
遺伝子改変動物の新規用途
【課題】膵再生因子の探索や、膵内分泌細胞の分化を促進し得る化合物のスクリーニング等に有用なツールとなり得る、膵内分泌細胞の再生モデル系の提供。
【解決手段】ATP感受性カリウムイオン(KATP)チャンネルKir6.2のドミナントネガティ
ブ体を膵β細胞で発現し得るトランスジェニック(Tg)非ヒト哺乳動物またはその生体の一部から実質的になる膵再生モデル系、それを用いた膵再生関連遺伝子(因子)の同定方法、膵再生調節物質のスクリーニング、並びにKATPチャンネルKir6.2のドミナントネガティブ体を膵β細胞で発現することができ、且つ1以上の他の遺伝子変異または改変を有する非ヒト哺乳動物またはその生体の一部。
【解決手段】ATP感受性カリウムイオン(KATP)チャンネルKir6.2のドミナントネガティ
ブ体を膵β細胞で発現し得るトランスジェニック(Tg)非ヒト哺乳動物またはその生体の一部から実質的になる膵再生モデル系、それを用いた膵再生関連遺伝子(因子)の同定方法、膵再生調節物質のスクリーニング、並びにKATPチャンネルKir6.2のドミナントネガティブ体を膵β細胞で発現することができ、且つ1以上の他の遺伝子変異または改変を有する非ヒト哺乳動物またはその生体の一部。
【発明の詳細な説明】
【技術分野】
【0001】
本発明は、ATP感受性カリウムイオンチャンネル遺伝子に改変を有するトランスジェニック非ヒト哺乳動物の新規用途に関する。詳細には、本発明は、ATP感受性カリウムイオンチャンネルのKir6.2のドミナントネガティブ体をコードする遺伝子を導入されたトランスジェニック非ヒト哺乳動物の膵再生モデルとしての用途等に関する。
【背景技術】
【0002】
糖尿病はインスリンの相対的もしくは絶対的不足によって起こり、その終末像は膵β細胞の破壊あるいは機能不全によるインスリン欠乏であり、インスリン注射が不可欠となる。しかし、インスリン注射では血糖値を生理的範囲内にコントロールすることは不可能であり、糖尿病性腎症や網膜症などの重篤な合併症の進展抑制は困難である。糖尿病の根本的な治療法としては、膵島細胞の移植が挙げられるが、多くの患者の治療に必要な数のヒト膵島を得ることは実際上不可能である。そこで、適当な前駆細胞をインビトロでβ細胞に分化させることができれば、移植用細胞の無限の供給源になると期待される。あるいは、膵β細胞の分化を誘導する内在因子や膵再生を促進する薬物の投与によってインビボで膵β細胞の再生(新生)を誘導することができれば、移植を必要としない薬物による糖尿病の根本的治療も不可能ではない。
【0003】
従って、膵臓の発生分化・β細胞の再生の機構を理解することは、糖尿病治療の観点上、極めて重要である。膵臓は成体ではほとんど分裂しないが、何らかの原因でβ細胞が損傷されると、膵再生が起こることが観察されていたので、分化増殖能を有する膵幹細胞の存在が考えられていた。例えば、ストレプトゾトシン(STZ)処理、膵切除、アロキサン投与、インターフェロン(IFN)γトランスジェニックマウス等で膵再生が観察されているが、これらの再生モデルでは、外分泌細胞は総じて再生能力が高いのに対して、内分泌細胞の再生能は低い。また、マウスやヒトのES細胞、肝細胞、膵導管細胞からインスリン分泌細胞を作製したとの報告もなされているが、その再現性、効率性には未だ問題が多く、確立した方法論として認められるには至っていない。
【0004】
ATP感受性カリウムイオン(以下、KATPと略記する場合がある)チャンネルは、種々の細胞において細胞の代謝状態を膜電位に連動させている中心的な分子であるが、膵β細胞においては、グルコース誘導性のインスリン分泌機構において中心的な役割を果たしている。即ち、グルコース代謝により生成したATPに応答してKATPチャンネルが閉じることによりβ細胞膜の脱分極を引き起こし、それに続いて、電位依存性のカルシウムチャンネルが開いてβ細胞内にカルシウムイオンが流入することにより、細胞内カルシウムイオン濃度が上昇してカルシウム依存性のインスリン分泌が起こる。
【0005】
膵β細胞のKATPチャンネルは、内向き整流特性を示すポア部分であるKir6.2の4量体と、調節サブユニットであるスルホニル尿素受容体SUR1の4量体とからなる複合体である。KATPチャンネルにおいて、カリウムイオン選択的性に寄与しているのはKir6.2である。本発明者らは、膵β細胞におけるKATPチャンネルの役割を直接的に検証するために、Kir6.2遺伝子を破壊したノックアウト(KO)マウスや(特許文献1および非特許文献1)、Kir6.2のドミナントネガティブ体を膵β細胞で特異的に発現するトランスジェニック(Tg)マウス(非特許文献2)を作製した。得られたマウスは、いずれも若週齢で膵島構造の破壊とβ細胞の減少およびα細胞の増加を認めたが、膵島構造の破壊やβ細胞数の減少はTgマウスにおいてより顕著であった(非特許文献3)。非特許文献4にもKir6.2のドミナントネガティブ体を導入したTgマウスが開示されているが、これらのKir6.2遺伝子改変動物において膵再生現象が観察されたという報告は皆無である。
【特許文献1】特開平11-346597号公報
【非特許文献1】三木(Miki, T.)ら, プロシーディングズ・オヴ・ナショナル・アカデミー・オヴ・サイエンシイズ・ユーエスエー(Proc. Natl. Acad. Sci. USA), 第95巻, pp. 10402-10406, 1998年
【非特許文献2】三木(Miki, T.)ら, プロシーディングズ・オヴ・ナショナル・アカデミー・オヴ・サイエンシイズ・ユーエスエー(Proc. Natl. Acad. Sci. USA), 第94巻, pp. 11969-11973, 1997年
【非特許文献3】清野(Seino, S.)ら, ダイアビーティズ(Diabetes), 第49巻, pp. 311-318, 2000年
【非特許文献4】コースター(Koster, J.C.)ら, プロシーディングズ・オヴ・ナショナル・アカデミー・オヴ・サイエンシイズ・ユーエスエー(Proc. Natl. Acad. Sci. USA), 第99巻, pp. 16992-16997, 2002年
【発明の開示】
【発明が解決しようとする課題】
【0006】
本発明の目的は、膵再生因子の探索や膵内分泌細胞の分化を促進し得る化合物のスクリーニング等に有用なツールとなり得る、膵内分泌細胞の再生モデルを提供することである。
【課題を解決するための手段】
【0007】
本発明者らは、上記の課題を解決するために鋭意研究を重ねた結果、意外にも、本発明者らが作製した上記Tgマウス(該マウスは若週齢で高血糖・低インスリン血症を発症し、膵β細胞の細胞死を認める)が、加齢とともにインスリン分泌が回復、血糖値が改善し、膵管近傍に膵β細胞の新生が見られることを見出した。
本発明者らは、これらの知見に基づいて、さらに検討を重ねた結果、本発明を完成するに至った。
【0008】
すなわち、本発明は、
[1]ATP感受性カリウムイオンチャンネルKir6.2のドミナントネガティブ体を膵β細胞で発現し得るトランスジェニック非ヒト哺乳動物またはその生体の一部から実質的になる膵再生モデル系、
[2]ドミナントネガティブ体が、チャンネルのカリウムイオン選択性に関与する領域中の1以上のアミノ酸が変異したものである上記[1]記載のモデル系、
[3]ATP感受性カリウムイオンチャンネルKir6.2のドミナントネガティブ体を膵β細胞で発現し得るトランスジェニック非ヒト哺乳動物における、膵内分泌細胞分化前後の膵臓もしくはその一部由来の単離されたRNAまたはそのcDNA、
[4]ATP感受性カリウムイオンチャンネルKir6.2のドミナントネガティブ体を膵β細胞で発現し得るトランスジェニック非ヒト哺乳動物の生体の一部における、膵内分泌細胞分化前後の該生体の一部由来の単離されたRNAまたはそのcDNA、
[5]ATP感受性カリウムイオンチャンネルKir6.2のドミナントネガティブ体を膵β細胞で発現し得るトランスジェニック非ヒト哺乳動物において、膵内分泌細胞分化前後での膵臓もしくはその一部における遺伝子発現プロファイルを比較することを特徴とする、膵再生関連遺伝子の同定方法、
[6]ATP感受性カリウムイオンチャンネルKir6.2のドミナントネガティブ体を膵β細胞
で発現し得るトランスジェニック非ヒト哺乳動物の生体の一部において、膵内分泌細胞分化前後での該生体の一部における遺伝子発現プロファイルを比較することを特徴とする、膵再生関連遺伝子の同定方法、
[7]ATP感受性カリウムイオンチャンネルKir6.2のドミナントネガティブ体を膵β細胞で発現し得るトランスジェニック非ヒト哺乳動物またはその生体の一部に被検物質を適用し、膵内分泌細胞分化に及ぼす該物質の効果を検定することを特徴とする、膵再生調節物質のスクリーニング方法、
[8]ATP感受性カリウムイオンチャンネルKir6.2のドミナントネガティブ体を膵β細胞で発現することができ、且つ1以上の他の遺伝子変異または改変を有する非ヒト哺乳動物またはその生体の一部、
[9]他の遺伝子変異または改変の少なくとも1つが、膵内分泌機能異常が関与する疾患と同一もしくは類似の病態を生じさせるものである、上記[8]記載の動物またはその生体の一部、および
[10]上記[9]記載の動物に被検物質を投与し、膵内分泌機能異常が関与する疾患と同一もしくは類似の病態に及ぼす該物質の効果を検定することを特徴とする、該疾患の予防・治療物質のスクリーニング方法を提供する。
【発明の効果】
【0009】
Kir6.2のドミナントネガティブ体を導入されたTg動物は、若週齢で糖尿病を発症し、インスリン欠乏・膵β細胞の細胞死が顕著となるが、加齢とともにインスリン分泌が増大し、膵管近傍に高頻度に膵内分泌細胞が出現することから、該動物およびその生体の一部は、優れた膵内分泌細胞の再生モデル系となり得る。
【発明を実施するための最良の形態】
【0010】
本発明の「ATP感受性カリウムイオン(KATP)チャンネルKir6.2のドミナントネガティブ体を膵β細胞で発現し得るトランスジェニック(Tg)非ヒト哺乳動物」(以下、「本発明のTg動物」と略記する場合がある)は、非ヒト哺乳動物の受精卵や、未受精卵、精子およびその前駆細胞(始原生殖細胞、卵原細胞、卵母細胞、卵細胞、精原細胞、精母細胞、精細胞等)などに、好ましくは受精卵の胚発生の初期段階(さらに好ましくは8細胞期以前)において、リン酸カルシウム共沈殿法、電気穿孔(エレクトロポレーション)法、リポフェクション法、凝集法、顕微注入(マイクロインジェクション)法、遺伝子銃(パーティクルガン)法、DEAE-デキストラン法などの遺伝子導入法によって、目的とするKATPチャンネルKir6.2のドミナントネガティブ体をコードするDNAを、膵β細胞において発現し得る形態で、目的とする細胞に導入することにより作出される。本明細書におけるTg動物とは、導入遺伝子を発現可能な状態で安定に保持する動物をいう。「安定に保持」するとは、該動物の細胞内で導入したDNAが発現可能な状態で永続的に存在することを意味し、該DNAが宿主染色体上に組み込まれていても、あるいは染色体外DNAとして安定に存在していてもよいが、好ましくは、該DNAは宿主染色体上に組み込まれた状態で保持される。
また、上記遺伝子導入法により、非ヒト哺乳動物の体細胞、組織、臓器などに目的とするDNAを導入し、細胞培養、組織培養などに利用することもでき、さらに、この細胞を上述の胚(もしくは生殖)細胞と公知の細胞融合法を用いて融合させることによりTg動物を作出することもできる。あるいは、ノックアウト(KO)動物を作製する場合と同様に、非ヒト哺乳動物の胚性幹細胞(ES細胞)に上記の遺伝子導入法を用いて目的とするDNAを導入し、予め該DNAが安定に組み込まれたクローンを選択した後に、該ES細胞を胚盤胞に注入するかあるいはES細胞塊と8細胞期胚とを凝集させてキメラマウスを作製し、生殖系列に導入遺伝子が伝達されたものを選択することによってもTg動物を得ることが可能である。
また、このようにして作製されたTg動物の生体の一部(例えば、(1)KATPチャンネルKir6.2のドミナントネガティブ体をコードするDNAを有する細胞、組織、臓器など、(2)これらに由来する細胞または組織を培養し、必要に応じて継代したものなど、(3)該Tg動物から単離し得る各種蛋白質またはDNAなど)も、本発明の「KATPチャンネルKir6.2のドミナントネガティブ体をコードするDNAを有する非ヒト哺乳動物の生体の一部」として、本発明のTg動物と同様の目的に用いることが出来る。
【0011】
本発明のTg動物の生体の一部としては、KATPチャンネルKir6.2のドミナントネガティブ体を発現し得る細胞・組織であって、膵内分泌細胞への分化能を有する細胞を含むものであれば、特に限定されないが、好ましくは、膵臓、肝臓等の臓器、膵島、膵管等の組織、膵前駆細胞、膵幹細胞、肝前駆細胞などの細胞、あるいはそれらより特定の培養条件下で誘導される細胞・組織、さらには、該Tg動物の初期胚より誘導されるES細胞などが好ましい。
【0012】
本発明で対象とし得る「非ヒト哺乳動物」としては、トランスジェニック系が確立されたヒト以外の哺乳動物であれば特に制限はなく、例えば、ウシ、サル、ブタ、ヒツジ、ヤギ、ウサギ、イヌ、ネコ、モルモット、ハムスター、ラット、マウスなどが挙げられる。好ましくは、ウサギ、イヌ、ネコ、モルモット、ハムスター、マウス、ラット等であり、なかでも疾患モデル動物作製の面から個体発生および生物サイクルが比較的短く、繁殖が容易な齧歯動物がより好ましく、とりわけマウス(例えば、純系としてC57BL/6系統、DBA2系統など、交雑系としてB6C3F1系統、BDF1系統、B6D2F1系統、BALB/c系統、ICR系統など)およびラット(例えば、Wistar、SDなど)が好ましい。
また、哺乳動物以外にもニワトリなどの鳥類が本発明で対象とする「非ヒト哺乳動物」と同様の目的に用いることができる。
【0013】
「KATPチャンネルKir6.2のドミナントネガティブ体」とは、KATPチャンネルKir6.2の1または2以上のアミノ酸が変異したことによって、該サブユニットの機能が欠損した蛋白質をいう。該サブユニットの機能の欠損は、KATPチャンネル電流の消失もしくは低下によって特徴付けられる。
野生型Kir6.2蛋白質としては、配列番号:2で表されるアミノ酸配列もしくはそれと実質的に同一のアミノ酸配列を有する蛋白質が挙げられる。「実質的に同一のアミノ酸配列を有する蛋白質」としては、配列番号:2で表わされるアミノ酸配列と約90%以上、好ましくは95%以上、より好ましくは約98%以上の相同性を有するアミノ酸配列を含み、配列番号:2で表わされるアミノ酸配列を有する蛋白質と実質的に同質の活性を有する蛋白質などが挙げられる。ここで「相同性」とは、当該技術分野において公知の数学的アルゴリズムを用いて2つのアミノ酸配列をアラインさせた場合の、最適なアラインメント(好ましくは、該アルゴリズムは最適なアラインメントのために配列の一方もしくは両方へのギャップの導入を考慮し得るものである)における、オーバーラップする全アミノ酸残基に対する同一アミノ酸および類似アミノ酸残基の割合(%)を意味する。「類似アミノ酸」とは物理化学的性質において類似したアミノ酸を意味し、例えば、芳香族アミノ酸(Phe、Trp、Tyr)、脂肪族アミノ酸(Ala、Leu、Ile、Val)、極性アミノ酸(Gln、Asn
)、塩基性アミノ酸(Lys、Arg、His)、酸性アミノ酸(Glu、Asp)、水酸基を有するア
ミノ酸(Ser、Thr)、側鎖の小さいアミノ酸(Gly、Ala、Ser、Thr、Met)などの同じグ
ループに分類されるアミノ酸が挙げられる。このような類似アミノ酸による置換は蛋白質の表現型に変化をもたらさない(即ち、保存的アミノ酸置換である)ことが予測される。保存的アミノ酸置換の具体例は当該技術分野で周知であり、種々の文献に記載されている(例えば、Bowieら,Science, 247: 1306-1310 (1990)を参照)。
本明細書におけるアミノ酸配列の相同性は、相同性計算アルゴリズムNCBI BLAST(National Center for Biotechnology Information Basic Local Alignment Search Tool)を用い、以下の条件(期待値=10;ギャップを許す;マトリクス=BLOSUM62;フィルタリング=OFF)にて計算することができる。
アミノ酸配列の相同性を決定するための他のアルゴリズムとしては、例えば、Karlinら, Proc. Natl. Acad. Sci. USA, 90: 5873-5877 (1993)に記載のアルゴリズム[該アルゴリズムはNBLASTおよびXBLASTプログラム(version 2.0)に組み込まれている(Altschulら, Nucleic Acids Res., 25: 3389-3402 (1997))]、Needlemanら, J. Mol. Biol., 48:444-453 (1970)に記載のアルゴリズム[該アルゴリズムはGCGソフトウェアパッケージ中のGAPプログラムに組み込まれている]、MyersおよびMiller, CABIOS, 4: 11-17 (1988)に記載のアルゴリズム[該アルゴリズムはCGC配列アラインメントソフトウェアパッケージの一部であるALIGNプログラム(version 2.0)に組み込まれている]、Pearsonら, Proc. Natl. Acad. Sci. USA, 85: 2444-2448 (1988)に記載のアルゴリズム[該アルゴリズムはGCGソフトウェアパッケージ中のFASTAプログラムに組み込まれている]等が挙げられ、それらも同様に好ましく用いられ得る。
