
遺伝子改変されたマウスおよび移植方法
mIL-3遺伝子およびmGM-CSF遺伝子のヒト化、mRAG遺伝子のノックアウト、ならびにmIl2rgサブユニット遺伝子のノックアウト、ならびに任意でTPO遺伝子のヒト化を伴うマウスを記載する。RAG/Il2rg KO/hTPOノックインマウスを記載する。ヒト造血幹細胞(HSC)由来のヒト免疫細胞(HIC)集団を維持し、かつヒト病原体により、例えばチフス菌(S. typhi)または結核菌(M. tuberculosis)により感染され得る、HSCを用いて移植されたマウスを記載する。マウスにおけるモデル化が不十分であるヒト病原体感染をモデル化するマウス、例えばヒトマイコバクテリア感染をモデル化するマウスを記載し、該マウスは、ヒト免疫細胞を含む1つまたは複数の肉芽腫を発症する。初期ヒト造血細胞から発生するヒト造血器悪性腫瘍、例えば骨髄性白血病または骨髄増殖性腫瘍を含むマウスを記載する。

【発明の詳細な説明】
【技術分野】
【0001】
発明の分野
本発明は、遺伝子改変された非ヒト動物の分野に関し、特に、RAG遺伝子ノックアウト、Il2rgIl2rg遺伝子ノックアウト、ならびにIL-3およびGM-CSF遺伝子のヒト化、ならびに任意でTPO遺伝子のヒト化を有する、免疫不全マウス;TPO遺伝子のヒト化を有するRAG/Il2rgIl2rgノックアウトマウス;ヒト造血細胞を用いて移植された、遺伝子改変されたマウス;およびヒト病原体、例えばチフス菌(Salmonella typhi)または結核菌(Mycobacterium tuberculosis)に感染する、移植されたマウスに関するものである。
【背景技術】
【0002】
背景
遺伝子改変されたマウス、改変および移植されたマウス、ならびに、例えば薬物検査の目的のための、ヒト疾患のモデル化におけるそれらの使用は、当技術分野で公知である。ヒト免疫システムをモデル化するために、遺伝子改変されたマウスを使用することが試みられてきた。その分野の概説は、Manz (2007) Human-Hemato-Lympoid-System Mice: Opportunities and Challenges, Immunity, 26:537-541(非特許文献1)に提供されており、それを参照により本明細書に組み入れる。
【0003】
これまで、特定のヒト病原体、例えばチフス菌(S. typhi)への感染性を示す遺伝子改変されたマウスが作製されたことはなかった。マウスモデルが存在する病原性感染症の場合でさえ、該モデルはヒトの特定の病態の適切にモデルとなることができないことがあり、例えば、結核菌(M. tuberculosis)のマウスモデルでは境界のはっきりした肉芽腫またはヒト免疫細胞を含む肉芽腫を形成できなかった。特定の病原体がヒトに及ぼす影響を研究するために、かつ特定の病原体に感染したヒトを治療するうえでの有効性について薬物を試験するために、そのような病原体、例えばチフス菌に感染しやすくなるように、および/または感染がヒト病態のより厳密なモデル、例えば結核菌のヒト感染症のより厳密なモデルとなるように、遺伝子改変されたマウスなどの非ヒト動物があることは有用であろう。
【0004】
一般的には、ヒト造血幹細胞の維持および増殖を支持することができる遺伝子改変されたマウスの必要性、ならびに、例えばヒト病原体に応答して、ヒト血液-リンパシステムの一部をモデル化するまたはそれに近づけることができる、移植に適したマウスの必要性が存在している。
【先行技術文献】
【非特許文献】
【0005】
【非特許文献1】Manz (2007) Human-Hemato-Lympoid-System Mice: Opportunities and Challenges, Immunity, 26:537-541
【発明の概要】
【0006】
概要
遺伝子改変された非ヒト動物が提供される。その非ヒト動物には、1つまたは複数の内在性遺伝子のノックアウトおよび1つまたは複数のヒト化遺伝子(すなわち、内在性遺伝子のヒトの類似体または相同体との、その内在性の遺伝子座での置換)を含むマウスが含まれる。
【0007】
免疫システムを取り除かれたかまたは損なわれた遺伝子改変されたマウス(例えば、照射による)、ならびにヒト造血細胞またはヒト造血幹細胞前駆細胞(hematopoietic stem and progenitor cell)(HSPC)を用いて移植されたマウスが提供される。ヒト造血細胞またはHSPCに由来するヒト細胞を含む遺伝子改変されたマウスが提供され、ヒト血液-リンパシステムを含むマウスも提供される。
【0008】
野生型マウスには感染しないヒト病原体に感染することができる、遺伝子改変、照射および移植されたマウスが提供される。ヒト病原体(例えば、結核菌)への曝露に応答して、野生型マウスには観察されない特徴(例えば、境界のはっきりした肉芽腫またはヒト免疫細胞を含む肉芽腫の形成)を有する免疫応答を開始するマウスが提供される。
【0009】
ヒト病原体の薬物耐性株を同定するための、ヒトワクチンを試験するための、および抗病原体薬物を開発して試験するための、遺伝子改変、照射および移植されたマウス、ならびにそれらを用いるための組成物および方法が提供される。
【0010】
ヒト免疫細胞を受け取りかつ増殖することができる遺伝子改変されたマウスが、ヒト造血器悪性腫瘍を保持できるマウスを含めて、提供される。
【0011】
一局面において、(a)マウスRAG遺伝子ノックアウト;(b)マウスIl2rgIl2rg遺伝子ノックアウト;および(c) (i)マウスIL-3(mIL-3)遺伝子、(ii)マウスGM-CSF(mGM-CSF)遺伝子、および(iii)マウストロンボポエチン(mTPO)遺伝子から選択される1つまたは複数の遺伝子のヒト化を含む、遺伝子改変されたマウスが提供される。
【0012】
一態様において、RAG遺伝子ノックアウトはRAG2遺伝子ノックアウトである。
【0013】
一態様において、ヒト化はmTPO遺伝子のhTPO遺伝子との置換を含む。特定の態様では、ヒト化は、mTPO遺伝子のhTPO遺伝子によるヒト化から本質的になる。
【0014】
一態様において、ヒト化はmIL-3遺伝子のヒトIL-3(hIL-3)遺伝子との置換およびmGM-CSF遺伝子のヒトGM-CSF(hGM-CSF)遺伝子との置換を含む。別の態様では、マウスがmTPO遺伝子のヒトTPO(hTPO)遺伝子との置換をさらに含む。特定の態様では、ヒト化は、mIL-3遺伝子のhIL-3遺伝子によるヒト化およびmGM-CSF遺伝子のhGM-CSF遺伝子によるヒト化から本質的になる。
【0015】
一態様において、ヒト化はmGM-CSF遺伝子のヒトGM-CSF遺伝子との置換を含み、マウスにおいてヒトGM-CSFが肝臓および循環において優勢に発現されない。一態様では、ヒトGM-CSFがマウス肺において優勢に発現される。一態様では、ヒトGM-CSFの発現が組織特異的であり、かつヒトにおける組織特異的発現を反映している。
【0016】
一態様において、遺伝子改変されたマウスは、マウスに存在しうる内在性造血細胞を排除するように処置される。一態様では、その処置が遺伝子改変されたマウスを照射することを含む。特定の態様では、遺伝子改変されたマウスの新生仔が亜致死的に照射される。特定の態様では、新生仔が2×200cGyを4時間間隔で照射される。
【0017】
一態様において、遺伝子改変および処置されたマウスは、遺伝子改変および移植されたマウスを形成するために、ヒト造血細胞またはヒト造血幹細胞(HPSC)を用いて移植される。一態様では、造血細胞がヒト臍帯血細胞およびヒト胎児肝細胞から選択される。一態様では、移植に約1〜2×105個のヒトCD34+細胞を用いる。
【0018】
一態様において、遺伝子改変および移植されたマウスは、以下から選択されるヒト細胞を生じさせる:CD34+細胞、造血幹細胞、造血細胞、骨髄前駆細胞、骨髄細胞、樹状細胞、単球、顆粒球、好中球、肥満細胞、胸腺細胞、T細胞、B細胞、血小板、およびこれらの組合せ。一態様では、ヒト細胞が移植後4ヶ月、5ヶ月、6ヶ月、7ヶ月、8ヶ月、9ヶ月、10ヶ月、11ヶ月、または12ヶ月の時点で存在している。
【0019】
一態様において、遺伝子改変および移植されたマウスは、ヒト造血幹細胞前駆細胞、ヒト骨髄前駆細胞、ヒト骨髄細胞、ヒト樹状細胞、ヒト単球、ヒト顆粒球、ヒト好中球、ヒト肥満細胞、ヒト胸腺細胞、ヒトT細胞、ヒトB細胞、およびヒト血小板を含むヒト血液-リンパシステムを生じさせる。一態様では、ヒト血液-リンパシステムが移植後4ヶ月、5ヶ月、6ヶ月、7ヶ月、8ヶ月、9ヶ月、10ヶ月、11ヶ月、または12ヶ月の時点で存在している。
【0020】
一態様において、遺伝子改変および移植されたマウスは、ヒト細胞が介在する炎症反応を示す。特定の態様では、そのヒト細胞がマクロファージである。特定の態様では、マクロファージ介在炎症反応が肺胞マクロファージによって介在される。特定の態様では、肺胞マクロファージが介在する炎症反応は肉芽腫の形成を含む。特定の態様では、肉芽腫がヒト免疫細胞を含む。特定の態様では、肉芽腫が明確に組織化された肉芽腫である。特定の態様では、肉芽腫がマイコバクテリア、例えば結核菌への曝露後に形成される。一態様では、マウスが2つまたはそれ以上の肉芽腫を含む免疫反応を示す。一態様では、遺伝子改変および移植されたマウスがヒト結核菌感染のためのモデルである。
【0021】
一態様において、遺伝子改変および移植されたマウスは、ヒト免疫細胞を含む肉芽腫の形成によって少なくとも一部特徴づけられる結核菌感染を含む。特定の態様では、その肉芽腫が明確に組織化された肉芽腫である。特定の態様では、結核菌がヒト集団に感染する結核菌の薬物耐性株または多剤耐性株である。一態様では、結核菌に感染したマウスが肺に肉芽腫を含む。特定の態様では、その肉芽腫が十分に発達した肉芽腫である。特定の態様では、肺の肉芽腫がヒト免疫細胞を含む。特定の態様では、肉芽腫のヒト免疫細胞が活性化ヒトマクロファージ、活性化ヒトT細胞、およびこれらの組合せから選択される。
【0022】
一態様において、遺伝子改変および移植されたマウスは、IL-3、GM-CSFおよびTPO遺伝子の1つまたは複数のヒト化を欠く移植されたマウスに比べて、増強された粘膜免疫を示す。特定の態様では、増強された粘膜免疫がインフルエンザA感染後にインターフェロンβ(IFNβ)の高度発現を含む。
【0023】
一態様において、遺伝子改変および移植されたマウスは、結核菌感染およびチフス菌感染から選択される感染を含む。一態様では、マウスは、チフス菌または結核菌を繁殖させる。一態様では、マウスはヒト病原性マイコバクテリアに対する抗マイコバクテリア免疫応答を開始し、該免疫応答は、ヒト免疫細胞によって媒介されかつヒト免疫細胞を含む肉芽腫の形成を含む。特定の態様では、その肉芽腫が十分に発達した肉芽腫である。
【0024】
一態様において、遺伝子改変および移植されたマウスはmTPO遺伝子のヒト化を含み、hTPOを移植されたマウスを形成する。一態様では、hTPOを移植されたマウスは、mTPO遺伝子を含むがhTPO遺伝子を含まない移植されたマウスに比べて、骨髄でのヒト骨髄細胞の増加を示す。特定の態様では、ヒト骨髄細胞がhTPO遺伝子を欠く移植されたマウスに対して1.5倍、2倍、2.5倍または3倍に増加する。特定の態様では、顆粒球の増加が約1.5倍、2倍、2.5倍または3倍である。別の態様では、hTPO遺伝子を欠く移植されたマウスに比して、末梢血単球の増加が認められ、この場合の末梢血単球の増加は約1.5倍、2倍、2.5倍または3倍である。一態様では、遺伝子改変され移植されたマウスは、mTPO遺伝子を置換するhTPO遺伝子から本質的になるヒト化を含み、該マウスはマウスTPOを発現せずにヒトTPOを発現する。
【0025】
一局面において、Rag遺伝子のノックアウト、Il2rgIl2rgノックアウト、およびTPOのヒト化を含む、遺伝子改変および移植されたマウスが提供され、該マウスはヒト造血幹細胞またはヒト免疫細胞を用いて移植され、かつ初期ヒト造血細胞から発生するヒト造血器悪性腫瘍を含む。特定の態様では、その悪性腫瘍が骨髄性白血病および骨髄増殖性腫瘍から選択される。
【0026】
一態様において、マウスは、ヒトIL-3遺伝子およびヒトGM-CSF遺伝子、ならびに内在性マウスIL-3遺伝子のノックアウトおよび内在性マウスGM-CSF遺伝子のノックアウトをさらに含む。
【0027】
一局面においては、RAG遺伝子ノックアウト、Il2rg遺伝子ノックアウト、およびマウス骨髄細胞に対して競合的優位性をもつヒト骨髄細胞を提供する遺伝子改変を含むマウスが提供される。一態様では、その遺伝子改変がマウス骨髄細胞の発達および/または維持に必要とされるマウス遺伝子の対応ヒト遺伝子との置換である。一態様では、遺伝子改変がマウスIL-3遺伝子のヒトIL-3遺伝子との置換、マウスGM-CSF遺伝子のヒトGM-CSF遺伝子との置換、およびこれらの組合せから選択される。一態様では、マウスは、内在性のマウス造血細胞を欠くかまたは実質的に欠き、かつヒト造血細胞を含む。
【0028】
一局面においては、ヒト病原体に感染可能なマウスの作製方法が提供され、該方法は、本明細書に記載した通りにマウスを遺伝子改変しかつ該マウスに移植する段階、遺伝子改変および移植されたマウスをヒト病原体に曝露する段階、ならびにヒト病原体が該マウスに感染するのに十分な条件下で該マウスを維持する段階を含む。一態様では、ヒト病原体が結核菌およびチフス菌から選択される。一態様では、ヒト病原体が遺伝子改変を欠くマウスには病原性でないヒト病原体である。一態様では、ヒト病原体が遺伝子改変を欠くマウスには感染しないヒト病原体である。
【0029】
一局面においては、ヒト病原体に対する薬物の効果を判定する方法が提供され、該方法は、本明細書に記載した通りに遺伝子改変および移植されたマウスをヒト病原体に曝露する段階、ならびに該病原体を該マウスに感染させて、薬物の存在下および非存在下で経時的に感染のパラメーターを測定する段階を含む。一態様では、ヒト病原体が遺伝子改変を欠くマウスには感染しない病原体である。一態様では、ヒト病原体が結核菌およびチフス菌から選択される。一態様では、マウスが既知の感染単位数のヒト病原体に曝露され、感染のパラメーターがマウスの体液中または組織中のヒト病原体の感染単位数である。
【0030】
一態様において、感染のパラメーターはマウスの体液中の力価である。一態様では、感染が結核菌感染およびチフス菌感染から選択される。特定の態様では、感染が結核菌感染であり、かつパラメーターが肉芽腫の形成である。特定の態様では、肉芽腫が肺肉芽腫である。別の特定の態様では、肉芽腫が境界のはっきりした肉芽腫である。
【0031】
一局面において、(a)マウスRAG遺伝子ノックアウト;(b)マウスIl2rg遺伝子ノックアウト;および(c) (i)マウスIL-3(mIL-3)遺伝子および(ii)マウスGM-CSF(mGM-CSF)遺伝子のヒト化を含む、遺伝子改変されたマウスが提供され;該マウスは、内在性マウス造血細胞を取り除くために照射された後およびヒト造血幹細胞を移植された後に、そのヒト造血幹細胞を維持し、分化した機能的ヒト免疫細胞を含むヒト免疫細胞集団をヒト造血幹細胞から発生させる。分化したヒト免疫細胞には、ヒト骨髄前駆細胞、ヒト骨髄細胞、ヒト樹状細胞、ヒト単球、ヒト顆粒球、ヒト好中球、ヒト肥満細胞、ヒト胸腺細胞、ヒトT細胞、ヒトB細胞、およびヒト血小板が含まれる。別の局面において、マウスは(iii)マウストロンボポエチン(mTPO)遺伝子のヒト化をさらに含む。
【0032】
一態様において、マウスは、ヒトの免疫細胞の集団と同程度細胞型が多様であるヒト免疫細胞の集団を維持する。一態様では、ヒト免疫細胞が移植後に少なくとも4ヶ月、5ヶ月、6ヶ月、7ヶ月、8ヶ月、9ヶ月、10ヶ月、11ヶ月、または12ヶ月間維持される。
【0033】
一態様において、マウスは、ヒト病原体またはヒト病原体の抗原に曝露されると、該病原体に曝露されたヒトの感染のモデルとなる細胞性および/または体液性の免疫応答を開始する。一態様では、ヒト病原体が野生型マウスには感染しない病原体である。別の態様では、ヒト病原体が野生型マウスに感染する病原体であり、感染後の野生型マウスは、ヒトがその病原体に応答して開始する免疫応答をモデル化したものではない。一態様では、病原体がウイルス、マイコバクテリア、真菌、または細菌である。特定の態様において、病原体はヒトまたはブタまたはトリのインフルエンザウイルス、チフス菌、または結核菌である。
【0034】
本発明のさらなる応用および態様は、本開示を読むことによって当業者には明らかになるであろう。
【図面の簡単な説明】
【0035】
【図1】感染後10日の、移植したRAG KO, Il2rg KOマウスとRAG KO, Il2rg KO, hIL-3/hGM-CSFマウスにおけるチフス菌感染を示す。実験グループ:(1)移植:n=9 (4m/m、5h/m);血液中の生着率=6.5〜16.7%;(2)対照:n=8;血液中の生着率=0.04〜0.4%(フローサイトメトリーのバックグラウンドを反映する。対照マウスには移植しなかった)。
【図2】1×103個のチフス菌の感染後1週の、移植したRAG KO, Il2rg KOマウスの脾臓におけるチフス菌感染を示す。
【図3】1×104個のチフス菌の感染後4週の、移植したRAG KO, Il2rg KOマウスの脾臓および肝臓におけるチフス菌感染を示し、該マウスはCD34陽性胎児肝細胞を用いて移植された。
【図4】1×104個のチフス菌の感染後4週の、RAG KO, Il2rg KOマウスの胆のうにおけるチフス菌感染を示し、該マウスはCD34陽性胎児肝細胞を用いて移植された。
【図5】図5(a)〜(d)は、非移植hIL-3/hGM-CSFマウスにおけるhGM-CSF発現の検証試験の結果を示す。図5(e)は、マウスIL-3/GM-CSF遺伝子座でのヒト化戦略を示す。
【図6】図6(a)〜(e)は、移植されたヒト化(hIL-3/hGM-CSF)マウスの肺検査の結果を示す。図6(f)、(g)は、活性化脾細胞によるマウスおよびヒトのIL-3(f)およびGM-CSF(g)産生のELISA結果を示す。
【図7】図7(a)は、非移植または移植m/mもしくはh/h KIマウスからの肺組織切片のPAS染色、図7(b)は、非移植(non)または移植h/h KIマウスもしくはm/m対照マウス(グループあたりn=6)からのBAL液中の総タンパク質の定量化を示す。
【図8】図8(a)は、マウスHprtに対して正規化されたヒトHprtの発現、図8(b)は、マウスHprtに対して正規化されたヒトIFNγの発現、図8(c)は、ヒトHprtに対して正規化されたヒトIFNγの発現を示す。図8(d)は、定常状態での移植hIL-3/GM-CSF h/m KIマウスからのヒト骨髄細胞のフローサイトメトリー解析、図8(e)は、LPSの2回腹腔内注射後72時間のCB移植したm/mまたはh/m KIマウスからのヒト血液細胞のフローサイトメトリー解析、図8(f)は、LPS注射後72時間の移植したm/mまたはh/m KIマウスにおけるヒトCD14+血液細胞の頻度、図8(g)は、1回目(上)および2回目(下)のLPS注射後2〜3時間の移植したm/mまたはh/m KIマウスからの血清中のヒトIL-6のELISAを示す。
【図9】図9(a)は、肺におけるヒトT細胞(hCD45+hCD3+)の頻度、図9(b)は、肺におけるヒトCD4およびCD8 T細胞の分布、図9(c)肺におけるヒトCD4対CD8 T細胞の比率、図9(d)は、BCG感染後4週のBALB/cマウス、移植m/mマウス、および移植h/m KIマウスからの脾細胞のフローサイトメトリー解析、図9(e)は、BCG感染後4週のBALB/cマウス、非移植(non)m/mマウス、移植m/mマウス、および移植h/m KIマウスからの肺組織におけるヒトIFNγ(左)およびTNFα(右)遺伝子発現の定量的RT-PCR解析を示す。図9(f)は、非移植m/mまたはh/h KIマウスからのBAL細胞のDiffQuick(商標)染色;倍率400x、図9(g)は、非移植m/mまたはh/h KIマウスからの肺組織切片のPAS染色;倍率400xを示す。
【図10】図10(a)は、BCG感染後4週の移植h/m KIマウスからの肺組織切片のヘマトキシリンおよびエオシン(H&E)染色;倍率100x(左)および200x(右)、図10(b)は、BCG感染後4週の移植h/m KIマウスからのヒトCD45、CD3またはCD68を染色した肺組織切片;倍率200xを示す。
【図11】図11(a)は、Rag2+/-γcY/- TPOh/mマウスの異なる組織におけるマウスTPO(mTpo)およびヒトTPO(hTPO)発現のRT-PCR解析、図11(b)は、Rag2-/-γc-/- TPOm/m、TPOh/mおよびTPOh/hマウスの肝臓、腎臓および間葉系多能性間質細胞(MSC)におけるmTpoおよびhTPO発現のRT-PCR解析、図11(c)は、TPOm/m、TPOh/mおよびTPOh/hマウスの血清中の、ELISAにより測定されたマウスおよびヒトTPOタンパク質の濃度を示す。図11(d)は、mTPO遺伝子をhTPO遺伝子と置き換えるためのターゲティング構築物を示す。
【図12】図12(a)は、ヒトCD34+細胞の移植後3〜4ヶ月のRag2-/-γc-/- TPOm/mおよびTPOh/hマウスの骨髄中のヒトおよびマウスCD45+細胞のFACS解析、図12(b)は、移植後3〜4ヶ月(左)または6〜7ヶ月(右)の骨髄中のヒトCD45+細胞のパーセンテージ、図12(c)は、図12(b)と同じ動物の骨髄中のヒトCD45+細胞の絶対数を示す。図12(d)は、臍帯血(CB)または胎児肝臓(FL)から分離したヒトCD34+細胞を用いて移植されたRag2-/-γc-/- TPOm/mおよびTPOh/hマウスの骨髄中のヒトCD45+細胞のパーセンテージを示す。
【図13】図13(a)は、非移植Rag2-/-γc-/- TPOm/m、TPOh/mおよびTPOh/h成体マウスの血液中の血小板数、図13(b)は、移植後3〜4ヶ月のRag2-/-γc-/- TPOm/mおよびTPOh/hマウスの血液中のマウス(mCD61+)およびヒト(hCD41a+)血小板の代表的FACS解析、図13(c)は、TPOm/mおよびTPOh/hマウスにおける、FACSにより測定されたヒト血小板のキメリズム、図13(d)、(e)は、TPOm/mおよびTPOh/hレシピエントの血液中のマウス(mCD61+, 20d)およびヒト(hCD41a+, 20e)血小板のカウント数、図13(f)は、骨髄中のヒトCD45+細胞のうちのヒト巨核球パーセンテージ(CD41a+)を示す。図13(g)、(h)は、Rag2-/-γc-/- TPOm/mおよびTPOh/hマウスの血液および脾臓中のヒトCD45+細胞のパーセンテージを示す。図13(i)は、移植TPOm/mおよびTPOh/hレシピエントの胸腺の全細胞量を提供する。
【図14】図14(a)は、移植後3〜4ヶ月のRag2-/-γc-/- TPOm/mおよびTPOh/hマウスの骨髄中および血液中のヒト骨髄細胞集団のFACS解析、図14(b)は、全骨髄集団(CD33+細胞)、図14(c)は、顆粒球(CD33+CD66hi)、図14(d)は、TPOh/hレシピエントの骨髄から精製されたhCD45+SSChiCD33+CD66hi細胞のDiffQuick(商標)染色、図14(e)は、単球(CD33+CD66loCD14+)、図14(f)、(g)は、Rag2-/-γc-/- TPOm/mおよびTPOh/hレシピエントの血液中の全ヒトCD45+細胞キメリズムに対するヒト骨髄細胞集団の解析を示す。図14(f) 顆粒球(CD66+)、図14(g) 単球(CD14+)。
【図15】図15(a)は、WT TPO (TPOm/m) Rag2-/-γc-/-マウスと比較した、非移植Rag2-/-γc-/- TPOh/mおよびTPOh/hマウスの骨髄中のマウスLin-Sca1+c-Kit+幹細胞前駆細胞のFACS解析、図15(b)は、図15(a)に示された結果の定量的解析、図15(c)は、移植後3〜4ヶ月のRag2-/-γc-/- TPOm/mおよびTPOh/hマウスの骨髄中のヒトCD34+CD38-細胞のFACS解析、図15(d)は、TPOm/mおよびTPOh/hレシピエントマウスにおけるヒトCD45+CD34+集団中のCD38-細胞のパーセンテージの定量的解析、図15(e)は、図15(d)と同じマウスの骨髄中のヒトCD34+CD38-細胞、図15(f)、(g)は、Rag2-/-γc-/- TPOm/mおよびTPOh/hレシピエントから精製されたヒトCD45+CD34+細胞を用いるメチルセルロースコロニー形成アッセイ法を示し、図15(f)は、CFU-GEMM、図15(g)は、BFU-E(黒)、CFU-G(白)、CFU-M(灰色)およびCFU-GM(破線)、図15(h)は、Rag2-/-γc-/- TPOm/mおよびTPOh/hマウス由来のヒトCD45+CD34+細胞の新生仔Rag2-/-γc-/-マウスへの二次移植におけるヒトCD45+キメリズムである。
【発明を実施するための形態】
【0036】
詳細な説明
本発明は、記載した特定の態様に限定されるものではなく、認められる特許請求の範囲によって説明されるものである。
【0037】
特に指定のない限り、本明細書中で用いるすべての技術用語および科学用語は、本発明が属する分野の当業者が一般に理解している意味と同じ意味を含む。記載したものと類似のまたは同等の方法および材料はどれも、本発明を実施または使用する際に用いることができるが、ここでは特定の態様、方法および材料が記載される。記載したすべての刊行物は参照により本明細書に組み入れられる。矛盾が存在する場合には、組み入れられた刊行物のあらゆる開示に本開示が優先する。
【0038】
単数形「1つの(a)」、「1つの(an)」および「その(the)」は、文脈上明確に別途示されない限り、複数形の照応を含むものである。したがって、例えば、「遺伝子(a gene)」への言及は複数のそのような遺伝子を包含し、また、「遺伝子ノックアウト(the gene knockout)」への言及は1つまたは複数のノックアウトおよびその同等物への言及を包含する。
【0039】
ヒト免疫細胞を支持する改変されたマウス:hIL-3/GM-CSFマウス
ヒト免疫システムの成分を含むマウス(HISマウス)は、インビボでのヒト免疫システムの研究に、ヒトワクチンの試験に、およびヒトの疾患および障害を治療するための薬物の試験と開発にかなり有望である。HISマウスは、重症免疫不全マウス株(例えば、組換え活性化遺伝子2(Rag2)ノックアウト(KO)インターロイキン2受容体γ(Il2rg)KOマウス)にヒト造血幹細胞前駆細胞を移植することによって作製される。非ヒト霊長類に比べて、HISマウスは小型動物モデルの利点を有しており、すなわち、それらはより多目的な実験を可能にし、研究団体にとってよりアクセス可能であり、また、ヒト被験者を用いて実験を行うより倫理的に許容される。最も重要なことは、HISマウスから得られた実験結果がヒトへの関連性が高く、ヒトに応用できる可能性があることである。なぜならば、ヒト特異的病原体を用いた感染、ならびにヒト特異的免疫応答および免疫病理学の研究が今や実現可能になりつつあるからである。
【0040】
多くの進展が近年なされてきたが、現在のHISマウスモデルには、ヒト骨髄細胞とT細胞の発達、維持および機能が低いといった、いくつかの主要な制限がある。その結果、粘膜表面でのヒト炎症および免疫反応、または遅延型過敏症(DTH)などの強いヒトT細胞性応答がめったに観察されていない。それゆえ、現在のHISマウスは、深刻なヒト病原体である結核菌に起因する感染症および病理の研究に適していない。実際、肉芽腫、特に、マイコバクテリアに対するヒト免疫応答の顕著な特徴である、ヒト細胞を含む肉芽腫は、これまでHISマウスにおいて報告されていない(例えば、Manz et al. (2009) Renaissance for mouse models of human hematopoiesis and immunobiology, Nat. Immunol. 10:1039-1042を参照されたい)。
【0041】
現在のHISマウス宿主は、少なくとも一部には現在のHISマウス宿主がヒト細胞に適した生理的環境を提示しないので、一部の感染症のモデル化には適していない。数種のマウスサイトカイン、例えばIL-3とGM-CSFはヒトコグネイト受容体に作用しない。さらに、Rag2 KO Il2rg KOマウスは完全なままのマウス骨髄区画を持っており、ヒト骨髄細胞は宿主細胞に対して競合的に不利である可能性がある。これらの制限を克服するために、本開示は、マウスのサイトカインをコードする遺伝子がヒトの対応遺伝子によって置き換えられたヒトサイトカインノックイン(knock-in)マウスの作製について記載する。サイトカイン置換の基準は次のとおりである。(1)マウスサイトカインがヒト細胞に作用しないかまたは弱く作用する。(2)ヒトサイトカインが、ヒト細胞に競合的優位性を与えるために、マウス細胞に作用しないかまたは弱く作用する。(3)ヒトサイトカインが造血(移植)細胞によって排他的に産生されない。(4)マウスサイトカインの欠乏がマウス宿主に致命的ではないか、またはヒトKIサイトカインがマウスKO表現型をレスキューするために十分に交差反応性である。KI戦略は適切な器官における生理学的濃度での忠実な発現を可能にすべきである。重要なことは、ホモ接合体KIマウスでは、ヒトコグネイト受容体を発現する細胞がそれぞれのマウス細胞に対して競合的優位性を獲得する必要があることである。
【0042】
IL-3およびGM-CSFは、骨髄細胞の発達と機能に欠かせない2種のサイトカインである。どちらのサイトカインもヒトとマウスの間で交差反応性がない。IL-3はインビトロで初期造血前駆細胞を刺激するが、インビボでの定常状態の造血にはなくても困らない。しかし、GM-CSFと一緒になって、それはインビボで効果的なDTH応答に必要である。IL-3はまた、インビトロで肺胞マクロファージ(AM)の増殖を特異的に刺激する。GM-CSFは肺で高度に発現されており、インビボでは肺の恒常性に重要である。このことは、GM-CSF KOマウスが、表面活性物質のクリアランスの欠陥のため肺にタンパク質が蓄積することによって特徴づけられる肺胞タンパク症(PAP)を発症するという事実により証明される。GM-CSF KOマウスからの肺胞マクロファージは最終分化において欠陥を有し、これは肺の病原体に対する先天性免疫の低下につながる。GM-CSFはまた、インビトロでヒトAMの増殖を刺激する。IL-3と同様に、GM-CSFは定常状態の造血にほとんど不必要であり、同じことが両サイトカインを欠くマウスにあてはまる。対照的に、GM-CSFはマクロファージによる炎症性サイトカインの産生および単球の動員と補充などの炎症反応に必要である。GM-CSFは結核菌などを含む広範な病原体に対する防御免疫のためにも不可欠である。特に、結核菌に感染したGM-CSF KOマウスは、結核の顕著な特徴である肉芽腫を発症することがない。
【0043】
本開示は、少なくとも一部には、hIL-3/GM-CSF KIマウスを作製することがヒト骨髄細胞の再構成および機能ならびにマウスでの病原体に対するヒト先天性免疫応答を支持する上で価値があるであろうという認識に基づいている。そのようなKIマウスを用いて得られた、本開示に記載された結果は、この戦略がヒト骨髄細胞の発達、ヒト肺粘膜免疫、かつさらにマイコバクテリア感染後の肉芽腫形成の観点からHISマウスの現在のモデルに比べて大幅な改善を与えることを実証する。そのようなマウスのこれらのおよび他の有益な特性は本開示の他の個所で説明される。
【0044】
マウスにおいてインビボ設定でヒト組織を研究できることは、広範囲の可能な研究手段を開放している。主要な制限事項がそのアプローチの適用を妨げており、それらのうち最も重要な欠陥の1つは、マウス因子がヒト細胞を支持できないことである。実際、免疫システムでは、ヒト免疫細胞の発達と機能に必要な多くの必須因子は種特異的であり、かつマウスによって効果的に提供され得ない。したがって、マウス遺伝子をそれらのヒト対応物と置き換えて、ヒト細胞の良好な発達と機能を可能にし、かつ対応するマウス細胞のそれを潜在的に不可能にするという戦略に従うことが決定された。この概念をヒトサイトカインKIマウスに適用することによって、マウス宿主の免疫遺伝子のヒト遺伝子との置換はHISマウスを改善するという概念の証明が本明細書に提供される。具体的には、本開示は、マウスとヒトの間の不適切なサイトカイン交差反応性、およびマウス細胞と競合せざるを得ないことが、現在のHISマウスにおけるヒト骨髄細胞の生着と機能を実際に制限しているという見解を支持する。
【0045】
ヒトサイトカインは、静脈内注射によってHISマウスに投与することができ、例えばIL-15/IL-15Rα複合体およびIL-7の注射によって、それぞれヒトNK細胞とT細胞の再構成を高めることができる。別のアプローチは、肝臓での一過性の発現につながるヒトサイトカインを発現するプラスミドDNAのハイドロダイナミックインジェクションである。このアプローチはごく最近、GM-CSFとIL-4のハイドロダイナミックデリバリーによってヒトDCの再構成を改善するために使用された(Chen et al. (2009) Expression of human cytokines dramatically improves reconstitution of specific human-blood lineage cells in humanized mice, Proc Natl Acad Sci USA, 106(51):21783-21788を参照されたい)。本開示とは対照的に、骨髄細胞の機能的応答または病原体に対するインビボ応答はこれらのマウスではまったく報告されなかった。最後に、ヒトサイトカインはまた、HISマウスにおいて導入遺伝子として過剰発現させることもできる。このアプローチは、ヒトIL-3/GM-CSF/幹細胞因子(SCF)トランスジェニック(tg)マウスを作製するために採用された(Nicolini et al. (2004) NOD/SCID mice engineered to express human IL-3, GM-CSF and Steel factor constitutively mobilize engrafted human progenitors and compromise human stem cell regeneration, Leukemia 18:341-347を参照されたい)。これらのマウスでは、ヒトサイトカインの発現が遍在的発現をもたらすサイトメガロウイルスプロモーターによって駆動される。しかしながら、hIL-3/GM-CSF/SCF tg HISマウスは、骨髄におけるヒト造血幹細胞の維持の低下および終末骨髄造血の増加によって妨げられる。この場合も、本開示とは相違して、改善された骨髄細胞の機能またはインビボ応答は記載されなかった。対照的に、本明細書に記載するシステムでは、定常状態および炎症での標的遺伝子の生理的発現が適切な細胞型のみの適切な発達と機能を可能にする。重要なことは、本開示で説明するアプローチが、高度に再現可能な条件下での維持および増殖が可能であり、かつ研究のために世界中で利用できる、マウスの系統を作製することである。
【0046】
本開示で説明するhIL-3/GM-CSF KIマウスは、以前のHISマウスおよび上述した代替アプローチと比べてかなりの改善を示す。第一に、本明細書に記載のKI戦略によるヒトIL-3およびGM-CSFの送達は長期的なサイトカインの発現につながり、これは高価なサイトカインの反復注射の必要性を回避する。第二に、IL-3とGM-CSFが通常発現されている器官での忠実な発現が達成される。生理的条件下では、GM-CSFは主に肺で発現される(図5a)。対照的に、ハイドロダイナミックデリバリーは肝臓および循環における優勢発現をもたらす。どちらの器官においても、GM-CSFは定常状態の条件下では発現されない。第三に、ハイドロダイナミックインジェクションによる送達またはhIL-3/GM-CSF/SCF tgマウスにおける遍在的な過剰発現とは対照的に、KIマウスではIL-3とGM-CSFの生理的量が発現される。GM-CSFの生理的レベルは結核菌に対する防御免疫応答に重要であることが実証されている。こうして、肺でのGM-CSFの局所過剰発現を有するトランスジェニックマウスは、肉芽腫形成不全および結核菌に対する感受性の増加を示す。同様に、GM-CSFの静脈内投与もまた、マウスにおける結核菌感染の防御低下をまねく。第四に、ホモ接合体hIL-3/GM-CSF KIマウスは、マウスIL-3およびGM-CSFがホモ接合体マウスでは発現されないので、マウス骨髄区画の同時損傷を可能にする。これは、本開示で示す通り、ヒト骨髄細胞に競合的優位性をもたらす。
【0047】
結核菌感染が原因で起こる結核では、年間170万人が死亡している。したがって、新規の有効な予防対策および治療対策が緊急に必要とされている。マウスは結核菌に感染し得るが、それらはヒト結核の理想的なモデルを表すものではない。それは結核菌に対する免疫応答の種特異的な差異のためである。例えば、感染したマウスは明確に組織化された肉芽腫を発症しない。肉芽腫は、結核に罹ったヒトの免疫反応の顕著な特徴であり、かつ融合して類上皮および多核巨細胞を形成する活性化マクロファージと活性化T細胞とを含む。肉芽腫は、細菌の複製を制限する上で、さらにマイコバクテリアの広がりを制御する上で重要な役割を果たしている。GM-CSFはインビトロでAMの多核巨細胞への分化を促進する。トランスジェニックマウスによる研究でも、インビボで多核巨細胞を形成するためのマクロファージの融合におけるGM-CSFの役割が明らかにされた。さらに、GM-CSFはマイコバクテリア感染後の肉芽腫形成に不可欠である。結核菌に感染したGM-CSF KOマウスにおける肉芽腫の不在は、細菌複製の増加と生存の低下に関連づけられる。最後に、GM-CSFシグナル伝達の欠陥に起因するPAPを患うヒトは、マイコバクテリア感染に対し増加した感受性を示す。
【0048】
