
遺伝子およびポリペプチド配列
本発明は、(i)(a)分泌プレ配列、および(b)以下のモチーフ:−X1−X2−X3−X4−X5−(式中、X1はフェニルアラニン、トリプトファンまたはチロシンであり、X2はイソロイシン、ロイシン、バリン、アラニンまたはメチオニンであり、X3はロイシン、バリン、アラニンまたはメチオニンであり、X4はセリンまたはトレオニンであり、かつ、X5はイソロイシン、バリン、アラニンまたはメチオニンである)を含んでなるリーダー配列、ならびに(ii)そのリーダー配列に対して異種の目的タンパク質を含んでなるポリペプチドを提供する。本発明のポリペプチドは、リーダー配列の一部として、分泌プロ配列をさらに含んでなってよい。本発明はまた、本発明のポリペプチドをコードする配列を含んでなるポリヌクレオチドおよび該ポリヌクレオチドを含んでなる細胞、好ましくは、酵母細胞を提供する。

【発明の詳細な説明】
【発明の背景】
【0001】
発明の分野
本発明は、宿主細胞からタンパク質を分泌させるためのポリペプチドおよびポリヌクレオチド配列に関する。
【0002】
背景技術
宿主細胞から目的ペプチド、ポリペプチドおよびタンパク質(以下、本明細書では、これらの用語は同義的に用いる)を分泌させることを目的に、多くの天然または人工ポリペプチドシグナル配列(分泌プレ領域とも呼ばれる)が使用または開発されている。シグナル配列は、初期のタンパク質を、タンパク質をその細胞から周囲の媒質へ、または、場合によっては、ペリプラスム間隙へと輸送する細胞の機構に導くものである。シグナル配列は、通常、必ずではないが、一次翻訳産物のN末端に位置しており、一般に、必ずではないが、分泌プロセス中にその配列が目的タンパク質から切り離されて、「成熟」タンパク質が得られる。
【0003】
いくつかの目的タンパク質の場合では、シグナル配列の除去後、最初に分泌される存在物はそのN末端に「プロ」配列と呼ばれる付加的アミノ酸を含んでおり、その中間存在物は「プロ−タンパク質」と呼ばれている。これらのプロ配列は、最終的なタンパク質の折り畳みおよび機能化を手助けし、通常、その後に切り離される。他の例では、そのプロ領域はプレ−プロ領域を切り離す酵素の切断部位を容易に提供するもので、別の機能は有していないと思われている。
【0004】
プロ配列は、細胞からの目的タンパク質の分泌中、またはその細胞からの周囲の媒質またはペリプラスム間隙への輸送後のいずれかに除去される可能性がある。
【0005】
タンパク質の分泌を導くポリペプチド配列はまた、それらがシグナル(すなわち、プレ)配列またはプレ−プロ分泌配列と似ていようがいよまいが、リーダー配列と呼ばれることもある。タンパク質の分泌は翻訳、転座および翻訳後プロセシングを伴う動的プロセスであり、これらの工程の1以上は、別のものが開始する前、または完了する前に、必ずしも完了しなくてよい。
【0006】
酵母、サッカロミセス・セレビシエ(Saccharomyces cerevisiae)およびピキア・パストリス(Pichia pastoris)などの真核生物種においてタンパク質を産生させるための既知のリーダー配列としては、S.セレビシエ酸性ホスファターゼタンパク質(Pho5p)(EP366400を参照)、転化酵素タンパク質(Suc2p)(Smith et al. (1985) Science, 229, 1219-1224を参照)および熱ショックタンパク質−150(Hsp150p)(WO95/33833を参照)のものが挙げられる。さらに、S.セレビシエ接合因子α−1タンパク質(MFα−1)のリーダー配列ならびにヒトリゾチームおよびヒト血清アルブミン(HSA)タンパク質のリーダー配列が使用されているが、限定されるものではないが、ヒトアルブミンの分泌に向けては後者のものが特に利用されている。WO90/01063は、MFα−1リーダー配列を使用する場合に比べて混入するヒトアルブミン断片の産生が有利に減少する、MFα−1とHSAリーダー配列との融合物を開示している。
【発明の概要】
【0007】
本発明者らは、予想外にも、分泌されたタンパク質の収量がアミノ酸配列モチーフの導入により、好ましくは、リーダー配列の改変により、増大し得ることを見出した。この改変は、完全な天然アルブミンリーダー配列に行うか、その変異体に行うか、またはWO90/01063にて開示されているMFα−1とHSAリーダー配列との融合物などのヒトアルブミンリーダー配列の関連部分を用いた他のリーダー配列に行うかを問わず有効である。後者の場合、アルブミンが分泌されるタンパク質であるならば、このようにして産生されたアルブミンは混入する断片が減少するという有利な特徴を保有し、同時に収量も増大する。
【0008】
WO90/01063の融合されたリーダー配列の保存的改変は 、WO90/01063に概要が開示されている(例えば、WO90/01063の8頁を参照)が、ここではおよそ8×1012ポリペプチドの群が得られることが明らかになっている。遺伝子コードの縮退に従い、典型的なリーダー配列についてポリヌクレオチドコード配列が示された。これはまた、多数の可能性を示すものである。WO90/01063では、本発明により提供される、改変されたリーダー配列の特定群が分泌されるタンパク質の発現に有利な特性を有しているという理解はなされていない。
【0009】
本発明の第1の態様において、下記(i)および(ii)を含んでなるポリペプチドが提供される:
(i)下記(a)および(b)を含んでなるリーダー配列
(a)分泌プレ配列、および
(b)以下のモチーフ:
−X1−X2−X3−X4−X5−
(式中、
X1はフェニルアラニン、トリプトファンまたはチロシンであり、
X2はイソロイシン、ロイシン、バリン、アラニンまたはメチオニンであり、
X3はロイシン、バリン、アラニンまたはメチオニンであり、
X4はセリンまたはトレオニンであり、かつ、
X5はイソロイシン、バリン、アラニンまたはメチオニンである)、および
(ii)そのリーダー配列に対して異種の目的タンパク質。
【0010】
言い換えれば、そのポリペプチドは下記配列番号1の配列を含む。
【化1】
配列番号1
【0011】
本発明の第1の態様の好ましい態様では、X1はフェニルアラニンである。よって、好ましいポリペプチドは下記配列番号2の配列を含む。
【化2】
配列番号2
【0012】
本発明の第1の態様のもう1つの好ましい態様では、X2はイソロイシンである。よって、もう1つの好ましいポリペプチドは下記配列番号3の配列を含む。
【化3】
配列番号3
【0013】
本発明の第1の態様のもう1つの好ましい態様では、X3はバリンである。よって、もう1つの好ましいポリペプチドは下記配列番号4の配列を含む。
【化4】
配列番号4
【0014】
もう1つの好ましいポリペプチドでは、X4はセリンであり、よって、下記配列番号5の配列を含む。
【化5】
配列番号5
【0015】
本発明の第1の態様のもう1つの好ましい態様では、X4はトレオニンである。よって、もう1つの好ましいポリペプチドが配列番号29の配列を含む。
【化6】
配列番号29
【0016】
本発明の第1の態様のもう1つの好ましい態様では、X5はイソロイシンである。よって、もう1つの好ましいポリペプチドは配列番号6の配列を含む。
【化7】
配列番号6
【0017】
より好ましくは、X1、X2、X3、X4およびX5のうちの少なくとも2つ、なおいっそう好ましくは、少なくとも3つ、さらに好ましくは、少なくとも4つが上記の好ましい態様で定義されたとおりである。
【0018】
そのモチーフはリーダー配列に(すなわち、付加物として)挿入してもよいし、または1個、2個、3個、4個、5個以上の連続したアミノ酸の代用としてリーダー配列内に含めることもできる。
【0019】
1つの好ましい態様では、そのモチーフが天然アミノ酸の代用としてリーダー配列に含められている。言い換えれば、そのモチーフのアミノ酸が、本発明によるその最適化を行う前にリーダー配列内に存在していたか、または存在していたであろう5個の連続したアミノ酸の代わりに含められている。これに関連して使用する「天然(の)」(nathurally occurring)とは、本発明を天然のリーダー配列の最適化に限定するものではないことは読む人にも理解できるであろう。それどころか、本発明はその最適化が本明細書において例示されているHSA/MFα−1リーダー配列融合物などの人工リーダー配列の最適化にも適用できる。
【0020】
そのモチーフを代用物としてリーダー配列に含める場合にはX4が天然アミノ酸またはその変異体であることが好ましい。言い換えれば、X1、X2、X3およびX5だけを置換して、X4を変えないでおくか、または、その位置にある天然アミノ酸の、好ましくは、以下で定義する保存的置換として、変異体に単に変えることが好ましい。
【0021】
本発明の第1の態様の特に好ましい態様では、X1はフェニルアラニンであり、X2はイソロイシンであり、X3はバリンであり、X4はセリンであり、かつ、X5はイソロイシンである。よって、本発明の第1の態様の特に好ましい態様では、下記配列番号7の配列を含むポリペプチドが提供される。
【化8】
配列番号7
【0022】
上記スキームにおいて、「N」および「C」はポリペプチド配列の方向を示し、それらの実際の末端についてのそれらの解釈を限定するものではない。言い換えれば、ポリペプチド配列は、N末端もしくはC末端のいずれか一方にて、または最も一般的にはそれら両方の末端において、1以上の他のポリペプチド配列と結合される(例えば、融合されるか、コンジュゲートされるか、または連結される)。
【発明の具体的説明】
【0023】
本発明の第1の態様のポリペプチドは、リーダー配列に対して異種の成熟した目的タンパク質の配列を含んでなる。成熟した目的タンパク質の配列は、本発明のポリペプチドが発現される発現系による翻訳後プロセシング後の発現産物に存在するであろう一次アミノ酸配列である。目的タンパク質は、好ましくは、本発明のポリペプチドが発現される細胞から分泌させるのに好適であるものである。
【0024】
目的タンパク質はリーダー配列に対して異種である。言い換えれば、本発明の第1の態様のポリペプチドは、それらのリーダー配列内に上記のモチーフ−X1−X2−X3−X4−X5−を有する天然のタンパク質を含まないものである。好ましい態様では、本発明の第1の態様のポリペプチドは、任意の位置に上記のモチーフ−X1−X2−X3−X4−X5−を有する天然のタンパク質を含まないものである。これに関連して、「天然(の)」とは、組換えテクノロジー、部位特異的突然変異誘発またはヒトの介入を必要とする同等の人工的な技術による改変を受けていない天然に存在する生物によりコードされるタンパク質を指す。
【0025】
目的タンパク質は、天然タンパク質(チモーゲン(zymogen)を含む)、ポリペプチドもしくはペプチド、または天然タンパク質、ポリペプチドもしくはペプチドの変異体もしくは断片(例えば、一ドメインであってよい);あるいは完全合成タンパク質、ポリペプチドもしくはペプチド;あるいは異なるタンパク質、ポリペプチドもしくはペプチド(天然または合成)の単一もしくは多重融合物であれば、いずれの配列を含んでなってもよい。このようなタンパク質は、限定されるものではないが、それらの開示内容が引用することにより本明細書の一部とされる、WO01/79258、WO01/79271、WO01/79442、WO01/79443、WO01/79444およびWO01/79480にて記載されているリスト、またはその変異体もしくは断片から選択することができる。これらの特許出願はアルブミンの融合パートナーという観点からタンパク質のリストを示しているが、本発明はそのように限定されず、本発明の目的では、本明細書に列挙するいかなるタンパク質も単独でまたはアルブミン、免疫グロブリンのFc領域、トランスフェリンもしくは目的ポリペプチドとしての他のタンパク質の融合パートナーとして与えてよい。
【0026】
本発明による発現に向けた目的タンパク質の好ましい例としては、アルブミン、トランスフェリン、ラクトフェリン、エンドスタチン、アンギオスタチン、コラーゲン、免疫グロブリン、Fab’フラグメント、F(ab’)2、ScAb、ScFv、インターフェロン、IL10、IL11、IL2、インターフェロンα種および亜種、インターフェロンβ種および亜種、インターフェロンγ種および亜種、IL1−受容体アンタゴニスト、EPO、TPO、プロサプチド(prosaptide)、シアノビリン−N、5−ヘリックス、T20ペプチド、T1249ペプチド、HIV gp41、HIV gp120、フィブリノーゲン、ウロキナーゼ、プロウロキナーゼ、tPA(組織プラスミノーゲン活性化因子)、ヒルジン、血小板由来増殖因子、副甲状腺ホルモン、プロインスリン、インスリン、インスリン様増殖因子、カルシトニン、成長ホルモン、形質転換増殖因子β、腫瘍壊死因子、G−CSF、GM−CSF、M−CSF、限定されるものではないが、プラスミノーゲン、フィブリノーゲン、トロンビン、プレトロンビン、プロトロンビン、フォン・ウィルブランド因子、α1−アンチトリプシン、プラスミノーゲン活性化因子、第VII因子、第VIII因子、第IX因子、第X因子および第XIII因子をはじめとするプレ型および活性型いずれもの凝固因子、神経成長因子、LACI(組織因子経路阻害剤または外因経路阻害剤としても知られているリポタンパク質由来凝固阻害剤)、血小板由来内皮細胞増殖因子(PD−ECGF)、グルコースオキシダーゼ、血清コリンエステラーゼ、アプロチニン、アミロイド前駆体、インターαトリプシン阻害剤、アンチトロンビンIII、アポ−リポタンパク質種、プロテインC、プロテインS、上記のものの変異体または断片が挙げられる。
【0027】
目的タンパク質に関連して、「変異体」とは、1箇所以上の位置にアミノ酸の挿入、欠失、または保存的置換もしくは非保存的置換のいずれかの置換が存在するタンパク質を指すが、そのような変化の結果として、タンパク質の基礎特性、例えば、酵素活性または受容体結合(種類および比活性度)、熱安定性、特定のpH範囲における活性(pH安定性)が有意に変わっていないタンパク質が得られるという条件においてである。これに関連して「有意に」とは、当業者によって、その変異体の特性はまだ異なっているかもしれないが、最初のタンパク質の特性に関しては自明であると思われることを意味する。
【0028】
「保存的置換」とは、Val、Ile、Leu、Ala、Met;Asp、Glu;Asn、Gln;Ser、Thr、Gly、Ala;Lys、Arg、His;およびPhe、Tyr、Trpなどの組合せを意図するものである。好ましい保存的置換としては、Gly、Ala;Val、Ile、Leu;Asp、Glu;Asn、Gln;Ser、Thr;Lys、Arg;およびPhe、Tyrが挙げられる。
【0029】
「変異体」は、その変異体が誘導されるポリペプチドと通常、少なくとも25%、少なくとも50%、少なくとも60%または少なくとも70%、好ましくは、少なくとも80%、より好ましくは、少なくとも90%、なおいっそう好ましくは、少なくとも95%、さらに好ましくは、少なくとも99%、最も好ましくは、少なくとも99.5%の配列同一性を有する。
【0030】
2ポリペプチド間の配列同一性パーセントは、好適なコンピュータープログラム、例えば、ウィスコンシン大学遺伝子コンピューティンググループ(University of Wisconsin Genetic Computing Group)のGAPプログラムを用いて決定することができ、当然のことではあるが、同一性パーセントは、最適に配列をアラインメントしたポリペプチドと比較して計算される。
【0031】
アラインメントは、また、Clustal Wプログラムを用いて実施してもよい(Thompson et al., (1994) Nucleic Acids Res., 22 (22), 4673-80)。使用されるパラメーターは以下の通りである:
・ファストペアワイズアラインメントパラメーター:K−tuple(ワード)サイズ;1、ウインドウサイズ;5、ギャップペナルティー;3、トップダイアゴナル(top diagonals)数;5。スコアリング法:xパーセント。
・マルチプルアラインメントパラメーター:ギャップオープンペナルティー;10、ギャップエクステンションペナルティー;0.05。
・スコアリングマトリックス:BLOSUM。
【0032】
このような変異体は天然のものであってよいし、または当技術分野で周知のタンパク質工学手法および部位特異的突然変異誘発手法を用いて作製されたものであってよい。
【0033】
目的タンパク質に関連して、「断片」とは、1箇所以上の位置に欠失が存在するタンパク質を指す。よって、その断片が全長成熟ポリペプチドの完全配列の最大限5、10、20、30、40または50%を含んでなると思われる。通常、一断片は、全長の目的タンパク質の完全配列の最大60%、より通常は最大70%、好ましくは、最大80%、より好ましくは、最大90%、なおいっそう好ましくは、最大95%、さらに好ましくは、最大99%を含んでなる。目的タンパク質の特に好ましい断片は、目的タンパク質の1以上の完全ドメインを含んでなる。例えば、目的タンパク質がアルブミンであってよい。アルブミンは3つのドメインを有している。アルブミンの特に好ましい断片は、1つまたは2つのドメインを含み得るため、通常、アルブミンの完全配列の少なくとも33%または少なくとも66%を含んでなる。
【0034】
アルブミンおよびトランスフェリン、またはその変異体もしくは断片が、とりわけそれらがヒト起源のものである場合、すなわち、それらが自然に産生されたヒトタンパク質において見つけられるものと同じ配列を有している場合には、目的タンパク質として特に好ましい。
【0035】
「ヒトアルブミン」とは、本明細書において、ヒト血清アルブミンと区別できない材料、またはその変異体もしくは断片である材料を示すために用いる。「変異体」とは、挿入、欠失、および保存的置換または非保存的置換のいずれかの置換を含み、そのような変化によってアルブミンの膨張特性、有用なリガンド結合特性または免疫原特性が実質的に変わらないものである。例えば、EP−A−322 094で開示されているヒトアルブミンまたはヒトアルブミン類似体の自然発生する多形変異体が挙げられる。一般に、ヒトアルブミンの変異体または断片はヒト血清アルブミンのリガンド結合活性(例えば、ビリルビン結合)の少なくとも10%(好ましくは、少なくとも50%、80%、90%または95%)およびヒト血清アルブミンの膨張活性の少なくとも50%(好ましくは、少なくとも80%、90%または95%)、w/wを有している。アルブミン、アルブミン変異体またはアルブミン断片の、膠質浸透圧としても知られている膨張活性は、Hoefs, J. C. (1992) Hepatology 16: 396-403により記載されている方法により決定してもよい。ビリルビン結合は、527nmでの蛍光増感により、HSAと比較して測定してもよい。ビリルビン(1.0mg)を50μLの1M NaOHに溶かし、脱塩水で1.0mLに希釈する。このビリルビンストックを100mM Tris−HCl pH8.5、1mM EDTAで希釈して、蛍光光度計キュベット中0.6nmolビリルビンmL−1とする。HSA:ビリルビン比 0〜5mol:molの範囲にわたるHSAでの滴定中においての448nmでの励起および527nmでの発光(10nm スリット幅)により蛍光を測定する。
【0036】
好ましい態様では、目的タンパク質がトランスフェリンであってよい。このようなものとしては、トランスフェリンファミリーのメンバー(Testa, Proteins of iron metabolism, CRC Press, 2002; Harris & Aisen, Iron carriers and iron proteins, Vol. 5, Physical Bioinorganic Chemistry, VCH, 1991)およびそれらの誘導体、例えば、トランスフェリン、突然変異型トランスフェリン(Mason et al, 1993, Biochemistry, 32, 5472; Mason et al, 1998, Biochem. J., 330 (1), 35)、末端切断型トランスフェリン、トランスフェリンローブ(Mason et al, 1996, Protein Expr. Purif., 8, 119; Mason et al, 1991, Protein Expr. Purif., 2, 214)、ラクトフェリン、突然変異型ラクトフェリン、末端切断型ラクトフェリン、ラクトフェリンローブまたは上記のものと他のペプチド、ポリペプチドもしくはタンパク質との融合物(Shin et al, 1995, Proc. Natl. Acad. Sci. USA, 92, 2820; Ali et al, 1999, J. Biol. Chem., 274, 24066; Mason et al, 2002, Biochemistry, 41, 9448)が挙げられる。トランスフェリンがヒトトランスフェリンであってよい。
【0037】
「ヒトトランスフェリン」とは、本明細書において、ヒト由来のトランスフェリンと区別できない材料、またはその変異体もしくは断片である材料を示すために用いる。「変異体」とは、挿入、欠失、および保存的置換または非保存的置換のいずれかの置換を含み、そのような変化によってトランスフェリンの有用なリガンド結合特性または免疫原特性が実質的に変わらないものである。例えば、ヒトトランスフェリンまたはヒトトランスフェリン類似体の自然発生する多形変異体が挙げられる。一般に、ヒトトランスフェリンの変異体または断片はヒトトランスフェリンのリガンド結合活性(例えば、鉄結合)の少なくとも50%(好ましくは、少なくとも80%、90%または95%)、w/wを有している。トランスフェリンまたは試験サンプルの鉄結合活性は、分光光度法で、鉄遊離状態および鉄飽和状態におけるそのタンパク質の470nm:280nm吸光度比により決定することができる。特に断りのない限り、試薬は鉄を含まないものであるべきである。鉄は、0.1Mクエン酸塩、0.1M酢酸塩、10mM EDTA pH4.5に対する透析によりトランスフェリンまたは試験サンプルから除去することができる。タンパク質は、100mM HEPES、10mM NaHCO3 pH8.0中、約20mg/mLにて存在するはずである。280nmでの吸光度を分光光度法で正確に決定することができるように(0%鉄結合)、水で希釈したアポトランスフェリン(Calbiochem, CN Biosciences, Nottingham, UK)の470nm:280nm吸光度比を測定する。191mgニトロ三酢酸を2mL 1M NaOHに溶かして20mM鉄−ニトリロ三酢酸(FeNTA)溶液を調製した後、2mL 0.5M塩化第二鉄を添加する。脱イオン水で50mLに希釈する。新たに調製した20mM FeNTAを十分に過剰に添加して、アポトランスフェリンを鉄飽和させ(100%鉄結合)た後、470nm:280nmでの吸光度比を測定する前に、ホロトランスフェリン調製物を100mM HEPES、10mM NaHCO3 pH8.0に対して完全に透析して残留しているFeNTAを除去する。最初は鉄を含まないものであるべき試験サンプルを用いてこの手順を繰り返し、最終的な比を対照と比較する。
【0038】
さらに、上記のものの単一もしくは多重異種融合物;またはアルブミン、トランスフェリンもしくは免疫グロビンあるいはこれらのものの変異体もしくは断片との単一もしくは多重異種融合物を使用してもよい。このような融合物としては、WO01/79271により例示されている、アルブミンN末端融合物、アルブミンC末端融合物およびN末端およびC末端アルブミン同時融合物、ならびにトランスフェリンN末端融合物、トランスフェリンC末端融合物、およびN末端およびC末端トランスフェリン同時融合物が挙げられる。
【0039】
好ましい態様では、本発明の第1の態様のポリペプチドが以上で定義されたX1−X5ペンタペプチドモチーフの少なくとも一部を含む分泌プレ配列を含んでなる。言い換えれば、成熟した目的ポリペプチドの分泌を行うように作用するリーダー配列の領域が、X1−X5ペンタペプチドモチーフのアミノ酸のうちの1個、2個、3個、4個または5個のアミノ酸を含有する。分泌プレ配列領域がX1−X5ペンタペプチドモチーフのうちの5個より少ないアミノ酸を含有する場合、そのプレ配列に含まれるモチーフのそれらのアミノ酸は、それらがX1−X5ペンタペプチドモチーフの残りのアミノ酸の近傍に存在するように、プレ配列領域の一方の境界に位置している。
【0040】
より好ましい態様では、本発明の第1の態様のポリペプチドが、本発明の第1の態様により以上で定義されたモチーフを含む分泌プレ配列を含むことを特徴とするリーダー配列を含んでなる。そのリーダー配列は、通常、必ずではないが、一次翻訳産物のN末端に位置しており、一般に、必ずではないが、分泌プロセス中にタンパク質から切り離されて、成熟した「目的」タンパク質が得られる。
【0041】
分泌リーダー配列は、通常、必ずではないが、N末端のアミノ酸配列であり、そのアミノ酸配列が、そのアミノ酸配列が一部を形成するポリペプチドを、それが産生される宿主細胞から分泌させる。分泌は、リン脂質二重層、一般に、限定されるものではないが、真核生物の小胞体または原核生物の原形質膜を通じた細胞質のコンパートメントからのタンパク質の翻訳後転座の共翻訳により定義される。分泌されたタンパク質は、細胞の範囲内(一般に、限定されるものではないが、小胞体、ゴルジ体、液胞、リソソームまたはペリプラスム間隙内)にとどまるか、または細胞から培養培地へと分泌される。ある配列が、分泌プレ配列を含まない同等のポリペプチドに比べて、いっそう多くのポリペプチドを、それが産生される宿主細胞から分泌させる場合には、その配列は分泌リーダー配列としての役割を果たしている。一般的に言えば、リーダー配列を有するポリペプチドは分泌されるが、リーダー配列を有さないポリペプチドは分泌されない。しかしながら、本発明は、異なるリーダー配列の効率レベルは異なるという状況を意図している。よって、リーダー配列は、細胞により産生された成熟タンパク質の少なくとも10%、20%、30%もしくは40%または50%、一般に、少なくとも60%または70%、好ましくは、少なくとも80%、より好ましくは、少なくとも90%、なおいっそう好ましくは、少なくとも95%、さらに好ましくは、少なくとも98%、最も好ましくは、少なくとも99%を細胞から分泌させ得る。成熟ポリペプチドの細胞からの分泌は、例えば、宿主細胞に好適なDNA構築物を提供し、分泌された成熟タンパク質(例えば、ヒトアルブミン)の量を細胞内に産生されたいずれもの成熟タンパク質と比較して測定することにより決定することができる。
【0042】
好ましい分泌リーダー配列は、宿主細胞が酵母細胞(例えば、サッカロミセス・セレビシエまたはピキア・パストリス)である場合には、上記レベルの分泌を導く。酵母宿主細胞からの成熟ポリペプチドの分泌は、例えば、以下の実施例にて記載するものなどの方法により決定することができる。
【0043】
従って、分泌は、試験リーダー配列を含んでなるタンパク質の分泌レベルを対照リーダー配列を含んでなるタンパク質の分泌レベルと比較することにより測定することができる。所与の配列(「試験配列」)が一定の分泌レベルを達成し得るかどうかを判定するために、LEU2遺伝子とrHAをコードするポリヌクレオチドとともに、PRB1プロモーターおよび以下で記載するADH1ターミネーターなどの機能的酵母調節領域に作動可能なように連結された配列番号26で定義される改変リーダー配列を有する「スターター」プラスミド、一般に、EP 0 286 422で記載されているタイプの酵母分解ベクターを、試験配列をコードするポリヌクレオチド配列を、リーダー配列の相当する領域の代わりに含めるよう改変し、そうすることによって、試験プラスミドを提供するといった第1のプロトコールを用いてよい。第1の対照として、WO90/01063で記載されているリーダー配列をコードする改変されていない「スターター」プラスミドを使用する。サッカロミセス・セレビシエ株AH22cir°(Hinnen et al, 1978, Proc. Natl. Acad. Sci. USA, 75 (4), 1929-33; Mead et al, 1986, Mol. Gen. Genet., 205, 417)、復帰変異したHis4を試験宿主として使用する。AH22(leu2、his4、can1)のHIS4復帰突然変異体(すなわち、His+)は、十分なAH22細胞を0.002%(w/v)ロイシンを補給したBMMD寒天でコロニーが現れるまで培養することによって得ることができる。そのコロニーを試験し、0.002%(w/v)ロイシンを補給したBMMD寒天(プレート1)、0.002%(w/v)ロイシンを補給し、0.002%(w/v)ヒスチジンを補給したBMMD寒天(プレート2)、およびBMMD寒天(プレート3)に播種することによりそれらがLeu−およびHis+(すなわち、AH22 His+(leu2、can1))であることを確認する。AH22 His+(leu2、can1)単離物はプレート1およびプレート2では増殖するが、プレート3では増殖しない。試験宿主を試験および対照プラスミドを用いてロイシンプロトトロフィー(prototrophy)に形質転換する。これらの形質転換体を2%(w/v)グルコースを含有する緩衝最小培地(BMM、Kerry-Williams, S. M. et al. (1998) Yeast 14, 161-169により記載されている)(BMMD)にパッチアウト(patched out)し、さらなる解析に向けて十分に増殖するまで30℃にてインキュベートする。形質転換体をフィル&ドロー手順による高細胞密度発酵にて、WO96/37515にてフェドバッチ手順に関して記載されているある培地で、制御パラメーターを使用して培養する:フェドバッチ培養手順供給期の終了後、培養物量の90%を発酵容器から取り出す。供給物の添加を開始する前に、残りの10%量の培養物に、WO96/37515に記載されている培地および制御パラメーターを使用してバッチ培地を添加する(pHを制御しながら)。フィル&ドロー手順は無制限に繰り返してもよい。試験および対照プラスミドを含有する形質転換体のヒトアルブミン生産性(YP/S)を細胞を含まない全培養物のSDS−PAGEのスキャンニングデンシトメトリーにより評価する。YP/Sは発酵中に培養物に添加したスクロース1g当たりのヒトアルブミンタンパク質(mg)の比率を表す。
