
遺伝子モールスコード
本発明は、供試巨大分子上の少なくとも1つの対象ドメインの存在を検出する方法に関し、該方法は、
a)対象ドメイン上の少なくとも2つの標的領域を予め決定し、各標的領域の対応する標識プローブ(対象ドメインのプローブのセットと呼ぶ)を設計および取得し、これらのプローブのいずれかの、他のものに対する位置が選択され、供試巨大分子上の対象ドメインの特異的サインを形成する工程;
b)工程a)で得られたプローブが結合されている供試巨大分子を展開した後、線状化巨大分子に結合されているプローブのいずれかの、他のものに対する位置を検出し、対象ドメインのサインが検出されれば、供試巨大分子上に対象ドメインが存在することを示し、逆に、対象ドメインのサインまたはサインの一部が検出されなければ、供試巨大分子上に対象ドメインまたはドメインの一部が存在しないことを示す工程
を含む。
a)対象ドメイン上の少なくとも2つの標的領域を予め決定し、各標的領域の対応する標識プローブ(対象ドメインのプローブのセットと呼ぶ)を設計および取得し、これらのプローブのいずれかの、他のものに対する位置が選択され、供試巨大分子上の対象ドメインの特異的サインを形成する工程;
b)工程a)で得られたプローブが結合されている供試巨大分子を展開した後、線状化巨大分子に結合されているプローブのいずれかの、他のものに対する位置を検出し、対象ドメインのサインが検出されれば、供試巨大分子上に対象ドメインが存在することを示し、逆に、対象ドメインのサインまたはサインの一部が検出されなければ、供試巨大分子上に対象ドメインまたはドメインの一部が存在しないことを示す工程
を含む。
【発明の詳細な説明】
【発明の分野】
【0001】
本発明は、供試巨大分子上の少なくとも1つの対象ドメインの存在を検出する方法に関し、該方法は、
a)対象ドメイン上の少なくとも2つの標的領域を予め決定し、各標的領域の対応する標識プローブ(対象ドメインのプローブのセットと呼ぶ)を設計および取得し、これらのプローブのいずれかの、他のものに対する位置が選択され、供試巨大分子上の対象ドメインの特異的サインを形成する工程;
b)工程a)で得られたプローブが結合されている供試巨大分子を展開(spreading)した後、線状化巨大分子に結合されているプローブのいずれかの、他のものに対する位置を検出し、対象ドメインのサイン(signature)が検出されれば、供試巨大分子上に対象ドメインが存在することを示し、逆に、対象ドメインのサインまたはサインの一部が検出されなければ、供試巨大分子上の対象ドメインまたはドメインの一部が存在しないことを示す工程
を含む。
【0002】
より詳しくは、本発明は、少なくとも2セットの3つのプローブを含み、各プローブセットは1つのドメイン上に特異的に結合することができ、巨大分子上での1セットの少なくとも3つのプローブの結合が、少なくとも2つの異なるコード群(例えば、「ドット(dot)」と「ダッシュ(dash)」)から選択される少なくとも2つのコードの配列を形成し、この群が総てのドメインで同一であり、このコード配列が1つのドメインに特異的であり、そのドメインのサインである、巨大分子上の少なくとも2つの対象ドメインの存在を検出するためのキットに関する。
【背景】
【0003】
巨大分子(macromolecule)、特に、生体巨大分子(より具体的には、DNA)の研究は、「地図作成」のため、すなわち、ドメインの空間構成を研究するため、または巨大分子上の、反応または一連の化学反応もしくは生化学反応の位置決定のためには、多くの場合、正確にいくつかのドメインに印をつける必要がある。
【0004】
DNA配列の空間構成の観察を可能とする方法(Fiber−FISH、分子コーミングなど)は、多くの適用において、いくつかの領域に目印が付けられる、すなわち、何らかの検出技術によって特定の領域の同定が可能な方法でマークされる必要がある。取り組む主題がいくつかの領域の相対的位置である地図作成での適用、ならびにある(いくつかの)特定の遺伝子座において生体現象が研究される適用の場合がそうである。そして、ドメインは特異的マーカー、実際には一般に、対象配列(対象ドメインと呼ぶ)に相補的なDNAのプローブにより同定することができる。これらのマーカーは例えば蛍光顕微鏡、オートラジオグラフィーなどによって検出することができる。
【0005】
用いる検出技術が何であれ、目印を付ける常法は、検出を可能とするエレメント(蛍光色素、放射性元素など)に結合されたプローブ、すなわち、対象領域に相補的な配列の合成からなる。いくつかの領域を識別するために、主要な方法は、それらの長さまたは検出可能なエレメントの性質(一般に、色の異なる蛍光色素)などのそれらの固有の特性に基づいて識別可能なプローブを設計することである。この場合、プローブは、個別に検出される場合であっても同定可能である(それらは異なる性質のものである)ことから識別可能である。
【0006】
このアプローチは、極めて多数の領域を識別するようになると、すぐにその限界に達し、プローブの長さは技術的制約となり、異なる検出可能なエレメントの数が制限される場合が多い。
【0007】
実際、いくつかのドメインを同時に区別可能な方法でマークする必要のある場合が多い。区別可能なマーカーを開発する努力は基本的に、異なる「性質」の、すなわち、用いる検出方法によって個々に区別することができるマーカーを併用し、蛍光を用いた顕微鏡においてスペクトルの異なる蛍光色素を結びつけ、蛍光およびオートラジオグラフィーを用いた顕微鏡において異なるプローブ長を用いることからなる。これらのアプローチに伴う主要な制限は、このようにして得られた異なるマーカーの数が限定されることである。さらに、性質の異なるマーカーの同時検出は一般に、実施するには繊細で時間のかかる獲得方法を用いざるを得ない。
【発明の概要】
【0008】
本発明は、供試巨大分子上の少なくとも1つの対象ドメインの存在および/または位置を検出する方法に関し、該方法は、
a)対象ドメイン上の少なくとも2つの標的領域を予め決定し、各標的領域の対応する標識プローブ(対象ドメインのプローブのセットと呼ぶ)を設計および取得し、これらのプローブのいずれかの、他のものに対する位置が選択され、供試巨大分子上の対象ドメインの特異的サインを形成する工程;
b)工程a)で得られたプローブが結合されている供試巨大分子を展開した後、線状化巨大分子に結合されているプローブのいずれかの、他のものに対する位置を検出し、対象ドメインのサインが検出されれば、供試巨大分子上に対象ドメインが存在することを示し、逆に、対象ドメインのサインまたはサインの一部が検出されなければ、供試巨大分子上に対象ドメインまたはドメインの一部が存在しないことを示す工程
を含む。
【0009】
一実施形態では、この巨大分子は核酸、特にDNA、より詳しくは、二本鎖DNAである。
さらなる実施形態では、プローブのセットは、さらに2つのプローブ(プローブ1またはプローブ2)を含み、各プローブは対象ドメインの異なる末端に結合可能であり、対象ドメインにおいてその連続プローブと関連する(開始部または終了部の末端プローブ結合と呼ぶ)該プローブ1またはプローブ2の一方のシグナル(signal)を読み取ることにより、読み取りの開始または終了に関する情報が得られる。
【0010】
さらなる実施形態では、本発明は、少なくとも2セットの3つのプローブを含み、各プローブセットは1つのドメイン上に特異的に結合することができ、巨大分子上での1セットの少なくとも3つのプローブの結合が、少なくとも2つの異なるコード群(例えば、「ドット」および「ダッシュ」)から選択される少なくとも2つのコードの配列を形成し、この群が総てのドメインで同一であり、このコード配列が1つのドメインに特異的であり、そのドメインのサインである、巨大分子上の少なくとも2つの対象ドメインの存在を検出するためのキットに関する。
【詳細な説明】
【0011】
本発明者らは今般、性質の異なる極めて少数のプローブを用いて、あるいはある単一の性質のプローブ用いても、事実上無限数の領域の識別が可能なプローブを設計するための新規なアプローチを発見した。このアプローチでは、目印は、各目印に独特な記号を一緒に作り出す数個のプローブからなり、それはコードの配列であり、1つのコードは連続する2つのプローブの読み取りによって得られる。このモチーフ(コード配列)はプローブの性質と位置を合わせたものであり得る。
【0012】
本発明は、このアプローチの適用のいくつかの例を示し、その1つはある1つの遺伝子座における複製の研究に首尾よく使用された。この場合、プローブのある単一の性質が用いられた。また、性質の異なる極めて少数のプローブを用い、全染色体の大きさを解像度の高い領域でいかにマークできるかも示される(例えば、3つの性質のプローブは解像度40kbで60Mbをマーク可能である)。
【0013】
我々の発明は、性質が1つだけであるが、各ドメインに関して単一の連続するプローブを形成する(各ドメインに関して単一のコード配列を形成する)ように対象ドメインにおいて空間的にレイアウトされたマーカーを用いることにより、異なる方法でドメインをマークすることからなる。この方法の可能性のある実施の1つは、ゲノムの一部のドメインによる文字(コード)を用いることにより、モールスコード(Morse code)を採用することからなるので、本発明者らはコードネームを遺伝子モールス(Genetic Morse)とし、モールスコードの「ドット」と「ダッシュ」をそれぞれ短い、または長い間隔で隔てられたマーカーに置き換える。
【0014】
あるドメインの特定のコード配列の開始部と終了部を一義的に位置決定することが必要であるか、またはより有効である場合がある。この位置は例えばマーカーの「ダブレット(doublet)」(開始部と終了部の末端プローブ対、これらのプローブは、マーカー(標識)、すなわち、あるドメインの特定のコード配列の中で用いられる総てのものより短い間隔で隔てられている2つのマーカーで標識される)を用いてコード配列の開始部と終了部をマークすることにより得ることができる。コード配列の開始部と終了部をマークする「ダブレット」における短い間隔は長さの異なるものであってよく、コード配列の配向が可能となる。
【0015】
一般的な方法では、本発明者らの方法は、空間的にレイアウトされた、性質が1つだけのある数のマーカーからなる目印の巨大分子に、各目印に特異的なコードの配列を形成するように区切りを入れることからなり、これらのマーカーは検討される巨大分子に直接関連するものである。
【0016】
これらのマーカーが総て同じである場合、これらのマーカーの検出の多くの様式が可能である(オートラジオグラフィー、シンチレーション、蛍光、化学発光など)。実際には無限数の、それぞれあるドメインに特異的なコードの配列を得ることができ、唯一の制限は使用する方法の空間的解像度とマーカーの特定の配置の緯度である。
【0017】
明らかに、本発明によれば、性質の異なるマーカーが使用可能であれば、制限される数においても、これらの異なるマーカーの組合せによりコード配列を得ることができる。従って、目印のコード化の可能性はいっそう多い。
【0018】
本願の実施例1は、これらの目印により境界が定められた正確な遺伝子座における生物現象(実際には、DNA複製)の研究に関する。1.5Mbのドメインは、モールスコードから直接示唆された4つのコード配列を用いてマークされている。これら4つのコード配列のそれぞれは、性質が1つだけ(1つだけの蛍光色素および同じ長さ)の、可変数の5または7プローブからなり、可変様式で間隔を置いた、蛍光により検出される(図5a)。対象ドメインの各開始部は、別個の「開始部末端プローブ対」によりマークされ、この対の各プローブは17kbの間隔があり、それぞれ対象ドメインの終了部は「終了部末端プローブ対」によりマークされ、この対の各プローブは6kbの間隔がある。これらのコードはモールスコードの最初の4文字に相当し、ドットは短い距離(およそ25〜30kb)離れたプローブからなり、ダッシュは長い距離(55〜70KB)離れたプローブからなる。
【0019】
また、規則的に間隔を置いた、2つの異なる性質のマーカーを用いることで、モールスコードに従いつつ、ビーコン(beacon)をコードすることもできる(図5b)。例えば、蛍光では、モールスの「ドット」に特定の色が与え、「ダッシュ」に別の特定の色を与えることができる。この方法はより「小型(compact)」であるという利点を有し、すなわち、形成するコード配列全体が全体としてより短い配列中に含まれ、これが有利となり得る。
【0020】
前記の場合と同様に、あるドメインに特異的なコードの配列の開始部と終了部をマークすることが望ましい場合がある。それぞれ異なる色でマークされた2つのプローブのダブレット(または対)が使用でき、これらは体系的に同じ方向に向かせることができる。そして、このダブレットはそのコード配列の読み取りの方向を示す。
【0021】
また、これまでに記載した2つのシステムを組み合わせることもでき(図5c)、「ドット」は、短いスペースで隔てられた同じ色の2つのプローブからなり、「ダッシュ」は、長いスペースで隔てられた、異なる色の2つのプローブからなる。この2つのコードシステムにより、より強固となる。
【0022】
特に本発明の地図作成適用、例えば分子診断では、配列の変動が予測され、そのような変動はプローブにより形成されるコード配列を改変し、改変されたプローブ間が無くなったり、繰り返されたり、スペースができたりする可能性がある。コード配列の1つがこのように改変されれば、コードシステムを1つだけ用いる場合、良い場合には、そのコード配列を解説することができなくなり、悪い場合では、別のものとの混同が起こるおそれがある。2つの冗長システムを用いる場合には、別のコード配列との混同が起こるおそれがなくなり、良い場合には、コード配列を同定することができ、配列変動が明らかに定義できる(図6)。
【0023】
本発明に記載されるプローブ設計のアプローチの利点の1つは、少数の異なる「色」の標識で極めて多数のプローブを作出できることである。
【0024】
この特徴を強調するいくつかの実施を記載する。本発明に従い、3色だけを用いて作出した7つのプローブを用い、均一な長さおよび間隔のプローブで、2187(=37)の異なるコードを作出することができる。さらに、これら2187のコードを、1つのコードをなす最後の6つのプローブが次のコードの最初の6つのプローブに相当するような順序に並べることができる。
【0025】
これらのコードを重ね合わせると、7つの連続するプローブが、独特なコード、よってこれらのプローブが位置している配列上の独特な部位、従って、巨大分子上の特定の対象ドメインを定義する2193の一連のプローブを得ることができる。
【0026】
よって、例えば、50kbごとにプローブを配置しつつ、100Mbを超えるものを作出することができる(例えば、3色のカラーまたは擬似カラーで5kbプローブ)。少なくとも7プローブ(従って、400kbを超える総てのフラグメント)を検出可能な各DNAフラグメントの元の位置(100Mb−配列における位置)が明らかに同定される。図7の配列は3色のプローブに対応するA、BおよびCの3文字、総数2193文字からなり、各連続7文字は1回である。
【0027】
この実施の利点は、一方で、それがプローブの数と利用可能なフラグメント(すなわち、全配列において明白な方法で記録可能なもの)の解像度と割合の間の関係を至適化することであり、他方で、この実施が容易に自動化でき、これら一連のプローブがデータ処理によって容易に発見できることである。
【0028】
対称配列のフラグメントの配向の問題に伴う解決策は、対称性とは無関係に上記の連続プローブに「一定の方向を向いた」プローブを周期的(例えば、7プローブごと)に導入することである。例えば、7プローブごとに、プローブの「色」とその配向を同時に与えるダブレットに置き換えることができる。ダブレットは2つの密に置かれたプローブからなる。よって、「A」を「AB」ダブレットに「B」を「BC」ダブレットに、「C」を「CA」ダブレットに置き換えることができる。
【0029】
本発明によって有利な多くの利点がある。実際に、先行技術では、これまでの開発努力は異なる検出可能なエレメントの数を掛け合わせることからなった。特に、これにより主要な検出法として蛍光検出を使用することになるが、それはプローブにタグ付けするために多くの異なる色を使用できるのは蛍光だけであるからである。これに関し、限定されたカラーセットを用いることで、獲得時間および分析時間を著しく軽減することができる。さらに、1つのみ、または2つの異なる検出可能なエレメントを保有することができれば、はるかに迅速かつより容易に自動化可能な検出技術が使用可能となる。最後に、蛍光検出であっても、プローブの異なる、潜在的性質の数にはなお限界があるが、本発明者らの方法は事実上無限数の目印を作出することができる。
【0030】
結果として、一態様において、本発明は、供試巨大分子上の少なくとも1つの対象ドメインの存在および/または位置を検出する方法に関し、該方法は、
a)対象ドメイン上の少なくとも2つの標的領域を予め決定し、各標的領域の対応する標識プローブ(対象ドメインのプローブのセットと呼ぶ)を設計および取得し、これらのプローブのいずれかの、他のものに対する位置が選択され、供試巨大分子上の対象ドメインの特異的サインを形成する工程;
b)工程a)で得られたプローブが結合されている供試巨大分子を展開した後、線状化巨大分子に結合されているプローブのいずれかの、他のものに対する位置を検出し、対象ドメインのサインが検出されれば、供試巨大分子上に対象ドメインが存在することを示し、逆に、対象ドメインのサインまたはサインの一部が検出されなければ、供試巨大分子上に対象ドメインまたはドメインの一部が存在しないことを示す工程
を含む。
【0031】
これらのプローブのいずれかの、他のものに対する位置は、供試巨大分子上の対象ドメインの特異的サインを形成するように任意に、かつ特異的に選択される。
【0032】
任意にとは、本発明によれば、当業者が、いずれの条件、特にドメイン配列に依存せずに対象ドメイン上のプローブの位置を選択することができることを意味する。
【0033】
特異的にとは、本発明によれば、当業者が、巨大分子上のその有無の決定を可能とする対象ドメインの特異的サインを形成するように、対象ドメイン上のプローブの位置を選択することができることを意味する。このサインは当業者により選択され、唯一の条件は異なる対象ドメインに対して異なるサインを与えることである。
【0034】
2つの連続するシグナル間の間隔(またはギャップ)は、例えば顕微鏡またはオートラジオグラフィーが用いられる場合には直接的測定、または検出方法に応じて他の好適ないずれかの方法により測定される。
【0035】
一実施形態では、対象ドメインの存在の検出は部分的サインの検出であってもよく、例えば、1以上のプローブが存在しないか、または誤った位置に結合している場合(例えば、対象ドメイン上のその連続するプローブとともにより長い、またはより小さい間隔がある場合)には、この部分的サインは巨大分子上の対象ドメインの改変の指標となる。巨大分子がDNAである場合、この改変は、巨大分子上の対象ドメインの1個または数個のヌクレオチドの欠失、挿入または置換などの突然変異であり得る。
【0036】
一実施形態では、本方法は好ましくは、少なくとも2つの対象ドメイン存在および/または位置を決定するためのものであり、工程a)において対象ドメイン上の少なくとも3つの標的領域を予め決定することを含む。
【0037】
一実施形態では、本発明の対象ドメインのサインは、連続するプローブ間の一連の間隔(spacing)から生じるものである。
【0038】
一実施形態では、本発明の対象ドメインの位置は、化学反応または生化学反応を位置決定するための参照として用いられる。
【0039】
一実施形態では、本発明の対象ドメインの位置は、標的領域を包含する巨大分子において物理的地図を確立するために用いられる。
【0040】
一実施形態では、本発明の対象ドメインのサインは、一連の異なる標識プローブからなる。
【0041】
一実施形態では、本発明の標的領域のプローブのいくつかは、巨大分子上の付近に位置する少なくとも1つの他の対象ドメインのサインの一部でもある。
【0042】
一実施形態では、巨大分子は核酸、タンパク質、ポリマーまたは炭水化物である。特に、巨大分子はDNA、二本鎖DNAまたは一本鎖DNA、より詳しくは、分子コーミングの場合の二本鎖DNAがDNAの線状化に用いられる。
【0043】
本明細書では互換的に用いられるが、「オリゴヌクレオチド」、「核酸」および「ポリヌクレオチド」とは、一本鎖型または二本鎖型のいずれかの1を超えるヌクレオチドのRNA、DNAまたはRNA/DNAハイブリッド配列を含む。
【0044】
本明細書において、「核酸」および「核酸分子」は、DNA分子(例えば、cDNAまたはゲノムDNA)およびRNA分子(例えばmRNA)ならびにヌクレオチド類似体を用いて作製されたDNAまたはRNAの類似体を含むものとする。核酸分子は一本鎖であっても二本鎖であってもよいが、好ましくは、二本鎖DNAである。本明細書を通じ、「ヌクレオチド配列」という表現は、ポリヌクレオチドまたは核酸を等しく表すために用い得る。より厳密には、「ヌクレオチド配列」という表現は核酸材料それ自体を包含し、よって、特定のDNAまたはRNA分子を生化学的に特徴付ける配列情報(すなわち、4つの塩基文字から選択される連続する文字)に限定されない。また、本明細書では、「核酸」、「オリゴヌクレオチド」および「ポリヌクレオチド」は互換的に用いられる。
【0045】
巨大分子が一本鎖DNAであり、プローブがオリゴヌクレオチドである場合、「結合する」とは「ハイブリダイズすること」を意味する。
【0046】
本明細書において、「ハイブリダイゼーション」、「とハイブリダイズする」または「ハイブリダイズしている」とは、中ストリンジェントまたは高ストリンジェントのハイブリダイゼーション条件、好ましくは、ハイブリダイゼーションおよび洗浄条件が互いに少なくとも60%相同なヌクレオチド配列が互いにハイブリダイズされたままとなる場合を表すものとする。
【0047】
好ましくは、これらの条件は、互いに少なくとも約70%、より好ましくは少なくとも約80%、いっそうより好ましくは少なくとも約85%、90%、95%または98%相同な配列が互いにハイブリダイズされたままとなるようなものである。ストリンジェント条件は当業者に公知であり、Current Protocols in Molecular Biology, John Wiley & Sons, N.Y. (1989), 6.3.1-6.3.6に見出せる。
【0048】
好ましい配列と最適に整列した後、少なくとも80%、好ましくは85%、90%、95%および98%の同一性%を有する核酸配列とは、参照核酸配列に対して、特に欠失、末端切断、延長、キメラ融合および/または置換、特に点置換などのある特定の修飾を有する核酸配列を示すものとする。それは好ましくは、配列が参照配列と同じアミノ酸配列をコードする場合の配列(これは遺伝コードの縮重に関連する)、または好ましくは特に以下に定義されるものなどの高ストリンジェント条件下で、参照配列と特異的にハイブリダイズし得る相補的配列に関する。