ヒト抗マイコバクテリア免疫反応、特にヒト細胞による肉芽腫の形成は、HISマウスで以前に報告されたことがなかった。それはおそらくヒトマクロファージとT細胞の弱い応答によるものである。本開示では、マイコバクテリアに対する抗原特異的T細胞応答が、ヒト細胞を用いて移植されたマウスのサブセットにおいて検出された。さらに、肉芽腫の生物学におけるGM-CSFの顕著な役割を考えると、移植したhIL-3/GM-CSF KIマウスは肉芽腫形成を支持するための優れた宿主であることが仮定される。これは確かに、少なくともBCGを感染させたマウスのサブセットではそうであった。重要なことは、これらのマウスの肺肉芽腫が、マウス肉芽腫に特有のゆるい構成を持っていたものの、その肉芽腫がヒトT細胞とヒトマクロファージを含んでいたことである。今後の取り組みは、HISマウスにおいてヒトT細胞およびマクロファージの応答をさらに高めることを目指すべきである。これは、インビボで結核菌に対するヒト免疫応答の研究を可能にする小型動物モデルの開発につながるはずである。hIL-3/GM-CSF KIマウスはまた、インビボでヒト免疫応答を研究するための他の設定でも有用である可能性がある。それには、肺病原体による感染、自己免疫、およびヒト癌が含まれる。要約すると、本開示で提供されるhIL-3/GM-CSF KIマウスは、今後の研究のための多目的ツールとして役に立ち得る、かなり改善されたHISマウスモデルに相当する。
【0049】
ヒト免疫細胞を支持する改変されたマウス:hTPO
造血幹細胞(HSC)は2つの主な特性によって特徴づけられる:生涯にわたる自己再生能、およびすべての成熟した造血系細胞への分化能。HSCプールの恒常性を確保するために、細胞分裂時に、HSCは一方の機能的HSCを生成するが、他方の子孫細胞は高度に組織化された分化と細胞増殖のプログラムを受け、その間に複数の系列に方向づけされた前駆細胞、および最後には最終分化細胞が生成されると考えられる。
【0050】
マウスの造血は過去数十年間にわたり広く研究されており、インビボで幹細胞と前駆細胞とに非常に富んでいる、免疫表現型的に定義された細胞集団の同定および機能解析につながった。しかし、ヒト造血の見込みのある実験的インビボ研究は、明白な実際上および倫理上の制約によって制限されている。
【0051】
この制限を回避するために、ヒト造血のインビボ研究用の異種移植モデルがいくつか開発されている。これらのうち、免疫不全マウスへのヒト造血細胞の移植が実験的造血研究室において広く確立されている。今日最もよく用いられるモデルは、BALB/c Rag2-/-γc-/-またはNOD-SCIDγc-/-系統のマウスに依存している。両系統ともB細胞、T細胞およびNK細胞を欠いていて高度に免疫不全であり、それらの遺伝的背景はヒト造血移植および分化に対して寛容である。ヒトCD34+造血幹細胞前駆細胞を移植すると、大部分のヒト造血集団(B細胞、T細胞、単球、樹状細胞、赤血球および血小板を含む)は発達することができ、これらのモデルにおいて検出可能である。しかし、こうしたキメラな動物では、一般にヒト細胞の最大80%に相当する初期に高いB細胞数を伴うリンパ球の発達への偏りが見られ、骨髄単球の発達はわずかである。かつ、生着レベルは通常、移植後4〜6ヶ月で下降し始める。さらに、ヒト細胞のマウスへの異種移植には、マウス造血幹細胞前駆細胞のマウスへの、またはヒト細胞のヒトへのそれぞれ最適な移植に十分な数と比較して、多数の細胞の移植が必要である。さらに、マウスレシピエントに移植されたマウスHSCとは対照的に、ヒトHSCは異種マウス環境では増えることがなく、維持もされない。したがって、マウスの背景はヒトHSCの生理学を研究するための最適な環境を提供しない。それは、HSCの機能と維持を支持するのに必要な、増殖因子の非存在または限られた交差反応性が原因である可能性がある。
【0052】
トロンボポエチン(TPO)は、巨核球および血小板の発達を促進する成長因子として最初に同定された。TPOは、肝臓と腎臓で構成的に産生され、かつ血液循環に放出される。TPOの受容体であるc-Mplは骨髄中の造血幹細胞前駆細胞によって発現される。c-Mplは循環している血小板にも発現される。しかしながら、血小板へのTPOの結合はいかなるシグナル伝達経路をも活性化しない。かくして、血小板はTPOのためのシンクまたはスカベンジャーとして作用し、この機構を介して血小板産生の負の調節に寄与している。その後、TPOはHSCの増殖と自己再生を支持するその重要な機能について認識されてきた。TPOの欠乏は成体マウスにおいてHSC数の減少につながり、かつTPOの存在は静止期の成体HSCの維持に必要である。さらに、TPOは、照射した宿主の造血区画を補充するのに必要な、HSCの移植後増殖を支持するために必要である。興味深いことに、骨髄でのHSCニッチの形成に関与する骨芽細胞は、HSCの機能と維持に重要なTPOを産生することが実証されている。
【0053】
マウスおよびヒトTPOは、インビトロにおいて超生理的用量で用いた場合、それぞれのコグネイト受容体に対して双方交差反応性であるが、そのサイトカインがインビボ環境において生理的制限用量で作用する場合には、親和性と生物学的活性が異なることがある。こうして、マウスTPOはインビボでヒトc-Mpl受容体に適切な刺激を与えない可能性があり、それゆえにマウス環境でヒトHSCの特性が損なわれる主な原因となり得る。この潜在的欠陥を修正するために、Rag2-/-γc-/-マウスにおいてマウスTPOをコードする遺伝子がそのヒト対応物により置き換えられた。
【0054】
本開示は、少なくとも一部には、RAG2-/-γc-/-背景でhTPO KIマウスを作製することが、該マウスそれ自体およびhIL-3/GM-CSFマウスと交配させた該マウスの子孫において、ヒト骨髄細胞の再構成と機能ならびにマウスでの病原体に対するヒト先天性免疫応答を支持する上で価値があるという認識に基づいている。RAG2-/-γc-/-背景でhTPO KIマウスから得られた結果が本開示に記載される。ホモ接合体hTPO KIマウスは骨髄中の有意に増加したヒト生着レベルを有し、かつ造血細胞の多系列分化がmTPOマウスよりも改善され、hTPO KIマウスは骨髄単球系列対リンパ球系列の比率の増加を示した。ヒト幹細胞前駆細胞の数および自己再生能は、連続移植によって実証される通り、両方とも改善された。したがって、いくつかある適用例の中で特に、hTPO KIマウスは連続移植によるヒト細胞の増殖に有用である。
【0055】
hIL-3/GM-CSFマウスとhTPOマウスの交配
本明細書に記載されるhIL-3/GM-CSFおよびhTPOマウスの子孫は、少なくとも同じ関連する特徴を有し、親系統(すなわち、hIL-3/GM-CSFマウスとhTPOマウス)と少なくとも同じ利点を示すと予想される。例えば、いずれか一方の親系統、または両方、またはその子孫からヒト細胞集団を単離し、かつhTPOマウスまたはhIL-3/GM-CSFおよびhTPOマウスの子孫に連続移植することが可能である。したがって、一局面において、本明細書に記載するhIL-3/GM-CSFマウスとhTPOマウスの子孫(各関連遺伝子に関してホモ接合性に交配させた子孫を含む)である遺伝子改変されたマウスが提供され、かつこの遺伝子改変されたマウスはhIL-3/GM-CSFマウスとhTPOマウスの両方の利点および特徴を示す。一局面においては、除去された免疫システムを含む(例えば、照射されたマウス)、任意の移植されたマウス(例えば、本明細書に記載する移植されたマウス)からの移植および/または連続移植に適している、そのような子孫マウスが提供される。
【0056】
遺伝子改変されたマウスの移植
本発明による遺伝子改変されたマウスは、移植したヒト造血細胞からヒト免疫細胞を発達させることができる、ヒト造血細胞のレシピエントとして使用される。一態様では、ヒト造血細胞またはヒト造血幹細胞前駆細胞(HSPC)が、本発明によって遺伝子改変および照射されたマウスに配置される(移植される)。ヒト造血細胞またはヒト造血幹細胞は、遺伝子改変されたマウスにおいて、以下から選択される細胞を生じさせる:ヒトCD34陽性細胞、ヒト造血幹細胞、ヒト造血細胞、骨髄前駆細胞、骨髄細胞、樹状細胞、単球、好中球、肥満細胞、およびヒト血液-リンパシステム(ヒト造血幹細胞前駆細胞、ヒト骨髄前駆細胞、ヒト骨髄細胞、ヒト樹状細胞、ヒト単球、ヒト顆粒球、ヒト好中球、ヒト肥満細胞、ヒト胸腺細胞、ヒトT細胞、ヒトB細胞、ヒト血小板を含む)、ならびにこれらの組合せ。
【0057】
遺伝子改変されたマウスは、存在しうる内在性の造血細胞を排除するために照射することができ、かつ該マウスは、任意の適した供給源のヒト造血細胞を用いて移植されることが可能である。造血細胞の当技術分野で公知である1つの適した供給源はヒト臍帯血細胞、特にCD34陽性細胞である。別の造血細胞源はヒト胎児肝臓である。
【0058】
一態様において、本発明によるマウスのヒト造血細胞を用いた移植は、TPO遺伝子のヒト化を欠くか、IL-3およびGM-CSF遺伝子のヒト化を欠くか、またはTPO遺伝子とIL-3およびGM-CSF遺伝子のヒト化を欠く免疫不全マウスより、増大した数のヒト造血細胞を示すマウスをもたらす。
【0059】
一態様において、本発明によるマウスのヒト造血細胞を用いた移植は、ヒト化を欠く免疫不全マウスと比べて、増大した数のヒト血液細胞(例えば、成熟造血細胞)を示すマウスをもたらす。特定の態様では、ヒト造血細胞が以下から選択される:ヒトCD34陽性細胞、造血幹細胞、造血細胞、骨髄前駆細胞、骨髄細胞、樹状細胞、単球、顆粒球、好中球、肥満細胞、およびヒト血液-リンパシステム(ヒト造血幹細胞前駆細胞、ヒト骨髄前駆細胞、ヒト骨髄細胞、ヒト樹状細胞、ヒト単球、ヒト顆粒球、ヒト好中球、ヒト肥満細胞、ヒト胸腺細胞、ヒトT細胞、ヒトB細胞、ヒト血小板を含む)、ならびにこれらの組合せ。
【0060】
遺伝子改変され移植されたマウスの非限定的な適用
ヒト造血細胞を用いて移植された、遺伝子改変されたマウスは、通常はマウスに感染しない病原体を研究するための有用動物である。そのような一例は腸チフスの原因病原体チフス菌である。
【0061】
腸チフスは、米国での約400症例/年を含めて、世界中で - 主に開発途上国で - 2100万人を超える人々を苦しめている。腸チフスは以下の薬物で治療されている:アモキシリン、アンピシリン、セフォタキシム、セフトリアキソン、セフタジジム、クロラムフェニコール、シプロフロキサシン、コトリモキサゾール、エルタペネム、イミペネム、フルオロキノロン類(例えば、シプロフロキサシン、ガチフロキサシン、オフロキサシン)、ストレプトマイシン、スルファジアジン、スルファメトキサゾール、テトラサイクリン、およびこれらの組合せ。反復性の感染が一般的であり、そのことが抗生物質療法による疾病管理を制限している。さらに、チフス菌感染症では多剤耐性もよく見られる。
【0062】
新規治療薬、新規ワクチン、および治療薬とワクチンの有効性を試験するための新規方法が必要とされている。チフス菌によって感染され得るマウスは、例えば、新規治療薬および新規ワクチンを同定するのに有用であろう。新規治療薬および新規ワクチンは、そのようなマウスにおいて、例えば推定上の抗チフス菌剤による治療に応答したマウス中(血液または所定の組織中)のチフス菌の量を測定することによって、あるいはマウスに推定ワクチンを接種し、続いてチフス菌の感染性投与に曝露し、推定ワクチンの接種による感染力の何らかの変化を、ワクチンを接種しないがチフス菌に感染させた対照と比較して観察することによって、試験することができる。
【0063】
本発明による遺伝子改変および移植されたマウスは、マウスに感染しないヒト病原体によって感染され得るマウスを作製するのに有用である。例えば、マウスは、チフス菌によって感染され得る非ヒト動物として有用である。一態様では、遺伝子改変および移植されたマウスは、遺伝子改変を欠く移植されたマウスに比べて、増強されたヒト細胞の生着を示し、増強はチフス菌感染を維持するのに十分である。特定の態様では、チフス菌感染の維持は、マウス中で繁殖するチフス菌の能力を含む。特定の態様では、チフス菌感染は、チフス菌を繁殖させる感染したマウスの能力を含む。特定の態様では、マウスは、チフス菌の初期導入または感染性曝露後に少なくとも1週間、10日間、2週間、3週間、または4週間にわたりチフス菌を繁殖させることができる。
【0064】
抗チフス菌剤を同定するための方法も提供され、その方法はチフス菌によって感染され得る本明細書に記載のマウスを用いるものである。野生型マウスおよび他の公知の免疫不全マウス(例えば、RAG1/RAG2遺伝子ノックアウトマウス)はチフス菌によって感染され得ない。
【0065】
Il2rg遺伝子ノックアウトとRAG遺伝子ノックアウト(例えば、RAG2遺伝子ノックアウト)を含み(第1のタイプ)、さらに内在性マウスIL-3遺伝子のヒトIL-3遺伝子との置換および内在性マウスGM-CSF遺伝子のヒトGM-CSF遺伝子との置換を含む(第2のタイプ)遺伝子改変されたマウスが提供され、該遺伝子改変されたマウスは、ヒト造血細胞を用いて移植された場合、チフス菌による感染が可能である。
【0066】
図1に示したデータは、第1および第2の両タイプのマウスを代表するものである。図1のマウスの遺伝子改変は、(a)マウスRAG遺伝子ノックアウト;および(b)マウスIl2rg遺伝子ノックアウトを含む。図1のマウスはまた、ヒト造血細胞の移植を含む。このマウスは第2のタイプのマウスを作製するために2つのさらなる改変によってさらに改変することができ、それらは、(c)内在性マウスIL-3遺伝子のヒトIL-3遺伝子との置換;および(d)マウスGM-CSF遺伝子のヒトGM-CSF遺伝子との置換である。
【0067】
図2、3および4は、第1のタイプの改変されたマウス((a)マウスRAG遺伝子ノックアウトおよび(b)マウスIl2rg遺伝子ノックアウトの改変、ならびにヒト造血細胞の移植を含む)のみを用いて得られたものである。
【0068】
さまざまな態様において、チフス菌感染した、遺伝子改変されたマウスは、チフス菌の増殖性感染を含む。一態様では、マウスは、その細胞の1個または複数においてチフス菌を保有および繁殖させることができる。一態様では、マウスは、チフス菌への感染性曝露後に少なくとも1週間、10日間、2週間、3週間、または4週間にわたりその血液中または少なくとも1つの組織中にチフス菌の力価またはレベルを維持することができる。
【0069】
一態様において、方法は、本発明による遺伝子改変されたマウスに剤を投与する段階であって、該遺伝子改変されたマウスがチフス菌に感染している段階;該剤の投与後に該マウスの血液中または組織中のチフス菌のレベルを検出する段階;および任意で、該剤の投与が該マウスの血液中または組織中のチフス菌のレベルを低下させるかどうかを判定する段階を含む。一態様では、剤がワクチンである。別の態様では、剤が抗生物質または抗生物質の性質をもつと予想される剤である。一態様では、剤が抗原結合タンパク質であり、特定の態様では抗体である。一態様では、剤がヒト用の承認された医薬である。
【0070】
一態様において、方法は、本発明による遺伝子改変および移植されたマウスに既知量のチフス菌を感染させる段階、感染させた該マウスに剤を投与する段階、ならびに該剤の投与後に該遺伝子改変および移植されたマウス中のチフス菌の量を測定する段階を含む。一態様では、剤がマウスの血液中または組織中のチフス菌の量を、1回の剤投与または2回またはそれ以上の投与後に所定の期間にわたり、少なくとも半分に減少させる場合に、その剤は抗チフス菌剤であると判定される。
【0071】
一局面においては、関心対象のチフス菌の分離株または株が薬物耐性または多剤耐性であるかどうかを判定するための方法が提供され、該方法は、本発明による遺伝子改変および移植されたマウスに、チフス菌の治療に用いる薬物または薬物の組合せを投与する段階を含み、該マウスは関心対象のチフス菌の分離株または株に感染している。方法は、(a)薬物または薬物の組合せの投与後のある時点でのマウスの血液中または組織中の関心対象のチフス菌の分離株または株の力価、(b)薬物または薬物の組合せの1回またはそれ以上の投与後に関心対象のチフス菌の分離株または株がマウスにおける感染またはマウスの組織中のチフス菌のレベルを維持する能力、あるいは(c)薬物または薬物の組合せの投与後のある時点で関心対象のチフス菌の分離株または株がマウス中で繁殖する能力に及ぼす薬物または薬物の組合せの、もしあれば、効果を測定する段階を含む。特定の態様において、薬物はアモキシリン、アンピシリン、セフォタキシム、セフトリアキソン、セフタジジム、クロラムフェニコール、シプロフロキサシン、コトリモキサゾール、エルタペネム、イミペネム、フルオロキノロン類(例えば、シプロフロキサシン、ガチフロキサシン、オフロキサシン)、ストレプトマイシン、スルファジアジン、スルファメトキサゾール、テトラサイクリン、およびこれらの組合せからなる群より選択される。特定の態様では、薬物または薬物の組合せの投与が、チフス菌への感染を生じる曝露後少なくとも1週間、10日間、2週間、3週間、または4週間である。
【0072】
さまざまな局面および態様において、血液中または組織中のチフス菌のレベルは、血液または組織の単位(例えば、重量または体積)あたりのコロニー形成単位の数を確認することによって測定される。
【0073】
一態様において、本発明による遺伝子改変されかつヒト造血細胞を移植されたマウスは、ヒト造血細胞を用いて移植されていないマウスと比べて少なくとも100倍、1,000倍、または10,000倍の、コロニー形成単位(cfu)で測定した、チフス菌レベルを有する。
【0074】
抗チフス菌ワクチンの有効性を確認するのに有用な方法および組成物を提供する。一局面において、抗チフス菌ワクチンの有効性を確認するための方法が提供され、該方法は、本発明による遺伝子改変および移植されたマウスを抗チフス菌ワクチンに曝露する段階、その後該遺伝子改変および移植されたマウスをチフス菌に曝露する段階、ならびに該遺伝子改変および移植されたマウスがチフス菌によって感染され得るかどうか、またはどの程度に感染され得るかを判定する段階を含む。
【0075】
一態様において、抗チフス菌ワクチンはチフス菌の細胞表面タンパク質またはその免疫原性断片を含む。一態様では、ワクチンがチフス菌株の膜画分を含む。一態様では、ワクチンが組換えチフス菌タンパク質またはその免疫原性断片を含む。一態様では、ワクチンがチフス菌タンパク質またはその免疫原性断片をコードする発現ベクターを含む。一態様では、ワクチンが不活化チフス菌株または不活化チフス菌株の混合物を含む。
【0076】
本開示に記載される遺伝子改変および移植されたマウスはまた、既存のマウスより厳密にヒト病原体感染をモデル化するのに有用である。例えば、結核菌による感染。本明細書に記載される遺伝子改変および移植されたマウスは、例えば、ヒト免疫細胞を含む肉芽腫および境界のはっきりした肉芽腫を含む肉芽腫を発生する結核菌マウスモデルを提供することによって、マイコバクテリアのヒト感染をモデル化するのに有用である。記載される遺伝子改変および移植されたマウスのチフス菌感染に関連して説明した薬物およびワクチンの試験方法はまた、結核菌への適用例、例えば、薬物耐性株の同定、結核菌ワクチンの有効性の試験、抗結核菌剤の試験、抗結核菌剤に応答するcfuの測定などにも適用可能である。
【0077】
本開示に記載される遺伝子改変および移植されたマウスはまた、初期ヒト造血細胞から、例えばヒト造血または前駆細胞から発生する、ヒト造血器悪性腫瘍をモデル化するのに有用である。本開示に記載される遺伝子改変および移植されたマウスのさらなる適用例は、この開示を読むことによって当業者には明らかであろう。
【0078】
トロンボポエチンおよび移植
トロンボポエチン(TPO)は、巨核球および血小板の発達を促進する成長因子として最初に同定された(Wendling, F. et al. (1994) cMpl ligand is a humoral regulator of megakaryocytopoiesis, Nature 369:571-574; Kaushansky, K. et al. (1994) Promotion of megakaryocyte progenitor expansion and differentiation by the c-Mpl ligand thrombopoietin, Nature 369:568-571; Lok, S. et al. (1994) Cloning and expression of murine thrombopoietin cDNA and stimulation of platelet production in vivo, Nature 369:565-568; de Sauvage, F.J. et al. (1994) Stimulation of megakaryocytopoiesis and thrombopoiesis by the c-Mpl ligand, Nature 369:533-538; Bartley, T.D. et al. (1994) Identification and cloning of a megakaryocyte growth and development factor that is a ligand for the cytokine receptor Mpl, Cell 77:1117-1124; Kaushansky, K. (1998) Thrombopoietin, N Engl J Med 339:746-754; Kaushansky, K. (2005) The molecular mechanisms that control thrombopoiesis, J Clin Invest 115:3339-3347; Kaushansky, K. (2008) Historical review: megakaryopoiesis and thrombopoiesis, Blood 111:981-986)。
【0079】
TPOは、肝臓および腎臓で構成的に産生され、かつ血液循環に放出される。TPOの受容体であるc-Mplは骨髄中の造血幹細胞前駆細胞によって発現される。c-Mplは循環している血小板にも発現される。しかしながら、血小板へのTPOの結合はいかなるシグナル伝達経路をも活性化しない。かくして、血小板はTPOのためのシンクまたはスカベンジャーとして作用し、この機構を介して血小板産生の負の調節に寄与している(Kuter, D.J.およびRosenberg, R.D. (1995) The reciprocal relationship of thrombopoietin (c-Mpl ligand) to changes in the platelet mass during busulfan-induced thrombocytopenia in the rabbit, Blood 85:2720-2730)。その後、TPOはHSCの増殖と自己再生を支持するその重要な機能について認識されてきた(Fox, N., et al. (2002) Thrombopoietin expands hematopoietic stem cells after transplantation, J Clin Invest 110, 389-3894; Kirito, K. et al. (2003) Thrombopoietin stimulates Hoxb4 expression: an explanation for the favorable effects of TPO on hematopoietic stem cells, Blood 102:3172-3178)。
【0080】
TPO欠乏は、成体マウスにおいてHSC数の減少につながり、かつTPOの存在は静止期の成体HSCの維持に必要である(Yoshihara, H. et al. (2007) Thrombopoietin/MPL signaling regulates hematopoietic stem cell quiescence and interaction with the osteoblastic niche, Cell Stem Cell 1, 685-697; Qian, H. et al. (2007) Critical role of thrombopoietin in maintaining adult quiescent hematopoietic stem cells, Cell Stem Cell 1:671-684)。さらに、TPOは、照射した宿主の造血区画を補充するのに必要な、HSCの移植後増殖を支持するために必要である。興味深いことに、骨髄でのHSC「ニッチ」の形成に関与する骨芽細胞は、HSCの機能と維持に重要なTPOを産生することが実証されている。
【0081】
マウスおよびヒトのTPOは、インビトロにおいて超生理的用量で用いた場合、それぞれのコグネイト受容体に対して双方交差反応性であるが、そのサイトカインがインビボ環境において生理的制限用量で作用する場合には、親和性と生物学的活性が異なることがある。本発明者らは、マウスTPOはインビボでヒトc-Mpl受容体に適切な刺激を与えない可能性があり、それゆえにマウス環境でヒトHSCの特性が損なわれる主な原因になり得ると仮定した。この潜在的欠陥を修正するために、本発明者らは、Rag2-/-γc-/-マウスにおいて、マウスTPOをコードする遺伝子をそのヒト対応物により置き換えた。そのようなマウスはヒト血液-リンパ球生成システムの分化と機能を維持するための改善された能力をもつと仮定された。
【0082】
ヒト血液-リンパ球生成システムの分化と機能を維持するマウスの開発では、20年以上前の最初のモデルの発表以来、著しい進展が達成されてきた。しかしながら、以下の(i)〜(iii)を含むいくつかの制限が残っている:(i)レシピエントマウスの寿命の間中継続しない、一過性のヒト細胞生着、(ii)リンパ球系への非生理的な偏りおよび骨髄細胞の不十分な分化、ならびに(iii)マウスのグループが1人のヒト提供者由来の細胞を移植される場合でさえの、異なる動物間での生着レベルのばらつき。こうした制限は、ヒト細胞の非生理的な位置、免疫不全宿主の残留する異種反応性、マウス種とヒト種における血液-リンパ系細胞の異なる組成、および/または優先的なマウス細胞の支持につながる、造血支持因子のマウスとヒトとの交差反応性の欠如もしくは不足が原因である可能性がある。したがって、ヒト成長因子の生理的レベルを提供すること、および宿主においてそれぞれのマウス相同体を欠失させることは、ヒト細胞集団の発達と生存にさらに有利にはたらく可能性がある。本明細書では、本発明者らが、造血幹細胞の維持と自己再生に重要な機能をもつサイトカインである、トロンボポエチンをコードする遺伝子をヒト化した、新規のレシピエントマウス株が記載される。
【0083】
これらのヒト化トロンボポエチンマウスにヒトCD34+造血幹細胞前駆細胞を移植すると、以前に利用可能なモデルと比較して、上で挙げた3つすべての制限に対して著しい改善が見られた。すなわち、骨髄キメリズムがより高く、少なくとも6ヶ月間維持された。多系列、特に骨髄系の分化が増強された。かつ生着レベルのばらつきが減った。
【0084】
マウスとヒトの免疫システムの主な相違は血液中に存在する顆粒球の割合である。マウスではリンパ球が優勢であるが、ヒト血液は顆粒球に富んでいて、種差はその意義がはっきりしない。興味深いことに、ヒトTPOの存在はヒト顆粒球の分化を改善した(図14)。したがって、レシピエントマウスにおけるヒトTPOの存在は、ヒト生理的状態をより良く反映する顆粒球とリンパ球のバランスに有利にはたらき、おそらくヒト骨髄前駆細胞の向上した維持および/または分化によると考えられる。
【0085】
さらに重要なこととして、これらの結果は、TPOヒト化がマウス環境でヒト造血幹細胞前駆細胞を再増殖する二次レシピエントの維持に有利にはたらくことを示す(図15)。それゆえ、Rag2-/-γc-/-TPOh/hマウスはインビボでヒト幹細胞前駆細胞の機能のさまざまな面を研究するための新規モデルに相当する。
【0086】
それにもかかわらず、血液中の骨髄系とリンパ系との良好なバランスが観察されたとはいえ、末梢リンパ組織(脾臓、血液および胸腺を含む)における全体的生着レベルに対するTPOヒト化の有意な効果は認められなかった(図13(g)〜(i))。それは多様な要因によって説明され得る。第一に、レシピエントマウスは移植前に亜致死的に照射されるが、マウス骨髄細胞の大集団がまだ存在している。これらの細胞のうち、マクロファージはヒト細胞の食細胞として機能することができ、末梢での全体的生着レベルを制限する。したがって、マウスマクロファージの遺伝的枯渇、またはそれらの機能的不活性化は、より高い末梢生着レベルを可能にする場合がある。第二に、ヒト細胞は、それらの最終分化、骨髄からの退出および/または末梢でのそれらの生存を有利にはこぶために、さらなるヒト成長因子を必要とすることがある。サイトカインの多様なパネルが各系列について考慮され得る。最後に、二次リンパ器官がヒト化マウスに形成されるものの、それらの構造はヒト組織に比較して最適ではない。その部分的に欠陥のある構造は、これらの器官で生存できるヒト細胞の数への制限を表す場合があるい。
【0087】
マウスレシピエントをさらに改善するために、さらなる遺伝子置換を採用することができる。それを達成するために、マウス遺伝子のそのヒト相同体によるノックイン置換に基づく、本研究で用いた手法は、古典的なトランスジェニックアプローチと比べて2つの主な利点を提供する。第一に、それはマウス由来の調節配列の大部分を維持しているので、ヒト化遺伝子がマウス宿主内で忠実に発現されることを保証する。第二に、ノックイン戦略はマウスサイトカインをそのヒト相同体と置き換えるので、ヒトサイトカインがマウス受容体に対し十分な交差反応性を示さない場合に、それはマウスサイトカインに依存するマウス由来の細胞の集団に影響を及ぼし得る。これは移植後のヒト細胞集団にさらなる競合的優位性を提供することができる。実際、それはヒトTPOの事例であると考えられる。なぜなら、TPOのホモ接合性置換は非移植動物においてマウス血小板とHSCの減少を招くからである(図13(a)、15(a)および15(b))。
【0088】
ヒトTPOノックインマウスの場合には、一般的にはヒト造血のインビボ生理学、特にヒト造血幹細胞前駆細胞を研究するのに有用であり得る改良モデルが提供される。さらに、これらのマウスは、例えば骨髄性白血病および骨髄増殖性腫瘍など、初期造血細胞から発生するヒト造血器悪性腫瘍をインビボで維持する。
【実施例】
【0089】
以下の実施例は、本発明者らが発明と見なすものの範囲を限定することを意図しない。特に断りのない限り、部分は重量による部分であり、分子量は重量平均分子量であり、温度は摂氏であり、かつ圧力は大気圧またはほぼ大気圧である。
【0090】
実施例1
ヒトIL-3/GM-CSFおよびヒトTPOマウスの作製
hIL-3/GM-CSFターゲティング
1回のターゲティング工程でマウスIL-3遺伝子をヒトIL-3遺伝子と置換し、マウスGM-CSF遺伝子をヒトGM-CSF遺伝子と置換するためのターゲティング構築物は、ギャップ修復クローニングを利用するVELOCIGENE(登録商標)技術(例えば、米国特許第6,586,251号およびValenzuela et al. (2003) "High-throughput engineering of the mouse genome coupled with high-resolution expression analysis," Nat Biot 21(6):652-659を参照されたい。参照により本明細書に組み入れる)を用いて構築した。
【0091】
マウス配列は細菌人工染色体(BAC)RPCI-23、クローン5E15から取得した。ヒト配列はCaltech Dライブラリー(CTD)、BACクローン2333J5から取得した。
【0092】
p15複製起点を含むギャップ修復ドナーベクターは、mIL-3 ATGのすぐ上流の5'マウスホモロジーアーム、hIL-3 ATGから約274ntまでhIL-3遺伝子にわたるヒト5'IL-3ホモロジーアーム、ポリリンカー、hGM-CSF遺伝子のポリA配列の約2.9kb下流で開始する3'hGM-CSF(約233塩基)、および薬物選択カセットと、これに続くmGM-CSFポリA配列の下流(約2.9kb下流)の配列を有するマウス3'ホモロジーアームをクローニングすることによって構築した。このギャップ修復ベクターを線状化し、かつヒトCTD BACクローン2333J5と組換え酵素ベクターを含む大腸菌(E. coli)DH10B株に、Valenzuelaらに記載される通りに挿入した。
【0093】
細胞を薬物選択培地で増殖させた。個々のクローンを増殖させて、ギャップ修復ドナーベクターDNAを抽出し、適切なマウス-ヒト接合点について該ベクターの部分を配列決定した。パルスフィールドゲル電気泳動を用いて、挿入物のサイズおよび予想される制限断片長を確認した。
【0094】
hIL-3遺伝子に隣接するマウス上流および下流ホモロジーボックス、hGM-CSF遺伝子、およびロキシド(loxed)薬物選択カセットを含む捕捉ドナー(captured donor)を修復ドナーベクターから得て、その捕捉ドナーを線状化し、線状化した捕捉ドナーを、RPCI23クローン5E15とpABGベクターを含む大腸菌DH10Bに導入した。細胞を薬物選択培地で増殖させた。RPCI23クローン5E15 DNA中に捕捉ドナーDNAを含む(ターゲティングベクターを形成する)個々のクローンを単離し、ターゲティングベクターDNAを抽出し、かつ適切なマウス-ヒト接合点について該ベクターの部分を配列決定した。パルスフィールドゲル電気泳動を用いて、挿入物のサイズおよび予想される制限断片長を確認した。
【0095】
エレクトロポレーション
ターゲティングベクターを線状化し、これを用いてValenzuelaらに記載される通りにマウスES細胞をエレクトロポレーションにかけた。エレクトロポレーション処理を行った、ターゲティングベクターを含むマウスES細胞は、ロキシド薬物選択カセットを除くために、一過性Cre発現ベクターを用いてさらにエレクトロポレーションにかけた。そのターゲティングベクターをRag2 HET Il2rg y/- ES細胞にエレクトロポレーションにより導入した。RAG2遺伝子とIl2rg遺伝子のノックアウトがなされた親ES細胞株は市販のV17 ES細胞(BALB/c×129ヘテロ接合体)であった。hIL-3およびhGM-CSF遺伝子でターゲティングされたES細胞は、マウス胚に導入するために使用した。
【0096】
hIL-3/GM-CSFマウス
ターゲティングされたドナーES細胞を、8細胞期のマウス胚にVELOCIMOUSE(登録商標)法により導入する(例えば、米国特許第7,294,754号およびPoueymirou et al. (2007) "F0 generation mice that are essentially fully derived from the donor gene-targeted ES cells allowing immediate phenotypic analyses," Nat Biot 25(1):91-99を参照されたい。参照により本明細書に組み入れる)。ヒト化IL-3およびGM-CSF構築物を保持するVELOCIMICE(登録商標)(完全にドナーES細胞に由来するF0マウス)は、対立遺伝子アッセイ法の変法(例えば、Valenzuelaら参照)を用いて、マウス対立遺伝子の喪失とヒト対立遺伝子の獲得について遺伝子型を同定することによって同定される。これらのマウスを最初にBALB/cAnNCRと交配させ、次にRag2とIl2rgについてヘテロ接合性で、ヒトIL-3/GM-CSF KIを含むマウスを、移植研究のためにRag2/Il2rg二重KOマウスと交配させる。
【0097】
hIL-3/GM-CSFマウスの表現型分類
ヒト化マウスは、hGM-CSF特異的プライマーを用いるRT-PCRによって、ヒトGM-CSFの産生について試験した。試験した組織のヒトGM-CSFの発現パターンはマウスGM-CSFのそれに一致した(主に肺で発現)。ConAとIL-2で48時間刺激した、ヒト化マウス由来の脾細胞のELISAを実施して、hIL-3とhGM-CSFの存在を検出した。脾細胞はhIL-3とhGM-CSFの両方の発現について陽性であった。
【0098】
hTPOターゲティング
1回のターゲティング工程でマウスTpo(mTpo)遺伝子をヒトTPO(hTPO)遺伝子と置換するためのターゲティング構築物(図11(d))は、ギャップ修復クローニングを利用するVELOCIGENE(登録商標)技術を用いて構築した(Valenzuela et al.)。このベクターは、Tpoのオープンリーディングフレームを含む配列を置換するが、マウス由来のプロモーターと5'UTRを維持するように設計された。マウス配列を細菌人工染色体(BAC) RPCI-23、クローン98H7から得た。ヒト配列をBAC RPCI-11、クローン63m3から得た。p15複製起点を含むギャップ修復ドナーベクターは、mTpo ATGのすぐ上流の5'マウスホモロジーアーム、hTPO ATGから約275ntまでhTPO遺伝子にわたるヒト5'TPOホモロジーアーム、ポリリンカー、hTPO遺伝子のポリA配列の約1.5kb下流で開始する3'hTPOホモロジーアーム、およびロキシド薬物選択カセットと、これに続くmTpoポリA配列の下流(約3.5kb下流)の配列を有するマウス3'ホモロジーアームをクローニングすることによって構築した。このギャップ修復ベクターを線状化し、かつヒトBACクローンRPCI-11、63m3と組換え酵素ベクターを含む大腸菌DH10B株に挿入した。細胞を薬物選択培地で増殖させた。個々のクローンを増殖させて、ギャップ修復ドナーベクターDNAを抽出し、適切なマウス-ヒト接合点について該ベクターの部分を配列決定した。パルスフィールドゲル電気泳動を用いて、挿入物のサイズおよび予想される制限断片長を確認した。hTPO遺伝子に隣接するマウス上流および下流ホモロジーボックスおよびロキシド薬物選択カセットを含む捕捉ドナーを修復ドナーベクターから得て、その捕捉ドナーを線状化し、線状化した捕捉ドナーを、RPCI-23クローン98H7とpABGベクターを含む大腸菌DH10Bに導入した。細胞を薬物選択培地で増殖させた。RPCI-23クローン98H7 DNA中に捕捉ドナーDNAを含む(ターゲティングベクターを形成する)個々のクローンを単離し、ターゲティングベクターDNAを抽出し、かつ適切なマウス-ヒト接合点について該ベクターの部分を配列決定した。パルスフィールドゲル電気泳動を用いて、挿入物のサイズおよび予想される制限断片長を確認した。ターゲティングベクターを線状化し、これを用いてマウス胚性幹(ES)細胞をエレクトロポレーションにかけた。ターゲティングベクターはRAG2+/-γcY/-ES細胞にエレクトロポレーションにより導入した。親RAG2+/-γcY/-ES細胞株は、市販されているV17 ES細胞株(BALB/c×129ヘテロ接合体)から作られた。正しくターゲティングされたES細胞は、ロキシド薬物選択カセットを除くために、一過性Cre発現ベクターを用いてさらにエレクトロポレーションにかけた。hTPOでターゲティングされた、選択カセットを含まないES細胞を、8細胞期のマウス胚にVELOCIMOUSE(登録商標)法により導入した(Poueymirou et al)。野生型Tpo(TPOm/m)、ヘテロ接合型(TPOh/m)またはホモ接合型(TPOh/h)TPO遺伝子置換をもつRag2-/-γc-/-マウスが得られた。
