapI)とによるベクター線状化の比較 その認識配列がポリ(A)配列の3’にあり、切断部位が24〜26 bp上流、従ってポリ(A)配列内にあるIIS型制限切断部位を導入することにより、プラスミドをポリ(A)配列内で線状化することができる。
【0115】
【図3】図3:in vitro転写のための鋳型として本発明に従って作製されるベクター 発現のレベルおよび持続時間に対する本発明のRNA修飾の効果を研究するため、いくつかのベクターを作製し、その後、in vitro転写のための鋳型として用いた。a.マスクされたポリ(A)配列とマスクされていないポリ(A)配列を有するベクター;b.長さの異なるポリ(A)配列を有するベクター;c.ヒトβ−グロビンの3’非翻訳領域を有するベクター;d.SIINFEKLおよびpp65ベクター;Cap−5’−キャッピング;eGFP−GFPレポーター遺伝子;β−グロビンの3’βg−3’非翻訳領域。A(x)−xは、ポリ(A)配列中のAヌクレオチドの数を表す。
【0116】
【図4】図4:示された表面マスクによる未成熟樹状細胞と成熟樹状細胞の成熟状態の決定 本発明のRNA修飾の効果をヒト樹状細胞(DC)にて、DC成熟プロセスを誘導する免疫刺激を用いて調べた。これらのDCを、特異的DC成熟マーカーを認識する抗CD80抗体、抗CD83抗体、抗CD86抗体および抗HLA−DR抗体で染色し、フローサイトメトリーで分析した。
【0117】
【図5】図5:翻訳効率および転写産物の安定性に対する遊離型およびマスク型のポリ(A)配列の影響 a.FACS−Kaliburにて平均蛍光強度[MFI]を測定することによる、K562細胞および樹状細胞におけるeGFP RNAの翻訳効率に対する遊離型およびマスク型のポリ(A)配列の影響。b.48時間後の未成熟樹状細胞におけるeGFP RNAの転写産物安定性に対する遊離型およびマスク型のポリ(A)配列の影響。腫瘍細胞系統および未成熟DCの双方で、オープンエンド型ポリ(A)配列を有するRNAはマスクエンド型ポリ(A)配列を有するRNAよりも有効かつ長期間翻訳される。DCにおける非マスクエンド型ポリ(A)配列の翻訳効率は、同じ長さのポリ(A)配列の1.5倍高い。さらにオープンエンド型ポリ(A)配列は、より高いRNA安定性をもたらす。
【0118】
【図6】図6:翻訳効率および転写産物の安定性に対するポリ(A)配列長の影響 a.K562細胞および樹状細胞におけるeGFP RNAの翻訳効率に対するポリ(A)配列長の影響。b.K562細胞および樹状細胞におけるd2eGFP RNAの翻訳効率に対するポリ(A)配列長の影響。c.エレクトロポレーション48時間後のK562細胞におけるeGFP RNAの転写産物安定性に対するポリ(A)配列長の影響。ポリ(A)配列を120Aヌクレオチドまで延長すると、転写産物の安定性および翻訳が高まる。これを超える延長に有効な効果は見られない。ポリ(A)配列を51〜120Aヌクレオチド延長すると、翻訳効率が1.5〜2倍高まる。この効果はRNAの安定性にも反映される。
【0119】
【図7】図7:未成熟および成熟DCにおける翻訳効率に対するヒトβ−グロビンの3’非翻訳領域(BgUTR)の影響 ヒトβ−グロビンの3’非翻訳領域を導入すると、RNA転写産物の発現が増強する。ヒトβ−グロビンの3’非翻訳領域が二重にあると、24時間後の発現レベルが高まり、このレベルは個々の2つのヒトβ−グロビンの3’非翻訳領域を合わせた効果を著しく超える。
【0120】
【図8】図8:未成熟および成熟DCにおける翻訳効率に対する本発明の修飾を組み合わせた効果 未成熟および成熟DCにおけるeGFPの翻訳効率は、本発明に記載のRNA転写産物修飾を組み合わせることで、5倍を上回る増強が可能である。
【0121】
【図9】図9:EL4細胞上のMHC分子によるペプチド提示に対する本発明の修飾を組み合わせた効果 本発明に従って修飾されたRNA構築物を用いると、高い翻訳効率のために細胞表面上でのペプチド−MHC複合体の提示が高まる。記載されているIVTベクターでは、eGFPをOVA257−264エピトープ(SIINFEKL)に置換し、EL4細胞(ネズミ、T細胞リンパ腫)をトランスフェクションの標的細胞として用いた。
【0122】
【図10】図10:本発明に従って安定化されたIVT RNA構築物を用いることによる、抗原特異的ペプチド/MHC複合体の増加 細胞にSec−SIINFEKL−A67−ACUAG RNAまたはSec−SIINFEKL−2BgUTR−A120 RNA(EL4細胞:10pmol、50pmol;C57B1/J6未成熟BMDC150pmol3反復)でエレクトロポレーションを行った。バッファーだけを用いるエレクトロポレーションを対照として用いた。細胞をSIINFEKL/Kb複合体に対する25D1.16抗体で染色した。SIINFEKLペプチド濃度を生細胞の平均蛍光値から、検量曲線としてペプチド滴定を用いて算出した。BMDCデータは3回の実験の平均値±SEMとして示す。
【0123】
【図11】図11:in vivoおよびin vitroでのT細胞刺激に対する、本発明に従って安定化されたIVT RNA構築物の効果 (A)安定化されたIVT RNA構築物の使用によるin vivoT細胞増殖の向上。1×105個のTCR−トランスジェニックCD8+ OT−I細胞をC57B1/J6マウスに養子免疫伝達した。C57B1/J6マウスのBMDCを50pmolのRNA(Sec−SIINFEKL−A67−ACUAG、Sec−SIINFEKL−2BgUTR−A120または対照RNA)でトランスフェクトし、ポリ(I:C)(50μg/ml)を用いて16時間成熟させ、T細胞移入の1日前に腹腔内注射した(n=3)。4日目に末梢血を採取し、SIINFEKL四量体陽性CD8+T細胞を染色した。ドットブロットはCD8+T細胞を表し、示されている数値は四量体陽性CD8+T細胞のパーセンテージを表す。 (B)安定化されたIVT RNA構築物を含むヒトT細胞のin vitro増殖の向上。HCMV血清反応陽性健常ドナーからのCD8+およびCD4+リンパ球をSec−pp65−A67−ACUAG RNA、Sec−pp65−2BGUTR−A120 RNA、または対照RNA(データは示されていない)でトランスフェクトされたか、あるいは陽性対照としてpp65ペプチドプール(1.75μg/ml)で刺激した自己DCと同時培養した。7日間増殖させた後、各エフェクター細胞集団(4×104/ウェル)を、pp65ペプチドプールか無関連のペプチドプール(1.75μg/ml)のいずれかを付加した自己DC(3×104/ウェル)を用い、IFN−γ ELISpotにてアッセイした。グラフは3回測定したスポットの平均値±SEMを表す。
【実施例1】
【0124】
実施例1:ベクターの作製とRNAのin vitro転写
発現のレベルおよび持続時間に対する本発明のRNA修飾の効果を検討するため、いくつかのIVTベクターを作製した後、in vitro転写のための鋳型として用いた(図3)。
【0125】
半減期(HL)の異なる2つの分子eGFPとd2eGFPのレポーター遺伝子をベクターに挿入し、これにより本発明のRNA修飾の影響を分析可能とした。蛍光は減少し、平均HLはeGFPでは17.3時間、d2eGFPでは2時間であった。これらの構築物をそれぞれin vitro転写eGFP RNAおよびd2eGFP RNAの作製に用いた。
【実施例2】
【0126】
実施例2:本発明に従って修飾されたin vitro転写RNAによる細胞のトランスフェクションおよびRNA翻訳と安定性に対する効果
in vitro転写eGFP RNAおよびd2eGFP RNAを用い、エレクトロポレーションによりK562細胞(ヒト白血病)をトランスフェクトした。トランスフェクション効率はK562細胞で90%を超えた。
【0127】
その後、免疫系の最も重要なモジュレーターであるヒト樹状細胞(DC)において、記載のRNA修飾の作用をアッセイした。このアプローチは、RNAでトランスフェクトされたDCが予防接種として意図され得ることから、免疫関連のものである。未成熟DCを皮膚および末梢器官に付加する。ここで、これらは、十分研究されている表面マーカーにより特徴付けられ、また、高いエンドサイトーシス活性により機能的に識別される未成熟状態にある。例えば病原体の感染などの免疫刺激はDCの成熟プロセスを誘導する。それと同時に、この刺激は局部リンパ節へのDCの移動を開始させるが、ここでDCはT細胞およびB細胞免疫応答の最も有効なインデューサーである。また、これらのDCの成熟状態は表面マーカー発現および詳細に研究されているサイトカインの発現によっても、また、DC形態の特徴によっても特徴付けられる。血液単球から未成熟ヒトDCを分化させるための培養系が確立されている。これらは種々の刺激により成熟が起こり得る。
【0128】
一次樹状細胞のトランスフェクション効率は70〜80%であった。これらのDCを、特異的DC成熟マーカーを認識する抗CD80抗体、抗CD83抗体、抗CD86抗体および抗HLA−DR抗体で染色し、フローサイトメトリーで分析した(図4)。
【0129】
発現のレベルおよび持続時間を、FACS−Kaliburを用い、eGFPの蛍光強度を測定することで測定した。細胞のRNA量は、定量的RT−PCRを用いて測定した。
【0130】
a.RNAの翻訳および安定性に対するオープンエンド型ポリ(A)配列の効果
腫瘍細胞系統K562と未成熟DC(iDC)は双方とも、オープンエンド型ポリ(A)配列を有するRNAを、マスクエンド型ポリ(A)配列を有するRNAよりも効率的にかつ長期間翻訳することが示された(図5a)。未成熟DCにおける非マスクエンド型ポリ(A)配列の翻訳効率は、同じ長さのポリ(A)配列では1.5倍高くなる。さらに、この修飾から、より高いRNA安定性が得られる(図5b)。非マスクポリ(A)配列を有するRNAでトランスフェクトされた未成熟DCでは、エレクトロポレーション48時間後に4〜5倍量のRNAを検出することができる。
【0131】
b.RNAの翻訳および安定性に対するポリ(A)配列長の効果
16bp、42bp、51bp、67bp、120bp、200bp、300bpおよび600bpの長さのポリ(A)配列を有するRNAを分析したところ、120Aヌクレオチドまでのポリ(A)配列の延長が転写産物の安定性と翻訳を高めること、また、それを超える延長には有効な効果がないことが明らかになった。この効果はK562細胞と未成熟DC(iDC)の双方に見られる(図6aおよび6b)。ポリ(A)配列を51〜120Aヌクレオチド延長すると、翻訳効率が1.5〜2倍高まる。この効果はRNAの安定性にも反映される(図6c)。
【0132】
c.RNAの翻訳および安定性に対する3’非翻訳領域の存在の効果
K562細胞および未成熟DCを用いた経時的推移により、ヒトβ−グロビンの3’非翻訳領域(UTR)の導入がRNA転写産物の発現の増強をもたらすことが確認された。さらに、ヒトβ−グロビンの3’非翻訳領域(UTR)が二重にあると、24時間後に高い発現レベルが得られ、個々の2つのUTRを合わせた効果を大きく超えることが実証された(図7)。
【0133】
d.RNAの翻訳および安定性に対する上記修飾の組合せの効果
本発明によれば、RNA転写産物における上記修飾の組合せは、未成熟DCまた成熟DCでも、eGFPの翻訳効率を5倍を超えて高めることが示された(図8)。
【実施例3】
【0134】
実施例3:安定性および翻訳効率の高められたin vitro転写RNAを介して発現されたペプチドの、MHC分子による提示
本発明によれば、本発明に従って修飾されたRNA構築物を用いると、細胞表面でのペプチド−MHCの提示が高まることが示された。この目的で、記載のベクター中のeGFPをコードする核酸配列を、OVA257−264エピトープ(SIINFEKL)をコードする核酸配列に置換し、これらの構築物を互いに比較した。トランスフェクションに用いた標的細胞はEL4細胞(マウス、T細胞リンパ腫)であった。
【0135】
MHC分子により提示されたSIINFEKLペプチドを定量するため、これらの細胞を、エレクトロポレーション後種々の時点で、抗H2−Kb OVA257−264抗体で染色し、二次抗体の蛍光強度を、FACS−Kaliburを用いて測定した(図9)。
【0136】
さらに、このSIINFEKLペプチドを、あらゆる最適化を反映したベクター(pST1−Sec−SIINFEKL−2BgUTR−A120−Sap1)に、また、標準的な特徴を有するベクター(pST1−Sec−SIINFEKL−A67−Spe1)にクローニングした。両ベクターに由来するIVT RNAで、EL4細胞およびBMDCに対してエレクトロポレーションを行った。OVA−ペプチド/Kb複合体は細胞表面実質的に多数見られ、本発明に従って修飾されたRNA、Sec−SIINFEKL−2−BgUTR−A120のエレクトロポレーション後、長期間維持された(図10)。
【実施例4】
【0137】
実施例4:抗原特異的T細胞の発現に対する、提示されるペプチドをコードするin vitro転写RNAによる細胞のトランスフェクションの効果
刺激能に対する効果を評価するため、SIINFEKLペプチドのMHCクラスI提示を検出するためのC57BL/J6(B6)バックグラウンドに集中的に用いられてきたOT−I−TCRを用いた。T細胞受容体(TCR)に関してトランスジェニックであり、ニワトリOVA由来のKb−特異的ペプチドSIINFEKL(OVA257−264)を認識するOT−I CD8+T細胞はH. Schild (Institute of Immunology, University of Mainz, Germany)から厚意により提供されたものである。
【0138】
0日目、OT−I−CD8+T細胞を用いて、動物に養子免疫伝達を行った。この目的で、TCR tg OT−Iマウスから脾細胞を調製し、C57BL/J6レシピエントマウスの尾の静脈に導入した。細胞数は1×105 TCR tg CD8+T細胞に調整した。翌日、50pmolのSIINFEKLコードRNA構築物変異体でエレクトロポレーションを行い、ポリ(I:C)で16時間成熟させたC57BL/J6マウスの1×106 BMDCをマウスに腹腔内投与した。4日目、四量体法を用い、末梢血においてOT−I−CD8+T細胞を測定した。この目的で、眼窩後方の血液サンプルを採取し、抗CD8(Caltag Laboratories, Burlingame, USA)およびSIINFEKL四量体(H−2Kb/SIINFEKL 257−264; Beckman Coulter, Fullerton, USA)で染色した。
【0139】
抗原特異的TCR−トランスジェニックCD8+T細胞のin vivo増殖は、抗原供給源としてSec−SIINFEKL−2BgUTR−A120 RNAを用いた場合には、Sec−SIINFEKL−A67−ACUAG RNAの場合と比べて実質的に向上することが分かった(図11A)。
【0140】
抗原供給源として安定されたIVT RNA構築物が、ヒトT細胞の抗原特異的刺激も向上させるかどうか評価するため、ポリエピトープ性のT細胞反応の自己刺激をバリデートするために用いられることが多いヒトサイトメガロウイルスの免疫優性抗原であるHCMV−pp65を用いた。抗体をコーティングしたマイクロビーズ(Miltenyi Biotec, Bergisch−Gladbach, Germany)を用いた正磁場セルソーティングにより、HCMV血清反応陽性健常ドナーから精製されたCD4+T細胞およびCD8+T細胞を、pp65をコードする対応するIVT RNA変異体でマイクロポレーションを行った自己DC2×105個とともに同時培養した。T細胞の増殖を、全pp65タンパク質配列にわたる重複ペプチドのプールで、または対照タンパク質で刺激した自己DCを用いたIFN−γ−ELISpotで7日目に測定したところ、Sec−pp65−2BgUTR−A120が優れていることが実証され、CD4+T細胞の増殖に対する効果が最も顕著であった(図11B)。
【0141】
引用文献
Bargmann, C.I., Hung, M.C., and Weinberg, R.A. (1986). The neu oncogene encodes an epidermal growth factor receptor−related protein. Nature 319, 226−230.
Boczkowski, D., Nair, S.K., Nam, J.H., Lyerly H.K., and Gilboa, E. (2000). Induction of tumor immunity and cytotoxic T lymphocyte responses using dendritic cells transfected with messenger RNA amplified from tumor cells. Cancer Res. 60, 1028−1034.
Carralot, J.P., Probst, J., Hoerr, I., Scheel, B., Teufel, R., Jung, G., Rammensee, H.G., and Pascolo, S. (2004). Polarization of immunity induced by direct injection of naked sequence−stabilized mRNA vaccines. Cell Mol. Life Sci. 61, 2418−2424.
Condon, C., Watkins, S.C., Celluzzi, C.M., Thompson, K., and Falo, L.D., Jr. (1996). DNA−based immunization by in vivo transfections of dendritic cells. Nat. Med. 2, 1122−1128.
Conry, R.M., LoBuglio, A.F., Kantor, J., Schlom, J., Loechel, F., Moore, S.E., Sumerel, L.A., Barlow, D.L., Abrams, S., and Curiel, D.T. (1994). Immune response to a carcinoembryonic antigen polynucleotide vaccine. Cancer Res. 54, 1164−1168.
Conry, R.M., LoBuglio, A.F., Loechel, F., Moore, S.E., Sumerel, L.A., Barlow, D.L., and Curiel, D.T. (1995a). A carcinoembryonic antigen polynucleotide vaccine has in vivo antitumor activity. Gene Ther. 2, 59−65.
Conry, R.M., LoBuglio, A.F., Wright, M., Sumerel, L., Pike, M.J., Johanning, F., Benjamin, R., Lu, D., and Curiel, D.T. (1995b). Characterization of a messenger RNA polynucleotide vaccine vector. Cancer Res. 55, 1397−1400.
