
選択的IFN−γ経路インヒビターとしてのヒト抗IFN−γ中和抗体
【課題】IFN−γ媒介性疾患を処置するための方法を提供する。
【解決手段】ヒトインターフェロン−ガンマ(IFN−γ)と相互作用するかまたは結合する抗体、ならびにIFN−γに対する抗体の薬学的に有効な量を投与することによってIFN−γ媒介性疾患を処置するための方法。IFN−γに対する抗体を用いてサンプル中のIFN−γの量を検出する方法。インターフェロンγ(IFN−γ)に結合するモノクローナル抗体およびそれらをコードするポリヌクレオチド。この抗体は、IFN−γの生物学的活性の少なくとも1つを阻害または調節し得、そしてIFN−γ媒介性疾患の影響を緩和するために有用であり得る。上記モノクローナル抗体を産生して細胞培養培地中に分泌し得るハイブリドーマ細胞。
【解決手段】ヒトインターフェロン−ガンマ(IFN−γ)と相互作用するかまたは結合する抗体、ならびにIFN−γに対する抗体の薬学的に有効な量を投与することによってIFN−γ媒介性疾患を処置するための方法。IFN−γに対する抗体を用いてサンプル中のIFN−γの量を検出する方法。インターフェロンγ(IFN−γ)に結合するモノクローナル抗体およびそれらをコードするポリヌクレオチド。この抗体は、IFN−γの生物学的活性の少なくとも1つを阻害または調節し得、そしてIFN−γ媒介性疾患の影響を緩和するために有用であり得る。上記モノクローナル抗体を産生して細胞培養培地中に分泌し得るハイブリドーマ細胞。
【発明の詳細な説明】
【技術分野】
【0001】
本出願は、2002年10月16日出願の米国仮出願番号60/419,057および2003年6月17日出願の米国仮出願番号60/479,241号の利益を請求する。
【0002】
(発明の分野)
本発明は、インターフェロンγ(IFN−γ)に結合するヒトモノクローナル抗体に関する。IFN−γによって媒介される疾患を処置するための組成物および方法もまた、記載される。
【背景技術】
【0003】
(背景)
インターフェロン(IFN)はもともと、宿主細胞のウイルス感染を妨害する能力に基づいて命名された(非特許文献1:IsaacsおよびLindenman,1957,Proc.R.Soc.147:28〜267)。それらの発見以来、インターフェロンファミリーの多数のメンバーが、抗ウイルス防御に加えて、細胞増殖および細胞免疫を含む種々の生物学的役割を有することが特定されている。インターフェロンの型であるIFN−α、IFN−β、IFN−ωおよびIFN−γは、I型インターフェロンであり、I型IFNレセプターに結合するが、IFN−γはII型インターフェロンであり、II型IFNレセプターに結合する(非特許文献2:Pfefferら、1998,Cancer Res.58:2489〜2499)。
【0004】
IFN−γのシグナル伝達は、少なくとも以下の5つの別個のタンパク質に依存する:IFNGR1およびIFNGR2(IFN−γレセプターのサブユニット)、Jak1、Jak2および転写因子STAT1(非特許文献3:SchindlerおよびDarnell,1995,Annu.Rev.Biochem.64:621〜651;非特許文献4:Bachら、1997,Annu.Rev.Immunol.15:563〜591)。IFN−γレセプターは、成熟赤血球を除くほとんどの細胞型で見出される(非特許文献5:FarrarおよびSchreiber,1993,Annu.Rev.Immunol.11:571〜611)。Jak1タンパク質、Jak2タンパク質およびSTAT1タンパク質は、IFN−γシグナル伝達を媒介する。
【0005】
IFN−γは、種々の生物学的機能(例えば、抗ウイルス応答、細胞増殖、免疫応答および腫瘍抑制)を調節し、そしてIFN−γは、種々のヒト疾患を媒介し得る。従って、IFN−γの生物学的活性を調節し得る因子が当該分野では必要である。
【非特許文献1】IsaacsおよびLindenman,1957,Proc.R.Soc.147:28〜267
【非特許文献2】Pfefferら、1998,Cancer Res.58:2489〜2499)
【非特許文献3】SchindlerおよびDarnell,1995,Annu.Rev.Biochem.64:621〜651
【非特許文献4】Bachら、1997,Annu.Rev.Immunol.15:563〜591
【非特許文献5】FarrarおよびSchreiber,1993,Annu.Rev.Immunol.11:571〜611
【発明の開示】
【課題を解決するための手段】
【0006】
(発明の要旨)
本発明は、インターフェロンγ(IFN−γ)に結合するモノクローナル抗体およびそれらをコードするポリヌクレオチドを提供する。この抗体は、IFN−γの生物学的活性の少なくとも1つを阻害または調節し得、そしてIFN−γ媒介性疾患の影響を緩和するために有用であり得る。本発明のモノクローナル抗体を産生して細胞培養培地中に分泌し得るハイブリドーマ細胞もまた、本発明によって提供される。本発明の抗体は、IFN−γによって媒介される疾患を処置するために有用であり得る。
【0007】
特定の局面では、本発明は、抗体(必要に応じて、モノクローナル抗体および/またはヒト抗体)を提供し、この抗体は、重鎖および軽鎖を含み、ここでこの重鎖は、配列番号2に示されるアミノ酸配列、またはその抗原結合フラグメントもしくは免疫学的に機能的な免疫グロブリンフラグメントを含む。
【0008】
特定の局面では、本発明は、抗体(必要に応じて、モノクローナル抗体)を提供し、この抗体は、重鎖および軽鎖を含むヒト抗体であり得、この重鎖は、IgG1、IgG2またはIgG4重鎖定常領域を含む。いくつかの実施形態では、本発明の抗体は、配列番号2に示されるIgG1重鎖定常領域のアミノ酸配列、またはその抗原結合フラグメントもしくは免疫学的に機能的な免疫グロブリンフラグメントを含む。
【0009】
特定の局面では、本発明の抗体は、重鎖および軽鎖を含み、この重鎖の可変領域は、配列番号6、配列番号10、配列番号14もしくは配列番号30のいずれかに示されるアミノ酸配列、またはその抗原結合フラグメントもしくは免疫学的に機能的な免疫グロブリンフラグメントを含む。他の局面では、この軽鎖可変領域は、配列番号8、配列番号12、配列番号16もしくは配列番号31のいずれかに示されるアミノ酸配列、またはその抗原結合フラグメントもしくは免疫学的に機能的な免疫グロブリンフラグメントを含む。さらなる局面では、この重鎖は、配列番号17、配列番号19、配列番号21もしくは配列番号32のいずれかに示されるアミノ酸配列、またはその抗原結合フラグメントもしくは免疫学的に機能的な免疫グロブリンフラグメントを含む。なおさらなる局面では、この軽鎖は、配列番号18、配列番号20、配列番号22もしくは配列番号33のいずれかに示されるアミノ酸配列、またはその抗原結合フラグメントもしくは免疫学的に機能的な免疫グロブリンフラグメントを含む。
【0010】
本発明はまた、IFN−γに対して特異的に結合する抗体を提供し、ここでこの重鎖は、重鎖可変領域を含み、この重鎖可変領域は、配列番号6に示されるアミノ酸配列、またはその抗原結合フラグメントもしくは免疫学的に機能的な免疫グロブリンフラグメントを含み、そしてこの軽鎖は、軽鎖可変領域を含み、この軽鎖可変領域は、配列番号8に示されるアミノ酸配列、またはその抗原結合フラグメントもしくは免疫学的に機能的な免疫グロブリンフラグメントを含む。
【0011】
特定の局面では、本発明はまた、IFN−γに特異的に結合し得、そして/またはその生物学的活性を阻害もしくは調節し得る、抗体およびその免疫学的に機能的な免疫グロブリンフラグメントを提供し、この抗体は、重鎖および軽鎖を含み、この重鎖は、重鎖可変領域を含み、この重鎖可変領域は、配列番号6に示されるアミノ酸配列に対して少なくとも80%、85%、90%、91%、92%、93%、94%、95%、96%、97%、98%もしくは約99%の同一性を有する配列を含み、そしてこの軽鎖は、軽鎖可変領域を含み、この軽鎖可変領域は、配列番号8に示されるアミノ酸配列に対して少なくとも80%、85%、90%、91%、92%、93%、94%、95%、96%、97%、98%もしくは約99%の同一性を有する配列を含み、この抗体は、IFN−γと相互作用する。
【0012】
本発明はさらに、IFN−γの生物学的活性を阻害し得るかもしくは調節し得、そして/またはIFN−γに特異的に結合し得る抗体を提供し、この重鎖は、配列番号10に示されるアミノ酸配列を含む重鎖可変領域、またはその抗原結合フラグメントもしくは免疫学的に機能的な免疫グロブリンフラグメントを含み、そしてこの軽鎖は、配列番号12に示されるアミノ酸配列を含む軽鎖可変領域、またはその抗原結合フラグメントもしくは免疫学的に機能的な免疫グロブリンフラグメントを含む。
【0013】
特定の局面では、本発明は、IFN−γの生物学的活性を阻害し得るかもしくは調節し得、そして/またはIFN−γに特異的に結合し得る抗体を提供し、この抗体は、重鎖および軽鎖を含み、この重鎖は、重鎖可変領域を含み、この重鎖可変領域は、配列番号10に示されるアミノ酸配列に対して少なくとも80%、85%、90%、91%、92%、93%、94%、95%、96%、97%、98%もしくは約99%の同一性を有する配列を含み、そしてこの軽鎖は、軽鎖可変領域を含み、この軽鎖可変領域は、配列番号12に示されるアミノ酸配列に対して少なくとも80%、85%、90%、91%、92%、93%、94%、95%、96%、97%、98%もしくは約99%の同一性を有する配列を含み、この抗体は、IFN−γと相互作用する。
【0014】
本発明はさらに、IFN−γの生物学的活性を阻害し得るかもしくは調節し得、そして/またはIFN−γに特異的に結合し得る抗体を提供し、この重鎖は、配列番号30に示されるアミノ酸配列を含む重鎖可変領域、またはその抗原結合フラグメントもしくは免疫学的に機能的な免疫グロブリンフラグメントを含み、そしてこの軽鎖は、配列番号12に示されるアミノ酸配列を含む軽鎖可変領域、またはその抗原結合フラグメントもしくは免疫学的に機能的な免疫グロブリンフラグメントを含む。
【0015】
特定の局面では、本発明は、IFN−γの生物学的活性を阻害し得るかもしくは調節し得、そして/またはIFN−γに特異的に結合し得る抗体を提供し、この抗体は、重鎖および軽鎖を含み、この重鎖は、重鎖可変領域を含み、この重鎖可変領域は、配列番号30のいずれかに示されるアミノ酸配列に対して少なくとも80%、85%、90%、91%、92%、93%、94%、95%、96%、97%、98%もしくは約99%の同一性を有する配列を含み、そしてこの軽鎖は、軽鎖可変領域を含み、この軽鎖可変領域は、配列番号12に示されるアミノ酸配列に対して少なくとも80%、85%、90%、91%、92%、93%、94%、95%、96%、97%、98%もしくは約99%の同一性を有する配列を含み、この抗体は、IFN−γと相互作用する。
【0016】
本発明はまた、IFN−γの生物学的活性を阻害し得るかもしくは調節し得、そして/またはIFN−γに特異的に結合し得る抗体を提供し、この重鎖は、配列番号14に示されるアミノ酸配列を含む重鎖可変領域、またはその抗原結合フラグメントもしくは免疫学的に機能的な免疫グロブリンフラグメントを含み、そしてこの軽鎖は、配列番号16に示されるアミノ酸配列を含む軽鎖可変領域、またはその抗原結合フラグメントもしくは免疫学的に機能的な免疫グロブリンフラグメントを含む。
【0017】
特定の局面では、本発明は、IFN−γに特異的に結合し得そして/またはIFN−γの生物学的活性を阻害し得るかもしくは調節し得る抗体を提供し、この抗体は、重鎖および軽鎖を含み、この重鎖は、重鎖可変領域を含み、この重鎖可変領域は、配列番号14に示されるアミノ酸配列に対して少なくとも80%、85%、90%、91%、92%、93%、94%、95%、96%、97%、98%もしくは約99%の同一性を有する配列を含み、そしてこの軽鎖は、軽鎖可変領域を含み、この軽鎖可変領域は、配列番号16に示されるアミノ酸配列に対して少なくとも80%、85%、90%、91%、92%、93%、94%、95%、96%、97%、98%もしくは約99%の同一性を有する配列を含み、この抗体は、IFN−γと相互作用する。
【0018】
本発明はまた、IFN−γの生物学的活性を阻害し得るかもしくは調節し得、そして/またはIFN−γに特異的に結合し得る抗体を提供し、この重鎖は、配列番号14に示されるアミノ酸配列を含む重鎖可変領域、またはその抗原結合フラグメントもしくは免疫学的に機能的な免疫グロブリンフラグメントを含み、そしてこの軽鎖は、配列番号31に示されるアミノ酸配列を含む軽鎖可変領域、またはその抗原結合フラグメントもしくは免疫学的に機能的な免疫グロブリンフラグメントを含む。
【0019】
特定の局面では、本発明は、IFN−γの生物学的活性を阻害し得るかもしくは調節し得、そして/またはIFN−γに特異的に結合し得る抗体を提供し、この抗体は、重鎖および軽鎖を含み、この重鎖は、重鎖可変領域を含み、この重鎖可変領域は、配列番号14に示されるアミノ酸配列に対して少なくとも80%、85%、90%、91%、92%、93%、94%、95%、96%、97%、98%もしくは約99%の同一性を有する配列を含み、そしてこの軽鎖は、軽鎖可変領域を含み、この軽鎖可変領域は、配列番号31に示されるアミノ酸配列に対して少なくとも80%、85%、90%、91%、92%、93%、94%、95%、96%、97%、98%もしくは約99%の同一性を有する配列を含み、この抗体は、IFN−γと相互作用する。
【0020】
本発明はまた、IFN−γの生物学的活性を阻害し得るかもしくは調節し得、そして/またはIFN−γに特異的に結合し得る抗体を提供し、この重鎖は、配列番号17に示されるアミノ酸配列を含む重鎖可変領域、またはその抗原結合フラグメントもしくは免疫学的に機能的な免疫グロブリンフラグメントを含み、そしてこの軽鎖は、配列番号18に示されるアミノ酸配列、またはその抗原結合フラグメントもしくは免疫学的に機能的な免疫グロブリンフラグメントを含む。
【0021】
特定の局面では、本発明は、IFN−γの生物学的活性を阻害し得るかもしくは調節し得、そして/またはIFN−γに特異的に結合し得る抗体を提供し、この抗体は、重鎖および軽鎖を含み、この重鎖は、重鎖可変領域を含み、この重鎖可変領域は、配列番号17に示されるアミノ酸配列に対して少なくとも80%、85%、90%、91%、92%、93%、94%、95%、96%、97%、98%もしくは約99%の同一性を有する配列を含み、そしてこの軽鎖は、軽鎖可変領域を含み、この軽鎖可変領域は、配列番号18に示されるアミノ酸配列に対して少なくとも80%、85%、90%、91%、92%、93%、94%、95%、96%、97%、98%もしくは約99%の同一性を有するアミノ酸配列を含み、この抗体は、IFN−γと相互作用する。
【0022】
本発明はさらに、IFN−γの生物学的活性を阻害し得るかもしくは調節し得、そして/またはIFN−γに特異的に結合し得る抗体を提供し、この重鎖は、配列番号19に示されるアミノ酸配列、またはその抗原結合フラグメントもしくは免疫学的に機能的な免疫グロブリンフラグメントを含み、そしてこの軽鎖は、配列番号20に示されるアミノ酸配列、またはその抗原結合フラグメントもしくは免疫学的に機能的な免疫グロブリンフラグメントを含む。
【0023】
特定の局面では、本発明は、IFN−γの生物学的活性を阻害し得るかもしくは調節し得、そして/またはIFN−γに特異的に結合し得る抗体を提供し、この抗体は、重鎖および軽鎖を含み、この重鎖は、重鎖可変領域を含み、この重鎖可変領域は、配列番号19のいずれかに示されるアミノ酸配列に対して少なくとも80%、85%、90%、91%、92%、93%、94%、95%、96%、97%、98%もしくは約99%の同一性を有する配列を含み、そしてこの軽鎖は、軽鎖可変領域を含み、この軽鎖可変領域は、配列番号20に示されるアミノ酸配列に対して少なくとも80%、85%、90%、91%、92%、93%、94%、95%、96%、97%、98%もしくは約99%の同一性を有する配列を含み、この抗体は、IFN−γと相互作用する。
【0024】
本発明はまた、IFN−γの生物学的活性を阻害し得るかもしくは調節し得、そして/またはIFN−γに特異的に結合し得る抗体を提供し、この重鎖は、配列番号21に示されるアミノ酸配列、またはその抗原結合フラグメントもしくは免疫学的に機能的な免疫グロブリンフラグメントを含み、そしてこの軽鎖は、配列番号22に示されるアミノ酸配列、またはその抗原結合フラグメントもしくは免疫学的に機能的な免疫グロブリンフラグメントを含む。
【0025】
特定の局面では、本発明は、IFN−γの生物学的活性を阻害し得るかもしくは調節し得、そして/またはIFN−γに特異的に結合し得る抗体を提供し、この抗体は、重鎖および軽鎖を含み、この重鎖は、重鎖可変領域を含み、この重鎖可変領域は、配列番号21のいずれかに示されるアミノ酸配列に対して少なくとも80%、85%、90%、91%、92%、93%、94%、95%、96%、97%、98%もしくは約99%の同一性を有する配列を含み、そしてこの軽鎖は、軽鎖可変領域を含み、この軽鎖可変領域は、配列番号22に示されるアミノ酸配列に対して少なくとも80%、85%、90%、91%、92%、93%、94%、95%、96%、97%、98%もしくは約99%の同一性を有するアミノ酸配列を含み、この抗体は、IFN−γと相互作用する。
【0026】
本発明はまた、IFN−γの生物学的活性を阻害し得るかもしくは調節し得、そして/またはIFN−γに特異的に結合し得る抗体を提供し、この重鎖は、配列番号32に示されるアミノ酸配列、またはその抗原結合フラグメントもしくは免疫学的に機能的な免疫グロブリンフラグメントを含み、そしてこの軽鎖は、配列番号20に示されるアミノ酸配列、またはその抗原結合フラグメントもしくは免疫学的に機能的な免疫グロブリンフラグメントを含む。
【0027】
特定の局面では、本発明は、IFN−γの生物学的活性を阻害し得るかもしくは調節し得、そして/またはIFN−γに特異的に結合し得る抗体を提供し、この抗体は、重鎖および軽鎖を含み、この重鎖は、重鎖可変領域を含み、この重鎖可変領域は、配列番号32に示されるアミノ酸配列に対して少なくとも80%、85%、90%、91%、92%、93%、94%、95%、96%、97%、98%もしくは約99%の同一性を有する配列を含み、そしてこの軽鎖は、軽鎖可変領域を含み、この軽鎖可変領域は、配列番号20に示されるアミノ酸配列に対して少なくとも80%、85%、90%、91%、92%、93%、94%、95%、96%、97%、98%もしくは約99%の同一性を有する配列を含み、この抗体は、IFN−γと相互作用する。
【0028】
本発明はまた、IFN−γの生物学的活性を阻害し得るかもしくは調節し得、そして/またはIFN−γに特異的に結合し得る抗体を提供し、この重鎖は、配列番号21に示されるアミノ酸配列、またはその抗原結合フラグメントもしくは免疫学的に機能的な免疫グロブリンフラグメントを含み、そしてこの軽鎖は、配列番号33に示されるアミノ酸配列、またはその抗原結合フラグメントもしくは免疫学的に機能的な免疫グロブリンフラグメントを含む。
【0029】
特定の局面では、本発明は、IFN−γの生物学的活性を阻害し得るかもしくは調節し得、そして/またはIFN−γに特異的に結合し得る抗体を提供し、この抗体は、重鎖および軽鎖を含み、この重鎖は、重鎖可変領域を含み、この重鎖可変領域は、配列番号21に示されるアミノ酸配列に対して少なくとも80%、85%、90%、91%、92%、93%、94%、95%、96%、97%、98%もしくは約99%の同一性を有する配列を含み、そしてこの軽鎖は、軽鎖可変領域を含み、この軽鎖可変領域は、配列番号33に示されるアミノ酸配列に対して少なくとも80%、85%、90%、91%、92%、93%、94%、95%、96%、97%、98%もしくは約99%の同一性を有するアミノ酸配列を含み、この抗体は、IFN−γと相互作用する。
【0030】
本発明はまた、単鎖抗体、単鎖Fv抗体、F(ab)抗体、F(ab)’抗体および(Fab’)2抗体を提供する。
【0031】
特定の局面では、本発明は、配列番号8、配列番号12、配列番号16、配列番号18、配列番号20、配列番号22、配列番号31もしくは配列番号33のいずれかに示されるアミノ酸配列、またはその抗原結合フラグメントもしくは免疫学的に機能的な免疫グロブリンフラグメントを含む、軽鎖を提供する。
【0032】
さらに本発明は、配列番号6、配列番号10、配列番号14、配列番号17、配列番号19、配列番号21、配列番号30もしくは配列番号32のいずれかに示されるアミノ酸配列、またはその抗原結合フラグメントもしくは免疫学的に機能的な免疫グロブリンフラグメントを含む、重鎖を提供する。
【0033】
本発明はまた、IFN−γに特異的に結合する単離されたヒト抗体に関し、この抗体は、(a)ヒト重鎖フレームワーク領域、ヒト重鎖CDR1領域、ヒト重鎖CDR2領域およびヒト重鎖CDR3領域;ならびに(b)ヒト軽鎖フレームワーク領域、ヒト軽鎖CDR1領域、ヒト軽鎖CDR2領域およびヒト軽鎖CDR3領域、を含む。特定の局面では、このヒト重鎖CDR1領域は、図12および配列番号34に示されるモノクローナル抗体(mAb)1119、1118、1118*もしくは1121の重鎖CDR1領域であり得、そしてこのヒト軽鎖CDR1領域は、図13および配列番号38、配列番号39もしくは配列番号40に示されるmAb 1119、1118、1121*もしくは1121の軽鎖CDR1領域であり得る。他の局面では、このヒト重鎖CDR2領域は、図12および配列番号35に示されるmAb 1119、1118、1118*もしくは1121の重鎖CDR2領域であり得、そしてこのヒト軽鎖CDR2領域は、図13および配列番号41もしくは配列番号42に示されるmAb 1119、1118、1121*もしくは1121の軽鎖CDR2領域であり得る。さらに他の局面では、このヒト重鎖CDR3領域は、図12および配列番号36または配列番号37に示されるmAb 1119、1118、1118*もしくは1121の重鎖CDR3領域であり、そしてこのヒト軽鎖CDR3領域は、図13および配列番号43もしくは配列番号44に示されるmAb 1119、1118、1121*もしくは1121の軽鎖CDR3領域である。
【0034】
さらに、本発明は、IFN−γの産生の増大もしくはIFN−γに対する感受性の増大に関連する疾患、および/またはIFN−γによって媒介される疾患を処置するための方法を提供し、この方法は、本発明の1つ以上のモノクローナル抗体、またはその抗原結合フラグメントもしくは免疫学的に機能的な免疫グロブリンフラグメントの、薬学的に有効な量を、それを必要とする個体に対して投与する工程を包含する。
【0035】
本発明はまた、生物学的サンプル中のIFN−γのレベルを決定するための方法を提供し、この方法は、本発明のモノクローナル抗体またはその抗原結合フラグメントとサンプルとを接触させる工程を包含する。
【0036】
本発明はまた、IFN−γに特異的に結合し得るか、またはその生物学的活性を阻害もしくは調節し得る単離された抗体を提供し、この抗体は、重鎖CDR3を含み、この重鎖CDR3は、(a)配列番号36に存在するのと同じ順序かつ間隔で、配列番号36のアミノ酸のうち少なくとも7つからなる、アミノ酸配列;または(b)配列番号37を含むアミノ酸配列、であるアミノ酸配列を有する。この抗体はさらに、軽鎖CDR3を含み得、この軽鎖CDR3は、(a)配列番号43に存在するのと同じ順序かつ間隔で、配列番号43のアミノ酸のうち少なくとも8つからなる、アミノ酸配列;または(b)配列番号44に存在するのと同じ順序かつ間隔で、配列番号44のアミノ酸のうち少なくとも9つからなるアミノ酸配列、であるアミノ酸配列を有する。この抗体はさらに、配列番号34、配列番号35、配列番号38、配列番号39、配列番号40、配列番号41および配列番号42からなる群より選択される1つ以上のCDRを含み得る。この重鎖CDR3は、配列番号36の少なくともアミノ酸からなってもよく、そして軽鎖CDR3は、配列番号43の少なくともアミノ酸からなってもよい。
【0037】
さらに、本発明は、IFN−γに特異的に結合し得るか、そして/またはその生物学的活性を阻害もしくは調節し得る単離された抗体を提供し、この抗体は、軽鎖CDR3を含み、この軽鎖CDR3は、アミノ酸配列((a)配列番号43に存在するのと同じ順序かつ間隔で、配列番号43のアミノ酸のうち少なくとも8つからなるアミノ酸配列;または(b)配列番号44に存在するのと同じ順序かつ間隔で、配列番号44のアミノ酸のうち少なくとも9つからなるアミノ酸配列)を有する。この抗体はさらに、配列番号35、配列番号38、配列番号39、配列番号40、配列番号41または配列番号42のアミノ酸配列を有するCDRを含み得る。
【0038】
IFN−γに特異的に結合し得るか、そして/またはその生物学的活性を阻害もしくは調節し得る、本発明の単離された抗体は、少なくとも:(a)配列番号34;(b)配列番号35;(c)配列番号36または配列番号37;(d)配列番号38、配列番号39または配列番号40;(e)配列番号41または配列番号42;および(f)配列番号43または配列番号44;のアミノ酸配列を有する6つのCDRを含み得る。
【0039】
IFN−γに特異的に結合し得るか、そして/またはその生物学的活性を阻害もしくは調節し得る、本発明の単離された抗体は、配列番号6、配列番号10、配列番号14または配列番号30に対して少なくとも80%、85%、90%、91%、92%、93%、94%、95%、96%、97%、98%もしくは約99%同一のアミノ酸配列(このアミノ酸配列と、配列番号6、配列番号10、配列番号14もしくは配列番号30との間のアラインメントが、少なくとも50アミノ酸にまたがる)、および/または配列番号8、配列番号12、配列番号16もしくは配列番号31に対して少なくとも80%、85%、90%、91%、92%、93%、94%、95%、96%、97%、98%もしくは約99%同一のアミノ酸配列(このアミノ酸配列と、配列番号8、配列番号12、配列番号16もしくは配列番号31との間のアラインメントが、少なくとも50アミノ酸にまたがる)を含み得る。
【0040】
別の局面では、IFN−γに特異的に結合し得る、そして/またはその生物学的活性を阻害もしくは調節し得る本発明の抗体は、配列番号17、配列番号19、配列番号21または配列番号32に対して少なくとも80%、85%、90%、91%、92%、93%、94%、95%、96%、97%、98%もしくは約99%同一のアミノ酸配列(このアミノ酸配列と、配列番号17、配列番号19、配列番号21もしくは配列番号32との間のアラインメントが、少なくとも50アミノ酸にまたがる)、および/または配列番号18、配列番号20、配列番号22もしくは配列番号33に対して少なくとも80%、85%、90%、91%、92%、93%、94%、95%、96%、97%、98%もしくは約99%同一のアミノ酸配列(このアミノ酸配列と、配列番号18、配列番号20、配列番号22もしくは配列番号33との間のアラインメントが、少なくとも50アミノ酸にまたがる)を含み得る。これらの抗体内の1つのアミノ酸配列は、配列番号36もしくは配列番号37のアミノ酸のうち少なくとも5つを、配列番号36もしくは配列番号37に存在するのと同じ順序かつ間隔で含み得、そして/またはこれらの抗体内の1つのアミノ酸配列は、配列番号43もしくは配列番号44のアミノ酸のうち少なくとも6つを、配列番号43もしくは配列番号44に存在するのと同じ順序かつ間隔で含み得る。
【0041】
1つの局面では、本発明は、単離された完全ヒト抗体であり得る抗体を提供し、この抗体は、ヒトIFN−γの生物学的活性を阻害または調節し得る。別の局面では、本発明の抗体は、単離された完全ヒト抗体であり得、カニクイザルおよびマウスのIFN−γの生物学的活性を阻害も調節もできない。さらに別の局面では、本発明の完全ヒト抗体は、ヒトおよびチンパンジーのIFN−γの生物学的活性を阻害または調節し得るが、カニクイザルおよび/またはマウスのIFN−γの生物学的活性を阻害も調節もできない。さらに別の局面では、ヒトIFN−γの残基19をアスパラギン酸で置換し、および/または残基20をプロリンで置換すれば、この抗体によるヒトIFN−γの生物学的活性の阻害を妨げるかまたは拮抗する。さらに、この抗体は、カニクイザルIFN−γの残基19、20および65にてそれぞれヒスチジン、セリンおよびセリンで置換された変異バージョンの生物学的活性を阻害し得る。
【0042】
なおさらなる局面では、IFN−γに特異的に結合し得、および/またはその生物学的活性を阻害もしくは調節し得る本発明の単離された抗体は、配列番号17、配列番号19、配列番号21または配列番号32に対して少なくとも80%、85%、90%、91%、92%、93%、94%、95%、96%、97%、98%もしくは約99%同一のアミノ酸配列を含み得、このアミノ酸配列と、配列番号17、配列番号19、配列番号21もしくは配列番号32との間のアラインメントは、少なくとも50アミノ酸にまたがり、そして/または上記抗体は、配列番号18、配列番号20、配列番号22もしくは配列番号33に対して少なくとも80%、85%、90%、91%、92%、93%、94%、95%、96%、97%、98%もしくは約99%同一のアミノ酸配列を含み得、このアミノ酸配列と、配列番号18、配列番号20、配列番号22もしくは配列番号33との間のアラインメントは、少なくとも50アミノ酸にまたがる。
【0043】
さらなる実施形態では、本発明は、IFN−γに特異的に結合し得る単離された抗体を含み、この抗体は、(a)配列番号6、配列番号10、配列番号14もしくは配列番号30を含むアミノ酸配列、またはこれらの配列の1つのフラグメント、および(b)配列番号8、配列番号12、配列番号16もしくは配列番号31を含むアミノ酸配列、またはこれらの配列の1つのフラグメント、を含む。この抗体は、重鎖および軽鎖を含み得る。この抗体は、(a)配列番号6、配列番号10、配列番号14または配列番号30、および(b)配列番号8、配列番号12、配列番号16または配列番号31を含み得る。
【0044】
さらに、本発明のいずれの抗体も、ヒト化抗体であっても完全ヒト抗体であってもよい。
【0045】
本発明はまた、本明細書に記載される本発明の抗体のいずれかまたはその一部をコードする、ポリヌクレオチド(単離されたポリヌクレオチドを含む)を提供し、この抗体は、CDR領域、重鎖可変領域、軽鎖可変領域、単鎖抗体、単鎖Fv抗体、F(ab)抗体、F(ab)’抗体および(Fab’)2抗体を含む。本発明はさらに、このようなポリヌクレオチドを含むベクター、ならびにこのようなポリヌクレオチドおよび/またはベクターを含む宿主細胞(必要に応じて、哺乳動物宿主細胞)を提供する。本発明の抗体は、このような宿主細胞を培養する工程によって生成され得る。
本発明によって、例えば、以下が提供される。
(項目1)
重鎖CDR3を含む単離された抗体であって、該重鎖CDR3がアミノ酸配列、すなわち:
(a)配列番号36のアミノ酸のうち少なくとも7つを配列番号36に存在するのと同じ順序でかつ間隔で含むアミノ酸配列;または
(b)配列番号37を含むアミノ酸配列;
を有し、ここで該抗体がIFN−γに特異的に結合し得る、単離された抗体。
(項目2)
軽鎖CDR3をさらに含む項目1に記載の抗体であって、該軽鎖CDR3がアミノ酸配列、すなわち:
(a)配列番号43のアミノ酸のうち少なくとも8つを配列番号43に存在するのと同じ順序でかつ間隔で含むアミノ酸配列;または
(b)配列番号44のアミノ酸のうち少なくとも9つを配列番号44に存在するのと同じ順序でかつ間隔で含むアミノ酸配列;
を有する、単離された抗体。
(項目3)
軽鎖CDR3を含む単離された抗体であって、該軽鎖CDR3がアミノ酸配列、すなわち:
(a)配列番号43のアミノ酸のうち少なくとも8つを配列番号43に存在するのと同じ順序でかつ間隔で含むアミノ酸配列;または
(b)配列番号44のアミノ酸のうち少なくとも9つを配列番号44に存在するのと同じ順序でかつ間隔で含むアミノ酸配列;
を有し、ここで該抗体がIFN−γに特異的に結合し得る、単離された抗体。
(項目4)
CDRをさらに含む項目3に記載の抗体であって、該CDRが配列番号34、配列番号35、配列番号36、配列番号37、配列番号38、配列番号39、配列番号40、配列番号41または配列番号42のアミノ酸配列を有する抗体。
(項目5)
前記重鎖CDR3が配列番号36のアミノ酸を含み、前記軽鎖CDR3が配列番号43のアミノ酸を含む項目2に記載の抗体。
(項目6)
配列番号6、配列番号10、配列番号14および/または配列番号30に対して少なくとも80%同一の第一のアミノ酸配列を含む、項目1〜5のいずれか1項に記載の抗体であって、
該第一のアミノ酸配列と配列番号6、配列番号10、配列番号14または配列番号30との間のアラインメントが少なくとも50アミノ酸にまたがる、抗体。
(項目7)
前記第一のアミノ酸配列が配列番号6、配列番号10、配列番号14および/または配列番号30に対して90%同一である、項目6に記載の抗体。
(項目8)
前記第一のアミノ酸配列が配列番号6、配列番号10、配列番号14または配列番号30に対して同一である、項目7に記載の抗体。
(項目9)
配列番号8、配列番号12、配列番号16および/または配列番号31に対して少なくとも80%同一の第二のアミノ酸配列を含む、項目1〜8のいずれか1項に記載の抗体であって、
該第二のアミノ酸配列と配列番号8、配列番号12、配列番号16および/または配列番号31との間のアラインメントが少なくとも50アミノ酸にまたがる、抗体。
(項目10)
前記第二のアミノ酸配列が配列番号8、配列番号12、配列番号16および/または配列番号31に対して少なくとも90%同一である、項目9に記載の抗体。
(項目11)
前記第二のアミノ酸配列が配列番号8、配列番号12、配列番号16または配列番号31に対して同一である、項目10に記載の抗体。
(項目12)
配列番号17、配列番号19、配列番号21および/または配列番号32に対して少なくとも80%同一の第一のアミノ酸配列を含む、項目1〜5のいずれか1項に記載の抗体であって、
該第一のアミノ酸配列と配列番号17、配列番号19、配列番号21および/または配列番号32との間のアラインメントが少なくとも50アミノ酸にまたがる、抗体。
(項目13)
前記第一のアミノ酸配列が配列番号17、配列番号19、配列番号21および/または配列番号32に対して少なくとも90%同一である、項目12に記載の抗体。
(項目14)
前記第一のアミノ酸配列が配列番号17、配列番号19、配列番号21または配列番号32に対して同一である、項目13に記載の抗体。
(項目15)
配列番号18、配列番号20、配列番号22および/または配列番号33に対して少なくとも80%同一の第二のアミノ酸配列を含む、項目1〜5および項目12〜14のいずれか1項に記載の抗体であって、
該第二のアミノ酸配列と配列番号18、配列番号20、配列番号22および/または配列番号33との間のアラインメントが少なくとも50アミノ酸にまたがる、抗体。
(項目16)
前記第二のアミノ酸配列が配列番号18、配列番号20、配列番号22および/または配列番号33に対して少なくとも90%同一である、項目15に記載の抗体。
(項目17)
前記第二のアミノ酸配列が配列番号18、配列番号20、配列番号22または配列番号33に対して同一である、項目16に記載の抗体。
(項目18)
単離された抗体であって、第一のアミノ酸配列、すなわち:
(a)配列番号6に対して少なくとも96%同一であるアミノ酸配列;
(b)配列番号10に対して少なくとも93%同一であるアミノ酸配列;
(c)配列番号14に対して少なくとも91%同一であるアミノ酸配列;または
(d)配列番号30に対して少なくとも98%同一であるアミノ酸配列;
を含み、
ここで該第一のアミノ酸配列と配列番号6、配列番号10、配列番号14または配列番号30との間のアラインメントが少なくとも50アミノ酸にまたがり、そして
IFN−γに特異的に結合可能である単離された抗体。
(項目19)
前記第一のアミノ酸配列が配列番号6に対して少なくとも96%同一であるか、配列番号10に95%同一であるか、配列番号14に対して95%同一であるか、または配列番号30に対して98%同一である、項目18に記載の抗体。
(項目20)
前記第一のアミノ酸配列が配列番号6、配列番号10、配列番号14または配列番号30に対して同一である、項目19に記載の抗体。
(項目21)
配列番号8、配列番号12、配列番号16および/または配列番号31に対して少なくとも80%同一である第二のアミノ酸配列をさらに含み、該第二のアミノ酸配列と配列番号8、配列番号12、配列番号16または配列番号31との間のアラインメントが少なくとも50アミノ酸にまたがる、項目18〜20のいずれか1項に記載の抗体。
(項目22)
前記第二のアミノ酸配列が配列番号8、配列番号12、配列番号16および/または配列番号31に対して少なくとも90%同一である、項目21に記載の抗体。
(項目23)
前記第一のアミノ酸配列が配列番号36および/または配列番号37のアミノ酸のうち少なくとも5つを配列番号36および/または配列番号37に存在するのと同じ順序でかつ間隔で含む、項目18〜22のいずれか1項に記載の抗体。
(項目24)
前記第二のアミノ酸配列が配列番号43または配列番号44のアミノ酸のうち少なくとも6つを、配列番号43または配列番号44に存在するのと同じ順序でかつ間隔で含む、項目21または22に記載の抗体。
(項目25)
単離された抗体であって、第一のアミノ酸配列、すなわち:
(a)配列番号8に対して少なくとも97%同一であるアミノ酸配列;
(b)配列番号12に対して少なくとも96%同一であるアミノ酸配列;
(c)配列番号16に対して少なくとも97%同一であるアミノ酸配列;または
(d)配列番号31に対して少なくとも95%同一であるアミノ酸配列;
を含み、
ここで該配列番号8、配列番号12、配列番号16または配列番号31と該第一のアミノ酸配列との間のアラインメントが少なくとも50アミノ酸にまたがり、そして
IFN−γに特異的に結合可能である単離された抗体。
(項目26)
前記第一のアミノ酸配列が配列番号8、配列番号12、配列番号16または配列番号31に対して少なくとも97%同一である、項目25に記載の抗体。
(項目27)
前記第一のアミノ酸配列が配列番号8、配列番号12、配列番号16または配列番号31に対して同一である、項目26に記載の抗体。
(項目28)
前記第一のアミノ酸配列が配列番号43または配列番号44のアミノ酸のうち少なくとも6つを、配列番号43または配列番号44に存在するのと同じ順序でかつ間隔で含む、項目25に記載の抗体。
(項目29)
配列番号6、配列番号10、配列番号14および/または配列番号30に対して少なくとも80%同一である第二のアミノ酸配列をさらに含む、項目25〜28のいずれか1項に記載の抗体であって、
該第二のアミノ酸配列と配列番号6、配列番号10、配列番号14および/または配列番号30との間のアラインメントが少なくとも50アミノ酸にまたがる、抗体。
(項目30)
前記第二のアミノ酸配列が配列番号6、配列番号10、配列番号14および/または配列番号30に対して少なくとも90%同一である、項目29に記載の抗体。
(項目31)
前記第二のアミノ酸配列が配列番号36および/または配列番号37のアミノ酸のうち少なくとも5つを配列番号36および/または配列番号37に存在するのと同じ順序でかつ間隔で含む、項目29または30に記載の抗体。
(項目32)
単離された完全ヒト抗体であって、
該抗体は、ヒトIFN−γおよびチンパンジーIFN−γの生物学的活性を阻害し得、そして該抗体はカニクイザルIFN−γおよびマウスIFN−γの生物学的活性を阻害できず、
該抗体は、VH領域、CH1領域、CH2領域およびCH3領域を含む重鎖、ならびにVL領域およびCL領域を含む軽鎖を含む、抗体。
(項目33)
項目32に記載の抗体であって、ヒトIFN−γの残基20をプロリンで置換することによって、該抗体によるヒトIFN−γの生物学的活性の阻害を拮抗する抗体。
(項目34)
項目32または33に記載の抗体であって、該抗体はカニクイザルIFN−γの変異バージョンの生物学的活性を阻害し得、該変異バージョンは19位にヒスチジン、20位にセリン、そして65位にセリンを含む、抗体。
(項目35)
ヒトIFN−γの生物学的活性を阻害し得る単離された抗体であって、ヒトIFN−γの残基20をプロリンで置換することによって、該抗体によるヒトIFN−γの生物学的活性の阻害を拮抗し、かつ完全ヒト抗体である、単離された抗体。
(項目36)
軽鎖CDR1、軽鎖CDR2、軽鎖CDR3、重鎖CDR1、重鎖CDR2、および重鎖CDR3を含む単離された抗体であって、
該重鎖CDR1は配列番号34のアミノ酸配列を有し;
該重鎖CDR2は配列番号35のアミノ酸配列を有し;
該重鎖CDR3は配列番号36または配列番号37のアミノ酸配列を有し;
該軽鎖CDR1は配列番号38、配列番号39または配列番号40のアミノ酸配列を有し;
該軽鎖CDR2は配列番号41または配列番号42のアミノ酸配列を有し;
該軽鎖CDR3は配列番号43または配列番号44のアミノ酸配列を有し;そして
該抗体はIFN−γに特異的に結合し得る、単離された抗体。
(項目37)
項目36に記載の抗体であって、
前記重鎖CDR3は配列番号36のアミノ酸配列を有し、
前記軽鎖CDR1は配列番号38のアミノ酸配列を有し、
前記軽鎖CDR2は配列番号41のアミノ酸配列を有し、そして
前記軽鎖CDR3は配列番号43のアミノ酸配列を有する、抗体。
(項目38)
項目36または37に記載の抗体であって、
配列番号17、配列番号19、配列番号21および/または配列番号32に対して少なくとも80%同一の第一のアミノ酸配列であって、ここで該第一のアミノ酸配列と配列番号17、配列番号19、配列番号21および/または配列番号32との間のアラインメントが少なくとも50アミノ酸にまたがり、該第一のアミノ酸配列が重鎖CDR1、CDR2およびCDR3のアミノ酸配列を含む、第一のアミノ酸配列、ならびに
配列番号18、配列番号20、配列番号22および/または配列番号33に対して少なくとも80%同一の第二のアミノ酸配列であって、ここで該第二のアミノ酸配列と配列番号18、配列番号20、配列番号22および/または配列番号33との間のアラインメントが少なくとも50アミノ酸にまたがり、該第二のアミノ酸配列が軽鎖CDR1、CDR2およびCDR3のアミノ酸配列を含む、第二のアミノ酸配列、
を含む抗体。
(項目39)
項目38に記載の抗体であって、
第一のアミノ酸配列が、配列番号17、配列番号19、配列番号21および/または配列番号32に対して少なくとも90%同一であり、そして、
第二のアミノ酸配列が、配列番号18、配列番号20、配列番号22および/または配列番号33に対して少なくとも90%同一である、
抗体。
(項目40)
項目39に記載の抗体であって、
前記第一のアミノ酸配列が、配列番号17、配列番号19、配列番号21および/または配列番号32に対して同一であり、そして、
前記第二のアミノ酸配列が、配列番号18、配列番号20、配列番号22および/または配列番号33に対して同一である、
抗体。
(項目41)
項目36または37に記載の抗体であって、
配列番号6、配列番号10、配列番号14および/または配列番号30に対して少なくとも80%同一の第一のアミノ酸配列であって、該第一のアミノ酸配列と配列番号6、配列番号10、配列番号14および/または配列番号30との間のアラインメントが少なくとも50アミノ酸にまたがる、第一のアミノ酸配列;ならびに
配列番号8、配列番号12、配列番号16および/または配列番号31に対して少なくとも80%同一の第二のアミノ酸配列であって、該第二のアミノ酸配列と配列番号8、配列番号12、配列番号16および/または配列番号31との間のアラインメントが少なくとも50アミノ酸にまたがる、第二のアミノ酸配列、
を含む抗体。
(項目42)
項目41に記載の抗体であって、
前記第一のアミノ酸配列が、配列番号6、配列番号10、配列番号14および/または配列番号30に対して少なくとも90%同一であり、そして
前記第二のアミノ酸配列が、配列番号8、配列番号12、配列番号16および/または配列番号31に対して少なくとも90%同一である、抗体。
(項目43)
項目42に記載の抗体であって、
前記第一のアミノ酸配列が、配列番号6、配列番号10、配列番号14または配列番号30に対して同一であり、そして
前記第二のアミノ酸配列が、配列番号8、配列番号12、配列番号16または配列番号31に対して同一である、抗体。
(項目44)
単離された抗体であって、
(a)配列番号6、配列番号10、配列番号14もしくは配列番号30、またはこれらの配列の1つのフラグメントを含むアミノ酸配列、および
(b)配列番号8、配列番号12、配列番号16もしくは配列番号31、またはこれらの配列の1つのフラグメントを含むアミノ酸配列、
を含み、IFN−γに特異的に結合可能である、単離された抗体。
(項目45)
重鎖および軽鎖を含む項目44に記載の抗体であって、該重鎖は、VH領域、CH1領域、CH2領域およびCH3領域を含み、該軽鎖はVL領域およびCL領域を含む、抗体。
(項目46)
項目44または45に記載の抗体であって、
(a)配列番号6、配列番号10、配列番号14または配列番号30、および
(b)配列番号8、配列番号12、配列番号16または配列番号31、
を含む抗体。
(項目47)
前記抗体が配列番号6および配列番号8を含む、項目46に記載の抗体。
(項目48)
前記抗体が単鎖抗体である、項目1〜31および項目36〜47のいずれか1項に記載の抗体。
(項目49)
前記抗体が完全ヒト抗体である、項目1〜31および項目36〜47のいずれか1項に記載の抗体。
(項目50)
前記抗体がヒト化抗体である、項目1〜31および項目36〜47のいずれか1項に記載の抗体。
(項目51)
項目1〜50のいずれか1項に記載の抗体をコードする単離されたポリヌクレオチド。
(項目52)
項目51に記載のポリヌクレオチドを含むベクター。
(項目53)
項目52に記載のベクターを含む宿主細胞。
(項目54)
項目53に記載の宿主細胞を培養する工程を包含する、抗体の産生方法。
(項目55)
IFN−γ媒介性疾患に罹患している被験体に対して、項目1〜50のいずれか1項に記載の抗体の治療上有効な量を投与する工程を包含する、方法。
(項目56)
薬学的に受容可能なキャリア、および項目1〜50のいずれか1項に記載の抗体の治療上有効な量を含む、組成物。
(項目57)
IFN−γ媒介性疾患に罹患している患者に対して項目56に記載の組成物を投与する工程を包含する、方法。
(項目58)
前記IFN−γ媒介性疾患が、AIDS、若年性関節リウマチを含む関節リウマチ、潰瘍性大腸炎およびクローン病を含む炎症性腸疾患、多発性硬化症、アジソン病、糖尿病(I型)、精巣上体炎、糸球体腎炎、グレーブス病、ギヤン・バレー症候群、橋本病、溶血性貧血、全身性エリテマトーデス(SLE)、ループス腎炎、重症筋無力症、天疱瘡、乾癬、乾癬性関節症、アテローム性動脈硬化症、エリスロポエチン耐性、宿主対移植片病、移植片拒絶、自己免疫性肝炎誘導性肝障害、胆汁性肝硬変、アルコール肝硬変を含むアルコール誘導性肝障害、リウマチ熱、サルコイドーシス、強皮症、シェーグレン症候群、強直性脊椎炎を含む脊椎関節症、甲状腺炎、または脈管炎である、項目55または57に記載の方法。
【0046】
本発明の特に好ましい実施形態は、特定の好ましい実施形態の以下のさらに詳細な説明および特許請求の範囲から明らかになる。
【0047】
(詳細な説明)
本明細書中で用いた節の表題は、構成上の目的でしかなく、記載された内容に限定されると解釈されるべきではない。本出願に引用された全ての参考文献は、任意の目的について本明細書中で参考として明確に援用される。
(定義)
組み換えDNA、オリゴヌクレオチド合成、ならびに組織培養および形質転換(例えば、エレクトロポレーション、リポフェクチン)については標準的な技術を用いてもよい。酵素反応および精製技術は、製造業者の説明書に従って、または当該分野で一般に完成されているとおり、または本明細書に記載されているように行なってもよい。前述の技術および手順は、当該分野で周知の従来法によって、そして本明細書を通じて引用および考察されている種々の一般的およびより詳細な参考文献に記載されているとおりに一般に行なうことができる。例えば、任意の目的について参考として本明細書中に援用されている、Sambrookら、2001、MOLECULAR CLONING:A LABORATORY MANUAL、第三版、Cold Spring Harbor Laboratory Press,Cold Spring Harbor,N.Y.を参照のこと。特に定義しない限り、本明細書に記載の分析化学、合成有機化学および医学および薬学的化学に関連して利用される命名法、そしてそれらの実験室手順および技術は、当該分野で周知でありかつ一般に用いられるものである。化学合成、化学分析、製剤、処方および送達、ならびに患者の処置には標準的な技術が用いられ得る。
【0048】
本開示に関して使用する場合、以下の用語は、他に示さない限り、以下の意味を有すると理解されるべきである。
【0049】
本明細書において用いる場合、「単離されたポリヌクレオチド」という用語は、ゲノムのポリヌクレオチド、cDNAまたは合成起源またはそれらのいくつかの組み合わせを意味しており、その起源のおかげで、この単離されたポリヌクレオチドは、(1)その単離されたポリヌクレオチドが、天然に見出されるポリヌクレオチドの全てまたは一部を伴わないか、(2)天然には連結されないポリヌクレオチドに連結されるか、または(3)より大きい配列の一部として天然には存在しないかのいずれかである。
【0050】
本明細書において言及される「単離されたタンパク質」という用語は、このタンパク質が、(1)天然に代表的に見出される少なくともいくつかの他のタンパク質を含まないか、(2)同じ供給源由来、例えば同じ種由来の他のタンパク質を本質的に含まないか、(3)異なる種由来の細胞によって発現されるか、(4)天然にはともなわない、少なくとも約50%のポリヌクレオチド、脂質、炭水化物もしくは他の物質から分離されているか、(5)この「単離されたタンパク質」が天然では会合しているタンパク質の一部と会合していない(共有的な相互作用によっても、または非共有的な相互作用によっても)か、(6)天然には会合していないポリペプチドと作動可能に会合している(共有的な相互作用または非共有的な相互作用によって)か、または(7)天然には存在しないということを意味する。このような単離されたタンパク質は、合成起源のゲノムDNA、cDNA、mRNAもしくは他のRNA、またはそれらの任意の組み合わせによってコードされ得る。好ましくは、単離されたタンパク質は、その用途(治療上、診断上、予測、研究その他)を妨げる、天然の環境で見出されるタンパク質またはポリペプチドまたは他の混入物を実質的に含まない。
【0051】
「ポリペプチド」または「タンパク質」という用語は、アミノ酸の1つ以上の鎖であって、各々の鎖がペプチド結合によって共有結合されたアミノ酸を含み、このポリペプチドまたはタンパク質は、非共有結合的におよび/またはペプチド結合によって共有結合的に一緒に連結された複数の鎖を含み得、天然のタンパク質、すなわち、天然に存在する細胞そして詳細には非組み換え細胞によって産生されたタンパク質、または遺伝子操作された細胞かもしくは組み換え細胞によって産生されたタンパク質の配列を有し、そしてこの天然のタンパク質のアミノ酸配列を有する分子、またはこの天然の配列の1つ以上のアミノ酸からの欠失、それに対する付加および/または置換を有する分子を含む鎖を意味する。「ポリペプチド」および「タンパク質」という用語は、特に抗IFNγ抗体、または抗IFNγ抗体の1つ以上のアミノ酸からの欠失、それに対する付加および/または置換を有する配列を含む。従って、「ポリペプチド」または「タンパク質」は、1つのアミノ酸鎖(「単量体」と命名される)または複数のアミノ酸鎖(「多量体」と命名される)を包含し得る。
【0052】
「ポリペプチドフラグメント」という用語は、単量体であっても多量体であってもよく、アミノ末端欠失、カルボキシ末端欠失、および/または天然に存在するかもしくは組み換え産生されたポリペプチドの内部欠失もしくは置換を有するポリペプチドをいう。特定の実施形態では、ポリペプチドフラグメントは、少なくとも5〜約500アミノ酸長のアミノ酸鎖を含んでもよい。特定の実施形態では、フラグメントは少なくとも5、6、8、10、14、20、50、70、100、110、150、200、250、300、350、400または450アミノ酸長であることが理解される。特に有用なポリペプチドフラグメントとしては、結合ドメインを含む機能的なドメインが挙げられる。抗IFN−γ抗体の場合、有用なフラグメントとしては、CDR領域、特に重鎖または軽鎖のCDR3領域;重鎖または軽鎖の可変ドメイン;抗体鎖の一部または2つのCDRを含むちょうどその可変領域;などが挙げられるがこれらに限定されない。
【0053】
本明細書において用いる場合「免疫学的に機能的な免疫グロブリンフラグメント」という用語は、免疫グロブリン重鎖および軽鎖の少なくともCDRを含むポリペプチドフラグメントをいう。本発明の免疫学的に機能的な免疫グロブリンフラグメントは、抗原に結合し得る。好ましい実施形態では、この抗原はレセプターに特異的に結合するリガンドである。これらの実施形態では、本発明の免疫学的に機能的な免疫グロブリンフラグメントの結合は、そのレセプターに対するリガンドの結合を防止または阻害し、このレセプターに対するリガンド結合から生じる生物学的応答を遮断する。好ましくは、本発明の免疫学的に機能的な免疫グロブリンフラグメントは、IFN−γに特異的に結合する。最も好ましくは、このフラグメントはヒトIFN−γに特異的に結合するか、そして/またはその生物学的活性を阻害もしくは調節する。
【0054】
「天然に存在する」という用語は、本明細書において用いて物体に対して適用する場合、その物体が天然に見出され得るという事実をいう。例えば、天然の供給源から単離可能であり、人によって意図的に改変されていない、生物体(ウイルスを含む)に存在するポリペプチドまたはポリヌクレオチドの配列は、天然に存在する。
【0055】
「作動可能に連結された」という用語は、その用語が適用された成分が、適切な条件下でその固有の機能を行なうことを可能にする関係であることを意味する。例えば、タンパク質コード配列に「作動可能に連結された」転写制御配列は、このタンパク質コード配列の発現が制御配列の転写活性に適合する条件下で達成されるようにそこに連結されている。
【0056】
「制御配列」という用語は、本明細書中で用いられる場合、それらが連結されるコード配列の発現、プロセシングまたは細胞内局在化に影響を及ぼし得るポリヌクレオチド配列をいう。このようなコントロール配列の性質は、宿主生物体に依存し得る。特定の実施形態では、原核生物の転写制御配列としては、プロモーター、リボソーム結合部位および転写終結配列が挙げられ得る。他の特定の実施形態では、真核生物の転写制御配列としては、転写因子の1つまたは複数の認識部位を含むプロモーター、転写エンハンサー配列、転写終結配列およびポリアデニル化配列が挙げられ得る。特定の実施形態では、「制御配列」とは、リーダー配列および/または融合パートナー配列を含んでもよい。
【0057】
「ポリヌクレオチド」という用語は、本明細書中で言及する場合、少なくとも10塩基長の一本鎖または二本鎖の核酸ポリマーを意味する。特定の実施形態では、ポリヌクレオチドを含むヌクレオチドは、リボヌクレオチドもしくはデオキシリボヌクレオチドまたはいずれかのタイプのヌクレオチドの修飾型であってもよい。このような修飾としては、ブロモウリジンのような塩基修飾、アラビノシドのようなリボース修飾、ならびに2’,3’−ジデオキシリボースおよびヌクレオチド間連結修飾(例えば、ホスホロチオエート、ホスホロジチオエート、ホスホロセレノエート、ホスホロジセレノエート、ホスホロアニロチオエート、ホスホロアニラデートおよびホスホロアミデート)が挙げられる。「ポリヌクレオチド」という用語は詳細には、DNAの一本鎖および二本鎖型を含む。
【0058】
本明細書で言及する「オリゴヌクレオチド」という用語は、天然に存在する、そして修飾されたヌクレオチドであって、天然に存在するか、および/または天然には存在しないオリゴヌクレオチド連結によって一緒に連結されたヌクレオチドを含む。オリゴヌクレオチドは、一般的に一本鎖であり、200塩基以下の長さを有するメンバーを含むポリヌクレオチドサブセットである。特定の実施形態では、オリゴヌクレオチドは、10〜60塩基長である。特定の実施形態では、オリゴヌクレオチドは12、13、14、15、16、17、18、19、または20〜40塩基長である。オリゴヌクレオチドは、例えば、遺伝子変異体の構築における使用のために、一本鎖であっても二本鎖であってもよい。本発明のオリゴヌクレオチドは、タンパク質コード配列に関連してセンスオリゴヌクレオチドであってもアンチセンスオリゴヌクレオチドであってもよい。
【0059】
他に特定しない限り、一本鎖ポリヌクレオチド配列の左側末端は、5’末端であり;二本鎖ポリヌクレオチド配列の左側方向は、5’方向と呼ばれる。初期RNA転写物の5’〜3’付加の方向は、転写方向と呼ばれる;RNAと同じ配列を有するDNA鎖上の配列領域であって、RNA転写物の5’〜5’末端への領域は「上流配列」と呼ばれる;RNAと同じ配列を有するDNA鎖上の配列領域であって、RNA転写物の3’〜3’末端への領域は「下流配列」と呼ばれる。
【0060】
「天然に存在するヌクレオチド」という用語は、デオキシリボヌクレオチドおよびリボヌクレオチドを含む。「修飾されたヌクレオチド」という用語は、修飾または置換された糖基などを有するヌクレオチドを含む。「オリゴヌクレオチド連結」という用語は、オリゴヌクレオチド連鎖、例えば、ホスホロチオエート、ホスホロジチオエート、ホスホロセレノエート、ホスホロジセレノエート、ホスホロアニロチオエート、ホスホルアニラデート、ホスホロアミデートなどを含む。例えば、LaPlancheら、1986、Nucl.Acids Res.14:9081;Stecら、1984,J.Am.Chem.Soc.106:6077;Steinら、1988,Nucl.Acids Res.16:3209;Zonら、1991,Anti−Cancer Drug Design 6:539;Zonら、1991,OLIGONUCLEOTIDES AND ANALOGOUES:A PRACTICAL APPROACH、87〜108頁(F.Eckstein編)、Oxford University Press、Oxford England);Stecら、米国特許第5,151,510号;UhlmannおよびPeyman、1990、 Chemical Reviews 90:543(これらの開示は任意の目的のために参考として本明細書に援用される)を参照のこと。オリゴヌクレオチドは、オリゴヌクレオチドまたはそのハイブリダイゼーションの検出を可能にする検出可能な標識を含んでもよい。
【0061】
「ベクター」という用語は、宿主細胞に対してコード情報を移入するために用いられる任意の分子(例えば、核酸、プラスミドまたはウイルス)をいうために、使用される。
【0062】
「発現ベクター」という用語は、宿主細胞の形質転換に適切であって、挿入された異種核酸配列の発現を指向および/または制御する核酸配列を含むベクターをいう。発現としては、これらに限定されないが、転写、翻訳およびイントロンが存在する場合、RNAスプライシングのようなプロセスが挙げられる。
【0063】
「宿主細胞」という用語は、核酸配列を導入されているか、または核酸配列を導入されることが可能な細胞であって、さらに目的の選択された遺伝子を発現するかまたは発現可能である細胞をいうために使用される。この用語は、親の細胞の子孫を、この子孫が形態学的に、またはもとの親に対する遺伝的作製において同一であるか否かにかわらず、この選択された遺伝子が存在する限り、包含する。
【0064】
「形質導入」という用語は、通常は、ファージによる1つの細菌から別の細菌への遺伝子の移入をいうために使用される。「形質導入」とはまた、レトロウイルスによる、真核生物細胞配列の獲得および移入をいう。
【0065】
「トランスフェクション」という用語は、細胞による外因性または内因性のDNAの取り込みをいうために使用され、細胞は、外因性のDNAがその細胞膜の内側に導入されている場合、「トランスフェクト」されている。多数のトランスフェクション技術が当該分野で周知であり、本明細書に開示されている。例えば、Grahamら、1973,Virology 52:456;Sambrookら,2001,MOLECULAR CLONING,A LABORATORY MANUAL,Cold Spring Harbor Laboratories;Davisら,1986,BASIC METHODS IN MOLECULAR BIOLOGY,Elsevier;およびChuら,1981,Gene 13:197を参照のこと。このような技術は、1つ以上の外因性DNA部分を適切な宿主細胞に導入するために用いることができる。
【0066】
「形質転換」という用語は、本明細書中で用いる場合、細胞の遺伝的特徴における変化をいい、細胞が新しいDNAを含むように改変されている場合、形質転換されている。例えば、細胞が天然の状態から遺伝的に改変されている場合、形質転換されている。トランスフェクションまたは形質導入の後、この形質転換DNAは、細胞の染色体への物理的な組みこみによって細胞のDNAと組み換えられてもよいし、または複製されることのないエピソームエレメントとして一時的に維持されてもよいし、またはプラスミドとして独立して複製されてもよい。細胞の分裂とともにDNAが複製される場合、細胞は安定に形質転換されていると考えられる。
【0067】
「天然に存在する」または「天然の」という用語は、核酸分子、ポリペプチド、宿主細胞などのような生物学的物質に関連して用いられる場合、天然に見出され、人によって操作されていない物質をいう。同様に、「天然に存在しない」または「非天然の」とは、本明細書において用いる場合、天然に見出されないか、または人によって構造的に改変されているかもしくは合成されている物質をいう。
【0068】
「抗原」という用語は、選択的な結合因子、例えば抗体によって結合され得、そしてさらに動物において使用され、その抗原のエピトープに対して結合できる抗体を生成することができる分子または分子の一部をいう。抗原は、1つ以上のエピトープを有してもよい。
【0069】
「エピトープ」という用語は、免疫グロブリンまたはT細胞レセプターに特異的に結合し得る任意の決定基、好ましくはポリペプチド決定基を含む。エピトープは、抗体によって結合される抗原の領域である。特定の実施形態では、エピトープ決定基としては、分子の化学的に活性な表面の基、例えば、アミノ酸、糖側鎖、ホスホリルまたはスルホニルが挙げられ、特定の実施形態では、特定の三次元構造特徴および/または特異的な荷電特徴を有してもよい。特定の実施形態では、抗体は、それが好ましくはその標的抗原を、タンパク質および/または高分子の複雑な混合物中で認識する場合、抗原に特異的に結合すると言われる。抗体は、平衡解離定数が10−7または10−8M以下である場合、抗原に特異的に結合すると言われる。いくつかの実施形態では、平衡解離定数は、10−9M以下または10−10M以下であってもよい。
【0070】
本明細書中で用いる場合、第一の配列が、例えば、配列RASQSVSSSY(配列番号56)という10のアミノ酸からなる場合、別の配列は、7アミノ酸が、その配列におけるアミノ酸と同一であり、それらが配列に存在するのと同じ相対的な位置で存在する場合、7アミノ酸を、この第一の配列にそれが存在するのと「同じ順序かつ間隔で」有する。例えば、配列RAAAAVSSSY(配列番号57)は、7アミノ酸をRASQSVSSSY(配列番号56)に存在するのと同じ順序かつ間隔で有する。対照的に、これは、配列RASSVSSSY(配列番号58)にはあてはまらない。なぜなら、この配列はRASQSVSSSY(配列番号56)に対して内部欠失を含み、これが、この欠失のいずれかの側に3および6アミノ酸を有するからである。従って、この配列は、第一の配列と同じ順序かつ間隔で、最大6アミノ酸を有する。RASQSVSSSY(配列番号56)と同じ順序でかつ間隔で7アミノ酸を有する可能性のある最短の可能な配列は、7アミノ酸長、例えば、SQSVSSS(配列番号59)である。
【0071】
「同一性」という用語は、当該分野で公知であるとおり、2つ以上のポリペプチド分子または2つ以上の核酸分子の配列の間の関係であって、その配列を比較することによって決定される関係をいう。当該分野では、「同一性」とはまた、核酸分子またはポリペプチドの間の配列の関連性の程度を意味し、場合によっては、2つ以上のヌクレオチドまたは2つ以上のアミノ酸配列の文字列の間の適合によって決定されてもよい。「同一性」とは、特定の数学的モデルまたはコンピュータープログラム(すなわち、「アルゴリズム」)によってアドレスされるギャップアラインメント(もしあれば)と2つ以上の配列のうちより小さい配列の間での同一性適合の割合を測定する。
【0072】
「類似性」という用語は、関連する概念に関して当該分野で用いられるが、「同一性」とは対照的に、「類似性」は、同一性適合および保存的置換適合の両方を含む、関連性の指標をいう。2つのポリペプチド配列が、例えば、アミノ酸20個のうち10個の同一アミノ酸を有し、この残りが全て非保存的置換であれば、同一性パーセントおよび類似性パーセントは、両方とも50%である。同じ例で、保存的置換がさらに5つの位置にあれば、同一性パーセントは、50%のままで、ただし類似性パーセントは、75%である(15/20)。従って、保存的置換がある場合、2つのポリペプチドの間の類似性パーセントは、これらの2つのポリペプチドの間の同一性パーセントよりも高い。
【0073】
関連の核酸およびポリペプチドの同一性および類似性は、公知の方法によって容易に算出され得る。このような方法としては、COMPUTATIONAL MOLECULAR BIOLOGY,(Lesk,A.M.編),1988,Oxford University Press,New York;BIOCOMPUTING:INFORMATICS AND GENOME PROJECTS,(Smith,D.W.編),1993,Academic Press,New York;COMPUTER ANALYSIS OF SEQUENCE DATA,Part 1,(Griffin,A.M.およびGriffin、H.G.編),1994,Humana Press,New Jersey;von Heinje,G.,SEQUENCE ANALYSIS IN MOLECULAR BIOLOGY,1987,Academic Press;SEQUENCE ANALYSIS PRIMER,(Gribskov,M.およびDevereux,J.,編),1991,M.Stockton Press,New York;Carilloら、1988,SIAM J.Applied Math.,48:1073;ならびにDurbinら,1998,BIOLOGICAL
SEQUENCE ANALYSIS,Cambridge University Pressに記載される方法が挙げられるがこれらに限定されない。
【0074】
同一性を決定するために好ましい方法は、試験された配列の間で最大の適合を得るように設計される。同一性を決定する方法は、公的に利用可能なコンピュータープログラムに記載される。2つの配列の間の同一性を決定する好ましいコンピュータープログラム法としては、GAPを含むGCGプログラムパッケージ(Devereuxら,1984,Nucl.Acid.Res.,12:387;Genetics Computer Group,University of Wisconsin,Madison,WI),BLASTP,BLASTN、およびFASTA(Altschulら,1990,J.Mol.Biol.,215:403〜410)が挙げられるがこれらに限定されない。BLASTXプログラムは、National Center for Biotechnology Information(NCBI)および他の供給源(BLAST Manual,Altschulら、NCB/NLM/NIH Bethesda,MD
20894;Altschulら,1990(前出))から公的に入手可能である。周知のSmith Watermanアルゴリズムも、同一性を決定するのに用いることができる。
【0075】
2つのアミノ酸またはポリヌクレオチド配列の整列のための特定のアラインメントスキームによって、2つの配列の短い領域のみの適合を生じ得、そしてこの短い整列された領域は、2つの全長配列の間で顕著な関係がなくても極めて高い配列同一性を有し得る。従って、特定の実施形態では、選択されたアラインメント法(GAPプログラム)によって、標的ポリペプチドの少なくとも50個の連続アミノ酸に及ぶアラインメントを生じる。いくつかの実施形態では、このアラインメントは、標的ポリペプチドのうち少なくとも60、70、80、90、100、110または120アミノ酸を含み得る。ポリヌクレオチドがGAPを用いて整列される場合、このアラインメントは、連続し得る少なくとも約100、150、または200ヌクレオチドに及び得る。
【0076】
例えば、コンピューターアルゴリズムGAP(Genetics Computer Group,University of Wisconsin,Madison,WI)を用いて、配列同一性パーセントを決定すべき2つのポリペプチドを、そのそれぞれのアミノ酸の最適適合について整列させる(アルゴリズムによって決定される場合「適合したスパン(matched span)」)。特定の実施形態において、ギャップオープニングペナルティー(これは、3回の平均対角として計算される;ここで、「平均対角(average diagonal)」は、用いられる比較マトリックスの対角の平均であり;「対角(diagonal)」は、特定の比較マトリックスにより各々の完全アミノ酸適合に対して割り当てられた点数または数である)およびギャップ伸長ペナルティー(通常、ギャップオープニングペナルティーの十分の一)、ならびに比較マトリックス(例えば、PAM250またはBLOSUM 62)をこのアルゴリズムと組み合わせて用いる。特定の実施形態では、標準比較マトリックス(PAM 250比較マトリックスについては、Dayhoffら,1978,Atlas of Protein Sequence and Structure,5:345〜352;BLOSUM 62比較マトリックスについてはHenikoffら、1992,Proc.Natl.Acad.Sci.USA 89:10915〜10919を参照のこと)も、アルゴリズムによって用いられる。
【0077】
特定の実施形態では、ポリペプチド配列比較についてのパラメーターとしては以下が挙げられる:
アルゴリズム:Needlemanら,1970,J.Mol.Biol.48:443〜453;
比較マトリックス:BLOSUM 62(Henikoffら、1992、前出より);
ギャップペナルティー:12
ギャップ長ペナルティー:4
類似性閾値:0
GAPプログラムは、上記のパラメーターとともに有用であり得る。ヌクレオチド配列については、パラメーターは、50のギャップペナルティーおよび3のギャップ長ペナルティーを含み得、すなわち、各々のギャップにおける各記号について3のペナルティーである。特定の実施形態では、前述のパラメーターは、GAPアルゴリズムを用いるポリペプチド比較(エンドギャップについてのペナルティーを伴わない)のためのデフォルトパラメーターである。
【0078】
本明細書において用いる場合、20個の従来のアミノ酸およびその省略形は従来の用法に従う。IMMUNOLOGY−−A SYNTHESIS,第2版(E.S.GolubおよびD.R.Gren編),Sinauer Associates、Sunderland、MA,1991を参照のこと(これは、任意の目的のために参考として本明細書に援用される)。20個の従来のアミノ酸の立体異性体(例えば、Dアミノ酸);非天然のアミノ酸、例えば、α−、α−二置換アミノ酸、N−アルキルアミノ酸、乳酸、および他の従来とは異なるアミノ酸もまた、本発明のポリペプチドに適切な成分であり得る。従来とは異なるアミノ酸の例としては、以下が挙げられる:4−ヒドロキシプロリン、γ−カルボキシグルタメート、ε−N,N,N−トリメチルリジン、ε−N−アセチルリジン、O−ホスホセリン、N−アセチルセリン、N−ホルミルメチオニン、3−メチルヒスチジン、5−ヒドロキシリジン、σ−N−メチルアルギニン、ならびに他の同様のアミノ酸およびイミノ酸(例えば、4−ヒドロキシプロリン)。本明細書において用いられるポリペプチドの表記法においては、標準的な用法および慣習に従って、左手側の方向がアミノ末端方向であり、そして右手側の方向がカルボキシ末端方向である。
【0079】
天然に存在する残基は、共通の側鎖特性に基づいたクラスに分けることができる:
1)疎水性:ノルロイシン(Nor)、Met、Ala、Val、Leu、Ile;
2)中性の親水性:Cys、Ser,Thr、Asn,Gln;
3)酸性:Asp、Glu;
4)塩基性:His、Lys、Arg;
5)鎖の配向に影響する残基:Gly、Pro;および
6)芳香族:Trp、Tyr、Phe。
【0080】
保存的アミノ酸置換は、これらのクラスのうちの1つのメンバーを同じクラスの別のメンバーで交換することを包含し得る。保存的アミノ酸置換基は、天然には存在しないアミノ酸残基を含み得るが、これは代表的には、生物学的な系における合成ではなく化学的なペプチド合成によって取り込まれる。これらとしては、ペプチド模倣物およびアミノ酸部分の他の反転型または逆位型が挙げられる。
【0081】
非保存的置換としては、これらのクラスのうちの1つのメンバーの別のクラス由来のメンバーへの交換を挙げることができる。このような置換残基は、非ヒト抗体と相同であるヒト抗体の領域に導入されても、またはその分子の相同でない領域に導入されてもよい。
【0082】
このような変化を作製する工程において、特定の実施形態に従うと、アミノ酸の疎水性親水性指標が考慮され得る。各々のアミノ酸は、その疎水性および荷電の特徴に基づいて疎水性親水性指標を割り当てられている。それらは、以下である:イソロイシン(+4.5);バリン(+4.2);ロイシン(+3.8);フェニルアラニン(+2.8);システイン/シスチン(+2.5);メチオニン(+1.9);アラニン(+1.8);グリシン(−0.4);スレオニン(−0.7);セリン(−0.8);トリプトファン(−0.9);チロシン(−1.3);プロリン(−1.6);ヒスチジン(−3.2);グルタミン酸(−3.5);グルタミン(−3.5);アスパラギン酸(−3.5);アスパラギン(−3.5);リジン(−3.9);およびアルギニン(−4.5)である。
【0083】
タンパク質に相互作用的な生物学的機能を付与する工程における、アミノ酸の疎水性親水性指標の重要性は当該分野で理解されている(例えば、Kyteら,1982,J.Mol.Biol.157:105〜131を参照のこと)。特定のアミノ酸が類似の疎水性親水性指標またはスコアを有する他のアミノ酸によって置換可能であり、そしてなお類似の生物学的活性を保持することが、公知である。疎水性親水性指標に基づいてこのような変化を作製する際に、特定の実施形態では、その疎水性親水性指標が±2以内であるアミノ酸の置換が含まれる。特定の実施形態では、疎水性親水性指標が±1以内であるアミノ酸の置換が含まれ、そして特定の実施形態では、±0.5以内である置換が含まれる。
【0084】
特に、生物学的に機能的なタンパク質またはそれによって作製されるペプチドが、本明細書に開示されるように免疫学的な実施形態における使用を意図される場合、同様なアミノ酸の置換が親水性に基づいて有効になされ得ることもまた、当該分野で理解されている。特定の実施形態では、あるタンパク質の最大の局所平均親水性は、その隣接するアミノ酸の親水性によって支配されるので、そのタンパク質の免疫原性および抗原性と、すなわち生物学的特性と相関する。
【0085】
以下の親水性値がこれらのアミノ酸残基に割り当てられている:アルギニン(+3.0);リジン(+3.0);アスパラギン酸(+3.0±1);グルタミン酸(+3.0±1);セリン(+0.3);アスパラギン(+0.2);グルタミン(+0.2);グリシン(0);スレオニン(−0.4);プロリン(−0.5±1);アラニン(−0.5);ヒスチジン(−0.5);システイン(−1.0);メチオニン(−1.3);バリン(−1.5);ロイシン(−1.8);イソロイシン(−1.8);チロシン(−2.3);フェニルアラニン(−2.5)およびトリプトファン(−3.4)。同様の親水性値に基づく変化を作製する工程において、特定の実施形態では、その親水性値が±2以内であるアミノ酸の置換が含まれ、特定の実施形態では、疎水性親水性指標が±1以内であるアミノ酸の置換が含まれ、そして特定の実施形態では、±0.5以内である置換が含まれる。親水性に基づいて一次アミノ酸配列からエピトープを同定することができる。これらの領域はまた、「エピトープコア領域(epitopic core regions」とも呼ばれる。
【0086】
例示的なアミノ酸置換を表1に示す。
【0087】
(表1)
【0088】
【表1−1】