【0099】
実施例2
hIL-3/GM-CSFマウス:移植
ヒト造血幹細胞の単離
ヒト臍帯血および胎児肝臓試料は、それぞれイェールニューヘブン病院(Yale-New Haven Hospital)およびアルバート・アインシュタイン医科大学ニューヨーク(Albert Einstein Medical College New York)から、イェール大学の人権擁護調査委員会(Human Investigation Committee)の承認の下で得られたものである。CD34+細胞は、ヒト臍帯血または胎児肝臓から、密度勾配遠心およびCD34マイクロビーズ(Miltenyi Biotec社)を用いる免疫磁気選別によって単離した。単離されたCD34陽性細胞の純度をフローサイトメトリーにより確認した。精製したヒトCD34陽性細胞は凍結保存され、使用前に液体窒素中で保存した。
【0100】
ヒト造血幹細胞を用いたマウスの移植
移植は以前に記載された通りに行った(Traggiai et al. (2004) Development of a human adaptive immune system in cord blood cell-transplanted mice, Science 304:104-107)。簡単に説明すると、出生日に、RAG2遺伝子ノックアウト/Il2rg遺伝子ノックアウトの背景(hIL-3/hGM-CSFを含むまたは含まない)からの仔マウスを亜致死的に照射した(4時間間隔で2×200cGy)。照射後、新生仔は30ゲージ針を用いる肝内注射によって1〜2×105個のヒトCD34+細胞(25μLのPBS中に再懸濁したもの)を受け取った。対照にはPBSのみを注射した。マウスは生後3〜4週間で離乳させ、特定病原体フリーの条件下で維持した。日和見感染を防ぐために、マウスに飲料水中の予防的抗生物質(サルファトリム(Sulfatrim))を投与した。すべての動物実験作業はイェール大学の動物管理使用委員会(Institutional Animal Care and Use Committee: IACUC)によって承認され、IACUCの規則に従って実施された。
【0101】
移植されたマウスの解析
ヒト造血細胞の生着は移植後8〜12週で判定した。血液試料を後眼窩洞(retro-orbital sinus)から採取し、ACK溶解緩衝液(Lonza社)を用いて赤血球の溶解を行った。次に、試料をマウスCD45、ヒトCD45、ヒトCD3、およびヒトCD14に対する蛍光標識モノクローナル抗体(すべてBD Biosciences社から)で染色し、FACScalibur(商標)(BD Biosciences社)でフローサイトメトリーにより解析した。感染実験に用いたマウスは、特に明記しない限り、>4%のhCD45+細胞の血中生着レベルを有していた。適合したマウス、すなわち同じバッチのCD34+細胞を用いて移植されたマウスを、実験に使用した。特に明記しない限り、実験はFL由来のCD34+細胞を用いて移植されたマウスを用いて行った。
【0102】
フローサイトメトリー
hIL-3/GM-CSF研究のため、移植後10〜14週のマウスの肺、BAL、骨髄、胸腺、脾臓および血液から細胞懸濁液を調製した。RBCの溶解はACK溶解緩衝液(Lonza社)を用いて行った。次に、マウスおよびヒト細胞表面抗原に対する蛍光色素標識モノクローナル抗体(mAb)で試料を染色した。次のmAbを使用した:(1)抗ヒト:CD3 (UCHT1)、CD4 (RPA-T4)、CD8 (HIT8a)、CD11c (B-ly6)、CD14 (MoP9)、CD19 (HIB19)、CD33 (WM53)、CD45 (HI30および2D1)、CD56 (NCAM 16.2)、CD66 (B1.1)、CD116 (4H1)、CD123 (9F5)。(2)抗マウス:CD45 (30-F11)、F4/80 (BM8)。CD116、CD45 (30-F11)およびF4/80 mAbはeBioscience社から購入した。他のすべてのmAbはBD Biosciences社から購入した。試料はFACSCalibur(商標)またはLSRII(商標)フローサイトメーター(BD Biosciences社)で解析した。
【0103】
メチルセルロースCFUアッセイ法
hIL-3/GM-CSF研究のため、移植されたマウスからのヒトCD34+骨髄細胞をセルソーティングにより精製した。ソーティングされた細胞(1〜1.5×105個)をイスコフ改変ダルベッコ培地(IMDM、GIBCO社)ベースのメチルセルロース培地(Methocult(商標)H4100、StemCell Technologies社)で培養した。該培地には20%FBS、1%BSA、2mM L-グルタミン、55μM 2-メルカプトエタノール、および次のヒトサイトカインを添加した:幹細胞因子(10ng/ml)、FLT3リガンド(10ng/ml)、トロンボポエチン(50ng/ml)、IL-3 (20ng/ml)、IL-6 (10ng/ml)、IL-11 (10ng/ml)、GM-CSF (50ng/ml)、およびエリトロポエチン(4U/ml)(すべてR&D Systems社)。細胞を60mmペトリ皿で37℃/5%C02にてインキュベートした。12〜14日後にコロニーの数を顕微鏡で測定した。
【0104】
LPSに対する炎症反応
マウスにUltrapure LPS E. coli 0111:B4 (Invivogen社)の2回腹腔内注射を48時間間隔で行った(35および17.5μg)。各注射の2〜3時間後に血清を採取した。ヒトIL-6の血清濃度をELISA (R&D Systems社)で測定した。最初のLPS注射の72時間後にマウスを屠殺し、心穿刺で血液を採取してフローサイトメトリーに使用した。
【0105】
細胞内サイトカイン染色
hIL-3/GM-CSF研究のため、全TB10.4タンパク質(Skjot et al. (2002) Epitope mapping of the immunodominant antigen TB10.4 and the two homologous proteins TB10.3 and TB12.9, which constitute a subfamily of the esat-6 gene family, Infect. Immun. 70:5446-5453)をカバーする重複ペプチドをイェール大学のW.M. Keck Facilityで合成した。BCG感染したマウス由来の脾細胞(2×106個/ウェル)を混合ペプチド(5μg/mlの各ペプチド)と共に、96ウェルのU底マイクロタイタープレート(Becton Dickinson社)において200μl/ウェルの全体積で37℃/5%C02にて5時間インキュベートした。細胞培養のために10%FCS、1%ペニシリン-ストレプトマイシン、1% L-グルタミン、および55μM 2-メルカプトエタノールを添加したRPMI 1640培地(Invitrogen社)を用いた。細胞内サイトカイン染色はCytofix/Cytoperm(商標)キット(BD Biosciences社)を用いてメーカーの指示に従って行った。次のmAbを細胞内染色のために使用した(すべてBD Biosciences社)。抗ヒトIFNγ(B27)、抗マウスIFNγ(XMG1.2)。アイソタイプの適合したmAbを対照として用いた。
【0106】
組織学および免疫組織化学
hIL-3/GM-CSF研究のため、器官を取り出し、かつ組織学的分析のために10%中性緩衝ホルマリンまたはZinc Fixative (BD Biosciences社)中に固定した。パラフィン包埋組織切片を調製し、H&EもしくはPASで染色するか、またはイェール病理組織サービス(Yale Pathology Tissue Services)で免疫組織化学のために処理した。次の抗ヒト抗体を免疫組織化学のために使用した(すべてDako社から):CD45 (2B11+PD7/26)、CD3 (F7.2.38)、CD68 (PG-M1)。肉芽腫の存在についての組織切片の評価は盲検で行った。
【0107】
統計解析
hIL-3/GM-CSF研究のため、ノンパラメトリックなMann-WhitneyのU検定を用いて、2グループ間の統計的有意性(a=0.05)を判定した。多重グループ比較のために、我々は一元配置ANOVAとともにTukeyの多重比較検定(a=0.05)を用いる事後検定(post hoc testing)を採用した。統計的に有意なP値(P<0.05)のみが示される。
【0108】
実施例3
hIL-3/GM-CSFを移植されたマウス:感染
チフス菌ISP2825 (Galan J.E.およびCurtiss, R. (1991) Distribution of the invA, -B, -C, and -D genes of S. thyphimurium among other S. serovars: invA mutants of S. typhi are deficient for entry into mammalian cells, Infect. Immun. 59(9):2901-2908;参照により本明細書に組み入れる)は、腸チフス患者からの臨床分離株であるが、このチフス菌をLBブロスで一晩増殖させた。翌日、40μLの細菌細胞培養物を、0.3M NaClを含む新鮮なLBブロス2mLに移し、かつその培養物が約0.9のOD600に達するまで37℃で約3時間増殖させた。その細菌培養物をスピンダウンし、緩衝生理食塩水に再懸濁させて、感染のために使用した。9〜12週齢のヒト化マウスと対照マウスに1×103または1×104または1×105個のチフス菌を0日目に腹腔内接種した。感染したマウスを念入りに監視し、かつ感染後4週で屠殺した。脾臓、肝臓および胆のうを無菌的に摘出し、0.05%デオキシコール酸ナトリウムを含む無菌PBS 3〜5mL中で機械的にホモジナイズした。組織ホモジネートを連続希釈し、LB寒天プレートにまき、コロニー数の計測のために37℃で一晩インキュベートした。コロニー数を計測し、かつ回収された全コロニー形成単位の数を算出した。マウスのデータは図8〜11に提供される。図8において、「対照」マウスは、移植されていない遺伝子改変されたマウス(RAG KO, Il2rg KO/hIL-3, hGM-CSF)である。図9〜11において、「対照」マウスは移植されていないRAG KO/Il2rg KOマウスである(すなわち、それらはIL-3とGM-CSFのヒト化を欠くが、代わりに内在性マウスIL-3と内在性マウスGM-CSFをもつ)。「対照」マウスには、ヒトCD34+細胞の代わりにPBSを注射した。
【0109】
図8に示す通り、脾臓におけるチフス菌感染は感染後10日で2匹の遺伝子改変されたマウス(RAG KO, Il2rg KO/hIL-3, hGM-CSF)に検出される。
【0110】
図9に示す通り、1×103個のチフス菌の感染後1週で、3.8および3%の生着率を有する遺伝子改変されたマウス(RAG KO, Il2rg KO)は、対照マウスより約1000倍高い、脾臓での感染を示した(p<0.01)。対照マウスとヒト化マウス間の差のp値はp<0.01であった。
【0111】
図10に示す通り、胎児肝由来のCD34陽性細胞を用いて移植され、かつ1×104個のチフス菌を感染させた、遺伝子改変されたマウス(RAG KO, Il2rg KO)は、感染後4週で、脾臓(平均して約1,000〜約10,000倍)と肝臓(平均して約1,000倍)の両方にチフス菌感染を示した。脾臓のチフス菌について試験したコホート中の各マウスは、(図10の左パネルの「ヒト化」コホートにおいて上から下に)23.5、40.1、16.5、50、26、および51.7のヒト細胞の生着率を有していた。肝臓のチフス菌について試験したコホート中の各マウスは、(図10の右パネルの「ヒト化」コホートにおいて上から下に)16.5、40.1、23.5、26、50、および51.7のヒト細胞の生着率を有していた。脾臓では対照マウスとヒト化マウス間の差のp値はp<0.01であった;肝臓ではp<0.03。
【0112】
図11に示す通り、胎児肝由来のCD34陽性細胞を用いて移植され、かつ1×104個のチフス菌を感染させた、遺伝子改変されたマウス(RAG KO, Il2rg KO)は、感染後4週で、胆のうにおけるチフス菌感染を示し、チフス菌cfuは平均して対照マウスより100万倍高かった。胆のうのチフス菌について試験したコホート中の各マウスは、(図10の「ヒト化」コホートにおいて上から下に)23.5、16.5、40.1、50、26、および51.7のヒト細胞の生着率を有していた。対照マウスとヒト化マウス間の差のp値はp<0.03であった。
【0113】
これらの結果は、遺伝子改変されたマウス(RAG KO, Il2rg KO/hIL-3, hGM-CSF)が全身感染後にチフス菌によってコロニー化され得ることを証明している。
【0114】
実施例4
hIL-3/GM-CSFを移植されたマウス:肺病原体に対するヒト炎症反応のためのマウスモデルの検証
GM-CSF(Csf2)およびIL-3をコードする遺伝子は、ヒトおよびマウスではそれぞれ5番および11番染色体上で密接に連鎖している(<10kb)。これは、hIL-3/GM-CSF KIマウスを作製するために、両遺伝子のマウス遺伝子座とヒト遺伝子座との置換を可能にした(図5(e))。ヒトIL-3 KI対立遺伝子はマウス調節エレメントの制御下にある一方で、ヒトCsf2 KI対立遺伝子はそのヒト調節エレメントの制御下のままである。マウスおよびヒトGM-CSF mRNAの発現は、各マウス遺伝子の一方の対立遺伝子および各ヒト遺伝子の一方の対立遺伝子を発現するhIL-3/GM-CSF KIマウス(IL-3/GM-CSF「ヒト/マウス」(h/m)マウスと呼ばれる)においてRT-PCRにより解析した。IL-3およびGM-CSFのマウス対立遺伝子を唯一持つ野生型マウスは、IL-3/GM-CSF「マウス/マウス」(m/m)マウスと呼ばれる。
【0115】
hIL-3/GM-CSF KIマウスのRT-PCRおよびELISA解析
全RNAはホモジナイズした組織からTRIzol(商標)試薬(Invitrogen社)を用いてメーカーの指示に従って抽出した。等量のDNase処理RNAをSuperScript(商標)First-Strand Synthesis System(Invitrogen社)によるcDNA合成のために使用した。従来型RT-PCRは、次のプライマーを用いて行った。
定量的RT-PCRは、ABI社から購入したプライマー-プローブセットを用いる7500 FastリアルタイムPCRシステムで行った。発現量は比較閾値サイクル法(comparative threshold cycle method)を用いて計算し、マウスまたはヒトHPRTに対して正規化された。マウスとヒトのIL-3およびGM-CSFタンパク質は、R&D Systems社からの種特異的ELISAキットを用いてメーカーの指示に従って検出した。脾細胞は5μg/mlコンカナバリンA(ConA)と100U/ml IL-2で活性化し、48時間の刺激の後に上清をELISAのために回収した。
【0116】
hIL-3/GM-CSF KIマウスにおける発現
ヒトGM-CSF mRNAは、肺で最高の発現を示して、そのマウス対応物と同様のパターンで発現された(図5(a))。IL-3は主に活性化T細胞によって発現され、活性化T細胞はまたGM-CSFをも産生する。したがって、ELISAはh/mマウスから単離された活性化脾細胞からの上清で実施した。ヒトIL-3およびGM-CSFタンパク質の両方を検出することができた(図6(f)f、6(g))。ヒト造血細胞に競合的優位性を付与するために、ヒトIL-3およびGM-CSFの2つの対立遺伝子を発現する、IL-3/GM-CSF「ヒト/ヒト」(h/h)マウスと呼ばれるホモ接合体KIマウスを作製した。肺組織の従来型RT-PCR解析および定量的RT-PCR解析は、h/hマウスがヒトGM-CSF mRNAのみを発現し、マウスGM-CSF mRNAを発現しないことを示した(図5(b)、5(c))。ヒトGM-CSFタンパク質はELISAでh/hマウスの気管支肺胞洗浄(BAL)液中に検出することができた(図5(d))。これらの結果は、hIL-3/GM-CSF KIマウスがヒトGM-CSF(およびIL-3)を忠実に発現することを示している。
【0117】
図5(a)〜(d)は、非移植hIL-3/GM-CSF KIマウスにおけるヒトGM-CSF発現の検証を示す。図5(a)は、ヒトCsf2の1つの対立遺伝子とマウスCsf2の1つの対立遺伝子をもつKIマウス(h/m)由来の各種組織におけるGM-CSF mRNA発現の代表的なRT-PCR解析を示す。Li, 肝臓;Br, 脳;Lu, 肺;Mu, 筋肉;Sp, 脾臓;Th, 胸腺;LN, リンパ節;BM, 骨髄。下部:ヒトGM-CSFを検出するためのプライマーの特異性を対照マウス(m/m)由来の組織のRT-PCR解析によって検証した。リボソームタンパク質L13(Rpl13)を内在性対照として用いた。図5(b)は、m/mマウスまたはヒトCsf2の2つの対立遺伝子を発現するホモ接合体KIマウス(h/h)(それぞれn=5)由来の肺におけるGM-CSF mRNA発現のRT-PCR解析を示す。Rpl13を内在性対照として用いた;NTC, テンプレートを含まない対照。図5(c)は、(b)と同じGM-CSF mRNA発現の定量的RT-PCR解析を示す。GM-CSF発現はマウスHprtに対して正規化された(それぞれn=5)。図5(d)は、m/mまたはh/h KIマウス(それぞれn=6)から回収されたBAL液中のヒトGM-CSFタンパク質のELISAを示す。結果は2つの独立した実験を代表する。各ドットは1匹のマウスを表す。水平バーは平均値を示す。
【0118】
図5(e)は、hIL-3/GM-CSF KIマウスを作製するための戦略を示す。マウス(上)とヒト(下)のIL3およびCsf2の遺伝子座のゲノム構成がそれぞれ11番および5番染色体上に示される。マウス遺伝子座が本開示に記載する通りにヒト遺伝子座と置き換えられた。
【0119】
図6(f)、(g)は、非移植hIL-3/GM-CSF KIマウスにおけるヒトIL-3およびGM-CSFの発現を示す。活性化脾細胞によるヒトIL-3(f)およびGM-CSF(g)産生のELISA結果が提示される。m/mマウス(白のバー)またはh/m KIマウス(黒のバー)由来の脾細胞はConAとIL-2で48時間刺激し、上清を回収した(それぞれn=1)。m/mマウスではヒトIL-3およびGM-CSFが検出されなかった(ND)。
【0120】
実施例5
hIL-3/GM-CSFを移植されたマウス:増強されたヒト炎症反応
hIL-3/GM-CSF KIマウスは、Rag2とIl2rgの両方の1つの対立遺伝子がすでに欠失されている胚性幹(ES)細胞から作製された。その後Rag2 KO Il2rg KO背景へと交配させることで、ヒトCD34+造血細胞を移植することが可能になった。全体的なヒトCD45+造血細胞のキメリズム、ならびに骨髄中、胸腺中、脾臓および血液中のT細胞、B細胞およびナチュラルキラー(NK)細胞の分布は、hIL-3/GM-CSF KIマウスにおいて有意に増加していなかった(データは示さず)。さらに、全ヒトCD33+骨髄細胞、CD66+顆粒球、CD14+単球/マクロファージ、CD14loCD16+非古典的単球、CD11c+樹状細胞(DC)、およびCD123+CD11c-形質細胞様DCの頻度は、hIL-3/GM-CSF KIマウスにおいて有意に増加していなかった(データは示さず)。これは定常状態の条件下のh/mマウスとh/hマウスの両方にあてはまった。最後に、移植したhIL-3/GM-CSF KIマウス由来のヒト骨髄細胞は同様の能力を有して、インビトロでメチルセルロース中に骨髄コロニーを形成した(データは示さず)。これらの知見は、IL-3とGM-CSFの両方が本明細書で解析した器官での定常状態の骨髄造血にほとんど不要であることを示すKOマウス研究からの結果と一致している。
【0121】
対照的に、GM-CSFは炎症反応を媒介する上で重要な役割を果たしている。GM-CSFの発現は炎症性刺激によって誘導され、これは単球/マクロファージによる炎症性サイトカイン(IL-6およびTNFαなど)の産生と、炎症部位へのそれらの動員とにつながる。移植hIL-3/GM-CSF KIマウス由来のヒトCD14+単球は、GM-CSF受容体α鎖(CD116)の最高発現を示した(図8(d))。したがって、移植hIL-3/GM-CSF KIマウスの解析はヒト単球/マクロファージに集中した。最初に、移植hIL-3/GM-CSF KIマウスにおけるヒト単球の炎症反応が解析された。リポ多糖(LPS)の腹腔内(i.p.)注射によって全身性の炎症を誘発させた。LPS注射後に、循環ヒトCD14+単球の頻度が対照のm/mマウスと比較してh/mマウスでは有意に増加していた(図8(e)、(f))。h/mマウスでのヒト単球動員の増加は、1回および2回のLPS注射後のヒトIL-6の血清濃度の上昇と関連していた(図8(g))。ヒトTNFαのLPS誘発産生もh/mマウスで増加していたが、この結果は統計的有意性に達しなかった。これらのデータは、ヒト造血細胞を用いて移植されたhIL-3/GM-CSF KIマウスがヒト骨髄単球細胞によって媒介されるヒト炎症反応の亢進を有することを示している。
【0122】
図8(d)〜(g)は、移植hIL-3/GM-CSF KIマウスにおけるヒト炎症反応の亢進を示す。図8(d)は、定常状態の移植hIL-3/GM-CSF h/m KIマウス由来のヒト骨髄細胞のフローサイトメトリー解析を示す。ドットプロット(左)はhCD45+mCD45-細胞にゲーティングされる。ヒストグラム(右)はCD14-細胞(集団1)、CD14mid/SSChi顆粒球(集団2)、およびCD14hi単球(集団3)上のGM-CSF受容体α(CD116)発現を示す。解析した合計12匹のマウスからの代表的な一例が示される。図8(e)は、2回のLPS腹腔内注射後72時間のCB移植m/mまたはh/m KIマウス由来のヒト血液細胞の代表的なフローサイトメトリー解析を含む。プロットはhCD45+mCD45-細胞にゲーティングされる。ボックス領域の隣にある数字はヒトCD14+細胞のパーセンテージを示す。図8(f)は、LPS注射後72時間の移植m/mマウス(n=4)またはh/m KIマウス(n=8)におけるヒトCD14+血液細胞の頻度を示す。図8(g)は、1回目(上)および2回目(下)のLPS注射後2〜3時間の移植m/mマウス(n=4〜5)またはh/m KIマウス(n=8)由来の血清中のヒトIL-6についてのELISA結果を示す。1匹のm/mマウスは1回目のLPS注射後に死亡した。各ドットは1匹のマウスを表す。水平バーは平均値を示す。結果は2つの独立した実験を代表する。
【0123】
実施例6
hIL-3/GM-CSFを移植されたマウス:肺での増強されたヒトマクロファージ生着
BAL解析
hIL-3/GM-CSF研究のための気管支肺胞解析は次の方法で行った。気管に挿入したカテーテルから1mlのPBSを入れて肺を膨らませた。これを2回繰り返し、回収した洗浄液をプールした。遠心分離後、細胞フリーの上清は、ELISAによるGM-CSFタンパク質濃度の測定のために、またはBCA Protein Assay Kit(Pierce社)をメーカーの指示に従って用いる総タンパク質含有量の測定のために保存した。赤血球(RBC)をACK溶解緩衝液(Lonza社)で溶解した後、細胞ペレット数を計測し、フローサイトメトリーのためにまたはサイトスピン標本作成のために使用した。細胞をスライド上にスピンさせ、Diff-Quik(商標)Stain Set (Dade Behring社)でメーカーの指示に従って染色した。
【0124】
増強されたマクロファージ生着
マウスGM-CSFの非存在はマウス肺胞マクロファージ(AM)の機能低下につながり、これはホモ接合体hIL-3/GM-CSF KIマウスにおいてヒトマクロファージによる再構成に有利にはたらくはずである。その裏付けとして、h/hマウスの肺およびBALではヒトGM-CSFが高度に発現しており、一方マウスGM-CSFは欠損している。
【0125】
非移植h/hマウスからのマウスAMは肥大し、かつ典型的な「泡沫状」外観(図9(f))を呈した。この泡沫状外観はGM-CSF KOマウス由来のAMに関して記載されている。GM-CSF KOマウスは、最終分化に障害があるAMによる表面活性物質のクリアランスの欠陥に起因するPAPを発症する。GM-CSF KOマウスに関して報告されたものと同様に、非移植h/hマウスは、過ヨウ素酸シッフ(PAS)陽性物質に満ちたAMの胸膜下蓄積など、PAPの特徴を現した(図9(g))。したがって、非移植h/hマウスはマウスAMの分化障害を示してPAPを発症し、それゆえにGM-CSF KOマウスと機能的に同等であると結論づけられた。
【0126】
図9(f)、(g)は、非移植ホモ接合体hIL-3/GM-CSF KIマウスにおけるPAPの発症を示す。図9(f)は、非移植m/mまたはh/h KIマウス由来のBAL細胞のDiff-Quick(商標)染色を示し(倍率400x);グループごとに解析した合計6匹のマウスの代表的な一例が示される。図9(g)は、非移植m/mまたはh/h KIマウス由来の肺組織切片のPAS染色を示し(倍率400x);グループごとに解析した合計12匹のマウスの代表的な一例が示される。
【0127】
次に、ヒト造血細胞を移植した後のh/hマウスの肺区画を検討した。FACS解析は、h/hマウスがBAL中にかなり多くのヒトCD45+細胞をもつことを示した(図6(a)、(b))。肺組織の定量的RT-PCRは、このヒト細胞の増加が主として、ヒト骨髄マーカーCD33、CD11b、CD11c、およびCD14のmRNAを発現する細胞で構成されることを明らかにした(図6(c))。さらに、主に細胞内に発現される成熟マクロファージマーカーである、ヒトCD68のmRNA発現が移植h/hマウスでは著しく増加していた(図6(d))。h/hマウスでのこの増加は、AMによって発現される2つの転写因子、すなわちPU.1(Spi1)およびペルオキシソーム増殖因子活性化受容体γ(PPARγ)のより高い発現と関連していた(図6(d))。PU.1は最終的に分化したAMにおいてGM-CSF依存的方法で高度に発現される。重要なことは、インビトロでGM-CSF KO AMにPU.1を形質導入すると、それらの機能障害が改善することである。PPARγもまたAMで高度に発現されており、GM-CSF KOマウスと同様に、PPARγ KOマウスはPAPを発症する。肺切片の免疫組織学的染色は、移植h/hマウスにおいて、ヒトAMと一致して、典型的な肺胞内位置での多数のhCD68+細胞の存在を明らかにした(図6(e))。対照的に、移植m/m対照マウスではごくわずかなヒトAMしか検出できなかった。要約すると、CD34+造血細胞を移植したh/hマウスの肺は、際立って改善されたヒトマクロファージ生着を示す。
【0128】
図6(a)〜(e)は、ホモ接合体hIL-3/GM-CSF KIマウスが肺での良好なヒトマクロファージ生着を有することを示す。図6(a)は、移植したm/mおよびh/h KIマウス由来のBAL細胞の代表的なフローサイトメトリー解析を示す。丸で囲った領域の隣にある数字はhCD45+およびmCD45+造血細胞のパーセンテージを示す。mCD45+hCD45+細胞は高い自己蛍光を有し、F4/80+マウスAMを構成する。図6(b)は、移植したm/mおよびh/h KIマウス由来のBAL中のヒト造血(hCD45+)細胞の数を提供する(結果は3つの独立した実験(グループあたり合計n=15)から組み合わされる)。図6(c)は、移植したm/mおよびh/h KIマウス(それぞれn=4)由来の肺組織におけるヒトリンパ球および骨髄遺伝子発現の定量的RT-PCR解析の結果を示す。発現はマウスHPRTに対して正規化された(*, P<0.05)。図6(d)は、移植したm/mおよびh/h KIマウス(それぞれn=4)由来の肺組織におけるヒトマクロファージ遺伝子発現の定量的RT-PCR解析を示す。発現はマウスHPRTに対して正規化された(*, P<0.05)。各ドットは1匹のマウスを表す。水平バーは平均値を示す。図6(e)は、移植したm/mおよびh/h KIマウス由来の、ヒトCD68について染色した肺組織切片の免疫組織化学を示す(倍率100x(上)および200x(下))。グループごとに解析した合計10匹のマウスの代表的な一例が示される。
【0129】
実施例7
hIL-3/GM-CSFを移植されたマウス:ヒト造血細胞によって軽減されるPAP
h/hマウスのヒトマクロファージ生着の増加が肺における良好なヒト免疫機能につながるかどうかを検討した。最初に、ヒトマクロファージが非移植h/hマウスに見られるPAP症候群をレスキューできるかどうかを検討した。II型肺胞上皮細胞とAMは両方ともGM-CSFに応答し得るが、PAPは骨髄移植によってレスキューすることができる。これは、造血細胞、特にAM、がPAPを改善できる主な細胞型であることを示している。したがって、ヒト造血細胞を用いて移植されたh/hマウスはPAPの重症度が低いはずであると仮定された。予想通り、非移植h/hマウスはPAS陽性物質の肺胞内蓄積を示し(図7(a))、これはPAPの顕著な特徴である。この仮説に一致して、移植したh/hマウスは肺に軽度のタンパク質蓄積を有し、一部のh/hマウスの肺は(非移植または移植)m/m対照マウスに似ていた(図7(a))。さらに、移植h/hマウスは、BAL液中の総タンパク質量が非移植h/hマウスより有意に低かった(図7(b))。これらの結果は、移植されたヒト造血細胞(おそらくAM)がホモ接合体hIL-3/GM-CSF KIマウスにおいてPAPを軽減できることを示している。
【0130】
図7(a)〜(e)は、ヒト造血細胞がホモ接合体hIL-3/GM-CSF KIマウスにおいてPAPを軽減することを示す。図7(a)は、非移植または移植m/mもしくはh/h KIマウス由来の肺組織切片のPAS染色を示す。2匹の異なる移植h/h KIマウス由来の肺切片が示される(倍率400x)。グループごとに解析した合計10〜12匹のマウスの代表的な例が示される。図7(b)は、非移植(non)または移植h/h KIマウスもしくはm/m対照マウス(グループあたりn=6)由来のBAL液中の総タンパク質の定量を示す。P<0.0001(一元配置ANOVA検定)。Tukeyの多重比較検定で決定されたP値は星印によって示される(**, P<0.01;***, P<0.001)。
【0131】
実施例8
hIL-3/GM-CSFを移植されたマウス:インフルエンザAに対する強いヒトI型IFN応答
インフルエンザA感染
マウス(9〜10週齢)に2×104プラーク形成単位のインフルエンザA/PR8(H1N1)ウイルスを鼻腔内経路で感染させた。感染は、Anafane(商標)(Ivesco社)で深く麻酔しておいたマウスに、PBSで希釈したウイルスストック液50μl(または対照として等量のPBS)を鼻腔内投与することによって行った。感染の24時間後に、上記のRNA抽出および定量的RT-PCR解析のために肺を回収した。
【0132】
肺の恒常性におけるそれらの役割に加えて、AMは肺での宿主防衛に不可欠である。多くの研究は、GM-CSF KOマウスが肺のさまざまな病原体の影響を受けやすくなることを示している。移植されたヒトAMの肺病原体に対する機能的応答を評価するために、移植h/hマウスにインフルエンザA/PR8(H1N1)ウイルスを鼻腔内経路で感染させた。AMは肺ウイルス感染後のI型インターフェロン(IFN)の主な産生細胞であり、かつAMはインフルエンザAへの有効な先天性応答に必要とされる。ヒトヒポキサンチンホスホリボシルトランスフェラーゼ(HPRT)mRNAの発現は、対照m/mマウスと比べて、移植h/hマウスの肺で有意に高かった(図8(a))が、これは良好なヒト免疫細胞のキメリズムを示している。移植m/mマウスは、PBSを鼻腔内投与された移植m/mマウスと比べた場合、インフルエンザA感染後にヒトIFNβ mRNA発現の有意な誘導を示さなかった(図8(b))。対照的に、移植h/hマウスは、非感染h/hマウスと感染m/mマウスの両方よりも有意に多いヒトIFNβ mRNAを発現した(図8(b))。m/mマウスと比較して、h/hマウスにおけるヒトIFNβ mRNAの発現増加は、ヒトHPRTに、すなわち肺中のヒト細胞の数に、正規化された場合、まだ有意であった(図8(c))。まとめると、ホモ接合体hIL-3/GM-CSF KIマウスは、良好なヒトマクロファージキメリズムと、ウイルス感染に対して増強されたヒト粘膜免疫につながる肺での機能とを可能にする。
【0133】
図8(a)〜(g)は、ホモ接合体hIL-3/GM-CSF KIマウスがインフルエンザA感染に対して強いヒトI型IFN応答を開始することを示す。図8(a)〜(c)は、インフルエンザA(PR8)の鼻腔内感染後24時間のm/mおよびh/h KI由来の肺組織における遺伝子発現の定量的RT-PCR解析を示す(それぞれn=8)。PBSの鼻腔内投与を対照(PBS)として用いた(それぞれn=4)。図8(a)は、マウスHprtに正規化されたヒトHprtの発現を示す。P<0.0001(一元配置ANOVA検定)。図8(b)は、マウスHprtに正規化されたヒトIFNγの発現を示す。P=0.0171(一元配置ANOVA検定)。図8(c)は、ヒトHprtに正規化されたヒトIFNγの発現を示す。P=0.0032(一元配置ANOVA検定)。Tukeyの多重比較検定で決定されたP値は星印によって示される(*, P<0.05;**, P<0.01;***, P<0.001)。各ドットは1匹のマウスを表す。水平バーは平均値を示す。結果は2つの独立した実験を代表する。
【0134】
実施例9
hIL-3/GM-CSFを移植されたマウス:マイコバクテリア感染後のヒト細胞を含む肉芽腫
マクロファージが病原体特異的免疫応答において中心的な役割を果たす肺に向性を有する第2の病原体に対するヒト炎症反応を支持するhIL-3/GM-CSF KIマウスの能力が検討された。マイコバクテリア感染後の肉芽腫の形成が選択された。肉芽腫は、マイコバクテリア感染の特徴である、特殊化した局所炎症反応を表している。それはDTH応答の古典的な例であり、その形成は活性化T細胞とマクロファージとの相互作用に依存している。最適なDTH応答にはIL-3とGM-CSFの両方が必要であり、重要なことは、GM-CSF KOマウスがマイコバクテリアに感染した場合肉芽腫を形成しないことである。
【0135】
移植したhIL-3/GM-CSF h/m KIマウスにBacillus Calmette-Guerin (BCG)を静脈内注射により感染させた。BCGはヒト結核に対するワクチンとして使用されるM. ボビス(M. bovis)の弱毒株である。BCG感染実験に用いたマウスは>20%のhCD45+細胞の血中生着レベルを有し、hCD45+細胞の>8%がT細胞(hCD3+)であった。マウスは感染時に9〜10週齢であった。1×105コロニー形成単位のBCG(コペンハーゲンのStatens Serum Institute社)を0.1ml量で尾静脈注射によりマウスに感染させた。
【0136】
肉芽腫の形成にはT細胞が不可欠であることから、最初に、感染4週後のBCGに対するヒトT細胞応答を調べた。フローサイトメトリーは、移植したm/mおよびh/mマウスの両方の肺にヒトT細胞が存在することをはっきりと示した(図9(a))。実際に、T細胞はBCG感染肺において主要なヒト造血細胞型であった。m/m対照マウスに比較して、BCGに感染したh/mマウスは肺でのヒトCD4対CD8 T細胞の平均比率がより高かったが、その差は統計的有意性に完全には達しなかった(図9(b)、9(c))。これら2つのマウスグループ間で脾臓hCD4/hCD8 T細胞の比率の差は認められなかった。
【0137】
次に、2種類のT細胞由来サイトカイン、すなわちIFNγとTNFαの発現を解析した。これらは両方ともマイコバクテリアに対する防御免疫応答に極めて重要である。感染したマウス由来の脾細胞を、免疫優性マイコバクテリア抗原TB10.4から誘導されたペプチドで再刺激した後の、細胞内サイトカイン染色によるBCG特異的IFNγ産生が検討された。予想されたとおり、IFNγを産生するマウスT細胞の集団がBALB/cマウス由来の脾細胞の中に検出された(図9(d))。さらに、ヒトBCG特異的T細胞応答が移植したh/mおよびm/mマウスのサブセットで見られた(図9(d))。最後に、移植したh/mおよびm/mマウスの大部分はBCG感染後に肺でヒトIFNγおよびTNFα mRNAを発現した(図9(e))。これらの結果は、移植されたマウスのサブセットがBCGに対する病原体特異的ヒトT細胞応答を開始することができるが、この応答はhIL-3/GM-CSF KIマウスにおいて増強されなかったことを示している。これに一致して、細菌負荷にはh/mおよびm/mマウス間で差がなかった。
【0138】
