
農作物の真菌病の処置のためのメチオニンシンターゼ阻害剤の使用
本発明は、農作物の真菌病の処置のためのメチオニンシンターゼ阻害剤の使用に関する。本発明はさらに、メチオニンシンターゼ阻害剤の施用を含む、真菌病に対する農作物の処置のための方法、また、メチオニンシンターゼ阻害剤の特定のための工程を含む、新規殺菌化合物の特定のための方法に関する。
【発明の詳細な説明】
【技術分野】
【0001】
本発明は、真菌病の処置、より詳細には農作物種の真菌病の処置のためのメチオニンシンターゼ阻害剤の使用に関する。
【背景技術】
【0002】
真菌は、様々な植物種の農作物の実質的な損失をもたらし得る壊滅的な伝染病の原因である。病原性真菌の酵素の阻害剤を使用すること、及びこれらの真菌に対して活性な新しい分子を特定するために試験においてこれらの酵素を使用することの原理は、それ自体公知である。しかし、単に真菌酵素の特性を決定するだけではこの目的を達成するのに十分ではなく、潜在的抗真菌分子のための標的として選択される酵素は、同時に真菌の生存にとって必須でなければならず、抗真菌分子によるこの阻害が真菌の死滅を生じさせるか、又は真菌の病原性にとって不可欠でなければならず、この場合、この阻害は真菌にとって致死的ではなく、単にこの病原能力を阻害する。真菌の病原性及び生存にとって必須の代謝経路及び酵素の特定が、従って、新規抗真菌製品の開発のために必要である。
【発明の開示】
【発明が解決しようとする課題】
【0003】
硫黄同化経路は、硫酸イオン(SO42−)の取り込み、この活性化、及び還元硫黄(S2−)へのこの還元を含む。これらの工程は、ATPスルフリラーゼ(EC2.7.7.4)、APSキナーゼ(EC2.7.1.25)、PAPSレダクターゼ(EC1.8.4.8)(光合成生物におけるAPSレダクターゼ、EC1.8.4.9)、及び(NADPH2)亜硫酸レダクターゼ(EC1.8.1.2)(光合成生物におけるフェレドキシン依存性酵素、EC1.8.7.1)によって連続的に触媒される。全ての独立栄養生物において、硫酸イオン同化、活性化及び還元経路は、この一般原則の観点から言えば保存されている;炭素骨格への還元硫黄の取り込みは生物によってかなりの変動を示す。細菌1(例えば大腸菌)、植物2(例えばシロイヌナズナ)、酵母(例えばサッカロミセス・セレビシエ(Saccharomyces cerevisiae)3)及び糸状菌4。実際に、植物及び細菌では、還元硫黄は、セリンに由来するC3の分子に取り込まれてシステインを形成する。次に硫黄は、ホモセリンに由来するC4の分子に転移してホモシステインを形成する。この一連の反応が直接トランススルフレーション経路を形成する。逆に、サッカロミセス・セレビシエ(S.cerevisiae)では、硫黄は、ホモセリンに由来するC4の分子に直接取り込まれてホモシステインを形成する(直接スルフヒドリル化)3。次にシステインは、逆トランススルフレーション経路を構成する一連の反応によってホモシステインから合成される。糸状菌では、ホモシステインの合成は植物における直接経路(直接トランススルフレーション)及びサッカロミセス・セレビシエの経路(直接スルフヒドリル化)の両方によって行われる。さらに、システインの合成は、セリンによって又は逆トランススルフレーション経路を通してホモシステインから実施される。これらの様々な代謝経路は、ニューロスポラ・クラッサ(Neurospora crassa)(N.crassa)5及びアセペルギルス・ニドゥランス(Aspergillus nidulans)(A.nidulnas)6におけるシステイン及びメチオニンについての栄養素要求性突然変異体の特性決定に従って定義された。このモデルは、植物(例えばマグナポルテ・グリセア(Magnaporthe grisea)、M.grisea)及び動物(例えばアスペルギルス・フミガツス(Aspergillus fumigatus)(A.fumigatus))の病原性真菌を含む全ての糸状菌類に当てはめることができる。イネ作物への多大の被害の原因である子嚢菌型病原体、マグナポルテ・グリセアは、このようなアプローチのための選択モデルである。糸状菌におけるメチオニン合成は、植物におけるようなビタミンB12非依存型のメチオニンシンターゼの作用を必要とする。ヒト病原菌、クリプトコッカス・ネオフォルマンス(Cryptococcus neoformans)28のメチオニンシンターゼ遺伝子の試験において記述されたアプローチは、本発明とは異なる。実際に、動物(ヒトを含む)はメチオニンを合成することができるが、この工程は、植物及び菌類のような他の真核生物のものとは大きく異なるビタミンB12依存型メチオニンシンターゼによって触媒される。植物メチオニンシンターゼは、タンパク質レベルでマグナポルテ・グリセアと強い相同性を示すが、同時に、実施されるモデリングに従って構造型の相違も示す9、12、27。従って、真菌阻害剤だけの特定を可能にする、この特異的性質を決定するためには、真菌酵素の特定及びこの特性決定が必要とされる。農作物を処置するための方法におけるこのような阻害剤の選択及び適用は特異的である。従って、本発明は、MET6遺伝子の突然変異体、より詳細にはマグナポルテ・グリセアのメチオニンシンターゼをコードするMET6遺伝子の欠失突然変異体がメチオニンに関して栄養素要求性であり、非病原性であるという事実を述べる。これらの突然変異体では、感染過程は、植物細胞への病原体の貫入期のレベルだけでなく、感染組織において進行するこの能力によっても達成される。マグナポルテ・グリセアメチオニンシンターゼ突然変異体の病原力は、感染の間にメチオニンを添加するとき部分的に回復される。これらの結果は、メチオニンシンターゼ活性の不在が感染の間の真菌にとって致死的であることを示す。同様の結果がトウモロコシ黒穂病菌(U.maydis)及びフィトフトラ・インフェスタンス(Phytophtora infestans)(P.infestans)でも得られた。
【0004】
配列表の説明
配列番号1:マグナポルテ・グリセアメチオニンシンターゼ遺伝子
配列番号2:マグナポルテ・グリセアメチオニンシンターゼcDNA
配列番号3:マグナポルテ・グリセアメチオニンシンターゼタンパク質配列
配列番号4:Met6−5プライマー
配列番号5:Met6−6プライマー
配列番号6:HphRP10プライマー
配列番号7:Met6−7プライマー
配列番号8:Met6−10プライマー
配列番号9:dCGS−hph−end(−)プライマー
配列番号10:Met6−8プライマー
配列番号11:Met6−9プライマー
配列番号12:Met6−1プライマー
配列番号13:Met6−2プライマー
配列番号14:Met6−3プライマー
配列番号15:Met6−4プライマー
配列番号16:トウモロコシ黒穂病菌メチオニンシンターゼ遺伝子
配列番号17:トウモロコシ黒穂病菌メチオニンシンターゼcDNA
配列番号18:トウモロコシ黒穂病菌メチオニンシンターゼタンパク質配列
配列番号19:フィトフトラ・インフェスタンス(Phytophtora infestans)メチオニンシンターゼEST配列
配列番号20:推定フィトフトラ・インフェスタンス(Phytophtora infestans)メチオニンシンターゼタンパク質配列
【課題を解決するための手段】
【0005】
本発明の1つの主題は、有効量のメチオニンシンターゼ阻害剤の施用によって真菌病に対して農作物を処置するための方法である。
【0006】
本発明に関して、真菌病は、子嚢菌、担子菌及び卵菌ファミリーに属する病原性植物真菌による病害と定義される。
【0007】
本発明の1つの主題は、有効(農業的に有効)な非植物毒性量のメチオニンシンターゼ阻害剤を、植物が成長する又は成長しやすい土壌、植物の葉及び/又は果実又は植物の種子に施用することを特徴とする、治療的又は予防的性能で、農作物の植物病原性菌類を防除するための方法である。「有効な非植物毒性量」という用語は、作物上に存在する又は出現し得る真菌の発生周期の制御又は破壊を可能にするのに十分であり、前記作物に対して植物毒性の著明な病徴を生じさせない阻害剤の量を意味することが意図されている。このような量は、防除すべき真菌ファミリー、作物の種類、気候条件及び本発明に従った抗真菌組成物に含まれる化合物に依存して、広い範囲内で変動し得る。
【0008】
この量は、当業者の技術範囲内である、体系的圃場試験によって決定することができる。
【発明を実施するための最良の形態】
【0009】
本発明に従った方法は、穀物(特に小麦、ライ麦、ライ小麦及び大麦)、ジャガイモ、綿、エンドウ豆、菜種、トウモロコシ又は亜麻の種子、選択的に林木の種子又はさもなければこれらの植物の遺伝的に改変された種子を処置するために有用である。本発明はまた、農作物への茎葉散布、すなわち当該植物の茎葉、葉、果実及び/又は茎への散布に関するが、また他のいかなる種類の施用にも関する。本発明に従った方法が標的する植物の中で、稲、トウモロコシ、小麦、大麦又はライ小麦などの穀物、果樹、特にリンゴの木、ナシの木、桃の木、ブドウの木、バナナの木、オレンジの木、レモンの木等、油生成作物、例えば菜種又はヒマワリ、市場向け菜園及び野菜作物、トマト、サラダ用作物、タンパク質生成作物、エンドウ豆、ナス科作物、例えばジャガイモ、ビートの根、亜麻、及び林木、及びまたこれらの作物の遺伝的に改変された同族体が挙げられる。
【0010】
本発明に従った方法が標的する植物の中で、
小麦、以下の種子病害を防除することに関して:フザリア(ミクロドキウム・ニバル(Microdochium nivale)及びフザリウム・ロゼウム(Fusarium roseum))、なまぐさ黒穂病(ティレティア・カリエス(Tilletia caries)、ティレティア・コントロバーサ(Tilletia controversa)又はティレティア・インディカ(Tilletia indica))、セプトリア病(セプトリア・ノドラム(Septoria nodorum));裸黒穂病(ウスチラゴ・トリティキ(Ustilago tritici));
小麦、植物の地上部分の以下の病害を防除することに関して:穀類眼紋病(タペシア・ヤルンダ(Tapesia yallundae)、タペシア・アクイフォルミス(Tapesia acuiformis))、立ち枯れ病(ゲウマノミセス・グラミニス(Gaeumannomyces graminis))、胴枯れ病(foot blight)(F.セルモラム(F. culmorum)、F.グラミネアラム(F. graminearum))、赤かび病(F.セルモラム、F.グラミネアラム、ミクロドキウム・ニバル)、黒斑病(black speck)(リゾクトニア・セレアリス(Rhizoctonia cerealis))、うどん粉病(トリティキ種エリシフェ・グラミニス・フォルマ(Erysiphe graminis forma specie tritici))、さび病(プキニア・ストリイフォルミス(Puccinia striiformis))及びプキニア・レコンディタ(Puccinia recondite))及びセプトリア病(セプトリア・トリティキ(Septoria tritici)及びセプトリア・ノドラム)、網斑病(ドレクスレラ・トリティキ−レペンティス(Drechslera tritici−repentis));
大麦、以下の種子病害を防除することに関して:網斑病(ピレノフォラ・グラミネア(Pyrenophora graminea)、ピレノフォラ・テレス(Pyrenophora teres)及びコクリオボラス・サティバス(Cochliobolus sativus))、裸黒穂病(ウスティラゴ・ヌーダ(Ustilago nuda))及びフザリウム病(ミクロドキウム・ニバル及びフザリウム・ロゼウム(Fusarium roseum));
大麦、植物の地上部分の以下の病害を防除することに関して:穀類眼紋病(タペシア・ヤルンダ)、網斑病(ピレノフォラ・テレス及びコクリオボラス・サティバス)、うどん粉病(ホルデイ種エリシフェ・グラミニス・フォルマ(Erysiphe graminis forma specie hordei))、小さび病(プキニア・ホルデイ(Puccinia hordei))及び葉枯れ病(リンコスポリウム・セカリス(Rhynchosporium secalis));
ジャガイモ、塊茎疾患(特にヘルミントスポリウム・ソラニ(Helminthosporium solani)、フォーマ・ツベローサ(Phoma tuberosa)、リゾクトニア・ソラニ(Rhizoctonia solani)、フザリウム・ソラニ(Fusarium solani))、及びうどん粉病(フィトフトラ・インフェスタンス(Phytophtora infestans))を防除することに関して;
ジャガイモ、以下の茎葉疾患を防除することに関して、夏疫病(アルテナリア・ソラニ(Alternaria solani))、うどん粉病(フィトフトラ・インフェスタンス);
綿、種子から成長した幼植物の以下の疾患を防除することに関して、立ち枯れ病及び斑点落葉病(collar rot)(リゾクトニア・ソラニ(Rhizoctonia solani)、フザリウム・オキシスポラム(Fusarium oxysporum))、黒根病(チエラビオプシス・バシコラ(Thielaviopsis basicola));
タンパク質生成作物、例えばエンドウ豆、以下の種子疾患を防除することに関して、炭疽病(アスコキタ・ピシ(Ascochyta pisi)、ミコスフェレラ・ピノデス(Mycosphaerella pinodes))、フザリウム病(フザリウム・オキシスポラム)、灰色かび病(ボツリティス・シネレア(Botrytis cinerea))、うどん粉病(ペロノスポラ・ピシ(Peronospora pisi));
油生成作物、例えば菜種、以下の種子疾患を防除することに関して、フォーマ・リンガム(Phoma lingam)、アルテルナリア・ブラシカ(Alternaria brassicae)及びスクレロティニア・スクレロティオラム(Sclerotinia);
トウモロコシ、種子疾患(リゾプス種(Rhizopus sp.)、ペニシリウム種(Penicillium sp.)、トリコデルマ種(Trichoderma sp.)、アスペルギルス種(Aspergillus sp.)及びギベレラ・フジクロイ(Gibberella fujikuroi))を防除することに関して;
亜麻、種子疾患を防除することに関して、アルテルナリア・リニコラ(Alternaria linicola);