【0089】
【表1−2】
当業者は、周知の技術を用いて本明細書に示すポリペプチドの適切な改変体を決定することができる。特定の実施形態では、当業者は、活性に重要であるとは考えられない領域を標的することによって活性を破壊することなく変化され得る、分子の適切な領域を同定することができる。他の実施形態では、当業者は、同様のポリペプチドの間で保存されている分子の残基および部分を同定することができる。さらなる実施形態では、生物学的活性のためにまたは構造のために重要であり得る領域でさえ、生物学的活性を破壊することなく、ポリペプチド構造に有害に影響することもなく、保存的なアミノ酸置換に供することができる。
【0090】
さらに当業者は、活性または構造に重要な類似のポリペプチドにおける残基を同定する構造−機能の研究を概説することができる。このような比較の観点では、当業者は、類似のタンパク質における活性または構造について重要なアミノ酸残基に相当する、タンパク質中のアミノ酸残基の重要性を推測することができる。当業者は、このように推測された重要なアミノ酸残基に対する化学的に類似のアミノ酸置換基を選択することができる。
【0091】
当業者はまた、三次元構造およびアミノ酸配列を、類似のポリペプチドにおけるその構造に関して解析することができる。このような情報の観点では、当業者は、抗体のアミノ酸残基のアラインメントをその三次元構造に関して推測することができる。特定の実施形態では、当業者は、タンパク質の表面にあることが推定されるアミノ酸残基に対してラジカルな変化を作製しないように選択し得る。なぜなら、このような残基は、他の分子との重要な相互作用に関与し得るからである。さらに、当業者は、各々の所望のアミノ酸残基で単一のアミノ酸置換を含む試験改変体を生成することができる。次いで、この改変体を、当業者に公知の活性アッセイを用いてスクリーニングし得る。このような改変体を用いて、適切な改変体についての情報を集めることができる。例えば、特定のアミノ酸残基に対する変化によって、活性の破壊、所望されない減少または不適切な活性を生じるということが発見されている場合、このような変化を有する改変体は、回避され得る。言い換えれば、このような慣用的実験から収集された情報に基づいて、当業者は、さらなる置換単独、または他の変異と組み合わせた置換が回避されるべきアミノ酸を容易に決定することができる。
【0092】
二次構造の予測に対して多数の化学的刊行物がでている。例えば、Moult,1996,Curr.Op.in Biotech.7:422〜427;Chouら,1974,Biochemistry 13:222〜245;Chouら,1974,Biochemistry 113:211〜222;Chouら,1978,Adv.Enzymol.Relat.Areas Mol.Biol.47:45〜148;Chouら、1979,Ann.Rev.Biochem.47:251〜276;およびChouら、1979,Biophys.J.26:367〜384を参照のこと。さらに、二次構造を予測することを補助するコンピュータープログラムが現在利用可能である。二次構造を予測する1方法は、ホモロジー(相同性)モデリングに基づく。例えば、30%より大きい配列同一性または40%より大きい配列類似性を有する2つのポリペプチドまたはタンパク質はしばしば、類似の構造的位相学を有する。タンパク質構造データベース(PDB)の近年の成長によって、ポリペプチドの構造またはタンパク質の構造内の潜在的な折り畳み回数を含む二次構造の予想性の向上が得られた。Holmら、1999,Nucl.Acid.Res.27:244〜247を参照のこと。所定のポリペプチドまたはタンパク質においては折り畳み回数は限定されていること、および一旦、構造の臨界数が解明されれば、構造的な予測は劇的にさらに正確になることが示唆されている(Brennerら,1997,Curr.Op.Struct.Biol.7:369〜376)。
【0093】
二次構造を予測するさらなる方法としては、「スレッディング(threading)」(Jones,1997,Curr.Opin.Struct.Biol.7:377〜87;Sipplら,1996,Structure 4:15〜19)、「プロフィール解析」(Bowieら,1991,Science 253:164〜170;Gribskovら,1990,Meth.Enzym.183:146〜159;Gribskovら,1987,Proc.Nat.Acad.Sci.84:4355〜4358)および「進化的連鎖(evolutionary linkage)」(Holm,1999,前出;およびBrenner,1997,前出を参照のこと)が挙げられる。
【0094】
特定の実施形態によれば、アミノ酸置換は:(1)タンパク質分解に対する感受性を低下するか、(2)酸化に対する感受性を低下するか、(3)タンパク質複合体を形成するための結合親和性を変更するか、(4)結合親和性を変更するか、そして/または(5)このようなポリペプチドに他の物理化学的特性または機能的特性を付与または改変する、アミノ酸置換である。特定の実施形態によれば、単一または複数のアミノ酸置換(特定の実施形態では、保存的アミノ酸置換)は、天然に存在する配列において(特定の実施形態では、分子間接触を形成するドメイン(単数または複数)の外側のポリペプチドの一部において)作製されてもよい。好ましい実施形態では保存的アミノ酸置換は代表的には、親の配列の構造的特徴を実質的に変化しない(例えば、置換アミノ酸は、親の配列に存在するヘリックスを破壊する傾向でも、親の配列を特徴付ける他のタイプの二次構造を破壊する傾向でもあってはならない)。当該分野で認識されるポリペプチドの二次構造および三次構造の例は、各々が参考として本明細書に援用される、PROTEINS、STRUCTURES AND MOLECULAR PRINCIPLES(Creighton編)、1984,W.H.Freeman and Company、New York;INTRODUCTION TO PROTEIN STRUCTURE(C.BrandenおよびJ.Tooze編),1991,Garland Publishing、New York、N.Y.;ならびにThorntonら,1991,Nature
354:105に記載されている。
【0095】
ペプチドアナログは一般に、鋳型のペプチドの特性と類似の特性を有する非ペプチド薬として製薬産業において用いられる。これらのタイプの非ペプチド化合物は、「ペプチド模倣物(peptide mimetics)」または「ペプチド模倣物(peptidomimetics)」と名付けられる。任意の目的のために参考として本明細書に援用される、Fauchere、1986,Adv.Drug Res.15:29;VeberおよびFreidinger,1985,TINS p.392;ならびにEvansら、1987,J.Med.Chem.30:1229)を参照のこと。このような化合物はしばしば、コンピューターによる分子モデリングの補助によって開発される。治療上有用なペプチドに対して構造的に類似であるペプチド模倣物を用いて、同様の治療効果または予防効果を得ることができる。一般には、ペプチド模倣物は、パラダイム(paradigm)ポリペプチド(すなわち、生化学的な特性または生理学的な活性を有するポリペプチド)、例えばヒト抗体に対して構造的に類似であるが、当該分野で周知の方法によって、以下から選択される連結により必要に応じて置換される1つ以上のペプチド連結を有する:−CH2NH−、−CH2S−、−CH2−CH2−、−CH=CH−(シスおよびトランス)、−COCH2−、−CH(OH)CH2−、および−CH2SO−。同じタイプのDアミノ酸を有するコンセンサス配列の1つ以上のアミノ酸の系統的な置換(例えば、L−リジンの代わりに、D−リジン)を用いて、特定の実施形態では、さらに安定なペプチドを生成することができる。さらにコンセンサス配列または実質的に同一のコンセンサス配列のバリエーションを含む拘束されたペプチドは、当該分野で公知の方法(任意の目的のために参考として本明細書に援用される、RizoおよびGierasch,1992,Ann.Rev.Biochem.61:387)によって;例えば、ペプチドを環化する分子間ジスルフィド架橋を形成し得る内部システイン残基を付加することによって、生成することができる。
【0096】
本明細書において用いる場合、「抗体(antibody)」または「抗体ペプチド(antibody peptid)(単数または複数)」という用語は、1つ以上のポリペプチド鎖を含む単量体または多量体のタンパク質をいう。抗体は、抗原に特異的に結合し得、そして抗原の生物学的活性を阻害または調節し得るかもしれない。「抗体」とは、以下に記載される、天然に存在する抗体を包含する。特定の実施形態では、抗体は、組み換えDNA技術によって生成される。さらなる実施形態では、抗体は、天然に存在する抗体の酵素的切断もしくは化学的切断によって、産生される。抗体としては、限定はしないが、F(ab)、F(ab’)、F(ab’)2、Fv、および単鎖Fvフラグメント、ならびに単鎖抗体、キメラ抗体、ヒト化抗体、完全ヒト抗体、ポリクローナル抗体およびモノクローナル抗体が挙げられる。最低限、抗体とは、本明細書において意味する場合、軽鎖または重鎖可変領域の全てまたは一部を含む抗原に特異的に結合し得るポリペプチドを包含する。
【0097】
可変領域は、フレームワーク領域内(Kabatら(前出)によってフレームワーク領域1−4、FR1、FR2、FR3、およびFR4と命名された;ChothiaおよびLesk(前出)も参照のこと)に組み込まれた、少なくとも3つの重鎖または軽鎖相補性決定領域(CDR、Kabatら、1991,Sequences of Proteins of Immunological Interest,Public Health Service N.I.H.,Bethesda、MDによってCDR1、CDR2およびCDR3と命名された超可変領域としても公知;ChothiaおよびLesk,1987、J.Mol.Biol.196:901〜17;Chothiaら、1989,Nature 342:877〜883も参照のこと)を含む。このCDRおよびフレームワークセグメントは、可変領域のアミノ末端に始まって以下のように分散される:FR1−CDR1−FR2−CDR2−FR3−CDR3−FR4。
【0098】
抗体可変領域のフレームワーク領域の一次配列は、門を横切って普遍的に保存されている少量の残基を有する。しかし、多くの残基は、門を横切って、ならびに/または種および/もしくは門のなかで、高度に保存されており、そして抗体内の多くの位置が通常アミノ酸の公知の群の1つによって占有される。Kabatら(前出)を参照のこと。あるいは、配列はその予測される三次構造によって抗体として認識され得る。グリーク・キーβバレル(Greek key β barrel)として公知の構造を形成する9つのβ鎖を含む、可変領域の三次構造は、極めてよく保存されており、この構造内のCDRの位置はまた高度に保存されている。例えば、Borkら,1994,J.Mol.Biol.242:309〜20;HunkapillerおよびHook,1989,Adv.Immunol.44:1〜63;WilliamsおよびBarclay,1988,Ann.Rev.Immunol.6:381〜405;ChothiaおよびLesk(前出);Kabatら(前出)を参照のこと。
【0099】
三次構造は、種々のコンピュータープログラム、例えば、GENEFOLD(登録商標)(Tripos,Inc.,St.Louis,MO;GodzikおよびSkolnik,1992,Proc.Natl.Acad.Sci.USA 89:12098〜12102;Godzikら,1992,J.Mol.Biol.227:227〜38;Godzikら,1993,Protein Engng.6:801〜10)、Protein Data Bank(PDB)の代表的構造に問い合わせタンパク質配列を重ねるプロテインスレッディングプログラム(protein threading program)(Bermanら、2000,Nucleic Acids Res 28:235〜242;Jaroszewskiら,1998,Prot Sci 7:1431〜1440)などによって予測できる。新規なアミノ酸配列を分類するためにGENEFOLD(登録商標)を用いるために、この配列をプログラムに入れて、これによってGENEFOLD(登録商標)データベースに存在する以前に公知のタンパク質構造(「鋳型」構造)上にそれがどのように折り畳まれるかを反映する確率スコアを割り当てる。スコア付けについて、GENEFOLD(登録商標)は、一次アミノ酸配列の類似性、残基のベリアルパターン、局所相互作用および二次構造比較に依存する。GENEFOLD(登録商標)プログラムは、多数の抗体についての解明された構造を含む、タンパク質折り畳みの既存のデータベースにおける全てのテンプレート構造にアミノ酸配列を折り畳む(またはスレッドする)。GENEFOLD(登録商標)の出力は、その三次構造が、入力のアミノ酸配列によって考えられる可能性が最も高い、このデータベース内に由来するタンパク質の3つのリストである。この3つのリストは、(i)配列のみ、(ii)配列と局所高次構造優先度とベリアル(burial)期間、および(iii)配列と局所高次構造優先度とベリアル期間と二次構造、に基づいて算出された3つの異なるスコアを含む。各々の場合に、プログラムによって最適アラインメントを決定し、アラインメントのこの程度が偶然によって生じる確率(P値)を算出して、あり得る最高のスコアである999.9(9.999×102)のスコアとしてP値の逆を報告する。従って、最高のスコアによって、このアラインメントが偶然生じる可能性が最低であることが示される。従ってこれらのスコアは、新規なタンパク質が種々の参照構造にマッチする程度を反映しており、タンパク質の公知のファミリーにおけるメンバーに対して新規なタンパク質を割り当てるのに有用である。例えば、抗体可変領域の構造を有する配列は、合理的に高いP値、例えば少なくとも約200、300、400、500、600、700、800またはそれ以上を有する少なくとも1つの公知の可変領域と整列されることが期待される。
【0100】
「重鎖」という用語は、IFN−γに対する結合特異性を付与するのに十分な可変領域配列を有する任意の免疫グロブリンポリペプチドを包含する。「軽鎖」という用語は、IFN−γに対する結合特異性を付与するのに十分な可変領域配列を有する任意の免疫グロブリンポリペプチドを包含する。このような重鎖または軽鎖は、それが重鎖の場合、軽鎖の、またはそれが軽鎖の場合、重鎖の非存在下でIFN−γに結合し得るが、その必要はない。全長重鎖は、可変領域ドメイン、VHおよび3つの定常領域ドメインCH1、CH2およびCH3を含む。このVHドメインは、ポリペプチドのアミノ末端であり、CH3ドメインは、カルボキシ末端である。「重鎖」という用語は、本明細書において用いる場合、全長重鎖およびそのフラグメントを包含する。全長軽鎖は、可変領域ドメインVL、および定常領域ドメインCLを含む。重鎖と同様、軽鎖の可変領域ドメインは、ポリペプチドのアミノ末端である。「軽鎖」という用語は、本明細書において用いる場合、全長軽鎖およびそのフラグメントを包含する。F(ab)フラグメントは、1つの軽鎖および1つの重鎖のCH1および可変領域からなる。F(ab)分子の重鎖は、別の重鎖分子とジスルフィド結合を形成できない。F(ab’)フラグメントは、1つの軽鎖および1つの重鎖を含み、この重鎖がCH1ドメインとCH2ドメインとの間にさらに定常領域を含み、この結果鎖間ジスルフィド結合が2つの重鎖の間で形成されてF(ab’)2分子が形成され得る。このFv領域は、重鎖および軽鎖の両方由来の可変領域を含むが、定常領域は欠く。単鎖抗体は、Fv分子であり、ここでは重鎖および軽鎖の可変領域が可撓性のリンカーによって接続されて、抗原結合領域を形成する単鎖ポリペプチドを形成している。単鎖抗体は国際特許出願公開番号WO88/01649、ならびに米国特許第4,946,778号および同第5,260,203号に詳細に考察されている。
【0101】
本発明はまた、完全ヒト抗体、ヒト化抗体およびキメラ抗体を包含する。本明細書において意味するように、完全ヒト抗体は、最終的にはヒト由来のポリヌクレオチドのみによってコードされるアミノ酸配列、またはこのような配列に同一であるアミノ酸配列を含む。本明細書において意味するとおり、トランスジェニックマウスにおいて生成されるマウスゲノムに挿入されたヒト免疫グロブリンコードDNAによってコードされる抗体は、完全ヒト抗体である。なぜなら、それらは最終的にヒト由来であるDNAによってコードされるからである。この状況では、ヒト免疫グロブリンコードDNAは、マウス内で(抗体をコードするように)再配列され得、そして体細胞変異がまた生じ得る。マウスにおいてこのような変化を受けている、もともとヒトDNAによってコードされる抗体は、本明細書において意味するとおり、完全ヒト抗体である。このようなトランスジェニックマウスの使用によって、ヒト抗原に対する完全ヒト抗体を選択することが可能になる。事実上、これはほとんどの場合不可能である。なぜなら、自己抗原に対するヒト免疫応答は、正常には生じないからである。当業者は、完全ヒト抗体が治療剤としての使用に、特に慢性疾患を処置するために有利であることを理解する。なぜなら完全ヒト抗体は、それ自体に対する免疫応答を引き起こす可能性が低いからである。対照的に、多くの非ヒト抗体は、ヒトで用いられた場合、それ自体に対する免疫応答を引き起こすことが公知であり、この状況によってヒトでのこのような抗体の慢性使用は勧められない。従って完全ヒト抗体は、ヒトの疾患を含む慢性状態を処置するための抗体を用いるのに直面した長年にわたる問題を解決する。例えば、Billiau,1988,Immunol.Today 9:37〜40;Horneffら,1991,Clin.Immunol.& Immunopathol.59:89〜103;Tjandraら,1990,Immunol &
Cell Biol.68:367〜76を参照のこと。従って、完全ヒトIFNγ抗体は特に、慢性ヒトIFNγ媒介性疾患、例えば自己免疫疾患の処置に十分に適している。
【0102】
ヒト化抗体では、CDRを除いて抗体全体が、ヒト由来のポリヌクレオチドでコードされているか、またはそのCDR内を除いてこのような抗体と同一である。非ヒト生物体に由来する核酸によってコードされるCDRは、ヒト抗体可変領域のβシートフレームワークに接合されて、抗体が作製され、その特異性はこの接合されたCDRによって決定される。このような抗体の作製は、例えば、WO92/11018,Jonesら,1986,Nature 321:522〜25,Verhoeyenら,1988,Science 239:1534〜36に記載される。この研究によって、抗原結合部位の形成におけるCDRの中心的な重要性が裏付けられる。キメラ抗体は、ヒト定常領域(ヒト由来のポリヌクレオチドによってコードされるか、またはこのようなヒト定常領域に同一である)および非ヒト可変領域を含む。このような抗体の作製は、例えば、米国特許第5,681,722号に記載される。
【0103】
特定の実施形態では、「多価特異的(multispecific)」抗体または「多機能性(multifunctional)」抗体以外の二価抗体は、同一の抗原特異性を有する結合部位を含むことが理解される。
【0104】
本発明に従って抗体結合および特異性を評価することにおいて、抗体は、IFN−γに特異的に結合するか、そして/または過剰の抗体がIFN−γに対して結合するそのレセプターの量を(または逆も同様であるが)少なくとも20%、40%、60%、80%、85%もしくはそれ以上まで(インビトロ競合結合アッセイで測定した場合)低下させる場合、IFN−γのそのレセプターへの結合を実質的に阻害する。特異的に結合する抗体は、IFN−γに対する結合について、10−8モル以下、状況に応じて10−9または10−10モル以下という平衡解離定数を有することが予想され得る。
【0105】
治療用途のために、抗IFN−γ抗体の重要な特徴は、IFN−γの生物学的活性を阻害し得るか、または調節し得るかである。IFN−γは多くの異なる生物学的効果を有し、それは異なる細胞タイプにおいて多くの異なるアッセイで測定できる。抗IFN−γ抗体がIFN−γの生物学的活性を阻害または調節する能力は、以下の実施例6に記載のA549アッセイを用いて、またはIFN−γの存在下で観察される細胞増殖の阻害を逆転する抗体の能力を測定する同様のアッセイを用いて測定することができる。意味のある結果を得るためのアッセイについては、アッセイで用いられる細胞の増殖は、このアッセイで用いられるIFN−γによって阻害されなければらなない。ヒトIFN−γは、A549細胞を含むいくつかの細胞タイプの増殖を阻害し得る(実施例6および7)。マウスIFN−γは、RAW 264.7細胞の増殖を阻害し得る(実施例7)が、A549細胞は阻害し得ない。ある抗体が非ヒトIFN−γの生物学的活性を阻害または調節する能力を試験する場合特に、A549細胞以外の細胞タイプを用いてもよい。なぜなら、非ヒトIFN−γは、A549細胞の増殖を阻害できるかもしれないし、阻害できないかもしれないからである。抗原に特異的に結合する抗体全てが、その正常なレセプターに対する抗原結合をブロックすることが可能なわけではなく、従ってそのレセプターに対する結合の際に抗原の生物学的効果を阻害または調節することが可能なわけでもない。当該分野で公知のとおり、このような効果は、抗原のどの部分が抗体に結合するか、ならびに抗原および抗体(この場合、IFN−γおよび抗IFN−γ抗体)の絶対的濃度および相対的濃度の両方に依存し得る。本明細書において意味するとおり、IFN−γの生物学的活性を阻害または調節できると考えられるには、A549アッセイ(実施例6)または同様のアッセイにおいて蛍光で測定する場合、IFN−γの濃度がある範囲内、例えば、およそEC80またはEC90であるとき、少なくとも約20%、40%、60%、80%、85%、100%以上まで、抗体は、IFN−γの存在下で観察される細胞増殖の阻害を逆転することができなければならず、この場合その生物学的活性を阻害する因子の効果が容易に明白に成り得る。EC80とは、本明細書において意味する場合、観察されるIFN−γの最大効果の80%に必要なIFN−γの量である。IFN−γ濃度がEC90を十分上回る場合、阻害因子の効果は、過剰のIFN−γに起因して明白でないかもしれない。IFN−γの生物学的活性を阻害または調節するのに必要な抗体の濃度は、広範に変化し得、そしてこの抗体がIFN−γにどの程度きつく結合するかに依存し得る。例えば、A549アッセイで生物学的活性を阻害または調節するのには、IFN−γの1分子あたり1分子以下の抗体で十分であり得る。いくつかの実施形態では、IFN−γ濃度がおよそEC50〜およそEC90である場合、IFN−γの生物学的活性を阻害または調節するためには、およそ2:1、1:1、1:2、1:4、1:6、1:8、1:10、1:20、1:40、1:60、1:100または1:50,000というIFN−γ抗体の比が必要であり得る。これらの値の間のIFN−γ抗体の比も可能である。
【0106】
さらなる実施形態では、抗体の改変体としては、改変されたFcフラグメントまたは改変された重鎖定常領域を含む抗体を挙げることができる。「結晶化するフラグメント(fragment that crystallizes)」を意味するFcフラグメント、または重鎖定常領域は、抗体に変化した特徴を付与する変異によって改変され得る。例えば、BurtonおよびWoof,1992,Advances in Immunology 51:1〜84;RavetchおよびBolland,2001,Annu.Rev.Immunol.19:275〜90;Sheildsら,2001,Journal of Biol.Chem.276:6591〜6604;TellemanおよびJunghans,2000,Immunology 100:245〜251;Madesanら,1998,Eur.J.Immunol.28:2092〜2100;その全てが本明細書において参考として援用される)を参照のこと。このような変異としては、置換、付加、欠失、またはその任意の組み合わせを挙げることが可能であり、そして代表的には、本明細書に記載の方法に従って、そして当該分野で公知の方法に従って1つ以上の変異原性オリゴヌクレオチド(単数または複数)を用いて部位特異性変異誘発によって生成される(例えば、本明細書において参考として援用される、Maniatisら、MOLECULAR CLONING:A LABORATORY MANUAL、第三版、2001,Cold Spring Harbor,N.Y.およびBergerおよびKimmel,METHODS IN ENZYMOLOGY,152巻、Guide to Molecular Cloning Techniques,1987,Academic Press,Inc.,San Diego,CA.,を参照のこと)。
【0107】
特定の実施形態では、抗体改変体としては、グリコシル化部位の数および/またはタイプが親のポリペプチドのアミノ酸配列に比べて変更されているグリコシル化改変体が挙げられる。特定の実施形態では、タンパク質改変体は、天然のタンパク質よりも多いかまたは少ない数のN連結グリコシル化部位を含む。N連結グリコシル化部位は、以下の配列:Asn−X−SerまたはAsn−X−Thrによって特徴付けられるが、ここでXと命名されたアミノ酸残基は、プロリン以外の任意のアミノ酸残基であってもよい。この配列を作製するためのアミノ酸残基の置換によって、N連結糖鎖の付加のための潜在的な新しい部位が得られる。あるいは、この配列を除く置換は、既存のN連結糖鎖を除去する。また、N連結糖鎖の再配列も得られるが、ここでは1つ以上のN連結グリコシル化部位(代表的には、天然に存在するもの)が、除かれて、1つ以上の新規なN連結部位が作製される。さらに好ましい抗体改変体としては、1つ以上のシステイン残基が、親のアミノ酸配列に比べて欠失されるかまたは別のアミノ酸(例えば、セリン)で置換されているシステイン改変体が挙げられる。抗体が、例えば、不溶性の封入体の単離後に生物学的に活性な高次構造に再折り畳みされなければならない場合に、システイン改変体が有用であり得る。システイン改変体は一般に、天然のタンパク質よりも少ないシステイン残基を有し、そして代表的には、不対のシステインから生じる相互作用を最小にするために偶数を有する。
【0108】
「因子(agent)」という用語は、本明細書において用いる場合、化合物、化合物の混合物、生物学的高分子、または生物学的物質から作製された抽出物を意味する。
【0109】
本明細書において用いる場合、「標識(label)」または「標識された(labeled)」という用語は、例えば、放射線標識されたアミノ酸の組み込み、または蛍光マーカーのポリペプチドもしくは核酸に対する結合、検出可能な活性を有する化学発光マーカーもしくは酵素、または標識されたアビジンによって検出可能であるビオチン部分のポリペプチドに対する結合による、検出可能なマーカー(例えば、とりわけ、光学的または比色定量的な方法によって検出可能である、蛍光マーカー、化学発光マーカーまたは酵素活性などの検出可能なマーカーを好ましくは含むストレプトアビジン)の組み込みをいう。特定の状況では、この標識はまた治療剤であってもよい。ポリペプチドおよび糖タンパク質を標識する種々の方法は、当該分野で公知であり、かつ本明細書に開示された方法において有利に用いることができる。ポリペプチドのための標識の例としては、以下が挙げられるがこれらに限定されない:放射性同位体または放射性核種(例えば、3H、14C、15N、35S、90Y、99mTc、111In、125I、131I)、蛍光標識(例えば、フルオレセインイソチオシアネートすなわちFITC、ローダミン、またはランタニド蛍光物質)、酵素標識(例えば、西洋ワサビペルオキシダーゼ、βガラクトシダーゼ、ルシフェラーゼ、アルカリホスファターゼ)、化学発光標識、ハプテン標識、例えば、ビオチニル基、および二次レポーターによって認識される予め決定されたポリペプチドエピトープ(例えば、ロイシンジッパー対配列、二次抗体の結合部位、金属結合ドメイン、またはエピトープタグ)。特定の実施形態において、標識は、種々の長さのスペーサーアーム(例えば、(CH2)n、ここでn<約20)によって結合されて、可能性のある立体障害を減らす。
【0110】
「生物学的サンプル(biological sample)」という用語は、本明細書において用いる場合、生存物または以前に生存していた物由来の任意の量の物質を包含するがこれらに限定されない。このような生存している物としては、ヒト、マウス、サル、ラット、ウサギおよび他の動物が挙げられるがこれらに限定されない。このような物質としては、血液、血清、尿、細胞、器官、組織、骨、骨髄、リンパ節および皮膚が挙げられるがこれらに限定されない。
【0111】
「薬学的因子または薬物(pharmaceutical agent or drug)」という用語は、本明細書において用いる場合、患者に適切に投与される場合、所望の治療効果を誘導し得る化合物または組成物をいう。
【0112】
「IFN−γ媒介性疾患(IFN−γ mediated disease)」という用語は、炎症性疾患、感染性疾患、および自己免疫疾患を包含するがこれらに限定されない。本明細書において用いる場合、「自己免疫疾患」とは、患者の免疫応答が患者自身の構成要素に向かう疾患状態および条件をいう。例えば、IFN−γ媒介性疾患としては、後天性免疫不全症候群(AIDS)、若年性関節リウマチを含む関節リウマチ、潰瘍性大腸炎およびクローン病を含む炎症性腸疾患、多発性硬化症、アジソン病、糖尿病(I型)、精巣上体炎、糸球体腎炎、グレーブス病、ギラン・バレー症候群、橋本病、溶血性貧血、全身性エリテマトーデス(SLE)、ループス腎炎、重症筋無力症、天疱瘡、乾癬、乾癬性関節症、アテローム性動脈硬化症、エリスロポエチン耐性、宿主対移植片病、移植片拒絶、自己免疫性肝炎誘導性肝障害、胆汁性肝硬変、アルコール肝硬変を含むアルコール誘導性肝障害、リウマチ熱、サルコイドーシス、強皮症、シェーグレン症候群、強直性脊椎炎を含む脊椎関節症、甲状腺炎、および脈管炎が挙げられるがこれらに限定されない。「IFN−γ媒介性疾患」という用語はまた、IFN−γのレベルの増大またはIFN−γの感度の増大に関連する任意の病状を包含する。
【0113】
自己免疫疾患を含むIFN−γ媒介性疾患の処置は、障害の少なくとも1つの症状の緩和、疾患の重篤度の低下、または処置された状態の後ある程度の頻度で生じるさらに重篤な疾患への進行の遅延もしくは予防を含む。処置とは疾患が完全に治癒することを意味する必要はない。有用な治療剤とは単に、ある疾患の重篤度を低下するか、この疾患またはその処置に伴う症状(単数または複数)の重篤度を低下するか、または患者のクオリティーオブライフを改善するか、または処置された状態の後、ある程度の頻度で生じ得るさらに重篤な疾患の発現を遅延することを必要とする。例えば、この疾患が関節リウマチである場合、治療剤は関節の腫脹を軽減するか、罹患する関節の数を減らすか、または骨の損失を遅延もしくは阻害し得る。SLE患者は、皮膚病変、熱、衰弱、関節炎、リンパ節腫脹、胸膜炎、心外膜炎および/または貧血などのような症状を有し得る。このような症状は、例えば、視覚的観察、写真、体温測定、握力もしくは関節サイズ、および/または赤血球の濃度を決定するための血液の顕微鏡検査を含む任意の多数の従来の技術によって評価できる。本発明は、IFN−γ媒介性疾患に罹患した患者に対して本発明のIFN−γ抗体を、特定の障害の重篤度もしくはこの障害によって生じる症状の重篤度を反映する指標の規準を上回って改善の維持を誘導するか、またはいくつかのもしくは全ての場合には処置される条件に伴うさらに重篤な疾患の発現を遅延もしくは予防するのに十分な量および時間で投与する工程を包含する、処置の方法を包含する。本発明は、IFN−γ抗体を用いた処置の前、後、および/または間の他の治療剤を用いた可能性のある処置を除外しない。
【0114】
本明細書において用いる場合、「実質的に純粋な(substantially pure)」または「実質的に精製された(substantially purified)」とは、化合物または種が存在する主な種であるである(すなわち、モル基準では、この組成物における任意の他の個々の種よりもそれが豊富である)ことを意味する。特定の実施形態では、実質的に精製された画分とは、この種が存在する全ての高分子種のうちの少なくとも約50パーセント(モル基準で)を含む組成物である。特定の実施形態では、実質的に純粋な組成物は、その組成物中に存在する全ての高分子種のうち約80%、85%、90%、95%または99%より多い。特定の実施形態では、この種は、本質的に均一まで精製され(混入する種は、従来の検出法ではこの組成物中で検出できない)、ここでこの組成物は本質的に、単一の高分子種からなる。
【0115】
「患者(patient)」という用語は、ヒトおよび獣医学の被験体を包含する。
【0116】
文脈が他を要することがない限り、単数の用語は複数を包含する。
【0117】
IFN−γは、ウイルス感染から身体を防御することおよび免疫応答のいくつかの局面を調節することを含む、複数の機能を有するサイトカインであり、IFN−γ活性の増大は、いくつかの生理学的な条件に寄与し得る。本発明の特定の実施形態によれば、IFN−γに対する抗体は、上述のものを含むがそれらに限定されない、IFN−γ媒介性疾患を処置するために用いられ得る。
【0118】
本発明の1局面では、ヒトIFN−γに対して惹起され、かつそれに対する特異的な結合のための生物学的および免疫学的な特異性を有する完全ヒトモノクローナル抗体が提供される。このような抗体に含まれる可変領域(配列番号6、配列番号8、配列番号10、配列番号12、配列番号14、配列番号16、配列番号30および配列番号31)は、このような抗体の完全な重鎖および軽鎖(配列番号17、配列番号18、配列番号19、配列番号20、配列番号21、および配列番号22)、そして特定のCDR(重鎖および軽鎖CDR1、CDR2、および/またはCDR3;配列番号34〜配列番号44)を含む抗体は、本発明に包含される。本発明のこの局面の特定の実施形態は、本発明によって提供される重鎖および軽鎖の、CDR、特にCDR1〜CDR3に対応する配列である。さらに、本発明は、本明細書に開示されるCDR3配列(配列番号36、配列番号37、配列番号43および/または配列番号44)を含む抗体を包含するが、この配列はまた、任意の可変領域配列または本明細書に開示される完全な重鎖もしくは軽鎖の配列に対して少なくとも80%、85%、90%、91%、92%、93%、94%、95%、96%、97%、98%もしくは約99%同一である配列を含んでもよく、この抗体は、IFN−γの生物学的活性を阻害または調節し得る。
【0119】
別の局面では、本発明は、本発明の抗体をコードする単離された核酸またはポリヌクレオチドを提供する。本発明の抗体は、IFN−γに特異的に結合するか、および/またはIFN−γの生物学的活性を阻害もしくは調節し得る。本発明により特異的に包含されるのは、アミノ酸配列である配列番号17、配列番号18、配列番号19、配列番号20、配列番号21、配列番号22、配列番号32、配列番号33、配列番号5、配列番号7、配列番号9、配列番号11、配列番号13、配列番号15、配列番号30、配列番号31および/またはこれらの配列に対して少なくとも80%、85%、90%、91%、92%、93%、94%、95%、96%、97%、98%もしくは約99%同一性である配列をコードするヌクレオチド配列を含むポリヌクレオチドであり、ここで上記の配列と試験配列との間のアラインメントは、少なくとも約50、60、70、80、90もしくは100アミノ酸にまたがる。本発明はさらに、配列番号45、配列番号46、配列番号47および/または配列番号48を含むポリヌクレオチドであって、IFN−γに特異的に結合するかおよび/またはその生物学的活性を阻害もしくは調節し得る抗体をコードするポリヌクレオチドを提供する。さらに、本発明は、配列番号5、配列番号7、配列番号9、配列番号11、配列番号13、配列番号15、配列番号30、配列番号31、配列番号17、配列番号18、配列番号19、配列番号20、配列番号21、配列番号22、配列番号32もしくは配列番号33に対して少なくとも80%、85%、90%、91%、92%、93%、94%、95%、96%、97%、98%もしくは約99%同一であるポリヌクレオチドを包含するが、ここでこれらのポリヌクレオチドの各々によって部分的にコードされる抗体は、IFN−γの生物学的を阻害もしくは調節するか、および/またはIFN−γに特異的に結合し得、すぐ上に挙げられたヌクレオチド配列と試験配列との間のアラインメントは、少なくとも100、150、または200ヌクレオチドにまたがる。
【0120】
表2には、その配列識別番号に関連付けて配列の簡単な説明を提供する。
【0121】
(表2)
【0122】
【表2−1】
【0123】
【表2−2】
さらに別の局面では、本発明は、本発明の免疫グロブリン分子および抗体、必要に応じてモノクローナル抗体を発現するハイブリドーマおよび細胞株を提供する。さらなる局面では、本発明の免疫グロブリン分子または抗体を発現および/または分泌する、ハイブリドーマ細胞または細胞株由来の細胞は、患者に移植され得、これによって本発明の抗体またはその免疫学的に機能的な免疫グロブリンフラグメントは、患者において発現および/または分泌されて、これによってIFN−γ活性が阻害または調節される。
【0124】
本発明はまた、生物学的におよび免疫学的に精製された抗体の調製物であって、好ましくはヒトIFN−γに対して惹起されて、それに対して特異的に結合するための生物学的および免疫学的特異性を有するモノクローナル抗体の調製物を提供する。
【0125】
酵母人工染色体(YAC)におけるメガベースサイズのヒト遺伝子座をクローニングおよび再構築して、マウス生殖系列にそれらを導入する能力によって、極めて大きいかまたはおおまかにマッピングされた遺伝子座の機能的成分を解明し、そしてヒト疾患の有用なモデルを生成するための有利なアプローチの開発が可能になる。さらに、ヒトの等価な産物によるマウス遺伝子座の置換のためのこのような技術の利用によって、発達の間のヒト遺伝子産物の発現および調節、他の系との関連、ならびに疾患誘導および進行におけるその関与に対する固有の洞察が得られる。
【0126】
このようなストラテジーの重要な実際的な適用は、内因性Ig遺伝子が不活性化されているマウスへのヒト免疫グロブリン(Ig)遺伝子座の導入によるマウス体液性免疫系の変更である。国際出願番号WO93/12227。この系によって抗体のプログラムされた発現およびアセンブリの背景にある機構、ならびにB細胞発達におけるその役割を研究する機会が得られる。さらに、このようなストラテジーによって、完全ヒトモノクローナル抗体(MAb)の産生の供給源が得られる。完全ヒトMAbは、マウスまたはマウス由来MAbに内在する免疫原性およびアレルギー性の応答を最小化すること、それによって投与された抗体の有効性および安全性を増大することが期待される。完全ヒト抗体を、慢性および再発性のヒト疾患、例えば、変形性関節炎、関節リウマチおよび他の炎症性状態の処置において用いることが可能であり、その処置には抗体投与を反復することが必要である。従って、本発明の抗IFN−γ抗体の1つの特定の利点は、この抗体が完全ヒト抗体であり、マウス抗ヒト抗体または他の以前に記載された不完全ヒト抗体もしくは非ヒト種由来の非ヒト抗体と一般に関連する有害反応を最小限にしながら非急性の方式で患者に投与できるということである。
【0127】
本明細書に記載される方法を用いて、当業者は、ヒトIg遺伝子座の大きいフラグメントを用いてマウス抗体産生において欠損するマウス系統を操作することが可能であり、その結果このようなマウスは、マウス抗体の非存在下でヒト抗体を生成する。それより大きいヒトIgフラグメントは、可変性の大きい遺伝子多様性、ならびに抗体産生および発現の適切な調節を保存し得る。抗体多様化および選択ならびにヒト抗体に対する免疫学的寛容の欠失のためにマウス細胞機構を開発することによって、これらのマウス系統において再生されたヒト抗体レパートリーにより、ヒト抗原を含む目的の任意の抗原に対する高親和性抗体を得る。ハイブリドーマ技術を用いて、所望の特異性を有する抗原特異的ヒトMAbを生成および選択することができる。
【0128】
特定の実施形態では、当業者は、ヒト以外の種由来の定常領域をヒト可変領域(単数または複数)とともにこのようなマウスで用いてキメラ抗体を産生することができる。
(天然に存在する抗体構築物)
最も天然に存在する抗体構築物単位は代表的には、四量体を含む。各々のこのような四量体は代表的には、ポリペプチド鎖の2つの同一の対から構成され、各々の対が1つの全長「軽」鎖(代表的には約25kDaの分子量を有する)および1つの全長「重」鎖(代表的には約50〜70kDaの分子量を有する)を有する。各々の軽鎖および重鎖のアミノ末端部分は代表的には、代表的に抗原認識を担う、約100〜110以上のアミノ酸の可変領域を含む。各々の鎖のカルボキシ末端部分は代表的には、エフェクター機能を担う、定常領域を規定する。ヒト軽鎖は代表的には、κ軽鎖およびλ軽鎖として分類される。重鎖は代表的には、μ、δ、γ、α、またはεとして分類され、そしてそれぞれIgM、IgD、IgG、IgA、およびIgEとして抗体のアイソタイプを規定する。IgGは、IgG1、IgG2、IgG3およびIgG4が挙げられるがこれらに限定されないいくつかのサブクラスを有する。IgMは、IgM1およびIgM2が挙げられるがこれらに限定されないサブクラスを有する。IgAは同様に、IgA1およびIgA2が挙げられるがこれらに限定されないサブクラスに分割される。全長の軽鎖および重鎖内では、代表的には可変領域および定常領域は、約12以上のアミノ酸の「J」領域によって連結され、ここでこの重鎖はまた、約10アミノ酸より多い「D」領域を含む。例えば、FUNDAMENTAL IMMUNOLOGY 第7章、第2版(Paul、W.編、1989,Raven Press、N.Y.(全ての目的のためにその全体が参考として援用される))を参照のこと。各々の軽鎖/重鎖対の可変領域は、代表的には、抗体結合部位を形成する。
【0129】
ラクダおよびラマに見出されているいくつかの天然に存在する抗体は、2つの重鎖からなる二量体であって、軽鎖を含まない。Muldermansら,2001,J.Biotechnol.74:277〜302;Desmyterら,2001,J.Biol.Chem.276:26285〜90。本発明は、IFN−γに結合するか、および/またはその生物学的活性を阻害し得る2つの重鎖からなる二量体抗体を包含する。ラクダ抗体の結晶学的研究によって、19アミノ酸長を有する重鎖CDR3が抗原と相互作用して2つの他の超可変領域をカバーする表面を形成することが明らかになっている。Desmyterら(前出)。従ってCDR3は、二量体ラクダ抗体において、そしてさらに代表的な四量体抗体において、抗原結合に重要である。
【0130】
可変領域は代表的には、相補性決定領域すなわちCDRとも呼ばれる、3つの超可変領域に連結された比較的保存されたフレームワーク領域(FR)の同じ一般的な構造を示す。各々の対の2つの鎖由来のCDRは代表的には、フレームワーク領域内に包埋され、これが特定のエピトープに結合し得る。N末端からC末端へと、軽鎖および重鎖の可変領域はともに、代表的には、ドメインFR1、CDR1、FR2、CDR2、FR3、CDR3およびFR4を含む。各々のドメインに対するアミノ酸の割り当ては代表的には、以下にさらに詳細に説明されているように、Kabatらの定義に従っている。Kabatら、Sequences of Proteins of Immunological Interest(1991,National Institutes of Health,Bethesda,Md);またChothiaおよびLesk,1987,J.Mol.Biol.196:901〜917;Chothiaら,1989,Nature 342:878〜883も参照のこと。CDRは、抗原結合のための主な表面接触ポイントを構成する。例えば、ChothiaおよびLesk(前出)を参照のこと。さらに軽鎖のCDR3および特に重鎖のCDR3は、軽鎖および重鎖の可変領域内の抗原結合における最も重要な決定基を構成し得る。例えば、ChothiaおよびLesk(前出);Desiderioら(2001),J.Mol.Biol.310:603〜15;XuおよびDavis(2000),Immunity 13(1):37〜45;Desmyterら(2001),J.Biol.Chem.276(28):26285〜90;ならびにMuyldermans(2001),J.Biotechnol.,74(4):277〜302を参照のこと。いくつかの抗体では、重鎖CDR3は、抗原と抗体との間の接触の主な領域を構成するようである。Desmyterら(前出)。CDR3のみを変化させるインビトロ選択スキームを用いることによって、抗体の結合特性を変化させることができる。Muyldermans(前出);Desiderio(前出)。
【0131】
CDRは、以下の方法で重鎖可変領域配列に配置されてもよい。CDR1は、成熟抗体のおよそ31残基で開始して、通常は約5〜7アミノ酸長であり、そしてこれには、ほぼ常にCys−Xxx−Xxx−Xxx−Xxx−Xxx−Xxx−Xxx−Xxx(配列番号48)が先行している(ここで、「Xxx」は任意のアミノ酸である)。重鎖CDR1の後ろの残基はほぼ常にトリプトファン、しばしばTyp−Val,Trp−IleまたはTrp−Alaである。CDR1の最後の残基とCDR2の最初の残基との間に14個のアミノ酸がほぼ常にあり、そしてCDR2は代表的には16〜19個のアミノ酸を含む。CDR2には、その直前にLeu−Glu−Trp−Ile−Gly(配列番号49)が先行してもよく、そしてその直後にLys/Arg−Leu/Ile/Val/Phe/Thr/Ala−Thr/Ser/Ile/Alaが続く。他のアミノ酸は、CDR2に先行しても続いてもよい。CDR2の最後の残基とCDR3の最初の残基との間に32個のアミノ酸がほぼ常にあり、そしてCDR3は、約3〜25残基長であり得る。Cys−Xxx−Xxxはほぼ常に、CDR3に先行しており、そしてTrp−Gly−Xxx−Gly(配列番号50)がほぼ常にCDR3に続く。
【0132】
軽鎖CDRは、以下の方法で軽鎖配列に配置されてもよい。CDR1は、成熟抗体のおよそ24残基で開始して、通常は約10〜17残基長であり、そしてこれには、ほぼ常にCysが先行している。CDR1の最終残基とCDR2の最初の残基との間にはほぼ常に15アミノ酸があり、CDR2はほぼ常に7残基長である。CDR2は代表的には、Ile−Tyr、Val−Tyr、Ile−LysまたはIle−Pheが先行する。軽鎖のCDR2とCDR3との間にはほぼ常に32個の残基があり、そしてCDR3は通常は約7〜10アミノ酸長である。CDR3にはほぼ常にCysが先行しており、そして通常はPhe−Gly−Xxx−Gly(配列番号51)が続く。
【0133】
CDRを囲むフレームワーク領域の長さが典型的であるものとは異なる長さを生む挿入を含んでもまたは欠失を含んでもよいことを当業者は理解する。本明細書において意味されるとおり、重鎖フレームワーク領域の長さは、以下の範囲内におさまる:FR1、0〜41アミノ酸;FR2、5〜24アミノ酸;FR3、13〜42アミノ酸;およびFR4、0〜21アミノ酸。さらに、本発明は、軽鎖フレームワーク領域の長さが以下の範囲内におさまることを意図する:FR1、6〜35アミノ酸;FR2、4〜25アミノ酸;FR3、2〜42アミノ酸;およびFR4、0〜23アミノ酸。
【0134】
天然に存在する抗体は代表的には、タンパク質分泌のための細胞経路に抗体を指向し、かつ成熟抗体には存在しない、シグナル配列を含む。本発明の抗体をコードするポリヌクレオチドは、下記のように、天然に存在するシグナル配列、または異種のシグナル配列をコードしてもよい。
(抗体のインビトロ成熟)
抗体は、抗原についてのさらに高い親和性またはさらに低い解離定数のような特性の変化した抗体を生成するためにインビトロで成熟され得る。CDR、特にCDR3内の残基のみのバリエーションによって、同じ抗原に結合するがさらに親和性の大きい変化した抗体が生じ得る。例えば、Schierら,1996,J.Mol.Biol.263:551〜67;Yangら,1995,J.Mol.Biol.254:392〜403を参照のこと。本発明は、抗体が有利な特性を有するように選択することができる、種々のインビトロ選択スキーム、例えば、アフィニティー成熟および/またはチェーンシャッフリング(Kangら,1991,Proc.Natl.Acad.Sci.88:11120〜23)、またはDNAシャッフリング(Stemmer,1994,Nature
370:389〜391)によって作製された抗体を包含する。多くのスキームでは、公知の抗体を特定の位置、しばしばCDR内でインビトロで無作為化して、選択プロセスに供して、これによって所望の特性、例えば特定の抗原に対する親和性の増大を有する抗体を単離することができる。例えば、van den Beuckenら,2001,J.Mol.Biol.310:591〜601;Desiderioら,2001,J.Mol.Biol.310:603〜15;Yangら,1995,J.Mol.Biol.254:392〜403;Schierら,1996,J.Mol.Biol.263:551〜67を参照のこと。代表的には、このような変異された抗体は、突然変異誘発および選択工程のデザインに依存して、1つ以上のCDRにいくつかの変更された残基を含んでもよい。例えば、van den Beuckenら(前出)を参照のこと。
(二重特異性または二機能性抗体)
二重特異性抗体または二機能性抗体は代表的には、2つの異なる重鎖/軽鎖の対および2つの異なる結合部位を有する人工的ハイブリッド抗体である。二重特異性抗体は、限定はしないが、ハイブリドーマの融合またはFab’フラグメントの連結を含む種々の方法によって生成することができる。例えば、SongsivilaiおよびLachmann,1990 Clin.Exp.Immunol.79:315〜321;Kostelnyら、1992,J.Immunol.148:1547〜1553を参照のこと。(抗体の調製)
本発明は、ヒトIFN−γに特異的に結合する抗体を提供する。これらの抗体は、全長IFN−γまたはそのフラグメントを用いた免疫によって産生され得る。本発明の抗体は、ポリクローナル抗体であっても、もしくはモノクローナル抗体であってもよく、そして/または組み換え抗体であってもよい。特定の実施形態では、本発明の完全ヒト抗体は、例えば、ヒト抗体を産生し得るトランスジェニック動物の免疫によって調製される(例えば、国際特許出願公開WO 93/12227を参照のこと)。
【0135】
本発明の抗IFNγ抗体の軽鎖および重鎖の可変領域のCDRは、同じ種または別の種由来のフレームワーク領域(FR)に移植され得る。特定の実施形態では、抗IFNγ抗体の軽鎖および重鎖の可変領域のCDRは、「ヒト化」抗体を作製するためにコンセンサスなヒトFRに移植されてもよい。このようなヒト化抗体は、本発明によって包含される。コンセンサスヒトFRを作製するために、いくつかのヒト重鎖または軽鎖のアミノ酸配列由来のFRを、整列させてコンセンサスアミノ酸配列を同定する。抗IFNγ抗体重鎖または軽鎖のFRは、異なる重鎖または軽鎖由来のFRで置換されてもよい。抗IFNγ抗体の重鎖および軽鎖のFRにおけるまれなアミノ酸は代表的には、置換されないが、残りのFRアミノ酸は置換されてもよい。まれなアミノ酸とは、FRにおいて通常はみいだされない位置にある特定のアミノ酸である。本発明の抗IFNγ抗体由来の移植された可変領域は、抗IFNγ抗体のもとの定常領域と異なる定常領域とともに用いられてもよい。あるいは、移植された可変領域は、単鎖Fv抗体の一部である。CDR移植は、任意の目的のために参考として本明細書に援用される、例えば、米国特許第6,180,370号、同第5,693,762号、同第5,693,761号、同第5,585,089号および同第5,530,101号に記載されている。
【0136】
本発明の抗体は、マウスの抗体産生細胞に挿入されたヒト抗体産生遺伝子座の実質的な一部を有しており、そして内因性マウス抗体を産生するにおいて欠失されるようにさらに操作されている、トランスジェニックマウスを用いて調製されてもよい。このようなマウスは、ヒト免疫グロブリン分子および抗体を産生可能であり、そしてマウスの免疫グロブリン分子および抗体を産生しないかまたは実質的に少ない量を産生する。この結果を達成するために利用される技術は、本明細書に開示された特許、出願および引用文献に開示されている。特定の実施形態では、当業者は、任意の目的のために参考として本明細書に援用される、国際特許出願公開番号WO98/24893に開示されるような方法を使用してもよい。任意の目的のために参考として本明細書に援用される、Mendezら,1997,Nature Genetics 15:146〜156も参照のこと。
【0137】
本発明のモノクローナル抗体(mAb)は、従来のモノクローナル抗体方法論、例えば、KohlerおよびMilstein(1975,Nature 256:495)の標準的体細胞ハイブリダイゼーション技術を含む、種々の技術によって生成されてもよい。体細胞ハイブリダイゼーション手順が好ましいが、原則的には、モノクローナル抗体を産生するための他の技術、例えば、Bリンパ球のウイルスまたは発癌性形質転換を用いることもできる。
【0138】
ハイブリドーマを調製するための可能性のある動物系の1つはマウスである。マウスにおけるハイブリドーマ産生はきわめてよく確立されており、そして融合のための免疫された脾細胞の単離のための免疫プロトコールおよび技術は当該分野で周知である。融合パートナー(例えば、マウス骨髄腫細胞)および融合手順はまた公知である。
【0139】
いくつかの実施形態では、IFN−γ、必要に応じてヒトIFN−γに対する完全ヒトモノクローナル抗体は、マウス系とは異なるヒト免疫系の一部を有するトランスジェニックマウスを用いて生成され得る。これらのトランスジェニックマウスは、本明細書において「HuMab」マウスと呼ばれるが、これは、再配列されていないヒト重鎖(μおよびγ)およびκ軽鎖免疫グロブリン配列をコードするヒト免疫グロブリン遺伝子ミニ遺伝子座を、内因性μおよびκ鎖遺伝子座を不活性化する標的化変異とともに含む(Lonbergら,1994,Nature 368:856〜859)。従って、マウスは、マウスIgMまたはκの発現の低下を示し、そして免疫に応答して、導入されたヒト重鎖および軽鎖導入遺伝子は、クラス切り替えおよび体細胞変異を受けて高い親和性のヒトIgGκモノクローナル抗体を生成する(Lonbergら、前出;LonbergおよびHuszar,1995,Intern.Rev.Immunol.13:65〜93;HardingおよびLonberg,1995,Ann.N.Y.Acad.Sci.764:536〜546)。HuMabマウスの調製は、その全ての内容が全体が参考として本明細書に援用されている、Taylorら,1992,Nucleic Acids Res.20:6287〜6295;Chenら,1993,International Immunology 5:647〜656;Tuaillonら,1994,J.Immunol.152:2912〜2920;Lonbergら,1994,Nature 368:856〜859;Lonberg,1994,Handbook of Exp.Pharmacology 113:49〜101;Taylorら,1994,International Immunology 6:579〜591;LonbergおよびHuszar,1995,Intern.Rev.Immunol.13:65〜93;HardingおよびLonberg,1995,Ann.N.Y.Acad.Sci 764:536〜546;Fishwildら,1996,Nature Biotechnology 14:845〜851に詳細に記載されている。さらに、全てがLonbergおよびKayに対する、米国特許第5,545,806号;同第5,569,825号;同第5,625,126号;同第5,633,425号;同第5,789,650号;同第5,877,397号;同第5,661,016号;同第5,814,318号;同第5,874,299号および同第5,770,429号、ならびにその全ての開示が全体が参考として本明細書に援用されている、1993年6月24日公開のSuraniらの米国特許第5,545,807号;1993年6月24日公開、国際特許出願公開番号WO93/1227;1992年12月23日公開WO92/22646;および1992年3月19日公開WO92/03918を参照のこと。あるいは、以下の実施例に記載されたHCo7およびHCo12のトランスジェニックマウス系統は、ヒト抗IFNγ抗体を生成するために用いられ得る。
【0140】
これらの実施形態では、本発明の抗体は、10−7M、10−8M、10−9M、または10−10M未満という平衡解離定数(KD)でIFN−γに特異的に結合する。本発明の特定の実施形態では、この抗体は、約10−8M〜10−12MというKDでIFN−γに結合する。
【0141】
好ましい実施形態では、本発明の抗体は、IgG1、IgG2またはIgG4アイソタイプの抗体である。この抗体は、IgG1アイソタイプの抗体であってもよい。他の実施形態では、本発明の抗体は、IgM、IgA、IgEまたはIgDアイソタイプの抗体である。本発明の好ましい実施形態では、この抗体は、ヒトκ軽鎖およびヒトIgG1重鎖を含む。IgG1重鎖定常領域を含む本発明の抗体の発現は、以下の実施例に記載されている。特定の実施形態では、この抗体の可変領域は、IgG1アイソタイプの定常領域以外の定常領域に連結される。特定の実施形態では、本発明の抗体は、哺乳動物細胞における発現のためにクローニングされている。