次に、肉芽腫の形成が感染4週後に肺と肝臓で組織学により評価された。非移植m/mマウス(T細胞を欠く)は肉芽腫を発生せず(表1)、これは肉芽腫形成のためのT細胞の必要性と一致する。同様に、ヒト細胞を用いて移植されたm/mマウスは肺にも肝臓にも肉芽腫を示さなかった(表1)。これに対して、h/mマウスの大部分には肺(図10(a))または肝臓または両方の臓器に小さな病変または肉芽腫があった(表1)。一般的に、観察された肉芽腫は小さく、ヒトよりもマウスの肉芽腫に特有のゆるい組織化を有していた。しかしながら、hIL-3/GM-CSF KIマウスの肺肉芽腫は、免疫組織化学によって証明される通りに、ヒト造血細胞(hCD45+)を含んでいた(図10(b))。これらの細胞の大部分はヒトT細胞(hCD3+)であり、中心部に数個のヒトマクロファージ(hCD68+)が存在していた(図10(b))。まとめると、移植されたhIL-3/GM-CSF KIマウスは、マイコバクテリア感染に応答してヒトT細胞とヒトマクロファージを含む肉芽腫を発症することができるが、こうしたことはHISマウスでは以前に報告されていない。表1は、BCG感染4週後のBALB/c、非移植m/m(non)、移植m/m、および移植hIL-3/GM-CSF h/m KIマウス由来の肝臓および肺組織切片に認められた病変/肉芽腫を示す。
【0139】
(表1)BCGに感染させた移植したhIL-3/GM-CSF KIにおける肉芽腫
【0140】
図9(a)〜(g)は、移植hIL-3/GM-CSF KIマウスにおけるBCGに対するヒトT細胞応答を示す。図9(a)〜(c)は、BCG感染4週後の移植したm/mおよびh/m KIマウス由来の肺細胞のフローサイトメトリー解析を示す。図9(a)は、肺におけるヒトT細胞(hCD45+hCD3+)の頻度を示す。ボックス領域の隣にある数字は細胞のパーセンテージを示す。図9(b)は、肺におけるヒトCD4およびCD8 T細胞の分布を示す。ドットプロットはhCD45+hCD3+細胞に対しゲーティングされる。象限内の数字は細胞のパーセンテージを示す。図9(c)は、肺におけるヒトCD4対CD8 T細胞の比率を示す(それぞれn=6)。各ドットは1匹のマウスを表す。水平バーは平均値を示す。図9(d)は、BCG感染4週後のBALB/cマウス、移植m/mマウス、および移植h/m KIマウス由来の脾細胞のフローサイトメトリー解析を示す。脾細胞は、本明細書に記載したTB10.4タンパク質をカバーする重複ペプチドのプールを用いてインビトロで再刺激された。ドットプロットは、細胞内サイトカイン染色により測定されたマウスIFNγ+ CD4 T細胞(mCD4+)またはヒトIFNγ+ T細胞(hCD3+)の頻度を示す。アイソタイプの適合した抗体による染色を対照として用いた。図9(e)は、BCG感染4週後のBALB/cマウス、非移植(non)m/mマウス、移植m/mマウス、および移植h/m KIマウス由来の肺組織におけるヒトIFNγ(左)およびTNFα(右)遺伝子発現の定量的RT-PCR解析の結果を示す(グループあたりn=4〜7)。各ドットは1匹のマウスを表す。水平バーは平均値を示す。
【0141】
図10(a)、(b)は、移植したhIL-3/GM-CSF KIマウスがBCG感染後にヒト細胞を含む肉芽腫を発症することを示す。図10(a)は、BCG感染4週後の移植h/m KIマウス由来の肺組織切片のヘマトキシリンおよびエオシン(H&E)染色を示す(倍率100x(左)および倍率200x(右))。図10(b)は、BCG感染4週間後の移植h/m KIマウス由来のヒトCD45、CD3、またはCD68を染色した肺組織切片の免疫組織化学を示す(倍率200x)。肺肉芽腫を有する2匹のマウスのうち代表的な一例が示される。
【0142】
実施例10
hTPOマウス:生着および解析
TPOマウスへの移植
レシピエントマウスは、Traggiaiらに記載される通りにヒト造血前駆細胞を用いて移植された。臍帯血試料は、イェール大学の人権擁護調査委員会(Department of Labor and Birth, Yale New Haven Hospital, New Haven, CT)からの承認の下で、健康な満期新生児から採取された。胎児肝臓試料は、アルバート・アインシュタイン医科大学(Bronx, NY)のヒト胎児組織リポジトリ(Human Fetal Tissue Repository)から、およびAdvance Biosciences Resources社(Alameda, CA)から取得された。
【0143】
胎児肝臓試料を小さな断片に切断し、コラゲナーゼD(100ng/ml、Roche社)を用いて37℃で45分間処理し、かつ細胞懸濁液を調製した。ヒトCD34+細胞は、密度勾配遠心(リンパ球分離溶液(Lymphocyte Separation Medium)、MP Biomedicals社)と、その後の抗ヒトCD34マイクロビーズを用いる陽性免疫磁気選別(positive immunomagnetic selection)によりメーカーの指示(Miltenyi Biotec社)に従って、胎児肝臓試料からまたは臍帯血から精製した。細胞は、FBSを含む10%DMSO中で凍結させるか、または直接注入した。
【0144】
新生仔(生後1日以内)を亜致死的に照射し(X線照射、4時間間隔で2×150cGy)、20μlのPBS中の100,000〜200,000個のCD34+細胞を、22ゲージ針(Hamilton社, Reno, NV)を使って肝臓に注入した。
【0145】
すべての実験は、イェール大学人権擁護調査委員会プロトコルおよびイェール大学動物管理使用委員会プロトコルに従って行った。
【0146】
TPO発現
マウスおよびヒトTPOタンパク質の血清濃度は、種特異的ELISA(RayBiotech社)によりメーカーのプロトコルに従って測定した。TPOをコードするマウスおよびヒトmRNAの発現を測定するために、組織を成体動物から分離し、全RNAを、TRIzol(Invitrogen社)を用いてメーカーの指示に従って精製した。混入しているゲノムDNAをRNase-Free DNase I(Roche社)で処理して除去し、そのRNAを、SuperScript II逆転写酵素(Invitrogen社)とオリゴdTプライマーを用いて逆転写した。PCR増幅のために次のプライマーを使用した。
【0147】
ヒトTPOがこれらのマウスで忠実に発現されるかどうかを調べるために、全RNAをTPOh/mマウス由来のさまざまな器官から抽出した。かつTPOをコードするマウスおよびヒトmRNAの両方について同様の発現パターンがRT-PCRにより観察された(図11(a))。次に、TPOm/m、TPOh/mおよびTPOh/hマウス由来の、TPOを発現することが公知である3種の組織または細胞型(肝臓、腎臓および間葉系多能性間質細胞)における発現を比較した。TPOm/mおよびTPOh/mマウスからの試料ではマウスTpoの発現が検出され、一方TPOh/mおよびTPOh/hマウスではヒトTPOが発現された(図11(b))。標的マウスの血清中のTPOタンパク質の濃度も測定された。マウスTPOはTPOm/mおよびTPOh/m動物で検出され、ヒトTPOはTPOh/mおよびTPOh/hで検出された(図11(c))。測定されたヒトTPOの濃度はマウスTPOより約10倍低かった。しかしながら、この差異は健康なヒトとマウスで報告された生理的濃度に適合するものであり(図11(c))、サイトカイン半減期の種特異的な差異に起因する可能性がある。
【0148】
図11は、(a)Rag2+/-γcY/- TPOh/mマウスの異なる組織におけるマウスTPO(mTpo)およびヒトTPO(hTPO)発現のRT-PCR解析(マウスRpl13aはハウスキーピング遺伝子として使用した);(b)Rag2-/-γc-/- TPOm/m、TPOh/mおよびTPOh/hマウスの肝臓、腎臓および間葉系多能性間質細胞(MSC)におけるmTpoおよびhTPO発現のRT-PCR解析;(c)TPOm/m、TPOh/mおよびTPOh/hマウスの血清中の、ELISAにより測定されたマウスおよびヒトTPOタンパク質の濃度(pg/ml、平均±S.D.、n=7〜9)を示す。ND:検出されず。示した正常範囲はR&D Systems社のThrombopoietin Quantikineキットからである。
【0149】
間葉系多能性間質細胞の単離
TPO研究のため、マウスの大腿骨と脛骨を採取し、かつ骨髄細胞を流し出した。骨を小片に切断し、コラゲナーゼPおよびD(10μg/ml)を用いて37℃で45分間消化した。骨関連細胞を反復ピペッティングによって収集した。刺激用サプリメント(stimulatory supplements)(Stemcell Technologies社)を加えたMSC培地の存在下で細胞を14日間培養した。その培養物から、CD45およびTer119抗体を用いる免疫磁気セルソーティング(MACS、Miltenyi Biotec社)により造血系細胞を除去した。非造血細胞(CD45-Ter119-)をもう5日間培養し、MSC表現型(CD45-Ter119-Sca1+CD90+)をFACSにより確認した(Diminici et al. (2006) Minimal criteria for defining multipotent mesenchymal stromal cells, The International Society for Cellular Therapy position statement, Cytotherapy 8:315-317)。
【0150】
TPOマウスにおける造血細胞集団の解析
移植後8〜12週でマウスを採血した。ACK(Lonza社)を用いて赤血球を3回溶解し、細胞を抗マウスCD45および抗ヒトCD45抗体で染色した。CD45+細胞の少なくとも1%がヒト由来であった動物をさらなる解析のために使用した。移植されたマウスの約80%がこの生着閾値に達しており、TPOm/mおよびTPOh/hグループの間で違いは認められなかった。
【0151】
マウスを移植後3〜4または6〜7ヶ月で屠殺した。単一細胞懸濁液を骨髄(2本の大腿骨と2本の脛骨から流した)、脾臓および胸腺から調製した。赤血球をACK溶解により排除し、FACS解析のため次の抗体を用いて細胞を染色した。全体的な造血移植のために:抗マウスCD45-eFluor450 (30-F11、eBioscience社)および抗ヒトCD45-APC-Cy7 (2D1)。ヒト造血幹細胞前駆細胞および造血系列のために:抗ヒトCD14-PerCP (MoP9)、CD19-APC (HIB19)、CD33-APC (WM53)、CD34-PE (AC136、Miltenyi Biotec社)、CD38-FITC (HIT2)、CD41a-APC (HIP8)およびCD66-FITC (B1.1)。
【0152】
マウス幹細胞前駆細胞の解析のため、抗細胞系列カクテル(anti-lineage cocktail)はCD3ε(145-2C11)、CD11b (M1/70)、CD11c (HL3)、CD19 (1D3)、Gr1 (RB6-8C5)およびLy-76 (Ter119)に対するビオチン化抗体を含んでいた。その後、細胞をストレプトアビジン-APC-Cy7、抗cKit-APC (2B8)および抗Sca1-PE-Cy7 (D7)で染色した。
【0153】
すべての抗体は、特に指定した場合を除き、BD Biosciences社から入手した。データはFACSCalibur(商標)またはLSRII(商標)フローサイトメーター(BD Biosciences社)で取得し、FlowJo(商標)ソフトウェアを用いて解析した。
【0154】
TPOマウスにおけるヒト造血幹細胞前駆細胞の機能的特性解析
3〜7匹の移植されたマウスから得られた骨髄細胞をプールし、ヒトCD34+細胞を、マウスCD45+細胞のMACS枯渇(Miltenyi Biotec社)とその後のFACSAria(商標)フローサイトメーター(BD Biosciences社)でのヒトCD45+CD34+細胞のFACSソーティングによって精製した。
【0155】
ヒトCD34+細胞のコロニー形成能を評価するために、IMDMに20%FCS、2mM L-グルタミン、55μM 2-メルカプトエタノール(試薬はすべてGIBCO社から)を添加し、Methocult(商標)H4100、1%BSA(Stemcell Technologies社)と混合し、次のヒトサイトカインを加えた:SCF (10ng/ml)、FLT3I (10ng/ml)、TPO (50ng/ml)、IL-3 (20ng/ml)、IL-6 (10ng/ml)、IL-11 (10ng/ml)、GM-CSF (50ng/ml)、EPO (4U/ml) (すべてR&D systems社から)。100,000〜150,000個のソーティングされた細胞を60mmペトリ皿にまき、37℃、5%C02で12〜14日間インキュベートした。12〜14日でコロニー数を計測し、顕微鏡で特定の骨髄系列に分類した。
【0156】
二次移植実験のため、TPOm/mまたはTPOh/h一次レシピエントから精製された100,000個のCD34+細胞を、上記の通りに、亜致死的に照射(2×200cGy)したRag2-/-γc-/- TPOm/m二次レシピエントに注入した。これらのマウスを8週間後に屠殺し、骨髄中のヒトCD45+細胞の割合をFACSで測定した。
【0157】
TPOマウスデータの統計解析
データは対応のない両側t検定を用いて比較された。2つより多い試料を比較する場合は、一元配置ANOVAと、その後にTukeyの事後検定を行った。二次移植実験において生着を示したマウスの割合はPearsonのカイ二乗検定を用いて比較された。p値が0.05を下回った場合は、差を有意と見なした。
【0158】
実施例11
hTPOを移植されたマウス:TPOh/hレシピエントマウスの骨髄における改善されたヒト生着レベル
ヒト化および移植されたマウスの骨髄の表現型分類
ヒト化マウスの骨髄から分離された細胞をフローサイトメトリーで解析したところ、非ヒト化マウス(すなわち、TPO遺伝子のヒト化を欠くRAGおよびIl2rgノックアウト)の生着と比べて、全ヒト造血細胞、ヒト造血幹細胞、ヒト骨髄細胞、およびヒト顆粒球の生着の統計的に有意な改善が認められた。図12および14参照。
【0159】
野生型Tpo(TPOm/m)、ヘテロ接合型(TPOh/m)またはホモ接合型(TPOh/h)TPO遺伝子置換を有するRag2-/-γc-/-マウスを、記載した通りに作製した。照射(2×1.5Gy)した新生仔Rag2-/-γc-/- TPOm/mおよびTPOh/hマウスは、臍帯血または胎児肝臓から精製されたヒトCD34+細胞を用いて移植され、3〜4ヶ月または6〜7ヶ月後に骨髄における生着を解析された。
【0160】
図12は生着研究の結果を示す。図12(a)は、ヒトCD34+細胞の移植後3〜4ヶ月のRag2-/-γc-/- TPOm/mおよびTPOh/hマウスの骨髄におけるヒトおよびマウスCD45+細胞の代表的なFACS解析を示す。2匹の代表的なマウスの結果が各遺伝子型について示される。全(マウス+ヒト)CD45+細胞集団中のマウスおよびヒトCD45+細胞のパーセンテージが示される。図12(b)は、移植後3〜4ヶ月(左、n=42〜53)または6〜7ヶ月(右、n=20〜25)の骨髄中のヒトCD45+細胞のパーセンテージを示す。各記号は個々のマウスを表し、水平バーは平均値を示す。図12(c)は、(b)と同じ動物の骨髄中のヒトCD45+細胞の絶対数を示す。p値は統計的有意性を示す。
【0161】
TPOm/mレシピエントと比較して、TPOh/hの骨髄中のヒト造血細胞(hCD45+)のパーセンテージ(図12(a)および12(b))と絶対数(図12(c))に両時点で有意な増加が観察された。さらに、TPOh/hレシピエントはより低い生着のばらつきを示し、3〜4ヶ月の時点でマウスの75%に少なくとも80%のヒトキメリズムが見られた(図12(b))。CD34+細胞の供給源はこの結果に影響を及ぼさなかったが、それは、TPOh/h宿主でのキメリズムの同様の増加が臍帯血由来細胞と胎児肝臓由来細胞で観察されたからである(図12(d))。興味深いことに、TPOm/m宿主ではヒト細胞の数が早い時点と遅い時点の間に減少したが、TPOh/h動物ではそれらが一定のままであった(図12(c))。これらの結果は、マウスにおけるTPOの先に記載した機能と一致している。第一に、TPOは照射レシピエントマウスへの移植後にHSCの増幅に有利にはたらいて、生着レベルの増加をもたらす。第二に、それは成体HSCの維持に有利にはたらき、成体期を通じて持続的な造血をもたらす。
【0162】
実施例12
hTPOを移植されたマウス:マウスおよびヒト血小板に対するTPOヒト化の効果
TPOマウスにおける血小板解析
末梢血中の血小板数はHemavet(商標)950FS装置(Drew Scientific社)を用いて測定した。次に血液試料を抗マウスCD61-PE (2C9.G2)および抗ヒトCD41a-APC (HIP8)で染色し、マウスおよびヒト血小板のパーセンテージを、細胞のサイズ(FSC)または顆粒性(granulosity)(SSC)にいかなるゲートも配置せずに、フローサイトメトリーで測定した。絶対マウスおよびヒト血小板数は、これらの各パーセンテージを絶対血小板数と乗じることによって算出した。
【0163】
TPOは血小板産生に及ぼすその重要な機能について周知されているので、TPOのヒト化が血小板の発生に影響を与えるかどうかを検討した。TPO遺伝子の両方の対立遺伝子のヒト化は、非移植Rag2-/-γc-/-マウスの血中血小板数の約2倍低下につながった(図13(a))。ヒト細胞を移植した後では、TPOh/hマウスにおけるマウス血小板数は正常値の25%未満にさらに低下した(図13(d))。ヒト対マウス血小板の比率(図13(b)、13(c))、ならびにヒト血小板の絶対数(図13(e))は、TPOm/mよりもTPOh/hマウスにおいて高くなる傾向にあったが、これらの差はどれも統計的有意性に達しなかった。さらに、ヒト細胞のうちの骨髄巨核球(CD41a+細胞)のパーセンテージは両系統で同等であった(図13(f))。これらの結果は、ノックイン戦略によって達成されたヒトTPOのレベルまたは生物活性がマウスTPO機能を完全に置き換えるには十分でないことを実証しており、さらに、ヒトTPOが単独ではマウス環境においてヒト血小板産生を支持するのに十分でないことを示唆している。
【0164】
図13(a)は、成体非移植Rag2-/-γc-/- TPOm/m、TPOh/mおよびTPOh/hマウスの血液中の血小板数を示す。p<0.0001 (一元配置ANOVA、n=7〜17;示したp値はTukeyの事後検定により算出した)。各記号は個々のマウスを表し、水平バーは平均値を示す。図13(b)移植後3〜4ヶ月のRag2-/-γc-/- TPOm/mおよびTPOh/hマウスの血液中のマウス(mCD61+)およびヒト(hCD41a+)血小板の代表的FACS解析。数字は全イベントのうちの比率を示す。図13(c)TPOm/mおよびTPOh/hマウス(n=19〜22)における、FACSにより測定されたヒト血小板のキメリズム。血中のヒトCD45+細胞のパーセンテージが5%より高いマウスだけをこの解析に含めた。(d)、(e)TPOm/mおよびTPOh/hレシピエントの血液中のマウス(mCD61+,(d))およびヒト(hCD41a+,(e))血小板数;(f)骨髄中のヒトCD45+細胞のうちのヒト巨核球(CD41a+)のパーセンテージ。
【0165】
図13(g)〜(i)は、二次リンパ器官におけるヒト生着レベルを示す。図13(g)、(h)は、Rag2-/-γc-/- TPOm/mおよびTPOh/hマウスの血液(20g;n=43〜53)および脾臓(13h;n=35〜36)中のヒトCD45+細胞のパーセンテージを提供する(各記号は個々のマウスを表し、水平バーは平均値を示す)。図13(i)は、移植したTPOm/mおよびTPOh/hレシピエント(n=24〜34)の胸腺の全細胞性を示す。胸腺に見られた細胞の90%超がヒト由来(hCD45+)であった。
【0166】
実施例13
hTPOを移植されたマウス:TPOヒト化マウスにおける多系列の造血
ヒト化および移植されたマウスの血液細胞の表現型分類
ヒト化マウスの血液から分離された細胞をフローサイトメトリーで解析したところ、非ヒト化マウス(すなわち、TPO遺伝子のヒト化を欠くRAGおよびIl2rgノックアウト)の移植と比べて、ヒト単球および顆粒球の移植において統計的に有意な改善が認められた。図14参照。
【0167】
ヒトTPOがインビボでヒト造血幹細胞前駆細胞の多系列分化に有利にはたらくことができるかどうかを検討した。以前に報告された通りに(Traggiai et al.; Ishikawa et al. (2005) Development of functional human blood and immune systems in NOD/SCID/IL2 receptor gamma chain(null) mice, Blood 106:1565-1573))、移植されたヒト細胞は野生型Rag2-/-γc-/-宿主において主にB細胞(CD19+)を生じさせ(脾臓中のヒト細胞の61.51±4.71%、平均±標準誤差、n=32)、骨髄細胞はほんのわずかしか含まれていなかった。TPOm/mレシピエントとTPOh/hレシピエントを比較した場合、TPOh/hマウスの骨髄中のCD33+骨髄細胞の頻度の有意な増加が観察された(図14(a)および14(b))。興味深いことに、この増加は主として顆粒球(CD33+CD66hiSSChi細胞)によるものであり、単球(CD33hiCD66loCD14+)の割合は両系統で類似していた(図14(a)、(c)、(d)、(e))。骨髄細胞(顆粒球と単球の両方)のパーセンテージもまた、TPOh/h動物の末梢血で有意に増加していた(図14(a)、(f)、および(g))。
【0168】
図14(a)〜(g)は、移植したTPOm/mおよびTPOh/hマウスのCD33+、CD66+、CD14+細胞で測定した場合の、hTPOマウスにおける改善された多系列造血を示す。図14(a)は、移植後3〜4ヶ月のRag2-/-γc-/- TPOm/mおよびTPOh/hマウスの骨髄中および血液中のヒト骨髄細胞集団の代表的なFACS解析を示す。数字は示したゲーティングされた細胞集団のうちのパーセンテージを示す。図14(b)〜(e)は、Rag2-/-γc-/- TPOm/mおよびTPOh/hレシピエント(n=19)の骨髄における全ヒトCD45+細胞キメリズムに対するヒト骨髄細胞集団の解析を示す。図14(b)は、全骨髄集団(CD33+細胞)を提供する。図14(d)は、顆粒球(CD33+CD66hi)を提供し、図14(c)は、TPOh/hレシピエントの骨髄から精製されたhCD45+SSChiCD33+CD66hi細胞のDiff-Quick(商標)染色を示す。図14(e)は、単球(CD33+CD66loCD14+)を示す。各記号は個々のマウスを表し、水平バーは平均値を示す。図14(f)および(g)は、Rag2-/-γc-/- TPOm/mおよびTPOh/hレシピエント(n=6〜7)の血液中の全ヒトCD45+細胞キメリズムに対するヒト骨髄細胞集団の解析を示す;図14(f):顆粒球(CD66+);図14(g):単球(CD14+)。
【0169】
実施例14
hTPOを移植されたマウス:マウスおよびヒト造血幹細胞前駆細胞に対するヒト化効果
HSCならびに前駆細胞それら自体の数および機能に及ぼすヒトTPOの効果が解析された。TPOの遺伝子欠失は成体マウスではHSCの減少につながる。TPOヒト化がマウスHSCを含むと免疫表現型的に定義されるマウス集団に影響を及ぼすことができるかどうかを調べるために、非移植TPOm/m、TPOh/mおよびTPOh/h成体マウスの骨髄中の系列陰性(lineage-negative)Sca1+c-Kit+細胞のパーセンテージを比較した。
【0170】
図15は、ヒトTPOノックインマウスの骨髄中のlin-c-Kit+Sca1+細胞の減少、ならびにヒト幹細胞前駆細胞の数および自己再生能の増加を示す。図15(a)は、WT TPO (TPOm/m) Rag2-/-γc-/-マウスと比較した、非移植Rag2-/-γc-/- TPOh/mおよびTPOh/hマウスの骨髄中のマウスLin-Sca1+c-Kit+幹細胞前駆細胞のFACS解析の代表的な結果を示す。数字はLin-集団のうちのSca1+c-Kit+細胞のパーセンテージを示す。図15(b)は、(a)に示された結果の定量的解析を示す。p=0.0006(一元配置ANOVA;示したp値はTukeyの事後検定により算出した;遺伝子型あたりn=5;示された結果は2つの独立した実験を代表する)。各記号は個々のマウスを表し、水平バーは平均値を示す。図15(c)は、移植後3〜4ヶ月のRag2-/-γc-/- TPOm/mおよびTPOh/hマウスの骨髄中のヒトCD34+CD38-細胞の代表的なFACS解析を示す。数字はヒトCD45+CD34+細胞のうちのCD38-細胞のパーセンテージを示す。図15(d)は、TPOm/mおよびTPOh/hレシピエントマウス(n=43〜53)におけるヒトCD45+CD34+集団中のCD38-細胞のパーセンテージの定量的解析を示す。図15(e)は、15(d)dと同じマウスの骨髄中のヒトCD34+CD38-細胞の絶対数を示す。図15(f)および(g)は、Rag2-/-γc-/- TPOm/mおよびTPOh/hレシピエントから精製されたヒトCD45+CD34+細胞を用いるメチルセルロースコロニー形成アッセイ法を示す。図15(f)はCFU-GEMMであり、図15(g)はBFU-E(黒)、CFU-G(白)、CFU-M(灰色)およびCFU-GM(破線)である。CD34+細胞は3〜4匹のマウスのグループからプールして、レシピエントマウスの遺伝子型あたり4つの独立したプールを得た。図15(h)では、Rag2-/-γc-/- TPOm/mおよびTPOh/h一次レシピエントマウスからヒトCD45+CD34+細胞を精製し、新生仔Rag2-/-γc-/-マウスに移植し(マウスあたり100,000個の細胞)、8週間後に二次レシピエントにおけるヒトCD45+キメリズムを測定した。結果は2つの独立した実験からプールされたものである(n=7〜12一次レシピエント、n=11〜19二次レシピエント)。
【0171】
TPOm/mと比較して、TPOh/mとTPOh/hの両マウスではこれらの細胞のパーセンテージの有意な減少が観察され(図15(b))、ヒトTPOはマウス受容体に完全には交差反応性でないか、このノックイン設定ではマウス細胞に十分な量で利用されないことを示唆している。
【0172】
移植したTPOm/mおよびTPOh/h宿主の骨髄中のヒトCD34+集団が特性解析された。長期再増殖能を有するヒトHSCはLin-CD34+CD38-細胞画分に含まれる。ヒトCD45+集団のうちのCD34+細胞のパーセンテージはTPOh/hマウスでわずかに増加していた(12.39±0.79%対10.00±0.81%、平均±標準誤差、n=43〜53、p=0.037)。TPOm/mレシピエントに比較して、TPOh/hではCD34+集団内のCD38-細胞のパーセンテージに、少ないながらも(1.5倍)、統計的に有意な増加が観察された(図15(c)、(d))。全体として、これはTPOヒト化マウスにおいてCD34+CD38-細胞の絶対数の有意な増加(約2.8倍)をもたらした(図15(e))。したがって、細胞表面の免疫表現型に基づいて、ヒトTPOはHSCに非常に富むことが公知である細胞の集団を支持している。
【0173】
この細胞集団の機能的特性を検討するために、ヒトCD34+細胞をTPOm/mおよびTPOh/hマウスの骨髄から精製し、かつインビトロでメチルセルロースコロニー形成アッセイ法により評価した。CFU-GEMMは、すべての赤芽球-巨核球および骨髄細胞への分化能を少なくとも含む未熟細胞に由来する多系列骨髄系コロニーである。CFU-GEMMの形成は、少数ではあるが、TPOh/hレシピエントマウスから単離されたCD34+細胞の4つすべての試料から検出されたのに対し、TPOm/mからは1つの試料のみがCFU-GEMMを生成したにすぎなかった(図15(f))。この結果は、TPOh/hレシピエントにおける未熟なヒト造血前駆細胞の改善された維持を示している。さらに、インビボで観察された増大した骨髄分化と一致して(図14)、CFU-Mの数もまた、TPOm/mマウスに比較して、TPOh/hから単離されたヒトCD34+細胞試料において有意に高かった(培養した150,000個のCD34+細胞あたり225.0±12.25対81.25±10.80コロニー、平均±標準誤差、p=0.0001;図15(g))。
【0174】
HSCの維持および/または自己再生は機能的には二次移植の成功によって最もよく実証される。マウスにおいて連続的に生着するSCID repopulating cell(SRC)は現在、ヒトHSC機能のための信頼できるサロゲート実験基準を表す。したがって、ヒトCD34+細胞をTPOm/mおよびTPOh/h一次レシピエントの骨髄から精製し、かつRag2-/-γc-/-新生仔マウスに同様に少数(動物あたり100,000個のCD34+細胞)を移植した。二次レシピエントの骨髄を8週間後に解析した(図15(h))。TPOm/m一次レシピエントから単離されたヒトCD34+細胞は、ヒトCD45+細胞が11匹のうち2匹の二次レシピエントにしか検出されなかったので、連続的に生着する能力が非常に低かった。対照的に、TPOh/h一次レシピエントから単離されたCD34+細胞を用いて移植された19匹のうち15匹のマウスの骨髄にヒトCD45+細胞が存在していた(p=0.0012)。二次レシピエントマウスの遺伝子型は両グループ(TPOm/m)で同じだったので、この結果は、一次レシピエントにおけるヒトTPOの存在が自己再生能を高められたヒト細胞の維持に有利にはたらくことを示している。
【0175】
まとめると、これらの結果は、ホモ接合体TPOヒト化マウスがヒト造血幹細胞前駆細胞の自己再生能および多系列分化能を維持するための良好な環境を提供することを実証している。
【0176】
当業者は、本開示で明示的に記載または表現されていないが、本発明を具体化しかつ本発明の精神および範囲に含まれる、さまざまな修正を考案することができる。すべての実施例は、読者が本発明の原理と概念を理解するのを助けるために提供されるものであり、記載された特定の実施例および態様に限定されずに用いられる。本発明のすべての原理、局面、態様および実施例は、等価物が現在公知であろうと将来開発されようと、それらの等価物を包含することを意図している。本発明の範囲は本開示で示したまたは説明した態様および実施例に限定されるものではない。
【技術分野】
【0001】
発明の分野
本発明は、遺伝子改変された非ヒト動物の分野に関し、特に、RAG遺伝子ノックアウト、Il2rgIl2rg遺伝子ノックアウト、ならびにIL-3およびGM-CSF遺伝子のヒト化、ならびに任意でTPO遺伝子のヒト化を有する、免疫不全マウス;TPO遺伝子のヒト化を有するRAG/Il2rgIl2rgノックアウトマウス;ヒト造血細胞を用いて移植された、遺伝子改変されたマウス;およびヒト病原体、例えばチフス菌(Salmonella typhi)または結核菌(Mycobacterium tuberculosis)に感染する、移植されたマウスに関するものである。
【背景技術】
【0002】
背景
遺伝子改変されたマウス、改変および移植されたマウス、ならびに、例えば薬物検査の目的のための、ヒト疾患のモデル化におけるそれらの使用は、当技術分野で公知である。ヒト免疫システムをモデル化するために、遺伝子改変されたマウスを使用することが試みられてきた。その分野の概説は、Manz (2007) Human-Hemato-Lympoid-System Mice: Opportunities and Challenges, Immunity, 26:537-541(非特許文献1)に提供されており、それを参照により本明細書に組み入れる。
【0003】
これまで、特定のヒト病原体、例えばチフス菌(S. typhi)への感染性を示す遺伝子改変されたマウスが作製されたことはなかった。マウスモデルが存在する病原性感染症の場合でさえ、該モデルはヒトの特定の病態の適切にモデルとなることができないことがあり、例えば、結核菌(M. tuberculosis)のマウスモデルでは境界のはっきりした肉芽腫またはヒト免疫細胞を含む肉芽腫を形成できなかった。特定の病原体がヒトに及ぼす影響を研究するために、かつ特定の病原体に感染したヒトを治療するうえでの有効性について薬物を試験するために、そのような病原体、例えばチフス菌に感染しやすくなるように、および/または感染がヒト病態のより厳密なモデル、例えば結核菌のヒト感染症のより厳密なモデルとなるように、遺伝子改変されたマウスなどの非ヒト動物があることは有用であろう。
【0004】
一般的には、ヒト造血幹細胞の維持および増殖を支持することができる遺伝子改変されたマウスの必要性、ならびに、例えばヒト病原体に応答して、ヒト血液-リンパシステムの一部をモデル化するまたはそれに近づけることができる、移植に適したマウスの必要性が存在している。
【先行技術文献】
【非特許文献】
【0005】
【非特許文献1】Manz (2007) Human-Hemato-Lympoid-System Mice: Opportunities and Challenges, Immunity, 26:537-541
【発明の概要】
【0006】
概要
遺伝子改変された非ヒト動物が提供される。その非ヒト動物には、1つまたは複数の内在性遺伝子のノックアウトおよび1つまたは複数のヒト化遺伝子(すなわち、内在性遺伝子のヒトの類似体または相同体との、その内在性の遺伝子座での置換)を含むマウスが含まれる。
【0007】
免疫システムを取り除かれたかまたは損なわれた遺伝子改変されたマウス(例えば、照射による)、ならびにヒト造血細胞またはヒト造血幹細胞前駆細胞(hematopoietic stem and progenitor cell)(HSPC)を用いて移植されたマウスが提供される。ヒト造血細胞またはHSPCに由来するヒト細胞を含む遺伝子改変されたマウスが提供され、ヒト血液-リンパシステムを含むマウスも提供される。
【0008】
野生型マウスには感染しないヒト病原体に感染することができる、遺伝子改変、照射および移植されたマウスが提供される。ヒト病原体(例えば、結核菌)への曝露に応答して、野生型マウスには観察されない特徴(例えば、境界のはっきりした肉芽腫またはヒト免疫細胞を含む肉芽腫の形成)を有する免疫応答を開始するマウスが提供される。
【0009】
ヒト病原体の薬物耐性株を同定するための、ヒトワクチンを試験するための、および抗病原体薬物を開発して試験するための、遺伝子改変、照射および移植されたマウス、ならびにそれらを用いるための組成物および方法が提供される。
【0010】
ヒト免疫細胞を受け取りかつ増殖することができる遺伝子改変されたマウスが、ヒト造血器悪性腫瘍を保持できるマウスを含めて、提供される。
【0011】
一局面において、(a)マウスRAG遺伝子ノックアウト;(b)マウスIl2rgIl2rg遺伝子ノックアウト;および(c) (i)マウスIL-3(mIL-3)遺伝子、(ii)マウスGM-CSF(mGM-CSF)遺伝子、および(iii)マウストロンボポエチン(mTPO)遺伝子から選択される1つまたは複数の遺伝子のヒト化を含む、遺伝子改変されたマウスが提供される。
【0012】
一態様において、RAG遺伝子ノックアウトはRAG2遺伝子ノックアウトである。
【0013】
一態様において、ヒト化はmTPO遺伝子のhTPO遺伝子との置換を含む。特定の態様では、ヒト化は、mTPO遺伝子のhTPO遺伝子によるヒト化から本質的になる。
【0014】
一態様において、ヒト化はmIL-3遺伝子のヒトIL-3(hIL-3)遺伝子との置換およびmGM-CSF遺伝子のヒトGM-CSF(hGM-CSF)遺伝子との置換を含む。別の態様では、マウスがmTPO遺伝子のヒトTPO(hTPO)遺伝子との置換をさらに含む。特定の態様では、ヒト化は、mIL-3遺伝子のhIL-3遺伝子によるヒト化およびmGM-CSF遺伝子のhGM-CSF遺伝子によるヒト化から本質的になる。
【0015】
一態様において、ヒト化はmGM-CSF遺伝子のヒトGM-CSF遺伝子との置換を含み、マウスにおいてヒトGM-CSFが肝臓および循環において優勢に発現されない。一態様では、ヒトGM-CSFがマウス肺において優勢に発現される。一態様では、ヒトGM-CSFの発現が組織特異的であり、かつヒトにおける組織特異的発現を反映している。
【0016】
一態様において、遺伝子改変されたマウスは、マウスに存在しうる内在性造血細胞を排除するように処置される。一態様では、その処置が遺伝子改変されたマウスを照射することを含む。特定の態様では、遺伝子改変されたマウスの新生仔が亜致死的に照射される。特定の態様では、新生仔が2×200cGyを4時間間隔で照射される。
【0017】
一態様において、遺伝子改変および処置されたマウスは、遺伝子改変および移植されたマウスを形成するために、ヒト造血細胞またはヒト造血幹細胞(HPSC)を用いて移植される。一態様では、造血細胞がヒト臍帯血細胞およびヒト胎児肝細胞から選択される。一態様では、移植に約1〜2×105個のヒトCD34+細胞を用いる。
【0018】
一態様において、遺伝子改変および移植されたマウスは、以下から選択されるヒト細胞を生じさせる:CD34+細胞、造血幹細胞、造血細胞、骨髄前駆細胞、骨髄細胞、樹状細胞、単球、顆粒球、好中球、肥満細胞、胸腺細胞、T細胞、B細胞、血小板、およびこれらの組合せ。一態様では、ヒト細胞が移植後4ヶ月、5ヶ月、6ヶ月、7ヶ月、8ヶ月、9ヶ月、10ヶ月、11ヶ月、または12ヶ月の時点で存在している。
【0019】
一態様において、遺伝子改変および移植されたマウスは、ヒト造血幹細胞前駆細胞、ヒト骨髄前駆細胞、ヒト骨髄細胞、ヒト樹状細胞、ヒト単球、ヒト顆粒球、ヒト好中球、ヒト肥満細胞、ヒト胸腺細胞、ヒトT細胞、ヒトB細胞、およびヒト血小板を含むヒト血液-リンパシステムを生じさせる。一態様では、ヒト血液-リンパシステムが移植後4ヶ月、5ヶ月、6ヶ月、7ヶ月、8ヶ月、9ヶ月、10ヶ月、11ヶ月、または12ヶ月の時点で存在している。