【0044】
試験および第1の対照形質転換体を比較できる発酵槽配置で比較できる時間培養するとき、本発明のリーダー配列は、上記の第1のプロトコールにより測定するYP/Sにより判定した場合、第1の対照により得られる分泌レベルよりも少なくとも10%、20%、30%、40%、50%、60%、70%、80%、90%、100%、110%、120%、130%、140%、150%、160%、170%、180%、190%、200%、210%、220%、230%、240%、250%、260%、270%、280%、290%、300%、310%、320%、330%、340%、350%、360%、370%、380%、390%、400%、410%、420%、430%、440%、450%、460%、470%、480%、490%または500%高い分泌レベルを獲得し得る。よって、本発明のリーダー配列が第1の対照よりも最大400%、450%、500%、550%、600%、650%、700%、750%、またはそれ以上に高い分泌レベルであることが証明し得る。本発明のリーダー配列が、第1の対照により得られる分泌レベルよりも少なくとも400%、例えば、408%、または少なくとも440%、例えば、442%高い分泌レベルを獲得することが特に好ましい。
【0045】
第1のプロトコールに代わる方法として、第2のプロトコールを用いて、所与の配列(「試験配列」)が一定の分泌レベルを達成し得るかどうかを判定してもよい。第2のプロトコールは第1のプロトコールと本質的に同じである。しかしながら、第2のプロトコールでは、プラスミドが、配列番号26の代わりに、配列番号22で定義されるポリヌクレオチド配列を有し、rHAをコードするポリヌクレオチドに連結されたWO90/01063で記載されているアミノ酸配列を有するリーダー配列もコードする(「第2のスターター」プラスミド)ことを除いて上記の通りの「スターター」プラスミドを利用する。試験プラスミドは第2のスタータープラスミドを、試験配列をコードするポリヌクレオチド配列を、第2のスタータープラスミドのリーダー配列の相当する領域の代わりに含めるよう改変することにより作製する。第2の対照として、改変されていない第2のスタータープラスミドを使用する。試験および第2の対照プラスミドを含む形質転換体を第1のプロトコールに関して以上で記述したように調製し、フェドバッチ手順による高細胞密度発酵にて、WO96/37515にて記載されているある培地で、制御パラメーターを使用して培養する。YP/Sは上記の通りに評価する。
【0046】
試験および第2の対照形質転換体を比較できる発酵槽配置で比較できる時間培養するとき、本発明のリーダー配列は、上記の第2のプロトコールにより測定するYP/Sにより判定した場合、第2の対照により得られる分泌レベルよりも少なくとも1%、2%、3%、4%、5%、6%、7%、8%、9%、10%、11%、12%、13%、14%、15%、16%,17%,18%、19%または20%高い分泌レベルを獲得し得る。よって、本発明のリーダー配列が第2の対照よりも最大20%、30%、40%、50%、60%、70%、80%、90%、100%またはそれ以上に高い分泌レベルであることが証明し得る。本発明のリーダー配列が、第2の対照により得られる分泌レベルよりも少なくとも5%、例えば、6%、または少なくとも10%、例えば、12%、13%、14%、15%または16%高い分泌レベルを獲得することが特に好ましい。
【0047】
1つの態様では、本発明のリーダー配列が第1のプロトコールに関して以上で記述した分泌レベルを獲得する。もう1つの態様では、本発明のリーダー配列が第2のプロトコールに関して以上で記述した分泌レベルを獲得する。特に好ましい態様では、本発明のリーダー配列が第1および第2のプロトコール両方に関して以上で記述した分泌レベルを獲得する。
【0048】
細胞バイオマス由来の可溶性タンパク質および培養物上清中に分泌されたタンパク質は:
1.ゲル透過高速液体クロマトグラフィー
2.SDS−PAGEのデンシトメトリー
3.ロケット免疫電気泳動
により分析することができる。
【0049】
分泌された目的タンパク質の量および細胞内の目的タンパク質の量を、目的タンパク質の標準曲線と比べて定量し、当業者ならば周知のようにバイオマスの量に標準化する。
【0050】
通常、リーダー配列が、その配列を付けるか、または付けることが意図される未熟型の成熟タンパク質から誘導される場合が好ましい。よって、例えば、成熟タンパク質がアルブミンである場合には、天然のアルブミン分泌プレ配列またはプロ配列あるいはプレ−プロ配列を含んでなる配列を使用することが好ましい。しかしながら、リーダー配列は、もう1つの方法として、成熟タンパク質のもの以外の起源から誘導されてもよい。
【0051】
よって、1つの好ましい態様では、本発明の第1の態様のポリペプチドのリーダー配列がアルブミン分泌プレ配列またはその変異体から誘導された分泌プレ配列を含んでなる。
【0052】
以上で用いたアルブミンプレ配列の「変異体」とは、以上のX1、X2、X3、X4またはX5により定義された位置以外の、1箇所以上の位置にアミノ酸の挿入、欠失、または保存的置換(上記の通り)もしくは非保存的置換のいずれかの置換が存在するアルブミンプレ配列を指すが、そのような変化によってもそのペプチドがプレ配列の機能を果たし得るという条件においてである。
【0053】
好ましくは、アルブミンプレ配列の「変異体」が、以上のX1〜X5と定義される残基以外に、天然のアルブミンプレ配列、最も好ましくは、図1のアルブミンプレ配列と少なくとも2個、少なくとも3個または少なくとも4個、好ましくは、少なくとも5個、より好ましくは、少なくとも6個、なおいっそう好ましくは、少なくとも7個、さらに好ましくは、少なくとも8個、最も好ましくは、少なくとも9個の同一アミノ酸を有する。
【0054】
なおいっそう好ましくは、分泌プレ配列がアルブミン分泌プレ配列から誘導される場合に、本発明の第1の態様のポリペプチドがそれぞれ−20、−19、−18、−17および−16番にある天然アミノ酸の代わりに、それらの位置にX1、X2、X3、X4およびX5を有する、ここで、そのナンバリングが天然のアルブミン分泌プロ配列のC末端アミノ酸を−1位の残基とするものであり、かつ、X1、X2、X3、X4およびX5が上記のようなアミノ酸である。
【0055】
例えば、上記のナンバリングをヒトアルブミン分泌プレ配列の配列に適用する(例えば、WO90/01063で開示されている通り)と、以下のものが得られる:
【化9】
【0056】
特に好ましい態様では、使用される分泌プレ−配列がヒトアルブミン分泌プレ配列の配列から誘導される。
【0057】
よって、例えば、X1−X5ペンタペプチドがそのN末端において、以下の配列番号8の配列:
【化10】
配列番号8
またはその保存的に置換されたその変異体、すなわち:
【化11】
配列番号33
のC末端と直接または間接的に融合され得る。
【0058】
さらに、またはあるいは、それがそのC末端において、以下の配列
【化12】
配列番号9
または保存的に置換されたその変異体、すなわち:
【化13】
配列番号10
もしくは
【化14】
配列番号11
もしくは
【化15】
配列番号30
の少なくとも1つのN末端と直接または間接的に融合され得る。
【0059】
配列番号9で示す配列は天然ヒトアルブミンプレ配列の最後の9個のアミノ酸を表している。配列番号11の場合、これがWO90/01063の2つの主要な融合リーダー配列のうちの一方の最後の6個のアミノ酸と融合され、配列番号30の場合、配列番号9が天然ヒトアルブミンプロ配列の最後の6個のアミノ酸と融合される。
【0060】
好ましくは、いずれの場合にも、X1はFであり、X2はIであり、X3はVであり、X4はSまたはTであり、かつ、X5はIである。
【0061】
好ましい態様では、ペンタペプチドがそのN末端において配列番号8の配列または保存的に置換されたその変異体のC末端と融合され、かつ、そのC末端において配列番号9の配列、保存的に置換されたその変異体、配列番号10、11もしくは30のN末端と融合され、その結果として、例えば、以下の配列:
【化16】
配列番号12
または
【化17】
配列番号13
または
【化18】
配列番号14
【化19】
配列番号31
(式中、X1〜X5は上記のとおりである)、
あるいは以上で定義された保存的に置換されたその変異体
のうちの1つを形成する。
【0062】
特に好ましい態様は分泌プレ配列として、配列番号28の配列:
【化20】
配列番号28
を有する、すなわち、プレ配列がヒト血清アルブミン分泌プレ配列から誘導されたものであり、X1、X2、X3、X4およびX5が、−20、−19、−18、−17および−16番に存在し、かつ、X1、X2、X3、X4およびX5が配列番号7により示されるとおりである。
【0063】
以上のことからわかるように、以上で定義された分泌プレ配列、例えば、配列番号12または28の配列を分泌プロ配列と組み合わせて、機能的プレ−プロ分泌配列を作製してもよい。好ましい態様では、プレ配列モチーフをそのC末端において分泌プロ配列モチーフのN末端アミノ酸とペプチド結合により融合し、その結果として、プレ−プロ配列モチーフを形成する。リーダー配列を付けるか、または付けることが意図される未熟型の成熟タンパク質から誘導されたプロ配列を使用することも好ましくある。関連リーダー由来の改変されていないプレ配列またはプロ配列、もしくはその一部と事実上結合するプロ配列を使用することもまた好ましくある。
【0064】
好ましくは、プロ配列がそのC末端において、二塩基性アミノ酸対、すなわち、各々がLysまたはArgである、で終結する。
【0065】
一般に、分泌プロ配列モチーフが、アルブミン分泌プロ配列または二塩基性アミノ酸対を含み、その他の位置には保存的置換しかを有さないようなその変異体、通常、ヒトアルブミン分泌プロ配列、すなわち、配列N−Arg−Gly−Val−Phe−Arg−Arg−Cを有するものまたはその変異体である。もう1つの好ましい態様では、プロ配列が酵母MFα−1分泌プロ配列、すなわち、N−Ser−Leu−Asp−Lys−Arg−Cまたはアルブミンプロ配列に関して定義されたその変異体の全てまたは一部の配列を含んでなる。
【0066】
WO90/01063に記載されているリーダーおよびヒトアルブミンリーダーの相当する部分と比べて、本発明のポリペプチドは少なくとも4個のアミノ酸、すなわち、Ser−20PheまたはTrpまたはTyr;Phe−19IleまたはLeuまたはValまたはAlaまたはMet;Ile−18LeuまたはValまたはAlaまたはMet;およびLeu−16IleまたはValまたはAlaまたはMetが変化している、この表記法では、一例として挙げると、最初に名前を挙げた変異は−20番(すなわち、リーダー配列を用いて分泌されるであろう成熟タンパク質のN末端に対してマイナス20)にあるセリン残基がフェニルアラニン残基に変化していることを意味している。これについては図1にて例示する。
【0067】
1つの好ましいプレ−プロ配列は、下記配列を含んでなる:
【化21】
(式中、Y1はGlyまたはSerであり、Y2はValまたはLeuであり、Y3はPheまたはAspであり、Y4はArgまたはLysであり、かつ、Y5はArgまたはLysである)。
【0068】
好ましい態様では、Y1はGlyであり、Y2はValであり、かつ、Y3はPheである。もう1つの好ましい態様では、Y1はSerであり、Y2はLeuであり、かつ、Y3はAspである。
【0069】
一般に、Y4はArgであり、かつ、Y5はArgである。あるいは、Y4がLysであり、かつ、Y5がArgである場合が好ましい。もう1つの好ましい別のものがY4がLysであり、かつ、Y5がLysである場合である。Y5がLysである場合にはY4はArgであってもよい。
【0070】
特に好ましい態様は分泌プレプロ配列として、下記配列番号32の配列を有する。
【化22】
配列番号32
【0071】
本発明の第2の態様は、本発明の第1の態様により定義されたモチーフをコードする配列を有する単離ポリヌクレオチドを提供する。
【0072】
本明細書において、「単離」とは、ポリヌクレオチドが、DNA分子の場合、それが自然に見つけられる染色体の少なくとも大部分から独立して存在していること、また、RNA分子の場合、それが自然に転写される無傷細胞から独立して存在しているという意味を含む。言い換えれば、ポリヌクレオチドが、それがそれまでに存在していた、例えば、自然界での形態にあるとは言えない。よって、本発明の第2の態様のポリヌクレオチドは、細菌もしくは真菌ベクター、例えば、プラスミド、またはウイルスベクター、例えば、バクテリオファージにクローニングされたポリヌクレオチドを含む。好ましくは、このようなクローンは、関連染色体のDNAライブラリーを構成しているクローンから独立して存在している。
【0073】
線形アミノ酸配列は、大部分のアミノ酸が2以上のトリヌクレオチドコドンによってコードされているという標準的な遺伝子コード(図2)の縮重を用いてDNA配列へと逆翻訳することができる。
【0074】
例えば、配列番号1と定義されたペプチドをコードするDNA配列が、下記配列であると推測される:
【化23】
配列番号15
(式中、「3’」および「5’」は実際の末端というよりもポリヌクレオチド配列の方向を示し;言い換えれば、ポリヌクレオチド配列はいずれかの末端において、または両方の末端において、他のポリヌクレオチド配列と結合され得(例えば、融合され得るか、または連結され得)、かつ、Y、R、HおよびNは図2で定義されたとおりである)。
【0075】
同じ変換手順を利用して、下記DNA配列が、配列番号7のポリペプチドをコードすると推測される。
【化24】
配列番号16
【0076】
ヒトアルブミンのような天然の成熟タンパク質をコードする配列を含んでなるポリヌクレオチド配列の場合では、これがヒトアルブミン遺伝子配列などの天然のコード配列、または相補DNA配列(cDNA)もしくは1以上のイントロンを含むcDNAのいずれかであり得る。
【0077】
さらなる配列の改変を、例えば、コード領域に導入してもよい。本発明のポリペプチドをコードするDNAを改変するための所望の方法は、Saiki et al (1988) Science 239, 487-491により開示されているポリメラーゼ鎖反応を利用することである。この方法では、酵素により増幅しようとするDNAの両側に2つの特定のオリゴヌクレオチドプライマーを置くが、それらのプライマー自体が増幅されるDNAに組み込まれることとなる。該特定のプライマーは制限エンドヌクレアーゼ認識部位を含み、これらの部位は当技術分野で公知の方法を利用した発現ベクターへのクローニングに使用することができる。
【0078】
本発明のリーダー配列をコードするポリヌクレオチドは、最も便宜には、当技術分野では周知であり、かつ、以下でより詳細に記述する方法により、オリゴヌクレオチドを化学合成し、続いて、遺伝子構築物の他のエレメントと連結することにより作製される。
【0079】
成熟アルブミンをコードするポリヌクレオチドを改変することが望まれる場合には、最も便宜には、天然のcDNA配列からの部位特異的突然変異誘発またはPCR突然変異誘発により、または合成オリゴヌクレオチドのアセンブリングからこれを行ってよい。さらに、このような技術は当技術分野では標準的なものであり、また、いずれにせよ以下でより詳細に記述する。
【0080】
コード配列に対する改変は、特定の生物において、高度に発現されるあるタンパク質をコードするポリヌクレオチド配列が特定のアミノ酸に対してあるコドンに偏って使用していること;これはコドンバイアスと呼ばれていることである、から有利である場合がある。本発明の第2の態様の好ましい態様では、標準的な遺伝子コードを最適な宿主生物に対する好ましいコドンに変えることができる。本発明の第2の態様の特に好ましい態様では、標準的な遺伝子コードを酵母の好ましいコドンに変えることができる。(Sharp and Crowe (1991) Yeast 7, 657-678の表4を参照)有利には、好ましい酵母コドンのこのリストはアスパラギンコドン5’−GAT−3’を含めることにより改変される(図3)。
【0081】
一例として、配列番号1のペプチド配列を用いて、酵母のこのペプチドをコードするコドンに偏りのあるDNA配列が下記のものであると推測され得る。
【化25】
配列番号17
【0082】
同じ変換手順を利用して、配列番号7の配列を有する特に好ましいポリペプチドモチーフについてのコドンに偏りのある縮重DNA配列:
【化26】
配列番号18
が推定されるが、配列番号7の配列を有するポリペプチドモチーフをコードする最も好ましいコドンに偏りのあるDNA配列は下記のものである。
【化27】
配列番号34
【0083】
図2で示す遺伝子コードまたは対象となる宿主に使用可能な好ましいコドンバイアス表もしくは図3で示す好ましいコドンバイアスを利用して、目的のアミノ酸配列を同じ変換手順により部分的に冗長なポリヌクレオチド配列に変換することができる。この方法によりDNA配列に変換するアミノ酸配列は、限定されるものではないが、本発明の第1の態様のポリペプチドから選択してよい。例えば、成熟ヒトアルブミンのコード領域の配列はこの方法により誘導することができる。EP308381はヒトアルブミンの酵母コドン最適化部分的コード配列を開示している。本明細書における配列番号20はそういった配列のさらなるものである。有利には、DNA配列の冗長性が許せば、コードされたアミノ酸配列(または図3を利用する場合にはコドンバイアス)を乱すことなく、ドメインおよびサブドメインの境界に制限部位を導入することができる。
【0084】
残るDNA配列の冗長性を解明し、別のコドンの発生頻度を冗長なDNA配列を有する各アミノ酸に対して均等化することができる。縮重コドンのDNA配列の冗長性を利用することにより可能であれば、可能性ある転写終結配列を示すDNA配列を除くか、または縮小することが有利である。最後に、冗長なDNA配列を有するアミノ酸の別のコドンのバランスを再度均等化することができるが、これまでの改変と矛盾することはない。
【0085】
本発明の第2の態様のポリヌクレオチドは、その5’および/または3’末端において、1以上の他のヌクレオチド配列と直接または間接的に融合し、その結果として、例えば、完全な遺伝子または発現カセットを形成することができる。よって、発現カセットは転写開始および終結部位、ならびに、転写された領域内に、翻訳開始のためのリボソーム結合部位も含むことが望ましい(1998年4月23日に公開されたHastings et al、WO98/16643)。
【0086】
従って、本発明の第2の態様は、異種タンパク質をコードするDNAと、本発明の第1の態様のポリペプチド、特に、目的タンパク質がアルブミンである場合、をコードするDNA配列またはその変異体もしくは断片のいずれかとの連続したまたは不連続な融合物であるDNA配列を含んでなるポリヌクレオチドを含む。これに関連して、「異種タンパク質」とは、それが「目的タンパク質」と同じではないこと、すなわち、ホモ二量体を形成しないことを意味する。
【0087】
従って、ポリヌクレオチドはその5’末端においてプロモーター(RNAポリメラーゼの結合および転写を起こさせるDNA配列により形成された発現制御エレメント)と、かつ/またはその3’末端において他の調節配列、例えば、翻訳終結配列と直接または間接的に融合されていてもよい。よって、ポリヌクレオチドが1以上の調節領域、通常、転写調節領域に作動可能なように連結されていてもよい。「作動可能なように連結された」とは、調節領域が、それがポリヌクレオチド配列に対して作用を及ぼし得るように連結されていることを意味する。どの調節領域を使用するかの選択は、想定される宿主(すなわち、意図される発現系)にある程度依存するが、好ましい配列の選択は当業者には知られている。
【0088】
数多くの発現系が知られているが、そのような系としては、例えば、組換えバクテリオファージ、プラスミドまたはコスミドDNA発現ベクターで形質転換された細菌(例えば、枯草菌(Bacillus subtilis)または大腸菌(Escherichia coli));例えば、酵母発現ベクターで形質転換された酵母(例えば、サッカロミセス・セレビシエまたはピキア・パストリス);例えば、ウイルス発現ベクター(例えば、バキュロウイルス)で形質転換された昆虫細胞系;例えば、ウイルスまたは細菌発現ベクターで形質転換された植物細胞系;例えば、アデノウイルス発現ベクターで形質転換された、動物細胞系の細胞培養物、動物細胞系トランスジェニック細胞、または遺伝子療法としての動物細胞系のいずれか、を使用する系が挙げられる。宿主細胞は、好ましくは、酵母(最も好ましくは、サッカロミセス属(Saccharomyces)種、例えば、S.セレビシエまたはピキア属(Pichia)種、例えば、P.パストリス)である。
【0089】
従って、本発明の第3の態様は、本発明の第2の態様のポリヌクレオチドで形質転換された宿主細胞を提供する。その宿主細胞は、原核生物細胞でも真核生物細胞でもよい。細菌細胞が、特に、バチルス属(Bacillus)およびエシェリキア属(Escherichia)のいくつかの種が可能なようにそれらがタンパク質を分泌し得るならば好ましい原核生物の宿主細胞である。好ましい真核生物宿主細胞としては、植物、真菌、酵母および動物細胞、好ましくは、脊椎動物細胞、より好ましくは、哺乳類細胞、例えば、マウス、ラット、ウシ、ヒツジ、ヤギ、ブタ、バッファロー、ヤク、ウマもしくは他の家畜、サルまたはヒト由来のものが挙げられる。好適なヒト細胞としては、ヒト繊維芽細胞細胞系の細胞が挙げられる。よって、宿主細胞がin situの哺乳類のトランスジェニック細胞であってよく、それゆえ、遺伝子療法アプローチまたはトランスジェニック個体の作製による産物であり得る。後者の場合では、個体が非ヒト哺乳類であることが好ましい。
【0090】
細菌宿主の典型的な種類としては、大腸菌および枯草菌が挙げられる。
【0091】
植物宿主の典型的な種類としては、種子植物類、シダ植物類(例えば、シダ類、ヒカゲノカズラ類、トクサ類)、コケ植物類(例えば、ゼニゴケ類およびセン類)、および藻類が挙げられる。一般に、植物宿主細胞は多細胞植物、通常、裸子植物または被子植物などの種子植物から誘導されたものである。好適な裸子植物としては、針葉樹類(例えば、マツ、カラマツ、モミ、トウヒおよびヒマラヤスギ)、ソテツ類、イチイ類およびイチョウ類が挙げられる。より一般には、植物宿主細胞が単子葉植物または双子葉植物、好ましくは、作物であり得る被子植物の細胞である。好ましい単子葉植物としては、トウモロコシ、コムギ、オオムギ、サトウモロコシ、タマネギ、カラスムギ、カモガヤおよび他のイチゴツナギ亜科(Pooideae)が挙げられる。好ましい双子葉植物の作物としては、トマト、ジャガイモ、テンサイ、タピオカノキ、アブラナ科の作物(アブラナを含む)、アマニ、タバコ、ヒマワリ、繊維作物、例えば、綿、ならびに豆科植物、例えば、エンドウ、インゲンマメ、特に、ダイズ、およびアルファルファが挙げられる。よって、宿主細胞は自律的細胞(autonomous cell)、例えば、単細胞植物の細胞または細胞培養で維持される細胞であってよいし、または多細胞植物のin situ細胞であってもよい。従って、本発明は、好ましくは、安定した遺伝性のトランスジェニック表現型を保持する完全トランスジェニック植物の生産を意図する。
【0092】
真菌宿主の典型的な種類としては、アスペルギルス属(Aspergillus)(例えば、A.ニガー(A. niger)およびA.オリゼー(A. oryzae))、ストレプトミセス属(Streptomyces)、ペニシリウム属(Penicillium)および酵母が挙げられる。本発明の実施において有用であると考えられる酵母の典型的な種類としては、ピキア属(ハンセヌラ属(Hansenula))、サッカロミセス属、クルイベロミセス属(Kluyveromyces)、カンジダ属(Candida)、トルロプシス属(Torulopsis)、トルラスポラ属(Torulaspora)、シゾサッカロミセス属(Schizosaccharomyces)、サイテロミセス属(Citeromyces)、パキソレン属(Pachysolen)、デバロミセス属(Debaromyces)、メチニコウィア属(Metschunikowia)、ロドスポリジウム属(Rhodosporidium)、ロイコスポリジウム属(Leucosporidium)、ボトリオアスクス属(Botryoascus)、スポリジオボルス属(Sporidiobolus)、エンドミコプシス属(Endomycopsis)などがある。好ましい属は、ピキア属(ハンセヌラ属)、サッカロミセス属、クルイベロミセス属およびヤロウィア属(Yarrowia)からなる群から選択されるものである。サッカロミセス属種の例として、S.セレビシエ、S.イタリクス(S. italicus)およびS.ルキシー(S. rouxii)がある。クルイベロミセス属種の例として、K.フラギリス(K. fragilis)およびK.ラクティス(K. lactis)がある。ピキア属(ハンセヌラ属)種の例として、P.パストリス、P.アノマラ(P. anomala)およびP.カプスラータ(P. capsulata)がある。好適なヤロウィア属種の例としては、Y.リポリチカ(Y. lipolytica)がある。酵母宿主細胞としては、一般に、Stratagene Cloning Systems, La Jolla, CA 92037, USAから市販されているYPH499、YPH500およびYPH501が挙げられる。
【0093】
好ましい哺乳類宿主細胞としては、ATCCからCCL61として入手可能であるチャイニーズハムスター卵巣(CHO)細胞、ATCCからCRL1658として入手可能であるNIH Swissマウス胚細胞NIH/3T3、およびATCCからCRL1650として入手可能であるサル腎臓由来のCOS−1細胞が挙げられる。好ましい昆虫細胞としては、バキュロウイルス発現ベクターでトランスフェクトすることができるSf9細胞がある。
【0094】
上述したように、ポリヌクレオチド調節領域の選択は、意図される宿主の性質にある程度依存する。
【0095】
細菌宿主細胞における使用に好適なプロモーターとしては、大腸菌lacIおよびlacZプロモーター、T3およびT7プロモーター、gptプロモーター、ファージλPRおよびPLプロモーター、phoAプロモーターおよびtrpプロモーターが挙げられる。本発明のDNAセグメントの挿入に便宜な制限部位を含むプラスミドベクターには、通常、典型的な細菌宿主に適合するプロモーター配列が提供される。
【0096】
真核生物のプロモーターとしては、CMV前初期プロモーター、HSVチミジンキナーゼプロモーター、初期および後期SV40プロモーターならびにレトロウイルスLTRのプロモーターが挙げられる。他の好適なプロモーターは、当業者には公知である。
【0097】
S.セレビシエに好適なプロモーターとしては、PGK1遺伝子、GAL1またはGAL10遺伝子、CYC1、PHO5、TRP1、ADH1、ADH2、グリセルアルデヒド−3−リン酸デヒドロゲナーゼ、ヘキソキナーゼ、ピルビン酸デカルボキシラーゼ、ホスホフルクトキナーゼ、トリオースリン酸イソメラーゼ、ホスホグルコースイソメラーゼ、グルコキナーゼ、α接合因子フェロモン、a−接合因子フェロモンに関する遺伝子に関連するもの、PRB1プロモーター、GPD1プロモーター、ならびに、5’調節領域の部分と、その他のプロモーターの5’調節領域の部分とのハイブリッド、あるいは、上流活性化部位とのハイブリッドを含むハイブリッドプロモーター(例えば、EP−A−258 067のプロモーター)が挙げられる。
【0098】
もう1つの好適な宿主細胞であるシゾサッカロミセス・ポンベ(Schizosaccharoonyces pombe)における使用に便宜な調節プロモーターとしては、Maundrell (1990) J. Biol. Chem. 265, 10857-10864に記載されているnmt遺伝子に由来するチアミン抑制プロモーター、ならびにHoffman & Winston (1990) Genetics 124, 807-816に記載されているグルコース抑制fbp1遺伝子プロモーターある。がある。
【0099】