【0049】
高ストリンジェント条件下でのハイブリダイゼーションは、温度条件およびイオン強度条件が相補的DNAの2つのフラグメント間でハイブリダイゼーションの維持が可能なように選択されることを意味する。例として、上記のポリヌクレオチド断片を定義することを目的とするハイブリダイゼーション工程の高ストリンジェンシーは有利には次の通りである。
【0050】
DNA−DNAまたはDNA−RNAハイブリダイゼーションは2工程で行われる:(1)42℃、3時間、5×SSC(1×SSCは0.15M NaCl+0.015Mクエン酸ナトリウム溶液に相当する)、50%のホルムアミド、7%のドデシル硫酸ナトリウム(SDS)、10×デンハート溶液、5%の硫酸デキストランおよび1%のサケ精子DNAを含有するリン酸バッファー(20mM、pH7.5)中でのプレハイブリダイゼーション;(2)プローブサイズに応じた温度(すなわち、プローブサイズ>100ヌクレオチドの場合は42℃)で20時間、実際のハイブリダイゼーションの後、20℃で20分、2×SSC+2%のSDS中での洗浄2回、20℃で20分、0.1×SSC+0.1%のSDS中での洗浄1回。最後の洗浄は、0.1×SSC+0.1%のSDS中、プローブサイズ>100ヌクレオチドの場合には60℃で30分行う。当業者であれば、Sambrook et al., (1989, Molecular cloning: a laboratory manual. 2nd Ed. Cold Spring Harbor)の教示に従い、所定サイズのポリヌクレオチドに関して上記された高ストリンジェントのハイブリダイゼーション条件を、より大きな、またはより小さなサイズのオリゴヌクレオチドにも適合させることができる。
【0051】
一実施形態では、これらのプローブは、少なくとも15ヌクレオチド、好ましくは少なくとも1Kb、より好ましくは1〜10kbの間、いっそうより好ましくは4〜10kbの間のオリゴヌクレオチドである。
【0052】
コーミングしたDNA(combed DNA)上での最大解像度が1〜4kbであることから、本発明のプローブは好ましくは少なくとも4kbである。
【0053】
一実施形態では、巨大分子の線状化は、巨大分子上にプローブが結合する前または結合した後に行われる。
【0054】
一実施形態では、巨大分子の線状化は分子コーミングまたはファイバーFishによって行われる。
【0055】
分子コーミングは公表されている方法に従って行う(Lebofsky and Bensimon, 2005)。分子コーミング技術を用い、大きなゲノム領域におよぶ単一ゲノムの物理的特性決定が可能である。単一のコームDNA分子のアレイは、後退エア−ウォーターメニスカスにより、シラン化ガラス面にそれらの末端で結合した分子を延長することにより作製される。コームDNA上で蛍光ハイブリダイゼーションを行うことにより、ゲノムプローブの位置を直接可視化することができ、物理的地図を構成するため、例えば、微小再配列を検出するための手段を提供する。単一分子DNA複製もまた、コームDNA分子上に組み込まれたヌクレオチド類似体の蛍光検出によりモニタリングすることができる。
【0056】
FISH(蛍光in situハイブリダイゼーション)は、染色体上のDNA配列を検出および位置決定するために使用可能な細胞遺伝学的技術である。それは、それらの染色体部分だけに結合する蛍光プローブを用い、それをもって高い配列類似性を示す。蛍光顕微鏡により、蛍光プローブが染色体に結合している場所を見つけることができる。
【0057】
FISH法では、まず、プローブを構築する。プローブはその標的と(ゲノム内の類似の配列とではなく)特異的にハイブリダイズするのに十分長くなければならないが、ハイブリダイゼーション法を妨げるほど大きくてはならず、蛍光団、抗体に対する標的またはビオチンで直接タグ付けすべきである。これは例えばニックトランスレーションおよびタグ付きヌクレオチドを用いたPCRなどの種々の方法で行うことができる。その後、染色体調製を行う。染色体は基板、通常はガラスに固定する。作製後、このプローブを染色体DNAに適用し、ハイブリダイズを開始する。数回の洗浄工程で、ハイブリダイズしていないか、または部分的にしかハイブリダイズしていないプローブは総て洗い流される。シグナル増幅が顕微鏡の検出閾値を超える必要がある場合には(プローブの標識効率、プローブの種類および蛍光色素などの多くの因子によって異なる)、蛍光タグ抗体またはストレプトアビジンをタグ分子と結合させて蛍光を増幅する。最後に、このサンプルは漂白防止剤に包埋し、蛍光顕微鏡で観察する。
【0058】
ファイバーFishでは、間期染色体を、従来のFISHの場合のように強固に巻きつける、あるいは間期FISHの場合のようにランダムコンフォメーションを採用するのではなく、直線上に引き延ばすような方法でスライドに付着させる。これは、スライドに固定した後に溶解させた細胞か、精製されたDNAの溶液のいずれかにスライドの長さに添って機械的剪断をかけることで行われる。これらの染色体の延長コンフォメーションは、数キロベースに引き下がったとしても、劇的に高い解像度をもたらす。しかし、ファイバーFishサンプルの調製は、概念的に単純ではあるが、むしろ熟練技術であり、専門の研究室だけが慣例的にその使用が可能であることを意味する。
【0059】
ファイバーFish法のプロトコールは上記されている。
【0060】
器具および試薬:
・リンパ芽球状細胞培養
・PBS
・血球計算板
・溶解溶液
・5部の70mM NaOH、2部の無水エタノール(Fidlerova et al. 1994)。この溶液は室温で数ヶ月保存可能である。
【0061】
方法
・健全な培養物から細胞懸濁液1〜2mlを採取する。
・5ml PBSで2回洗浄する。
・1ml PBSに再懸濁させる。
・血球計算板を用いて細胞のアリコートを計数する。
・細胞を追加のPBSで希釈し、終濃度をおよそ2×106/mlとする。
・10μlの細胞懸濁液を清浄な顕微鏡スライドの上部に1cmの面積に広げる。
・風乾する。
・スライドをプラスチック製のCadenza(Shandon Southern)チャンバーおよびクランプにほぼ垂直の位置に取り付ける。
・このcadenzaの上に150μlの溶解溶液を適用する。
・レベルがスライドのつや消しのエッジより下に下がれば、200μlのエタノールを加える。
・軽く排水する。
・エッジを持って、スライドとcadenzaユニットをクランプの外へ注意深く持ち上げる。
・cadenzaからスライドの背面の上部を抜き、メニスカスをスライドの下へ移動させる。
・アングルで風乾させる。
・アセトン中で10分固定する。スライドは室温で数ヶ月、問題なく保存することができる。
【0062】
一実施形態では、総てのプローブは同じ標識で標識される。
【0063】
一実施形態では、プローブは少なくとも2つに異なる標識で標識される。一実施形態では、プローブは3つの標識で標識される。
【0064】
一実施形態では、結合またはハイブリダイズしたプローブは、プローブと結合している1以上の標識を検出することにより検出される。これらの標識は当業者に周知のいくつかの手段のいずれによって組み込んでもよい。しかしながら、好ましい実施形態では、標識は、プローブ調製時の増幅工程中に同時に組み込まれる。例えば、標識プライマーまたは標識ヌクレオチドを用いたポリメラーゼ連鎖反応(PCR)により、標識された増幅産物が得られる。プローブ(例えばDNA)は標識デオキシヌクレオチド三リン酸(dNTP)の存在下で増幅される。
【0065】
好ましい実施形態では、標識ヌクレオチド(例えば、蛍光標識UTPおよび/またはCTP)を用いた上記のような転写増幅が転写された核酸に標識を組み込む。
【0066】
あるいは、標識は、ものとプローブ(例えば、mRNA、ポリA mRNA、cDNAなど)または増幅が完了した後の増幅産物に直接加えてもよい。このような標識により、高収量の増幅産物が得られ、増幅反応に必要な時間が短縮される。プローブに標識を付着させる手段としては、例えば、ニックトランスレーションまたは核酸のキナーゼ処理の後に標識(例えば蛍光団)にプローブを結びつける核酸リンカーを付着(連結)させることによる末端標識(例えば、標識RNAを用いる)が挙げられる。
【0067】
好ましくは、本発明の標識ヌクレオチドは、クロロデオキシウリジン(CIdU)、ブロモデオキシウリジン(BrdU)および/またはヨードデオキシウリジン(IdU)である。
【0068】
本発明に用いるのに好適な検出可能な標識としては、顕微鏡的手段、光化学的手段、生化学的手段、免疫化学的手段、電気的手段、光学的手段または化学的手段によって検出可能ないずれの組成物も含む。本発明において有用な標識としては、標識ストレプトアビジンコンジュゲートで染色するためのビオチン、磁性ビーズ(例えば、Dynabeads(商標))、蛍光色素(例えば、フルオレセイン、テキサスレッド、ローダミン、緑色蛍光タンパク質など。例えば、Molecular Probes, Eugene, Oreg., USA参照)、放射性標識(例えば、3H、125I、35S、14Cまたは32P)、酵素(例えば、ホースラディッシュペルオキシダーゼ、アルカリ性ホスファターゼおよびその他ELISAに慣用されるもの)、ならびに金コロイド(例えば、直径40〜80nm範囲の、高効率で緑色光を発する金粒子)またはカラーガラスもしくはプラスチック(例えば、ポリスチレン、ポリプロピレン、ラテックスなど)ビーズのような比色定量標識が挙げられる。このような標識の使用を教示する特許としては、米国特許第3,817,837号、同第3,850,752号、同第3,939,350号、同第3,996,345号、同第4,277,437号、同第4,275,149号および同第4,366,241号がある。
【0069】
蛍光標識は、低いバックグラウンドで極めて強いシグナルを提供するので好ましい。また、迅速走査法によって高い解像度および感度で光学的に検出することもできる。
【0070】
プローブは総て、単一の標識、例えば単一の蛍光標識で標識することができる。あるいは、別の実施形態では、異なるプローブを、各プローブが異なる標識を有するように同時にハイブリダイズさせることもできる。例えば、ある標的は緑色蛍光標識を有し、別の標的が赤色蛍光標識を有してもよい。走査工程は緑色蛍光標識の結合部位から赤色標識の結合部位を識別する。各プローブ(標的核酸)は互いに独立に分析することができる。
【0071】
使用可能な好適な色素原としては、色が観察できるように識別的な波長域の光を吸収するか、あるいはまた例えば蛍光剤など、特定の波長または波長域の放射線を照射した際に発光する分子および化合物が挙げられる。
【0072】
広範囲の好適な色素が利用可能であり、基本的に、それらの周囲による吸収が最小で強い色をもたらすように選択される。例示的な色素種としては、キノリン色素、トリアリールメタン色素、アクリジン色素、アリザリン色素、フタレイン、昆虫色素、アゾ色素、アントラキノイド色素、シアニン色素、フェンアザチオニウム色素およびフェンアゾキソニウム色素が挙げられる。
【0073】
広範な蛍光剤が単独、あるいはまた消光分子と組み合わせて使用可能である。対象蛍光剤はある主要な官能基を有する多様なカテゴリーに分類される。これらの主要な官能基としては、1−および2−アミノナフタレン、p,p’−ジアミノスチルベン、ピレン、第四級フェナントリジン塩、9−アミノアクリジン、p,p’−ジアミノベンゾフェノンイミン、アントラセン、オキサカルボシアニン、マロシアニン、3−アミノエキレニン、ペリレン、ビスベンズオキサゾール、ビス−p−オキサゾリルベンゼン、1,2−ベンゾフェナジン、レチノール、ビス−3−アミノピリジニウム塩、ヘレブリゲニン、テトラサイクリン、ステロフェノール、ベンズイミダゾリルフェニルアミン、2−オキソ−3−クロメン、インドール、キサンテン、7−ヒドロキシクマリン、フェノキサジン、サリチレート、ストロファンチジン、ポルフィリン、トリアリールメタンおよびフラビンが挙げられる。
【0074】
連結のための官能基を有するか、またはこのような官能基を組み込むよう修飾可能な個々の蛍光化合物としては、例えば、塩化ダンシル;3,6−ジヒドロキシ−9−フェニルキサントヒドロールなどのフルオレセイン;ローダミンイソチオシアネート;N−フェニル1−アミノ−8−スルホナトナフタレン;N−フェニル2−アミノ−6−スルホナトナフタレン:4−アセトアミド−4−イソチオシアナト−スチルベン−2,2’−ジスルホン酸;ピレン−3−スルホン酸;2−トルイジノナフタレン−6−スルホン酸;N−フェニル、N−メチル2−アミノナフタレン(aminoaphthalene)−6−スルホン酸;臭化エチジウム;ステブリン;オーラミン(auromine)−0,2−(9’−アンスロイル)パルミチン酸;ダンシルホスファチジルエタノールアミン;N,N’−ジオクタデシルオキサカルボシアニン;N,N’−ジヘキシルオキサカルボシアニン;メロシアニン、4(3’ピレニル)酪酸;d−3−アミノデソキシ−エキレニン;12−(9’−アンスロイル)ステアリン酸;2−メチルアントラセン;9−ビニルアントラセン;2,2’(ビニレン−p−フェニレン)ビスベンゾオキサゾール;p−ビス[2−(4−メチル−5−フェニルオキサゾリル)]ベンゼン;6−ジメチルアミノ−1,2−ベンゾフェナジン;レチノール;ビス(3’−アミノピリジニウム)−1,10−デカンジイルジヨージド;ヘリブリエニンのスルホナフチルヒドラゾン;クロロテトラサイクリン;N(7−ジメチルアミノ−4−メチル−2−オキソ−3−クロメニル)マレイミド;N−[p−(2−ベンズイミダゾリル)−フェニル]マレイミド;N−(4−フルオランチル)マレイミド;ビス(ホモバニリン酸);レサザリン;4−クロロ−7−ニトロ−2,1,3ベンゾオキサジアゾール;メロシアニン540;レゾルフィン;ローズベンガル;および2,4−ジフェニル−3(2H)−フラノンが挙げられる。
特に、本発明の蛍光標識は、1−クロロ−9,10−ビス(フェニルエチニル)アントラセン、5,12−ビス(フェニルエチニル)ナフタセン、9,10−ビス(フェニルエチニル)アントラセン、アクリジンオレンジ、オーラミンO、ベンズアントロン、クマリン、4’,6−ジアミジノ−2−フェニルインドール(DAPI)、臭化エチジウム、フルオレセイン、緑色蛍光タンパク質、ヘキストステイン(Hoechst stain)、インディアンイエロー、ルシフェリン、フィコビリン、フィコエリトリン、ローダミン、ルブレン、スチルベン、TSQ、テキサスレッドおよびウンベリフェロンである。
【0075】
望ましくは、蛍光剤は約300nm、好ましくは約350nmより上、より好ましくは約400nmより上の光を吸収すべきであり、通常には、吸収される光の波長の約10nmを超える高い波長で発光する。結合色素の吸収および発光特性は結合していない色素とは異なることに注意すべきである。よって、種々の波長範囲および色素の特徴に対して言及する場合、コンジュゲートされておらず、任意の溶媒中で特徴付けられる色素ではなく、使用されるような色素を示すものとする。
【0076】
蛍光剤は一般に、蛍光剤に光を照射することで複数の発光を得ることができるので好ましい。よって、単一の標識で複数の測定可能な事象を得ることができる。
【0077】
本発明によれば、蛍光標識で標識が行われる場合、シグナルの読み取りは蛍光検出によりなされ、蛍光標識されたプローブは光により励起され、その後、その励起の発光が、デジタル画像を取り込み、さらなるデータ解析を可能とする適当な発光フィルターを装備したCCDカメラなどの光センサーにより検出される。
【0078】
また、化学発光源および生物発光源によっても検出可能なシグナルが得られる。化学発光源としては、化学反応により電気的に励起されるようになり、その後、検出可能なシグナルとして働くか、または蛍光アクセプターにエネルギーを与える光を発し得る化合物が含まれる。多様な数の化合物群が多様な条件下で化学発光を与えることが見出されている。1つの化合物群が2,3−ジヒドロ−1,−4−フタラジンジオンである。最も知られている化合物はルミノールであり、これは5−アミノ化合物である。この群の他のものとしては、5−アミノ−6,7,8−トリメトキシ−およびジメチルアミノ[ca]ベンズ類似体がある。これらの化合物はアルカリ性過酸化水素または次亜塩素酸カルシウムおよび塩基で発光をもたらし得る。別の化合物群としては、2,4,5−トリフェニルイミダゾールがあり、親生成物の一般名はロフィンである。化学発光類似体としては、パラ−ジメチルアミノおよび−メトキシ置換基が挙げられる。化学発光はまた、塩基性条件下でシュウ酸塩、通常にはオキサリル活性エステル、例えばp−ニトロフェニルおよび過酸化物、例えば過酸化水素で得ることもできる。あるいは、生物発光を得るために、ルシフェリンをルシフェラーゼまたはルシゲニンと組み合わせて使用することもできる。
【0079】
スピン標識は、電子スピン共鳴(ESR)分光法によって検出可能な不対電子スピンを伴うリポーター分子により得られる。スピン標識の例としては、有機フリーラジカル、遷移金属錯体、特に、バナジウム、銅、鉄およびマンガンなどが挙げられる。スピン標識の例としては、ニトロキシドフリーラジカルが挙げられる。
【0080】
標識はハイブリダイゼーション前または後にプローブ(または標的、これは特に核酸である)に加えることができる。いわゆる「直接標識」は、ハイブリダイゼーション前にプローブに直接結合されるか、組み込まれる検出可能な標識である。これに対し、いわゆる「間接標識」は、ハイブリダイゼーション後にハイブリッド二本鎖に連結される。多くの場合、間接標識は、ハイブリダイゼーション前にプローブに結合されている結合部分に結合される。よって、例えば、プローブはハイブリダイゼーション前にビオチン化することができる。ハイブリダイゼーション後、アビジンとコンジュゲートされた蛍光団を、容易に検出される標識をもたらすビオチン担持ハイブリッド二本鎖に結合させる。核酸を標識し、標識されたハイブリダイズ核酸を検出する方法に関する詳細な総説としては、Laboratory Techniques in Biochemistry and Molecular Biology, Vol. 24: Hybridization With Nucleic Acid Probs, P. Tijssen, ed. Elsevier, N.Y., (1993)を参照。
【0081】
これらの標識は直接、またはリンカー部分を介して結合させることができる。一般に、標識またはリンカー−標識結合部位は特定の位置に限定されない。例えば、標識は、所望により、検出またはハイブリダイゼーションを妨げないいずれの位置でヌクレオシド、ヌクレオチドまたは類似体と結合させてもよい。例えば、Clontech (Palo Alto, Calif.)からのある種のLabel−ON試薬は、オリゴヌクレオチドのリン酸主鎖に散在した標識と3’および5’末端における末端標識を与える。例えば本明細書に示されるように、標識はリボース環上の位置に結合可能であり、このリボースは所望により修飾、さらには排除することもできる。有用な標識試薬の塩基部分は、天然に存在するもの、またはそれらが置かれている目的を妨げない様式で修飾されたものを含み得る。修飾塩基としては、限定されるものではないが、7−デアザAおよびG、7−デアザ−8−アザAおよびGおよび他の複素環式部分が挙げられる。
【0082】
末端標識プローブに関して、多くの適用では、増幅、転写またはその他の変換工程を介して行う必要のない直接的標識プローブに有用である。一般に、末端標識法は、標識される核酸のサイズの至適化を可能とする。末端標識法はまた、ポリメラーゼ補助標識法に伴うことがある配列の偏りを軽減する。末端標識は末端トランスフェラーゼ(TdT)を用いて行うことができる。
【0083】
末端標識はまた、プローブの末端に標識オリゴヌクレオチドまたはその類似体を連結することにより行うこともできる。他の末端標識法としては、例えばリガーゼまたは末端トランスフェラーゼを用いて核酸の標識または非標識「テール」を作出することを含む。次に、このテールを付けた核酸を、そのテールと優先的に結合する標識部分に曝す。このテールとそのテールと優先的に結合する部分は、核酸、ペプチドまたは炭水化物などのポリマーであり得る。このテールとその認識部分は両者の間の認識が可能ないずれのものであってもよく、ハプテン、エピトープ、抗体、酵素とそれらの基質、ならびに相補的核酸およびその類似体などの、リガンド−基質の関係を有する分子を含む。
【0084】
テールまたはテール認識部分と結合される標識は検出可能な部分を含む。テールおよびその認識部分は双方とも標識され、それぞれと結合される個々の標識はそれ自体リガンド−基質の関係を有する。個々の標識はまた、異なる分光光度的特徴を有する色素などのエネルギー移動試薬も含み得る。エネルギー移動対は所望の組合せのスペクトル的特徴が得られるように選択することができる。例えば、第二の色素によって吸収されるよりも短い波長で吸収する第一の色素は、そのより短い波長で吸収する際に、第二の色素へエネルギーを移動する。その後、第二の色素は、第一の色素単独で発せられたものよりも長い波長で電磁線を発する。エネルギー移動試薬は、1995年9月15日出願の米国出願番号08/529,115の一部継続である、1996年12月23日に出願の同時係属中米国特許出願、および1990年12月6日出願の米国出願番号07/624,114の継続出願である、1993年12月15日出願の米国出願番号08/168,904の分割出願である、1996年6月25日出願の米国出願番号08/670,118の一部継続出願でもある国際出願WO96/14839に示されているものなどの2色標識スキームに特に有用であり得る。米国出願番号07/624,114は、参照することにより本明細書の一部とされる1990年6月7日出願の米国出願番号07/362,901の一部継続出願である。
【0085】
一実施形態では、標識が放射性標識で行われる場合、シグナルの読み取りは放射性検出により行われる。
【0086】
放射性検出はX線フィルムまたはホスホルイメージャーを用いて行うことができる。
【0087】
本発明の放射性標識の例は3H、125I、35S、14Cまたは32Pである。
【0088】
好ましい実施形態では、プローブは蛍光標識で標識される。
【0089】
好ましい実施形態では、プローブは放射性標識で標識される。
【0090】
本発明によれば、プローブが少なくとも2つの異なる標識で標識される場合、対象ドメインのサインは連続する標識を生じる。
【0091】
一実施形態では、巨大分子上の対象ドメインに対応する少なくとも3つのプローブの結合は、少なくとも2つの異なるスペース(例えば、「ショート」と「ラージ」)の群(この群は各対象ドメインで同一である)から選択される少なくとも2つのスペースの配列を形成する。
【0092】
一実施形態では、プローブセットはさらに2つのプローブ(プローブ1またはプローブ2)を含み、各プローブは対象ドメインの異なる末端に結合することができ、対象ドメインにおけるその連続するプローブと関連する前記プローブ1またはプローブ2(開始部または終了部の末端(端部)プローブ対(extremity probe couple)と呼ぶ)の1つのシグナルを読み取ることによって、読み取りの開始または終了に関する情報を得ることができる。