【0142】
特定の実施形態では、抗IFNγ抗体の重鎖および軽鎖に対する(そしてそのコードするヌクレオチドに対する改変に相当する)保存的改変は、抗IFNγ抗体の特徴と類似の機能的および化学的特徴を有する抗IFNγ抗体を生成する。対照的に、抗IFNγ抗体の機能および/または化学的特徴における実質的な改変は、(a)置換の領域における分子骨格の構造、例えばβシートまたはらせん高次構造、(b)標的部位における分子の変化もしくは疎水性、または(c)側鎖のバルク、の維持に対する効果が有意に異なる重鎖および軽鎖のアミノ酸配列の置換を選択することによって達成され得る。
【0143】
例えば、「保存的アミノ酸置換」とは、その位置でのアミノ酸残基の極性または荷電に対して影響がほとんどないかまたは全くないような、天然ではない残基による天然のアミノ酸残基の置換を包含し得る。さらに、ポリペプチドにおける任意の天然の残基はまた、「アラニンスキャニング突然変異誘発(alanine scanning mutagenesis)」について以前に記載されているとおり、アラニンで置換されてもよい。
【0144】
所望のアミノ酸置換(保存的であっても、または非保存的であっても)は、このような置換が所望される時点で当業者によって決定され得る。特定の実施形態において、アミノ酸置換を用いて、抗IFNγ抗体の重要な残基を同定すること、または本明細書に記載される抗IFNγ抗体の親和性を増大もしくは低下させることができる。
【0145】
別の実施形態では、本発明の抗体は、ハイブリドーマ細胞株とは異なる細胞株で発現され得る。これらの実施形態では、適切な哺乳動物宿主細胞の形質転換のために特定の抗体をコードする配列を用いてもよい。これらの実施形態によれば、米国特許第4,399,216号、同第4,912,040号、同第4,740,461号および同第4,959,455号(その全てが任意の目的のために参考として本明細書に援用される)に例示されているとおり、例えばウイルスへ(またはウイルスベクターへ)ポリヌクレオチドをパッケージングすること、およびこのウイルス(またはベクター)によって宿主細胞を形質導入すること、または当該分野で公知のトランスフェクション手順によって、などを含む、宿主細胞にポリヌクレオチドを導入するための任意の公知の方法を用いて、形質転換を達成することができる。一般的に、用いられる形質転換手順は、形質転換されるべき宿主に依存し得る。哺乳動物細胞へ異種ポリヌクレオチドを導入するための方法は、当該分野で周知であり、そしてこれには、デキストラン媒介性トランスフェクション、リン酸カルシウム沈殿、ポリブレン媒介性トランスフェクション、プロトプラスト融合、エレクトロポレーション、リポソームへのポリヌクレオチド(単数または複数)のカプセル化、および核へのDNAの直接マイクロインジェクションが挙げられるがこれらに限定されない。
【0146】
本発明の抗IFNγ抗体の重鎖定常領域、重鎖可変領域、軽鎖定常領域、または軽鎖可変領域のアミノ酸配列をコードする核酸分子(またはポリヌクレオチド)は、本発明によって包含される。このようなポリヌクレオチドは、標準的な連結技術を用いて適切な発現ベクター中に挿入されてもよい。好ましい実施形態では、抗IFNγ抗体重鎖または軽鎖の定常領域をコードするポリヌクレオチドは、適切な可変領域をコードするポリヌクレオチドの下流に付加され、そして発現ベクター中に連結される。このベクターは代表的には、使用される特定の宿主細胞において機能的である(すなわち、このベクターは、遺伝子の増幅および/または遺伝子の発現が生じ得るように宿主細胞の機構に適切である)ように選択される。発現ベクターの概説に関しては、METH.ENZ.185(Goeddel、編)1990,Academic Pressを参照のこと。
【0147】
代表的には、任意の宿主細胞において用いられる発現ベクターは、プラスミド維持のため、ならびに内因性ヌクレオチド配列のクローニングおよび発現のための配列を含む。このような配列は総称的に、「隣接配列(flanking sequences)」と呼ばれるが、特定の実施形態では、代表的には、以下のヌクレオチド配列のうちの1つ以上を含む:プロモーター、1つ以上のエンハンサー配列、複製起点、転写終結配列、ドナーおよびアクセプタースプライス部位を含む完全イントロン配列、ポリペプチド分泌のためのリーダー配列をコードする配列、リボソーム結合部位、ポリアデニル化配列、発現されるべきポリペプチドをコードする核酸を挿入するためのポリリンカー領域、および選択可能なマーカーエレメント。これらの配列の各々は以下に考察される。
【0148】
必要に応じて、ベクターは、「タグ(tag)」コード配列、すなわち、抗IFNγ抗体ポリペプチドコード配列の5’または3’末端に位置するオリゴヌクレオチド分子を含んでもよい;このオリゴヌクレオチド配列は、ポリHis(例えば、hexaHis)、または別の「タグ」、例えばFLAG、HA(インフルエンザウイルス血球凝集素)、またはmyc(これについては、市販の抗体が存在する)をコードする。このタグは代表的には、ポリペプチドの発現の際にポリペプチドに融合され、そして宿主細胞からのIFN−γ抗体のアフィニティー精製または検出のための手段として機能し得る。アフィニティー精製は、例えば、アフィニティーマトリックスとしてタグに対する抗体を用いるカラムクロマトグラフィーによって達成され得る。必要に応じて、タグは、種々の手段によって、例えば、切断のための特定のペプチダーゼを用いて、精製された抗IFNγ抗体ポリペプチドから引き続いて除去されてもよい。
【0149】
隣接配列は、相同(すなわち、宿主細胞と同じ種および/または株由来)、異種(すなわち、宿主細胞種または株以外の種由来)、ハイブリッド(すなわち、2つ以上の供給源由来の隣接配列の組み合わせ)、合成または天然であってもよい。従って、隣接配列の供給源は、この隣接配列が機能的であり、宿主細胞の機構によって活性化され得る条件下では、任意の原核生物生物体であっても、または真核生物生物体であっても、任意の脊椎動物もしくは無脊椎動物であっても、または任意の植物であってもよい。
【0150】
本発明のベクターにおいて有用な隣接配列は、当該分野で周知の任意のいくつかの方法によって達成され得る。代表的には、本明細書において有用な隣接配列は、マッピングによって、および/または制限エンドヌクレアーゼ消化によって以前に同定されており、従って適切な制限エンドヌクレアーゼを用いて適切な組織供給源から単離可能である。ある場合には、隣接配列の全ヌクレオチド配列が公知であり得る。ここでは、隣接配列は、核酸合成またはクローニングのために本明細書において記載された方法を用いて合成され得る。
【0151】
隣接配列の全てが既知であるか一部のみが既知であるかにかかわらず、これは、ポリメラーゼ連鎖反応(PCR)を用いて、ならびに/または適切なプローブ、例えば、同じもしくは別の種由来のオリゴヌクレオチドおよび/もしくは隣接配列のフラグメントを用いてゲノムライブラリーをスクリーニングすることによって、得ることができる。隣接配列が既知でない場合、隣接配列を含むDNAのフラグメントは、例えば、コード配列またはさらに別の遺伝子(単数または複数)を含み得る、DNAのさらに大きい小片から単離することができる。単離は、適切なDNAフラグメントを生成するための制限エンドヌクレアーゼ消化によって、その後のアガロースゲル精製、Qiagen(登録商標)カラムクロマトグラフィー(Chatsworth,CA)または当業者に公知の他の方法を用いる単離によって達成できる。この目的を達成するための適切な酵素の選択は、当業者に容易に明らかである。
【0152】
複製起点は代表的には、市販の原核生物発現ベクターの一部であり、この起点は宿主細胞においてベクターの増幅を補助する。選択したベクターが複製起点の部位を含まない場合、既知の配列に基づいて化学的に合成して、ベクターに連結してもよい。例えば、プラスミドpBR322(New England Biolabs,Beverly,MA)由来の複製起点は、ほとんどのグラム陰性細菌に適切であり、種々のウイルス起点(例えば、SV40、ポリオーマ、アデノウイルス、水疱性口内炎ウイルス(VSV)、またはパピローマウイルス、例えば、HPVまたはBPV)は、哺乳動物細胞におけるクローニングベクターに有用である。一般的に、複製起点の成分は、哺乳動物発現ベクターには必要ではない(例えば、SV40起点は、ウイルス初期プロモーターをも含むので、しばしば単独で用いられる)。
【0153】
転写終結配列は代表的には、ポリペプチドコード領域の末端に対して3’に位置して、転写を集結するように機能する。通常、原核生物細胞における転写終結配列は、G−Cリッチフラグメントであり、ポリT配列が続く。この配列はライブラリーから容易にクローニングされるかまたはベクターの一部として市販されてさえいるが、本明細書に記載の方法のような核酸合成の方法を用いて容易に合成することもできる。
【0154】
選択マーカー遺伝子は、選択性の増殖培地中で増殖される宿主細胞増殖の生存および増殖に必要なタンパク質をコードする。代表的な選択マーカー遺伝子は、(a)原核生物宿主細胞については、抗生物質または他の毒素、例えば、アンピシリン、テトラサイクリンもしくはカナマイシンに対する耐性を付与するか;(b)細胞の栄養要求欠損を補完するか;あるいは(c)複合培地または規定の培地から利用できない重要な栄養物を供給する、タンパク質をコードする。好ましい選択マーカーは、カナマイシン耐性遺伝子、アンピシリン耐性遺伝子およびテトラマイシン耐性遺伝子である。有利には、ネオマイシン耐性遺伝子もまた、原核生物および真核生物の宿主細胞の両方における選択のために用いることができる。
【0155】
発現される遺伝子を増幅するために、他の選択遺伝子が用いられてもよい。増幅とは、増殖または細胞の生存に重要なタンパク質の産生に必要な遺伝子が、組み換え細胞の連続的世代の染色体内において一列で反復されているプロセスである。哺乳動物細胞のための適切な選択マーカーの例としては、ジヒドロ葉酸還元酵素(DHFR)およびプロモーターなしのチミジンキナーゼ遺伝子が挙げられる。哺乳動物細胞形質転換体は、ベクターに存在する選択遺伝子のおかげで形質転換体のみが生存するように固有に適合されるという選択圧下に置かれる。選択圧は、培地中の選択因子の濃度が連続的に増大される条件下で形質転換細胞を培養することによって課され、これによって選択遺伝子および別の遺伝子、例えばIFNγポリペプチドに結合する抗体をコードするDNAの両方の増幅がもたらされる。結果として、抗IFNγ抗体のようなポリペプチドの増大した量は、増幅されたDNAから合成される。
【0156】
リボソーム結合部位は通常、mRNAの翻訳開始に必要であり、シャイン・ダルガーノ(Shine−Dalgarno)配列(原核生物)またはKozak配列(真核生物)によって特徴付けられる。このエレメントは代表的には、プロモーターの3’側および発現されるべきポリペプチドのコード配列の5’側に位置する。
【0157】
真核生物の宿主細胞発現系においてグリコシル化が所望される場合のような、ある場合には、グリコシル化または収率を改善するための種々のプレ配列またはプロ配列を操作してもよい。例えば、特定のシグナルペプチドのペプチダーゼ切断部位を変更してもよいし、またはグリコシル化に影響し得るプロ配列を付加してもよい。最終タンパク質産物は、−1位(成熟タンパク質の第一アミノ酸に対する位置)に、発現に付随する1つ以上のさらなるアミノ酸付加を有してもよく、これは完全に取り除かれていなくてもよい。例えば、最終タンパク質産物は、アミノ末端に結合されたペプチダーゼ切断部位に見出される2つ以上のアミノ酸残基を有し得る。あるいは、いくつかの酵素切断部位の使用によって、酵素が成熟ポリペプチド内のこのような領域で切断する場合、所望のポリペプチドのわずかに短縮された形態が生じ得る。
【0158】
本発明の発現ベクターおよびクローニングベクターは代表的には、宿主生物体によって認識され、かつ抗IFNγ抗体をコードする分子に作動可能に連結されているプロモーターを含む。プロモーターは、構造遺伝子の転写を制御する構造遺伝子(一般的には約100〜1000bp内)の開始コドンに対して上流(すなわち、5’側)に位置する非翻訳配列である。プロモーターは以下の2つのクラスのうちの1つに慣用的に分類される:誘導性プロモーターおよび構成的プロモーター。誘導性プロモーターは、栄養物の有無または温度の変化のような培養条件におけるいくつかの変化に応答してそれらの制御下でDNAから増大したレベルの転写を開始する。構成的プロモーターは他方では、それらが作動可能に連結されている遺伝子を均一に転写し、すなわち、遺伝子発現に対する制御はほとんどないかまたは全くない。種々の潜在的な宿主細胞によって認識される多数のプロモーターは周知である。適切なプロモーターは、制限酵素消化によって供給源のDNAからプロモーターを除去すること、およびこのベクターに所望のプロモーター配列を挿入することによって、本発明の抗IFNγ抗体を含む重鎖または軽鎖をコードするDNAに作動可能に連結される。
【0159】
酵母宿主との使用のための適切なプロモーターもまた、当該分野で周知である。酵母エンハンサーは、酵母プロモーターとともに有利に用いられる。哺乳動物宿主細胞とともに使用するための適切なプロモーターは周知であり、そしてウイルス、例えば、ポリオーマウイルス、鶏痘ウイルス、アデノウイルス(例えば、アデノウイルス2)、ウシパピローマウイルス、トリ肉腫ウイルス、サイトメガロウイルス、レトロウイルス、B型肝炎ウイルスおよび最も好ましくはシミアンウイルス40(SV40)のゲノムから得られたプロモーターが挙げられるがこれらに限定されない。他の適切な哺乳動物プロモーターとしては、異種哺乳動物プロモーター、例えば、熱ショックプロモーターおよびアクチンプロモーターが挙げられる。
【0160】
目的であり得るさらなるプロモーターとしては以下が挙げられるがこれらに限定されない:SV40初期プロモーター(BenoistおよびChambon,1981,Nature 290:304〜10);CMVプロモーター(Thomsenら、1984,Proc.Natl.Acad.USA 81:659〜663);ラウス肉腫ウイルスの3’長末端反復配列に含まれるプロモーター(Yamamotoら,1980,Cell 22:787〜97);ヘルペスチミジンキナーゼプロモーター(Wagnerら、1981,Proc.Natl.Acad.Sci.U.S.A.78:1444〜45);メタトリオネイン遺伝子由来のプロモーターおよび調節性配列(Brinsterら、1982,Nature 296:39〜42);ならびに原核生物プロモーター、例えば、βラクタマーゼプロモーター(Villa−Kamaroffら、1978,Proc.Natl.Acad.Sci.U.S.A.,75:3727〜31);またはtacプロモーター(DeBoerら、1983,Proc.Natl.Acad.Sci.U.S.A.,80:21〜25)。また目的のものは、組織特異性を示して、トランスジェニック動物において利用されている、以下の動物転写制御領域である:膵臓の腺房細胞において活性であるエラスターゼI遺伝子制御領域(Swiftら,1984,Cell 38:639〜46;Ornitzら,1986,Cold Spring Harbor Symp.Quant.Biol.50:399〜409(1986);MacDonald,1987,Hepatology 7:425〜515);膵臓β細胞において活性なインスリン遺伝子制御領域(Hanahan,1985,Nature
315:115〜22);リンパ細胞において活性な免疫グロブリン遺伝子制御領域(Grosschedlら、1984,Cell 38:647〜58;Adamesら,1985,Nature 318:533〜38;Alexanderら,1987,Mol.Cell.Biol.,7:1436〜44);精巣、乳房、リンパ系細胞および肥満細胞において活性なマウス乳腺腫瘍ウイルス制御領域(Lederら、1986,Cell 45:485〜95);肝臓で活性なアルブミン遺伝子制御領域(Pinkertら、1987,Genes and Devel.1:268〜76);肝臓においおいて活性なαフェトプロテイン遺伝子制御領域(Krumlaufら、1985,Mol.Cell.Biol.,5:1639〜48;Hammerら、1987,Science 235:53〜58);肝臓において活性なα1−抗トリプシン遺伝子制御領域(Kelseyら、1987,Genes and Devel.1:161〜71);骨髄性細胞において活性なβグロビン遺伝子制御領域(Mogramら、1985,Nature 315:338〜40;Kolliasら、1986,Cell 46:89〜94);脳における稀突起膠細胞において活性なミエリン塩基性タンパク質遺伝子制御領域(Readheadら、1987,Cell 48:703〜12);骨格筋において活性であるミオシン軽鎖2遺伝子制御領域(Sani,1985,Nature 314:283〜86);ならびに視床下部で活性な生殖腺刺激ホルモン放出ホルモン遺伝子制御領域(Masonら、1986,Science 234:1372〜78)。
【0161】
エンハンサー配列をベクターに挿入して、高等な真核生物によって本発明の抗IFNγ抗体を含む軽鎖または重鎖をコードするDNAの転写を増大する。エンハンサーは、通常約10〜300bp長のDNAのシス作用性エレメントであり、これはプロモーター上で転写を増大するように働く。エンハンサーは比較的方向および位置に依存せず、転写単位の5’および3’の両方の位置で見出されている。哺乳動物遺伝子から利用可能ないくつかのエンハンサー配列は公知である(例えば、グロビン、エラスターゼ、アルブミン、αフェトプロテインおよびインスリン)。しかし、代表的には、ウイルス由来のエンハンサーを用いる。当該分野で公知のSV40エンハンサー、サイトメガロウイルス初期プロモーターエンハンサー、ポリオーマエンハンサー、およびアデノウイルスエンハンサーは、真核生物プロモーターの活性化のための例示的な増強エレメントである。エンハンサーは、ベクター中でコード配列の5’または3’のいずれに位置してもよいが、代表的にはプロモーターから5’部位に位置する。
【0162】
適切な天然のシグナル配列または異種のシグナル配列をコードする配列(リーダー配列またはシグナルペプチド)を発現ベクターに組み込んで、抗体の細胞外分泌を促進することが可能である。シグナルペプチドまたはリーダーの選択は、抗体が産生される宿主細胞のタイプに依存し、そして異種シグナル配列は、天然のシグナル配列を置き換え得る。哺乳動物宿主細胞で機能的であるシグナルペプチドの例としては、以下が挙げられる:米国特許第4,965,195号に記載されるインターロイキン7(IL−7)のシグナル配列;Cosmanらに記載のインターロイキン2レセプターのシグナル配列(1984,Nature 312:768);欧州特許第0 367 566号に記載のインターロイキン−4レセプターシグナルペプチド;米国特許第4,968,607号に記載のI型インターロイキン−1レセプターシグナルペプチド;欧州特許第0 460 846号に記載のII型インターロイキン−1レセプターシグナルペプチド;ヒトIgKのシグナル配列(METDTLLLWVLLLWVPGSTG(配列番号52)である);ヒト成長ホルモンのシグナル配列(MATGSRTSLLLAFGLLCLPWLQEGSA;配列番号53である);ならびに実施例3において単離された重鎖および軽鎖をコードするcDNAによってコードされた、ヒトシグナル配列MGSTAILALLLAVLQGVCA(配列番号54)およびMETPAQLLFLLLLWLPDTTG(配列番号55)。
【0163】
本発明の発現ベクターは、市販のベクターのような開始ベクターから構築されてもよい。このようなベクターは、所望の隣接配列の全てを含んでも含まなくてもよい。本明細書に記載される隣接配列の1つ以上は既にベクターには存在しないが、それらを個々に入手してベクターに連結してもよい。隣接配列の各々を得るために用いた方法は、当業者に周知である。
【0164】
ベクターが構築され、そして軽鎖、重鎖、または抗IFNγ抗体を含む軽鎖および重鎖をコードする核酸分子がこのベクターの適切な部位に挿入された後、この完成したベクターを増幅および/またはポリペプチド発現のために適切な宿主細胞中に挿入してもよい。選択された宿主細胞への抗IFNγ抗体のための発現ベクターの形質転換は、トランスフェクション、感染、リン酸カルシウム共沈殿、エレクトロポレーション、マイクロインジェクション、リポフェクション、DEAEデキストラン媒介性トランスフェクション、または他の公知の技術を含む周知の方法によって達成され得る。選択される方法は一部は、用いられる宿主細胞のタイプに関連する。これらの方法および他の適切な方法は、当業者に周知であり、そして例えば、Sambrookら(前出)に記載されている。
【0165】
宿主細胞は、適切な条件下で培養された場合、抗IFNγ抗体を合成し、この抗体は引き続いて、培養培地から(宿主細胞が培地中にこの抗体を分泌する場合)、またはこの抗体を産生する宿主細胞から直接(この抗体が分泌されない場合)、収集できる。適切な宿主細胞の選択は、種々の要因、例えば、所望の発現レベル、活性のために所望されるかまたは必要であるポリペプチド改変(例えば、グリコシル化またはリン酸化)および生物学的に活性な分子への折り畳みの容易さに依存する。
【0166】
発現のための宿主として利用可能な哺乳動物細胞株は、当該分野で周知であり、そしてこれらとしては、限定はしないがAmerican Type Culture Collection(ATCC)から入手可能な不死化細胞株が挙げられ、これらとしては、限定はしないが、チャイニーズハムスター卵巣(CHO)細胞、HeLa細胞、ベビーハムスター腎臓(BHK)細胞、サル腎臓細胞(COS)、ヒト肝細胞癌腫細胞(例えば、Hep G2)および多数の他の細胞株が挙げられる。特定の実施形態では、どの細胞株が高い発現レベルを有し、IFNγ結合特性を有する抗体を構成的に生成するかを決定することを通じて細胞株を選択することができる。別の実施形態では、それ自体の抗体を作製しないが、異種の抗体を作製および分泌する能力を有するB細胞系列由来の細胞株を選択することができる。
【0167】
本発明の抗体は、生物学的サンプル中のIFNγを検出するため、およびIFNγタンパク質を産生する細胞または組織の同定のために有用である。IFNγに特異的に結合する本発明の抗体は、IFNγ媒介性疾患の処置において有用であり得る。このような抗体は、IFNγを検出するため、およびIFNγがIFNγレセプターとの複合体を形成することを阻害するための結合アッセイにおいて用いられ得る。IFNγに結合して、他の結合化合物との相互作用をブロックするこのような抗体は、IFNγ媒介性疾患を調節するのにおいて治療的な用途を有し得る。好ましい実施形態では、IFNγに対する抗体は、そのレセプターに対するIFNγ結合をブロックし得、これがIFNγ誘導性シグナル伝達カスケードの破壊を生じ得る。
【0168】
いくつかの実施形態では、本発明は薬学的組成物であって、治療上有効な量の1つまたは複数の本発明の抗体を薬学的に受容可能な希釈剤、キャリア、可溶化剤、乳化剤、保存剤および/またはアジュバントとともに含む薬学的組成物を提供する。好ましくは、受容可能な処方物質は、使用される投薬量および濃度でレシピエントに対して非毒性である。好ましい実施形態では、治療上有効な量の抗IFNγ抗体を含む薬学的組成物が提供される。
【0169】
特定の実施形態では、受容可能な処方物質は好ましくは、使用される投薬量および濃度でレシピエントに対して非毒性である。
【0170】
特定の実施形態では、薬学的組成物は、例えば、pH、浸透圧、粘度、清澄度、色調、等張性、臭気、無菌性、安定性、分解もしくは放出の速度、組成物の吸着もしくは浸透を改変、維持または保存するための処方物質を含んでもよい。このような実施形態では、適切な処方物質としては、限定はしないが、アミノ酸(例えば、グリシン、グルタミン、アスパラギン、アルギニンまたはリジン);抗菌剤;抗酸化剤(例えば、アスコルビン酸、亜硫酸ナトリウムまたは亜硫酸水素ナトリウム);緩衝液(例えば、ホウ酸塩、炭酸水素塩、Tris−HCl、クエン酸、リン酸または他の有機酸);充填剤(例えば、マンニトールまたはグリシン);キレート剤(例えば、エチレンジアミン四酢酸(EDTA));錯化剤(例えば、カフェイン、ポリビニルピロリドン、βサイクロデキストリンまたはヒドロキシプロピルβシクロデキストリン);賦形剤(filler);単糖類;二糖類;ならびに他の炭水化物(例えば、グルコース、マンノースまたはデキストリン);タンパク質(例えば、血清アルブミン、ゼラチンまたは免疫グロブリン);着色剤、香味料および希釈剤;乳化剤;親水性のポリマー(例えば、ポリビニルピロリドン);低分子量ポリペプチド;塩形成対イオン(例えば、ナトリウム);防腐剤(例えば、塩化ベンザルコニウム、安息香酸、サリチル酸、チメロサール、フェネチルアルコール、メチルパラベン、プロピルパラベン、クロルヘキシジン、ソルビン酸または過酸化水素);溶媒(例えば、グリセリン、プロピレングリコールまたはポリエチレングリコール);糖アルコール(例えば、マンニトールまたはソルビトール);懸濁剤;界面活性剤または湿潤剤(例えば、プルロニック、PEG、ソルビタンエステル、ポリソルベート、例えば、ポリソルベート20、ポリソルベート80、トリトン(triton)、トロメタミン、レシチン、コレステロール、チロキサポール);安定性強化剤(例えば、スクロースまたはソルビトール);等張化剤(例えば、ハロゲン化アルカリ金属、好ましくは塩化ナトリウムまたは塩化カリウム、マンニトール ソルビトール);送達ビヒクル;希釈剤;賦形剤(excipient)および/または薬学的アジュバントが挙げられる。REMINGTON’S
PHARMACEUTICAL SCIENCES,第18版(A.R.Gennaro編),1990,Mack Publishing Companyを参照のこと。
【0171】
特定の実施形態では、最適の薬学的組成物は、例えば、意図される投与経路、送達様式および所望の投薬量に依存して当業者によって決定される。例えば、REMINGTON’S PHARMACEUTICAL SCIENCES,前出を参照のこと。特定の実施形態では、このような組成物は、本発明の抗体の物理的状態、安定性、インビボでの放出の速度、およびインビボでのクリアランスの速度に影響し得る。
【0172】
特定の実施形態では、薬学的組成物における主なビヒクルまたはキャリアは、天然には水性であっても非水性であってもよい。例えば、適切なビヒクルまたはキャリアは、(おそらくは非経口投与のための組成物中に共通の他の物質で補充された)注射用水、生理学的生理食塩水または人工的な脳脊髄液であってもよい。中性の緩衝化生理食塩水、または血清アルブミンと混合された生理食塩水はさらに、例示的なビヒクルである。好ましい実施形態では、薬学的組成物は、約pH7.0〜8.5のTris緩衝液、または約pH4.0〜5.5の酢酸塩緩衝液を含み、そしてさらにソルビトールまたはその適切な代替物を含んでもよい。本発明の特定の実施形態では、抗IFNγ抗体組成物は、所望の程度の純度を有する選択された組成物を最適の処方剤(REMINGTON’S PHARMACEUTICAL SCIENCES、前出)とともに、凍結乾燥ケーキまたは水溶液の形態で混合することによって貯蔵のために調製され得る。さらに、特定の実施形態では、抗IFNγ抗体産物は、スクロースのような適切な賦形剤を用いて凍結乾燥剤として処方されてもよい。
【0173】
本発明の薬学的組成物は、非経口送達のために選択され得る。あるいは、この組成物は、吸入のために、または経口のような消化管を通じた送達のために選択され得る。このような薬学的に受容可能な組成物の調製は、当該分野の技術の範囲内である。
【0174】
処方成分は、投与の部位に受容可能な濃度で存在することが好ましい。特定の実施形態では、緩衝液を用いて、この組成物を生理学的なpHで、またはわずかに低いpHで、代表的には約5〜約8のpH範囲内で維持する。
【0175】
非経口的投与を考慮する場合、本発明における使用のための治療組成物は、所望の抗IFNγ抗体を薬学的に受容可能なビヒクル中に含む、発熱物質なしの、非経口的に受容可能な水溶液の形態で提供されてもよい。非経口注射のために特に適切なビヒクルは、抗IFNγ抗体が、適切に保存された無菌の等張性溶液として処方されている、滅菌蒸留水である。特定の実施形態では、この調製物は、所望の分子の処方物を、デポ注射を介して送達できる産物の制御放出または徐放性放出を提供し得る、ある因子、例えば、注射用マイクロスフェア、生体侵食性粒子、ポリマー化合物(例えば、ポリ乳酸またはポリグリコール酸)、ビーズまたはリポソームとともに含んでもよい。特定の実施形態では、循環中の持続期間を延長する効果を有する、ヒアルロン酸も用いることができる。特定の実施形態では、移植可能な薬物送達デバイスを用いて所望の抗体分子を導入することができる。
【0176】
本発明の薬学的組成物は、吸入のために処方され得る。これらの実施形態では、抗IFNγ抗体は、乾燥した吸入可能な粉末として有利に処方される。好ましい実施形態では、抗IFNγ抗体吸入溶液はまた、エアロゾル送達のための噴霧剤とともに処方され得る。特定の実施形態では、溶液は噴霧され得る。従って、肺投与および処方の方法はさらに、参考として援用されており、化学的に修飾されたタンパク質の肺送達を記載する、国際特許出願番号PCT/US94/001875に記載されている。
【0177】
処方物は経口的に投与できることも考えられる。この様式で投与される抗IFNγ抗体は、錠剤およびカプセルのような固体投薬形態の配合に習慣的に用いられるキャリアの有無とともに処方され得る。特定の実施形態では、カプセルは、バイオアベイラビリティーが最大化されて予備的な全身性の分解が最小化される胃腸管におけるポイントでこの処方物の活性部位を遊離するように設計され得る。抗IFNγ抗体の吸着を促進するためにさらなる因子が含まれてもよい。希釈剤、香味料、低融点ワックス、植物油、潤滑剤、懸濁剤、錠剤崩壊剤および結合剤も使用され得る。
【0178】
本発明の薬学的組成物は好ましくは、1または複数の抗IFNγ抗体の有効量を錠剤の製造業者に適切な非毒性賦形剤と混合して含むように提供される。滅菌水、または別の適切なビヒクルに錠剤を溶解することによって、単位用量形態で溶液を調製し得る。適切な賦形剤としては、限定はしないが、不活性希釈剤、例えば、炭酸カルシウム、炭酸ナトリウムまたは重炭酸ナトリウム、ラクトースもしくはリン酸カルシウム;または結合剤、例えばデンプン、ゼラチンもしくはアラビアゴム;または潤滑剤、例えば、ステアリン酸マグネシウム、ステアリン酸、もしくは滑石が挙げられる。
【0179】
さらなる薬学的組成物は、当業者には明白であり、これらとしては、徐放性または制御送達の処方物に含まれた抗IFNγ抗体を含む処方物が挙げられる。種々の他の徐放性または制御送達の手段、例えば、リポソームキャリア、生体侵食性微粒子または多孔性ビーズおよびデポ注射剤を処方するための技術もまた、当業者には公知である。例えば、参考として援用されており、薬学的組成物の送達のための多孔性ポリマー微粒子の制御放出を記載している、国際特許出願番号PCT/US93/00829を参照のこと。徐放性調製物は、成形された物質の形態で半透過性ポリマー物質、例えばフィルムまたはマイクロカプセルを含んでもよい。徐放性マトリックスは、ポリエステル、ヒドロゲル、ポリラクチド(参考として各々が援用される、米国特許第3,773,919号および欧州特許出願公開番号EP058481に開示される)、Lグルタミン酸およびγエチル−L−グルタメートのコポリマー(Sidmanら、1983,Biopolymers 22:547〜556)、ポリ(2−ヒドロキシエチル−メタクリレート)(Langerら、1981,J.Biomed.Mater.Res.15:167〜277およびLanger,1982,Chem.Tech.12:98〜105)、エチレン酢酸ビニル(Langerら、前出)またはポリ−D(−)−3−ヒドロキシ酪酸(欧州特許出願公開番号EP133,988)を含んでもよい。徐放性組成物はまた、当該分野で公知の任意のいくつかの方法によって調製され得るリポソームを含んでもよい。例えば、参考として援用される、Eppsteinら、1985,Proc.Natl.Acad.Sci.USA 82:3688〜3692;欧州特許出願公開番号EP 036,676;EP 088,046およびEP 143,949を参照のこと。
【0180】
インビボ投与のために用いられる薬学的組成物は代表的には、滅菌調製物として提供される。滅菌は、滅菌濾過膜を通した濾過によって達成され得る。組成物が凍結乾燥される場合、この方法を用いる滅菌は、凍結乾燥および再構成の前または後のいずれに行なわれてもよい。非経口的な投与のための組成物は、凍結乾燥型または溶液として貯蔵されてもよい。非経口的な組成物は一般には、無菌アクセスポートを有する容器、例えば静脈内溶液バッグ、または皮下注射針によって穿通可能な栓を有するバイアルに入れられる。
【0181】
一旦薬学的組成物が処方されれば、これは、滅菌バイアル中に、溶液、懸濁液、ゲル、エマルジョン、固体、結晶として、または脱水もしくは凍結乾燥された粉末として貯蔵され得る。このような処方物は、レディーツーユースの形態、または投与の前に再構成される形態(例えば、凍結乾燥された形態)のいずれで貯蔵されてもよい。
【0182】
本発明はまた、単回用量投与単位を生成するためのキットを提供する。本発明のキットは各々が、乾燥タンパク質を有する第一の容器、および水性処方物を有する第二の容器の両方を含む。本発明の特定の実施形態では、単一および複数のチャンバを有する事前充填(pre−filled)シリンジ(例えば、液体シリンジおよびライオシリンジ(lyosyringe))を備えるキットが提供される。
【0183】
使用されるべき、抗IFNγ抗体を含有する薬学的組成物の治療的有効量は、例えば、治療の状況および目的に依存する。処置のために適切な投薬レベルは部分的には送達される分子、抗IFNγ抗体が用いられている徴候、投与経路、ならびに患者のサイズ(体重、体表面または器官サイズ)および/または状態(年齢および全身健康状態)に依存して変化することが当業者には理解される。特定の実施形態では、最適の治療効果を得るために、臨床医が投薬量を滴定して、投与経路を改変してもよい。代表的な投薬量は、上述の要因に依存して、約0.1μg/kg〜約30mg/kg以上までに及んでもよい。好ましい実施形態では、投薬量は0.1μg/kg〜約30mg/kg、必要に応じて1μg/kg〜約30mg/kgまたは10μg/kg〜約5mg/kgに及んでもよい。
【0184】
投与頻度は、用いられる処方物中の特定の抗IFNγ抗体の薬物動態的パラメーターに依存する。代表的には、所望の効果を達成する投薬量が得られるまで、臨床医がこの組成物を投与する。従って、この組成物は単回用量として、または2回以上の用量(これは同じ量の所望の分子を含んでも含まなくてもよい)として、または移植デバイスもしくはカテーテルを介した連続注入として、経時的に投与されてもよい。適切な投薬量のさらなる工夫は当業者によって慣用的に行なわれ、そしてそれらによって慣用的に行なわれる課題の範囲内である。適切な投薬量は、適切な用量応答データの使用を通じて得ることができる。特定の実施形態では、本発明の抗体は、長期間にわたって患者に投与されてもよい。本発明の抗体の慢性投与によって、非ヒト動物においてヒト抗原に対して惹起された抗体、例えば、ヒト以外の種で産生された不完全ヒト抗体または非ヒト抗体に一般に関連する有害な免疫応答またはアレルギー応答が最小化される。
【0185】
薬学的組成物の投与経路は、徐放性システムによるかまたは移植デバイスによる、公知の方法、例えば、経口、静脈内、腹腔内、脳内(実質内)、脳室内、筋肉内、眼内、動脈内、門脈内または病巣内の経路による注射を通じた公知の方法に従う。特定の実施形態では、組成物は、ボーラス注射によって投与されても、もしくは注入によって連続的に投与されても、または移植デバイスによって投与されてもよい。
【0186】
この組成物はまた、所望の分子がその上に吸着されるかまたはカプセル化されている、膜、スポンジまたは別の適切な物質の移植によって局所投与され得る。移植デバイスを用いる特定の実施形態では、デバイスは任意の適切な組織または器官に移植されてもよく、そして所望の分子の送達は、拡散、時限放出ボーラス、または連続的投与を介してもよい。
【0187】
本発明による抗IFNγ抗体の薬学的組成物をエキソビボで用いることもまた所望され得る。このような場合、患者から除去されている細胞、組織または器官を、抗IFNγ抗体の薬学的組成物に曝露して、その後に細胞、組織および/または器官を患者に引き続き移植して戻す。
【0188】
詳細には、抗IFNγ抗体は、本明細書において記載される方法のような方法を用いて、ポリペプチドを発現および分泌するように遺伝子操作されている特定の細胞を移植することによって送達できる。特定の実施形態では、このような細胞は、動物細胞であってもまたはヒト細胞であってもよく、そして自家性、非相同性または異種性であってもよい。特定の実施形態では、細胞は不死化されてもよい。他の実施形態では、免疫学的応答の機会を低下させるために、細胞は、周囲の組織の浸潤を回避するためにカプセル化されてもよい。さらなる実施形態では、カプセル化物質は代表的には、タンパク質産物(単数または複数)の放出を可能にするが、患者の免疫系によるか、または周囲の組織からの他の有害な要因による、細胞の破壊を妨げる生体適合性、半透過性ポリマー同封物または膜である。
【実施例】
【0189】
以下の実施例は、行なった実験および得られた結果を含んでいるが、これは、例示目的のためにのみ提供されており、本発明を限定すると解釈されるべきではない。
【0190】
(実施例1)
(CHO細胞からのヒトIFNγタンパク質の生成)
テンプレートとしてヒト脾臓Marathon−Ready cDNA(Clontech)を用いて、全長ヒトIFNγ cDNAをPCR(標準的条件下)によって増幅した。この配列をpDSRα2プラスミド中にサブクローニングした。DH10B(Escherichia coli)細胞をpDSRα2プラスミドを用いて形質転換した。標準的技術を用いてDNAを調製して、CHO細胞をリン酸カルシウム法(Speciality Media,Inc)によってトランスフェクトした。高発現細胞株クローンを用いて、無血清馴化培地を生成した。
【0191】
ヒトIFNγ(hu−IFN−γ)を含むCHO細胞馴化培地を濃縮して、透析して、数段階のクロマトグラフィー工程を通じて精製した。第一の工程は、グリコシル化されていないhu−IFN−γ形態から高度にグリコシル化された形態を分離するために、標準的なNaCl勾配を用いるQ−HP(Pharmacia)クロマトグラフィーであった。Q−HPプールをさらに、コムギ胚芽凝集素クロマトグラフィー(EY Laboratories)を通じて精製した。この精製された物質をSDS−ポリアクリルアミドゲル電気泳動(SDS−PAGE)によって分離して、クマーシー・ブルーおよび銀染色によって分析した。精製された物質は、クマーシー・ブルーおよび銀の両方で染色されたSDS−PAGEによって決定した場合95%を超える純度であった。この物質をゲル凝固(gel−clot)法(Limulus Amebocyte Lysate)によってもアッセイして、内毒素レベルが低いことを示した。hu−IFN−γの同一性は、R&D Systemsの抗AF−285 NA抗体を用いてウエスタンブロットによって確認した。吸光係数方法を用いて、吸光度(A280)から最終のタンパク質濃度を決定したが、ここでA280読み取り/吸光係数=濃度(g/L)(吸光係数=0.66)である。
【0192】
(実施例2)
(IFNγに対するヒトモノクローナル抗体の産生)
(トランスジェニックHuMabマウス)
IFN−γに対する完全ヒトモノクローナル抗体は、ヒト抗体遺伝子を各々が発現する、トランスジェニックマウスのHCo7株、HCo12株およびHCo7+HCo12株を用いて調製された。これらの株の各々において、内因性マウスκ軽鎖遺伝子は、Chenらに記載されたとおりホモ接合性に分裂されており(1993,EMBO J.12:811〜820)、そして内因性マウス重鎖遺伝子は、国際特許出願公開番号WO 01/09187(参考として援用される)の実施例1に記載のようにホモ接合性に分裂されていた。各々の株は、Fishwildら(1996,Nature Biotechnology 14:845〜851)に記載されているとおり、ヒトκ軽鎖導入遺伝子KCo5を担持した。HCo7株は、米国特許第5,545,806号、同第5,625,825号および同第5,545,807号(参考として援用される)に記載のとおりHCo7ヒト重鎖導入遺伝子を担持する。HCo12株は、国際特許出願番号WO 01/09187(参考として援用される)の実施例2に記載のとおりHCo12ヒト重鎖導入遺伝子を担持した。HCo7+HCo12株は、HCo7およびHCo12の両方の重鎖導入遺伝子を担持し、そして各々の導入遺伝子について半接合性であった。これらの株の全てが本明細書においてHuMabマウスと呼ばれる。
【0193】
(HuMab免疫:)
IFN−γに対する完全ヒトモノクローナル抗体を生成するため、HuMabマウスを、抗原としてE.coliまたはCHO細胞由来の精製組み換えヒトIFN−γを用いて免疫した。HuMabマウスの一般的な免疫スキームは、Lonbergらに記載されている(各々の教示が参考として援用されている、1994,Nature 368:856〜859;Fishwildら(前出)および国際特許出願公開番号WO98/24884)。マウスは、抗原の第一回の注入の際に6〜16週齢であった。IFN−γ抗原(例えば、IFN−γを発現する、トランスフェクトされたE.coliまたはCHO細胞から精製された)の精製組み換え調製物(25〜100μg)を用いて、HuMabマウスを腹腔内(IP)または皮下(Sc)で免疫した。
【0194】
HuMabトランスジェニックマウスの免疫は、完全フロイントアジュバントおよび2つの注射物中の抗原を用いて、続いて、不完全フロイントアジュバント中の抗原を用いた2〜4週のIP免疫(全部で9免疫まで)によって行なった。数ダースのマウスを各々の抗原について免疫した(E.coliまたはCHO細胞のいずれかで生成したヒトIFN−γ)。HCo7株、HCo12株、およびHCo7+HCo12株の全部で91匹のマウスをIFN−γで免疫した。免疫応答を後眼窩採血によってモニターした。
【0195】
IFN−γに結合する抗体を産生するHuMabマウスを選択するため、免疫したマウス由来の血清をFishwildら(前出)に記載されるとおりELISAによって試験した。要するに、マイクロタイタープレートを、CHO細胞またはE.coli由来の精製された組み換えIFN−γをPBS中に1〜2μL/mLで用いてコーティングして、50μL/ウェルで、4℃で一晩インキュベートし、次いで5%ニワトリ血清を含むPBS/Tween(0.05%)の200μL/ウェルを用いてブロックした。IFN−γ免疫したマウスからの血漿の希釈物を、各々ウェルに添加して、環境温度で1〜2時間インキュベートした。このプレートをPBS/Tweenを用いて洗浄して、次いで西洋ワサビペルオキシダーゼ(HRP)と結合体化したヤギ抗ヒトIgG Fc特異的ポリクローナル試薬とともに室温で1時間インキュベートした。プレートをPBS/Tweenを用いて洗浄して、西洋ワサビペルオキシダーゼ(HRP)と結合体化したヤギ抗ヒトIgG Fc特異的ポリクローナル試薬とともに室温で1時間インキュベートした。洗浄後、このプレートをABTS基質(Sigma Chemical Co.,St.Louis,MO,カタログ番号A−1888,0.22mg/mL)を用いて発色させて、415〜495nmの波長で光学密度(OD)を決定することによって分光光度的に分析した。抗IFN−γヒト免疫グロブリンの十分な力価を有するマウスを用いて下記のとおりモノクローナル抗体を産生した。
【0196】
(IFN−γに対するヒトモノクローナル抗体を産生するハイブリドーマの生成)
屠殺の2日前に抗原を用いて静脈内にブースト(追加免疫)することによりモノクローナル抗体産生のためにマウスを準備して、その後に脾臓を取り出した。HuMabマウスからマウス脾細胞を単離して、標準的なプロトコールを用いてPEGとともにマウス骨髄腫細胞株に融合した。代表的には、各々の抗原について10〜20の融合を行なった。
【0197】
要するに、免疫されたマウス由来の脾臓リンパ球の単一細胞懸濁物を、50%PEG(Sigma)を用いてP3X63−Ag8.653非分泌マウス骨髄腫細胞(ATCC,登録番号CRL1580)の数の四分の一に融合した。細胞を平底マイクロタイタープレート中で約1×105/ウェルでプレートして、続いて10%ウシ胎仔血清、10%P388D1−(ATCC,登録番号CRL TIB−63)馴化培地、3〜5%ORIGEN(登録商標)Hybridoma Cloning Factor(IGEN)、マウスマクロファージ様細胞株を培養するために用いた培地由来の部分的に精製されたハイブリドーマ増殖培地補充物をDMEM(Mediatech,カタログ番号CRL10013、高いグルコース、Lグルタミンおよびピルビン酸ナトリウムを含む)中に、さらに5mM HEPES、0.055mM 2−メルカプトエタノール、50mg/mLゲンタマイシンおよび1×HAT(Sigma,カタログ番号、CRL P−7185)とともに含有する選択培地中での約2週間のインキュベーションを続けた。1〜2週後、HATをHTに置き換えた培地中で細胞を培養した。
【0198】
得られたハイブリドーマを抗原特異的抗体の産生についてスクリーニングした。個々のウェルをELISA(上記)によって、ヒト抗IFN−γモノクローナルIgG抗体についてスクリーニングした。一旦、過剰のハイブリドーマ増殖が生じれば、通常10〜14日後に培地をモニターした。抗体分泌するハイブリドーマを再度プレートして、再度スクリーニングして、依然としてヒトIgGについて陽性である場合、抗IFN−γモノクローナル抗体を少なくとも2回、限界希釈によってサブクローニングした。次いで、安定なサブクローンをインビトロで培養して、精製および特徴づけのために組織培養培地中で少量の抗体を生成した。
【0199】
(IFN−γに結合するヒトモノクローナル抗体の選択)
上記のようなELISAアッセイを用いて、IFN−γ免疫原と正の反応性を示したハイブリドーマをスクリーニングした。IFN−γに対して高い結合活性で結合するモノクローナル抗体を分泌するハイブリドーマをサブクローニングしてさらに特徴付けた。親細胞の反応性を保持している(ELISAによって決定した場合)、各々のハイブリドーマ由来の1つのクローンを、液体窒素中に保持した5〜10バイアルの細胞バンクを作製するために選択した。
【0200】
アイソタイプ特異的ELISAを行なって、本明細書に開示されたとおり産生したモノクローナル抗体のアイソタイプを決定した。これらの実験では、マイクロタイタープレートウェルを、PBS中1μg/mLのマウス抗ヒトκ軽鎖溶液を50μL/ウェルで用いてコーティングして、4℃で一晩インキュベートした。5%ニワトリ血清を用いたブロッキング後、このプレートを各々の試験したモノクローナル抗体由来の上清および精製されたアイソタイプコントロールと反応させた。プレートを環境温度で1〜2時間インキュベートした。次いで、このウェルをヒトIgG1またはIgG3特異的な西洋ワサビペルオキシダーゼ結合体化ヤギ抗ヒトポリクローナル抗血清のいずれかと反応させて、プレートを発色させ、上記のように分析した。
【0201】
ELISAによって検出した場合IFN−γに対する有意な結合を示したハイブリドーマ上清から精製したモノクローナル抗体をさらに、下記のような種々のバイオアッセイを用いて生物学的活性について試験した。選択された抗体を1119、1121、1118*、1121*および1118と命名した。
【0202】
(実施例3)
(抗IFN−γ抗体重鎖および軽鎖のクローニング)
上記の実施例2で同定されたIFN−γ結合モノクローナル抗体1119、1121、1118*、1121*および1118を発現するハイブリドーマを供給源として用いて、総RNA、DNAおよびタンパク質を単離するのに適切なフェノールおよびグアニジンイソチオシアネートの単層溶液であるTRIzol(登録商標)試薬(Invitrogen)を用いて総RNAを単離した。第一鎖のcDNAを、伸長アダプター(5’−GGC CGG ATA GGC CTC CAN NNN NNT−3’)(配列番号23)とともにランダムプライマーを用いて合成し、そして5’RACE(cDNA末端の迅速増幅)分取アッセイを、有効性が改善されているcDNA末端の迅速増幅(RACE)のためのキットであるGENERACERTMKit(Invitrogen)を用いて、製造業者の指示に従って行った。完全な軽鎖コードcDNAを調製するために、フォワードプライマーは、GENERACERTMネスト化プライマーであって、リバースプライマーは5’−GGG GTC AGG CTG GAA CTG AGG−3’(配列番号24)であった。リバースプライマーは、ヒトκ鎖の3’非翻訳領域において見出されるcDNA配列の保存領域を認識するように設計した。重鎖の可変領域をコードするcDNAを調製するために、フォワードプライマーはGENERACERTMネスト化プライマーであって、リバースプライマーは5’−TGA GGA CGC TGA CCA
CAC G−3’(配列番号25)であって、これは、ヒトIgG鎖のFc領域におけるコード配列中の保存領域を認識するように設計した。RACE産物をpCR4−TOPO(Invitrogen)中にクローニングして、その配列を決定した。コンセンサス配列を用いて全長抗体鎖PCR増幅のプライマーを設計した。
【0203】
抗IFN−γκ軽鎖をコードするcDNAを調製するために、5’PCRプライマーは、シグナル配列のアミノ末端、XbaI制限酵素部位、および最適化Kozak配列(5’−ACA ACA AAG CTT CTA GAC CAC CAT GGA AAC CCC AGC TCA GCT TCT CTT−3’;配列番号26)をコードした。3’プライマーは、カルボキシ末端および終止コドン、ならびにSalI制限部位(5’−CTT GTC GAC TCA ACA CTC TCC CCT GTT GAA GCT−3’;配列番号27)をコードした。得られたPCR産物のフラグメントを精製してXbaIおよびSalIを用いて消化し、次いでゲル単離して、哺乳動物発現ベクターpDSRα19に連結した(任意の目的のために参考として本明細書に援用される国際出願公開番号WO 90/14363を参照のこと)。
【0204】
抗IFNγ重鎖をコードするcDNAを調製するため、5’PCRプライマーは、シグナル配列のアミノ末端、XbaI制限酵素部位、および最適化Kozak配列(5’−CAG CAG AAG CTT CTA GAC CAC CAT GGG GTC AAC CGC CAT CCT CG−3’;配列番号28)をコードした。3’プライマーは、天然に存在するセンス鎖BsmBI部位(5’−CTT GGT GGA GGC ACT AGA GAC GGT GAC CAG GGT GCC ACG GCC−3’;配列番号29)を含む可変領域のカルボキシル末端をコードした。得られた産物を精製して、XbaIおよびBsmBIを用いて消化し、ゲル単離して、ヒトIgG1定常領域を含むpDSRα19ベクターに連結した。
【0205】
(実施例4)
(チャイニーズハムスター卵巣(CHO)細胞における抗IFNγ抗体の発現)
リン酸カルシウム法を用いて、1119−重鎖/pDSRα19および1119κ鎖/pDSRα19プラスミドを、ジヒドロ葉酸還元酵素欠損(DHFR−)無血清適合チャイニーズハムスター卵巣(CHO)細胞中に同時トランスフェクションすることによって、1119抗IFN−γ mAbの安定な発現を得た。トランスフェクトされた細胞を透析血清を含むが、ヒポキサンチン−チミジンを含まない培地中で選択して、DHFR酵素を発現する細胞の増殖を確保した。トランスフェクトされたクローンをELISAのようなアッセイを用いてスクリーニングして、馴化培地中で1119抗IFN−γ mAbの発現を検出した。1119発現細胞株をメトトレキセート増幅に供した。増幅の際の最高発現クローンを単細胞クローニングおよび細胞バンクの作製のために選択した。
【0206】
本発明の任意の組み換え抗IFN−γ抗体は、1119MAbについて上記と同じプロトコールを用いて、DHFRの欠損したチャイニーズハムスター卵巣細胞において生成し得る。本発明の各々の抗IFN−γ抗体の完全重鎖または軽鎖をコードするDNA配列を発現ベクター中にクローニングする。DHFRを欠くCHO細胞を、適切な抗IFN−γ抗体の完全重鎖を発現できる発現ベクターおよび完全軽鎖を発現できる発現ベクターを用いて同時トランスフェクトする。例えば、1118抗体を生成するために、配列番号19に示されるアミノ酸配列を含む完全重鎖を発現できるベクターおよび配列番号20に示されるアミノ酸配列を含む完全軽鎖を発現できるベクターを用いて細胞を同時トランスフェクトする。1121抗体を生成するために、配列番号21に示されるアミノ酸配列を含む完全重鎖を発現できるベクター、および配列番号22に示されるアミノ酸配列を含む完全軽鎖を発現できるベクターを用いて細胞を同時トランスフェクトする。1118*抗体を生成するために、配列番号32に示されるアミノ酸配列を含む完全重鎖を発現できるベクター、および配列番号20に示されるアミノ酸配列を含む完全軽鎖を発現できるベクターを用いて細胞を同時トランスフェクトする。1121*抗体を生成するために、配列番号21に示されるアミノ酸配列を含む完全重鎖を発現できるベクター、および配列番号33に示されるアミノ酸配列を含む完全軽鎖を発現できるベクターを用いて細胞を同時トランスフェクトする。表3は、種々のIFNγ抗体についての完全重鎖および完全軽鎖をまとめる。
【0207】
【表3】
(実施例5)
(抗IFN−γ抗体の産生)
1119抗体を発現するCHO細胞のクローン株における発現によって1119抗体を産生した。産生を行なうために、単一のバイアルから細胞を無血清細胞培養培地中に溶かした。この細胞を250mLの振盪フラスコ中で最初に、次いでスピナーフラスコ中で、そして最後にステンレス鋼リアクター中で2000Lのバイオリアクターまでスケールアップして増殖させた。流加培養法を用いて2000Lのバイオリアクター中で産生を行ない、ここには濃縮された媒体成分を含む栄養供給原を加えて、細胞の増殖および培養生存度を維持した。産生を約2週間続けて、この間に細胞によって1119抗体を構成的に産生して、細胞培養培地に分泌させた。
【0208】
産生リアクターを予め決定したpH、温度および溶存酸素レベルで制御した。このpHを二酸化炭素ガスおよび炭酸ナトリウムの添加によって制御した。溶存酸素を、空気、窒素および酸素のガスフローによって制御した。
【0209】
産生の終わりに、細胞ブロスをディスクスタック遠心分離に供して、培養上清を細胞から分離した。さらにこの濃縮物をデプスフィルター、続いて0.2μmのフィルターを通して清澄化する。次いで、清澄化した馴化培地を接線流限外濾過によって濃縮した。馴化培地を15〜30倍に濃縮した。次いで、得られた濃縮された馴化培地を処理して抗体を精製したが、これは後日精製するために凍結してもよい。本明細書に記載される任意の他の抗体は、同様の様式で産生できる。
【0210】
(実施例6)
(抗IFNγ抗体の活性の特徴づけ)
IFN−γは多数の生物学的効果を有するので、いくつかの異なるバイオアッセイを用いて種々のIFN−γ抗体の力価を比較する。以下で記載されたA549アッセイを、そのアッセイでの選択された候補物の能力に基づいたさらなる分析のためにそれらの選択された候補物を用いる一次スクリーニングに使用した。選択された候補物は、1119抗体、1118抗体および1121抗体を含んだ。
【0211】
(A549 バイオアッセイ)
IFN−γの確立された特性の1つは種々の細胞集団に対するその抗増殖効果である。例えば、AuneおよびPogue,1989,J.Clin.Invest.84:863〜75を参照のこと。ヒト肺細胞株A549は、IFN−γの生物活性を記載している刊行物では頻繁に用いられている。例えば、AuneおよびPogue、前出;Hillら、1993,Immunology 79:236〜40を参照のこと。一般には、用量のわずかな変化が応答の変化を生じる、用量応答曲線の一部におさまる刺激物質の濃度でインヒビターの活性を試験する。当業者は、過剰な用量の刺激物質が用いられる場合、応答の変化を観察するには極めて高用量のインヒビターが必要であり得ることを認識する。物質を刺激するために一般に用いられる濃度は、EC80およびEC90(最大応答のそれぞれ80%または90%が達成される濃度)である。
【0212】
IFN−γ用量応答曲線を作成して肺上皮癌細胞株A549のEC90を決定した(約30pM)。引き続く実験では、異なる濃度の精製された抗体を固定用量のIFN−γ(30pM)と混合して、この抗体がIFN−γの抗増殖性効果の生物学的活性を阻害する能力を決定した。このアッセイを5日間行なって、代謝的に活性な、すなわち増殖している細胞による、細胞増殖を示すために用いられる色素であるALAMARBLUETM(AccuMed International,Inc.,Chicago,Illinois)の減少によって生じた蛍光を決定することによって増殖を測定した。例えば、de FriesおよびMitsuhashi,1995,J.Clin.Lab.Analysis 9(2):89〜95;Ahmedら、1994,J.Immunol.Methods 170(2):211〜24を参照のこと。