【0014】
「実質的に同質の活性」とは、例えば、KATPチャンネル活性が性質的に同質であることを示す。したがって、KATPチャンネル活性が同等(例、約0.5〜約2倍)であることが好ましいが、活性の程度や蛋白質の分子量などの量的要素は異なっていてもよい。KATPチャンネル活性の測定は、自体公知の方法に準じて行なうことができるが、例えば、該蛋白質を発現する細胞における標識Rb+の流出を検出する方法、パッチ−クランプ法でカリウムイオン電流を測定する方法等が挙げられる。
【0015】
また、Kir6.2蛋白質としては、例えば、(1)配列番号:2で表されるアミノ酸配列中の1または2個以上(好ましくは、1〜30個程度、好ましくは1〜10個程度、さらに好ましくは1〜5個)のアミノ酸が欠失したアミノ酸配列、(2)配列番号:2で表されるアミノ酸配列に1または2個以上(好ましくは、1〜30個程度、好ましくは1〜10個程度、さらに好ましくは1〜5個)のアミノ酸が付加したアミノ酸配列、(3)配列番号:2で表されるアミノ酸配列に1または2個以上(好ましくは、1〜30個程度、好ましくは1〜10個程度、さらに好ましくは1〜5個)のアミノ酸が挿入されたアミノ酸配列、(4)配列番号:2で表されるアミノ酸配列中の1または2個以上(好ましくは、1〜30個程度、好ましくは1〜10個程度、さらに好ましくは1〜5個)のアミノ酸が他のアミノ酸で置換されたアミノ酸配列、または(5)それらを組み合わせたアミノ酸配列を含有する蛋白質であって、配列番号:2で表されるアミノ酸配列を含有する蛋白質と実質的に同質の活性を有する蛋白質も含まれる。
上記のようにアミノ酸配列が挿入、欠失または置換されている場合、その挿入、欠失または置換の位置は、当該蛋白質の活性を損なわない限り、特に限定されない。
【0016】
KATPチャンネルKir6.2のドミナントネガティブ体としては、上述のように、膵β細胞で発現した際にKATPチャンネルのポア成分を構成して該チャンネル電流を消失もしくは低下させ得る限り、いかなるものであってもよいが、好ましくは、配列番号:2で表されるアミノ酸配列もしくはそれと実質的に同一のアミノ酸配列において、(1)1または2個以上(好ましくは、1〜30個程度、好ましくは1〜10個程度、さらに好ましくは1〜5個)のアミノ酸が欠失したアミノ酸配列、(2)1または2個以上(好ましくは、1〜30個程度、好ましくは1〜10個程度、さらに好ましくは1〜5個)のアミノ酸が付加したアミノ酸配列、(3)1または2個以上(好ましくは、1〜30個程度、好ましくは1〜10個程度、さらに好ましくは1〜5個)のアミノ酸が挿入されたアミノ酸配列、(4)1または2個以上(好ましくは、1〜30個程度、好ましくは1〜10個程度、さらに好ましくは1〜5個)のアミノ酸が他のアミノ酸で置換されたアミノ酸配列、または(5)それらを組み合わせたアミノ酸配列を含有する蛋白質であって、膵β細胞のKATPチャンネル電流を消失もしくは低下させ得る蛋白質である。
【0017】
より好ましくは、本発明で用いられるドミナントネガティブ体は、KATPチャンネルKir6.2のカリウムイオン選択性に関与する領域中の1以上のアミノ酸が変異(例えば、置換、欠失、挿入、付加など)したものである。「カリウムイオン選択性に関与する領域」としては、例えば、2つの膜貫通ドメイン(TM1およびTM2)の間の高度に保存されたH5領域、好ましくは、H5領域中のGly-Tyr(又はPhe)-Glyのモチーフ(以下、GY(F)Gモチーフという)が挙げられる。特に好ましくは、本発明で用いられるドミナントネガティブ体は、上記GY(F)Gモチーフの1以上のアミノ酸が他のアミノ酸で置換されたものである。他のアミノ酸としては、保存的アミノ酸以外のアミノ酸が挙げられる。例えば、配列番号:2で表されるアミノ酸配列においては、アミノ酸番号132〜134で示されるGFGが該Kir6.2蛋白質におけるGFGモチーフであるので、これらのアミノ酸のうちの1以上を他のアミノ酸で置換したものが挙げられる。とりわけ、アミノ酸番号132のGlyをSerで置換したアミノ酸配列(以下、「Kir6.2G132S」と略記する場合がある)もしくはそれと実質的に同一のアミノ酸配列を有する蛋白質が好ましい。ここで「実質的に同一のアミノ酸配列」とは上記と同義であるが、この場合の「同質の活性」とは、ドミナントネガティブ活性(即ち、KATPチャンネル電流を消失もしくは低下させる活性)が同質であることを意味する。
【0018】
KATPチャンネルKir6.2のドミナントネガティブ体をコードするDNAとしては、好ましくは、KATPチャンネルKir6.2のカリウムイオン選択性に関与する領域中の1以上のアミノ酸が変異(例えば、置換、欠失、挿入、付加など)した蛋白質をコードするDNAであり、より好ましくは、該サブユニット蛋白質のH5領域、さらに好ましくはGY(F)Gモチーフ内の1以上のアミノ酸が他のアミノ酸で置換された蛋白質、特に好ましくは、配列番号:2で表されるアミノ酸配列中アミノ酸番号132〜134で示されるGFGのうちの1以上のアミノ酸が他のアミノ酸で置換されたアミノ酸配列もしくはそれと実質的に同一のアミノ酸配列を有する蛋白質、就中、Kir6.2G132Sもしくはそれと実質的に同一のアミノ酸配列を有する蛋白質をコードするDNAである。
【0019】
配列番号:2で表されるアミノ酸配列をコードするDNAとしては、例えば、配列番号:1で表される塩基配列もしくは該塩基配列の相補鎖配列とストリンジェントな条件下でハイブリダイズし得る塩基配列を含み、且つ配列番号:2で表されるアミノ酸配列を有する蛋白質と実質的に同質の活性を有する蛋白質をコードするDNAが挙げられる。従って、配列番号:2で表されるアミノ酸配列を有する蛋白質のドミナントネガティブ体をコードするDNAとしては、好ましくは、配列番号:1で表される塩基配列中H5領域をコードする塩基配列が、該領域中の1以上のアミノ酸が他のアミノ酸で置換されたアミノ酸配列をコードする配列で置換された塩基配列、より好ましくは、配列番号:1で表される塩基配列中塩基番号394〜402で示される塩基配列が、GFGのうちの1以上のアミノ酸が他のアミノ酸で置換されたアミノ酸配列をコードする配列で置換された塩基配列、特に好ましくは、配列番号:1で表される塩基配列中塩基番号394〜396で示される塩基配列が、Serをコードする配列で置換された塩基配列を含むDNA、あるいはそれらの相補鎖配列とストリンジェントな条件下でハイブリダイズし得る塩基配列を含み、且つ上記いずれかのDNAにコードされる蛋白質と実質的に同質の活性(ドミナントネガティブ活性)を有する蛋白質をコードするDNAが挙げられる。「ストリンジェントな条件」としては、例えば、ナトリウム塩濃度が約19〜約40mM、好ましくは約19〜約20mMで、温度が約50〜約70℃、好ましくは約60〜約65℃の条件等が挙げられる。特に、ナトリウム塩濃度が約19mMで温度が約65℃の場合が好ましい。当業者は、ハイブリダイゼーション溶液の塩濃度、ハイブリダゼーション反応の温度、プローブ濃度、プローブの長さ、ミスマッチの数、ハイブリダイゼーション反応の時間、洗浄液の塩濃度、洗浄の温度等を適宜変更することにより、所望のストリンジェンシーに容易に調節することができる。
【0020】
KATPチャンネルKir6.2のドミナントネガティブ体をコードするDNAは、各種哺乳動物(ヒト、ウシ、サル、ブタ、ヒツジ、ヤギ、ウサギ、イヌ、ネコ、モルモット、ハムスター、ラット、マウスなど)の膵臓、膵島、膵β細胞、骨格筋、心筋、平滑筋など、好ましくは、膵臓、膵島、膵β細胞等に由来するDNAおよび市販の各種ゲノムDNAライブラリーに由来するゲノムDNAの全てあるいは一部を原料として用い、あるいは各種哺乳動物の膵臓、膵島、膵β細胞、骨格筋、心筋、平滑筋など、好ましくは、膵臓、膵島、膵β細胞等に由来するRNAから公知の方法により調製されたcDNAを原料として用い、公知の該サブユニット遺伝子の配列をもとに作製したオリゴヌクレオチドをプローブもしくはプライマーとして、ハイブリダイゼーション法もしくはPCR法などにより単離したKATPチャンネルKir6.2をコードするDNAを鋳型として用い、従来公知の部位特異的変異誘発により、所望の部位(例えば、上記H5領域、好ましくはGY(F)Gモチーフ)に変異を導入することにより調製することができる。具体的には、例えば、上記非特許文献2に記載の方法などが挙げられるが、それに限定されない。
【0021】
本発明のTg動物は、KATPチャンネルKir6.2のドミナントネガティブ体をコードするDNAを膵β細胞で「発現可能な状態で」保持している。したがって、当該DNAを対象動物に導入するにあたっては、当該DNAを対象動物の膵β細胞内で機能し得るプロモーターの下流に連結した発現カセットを含む形態(例、発現ベクターなど)で用いるのが一般に有利である。
【0022】
KATPチャンネルKir6.2のドミナントネガティブ体をコードするDNAを担持するベクターとしては、大腸菌由来のプラスミド、枯草菌由来のプラスミド、酵母由来のプラスミド、λファージなどのバクテリオファージ、モロニー白血病ウイルスなどのレトロウイルス、ワクシニアウイルスまたはバキュロウイルスなどの動物もしくは昆虫ウイルスなどが用いられる。なかでも、大腸菌由来のプラスミド、枯草菌由来のプラスミドまたは酵母由来のプラスミドなどが好ましく用いられ、特に大腸菌由来のプラスミドが好ましい。
【0023】
KATPチャンネルKir6.2のドミナントネガティブ体の遺伝子発現調節を行うプロモーターとしては、少なくとも膵β細胞内でプロモーター活性を発揮し得るものであれば特に制限されず、例えばウイルス(サイトメガロウイルス、モロニー白血病ウイルス、JCウイルス、乳癌ウイルスなど)に由来する遺伝子のプロモーター、各種哺乳動物(ヒト、ウシ、サル、ブタ、ヒツジ、ヤギ、ウサギ、イヌ、ネコ、モルモット、ハムスター、ラット、マウスなど)および鳥類(ニワトリなど)に由来する遺伝子[例えば、アルブミン、エンドセリン、オステオカルシン、筋クレアチンキナーゼ、I型およびII型コラーゲン、サイクリックAMP依存蛋白キナーゼβIサブユニット、心房ナトリウム利尿性因子、ドーパミンβ−水酸化酵素、ニューロフィラメント軽鎖、メタロチオネインIおよびIIA、メタロプロティナーゼ1組織インヒビター、平滑筋αアクチン、ポリペプチド鎖伸長因子1α(EF1−α)、βアクチン、αおよびβミオシン重鎖、ミオシン軽鎖1および2、ミエリン塩基性蛋白、血清アミロイドPコンポーネント、レニンなど]のプロモーターなども挙げられるが、好ましくは、膵β細胞以外にプロモーター活性を発揮する細胞種が限定されるプロモーター(例えば、アミリンプロモーター、pdx-1プロモーター等)、より好ましくは膵β細胞特異的プロモーター(例えば、インスリンIもしくはインスリンIIプロモーター等)である。
【0024】
KATPチャンネルKir6.2のドミナントネガティブ体をコードするDNAの下流には、Tg動物において目的とするmRNAの転写を終結させる配列(ポリアデニレーション(polyA)シグナル、ターミネーターとも呼ばれる)を有していることが好ましく、例えば、ウイルス遺伝子由来、あるいは各種哺乳動物または鳥類の遺伝子由来のターミネーター配列を用いて、効率よい導入遺伝子の発現を達成することができる。好ましくは、シミアンウイルスのSV40ターミネーターなどが用いられる。その他、目的の遺伝子をさらに高発現させる目的で、各遺伝子のスプライシングシグナル、エンハンサー領域、真核遺伝子のイントロンの一部を、プロモーター領域の5’上流、プロモーター領域と翻訳領域間あるいは翻訳領域の3’下流に連結することも目的により可能である。
【0025】
また、胚性幹細胞(ES細胞)を用いてTg動物を作製する場合、上記のベクターは、導入DNAが安定に組み込まれたクローンを選択するための選択マーカー遺伝子(例:ネオマイシン耐性遺伝子、ハイグロマイシン耐性遺伝子などの薬剤耐性遺伝子)をさらに含むことが好ましい。
【0026】
上記のプロモーター、KATPチャンネルKir6.2のドミナントネガティブ体をコードするDNA、ターミネーターなどは、適当な制限酵素およびDNAリガーゼ等を用いた通常の遺伝子工学的手法により、上記のベクター中に正しい配置で、即ち、Tg動物においてKATPチャンネルKir6.2のドミナントネガティブ体を発現可能な配置で、挿入することができる。
【0027】
好ましい一実施態様においては、上記のようにして得られるKATPチャンネルKir6.2のドミナントネガティブ体をコードするDNAを含む発現ベクターは、マイクロインジェクション法により対象となる非ヒト哺乳動物の初期胚に導入される。
【0028】
対象非ヒト哺乳動物の初期胚は、同種の非ヒト哺乳動物の雌雄を交配させて得られる体内受精卵を採取するか、あるいは同種の非ヒト哺乳動物の雌雄からそれぞれ採取した卵と精子を体外受精させることにより得ることができる。
用いる非ヒト哺乳動物の齢や飼育条件等は動物種によってそれぞれ異なるが、例えばマウス(好ましくはC57BL/6J(B6)などの近交系マウス、B6と他の近交系とのF1など)を用いる場合は、雌が約4〜約6週齢、雄が約2〜約8月齢程度のものが好ましく、また、約12時間明期条件(例えば7:00−19:00)で約1週間飼育したものが好ましい。
体内受精は自然交配によってもよいが、性周期の調節と1個体から多数の初期胚を得ることを目的として、雌非ヒト哺乳動物に性腺刺激ホルモンを投与して過剰排卵を誘起した後、雄非ヒト哺乳動物と交配させる方法が好ましい。雌非ヒト哺乳動物の排卵誘発法としては、例えば初めに卵胞刺激ホルモン(妊馬血清性性腺刺激ホルモン、一般にPMSGと略する)、次いで黄体形成ホルモン(ヒト絨毛性性腺刺激ホルモン、一般にhCGと略する)を、例えば腹腔内注射などにより投与する方法が好ましいが、好ましいホルモンの投与量、投与間隔は非ヒト哺乳動物の種類によりそれぞれ異なる。例えば、非ヒト哺乳動物がマウス(好ましくはC57BL/6J(B6)などの近交系マウス、B6と他の近交系とのF1など)の場合は、通常、卵胞刺激ホルモン投与後、約48時間後に黄体形成ホルモンを投与し、直ちに雄マウスと交配させることにより受精卵を得る方法が好ましく、卵胞刺激ホルモンの投与量は約20〜約50IU/個体、好ましくは約30IU/個体、黄体形成ホルモンの投与量は約0〜約10IU/個体、好ましくは約5IU/個体である。
一定時間経過後、膣栓の検査等により交配を確認した雌非ヒト哺乳動物の腹腔を開き、卵管から受精卵を取り出して胚培養用培地(例:M16培地、修正Whitten培地、BWW培地、M2培地、WM−HEPES培地、BWW−HEPES培地等)中で洗って卵丘細胞を除き、微小滴培養法等により5%炭酸ガス/95%大気下でDNA顕微注入まで培養する。直ちに顕微注入を行わない場合、採取した受精卵を緩慢法または超急速法等で凍結保存することも可能である。
【0029】
一方、体外受精の場合は、採卵用雌非ヒト哺乳動物(体内受精の場合と同様のものが好ましく用いられる)に上記と同様に卵胞刺激ホルモンおよび黄体形成ホルモンを投与して排卵を誘発させた後、卵子を採取して受精用培地(例:TYH培地)中で体外受精時まで微小滴培養法等により5%炭酸ガス/95%大気下で培養する。他方、同種の雄非ヒト哺乳動物(体内受精の場合と同様のものが好ましく用いられる)から精巣上体尾部を取り出し、精子塊を採取して受精用培地中で前培養する。前培養終了後の精子を卵子を含む受精用培地に添加し、微小滴培養法等により5%炭酸ガス/95%大気下で培養した後、2個の前核を有する受精卵を顕微鏡下で選抜する。直ちにDNAの顕微注入を行わない場合は、得られた受精卵を緩慢法または超急速法等で凍結保存することも可能である。
【0030】
受精卵へのDNAの顕微注入は、マイクロマニピュレーター等の公知の装置を用いて常法
に従って実施することができる。簡潔に言えば、胚培養用培地の微小滴中に入れた受精卵をホールディングピペットで吸引して固定し、インジェクションピペットを用いてDNA溶液を雄性もしくは雌性前核、好ましくは雄性前核内に直接注入する。導入DNAはCsCl密度勾配超遠心等で高度に精製したものを用いることが好ましい。また、導入DNAは制限酵素を用いてベクター部分を切断し、直鎖状にしておくことが好ましい。
【0031】
DNA導入後の受精卵は胚培養用培地中で微小滴培養法等により5%炭酸ガス/95%大気下で1細胞期〜胚盤胞期まで培養した後、偽妊娠させた受胚用雌非ヒト哺乳動物の卵管または子宮内に移植される。受胚用雌非ヒト哺乳動物は移植される初期胚が由来する動物と同種のものであればよく、例えば、マウス初期胚を移植する場合は、ICR系の雌マウス(好ましくは約8〜約10週齢)などが好ましく用いられる。受胚用雌非ヒト哺乳動物を偽妊娠状態にする方法としては、例えば、同種の精管切除(結紮)雄非ヒト哺乳動物(例えば、マウスの場合、ICR系の雄マウス(好ましくは約2月齢以上))と交配させて、膣栓の存在が確認されたものを選択する方法が知られている。