林木、立ち枯れ病(フザリウム・オキシスポラム、リゾクトニア・ソラニ)を防除することに関して;
イネ、地上部分の以下の病害を防除することに関して、いもち病(マグナポルテ・グリセア)、黒斑病(リゾクトニア・ソラニ);
野菜作物、種子又は種子から成長した幼植物の以下の疾患を防除することに関して、立ち枯れ病及び斑点落葉病(フザリウム・オキシスポラム、フザリウム・ロゼウム、リゾクトニア・ソラニ、ピチウム種(Pythium sp.));
野菜作物、地上部分の以下の病害を防除することに関して、灰色かび病(ボツリティス種(Botrytis sp.))、うどん粉病(特にエリシフェ・シコラセアラム(Erysiphe cichoracearum)、スフェロテカ・フリギニエ(Sphaerotheca fuliginea)、レヴェイルラ・タウリカ(Leveillula taurica))、フザリウム病(フザリウム・オキシスポラム、フザリウム・ロゼウム)、葉枯れ病(クラドスポリウム種(Cladosporium sp.))、アルテルナリア斑点病(アルテルナリア種(Alternaria))、炭疽病(コレトトリカム種(Colletotrichum sp))、セプトリア葉枯れ病(セプトリア種(Septoria sp.))、黒斑病(リゾクトニア・ソラニ)、うどん粉病(例えばブレミア・ラクチュカ(Bremia lactucae)、ペロノスポラ種(Peronospora sp.)、シュードペルノスポラ種(Pseudoperonospora sp.)、フィトフトラ種(Phytophthora sp.));
果樹、地上部分の病害に関して、モニリア病(モニリア・フルクチゲナ(Monilia fructigenae)、M.ラクサ(M.laxa))、そうか病(ベンチュリア・イナクアリス(Venturia inaequalis))、うどん粉病(ポドスファエラ・リューコトリカ(Podosphaera leucotricha));
ブドウ、茎葉病害に関して、特に灰色かび病(ボツリティス・シネレア(Botrytis cinerea))、うどん粉病(ウンキヌラ・ネカトル(Uncinula necator))、胴枯れ病(ギグナルディア・ビウェリ(Guignardia biwelli))、うどん粉病(プラスモパラ・ヴィティコラ(Plasmopara viticola));
ビートの根、地上部分の以下の病害に関して、セルコスポラ病(セルコスポラ・ベティコラ(Cercospora beticola))、うどん粉病(エリシフェ・ベティコラ(Erysiphe beticola))、葉枯れ病(ラムラリア・ベティコラ(Ramularia beticola))
が挙げられる。
【0011】
メチオニンシンターゼは、植物及び微生物(細菌、酵母、真菌)において認められる広く特性付けられた酵素である。本発明の方法はメチオニンシンターゼ阻害剤を使用する。最初の実施形態では、本発明は、農作物の真菌病の処置のための、真菌メチオニンシンターゼの阻害剤、より好ましくは植物性病原真菌のメチオニンシンターゼの阻害剤の使用に関する。
【0012】
好ましくは、メチオニンシンターゼはこの天然環境から単離、精製又は部分精製される。メチオニンシンターゼは様々な方法によって調製することができる。これらの方法は、特に、これらのポリペプチドを天然に発現する細胞などの天然ソースからの精製、適切な宿主細胞による組換えポリペプチドの生産とその後の精製、化学合成による生産又は、最後に、これらの様々なアプローチの組合せである。これらの様々な生産方法は当業者に周知である。
【0013】
本発明の実施形態の1つでは、この酵素を天然に生産する生物、例えば大腸菌のような細菌、S.セレビシエのような酵母、又はニューロスポラ・クラッサ又はマグナポルテ・グリセアのような真菌からメチオニンシンターゼを精製する。
【0014】
本発明の好ましい実施形態では、メチオニンシンターゼは組換え宿主生物において過剰発現される。DNAフラグメントを工作する方法及び宿主細胞におけるポリペプチドの発現は当業者に周知であり、例えばGreene Publishing Associates and Wiley−Interscience発行の「Current Protocols in Molecular Biology」Volumes 1 and 2(1989)、 Ausubel F.M.ら、又はMolecular Cloning,T.Maniatis,E.F.Fritsch,J.Sambrook(1982)に述べられている。
【0015】
本発明の特定実施形態では、メチオニンシンターゼ阻害剤はマグナポルテ・グリセアの、トウモロコシ黒穂病菌の、及びより詳細には配列表の配列番号18を含む配列によって表わされるメチオニンシンターゼ、又はさもなければフィトフトラ・インフェスタンス(Phytophtora infestans)の、特に配列表の配列番号20を含む配列によって表わされるメチオニンシンターゼを阻害する;前記メチオニンシンターゼは、配列表の配列番号1を含む配列によって表わされるマグナポルテ・グリセアの遺伝子によって、又は配列表の配列番号2を含む配列によって表わされるcDNAによって、配列表の配列番号16を含む配列によって表わされるトウモロコシ黒穂病菌の遺伝子によって、又は配列表の配列番号17を含む配列によって表わされるcDNAによって、又はさもなければ配列表の配列番号19を含む配列によって表わされるフィトフトラ・インフェスタンス(Phytophtora infestans)の遺伝子によってコードされ得る。
【0016】
本発明の1つの主題はまた、メチオニンシンターゼ阻害剤ともう1つの抗真菌化合物を含む抗真菌組成物である。他の抗真菌化合物との混合物、特にアシベンゾラル−S−メチル、アゾキシストロビン、ベナラキシル、ベノミル、ブラスチシジン−S、ブロムコナゾール、カプタフォル、カプタン、カルベンダジム、カルボキシン、カルプロパミド、クロロタロニル、水酸化銅又はオキシ塩化銅のような銅又は銅誘導体に基づく抗真菌組成物、シアゾファミド、シモキサニル、シプロコナゾール、シプロジニル、ジクロラン、ジクロシメット、ジクロラン、ジエトフェンカルブ、ジフェノコナゾール、ジフルメトリム、ジメトモルフ、ジニコナゾール、ジスコストロビン、ドデモルフ、ドジン、エジフェンフォス、エポキシコナゾール、エタボキサム、エチリモール、ファモキサドン、フェナミドン、フェナリモール、フェンブコナゾール、フェンヘキサミド、フェンピクロニル、フェンプロピジン、フェンプロピモルフ、フェリムゾン、フルアジナム、フルジオキソニル、フルメトバー、フルキンコナゾール、フルシラゾール、フルスルファミド、フルトラニル、フルトリアフォル、フォルペット、フララキシル、フラメトピル、グアザチン、ヘキサコナゾール、ヒメキサゾル、イマザリル、イプロベンフォス、イプロジオン、イソプロチオラン、カスガマイシン、クレソキシム−メチル、マンコゼブ、マネブ、メフェノキサム、メパニピリム、メタラキシル及びメタラキシル−Mのようなこの鏡像異性体、メトコナゾール、メチラム−亜鉛、メトミノストロビン、オキサジキシル、ペフラゾエート、ペンコナゾール、ペンシクロン、リン酸及びフォセチル−Alのようなこの誘導体、フタリド、ピコキシストロビン、プロベナゾール、プロクロラズ、プロシミドン、プロパモカルブ、プロピコナゾール、ピラクロストロビン、ピリメタニル、ピロキロン、キノキシフェン、シルチオファム、シメコナゾール、スピロキサミン、テブコナゾール、テトラコナゾール、チアベンダゾール、チフルザミド、チオファネート、例えばチオファネート−メチル、チラム、トリアジメフォン、トリアジメノール、トリシクラゾール、トリデモルフ、トリフロキシストロビン、トリチコナゾール、バリンアミド誘導体、例えばイプロバリカルブ、ビンクロゾリン、ジネブ及びゾキサミドとの混合物は、特に有益である。このようにして得られる混合物はより広い活性スペクトルを有する。本発明に従った組成物はまた、1以上の殺虫剤、殺菌剤、殺ダニ剤又はフェロモン、又は生物活性を有する他の化合物を含み得る。
【0017】
本発明の1つの主題はまた、メチオニンシンターゼ阻害剤を使用して抗真菌組成物を生産するための方法である。
【0018】
本発明の1つの主題はまた、メチオニンシンターゼの酵素活性を阻害する化合物の特定を含む、抗真菌化合物を調製するための方法である。
【0019】
メチオニンシンターゼの酵素活性の阻害を測定するために試験化合物の存在下で酵素反応を実施する。メチオニンシンターゼの酵素活性を測定するため、及び従って、この酵素活性を阻害する化合物を特定するための全ての生化学試験が、本発明に従った方法において使用できる。
【0020】
この酵素の特異的阻害剤をスクリーニングするために高速(high−throughput)生化学アッセイを提案する。
【0021】
好ましくは、メチオニンシンターゼの活性を阻害する化合物を特定するための方法は、これらの化合物を、この基質、ホモシステイン、メチルテトラヒドロ葉酸又はメチルテトラヒドロ葉酸のポリグルタミン酸誘導体((CH3−H4)PteGlun)、及びリン酸塩、マグネシウム及び亜鉛のような様々な補因子の存在下でメチオニンシンターゼと接触させること;及び酵素活性を測定することを含む。
【0022】
メチオニンシンターゼの酵素活性を測定することは、メチオニン、テトラヒドロ葉酸又はさもなければメテニルテトラヒドロ葉酸又はこのようにして得られる何らかの産物の形成を測定すること、又はさもなければ他の何らかの化学又は酵素反応によって前記活性を測定することに結びつけることができる。
【0023】
メチオニンシンターゼの酵素活性の測定はまた、共役酵素の存在下で実施することもできる。S−アデノシルメチオニンシンターゼ(AdoMetS)はそれ自体で使用できる;これは、メチオニン、ATP及びマグネシウムの存在下でS−アデノシルメチオニン(S−AdoMet)の形成を触媒する。メチオニンシンターゼの酵素活性の測定は、従って、S−アデノシルメチオニン、リン酸塩又はピロリン酸塩の形成の測定と結びつけることができる。
【0024】
本発明のもう1つの態様によれば、メチオニンシンターゼの酵素活性を阻害する化合物の特定は、宿主生物においてメチオニンシンターゼを発現させること、宿主生物によって生産されたメチオニンシンターゼを精製すること、これらの化合物を精製メチオニンシンターゼ及びこの基質と接触させること、及び酵素活性を測定することを含む。
【0025】
好ましい実施形態では、これらの方法は全て、メチオニンシンターゼの酵素活性を阻害する前記化合物が、真菌の増殖及び/又は病原性を阻害するかどうかを測定する追加工程を含む。
【0026】
本発明は、従って、メチオニンシンターゼの酵素活性を阻害することによって真菌の増殖及び/又は病原性を阻害する化合物を特定するための方法に関する。これらの方法は、化合物又は化合物の混合物を、メチオニンシンターゼを阻害する化合物を特定するための適切なアッセイに供すること、及び前記アッセイに対して陽性に反応する化合物を選択すること、適宜にこれらを単離すること、及びその後これらを特定することから成る。
【0027】
好ましくは、適切なアッセイは、上記で定義したようなメチオニンシンターゼの酵素活性のアッセイである。
【0028】
好ましくは、これらの方法に従って特定される化合物を、その後、当業者に公知の方法に従ってこれらの抗真菌特性に関して試験する。好ましくは、切断葉又は植物全体に関する病原性アッセイのような表現型試験によって化合物を評価する。
【0029】
「化合物」という用語は、本発明によれば、ペプチド及びタンパク質を含む、何らかの化合物又は化合物の混合物を意味することが意図されている。
【0030】
「化合物の混合物」という用語は、本発明によれば、例えば分子の立体異性体、生物学的材料(植物、植物組織、細菌培養物、酵母又は真菌の培養物、昆虫、動物組織等)の抽出に由来する天然起源の混合物、又は精製されていないか又は完全に又は部分的に精製された反応混合物、又はコンビナトリアル化学手法に由来する生成物の混合物のような、少なくとも2つの異なる化合物を意味すると理解される。
【0031】
最後に、本発明は、メチオニンシンターゼの酵素活性を阻害する新規真菌病原性阻害化合物、特に本発明に従った方法によって特定される化合物及び/又は本発明に従った方法によって特定される化合物に由来する化合物に関する。
【0032】
好ましくは、メチオニンシンターゼの酵素活性を阻害する真菌病原性阻害化合物は、酵素の一般的阻害剤ではない。また好ましくは、本発明に従った化合物は、抗真菌活性及び/又は真菌病原性への活性を有することが既に公知の化合物ではない。
【実施例1】
【0033】
真菌におけるメチオニンシンターゼ遺伝子の特性決定
A.ニデュランス7(NCBI、アクセッション番号:AAF82115)のメチオニンシンターゼのタンパク質配列をモデルとして使用して、マグナポルテ・グリセアV2型のゲノムにおいてメチオニンシンターゼ遺伝子を特定した。コンティグ2.150(MG_コンティグ_2.150、6196−8629位、相補鎖、配列番号1)に位置するメチオニンシンターゼ遺伝子の完全なヌクレオチド配列は、766アミノ酸のポリペプチド(配列番号3)をコードする2301bpのcDNA(配列番号2)に対応する3個のエクソンを含む。この遺伝子の配列及び最終的メッセンジャーを生じさせるスプライシングは、様々な公的及び私的ベースで特定される数多くのESTの効力によって確認することができる。マグナポルテ・グリセアメチオニンシンターゼは、A.ニデュランスにおけるように71個の遺伝子によってコードされる。推定cDNAから推測される一次タンパク質配列の分析は、S.セレビシエ(P05694)、A.ニデュランス(AAF82115)、大腸菌(P13009)及びシロイヌナズナ植物(AAF00639)のビタミンB12非依存性メチオニンシンターゼと48%から79%の相同性を示す。
【0034】
マグナポルテ・グリセアメチオニンシンターゼの一次配列は、この酵素の特徴であるメチオニンシンターゼドメイン(334残基、E=4e−116、pfam01717)に対応する2個の保存されたドメインを有する。このドメインは、メチルテトラヒドロ葉酸トリグルタメートからホモシステインへのメチル基の移動によるメチオニンの生産を可能にする。この領域は、タンパク質のC末端部分に位置する。2番目のドメイン、COG0620又はメチオニンシンターゼII(メチルトランスフェラーゼ)は、タンパク質のN末端部分に関する(330アミノ酸)8。最近、シロイヌナズナから結晶化した酵素に関して9、この酵素の基質であるホモシステイン及びメチルテトラヒドロ葉酸、及び反応産物であるメチオニンに関するこれらのドメインの各々の特異性を決定することが可能となった。