【0213】
図9に示されるとおり、1119抗体は、14pMというIC50(IFN−γの効果の50%阻害を達成した濃度)を有する最も強力な抗体であって、1121(46pM)および1118(97pM)がそれに続く。
【0214】
(HLA DRバイオアッセイ)
IFN−γの確立された別の特性は、種々の細胞タイプにおけるMHCクラスIおよびクラスII遺伝子の発現を上方制御する能力である。この活性は特に、ループス腎炎に関連し得る(Yokoyamaら、1992,Kidney Int.42:755〜63)。THP−1ヒト単球細胞株は、IFN−γのこの生物活性について述べている刊行物では頻繁に用いられている。IFN−γ用量応答曲線を作成して、この実験に用いられる特定のTHP−1細胞株のEC80を決定した(約21pM)。引き続く実験では、種々の濃度の精製された抗体を固定用量のIFN−γ(21pM)と混合して、この抗体が細胞表面上でのHLA DR発現のIFN−γ誘導性上方制御を中和または阻害する能力を決定した。このアッセイを24時間行ない、測定した指標は、細胞に対するFITC標識抗HLA DR抗体の結合を検出するためのFACS解析によって決定された平均蛍光強度であった。
【0215】
図10に示されるとおり、1119抗体は、14pMというIC50を有する最も強力な抗体であって、1121抗体(60pM)および1118抗体(86pM)がそれに続く。
【0216】
(全血のバイオアッセイ)
いくつかの異なる細胞株におけるIP−10ケモカインの産生をIFN−γが上方制御するという公表された観察に基づいてヒト全血のアッセイを行なった。この活性は特に、ループス腎炎に関連し得る(Narumiら、2000,Cytokine 12:1561〜1565)。多数の正常ヒトドナー由来の全血を、IFN−γがIP−10産生を増大する能力について試験した。IFN−γ用量応答曲線を作成して、個々のドナーについてのEC50を決定した。予想どおり、いくつかの変動がドナーの間で観察された。一般に、50〜100pMのEC50を再現可能に示すと考えられるドナーを用いた。全血を固定濃度のIFN−γおよび種々の濃度の抗体と混合して、18.5時間インキュベートして、次いでELISAによってIP−10レベルを決定した。全血アッセイからの2つの異なるドナーについての代表的な結果を図11に示す。これらの2つのドナーからのIC50は17pMおよび14pMであった。現在まで、外因性IFN−γの添加の必要なしに全血アッセイにおいて自然に上昇したIP−10レベルを有する1つのドナーが同定されている。抗IFNγ抗体は、おそらく内因性に生成されたIFN−γをブロックすることによって、IP−10のこの自然な産生をブロックできた。
【0217】
(生化学的アッセイ)
IFN−γに対するいくつかの抗体についての結合動態を、BIAコア分析によって測定した。最初の結果では、この抗体が、分子間の結合を測定するために表面プラズモン共鳴を用いる装置である、BIACORETM(Pharmacis Biosensor
AB Corporation,Uppsala,Sweden)での信頼性のある測定の限界にアプローチするオフレート(off−rate)を有することが示唆された。従って、平衡結合アッセイを行なって用いた。平衡に達するように、固定量の抗体を種々の濃度のIFN−γとともに5時間を超えてインキュベートし、次いでIFN−γ結合ビーズと極めて短時間接触させて、このビーズに結合した遊離の抗体の量を、蛍光に基づく免疫アッセイ装置であるKINEXATM装置(Sapidyne Instruments Inc.,Boise,ID)で測定した。1119抗体では、最低の平衡解離定数である約24pMが得られた。
【0218】
(実施例7)
(種交差反応性)
上記の抗体がいくつかの異なる種由来の組み換えIFN−γタンパク質を中和または阻害する能力について、それらの抗体を試験した。マウスIFN−γタンパク質は市販されているが、ヒト、カニクイザルおよびチンパンジーのIFN−γタンパク質は、クローニングして、ヒト293細胞のような従来の哺乳動物発現系において発現した。ヒト、カニクイザルおよびチンパンジーのIFN−γタンパク質は全てが以前に記載されたA549アッセイで活性であるが、マウスタンパク質は、このアッセイでは活性ではない。マウスタンパク質は、マウス細胞株の置換について以外は以前に記載されたA549アッセイと本質的に同一である、RAW 264.7細胞株に基づくアッセイにおいて活性であった。RAW 264.7はマウス単球マクロファージ細胞株であり、そして例えば、American Type Culture Collectionから得ることができる。表4に示されるとおり、3つ全ての抗体がヒトおよびチンパンジーのIFN−γを中和できたが、3つのうちどれもカニクイザルまたはマウス由来のIFN−γの生物学的活性を中和も阻害もできなかった。
【0219】
【表4】
(実施例8)
(抗IFN−γ抗体のエピトープの同定)
成熟ヒトおよびカニクイザルのIFN−γのアミノ酸配列の比較によって、それらの間に9つのアミノ酸の相違が、ヒトIFN−γ配列の19、20、31、34、65、77、103、110および126位置において存在することが示された。ヒトおよびチンパンジーのIFN−γの配列は、ThakurおよびLandolfi(1999),Molecular Immunology 36:1107〜15に開示されている。カニクイザルIFN−γ配列は、TatsumiおよびSata(1997)、Int.Arch.Allergy Immunol.114(3):229〜36に開示されている;そしてマウスIFN−γ配列は、例えば、National Center for Biotechnology Information(NCBI)アクセッション番号NP_032363に開示されている。市販のキットを用いる部位特異的突然変異誘発を用いて、ヒトIFN−γ内の相違するヒトアミノ酸の各々を、カニクイザルタンパク質由来の対応するアミノ酸によって個々に置換した。各々の置換されたIFN−γは、ヒトIFNγ配列に存在するアミノ酸を置換するために用いたアミノ酸についての記号、およびこの置換が生じた成熟ヒトIFN−γ配列における位置に従って「huIFNγ」と命名した。例えば、「huIFNγD19」は、19位置を除いてヒトIFN−γに同一のIFN−γのバージョンを表し、ここでは正常にはこの位置を占めるヒスチジンにアスパラギン酸が置き換わっている。同様の実験を行ない、ここではカニクイザルIFN−γ内で開始して、各々の相違するアミノ酸をヒトIFN−γに存在するアミノ酸で置換した。例えば、「cynoIFNγL103」は、103位置を除いてカニクイザルIFN−γに同一のIFN−γのバージョンを表し、ここでは正常にはこの位置を占めるセリンにロイシン酸が置き換わっている。表5および6を参照のこと。これらの変異タンパク質を、ヒト293細胞のような従来の哺乳動物発現系で発現させた。全ての変異体IFN−γタンパク質が、A549アッセイにおいて決定された活性を保持していた。1119抗体が種々の変異体IFN−γタンパク質の生物学的活性を中和または阻害する能力について、A549バイオアッセイを用いて決定して、この抗体を試験した。1119抗体がヒトIFN−γ抗増殖活性を中和または阻害する能力を、上記のような蛍光を測定することによって決定して、これを1119抗体がIFN−γの改変型の各活性を中和する能力を比較するためのベースラインとして用いた。ヒトIFN−γで開始した構築物について、IFN−γ活性の阻害をパーセンテージとして測定し、ここではヒトIFN−γおよび1119抗体の存在下で観察された蛍光の最大レベル(増殖している細胞によるALAMARBLUETMの減少に起因する)を100%に設定して、IFN−γの各変化型に加えて1119抗体の存在下で測定した最大レベルとこれとを比較した。あるいは、カニクイザルIFN−γで開始する構築物については、観察された蛍光に基づいて阻害を定量的にスコア付けした。
【0220】
表5にまとめたとおり、1119抗体は、ヒトIFN−γ、ならびにhuIFNγD19およびhuIFNγP20を除くヒトIFNγの全ての置換変異体の生物学的活性を中和または阻害することができた。前の実施例のように、カニクイザルIFN−γタンパク質はまた、1119抗体では阻害されなかった。この解析によって、残基19および20が1119抗体とIFN−γとの間の相互作用に特に重要であり、そして1119抗体とヒトIFN−γとの間の接触のポイントとして機能し得ることが示される。
【0221】
【表5】
表6は、19位置および20位置でヒト配列に適合するカニクイザルIFNγ配列を作成する置換を含むカニクイザルIFNγの変更バージョンが、1119抗体によって中和されなかったことを示す。しかし、19、20および65、または19、20および103位置にヒト配列置換を有するカニクイザルIFNγのバージョンは、1119抗体によって中和され、これはこれらのいずれかに加えてさらに置換を含むバージョンと同様であった。
【0222】
【表6】
(実施例9)
(チンパンジーに対する投与の際の抗IFN−γ抗体の生物学的活性)
3週間の間、毎週20mg/kgの用量で2匹のチンパンジーに1119抗体を投与した。抗体の投与の2週前(−2)または1週前(−1)にチンパンジーから血液を採取し、さらに抗体の初回用量の投与の2、8、15、29および36日後に血液を採取した。実施例6に記載のヒト全血アッセイにおいて用いたのと本質的に同じ方法を用いて、この採取した血液でチンパンジーの全血アッセイを行なったが、重要な相違は、この抗体は全血を外因性に加えたのではなくこのチンパンジーに事前にインビボで投与したということであった。IFN−γを種々の濃度で血液に加えて(0.01ng/ml、3.9ng/mlまたは1μg/ml)、この血液を37℃で20〜24時間インキュベートして、次いでビーズに基づくELISAによってIP−10産生を測定した。
【0223】
図14および15でわかるように、抗体投与の前に動物から採取された血液は、IFN−γと反応性であって、IP−10産生が濃度依存性に増大した。対照的に、抗体投与後に採取された血液は、試験したどの濃度のIFN−γとも反応せず、IP−10の産生が増大した。これらデータによって、この抗体はインビボでの投与後でさえ、IFN−γを中和する能力を保持していたことが示される。さらにこれらの培養物に添加されたIFN−γの量は、チンパンジーでのIFN−γの内因性のレベルを大きく超えており、これによって、投与された抗体がインビボで内因性のIFN−γを中和できることが示唆された。
【0224】
前述の開示によって本発明の特定の実施形態が強調されること、およびその全ての改変または変更が添付の特許請求の範囲に示される本発明と等価であり、そのうえ本発明の趣旨および範囲内であることが理解されるべきである。本明細書に引用される全ての参考文献はその全体が参考として援用される。
【図面の簡単な説明】
【0225】
【図1A】図1A〜1Bは、1118、1118*、1119、1121および1121*の抗IFN−γ抗体の重鎖定常領域のアミノ酸配列(図1B;配列番号2)をコードするcDNA(図1A;配列番号1)の一部のヌクレオチド配列を示す。
【図1B】図1A〜1Bは、1118、1118*、1119、1121および1121*の抗IFN−γ抗体の重鎖定常領域のアミノ酸配列(図1B;配列番号2)をコードするcDNA(図1A;配列番号1)の一部のヌクレオチド配列を示す。
【図2A】図2A〜2Bは、1118、1118*、1119、1121および1121*の抗IFN−γ抗体の軽鎖定常領域のアミノ酸配列(図2B;配列番号4)をコードするcDNA(図2A;配列番号3)の一部のヌクレオチド配列を示す。
【図2B】図2A〜2Bは、1118、1118*、1119、1121および1121*の抗IFN−γ抗体の軽鎖定常領域のアミノ酸配列(図2B;配列番号4)をコードするcDNA(図2A;配列番号3)の一部のヌクレオチド配列を示す。
【図3A】図3A〜3Bは、1119の抗IFN−γ抗体の重鎖可変領域のアミノ酸配列(図3B;配列番号6)をコードするcDNA(図3A;配列番号5)の一部のヌクレオチド配列を示す。
【図3B】図3A〜3Bは、1119の抗IFN−γ抗体の重鎖可変領域のアミノ酸配列(図3B;配列番号6)をコードするcDNA(図3A;配列番号5)の一部のヌクレオチド配列を示す。
【図4A】図4A〜4Bは、1119の抗IFN−γ抗体の軽鎖可変領域のアミノ酸配列(図4B;配列番号8)をコードするcDNA(図4A;配列番号7)の一部のヌクレオチド配列を示す。
【図4B】図4A〜4Bは、1119の抗IFN−γ抗体の軽鎖可変領域のアミノ酸配列(図4B;配列番号8)をコードするcDNA(図4A;配列番号7)の一部のヌクレオチド配列を示す。
【図5A】図5A〜5Bは、1118の抗IFN−γ抗体の重鎖可変領域のアミノ酸配列(図5B;配列番号10)をコードするcDNA(図5A;配列番号9)の一部のヌクレオチド配列を示す。図5Cは、1118*の抗IFN−γ抗体の重鎖可変領域のアミノ酸配列(配列番号30)を示す。図5Dは、1118*の抗IFN−γ抗体の重鎖可変領域のヌクレオチド配列(配列番号56)を示す。
【図5B】図5A〜5Bは、1118の抗IFN−γ抗体の重鎖可変領域のアミノ酸配列(図5B;配列番号10)をコードするcDNA(図5A;配列番号9)の一部のヌクレオチド配列を示す。図5Cは、1118*の抗IFN−γ抗体の重鎖可変領域のアミノ酸配列(配列番号30)を示す。図5Dは、1118*の抗IFN−γ抗体の重鎖可変領域のヌクレオチド配列(配列番号56)を示す。
【図5C】図5A〜5Bは、1118の抗IFN−γ抗体の重鎖可変領域のアミノ酸配列(図5B;配列番号10)をコードするcDNA(図5A;配列番号9)の一部のヌクレオチド配列を示す。図5Cは、1118*の抗IFN−γ抗体の重鎖可変領域のアミノ酸配列(配列番号30)を示す。図5Dは、1118*の抗IFN−γ抗体の重鎖可変領域のヌクレオチド配列(配列番号56)を示す。
【図5D】図5A〜5Bは、1118の抗IFN−γ抗体の重鎖可変領域のアミノ酸配列(図5B;配列番号10)をコードするcDNA(図5A;配列番号9)の一部のヌクレオチド配列を示す。図5Cは、1118*の抗IFN−γ抗体の重鎖可変領域のアミノ酸配列(配列番号30)を示す。図5Dは、1118*の抗IFN−γ抗体の重鎖可変領域のヌクレオチド配列(配列番号56)を示す。
【図6A】図6A〜6Bは、1118または1118*の抗IFN−γ抗体の軽鎖可変領域のアミノ酸(図6B;配列番号12)をコードするcDNA(図6A;配列番号11)の一部のヌクレオチド配列を示す。
【図6B】図6A〜6Bは、1118または1118*の抗IFN−γ抗体の軽鎖可変領域のアミノ酸(図6B;配列番号12)をコードするcDNA(図6A;配列番号11)の一部のヌクレオチド配列を示す。
【図7A】図7A〜7Bは、1121、または1121*の抗IFN−γ抗体の重鎖定常領域のアミノ酸配列(図7B;配列番号14)をコードするcDNA(図7A;配列番号13)の一部のヌクレオチド配列を示す。
【図7B】図7A〜7Bは、1121、または1121*の抗IFN−γ抗体の重鎖定常領域のアミノ酸配列(図7B;配列番号14)をコードするcDNA(図7A;配列番号13)の一部のヌクレオチド配列を示す。
【図8A】図8A〜8Bは、1121の抗IFN−γ抗体の軽鎖可変領域のアミノ酸配列(図8B;配列番号16)をコードするcDNA(図8A;配列番号15)の一部のヌクレオチド配列を示す。図8Cは、1121*の抗IFN−γ抗体の軽鎖可変領域のアミノ酸配列(図31)を示す。図8Dは、1121*の抗IFN−γ抗体の軽鎖可変領域のヌクレオチド配列(配列番号57)を示す。
【図8B】図8A〜8Bは、1121の抗IFN−γ抗体の軽鎖可変領域のアミノ酸配列(図8B;配列番号16)をコードするcDNA(図8A;配列番号15)の一部のヌクレオチド配列を示す。図8Cは、1121*の抗IFN−γ抗体の軽鎖可変領域のアミノ酸配列(図31)を示す。図8Dは、1121*の抗IFN−γ抗体の軽鎖可変領域のヌクレオチド配列(配列番号57)を示す。
【図8C】図8A〜8Bは、1121の抗IFN−γ抗体の軽鎖可変領域のアミノ酸配列(図8B;配列番号16)をコードするcDNA(図8A;配列番号15)の一部のヌクレオチド配列を示す。図8Cは、1121*の抗IFN−γ抗体の軽鎖可変領域のアミノ酸配列(図31)を示す。図8Dは、1121*の抗IFN−γ抗体の軽鎖可変領域のヌクレオチド配列(配列番号57)を示す。
【図8D】図8A〜8Bは、1121の抗IFN−γ抗体の軽鎖可変領域のアミノ酸配列(図8B;配列番号16)をコードするcDNA(図8A;配列番号15)の一部のヌクレオチド配列を示す。図8Cは、1121*の抗IFN−γ抗体の軽鎖可変領域のアミノ酸配列(図31)を示す。図8Dは、1121*の抗IFN−γ抗体の軽鎖可変領域のヌクレオチド配列(配列番号57)を示す。
【図9】図9は、1119、1118および1121のモノクローナル抗体を用いたA549バイオアッセイにおけるIFN−γの生物学的活性の中和または阻害を示すグラフを含む。
【図10】図10は、1119、1118および1121のモノクローナル抗体によるTHP−1/HLA DRバイオアッセイにおけるIFN−γの生物学的活性の中和または阻害を示すグラフを含む。
【図11】図11は、1119のモノクローナル抗体による全血バイオアッセイ(2つのドナー)におけるIFN−γの生物学的活性の中和または阻害を示すグラフを含む。
【図12】図12は、1118、1118*、1121および1119と命名された抗IFNモノクローナル抗体の重鎖のアミノ末端部分(可変領域を含む)のアラインメントを示す。この配列は、実施例3において単離されたcDNA上にコードされるシグナル配列を含む。このシグナル配列は、1位〜19位に延びる。CDRには下線を付している。図に示すとおり、CDR1は、アミノ酸50〜54にまたがり、CDR2は、69〜85にまたがり、そしてCDR3は、118〜125にまたがる。Kabatら(1991、Sequences of Proteins of Immunological Interest,Public Health Service N.I.H.,Bethesda,MD)の番号付けシステムは、成熟抗体の最初のアミノ酸で開始して、シグナル配列は除く。従って、この図の20位は、Kabatら(前出)の1位に相当する。
【図13】図13は、1118、1121、1121*および1119と命名された抗IFNモノクローナル抗体の軽鎖のアミノ末端部分(可変領域を含む)のアラインメントを示す。この配列は、実施例3において単離されたcDNA上にコードされるシグナル配列を含む。このシグナル配列は、1〜20位に延びる。CDRには下線を付している。図に示すとおり、CDR1は、アミノ酸44〜55にまたがり、CDR2は、71〜77にまたがり、そしてCDR3は、110〜118にまたがる。Kabatら(前出)の番号付けシステムは、シグナル配列を除くので、この図の21位は、Kabatらの1位に相当する。
【図14】図14は、週に1回行なった、抗IFNγ抗体の3回の注射の開始の2週間前もしくは1週間前(図14では「−2」および「−1」と表示した線)、または2日間後、8日間後、15日間後、29日間後もしくは36日後(図14では「2」、「8」、「15」、「29」もしくは「36」と表示した線)においてチンパンジーから採取した全血による、IFN−γに応答したIP−10の産生を示す。
【図15】図15は、異なるチンパンジーの血液を用いた以外は、図14と同様である。
【技術分野】
【0001】
本出願は、2002年10月16日出願の米国仮出願番号60/419,057および2003年6月17日出願の米国仮出願番号60/479,241号の利益を請求する。
【0002】
(発明の分野)
本発明は、インターフェロンγ(IFN−γ)に結合するヒトモノクローナル抗体に関する。IFN−γによって媒介される疾患を処置するための組成物および方法もまた、記載される。
【背景技術】
【0003】
(背景)
インターフェロン(IFN)はもともと、宿主細胞のウイルス感染を妨害する能力に基づいて命名された(非特許文献1:IsaacsおよびLindenman,1957,Proc.R.Soc.147:28〜267)。それらの発見以来、インターフェロンファミリーの多数のメンバーが、抗ウイルス防御に加えて、細胞増殖および細胞免疫を含む種々の生物学的役割を有することが特定されている。インターフェロンの型であるIFN−α、IFN−β、IFN−ωおよびIFN−γは、I型インターフェロンであり、I型IFNレセプターに結合するが、IFN−γはII型インターフェロンであり、II型IFNレセプターに結合する(非特許文献2:Pfefferら、1998,Cancer Res.58:2489〜2499)。
【0004】
IFN−γのシグナル伝達は、少なくとも以下の5つの別個のタンパク質に依存する:IFNGR1およびIFNGR2(IFN−γレセプターのサブユニット)、Jak1、Jak2および転写因子STAT1(非特許文献3:SchindlerおよびDarnell,1995,Annu.Rev.Biochem.64:621〜651;非特許文献4:Bachら、1997,Annu.Rev.Immunol.15:563〜591)。IFN−γレセプターは、成熟赤血球を除くほとんどの細胞型で見出される(非特許文献5:FarrarおよびSchreiber,1993,Annu.Rev.Immunol.11:571〜611)。Jak1タンパク質、Jak2タンパク質およびSTAT1タンパク質は、IFN−γシグナル伝達を媒介する。
【0005】
IFN−γは、種々の生物学的機能(例えば、抗ウイルス応答、細胞増殖、免疫応答および腫瘍抑制)を調節し、そしてIFN−γは、種々のヒト疾患を媒介し得る。従って、IFN−γの生物学的活性を調節し得る因子が当該分野では必要である。
【非特許文献1】IsaacsおよびLindenman,1957,Proc.R.Soc.147:28〜267
【非特許文献2】Pfefferら、1998,Cancer Res.58:2489〜2499)
【非特許文献3】SchindlerおよびDarnell,1995,Annu.Rev.Biochem.64:621〜651
【非特許文献4】Bachら、1997,Annu.Rev.Immunol.15:563〜591
【非特許文献5】FarrarおよびSchreiber,1993,Annu.Rev.Immunol.11:571〜611
【発明の開示】
【課題を解決するための手段】
【0006】
(発明の要旨)
本発明は、インターフェロンγ(IFN−γ)に結合するモノクローナル抗体およびそれらをコードするポリヌクレオチドを提供する。この抗体は、IFN−γの生物学的活性の少なくとも1つを阻害または調節し得、そしてIFN−γ媒介性疾患の影響を緩和するために有用であり得る。本発明のモノクローナル抗体を産生して細胞培養培地中に分泌し得るハイブリドーマ細胞もまた、本発明によって提供される。本発明の抗体は、IFN−γによって媒介される疾患を処置するために有用であり得る。
【0007】
特定の局面では、本発明は、抗体(必要に応じて、モノクローナル抗体および/またはヒト抗体)を提供し、この抗体は、重鎖および軽鎖を含み、ここでこの重鎖は、配列番号2に示されるアミノ酸配列、またはその抗原結合フラグメントもしくは免疫学的に機能的な免疫グロブリンフラグメントを含む。
【0008】
特定の局面では、本発明は、抗体(必要に応じて、モノクローナル抗体)を提供し、この抗体は、重鎖および軽鎖を含むヒト抗体であり得、この重鎖は、IgG1、IgG2またはIgG4重鎖定常領域を含む。いくつかの実施形態では、本発明の抗体は、配列番号2に示されるIgG1重鎖定常領域のアミノ酸配列、またはその抗原結合フラグメントもしくは免疫学的に機能的な免疫グロブリンフラグメントを含む。
【0009】
特定の局面では、本発明の抗体は、重鎖および軽鎖を含み、この重鎖の可変領域は、配列番号6、配列番号10、配列番号14もしくは配列番号30のいずれかに示されるアミノ酸配列、またはその抗原結合フラグメントもしくは免疫学的に機能的な免疫グロブリンフラグメントを含む。他の局面では、この軽鎖可変領域は、配列番号8、配列番号12、配列番号16もしくは配列番号31のいずれかに示されるアミノ酸配列、またはその抗原結合フラグメントもしくは免疫学的に機能的な免疫グロブリンフラグメントを含む。さらなる局面では、この重鎖は、配列番号17、配列番号19、配列番号21もしくは配列番号32のいずれかに示されるアミノ酸配列、またはその抗原結合フラグメントもしくは免疫学的に機能的な免疫グロブリンフラグメントを含む。なおさらなる局面では、この軽鎖は、配列番号18、配列番号20、配列番号22もしくは配列番号33のいずれかに示されるアミノ酸配列、またはその抗原結合フラグメントもしくは免疫学的に機能的な免疫グロブリンフラグメントを含む。
【0010】
本発明はまた、IFN−γに対して特異的に結合する抗体を提供し、ここでこの重鎖は、重鎖可変領域を含み、この重鎖可変領域は、配列番号6に示されるアミノ酸配列、またはその抗原結合フラグメントもしくは免疫学的に機能的な免疫グロブリンフラグメントを含み、そしてこの軽鎖は、軽鎖可変領域を含み、この軽鎖可変領域は、配列番号8に示されるアミノ酸配列、またはその抗原結合フラグメントもしくは免疫学的に機能的な免疫グロブリンフラグメントを含む。
【0011】
特定の局面では、本発明はまた、IFN−γに特異的に結合し得、そして/またはその生物学的活性を阻害もしくは調節し得る、抗体およびその免疫学的に機能的な免疫グロブリンフラグメントを提供し、この抗体は、重鎖および軽鎖を含み、この重鎖は、重鎖可変領域を含み、この重鎖可変領域は、配列番号6に示されるアミノ酸配列に対して少なくとも80%、85%、90%、91%、92%、93%、94%、95%、96%、97%、98%もしくは約99%の同一性を有する配列を含み、そしてこの軽鎖は、軽鎖可変領域を含み、この軽鎖可変領域は、配列番号8に示されるアミノ酸配列に対して少なくとも80%、85%、90%、91%、92%、93%、94%、95%、96%、97%、98%もしくは約99%の同一性を有する配列を含み、この抗体は、IFN−γと相互作用する。
【0012】
本発明はさらに、IFN−γの生物学的活性を阻害し得るかもしくは調節し得、そして/またはIFN−γに特異的に結合し得る抗体を提供し、この重鎖は、配列番号10に示されるアミノ酸配列を含む重鎖可変領域、またはその抗原結合フラグメントもしくは免疫学的に機能的な免疫グロブリンフラグメントを含み、そしてこの軽鎖は、配列番号12に示されるアミノ酸配列を含む軽鎖可変領域、またはその抗原結合フラグメントもしくは免疫学的に機能的な免疫グロブリンフラグメントを含む。
【0013】
特定の局面では、本発明は、IFN−γの生物学的活性を阻害し得るかもしくは調節し得、そして/またはIFN−γに特異的に結合し得る抗体を提供し、この抗体は、重鎖および軽鎖を含み、この重鎖は、重鎖可変領域を含み、この重鎖可変領域は、配列番号10に示されるアミノ酸配列に対して少なくとも80%、85%、90%、91%、92%、93%、94%、95%、96%、97%、98%もしくは約99%の同一性を有する配列を含み、そしてこの軽鎖は、軽鎖可変領域を含み、この軽鎖可変領域は、配列番号12に示されるアミノ酸配列に対して少なくとも80%、85%、90%、91%、92%、93%、94%、95%、96%、97%、98%もしくは約99%の同一性を有する配列を含み、この抗体は、IFN−γと相互作用する。
【0014】
本発明はさらに、IFN−γの生物学的活性を阻害し得るかもしくは調節し得、そして/またはIFN−γに特異的に結合し得る抗体を提供し、この重鎖は、配列番号30に示されるアミノ酸配列を含む重鎖可変領域、またはその抗原結合フラグメントもしくは免疫学的に機能的な免疫グロブリンフラグメントを含み、そしてこの軽鎖は、配列番号12に示されるアミノ酸配列を含む軽鎖可変領域、またはその抗原結合フラグメントもしくは免疫学的に機能的な免疫グロブリンフラグメントを含む。
【0015】
特定の局面では、本発明は、IFN−γの生物学的活性を阻害し得るかもしくは調節し得、そして/またはIFN−γに特異的に結合し得る抗体を提供し、この抗体は、重鎖および軽鎖を含み、この重鎖は、重鎖可変領域を含み、この重鎖可変領域は、配列番号30のいずれかに示されるアミノ酸配列に対して少なくとも80%、85%、90%、91%、92%、93%、94%、95%、96%、97%、98%もしくは約99%の同一性を有する配列を含み、そしてこの軽鎖は、軽鎖可変領域を含み、この軽鎖可変領域は、配列番号12に示されるアミノ酸配列に対して少なくとも80%、85%、90%、91%、92%、93%、94%、95%、96%、97%、98%もしくは約99%の同一性を有する配列を含み、この抗体は、IFN−γと相互作用する。
【0016】
本発明はまた、IFN−γの生物学的活性を阻害し得るかもしくは調節し得、そして/またはIFN−γに特異的に結合し得る抗体を提供し、この重鎖は、配列番号14に示されるアミノ酸配列を含む重鎖可変領域、またはその抗原結合フラグメントもしくは免疫学的に機能的な免疫グロブリンフラグメントを含み、そしてこの軽鎖は、配列番号16に示されるアミノ酸配列を含む軽鎖可変領域、またはその抗原結合フラグメントもしくは免疫学的に機能的な免疫グロブリンフラグメントを含む。
【0017】
特定の局面では、本発明は、IFN−γに特異的に結合し得そして/またはIFN−γの生物学的活性を阻害し得るかもしくは調節し得る抗体を提供し、この抗体は、重鎖および軽鎖を含み、この重鎖は、重鎖可変領域を含み、この重鎖可変領域は、配列番号14に示されるアミノ酸配列に対して少なくとも80%、85%、90%、91%、92%、93%、94%、95%、96%、97%、98%もしくは約99%の同一性を有する配列を含み、そしてこの軽鎖は、軽鎖可変領域を含み、この軽鎖可変領域は、配列番号16に示されるアミノ酸配列に対して少なくとも80%、85%、90%、91%、92%、93%、94%、95%、96%、97%、98%もしくは約99%の同一性を有する配列を含み、この抗体は、IFN−γと相互作用する。
【0018】
本発明はまた、IFN−γの生物学的活性を阻害し得るかもしくは調節し得、そして/またはIFN−γに特異的に結合し得る抗体を提供し、この重鎖は、配列番号14に示されるアミノ酸配列を含む重鎖可変領域、またはその抗原結合フラグメントもしくは免疫学的に機能的な免疫グロブリンフラグメントを含み、そしてこの軽鎖は、配列番号31に示されるアミノ酸配列を含む軽鎖可変領域、またはその抗原結合フラグメントもしくは免疫学的に機能的な免疫グロブリンフラグメントを含む。
【0019】
特定の局面では、本発明は、IFN−γの生物学的活性を阻害し得るかもしくは調節し得、そして/またはIFN−γに特異的に結合し得る抗体を提供し、この抗体は、重鎖および軽鎖を含み、この重鎖は、重鎖可変領域を含み、この重鎖可変領域は、配列番号14に示されるアミノ酸配列に対して少なくとも80%、85%、90%、91%、92%、93%、94%、95%、96%、97%、98%もしくは約99%の同一性を有する配列を含み、そしてこの軽鎖は、軽鎖可変領域を含み、この軽鎖可変領域は、配列番号31に示されるアミノ酸配列に対して少なくとも80%、85%、90%、91%、92%、93%、94%、95%、96%、97%、98%もしくは約99%の同一性を有する配列を含み、この抗体は、IFN−γと相互作用する。
【0020】
本発明はまた、IFN−γの生物学的活性を阻害し得るかもしくは調節し得、そして/またはIFN−γに特異的に結合し得る抗体を提供し、この重鎖は、配列番号17に示されるアミノ酸配列を含む重鎖可変領域、またはその抗原結合フラグメントもしくは免疫学的に機能的な免疫グロブリンフラグメントを含み、そしてこの軽鎖は、配列番号18に示されるアミノ酸配列、またはその抗原結合フラグメントもしくは免疫学的に機能的な免疫グロブリンフラグメントを含む。
【0021】
特定の局面では、本発明は、IFN−γの生物学的活性を阻害し得るかもしくは調節し得、そして/またはIFN−γに特異的に結合し得る抗体を提供し、この抗体は、重鎖および軽鎖を含み、この重鎖は、重鎖可変領域を含み、この重鎖可変領域は、配列番号17に示されるアミノ酸配列に対して少なくとも80%、85%、90%、91%、92%、93%、94%、95%、96%、97%、98%もしくは約99%の同一性を有する配列を含み、そしてこの軽鎖は、軽鎖可変領域を含み、この軽鎖可変領域は、配列番号18に示されるアミノ酸配列に対して少なくとも80%、85%、90%、91%、92%、93%、94%、95%、96%、97%、98%もしくは約99%の同一性を有するアミノ酸配列を含み、この抗体は、IFN−γと相互作用する。
【0022】
本発明はさらに、IFN−γの生物学的活性を阻害し得るかもしくは調節し得、そして/またはIFN−γに特異的に結合し得る抗体を提供し、この重鎖は、配列番号19に示されるアミノ酸配列、またはその抗原結合フラグメントもしくは免疫学的に機能的な免疫グロブリンフラグメントを含み、そしてこの軽鎖は、配列番号20に示されるアミノ酸配列、またはその抗原結合フラグメントもしくは免疫学的に機能的な免疫グロブリンフラグメントを含む。
【0023】
特定の局面では、本発明は、IFN−γの生物学的活性を阻害し得るかもしくは調節し得、そして/またはIFN−γに特異的に結合し得る抗体を提供し、この抗体は、重鎖および軽鎖を含み、この重鎖は、重鎖可変領域を含み、この重鎖可変領域は、配列番号19のいずれかに示されるアミノ酸配列に対して少なくとも80%、85%、90%、91%、92%、93%、94%、95%、96%、97%、98%もしくは約99%の同一性を有する配列を含み、そしてこの軽鎖は、軽鎖可変領域を含み、この軽鎖可変領域は、配列番号20に示されるアミノ酸配列に対して少なくとも80%、85%、90%、91%、92%、93%、94%、95%、96%、97%、98%もしくは約99%の同一性を有する配列を含み、この抗体は、IFN−γと相互作用する。
【0024】
本発明はまた、IFN−γの生物学的活性を阻害し得るかもしくは調節し得、そして/またはIFN−γに特異的に結合し得る抗体を提供し、この重鎖は、配列番号21に示されるアミノ酸配列、またはその抗原結合フラグメントもしくは免疫学的に機能的な免疫グロブリンフラグメントを含み、そしてこの軽鎖は、配列番号22に示されるアミノ酸配列、またはその抗原結合フラグメントもしくは免疫学的に機能的な免疫グロブリンフラグメントを含む。
【0025】
特定の局面では、本発明は、IFN−γの生物学的活性を阻害し得るかもしくは調節し得、そして/またはIFN−γに特異的に結合し得る抗体を提供し、この抗体は、重鎖および軽鎖を含み、この重鎖は、重鎖可変領域を含み、この重鎖可変領域は、配列番号21のいずれかに示されるアミノ酸配列に対して少なくとも80%、85%、90%、91%、92%、93%、94%、95%、96%、97%、98%もしくは約99%の同一性を有する配列を含み、そしてこの軽鎖は、軽鎖可変領域を含み、この軽鎖可変領域は、配列番号22に示されるアミノ酸配列に対して少なくとも80%、85%、90%、91%、92%、93%、94%、95%、96%、97%、98%もしくは約99%の同一性を有するアミノ酸配列を含み、この抗体は、IFN−γと相互作用する。
【0026】
本発明はまた、IFN−γの生物学的活性を阻害し得るかもしくは調節し得、そして/またはIFN−γに特異的に結合し得る抗体を提供し、この重鎖は、配列番号32に示されるアミノ酸配列、またはその抗原結合フラグメントもしくは免疫学的に機能的な免疫グロブリンフラグメントを含み、そしてこの軽鎖は、配列番号20に示されるアミノ酸配列、またはその抗原結合フラグメントもしくは免疫学的に機能的な免疫グロブリンフラグメントを含む。
【0027】
特定の局面では、本発明は、IFN−γの生物学的活性を阻害し得るかもしくは調節し得、そして/またはIFN−γに特異的に結合し得る抗体を提供し、この抗体は、重鎖および軽鎖を含み、この重鎖は、重鎖可変領域を含み、この重鎖可変領域は、配列番号32に示されるアミノ酸配列に対して少なくとも80%、85%、90%、91%、92%、93%、94%、95%、96%、97%、98%もしくは約99%の同一性を有する配列を含み、そしてこの軽鎖は、軽鎖可変領域を含み、この軽鎖可変領域は、配列番号20に示されるアミノ酸配列に対して少なくとも80%、85%、90%、91%、92%、93%、94%、95%、96%、97%、98%もしくは約99%の同一性を有する配列を含み、この抗体は、IFN−γと相互作用する。
【0028】
本発明はまた、IFN−γの生物学的活性を阻害し得るかもしくは調節し得、そして/またはIFN−γに特異的に結合し得る抗体を提供し、この重鎖は、配列番号21に示されるアミノ酸配列、またはその抗原結合フラグメントもしくは免疫学的に機能的な免疫グロブリンフラグメントを含み、そしてこの軽鎖は、配列番号33に示されるアミノ酸配列、またはその抗原結合フラグメントもしくは免疫学的に機能的な免疫グロブリンフラグメントを含む。
【0029】
特定の局面では、本発明は、IFN−γの生物学的活性を阻害し得るかもしくは調節し得、そして/またはIFN−γに特異的に結合し得る抗体を提供し、この抗体は、重鎖および軽鎖を含み、この重鎖は、重鎖可変領域を含み、この重鎖可変領域は、配列番号21に示されるアミノ酸配列に対して少なくとも80%、85%、90%、91%、92%、93%、94%、95%、96%、97%、98%もしくは約99%の同一性を有する配列を含み、そしてこの軽鎖は、軽鎖可変領域を含み、この軽鎖可変領域は、配列番号33に示されるアミノ酸配列に対して少なくとも80%、85%、90%、91%、92%、93%、94%、95%、96%、97%、98%もしくは約99%の同一性を有するアミノ酸配列を含み、この抗体は、IFN−γと相互作用する。
【0030】
本発明はまた、単鎖抗体、単鎖Fv抗体、F(ab)抗体、F(ab)’抗体および(Fab’)2抗体を提供する。
【0031】
特定の局面では、本発明は、配列番号8、配列番号12、配列番号16、配列番号18、配列番号20、配列番号22、配列番号31もしくは配列番号33のいずれかに示されるアミノ酸配列、またはその抗原結合フラグメントもしくは免疫学的に機能的な免疫グロブリンフラグメントを含む、軽鎖を提供する。
【0032】
さらに本発明は、配列番号6、配列番号10、配列番号14、配列番号17、配列番号19、配列番号21、配列番号30もしくは配列番号32のいずれかに示されるアミノ酸配列、またはその抗原結合フラグメントもしくは免疫学的に機能的な免疫グロブリンフラグメントを含む、重鎖を提供する。
【0033】
本発明はまた、IFN−γに特異的に結合する単離されたヒト抗体に関し、この抗体は、(a)ヒト重鎖フレームワーク領域、ヒト重鎖CDR1領域、ヒト重鎖CDR2領域およびヒト重鎖CDR3領域;ならびに(b)ヒト軽鎖フレームワーク領域、ヒト軽鎖CDR1領域、ヒト軽鎖CDR2領域およびヒト軽鎖CDR3領域、を含む。特定の局面では、このヒト重鎖CDR1領域は、図12および配列番号34に示されるモノクローナル抗体(mAb)1119、1118、1118*もしくは1121の重鎖CDR1領域であり得、そしてこのヒト軽鎖CDR1領域は、図13および配列番号38、配列番号39もしくは配列番号40に示されるmAb 1119、1118、1121*もしくは1121の軽鎖CDR1領域であり得る。他の局面では、このヒト重鎖CDR2領域は、図12および配列番号35に示されるmAb 1119、1118、1118*もしくは1121の重鎖CDR2領域であり得、そしてこのヒト軽鎖CDR2領域は、図13および配列番号41もしくは配列番号42に示されるmAb 1119、1118、1121*もしくは1121の軽鎖CDR2領域であり得る。さらに他の局面では、このヒト重鎖CDR3領域は、図12および配列番号36または配列番号37に示されるmAb 1119、1118、1118*もしくは1121の重鎖CDR3領域であり、そしてこのヒト軽鎖CDR3領域は、図13および配列番号43もしくは配列番号44に示されるmAb 1119、1118、1121*もしくは1121の軽鎖CDR3領域である。
【0034】
さらに、本発明は、IFN−γの産生の増大もしくはIFN−γに対する感受性の増大に関連する疾患、および/またはIFN−γによって媒介される疾患を処置するための方法を提供し、この方法は、本発明の1つ以上のモノクローナル抗体、またはその抗原結合フラグメントもしくは免疫学的に機能的な免疫グロブリンフラグメントの、薬学的に有効な量を、それを必要とする個体に対して投与する工程を包含する。
【0035】
本発明はまた、生物学的サンプル中のIFN−γのレベルを決定するための方法を提供し、この方法は、本発明のモノクローナル抗体またはその抗原結合フラグメントとサンプルとを接触させる工程を包含する。
【0036】
本発明はまた、IFN−γに特異的に結合し得るか、またはその生物学的活性を阻害もしくは調節し得る単離された抗体を提供し、この抗体は、重鎖CDR3を含み、この重鎖CDR3は、(a)配列番号36に存在するのと同じ順序かつ間隔で、配列番号36のアミノ酸のうち少なくとも7つからなる、アミノ酸配列;または(b)配列番号37を含むアミノ酸配列、であるアミノ酸配列を有する。この抗体はさらに、軽鎖CDR3を含み得、この軽鎖CDR3は、(a)配列番号43に存在するのと同じ順序かつ間隔で、配列番号43のアミノ酸のうち少なくとも8つからなる、アミノ酸配列;または(b)配列番号44に存在するのと同じ順序かつ間隔で、配列番号44のアミノ酸のうち少なくとも9つからなるアミノ酸配列、であるアミノ酸配列を有する。この抗体はさらに、配列番号34、配列番号35、配列番号38、配列番号39、配列番号40、配列番号41および配列番号42からなる群より選択される1つ以上のCDRを含み得る。この重鎖CDR3は、配列番号36の少なくともアミノ酸からなってもよく、そして軽鎖CDR3は、配列番号43の少なくともアミノ酸からなってもよい。
【0037】
さらに、本発明は、IFN−γに特異的に結合し得るか、そして/またはその生物学的活性を阻害もしくは調節し得る単離された抗体を提供し、この抗体は、軽鎖CDR3を含み、この軽鎖CDR3は、アミノ酸配列((a)配列番号43に存在するのと同じ順序かつ間隔で、配列番号43のアミノ酸のうち少なくとも8つからなるアミノ酸配列;または(b)配列番号44に存在するのと同じ順序かつ間隔で、配列番号44のアミノ酸のうち少なくとも9つからなるアミノ酸配列)を有する。この抗体はさらに、配列番号35、配列番号38、配列番号39、配列番号40、配列番号41または配列番号42のアミノ酸配列を有するCDRを含み得る。
【0038】
IFN−γに特異的に結合し得るか、そして/またはその生物学的活性を阻害もしくは調節し得る、本発明の単離された抗体は、少なくとも:(a)配列番号34;(b)配列番号35;(c)配列番号36または配列番号37;(d)配列番号38、配列番号39または配列番号40;(e)配列番号41または配列番号42;および(f)配列番号43または配列番号44;のアミノ酸配列を有する6つのCDRを含み得る。
【0039】
IFN−γに特異的に結合し得るか、そして/またはその生物学的活性を阻害もしくは調節し得る、本発明の単離された抗体は、配列番号6、配列番号10、配列番号14または配列番号30に対して少なくとも80%、85%、90%、91%、92%、93%、94%、95%、96%、97%、98%もしくは約99%同一のアミノ酸配列(このアミノ酸配列と、配列番号6、配列番号10、配列番号14もしくは配列番号30との間のアラインメントが、少なくとも50アミノ酸にまたがる)、および/または配列番号8、配列番号12、配列番号16もしくは配列番号31に対して少なくとも80%、85%、90%、91%、92%、93%、94%、95%、96%、97%、98%もしくは約99%同一のアミノ酸配列(このアミノ酸配列と、配列番号8、配列番号12、配列番号16もしくは配列番号31との間のアラインメントが、少なくとも50アミノ酸にまたがる)を含み得る。
【0040】
別の局面では、IFN−γに特異的に結合し得る、そして/またはその生物学的活性を阻害もしくは調節し得る本発明の抗体は、配列番号17、配列番号19、配列番号21または配列番号32に対して少なくとも80%、85%、90%、91%、92%、93%、94%、95%、96%、97%、98%もしくは約99%同一のアミノ酸配列(このアミノ酸配列と、配列番号17、配列番号19、配列番号21もしくは配列番号32との間のアラインメントが、少なくとも50アミノ酸にまたがる)、および/または配列番号18、配列番号20、配列番号22もしくは配列番号33に対して少なくとも80%、85%、90%、91%、92%、93%、94%、95%、96%、97%、98%もしくは約99%同一のアミノ酸配列(このアミノ酸配列と、配列番号18、配列番号20、配列番号22もしくは配列番号33との間のアラインメントが、少なくとも50アミノ酸にまたがる)を含み得る。これらの抗体内の1つのアミノ酸配列は、配列番号36もしくは配列番号37のアミノ酸のうち少なくとも5つを、配列番号36もしくは配列番号37に存在するのと同じ順序かつ間隔で含み得、そして/またはこれらの抗体内の1つのアミノ酸配列は、配列番号43もしくは配列番号44のアミノ酸のうち少なくとも6つを、配列番号43もしくは配列番号44に存在するのと同じ順序かつ間隔で含み得る。
【0041】
1つの局面では、本発明は、単離された完全ヒト抗体であり得る抗体を提供し、この抗体は、ヒトIFN−γの生物学的活性を阻害または調節し得る。別の局面では、本発明の抗体は、単離された完全ヒト抗体であり得、カニクイザルおよびマウスのIFN−γの生物学的活性を阻害も調節もできない。さらに別の局面では、本発明の完全ヒト抗体は、ヒトおよびチンパンジーのIFN−γの生物学的活性を阻害または調節し得るが、カニクイザルおよび/またはマウスのIFN−γの生物学的活性を阻害も調節もできない。さらに別の局面では、ヒトIFN−γの残基19をアスパラギン酸で置換し、および/または残基20をプロリンで置換すれば、この抗体によるヒトIFN−γの生物学的活性の阻害を妨げるかまたは拮抗する。さらに、この抗体は、カニクイザルIFN−γの残基19、20および65にてそれぞれヒスチジン、セリンおよびセリンで置換された変異バージョンの生物学的活性を阻害し得る。
【0042】
なおさらなる局面では、IFN−γに特異的に結合し得、および/またはその生物学的活性を阻害もしくは調節し得る本発明の単離された抗体は、配列番号17、配列番号19、配列番号21または配列番号32に対して少なくとも80%、85%、90%、91%、92%、93%、94%、95%、96%、97%、98%もしくは約99%同一のアミノ酸配列を含み得、このアミノ酸配列と、配列番号17、配列番号19、配列番号21もしくは配列番号32との間のアラインメントは、少なくとも50アミノ酸にまたがり、そして/または上記抗体は、配列番号18、配列番号20、配列番号22もしくは配列番号33に対して少なくとも80%、85%、90%、91%、92%、93%、94%、95%、96%、97%、98%もしくは約99%同一のアミノ酸配列を含み得、このアミノ酸配列と、配列番号18、配列番号20、配列番号22もしくは配列番号33との間のアラインメントは、少なくとも50アミノ酸にまたがる。
【0043】
さらなる実施形態では、本発明は、IFN−γに特異的に結合し得る単離された抗体を含み、この抗体は、(a)配列番号6、配列番号10、配列番号14もしくは配列番号30を含むアミノ酸配列、またはこれらの配列の1つのフラグメント、および(b)配列番号8、配列番号12、配列番号16もしくは配列番号31を含むアミノ酸配列、またはこれらの配列の1つのフラグメント、を含む。この抗体は、重鎖および軽鎖を含み得る。この抗体は、(a)配列番号6、配列番号10、配列番号14または配列番号30、および(b)配列番号8、配列番号12、配列番号16または配列番号31を含み得る。
【0044】
さらに、本発明のいずれの抗体も、ヒト化抗体であっても完全ヒト抗体であってもよい。
【0045】
本発明はまた、本明細書に記載される本発明の抗体のいずれかまたはその一部をコードする、ポリヌクレオチド(単離されたポリヌクレオチドを含む)を提供し、この抗体は、CDR領域、重鎖可変領域、軽鎖可変領域、単鎖抗体、単鎖Fv抗体、F(ab)抗体、F(ab)’抗体および(Fab’)2抗体を含む。本発明はさらに、このようなポリヌクレオチドを含むベクター、ならびにこのようなポリヌクレオチドおよび/またはベクターを含む宿主細胞(必要に応じて、哺乳動物宿主細胞)を提供する。本発明の抗体は、このような宿主細胞を培養する工程によって生成され得る。
本発明によって、例えば、以下が提供される。
(項目1)
重鎖CDR3を含む単離された抗体であって、該重鎖CDR3がアミノ酸配列、すなわち:
(a)配列番号36のアミノ酸のうち少なくとも7つを配列番号36に存在するのと同じ順序でかつ間隔で含むアミノ酸配列;または
(b)配列番号37を含むアミノ酸配列;
を有し、ここで該抗体がIFN−γに特異的に結合し得る、単離された抗体。
(項目2)
軽鎖CDR3をさらに含む項目1に記載の抗体であって、該軽鎖CDR3がアミノ酸配列、すなわち:
(a)配列番号43のアミノ酸のうち少なくとも8つを配列番号43に存在するのと同じ順序でかつ間隔で含むアミノ酸配列;または
(b)配列番号44のアミノ酸のうち少なくとも9つを配列番号44に存在するのと同じ順序でかつ間隔で含むアミノ酸配列;
を有する、単離された抗体。
(項目3)
軽鎖CDR3を含む単離された抗体であって、該軽鎖CDR3がアミノ酸配列、すなわち:
(a)配列番号43のアミノ酸のうち少なくとも8つを配列番号43に存在するのと同じ順序でかつ間隔で含むアミノ酸配列;または
(b)配列番号44のアミノ酸のうち少なくとも9つを配列番号44に存在するのと同じ順序でかつ間隔で含むアミノ酸配列;
を有し、ここで該抗体がIFN−γに特異的に結合し得る、単離された抗体。
(項目4)
CDRをさらに含む項目3に記載の抗体であって、該CDRが配列番号34、配列番号35、配列番号36、配列番号37、配列番号38、配列番号39、配列番号40、配列番号41または配列番号42のアミノ酸配列を有する抗体。
(項目5)
前記重鎖CDR3が配列番号36のアミノ酸を含み、前記軽鎖CDR3が配列番号43のアミノ酸を含む項目2に記載の抗体。
(項目6)
配列番号6、配列番号10、配列番号14および/または配列番号30に対して少なくとも80%同一の第一のアミノ酸配列を含む、項目1〜5のいずれか1項に記載の抗体であって、
該第一のアミノ酸配列と配列番号6、配列番号10、配列番号14または配列番号30との間のアラインメントが少なくとも50アミノ酸にまたがる、抗体。
(項目7)
前記第一のアミノ酸配列が配列番号6、配列番号10、配列番号14および/または配列番号30に対して90%同一である、項目6に記載の抗体。
(項目8)
前記第一のアミノ酸配列が配列番号6、配列番号10、配列番号14または配列番号30に対して同一である、項目7に記載の抗体。
(項目9)
配列番号8、配列番号12、配列番号16および/または配列番号31に対して少なくとも80%同一の第二のアミノ酸配列を含む、項目1〜8のいずれか1項に記載の抗体であって、
該第二のアミノ酸配列と配列番号8、配列番号12、配列番号16および/または配列番号31との間のアラインメントが少なくとも50アミノ酸にまたがる、抗体。
(項目10)
前記第二のアミノ酸配列が配列番号8、配列番号12、配列番号16および/または配列番号31に対して少なくとも90%同一である、項目9に記載の抗体。
(項目11)
前記第二のアミノ酸配列が配列番号8、配列番号12、配列番号16または配列番号31に対して同一である、項目10に記載の抗体。
(項目12)
配列番号17、配列番号19、配列番号21および/または配列番号32に対して少なくとも80%同一の第一のアミノ酸配列を含む、項目1〜5のいずれか1項に記載の抗体であって、
該第一のアミノ酸配列と配列番号17、配列番号19、配列番号21および/または配列番号32との間のアラインメントが少なくとも50アミノ酸にまたがる、抗体。
(項目13)
前記第一のアミノ酸配列が配列番号17、配列番号19、配列番号21および/または配列番号32に対して少なくとも90%同一である、項目12に記載の抗体。
(項目14)
前記第一のアミノ酸配列が配列番号17、配列番号19、配列番号21または配列番号32に対して同一である、項目13に記載の抗体。
(項目15)
配列番号18、配列番号20、配列番号22および/または配列番号33に対して少なくとも80%同一の第二のアミノ酸配列を含む、項目1〜5および項目12〜14のいずれか1項に記載の抗体であって、
該第二のアミノ酸配列と配列番号18、配列番号20、配列番号22および/または配列番号33との間のアラインメントが少なくとも50アミノ酸にまたがる、抗体。
(項目16)
前記第二のアミノ酸配列が配列番号18、配列番号20、配列番号22および/または配列番号33に対して少なくとも90%同一である、項目15に記載の抗体。
(項目17)
前記第二のアミノ酸配列が配列番号18、配列番号20、配列番号22または配列番号33に対して同一である、項目16に記載の抗体。
(項目18)
単離された抗体であって、第一のアミノ酸配列、すなわち:
(a)配列番号6に対して少なくとも96%同一であるアミノ酸配列;
(b)配列番号10に対して少なくとも93%同一であるアミノ酸配列;
(c)配列番号14に対して少なくとも91%同一であるアミノ酸配列;または
(d)配列番号30に対して少なくとも98%同一であるアミノ酸配列;
を含み、
ここで該第一のアミノ酸配列と配列番号6、配列番号10、配列番号14または配列番号30との間のアラインメントが少なくとも50アミノ酸にまたがり、そして
IFN−γに特異的に結合可能である単離された抗体。
(項目19)
前記第一のアミノ酸配列が配列番号6に対して少なくとも96%同一であるか、配列番号10に95%同一であるか、配列番号14に対して95%同一であるか、または配列番号30に対して98%同一である、項目18に記載の抗体。
(項目20)
前記第一のアミノ酸配列が配列番号6、配列番号10、配列番号14または配列番号30に対して同一である、項目19に記載の抗体。
(項目21)
配列番号8、配列番号12、配列番号16および/または配列番号31に対して少なくとも80%同一である第二のアミノ酸配列をさらに含み、該第二のアミノ酸配列と配列番号8、配列番号12、配列番号16または配列番号31との間のアラインメントが少なくとも50アミノ酸にまたがる、項目18〜20のいずれか1項に記載の抗体。
(項目22)
前記第二のアミノ酸配列が配列番号8、配列番号12、配列番号16および/または配列番号31に対して少なくとも90%同一である、項目21に記載の抗体。
(項目23)
前記第一のアミノ酸配列が配列番号36および/または配列番号37のアミノ酸のうち少なくとも5つを配列番号36および/または配列番号37に存在するのと同じ順序でかつ間隔で含む、項目18〜22のいずれか1項に記載の抗体。
(項目24)
前記第二のアミノ酸配列が配列番号43または配列番号44のアミノ酸のうち少なくとも6つを、配列番号43または配列番号44に存在するのと同じ順序でかつ間隔で含む、項目21または22に記載の抗体。
(項目25)
単離された抗体であって、第一のアミノ酸配列、すなわち:
(a)配列番号8に対して少なくとも97%同一であるアミノ酸配列;
(b)配列番号12に対して少なくとも96%同一であるアミノ酸配列;
(c)配列番号16に対して少なくとも97%同一であるアミノ酸配列;または
(d)配列番号31に対して少なくとも95%同一であるアミノ酸配列;
を含み、
ここで該配列番号8、配列番号12、配列番号16または配列番号31と該第一のアミノ酸配列との間のアラインメントが少なくとも50アミノ酸にまたがり、そして
IFN−γに特異的に結合可能である単離された抗体。
(項目26)
前記第一のアミノ酸配列が配列番号8、配列番号12、配列番号16または配列番号31に対して少なくとも97%同一である、項目25に記載の抗体。
(項目27)
前記第一のアミノ酸配列が配列番号8、配列番号12、配列番号16または配列番号31に対して同一である、項目26に記載の抗体。
(項目28)
前記第一のアミノ酸配列が配列番号43または配列番号44のアミノ酸のうち少なくとも6つを、配列番号43または配列番号44に存在するのと同じ順序でかつ間隔で含む、項目25に記載の抗体。
(項目29)
配列番号6、配列番号10、配列番号14および/または配列番号30に対して少なくとも80%同一である第二のアミノ酸配列をさらに含む、項目25〜28のいずれか1項に記載の抗体であって、
該第二のアミノ酸配列と配列番号6、配列番号10、配列番号14および/または配列番号30との間のアラインメントが少なくとも50アミノ酸にまたがる、抗体。
(項目30)
前記第二のアミノ酸配列が配列番号6、配列番号10、配列番号14および/または配列番号30に対して少なくとも90%同一である、項目29に記載の抗体。
(項目31)
前記第二のアミノ酸配列が配列番号36および/または配列番号37のアミノ酸のうち少なくとも5つを配列番号36および/または配列番号37に存在するのと同じ順序でかつ間隔で含む、項目29または30に記載の抗体。
(項目32)
単離された完全ヒト抗体であって、