【0093】
一実施形態では、読み取りの開始に関する情報は、開始部の末端プローブ対の2つの連続するプローブ間の間隔の読み取りの結果である。
【0094】
一実施形態では、読み取りの終了に関する情報は、終了部の末端プローブ対の2つの連続するプローブ間の間隔の読み取りの結果である。
【0095】
一実施形態では、読み取りの開始に関する情報は、開始部の末端プローブ対の2つの連続するプローブ間の間隔の読み取りの結果であり、読み取りの終了に関する情報は、終了部の末端プローブ対の2つの連続するプローブ間の間隔の読み取りの結果であり、この間隔は、開始と終了の情報を区別するために、開始部の末端プローブ対と終了部の末端プローブ対では異なる。
【0096】
本発明の別の態様は、巨大分子上の少なくとも1つの対象ドメインの存在を同定する方法に関し、前記ドメイン上に特異的に結合し得る少なくとも3つのプローブのセットが設計され、巨大分子上の1セットの少なくとも3つのプローブの結合が少なくとも2つの異なるコードの群((例えば、「ドット」と「ダッシュ」)から選択される少なくとも2つのコードの配列を形成し、この群が各ドメインで同一であり、このコードの配列がそのドメインに特異的であり、そのドメインのサインであり、該方法は、
a)少なくとも3つのプローブの少なくとも2セットを巨大分子と接触させる工程(各セットは各対象ドメインに特異的である);
b)シグナルを読み取ること(1つのシグナルは1つのプローブに相当する);
c)該シグナルをコードで転写する工程;
d)対象ドメインに特異的なコード配列の存在の有無を検出する工程(該コード配列の存在が存在に相当し、巨大分子上のドメインの位置を示す);および
所望により、工程a)の前または工程a)とb)の間に巨大分子を線状化する工程
を含む。
【0097】
好ましい実施形態では、方法は、巨大分子上の少なくとも2つの対象ドメインの存在を同定する。
【0098】
好ましい実施形態では、得られたコードは2つの連続シグナルの特徴の読み取りの結果である。
【0099】
一実施形態では、2つの連続シグナルの特徴はプローブの標識により得られる。
【0100】
一実施形態では、標識は蛍光標識で行われ、特定の蛍光標識は特定のコードに相当する。
【0101】
一実施形態では、標識は放射性標識で行われ、特定の放射性標識は特定のコードに相当する。
【0102】
一実施形態では、得られたコードは2つの連続シグナル間の間隔の読み取りの結果である。
【0103】
一実施形態では、2つのシグナル間の特定の間隔は特定のコードに相当する。
【0104】
例えば、本発明によれば、小さなギャップはドットと呼ばれるコードに相当し、大きなギャップはダッシュと呼ばれるコードに相当する。小さなコードは例えば25〜30kbを測定するように設計することができ、大きなコードは例えば55〜70kbを測定するように設計することができる。
【0105】
一実施形態では、2つの連続シグナルの特徴はプローブの長さによって与えられ、各長さが特定のコードに相当する。
【0106】
本発明は、2つのコードにまで減少されず、3つ、4つまたは4つを超えるコードを含むことができ、コードを定義するためにいずれのサイズのギャップが選択されてもよく、このサイズは任意である。
【0107】
本発明によれば、これらのプローブセットはさらに2つのプローブ(プローブ1またはプローブ2)を含み、各プローブは対象ドメインの異なる末端に結合することができ、対象ドメインにおけるその連続プローブと関連する該プローブ1またはプローブ2(「開始または終了の末端プローブ対」と呼ばれる)の1つのシグナルを読み取ることで、読み取りの開始または終了に関する情報が得られる。
【0108】
一実施形態では、本発明のサインはプローブセットにおける第一のプローブと第二のプローブの間にスペースを含み、このスペースはそのサインにおいて他の総てのスペースと異なり、このスペースはサインの開始に関する情報を得るために使用することができる。
【0109】
一実施形態では、本発明のサインは、プローブセットにおいて最後の手前のプローブと最後のプローブの間にスペースを含み、このスペースはそのサインにおいて他の総てのスペースと異なり、このスペースはサインの終了に関する情報を得るために使用することができる。
【0110】
さらなる実施形態では、本発明は、少なくとも2セットの3つのプローブを含み、各プローブセットは1つのドメイン上に特異的に結合することができ、巨大分子上での1セットの少なくとも3つのプローブの結合が、少なくとも2つの異なるコード群(例えば、「ドット」と「ダッシュ」)から選択される少なくとも2つのコードの配列を形成し、この群が総てのドメインで同一であり、このコード配列が1つのドメインに特異的であり、そのドメインのサインである、巨大分子上の少なくとも2つの対象ドメインの存在を検出するためのキットに関する。
【0111】
本発明は、本出願者が行った研究に関して遂行された実験検討の記載を読めばより明確に理解される。これらは何ら限定と解釈されるべきでない。
【実施例】
【0112】
哺乳類DNA複製開始点は数bp〜数十kbの範囲の部位に位置する。個々の細胞周期における開始の規則的分布は、これら多くの潜在的開始部位のうち限られた数だけが活性化開始点へと変換されることを示唆する。開始点干渉は冗長開始点を封じることができるが、干渉が機能的ヒト開始事象のスペーシングに関与するかどうかは現在のところ知られていない。一次ケラチノサイト由来の単一にコーミングされたDNA分子に対して新規なハイブリダイゼーション戦略である遺伝子モールスコードを用いることにより、本発明者らは1.5Mbのヒト染色体14q11.2上に存在する開始部位を報告する。本発明者らは、開始ゾーンがヒト細胞に拡散していることを確認した。個々の細胞周期において用いられる開始点は、存在し得る開始部位よりも少なく、それらの限定された使用は規則的な開始点間発火距離を作り出す。
【0113】
ゾーン間干渉は活性開始点からの距離に比例して減少するが、ゾーン内干渉は100%有効である。これらの結果によってヒト細胞において有効な開始点の階層的構成が確認される。機能的開始点は、その付近で開始点が発火する確率を支配し、これはゾーン間干渉およびゾーン内干渉によって媒介される。これにより、DNA複製の複数の複雑な、潜在的開始部位に関して、開始事象がヒト染色体の長さに添って一様に分布していることが確認される。
【0114】
導入
真核細胞は、S期の長さによって規定され、ゲノム倍加回数が限定されている。これは、複数の開始部位から伸びる複製分岐においてDNAを合成することにより果たされる。分岐(fork)の速度はS期の長さに応じて尺度化されるのではないので、各個の染色体の開始頻度の調節には有糸分裂の開始前に非複製ギャップを阻害する必要がある(Hand and Tamm, 1973; Edenberg and Huberman, 1975)。例外もあるが、一般的な見解では、体細胞哺乳類開始点は50〜300kb間隔で始まっている(Edenberg and Huberman, 1975; Berezney et al., 2000)。このことは、後生動物が開始事象を一様に分布させる機構を有していることを示唆する。一定の距離に強力なレプリケーター配列を配置することは、発芽酵母サッカロミセス・セレビシエ(Saccharomyces cerevisiae)に用いられているこのような機構の1つである(Newlon et al., 1991; Shirahige et al., 1993)。もっと高等な真核生物では、遺伝エレメントは開始点の活性化に役割を果たすが、それらはそれ自体開始を駆動するには十分なものでない(Gilbert, 2004)。さらに、後生動物の開始点には数bpという限局化された部位に存在するものがあるが、大多数は数十kbまでの範囲のより分散した開始ゾーンに存在する(DePamphilis, 1999)。これにより内因的効率の低い、ある範囲の潜在的部位からいかにして活性化開始点の規則的分布を果たすかという問題が持ち上がる。
【0115】
開始点活性を調節する1つの方法は、それが受動的に複製される確率を変化させることである。開始点付近から伸張している分岐はこの抑制を媒介するので、この形式の開始点の不活性化は「開始点干渉」と呼ばれている(Brewer and Fangman, 1993)。開始点干渉に関する本発明者らの理解の大部分は、S.セレビシエでの研究により得られたものである。発芽酵母では、複製の完了に必要とされるか、または用いられるものよりも多くのより集合した複製前駆複合体(pre−RC)がある(Dershowitz and Newlon, 1993; Raghuraman et al., 2001; Wyrick et al., 2001; Pasero et al., 2002)。酵母染色体IIIおよびVIに対して開始点効率を分析したところ、開始点が細胞周期の5〜90%の間で用いられていることが明らかになった(Friedman et al., 1997; Yamashita et al., 1997; Poloumienko et al., 2001)。許可された開始点は、その近傍の他の開始点に比べて、予定されたS期のタイミングが遅いか、比較的遅いために効果的でない(Santocanale and Diffley, 1996; Vujcic et al., 1999)。結果として、これらの適格開始点は、隣接する開始部位から伸びている分岐により受動的に複製される(Santocanale et al., 1999)。
【0116】
酵母から得られたデータによれば、開始点が互いに干渉すべきであれば、開始点付近がS期の異なる時間に発火する(fire)ようにG1期において予めプログラムされていなければならない(Raghuraman et al., 1997)。しかし、この必要条件は、より高等な真核生物において、1)タイミングの制御が〜100kbという拡大された領域にわたって発揮され(MacAlpine et al., 2004; Norio et al., 2005)、かつ、2)クラスターにおいて互いに次に位置する開始点が同時に発する場合には満足のいくものでない可能性がある(Berezney et al., 2000)。
【0117】
実際、CHO細胞の増幅されたAMPD2遺伝子座では、近傍の開始点間の有意な予め定義されたタイミングの違いは見られなかった(Anglana et al., 2003)。やはり隣接する開始点の、よく定義されたbp部位での同時活性化は遮断された(Anglana et al., 2003)。これが広い開始ゾーンに当てはまるかどうか依然として判定すべきである。
【0118】
規則的な開始間隔がヒト細胞でいかにして果たされるか、また、開始点干渉がこのプロセスに寄与するかどうかを理解するため、本発明者らは開始点活性に関して、一次ケラチノサイト由来のヒト染色体14q11.2の1.5Mb領域に疑問を持った。次のような理由で、分子コーミング技術を採用した一分子アプローチを選択した。第一に、特定の細胞種におけるDNA複製の総ての潜在的開始部位の位置を特定するために十分な開始点発火事象が得られる。第二に、本発明者らは、単一の細胞が個々のS期においてどの開始点を用いるかということと、それらの互いに対する活性化のタイミングを決定することができる。これには開始事象の時空間分布を確認する必要がある。これらのデータを組み合わせて、既に発火した開始点が下流の、潜在的開始部位の使用を調節するかどうかを評価した。本発明者らは、開始点が活性開始点によって確立された階層に従って互いに自己調節し、予め定義されたタイミングの優先なく、確率論的に選択されることを見出した。さらに、開始点干渉により、開始事象の間隔の保存がもたらされる。その理由およびヒト開始点干渉の実行に用いられる機構が議論される。
【0119】
材料および方法
DNA調製
正常なヒト一次ケラチノサイトを皮膚生検から誘導した。細胞を標準的なケラチノサイトSFM中で培養した。その後、新生DNAをそれぞれ記載されているように(Lebofsky and Bensimon, 2005)、IdUおよびCIdUで20分間標識した。DNA抽出およびコーミングは標準的なプロトコールに従って行った。
【0120】
ハイブリダイゼーションおよび蛍光呈示
表S1に示されているプライマー対を用い、広範囲PCRにより、25GMCプローブを作製した。5〜7kbのプローブの増幅を助けるため、TaKaRa LA Taqを用いた(Takara Bio)。PCR産物の鋳型として用いたBACを表S2に示す。プローブをそれらの記号(A、B、CおよびD)に従い、終濃度〜20ng/μlでプールした。プローブのビオチン化は、個別に4つの記号のランダムプライミング(Invitrogen)を行うことにより達成した。個々のスライドアッセイについては、〜250ngの各ビオチン化プローブを10μgのヒトcot−1 DNA(Gibco BRL)と合わせた。その後のコームDNAに対するハイブリダイゼーションは公表されている方法(Lebofsky and Bensimon, 2005)に従った。プローブIdUおよびCIdUを検出する免疫蛍光工程は次の通り:(i)Alexa 488コンジュゲートストレプトアビジン(Moleccular Probs)、マウス抗ブロモデオキシウリジン(Becton Dickinson)およびラット抗ブロモデオキシウリジン(Harlan Seralab);(ii)ビオチンコンジュゲートウサギ抗ストレプトアビジン(Rockland)、Alexa 350コンジュゲートヤギ抗マウス(Moleccular Probs)およびテキサスレッドコンジュゲートロバ抗ラット(Jackson);(iii)Alexa 488コンジュゲートストレプトアビジン(Moleccular Probs)およびAlexa 350コンジュゲートロバ抗ヤギ(Moleccular Probs)。抗体のインキュベーション、洗浄およびスライドのマウントはこれまでに報告されている通りに行った(Lebofsky and Bensimon, 2005)。
【0121】
画像の採取
HQ CCDカメラを装備したZeiss Axioplan 2顕微鏡(Photometries)を用いて画像の半分を取り込んだ。あとの半分をCytoscoutハイスループットスキャニング装置(Upper Austrian Research)を用いて採取した。バックグラウンド蛍光のドットを、Photoshop(Adobe)を用いて除去し、対象分子を強調した。
【0122】
結果
ゲノムモールスコードハイブリダイゼーション戦略
ある細胞周期の中でどの開始点が活性、またサイレントなのかを分析するための第一段階として、ある領域内の総ての可能性ある開始部位を得た。このために、本発明者らは分子コーミングに基づく単一分子アプローチを用いた(Bensimon et al, 1994)。正常な核型を示す非同期性ヒト一次ケラチノサイトに20分間ヨードデオキシウリジン(IdU)、次いでクロロデオキシウリジン(CIdU)のパルスを与えた。この細胞集団からDNAを抽出し、コーミングした。新たに合成されたDNAにおけるBrdU類似体の組み込みをIdU(青)およびCIdU(赤)の免疫検出によって可視化した。この実験パラダイムは、二方向性複製の開始部位が推察できる3種類のシグナルを生じる(図1A;詳細はAnglana et al, 2003; Lebofsky and Bensimon, 2005を参照)。コーミングBrdUのFISHによる呈示により、複製トラックがプローブシグナルとどこで重複しようと、開始を特異的配列トラックに帰すことができる。
【0123】
慣例的に、スライド上のゲノム領域を可視化するためには色が異なる(図1Bi)か、サイズが異なる(図1Bii)プローブ対が用いられる。しかし、DNAの破断が、それらがカバーできる距離を制限する。結果として、このハイブリダイゼーション戦略を用いた染色体のウォーキングダウンは、実験がプローブ対の数に比例して増えるので、時間がかかる方法である。理想的には、数色および/または大きな領域をカバーするために様々な長さのプローブを使用することができるが、これらの戦略はそれぞれスペクトルの重複および繰り返し配列の非特異的ハイブリダイゼーションのために適切なものではない。
【0124】
本発明者ら独自の解決策の第一の部分は、サイズの異なるギャップが、色またはサイズの異なるプローブと同じ情報を与えることを実現したことによる。図1Biiiに示される例では、ギャップ1はあるプローブセットにより定義され、ギャップ2は別のプローブセットにより定義される。また、この2つのプローセット間のギャップサイズはギャップ1および2とは明瞭に異なる。ギャップ3はDNA破断の際に有用となる。その助けで、その分子は完全なプローブセットが可視化されなくてもなお一定方向を向くことができる(図1Bivおよびv)。ギャップは位置情報を与えるので、それらの数にもはや制限はなく、すなわち、スペクトルの重複やハイブリダイゼーションの際の繰り返し配列はもはや問題でない。サイズの異なるギャップを用いることで、ヒト染色体14q11.2において〜1.5Mbに及ぶ遺伝子モールスコード(GMC)が作製された(図1C)。この全GMCを個々のアッセイでハイブリダイズさせ、総てのプローブが緑で検出された。分子コーミングに先立つDNA操作は繊維をランダムな位置で破断させる。しかし、繊維のサイズは個々の分子上で複数の記号を可視化するに十分なものであった。これに対し、繊維破断のため、記号から数プローブだけしか検出されない場合もあった。開始点は、複製トラックが解読可能なGMCプローブセットと同時に局在した開始点を示す限りマッピングされた(図1D)。よって、新規なハイブリダイゼーション戦略GMCにより、限られた数の実験で、大きな領域の開始点マッピングが可能であった。
【0125】
1.5Mbのヒト染色体14q11.2に対する開始点マッピング
この実験パラダイムを用い、本発明者らは、GMC領域において232の単一DNA分子に対し、307の開始事象を検出した。優先的開始ゾーンを客観的に確立するために、データのクラスター化を行った。まず、本発明者らは、階層的クラスタリングツリーを作出した。最適な区切りを設けるためには、クラスター内のデータの分散を最小化し、クラスター間の分離は最大にすべきである。これら2つの特徴を級内分散(W)および級間分散(B)と呼ぶ(図2A)。従って、望ましいクラスターセットは小さなWと大きなBまたはB−Wの最大値を持たなければならない。これらのデータを9、22および45のクラスターに分けたところ、比較的高いB−W値が得られた(図2A)。図2Bは、開始事象がこれらのクラスターセットに従ってどのように分割されるかを示している。データを45のクラスターに分けると、最も狭い開始領域が得られる。本発明者らは、これら45のクラスターを個々の開始ゾーンを示すものと考えた(図2C)。これらのクラスターの中には極めて少ない開始事象しか含まないものがあり、これはバックグラウンドのノイズによるものでる可能性がある。よって、3つを超える開始事象を有するクラスターだけを次の分析に用いた。このようにして、38の開始ゾーンを同定した。
【0126】
開始ゾーンのサイズは2.6kb(最小)〜21.6kb(最大)の間で様々であり、平均は13.5+/−5.2kbであった。これらの値は哺乳類細胞に関して報告されている(DePamphilis, 1999)他の開始ゾーンの範囲内にある。このゾーン内で、1〜2kbの特定の開始部位は見られなかった。
【0127】
これは、開始点ピークを識別するためのデータ数が不十分なためである可能性があった。コーミングされた分子上の最大解像度は1〜4kbの間である。よって、これらの開始の優先性が、マッピングされた各開始点に固有の標準偏差の結果として取り払われた可能性もある。
【0128】
あるいは、散在した発火部位を含む開始ゾーンがこれまでに考えられていたものよりも優勢である可能性がある。本発明者らのデータは、他のもっと解像度の高い技術でプローブ可能な標的領域を提供する(Todorovic et al, 2005)。よって、これらの可能性を区別することができる。
【0129】
活性化された開始点近傍の時空間分析
本発明者らは次に、開始ゾーンが互いに対してどのように分布するかということに注意を向けた。ゾーン間の距離を測定したところ、ゾーン間平均は40.6+/−20.7kb(最小値=14.3kb、最大値=93.1kb)であることが明らかになった。哺乳類細胞の開始点間距離が一般に100〜150kbであること(Berezney et al, 2000)を考えれば、これは驚くべきことである。この相違は、細胞周期ごとにゾーンのサブセットだけが活性化されるとすると説明がつく。この可能性を調べるため、本発明者らは個々の繊維上の複数の開始点間の間隔を分析した(図3A)。
【0130】
本発明者らの分析は一分子レベルであるため、これらの開始点は、1回のS期において1個の細胞によって実際に用いられるものに相当する。DNA破断は、観察された307の開始事象のうち173に関して、隣接する開始点の可視化を妨げた。残りの場合は、活性のある開始点近傍が存在する場合に見られた(134/307)。この2つの最も近い機能的開始点と最も遠い機能的開始点はそれぞれ31.4kbおよび390.8kb離れていた。興味深いことに、平均開始点間距離は113+/−66.4kbであると算出された(図3B)。ゾーン間距離(〜40kb)に比べて、この結果は、ある細胞周期において3つの潜在的ゾーンのうち平均して1つだけの開始点から発火することを示唆する(図3C)。
【0131】
特定のゾーンに由来する開始点が互いに対して早くまたは遅れて再現性よく発火されたかどうかを検討するため、活性化時間を調べた。開始点を示す複製シグナルの種類に基づき(図1A)、開始時間を標識期間に対して関連づけることができる。これは、IdUまたはCIdUパルスのいずれかの際に発火した開始点にも当てはまり、各20分の時間であった(図1Aiiおよびiii)。これは同様に、標識期間前に発火した開始点にも当てはまり、先行する分岐が修飾されたヌクレオチドの組み込みによって可視化できることが示された(図1Ai)。これが生じるためには、開始点の活性化の時間が平均20分を超えてIdU/CldUパルスに先行することはできなかった(例えば、図3Ciの2番目の分子を参照)。従って、分析枠は、パルス20分前、IdUパルス中20分およびCIdUパルス中20分を含む、全〜60分に及ぶ。本発明者らは、これら38の開始ゾーンのいずれにもタイミング的に優先されるものを見出すことができなかった。さらに、隣接する開始点は同時には発火しなかった(図3C)。個々の繊維上の複数の開始部が本発明者らの実験パラダイムによって与えられる60分以内に検出可能であったことに注目すべきである。従って、開始事象間の厳密な同期性は見られなかったが、任意の2つの活性化開始点の間のタイミングの差はおよそ1時間に限定される。
【0132】