ピキア属に好適なプロモーター、形質転換プロトコールおよび培養条件については、米国特許第5986062号(引用することにより本明細書の一部とされる)で見つけることができる。例えば、HSA産生宿主(またはHSA産生株)の作製は、組換えプラスミドを染色体に導入するという方法(EP−A−399455に対応するJP−A−3−72889)、HSAを酵母で発現させるという方法(EP−A−123544に対応するJP−A−60−41487、EP−A−248657に対応するJP−A−63−39576およびEP−A−251744に対応するJP−A−63−74493)、およびHSAをピキア属で発現させるという方法(EP−A−344459に対応するJP−A−2−104290)を利用して行い得る。HSA産生宿主の培養(HSA産生プロセス)は、米国特許第5,986,062号に記載されている方法などの既知の方法を利用して、例えば、JP−A−3−83595またはJP−A−4−293495(EP−A−504823に対応)に開示されている方法により実施することができる。形質転換された宿主を培養するための培地は、米国特許第5,986,062号に従って調製し得、宿主の培養は、米国特許第5,986,062号の開示内容に従って、静置もしくは振盪培養または攪拌および通気下でのバッチ式、半バッチ式もしくは連続培養により、好ましくは、15〜43℃(より好ましくは、20〜30℃)にて1〜1,000時間実施することができる。
【0100】
好適な転写終結シグナルは当技術分野では周知である。宿主細胞が真核生物である場合、転写終結シグナルは、好ましくは、適正な転写終結シグナルおよびポリアデニル化シグナルを含む真核生物の遺伝子の3’フランキング配列由来のものである。好適な3’フランキング配列が、例えば、使用される発現制御配列に自然に連結された遺伝子のものであってよい、すなわち、それらがプロモーターと一致したものであってもよい。あるいは、それらが異なっていてもよい。その場合には、宿主が酵母、好ましくは、S.セレビシエであれば、S.セレビシエADH1遺伝子の終結シグナルが好ましい。
【0101】
よって、本発明の第2の態様のポリヌクレオチドは、以上で記述したような方法を利用することによりいかなる所望の宿主に対しても開発することができる。
【0102】
成熟ヒトアルブミンをコードするDNA配列は、以上で記述した、天然の遺伝子、cDNAまたは1以上のイントロンを含むcDNAと、以上で記述した方法により誘導したコドンに偏りのあるヒトアルブミンDNA配列とのDNA融合物から開発することができる。
【0103】
配列番号19は、翻訳開始部位の5’に22ヌクレオチド、分泌リーダー配列 配列番号32の好ましいポリヌクレオチドコード配列、および成熟ヒトアルブミンコード領域 配列番号20を含んでなるポリヌクレオチド配列である。コード配列は翻訳終止コドンで終わる。このコドンは、一般には、TGA、TAGまたはTAAであるが、酵母ではTAAが最も有効である。好ましくは、さらなる翻訳終止コドン(好ましくは、各々TAAである)、通常1つまたは2つが、好ましくは、互いに隣接してまたは終止コドンの各対間3塩基対以下で含まれる。配列番号19は両末端に好適なクローニング部位がフランキングする。
【0104】
本発明の第2の態様のポリヌクレオチドはまた、好適な宿主への導入に向けて多くの種類の他のDNA配列と結合し得る。コンパニオン配列は、宿主の性質、ポリヌクレオチドの宿主への導入方法、およびエピソーム維持または組込みが所望か否かに依存する。例えば、ベクターを他の非原核生物細胞型での発現に使用する場合でも、ベクターが原核生物における増殖のためのCol E1 oriなどの原核生物レプリコンを含むことがある。
【0105】
一般に、本発明の第2の態様のポリヌクレオチドは、プラスミドなどの発現ベクターに、適正な方向および正確な発現用リーディングフレーム内に挿入される。
【0106】
よって、ポリヌクレオチドを本明細書に記載の教示に照らして好適に改変された既知の技術に従って使用して、限定されるものではないが、組込みベクター、動原体ベクターおよびエピソームベクターをはじめとする発現ベクターを構築し得る。
【0107】
よって、本発明の第2の態様の1つの態様では、ポリヌクレオチドがベクターである。
【0108】
一般的な原核生物ベクタープラスミドとしては、Biorad Laboratories(Richmond, CA, USA)から市販されているpUC18、pUC19、pBR322およびpBR329;Pharmacia(Piscataway, NJ, USA)から市販されているpTrc99A、pKK223−3、pKK233−3、pDR540およびpRIT5;Stratagene Cloning Systems(La Jolla, CA 92037, USA)から市販されているpBSベクター、Phagescriptベクター、Bluescriptベクター、pNH8A、pNH16A、pNH18A、pNH46Aがある。
【0109】
一般的な哺乳類細胞ベクタープラスミドとしては、Pharmacia(Piscataway, NJ, USA)から市販されているpSVLがある。このベクターはSV40後期プロモーターを使用してクローニングされた遺伝子の発現を推進するが、COS−1細胞などのT抗原産生細胞において最高レベルの発現が認められる。誘導性哺乳類発現ベクターの例としては、Pharmacia(Piscataway, NJ, USA)からも市販されているpMSGがある。このベクターはマウス乳癌ウイルスの長い末端反復配列のグルココルチコイド誘導性プロモーターを使用してクローニングされた遺伝子の発現を推進する。
【0110】
有用な酵母エピソームプラスミドベクターとしては、pRS403−406およびpRS413−416、これらは、一般に、Stratagene Cloning Systems(La Jolla, CA 92037, USA)から市販されている、YEp24(Botstein, D., et al. (1979) Gene 8, 17-24)、ならびにYEplac122、YEplac195およびYEplac181(Gietz, R. D. and Sugino. A. (1988) Gene 74, 527-534)がある。他の酵母プラスミドは、EP−A−286 424の「分解ベクター」に加えてさらに、WO90/01063およびEP424117で記載されている。プラスミドpRS403、pRS404、pRS405およびpRS406は、YIplac204、YIplac211およびYIplac128(Gietz, R. D. and Sugino. A. (1988) Gene 74, 527-534)がそうであるように、酵母組込み型プラスミド(YIp)であり、酵母選択マーカーHIS3、TRP1、LEU2およびURA3を組み込んでいる。プラスミドpRS413−416は、YCplac22、YCplac33およびYCplac111(Gietz, R. D. and Sugino. A. (1988) Gene 74, 527-534)がそうであるように、酵母動原体プラスミド(YCp)である。
【0111】
当業者には周知の方法を利用して、コード配列と、例えば、好適な転写または翻訳制御物を含む発現ベクターを構築することができる。このような1つの方法では、突出末端を介した連結を必要とする。好適な制限酵素の作用によりDNA断片およびベクターに適合する突出末端を作成することができる。これらの末端を相補塩基対合により迅速にアニーリングし、残るニックをDNAリガーゼの作用により閉じることができる。
【0112】
さらなる方法では、合成二本鎖オリゴヌクレオチドリンカーとアダプターを使用する。平滑末端を有するDNA断片を、凸部3’末端を除去し、凹部3’末端を埋めるバクテリオファージ T4 DNAポリメラーゼまたは大腸菌DNAポリメラーゼIにより作製する。合成リンカーおよび特定の制限酵素の認識配列を含む平滑末端を有する二本鎖DNAの断片をT4 DNAリガーゼにより平滑末端を有するDNA断片に連結することができる。続いて、それらを好適な制限酵素で消化して突出末端を作り出し、適合する末端を有する発現ベクターに連結する。また、アダプターが連結に用いる1つの平滑末端を含み、また、1つの成形した突出末端も保有する化学合成されたDNA断片でもある。あるいは、1DNA断片または複数のDNA断片を、所望により、突出末端を有していてもよい1以上の合成二本鎖オリゴヌクレオチドの存在または不在下で、DNAリガーゼの作用により一緒に連結してもよい。
【0113】
種々の制限エンドヌクレアーゼ部位を含む合成リンカーが、Sigma-Genosys Ltd, London Road, Pampisford, Cambridge, United Kingdomをはじめとする多数の供給源から市販されている。
【0114】
このようにして作製される本発明のベクターを、本発明の第1の態様で定義されたとおりの配列を含んでなるポリペプチドの発現および産生に向けて好適な宿主細胞を形質転換するために使用してもよい。このような技術としては、その全てが引用することにより本明細書の一部とされる、1984年4月3日に発行されたRutter et alの米国特許第4,440,859号、1985年7月23日に発行されたWeissmanの同第4,530,901号、1986年4月15日に発行されたCrowlの同第4,582,800号、1987年6月30日に発行されたMark et alの同第4,677,063号、1987年7月7日に発行されたGoeddelの同第4,678,751号、1987年11月3日に発行されたItakura et alの同第4,704,362号、1987年12月1日に発行されたMurrayの同第4,710,463号、1988年7月12日に発行されたToole, Jr. et alの同第4,757,006号、1988年8月23日に発行されたGoeddel et alの同第4,766,075号、および1989年3月7日に発行されたStalkerの同第4,810,648号で開示されているものが挙げられる。
【0115】
本発明のDNA構築物での好適な細胞宿主の形質転換は、周知の方法により実施されるが、一般に、使用するベクターのタイプよって異なる。原核生物宿主細胞の形質転換に関しては、例えば、Cohen et al (1972) Proc. Natl. Acad. Sci. USA 69, 2110およびSambrook et al (2001) Molecular Cloning, A Laboratory Manual, 3'd Ed. Cold Spring Harbor Laboratory, Cold Spring Harbor, NYを参照のこと。酵母細胞の形質転換については、Sherman et al (1986) Methods In Yeast Genetics, A Laboratory Manual, Cold Spring Harbor, NYで記載されている。Beggs (1978) Nature 275, 104-109の方法もまた有用である。S.セレビシエを形質転換する方法については、その全てが引用することにより本明細書の一部とされる、EP251744、EP258067およびWO90/01063で一般的に教示されている。脊椎動物細胞に関し、このような細胞をトランスフェクトするのに有用な試薬、例えば、リン酸カルシウムおよびDEAE−デキストランまたはリポソーム製剤は、Stratagene Cloning Systems、またはLife Technologies Inc., Gaithersburg, MD 20877, USAから市販されている。
【0116】
エレクトロポレーションもまた、細胞の形質転換に有用であり、酵母細胞、細菌細胞および脊椎動物細胞を形質転換する技術分野では周知のことである。エレクトロポレーションにより酵母を形質転換する方法については、Becker & Guarente (1990) Methods Enzymol. 194, 182で開示されている。
【0117】
DNAを動物および植物細胞に導入するために物質的な方法を使用してもよい。例えば、マイクロインジェクションでは、極細のピペットを使用して、DNA分子を形質転換しようとする細胞の核に直接注入する。もう1つの例では、高速マイクロプロジェクタイル、通常、DNAでコーティングした金またはタングステンの粒子の細胞への撃ち込みを使用する。
【0118】
植物は多くの当技術分野において承認されている方法により形質転換し得る。当業者ならば、その選択する方法が形質転換の対象となる植物のタイプに依存することが分かるであろう。植物細胞を形質転換する好適な方法の例としては、マイクロインジェクション(Crossway et al., BioTechniques 4: 320-334 (1986))、エレクトロポレーション(Riggs et al., Proc. Natl. Acad. Sci. USA 83: 5602-5606 (1986)、アグロバクテリウム(Agrobacterium)媒介形質転換(Hinchee et al, Biotechnology 6: 915-921 (1988);トウモロコシの形質転換についてはIshida et al., Nature Biotechnology 14: 745-750 (1996)も参照のこと)、直接遺伝子導入(Paszkowski et al., EMBO J. 3: 2717-2722 (1984);Hayashimoto et al., Plant Physiol. 93: 857-863 (1990)(イネ))、およびAgracetus, Inc., Madison, Wisconsin and Dupont, Inc., Wilmington, Delawareから市販されている装置を使用した弾道学的粒子加速(例えば、Sanford et al., 米国特許第4,945,050号;およびMcCabe et al., Biotechnology 6: 923-926 (1988)を参照)が挙げられる。また、Weissinger et al., Annual Rev. Genet. 22: 421-477 (1988);Sanford et al., Particulate Science and Technology 5: 27-37 91987)(タマネギ);Svab et al., Proc. Natl. Acad. Sci. USA 87: 8526-8530 (1990)(タバコ葉緑体);Christou et al., Plant Physiol. 87: 671-674 (1988)(ダイズ);McCabe et al., Bio/Technology 6: 923-926 (1988)(ダイズ);Klein et al., Proc. Natl. Acad. Sci. USA, 85: 4305-4309 (1988)(トウモロコシ);Klein et al., Bio/Technology 6: 559-563 (1988)(トウモロコシ);Klein et al., Plant Physiol. 91: 440-444 (1988)(トウモロコシ);Fromm et al., Bio/Technology 8: 833-839 (1990);およびGordon-Kamm et al., Plant Cell 2: 603-618 (1990)(トウモロコシ);Koziel et al., Biotechnology 11: 194-200 (1993)(トウモロコシ);Shimamoto et al., Nature 338: 274-277 (1989)(イネ);Christou et al., Biotechnology 9: 957-962 (1991)(イネ);Datta et al., Bio/Technology 8: 736-740 (1990)(イネ);欧州特許出願EP−A−332 581(カモガヤおよび他のイチゴツナギ亜科);Vasil et al., Biotechnology 11: 1553-1558 (1993)(コムギ);Weeks et al., Plant Physiol. 102: 1077-1084 (1993)(コムギ);Wan et al., Plant Physiol. 104: 37-48 (1994)(オオムギ);Jahne et al., Theor. Appl. Genet. 89: 525-533 (1994)(オオムギ);Umbeck et al., Bio/Technology 5: 263-266 (1987)(綿);Casas et al., Proc.Natl. Acad. Sci. USA 90: 11212-11216 (1993)(サトウモロコシ);Somers et al., Bio/Technology 10: 1589-1594 (1992)(カラスムギ);Torbert et al., Plant Cell Reports 14: 6
35-640 (1995)(カラスムギ);Weeks et al., Plant Physiol. 102. 1077-1084 (1993)(コムギ);Chang et al., WO94/13822(コムギ)ならびにNehra et al., The Plant Journal 5: 285-297 (1994)(コムギ)も参照のこと。アグロバクテリウム媒介形質転換は、一般的には、単子葉植物に対して効果がなく、この種の植物には以上で記述した他の方法が好ましい。
【0119】
一般に、ベクターが宿主の全てを形質転換するのではないため、形質転換する宿主細胞に対してベクターを選択する必要がある。1つの選択手法では、形質転換される細胞において選択可能な特徴をコードする、必要な制御エレメントを含むDNA配列マーカーの発現ベクターへの組込みを利用する。これらのマーカーとしては、ジヒドロ葉酸レダクターゼ、G418または真核細胞培養に関するネオマイシン耐性、ならびに大腸菌および他の細菌での培養に関するテトラサイクリン、カナマイシンまたはアンピシリン耐性遺伝子が挙げられる。あるいは、このような選択可能な特徴に関連する遺伝子が、所望の宿主細胞を同時に形質転換するために使用されるもう1つのベクターにあってもよい。
【0120】
マーカー遺伝子を使用して形質転換体を同定することができるが、どの細胞が組換えDNA分子を含んでいるのか、また、どれが自己連結したベクター分子を含んでいるのかが判定できることが望ましい。これは、DNA断片を挿入することによってその分子に存在する遺伝子のうちの1つの完全性を破壊するクローニングベクターを利用することによって行うことができる。よって、組換え体はその遺伝子の機能の喪失によって同定することができる。
【0121】
首尾よく形質転換された細胞を同定するもう1つの方法では、本発明のポリペプチドを産生させるための本発明の発現構築物の導入により得られた細胞を増殖させる。細胞を回収し、それらを溶解し、それらのDNA含量をDNAの存在について、Southern (1975) J. Mol. Biol. 98, 503またはBerent et al (1985) Biotech. 3, 208により記載されているような方法を利用して調べてもよい。あるいは、形質転換細胞の培養物上清中の成熟タンパク質の存在を抗体を用いて検出してもよい。
【0122】
組換えDNAの存在について直接アッセイすることに加えて、組換えDNAがタンパク質の発現を誘導し得る場合には周知の免疫学的方法により形質転換の成功を確認することができる。例えば、発現ベクターで首尾よく形質転換された細胞は、好適な抗原性を提示するタンパク質を産生する。形質転換されたと思われる細胞のサンプルを回収し、好適な抗体を用いてそのタンパク質についてアッセイする。
【0123】
よって、形質転換された宿主細胞自体に加えて、本発明はまた、これらの細胞の培養物、好ましくは、モノクローナル(クローン的に同質な)培養物、または栄養培地においてモノクローナル培養物から誘導された培養物も意図する。
【0124】
従って、本発明の第4の態様では、本発明の第3の態様の細胞と培養培地を含んでなる細胞培養物を提供する。一般に、培養培地は本発明の第1の態様のポリペプチドの発現系内での発現により、通常、プレおよび/またはプロ配列の除去などのさらなる翻訳プロセシングにより生じる成熟ポリペプチドを含有する。
【0125】
大腸菌などの原核生物宿主細胞、ならびに哺乳類細胞などの真核生物宿主細胞の培養方法は当技術分野では周知である。酵母の培養方法はEP330451およびEP361991で一般的に教示されている。
【0126】
本発明の組換えDNAにより形質転換された宿主細胞を本明細書に開示されている教示に照らして、十分な時間、当業者には公知の好適な条件下で培養することにより、本発明の第1の態様のポリペプチドの発現が可能になる。このようにして産生されたポリペプチドに、プレおよび/またはプロ配列が除去されるように宿主細胞によるプロセシングをさらに行ってもよい。従って、「成熟した」目的タンパク質が最初に翻訳されたタンパク質とは異なっている場合がある。
【0127】
よって、本発明はまた、第5の態様として、以上で定義された成熟した目的タンパク質を作製する方法を提供する。この方法は、本発明の第3の態様の細胞を培養培地で培養する工程(ここで、その細胞は、本発明の第1の態様で定義されたとおりのポリペプチドの発現の結果として、成熟した目的タンパク質を分泌し、その際、そのタンパク質はペリプラスム間隙、その培養培地またはそれらの両方に蓄積するが、好ましくは、その培養培地に蓄積する)を含んでなる。次いで、分泌された目的タンパク質を含有する培養培地を細胞培養物中の細胞から分離する。細胞壁と結合した分泌されたタンパク質は、一般に、浸透性補助(例えば、ソルビトール)条件下(選択的に分泌されたタンパク質を徐々に放出する)、細胞壁融解酵素を用いて解離させることができる。Elango et al., J. Biol. Chem. 257: 1398-1400 (1982)を参照のこと。この目的に好適な細胞壁融解酵素の例としては、lyticase、Zymolyase-60,000およびGlusulaseが挙げられ、それらの全てが市販されており、例えば、後の2つの場合では、Seikagaku KogyoまたはKirin Breweryから、また、各々Boehringer Mannheimから市販されている。
【0128】
好ましくは、培養培地の単離後、成熟した目的タンパク質をその培地から分離する。なおいっそう好ましくは、このようにして得られた成熟した目的タンパク質をさらに精製する。
【0129】
目的の成熟タンパク質は当技術分野で公知の多くの方法により培養培地から抽出し得る。例えば、組換えにより発現されるアルブミンの回収に関する精製技術は、WO92/04367、基質由来の色素の除去;EP464590、酵母由来の染料の除去;EP319067、アルカリ沈殿およびその後のアルブミンの親油相への適用;ならびに全精製方法を記述しているWO96/37515、米国特許第5728553号およびWO00/44772で開示されており、その全てが引用することにより本明細書の一部とされる。本発明の改変リーダー配列は成熟タンパク質それ自体に影響を及ぼさないため、アルブミン以外のタンパク質を、このようなタンパク質の精製に有用であることが分かっているいずれかの技術により培養培地から精製してもよい。
【0130】
このような周知の方法としては、硫酸アンモニウムまたはエタノール沈降、酸抽出、陰イオンまたは陽イオン交換クロマトグラフィー、ホスホセルロースクロマトグラフィー、疎水性相互作用クロマトグラフィー、アフィニティークロマトグラフィー、ヒドロキシルアパタイトクロマトグラフィーおよびレクチンクロマトグラフィーが挙げられる。最も好ましくは、高速液体クロマトグラフィー(「HPLC」)を精製に用いる。
【0131】
得られたタンパク質は、その知られた有用性のいずれかに向けて使用することができ、アルブミンの場合では、重度の火傷、ショック状態および失血を治療するための患者へのi.v.投与物、培養培地への補給物、ならびに他のタンパク質製剤の賦形剤としての使用を含む。
【0132】
本発明の方法により得られる治療上有用な目的タンパク質は、単独で投与することが可能であるが、そのタンパク質を1種以上の許容される担体または希釈剤とともに医薬製剤として提供することが好ましい。担体または希釈剤は、目的タンパク質と適合し、かつ、その受容者に対して有害ではないという意味において「許容される」ものでなければならない。一般に、担体または希釈剤は、無菌かつ発熱物質を含まない水または生理食塩水である。
【0133】
よって、本発明の第6の態様は、本発明の第5の態様の方法により得られる目的タンパク質を治療上許容される担体または希釈剤とともに製剤し、そうすることによって、ヒトまたは動物への投与に好適な治療用製品を生産する方法を提供する。
【0134】
治療用製品は、単位投与形で提供することが便宜であり、製薬分野では周知の方法のいずれによっても調製し得る。好ましい単位投与形製剤は、有効成分の、1日用量もしくは単位、1日用量以下の量またはその好適な分割量を含むものである。
【0135】
当然のことながら、治療用製品は、以上で特記した成分に加えて、問題の製品のタイプを考慮し、当技術分野で常用の他の薬剤を含んでいてもよい。
【0136】
以下、本発明を添付の限定されない図面および実施例を参照してより詳細に説明する。
【実施例】
【0137】
実施例1
下記の2種類の一本鎖オリゴヌクレオチドPRBJM1およびPRBJM2を用いたPCRにより、サッカロミセス・セレビシエ PRBIプロモーターを酵母ゲノムDNAから単離した。
PRBJM1:
【化28】
配列番号35
PRBJM2:
【化29】
配列番号36
【0138】
PCR条件 94℃で30秒間、50℃で40秒間、72℃で120秒間、次いで、72℃で600秒間、その後、4℃で維持を40サイクル。0.85kb DNA断片をNotIとHindIIIの両方で消化し、WO97/24445で記載されている、NotIとHindIIIで同様に消化したpBST+に連結して、プラスミドpAYE438(図4)を作出した。プラスミドpAYE438をHindIIIとBamHIで消化し、WO00/44772で従前に開示されているpAYE440由来の0.48kb HindIII/BamHI ADHIターミネーターDNA断片と連結して、プラスミドpAYE441(図5)を作出した。プラスミドpAYE441を独自のHindIII部位にて線状化し、従前に開示されている (Sleep, D. et al. (1991) Bio/Technology 9, 183-187およびEP−A−0431880)pAYE309(図6)由来の1.8kb HindIII/Bsu36I断片および二本鎖オリゴヌクレオチドリンカー:
【化30】
配列番号37
【化31】
配列番号38
と連結して、pAYE467(図7)を作出した。pAYE467由来の3.2kb NotI、発現カセットを、仔ウシ小腸ホスファターゼ(CIP)で予め処理しておいたNotI線状化pSAC35(Sleep et al. (1991), Bio/Technology 9: 183-187)に連結して、プラスミドpAYE443(図8)を作出した。配列番号22は、pAYE467およびpAYE443両方のDNA配列内に見られるHSA/MFα−1融合リーダー配列のコード領域と成熟ヒトアルブミンコード領域を含んでなるポリヌクレオチドを示すものである。HSA/MFα−1融合リーダー配列をコードするポリヌクレオチド配列を、DNA配列:
【化32】
配列番号23
を有するCPK1と呼ばれる一本鎖オリゴヌクレオチドを用いた部位特異的突然変異誘発により改変した。
【0139】
部位特異的突然変異誘発(SDM)を標準的なプロトコール(引用することにより本明細書の一部とされるBotstein and Shortle, "Strategies and Applications of In Vitro Mutagenesis," Science, 229: 193-1210 (1985))に従って実施したが、他の好適な技術を利用してもよい。CPK1のヌクレオチド配列は、HSA/MFα−1融合リーダー配列のアミノ酸配列を改変して、以下の変異Thr−20Phe、Phe−19Ile、Ile−18ValおよびLeu−16Ileを導入するよう設計されたものである、ここで、そのナンバリング(−20など)がHSA/MFα−1融合リーダー配列のC末端アミノ酸を−1位の残基とするものである。
【0140】
突然変異を誘発させたプラスミドのDNA配列をジデオキシヌクレオチド配列決定法により確認し、ポリヌクレオチド配列が突然変異誘発を受けて目的の配列となったことおよび他のDNA配列の変化が導入されていないことをこの方法により確認した。この新規プラスミドをpAYE653(図9)と名づけた。配列番号24は、改変されたHSA/MFα−1融合リーダー配列のコード領域を含んでなるポリヌクレオチド配列を示し、配列番号25は、pAYE653のポリヌクレオチド配列内に見られる改変されたHSA/MFα−1融合リーダー配列のコード領域と成熟ヒトアルブミンコード領域を含んでなるポリヌクレオチド配列を示すものである。
【0141】
pAYE653からNotIヒトアルブミン発現カセットを単離し、プラスミドpSAC35の独自のNotI部位に連結して、プラスミドpAYE655(図10)を作製した。
【0142】
実施例2
配列番号19は、サッカロミセス・セレビシエPRBIプロモーター由来の5’UTRを含む非コード領域;本発明の改変されたHSA/MFα−1融合リーダー配列をコードするポリヌクレオチド領域;成熟ヒトアルブミンのコドン最適化コード領域および翻訳終結部位を含んでなるDNA配列を示すものである。
【0143】
配列番号19のリーダー配列にもたらされる配列の改変の効果を比較するための対照として、配列番号40が、配列番号40のDNA配列が、本発明の第2の態様を示す15ポリヌクレオチド領域の代わりに、改変されていないHSA/MFα−1融合リーダー配列、すなわち、SFISLの5個のアミノ酸をコードする15ポリヌクレオチド領域を含んでなることを除いて配列番号19と本質的に同じであるDNA配列を示すものである。
【0144】