該抗体は、ヒトIFN−γおよびチンパンジーIFN−γの生物学的活性を阻害し得、そして該抗体はカニクイザルIFN−γおよびマウスIFN−γの生物学的活性を阻害できず、
該抗体は、VH領域、CH1領域、CH2領域およびCH3領域を含む重鎖、ならびにVL領域およびCL領域を含む軽鎖を含む、抗体。
(項目33)
項目32に記載の抗体であって、ヒトIFN−γの残基20をプロリンで置換することによって、該抗体によるヒトIFN−γの生物学的活性の阻害を拮抗する抗体。
(項目34)
項目32または33に記載の抗体であって、該抗体はカニクイザルIFN−γの変異バージョンの生物学的活性を阻害し得、該変異バージョンは19位にヒスチジン、20位にセリン、そして65位にセリンを含む、抗体。
(項目35)
ヒトIFN−γの生物学的活性を阻害し得る単離された抗体であって、ヒトIFN−γの残基20をプロリンで置換することによって、該抗体によるヒトIFN−γの生物学的活性の阻害を拮抗し、かつ完全ヒト抗体である、単離された抗体。
(項目36)
軽鎖CDR1、軽鎖CDR2、軽鎖CDR3、重鎖CDR1、重鎖CDR2、および重鎖CDR3を含む単離された抗体であって、
該重鎖CDR1は配列番号34のアミノ酸配列を有し;
該重鎖CDR2は配列番号35のアミノ酸配列を有し;
該重鎖CDR3は配列番号36または配列番号37のアミノ酸配列を有し;
該軽鎖CDR1は配列番号38、配列番号39または配列番号40のアミノ酸配列を有し;
該軽鎖CDR2は配列番号41または配列番号42のアミノ酸配列を有し;
該軽鎖CDR3は配列番号43または配列番号44のアミノ酸配列を有し;そして
該抗体はIFN−γに特異的に結合し得る、単離された抗体。
(項目37)
項目36に記載の抗体であって、
前記重鎖CDR3は配列番号36のアミノ酸配列を有し、
前記軽鎖CDR1は配列番号38のアミノ酸配列を有し、
前記軽鎖CDR2は配列番号41のアミノ酸配列を有し、そして
前記軽鎖CDR3は配列番号43のアミノ酸配列を有する、抗体。
(項目38)
項目36または37に記載の抗体であって、
配列番号17、配列番号19、配列番号21および/または配列番号32に対して少なくとも80%同一の第一のアミノ酸配列であって、ここで該第一のアミノ酸配列と配列番号17、配列番号19、配列番号21および/または配列番号32との間のアラインメントが少なくとも50アミノ酸にまたがり、該第一のアミノ酸配列が重鎖CDR1、CDR2およびCDR3のアミノ酸配列を含む、第一のアミノ酸配列、ならびに
配列番号18、配列番号20、配列番号22および/または配列番号33に対して少なくとも80%同一の第二のアミノ酸配列であって、ここで該第二のアミノ酸配列と配列番号18、配列番号20、配列番号22および/または配列番号33との間のアラインメントが少なくとも50アミノ酸にまたがり、該第二のアミノ酸配列が軽鎖CDR1、CDR2およびCDR3のアミノ酸配列を含む、第二のアミノ酸配列、
を含む抗体。
(項目39)
項目38に記載の抗体であって、
第一のアミノ酸配列が、配列番号17、配列番号19、配列番号21および/または配列番号32に対して少なくとも90%同一であり、そして、
第二のアミノ酸配列が、配列番号18、配列番号20、配列番号22および/または配列番号33に対して少なくとも90%同一である、
抗体。
(項目40)
項目39に記載の抗体であって、
前記第一のアミノ酸配列が、配列番号17、配列番号19、配列番号21および/または配列番号32に対して同一であり、そして、
前記第二のアミノ酸配列が、配列番号18、配列番号20、配列番号22および/または配列番号33に対して同一である、
抗体。
(項目41)
項目36または37に記載の抗体であって、
配列番号6、配列番号10、配列番号14および/または配列番号30に対して少なくとも80%同一の第一のアミノ酸配列であって、該第一のアミノ酸配列と配列番号6、配列番号10、配列番号14および/または配列番号30との間のアラインメントが少なくとも50アミノ酸にまたがる、第一のアミノ酸配列;ならびに
配列番号8、配列番号12、配列番号16および/または配列番号31に対して少なくとも80%同一の第二のアミノ酸配列であって、該第二のアミノ酸配列と配列番号8、配列番号12、配列番号16および/または配列番号31との間のアラインメントが少なくとも50アミノ酸にまたがる、第二のアミノ酸配列、
を含む抗体。
(項目42)
項目41に記載の抗体であって、
前記第一のアミノ酸配列が、配列番号6、配列番号10、配列番号14および/または配列番号30に対して少なくとも90%同一であり、そして
前記第二のアミノ酸配列が、配列番号8、配列番号12、配列番号16および/または配列番号31に対して少なくとも90%同一である、抗体。
(項目43)
項目42に記載の抗体であって、
前記第一のアミノ酸配列が、配列番号6、配列番号10、配列番号14または配列番号30に対して同一であり、そして
前記第二のアミノ酸配列が、配列番号8、配列番号12、配列番号16または配列番号31に対して同一である、抗体。
(項目44)
単離された抗体であって、
(a)配列番号6、配列番号10、配列番号14もしくは配列番号30、またはこれらの配列の1つのフラグメントを含むアミノ酸配列、および
(b)配列番号8、配列番号12、配列番号16もしくは配列番号31、またはこれらの配列の1つのフラグメントを含むアミノ酸配列、
を含み、IFN−γに特異的に結合可能である、単離された抗体。
(項目45)
重鎖および軽鎖を含む項目44に記載の抗体であって、該重鎖は、VH領域、CH1領域、CH2領域およびCH3領域を含み、該軽鎖はVL領域およびCL領域を含む、抗体。
(項目46)
項目44または45に記載の抗体であって、
(a)配列番号6、配列番号10、配列番号14または配列番号30、および
(b)配列番号8、配列番号12、配列番号16または配列番号31、
を含む抗体。
(項目47)
前記抗体が配列番号6および配列番号8を含む、項目46に記載の抗体。
(項目48)
前記抗体が単鎖抗体である、項目1〜31および項目36〜47のいずれか1項に記載の抗体。
(項目49)
前記抗体が完全ヒト抗体である、項目1〜31および項目36〜47のいずれか1項に記載の抗体。
(項目50)
前記抗体がヒト化抗体である、項目1〜31および項目36〜47のいずれか1項に記載の抗体。
(項目51)
項目1〜50のいずれか1項に記載の抗体をコードする単離されたポリヌクレオチド。
(項目52)
項目51に記載のポリヌクレオチドを含むベクター。
(項目53)
項目52に記載のベクターを含む宿主細胞。
(項目54)
項目53に記載の宿主細胞を培養する工程を包含する、抗体の産生方法。
(項目55)
IFN−γ媒介性疾患に罹患している被験体に対して、項目1〜50のいずれか1項に記載の抗体の治療上有効な量を投与する工程を包含する、方法。
(項目56)
薬学的に受容可能なキャリア、および項目1〜50のいずれか1項に記載の抗体の治療上有効な量を含む、組成物。
(項目57)
IFN−γ媒介性疾患に罹患している患者に対して項目56に記載の組成物を投与する工程を包含する、方法。
(項目58)
前記IFN−γ媒介性疾患が、AIDS、若年性関節リウマチを含む関節リウマチ、潰瘍性大腸炎およびクローン病を含む炎症性腸疾患、多発性硬化症、アジソン病、糖尿病(I型)、精巣上体炎、糸球体腎炎、グレーブス病、ギヤン・バレー症候群、橋本病、溶血性貧血、全身性エリテマトーデス(SLE)、ループス腎炎、重症筋無力症、天疱瘡、乾癬、乾癬性関節症、アテローム性動脈硬化症、エリスロポエチン耐性、宿主対移植片病、移植片拒絶、自己免疫性肝炎誘導性肝障害、胆汁性肝硬変、アルコール肝硬変を含むアルコール誘導性肝障害、リウマチ熱、サルコイドーシス、強皮症、シェーグレン症候群、強直性脊椎炎を含む脊椎関節症、甲状腺炎、または脈管炎である、項目55または57に記載の方法。
【0046】
本発明の特に好ましい実施形態は、特定の好ましい実施形態の以下のさらに詳細な説明および特許請求の範囲から明らかになる。
【0047】
(詳細な説明)
本明細書中で用いた節の表題は、構成上の目的でしかなく、記載された内容に限定されると解釈されるべきではない。本出願に引用された全ての参考文献は、任意の目的について本明細書中で参考として明確に援用される。
(定義)
組み換えDNA、オリゴヌクレオチド合成、ならびに組織培養および形質転換(例えば、エレクトロポレーション、リポフェクチン)については標準的な技術を用いてもよい。酵素反応および精製技術は、製造業者の説明書に従って、または当該分野で一般に完成されているとおり、または本明細書に記載されているように行なってもよい。前述の技術および手順は、当該分野で周知の従来法によって、そして本明細書を通じて引用および考察されている種々の一般的およびより詳細な参考文献に記載されているとおりに一般に行なうことができる。例えば、任意の目的について参考として本明細書中に援用されている、Sambrookら、2001、MOLECULAR CLONING:A LABORATORY MANUAL、第三版、Cold Spring Harbor Laboratory Press,Cold Spring Harbor,N.Y.を参照のこと。特に定義しない限り、本明細書に記載の分析化学、合成有機化学および医学および薬学的化学に関連して利用される命名法、そしてそれらの実験室手順および技術は、当該分野で周知でありかつ一般に用いられるものである。化学合成、化学分析、製剤、処方および送達、ならびに患者の処置には標準的な技術が用いられ得る。
【0048】
本開示に関して使用する場合、以下の用語は、他に示さない限り、以下の意味を有すると理解されるべきである。
【0049】
本明細書において用いる場合、「単離されたポリヌクレオチド」という用語は、ゲノムのポリヌクレオチド、cDNAまたは合成起源またはそれらのいくつかの組み合わせを意味しており、その起源のおかげで、この単離されたポリヌクレオチドは、(1)その単離されたポリヌクレオチドが、天然に見出されるポリヌクレオチドの全てまたは一部を伴わないか、(2)天然には連結されないポリヌクレオチドに連結されるか、または(3)より大きい配列の一部として天然には存在しないかのいずれかである。
【0050】
本明細書において言及される「単離されたタンパク質」という用語は、このタンパク質が、(1)天然に代表的に見出される少なくともいくつかの他のタンパク質を含まないか、(2)同じ供給源由来、例えば同じ種由来の他のタンパク質を本質的に含まないか、(3)異なる種由来の細胞によって発現されるか、(4)天然にはともなわない、少なくとも約50%のポリヌクレオチド、脂質、炭水化物もしくは他の物質から分離されているか、(5)この「単離されたタンパク質」が天然では会合しているタンパク質の一部と会合していない(共有的な相互作用によっても、または非共有的な相互作用によっても)か、(6)天然には会合していないポリペプチドと作動可能に会合している(共有的な相互作用または非共有的な相互作用によって)か、または(7)天然には存在しないということを意味する。このような単離されたタンパク質は、合成起源のゲノムDNA、cDNA、mRNAもしくは他のRNA、またはそれらの任意の組み合わせによってコードされ得る。好ましくは、単離されたタンパク質は、その用途(治療上、診断上、予測、研究その他)を妨げる、天然の環境で見出されるタンパク質またはポリペプチドまたは他の混入物を実質的に含まない。
【0051】
「ポリペプチド」または「タンパク質」という用語は、アミノ酸の1つ以上の鎖であって、各々の鎖がペプチド結合によって共有結合されたアミノ酸を含み、このポリペプチドまたはタンパク質は、非共有結合的におよび/またはペプチド結合によって共有結合的に一緒に連結された複数の鎖を含み得、天然のタンパク質、すなわち、天然に存在する細胞そして詳細には非組み換え細胞によって産生されたタンパク質、または遺伝子操作された細胞かもしくは組み換え細胞によって産生されたタンパク質の配列を有し、そしてこの天然のタンパク質のアミノ酸配列を有する分子、またはこの天然の配列の1つ以上のアミノ酸からの欠失、それに対する付加および/または置換を有する分子を含む鎖を意味する。「ポリペプチド」および「タンパク質」という用語は、特に抗IFNγ抗体、または抗IFNγ抗体の1つ以上のアミノ酸からの欠失、それに対する付加および/または置換を有する配列を含む。従って、「ポリペプチド」または「タンパク質」は、1つのアミノ酸鎖(「単量体」と命名される)または複数のアミノ酸鎖(「多量体」と命名される)を包含し得る。
【0052】
「ポリペプチドフラグメント」という用語は、単量体であっても多量体であってもよく、アミノ末端欠失、カルボキシ末端欠失、および/または天然に存在するかもしくは組み換え産生されたポリペプチドの内部欠失もしくは置換を有するポリペプチドをいう。特定の実施形態では、ポリペプチドフラグメントは、少なくとも5〜約500アミノ酸長のアミノ酸鎖を含んでもよい。特定の実施形態では、フラグメントは少なくとも5、6、8、10、14、20、50、70、100、110、150、200、250、300、350、400または450アミノ酸長であることが理解される。特に有用なポリペプチドフラグメントとしては、結合ドメインを含む機能的なドメインが挙げられる。抗IFN−γ抗体の場合、有用なフラグメントとしては、CDR領域、特に重鎖または軽鎖のCDR3領域;重鎖または軽鎖の可変ドメイン;抗体鎖の一部または2つのCDRを含むちょうどその可変領域;などが挙げられるがこれらに限定されない。
【0053】
本明細書において用いる場合「免疫学的に機能的な免疫グロブリンフラグメント」という用語は、免疫グロブリン重鎖および軽鎖の少なくともCDRを含むポリペプチドフラグメントをいう。本発明の免疫学的に機能的な免疫グロブリンフラグメントは、抗原に結合し得る。好ましい実施形態では、この抗原はレセプターに特異的に結合するリガンドである。これらの実施形態では、本発明の免疫学的に機能的な免疫グロブリンフラグメントの結合は、そのレセプターに対するリガンドの結合を防止または阻害し、このレセプターに対するリガンド結合から生じる生物学的応答を遮断する。好ましくは、本発明の免疫学的に機能的な免疫グロブリンフラグメントは、IFN−γに特異的に結合する。最も好ましくは、このフラグメントはヒトIFN−γに特異的に結合するか、そして/またはその生物学的活性を阻害もしくは調節する。
【0054】
「天然に存在する」という用語は、本明細書において用いて物体に対して適用する場合、その物体が天然に見出され得るという事実をいう。例えば、天然の供給源から単離可能であり、人によって意図的に改変されていない、生物体(ウイルスを含む)に存在するポリペプチドまたはポリヌクレオチドの配列は、天然に存在する。
【0055】
「作動可能に連結された」という用語は、その用語が適用された成分が、適切な条件下でその固有の機能を行なうことを可能にする関係であることを意味する。例えば、タンパク質コード配列に「作動可能に連結された」転写制御配列は、このタンパク質コード配列の発現が制御配列の転写活性に適合する条件下で達成されるようにそこに連結されている。
【0056】
「制御配列」という用語は、本明細書中で用いられる場合、それらが連結されるコード配列の発現、プロセシングまたは細胞内局在化に影響を及ぼし得るポリヌクレオチド配列をいう。このようなコントロール配列の性質は、宿主生物体に依存し得る。特定の実施形態では、原核生物の転写制御配列としては、プロモーター、リボソーム結合部位および転写終結配列が挙げられ得る。他の特定の実施形態では、真核生物の転写制御配列としては、転写因子の1つまたは複数の認識部位を含むプロモーター、転写エンハンサー配列、転写終結配列およびポリアデニル化配列が挙げられ得る。特定の実施形態では、「制御配列」とは、リーダー配列および/または融合パートナー配列を含んでもよい。
【0057】
「ポリヌクレオチド」という用語は、本明細書中で言及する場合、少なくとも10塩基長の一本鎖または二本鎖の核酸ポリマーを意味する。特定の実施形態では、ポリヌクレオチドを含むヌクレオチドは、リボヌクレオチドもしくはデオキシリボヌクレオチドまたはいずれかのタイプのヌクレオチドの修飾型であってもよい。このような修飾としては、ブロモウリジンのような塩基修飾、アラビノシドのようなリボース修飾、ならびに2’,3’−ジデオキシリボースおよびヌクレオチド間連結修飾(例えば、ホスホロチオエート、ホスホロジチオエート、ホスホロセレノエート、ホスホロジセレノエート、ホスホロアニロチオエート、ホスホロアニラデートおよびホスホロアミデート)が挙げられる。「ポリヌクレオチド」という用語は詳細には、DNAの一本鎖および二本鎖型を含む。
【0058】
本明細書で言及する「オリゴヌクレオチド」という用語は、天然に存在する、そして修飾されたヌクレオチドであって、天然に存在するか、および/または天然には存在しないオリゴヌクレオチド連結によって一緒に連結されたヌクレオチドを含む。オリゴヌクレオチドは、一般的に一本鎖であり、200塩基以下の長さを有するメンバーを含むポリヌクレオチドサブセットである。特定の実施形態では、オリゴヌクレオチドは、10〜60塩基長である。特定の実施形態では、オリゴヌクレオチドは12、13、14、15、16、17、18、19、または20〜40塩基長である。オリゴヌクレオチドは、例えば、遺伝子変異体の構築における使用のために、一本鎖であっても二本鎖であってもよい。本発明のオリゴヌクレオチドは、タンパク質コード配列に関連してセンスオリゴヌクレオチドであってもアンチセンスオリゴヌクレオチドであってもよい。
【0059】
他に特定しない限り、一本鎖ポリヌクレオチド配列の左側末端は、5’末端であり;二本鎖ポリヌクレオチド配列の左側方向は、5’方向と呼ばれる。初期RNA転写物の5’〜3’付加の方向は、転写方向と呼ばれる;RNAと同じ配列を有するDNA鎖上の配列領域であって、RNA転写物の5’〜5’末端への領域は「上流配列」と呼ばれる;RNAと同じ配列を有するDNA鎖上の配列領域であって、RNA転写物の3’〜3’末端への領域は「下流配列」と呼ばれる。
【0060】
「天然に存在するヌクレオチド」という用語は、デオキシリボヌクレオチドおよびリボヌクレオチドを含む。「修飾されたヌクレオチド」という用語は、修飾または置換された糖基などを有するヌクレオチドを含む。「オリゴヌクレオチド連結」という用語は、オリゴヌクレオチド連鎖、例えば、ホスホロチオエート、ホスホロジチオエート、ホスホロセレノエート、ホスホロジセレノエート、ホスホロアニロチオエート、ホスホルアニラデート、ホスホロアミデートなどを含む。例えば、LaPlancheら、1986、Nucl.Acids Res.14:9081;Stecら、1984,J.Am.Chem.Soc.106:6077;Steinら、1988,Nucl.Acids Res.16:3209;Zonら、1991,Anti−Cancer Drug Design 6:539;Zonら、1991,OLIGONUCLEOTIDES AND ANALOGOUES:A PRACTICAL APPROACH、87〜108頁(F.Eckstein編)、Oxford University Press、Oxford England);Stecら、米国特許第5,151,510号;UhlmannおよびPeyman、1990、 Chemical Reviews 90:543(これらの開示は任意の目的のために参考として本明細書に援用される)を参照のこと。オリゴヌクレオチドは、オリゴヌクレオチドまたはそのハイブリダイゼーションの検出を可能にする検出可能な標識を含んでもよい。
【0061】
「ベクター」という用語は、宿主細胞に対してコード情報を移入するために用いられる任意の分子(例えば、核酸、プラスミドまたはウイルス)をいうために、使用される。
【0062】
「発現ベクター」という用語は、宿主細胞の形質転換に適切であって、挿入された異種核酸配列の発現を指向および/または制御する核酸配列を含むベクターをいう。発現としては、これらに限定されないが、転写、翻訳およびイントロンが存在する場合、RNAスプライシングのようなプロセスが挙げられる。
【0063】
「宿主細胞」という用語は、核酸配列を導入されているか、または核酸配列を導入されることが可能な細胞であって、さらに目的の選択された遺伝子を発現するかまたは発現可能である細胞をいうために使用される。この用語は、親の細胞の子孫を、この子孫が形態学的に、またはもとの親に対する遺伝的作製において同一であるか否かにかわらず、この選択された遺伝子が存在する限り、包含する。
【0064】
「形質導入」という用語は、通常は、ファージによる1つの細菌から別の細菌への遺伝子の移入をいうために使用される。「形質導入」とはまた、レトロウイルスによる、真核生物細胞配列の獲得および移入をいう。
【0065】
「トランスフェクション」という用語は、細胞による外因性または内因性のDNAの取り込みをいうために使用され、細胞は、外因性のDNAがその細胞膜の内側に導入されている場合、「トランスフェクト」されている。多数のトランスフェクション技術が当該分野で周知であり、本明細書に開示されている。例えば、Grahamら、1973,Virology 52:456;Sambrookら,2001,MOLECULAR CLONING,A LABORATORY MANUAL,Cold Spring Harbor Laboratories;Davisら,1986,BASIC METHODS IN MOLECULAR BIOLOGY,Elsevier;およびChuら,1981,Gene 13:197を参照のこと。このような技術は、1つ以上の外因性DNA部分を適切な宿主細胞に導入するために用いることができる。
【0066】
「形質転換」という用語は、本明細書中で用いる場合、細胞の遺伝的特徴における変化をいい、細胞が新しいDNAを含むように改変されている場合、形質転換されている。例えば、細胞が天然の状態から遺伝的に改変されている場合、形質転換されている。トランスフェクションまたは形質導入の後、この形質転換DNAは、細胞の染色体への物理的な組みこみによって細胞のDNAと組み換えられてもよいし、または複製されることのないエピソームエレメントとして一時的に維持されてもよいし、またはプラスミドとして独立して複製されてもよい。細胞の分裂とともにDNAが複製される場合、細胞は安定に形質転換されていると考えられる。
【0067】
「天然に存在する」または「天然の」という用語は、核酸分子、ポリペプチド、宿主細胞などのような生物学的物質に関連して用いられる場合、天然に見出され、人によって操作されていない物質をいう。同様に、「天然に存在しない」または「非天然の」とは、本明細書において用いる場合、天然に見出されないか、または人によって構造的に改変されているかもしくは合成されている物質をいう。
【0068】
「抗原」という用語は、選択的な結合因子、例えば抗体によって結合され得、そしてさらに動物において使用され、その抗原のエピトープに対して結合できる抗体を生成することができる分子または分子の一部をいう。抗原は、1つ以上のエピトープを有してもよい。
【0069】
「エピトープ」という用語は、免疫グロブリンまたはT細胞レセプターに特異的に結合し得る任意の決定基、好ましくはポリペプチド決定基を含む。エピトープは、抗体によって結合される抗原の領域である。特定の実施形態では、エピトープ決定基としては、分子の化学的に活性な表面の基、例えば、アミノ酸、糖側鎖、ホスホリルまたはスルホニルが挙げられ、特定の実施形態では、特定の三次元構造特徴および/または特異的な荷電特徴を有してもよい。特定の実施形態では、抗体は、それが好ましくはその標的抗原を、タンパク質および/または高分子の複雑な混合物中で認識する場合、抗原に特異的に結合すると言われる。抗体は、平衡解離定数が10−7または10−8M以下である場合、抗原に特異的に結合すると言われる。いくつかの実施形態では、平衡解離定数は、10−9M以下または10−10M以下であってもよい。
【0070】
本明細書中で用いる場合、第一の配列が、例えば、配列RASQSVSSSY(配列番号56)という10のアミノ酸からなる場合、別の配列は、7アミノ酸が、その配列におけるアミノ酸と同一であり、それらが配列に存在するのと同じ相対的な位置で存在する場合、7アミノ酸を、この第一の配列にそれが存在するのと「同じ順序かつ間隔で」有する。例えば、配列RAAAAVSSSY(配列番号57)は、7アミノ酸をRASQSVSSSY(配列番号56)に存在するのと同じ順序かつ間隔で有する。対照的に、これは、配列RASSVSSSY(配列番号58)にはあてはまらない。なぜなら、この配列はRASQSVSSSY(配列番号56)に対して内部欠失を含み、これが、この欠失のいずれかの側に3および6アミノ酸を有するからである。従って、この配列は、第一の配列と同じ順序かつ間隔で、最大6アミノ酸を有する。RASQSVSSSY(配列番号56)と同じ順序でかつ間隔で7アミノ酸を有する可能性のある最短の可能な配列は、7アミノ酸長、例えば、SQSVSSS(配列番号59)である。
【0071】
「同一性」という用語は、当該分野で公知であるとおり、2つ以上のポリペプチド分子または2つ以上の核酸分子の配列の間の関係であって、その配列を比較することによって決定される関係をいう。当該分野では、「同一性」とはまた、核酸分子またはポリペプチドの間の配列の関連性の程度を意味し、場合によっては、2つ以上のヌクレオチドまたは2つ以上のアミノ酸配列の文字列の間の適合によって決定されてもよい。「同一性」とは、特定の数学的モデルまたはコンピュータープログラム(すなわち、「アルゴリズム」)によってアドレスされるギャップアラインメント(もしあれば)と2つ以上の配列のうちより小さい配列の間での同一性適合の割合を測定する。
【0072】
「類似性」という用語は、関連する概念に関して当該分野で用いられるが、「同一性」とは対照的に、「類似性」は、同一性適合および保存的置換適合の両方を含む、関連性の指標をいう。2つのポリペプチド配列が、例えば、アミノ酸20個のうち10個の同一アミノ酸を有し、この残りが全て非保存的置換であれば、同一性パーセントおよび類似性パーセントは、両方とも50%である。同じ例で、保存的置換がさらに5つの位置にあれば、同一性パーセントは、50%のままで、ただし類似性パーセントは、75%である(15/20)。従って、保存的置換がある場合、2つのポリペプチドの間の類似性パーセントは、これらの2つのポリペプチドの間の同一性パーセントよりも高い。
【0073】
関連の核酸およびポリペプチドの同一性および類似性は、公知の方法によって容易に算出され得る。このような方法としては、COMPUTATIONAL MOLECULAR BIOLOGY,(Lesk,A.M.編),1988,Oxford University Press,New York;BIOCOMPUTING:INFORMATICS AND GENOME PROJECTS,(Smith,D.W.編),1993,Academic Press,New York;COMPUTER ANALYSIS OF SEQUENCE DATA,Part 1,(Griffin,A.M.およびGriffin、H.G.編),1994,Humana Press,New Jersey;von Heinje,G.,SEQUENCE ANALYSIS IN MOLECULAR BIOLOGY,1987,Academic Press;SEQUENCE ANALYSIS PRIMER,(Gribskov,M.およびDevereux,J.,編),1991,M.Stockton Press,New York;Carilloら、1988,SIAM J.Applied Math.,48:1073;ならびにDurbinら,1998,BIOLOGICAL
SEQUENCE ANALYSIS,Cambridge University Pressに記載される方法が挙げられるがこれらに限定されない。
【0074】
同一性を決定するために好ましい方法は、試験された配列の間で最大の適合を得るように設計される。同一性を決定する方法は、公的に利用可能なコンピュータープログラムに記載される。2つの配列の間の同一性を決定する好ましいコンピュータープログラム法としては、GAPを含むGCGプログラムパッケージ(Devereuxら,1984,Nucl.Acid.Res.,12:387;Genetics Computer Group,University of Wisconsin,Madison,WI),BLASTP,BLASTN、およびFASTA(Altschulら,1990,J.Mol.Biol.,215:403〜410)が挙げられるがこれらに限定されない。BLASTXプログラムは、National Center for Biotechnology Information(NCBI)および他の供給源(BLAST Manual,Altschulら、NCB/NLM/NIH Bethesda,MD
20894;Altschulら,1990(前出))から公的に入手可能である。周知のSmith Watermanアルゴリズムも、同一性を決定するのに用いることができる。
【0075】
2つのアミノ酸またはポリヌクレオチド配列の整列のための特定のアラインメントスキームによって、2つの配列の短い領域のみの適合を生じ得、そしてこの短い整列された領域は、2つの全長配列の間で顕著な関係がなくても極めて高い配列同一性を有し得る。従って、特定の実施形態では、選択されたアラインメント法(GAPプログラム)によって、標的ポリペプチドの少なくとも50個の連続アミノ酸に及ぶアラインメントを生じる。いくつかの実施形態では、このアラインメントは、標的ポリペプチドのうち少なくとも60、70、80、90、100、110または120アミノ酸を含み得る。ポリヌクレオチドがGAPを用いて整列される場合、このアラインメントは、連続し得る少なくとも約100、150、または200ヌクレオチドに及び得る。
【0076】
例えば、コンピューターアルゴリズムGAP(Genetics Computer Group,University of Wisconsin,Madison,WI)を用いて、配列同一性パーセントを決定すべき2つのポリペプチドを、そのそれぞれのアミノ酸の最適適合について整列させる(アルゴリズムによって決定される場合「適合したスパン(matched span)」)。特定の実施形態において、ギャップオープニングペナルティー(これは、3回の平均対角として計算される;ここで、「平均対角(average diagonal)」は、用いられる比較マトリックスの対角の平均であり;「対角(diagonal)」は、特定の比較マトリックスにより各々の完全アミノ酸適合に対して割り当てられた点数または数である)およびギャップ伸長ペナルティー(通常、ギャップオープニングペナルティーの十分の一)、ならびに比較マトリックス(例えば、PAM250またはBLOSUM 62)をこのアルゴリズムと組み合わせて用いる。特定の実施形態では、標準比較マトリックス(PAM 250比較マトリックスについては、Dayhoffら,1978,Atlas of Protein Sequence and Structure,5:345〜352;BLOSUM 62比較マトリックスについてはHenikoffら、1992,Proc.Natl.Acad.Sci.USA 89:10915〜10919を参照のこと)も、アルゴリズムによって用いられる。
【0077】
特定の実施形態では、ポリペプチド配列比較についてのパラメーターとしては以下が挙げられる:
アルゴリズム:Needlemanら,1970,J.Mol.Biol.48:443〜453;
比較マトリックス:BLOSUM 62(Henikoffら、1992、前出より);
ギャップペナルティー:12
ギャップ長ペナルティー:4
類似性閾値:0
GAPプログラムは、上記のパラメーターとともに有用であり得る。ヌクレオチド配列については、パラメーターは、50のギャップペナルティーおよび3のギャップ長ペナルティーを含み得、すなわち、各々のギャップにおける各記号について3のペナルティーである。特定の実施形態では、前述のパラメーターは、GAPアルゴリズムを用いるポリペプチド比較(エンドギャップについてのペナルティーを伴わない)のためのデフォルトパラメーターである。
【0078】
本明細書において用いる場合、20個の従来のアミノ酸およびその省略形は従来の用法に従う。IMMUNOLOGY−−A SYNTHESIS,第2版(E.S.GolubおよびD.R.Gren編),Sinauer Associates、Sunderland、MA,1991を参照のこと(これは、任意の目的のために参考として本明細書に援用される)。20個の従来のアミノ酸の立体異性体(例えば、Dアミノ酸);非天然のアミノ酸、例えば、α−、α−二置換アミノ酸、N−アルキルアミノ酸、乳酸、および他の従来とは異なるアミノ酸もまた、本発明のポリペプチドに適切な成分であり得る。従来とは異なるアミノ酸の例としては、以下が挙げられる:4−ヒドロキシプロリン、γ−カルボキシグルタメート、ε−N,N,N−トリメチルリジン、ε−N−アセチルリジン、O−ホスホセリン、N−アセチルセリン、N−ホルミルメチオニン、3−メチルヒスチジン、5−ヒドロキシリジン、σ−N−メチルアルギニン、ならびに他の同様のアミノ酸およびイミノ酸(例えば、4−ヒドロキシプロリン)。本明細書において用いられるポリペプチドの表記法においては、標準的な用法および慣習に従って、左手側の方向がアミノ末端方向であり、そして右手側の方向がカルボキシ末端方向である。
【0079】
天然に存在する残基は、共通の側鎖特性に基づいたクラスに分けることができる:
1)疎水性:ノルロイシン(Nor)、Met、Ala、Val、Leu、Ile;
2)中性の親水性:Cys、Ser,Thr、Asn,Gln;
3)酸性:Asp、Glu;
4)塩基性:His、Lys、Arg;
5)鎖の配向に影響する残基:Gly、Pro;および
6)芳香族:Trp、Tyr、Phe。
【0080】
保存的アミノ酸置換は、これらのクラスのうちの1つのメンバーを同じクラスの別のメンバーで交換することを包含し得る。保存的アミノ酸置換基は、天然には存在しないアミノ酸残基を含み得るが、これは代表的には、生物学的な系における合成ではなく化学的なペプチド合成によって取り込まれる。これらとしては、ペプチド模倣物およびアミノ酸部分の他の反転型または逆位型が挙げられる。
【0081】
非保存的置換としては、これらのクラスのうちの1つのメンバーの別のクラス由来のメンバーへの交換を挙げることができる。このような置換残基は、非ヒト抗体と相同であるヒト抗体の領域に導入されても、またはその分子の相同でない領域に導入されてもよい。
【0082】
このような変化を作製する工程において、特定の実施形態に従うと、アミノ酸の疎水性親水性指標が考慮され得る。各々のアミノ酸は、その疎水性および荷電の特徴に基づいて疎水性親水性指標を割り当てられている。それらは、以下である:イソロイシン(+4.5);バリン(+4.2);ロイシン(+3.8);フェニルアラニン(+2.8);システイン/シスチン(+2.5);メチオニン(+1.9);アラニン(+1.8);グリシン(−0.4);スレオニン(−0.7);セリン(−0.8);トリプトファン(−0.9);チロシン(−1.3);プロリン(−1.6);ヒスチジン(−3.2);グルタミン酸(−3.5);グルタミン(−3.5);アスパラギン酸(−3.5);アスパラギン(−3.5);リジン(−3.9);およびアルギニン(−4.5)である。
【0083】
タンパク質に相互作用的な生物学的機能を付与する工程における、アミノ酸の疎水性親水性指標の重要性は当該分野で理解されている(例えば、Kyteら,1982,J.Mol.Biol.157:105〜131を参照のこと)。特定のアミノ酸が類似の疎水性親水性指標またはスコアを有する他のアミノ酸によって置換可能であり、そしてなお類似の生物学的活性を保持することが、公知である。疎水性親水性指標に基づいてこのような変化を作製する際に、特定の実施形態では、その疎水性親水性指標が±2以内であるアミノ酸の置換が含まれる。特定の実施形態では、疎水性親水性指標が±1以内であるアミノ酸の置換が含まれ、そして特定の実施形態では、±0.5以内である置換が含まれる。
【0084】
特に、生物学的に機能的なタンパク質またはそれによって作製されるペプチドが、本明細書に開示されるように免疫学的な実施形態における使用を意図される場合、同様なアミノ酸の置換が親水性に基づいて有効になされ得ることもまた、当該分野で理解されている。特定の実施形態では、あるタンパク質の最大の局所平均親水性は、その隣接するアミノ酸の親水性によって支配されるので、そのタンパク質の免疫原性および抗原性と、すなわち生物学的特性と相関する。
【0085】
以下の親水性値がこれらのアミノ酸残基に割り当てられている:アルギニン(+3.0);リジン(+3.0);アスパラギン酸(+3.0±1);グルタミン酸(+3.0±1);セリン(+0.3);アスパラギン(+0.2);グルタミン(+0.2);グリシン(0);スレオニン(−0.4);プロリン(−0.5±1);アラニン(−0.5);ヒスチジン(−0.5);システイン(−1.0);メチオニン(−1.3);バリン(−1.5);ロイシン(−1.8);イソロイシン(−1.8);チロシン(−2.3);フェニルアラニン(−2.5)およびトリプトファン(−3.4)。同様の親水性値に基づく変化を作製する工程において、特定の実施形態では、その親水性値が±2以内であるアミノ酸の置換が含まれ、特定の実施形態では、疎水性親水性指標が±1以内であるアミノ酸の置換が含まれ、そして特定の実施形態では、±0.5以内である置換が含まれる。親水性に基づいて一次アミノ酸配列からエピトープを同定することができる。これらの領域はまた、「エピトープコア領域(epitopic core regions」とも呼ばれる。
【0086】
例示的なアミノ酸置換を表1に示す。
【0087】
(表1)
【0088】
【表1−1】
【0089】
【表1−2】
当業者は、周知の技術を用いて本明細書に示すポリペプチドの適切な改変体を決定することができる。特定の実施形態では、当業者は、活性に重要であるとは考えられない領域を標的することによって活性を破壊することなく変化され得る、分子の適切な領域を同定することができる。他の実施形態では、当業者は、同様のポリペプチドの間で保存されている分子の残基および部分を同定することができる。さらなる実施形態では、生物学的活性のためにまたは構造のために重要であり得る領域でさえ、生物学的活性を破壊することなく、ポリペプチド構造に有害に影響することもなく、保存的なアミノ酸置換に供することができる。
【0090】
さらに当業者は、活性または構造に重要な類似のポリペプチドにおける残基を同定する構造−機能の研究を概説することができる。このような比較の観点では、当業者は、類似のタンパク質における活性または構造について重要なアミノ酸残基に相当する、タンパク質中のアミノ酸残基の重要性を推測することができる。当業者は、このように推測された重要なアミノ酸残基に対する化学的に類似のアミノ酸置換基を選択することができる。
【0091】
当業者はまた、三次元構造およびアミノ酸配列を、類似のポリペプチドにおけるその構造に関して解析することができる。このような情報の観点では、当業者は、抗体のアミノ酸残基のアラインメントをその三次元構造に関して推測することができる。特定の実施形態では、当業者は、タンパク質の表面にあることが推定されるアミノ酸残基に対してラジカルな変化を作製しないように選択し得る。なぜなら、このような残基は、他の分子との重要な相互作用に関与し得るからである。さらに、当業者は、各々の所望のアミノ酸残基で単一のアミノ酸置換を含む試験改変体を生成することができる。次いで、この改変体を、当業者に公知の活性アッセイを用いてスクリーニングし得る。このような改変体を用いて、適切な改変体についての情報を集めることができる。例えば、特定のアミノ酸残基に対する変化によって、活性の破壊、所望されない減少または不適切な活性を生じるということが発見されている場合、このような変化を有する改変体は、回避され得る。言い換えれば、このような慣用的実験から収集された情報に基づいて、当業者は、さらなる置換単独、または他の変異と組み合わせた置換が回避されるべきアミノ酸を容易に決定することができる。
【0092】
二次構造の予測に対して多数の化学的刊行物がでている。例えば、Moult,1996,Curr.Op.in Biotech.7:422〜427;Chouら,1974,Biochemistry 13:222〜245;Chouら,1974,Biochemistry 113:211〜222;Chouら,1978,Adv.Enzymol.Relat.Areas Mol.Biol.47:45〜148;Chouら、1979,Ann.Rev.Biochem.47:251〜276;およびChouら、1979,Biophys.J.26:367〜384を参照のこと。さらに、二次構造を予測することを補助するコンピュータープログラムが現在利用可能である。二次構造を予測する1方法は、ホモロジー(相同性)モデリングに基づく。例えば、30%より大きい配列同一性または40%より大きい配列類似性を有する2つのポリペプチドまたはタンパク質はしばしば、類似の構造的位相学を有する。タンパク質構造データベース(PDB)の近年の成長によって、ポリペプチドの構造またはタンパク質の構造内の潜在的な折り畳み回数を含む二次構造の予想性の向上が得られた。Holmら、1999,Nucl.Acid.Res.27:244〜247を参照のこと。所定のポリペプチドまたはタンパク質においては折り畳み回数は限定されていること、および一旦、構造の臨界数が解明されれば、構造的な予測は劇的にさらに正確になることが示唆されている(Brennerら,1997,Curr.Op.Struct.Biol.7:369〜376)。
【0093】
二次構造を予測するさらなる方法としては、「スレッディング(threading)」(Jones,1997,Curr.Opin.Struct.Biol.7:377〜87;Sipplら,1996,Structure 4:15〜19)、「プロフィール解析」(Bowieら,1991,Science 253:164〜170;Gribskovら,1990,Meth.Enzym.183:146〜159;Gribskovら,1987,Proc.Nat.Acad.Sci.84:4355〜4358)および「進化的連鎖(evolutionary linkage)」(Holm,1999,前出;およびBrenner,1997,前出を参照のこと)が挙げられる。
【0094】
特定の実施形態によれば、アミノ酸置換は:(1)タンパク質分解に対する感受性を低下するか、(2)酸化に対する感受性を低下するか、(3)タンパク質複合体を形成するための結合親和性を変更するか、(4)結合親和性を変更するか、そして/または(5)このようなポリペプチドに他の物理化学的特性または機能的特性を付与または改変する、アミノ酸置換である。特定の実施形態によれば、単一または複数のアミノ酸置換(特定の実施形態では、保存的アミノ酸置換)は、天然に存在する配列において(特定の実施形態では、分子間接触を形成するドメイン(単数または複数)の外側のポリペプチドの一部において)作製されてもよい。好ましい実施形態では保存的アミノ酸置換は代表的には、親の配列の構造的特徴を実質的に変化しない(例えば、置換アミノ酸は、親の配列に存在するヘリックスを破壊する傾向でも、親の配列を特徴付ける他のタイプの二次構造を破壊する傾向でもあってはならない)。当該分野で認識されるポリペプチドの二次構造および三次構造の例は、各々が参考として本明細書に援用される、PROTEINS、STRUCTURES AND MOLECULAR PRINCIPLES(Creighton編)、1984,W.H.Freeman and Company、New York;INTRODUCTION TO PROTEIN STRUCTURE(C.BrandenおよびJ.Tooze編),1991,Garland Publishing、New York、N.Y.;ならびにThorntonら,1991,Nature
354:105に記載されている。
【0095】
ペプチドアナログは一般に、鋳型のペプチドの特性と類似の特性を有する非ペプチド薬として製薬産業において用いられる。これらのタイプの非ペプチド化合物は、「ペプチド模倣物(peptide mimetics)」または「ペプチド模倣物(peptidomimetics)」と名付けられる。任意の目的のために参考として本明細書に援用される、Fauchere、1986,Adv.Drug Res.15:29;VeberおよびFreidinger,1985,TINS p.392;ならびにEvansら、1987,J.Med.Chem.30:1229)を参照のこと。このような化合物はしばしば、コンピューターによる分子モデリングの補助によって開発される。治療上有用なペプチドに対して構造的に類似であるペプチド模倣物を用いて、同様の治療効果または予防効果を得ることができる。一般には、ペプチド模倣物は、パラダイム(paradigm)ポリペプチド(すなわち、生化学的な特性または生理学的な活性を有するポリペプチド)、例えばヒト抗体に対して構造的に類似であるが、当該分野で周知の方法によって、以下から選択される連結により必要に応じて置換される1つ以上のペプチド連結を有する:−CH2NH−、−CH2S−、−CH2−CH2−、−CH=CH−(シスおよびトランス)、−COCH2−、−CH(OH)CH2−、および−CH2SO−。同じタイプのDアミノ酸を有するコンセンサス配列の1つ以上のアミノ酸の系統的な置換(例えば、L−リジンの代わりに、D−リジン)を用いて、特定の実施形態では、さらに安定なペプチドを生成することができる。さらにコンセンサス配列または実質的に同一のコンセンサス配列のバリエーションを含む拘束されたペプチドは、当該分野で公知の方法(任意の目的のために参考として本明細書に援用される、RizoおよびGierasch,1992,Ann.Rev.Biochem.61:387)によって;例えば、ペプチドを環化する分子間ジスルフィド架橋を形成し得る内部システイン残基を付加することによって、生成することができる。
【0096】
本明細書において用いる場合、「抗体(antibody)」または「抗体ペプチド(antibody peptid)(単数または複数)」という用語は、1つ以上のポリペプチド鎖を含む単量体または多量体のタンパク質をいう。抗体は、抗原に特異的に結合し得、そして抗原の生物学的活性を阻害または調節し得るかもしれない。「抗体」とは、以下に記載される、天然に存在する抗体を包含する。特定の実施形態では、抗体は、組み換えDNA技術によって生成される。さらなる実施形態では、抗体は、天然に存在する抗体の酵素的切断もしくは化学的切断によって、産生される。抗体としては、限定はしないが、F(ab)、F(ab’)、F(ab’)2、Fv、および単鎖Fvフラグメント、ならびに単鎖抗体、キメラ抗体、ヒト化抗体、完全ヒト抗体、ポリクローナル抗体およびモノクローナル抗体が挙げられる。最低限、抗体とは、本明細書において意味する場合、軽鎖または重鎖可変領域の全てまたは一部を含む抗原に特異的に結合し得るポリペプチドを包含する。
【0097】
可変領域は、フレームワーク領域内(Kabatら(前出)によってフレームワーク領域1−4、FR1、FR2、FR3、およびFR4と命名された;ChothiaおよびLesk(前出)も参照のこと)に組み込まれた、少なくとも3つの重鎖または軽鎖相補性決定領域(CDR、Kabatら、1991,Sequences of Proteins of Immunological Interest,Public Health Service N.I.H.,Bethesda、MDによってCDR1、CDR2およびCDR3と命名された超可変領域としても公知;ChothiaおよびLesk,1987、J.Mol.Biol.196:901〜17;Chothiaら、1989,Nature 342:877〜883も参照のこと)を含む。このCDRおよびフレームワークセグメントは、可変領域のアミノ末端に始まって以下のように分散される:FR1−CDR1−FR2−CDR2−FR3−CDR3−FR4。
【0098】
抗体可変領域のフレームワーク領域の一次配列は、門を横切って普遍的に保存されている少量の残基を有する。しかし、多くの残基は、門を横切って、ならびに/または種および/もしくは門のなかで、高度に保存されており、そして抗体内の多くの位置が通常アミノ酸の公知の群の1つによって占有される。Kabatら(前出)を参照のこと。あるいは、配列はその予測される三次構造によって抗体として認識され得る。グリーク・キーβバレル(Greek key β barrel)として公知の構造を形成する9つのβ鎖を含む、可変領域の三次構造は、極めてよく保存されており、この構造内のCDRの位置はまた高度に保存されている。例えば、Borkら,1994,J.Mol.Biol.242:309〜20;HunkapillerおよびHook,1989,Adv.Immunol.44:1〜63;WilliamsおよびBarclay,1988,Ann.Rev.Immunol.6:381〜405;ChothiaおよびLesk(前出);Kabatら(前出)を参照のこと。
【0099】
三次構造は、種々のコンピュータープログラム、例えば、GENEFOLD(登録商標)(Tripos,Inc.,St.Louis,MO;GodzikおよびSkolnik,1992,Proc.Natl.Acad.Sci.USA 89:12098〜12102;Godzikら,1992,J.Mol.Biol.227:227〜38;Godzikら,1993,Protein Engng.6:801〜10)、Protein Data Bank(PDB)の代表的構造に問い合わせタンパク質配列を重ねるプロテインスレッディングプログラム(protein threading program)(Bermanら、2000,Nucleic Acids Res 28:235〜242;Jaroszewskiら,1998,Prot Sci 7:1431〜1440)などによって予測できる。新規なアミノ酸配列を分類するためにGENEFOLD(登録商標)を用いるために、この配列をプログラムに入れて、これによってGENEFOLD(登録商標)データベースに存在する以前に公知のタンパク質構造(「鋳型」構造)上にそれがどのように折り畳まれるかを反映する確率スコアを割り当てる。スコア付けについて、GENEFOLD(登録商標)は、一次アミノ酸配列の類似性、残基のベリアルパターン、局所相互作用および二次構造比較に依存する。GENEFOLD(登録商標)プログラムは、多数の抗体についての解明された構造を含む、タンパク質折り畳みの既存のデータベースにおける全てのテンプレート構造にアミノ酸配列を折り畳む(またはスレッドする)。GENEFOLD(登録商標)の出力は、その三次構造が、入力のアミノ酸配列によって考えられる可能性が最も高い、このデータベース内に由来するタンパク質の3つのリストである。この3つのリストは、(i)配列のみ、(ii)配列と局所高次構造優先度とベリアル(burial)期間、および(iii)配列と局所高次構造優先度とベリアル期間と二次構造、に基づいて算出された3つの異なるスコアを含む。各々の場合に、プログラムによって最適アラインメントを決定し、アラインメントのこの程度が偶然によって生じる確率(P値)を算出して、あり得る最高のスコアである999.9(9.999×102)のスコアとしてP値の逆を報告する。従って、最高のスコアによって、このアラインメントが偶然生じる可能性が最低であることが示される。従ってこれらのスコアは、新規なタンパク質が種々の参照構造にマッチする程度を反映しており、タンパク質の公知のファミリーにおけるメンバーに対して新規なタンパク質を割り当てるのに有用である。例えば、抗体可変領域の構造を有する配列は、合理的に高いP値、例えば少なくとも約200、300、400、500、600、700、800またはそれ以上を有する少なくとも1つの公知の可変領域と整列されることが期待される。
【0100】
「重鎖」という用語は、IFN−γに対する結合特異性を付与するのに十分な可変領域配列を有する任意の免疫グロブリンポリペプチドを包含する。「軽鎖」という用語は、IFN−γに対する結合特異性を付与するのに十分な可変領域配列を有する任意の免疫グロブリンポリペプチドを包含する。このような重鎖または軽鎖は、それが重鎖の場合、軽鎖の、またはそれが軽鎖の場合、重鎖の非存在下でIFN−γに結合し得るが、その必要はない。全長重鎖は、可変領域ドメイン、VHおよび3つの定常領域ドメインCH1、CH2およびCH3を含む。このVHドメインは、ポリペプチドのアミノ末端であり、CH3ドメインは、カルボキシ末端である。「重鎖」という用語は、本明細書において用いる場合、全長重鎖およびそのフラグメントを包含する。全長軽鎖は、可変領域ドメインVL、および定常領域ドメインCLを含む。重鎖と同様、軽鎖の可変領域ドメインは、ポリペプチドのアミノ末端である。「軽鎖」という用語は、本明細書において用いる場合、全長軽鎖およびそのフラグメントを包含する。F(ab)フラグメントは、1つの軽鎖および1つの重鎖のCH1および可変領域からなる。F(ab)分子の重鎖は、別の重鎖分子とジスルフィド結合を形成できない。F(ab’)フラグメントは、1つの軽鎖および1つの重鎖を含み、この重鎖がCH1ドメインとCH2ドメインとの間にさらに定常領域を含み、この結果鎖間ジスルフィド結合が2つの重鎖の間で形成されてF(ab’)2分子が形成され得る。このFv領域は、重鎖および軽鎖の両方由来の可変領域を含むが、定常領域は欠く。単鎖抗体は、Fv分子であり、ここでは重鎖および軽鎖の可変領域が可撓性のリンカーによって接続されて、抗原結合領域を形成する単鎖ポリペプチドを形成している。単鎖抗体は国際特許出願公開番号WO88/01649、ならびに米国特許第4,946,778号および同第5,260,203号に詳細に考察されている。
【0101】
本発明はまた、完全ヒト抗体、ヒト化抗体およびキメラ抗体を包含する。本明細書において意味するように、完全ヒト抗体は、最終的にはヒト由来のポリヌクレオチドのみによってコードされるアミノ酸配列、またはこのような配列に同一であるアミノ酸配列を含む。本明細書において意味するとおり、トランスジェニックマウスにおいて生成されるマウスゲノムに挿入されたヒト免疫グロブリンコードDNAによってコードされる抗体は、完全ヒト抗体である。なぜなら、それらは最終的にヒト由来であるDNAによってコードされるからである。この状況では、ヒト免疫グロブリンコードDNAは、マウス内で(抗体をコードするように)再配列され得、そして体細胞変異がまた生じ得る。マウスにおいてこのような変化を受けている、もともとヒトDNAによってコードされる抗体は、本明細書において意味するとおり、完全ヒト抗体である。このようなトランスジェニックマウスの使用によって、ヒト抗原に対する完全ヒト抗体を選択することが可能になる。事実上、これはほとんどの場合不可能である。なぜなら、自己抗原に対するヒト免疫応答は、正常には生じないからである。当業者は、完全ヒト抗体が治療剤としての使用に、特に慢性疾患を処置するために有利であることを理解する。なぜなら完全ヒト抗体は、それ自体に対する免疫応答を引き起こす可能性が低いからである。対照的に、多くの非ヒト抗体は、ヒトで用いられた場合、それ自体に対する免疫応答を引き起こすことが公知であり、この状況によってヒトでのこのような抗体の慢性使用は勧められない。従って完全ヒト抗体は、ヒトの疾患を含む慢性状態を処置するための抗体を用いるのに直面した長年にわたる問題を解決する。例えば、Billiau,1988,Immunol.Today 9:37〜40;Horneffら,1991,Clin.Immunol.& Immunopathol.59:89〜103;Tjandraら,1990,Immunol &
Cell Biol.68:367〜76を参照のこと。従って、完全ヒトIFNγ抗体は特に、慢性ヒトIFNγ媒介性疾患、例えば自己免疫疾患の処置に十分に適している。
【0102】
ヒト化抗体では、CDRを除いて抗体全体が、ヒト由来のポリヌクレオチドでコードされているか、またはそのCDR内を除いてこのような抗体と同一である。非ヒト生物体に由来する核酸によってコードされるCDRは、ヒト抗体可変領域のβシートフレームワークに接合されて、抗体が作製され、その特異性はこの接合されたCDRによって決定される。このような抗体の作製は、例えば、WO92/11018,Jonesら,1986,Nature 321:522〜25,Verhoeyenら,1988,Science 239:1534〜36に記載される。この研究によって、抗原結合部位の形成におけるCDRの中心的な重要性が裏付けられる。キメラ抗体は、ヒト定常領域(ヒト由来のポリヌクレオチドによってコードされるか、またはこのようなヒト定常領域に同一である)および非ヒト可変領域を含む。このような抗体の作製は、例えば、米国特許第5,681,722号に記載される。
【0103】
特定の実施形態では、「多価特異的(multispecific)」抗体または「多機能性(multifunctional)」抗体以外の二価抗体は、同一の抗原特異性を有する結合部位を含むことが理解される。
【0104】
本発明に従って抗体結合および特異性を評価することにおいて、抗体は、IFN−γに特異的に結合するか、そして/または過剰の抗体がIFN−γに対して結合するそのレセプターの量を(または逆も同様であるが)少なくとも20%、40%、60%、80%、85%もしくはそれ以上まで(インビトロ競合結合アッセイで測定した場合)低下させる場合、IFN−γのそのレセプターへの結合を実質的に阻害する。特異的に結合する抗体は、IFN−γに対する結合について、10−8モル以下、状況に応じて10−9または10−10モル以下という平衡解離定数を有することが予想され得る。