Cox, G.J., Zamb, T.J., and Babiuk, L.A. (1993). Bovine herpesvirus 1: immune responses in mice and cattle injected with plasmid DNA. J. Virol. 67, 5664−5667.
Davis, H.L., Michel, M.L., and Whalen, R.G. (1993). DNA−based immunization induces continuous secretion of hepatitis B surface antigen and high levels of circulating antibody. Hum. Mol. Genet. 2, 1847−1851.
Gallie, D.R. (1991). The cap and poly(A) tail function synergistically to regulate mRNA translational efficiency. Genes Dev. 5, 2108−2116.
Gilkeson, G.S., Pippen, A.M., and Pisetsky, D.S. (1995). Induction of cross−reactive anti−dsDNA antibodies in preautoimmune NZB/NZW mice by immunization with bacterial DNA. J. Clin. Invest. 95, 1398−1402.
Greenblatt, M.S., Bennett, W.P., Hollstein, M., and Harris, C.C. (1994). Mutations in the p53 tumor suppressor gene: clues to cancer etiology and molecular pathogenesis. Cancer Res. 54, 4855−4878.
Heiser, A., Coleman, D., Dannull, J., Yancey, D., Maurice, M.A., Lallas, C.D., Dahm, P., Niedzwiecki, D., Gilboa, E., and Vieweg, J. (2002). Autologous dendritic cells transfected with prostate−specific antigen RNA stimulate CTL responses against metastatic prostate tumors. J. Clin. Invest 109, 409−417.
Heiser, A., Dahm, P., Yancey, D.R., Maurice, M.A., Boczkowski, D., Nair, S.K., Gilboa, E., and Vieweg, J. (2000). Human dendritic cells transfected with RNA encoding prostate−specific antigen stimulate prostate−specific CTL responses in vitro. J. Immunol. 164, 5508−5514.
Hoerr, I., Obst, R., Rammensee, H.G., and Jung, G. (2000). In vivo application of RNA leads to induction of specific cytotoxic T lymphocytes and antibodies. Eur. J. Immunol. 30, 1−7.
Malone, R.W., Felgner, P.L., and Verma, I.M. (1989). Cationic liposome−mediated RNA transfection. Proc. Natl. Acad. Sci. USA 86, 6077−6081.
Preiss, T. and Hentze, M.W. (1998). Dual function of the messenger RNA cap structure in poly(A)−tail promoted translation in yeast. Nature 392, 516−520.
Spooner, R.A., Deonarain, M.P., and Epenetos, A.A. (1995). DNA vaccination for cancer treatment. Gene Ther. 2, 173−180.
Strong, T.V., Hampton, T.A., Louro, I., Bilbao, G., Conry, R.M., and Curiel, D.T. (1997). Incorporation of beta−globin untranslated regions into a Sindbis virus vector for augmentation of heterologous mRNA expression. Gene Ther. 4, 624−627.
Su, Z., Dannull, J., Heiser, A., Yancey, D., Pruitt, S., Madden, J., Coleman, D., Niedzwiecki, D., Gilboa, E., and Vieweg, J. (2003). Immunological and clinical responses in metastatic renal cancer patients vaccinated with tumor RNA−transfected dendritic cells. Cancer Res. 63, 2127−2133.
Tang, D.C., DeVit, M., and Johnston, S.A. (1992). Genetic immunization is a simple method for eliciting an immune response. Nature 356, 152−154.
Teufel, R., Carralot, J.P., Scheel, B., Probst, J., Walter, S., Jung, G., Hoerr, I., Rammensee, H.G., and Pascolo, S. (2005). Human peripheral blood monuclear cells transfected with messenger RNA stimulate antigen−specific cytotoxic T−lymphocytes in vitro. Cell Mol. Life Sci. 62, 1755−1762.
Ulmer, J.B., Donnelly, J.J., Parker, S.E., Rhodes, G.H., Felgner, P.L., Dwarki, V.J., Gromskowski, S.H., Deck, R.R., DeWitt, C.M., Friedman, A., et al. (1993). Heterologous protection against influenza by injection of DNA encoding a viral protein. Science 259, 1745−1749.
Wang, B., Merva, M., Dang, K., Ugen, K.E., Williams, W.V., and Weiner, D.B. (1995). Immunization by direct DNA inoculation induces rejection of tumor cell challenge. Hum. Gene Ther. 6, 407−418.
Wang, B., Ugen, K.E., Srikantan, V., Agadjanyan, M.G., Dang, K., Refaeli, Y., Sato, A.I., Boyer, J., Williams, W.V., and Weiner, D.B. (1993). Gene inoculation generates immune responses against human immunodeficiency virus type 1. Proc. Natl. Acad. Sci. USA 90, 4156−4160.
Wolff, J.A., Malone, R.W., Williams, P., Chong, W., Acsadi, G., Jani, A., and Felgner, P.L. (1990). Direct gene transfer into mouse muscle in vivo. Science 247, 1465−1468.
Ying, H., Zaks, T.Z., Wang, R.F., Irvine, K.R., Kammula, U.S., Marincola, F.M., Leitner, W.W., and Restifo, N.P. (1999). Cancer therapy using a self−replicating RNA vaccine. Nat. Med. 5, 823−827.
【技術分野】
【0001】
弱毒または不活性化された病原体を含む従来のワクチンは多くの分野で有効であるものの、いくつかの感染性病原体や腫瘍に対しては有効な防御免疫を付与しない。これには有効で、用途が広く、生産が容易かつコスト効果があり、保存が容易なワクチンが必要である。
【背景技術】
【0002】
プラスミドDNAを直接筋肉注射した後では、コードされている遺伝子の細胞表面での発現が持続することが示されており(Wolff et al., 1990)、DNAに基づくワクチンが新たな有望な免疫戦略としてみなされている。これは核酸に基づくワクチンを開発するための重要な動機となった。当初、感染性病原体に対するDNAに基づくワクチンが試験されたが(Cox et al., 1993; Davis et al., 1993; Ulmer et al., 1993; Wang et al., 1993)、すぐに、特異的抗腫瘍免疫を誘導することを目的とする対腫瘍遺伝子療法においてさらに詳細に研究されるようになった(Conry et al., 1994; Conry et al., 1995a; Spooner et al., 1995; Wang et al., 1995)。この腫瘍免疫の戦略には重要な利点がいくつかある。核酸に基づくワクチンは製造が容易で、比較的安価である。さらに、この核酸に基づくワクチンは少数の細胞から増幅させることができる。
【0003】
DNAはRNAよりも安定であるが、抗DNA抗体の誘導(Gilkeson et al., 1995)および宿主ゲノムへの導入遺伝子(transgen)の組込みなど、安全性に対する潜在的なリスクがいくつかある。これは細胞の遺伝子を不活性化し、該導入遺伝子(transgen)の制御できない長期的発現または発癌を招き、従って、例えばerb−B2(Bargmann et al., 1986)やp53(Greenblatt et al., 1994)などの発癌能を有する腫瘍関連抗原には通常適用できない。RNAの使用はこれらの潜在的リスクを回避するための魅力的な代替法となる。
【0004】
可逆的な遺伝子療法の一種としてRNAを用いることの利点には、一時的発現および非形質転換特性が含まれる。RNAはトランスジェニック的に発現させるために核に入る必要がなく、もっと言えば、宿主ゲノムに組み込まれることはなく、それにより発癌のリスクがなくなる。DNAを用いる場合 (Condon et al., 1996; Tang et al., 1992)と同様、RNAの注射もまた、in vivoで細胞性免疫応答と体液性免疫応答の双方を誘導し得る(Hoerr et al., 2000; Ying et al., 1999)。
【0005】
in vitro転写されたRNA(IVT−RNA)による免疫療法は、双方とも種々の動物モデルにおいて引き続いて試験されている2つの異なる戦略の使用に至らせる。RNAは種々の免疫経路を介して直接注入される(Hoerr et al., 2000)か、または樹状細胞(DC)がリポフェクションもしくはエレクトロポレーションにより、in vitro転写されたRNAでトランスフェクトされ、その後投与される(Heiser et al., 2000)。最近発表された研究では、RNAでトランスフェクトされたDCによる免疫化は、in vitroおよびin vivoで抗原特異的細胞傷害性Tリンパ球(CTL)を誘導ことが示されている(Su et al., 2003; Heiser et al., 2002)。T細胞介在性の免疫応答の最適な誘導の中枢的に重要な因子はとりわけ用量、すなわち、DCに対する抗原提示の密度である。導入されたIVT−RNAの長期発現を達成するためには、種々の修飾によりIVT−RNAを安定化させる、それにより、DCに対する抗原提示を高めることが試みられている。翻訳の基本要件は、その翻訳効率がポリ(A)の長さに相関する3’ポリ(A)配列が存在することである(Preiss and Hentze, 1998)。5’キャップおよび3’ポリ(A)配列は相乗作用的にin vivo翻訳を活性化する(Gallie, 1991)。グロビン遺伝子の非翻訳領域(UTR)は、RNAの安定化および翻訳効率の増強に寄与し得る他の公知のエレメントである(Malone et al., 1989)。
【0006】
いくつかのIVTベクターは文献で知られ、in vitro転写の鋳型として標準的な方法で使用され、安定化されたRNA転写産物が生成されるように遺伝的に修飾されている。これまでに文献に記載されているプロトコール(Conry et al., 1995b; Teufel et al., 2005; Strong et al., 1997; Carralot et al., 2004; Boczkowski et al., 2000)は、次の構造:RNA転写を可能とする5’RNAポリメラーゼプロモーター、それに続く、3’および/または5’のいずれかで非翻訳領域(UTR)にフランキングされた目的遺伝子、および50〜70Aヌクレオチドを含む3’ポリアデニルカセットを有するプラスミドベクターに基づく。in vitro転写前に、この環状プラスミドは、II型制限酵素(認識配列は切断部位に相当する)によりポリアデニルカセットの下流で線状化される。よって、このポリアデニルカセットは、その転写産物において後期ポリ(A)配列に相当する。この手順の結果として、いくつかのヌクレオチドが線状化後に酵素切断部位の一部として残り、その3’末端においてポリ(A)配列を延長するか、またはマスクする。この非生理学的オーバーハングがこのような構築物から細胞内で産生されるタンパク質の量に影響を及ぼすかどうかは明らかでない。
【0007】
よって、RNAは臨床適用に特に好適であると思われる。しかしながら、遺伝子療法におけるRNAの利用は、特に細胞質ではRNAの短い半減期により著しく制限され、その結果、タンパク質発現は低くなる。
【発明の概要】
【発明が解決しようとする課題】
【0008】
本発明の目的は、安定性および翻訳効率が向上されたRNAおよびこのようなRNAを得るための手段を提供することである。該RNAを遺伝子療法アプローチに用いることで、高いグレードの発現が得られるに違いない。
【0009】
この目的は特許請求の範囲の主題により本発明に従い達成される。
【課題を解決するための手段】
【0010】
本発明は、RNA、特にmRNAの安定化およびmRNA翻訳の増強に関する。本発明は特に、転写産物の安定性および翻訳効率の向上をもたらすRNA、特にin vitro転写されたRNAの3つの修飾に関する。
【発明の効果】
【0011】
本発明によれば、オープンエンド型ポリ(A)配列を有するRNAは、末端がマスクされたポリ(A)配列を有するRNAより効率的に翻訳されることが分かった。特に約120 bpの長いポリ(A)配列は最適なRNA転写産物の安定性と翻訳効率をもたらすことが分かった。本発明はまた、RNA分子において、特にヒトβ−グロビン遺伝子の二重の3’非翻訳領域(UTR)が、個々の2つのUTRを用いた場合に予測されるトータル効果を明らかに超えるほどに翻訳効率を向上させることが実証された。本発明によれば、上記の修飾の組合せは、RNAの安定化および翻訳の増強に相乗作用を有することが分かった。
【0012】
転写産物の量およびタンパク質収量を測定するための定量的RT−PCRおよびeGFP変異体を用い、本発明は、本発明のRNA修飾が樹状細胞(DC)のトランスフェクションにおいて独立にRNAの安定性と翻訳効率増強することを実証した。よって、トランスフェクトされた細胞上での抗原特異的ペプチド/MHC複合体の密度ならびに抗原特異的CD4+およびCD8+T細胞を刺激および拡大するそれらの能力を増強することができた。よって、本発明は、本発明に記載のRNA修飾により修飾されたRNAを用いることでRNAトランスフェクトDCワクチンを至適化するための戦略に関する。
【0013】
本発明によれば、修飾、およびそれによるRNAの安定化および/または翻訳効率の向上は好ましくは、in vitroにおいてRNA転写の鋳型として好ましく働く発現ベクターを遺伝的に修飾することにより達成される。
【0014】
この種のベクターは特に、好ましくは該RNAにオープンエンドを有する、すなわち、Aヌクレオチド以外のヌクレオチドが該ポリ(A)配列の3’末端にフランキングしていないポリ(A)配列を有するRNAを転写させることを意図する。該RNAにおけるオープンエンド型ポリ(A)配列は、5’RNAポリメラーゼプロモーターの制御下でRNAを転写させ、ポリアデニルカセット(ポリ(A)配列)を含む発現ベクターにIIS型制限切断部位を導入することにより達成することができ、この認識配列はポリ(A)配列の3’に位置し、切断部位は上流、従ってポリ(A)配列内に位置する。このIIS型制限切断部位における制限切断により、プラスミドをポリ(A)配列内で線状化することができる(図2)。その後、線状化プラスミドをin vitro転写のための鋳型として用い、マスクされていないポリ(A)配列で終わる転写産物を得ることができる。
【0015】
その上、またはその代わりに、修飾、ならびにそれによるRNAの安定化および/または翻訳効率の向上は、本発明に従い、その3’末端に、好ましくはペプチドまたはタンパク質をコードする配列(オープンリーディングフレーム)とポリ(A)配列との間に2以上の3’非翻訳領域を有するRNAを転写させるように発現ベクターを遺伝的に修飾することで達成することができる。
【0016】
一態様では、本発明は、5’→3’転写方向に、(a)プロモーター;(b)転写可能な核酸配列または転写可能な核酸配列を導入するための核酸配列;(c−1)遺伝子の3’非翻訳領域に相当する、またはそれに由来する第一の核酸配列;および(c−2)遺伝子の3’非翻訳領域に相当する、またはそれに由来する第二の核酸配列を含む核酸分子に関する。
【0017】
一実施形態では、本発明の核酸分子は、(c−3)遺伝子の3’ 非翻訳領域に相当する、またはそれに由来する少なくとも1つのさらなる核酸配列をさらに含む。
【0018】