【0035】
マグナポルテ・グリセアメチオニンシンターゼの一次配列を、このゲノムが部分的に又は完全に配列決定されている様々な真菌種におけるオーソログに関して検索するために使用した。これらの様々な一次配列を、その後、植物、細菌及び動物のような様々な生物において記述されているメチオニンシンターゼと比較した。適切なプログラム(tblastn;FGENSH;PSI−PHI−BLAST)を使用して遺伝子の構造(イントロン+エクソン)及び一次アミノ酸配列の特性決定を実施した。この手順に従って、いくつかの真菌(子嚢菌類及び担子菌類)及び植物病原性卵菌類(P.ソージャ及びP.インフェスタンス)においてメチオニンシンターゼを特性決定することができた。メチオニンシンターゼについての系統樹を確立することができ、得られた表示は、マグナポルテ・グリセアメチオニンシンターゼが子嚢菌類のメチオニンシンターゼに属すること及び担子菌類のメチオニンシンターゼからは遠いことを示す。全体として、得られた系統樹は、これらの生物の系統発生起源10を遡るものと一致する。
【実施例2】
【0036】
マグナポルテ・グリセアメチオニンシンターゼ遺伝子の欠失
マグナポルテ・グリセアの発生及び感染過程におけるメチオニンシンターゼ遺伝子の役割の検討を、この遺伝子の欠失突然変異体の表現型を試験することによって実施した。欠失突然変異体を得るための戦略は、MET6遺伝子を、MET6オープンリーディングフレームが形質転換体の選択のための抗生物質(ハイグロマイシン)に対する耐性のためのカセットで置換された突然変異型対立遺伝子で置換することに基づく。
【0037】
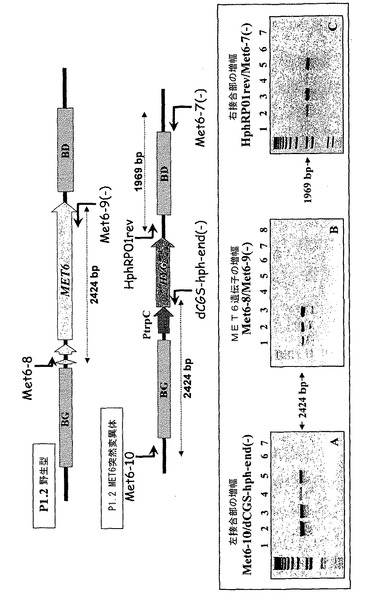
マグナポルテ・グリセアMET6遺伝子を置換するためのこのベクターの構築は、2工程で実施される。(i)この遺伝子に隣接し、それぞれMET6のいずれかの側に位置する約1kbのゲノムの領域に対応する領域のPCR増幅、(ii)形質転換体の選択を可能にする、抗生物質に対する耐性のための遺伝子へのこれらのゲノムDNAフラグメントの連結。従って、遺伝子を置換するために使用されるPCRフラグメントは、検討する遺伝子の左境界及び右境界と称される2つの領域から成る(図1)。我々は、選択遺伝子としてハイグロマイシン耐性遺伝子(HYG、PtrpCプロモーター、ハイグロマイシン耐性遺伝子hphのコード部分を含む)を選択した。HYG遺伝子の連結は、最初、MET6遺伝子の左境界のSacII/BglII部位及びEcoRI/SacII部位(Met6−1/Met6−2プライマー)によって実施する。MET6遺伝子の右境界(Met6−3/Met6−4プライマー)を、次に、hph遺伝子の下流のPmeI部位を通して導入する(図1)。置換ベクターは、従って、MET6遺伝子の左境界(BG)(1475bpのプロモーター領域)、HYGカセット(1400bp)及びMET6遺伝子の右境界(BD)(1251bpのターミネーター領域)を含む。その後、連結産物(BG−hph−BD)をプラスミドベクターにクローニングした。MET6遺伝子置換カセットを、その後、MET6遺伝子の両境界の末端に特異的なプライマー(Met6−5/Met6−6プライマー)を用いたPCRによって対応するプラスミドから増幅した。境界の間の接合部及び置換ベクターのHYGカセットの配列決定によって構築を確認することができた。アガロースゲル電気泳動によって精製したPCR産物(1μg)を、次に、実験室において開発された従来の手法に従ってマグナポルテ・グリセアP1.2野生型株のプロトプラストを形質転換するために使用する。形質転換に由来する産物を、対応する抗生物質(ハイグロマイシン)を含む培地で選択する。
【実施例3】
【0038】
遺伝子置換によって得られるmet6Δ::hph欠失突然変異体の特定及び栄養特性決定
一次形質転換体を、ハイグロマイシンの存在下で発生するこれらの能力に関して選択する。1mMメチオニンを添加した又は添加していない、ハイグロマイシンを含む最小培地での形質転換体の異なる増殖を測定することにより、met6Δ::hph突然変異体の特定を実施した。met6Δ::hph突然変異体はこの最小培地で発生することができないが、メチオニンを添加したこの最小培地では正常に増殖する。突然変異体の発生率は、我々の実験条件下で分析した一次形質転換体の約20%である。これらの突然変異体を、その後、単胞子(monosphores)の単離によって遺伝的に精製する。
【0039】
得られた5個のmet6Δ::hph突然変異体(4.1、15.1、22.1及び23.1)は、P1.2野生型株の増殖を可能にする最小培地で発生することができない。最小培地へのメチオニンの添加は突然変異体の増殖を回復する。メチオニンの添加によるmet6Δ::hph突然変異体の栄養補完は、メチオニンシンターゼがMET6遺伝子欠失によって影響されることを指示する。
【0040】
システイン又はグルタチオン(メチオニンシンターゼからの基質であるホモシステインの前駆体)のような硫黄供与体の添加は、マグナポルテ・グリセアmet6Δ::hph突然変異体の増殖を回復するのに十分ではない。従って、この真菌においてメチオニン合成のためにメチルテトラヒドロ葉酸又はこのポリグルタミン酸誘導体を使用する平行経路は存在しない。従って、メチオニンの新たな合成は専らメチオニンシンターゼによってのみ触媒される。この酵素の活性は、従って、真菌の発生にとって必須である。
【0041】
他方で、直接メチオニンに由来し、細胞周期に必須である化合物、S−アデノシルメチオニン(SAM又はAdoMet)、又は植物において合成される化合物、S−メチルメチオニン(SMM)の存在下で、マグナポルテ・グリセアmet6Δ::hph突然変異体は発生することができるが、P1.2野生型株に比べて増殖率は低い。SAM及びSMMは、浸透することができ、マグナポルテ・グリセアによってメチオニンに代謝され得る。この機構はおそらく、酵母S.セレビシエにおいて記述されているものと同様である。実際に、この子嚢菌では、SAMとSMMは輸送体SAM3及びMPP1によって細胞内に取り込まれ、その後ホモシステインの存在下で、SAM又はSMMをそれぞれメチル基供与体として使用するホモシステイン−S−メチルトランスフェラーゼ(SAM4及びMHT1)によってメチオニンに変換される(S.セレビシエ)11。我々の実験条件によれば、最小培地へのSAM(1mM)の添加は、met6Δ::hph突然変異体の増殖を回復する上でSMMの添加よりも有効である。これらの結果は、マグナポルテ・グリセアが、S.セレビシエにおいて記述されているのと同様の輸送体及びホモシステインメチルトランスフェラーゼを有することを示唆する。酵母SAM4及びMHT1タンパク質に関する配列相同性検索(tBlastN)によるマグナポルテ・グリセアゲノムの分析は、マグナポルテ・グリセアにおけるSAM4のオーソログである遺伝子を明らかにすることを可能にする。糸状菌は1個の遺伝子だけを有する(MHT1のオーソログなし)と思われる。我々の結果によれば(SAMでのSMMよりも良好な増殖)、このタンパク質はSMMよりもSAMに対してより大きな親和性を有すると考えられる。
【0042】
MET6遺伝子以外の遺伝子座に置換ベクターBG−hph−BDを組み込んだ形質転換体に対応する、いくつかの「異所性」形質転換体も分析した。これらのハイグロマイシン耐性異所性形質転換体は最小培地で発生することができる。メチオニンシンターゼ遺伝子は、従って、これらの異所性形質転換体において機能性であり、ベクターは、我々の実験条件下で病原体の発生(最小又は完全培地で発生する能力、胞子形成)に影響を及ぼさないゲノムの遺伝子座に挿入された。
【実施例4】
【0043】
met6Δ::hph突然変異体の分子特性決定
PCR及びサザンハイブリダイゼーションによるMET6遺伝子の分子分析を実施するために使用するゲノムDNAを抽出するために、メチオニン(1mM)を含む培地でmet6Δ::hph突然変異体を培養する。
【0044】
形質転換体の分子分析を、P1.2のMET6遺伝子の野生型対立遺伝子を突然変異型対立遺伝子met6Δ::hphで置換するための様々な特異的プライマーを使用して、MET6遺伝子座のゲノム領域の増幅によって実施する。これらのPCRは、特異的オリゴヌクレオチドを用いて各々の突然変異体に関して実施する。反応は、最初にハイグロマイシン耐性遺伝子hphと、2番目に、置換ベクターを構築するために使用したMET6領域の外側に位置するMET6遺伝子座のゲノム配列と(左接合物及び右接合部);MET6遺伝子に相同な配列と、ハイブリダイズするオリゴヌクレオチドを使用する。従って、1969bp(左接合部)又は2447bp(右接合部)のフラグメントの増幅は、構築物のmet6Δ::hph遺伝子による野生型遺伝子の置換の場合にのみ起こる(プライマーHphRP01rev(配列番号6)/Met6−7(−)(配列番号7)及びMet6−10(配列番号8)/dCGS−hph−end(−)(配列番号9)(図1))。P1.2野生型株又は異所性形質転換体の場合には増幅は得られない。同様に、met6Δ::hph突然変異体内にMET6遺伝子の増幅が存在しないことは(プライマーMet6−8、配列番号10/Met6−9、配列番号11)(図1)、MET6遺伝子が実際にこれらの形質転換体に存在しないことの指標である。他方で、これらのプライマーにより、2424bpのフラグメントがP1.2野生型株及び異所性形質転換体において増幅される。突然変異体22.2及び23.1に関して実施したPCRだけが、これら3つのタイプのPCRに関して予想された結果を与える。PCR Met6−10(配列番号8)/dCGS−hph−end(配列番号9)及びhphRP01(配列番号6)/Met6−7(−)、配列番号7)では、説明のつかない理由で、増幅が得られなかった。
【0045】
BamHI制限酵素によるゲノムDNAの消化後、サザンハイブリダイゼーションによる分析を実施し、4個の突然変異体に関して得られたハイブリダイゼーションシグナルをP1.2野生型株及び異所性形質転換体(19.1)で得られたものと比較した。置換ベクター内に存在するMET6左境界(MET6プロモーター領域)に対応する(Met6−1(配列番号12)/Met6−2(配列番号13))PCRフラグメントをプローブとして使用して、大きさがP1.2野生型株及び異所性形質転換体(19.1)のものとは異なる2つのバンドが突然変異体に関して認められる。シグナルは、P1.2野生型株及び異所性形質転換体19.1内のMET6プロモーター領域に対応する。後者はまた、別のゲノム領域に挿入された置換ベクターに対応するハイブリダイゼーションシグナルも示す。同様の結果が、置換ベクター内に存在するMET6右境界(MET6ターミネーター領域)に対応する(Met6−3(配列番号14)/Met6−4(配列番号15))PCRフラグメントを使用して得られる。挿入遺伝子(hph)に特異的なプローブにより、met6Δ::hph突然変異体及び異所性形質転換体19.1だけが、MET6遺伝子座におけるhphの存在(突然変異体)又は置換ベクターBG−met6Δ::hph−BD(異所性)のいずれかに対応するハイブリダイゼーションシグナルを示す。後者の結果は、分析した様々な突然変異体が分子レベルで同一であり、MET6コード相(coding phase)の代わりにMET6遺伝子座として挿入されたhph遺伝子の1コピーだけを含むことを指示する。
【実施例5】
【0046】
マグナポルテ・グリセアmet6Δ::hph突然変異体の病原力の分析
マグナポルテ・グリセアmet6Δ::hph突然変異体の病原力を、生存条件下の大麦の葉に関して及び大麦及びイネ植物全体に関する感染試験によって評価した。この分析の後に、胞子発芽率、付着器分化及び大麦葉への貫入測定を実施した。P1.2野生型株及びmet6Δ::hph突然変異体4.1、15.1、22.1、23.1及び24.1の胞子を、1mMを含む米粉ベースの培地での14日間の増殖後に採取する。使用する植物材料は、大麦(cv.Express)及びイネ(cv.Sariceltik)である。
【0047】
生存条件下で大麦の葉に関して実施する実験では、高湿度(100%)及び100マイクロアインシュタインの光下で温帯気候室(26℃)において、キネチン(2mg/ml−1)を含む寒天培地(1%寒天−H2O)で葉をインキュベートする。感染の間、胞子を液滴の形態で(35μl)又は綿毛芽を用いて胞子の懸濁液で葉の表面を被覆することによって葉に付着させる。これらの実験を、インキュベーションの間を通じて又は24時間だけ、1mMメチオニンの不在下又は存在下で実施する。胞子濃度は、水中3×104胞子/mlから1×105胞子/mlである。次に、マグナポルテ・グリセアによって引き起こされる病徴の出現を少なくともインキュベーションの7日間観察する。マグナポルテ・グリセア野生型株P1.2の胞子による生存条件下での大麦葉の接種は、感染葉における真菌の発生の特徴である壊死(胞子形成病変)を生じさせる。反対に、met6Δ::hph突然変異体4.1、15.1、22.1、23.1及び24.1は、接種の方法に関わらず、大麦葉にいかなる病徴も生じさせず、従って非病原性である。met6Δ::hph突然変異体胞子へのメチオニンの添加は、これらの病原力を部分的に回復することを可能にする(特徴的であるが非胞子形成性病変の発現)。さらに、我々の実験条件下では、損傷葉又は非損傷葉に関して実施した感染の間で著明な差を認めなかった。これらの所見は、met6Δ::hph突然変異体が、損傷した大麦葉でさえも、大麦葉に貫入できないことを示唆する。被覆による胞子の分散に関して実施した実験で認められた葉の黄化は、おそらく我々の実験条件下での葉の早期老化によるものである。結果を以下の表に要約する。
【0048】
【表1】

【0049】
感染の様々な段階(発芽、付着器の形成及び貫入)の観察及び定量を、胞子の懸濁液(3×105胞子/ml)35μlの液滴による生存条件下での大麦葉の接種に基づいて実施する。24時間後、付着器を分化した発芽胞子の数及び表皮細胞への貫入事象の数を顕微鏡下で観察するために葉の表皮をはがす。0.01%のカルコフロールの溶液は、植物の表面に位置する真菌細胞の壁の強い蛍光を生じさせることを可能にする(表皮細胞内に位置する菌糸は着色されない)。これらの観察(表II)は、met6Δ::hph突然変異体がわずかに低い発芽を有する(野生型株と比較して−10%から−40%)ことを明らかにする。これらの付着器の分化率もわずかに低い(野生型株と比較して0%から−30%)。他方で、これらの突然変異型付着器は茎葉組織に貫入することができない。