【0020】
一態様において、遺伝子改変および移植されたマウスは、ヒト細胞が介在する炎症反応を示す。特定の態様では、そのヒト細胞がマクロファージである。特定の態様では、マクロファージ介在炎症反応が肺胞マクロファージによって介在される。特定の態様では、肺胞マクロファージが介在する炎症反応は肉芽腫の形成を含む。特定の態様では、肉芽腫がヒト免疫細胞を含む。特定の態様では、肉芽腫が明確に組織化された肉芽腫である。特定の態様では、肉芽腫がマイコバクテリア、例えば結核菌への曝露後に形成される。一態様では、マウスが2つまたはそれ以上の肉芽腫を含む免疫反応を示す。一態様では、遺伝子改変および移植されたマウスがヒト結核菌感染のためのモデルである。
【0021】
一態様において、遺伝子改変および移植されたマウスは、ヒト免疫細胞を含む肉芽腫の形成によって少なくとも一部特徴づけられる結核菌感染を含む。特定の態様では、その肉芽腫が明確に組織化された肉芽腫である。特定の態様では、結核菌がヒト集団に感染する結核菌の薬物耐性株または多剤耐性株である。一態様では、結核菌に感染したマウスが肺に肉芽腫を含む。特定の態様では、その肉芽腫が十分に発達した肉芽腫である。特定の態様では、肺の肉芽腫がヒト免疫細胞を含む。特定の態様では、肉芽腫のヒト免疫細胞が活性化ヒトマクロファージ、活性化ヒトT細胞、およびこれらの組合せから選択される。
【0022】
一態様において、遺伝子改変および移植されたマウスは、IL-3、GM-CSFおよびTPO遺伝子の1つまたは複数のヒト化を欠く移植されたマウスに比べて、増強された粘膜免疫を示す。特定の態様では、増強された粘膜免疫がインフルエンザA感染後にインターフェロンβ(IFNβ)の高度発現を含む。
【0023】
一態様において、遺伝子改変および移植されたマウスは、結核菌感染およびチフス菌感染から選択される感染を含む。一態様では、マウスは、チフス菌または結核菌を繁殖させる。一態様では、マウスはヒト病原性マイコバクテリアに対する抗マイコバクテリア免疫応答を開始し、該免疫応答は、ヒト免疫細胞によって媒介されかつヒト免疫細胞を含む肉芽腫の形成を含む。特定の態様では、その肉芽腫が十分に発達した肉芽腫である。
【0024】
一態様において、遺伝子改変および移植されたマウスはmTPO遺伝子のヒト化を含み、hTPOを移植されたマウスを形成する。一態様では、hTPOを移植されたマウスは、mTPO遺伝子を含むがhTPO遺伝子を含まない移植されたマウスに比べて、骨髄でのヒト骨髄細胞の増加を示す。特定の態様では、ヒト骨髄細胞がhTPO遺伝子を欠く移植されたマウスに対して1.5倍、2倍、2.5倍または3倍に増加する。特定の態様では、顆粒球の増加が約1.5倍、2倍、2.5倍または3倍である。別の態様では、hTPO遺伝子を欠く移植されたマウスに比して、末梢血単球の増加が認められ、この場合の末梢血単球の増加は約1.5倍、2倍、2.5倍または3倍である。一態様では、遺伝子改変され移植されたマウスは、mTPO遺伝子を置換するhTPO遺伝子から本質的になるヒト化を含み、該マウスはマウスTPOを発現せずにヒトTPOを発現する。
【0025】
一局面において、Rag遺伝子のノックアウト、Il2rgIl2rgノックアウト、およびTPOのヒト化を含む、遺伝子改変および移植されたマウスが提供され、該マウスはヒト造血幹細胞またはヒト免疫細胞を用いて移植され、かつ初期ヒト造血細胞から発生するヒト造血器悪性腫瘍を含む。特定の態様では、その悪性腫瘍が骨髄性白血病および骨髄増殖性腫瘍から選択される。
【0026】
一態様において、マウスは、ヒトIL-3遺伝子およびヒトGM-CSF遺伝子、ならびに内在性マウスIL-3遺伝子のノックアウトおよび内在性マウスGM-CSF遺伝子のノックアウトをさらに含む。
【0027】
一局面においては、RAG遺伝子ノックアウト、Il2rg遺伝子ノックアウト、およびマウス骨髄細胞に対して競合的優位性をもつヒト骨髄細胞を提供する遺伝子改変を含むマウスが提供される。一態様では、その遺伝子改変がマウス骨髄細胞の発達および/または維持に必要とされるマウス遺伝子の対応ヒト遺伝子との置換である。一態様では、遺伝子改変がマウスIL-3遺伝子のヒトIL-3遺伝子との置換、マウスGM-CSF遺伝子のヒトGM-CSF遺伝子との置換、およびこれらの組合せから選択される。一態様では、マウスは、内在性のマウス造血細胞を欠くかまたは実質的に欠き、かつヒト造血細胞を含む。
【0028】
一局面においては、ヒト病原体に感染可能なマウスの作製方法が提供され、該方法は、本明細書に記載した通りにマウスを遺伝子改変しかつ該マウスに移植する段階、遺伝子改変および移植されたマウスをヒト病原体に曝露する段階、ならびにヒト病原体が該マウスに感染するのに十分な条件下で該マウスを維持する段階を含む。一態様では、ヒト病原体が結核菌およびチフス菌から選択される。一態様では、ヒト病原体が遺伝子改変を欠くマウスには病原性でないヒト病原体である。一態様では、ヒト病原体が遺伝子改変を欠くマウスには感染しないヒト病原体である。
【0029】
一局面においては、ヒト病原体に対する薬物の効果を判定する方法が提供され、該方法は、本明細書に記載した通りに遺伝子改変および移植されたマウスをヒト病原体に曝露する段階、ならびに該病原体を該マウスに感染させて、薬物の存在下および非存在下で経時的に感染のパラメーターを測定する段階を含む。一態様では、ヒト病原体が遺伝子改変を欠くマウスには感染しない病原体である。一態様では、ヒト病原体が結核菌およびチフス菌から選択される。一態様では、マウスが既知の感染単位数のヒト病原体に曝露され、感染のパラメーターがマウスの体液中または組織中のヒト病原体の感染単位数である。
【0030】
一態様において、感染のパラメーターはマウスの体液中の力価である。一態様では、感染が結核菌感染およびチフス菌感染から選択される。特定の態様では、感染が結核菌感染であり、かつパラメーターが肉芽腫の形成である。特定の態様では、肉芽腫が肺肉芽腫である。別の特定の態様では、肉芽腫が境界のはっきりした肉芽腫である。
【0031】
一局面において、(a)マウスRAG遺伝子ノックアウト;(b)マウスIl2rg遺伝子ノックアウト;および(c) (i)マウスIL-3(mIL-3)遺伝子および(ii)マウスGM-CSF(mGM-CSF)遺伝子のヒト化を含む、遺伝子改変されたマウスが提供され;該マウスは、内在性マウス造血細胞を取り除くために照射された後およびヒト造血幹細胞を移植された後に、そのヒト造血幹細胞を維持し、分化した機能的ヒト免疫細胞を含むヒト免疫細胞集団をヒト造血幹細胞から発生させる。分化したヒト免疫細胞には、ヒト骨髄前駆細胞、ヒト骨髄細胞、ヒト樹状細胞、ヒト単球、ヒト顆粒球、ヒト好中球、ヒト肥満細胞、ヒト胸腺細胞、ヒトT細胞、ヒトB細胞、およびヒト血小板が含まれる。別の局面において、マウスは(iii)マウストロンボポエチン(mTPO)遺伝子のヒト化をさらに含む。
【0032】
一態様において、マウスは、ヒトの免疫細胞の集団と同程度細胞型が多様であるヒト免疫細胞の集団を維持する。一態様では、ヒト免疫細胞が移植後に少なくとも4ヶ月、5ヶ月、6ヶ月、7ヶ月、8ヶ月、9ヶ月、10ヶ月、11ヶ月、または12ヶ月間維持される。
【0033】
一態様において、マウスは、ヒト病原体またはヒト病原体の抗原に曝露されると、該病原体に曝露されたヒトの感染のモデルとなる細胞性および/または体液性の免疫応答を開始する。一態様では、ヒト病原体が野生型マウスには感染しない病原体である。別の態様では、ヒト病原体が野生型マウスに感染する病原体であり、感染後の野生型マウスは、ヒトがその病原体に応答して開始する免疫応答をモデル化したものではない。一態様では、病原体がウイルス、マイコバクテリア、真菌、または細菌である。特定の態様において、病原体はヒトまたはブタまたはトリのインフルエンザウイルス、チフス菌、または結核菌である。
【0034】
本発明のさらなる応用および態様は、本開示を読むことによって当業者には明らかになるであろう。
【図面の簡単な説明】
【0035】
【図1】感染後10日の、移植したRAG KO, Il2rg KOマウスとRAG KO, Il2rg KO, hIL-3/hGM-CSFマウスにおけるチフス菌感染を示す。実験グループ:(1)移植:n=9 (4m/m、5h/m);血液中の生着率=6.5〜16.7%;(2)対照:n=8;血液中の生着率=0.04〜0.4%(フローサイトメトリーのバックグラウンドを反映する。対照マウスには移植しなかった)。
【図2】1×103個のチフス菌の感染後1週の、移植したRAG KO, Il2rg KOマウスの脾臓におけるチフス菌感染を示す。
【図3】1×104個のチフス菌の感染後4週の、移植したRAG KO, Il2rg KOマウスの脾臓および肝臓におけるチフス菌感染を示し、該マウスはCD34陽性胎児肝細胞を用いて移植された。
【図4】1×104個のチフス菌の感染後4週の、RAG KO, Il2rg KOマウスの胆のうにおけるチフス菌感染を示し、該マウスはCD34陽性胎児肝細胞を用いて移植された。
【図5】図5(a)〜(d)は、非移植hIL-3/hGM-CSFマウスにおけるhGM-CSF発現の検証試験の結果を示す。図5(e)は、マウスIL-3/GM-CSF遺伝子座でのヒト化戦略を示す。
【図6】図6(a)〜(e)は、移植されたヒト化(hIL-3/hGM-CSF)マウスの肺検査の結果を示す。図6(f)、(g)は、活性化脾細胞によるマウスおよびヒトのIL-3(f)およびGM-CSF(g)産生のELISA結果を示す。
【図7】図7(a)は、非移植または移植m/mもしくはh/h KIマウスからの肺組織切片のPAS染色、図7(b)は、非移植(non)または移植h/h KIマウスもしくはm/m対照マウス(グループあたりn=6)からのBAL液中の総タンパク質の定量化を示す。
【図8】図8(a)は、マウスHprtに対して正規化されたヒトHprtの発現、図8(b)は、マウスHprtに対して正規化されたヒトIFNγの発現、図8(c)は、ヒトHprtに対して正規化されたヒトIFNγの発現を示す。図8(d)は、定常状態での移植hIL-3/GM-CSF h/m KIマウスからのヒト骨髄細胞のフローサイトメトリー解析、図8(e)は、LPSの2回腹腔内注射後72時間のCB移植したm/mまたはh/m KIマウスからのヒト血液細胞のフローサイトメトリー解析、図8(f)は、LPS注射後72時間の移植したm/mまたはh/m KIマウスにおけるヒトCD14+血液細胞の頻度、図8(g)は、1回目(上)および2回目(下)のLPS注射後2〜3時間の移植したm/mまたはh/m KIマウスからの血清中のヒトIL-6のELISAを示す。
【図9】図9(a)は、肺におけるヒトT細胞(hCD45+hCD3+)の頻度、図9(b)は、肺におけるヒトCD4およびCD8 T細胞の分布、図9(c)肺におけるヒトCD4対CD8 T細胞の比率、図9(d)は、BCG感染後4週のBALB/cマウス、移植m/mマウス、および移植h/m KIマウスからの脾細胞のフローサイトメトリー解析、図9(e)は、BCG感染後4週のBALB/cマウス、非移植(non)m/mマウス、移植m/mマウス、および移植h/m KIマウスからの肺組織におけるヒトIFNγ(左)およびTNFα(右)遺伝子発現の定量的RT-PCR解析を示す。図9(f)は、非移植m/mまたはh/h KIマウスからのBAL細胞のDiffQuick(商標)染色;倍率400x、図9(g)は、非移植m/mまたはh/h KIマウスからの肺組織切片のPAS染色;倍率400xを示す。
【図10】図10(a)は、BCG感染後4週の移植h/m KIマウスからの肺組織切片のヘマトキシリンおよびエオシン(H&E)染色;倍率100x(左)および200x(右)、図10(b)は、BCG感染後4週の移植h/m KIマウスからのヒトCD45、CD3またはCD68を染色した肺組織切片;倍率200xを示す。
【図11】図11(a)は、Rag2+/-γcY/- TPOh/mマウスの異なる組織におけるマウスTPO(mTpo)およびヒトTPO(hTPO)発現のRT-PCR解析、図11(b)は、Rag2-/-γc-/- TPOm/m、TPOh/mおよびTPOh/hマウスの肝臓、腎臓および間葉系多能性間質細胞(MSC)におけるmTpoおよびhTPO発現のRT-PCR解析、図11(c)は、TPOm/m、TPOh/mおよびTPOh/hマウスの血清中の、ELISAにより測定されたマウスおよびヒトTPOタンパク質の濃度を示す。図11(d)は、mTPO遺伝子をhTPO遺伝子と置き換えるためのターゲティング構築物を示す。
【図12】図12(a)は、ヒトCD34+細胞の移植後3〜4ヶ月のRag2-/-γc-/- TPOm/mおよびTPOh/hマウスの骨髄中のヒトおよびマウスCD45+細胞のFACS解析、図12(b)は、移植後3〜4ヶ月(左)または6〜7ヶ月(右)の骨髄中のヒトCD45+細胞のパーセンテージ、図12(c)は、図12(b)と同じ動物の骨髄中のヒトCD45+細胞の絶対数を示す。図12(d)は、臍帯血(CB)または胎児肝臓(FL)から分離したヒトCD34+細胞を用いて移植されたRag2-/-γc-/- TPOm/mおよびTPOh/hマウスの骨髄中のヒトCD45+細胞のパーセンテージを示す。
【図13】図13(a)は、非移植Rag2-/-γc-/- TPOm/m、TPOh/mおよびTPOh/h成体マウスの血液中の血小板数、図13(b)は、移植後3〜4ヶ月のRag2-/-γc-/- TPOm/mおよびTPOh/hマウスの血液中のマウス(mCD61+)およびヒト(hCD41a+)血小板の代表的FACS解析、図13(c)は、TPOm/mおよびTPOh/hマウスにおける、FACSにより測定されたヒト血小板のキメリズム、図13(d)、(e)は、TPOm/mおよびTPOh/hレシピエントの血液中のマウス(mCD61+, 20d)およびヒト(hCD41a+, 20e)血小板のカウント数、図13(f)は、骨髄中のヒトCD45+細胞のうちのヒト巨核球パーセンテージ(CD41a+)を示す。図13(g)、(h)は、Rag2-/-γc-/- TPOm/mおよびTPOh/hマウスの血液および脾臓中のヒトCD45+細胞のパーセンテージを示す。図13(i)は、移植TPOm/mおよびTPOh/hレシピエントの胸腺の全細胞量を提供する。
【図14】図14(a)は、移植後3〜4ヶ月のRag2-/-γc-/- TPOm/mおよびTPOh/hマウスの骨髄中および血液中のヒト骨髄細胞集団のFACS解析、図14(b)は、全骨髄集団(CD33+細胞)、図14(c)は、顆粒球(CD33+CD66hi)、図14(d)は、TPOh/hレシピエントの骨髄から精製されたhCD45+SSChiCD33+CD66hi細胞のDiffQuick(商標)染色、図14(e)は、単球(CD33+CD66loCD14+)、図14(f)、(g)は、Rag2-/-γc-/- TPOm/mおよびTPOh/hレシピエントの血液中の全ヒトCD45+細胞キメリズムに対するヒト骨髄細胞集団の解析を示す。図14(f) 顆粒球(CD66+)、図14(g) 単球(CD14+)。
【図15】図15(a)は、WT TPO (TPOm/m) Rag2-/-γc-/-マウスと比較した、非移植Rag2-/-γc-/- TPOh/mおよびTPOh/hマウスの骨髄中のマウスLin-Sca1+c-Kit+幹細胞前駆細胞のFACS解析、図15(b)は、図15(a)に示された結果の定量的解析、図15(c)は、移植後3〜4ヶ月のRag2-/-γc-/- TPOm/mおよびTPOh/hマウスの骨髄中のヒトCD34+CD38-細胞のFACS解析、図15(d)は、TPOm/mおよびTPOh/hレシピエントマウスにおけるヒトCD45+CD34+集団中のCD38-細胞のパーセンテージの定量的解析、図15(e)は、図15(d)と同じマウスの骨髄中のヒトCD34+CD38-細胞、図15(f)、(g)は、Rag2-/-γc-/- TPOm/mおよびTPOh/hレシピエントから精製されたヒトCD45+CD34+細胞を用いるメチルセルロースコロニー形成アッセイ法を示し、図15(f)は、CFU-GEMM、図15(g)は、BFU-E(黒)、CFU-G(白)、CFU-M(灰色)およびCFU-GM(破線)、図15(h)は、Rag2-/-γc-/- TPOm/mおよびTPOh/hマウス由来のヒトCD45+CD34+細胞の新生仔Rag2-/-γc-/-マウスへの二次移植におけるヒトCD45+キメリズムである。
【発明を実施するための形態】
【0036】
詳細な説明
本発明は、記載した特定の態様に限定されるものではなく、認められる特許請求の範囲によって説明されるものである。
【0037】
特に指定のない限り、本明細書中で用いるすべての技術用語および科学用語は、本発明が属する分野の当業者が一般に理解している意味と同じ意味を含む。記載したものと類似のまたは同等の方法および材料はどれも、本発明を実施または使用する際に用いることができるが、ここでは特定の態様、方法および材料が記載される。記載したすべての刊行物は参照により本明細書に組み入れられる。矛盾が存在する場合には、組み入れられた刊行物のあらゆる開示に本開示が優先する。
【0038】
単数形「1つの(a)」、「1つの(an)」および「その(the)」は、文脈上明確に別途示されない限り、複数形の照応を含むものである。したがって、例えば、「遺伝子(a gene)」への言及は複数のそのような遺伝子を包含し、また、「遺伝子ノックアウト(the gene knockout)」への言及は1つまたは複数のノックアウトおよびその同等物への言及を包含する。
【0039】
ヒト免疫細胞を支持する改変されたマウス:hIL-3/GM-CSFマウス
ヒト免疫システムの成分を含むマウス(HISマウス)は、インビボでのヒト免疫システムの研究に、ヒトワクチンの試験に、およびヒトの疾患および障害を治療するための薬物の試験と開発にかなり有望である。HISマウスは、重症免疫不全マウス株(例えば、組換え活性化遺伝子2(Rag2)ノックアウト(KO)インターロイキン2受容体γ(Il2rg)KOマウス)にヒト造血幹細胞前駆細胞を移植することによって作製される。非ヒト霊長類に比べて、HISマウスは小型動物モデルの利点を有しており、すなわち、それらはより多目的な実験を可能にし、研究団体にとってよりアクセス可能であり、また、ヒト被験者を用いて実験を行うより倫理的に許容される。最も重要なことは、HISマウスから得られた実験結果がヒトへの関連性が高く、ヒトに応用できる可能性があることである。なぜならば、ヒト特異的病原体を用いた感染、ならびにヒト特異的免疫応答および免疫病理学の研究が今や実現可能になりつつあるからである。
【0040】
多くの進展が近年なされてきたが、現在のHISマウスモデルには、ヒト骨髄細胞とT細胞の発達、維持および機能が低いといった、いくつかの主要な制限がある。その結果、粘膜表面でのヒト炎症および免疫反応、または遅延型過敏症(DTH)などの強いヒトT細胞性応答がめったに観察されていない。それゆえ、現在のHISマウスは、深刻なヒト病原体である結核菌に起因する感染症および病理の研究に適していない。実際、肉芽腫、特に、マイコバクテリアに対するヒト免疫応答の顕著な特徴である、ヒト細胞を含む肉芽腫は、これまでHISマウスにおいて報告されていない(例えば、Manz et al. (2009) Renaissance for mouse models of human hematopoiesis and immunobiology, Nat. Immunol. 10:1039-1042を参照されたい)。
【0041】
現在のHISマウス宿主は、少なくとも一部には現在のHISマウス宿主がヒト細胞に適した生理的環境を提示しないので、一部の感染症のモデル化には適していない。数種のマウスサイトカイン、例えばIL-3とGM-CSFはヒトコグネイト受容体に作用しない。さらに、Rag2 KO Il2rg KOマウスは完全なままのマウス骨髄区画を持っており、ヒト骨髄細胞は宿主細胞に対して競合的に不利である可能性がある。これらの制限を克服するために、本開示は、マウスのサイトカインをコードする遺伝子がヒトの対応遺伝子によって置き換えられたヒトサイトカインノックイン(knock-in)マウスの作製について記載する。サイトカイン置換の基準は次のとおりである。(1)マウスサイトカインがヒト細胞に作用しないかまたは弱く作用する。(2)ヒトサイトカインが、ヒト細胞に競合的優位性を与えるために、マウス細胞に作用しないかまたは弱く作用する。(3)ヒトサイトカインが造血(移植)細胞によって排他的に産生されない。(4)マウスサイトカインの欠乏がマウス宿主に致命的ではないか、またはヒトKIサイトカインがマウスKO表現型をレスキューするために十分に交差反応性である。KI戦略は適切な器官における生理学的濃度での忠実な発現を可能にすべきである。重要なことは、ホモ接合体KIマウスでは、ヒトコグネイト受容体を発現する細胞がそれぞれのマウス細胞に対して競合的優位性を獲得する必要があることである。
【0042】
IL-3およびGM-CSFは、骨髄細胞の発達と機能に欠かせない2種のサイトカインである。どちらのサイトカインもヒトとマウスの間で交差反応性がない。IL-3はインビトロで初期造血前駆細胞を刺激するが、インビボでの定常状態の造血にはなくても困らない。しかし、GM-CSFと一緒になって、それはインビボで効果的なDTH応答に必要である。IL-3はまた、インビトロで肺胞マクロファージ(AM)の増殖を特異的に刺激する。GM-CSFは肺で高度に発現されており、インビボでは肺の恒常性に重要である。このことは、GM-CSF KOマウスが、表面活性物質のクリアランスの欠陥のため肺にタンパク質が蓄積することによって特徴づけられる肺胞タンパク症(PAP)を発症するという事実により証明される。GM-CSF KOマウスからの肺胞マクロファージは最終分化において欠陥を有し、これは肺の病原体に対する先天性免疫の低下につながる。GM-CSFはまた、インビトロでヒトAMの増殖を刺激する。IL-3と同様に、GM-CSFは定常状態の造血にほとんど不必要であり、同じことが両サイトカインを欠くマウスにあてはまる。対照的に、GM-CSFはマクロファージによる炎症性サイトカインの産生および単球の動員と補充などの炎症反応に必要である。GM-CSFは結核菌などを含む広範な病原体に対する防御免疫のためにも不可欠である。特に、結核菌に感染したGM-CSF KOマウスは、結核の顕著な特徴である肉芽腫を発症することがない。
【0043】
本開示は、少なくとも一部には、hIL-3/GM-CSF KIマウスを作製することがヒト骨髄細胞の再構成および機能ならびにマウスでの病原体に対するヒト先天性免疫応答を支持する上で価値があるであろうという認識に基づいている。そのようなKIマウスを用いて得られた、本開示に記載された結果は、この戦略がヒト骨髄細胞の発達、ヒト肺粘膜免疫、かつさらにマイコバクテリア感染後の肉芽腫形成の観点からHISマウスの現在のモデルに比べて大幅な改善を与えることを実証する。そのようなマウスのこれらのおよび他の有益な特性は本開示の他の個所で説明される。
【0044】
マウスにおいてインビボ設定でヒト組織を研究できることは、広範囲の可能な研究手段を開放している。主要な制限事項がそのアプローチの適用を妨げており、それらのうち最も重要な欠陥の1つは、マウス因子がヒト細胞を支持できないことである。実際、免疫システムでは、ヒト免疫細胞の発達と機能に必要な多くの必須因子は種特異的であり、かつマウスによって効果的に提供され得ない。したがって、マウス遺伝子をそれらのヒト対応物と置き換えて、ヒト細胞の良好な発達と機能を可能にし、かつ対応するマウス細胞のそれを潜在的に不可能にするという戦略に従うことが決定された。この概念をヒトサイトカインKIマウスに適用することによって、マウス宿主の免疫遺伝子のヒト遺伝子との置換はHISマウスを改善するという概念の証明が本明細書に提供される。具体的には、本開示は、マウスとヒトの間の不適切なサイトカイン交差反応性、およびマウス細胞と競合せざるを得ないことが、現在のHISマウスにおけるヒト骨髄細胞の生着と機能を実際に制限しているという見解を支持する。
【0045】
ヒトサイトカインは、静脈内注射によってHISマウスに投与することができ、例えばIL-15/IL-15Rα複合体およびIL-7の注射によって、それぞれヒトNK細胞とT細胞の再構成を高めることができる。別のアプローチは、肝臓での一過性の発現につながるヒトサイトカインを発現するプラスミドDNAのハイドロダイナミックインジェクションである。このアプローチはごく最近、GM-CSFとIL-4のハイドロダイナミックデリバリーによってヒトDCの再構成を改善するために使用された(Chen et al. (2009) Expression of human cytokines dramatically improves reconstitution of specific human-blood lineage cells in humanized mice, Proc Natl Acad Sci USA, 106(51):21783-21788を参照されたい)。本開示とは対照的に、骨髄細胞の機能的応答または病原体に対するインビボ応答はこれらのマウスではまったく報告されなかった。最後に、ヒトサイトカインはまた、HISマウスにおいて導入遺伝子として過剰発現させることもできる。このアプローチは、ヒトIL-3/GM-CSF/幹細胞因子(SCF)トランスジェニック(tg)マウスを作製するために採用された(Nicolini et al. (2004) NOD/SCID mice engineered to express human IL-3, GM-CSF and Steel factor constitutively mobilize engrafted human progenitors and compromise human stem cell regeneration, Leukemia 18:341-347を参照されたい)。これらのマウスでは、ヒトサイトカインの発現が遍在的発現をもたらすサイトメガロウイルスプロモーターによって駆動される。しかしながら、hIL-3/GM-CSF/SCF tg HISマウスは、骨髄におけるヒト造血幹細胞の維持の低下および終末骨髄造血の増加によって妨げられる。この場合も、本開示とは相違して、改善された骨髄細胞の機能またはインビボ応答は記載されなかった。対照的に、本明細書に記載するシステムでは、定常状態および炎症での標的遺伝子の生理的発現が適切な細胞型のみの適切な発達と機能を可能にする。重要なことは、本開示で説明するアプローチが、高度に再現可能な条件下での維持および増殖が可能であり、かつ研究のために世界中で利用できる、マウスの系統を作製することである。
【0046】
本開示で説明するhIL-3/GM-CSF KIマウスは、以前のHISマウスおよび上述した代替アプローチと比べてかなりの改善を示す。第一に、本明細書に記載のKI戦略によるヒトIL-3およびGM-CSFの送達は長期的なサイトカインの発現につながり、これは高価なサイトカインの反復注射の必要性を回避する。第二に、IL-3とGM-CSFが通常発現されている器官での忠実な発現が達成される。生理的条件下では、GM-CSFは主に肺で発現される(図5a)。対照的に、ハイドロダイナミックデリバリーは肝臓および循環における優勢発現をもたらす。どちらの器官においても、GM-CSFは定常状態の条件下では発現されない。第三に、ハイドロダイナミックインジェクションによる送達またはhIL-3/GM-CSF/SCF tgマウスにおける遍在的な過剰発現とは対照的に、KIマウスではIL-3とGM-CSFの生理的量が発現される。GM-CSFの生理的レベルは結核菌に対する防御免疫応答に重要であることが実証されている。こうして、肺でのGM-CSFの局所過剰発現を有するトランスジェニックマウスは、肉芽腫形成不全および結核菌に対する感受性の増加を示す。同様に、GM-CSFの静脈内投与もまた、マウスにおける結核菌感染の防御低下をまねく。第四に、ホモ接合体hIL-3/GM-CSF KIマウスは、マウスIL-3およびGM-CSFがホモ接合体マウスでは発現されないので、マウス骨髄区画の同時損傷を可能にする。これは、本開示で示す通り、ヒト骨髄細胞に競合的優位性をもたらす。
【0047】
結核菌感染が原因で起こる結核では、年間170万人が死亡している。したがって、新規の有効な予防対策および治療対策が緊急に必要とされている。マウスは結核菌に感染し得るが、それらはヒト結核の理想的なモデルを表すものではない。それは結核菌に対する免疫応答の種特異的な差異のためである。例えば、感染したマウスは明確に組織化された肉芽腫を発症しない。肉芽腫は、結核に罹ったヒトの免疫反応の顕著な特徴であり、かつ融合して類上皮および多核巨細胞を形成する活性化マクロファージと活性化T細胞とを含む。肉芽腫は、細菌の複製を制限する上で、さらにマイコバクテリアの広がりを制御する上で重要な役割を果たしている。GM-CSFはインビトロでAMの多核巨細胞への分化を促進する。トランスジェニックマウスによる研究でも、インビボで多核巨細胞を形成するためのマクロファージの融合におけるGM-CSFの役割が明らかにされた。さらに、GM-CSFはマイコバクテリア感染後の肉芽腫形成に不可欠である。結核菌に感染したGM-CSF KOマウスにおける肉芽腫の不在は、細菌複製の増加と生存の低下に関連づけられる。最後に、GM-CSFシグナル伝達の欠陥に起因するPAPを患うヒトは、マイコバクテリア感染に対し増加した感受性を示す。
【0048】
ヒト抗マイコバクテリア免疫反応、特にヒト細胞による肉芽腫の形成は、HISマウスで以前に報告されたことがなかった。それはおそらくヒトマクロファージとT細胞の弱い応答によるものである。本開示では、マイコバクテリアに対する抗原特異的T細胞応答が、ヒト細胞を用いて移植されたマウスのサブセットにおいて検出された。さらに、肉芽腫の生物学におけるGM-CSFの顕著な役割を考えると、移植したhIL-3/GM-CSF KIマウスは肉芽腫形成を支持するための優れた宿主であることが仮定される。これは確かに、少なくともBCGを感染させたマウスのサブセットではそうであった。重要なことは、これらのマウスの肺肉芽腫が、マウス肉芽腫に特有のゆるい構成を持っていたものの、その肉芽腫がヒトT細胞とヒトマクロファージを含んでいたことである。今後の取り組みは、HISマウスにおいてヒトT細胞およびマクロファージの応答をさらに高めることを目指すべきである。これは、インビボで結核菌に対するヒト免疫応答の研究を可能にする小型動物モデルの開発につながるはずである。hIL-3/GM-CSF KIマウスはまた、インビボでヒト免疫応答を研究するための他の設定でも有用である可能性がある。それには、肺病原体による感染、自己免疫、およびヒト癌が含まれる。要約すると、本開示で提供されるhIL-3/GM-CSF KIマウスは、今後の研究のための多目的ツールとして役に立ち得る、かなり改善されたHISマウスモデルに相当する。
【0049】
ヒト免疫細胞を支持する改変されたマウス:hTPO
造血幹細胞(HSC)は2つの主な特性によって特徴づけられる:生涯にわたる自己再生能、およびすべての成熟した造血系細胞への分化能。HSCプールの恒常性を確保するために、細胞分裂時に、HSCは一方の機能的HSCを生成するが、他方の子孫細胞は高度に組織化された分化と細胞増殖のプログラムを受け、その間に複数の系列に方向づけされた前駆細胞、および最後には最終分化細胞が生成されると考えられる。
【0050】
マウスの造血は過去数十年間にわたり広く研究されており、インビボで幹細胞と前駆細胞とに非常に富んでいる、免疫表現型的に定義された細胞集団の同定および機能解析につながった。しかし、ヒト造血の見込みのある実験的インビボ研究は、明白な実際上および倫理上の制約によって制限されている。
【0051】
この制限を回避するために、ヒト造血のインビボ研究用の異種移植モデルがいくつか開発されている。これらのうち、免疫不全マウスへのヒト造血細胞の移植が実験的造血研究室において広く確立されている。今日最もよく用いられるモデルは、BALB/c Rag2-/-γc-/-またはNOD-SCIDγc-/-系統のマウスに依存している。両系統ともB細胞、T細胞およびNK細胞を欠いていて高度に免疫不全であり、それらの遺伝的背景はヒト造血移植および分化に対して寛容である。ヒトCD34+造血幹細胞前駆細胞を移植すると、大部分のヒト造血集団(B細胞、T細胞、単球、樹状細胞、赤血球および血小板を含む)は発達することができ、これらのモデルにおいて検出可能である。しかし、こうしたキメラな動物では、一般にヒト細胞の最大80%に相当する初期に高いB細胞数を伴うリンパ球の発達への偏りが見られ、骨髄単球の発達はわずかである。かつ、生着レベルは通常、移植後4〜6ヶ月で下降し始める。さらに、ヒト細胞のマウスへの異種移植には、マウス造血幹細胞前駆細胞のマウスへの、またはヒト細胞のヒトへのそれぞれ最適な移植に十分な数と比較して、多数の細胞の移植が必要である。さらに、マウスレシピエントに移植されたマウスHSCとは対照的に、ヒトHSCは異種マウス環境では増えることがなく、維持もされない。したがって、マウスの背景はヒトHSCの生理学を研究するための最適な環境を提供しない。それは、HSCの機能と維持を支持するのに必要な、増殖因子の非存在または限られた交差反応性が原因である可能性がある。
【0052】
トロンボポエチン(TPO)は、巨核球および血小板の発達を促進する成長因子として最初に同定された。TPOは、肝臓と腎臓で構成的に産生され、かつ血液循環に放出される。TPOの受容体であるc-Mplは骨髄中の造血幹細胞前駆細胞によって発現される。c-Mplは循環している血小板にも発現される。しかしながら、血小板へのTPOの結合はいかなるシグナル伝達経路をも活性化しない。かくして、血小板はTPOのためのシンクまたはスカベンジャーとして作用し、この機構を介して血小板産生の負の調節に寄与している。その後、TPOはHSCの増殖と自己再生を支持するその重要な機能について認識されてきた。TPOの欠乏は成体マウスにおいてHSC数の減少につながり、かつTPOの存在は静止期の成体HSCの維持に必要である。さらに、TPOは、照射した宿主の造血区画を補充するのに必要な、HSCの移植後増殖を支持するために必要である。興味深いことに、骨髄でのHSCニッチの形成に関与する骨芽細胞は、HSCの機能と維持に重要なTPOを産生することが実証されている。
【0053】
マウスおよびヒトTPOは、インビトロにおいて超生理的用量で用いた場合、それぞれのコグネイト受容体に対して双方交差反応性であるが、そのサイトカインがインビボ環境において生理的制限用量で作用する場合には、親和性と生物学的活性が異なることがある。こうして、マウスTPOはインビボでヒトc-Mpl受容体に適切な刺激を与えない可能性があり、それゆえにマウス環境でヒトHSCの特性が損なわれる主な原因となり得る。この潜在的欠陥を修正するために、Rag2-/-γc-/-マウスにおいてマウスTPOをコードする遺伝子がそのヒト対応物により置き換えられた。
【0054】
本開示は、少なくとも一部には、RAG2-/-γc-/-背景でhTPO KIマウスを作製することが、該マウスそれ自体およびhIL-3/GM-CSFマウスと交配させた該マウスの子孫において、ヒト骨髄細胞の再構成と機能ならびにマウスでの病原体に対するヒト先天性免疫応答を支持する上で価値があるという認識に基づいている。RAG2-/-γc-/-背景でhTPO KIマウスから得られた結果が本開示に記載される。ホモ接合体hTPO KIマウスは骨髄中の有意に増加したヒト生着レベルを有し、かつ造血細胞の多系列分化がmTPOマウスよりも改善され、hTPO KIマウスは骨髄単球系列対リンパ球系列の比率の増加を示した。ヒト幹細胞前駆細胞の数および自己再生能は、連続移植によって実証される通り、両方とも改善された。したがって、いくつかある適用例の中で特に、hTPO KIマウスは連続移植によるヒト細胞の増殖に有用である。
【0055】
hIL-3/GM-CSFマウスとhTPOマウスの交配
本明細書に記載されるhIL-3/GM-CSFおよびhTPOマウスの子孫は、少なくとも同じ関連する特徴を有し、親系統(すなわち、hIL-3/GM-CSFマウスとhTPOマウス)と少なくとも同じ利点を示すと予想される。例えば、いずれか一方の親系統、または両方、またはその子孫からヒト細胞集団を単離し、かつhTPOマウスまたはhIL-3/GM-CSFおよびhTPOマウスの子孫に連続移植することが可能である。したがって、一局面において、本明細書に記載するhIL-3/GM-CSFマウスとhTPOマウスの子孫(各関連遺伝子に関してホモ接合性に交配させた子孫を含む)である遺伝子改変されたマウスが提供され、かつこの遺伝子改変されたマウスはhIL-3/GM-CSFマウスとhTPOマウスの両方の利点および特徴を示す。一局面においては、除去された免疫システムを含む(例えば、照射されたマウス)、任意の移植されたマウス(例えば、本明細書に記載する移植されたマウス)からの移植および/または連続移植に適している、そのような子孫マウスが提供される。
【0056】
遺伝子改変されたマウスの移植
本発明による遺伝子改変されたマウスは、移植したヒト造血細胞からヒト免疫細胞を発達させることができる、ヒト造血細胞のレシピエントとして使用される。一態様では、ヒト造血細胞またはヒト造血幹細胞前駆細胞(HSPC)が、本発明によって遺伝子改変および照射されたマウスに配置される(移植される)。ヒト造血細胞またはヒト造血幹細胞は、遺伝子改変されたマウスにおいて、以下から選択される細胞を生じさせる:ヒトCD34陽性細胞、ヒト造血幹細胞、ヒト造血細胞、骨髄前駆細胞、骨髄細胞、樹状細胞、単球、好中球、肥満細胞、およびヒト血液-リンパシステム(ヒト造血幹細胞前駆細胞、ヒト骨髄前駆細胞、ヒト骨髄細胞、ヒト樹状細胞、ヒト単球、ヒト顆粒球、ヒト好中球、ヒト肥満細胞、ヒト胸腺細胞、ヒトT細胞、ヒトB細胞、ヒト血小板を含む)、ならびにこれらの組合せ。
【0057】
遺伝子改変されたマウスは、存在しうる内在性の造血細胞を排除するために照射することができ、かつ該マウスは、任意の適した供給源のヒト造血細胞を用いて移植されることが可能である。造血細胞の当技術分野で公知である1つの適した供給源はヒト臍帯血細胞、特にCD34陽性細胞である。別の造血細胞源はヒト胎児肝臓である。