本発明の核酸分子では、プロモーター(a)の制御下の核酸配列(b)、(c−1)、(c−2)および適当であれば(c−3)は転写されて、核酸配列(c−1)および/または(c−2)および/または適当であれば(c−3)から転写された核酸配列に、好ましくは転写可能な核酸配列(b)から転写された核酸配列の翻訳効率および/または安定性を高めるような活性がある共通転写産物を生じ得ることが好ましい。
【0019】
核酸配列(c−1)、(c−2)および適当であれば(c−3)は同一であっても異なっていてもよい。
【0020】
一実施形態では、該核酸配列は、(d)プロモーター(a)の制御下で転写された際に、その転写産物に少なくとも20の連続するAヌクレオチドのヌクレオチド配列をコードする核酸配列をさらに含む。
【0021】
プロモーター(a)の制御下の核酸配列(b)、(c−1)、(c−2)、適当であれば(c−3)および(d)は転写されて、核酸配列(c−1)および/または(c−2)および/または適当であれば(c−3)および/または(d)から転写された核酸配列に、好ましくは転写可能な核酸配列(b)から転写された核酸配列の翻訳効率および/または安定性を高めるような活性がある共通転写産物を生じ得ることが好ましい。
【0022】
特定の実施形態では、核酸配列(d)は、プロモーター(a)の制御下で転写された際に、その転写産物に少なくとも40、好ましくは少なくとも80、好ましくは少なくとも100、特に約120の連続するAヌクレオチドのヌクレオチド配列をコードする。核酸配列(d)は、プロモーター(a)の制御下で転写された際に、その転写産物に500まで、好ましくは400まで、好ましくは300まで、好ましくは200まで、特に150までの連続するAヌクレオチドのヌクレオチド配列をコードするのが好ましい。
【0023】
一実施形態では、該核酸分子は、切断が5’→3’転写方向に、プロモーター(a)、核酸配列(b)、核酸配列(c−1)、(c−2)および適当であれば(c−3)、ならびに核酸配列(d)の少なくとも一部(この核酸配列(d)の少なくとも一部は、プロモーター(a)の制御下で転写された際に、その転写産物に少なくとも20の連続するAヌクレオチドのヌクレオチド配列をコードし、その転写産物において3’末端ヌクレオチドは少なくとも20の連続するAヌクレオチドの該ヌクレオチド配列のAヌクレオチドである)を含む核酸分子を生じるように、核酸配列(d)内で、好ましくは酵素的にまたは別の生化学的方法で切断され得ることを特徴とする。
【0024】
好ましくは、切断後、該核酸分子は、少なくとも20の連続するAヌクレオチドのヌクレオチド配列の鋳型として働く鎖の末端に、その転写産物において少なくとも20の連続するAヌクレオチドのヌクレオチド配列の鋳型として働くヌクレオチド配列の一部であるTヌクレオチドを有する。
【0025】
特定の実施形態では、核酸配列(d)の少なくとも一部は、プロモーター(a)の制御下で転写された際に、その転写産物に少なくとも40、好ましくは少なくとも80、好ましくは少なくとも100、特に約120の連続するAヌクレオチドのヌクレオチド配列をコードする。核酸配列(d)は、プロモーター(a)の制御下で転写された際に、その転写産物に500まで、好ましくは400まで、好ましくは300まで、好ましくは200まで、特に150までの連続するAヌクレオチドのヌクレオチド配列をコードするのが好ましい。
【0026】
本発明の核酸分子は好ましくは、切断前には閉環分子であり、切断後には直鎖分子である。
【0027】
好ましくはIIS型制限エンドヌクレアーゼの制限切断部位である制限切断部位の助けで切断が行われるのが好ましい。
【0028】
一実施形態では、IIS型制限エンドヌクレアーゼの認識配列は核酸配列(d)の3’末端の5〜26塩基対、好ましくは24〜26塩基対下流に位置する。
【0029】
好ましい実施形態では、核酸配列(c−1)、(c−2)および適当であれば(c−3)は互いに独立にα2−グロビン、α1−グロビン、β−グロビンおよび成長ホルモン、好ましくは、ヒトβ−グロビンなどのグロビン遺伝子からなる群から選択される遺伝子に由来し、特に好ましい実施形態では、配列表の配列番号1の核酸配列またはそれに由来する核酸配列に相当する。
【0030】
さらなる態様において、本発明は、5’→3’転写方向に、(a)プロモーター;(b)転写可能な核酸配列または転写可能な核酸配列を導入するための核酸配列;および(c)プロモーター(a)の制御下で転写された際に、その転写産物に少なくとも20の連続するAヌクレオチドのヌクレオチド配列をコードする核酸配列を含む核酸分子に関する。
【0031】
プロモーター(a)の制御下の核酸配列(b)および(c)は転写されて、核酸配列(c)から転写された核酸配列に、好ましくは転写可能な核酸配列(b)から転写された核酸配列の翻訳効率および/または安定性を高めるような活性がある共通転写産物を生じ得ることが好ましい。
【0032】
特定の実施形態では、核酸配列(c)は、プロモーター(a)の制御下で転写された際に、その転写産物に少なくとも40、好ましくは少なくとも80、好ましくは少なくとも100、特に約120の連続するAヌクレオチドのヌクレオチド配列をコードする。核酸配列(c)は、プロモーター(a)の制御下で転写された際に、その転写産物に500まで、好ましくは400まで、好ましくは300まで、好ましくは200まで、特に150までの連続するAヌクレオチドのヌクレオチド配列をコードするのが好ましい。
【0033】
一実施形態では、該核酸分子は、切断が5’→3’転写方向に、プロモーター(a)、核酸配列(b)、および核酸配列(c)の少なくとも一部(この核酸配列(c)の少なくとも一部は、プロモーター(a)の制御下で転写された際に、その転写産物に少なくとも20の連続するAヌクレオチドのヌクレオチド配列をコードし、その転写産物において3’末端ヌクレオチドは少なくとも20の連続するAヌクレオチドの該ヌクレオチド配列のAヌクレオチドである)を含む核酸分子を生じるように、核酸配列(c)内で、好ましくは酵素的にまたは別の生化学的方法で切断され得る。
【0034】
好ましくは、切断後、該核酸分子は、少なくとも20の連続するAヌクレオチドのヌクレオチド配列の鋳型として働く鎖の末端に、その転写産物において少なくとも20の連続するAヌクレオチドのヌクレオチド配列の鋳型として働くヌクレオチド配列の一部であるTヌクレオチドを有する。
【0035】
特定の実施形態では、核酸配列(c)の少なくとも一部は、プロモーター(a)の制御下で転写された際に、その転写産物に少なくとも40、好ましくは少なくとも80、好ましくは少なくとも100、特に約120の連続するAヌクレオチドのヌクレオチド配列をコードする。核酸配列(c)は、プロモーター(a)の制御下で転写された際に、その転写産物に500まで、好ましくは400まで、好ましくは300まで、好ましくは200まで、特に150までの連続するAヌクレオチドのヌクレオチド配列をコードするのが好ましい。
【0036】
該核酸分子は好ましくは、切断前には閉環分子であり、切断後には直鎖分子である。
【0037】
好ましくはIIS型制限エンドヌクレアーゼの制限切断部位である制限切断部位の助けで切断が行われるのが好ましい。
【0038】
一実施形態では、IIS型制限エンドヌクレアーゼの認識配列は核酸配列(c)の3’末端の5〜26塩基対、好ましくは24〜26塩基対下流に位置する。
【0039】
本発明の1つの核酸分子では、転写可能な核酸配列は好ましくはペプチドまたはタンパク質をコードする核酸配列を含み、転写可能な核酸配列を導入するための核酸配列は好ましくは多重クローニング部位である。
【0040】
本発明の核酸分子は、(i)レポーター遺伝子;(ii)選択マーカー;および(iii)複製起点からなる群から選択される1以上のメンバーをさらに含み得る。
【0041】
一実施形態では、本発明の核酸分子は閉環コンフォメーションであり、特に線状化後に、RNA、特にmRNAのin vitro転写に好適であるのが好ましい。
【0042】
さらなる態様では、本発明は、上記の核酸分子の線状化により、好ましくは少なくとも20の連続するAヌクレオチドのヌクレオチド配列をコードする核酸配列内での切断により得られる核酸分子、ならびにプロモーター(a)の制御下、上記の核酸分子との転写、好ましくはin vitro転写により得られるRNAに関する。
【0043】
さらなる態様では、本発明は、その安定性および/または翻訳効率を高めるよう選択されたRNA分子をin vitroで転写させる方法であって、(i)遺伝子の3’非翻訳領域に相当する、またはそれに由来する第一の核酸配列(b−1)を、転写されて該RNA分子を生じ得る核酸配列(a)の3’末端に結合させること、(ii)遺伝子の3’非翻訳領域に相当する、またはそれに由来する第二の核酸配列(b−2)を、該第一の核酸配列(b−1)の3’末端に結合させること、および(iii)得られた核酸をin vitroで転写させることを含む方法に関する。
【0044】
さらなる態様において、本発明は、その発現を増強するよう選択されたmRNA分子を転写させる方法であって、(i)遺伝子の3’非翻訳領域に相当する、またはそれに由来する第一の核酸配列(b−1)を、転写されて該RNA分子を生じ得る核酸配列(a)の3’末端に結合させること、(ii)遺伝子の3’非翻訳領域に相当する、またはそれに由来する第二の核酸配列(b−2)を、該第一の核酸配列(b−1)の3’末端に結合させること、および(iii)得られた核酸を転写させることにより得ることができるmRNAを翻訳することを含む方法に関する。転写は好ましくはin vitroで行われる。
【0045】
本発明おいて「核酸配列を核酸配列の3’末端に結合させる」とは、第一の核酸配列が第二の核酸配列の下流になり、付加的核酸配列により後者から離されてもよいような、2つの核酸配列の共有結合に関していう。
【0046】
一実施形態では、本発明の方法は、遺伝子の3’非翻訳領域に相当する、またはそれに由来する少なくとも1つのさらなる核酸配列(b−3)を第二の核酸配列(b−2)の3’末端に結合させることをさらに含む。
【0047】
核酸配列(a)、(b−1)、(b−2)および適当であれば(b−3)は転写されて、核酸配列(b−1)および/または(b−2)および/または適当であれば(b−3)から転写された核酸配列に、好ましくは核酸配列(a)により転写された核酸配列の翻訳効率および/または安定性を高めるような活性がある共通転写産物を生じ得ることが好ましい。
【0048】
さらなる実施形態では、本発明の方法は、転写された際に、少なくとも20の連続するAヌクレオチドのヌクレオチド配列をコードする核酸配列(c)を核酸配列(b−2)の、または適当であれば核酸配列(b−3)の3’末端に結合させることをさらに含む。
【0049】
核酸配列(a)、(b−1)、(b−2)および適当であれば(b−3)ならびに(c)は転写されて、核酸配列(b−1)および/または(b−2)および/または適当であれば(b−3)および/または(c)から転写された核酸配列に、好ましくは核酸配列(a)から転写された核酸配列の翻訳効率および/または安定性を高めるような活性がある共通転写産物を生じ得ることが好ましい。
【0050】
特定の実施形態では、核酸配列(c)は、その転写産物に少なくとも40、好ましくは少なくとも80、好ましくは少なくとも100、特に約120の連続するAヌクレオチドのヌクレオチド配列をコードする。核酸配列(c)は、転写された際に、その転写産物に500まで、好ましくは400まで、好ましくは300まで、好ましくは200まで、特に150までの連続するAヌクレオチドのヌクレオチド配列をコードするのが好ましい。
【0051】
特定の実施形態では、本発明の方法は、得られた核酸の転写の前に、このようにして得られた核酸の転写が、核酸配列(a)、(b−1)、(b−2)および適当であれば(b−3)ならびに少なくとも20の連続するAヌクレオチドの3’末端ヌクレオチド配列から転写された核酸配列を有する転写産物(該転写産物の3’末端ヌクレオチドは少なくとも20の連続するAヌクレオチドのヌクレオチド配列のAヌクレオチドである)を生じるように核酸配列(c)内で切断することをさらに含む。
【0052】
特定の実施形態では、該転写産物は、その3’末端に少なくとも40、好ましくは少なくとも80、好ましくは少なくとも100、特に約120の連続するAヌクレオチドのヌクレオチド配列を有する。該転写産物は、その3’末端に500まで、好ましくは400まで、好ましくは300まで、好ましくは200まで、特に150までの連続するAヌクレオチドのヌクレオチド配列を有するのが好ましい。
【0053】
好ましい実施形態では、核酸配列(b−1)、(b−2)および適当であれば(b−3)は互いに独立にα2−グロビン、α1−グロビン、β−グロビンおよび成長ホルモン、好ましくはヒトβ−グロビンなどのグロビン遺伝子からなる群から選択される遺伝子に由来し、特に好ましい実施形態では、配列表の配列番号1の核酸配列またはそれに由来する核酸配列に相当する。
【0054】
さらなる態様において、本発明は、その安定性および/または翻訳効率を高めるよう選択されたRNA分子をin vitroで転写させる方法であって、(i)転写された際に、少なくとも20の連続するAヌクレオチドのヌクレオチド配列をコードする核酸配列(b)を、転写されて前記RNA分子を生じ得る核酸配列(a)の3’末端に結合させること、および(ii)得られた核酸をin vitroで転写させることを含む方法に関する。
【0055】
さらなる態様において、本発明は、その発現を増強するよう選択されたmRNA分子を翻訳する方法であって、(i)転写された際に、少なくとも20の連続するAヌクレオチドのヌクレオチド配列をコードする核酸配列(b)を、転写されて前記mRNA分子を生じ得る核酸配列(a)の3’末端に結合させること、および(ii)得られた核酸を転写させることにより得ることができるmRNAを翻訳することを含む方法に関する。転写はin vitroで行われるのが好ましい。
【0056】
核酸配列(a)および(b)は転写されて、核酸配列(b)から転写された核酸配列に、好ましくは核酸配列(a)から転写された核酸配列の翻訳効率および/または安定性を高めるような活性がある共通転写産物を生じ得ることが好ましい。
【0057】
特定の実施形態では、核酸配列(b)は、その転写産物に少なくとも40、好ましくは少なくとも80、好ましくは少なくとも100、特に約120の連続するAヌクレオチドのヌクレオチド配列をコードする。核酸配列(b)は、転写された際に、その転写産物に500まで、好ましくは400まで、好ましくは300まで、好ましくは200まで、特に150までの連続するAヌクレオチドのヌクレオチド配列をコードするのが好ましい。
【0058】
特定の実施形態では、本発明の方法は、得られた核酸の転写の前に、このようにして得られた核酸の転写が、核酸配列(a)および少なくとも20の連続するAヌクレオチドの3’末端ヌクレオチド配列から転写された核酸配列を有する転写産物(該転写産物の3’末端ヌクレオチドは少なくとも20の連続するAヌクレオチドのヌクレオチド配列のAヌクレオチドである)を生じるように核酸配列(b)内で切断することをさらに含む。
【0059】
特定の実施形態では、該転写産物は、その3’末端に少なくとも40、好ましくは少なくとも80、好ましくは少なくとも100、特に約120の連続するAヌクレオチドを有する。該転写産物は、その3’末端に500まで、好ましくは400まで、好ましくは300まで、好ましくは200まで、特に150までの連続するAヌクレオチドを有するのが好ましい。
【0060】
本発明の方法の全ての態様において、切断は、好ましくはIIS型制限エンドヌクレアーゼの制限切断部位である制限切断部位の助けで行われるのが好ましい。
【0061】
一実施形態では、IIS型制限エンドヌクレアーゼの認識配列は、転写された際に、少なくとも20の連続するAヌクレオチドのヌクレオチド配列をコードする核酸配列の3’末端の5〜26塩基対、好ましくは24〜26塩基対下流に位置する。
【0062】
本発明はまた、選択されたRNA分子をin vitroで転写させる本発明の方法により得ることができるRNAに関する。本発明の核酸分子から、選択されたRNA分子をin vitroで転写させる本発明の方法により得ることができる、鋳型としてのRNA調製物は、そのRNAのポリ(A)配列の長さに関して好ましくは均質または本質的に均質であり、すなわち、このポリ(A)配列の長さは、10個だけ、好ましくは5個、4個、3個、2個または1個のAヌクレオチドだけ異なるRNA分子が調製物中に90%以下、好ましくは95%以下、好ましくは98%または99%以下である。
【0063】
本発明は例えば、細胞内での転写および発現において組換えタンパク質の発現を増強するために利用可能である。より具体的には、組換えタンパク質を産生する場合、本発明に記載の修飾およびその組合せを発現ベクターに導入し、細胞系において組換え核酸の転写および組換えタンパク質の発現を増強する目的でそれらを利用することができる。これには例えば、組換え抗体、ホルモン、サイトカインおよび酵素などの生産が含まれる。これによりとりわけ生産コストの低減が可能となる。
【0064】
また、遺伝子療法適用で、本発明に記載の修飾およびその組合せを利用することもできる。該修飾は遺伝子療法ベクターに導入し、それにより導入遺伝子(transgen)の発現を増強させるために利用することができる。この目的で、いずれの核酸(DNA/RNA)に基づくベクター系(例えば、プラスミド、アデノウイルス、ポックスウイルスベクター、インフルエンザウイルスベクター、およびαウイルスベクターなど)も使用可能である。例えばリンパ球または樹状細胞では、in vitroにて細胞をこれらのベクターでトランスフェクトすることができ、あるいは直接in vivoに投与することもできる。
【0065】
さらに、本発明に記載の修飾およびその組合せは、リボ核酸の安定性および/または発現を、そしてそれにより該リボ核酸によりコードされるペプチドまたはタンパク質の量を増強させることができる。コードリボ核酸は例えば遺伝子の一時的発現ために利用可能であり、可能性のある適用分野としては、in vitroにて細胞にトランスフェクトされる、または直接in vivoに投与されるRNAに基づくワクチン、例えば細胞において分化プロセスを開始させるため、またはタンパク質の機能を研究するためのin vitroにおける機能的組換えタンパク質の一時的発現、ならびにin vivoにおける、特に医薬としての、エリスロポエチン、ホルモン、血液凝固阻害剤などのような機能的組換えタンパク質の一時的発現がある。
【0066】
本発明に記載の修飾により修飾されたRNA、特にin vitroで転写されたRNAは特に抗原提示細胞をトラスフェクトするために、従って提示されるべき抗原を送達するため、また、抗原提示細胞に、該RNAから発現されたまたはそれに由来するペプチドまたはタンパク質に相当する該提示抗原を、特に切断などの細胞内プロセシングにより付加するため(すなわち、この提示抗原は例えばRNAから発現されたペプチドまたはタンパク質の断片である)のツールとして使用することができる。このような抗原提示細胞はT細胞、特にCD4+および/またはCD8+T細胞を刺激するために使用可能である。
【0067】
発明の詳細な説明
本発明によれば、組換え核酸の作製、細胞の培養および核酸、特にRNAの細胞への導入、特にエレクトロポレーションおよびリポフェクションには、標準的な方法が使用可能である。酵素反応は製造業者の使用説明書に従い、またはそれ自体公知の方法で行われる。
【0068】