隣接する開始点間の活性化時間はわずかに違いがあることから、2つの接近する分岐間の非複製領域における潜在的開始点は、なお、いくらか後の時間に活性化された可能性がある(例えば、図3Ciiの3番目および5番目の分子を参照)。これらの領域から発火する開始点はより短い開始点間距離をもたらす。隣接する開始点の大部分は互いに30分以内に発火すると考えられる(Berezney et al, 2000)。本発明者らの分析枠は60分(前段参照)であるので、クラスター内のほとんど総ての開始点が活性化されると予測される。
【0133】
従って、開始点の発火の遅れが開始点間距離の測定値を有意に変化させる可能性は低い。
【0134】
潜在的開始部位にわたる分岐の伸張
これまでに、開始部位または開始部を意味するためには、唯一、複製トラックが用いられてきた。しかしながら、それらの周囲領域への二方向性の伸張は、データの別の重要な部分を提供する。隣接するゾーンと重複する1つの開始ゾーンから生じるシグナルは、後者の場合には、複製分岐の先行通過および開始点の許可の剥奪を意味する。
【0135】
これは、受動的に複製されたゾーンをS期のいくらか遅い時間における発火に不応とする。
【0136】
ヒト細胞における総ての潜在的開始点が、それらが酵母である場合(Santocanale and Diffley, 1996)と同様に許可される限り、この観察は開始点干渉の証拠となる(図4A)。開始ゾーンへ向かっていくらか伸張する分岐では、ゾーンは重心が達せられた場合にのみ抑制されると考えられる。本発明者らは、伸張中の分岐からのシグナルを用い、活性開始点干渉からどれくらい離れて起こるのか分析した(図4B)。
【0137】
全部で528の開始ゾーンが抑制されることが分かった。末端小粒方向へ移動する分岐を表す標識(55.1+/−38.4kb、N=262;図4C)に対して動原体方向へ移動する分岐を表す標識(56.5+/−37.7kb、N=266)を考えると、それらの開始部位からの距離は有意には異なっていなかった。結果として、以下の分析のため、両方向に移動する分岐により媒介される開始点干渉を合わせた。活性開始点からの複製トラックは、平均して55.8+/−38kb離れたゾーンと重複していた。最も近い、また最も遠い抑制ゾーンは機能的開始点からそれぞれ7kbおよび284.6kbの位置にあった。これらのデータは次のように解釈される。開始部位のすぐ隣に位置するゾーンは314回抑制された。2番目のゾーンは137回抑制され、その後(3番目以降)のゾーンは77回抑制された。528の抑制された開始ゾーンに基づくこれらのデータを考え合わせると、開始点干渉が1〜2の隣接する開始ゾーンのほとんどの部分に及んでいることが示唆される。
【0138】
ゾーン間干渉の他、本発明者らはゾーン内干渉も分析した。
【0139】
活性開始点からの分岐は、100%の時間、その固有の開始ゾーンの境界を越えて伸張した(例えば、図4B参照)。この形態の干渉が多ければ、与えられた任意の細胞周期において開始ゾーンにつき1を超える開始点が発火する確率は低いはずである。この分析を行うことは、比較的小さな開始ゾーンにおいて密に配置された開始点に代表的な短い複製トラックを識別する本発明者らの能力にかかっている。コームDNA上の線状蛍光セグメントの最大解像度は1〜4kbである。これは平均13.5kbより小さいゾーンにおける複製の開始点の可視化を面倒にする。しかしながら、より大きな開始ゾーンでは、いくつかの開始点の観察を分子コーミングの解像度により制限されない。開始ゾーンのサイズにかかわらず、個々のS期において同じ開始ゾーン内から2以上の開始点が発火するということは見られなかった。よって、開始事象からの距離によって低下するゾーン間干渉とは対照的に、ゾーン内干渉は極めて効率的であり、分岐が移動しなければならない距離には依存しない。
【0140】
結論
結論として、本発明者らは、一分子アプローチを用い、ヒト染色体14q11.2の1.5Mb領域の総ての潜在的開始ゾーンをマッピングした(図1および2)。本発明者らはまた、それらの一部だけが実際に個々の細胞周期に用いられることも実証した(図3)。
【0141】
最後に、限定された開始点の活性化は、規則的な開始点間発火距離をもたらす(図3)。
【0142】
従って、本発明者らは、保存された開始事象間隔が広範な哺乳類開始ゾーンに関して維持されていることを初めて示す。
【0143】
酵母およびCHO細胞では、高い潜在的開始点/活性開始点比が記載されている(Raghuraman et al, 2001; Wyrick et al, 2001; Pasero et al, 2002; Anglana et al, 2003)。本発明者らのデータは、この比がヒト細胞においても保存された特徴であることを示唆する。これにより、真核細胞において繰り返されるテーマである、なぜ開始点が冗長なのかという重要な疑問が持ち上がる。酵母染色体の1つの腕におけるいくつかの開始点を欠失させても、ゲノムの安定性に無視できるほどの影響しかなかった(Dershowitz and Newlon, 1993)。このことは、それほど多くの開始点が必要でないことを示唆する。
【0144】
しかしながら、さらに最近では、組み立てられたpre−RCの完全な相補物を阻害すると染色体の再配列が起こった(Lengronne and Schwob, 2002; Tanaka and Diffley, 2002)。この理由は分かっていないが、いくつかの提案が、余分な潜在的開始点が、混乱したDNA複製の事象において安全ネットを提供するという観点に収束している(Schwob, 2004)。第一に、分岐が遮断されれば、それは組換えの基質へと変換することができる(Rothstein et al, 2000)。下流の「余分な」開始点の活性化は接近する分岐を生じる。この分岐は遮断された分岐と合流し、それによりそれを組換えから救い出す。
【0145】
第二に、いくつかの開始点が発火できない場合、細胞は複製されないDNAとともに有糸分裂を受け得る。このフラグメントは動原体が引き離される際に破断する。潜在的開始点の供給過剰は偶発事象の確率を減らす。最後に、S期のチェックポイントによる最適な細胞周期の休止には、十分な数の分岐が必要である(Shimada et al, 2002)。開始の試みに失敗すると、分岐が失われる。バックアップ開始点の発火が、補償のための2つの付加的な分岐を生成し、これによりチェックポイントが作動可能となる。これらのモデルのいずれかが適用可能であるかを評価するには、明らかにさらなる研究が必要である。
【0146】
開始点干渉は、高い潜在的開始点/活性開始点比が真核生物でいかに達成されるかを説明する機構として取り上げられてきた。それは、早期に活性化された開始点から進行する分岐により、許可された開始点を表すpre−RCの除去を含む(Brewer and Fangman, 1993)。
【0147】
開始点干渉は酵母、ツメガエルおよびCHO細胞で見られている(Brewer and Fangman, 1993; Lucas et al, 2000; Anglana et al, 2003)。ここで、本発明者らは、開始点干渉がヒト細胞における開始点機能の調節に重要な役割を果たすこと、さらにはこれが開始ゾーンに関して起こることを初めて示す(図4)。分子コーミングに先立ち、DNAは除タンパクされる。従って、どの開始ゾーンが許可された開始点を含んでいたか確認することはできなかった。実際に、受動的に複製されたゾーン(抑制されていると解釈される)は単にそれから始まることが許可されていないということかもしれない。今後の研究で、開始ゾーン間のpre−RCの会合がアッセイされる。これにより、本発明者らは、開始点干渉がこの用語の正式な定義に従って起こるかどうかを決定することができる。
【0148】
開始点干渉を説明するため、酵母から得られたデータを挙げられており、これによれば、G1において確立された種々の活性化時間が後の開始点を先の開始点からの分岐により受動的に複製させる(Lucas and Raghuraman, 2003)。CHO細胞で行われた研究(Anglana et al., 2003)と一致して、本発明者らは隣接する開始点に関して強い、予めプログラムされたタイミングの違いを見出せなかった(図3)。よって、本発明者らのデータは、ヒト細胞では酵母に存在するものとは異なる機構が作動していることを示唆する。
【0149】
本発明者らは、ゾーン間干渉が活性開始点からの距離に応じて徐々に低下することを見出した(図4)。開始点の発火の確率が限定された開始因子のために低い場合(Walter and Newport, 1997)には、本明細書で報告されている開始点干渉はこの低い確率および結果としての受動的な現象の直接的結果であり得る。他方で、開始点発火確率が高ければ、開始点干渉は能動的に調節されるはずである。例えば、混乱のない伸張分岐に存在するチェックポイントタンパク質は遠位開始点の発火を抑制し得る。
【0150】
これにより、遅れた開始点が受動的に複製され、従って抑制される機会が能動的に増える(Marheineke and Hyrien, 2004; Shechter et al., 2004; Sorensen et al., 2004; Syljuasen et al., 2005)。今後の研究で、いずれのモデルがゾーン間干渉を担うのかが明らかになるであろう。
【0151】
最近、数学的研究により、複製中心においてともに隔離され、従って同時に活性化され得るのはわずかに11kb離れた潜在的開始点だけであるということが提示された(Jun et al., 2004)。この制限は、DNAの屈曲を制限するDNAの持続長により決定される。持続長はゾーン内干渉を説明することができ、DNAの堅さが、あるゾーンに由来する2つの潜在的開始部位が複製中心内に集中することを妨げ、従って、それらの同時活性化を妨げる。DNAの物理的特性に基づく機構が堅牢なため、本明細書で報告される高効率のゾーン内干渉が生じ得る。
【0152】
哺乳類開始ゾーン内およびゾーン間の開始点干渉機構は、これらの領域の基礎にある分子決定基に依存する。許可の過程で、複数のミニ染色体維持(MiniChromosome Maintenance)(MCM)複合体はpre−RCから離れて拡散する(Ritzi et al, 1998; Edwards et al, 2002)。これらのMCM部位の1つで発火する開始点は哺乳類細胞における開始ゾーンの存在を説明することが示唆されている(Hyrien et al, 2003; Blow and Dutta, 2005; Cvetic and Walter, 2005)。よって、本明細書で報告される開始ゾーン(図2)は反復MCMローディングのために生じ得る。拡散MCMまたは他の未知の因子が哺乳類開始ゾーンを担っているかどうかの判定は、我々が、ヒト開始点干渉がいかにして実行され、開始事象間隔が調節されるかを理解する助けとなる。
【参考文献】
【0153】

【図面の簡単な説明】
【0154】
【図1】ヒト染色体14q11.2における1.5Mbに対するDNA複製開始マッピングを示す。 (A)開始点を示すコーミングされたDNA上の3種類の複製シグナル。開始は、標識前、IdUパルス(青)中、およびCIdUパルス(赤)中に起こり、それぞれ(i)、(ii)および(iii)に示されるシグナルを生じる。3つの場合の総てで、これらのトラックの中点が開始部位であると推定される。 (B)コーミングされたDNAに対するハイブリダイゼーション戦略。長さが等しいが色の違いで検出される2つのプローブ(i)または同じ色で検出される、長さが異なる2つのプローブ(ii)をハイブリダイズさせ、対象ゲノム領域を可視化する。あるいは、同じ情報を得るためにプローブセット間のギャップを使用することもできる。(iii)では、4つの短いプローブをハイブリダイズさせ、情報が得られる3つのギャップを生じさせる。ギャップ3は、分解の際に分子を一定の方向に向かせる(iv、v)。DNA分解は一対の垂直実線により表される。 (C)ヒト染色体14q11.2における1.356Mbにわたる遺伝子モールスコード(GMC)。モールスコードA、B、CおよびDで、最初の4つの記号の直線パターンを示す。GMCはこれら4つの記号を含み、各記号は1セットのプローブで表される。プローブは緑で示される。コードギャップは短ギャップと長ギャップである。開始ギャップおよび終了ギャップは、DNA破断の際の記号の配向を助けるために含めた。記号はスペースギャップにより隔てられている。プローブおよびギャップのサイズ(kb)がそれぞれ上に示されている。 (D)GMC領域において観察される開始事象の例。白い矢印は開始部位を示す。ファイバー2、7、8および13では、GMCは、ある記号に属す総てのプローブが存在しない場合でもなお解読可能である。それらの記号と隣接する開始事象は、隣接する記号の1つが解読されるか(分子1、5、11および14)、スペースギャップ情報が入手可能な場合(分子4および10)にマッピングされた(バー=100kb)。
【図2】クラスター分析による開始ゾーンの同定。 (A)理想的クラスターの定義。分散内(W)および分散間(B)の方程式を挿入枠内に示す。Wに関して、Nはクラスター数であり。Viはクラスターiの分散である。Bに関して、Ciはクラスターiの重心であり、Cはクラスターセットにおける総ての重心の平均である。クラスター数に関数としてB−Wをプロットしたところ、データを9群(青丸)、22群(緑丸)および45群(赤丸)に分けた際の最大値が明らかになった。 (B)9(青線)、22(緑線)および45(赤線)の区分がデータに適用される際の、あるクラスターの、その成分への分解。赤線の下の領域は開始ゾーンを表し、分子の上の白四角により示される。白い矢印は開始部位を示す(バー=100kb)。 (C)GMC領域における開始ゾーンの分布。水平の赤線はゾーンのサイズと位置を表す。垂直の白線はマッピングされた307の開始部位の総ての位置を表す。4未満のデータ点を含むゾーンは、赤線の代わりに水平のグレーの線でマークされ、その後の分析では考慮されていない。この図面のパネルBにおけるクラスターはアスタリスクで示される(バー=100kb)。
【図3】機能的開始点の時空間分析を示す。 (A)開始点間距離(X)を示す複製シグナル。(i)では、2つの開始部位からの複製トラックには隔たりがあるままである。(ii)および(iii)では、それぞれIdUパルスおよびCIdUパルス中に、接近する分岐が合流する。(B)測定された開始点間距離の頻度を示すヒストグラム。 (C)記号A、B(i)およびC、D(ii)において少なくとも2つの開始事象を有する分子の例。白い矢印は開始部位を示す。開始ゾーンは水平の赤線でマークされている。個々の分子について、開始点が発火する開始ゾーンは白四角で示されている。影付きの四角はサイレント開始ゾーンを表す(バー=100kb)。
【図4】分岐の伸張に基づく開始点干渉を示す。 (A)ゾーン(v)の活性開始点から伸びている分岐は垂線対に接している領域を含んでいる。左に移動している分岐の終点が見られる(白ぬきの逆三角)。着目する開始点からの分岐はその境界部に達しない限り、開始ゾーン(i)および(ii)は干渉を受けない。その分岐が受動的にその全長を複製すれば、ゾーン(iii)、(iv)および(vi)は抑制される。右へ移動している分岐はゾーン(vii)を通り抜けるが、その重心(黒いドット)には達しない。このゾーンは開始点干渉データには含まれていない。グレーの長方形は開始ゾーンを表し、濃いグレーの長方形は抑制された開始ゾーンを表す。 (B)開始点干渉を示す分子の例。この図の開始ゾーンは水平の赤線で示される。白四角は開始点が発火する(白い矢印)開始ゾーンを示す。影付きの四角は、分岐の伸張によって抑制されている開始ゾーンを示す(バー=100kb)。 (C)開始事象と干渉を受けているゾーンの間の距離の頻度を示すヒストグラム。動原体方向へ移動している分岐および末端小粒方向へ移動している分岐により抑制されたゾーンはそれぞれ、負の値および正の値で表される。
【図5】3種類のコードを示す。 A:連続するプローブ間の間隔に基づくコード。 B:標識(より詳しくは、色)に基づくコード。 C:標識(色)+連続するプローブ間の間隔に基づく冗長コード。 3種類のコード法。プローブはカラーの短いセグメントで示される。これらのプローブは例えば5〜10Kbを測定することができる。ここで示される「色」は、性質の異なるプローブと同様に含め/理解しなければならず、それは結びついている蛍光色素で異なる場合もあるし、大きさまたは他のいずれかの特徴または特徴の組合せで異なる場合もあり、特にそれ自体蛍光プローブの問題ではない。プローブ間の間隔は例えば短いスペースで約20KB、長いものでは40KB、ダブレットの場合では10KB未満(最初の場合)であり得る。冗長コードは、その他の双方の組合せである。
【図6】冗長コードの興味深い点を示す。 冗長コードの興味深い点。記載されている3種類のコード1に関して、青い矢印のマークされている配列の一部における欠失が示されている。最初の両方の場合では、この欠失に関する理由の修正点として、別のコード配列との混同が含まれる。3番目の場合では、得られたコード配列には矛盾があり、従って、再配列があったことを明らかに示す。この再配列(欠失)の性質は本明細書によれば同定可能である。
【図7】重複コードを示す。 重複コード。プローブの3つの性質はA、BおよびCの文字で表される。上段では、あるものが、7つの連続するプローブからなる種々のコード配列を、その最後の6つのプローブが次にくるものの最初の6つのプローブに相当するように並べることによりどのようなものかを示し、7文字の連続が1回である連続プローブを得ることができる。下段では、7文字のあり得る総ての前提など、連続する2193文字のA、BおよびCが一度だけ表される。従って、例えばコード配列abaaccbを含む/理解するフラグメントが検出されれば、それは赤色の領域から得られるものであることが分かる。
【発明の分野】
【0001】
本発明は、供試巨大分子上の少なくとも1つの対象ドメインの存在を検出する方法に関し、該方法は、
a)対象ドメイン上の少なくとも2つの標的領域を予め決定し、各標的領域の対応する標識プローブ(対象ドメインのプローブのセットと呼ぶ)を設計および取得し、これらのプローブのいずれかの、他のものに対する位置が選択され、供試巨大分子上の対象ドメインの特異的サインを形成する工程;
b)工程a)で得られたプローブが結合されている供試巨大分子を展開(spreading)した後、線状化巨大分子に結合されているプローブのいずれかの、他のものに対する位置を検出し、対象ドメインのサイン(signature)が検出されれば、供試巨大分子上に対象ドメインが存在することを示し、逆に、対象ドメインのサインまたはサインの一部が検出されなければ、供試巨大分子上の対象ドメインまたはドメインの一部が存在しないことを示す工程
を含む。
【0002】
より詳しくは、本発明は、少なくとも2セットの3つのプローブを含み、各プローブセットは1つのドメイン上に特異的に結合することができ、巨大分子上での1セットの少なくとも3つのプローブの結合が、少なくとも2つの異なるコード群(例えば、「ドット(dot)」と「ダッシュ(dash)」)から選択される少なくとも2つのコードの配列を形成し、この群が総てのドメインで同一であり、このコード配列が1つのドメインに特異的であり、そのドメインのサインである、巨大分子上の少なくとも2つの対象ドメインの存在を検出するためのキットに関する。
【背景】
【0003】
巨大分子(macromolecule)、特に、生体巨大分子(より具体的には、DNA)の研究は、「地図作成」のため、すなわち、ドメインの空間構成を研究するため、または巨大分子上の、反応または一連の化学反応もしくは生化学反応の位置決定のためには、多くの場合、正確にいくつかのドメインに印をつける必要がある。
【0004】
DNA配列の空間構成の観察を可能とする方法(Fiber−FISH、分子コーミングなど)は、多くの適用において、いくつかの領域に目印が付けられる、すなわち、何らかの検出技術によって特定の領域の同定が可能な方法でマークされる必要がある。取り組む主題がいくつかの領域の相対的位置である地図作成での適用、ならびにある(いくつかの)特定の遺伝子座において生体現象が研究される適用の場合がそうである。そして、ドメインは特異的マーカー、実際には一般に、対象配列(対象ドメインと呼ぶ)に相補的なDNAのプローブにより同定することができる。これらのマーカーは例えば蛍光顕微鏡、オートラジオグラフィーなどによって検出することができる。
【0005】
用いる検出技術が何であれ、目印を付ける常法は、検出を可能とするエレメント(蛍光色素、放射性元素など)に結合されたプローブ、すなわち、対象領域に相補的な配列の合成からなる。いくつかの領域を識別するために、主要な方法は、それらの長さまたは検出可能なエレメントの性質(一般に、色の異なる蛍光色素)などのそれらの固有の特性に基づいて識別可能なプローブを設計することである。この場合、プローブは、個別に検出される場合であっても同定可能である(それらは異なる性質のものである)ことから識別可能である。
【0006】
このアプローチは、極めて多数の領域を識別するようになると、すぐにその限界に達し、プローブの長さは技術的制約となり、異なる検出可能なエレメントの数が制限される場合が多い。
【0007】
実際、いくつかのドメインを同時に区別可能な方法でマークする必要のある場合が多い。区別可能なマーカーを開発する努力は基本的に、異なる「性質」の、すなわち、用いる検出方法によって個々に区別することができるマーカーを併用し、蛍光を用いた顕微鏡においてスペクトルの異なる蛍光色素を結びつけ、蛍光およびオートラジオグラフィーを用いた顕微鏡において異なるプローブ長を用いることからなる。これらのアプローチに伴う主要な制限は、このようにして得られた異なるマーカーの数が限定されることである。さらに、性質の異なるマーカーの同時検出は一般に、実施するには繊細で時間のかかる獲得方法を用いざるを得ない。
【発明の概要】
【0008】
本発明は、供試巨大分子上の少なくとも1つの対象ドメインの存在および/または位置を検出する方法に関し、該方法は、
a)対象ドメイン上の少なくとも2つの標的領域を予め決定し、各標的領域の対応する標識プローブ(対象ドメインのプローブのセットと呼ぶ)を設計および取得し、これらのプローブのいずれかの、他のものに対する位置が選択され、供試巨大分子上の対象ドメインの特異的サインを形成する工程;
b)工程a)で得られたプローブが結合されている供試巨大分子を展開した後、線状化巨大分子に結合されているプローブのいずれかの、他のものに対する位置を検出し、対象ドメインのサインが検出されれば、供試巨大分子上に対象ドメインが存在することを示し、逆に、対象ドメインのサインまたはサインの一部が検出されなければ、供試巨大分子上に対象ドメインまたはドメインの一部が存在しないことを示す工程
を含む。
【0009】
一実施形態では、この巨大分子は核酸、特にDNA、より詳しくは、二本鎖DNAである。
さらなる実施形態では、プローブのセットは、さらに2つのプローブ(プローブ1またはプローブ2)を含み、各プローブは対象ドメインの異なる末端に結合可能であり、対象ドメインにおいてその連続プローブと関連する(開始部または終了部の末端プローブ結合と呼ぶ)該プローブ1またはプローブ2の一方のシグナル(signal)を読み取ることにより、読み取りの開始または終了に関する情報が得られる。
【0010】