【0058】
一態様において、本発明によるマウスのヒト造血細胞を用いた移植は、TPO遺伝子のヒト化を欠くか、IL-3およびGM-CSF遺伝子のヒト化を欠くか、またはTPO遺伝子とIL-3およびGM-CSF遺伝子のヒト化を欠く免疫不全マウスより、増大した数のヒト造血細胞を示すマウスをもたらす。
【0059】
一態様において、本発明によるマウスのヒト造血細胞を用いた移植は、ヒト化を欠く免疫不全マウスと比べて、増大した数のヒト血液細胞(例えば、成熟造血細胞)を示すマウスをもたらす。特定の態様では、ヒト造血細胞が以下から選択される:ヒトCD34陽性細胞、造血幹細胞、造血細胞、骨髄前駆細胞、骨髄細胞、樹状細胞、単球、顆粒球、好中球、肥満細胞、およびヒト血液-リンパシステム(ヒト造血幹細胞前駆細胞、ヒト骨髄前駆細胞、ヒト骨髄細胞、ヒト樹状細胞、ヒト単球、ヒト顆粒球、ヒト好中球、ヒト肥満細胞、ヒト胸腺細胞、ヒトT細胞、ヒトB細胞、ヒト血小板を含む)、ならびにこれらの組合せ。
【0060】
遺伝子改変され移植されたマウスの非限定的な適用
ヒト造血細胞を用いて移植された、遺伝子改変されたマウスは、通常はマウスに感染しない病原体を研究するための有用動物である。そのような一例は腸チフスの原因病原体チフス菌である。
【0061】
腸チフスは、米国での約400症例/年を含めて、世界中で - 主に開発途上国で - 2100万人を超える人々を苦しめている。腸チフスは以下の薬物で治療されている:アモキシリン、アンピシリン、セフォタキシム、セフトリアキソン、セフタジジム、クロラムフェニコール、シプロフロキサシン、コトリモキサゾール、エルタペネム、イミペネム、フルオロキノロン類(例えば、シプロフロキサシン、ガチフロキサシン、オフロキサシン)、ストレプトマイシン、スルファジアジン、スルファメトキサゾール、テトラサイクリン、およびこれらの組合せ。反復性の感染が一般的であり、そのことが抗生物質療法による疾病管理を制限している。さらに、チフス菌感染症では多剤耐性もよく見られる。
【0062】
新規治療薬、新規ワクチン、および治療薬とワクチンの有効性を試験するための新規方法が必要とされている。チフス菌によって感染され得るマウスは、例えば、新規治療薬および新規ワクチンを同定するのに有用であろう。新規治療薬および新規ワクチンは、そのようなマウスにおいて、例えば推定上の抗チフス菌剤による治療に応答したマウス中(血液または所定の組織中)のチフス菌の量を測定することによって、あるいはマウスに推定ワクチンを接種し、続いてチフス菌の感染性投与に曝露し、推定ワクチンの接種による感染力の何らかの変化を、ワクチンを接種しないがチフス菌に感染させた対照と比較して観察することによって、試験することができる。
【0063】
本発明による遺伝子改変および移植されたマウスは、マウスに感染しないヒト病原体によって感染され得るマウスを作製するのに有用である。例えば、マウスは、チフス菌によって感染され得る非ヒト動物として有用である。一態様では、遺伝子改変および移植されたマウスは、遺伝子改変を欠く移植されたマウスに比べて、増強されたヒト細胞の生着を示し、増強はチフス菌感染を維持するのに十分である。特定の態様では、チフス菌感染の維持は、マウス中で繁殖するチフス菌の能力を含む。特定の態様では、チフス菌感染は、チフス菌を繁殖させる感染したマウスの能力を含む。特定の態様では、マウスは、チフス菌の初期導入または感染性曝露後に少なくとも1週間、10日間、2週間、3週間、または4週間にわたりチフス菌を繁殖させることができる。
【0064】
抗チフス菌剤を同定するための方法も提供され、その方法はチフス菌によって感染され得る本明細書に記載のマウスを用いるものである。野生型マウスおよび他の公知の免疫不全マウス(例えば、RAG1/RAG2遺伝子ノックアウトマウス)はチフス菌によって感染され得ない。
【0065】
Il2rg遺伝子ノックアウトとRAG遺伝子ノックアウト(例えば、RAG2遺伝子ノックアウト)を含み(第1のタイプ)、さらに内在性マウスIL-3遺伝子のヒトIL-3遺伝子との置換および内在性マウスGM-CSF遺伝子のヒトGM-CSF遺伝子との置換を含む(第2のタイプ)遺伝子改変されたマウスが提供され、該遺伝子改変されたマウスは、ヒト造血細胞を用いて移植された場合、チフス菌による感染が可能である。
【0066】
図1に示したデータは、第1および第2の両タイプのマウスを代表するものである。図1のマウスの遺伝子改変は、(a)マウスRAG遺伝子ノックアウト;および(b)マウスIl2rg遺伝子ノックアウトを含む。図1のマウスはまた、ヒト造血細胞の移植を含む。このマウスは第2のタイプのマウスを作製するために2つのさらなる改変によってさらに改変することができ、それらは、(c)内在性マウスIL-3遺伝子のヒトIL-3遺伝子との置換;および(d)マウスGM-CSF遺伝子のヒトGM-CSF遺伝子との置換である。
【0067】
図2、3および4は、第1のタイプの改変されたマウス((a)マウスRAG遺伝子ノックアウトおよび(b)マウスIl2rg遺伝子ノックアウトの改変、ならびにヒト造血細胞の移植を含む)のみを用いて得られたものである。
【0068】
さまざまな態様において、チフス菌感染した、遺伝子改変されたマウスは、チフス菌の増殖性感染を含む。一態様では、マウスは、その細胞の1個または複数においてチフス菌を保有および繁殖させることができる。一態様では、マウスは、チフス菌への感染性曝露後に少なくとも1週間、10日間、2週間、3週間、または4週間にわたりその血液中または少なくとも1つの組織中にチフス菌の力価またはレベルを維持することができる。
【0069】
一態様において、方法は、本発明による遺伝子改変されたマウスに剤を投与する段階であって、該遺伝子改変されたマウスがチフス菌に感染している段階;該剤の投与後に該マウスの血液中または組織中のチフス菌のレベルを検出する段階;および任意で、該剤の投与が該マウスの血液中または組織中のチフス菌のレベルを低下させるかどうかを判定する段階を含む。一態様では、剤がワクチンである。別の態様では、剤が抗生物質または抗生物質の性質をもつと予想される剤である。一態様では、剤が抗原結合タンパク質であり、特定の態様では抗体である。一態様では、剤がヒト用の承認された医薬である。
【0070】
一態様において、方法は、本発明による遺伝子改変および移植されたマウスに既知量のチフス菌を感染させる段階、感染させた該マウスに剤を投与する段階、ならびに該剤の投与後に該遺伝子改変および移植されたマウス中のチフス菌の量を測定する段階を含む。一態様では、剤がマウスの血液中または組織中のチフス菌の量を、1回の剤投与または2回またはそれ以上の投与後に所定の期間にわたり、少なくとも半分に減少させる場合に、その剤は抗チフス菌剤であると判定される。
【0071】
一局面においては、関心対象のチフス菌の分離株または株が薬物耐性または多剤耐性であるかどうかを判定するための方法が提供され、該方法は、本発明による遺伝子改変および移植されたマウスに、チフス菌の治療に用いる薬物または薬物の組合せを投与する段階を含み、該マウスは関心対象のチフス菌の分離株または株に感染している。方法は、(a)薬物または薬物の組合せの投与後のある時点でのマウスの血液中または組織中の関心対象のチフス菌の分離株または株の力価、(b)薬物または薬物の組合せの1回またはそれ以上の投与後に関心対象のチフス菌の分離株または株がマウスにおける感染またはマウスの組織中のチフス菌のレベルを維持する能力、あるいは(c)薬物または薬物の組合せの投与後のある時点で関心対象のチフス菌の分離株または株がマウス中で繁殖する能力に及ぼす薬物または薬物の組合せの、もしあれば、効果を測定する段階を含む。特定の態様において、薬物はアモキシリン、アンピシリン、セフォタキシム、セフトリアキソン、セフタジジム、クロラムフェニコール、シプロフロキサシン、コトリモキサゾール、エルタペネム、イミペネム、フルオロキノロン類(例えば、シプロフロキサシン、ガチフロキサシン、オフロキサシン)、ストレプトマイシン、スルファジアジン、スルファメトキサゾール、テトラサイクリン、およびこれらの組合せからなる群より選択される。特定の態様では、薬物または薬物の組合せの投与が、チフス菌への感染を生じる曝露後少なくとも1週間、10日間、2週間、3週間、または4週間である。
【0072】
さまざまな局面および態様において、血液中または組織中のチフス菌のレベルは、血液または組織の単位(例えば、重量または体積)あたりのコロニー形成単位の数を確認することによって測定される。
【0073】
一態様において、本発明による遺伝子改変されかつヒト造血細胞を移植されたマウスは、ヒト造血細胞を用いて移植されていないマウスと比べて少なくとも100倍、1,000倍、または10,000倍の、コロニー形成単位(cfu)で測定した、チフス菌レベルを有する。
【0074】
抗チフス菌ワクチンの有効性を確認するのに有用な方法および組成物を提供する。一局面において、抗チフス菌ワクチンの有効性を確認するための方法が提供され、該方法は、本発明による遺伝子改変および移植されたマウスを抗チフス菌ワクチンに曝露する段階、その後該遺伝子改変および移植されたマウスをチフス菌に曝露する段階、ならびに該遺伝子改変および移植されたマウスがチフス菌によって感染され得るかどうか、またはどの程度に感染され得るかを判定する段階を含む。
【0075】
一態様において、抗チフス菌ワクチンはチフス菌の細胞表面タンパク質またはその免疫原性断片を含む。一態様では、ワクチンがチフス菌株の膜画分を含む。一態様では、ワクチンが組換えチフス菌タンパク質またはその免疫原性断片を含む。一態様では、ワクチンがチフス菌タンパク質またはその免疫原性断片をコードする発現ベクターを含む。一態様では、ワクチンが不活化チフス菌株または不活化チフス菌株の混合物を含む。
【0076】
本開示に記載される遺伝子改変および移植されたマウスはまた、既存のマウスより厳密にヒト病原体感染をモデル化するのに有用である。例えば、結核菌による感染。本明細書に記載される遺伝子改変および移植されたマウスは、例えば、ヒト免疫細胞を含む肉芽腫および境界のはっきりした肉芽腫を含む肉芽腫を発生する結核菌マウスモデルを提供することによって、マイコバクテリアのヒト感染をモデル化するのに有用である。記載される遺伝子改変および移植されたマウスのチフス菌感染に関連して説明した薬物およびワクチンの試験方法はまた、結核菌への適用例、例えば、薬物耐性株の同定、結核菌ワクチンの有効性の試験、抗結核菌剤の試験、抗結核菌剤に応答するcfuの測定などにも適用可能である。
【0077】
本開示に記載される遺伝子改変および移植されたマウスはまた、初期ヒト造血細胞から、例えばヒト造血または前駆細胞から発生する、ヒト造血器悪性腫瘍をモデル化するのに有用である。本開示に記載される遺伝子改変および移植されたマウスのさらなる適用例は、この開示を読むことによって当業者には明らかであろう。
【0078】
トロンボポエチンおよび移植
トロンボポエチン(TPO)は、巨核球および血小板の発達を促進する成長因子として最初に同定された(Wendling, F. et al. (1994) cMpl ligand is a humoral regulator of megakaryocytopoiesis, Nature 369:571-574; Kaushansky, K. et al. (1994) Promotion of megakaryocyte progenitor expansion and differentiation by the c-Mpl ligand thrombopoietin, Nature 369:568-571; Lok, S. et al. (1994) Cloning and expression of murine thrombopoietin cDNA and stimulation of platelet production in vivo, Nature 369:565-568; de Sauvage, F.J. et al. (1994) Stimulation of megakaryocytopoiesis and thrombopoiesis by the c-Mpl ligand, Nature 369:533-538; Bartley, T.D. et al. (1994) Identification and cloning of a megakaryocyte growth and development factor that is a ligand for the cytokine receptor Mpl, Cell 77:1117-1124; Kaushansky, K. (1998) Thrombopoietin, N Engl J Med 339:746-754; Kaushansky, K. (2005) The molecular mechanisms that control thrombopoiesis, J Clin Invest 115:3339-3347; Kaushansky, K. (2008) Historical review: megakaryopoiesis and thrombopoiesis, Blood 111:981-986)。
【0079】
TPOは、肝臓および腎臓で構成的に産生され、かつ血液循環に放出される。TPOの受容体であるc-Mplは骨髄中の造血幹細胞前駆細胞によって発現される。c-Mplは循環している血小板にも発現される。しかしながら、血小板へのTPOの結合はいかなるシグナル伝達経路をも活性化しない。かくして、血小板はTPOのためのシンクまたはスカベンジャーとして作用し、この機構を介して血小板産生の負の調節に寄与している(Kuter, D.J.およびRosenberg, R.D. (1995) The reciprocal relationship of thrombopoietin (c-Mpl ligand) to changes in the platelet mass during busulfan-induced thrombocytopenia in the rabbit, Blood 85:2720-2730)。その後、TPOはHSCの増殖と自己再生を支持するその重要な機能について認識されてきた(Fox, N., et al. (2002) Thrombopoietin expands hematopoietic stem cells after transplantation, J Clin Invest 110, 389-3894; Kirito, K. et al. (2003) Thrombopoietin stimulates Hoxb4 expression: an explanation for the favorable effects of TPO on hematopoietic stem cells, Blood 102:3172-3178)。
【0080】
TPO欠乏は、成体マウスにおいてHSC数の減少につながり、かつTPOの存在は静止期の成体HSCの維持に必要である(Yoshihara, H. et al. (2007) Thrombopoietin/MPL signaling regulates hematopoietic stem cell quiescence and interaction with the osteoblastic niche, Cell Stem Cell 1, 685-697; Qian, H. et al. (2007) Critical role of thrombopoietin in maintaining adult quiescent hematopoietic stem cells, Cell Stem Cell 1:671-684)。さらに、TPOは、照射した宿主の造血区画を補充するのに必要な、HSCの移植後増殖を支持するために必要である。興味深いことに、骨髄でのHSC「ニッチ」の形成に関与する骨芽細胞は、HSCの機能と維持に重要なTPOを産生することが実証されている。
【0081】
マウスおよびヒトのTPOは、インビトロにおいて超生理的用量で用いた場合、それぞれのコグネイト受容体に対して双方交差反応性であるが、そのサイトカインがインビボ環境において生理的制限用量で作用する場合には、親和性と生物学的活性が異なることがある。本発明者らは、マウスTPOはインビボでヒトc-Mpl受容体に適切な刺激を与えない可能性があり、それゆえにマウス環境でヒトHSCの特性が損なわれる主な原因になり得ると仮定した。この潜在的欠陥を修正するために、本発明者らは、Rag2-/-γc-/-マウスにおいて、マウスTPOをコードする遺伝子をそのヒト対応物により置き換えた。そのようなマウスはヒト血液-リンパ球生成システムの分化と機能を維持するための改善された能力をもつと仮定された。
【0082】
ヒト血液-リンパ球生成システムの分化と機能を維持するマウスの開発では、20年以上前の最初のモデルの発表以来、著しい進展が達成されてきた。しかしながら、以下の(i)〜(iii)を含むいくつかの制限が残っている:(i)レシピエントマウスの寿命の間中継続しない、一過性のヒト細胞生着、(ii)リンパ球系への非生理的な偏りおよび骨髄細胞の不十分な分化、ならびに(iii)マウスのグループが1人のヒト提供者由来の細胞を移植される場合でさえの、異なる動物間での生着レベルのばらつき。こうした制限は、ヒト細胞の非生理的な位置、免疫不全宿主の残留する異種反応性、マウス種とヒト種における血液-リンパ系細胞の異なる組成、および/または優先的なマウス細胞の支持につながる、造血支持因子のマウスとヒトとの交差反応性の欠如もしくは不足が原因である可能性がある。したがって、ヒト成長因子の生理的レベルを提供すること、および宿主においてそれぞれのマウス相同体を欠失させることは、ヒト細胞集団の発達と生存にさらに有利にはたらく可能性がある。本明細書では、本発明者らが、造血幹細胞の維持と自己再生に重要な機能をもつサイトカインである、トロンボポエチンをコードする遺伝子をヒト化した、新規のレシピエントマウス株が記載される。
【0083】
これらのヒト化トロンボポエチンマウスにヒトCD34+造血幹細胞前駆細胞を移植すると、以前に利用可能なモデルと比較して、上で挙げた3つすべての制限に対して著しい改善が見られた。すなわち、骨髄キメリズムがより高く、少なくとも6ヶ月間維持された。多系列、特に骨髄系の分化が増強された。かつ生着レベルのばらつきが減った。
【0084】
マウスとヒトの免疫システムの主な相違は血液中に存在する顆粒球の割合である。マウスではリンパ球が優勢であるが、ヒト血液は顆粒球に富んでいて、種差はその意義がはっきりしない。興味深いことに、ヒトTPOの存在はヒト顆粒球の分化を改善した(図14)。したがって、レシピエントマウスにおけるヒトTPOの存在は、ヒト生理的状態をより良く反映する顆粒球とリンパ球のバランスに有利にはたらき、おそらくヒト骨髄前駆細胞の向上した維持および/または分化によると考えられる。
【0085】
さらに重要なこととして、これらの結果は、TPOヒト化がマウス環境でヒト造血幹細胞前駆細胞を再増殖する二次レシピエントの維持に有利にはたらくことを示す(図15)。それゆえ、Rag2-/-γc-/-TPOh/hマウスはインビボでヒト幹細胞前駆細胞の機能のさまざまな面を研究するための新規モデルに相当する。
【0086】
それにもかかわらず、血液中の骨髄系とリンパ系との良好なバランスが観察されたとはいえ、末梢リンパ組織(脾臓、血液および胸腺を含む)における全体的生着レベルに対するTPOヒト化の有意な効果は認められなかった(図13(g)〜(i))。それは多様な要因によって説明され得る。第一に、レシピエントマウスは移植前に亜致死的に照射されるが、マウス骨髄細胞の大集団がまだ存在している。これらの細胞のうち、マクロファージはヒト細胞の食細胞として機能することができ、末梢での全体的生着レベルを制限する。したがって、マウスマクロファージの遺伝的枯渇、またはそれらの機能的不活性化は、より高い末梢生着レベルを可能にする場合がある。第二に、ヒト細胞は、それらの最終分化、骨髄からの退出および/または末梢でのそれらの生存を有利にはこぶために、さらなるヒト成長因子を必要とすることがある。サイトカインの多様なパネルが各系列について考慮され得る。最後に、二次リンパ器官がヒト化マウスに形成されるものの、それらの構造はヒト組織に比較して最適ではない。その部分的に欠陥のある構造は、これらの器官で生存できるヒト細胞の数への制限を表す場合があるい。
【0087】
マウスレシピエントをさらに改善するために、さらなる遺伝子置換を採用することができる。それを達成するために、マウス遺伝子のそのヒト相同体によるノックイン置換に基づく、本研究で用いた手法は、古典的なトランスジェニックアプローチと比べて2つの主な利点を提供する。第一に、それはマウス由来の調節配列の大部分を維持しているので、ヒト化遺伝子がマウス宿主内で忠実に発現されることを保証する。第二に、ノックイン戦略はマウスサイトカインをそのヒト相同体と置き換えるので、ヒトサイトカインがマウス受容体に対し十分な交差反応性を示さない場合に、それはマウスサイトカインに依存するマウス由来の細胞の集団に影響を及ぼし得る。これは移植後のヒト細胞集団にさらなる競合的優位性を提供することができる。実際、それはヒトTPOの事例であると考えられる。なぜなら、TPOのホモ接合性置換は非移植動物においてマウス血小板とHSCの減少を招くからである(図13(a)、15(a)および15(b))。
【0088】
ヒトTPOノックインマウスの場合には、一般的にはヒト造血のインビボ生理学、特にヒト造血幹細胞前駆細胞を研究するのに有用であり得る改良モデルが提供される。さらに、これらのマウスは、例えば骨髄性白血病および骨髄増殖性腫瘍など、初期造血細胞から発生するヒト造血器悪性腫瘍をインビボで維持する。
【実施例】
【0089】
以下の実施例は、本発明者らが発明と見なすものの範囲を限定することを意図しない。特に断りのない限り、部分は重量による部分であり、分子量は重量平均分子量であり、温度は摂氏であり、かつ圧力は大気圧またはほぼ大気圧である。
【0090】
実施例1
ヒトIL-3/GM-CSFおよびヒトTPOマウスの作製
hIL-3/GM-CSFターゲティング
1回のターゲティング工程でマウスIL-3遺伝子をヒトIL-3遺伝子と置換し、マウスGM-CSF遺伝子をヒトGM-CSF遺伝子と置換するためのターゲティング構築物は、ギャップ修復クローニングを利用するVELOCIGENE(登録商標)技術(例えば、米国特許第6,586,251号およびValenzuela et al. (2003) "High-throughput engineering of the mouse genome coupled with high-resolution expression analysis," Nat Biot 21(6):652-659を参照されたい。参照により本明細書に組み入れる)を用いて構築した。
【0091】
マウス配列は細菌人工染色体(BAC)RPCI-23、クローン5E15から取得した。ヒト配列はCaltech Dライブラリー(CTD)、BACクローン2333J5から取得した。
【0092】
p15複製起点を含むギャップ修復ドナーベクターは、mIL-3 ATGのすぐ上流の5'マウスホモロジーアーム、hIL-3 ATGから約274ntまでhIL-3遺伝子にわたるヒト5'IL-3ホモロジーアーム、ポリリンカー、hGM-CSF遺伝子のポリA配列の約2.9kb下流で開始する3'hGM-CSF(約233塩基)、および薬物選択カセットと、これに続くmGM-CSFポリA配列の下流(約2.9kb下流)の配列を有するマウス3'ホモロジーアームをクローニングすることによって構築した。このギャップ修復ベクターを線状化し、かつヒトCTD BACクローン2333J5と組換え酵素ベクターを含む大腸菌(E. coli)DH10B株に、Valenzuelaらに記載される通りに挿入した。
【0093】
細胞を薬物選択培地で増殖させた。個々のクローンを増殖させて、ギャップ修復ドナーベクターDNAを抽出し、適切なマウス-ヒト接合点について該ベクターの部分を配列決定した。パルスフィールドゲル電気泳動を用いて、挿入物のサイズおよび予想される制限断片長を確認した。
【0094】
hIL-3遺伝子に隣接するマウス上流および下流ホモロジーボックス、hGM-CSF遺伝子、およびロキシド(loxed)薬物選択カセットを含む捕捉ドナー(captured donor)を修復ドナーベクターから得て、その捕捉ドナーを線状化し、線状化した捕捉ドナーを、RPCI23クローン5E15とpABGベクターを含む大腸菌DH10Bに導入した。細胞を薬物選択培地で増殖させた。RPCI23クローン5E15 DNA中に捕捉ドナーDNAを含む(ターゲティングベクターを形成する)個々のクローンを単離し、ターゲティングベクターDNAを抽出し、かつ適切なマウス-ヒト接合点について該ベクターの部分を配列決定した。パルスフィールドゲル電気泳動を用いて、挿入物のサイズおよび予想される制限断片長を確認した。
【0095】
エレクトロポレーション
ターゲティングベクターを線状化し、これを用いてValenzuelaらに記載される通りにマウスES細胞をエレクトロポレーションにかけた。エレクトロポレーション処理を行った、ターゲティングベクターを含むマウスES細胞は、ロキシド薬物選択カセットを除くために、一過性Cre発現ベクターを用いてさらにエレクトロポレーションにかけた。そのターゲティングベクターをRag2 HET Il2rg y/- ES細胞にエレクトロポレーションにより導入した。RAG2遺伝子とIl2rg遺伝子のノックアウトがなされた親ES細胞株は市販のV17 ES細胞(BALB/c×129ヘテロ接合体)であった。hIL-3およびhGM-CSF遺伝子でターゲティングされたES細胞は、マウス胚に導入するために使用した。
【0096】
hIL-3/GM-CSFマウス
ターゲティングされたドナーES細胞を、8細胞期のマウス胚にVELOCIMOUSE(登録商標)法により導入する(例えば、米国特許第7,294,754号およびPoueymirou et al. (2007) "F0 generation mice that are essentially fully derived from the donor gene-targeted ES cells allowing immediate phenotypic analyses," Nat Biot 25(1):91-99を参照されたい。参照により本明細書に組み入れる)。ヒト化IL-3およびGM-CSF構築物を保持するVELOCIMICE(登録商標)(完全にドナーES細胞に由来するF0マウス)は、対立遺伝子アッセイ法の変法(例えば、Valenzuelaら参照)を用いて、マウス対立遺伝子の喪失とヒト対立遺伝子の獲得について遺伝子型を同定することによって同定される。これらのマウスを最初にBALB/cAnNCRと交配させ、次にRag2とIl2rgについてヘテロ接合性で、ヒトIL-3/GM-CSF KIを含むマウスを、移植研究のためにRag2/Il2rg二重KOマウスと交配させる。
【0097】
hIL-3/GM-CSFマウスの表現型分類
ヒト化マウスは、hGM-CSF特異的プライマーを用いるRT-PCRによって、ヒトGM-CSFの産生について試験した。試験した組織のヒトGM-CSFの発現パターンはマウスGM-CSFのそれに一致した(主に肺で発現)。ConAとIL-2で48時間刺激した、ヒト化マウス由来の脾細胞のELISAを実施して、hIL-3とhGM-CSFの存在を検出した。脾細胞はhIL-3とhGM-CSFの両方の発現について陽性であった。
【0098】
hTPOターゲティング
1回のターゲティング工程でマウスTpo(mTpo)遺伝子をヒトTPO(hTPO)遺伝子と置換するためのターゲティング構築物(図11(d))は、ギャップ修復クローニングを利用するVELOCIGENE(登録商標)技術を用いて構築した(Valenzuela et al.)。このベクターは、Tpoのオープンリーディングフレームを含む配列を置換するが、マウス由来のプロモーターと5'UTRを維持するように設計された。マウス配列を細菌人工染色体(BAC) RPCI-23、クローン98H7から得た。ヒト配列をBAC RPCI-11、クローン63m3から得た。p15複製起点を含むギャップ修復ドナーベクターは、mTpo ATGのすぐ上流の5'マウスホモロジーアーム、hTPO ATGから約275ntまでhTPO遺伝子にわたるヒト5'TPOホモロジーアーム、ポリリンカー、hTPO遺伝子のポリA配列の約1.5kb下流で開始する3'hTPOホモロジーアーム、およびロキシド薬物選択カセットと、これに続くmTpoポリA配列の下流(約3.5kb下流)の配列を有するマウス3'ホモロジーアームをクローニングすることによって構築した。このギャップ修復ベクターを線状化し、かつヒトBACクローンRPCI-11、63m3と組換え酵素ベクターを含む大腸菌DH10B株に挿入した。細胞を薬物選択培地で増殖させた。個々のクローンを増殖させて、ギャップ修復ドナーベクターDNAを抽出し、適切なマウス-ヒト接合点について該ベクターの部分を配列決定した。パルスフィールドゲル電気泳動を用いて、挿入物のサイズおよび予想される制限断片長を確認した。hTPO遺伝子に隣接するマウス上流および下流ホモロジーボックスおよびロキシド薬物選択カセットを含む捕捉ドナーを修復ドナーベクターから得て、その捕捉ドナーを線状化し、線状化した捕捉ドナーを、RPCI-23クローン98H7とpABGベクターを含む大腸菌DH10Bに導入した。細胞を薬物選択培地で増殖させた。RPCI-23クローン98H7 DNA中に捕捉ドナーDNAを含む(ターゲティングベクターを形成する)個々のクローンを単離し、ターゲティングベクターDNAを抽出し、かつ適切なマウス-ヒト接合点について該ベクターの部分を配列決定した。パルスフィールドゲル電気泳動を用いて、挿入物のサイズおよび予想される制限断片長を確認した。ターゲティングベクターを線状化し、これを用いてマウス胚性幹(ES)細胞をエレクトロポレーションにかけた。ターゲティングベクターはRAG2+/-γcY/-ES細胞にエレクトロポレーションにより導入した。親RAG2+/-γcY/-ES細胞株は、市販されているV17 ES細胞株(BALB/c×129ヘテロ接合体)から作られた。正しくターゲティングされたES細胞は、ロキシド薬物選択カセットを除くために、一過性Cre発現ベクターを用いてさらにエレクトロポレーションにかけた。hTPOでターゲティングされた、選択カセットを含まないES細胞を、8細胞期のマウス胚にVELOCIMOUSE(登録商標)法により導入した(Poueymirou et al)。野生型Tpo(TPOm/m)、ヘテロ接合型(TPOh/m)またはホモ接合型(TPOh/h)TPO遺伝子置換をもつRag2-/-γc-/-マウスが得られた。
【0099】
実施例2
hIL-3/GM-CSFマウス:移植
ヒト造血幹細胞の単離
ヒト臍帯血および胎児肝臓試料は、それぞれイェールニューヘブン病院(Yale-New Haven Hospital)およびアルバート・アインシュタイン医科大学ニューヨーク(Albert Einstein Medical College New York)から、イェール大学の人権擁護調査委員会(Human Investigation Committee)の承認の下で得られたものである。CD34+細胞は、ヒト臍帯血または胎児肝臓から、密度勾配遠心およびCD34マイクロビーズ(Miltenyi Biotec社)を用いる免疫磁気選別によって単離した。単離されたCD34陽性細胞の純度をフローサイトメトリーにより確認した。精製したヒトCD34陽性細胞は凍結保存され、使用前に液体窒素中で保存した。
【0100】
ヒト造血幹細胞を用いたマウスの移植
移植は以前に記載された通りに行った(Traggiai et al. (2004) Development of a human adaptive immune system in cord blood cell-transplanted mice, Science 304:104-107)。簡単に説明すると、出生日に、RAG2遺伝子ノックアウト/Il2rg遺伝子ノックアウトの背景(hIL-3/hGM-CSFを含むまたは含まない)からの仔マウスを亜致死的に照射した(4時間間隔で2×200cGy)。照射後、新生仔は30ゲージ針を用いる肝内注射によって1〜2×105個のヒトCD34+細胞(25μLのPBS中に再懸濁したもの)を受け取った。対照にはPBSのみを注射した。マウスは生後3〜4週間で離乳させ、特定病原体フリーの条件下で維持した。日和見感染を防ぐために、マウスに飲料水中の予防的抗生物質(サルファトリム(Sulfatrim))を投与した。すべての動物実験作業はイェール大学の動物管理使用委員会(Institutional Animal Care and Use Committee: IACUC)によって承認され、IACUCの規則に従って実施された。
【0101】
移植されたマウスの解析
ヒト造血細胞の生着は移植後8〜12週で判定した。血液試料を後眼窩洞(retro-orbital sinus)から採取し、ACK溶解緩衝液(Lonza社)を用いて赤血球の溶解を行った。次に、試料をマウスCD45、ヒトCD45、ヒトCD3、およびヒトCD14に対する蛍光標識モノクローナル抗体(すべてBD Biosciences社から)で染色し、FACScalibur(商標)(BD Biosciences社)でフローサイトメトリーにより解析した。感染実験に用いたマウスは、特に明記しない限り、>4%のhCD45+細胞の血中生着レベルを有していた。適合したマウス、すなわち同じバッチのCD34+細胞を用いて移植されたマウスを、実験に使用した。特に明記しない限り、実験はFL由来のCD34+細胞を用いて移植されたマウスを用いて行った。
【0102】
フローサイトメトリー
hIL-3/GM-CSF研究のため、移植後10〜14週のマウスの肺、BAL、骨髄、胸腺、脾臓および血液から細胞懸濁液を調製した。RBCの溶解はACK溶解緩衝液(Lonza社)を用いて行った。次に、マウスおよびヒト細胞表面抗原に対する蛍光色素標識モノクローナル抗体(mAb)で試料を染色した。次のmAbを使用した:(1)抗ヒト:CD3 (UCHT1)、CD4 (RPA-T4)、CD8 (HIT8a)、CD11c (B-ly6)、CD14 (MoP9)、CD19 (HIB19)、CD33 (WM53)、CD45 (HI30および2D1)、CD56 (NCAM 16.2)、CD66 (B1.1)、CD116 (4H1)、CD123 (9F5)。(2)抗マウス:CD45 (30-F11)、F4/80 (BM8)。CD116、CD45 (30-F11)およびF4/80 mAbはeBioscience社から購入した。他のすべてのmAbはBD Biosciences社から購入した。試料はFACSCalibur(商標)またはLSRII(商標)フローサイトメーター(BD Biosciences社)で解析した。
【0103】
メチルセルロースCFUアッセイ法
hIL-3/GM-CSF研究のため、移植されたマウスからのヒトCD34+骨髄細胞をセルソーティングにより精製した。ソーティングされた細胞(1〜1.5×105個)をイスコフ改変ダルベッコ培地(IMDM、GIBCO社)ベースのメチルセルロース培地(Methocult(商標)H4100、StemCell Technologies社)で培養した。該培地には20%FBS、1%BSA、2mM L-グルタミン、55μM 2-メルカプトエタノール、および次のヒトサイトカインを添加した:幹細胞因子(10ng/ml)、FLT3リガンド(10ng/ml)、トロンボポエチン(50ng/ml)、IL-3 (20ng/ml)、IL-6 (10ng/ml)、IL-11 (10ng/ml)、GM-CSF (50ng/ml)、およびエリトロポエチン(4U/ml)(すべてR&D Systems社)。細胞を60mmペトリ皿で37℃/5%C02にてインキュベートした。12〜14日後にコロニーの数を顕微鏡で測定した。
【0104】
LPSに対する炎症反応
マウスにUltrapure LPS E. coli 0111:B4 (Invivogen社)の2回腹腔内注射を48時間間隔で行った(35および17.5μg)。各注射の2〜3時間後に血清を採取した。ヒトIL-6の血清濃度をELISA (R&D Systems社)で測定した。最初のLPS注射の72時間後にマウスを屠殺し、心穿刺で血液を採取してフローサイトメトリーに使用した。
【0105】
細胞内サイトカイン染色
hIL-3/GM-CSF研究のため、全TB10.4タンパク質(Skjot et al. (2002) Epitope mapping of the immunodominant antigen TB10.4 and the two homologous proteins TB10.3 and TB12.9, which constitute a subfamily of the esat-6 gene family, Infect. Immun. 70:5446-5453)をカバーする重複ペプチドをイェール大学のW.M. Keck Facilityで合成した。BCG感染したマウス由来の脾細胞(2×106個/ウェル)を混合ペプチド(5μg/mlの各ペプチド)と共に、96ウェルのU底マイクロタイタープレート(Becton Dickinson社)において200μl/ウェルの全体積で37℃/5%C02にて5時間インキュベートした。細胞培養のために10%FCS、1%ペニシリン-ストレプトマイシン、1% L-グルタミン、および55μM 2-メルカプトエタノールを添加したRPMI 1640培地(Invitrogen社)を用いた。細胞内サイトカイン染色はCytofix/Cytoperm(商標)キット(BD Biosciences社)を用いてメーカーの指示に従って行った。次のmAbを細胞内染色のために使用した(すべてBD Biosciences社)。抗ヒトIFNγ(B27)、抗マウスIFNγ(XMG1.2)。アイソタイプの適合したmAbを対照として用いた。