いずれのDNA配列もオーバーラッピング一本鎖オリゴヌクレオチドからGenosys, Inc (Cambridge, UK)により合成した。
【0145】
プラスミドpAYE639(図11)では、配列番号40を1.865kb SacI−HindIII DNA断片として合成し、プラスミドpBSSK−(Stratagene Europe, P. O. Box 12085, Amsterdam, The Netherlands)のSacI−HindIII部位にクローニングした。
【0146】
2種類の一本鎖オリゴヌクレオチドPRBJM1およびPRBJM3(下記)を用いたPCRによりサッカロミセス・セレビシエPRBIプロモーターを酵母ゲノムDNAから単離した。
PRBJM3:
【化33】
配列番号39
【0147】
PCR条件 94℃で30秒間、50℃で40秒間、72℃で120秒間、次いで、72℃で600秒間、その後、4℃で維持を40サイクル。0.81kb DNA断片をNotIとHindIIIの両方で消化し、WO 97/24445で記載されている、NotIとHindIIIで同様に消化したpBST+に連結して、プラスミドpAYE439(図12)を作出した。プラスミドpAYE439をHindIIIとBa7nHIで消化し、WO00/44772で従前に開示されているpAYE440由来の0.48kb HindIII/BamHI ADHIターミネーターDNA断片と連結して、プラスミドpAYE466(図13)を作出した。
【0148】
配列番号40の1.865kb HindIII DNA断片をプラスミドpAYE466の独自のHindIII部位にクローニングして、プラスミドpAYE640を作出した。このプラスミドが配列番号40の1.865kb HindIII DNA断片をPRB1プロモーターとADH1ターミネーターとの間に、PRB1プロモーターからの発現に対して適正な方向で含むことが分かった(図14)。
【0149】
プラスミドpAYE640をNotI/PvuIで消化完了し、NotI 3.2kb、PRBIプロモーター/配列番号40のHindIII DNA断片遺伝子/ADHIターミネーター発現カセットを精製した。発現カセット(3.2kb)とpBST+プラスミドバックボーン(3.15kb)は大きさが同じであるため、pAYE640のNotI/PvuI二重消化がNotI消化だけの場合よりも望ましい。pAYE640由来の3.2kb NotI、発現カセットを、仔ウシ小腸ホスファターゼ(CIP)で予め処理しておいたNotI線状化pSAC35(Sleep et al. (1991), Bio/Technology 9: 183-187)に連結して、プラスミド pAYE638(図15)を作出した。プラスミドpAYE638が、pSAC35のNotI部位に挿入され、HSA遺伝子の発現がLEU2栄養要求性マーカーから離れ、2μm複製起点に向かうように方向付けられたNotI HSA発現カセットを含むことが分かった。プラスミドpAYE642は同じHSA発現カセットを含むが、正反対の方向に配列されている(図15)。
【0150】
プラスミドpAYE643(図16)では、配列番号19を1.865kb SacI−HindIII DNA断片として合成し、pBSSK−(Stratagene Europe, P. O. Box 12085, Amsterdam, The Netherlands)にクローニングした。pAYE643内のHSA/MFα−1融合リーダー配列−アルブミン融合物をコードするDNA配列を配列番号27で示す。pAYE643から配列番号19の1.865kb HindIII断片を単離し、pAYE466の独自のHindIII部位に連結して、プラスミドpAYE645(図17)を作出した。NotI/PvuIでの消化によりpAYE645からNotI PRBI rHA発現カセットを単離し、pSAC35の独自のNotI部位に連結して、プラスミドpAYE646(図18)およびpAYE647(図19)を作製した。プラスミドpAYE646内のNotI 発現カセットはプラスミドpAYE638およびpAYE443と同じ方向に配向したが、プラスミドpAYE647内のNotI発現カセットは逆方向に配向し、プラスミドpAYE642と同じにした。
【0151】
実施例3
3種の異なる酵母株、A、BおよびCをプラスミドpAYE443、pAYE638、pAYE646およびpAYE655を用いてロイシンプロトトロフィーに形質転換した。これらの形質転換体を2%(w/v)グルコースを含有する緩衝最小培地(BMM、Kerry-Williams, S. M. et al. (1998) Yeast 14, 161-169により記載されている)(BMMD)にパッチアウトし、さらなる解析のために十分に増殖するまで30℃にてインキュベートした。2%(W/V)グルコースを含有する10mL YEP(1%(w/v)酵母抽出物;2%(W/V)バクトペプトン)(YEPD)およびBMMD振盪フラスコ培養(30℃、200rpm、72時間)から細胞を含まない培養上清のロケット免疫電気泳動により形質転換体のヒトアルブミン生産性を解析した(図20)。
【0152】
これらの結果から、pAYE638で形質転換された3種類全ての株のヒトアルブミン生産性が富栄養培地および規定培地の両方においてpAYE443で形質転換された同じ株で観察された生産性よりも約4〜5倍低いことが分かった(それらのいずれもがHSA/MFα−1融合リーダー配列を含むが、それらは異なるポリヌクレオチド配列によってコードされている)。予想外にも、pAYE646またはpAYE655で形質転換された3種類全ての株のヒトアルブミン生産性はpAYE638で観察された生産性よりもかなり高く、pAYE443で形質転換された同じ株で観察された生産性と同等か、またはそれよりも少し高かった。
【0153】
実施例4
酵母株C[pAYE443]、株C[pAYE655]、株C[pAYE638]および株C[pAYE646]、ならびに株B[pAYE443]および株B[pAYE646]をフェドバッチおよびフィル&ドロー両手順による高細胞密度発酵にて培養した。フェドバッチ手順では、WO96/37515に記載されている培地および制御パラメーターを使用した。フィル&ドロー手順では、以上で記述したフェドバッチ手順を使用するが、以下の工程もさらに含めた:フェドバッチ培養手順供給期の終了後、培養物量の90%を発酵容器から取り出し、供給物の添加を開始する前に、残りの10%量の培養物に、WO 96/37515に記載されている培地および制御パラメーターを使用してバッチ培地を添加した(pHを制御しながら)。ヒトアルブミン生産性(YP/S)およびヒトアルブミン濃度(g/L)を細胞を含まない全培養物のSDS−PAGEのスキャンニングデンシトメトリーにより評価した。重量測定によりバイオマス収率(YX/S)も計算した。これらの結果により(図21)、実施例3ですでに分かったように、HSA/MFα−1融合リーダー配列と成熟ヒトアルブミの両方のアミノ酸配列が同一であっても、ヒトアルブミン発現プラスミド pAYE638(天然のポリペプチド配列で酵母の偏りのあるコドン)を含有する酵母株のヒトアルブミン生産性(YP/S)およびヒトアルブミン濃度(g/L)が、ヒトアルブミン発現プラスミド pAYE443(天然のポリペプチド配列でリーダーおよび成熟アルブミンの自然コドンバイアス)を含有する同じ株よりもかなり低い生産性を有することが示された。
【0154】
株Cの発酵をフェドバッチ様式で行った場合、株C[pAYE655]および株C[pAYE646](本発明に従って改変されたリーダー配列を組み込んだヒトアルブミン発現プラスミド)をそれぞれ、同等の時間培養すると、ヒトアルブミン生産性(YP/S)の、株C[pAYE443]の生産性に比べて16%および12%の上昇が観察された。株Bの発酵をフェドバッチ様式で行った場合、株B[pAYE646](本発明に従って改変されたリーダー配列を組み込んだヒトアルブミン発現プラスミド)を同等の時間培養すると、ヒトアルブミン生産性(YP/S)の、株B[pAYE443]の生産性に比べて24%の上昇が観察された。
【0155】
株Cの発酵をフィルアンドドロー様式で行った場合、株C[pAYE655]および株C[pAYE646](本発明に従って改変されたリーダー配列を組み込んだヒトアルブミン発現プラスミド)をそれぞれ、同等の時間培養すると、ヒトアルブミン生産性(YP/S)の、株C[pAYE443]の生産性に比べて13%および6%の上昇が観察された。株C[pAYE655]および株C[pAYE646](本発明に従って改変されたリーダー配列を組み込んだヒトアルブミン発現プラスミド)をそれぞれ、同等の時間培養すると、この生産性は株C[pAYE638]の生産性に比べて442%および408%まで上昇した。
【0156】
概要
プラスミドpAYE443およびpAYE638はいずれもHSA/MFα−1融合リーダー配列由来のリーダー配列を有するヒトアルブミンをコードするが、前者は天然のポリヌクレオチド配列の自然のコドンバイアスを用い、一方、後者は酵母発現に向けて十分にコドン最適化されたポリヌクレオチド配列を用いる。pAYE638から得られたヒトアルブミンの発現は、pAYE443を用いて得られたものより4〜5倍低い。本発明に従って改変されたリーダー配列をコードするポリヌクレオチド配列を、pAYE443およびpAYE638の両方のHSA/MFα−1融合リーダー配列をコードするポリヌクレオチド配列の代わりに使用して、各々、ヒトアルブミン発現プラスミドpAYE665およびpAYE646を作出した。本発明のポリペプチド配列を導入することによって、目的ポリペプチドの産生におけるかなりの改善がもたらされた。
【0157】
【化34】
【図面の簡単な説明】
【0158】
【図1】天然のHSAリーダー(プレ領域およびプロ領域を有する)(上列)とWO90/01063で開示されているHSA/MFα−1融合リーダー配列(第2列)および本発明の好ましい改変リーダー配列(第3列)との比較。
【図2】標準的な遺伝子コード。
【図3】好ましいS.セレビシエコドンの改変リスト。
【図4】pAYE438のプラスミドマップ。
【図5】pAYE441のプラスミドマップ。
【図6】pAYE309のプラスミドマップ。
【図7】pAYE467のプラスミドマップ。
【図8】pAYE443のプラスミドマップ。
【図9】pAYE653のプラスミドマップ。
【図10】pAYE655のプラスミドマップ。
【図11】pAYE639のプラスミドマップ。
【図12】pAYE439のプラスミドマップ。
【図13】pAYE466のプラスミドマップ。
【図14】pAYE640のプラスミドマップ。
【図15】pAYE638およびpAYE642のプラスミドマップ。
【図16】pAYE643のプラスミドマップ。
【図17】pAYE645のプラスミドマップ。
【図18】pAYE646のプラスミドマップ。
【図19】pAYE647のプラスミドマップ。
【図20】ロケット免疫電気泳動によるrHA生産性の解析。酵母をYEP、2%(W/V)スクロースまたはB/MM、2%(W/V)スクロースにおいて30℃にて200rpm、72時間培養した。定量はHSA標品(mg.L−1)を参照することにより行った。
【図21】高細胞密度発酵におけるアルブミン生産性。*ヒトアルブミンレベルが低すぎて定量不可能であることを意味する。
【図22】実施例にて使用する構築物の特性の要約。
【発明の背景】
【0001】
発明の分野
本発明は、宿主細胞からタンパク質を分泌させるためのポリペプチドおよびポリヌクレオチド配列に関する。
【0002】
背景技術
宿主細胞から目的ペプチド、ポリペプチドおよびタンパク質(以下、本明細書では、これらの用語は同義的に用いる)を分泌させることを目的に、多くの天然または人工ポリペプチドシグナル配列(分泌プレ領域とも呼ばれる)が使用または開発されている。シグナル配列は、初期のタンパク質を、タンパク質をその細胞から周囲の媒質へ、または、場合によっては、ペリプラスム間隙へと輸送する細胞の機構に導くものである。シグナル配列は、通常、必ずではないが、一次翻訳産物のN末端に位置しており、一般に、必ずではないが、分泌プロセス中にその配列が目的タンパク質から切り離されて、「成熟」タンパク質が得られる。
【0003】
いくつかの目的タンパク質の場合では、シグナル配列の除去後、最初に分泌される存在物はそのN末端に「プロ」配列と呼ばれる付加的アミノ酸を含んでおり、その中間存在物は「プロ−タンパク質」と呼ばれている。これらのプロ配列は、最終的なタンパク質の折り畳みおよび機能化を手助けし、通常、その後に切り離される。他の例では、そのプロ領域はプレ−プロ領域を切り離す酵素の切断部位を容易に提供するもので、別の機能は有していないと思われている。
【0004】
プロ配列は、細胞からの目的タンパク質の分泌中、またはその細胞からの周囲の媒質またはペリプラスム間隙への輸送後のいずれかに除去される可能性がある。
【0005】
タンパク質の分泌を導くポリペプチド配列はまた、それらがシグナル(すなわち、プレ)配列またはプレ−プロ分泌配列と似ていようがいよまいが、リーダー配列と呼ばれることもある。タンパク質の分泌は翻訳、転座および翻訳後プロセシングを伴う動的プロセスであり、これらの工程の1以上は、別のものが開始する前、または完了する前に、必ずしも完了しなくてよい。
【0006】
酵母、サッカロミセス・セレビシエ(Saccharomyces cerevisiae)およびピキア・パストリス(Pichia pastoris)などの真核生物種においてタンパク質を産生させるための既知のリーダー配列としては、S.セレビシエ酸性ホスファターゼタンパク質(Pho5p)(EP366400を参照)、転化酵素タンパク質(Suc2p)(Smith et al. (1985) Science, 229, 1219-1224を参照)および熱ショックタンパク質−150(Hsp150p)(WO95/33833を参照)のものが挙げられる。さらに、S.セレビシエ接合因子α−1タンパク質(MFα−1)のリーダー配列ならびにヒトリゾチームおよびヒト血清アルブミン(HSA)タンパク質のリーダー配列が使用されているが、限定されるものではないが、ヒトアルブミンの分泌に向けては後者のものが特に利用されている。WO90/01063は、MFα−1リーダー配列を使用する場合に比べて混入するヒトアルブミン断片の産生が有利に減少する、MFα−1とHSAリーダー配列との融合物を開示している。
【発明の概要】
【0007】
本発明者らは、予想外にも、分泌されたタンパク質の収量がアミノ酸配列モチーフの導入により、好ましくは、リーダー配列の改変により、増大し得ることを見出した。この改変は、完全な天然アルブミンリーダー配列に行うか、その変異体に行うか、またはWO90/01063にて開示されているMFα−1とHSAリーダー配列との融合物などのヒトアルブミンリーダー配列の関連部分を用いた他のリーダー配列に行うかを問わず有効である。後者の場合、アルブミンが分泌されるタンパク質であるならば、このようにして産生されたアルブミンは混入する断片が減少するという有利な特徴を保有し、同時に収量も増大する。
【0008】
WO90/01063の融合されたリーダー配列の保存的改変は 、WO90/01063に概要が開示されている(例えば、WO90/01063の8頁を参照)が、ここではおよそ8×1012ポリペプチドの群が得られることが明らかになっている。遺伝子コードの縮退に従い、典型的なリーダー配列についてポリヌクレオチドコード配列が示された。これはまた、多数の可能性を示すものである。WO90/01063では、本発明により提供される、改変されたリーダー配列の特定群が分泌されるタンパク質の発現に有利な特性を有しているという理解はなされていない。
【0009】
本発明の第1の態様において、下記(i)および(ii)を含んでなるポリペプチドが提供される:
(i)下記(a)および(b)を含んでなるリーダー配列
(a)分泌プレ配列、および
(b)以下のモチーフ:
−X1−X2−X3−X4−X5−
(式中、
X1はフェニルアラニン、トリプトファンまたはチロシンであり、
X2はイソロイシン、ロイシン、バリン、アラニンまたはメチオニンであり、
X3はロイシン、バリン、アラニンまたはメチオニンであり、
X4はセリンまたはトレオニンであり、かつ、
X5はイソロイシン、バリン、アラニンまたはメチオニンである)、および
(ii)そのリーダー配列に対して異種の目的タンパク質。
【0010】
言い換えれば、そのポリペプチドは下記配列番号1の配列を含む。
【化1】
配列番号1
【0011】
本発明の第1の態様の好ましい態様では、X1はフェニルアラニンである。よって、好ましいポリペプチドは下記配列番号2の配列を含む。
【化2】
配列番号2
【0012】
本発明の第1の態様のもう1つの好ましい態様では、X2はイソロイシンである。よって、もう1つの好ましいポリペプチドは下記配列番号3の配列を含む。
【化3】
配列番号3
【0013】
本発明の第1の態様のもう1つの好ましい態様では、X3はバリンである。よって、もう1つの好ましいポリペプチドは下記配列番号4の配列を含む。
【化4】
配列番号4
【0014】
もう1つの好ましいポリペプチドでは、X4はセリンであり、よって、下記配列番号5の配列を含む。
【化5】
配列番号5
【0015】
本発明の第1の態様のもう1つの好ましい態様では、X4はトレオニンである。よって、もう1つの好ましいポリペプチドが配列番号29の配列を含む。
【化6】
配列番号29
【0016】
本発明の第1の態様のもう1つの好ましい態様では、X5はイソロイシンである。よって、もう1つの好ましいポリペプチドは配列番号6の配列を含む。
【化7】
配列番号6
【0017】
より好ましくは、X1、X2、X3、X4およびX5のうちの少なくとも2つ、なおいっそう好ましくは、少なくとも3つ、さらに好ましくは、少なくとも4つが上記の好ましい態様で定義されたとおりである。
【0018】
そのモチーフはリーダー配列に(すなわち、付加物として)挿入してもよいし、または1個、2個、3個、4個、5個以上の連続したアミノ酸の代用としてリーダー配列内に含めることもできる。
【0019】
1つの好ましい態様では、そのモチーフが天然アミノ酸の代用としてリーダー配列に含められている。言い換えれば、そのモチーフのアミノ酸が、本発明によるその最適化を行う前にリーダー配列内に存在していたか、または存在していたであろう5個の連続したアミノ酸の代わりに含められている。これに関連して使用する「天然(の)」(nathurally occurring)とは、本発明を天然のリーダー配列の最適化に限定するものではないことは読む人にも理解できるであろう。それどころか、本発明はその最適化が本明細書において例示されているHSA/MFα−1リーダー配列融合物などの人工リーダー配列の最適化にも適用できる。
【0020】
そのモチーフを代用物としてリーダー配列に含める場合にはX4が天然アミノ酸またはその変異体であることが好ましい。言い換えれば、X1、X2、X3およびX5だけを置換して、X4を変えないでおくか、または、その位置にある天然アミノ酸の、好ましくは、以下で定義する保存的置換として、変異体に単に変えることが好ましい。
【0021】
本発明の第1の態様の特に好ましい態様では、X1はフェニルアラニンであり、X2はイソロイシンであり、X3はバリンであり、X4はセリンであり、かつ、X5はイソロイシンである。よって、本発明の第1の態様の特に好ましい態様では、下記配列番号7の配列を含むポリペプチドが提供される。
【化8】
配列番号7
【0022】
上記スキームにおいて、「N」および「C」はポリペプチド配列の方向を示し、それらの実際の末端についてのそれらの解釈を限定するものではない。言い換えれば、ポリペプチド配列は、N末端もしくはC末端のいずれか一方にて、または最も一般的にはそれら両方の末端において、1以上の他のポリペプチド配列と結合される(例えば、融合されるか、コンジュゲートされるか、または連結される)。
【発明の具体的説明】
【0023】
本発明の第1の態様のポリペプチドは、リーダー配列に対して異種の成熟した目的タンパク質の配列を含んでなる。成熟した目的タンパク質の配列は、本発明のポリペプチドが発現される発現系による翻訳後プロセシング後の発現産物に存在するであろう一次アミノ酸配列である。目的タンパク質は、好ましくは、本発明のポリペプチドが発現される細胞から分泌させるのに好適であるものである。
【0024】
目的タンパク質はリーダー配列に対して異種である。言い換えれば、本発明の第1の態様のポリペプチドは、それらのリーダー配列内に上記のモチーフ−X1−X2−X3−X4−X5−を有する天然のタンパク質を含まないものである。好ましい態様では、本発明の第1の態様のポリペプチドは、任意の位置に上記のモチーフ−X1−X2−X3−X4−X5−を有する天然のタンパク質を含まないものである。これに関連して、「天然(の)」とは、組換えテクノロジー、部位特異的突然変異誘発またはヒトの介入を必要とする同等の人工的な技術による改変を受けていない天然に存在する生物によりコードされるタンパク質を指す。
【0025】
目的タンパク質は、天然タンパク質(チモーゲン(zymogen)を含む)、ポリペプチドもしくはペプチド、または天然タンパク質、ポリペプチドもしくはペプチドの変異体もしくは断片(例えば、一ドメインであってよい);あるいは完全合成タンパク質、ポリペプチドもしくはペプチド;あるいは異なるタンパク質、ポリペプチドもしくはペプチド(天然または合成)の単一もしくは多重融合物であれば、いずれの配列を含んでなってもよい。このようなタンパク質は、限定されるものではないが、それらの開示内容が引用することにより本明細書の一部とされる、WO01/79258、WO01/79271、WO01/79442、WO01/79443、WO01/79444およびWO01/79480にて記載されているリスト、またはその変異体もしくは断片から選択することができる。これらの特許出願はアルブミンの融合パートナーという観点からタンパク質のリストを示しているが、本発明はそのように限定されず、本発明の目的では、本明細書に列挙するいかなるタンパク質も単独でまたはアルブミン、免疫グロブリンのFc領域、トランスフェリンもしくは目的ポリペプチドとしての他のタンパク質の融合パートナーとして与えてよい。
【0026】
本発明による発現に向けた目的タンパク質の好ましい例としては、アルブミン、トランスフェリン、ラクトフェリン、エンドスタチン、アンギオスタチン、コラーゲン、免疫グロブリン、Fab’フラグメント、F(ab’)2、ScAb、ScFv、インターフェロン、IL10、IL11、IL2、インターフェロンα種および亜種、インターフェロンβ種および亜種、インターフェロンγ種および亜種、IL1−受容体アンタゴニスト、EPO、TPO、プロサプチド(prosaptide)、シアノビリン−N、5−ヘリックス、T20ペプチド、T1249ペプチド、HIV gp41、HIV gp120、フィブリノーゲン、ウロキナーゼ、プロウロキナーゼ、tPA(組織プラスミノーゲン活性化因子)、ヒルジン、血小板由来増殖因子、副甲状腺ホルモン、プロインスリン、インスリン、インスリン様増殖因子、カルシトニン、成長ホルモン、形質転換増殖因子β、腫瘍壊死因子、G−CSF、GM−CSF、M−CSF、限定されるものではないが、プラスミノーゲン、フィブリノーゲン、トロンビン、プレトロンビン、プロトロンビン、フォン・ウィルブランド因子、α1−アンチトリプシン、プラスミノーゲン活性化因子、第VII因子、第VIII因子、第IX因子、第X因子および第XIII因子をはじめとするプレ型および活性型いずれもの凝固因子、神経成長因子、LACI(組織因子経路阻害剤または外因経路阻害剤としても知られているリポタンパク質由来凝固阻害剤)、血小板由来内皮細胞増殖因子(PD−ECGF)、グルコースオキシダーゼ、血清コリンエステラーゼ、アプロチニン、アミロイド前駆体、インターαトリプシン阻害剤、アンチトロンビンIII、アポ−リポタンパク質種、プロテインC、プロテインS、上記のものの変異体または断片が挙げられる。
【0027】
目的タンパク質に関連して、「変異体」とは、1箇所以上の位置にアミノ酸の挿入、欠失、または保存的置換もしくは非保存的置換のいずれかの置換が存在するタンパク質を指すが、そのような変化の結果として、タンパク質の基礎特性、例えば、酵素活性または受容体結合(種類および比活性度)、熱安定性、特定のpH範囲における活性(pH安定性)が有意に変わっていないタンパク質が得られるという条件においてである。これに関連して「有意に」とは、当業者によって、その変異体の特性はまだ異なっているかもしれないが、最初のタンパク質の特性に関しては自明であると思われることを意味する。
【0028】
「保存的置換」とは、Val、Ile、Leu、Ala、Met;Asp、Glu;Asn、Gln;Ser、Thr、Gly、Ala;Lys、Arg、His;およびPhe、Tyr、Trpなどの組合せを意図するものである。好ましい保存的置換としては、Gly、Ala;Val、Ile、Leu;Asp、Glu;Asn、Gln;Ser、Thr;Lys、Arg;およびPhe、Tyrが挙げられる。
【0029】
「変異体」は、その変異体が誘導されるポリペプチドと通常、少なくとも25%、少なくとも50%、少なくとも60%または少なくとも70%、好ましくは、少なくとも80%、より好ましくは、少なくとも90%、なおいっそう好ましくは、少なくとも95%、さらに好ましくは、少なくとも99%、最も好ましくは、少なくとも99.5%の配列同一性を有する。
【0030】
2ポリペプチド間の配列同一性パーセントは、好適なコンピュータープログラム、例えば、ウィスコンシン大学遺伝子コンピューティンググループ(University of Wisconsin Genetic Computing Group)のGAPプログラムを用いて決定することができ、当然のことではあるが、同一性パーセントは、最適に配列をアラインメントしたポリペプチドと比較して計算される。
【0031】
アラインメントは、また、Clustal Wプログラムを用いて実施してもよい(Thompson et al., (1994) Nucleic Acids Res., 22 (22), 4673-80)。使用されるパラメーターは以下の通りである:
・ファストペアワイズアラインメントパラメーター:K−tuple(ワード)サイズ;1、ウインドウサイズ;5、ギャップペナルティー;3、トップダイアゴナル(top diagonals)数;5。スコアリング法:xパーセント。
・マルチプルアラインメントパラメーター:ギャップオープンペナルティー;10、ギャップエクステンションペナルティー;0.05。
・スコアリングマトリックス:BLOSUM。
【0032】
このような変異体は天然のものであってよいし、または当技術分野で周知のタンパク質工学手法および部位特異的突然変異誘発手法を用いて作製されたものであってよい。
【0033】
目的タンパク質に関連して、「断片」とは、1箇所以上の位置に欠失が存在するタンパク質を指す。よって、その断片が全長成熟ポリペプチドの完全配列の最大限5、10、20、30、40または50%を含んでなると思われる。通常、一断片は、全長の目的タンパク質の完全配列の最大60%、より通常は最大70%、好ましくは、最大80%、より好ましくは、最大90%、なおいっそう好ましくは、最大95%、さらに好ましくは、最大99%を含んでなる。目的タンパク質の特に好ましい断片は、目的タンパク質の1以上の完全ドメインを含んでなる。例えば、目的タンパク質がアルブミンであってよい。アルブミンは3つのドメインを有している。アルブミンの特に好ましい断片は、1つまたは2つのドメインを含み得るため、通常、アルブミンの完全配列の少なくとも33%または少なくとも66%を含んでなる。
【0034】
アルブミンおよびトランスフェリン、またはその変異体もしくは断片が、とりわけそれらがヒト起源のものである場合、すなわち、それらが自然に産生されたヒトタンパク質において見つけられるものと同じ配列を有している場合には、目的タンパク質として特に好ましい。
【0035】
「ヒトアルブミン」とは、本明細書において、ヒト血清アルブミンと区別できない材料、またはその変異体もしくは断片である材料を示すために用いる。「変異体」とは、挿入、欠失、および保存的置換または非保存的置換のいずれかの置換を含み、そのような変化によってアルブミンの膨張特性、有用なリガンド結合特性または免疫原特性が実質的に変わらないものである。例えば、EP−A−322 094で開示されているヒトアルブミンまたはヒトアルブミン類似体の自然発生する多形変異体が挙げられる。一般に、ヒトアルブミンの変異体または断片はヒト血清アルブミンのリガンド結合活性(例えば、ビリルビン結合)の少なくとも10%(好ましくは、少なくとも50%、80%、90%または95%)およびヒト血清アルブミンの膨張活性の少なくとも50%(好ましくは、少なくとも80%、90%または95%)、w/wを有している。アルブミン、アルブミン変異体またはアルブミン断片の、膠質浸透圧としても知られている膨張活性は、Hoefs, J. C. (1992) Hepatology 16: 396-403により記載されている方法により決定してもよい。ビリルビン結合は、527nmでの蛍光増感により、HSAと比較して測定してもよい。ビリルビン(1.0mg)を50μLの1M NaOHに溶かし、脱塩水で1.0mLに希釈する。このビリルビンストックを100mM Tris−HCl pH8.5、1mM EDTAで希釈して、蛍光光度計キュベット中0.6nmolビリルビンmL−1とする。HSA:ビリルビン比 0〜5mol:molの範囲にわたるHSAでの滴定中においての448nmでの励起および527nmでの発光(10nm スリット幅)により蛍光を測定する。