さらなる実施形態では、本発明は、少なくとも2セットの3つのプローブを含み、各プローブセットは1つのドメイン上に特異的に結合することができ、巨大分子上での1セットの少なくとも3つのプローブの結合が、少なくとも2つの異なるコード群(例えば、「ドット」および「ダッシュ」)から選択される少なくとも2つのコードの配列を形成し、この群が総てのドメインで同一であり、このコード配列が1つのドメインに特異的であり、そのドメインのサインである、巨大分子上の少なくとも2つの対象ドメインの存在を検出するためのキットに関する。
【詳細な説明】
【0011】
本発明者らは今般、性質の異なる極めて少数のプローブを用いて、あるいはある単一の性質のプローブ用いても、事実上無限数の領域の識別が可能なプローブを設計するための新規なアプローチを発見した。このアプローチでは、目印は、各目印に独特な記号を一緒に作り出す数個のプローブからなり、それはコードの配列であり、1つのコードは連続する2つのプローブの読み取りによって得られる。このモチーフ(コード配列)はプローブの性質と位置を合わせたものであり得る。
【0012】
本発明は、このアプローチの適用のいくつかの例を示し、その1つはある1つの遺伝子座における複製の研究に首尾よく使用された。この場合、プローブのある単一の性質が用いられた。また、性質の異なる極めて少数のプローブを用い、全染色体の大きさを解像度の高い領域でいかにマークできるかも示される(例えば、3つの性質のプローブは解像度40kbで60Mbをマーク可能である)。
【0013】
我々の発明は、性質が1つだけであるが、各ドメインに関して単一の連続するプローブを形成する(各ドメインに関して単一のコード配列を形成する)ように対象ドメインにおいて空間的にレイアウトされたマーカーを用いることにより、異なる方法でドメインをマークすることからなる。この方法の可能性のある実施の1つは、ゲノムの一部のドメインによる文字(コード)を用いることにより、モールスコード(Morse code)を採用することからなるので、本発明者らはコードネームを遺伝子モールス(Genetic Morse)とし、モールスコードの「ドット」と「ダッシュ」をそれぞれ短い、または長い間隔で隔てられたマーカーに置き換える。
【0014】
あるドメインの特定のコード配列の開始部と終了部を一義的に位置決定することが必要であるか、またはより有効である場合がある。この位置は例えばマーカーの「ダブレット(doublet)」(開始部と終了部の末端プローブ対、これらのプローブは、マーカー(標識)、すなわち、あるドメインの特定のコード配列の中で用いられる総てのものより短い間隔で隔てられている2つのマーカーで標識される)を用いてコード配列の開始部と終了部をマークすることにより得ることができる。コード配列の開始部と終了部をマークする「ダブレット」における短い間隔は長さの異なるものであってよく、コード配列の配向が可能となる。
【0015】
一般的な方法では、本発明者らの方法は、空間的にレイアウトされた、性質が1つだけのある数のマーカーからなる目印の巨大分子に、各目印に特異的なコードの配列を形成するように区切りを入れることからなり、これらのマーカーは検討される巨大分子に直接関連するものである。
【0016】
これらのマーカーが総て同じである場合、これらのマーカーの検出の多くの様式が可能である(オートラジオグラフィー、シンチレーション、蛍光、化学発光など)。実際には無限数の、それぞれあるドメインに特異的なコードの配列を得ることができ、唯一の制限は使用する方法の空間的解像度とマーカーの特定の配置の緯度である。
【0017】
明らかに、本発明によれば、性質の異なるマーカーが使用可能であれば、制限される数においても、これらの異なるマーカーの組合せによりコード配列を得ることができる。従って、目印のコード化の可能性はいっそう多い。
【0018】
本願の実施例1は、これらの目印により境界が定められた正確な遺伝子座における生物現象(実際には、DNA複製)の研究に関する。1.5Mbのドメインは、モールスコードから直接示唆された4つのコード配列を用いてマークされている。これら4つのコード配列のそれぞれは、性質が1つだけ(1つだけの蛍光色素および同じ長さ)の、可変数の5または7プローブからなり、可変様式で間隔を置いた、蛍光により検出される(図5a)。対象ドメインの各開始部は、別個の「開始部末端プローブ対」によりマークされ、この対の各プローブは17kbの間隔があり、それぞれ対象ドメインの終了部は「終了部末端プローブ対」によりマークされ、この対の各プローブは6kbの間隔がある。これらのコードはモールスコードの最初の4文字に相当し、ドットは短い距離(およそ25〜30kb)離れたプローブからなり、ダッシュは長い距離(55〜70KB)離れたプローブからなる。
【0019】
また、規則的に間隔を置いた、2つの異なる性質のマーカーを用いることで、モールスコードに従いつつ、ビーコン(beacon)をコードすることもできる(図5b)。例えば、蛍光では、モールスの「ドット」に特定の色が与え、「ダッシュ」に別の特定の色を与えることができる。この方法はより「小型(compact)」であるという利点を有し、すなわち、形成するコード配列全体が全体としてより短い配列中に含まれ、これが有利となり得る。
【0020】
前記の場合と同様に、あるドメインに特異的なコードの配列の開始部と終了部をマークすることが望ましい場合がある。それぞれ異なる色でマークされた2つのプローブのダブレット(または対)が使用でき、これらは体系的に同じ方向に向かせることができる。そして、このダブレットはそのコード配列の読み取りの方向を示す。
【0021】
また、これまでに記載した2つのシステムを組み合わせることもでき(図5c)、「ドット」は、短いスペースで隔てられた同じ色の2つのプローブからなり、「ダッシュ」は、長いスペースで隔てられた、異なる色の2つのプローブからなる。この2つのコードシステムにより、より強固となる。
【0022】
特に本発明の地図作成適用、例えば分子診断では、配列の変動が予測され、そのような変動はプローブにより形成されるコード配列を改変し、改変されたプローブ間が無くなったり、繰り返されたり、スペースができたりする可能性がある。コード配列の1つがこのように改変されれば、コードシステムを1つだけ用いる場合、良い場合には、そのコード配列を解説することができなくなり、悪い場合では、別のものとの混同が起こるおそれがある。2つの冗長システムを用いる場合には、別のコード配列との混同が起こるおそれがなくなり、良い場合には、コード配列を同定することができ、配列変動が明らかに定義できる(図6)。
【0023】
本発明に記載されるプローブ設計のアプローチの利点の1つは、少数の異なる「色」の標識で極めて多数のプローブを作出できることである。
【0024】
この特徴を強調するいくつかの実施を記載する。本発明に従い、3色だけを用いて作出した7つのプローブを用い、均一な長さおよび間隔のプローブで、2187(=37)の異なるコードを作出することができる。さらに、これら2187のコードを、1つのコードをなす最後の6つのプローブが次のコードの最初の6つのプローブに相当するような順序に並べることができる。
【0025】
これらのコードを重ね合わせると、7つの連続するプローブが、独特なコード、よってこれらのプローブが位置している配列上の独特な部位、従って、巨大分子上の特定の対象ドメインを定義する2193の一連のプローブを得ることができる。
【0026】
よって、例えば、50kbごとにプローブを配置しつつ、100Mbを超えるものを作出することができる(例えば、3色のカラーまたは擬似カラーで5kbプローブ)。少なくとも7プローブ(従って、400kbを超える総てのフラグメント)を検出可能な各DNAフラグメントの元の位置(100Mb−配列における位置)が明らかに同定される。図7の配列は3色のプローブに対応するA、BおよびCの3文字、総数2193文字からなり、各連続7文字は1回である。
【0027】
この実施の利点は、一方で、それがプローブの数と利用可能なフラグメント(すなわち、全配列において明白な方法で記録可能なもの)の解像度と割合の間の関係を至適化することであり、他方で、この実施が容易に自動化でき、これら一連のプローブがデータ処理によって容易に発見できることである。
【0028】
対称配列のフラグメントの配向の問題に伴う解決策は、対称性とは無関係に上記の連続プローブに「一定の方向を向いた」プローブを周期的(例えば、7プローブごと)に導入することである。例えば、7プローブごとに、プローブの「色」とその配向を同時に与えるダブレットに置き換えることができる。ダブレットは2つの密に置かれたプローブからなる。よって、「A」を「AB」ダブレットに「B」を「BC」ダブレットに、「C」を「CA」ダブレットに置き換えることができる。
【0029】
本発明によって有利な多くの利点がある。実際に、先行技術では、これまでの開発努力は異なる検出可能なエレメントの数を掛け合わせることからなった。特に、これにより主要な検出法として蛍光検出を使用することになるが、それはプローブにタグ付けするために多くの異なる色を使用できるのは蛍光だけであるからである。これに関し、限定されたカラーセットを用いることで、獲得時間および分析時間を著しく軽減することができる。さらに、1つのみ、または2つの異なる検出可能なエレメントを保有することができれば、はるかに迅速かつより容易に自動化可能な検出技術が使用可能となる。最後に、蛍光検出であっても、プローブの異なる、潜在的性質の数にはなお限界があるが、本発明者らの方法は事実上無限数の目印を作出することができる。
【0030】
結果として、一態様において、本発明は、供試巨大分子上の少なくとも1つの対象ドメインの存在および/または位置を検出する方法に関し、該方法は、
a)対象ドメイン上の少なくとも2つの標的領域を予め決定し、各標的領域の対応する標識プローブ(対象ドメインのプローブのセットと呼ぶ)を設計および取得し、これらのプローブのいずれかの、他のものに対する位置が選択され、供試巨大分子上の対象ドメインの特異的サインを形成する工程;
b)工程a)で得られたプローブが結合されている供試巨大分子を展開した後、線状化巨大分子に結合されているプローブのいずれかの、他のものに対する位置を検出し、対象ドメインのサインが検出されれば、供試巨大分子上に対象ドメインが存在することを示し、逆に、対象ドメインのサインまたはサインの一部が検出されなければ、供試巨大分子上に対象ドメインまたはドメインの一部が存在しないことを示す工程
を含む。
【0031】
これらのプローブのいずれかの、他のものに対する位置は、供試巨大分子上の対象ドメインの特異的サインを形成するように任意に、かつ特異的に選択される。
【0032】
任意にとは、本発明によれば、当業者が、いずれの条件、特にドメイン配列に依存せずに対象ドメイン上のプローブの位置を選択することができることを意味する。
【0033】
特異的にとは、本発明によれば、当業者が、巨大分子上のその有無の決定を可能とする対象ドメインの特異的サインを形成するように、対象ドメイン上のプローブの位置を選択することができることを意味する。このサインは当業者により選択され、唯一の条件は異なる対象ドメインに対して異なるサインを与えることである。
【0034】
2つの連続するシグナル間の間隔(またはギャップ)は、例えば顕微鏡またはオートラジオグラフィーが用いられる場合には直接的測定、または検出方法に応じて他の好適ないずれかの方法により測定される。
【0035】
一実施形態では、対象ドメインの存在の検出は部分的サインの検出であってもよく、例えば、1以上のプローブが存在しないか、または誤った位置に結合している場合(例えば、対象ドメイン上のその連続するプローブとともにより長い、またはより小さい間隔がある場合)には、この部分的サインは巨大分子上の対象ドメインの改変の指標となる。巨大分子がDNAである場合、この改変は、巨大分子上の対象ドメインの1個または数個のヌクレオチドの欠失、挿入または置換などの突然変異であり得る。
【0036】
一実施形態では、本方法は好ましくは、少なくとも2つの対象ドメイン存在および/または位置を決定するためのものであり、工程a)において対象ドメイン上の少なくとも3つの標的領域を予め決定することを含む。
【0037】
一実施形態では、本発明の対象ドメインのサインは、連続するプローブ間の一連の間隔(spacing)から生じるものである。
【0038】
一実施形態では、本発明の対象ドメインの位置は、化学反応または生化学反応を位置決定するための参照として用いられる。
【0039】
一実施形態では、本発明の対象ドメインの位置は、標的領域を包含する巨大分子において物理的地図を確立するために用いられる。
【0040】
一実施形態では、本発明の対象ドメインのサインは、一連の異なる標識プローブからなる。
【0041】
一実施形態では、本発明の標的領域のプローブのいくつかは、巨大分子上の付近に位置する少なくとも1つの他の対象ドメインのサインの一部でもある。
【0042】
一実施形態では、巨大分子は核酸、タンパク質、ポリマーまたは炭水化物である。特に、巨大分子はDNA、二本鎖DNAまたは一本鎖DNA、より詳しくは、分子コーミングの場合の二本鎖DNAがDNAの線状化に用いられる。
【0043】
本明細書では互換的に用いられるが、「オリゴヌクレオチド」、「核酸」および「ポリヌクレオチド」とは、一本鎖型または二本鎖型のいずれかの1を超えるヌクレオチドのRNA、DNAまたはRNA/DNAハイブリッド配列を含む。
【0044】
本明細書において、「核酸」および「核酸分子」は、DNA分子(例えば、cDNAまたはゲノムDNA)およびRNA分子(例えばmRNA)ならびにヌクレオチド類似体を用いて作製されたDNAまたはRNAの類似体を含むものとする。核酸分子は一本鎖であっても二本鎖であってもよいが、好ましくは、二本鎖DNAである。本明細書を通じ、「ヌクレオチド配列」という表現は、ポリヌクレオチドまたは核酸を等しく表すために用い得る。より厳密には、「ヌクレオチド配列」という表現は核酸材料それ自体を包含し、よって、特定のDNAまたはRNA分子を生化学的に特徴付ける配列情報(すなわち、4つの塩基文字から選択される連続する文字)に限定されない。また、本明細書では、「核酸」、「オリゴヌクレオチド」および「ポリヌクレオチド」は互換的に用いられる。
【0045】
巨大分子が一本鎖DNAであり、プローブがオリゴヌクレオチドである場合、「結合する」とは「ハイブリダイズすること」を意味する。
【0046】
本明細書において、「ハイブリダイゼーション」、「とハイブリダイズする」または「ハイブリダイズしている」とは、中ストリンジェントまたは高ストリンジェントのハイブリダイゼーション条件、好ましくは、ハイブリダイゼーションおよび洗浄条件が互いに少なくとも60%相同なヌクレオチド配列が互いにハイブリダイズされたままとなる場合を表すものとする。
【0047】
好ましくは、これらの条件は、互いに少なくとも約70%、より好ましくは少なくとも約80%、いっそうより好ましくは少なくとも約85%、90%、95%または98%相同な配列が互いにハイブリダイズされたままとなるようなものである。ストリンジェント条件は当業者に公知であり、Current Protocols in Molecular Biology, John Wiley & Sons, N.Y. (1989), 6.3.1-6.3.6に見出せる。
【0048】
好ましい配列と最適に整列した後、少なくとも80%、好ましくは85%、90%、95%および98%の同一性%を有する核酸配列とは、参照核酸配列に対して、特に欠失、末端切断、延長、キメラ融合および/または置換、特に点置換などのある特定の修飾を有する核酸配列を示すものとする。それは好ましくは、配列が参照配列と同じアミノ酸配列をコードする場合の配列(これは遺伝コードの縮重に関連する)、または好ましくは特に以下に定義されるものなどの高ストリンジェント条件下で、参照配列と特異的にハイブリダイズし得る相補的配列に関する。
【0049】
高ストリンジェント条件下でのハイブリダイゼーションは、温度条件およびイオン強度条件が相補的DNAの2つのフラグメント間でハイブリダイゼーションの維持が可能なように選択されることを意味する。例として、上記のポリヌクレオチド断片を定義することを目的とするハイブリダイゼーション工程の高ストリンジェンシーは有利には次の通りである。
【0050】
DNA−DNAまたはDNA−RNAハイブリダイゼーションは2工程で行われる:(1)42℃、3時間、5×SSC(1×SSCは0.15M NaCl+0.015Mクエン酸ナトリウム溶液に相当する)、50%のホルムアミド、7%のドデシル硫酸ナトリウム(SDS)、10×デンハート溶液、5%の硫酸デキストランおよび1%のサケ精子DNAを含有するリン酸バッファー(20mM、pH7.5)中でのプレハイブリダイゼーション;(2)プローブサイズに応じた温度(すなわち、プローブサイズ>100ヌクレオチドの場合は42℃)で20時間、実際のハイブリダイゼーションの後、20℃で20分、2×SSC+2%のSDS中での洗浄2回、20℃で20分、0.1×SSC+0.1%のSDS中での洗浄1回。最後の洗浄は、0.1×SSC+0.1%のSDS中、プローブサイズ>100ヌクレオチドの場合には60℃で30分行う。当業者であれば、Sambrook et al., (1989, Molecular cloning: a laboratory manual. 2nd Ed. Cold Spring Harbor)の教示に従い、所定サイズのポリヌクレオチドに関して上記された高ストリンジェントのハイブリダイゼーション条件を、より大きな、またはより小さなサイズのオリゴヌクレオチドにも適合させることができる。
【0051】
一実施形態では、これらのプローブは、少なくとも15ヌクレオチド、好ましくは少なくとも1Kb、より好ましくは1〜10kbの間、いっそうより好ましくは4〜10kbの間のオリゴヌクレオチドである。
【0052】
コーミングしたDNA(combed DNA)上での最大解像度が1〜4kbであることから、本発明のプローブは好ましくは少なくとも4kbである。
【0053】
一実施形態では、巨大分子の線状化は、巨大分子上にプローブが結合する前または結合した後に行われる。
【0054】
一実施形態では、巨大分子の線状化は分子コーミングまたはファイバーFishによって行われる。
【0055】
分子コーミングは公表されている方法に従って行う(Lebofsky and Bensimon, 2005)。分子コーミング技術を用い、大きなゲノム領域におよぶ単一ゲノムの物理的特性決定が可能である。単一のコームDNA分子のアレイは、後退エア−ウォーターメニスカスにより、シラン化ガラス面にそれらの末端で結合した分子を延長することにより作製される。コームDNA上で蛍光ハイブリダイゼーションを行うことにより、ゲノムプローブの位置を直接可視化することができ、物理的地図を構成するため、例えば、微小再配列を検出するための手段を提供する。単一分子DNA複製もまた、コームDNA分子上に組み込まれたヌクレオチド類似体の蛍光検出によりモニタリングすることができる。
【0056】
FISH(蛍光in situハイブリダイゼーション)は、染色体上のDNA配列を検出および位置決定するために使用可能な細胞遺伝学的技術である。それは、それらの染色体部分だけに結合する蛍光プローブを用い、それをもって高い配列類似性を示す。蛍光顕微鏡により、蛍光プローブが染色体に結合している場所を見つけることができる。
【0057】
FISH法では、まず、プローブを構築する。プローブはその標的と(ゲノム内の類似の配列とではなく)特異的にハイブリダイズするのに十分長くなければならないが、ハイブリダイゼーション法を妨げるほど大きくてはならず、蛍光団、抗体に対する標的またはビオチンで直接タグ付けすべきである。これは例えばニックトランスレーションおよびタグ付きヌクレオチドを用いたPCRなどの種々の方法で行うことができる。その後、染色体調製を行う。染色体は基板、通常はガラスに固定する。作製後、このプローブを染色体DNAに適用し、ハイブリダイズを開始する。数回の洗浄工程で、ハイブリダイズしていないか、または部分的にしかハイブリダイズしていないプローブは総て洗い流される。シグナル増幅が顕微鏡の検出閾値を超える必要がある場合には(プローブの標識効率、プローブの種類および蛍光色素などの多くの因子によって異なる)、蛍光タグ抗体またはストレプトアビジンをタグ分子と結合させて蛍光を増幅する。最後に、このサンプルは漂白防止剤に包埋し、蛍光顕微鏡で観察する。
【0058】
ファイバーFishでは、間期染色体を、従来のFISHの場合のように強固に巻きつける、あるいは間期FISHの場合のようにランダムコンフォメーションを採用するのではなく、直線上に引き延ばすような方法でスライドに付着させる。これは、スライドに固定した後に溶解させた細胞か、精製されたDNAの溶液のいずれかにスライドの長さに添って機械的剪断をかけることで行われる。これらの染色体の延長コンフォメーションは、数キロベースに引き下がったとしても、劇的に高い解像度をもたらす。しかし、ファイバーFishサンプルの調製は、概念的に単純ではあるが、むしろ熟練技術であり、専門の研究室だけが慣例的にその使用が可能であることを意味する。
【0059】
ファイバーFish法のプロトコールは上記されている。
【0060】
器具および試薬:
・リンパ芽球状細胞培養
・PBS
・血球計算板
・溶解溶液
・5部の70mM NaOH、2部の無水エタノール(Fidlerova et al. 1994)。この溶液は室温で数ヶ月保存可能である。
【0061】
方法
・健全な培養物から細胞懸濁液1〜2mlを採取する。
・5ml PBSで2回洗浄する。
・1ml PBSに再懸濁させる。
・血球計算板を用いて細胞のアリコートを計数する。
・細胞を追加のPBSで希釈し、終濃度をおよそ2×106/mlとする。
・10μlの細胞懸濁液を清浄な顕微鏡スライドの上部に1cmの面積に広げる。
・風乾する。
・スライドをプラスチック製のCadenza(Shandon Southern)チャンバーおよびクランプにほぼ垂直の位置に取り付ける。
・このcadenzaの上に150μlの溶解溶液を適用する。
・レベルがスライドのつや消しのエッジより下に下がれば、200μlのエタノールを加える。
・軽く排水する。
・エッジを持って、スライドとcadenzaユニットをクランプの外へ注意深く持ち上げる。
・cadenzaからスライドの背面の上部を抜き、メニスカスをスライドの下へ移動させる。
・アングルで風乾させる。
・アセトン中で10分固定する。スライドは室温で数ヶ月、問題なく保存することができる。
【0062】
一実施形態では、総てのプローブは同じ標識で標識される。
【0063】
一実施形態では、プローブは少なくとも2つに異なる標識で標識される。一実施形態では、プローブは3つの標識で標識される。
【0064】
