
過剰発現で種子を大きくする遺伝子
【課題】イネにおいてBRのシグナル伝達に関与する遺伝子を単離することを課題とする。
【解決手段】BR内生量の低いbrd1変異体に代表的なブラシノステロイドであるブラシノライド(BL)を添加し、3時間及び24時間後に発現レベルが上昇する遺伝子の単離を試みた。マイクロアレイ解析の結果、3時間後、24時間後共にブラシノライド非添加コントロールに比べて2倍以上発現の増加している遺伝子AK071601を見出した。AK071601を過剰発現するイネを作製し、形態等に変化が見られるかどうか確認を行った結果、T0世代のOsBU3過剰発現カルスの生育が、コントロールと比較して顕著に促進することが確認された。またT1種子はOsBU3の転写量に応じて長さ、幅共に大きくなった。更に生育後期では、OsBU3過剰発現イネにおいて節間の異常な伸長が認められた。
【解決手段】BR内生量の低いbrd1変異体に代表的なブラシノステロイドであるブラシノライド(BL)を添加し、3時間及び24時間後に発現レベルが上昇する遺伝子の単離を試みた。マイクロアレイ解析の結果、3時間後、24時間後共にブラシノライド非添加コントロールに比べて2倍以上発現の増加している遺伝子AK071601を見出した。AK071601を過剰発現するイネを作製し、形態等に変化が見られるかどうか確認を行った結果、T0世代のOsBU3過剰発現カルスの生育が、コントロールと比較して顕著に促進することが確認された。またT1種子はOsBU3の転写量に応じて長さ、幅共に大きくなった。更に生育後期では、OsBU3過剰発現イネにおいて節間の異常な伸長が認められた。
【発明の詳細な説明】
【技術分野】
【0001】
本発明は、種子の大型化、カルスの生育促進、節間の異常な伸長からなる群より選択される少なくとも1つの形質を有するカルス又は植物体の製造方法及びこれを用いて得られるカルス及び植物体、並びにその利用に関する。
【背景技術】
【0002】
植物ホルモンのブラシノステロイド(BR)は植物の形態形成や、環境ストレスへの応答等に関与することが知られている。化学構造的にはステロイド骨格を有している点がBRの特徴で、被子植物、裸子植物、藻類およびシダ類を含む40種類以上の植物から、ブラシノライド(BL)、カスタステロンを代表に40種類以上の遊離型が単離・同定されている(Fujioka and Sakurai, 1997)。中でもBLは植物界における分布が広く、かつ天然のBRのなかで最も強い活性を示すことから、最も重要な役割を果たしていると考えられている。
【0003】
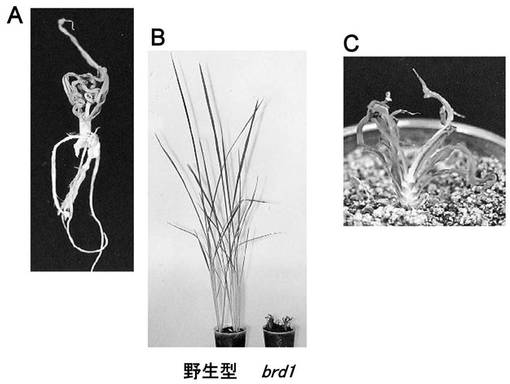
本発明者らは既に、イネでBRの生合成変異体brd1(brassinosteroid-dpendent 1)を単離しており、brd1変異体は極わい性で葉鞘が極めて短く葉身は巻いていることを示している(Mori et al. 2002, 図1)。また本発明者らは、BRはイネにおいても、葉身の伸長や展開、分げつ形成、根の発達、生殖成長等の多くの形態的及び生理的プロセスに関与していることを示している(Mori et al. 2002)。
【0004】
本発明の先行技術文献を以下に示す。
【非特許文献1】Fujioka, S. and Sakurai, A. (1997) Natural Products Report,14,1-10
【非特許文献2】Livak, KJ, Schmittgen, TD (2001) Methods 25:402-408
【非特許文献3】Mori, M et al., (2002) Plant Physiol. 130:1152-1161.
【非特許文献4】Nakamura, H et al., (2007) Plant Mol Biol. 65:357-371
【非特許文献5】Sato, H et al., (1996) Plant Sci. 119:39-47.
【非特許文献6】Toki, S et al., (2006) Plant J. 47:969-76.
【非特許文献7】Yazaki, J et al.,(2004) Physiol Genomics. 17(2):87-100.
【発明の開示】
【発明が解決しようとする課題】
【0005】
上述のように、BRは植物の形態形成や、環境ストレスへの応答等に関与することが知られている。そこで本発明は、BRの有用な作用を農業等に利用することを目的として、イネにおいてBRのシグナル伝達に関与する遺伝子を単離することを課題とした。
【課題を解決するための手段】
【0006】
上記課題を解決するにあたり本発明者らは、BRのシグナル伝達に関与する遺伝子は、BR内生量の低いbrd1変異体では発現が抑制されており、brd1にBLを添加すると添加後初期段階で発現が増大するのではないかと考えた。そこで本発明者らは、brd1変異体に代表的なブラシノステロイドであるブラシノライド(BL)を添加し、3時間及び24時間後に発現レベルが上昇する遺伝子の単離を試みた。マイクロアレイ解析の結果本発明者らは、3時間後、24時間後共にブラシノライド非添加コントロールに比べて2倍以上発現の増加している遺伝子(DDBJ Accession No. AK071601、OsBU3 (Oryza sative Brassinosteroid Upregulated 3)と命名)を単離することに成功した。
【0007】
次に本発明者らは、AK071601を過剰発現するイネを作製し、形態等に変化が見られるかどうか確認を行った。その結果、T0世代のOsBU3過剰発現カルスの生育が、コントロールと比較して顕著に促進することが確認された。
また再分化した植物体を野生型(WT)と比較すると、ラミナジョイントの顕著な屈曲が認められた。また種子はOsBU3の転写量に応じて長さ、幅共に大きくなった。更に生育後期では、OsBU3過剰発現イネにおいて節間の異常な伸長が認められた。
このようなイネカルスの生育促進、種子の大型化、明所における異常な節間伸長と、BRシグナルの増強の関連が見出されたのは、今回が初めてである。
【0008】
本発明は、ラミナジョイントの屈曲に加え、種子の大型化、カルスの生育促進、節間の異常な伸長作用を有する遺伝子、及び当該遺伝子が導入されたカルス及び植物体に関するものであり、より詳しくは、以下の〔1〕〜〔6〕を提供する。
〔1〕以下(a)及び(b)の工程を含む、種子の大型化、カルスの生育促進、節間の異常な伸長からなる群より選択される少なくとも1つの形質を植物に付与する方法;
(a)下記(i)から(iv)からなる群より選択されるDNA又は該DNAを含むベクターを植物細胞に導入する工程、及び
(b)工程(a)においてDNA又はベクターが導入された植物細胞からカルスを形成又は植物体を再生する工程、
(i)配列番号:2に記載のアミノ酸配列を含むタンパク質をコードするDNA、
(ii)配列番号:1に記載の塩基配列のコード領域を含むDNA、
(iii)配列番号:2に記載のアミノ酸配列において1もしくは複数のアミノ酸が置換、欠失、付加、及び/又は挿入されたアミノ酸配列を含むタンパク質をコードするDNA、及び
(iv)配列番号:1に記載の塩基配列を含むDNAとストリンジェントな条件下でハイブリダイズするDNA、
〔2〕以下(a)から(d)のいずれかに記載のDNAまたは該DNAを含むベクターが導入された植物細胞であって、種子の大型化、カルスの生育促進、節間の異常な伸長からなる群より選択される少なくとも1つの形質を有するカルスを形成又は植物体を再生しうる植物細胞;
(a)配列番号:2に記載のアミノ酸配列を含むタンパク質をコードするDNA、
(b)配列番号:1に記載の塩基配列のコード領域を含むDNA、
(c)配列番号:2に記載のアミノ酸配列において1もしくは複数のアミノ酸が置換、欠失、付加、及び/又は挿入されたアミノ酸配列を含むタンパク質をコードするDNA、及び
(d)配列番号:1に記載の塩基配列を含むDNAとストリンジェントな条件下でハイブリダイズするDNA、
〔3〕〔2〕に記載の植物細胞から形成されたカルス又は再生された植物体であって、種子の大型化、カルスの生育促進、節間の異常な伸長からなる群より選択される少なくとも1つの形質を有するカルス又は植物体、
〔4〕〔3〕に記載のカルス又は植物体の子孫またはクローンである、種子の大型化、カルスの生育促進、節間の異常な伸長からなる群より選択される少なくとも1つの形質を有するカルス又は植物体、
〔5〕〔3〕または〔4〕に記載の植物体の繁殖材料、
〔6〕下記(a)及び(b)の工程を含む、種子の大型化、カルスの生育促進、節間の異常な伸長からなる群より選択される少なくとも1つの形質を有するカルス又は植物体の製造方法;
(a)下記(i)から(iv)からなる群より選択されるDNA又は該DNAを含むベクターを植物細胞に導入する工程、及び
(b)工程(a)においてDNA又はベクターが導入された植物細胞からカルスを形成又は植物体を再生する工程、
(i)配列番号:2に記載のアミノ酸配列を含むタンパク質をコードするDNA、
(ii)配列番号:1に記載の塩基配列のコード領域を含むDNA、
(iii)配列番号:2に記載のアミノ酸配列において1もしくは複数のアミノ酸が置換、欠失、付加、及び/又は挿入されたアミノ酸配列を含むタンパク質をコードするDNA、及び