【0106】
組織学および免疫組織化学
hIL-3/GM-CSF研究のため、器官を取り出し、かつ組織学的分析のために10%中性緩衝ホルマリンまたはZinc Fixative (BD Biosciences社)中に固定した。パラフィン包埋組織切片を調製し、H&EもしくはPASで染色するか、またはイェール病理組織サービス(Yale Pathology Tissue Services)で免疫組織化学のために処理した。次の抗ヒト抗体を免疫組織化学のために使用した(すべてDako社から):CD45 (2B11+PD7/26)、CD3 (F7.2.38)、CD68 (PG-M1)。肉芽腫の存在についての組織切片の評価は盲検で行った。
【0107】
統計解析
hIL-3/GM-CSF研究のため、ノンパラメトリックなMann-WhitneyのU検定を用いて、2グループ間の統計的有意性(a=0.05)を判定した。多重グループ比較のために、我々は一元配置ANOVAとともにTukeyの多重比較検定(a=0.05)を用いる事後検定(post hoc testing)を採用した。統計的に有意なP値(P<0.05)のみが示される。
【0108】
実施例3
hIL-3/GM-CSFを移植されたマウス:感染
チフス菌ISP2825 (Galan J.E.およびCurtiss, R. (1991) Distribution of the invA, -B, -C, and -D genes of S. thyphimurium among other S. serovars: invA mutants of S. typhi are deficient for entry into mammalian cells, Infect. Immun. 59(9):2901-2908;参照により本明細書に組み入れる)は、腸チフス患者からの臨床分離株であるが、このチフス菌をLBブロスで一晩増殖させた。翌日、40μLの細菌細胞培養物を、0.3M NaClを含む新鮮なLBブロス2mLに移し、かつその培養物が約0.9のOD600に達するまで37℃で約3時間増殖させた。その細菌培養物をスピンダウンし、緩衝生理食塩水に再懸濁させて、感染のために使用した。9〜12週齢のヒト化マウスと対照マウスに1×103または1×104または1×105個のチフス菌を0日目に腹腔内接種した。感染したマウスを念入りに監視し、かつ感染後4週で屠殺した。脾臓、肝臓および胆のうを無菌的に摘出し、0.05%デオキシコール酸ナトリウムを含む無菌PBS 3〜5mL中で機械的にホモジナイズした。組織ホモジネートを連続希釈し、LB寒天プレートにまき、コロニー数の計測のために37℃で一晩インキュベートした。コロニー数を計測し、かつ回収された全コロニー形成単位の数を算出した。マウスのデータは図8〜11に提供される。図8において、「対照」マウスは、移植されていない遺伝子改変されたマウス(RAG KO, Il2rg KO/hIL-3, hGM-CSF)である。図9〜11において、「対照」マウスは移植されていないRAG KO/Il2rg KOマウスである(すなわち、それらはIL-3とGM-CSFのヒト化を欠くが、代わりに内在性マウスIL-3と内在性マウスGM-CSFをもつ)。「対照」マウスには、ヒトCD34+細胞の代わりにPBSを注射した。
【0109】
図8に示す通り、脾臓におけるチフス菌感染は感染後10日で2匹の遺伝子改変されたマウス(RAG KO, Il2rg KO/hIL-3, hGM-CSF)に検出される。
【0110】
図9に示す通り、1×103個のチフス菌の感染後1週で、3.8および3%の生着率を有する遺伝子改変されたマウス(RAG KO, Il2rg KO)は、対照マウスより約1000倍高い、脾臓での感染を示した(p<0.01)。対照マウスとヒト化マウス間の差のp値はp<0.01であった。
【0111】
図10に示す通り、胎児肝由来のCD34陽性細胞を用いて移植され、かつ1×104個のチフス菌を感染させた、遺伝子改変されたマウス(RAG KO, Il2rg KO)は、感染後4週で、脾臓(平均して約1,000〜約10,000倍)と肝臓(平均して約1,000倍)の両方にチフス菌感染を示した。脾臓のチフス菌について試験したコホート中の各マウスは、(図10の左パネルの「ヒト化」コホートにおいて上から下に)23.5、40.1、16.5、50、26、および51.7のヒト細胞の生着率を有していた。肝臓のチフス菌について試験したコホート中の各マウスは、(図10の右パネルの「ヒト化」コホートにおいて上から下に)16.5、40.1、23.5、26、50、および51.7のヒト細胞の生着率を有していた。脾臓では対照マウスとヒト化マウス間の差のp値はp<0.01であった;肝臓ではp<0.03。
【0112】
図11に示す通り、胎児肝由来のCD34陽性細胞を用いて移植され、かつ1×104個のチフス菌を感染させた、遺伝子改変されたマウス(RAG KO, Il2rg KO)は、感染後4週で、胆のうにおけるチフス菌感染を示し、チフス菌cfuは平均して対照マウスより100万倍高かった。胆のうのチフス菌について試験したコホート中の各マウスは、(図10の「ヒト化」コホートにおいて上から下に)23.5、16.5、40.1、50、26、および51.7のヒト細胞の生着率を有していた。対照マウスとヒト化マウス間の差のp値はp<0.03であった。
【0113】
これらの結果は、遺伝子改変されたマウス(RAG KO, Il2rg KO/hIL-3, hGM-CSF)が全身感染後にチフス菌によってコロニー化され得ることを証明している。
【0114】
実施例4
hIL-3/GM-CSFを移植されたマウス:肺病原体に対するヒト炎症反応のためのマウスモデルの検証
GM-CSF(Csf2)およびIL-3をコードする遺伝子は、ヒトおよびマウスではそれぞれ5番および11番染色体上で密接に連鎖している(<10kb)。これは、hIL-3/GM-CSF KIマウスを作製するために、両遺伝子のマウス遺伝子座とヒト遺伝子座との置換を可能にした(図5(e))。ヒトIL-3 KI対立遺伝子はマウス調節エレメントの制御下にある一方で、ヒトCsf2 KI対立遺伝子はそのヒト調節エレメントの制御下のままである。マウスおよびヒトGM-CSF mRNAの発現は、各マウス遺伝子の一方の対立遺伝子および各ヒト遺伝子の一方の対立遺伝子を発現するhIL-3/GM-CSF KIマウス(IL-3/GM-CSF「ヒト/マウス」(h/m)マウスと呼ばれる)においてRT-PCRにより解析した。IL-3およびGM-CSFのマウス対立遺伝子を唯一持つ野生型マウスは、IL-3/GM-CSF「マウス/マウス」(m/m)マウスと呼ばれる。
【0115】
hIL-3/GM-CSF KIマウスのRT-PCRおよびELISA解析
全RNAはホモジナイズした組織からTRIzol(商標)試薬(Invitrogen社)を用いてメーカーの指示に従って抽出した。等量のDNase処理RNAをSuperScript(商標)First-Strand Synthesis System(Invitrogen社)によるcDNA合成のために使用した。従来型RT-PCRは、次のプライマーを用いて行った。
定量的RT-PCRは、ABI社から購入したプライマー-プローブセットを用いる7500 FastリアルタイムPCRシステムで行った。発現量は比較閾値サイクル法(comparative threshold cycle method)を用いて計算し、マウスまたはヒトHPRTに対して正規化された。マウスとヒトのIL-3およびGM-CSFタンパク質は、R&D Systems社からの種特異的ELISAキットを用いてメーカーの指示に従って検出した。脾細胞は5μg/mlコンカナバリンA(ConA)と100U/ml IL-2で活性化し、48時間の刺激の後に上清をELISAのために回収した。
【0116】
hIL-3/GM-CSF KIマウスにおける発現
ヒトGM-CSF mRNAは、肺で最高の発現を示して、そのマウス対応物と同様のパターンで発現された(図5(a))。IL-3は主に活性化T細胞によって発現され、活性化T細胞はまたGM-CSFをも産生する。したがって、ELISAはh/mマウスから単離された活性化脾細胞からの上清で実施した。ヒトIL-3およびGM-CSFタンパク質の両方を検出することができた(図6(f)f、6(g))。ヒト造血細胞に競合的優位性を付与するために、ヒトIL-3およびGM-CSFの2つの対立遺伝子を発現する、IL-3/GM-CSF「ヒト/ヒト」(h/h)マウスと呼ばれるホモ接合体KIマウスを作製した。肺組織の従来型RT-PCR解析および定量的RT-PCR解析は、h/hマウスがヒトGM-CSF mRNAのみを発現し、マウスGM-CSF mRNAを発現しないことを示した(図5(b)、5(c))。ヒトGM-CSFタンパク質はELISAでh/hマウスの気管支肺胞洗浄(BAL)液中に検出することができた(図5(d))。これらの結果は、hIL-3/GM-CSF KIマウスがヒトGM-CSF(およびIL-3)を忠実に発現することを示している。
【0117】
図5(a)〜(d)は、非移植hIL-3/GM-CSF KIマウスにおけるヒトGM-CSF発現の検証を示す。図5(a)は、ヒトCsf2の1つの対立遺伝子とマウスCsf2の1つの対立遺伝子をもつKIマウス(h/m)由来の各種組織におけるGM-CSF mRNA発現の代表的なRT-PCR解析を示す。Li, 肝臓;Br, 脳;Lu, 肺;Mu, 筋肉;Sp, 脾臓;Th, 胸腺;LN, リンパ節;BM, 骨髄。下部:ヒトGM-CSFを検出するためのプライマーの特異性を対照マウス(m/m)由来の組織のRT-PCR解析によって検証した。リボソームタンパク質L13(Rpl13)を内在性対照として用いた。図5(b)は、m/mマウスまたはヒトCsf2の2つの対立遺伝子を発現するホモ接合体KIマウス(h/h)(それぞれn=5)由来の肺におけるGM-CSF mRNA発現のRT-PCR解析を示す。Rpl13を内在性対照として用いた;NTC, テンプレートを含まない対照。図5(c)は、(b)と同じGM-CSF mRNA発現の定量的RT-PCR解析を示す。GM-CSF発現はマウスHprtに対して正規化された(それぞれn=5)。図5(d)は、m/mまたはh/h KIマウス(それぞれn=6)から回収されたBAL液中のヒトGM-CSFタンパク質のELISAを示す。結果は2つの独立した実験を代表する。各ドットは1匹のマウスを表す。水平バーは平均値を示す。
【0118】
図5(e)は、hIL-3/GM-CSF KIマウスを作製するための戦略を示す。マウス(上)とヒト(下)のIL3およびCsf2の遺伝子座のゲノム構成がそれぞれ11番および5番染色体上に示される。マウス遺伝子座が本開示に記載する通りにヒト遺伝子座と置き換えられた。
【0119】
図6(f)、(g)は、非移植hIL-3/GM-CSF KIマウスにおけるヒトIL-3およびGM-CSFの発現を示す。活性化脾細胞によるヒトIL-3(f)およびGM-CSF(g)産生のELISA結果が提示される。m/mマウス(白のバー)またはh/m KIマウス(黒のバー)由来の脾細胞はConAとIL-2で48時間刺激し、上清を回収した(それぞれn=1)。m/mマウスではヒトIL-3およびGM-CSFが検出されなかった(ND)。
【0120】
実施例5
hIL-3/GM-CSFを移植されたマウス:増強されたヒト炎症反応
hIL-3/GM-CSF KIマウスは、Rag2とIl2rgの両方の1つの対立遺伝子がすでに欠失されている胚性幹(ES)細胞から作製された。その後Rag2 KO Il2rg KO背景へと交配させることで、ヒトCD34+造血細胞を移植することが可能になった。全体的なヒトCD45+造血細胞のキメリズム、ならびに骨髄中、胸腺中、脾臓および血液中のT細胞、B細胞およびナチュラルキラー(NK)細胞の分布は、hIL-3/GM-CSF KIマウスにおいて有意に増加していなかった(データは示さず)。さらに、全ヒトCD33+骨髄細胞、CD66+顆粒球、CD14+単球/マクロファージ、CD14loCD16+非古典的単球、CD11c+樹状細胞(DC)、およびCD123+CD11c-形質細胞様DCの頻度は、hIL-3/GM-CSF KIマウスにおいて有意に増加していなかった(データは示さず)。これは定常状態の条件下のh/mマウスとh/hマウスの両方にあてはまった。最後に、移植したhIL-3/GM-CSF KIマウス由来のヒト骨髄細胞は同様の能力を有して、インビトロでメチルセルロース中に骨髄コロニーを形成した(データは示さず)。これらの知見は、IL-3とGM-CSFの両方が本明細書で解析した器官での定常状態の骨髄造血にほとんど不要であることを示すKOマウス研究からの結果と一致している。
【0121】
対照的に、GM-CSFは炎症反応を媒介する上で重要な役割を果たしている。GM-CSFの発現は炎症性刺激によって誘導され、これは単球/マクロファージによる炎症性サイトカイン(IL-6およびTNFαなど)の産生と、炎症部位へのそれらの動員とにつながる。移植hIL-3/GM-CSF KIマウス由来のヒトCD14+単球は、GM-CSF受容体α鎖(CD116)の最高発現を示した(図8(d))。したがって、移植hIL-3/GM-CSF KIマウスの解析はヒト単球/マクロファージに集中した。最初に、移植hIL-3/GM-CSF KIマウスにおけるヒト単球の炎症反応が解析された。リポ多糖(LPS)の腹腔内(i.p.)注射によって全身性の炎症を誘発させた。LPS注射後に、循環ヒトCD14+単球の頻度が対照のm/mマウスと比較してh/mマウスでは有意に増加していた(図8(e)、(f))。h/mマウスでのヒト単球動員の増加は、1回および2回のLPS注射後のヒトIL-6の血清濃度の上昇と関連していた(図8(g))。ヒトTNFαのLPS誘発産生もh/mマウスで増加していたが、この結果は統計的有意性に達しなかった。これらのデータは、ヒト造血細胞を用いて移植されたhIL-3/GM-CSF KIマウスがヒト骨髄単球細胞によって媒介されるヒト炎症反応の亢進を有することを示している。
【0122】
図8(d)〜(g)は、移植hIL-3/GM-CSF KIマウスにおけるヒト炎症反応の亢進を示す。図8(d)は、定常状態の移植hIL-3/GM-CSF h/m KIマウス由来のヒト骨髄細胞のフローサイトメトリー解析を示す。ドットプロット(左)はhCD45+mCD45-細胞にゲーティングされる。ヒストグラム(右)はCD14-細胞(集団1)、CD14mid/SSChi顆粒球(集団2)、およびCD14hi単球(集団3)上のGM-CSF受容体α(CD116)発現を示す。解析した合計12匹のマウスからの代表的な一例が示される。図8(e)は、2回のLPS腹腔内注射後72時間のCB移植m/mまたはh/m KIマウス由来のヒト血液細胞の代表的なフローサイトメトリー解析を含む。プロットはhCD45+mCD45-細胞にゲーティングされる。ボックス領域の隣にある数字はヒトCD14+細胞のパーセンテージを示す。図8(f)は、LPS注射後72時間の移植m/mマウス(n=4)またはh/m KIマウス(n=8)におけるヒトCD14+血液細胞の頻度を示す。図8(g)は、1回目(上)および2回目(下)のLPS注射後2〜3時間の移植m/mマウス(n=4〜5)またはh/m KIマウス(n=8)由来の血清中のヒトIL-6についてのELISA結果を示す。1匹のm/mマウスは1回目のLPS注射後に死亡した。各ドットは1匹のマウスを表す。水平バーは平均値を示す。結果は2つの独立した実験を代表する。
【0123】
実施例6
hIL-3/GM-CSFを移植されたマウス:肺での増強されたヒトマクロファージ生着
BAL解析
hIL-3/GM-CSF研究のための気管支肺胞解析は次の方法で行った。気管に挿入したカテーテルから1mlのPBSを入れて肺を膨らませた。これを2回繰り返し、回収した洗浄液をプールした。遠心分離後、細胞フリーの上清は、ELISAによるGM-CSFタンパク質濃度の測定のために、またはBCA Protein Assay Kit(Pierce社)をメーカーの指示に従って用いる総タンパク質含有量の測定のために保存した。赤血球(RBC)をACK溶解緩衝液(Lonza社)で溶解した後、細胞ペレット数を計測し、フローサイトメトリーのためにまたはサイトスピン標本作成のために使用した。細胞をスライド上にスピンさせ、Diff-Quik(商標)Stain Set (Dade Behring社)でメーカーの指示に従って染色した。
【0124】
増強されたマクロファージ生着
マウスGM-CSFの非存在はマウス肺胞マクロファージ(AM)の機能低下につながり、これはホモ接合体hIL-3/GM-CSF KIマウスにおいてヒトマクロファージによる再構成に有利にはたらくはずである。その裏付けとして、h/hマウスの肺およびBALではヒトGM-CSFが高度に発現しており、一方マウスGM-CSFは欠損している。
【0125】
非移植h/hマウスからのマウスAMは肥大し、かつ典型的な「泡沫状」外観(図9(f))を呈した。この泡沫状外観はGM-CSF KOマウス由来のAMに関して記載されている。GM-CSF KOマウスは、最終分化に障害があるAMによる表面活性物質のクリアランスの欠陥に起因するPAPを発症する。GM-CSF KOマウスに関して報告されたものと同様に、非移植h/hマウスは、過ヨウ素酸シッフ(PAS)陽性物質に満ちたAMの胸膜下蓄積など、PAPの特徴を現した(図9(g))。したがって、非移植h/hマウスはマウスAMの分化障害を示してPAPを発症し、それゆえにGM-CSF KOマウスと機能的に同等であると結論づけられた。
【0126】
図9(f)、(g)は、非移植ホモ接合体hIL-3/GM-CSF KIマウスにおけるPAPの発症を示す。図9(f)は、非移植m/mまたはh/h KIマウス由来のBAL細胞のDiff-Quick(商標)染色を示し(倍率400x);グループごとに解析した合計6匹のマウスの代表的な一例が示される。図9(g)は、非移植m/mまたはh/h KIマウス由来の肺組織切片のPAS染色を示し(倍率400x);グループごとに解析した合計12匹のマウスの代表的な一例が示される。
【0127】
次に、ヒト造血細胞を移植した後のh/hマウスの肺区画を検討した。FACS解析は、h/hマウスがBAL中にかなり多くのヒトCD45+細胞をもつことを示した(図6(a)、(b))。肺組織の定量的RT-PCRは、このヒト細胞の増加が主として、ヒト骨髄マーカーCD33、CD11b、CD11c、およびCD14のmRNAを発現する細胞で構成されることを明らかにした(図6(c))。さらに、主に細胞内に発現される成熟マクロファージマーカーである、ヒトCD68のmRNA発現が移植h/hマウスでは著しく増加していた(図6(d))。h/hマウスでのこの増加は、AMによって発現される2つの転写因子、すなわちPU.1(Spi1)およびペルオキシソーム増殖因子活性化受容体γ(PPARγ)のより高い発現と関連していた(図6(d))。PU.1は最終的に分化したAMにおいてGM-CSF依存的方法で高度に発現される。重要なことは、インビトロでGM-CSF KO AMにPU.1を形質導入すると、それらの機能障害が改善することである。PPARγもまたAMで高度に発現されており、GM-CSF KOマウスと同様に、PPARγ KOマウスはPAPを発症する。肺切片の免疫組織学的染色は、移植h/hマウスにおいて、ヒトAMと一致して、典型的な肺胞内位置での多数のhCD68+細胞の存在を明らかにした(図6(e))。対照的に、移植m/m対照マウスではごくわずかなヒトAMしか検出できなかった。要約すると、CD34+造血細胞を移植したh/hマウスの肺は、際立って改善されたヒトマクロファージ生着を示す。
【0128】
図6(a)〜(e)は、ホモ接合体hIL-3/GM-CSF KIマウスが肺での良好なヒトマクロファージ生着を有することを示す。図6(a)は、移植したm/mおよびh/h KIマウス由来のBAL細胞の代表的なフローサイトメトリー解析を示す。丸で囲った領域の隣にある数字はhCD45+およびmCD45+造血細胞のパーセンテージを示す。mCD45+hCD45+細胞は高い自己蛍光を有し、F4/80+マウスAMを構成する。図6(b)は、移植したm/mおよびh/h KIマウス由来のBAL中のヒト造血(hCD45+)細胞の数を提供する(結果は3つの独立した実験(グループあたり合計n=15)から組み合わされる)。図6(c)は、移植したm/mおよびh/h KIマウス(それぞれn=4)由来の肺組織におけるヒトリンパ球および骨髄遺伝子発現の定量的RT-PCR解析の結果を示す。発現はマウスHPRTに対して正規化された(*, P<0.05)。図6(d)は、移植したm/mおよびh/h KIマウス(それぞれn=4)由来の肺組織におけるヒトマクロファージ遺伝子発現の定量的RT-PCR解析を示す。発現はマウスHPRTに対して正規化された(*, P<0.05)。各ドットは1匹のマウスを表す。水平バーは平均値を示す。図6(e)は、移植したm/mおよびh/h KIマウス由来の、ヒトCD68について染色した肺組織切片の免疫組織化学を示す(倍率100x(上)および200x(下))。グループごとに解析した合計10匹のマウスの代表的な一例が示される。
【0129】
実施例7
hIL-3/GM-CSFを移植されたマウス:ヒト造血細胞によって軽減されるPAP
h/hマウスのヒトマクロファージ生着の増加が肺における良好なヒト免疫機能につながるかどうかを検討した。最初に、ヒトマクロファージが非移植h/hマウスに見られるPAP症候群をレスキューできるかどうかを検討した。II型肺胞上皮細胞とAMは両方ともGM-CSFに応答し得るが、PAPは骨髄移植によってレスキューすることができる。これは、造血細胞、特にAM、がPAPを改善できる主な細胞型であることを示している。したがって、ヒト造血細胞を用いて移植されたh/hマウスはPAPの重症度が低いはずであると仮定された。予想通り、非移植h/hマウスはPAS陽性物質の肺胞内蓄積を示し(図7(a))、これはPAPの顕著な特徴である。この仮説に一致して、移植したh/hマウスは肺に軽度のタンパク質蓄積を有し、一部のh/hマウスの肺は(非移植または移植)m/m対照マウスに似ていた(図7(a))。さらに、移植h/hマウスは、BAL液中の総タンパク質量が非移植h/hマウスより有意に低かった(図7(b))。これらの結果は、移植されたヒト造血細胞(おそらくAM)がホモ接合体hIL-3/GM-CSF KIマウスにおいてPAPを軽減できることを示している。
【0130】
図7(a)〜(e)は、ヒト造血細胞がホモ接合体hIL-3/GM-CSF KIマウスにおいてPAPを軽減することを示す。図7(a)は、非移植または移植m/mもしくはh/h KIマウス由来の肺組織切片のPAS染色を示す。2匹の異なる移植h/h KIマウス由来の肺切片が示される(倍率400x)。グループごとに解析した合計10〜12匹のマウスの代表的な例が示される。図7(b)は、非移植(non)または移植h/h KIマウスもしくはm/m対照マウス(グループあたりn=6)由来のBAL液中の総タンパク質の定量を示す。P<0.0001(一元配置ANOVA検定)。Tukeyの多重比較検定で決定されたP値は星印によって示される(**, P<0.01;***, P<0.001)。
【0131】
実施例8
hIL-3/GM-CSFを移植されたマウス:インフルエンザAに対する強いヒトI型IFN応答
インフルエンザA感染
マウス(9〜10週齢)に2×104プラーク形成単位のインフルエンザA/PR8(H1N1)ウイルスを鼻腔内経路で感染させた。感染は、Anafane(商標)(Ivesco社)で深く麻酔しておいたマウスに、PBSで希釈したウイルスストック液50μl(または対照として等量のPBS)を鼻腔内投与することによって行った。感染の24時間後に、上記のRNA抽出および定量的RT-PCR解析のために肺を回収した。
【0132】
肺の恒常性におけるそれらの役割に加えて、AMは肺での宿主防衛に不可欠である。多くの研究は、GM-CSF KOマウスが肺のさまざまな病原体の影響を受けやすくなることを示している。移植されたヒトAMの肺病原体に対する機能的応答を評価するために、移植h/hマウスにインフルエンザA/PR8(H1N1)ウイルスを鼻腔内経路で感染させた。AMは肺ウイルス感染後のI型インターフェロン(IFN)の主な産生細胞であり、かつAMはインフルエンザAへの有効な先天性応答に必要とされる。ヒトヒポキサンチンホスホリボシルトランスフェラーゼ(HPRT)mRNAの発現は、対照m/mマウスと比べて、移植h/hマウスの肺で有意に高かった(図8(a))が、これは良好なヒト免疫細胞のキメリズムを示している。移植m/mマウスは、PBSを鼻腔内投与された移植m/mマウスと比べた場合、インフルエンザA感染後にヒトIFNβ mRNA発現の有意な誘導を示さなかった(図8(b))。対照的に、移植h/hマウスは、非感染h/hマウスと感染m/mマウスの両方よりも有意に多いヒトIFNβ mRNAを発現した(図8(b))。m/mマウスと比較して、h/hマウスにおけるヒトIFNβ mRNAの発現増加は、ヒトHPRTに、すなわち肺中のヒト細胞の数に、正規化された場合、まだ有意であった(図8(c))。まとめると、ホモ接合体hIL-3/GM-CSF KIマウスは、良好なヒトマクロファージキメリズムと、ウイルス感染に対して増強されたヒト粘膜免疫につながる肺での機能とを可能にする。
【0133】
図8(a)〜(g)は、ホモ接合体hIL-3/GM-CSF KIマウスがインフルエンザA感染に対して強いヒトI型IFN応答を開始することを示す。図8(a)〜(c)は、インフルエンザA(PR8)の鼻腔内感染後24時間のm/mおよびh/h KI由来の肺組織における遺伝子発現の定量的RT-PCR解析を示す(それぞれn=8)。PBSの鼻腔内投与を対照(PBS)として用いた(それぞれn=4)。図8(a)は、マウスHprtに正規化されたヒトHprtの発現を示す。P<0.0001(一元配置ANOVA検定)。図8(b)は、マウスHprtに正規化されたヒトIFNγの発現を示す。P=0.0171(一元配置ANOVA検定)。図8(c)は、ヒトHprtに正規化されたヒトIFNγの発現を示す。P=0.0032(一元配置ANOVA検定)。Tukeyの多重比較検定で決定されたP値は星印によって示される(*, P<0.05;**, P<0.01;***, P<0.001)。各ドットは1匹のマウスを表す。水平バーは平均値を示す。結果は2つの独立した実験を代表する。
【0134】
実施例9
hIL-3/GM-CSFを移植されたマウス:マイコバクテリア感染後のヒト細胞を含む肉芽腫
マクロファージが病原体特異的免疫応答において中心的な役割を果たす肺に向性を有する第2の病原体に対するヒト炎症反応を支持するhIL-3/GM-CSF KIマウスの能力が検討された。マイコバクテリア感染後の肉芽腫の形成が選択された。肉芽腫は、マイコバクテリア感染の特徴である、特殊化した局所炎症反応を表している。それはDTH応答の古典的な例であり、その形成は活性化T細胞とマクロファージとの相互作用に依存している。最適なDTH応答にはIL-3とGM-CSFの両方が必要であり、重要なことは、GM-CSF KOマウスがマイコバクテリアに感染した場合肉芽腫を形成しないことである。
【0135】
移植したhIL-3/GM-CSF h/m KIマウスにBacillus Calmette-Guerin (BCG)を静脈内注射により感染させた。BCGはヒト結核に対するワクチンとして使用されるM. ボビス(M. bovis)の弱毒株である。BCG感染実験に用いたマウスは>20%のhCD45+細胞の血中生着レベルを有し、hCD45+細胞の>8%がT細胞(hCD3+)であった。マウスは感染時に9〜10週齢であった。1×105コロニー形成単位のBCG(コペンハーゲンのStatens Serum Institute社)を0.1ml量で尾静脈注射によりマウスに感染させた。
【0136】
肉芽腫の形成にはT細胞が不可欠であることから、最初に、感染4週後のBCGに対するヒトT細胞応答を調べた。フローサイトメトリーは、移植したm/mおよびh/mマウスの両方の肺にヒトT細胞が存在することをはっきりと示した(図9(a))。実際に、T細胞はBCG感染肺において主要なヒト造血細胞型であった。m/m対照マウスに比較して、BCGに感染したh/mマウスは肺でのヒトCD4対CD8 T細胞の平均比率がより高かったが、その差は統計的有意性に完全には達しなかった(図9(b)、9(c))。これら2つのマウスグループ間で脾臓hCD4/hCD8 T細胞の比率の差は認められなかった。
【0137】
次に、2種類のT細胞由来サイトカイン、すなわちIFNγとTNFαの発現を解析した。これらは両方ともマイコバクテリアに対する防御免疫応答に極めて重要である。感染したマウス由来の脾細胞を、免疫優性マイコバクテリア抗原TB10.4から誘導されたペプチドで再刺激した後の、細胞内サイトカイン染色によるBCG特異的IFNγ産生が検討された。予想されたとおり、IFNγを産生するマウスT細胞の集団がBALB/cマウス由来の脾細胞の中に検出された(図9(d))。さらに、ヒトBCG特異的T細胞応答が移植したh/mおよびm/mマウスのサブセットで見られた(図9(d))。最後に、移植したh/mおよびm/mマウスの大部分はBCG感染後に肺でヒトIFNγおよびTNFα mRNAを発現した(図9(e))。これらの結果は、移植されたマウスのサブセットがBCGに対する病原体特異的ヒトT細胞応答を開始することができるが、この応答はhIL-3/GM-CSF KIマウスにおいて増強されなかったことを示している。これに一致して、細菌負荷にはh/mおよびm/mマウス間で差がなかった。
【0138】
次に、肉芽腫の形成が感染4週後に肺と肝臓で組織学により評価された。非移植m/mマウス(T細胞を欠く)は肉芽腫を発生せず(表1)、これは肉芽腫形成のためのT細胞の必要性と一致する。同様に、ヒト細胞を用いて移植されたm/mマウスは肺にも肝臓にも肉芽腫を示さなかった(表1)。これに対して、h/mマウスの大部分には肺(図10(a))または肝臓または両方の臓器に小さな病変または肉芽腫があった(表1)。一般的に、観察された肉芽腫は小さく、ヒトよりもマウスの肉芽腫に特有のゆるい組織化を有していた。しかしながら、hIL-3/GM-CSF KIマウスの肺肉芽腫は、免疫組織化学によって証明される通りに、ヒト造血細胞(hCD45+)を含んでいた(図10(b))。これらの細胞の大部分はヒトT細胞(hCD3+)であり、中心部に数個のヒトマクロファージ(hCD68+)が存在していた(図10(b))。まとめると、移植されたhIL-3/GM-CSF KIマウスは、マイコバクテリア感染に応答してヒトT細胞とヒトマクロファージを含む肉芽腫を発症することができるが、こうしたことはHISマウスでは以前に報告されていない。表1は、BCG感染4週後のBALB/c、非移植m/m(non)、移植m/m、および移植hIL-3/GM-CSF h/m KIマウス由来の肝臓および肺組織切片に認められた病変/肉芽腫を示す。
【0139】
(表1)BCGに感染させた移植したhIL-3/GM-CSF KIにおける肉芽腫
【0140】
図9(a)〜(g)は、移植hIL-3/GM-CSF KIマウスにおけるBCGに対するヒトT細胞応答を示す。図9(a)〜(c)は、BCG感染4週後の移植したm/mおよびh/m KIマウス由来の肺細胞のフローサイトメトリー解析を示す。図9(a)は、肺におけるヒトT細胞(hCD45+hCD3+)の頻度を示す。ボックス領域の隣にある数字は細胞のパーセンテージを示す。図9(b)は、肺におけるヒトCD4およびCD8 T細胞の分布を示す。ドットプロットはhCD45+hCD3+細胞に対しゲーティングされる。象限内の数字は細胞のパーセンテージを示す。図9(c)は、肺におけるヒトCD4対CD8 T細胞の比率を示す(それぞれn=6)。各ドットは1匹のマウスを表す。水平バーは平均値を示す。図9(d)は、BCG感染4週後のBALB/cマウス、移植m/mマウス、および移植h/m KIマウス由来の脾細胞のフローサイトメトリー解析を示す。脾細胞は、本明細書に記載したTB10.4タンパク質をカバーする重複ペプチドのプールを用いてインビトロで再刺激された。ドットプロットは、細胞内サイトカイン染色により測定されたマウスIFNγ+ CD4 T細胞(mCD4+)またはヒトIFNγ+ T細胞(hCD3+)の頻度を示す。アイソタイプの適合した抗体による染色を対照として用いた。図9(e)は、BCG感染4週後のBALB/cマウス、非移植(non)m/mマウス、移植m/mマウス、および移植h/m KIマウス由来の肺組織におけるヒトIFNγ(左)およびTNFα(右)遺伝子発現の定量的RT-PCR解析の結果を示す(グループあたりn=4〜7)。各ドットは1匹のマウスを表す。水平バーは平均値を示す。
【0141】
図10(a)、(b)は、移植したhIL-3/GM-CSF KIマウスがBCG感染後にヒト細胞を含む肉芽腫を発症することを示す。図10(a)は、BCG感染4週後の移植h/m KIマウス由来の肺組織切片のヘマトキシリンおよびエオシン(H&E)染色を示す(倍率100x(左)および倍率200x(右))。図10(b)は、BCG感染4週間後の移植h/m KIマウス由来のヒトCD45、CD3、またはCD68を染色した肺組織切片の免疫組織化学を示す(倍率200x)。肺肉芽腫を有する2匹のマウスのうち代表的な一例が示される。
【0142】
実施例10
hTPOマウス:生着および解析
TPOマウスへの移植
レシピエントマウスは、Traggiaiらに記載される通りにヒト造血前駆細胞を用いて移植された。臍帯血試料は、イェール大学の人権擁護調査委員会(Department of Labor and Birth, Yale New Haven Hospital, New Haven, CT)からの承認の下で、健康な満期新生児から採取された。胎児肝臓試料は、アルバート・アインシュタイン医科大学(Bronx, NY)のヒト胎児組織リポジトリ(Human Fetal Tissue Repository)から、およびAdvance Biosciences Resources社(Alameda, CA)から取得された。
【0143】
胎児肝臓試料を小さな断片に切断し、コラゲナーゼD(100ng/ml、Roche社)を用いて37℃で45分間処理し、かつ細胞懸濁液を調製した。ヒトCD34+細胞は、密度勾配遠心(リンパ球分離溶液(Lymphocyte Separation Medium)、MP Biomedicals社)と、その後の抗ヒトCD34マイクロビーズを用いる陽性免疫磁気選別(positive immunomagnetic selection)によりメーカーの指示(Miltenyi Biotec社)に従って、胎児肝臓試料からまたは臍帯血から精製した。細胞は、FBSを含む10%DMSO中で凍結させるか、または直接注入した。
【0144】
新生仔(生後1日以内)を亜致死的に照射し(X線照射、4時間間隔で2×150cGy)、20μlのPBS中の100,000〜200,000個のCD34+細胞を、22ゲージ針(Hamilton社, Reno, NV)を使って肝臓に注入した。
【0145】
すべての実験は、イェール大学人権擁護調査委員会プロトコルおよびイェール大学動物管理使用委員会プロトコルに従って行った。
【0146】
TPO発現
マウスおよびヒトTPOタンパク質の血清濃度は、種特異的ELISA(RayBiotech社)によりメーカーのプロトコルに従って測定した。TPOをコードするマウスおよびヒトmRNAの発現を測定するために、組織を成体動物から分離し、全RNAを、TRIzol(Invitrogen社)を用いてメーカーの指示に従って精製した。混入しているゲノムDNAをRNase-Free DNase I(Roche社)で処理して除去し、そのRNAを、SuperScript II逆転写酵素(Invitrogen社)とオリゴdTプライマーを用いて逆転写した。PCR増幅のために次のプライマーを使用した。
【0147】
ヒトTPOがこれらのマウスで忠実に発現されるかどうかを調べるために、全RNAをTPOh/mマウス由来のさまざまな器官から抽出した。かつTPOをコードするマウスおよびヒトmRNAの両方について同様の発現パターンがRT-PCRにより観察された(図11(a))。次に、TPOm/m、TPOh/mおよびTPOh/hマウス由来の、TPOを発現することが公知である3種の組織または細胞型(肝臓、腎臓および間葉系多能性間質細胞)における発現を比較した。TPOm/mおよびTPOh/mマウスからの試料ではマウスTpoの発現が検出され、一方TPOh/mおよびTPOh/hマウスではヒトTPOが発現された(図11(b))。標的マウスの血清中のTPOタンパク質の濃度も測定された。マウスTPOはTPOm/mおよびTPOh/m動物で検出され、ヒトTPOはTPOh/mおよびTPOh/hで検出された(図11(c))。測定されたヒトTPOの濃度はマウスTPOより約10倍低かった。しかしながら、この差異は健康なヒトとマウスで報告された生理的濃度に適合するものであり(図11(c))、サイトカイン半減期の種特異的な差異に起因する可能性がある。
【0148】
図11は、(a)Rag2+/-γcY/- TPOh/mマウスの異なる組織におけるマウスTPO(mTpo)およびヒトTPO(hTPO)発現のRT-PCR解析(マウスRpl13aはハウスキーピング遺伝子として使用した);(b)Rag2-/-γc-/- TPOm/m、TPOh/mおよびTPOh/hマウスの肝臓、腎臓および間葉系多能性間質細胞(MSC)におけるmTpoおよびhTPO発現のRT-PCR解析;(c)TPOm/m、TPOh/mおよびTPOh/hマウスの血清中の、ELISAにより測定されたマウスおよびヒトTPOタンパク質の濃度(pg/ml、平均±S.D.、n=7〜9)を示す。ND:検出されず。示した正常範囲はR&D Systems社のThrombopoietin Quantikineキットからである。
【0149】
間葉系多能性間質細胞の単離