一実施形態では、結合またはハイブリダイズしたプローブは、プローブと結合している1以上の標識を検出することにより検出される。これらの標識は当業者に周知のいくつかの手段のいずれによって組み込んでもよい。しかしながら、好ましい実施形態では、標識は、プローブ調製時の増幅工程中に同時に組み込まれる。例えば、標識プライマーまたは標識ヌクレオチドを用いたポリメラーゼ連鎖反応(PCR)により、標識された増幅産物が得られる。プローブ(例えばDNA)は標識デオキシヌクレオチド三リン酸(dNTP)の存在下で増幅される。
【0065】
好ましい実施形態では、標識ヌクレオチド(例えば、蛍光標識UTPおよび/またはCTP)を用いた上記のような転写増幅が転写された核酸に標識を組み込む。
【0066】
あるいは、標識は、ものとプローブ(例えば、mRNA、ポリA mRNA、cDNAなど)または増幅が完了した後の増幅産物に直接加えてもよい。このような標識により、高収量の増幅産物が得られ、増幅反応に必要な時間が短縮される。プローブに標識を付着させる手段としては、例えば、ニックトランスレーションまたは核酸のキナーゼ処理の後に標識(例えば蛍光団)にプローブを結びつける核酸リンカーを付着(連結)させることによる末端標識(例えば、標識RNAを用いる)が挙げられる。
【0067】
好ましくは、本発明の標識ヌクレオチドは、クロロデオキシウリジン(CIdU)、ブロモデオキシウリジン(BrdU)および/またはヨードデオキシウリジン(IdU)である。
【0068】
本発明に用いるのに好適な検出可能な標識としては、顕微鏡的手段、光化学的手段、生化学的手段、免疫化学的手段、電気的手段、光学的手段または化学的手段によって検出可能ないずれの組成物も含む。本発明において有用な標識としては、標識ストレプトアビジンコンジュゲートで染色するためのビオチン、磁性ビーズ(例えば、Dynabeads(商標))、蛍光色素(例えば、フルオレセイン、テキサスレッド、ローダミン、緑色蛍光タンパク質など。例えば、Molecular Probes, Eugene, Oreg., USA参照)、放射性標識(例えば、3H、125I、35S、14Cまたは32P)、酵素(例えば、ホースラディッシュペルオキシダーゼ、アルカリ性ホスファターゼおよびその他ELISAに慣用されるもの)、ならびに金コロイド(例えば、直径40〜80nm範囲の、高効率で緑色光を発する金粒子)またはカラーガラスもしくはプラスチック(例えば、ポリスチレン、ポリプロピレン、ラテックスなど)ビーズのような比色定量標識が挙げられる。このような標識の使用を教示する特許としては、米国特許第3,817,837号、同第3,850,752号、同第3,939,350号、同第3,996,345号、同第4,277,437号、同第4,275,149号および同第4,366,241号がある。
【0069】
蛍光標識は、低いバックグラウンドで極めて強いシグナルを提供するので好ましい。また、迅速走査法によって高い解像度および感度で光学的に検出することもできる。
【0070】
プローブは総て、単一の標識、例えば単一の蛍光標識で標識することができる。あるいは、別の実施形態では、異なるプローブを、各プローブが異なる標識を有するように同時にハイブリダイズさせることもできる。例えば、ある標的は緑色蛍光標識を有し、別の標的が赤色蛍光標識を有してもよい。走査工程は緑色蛍光標識の結合部位から赤色標識の結合部位を識別する。各プローブ(標的核酸)は互いに独立に分析することができる。
【0071】
使用可能な好適な色素原としては、色が観察できるように識別的な波長域の光を吸収するか、あるいはまた例えば蛍光剤など、特定の波長または波長域の放射線を照射した際に発光する分子および化合物が挙げられる。
【0072】
広範囲の好適な色素が利用可能であり、基本的に、それらの周囲による吸収が最小で強い色をもたらすように選択される。例示的な色素種としては、キノリン色素、トリアリールメタン色素、アクリジン色素、アリザリン色素、フタレイン、昆虫色素、アゾ色素、アントラキノイド色素、シアニン色素、フェンアザチオニウム色素およびフェンアゾキソニウム色素が挙げられる。
【0073】
広範な蛍光剤が単独、あるいはまた消光分子と組み合わせて使用可能である。対象蛍光剤はある主要な官能基を有する多様なカテゴリーに分類される。これらの主要な官能基としては、1−および2−アミノナフタレン、p,p’−ジアミノスチルベン、ピレン、第四級フェナントリジン塩、9−アミノアクリジン、p,p’−ジアミノベンゾフェノンイミン、アントラセン、オキサカルボシアニン、マロシアニン、3−アミノエキレニン、ペリレン、ビスベンズオキサゾール、ビス−p−オキサゾリルベンゼン、1,2−ベンゾフェナジン、レチノール、ビス−3−アミノピリジニウム塩、ヘレブリゲニン、テトラサイクリン、ステロフェノール、ベンズイミダゾリルフェニルアミン、2−オキソ−3−クロメン、インドール、キサンテン、7−ヒドロキシクマリン、フェノキサジン、サリチレート、ストロファンチジン、ポルフィリン、トリアリールメタンおよびフラビンが挙げられる。
【0074】
連結のための官能基を有するか、またはこのような官能基を組み込むよう修飾可能な個々の蛍光化合物としては、例えば、塩化ダンシル;3,6−ジヒドロキシ−9−フェニルキサントヒドロールなどのフルオレセイン;ローダミンイソチオシアネート;N−フェニル1−アミノ−8−スルホナトナフタレン;N−フェニル2−アミノ−6−スルホナトナフタレン:4−アセトアミド−4−イソチオシアナト−スチルベン−2,2’−ジスルホン酸;ピレン−3−スルホン酸;2−トルイジノナフタレン−6−スルホン酸;N−フェニル、N−メチル2−アミノナフタレン(aminoaphthalene)−6−スルホン酸;臭化エチジウム;ステブリン;オーラミン(auromine)−0,2−(9’−アンスロイル)パルミチン酸;ダンシルホスファチジルエタノールアミン;N,N’−ジオクタデシルオキサカルボシアニン;N,N’−ジヘキシルオキサカルボシアニン;メロシアニン、4(3’ピレニル)酪酸;d−3−アミノデソキシ−エキレニン;12−(9’−アンスロイル)ステアリン酸;2−メチルアントラセン;9−ビニルアントラセン;2,2’(ビニレン−p−フェニレン)ビスベンゾオキサゾール;p−ビス[2−(4−メチル−5−フェニルオキサゾリル)]ベンゼン;6−ジメチルアミノ−1,2−ベンゾフェナジン;レチノール;ビス(3’−アミノピリジニウム)−1,10−デカンジイルジヨージド;ヘリブリエニンのスルホナフチルヒドラゾン;クロロテトラサイクリン;N(7−ジメチルアミノ−4−メチル−2−オキソ−3−クロメニル)マレイミド;N−[p−(2−ベンズイミダゾリル)−フェニル]マレイミド;N−(4−フルオランチル)マレイミド;ビス(ホモバニリン酸);レサザリン;4−クロロ−7−ニトロ−2,1,3ベンゾオキサジアゾール;メロシアニン540;レゾルフィン;ローズベンガル;および2,4−ジフェニル−3(2H)−フラノンが挙げられる。
特に、本発明の蛍光標識は、1−クロロ−9,10−ビス(フェニルエチニル)アントラセン、5,12−ビス(フェニルエチニル)ナフタセン、9,10−ビス(フェニルエチニル)アントラセン、アクリジンオレンジ、オーラミンO、ベンズアントロン、クマリン、4’,6−ジアミジノ−2−フェニルインドール(DAPI)、臭化エチジウム、フルオレセイン、緑色蛍光タンパク質、ヘキストステイン(Hoechst stain)、インディアンイエロー、ルシフェリン、フィコビリン、フィコエリトリン、ローダミン、ルブレン、スチルベン、TSQ、テキサスレッドおよびウンベリフェロンである。
【0075】
望ましくは、蛍光剤は約300nm、好ましくは約350nmより上、より好ましくは約400nmより上の光を吸収すべきであり、通常には、吸収される光の波長の約10nmを超える高い波長で発光する。結合色素の吸収および発光特性は結合していない色素とは異なることに注意すべきである。よって、種々の波長範囲および色素の特徴に対して言及する場合、コンジュゲートされておらず、任意の溶媒中で特徴付けられる色素ではなく、使用されるような色素を示すものとする。
【0076】
蛍光剤は一般に、蛍光剤に光を照射することで複数の発光を得ることができるので好ましい。よって、単一の標識で複数の測定可能な事象を得ることができる。
【0077】
本発明によれば、蛍光標識で標識が行われる場合、シグナルの読み取りは蛍光検出によりなされ、蛍光標識されたプローブは光により励起され、その後、その励起の発光が、デジタル画像を取り込み、さらなるデータ解析を可能とする適当な発光フィルターを装備したCCDカメラなどの光センサーにより検出される。
【0078】
また、化学発光源および生物発光源によっても検出可能なシグナルが得られる。化学発光源としては、化学反応により電気的に励起されるようになり、その後、検出可能なシグナルとして働くか、または蛍光アクセプターにエネルギーを与える光を発し得る化合物が含まれる。多様な数の化合物群が多様な条件下で化学発光を与えることが見出されている。1つの化合物群が2,3−ジヒドロ−1,−4−フタラジンジオンである。最も知られている化合物はルミノールであり、これは5−アミノ化合物である。この群の他のものとしては、5−アミノ−6,7,8−トリメトキシ−およびジメチルアミノ[ca]ベンズ類似体がある。これらの化合物はアルカリ性過酸化水素または次亜塩素酸カルシウムおよび塩基で発光をもたらし得る。別の化合物群としては、2,4,5−トリフェニルイミダゾールがあり、親生成物の一般名はロフィンである。化学発光類似体としては、パラ−ジメチルアミノおよび−メトキシ置換基が挙げられる。化学発光はまた、塩基性条件下でシュウ酸塩、通常にはオキサリル活性エステル、例えばp−ニトロフェニルおよび過酸化物、例えば過酸化水素で得ることもできる。あるいは、生物発光を得るために、ルシフェリンをルシフェラーゼまたはルシゲニンと組み合わせて使用することもできる。
【0079】
スピン標識は、電子スピン共鳴(ESR)分光法によって検出可能な不対電子スピンを伴うリポーター分子により得られる。スピン標識の例としては、有機フリーラジカル、遷移金属錯体、特に、バナジウム、銅、鉄およびマンガンなどが挙げられる。スピン標識の例としては、ニトロキシドフリーラジカルが挙げられる。
【0080】
標識はハイブリダイゼーション前または後にプローブ(または標的、これは特に核酸である)に加えることができる。いわゆる「直接標識」は、ハイブリダイゼーション前にプローブに直接結合されるか、組み込まれる検出可能な標識である。これに対し、いわゆる「間接標識」は、ハイブリダイゼーション後にハイブリッド二本鎖に連結される。多くの場合、間接標識は、ハイブリダイゼーション前にプローブに結合されている結合部分に結合される。よって、例えば、プローブはハイブリダイゼーション前にビオチン化することができる。ハイブリダイゼーション後、アビジンとコンジュゲートされた蛍光団を、容易に検出される標識をもたらすビオチン担持ハイブリッド二本鎖に結合させる。核酸を標識し、標識されたハイブリダイズ核酸を検出する方法に関する詳細な総説としては、Laboratory Techniques in Biochemistry and Molecular Biology, Vol. 24: Hybridization With Nucleic Acid Probs, P. Tijssen, ed. Elsevier, N.Y., (1993)を参照。
【0081】
これらの標識は直接、またはリンカー部分を介して結合させることができる。一般に、標識またはリンカー−標識結合部位は特定の位置に限定されない。例えば、標識は、所望により、検出またはハイブリダイゼーションを妨げないいずれの位置でヌクレオシド、ヌクレオチドまたは類似体と結合させてもよい。例えば、Clontech (Palo Alto, Calif.)からのある種のLabel−ON試薬は、オリゴヌクレオチドのリン酸主鎖に散在した標識と3’および5’末端における末端標識を与える。例えば本明細書に示されるように、標識はリボース環上の位置に結合可能であり、このリボースは所望により修飾、さらには排除することもできる。有用な標識試薬の塩基部分は、天然に存在するもの、またはそれらが置かれている目的を妨げない様式で修飾されたものを含み得る。修飾塩基としては、限定されるものではないが、7−デアザAおよびG、7−デアザ−8−アザAおよびGおよび他の複素環式部分が挙げられる。
【0082】
末端標識プローブに関して、多くの適用では、増幅、転写またはその他の変換工程を介して行う必要のない直接的標識プローブに有用である。一般に、末端標識法は、標識される核酸のサイズの至適化を可能とする。末端標識法はまた、ポリメラーゼ補助標識法に伴うことがある配列の偏りを軽減する。末端標識は末端トランスフェラーゼ(TdT)を用いて行うことができる。
【0083】
末端標識はまた、プローブの末端に標識オリゴヌクレオチドまたはその類似体を連結することにより行うこともできる。他の末端標識法としては、例えばリガーゼまたは末端トランスフェラーゼを用いて核酸の標識または非標識「テール」を作出することを含む。次に、このテールを付けた核酸を、そのテールと優先的に結合する標識部分に曝す。このテールとそのテールと優先的に結合する部分は、核酸、ペプチドまたは炭水化物などのポリマーであり得る。このテールとその認識部分は両者の間の認識が可能ないずれのものであってもよく、ハプテン、エピトープ、抗体、酵素とそれらの基質、ならびに相補的核酸およびその類似体などの、リガンド−基質の関係を有する分子を含む。
【0084】
テールまたはテール認識部分と結合される標識は検出可能な部分を含む。テールおよびその認識部分は双方とも標識され、それぞれと結合される個々の標識はそれ自体リガンド−基質の関係を有する。個々の標識はまた、異なる分光光度的特徴を有する色素などのエネルギー移動試薬も含み得る。エネルギー移動対は所望の組合せのスペクトル的特徴が得られるように選択することができる。例えば、第二の色素によって吸収されるよりも短い波長で吸収する第一の色素は、そのより短い波長で吸収する際に、第二の色素へエネルギーを移動する。その後、第二の色素は、第一の色素単独で発せられたものよりも長い波長で電磁線を発する。エネルギー移動試薬は、1995年9月15日出願の米国出願番号08/529,115の一部継続である、1996年12月23日に出願の同時係属中米国特許出願、および1990年12月6日出願の米国出願番号07/624,114の継続出願である、1993年12月15日出願の米国出願番号08/168,904の分割出願である、1996年6月25日出願の米国出願番号08/670,118の一部継続出願でもある国際出願WO96/14839に示されているものなどの2色標識スキームに特に有用であり得る。米国出願番号07/624,114は、参照することにより本明細書の一部とされる1990年6月7日出願の米国出願番号07/362,901の一部継続出願である。
【0085】
一実施形態では、標識が放射性標識で行われる場合、シグナルの読み取りは放射性検出により行われる。
【0086】
放射性検出はX線フィルムまたはホスホルイメージャーを用いて行うことができる。
【0087】
本発明の放射性標識の例は3H、125I、35S、14Cまたは32Pである。
【0088】
好ましい実施形態では、プローブは蛍光標識で標識される。
【0089】
好ましい実施形態では、プローブは放射性標識で標識される。
【0090】
本発明によれば、プローブが少なくとも2つの異なる標識で標識される場合、対象ドメインのサインは連続する標識を生じる。
【0091】
一実施形態では、巨大分子上の対象ドメインに対応する少なくとも3つのプローブの結合は、少なくとも2つの異なるスペース(例えば、「ショート」と「ラージ」)の群(この群は各対象ドメインで同一である)から選択される少なくとも2つのスペースの配列を形成する。
【0092】
一実施形態では、プローブセットはさらに2つのプローブ(プローブ1またはプローブ2)を含み、各プローブは対象ドメインの異なる末端に結合することができ、対象ドメインにおけるその連続するプローブと関連する前記プローブ1またはプローブ2(開始部または終了部の末端(端部)プローブ対(extremity probe couple)と呼ぶ)の1つのシグナルを読み取ることによって、読み取りの開始または終了に関する情報を得ることができる。
【0093】
一実施形態では、読み取りの開始に関する情報は、開始部の末端プローブ対の2つの連続するプローブ間の間隔の読み取りの結果である。
【0094】
一実施形態では、読み取りの終了に関する情報は、終了部の末端プローブ対の2つの連続するプローブ間の間隔の読み取りの結果である。
【0095】
一実施形態では、読み取りの開始に関する情報は、開始部の末端プローブ対の2つの連続するプローブ間の間隔の読み取りの結果であり、読み取りの終了に関する情報は、終了部の末端プローブ対の2つの連続するプローブ間の間隔の読み取りの結果であり、この間隔は、開始と終了の情報を区別するために、開始部の末端プローブ対と終了部の末端プローブ対では異なる。
【0096】
本発明の別の態様は、巨大分子上の少なくとも1つの対象ドメインの存在を同定する方法に関し、前記ドメイン上に特異的に結合し得る少なくとも3つのプローブのセットが設計され、巨大分子上の1セットの少なくとも3つのプローブの結合が少なくとも2つの異なるコードの群((例えば、「ドット」と「ダッシュ」)から選択される少なくとも2つのコードの配列を形成し、この群が各ドメインで同一であり、このコードの配列がそのドメインに特異的であり、そのドメインのサインであり、該方法は、
a)少なくとも3つのプローブの少なくとも2セットを巨大分子と接触させる工程(各セットは各対象ドメインに特異的である);
b)シグナルを読み取ること(1つのシグナルは1つのプローブに相当する);
c)該シグナルをコードで転写する工程;
d)対象ドメインに特異的なコード配列の存在の有無を検出する工程(該コード配列の存在が存在に相当し、巨大分子上のドメインの位置を示す);および
所望により、工程a)の前または工程a)とb)の間に巨大分子を線状化する工程
を含む。
【0097】
好ましい実施形態では、方法は、巨大分子上の少なくとも2つの対象ドメインの存在を同定する。
【0098】
好ましい実施形態では、得られたコードは2つの連続シグナルの特徴の読み取りの結果である。
【0099】
一実施形態では、2つの連続シグナルの特徴はプローブの標識により得られる。
【0100】
一実施形態では、標識は蛍光標識で行われ、特定の蛍光標識は特定のコードに相当する。
【0101】
一実施形態では、標識は放射性標識で行われ、特定の放射性標識は特定のコードに相当する。
【0102】
一実施形態では、得られたコードは2つの連続シグナル間の間隔の読み取りの結果である。
【0103】
一実施形態では、2つのシグナル間の特定の間隔は特定のコードに相当する。
【0104】
例えば、本発明によれば、小さなギャップはドットと呼ばれるコードに相当し、大きなギャップはダッシュと呼ばれるコードに相当する。小さなコードは例えば25〜30kbを測定するように設計することができ、大きなコードは例えば55〜70kbを測定するように設計することができる。
【0105】
一実施形態では、2つの連続シグナルの特徴はプローブの長さによって与えられ、各長さが特定のコードに相当する。
【0106】
本発明は、2つのコードにまで減少されず、3つ、4つまたは4つを超えるコードを含むことができ、コードを定義するためにいずれのサイズのギャップが選択されてもよく、このサイズは任意である。
【0107】
本発明によれば、これらのプローブセットはさらに2つのプローブ(プローブ1またはプローブ2)を含み、各プローブは対象ドメインの異なる末端に結合することができ、対象ドメインにおけるその連続プローブと関連する該プローブ1またはプローブ2(「開始または終了の末端プローブ対」と呼ばれる)の1つのシグナルを読み取ることで、読み取りの開始または終了に関する情報が得られる。
【0108】
一実施形態では、本発明のサインはプローブセットにおける第一のプローブと第二のプローブの間にスペースを含み、このスペースはそのサインにおいて他の総てのスペースと異なり、このスペースはサインの開始に関する情報を得るために使用することができる。
【0109】
一実施形態では、本発明のサインは、プローブセットにおいて最後の手前のプローブと最後のプローブの間にスペースを含み、このスペースはそのサインにおいて他の総てのスペースと異なり、このスペースはサインの終了に関する情報を得るために使用することができる。
【0110】
さらなる実施形態では、本発明は、少なくとも2セットの3つのプローブを含み、各プローブセットは1つのドメイン上に特異的に結合することができ、巨大分子上での1セットの少なくとも3つのプローブの結合が、少なくとも2つの異なるコード群(例えば、「ドット」と「ダッシュ」)から選択される少なくとも2つのコードの配列を形成し、この群が総てのドメインで同一であり、このコード配列が1つのドメインに特異的であり、そのドメインのサインである、巨大分子上の少なくとも2つの対象ドメインの存在を検出するためのキットに関する。
【0111】
本発明は、本出願者が行った研究に関して遂行された実験検討の記載を読めばより明確に理解される。これらは何ら限定と解釈されるべきでない。
【実施例】
【0112】
哺乳類DNA複製開始点は数bp〜数十kbの範囲の部位に位置する。個々の細胞周期における開始の規則的分布は、これら多くの潜在的開始部位のうち限られた数だけが活性化開始点へと変換されることを示唆する。開始点干渉は冗長開始点を封じることができるが、干渉が機能的ヒト開始事象のスペーシングに関与するかどうかは現在のところ知られていない。一次ケラチノサイト由来の単一にコーミングされたDNA分子に対して新規なハイブリダイゼーション戦略である遺伝子モールスコードを用いることにより、本発明者らは1.5Mbのヒト染色体14q11.2上に存在する開始部位を報告する。本発明者らは、開始ゾーンがヒト細胞に拡散していることを確認した。個々の細胞周期において用いられる開始点は、存在し得る開始部位よりも少なく、それらの限定された使用は規則的な開始点間発火距離を作り出す。
【0113】
ゾーン間干渉は活性開始点からの距離に比例して減少するが、ゾーン内干渉は100%有効である。これらの結果によってヒト細胞において有効な開始点の階層的構成が確認される。機能的開始点は、その付近で開始点が発火する確率を支配し、これはゾーン間干渉およびゾーン内干渉によって媒介される。これにより、DNA複製の複数の複雑な、潜在的開始部位に関して、開始事象がヒト染色体の長さに添って一様に分布していることが確認される。
【0114】
導入