(iv)配列番号:1に記載の塩基配列を含むDNAとストリンジェントな条件下でハイブリダイズするDNA。
【発明の効果】
【0009】
本発明により、カルスの生育を促進し、種子を大型化し、かつ明所で節間の異常な伸長作用を有する遺伝子(OsBU3)が提供された。本発明にて同定された遺伝子を過剰発現させたイネは、ラミナジョイントの屈曲に加え、種子の大型化、カルスの生育促進、節間の異常な伸長が認められた。
OsBU3あるいはそのホモログ遺伝子の発現レベルを変化させ、種子サイズを変化させることにより、主に種子が可食部の植物(穀物等)で新たな商品価値を付与することが可能である。また、過剰発現や発現抑制により草型や節間長を変化させることにより、作物や観葉植物で新たな商品価値を付与することが可能となった。またOsBU3を過剰発現することによりカルスの増殖が促進するため、OsBU3が導入されたカルスを色素やアルカロイド、薬品などの物質生産に利用できる可能性がある。
【0010】
〔発明の実施の形態〕
本発明は、OsBU3(Oryza sative Brassinosteroid Upregulated 3、DDBJ Accession No. AK071601)を用いた、種子の大型化、カルスの生育促進、節間の異常な伸長からなる群より選択される少なくとも1つの形質を植物に付与する方法を提供する。
本発明において「種子の大型化」とは、野生型の植物体と比較して種子の大きさ(長さや幅)がわずかでも増加することを意味する。本発明の方法によって得られた植物体の種子の大きさが野生型と比較して増加したか否かは、例えば種子の長さや幅等を測定することによって判定することが出来るが、これに限定されない。
また本発明において「カルスの生育促進」とは、野生型のカルスと比較して生育の速度がわずかでも増加することを意味する。
また本発明において「節間の異常な伸長」とは、穂首下の上位4節間以外の節間がWTより長く伸長することを意味する。また通常は長く伸長した節間の節から分げつができることはほとんどないが、伸長した節間の節から分げつができ易くなる点も、本発明の「節間の異常な伸長」に含まれる。
【0011】
本発明に用いるDNA(以下「本発明のDNA」と表現する場合あり)の形態に特に制限はなく、cDNAであってもゲノムDNAであってもよい。ゲノムDNAおよびcDNAの調製は、当業者にとって常套手段を利用して行うことが可能である。例えば、ゲノムDNAはOsBU3(DDBJ Accession No. AK071601)の公知の塩基配列情報(配列番号:1)から適当なプライマー対を設計して、目的の植物から調製したゲノムDNAを鋳型にPCRを行い、得られる増幅DNA断片をプローブとしてゲノミックライブラリーをスクリーニングすることによって調製することができる。また、同様にプライマー対を設計して、目的の植物から調製したcDNAまたはmRNAを鋳型にPCRを行い、得られる増幅DNA断片をプローブとして用いてcDNAライブラリーをスクリーニングすることにより、本発明のDNAを調製することができる。さらに市販のDNA合成機を用いれば、目的のDNAを合成により調製することも可能である。
【0012】
本発明のDNAは、種子の大型化、カルスの生育促進、節間の異常な伸長からなる群より選択される少なくとも1つの形質を植物に付与する機能を有している限り、イネ由来のOsBU3タンパク質(配列番号:2)をコードするDNAのみならず、該タンパク質に構造的に類似したタンパク質をコードするDNA(例えば、変異体、誘導体、アレル、バリアントおよびホモログ)を用いることもできる。このようなDNAには、例えば、配列番号:2に記載のアミノ酸配列において1もしくは複数のアミノ酸が置換、欠失、付加、および/または挿入されたアミノ酸配列からなるタンパク質をコードするDNAが含まれる。
【0013】
アミノ酸配列が改変されたタンパク質をコードするDNAを調製するための当業者によく知られた方法としては、例えば、site-directed mutagenesis法(Kramer, W. and Fritz, H.J. Oligonucleotide-directed construction of mutagenesis via gapped duplex DNA. Methods in Enzymology. 154, 1987, 350-367.)が挙げられる。また、塩基配列の変異によりコードするタンパク質のアミノ酸配列が変異することは、自然界においても生じ得る。このように天然型のOsBU3タンパク質のアミノ酸配列(配列番号:2)において1もしくは複数のアミノ酸が置換、欠失もしくは付加したアミノ酸配列を有するタンパク質をコードするDNAであっても、種子の大型化、カルスの生育促進、節間の異常な伸長からなる群より選択される少なくとも1つの形質を植物に付与する機能を有している限り、本発明のDNAに含まれる。
【0014】
改変されるアミノ酸の数は、特に制限はないが、一般的には、50アミノ酸以内、好ましくは30アミノ酸以内、より好ましくは10アミノ酸以内(例えば、5アミノ酸以内、3アミノ酸以内)である。アミノ酸の改変は、好ましくは保存的置換である。改変前と改変後の各アミノ酸についてのhydropathic index(Kyte, J. and Doolittle, R.F. J Mol Biol. 157(1), 1982, 105-132.)やHydrophilicity value(米国特許第4,554,101号)の数値は、±2以内が好ましく、さらに好ましくは±1以内であり、最も好ましくは±0.5以内である。
【0015】
また、たとえ、塩基配列が変異した場合でも、それがタンパク質中のアミノ酸の変異を伴わない場合(縮重変異)もあり、このような縮重変異体も本発明のDNAに含まれる。
【0016】
イネ由来のOsBU3タンパク質(配列番号:2)に構造的に類似したタンパク質をコードするDNAとしては、ハイブリダイゼーション技術(Southern, E.M. Journal of Molecular Biology. 98, 1975, 503.)やポリメラーゼ連鎖反応(PCR)技術(Saiki, R.K. et al. Science. 230, 1985, 1350-1354.; Saiki, R.K. et al. Science, 239, 1988, 487-491.)を利用して調製したものを用いることも可能である。即ち、本発明のDNAには、配列番号:1に記載の塩基配列からなるDNAとストリンジェントな条件でハイブリダイズするDNAが含まれる。このようなDNAを単離するためには、好ましくはストリンジェントな条件下でハイブリダイゼーション反応を行なう。本発明において「ストリンジェントな条件」とは、6M尿素、0.4%SDS、0.5xSSCの条件またはこれと同等のストリンジェンシーのハイブリダイゼーション条件を指すが、特にこれらの条件に限定されるものではない。よりストリンジェンシーの高い条件、例えば、6M尿素、0.4%SDS、0.1xSSCの条件を用いれば、より相同性の高いDNAの単離を期待することができる。
【0017】
ハイブリダイゼーションのストリンジェンシーに影響する要素としては温度や塩濃度など複数の要素が考えられるが、当業者であればこれら要素を適宜選択することで最適なストリンジェンシーを実現することが可能である。これにより単離されたDNAは、アミノ酸レベルにおいて、イネ由来のOsBU3タンパク質のアミノ酸配列(配列番号:2)と高い相同性を有すると考えられる。高い相同性とは、アミノ酸配列全体で、少なくとも50%以上、さらに好ましくは70%以上、さらに好ましくは90%以上(例えば、95%,96%,97%,98%,99%以上)の配列の同一性を指す。アミノ酸配列や塩基配列の同一性は、カーリンおよびアルチュールによるアルゴリズムBLAST(Karlin, S. and Altschul, S.F. Proc Natl Acad Sci U S A. 87(6), 1990, 2264-2268.; Karlin, S. and Altschul, S.F. Proc Natl Acad Sci U S A. 90(12), 1993, 5873-5877.)を用いて決定できる。BLASTのアルゴリズムに基づいたBLASTNやBLASTXと呼ばれるプログラムが開発されている(Altschul, S.F. et al. J Mol Biol. 215(3), 1990, 403-410.)。BLASTNを用いて塩基配列を解析する場合は、パラメーターは、例えばscore=100、wordlength=12とする。また、BLASTXを用いてアミノ酸配列を解析する場合は、パラメーターは、例えばscore=50、wordlength=3とする。BLASTとGapped BLASTプログラムを用いる場合は、各プログラムのデフォルトパラメーターを用いる。これらの解析方法の具体的な手法は公知である。
【0018】