【0050】
【表2】
【0051】
付着器の形成はまた、メチオニン(我々の実験条件下で1mM濃度)を添加した又は添加していないテフロン(登録商標)膜上で胞子が発芽する、人工条件下でも認められる。これらの非常に疎水性の膜は葉の表面を模倣し、これによって付着器の形成を誘導すること及び付着器分化率を容易に測定することを可能にする。
【0052】
結論として、マグナポルテ・グリセアmet6Δ::hph突然変異体は、従って、付着器を分化するが、植物に貫入することはできない。これらの結果は、これらの突然変異体が非機能性付着器を有することを示す。
【0053】
3週齢の大麦植物(第二葉の出芽に相当する)を含む鉢を、マグナポルテ・グリセア胞子の懸濁液(3×104胞子/mlの濃度の胞子及び水滴の葉への付着のための0.3%ゼラチンを含む水10ml)による散布に供する。散布後少なくとも8日間病徴の観察を実施する。野生型株(P1.2)及び異所性形質転換体(19.1及び20.1)で処置した大麦植物は、感染葉における真菌の発生によって引き起こされる壊死病変を示す。他方で、マグナポルテ・グリセアmet6Δ::hph突然変異体を接種した場合は病害の症状を認めなかった。これらの突然変異体は、従って、非病原性であると考えられる。さらに、この試験は、生存条件下で大麦葉に関して実施した分析と同様に、葉の組織中のメチオニン(又はSAM及びSMMのようなメチオニンに由来する他の化合物)の量が、met6Δ::hph突然変異体のメチオニン合成の欠損を補うには不十分であることを指示する。従って、インビトロ実験で明らかにされた経路(SAM及びSMMの存在下での補完)は、真菌が植物において成長している間、真菌において機能性ではないと思われる。
【0054】
得られたmet6Δ::hph突然変異体は、ハイグロマイシン耐性を付与する、hph遺伝子によって置換されるMET6遺伝子のコード相の欠失に相当する。これらの突然変異体はホモシステインからメチオニンを合成することができず、従って最小培地で増殖することができない。この最小培地へのメチオニンの添加はこれらの突然変異体の発生のために必須である。met6Δ::hph突然変異体は、生存条件下の大麦葉又は全植物に接種するために使用したとき、高い胞子濃度であっても、病害の症状を生じさせない。従って、メチオニンシンターゼによるメチオニン合成は、インビトロで及び植物において、真菌の発生に必須である。
【0055】
met6Δ::hph突然変異体はメチオニンの不在下で付着器に分化することができる。この態様は、胞子が、この細胞の発生のために必要なタンパク質及び代謝産物の合成を可能にするメチオニンの少なからぬ貯蔵を有し得ることを指示する。この貯蔵は、使用した米粉ベースの培地(1mMメチオニンを含む)での突然変異体の増殖とこの胞子形成を可能にするために突然変異体に供給されたメチオニンに由来するはずである。他方で、met6Δ::hph突然変異体の葉への貫入が存在しないことは、前記突然変異体が、真菌の植物への貫入を指令することができない非機能性付着器に分化することを指示する。おそらく、突然変異型付着器はこれらのメチオニン貯蔵を急速に使い果たしたと考えられる。実際に、突然変異体胞子へのメチオニンの添加は植物への貫入と葉における発生の開始を可能にする。しかし、前記発生は完全ではなく、胞子形成病変の形成の不在は、胞子に供給されるメチオニン貯蔵がひとたび使い果たされると、植物は突然変異体のメチオニン欠乏を補うことができない。
【実施例6】
【0056】
メチオニンシンターゼの酵素活性を阻害する分子を検定し、特性決定するための方法
本発明の方法は、この作用が、直接又は間接的手法(メチオニンシンターゼの活性を測定するための「共役」酵素の使用を含む)に従って定量される基質の消費又は生成物の形成を阻害する全ての分子の特性決定を含む。メチオニンシンターゼは、文献12、13、14、15、27に記述されている公知のアッセイに従って様々な補因子(リン酸塩、マグネシウム、亜鉛)の存在下で、ホモシステイン、メチルテトラヒドロ葉酸(n=1)又はこのポリグルタミン酸誘導体(n≧3)の存在下で不可逆的反応を触媒する。この方法は、逆相HPLCクロマトグラフィーによって化合物を分離するための手法16により、生産されるメチオニン又は葉酸誘導体を定量することを含む。メチルテトラヒドロ葉酸基質は手順17に述べられている実験条件下で検出可能ではないので、テトラヒドロ葉酸がメテニルテトラヒドロ葉酸に変換された後、反応の間に生産されたテトラヒドロ葉酸の検定を350nmの分光光度計で実施することができる。提案される代替法は、S−アデノシルメチオニンシンテターゼの存在下でメチオニンシンターゼ活性を検定することである。このアッセイでは、マグナポルテ・グリセアのS−アデノシルメチオニンシンテターゼ(AdoMetS)が好ましく使用されるが、いかなるAdoMetSも「共役」酵素として使用できる。AdoMetS酵素は、メチオニン、ATP及びマグネシウムの存在下で、S−アデノシルメチオニン(SAM)、リン酸塩及びピロリン酸塩を生産する不可逆的反応を触媒する。
【0057】
メチオニンシンターゼ活性は、結局、共役酵素の存在下での生成物の変換後に比色法によって及び/又は分光光度計で又はメチオニンシンターゼ活性を測定するための他の何らかの化学又は酵素反応によって、生産されるSAM、リン酸塩又はピロリン酸塩の量を通して検定される。これらの様々な方法は文献における多くの記述の対象であり、実験者に従って適合させることができる18、19、20、21、22、23。例えばSAMシンテターゼの存在下でメチオニンシンターゼ活性を検定するための方法の感受性は、ピロリン酸塩を2モルのリン酸塩に変換するピロホスファターゼの添加を通して改善することができる。従って、合成されるメチオニンの各モルについて、前記方法は3モルのリン酸塩を生産する。
【0058】
マグナポルテ・グリセアメチオニンシンターゼの精製
大量のメチオニンシンターゼの生産は、細菌又は酵母におけるタンパク質の過剰生産のための発現ベクターを使用する手法を用いて実施される。この手法は、好ましくはタンパク質のN末端又はC末端にヒスチジンタグ延長部を組み込むことを可能にする発現ベクターへのcDNAのクローニングを用いる。例えばpET−28b(+)ベクター(Novagen)24を選択するとき、従来の分子生物学手法に従って2301bpのcDNAをNdEI及びEcoRIにクローニングする。pET−28−MgMET6と呼ばれる、得られた構築物を大腸菌BL21型DE3(pLysS)菌株に導入し、IPTG(0.5mM)による誘導後に発現を生じさせる。組換え細菌を28℃で4時間培養する。その後遠心分離によって細胞を収集し、得られたペレットをタンパク質の安定性に適した溶解緩衝液に再懸濁する。試料の超音波処理後、遠心分離後に得られる、組換えタンパク質を含む可溶性分画をNi−NTAアガロース型のカラムに負荷する。次に、マトリックスをイミダゾール溶液で数回連続的に洗浄した後、酵素の精製及び溶出を実施する。手順はQiagenによって定義されたプロトコール25に従う。
【0059】
溶出後、組換えメチオニンシンターゼを含むタンパク質分画を限外ろ過によって濃縮し、イミダゾールの全ての痕跡を除去するためにPD10(Pharmacia)27での分子ろ過に供する。組換えタンパク質の精製は、Superdex S200(Pharmacia)26でのクロマトグラフィーによる分子ろ過又はMonoQ HR10/10(Pharmacia)26でのイオン交換クロマトグラフィーから成る第二工程を伴い得る。適切な直接測定アッセイを使用して、精製の間メチオニンシンターゼの活性を追跡する。
【0060】
【表3】
【図面の簡単な説明】
【0061】
【図1】遺伝子置換のためのフラグメント、ならびに増幅の概略図。
【技術分野】
【0001】
本発明は、真菌病の処置、より詳細には農作物種の真菌病の処置のためのメチオニンシンターゼ阻害剤の使用に関する。
【背景技術】
【0002】
真菌は、様々な植物種の農作物の実質的な損失をもたらし得る壊滅的な伝染病の原因である。病原性真菌の酵素の阻害剤を使用すること、及びこれらの真菌に対して活性な新しい分子を特定するために試験においてこれらの酵素を使用することの原理は、それ自体公知である。しかし、単に真菌酵素の特性を決定するだけではこの目的を達成するのに十分ではなく、潜在的抗真菌分子のための標的として選択される酵素は、同時に真菌の生存にとって必須でなければならず、抗真菌分子によるこの阻害が真菌の死滅を生じさせるか、又は真菌の病原性にとって不可欠でなければならず、この場合、この阻害は真菌にとって致死的ではなく、単にこの病原能力を阻害する。真菌の病原性及び生存にとって必須の代謝経路及び酵素の特定が、従って、新規抗真菌製品の開発のために必要である。
【発明の開示】
【発明が解決しようとする課題】
【0003】
硫黄同化経路は、硫酸イオン(SO42−)の取り込み、この活性化、及び還元硫黄(S2−)へのこの還元を含む。これらの工程は、ATPスルフリラーゼ(EC2.7.7.4)、APSキナーゼ(EC2.7.1.25)、PAPSレダクターゼ(EC1.8.4.8)(光合成生物におけるAPSレダクターゼ、EC1.8.4.9)、及び(NADPH2)亜硫酸レダクターゼ(EC1.8.1.2)(光合成生物におけるフェレドキシン依存性酵素、EC1.8.7.1)によって連続的に触媒される。全ての独立栄養生物において、硫酸イオン同化、活性化及び還元経路は、この一般原則の観点から言えば保存されている;炭素骨格への還元硫黄の取り込みは生物によってかなりの変動を示す。細菌1(例えば大腸菌)、植物2(例えばシロイヌナズナ)、酵母(例えばサッカロミセス・セレビシエ(Saccharomyces cerevisiae)3)及び糸状菌4。実際に、植物及び細菌では、還元硫黄は、セリンに由来するC3の分子に取り込まれてシステインを形成する。次に硫黄は、ホモセリンに由来するC4の分子に転移してホモシステインを形成する。この一連の反応が直接トランススルフレーション経路を形成する。逆に、サッカロミセス・セレビシエ(S.cerevisiae)では、硫黄は、ホモセリンに由来するC4の分子に直接取り込まれてホモシステインを形成する(直接スルフヒドリル化)3。次にシステインは、逆トランススルフレーション経路を構成する一連の反応によってホモシステインから合成される。糸状菌では、ホモシステインの合成は植物における直接経路(直接トランススルフレーション)及びサッカロミセス・セレビシエの経路(直接スルフヒドリル化)の両方によって行われる。さらに、システインの合成は、セリンによって又は逆トランススルフレーション経路を通してホモシステインから実施される。これらの様々な代謝経路は、ニューロスポラ・クラッサ(Neurospora crassa)(N.crassa)5及びアセペルギルス・ニドゥランス(Aspergillus nidulans)(A.nidulnas)6におけるシステイン及びメチオニンについての栄養素要求性突然変異体の特性決定に従って定義された。このモデルは、植物(例えばマグナポルテ・グリセア(Magnaporthe grisea)、M.grisea)及び動物(例えばアスペルギルス・フミガツス(Aspergillus fumigatus)(A.fumigatus))の病原性真菌を含む全ての糸状菌類に当てはめることができる。イネ作物への多大の被害の原因である子嚢菌型病原体、マグナポルテ・グリセアは、このようなアプローチのための選択モデルである。糸状菌におけるメチオニン合成は、植物におけるようなビタミンB12非依存型のメチオニンシンターゼの作用を必要とする。ヒト病原菌、クリプトコッカス・ネオフォルマンス(Cryptococcus neoformans)28のメチオニンシンターゼ遺伝子の試験において記述されたアプローチは、本発明とは異なる。実際に、動物(ヒトを含む)はメチオニンを合成することができるが、この工程は、植物及び菌類のような他の真核生物のものとは大きく異なるビタミンB12依存型メチオニンシンターゼによって触媒される。植物メチオニンシンターゼは、タンパク質レベルでマグナポルテ・グリセアと強い相同性を示すが、同時に、実施されるモデリングに従って構造型の相違も示す9、12、27。従って、真菌阻害剤だけの特定を可能にする、この特異的性質を決定するためには、真菌酵素の特定及びこの特性決定が必要とされる。農作物を処置するための方法におけるこのような阻害剤の選択及び適用は特異的である。従って、本発明は、MET6遺伝子の突然変異体、より詳細にはマグナポルテ・グリセアのメチオニンシンターゼをコードするMET6遺伝子の欠失突然変異体がメチオニンに関して栄養素要求性であり、非病原性であるという事実を述べる。これらの突然変異体では、感染過程は、植物細胞への病原体の貫入期のレベルだけでなく、感染組織において進行するこの能力によっても達成される。マグナポルテ・グリセアメチオニンシンターゼ突然変異体の病原力は、感染の間にメチオニンを添加するとき部分的に回復される。これらの結果は、メチオニンシンターゼ活性の不在が感染の間の真菌にとって致死的であることを示す。同様の結果がトウモロコシ黒穂病菌(U.maydis)及びフィトフトラ・インフェスタンス(Phytophtora infestans)(P.infestans)でも得られた。
【0004】
配列表の説明
配列番号1:マグナポルテ・グリセアメチオニンシンターゼ遺伝子
配列番号2:マグナポルテ・グリセアメチオニンシンターゼcDNA
配列番号3:マグナポルテ・グリセアメチオニンシンターゼタンパク質配列
配列番号4:Met6−5プライマー
配列番号5:Met6−6プライマー
配列番号6:HphRP10プライマー
配列番号7:Met6−7プライマー
配列番号8:Met6−10プライマー
配列番号9:dCGS−hph−end(−)プライマー
配列番号10:Met6−8プライマー
配列番号11:Met6−9プライマー
配列番号12:Met6−1プライマー
配列番号13:Met6−2プライマー
配列番号14:Met6−3プライマー
配列番号15:Met6−4プライマー
配列番号16:トウモロコシ黒穂病菌メチオニンシンターゼ遺伝子
配列番号17:トウモロコシ黒穂病菌メチオニンシンターゼcDNA
配列番号18:トウモロコシ黒穂病菌メチオニンシンターゼタンパク質配列