【0105】
治療用途のために、抗IFN−γ抗体の重要な特徴は、IFN−γの生物学的活性を阻害し得るか、または調節し得るかである。IFN−γは多くの異なる生物学的効果を有し、それは異なる細胞タイプにおいて多くの異なるアッセイで測定できる。抗IFN−γ抗体がIFN−γの生物学的活性を阻害または調節する能力は、以下の実施例6に記載のA549アッセイを用いて、またはIFN−γの存在下で観察される細胞増殖の阻害を逆転する抗体の能力を測定する同様のアッセイを用いて測定することができる。意味のある結果を得るためのアッセイについては、アッセイで用いられる細胞の増殖は、このアッセイで用いられるIFN−γによって阻害されなければらなない。ヒトIFN−γは、A549細胞を含むいくつかの細胞タイプの増殖を阻害し得る(実施例6および7)。マウスIFN−γは、RAW 264.7細胞の増殖を阻害し得る(実施例7)が、A549細胞は阻害し得ない。ある抗体が非ヒトIFN−γの生物学的活性を阻害または調節する能力を試験する場合特に、A549細胞以外の細胞タイプを用いてもよい。なぜなら、非ヒトIFN−γは、A549細胞の増殖を阻害できるかもしれないし、阻害できないかもしれないからである。抗原に特異的に結合する抗体全てが、その正常なレセプターに対する抗原結合をブロックすることが可能なわけではなく、従ってそのレセプターに対する結合の際に抗原の生物学的効果を阻害または調節することが可能なわけでもない。当該分野で公知のとおり、このような効果は、抗原のどの部分が抗体に結合するか、ならびに抗原および抗体(この場合、IFN−γおよび抗IFN−γ抗体)の絶対的濃度および相対的濃度の両方に依存し得る。本明細書において意味するとおり、IFN−γの生物学的活性を阻害または調節できると考えられるには、A549アッセイ(実施例6)または同様のアッセイにおいて蛍光で測定する場合、IFN−γの濃度がある範囲内、例えば、およそEC80またはEC90であるとき、少なくとも約20%、40%、60%、80%、85%、100%以上まで、抗体は、IFN−γの存在下で観察される細胞増殖の阻害を逆転することができなければならず、この場合その生物学的活性を阻害する因子の効果が容易に明白に成り得る。EC80とは、本明細書において意味する場合、観察されるIFN−γの最大効果の80%に必要なIFN−γの量である。IFN−γ濃度がEC90を十分上回る場合、阻害因子の効果は、過剰のIFN−γに起因して明白でないかもしれない。IFN−γの生物学的活性を阻害または調節するのに必要な抗体の濃度は、広範に変化し得、そしてこの抗体がIFN−γにどの程度きつく結合するかに依存し得る。例えば、A549アッセイで生物学的活性を阻害または調節するのには、IFN−γの1分子あたり1分子以下の抗体で十分であり得る。いくつかの実施形態では、IFN−γ濃度がおよそEC50〜およそEC90である場合、IFN−γの生物学的活性を阻害または調節するためには、およそ2:1、1:1、1:2、1:4、1:6、1:8、1:10、1:20、1:40、1:60、1:100または1:50,000というIFN−γ抗体の比が必要であり得る。これらの値の間のIFN−γ抗体の比も可能である。
【0106】
さらなる実施形態では、抗体の改変体としては、改変されたFcフラグメントまたは改変された重鎖定常領域を含む抗体を挙げることができる。「結晶化するフラグメント(fragment that crystallizes)」を意味するFcフラグメント、または重鎖定常領域は、抗体に変化した特徴を付与する変異によって改変され得る。例えば、BurtonおよびWoof,1992,Advances in Immunology 51:1〜84;RavetchおよびBolland,2001,Annu.Rev.Immunol.19:275〜90;Sheildsら,2001,Journal of Biol.Chem.276:6591〜6604;TellemanおよびJunghans,2000,Immunology 100:245〜251;Madesanら,1998,Eur.J.Immunol.28:2092〜2100;その全てが本明細書において参考として援用される)を参照のこと。このような変異としては、置換、付加、欠失、またはその任意の組み合わせを挙げることが可能であり、そして代表的には、本明細書に記載の方法に従って、そして当該分野で公知の方法に従って1つ以上の変異原性オリゴヌクレオチド(単数または複数)を用いて部位特異性変異誘発によって生成される(例えば、本明細書において参考として援用される、Maniatisら、MOLECULAR CLONING:A LABORATORY MANUAL、第三版、2001,Cold Spring Harbor,N.Y.およびBergerおよびKimmel,METHODS IN ENZYMOLOGY,152巻、Guide to Molecular Cloning Techniques,1987,Academic Press,Inc.,San Diego,CA.,を参照のこと)。
【0107】
特定の実施形態では、抗体改変体としては、グリコシル化部位の数および/またはタイプが親のポリペプチドのアミノ酸配列に比べて変更されているグリコシル化改変体が挙げられる。特定の実施形態では、タンパク質改変体は、天然のタンパク質よりも多いかまたは少ない数のN連結グリコシル化部位を含む。N連結グリコシル化部位は、以下の配列:Asn−X−SerまたはAsn−X−Thrによって特徴付けられるが、ここでXと命名されたアミノ酸残基は、プロリン以外の任意のアミノ酸残基であってもよい。この配列を作製するためのアミノ酸残基の置換によって、N連結糖鎖の付加のための潜在的な新しい部位が得られる。あるいは、この配列を除く置換は、既存のN連結糖鎖を除去する。また、N連結糖鎖の再配列も得られるが、ここでは1つ以上のN連結グリコシル化部位(代表的には、天然に存在するもの)が、除かれて、1つ以上の新規なN連結部位が作製される。さらに好ましい抗体改変体としては、1つ以上のシステイン残基が、親のアミノ酸配列に比べて欠失されるかまたは別のアミノ酸(例えば、セリン)で置換されているシステイン改変体が挙げられる。抗体が、例えば、不溶性の封入体の単離後に生物学的に活性な高次構造に再折り畳みされなければならない場合に、システイン改変体が有用であり得る。システイン改変体は一般に、天然のタンパク質よりも少ないシステイン残基を有し、そして代表的には、不対のシステインから生じる相互作用を最小にするために偶数を有する。
【0108】
「因子(agent)」という用語は、本明細書において用いる場合、化合物、化合物の混合物、生物学的高分子、または生物学的物質から作製された抽出物を意味する。
【0109】
本明細書において用いる場合、「標識(label)」または「標識された(labeled)」という用語は、例えば、放射線標識されたアミノ酸の組み込み、または蛍光マーカーのポリペプチドもしくは核酸に対する結合、検出可能な活性を有する化学発光マーカーもしくは酵素、または標識されたアビジンによって検出可能であるビオチン部分のポリペプチドに対する結合による、検出可能なマーカー(例えば、とりわけ、光学的または比色定量的な方法によって検出可能である、蛍光マーカー、化学発光マーカーまたは酵素活性などの検出可能なマーカーを好ましくは含むストレプトアビジン)の組み込みをいう。特定の状況では、この標識はまた治療剤であってもよい。ポリペプチドおよび糖タンパク質を標識する種々の方法は、当該分野で公知であり、かつ本明細書に開示された方法において有利に用いることができる。ポリペプチドのための標識の例としては、以下が挙げられるがこれらに限定されない:放射性同位体または放射性核種(例えば、3H、14C、15N、35S、90Y、99mTc、111In、125I、131I)、蛍光標識(例えば、フルオレセインイソチオシアネートすなわちFITC、ローダミン、またはランタニド蛍光物質)、酵素標識(例えば、西洋ワサビペルオキシダーゼ、βガラクトシダーゼ、ルシフェラーゼ、アルカリホスファターゼ)、化学発光標識、ハプテン標識、例えば、ビオチニル基、および二次レポーターによって認識される予め決定されたポリペプチドエピトープ(例えば、ロイシンジッパー対配列、二次抗体の結合部位、金属結合ドメイン、またはエピトープタグ)。特定の実施形態において、標識は、種々の長さのスペーサーアーム(例えば、(CH2)n、ここでn<約20)によって結合されて、可能性のある立体障害を減らす。
【0110】
「生物学的サンプル(biological sample)」という用語は、本明細書において用いる場合、生存物または以前に生存していた物由来の任意の量の物質を包含するがこれらに限定されない。このような生存している物としては、ヒト、マウス、サル、ラット、ウサギおよび他の動物が挙げられるがこれらに限定されない。このような物質としては、血液、血清、尿、細胞、器官、組織、骨、骨髄、リンパ節および皮膚が挙げられるがこれらに限定されない。
【0111】
「薬学的因子または薬物(pharmaceutical agent or drug)」という用語は、本明細書において用いる場合、患者に適切に投与される場合、所望の治療効果を誘導し得る化合物または組成物をいう。
【0112】
「IFN−γ媒介性疾患(IFN−γ mediated disease)」という用語は、炎症性疾患、感染性疾患、および自己免疫疾患を包含するがこれらに限定されない。本明細書において用いる場合、「自己免疫疾患」とは、患者の免疫応答が患者自身の構成要素に向かう疾患状態および条件をいう。例えば、IFN−γ媒介性疾患としては、後天性免疫不全症候群(AIDS)、若年性関節リウマチを含む関節リウマチ、潰瘍性大腸炎およびクローン病を含む炎症性腸疾患、多発性硬化症、アジソン病、糖尿病(I型)、精巣上体炎、糸球体腎炎、グレーブス病、ギラン・バレー症候群、橋本病、溶血性貧血、全身性エリテマトーデス(SLE)、ループス腎炎、重症筋無力症、天疱瘡、乾癬、乾癬性関節症、アテローム性動脈硬化症、エリスロポエチン耐性、宿主対移植片病、移植片拒絶、自己免疫性肝炎誘導性肝障害、胆汁性肝硬変、アルコール肝硬変を含むアルコール誘導性肝障害、リウマチ熱、サルコイドーシス、強皮症、シェーグレン症候群、強直性脊椎炎を含む脊椎関節症、甲状腺炎、および脈管炎が挙げられるがこれらに限定されない。「IFN−γ媒介性疾患」という用語はまた、IFN−γのレベルの増大またはIFN−γの感度の増大に関連する任意の病状を包含する。
【0113】
自己免疫疾患を含むIFN−γ媒介性疾患の処置は、障害の少なくとも1つの症状の緩和、疾患の重篤度の低下、または処置された状態の後ある程度の頻度で生じるさらに重篤な疾患への進行の遅延もしくは予防を含む。処置とは疾患が完全に治癒することを意味する必要はない。有用な治療剤とは単に、ある疾患の重篤度を低下するか、この疾患またはその処置に伴う症状(単数または複数)の重篤度を低下するか、または患者のクオリティーオブライフを改善するか、または処置された状態の後、ある程度の頻度で生じ得るさらに重篤な疾患の発現を遅延することを必要とする。例えば、この疾患が関節リウマチである場合、治療剤は関節の腫脹を軽減するか、罹患する関節の数を減らすか、または骨の損失を遅延もしくは阻害し得る。SLE患者は、皮膚病変、熱、衰弱、関節炎、リンパ節腫脹、胸膜炎、心外膜炎および/または貧血などのような症状を有し得る。このような症状は、例えば、視覚的観察、写真、体温測定、握力もしくは関節サイズ、および/または赤血球の濃度を決定するための血液の顕微鏡検査を含む任意の多数の従来の技術によって評価できる。本発明は、IFN−γ媒介性疾患に罹患した患者に対して本発明のIFN−γ抗体を、特定の障害の重篤度もしくはこの障害によって生じる症状の重篤度を反映する指標の規準を上回って改善の維持を誘導するか、またはいくつかのもしくは全ての場合には処置される条件に伴うさらに重篤な疾患の発現を遅延もしくは予防するのに十分な量および時間で投与する工程を包含する、処置の方法を包含する。本発明は、IFN−γ抗体を用いた処置の前、後、および/または間の他の治療剤を用いた可能性のある処置を除外しない。
【0114】
本明細書において用いる場合、「実質的に純粋な(substantially pure)」または「実質的に精製された(substantially purified)」とは、化合物または種が存在する主な種であるである(すなわち、モル基準では、この組成物における任意の他の個々の種よりもそれが豊富である)ことを意味する。特定の実施形態では、実質的に精製された画分とは、この種が存在する全ての高分子種のうちの少なくとも約50パーセント(モル基準で)を含む組成物である。特定の実施形態では、実質的に純粋な組成物は、その組成物中に存在する全ての高分子種のうち約80%、85%、90%、95%または99%より多い。特定の実施形態では、この種は、本質的に均一まで精製され(混入する種は、従来の検出法ではこの組成物中で検出できない)、ここでこの組成物は本質的に、単一の高分子種からなる。
【0115】
「患者(patient)」という用語は、ヒトおよび獣医学の被験体を包含する。
【0116】
文脈が他を要することがない限り、単数の用語は複数を包含する。
【0117】
IFN−γは、ウイルス感染から身体を防御することおよび免疫応答のいくつかの局面を調節することを含む、複数の機能を有するサイトカインであり、IFN−γ活性の増大は、いくつかの生理学的な条件に寄与し得る。本発明の特定の実施形態によれば、IFN−γに対する抗体は、上述のものを含むがそれらに限定されない、IFN−γ媒介性疾患を処置するために用いられ得る。
【0118】
本発明の1局面では、ヒトIFN−γに対して惹起され、かつそれに対する特異的な結合のための生物学的および免疫学的な特異性を有する完全ヒトモノクローナル抗体が提供される。このような抗体に含まれる可変領域(配列番号6、配列番号8、配列番号10、配列番号12、配列番号14、配列番号16、配列番号30および配列番号31)は、このような抗体の完全な重鎖および軽鎖(配列番号17、配列番号18、配列番号19、配列番号20、配列番号21、および配列番号22)、そして特定のCDR(重鎖および軽鎖CDR1、CDR2、および/またはCDR3;配列番号34〜配列番号44)を含む抗体は、本発明に包含される。本発明のこの局面の特定の実施形態は、本発明によって提供される重鎖および軽鎖の、CDR、特にCDR1〜CDR3に対応する配列である。さらに、本発明は、本明細書に開示されるCDR3配列(配列番号36、配列番号37、配列番号43および/または配列番号44)を含む抗体を包含するが、この配列はまた、任意の可変領域配列または本明細書に開示される完全な重鎖もしくは軽鎖の配列に対して少なくとも80%、85%、90%、91%、92%、93%、94%、95%、96%、97%、98%もしくは約99%同一である配列を含んでもよく、この抗体は、IFN−γの生物学的活性を阻害または調節し得る。
【0119】
別の局面では、本発明は、本発明の抗体をコードする単離された核酸またはポリヌクレオチドを提供する。本発明の抗体は、IFN−γに特異的に結合するか、および/またはIFN−γの生物学的活性を阻害もしくは調節し得る。本発明により特異的に包含されるのは、アミノ酸配列である配列番号17、配列番号18、配列番号19、配列番号20、配列番号21、配列番号22、配列番号32、配列番号33、配列番号5、配列番号7、配列番号9、配列番号11、配列番号13、配列番号15、配列番号30、配列番号31および/またはこれらの配列に対して少なくとも80%、85%、90%、91%、92%、93%、94%、95%、96%、97%、98%もしくは約99%同一性である配列をコードするヌクレオチド配列を含むポリヌクレオチドであり、ここで上記の配列と試験配列との間のアラインメントは、少なくとも約50、60、70、80、90もしくは100アミノ酸にまたがる。本発明はさらに、配列番号45、配列番号46、配列番号47および/または配列番号48を含むポリヌクレオチドであって、IFN−γに特異的に結合するかおよび/またはその生物学的活性を阻害もしくは調節し得る抗体をコードするポリヌクレオチドを提供する。さらに、本発明は、配列番号5、配列番号7、配列番号9、配列番号11、配列番号13、配列番号15、配列番号30、配列番号31、配列番号17、配列番号18、配列番号19、配列番号20、配列番号21、配列番号22、配列番号32もしくは配列番号33に対して少なくとも80%、85%、90%、91%、92%、93%、94%、95%、96%、97%、98%もしくは約99%同一であるポリヌクレオチドを包含するが、ここでこれらのポリヌクレオチドの各々によって部分的にコードされる抗体は、IFN−γの生物学的を阻害もしくは調節するか、および/またはIFN−γに特異的に結合し得、すぐ上に挙げられたヌクレオチド配列と試験配列との間のアラインメントは、少なくとも100、150、または200ヌクレオチドにまたがる。
【0120】
表2には、その配列識別番号に関連付けて配列の簡単な説明を提供する。
【0121】
(表2)
【0122】
【表2−1】
【0123】
【表2−2】
さらに別の局面では、本発明は、本発明の免疫グロブリン分子および抗体、必要に応じてモノクローナル抗体を発現するハイブリドーマおよび細胞株を提供する。さらなる局面では、本発明の免疫グロブリン分子または抗体を発現および/または分泌する、ハイブリドーマ細胞または細胞株由来の細胞は、患者に移植され得、これによって本発明の抗体またはその免疫学的に機能的な免疫グロブリンフラグメントは、患者において発現および/または分泌されて、これによってIFN−γ活性が阻害または調節される。
【0124】
本発明はまた、生物学的におよび免疫学的に精製された抗体の調製物であって、好ましくはヒトIFN−γに対して惹起されて、それに対して特異的に結合するための生物学的および免疫学的特異性を有するモノクローナル抗体の調製物を提供する。
【0125】
酵母人工染色体(YAC)におけるメガベースサイズのヒト遺伝子座をクローニングおよび再構築して、マウス生殖系列にそれらを導入する能力によって、極めて大きいかまたはおおまかにマッピングされた遺伝子座の機能的成分を解明し、そしてヒト疾患の有用なモデルを生成するための有利なアプローチの開発が可能になる。さらに、ヒトの等価な産物によるマウス遺伝子座の置換のためのこのような技術の利用によって、発達の間のヒト遺伝子産物の発現および調節、他の系との関連、ならびに疾患誘導および進行におけるその関与に対する固有の洞察が得られる。
【0126】
このようなストラテジーの重要な実際的な適用は、内因性Ig遺伝子が不活性化されているマウスへのヒト免疫グロブリン(Ig)遺伝子座の導入によるマウス体液性免疫系の変更である。国際出願番号WO93/12227。この系によって抗体のプログラムされた発現およびアセンブリの背景にある機構、ならびにB細胞発達におけるその役割を研究する機会が得られる。さらに、このようなストラテジーによって、完全ヒトモノクローナル抗体(MAb)の産生の供給源が得られる。完全ヒトMAbは、マウスまたはマウス由来MAbに内在する免疫原性およびアレルギー性の応答を最小化すること、それによって投与された抗体の有効性および安全性を増大することが期待される。完全ヒト抗体を、慢性および再発性のヒト疾患、例えば、変形性関節炎、関節リウマチおよび他の炎症性状態の処置において用いることが可能であり、その処置には抗体投与を反復することが必要である。従って、本発明の抗IFN−γ抗体の1つの特定の利点は、この抗体が完全ヒト抗体であり、マウス抗ヒト抗体または他の以前に記載された不完全ヒト抗体もしくは非ヒト種由来の非ヒト抗体と一般に関連する有害反応を最小限にしながら非急性の方式で患者に投与できるということである。
【0127】
本明細書に記載される方法を用いて、当業者は、ヒトIg遺伝子座の大きいフラグメントを用いてマウス抗体産生において欠損するマウス系統を操作することが可能であり、その結果このようなマウスは、マウス抗体の非存在下でヒト抗体を生成する。それより大きいヒトIgフラグメントは、可変性の大きい遺伝子多様性、ならびに抗体産生および発現の適切な調節を保存し得る。抗体多様化および選択ならびにヒト抗体に対する免疫学的寛容の欠失のためにマウス細胞機構を開発することによって、これらのマウス系統において再生されたヒト抗体レパートリーにより、ヒト抗原を含む目的の任意の抗原に対する高親和性抗体を得る。ハイブリドーマ技術を用いて、所望の特異性を有する抗原特異的ヒトMAbを生成および選択することができる。
【0128】
特定の実施形態では、当業者は、ヒト以外の種由来の定常領域をヒト可変領域(単数または複数)とともにこのようなマウスで用いてキメラ抗体を産生することができる。
(天然に存在する抗体構築物)
最も天然に存在する抗体構築物単位は代表的には、四量体を含む。各々のこのような四量体は代表的には、ポリペプチド鎖の2つの同一の対から構成され、各々の対が1つの全長「軽」鎖(代表的には約25kDaの分子量を有する)および1つの全長「重」鎖(代表的には約50〜70kDaの分子量を有する)を有する。各々の軽鎖および重鎖のアミノ末端部分は代表的には、代表的に抗原認識を担う、約100〜110以上のアミノ酸の可変領域を含む。各々の鎖のカルボキシ末端部分は代表的には、エフェクター機能を担う、定常領域を規定する。ヒト軽鎖は代表的には、κ軽鎖およびλ軽鎖として分類される。重鎖は代表的には、μ、δ、γ、α、またはεとして分類され、そしてそれぞれIgM、IgD、IgG、IgA、およびIgEとして抗体のアイソタイプを規定する。IgGは、IgG1、IgG2、IgG3およびIgG4が挙げられるがこれらに限定されないいくつかのサブクラスを有する。IgMは、IgM1およびIgM2が挙げられるがこれらに限定されないサブクラスを有する。IgAは同様に、IgA1およびIgA2が挙げられるがこれらに限定されないサブクラスに分割される。全長の軽鎖および重鎖内では、代表的には可変領域および定常領域は、約12以上のアミノ酸の「J」領域によって連結され、ここでこの重鎖はまた、約10アミノ酸より多い「D」領域を含む。例えば、FUNDAMENTAL IMMUNOLOGY 第7章、第2版(Paul、W.編、1989,Raven Press、N.Y.(全ての目的のためにその全体が参考として援用される))を参照のこと。各々の軽鎖/重鎖対の可変領域は、代表的には、抗体結合部位を形成する。
【0129】
ラクダおよびラマに見出されているいくつかの天然に存在する抗体は、2つの重鎖からなる二量体であって、軽鎖を含まない。Muldermansら,2001,J.Biotechnol.74:277〜302;Desmyterら,2001,J.Biol.Chem.276:26285〜90。本発明は、IFN−γに結合するか、および/またはその生物学的活性を阻害し得る2つの重鎖からなる二量体抗体を包含する。ラクダ抗体の結晶学的研究によって、19アミノ酸長を有する重鎖CDR3が抗原と相互作用して2つの他の超可変領域をカバーする表面を形成することが明らかになっている。Desmyterら(前出)。従ってCDR3は、二量体ラクダ抗体において、そしてさらに代表的な四量体抗体において、抗原結合に重要である。
【0130】
可変領域は代表的には、相補性決定領域すなわちCDRとも呼ばれる、3つの超可変領域に連結された比較的保存されたフレームワーク領域(FR)の同じ一般的な構造を示す。各々の対の2つの鎖由来のCDRは代表的には、フレームワーク領域内に包埋され、これが特定のエピトープに結合し得る。N末端からC末端へと、軽鎖および重鎖の可変領域はともに、代表的には、ドメインFR1、CDR1、FR2、CDR2、FR3、CDR3およびFR4を含む。各々のドメインに対するアミノ酸の割り当ては代表的には、以下にさらに詳細に説明されているように、Kabatらの定義に従っている。Kabatら、Sequences of Proteins of Immunological Interest(1991,National Institutes of Health,Bethesda,Md);またChothiaおよびLesk,1987,J.Mol.Biol.196:901〜917;Chothiaら,1989,Nature 342:878〜883も参照のこと。CDRは、抗原結合のための主な表面接触ポイントを構成する。例えば、ChothiaおよびLesk(前出)を参照のこと。さらに軽鎖のCDR3および特に重鎖のCDR3は、軽鎖および重鎖の可変領域内の抗原結合における最も重要な決定基を構成し得る。例えば、ChothiaおよびLesk(前出);Desiderioら(2001),J.Mol.Biol.310:603〜15;XuおよびDavis(2000),Immunity 13(1):37〜45;Desmyterら(2001),J.Biol.Chem.276(28):26285〜90;ならびにMuyldermans(2001),J.Biotechnol.,74(4):277〜302を参照のこと。いくつかの抗体では、重鎖CDR3は、抗原と抗体との間の接触の主な領域を構成するようである。Desmyterら(前出)。CDR3のみを変化させるインビトロ選択スキームを用いることによって、抗体の結合特性を変化させることができる。Muyldermans(前出);Desiderio(前出)。
【0131】
CDRは、以下の方法で重鎖可変領域配列に配置されてもよい。CDR1は、成熟抗体のおよそ31残基で開始して、通常は約5〜7アミノ酸長であり、そしてこれには、ほぼ常にCys−Xxx−Xxx−Xxx−Xxx−Xxx−Xxx−Xxx−Xxx(配列番号48)が先行している(ここで、「Xxx」は任意のアミノ酸である)。重鎖CDR1の後ろの残基はほぼ常にトリプトファン、しばしばTyp−Val,Trp−IleまたはTrp−Alaである。CDR1の最後の残基とCDR2の最初の残基との間に14個のアミノ酸がほぼ常にあり、そしてCDR2は代表的には16〜19個のアミノ酸を含む。CDR2には、その直前にLeu−Glu−Trp−Ile−Gly(配列番号49)が先行してもよく、そしてその直後にLys/Arg−Leu/Ile/Val/Phe/Thr/Ala−Thr/Ser/Ile/Alaが続く。他のアミノ酸は、CDR2に先行しても続いてもよい。CDR2の最後の残基とCDR3の最初の残基との間に32個のアミノ酸がほぼ常にあり、そしてCDR3は、約3〜25残基長であり得る。Cys−Xxx−Xxxはほぼ常に、CDR3に先行しており、そしてTrp−Gly−Xxx−Gly(配列番号50)がほぼ常にCDR3に続く。
【0132】
軽鎖CDRは、以下の方法で軽鎖配列に配置されてもよい。CDR1は、成熟抗体のおよそ24残基で開始して、通常は約10〜17残基長であり、そしてこれには、ほぼ常にCysが先行している。CDR1の最終残基とCDR2の最初の残基との間にはほぼ常に15アミノ酸があり、CDR2はほぼ常に7残基長である。CDR2は代表的には、Ile−Tyr、Val−Tyr、Ile−LysまたはIle−Pheが先行する。軽鎖のCDR2とCDR3との間にはほぼ常に32個の残基があり、そしてCDR3は通常は約7〜10アミノ酸長である。CDR3にはほぼ常にCysが先行しており、そして通常はPhe−Gly−Xxx−Gly(配列番号51)が続く。
【0133】
CDRを囲むフレームワーク領域の長さが典型的であるものとは異なる長さを生む挿入を含んでもまたは欠失を含んでもよいことを当業者は理解する。本明細書において意味されるとおり、重鎖フレームワーク領域の長さは、以下の範囲内におさまる:FR1、0〜41アミノ酸;FR2、5〜24アミノ酸;FR3、13〜42アミノ酸;およびFR4、0〜21アミノ酸。さらに、本発明は、軽鎖フレームワーク領域の長さが以下の範囲内におさまることを意図する:FR1、6〜35アミノ酸;FR2、4〜25アミノ酸;FR3、2〜42アミノ酸;およびFR4、0〜23アミノ酸。
【0134】
天然に存在する抗体は代表的には、タンパク質分泌のための細胞経路に抗体を指向し、かつ成熟抗体には存在しない、シグナル配列を含む。本発明の抗体をコードするポリヌクレオチドは、下記のように、天然に存在するシグナル配列、または異種のシグナル配列をコードしてもよい。
(抗体のインビトロ成熟)
抗体は、抗原についてのさらに高い親和性またはさらに低い解離定数のような特性の変化した抗体を生成するためにインビトロで成熟され得る。CDR、特にCDR3内の残基のみのバリエーションによって、同じ抗原に結合するがさらに親和性の大きい変化した抗体が生じ得る。例えば、Schierら,1996,J.Mol.Biol.263:551〜67;Yangら,1995,J.Mol.Biol.254:392〜403を参照のこと。本発明は、抗体が有利な特性を有するように選択することができる、種々のインビトロ選択スキーム、例えば、アフィニティー成熟および/またはチェーンシャッフリング(Kangら,1991,Proc.Natl.Acad.Sci.88:11120〜23)、またはDNAシャッフリング(Stemmer,1994,Nature
370:389〜391)によって作製された抗体を包含する。多くのスキームでは、公知の抗体を特定の位置、しばしばCDR内でインビトロで無作為化して、選択プロセスに供して、これによって所望の特性、例えば特定の抗原に対する親和性の増大を有する抗体を単離することができる。例えば、van den Beuckenら,2001,J.Mol.Biol.310:591〜601;Desiderioら,2001,J.Mol.Biol.310:603〜15;Yangら,1995,J.Mol.Biol.254:392〜403;Schierら,1996,J.Mol.Biol.263:551〜67を参照のこと。代表的には、このような変異された抗体は、突然変異誘発および選択工程のデザインに依存して、1つ以上のCDRにいくつかの変更された残基を含んでもよい。例えば、van den Beuckenら(前出)を参照のこと。
(二重特異性または二機能性抗体)
二重特異性抗体または二機能性抗体は代表的には、2つの異なる重鎖/軽鎖の対および2つの異なる結合部位を有する人工的ハイブリッド抗体である。二重特異性抗体は、限定はしないが、ハイブリドーマの融合またはFab’フラグメントの連結を含む種々の方法によって生成することができる。例えば、SongsivilaiおよびLachmann,1990 Clin.Exp.Immunol.79:315〜321;Kostelnyら、1992,J.Immunol.148:1547〜1553を参照のこと。(抗体の調製)
本発明は、ヒトIFN−γに特異的に結合する抗体を提供する。これらの抗体は、全長IFN−γまたはそのフラグメントを用いた免疫によって産生され得る。本発明の抗体は、ポリクローナル抗体であっても、もしくはモノクローナル抗体であってもよく、そして/または組み換え抗体であってもよい。特定の実施形態では、本発明の完全ヒト抗体は、例えば、ヒト抗体を産生し得るトランスジェニック動物の免疫によって調製される(例えば、国際特許出願公開WO 93/12227を参照のこと)。
【0135】
本発明の抗IFNγ抗体の軽鎖および重鎖の可変領域のCDRは、同じ種または別の種由来のフレームワーク領域(FR)に移植され得る。特定の実施形態では、抗IFNγ抗体の軽鎖および重鎖の可変領域のCDRは、「ヒト化」抗体を作製するためにコンセンサスなヒトFRに移植されてもよい。このようなヒト化抗体は、本発明によって包含される。コンセンサスヒトFRを作製するために、いくつかのヒト重鎖または軽鎖のアミノ酸配列由来のFRを、整列させてコンセンサスアミノ酸配列を同定する。抗IFNγ抗体重鎖または軽鎖のFRは、異なる重鎖または軽鎖由来のFRで置換されてもよい。抗IFNγ抗体の重鎖および軽鎖のFRにおけるまれなアミノ酸は代表的には、置換されないが、残りのFRアミノ酸は置換されてもよい。まれなアミノ酸とは、FRにおいて通常はみいだされない位置にある特定のアミノ酸である。本発明の抗IFNγ抗体由来の移植された可変領域は、抗IFNγ抗体のもとの定常領域と異なる定常領域とともに用いられてもよい。あるいは、移植された可変領域は、単鎖Fv抗体の一部である。CDR移植は、任意の目的のために参考として本明細書に援用される、例えば、米国特許第6,180,370号、同第5,693,762号、同第5,693,761号、同第5,585,089号および同第5,530,101号に記載されている。
【0136】
本発明の抗体は、マウスの抗体産生細胞に挿入されたヒト抗体産生遺伝子座の実質的な一部を有しており、そして内因性マウス抗体を産生するにおいて欠失されるようにさらに操作されている、トランスジェニックマウスを用いて調製されてもよい。このようなマウスは、ヒト免疫グロブリン分子および抗体を産生可能であり、そしてマウスの免疫グロブリン分子および抗体を産生しないかまたは実質的に少ない量を産生する。この結果を達成するために利用される技術は、本明細書に開示された特許、出願および引用文献に開示されている。特定の実施形態では、当業者は、任意の目的のために参考として本明細書に援用される、国際特許出願公開番号WO98/24893に開示されるような方法を使用してもよい。任意の目的のために参考として本明細書に援用される、Mendezら,1997,Nature Genetics 15:146〜156も参照のこと。
【0137】
本発明のモノクローナル抗体(mAb)は、従来のモノクローナル抗体方法論、例えば、KohlerおよびMilstein(1975,Nature 256:495)の標準的体細胞ハイブリダイゼーション技術を含む、種々の技術によって生成されてもよい。体細胞ハイブリダイゼーション手順が好ましいが、原則的には、モノクローナル抗体を産生するための他の技術、例えば、Bリンパ球のウイルスまたは発癌性形質転換を用いることもできる。
【0138】
ハイブリドーマを調製するための可能性のある動物系の1つはマウスである。マウスにおけるハイブリドーマ産生はきわめてよく確立されており、そして融合のための免疫された脾細胞の単離のための免疫プロトコールおよび技術は当該分野で周知である。融合パートナー(例えば、マウス骨髄腫細胞)および融合手順はまた公知である。
【0139】
いくつかの実施形態では、IFN−γ、必要に応じてヒトIFN−γに対する完全ヒトモノクローナル抗体は、マウス系とは異なるヒト免疫系の一部を有するトランスジェニックマウスを用いて生成され得る。これらのトランスジェニックマウスは、本明細書において「HuMab」マウスと呼ばれるが、これは、再配列されていないヒト重鎖(μおよびγ)およびκ軽鎖免疫グロブリン配列をコードするヒト免疫グロブリン遺伝子ミニ遺伝子座を、内因性μおよびκ鎖遺伝子座を不活性化する標的化変異とともに含む(Lonbergら,1994,Nature 368:856〜859)。従って、マウスは、マウスIgMまたはκの発現の低下を示し、そして免疫に応答して、導入されたヒト重鎖および軽鎖導入遺伝子は、クラス切り替えおよび体細胞変異を受けて高い親和性のヒトIgGκモノクローナル抗体を生成する(Lonbergら、前出;LonbergおよびHuszar,1995,Intern.Rev.Immunol.13:65〜93;HardingおよびLonberg,1995,Ann.N.Y.Acad.Sci.764:536〜546)。HuMabマウスの調製は、その全ての内容が全体が参考として本明細書に援用されている、Taylorら,1992,Nucleic Acids Res.20:6287〜6295;Chenら,1993,International Immunology 5:647〜656;Tuaillonら,1994,J.Immunol.152:2912〜2920;Lonbergら,1994,Nature 368:856〜859;Lonberg,1994,Handbook of Exp.Pharmacology 113:49〜101;Taylorら,1994,International Immunology 6:579〜591;LonbergおよびHuszar,1995,Intern.Rev.Immunol.13:65〜93;HardingおよびLonberg,1995,Ann.N.Y.Acad.Sci 764:536〜546;Fishwildら,1996,Nature Biotechnology 14:845〜851に詳細に記載されている。さらに、全てがLonbergおよびKayに対する、米国特許第5,545,806号;同第5,569,825号;同第5,625,126号;同第5,633,425号;同第5,789,650号;同第5,877,397号;同第5,661,016号;同第5,814,318号;同第5,874,299号および同第5,770,429号、ならびにその全ての開示が全体が参考として本明細書に援用されている、1993年6月24日公開のSuraniらの米国特許第5,545,807号;1993年6月24日公開、国際特許出願公開番号WO93/1227;1992年12月23日公開WO92/22646;および1992年3月19日公開WO92/03918を参照のこと。あるいは、以下の実施例に記載されたHCo7およびHCo12のトランスジェニックマウス系統は、ヒト抗IFNγ抗体を生成するために用いられ得る。
【0140】
これらの実施形態では、本発明の抗体は、10−7M、10−8M、10−9M、または10−10M未満という平衡解離定数(KD)でIFN−γに特異的に結合する。本発明の特定の実施形態では、この抗体は、約10−8M〜10−12MというKDでIFN−γに結合する。
【0141】
好ましい実施形態では、本発明の抗体は、IgG1、IgG2またはIgG4アイソタイプの抗体である。この抗体は、IgG1アイソタイプの抗体であってもよい。他の実施形態では、本発明の抗体は、IgM、IgA、IgEまたはIgDアイソタイプの抗体である。本発明の好ましい実施形態では、この抗体は、ヒトκ軽鎖およびヒトIgG1重鎖を含む。IgG1重鎖定常領域を含む本発明の抗体の発現は、以下の実施例に記載されている。特定の実施形態では、この抗体の可変領域は、IgG1アイソタイプの定常領域以外の定常領域に連結される。特定の実施形態では、本発明の抗体は、哺乳動物細胞における発現のためにクローニングされている。
【0142】
特定の実施形態では、抗IFNγ抗体の重鎖および軽鎖に対する(そしてそのコードするヌクレオチドに対する改変に相当する)保存的改変は、抗IFNγ抗体の特徴と類似の機能的および化学的特徴を有する抗IFNγ抗体を生成する。対照的に、抗IFNγ抗体の機能および/または化学的特徴における実質的な改変は、(a)置換の領域における分子骨格の構造、例えばβシートまたはらせん高次構造、(b)標的部位における分子の変化もしくは疎水性、または(c)側鎖のバルク、の維持に対する効果が有意に異なる重鎖および軽鎖のアミノ酸配列の置換を選択することによって達成され得る。
【0143】
例えば、「保存的アミノ酸置換」とは、その位置でのアミノ酸残基の極性または荷電に対して影響がほとんどないかまたは全くないような、天然ではない残基による天然のアミノ酸残基の置換を包含し得る。さらに、ポリペプチドにおける任意の天然の残基はまた、「アラニンスキャニング突然変異誘発(alanine scanning mutagenesis)」について以前に記載されているとおり、アラニンで置換されてもよい。
【0144】
所望のアミノ酸置換(保存的であっても、または非保存的であっても)は、このような置換が所望される時点で当業者によって決定され得る。特定の実施形態において、アミノ酸置換を用いて、抗IFNγ抗体の重要な残基を同定すること、または本明細書に記載される抗IFNγ抗体の親和性を増大もしくは低下させることができる。
【0145】
別の実施形態では、本発明の抗体は、ハイブリドーマ細胞株とは異なる細胞株で発現され得る。これらの実施形態では、適切な哺乳動物宿主細胞の形質転換のために特定の抗体をコードする配列を用いてもよい。これらの実施形態によれば、米国特許第4,399,216号、同第4,912,040号、同第4,740,461号および同第4,959,455号(その全てが任意の目的のために参考として本明細書に援用される)に例示されているとおり、例えばウイルスへ(またはウイルスベクターへ)ポリヌクレオチドをパッケージングすること、およびこのウイルス(またはベクター)によって宿主細胞を形質導入すること、または当該分野で公知のトランスフェクション手順によって、などを含む、宿主細胞にポリヌクレオチドを導入するための任意の公知の方法を用いて、形質転換を達成することができる。一般的に、用いられる形質転換手順は、形質転換されるべき宿主に依存し得る。哺乳動物細胞へ異種ポリヌクレオチドを導入するための方法は、当該分野で周知であり、そしてこれには、デキストラン媒介性トランスフェクション、リン酸カルシウム沈殿、ポリブレン媒介性トランスフェクション、プロトプラスト融合、エレクトロポレーション、リポソームへのポリヌクレオチド(単数または複数)のカプセル化、および核へのDNAの直接マイクロインジェクションが挙げられるがこれらに限定されない。
【0146】
本発明の抗IFNγ抗体の重鎖定常領域、重鎖可変領域、軽鎖定常領域、または軽鎖可変領域のアミノ酸配列をコードする核酸分子(またはポリヌクレオチド)は、本発明によって包含される。このようなポリヌクレオチドは、標準的な連結技術を用いて適切な発現ベクター中に挿入されてもよい。好ましい実施形態では、抗IFNγ抗体重鎖または軽鎖の定常領域をコードするポリヌクレオチドは、適切な可変領域をコードするポリヌクレオチドの下流に付加され、そして発現ベクター中に連結される。このベクターは代表的には、使用される特定の宿主細胞において機能的である(すなわち、このベクターは、遺伝子の増幅および/または遺伝子の発現が生じ得るように宿主細胞の機構に適切である)ように選択される。発現ベクターの概説に関しては、METH.ENZ.185(Goeddel、編)1990,Academic Pressを参照のこと。
【0147】
代表的には、任意の宿主細胞において用いられる発現ベクターは、プラスミド維持のため、ならびに内因性ヌクレオチド配列のクローニングおよび発現のための配列を含む。このような配列は総称的に、「隣接配列(flanking sequences)」と呼ばれるが、特定の実施形態では、代表的には、以下のヌクレオチド配列のうちの1つ以上を含む:プロモーター、1つ以上のエンハンサー配列、複製起点、転写終結配列、ドナーおよびアクセプタースプライス部位を含む完全イントロン配列、ポリペプチド分泌のためのリーダー配列をコードする配列、リボソーム結合部位、ポリアデニル化配列、発現されるべきポリペプチドをコードする核酸を挿入するためのポリリンカー領域、および選択可能なマーカーエレメント。これらの配列の各々は以下に考察される。
【0148】
必要に応じて、ベクターは、「タグ(tag)」コード配列、すなわち、抗IFNγ抗体ポリペプチドコード配列の5’または3’末端に位置するオリゴヌクレオチド分子を含んでもよい;このオリゴヌクレオチド配列は、ポリHis(例えば、hexaHis)、または別の「タグ」、例えばFLAG、HA(インフルエンザウイルス血球凝集素)、またはmyc(これについては、市販の抗体が存在する)をコードする。このタグは代表的には、ポリペプチドの発現の際にポリペプチドに融合され、そして宿主細胞からのIFN−γ抗体のアフィニティー精製または検出のための手段として機能し得る。アフィニティー精製は、例えば、アフィニティーマトリックスとしてタグに対する抗体を用いるカラムクロマトグラフィーによって達成され得る。必要に応じて、タグは、種々の手段によって、例えば、切断のための特定のペプチダーゼを用いて、精製された抗IFNγ抗体ポリペプチドから引き続いて除去されてもよい。
【0149】
隣接配列は、相同(すなわち、宿主細胞と同じ種および/または株由来)、異種(すなわち、宿主細胞種または株以外の種由来)、ハイブリッド(すなわち、2つ以上の供給源由来の隣接配列の組み合わせ)、合成または天然であってもよい。従って、隣接配列の供給源は、この隣接配列が機能的であり、宿主細胞の機構によって活性化され得る条件下では、任意の原核生物生物体であっても、または真核生物生物体であっても、任意の脊椎動物もしくは無脊椎動物であっても、または任意の植物であってもよい。
【0150】
本発明のベクターにおいて有用な隣接配列は、当該分野で周知の任意のいくつかの方法によって達成され得る。代表的には、本明細書において有用な隣接配列は、マッピングによって、および/または制限エンドヌクレアーゼ消化によって以前に同定されており、従って適切な制限エンドヌクレアーゼを用いて適切な組織供給源から単離可能である。ある場合には、隣接配列の全ヌクレオチド配列が公知であり得る。ここでは、隣接配列は、核酸合成またはクローニングのために本明細書において記載された方法を用いて合成され得る。
【0151】
隣接配列の全てが既知であるか一部のみが既知であるかにかかわらず、これは、ポリメラーゼ連鎖反応(PCR)を用いて、ならびに/または適切なプローブ、例えば、同じもしくは別の種由来のオリゴヌクレオチドおよび/もしくは隣接配列のフラグメントを用いてゲノムライブラリーをスクリーニングすることによって、得ることができる。隣接配列が既知でない場合、隣接配列を含むDNAのフラグメントは、例えば、コード配列またはさらに別の遺伝子(単数または複数)を含み得る、DNAのさらに大きい小片から単離することができる。単離は、適切なDNAフラグメントを生成するための制限エンドヌクレアーゼ消化によって、その後のアガロースゲル精製、Qiagen(登録商標)カラムクロマトグラフィー(Chatsworth,CA)または当業者に公知の他の方法を用いる単離によって達成できる。この目的を達成するための適切な酵素の選択は、当業者に容易に明らかである。
【0152】
複製起点は代表的には、市販の原核生物発現ベクターの一部であり、この起点は宿主細胞においてベクターの増幅を補助する。選択したベクターが複製起点の部位を含まない場合、既知の配列に基づいて化学的に合成して、ベクターに連結してもよい。例えば、プラスミドpBR322(New England Biolabs,Beverly,MA)由来の複製起点は、ほとんどのグラム陰性細菌に適切であり、種々のウイルス起点(例えば、SV40、ポリオーマ、アデノウイルス、水疱性口内炎ウイルス(VSV)、またはパピローマウイルス、例えば、HPVまたはBPV)は、哺乳動物細胞におけるクローニングベクターに有用である。一般的に、複製起点の成分は、哺乳動物発現ベクターには必要ではない(例えば、SV40起点は、ウイルス初期プロモーターをも含むので、しばしば単独で用いられる)。
【0153】
転写終結配列は代表的には、ポリペプチドコード領域の末端に対して3’に位置して、転写を集結するように機能する。通常、原核生物細胞における転写終結配列は、G−Cリッチフラグメントであり、ポリT配列が続く。この配列はライブラリーから容易にクローニングされるかまたはベクターの一部として市販されてさえいるが、本明細書に記載の方法のような核酸合成の方法を用いて容易に合成することもできる。
【0154】
選択マーカー遺伝子は、選択性の増殖培地中で増殖される宿主細胞増殖の生存および増殖に必要なタンパク質をコードする。代表的な選択マーカー遺伝子は、(a)原核生物宿主細胞については、抗生物質または他の毒素、例えば、アンピシリン、テトラサイクリンもしくはカナマイシンに対する耐性を付与するか;(b)細胞の栄養要求欠損を補完するか;あるいは(c)複合培地または規定の培地から利用できない重要な栄養物を供給する、タンパク質をコードする。好ましい選択マーカーは、カナマイシン耐性遺伝子、アンピシリン耐性遺伝子およびテトラマイシン耐性遺伝子である。有利には、ネオマイシン耐性遺伝子もまた、原核生物および真核生物の宿主細胞の両方における選択のために用いることができる。
【0155】
発現される遺伝子を増幅するために、他の選択遺伝子が用いられてもよい。増幅とは、増殖または細胞の生存に重要なタンパク質の産生に必要な遺伝子が、組み換え細胞の連続的世代の染色体内において一列で反復されているプロセスである。哺乳動物細胞のための適切な選択マーカーの例としては、ジヒドロ葉酸還元酵素(DHFR)およびプロモーターなしのチミジンキナーゼ遺伝子が挙げられる。哺乳動物細胞形質転換体は、ベクターに存在する選択遺伝子のおかげで形質転換体のみが生存するように固有に適合されるという選択圧下に置かれる。選択圧は、培地中の選択因子の濃度が連続的に増大される条件下で形質転換細胞を培養することによって課され、これによって選択遺伝子および別の遺伝子、例えばIFNγポリペプチドに結合する抗体をコードするDNAの両方の増幅がもたらされる。結果として、抗IFNγ抗体のようなポリペプチドの増大した量は、増幅されたDNAから合成される。
【0156】
リボソーム結合部位は通常、mRNAの翻訳開始に必要であり、シャイン・ダルガーノ(Shine−Dalgarno)配列(原核生物)またはKozak配列(真核生物)によって特徴付けられる。このエレメントは代表的には、プロモーターの3’側および発現されるべきポリペプチドのコード配列の5’側に位置する。
【0157】
真核生物の宿主細胞発現系においてグリコシル化が所望される場合のような、ある場合には、グリコシル化または収率を改善するための種々のプレ配列またはプロ配列を操作してもよい。例えば、特定のシグナルペプチドのペプチダーゼ切断部位を変更してもよいし、またはグリコシル化に影響し得るプロ配列を付加してもよい。最終タンパク質産物は、−1位(成熟タンパク質の第一アミノ酸に対する位置)に、発現に付随する1つ以上のさらなるアミノ酸付加を有してもよく、これは完全に取り除かれていなくてもよい。例えば、最終タンパク質産物は、アミノ末端に結合されたペプチダーゼ切断部位に見出される2つ以上のアミノ酸残基を有し得る。あるいは、いくつかの酵素切断部位の使用によって、酵素が成熟ポリペプチド内のこのような領域で切断する場合、所望のポリペプチドのわずかに短縮された形態が生じ得る。
【0158】
本発明の発現ベクターおよびクローニングベクターは代表的には、宿主生物体によって認識され、かつ抗IFNγ抗体をコードする分子に作動可能に連結されているプロモーターを含む。プロモーターは、構造遺伝子の転写を制御する構造遺伝子(一般的には約100〜1000bp内)の開始コドンに対して上流(すなわち、5’側)に位置する非翻訳配列である。プロモーターは以下の2つのクラスのうちの1つに慣用的に分類される:誘導性プロモーターおよび構成的プロモーター。誘導性プロモーターは、栄養物の有無または温度の変化のような培養条件におけるいくつかの変化に応答してそれらの制御下でDNAから増大したレベルの転写を開始する。構成的プロモーターは他方では、それらが作動可能に連結されている遺伝子を均一に転写し、すなわち、遺伝子発現に対する制御はほとんどないかまたは全くない。種々の潜在的な宿主細胞によって認識される多数のプロモーターは周知である。適切なプロモーターは、制限酵素消化によって供給源のDNAからプロモーターを除去すること、およびこのベクターに所望のプロモーター配列を挿入することによって、本発明の抗IFNγ抗体を含む重鎖または軽鎖をコードするDNAに作動可能に連結される。
【0159】
酵母宿主との使用のための適切なプロモーターもまた、当該分野で周知である。酵母エンハンサーは、酵母プロモーターとともに有利に用いられる。哺乳動物宿主細胞とともに使用するための適切なプロモーターは周知であり、そしてウイルス、例えば、ポリオーマウイルス、鶏痘ウイルス、アデノウイルス(例えば、アデノウイルス2)、ウシパピローマウイルス、トリ肉腫ウイルス、サイトメガロウイルス、レトロウイルス、B型肝炎ウイルスおよび最も好ましくはシミアンウイルス40(SV40)のゲノムから得られたプロモーターが挙げられるがこれらに限定されない。他の適切な哺乳動物プロモーターとしては、異種哺乳動物プロモーター、例えば、熱ショックプロモーターおよびアクチンプロモーターが挙げられる。
【0160】
目的であり得るさらなるプロモーターとしては以下が挙げられるがこれらに限定されない:SV40初期プロモーター(BenoistおよびChambon,1981,Nature 290:304〜10);CMVプロモーター(Thomsenら、1984,Proc.Natl.Acad.USA 81:659〜663);ラウス肉腫ウイルスの3’長末端反復配列に含まれるプロモーター(Yamamotoら,1980,Cell 22:787〜97);ヘルペスチミジンキナーゼプロモーター(Wagnerら、1981,Proc.Natl.Acad.Sci.U.S.A.78:1444〜45);メタトリオネイン遺伝子由来のプロモーターおよび調節性配列(Brinsterら、1982,Nature 296:39〜42);ならびに原核生物プロモーター、例えば、βラクタマーゼプロモーター(Villa−Kamaroffら、1978,Proc.Natl.Acad.Sci.U.S.A.,75:3727〜31);またはtacプロモーター(DeBoerら、1983,Proc.Natl.Acad.Sci.U.S.A.,80:21〜25)。また目的のものは、組織特異性を示して、トランスジェニック動物において利用されている、以下の動物転写制御領域である:膵臓の腺房細胞において活性であるエラスターゼI遺伝子制御領域(Swiftら,1984,Cell 38:639〜46;Ornitzら,1986,Cold Spring Harbor Symp.Quant.Biol.50:399〜409(1986);MacDonald,1987,Hepatology 7:425〜515);膵臓β細胞において活性なインスリン遺伝子制御領域(Hanahan,1985,Nature
315:115〜22);リンパ細胞において活性な免疫グロブリン遺伝子制御領域(Grosschedlら、1984,Cell 38:647〜58;Adamesら,1985,Nature 318:533〜38;Alexanderら,1987,Mol.Cell.Biol.,7:1436〜44);精巣、乳房、リンパ系細胞および肥満細胞において活性なマウス乳腺腫瘍ウイルス制御領域(Lederら、1986,Cell 45:485〜95);肝臓で活性なアルブミン遺伝子制御領域(Pinkertら、1987,Genes and Devel.1:268〜76);肝臓においおいて活性なαフェトプロテイン遺伝子制御領域(Krumlaufら、1985,Mol.Cell.Biol.,5:1639〜48;Hammerら、1987,Science 235:53〜58);肝臓において活性なα1−抗トリプシン遺伝子制御領域(Kelseyら、1987,Genes and Devel.1:161〜71);骨髄性細胞において活性なβグロビン遺伝子制御領域(Mogramら、1985,Nature 315:338〜40;Kolliasら、1986,Cell 46:89〜94);脳における稀突起膠細胞において活性なミエリン塩基性タンパク質遺伝子制御領域(Readheadら、1987,Cell 48:703〜12);骨格筋において活性であるミオシン軽鎖2遺伝子制御領域(Sani,1985,Nature 314:283〜86);ならびに視床下部で活性な生殖腺刺激ホルモン放出ホルモン遺伝子制御領域(Masonら、1986,Science 234:1372〜78)。
【0161】
エンハンサー配列をベクターに挿入して、高等な真核生物によって本発明の抗IFNγ抗体を含む軽鎖または重鎖をコードするDNAの転写を増大する。エンハンサーは、通常約10〜300bp長のDNAのシス作用性エレメントであり、これはプロモーター上で転写を増大するように働く。エンハンサーは比較的方向および位置に依存せず、転写単位の5’および3’の両方の位置で見出されている。哺乳動物遺伝子から利用可能ないくつかのエンハンサー配列は公知である(例えば、グロビン、エラスターゼ、アルブミン、αフェトプロテインおよびインスリン)。しかし、代表的には、ウイルス由来のエンハンサーを用いる。当該分野で公知のSV40エンハンサー、サイトメガロウイルス初期プロモーターエンハンサー、ポリオーマエンハンサー、およびアデノウイルスエンハンサーは、真核生物プロモーターの活性化のための例示的な増強エレメントである。エンハンサーは、ベクター中でコード配列の5’または3’のいずれに位置してもよいが、代表的にはプロモーターから5’部位に位置する。
【0162】
適切な天然のシグナル配列または異種のシグナル配列をコードする配列(リーダー配列またはシグナルペプチド)を発現ベクターに組み込んで、抗体の細胞外分泌を促進することが可能である。シグナルペプチドまたはリーダーの選択は、抗体が産生される宿主細胞のタイプに依存し、そして異種シグナル配列は、天然のシグナル配列を置き換え得る。哺乳動物宿主細胞で機能的であるシグナルペプチドの例としては、以下が挙げられる:米国特許第4,965,195号に記載されるインターロイキン7(IL−7)のシグナル配列;Cosmanらに記載のインターロイキン2レセプターのシグナル配列(1984,Nature 312:768);欧州特許第0 367 566号に記載のインターロイキン−4レセプターシグナルペプチド;米国特許第4,968,607号に記載のI型インターロイキン−1レセプターシグナルペプチド;欧州特許第0 460 846号に記載のII型インターロイキン−1レセプターシグナルペプチド;ヒトIgKのシグナル配列(METDTLLLWVLLLWVPGSTG(配列番号52)である);ヒト成長ホルモンのシグナル配列(MATGSRTSLLLAFGLLCLPWLQEGSA;配列番号53である);ならびに実施例3において単離された重鎖および軽鎖をコードするcDNAによってコードされた、ヒトシグナル配列MGSTAILALLLAVLQGVCA(配列番号54)およびMETPAQLLFLLLLWLPDTTG(配列番号55)。
【0163】