受胚用雌は自然排卵のものを用いてもよいし、あるいは精管切除(結紮)雄との交配に先立って、黄体形成ホルモン放出ホルモン(一般にLHRHと略する)もしくはその類縁体を投与し、受精能を誘起させたものを用いてもよい。LHRH類縁体としては、例えば、[3,5-DiI-Tyr5]-LH-RH、[Gln8]-LH-RH、[D-Ala6]-LH-RH、[des-Gly10]-LH-RH、[D-His(Bzl)6]-LH-RHおよびそれらのEthylamideなどが挙げられる。LHRHもしくはその類縁体の投与量、ならびにその投与後に雄非ヒト哺乳動物と交配させる時期は、非ヒト哺乳動物の種類によりそれぞれ異なる。例えば、非ヒト哺乳動物がマウス(好ましくはICR系のマウスなど)の場合には、通常、LHRHもしくはその類縁体を投与した後、約4日目に雄マウスと交配させることが好ましく、LHRHあるいはその類縁体の投与量は、通常、約10〜60μg/個体、好ましくは約40μg/個体である。
【0032】
通常、移植される初期胚が桑実胚期以後の場合は受胚用雌の子宮に、それより前(例えば、1細胞期〜8細胞期胚)であれば卵管に胚移植される。受胚用雌は、移植胚の発生段階に応じて偽妊娠からある日数が経過したものが適宜使用される。例えばマウスの場合、2細胞期胚を移植するには偽妊娠後約0.5日の雌マウスが、胚盤胞期胚を移植するには偽妊娠後約2.5日の雌マウスが好ましい。受胚用雌を麻酔(好ましくはAvertin、ネンブタール等が使用される)後、切開して卵巣を引き出し、胚培養用培地に懸濁した初期胚(約5〜約10個)を胚移植用ピペットを用いて、卵管腹腔口もしくは子宮角の卵管接合部付近に注入する。
【0033】
移植胚が首尾よく着床し受胚雌が妊娠すれば、自然分娩もしくは帝王切開により仔非ヒト哺乳動物が得られる。自然分娩した受胚雌にはそのまま哺乳を継続させればよく、帝王切開により出産した場合は、産仔は別途用意した哺乳用雌(例えばマウスの場合、通常に交配・分娩した雌マウス(好ましくはICR系の雌マウス等))に哺乳させることができる。
【0034】
受精卵細胞段階におけるKATPチャンネルKir6.2のドミナントネガティブ体をコードするDNAの導入は、導入DNAが対象非ヒト哺乳動物の生殖系列細胞および体細胞のすべてに存在するように確保される。導入DNAが染色体DNAに組み込まれているか否かは、例えば、産仔の尾部より分離抽出した染色体DNAをサザンハイブリダイゼーションまたはPCR法によりスクリーニングすることにより検定することができる。上記のようにして得られる仔非ヒト哺乳動物(F0)の生殖系列細胞においてKATPチャンネルKir6.2のドミナントネガティブ体をコードするDNAが存在することは、その後代(F1)の動物全てが、その生殖系列細胞および体細胞のすべてにKATPチャンネルKir6.2のドミナントネガティブ体をコードするDNAが存在することを意味する。
通常、F0動物は相同染色体の一方にのみ導入DNAを有するヘテロ接合体として得られる。また、個々のF0個体は相同組換えによらない限り異なる染色体上にランダムに挿入される。相同染色体の両方にKATPチャンネルKir6.2のドミナントネガティブ体をコードするDNAを有するホモ接合体を得るためには、F0動物と非トランスジェニック動物とを交雑してF1動物を作製し、相同染色体の一方にのみ導入DNAを有するヘテロ接合体の兄妹同士を交雑すればよい。1遺伝子座にのみ導入DNAが組み込まれていれば、得られるF2動物の1/4がホモ接合体となる。
【0035】
別の好ましい一実施態様においては、KATPチャンネルKir6.2のドミナントネガティブ体をコードするDNAを含む発現ベクターは、エレクトロポレーション法等の公知の遺伝子導入法により対象となる非ヒト哺乳動物のES細胞に導入される。
【0036】
ES細胞は胚盤胞期の受精卵の内部細胞塊(ICM)に由来し、インビトロで未分化状態を保ったまま培養維持できる細胞をいう。ICMの細胞は将来、胚本体を形成する細胞であり、生殖細胞を含むすべての組織の基になる幹細胞である。ES細胞としては、既に樹立された細胞株を用いてもよく、また、EvansとKaufmanの方法(ネイチャー(Nature)第292巻、154頁、1981年)に準じて新しく樹立したものでもよい。例えば、マウスES細胞の場合、現在、一般的には129系マウス由来のES細胞が使用されているが、免疫学的背景がはっきりしていないので、これに代わる純系で免疫学的に遺伝的背景が明らかなES細胞を取得するなどの目的で、例えば、C57BL/6マウスやC57BL/6の採卵数の少なさをDBA/2との交雑により改善したBDF1マウス(C57BL/6とDBA/2とのF1)から樹立されるES細胞なども良好に用いることができる。BDF1マウスは、採卵数が多く、かつ卵が丈夫であるという利点に加えて、C57BL/6マウスを背景に持つので、これ由来のES細胞は疾患モデルマウスを作製したとき、C57BL/6マウスと戻し交雑することでその遺伝的背景をC57BL/6マウスに代えることが可能である点で有利に用い得る。
【0037】
ES細胞の調製は、例えば以下のようにして行うことができる。交配後の雌非ヒト哺乳動物[例えばマウス(好ましくはC57BL/6J(B6)などの近交系マウス、B6と他の近交系とのF1など)を用いる場合は、約2月齢以上の雄マウスと交配させた約8〜約10週齢程度の雌マウス(妊娠約3.5日)が好ましく用いられる]の子宮から胚盤胞期胚を採取して(あるいは桑実胚期以前の初期胚を卵管から採取した後、胚培養用培地中で上記と同様にして胚盤胞期まで培養してもよい)、適当なフィーダー細胞(例えばマウスの場合、マウス胎仔から調製される初代繊維芽細胞や公知のSTO繊維芽細胞株等)層上で培養すると、胚盤胞の一部の細胞が集合して将来胚に分化するICMを形成する。この内部細胞塊をトリプシン処理して単細胞を解離させ、適切な細胞密度を保ち、培地交換を行いながら、解離と継代を繰り返すことによりES細胞が得られる。
【0038】
ES細胞は雌雄いずれを用いてもよいが、通常雄のES細胞の方が生殖系列キメラを作製するのに都合が良い。また、煩雑な培養の手間を削減するためにもできるだけ早く雌雄の判別を行なうことが望ましい。ES細胞の雌雄の判定方法としては、例えば、PCR法によりY染色体上の性決定領域の遺伝子を増幅、検出する方法が、その1例としてあげることができる。この方法を使用すれば、従来、核型分析をするのに約106個の細胞数を要していたのに対して、1コロニー程度のES細胞数(約50個)で済むので、培養初期におけるES細胞の第一次セレクションを雌雄の判別で行なうことが可能であり、早期に雄細胞の選定を可能にしたことにより培養初期の手間は大幅に削減できる。
また、第二次セレクションとして、例えば、G−バンディング法による染色体数の確認等により行うことができる。得られるES細胞の染色体数は正常数の100%が望ましいが、細胞株樹立の際の物理的操作等の関係上困難な場合は、ES細胞への遺伝子導入の後、正常細胞(例えば、マウスでは染色体数が2n=40である細胞)に再びクローニングすることが望ましい。
【0039】
このようにして得られるES細胞株は、未分化幹細胞の性質を維持するために注意深く継代培養することが必要である。例えば、STO繊維芽細胞のような適当なフィーダー細胞上で、分化抑制因子として知られるLIF(1〜10,000U/ml)存在下に炭酸ガス培養器内(好ましくは、5%炭酸ガス/95%空気または5%酸素/5%炭酸ガス/90%空気)で約37℃で培養するなどの方法で培養し、継代時には、例えば、トリプシン/EDTA溶液(通常0.001〜0.5%トリプシン/0.1〜5mM EDTA、好ましくは約0.1%トリプシン/1mM EDTA)処理により単細胞化し、新たに用意したフィーダー細胞上に播種する方法などがとられる。このような継代は、通常1〜3日毎に行なうが、この際に細胞の観察を行い、形態的に異常な細胞が見受けられた場合はその培養細胞は放棄することが望まれる。
ES細胞は、適当な条件により、高密度に至るまで単層培養するか、または細胞集塊を形成するまで浮遊培養することにより、頭頂筋、内臓筋、心筋などの種々のタイプの細胞に分化させることが可能であり〔M. J. Evans及びM. H. Kaufman, ネイチャー(Nature)第292巻、154頁、1981年;G. R. Martin プロシーディングス・オブ・ナショナル・アカデミー・オブ・サイエンス・ユーエスエー(Proc. Natl. Acad. Sci. USA)第78巻、7634頁、1981年;T. C. Doetschman ら、ジャーナル・オブ・エンブリオロジー・アンド・エクスペリメンタル・モルフォロジー、第87巻、27頁、1985年〕、本発明のKATPチャンネルKir6.2のドミナントネガティブ体をコードするDNAを導入されたES細胞を分化させて得られる該ドミナントネガティブ体発現非ヒト哺乳動物細胞は、インビトロにおけるKATPチャンネルKir6.2の細胞生物学的検討において有用である。
【0040】
ES細胞への遺伝子導入には、リン酸カルシウム共沈殿法、電気穿孔(エレクトロポレーション)法、リポフェクション法、レトロウイルス感染法、凝集法、顕微注入(マイクロインジェクション)法、遺伝子銃(パーティクルガン)法、DEAE−デキストラン法などのいずれも用いることができるが、簡便に多数の細胞を処理できること等の点からエレクトロポレーション法が一般的に選択されている。エレクトロポレーションには通常の動物細胞への遺伝子導入に使用されている条件をそのまま用いればよく、例えば、対数増殖期にあるES細胞をトリプシン処理して単一細胞に分散させた後、106〜108細胞/mlとなるように培地に懸濁してキュベットに移し、KATPチャンネルKir6.2のドミナントネガティブ体をコードするDNAを含むベクターを10〜100μg添加し、200〜600V/cmの電気パルスを印加することにより行なうことができる。
【0041】
導入DNAが組み込まれたES細胞は、単一細胞をフィーダー細胞上で培養して得られるコロニーから分離抽出した染色体DNAをサザンハイブリダイゼーションまたはPCR法によりスクリーニングすることによっても検定することができるが、ES細胞を用いるトランスジェニック系の最大の長所は、薬剤耐性遺伝子やレポーター遺伝子の発現を指標として細胞段階で形質転換体を選択できることである。したがって、ここで使用される導入ベクターは、KATPチャンネルKir6.2のドミナントネガティブ体をコードするDNAを含む発現カセットに加えて、薬剤耐性遺伝子(例:ネオマイシンホスホトランスフェラーゼII(nptII)遺伝子、ハイグロマイシンホスホトランスフェラーゼ(hpt)遺伝子など)やレポーター遺伝子(例:β−ガラクトシダーゼ(lacZ)遺伝子、クロラムフェニコールアセチルトランスフェラーゼ(cat)遺伝子など)等の選択マーカー遺伝子をさらに含むことが望ましい。例えば、選択マーカー遺伝子としてnptII遺伝子を含むベクターを用いた場合、遺伝子導入処理後のES細胞をG418などのネオマイシン系抗生物質を含有する培地中で培養し、出現した耐性コロニーをそれぞれ培養プレートに移してトリプシン処理、培地交換を繰り返した後、一部を培養用として残し、残りをPCRもしくはサザンハイブリダイゼーションにかけて導入DNAの存在を確認する。
【0042】
導入DNAの組込みが確認されたES細胞を同種の非ヒト哺乳動物由来の胚内に戻すと、宿主胚のICMに組み込まれてキメラ胚が形成される。これを仮親(受胚用雌)に移植してさらに発生を続けさせることにより、キメラトランスジェニック動物が得られる。キメラ動物の中でES細胞が将来卵や精子に分化する始原生殖細胞の形成に寄与した場合には、生殖系列キメラが得られることとなり、これを交配することにより導入DNAが遺伝的に固定された遺伝子導入非ヒト哺乳動物を作製することができる。
【0043】
キメラ胚の作製方法としては、桑実胚期までの初期胚同士を接着させて集合させる方法(集合キメラ法)と、胚盤胞の割腔内に細胞を顕微注入する方法(注入キメラ法)とがある。ES細胞によるキメラ胚の作製においては従来より後者が広く行なわれているが、最近では、8細胞期胚の透明帯内へのES細胞の注入により集合キメラを作る方法や、マイクロマニピュレーターが不要で操作が容易な方法として、ES細胞塊と透明帯を除去した8細胞期胚とを共培養して凝集させることによって集合キメラを作製する方法も行われている。
いずれの場合も、宿主胚は受精卵への遺伝子導入における採卵用雌として使用され得る非ヒト哺乳動物から同様にして採取することができるが、例えばマウスの場合、キメラマウス形成へのES細胞の寄与率を毛色(コートカラー)で判定し得るように、ES細胞の由来する系統とは毛色の異なる系統のマウスから宿主胚を採取することが好ましい。例えば、ES細胞が129系マウス(毛色:アグーチ)由来であれば、採卵用雌としてC57BL/6マウス(毛色:ブラック)やICRマウス(毛色:アルビノ)を用い、ES細胞がC57BL/6もしくはDBF1マウス(毛色:ブラック)由来やTT2細胞(C57BL/6とCBAとのF1(毛色:アグーチ)由来)であれば、採卵用雌としてICRマウスやBALB/cマウス(毛色:アルビノ)を用いることができる。
また、生殖系列キメラ形成能はES細胞と宿主胚との組み合わせに大きく依存するので、生殖系列キメラ形成能の高い組み合わせを選択することがより好ましい。例えばマウスの場合、129系統由来のES細胞に対してはC57BL/6系統由来の宿主胚等を用いることが好ましく、C57BL/6系統由来のES細胞に対してはBALB/c系統由来の宿主胚等が好ましい。
採卵用雌マウスは約4〜約6週齢程度が好ましく、交配用の雄マウスとしては約2〜約8月齢程度の同系統のものが好ましい。交配は自然交配によってもよいが、好ましくは性腺刺激ホルモン(卵胞刺激ホルモン、次いで黄体形成ホルモン)を投与して過剰排卵を誘起した後に行なわれる。
【0044】
胚盤注入法による場合は、胚盤胞期胚(例えばマウスの場合、交配後約3.5日)を採卵用雌の子宮から採取し(あるいは桑実胚期以前の初期胚を卵管から採取した後、上述の胚培養用培地中で胚盤胞期まで培養してもよい)、マイクロマニピュレーターを用いて胚盤胞の割腔内にKATPチャンネルKir6.2のドミナントネガティブ体をコードするDNAが導入されたES細胞(約10〜約15個)を注入した後、偽妊娠させた受胚用雌非ヒト哺乳動物の子宮内に移植する。受胚用雌非ヒト哺乳動物は受精卵への遺伝子導入における受胚用雌として使用され得る非ヒト哺乳動物を同様に用いることができる。
共培養法による場合は、8細胞期胚および桑実胚(例えばマウスの場合、交配後約2.5日)を採卵用雌の卵管および子宮から採取して(あるいは8細胞期以前の初期胚を卵管から採取した後、上述の胚培養用培地中で8細胞期または桑実胚期まで培養してもよい)酸性タイロード液中で透明帯を溶解した後、ミネラルオイルを重層した胚培養用培地の微小滴中にKATPチャンネルKir6.2のドミナントネガティブ体をコードするDNAが導入されたES細胞塊(細胞数約10〜約15個)を入れ、さらに上記8細胞期胚または桑実胚(好ましくは2個)を入れて一晩共培養する。得られた桑実胚または胚盤胞を上記と同様にして受胚用雌非ヒト哺乳動物の子宮内に移植する。
【0045】
移植胚が首尾よく着床し受胚雌が妊娠すれば、自然分娩もしくは帝王切開によりキメラ非ヒト哺乳動物が得られる。自然分娩した受胚雌にはそのまま哺乳を継続させればよく、帝王切開により出産した場合は、産仔は別途用意した哺乳用雌(通常に交配・分娩した雌非ヒト哺乳動物)に哺乳させることができる。
生殖系列キメラの選択は、まずES細胞の雌雄が予め判別されている場合はES細胞と同じ性別のキメラマウスを選択し(通常は雄性ES細胞が使用されるので、雄キメラマウスが選択される)、次いで毛色等の表現型からES細胞の寄与率が高いキメラマウス(例えば、50%以上)を選択する。例えば、129系マウス由来の雄性ES細胞であるD3細胞とC57BL/6マウス由来の宿主胚とのキメラ胚から得られるキメラマウスの場合、アグーチの毛色の占める割合の高い雄マウスを選択するのが好ましい。選択されたキメラ非ヒト哺乳動物が生殖系列キメラであるか否かの確認は、適当な系統の同種動物との交雑により得られるF1動物の表現型に基づいて行なうことができる。例えば、上記キメラマウスの場合、アグーチはブラックに対して優性であるので、雌C57BL/6マウスと交雑すると、選択された雄マウスが生殖系列キメラであれば得られるF1の毛色はアグーチとなる。
【0046】