本発明のDNAは、ベクターに挿入された形態であってもよい。ベクターとしては、植物細胞内で挿入遺伝子を発現させることが可能なものであれば特に制限はない。例えば、植物細胞内での恒常的な遺伝子発現を行うためのプロモーター(例えば、ジャガイモ・キチナーゼ遺伝子SK2のプロモーター、カリフラワーモザイクウイルスの35Sプロモーター等)を有するベクターや外的な刺激により誘導的に活性化されるプロモーターを有するベクターを用いることも可能である。
【0019】
なお、配列番号:1に記載の塩基配列におけるタンパク質のコード領域は、107番目の塩基から367番目の塩基である。配列番号:1に記載の塩基配列のうち107番目の塩基から367番目の塩基から生成されるアミノ酸配列は、配列番号:2に示される。
【0020】
種子の大型化、カルスの生育促進、節間の異常な伸長からなる群より選択される少なくとも1つの形質を植物に付与する機能を有する上記のDNAや該DNAを含むベクターを植物細胞に導入し、該植物細胞からカルスを形成又は植物体を再生することにより、種子の大型化、カルスの生育促進、節間の異常な伸長からなる群より選択される少なくとも1つの形質を有するカルス又は植物体を製造することができる。従って本発明は、種子の大型化、カルスの生育促進、節間の異常な伸長からなる群より選択される少なくとも1つの形質を有するカルス又は植物体の製造方法を提供する。
【0021】
上記DNAやベクターを導入する植物細胞の種類としては、例えば、イネ、小麦、大麦、トウモロコシ、ソルガム等の単子葉植物、シロイヌナズナ、ナタネ、トマト、大豆、ジャガイモ、タバコ、ムラサキ等の双子葉植物等があげられるがこれらに限定されない。
【0022】
上記DNAやベクターが導入される植物細胞の形態は、植物体を再生しうるものであれば、特に制限はなく、例えば、懸濁培養細胞、プロトプラスト、葉の切片、カルスなどが含まれる。
【0023】
上記DNAやベクターの植物細胞への導入は、例えば、ポリエチレングリコール法、電気穿孔法(エレクトロポーレーション)、アグロバクテリウムを介する方法、パーティクルガン法等の当業者に公知の方法によって実施することができる。アグロバクテリウムを介する方法においては、例えばNagelらの方法(Nagel, R. et al. FEMS Microbiol Lett. 67, 1990, 325-328.)にしたがって、上記DNAが挿入された発現ベクターをアグロバクテリウムに導入し、このアグロバクテリウムを直接感染法やリーフディスク法で植物細胞に感染させることにより、上記DNAを植物細胞に導入することができる。
【0024】
植物細胞からの植物体の再生は、植物の種類に応じて当業者に公知の方法で行うことが可能である。例えば、イネであればFujimuraら(Fujimura. et al. Tissue Culture Lett. 2, 1995, 74.)の方法が挙げられ、小麦であればHarrisら(Harris, R. et al. Plant Cell Reports. 7, 1988, 337-340)の方法やOzgenら(Ozgen, M. et al. Plant Cell Reports. 18, 1998, 331-335)の方法が挙げられ、大麦であればKiharaとFunatsuki(Kihara, M. and Funatsuki, H. Breeding Sci. 44, 1994, 157-160.)の方法やLursとLorz(Lurs, R. and Lorz, H. Theor. Appl. Genet. 75, 1987, 16-25.)の方法が挙げられ、トウモロコシであればShillitoら(Shillito, R.D., et al. Bio/Technology, 7, 1989, 581-587.)の方法やGordon-Kammら(Gordon-Kamm, W.J. et al. Plant Cell. 2(7), 1990, 603-618.)の方法が挙げられ、ソルガムであればWenら(Wen, F.S., et al. Euphytica. 52, 1991, 177-181.)の方法やHagio(Hagio, T. Breeding Sci. 44, 1994, 121-126.)の方法が挙げられるが、これらに限定されない。
またシロイヌナズナであればAkamaら(Akama. et al. Plant Cell Reports. 12, 1992, 7-11.)の方法が挙げられ、ナタネであればWangら(Wang, Y.P. et al. Plant Breeding. 124, 2005, 1-4.)の方法が挙げられ、トマトであればKoblitzとKoblitz(Koblitz, H and Koblitz, D. Plant Cell Reports. 1, 1982, 143-146.)の方法やMorganとCocking(Morgan, A. and Cocking, E.C. Z.Pflanzenpysiol. 106, 1982, 97-104.)の方法が挙げられ、大豆であればLazzeriら(Lazzeri, P.A. et al., Plant Mol. Biol. Rep. 3, 1985, 160-167.)の方法やRanchら(Ranch, J.P. et al., In Vitro Cell Dev. Biol. 21, 1985, 653-658.)の方法が挙げられ、ジャガイモであればVisserら(Visser, R.G.F. et al. Theor. Appl. Genet. 78, 1989, 594-600.)の方法が挙げられ、タバコであればHorschら (Horsch, R. B. et al., Science, 227, 1985, 1229-1231.)の方法が挙げられるが、これらに限定されない。
【0025】
一旦、ゲノム内に上記DNAやベクターが導入された形質転換植物体が得られれば、該植物体から有性生殖または無性生殖により子孫あるいはクローンを得ることが可能である。また、該植物体やその子孫あるいはクローンから繁殖材料(例えば、種子、果実、切穂、塊茎、塊根、株、カルス、プロトプラスト等)を得て、それらを基に該植物体を量産することも可能である。
【0026】
カルス又は植物体が、種子の大型化、カルスの生育促進、節間の異常な伸長からなる群より選択される少なくとも1つの形質を有するか否かは、対照と比較することによって判断することが出来る。本発明において対照とは、本発明のカルス又は植物体と同じ種のカルス又は植物体であって、本発明のDNAが過剰発現していないカルス又は植物体を意味する。本発明における対照は、本発明のカルス又は植物体と同じ種の植物体であって本発明のDNAが過剰発現していないものである限り何ら限定されない。従って本発明の対照には、例えば本発明のイネ由来のOsBU3以外のDNAが導入されたカルス又は植物体も含まれる。このような植物体の例としては、例えば、本発明の形質転換植物体と同じ種のカルス又は植物体であって、本発明のDNA以外のDNAで形質転換されたカルス又は植物体、本発明のDNAの機能欠失変異導入のDNAで形質転換されたカルス又は植物体、本発明のDNAの機能抑制型に変換されたDNAで形質転換されたカルス又は植物体、本発明のDNAの機能発現に不十分な領域のDNA断片で形質転換されたカルス又は植物体などが挙げられるが、これらに制限されない。
【0027】
このように本発明は、種子の大型化、カルスの生育促進、節間の異常な伸長からなる群より選択される少なくとも1つの形質を有するカルス又は植物体、該カルス形成又は植物体を再生しうる植物細胞、該カルス又は植物体の子孫あるいはクローンであるカルス又は植物体、および上記カルス又は植物体の繁殖材料をも提供する。
なお本発明のカルス又は植物体、植物細胞、子孫あるいはクローンである植物体、繁殖材料は、種子の大型化、カルスの生育促進、節間の異常な伸長の全ての形質を有していてもよい。
【実施例】
【0028】
次に実施例を挙げて本発明を詳細に説明するが、本発明は以下の実施例になんら制限されるものではない。
【0029】
1.材料と方法
1−1.イネの水耕栽培
1/2ムラシゲ・スクーグ培地(含3%ショ糖、0.4%ゲルライト)に播種後2週間後の苗を、水耕栽培に移行し、グロスチャンバー内では16時間明条件下で生育させた。なお、水耕液は木村氏B液(Sato et al. 1996)を用いた。
【0030】
1−2.BL処理およびサンプリング
播種50日後にBL(和光)処理を行った。光照射開始約3時間後に一方の水耕液内にBL(40nM)を添加し、もう一方はそのままの状態でグロスチャンバー内に静置した。BL添加、非添加(コントロール)の植物体(若い葉身および葉鞘各5枚)のサンプリングをBL処理3時間および24時間後に行い、液体窒素ですばやく凍結した。
【0031】
1−3.total RNAの抽出および精製
ISOGEN(ニッポンジーン)を用いて抽出した後、RNeasy Mini kit(QIAGEN)を用いて精製したものをRNAとして使用した。
【0032】
1−4.マイクロアレイ解析