【0036】
好ましい態様では、目的タンパク質がトランスフェリンであってよい。このようなものとしては、トランスフェリンファミリーのメンバー(Testa, Proteins of iron metabolism, CRC Press, 2002; Harris & Aisen, Iron carriers and iron proteins, Vol. 5, Physical Bioinorganic Chemistry, VCH, 1991)およびそれらの誘導体、例えば、トランスフェリン、突然変異型トランスフェリン(Mason et al, 1993, Biochemistry, 32, 5472; Mason et al, 1998, Biochem. J., 330 (1), 35)、末端切断型トランスフェリン、トランスフェリンローブ(Mason et al, 1996, Protein Expr. Purif., 8, 119; Mason et al, 1991, Protein Expr. Purif., 2, 214)、ラクトフェリン、突然変異型ラクトフェリン、末端切断型ラクトフェリン、ラクトフェリンローブまたは上記のものと他のペプチド、ポリペプチドもしくはタンパク質との融合物(Shin et al, 1995, Proc. Natl. Acad. Sci. USA, 92, 2820; Ali et al, 1999, J. Biol. Chem., 274, 24066; Mason et al, 2002, Biochemistry, 41, 9448)が挙げられる。トランスフェリンがヒトトランスフェリンであってよい。
【0037】
「ヒトトランスフェリン」とは、本明細書において、ヒト由来のトランスフェリンと区別できない材料、またはその変異体もしくは断片である材料を示すために用いる。「変異体」とは、挿入、欠失、および保存的置換または非保存的置換のいずれかの置換を含み、そのような変化によってトランスフェリンの有用なリガンド結合特性または免疫原特性が実質的に変わらないものである。例えば、ヒトトランスフェリンまたはヒトトランスフェリン類似体の自然発生する多形変異体が挙げられる。一般に、ヒトトランスフェリンの変異体または断片はヒトトランスフェリンのリガンド結合活性(例えば、鉄結合)の少なくとも50%(好ましくは、少なくとも80%、90%または95%)、w/wを有している。トランスフェリンまたは試験サンプルの鉄結合活性は、分光光度法で、鉄遊離状態および鉄飽和状態におけるそのタンパク質の470nm:280nm吸光度比により決定することができる。特に断りのない限り、試薬は鉄を含まないものであるべきである。鉄は、0.1Mクエン酸塩、0.1M酢酸塩、10mM EDTA pH4.5に対する透析によりトランスフェリンまたは試験サンプルから除去することができる。タンパク質は、100mM HEPES、10mM NaHCO3 pH8.0中、約20mg/mLにて存在するはずである。280nmでの吸光度を分光光度法で正確に決定することができるように(0%鉄結合)、水で希釈したアポトランスフェリン(Calbiochem, CN Biosciences, Nottingham, UK)の470nm:280nm吸光度比を測定する。191mgニトロ三酢酸を2mL 1M NaOHに溶かして20mM鉄−ニトリロ三酢酸(FeNTA)溶液を調製した後、2mL 0.5M塩化第二鉄を添加する。脱イオン水で50mLに希釈する。新たに調製した20mM FeNTAを十分に過剰に添加して、アポトランスフェリンを鉄飽和させ(100%鉄結合)た後、470nm:280nmでの吸光度比を測定する前に、ホロトランスフェリン調製物を100mM HEPES、10mM NaHCO3 pH8.0に対して完全に透析して残留しているFeNTAを除去する。最初は鉄を含まないものであるべき試験サンプルを用いてこの手順を繰り返し、最終的な比を対照と比較する。
【0038】
さらに、上記のものの単一もしくは多重異種融合物;またはアルブミン、トランスフェリンもしくは免疫グロビンあるいはこれらのものの変異体もしくは断片との単一もしくは多重異種融合物を使用してもよい。このような融合物としては、WO01/79271により例示されている、アルブミンN末端融合物、アルブミンC末端融合物およびN末端およびC末端アルブミン同時融合物、ならびにトランスフェリンN末端融合物、トランスフェリンC末端融合物、およびN末端およびC末端トランスフェリン同時融合物が挙げられる。
【0039】
好ましい態様では、本発明の第1の態様のポリペプチドが以上で定義されたX1−X5ペンタペプチドモチーフの少なくとも一部を含む分泌プレ配列を含んでなる。言い換えれば、成熟した目的ポリペプチドの分泌を行うように作用するリーダー配列の領域が、X1−X5ペンタペプチドモチーフのアミノ酸のうちの1個、2個、3個、4個または5個のアミノ酸を含有する。分泌プレ配列領域がX1−X5ペンタペプチドモチーフのうちの5個より少ないアミノ酸を含有する場合、そのプレ配列に含まれるモチーフのそれらのアミノ酸は、それらがX1−X5ペンタペプチドモチーフの残りのアミノ酸の近傍に存在するように、プレ配列領域の一方の境界に位置している。
【0040】
より好ましい態様では、本発明の第1の態様のポリペプチドが、本発明の第1の態様により以上で定義されたモチーフを含む分泌プレ配列を含むことを特徴とするリーダー配列を含んでなる。そのリーダー配列は、通常、必ずではないが、一次翻訳産物のN末端に位置しており、一般に、必ずではないが、分泌プロセス中にタンパク質から切り離されて、成熟した「目的」タンパク質が得られる。
【0041】
分泌リーダー配列は、通常、必ずではないが、N末端のアミノ酸配列であり、そのアミノ酸配列が、そのアミノ酸配列が一部を形成するポリペプチドを、それが産生される宿主細胞から分泌させる。分泌は、リン脂質二重層、一般に、限定されるものではないが、真核生物の小胞体または原核生物の原形質膜を通じた細胞質のコンパートメントからのタンパク質の翻訳後転座の共翻訳により定義される。分泌されたタンパク質は、細胞の範囲内(一般に、限定されるものではないが、小胞体、ゴルジ体、液胞、リソソームまたはペリプラスム間隙内)にとどまるか、または細胞から培養培地へと分泌される。ある配列が、分泌プレ配列を含まない同等のポリペプチドに比べて、いっそう多くのポリペプチドを、それが産生される宿主細胞から分泌させる場合には、その配列は分泌リーダー配列としての役割を果たしている。一般的に言えば、リーダー配列を有するポリペプチドは分泌されるが、リーダー配列を有さないポリペプチドは分泌されない。しかしながら、本発明は、異なるリーダー配列の効率レベルは異なるという状況を意図している。よって、リーダー配列は、細胞により産生された成熟タンパク質の少なくとも10%、20%、30%もしくは40%または50%、一般に、少なくとも60%または70%、好ましくは、少なくとも80%、より好ましくは、少なくとも90%、なおいっそう好ましくは、少なくとも95%、さらに好ましくは、少なくとも98%、最も好ましくは、少なくとも99%を細胞から分泌させ得る。成熟ポリペプチドの細胞からの分泌は、例えば、宿主細胞に好適なDNA構築物を提供し、分泌された成熟タンパク質(例えば、ヒトアルブミン)の量を細胞内に産生されたいずれもの成熟タンパク質と比較して測定することにより決定することができる。
【0042】
好ましい分泌リーダー配列は、宿主細胞が酵母細胞(例えば、サッカロミセス・セレビシエまたはピキア・パストリス)である場合には、上記レベルの分泌を導く。酵母宿主細胞からの成熟ポリペプチドの分泌は、例えば、以下の実施例にて記載するものなどの方法により決定することができる。
【0043】
従って、分泌は、試験リーダー配列を含んでなるタンパク質の分泌レベルを対照リーダー配列を含んでなるタンパク質の分泌レベルと比較することにより測定することができる。所与の配列(「試験配列」)が一定の分泌レベルを達成し得るかどうかを判定するために、LEU2遺伝子とrHAをコードするポリヌクレオチドとともに、PRB1プロモーターおよび以下で記載するADH1ターミネーターなどの機能的酵母調節領域に作動可能なように連結された配列番号26で定義される改変リーダー配列を有する「スターター」プラスミド、一般に、EP 0 286 422で記載されているタイプの酵母分解ベクターを、試験配列をコードするポリヌクレオチド配列を、リーダー配列の相当する領域の代わりに含めるよう改変し、そうすることによって、試験プラスミドを提供するといった第1のプロトコールを用いてよい。第1の対照として、WO90/01063で記載されているリーダー配列をコードする改変されていない「スターター」プラスミドを使用する。サッカロミセス・セレビシエ株AH22cir°(Hinnen et al, 1978, Proc. Natl. Acad. Sci. USA, 75 (4), 1929-33; Mead et al, 1986, Mol. Gen. Genet., 205, 417)、復帰変異したHis4を試験宿主として使用する。AH22(leu2、his4、can1)のHIS4復帰突然変異体(すなわち、His+)は、十分なAH22細胞を0.002%(w/v)ロイシンを補給したBMMD寒天でコロニーが現れるまで培養することによって得ることができる。そのコロニーを試験し、0.002%(w/v)ロイシンを補給したBMMD寒天(プレート1)、0.002%(w/v)ロイシンを補給し、0.002%(w/v)ヒスチジンを補給したBMMD寒天(プレート2)、およびBMMD寒天(プレート3)に播種することによりそれらがLeu−およびHis+(すなわち、AH22 His+(leu2、can1))であることを確認する。AH22 His+(leu2、can1)単離物はプレート1およびプレート2では増殖するが、プレート3では増殖しない。試験宿主を試験および対照プラスミドを用いてロイシンプロトトロフィー(prototrophy)に形質転換する。これらの形質転換体を2%(w/v)グルコースを含有する緩衝最小培地(BMM、Kerry-Williams, S. M. et al. (1998) Yeast 14, 161-169により記載されている)(BMMD)にパッチアウト(patched out)し、さらなる解析に向けて十分に増殖するまで30℃にてインキュベートする。形質転換体をフィル&ドロー手順による高細胞密度発酵にて、WO96/37515にてフェドバッチ手順に関して記載されているある培地で、制御パラメーターを使用して培養する:フェドバッチ培養手順供給期の終了後、培養物量の90%を発酵容器から取り出す。供給物の添加を開始する前に、残りの10%量の培養物に、WO96/37515に記載されている培地および制御パラメーターを使用してバッチ培地を添加する(pHを制御しながら)。フィル&ドロー手順は無制限に繰り返してもよい。試験および対照プラスミドを含有する形質転換体のヒトアルブミン生産性(YP/S)を細胞を含まない全培養物のSDS−PAGEのスキャンニングデンシトメトリーにより評価する。YP/Sは発酵中に培養物に添加したスクロース1g当たりのヒトアルブミンタンパク質(mg)の比率を表す。
【0044】
試験および第1の対照形質転換体を比較できる発酵槽配置で比較できる時間培養するとき、本発明のリーダー配列は、上記の第1のプロトコールにより測定するYP/Sにより判定した場合、第1の対照により得られる分泌レベルよりも少なくとも10%、20%、30%、40%、50%、60%、70%、80%、90%、100%、110%、120%、130%、140%、150%、160%、170%、180%、190%、200%、210%、220%、230%、240%、250%、260%、270%、280%、290%、300%、310%、320%、330%、340%、350%、360%、370%、380%、390%、400%、410%、420%、430%、440%、450%、460%、470%、480%、490%または500%高い分泌レベルを獲得し得る。よって、本発明のリーダー配列が第1の対照よりも最大400%、450%、500%、550%、600%、650%、700%、750%、またはそれ以上に高い分泌レベルであることが証明し得る。本発明のリーダー配列が、第1の対照により得られる分泌レベルよりも少なくとも400%、例えば、408%、または少なくとも440%、例えば、442%高い分泌レベルを獲得することが特に好ましい。
【0045】
第1のプロトコールに代わる方法として、第2のプロトコールを用いて、所与の配列(「試験配列」)が一定の分泌レベルを達成し得るかどうかを判定してもよい。第2のプロトコールは第1のプロトコールと本質的に同じである。しかしながら、第2のプロトコールでは、プラスミドが、配列番号26の代わりに、配列番号22で定義されるポリヌクレオチド配列を有し、rHAをコードするポリヌクレオチドに連結されたWO90/01063で記載されているアミノ酸配列を有するリーダー配列もコードする(「第2のスターター」プラスミド)ことを除いて上記の通りの「スターター」プラスミドを利用する。試験プラスミドは第2のスタータープラスミドを、試験配列をコードするポリヌクレオチド配列を、第2のスタータープラスミドのリーダー配列の相当する領域の代わりに含めるよう改変することにより作製する。第2の対照として、改変されていない第2のスタータープラスミドを使用する。試験および第2の対照プラスミドを含む形質転換体を第1のプロトコールに関して以上で記述したように調製し、フェドバッチ手順による高細胞密度発酵にて、WO96/37515にて記載されているある培地で、制御パラメーターを使用して培養する。YP/Sは上記の通りに評価する。
【0046】
試験および第2の対照形質転換体を比較できる発酵槽配置で比較できる時間培養するとき、本発明のリーダー配列は、上記の第2のプロトコールにより測定するYP/Sにより判定した場合、第2の対照により得られる分泌レベルよりも少なくとも1%、2%、3%、4%、5%、6%、7%、8%、9%、10%、11%、12%、13%、14%、15%、16%,17%,18%、19%または20%高い分泌レベルを獲得し得る。よって、本発明のリーダー配列が第2の対照よりも最大20%、30%、40%、50%、60%、70%、80%、90%、100%またはそれ以上に高い分泌レベルであることが証明し得る。本発明のリーダー配列が、第2の対照により得られる分泌レベルよりも少なくとも5%、例えば、6%、または少なくとも10%、例えば、12%、13%、14%、15%または16%高い分泌レベルを獲得することが特に好ましい。
【0047】
1つの態様では、本発明のリーダー配列が第1のプロトコールに関して以上で記述した分泌レベルを獲得する。もう1つの態様では、本発明のリーダー配列が第2のプロトコールに関して以上で記述した分泌レベルを獲得する。特に好ましい態様では、本発明のリーダー配列が第1および第2のプロトコール両方に関して以上で記述した分泌レベルを獲得する。
【0048】
細胞バイオマス由来の可溶性タンパク質および培養物上清中に分泌されたタンパク質は:
1.ゲル透過高速液体クロマトグラフィー
2.SDS−PAGEのデンシトメトリー
3.ロケット免疫電気泳動
により分析することができる。
【0049】
分泌された目的タンパク質の量および細胞内の目的タンパク質の量を、目的タンパク質の標準曲線と比べて定量し、当業者ならば周知のようにバイオマスの量に標準化する。
【0050】
通常、リーダー配列が、その配列を付けるか、または付けることが意図される未熟型の成熟タンパク質から誘導される場合が好ましい。よって、例えば、成熟タンパク質がアルブミンである場合には、天然のアルブミン分泌プレ配列またはプロ配列あるいはプレ−プロ配列を含んでなる配列を使用することが好ましい。しかしながら、リーダー配列は、もう1つの方法として、成熟タンパク質のもの以外の起源から誘導されてもよい。
【0051】
よって、1つの好ましい態様では、本発明の第1の態様のポリペプチドのリーダー配列がアルブミン分泌プレ配列またはその変異体から誘導された分泌プレ配列を含んでなる。
【0052】
以上で用いたアルブミンプレ配列の「変異体」とは、以上のX1、X2、X3、X4またはX5により定義された位置以外の、1箇所以上の位置にアミノ酸の挿入、欠失、または保存的置換(上記の通り)もしくは非保存的置換のいずれかの置換が存在するアルブミンプレ配列を指すが、そのような変化によってもそのペプチドがプレ配列の機能を果たし得るという条件においてである。
【0053】
好ましくは、アルブミンプレ配列の「変異体」が、以上のX1〜X5と定義される残基以外に、天然のアルブミンプレ配列、最も好ましくは、図1のアルブミンプレ配列と少なくとも2個、少なくとも3個または少なくとも4個、好ましくは、少なくとも5個、より好ましくは、少なくとも6個、なおいっそう好ましくは、少なくとも7個、さらに好ましくは、少なくとも8個、最も好ましくは、少なくとも9個の同一アミノ酸を有する。
【0054】
なおいっそう好ましくは、分泌プレ配列がアルブミン分泌プレ配列から誘導される場合に、本発明の第1の態様のポリペプチドがそれぞれ−20、−19、−18、−17および−16番にある天然アミノ酸の代わりに、それらの位置にX1、X2、X3、X4およびX5を有する、ここで、そのナンバリングが天然のアルブミン分泌プロ配列のC末端アミノ酸を−1位の残基とするものであり、かつ、X1、X2、X3、X4およびX5が上記のようなアミノ酸である。
【0055】
例えば、上記のナンバリングをヒトアルブミン分泌プレ配列の配列に適用する(例えば、WO90/01063で開示されている通り)と、以下のものが得られる:
【化9】
【0056】
特に好ましい態様では、使用される分泌プレ−配列がヒトアルブミン分泌プレ配列の配列から誘導される。
【0057】
よって、例えば、X1−X5ペンタペプチドがそのN末端において、以下の配列番号8の配列:
【化10】
配列番号8
またはその保存的に置換されたその変異体、すなわち:
【化11】
配列番号33
のC末端と直接または間接的に融合され得る。
【0058】
さらに、またはあるいは、それがそのC末端において、以下の配列
【化12】
配列番号9
または保存的に置換されたその変異体、すなわち:
【化13】
配列番号10
もしくは
【化14】
配列番号11
もしくは
【化15】
配列番号30
の少なくとも1つのN末端と直接または間接的に融合され得る。
【0059】
配列番号9で示す配列は天然ヒトアルブミンプレ配列の最後の9個のアミノ酸を表している。配列番号11の場合、これがWO90/01063の2つの主要な融合リーダー配列のうちの一方の最後の6個のアミノ酸と融合され、配列番号30の場合、配列番号9が天然ヒトアルブミンプロ配列の最後の6個のアミノ酸と融合される。
【0060】
好ましくは、いずれの場合にも、X1はFであり、X2はIであり、X3はVであり、X4はSまたはTであり、かつ、X5はIである。
【0061】
好ましい態様では、ペンタペプチドがそのN末端において配列番号8の配列または保存的に置換されたその変異体のC末端と融合され、かつ、そのC末端において配列番号9の配列、保存的に置換されたその変異体、配列番号10、11もしくは30のN末端と融合され、その結果として、例えば、以下の配列:
【化16】
配列番号12
または
【化17】
配列番号13
または
【化18】
配列番号14
【化19】
配列番号31
(式中、X1〜X5は上記のとおりである)、
あるいは以上で定義された保存的に置換されたその変異体
のうちの1つを形成する。
【0062】
特に好ましい態様は分泌プレ配列として、配列番号28の配列:
【化20】
配列番号28
を有する、すなわち、プレ配列がヒト血清アルブミン分泌プレ配列から誘導されたものであり、X1、X2、X3、X4およびX5が、−20、−19、−18、−17および−16番に存在し、かつ、X1、X2、X3、X4およびX5が配列番号7により示されるとおりである。
【0063】
以上のことからわかるように、以上で定義された分泌プレ配列、例えば、配列番号12または28の配列を分泌プロ配列と組み合わせて、機能的プレ−プロ分泌配列を作製してもよい。好ましい態様では、プレ配列モチーフをそのC末端において分泌プロ配列モチーフのN末端アミノ酸とペプチド結合により融合し、その結果として、プレ−プロ配列モチーフを形成する。リーダー配列を付けるか、または付けることが意図される未熟型の成熟タンパク質から誘導されたプロ配列を使用することも好ましくある。関連リーダー由来の改変されていないプレ配列またはプロ配列、もしくはその一部と事実上結合するプロ配列を使用することもまた好ましくある。
【0064】
好ましくは、プロ配列がそのC末端において、二塩基性アミノ酸対、すなわち、各々がLysまたはArgである、で終結する。
【0065】
一般に、分泌プロ配列モチーフが、アルブミン分泌プロ配列または二塩基性アミノ酸対を含み、その他の位置には保存的置換しかを有さないようなその変異体、通常、ヒトアルブミン分泌プロ配列、すなわち、配列N−Arg−Gly−Val−Phe−Arg−Arg−Cを有するものまたはその変異体である。もう1つの好ましい態様では、プロ配列が酵母MFα−1分泌プロ配列、すなわち、N−Ser−Leu−Asp−Lys−Arg−Cまたはアルブミンプロ配列に関して定義されたその変異体の全てまたは一部の配列を含んでなる。
【0066】
WO90/01063に記載されているリーダーおよびヒトアルブミンリーダーの相当する部分と比べて、本発明のポリペプチドは少なくとも4個のアミノ酸、すなわち、Ser−20PheまたはTrpまたはTyr;Phe−19IleまたはLeuまたはValまたはAlaまたはMet;Ile−18LeuまたはValまたはAlaまたはMet;およびLeu−16IleまたはValまたはAlaまたはMetが変化している、この表記法では、一例として挙げると、最初に名前を挙げた変異は−20番(すなわち、リーダー配列を用いて分泌されるであろう成熟タンパク質のN末端に対してマイナス20)にあるセリン残基がフェニルアラニン残基に変化していることを意味している。これについては図1にて例示する。
【0067】
1つの好ましいプレ−プロ配列は、下記配列を含んでなる:
【化21】
(式中、Y1はGlyまたはSerであり、Y2はValまたはLeuであり、Y3はPheまたはAspであり、Y4はArgまたはLysであり、かつ、Y5はArgまたはLysである)。
【0068】
好ましい態様では、Y1はGlyであり、Y2はValであり、かつ、Y3はPheである。もう1つの好ましい態様では、Y1はSerであり、Y2はLeuであり、かつ、Y3はAspである。
【0069】
一般に、Y4はArgであり、かつ、Y5はArgである。あるいは、Y4がLysであり、かつ、Y5がArgである場合が好ましい。もう1つの好ましい別のものがY4がLysであり、かつ、Y5がLysである場合である。Y5がLysである場合にはY4はArgであってもよい。
【0070】
特に好ましい態様は分泌プレプロ配列として、下記配列番号32の配列を有する。
【化22】
配列番号32
【0071】
本発明の第2の態様は、本発明の第1の態様により定義されたモチーフをコードする配列を有する単離ポリヌクレオチドを提供する。
【0072】
本明細書において、「単離」とは、ポリヌクレオチドが、DNA分子の場合、それが自然に見つけられる染色体の少なくとも大部分から独立して存在していること、また、RNA分子の場合、それが自然に転写される無傷細胞から独立して存在しているという意味を含む。言い換えれば、ポリヌクレオチドが、それがそれまでに存在していた、例えば、自然界での形態にあるとは言えない。よって、本発明の第2の態様のポリヌクレオチドは、細菌もしくは真菌ベクター、例えば、プラスミド、またはウイルスベクター、例えば、バクテリオファージにクローニングされたポリヌクレオチドを含む。好ましくは、このようなクローンは、関連染色体のDNAライブラリーを構成しているクローンから独立して存在している。
【0073】
線形アミノ酸配列は、大部分のアミノ酸が2以上のトリヌクレオチドコドンによってコードされているという標準的な遺伝子コード(図2)の縮重を用いてDNA配列へと逆翻訳することができる。
【0074】
例えば、配列番号1と定義されたペプチドをコードするDNA配列が、下記配列であると推測される:
【化23】
配列番号15
(式中、「3’」および「5’」は実際の末端というよりもポリヌクレオチド配列の方向を示し;言い換えれば、ポリヌクレオチド配列はいずれかの末端において、または両方の末端において、他のポリヌクレオチド配列と結合され得(例えば、融合され得るか、または連結され得)、かつ、Y、R、HおよびNは図2で定義されたとおりである)。
【0075】
同じ変換手順を利用して、下記DNA配列が、配列番号7のポリペプチドをコードすると推測される。
【化24】
配列番号16
【0076】
ヒトアルブミンのような天然の成熟タンパク質をコードする配列を含んでなるポリヌクレオチド配列の場合では、これがヒトアルブミン遺伝子配列などの天然のコード配列、または相補DNA配列(cDNA)もしくは1以上のイントロンを含むcDNAのいずれかであり得る。
【0077】
さらなる配列の改変を、例えば、コード領域に導入してもよい。本発明のポリペプチドをコードするDNAを改変するための所望の方法は、Saiki et al (1988) Science 239, 487-491により開示されているポリメラーゼ鎖反応を利用することである。この方法では、酵素により増幅しようとするDNAの両側に2つの特定のオリゴヌクレオチドプライマーを置くが、それらのプライマー自体が増幅されるDNAに組み込まれることとなる。該特定のプライマーは制限エンドヌクレアーゼ認識部位を含み、これらの部位は当技術分野で公知の方法を利用した発現ベクターへのクローニングに使用することができる。
【0078】
本発明のリーダー配列をコードするポリヌクレオチドは、最も便宜には、当技術分野では周知であり、かつ、以下でより詳細に記述する方法により、オリゴヌクレオチドを化学合成し、続いて、遺伝子構築物の他のエレメントと連結することにより作製される。
【0079】
成熟アルブミンをコードするポリヌクレオチドを改変することが望まれる場合には、最も便宜には、天然のcDNA配列からの部位特異的突然変異誘発またはPCR突然変異誘発により、または合成オリゴヌクレオチドのアセンブリングからこれを行ってよい。さらに、このような技術は当技術分野では標準的なものであり、また、いずれにせよ以下でより詳細に記述する。
【0080】
コード配列に対する改変は、特定の生物において、高度に発現されるあるタンパク質をコードするポリヌクレオチド配列が特定のアミノ酸に対してあるコドンに偏って使用していること;これはコドンバイアスと呼ばれていることである、から有利である場合がある。本発明の第2の態様の好ましい態様では、標準的な遺伝子コードを最適な宿主生物に対する好ましいコドンに変えることができる。本発明の第2の態様の特に好ましい態様では、標準的な遺伝子コードを酵母の好ましいコドンに変えることができる。(Sharp and Crowe (1991) Yeast 7, 657-678の表4を参照)有利には、好ましい酵母コドンのこのリストはアスパラギンコドン5’−GAT−3’を含めることにより改変される(図3)。
【0081】
一例として、配列番号1のペプチド配列を用いて、酵母のこのペプチドをコードするコドンに偏りのあるDNA配列が下記のものであると推測され得る。
【化25】
配列番号17
【0082】
同じ変換手順を利用して、配列番号7の配列を有する特に好ましいポリペプチドモチーフについてのコドンに偏りのある縮重DNA配列:
【化26】
配列番号18
が推定されるが、配列番号7の配列を有するポリペプチドモチーフをコードする最も好ましいコドンに偏りのあるDNA配列は下記のものである。
【化27】
配列番号34
【0083】
図2で示す遺伝子コードまたは対象となる宿主に使用可能な好ましいコドンバイアス表もしくは図3で示す好ましいコドンバイアスを利用して、目的のアミノ酸配列を同じ変換手順により部分的に冗長なポリヌクレオチド配列に変換することができる。この方法によりDNA配列に変換するアミノ酸配列は、限定されるものではないが、本発明の第1の態様のポリペプチドから選択してよい。例えば、成熟ヒトアルブミンのコード領域の配列はこの方法により誘導することができる。EP308381はヒトアルブミンの酵母コドン最適化部分的コード配列を開示している。本明細書における配列番号20はそういった配列のさらなるものである。有利には、DNA配列の冗長性が許せば、コードされたアミノ酸配列(または図3を利用する場合にはコドンバイアス)を乱すことなく、ドメインおよびサブドメインの境界に制限部位を導入することができる。
【0084】
残るDNA配列の冗長性を解明し、別のコドンの発生頻度を冗長なDNA配列を有する各アミノ酸に対して均等化することができる。縮重コドンのDNA配列の冗長性を利用することにより可能であれば、可能性ある転写終結配列を示すDNA配列を除くか、または縮小することが有利である。最後に、冗長なDNA配列を有するアミノ酸の別のコドンのバランスを再度均等化することができるが、これまでの改変と矛盾することはない。
【0085】
本発明の第2の態様のポリヌクレオチドは、その5’および/または3’末端において、1以上の他のヌクレオチド配列と直接または間接的に融合し、その結果として、例えば、完全な遺伝子または発現カセットを形成することができる。よって、発現カセットは転写開始および終結部位、ならびに、転写された領域内に、翻訳開始のためのリボソーム結合部位も含むことが望ましい(1998年4月23日に公開されたHastings et al、WO98/16643)。