上記のようにして得られるKATPチャンネルKir6.2のドミナントネガティブ体をコードするDNAが導入された生殖系列キメラ非ヒト哺乳動物(ファウンダー)は、通常、相同染色体の一方にのみ導入DNAを有するヘテロ接合体として得られる。また、個々のファウンダーは相同組換えによらない限り異なる染色体上にランダムに挿入される。相同染色体の両方にKATPチャンネルKir6.2のドミナントネガティブ体をコードするDNAを有するホモ接合体を得るためには、上記のようにして得られるF1動物のうち相同染色体の一方にのみ導入DNAを有するヘテロ接合体の兄妹同士を交雑すればよい。ヘテロ接合体の選択は、例えばF1動物の尾部より分離抽出した染色体DNAをサザンハイブリダイゼーションまたはPCR法によりスクリーニングすることにより検定することができる。1遺伝子座にのみ導入DNAが組み込まれていれば、得られるF2動物の1/4がホモ接合体となる。
【0047】
上記のようにして得られうる、KATPチャンネルKir6.2のドミナントネガティブ体を膵β細胞で発現し得る本発明のTg動物は、以下の特性:
(a)血糖値およびインスリン分泌
(1)新生仔で低血糖・高インスリン血症を呈する
(2)若週(約4〜約8週)齢で高血糖・インスリン欠乏症を呈する
(3)加齢(約9〜約12週齢以降)とともに血糖低下・インスリン分泌増加を呈する
(b)膵β細胞・膵島構造
(1)若週齢でβ細胞の細胞死・α細胞の増加および正常な膵島構造(中心部にβ細胞が存
在し、α細胞は周縁部に限局される)の破壊を認める
(2)加齢とともに膵管近傍に膵β細胞の新生を認める
を有することから、該Tg動物、あるいは膵内分泌細胞に分化し得るその生体の一部は、膵再生、特に膵内分泌細胞再生の有用なモデル系となり得る。ここで「モデル系」なる用語は、再生モデルは再生現象の前後において物質としての性状が変化することから、経時的要素を含むことを意味する目的で用いられる。
【0048】
本発明の膵再生モデル系の好ましい一実施態様は、本発明のTg動物の個体全体からなるものである。即ち、本発明のTg動物を、通常の飼育条件下(例えば、約12時間明期条件(例えば7:00−19:00)下で、普通食(必要に応じて高脂肪食等を用いることもできる)および水を自由摂取の状態で飼育する(SPF条件下が望ましいが、コンベンショナルな条件でもよい)など)で、少なくとも膵内分泌細胞の分化が起こるまで、例えば、少なくとも約9〜約12週齢以降まで飼育し続けることにより、膵再生現象を観察することができる。
【0049】
本発明の膵再生モデル系の別の好ましい一実施態様は、本発明のTg動物の生体の一部から実質的になるものである。ここで「実質的になる」とは、例えば、培地成分や分化誘導物質などの、膵再生現象の主体ではないが該生体の一部における膵再生を誘導するために必要もしくは望ましい他の要素を含んでいてもよいことを意味する。該生体の一部は、少なくとも膵内分泌細胞に分化し得る細胞を含むものであり、例えば、膵臓、肝臓等の臓器、膵島、膵管等の組織、膵前駆細胞、膵幹細胞、肝前駆細胞などの細胞、あるいはそれらより特定の培養条件下で誘導される細胞・組織、さらには、該Tg動物の初期胚より誘導されるES細胞などが挙げられるが、それらに限定されない。
【0050】
本発明のTg動物の生体の一部から実質的になる膵再生モデル系に含有され得る培地としては、該生体の一部の維持・増殖・分化に適したものであれば特に制限はなく、例えば、最小必須培地(MEM)、ダルベッコ改変イーグル培地(DMEM)、ハムF12培地、F10培地、RPMI1640培地等が挙げられる。これらの培地には、インスリン、トランスフェリン等の成分が補充されていることが望ましい。
本発明のTg動物の生体の一部から実質的になる膵再生モデル系に含有され得る分化誘導剤としては、例えば、FGFファミリー(FGF1、FGF2、FGF7、FGF10など)、EGFファミリー(EGF、TGF-α、βセルリン、βセルリンδ4、amphiregulin、Heparin-binding EGF)、keratinocyte growth factor、アクチビン、TGFβ1-3、HGF、GLP-1、CCK、ガストリン、脂肪酸、ニコチンアミド、セレニウム、プロゲステロン、プトレッシン等が挙げられるが、これらに限定されない。
【0051】
本発明は、本発明のTg動物またはその生体の一部を用いた膵再生関連遺伝子の同定方法を提供する。本方法は、膵内分泌細胞の分化前後(ここで「分化前後」とは、分化が起こる前と分化が完了した後との比較だけを意味するのではなく、分化前と今まさに分化が起こっている最中や分化誘導のスイッチがオンされた直後等との比較をも包含する意味で用いられる)で、該Tg動物から膵臓もしくはその一部(例えば、膵島、膵管など)を摘出し、あるいは該Tg動物の生体の一部を膵内分泌細胞への分化が誘導され得る条件下で培養し、それらから常法に従って時系列的に全RNAもしくはpolyA RNA (mRNA)を抽出して、例えば、膵島特異的なcDNAマイクロアレイ等を用いて、網羅的に遺伝子発現プロファイルの変動を解析することにより行うことができる。RNA画分の調製は、グアニジン−CsCl超遠心法、AGPC法など公知の手法を用いて行うことができるが、市販のRNA抽出用キット(例:RNeasy Mini Kit; QIAGEN製等)を用いて、微量試料から迅速且つ簡便に高純度の全RNAを調製することができる。
本発明のTg動物の生体の一部を膵内分泌細胞への分化が誘導される条件下で培養する方法としては、例えば、該Tg動物から膵導管細胞画分を採取して培養する方法(Proc. Natl. Acad. Sci. USA, 97: 7999-8004 (2000))、該Tg動物とNODマウスとの交配により得られた動物の膵導管上皮を長期培養してインスリン産生細胞を取得する方法(Nat.Med., 6: 278-282 (2000))、膵島に存在するネスチン陽性細胞で、終分化マーカーと共発現しない細胞を分取し、インビトロで培養することにより、Pdx-1、インスリン、グルカゴン、GLP-1陽性細胞を生じさせる方法(Diabetes, 50: 521-533 (2001))、肝前駆細胞をニコチンアミド含有培地で長期培養することによりインスリンIおよびII遺伝子を発現する細胞を得る方法(J. Cell Biol., 156: 173-184 (2002))等が挙げられるが、それらに限定されない。膵内分泌細胞への分化は、例えば、膵再生への関連が示されている各種転写因子(例:Ngn3、Nkx2、NeuroD、Nkx6.1、PAX-4等)の発現をRT-PCR、in situハイブリダイゼーション、免疫染色等で検出するか、膵再生への関連が示されている因子に対する蛍光抗体を用いたセルソーティング、後述する本発明のTg動物にさらに他の1つ以上の遺伝子改変を有する動物において、該他の1つの遺伝子改変がβ細胞特異的にレポーター遺伝子を発現させるものであるTg動物を用いて、該レポーター遺伝子の発現を検出する等の手法によって確認することができる。
【0052】
マイクロアレイ解析は、例えば、上記のようにして調製したRNA画分から、逆転写反応によりT7プロモーター等の適当なプロモーターを導入したcDNAを合成し、さらにRNAポリメラーゼを用いてcRNAを合成する(この時ビオチンなどで標識したモノヌクレオチドを基質として用いることにより、標識されたcRNAが得られる)。この標識cRNAを、例えば、膵島特異的cDNAマイクロアレイ(正常膵島由来のcDNAを、適当な固相、例えば、ガラス、シリコン、プラスチック、ニトロセルロース、ナイロン、ポリビニリデンジフロリド等に固定化したもの。固定化手段としては、予め核酸にアミノ基、アルデヒド基、SH基、ビオチンなどの官能基を導入しておき、一方、固相上にも該核酸と反応し得る官能基(例:アルデヒド基、アミノ基、SH基、ストレプトアビジンなど)を導入し、両官能基間の共有結合で固相と核酸を架橋したり、ポリアニオン性の核酸に対して、固相をポリカチオンコーティングして静電結合を利用して核酸を固定化するなどの方法が挙げられるが、これらに限定されない)と接触させてハイブリダイゼーション反応させ、マイクロアレイ上の各プローブに結合した標識量を測定することにより、各時点でのTg動物の膵臓もしくはその一部、あるいは該Tg動物由来の細胞・組織における遺伝子の発現量を測定することができる。これらの各遺伝子の発現量を分化誘導前後で比較した結果、分化誘導前後で発現量が変動した遺伝子を膵(内分泌細胞)再生関連(促進もしくは抑制)遺伝子の候補として選択することができる。
【0053】
本発明はまた、上記の膵再生関連遺伝子の同定方法を迅速且つ簡便に行うためのツールとして有用な、本発明のTg動物の膵内分泌細胞分化前後の膵臓もしくはその一部(例えば、膵島、膵管など)由来の単離されたRNAまたはそれを鋳型として逆転写反応によって調製されるcDNA(さらにRNAポリメラーゼの反応によって合成され得るcRNAも包含される)、あるいは該Tg動物の生体の一部における膵内分泌細胞分化前後の該生体の一部由来の単離されたRNAまたはそのcDNAを提供する。ここで「膵内分泌細胞分化前後」とは、分化前、分化誘導直後、分化の途中、分化終了後のいかなる段階であってもよく、それらのいずれか1つであっても、2つ以上であってもよい。RNA画分の調製法としては、上記と同様の方法が例示される。
【0054】
本発明の膵再生モデル系を用いた膵再生因子の別の同定方法としては、該モデル系より常法に従って蛋白質画分を調製し、例えば、サイファージェン・バイオシステムズ社のSELDI技術等を用いて、蛋白質を特異的に質量分析用プローブの表面上に捕捉させ、質量分析、好ましくはMALDI-TOFMS法などによりプローブに捕捉された蛋白質の分子量を測定、膵内分泌細胞分化前後でピーク強度の変化した分子量を示した蛋白質について、例えばタンデム質量分析(MS/MS)等を実施し、そのアミノ酸配列を同定する方法などが挙げられる。
【0055】
本発明はまた、本発明のTg動物に被検物質を投与、あるいは該Tg動物の生体の一部に被検物質を添加し、膵内分泌細胞分化に及ぼす該物質の効果を検定することによる、膵再生調節物質のスクリーニング方法を提供する。即ち、被検物質を投与(添加)しなかった場合と比較して、膵内分泌細胞分化が促進(もしくは抑制)されたか否かを判定することにより該物質の効果を検定する。膵内分泌細胞分化は、上記と同様に、例えば、膵再生への関連が示されている各種転写因子(例:Ngn3、Nkx2、NeuroD、Nkx6.1、PAX-4等)の発現をRT-PCR、in situハイブリダイゼーション、免疫染色等で検出するか、膵再生への関連が示されている因子に対する蛍光抗体を用いたセルソーティング、後述する本発明のTg動物にさらに他の1つ以上の遺伝子改変を有する動物において、該他の1つの遺伝子改変がβ細胞特異的にレポーター遺伝子を発現させるものであるTg動物を用いて、該レポーター遺伝子の発現を検出する等の手法によって確認することができる。
その結果、被検物質を投与(添加)しなかった場合に比べて、該Tg動物もしくはその生体の一部における内分泌細胞分化の開始が早まった場合(あるいは、同期間で分化がより進んだ場合)、該被検物質を膵再生促進物質として、内分泌細胞分化の開始が遅くなった場合(あるいは、同期間で分化がより進まなかった場合)、該被検物質を膵再生抑制物質として、それぞれ選択することができる。
【0056】
上記スクリーニング方法において、膵内分泌細胞分化を、上記の同定方法によって同定された膵再生関連遺伝子(因子)の発現を指標にして測定することもできる。従って、本発明はまた、本発明のTg動物に被検物質を投与、あるいは該Tg動物の生体の一部に被検物質を添加し、膵再生関連遺伝子(因子)の発現に及ぼす該物質の効果を検定することによる、膵再生関連遺伝子(因子)の発現調節物質のスクリーニング方法を提供する。
【0057】
上記スクリーニング方法によって得られうる、膵再生促進物質または膵再生促進遺伝子(因子)の発現促進物質もしくは膵再生抑制遺伝子(因子)の発現抑制物質(以下、包括して「膵再生促進薬」という)は、膵疲弊・膵傷害を伴う疾患、例えば、1型および2型糖尿病、糖尿病合併症(糖尿病性腎症、糖尿病性網膜症、糖尿病性神経障害等)、耐糖能異常、低インスリン血症等の予防・治療薬として安全かつ有効に用いることができる。
【0058】
膵再生促進薬は、例えば、必要に応じて糖衣を施した錠剤、カプセル剤、エリキシル剤、マイクロカプセル剤などとして経口的に、あるいは水もしくはそれ以外の薬学的に許容し得る液との無菌性溶液、または懸濁液剤などの注射剤の形で非経口的に使用できる。該作動薬は、生理学的に認められる担体、香味剤、賦形剤、ベヒクル、防腐剤、安定剤、結合剤などとともに一般に認められた製剤実施に要求される単位用量形態で混和することによって製剤化することができる。これら製剤における有効成分量は後述する投与量を考慮して適宜選択される。
【0059】
錠剤、カプセル剤などに混和することができる添加剤としては、例えば、ゼラチン、コーンスターチ、トラガント、アラビアゴムのような結合剤、結晶性セルロースのような賦形剤、コーンスターチ、ゼラチン、アルギン酸などのような膨化剤、ステアリン酸マグネシウムのような潤滑剤、ショ糖、乳糖またはサッカリンのような甘味剤、ペパーミント、アカモノ油またはチェリーのような香味剤などが用いられる。調剤単位形態がカプセルである場合には、前記タイプの材料にさらに油脂のような液状担体を含有することができる。注射のための無菌組成物は注射用水のようなベヒクル中の活性物質、胡麻油、椰子油などのような天然産出植物油などを溶解または懸濁させるなどの通常の製剤実施に従って処方することができる。
【0060】
注射用の水性液としては、例えば、生理食塩水、ブドウ糖やその他の補助薬を含む等張液(例えば、D−ソルビトール、D−マンニトール、塩化ナトリウムなど)などが挙げられ、適当な溶解補助剤、例えば、アルコール(例えば、エタノールなど)、ポリアルコール(例えば、プロピレングリコール、ポリエチレングリコールなど)、非イオン性界面活性剤(例えば、ポリソルベート80TM、HCO−50など)などと併用してもよい。油性液としては、例えば、ゴマ油、大豆油などが挙げられ、溶解補助剤として安息香酸ベンジル、ベンジルアルコールなどと併用してもよい。また、緩衝剤(例えば、リン酸塩緩衝液、酢酸ナトリウム緩衝液など)、無痛化剤(例えば、塩化ベンザルコニウム、塩酸プロカインなど)、安定剤(例えば、ヒト血清アルブミン、ポリエチレングリコールなど)、保存剤(例えば、ベンジルアルコール、フェノールなど)、酸化防止剤などと配合してもよい。調製された注射液は、通常、適当なアンプルに充填される。
【0061】
また、膵再生促進薬がDNA、RNAなどの核酸である場合、当該核酸を単独で、あるいは当該DNA(または当該RNAに対応するDNA)をレトロウイルスベクター、アデノウイルスベクター、アデノウイルスアソシエーテッドウイルスベクターなどの適当なベクターに挿入した後、常套手段に従って、ヒトまたは他の哺乳動物に投与することができる。当該核酸は、そのままで、あるいは摂取促進のための補助剤などの生理学的に認められる担体とともに製剤化した後、遺伝子銃やハイドロゲルカテーテルのようなカテーテルによって投与できる。
【0062】
膵再生促進薬の投与量は、対象疾患、投与対象、投与ルートなどにより異なるが、例えば、糖尿病の治療目的で経口投与する場合、一般的に成人(体重60kg)においては、一日につき約0.1mg〜100mg、好ましくは約1.0〜50mg、より好ましくは約1.0〜20mgである。非経口投与の場合、当該薬物の投与量は投与対象、対象疾患などによっても異なるが、例えば、糖尿病の治療目的で注射剤として成人(体重60kg)に投与する場合、一日につき約0.01〜30mg程度、好ましくは約0.1〜20mg程度、より好ましくは約0.1〜10mg程度である。投与対象がヒト以外の動物の場合も、体重60kg当たりに換算した量を投与することができる。
【0063】
本発明のTg動物は、KATPチャンネルKir6.2のドミナントネガティブ体をコードするDNAを膵β細胞で発現可能な状態で安定に保持することに加えて、1以上の他の遺伝子変異または改変を有することをさらなる特徴とする。「他の遺伝子変異」とは、自然突然変異により内因性遺伝子が欠損、機能低下、あるいは機能亢進された自然発症疾患モデル動物をいい、「他の遺伝子改変」とは、KATPチャンネルKir6.2のドミナントネガティブ体をコードするDNAが導入される以外の遺伝子改変、即ち、他の遺伝子をさらに導入されたトランスジェニック動物、内因性遺伝子を不活化されたノックアウト動物(挿入突然変異等による遺伝子破壊のほか、アンチセンスDNAや中和抗体をコードするDNAの導入により遺伝子発現が検出不可能もしくは無視し得る程度にまで低下したトランスジェニック動物を含む)、変異内因性遺伝子が導入されたドミナントネガティブ変異体などが含まれる。
【0064】
「他の遺伝子変異または改変」は、膵再生関連遺伝子(因子)の探索、膵再生促進薬の探索、並びに膵再生促進薬の薬効評価用としての用途に有利なものであれば特に制限はないが、例えば、膵内分泌機能異常が関与する疾患と同一もしくは類似の病態を生じさせるような遺伝子変異または改変であることが好ましい。
「膵内分泌機能異常が関与する疾患」とは、膵内分泌機能の異常に起因するかもしくは結果的に膵内分泌機能の異常を生じる疾患だけでなく、膵内分泌機能を調節することにより予防および/または治療効果が得られ得る疾患をも含めた概念として把握されるべきである。例えば、1型および2型糖尿病、糖尿病合併症(糖尿病性腎症、糖尿病性網膜症、糖尿病性神経障害等)、耐糖能異常、低インスリン血症等が挙げられる。