BL添加および非添加(コントロール)のbrd1より抽出したtotal RNAからcDNAを合成、増幅し、cRNAへの逆転写合成と同時にCy3およびCy5で標識した。それぞれ標識したcRNA(500ng)を22Kイネアレイ(Yazaki et al. 2004)に対してハイブリダイズ(60℃、17時間)し、シグナルの強度をスキャナーで読み取った。実験は、BL添加(Cy3) vs コントロール(Cy5)およびBL添加(Cy5) vs コントロール(Cy3)について同total RNAサンプルを蛍光色素の標識を変え、Dye-swap実験により比較した。なお、Dye-swap実験より、2slide双方において蛍光強度の比が2以上の遺伝子を増加、0.5以下の遺伝子を減少したとみなした。
【0033】
1−5.Real time-PCR
1st -strand合成にはtotal RNAを4μg用い、1st-strand cDNA Synthesis Kit(Amersham Biosciences)を用いて行った。Real time-PCRにはiCycle iQ リアルタイム PCR 解析 システム MC (Bio-Rad)を用いた。サイバーグリーンはiQ SYBR Green Supermix(Bio-Rad)を用いた。スタンダードとしてpolyubiquitin(RUBQ2, Genbank Accession No. AF184280)を用いた。なお、RUBQ2のプライマーペアには5’-GGTCGTCCCGAGCCTCTGTT-3’(配列番号:3)と5’-GCAAATGAGCAAATTGAGCA-3’(配列番号:4)をOsBU3(DDBJ Accession No. AK071601)のプライマーペアには 5’-ggaggagatcaacgagctcat-3’(配列番号:5)と 5’- cggtgcaggctcttgatgta-3’(配列番号:6)を使用した。PCRの温度条件と反応時間・サイクル数は、95℃15分を1サイクル、94℃15秒、60℃30秒、72℃30秒を45サイクル、72℃7分を1サイクル行った。OsBU3の発現レベルはcomparative CT法(Livak and Schmittegen, 2001)により計算した。
【0034】
1−6.形質転換イネの作製
イネの品種は日本晴を用いた。イネの発現ベクターはpRiceFOX (Nakamura et al. 2007)を用いた。農業生物資源研究所のイネゲノムリソースセンターから得た完全長cDNA(AK071601)をSfiIで切断後pRiceFOXベクターに連結した。イネの形質転換はアグロバクテリウムEH105株を用いて高速形質転換法(Toki et al. 2006)で行った。形質転換イネは隔離温室で28℃、13.5時間日長で栽培した。
【0035】
2.結果
2−1.マイクロアレイ及びReal-time PCRによる遺伝子発現解析
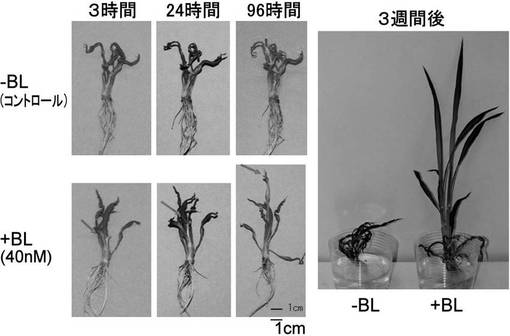
水耕栽培したbrd1変異体にブラシノライド(BL)を添加後経時的に形態を観察すると、3時間及び24時間後はほとんど変化がみられないが、96時間後には最も若い葉の伸長が認められた(図2)。BL添加後初期の遺伝子発現を調べるために、添加後3時間、24時間の葉をサンプリングしRNAを抽出し、22Kマイクロアレイを用いてBL非添加サンプルとの発現を網羅的に比較した。その中で3時間後、24時間後共に非添加コントロールに比べて2倍以上発現の増加している遺伝子を選抜した。選抜された遺伝子の1つでbHLHモチーフを持つ、DDBJ Accession No. AK071601の遺伝子をOsBU3 (Oryza sative Brassinosteroid Upregulated 3)と命名した。Real-time PCRでOsBU3の発現を確認すると、少なくともBL処理後24時間、96時間で顕著に発現が増加していることが示された。
【0036】
(表1の説明)
Real-time PCRで調べた。brd1変異体にBL処理した場合にOsBU3の転写量が、無処理サンプルに比べ何倍増加したかを示す。(1)(2)は栽培、BL処理、RNA調製全てのステップを別々に行ったbiological repeatを示す。
【0037】
【表1】

【0038】
2−2.過剰発現イネカルスにおける表現型
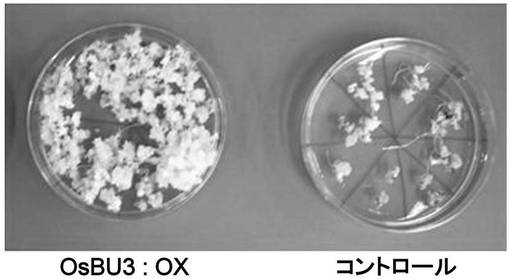
OsBU3はBL添加後初期に発現が増加しているため、BRシグナル伝達に関与している可能性が考えられた。よってOsBU3を過剰発現するイネを作製し、形態等に変化が見られるかどうか調べることにした。すると、T0世代のOsBU3過剰発現カルスの段階で、コントロールと比較して顕著な生育促進が認められた(図3)。
【0039】
2−3.過剰発現イネ植物体における表現型
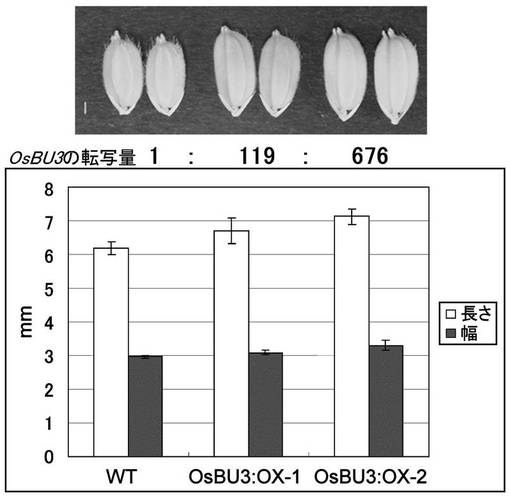
再分化した植物体でWTと比較すると、ラミナジョイントの顕著な屈曲が認められ、葉身が開いた草型を示した(図4)。また種子はOsBU3の転写量に比例して長さ、幅共に大きくなった。高レベルで発現しているOsBU3:OX-2の種子は約15%大型化していた(図5)。しかしながら稔性はWTに比べ低下していた。更に生育後期では、OsBU3過剰発現イネにおいて節間の異常な伸長が認められた(図6)。
【0040】
本発明者らはbrd1変異体を用いてBL誘導性遺伝子OsBU3を同定した。OsBU3を日本晴(WT)で過剰発現させると、BL添加の際に認められる典型的な反応であるラミナジョイントの屈曲が起こり、葉身が開いた草型になった。それ以外に、種子の大型化、カルスの生育促進、節間の異常な伸長が認められた。以上の結果から本遺伝子は、遺伝子組み換え技術を用いて過剰発現させることにより、種々の植物に同様の性質を付与するための素材となりうる。
【図面の簡単な説明】
【0041】
【図1】イネのbrd1変異体(Mori et al., 2002)を示す写真である。A.発芽後3週間の植物体、B.発芽後80日の植物体。左はWT(日本晴)、右はbrd1 C.B.のbrd1の拡大図。
【図2】ブラシノライド(BL)添加による表現型の回復を示す写真である。brd1変異体を水耕栽培しBL処理すると、若い葉(矢印)で表現型の回復がみられる。96時間後にはかなり回復して葉の伸長が認められる。3週間後にはほぼ正常な形態に回復する。
【図3】OsBU3過剰発現カルスの表現型を示す写真である。T0世代のカルス。再分化培地上。OsBU3:OXはOsBU3の過剰発現カルス。
【図4】OsBU3過剰発現イネ植物体の表現型を示す写真である。OsBU3:OXはOsBU3の過剰発現体。ラミナジョイントがWT(日本晴)に比べ屈曲しており葉身が開いた草型を示す。転写量はReal-time PCRで調べた。
【図5】OsBU3過剰発現体における種子の表現型を示す写真およびグラフである。上の写真は左からWT(日本晴)、OsBU3:OX-1, OsBU3:OX-2の順。OsBU3:OX-1, OsBU3:OX-2はそれぞれ独立の過剰発現体。転写量はReal-time PCRで調べた。下のグラフはそれぞれの種子の長さ、幅を示した。
【図6】OsBU3過剰発現体における節間の表現型を示す写真である。OsBU3:OX-2(図5参照)の節間を示した。矢印は節を示す。伸長した節間の節から分げつが伸長している様子がわかる。
【技術分野】
【0001】
本発明は、種子の大型化、カルスの生育促進、節間の異常な伸長からなる群より選択される少なくとも1つの形質を有するカルス又は植物体の製造方法及びこれを用いて得られるカルス及び植物体、並びにその利用に関する。
【背景技術】
【0002】
植物ホルモンのブラシノステロイド(BR)は植物の形態形成や、環境ストレスへの応答等に関与することが知られている。化学構造的にはステロイド骨格を有している点がBRの特徴で、被子植物、裸子植物、藻類およびシダ類を含む40種類以上の植物から、ブラシノライド(BL)、カスタステロンを代表に40種類以上の遊離型が単離・同定されている(Fujioka and Sakurai, 1997)。中でもBLは植物界における分布が広く、かつ天然のBRのなかで最も強い活性を示すことから、最も重要な役割を果たしていると考えられている。
【0003】
本発明者らは既に、イネでBRの生合成変異体brd1(brassinosteroid-dpendent 1)を単離しており、brd1変異体は極わい性で葉鞘が極めて短く葉身は巻いていることを示している(Mori et al. 2002, 図1)。また本発明者らは、BRはイネにおいても、葉身の伸長や展開、分げつ形成、根の発達、生殖成長等の多くの形態的及び生理的プロセスに関与していることを示している(Mori et al. 2002)。
【0004】
本発明の先行技術文献を以下に示す。
【非特許文献1】Fujioka, S. and Sakurai, A. (1997) Natural Products Report,14,1-10
【非特許文献2】Livak, KJ, Schmittgen, TD (2001) Methods 25:402-408
【非特許文献3】Mori, M et al., (2002) Plant Physiol. 130:1152-1161.
【非特許文献4】Nakamura, H et al., (2007) Plant Mol Biol. 65:357-371
【非特許文献5】Sato, H et al., (1996) Plant Sci. 119:39-47.