本発明によれば、核酸分子または核酸配列とは、好ましくはデオキシリボ核酸(DNA)またはリボ核酸(RNA)である核酸を指す。本発明によれば、核酸はゲノムDNA、cDNA、mRNA、組換え生産された分子、および化学的に合成された分子を含む。本発明によれば、核酸は一本鎖または二本鎖、および直鎖または共有結合的に閉環した分子の形態であってよい。
【0069】
「mRNA」とは「メッセンジャーRNA」を意味し、DNAを鋳型として生産され、それ自体ペプチドまたはタンパク質をコードする「転写産物」を指す。mRNAは一般に5’非翻訳領域、タンパク質コード領域および3’非翻訳領域を含む。mRNAは、細胞内でもin vitroでも限られた半減期を有する。本発明によれば、mRNAはDNA鋳型からin vitro転写により作製することができる。このmRNAは、本発明の修飾に加え、さらなる安定化修飾およびキャッピングにより修飾してもよい。
【0070】
「核酸」とはさらにまた、ヌクレオチド塩基、糖またはリン酸基に対する核酸の化学誘導体、ならびに非天然ヌクレオチドおよびヌクレオチド類似体を含有する核酸を含む。
【0071】
本発明によれば「核酸配列に由来する核酸配列」とは、それが由来する核酸に比べて、単一または複数のヌクレオチド置換、欠失および/または付加を含み、好ましくはそれが由来する核酸と相補的である、すなわち、該核酸間にある程度の相同性があり、該核酸のヌクレオチド配列が有意な直接的または相補的様式で一致する、核酸配列を指す。本発明によれば、核酸に由来する核酸は、それが由来する核酸の機能的特性を有している。このような機能的特性としては、特に、RNAへと転写し得る核酸(転写可能な核酸配列)との機能的連結において、完全なRNA分子においてこの核酸から産生されるRNAの安定性および/または翻訳効率を高める能力が含まれる。
【0072】
本発明によれば「機能的連結」または「機能的に連結された」とは、機能的関係の範囲内の連結に関していう。核酸は、別の核酸配列と機能的に関連している場合に、「機能的に連結されている」という。例えば、プロモーターは、コード配列の転写に影響を及ぼす場合に、該コード配列に機能的に連結されているという。機能的に連結されている核酸は一般に、別の核酸と、適当であればさらなる核酸配列により離されて隣接しており、特定の実施形態では、RNAポリメラーゼにより転写されて、単一のRNA分子(共通転写産物)が得られる。
【0073】
本発明に記載される核酸は単離されていることが好ましい。「単離された核酸」とは、本発明によれば、その核酸が(i)例えばポリメラーゼ連鎖反応(PCR)によりin vitroで増幅されたものであるか、(ii)クローニングにより組換え生産されたものであるか、(iii)例えば切断およびゲル濾過分画により精製されたものであるか、または(iv)例えば化学合成により合成されたものであることを意味する。単離された核酸は組換えDNA技術による操作に利用可能な核酸である。
【0074】
核酸は、その2つの配列が互いにハイブリダイズして安定な二重らせんを形成するこができる場合に別の核酸と「相補的」であるといい、該ハイブリダイゼーションは好ましくはポリヌクレオチド間の特異的ハイブリダイゼーションを可能とする条件(ストリンジェント条件)下で行われる。ストリンジェント条件は、例えば、Molecular Cloning: A Laboratory Manual, J. Sambrook et al., eds., 2nd edition, Cold Spring Harbor Laboratory Press, Cold Spring Harbor, New York 1989 or Current Protocols in Molecular Biology, F.M. Ausubel et al., eds., John Wiley & Sons, Inc., New Yorkに記載されており、例えば、65℃、ハイブリダイゼーションバッファー(3.5×SSC、0.02%フィコール、0.02%ポリビニルピロリドン、0.02%ウシ血清アルブミン、2.5mM NaH2PO4(pH7)、0.5%SDS、2mM EDTA)中でのハイブリダイゼーションを指す。SSCは0.15M塩化ナトリウム/0.15Mクエン酸ナトリウム、pH 7である。ハイブリダイゼーション後、DNAを写し取ったメンブランを、例えば2×SSC中、室温で洗浄した後、0.1〜0.5×SSC/0.1×SDS中、68℃までの温度で洗浄する。
【0075】
本発明によれば、相補的核酸は、少なくとも60%、少なくとも70%、少なくとも80%、少なくとも90%、好ましくは少なくとも95%、少なくとも98%または少なくとも99%同一であるヌクレオチドを有する。
【0076】
「同一性%」とは、比較する2つの配列間で最適なアライメントにおいて同一であるヌクレオチドのパーセンテージを指し、該パーセンテージは単に統計学的なものであり、この2配列間の違いは、配列の全長にわたってランダムに分布している場合もあるし、また、2配列間の最適アライメントを得るために、比較配列は参照配列に対して付加もしくは欠失を含んでいる場合もある。2配列の比較は通常、対応する配列の局部領域を特定するために、最適アライメントの後にこれらの配列をセグメントまたは「比較枠」に関して比較することにより行う。比較のための最適アライメントは手動で、またはSmith and Waterman, 1981, Ads App. Math. 2, 482によるローカルホモロジーアルゴリズム、Neddleman and Wunsch, 1970, J. Mol. Biol. 48, 443によるローカルホモロジーアルゴリズム、およびPearson and Lipman, 1988, Proc. Natl Acad. Sci. USA 85, 2444による類似性検索アルゴリズムまたは前記アルゴリズムを用いたコンピュータープログラム(GAP, BESTFIT, FASTA, BLAST P, BLAST N and TFASTA in Wisconsin Genetics Software Package, Genetics Computer Group, 575 Science Drive, Madison, Wis.)の助けで行うことができる。
【0077】
同一性パーセンテージは、比較配列が一致する位置の数を求め、この数を比較した位置の数で割り、この解に100を掛けることにより得られる。
【0078】
例えば、ウェブサイトhttp://www.ncbi.nlm.nih.gov/blast/bl2seq/wblast2.cgiで入手できるBLASTプログラム「BLAST2配列」が使用可能である。
【0079】
「核酸の3’末端」とは、本発明では、遊離ヒドロキシ基を有する末端を指す。二本鎖核酸、特にDNAの模式図では、3’末端は常に右側にある。「核酸の5’末端」とは、本発明では、遊離リン酸基を有する末端を指す。二本鎖核酸、特にDNAの模式図では、5’末端は常に左側にある。
5’末端 5’−−P−NNNNNNN−OH−3’
3’末端 3’−HO−NNNNNNN−P−−5’
【0080】
特定の実施形態では、核酸は、本発明に従い、その核酸に対して同種であっても異種であってもよい発現制御配列と機能的に連結されている。
【0081】
転写可能な核酸、特にペプチドまたはタンパク質をコードする核酸および発現制御配列は、それらが、転写可能な、特にコード核酸の転写または発現がその発現制御配列の制御下または影響下にあるように互いに共有結合されている場合に、互いに「機能的に」連結されているという。この核酸が機能的ペプチドまたはタンパク質に翻訳されるならば、そのコード配列に機能的に連結されている発現制御配列の誘導は、コード配列にフレームシフトを起こすことなく、またはコード配列が所望のペプチドまたはタンパク質の翻訳不能となることなく、コード配列の転写を起こす。
【0082】
「発現制御配列」は、本発明によれば、プロモーター、リボソーム結合配列および遺伝子の転写または誘導されるRNAの翻訳を制御する他の制御エレメントを含む。本発明の特定の実施形態では、発現制御配列は調節可能である。発現制御配列の厳密な構造は、種または細胞種によって異なるが、通常、TATAボックス、キャッピング配列およびCAAT配列など、それぞれ転写および翻訳の開始に関与する5’非転写配列ならびに5’および3’非翻訳配列を含む。より具体的には、5’非転写発現制御配列は、機能的に連結された遺伝子の転写制御のためのプロモーター配列を包含するプロモーター領域を含む。発現制御配列はまたエンハンサー配列または上流アクチベーター配列も含み得る。
【0083】
本明細書に明示される核酸、特に転写可能なコード核酸は、その核酸に対して同種であっても異種であってもよい発現制御配列、特にプロモーターのいずれと組み合わせてもよく、「同種」とは、核酸がまた発現制御配列に天然状態で機能的に連結されているということを表し、「異種」とは、核酸が発現制御配列に天然状態では機能的に連結されていないということを表す。
【0084】
「プロモーター」または「プロモーター領域」とは、RNAポリメラーゼの認識および結合部位を提供することでコード配列の発現を制御する、遺伝子のコード配列の上流(5’)DNA配列を指す。このプロモーター領域は、該遺伝子の転写の調節に関与するさらなる因子のためのさらなる認識または結合部位を含んでいてもよい。プロモーターは原核生物または真核生物の遺伝子の転写を制御し得る。プロモーターはインデューサーに応答して転写を開始する「誘導型」であるか、転写がインデューサーによって制御されなければ「構成型」である。誘導型プロモーターは、インデューサーが存在しなければ、極めて小さな程度でしか発現しないか、または全く発現しない。インデューサーの存在下では、この遺伝子は「スイッチオン」となるか、または転写レベルが高まる。これは通常、特定の転写因子の結合によって媒介される。
【0085】
本発明に従う好ましいプロモーターの例としては、SP6、T3またはT7ポリメラーゼのプロモーターがある。
【0086】
本発明によれば、「発現」とは、その最も一般的に意味で用いられ、RNAまたはRNAとタンパク質の産生を含む。また、核酸の部分的発現も含まれる。さらに、発現は一時的なものであっても安定したものであってもよい。RNAに関して「発現」または「翻訳」とは、特にペプチドまたはタンパク質の産生を指す。
【0087】
「転写されて共通転写産物を生じ得る核酸」とは、その核酸が、適当であれば該核酸を含む核酸分子の、特に、閉環核酸分子の制限酵素切断などの線状化の後、プロモーターの制御下での転写が、適当であればその間に位置する配列により離されて、互いに共有結合された該核酸の転写産物を含むRNA分子を生じるように、互いに機能的に連結されていることを意味する。
【0088】
本発明によれば、「転写」には「in vitro転写」が含まれ、この「in vitro転写」とは、RNA、特にmRNAが無細胞様式で、すなわち、好ましくは適宜調製された細胞抽出液を用いることでin vitro合成される方法に関していう。転写産物の調製には、一般に転写ベクターと呼ばれ、本発明によれば「ベクター」という用語に含まれる、クローニングベクターを用いるのが好ましい。
【0089】
「核酸配列から転写された核酸配列」とは、適当であれば後者の核酸配列の転写産物である完全RNA分子の一部としてのRNAを指す。
【0090】
「核酸配列の翻訳効率および/または安定性を高めるために活性である核酸配列」とは、第一の核酸が第二の核酸との共通転写産物において、第二の核酸の翻訳効率および/または安定性を、その翻訳効率および/または安定性が第一の核酸を用いない場合の第二の核酸の翻訳効率および/または安定性に比べて高まるように修飾することができることを意味する。これに関して、「翻訳効率」とは、特定の時間内にRNA分子がもたらす翻訳産物の量に関し、「安定性」とは、RNA分子の半減期に関していう。
【0091】
3’非翻訳領域は、タンパク質コード領域の末端コドンの下流の、遺伝子の3’末端に位置し、転写されるがアミノ酸配列へ翻訳されない領域に関していう。
【0092】
本発明によれば、第一のポリヌクレオチド領域の5’末端が第二のポリヌクレオチド領域の3’末端に最も近い第一のポリヌクレオチド領域の一部である場合に、第一のポリヌクレオチド領域は、第二のポリヌクレオチドの下流に位置するとみなされる。
【0093】
3’非翻訳領域は一般に、翻訳産物の末端コドンから、通常転写工程後に付着されるポリ(A)配列へと伸びる。哺乳類mRNAの3’非翻訳領域は一般に、AAUAAAヘキサヌクレオチド配列として知られる相同性領域を有する。この配列はおそらくポリ(A)付着シグナルであり、ポリ(A)付着部位の10〜30塩基上流に位置している場合が多い。
【0094】
3’非翻訳領域は、エキソリボヌクレアーゼに対するバリアとして働く、またはRNAの安定性を高めることが知られているタンパク質(例えば、RNA結合タンパク質)と相互作用するステム−ループ構造が得られるように折り畳み可能な1以上の逆方向反復を含み得る。
【0095】
本発明によれば5’および/または3’非翻訳領域は転写可能な、特にコード核酸と機能的に連結させることができ、これによりこれらの領域を、転写可能な核酸から転写されたRNAの安定性および/または翻訳効率が高まるように核酸と結合させる。
【0096】
免疫グロブリンmRNAの3’非翻訳領域は比較的短く(約300ヌクレオチドより短い)が、他の遺伝子の3’非翻訳領域は比較的長い。例えば、tPAの3’非翻訳領域は約800ヌクレオチドの長さであり、第VIII因子の3’非翻訳領域は約1800ヌクレオチドの長さであり、エリスロポエチンの3’非翻訳領域は約560ヌクレオチドの長さである。
【0097】
本発明によれば、3’非翻訳領域またはそれに由来する核酸配列がRNAの安定性および/または翻訳効率を高めるかどうかは、その3’非翻訳領域またはそれに由来する核酸配列を遺伝子の3’非翻訳領域に組み込み、その組み込みによりタンパク質の合成量が高まるかどうかを測定することによって判定することができる。
【0098】
上記のことは本発明において核酸が2以上の3’非翻訳領域を含む場合にも適用され、この場合には、間に好ましくは「ヘッド・トゥ・テール関係」(すなわち、これらの3’非翻訳領域は同じ配向、好ましくは核酸において天然の配向を持つ)のリンカーを用いて、または用いずに順次結合されていることが好ましい。
【0099】
本発明によれば「遺伝子」とは、1以上の細胞産物の産生および/または1以上の細胞間もしくは細胞内機能の達成を担う特定の核酸配列に関していう。より具体的には、この用語は、特定のタンパク質または機能的もしくは構造的RNA分子をコードする核酸を含むDNA部分に関する。
【0100】
「ポリアデニルカセット」または「ポリ(A)配列」は、一般にRNA分子の3’末端に位置するアデニル残基の配列を指す。本発明は、コード鎖に相補的な鎖中の反復チミジル残基に基づくDNA鋳型を介したRNA転写中に付着されるこのような配列を提供するが、このような配列は通常そのDNAにはコードされておらず、核内で転写後に鋳型依存性のRNAポリメラーゼによりRNAの空いている3’末端に付着される。本発明によれば、この種のポリ(A)配列は少なくとも20、好ましくは少なくとも40、好ましくは少なくとも80、好ましくは少なくとも100で、好ましくは500まで、好ましくは400まで、好ましくは300まで、好ましくは200まで、特に150までの連続するAヌクレオチド、特には約120の連続するAヌクレオチドのヌクレオチド配列を意味するものと理解され、「Aヌクレオチド」とはアデニル残基を指す。
【0101】
好ましい実施形態では、本発明の核酸分子はベクターである。「ベクター」とは、ここではその最も一般的な意味で用いられ、例えば核酸を原核生物および/または真核生物の宿主細胞へ導入可能とし、および適当であればゲノムへ導入可能とする核酸のいずれの中間ビヒクルも含む。このようなベクターは細胞内で複製および/または発現されることが好ましい。ベクターはプラスミド、ファージミドまたはウイルスゲノムを含む。「プラスミド」とは本明細書において一般に、染色体DNAとは独立に複製可能な染色体外遺伝物質の構築物、通常、環状DNA二重らせんに関していう。
【0102】
本発明によれば、「宿主細胞」とは、外来核酸で形質転換またはトランスフェクション可能ないずれの細胞も指す。「宿主細胞」とは、本発明によれば、原核生物(例えば、大腸菌(E. coli))または真核細胞(例えば、酵母細胞および昆虫細胞)を含む。ヒト、マウス、ハムスター、ブタ、ヤギ、霊長類などの哺乳類細胞が特に好ましい。これらの細胞は多数の細胞種に由来し、一次細胞と細胞系統を含む。特定の例としては、ケラチノサイト、末梢血白血球、骨髄幹細胞および胚幹細胞が挙げられる。他の実施形態では、宿主細胞は抗原提示細胞、特に樹状細胞、単球またはマクロファージである。核酸は一コピーまたは数個ピーで宿主細胞に存在してもよく、一実施形態では宿主細胞内で発現される。
【0103】
本発明によれば、核酸によりコードされるペプチドまたはタンパク質は、細胞質、核、膜、オルガネラに存在するペプチドまたはタンパク質であってもよいし、あるいは分泌されるものでもよい。これらのものとして構造タンパク質、調節タンパク質、ホルモン、神経伝達物質、成長調節因子、分化因子、遺伝子発現調節因子、DNA会合タンパク質、酵素、血清タンパク質、受容体、薬剤、免疫調節剤、癌遺伝子、毒素、腫瘍抗原または抗原が挙げられる。これらのペプチドまたはタンパク質は天然配列を持っていてもよいし、あるいはそれらの生物活性を増強、阻害、調節または排除するために変異された配列を持っていてもよい。
【0104】
「ペプチド」とは、ペプチド結合を介して互いに連結された2以上、好ましくは3以上、好ましくは4以上、好ましくは6以上、好ましくは8以上、好ましくは10以上、好ましくは13以上、好ましくは16以上、好ましくは20以上で、好ましくは50まで、好ましくは100までまたは好ましくは150までの連続するアミノ酸を含む物質を指す。「タンパク質」とは、大きなペプチド、好ましくは少なくとも151のアミノ酸を有するペプチドを指すが、本明細書では「ペプチド」と「タンパク質」は通常同義で用いる。本発明において「ペプチド」および「タンパク質」は、アミノ酸成分だけでなく、糖およびリン酸構造などの非アミノ酸成分も含む物質を含み、またエステル、チオエーテルまたはジスルフィド結合などの結合を含む物質も含む。
【0105】
本発明は、患者に投与する核酸、特にRNAを提供する。一実施形態では、核酸はex vivo法、すなわち、患者から細胞を取り出し、それらの細胞を遺伝的に修飾し、修飾された細胞を患者に再び導入することにより投与される。トランスフェクションおよび形質導入法は当業者に公知である。本発明はまた、in vivo投与される核酸も提供する。
【0106】
本発明によれば、「トランスフェクション」とは、1以上の核酸を生物体または宿主細胞に導入することを指す。本発明に従いin vitroまたはin vivoで核酸を細胞に導入するためには、様々な方法が使用できる。このような方法としては、核酸 CaPO4沈降物のトランスフェクション、DEAEと会合した核酸のトランスフェクション、目的の核酸を運ぶウイルスによるトランスフェクションまたは感染、およびリポソームに媒介されるトランスフェクションなどが挙げられる。特定の実施形態では、核酸を特定の細胞に向けることが好ましい。このような実施形態では、核酸を細胞へ投与するために用いる担体(例えば、レトロウイルスまたはリポソーム)にターゲティング分子を結合させてもよい。例えば、標的細胞上の表面膜タンパク質に特異的な抗体または標的細胞上の受容体に対するリガンドなどの分子を、核酸担体に組み込むか結合させればよい。リポソームによる核酸の投与が望ましければ、ターゲティングおよび/または吸収を可能とするためにエンドサイトーシスに関連する表面膜タンパク質に結合するタンパク質をリポソーム製剤に組み込めばよい。このようなタンパク質としては、特定の細胞種に特異的なキャプシドタンパク質またはその断片、インターナライズされるタンパク質に対する抗体、および細胞内部位をターゲティングするタンパク質などが挙げられる。