【0065】
「膵内分泌機能異常が関与する疾患と同一もしくは類似の病態を生じさせる1以上の他の遺伝子変異または改変を有する疾患モデル」としては、例えば、自然発症糖尿病モデルマウスであるKKマウス、KKAyマウス、db/dbマウス、ob/obマウス、Akitaマウス等、あるいはヒトIAPP遺伝子を導入したTgマウス等の膵疲弊または膵傷害を示す各種Tg/KOマウスが挙げられる。
これらの「他の遺伝子変異または改変を有する疾患モデル」は、例えば、米国のJackson研究所などから購入可能であるか、あるいは周知の遺伝子改変技術を用いて容易に作製することができる。
【0066】
本発明のTg動物は、「膵内分泌機能異常が関与する疾患と同一もしくは類似の病態を生じさせる1以上の他の遺伝子変異または改変」に加えて、同一もしくは他の疾患モデルを作製し得る非遺伝的処理を施されていてもよい。「非遺伝的処理」とは対象非ヒト哺乳動物における遺伝子変異または改変を生じさせない処理を意味する。このような処理としては、例えば、高脂肪食負荷処理、糖負荷処理、飢餓処理、膵管結紮、STZ処理、膵切除等が挙げられる。
【0067】
膵再生促進薬の薬効評価用としての本発明のTg動物の用途に有利な「他の遺伝子改変」の別の好ましい態様としては、膵β細胞特異的なプロモーターの制御下にあるレポーター遺伝子の導入が挙げられる。該プロモーターの制御下にあるレポーター遺伝子を導入された非ヒト哺乳動物では、膵β細胞の再生によりβ細胞数が増加すると該レポーターの発現量が増大するので、該レポーターの発現に及ぼす被検物質の効果を検定することにより、該物質の膵再生促進活性を評価することができる。膵β細胞特異的プロモーターとしては、例えばインスリンIもしくはIIプロモーター等が挙げられる。また、レポーター遺伝子としては、例えば、β−ガラクトシダーゼ遺伝子、ルシフェラーゼ遺伝子、クロラムフェニコールアセチルトランスフェラーゼ遺伝子、アルカリホスファターゼ遺伝子、ペルオキシダーゼ遺伝子等が挙げられる。
【0068】
KATPチャンネルKir6.2のドミナントネガティブ体をコードするDNAを導入された非ヒト哺乳動物に、膵内分泌機能異常が関与する疾患と同一もしくは類似の病態を生じさせる1以上の他の遺伝子変異または改変、あるいは膵β細胞特異的プロモーターの制御下にあるレポーター遺伝子を導入する方法は特に制限はなく、例えば、KATPチャンネルKir6.2のドミナントネガティブ体をコードするDNAを導入された非ヒト哺乳動物と、膵内分泌機能異常が関与する疾患と同一もしくは類似の病態を生じさせる1以上の他の遺伝子変異または改変、あるいは膵β細胞特異的プロモーターの制御下にあるレポーター遺伝子を有する同種の非ヒト哺乳動物とを交雑する方法;膵内分泌機能異常が関与する疾患と同一もしくは類似の病態を生じさせる1以上の他の遺伝子変異または改変、あるいは膵β細胞特異的プロモーターの制御下にあるレポーター遺伝子を有する非ヒト哺乳動物の初期胚やES細胞に、上述の方法によりKATPチャンネルKir6.2のドミナントネガティブ体をコードするDNAを導入してトランスジェニック動物を得る方法;KATPチャンネルKir6.2のドミナントネガティブ体をコードするDNAを導入された非ヒト哺乳動物の初期胚やES細胞に、上述の方法により、あるいはノックアウト技術により、膵内分泌機能異常が関与する疾患と同一もしくは類似の病態を生じさせる1以上の他の遺伝子改変、あるいは膵β細胞特異的プロモーターの制御下にあるレポーター遺伝子を導入する方法等が挙げられる。また、膵内分泌機能異常が関与する疾患と同一もしくは類似の病態を生じさせる1以上の他の遺伝子改変が外来遺伝子やドミナント変異遺伝子の導入による場合、野生型非ヒト哺乳動物の初期胚やES細胞に、該外来遺伝子等とKATPチャンネルKir6.2のドミナントネガティブ体をコードするDNAとを同時にもしくは順次導入してトランスジェニック動物を得てもよい。他の遺伝子改変が膵β細胞特異的プロモーターの制御下にあるレポーター遺伝子の導入である場合も同様に、該レポーター遺伝子とKATPチャンネルKir6.2のドミナントネガティブ体をコードするDNAとを同時にもしくは順次導入してトランスジェニック動物を得ることができる。さらに、膵内分泌機能異常が関与する疾患と同一もしくは類似の病態を生じさせる1以上の他の遺伝子改変が内因性遺伝子の破壊による場合は、KATPチャンネルKir6.2のドミナントネガティブ体をコードするDNAを破壊すべき内因性遺伝子にターゲッティングされ得るようにデザインして野生型非ヒト哺乳動物のES細胞に導入してもよい。この場合、従来公知のノックイン動物作製技術が好ましく使用され得る。
【0069】
KATPチャンネルKir6.2のドミナントネガティブ体をコードするDNAを導入された非ヒト哺乳動物と、膵内分泌機能異常が関与する疾患と同一もしくは類似の病態を生じさせる1以上の他の遺伝子変異または改変(あるいは膵β細胞特異的プロモーターの制御下にあるレポーター遺伝子)を有する同種の疾患モデル非ヒト哺乳動物とを交雑する場合、ホモ接合体同士を交雑することが望ましい。例えば、KATPチャンネルKir6.2のドミナントネガティブ体をコードするDNAが1遺伝子座に組み込まれたホモ接合体と、db/dbマウスとを交雑して得られるF1は両遺伝子についてヘテロである。このF1同士を兄妹交配して得られるF2個体の1/16はドミナントネガティブホモ導入・dbホモとなる。
【0070】
KATPチャンネルKir6.2のドミナントネガティブ体をコードするDNAを導入され、且つ膵内分泌機能異常が関与する疾患と同一もしくは類似の病態を生じさせる1以上の他の遺伝子変異または改変を有する本発明のTg動物は、膵内分泌機能異常が関与する疾患と同一もしくは類似の病態を示すので、該動物に被検物質を投与してその病態に及ぼす該物質の効果を検定することによって、膵内分泌機能異常が関与する疾患に対して予防・治療活性を有する物質をスクリーニングすることができる。指標となる病態は疾患モデルの種類に応じて適宜選択され得るが、例えば、該Tg動物が糖尿病モデルであれば、血糖値・インスリン分泌の改善、膵内分泌細胞の増殖・分化促進などを指標として、被検物質の疾患予防・治療効果を評価することができる。
【0071】
本願明細書において、塩基やアミノ酸などを略号で表示する場合、IUPAC-IUB Commission on Biochemical Nomenclatureによる略号あるいは当該分野における慣用略号に基づくものであり、その例を次に挙げる。
DNA :デオキシリボ核酸
cDNA :相補的デオキシリボ核酸
A :アデニン
T :チミン
G :グアニン
C :シトシン
RNA :リボ核酸
mRNA :メッセンジャーリボ核酸
dATP :デオキシアデノシン三リン酸
dTTP :デオキシチミジン三リン酸
dGTP :デオキシグアノシン三リン酸
dCTP :デオキシシチジン三リン酸
ATP :アデノシン三リン酸
EDTA :エチレンジアミン四酢酸
SDS :ドデシル硫酸ナトリウム
Gly :グリシン
Ala :アラニン
Val :バリン
Leu :ロイシン
Ile :イソロイシン
Ser :セリン
Thr :スレオニン
Cys :システイン
Met :メチオニン
Glu :グルタミン酸
Asp :アスパラギン酸
Lys :リジン
Arg :アルギニン
His :ヒスチジン
Phe :フェニルアラニン
Tyr :チロシン
Trp :トリプトファン
Pro :プロリン
Asn :アスパラギン
Gln :グルタミン
【実施例】
【0072】
以下に実施例を挙げて本発明をより具体的に説明するが、本発明がこれらに限定されないことは言うまでもない。
【0073】
実施例1 Kir6.2G132S Tgマウスにおける血糖値の推移
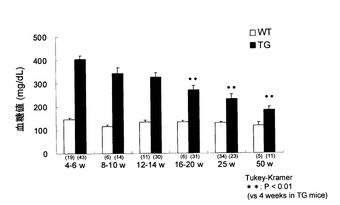
上記非特許文献2に記載される方法により作製されたKir6.2G132S Tgマウス(line M45)について、同文献と同様の方法により、4-6(n=43)、8-10(n=14)、12-14(n=30)、16-20(n=31)、25(n=23)および50(n=11)週齢における血糖値をそれぞれ測定した。結果を図1に示す。経時的に血糖値を観察したところ、4-6週齢に認められた高血糖が経時的に低下し、16-20週齢以降、4-6週齢に比べて有意な血糖改善が認められ、50週齢ではほぼ正常値を示すまでに至った。
【0074】
実施例2 Kir6.2G132S Tgマウスにおける血清インスリン値の推移
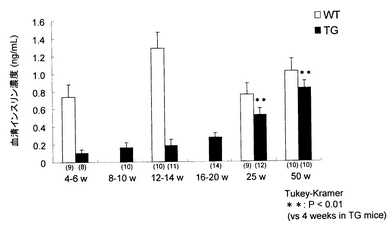
Kir6.2G132S Tgマウス(line M45)について、上記非特許文献2と同様の方法により、4-6(n=8)、8-10(n=10)、12-14(n=11)、16-20(n=14)、25(n=12)および50(n=10)週齢における血清インスリン値をそれぞれ測定した。結果を図2に示す。経時的に血清インスリン値を観察したところ、4-6週齢に認められたインスリン欠乏状態が経時的に改善された。25週齢以降は有意な改善が認められ、50週齢ではほぼ正常値を示すまでに至った。
【0075】
実施例3 Kir6.2G132S Tgマウスにおける膵組織像
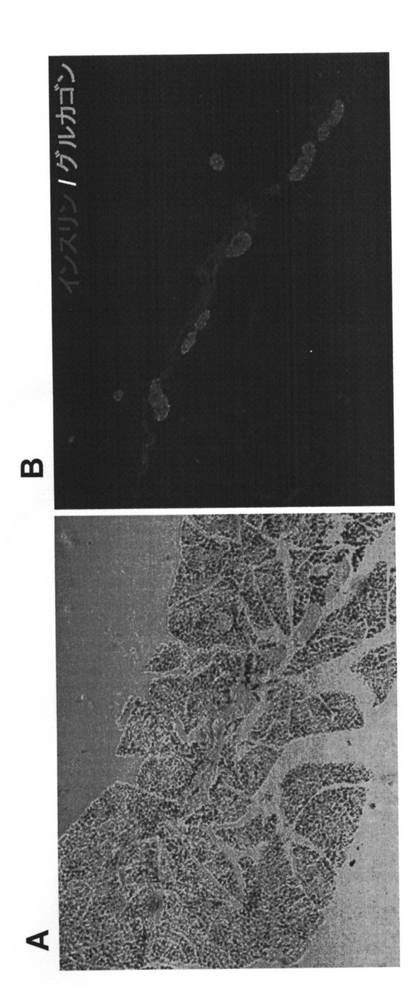
Kir6.2G132S Tgマウス(line M45;11週齢)より膵臓を摘出し、膵管近傍組織切片を作製した。抗インスリン抗体および抗グルカゴン抗体をそれぞれ蛍光色素Cy3(赤)およびCy2(緑)で標識し、二重免疫染色を行った。結果を図3に示す。血糖値が改善し始める12週齢の前後では、膵管近傍において膵β細胞の新生像が観察された(図3B)。
【0076】
実施例4 Kir6.2G132S Tgマウスにおけるβ細胞組織像
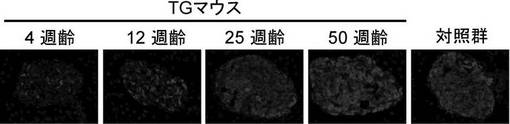
Kir6.2G132S Tgマウス(line M45)について、4、12、25及び50週齢時に膵臓を摘出し、β細胞組織切片を作製した。対照群(正常マウス)についても膵臓を摘出し、β細胞組織切片を作製した。抗インスリン抗体を蛍光色素Cy3(赤)で標識し、免疫染色を行った。結果を図4に示す。Kir6.2G132S Tgマウスのβ細胞において、4週齢に認められたインスリン含量の低下は経時的に改善され、50週齢ではほぼ正常化するに至った。
【産業上の利用可能性】
【0077】
Kir6.2のドミナントネガティブ体を導入されたTg動物は、若週齢で糖尿病を発症し、インスリン欠乏・膵β細胞の細胞死が顕著となるが、加齢とともにインスリン分泌が増大し、膵管近傍に高頻度に膵内分泌細胞が出現することから、該動物およびその生体の一部は、優れた膵内分泌細胞の再生モデル系として、膵再生関連遺伝子(因子)の探索、膵再生促進薬の探索、膵再生促進薬の薬効評価に利用することができる。
【図面の簡単な説明】
【0078】
【図1】Kir6.2G132S Tgマウス(TG)および野生型マウス(WT)における血糖値の推移を示す図である。グラフの各棒の下の括弧内の数字は例数(匹)を示す。
【図2】Kir6.2G132S Tgマウス(TG)および野生型マウス(WT)における血清インスリン値の推移を示す図である。グラフの各棒の下の括弧内の数字は例数(匹)を示す。
【図3】Kir6.2G132S Tgマウス(11週齢)の膵管近傍における組織像を示す図である。図3Aは位相差顕微鏡像を、図3Bは蛍光色素によるインスリン/グルカゴン共染色像をそれぞれ示す。
【図4】Kir6.2G132S Tgマウスのβ細胞組織におけるインスリン含量の経時変化を示す図である。抗インスリン抗体を蛍光色素Cy3(赤)で標識し、免疫染色を行った。
【技術分野】
【0001】
本発明は、ATP感受性カリウムイオンチャンネル遺伝子に改変を有するトランスジェニック非ヒト哺乳動物の新規用途に関する。詳細には、本発明は、ATP感受性カリウムイオンチャンネルのKir6.2のドミナントネガティブ体をコードする遺伝子を導入されたトランスジェニック非ヒト哺乳動物の膵再生モデルとしての用途等に関する。
【背景技術】
【0002】
糖尿病はインスリンの相対的もしくは絶対的不足によって起こり、その終末像は膵β細胞の破壊あるいは機能不全によるインスリン欠乏であり、インスリン注射が不可欠となる。しかし、インスリン注射では血糖値を生理的範囲内にコントロールすることは不可能であり、糖尿病性腎症や網膜症などの重篤な合併症の進展抑制は困難である。糖尿病の根本的な治療法としては、膵島細胞の移植が挙げられるが、多くの患者の治療に必要な数のヒト膵島を得ることは実際上不可能である。そこで、適当な前駆細胞をインビトロでβ細胞に分化させることができれば、移植用細胞の無限の供給源になると期待される。あるいは、膵β細胞の分化を誘導する内在因子や膵再生を促進する薬物の投与によってインビボで膵β細胞の再生(新生)を誘導することができれば、移植を必要としない薬物による糖尿病の根本的治療も不可能ではない。
【0003】
従って、膵臓の発生分化・β細胞の再生の機構を理解することは、糖尿病治療の観点上、極めて重要である。膵臓は成体ではほとんど分裂しないが、何らかの原因でβ細胞が損傷されると、膵再生が起こることが観察されていたので、分化増殖能を有する膵幹細胞の存在が考えられていた。例えば、ストレプトゾトシン(STZ)処理、膵切除、アロキサン投与、インターフェロン(IFN)γトランスジェニックマウス等で膵再生が観察されているが、これらの再生モデルでは、外分泌細胞は総じて再生能力が高いのに対して、内分泌細胞の再生能は低い。また、マウスやヒトのES細胞、肝細胞、膵導管細胞からインスリン分泌細胞を作製したとの報告もなされているが、その再現性、効率性には未だ問題が多く、確立した方法論として認められるには至っていない。
【0004】
ATP感受性カリウムイオン(以下、KATPと略記する場合がある)チャンネルは、種々の細胞において細胞の代謝状態を膜電位に連動させている中心的な分子であるが、膵β細胞においては、グルコース誘導性のインスリン分泌機構において中心的な役割を果たしている。即ち、グルコース代謝により生成したATPに応答してKATPチャンネルが閉じることによりβ細胞膜の脱分極を引き起こし、それに続いて、電位依存性のカルシウムチャンネルが開いてβ細胞内にカルシウムイオンが流入することにより、細胞内カルシウムイオン濃度が上昇してカルシウム依存性のインスリン分泌が起こる。
【0005】
膵β細胞のKATPチャンネルは、内向き整流特性を示すポア部分であるKir6.2の4量体と、調節サブユニットであるスルホニル尿素受容体SUR1の4量体とからなる複合体である。KATPチャンネルにおいて、カリウムイオン選択的性に寄与しているのはKir6.2である。本発明者らは、膵β細胞におけるKATPチャンネルの役割を直接的に検証するために、Kir6.2遺伝子を破壊したノックアウト(KO)マウスや(特許文献1および非特許文献1)、Kir6.2のドミナントネガティブ体を膵β細胞で特異的に発現するトランスジェニック(Tg)マウス(非特許文献2)を作製した。得られたマウスは、いずれも若週齢で膵島構造の破壊とβ細胞の減少およびα細胞の増加を認めたが、膵島構造の破壊やβ細胞数の減少はTgマウスにおいてより顕著であった(非特許文献3)。非特許文献4にもKir6.2のドミナントネガティブ体を導入したTgマウスが開示されているが、これらのKir6.2遺伝子改変動物において膵再生現象が観察されたという報告は皆無である。
【特許文献1】特開平11-346597号公報
【非特許文献1】三木(Miki, T.)ら, プロシーディングズ・オヴ・ナショナル・アカデミー・オヴ・サイエンシイズ・ユーエスエー(Proc. Natl. Acad. Sci. USA), 第95巻, pp. 10402-10406, 1998年
【非特許文献2】三木(Miki, T.)ら, プロシーディングズ・オヴ・ナショナル・アカデミー・オヴ・サイエンシイズ・ユーエスエー(Proc. Natl. Acad. Sci. USA), 第94巻, pp. 11969-11973, 1997年
【非特許文献3】清野(Seino, S.)ら, ダイアビーティズ(Diabetes), 第49巻, pp. 311-318, 2000年
【非特許文献4】コースター(Koster, J.C.)ら, プロシーディングズ・オヴ・ナショナル・アカデミー・オヴ・サイエンシイズ・ユーエスエー(Proc. Natl. Acad. Sci. USA), 第99巻, pp. 16992-16997, 2002年
【発明の開示】
【発明が解決しようとする課題】
【0006】
本発明の目的は、膵再生因子の探索や膵内分泌細胞の分化を促進し得る化合物のスクリーニング等に有用なツールとなり得る、膵内分泌細胞の再生モデルを提供することである。
【課題を解決するための手段】
【0007】