【非特許文献6】Toki, S et al., (2006) Plant J. 47:969-76.
【非特許文献7】Yazaki, J et al.,(2004) Physiol Genomics. 17(2):87-100.
【発明の開示】
【発明が解決しようとする課題】
【0005】
上述のように、BRは植物の形態形成や、環境ストレスへの応答等に関与することが知られている。そこで本発明は、BRの有用な作用を農業等に利用することを目的として、イネにおいてBRのシグナル伝達に関与する遺伝子を単離することを課題とした。
【課題を解決するための手段】
【0006】
上記課題を解決するにあたり本発明者らは、BRのシグナル伝達に関与する遺伝子は、BR内生量の低いbrd1変異体では発現が抑制されており、brd1にBLを添加すると添加後初期段階で発現が増大するのではないかと考えた。そこで本発明者らは、brd1変異体に代表的なブラシノステロイドであるブラシノライド(BL)を添加し、3時間及び24時間後に発現レベルが上昇する遺伝子の単離を試みた。マイクロアレイ解析の結果本発明者らは、3時間後、24時間後共にブラシノライド非添加コントロールに比べて2倍以上発現の増加している遺伝子(DDBJ Accession No. AK071601、OsBU3 (Oryza sative Brassinosteroid Upregulated 3)と命名)を単離することに成功した。
【0007】
次に本発明者らは、AK071601を過剰発現するイネを作製し、形態等に変化が見られるかどうか確認を行った。その結果、T0世代のOsBU3過剰発現カルスの生育が、コントロールと比較して顕著に促進することが確認された。
また再分化した植物体を野生型(WT)と比較すると、ラミナジョイントの顕著な屈曲が認められた。また種子はOsBU3の転写量に応じて長さ、幅共に大きくなった。更に生育後期では、OsBU3過剰発現イネにおいて節間の異常な伸長が認められた。
このようなイネカルスの生育促進、種子の大型化、明所における異常な節間伸長と、BRシグナルの増強の関連が見出されたのは、今回が初めてである。
【0008】
本発明は、ラミナジョイントの屈曲に加え、種子の大型化、カルスの生育促進、節間の異常な伸長作用を有する遺伝子、及び当該遺伝子が導入されたカルス及び植物体に関するものであり、より詳しくは、以下の〔1〕〜〔6〕を提供する。
〔1〕以下(a)及び(b)の工程を含む、種子の大型化、カルスの生育促進、節間の異常な伸長からなる群より選択される少なくとも1つの形質を植物に付与する方法;
(a)下記(i)から(iv)からなる群より選択されるDNA又は該DNAを含むベクターを植物細胞に導入する工程、及び
(b)工程(a)においてDNA又はベクターが導入された植物細胞からカルスを形成又は植物体を再生する工程、
(i)配列番号:2に記載のアミノ酸配列を含むタンパク質をコードするDNA、
(ii)配列番号:1に記載の塩基配列のコード領域を含むDNA、
(iii)配列番号:2に記載のアミノ酸配列において1もしくは複数のアミノ酸が置換、欠失、付加、及び/又は挿入されたアミノ酸配列を含むタンパク質をコードするDNA、及び
(iv)配列番号:1に記載の塩基配列を含むDNAとストリンジェントな条件下でハイブリダイズするDNA、
〔2〕以下(a)から(d)のいずれかに記載のDNAまたは該DNAを含むベクターが導入された植物細胞であって、種子の大型化、カルスの生育促進、節間の異常な伸長からなる群より選択される少なくとも1つの形質を有するカルスを形成又は植物体を再生しうる植物細胞;
(a)配列番号:2に記載のアミノ酸配列を含むタンパク質をコードするDNA、
(b)配列番号:1に記載の塩基配列のコード領域を含むDNA、
(c)配列番号:2に記載のアミノ酸配列において1もしくは複数のアミノ酸が置換、欠失、付加、及び/又は挿入されたアミノ酸配列を含むタンパク質をコードするDNA、及び
(d)配列番号:1に記載の塩基配列を含むDNAとストリンジェントな条件下でハイブリダイズするDNA、
〔3〕〔2〕に記載の植物細胞から形成されたカルス又は再生された植物体であって、種子の大型化、カルスの生育促進、節間の異常な伸長からなる群より選択される少なくとも1つの形質を有するカルス又は植物体、
〔4〕〔3〕に記載のカルス又は植物体の子孫またはクローンである、種子の大型化、カルスの生育促進、節間の異常な伸長からなる群より選択される少なくとも1つの形質を有するカルス又は植物体、
〔5〕〔3〕または〔4〕に記載の植物体の繁殖材料、
〔6〕下記(a)及び(b)の工程を含む、種子の大型化、カルスの生育促進、節間の異常な伸長からなる群より選択される少なくとも1つの形質を有するカルス又は植物体の製造方法;
(a)下記(i)から(iv)からなる群より選択されるDNA又は該DNAを含むベクターを植物細胞に導入する工程、及び
(b)工程(a)においてDNA又はベクターが導入された植物細胞からカルスを形成又は植物体を再生する工程、
(i)配列番号:2に記載のアミノ酸配列を含むタンパク質をコードするDNA、
(ii)配列番号:1に記載の塩基配列のコード領域を含むDNA、
(iii)配列番号:2に記載のアミノ酸配列において1もしくは複数のアミノ酸が置換、欠失、付加、及び/又は挿入されたアミノ酸配列を含むタンパク質をコードするDNA、及び
(iv)配列番号:1に記載の塩基配列を含むDNAとストリンジェントな条件下でハイブリダイズするDNA。
【発明の効果】
【0009】
本発明により、カルスの生育を促進し、種子を大型化し、かつ明所で節間の異常な伸長作用を有する遺伝子(OsBU3)が提供された。本発明にて同定された遺伝子を過剰発現させたイネは、ラミナジョイントの屈曲に加え、種子の大型化、カルスの生育促進、節間の異常な伸長が認められた。
OsBU3あるいはそのホモログ遺伝子の発現レベルを変化させ、種子サイズを変化させることにより、主に種子が可食部の植物(穀物等)で新たな商品価値を付与することが可能である。また、過剰発現や発現抑制により草型や節間長を変化させることにより、作物や観葉植物で新たな商品価値を付与することが可能となった。またOsBU3を過剰発現することによりカルスの増殖が促進するため、OsBU3が導入されたカルスを色素やアルカロイド、薬品などの物質生産に利用できる可能性がある。
【0010】
〔発明の実施の形態〕
本発明は、OsBU3(Oryza sative Brassinosteroid Upregulated 3、DDBJ Accession No. AK071601)を用いた、種子の大型化、カルスの生育促進、節間の異常な伸長からなる群より選択される少なくとも1つの形質を植物に付与する方法を提供する。
本発明において「種子の大型化」とは、野生型の植物体と比較して種子の大きさ(長さや幅)がわずかでも増加することを意味する。本発明の方法によって得られた植物体の種子の大きさが野生型と比較して増加したか否かは、例えば種子の長さや幅等を測定することによって判定することが出来るが、これに限定されない。
また本発明において「カルスの生育促進」とは、野生型のカルスと比較して生育の速度がわずかでも増加することを意味する。
また本発明において「節間の異常な伸長」とは、穂首下の上位4節間以外の節間がWTより長く伸長することを意味する。また通常は長く伸長した節間の節から分げつができることはほとんどないが、伸長した節間の節から分げつができ易くなる点も、本発明の「節間の異常な伸長」に含まれる。
【0011】
本発明に用いるDNA(以下「本発明のDNA」と表現する場合あり)の形態に特に制限はなく、cDNAであってもゲノムDNAであってもよい。ゲノムDNAおよびcDNAの調製は、当業者にとって常套手段を利用して行うことが可能である。例えば、ゲノムDNAはOsBU3(DDBJ Accession No. AK071601)の公知の塩基配列情報(配列番号:1)から適当なプライマー対を設計して、目的の植物から調製したゲノムDNAを鋳型にPCRを行い、得られる増幅DNA断片をプローブとしてゲノミックライブラリーをスクリーニングすることによって調製することができる。また、同様にプライマー対を設計して、目的の植物から調製したcDNAまたはmRNAを鋳型にPCRを行い、得られる増幅DNA断片をプローブとして用いてcDNAライブラリーをスクリーニングすることにより、本発明のDNAを調製することができる。さらに市販のDNA合成機を用いれば、目的のDNAを合成により調製することも可能である。
【0012】
本発明のDNAは、種子の大型化、カルスの生育促進、節間の異常な伸長からなる群より選択される少なくとも1つの形質を植物に付与する機能を有している限り、イネ由来のOsBU3タンパク質(配列番号:2)をコードするDNAのみならず、該タンパク質に構造的に類似したタンパク質をコードするDNA(例えば、変異体、誘導体、アレル、バリアントおよびホモログ)を用いることもできる。このようなDNAには、例えば、配列番号:2に記載のアミノ酸配列において1もしくは複数のアミノ酸が置換、欠失、付加、および/または挿入されたアミノ酸配列からなるタンパク質をコードするDNAが含まれる。
【0013】