真核細胞は、S期の長さによって規定され、ゲノム倍加回数が限定されている。これは、複数の開始部位から伸びる複製分岐においてDNAを合成することにより果たされる。分岐(fork)の速度はS期の長さに応じて尺度化されるのではないので、各個の染色体の開始頻度の調節には有糸分裂の開始前に非複製ギャップを阻害する必要がある(Hand and Tamm, 1973; Edenberg and Huberman, 1975)。例外もあるが、一般的な見解では、体細胞哺乳類開始点は50〜300kb間隔で始まっている(Edenberg and Huberman, 1975; Berezney et al., 2000)。このことは、後生動物が開始事象を一様に分布させる機構を有していることを示唆する。一定の距離に強力なレプリケーター配列を配置することは、発芽酵母サッカロミセス・セレビシエ(Saccharomyces cerevisiae)に用いられているこのような機構の1つである(Newlon et al., 1991; Shirahige et al., 1993)。もっと高等な真核生物では、遺伝エレメントは開始点の活性化に役割を果たすが、それらはそれ自体開始を駆動するには十分なものでない(Gilbert, 2004)。さらに、後生動物の開始点には数bpという限局化された部位に存在するものがあるが、大多数は数十kbまでの範囲のより分散した開始ゾーンに存在する(DePamphilis, 1999)。これにより内因的効率の低い、ある範囲の潜在的部位からいかにして活性化開始点の規則的分布を果たすかという問題が持ち上がる。
【0115】
開始点活性を調節する1つの方法は、それが受動的に複製される確率を変化させることである。開始点付近から伸張している分岐はこの抑制を媒介するので、この形式の開始点の不活性化は「開始点干渉」と呼ばれている(Brewer and Fangman, 1993)。開始点干渉に関する本発明者らの理解の大部分は、S.セレビシエでの研究により得られたものである。発芽酵母では、複製の完了に必要とされるか、または用いられるものよりも多くのより集合した複製前駆複合体(pre−RC)がある(Dershowitz and Newlon, 1993; Raghuraman et al., 2001; Wyrick et al., 2001; Pasero et al., 2002)。酵母染色体IIIおよびVIに対して開始点効率を分析したところ、開始点が細胞周期の5〜90%の間で用いられていることが明らかになった(Friedman et al., 1997; Yamashita et al., 1997; Poloumienko et al., 2001)。許可された開始点は、その近傍の他の開始点に比べて、予定されたS期のタイミングが遅いか、比較的遅いために効果的でない(Santocanale and Diffley, 1996; Vujcic et al., 1999)。結果として、これらの適格開始点は、隣接する開始部位から伸びている分岐により受動的に複製される(Santocanale et al., 1999)。
【0116】
酵母から得られたデータによれば、開始点が互いに干渉すべきであれば、開始点付近がS期の異なる時間に発火する(fire)ようにG1期において予めプログラムされていなければならない(Raghuraman et al., 1997)。しかし、この必要条件は、より高等な真核生物において、1)タイミングの制御が〜100kbという拡大された領域にわたって発揮され(MacAlpine et al., 2004; Norio et al., 2005)、かつ、2)クラスターにおいて互いに次に位置する開始点が同時に発する場合には満足のいくものでない可能性がある(Berezney et al., 2000)。
【0117】
実際、CHO細胞の増幅されたAMPD2遺伝子座では、近傍の開始点間の有意な予め定義されたタイミングの違いは見られなかった(Anglana et al., 2003)。やはり隣接する開始点の、よく定義されたbp部位での同時活性化は遮断された(Anglana et al., 2003)。これが広い開始ゾーンに当てはまるかどうか依然として判定すべきである。
【0118】
規則的な開始間隔がヒト細胞でいかにして果たされるか、また、開始点干渉がこのプロセスに寄与するかどうかを理解するため、本発明者らは開始点活性に関して、一次ケラチノサイト由来のヒト染色体14q11.2の1.5Mb領域に疑問を持った。次のような理由で、分子コーミング技術を採用した一分子アプローチを選択した。第一に、特定の細胞種におけるDNA複製の総ての潜在的開始部位の位置を特定するために十分な開始点発火事象が得られる。第二に、本発明者らは、単一の細胞が個々のS期においてどの開始点を用いるかということと、それらの互いに対する活性化のタイミングを決定することができる。これには開始事象の時空間分布を確認する必要がある。これらのデータを組み合わせて、既に発火した開始点が下流の、潜在的開始部位の使用を調節するかどうかを評価した。本発明者らは、開始点が活性開始点によって確立された階層に従って互いに自己調節し、予め定義されたタイミングの優先なく、確率論的に選択されることを見出した。さらに、開始点干渉により、開始事象の間隔の保存がもたらされる。その理由およびヒト開始点干渉の実行に用いられる機構が議論される。
【0119】
材料および方法
DNA調製
正常なヒト一次ケラチノサイトを皮膚生検から誘導した。細胞を標準的なケラチノサイトSFM中で培養した。その後、新生DNAをそれぞれ記載されているように(Lebofsky and Bensimon, 2005)、IdUおよびCIdUで20分間標識した。DNA抽出およびコーミングは標準的なプロトコールに従って行った。
【0120】
ハイブリダイゼーションおよび蛍光呈示
表S1に示されているプライマー対を用い、広範囲PCRにより、25GMCプローブを作製した。5〜7kbのプローブの増幅を助けるため、TaKaRa LA Taqを用いた(Takara Bio)。PCR産物の鋳型として用いたBACを表S2に示す。プローブをそれらの記号(A、B、CおよびD)に従い、終濃度〜20ng/μlでプールした。プローブのビオチン化は、個別に4つの記号のランダムプライミング(Invitrogen)を行うことにより達成した。個々のスライドアッセイについては、〜250ngの各ビオチン化プローブを10μgのヒトcot−1 DNA(Gibco BRL)と合わせた。その後のコームDNAに対するハイブリダイゼーションは公表されている方法(Lebofsky and Bensimon, 2005)に従った。プローブIdUおよびCIdUを検出する免疫蛍光工程は次の通り:(i)Alexa 488コンジュゲートストレプトアビジン(Moleccular Probs)、マウス抗ブロモデオキシウリジン(Becton Dickinson)およびラット抗ブロモデオキシウリジン(Harlan Seralab);(ii)ビオチンコンジュゲートウサギ抗ストレプトアビジン(Rockland)、Alexa 350コンジュゲートヤギ抗マウス(Moleccular Probs)およびテキサスレッドコンジュゲートロバ抗ラット(Jackson);(iii)Alexa 488コンジュゲートストレプトアビジン(Moleccular Probs)およびAlexa 350コンジュゲートロバ抗ヤギ(Moleccular Probs)。抗体のインキュベーション、洗浄およびスライドのマウントはこれまでに報告されている通りに行った(Lebofsky and Bensimon, 2005)。
【0121】
画像の採取
HQ CCDカメラを装備したZeiss Axioplan 2顕微鏡(Photometries)を用いて画像の半分を取り込んだ。あとの半分をCytoscoutハイスループットスキャニング装置(Upper Austrian Research)を用いて採取した。バックグラウンド蛍光のドットを、Photoshop(Adobe)を用いて除去し、対象分子を強調した。
【0122】
結果
ゲノムモールスコードハイブリダイゼーション戦略
ある細胞周期の中でどの開始点が活性、またサイレントなのかを分析するための第一段階として、ある領域内の総ての可能性ある開始部位を得た。このために、本発明者らは分子コーミングに基づく単一分子アプローチを用いた(Bensimon et al, 1994)。正常な核型を示す非同期性ヒト一次ケラチノサイトに20分間ヨードデオキシウリジン(IdU)、次いでクロロデオキシウリジン(CIdU)のパルスを与えた。この細胞集団からDNAを抽出し、コーミングした。新たに合成されたDNAにおけるBrdU類似体の組み込みをIdU(青)およびCIdU(赤)の免疫検出によって可視化した。この実験パラダイムは、二方向性複製の開始部位が推察できる3種類のシグナルを生じる(図1A;詳細はAnglana et al, 2003; Lebofsky and Bensimon, 2005を参照)。コーミングBrdUのFISHによる呈示により、複製トラックがプローブシグナルとどこで重複しようと、開始を特異的配列トラックに帰すことができる。
【0123】
慣例的に、スライド上のゲノム領域を可視化するためには色が異なる(図1Bi)か、サイズが異なる(図1Bii)プローブ対が用いられる。しかし、DNAの破断が、それらがカバーできる距離を制限する。結果として、このハイブリダイゼーション戦略を用いた染色体のウォーキングダウンは、実験がプローブ対の数に比例して増えるので、時間がかかる方法である。理想的には、数色および/または大きな領域をカバーするために様々な長さのプローブを使用することができるが、これらの戦略はそれぞれスペクトルの重複および繰り返し配列の非特異的ハイブリダイゼーションのために適切なものではない。
【0124】
本発明者ら独自の解決策の第一の部分は、サイズの異なるギャップが、色またはサイズの異なるプローブと同じ情報を与えることを実現したことによる。図1Biiiに示される例では、ギャップ1はあるプローブセットにより定義され、ギャップ2は別のプローブセットにより定義される。また、この2つのプローセット間のギャップサイズはギャップ1および2とは明瞭に異なる。ギャップ3はDNA破断の際に有用となる。その助けで、その分子は完全なプローブセットが可視化されなくてもなお一定方向を向くことができる(図1Bivおよびv)。ギャップは位置情報を与えるので、それらの数にもはや制限はなく、すなわち、スペクトルの重複やハイブリダイゼーションの際の繰り返し配列はもはや問題でない。サイズの異なるギャップを用いることで、ヒト染色体14q11.2において〜1.5Mbに及ぶ遺伝子モールスコード(GMC)が作製された(図1C)。この全GMCを個々のアッセイでハイブリダイズさせ、総てのプローブが緑で検出された。分子コーミングに先立つDNA操作は繊維をランダムな位置で破断させる。しかし、繊維のサイズは個々の分子上で複数の記号を可視化するに十分なものであった。これに対し、繊維破断のため、記号から数プローブだけしか検出されない場合もあった。開始点は、複製トラックが解読可能なGMCプローブセットと同時に局在した開始点を示す限りマッピングされた(図1D)。よって、新規なハイブリダイゼーション戦略GMCにより、限られた数の実験で、大きな領域の開始点マッピングが可能であった。
【0125】
1.5Mbのヒト染色体14q11.2に対する開始点マッピング
この実験パラダイムを用い、本発明者らは、GMC領域において232の単一DNA分子に対し、307の開始事象を検出した。優先的開始ゾーンを客観的に確立するために、データのクラスター化を行った。まず、本発明者らは、階層的クラスタリングツリーを作出した。最適な区切りを設けるためには、クラスター内のデータの分散を最小化し、クラスター間の分離は最大にすべきである。これら2つの特徴を級内分散(W)および級間分散(B)と呼ぶ(図2A)。従って、望ましいクラスターセットは小さなWと大きなBまたはB−Wの最大値を持たなければならない。これらのデータを9、22および45のクラスターに分けたところ、比較的高いB−W値が得られた(図2A)。図2Bは、開始事象がこれらのクラスターセットに従ってどのように分割されるかを示している。データを45のクラスターに分けると、最も狭い開始領域が得られる。本発明者らは、これら45のクラスターを個々の開始ゾーンを示すものと考えた(図2C)。これらのクラスターの中には極めて少ない開始事象しか含まないものがあり、これはバックグラウンドのノイズによるものでる可能性がある。よって、3つを超える開始事象を有するクラスターだけを次の分析に用いた。このようにして、38の開始ゾーンを同定した。
【0126】
開始ゾーンのサイズは2.6kb(最小)〜21.6kb(最大)の間で様々であり、平均は13.5+/−5.2kbであった。これらの値は哺乳類細胞に関して報告されている(DePamphilis, 1999)他の開始ゾーンの範囲内にある。このゾーン内で、1〜2kbの特定の開始部位は見られなかった。
【0127】
これは、開始点ピークを識別するためのデータ数が不十分なためである可能性があった。コーミングされた分子上の最大解像度は1〜4kbの間である。よって、これらの開始の優先性が、マッピングされた各開始点に固有の標準偏差の結果として取り払われた可能性もある。
【0128】
あるいは、散在した発火部位を含む開始ゾーンがこれまでに考えられていたものよりも優勢である可能性がある。本発明者らのデータは、他のもっと解像度の高い技術でプローブ可能な標的領域を提供する(Todorovic et al, 2005)。よって、これらの可能性を区別することができる。
【0129】
活性化された開始点近傍の時空間分析
本発明者らは次に、開始ゾーンが互いに対してどのように分布するかということに注意を向けた。ゾーン間の距離を測定したところ、ゾーン間平均は40.6+/−20.7kb(最小値=14.3kb、最大値=93.1kb)であることが明らかになった。哺乳類細胞の開始点間距離が一般に100〜150kbであること(Berezney et al, 2000)を考えれば、これは驚くべきことである。この相違は、細胞周期ごとにゾーンのサブセットだけが活性化されるとすると説明がつく。この可能性を調べるため、本発明者らは個々の繊維上の複数の開始点間の間隔を分析した(図3A)。
【0130】
本発明者らの分析は一分子レベルであるため、これらの開始点は、1回のS期において1個の細胞によって実際に用いられるものに相当する。DNA破断は、観察された307の開始事象のうち173に関して、隣接する開始点の可視化を妨げた。残りの場合は、活性のある開始点近傍が存在する場合に見られた(134/307)。この2つの最も近い機能的開始点と最も遠い機能的開始点はそれぞれ31.4kbおよび390.8kb離れていた。興味深いことに、平均開始点間距離は113+/−66.4kbであると算出された(図3B)。ゾーン間距離(〜40kb)に比べて、この結果は、ある細胞周期において3つの潜在的ゾーンのうち平均して1つだけの開始点から発火することを示唆する(図3C)。
【0131】
特定のゾーンに由来する開始点が互いに対して早くまたは遅れて再現性よく発火されたかどうかを検討するため、活性化時間を調べた。開始点を示す複製シグナルの種類に基づき(図1A)、開始時間を標識期間に対して関連づけることができる。これは、IdUまたはCIdUパルスのいずれかの際に発火した開始点にも当てはまり、各20分の時間であった(図1Aiiおよびiii)。これは同様に、標識期間前に発火した開始点にも当てはまり、先行する分岐が修飾されたヌクレオチドの組み込みによって可視化できることが示された(図1Ai)。これが生じるためには、開始点の活性化の時間が平均20分を超えてIdU/CldUパルスに先行することはできなかった(例えば、図3Ciの2番目の分子を参照)。従って、分析枠は、パルス20分前、IdUパルス中20分およびCIdUパルス中20分を含む、全〜60分に及ぶ。本発明者らは、これら38の開始ゾーンのいずれにもタイミング的に優先されるものを見出すことができなかった。さらに、隣接する開始点は同時には発火しなかった(図3C)。個々の繊維上の複数の開始部が本発明者らの実験パラダイムによって与えられる60分以内に検出可能であったことに注目すべきである。従って、開始事象間の厳密な同期性は見られなかったが、任意の2つの活性化開始点の間のタイミングの差はおよそ1時間に限定される。
【0132】
隣接する開始点間の活性化時間はわずかに違いがあることから、2つの接近する分岐間の非複製領域における潜在的開始点は、なお、いくらか後の時間に活性化された可能性がある(例えば、図3Ciiの3番目および5番目の分子を参照)。これらの領域から発火する開始点はより短い開始点間距離をもたらす。隣接する開始点の大部分は互いに30分以内に発火すると考えられる(Berezney et al, 2000)。本発明者らの分析枠は60分(前段参照)であるので、クラスター内のほとんど総ての開始点が活性化されると予測される。
【0133】
従って、開始点の発火の遅れが開始点間距離の測定値を有意に変化させる可能性は低い。
【0134】
潜在的開始部位にわたる分岐の伸張
これまでに、開始部位または開始部を意味するためには、唯一、複製トラックが用いられてきた。しかしながら、それらの周囲領域への二方向性の伸張は、データの別の重要な部分を提供する。隣接するゾーンと重複する1つの開始ゾーンから生じるシグナルは、後者の場合には、複製分岐の先行通過および開始点の許可の剥奪を意味する。
【0135】
これは、受動的に複製されたゾーンをS期のいくらか遅い時間における発火に不応とする。
【0136】
ヒト細胞における総ての潜在的開始点が、それらが酵母である場合(Santocanale and Diffley, 1996)と同様に許可される限り、この観察は開始点干渉の証拠となる(図4A)。開始ゾーンへ向かっていくらか伸張する分岐では、ゾーンは重心が達せられた場合にのみ抑制されると考えられる。本発明者らは、伸張中の分岐からのシグナルを用い、活性開始点干渉からどれくらい離れて起こるのか分析した(図4B)。
【0137】
全部で528の開始ゾーンが抑制されることが分かった。末端小粒方向へ移動する分岐を表す標識(55.1+/−38.4kb、N=262;図4C)に対して動原体方向へ移動する分岐を表す標識(56.5+/−37.7kb、N=266)を考えると、それらの開始部位からの距離は有意には異なっていなかった。結果として、以下の分析のため、両方向に移動する分岐により媒介される開始点干渉を合わせた。活性開始点からの複製トラックは、平均して55.8+/−38kb離れたゾーンと重複していた。最も近い、また最も遠い抑制ゾーンは機能的開始点からそれぞれ7kbおよび284.6kbの位置にあった。これらのデータは次のように解釈される。開始部位のすぐ隣に位置するゾーンは314回抑制された。2番目のゾーンは137回抑制され、その後(3番目以降)のゾーンは77回抑制された。528の抑制された開始ゾーンに基づくこれらのデータを考え合わせると、開始点干渉が1〜2の隣接する開始ゾーンのほとんどの部分に及んでいることが示唆される。
【0138】
ゾーン間干渉の他、本発明者らはゾーン内干渉も分析した。
【0139】
活性開始点からの分岐は、100%の時間、その固有の開始ゾーンの境界を越えて伸張した(例えば、図4B参照)。この形態の干渉が多ければ、与えられた任意の細胞周期において開始ゾーンにつき1を超える開始点が発火する確率は低いはずである。この分析を行うことは、比較的小さな開始ゾーンにおいて密に配置された開始点に代表的な短い複製トラックを識別する本発明者らの能力にかかっている。コームDNA上の線状蛍光セグメントの最大解像度は1〜4kbである。これは平均13.5kbより小さいゾーンにおける複製の開始点の可視化を面倒にする。しかしながら、より大きな開始ゾーンでは、いくつかの開始点の観察を分子コーミングの解像度により制限されない。開始ゾーンのサイズにかかわらず、個々のS期において同じ開始ゾーン内から2以上の開始点が発火するということは見られなかった。よって、開始事象からの距離によって低下するゾーン間干渉とは対照的に、ゾーン内干渉は極めて効率的であり、分岐が移動しなければならない距離には依存しない。
【0140】
結論
結論として、本発明者らは、一分子アプローチを用い、ヒト染色体14q11.2の1.5Mb領域の総ての潜在的開始ゾーンをマッピングした(図1および2)。本発明者らはまた、それらの一部だけが実際に個々の細胞周期に用いられることも実証した(図3)。
【0141】
最後に、限定された開始点の活性化は、規則的な開始点間発火距離をもたらす(図3)。
【0142】
従って、本発明者らは、保存された開始事象間隔が広範な哺乳類開始ゾーンに関して維持されていることを初めて示す。
【0143】
酵母およびCHO細胞では、高い潜在的開始点/活性開始点比が記載されている(Raghuraman et al, 2001; Wyrick et al, 2001; Pasero et al, 2002; Anglana et al, 2003)。本発明者らのデータは、この比がヒト細胞においても保存された特徴であることを示唆する。これにより、真核細胞において繰り返されるテーマである、なぜ開始点が冗長なのかという重要な疑問が持ち上がる。酵母染色体の1つの腕におけるいくつかの開始点を欠失させても、ゲノムの安定性に無視できるほどの影響しかなかった(Dershowitz and Newlon, 1993)。このことは、それほど多くの開始点が必要でないことを示唆する。
【0144】
しかしながら、さらに最近では、組み立てられたpre−RCの完全な相補物を阻害すると染色体の再配列が起こった(Lengronne and Schwob, 2002; Tanaka and Diffley, 2002)。この理由は分かっていないが、いくつかの提案が、余分な潜在的開始点が、混乱したDNA複製の事象において安全ネットを提供するという観点に収束している(Schwob, 2004)。第一に、分岐が遮断されれば、それは組換えの基質へと変換することができる(Rothstein et al, 2000)。下流の「余分な」開始点の活性化は接近する分岐を生じる。この分岐は遮断された分岐と合流し、それによりそれを組換えから救い出す。
【0145】
第二に、いくつかの開始点が発火できない場合、細胞は複製されないDNAとともに有糸分裂を受け得る。このフラグメントは動原体が引き離される際に破断する。潜在的開始点の供給過剰は偶発事象の確率を減らす。最後に、S期のチェックポイントによる最適な細胞周期の休止には、十分な数の分岐が必要である(Shimada et al, 2002)。開始の試みに失敗すると、分岐が失われる。バックアップ開始点の発火が、補償のための2つの付加的な分岐を生成し、これによりチェックポイントが作動可能となる。これらのモデルのいずれかが適用可能であるかを評価するには、明らかにさらなる研究が必要である。
【0146】
開始点干渉は、高い潜在的開始点/活性開始点比が真核生物でいかに達成されるかを説明する機構として取り上げられてきた。それは、早期に活性化された開始点から進行する分岐により、許可された開始点を表すpre−RCの除去を含む(Brewer and Fangman, 1993)。
【0147】
開始点干渉は酵母、ツメガエルおよびCHO細胞で見られている(Brewer and Fangman, 1993; Lucas et al, 2000; Anglana et al, 2003)。ここで、本発明者らは、開始点干渉がヒト細胞における開始点機能の調節に重要な役割を果たすこと、さらにはこれが開始ゾーンに関して起こることを初めて示す(図4)。分子コーミングに先立ち、DNAは除タンパクされる。従って、どの開始ゾーンが許可された開始点を含んでいたか確認することはできなかった。実際に、受動的に複製されたゾーン(抑制されていると解釈される)は単にそれから始まることが許可されていないということかもしれない。今後の研究で、開始ゾーン間のpre−RCの会合がアッセイされる。これにより、本発明者らは、開始点干渉がこの用語の正式な定義に従って起こるかどうかを決定することができる。