【0107】
「レポーター」とは、レポーター遺伝子によりコードされ、レポーターアッセイで測定される分子、一般にペプチドまたはタンパク質に関していう。慣例の系としては、通常、酵素レポーターを用い、そのレポーターの活性を測定する。
【0108】
「多重クローニング部位」とは、そのいずれか1つが例えばベクターの切断および核酸の挿入に使用できる制限酵素部位を含む核酸領域に関していう。
【0109】
本発明によれば、ヌクレオチドまたはアミノ酸などの2つのエレメントは、互いに直接隣接していれば連続のものである。例えば、x個の連続するヌクレオチドNの配列は、配列(N)xと表す。
【0110】
「制限エンドヌクレアーゼ」または「制限酵素」とは、DNA分子の両鎖のホスホジエステル結合を特定の塩基配列内で切断する酵素種を指す。これらの酵素は、二本鎖DNA分子上の、認識配列と呼ばれる特定の結合部位を認識する。DNA中のホスホジエステル結合がこのような酵素により切断される部位は切断部位と呼ばれる。IIS型酵素の場合、この切断部位はDNA結合部位から所定の距離離れたところに存在する。本発明によれば、「制限エンドヌクレアーゼ」には、例えば、酵素SapI、EciI、BpiI、AarI、AloI、BaeI、BbvCI、PpiIおよびPsrI、BsrD1、BtsI、EarI、BmrI、BsaI、BsmBI、FauI、BbsI、BciVI、BfuAI、BspMI、BseRI、EciI、BtgZI、BpuEI、BsgI、MmeI、CspCI、BaeI、BsaMI、Mva1269I、PctI、Bse3DI、BseMI、Bst6I、Eam1104I、Ksp632I、BfiI、Bso31I、BspTNI、Eco31I、Esp3I、BfuI、Acc36I、AarI、Eco57I、Eco57MI、GsuI、AloI、Hin4I、PpiI、およびPsrIが含まれる。
【0111】
「半減期」とは、分子の活性、量または数の半分が消失するのにかかる時間を指す。
【0112】
以下、本発明を図面および実施例により詳しく説明するが、これらは単に例示であって、限定を示すものではない。これらの説明および実施例に基づいて当業者ならばさらなる実施例を利用することができるが、それらも同様に本発明の範囲内にある。
【図面の簡単な説明】
【0113】
【図1】図1:さらなるクローニングのため本発明に従って用いられる基本ベクター これらのベクターはRNAポリメラーゼ5’プロモーターの制御下でRNA転写が可能であり、ポリアデニルカセットを含む。
【0114】
【図2】図2:II型制限酵素(例えば、SpeI)とIIS型制限酵素(例えば、SapI)とによるベクター線状化の比較 その認識配列がポリ(A)配列の3’にあり、切断部位が24〜26 bp上流、従ってポリ(A)配列内にあるIIS型制限切断部位を導入することにより、プラスミドをポリ(A)配列内で線状化することができる。
【0115】
【図3】図3:in vitro転写のための鋳型として本発明に従って作製されるベクター 発現のレベルおよび持続時間に対する本発明のRNA修飾の効果を研究するため、いくつかのベクターを作製し、その後、in vitro転写のための鋳型として用いた。a.マスクされたポリ(A)配列とマスクされていないポリ(A)配列を有するベクター;b.長さの異なるポリ(A)配列を有するベクター;c.ヒトβ−グロビンの3’非翻訳領域を有するベクター;d.SIINFEKLおよびpp65ベクター;Cap−5’−キャッピング;eGFP−GFPレポーター遺伝子;β−グロビンの3’βg−3’非翻訳領域。A(x)−xは、ポリ(A)配列中のAヌクレオチドの数を表す。
【0116】
【図4】図4:示された表面マスクによる未成熟樹状細胞と成熟樹状細胞の成熟状態の決定 本発明のRNA修飾の効果をヒト樹状細胞(DC)にて、DC成熟プロセスを誘導する免疫刺激を用いて調べた。これらのDCを、特異的DC成熟マーカーを認識する抗CD80抗体、抗CD83抗体、抗CD86抗体および抗HLA−DR抗体で染色し、フローサイトメトリーで分析した。
【0117】
【図5】図5:翻訳効率および転写産物の安定性に対する遊離型およびマスク型のポリ(A)配列の影響 a.FACS−Kaliburにて平均蛍光強度[MFI]を測定することによる、K562細胞および樹状細胞におけるeGFP RNAの翻訳効率に対する遊離型およびマスク型のポリ(A)配列の影響。b.48時間後の未成熟樹状細胞におけるeGFP RNAの転写産物安定性に対する遊離型およびマスク型のポリ(A)配列の影響。腫瘍細胞系統および未成熟DCの双方で、オープンエンド型ポリ(A)配列を有するRNAはマスクエンド型ポリ(A)配列を有するRNAよりも有効かつ長期間翻訳される。DCにおける非マスクエンド型ポリ(A)配列の翻訳効率は、同じ長さのポリ(A)配列の1.5倍高い。さらにオープンエンド型ポリ(A)配列は、より高いRNA安定性をもたらす。
【0118】
【図6】図6:翻訳効率および転写産物の安定性に対するポリ(A)配列長の影響 a.K562細胞および樹状細胞におけるeGFP RNAの翻訳効率に対するポリ(A)配列長の影響。b.K562細胞および樹状細胞におけるd2eGFP RNAの翻訳効率に対するポリ(A)配列長の影響。c.エレクトロポレーション48時間後のK562細胞におけるeGFP RNAの転写産物安定性に対するポリ(A)配列長の影響。ポリ(A)配列を120Aヌクレオチドまで延長すると、転写産物の安定性および翻訳が高まる。これを超える延長に有効な効果は見られない。ポリ(A)配列を51〜120Aヌクレオチド延長すると、翻訳効率が1.5〜2倍高まる。この効果はRNAの安定性にも反映される。
【0119】
【図7】図7:未成熟および成熟DCにおける翻訳効率に対するヒトβ−グロビンの3’非翻訳領域(BgUTR)の影響 ヒトβ−グロビンの3’非翻訳領域を導入すると、RNA転写産物の発現が増強する。ヒトβ−グロビンの3’非翻訳領域が二重にあると、24時間後の発現レベルが高まり、このレベルは個々の2つのヒトβ−グロビンの3’非翻訳領域を合わせた効果を著しく超える。
【0120】
【図8】図8:未成熟および成熟DCにおける翻訳効率に対する本発明の修飾を組み合わせた効果 未成熟および成熟DCにおけるeGFPの翻訳効率は、本発明に記載のRNA転写産物修飾を組み合わせることで、5倍を上回る増強が可能である。
【0121】
【図9】図9:EL4細胞上のMHC分子によるペプチド提示に対する本発明の修飾を組み合わせた効果 本発明に従って修飾されたRNA構築物を用いると、高い翻訳効率のために細胞表面上でのペプチド−MHC複合体の提示が高まる。記載されているIVTベクターでは、eGFPをOVA257−264エピトープ(SIINFEKL)に置換し、EL4細胞(ネズミ、T細胞リンパ腫)をトランスフェクションの標的細胞として用いた。
【0122】
【図10】図10:本発明に従って安定化されたIVT RNA構築物を用いることによる、抗原特異的ペプチド/MHC複合体の増加 細胞にSec−SIINFEKL−A67−ACUAG RNAまたはSec−SIINFEKL−2BgUTR−A120 RNA(EL4細胞:10pmol、50pmol;C57B1/J6未成熟BMDC150pmol3反復)でエレクトロポレーションを行った。バッファーだけを用いるエレクトロポレーションを対照として用いた。細胞をSIINFEKL/Kb複合体に対する25D1.16抗体で染色した。SIINFEKLペプチド濃度を生細胞の平均蛍光値から、検量曲線としてペプチド滴定を用いて算出した。BMDCデータは3回の実験の平均値±SEMとして示す。
【0123】
【図11】図11:in vivoおよびin vitroでのT細胞刺激に対する、本発明に従って安定化されたIVT RNA構築物の効果 (A)安定化されたIVT RNA構築物の使用によるin vivoT細胞増殖の向上。1×105個のTCR−トランスジェニックCD8+ OT−I細胞をC57B1/J6マウスに養子免疫伝達した。C57B1/J6マウスのBMDCを50pmolのRNA(Sec−SIINFEKL−A67−ACUAG、Sec−SIINFEKL−2BgUTR−A120または対照RNA)でトランスフェクトし、ポリ(I:C)(50μg/ml)を用いて16時間成熟させ、T細胞移入の1日前に腹腔内注射した(n=3)。4日目に末梢血を採取し、SIINFEKL四量体陽性CD8+T細胞を染色した。ドットブロットはCD8+T細胞を表し、示されている数値は四量体陽性CD8+T細胞のパーセンテージを表す。 (B)安定化されたIVT RNA構築物を含むヒトT細胞のin vitro増殖の向上。HCMV血清反応陽性健常ドナーからのCD8+およびCD4+リンパ球をSec−pp65−A67−ACUAG RNA、Sec−pp65−2BGUTR−A120 RNA、または対照RNA(データは示されていない)でトランスフェクトされたか、あるいは陽性対照としてpp65ペプチドプール(1.75μg/ml)で刺激した自己DCと同時培養した。7日間増殖させた後、各エフェクター細胞集団(4×104/ウェル)を、pp65ペプチドプールか無関連のペプチドプール(1.75μg/ml)のいずれかを付加した自己DC(3×104/ウェル)を用い、IFN−γ ELISpotにてアッセイした。グラフは3回測定したスポットの平均値±SEMを表す。
【実施例1】
【0124】
実施例1:ベクターの作製とRNAのin vitro転写
発現のレベルおよび持続時間に対する本発明のRNA修飾の効果を検討するため、いくつかのIVTベクターを作製した後、in vitro転写のための鋳型として用いた(図3)。
【0125】
半減期(HL)の異なる2つの分子eGFPとd2eGFPのレポーター遺伝子をベクターに挿入し、これにより本発明のRNA修飾の影響を分析可能とした。蛍光は減少し、平均HLはeGFPでは17.3時間、d2eGFPでは2時間であった。これらの構築物をそれぞれin vitro転写eGFP RNAおよびd2eGFP RNAの作製に用いた。
【実施例2】
【0126】
実施例2:本発明に従って修飾されたin vitro転写RNAによる細胞のトランスフェクションおよびRNA翻訳と安定性に対する効果
in vitro転写eGFP RNAおよびd2eGFP RNAを用い、エレクトロポレーションによりK562細胞(ヒト白血病)をトランスフェクトした。トランスフェクション効率はK562細胞で90%を超えた。
【0127】
その後、免疫系の最も重要なモジュレーターであるヒト樹状細胞(DC)において、記載のRNA修飾の作用をアッセイした。このアプローチは、RNAでトランスフェクトされたDCが予防接種として意図され得ることから、免疫関連のものである。未成熟DCを皮膚および末梢器官に付加する。ここで、これらは、十分研究されている表面マーカーにより特徴付けられ、また、高いエンドサイトーシス活性により機能的に識別される未成熟状態にある。例えば病原体の感染などの免疫刺激はDCの成熟プロセスを誘導する。それと同時に、この刺激は局部リンパ節へのDCの移動を開始させるが、ここでDCはT細胞およびB細胞免疫応答の最も有効なインデューサーである。また、これらのDCの成熟状態は表面マーカー発現および詳細に研究されているサイトカインの発現によっても、また、DC形態の特徴によっても特徴付けられる。血液単球から未成熟ヒトDCを分化させるための培養系が確立されている。これらは種々の刺激により成熟が起こり得る。
【0128】
一次樹状細胞のトランスフェクション効率は70〜80%であった。これらのDCを、特異的DC成熟マーカーを認識する抗CD80抗体、抗CD83抗体、抗CD86抗体および抗HLA−DR抗体で染色し、フローサイトメトリーで分析した(図4)。
【0129】
発現のレベルおよび持続時間を、FACS−Kaliburを用い、eGFPの蛍光強度を測定することで測定した。細胞のRNA量は、定量的RT−PCRを用いて測定した。
【0130】
a.RNAの翻訳および安定性に対するオープンエンド型ポリ(A)配列の効果
腫瘍細胞系統K562と未成熟DC(iDC)は双方とも、オープンエンド型ポリ(A)配列を有するRNAを、マスクエンド型ポリ(A)配列を有するRNAよりも効率的にかつ長期間翻訳することが示された(図5a)。未成熟DCにおける非マスクエンド型ポリ(A)配列の翻訳効率は、同じ長さのポリ(A)配列では1.5倍高くなる。さらに、この修飾から、より高いRNA安定性が得られる(図5b)。非マスクポリ(A)配列を有するRNAでトランスフェクトされた未成熟DCでは、エレクトロポレーション48時間後に4〜5倍量のRNAを検出することができる。
【0131】
b.RNAの翻訳および安定性に対するポリ(A)配列長の効果
16bp、42bp、51bp、67bp、120bp、200bp、300bpおよび600bpの長さのポリ(A)配列を有するRNAを分析したところ、120Aヌクレオチドまでのポリ(A)配列の延長が転写産物の安定性と翻訳を高めること、また、それを超える延長には有効な効果がないことが明らかになった。この効果はK562細胞と未成熟DC(iDC)の双方に見られる(図6aおよび6b)。ポリ(A)配列を51〜120Aヌクレオチド延長すると、翻訳効率が1.5〜2倍高まる。この効果はRNAの安定性にも反映される(図6c)。
【0132】
c.RNAの翻訳および安定性に対する3’非翻訳領域の存在の効果
K562細胞および未成熟DCを用いた経時的推移により、ヒトβ−グロビンの3’非翻訳領域(UTR)の導入がRNA転写産物の発現の増強をもたらすことが確認された。さらに、ヒトβ−グロビンの3’非翻訳領域(UTR)が二重にあると、24時間後に高い発現レベルが得られ、個々の2つのUTRを合わせた効果を大きく超えることが実証された(図7)。
【0133】
d.RNAの翻訳および安定性に対する上記修飾の組合せの効果
本発明によれば、RNA転写産物における上記修飾の組合せは、未成熟DCまた成熟DCでも、eGFPの翻訳効率を5倍を超えて高めることが示された(図8)。
【実施例3】
【0134】
実施例3:安定性および翻訳効率の高められたin vitro転写RNAを介して発現されたペプチドの、MHC分子による提示
本発明によれば、本発明に従って修飾されたRNA構築物を用いると、細胞表面でのペプチド−MHCの提示が高まることが示された。この目的で、記載のベクター中のeGFPをコードする核酸配列を、OVA257−264エピトープ(SIINFEKL)をコードする核酸配列に置換し、これらの構築物を互いに比較した。トランスフェクションに用いた標的細胞はEL4細胞(マウス、T細胞リンパ腫)であった。
【0135】
MHC分子により提示されたSIINFEKLペプチドを定量するため、これらの細胞を、エレクトロポレーション後種々の時点で、抗H2−Kb OVA257−264抗体で染色し、二次抗体の蛍光強度を、FACS−Kaliburを用いて測定した(図9)。
【0136】
さらに、このSIINFEKLペプチドを、あらゆる最適化を反映したベクター(pST1−Sec−SIINFEKL−2BgUTR−A120−Sap1)に、また、標準的な特徴を有するベクター(pST1−Sec−SIINFEKL−A67−Spe1)にクローニングした。両ベクターに由来するIVT RNAで、EL4細胞およびBMDCに対してエレクトロポレーションを行った。OVA−ペプチド/Kb複合体は細胞表面実質的に多数見られ、本発明に従って修飾されたRNA、Sec−SIINFEKL−2−BgUTR−A120のエレクトロポレーション後、長期間維持された(図10)。
【実施例4】
【0137】
実施例4:抗原特異的T細胞の発現に対する、提示されるペプチドをコードするin vitro転写RNAによる細胞のトランスフェクションの効果
刺激能に対する効果を評価するため、SIINFEKLペプチドのMHCクラスI提示を検出するためのC57BL/J6(B6)バックグラウンドに集中的に用いられてきたOT−I−TCRを用いた。T細胞受容体(TCR)に関してトランスジェニックであり、ニワトリOVA由来のKb−特異的ペプチドSIINFEKL(OVA257−264)を認識するOT−I CD8+T細胞はH. Schild (Institute of Immunology, University of Mainz, Germany)から厚意により提供されたものである。
【0138】
0日目、OT−I−CD8+T細胞を用いて、動物に養子免疫伝達を行った。この目的で、TCR tg OT−Iマウスから脾細胞を調製し、C57BL/J6レシピエントマウスの尾の静脈に導入した。細胞数は1×105 TCR tg CD8+T細胞に調整した。翌日、50pmolのSIINFEKLコードRNA構築物変異体でエレクトロポレーションを行い、ポリ(I:C)で16時間成熟させたC57BL/J6マウスの1×106 BMDCをマウスに腹腔内投与した。4日目、四量体法を用い、末梢血においてOT−I−CD8+T細胞を測定した。この目的で、眼窩後方の血液サンプルを採取し、抗CD8(Caltag Laboratories, Burlingame, USA)およびSIINFEKL四量体(H−2Kb/SIINFEKL 257−264; Beckman Coulter, Fullerton, USA)で染色した。
【0139】
抗原特異的TCR−トランスジェニックCD8+T細胞のin vivo増殖は、抗原供給源としてSec−SIINFEKL−2BgUTR−A120 RNAを用いた場合には、Sec−SIINFEKL−A67−ACUAG RNAの場合と比べて実質的に向上することが分かった(図11A)。
【0140】
抗原供給源として安定されたIVT RNA構築物が、ヒトT細胞の抗原特異的刺激も向上させるかどうか評価するため、ポリエピトープ性のT細胞反応の自己刺激をバリデートするために用いられることが多いヒトサイトメガロウイルスの免疫優性抗原であるHCMV−pp65を用いた。抗体をコーティングしたマイクロビーズ(Miltenyi Biotec, Bergisch−Gladbach, Germany)を用いた正磁場セルソーティングにより、HCMV血清反応陽性健常ドナーから精製されたCD4+T細胞およびCD8+T細胞を、pp65をコードする対応するIVT RNA変異体でマイクロポレーションを行った自己DC2×105個とともに同時培養した。T細胞の増殖を、全pp65タンパク質配列にわたる重複ペプチドのプールで、または対照タンパク質で刺激した自己DCを用いたIFN−γ−ELISpotで7日目に測定したところ、Sec−pp65−2BgUTR−A120が優れていることが実証され、CD4+T細胞の増殖に対する効果が最も顕著であった(図11B)。
【0141】
引用文献
Bargmann, C.I., Hung, M.C., and Weinberg, R.A. (1986). The neu oncogene encodes an epidermal growth factor receptor−related protein. Nature 319, 226−230.