配列番号19:フィトフトラ・インフェスタンス(Phytophtora infestans)メチオニンシンターゼEST配列
配列番号20:推定フィトフトラ・インフェスタンス(Phytophtora infestans)メチオニンシンターゼタンパク質配列
【課題を解決するための手段】
【0005】
本発明の1つの主題は、有効量のメチオニンシンターゼ阻害剤の施用によって真菌病に対して農作物を処置するための方法である。
【0006】
本発明に関して、真菌病は、子嚢菌、担子菌及び卵菌ファミリーに属する病原性植物真菌による病害と定義される。
【0007】
本発明の1つの主題は、有効(農業的に有効)な非植物毒性量のメチオニンシンターゼ阻害剤を、植物が成長する又は成長しやすい土壌、植物の葉及び/又は果実又は植物の種子に施用することを特徴とする、治療的又は予防的性能で、農作物の植物病原性菌類を防除するための方法である。「有効な非植物毒性量」という用語は、作物上に存在する又は出現し得る真菌の発生周期の制御又は破壊を可能にするのに十分であり、前記作物に対して植物毒性の著明な病徴を生じさせない阻害剤の量を意味することが意図されている。このような量は、防除すべき真菌ファミリー、作物の種類、気候条件及び本発明に従った抗真菌組成物に含まれる化合物に依存して、広い範囲内で変動し得る。
【0008】
この量は、当業者の技術範囲内である、体系的圃場試験によって決定することができる。
【発明を実施するための最良の形態】
【0009】
本発明に従った方法は、穀物(特に小麦、ライ麦、ライ小麦及び大麦)、ジャガイモ、綿、エンドウ豆、菜種、トウモロコシ又は亜麻の種子、選択的に林木の種子又はさもなければこれらの植物の遺伝的に改変された種子を処置するために有用である。本発明はまた、農作物への茎葉散布、すなわち当該植物の茎葉、葉、果実及び/又は茎への散布に関するが、また他のいかなる種類の施用にも関する。本発明に従った方法が標的する植物の中で、稲、トウモロコシ、小麦、大麦又はライ小麦などの穀物、果樹、特にリンゴの木、ナシの木、桃の木、ブドウの木、バナナの木、オレンジの木、レモンの木等、油生成作物、例えば菜種又はヒマワリ、市場向け菜園及び野菜作物、トマト、サラダ用作物、タンパク質生成作物、エンドウ豆、ナス科作物、例えばジャガイモ、ビートの根、亜麻、及び林木、及びまたこれらの作物の遺伝的に改変された同族体が挙げられる。
【0010】
本発明に従った方法が標的する植物の中で、
小麦、以下の種子病害を防除することに関して:フザリア(ミクロドキウム・ニバル(Microdochium nivale)及びフザリウム・ロゼウム(Fusarium roseum))、なまぐさ黒穂病(ティレティア・カリエス(Tilletia caries)、ティレティア・コントロバーサ(Tilletia controversa)又はティレティア・インディカ(Tilletia indica))、セプトリア病(セプトリア・ノドラム(Septoria nodorum));裸黒穂病(ウスチラゴ・トリティキ(Ustilago tritici));
小麦、植物の地上部分の以下の病害を防除することに関して:穀類眼紋病(タペシア・ヤルンダ(Tapesia yallundae)、タペシア・アクイフォルミス(Tapesia acuiformis))、立ち枯れ病(ゲウマノミセス・グラミニス(Gaeumannomyces graminis))、胴枯れ病(foot blight)(F.セルモラム(F. culmorum)、F.グラミネアラム(F. graminearum))、赤かび病(F.セルモラム、F.グラミネアラム、ミクロドキウム・ニバル)、黒斑病(black speck)(リゾクトニア・セレアリス(Rhizoctonia cerealis))、うどん粉病(トリティキ種エリシフェ・グラミニス・フォルマ(Erysiphe graminis forma specie tritici))、さび病(プキニア・ストリイフォルミス(Puccinia striiformis))及びプキニア・レコンディタ(Puccinia recondite))及びセプトリア病(セプトリア・トリティキ(Septoria tritici)及びセプトリア・ノドラム)、網斑病(ドレクスレラ・トリティキ−レペンティス(Drechslera tritici−repentis));
大麦、以下の種子病害を防除することに関して:網斑病(ピレノフォラ・グラミネア(Pyrenophora graminea)、ピレノフォラ・テレス(Pyrenophora teres)及びコクリオボラス・サティバス(Cochliobolus sativus))、裸黒穂病(ウスティラゴ・ヌーダ(Ustilago nuda))及びフザリウム病(ミクロドキウム・ニバル及びフザリウム・ロゼウム(Fusarium roseum));
大麦、植物の地上部分の以下の病害を防除することに関して:穀類眼紋病(タペシア・ヤルンダ)、網斑病(ピレノフォラ・テレス及びコクリオボラス・サティバス)、うどん粉病(ホルデイ種エリシフェ・グラミニス・フォルマ(Erysiphe graminis forma specie hordei))、小さび病(プキニア・ホルデイ(Puccinia hordei))及び葉枯れ病(リンコスポリウム・セカリス(Rhynchosporium secalis));
ジャガイモ、塊茎疾患(特にヘルミントスポリウム・ソラニ(Helminthosporium solani)、フォーマ・ツベローサ(Phoma tuberosa)、リゾクトニア・ソラニ(Rhizoctonia solani)、フザリウム・ソラニ(Fusarium solani))、及びうどん粉病(フィトフトラ・インフェスタンス(Phytophtora infestans))を防除することに関して;
ジャガイモ、以下の茎葉疾患を防除することに関して、夏疫病(アルテナリア・ソラニ(Alternaria solani))、うどん粉病(フィトフトラ・インフェスタンス);
綿、種子から成長した幼植物の以下の疾患を防除することに関して、立ち枯れ病及び斑点落葉病(collar rot)(リゾクトニア・ソラニ(Rhizoctonia solani)、フザリウム・オキシスポラム(Fusarium oxysporum))、黒根病(チエラビオプシス・バシコラ(Thielaviopsis basicola));
タンパク質生成作物、例えばエンドウ豆、以下の種子疾患を防除することに関して、炭疽病(アスコキタ・ピシ(Ascochyta pisi)、ミコスフェレラ・ピノデス(Mycosphaerella pinodes))、フザリウム病(フザリウム・オキシスポラム)、灰色かび病(ボツリティス・シネレア(Botrytis cinerea))、うどん粉病(ペロノスポラ・ピシ(Peronospora pisi));
油生成作物、例えば菜種、以下の種子疾患を防除することに関して、フォーマ・リンガム(Phoma lingam)、アルテルナリア・ブラシカ(Alternaria brassicae)及びスクレロティニア・スクレロティオラム(Sclerotinia);
トウモロコシ、種子疾患(リゾプス種(Rhizopus sp.)、ペニシリウム種(Penicillium sp.)、トリコデルマ種(Trichoderma sp.)、アスペルギルス種(Aspergillus sp.)及びギベレラ・フジクロイ(Gibberella fujikuroi))を防除することに関して;
亜麻、種子疾患を防除することに関して、アルテルナリア・リニコラ(Alternaria linicola);
林木、立ち枯れ病(フザリウム・オキシスポラム、リゾクトニア・ソラニ)を防除することに関して;
イネ、地上部分の以下の病害を防除することに関して、いもち病(マグナポルテ・グリセア)、黒斑病(リゾクトニア・ソラニ);
野菜作物、種子又は種子から成長した幼植物の以下の疾患を防除することに関して、立ち枯れ病及び斑点落葉病(フザリウム・オキシスポラム、フザリウム・ロゼウム、リゾクトニア・ソラニ、ピチウム種(Pythium sp.));
野菜作物、地上部分の以下の病害を防除することに関して、灰色かび病(ボツリティス種(Botrytis sp.))、うどん粉病(特にエリシフェ・シコラセアラム(Erysiphe cichoracearum)、スフェロテカ・フリギニエ(Sphaerotheca fuliginea)、レヴェイルラ・タウリカ(Leveillula taurica))、フザリウム病(フザリウム・オキシスポラム、フザリウム・ロゼウム)、葉枯れ病(クラドスポリウム種(Cladosporium sp.))、アルテルナリア斑点病(アルテルナリア種(Alternaria))、炭疽病(コレトトリカム種(Colletotrichum sp))、セプトリア葉枯れ病(セプトリア種(Septoria sp.))、黒斑病(リゾクトニア・ソラニ)、うどん粉病(例えばブレミア・ラクチュカ(Bremia lactucae)、ペロノスポラ種(Peronospora sp.)、シュードペルノスポラ種(Pseudoperonospora sp.)、フィトフトラ種(Phytophthora sp.));
果樹、地上部分の病害に関して、モニリア病(モニリア・フルクチゲナ(Monilia fructigenae)、M.ラクサ(M.laxa))、そうか病(ベンチュリア・イナクアリス(Venturia inaequalis))、うどん粉病(ポドスファエラ・リューコトリカ(Podosphaera leucotricha));
ブドウ、茎葉病害に関して、特に灰色かび病(ボツリティス・シネレア(Botrytis cinerea))、うどん粉病(ウンキヌラ・ネカトル(Uncinula necator))、胴枯れ病(ギグナルディア・ビウェリ(Guignardia biwelli))、うどん粉病(プラスモパラ・ヴィティコラ(Plasmopara viticola));
ビートの根、地上部分の以下の病害に関して、セルコスポラ病(セルコスポラ・ベティコラ(Cercospora beticola))、うどん粉病(エリシフェ・ベティコラ(Erysiphe beticola))、葉枯れ病(ラムラリア・ベティコラ(Ramularia beticola))
が挙げられる。
【0011】
メチオニンシンターゼは、植物及び微生物(細菌、酵母、真菌)において認められる広く特性付けられた酵素である。本発明の方法はメチオニンシンターゼ阻害剤を使用する。最初の実施形態では、本発明は、農作物の真菌病の処置のための、真菌メチオニンシンターゼの阻害剤、より好ましくは植物性病原真菌のメチオニンシンターゼの阻害剤の使用に関する。
【0012】
好ましくは、メチオニンシンターゼはこの天然環境から単離、精製又は部分精製される。メチオニンシンターゼは様々な方法によって調製することができる。これらの方法は、特に、これらのポリペプチドを天然に発現する細胞などの天然ソースからの精製、適切な宿主細胞による組換えポリペプチドの生産とその後の精製、化学合成による生産又は、最後に、これらの様々なアプローチの組合せである。これらの様々な生産方法は当業者に周知である。
【0013】
本発明の実施形態の1つでは、この酵素を天然に生産する生物、例えば大腸菌のような細菌、S.セレビシエのような酵母、又はニューロスポラ・クラッサ又はマグナポルテ・グリセアのような真菌からメチオニンシンターゼを精製する。
【0014】
本発明の好ましい実施形態では、メチオニンシンターゼは組換え宿主生物において過剰発現される。DNAフラグメントを工作する方法及び宿主細胞におけるポリペプチドの発現は当業者に周知であり、例えばGreene Publishing Associates and Wiley−Interscience発行の「Current Protocols in Molecular Biology」Volumes 1 and 2(1989)、 Ausubel F.M.ら、又はMolecular Cloning,T.Maniatis,E.F.Fritsch,J.Sambrook(1982)に述べられている。
【0015】
本発明の特定実施形態では、メチオニンシンターゼ阻害剤はマグナポルテ・グリセアの、トウモロコシ黒穂病菌の、及びより詳細には配列表の配列番号18を含む配列によって表わされるメチオニンシンターゼ、又はさもなければフィトフトラ・インフェスタンス(Phytophtora infestans)の、特に配列表の配列番号20を含む配列によって表わされるメチオニンシンターゼを阻害する;前記メチオニンシンターゼは、配列表の配列番号1を含む配列によって表わされるマグナポルテ・グリセアの遺伝子によって、又は配列表の配列番号2を含む配列によって表わされるcDNAによって、配列表の配列番号16を含む配列によって表わされるトウモロコシ黒穂病菌の遺伝子によって、又は配列表の配列番号17を含む配列によって表わされるcDNAによって、又はさもなければ配列表の配列番号19を含む配列によって表わされるフィトフトラ・インフェスタンス(Phytophtora infestans)の遺伝子によってコードされ得る。
【0016】
本発明の1つの主題はまた、メチオニンシンターゼ阻害剤ともう1つの抗真菌化合物を含む抗真菌組成物である。他の抗真菌化合物との混合物、特にアシベンゾラル−S−メチル、アゾキシストロビン、ベナラキシル、ベノミル、ブラスチシジン−S、ブロムコナゾール、カプタフォル、カプタン、カルベンダジム、カルボキシン、カルプロパミド、クロロタロニル、水酸化銅又はオキシ塩化銅のような銅又は銅誘導体に基づく抗真菌組成物、シアゾファミド、シモキサニル、シプロコナゾール、シプロジニル、ジクロラン、ジクロシメット、ジクロラン、ジエトフェンカルブ、ジフェノコナゾール、ジフルメトリム、ジメトモルフ、ジニコナゾール、ジスコストロビン、ドデモルフ、ドジン、エジフェンフォス、エポキシコナゾール、エタボキサム、エチリモール、ファモキサドン、フェナミドン、フェナリモール、フェンブコナゾール、フェンヘキサミド、フェンピクロニル、フェンプロピジン、フェンプロピモルフ、フェリムゾン、フルアジナム、フルジオキソニル、フルメトバー、フルキンコナゾール、フルシラゾール、フルスルファミド、フルトラニル、フルトリアフォル、フォルペット、フララキシル、フラメトピル、グアザチン、ヘキサコナゾール、ヒメキサゾル、イマザリル、イプロベンフォス、イプロジオン、イソプロチオラン、カスガマイシン、クレソキシム−メチル、マンコゼブ、マネブ、メフェノキサム、メパニピリム、メタラキシル及びメタラキシル−Mのようなこの鏡像異性体、メトコナゾール、メチラム−亜鉛、メトミノストロビン、オキサジキシル、ペフラゾエート、ペンコナゾール、ペンシクロン、リン酸及びフォセチル−Alのようなこの誘導体、フタリド、ピコキシストロビン、プロベナゾール、プロクロラズ、プロシミドン、プロパモカルブ、プロピコナゾール、ピラクロストロビン、ピリメタニル、ピロキロン、キノキシフェン、シルチオファム、シメコナゾール、スピロキサミン、テブコナゾール、テトラコナゾール、チアベンダゾール、チフルザミド、チオファネート、例えばチオファネート−メチル、チラム、トリアジメフォン、トリアジメノール、トリシクラゾール、トリデモルフ、トリフロキシストロビン、トリチコナゾール、バリンアミド誘導体、例えばイプロバリカルブ、ビンクロゾリン、ジネブ及びゾキサミドとの混合物は、特に有益である。このようにして得られる混合物はより広い活性スペクトルを有する。本発明に従った組成物はまた、1以上の殺虫剤、殺菌剤、殺ダニ剤又はフェロモン、又は生物活性を有する他の化合物を含み得る。