本発明者らは、上記の課題を解決するために鋭意研究を重ねた結果、意外にも、本発明者らが作製した上記Tgマウス(該マウスは若週齢で高血糖・低インスリン血症を発症し、膵β細胞の細胞死を認める)が、加齢とともにインスリン分泌が回復、血糖値が改善し、膵管近傍に膵β細胞の新生が見られることを見出した。
本発明者らは、これらの知見に基づいて、さらに検討を重ねた結果、本発明を完成するに至った。
【0008】
すなわち、本発明は、
[1]ATP感受性カリウムイオンチャンネルKir6.2のドミナントネガティブ体を膵β細胞で発現し得るトランスジェニック非ヒト哺乳動物またはその生体の一部から実質的になる膵再生モデル系、
[2]ドミナントネガティブ体が、チャンネルのカリウムイオン選択性に関与する領域中の1以上のアミノ酸が変異したものである上記[1]記載のモデル系、
[3]ATP感受性カリウムイオンチャンネルKir6.2のドミナントネガティブ体を膵β細胞で発現し得るトランスジェニック非ヒト哺乳動物における、膵内分泌細胞分化前後の膵臓もしくはその一部由来の単離されたRNAまたはそのcDNA、
[4]ATP感受性カリウムイオンチャンネルKir6.2のドミナントネガティブ体を膵β細胞で発現し得るトランスジェニック非ヒト哺乳動物の生体の一部における、膵内分泌細胞分化前後の該生体の一部由来の単離されたRNAまたはそのcDNA、
[5]ATP感受性カリウムイオンチャンネルKir6.2のドミナントネガティブ体を膵β細胞で発現し得るトランスジェニック非ヒト哺乳動物において、膵内分泌細胞分化前後での膵臓もしくはその一部における遺伝子発現プロファイルを比較することを特徴とする、膵再生関連遺伝子の同定方法、
[6]ATP感受性カリウムイオンチャンネルKir6.2のドミナントネガティブ体を膵β細胞
で発現し得るトランスジェニック非ヒト哺乳動物の生体の一部において、膵内分泌細胞分化前後での該生体の一部における遺伝子発現プロファイルを比較することを特徴とする、膵再生関連遺伝子の同定方法、
[7]ATP感受性カリウムイオンチャンネルKir6.2のドミナントネガティブ体を膵β細胞で発現し得るトランスジェニック非ヒト哺乳動物またはその生体の一部に被検物質を適用し、膵内分泌細胞分化に及ぼす該物質の効果を検定することを特徴とする、膵再生調節物質のスクリーニング方法、
[8]ATP感受性カリウムイオンチャンネルKir6.2のドミナントネガティブ体を膵β細胞で発現することができ、且つ1以上の他の遺伝子変異または改変を有する非ヒト哺乳動物またはその生体の一部、
[9]他の遺伝子変異または改変の少なくとも1つが、膵内分泌機能異常が関与する疾患と同一もしくは類似の病態を生じさせるものである、上記[8]記載の動物またはその生体の一部、および
[10]上記[9]記載の動物に被検物質を投与し、膵内分泌機能異常が関与する疾患と同一もしくは類似の病態に及ぼす該物質の効果を検定することを特徴とする、該疾患の予防・治療物質のスクリーニング方法を提供する。
【発明の効果】
【0009】
Kir6.2のドミナントネガティブ体を導入されたTg動物は、若週齢で糖尿病を発症し、インスリン欠乏・膵β細胞の細胞死が顕著となるが、加齢とともにインスリン分泌が増大し、膵管近傍に高頻度に膵内分泌細胞が出現することから、該動物およびその生体の一部は、優れた膵内分泌細胞の再生モデル系となり得る。
【発明を実施するための最良の形態】
【0010】
本発明の「ATP感受性カリウムイオン(KATP)チャンネルKir6.2のドミナントネガティブ体を膵β細胞で発現し得るトランスジェニック(Tg)非ヒト哺乳動物」(以下、「本発明のTg動物」と略記する場合がある)は、非ヒト哺乳動物の受精卵や、未受精卵、精子およびその前駆細胞(始原生殖細胞、卵原細胞、卵母細胞、卵細胞、精原細胞、精母細胞、精細胞等)などに、好ましくは受精卵の胚発生の初期段階(さらに好ましくは8細胞期以前)において、リン酸カルシウム共沈殿法、電気穿孔(エレクトロポレーション)法、リポフェクション法、凝集法、顕微注入(マイクロインジェクション)法、遺伝子銃(パーティクルガン)法、DEAE-デキストラン法などの遺伝子導入法によって、目的とするKATPチャンネルKir6.2のドミナントネガティブ体をコードするDNAを、膵β細胞において発現し得る形態で、目的とする細胞に導入することにより作出される。本明細書におけるTg動物とは、導入遺伝子を発現可能な状態で安定に保持する動物をいう。「安定に保持」するとは、該動物の細胞内で導入したDNAが発現可能な状態で永続的に存在することを意味し、該DNAが宿主染色体上に組み込まれていても、あるいは染色体外DNAとして安定に存在していてもよいが、好ましくは、該DNAは宿主染色体上に組み込まれた状態で保持される。
また、上記遺伝子導入法により、非ヒト哺乳動物の体細胞、組織、臓器などに目的とするDNAを導入し、細胞培養、組織培養などに利用することもでき、さらに、この細胞を上述の胚(もしくは生殖)細胞と公知の細胞融合法を用いて融合させることによりTg動物を作出することもできる。あるいは、ノックアウト(KO)動物を作製する場合と同様に、非ヒト哺乳動物の胚性幹細胞(ES細胞)に上記の遺伝子導入法を用いて目的とするDNAを導入し、予め該DNAが安定に組み込まれたクローンを選択した後に、該ES細胞を胚盤胞に注入するかあるいはES細胞塊と8細胞期胚とを凝集させてキメラマウスを作製し、生殖系列に導入遺伝子が伝達されたものを選択することによってもTg動物を得ることが可能である。
また、このようにして作製されたTg動物の生体の一部(例えば、(1)KATPチャンネルKir6.2のドミナントネガティブ体をコードするDNAを有する細胞、組織、臓器など、(2)これらに由来する細胞または組織を培養し、必要に応じて継代したものなど、(3)該Tg動物から単離し得る各種蛋白質またはDNAなど)も、本発明の「KATPチャンネルKir6.2のドミナントネガティブ体をコードするDNAを有する非ヒト哺乳動物の生体の一部」として、本発明のTg動物と同様の目的に用いることが出来る。
【0011】
本発明のTg動物の生体の一部としては、KATPチャンネルKir6.2のドミナントネガティブ体を発現し得る細胞・組織であって、膵内分泌細胞への分化能を有する細胞を含むものであれば、特に限定されないが、好ましくは、膵臓、肝臓等の臓器、膵島、膵管等の組織、膵前駆細胞、膵幹細胞、肝前駆細胞などの細胞、あるいはそれらより特定の培養条件下で誘導される細胞・組織、さらには、該Tg動物の初期胚より誘導されるES細胞などが好ましい。
【0012】
本発明で対象とし得る「非ヒト哺乳動物」としては、トランスジェニック系が確立されたヒト以外の哺乳動物であれば特に制限はなく、例えば、ウシ、サル、ブタ、ヒツジ、ヤギ、ウサギ、イヌ、ネコ、モルモット、ハムスター、ラット、マウスなどが挙げられる。好ましくは、ウサギ、イヌ、ネコ、モルモット、ハムスター、マウス、ラット等であり、なかでも疾患モデル動物作製の面から個体発生および生物サイクルが比較的短く、繁殖が容易な齧歯動物がより好ましく、とりわけマウス(例えば、純系としてC57BL/6系統、DBA2系統など、交雑系としてB6C3F1系統、BDF1系統、B6D2F1系統、BALB/c系統、ICR系統など)およびラット(例えば、Wistar、SDなど)が好ましい。
また、哺乳動物以外にもニワトリなどの鳥類が本発明で対象とする「非ヒト哺乳動物」と同様の目的に用いることができる。
【0013】
「KATPチャンネルKir6.2のドミナントネガティブ体」とは、KATPチャンネルKir6.2の1または2以上のアミノ酸が変異したことによって、該サブユニットの機能が欠損した蛋白質をいう。該サブユニットの機能の欠損は、KATPチャンネル電流の消失もしくは低下によって特徴付けられる。
野生型Kir6.2蛋白質としては、配列番号:2で表されるアミノ酸配列もしくはそれと実質的に同一のアミノ酸配列を有する蛋白質が挙げられる。「実質的に同一のアミノ酸配列を有する蛋白質」としては、配列番号:2で表わされるアミノ酸配列と約90%以上、好ましくは95%以上、より好ましくは約98%以上の相同性を有するアミノ酸配列を含み、配列番号:2で表わされるアミノ酸配列を有する蛋白質と実質的に同質の活性を有する蛋白質などが挙げられる。ここで「相同性」とは、当該技術分野において公知の数学的アルゴリズムを用いて2つのアミノ酸配列をアラインさせた場合の、最適なアラインメント(好ましくは、該アルゴリズムは最適なアラインメントのために配列の一方もしくは両方へのギャップの導入を考慮し得るものである)における、オーバーラップする全アミノ酸残基に対する同一アミノ酸および類似アミノ酸残基の割合(%)を意味する。「類似アミノ酸」とは物理化学的性質において類似したアミノ酸を意味し、例えば、芳香族アミノ酸(Phe、Trp、Tyr)、脂肪族アミノ酸(Ala、Leu、Ile、Val)、極性アミノ酸(Gln、Asn
)、塩基性アミノ酸(Lys、Arg、His)、酸性アミノ酸(Glu、Asp)、水酸基を有するア
ミノ酸(Ser、Thr)、側鎖の小さいアミノ酸(Gly、Ala、Ser、Thr、Met)などの同じグ
ループに分類されるアミノ酸が挙げられる。このような類似アミノ酸による置換は蛋白質の表現型に変化をもたらさない(即ち、保存的アミノ酸置換である)ことが予測される。保存的アミノ酸置換の具体例は当該技術分野で周知であり、種々の文献に記載されている(例えば、Bowieら,Science, 247: 1306-1310 (1990)を参照)。
本明細書におけるアミノ酸配列の相同性は、相同性計算アルゴリズムNCBI BLAST(National Center for Biotechnology Information Basic Local Alignment Search Tool)を用い、以下の条件(期待値=10;ギャップを許す;マトリクス=BLOSUM62;フィルタリング=OFF)にて計算することができる。
アミノ酸配列の相同性を決定するための他のアルゴリズムとしては、例えば、Karlinら, Proc. Natl. Acad. Sci. USA, 90: 5873-5877 (1993)に記載のアルゴリズム[該アルゴリズムはNBLASTおよびXBLASTプログラム(version 2.0)に組み込まれている(Altschulら, Nucleic Acids Res., 25: 3389-3402 (1997))]、Needlemanら, J. Mol. Biol., 48:444-453 (1970)に記載のアルゴリズム[該アルゴリズムはGCGソフトウェアパッケージ中のGAPプログラムに組み込まれている]、MyersおよびMiller, CABIOS, 4: 11-17 (1988)に記載のアルゴリズム[該アルゴリズムはCGC配列アラインメントソフトウェアパッケージの一部であるALIGNプログラム(version 2.0)に組み込まれている]、Pearsonら, Proc. Natl. Acad. Sci. USA, 85: 2444-2448 (1988)に記載のアルゴリズム[該アルゴリズムはGCGソフトウェアパッケージ中のFASTAプログラムに組み込まれている]等が挙げられ、それらも同様に好ましく用いられ得る。
【0014】
「実質的に同質の活性」とは、例えば、KATPチャンネル活性が性質的に同質であることを示す。したがって、KATPチャンネル活性が同等(例、約0.5〜約2倍)であることが好ましいが、活性の程度や蛋白質の分子量などの量的要素は異なっていてもよい。KATPチャンネル活性の測定は、自体公知の方法に準じて行なうことができるが、例えば、該蛋白質を発現する細胞における標識Rb+の流出を検出する方法、パッチ−クランプ法でカリウムイオン電流を測定する方法等が挙げられる。
【0015】
また、Kir6.2蛋白質としては、例えば、(1)配列番号:2で表されるアミノ酸配列中の1または2個以上(好ましくは、1〜30個程度、好ましくは1〜10個程度、さらに好ましくは1〜5個)のアミノ酸が欠失したアミノ酸配列、(2)配列番号:2で表されるアミノ酸配列に1または2個以上(好ましくは、1〜30個程度、好ましくは1〜10個程度、さらに好ましくは1〜5個)のアミノ酸が付加したアミノ酸配列、(3)配列番号:2で表されるアミノ酸配列に1または2個以上(好ましくは、1〜30個程度、好ましくは1〜10個程度、さらに好ましくは1〜5個)のアミノ酸が挿入されたアミノ酸配列、(4)配列番号:2で表されるアミノ酸配列中の1または2個以上(好ましくは、1〜30個程度、好ましくは1〜10個程度、さらに好ましくは1〜5個)のアミノ酸が他のアミノ酸で置換されたアミノ酸配列、または(5)それらを組み合わせたアミノ酸配列を含有する蛋白質であって、配列番号:2で表されるアミノ酸配列を含有する蛋白質と実質的に同質の活性を有する蛋白質も含まれる。
上記のようにアミノ酸配列が挿入、欠失または置換されている場合、その挿入、欠失または置換の位置は、当該蛋白質の活性を損なわない限り、特に限定されない。
【0016】
KATPチャンネルKir6.2のドミナントネガティブ体としては、上述のように、膵β細胞で発現した際にKATPチャンネルのポア成分を構成して該チャンネル電流を消失もしくは低下させ得る限り、いかなるものであってもよいが、好ましくは、配列番号:2で表されるアミノ酸配列もしくはそれと実質的に同一のアミノ酸配列において、(1)1または2個以上(好ましくは、1〜30個程度、好ましくは1〜10個程度、さらに好ましくは1〜5個)のアミノ酸が欠失したアミノ酸配列、(2)1または2個以上(好ましくは、1〜30個程度、好ましくは1〜10個程度、さらに好ましくは1〜5個)のアミノ酸が付加したアミノ酸配列、(3)1または2個以上(好ましくは、1〜30個程度、好ましくは1〜10個程度、さらに好ましくは1〜5個)のアミノ酸が挿入されたアミノ酸配列、(4)1または2個以上(好ましくは、1〜30個程度、好ましくは1〜10個程度、さらに好ましくは1〜5個)のアミノ酸が他のアミノ酸で置換されたアミノ酸配列、または(5)それらを組み合わせたアミノ酸配列を含有する蛋白質であって、膵β細胞のKATPチャンネル電流を消失もしくは低下させ得る蛋白質である。
【0017】
より好ましくは、本発明で用いられるドミナントネガティブ体は、KATPチャンネルKir6.2のカリウムイオン選択性に関与する領域中の1以上のアミノ酸が変異(例えば、置換、欠失、挿入、付加など)したものである。「カリウムイオン選択性に関与する領域」としては、例えば、2つの膜貫通ドメイン(TM1およびTM2)の間の高度に保存されたH5領域、好ましくは、H5領域中のGly-Tyr(又はPhe)-Glyのモチーフ(以下、GY(F)Gモチーフという)が挙げられる。特に好ましくは、本発明で用いられるドミナントネガティブ体は、上記GY(F)Gモチーフの1以上のアミノ酸が他のアミノ酸で置換されたものである。他のアミノ酸としては、保存的アミノ酸以外のアミノ酸が挙げられる。例えば、配列番号:2で表されるアミノ酸配列においては、アミノ酸番号132〜134で示されるGFGが該Kir6.2蛋白質におけるGFGモチーフであるので、これらのアミノ酸のうちの1以上を他のアミノ酸で置換したものが挙げられる。とりわけ、アミノ酸番号132のGlyをSerで置換したアミノ酸配列(以下、「Kir6.2G132S」と略記する場合がある)もしくはそれと実質的に同一のアミノ酸配列を有する蛋白質が好ましい。ここで「実質的に同一のアミノ酸配列」とは上記と同義であるが、この場合の「同質の活性」とは、ドミナントネガティブ活性(即ち、KATPチャンネル電流を消失もしくは低下させる活性)が同質であることを意味する。
【0018】
KATPチャンネルKir6.2のドミナントネガティブ体をコードするDNAとしては、好ましくは、KATPチャンネルKir6.2のカリウムイオン選択性に関与する領域中の1以上のアミノ酸が変異(例えば、置換、欠失、挿入、付加など)した蛋白質をコードするDNAであり、より好ましくは、該サブユニット蛋白質のH5領域、さらに好ましくはGY(F)Gモチーフ内の1以上のアミノ酸が他のアミノ酸で置換された蛋白質、特に好ましくは、配列番号:2で表されるアミノ酸配列中アミノ酸番号132〜134で示されるGFGのうちの1以上のアミノ酸が他のアミノ酸で置換されたアミノ酸配列もしくはそれと実質的に同一のアミノ酸配列を有する蛋白質、就中、Kir6.2G132Sもしくはそれと実質的に同一のアミノ酸配列を有する蛋白質をコードするDNAである。
【0019】