Boczkowski, D., Nair, S.K., Nam, J.H., Lyerly H.K., and Gilboa, E. (2000). Induction of tumor immunity and cytotoxic T lymphocyte responses using dendritic cells transfected with messenger RNA amplified from tumor cells. Cancer Res. 60, 1028−1034.
Carralot, J.P., Probst, J., Hoerr, I., Scheel, B., Teufel, R., Jung, G., Rammensee, H.G., and Pascolo, S. (2004). Polarization of immunity induced by direct injection of naked sequence−stabilized mRNA vaccines. Cell Mol. Life Sci. 61, 2418−2424.
Condon, C., Watkins, S.C., Celluzzi, C.M., Thompson, K., and Falo, L.D., Jr. (1996). DNA−based immunization by in vivo transfections of dendritic cells. Nat. Med. 2, 1122−1128.
Conry, R.M., LoBuglio, A.F., Kantor, J., Schlom, J., Loechel, F., Moore, S.E., Sumerel, L.A., Barlow, D.L., Abrams, S., and Curiel, D.T. (1994). Immune response to a carcinoembryonic antigen polynucleotide vaccine. Cancer Res. 54, 1164−1168.
Conry, R.M., LoBuglio, A.F., Loechel, F., Moore, S.E., Sumerel, L.A., Barlow, D.L., and Curiel, D.T. (1995a). A carcinoembryonic antigen polynucleotide vaccine has in vivo antitumor activity. Gene Ther. 2, 59−65.
Conry, R.M., LoBuglio, A.F., Wright, M., Sumerel, L., Pike, M.J., Johanning, F., Benjamin, R., Lu, D., and Curiel, D.T. (1995b). Characterization of a messenger RNA polynucleotide vaccine vector. Cancer Res. 55, 1397−1400.
Cox, G.J., Zamb, T.J., and Babiuk, L.A. (1993). Bovine herpesvirus 1: immune responses in mice and cattle injected with plasmid DNA. J. Virol. 67, 5664−5667.
Davis, H.L., Michel, M.L., and Whalen, R.G. (1993). DNA−based immunization induces continuous secretion of hepatitis B surface antigen and high levels of circulating antibody. Hum. Mol. Genet. 2, 1847−1851.
Gallie, D.R. (1991). The cap and poly(A) tail function synergistically to regulate mRNA translational efficiency. Genes Dev. 5, 2108−2116.
Gilkeson, G.S., Pippen, A.M., and Pisetsky, D.S. (1995). Induction of cross−reactive anti−dsDNA antibodies in preautoimmune NZB/NZW mice by immunization with bacterial DNA. J. Clin. Invest. 95, 1398−1402.
Greenblatt, M.S., Bennett, W.P., Hollstein, M., and Harris, C.C. (1994). Mutations in the p53 tumor suppressor gene: clues to cancer etiology and molecular pathogenesis. Cancer Res. 54, 4855−4878.
Heiser, A., Coleman, D., Dannull, J., Yancey, D., Maurice, M.A., Lallas, C.D., Dahm, P., Niedzwiecki, D., Gilboa, E., and Vieweg, J. (2002). Autologous dendritic cells transfected with prostate−specific antigen RNA stimulate CTL responses against metastatic prostate tumors. J. Clin. Invest 109, 409−417.
Heiser, A., Dahm, P., Yancey, D.R., Maurice, M.A., Boczkowski, D., Nair, S.K., Gilboa, E., and Vieweg, J. (2000). Human dendritic cells transfected with RNA encoding prostate−specific antigen stimulate prostate−specific CTL responses in vitro. J. Immunol. 164, 5508−5514.
Hoerr, I., Obst, R., Rammensee, H.G., and Jung, G. (2000). In vivo application of RNA leads to induction of specific cytotoxic T lymphocytes and antibodies. Eur. J. Immunol. 30, 1−7.
Malone, R.W., Felgner, P.L., and Verma, I.M. (1989). Cationic liposome−mediated RNA transfection. Proc. Natl. Acad. Sci. USA 86, 6077−6081.
Preiss, T. and Hentze, M.W. (1998). Dual function of the messenger RNA cap structure in poly(A)−tail promoted translation in yeast. Nature 392, 516−520.
Spooner, R.A., Deonarain, M.P., and Epenetos, A.A. (1995). DNA vaccination for cancer treatment. Gene Ther. 2, 173−180.
Strong, T.V., Hampton, T.A., Louro, I., Bilbao, G., Conry, R.M., and Curiel, D.T. (1997). Incorporation of beta−globin untranslated regions into a Sindbis virus vector for augmentation of heterologous mRNA expression. Gene Ther. 4, 624−627.
Su, Z., Dannull, J., Heiser, A., Yancey, D., Pruitt, S., Madden, J., Coleman, D., Niedzwiecki, D., Gilboa, E., and Vieweg, J. (2003). Immunological and clinical responses in metastatic renal cancer patients vaccinated with tumor RNA−transfected dendritic cells. Cancer Res. 63, 2127−2133.
Tang, D.C., DeVit, M., and Johnston, S.A. (1992). Genetic immunization is a simple method for eliciting an immune response. Nature 356, 152−154.
Teufel, R., Carralot, J.P., Scheel, B., Probst, J., Walter, S., Jung, G., Hoerr, I., Rammensee, H.G., and Pascolo, S. (2005). Human peripheral blood monuclear cells transfected with messenger RNA stimulate antigen−specific cytotoxic T−lymphocytes in vitro. Cell Mol. Life Sci. 62, 1755−1762.
Ulmer, J.B., Donnelly, J.J., Parker, S.E., Rhodes, G.H., Felgner, P.L., Dwarki, V.J., Gromskowski, S.H., Deck, R.R., DeWitt, C.M., Friedman, A., et al. (1993). Heterologous protection against influenza by injection of DNA encoding a viral protein. Science 259, 1745−1749.
Wang, B., Merva, M., Dang, K., Ugen, K.E., Williams, W.V., and Weiner, D.B. (1995). Immunization by direct DNA inoculation induces rejection of tumor cell challenge. Hum. Gene Ther. 6, 407−418.
Wang, B., Ugen, K.E., Srikantan, V., Agadjanyan, M.G., Dang, K., Refaeli, Y., Sato, A.I., Boyer, J., Williams, W.V., and Weiner, D.B. (1993). Gene inoculation generates immune responses against human immunodeficiency virus type 1. Proc. Natl. Acad. Sci. USA 90, 4156−4160.
Wolff, J.A., Malone, R.W., Williams, P., Chong, W., Acsadi, G., Jani, A., and Felgner, P.L. (1990). Direct gene transfer into mouse muscle in vivo. Science 247, 1465−1468.
Ying, H., Zaks, T.Z., Wang, R.F., Irvine, K.R., Kammula, U.S., Marincola, F.M., Leitner, W.W., and Restifo, N.P. (1999). Cancer therapy using a self−replicating RNA vaccine. Nat. Med. 5, 823−827.
【特許請求の範囲】
【請求項1】
5’→3’転写方向に、
(a)プロモーター;
(b)転写可能な核酸配列または転写可能な核酸配列を導入するための核酸配列;
(c−1)遺伝子の3’非翻訳領域に相当する、またはそれに由来する第一の核酸配列;および
(c−2)遺伝子の3’非翻訳領域に相当する、またはそれに由来する第二の核酸配列
を含む、核酸分子。
【請求項2】
(c−3)遺伝子の3’非翻訳領域に相当する、またはそれに由来する少なくとも1つのさらなる核酸配列をさらに含む、請求項1に記載の核酸分子。
【請求項3】
プロモーター(a)の制御下の核酸配列(b)、(c−1)、(c−2)および適当であれば(c−3)が転写されて、核酸配列(c−1)および/または(c−2)および/または適当であれば(c−3)から転写された核酸配列に、転写可能な核酸配列(b)から転写された核酸配列の翻訳効率および/または安定性を高めるような活性がある共通転写産物を生じ得る、請求項1または2に記載の核酸分子。
【請求項4】
核酸配列(c−1)、(c−2)および適当であれば(c−3)が同一であっても異なっていてもよい、請求項1〜3のいずれか一項に記載の核酸分子。
【請求項5】
(d)プロモーター(a)の制御下で転写された際に、その転写産物に少なくとも20の連続するAヌクレオチドのヌクレオチド配列をコードする核酸配列をさらに含む、請求項1〜4のいずれか一項に記載の核酸分子。
【請求項6】
プロモーター(a)の制御下の核酸配列(b)、(c−1)、(c−2)、適当であれば(c−3)および(d)が転写されて、核酸配列(c−1)および/または(c−2)および/または適当であれば(c−3)および/または(d)から転写された核酸配列に、転写可能な核酸配列(b)から転写された核酸配列の翻訳効率および/または安定性を高めるような活性がある共通転写産物を生じ得る、請求項5に記載の核酸分子。
【請求項7】
核酸配列(d)が、プロモーター(a)の制御下で転写された際に、その転写産物に少なくとも40の連続するAヌクレオチドのヌクレオチド配列をコードする、請求項5または6に記載の核酸分子。
【請求項8】
核酸配列(d)が、プロモーター(a)の制御下で転写された際に、その転写産物に少なくとも80の連続するAヌクレオチドのヌクレオチド配列をコードする、請求項7に記載の核酸分子。
【請求項9】
核酸配列(d)が、プロモーター(a)の制御下で転写された際に、その転写産物に少なくとも100の連続するAヌクレオチドのヌクレオチド配列をコードする、請求項8に記載の核酸分子。
【請求項10】
核酸配列(d)が、プロモーター(a)の制御下で転写された際に、その転写産物に約120の連続するAヌクレオチドのヌクレオチド配列をコードする、請求項9に記載の核酸分子。
【請求項11】
切断が5’→3’転写方向に、プロモーター(a)、核酸配列(b)、核酸配列(c−1)、(c−2)および適当であれば(c−3)、ならびに核酸配列(d)の少なくとも一部(この核酸配列(d)の少なくとも一部は、プロモーター(a)の制御下で転写された際に、その転写産物に少なくとも20の連続するAヌクレオチドのヌクレオチド配列をコードし、その転写産物において3’末端ヌクレオチドは少なくとも20の連続するAヌクレオチドの該ヌクレオチド配列のAヌクレオチドである)を含む核酸分子を生じるように、核酸配列(d)内で、好ましくは酵素的にまたは別の生化学的方法で切断され得ることを特徴とする、請求項5〜10のいずれか一項に記載の核酸分子。
【請求項12】
切断後、前記核酸分子が、少なくとも20の連続するAヌクレオチドのヌクレオチド配列の鋳型として働く鎖の末端に、その転写産物において少なくとも20の連続するAヌクレオチドのヌクレオチド配列の鋳型として働くヌクレオチド配列の一部であるTヌクレオチドを有する、請求項11に記載の核酸分子。
【請求項13】
前記核酸配列(d)の少なくとも一部が、プロモーター(a)の制御下で転写された際に、その転写産物に少なくとも40の連続するAヌクレオチドのヌクレオチド配列をコードする、請求項11または12に記載の核酸分子。
【請求項14】
前記核酸配列(d)の少なくとも一部が、プロモーター(a)の制御下で転写された際に、その転写産物に少なくとも80の連続するAヌクレオチドのヌクレオチド配列をコードする、請求項13に記載の核酸分子。
【請求項15】