【0086】
従って、本発明の第2の態様は、異種タンパク質をコードするDNAと、本発明の第1の態様のポリペプチド、特に、目的タンパク質がアルブミンである場合、をコードするDNA配列またはその変異体もしくは断片のいずれかとの連続したまたは不連続な融合物であるDNA配列を含んでなるポリヌクレオチドを含む。これに関連して、「異種タンパク質」とは、それが「目的タンパク質」と同じではないこと、すなわち、ホモ二量体を形成しないことを意味する。
【0087】
従って、ポリヌクレオチドはその5’末端においてプロモーター(RNAポリメラーゼの結合および転写を起こさせるDNA配列により形成された発現制御エレメント)と、かつ/またはその3’末端において他の調節配列、例えば、翻訳終結配列と直接または間接的に融合されていてもよい。よって、ポリヌクレオチドが1以上の調節領域、通常、転写調節領域に作動可能なように連結されていてもよい。「作動可能なように連結された」とは、調節領域が、それがポリヌクレオチド配列に対して作用を及ぼし得るように連結されていることを意味する。どの調節領域を使用するかの選択は、想定される宿主(すなわち、意図される発現系)にある程度依存するが、好ましい配列の選択は当業者には知られている。
【0088】
数多くの発現系が知られているが、そのような系としては、例えば、組換えバクテリオファージ、プラスミドまたはコスミドDNA発現ベクターで形質転換された細菌(例えば、枯草菌(Bacillus subtilis)または大腸菌(Escherichia coli));例えば、酵母発現ベクターで形質転換された酵母(例えば、サッカロミセス・セレビシエまたはピキア・パストリス);例えば、ウイルス発現ベクター(例えば、バキュロウイルス)で形質転換された昆虫細胞系;例えば、ウイルスまたは細菌発現ベクターで形質転換された植物細胞系;例えば、アデノウイルス発現ベクターで形質転換された、動物細胞系の細胞培養物、動物細胞系トランスジェニック細胞、または遺伝子療法としての動物細胞系のいずれか、を使用する系が挙げられる。宿主細胞は、好ましくは、酵母(最も好ましくは、サッカロミセス属(Saccharomyces)種、例えば、S.セレビシエまたはピキア属(Pichia)種、例えば、P.パストリス)である。
【0089】
従って、本発明の第3の態様は、本発明の第2の態様のポリヌクレオチドで形質転換された宿主細胞を提供する。その宿主細胞は、原核生物細胞でも真核生物細胞でもよい。細菌細胞が、特に、バチルス属(Bacillus)およびエシェリキア属(Escherichia)のいくつかの種が可能なようにそれらがタンパク質を分泌し得るならば好ましい原核生物の宿主細胞である。好ましい真核生物宿主細胞としては、植物、真菌、酵母および動物細胞、好ましくは、脊椎動物細胞、より好ましくは、哺乳類細胞、例えば、マウス、ラット、ウシ、ヒツジ、ヤギ、ブタ、バッファロー、ヤク、ウマもしくは他の家畜、サルまたはヒト由来のものが挙げられる。好適なヒト細胞としては、ヒト繊維芽細胞細胞系の細胞が挙げられる。よって、宿主細胞がin situの哺乳類のトランスジェニック細胞であってよく、それゆえ、遺伝子療法アプローチまたはトランスジェニック個体の作製による産物であり得る。後者の場合では、個体が非ヒト哺乳類であることが好ましい。
【0090】
細菌宿主の典型的な種類としては、大腸菌および枯草菌が挙げられる。
【0091】
植物宿主の典型的な種類としては、種子植物類、シダ植物類(例えば、シダ類、ヒカゲノカズラ類、トクサ類)、コケ植物類(例えば、ゼニゴケ類およびセン類)、および藻類が挙げられる。一般に、植物宿主細胞は多細胞植物、通常、裸子植物または被子植物などの種子植物から誘導されたものである。好適な裸子植物としては、針葉樹類(例えば、マツ、カラマツ、モミ、トウヒおよびヒマラヤスギ)、ソテツ類、イチイ類およびイチョウ類が挙げられる。より一般には、植物宿主細胞が単子葉植物または双子葉植物、好ましくは、作物であり得る被子植物の細胞である。好ましい単子葉植物としては、トウモロコシ、コムギ、オオムギ、サトウモロコシ、タマネギ、カラスムギ、カモガヤおよび他のイチゴツナギ亜科(Pooideae)が挙げられる。好ましい双子葉植物の作物としては、トマト、ジャガイモ、テンサイ、タピオカノキ、アブラナ科の作物(アブラナを含む)、アマニ、タバコ、ヒマワリ、繊維作物、例えば、綿、ならびに豆科植物、例えば、エンドウ、インゲンマメ、特に、ダイズ、およびアルファルファが挙げられる。よって、宿主細胞は自律的細胞(autonomous cell)、例えば、単細胞植物の細胞または細胞培養で維持される細胞であってよいし、または多細胞植物のin situ細胞であってもよい。従って、本発明は、好ましくは、安定した遺伝性のトランスジェニック表現型を保持する完全トランスジェニック植物の生産を意図する。
【0092】
真菌宿主の典型的な種類としては、アスペルギルス属(Aspergillus)(例えば、A.ニガー(A. niger)およびA.オリゼー(A. oryzae))、ストレプトミセス属(Streptomyces)、ペニシリウム属(Penicillium)および酵母が挙げられる。本発明の実施において有用であると考えられる酵母の典型的な種類としては、ピキア属(ハンセヌラ属(Hansenula))、サッカロミセス属、クルイベロミセス属(Kluyveromyces)、カンジダ属(Candida)、トルロプシス属(Torulopsis)、トルラスポラ属(Torulaspora)、シゾサッカロミセス属(Schizosaccharomyces)、サイテロミセス属(Citeromyces)、パキソレン属(Pachysolen)、デバロミセス属(Debaromyces)、メチニコウィア属(Metschunikowia)、ロドスポリジウム属(Rhodosporidium)、ロイコスポリジウム属(Leucosporidium)、ボトリオアスクス属(Botryoascus)、スポリジオボルス属(Sporidiobolus)、エンドミコプシス属(Endomycopsis)などがある。好ましい属は、ピキア属(ハンセヌラ属)、サッカロミセス属、クルイベロミセス属およびヤロウィア属(Yarrowia)からなる群から選択されるものである。サッカロミセス属種の例として、S.セレビシエ、S.イタリクス(S. italicus)およびS.ルキシー(S. rouxii)がある。クルイベロミセス属種の例として、K.フラギリス(K. fragilis)およびK.ラクティス(K. lactis)がある。ピキア属(ハンセヌラ属)種の例として、P.パストリス、P.アノマラ(P. anomala)およびP.カプスラータ(P. capsulata)がある。好適なヤロウィア属種の例としては、Y.リポリチカ(Y. lipolytica)がある。酵母宿主細胞としては、一般に、Stratagene Cloning Systems, La Jolla, CA 92037, USAから市販されているYPH499、YPH500およびYPH501が挙げられる。
【0093】
好ましい哺乳類宿主細胞としては、ATCCからCCL61として入手可能であるチャイニーズハムスター卵巣(CHO)細胞、ATCCからCRL1658として入手可能であるNIH Swissマウス胚細胞NIH/3T3、およびATCCからCRL1650として入手可能であるサル腎臓由来のCOS−1細胞が挙げられる。好ましい昆虫細胞としては、バキュロウイルス発現ベクターでトランスフェクトすることができるSf9細胞がある。
【0094】
上述したように、ポリヌクレオチド調節領域の選択は、意図される宿主の性質にある程度依存する。
【0095】
細菌宿主細胞における使用に好適なプロモーターとしては、大腸菌lacIおよびlacZプロモーター、T3およびT7プロモーター、gptプロモーター、ファージλPRおよびPLプロモーター、phoAプロモーターおよびtrpプロモーターが挙げられる。本発明のDNAセグメントの挿入に便宜な制限部位を含むプラスミドベクターには、通常、典型的な細菌宿主に適合するプロモーター配列が提供される。
【0096】
真核生物のプロモーターとしては、CMV前初期プロモーター、HSVチミジンキナーゼプロモーター、初期および後期SV40プロモーターならびにレトロウイルスLTRのプロモーターが挙げられる。他の好適なプロモーターは、当業者には公知である。
【0097】
S.セレビシエに好適なプロモーターとしては、PGK1遺伝子、GAL1またはGAL10遺伝子、CYC1、PHO5、TRP1、ADH1、ADH2、グリセルアルデヒド−3−リン酸デヒドロゲナーゼ、ヘキソキナーゼ、ピルビン酸デカルボキシラーゼ、ホスホフルクトキナーゼ、トリオースリン酸イソメラーゼ、ホスホグルコースイソメラーゼ、グルコキナーゼ、α接合因子フェロモン、a−接合因子フェロモンに関する遺伝子に関連するもの、PRB1プロモーター、GPD1プロモーター、ならびに、5’調節領域の部分と、その他のプロモーターの5’調節領域の部分とのハイブリッド、あるいは、上流活性化部位とのハイブリッドを含むハイブリッドプロモーター(例えば、EP−A−258 067のプロモーター)が挙げられる。
【0098】
もう1つの好適な宿主細胞であるシゾサッカロミセス・ポンベ(Schizosaccharoonyces pombe)における使用に便宜な調節プロモーターとしては、Maundrell (1990) J. Biol. Chem. 265, 10857-10864に記載されているnmt遺伝子に由来するチアミン抑制プロモーター、ならびにHoffman & Winston (1990) Genetics 124, 807-816に記載されているグルコース抑制fbp1遺伝子プロモーターある。がある。
【0099】
ピキア属に好適なプロモーター、形質転換プロトコールおよび培養条件については、米国特許第5986062号(引用することにより本明細書の一部とされる)で見つけることができる。例えば、HSA産生宿主(またはHSA産生株)の作製は、組換えプラスミドを染色体に導入するという方法(EP−A−399455に対応するJP−A−3−72889)、HSAを酵母で発現させるという方法(EP−A−123544に対応するJP−A−60−41487、EP−A−248657に対応するJP−A−63−39576およびEP−A−251744に対応するJP−A−63−74493)、およびHSAをピキア属で発現させるという方法(EP−A−344459に対応するJP−A−2−104290)を利用して行い得る。HSA産生宿主の培養(HSA産生プロセス)は、米国特許第5,986,062号に記載されている方法などの既知の方法を利用して、例えば、JP−A−3−83595またはJP−A−4−293495(EP−A−504823に対応)に開示されている方法により実施することができる。形質転換された宿主を培養するための培地は、米国特許第5,986,062号に従って調製し得、宿主の培養は、米国特許第5,986,062号の開示内容に従って、静置もしくは振盪培養または攪拌および通気下でのバッチ式、半バッチ式もしくは連続培養により、好ましくは、15〜43℃(より好ましくは、20〜30℃)にて1〜1,000時間実施することができる。
【0100】
好適な転写終結シグナルは当技術分野では周知である。宿主細胞が真核生物である場合、転写終結シグナルは、好ましくは、適正な転写終結シグナルおよびポリアデニル化シグナルを含む真核生物の遺伝子の3’フランキング配列由来のものである。好適な3’フランキング配列が、例えば、使用される発現制御配列に自然に連結された遺伝子のものであってよい、すなわち、それらがプロモーターと一致したものであってもよい。あるいは、それらが異なっていてもよい。その場合には、宿主が酵母、好ましくは、S.セレビシエであれば、S.セレビシエADH1遺伝子の終結シグナルが好ましい。
【0101】
よって、本発明の第2の態様のポリヌクレオチドは、以上で記述したような方法を利用することによりいかなる所望の宿主に対しても開発することができる。
【0102】
成熟ヒトアルブミンをコードするDNA配列は、以上で記述した、天然の遺伝子、cDNAまたは1以上のイントロンを含むcDNAと、以上で記述した方法により誘導したコドンに偏りのあるヒトアルブミンDNA配列とのDNA融合物から開発することができる。
【0103】
配列番号19は、翻訳開始部位の5’に22ヌクレオチド、分泌リーダー配列 配列番号32の好ましいポリヌクレオチドコード配列、および成熟ヒトアルブミンコード領域 配列番号20を含んでなるポリヌクレオチド配列である。コード配列は翻訳終止コドンで終わる。このコドンは、一般には、TGA、TAGまたはTAAであるが、酵母ではTAAが最も有効である。好ましくは、さらなる翻訳終止コドン(好ましくは、各々TAAである)、通常1つまたは2つが、好ましくは、互いに隣接してまたは終止コドンの各対間3塩基対以下で含まれる。配列番号19は両末端に好適なクローニング部位がフランキングする。
【0104】
本発明の第2の態様のポリヌクレオチドはまた、好適な宿主への導入に向けて多くの種類の他のDNA配列と結合し得る。コンパニオン配列は、宿主の性質、ポリヌクレオチドの宿主への導入方法、およびエピソーム維持または組込みが所望か否かに依存する。例えば、ベクターを他の非原核生物細胞型での発現に使用する場合でも、ベクターが原核生物における増殖のためのCol E1 oriなどの原核生物レプリコンを含むことがある。
【0105】
一般に、本発明の第2の態様のポリヌクレオチドは、プラスミドなどの発現ベクターに、適正な方向および正確な発現用リーディングフレーム内に挿入される。
【0106】
よって、ポリヌクレオチドを本明細書に記載の教示に照らして好適に改変された既知の技術に従って使用して、限定されるものではないが、組込みベクター、動原体ベクターおよびエピソームベクターをはじめとする発現ベクターを構築し得る。
【0107】
よって、本発明の第2の態様の1つの態様では、ポリヌクレオチドがベクターである。
【0108】
一般的な原核生物ベクタープラスミドとしては、Biorad Laboratories(Richmond, CA, USA)から市販されているpUC18、pUC19、pBR322およびpBR329;Pharmacia(Piscataway, NJ, USA)から市販されているpTrc99A、pKK223−3、pKK233−3、pDR540およびpRIT5;Stratagene Cloning Systems(La Jolla, CA 92037, USA)から市販されているpBSベクター、Phagescriptベクター、Bluescriptベクター、pNH8A、pNH16A、pNH18A、pNH46Aがある。
【0109】
一般的な哺乳類細胞ベクタープラスミドとしては、Pharmacia(Piscataway, NJ, USA)から市販されているpSVLがある。このベクターはSV40後期プロモーターを使用してクローニングされた遺伝子の発現を推進するが、COS−1細胞などのT抗原産生細胞において最高レベルの発現が認められる。誘導性哺乳類発現ベクターの例としては、Pharmacia(Piscataway, NJ, USA)からも市販されているpMSGがある。このベクターはマウス乳癌ウイルスの長い末端反復配列のグルココルチコイド誘導性プロモーターを使用してクローニングされた遺伝子の発現を推進する。
【0110】
有用な酵母エピソームプラスミドベクターとしては、pRS403−406およびpRS413−416、これらは、一般に、Stratagene Cloning Systems(La Jolla, CA 92037, USA)から市販されている、YEp24(Botstein, D., et al. (1979) Gene 8, 17-24)、ならびにYEplac122、YEplac195およびYEplac181(Gietz, R. D. and Sugino. A. (1988) Gene 74, 527-534)がある。他の酵母プラスミドは、EP−A−286 424の「分解ベクター」に加えてさらに、WO90/01063およびEP424117で記載されている。プラスミドpRS403、pRS404、pRS405およびpRS406は、YIplac204、YIplac211およびYIplac128(Gietz, R. D. and Sugino. A. (1988) Gene 74, 527-534)がそうであるように、酵母組込み型プラスミド(YIp)であり、酵母選択マーカーHIS3、TRP1、LEU2およびURA3を組み込んでいる。プラスミドpRS413−416は、YCplac22、YCplac33およびYCplac111(Gietz, R. D. and Sugino. A. (1988) Gene 74, 527-534)がそうであるように、酵母動原体プラスミド(YCp)である。
【0111】
当業者には周知の方法を利用して、コード配列と、例えば、好適な転写または翻訳制御物を含む発現ベクターを構築することができる。このような1つの方法では、突出末端を介した連結を必要とする。好適な制限酵素の作用によりDNA断片およびベクターに適合する突出末端を作成することができる。これらの末端を相補塩基対合により迅速にアニーリングし、残るニックをDNAリガーゼの作用により閉じることができる。
【0112】
さらなる方法では、合成二本鎖オリゴヌクレオチドリンカーとアダプターを使用する。平滑末端を有するDNA断片を、凸部3’末端を除去し、凹部3’末端を埋めるバクテリオファージ T4 DNAポリメラーゼまたは大腸菌DNAポリメラーゼIにより作製する。合成リンカーおよび特定の制限酵素の認識配列を含む平滑末端を有する二本鎖DNAの断片をT4 DNAリガーゼにより平滑末端を有するDNA断片に連結することができる。続いて、それらを好適な制限酵素で消化して突出末端を作り出し、適合する末端を有する発現ベクターに連結する。また、アダプターが連結に用いる1つの平滑末端を含み、また、1つの成形した突出末端も保有する化学合成されたDNA断片でもある。あるいは、1DNA断片または複数のDNA断片を、所望により、突出末端を有していてもよい1以上の合成二本鎖オリゴヌクレオチドの存在または不在下で、DNAリガーゼの作用により一緒に連結してもよい。
【0113】
種々の制限エンドヌクレアーゼ部位を含む合成リンカーが、Sigma-Genosys Ltd, London Road, Pampisford, Cambridge, United Kingdomをはじめとする多数の供給源から市販されている。
【0114】
このようにして作製される本発明のベクターを、本発明の第1の態様で定義されたとおりの配列を含んでなるポリペプチドの発現および産生に向けて好適な宿主細胞を形質転換するために使用してもよい。このような技術としては、その全てが引用することにより本明細書の一部とされる、1984年4月3日に発行されたRutter et alの米国特許第4,440,859号、1985年7月23日に発行されたWeissmanの同第4,530,901号、1986年4月15日に発行されたCrowlの同第4,582,800号、1987年6月30日に発行されたMark et alの同第4,677,063号、1987年7月7日に発行されたGoeddelの同第4,678,751号、1987年11月3日に発行されたItakura et alの同第4,704,362号、1987年12月1日に発行されたMurrayの同第4,710,463号、1988年7月12日に発行されたToole, Jr. et alの同第4,757,006号、1988年8月23日に発行されたGoeddel et alの同第4,766,075号、および1989年3月7日に発行されたStalkerの同第4,810,648号で開示されているものが挙げられる。
【0115】
本発明のDNA構築物での好適な細胞宿主の形質転換は、周知の方法により実施されるが、一般に、使用するベクターのタイプよって異なる。原核生物宿主細胞の形質転換に関しては、例えば、Cohen et al (1972) Proc. Natl. Acad. Sci. USA 69, 2110およびSambrook et al (2001) Molecular Cloning, A Laboratory Manual, 3'd Ed. Cold Spring Harbor Laboratory, Cold Spring Harbor, NYを参照のこと。酵母細胞の形質転換については、Sherman et al (1986) Methods In Yeast Genetics, A Laboratory Manual, Cold Spring Harbor, NYで記載されている。Beggs (1978) Nature 275, 104-109の方法もまた有用である。S.セレビシエを形質転換する方法については、その全てが引用することにより本明細書の一部とされる、EP251744、EP258067およびWO90/01063で一般的に教示されている。脊椎動物細胞に関し、このような細胞をトランスフェクトするのに有用な試薬、例えば、リン酸カルシウムおよびDEAE−デキストランまたはリポソーム製剤は、Stratagene Cloning Systems、またはLife Technologies Inc., Gaithersburg, MD 20877, USAから市販されている。
【0116】
エレクトロポレーションもまた、細胞の形質転換に有用であり、酵母細胞、細菌細胞および脊椎動物細胞を形質転換する技術分野では周知のことである。エレクトロポレーションにより酵母を形質転換する方法については、Becker & Guarente (1990) Methods Enzymol. 194, 182で開示されている。
【0117】
DNAを動物および植物細胞に導入するために物質的な方法を使用してもよい。例えば、マイクロインジェクションでは、極細のピペットを使用して、DNA分子を形質転換しようとする細胞の核に直接注入する。もう1つの例では、高速マイクロプロジェクタイル、通常、DNAでコーティングした金またはタングステンの粒子の細胞への撃ち込みを使用する。
【0118】
植物は多くの当技術分野において承認されている方法により形質転換し得る。当業者ならば、その選択する方法が形質転換の対象となる植物のタイプに依存することが分かるであろう。植物細胞を形質転換する好適な方法の例としては、マイクロインジェクション(Crossway et al., BioTechniques 4: 320-334 (1986))、エレクトロポレーション(Riggs et al., Proc. Natl. Acad. Sci. USA 83: 5602-5606 (1986)、アグロバクテリウム(Agrobacterium)媒介形質転換(Hinchee et al, Biotechnology 6: 915-921 (1988);トウモロコシの形質転換についてはIshida et al., Nature Biotechnology 14: 745-750 (1996)も参照のこと)、直接遺伝子導入(Paszkowski et al., EMBO J. 3: 2717-2722 (1984);Hayashimoto et al., Plant Physiol. 93: 857-863 (1990)(イネ))、およびAgracetus, Inc., Madison, Wisconsin and Dupont, Inc., Wilmington, Delawareから市販されている装置を使用した弾道学的粒子加速(例えば、Sanford et al., 米国特許第4,945,050号;およびMcCabe et al., Biotechnology 6: 923-926 (1988)を参照)が挙げられる。また、Weissinger et al., Annual Rev. Genet. 22: 421-477 (1988);Sanford et al., Particulate Science and Technology 5: 27-37 91987)(タマネギ);Svab et al., Proc. Natl. Acad. Sci. USA 87: 8526-8530 (1990)(タバコ葉緑体);Christou et al., Plant Physiol. 87: 671-674 (1988)(ダイズ);McCabe et al., Bio/Technology 6: 923-926 (1988)(ダイズ);Klein et al., Proc. Natl. Acad. Sci. USA, 85: 4305-4309 (1988)(トウモロコシ);Klein et al., Bio/Technology 6: 559-563 (1988)(トウモロコシ);Klein et al., Plant Physiol. 91: 440-444 (1988)(トウモロコシ);Fromm et al., Bio/Technology 8: 833-839 (1990);およびGordon-Kamm et al., Plant Cell 2: 603-618 (1990)(トウモロコシ);Koziel et al., Biotechnology 11: 194-200 (1993)(トウモロコシ);Shimamoto et al., Nature 338: 274-277 (1989)(イネ);Christou et al., Biotechnology 9: 957-962 (1991)(イネ);Datta et al., Bio/Technology 8: 736-740 (1990)(イネ);欧州特許出願EP−A−332 581(カモガヤおよび他のイチゴツナギ亜科);Vasil et al., Biotechnology 11: 1553-1558 (1993)(コムギ);Weeks et al., Plant Physiol. 102: 1077-1084 (1993)(コムギ);Wan et al., Plant Physiol. 104: 37-48 (1994)(オオムギ);Jahne et al., Theor. Appl. Genet. 89: 525-533 (1994)(オオムギ);Umbeck et al., Bio/Technology 5: 263-266 (1987)(綿);Casas et al., Proc.Natl. Acad. Sci. USA 90: 11212-11216 (1993)(サトウモロコシ);Somers et al., Bio/Technology 10: 1589-1594 (1992)(カラスムギ);Torbert et al., Plant Cell Reports 14: 6
35-640 (1995)(カラスムギ);Weeks et al., Plant Physiol. 102. 1077-1084 (1993)(コムギ);Chang et al., WO94/13822(コムギ)ならびにNehra et al., The Plant Journal 5: 285-297 (1994)(コムギ)も参照のこと。アグロバクテリウム媒介形質転換は、一般的には、単子葉植物に対して効果がなく、この種の植物には以上で記述した他の方法が好ましい。
【0119】
一般に、ベクターが宿主の全てを形質転換するのではないため、形質転換する宿主細胞に対してベクターを選択する必要がある。1つの選択手法では、形質転換される細胞において選択可能な特徴をコードする、必要な制御エレメントを含むDNA配列マーカーの発現ベクターへの組込みを利用する。これらのマーカーとしては、ジヒドロ葉酸レダクターゼ、G418または真核細胞培養に関するネオマイシン耐性、ならびに大腸菌および他の細菌での培養に関するテトラサイクリン、カナマイシンまたはアンピシリン耐性遺伝子が挙げられる。あるいは、このような選択可能な特徴に関連する遺伝子が、所望の宿主細胞を同時に形質転換するために使用されるもう1つのベクターにあってもよい。
【0120】
マーカー遺伝子を使用して形質転換体を同定することができるが、どの細胞が組換えDNA分子を含んでいるのか、また、どれが自己連結したベクター分子を含んでいるのかが判定できることが望ましい。これは、DNA断片を挿入することによってその分子に存在する遺伝子のうちの1つの完全性を破壊するクローニングベクターを利用することによって行うことができる。よって、組換え体はその遺伝子の機能の喪失によって同定することができる。
【0121】
首尾よく形質転換された細胞を同定するもう1つの方法では、本発明のポリペプチドを産生させるための本発明の発現構築物の導入により得られた細胞を増殖させる。細胞を回収し、それらを溶解し、それらのDNA含量をDNAの存在について、Southern (1975) J. Mol. Biol. 98, 503またはBerent et al (1985) Biotech. 3, 208により記載されているような方法を利用して調べてもよい。あるいは、形質転換細胞の培養物上清中の成熟タンパク質の存在を抗体を用いて検出してもよい。
【0122】
組換えDNAの存在について直接アッセイすることに加えて、組換えDNAがタンパク質の発現を誘導し得る場合には周知の免疫学的方法により形質転換の成功を確認することができる。例えば、発現ベクターで首尾よく形質転換された細胞は、好適な抗原性を提示するタンパク質を産生する。形質転換されたと思われる細胞のサンプルを回収し、好適な抗体を用いてそのタンパク質についてアッセイする。
【0123】