【0017】
本発明の1つの主題はまた、メチオニンシンターゼ阻害剤を使用して抗真菌組成物を生産するための方法である。
【0018】
本発明の1つの主題はまた、メチオニンシンターゼの酵素活性を阻害する化合物の特定を含む、抗真菌化合物を調製するための方法である。
【0019】
メチオニンシンターゼの酵素活性の阻害を測定するために試験化合物の存在下で酵素反応を実施する。メチオニンシンターゼの酵素活性を測定するため、及び従って、この酵素活性を阻害する化合物を特定するための全ての生化学試験が、本発明に従った方法において使用できる。
【0020】
この酵素の特異的阻害剤をスクリーニングするために高速(high−throughput)生化学アッセイを提案する。
【0021】
好ましくは、メチオニンシンターゼの活性を阻害する化合物を特定するための方法は、これらの化合物を、この基質、ホモシステイン、メチルテトラヒドロ葉酸又はメチルテトラヒドロ葉酸のポリグルタミン酸誘導体((CH3−H4)PteGlun)、及びリン酸塩、マグネシウム及び亜鉛のような様々な補因子の存在下でメチオニンシンターゼと接触させること;及び酵素活性を測定することを含む。
【0022】
メチオニンシンターゼの酵素活性を測定することは、メチオニン、テトラヒドロ葉酸又はさもなければメテニルテトラヒドロ葉酸又はこのようにして得られる何らかの産物の形成を測定すること、又はさもなければ他の何らかの化学又は酵素反応によって前記活性を測定することに結びつけることができる。
【0023】
メチオニンシンターゼの酵素活性の測定はまた、共役酵素の存在下で実施することもできる。S−アデノシルメチオニンシンターゼ(AdoMetS)はそれ自体で使用できる;これは、メチオニン、ATP及びマグネシウムの存在下でS−アデノシルメチオニン(S−AdoMet)の形成を触媒する。メチオニンシンターゼの酵素活性の測定は、従って、S−アデノシルメチオニン、リン酸塩又はピロリン酸塩の形成の測定と結びつけることができる。
【0024】
本発明のもう1つの態様によれば、メチオニンシンターゼの酵素活性を阻害する化合物の特定は、宿主生物においてメチオニンシンターゼを発現させること、宿主生物によって生産されたメチオニンシンターゼを精製すること、これらの化合物を精製メチオニンシンターゼ及びこの基質と接触させること、及び酵素活性を測定することを含む。
【0025】
好ましい実施形態では、これらの方法は全て、メチオニンシンターゼの酵素活性を阻害する前記化合物が、真菌の増殖及び/又は病原性を阻害するかどうかを測定する追加工程を含む。
【0026】
本発明は、従って、メチオニンシンターゼの酵素活性を阻害することによって真菌の増殖及び/又は病原性を阻害する化合物を特定するための方法に関する。これらの方法は、化合物又は化合物の混合物を、メチオニンシンターゼを阻害する化合物を特定するための適切なアッセイに供すること、及び前記アッセイに対して陽性に反応する化合物を選択すること、適宜にこれらを単離すること、及びその後これらを特定することから成る。
【0027】
好ましくは、適切なアッセイは、上記で定義したようなメチオニンシンターゼの酵素活性のアッセイである。
【0028】
好ましくは、これらの方法に従って特定される化合物を、その後、当業者に公知の方法に従ってこれらの抗真菌特性に関して試験する。好ましくは、切断葉又は植物全体に関する病原性アッセイのような表現型試験によって化合物を評価する。
【0029】
「化合物」という用語は、本発明によれば、ペプチド及びタンパク質を含む、何らかの化合物又は化合物の混合物を意味することが意図されている。
【0030】
「化合物の混合物」という用語は、本発明によれば、例えば分子の立体異性体、生物学的材料(植物、植物組織、細菌培養物、酵母又は真菌の培養物、昆虫、動物組織等)の抽出に由来する天然起源の混合物、又は精製されていないか又は完全に又は部分的に精製された反応混合物、又はコンビナトリアル化学手法に由来する生成物の混合物のような、少なくとも2つの異なる化合物を意味すると理解される。
【0031】
最後に、本発明は、メチオニンシンターゼの酵素活性を阻害する新規真菌病原性阻害化合物、特に本発明に従った方法によって特定される化合物及び/又は本発明に従った方法によって特定される化合物に由来する化合物に関する。
【0032】
好ましくは、メチオニンシンターゼの酵素活性を阻害する真菌病原性阻害化合物は、酵素の一般的阻害剤ではない。また好ましくは、本発明に従った化合物は、抗真菌活性及び/又は真菌病原性への活性を有することが既に公知の化合物ではない。
【実施例1】
【0033】
真菌におけるメチオニンシンターゼ遺伝子の特性決定
A.ニデュランス7(NCBI、アクセッション番号:AAF82115)のメチオニンシンターゼのタンパク質配列をモデルとして使用して、マグナポルテ・グリセアV2型のゲノムにおいてメチオニンシンターゼ遺伝子を特定した。コンティグ2.150(MG_コンティグ_2.150、6196−8629位、相補鎖、配列番号1)に位置するメチオニンシンターゼ遺伝子の完全なヌクレオチド配列は、766アミノ酸のポリペプチド(配列番号3)をコードする2301bpのcDNA(配列番号2)に対応する3個のエクソンを含む。この遺伝子の配列及び最終的メッセンジャーを生じさせるスプライシングは、様々な公的及び私的ベースで特定される数多くのESTの効力によって確認することができる。マグナポルテ・グリセアメチオニンシンターゼは、A.ニデュランスにおけるように71個の遺伝子によってコードされる。推定cDNAから推測される一次タンパク質配列の分析は、S.セレビシエ(P05694)、A.ニデュランス(AAF82115)、大腸菌(P13009)及びシロイヌナズナ植物(AAF00639)のビタミンB12非依存性メチオニンシンターゼと48%から79%の相同性を示す。
【0034】
マグナポルテ・グリセアメチオニンシンターゼの一次配列は、この酵素の特徴であるメチオニンシンターゼドメイン(334残基、E=4e−116、pfam01717)に対応する2個の保存されたドメインを有する。このドメインは、メチルテトラヒドロ葉酸トリグルタメートからホモシステインへのメチル基の移動によるメチオニンの生産を可能にする。この領域は、タンパク質のC末端部分に位置する。2番目のドメイン、COG0620又はメチオニンシンターゼII(メチルトランスフェラーゼ)は、タンパク質のN末端部分に関する(330アミノ酸)8。最近、シロイヌナズナから結晶化した酵素に関して9、この酵素の基質であるホモシステイン及びメチルテトラヒドロ葉酸、及び反応産物であるメチオニンに関するこれらのドメインの各々の特異性を決定することが可能となった。
【0035】
マグナポルテ・グリセアメチオニンシンターゼの一次配列を、このゲノムが部分的に又は完全に配列決定されている様々な真菌種におけるオーソログに関して検索するために使用した。これらの様々な一次配列を、その後、植物、細菌及び動物のような様々な生物において記述されているメチオニンシンターゼと比較した。適切なプログラム(tblastn;FGENSH;PSI−PHI−BLAST)を使用して遺伝子の構造(イントロン+エクソン)及び一次アミノ酸配列の特性決定を実施した。この手順に従って、いくつかの真菌(子嚢菌類及び担子菌類)及び植物病原性卵菌類(P.ソージャ及びP.インフェスタンス)においてメチオニンシンターゼを特性決定することができた。メチオニンシンターゼについての系統樹を確立することができ、得られた表示は、マグナポルテ・グリセアメチオニンシンターゼが子嚢菌類のメチオニンシンターゼに属すること及び担子菌類のメチオニンシンターゼからは遠いことを示す。全体として、得られた系統樹は、これらの生物の系統発生起源10を遡るものと一致する。
【実施例2】
【0036】
マグナポルテ・グリセアメチオニンシンターゼ遺伝子の欠失
マグナポルテ・グリセアの発生及び感染過程におけるメチオニンシンターゼ遺伝子の役割の検討を、この遺伝子の欠失突然変異体の表現型を試験することによって実施した。欠失突然変異体を得るための戦略は、MET6遺伝子を、MET6オープンリーディングフレームが形質転換体の選択のための抗生物質(ハイグロマイシン)に対する耐性のためのカセットで置換された突然変異型対立遺伝子で置換することに基づく。
【0037】
マグナポルテ・グリセアMET6遺伝子を置換するためのこのベクターの構築は、2工程で実施される。(i)この遺伝子に隣接し、それぞれMET6のいずれかの側に位置する約1kbのゲノムの領域に対応する領域のPCR増幅、(ii)形質転換体の選択を可能にする、抗生物質に対する耐性のための遺伝子へのこれらのゲノムDNAフラグメントの連結。従って、遺伝子を置換するために使用されるPCRフラグメントは、検討する遺伝子の左境界及び右境界と称される2つの領域から成る(図1)。我々は、選択遺伝子としてハイグロマイシン耐性遺伝子(HYG、PtrpCプロモーター、ハイグロマイシン耐性遺伝子hphのコード部分を含む)を選択した。HYG遺伝子の連結は、最初、MET6遺伝子の左境界のSacII/BglII部位及びEcoRI/SacII部位(Met6−1/Met6−2プライマー)によって実施する。MET6遺伝子の右境界(Met6−3/Met6−4プライマー)を、次に、hph遺伝子の下流のPmeI部位を通して導入する(図1)。置換ベクターは、従って、MET6遺伝子の左境界(BG)(1475bpのプロモーター領域)、HYGカセット(1400bp)及びMET6遺伝子の右境界(BD)(1251bpのターミネーター領域)を含む。その後、連結産物(BG−hph−BD)をプラスミドベクターにクローニングした。MET6遺伝子置換カセットを、その後、MET6遺伝子の両境界の末端に特異的なプライマー(Met6−5/Met6−6プライマー)を用いたPCRによって対応するプラスミドから増幅した。境界の間の接合部及び置換ベクターのHYGカセットの配列決定によって構築を確認することができた。アガロースゲル電気泳動によって精製したPCR産物(1μg)を、次に、実験室において開発された従来の手法に従ってマグナポルテ・グリセアP1.2野生型株のプロトプラストを形質転換するために使用する。形質転換に由来する産物を、対応する抗生物質(ハイグロマイシン)を含む培地で選択する。
【実施例3】
【0038】
遺伝子置換によって得られるmet6Δ::hph欠失突然変異体の特定及び栄養特性決定
一次形質転換体を、ハイグロマイシンの存在下で発生するこれらの能力に関して選択する。1mMメチオニンを添加した又は添加していない、ハイグロマイシンを含む最小培地での形質転換体の異なる増殖を測定することにより、met6Δ::hph突然変異体の特定を実施した。met6Δ::hph突然変異体はこの最小培地で発生することができないが、メチオニンを添加したこの最小培地では正常に増殖する。突然変異体の発生率は、我々の実験条件下で分析した一次形質転換体の約20%である。これらの突然変異体を、その後、単胞子(monosphores)の単離によって遺伝的に精製する。
【0039】
得られた5個のmet6Δ::hph突然変異体(4.1、15.1、22.1及び23.1)は、P1.2野生型株の増殖を可能にする最小培地で発生することができない。最小培地へのメチオニンの添加は突然変異体の増殖を回復する。メチオニンの添加によるmet6Δ::hph突然変異体の栄養補完は、メチオニンシンターゼがMET6遺伝子欠失によって影響されることを指示する。
【0040】
システイン又はグルタチオン(メチオニンシンターゼからの基質であるホモシステインの前駆体)のような硫黄供与体の添加は、マグナポルテ・グリセアmet6Δ::hph突然変異体の増殖を回復するのに十分ではない。従って、この真菌においてメチオニン合成のためにメチルテトラヒドロ葉酸又はこのポリグルタミン酸誘導体を使用する平行経路は存在しない。従って、メチオニンの新たな合成は専らメチオニンシンターゼによってのみ触媒される。この酵素の活性は、従って、真菌の発生にとって必須である。
【0041】
他方で、直接メチオニンに由来し、細胞周期に必須である化合物、S−アデノシルメチオニン(SAM又はAdoMet)、又は植物において合成される化合物、S−メチルメチオニン(SMM)の存在下で、マグナポルテ・グリセアmet6Δ::hph突然変異体は発生することができるが、P1.2野生型株に比べて増殖率は低い。SAM及びSMMは、浸透することができ、マグナポルテ・グリセアによってメチオニンに代謝され得る。この機構はおそらく、酵母S.セレビシエにおいて記述されているものと同様である。実際に、この子嚢菌では、SAMとSMMは輸送体SAM3及びMPP1によって細胞内に取り込まれ、その後ホモシステインの存在下で、SAM又はSMMをそれぞれメチル基供与体として使用するホモシステイン−S−メチルトランスフェラーゼ(SAM4及びMHT1)によってメチオニンに変換される(S.セレビシエ)11。我々の実験条件によれば、最小培地へのSAM(1mM)の添加は、met6Δ::hph突然変異体の増殖を回復する上でSMMの添加よりも有効である。これらの結果は、マグナポルテ・グリセアが、S.セレビシエにおいて記述されているのと同様の輸送体及びホモシステインメチルトランスフェラーゼを有することを示唆する。酵母SAM4及びMHT1タンパク質に関する配列相同性検索(tBlastN)によるマグナポルテ・グリセアゲノムの分析は、マグナポルテ・グリセアにおけるSAM4のオーソログである遺伝子を明らかにすることを可能にする。糸状菌は1個の遺伝子だけを有する(MHT1のオーソログなし)と思われる。我々の結果によれば(SAMでのSMMよりも良好な増殖)、このタンパク質はSMMよりもSAMに対してより大きな親和性を有すると考えられる。