前記核酸配列(d)の少なくとも一部が、プロモーター(a)の制御下で転写された際に、その転写産物に少なくとも100の連続するAヌクレオチドのヌクレオチド配列をコードする、請求項14に記載の核酸分子。
【請求項16】
前記核酸配列(d)の少なくとも一部が、プロモーター(a)の制御下で転写された際に、その転写産物に約120の連続するAヌクレオチドのヌクレオチド配列をコードする、請求項15に記載の核酸分子。
【請求項17】
切断前には閉環分子であり、切断後には直鎖分子であることを特徴とする、請求項11〜16のいずれか一項に記載の核酸分子。
【請求項18】
前記切断が制限切断部位の助けで行われる、請求項11〜17のいずれか一項に記載の核酸分子。
【請求項19】
前記制限切断部位がIIS型制限エンドヌクレアーゼの制限切断部位である、請求項18に記載の核酸分子。
【請求項20】
前記IIS型制限エンドヌクレアーゼの認識配列が、核酸配列(d)の3’末端の5〜26塩基対下流に位置する、請求項19に記載の核酸分子。
【請求項21】
前記IIS型制限エンドヌクレアーゼの認識配列が、核酸配列(d)の3’末端の24〜26塩基対下流に位置する、請求項20に記載の核酸分子。
【請求項22】
核酸配列(c−1)、(c−2)および適当であれば(c−3)が互いに独立にα2−グロビン、α1−グロビン、β−グロビンおよび成長ホルモン、好ましくは、ヒトβ−グロビンからなる群から選択される遺伝子に由来する、請求項1〜21のいずれか一項に記載の核酸分子。
【請求項23】
5’→3’転写方向に、
(a)プロモーター;
(b)転写可能な核酸配列または転写可能な核酸配列を導入するための核酸配列;および
(c)プロモーター(a)の制御下で転写された際に、その転写産物に少なくとも20の連続するAヌクレオチドのヌクレオチド配列をコードする核酸配列
を含む、核酸分子。
【請求項24】
プロモーター(a)の制御下の核酸配列(b)および(c)が転写されて、核酸配列(c)から転写された核酸配列に、転写可能な核酸配列(b)から転写された核酸配列の翻訳効率および/または安定性を高めるような活性がある共通転写産物を生じ得る、請求項23に記載の核酸分子。
【請求項25】
核酸配列(c)が、プロモーター(a)の制御下で転写された際に、その転写産物に少なくとも40の連続するAヌクレオチドのヌクレオチド配列をコードする、請求項23または24に記載の核酸分子。
【請求項26】
核酸配列(c)が、プロモーター(a)の制御下で転写された際に、その転写産物に少なくとも80の連続するAヌクレオチドのヌクレオチド配列をコードする、請求項25に記載の核酸分子。
【請求項27】
核酸配列(c)が、プロモーター(a)の制御下で転写された際に、その転写産物に少なくとも100の連続するAヌクレオチドのヌクレオチド配列をコードする、請求項26に記載の核酸分子。
【請求項28】
核酸配列(c)が、プロモーター(a)の制御下で転写された際に、その転写産物に約120の連続するAヌクレオチドのヌクレオチド配列をコードする、請求項27に記載の核酸分子。
【請求項29】
切断が5’→3’転写方向に、プロモーター(a)、核酸配列(b)、および核酸配列(c)の少なくとも一部(この核酸配列(c)の少なくとも一部は、プロモーター(a)の制御下で転写された際に、その転写産物に少なくとも20の連続するAヌクレオチドのヌクレオチド配列をコードし、その転写産物において3’末端ヌクレオチドは少なくとも20の連続するAヌクレオチドの該ヌクレオチド配列のAヌクレオチドである)を含む核酸分子を生じるように、核酸配列(c)内で、好ましくは酵素的にまたは別の生化学的方法で切断され得ることを特徴とする、請求項23〜28のいずれか一項に記載の核酸分子。
【請求項30】
切断後、前記核酸分子が、少なくとも20の連続するAヌクレオチドのヌクレオチド配列の鋳型として働く鎖の末端に、その転写産物において少なくとも20の連続するAヌクレオチドのヌクレオチド配列の鋳型として働くヌクレオチド配列の一部であるTヌクレオチドを有する、請求項29に記載の核酸分子。
【請求項31】
前記核酸配列(c)の少なくとも一部が、プロモーター(a)の制御下で転写された際に、その転写産物に少なくとも40の連続するAヌクレオチドのヌクレオチド配列をコードする、請求項29または30に記載の核酸分子。
【請求項32】
前記核酸配列(c)の少なくとも一部が、プロモーター(a)の制御下で転写された際に、その転写産物に少なくとも80の連続するAヌクレオチドのヌクレオチド配列をコードする、請求項31に記載の核酸分子。
【請求項33】
前記核酸配列(c)の少なくとも一部が、プロモーター(a)の制御下で転写された際に、その転写産物に少なくとも100の連続するAヌクレオチドのヌクレオチド配列をコードする、請求項32に記載の核酸分子。
【請求項34】
前記核酸配列(c)の少なくとも一部が、プロモーター(a)の制御下で転写された際に、その転写産物に約120連続するAヌクレオチドのヌクレオチド配列をコードする、請求項33に記載の核酸分子。
【請求項35】
切断前には閉環分子であり、切断後には直鎖分子であることを特徴とする、請求項29〜34のいずれか一項に記載の核酸分子。
【請求項36】
切断が制限切断部位の助けで行われる、請求項29〜35のいずれか一項に記載の核酸分子。
【請求項37】
前記制限切断部位がIIS型制限エンドヌクレアーゼの制限切断部位である、請求項36に記載の核酸分子。
【請求項38】
前記IIS型制限エンドヌクレアーゼの認識配列が、核酸配列(c)の3’末端の5〜26塩基対下流に位置する、請求項37に記載の核酸分子。
【請求項39】
前記IIS型制限エンドヌクレアーゼの認識配列が、核酸配列(c)の3’末端の24〜26塩基対下流に位置する、請求項38に記載の核酸分子。
【請求項40】
前記の転写可能な核酸配列がペプチドまたはタンパク質をコードする核酸配列を含み、前記の転写可能な核酸配列を導入するための核酸配列が多重クローニング部位である、請求項1〜39のいずれか一項に記載の核酸分子。
【請求項41】
(i)レポーター遺伝子;(ii)選択マーカー;および(iii)複製起点からなる群から選択される1以上のメンバーをさらに含む、請求項1〜40のいずれか一項に記載の核酸分子。
【請求項42】
前記核酸分子が閉環コンフォメーションである、請求項1〜41のいずれか一項に記載の核酸分子。
【請求項43】
特に線状化後に、RNA、特にmRNAのin vitro転写に好適である、請求項1〜42のいずれか一項に記載の核酸分子。
【請求項44】
請求項1〜43のいずれか一項に記載の核酸分子の線状化により得られる、核酸分子。
【請求項45】
プロモーター(a)の制御下、請求項1〜44のいずれか一項に記載の核酸分子とともに転写される、好ましくはin vitro転写させることにより得られる、RNA。
【請求項46】
その安定性および/または翻訳効率を高めるよう選択されたRNA分子をin vitroで転写させる方法であって、
(i)遺伝子の3’非翻訳領域に相当する、またはそれに由来する第一の核酸配列(b−1)を、転写されて該RNA分子を生じ得る核酸配列(a)の3’末端に結合させること、
(ii)遺伝子の3’非翻訳領域に相当する、またはそれに由来する第二の核酸配列(b−2)を、該第一の核酸配列(b−1)の3’末端に結合させること、および
(iii)得られた核酸をin vitroで転写させること
を含む、方法。
【請求項47】
その発現を増強するよう選択されたmRNA分子を転写させる方法であって、
(i)遺伝子の3’非翻訳領域に相当する、またはそれに由来する第一の核酸配列(b−1)を、転写されて該RNA分子を生じ得る核酸配列(a)の3’末端に結合させること、
(ii)遺伝子の3’非翻訳領域に相当する、またはそれに由来する第二の核酸配列(b−2)を、該第一の核酸配列(b−1)の3’末端に結合させること、および
(iii)得られた核酸を転写させることにより得ることができるmRNAを翻訳すること
を含む、方法。
【請求項48】
転写がin vitroで行われる、請求項47に記載の方法。
【請求項49】
遺伝子の3’非翻訳領域に相当する、またはそれに由来する少なくとも1つのさらなる核酸配列(b−3)を第二の核酸配列(b−2)の3’末端に結合させることをさらに含む、請求項46〜48のいずれか一項に記載の方法。
【請求項50】
核酸配列(a)、(b−1)、(b−2)および適当であれば(b−3)が転写されて、核酸配列(b−1)および/または(b−2)および/または適当であれば(b−3)から転写された核酸配列に、核酸配列(a)から転写された核酸配列の翻訳効率および/または安定性を高めるような活性がある共通転写産物を生じ得る、請求項46〜49のいずれか一項に記載の方法。
【請求項51】
転写された際に、少なくとも20の連続するAヌクレオチドのヌクレオチド配列をコードする核酸配列(c)を核酸配列(b−2)の、または適当であれば核酸配列(b−3)の3’末端に結合させることをさらに含む、請求項46〜50のいずれか一項に記載の方法。
【請求項52】
核酸配列(a)、(b−1)、(b−2)および適当であれば(b−3)ならびに(c)が転写されて、核酸配列(b−1)および/または(b−2)および/または適当であれば(b−3)および/または(c)から転写された核酸配列に、核酸配列(a)から転写された核酸配列の翻訳効率および/または安定性を高めるような活性がある共通転写産物を生じ得る、請求項51に記載の方法。
【請求項53】
得られた核酸の転写の前に、このようにして得られた核酸の転写が、核酸配列(a)、(b−1)、(b−2)および適当であれば(b−3)ならびに少なくとも20の連続するAヌクレオチドの3’末端ヌクレオチド配列から転写された核酸配列を有する転写産物(該転写産物の3’末端ヌクレオチドは少なくとも20の連続するAヌクレオチドのヌクレオチド配列のAヌクレオチドである)を生じるように核酸配列(c)内で切断することをさらに含むことを特徴とする、請求項51または52に記載の方法。
【請求項54】
核酸配列(b−1)、(b−2)および適当であれば(b−3)が互いに独立にα2−グロビン、α1−グロビン、β−グロビンおよび成長ホルモン、好ましくはヒトβ−グロビンからなる群から選択される遺伝子に由来する、請求項46〜53のいずれか一項に記載の方法。
【請求項55】
その安定性および/または翻訳効率を高めるよう選択されたRNA分子をin vitroで転写させる方法であって、
(i)転写された際に、少なくとも20の連続するAヌクレオチドのヌクレオチド配列をコードする核酸配列(b)を、転写されて前記RNA分子を生じ得る核酸配列(a)の3’末端に結合させること、および
(ii)得られた核酸をin vitroで転写させること
を含む、方法。
【請求項56】
その発現を増強するよう選択されたmRNA分子を翻訳する方法であって、
(i)転写された際に、少なくとも20の連続するAヌクレオチドのヌクレオチド配列をコードする核酸配列(b)を、転写されて前記mRNA分子を生じ得る核酸配列(a)の3’末端に結合させること、および
(ii)得られた核酸を転写させることにより得ることができるmRNAを翻訳すること
を含む、方法。
【請求項57】
転写がin vitroで行われる、請求項56に記載の方法。
【請求項58】
核酸配列(a)および(b)が転写されて、核酸配列(b)から転写された核酸配列に、核酸配列(a)から転写された核酸配列の翻訳効率および/または安定性を高めるような活性がある共通転写産物を生じ得る、請求項55〜57のいずれか一項に記載の方法。
【請求項59】
得られた核酸の転写の前に、このようにして得られた核酸の転写が、核酸配列(a)、および少なくとも20の連続するAヌクレオチドの3’末端ヌクレオチド配列から転写された核酸配列を有する転写産物(該転写産物の3’末端ヌクレオチドは少なくとも20の連続するAヌクレオチドのヌクレオチド配列のAヌクレオチドである)を生じるように核酸配列(b)内で切断することをさらに含むことを特徴とする、請求項55〜58のいずれか一項に記載の方法。
【請求項60】
切断が制限切断部位の助けで行われる、請求項53または59に記載の方法。
【請求項61】
前記制限切断部位がIIS型制限エンドヌクレアーゼの制限切断部位である、請求項60に記載の方法。
【請求項62】
前記IIS型制限エンドヌクレアーゼの認識配列が、転写された際に、少なくとも20の連続するAヌクレオチドのヌクレオチド配列をコードする核酸配列の3’末端の5〜26塩基対下流に位置する、請求項61に記載の方法。
【請求項63】
前記IIS型制限エンドヌクレアーゼの認識配列が、転写された際に、少なくとも20の連続するAヌクレオチドのヌクレオチド配列をコードする核酸配列の3’末端の24〜26塩基対下流に位置する、請求項62に記載の方法。
【請求項64】
請求項46、49〜55および58〜63のいずれか一項に記載の選択されたRNA分子をin vitroで転写させる方法により得ることができるRNA。
【請求項65】
宿主細胞をトランスフェクトするための請求項45または64に記載のRNAの使用。
【請求項66】
前記宿主細胞が抗原提示細胞、特に、樹状細胞、単球またはマクロファージである、請求項65に記載の使用。
【請求項1】
5’→3’転写方向に、
(a)プロモーター;
(b)転写可能な核酸配列または転写可能な核酸配列を導入するための核酸配列;
(c−1)遺伝子の3’非翻訳領域に相当する、またはそれに由来する第一の核酸配列;および
(c−2)遺伝子の3’非翻訳領域に相当する、またはそれに由来する第二の核酸配列
を含む、核酸分子。
【請求項2】
(c−3)遺伝子の3’非翻訳領域に相当する、またはそれに由来する少なくとも1つのさらなる核酸配列をさらに含む、請求項1に記載の核酸分子。
【請求項3】
プロモーター(a)の制御下の核酸配列(b)、(c−1)、(c−2)および適当であれば(c−3)が転写されて、核酸配列(c−1)および/または(c−2)および/または適当であれば(c−3)から転写された核酸配列に、転写可能な核酸配列(b)から転写された核酸配列の翻訳効率および/または安定性を高めるような活性がある共通転写産物を生じ得る、請求項1または2に記載の核酸分子。
【請求項4】
核酸配列(c−1)、(c−2)および適当であれば(c−3)が同一であっても異なっていてもよい、請求項1〜3のいずれか一項に記載の核酸分子。
【請求項5】
(d)プロモーター(a)の制御下で転写された際に、その転写産物に少なくとも20の連続するAヌクレオチドのヌクレオチド配列をコードする核酸配列をさらに含む、請求項1〜4のいずれか一項に記載の核酸分子。
【請求項6】
プロモーター(a)の制御下の核酸配列(b)、(c−1)、(c−2)、適当であれば(c−3)および(d)が転写されて、核酸配列(c−1)および/または(c−2)および/または適当であれば(c−3)および/または(d)から転写された核酸配列に、転写可能な核酸配列(b)から転写された核酸配列の翻訳効率および/または安定性を高めるような活性がある共通転写産物を生じ得る、請求項5に記載の核酸分子。
【請求項7】
核酸配列(d)が、プロモーター(a)の制御下で転写された際に、その転写産物に少なくとも40の連続するAヌクレオチドのヌクレオチド配列をコードする、請求項5または6に記載の核酸分子。
【請求項8】
核酸配列(d)が、プロモーター(a)の制御下で転写された際に、その転写産物に少なくとも80の連続するAヌクレオチドのヌクレオチド配列をコードする、請求項7に記載の核酸分子。
【請求項9】
核酸配列(d)が、プロモーター(a)の制御下で転写された際に、その転写産物に少なくとも100の連続するAヌクレオチドのヌクレオチド配列をコードする、請求項8に記載の核酸分子。
【請求項10】
核酸配列(d)が、プロモーター(a)の制御下で転写された際に、その転写産物に約120の連続するAヌクレオチドのヌクレオチド配列をコードする、請求項9に記載の核酸分子。
【請求項11】
切断が5’→3’転写方向に、プロモーター(a)、核酸配列(b)、核酸配列(c−1)、(c−2)および適当であれば(c−3)、ならびに核酸配列(d)の少なくとも一部(この核酸配列(d)の少なくとも一部は、プロモーター(a)の制御下で転写された際に、その転写産物に少なくとも20の連続するAヌクレオチドのヌクレオチド配列をコードし、その転写産物において3’末端ヌクレオチドは少なくとも20の連続するAヌクレオチドの該ヌクレオチド配列のAヌクレオチドである)を含む核酸分子を生じるように、核酸配列(d)内で、好ましくは酵素的にまたは別の生化学的方法で切断され得ることを特徴とする、請求項5〜10のいずれか一項に記載の核酸分子。
【請求項12】
切断後、前記核酸分子が、少なくとも20の連続するAヌクレオチドのヌクレオチド配列の鋳型として働く鎖の末端に、その転写産物において少なくとも20の連続するAヌクレオチドのヌクレオチド配列の鋳型として働くヌクレオチド配列の一部であるTヌクレオチドを有する、請求項11に記載の核酸分子。