配列番号:2で表されるアミノ酸配列をコードするDNAとしては、例えば、配列番号:1で表される塩基配列もしくは該塩基配列の相補鎖配列とストリンジェントな条件下でハイブリダイズし得る塩基配列を含み、且つ配列番号:2で表されるアミノ酸配列を有する蛋白質と実質的に同質の活性を有する蛋白質をコードするDNAが挙げられる。従って、配列番号:2で表されるアミノ酸配列を有する蛋白質のドミナントネガティブ体をコードするDNAとしては、好ましくは、配列番号:1で表される塩基配列中H5領域をコードする塩基配列が、該領域中の1以上のアミノ酸が他のアミノ酸で置換されたアミノ酸配列をコードする配列で置換された塩基配列、より好ましくは、配列番号:1で表される塩基配列中塩基番号394〜402で示される塩基配列が、GFGのうちの1以上のアミノ酸が他のアミノ酸で置換されたアミノ酸配列をコードする配列で置換された塩基配列、特に好ましくは、配列番号:1で表される塩基配列中塩基番号394〜396で示される塩基配列が、Serをコードする配列で置換された塩基配列を含むDNA、あるいはそれらの相補鎖配列とストリンジェントな条件下でハイブリダイズし得る塩基配列を含み、且つ上記いずれかのDNAにコードされる蛋白質と実質的に同質の活性(ドミナントネガティブ活性)を有する蛋白質をコードするDNAが挙げられる。「ストリンジェントな条件」としては、例えば、ナトリウム塩濃度が約19〜約40mM、好ましくは約19〜約20mMで、温度が約50〜約70℃、好ましくは約60〜約65℃の条件等が挙げられる。特に、ナトリウム塩濃度が約19mMで温度が約65℃の場合が好ましい。当業者は、ハイブリダイゼーション溶液の塩濃度、ハイブリダゼーション反応の温度、プローブ濃度、プローブの長さ、ミスマッチの数、ハイブリダイゼーション反応の時間、洗浄液の塩濃度、洗浄の温度等を適宜変更することにより、所望のストリンジェンシーに容易に調節することができる。
【0020】
KATPチャンネルKir6.2のドミナントネガティブ体をコードするDNAは、各種哺乳動物(ヒト、ウシ、サル、ブタ、ヒツジ、ヤギ、ウサギ、イヌ、ネコ、モルモット、ハムスター、ラット、マウスなど)の膵臓、膵島、膵β細胞、骨格筋、心筋、平滑筋など、好ましくは、膵臓、膵島、膵β細胞等に由来するDNAおよび市販の各種ゲノムDNAライブラリーに由来するゲノムDNAの全てあるいは一部を原料として用い、あるいは各種哺乳動物の膵臓、膵島、膵β細胞、骨格筋、心筋、平滑筋など、好ましくは、膵臓、膵島、膵β細胞等に由来するRNAから公知の方法により調製されたcDNAを原料として用い、公知の該サブユニット遺伝子の配列をもとに作製したオリゴヌクレオチドをプローブもしくはプライマーとして、ハイブリダイゼーション法もしくはPCR法などにより単離したKATPチャンネルKir6.2をコードするDNAを鋳型として用い、従来公知の部位特異的変異誘発により、所望の部位(例えば、上記H5領域、好ましくはGY(F)Gモチーフ)に変異を導入することにより調製することができる。具体的には、例えば、上記非特許文献2に記載の方法などが挙げられるが、それに限定されない。
【0021】
本発明のTg動物は、KATPチャンネルKir6.2のドミナントネガティブ体をコードするDNAを膵β細胞で「発現可能な状態で」保持している。したがって、当該DNAを対象動物に導入するにあたっては、当該DNAを対象動物の膵β細胞内で機能し得るプロモーターの下流に連結した発現カセットを含む形態(例、発現ベクターなど)で用いるのが一般に有利である。
【0022】
KATPチャンネルKir6.2のドミナントネガティブ体をコードするDNAを担持するベクターとしては、大腸菌由来のプラスミド、枯草菌由来のプラスミド、酵母由来のプラスミド、λファージなどのバクテリオファージ、モロニー白血病ウイルスなどのレトロウイルス、ワクシニアウイルスまたはバキュロウイルスなどの動物もしくは昆虫ウイルスなどが用いられる。なかでも、大腸菌由来のプラスミド、枯草菌由来のプラスミドまたは酵母由来のプラスミドなどが好ましく用いられ、特に大腸菌由来のプラスミドが好ましい。
【0023】
KATPチャンネルKir6.2のドミナントネガティブ体の遺伝子発現調節を行うプロモーターとしては、少なくとも膵β細胞内でプロモーター活性を発揮し得るものであれば特に制限されず、例えばウイルス(サイトメガロウイルス、モロニー白血病ウイルス、JCウイルス、乳癌ウイルスなど)に由来する遺伝子のプロモーター、各種哺乳動物(ヒト、ウシ、サル、ブタ、ヒツジ、ヤギ、ウサギ、イヌ、ネコ、モルモット、ハムスター、ラット、マウスなど)および鳥類(ニワトリなど)に由来する遺伝子[例えば、アルブミン、エンドセリン、オステオカルシン、筋クレアチンキナーゼ、I型およびII型コラーゲン、サイクリックAMP依存蛋白キナーゼβIサブユニット、心房ナトリウム利尿性因子、ドーパミンβ−水酸化酵素、ニューロフィラメント軽鎖、メタロチオネインIおよびIIA、メタロプロティナーゼ1組織インヒビター、平滑筋αアクチン、ポリペプチド鎖伸長因子1α(EF1−α)、βアクチン、αおよびβミオシン重鎖、ミオシン軽鎖1および2、ミエリン塩基性蛋白、血清アミロイドPコンポーネント、レニンなど]のプロモーターなども挙げられるが、好ましくは、膵β細胞以外にプロモーター活性を発揮する細胞種が限定されるプロモーター(例えば、アミリンプロモーター、pdx-1プロモーター等)、より好ましくは膵β細胞特異的プロモーター(例えば、インスリンIもしくはインスリンIIプロモーター等)である。
【0024】
KATPチャンネルKir6.2のドミナントネガティブ体をコードするDNAの下流には、Tg動物において目的とするmRNAの転写を終結させる配列(ポリアデニレーション(polyA)シグナル、ターミネーターとも呼ばれる)を有していることが好ましく、例えば、ウイルス遺伝子由来、あるいは各種哺乳動物または鳥類の遺伝子由来のターミネーター配列を用いて、効率よい導入遺伝子の発現を達成することができる。好ましくは、シミアンウイルスのSV40ターミネーターなどが用いられる。その他、目的の遺伝子をさらに高発現させる目的で、各遺伝子のスプライシングシグナル、エンハンサー領域、真核遺伝子のイントロンの一部を、プロモーター領域の5’上流、プロモーター領域と翻訳領域間あるいは翻訳領域の3’下流に連結することも目的により可能である。
【0025】
また、胚性幹細胞(ES細胞)を用いてTg動物を作製する場合、上記のベクターは、導入DNAが安定に組み込まれたクローンを選択するための選択マーカー遺伝子(例:ネオマイシン耐性遺伝子、ハイグロマイシン耐性遺伝子などの薬剤耐性遺伝子)をさらに含むことが好ましい。
【0026】
上記のプロモーター、KATPチャンネルKir6.2のドミナントネガティブ体をコードするDNA、ターミネーターなどは、適当な制限酵素およびDNAリガーゼ等を用いた通常の遺伝子工学的手法により、上記のベクター中に正しい配置で、即ち、Tg動物においてKATPチャンネルKir6.2のドミナントネガティブ体を発現可能な配置で、挿入することができる。
【0027】
好ましい一実施態様においては、上記のようにして得られるKATPチャンネルKir6.2のドミナントネガティブ体をコードするDNAを含む発現ベクターは、マイクロインジェクション法により対象となる非ヒト哺乳動物の初期胚に導入される。
【0028】
対象非ヒト哺乳動物の初期胚は、同種の非ヒト哺乳動物の雌雄を交配させて得られる体内受精卵を採取するか、あるいは同種の非ヒト哺乳動物の雌雄からそれぞれ採取した卵と精子を体外受精させることにより得ることができる。
用いる非ヒト哺乳動物の齢や飼育条件等は動物種によってそれぞれ異なるが、例えばマウス(好ましくはC57BL/6J(B6)などの近交系マウス、B6と他の近交系とのF1など)を用いる場合は、雌が約4〜約6週齢、雄が約2〜約8月齢程度のものが好ましく、また、約12時間明期条件(例えば7:00−19:00)で約1週間飼育したものが好ましい。
体内受精は自然交配によってもよいが、性周期の調節と1個体から多数の初期胚を得ることを目的として、雌非ヒト哺乳動物に性腺刺激ホルモンを投与して過剰排卵を誘起した後、雄非ヒト哺乳動物と交配させる方法が好ましい。雌非ヒト哺乳動物の排卵誘発法としては、例えば初めに卵胞刺激ホルモン(妊馬血清性性腺刺激ホルモン、一般にPMSGと略する)、次いで黄体形成ホルモン(ヒト絨毛性性腺刺激ホルモン、一般にhCGと略する)を、例えば腹腔内注射などにより投与する方法が好ましいが、好ましいホルモンの投与量、投与間隔は非ヒト哺乳動物の種類によりそれぞれ異なる。例えば、非ヒト哺乳動物がマウス(好ましくはC57BL/6J(B6)などの近交系マウス、B6と他の近交系とのF1など)の場合は、通常、卵胞刺激ホルモン投与後、約48時間後に黄体形成ホルモンを投与し、直ちに雄マウスと交配させることにより受精卵を得る方法が好ましく、卵胞刺激ホルモンの投与量は約20〜約50IU/個体、好ましくは約30IU/個体、黄体形成ホルモンの投与量は約0〜約10IU/個体、好ましくは約5IU/個体である。
一定時間経過後、膣栓の検査等により交配を確認した雌非ヒト哺乳動物の腹腔を開き、卵管から受精卵を取り出して胚培養用培地(例:M16培地、修正Whitten培地、BWW培地、M2培地、WM−HEPES培地、BWW−HEPES培地等)中で洗って卵丘細胞を除き、微小滴培養法等により5%炭酸ガス/95%大気下でDNA顕微注入まで培養する。直ちに顕微注入を行わない場合、採取した受精卵を緩慢法または超急速法等で凍結保存することも可能である。
【0029】
一方、体外受精の場合は、採卵用雌非ヒト哺乳動物(体内受精の場合と同様のものが好ましく用いられる)に上記と同様に卵胞刺激ホルモンおよび黄体形成ホルモンを投与して排卵を誘発させた後、卵子を採取して受精用培地(例:TYH培地)中で体外受精時まで微小滴培養法等により5%炭酸ガス/95%大気下で培養する。他方、同種の雄非ヒト哺乳動物(体内受精の場合と同様のものが好ましく用いられる)から精巣上体尾部を取り出し、精子塊を採取して受精用培地中で前培養する。前培養終了後の精子を卵子を含む受精用培地に添加し、微小滴培養法等により5%炭酸ガス/95%大気下で培養した後、2個の前核を有する受精卵を顕微鏡下で選抜する。直ちにDNAの顕微注入を行わない場合は、得られた受精卵を緩慢法または超急速法等で凍結保存することも可能である。
【0030】
受精卵へのDNAの顕微注入は、マイクロマニピュレーター等の公知の装置を用いて常法
に従って実施することができる。簡潔に言えば、胚培養用培地の微小滴中に入れた受精卵をホールディングピペットで吸引して固定し、インジェクションピペットを用いてDNA溶液を雄性もしくは雌性前核、好ましくは雄性前核内に直接注入する。導入DNAはCsCl密度勾配超遠心等で高度に精製したものを用いることが好ましい。また、導入DNAは制限酵素を用いてベクター部分を切断し、直鎖状にしておくことが好ましい。
【0031】
DNA導入後の受精卵は胚培養用培地中で微小滴培養法等により5%炭酸ガス/95%大気下で1細胞期〜胚盤胞期まで培養した後、偽妊娠させた受胚用雌非ヒト哺乳動物の卵管または子宮内に移植される。受胚用雌非ヒト哺乳動物は移植される初期胚が由来する動物と同種のものであればよく、例えば、マウス初期胚を移植する場合は、ICR系の雌マウス(好ましくは約8〜約10週齢)などが好ましく用いられる。受胚用雌非ヒト哺乳動物を偽妊娠状態にする方法としては、例えば、同種の精管切除(結紮)雄非ヒト哺乳動物(例えば、マウスの場合、ICR系の雄マウス(好ましくは約2月齢以上))と交配させて、膣栓の存在が確認されたものを選択する方法が知られている。
受胚用雌は自然排卵のものを用いてもよいし、あるいは精管切除(結紮)雄との交配に先立って、黄体形成ホルモン放出ホルモン(一般にLHRHと略する)もしくはその類縁体を投与し、受精能を誘起させたものを用いてもよい。LHRH類縁体としては、例えば、[3,5-DiI-Tyr5]-LH-RH、[Gln8]-LH-RH、[D-Ala6]-LH-RH、[des-Gly10]-LH-RH、[D-His(Bzl)6]-LH-RHおよびそれらのEthylamideなどが挙げられる。LHRHもしくはその類縁体の投与量、ならびにその投与後に雄非ヒト哺乳動物と交配させる時期は、非ヒト哺乳動物の種類によりそれぞれ異なる。例えば、非ヒト哺乳動物がマウス(好ましくはICR系のマウスなど)の場合には、通常、LHRHもしくはその類縁体を投与した後、約4日目に雄マウスと交配させることが好ましく、LHRHあるいはその類縁体の投与量は、通常、約10〜60μg/個体、好ましくは約40μg/個体である。
【0032】
通常、移植される初期胚が桑実胚期以後の場合は受胚用雌の子宮に、それより前(例えば、1細胞期〜8細胞期胚)であれば卵管に胚移植される。受胚用雌は、移植胚の発生段階に応じて偽妊娠からある日数が経過したものが適宜使用される。例えばマウスの場合、2細胞期胚を移植するには偽妊娠後約0.5日の雌マウスが、胚盤胞期胚を移植するには偽妊娠後約2.5日の雌マウスが好ましい。受胚用雌を麻酔(好ましくはAvertin、ネンブタール等が使用される)後、切開して卵巣を引き出し、胚培養用培地に懸濁した初期胚(約5〜約10個)を胚移植用ピペットを用いて、卵管腹腔口もしくは子宮角の卵管接合部付近に注入する。
【0033】
移植胚が首尾よく着床し受胚雌が妊娠すれば、自然分娩もしくは帝王切開により仔非ヒト哺乳動物が得られる。自然分娩した受胚雌にはそのまま哺乳を継続させればよく、帝王切開により出産した場合は、産仔は別途用意した哺乳用雌(例えばマウスの場合、通常に交配・分娩した雌マウス(好ましくはICR系の雌マウス等))に哺乳させることができる。
【0034】
受精卵細胞段階におけるKATPチャンネルKir6.2のドミナントネガティブ体をコードするDNAの導入は、導入DNAが対象非ヒト哺乳動物の生殖系列細胞および体細胞のすべてに存在するように確保される。導入DNAが染色体DNAに組み込まれているか否かは、例えば、産仔の尾部より分離抽出した染色体DNAをサザンハイブリダイゼーションまたはPCR法によりスクリーニングすることにより検定することができる。上記のようにして得られる仔非ヒト哺乳動物(F0)の生殖系列細胞においてKATPチャンネルKir6.2のドミナントネガティブ体をコードするDNAが存在することは、その後代(F1)の動物全てが、その生殖系列細胞および体細胞のすべてにKATPチャンネルKir6.2のドミナントネガティブ体をコードするDNAが存在することを意味する。
通常、F0動物は相同染色体の一方にのみ導入DNAを有するヘテロ接合体として得られる。また、個々のF0個体は相同組換えによらない限り異なる染色体上にランダムに挿入される。相同染色体の両方にKATPチャンネルKir6.2のドミナントネガティブ体をコードするDNAを有するホモ接合体を得るためには、F0動物と非トランスジェニック動物とを交雑してF1動物を作製し、相同染色体の一方にのみ導入DNAを有するヘテロ接合体の兄妹同士を交雑すればよい。1遺伝子座にのみ導入DNAが組み込まれていれば、得られるF2動物の1/4がホモ接合体となる。
【0035】
別の好ましい一実施態様においては、KATPチャンネルKir6.2のドミナントネガティブ体をコードするDNAを含む発現ベクターは、エレクトロポレーション法等の公知の遺伝子導入法により対象となる非ヒト哺乳動物のES細胞に導入される。
【0036】
ES細胞は胚盤胞期の受精卵の内部細胞塊(ICM)に由来し、インビトロで未分化状態を保ったまま培養維持できる細胞をいう。ICMの細胞は将来、胚本体を形成する細胞であり、生殖細胞を含むすべての組織の基になる幹細胞である。ES細胞としては、既に樹立された細胞株を用いてもよく、また、EvansとKaufmanの方法(ネイチャー(Nature)第292巻、154頁、1981年)に準じて新しく樹立したものでもよい。例えば、マウスES細胞の場合、現在、一般的には129系マウス由来のES細胞が使用されているが、免疫学的背景がはっきりしていないので、これに代わる純系で免疫学的に遺伝的背景が明らかなES細胞を取得するなどの目的で、例えば、C57BL/6マウスやC57BL/6の採卵数の少なさをDBA/2との交雑により改善したBDF1マウス(C57BL/6とDBA/2とのF1)から樹立されるES細胞なども良好に用いることができる。BDF1マウスは、採卵数が多く、かつ卵が丈夫であるという利点に加えて、C57BL/6マウスを背景に持つので、これ由来のES細胞は疾患モデルマウスを作製したとき、C57BL/6マウスと戻し交雑することでその遺伝的背景をC57BL/6マウスに代えることが可能である点で有利に用い得る。
【0037】
ES細胞の調製は、例えば以下のようにして行うことができる。交配後の雌非ヒト哺乳動物[例えばマウス(好ましくはC57BL/6J(B6)などの近交系マウス、B6と他の近交系とのF1など)を用いる場合は、約2月齢以上の雄マウスと交配させた約8〜約10週齢程度の雌マウス(妊娠約3.5日)が好ましく用いられる]の子宮から胚盤胞期胚を採取して(あるいは桑実胚期以前の初期胚を卵管から採取した後、胚培養用培地中で上記と同様にして胚盤胞期まで培養してもよい)、適当なフィーダー細胞(例えばマウスの場合、マウス胎仔から調製される初代繊維芽細胞や公知のSTO繊維芽細胞株等)層上で培養すると、胚盤胞の一部の細胞が集合して将来胚に分化するICMを形成する。この内部細胞塊をトリプシン処理して単細胞を解離させ、適切な細胞密度を保ち、培地交換を行いながら、解離と継代を繰り返すことによりES細胞が得られる。
【0038】