アミノ酸配列が改変されたタンパク質をコードするDNAを調製するための当業者によく知られた方法としては、例えば、site-directed mutagenesis法(Kramer, W. and Fritz, H.J. Oligonucleotide-directed construction of mutagenesis via gapped duplex DNA. Methods in Enzymology. 154, 1987, 350-367.)が挙げられる。また、塩基配列の変異によりコードするタンパク質のアミノ酸配列が変異することは、自然界においても生じ得る。このように天然型のOsBU3タンパク質のアミノ酸配列(配列番号:2)において1もしくは複数のアミノ酸が置換、欠失もしくは付加したアミノ酸配列を有するタンパク質をコードするDNAであっても、種子の大型化、カルスの生育促進、節間の異常な伸長からなる群より選択される少なくとも1つの形質を植物に付与する機能を有している限り、本発明のDNAに含まれる。
【0014】
改変されるアミノ酸の数は、特に制限はないが、一般的には、50アミノ酸以内、好ましくは30アミノ酸以内、より好ましくは10アミノ酸以内(例えば、5アミノ酸以内、3アミノ酸以内)である。アミノ酸の改変は、好ましくは保存的置換である。改変前と改変後の各アミノ酸についてのhydropathic index(Kyte, J. and Doolittle, R.F. J Mol Biol. 157(1), 1982, 105-132.)やHydrophilicity value(米国特許第4,554,101号)の数値は、±2以内が好ましく、さらに好ましくは±1以内であり、最も好ましくは±0.5以内である。
【0015】
また、たとえ、塩基配列が変異した場合でも、それがタンパク質中のアミノ酸の変異を伴わない場合(縮重変異)もあり、このような縮重変異体も本発明のDNAに含まれる。
【0016】
イネ由来のOsBU3タンパク質(配列番号:2)に構造的に類似したタンパク質をコードするDNAとしては、ハイブリダイゼーション技術(Southern, E.M. Journal of Molecular Biology. 98, 1975, 503.)やポリメラーゼ連鎖反応(PCR)技術(Saiki, R.K. et al. Science. 230, 1985, 1350-1354.; Saiki, R.K. et al. Science, 239, 1988, 487-491.)を利用して調製したものを用いることも可能である。即ち、本発明のDNAには、配列番号:1に記載の塩基配列からなるDNAとストリンジェントな条件でハイブリダイズするDNAが含まれる。このようなDNAを単離するためには、好ましくはストリンジェントな条件下でハイブリダイゼーション反応を行なう。本発明において「ストリンジェントな条件」とは、6M尿素、0.4%SDS、0.5xSSCの条件またはこれと同等のストリンジェンシーのハイブリダイゼーション条件を指すが、特にこれらの条件に限定されるものではない。よりストリンジェンシーの高い条件、例えば、6M尿素、0.4%SDS、0.1xSSCの条件を用いれば、より相同性の高いDNAの単離を期待することができる。
【0017】
ハイブリダイゼーションのストリンジェンシーに影響する要素としては温度や塩濃度など複数の要素が考えられるが、当業者であればこれら要素を適宜選択することで最適なストリンジェンシーを実現することが可能である。これにより単離されたDNAは、アミノ酸レベルにおいて、イネ由来のOsBU3タンパク質のアミノ酸配列(配列番号:2)と高い相同性を有すると考えられる。高い相同性とは、アミノ酸配列全体で、少なくとも50%以上、さらに好ましくは70%以上、さらに好ましくは90%以上(例えば、95%,96%,97%,98%,99%以上)の配列の同一性を指す。アミノ酸配列や塩基配列の同一性は、カーリンおよびアルチュールによるアルゴリズムBLAST(Karlin, S. and Altschul, S.F. Proc Natl Acad Sci U S A. 87(6), 1990, 2264-2268.; Karlin, S. and Altschul, S.F. Proc Natl Acad Sci U S A. 90(12), 1993, 5873-5877.)を用いて決定できる。BLASTのアルゴリズムに基づいたBLASTNやBLASTXと呼ばれるプログラムが開発されている(Altschul, S.F. et al. J Mol Biol. 215(3), 1990, 403-410.)。BLASTNを用いて塩基配列を解析する場合は、パラメーターは、例えばscore=100、wordlength=12とする。また、BLASTXを用いてアミノ酸配列を解析する場合は、パラメーターは、例えばscore=50、wordlength=3とする。BLASTとGapped BLASTプログラムを用いる場合は、各プログラムのデフォルトパラメーターを用いる。これらの解析方法の具体的な手法は公知である。
【0018】
本発明のDNAは、ベクターに挿入された形態であってもよい。ベクターとしては、植物細胞内で挿入遺伝子を発現させることが可能なものであれば特に制限はない。例えば、植物細胞内での恒常的な遺伝子発現を行うためのプロモーター(例えば、ジャガイモ・キチナーゼ遺伝子SK2のプロモーター、カリフラワーモザイクウイルスの35Sプロモーター等)を有するベクターや外的な刺激により誘導的に活性化されるプロモーターを有するベクターを用いることも可能である。
【0019】
なお、配列番号:1に記載の塩基配列におけるタンパク質のコード領域は、107番目の塩基から367番目の塩基である。配列番号:1に記載の塩基配列のうち107番目の塩基から367番目の塩基から生成されるアミノ酸配列は、配列番号:2に示される。
【0020】
種子の大型化、カルスの生育促進、節間の異常な伸長からなる群より選択される少なくとも1つの形質を植物に付与する機能を有する上記のDNAや該DNAを含むベクターを植物細胞に導入し、該植物細胞からカルスを形成又は植物体を再生することにより、種子の大型化、カルスの生育促進、節間の異常な伸長からなる群より選択される少なくとも1つの形質を有するカルス又は植物体を製造することができる。従って本発明は、種子の大型化、カルスの生育促進、節間の異常な伸長からなる群より選択される少なくとも1つの形質を有するカルス又は植物体の製造方法を提供する。
【0021】
上記DNAやベクターを導入する植物細胞の種類としては、例えば、イネ、小麦、大麦、トウモロコシ、ソルガム等の単子葉植物、シロイヌナズナ、ナタネ、トマト、大豆、ジャガイモ、タバコ、ムラサキ等の双子葉植物等があげられるがこれらに限定されない。
【0022】
上記DNAやベクターが導入される植物細胞の形態は、植物体を再生しうるものであれば、特に制限はなく、例えば、懸濁培養細胞、プロトプラスト、葉の切片、カルスなどが含まれる。
【0023】
上記DNAやベクターの植物細胞への導入は、例えば、ポリエチレングリコール法、電気穿孔法(エレクトロポーレーション)、アグロバクテリウムを介する方法、パーティクルガン法等の当業者に公知の方法によって実施することができる。アグロバクテリウムを介する方法においては、例えばNagelらの方法(Nagel, R. et al. FEMS Microbiol Lett. 67, 1990, 325-328.)にしたがって、上記DNAが挿入された発現ベクターをアグロバクテリウムに導入し、このアグロバクテリウムを直接感染法やリーフディスク法で植物細胞に感染させることにより、上記DNAを植物細胞に導入することができる。
【0024】
植物細胞からの植物体の再生は、植物の種類に応じて当業者に公知の方法で行うことが可能である。例えば、イネであればFujimuraら(Fujimura. et al. Tissue Culture Lett. 2, 1995, 74.)の方法が挙げられ、小麦であればHarrisら(Harris, R. et al. Plant Cell Reports. 7, 1988, 337-340)の方法やOzgenら(Ozgen, M. et al. Plant Cell Reports. 18, 1998, 331-335)の方法が挙げられ、大麦であればKiharaとFunatsuki(Kihara, M. and Funatsuki, H. Breeding Sci. 44, 1994, 157-160.)の方法やLursとLorz(Lurs, R. and Lorz, H. Theor. Appl. Genet. 75, 1987, 16-25.)の方法が挙げられ、トウモロコシであればShillitoら(Shillito, R.D., et al. Bio/Technology, 7, 1989, 581-587.)の方法やGordon-Kammら(Gordon-Kamm, W.J. et al. Plant Cell. 2(7), 1990, 603-618.)の方法が挙げられ、ソルガムであればWenら(Wen, F.S., et al. Euphytica. 52, 1991, 177-181.)の方法やHagio(Hagio, T. Breeding Sci. 44, 1994, 121-126.)の方法が挙げられるが、これらに限定されない。