よって、形質転換された宿主細胞自体に加えて、本発明はまた、これらの細胞の培養物、好ましくは、モノクローナル(クローン的に同質な)培養物、または栄養培地においてモノクローナル培養物から誘導された培養物も意図する。
【0124】
従って、本発明の第4の態様では、本発明の第3の態様の細胞と培養培地を含んでなる細胞培養物を提供する。一般に、培養培地は本発明の第1の態様のポリペプチドの発現系内での発現により、通常、プレおよび/またはプロ配列の除去などのさらなる翻訳プロセシングにより生じる成熟ポリペプチドを含有する。
【0125】
大腸菌などの原核生物宿主細胞、ならびに哺乳類細胞などの真核生物宿主細胞の培養方法は当技術分野では周知である。酵母の培養方法はEP330451およびEP361991で一般的に教示されている。
【0126】
本発明の組換えDNAにより形質転換された宿主細胞を本明細書に開示されている教示に照らして、十分な時間、当業者には公知の好適な条件下で培養することにより、本発明の第1の態様のポリペプチドの発現が可能になる。このようにして産生されたポリペプチドに、プレおよび/またはプロ配列が除去されるように宿主細胞によるプロセシングをさらに行ってもよい。従って、「成熟した」目的タンパク質が最初に翻訳されたタンパク質とは異なっている場合がある。
【0127】
よって、本発明はまた、第5の態様として、以上で定義された成熟した目的タンパク質を作製する方法を提供する。この方法は、本発明の第3の態様の細胞を培養培地で培養する工程(ここで、その細胞は、本発明の第1の態様で定義されたとおりのポリペプチドの発現の結果として、成熟した目的タンパク質を分泌し、その際、そのタンパク質はペリプラスム間隙、その培養培地またはそれらの両方に蓄積するが、好ましくは、その培養培地に蓄積する)を含んでなる。次いで、分泌された目的タンパク質を含有する培養培地を細胞培養物中の細胞から分離する。細胞壁と結合した分泌されたタンパク質は、一般に、浸透性補助(例えば、ソルビトール)条件下(選択的に分泌されたタンパク質を徐々に放出する)、細胞壁融解酵素を用いて解離させることができる。Elango et al., J. Biol. Chem. 257: 1398-1400 (1982)を参照のこと。この目的に好適な細胞壁融解酵素の例としては、lyticase、Zymolyase-60,000およびGlusulaseが挙げられ、それらの全てが市販されており、例えば、後の2つの場合では、Seikagaku KogyoまたはKirin Breweryから、また、各々Boehringer Mannheimから市販されている。
【0128】
好ましくは、培養培地の単離後、成熟した目的タンパク質をその培地から分離する。なおいっそう好ましくは、このようにして得られた成熟した目的タンパク質をさらに精製する。
【0129】
目的の成熟タンパク質は当技術分野で公知の多くの方法により培養培地から抽出し得る。例えば、組換えにより発現されるアルブミンの回収に関する精製技術は、WO92/04367、基質由来の色素の除去;EP464590、酵母由来の染料の除去;EP319067、アルカリ沈殿およびその後のアルブミンの親油相への適用;ならびに全精製方法を記述しているWO96/37515、米国特許第5728553号およびWO00/44772で開示されており、その全てが引用することにより本明細書の一部とされる。本発明の改変リーダー配列は成熟タンパク質それ自体に影響を及ぼさないため、アルブミン以外のタンパク質を、このようなタンパク質の精製に有用であることが分かっているいずれかの技術により培養培地から精製してもよい。
【0130】
このような周知の方法としては、硫酸アンモニウムまたはエタノール沈降、酸抽出、陰イオンまたは陽イオン交換クロマトグラフィー、ホスホセルロースクロマトグラフィー、疎水性相互作用クロマトグラフィー、アフィニティークロマトグラフィー、ヒドロキシルアパタイトクロマトグラフィーおよびレクチンクロマトグラフィーが挙げられる。最も好ましくは、高速液体クロマトグラフィー(「HPLC」)を精製に用いる。
【0131】
得られたタンパク質は、その知られた有用性のいずれかに向けて使用することができ、アルブミンの場合では、重度の火傷、ショック状態および失血を治療するための患者へのi.v.投与物、培養培地への補給物、ならびに他のタンパク質製剤の賦形剤としての使用を含む。
【0132】
本発明の方法により得られる治療上有用な目的タンパク質は、単独で投与することが可能であるが、そのタンパク質を1種以上の許容される担体または希釈剤とともに医薬製剤として提供することが好ましい。担体または希釈剤は、目的タンパク質と適合し、かつ、その受容者に対して有害ではないという意味において「許容される」ものでなければならない。一般に、担体または希釈剤は、無菌かつ発熱物質を含まない水または生理食塩水である。
【0133】
よって、本発明の第6の態様は、本発明の第5の態様の方法により得られる目的タンパク質を治療上許容される担体または希釈剤とともに製剤し、そうすることによって、ヒトまたは動物への投与に好適な治療用製品を生産する方法を提供する。
【0134】
治療用製品は、単位投与形で提供することが便宜であり、製薬分野では周知の方法のいずれによっても調製し得る。好ましい単位投与形製剤は、有効成分の、1日用量もしくは単位、1日用量以下の量またはその好適な分割量を含むものである。
【0135】
当然のことながら、治療用製品は、以上で特記した成分に加えて、問題の製品のタイプを考慮し、当技術分野で常用の他の薬剤を含んでいてもよい。
【0136】
以下、本発明を添付の限定されない図面および実施例を参照してより詳細に説明する。
【実施例】
【0137】
実施例1
下記の2種類の一本鎖オリゴヌクレオチドPRBJM1およびPRBJM2を用いたPCRにより、サッカロミセス・セレビシエ PRBIプロモーターを酵母ゲノムDNAから単離した。
PRBJM1:
【化28】
配列番号35
PRBJM2:
【化29】
配列番号36
【0138】
PCR条件 94℃で30秒間、50℃で40秒間、72℃で120秒間、次いで、72℃で600秒間、その後、4℃で維持を40サイクル。0.85kb DNA断片をNotIとHindIIIの両方で消化し、WO97/24445で記載されている、NotIとHindIIIで同様に消化したpBST+に連結して、プラスミドpAYE438(図4)を作出した。プラスミドpAYE438をHindIIIとBamHIで消化し、WO00/44772で従前に開示されているpAYE440由来の0.48kb HindIII/BamHI ADHIターミネーターDNA断片と連結して、プラスミドpAYE441(図5)を作出した。プラスミドpAYE441を独自のHindIII部位にて線状化し、従前に開示されている (Sleep, D. et al. (1991) Bio/Technology 9, 183-187およびEP−A−0431880)pAYE309(図6)由来の1.8kb HindIII/Bsu36I断片および二本鎖オリゴヌクレオチドリンカー:
【化30】
配列番号37
【化31】
配列番号38
と連結して、pAYE467(図7)を作出した。pAYE467由来の3.2kb NotI、発現カセットを、仔ウシ小腸ホスファターゼ(CIP)で予め処理しておいたNotI線状化pSAC35(Sleep et al. (1991), Bio/Technology 9: 183-187)に連結して、プラスミドpAYE443(図8)を作出した。配列番号22は、pAYE467およびpAYE443両方のDNA配列内に見られるHSA/MFα−1融合リーダー配列のコード領域と成熟ヒトアルブミンコード領域を含んでなるポリヌクレオチドを示すものである。HSA/MFα−1融合リーダー配列をコードするポリヌクレオチド配列を、DNA配列:
【化32】
配列番号23
を有するCPK1と呼ばれる一本鎖オリゴヌクレオチドを用いた部位特異的突然変異誘発により改変した。
【0139】
部位特異的突然変異誘発(SDM)を標準的なプロトコール(引用することにより本明細書の一部とされるBotstein and Shortle, "Strategies and Applications of In Vitro Mutagenesis," Science, 229: 193-1210 (1985))に従って実施したが、他の好適な技術を利用してもよい。CPK1のヌクレオチド配列は、HSA/MFα−1融合リーダー配列のアミノ酸配列を改変して、以下の変異Thr−20Phe、Phe−19Ile、Ile−18ValおよびLeu−16Ileを導入するよう設計されたものである、ここで、そのナンバリング(−20など)がHSA/MFα−1融合リーダー配列のC末端アミノ酸を−1位の残基とするものである。
【0140】
突然変異を誘発させたプラスミドのDNA配列をジデオキシヌクレオチド配列決定法により確認し、ポリヌクレオチド配列が突然変異誘発を受けて目的の配列となったことおよび他のDNA配列の変化が導入されていないことをこの方法により確認した。この新規プラスミドをpAYE653(図9)と名づけた。配列番号24は、改変されたHSA/MFα−1融合リーダー配列のコード領域を含んでなるポリヌクレオチド配列を示し、配列番号25は、pAYE653のポリヌクレオチド配列内に見られる改変されたHSA/MFα−1融合リーダー配列のコード領域と成熟ヒトアルブミンコード領域を含んでなるポリヌクレオチド配列を示すものである。
【0141】
pAYE653からNotIヒトアルブミン発現カセットを単離し、プラスミドpSAC35の独自のNotI部位に連結して、プラスミドpAYE655(図10)を作製した。
【0142】
実施例2
配列番号19は、サッカロミセス・セレビシエPRBIプロモーター由来の5’UTRを含む非コード領域;本発明の改変されたHSA/MFα−1融合リーダー配列をコードするポリヌクレオチド領域;成熟ヒトアルブミンのコドン最適化コード領域および翻訳終結部位を含んでなるDNA配列を示すものである。
【0143】
配列番号19のリーダー配列にもたらされる配列の改変の効果を比較するための対照として、配列番号40が、配列番号40のDNA配列が、本発明の第2の態様を示す15ポリヌクレオチド領域の代わりに、改変されていないHSA/MFα−1融合リーダー配列、すなわち、SFISLの5個のアミノ酸をコードする15ポリヌクレオチド領域を含んでなることを除いて配列番号19と本質的に同じであるDNA配列を示すものである。
【0144】
いずれのDNA配列もオーバーラッピング一本鎖オリゴヌクレオチドからGenosys, Inc (Cambridge, UK)により合成した。
【0145】
プラスミドpAYE639(図11)では、配列番号40を1.865kb SacI−HindIII DNA断片として合成し、プラスミドpBSSK−(Stratagene Europe, P. O. Box 12085, Amsterdam, The Netherlands)のSacI−HindIII部位にクローニングした。
【0146】
2種類の一本鎖オリゴヌクレオチドPRBJM1およびPRBJM3(下記)を用いたPCRによりサッカロミセス・セレビシエPRBIプロモーターを酵母ゲノムDNAから単離した。
PRBJM3:
【化33】
配列番号39
【0147】
PCR条件 94℃で30秒間、50℃で40秒間、72℃で120秒間、次いで、72℃で600秒間、その後、4℃で維持を40サイクル。0.81kb DNA断片をNotIとHindIIIの両方で消化し、WO 97/24445で記載されている、NotIとHindIIIで同様に消化したpBST+に連結して、プラスミドpAYE439(図12)を作出した。プラスミドpAYE439をHindIIIとBa7nHIで消化し、WO00/44772で従前に開示されているpAYE440由来の0.48kb HindIII/BamHI ADHIターミネーターDNA断片と連結して、プラスミドpAYE466(図13)を作出した。
【0148】
配列番号40の1.865kb HindIII DNA断片をプラスミドpAYE466の独自のHindIII部位にクローニングして、プラスミドpAYE640を作出した。このプラスミドが配列番号40の1.865kb HindIII DNA断片をPRB1プロモーターとADH1ターミネーターとの間に、PRB1プロモーターからの発現に対して適正な方向で含むことが分かった(図14)。
【0149】
プラスミドpAYE640をNotI/PvuIで消化完了し、NotI 3.2kb、PRBIプロモーター/配列番号40のHindIII DNA断片遺伝子/ADHIターミネーター発現カセットを精製した。発現カセット(3.2kb)とpBST+プラスミドバックボーン(3.15kb)は大きさが同じであるため、pAYE640のNotI/PvuI二重消化がNotI消化だけの場合よりも望ましい。pAYE640由来の3.2kb NotI、発現カセットを、仔ウシ小腸ホスファターゼ(CIP)で予め処理しておいたNotI線状化pSAC35(Sleep et al. (1991), Bio/Technology 9: 183-187)に連結して、プラスミド pAYE638(図15)を作出した。プラスミドpAYE638が、pSAC35のNotI部位に挿入され、HSA遺伝子の発現がLEU2栄養要求性マーカーから離れ、2μm複製起点に向かうように方向付けられたNotI HSA発現カセットを含むことが分かった。プラスミドpAYE642は同じHSA発現カセットを含むが、正反対の方向に配列されている(図15)。
【0150】
プラスミドpAYE643(図16)では、配列番号19を1.865kb SacI−HindIII DNA断片として合成し、pBSSK−(Stratagene Europe, P. O. Box 12085, Amsterdam, The Netherlands)にクローニングした。pAYE643内のHSA/MFα−1融合リーダー配列−アルブミン融合物をコードするDNA配列を配列番号27で示す。pAYE643から配列番号19の1.865kb HindIII断片を単離し、pAYE466の独自のHindIII部位に連結して、プラスミドpAYE645(図17)を作出した。NotI/PvuIでの消化によりpAYE645からNotI PRBI rHA発現カセットを単離し、pSAC35の独自のNotI部位に連結して、プラスミドpAYE646(図18)およびpAYE647(図19)を作製した。プラスミドpAYE646内のNotI 発現カセットはプラスミドpAYE638およびpAYE443と同じ方向に配向したが、プラスミドpAYE647内のNotI発現カセットは逆方向に配向し、プラスミドpAYE642と同じにした。
【0151】
実施例3
3種の異なる酵母株、A、BおよびCをプラスミドpAYE443、pAYE638、pAYE646およびpAYE655を用いてロイシンプロトトロフィーに形質転換した。これらの形質転換体を2%(w/v)グルコースを含有する緩衝最小培地(BMM、Kerry-Williams, S. M. et al. (1998) Yeast 14, 161-169により記載されている)(BMMD)にパッチアウトし、さらなる解析のために十分に増殖するまで30℃にてインキュベートした。2%(W/V)グルコースを含有する10mL YEP(1%(w/v)酵母抽出物;2%(W/V)バクトペプトン)(YEPD)およびBMMD振盪フラスコ培養(30℃、200rpm、72時間)から細胞を含まない培養上清のロケット免疫電気泳動により形質転換体のヒトアルブミン生産性を解析した(図20)。
【0152】
これらの結果から、pAYE638で形質転換された3種類全ての株のヒトアルブミン生産性が富栄養培地および規定培地の両方においてpAYE443で形質転換された同じ株で観察された生産性よりも約4〜5倍低いことが分かった(それらのいずれもがHSA/MFα−1融合リーダー配列を含むが、それらは異なるポリヌクレオチド配列によってコードされている)。予想外にも、pAYE646またはpAYE655で形質転換された3種類全ての株のヒトアルブミン生産性はpAYE638で観察された生産性よりもかなり高く、pAYE443で形質転換された同じ株で観察された生産性と同等か、またはそれよりも少し高かった。
【0153】
実施例4
酵母株C[pAYE443]、株C[pAYE655]、株C[pAYE638]および株C[pAYE646]、ならびに株B[pAYE443]および株B[pAYE646]をフェドバッチおよびフィル&ドロー両手順による高細胞密度発酵にて培養した。フェドバッチ手順では、WO96/37515に記載されている培地および制御パラメーターを使用した。フィル&ドロー手順では、以上で記述したフェドバッチ手順を使用するが、以下の工程もさらに含めた:フェドバッチ培養手順供給期の終了後、培養物量の90%を発酵容器から取り出し、供給物の添加を開始する前に、残りの10%量の培養物に、WO 96/37515に記載されている培地および制御パラメーターを使用してバッチ培地を添加した(pHを制御しながら)。ヒトアルブミン生産性(YP/S)およびヒトアルブミン濃度(g/L)を細胞を含まない全培養物のSDS−PAGEのスキャンニングデンシトメトリーにより評価した。重量測定によりバイオマス収率(YX/S)も計算した。これらの結果により(図21)、実施例3ですでに分かったように、HSA/MFα−1融合リーダー配列と成熟ヒトアルブミの両方のアミノ酸配列が同一であっても、ヒトアルブミン発現プラスミド pAYE638(天然のポリペプチド配列で酵母の偏りのあるコドン)を含有する酵母株のヒトアルブミン生産性(YP/S)およびヒトアルブミン濃度(g/L)が、ヒトアルブミン発現プラスミド pAYE443(天然のポリペプチド配列でリーダーおよび成熟アルブミンの自然コドンバイアス)を含有する同じ株よりもかなり低い生産性を有することが示された。
【0154】
株Cの発酵をフェドバッチ様式で行った場合、株C[pAYE655]および株C[pAYE646](本発明に従って改変されたリーダー配列を組み込んだヒトアルブミン発現プラスミド)をそれぞれ、同等の時間培養すると、ヒトアルブミン生産性(YP/S)の、株C[pAYE443]の生産性に比べて16%および12%の上昇が観察された。株Bの発酵をフェドバッチ様式で行った場合、株B[pAYE646](本発明に従って改変されたリーダー配列を組み込んだヒトアルブミン発現プラスミド)を同等の時間培養すると、ヒトアルブミン生産性(YP/S)の、株B[pAYE443]の生産性に比べて24%の上昇が観察された。
【0155】
株Cの発酵をフィルアンドドロー様式で行った場合、株C[pAYE655]および株C[pAYE646](本発明に従って改変されたリーダー配列を組み込んだヒトアルブミン発現プラスミド)をそれぞれ、同等の時間培養すると、ヒトアルブミン生産性(YP/S)の、株C[pAYE443]の生産性に比べて13%および6%の上昇が観察された。株C[pAYE655]および株C[pAYE646](本発明に従って改変されたリーダー配列を組み込んだヒトアルブミン発現プラスミド)をそれぞれ、同等の時間培養すると、この生産性は株C[pAYE638]の生産性に比べて442%および408%まで上昇した。
【0156】
概要
プラスミドpAYE443およびpAYE638はいずれもHSA/MFα−1融合リーダー配列由来のリーダー配列を有するヒトアルブミンをコードするが、前者は天然のポリヌクレオチド配列の自然のコドンバイアスを用い、一方、後者は酵母発現に向けて十分にコドン最適化されたポリヌクレオチド配列を用いる。pAYE638から得られたヒトアルブミンの発現は、pAYE443を用いて得られたものより4〜5倍低い。本発明に従って改変されたリーダー配列をコードするポリヌクレオチド配列を、pAYE443およびpAYE638の両方のHSA/MFα−1融合リーダー配列をコードするポリヌクレオチド配列の代わりに使用して、各々、ヒトアルブミン発現プラスミドpAYE665およびpAYE646を作出した。本発明のポリペプチド配列を導入することによって、目的ポリペプチドの産生におけるかなりの改善がもたらされた。
【0157】
【化34】
【図面の簡単な説明】
【0158】
【図1】天然のHSAリーダー(プレ領域およびプロ領域を有する)(上列)とWO90/01063で開示されているHSA/MFα−1融合リーダー配列(第2列)および本発明の好ましい改変リーダー配列(第3列)との比較。
【図2】標準的な遺伝子コード。
【図3】好ましいS.セレビシエコドンの改変リスト。
【図4】pAYE438のプラスミドマップ。
【図5】pAYE441のプラスミドマップ。
【図6】pAYE309のプラスミドマップ。
【図7】pAYE467のプラスミドマップ。
【図8】pAYE443のプラスミドマップ。
【図9】pAYE653のプラスミドマップ。
【図10】pAYE655のプラスミドマップ。
【図11】pAYE639のプラスミドマップ。
【図12】pAYE439のプラスミドマップ。
【図13】pAYE466のプラスミドマップ。
【図14】pAYE640のプラスミドマップ。
【図15】pAYE638およびpAYE642のプラスミドマップ。
【図16】pAYE643のプラスミドマップ。
【図17】pAYE645のプラスミドマップ。
【図18】pAYE646のプラスミドマップ。
【図19】pAYE647のプラスミドマップ。
【図20】ロケット免疫電気泳動によるrHA生産性の解析。酵母をYEP、2%(W/V)スクロースまたはB/MM、2%(W/V)スクロースにおいて30℃にて200rpm、72時間培養した。定量はHSA標品(mg.L−1)を参照することにより行った。
【図21】高細胞密度発酵におけるアルブミン生産性。*ヒトアルブミンレベルが低すぎて定量不可能であることを意味する。
【図22】実施例にて使用する構築物の特性の要約。
【特許請求の範囲】
【請求項1】
下記(i)および(ii)を含んでなる、ポリペプチド:
(i)次の(a)および(b)を含んでなる、リーダー配列:
(a)分泌プレ配列、および
(b)以下のモチーフ:
−X1−X2−X3−X4−X5−
(式中、
X1はフェニルアラニン、トリプトファンまたはチロシンであり、
X2はイソロイシン、ロイシン、バリン、アラニンまたはメチオニンであり、
X3はロイシン、バリン、アラニンまたはメチオニンであり、
X4はセリンまたはトレオニンであり、かつ、
X5はイソロイシン、バリン、アラニンまたはメチオニンである)、および
(ii)そのリーダー配列に対して異種の目的タンパク質。
【請求項2】
X1がフェニルアラニンである、請求項1に記載のポリペプチド。
【請求項3】
X2がイソロイシンである、請求項1または2に記載のポリペプチド。
【請求項4】
X3がバリンである、請求項1〜3のいずれか一項に記載のポリペプチド。
【請求項5】
モチーフのアミノ酸を天然アミノ酸の代用としてポリペプチド中に含む、請求項1〜4のいずれか一項に記載のポリペプチド。
【請求項6】
X5がイソロイシンである、請求項1〜5のいずれか一項に記載のポリペプチド。
【請求項7】
モチーフが−Phe−Ile−Val−Ser−Ile−である、請求項1〜6のいずれか一項に記載のポリペプチド。
【請求項8】
分泌プレ配列がアルブミン分泌プレ配列またはその変異体である、請求項1〜7のいずれか一項に記載のポリペプチド。
【請求項9】
X1、X2、X3、X4およびX5がそれぞれ−20、−19、−18、−17および−16位に、それらの位置の天然アミノ酸の代わりに存在し、そのナンバリングが天然のアルブミン分泌プロ配列のC末端アミノ酸を−1番残基とするものであり、かつ、X1、X2、X3、X4およびX5が請求項1〜7のいずれか一項により定義されるアミノ酸である、請求項8に記載のポリペプチド。
【請求項10】
アルブミン分泌プレ配列またはその変異体がヒトアルブミン分泌プレ配列またはその変異体である、請求項8または9に記載のポリペプチド。
【請求項11】
分泌プレ配列MKWVFIVSILFLFSSAYSを含んでなる、請求項10に記載のポリペプチド。
【請求項12】
リーダー配列が分泌プロ配列を含んでなる、請求項1〜11のいずれか一項に記載のポリペプチド。
【請求項13】
アルブミン分泌プレ配列またはその変異体が、そのC末端において分泌プロ配列またはその変異体のN末端アミノ酸とペプチド結合により融合され、それにより、プレ−プロ配列を形成する、請求項12に記載のポリペプチド。
【請求項14】
分泌プロ配列がアルブミン分泌プロ配列またはその変異体である、請求項12または13に記載のポリペプチド。
【請求項15】
アルブミン分泌プロ配列がヒト血清アルブミン分泌プロ配列またはその変異体である、請求項14に記載のポリペプチド。
【請求項16】
分泌プロ配列モチーフが酵母MFα−1分泌プロ配列またはその変異体である、請求項14または15に記載のポリペプチド。
【請求項17】
下記配列、またはその変異体を含んでなる、請求項12に記載のポリペプチド:
【化1】
(式中、
Y1がGlyまたはSerであり、
Y2がValまたはLeuであり、
Y3がPheまたはAspであり、
Y4がArgまたはLysであり、かつ、
Y5がArgまたはLysである)。
【請求項18】
Y1がGlyであり、Y2がValであり、かつ、Y3がPheであるか、または
Y1がSerであり、Y2がLeuであり、かつ、Y3がAspである、請求項17に記載のポリペプチド。
【請求項19】
Y4がArgであり、かつ、Y5がArgであるか、
Y4がLysであり、かつ、Y5がArgであるか、
Y4がLysであり、かつ、YがLysであるか、または、
Y4がArgであり、かつ、Y5がLysである、請求項17または18に記載のポリペプチド。
【請求項20】
該モチーフの少なくとも一部が分泌プレ−配列中に存在する、請求項1〜7のいずれか一項に記載のポリペプチド。
【請求項21】
目的タンパク質の配列がそのN末端においてリーダー配列のC末端アミノ酸と融合される、請求項1〜20のいずれか一項に記載のポリペプチド。
【請求項22】
目的タンパク質がアルブミンまたはその変異体、断片もしくは融合物である、請求項1〜21のいずれか一項に記載のポリペプチド。
【請求項23】
アルブミンがヒトアルブミンである、請求項22に記載のポリペプチド。
【請求項24】
成熟ポリペプチドがトランスフェリンまたはその変異体、断片もしくは融合物である、請求項1〜21のいずれか一項に記載のポリペプチド。
【請求項25】
トランスフェリンがヒトトランスフェリンである、請求項24に記載のポリペプチド。
【請求項26】
請求項1〜25のいずれか一項に記載モチーフをコードする配列を含んでなる、単離ポリヌクレオチド。
【請求項27】
配列番号15の配列を含んでなる、請求項26に記載のポリヌクレオチド。
【請求項28】
配列番号16の配列を含んでなる、請求項26に記載のポリヌクレオチド。
【請求項29】
配列番号17の配列を含んでなる、請求項26に記載のポリヌクレオチド。
【請求項30】
配列番号18の配列を含んでなる、請求項26に記載のポリヌクレオチド。
【請求項31】
配列番号34の配列を含んでなる、請求項26に記載のポリヌクレオチド。
【請求項32】
配列番号24の配列を含んでなる、請求項30または31に記載のポリヌクレオチド。
【請求項33】
配列番号25の配列、またはその変異体を含んでなる、請求項32に記載のポリヌクレオチドであって、
該変異体が、配列番号24のリーダー配列を有し、かつ、配列番号25によりコードされるアルブミンの変異体もしくは断片をコードするものである、ポリヌクレオチド。
【請求項34】
配列番号27の配列を含んでなる、請求項30または31に記載のポリヌクレオチド。
【請求項35】
配列番号21の配列、またはその変異体を含んでなる、請求項34に記載のポリヌクレオチドであって、
該変異体が、配列番号27のリーダー配列を有し、かつ、配列番号21によりコードされるアルブミンの変異体もしくは断片をコードするものである、ポリヌクレオチド。
【請求項36】
配列番号21の配列またはその断片を含んでなる、ポリヌクレオチド。
【請求項37】
ポリヌクレオチドが、異種タンパク質をコードしているDNA配列と、配列番号25のDNA配列または配列番号21のDNA配列のいずれかとの、連続したまたは不連続な融合物であるDNA配列を含んでなる、請求項33、35または36のいずれか一項に記載のポリヌクレオチド。
【請求項38】
請求項26〜37のいずれか一項に記載のポリヌクレオチドの相補鎖である、ポリヌクレオチド。
【請求項39】
作動可能なように連結された転写調節領域を含んでなる、請求項26〜38のいずれか一項に記載のポリヌクレオチド。
【請求項40】
転写調節領域が転写プロモーターを含んでなる、請求項39に記載のポリヌクレオチド。
【請求項41】
請求項26〜40のいずれか一項に記載のポリヌクレオチドを含んでなる、自己複製可能なポリヌクレオチド配列。
【請求項42】
請求項26〜41のいずれか一項に記載のポリヌクレオチドを含んでなる、細胞。
【請求項43】
真核生物細胞である、請求項42に記載の細胞。
【請求項44】
真菌細胞である、請求項43に記載の細胞。
【請求項45】
アスペルギルス属の細胞である、請求項44に記載の細胞。
【請求項46】
酵母細胞である、請求項44に記載の細胞。
【請求項47】
サッカロミセス属、クルイベロミセス属、シゾサッカロミセス属またはピキア属の細胞である、請求項46に記載の細胞。
【請求項48】
請求項42〜47のいずれか一項に記載の細胞および培養培地を含んでなる、細胞培養物。
【請求項49】
その培地が、請求項1〜22のいずれか一項に記載のポリペプチドの産生の結果として、成熟した目的タンパク質を含有する、請求項48に記載の細胞培養物。