本発明の発現ベクターは、市販のベクターのような開始ベクターから構築されてもよい。このようなベクターは、所望の隣接配列の全てを含んでも含まなくてもよい。本明細書に記載される隣接配列の1つ以上は既にベクターには存在しないが、それらを個々に入手してベクターに連結してもよい。隣接配列の各々を得るために用いた方法は、当業者に周知である。
【0164】
ベクターが構築され、そして軽鎖、重鎖、または抗IFNγ抗体を含む軽鎖および重鎖をコードする核酸分子がこのベクターの適切な部位に挿入された後、この完成したベクターを増幅および/またはポリペプチド発現のために適切な宿主細胞中に挿入してもよい。選択された宿主細胞への抗IFNγ抗体のための発現ベクターの形質転換は、トランスフェクション、感染、リン酸カルシウム共沈殿、エレクトロポレーション、マイクロインジェクション、リポフェクション、DEAEデキストラン媒介性トランスフェクション、または他の公知の技術を含む周知の方法によって達成され得る。選択される方法は一部は、用いられる宿主細胞のタイプに関連する。これらの方法および他の適切な方法は、当業者に周知であり、そして例えば、Sambrookら(前出)に記載されている。
【0165】
宿主細胞は、適切な条件下で培養された場合、抗IFNγ抗体を合成し、この抗体は引き続いて、培養培地から(宿主細胞が培地中にこの抗体を分泌する場合)、またはこの抗体を産生する宿主細胞から直接(この抗体が分泌されない場合)、収集できる。適切な宿主細胞の選択は、種々の要因、例えば、所望の発現レベル、活性のために所望されるかまたは必要であるポリペプチド改変(例えば、グリコシル化またはリン酸化)および生物学的に活性な分子への折り畳みの容易さに依存する。
【0166】
発現のための宿主として利用可能な哺乳動物細胞株は、当該分野で周知であり、そしてこれらとしては、限定はしないがAmerican Type Culture Collection(ATCC)から入手可能な不死化細胞株が挙げられ、これらとしては、限定はしないが、チャイニーズハムスター卵巣(CHO)細胞、HeLa細胞、ベビーハムスター腎臓(BHK)細胞、サル腎臓細胞(COS)、ヒト肝細胞癌腫細胞(例えば、Hep G2)および多数の他の細胞株が挙げられる。特定の実施形態では、どの細胞株が高い発現レベルを有し、IFNγ結合特性を有する抗体を構成的に生成するかを決定することを通じて細胞株を選択することができる。別の実施形態では、それ自体の抗体を作製しないが、異種の抗体を作製および分泌する能力を有するB細胞系列由来の細胞株を選択することができる。
【0167】
本発明の抗体は、生物学的サンプル中のIFNγを検出するため、およびIFNγタンパク質を産生する細胞または組織の同定のために有用である。IFNγに特異的に結合する本発明の抗体は、IFNγ媒介性疾患の処置において有用であり得る。このような抗体は、IFNγを検出するため、およびIFNγがIFNγレセプターとの複合体を形成することを阻害するための結合アッセイにおいて用いられ得る。IFNγに結合して、他の結合化合物との相互作用をブロックするこのような抗体は、IFNγ媒介性疾患を調節するのにおいて治療的な用途を有し得る。好ましい実施形態では、IFNγに対する抗体は、そのレセプターに対するIFNγ結合をブロックし得、これがIFNγ誘導性シグナル伝達カスケードの破壊を生じ得る。
【0168】
いくつかの実施形態では、本発明は薬学的組成物であって、治療上有効な量の1つまたは複数の本発明の抗体を薬学的に受容可能な希釈剤、キャリア、可溶化剤、乳化剤、保存剤および/またはアジュバントとともに含む薬学的組成物を提供する。好ましくは、受容可能な処方物質は、使用される投薬量および濃度でレシピエントに対して非毒性である。好ましい実施形態では、治療上有効な量の抗IFNγ抗体を含む薬学的組成物が提供される。
【0169】
特定の実施形態では、受容可能な処方物質は好ましくは、使用される投薬量および濃度でレシピエントに対して非毒性である。
【0170】
特定の実施形態では、薬学的組成物は、例えば、pH、浸透圧、粘度、清澄度、色調、等張性、臭気、無菌性、安定性、分解もしくは放出の速度、組成物の吸着もしくは浸透を改変、維持または保存するための処方物質を含んでもよい。このような実施形態では、適切な処方物質としては、限定はしないが、アミノ酸(例えば、グリシン、グルタミン、アスパラギン、アルギニンまたはリジン);抗菌剤;抗酸化剤(例えば、アスコルビン酸、亜硫酸ナトリウムまたは亜硫酸水素ナトリウム);緩衝液(例えば、ホウ酸塩、炭酸水素塩、Tris−HCl、クエン酸、リン酸または他の有機酸);充填剤(例えば、マンニトールまたはグリシン);キレート剤(例えば、エチレンジアミン四酢酸(EDTA));錯化剤(例えば、カフェイン、ポリビニルピロリドン、βサイクロデキストリンまたはヒドロキシプロピルβシクロデキストリン);賦形剤(filler);単糖類;二糖類;ならびに他の炭水化物(例えば、グルコース、マンノースまたはデキストリン);タンパク質(例えば、血清アルブミン、ゼラチンまたは免疫グロブリン);着色剤、香味料および希釈剤;乳化剤;親水性のポリマー(例えば、ポリビニルピロリドン);低分子量ポリペプチド;塩形成対イオン(例えば、ナトリウム);防腐剤(例えば、塩化ベンザルコニウム、安息香酸、サリチル酸、チメロサール、フェネチルアルコール、メチルパラベン、プロピルパラベン、クロルヘキシジン、ソルビン酸または過酸化水素);溶媒(例えば、グリセリン、プロピレングリコールまたはポリエチレングリコール);糖アルコール(例えば、マンニトールまたはソルビトール);懸濁剤;界面活性剤または湿潤剤(例えば、プルロニック、PEG、ソルビタンエステル、ポリソルベート、例えば、ポリソルベート20、ポリソルベート80、トリトン(triton)、トロメタミン、レシチン、コレステロール、チロキサポール);安定性強化剤(例えば、スクロースまたはソルビトール);等張化剤(例えば、ハロゲン化アルカリ金属、好ましくは塩化ナトリウムまたは塩化カリウム、マンニトール ソルビトール);送達ビヒクル;希釈剤;賦形剤(excipient)および/または薬学的アジュバントが挙げられる。REMINGTON’S
PHARMACEUTICAL SCIENCES,第18版(A.R.Gennaro編),1990,Mack Publishing Companyを参照のこと。
【0171】
特定の実施形態では、最適の薬学的組成物は、例えば、意図される投与経路、送達様式および所望の投薬量に依存して当業者によって決定される。例えば、REMINGTON’S PHARMACEUTICAL SCIENCES,前出を参照のこと。特定の実施形態では、このような組成物は、本発明の抗体の物理的状態、安定性、インビボでの放出の速度、およびインビボでのクリアランスの速度に影響し得る。
【0172】
特定の実施形態では、薬学的組成物における主なビヒクルまたはキャリアは、天然には水性であっても非水性であってもよい。例えば、適切なビヒクルまたはキャリアは、(おそらくは非経口投与のための組成物中に共通の他の物質で補充された)注射用水、生理学的生理食塩水または人工的な脳脊髄液であってもよい。中性の緩衝化生理食塩水、または血清アルブミンと混合された生理食塩水はさらに、例示的なビヒクルである。好ましい実施形態では、薬学的組成物は、約pH7.0〜8.5のTris緩衝液、または約pH4.0〜5.5の酢酸塩緩衝液を含み、そしてさらにソルビトールまたはその適切な代替物を含んでもよい。本発明の特定の実施形態では、抗IFNγ抗体組成物は、所望の程度の純度を有する選択された組成物を最適の処方剤(REMINGTON’S PHARMACEUTICAL SCIENCES、前出)とともに、凍結乾燥ケーキまたは水溶液の形態で混合することによって貯蔵のために調製され得る。さらに、特定の実施形態では、抗IFNγ抗体産物は、スクロースのような適切な賦形剤を用いて凍結乾燥剤として処方されてもよい。
【0173】
本発明の薬学的組成物は、非経口送達のために選択され得る。あるいは、この組成物は、吸入のために、または経口のような消化管を通じた送達のために選択され得る。このような薬学的に受容可能な組成物の調製は、当該分野の技術の範囲内である。
【0174】
処方成分は、投与の部位に受容可能な濃度で存在することが好ましい。特定の実施形態では、緩衝液を用いて、この組成物を生理学的なpHで、またはわずかに低いpHで、代表的には約5〜約8のpH範囲内で維持する。
【0175】
非経口的投与を考慮する場合、本発明における使用のための治療組成物は、所望の抗IFNγ抗体を薬学的に受容可能なビヒクル中に含む、発熱物質なしの、非経口的に受容可能な水溶液の形態で提供されてもよい。非経口注射のために特に適切なビヒクルは、抗IFNγ抗体が、適切に保存された無菌の等張性溶液として処方されている、滅菌蒸留水である。特定の実施形態では、この調製物は、所望の分子の処方物を、デポ注射を介して送達できる産物の制御放出または徐放性放出を提供し得る、ある因子、例えば、注射用マイクロスフェア、生体侵食性粒子、ポリマー化合物(例えば、ポリ乳酸またはポリグリコール酸)、ビーズまたはリポソームとともに含んでもよい。特定の実施形態では、循環中の持続期間を延長する効果を有する、ヒアルロン酸も用いることができる。特定の実施形態では、移植可能な薬物送達デバイスを用いて所望の抗体分子を導入することができる。
【0176】
本発明の薬学的組成物は、吸入のために処方され得る。これらの実施形態では、抗IFNγ抗体は、乾燥した吸入可能な粉末として有利に処方される。好ましい実施形態では、抗IFNγ抗体吸入溶液はまた、エアロゾル送達のための噴霧剤とともに処方され得る。特定の実施形態では、溶液は噴霧され得る。従って、肺投与および処方の方法はさらに、参考として援用されており、化学的に修飾されたタンパク質の肺送達を記載する、国際特許出願番号PCT/US94/001875に記載されている。
【0177】
処方物は経口的に投与できることも考えられる。この様式で投与される抗IFNγ抗体は、錠剤およびカプセルのような固体投薬形態の配合に習慣的に用いられるキャリアの有無とともに処方され得る。特定の実施形態では、カプセルは、バイオアベイラビリティーが最大化されて予備的な全身性の分解が最小化される胃腸管におけるポイントでこの処方物の活性部位を遊離するように設計され得る。抗IFNγ抗体の吸着を促進するためにさらなる因子が含まれてもよい。希釈剤、香味料、低融点ワックス、植物油、潤滑剤、懸濁剤、錠剤崩壊剤および結合剤も使用され得る。
【0178】
本発明の薬学的組成物は好ましくは、1または複数の抗IFNγ抗体の有効量を錠剤の製造業者に適切な非毒性賦形剤と混合して含むように提供される。滅菌水、または別の適切なビヒクルに錠剤を溶解することによって、単位用量形態で溶液を調製し得る。適切な賦形剤としては、限定はしないが、不活性希釈剤、例えば、炭酸カルシウム、炭酸ナトリウムまたは重炭酸ナトリウム、ラクトースもしくはリン酸カルシウム;または結合剤、例えばデンプン、ゼラチンもしくはアラビアゴム;または潤滑剤、例えば、ステアリン酸マグネシウム、ステアリン酸、もしくは滑石が挙げられる。
【0179】
さらなる薬学的組成物は、当業者には明白であり、これらとしては、徐放性または制御送達の処方物に含まれた抗IFNγ抗体を含む処方物が挙げられる。種々の他の徐放性または制御送達の手段、例えば、リポソームキャリア、生体侵食性微粒子または多孔性ビーズおよびデポ注射剤を処方するための技術もまた、当業者には公知である。例えば、参考として援用されており、薬学的組成物の送達のための多孔性ポリマー微粒子の制御放出を記載している、国際特許出願番号PCT/US93/00829を参照のこと。徐放性調製物は、成形された物質の形態で半透過性ポリマー物質、例えばフィルムまたはマイクロカプセルを含んでもよい。徐放性マトリックスは、ポリエステル、ヒドロゲル、ポリラクチド(参考として各々が援用される、米国特許第3,773,919号および欧州特許出願公開番号EP058481に開示される)、Lグルタミン酸およびγエチル−L−グルタメートのコポリマー(Sidmanら、1983,Biopolymers 22:547〜556)、ポリ(2−ヒドロキシエチル−メタクリレート)(Langerら、1981,J.Biomed.Mater.Res.15:167〜277およびLanger,1982,Chem.Tech.12:98〜105)、エチレン酢酸ビニル(Langerら、前出)またはポリ−D(−)−3−ヒドロキシ酪酸(欧州特許出願公開番号EP133,988)を含んでもよい。徐放性組成物はまた、当該分野で公知の任意のいくつかの方法によって調製され得るリポソームを含んでもよい。例えば、参考として援用される、Eppsteinら、1985,Proc.Natl.Acad.Sci.USA 82:3688〜3692;欧州特許出願公開番号EP 036,676;EP 088,046およびEP 143,949を参照のこと。
【0180】
インビボ投与のために用いられる薬学的組成物は代表的には、滅菌調製物として提供される。滅菌は、滅菌濾過膜を通した濾過によって達成され得る。組成物が凍結乾燥される場合、この方法を用いる滅菌は、凍結乾燥および再構成の前または後のいずれに行なわれてもよい。非経口的な投与のための組成物は、凍結乾燥型または溶液として貯蔵されてもよい。非経口的な組成物は一般には、無菌アクセスポートを有する容器、例えば静脈内溶液バッグ、または皮下注射針によって穿通可能な栓を有するバイアルに入れられる。
【0181】
一旦薬学的組成物が処方されれば、これは、滅菌バイアル中に、溶液、懸濁液、ゲル、エマルジョン、固体、結晶として、または脱水もしくは凍結乾燥された粉末として貯蔵され得る。このような処方物は、レディーツーユースの形態、または投与の前に再構成される形態(例えば、凍結乾燥された形態)のいずれで貯蔵されてもよい。
【0182】
本発明はまた、単回用量投与単位を生成するためのキットを提供する。本発明のキットは各々が、乾燥タンパク質を有する第一の容器、および水性処方物を有する第二の容器の両方を含む。本発明の特定の実施形態では、単一および複数のチャンバを有する事前充填(pre−filled)シリンジ(例えば、液体シリンジおよびライオシリンジ(lyosyringe))を備えるキットが提供される。
【0183】
使用されるべき、抗IFNγ抗体を含有する薬学的組成物の治療的有効量は、例えば、治療の状況および目的に依存する。処置のために適切な投薬レベルは部分的には送達される分子、抗IFNγ抗体が用いられている徴候、投与経路、ならびに患者のサイズ(体重、体表面または器官サイズ)および/または状態(年齢および全身健康状態)に依存して変化することが当業者には理解される。特定の実施形態では、最適の治療効果を得るために、臨床医が投薬量を滴定して、投与経路を改変してもよい。代表的な投薬量は、上述の要因に依存して、約0.1μg/kg〜約30mg/kg以上までに及んでもよい。好ましい実施形態では、投薬量は0.1μg/kg〜約30mg/kg、必要に応じて1μg/kg〜約30mg/kgまたは10μg/kg〜約5mg/kgに及んでもよい。
【0184】
投与頻度は、用いられる処方物中の特定の抗IFNγ抗体の薬物動態的パラメーターに依存する。代表的には、所望の効果を達成する投薬量が得られるまで、臨床医がこの組成物を投与する。従って、この組成物は単回用量として、または2回以上の用量(これは同じ量の所望の分子を含んでも含まなくてもよい)として、または移植デバイスもしくはカテーテルを介した連続注入として、経時的に投与されてもよい。適切な投薬量のさらなる工夫は当業者によって慣用的に行なわれ、そしてそれらによって慣用的に行なわれる課題の範囲内である。適切な投薬量は、適切な用量応答データの使用を通じて得ることができる。特定の実施形態では、本発明の抗体は、長期間にわたって患者に投与されてもよい。本発明の抗体の慢性投与によって、非ヒト動物においてヒト抗原に対して惹起された抗体、例えば、ヒト以外の種で産生された不完全ヒト抗体または非ヒト抗体に一般に関連する有害な免疫応答またはアレルギー応答が最小化される。
【0185】
薬学的組成物の投与経路は、徐放性システムによるかまたは移植デバイスによる、公知の方法、例えば、経口、静脈内、腹腔内、脳内(実質内)、脳室内、筋肉内、眼内、動脈内、門脈内または病巣内の経路による注射を通じた公知の方法に従う。特定の実施形態では、組成物は、ボーラス注射によって投与されても、もしくは注入によって連続的に投与されても、または移植デバイスによって投与されてもよい。
【0186】
この組成物はまた、所望の分子がその上に吸着されるかまたはカプセル化されている、膜、スポンジまたは別の適切な物質の移植によって局所投与され得る。移植デバイスを用いる特定の実施形態では、デバイスは任意の適切な組織または器官に移植されてもよく、そして所望の分子の送達は、拡散、時限放出ボーラス、または連続的投与を介してもよい。
【0187】
本発明による抗IFNγ抗体の薬学的組成物をエキソビボで用いることもまた所望され得る。このような場合、患者から除去されている細胞、組織または器官を、抗IFNγ抗体の薬学的組成物に曝露して、その後に細胞、組織および/または器官を患者に引き続き移植して戻す。
【0188】
詳細には、抗IFNγ抗体は、本明細書において記載される方法のような方法を用いて、ポリペプチドを発現および分泌するように遺伝子操作されている特定の細胞を移植することによって送達できる。特定の実施形態では、このような細胞は、動物細胞であってもまたはヒト細胞であってもよく、そして自家性、非相同性または異種性であってもよい。特定の実施形態では、細胞は不死化されてもよい。他の実施形態では、免疫学的応答の機会を低下させるために、細胞は、周囲の組織の浸潤を回避するためにカプセル化されてもよい。さらなる実施形態では、カプセル化物質は代表的には、タンパク質産物(単数または複数)の放出を可能にするが、患者の免疫系によるか、または周囲の組織からの他の有害な要因による、細胞の破壊を妨げる生体適合性、半透過性ポリマー同封物または膜である。
【実施例】
【0189】
以下の実施例は、行なった実験および得られた結果を含んでいるが、これは、例示目的のためにのみ提供されており、本発明を限定すると解釈されるべきではない。
【0190】
(実施例1)
(CHO細胞からのヒトIFNγタンパク質の生成)
テンプレートとしてヒト脾臓Marathon−Ready cDNA(Clontech)を用いて、全長ヒトIFNγ cDNAをPCR(標準的条件下)によって増幅した。この配列をpDSRα2プラスミド中にサブクローニングした。DH10B(Escherichia coli)細胞をpDSRα2プラスミドを用いて形質転換した。標準的技術を用いてDNAを調製して、CHO細胞をリン酸カルシウム法(Speciality Media,Inc)によってトランスフェクトした。高発現細胞株クローンを用いて、無血清馴化培地を生成した。
【0191】
ヒトIFNγ(hu−IFN−γ)を含むCHO細胞馴化培地を濃縮して、透析して、数段階のクロマトグラフィー工程を通じて精製した。第一の工程は、グリコシル化されていないhu−IFN−γ形態から高度にグリコシル化された形態を分離するために、標準的なNaCl勾配を用いるQ−HP(Pharmacia)クロマトグラフィーであった。Q−HPプールをさらに、コムギ胚芽凝集素クロマトグラフィー(EY Laboratories)を通じて精製した。この精製された物質をSDS−ポリアクリルアミドゲル電気泳動(SDS−PAGE)によって分離して、クマーシー・ブルーおよび銀染色によって分析した。精製された物質は、クマーシー・ブルーおよび銀の両方で染色されたSDS−PAGEによって決定した場合95%を超える純度であった。この物質をゲル凝固(gel−clot)法(Limulus Amebocyte Lysate)によってもアッセイして、内毒素レベルが低いことを示した。hu−IFN−γの同一性は、R&D Systemsの抗AF−285 NA抗体を用いてウエスタンブロットによって確認した。吸光係数方法を用いて、吸光度(A280)から最終のタンパク質濃度を決定したが、ここでA280読み取り/吸光係数=濃度(g/L)(吸光係数=0.66)である。
【0192】
(実施例2)
(IFNγに対するヒトモノクローナル抗体の産生)
(トランスジェニックHuMabマウス)
IFN−γに対する完全ヒトモノクローナル抗体は、ヒト抗体遺伝子を各々が発現する、トランスジェニックマウスのHCo7株、HCo12株およびHCo7+HCo12株を用いて調製された。これらの株の各々において、内因性マウスκ軽鎖遺伝子は、Chenらに記載されたとおりホモ接合性に分裂されており(1993,EMBO J.12:811〜820)、そして内因性マウス重鎖遺伝子は、国際特許出願公開番号WO 01/09187(参考として援用される)の実施例1に記載のようにホモ接合性に分裂されていた。各々の株は、Fishwildら(1996,Nature Biotechnology 14:845〜851)に記載されているとおり、ヒトκ軽鎖導入遺伝子KCo5を担持した。HCo7株は、米国特許第5,545,806号、同第5,625,825号および同第5,545,807号(参考として援用される)に記載のとおりHCo7ヒト重鎖導入遺伝子を担持する。HCo12株は、国際特許出願番号WO 01/09187(参考として援用される)の実施例2に記載のとおりHCo12ヒト重鎖導入遺伝子を担持した。HCo7+HCo12株は、HCo7およびHCo12の両方の重鎖導入遺伝子を担持し、そして各々の導入遺伝子について半接合性であった。これらの株の全てが本明細書においてHuMabマウスと呼ばれる。
【0193】
(HuMab免疫:)
IFN−γに対する完全ヒトモノクローナル抗体を生成するため、HuMabマウスを、抗原としてE.coliまたはCHO細胞由来の精製組み換えヒトIFN−γを用いて免疫した。HuMabマウスの一般的な免疫スキームは、Lonbergらに記載されている(各々の教示が参考として援用されている、1994,Nature 368:856〜859;Fishwildら(前出)および国際特許出願公開番号WO98/24884)。マウスは、抗原の第一回の注入の際に6〜16週齢であった。IFN−γ抗原(例えば、IFN−γを発現する、トランスフェクトされたE.coliまたはCHO細胞から精製された)の精製組み換え調製物(25〜100μg)を用いて、HuMabマウスを腹腔内(IP)または皮下(Sc)で免疫した。
【0194】
HuMabトランスジェニックマウスの免疫は、完全フロイントアジュバントおよび2つの注射物中の抗原を用いて、続いて、不完全フロイントアジュバント中の抗原を用いた2〜4週のIP免疫(全部で9免疫まで)によって行なった。数ダースのマウスを各々の抗原について免疫した(E.coliまたはCHO細胞のいずれかで生成したヒトIFN−γ)。HCo7株、HCo12株、およびHCo7+HCo12株の全部で91匹のマウスをIFN−γで免疫した。免疫応答を後眼窩採血によってモニターした。
【0195】
IFN−γに結合する抗体を産生するHuMabマウスを選択するため、免疫したマウス由来の血清をFishwildら(前出)に記載されるとおりELISAによって試験した。要するに、マイクロタイタープレートを、CHO細胞またはE.coli由来の精製された組み換えIFN−γをPBS中に1〜2μL/mLで用いてコーティングして、50μL/ウェルで、4℃で一晩インキュベートし、次いで5%ニワトリ血清を含むPBS/Tween(0.05%)の200μL/ウェルを用いてブロックした。IFN−γ免疫したマウスからの血漿の希釈物を、各々ウェルに添加して、環境温度で1〜2時間インキュベートした。このプレートをPBS/Tweenを用いて洗浄して、次いで西洋ワサビペルオキシダーゼ(HRP)と結合体化したヤギ抗ヒトIgG Fc特異的ポリクローナル試薬とともに室温で1時間インキュベートした。プレートをPBS/Tweenを用いて洗浄して、西洋ワサビペルオキシダーゼ(HRP)と結合体化したヤギ抗ヒトIgG Fc特異的ポリクローナル試薬とともに室温で1時間インキュベートした。洗浄後、このプレートをABTS基質(Sigma Chemical Co.,St.Louis,MO,カタログ番号A−1888,0.22mg/mL)を用いて発色させて、415〜495nmの波長で光学密度(OD)を決定することによって分光光度的に分析した。抗IFN−γヒト免疫グロブリンの十分な力価を有するマウスを用いて下記のとおりモノクローナル抗体を産生した。
【0196】
(IFN−γに対するヒトモノクローナル抗体を産生するハイブリドーマの生成)
屠殺の2日前に抗原を用いて静脈内にブースト(追加免疫)することによりモノクローナル抗体産生のためにマウスを準備して、その後に脾臓を取り出した。HuMabマウスからマウス脾細胞を単離して、標準的なプロトコールを用いてPEGとともにマウス骨髄腫細胞株に融合した。代表的には、各々の抗原について10〜20の融合を行なった。
【0197】
要するに、免疫されたマウス由来の脾臓リンパ球の単一細胞懸濁物を、50%PEG(Sigma)を用いてP3X63−Ag8.653非分泌マウス骨髄腫細胞(ATCC,登録番号CRL1580)の数の四分の一に融合した。細胞を平底マイクロタイタープレート中で約1×105/ウェルでプレートして、続いて10%ウシ胎仔血清、10%P388D1−(ATCC,登録番号CRL TIB−63)馴化培地、3〜5%ORIGEN(登録商標)Hybridoma Cloning Factor(IGEN)、マウスマクロファージ様細胞株を培養するために用いた培地由来の部分的に精製されたハイブリドーマ増殖培地補充物をDMEM(Mediatech,カタログ番号CRL10013、高いグルコース、Lグルタミンおよびピルビン酸ナトリウムを含む)中に、さらに5mM HEPES、0.055mM 2−メルカプトエタノール、50mg/mLゲンタマイシンおよび1×HAT(Sigma,カタログ番号、CRL P−7185)とともに含有する選択培地中での約2週間のインキュベーションを続けた。1〜2週後、HATをHTに置き換えた培地中で細胞を培養した。
【0198】
得られたハイブリドーマを抗原特異的抗体の産生についてスクリーニングした。個々のウェルをELISA(上記)によって、ヒト抗IFN−γモノクローナルIgG抗体についてスクリーニングした。一旦、過剰のハイブリドーマ増殖が生じれば、通常10〜14日後に培地をモニターした。抗体分泌するハイブリドーマを再度プレートして、再度スクリーニングして、依然としてヒトIgGについて陽性である場合、抗IFN−γモノクローナル抗体を少なくとも2回、限界希釈によってサブクローニングした。次いで、安定なサブクローンをインビトロで培養して、精製および特徴づけのために組織培養培地中で少量の抗体を生成した。
【0199】
(IFN−γに結合するヒトモノクローナル抗体の選択)
上記のようなELISAアッセイを用いて、IFN−γ免疫原と正の反応性を示したハイブリドーマをスクリーニングした。IFN−γに対して高い結合活性で結合するモノクローナル抗体を分泌するハイブリドーマをサブクローニングしてさらに特徴付けた。親細胞の反応性を保持している(ELISAによって決定した場合)、各々のハイブリドーマ由来の1つのクローンを、液体窒素中に保持した5〜10バイアルの細胞バンクを作製するために選択した。
【0200】
アイソタイプ特異的ELISAを行なって、本明細書に開示されたとおり産生したモノクローナル抗体のアイソタイプを決定した。これらの実験では、マイクロタイタープレートウェルを、PBS中1μg/mLのマウス抗ヒトκ軽鎖溶液を50μL/ウェルで用いてコーティングして、4℃で一晩インキュベートした。5%ニワトリ血清を用いたブロッキング後、このプレートを各々の試験したモノクローナル抗体由来の上清および精製されたアイソタイプコントロールと反応させた。プレートを環境温度で1〜2時間インキュベートした。次いで、このウェルをヒトIgG1またはIgG3特異的な西洋ワサビペルオキシダーゼ結合体化ヤギ抗ヒトポリクローナル抗血清のいずれかと反応させて、プレートを発色させ、上記のように分析した。
【0201】
ELISAによって検出した場合IFN−γに対する有意な結合を示したハイブリドーマ上清から精製したモノクローナル抗体をさらに、下記のような種々のバイオアッセイを用いて生物学的活性について試験した。選択された抗体を1119、1121、1118*、1121*および1118と命名した。
【0202】
(実施例3)
(抗IFN−γ抗体重鎖および軽鎖のクローニング)
上記の実施例2で同定されたIFN−γ結合モノクローナル抗体1119、1121、1118*、1121*および1118を発現するハイブリドーマを供給源として用いて、総RNA、DNAおよびタンパク質を単離するのに適切なフェノールおよびグアニジンイソチオシアネートの単層溶液であるTRIzol(登録商標)試薬(Invitrogen)を用いて総RNAを単離した。第一鎖のcDNAを、伸長アダプター(5’−GGC CGG ATA GGC CTC CAN NNN NNT−3’)(配列番号23)とともにランダムプライマーを用いて合成し、そして5’RACE(cDNA末端の迅速増幅)分取アッセイを、有効性が改善されているcDNA末端の迅速増幅(RACE)のためのキットであるGENERACERTMKit(Invitrogen)を用いて、製造業者の指示に従って行った。完全な軽鎖コードcDNAを調製するために、フォワードプライマーは、GENERACERTMネスト化プライマーであって、リバースプライマーは5’−GGG GTC AGG CTG GAA CTG AGG−3’(配列番号24)であった。リバースプライマーは、ヒトκ鎖の3’非翻訳領域において見出されるcDNA配列の保存領域を認識するように設計した。重鎖の可変領域をコードするcDNAを調製するために、フォワードプライマーはGENERACERTMネスト化プライマーであって、リバースプライマーは5’−TGA GGA CGC TGA CCA
CAC G−3’(配列番号25)であって、これは、ヒトIgG鎖のFc領域におけるコード配列中の保存領域を認識するように設計した。RACE産物をpCR4−TOPO(Invitrogen)中にクローニングして、その配列を決定した。コンセンサス配列を用いて全長抗体鎖PCR増幅のプライマーを設計した。
【0203】
抗IFN−γκ軽鎖をコードするcDNAを調製するために、5’PCRプライマーは、シグナル配列のアミノ末端、XbaI制限酵素部位、および最適化Kozak配列(5’−ACA ACA AAG CTT CTA GAC CAC CAT GGA AAC CCC AGC TCA GCT TCT CTT−3’;配列番号26)をコードした。3’プライマーは、カルボキシ末端および終止コドン、ならびにSalI制限部位(5’−CTT GTC GAC TCA ACA CTC TCC CCT GTT GAA GCT−3’;配列番号27)をコードした。得られたPCR産物のフラグメントを精製してXbaIおよびSalIを用いて消化し、次いでゲル単離して、哺乳動物発現ベクターpDSRα19に連結した(任意の目的のために参考として本明細書に援用される国際出願公開番号WO 90/14363を参照のこと)。
【0204】
抗IFNγ重鎖をコードするcDNAを調製するため、5’PCRプライマーは、シグナル配列のアミノ末端、XbaI制限酵素部位、および最適化Kozak配列(5’−CAG CAG AAG CTT CTA GAC CAC CAT GGG GTC AAC CGC CAT CCT CG−3’;配列番号28)をコードした。3’プライマーは、天然に存在するセンス鎖BsmBI部位(5’−CTT GGT GGA GGC ACT AGA GAC GGT GAC CAG GGT GCC ACG GCC−3’;配列番号29)を含む可変領域のカルボキシル末端をコードした。得られた産物を精製して、XbaIおよびBsmBIを用いて消化し、ゲル単離して、ヒトIgG1定常領域を含むpDSRα19ベクターに連結した。
【0205】
(実施例4)
(チャイニーズハムスター卵巣(CHO)細胞における抗IFNγ抗体の発現)
リン酸カルシウム法を用いて、1119−重鎖/pDSRα19および1119κ鎖/pDSRα19プラスミドを、ジヒドロ葉酸還元酵素欠損(DHFR−)無血清適合チャイニーズハムスター卵巣(CHO)細胞中に同時トランスフェクションすることによって、1119抗IFN−γ mAbの安定な発現を得た。トランスフェクトされた細胞を透析血清を含むが、ヒポキサンチン−チミジンを含まない培地中で選択して、DHFR酵素を発現する細胞の増殖を確保した。トランスフェクトされたクローンをELISAのようなアッセイを用いてスクリーニングして、馴化培地中で1119抗IFN−γ mAbの発現を検出した。1119発現細胞株をメトトレキセート増幅に供した。増幅の際の最高発現クローンを単細胞クローニングおよび細胞バンクの作製のために選択した。
【0206】
本発明の任意の組み換え抗IFN−γ抗体は、1119MAbについて上記と同じプロトコールを用いて、DHFRの欠損したチャイニーズハムスター卵巣細胞において生成し得る。本発明の各々の抗IFN−γ抗体の完全重鎖または軽鎖をコードするDNA配列を発現ベクター中にクローニングする。DHFRを欠くCHO細胞を、適切な抗IFN−γ抗体の完全重鎖を発現できる発現ベクターおよび完全軽鎖を発現できる発現ベクターを用いて同時トランスフェクトする。例えば、1118抗体を生成するために、配列番号19に示されるアミノ酸配列を含む完全重鎖を発現できるベクターおよび配列番号20に示されるアミノ酸配列を含む完全軽鎖を発現できるベクターを用いて細胞を同時トランスフェクトする。1121抗体を生成するために、配列番号21に示されるアミノ酸配列を含む完全重鎖を発現できるベクター、および配列番号22に示されるアミノ酸配列を含む完全軽鎖を発現できるベクターを用いて細胞を同時トランスフェクトする。1118*抗体を生成するために、配列番号32に示されるアミノ酸配列を含む完全重鎖を発現できるベクター、および配列番号20に示されるアミノ酸配列を含む完全軽鎖を発現できるベクターを用いて細胞を同時トランスフェクトする。1121*抗体を生成するために、配列番号21に示されるアミノ酸配列を含む完全重鎖を発現できるベクター、および配列番号33に示されるアミノ酸配列を含む完全軽鎖を発現できるベクターを用いて細胞を同時トランスフェクトする。表3は、種々のIFNγ抗体についての完全重鎖および完全軽鎖をまとめる。
【0207】
【表3】
(実施例5)
(抗IFN−γ抗体の産生)
1119抗体を発現するCHO細胞のクローン株における発現によって1119抗体を産生した。産生を行なうために、単一のバイアルから細胞を無血清細胞培養培地中に溶かした。この細胞を250mLの振盪フラスコ中で最初に、次いでスピナーフラスコ中で、そして最後にステンレス鋼リアクター中で2000Lのバイオリアクターまでスケールアップして増殖させた。流加培養法を用いて2000Lのバイオリアクター中で産生を行ない、ここには濃縮された媒体成分を含む栄養供給原を加えて、細胞の増殖および培養生存度を維持した。産生を約2週間続けて、この間に細胞によって1119抗体を構成的に産生して、細胞培養培地に分泌させた。
【0208】
産生リアクターを予め決定したpH、温度および溶存酸素レベルで制御した。このpHを二酸化炭素ガスおよび炭酸ナトリウムの添加によって制御した。溶存酸素を、空気、窒素および酸素のガスフローによって制御した。
【0209】
産生の終わりに、細胞ブロスをディスクスタック遠心分離に供して、培養上清を細胞から分離した。さらにこの濃縮物をデプスフィルター、続いて0.2μmのフィルターを通して清澄化する。次いで、清澄化した馴化培地を接線流限外濾過によって濃縮した。馴化培地を15〜30倍に濃縮した。次いで、得られた濃縮された馴化培地を処理して抗体を精製したが、これは後日精製するために凍結してもよい。本明細書に記載される任意の他の抗体は、同様の様式で産生できる。
【0210】
(実施例6)
(抗IFNγ抗体の活性の特徴づけ)
IFN−γは多数の生物学的効果を有するので、いくつかの異なるバイオアッセイを用いて種々のIFN−γ抗体の力価を比較する。以下で記載されたA549アッセイを、そのアッセイでの選択された候補物の能力に基づいたさらなる分析のためにそれらの選択された候補物を用いる一次スクリーニングに使用した。選択された候補物は、1119抗体、1118抗体および1121抗体を含んだ。
【0211】
(A549 バイオアッセイ)
IFN−γの確立された特性の1つは種々の細胞集団に対するその抗増殖効果である。例えば、AuneおよびPogue,1989,J.Clin.Invest.84:863〜75を参照のこと。ヒト肺細胞株A549は、IFN−γの生物活性を記載している刊行物では頻繁に用いられている。例えば、AuneおよびPogue、前出;Hillら、1993,Immunology 79:236〜40を参照のこと。一般には、用量のわずかな変化が応答の変化を生じる、用量応答曲線の一部におさまる刺激物質の濃度でインヒビターの活性を試験する。当業者は、過剰な用量の刺激物質が用いられる場合、応答の変化を観察するには極めて高用量のインヒビターが必要であり得ることを認識する。物質を刺激するために一般に用いられる濃度は、EC80およびEC90(最大応答のそれぞれ80%または90%が達成される濃度)である。
【0212】
IFN−γ用量応答曲線を作成して肺上皮癌細胞株A549のEC90を決定した(約30pM)。引き続く実験では、異なる濃度の精製された抗体を固定用量のIFN−γ(30pM)と混合して、この抗体がIFN−γの抗増殖性効果の生物学的活性を阻害する能力を決定した。このアッセイを5日間行なって、代謝的に活性な、すなわち増殖している細胞による、細胞増殖を示すために用いられる色素であるALAMARBLUETM(AccuMed International,Inc.,Chicago,Illinois)の減少によって生じた蛍光を決定することによって増殖を測定した。例えば、de FriesおよびMitsuhashi,1995,J.Clin.Lab.Analysis 9(2):89〜95;Ahmedら、1994,J.Immunol.Methods 170(2):211〜24を参照のこと。
【0213】
図9に示されるとおり、1119抗体は、14pMというIC50(IFN−γの効果の50%阻害を達成した濃度)を有する最も強力な抗体であって、1121(46pM)および1118(97pM)がそれに続く。
【0214】
(HLA DRバイオアッセイ)
IFN−γの確立された別の特性は、種々の細胞タイプにおけるMHCクラスIおよびクラスII遺伝子の発現を上方制御する能力である。この活性は特に、ループス腎炎に関連し得る(Yokoyamaら、1992,Kidney Int.42:755〜63)。THP−1ヒト単球細胞株は、IFN−γのこの生物活性について述べている刊行物では頻繁に用いられている。IFN−γ用量応答曲線を作成して、この実験に用いられる特定のTHP−1細胞株のEC80を決定した(約21pM)。引き続く実験では、種々の濃度の精製された抗体を固定用量のIFN−γ(21pM)と混合して、この抗体が細胞表面上でのHLA DR発現のIFN−γ誘導性上方制御を中和または阻害する能力を決定した。このアッセイを24時間行ない、測定した指標は、細胞に対するFITC標識抗HLA DR抗体の結合を検出するためのFACS解析によって決定された平均蛍光強度であった。
【0215】
図10に示されるとおり、1119抗体は、14pMというIC50を有する最も強力な抗体であって、1121抗体(60pM)および1118抗体(86pM)がそれに続く。
【0216】
(全血のバイオアッセイ)
いくつかの異なる細胞株におけるIP−10ケモカインの産生をIFN−γが上方制御するという公表された観察に基づいてヒト全血のアッセイを行なった。この活性は特に、ループス腎炎に関連し得る(Narumiら、2000,Cytokine 12:1561〜1565)。多数の正常ヒトドナー由来の全血を、IFN−γがIP−10産生を増大する能力について試験した。IFN−γ用量応答曲線を作成して、個々のドナーについてのEC50を決定した。予想どおり、いくつかの変動がドナーの間で観察された。一般に、50〜100pMのEC50を再現可能に示すと考えられるドナーを用いた。全血を固定濃度のIFN−γおよび種々の濃度の抗体と混合して、18.5時間インキュベートして、次いでELISAによってIP−10レベルを決定した。全血アッセイからの2つの異なるドナーについての代表的な結果を図11に示す。これらの2つのドナーからのIC50は17pMおよび14pMであった。現在まで、外因性IFN−γの添加の必要なしに全血アッセイにおいて自然に上昇したIP−10レベルを有する1つのドナーが同定されている。抗IFNγ抗体は、おそらく内因性に生成されたIFN−γをブロックすることによって、IP−10のこの自然な産生をブロックできた。
【0217】
(生化学的アッセイ)
IFN−γに対するいくつかの抗体についての結合動態を、BIAコア分析によって測定した。最初の結果では、この抗体が、分子間の結合を測定するために表面プラズモン共鳴を用いる装置である、BIACORETM(Pharmacis Biosensor
AB Corporation,Uppsala,Sweden)での信頼性のある測定の限界にアプローチするオフレート(off−rate)を有することが示唆された。従って、平衡結合アッセイを行なって用いた。平衡に達するように、固定量の抗体を種々の濃度のIFN−γとともに5時間を超えてインキュベートし、次いでIFN−γ結合ビーズと極めて短時間接触させて、このビーズに結合した遊離の抗体の量を、蛍光に基づく免疫アッセイ装置であるKINEXATM装置(Sapidyne Instruments Inc.,Boise,ID)で測定した。1119抗体では、最低の平衡解離定数である約24pMが得られた。
【0218】
(実施例7)
(種交差反応性)
上記の抗体がいくつかの異なる種由来の組み換えIFN−γタンパク質を中和または阻害する能力について、それらの抗体を試験した。マウスIFN−γタンパク質は市販されているが、ヒト、カニクイザルおよびチンパンジーのIFN−γタンパク質は、クローニングして、ヒト293細胞のような従来の哺乳動物発現系において発現した。ヒト、カニクイザルおよびチンパンジーのIFN−γタンパク質は全てが以前に記載されたA549アッセイで活性であるが、マウスタンパク質は、このアッセイでは活性ではない。マウスタンパク質は、マウス細胞株の置換について以外は以前に記載されたA549アッセイと本質的に同一である、RAW 264.7細胞株に基づくアッセイにおいて活性であった。RAW 264.7はマウス単球マクロファージ細胞株であり、そして例えば、American Type Culture Collectionから得ることができる。表4に示されるとおり、3つ全ての抗体がヒトおよびチンパンジーのIFN−γを中和できたが、3つのうちどれもカニクイザルまたはマウス由来のIFN−γの生物学的活性を中和も阻害もできなかった。
【0219】
【表4】
(実施例8)
(抗IFN−γ抗体のエピトープの同定)
成熟ヒトおよびカニクイザルのIFN−γのアミノ酸配列の比較によって、それらの間に9つのアミノ酸の相違が、ヒトIFN−γ配列の19、20、31、34、65、77、103、110および126位置において存在することが示された。ヒトおよびチンパンジーのIFN−γの配列は、ThakurおよびLandolfi(1999),Molecular Immunology 36:1107〜15に開示されている。カニクイザルIFN−γ配列は、TatsumiおよびSata(1997)、Int.Arch.Allergy Immunol.114(3):229〜36に開示されている;そしてマウスIFN−γ配列は、例えば、National Center for Biotechnology Information(NCBI)アクセッション番号NP_032363に開示されている。市販のキットを用いる部位特異的突然変異誘発を用いて、ヒトIFN−γ内の相違するヒトアミノ酸の各々を、カニクイザルタンパク質由来の対応するアミノ酸によって個々に置換した。各々の置換されたIFN−γは、ヒトIFNγ配列に存在するアミノ酸を置換するために用いたアミノ酸についての記号、およびこの置換が生じた成熟ヒトIFN−γ配列における位置に従って「huIFNγ」と命名した。例えば、「huIFNγD19」は、19位置を除いてヒトIFN−γに同一のIFN−γのバージョンを表し、ここでは正常にはこの位置を占めるヒスチジンにアスパラギン酸が置き換わっている。同様の実験を行ない、ここではカニクイザルIFN−γ内で開始して、各々の相違するアミノ酸をヒトIFN−γに存在するアミノ酸で置換した。例えば、「cynoIFNγL103」は、103位置を除いてカニクイザルIFN−γに同一のIFN−γのバージョンを表し、ここでは正常にはこの位置を占めるセリンにロイシン酸が置き換わっている。表5および6を参照のこと。これらの変異タンパク質を、ヒト293細胞のような従来の哺乳動物発現系で発現させた。全ての変異体IFN−γタンパク質が、A549アッセイにおいて決定された活性を保持していた。1119抗体が種々の変異体IFN−γタンパク質の生物学的活性を中和または阻害する能力について、A549バイオアッセイを用いて決定して、この抗体を試験した。1119抗体がヒトIFN−γ抗増殖活性を中和または阻害する能力を、上記のような蛍光を測定することによって決定して、これを1119抗体がIFN−γの改変型の各活性を中和する能力を比較するためのベースラインとして用いた。ヒトIFN−γで開始した構築物について、IFN−γ活性の阻害をパーセンテージとして測定し、ここではヒトIFN−γおよび1119抗体の存在下で観察された蛍光の最大レベル(増殖している細胞によるALAMARBLUETMの減少に起因する)を100%に設定して、IFN−γの各変化型に加えて1119抗体の存在下で測定した最大レベルとこれとを比較した。あるいは、カニクイザルIFN−γで開始する構築物については、観察された蛍光に基づいて阻害を定量的にスコア付けした。
【0220】
表5にまとめたとおり、1119抗体は、ヒトIFN−γ、ならびにhuIFNγD19およびhuIFNγP20を除くヒトIFNγの全ての置換変異体の生物学的活性を中和または阻害することができた。前の実施例のように、カニクイザルIFN−γタンパク質はまた、1119抗体では阻害されなかった。この解析によって、残基19および20が1119抗体とIFN−γとの間の相互作用に特に重要であり、そして1119抗体とヒトIFN−γとの間の接触のポイントとして機能し得ることが示される。
【0221】
【表5】
表6は、19位置および20位置でヒト配列に適合するカニクイザルIFNγ配列を作成する置換を含むカニクイザルIFNγの変更バージョンが、1119抗体によって中和されなかったことを示す。しかし、19、20および65、または19、20および103位置にヒト配列置換を有するカニクイザルIFNγのバージョンは、1119抗体によって中和され、これはこれらのいずれかに加えてさらに置換を含むバージョンと同様であった。
【0222】
【表6】
(実施例9)
(チンパンジーに対する投与の際の抗IFN−γ抗体の生物学的活性)
3週間の間、毎週20mg/kgの用量で2匹のチンパンジーに1119抗体を投与した。抗体の投与の2週前(−2)または1週前(−1)にチンパンジーから血液を採取し、さらに抗体の初回用量の投与の2、8、15、29および36日後に血液を採取した。実施例6に記載のヒト全血アッセイにおいて用いたのと本質的に同じ方法を用いて、この採取した血液でチンパンジーの全血アッセイを行なったが、重要な相違は、この抗体は全血を外因性に加えたのではなくこのチンパンジーに事前にインビボで投与したということであった。IFN−γを種々の濃度で血液に加えて(0.01ng/ml、3.9ng/mlまたは1μg/ml)、この血液を37℃で20〜24時間インキュベートして、次いでビーズに基づくELISAによってIP−10産生を測定した。
【0223】
図14および15でわかるように、抗体投与の前に動物から採取された血液は、IFN−γと反応性であって、IP−10産生が濃度依存性に増大した。対照的に、抗体投与後に採取された血液は、試験したどの濃度のIFN−γとも反応せず、IP−10の産生が増大した。これらデータによって、この抗体はインビボでの投与後でさえ、IFN−γを中和する能力を保持していたことが示される。さらにこれらの培養物に添加されたIFN−γの量は、チンパンジーでのIFN−γの内因性のレベルを大きく超えており、これによって、投与された抗体がインビボで内因性のIFN−γを中和できることが示唆された。
【0224】
前述の開示によって本発明の特定の実施形態が強調されること、およびその全ての改変または変更が添付の特許請求の範囲に示される本発明と等価であり、そのうえ本発明の趣旨および範囲内であることが理解されるべきである。本明細書に引用される全ての参考文献はその全体が参考として援用される。
【図面の簡単な説明】
【0225】
【図1A】図1A〜1Bは、1118、1118*、1119、1121および1121*の抗IFN−γ抗体の重鎖定常領域のアミノ酸配列(図1B;配列番号2)をコードするcDNA(図1A;配列番号1)の一部のヌクレオチド配列を示す。
【図1B】図1A〜1Bは、1118、1118*、1119、1121および1121*の抗IFN−γ抗体の重鎖定常領域のアミノ酸配列(図1B;配列番号2)をコードするcDNA(図1A;配列番号1)の一部のヌクレオチド配列を示す。
【図2A】図2A〜2Bは、1118、1118*、1119、1121および1121*の抗IFN−γ抗体の軽鎖定常領域のアミノ酸配列(図2B;配列番号4)をコードするcDNA(図2A;配列番号3)の一部のヌクレオチド配列を示す。
【図2B】図2A〜2Bは、1118、1118*、1119、1121および1121*の抗IFN−γ抗体の軽鎖定常領域のアミノ酸配列(図2B;配列番号4)をコードするcDNA(図2A;配列番号3)の一部のヌクレオチド配列を示す。
【図3A】図3A〜3Bは、1119の抗IFN−γ抗体の重鎖可変領域のアミノ酸配列(図3B;配列番号6)をコードするcDNA(図3A;配列番号5)の一部のヌクレオチド配列を示す。
【図3B】図3A〜3Bは、1119の抗IFN−γ抗体の重鎖可変領域のアミノ酸配列(図3B;配列番号6)をコードするcDNA(図3A;配列番号5)の一部のヌクレオチド配列を示す。
【図4A】図4A〜4Bは、1119の抗IFN−γ抗体の軽鎖可変領域のアミノ酸配列(図4B;配列番号8)をコードするcDNA(図4A;配列番号7)の一部のヌクレオチド配列を示す。
【図4B】図4A〜4Bは、1119の抗IFN−γ抗体の軽鎖可変領域のアミノ酸配列(図4B;配列番号8)をコードするcDNA(図4A;配列番号7)の一部のヌクレオチド配列を示す。
【図5A】図5A〜5Bは、1118の抗IFN−γ抗体の重鎖可変領域のアミノ酸配列(図5B;配列番号10)をコードするcDNA(図5A;配列番号9)の一部のヌクレオチド配列を示す。図5Cは、1118*の抗IFN−γ抗体の重鎖可変領域のアミノ酸配列(配列番号30)を示す。図5Dは、1118*の抗IFN−γ抗体の重鎖可変領域のヌクレオチド配列(配列番号56)を示す。
【図5B】図5A〜5Bは、1118の抗IFN−γ抗体の重鎖可変領域のアミノ酸配列(図5B;配列番号10)をコードするcDNA(図5A;配列番号9)の一部のヌクレオチド配列を示す。図5Cは、1118*の抗IFN−γ抗体の重鎖可変領域のアミノ酸配列(配列番号30)を示す。図5Dは、1118*の抗IFN−γ抗体の重鎖可変領域のヌクレオチド配列(配列番号56)を示す。
【図5C】図5A〜5Bは、1118の抗IFN−γ抗体の重鎖可変領域のアミノ酸配列(図5B;配列番号10)をコードするcDNA(図5A;配列番号9)の一部のヌクレオチド配列を示す。図5Cは、1118*の抗IFN−γ抗体の重鎖可変領域のアミノ酸配列(配列番号30)を示す。図5Dは、1118*の抗IFN−γ抗体の重鎖可変領域のヌクレオチド配列(配列番号56)を示す。
【図5D】図5A〜5Bは、1118の抗IFN−γ抗体の重鎖可変領域のアミノ酸配列(図5B;配列番号10)をコードするcDNA(図5A;配列番号9)の一部のヌクレオチド配列を示す。図5Cは、1118*の抗IFN−γ抗体の重鎖可変領域のアミノ酸配列(配列番号30)を示す。図5Dは、1118*の抗IFN−γ抗体の重鎖可変領域のヌクレオチド配列(配列番号56)を示す。
【図6A】図6A〜6Bは、1118または1118*の抗IFN−γ抗体の軽鎖可変領域のアミノ酸(図6B;配列番号12)をコードするcDNA(図6A;配列番号11)の一部のヌクレオチド配列を示す。
【図6B】図6A〜6Bは、1118または1118*の抗IFN−γ抗体の軽鎖可変領域のアミノ酸(図6B;配列番号12)をコードするcDNA(図6A;配列番号11)の一部のヌクレオチド配列を示す。
【図7A】図7A〜7Bは、1121、または1121*の抗IFN−γ抗体の重鎖定常領域のアミノ酸配列(図7B;配列番号14)をコードするcDNA(図7A;配列番号13)の一部のヌクレオチド配列を示す。
【図7B】図7A〜7Bは、1121、または1121*の抗IFN−γ抗体の重鎖定常領域のアミノ酸配列(図7B;配列番号14)をコードするcDNA(図7A;配列番号13)の一部のヌクレオチド配列を示す。
【図8A】図8A〜8Bは、1121の抗IFN−γ抗体の軽鎖可変領域のアミノ酸配列(図8B;配列番号16)をコードするcDNA(図8A;配列番号15)の一部のヌクレオチド配列を示す。図8Cは、1121*の抗IFN−γ抗体の軽鎖可変領域のアミノ酸配列(図31)を示す。図8Dは、1121*の抗IFN−γ抗体の軽鎖可変領域のヌクレオチド配列(配列番号57)を示す。
【図8B】図8A〜8Bは、1121の抗IFN−γ抗体の軽鎖可変領域のアミノ酸配列(図8B;配列番号16)をコードするcDNA(図8A;配列番号15)の一部のヌクレオチド配列を示す。図8Cは、1121*の抗IFN−γ抗体の軽鎖可変領域のアミノ酸配列(図31)を示す。図8Dは、1121*の抗IFN−γ抗体の軽鎖可変領域のヌクレオチド配列(配列番号57)を示す。
【図8C】図8A〜8Bは、1121の抗IFN−γ抗体の軽鎖可変領域のアミノ酸配列(図8B;配列番号16)をコードするcDNA(図8A;配列番号15)の一部のヌクレオチド配列を示す。図8Cは、1121*の抗IFN−γ抗体の軽鎖可変領域のアミノ酸配列(図31)を示す。図8Dは、1121*の抗IFN−γ抗体の軽鎖可変領域のヌクレオチド配列(配列番号57)を示す。
【図8D】図8A〜8Bは、1121の抗IFN−γ抗体の軽鎖可変領域のアミノ酸配列(図8B;配列番号16)をコードするcDNA(図8A;配列番号15)の一部のヌクレオチド配列を示す。図8Cは、1121*の抗IFN−γ抗体の軽鎖可変領域のアミノ酸配列(図31)を示す。図8Dは、1121*の抗IFN−γ抗体の軽鎖可変領域のヌクレオチド配列(配列番号57)を示す。
【図9】図9は、1119、1118および1121のモノクローナル抗体を用いたA549バイオアッセイにおけるIFN−γの生物学的活性の中和または阻害を示すグラフを含む。
【図10】図10は、1119、1118および1121のモノクローナル抗体によるTHP−1/HLA DRバイオアッセイにおけるIFN−γの生物学的活性の中和または阻害を示すグラフを含む。
【図11】図11は、1119のモノクローナル抗体による全血バイオアッセイ(2つのドナー)におけるIFN−γの生物学的活性の中和または阻害を示すグラフを含む。
【図12】図12は、1118、1118*、1121および1119と命名された抗IFNモノクローナル抗体の重鎖のアミノ末端部分(可変領域を含む)のアラインメントを示す。この配列は、実施例3において単離されたcDNA上にコードされるシグナル配列を含む。このシグナル配列は、1位〜19位に延びる。CDRには下線を付している。図に示すとおり、CDR1は、アミノ酸50〜54にまたがり、CDR2は、69〜85にまたがり、そしてCDR3は、118〜125にまたがる。Kabatら(1991、Sequences of Proteins of Immunological Interest,Public Health Service N.I.H.,Bethesda,MD)の番号付けシステムは、成熟抗体の最初のアミノ酸で開始して、シグナル配列は除く。従って、この図の20位は、Kabatら(前出)の1位に相当する。
【図13】図13は、1118、1121、1121*および1119と命名された抗IFNモノクローナル抗体の軽鎖のアミノ末端部分(可変領域を含む)のアラインメントを示す。この配列は、実施例3において単離されたcDNA上にコードされるシグナル配列を含む。このシグナル配列は、1〜20位に延びる。CDRには下線を付している。図に示すとおり、CDR1は、アミノ酸44〜55にまたがり、CDR2は、71〜77にまたがり、そしてCDR3は、110〜118にまたがる。Kabatら(前出)の番号付けシステムは、シグナル配列を除くので、この図の21位は、Kabatらの1位に相当する。
【図14】図14は、週に1回行なった、抗IFNγ抗体の3回の注射の開始の2週間前もしくは1週間前(図14では「−2」および「−1」と表示した線)、または2日間後、8日間後、15日間後、29日間後もしくは36日後(図14では「2」、「8」、「15」、「29」もしくは「36」と表示した線)においてチンパンジーから採取した全血による、IFN−γに応答したIP−10の産生を示す。
【図15】図15は、異なるチンパンジーの血液を用いた以外は、図14と同様である。
【特許請求の範囲】
【請求項1】
本明細書に記載される発明。
【請求項1】
本明細書に記載される発明。
【図1A】