またシロイヌナズナであればAkamaら(Akama. et al. Plant Cell Reports. 12, 1992, 7-11.)の方法が挙げられ、ナタネであればWangら(Wang, Y.P. et al. Plant Breeding. 124, 2005, 1-4.)の方法が挙げられ、トマトであればKoblitzとKoblitz(Koblitz, H and Koblitz, D. Plant Cell Reports. 1, 1982, 143-146.)の方法やMorganとCocking(Morgan, A. and Cocking, E.C. Z.Pflanzenpysiol. 106, 1982, 97-104.)の方法が挙げられ、大豆であればLazzeriら(Lazzeri, P.A. et al., Plant Mol. Biol. Rep. 3, 1985, 160-167.)の方法やRanchら(Ranch, J.P. et al., In Vitro Cell Dev. Biol. 21, 1985, 653-658.)の方法が挙げられ、ジャガイモであればVisserら(Visser, R.G.F. et al. Theor. Appl. Genet. 78, 1989, 594-600.)の方法が挙げられ、タバコであればHorschら (Horsch, R. B. et al., Science, 227, 1985, 1229-1231.)の方法が挙げられるが、これらに限定されない。
【0025】
一旦、ゲノム内に上記DNAやベクターが導入された形質転換植物体が得られれば、該植物体から有性生殖または無性生殖により子孫あるいはクローンを得ることが可能である。また、該植物体やその子孫あるいはクローンから繁殖材料(例えば、種子、果実、切穂、塊茎、塊根、株、カルス、プロトプラスト等)を得て、それらを基に該植物体を量産することも可能である。
【0026】
カルス又は植物体が、種子の大型化、カルスの生育促進、節間の異常な伸長からなる群より選択される少なくとも1つの形質を有するか否かは、対照と比較することによって判断することが出来る。本発明において対照とは、本発明のカルス又は植物体と同じ種のカルス又は植物体であって、本発明のDNAが過剰発現していないカルス又は植物体を意味する。本発明における対照は、本発明のカルス又は植物体と同じ種の植物体であって本発明のDNAが過剰発現していないものである限り何ら限定されない。従って本発明の対照には、例えば本発明のイネ由来のOsBU3以外のDNAが導入されたカルス又は植物体も含まれる。このような植物体の例としては、例えば、本発明の形質転換植物体と同じ種のカルス又は植物体であって、本発明のDNA以外のDNAで形質転換されたカルス又は植物体、本発明のDNAの機能欠失変異導入のDNAで形質転換されたカルス又は植物体、本発明のDNAの機能抑制型に変換されたDNAで形質転換されたカルス又は植物体、本発明のDNAの機能発現に不十分な領域のDNA断片で形質転換されたカルス又は植物体などが挙げられるが、これらに制限されない。
【0027】
このように本発明は、種子の大型化、カルスの生育促進、節間の異常な伸長からなる群より選択される少なくとも1つの形質を有するカルス又は植物体、該カルス形成又は植物体を再生しうる植物細胞、該カルス又は植物体の子孫あるいはクローンであるカルス又は植物体、および上記カルス又は植物体の繁殖材料をも提供する。
なお本発明のカルス又は植物体、植物細胞、子孫あるいはクローンである植物体、繁殖材料は、種子の大型化、カルスの生育促進、節間の異常な伸長の全ての形質を有していてもよい。
【実施例】
【0028】
次に実施例を挙げて本発明を詳細に説明するが、本発明は以下の実施例になんら制限されるものではない。
【0029】
1.材料と方法
1−1.イネの水耕栽培
1/2ムラシゲ・スクーグ培地(含3%ショ糖、0.4%ゲルライト)に播種後2週間後の苗を、水耕栽培に移行し、グロスチャンバー内では16時間明条件下で生育させた。なお、水耕液は木村氏B液(Sato et al. 1996)を用いた。
【0030】
1−2.BL処理およびサンプリング
播種50日後にBL(和光)処理を行った。光照射開始約3時間後に一方の水耕液内にBL(40nM)を添加し、もう一方はそのままの状態でグロスチャンバー内に静置した。BL添加、非添加(コントロール)の植物体(若い葉身および葉鞘各5枚)のサンプリングをBL処理3時間および24時間後に行い、液体窒素ですばやく凍結した。
【0031】
1−3.total RNAの抽出および精製
ISOGEN(ニッポンジーン)を用いて抽出した後、RNeasy Mini kit(QIAGEN)を用いて精製したものをRNAとして使用した。
【0032】
1−4.マイクロアレイ解析
BL添加および非添加(コントロール)のbrd1より抽出したtotal RNAからcDNAを合成、増幅し、cRNAへの逆転写合成と同時にCy3およびCy5で標識した。それぞれ標識したcRNA(500ng)を22Kイネアレイ(Yazaki et al. 2004)に対してハイブリダイズ(60℃、17時間)し、シグナルの強度をスキャナーで読み取った。実験は、BL添加(Cy3) vs コントロール(Cy5)およびBL添加(Cy5) vs コントロール(Cy3)について同total RNAサンプルを蛍光色素の標識を変え、Dye-swap実験により比較した。なお、Dye-swap実験より、2slide双方において蛍光強度の比が2以上の遺伝子を増加、0.5以下の遺伝子を減少したとみなした。
【0033】
1−5.Real time-PCR
1st -strand合成にはtotal RNAを4μg用い、1st-strand cDNA Synthesis Kit(Amersham Biosciences)を用いて行った。Real time-PCRにはiCycle iQ リアルタイム PCR 解析 システム MC (Bio-Rad)を用いた。サイバーグリーンはiQ SYBR Green Supermix(Bio-Rad)を用いた。スタンダードとしてpolyubiquitin(RUBQ2, Genbank Accession No. AF184280)を用いた。なお、RUBQ2のプライマーペアには5’-GGTCGTCCCGAGCCTCTGTT-3’(配列番号:3)と5’-GCAAATGAGCAAATTGAGCA-3’(配列番号:4)をOsBU3(DDBJ Accession No. AK071601)のプライマーペアには 5’-ggaggagatcaacgagctcat-3’(配列番号:5)と 5’- cggtgcaggctcttgatgta-3’(配列番号:6)を使用した。PCRの温度条件と反応時間・サイクル数は、95℃15分を1サイクル、94℃15秒、60℃30秒、72℃30秒を45サイクル、72℃7分を1サイクル行った。OsBU3の発現レベルはcomparative CT法(Livak and Schmittegen, 2001)により計算した。
【0034】
1−6.形質転換イネの作製
イネの品種は日本晴を用いた。イネの発現ベクターはpRiceFOX (Nakamura et al. 2007)を用いた。農業生物資源研究所のイネゲノムリソースセンターから得た完全長cDNA(AK071601)をSfiIで切断後pRiceFOXベクターに連結した。イネの形質転換はアグロバクテリウムEH105株を用いて高速形質転換法(Toki et al. 2006)で行った。形質転換イネは隔離温室で28℃、13.5時間日長で栽培した。
【0035】
2.結果
2−1.マイクロアレイ及びReal-time PCRによる遺伝子発現解析
水耕栽培したbrd1変異体にブラシノライド(BL)を添加後経時的に形態を観察すると、3時間及び24時間後はほとんど変化がみられないが、96時間後には最も若い葉の伸長が認められた(図2)。BL添加後初期の遺伝子発現を調べるために、添加後3時間、24時間の葉をサンプリングしRNAを抽出し、22Kマイクロアレイを用いてBL非添加サンプルとの発現を網羅的に比較した。その中で3時間後、24時間後共に非添加コントロールに比べて2倍以上発現の増加している遺伝子を選抜した。選抜された遺伝子の1つでbHLHモチーフを持つ、DDBJ Accession No. AK071601の遺伝子をOsBU3 (Oryza sative Brassinosteroid Upregulated 3)と命名した。Real-time PCRでOsBU3の発現を確認すると、少なくともBL処理後24時間、96時間で顕著に発現が増加していることが示された。
【0036】
(表1の説明)
Real-time PCRで調べた。brd1変異体にBL処理した場合にOsBU3の転写量が、無処理サンプルに比べ何倍増加したかを示す。(1)(2)は栽培、BL処理、RNA調製全てのステップを別々に行ったbiological repeatを示す。
【0037】
【表1】
【0038】
2−2.過剰発現イネカルスにおける表現型
OsBU3はBL添加後初期に発現が増加しているため、BRシグナル伝達に関与している可能性が考えられた。よってOsBU3を過剰発現するイネを作製し、形態等に変化が見られるかどうか調べることにした。すると、T0世代のOsBU3過剰発現カルスの段階で、コントロールと比較して顕著な生育促進が認められた(図3)。
【0039】
2−3.過剰発現イネ植物体における表現型
再分化した植物体でWTと比較すると、ラミナジョイントの顕著な屈曲が認められ、葉身が開いた草型を示した(図4)。また種子はOsBU3の転写量に比例して長さ、幅共に大きくなった。高レベルで発現しているOsBU3:OX-2の種子は約15%大型化していた(図5)。しかしながら稔性はWTに比べ低下していた。更に生育後期では、OsBU3過剰発現イネにおいて節間の異常な伸長が認められた(図6)。
【0040】