【0148】
開始点干渉を説明するため、酵母から得られたデータを挙げられており、これによれば、G1において確立された種々の活性化時間が後の開始点を先の開始点からの分岐により受動的に複製させる(Lucas and Raghuraman, 2003)。CHO細胞で行われた研究(Anglana et al., 2003)と一致して、本発明者らは隣接する開始点に関して強い、予めプログラムされたタイミングの違いを見出せなかった(図3)。よって、本発明者らのデータは、ヒト細胞では酵母に存在するものとは異なる機構が作動していることを示唆する。
【0149】
本発明者らは、ゾーン間干渉が活性開始点からの距離に応じて徐々に低下することを見出した(図4)。開始点の発火の確率が限定された開始因子のために低い場合(Walter and Newport, 1997)には、本明細書で報告されている開始点干渉はこの低い確率および結果としての受動的な現象の直接的結果であり得る。他方で、開始点発火確率が高ければ、開始点干渉は能動的に調節されるはずである。例えば、混乱のない伸張分岐に存在するチェックポイントタンパク質は遠位開始点の発火を抑制し得る。
【0150】
これにより、遅れた開始点が受動的に複製され、従って抑制される機会が能動的に増える(Marheineke and Hyrien, 2004; Shechter et al., 2004; Sorensen et al., 2004; Syljuasen et al., 2005)。今後の研究で、いずれのモデルがゾーン間干渉を担うのかが明らかになるであろう。
【0151】
最近、数学的研究により、複製中心においてともに隔離され、従って同時に活性化され得るのはわずかに11kb離れた潜在的開始点だけであるということが提示された(Jun et al., 2004)。この制限は、DNAの屈曲を制限するDNAの持続長により決定される。持続長はゾーン内干渉を説明することができ、DNAの堅さが、あるゾーンに由来する2つの潜在的開始部位が複製中心内に集中することを妨げ、従って、それらの同時活性化を妨げる。DNAの物理的特性に基づく機構が堅牢なため、本明細書で報告される高効率のゾーン内干渉が生じ得る。
【0152】
哺乳類開始ゾーン内およびゾーン間の開始点干渉機構は、これらの領域の基礎にある分子決定基に依存する。許可の過程で、複数のミニ染色体維持(MiniChromosome Maintenance)(MCM)複合体はpre−RCから離れて拡散する(Ritzi et al, 1998; Edwards et al, 2002)。これらのMCM部位の1つで発火する開始点は哺乳類細胞における開始ゾーンの存在を説明することが示唆されている(Hyrien et al, 2003; Blow and Dutta, 2005; Cvetic and Walter, 2005)。よって、本明細書で報告される開始ゾーン(図2)は反復MCMローディングのために生じ得る。拡散MCMまたは他の未知の因子が哺乳類開始ゾーンを担っているかどうかの判定は、我々が、ヒト開始点干渉がいかにして実行され、開始事象間隔が調節されるかを理解する助けとなる。
【参考文献】
【0153】
【図面の簡単な説明】
【0154】
【図1】ヒト染色体14q11.2における1.5Mbに対するDNA複製開始マッピングを示す。 (A)開始点を示すコーミングされたDNA上の3種類の複製シグナル。開始は、標識前、IdUパルス(青)中、およびCIdUパルス(赤)中に起こり、それぞれ(i)、(ii)および(iii)に示されるシグナルを生じる。3つの場合の総てで、これらのトラックの中点が開始部位であると推定される。 (B)コーミングされたDNAに対するハイブリダイゼーション戦略。長さが等しいが色の違いで検出される2つのプローブ(i)または同じ色で検出される、長さが異なる2つのプローブ(ii)をハイブリダイズさせ、対象ゲノム領域を可視化する。あるいは、同じ情報を得るためにプローブセット間のギャップを使用することもできる。(iii)では、4つの短いプローブをハイブリダイズさせ、情報が得られる3つのギャップを生じさせる。ギャップ3は、分解の際に分子を一定の方向に向かせる(iv、v)。DNA分解は一対の垂直実線により表される。 (C)ヒト染色体14q11.2における1.356Mbにわたる遺伝子モールスコード(GMC)。モールスコードA、B、CおよびDで、最初の4つの記号の直線パターンを示す。GMCはこれら4つの記号を含み、各記号は1セットのプローブで表される。プローブは緑で示される。コードギャップは短ギャップと長ギャップである。開始ギャップおよび終了ギャップは、DNA破断の際の記号の配向を助けるために含めた。記号はスペースギャップにより隔てられている。プローブおよびギャップのサイズ(kb)がそれぞれ上に示されている。 (D)GMC領域において観察される開始事象の例。白い矢印は開始部位を示す。ファイバー2、7、8および13では、GMCは、ある記号に属す総てのプローブが存在しない場合でもなお解読可能である。それらの記号と隣接する開始事象は、隣接する記号の1つが解読されるか(分子1、5、11および14)、スペースギャップ情報が入手可能な場合(分子4および10)にマッピングされた(バー=100kb)。
【図2】クラスター分析による開始ゾーンの同定。 (A)理想的クラスターの定義。分散内(W)および分散間(B)の方程式を挿入枠内に示す。Wに関して、Nはクラスター数であり。Viはクラスターiの分散である。Bに関して、Ciはクラスターiの重心であり、Cはクラスターセットにおける総ての重心の平均である。クラスター数に関数としてB−Wをプロットしたところ、データを9群(青丸)、22群(緑丸)および45群(赤丸)に分けた際の最大値が明らかになった。 (B)9(青線)、22(緑線)および45(赤線)の区分がデータに適用される際の、あるクラスターの、その成分への分解。赤線の下の領域は開始ゾーンを表し、分子の上の白四角により示される。白い矢印は開始部位を示す(バー=100kb)。 (C)GMC領域における開始ゾーンの分布。水平の赤線はゾーンのサイズと位置を表す。垂直の白線はマッピングされた307の開始部位の総ての位置を表す。4未満のデータ点を含むゾーンは、赤線の代わりに水平のグレーの線でマークされ、その後の分析では考慮されていない。この図面のパネルBにおけるクラスターはアスタリスクで示される(バー=100kb)。
【図3】機能的開始点の時空間分析を示す。 (A)開始点間距離(X)を示す複製シグナル。(i)では、2つの開始部位からの複製トラックには隔たりがあるままである。(ii)および(iii)では、それぞれIdUパルスおよびCIdUパルス中に、接近する分岐が合流する。(B)測定された開始点間距離の頻度を示すヒストグラム。 (C)記号A、B(i)およびC、D(ii)において少なくとも2つの開始事象を有する分子の例。白い矢印は開始部位を示す。開始ゾーンは水平の赤線でマークされている。個々の分子について、開始点が発火する開始ゾーンは白四角で示されている。影付きの四角はサイレント開始ゾーンを表す(バー=100kb)。
【図4】分岐の伸張に基づく開始点干渉を示す。 (A)ゾーン(v)の活性開始点から伸びている分岐は垂線対に接している領域を含んでいる。左に移動している分岐の終点が見られる(白ぬきの逆三角)。着目する開始点からの分岐はその境界部に達しない限り、開始ゾーン(i)および(ii)は干渉を受けない。その分岐が受動的にその全長を複製すれば、ゾーン(iii)、(iv)および(vi)は抑制される。右へ移動している分岐はゾーン(vii)を通り抜けるが、その重心(黒いドット)には達しない。このゾーンは開始点干渉データには含まれていない。グレーの長方形は開始ゾーンを表し、濃いグレーの長方形は抑制された開始ゾーンを表す。 (B)開始点干渉を示す分子の例。この図の開始ゾーンは水平の赤線で示される。白四角は開始点が発火する(白い矢印)開始ゾーンを示す。影付きの四角は、分岐の伸張によって抑制されている開始ゾーンを示す(バー=100kb)。 (C)開始事象と干渉を受けているゾーンの間の距離の頻度を示すヒストグラム。動原体方向へ移動している分岐および末端小粒方向へ移動している分岐により抑制されたゾーンはそれぞれ、負の値および正の値で表される。
【図5】3種類のコードを示す。 A:連続するプローブ間の間隔に基づくコード。 B:標識(より詳しくは、色)に基づくコード。 C:標識(色)+連続するプローブ間の間隔に基づく冗長コード。 3種類のコード法。プローブはカラーの短いセグメントで示される。これらのプローブは例えば5〜10Kbを測定することができる。ここで示される「色」は、性質の異なるプローブと同様に含め/理解しなければならず、それは結びついている蛍光色素で異なる場合もあるし、大きさまたは他のいずれかの特徴または特徴の組合せで異なる場合もあり、特にそれ自体蛍光プローブの問題ではない。プローブ間の間隔は例えば短いスペースで約20KB、長いものでは40KB、ダブレットの場合では10KB未満(最初の場合)であり得る。冗長コードは、その他の双方の組合せである。
【図6】冗長コードの興味深い点を示す。 冗長コードの興味深い点。記載されている3種類のコード1に関して、青い矢印のマークされている配列の一部における欠失が示されている。最初の両方の場合では、この欠失に関する理由の修正点として、別のコード配列との混同が含まれる。3番目の場合では、得られたコード配列には矛盾があり、従って、再配列があったことを明らかに示す。この再配列(欠失)の性質は本明細書によれば同定可能である。
【図7】重複コードを示す。 重複コード。プローブの3つの性質はA、BおよびCの文字で表される。上段では、あるものが、7つの連続するプローブからなる種々のコード配列を、その最後の6つのプローブが次にくるものの最初の6つのプローブに相当するように並べることによりどのようなものかを示し、7文字の連続が1回である連続プローブを得ることができる。下段では、7文字のあり得る総ての前提など、連続する2193文字のA、BおよびCが一度だけ表される。従って、例えばコード配列abaaccbを含む/理解するフラグメントが検出されれば、それは赤色の領域から得られるものであることが分かる。
【特許請求の範囲】
【請求項1】
供試巨大分子上の少なくとも1つの対象ドメインの存在を検出する方法であって、
a)対象ドメイン上の少なくとも2つの標的領域を予め決定し、各標的領域の対応する標識プローブ(対象ドメインのプローブのセットと呼ぶ)を設計および取得し、これらのプローブのいずれかの、他のものに対する位置が選択され、供試巨大分子上の対象ドメインの特異的サインを形成する工程;
b)工程a)で得られたプローブが結合されている供試巨大分子を展開した後、線状化巨大分子に結合されているプローブのいずれかの、他のものに対する位置を検出し、対象ドメインのサインが検出されれば、供試巨大分子上に対象ドメインが存在することを示し、逆に、対象ドメインのサインまたはサインの一部が検出されなければ、供試巨大分子上に対象ドメインまたはドメインの一部が存在しないことを示す工程
を含む、方法。
【請求項2】
少なくとも2つの対象ドメインの存在を同定するためであって、工程a)において各対象ドメイン上の少なくとも3つの標的領域を予め決定することを含む、請求項1に記載の方法。
【請求項3】
対象ドメインのサインが、連続するプローブ間の一連の間隔から生じるものである、請求項1に記載の方法。
【請求項4】
対象ドメインの位置が、化学反応または生化学反応を位置決定するための参照として用いられる、請求項1に記載の方法。
【請求項5】
対象ドメインの位置が、標的領域を包含する巨大分子における物理的地図を確立するために用いられる、請求項1に記載の方法。
【請求項6】
対象ドメインのサインが、一連の異なる標識プローブからなる、請求項1に記載の方法。
【請求項7】
標的領域のプローブのいくつかが、巨大分子上付近に位置した少なくとも1つの他の対象ドメインのサインの一部でもある、請求項1に記載の方法。
【請求項8】
総てのプローブが、同じ標識で標識される、請求項1に記載の方法。
【請求項9】
プローブが、少なくとも2つに異なる標識で標識される、請求項1に記載の方法。
【請求項10】
対象ドメインのサインが、標識の連続をもたらす、請求項9に記載の方法。
【請求項11】
巨大分子が、核酸、特にDNA、より詳しくは、二本鎖DNAである、請求項1に記載の方法。
【請求項12】
プローブが、少なくとも1kbのオリゴヌクレオチドである、請求項11に記載の方法。
【請求項13】
巨大分子の展開が、線状化によりなされる、請求項1に記載の方法。
【請求項14】
巨大分子の線状化が、巨大分子上にプローブが結合する前または結合した後に行われる、請求項13に記載の方法。
【請求項15】
巨大分子の線状化が分子コーミングまたはファイバーFishによって行われる、請求項13に記載の方法。
【請求項16】
巨大分子上の対象ドメインに相当する少なくとも3つのプローブの結合が、少なくとも2つの異なるスペース(例えば、「ショート」と「ラージ」)の群(この群は各対象ドメインで同一である)から選択される少なくとも2つのスペースの配列を形成する、請求項2に記載の方法。
【請求項17】
プローブセットがさらに2つのプローブ(プローブ1またはプローブ2)を含み、各プローブが対象ドメインの異なる末端に結合することができ、対象ドメインにおいてその連続するプローブと関連する前記プローブ1またはプローブ2(開始部または終了部の末端プローブ対と呼ぶ)の1つのシグナルを読み取ることによって、読み取りの開始または終了に関する情報が得られる、請求項2に記載の方法。
【請求項18】
読み取りの開始に関する情報が、開始部の末端プローブ対の2つの連続するプローブ間の間隔の読み取りの結果である、請求項17に記載の方法。
【請求項19】
読み取りの終了に関する情報が、終了部の末端プローブ対の2つの連続するプローブ間の間隔の読み取りの結果である、請求項17に記載の方法。
【請求項20】
読み取りの開始に関する情報が、開始部の末端プローブ対の2つの連続するプローブ間の間隔の読み取りの結果であり、読み取りの終了に関する情報が終了部の末端プローブ対の2つの連続するプローブ間の間隔の読み取りの結果であり、この間隔が、開始と終了の情報を区別するために開始部の末端プローブ対と終了部の末端プローブ対では異なる、請求項17に記載の方法。
【請求項21】
プローブが蛍光標識で標識される、請求項1に記載の方法。
【請求項22】
プローブが放射性標識で標識される、請求項1に記載の方法。
【請求項23】
前記サインが、プローブセットにおける第一プローブと第二プローブの間のスペースを含み、このスペースがそのサインにおける他の総てのスペースと異なり、このスペースがこのサインの開始についての情報を得るために使用可能である、請求項1に記載の方法。
【請求項24】
前記サインが、プローブセットにおいて最後の手前のプローブと最後のプローブの間にスペースを含み、このスペースがそのサインにおける他の総てのスペースと異なり、このスペースがサインの終了に関する情報を得るために使用可能である、請求項1に記載の方法。
【請求項25】
少なくとも2セットの3つのプローブを含み、各プローブセットは1つのドメイン上に特異的に結合することができ、巨大分子上での1セットの少なくとも3つのプローブの結合が、少なくとも2つの異なるコード群(例えば、「ドット」と「ダッシュ」)から選択される少なくとも2つのコードの配列を形成し、この群が総てのドメインで同一であり、このコード配列が1つのドメインに特異的であり、そのドメインのサインである、巨大分子上の少なくとも2つの対象ドメインの存在を検出するためのキット。
【請求項1】
供試巨大分子上の少なくとも1つの対象ドメインの存在を検出する方法であって、
a)対象ドメイン上の少なくとも2つの標的領域を予め決定し、各標的領域の対応する標識プローブ(対象ドメインのプローブのセットと呼ぶ)を設計および取得し、これらのプローブのいずれかの、他のものに対する位置が選択され、供試巨大分子上の対象ドメインの特異的サインを形成する工程;
b)工程a)で得られたプローブが結合されている供試巨大分子を展開した後、線状化巨大分子に結合されているプローブのいずれかの、他のものに対する位置を検出し、対象ドメインのサインが検出されれば、供試巨大分子上に対象ドメインが存在することを示し、逆に、対象ドメインのサインまたはサインの一部が検出されなければ、供試巨大分子上に対象ドメインまたはドメインの一部が存在しないことを示す工程
を含む、方法。
【請求項2】
少なくとも2つの対象ドメインの存在を同定するためであって、工程a)において各対象ドメイン上の少なくとも3つの標的領域を予め決定することを含む、請求項1に記載の方法。
【請求項3】
対象ドメインのサインが、連続するプローブ間の一連の間隔から生じるものである、請求項1に記載の方法。
【請求項4】
対象ドメインの位置が、化学反応または生化学反応を位置決定するための参照として用いられる、請求項1に記載の方法。
【請求項5】
対象ドメインの位置が、標的領域を包含する巨大分子における物理的地図を確立するために用いられる、請求項1に記載の方法。
【請求項6】
対象ドメインのサインが、一連の異なる標識プローブからなる、請求項1に記載の方法。
【請求項7】
標的領域のプローブのいくつかが、巨大分子上付近に位置した少なくとも1つの他の対象ドメインのサインの一部でもある、請求項1に記載の方法。
【請求項8】
総てのプローブが、同じ標識で標識される、請求項1に記載の方法。
【請求項9】
プローブが、少なくとも2つに異なる標識で標識される、請求項1に記載の方法。
【請求項10】
対象ドメインのサインが、標識の連続をもたらす、請求項9に記載の方法。
【請求項11】
巨大分子が、核酸、特にDNA、より詳しくは、二本鎖DNAである、請求項1に記載の方法。
【請求項12】
プローブが、少なくとも1kbのオリゴヌクレオチドである、請求項11に記載の方法。
【請求項13】
巨大分子の展開が、線状化によりなされる、請求項1に記載の方法。
【請求項14】
巨大分子の線状化が、巨大分子上にプローブが結合する前または結合した後に行われる、請求項13に記載の方法。
【請求項15】
巨大分子の線状化が分子コーミングまたはファイバーFishによって行われる、請求項13に記載の方法。
【請求項16】
巨大分子上の対象ドメインに相当する少なくとも3つのプローブの結合が、少なくとも2つの異なるスペース(例えば、「ショート」と「ラージ」)の群(この群は各対象ドメインで同一である)から選択される少なくとも2つのスペースの配列を形成する、請求項2に記載の方法。
【請求項17】
プローブセットがさらに2つのプローブ(プローブ1またはプローブ2)を含み、各プローブが対象ドメインの異なる末端に結合することができ、対象ドメインにおいてその連続するプローブと関連する前記プローブ1またはプローブ2(開始部または終了部の末端プローブ対と呼ぶ)の1つのシグナルを読み取ることによって、読み取りの開始または終了に関する情報が得られる、請求項2に記載の方法。
【請求項18】
読み取りの開始に関する情報が、開始部の末端プローブ対の2つの連続するプローブ間の間隔の読み取りの結果である、請求項17に記載の方法。
【請求項19】
読み取りの終了に関する情報が、終了部の末端プローブ対の2つの連続するプローブ間の間隔の読み取りの結果である、請求項17に記載の方法。
【請求項20】
読み取りの開始に関する情報が、開始部の末端プローブ対の2つの連続するプローブ間の間隔の読み取りの結果であり、読み取りの終了に関する情報が終了部の末端プローブ対の2つの連続するプローブ間の間隔の読み取りの結果であり、この間隔が、開始と終了の情報を区別するために開始部の末端プローブ対と終了部の末端プローブ対では異なる、請求項17に記載の方法。
【請求項21】
プローブが蛍光標識で標識される、請求項1に記載の方法。
【請求項22】
プローブが放射性標識で標識される、請求項1に記載の方法。
【請求項23】
前記サインが、プローブセットにおける第一プローブと第二プローブの間のスペースを含み、このスペースがそのサインにおける他の総てのスペースと異なり、このスペースがこのサインの開始についての情報を得るために使用可能である、請求項1に記載の方法。
【請求項24】
前記サインが、プローブセットにおいて最後の手前のプローブと最後のプローブの間にスペースを含み、このスペースがそのサインにおける他の総てのスペースと異なり、このスペースがサインの終了に関する情報を得るために使用可能である、請求項1に記載の方法。
【請求項25】
少なくとも2セットの3つのプローブを含み、各プローブセットは1つのドメイン上に特異的に結合することができ、巨大分子上での1セットの少なくとも3つのプローブの結合が、少なくとも2つの異なるコード群(例えば、「ドット」と「ダッシュ」)から選択される少なくとも2つのコードの配列を形成し、この群が総てのドメインで同一であり、このコード配列が1つのドメインに特異的であり、そのドメインのサインである、巨大分子上の少なくとも2つの対象ドメインの存在を検出するためのキット。
【図5】

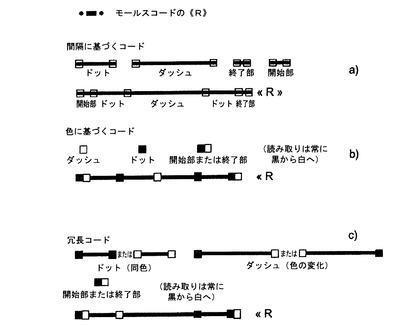
【図6】
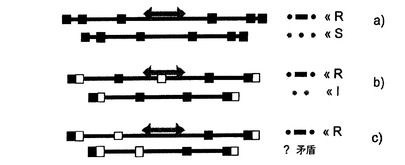
【図7】

【図1】

【図2】
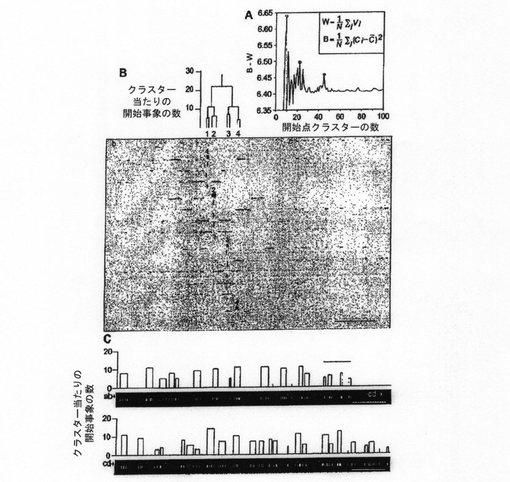
【図3】
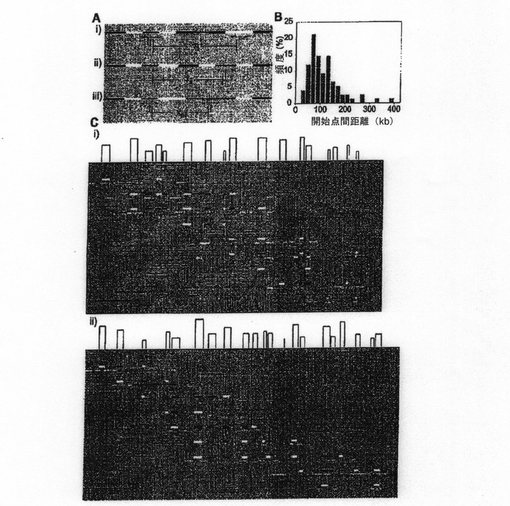
【図4】
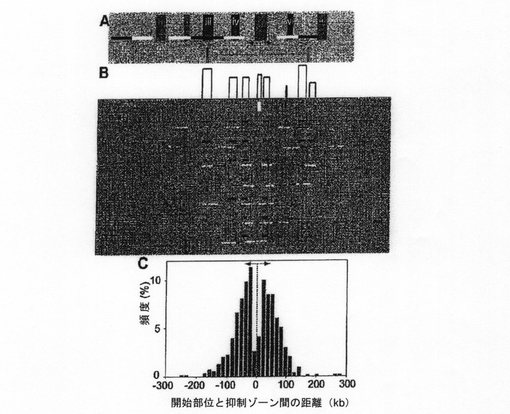
【図6】
【図7】
【図1】
【図2】
【図3】
【図4】
【公表番号】特表2010−502206(P2010−502206A)
【公表日】平成22年1月28日(2010.1.28)
【国際特許分類】
【出願番号】特願2009−527137(P2009−527137)
【出願日】平成19年9月5日(2007.9.5)
【国際出願番号】PCT/EP2007/059299
【国際公開番号】WO2008/028931
【国際公開日】平成20年3月13日(2008.3.13)
【出願人】(501173391)
【氏名又は名称原語表記】INSTITUT PASTEUR
【出願人】(509066330)
【氏名又は名称原語表記】GENOMIC VISION
【Fターム(参考)】
【公表日】平成22年1月28日(2010.1.28)
【国際特許分類】
【出願日】平成19年9月5日(2007.9.5)
【国際出願番号】PCT/EP2007/059299
【国際公開番号】WO2008/028931
【国際公開日】平成20年3月13日(2008.3.13)
【出願人】(501173391)
【氏名又は名称原語表記】INSTITUT PASTEUR
【出願人】(509066330)
【氏名又は名称原語表記】GENOMIC VISION
【Fターム(参考)】
[ Back to top ]