【請求項13】
前記核酸配列(d)の少なくとも一部が、プロモーター(a)の制御下で転写された際に、その転写産物に少なくとも40の連続するAヌクレオチドのヌクレオチド配列をコードする、請求項11または12に記載の核酸分子。
【請求項14】
前記核酸配列(d)の少なくとも一部が、プロモーター(a)の制御下で転写された際に、その転写産物に少なくとも80の連続するAヌクレオチドのヌクレオチド配列をコードする、請求項13に記載の核酸分子。
【請求項15】
前記核酸配列(d)の少なくとも一部が、プロモーター(a)の制御下で転写された際に、その転写産物に少なくとも100の連続するAヌクレオチドのヌクレオチド配列をコードする、請求項14に記載の核酸分子。
【請求項16】
前記核酸配列(d)の少なくとも一部が、プロモーター(a)の制御下で転写された際に、その転写産物に約120の連続するAヌクレオチドのヌクレオチド配列をコードする、請求項15に記載の核酸分子。
【請求項17】
切断前には閉環分子であり、切断後には直鎖分子であることを特徴とする、請求項11〜16のいずれか一項に記載の核酸分子。
【請求項18】
前記切断が制限切断部位の助けで行われる、請求項11〜17のいずれか一項に記載の核酸分子。
【請求項19】
前記制限切断部位がIIS型制限エンドヌクレアーゼの制限切断部位である、請求項18に記載の核酸分子。
【請求項20】
前記IIS型制限エンドヌクレアーゼの認識配列が、核酸配列(d)の3’末端の5〜26塩基対下流に位置する、請求項19に記載の核酸分子。
【請求項21】
前記IIS型制限エンドヌクレアーゼの認識配列が、核酸配列(d)の3’末端の24〜26塩基対下流に位置する、請求項20に記載の核酸分子。
【請求項22】
核酸配列(c−1)、(c−2)および適当であれば(c−3)が互いに独立にα2−グロビン、α1−グロビン、β−グロビンおよび成長ホルモン、好ましくは、ヒトβ−グロビンからなる群から選択される遺伝子に由来する、請求項1〜21のいずれか一項に記載の核酸分子。
【請求項23】
5’→3’転写方向に、
(a)プロモーター;
(b)転写可能な核酸配列または転写可能な核酸配列を導入するための核酸配列;および
(c)プロモーター(a)の制御下で転写された際に、その転写産物に少なくとも20の連続するAヌクレオチドのヌクレオチド配列をコードする核酸配列
を含む、核酸分子。
【請求項24】
プロモーター(a)の制御下の核酸配列(b)および(c)が転写されて、核酸配列(c)から転写された核酸配列に、転写可能な核酸配列(b)から転写された核酸配列の翻訳効率および/または安定性を高めるような活性がある共通転写産物を生じ得る、請求項23に記載の核酸分子。
【請求項25】
核酸配列(c)が、プロモーター(a)の制御下で転写された際に、その転写産物に少なくとも40の連続するAヌクレオチドのヌクレオチド配列をコードする、請求項23または24に記載の核酸分子。
【請求項26】
核酸配列(c)が、プロモーター(a)の制御下で転写された際に、その転写産物に少なくとも80の連続するAヌクレオチドのヌクレオチド配列をコードする、請求項25に記載の核酸分子。
【請求項27】
核酸配列(c)が、プロモーター(a)の制御下で転写された際に、その転写産物に少なくとも100の連続するAヌクレオチドのヌクレオチド配列をコードする、請求項26に記載の核酸分子。
【請求項28】
核酸配列(c)が、プロモーター(a)の制御下で転写された際に、その転写産物に約120の連続するAヌクレオチドのヌクレオチド配列をコードする、請求項27に記載の核酸分子。
【請求項29】
切断が5’→3’転写方向に、プロモーター(a)、核酸配列(b)、および核酸配列(c)の少なくとも一部(この核酸配列(c)の少なくとも一部は、プロモーター(a)の制御下で転写された際に、その転写産物に少なくとも20の連続するAヌクレオチドのヌクレオチド配列をコードし、その転写産物において3’末端ヌクレオチドは少なくとも20の連続するAヌクレオチドの該ヌクレオチド配列のAヌクレオチドである)を含む核酸分子を生じるように、核酸配列(c)内で、好ましくは酵素的にまたは別の生化学的方法で切断され得ることを特徴とする、請求項23〜28のいずれか一項に記載の核酸分子。
【請求項30】
切断後、前記核酸分子が、少なくとも20の連続するAヌクレオチドのヌクレオチド配列の鋳型として働く鎖の末端に、その転写産物において少なくとも20の連続するAヌクレオチドのヌクレオチド配列の鋳型として働くヌクレオチド配列の一部であるTヌクレオチドを有する、請求項29に記載の核酸分子。
【請求項31】
前記核酸配列(c)の少なくとも一部が、プロモーター(a)の制御下で転写された際に、その転写産物に少なくとも40の連続するAヌクレオチドのヌクレオチド配列をコードする、請求項29または30に記載の核酸分子。
【請求項32】
前記核酸配列(c)の少なくとも一部が、プロモーター(a)の制御下で転写された際に、その転写産物に少なくとも80の連続するAヌクレオチドのヌクレオチド配列をコードする、請求項31に記載の核酸分子。
【請求項33】
前記核酸配列(c)の少なくとも一部が、プロモーター(a)の制御下で転写された際に、その転写産物に少なくとも100の連続するAヌクレオチドのヌクレオチド配列をコードする、請求項32に記載の核酸分子。
【請求項34】
前記核酸配列(c)の少なくとも一部が、プロモーター(a)の制御下で転写された際に、その転写産物に約120連続するAヌクレオチドのヌクレオチド配列をコードする、請求項33に記載の核酸分子。
【請求項35】
切断前には閉環分子であり、切断後には直鎖分子であることを特徴とする、請求項29〜34のいずれか一項に記載の核酸分子。
【請求項36】
切断が制限切断部位の助けで行われる、請求項29〜35のいずれか一項に記載の核酸分子。
【請求項37】
前記制限切断部位がIIS型制限エンドヌクレアーゼの制限切断部位である、請求項36に記載の核酸分子。
【請求項38】
前記IIS型制限エンドヌクレアーゼの認識配列が、核酸配列(c)の3’末端の5〜26塩基対下流に位置する、請求項37に記載の核酸分子。
【請求項39】
前記IIS型制限エンドヌクレアーゼの認識配列が、核酸配列(c)の3’末端の24〜26塩基対下流に位置する、請求項38に記載の核酸分子。
【請求項40】
前記の転写可能な核酸配列がペプチドまたはタンパク質をコードする核酸配列を含み、前記の転写可能な核酸配列を導入するための核酸配列が多重クローニング部位である、請求項1〜39のいずれか一項に記載の核酸分子。
【請求項41】
(i)レポーター遺伝子;(ii)選択マーカー;および(iii)複製起点からなる群から選択される1以上のメンバーをさらに含む、請求項1〜40のいずれか一項に記載の核酸分子。
【請求項42】
前記核酸分子が閉環コンフォメーションである、請求項1〜41のいずれか一項に記載の核酸分子。
【請求項43】
特に線状化後に、RNA、特にmRNAのin vitro転写に好適である、請求項1〜42のいずれか一項に記載の核酸分子。
【請求項44】
請求項1〜43のいずれか一項に記載の核酸分子の線状化により得られる、核酸分子。
【請求項45】
プロモーター(a)の制御下、請求項1〜44のいずれか一項に記載の核酸分子とともに転写される、好ましくはin vitro転写させることにより得られる、RNA。
【請求項46】
その安定性および/または翻訳効率を高めるよう選択されたRNA分子をin vitroで転写させる方法であって、
(i)遺伝子の3’非翻訳領域に相当する、またはそれに由来する第一の核酸配列(b−1)を、転写されて該RNA分子を生じ得る核酸配列(a)の3’末端に結合させること、
(ii)遺伝子の3’非翻訳領域に相当する、またはそれに由来する第二の核酸配列(b−2)を、該第一の核酸配列(b−1)の3’末端に結合させること、および
(iii)得られた核酸をin vitroで転写させること
を含む、方法。
【請求項47】
その発現を増強するよう選択されたmRNA分子を転写させる方法であって、
(i)遺伝子の3’非翻訳領域に相当する、またはそれに由来する第一の核酸配列(b−1)を、転写されて該RNA分子を生じ得る核酸配列(a)の3’末端に結合させること、
(ii)遺伝子の3’非翻訳領域に相当する、またはそれに由来する第二の核酸配列(b−2)を、該第一の核酸配列(b−1)の3’末端に結合させること、および
(iii)得られた核酸を転写させることにより得ることができるmRNAを翻訳すること
を含む、方法。
【請求項48】
転写がin vitroで行われる、請求項47に記載の方法。
【請求項49】
遺伝子の3’非翻訳領域に相当する、またはそれに由来する少なくとも1つのさらなる核酸配列(b−3)を第二の核酸配列(b−2)の3’末端に結合させることをさらに含む、請求項46〜48のいずれか一項に記載の方法。
【請求項50】
核酸配列(a)、(b−1)、(b−2)および適当であれば(b−3)が転写されて、核酸配列(b−1)および/または(b−2)および/または適当であれば(b−3)から転写された核酸配列に、核酸配列(a)から転写された核酸配列の翻訳効率および/または安定性を高めるような活性がある共通転写産物を生じ得る、請求項46〜49のいずれか一項に記載の方法。
【請求項51】
転写された際に、少なくとも20の連続するAヌクレオチドのヌクレオチド配列をコードする核酸配列(c)を核酸配列(b−2)の、または適当であれば核酸配列(b−3)の3’末端に結合させることをさらに含む、請求項46〜50のいずれか一項に記載の方法。
【請求項52】
核酸配列(a)、(b−1)、(b−2)および適当であれば(b−3)ならびに(c)が転写されて、核酸配列(b−1)および/または(b−2)および/または適当であれば(b−3)および/または(c)から転写された核酸配列に、核酸配列(a)から転写された核酸配列の翻訳効率および/または安定性を高めるような活性がある共通転写産物を生じ得る、請求項51に記載の方法。
【請求項53】
得られた核酸の転写の前に、このようにして得られた核酸の転写が、核酸配列(a)、(b−1)、(b−2)および適当であれば(b−3)ならびに少なくとも20の連続するAヌクレオチドの3’末端ヌクレオチド配列から転写された核酸配列を有する転写産物(該転写産物の3’末端ヌクレオチドは少なくとも20の連続するAヌクレオチドのヌクレオチド配列のAヌクレオチドである)を生じるように核酸配列(c)内で切断することをさらに含むことを特徴とする、請求項51または52に記載の方法。
【請求項54】
核酸配列(b−1)、(b−2)および適当であれば(b−3)が互いに独立にα2−グロビン、α1−グロビン、β−グロビンおよび成長ホルモン、好ましくはヒトβ−グロビンからなる群から選択される遺伝子に由来する、請求項46〜53のいずれか一項に記載の方法。
【請求項55】
その安定性および/または翻訳効率を高めるよう選択されたRNA分子をin vitroで転写させる方法であって、
(i)転写された際に、少なくとも20の連続するAヌクレオチドのヌクレオチド配列をコードする核酸配列(b)を、転写されて前記RNA分子を生じ得る核酸配列(a)の3’末端に結合させること、および
(ii)得られた核酸をin vitroで転写させること
を含む、方法。
【請求項56】
その発現を増強するよう選択されたmRNA分子を翻訳する方法であって、
(i)転写された際に、少なくとも20の連続するAヌクレオチドのヌクレオチド配列をコードする核酸配列(b)を、転写されて前記mRNA分子を生じ得る核酸配列(a)の3’末端に結合させること、および
(ii)得られた核酸を転写させることにより得ることができるmRNAを翻訳すること
を含む、方法。
【請求項57】
転写がin vitroで行われる、請求項56に記載の方法。
【請求項58】
核酸配列(a)および(b)が転写されて、核酸配列(b)から転写された核酸配列に、核酸配列(a)から転写された核酸配列の翻訳効率および/または安定性を高めるような活性がある共通転写産物を生じ得る、請求項55〜57のいずれか一項に記載の方法。
【請求項59】
得られた核酸の転写の前に、このようにして得られた核酸の転写が、核酸配列(a)、および少なくとも20の連続するAヌクレオチドの3’末端ヌクレオチド配列から転写された核酸配列を有する転写産物(該転写産物の3’末端ヌクレオチドは少なくとも20の連続するAヌクレオチドのヌクレオチド配列のAヌクレオチドである)を生じるように核酸配列(b)内で切断することをさらに含むことを特徴とする、請求項55〜58のいずれか一項に記載の方法。
【請求項60】
切断が制限切断部位の助けで行われる、請求項53または59に記載の方法。
【請求項61】
前記制限切断部位がIIS型制限エンドヌクレアーゼの制限切断部位である、請求項60に記載の方法。
【請求項62】
前記IIS型制限エンドヌクレアーゼの認識配列が、転写された際に、少なくとも20の連続するAヌクレオチドのヌクレオチド配列をコードする核酸配列の3’末端の5〜26塩基対下流に位置する、請求項61に記載の方法。
【請求項63】
前記IIS型制限エンドヌクレアーゼの認識配列が、転写された際に、少なくとも20の連続するAヌクレオチドのヌクレオチド配列をコードする核酸配列の3’末端の24〜26塩基対下流に位置する、請求項62に記載の方法。
【請求項64】
請求項46、49〜55および58〜63のいずれか一項に記載の選択されたRNA分子をin vitroで転写させる方法により得ることができるRNA。
【請求項65】
宿主細胞をトランスフェクトするための請求項45または64に記載のRNAの使用。
【請求項66】
前記宿主細胞が抗原提示細胞、特に、樹状細胞、単球またはマクロファージである、請求項65に記載の使用。
【図1】


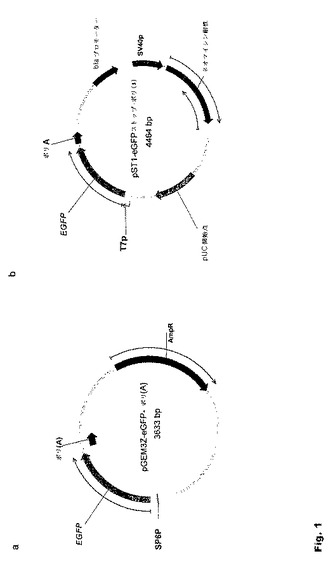
【図2】
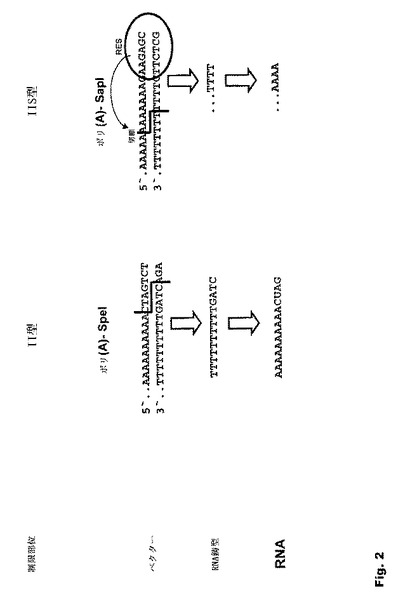
【図3−1】
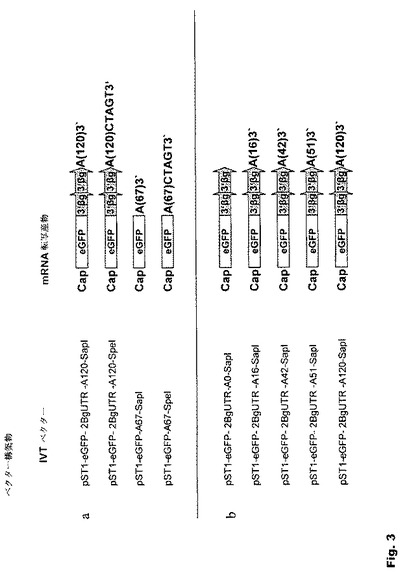
【図3−2】

【図4】
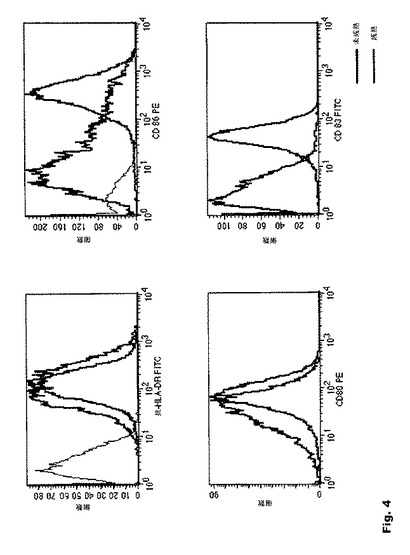
【図5(a)】
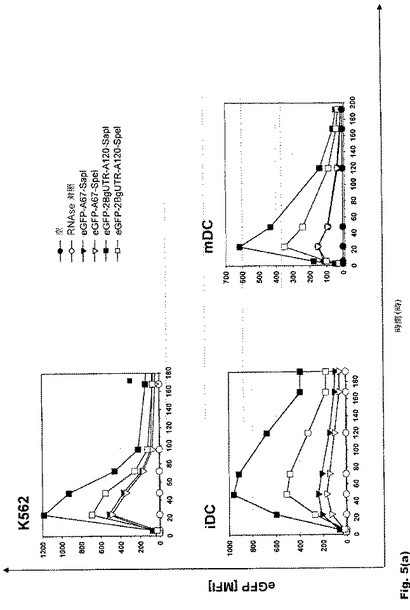
【図5(b)】
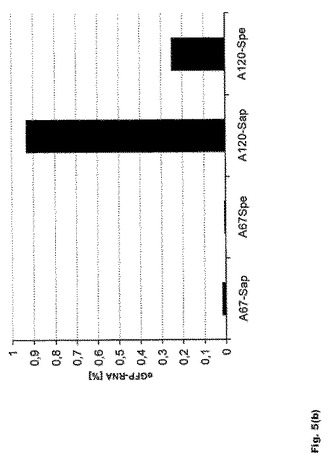
【図6(a)】
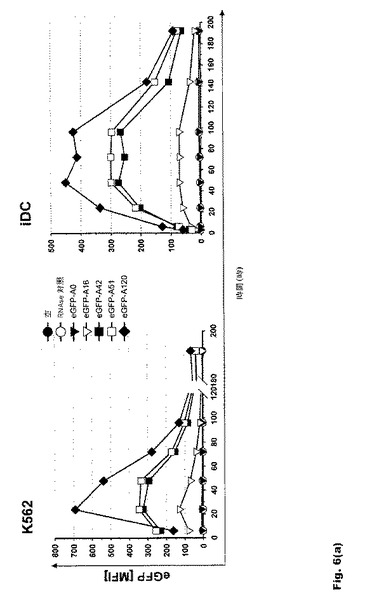
【図6(b)】
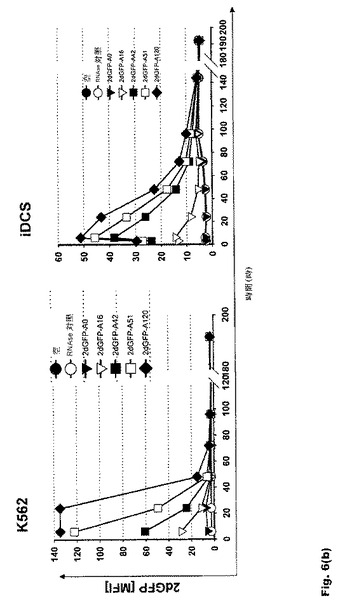
【図6(c)】
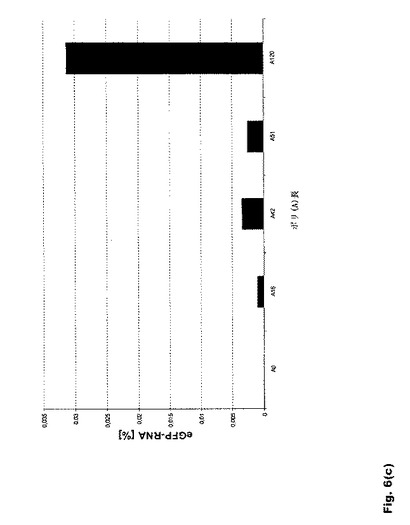
【図7】
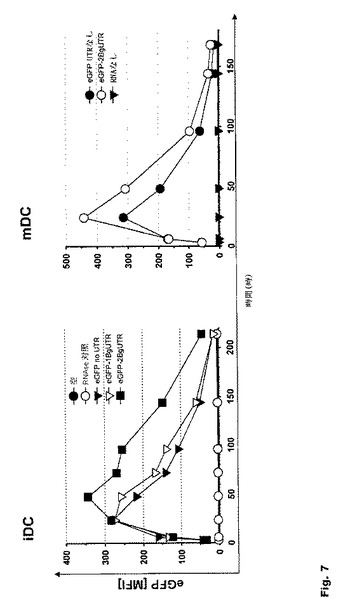
【図8】
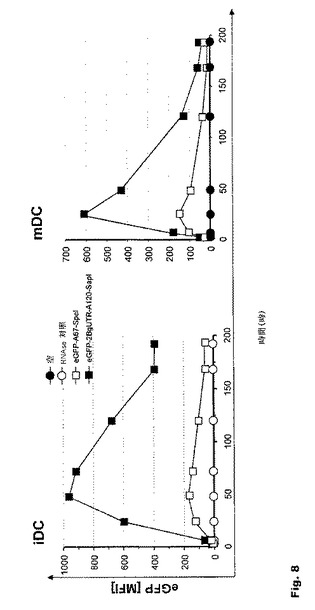
【図9】
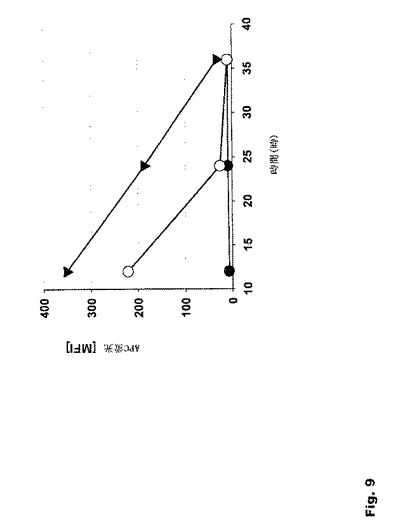
【図10】
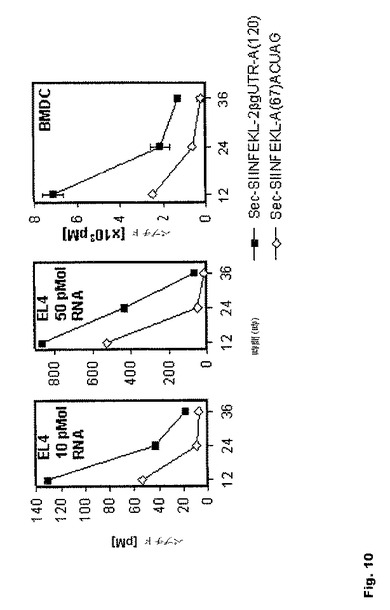
【図11】
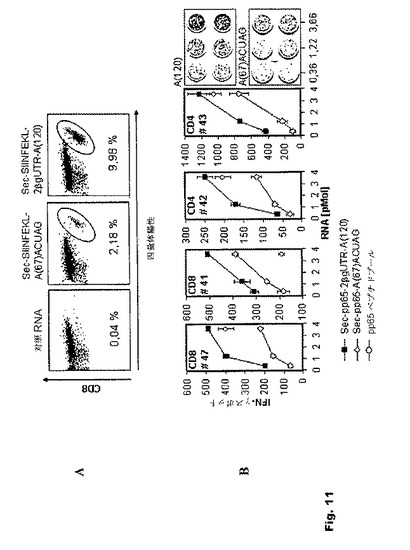
【図2】
【図3−1】
【図3−2】
【図4】
【図5(a)】
【図5(b)】
【図6(a)】
【図6(b)】
【図6(c)】
【図7】
【図8】
【図9】
【図10】
【図11】
【公開番号】特開2012−235786(P2012−235786A)
【公開日】平成24年12月6日(2012.12.6)
【国際特許分類】
【外国語出願】
【出願番号】特願2012−154852(P2012−154852)
【出願日】平成24年7月10日(2012.7.10)
【分割の表示】特願2008−532678(P2008−532678)の分割
【原出願日】平成18年9月28日(2006.9.28)
【出願人】(509162551)ビオエンテッヒ・アクチェンゲゼルシャフト (1)
【氏名又は名称原語表記】BioNTech AG
【Fターム(参考)】
【公開日】平成24年12月6日(2012.12.6)
【国際特許分類】
【出願番号】特願2012−154852(P2012−154852)
【出願日】平成24年7月10日(2012.7.10)
【分割の表示】特願2008−532678(P2008−532678)の分割
【原出願日】平成18年9月28日(2006.9.28)
【出願人】(509162551)ビオエンテッヒ・アクチェンゲゼルシャフト (1)
【氏名又は名称原語表記】BioNTech AG
【Fターム(参考)】
[ Back to top ]