【0042】
MET6遺伝子以外の遺伝子座に置換ベクターBG−hph−BDを組み込んだ形質転換体に対応する、いくつかの「異所性」形質転換体も分析した。これらのハイグロマイシン耐性異所性形質転換体は最小培地で発生することができる。メチオニンシンターゼ遺伝子は、従って、これらの異所性形質転換体において機能性であり、ベクターは、我々の実験条件下で病原体の発生(最小又は完全培地で発生する能力、胞子形成)に影響を及ぼさないゲノムの遺伝子座に挿入された。
【実施例4】
【0043】
met6Δ::hph突然変異体の分子特性決定
PCR及びサザンハイブリダイゼーションによるMET6遺伝子の分子分析を実施するために使用するゲノムDNAを抽出するために、メチオニン(1mM)を含む培地でmet6Δ::hph突然変異体を培養する。
【0044】
形質転換体の分子分析を、P1.2のMET6遺伝子の野生型対立遺伝子を突然変異型対立遺伝子met6Δ::hphで置換するための様々な特異的プライマーを使用して、MET6遺伝子座のゲノム領域の増幅によって実施する。これらのPCRは、特異的オリゴヌクレオチドを用いて各々の突然変異体に関して実施する。反応は、最初にハイグロマイシン耐性遺伝子hphと、2番目に、置換ベクターを構築するために使用したMET6領域の外側に位置するMET6遺伝子座のゲノム配列と(左接合物及び右接合部);MET6遺伝子に相同な配列と、ハイブリダイズするオリゴヌクレオチドを使用する。従って、1969bp(左接合部)又は2447bp(右接合部)のフラグメントの増幅は、構築物のmet6Δ::hph遺伝子による野生型遺伝子の置換の場合にのみ起こる(プライマーHphRP01rev(配列番号6)/Met6−7(−)(配列番号7)及びMet6−10(配列番号8)/dCGS−hph−end(−)(配列番号9)(図1))。P1.2野生型株又は異所性形質転換体の場合には増幅は得られない。同様に、met6Δ::hph突然変異体内にMET6遺伝子の増幅が存在しないことは(プライマーMet6−8、配列番号10/Met6−9、配列番号11)(図1)、MET6遺伝子が実際にこれらの形質転換体に存在しないことの指標である。他方で、これらのプライマーにより、2424bpのフラグメントがP1.2野生型株及び異所性形質転換体において増幅される。突然変異体22.2及び23.1に関して実施したPCRだけが、これら3つのタイプのPCRに関して予想された結果を与える。PCR Met6−10(配列番号8)/dCGS−hph−end(配列番号9)及びhphRP01(配列番号6)/Met6−7(−)、配列番号7)では、説明のつかない理由で、増幅が得られなかった。
【0045】
BamHI制限酵素によるゲノムDNAの消化後、サザンハイブリダイゼーションによる分析を実施し、4個の突然変異体に関して得られたハイブリダイゼーションシグナルをP1.2野生型株及び異所性形質転換体(19.1)で得られたものと比較した。置換ベクター内に存在するMET6左境界(MET6プロモーター領域)に対応する(Met6−1(配列番号12)/Met6−2(配列番号13))PCRフラグメントをプローブとして使用して、大きさがP1.2野生型株及び異所性形質転換体(19.1)のものとは異なる2つのバンドが突然変異体に関して認められる。シグナルは、P1.2野生型株及び異所性形質転換体19.1内のMET6プロモーター領域に対応する。後者はまた、別のゲノム領域に挿入された置換ベクターに対応するハイブリダイゼーションシグナルも示す。同様の結果が、置換ベクター内に存在するMET6右境界(MET6ターミネーター領域)に対応する(Met6−3(配列番号14)/Met6−4(配列番号15))PCRフラグメントを使用して得られる。挿入遺伝子(hph)に特異的なプローブにより、met6Δ::hph突然変異体及び異所性形質転換体19.1だけが、MET6遺伝子座におけるhphの存在(突然変異体)又は置換ベクターBG−met6Δ::hph−BD(異所性)のいずれかに対応するハイブリダイゼーションシグナルを示す。後者の結果は、分析した様々な突然変異体が分子レベルで同一であり、MET6コード相(coding phase)の代わりにMET6遺伝子座として挿入されたhph遺伝子の1コピーだけを含むことを指示する。
【実施例5】
【0046】
マグナポルテ・グリセアmet6Δ::hph突然変異体の病原力の分析
マグナポルテ・グリセアmet6Δ::hph突然変異体の病原力を、生存条件下の大麦の葉に関して及び大麦及びイネ植物全体に関する感染試験によって評価した。この分析の後に、胞子発芽率、付着器分化及び大麦葉への貫入測定を実施した。P1.2野生型株及びmet6Δ::hph突然変異体4.1、15.1、22.1、23.1及び24.1の胞子を、1mMを含む米粉ベースの培地での14日間の増殖後に採取する。使用する植物材料は、大麦(cv.Express)及びイネ(cv.Sariceltik)である。
【0047】
生存条件下で大麦の葉に関して実施する実験では、高湿度(100%)及び100マイクロアインシュタインの光下で温帯気候室(26℃)において、キネチン(2mg/ml−1)を含む寒天培地(1%寒天−H2O)で葉をインキュベートする。感染の間、胞子を液滴の形態で(35μl)又は綿毛芽を用いて胞子の懸濁液で葉の表面を被覆することによって葉に付着させる。これらの実験を、インキュベーションの間を通じて又は24時間だけ、1mMメチオニンの不在下又は存在下で実施する。胞子濃度は、水中3×104胞子/mlから1×105胞子/mlである。次に、マグナポルテ・グリセアによって引き起こされる病徴の出現を少なくともインキュベーションの7日間観察する。マグナポルテ・グリセア野生型株P1.2の胞子による生存条件下での大麦葉の接種は、感染葉における真菌の発生の特徴である壊死(胞子形成病変)を生じさせる。反対に、met6Δ::hph突然変異体4.1、15.1、22.1、23.1及び24.1は、接種の方法に関わらず、大麦葉にいかなる病徴も生じさせず、従って非病原性である。met6Δ::hph突然変異体胞子へのメチオニンの添加は、これらの病原力を部分的に回復することを可能にする(特徴的であるが非胞子形成性病変の発現)。さらに、我々の実験条件下では、損傷葉又は非損傷葉に関して実施した感染の間で著明な差を認めなかった。これらの所見は、met6Δ::hph突然変異体が、損傷した大麦葉でさえも、大麦葉に貫入できないことを示唆する。被覆による胞子の分散に関して実施した実験で認められた葉の黄化は、おそらく我々の実験条件下での葉の早期老化によるものである。結果を以下の表に要約する。
【0048】
【表1】
【0049】
感染の様々な段階(発芽、付着器の形成及び貫入)の観察及び定量を、胞子の懸濁液(3×105胞子/ml)35μlの液滴による生存条件下での大麦葉の接種に基づいて実施する。24時間後、付着器を分化した発芽胞子の数及び表皮細胞への貫入事象の数を顕微鏡下で観察するために葉の表皮をはがす。0.01%のカルコフロールの溶液は、植物の表面に位置する真菌細胞の壁の強い蛍光を生じさせることを可能にする(表皮細胞内に位置する菌糸は着色されない)。これらの観察(表II)は、met6Δ::hph突然変異体がわずかに低い発芽を有する(野生型株と比較して−10%から−40%)ことを明らかにする。これらの付着器の分化率もわずかに低い(野生型株と比較して0%から−30%)。他方で、これらの突然変異型付着器は茎葉組織に貫入することができない。
【0050】
【表2】
【0051】
付着器の形成はまた、メチオニン(我々の実験条件下で1mM濃度)を添加した又は添加していないテフロン(登録商標)膜上で胞子が発芽する、人工条件下でも認められる。これらの非常に疎水性の膜は葉の表面を模倣し、これによって付着器の形成を誘導すること及び付着器分化率を容易に測定することを可能にする。
【0052】
結論として、マグナポルテ・グリセアmet6Δ::hph突然変異体は、従って、付着器を分化するが、植物に貫入することはできない。これらの結果は、これらの突然変異体が非機能性付着器を有することを示す。
【0053】
3週齢の大麦植物(第二葉の出芽に相当する)を含む鉢を、マグナポルテ・グリセア胞子の懸濁液(3×104胞子/mlの濃度の胞子及び水滴の葉への付着のための0.3%ゼラチンを含む水10ml)による散布に供する。散布後少なくとも8日間病徴の観察を実施する。野生型株(P1.2)及び異所性形質転換体(19.1及び20.1)で処置した大麦植物は、感染葉における真菌の発生によって引き起こされる壊死病変を示す。他方で、マグナポルテ・グリセアmet6Δ::hph突然変異体を接種した場合は病害の症状を認めなかった。これらの突然変異体は、従って、非病原性であると考えられる。さらに、この試験は、生存条件下で大麦葉に関して実施した分析と同様に、葉の組織中のメチオニン(又はSAM及びSMMのようなメチオニンに由来する他の化合物)の量が、met6Δ::hph突然変異体のメチオニン合成の欠損を補うには不十分であることを指示する。従って、インビトロ実験で明らかにされた経路(SAM及びSMMの存在下での補完)は、真菌が植物において成長している間、真菌において機能性ではないと思われる。
【0054】
得られたmet6Δ::hph突然変異体は、ハイグロマイシン耐性を付与する、hph遺伝子によって置換されるMET6遺伝子のコード相の欠失に相当する。これらの突然変異体はホモシステインからメチオニンを合成することができず、従って最小培地で増殖することができない。この最小培地へのメチオニンの添加はこれらの突然変異体の発生のために必須である。met6Δ::hph突然変異体は、生存条件下の大麦葉又は全植物に接種するために使用したとき、高い胞子濃度であっても、病害の症状を生じさせない。従って、メチオニンシンターゼによるメチオニン合成は、インビトロで及び植物において、真菌の発生に必須である。
【0055】
met6Δ::hph突然変異体はメチオニンの不在下で付着器に分化することができる。この態様は、胞子が、この細胞の発生のために必要なタンパク質及び代謝産物の合成を可能にするメチオニンの少なからぬ貯蔵を有し得ることを指示する。この貯蔵は、使用した米粉ベースの培地(1mMメチオニンを含む)での突然変異体の増殖とこの胞子形成を可能にするために突然変異体に供給されたメチオニンに由来するはずである。他方で、met6Δ::hph突然変異体の葉への貫入が存在しないことは、前記突然変異体が、真菌の植物への貫入を指令することができない非機能性付着器に分化することを指示する。おそらく、突然変異型付着器はこれらのメチオニン貯蔵を急速に使い果たしたと考えられる。実際に、突然変異体胞子へのメチオニンの添加は植物への貫入と葉における発生の開始を可能にする。しかし、前記発生は完全ではなく、胞子形成病変の形成の不在は、胞子に供給されるメチオニン貯蔵がひとたび使い果たされると、植物は突然変異体のメチオニン欠乏を補うことができない。
【実施例6】
【0056】
メチオニンシンターゼの酵素活性を阻害する分子を検定し、特性決定するための方法
本発明の方法は、この作用が、直接又は間接的手法(メチオニンシンターゼの活性を測定するための「共役」酵素の使用を含む)に従って定量される基質の消費又は生成物の形成を阻害する全ての分子の特性決定を含む。メチオニンシンターゼは、文献12、13、14、15、27に記述されている公知のアッセイに従って様々な補因子(リン酸塩、マグネシウム、亜鉛)の存在下で、ホモシステイン、メチルテトラヒドロ葉酸(n=1)又はこのポリグルタミン酸誘導体(n≧3)の存在下で不可逆的反応を触媒する。この方法は、逆相HPLCクロマトグラフィーによって化合物を分離するための手法16により、生産されるメチオニン又は葉酸誘導体を定量することを含む。メチルテトラヒドロ葉酸基質は手順17に述べられている実験条件下で検出可能ではないので、テトラヒドロ葉酸がメテニルテトラヒドロ葉酸に変換された後、反応の間に生産されたテトラヒドロ葉酸の検定を350nmの分光光度計で実施することができる。提案される代替法は、S−アデノシルメチオニンシンテターゼの存在下でメチオニンシンターゼ活性を検定することである。このアッセイでは、マグナポルテ・グリセアのS−アデノシルメチオニンシンテターゼ(AdoMetS)が好ましく使用されるが、いかなるAdoMetSも「共役」酵素として使用できる。AdoMetS酵素は、メチオニン、ATP及びマグネシウムの存在下で、S−アデノシルメチオニン(SAM)、リン酸塩及びピロリン酸塩を生産する不可逆的反応を触媒する。
【0057】
メチオニンシンターゼ活性は、結局、共役酵素の存在下での生成物の変換後に比色法によって及び/又は分光光度計で又はメチオニンシンターゼ活性を測定するための他の何らかの化学又は酵素反応によって、生産されるSAM、リン酸塩又はピロリン酸塩の量を通して検定される。これらの様々な方法は文献における多くの記述の対象であり、実験者に従って適合させることができる18、19、20、21、22、23。例えばSAMシンテターゼの存在下でメチオニンシンターゼ活性を検定するための方法の感受性は、ピロリン酸塩を2モルのリン酸塩に変換するピロホスファターゼの添加を通して改善することができる。従って、合成されるメチオニンの各モルについて、前記方法は3モルのリン酸塩を生産する。
【0058】
マグナポルテ・グリセアメチオニンシンターゼの精製
大量のメチオニンシンターゼの生産は、細菌又は酵母におけるタンパク質の過剰生産のための発現ベクターを使用する手法を用いて実施される。この手法は、好ましくはタンパク質のN末端又はC末端にヒスチジンタグ延長部を組み込むことを可能にする発現ベクターへのcDNAのクローニングを用いる。例えばpET−28b(+)ベクター(Novagen)24を選択するとき、従来の分子生物学手法に従って2301bpのcDNAをNdEI及びEcoRIにクローニングする。pET−28−MgMET6と呼ばれる、得られた構築物を大腸菌BL21型DE3(pLysS)菌株に導入し、IPTG(0.5mM)による誘導後に発現を生じさせる。組換え細菌を28℃で4時間培養する。その後遠心分離によって細胞を収集し、得られたペレットをタンパク質の安定性に適した溶解緩衝液に再懸濁する。試料の超音波処理後、遠心分離後に得られる、組換えタンパク質を含む可溶性分画をNi−NTAアガロース型のカラムに負荷する。次に、マトリックスをイミダゾール溶液で数回連続的に洗浄した後、酵素の精製及び溶出を実施する。手順はQiagenによって定義されたプロトコール25に従う。