本発明者らはbrd1変異体を用いてBL誘導性遺伝子OsBU3を同定した。OsBU3を日本晴(WT)で過剰発現させると、BL添加の際に認められる典型的な反応であるラミナジョイントの屈曲が起こり、葉身が開いた草型になった。それ以外に、種子の大型化、カルスの生育促進、節間の異常な伸長が認められた。以上の結果から本遺伝子は、遺伝子組み換え技術を用いて過剰発現させることにより、種々の植物に同様の性質を付与するための素材となりうる。
【図面の簡単な説明】
【0041】
【図1】イネのbrd1変異体(Mori et al., 2002)を示す写真である。A.発芽後3週間の植物体、B.発芽後80日の植物体。左はWT(日本晴)、右はbrd1 C.B.のbrd1の拡大図。
【図2】ブラシノライド(BL)添加による表現型の回復を示す写真である。brd1変異体を水耕栽培しBL処理すると、若い葉(矢印)で表現型の回復がみられる。96時間後にはかなり回復して葉の伸長が認められる。3週間後にはほぼ正常な形態に回復する。
【図3】OsBU3過剰発現カルスの表現型を示す写真である。T0世代のカルス。再分化培地上。OsBU3:OXはOsBU3の過剰発現カルス。
【図4】OsBU3過剰発現イネ植物体の表現型を示す写真である。OsBU3:OXはOsBU3の過剰発現体。ラミナジョイントがWT(日本晴)に比べ屈曲しており葉身が開いた草型を示す。転写量はReal-time PCRで調べた。
【図5】OsBU3過剰発現体における種子の表現型を示す写真およびグラフである。上の写真は左からWT(日本晴)、OsBU3:OX-1, OsBU3:OX-2の順。OsBU3:OX-1, OsBU3:OX-2はそれぞれ独立の過剰発現体。転写量はReal-time PCRで調べた。下のグラフはそれぞれの種子の長さ、幅を示した。
【図6】OsBU3過剰発現体における節間の表現型を示す写真である。OsBU3:OX-2(図5参照)の節間を示した。矢印は節を示す。伸長した節間の節から分げつが伸長している様子がわかる。
【特許請求の範囲】
【請求項1】
以下(a)及び(b)の工程を含む、種子の大型化、カルスの生育促進、節間の異常な伸長からなる群より選択される少なくとも1つの形質を植物に付与する方法;
(a)以下(i)から(iv)からなる群より選択されるDNA又は該DNAを含むベクターを植物細胞に導入する工程、及び
(b)工程(a)においてDNA又はベクターが導入された植物細胞からカルスを形成又は植物体を再生する工程、
(i)配列番号:2に記載のアミノ酸配列を含むタンパク質をコードするDNA、
(ii)配列番号:1に記載の塩基配列のコード領域を含むDNA、
(iii)配列番号:2に記載のアミノ酸配列において1もしくは複数のアミノ酸が置換、欠失、付加、及び/又は挿入されたアミノ酸配列を含むタンパク質をコードするDNA、及び
(iv)配列番号:1に記載の塩基配列を含むDNAとストリンジェントな条件下でハイブリダイズするDNA。
【請求項2】
以下(a)から(d)のいずれかに記載のDNAまたは該DNAを含むベクターが導入された植物細胞であって、種子の大型化、カルスの生育促進、節間の異常な伸長からなる群より選択される少なくとも1つの形質を有するカルスを形成又は植物体を再生しうる植物細胞;
(a)配列番号:2に記載のアミノ酸配列を含むタンパク質をコードするDNA、
(b)配列番号:1に記載の塩基配列のコード領域を含むDNA、
(c)配列番号:2に記載のアミノ酸配列において1もしくは複数のアミノ酸が置換、欠失、付加、及び/又は挿入されたアミノ酸配列を含むタンパク質をコードするDNA、及び
(d)配列番号:1に記載の塩基配列を含むDNAとストリンジェントな条件下でハイブリダイズするDNA。
【請求項3】
請求項2に記載の植物細胞から形成されたカルス又は再生された植物体であって、種子の大型化、カルスの生育促進、節間の異常な伸長からなる群より選択される少なくとも1つの形質を有するカルス又は植物体。
【請求項4】
請求項3に記載のカルス又は植物体の子孫またはクローンである、種子の大型化、カルスの生育促進、節間の異常な伸長からなる群より選択される少なくとも1つの形質を有するカルス又は植物体。
【請求項5】
請求項3または4に記載のカルス又は植物体の繁殖材料。
【請求項6】
以下(a)及び(b)の工程を含む、種子の大型化、カルスの生育促進、節間の異常な伸長からなる群より選択される少なくとも1つの形質を有するカルス又は植物体の製造方法;
(a)以下(i)から(iv)からなる群より選択されるDNA又は該DNAを含むベクターを植物細胞に導入する工程、及び
(b)工程(a)においてDNA又はベクターが導入された植物細胞からカルスを形成又は植物体を再生する工程、
(i)配列番号:2に記載のアミノ酸配列を含むタンパク質をコードするDNA、
(ii)配列番号:1に記載の塩基配列のコード領域を含むDNA、
(iii)配列番号:2に記載のアミノ酸配列において1もしくは複数のアミノ酸が置換、欠失、付加、及び/又は挿入されたアミノ酸配列を含むタンパク質をコードするDNA、及び
(iv)配列番号:1に記載の塩基配列を含むDNAとストリンジェントな条件下でハイブリダイズするDNA。
【請求項1】
以下(a)及び(b)の工程を含む、種子の大型化、カルスの生育促進、節間の異常な伸長からなる群より選択される少なくとも1つの形質を植物に付与する方法;
(a)以下(i)から(iv)からなる群より選択されるDNA又は該DNAを含むベクターを植物細胞に導入する工程、及び
(b)工程(a)においてDNA又はベクターが導入された植物細胞からカルスを形成又は植物体を再生する工程、
(i)配列番号:2に記載のアミノ酸配列を含むタンパク質をコードするDNA、
(ii)配列番号:1に記載の塩基配列のコード領域を含むDNA、
(iii)配列番号:2に記載のアミノ酸配列において1もしくは複数のアミノ酸が置換、欠失、付加、及び/又は挿入されたアミノ酸配列を含むタンパク質をコードするDNA、及び
(iv)配列番号:1に記載の塩基配列を含むDNAとストリンジェントな条件下でハイブリダイズするDNA。
【請求項2】
以下(a)から(d)のいずれかに記載のDNAまたは該DNAを含むベクターが導入された植物細胞であって、種子の大型化、カルスの生育促進、節間の異常な伸長からなる群より選択される少なくとも1つの形質を有するカルスを形成又は植物体を再生しうる植物細胞;
(a)配列番号:2に記載のアミノ酸配列を含むタンパク質をコードするDNA、
(b)配列番号:1に記載の塩基配列のコード領域を含むDNA、
(c)配列番号:2に記載のアミノ酸配列において1もしくは複数のアミノ酸が置換、欠失、付加、及び/又は挿入されたアミノ酸配列を含むタンパク質をコードするDNA、及び
(d)配列番号:1に記載の塩基配列を含むDNAとストリンジェントな条件下でハイブリダイズするDNA。
【請求項3】
請求項2に記載の植物細胞から形成されたカルス又は再生された植物体であって、種子の大型化、カルスの生育促進、節間の異常な伸長からなる群より選択される少なくとも1つの形質を有するカルス又は植物体。
【請求項4】
請求項3に記載のカルス又は植物体の子孫またはクローンである、種子の大型化、カルスの生育促進、節間の異常な伸長からなる群より選択される少なくとも1つの形質を有するカルス又は植物体。
【請求項5】
請求項3または4に記載のカルス又は植物体の繁殖材料。
【請求項6】
以下(a)及び(b)の工程を含む、種子の大型化、カルスの生育促進、節間の異常な伸長からなる群より選択される少なくとも1つの形質を有するカルス又は植物体の製造方法;
(a)以下(i)から(iv)からなる群より選択されるDNA又は該DNAを含むベクターを植物細胞に導入する工程、及び
(b)工程(a)においてDNA又はベクターが導入された植物細胞からカルスを形成又は植物体を再生する工程、
(i)配列番号:2に記載のアミノ酸配列を含むタンパク質をコードするDNA、
(ii)配列番号:1に記載の塩基配列のコード領域を含むDNA、
(iii)配列番号:2に記載のアミノ酸配列において1もしくは複数のアミノ酸が置換、欠失、付加、及び/又は挿入されたアミノ酸配列を含むタンパク質をコードするDNA、及び
(iv)配列番号:1に記載の塩基配列を含むDNAとストリンジェントな条件下でハイブリダイズするDNA。
【図1】

【図2】
【図3】
【図4】

【図5】
【図6】

【図2】
【図3】
【図4】
【図5】
【図6】
【公開番号】特開2010−81839(P2010−81839A)
【公開日】平成22年4月15日(2010.4.15)
【国際特許分類】
【出願番号】特願2008−253289(P2008−253289)
【出願日】平成20年9月30日(2008.9.30)
【国等の委託研究の成果に係る記載事項】(出願人による申告)平成19年度農林水産省、生物系特定産業技術研究支援センター「植物ホルモン関連新規遺伝子のイネにおける機能解明と有用性の検討」委託事業、産業技術力強化法第19条の適用を受けるもの
【出願人】(501167644)独立行政法人農業生物資源研究所 (200)
【Fターム(参考)】
【公開日】平成22年4月15日(2010.4.15)
【国際特許分類】
【出願日】平成20年9月30日(2008.9.30)
【国等の委託研究の成果に係る記載事項】(出願人による申告)平成19年度農林水産省、生物系特定産業技術研究支援センター「植物ホルモン関連新規遺伝子のイネにおける機能解明と有用性の検討」委託事業、産業技術力強化法第19条の適用を受けるもの
【出願人】(501167644)独立行政法人農業生物資源研究所 (200)
【Fターム(参考)】
[ Back to top ]