【図1B】

【図2A】

【図2B】

【図3A】

【図3B】

【図4A】

【図4B】

【図5A】

【図5B】

【図5C】

【図5D】

【図6A】

【図6B】

【図7A】

【図7B】

【図8A】

【図8B】

【図8C】

【図8D】

【図9】
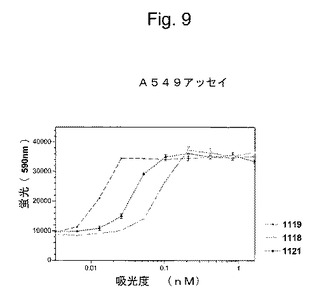
【図10】
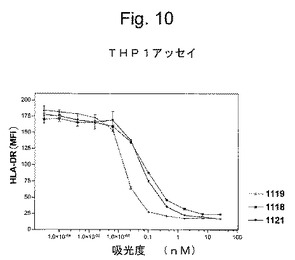
【図11】
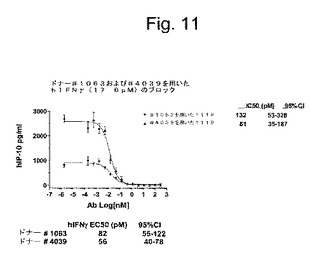
【図12】

【図13】
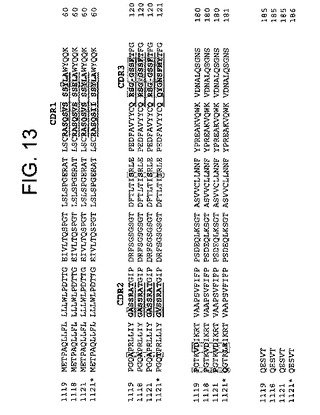
【図14】
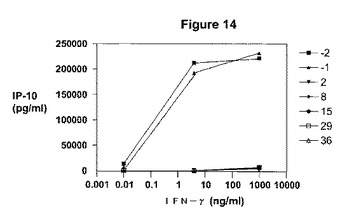
【図15】
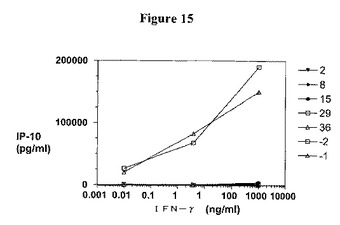
【図1B】
【図2A】
【図2B】
【図3A】
【図3B】
【図4A】
【図4B】
【図5A】
【図5B】
【図5C】
【図5D】
【図6A】
【図6B】
【図7A】
【図7B】
【図8A】
【図8B】
【図8C】
【図8D】
【図9】
【図10】
【図11】
【図12】
【図13】
【図14】
【図15】
【公開番号】特開2009−82133(P2009−82133A)
【公開日】平成21年4月23日(2009.4.23)
【国際特許分類】
【出願番号】特願2008−265800(P2008−265800)
【出願日】平成20年10月14日(2008.10.14)
【分割の表示】特願2005−501441(P2005−501441)の分割
【原出願日】平成15年10月16日(2003.10.16)
【出願人】(500203709)アムジェン インコーポレイテッド (76)
【出願人】(502066649)メダレックス, インコーポレイテッド (13)
【Fターム(参考)】
【公開日】平成21年4月23日(2009.4.23)
【国際特許分類】
【出願日】平成20年10月14日(2008.10.14)
【分割の表示】特願2005−501441(P2005−501441)の分割
【原出願日】平成15年10月16日(2003.10.16)
【出願人】(500203709)アムジェン インコーポレイテッド (76)
【出願人】(502066649)メダレックス, インコーポレイテッド (13)
【Fターム(参考)】
[ Back to top ]