【0059】
溶出後、組換えメチオニンシンターゼを含むタンパク質分画を限外ろ過によって濃縮し、イミダゾールの全ての痕跡を除去するためにPD10(Pharmacia)27での分子ろ過に供する。組換えタンパク質の精製は、Superdex S200(Pharmacia)26でのクロマトグラフィーによる分子ろ過又はMonoQ HR10/10(Pharmacia)26でのイオン交換クロマトグラフィーから成る第二工程を伴い得る。適切な直接測定アッセイを使用して、精製の間メチオニンシンターゼの活性を追跡する。
【0060】
【表3】
【図面の簡単な説明】
【0061】
【図1】遺伝子置換のためのフラグメント、ならびに増幅の概略図。
【特許請求の範囲】
【請求項1】
有効量のメチオニンシンターゼ阻害剤を施用することを特徴とする、真菌病に対して作物を処置するための方法。
【請求項2】
メチオニンシンターゼが真菌由来であることを特徴とする、請求項1に記載の方法。
【請求項3】
メチオニンシンターゼがマグナポルテ・グリセア(Magnaporthe grisea)に由来することを特徴とする、請求項2に記載の方法。
【請求項4】
メチオニンシンターゼが配列番号3を含む、請求項3に記載の方法。
【請求項5】
メチオニンシンターゼが、配列番号1又は配列番号2を含む配列によってコードされる、請求項3に記載の方法。
【請求項6】
メチオニンシンターゼがトウモロコシ黒穂病菌(Ustilago maydis)に由来することを特徴とする、請求項2に記載の方法。
【請求項7】
メチオニンシンターゼが配列番号18を含む、請求項6に記載の方法。
【請求項8】
メチオニンシンターゼが、配列番号16又は配列番号17を含む配列によってコードされる、請求項6に記載の方法。
【請求項9】
メチオニンシンターゼがフィトフトラ・インフェスタンス(Phytophthora infestans)に由来することを特徴とする、請求項2に記載の方法。
【請求項10】
メチオニンシンターゼが配列番号20を含む、請求項9に記載の方法。
【請求項11】
メチオニンシンターゼが、配列番号19を含む配列によってコードされる、請求項9に記載の方法。
【請求項12】
メチオニンシンターゼ阻害剤が抗真菌組成物の形態である、請求項1から11のいずれか一項に記載の方法。
【請求項13】
メチオニンシンターゼ阻害剤と抗真菌化合物を含むことを特徴とする、抗真菌組成物。
【請求項14】
メチオニンシンターゼ阻害剤を使用することを特徴とする、抗真菌組成物を生産するための方法。
【請求項15】
メチオニンシンターゼの酵素活性を阻害する化合物の特定を含む、抗真菌化合物を調製するための方法。
【請求項16】
メチオニンシンターゼの酵素活性を阻害する化合物の特定が、以下の工程、
前記化合物を、ホモシステイン及びメチルテトラグルタメート又はこのポリグルタミン酸誘導体、及び補因子の存在下でメチオニンシンターゼと接触させる工程;及び
前記酵素活性を測定する工程
を含む、請求項15に記載の方法。
【請求項17】
メチオニンシンターゼの酵素活性を阻害する化合物の特定が、以下の工程、
前記化合物を、ホモシステイン及びメチルテトラグルタメート又はこのポリグルタミン酸誘導体、及び補因子の存在下でメチオニンシンターゼと接触させる工程;及び
メチオニンの形成を測定する工程
を含む、請求項16に記載の方法。
【請求項18】
メチオニンシンターゼの酵素活性を阻害する化合物の特定が、以下の工程、
前記化合物を、ホモシステイン、メチルテトラヒドロ葉酸及びリン酸塩、マグネシウム及び亜鉛の存在下でメチオニンシンターゼと接触させる工程;及び
メチオニンの形成を測定する工程
を含む、請求項17に記載の方法。
【請求項19】
メチオニンシンターゼの酵素活性を阻害する化合物の特定が、以下の工程、
前記化合物を、ホモシステイン、メチルテトラヒドロ葉酸又はこのポリグルタミン酸誘導体、S−アデノシル−メチオニンシンテターゼ、ATP及びMg、及び補因子の存在下でメチオニンシンターゼと接触させる工程;及び
S−アデノシル−メチオニン、リン酸塩又はピロリン酸塩の形成を測定する工程
を含む、請求項15に記載の方法。
【請求項20】
メチオニンシンターゼの酵素活性を阻害する化合物の特定が、以下の工程、
前記化合物を、ホモシステイン、メチルテトラヒドロ葉酸又はこのポリグルタミン酸誘導体、S−アデノシル−メチオニンシンテターゼ、ATP及びMg、及び補因子の存在下でメチオニンシンターゼと接触させる工程;及び
リン酸塩の形成を測定する工程
を含む、請求項19に記載の方法。
【請求項21】
メチオニンシンターゼの酵素活性を阻害する化合物の特定が、以下の工程、
宿主生物においてメチオニンシンターゼを発現させる工程;
前記宿主生物によって生産されたメチオニンシンターゼを精製する工程;
前記化合物を、ホモシステイン及びメチルテトラヒドロ葉酸又はこのポリグルタミン酸誘導体、及び補因子の存在下で前記精製メチオニンシンターゼと接触させる工程;及び
酵素活性を測定する工程
を含む、請求項15に記載の方法。
【請求項22】
メチオニンシンターゼの酵素活性を阻害する化合物の特定が、以下の工程、
宿主生物においてメチオニンシンターゼを発現させる工程;
前記宿主生物によって生産されたメチオニンシンターゼを精製する工程;
前記化合物を、ホモシステイン、メチルテトラヒドロ葉酸及びリン酸塩、マグネシウム及び亜鉛の存在下で前記精製メチオニンシンターゼと接触させる工程;及び
メチオニンの形成を測定する工程
を含む、請求項21に記載の方法。
【請求項23】
メチオニンシンターゼの酵素活性を阻害する化合物の特定が、以下の工程、
宿主生物においてメチオニンシンターゼを発現させる工程;
前記宿主生物によって生産されたメチオニンシンターゼを精製する工程;
前記化合物を、ホモシステイン及びメチルテトラヒドロ葉酸又はこのポリグルタミン酸誘導体、S−アデノシル−メチオニンシンテターゼ、ATP及びMg、及び補因子の存在下で前記精製メチオニンシンターゼと接触させる工程;及び
S−アデノシル−メチオニン、リン酸塩又はピロリン酸塩の形成を測定する工程
を含む、請求項15に記載の方法。
【請求項24】
メチオニンシンターゼが真菌由来であることを特徴とする、請求項15から23のいずれか一項に記載の方法。
【請求項25】
発現されるメチオニンシンターゼが、配列番号1、配列番号2、配列番号16、配列番号17及び/又は配列番号19を含む配列から選択されるポリヌクレオチドによってコードされる、請求項24に記載の方法。
【請求項26】
発現されるメチオニンシンターゼが、配列番号3、配列番号18及び/又は配列番号20を含む配列から選択される、請求項24に記載の方法。
【請求項27】
MSの酵素活性を阻害する前記化合物が、真菌の増殖及び/又は病原性を阻害するかどうかを測定する追加工程を含む、請求項15から26のいずれか一項に記載の方法。
【請求項28】
請求項15から27のいずれか一項に記載の方法の一つによって特定されるメチオニンシンターゼ阻害剤。
【請求項1】
有効量のメチオニンシンターゼ阻害剤を施用することを特徴とする、真菌病に対して作物を処置するための方法。
【請求項2】
メチオニンシンターゼが真菌由来であることを特徴とする、請求項1に記載の方法。
【請求項3】
メチオニンシンターゼがマグナポルテ・グリセア(Magnaporthe grisea)に由来することを特徴とする、請求項2に記載の方法。
【請求項4】
メチオニンシンターゼが配列番号3を含む、請求項3に記載の方法。
【請求項5】
メチオニンシンターゼが、配列番号1又は配列番号2を含む配列によってコードされる、請求項3に記載の方法。
【請求項6】
メチオニンシンターゼがトウモロコシ黒穂病菌(Ustilago maydis)に由来することを特徴とする、請求項2に記載の方法。
【請求項7】
メチオニンシンターゼが配列番号18を含む、請求項6に記載の方法。
【請求項8】
メチオニンシンターゼが、配列番号16又は配列番号17を含む配列によってコードされる、請求項6に記載の方法。
【請求項9】
メチオニンシンターゼがフィトフトラ・インフェスタンス(Phytophthora infestans)に由来することを特徴とする、請求項2に記載の方法。
【請求項10】
メチオニンシンターゼが配列番号20を含む、請求項9に記載の方法。
【請求項11】
メチオニンシンターゼが、配列番号19を含む配列によってコードされる、請求項9に記載の方法。
【請求項12】
メチオニンシンターゼ阻害剤が抗真菌組成物の形態である、請求項1から11のいずれか一項に記載の方法。
【請求項13】
メチオニンシンターゼ阻害剤と抗真菌化合物を含むことを特徴とする、抗真菌組成物。
【請求項14】
メチオニンシンターゼ阻害剤を使用することを特徴とする、抗真菌組成物を生産するための方法。
【請求項15】
メチオニンシンターゼの酵素活性を阻害する化合物の特定を含む、抗真菌化合物を調製するための方法。
【請求項16】
メチオニンシンターゼの酵素活性を阻害する化合物の特定が、以下の工程、
前記化合物を、ホモシステイン及びメチルテトラグルタメート又はこのポリグルタミン酸誘導体、及び補因子の存在下でメチオニンシンターゼと接触させる工程;及び
前記酵素活性を測定する工程
を含む、請求項15に記載の方法。
【請求項17】
メチオニンシンターゼの酵素活性を阻害する化合物の特定が、以下の工程、
前記化合物を、ホモシステイン及びメチルテトラグルタメート又はこのポリグルタミン酸誘導体、及び補因子の存在下でメチオニンシンターゼと接触させる工程;及び
メチオニンの形成を測定する工程
を含む、請求項16に記載の方法。
【請求項18】
メチオニンシンターゼの酵素活性を阻害する化合物の特定が、以下の工程、
前記化合物を、ホモシステイン、メチルテトラヒドロ葉酸及びリン酸塩、マグネシウム及び亜鉛の存在下でメチオニンシンターゼと接触させる工程;及び
メチオニンの形成を測定する工程
を含む、請求項17に記載の方法。
【請求項19】
メチオニンシンターゼの酵素活性を阻害する化合物の特定が、以下の工程、
前記化合物を、ホモシステイン、メチルテトラヒドロ葉酸又はこのポリグルタミン酸誘導体、S−アデノシル−メチオニンシンテターゼ、ATP及びMg、及び補因子の存在下でメチオニンシンターゼと接触させる工程;及び
S−アデノシル−メチオニン、リン酸塩又はピロリン酸塩の形成を測定する工程
を含む、請求項15に記載の方法。
【請求項20】
メチオニンシンターゼの酵素活性を阻害する化合物の特定が、以下の工程、
前記化合物を、ホモシステイン、メチルテトラヒドロ葉酸又はこのポリグルタミン酸誘導体、S−アデノシル−メチオニンシンテターゼ、ATP及びMg、及び補因子の存在下でメチオニンシンターゼと接触させる工程;及び
リン酸塩の形成を測定する工程
を含む、請求項19に記載の方法。
【請求項21】
メチオニンシンターゼの酵素活性を阻害する化合物の特定が、以下の工程、
宿主生物においてメチオニンシンターゼを発現させる工程;
前記宿主生物によって生産されたメチオニンシンターゼを精製する工程;
前記化合物を、ホモシステイン及びメチルテトラヒドロ葉酸又はこのポリグルタミン酸誘導体、及び補因子の存在下で前記精製メチオニンシンターゼと接触させる工程;及び
酵素活性を測定する工程
を含む、請求項15に記載の方法。
【請求項22】
メチオニンシンターゼの酵素活性を阻害する化合物の特定が、以下の工程、
宿主生物においてメチオニンシンターゼを発現させる工程;
前記宿主生物によって生産されたメチオニンシンターゼを精製する工程;
前記化合物を、ホモシステイン、メチルテトラヒドロ葉酸及びリン酸塩、マグネシウム及び亜鉛の存在下で前記精製メチオニンシンターゼと接触させる工程;及び
メチオニンの形成を測定する工程
を含む、請求項21に記載の方法。
【請求項23】
メチオニンシンターゼの酵素活性を阻害する化合物の特定が、以下の工程、
宿主生物においてメチオニンシンターゼを発現させる工程;
前記宿主生物によって生産されたメチオニンシンターゼを精製する工程;
前記化合物を、ホモシステイン及びメチルテトラヒドロ葉酸又はこのポリグルタミン酸誘導体、S−アデノシル−メチオニンシンテターゼ、ATP及びMg、及び補因子の存在下で前記精製メチオニンシンターゼと接触させる工程;及び
S−アデノシル−メチオニン、リン酸塩又はピロリン酸塩の形成を測定する工程
を含む、請求項15に記載の方法。
【請求項24】
メチオニンシンターゼが真菌由来であることを特徴とする、請求項15から23のいずれか一項に記載の方法。
【請求項25】
発現されるメチオニンシンターゼが、配列番号1、配列番号2、配列番号16、配列番号17及び/又は配列番号19を含む配列から選択されるポリヌクレオチドによってコードされる、請求項24に記載の方法。
【請求項26】
発現されるメチオニンシンターゼが、配列番号3、配列番号18及び/又は配列番号20を含む配列から選択される、請求項24に記載の方法。
【請求項27】
MSの酵素活性を阻害する前記化合物が、真菌の増殖及び/又は病原性を阻害するかどうかを測定する追加工程を含む、請求項15から26のいずれか一項に記載の方法。
【請求項28】
請求項15から27のいずれか一項に記載の方法の一つによって特定されるメチオニンシンターゼ阻害剤。
【図1】

【公表番号】特表2008−523791(P2008−523791A)
【公表日】平成20年7月10日(2008.7.10)
【国際特許分類】
【出願番号】特願2007−545989(P2007−545989)
【出願日】平成17年12月20日(2005.12.20)
【国際出願番号】PCT/EP2005/014209
【国際公開番号】WO2006/066974
【国際公開日】平成18年6月29日(2006.6.29)
【出願人】(503325538)バイエル・クロツプサイエンス・エス・アー (73)
【Fターム(参考)】
【公表日】平成20年7月10日(2008.7.10)
【国際特許分類】
【出願日】平成17年12月20日(2005.12.20)
【国際出願番号】PCT/EP2005/014209
【国際公開番号】WO2006/066974
【国際公開日】平成18年6月29日(2006.6.29)
【出願人】(503325538)バイエル・クロツプサイエンス・エス・アー (73)
【Fターム(参考)】
[ Back to top ]