TPO研究のため、マウスの大腿骨と脛骨を採取し、かつ骨髄細胞を流し出した。骨を小片に切断し、コラゲナーゼPおよびD(10μg/ml)を用いて37℃で45分間消化した。骨関連細胞を反復ピペッティングによって収集した。刺激用サプリメント(stimulatory supplements)(Stemcell Technologies社)を加えたMSC培地の存在下で細胞を14日間培養した。その培養物から、CD45およびTer119抗体を用いる免疫磁気セルソーティング(MACS、Miltenyi Biotec社)により造血系細胞を除去した。非造血細胞(CD45-Ter119-)をもう5日間培養し、MSC表現型(CD45-Ter119-Sca1+CD90+)をFACSにより確認した(Diminici et al. (2006) Minimal criteria for defining multipotent mesenchymal stromal cells, The International Society for Cellular Therapy position statement, Cytotherapy 8:315-317)。
【0150】
TPOマウスにおける造血細胞集団の解析
移植後8〜12週でマウスを採血した。ACK(Lonza社)を用いて赤血球を3回溶解し、細胞を抗マウスCD45および抗ヒトCD45抗体で染色した。CD45+細胞の少なくとも1%がヒト由来であった動物をさらなる解析のために使用した。移植されたマウスの約80%がこの生着閾値に達しており、TPOm/mおよびTPOh/hグループの間で違いは認められなかった。
【0151】
マウスを移植後3〜4または6〜7ヶ月で屠殺した。単一細胞懸濁液を骨髄(2本の大腿骨と2本の脛骨から流した)、脾臓および胸腺から調製した。赤血球をACK溶解により排除し、FACS解析のため次の抗体を用いて細胞を染色した。全体的な造血移植のために:抗マウスCD45-eFluor450 (30-F11、eBioscience社)および抗ヒトCD45-APC-Cy7 (2D1)。ヒト造血幹細胞前駆細胞および造血系列のために:抗ヒトCD14-PerCP (MoP9)、CD19-APC (HIB19)、CD33-APC (WM53)、CD34-PE (AC136、Miltenyi Biotec社)、CD38-FITC (HIT2)、CD41a-APC (HIP8)およびCD66-FITC (B1.1)。
【0152】
マウス幹細胞前駆細胞の解析のため、抗細胞系列カクテル(anti-lineage cocktail)はCD3ε(145-2C11)、CD11b (M1/70)、CD11c (HL3)、CD19 (1D3)、Gr1 (RB6-8C5)およびLy-76 (Ter119)に対するビオチン化抗体を含んでいた。その後、細胞をストレプトアビジン-APC-Cy7、抗cKit-APC (2B8)および抗Sca1-PE-Cy7 (D7)で染色した。
【0153】
すべての抗体は、特に指定した場合を除き、BD Biosciences社から入手した。データはFACSCalibur(商標)またはLSRII(商標)フローサイトメーター(BD Biosciences社)で取得し、FlowJo(商標)ソフトウェアを用いて解析した。
【0154】
TPOマウスにおけるヒト造血幹細胞前駆細胞の機能的特性解析
3〜7匹の移植されたマウスから得られた骨髄細胞をプールし、ヒトCD34+細胞を、マウスCD45+細胞のMACS枯渇(Miltenyi Biotec社)とその後のFACSAria(商標)フローサイトメーター(BD Biosciences社)でのヒトCD45+CD34+細胞のFACSソーティングによって精製した。
【0155】
ヒトCD34+細胞のコロニー形成能を評価するために、IMDMに20%FCS、2mM L-グルタミン、55μM 2-メルカプトエタノール(試薬はすべてGIBCO社から)を添加し、Methocult(商標)H4100、1%BSA(Stemcell Technologies社)と混合し、次のヒトサイトカインを加えた:SCF (10ng/ml)、FLT3I (10ng/ml)、TPO (50ng/ml)、IL-3 (20ng/ml)、IL-6 (10ng/ml)、IL-11 (10ng/ml)、GM-CSF (50ng/ml)、EPO (4U/ml) (すべてR&D systems社から)。100,000〜150,000個のソーティングされた細胞を60mmペトリ皿にまき、37℃、5%C02で12〜14日間インキュベートした。12〜14日でコロニー数を計測し、顕微鏡で特定の骨髄系列に分類した。
【0156】
二次移植実験のため、TPOm/mまたはTPOh/h一次レシピエントから精製された100,000個のCD34+細胞を、上記の通りに、亜致死的に照射(2×200cGy)したRag2-/-γc-/- TPOm/m二次レシピエントに注入した。これらのマウスを8週間後に屠殺し、骨髄中のヒトCD45+細胞の割合をFACSで測定した。
【0157】
TPOマウスデータの統計解析
データは対応のない両側t検定を用いて比較された。2つより多い試料を比較する場合は、一元配置ANOVAと、その後にTukeyの事後検定を行った。二次移植実験において生着を示したマウスの割合はPearsonのカイ二乗検定を用いて比較された。p値が0.05を下回った場合は、差を有意と見なした。
【0158】
実施例11
hTPOを移植されたマウス:TPOh/hレシピエントマウスの骨髄における改善されたヒト生着レベル
ヒト化および移植されたマウスの骨髄の表現型分類
ヒト化マウスの骨髄から分離された細胞をフローサイトメトリーで解析したところ、非ヒト化マウス(すなわち、TPO遺伝子のヒト化を欠くRAGおよびIl2rgノックアウト)の生着と比べて、全ヒト造血細胞、ヒト造血幹細胞、ヒト骨髄細胞、およびヒト顆粒球の生着の統計的に有意な改善が認められた。図12および14参照。
【0159】
野生型Tpo(TPOm/m)、ヘテロ接合型(TPOh/m)またはホモ接合型(TPOh/h)TPO遺伝子置換を有するRag2-/-γc-/-マウスを、記載した通りに作製した。照射(2×1.5Gy)した新生仔Rag2-/-γc-/- TPOm/mおよびTPOh/hマウスは、臍帯血または胎児肝臓から精製されたヒトCD34+細胞を用いて移植され、3〜4ヶ月または6〜7ヶ月後に骨髄における生着を解析された。
【0160】
図12は生着研究の結果を示す。図12(a)は、ヒトCD34+細胞の移植後3〜4ヶ月のRag2-/-γc-/- TPOm/mおよびTPOh/hマウスの骨髄におけるヒトおよびマウスCD45+細胞の代表的なFACS解析を示す。2匹の代表的なマウスの結果が各遺伝子型について示される。全(マウス+ヒト)CD45+細胞集団中のマウスおよびヒトCD45+細胞のパーセンテージが示される。図12(b)は、移植後3〜4ヶ月(左、n=42〜53)または6〜7ヶ月(右、n=20〜25)の骨髄中のヒトCD45+細胞のパーセンテージを示す。各記号は個々のマウスを表し、水平バーは平均値を示す。図12(c)は、(b)と同じ動物の骨髄中のヒトCD45+細胞の絶対数を示す。p値は統計的有意性を示す。
【0161】
TPOm/mレシピエントと比較して、TPOh/hの骨髄中のヒト造血細胞(hCD45+)のパーセンテージ(図12(a)および12(b))と絶対数(図12(c))に両時点で有意な増加が観察された。さらに、TPOh/hレシピエントはより低い生着のばらつきを示し、3〜4ヶ月の時点でマウスの75%に少なくとも80%のヒトキメリズムが見られた(図12(b))。CD34+細胞の供給源はこの結果に影響を及ぼさなかったが、それは、TPOh/h宿主でのキメリズムの同様の増加が臍帯血由来細胞と胎児肝臓由来細胞で観察されたからである(図12(d))。興味深いことに、TPOm/m宿主ではヒト細胞の数が早い時点と遅い時点の間に減少したが、TPOh/h動物ではそれらが一定のままであった(図12(c))。これらの結果は、マウスにおけるTPOの先に記載した機能と一致している。第一に、TPOは照射レシピエントマウスへの移植後にHSCの増幅に有利にはたらいて、生着レベルの増加をもたらす。第二に、それは成体HSCの維持に有利にはたらき、成体期を通じて持続的な造血をもたらす。
【0162】
実施例12
hTPOを移植されたマウス:マウスおよびヒト血小板に対するTPOヒト化の効果
TPOマウスにおける血小板解析
末梢血中の血小板数はHemavet(商標)950FS装置(Drew Scientific社)を用いて測定した。次に血液試料を抗マウスCD61-PE (2C9.G2)および抗ヒトCD41a-APC (HIP8)で染色し、マウスおよびヒト血小板のパーセンテージを、細胞のサイズ(FSC)または顆粒性(granulosity)(SSC)にいかなるゲートも配置せずに、フローサイトメトリーで測定した。絶対マウスおよびヒト血小板数は、これらの各パーセンテージを絶対血小板数と乗じることによって算出した。
【0163】
TPOは血小板産生に及ぼすその重要な機能について周知されているので、TPOのヒト化が血小板の発生に影響を与えるかどうかを検討した。TPO遺伝子の両方の対立遺伝子のヒト化は、非移植Rag2-/-γc-/-マウスの血中血小板数の約2倍低下につながった(図13(a))。ヒト細胞を移植した後では、TPOh/hマウスにおけるマウス血小板数は正常値の25%未満にさらに低下した(図13(d))。ヒト対マウス血小板の比率(図13(b)、13(c))、ならびにヒト血小板の絶対数(図13(e))は、TPOm/mよりもTPOh/hマウスにおいて高くなる傾向にあったが、これらの差はどれも統計的有意性に達しなかった。さらに、ヒト細胞のうちの骨髄巨核球(CD41a+細胞)のパーセンテージは両系統で同等であった(図13(f))。これらの結果は、ノックイン戦略によって達成されたヒトTPOのレベルまたは生物活性がマウスTPO機能を完全に置き換えるには十分でないことを実証しており、さらに、ヒトTPOが単独ではマウス環境においてヒト血小板産生を支持するのに十分でないことを示唆している。
【0164】
図13(a)は、成体非移植Rag2-/-γc-/- TPOm/m、TPOh/mおよびTPOh/hマウスの血液中の血小板数を示す。p<0.0001 (一元配置ANOVA、n=7〜17;示したp値はTukeyの事後検定により算出した)。各記号は個々のマウスを表し、水平バーは平均値を示す。図13(b)移植後3〜4ヶ月のRag2-/-γc-/- TPOm/mおよびTPOh/hマウスの血液中のマウス(mCD61+)およびヒト(hCD41a+)血小板の代表的FACS解析。数字は全イベントのうちの比率を示す。図13(c)TPOm/mおよびTPOh/hマウス(n=19〜22)における、FACSにより測定されたヒト血小板のキメリズム。血中のヒトCD45+細胞のパーセンテージが5%より高いマウスだけをこの解析に含めた。(d)、(e)TPOm/mおよびTPOh/hレシピエントの血液中のマウス(mCD61+,(d))およびヒト(hCD41a+,(e))血小板数;(f)骨髄中のヒトCD45+細胞のうちのヒト巨核球(CD41a+)のパーセンテージ。
【0165】
図13(g)〜(i)は、二次リンパ器官におけるヒト生着レベルを示す。図13(g)、(h)は、Rag2-/-γc-/- TPOm/mおよびTPOh/hマウスの血液(20g;n=43〜53)および脾臓(13h;n=35〜36)中のヒトCD45+細胞のパーセンテージを提供する(各記号は個々のマウスを表し、水平バーは平均値を示す)。図13(i)は、移植したTPOm/mおよびTPOh/hレシピエント(n=24〜34)の胸腺の全細胞性を示す。胸腺に見られた細胞の90%超がヒト由来(hCD45+)であった。
【0166】
実施例13
hTPOを移植されたマウス:TPOヒト化マウスにおける多系列の造血
ヒト化および移植されたマウスの血液細胞の表現型分類
ヒト化マウスの血液から分離された細胞をフローサイトメトリーで解析したところ、非ヒト化マウス(すなわち、TPO遺伝子のヒト化を欠くRAGおよびIl2rgノックアウト)の移植と比べて、ヒト単球および顆粒球の移植において統計的に有意な改善が認められた。図14参照。
【0167】
ヒトTPOがインビボでヒト造血幹細胞前駆細胞の多系列分化に有利にはたらくことができるかどうかを検討した。以前に報告された通りに(Traggiai et al.; Ishikawa et al. (2005) Development of functional human blood and immune systems in NOD/SCID/IL2 receptor gamma chain(null) mice, Blood 106:1565-1573))、移植されたヒト細胞は野生型Rag2-/-γc-/-宿主において主にB細胞(CD19+)を生じさせ(脾臓中のヒト細胞の61.51±4.71%、平均±標準誤差、n=32)、骨髄細胞はほんのわずかしか含まれていなかった。TPOm/mレシピエントとTPOh/hレシピエントを比較した場合、TPOh/hマウスの骨髄中のCD33+骨髄細胞の頻度の有意な増加が観察された(図14(a)および14(b))。興味深いことに、この増加は主として顆粒球(CD33+CD66hiSSChi細胞)によるものであり、単球(CD33hiCD66loCD14+)の割合は両系統で類似していた(図14(a)、(c)、(d)、(e))。骨髄細胞(顆粒球と単球の両方)のパーセンテージもまた、TPOh/h動物の末梢血で有意に増加していた(図14(a)、(f)、および(g))。
【0168】
図14(a)〜(g)は、移植したTPOm/mおよびTPOh/hマウスのCD33+、CD66+、CD14+細胞で測定した場合の、hTPOマウスにおける改善された多系列造血を示す。図14(a)は、移植後3〜4ヶ月のRag2-/-γc-/- TPOm/mおよびTPOh/hマウスの骨髄中および血液中のヒト骨髄細胞集団の代表的なFACS解析を示す。数字は示したゲーティングされた細胞集団のうちのパーセンテージを示す。図14(b)〜(e)は、Rag2-/-γc-/- TPOm/mおよびTPOh/hレシピエント(n=19)の骨髄における全ヒトCD45+細胞キメリズムに対するヒト骨髄細胞集団の解析を示す。図14(b)は、全骨髄集団(CD33+細胞)を提供する。図14(d)は、顆粒球(CD33+CD66hi)を提供し、図14(c)は、TPOh/hレシピエントの骨髄から精製されたhCD45+SSChiCD33+CD66hi細胞のDiff-Quick(商標)染色を示す。図14(e)は、単球(CD33+CD66loCD14+)を示す。各記号は個々のマウスを表し、水平バーは平均値を示す。図14(f)および(g)は、Rag2-/-γc-/- TPOm/mおよびTPOh/hレシピエント(n=6〜7)の血液中の全ヒトCD45+細胞キメリズムに対するヒト骨髄細胞集団の解析を示す;図14(f):顆粒球(CD66+);図14(g):単球(CD14+)。
【0169】
実施例14
hTPOを移植されたマウス:マウスおよびヒト造血幹細胞前駆細胞に対するヒト化効果
HSCならびに前駆細胞それら自体の数および機能に及ぼすヒトTPOの効果が解析された。TPOの遺伝子欠失は成体マウスではHSCの減少につながる。TPOヒト化がマウスHSCを含むと免疫表現型的に定義されるマウス集団に影響を及ぼすことができるかどうかを調べるために、非移植TPOm/m、TPOh/mおよびTPOh/h成体マウスの骨髄中の系列陰性(lineage-negative)Sca1+c-Kit+細胞のパーセンテージを比較した。
【0170】
図15は、ヒトTPOノックインマウスの骨髄中のlin-c-Kit+Sca1+細胞の減少、ならびにヒト幹細胞前駆細胞の数および自己再生能の増加を示す。図15(a)は、WT TPO (TPOm/m) Rag2-/-γc-/-マウスと比較した、非移植Rag2-/-γc-/- TPOh/mおよびTPOh/hマウスの骨髄中のマウスLin-Sca1+c-Kit+幹細胞前駆細胞のFACS解析の代表的な結果を示す。数字はLin-集団のうちのSca1+c-Kit+細胞のパーセンテージを示す。図15(b)は、(a)に示された結果の定量的解析を示す。p=0.0006(一元配置ANOVA;示したp値はTukeyの事後検定により算出した;遺伝子型あたりn=5;示された結果は2つの独立した実験を代表する)。各記号は個々のマウスを表し、水平バーは平均値を示す。図15(c)は、移植後3〜4ヶ月のRag2-/-γc-/- TPOm/mおよびTPOh/hマウスの骨髄中のヒトCD34+CD38-細胞の代表的なFACS解析を示す。数字はヒトCD45+CD34+細胞のうちのCD38-細胞のパーセンテージを示す。図15(d)は、TPOm/mおよびTPOh/hレシピエントマウス(n=43〜53)におけるヒトCD45+CD34+集団中のCD38-細胞のパーセンテージの定量的解析を示す。図15(e)は、15(d)dと同じマウスの骨髄中のヒトCD34+CD38-細胞の絶対数を示す。図15(f)および(g)は、Rag2-/-γc-/- TPOm/mおよびTPOh/hレシピエントから精製されたヒトCD45+CD34+細胞を用いるメチルセルロースコロニー形成アッセイ法を示す。図15(f)はCFU-GEMMであり、図15(g)はBFU-E(黒)、CFU-G(白)、CFU-M(灰色)およびCFU-GM(破線)である。CD34+細胞は3〜4匹のマウスのグループからプールして、レシピエントマウスの遺伝子型あたり4つの独立したプールを得た。図15(h)では、Rag2-/-γc-/- TPOm/mおよびTPOh/h一次レシピエントマウスからヒトCD45+CD34+細胞を精製し、新生仔Rag2-/-γc-/-マウスに移植し(マウスあたり100,000個の細胞)、8週間後に二次レシピエントにおけるヒトCD45+キメリズムを測定した。結果は2つの独立した実験からプールされたものである(n=7〜12一次レシピエント、n=11〜19二次レシピエント)。
【0171】
TPOm/mと比較して、TPOh/mとTPOh/hの両マウスではこれらの細胞のパーセンテージの有意な減少が観察され(図15(b))、ヒトTPOはマウス受容体に完全には交差反応性でないか、このノックイン設定ではマウス細胞に十分な量で利用されないことを示唆している。
【0172】
移植したTPOm/mおよびTPOh/h宿主の骨髄中のヒトCD34+集団が特性解析された。長期再増殖能を有するヒトHSCはLin-CD34+CD38-細胞画分に含まれる。ヒトCD45+集団のうちのCD34+細胞のパーセンテージはTPOh/hマウスでわずかに増加していた(12.39±0.79%対10.00±0.81%、平均±標準誤差、n=43〜53、p=0.037)。TPOm/mレシピエントに比較して、TPOh/hではCD34+集団内のCD38-細胞のパーセンテージに、少ないながらも(1.5倍)、統計的に有意な増加が観察された(図15(c)、(d))。全体として、これはTPOヒト化マウスにおいてCD34+CD38-細胞の絶対数の有意な増加(約2.8倍)をもたらした(図15(e))。したがって、細胞表面の免疫表現型に基づいて、ヒトTPOはHSCに非常に富むことが公知である細胞の集団を支持している。
【0173】
この細胞集団の機能的特性を検討するために、ヒトCD34+細胞をTPOm/mおよびTPOh/hマウスの骨髄から精製し、かつインビトロでメチルセルロースコロニー形成アッセイ法により評価した。CFU-GEMMは、すべての赤芽球-巨核球および骨髄細胞への分化能を少なくとも含む未熟細胞に由来する多系列骨髄系コロニーである。CFU-GEMMの形成は、少数ではあるが、TPOh/hレシピエントマウスから単離されたCD34+細胞の4つすべての試料から検出されたのに対し、TPOm/mからは1つの試料のみがCFU-GEMMを生成したにすぎなかった(図15(f))。この結果は、TPOh/hレシピエントにおける未熟なヒト造血前駆細胞の改善された維持を示している。さらに、インビボで観察された増大した骨髄分化と一致して(図14)、CFU-Mの数もまた、TPOm/mマウスに比較して、TPOh/hから単離されたヒトCD34+細胞試料において有意に高かった(培養した150,000個のCD34+細胞あたり225.0±12.25対81.25±10.80コロニー、平均±標準誤差、p=0.0001;図15(g))。
【0174】
HSCの維持および/または自己再生は機能的には二次移植の成功によって最もよく実証される。マウスにおいて連続的に生着するSCID repopulating cell(SRC)は現在、ヒトHSC機能のための信頼できるサロゲート実験基準を表す。したがって、ヒトCD34+細胞をTPOm/mおよびTPOh/h一次レシピエントの骨髄から精製し、かつRag2-/-γc-/-新生仔マウスに同様に少数(動物あたり100,000個のCD34+細胞)を移植した。二次レシピエントの骨髄を8週間後に解析した(図15(h))。TPOm/m一次レシピエントから単離されたヒトCD34+細胞は、ヒトCD45+細胞が11匹のうち2匹の二次レシピエントにしか検出されなかったので、連続的に生着する能力が非常に低かった。対照的に、TPOh/h一次レシピエントから単離されたCD34+細胞を用いて移植された19匹のうち15匹のマウスの骨髄にヒトCD45+細胞が存在していた(p=0.0012)。二次レシピエントマウスの遺伝子型は両グループ(TPOm/m)で同じだったので、この結果は、一次レシピエントにおけるヒトTPOの存在が自己再生能を高められたヒト細胞の維持に有利にはたらくことを示している。
【0175】
まとめると、これらの結果は、ホモ接合体TPOヒト化マウスがヒト造血幹細胞前駆細胞の自己再生能および多系列分化能を維持するための良好な環境を提供することを実証している。
【0176】
当業者は、本開示で明示的に記載または表現されていないが、本発明を具体化しかつ本発明の精神および範囲に含まれる、さまざまな修正を考案することができる。すべての実施例は、読者が本発明の原理と概念を理解するのを助けるために提供されるものであり、記載された特定の実施例および態様に限定されずに用いられる。本発明のすべての原理、局面、態様および実施例は、等価物が現在公知であろうと将来開発されようと、それらの等価物を包含することを意図している。本発明の範囲は本開示で示したまたは説明した態様および実施例に限定されるものではない。
【特許請求の範囲】
【請求項1】
(a) マウスRAG遺伝子ノックアウト;
(b) マウスIl2rg遺伝子ノックアウト;
(c) ヒトIL-3遺伝子がマウスIL-3遺伝子座に存在する、マウスIL-3遺伝子の該ヒトIL-3遺伝子との置換;および
(d) ヒトGM-CSF遺伝子がマウスGM-CSF遺伝子座に存在する、マウスGM-CSF遺伝子の該ヒトGM-CSF遺伝子との置換
を含む、遺伝子改変されたマウス。
【請求項2】
ヒトトロンボポエチン遺伝子がマウストロンボポエチン遺伝子座に存在する、マウストロンボポエチン遺伝子の該ヒトトロンボポエチン遺伝子との置換
をさらに含む、請求項1記載のマウス。
【請求項3】
前記RAG遺伝子ノックアウトが、RAG1遺伝子ノックアウト、RAG2遺伝子ノックアウト、およびこれらの組合せから選択される、請求項1記載のマウス。
【請求項4】
ヒト造血細胞の移植をさらに含む、請求項1記載のマウス。
【請求項5】
前記ヒト造血細胞が、臍帯血細胞および胎児肝細胞からなる群より選択される、請求項4記載のマウス。
【請求項6】
前記ヒト造血細胞が、CD34+細胞であるヒト臍帯血細胞である、請求項5記載のマウス。
【請求項7】
CD34陽性細胞、造血幹細胞、造血細胞、骨髄前駆細胞、骨髄細胞、樹状細胞、単球、顆粒球、好中球、肥満細胞、またはこれらの組合せであるヒト細胞を含む、請求項3記載のマウス。
【請求項8】
ヒト造血幹細胞前駆細胞(hematopoietic stem and progenitor cell)、ヒト骨髄前駆細胞、ヒト骨髄細胞、ヒト樹状細胞、ヒト単球、ヒト顆粒球、ヒト好中球、ヒト肥満細胞、ヒト胸腺細胞、ヒトT細胞、ヒトB細胞、ヒト血小板を含む、請求項3記載のマウス。
【請求項9】
チフス菌(Salmonella typhi)感染を含む、請求項4記載のマウス。
【請求項10】
前記チフス菌感染が、全身的なチフス菌感染である、請求項9記載のマウス。
【請求項11】
結核菌(Mycobacterium tuberculosis)感染を含む、請求項4記載のマウス。
【請求項12】
ヒト免疫細胞を含む肺肉芽腫を含む、請求項11記載のマウス。
【請求項13】
(a) 遺伝子改変され、移植され、かつヒト病原体に感染したマウスに剤を投与する段階であって、
該マウスが、
(i) RAG1遺伝子ノックアウトもしくはRAG2遺伝子ノックアウト、またはこれらの組合せ、
(ii) Il2rg遺伝子ノックアウト、
(iii) ヒトIL-3遺伝子がマウスIL-3遺伝子座に存在する、マウスIL-3遺伝子の該ヒトIL-3遺伝子との置換、および
(iv) ヒトGM-CSF遺伝子がマウスGM-CSF遺伝子座に存在する、マウスGM-CSF遺伝子の該ヒトGM-CSF遺伝子との置換
を含み、
該マウスが、CD34陽性ヒト臍帯血細胞またはCD34陽性ヒト胎児肝細胞を用いて移植される、段階;ならびに
(b) 該剤がヒト病原体に感染した該マウスにおいて該ヒト病原体の量を低減するかどうかを判定する段階
を含む、ヒト病原体感染を抑制する剤を同定するための方法。
【請求項14】
前記マウスが、マウストロンボポエチン遺伝子のヒトトロンボポエチン遺伝子との置換をさらに含む、請求項13記載の方法。
【請求項15】
前記ヒト病原体がチフス菌である、請求項13記載の方法。
【請求項16】
前記ヒト病原体がマイコバクテリアであり、かつ該マイコバクテリアが、ヒト免疫細胞を含む肉芽腫を前記マウスにおいて引き起こす、請求項13記載の方法。
【請求項17】
前記マイコバクテリアが結核菌である、請求項16記載の方法。
【請求項1】
(a) マウスRAG遺伝子ノックアウト;
(b) マウスIl2rg遺伝子ノックアウト;
(c) ヒトIL-3遺伝子がマウスIL-3遺伝子座に存在する、マウスIL-3遺伝子の該ヒトIL-3遺伝子との置換;および
(d) ヒトGM-CSF遺伝子がマウスGM-CSF遺伝子座に存在する、マウスGM-CSF遺伝子の該ヒトGM-CSF遺伝子との置換
を含む、遺伝子改変されたマウス。
【請求項2】
ヒトトロンボポエチン遺伝子がマウストロンボポエチン遺伝子座に存在する、マウストロンボポエチン遺伝子の該ヒトトロンボポエチン遺伝子との置換
をさらに含む、請求項1記載のマウス。
【請求項3】
前記RAG遺伝子ノックアウトが、RAG1遺伝子ノックアウト、RAG2遺伝子ノックアウト、およびこれらの組合せから選択される、請求項1記載のマウス。
【請求項4】
ヒト造血細胞の移植をさらに含む、請求項1記載のマウス。
【請求項5】
前記ヒト造血細胞が、臍帯血細胞および胎児肝細胞からなる群より選択される、請求項4記載のマウス。
【請求項6】
前記ヒト造血細胞が、CD34+細胞であるヒト臍帯血細胞である、請求項5記載のマウス。
【請求項7】
CD34陽性細胞、造血幹細胞、造血細胞、骨髄前駆細胞、骨髄細胞、樹状細胞、単球、顆粒球、好中球、肥満細胞、またはこれらの組合せであるヒト細胞を含む、請求項3記載のマウス。
【請求項8】
ヒト造血幹細胞前駆細胞(hematopoietic stem and progenitor cell)、ヒト骨髄前駆細胞、ヒト骨髄細胞、ヒト樹状細胞、ヒト単球、ヒト顆粒球、ヒト好中球、ヒト肥満細胞、ヒト胸腺細胞、ヒトT細胞、ヒトB細胞、ヒト血小板を含む、請求項3記載のマウス。
【請求項9】
チフス菌(Salmonella typhi)感染を含む、請求項4記載のマウス。
【請求項10】
前記チフス菌感染が、全身的なチフス菌感染である、請求項9記載のマウス。
【請求項11】
結核菌(Mycobacterium tuberculosis)感染を含む、請求項4記載のマウス。
【請求項12】
ヒト免疫細胞を含む肺肉芽腫を含む、請求項11記載のマウス。
【請求項13】
(a) 遺伝子改変され、移植され、かつヒト病原体に感染したマウスに剤を投与する段階であって、
該マウスが、
(i) RAG1遺伝子ノックアウトもしくはRAG2遺伝子ノックアウト、またはこれらの組合せ、
(ii) Il2rg遺伝子ノックアウト、
(iii) ヒトIL-3遺伝子がマウスIL-3遺伝子座に存在する、マウスIL-3遺伝子の該ヒトIL-3遺伝子との置換、および
(iv) ヒトGM-CSF遺伝子がマウスGM-CSF遺伝子座に存在する、マウスGM-CSF遺伝子の該ヒトGM-CSF遺伝子との置換
を含み、
該マウスが、CD34陽性ヒト臍帯血細胞またはCD34陽性ヒト胎児肝細胞を用いて移植される、段階;ならびに
(b) 該剤がヒト病原体に感染した該マウスにおいて該ヒト病原体の量を低減するかどうかを判定する段階
を含む、ヒト病原体感染を抑制する剤を同定するための方法。
【請求項14】
前記マウスが、マウストロンボポエチン遺伝子のヒトトロンボポエチン遺伝子との置換をさらに含む、請求項13記載の方法。
【請求項15】
前記ヒト病原体がチフス菌である、請求項13記載の方法。
【請求項16】
前記ヒト病原体がマイコバクテリアであり、かつ該マイコバクテリアが、ヒト免疫細胞を含む肉芽腫を前記マウスにおいて引き起こす、請求項13記載の方法。
【請求項17】
前記マイコバクテリアが結核菌である、請求項16記載の方法。
【図1】

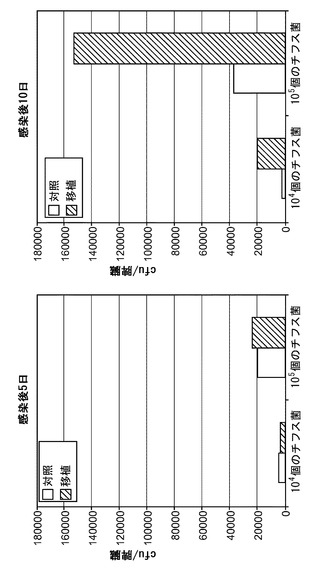
【図2】
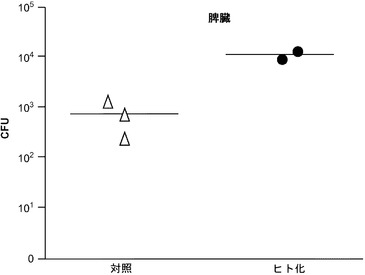
【図3】
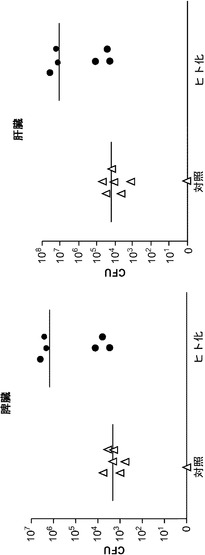
【図4】
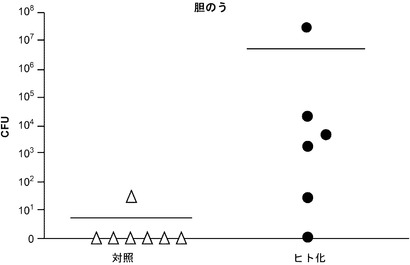
【図5A】
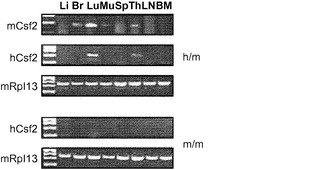
【図5B】
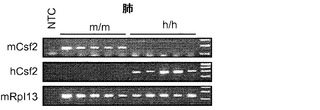
【図5C】
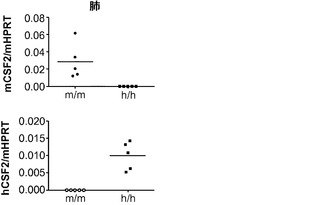
【図5D】
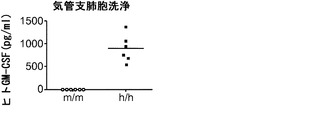
【図5E】
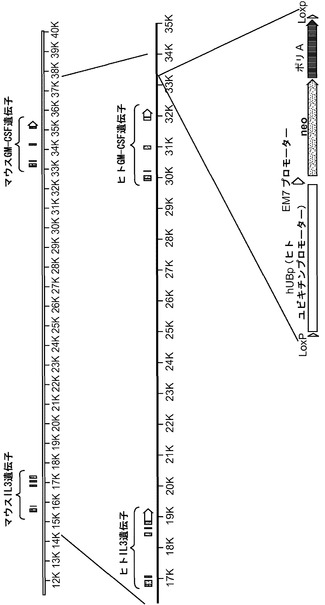
【図6A】
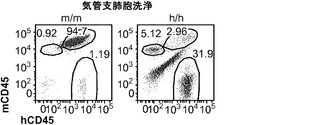
【図6B】
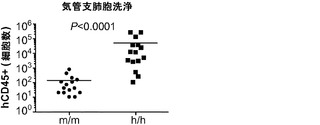
【図6C】
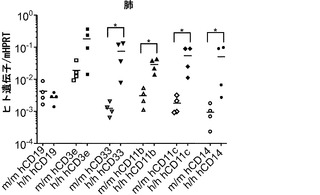
【図6D】
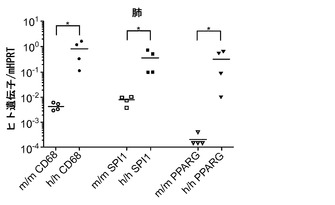
【図6E】
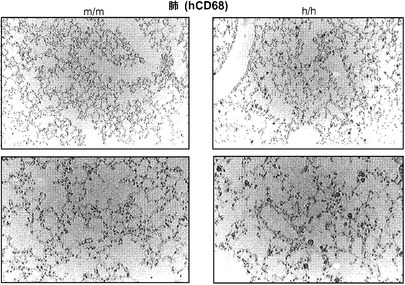
【図6F】
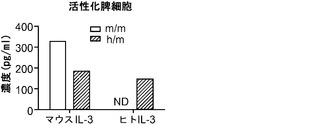
【図6G】
【図7A】
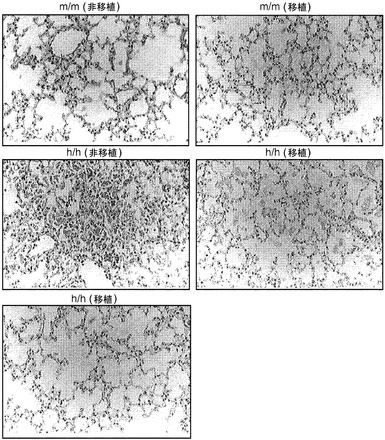
【図7B】
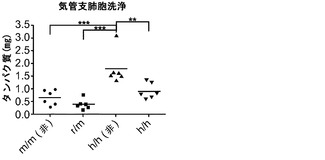
【図8A】
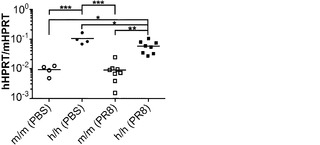
【図8B】
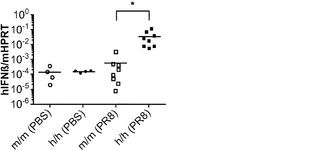
【図8C】
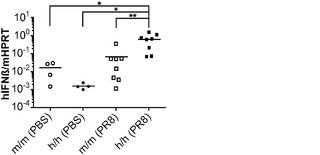
【図8D】
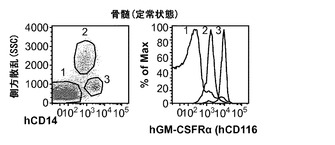
【図8E】
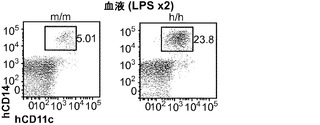
【図8F】
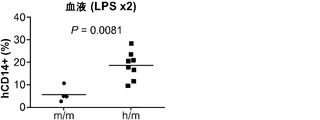
【図8G】
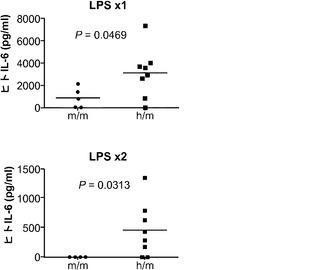
【図9A】
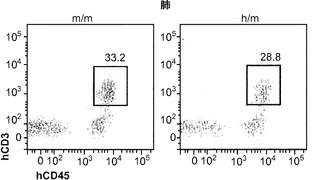
【図9B】
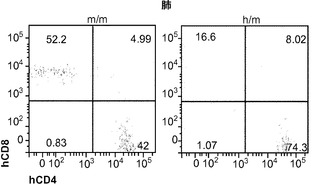
【図9C】
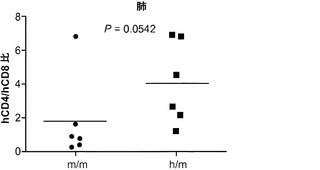
【図9D】
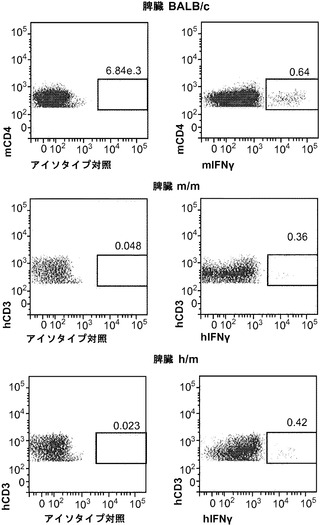
【図9E】
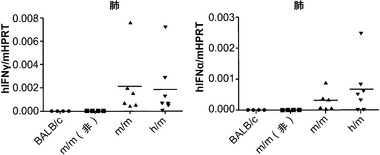
【図9F】
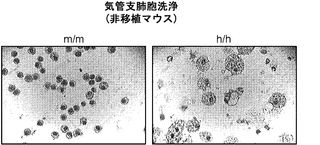
【図9G】
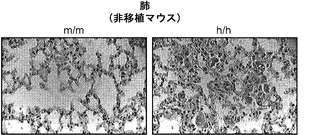
【図10A】
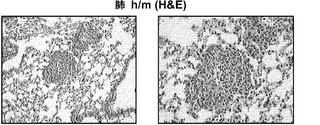
【図10B】
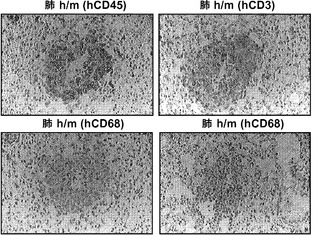
【図11A】
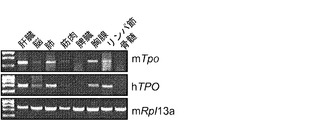
【図11B】

【図11C】

【図11D】
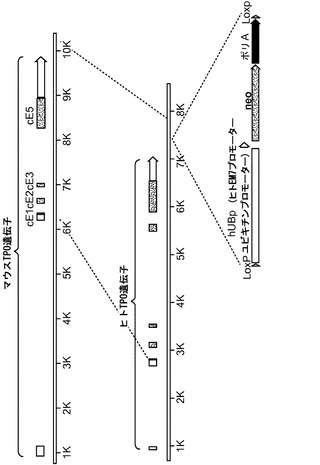
【図12A】
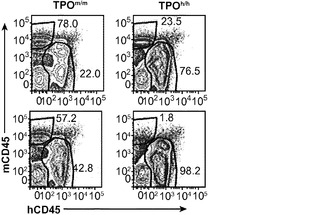
【図12B】
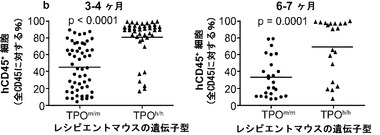
【図12C】
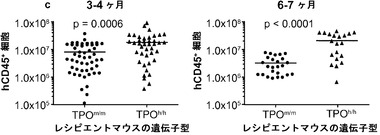
【図12D】
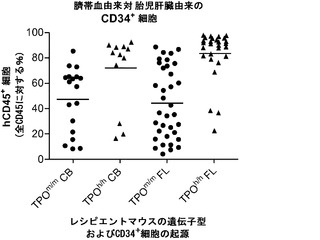
【図13A】
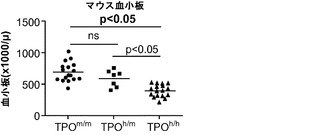
【図13B】
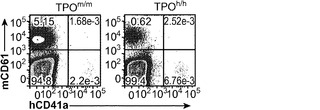
【図13C】
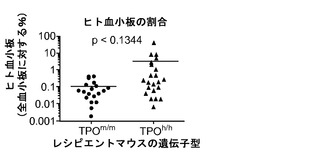
【図13D】
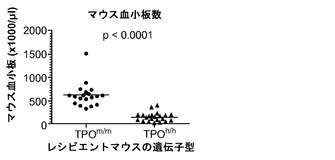
【図13E】
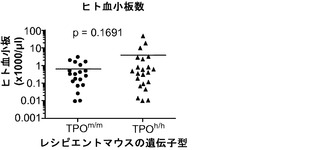
【図13F】
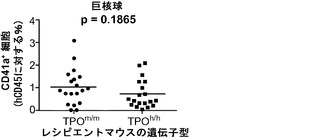
【図13G】

【図13H】

【図13I】
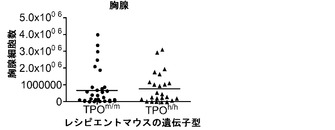
【図14A】
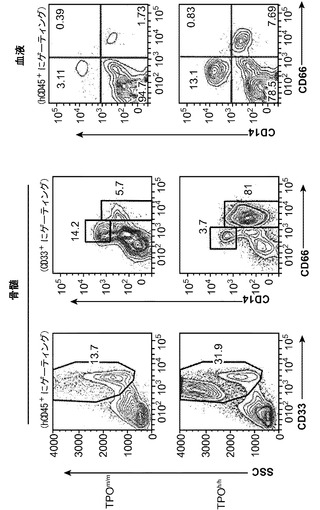
【図14B】
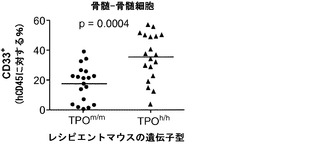
【図14C】
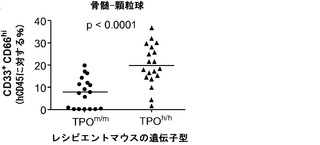
【図14D】

【図14E】
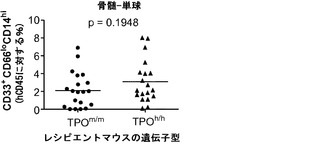
【図14F】
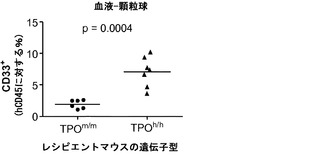
【図14G】
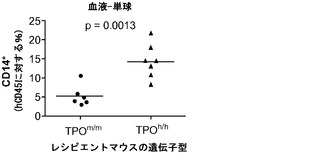
【図15A】
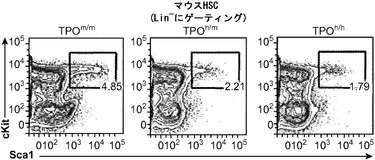
【図15B】
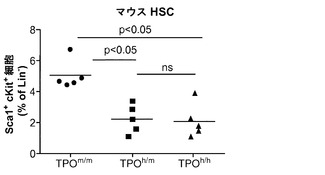
【図15C】
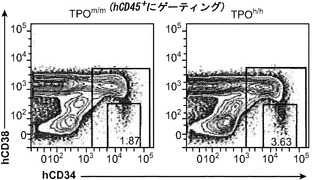
【図15D】
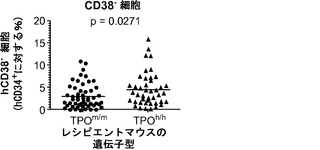
【図15E】
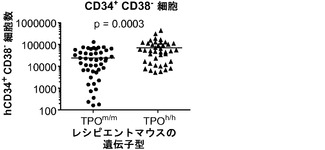
【図15F】

【図15G】
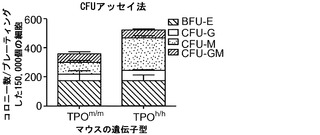
【図15H】
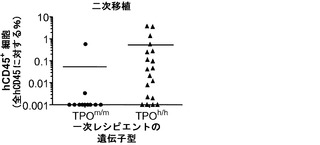
【図2】
【図3】
【図4】
【図5A】
【図5B】
【図5C】
【図5D】
【図5E】
【図6A】
【図6B】
【図6C】
【図6D】
【図6E】
【図6F】
【図6G】
【図7A】
【図7B】
【図8A】
【図8B】
【図8C】
【図8D】
【図8E】
【図8F】
【図8G】
【図9A】
【図9B】
【図9C】
【図9D】
【図9E】
【図9F】
【図9G】
【図10A】
【図10B】
【図11A】
【図11B】
【図11C】
【図11D】
【図12A】
【図12B】
【図12C】
【図12D】
【図13A】
【図13B】
【図13C】
【図13D】
【図13E】
【図13F】
【図13G】
【図13H】
【図13I】
【図14A】
【図14B】
【図14C】
【図14D】
【図14E】
【図14F】
【図14G】
【図15A】
【図15B】
【図15C】
【図15D】
【図15E】
【図15F】
【図15G】
【図15H】
【公表番号】特表2013−506433(P2013−506433A)
【公表日】平成25年2月28日(2013.2.28)
【国際特許分類】
【出願番号】特願2012−533235(P2012−533235)
【出願日】平成22年10月4日(2010.10.4)
【国際出願番号】PCT/US2010/051339
【国際公開番号】WO2011/044050
【国際公開日】平成23年4月14日(2011.4.14)
【出願人】(507302748)リジェネロン・ファーマシューティカルズ・インコーポレイテッド (9)
【出願人】(503469393)イエール ユニバーシティ (11)
【出願人】(512089900)インスティテュート フォー リサーチ イン バイオメディシン (アイアールビー) (1)
【Fターム(参考)】
【公表日】平成25年2月28日(2013.2.28)
【国際特許分類】
【出願日】平成22年10月4日(2010.10.4)
【国際出願番号】PCT/US2010/051339
【国際公開番号】WO2011/044050
【国際公開日】平成23年4月14日(2011.4.14)
【出願人】(507302748)リジェネロン・ファーマシューティカルズ・インコーポレイテッド (9)
【出願人】(503469393)イエール ユニバーシティ (11)
【出願人】(512089900)インスティテュート フォー リサーチ イン バイオメディシン (アイアールビー) (1)
【Fターム(参考)】
[ Back to top ]