【請求項50】
成熟した目的タンパク質を作製する方法であって、
(1)請求項42〜47のいずれか一項に記載の細胞を培養培地で培養し、ここで、その細胞が請求項1〜25のいずれか一項に記載のポリペプチドの産生の結果として、成熟した目的タンパク質を培養培地中に分泌し、かつ
(2)分泌された成熟タンパク質を含有する培養培地を細胞から分離する
ことを含んでなる、方法。
【請求項51】
成熟した目的タンパク質を培地から分離し、必要に応じて、成熟した目的タンパク質をさらに精製する工程をさらに含んでなる、請求項50に記載の方法。
【請求項52】
このようにして分離および/または精製された、成熟した目的タンパク質を、治療上許容されうる担体または希釈剤とともに製剤し、それにより、ヒトまたは動物への投与に好適な治療用製品を生産する工程をさらに含んでなる、請求項51に記載の方法。
【特許請求の範囲】
【請求項1】
下記(i)および(ii)を含んでなる、ポリペプチド:
(i)次の(a)および(b)を含んでなる、リーダー配列:
(a)分泌プレ配列、および
(b)以下のモチーフ:
−X1−X2−X3−X4−X5−
(式中、
X1はフェニルアラニン、トリプトファンまたはチロシンであり、
X2はイソロイシン、ロイシン、バリン、アラニンまたはメチオニンであり、
X3はロイシン、バリン、アラニンまたはメチオニンであり、
X4はセリンまたはトレオニンであり、かつ、
X5はイソロイシン、バリン、アラニンまたはメチオニンである)、および
(ii)そのリーダー配列に対して異種の目的タンパク質。
【請求項2】
X1がフェニルアラニンである、請求項1に記載のポリペプチド。
【請求項3】
X2がイソロイシンである、請求項1または2に記載のポリペプチド。
【請求項4】
X3がバリンである、請求項1〜3のいずれか一項に記載のポリペプチド。
【請求項5】
モチーフのアミノ酸を天然アミノ酸の代用としてポリペプチド中に含む、請求項1〜4のいずれか一項に記載のポリペプチド。
【請求項6】
X5がイソロイシンである、請求項1〜5のいずれか一項に記載のポリペプチド。
【請求項7】
モチーフが−Phe−Ile−Val−Ser−Ile−である、請求項1〜6のいずれか一項に記載のポリペプチド。
【請求項8】
分泌プレ配列がアルブミン分泌プレ配列またはその変異体である、請求項1〜7のいずれか一項に記載のポリペプチド。
【請求項9】
X1、X2、X3、X4およびX5がそれぞれ−20、−19、−18、−17および−16位に、それらの位置の天然アミノ酸の代わりに存在し、そのナンバリングが天然のアルブミン分泌プロ配列のC末端アミノ酸を−1番残基とするものであり、かつ、X1、X2、X3、X4およびX5が請求項1〜7のいずれか一項により定義されるアミノ酸である、請求項8に記載のポリペプチド。
【請求項10】
アルブミン分泌プレ配列またはその変異体がヒトアルブミン分泌プレ配列またはその変異体である、請求項8または9に記載のポリペプチド。
【請求項11】
分泌プレ配列MKWVFIVSILFLFSSAYSを含んでなる、請求項10に記載のポリペプチド。
【請求項12】
リーダー配列が分泌プロ配列を含んでなる、請求項1〜11のいずれか一項に記載のポリペプチド。
【請求項13】
アルブミン分泌プレ配列またはその変異体が、そのC末端において分泌プロ配列またはその変異体のN末端アミノ酸とペプチド結合により融合され、それにより、プレ−プロ配列を形成する、請求項12に記載のポリペプチド。
【請求項14】
分泌プロ配列がアルブミン分泌プロ配列またはその変異体である、請求項12または13に記載のポリペプチド。
【請求項15】
アルブミン分泌プロ配列がヒト血清アルブミン分泌プロ配列またはその変異体である、請求項14に記載のポリペプチド。
【請求項16】
分泌プロ配列モチーフが酵母MFα−1分泌プロ配列またはその変異体である、請求項14または15に記載のポリペプチド。
【請求項17】
下記配列、またはその変異体を含んでなる、請求項12に記載のポリペプチド:
【化1】
(式中、
Y1がGlyまたはSerであり、
Y2がValまたはLeuであり、
Y3がPheまたはAspであり、
Y4がArgまたはLysであり、かつ、
Y5がArgまたはLysである)。
【請求項18】
Y1がGlyであり、Y2がValであり、かつ、Y3がPheであるか、または
Y1がSerであり、Y2がLeuであり、かつ、Y3がAspである、請求項17に記載のポリペプチド。
【請求項19】
Y4がArgであり、かつ、Y5がArgであるか、
Y4がLysであり、かつ、Y5がArgであるか、
Y4がLysであり、かつ、YがLysであるか、または、
Y4がArgであり、かつ、Y5がLysである、請求項17または18に記載のポリペプチド。
【請求項20】
該モチーフの少なくとも一部が分泌プレ−配列中に存在する、請求項1〜7のいずれか一項に記載のポリペプチド。
【請求項21】
目的タンパク質の配列がそのN末端においてリーダー配列のC末端アミノ酸と融合される、請求項1〜20のいずれか一項に記載のポリペプチド。
【請求項22】
目的タンパク質がアルブミンまたはその変異体、断片もしくは融合物である、請求項1〜21のいずれか一項に記載のポリペプチド。
【請求項23】
アルブミンがヒトアルブミンである、請求項22に記載のポリペプチド。
【請求項24】
成熟ポリペプチドがトランスフェリンまたはその変異体、断片もしくは融合物である、請求項1〜21のいずれか一項に記載のポリペプチド。
【請求項25】
トランスフェリンがヒトトランスフェリンである、請求項24に記載のポリペプチド。
【請求項26】
請求項1〜25のいずれか一項に記載モチーフをコードする配列を含んでなる、単離ポリヌクレオチド。
【請求項27】
配列番号15の配列を含んでなる、請求項26に記載のポリヌクレオチド。
【請求項28】
配列番号16の配列を含んでなる、請求項26に記載のポリヌクレオチド。
【請求項29】
配列番号17の配列を含んでなる、請求項26に記載のポリヌクレオチド。
【請求項30】
配列番号18の配列を含んでなる、請求項26に記載のポリヌクレオチド。
【請求項31】
配列番号34の配列を含んでなる、請求項26に記載のポリヌクレオチド。
【請求項32】
配列番号24の配列を含んでなる、請求項30または31に記載のポリヌクレオチド。
【請求項33】
配列番号25の配列、またはその変異体を含んでなる、請求項32に記載のポリヌクレオチドであって、
該変異体が、配列番号24のリーダー配列を有し、かつ、配列番号25によりコードされるアルブミンの変異体もしくは断片をコードするものである、ポリヌクレオチド。
【請求項34】
配列番号27の配列を含んでなる、請求項30または31に記載のポリヌクレオチド。
【請求項35】
配列番号21の配列、またはその変異体を含んでなる、請求項34に記載のポリヌクレオチドであって、
該変異体が、配列番号27のリーダー配列を有し、かつ、配列番号21によりコードされるアルブミンの変異体もしくは断片をコードするものである、ポリヌクレオチド。
【請求項36】
配列番号21の配列を含んでなる、ポリヌクレオチド。
【請求項37】
ポリヌクレオチドが、異種タンパク質をコードしているDNA配列と、配列番号25のDNA配列または配列番号21のDNA配列のいずれかとの、連続したまたは不連続な融合物であるDNA配列を含んでなる、請求項33、35または36のいずれか一項に記載のポリヌクレオチド。
【請求項38】
請求項26〜37のいずれか一項に記載のポリヌクレオチドの相補鎖である、ポリヌクレオチド。
【請求項39】
作動可能なように連結された転写調節領域を含んでなる、請求項26〜38のいずれか一項に記載のポリヌクレオチド。
【請求項40】
転写調節領域が転写プロモーターを含んでなる、請求項39に記載のポリヌクレオチド。
【請求項41】
請求項26〜40のいずれか一項に記載のポリヌクレオチドを含んでなる、自己複製可能なポリヌクレオチド配列。
【請求項42】
請求項26〜41のいずれか一項に記載のポリヌクレオチドを含んでなる、細胞。
【請求項43】
真核生物細胞である、請求項42に記載の細胞。
【請求項44】
真菌細胞である、請求項43に記載の細胞。
【請求項45】
アスペルギルス属の細胞である、請求項44に記載の細胞。
【請求項46】
酵母細胞である、請求項44に記載の細胞。
【請求項47】
サッカロミセス属、クルイベロミセス属、シゾサッカロミセス属またはピキア属の細胞である、請求項46に記載の細胞。
【請求項48】
請求項42〜47のいずれか一項に記載の細胞および培養培地を含んでなる、細胞培養物。
【請求項49】
その培地が、請求項1〜22のいずれか一項に記載のポリペプチドの産生の結果として、成熟した目的タンパク質を含有する、請求項48に記載の細胞培養物。
【請求項50】
成熟した目的タンパク質を作製する方法であって、
(1)請求項42〜47のいずれか一項に記載の細胞を培養培地で培養し、ここで、その細胞が請求項1〜25のいずれか一項に記載のポリペプチドの産生の結果として、成熟した目的タンパク質を培養培地中に分泌し、かつ
(2)分泌された成熟タンパク質を含有する培養培地を細胞から分離する
ことを含んでなる、方法。
【請求項51】
成熟した目的タンパク質を培地から分離し、必要に応じて、成熟した目的タンパク質をさらに精製する工程をさらに含んでなる、請求項50に記載の方法。
【請求項52】
このようにして分離および/または精製された、成熟した目的タンパク質を、治療上許容されうる担体または希釈剤とともに製剤し、それにより、ヒトまたは動物への投与に好適な治療用製品を生産する工程をさらに含んでなる、請求項51に記載の方法。
【請求項1】
下記(i)および(ii)を含んでなる、ポリペプチド:
(i)次の(a)および(b)を含んでなる、リーダー配列:
(a)分泌プレ配列、および
(b)以下のモチーフ:
−X1−X2−X3−X4−X5−
(式中、
X1はフェニルアラニン、トリプトファンまたはチロシンであり、
X2はイソロイシン、ロイシン、バリン、アラニンまたはメチオニンであり、
X3はロイシン、バリン、アラニンまたはメチオニンであり、
X4はセリンまたはトレオニンであり、かつ、
X5はイソロイシン、バリン、アラニンまたはメチオニンである)、および
(ii)そのリーダー配列に対して異種の目的タンパク質。
【請求項2】
X1がフェニルアラニンである、請求項1に記載のポリペプチド。
【請求項3】
X2がイソロイシンである、請求項1または2に記載のポリペプチド。
【請求項4】
X3がバリンである、請求項1〜3のいずれか一項に記載のポリペプチド。
【請求項5】
モチーフのアミノ酸を天然アミノ酸の代用としてポリペプチド中に含む、請求項1〜4のいずれか一項に記載のポリペプチド。
【請求項6】
X5がイソロイシンである、請求項1〜5のいずれか一項に記載のポリペプチド。
【請求項7】
モチーフが−Phe−Ile−Val−Ser−Ile−である、請求項1〜6のいずれか一項に記載のポリペプチド。
【請求項8】
分泌プレ配列がアルブミン分泌プレ配列またはその変異体である、請求項1〜7のいずれか一項に記載のポリペプチド。
【請求項9】
X1、X2、X3、X4およびX5がそれぞれ−20、−19、−18、−17および−16位に、それらの位置の天然アミノ酸の代わりに存在し、そのナンバリングが天然のアルブミン分泌プロ配列のC末端アミノ酸を−1番残基とするものであり、かつ、X1、X2、X3、X4およびX5が請求項1〜7のいずれか一項により定義されるアミノ酸である、請求項8に記載のポリペプチド。
【請求項10】
アルブミン分泌プレ配列またはその変異体がヒトアルブミン分泌プレ配列またはその変異体である、請求項8または9に記載のポリペプチド。
【請求項11】
分泌プレ配列MKWVFIVSILFLFSSAYSを含んでなる、請求項10に記載のポリペプチド。
【請求項12】
リーダー配列が分泌プロ配列を含んでなる、請求項1〜11のいずれか一項に記載のポリペプチド。
【請求項13】
アルブミン分泌プレ配列またはその変異体が、そのC末端において分泌プロ配列またはその変異体のN末端アミノ酸とペプチド結合により融合され、それにより、プレ−プロ配列を形成する、請求項12に記載のポリペプチド。
【請求項14】
分泌プロ配列がアルブミン分泌プロ配列またはその変異体である、請求項12または13に記載のポリペプチド。
【請求項15】
アルブミン分泌プロ配列がヒト血清アルブミン分泌プロ配列またはその変異体である、請求項14に記載のポリペプチド。
【請求項16】
分泌プロ配列モチーフが酵母MFα−1分泌プロ配列またはその変異体である、請求項14または15に記載のポリペプチド。
【請求項17】
下記配列、またはその変異体を含んでなる、請求項12に記載のポリペプチド:
【化1】
(式中、
Y1がGlyまたはSerであり、
Y2がValまたはLeuであり、
Y3がPheまたはAspであり、
Y4がArgまたはLysであり、かつ、
Y5がArgまたはLysである)。
【請求項18】
Y1がGlyであり、Y2がValであり、かつ、Y3がPheであるか、または
Y1がSerであり、Y2がLeuであり、かつ、Y3がAspである、請求項17に記載のポリペプチド。
【請求項19】
Y4がArgであり、かつ、Y5がArgであるか、
Y4がLysであり、かつ、Y5がArgであるか、
Y4がLysであり、かつ、YがLysであるか、または、
Y4がArgであり、かつ、Y5がLysである、請求項17または18に記載のポリペプチド。
【請求項20】
該モチーフの少なくとも一部が分泌プレ−配列中に存在する、請求項1〜7のいずれか一項に記載のポリペプチド。
【請求項21】
目的タンパク質の配列がそのN末端においてリーダー配列のC末端アミノ酸と融合される、請求項1〜20のいずれか一項に記載のポリペプチド。
【請求項22】
目的タンパク質がアルブミンまたはその変異体、断片もしくは融合物である、請求項1〜21のいずれか一項に記載のポリペプチド。
【請求項23】
アルブミンがヒトアルブミンである、請求項22に記載のポリペプチド。
【請求項24】
成熟ポリペプチドがトランスフェリンまたはその変異体、断片もしくは融合物である、請求項1〜21のいずれか一項に記載のポリペプチド。
【請求項25】
トランスフェリンがヒトトランスフェリンである、請求項24に記載のポリペプチド。
【請求項26】
請求項1〜25のいずれか一項に記載モチーフをコードする配列を含んでなる、単離ポリヌクレオチド。
【請求項27】
配列番号15の配列を含んでなる、請求項26に記載のポリヌクレオチド。
【請求項28】
配列番号16の配列を含んでなる、請求項26に記載のポリヌクレオチド。
【請求項29】
配列番号17の配列を含んでなる、請求項26に記載のポリヌクレオチド。
【請求項30】
配列番号18の配列を含んでなる、請求項26に記載のポリヌクレオチド。
【請求項31】
配列番号34の配列を含んでなる、請求項26に記載のポリヌクレオチド。
【請求項32】
配列番号24の配列を含んでなる、請求項30または31に記載のポリヌクレオチド。
【請求項33】
配列番号25の配列、またはその変異体を含んでなる、請求項32に記載のポリヌクレオチドであって、
該変異体が、配列番号24のリーダー配列を有し、かつ、配列番号25によりコードされるアルブミンの変異体もしくは断片をコードするものである、ポリヌクレオチド。
【請求項34】
配列番号27の配列を含んでなる、請求項30または31に記載のポリヌクレオチド。
【請求項35】
配列番号21の配列、またはその変異体を含んでなる、請求項34に記載のポリヌクレオチドであって、
該変異体が、配列番号27のリーダー配列を有し、かつ、配列番号21によりコードされるアルブミンの変異体もしくは断片をコードするものである、ポリヌクレオチド。
【請求項36】
配列番号21の配列またはその断片を含んでなる、ポリヌクレオチド。
【請求項37】
ポリヌクレオチドが、異種タンパク質をコードしているDNA配列と、配列番号25のDNA配列または配列番号21のDNA配列のいずれかとの、連続したまたは不連続な融合物であるDNA配列を含んでなる、請求項33、35または36のいずれか一項に記載のポリヌクレオチド。
【請求項38】
請求項26〜37のいずれか一項に記載のポリヌクレオチドの相補鎖である、ポリヌクレオチド。
【請求項39】
作動可能なように連結された転写調節領域を含んでなる、請求項26〜38のいずれか一項に記載のポリヌクレオチド。
【請求項40】
転写調節領域が転写プロモーターを含んでなる、請求項39に記載のポリヌクレオチド。
【請求項41】
請求項26〜40のいずれか一項に記載のポリヌクレオチドを含んでなる、自己複製可能なポリヌクレオチド配列。
【請求項42】
請求項26〜41のいずれか一項に記載のポリヌクレオチドを含んでなる、細胞。
【請求項43】
真核生物細胞である、請求項42に記載の細胞。
【請求項44】
真菌細胞である、請求項43に記載の細胞。
【請求項45】
アスペルギルス属の細胞である、請求項44に記載の細胞。
【請求項46】
酵母細胞である、請求項44に記載の細胞。
【請求項47】
サッカロミセス属、クルイベロミセス属、シゾサッカロミセス属またはピキア属の細胞である、請求項46に記載の細胞。
【請求項48】
請求項42〜47のいずれか一項に記載の細胞および培養培地を含んでなる、細胞培養物。
【請求項49】
その培地が、請求項1〜22のいずれか一項に記載のポリペプチドの産生の結果として、成熟した目的タンパク質を含有する、請求項48に記載の細胞培養物。
【請求項50】
成熟した目的タンパク質を作製する方法であって、
(1)請求項42〜47のいずれか一項に記載の細胞を培養培地で培養し、ここで、その細胞が請求項1〜25のいずれか一項に記載のポリペプチドの産生の結果として、成熟した目的タンパク質を培養培地中に分泌し、かつ
(2)分泌された成熟タンパク質を含有する培養培地を細胞から分離する
ことを含んでなる、方法。
【請求項51】
成熟した目的タンパク質を培地から分離し、必要に応じて、成熟した目的タンパク質をさらに精製する工程をさらに含んでなる、請求項50に記載の方法。
【請求項52】
このようにして分離および/または精製された、成熟した目的タンパク質を、治療上許容されうる担体または希釈剤とともに製剤し、それにより、ヒトまたは動物への投与に好適な治療用製品を生産する工程をさらに含んでなる、請求項51に記載の方法。
【特許請求の範囲】
【請求項1】
下記(i)および(ii)を含んでなる、ポリペプチド:
(i)次の(a)および(b)を含んでなる、リーダー配列:
(a)分泌プレ配列、および
(b)以下のモチーフ:
−X1−X2−X3−X4−X5−
(式中、
X1はフェニルアラニン、トリプトファンまたはチロシンであり、
X2はイソロイシン、ロイシン、バリン、アラニンまたはメチオニンであり、
X3はロイシン、バリン、アラニンまたはメチオニンであり、
X4はセリンまたはトレオニンであり、かつ、
X5はイソロイシン、バリン、アラニンまたはメチオニンである)、および
(ii)そのリーダー配列に対して異種の目的タンパク質。
【請求項2】
X1がフェニルアラニンである、請求項1に記載のポリペプチド。
【請求項3】
X2がイソロイシンである、請求項1または2に記載のポリペプチド。
【請求項4】
X3がバリンである、請求項1〜3のいずれか一項に記載のポリペプチド。
【請求項5】
モチーフのアミノ酸を天然アミノ酸の代用としてポリペプチド中に含む、請求項1〜4のいずれか一項に記載のポリペプチド。
【請求項6】
X5がイソロイシンである、請求項1〜5のいずれか一項に記載のポリペプチド。
【請求項7】
モチーフが−Phe−Ile−Val−Ser−Ile−である、請求項1〜6のいずれか一項に記載のポリペプチド。
【請求項8】
分泌プレ配列がアルブミン分泌プレ配列またはその変異体である、請求項1〜7のいずれか一項に記載のポリペプチド。
【請求項9】
X1、X2、X3、X4およびX5がそれぞれ−20、−19、−18、−17および−16位に、それらの位置の天然アミノ酸の代わりに存在し、そのナンバリングが天然のアルブミン分泌プロ配列のC末端アミノ酸を−1番残基とするものであり、かつ、X1、X2、X3、X4およびX5が請求項1〜7のいずれか一項により定義されるアミノ酸である、請求項8に記載のポリペプチド。
【請求項10】
アルブミン分泌プレ配列またはその変異体がヒトアルブミン分泌プレ配列またはその変異体である、請求項8または9に記載のポリペプチド。
【請求項11】
分泌プレ配列MKWVFIVSILFLFSSAYSを含んでなる、請求項10に記載のポリペプチド。
【請求項12】
リーダー配列が分泌プロ配列を含んでなる、請求項1〜11のいずれか一項に記載のポリペプチド。
【請求項13】
アルブミン分泌プレ配列またはその変異体が、そのC末端において分泌プロ配列またはその変異体のN末端アミノ酸とペプチド結合により融合され、それにより、プレ−プロ配列を形成する、請求項12に記載のポリペプチド。
【請求項14】
分泌プロ配列がアルブミン分泌プロ配列またはその変異体である、請求項12または13に記載のポリペプチド。
【請求項15】
アルブミン分泌プロ配列がヒト血清アルブミン分泌プロ配列またはその変異体である、請求項14に記載のポリペプチド。
【請求項16】
分泌プロ配列モチーフが酵母MFα−1分泌プロ配列またはその変異体である、請求項14または15に記載のポリペプチド。
【請求項17】
下記配列、またはその変異体を含んでなる、請求項12に記載のポリペプチド:
【化1】
(式中、
Y1がGlyまたはSerであり、
Y2がValまたはLeuであり、
Y3がPheまたはAspであり、
Y4がArgまたはLysであり、かつ、
Y5がArgまたはLysである)。
【請求項18】
Y1がGlyであり、Y2がValであり、かつ、Y3がPheであるか、または
Y1がSerであり、Y2がLeuであり、かつ、Y3がAspである、請求項17に記載のポリペプチド。
【請求項19】
Y4がArgであり、かつ、Y5がArgであるか、
Y4がLysであり、かつ、Y5がArgであるか、
Y4がLysであり、かつ、YがLysであるか、または、
Y4がArgであり、かつ、Y5がLysである、請求項17または18に記載のポリペプチド。
【請求項20】
該モチーフの少なくとも一部が分泌プレ−配列中に存在する、請求項1〜7のいずれか一項に記載のポリペプチド。
【請求項21】
目的タンパク質の配列がそのN末端においてリーダー配列のC末端アミノ酸と融合される、請求項1〜20のいずれか一項に記載のポリペプチド。
【請求項22】
目的タンパク質がアルブミンまたはその変異体、断片もしくは融合物である、請求項1〜21のいずれか一項に記載のポリペプチド。
【請求項23】
アルブミンがヒトアルブミンである、請求項22に記載のポリペプチド。
【請求項24】
成熟ポリペプチドがトランスフェリンまたはその変異体、断片もしくは融合物である、請求項1〜21のいずれか一項に記載のポリペプチド。
【請求項25】
トランスフェリンがヒトトランスフェリンである、請求項24に記載のポリペプチド。
【請求項26】
請求項1〜25のいずれか一項に記載モチーフをコードする配列を含んでなる、単離ポリヌクレオチド。
【請求項27】
配列番号15の配列を含んでなる、請求項26に記載のポリヌクレオチド。
【請求項28】
配列番号16の配列を含んでなる、請求項26に記載のポリヌクレオチド。
【請求項29】
配列番号17の配列を含んでなる、請求項26に記載のポリヌクレオチド。
【請求項30】
配列番号18の配列を含んでなる、請求項26に記載のポリヌクレオチド。
【請求項31】
配列番号34の配列を含んでなる、請求項26に記載のポリヌクレオチド。
【請求項32】
配列番号24の配列を含んでなる、請求項30または31に記載のポリヌクレオチド。
【請求項33】
配列番号25の配列、またはその変異体を含んでなる、請求項32に記載のポリヌクレオチドであって、
該変異体が、配列番号24のリーダー配列を有し、かつ、配列番号25によりコードされるアルブミンの変異体もしくは断片をコードするものである、ポリヌクレオチド。
【請求項34】
配列番号27の配列を含んでなる、請求項30または31に記載のポリヌクレオチド。
【請求項35】
配列番号21の配列、またはその変異体を含んでなる、請求項34に記載のポリヌクレオチドであって、
該変異体が、配列番号27のリーダー配列を有し、かつ、配列番号21によりコードされるアルブミンの変異体もしくは断片をコードするものである、ポリヌクレオチド。
【請求項36】
配列番号21の配列を含んでなる、ポリヌクレオチド。
【請求項37】
ポリヌクレオチドが、異種タンパク質をコードしているDNA配列と、配列番号25のDNA配列または配列番号21のDNA配列のいずれかとの、連続したまたは不連続な融合物であるDNA配列を含んでなる、請求項33、35または36のいずれか一項に記載のポリヌクレオチド。
【請求項38】
請求項26〜37のいずれか一項に記載のポリヌクレオチドの相補鎖である、ポリヌクレオチド。
【請求項39】
作動可能なように連結された転写調節領域を含んでなる、請求項26〜38のいずれか一項に記載のポリヌクレオチド。
【請求項40】
転写調節領域が転写プロモーターを含んでなる、請求項39に記載のポリヌクレオチド。
【請求項41】
請求項26〜40のいずれか一項に記載のポリヌクレオチドを含んでなる、自己複製可能なポリヌクレオチド配列。
【請求項42】
請求項26〜41のいずれか一項に記載のポリヌクレオチドを含んでなる、細胞。
【請求項43】
真核生物細胞である、請求項42に記載の細胞。
【請求項44】
真菌細胞である、請求項43に記載の細胞。
【請求項45】
アスペルギルス属の細胞である、請求項44に記載の細胞。
【請求項46】
酵母細胞である、請求項44に記載の細胞。
【請求項47】
サッカロミセス属、クルイベロミセス属、シゾサッカロミセス属またはピキア属の細胞である、請求項46に記載の細胞。
【請求項48】
請求項42〜47のいずれか一項に記載の細胞および培養培地を含んでなる、細胞培養物。
【請求項49】
その培地が、請求項1〜22のいずれか一項に記載のポリペプチドの産生の結果として、成熟した目的タンパク質を含有する、請求項48に記載の細胞培養物。
【請求項50】
成熟した目的タンパク質を作製する方法であって、
(1)請求項42〜47のいずれか一項に記載の細胞を培養培地で培養し、ここで、その細胞が請求項1〜25のいずれか一項に記載のポリペプチドの産生の結果として、成熟した目的タンパク質を培養培地中に分泌し、かつ
(2)分泌された成熟タンパク質を含有する培養培地を細胞から分離する
ことを含んでなる、方法。
【請求項51】
成熟した目的タンパク質を培地から分離し、必要に応じて、成熟した目的タンパク質をさらに精製する工程をさらに含んでなる、請求項50に記載の方法。
【請求項52】
このようにして分離および/または精製された、成熟した目的タンパク質を、治療上許容されうる担体または希釈剤とともに製剤し、それにより、ヒトまたは動物への投与に好適な治療用製品を生産する工程をさらに含んでなる、請求項51に記載の方法。
【図1】


【図2】

【図3】
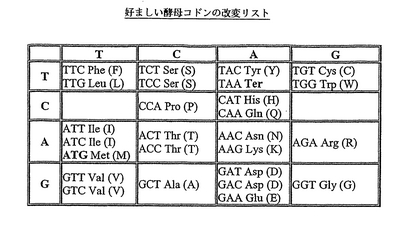
【図4】
【図5】
【図6】
【図7】
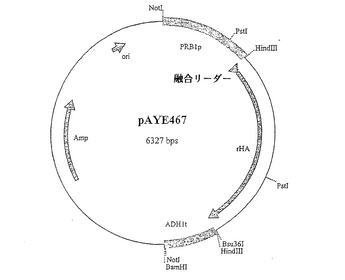
【図8】
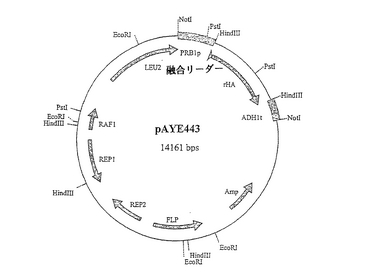
【図9】
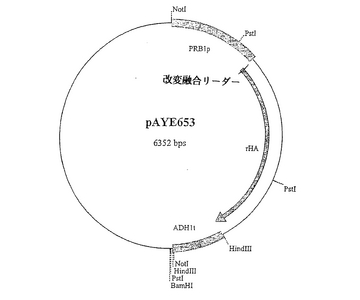
【図10】
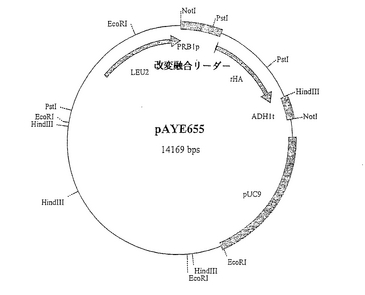
【図11】
【図12】
【図13】
【図14】
【図15】
【図16】
【図17】
【図18】
【図19】
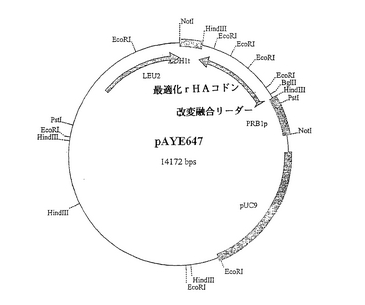
【図20】
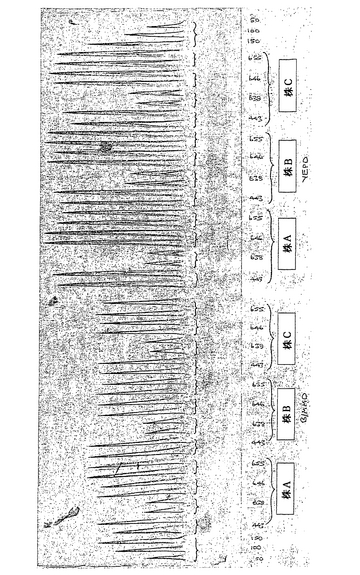
【図21】
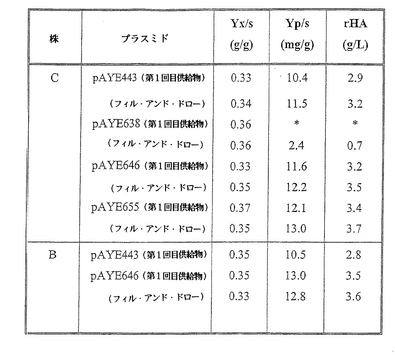
【図22】
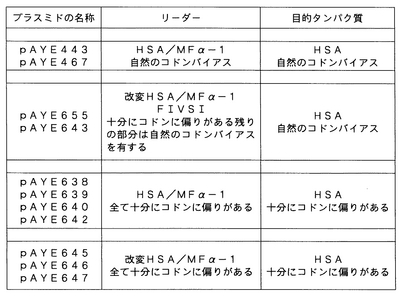
【図2】
【図3】
【図4】
【図5】
【図6】
【図7】
【図8】
【図9】
【図10】
【図11】
【図12】
【図13】
【図14】
【図15】
【図16】
【図17】
【図18】
【図19】
【図20】
【図21】
【図22】
【公表番号】特表2006−501818(P2006−501818A)
【公表日】平成18年1月19日(2006.1.19)
【国際特許分類】
【出願番号】特願2004−522368(P2004−522368)
【出願日】平成15年7月23日(2003.7.23)
【国際出願番号】PCT/GB2003/003273
【国際公開番号】WO2004/009819
【国際公開日】平成16年1月29日(2004.1.29)
【出願人】(591131523)デルタ バイオテクノロジー リミテッド (9)
【氏名又は名称原語表記】DELTA BIOTECHNOLOGY LIMITED
【Fターム(参考)】
【公表日】平成18年1月19日(2006.1.19)
【国際特許分類】
【出願日】平成15年7月23日(2003.7.23)
【国際出願番号】PCT/GB2003/003273
【国際公開番号】WO2004/009819
【国際公開日】平成16年1月29日(2004.1.29)
【出願人】(591131523)デルタ バイオテクノロジー リミテッド (9)
【氏名又は名称原語表記】DELTA BIOTECHNOLOGY LIMITED
【Fターム(参考)】
[ Back to top ]