ES細胞は雌雄いずれを用いてもよいが、通常雄のES細胞の方が生殖系列キメラを作製するのに都合が良い。また、煩雑な培養の手間を削減するためにもできるだけ早く雌雄の判別を行なうことが望ましい。ES細胞の雌雄の判定方法としては、例えば、PCR法によりY染色体上の性決定領域の遺伝子を増幅、検出する方法が、その1例としてあげることができる。この方法を使用すれば、従来、核型分析をするのに約106個の細胞数を要していたのに対して、1コロニー程度のES細胞数(約50個)で済むので、培養初期におけるES細胞の第一次セレクションを雌雄の判別で行なうことが可能であり、早期に雄細胞の選定を可能にしたことにより培養初期の手間は大幅に削減できる。
また、第二次セレクションとして、例えば、G−バンディング法による染色体数の確認等により行うことができる。得られるES細胞の染色体数は正常数の100%が望ましいが、細胞株樹立の際の物理的操作等の関係上困難な場合は、ES細胞への遺伝子導入の後、正常細胞(例えば、マウスでは染色体数が2n=40である細胞)に再びクローニングすることが望ましい。
【0039】
このようにして得られるES細胞株は、未分化幹細胞の性質を維持するために注意深く継代培養することが必要である。例えば、STO繊維芽細胞のような適当なフィーダー細胞上で、分化抑制因子として知られるLIF(1〜10,000U/ml)存在下に炭酸ガス培養器内(好ましくは、5%炭酸ガス/95%空気または5%酸素/5%炭酸ガス/90%空気)で約37℃で培養するなどの方法で培養し、継代時には、例えば、トリプシン/EDTA溶液(通常0.001〜0.5%トリプシン/0.1〜5mM EDTA、好ましくは約0.1%トリプシン/1mM EDTA)処理により単細胞化し、新たに用意したフィーダー細胞上に播種する方法などがとられる。このような継代は、通常1〜3日毎に行なうが、この際に細胞の観察を行い、形態的に異常な細胞が見受けられた場合はその培養細胞は放棄することが望まれる。
ES細胞は、適当な条件により、高密度に至るまで単層培養するか、または細胞集塊を形成するまで浮遊培養することにより、頭頂筋、内臓筋、心筋などの種々のタイプの細胞に分化させることが可能であり〔M. J. Evans及びM. H. Kaufman, ネイチャー(Nature)第292巻、154頁、1981年;G. R. Martin プロシーディングス・オブ・ナショナル・アカデミー・オブ・サイエンス・ユーエスエー(Proc. Natl. Acad. Sci. USA)第78巻、7634頁、1981年;T. C. Doetschman ら、ジャーナル・オブ・エンブリオロジー・アンド・エクスペリメンタル・モルフォロジー、第87巻、27頁、1985年〕、本発明のKATPチャンネルKir6.2のドミナントネガティブ体をコードするDNAを導入されたES細胞を分化させて得られる該ドミナントネガティブ体発現非ヒト哺乳動物細胞は、インビトロにおけるKATPチャンネルKir6.2の細胞生物学的検討において有用である。
【0040】
ES細胞への遺伝子導入には、リン酸カルシウム共沈殿法、電気穿孔(エレクトロポレーション)法、リポフェクション法、レトロウイルス感染法、凝集法、顕微注入(マイクロインジェクション)法、遺伝子銃(パーティクルガン)法、DEAE−デキストラン法などのいずれも用いることができるが、簡便に多数の細胞を処理できること等の点からエレクトロポレーション法が一般的に選択されている。エレクトロポレーションには通常の動物細胞への遺伝子導入に使用されている条件をそのまま用いればよく、例えば、対数増殖期にあるES細胞をトリプシン処理して単一細胞に分散させた後、106〜108細胞/mlとなるように培地に懸濁してキュベットに移し、KATPチャンネルKir6.2のドミナントネガティブ体をコードするDNAを含むベクターを10〜100μg添加し、200〜600V/cmの電気パルスを印加することにより行なうことができる。
【0041】
導入DNAが組み込まれたES細胞は、単一細胞をフィーダー細胞上で培養して得られるコロニーから分離抽出した染色体DNAをサザンハイブリダイゼーションまたはPCR法によりスクリーニングすることによっても検定することができるが、ES細胞を用いるトランスジェニック系の最大の長所は、薬剤耐性遺伝子やレポーター遺伝子の発現を指標として細胞段階で形質転換体を選択できることである。したがって、ここで使用される導入ベクターは、KATPチャンネルKir6.2のドミナントネガティブ体をコードするDNAを含む発現カセットに加えて、薬剤耐性遺伝子(例:ネオマイシンホスホトランスフェラーゼII(nptII)遺伝子、ハイグロマイシンホスホトランスフェラーゼ(hpt)遺伝子など)やレポーター遺伝子(例:β−ガラクトシダーゼ(lacZ)遺伝子、クロラムフェニコールアセチルトランスフェラーゼ(cat)遺伝子など)等の選択マーカー遺伝子をさらに含むことが望ましい。例えば、選択マーカー遺伝子としてnptII遺伝子を含むベクターを用いた場合、遺伝子導入処理後のES細胞をG418などのネオマイシン系抗生物質を含有する培地中で培養し、出現した耐性コロニーをそれぞれ培養プレートに移してトリプシン処理、培地交換を繰り返した後、一部を培養用として残し、残りをPCRもしくはサザンハイブリダイゼーションにかけて導入DNAの存在を確認する。
【0042】
導入DNAの組込みが確認されたES細胞を同種の非ヒト哺乳動物由来の胚内に戻すと、宿主胚のICMに組み込まれてキメラ胚が形成される。これを仮親(受胚用雌)に移植してさらに発生を続けさせることにより、キメラトランスジェニック動物が得られる。キメラ動物の中でES細胞が将来卵や精子に分化する始原生殖細胞の形成に寄与した場合には、生殖系列キメラが得られることとなり、これを交配することにより導入DNAが遺伝的に固定された遺伝子導入非ヒト哺乳動物を作製することができる。
【0043】
キメラ胚の作製方法としては、桑実胚期までの初期胚同士を接着させて集合させる方法(集合キメラ法)と、胚盤胞の割腔内に細胞を顕微注入する方法(注入キメラ法)とがある。ES細胞によるキメラ胚の作製においては従来より後者が広く行なわれているが、最近では、8細胞期胚の透明帯内へのES細胞の注入により集合キメラを作る方法や、マイクロマニピュレーターが不要で操作が容易な方法として、ES細胞塊と透明帯を除去した8細胞期胚とを共培養して凝集させることによって集合キメラを作製する方法も行われている。
いずれの場合も、宿主胚は受精卵への遺伝子導入における採卵用雌として使用され得る非ヒト哺乳動物から同様にして採取することができるが、例えばマウスの場合、キメラマウス形成へのES細胞の寄与率を毛色(コートカラー)で判定し得るように、ES細胞の由来する系統とは毛色の異なる系統のマウスから宿主胚を採取することが好ましい。例えば、ES細胞が129系マウス(毛色:アグーチ)由来であれば、採卵用雌としてC57BL/6マウス(毛色:ブラック)やICRマウス(毛色:アルビノ)を用い、ES細胞がC57BL/6もしくはDBF1マウス(毛色:ブラック)由来やTT2細胞(C57BL/6とCBAとのF1(毛色:アグーチ)由来)であれば、採卵用雌としてICRマウスやBALB/cマウス(毛色:アルビノ)を用いることができる。
また、生殖系列キメラ形成能はES細胞と宿主胚との組み合わせに大きく依存するので、生殖系列キメラ形成能の高い組み合わせを選択することがより好ましい。例えばマウスの場合、129系統由来のES細胞に対してはC57BL/6系統由来の宿主胚等を用いることが好ましく、C57BL/6系統由来のES細胞に対してはBALB/c系統由来の宿主胚等が好ましい。
採卵用雌マウスは約4〜約6週齢程度が好ましく、交配用の雄マウスとしては約2〜約8月齢程度の同系統のものが好ましい。交配は自然交配によってもよいが、好ましくは性腺刺激ホルモン(卵胞刺激ホルモン、次いで黄体形成ホルモン)を投与して過剰排卵を誘起した後に行なわれる。
【0044】
胚盤注入法による場合は、胚盤胞期胚(例えばマウスの場合、交配後約3.5日)を採卵用雌の子宮から採取し(あるいは桑実胚期以前の初期胚を卵管から採取した後、上述の胚培養用培地中で胚盤胞期まで培養してもよい)、マイクロマニピュレーターを用いて胚盤胞の割腔内にKATPチャンネルKir6.2のドミナントネガティブ体をコードするDNAが導入されたES細胞(約10〜約15個)を注入した後、偽妊娠させた受胚用雌非ヒト哺乳動物の子宮内に移植する。受胚用雌非ヒト哺乳動物は受精卵への遺伝子導入における受胚用雌として使用され得る非ヒト哺乳動物を同様に用いることができる。
共培養法による場合は、8細胞期胚および桑実胚(例えばマウスの場合、交配後約2.5日)を採卵用雌の卵管および子宮から採取して(あるいは8細胞期以前の初期胚を卵管から採取した後、上述の胚培養用培地中で8細胞期または桑実胚期まで培養してもよい)酸性タイロード液中で透明帯を溶解した後、ミネラルオイルを重層した胚培養用培地の微小滴中にKATPチャンネルKir6.2のドミナントネガティブ体をコードするDNAが導入されたES細胞塊(細胞数約10〜約15個)を入れ、さらに上記8細胞期胚または桑実胚(好ましくは2個)を入れて一晩共培養する。得られた桑実胚または胚盤胞を上記と同様にして受胚用雌非ヒト哺乳動物の子宮内に移植する。
【0045】
移植胚が首尾よく着床し受胚雌が妊娠すれば、自然分娩もしくは帝王切開によりキメラ非ヒト哺乳動物が得られる。自然分娩した受胚雌にはそのまま哺乳を継続させればよく、帝王切開により出産した場合は、産仔は別途用意した哺乳用雌(通常に交配・分娩した雌非ヒト哺乳動物)に哺乳させることができる。
生殖系列キメラの選択は、まずES細胞の雌雄が予め判別されている場合はES細胞と同じ性別のキメラマウスを選択し(通常は雄性ES細胞が使用されるので、雄キメラマウスが選択される)、次いで毛色等の表現型からES細胞の寄与率が高いキメラマウス(例えば、50%以上)を選択する。例えば、129系マウス由来の雄性ES細胞であるD3細胞とC57BL/6マウス由来の宿主胚とのキメラ胚から得られるキメラマウスの場合、アグーチの毛色の占める割合の高い雄マウスを選択するのが好ましい。選択されたキメラ非ヒト哺乳動物が生殖系列キメラであるか否かの確認は、適当な系統の同種動物との交雑により得られるF1動物の表現型に基づいて行なうことができる。例えば、上記キメラマウスの場合、アグーチはブラックに対して優性であるので、雌C57BL/6マウスと交雑すると、選択された雄マウスが生殖系列キメラであれば得られるF1の毛色はアグーチとなる。
【0046】
上記のようにして得られるKATPチャンネルKir6.2のドミナントネガティブ体をコードするDNAが導入された生殖系列キメラ非ヒト哺乳動物(ファウンダー)は、通常、相同染色体の一方にのみ導入DNAを有するヘテロ接合体として得られる。また、個々のファウンダーは相同組換えによらない限り異なる染色体上にランダムに挿入される。相同染色体の両方にKATPチャンネルKir6.2のドミナントネガティブ体をコードするDNAを有するホモ接合体を得るためには、上記のようにして得られるF1動物のうち相同染色体の一方にのみ導入DNAを有するヘテロ接合体の兄妹同士を交雑すればよい。ヘテロ接合体の選択は、例えばF1動物の尾部より分離抽出した染色体DNAをサザンハイブリダイゼーションまたはPCR法によりスクリーニングすることにより検定することができる。1遺伝子座にのみ導入DNAが組み込まれていれば、得られるF2動物の1/4がホモ接合体となる。
【0047】
上記のようにして得られうる、KATPチャンネルKir6.2のドミナントネガティブ体を膵β細胞で発現し得る本発明のTg動物は、以下の特性:
(a)血糖値およびインスリン分泌
(1)新生仔で低血糖・高インスリン血症を呈する
(2)若週(約4〜約8週)齢で高血糖・インスリン欠乏症を呈する
(3)加齢(約9〜約12週齢以降)とともに血糖低下・インスリン分泌増加を呈する
(b)膵β細胞・膵島構造
(1)若週齢でβ細胞の細胞死・α細胞の増加および正常な膵島構造(中心部にβ細胞が存
在し、α細胞は周縁部に限局される)の破壊を認める
(2)加齢とともに膵管近傍に膵β細胞の新生を認める
を有することから、該Tg動物、あるいは膵内分泌細胞に分化し得るその生体の一部は、膵再生、特に膵内分泌細胞再生の有用なモデル系となり得る。ここで「モデル系」なる用語は、再生モデルは再生現象の前後において物質としての性状が変化することから、経時的要素を含むことを意味する目的で用いられる。
【0048】
本発明の膵再生モデル系の好ましい一実施態様は、本発明のTg動物の個体全体からなるものである。即ち、本発明のTg動物を、通常の飼育条件下(例えば、約12時間明期条件(例えば7:00−19:00)下で、普通食(必要に応じて高脂肪食等を用いることもできる)および水を自由摂取の状態で飼育する(SPF条件下が望ましいが、コンベンショナルな条件でもよい)など)で、少なくとも膵内分泌細胞の分化が起こるまで、例えば、少なくとも約9〜約12週齢以降まで飼育し続けることにより、膵再生現象を観察することができる。
【0049】
本発明の膵再生モデル系の別の好ましい一実施態様は、本発明のTg動物の生体の一部から実質的になるものである。ここで「実質的になる」とは、例えば、培地成分や分化誘導物質などの、膵再生現象の主体ではないが該生体の一部における膵再生を誘導するために必要もしくは望ましい他の要素を含んでいてもよいことを意味する。該生体の一部は、少なくとも膵内分泌細胞に分化し得る細胞を含むものであり、例えば、膵臓、肝臓等の臓器、膵島、膵管等の組織、膵前駆細胞、膵幹細胞、肝前駆細胞などの細胞、あるいはそれらより特定の培養条件下で誘導される細胞・組織、さらには、該Tg動物の初期胚より誘導されるES細胞などが挙げられるが、それらに限定されない。
【0050】
本発明のTg動物の生体の一部から実質的になる膵再生モデル系に含有され得る培地としては、該生体の一部の維持・増殖・分化に適したものであれば特に制限はなく、例えば、最小必須培地(MEM)、ダルベッコ改変イーグル培地(DMEM)、ハムF12培地、F10培地、RPMI1640培地等が挙げられる。これらの培地には、インスリン、トランスフェリン等の成分が補充されていることが望ましい。
本発明のTg動物の生体の一部から実質的になる膵再生モデル系に含有され得る分化誘導剤としては、例えば、FGFファミリー(FGF1、FGF2、FGF7、FGF10など)、EGFファミリー(EGF、TGF-α、βセルリン、βセルリンδ4、amphiregulin、Heparin-binding EGF)、keratinocyte growth factor、アクチビン、TGFβ1-3、HGF、GLP-1、CCK、ガストリン、脂肪酸、ニコチンアミド、セレニウム、プロゲステロン、プトレッシン等が挙げられるが、これらに限定されない。
【0051】
本発明は、本発明のTg動物またはその生体の一部を用いた膵再生関連遺伝子の同定方法を提供する。本方法は、膵内分泌細胞の分化前後(ここで「分化前後」とは、分化が起こる前と分化が完了した後との比較だけを意味するのではなく、分化前と今まさに分化が起こっている最中や分化誘導のスイッチがオンされた直後等との比較をも包含する意味で用いられる)で、該Tg動物から膵臓もしくはその一部(例えば、膵島、膵管など)を摘出し、あるいは該Tg動物の生体の一部を膵内分泌細胞への分化が誘導され得る条件下で培養し、それらから常法に従って時系列的に全RNAもしくはpolyA RNA (mRNA)を抽出して、例えば、膵島特異的なcDNAマイクロアレイ等を用いて、網羅的に遺伝子発現プロファイルの変動を解析することにより行うことができる。RNA画分の調製は、グアニジン−CsCl超遠心法、AGPC法など公知の手法を用いて行うことができるが、市販のRNA抽出用キット(例:RNeasy Mini Kit; QIAGEN製等)を用いて、微量試料から迅速且つ簡便に高純度の全RNAを調製することができる。
本発明のTg動物の生体の一部を膵内分泌細胞への分化が誘導される条件下で培養する方法としては、例えば、該Tg動物から膵導管細胞画分を採取して培養する方法(Proc. Natl. Acad. Sci. USA, 97: 7999-8004 (2000))、該Tg動物とNODマウスとの交配により得られた動物の膵導管上皮を長期培養してインスリン産生細胞を取得する方法(Nat.Med., 6: 278-282 (2000))、膵島に存在するネスチン陽性細胞で、終分化マーカーと共発現しない細胞を分取し、インビトロで培養することにより、Pdx-1、インスリン、グルカゴン、GLP-1陽性細胞を生じさせる方法(Diabetes, 50: 521-533 (2001))、肝前駆細胞をニコチンアミド含有培地で長期培養することによりインスリンIおよびII遺伝子を発現する細胞を得る方法(J. Cell Biol., 156: 173-184 (2002))等が挙げられるが、それらに限定されない。膵内分泌細胞への分化は、例えば、膵再生への関連が示されている各種転写因子(例:Ngn3、Nkx2、NeuroD、Nkx6.1、PAX-4等)の発現をRT-PCR、in situハイブリダイゼーション、免疫染色等で検出するか、膵再生への関連が示されている因子に対する蛍光抗体を用いたセルソーティング、後述する本発明のTg動物にさらに他の1つ以上の遺伝子改変を有する動物において、該他の1つの遺伝子改変がβ細胞特異的にレポーター遺伝子を発現させるものであるTg動物を用いて、該レポーター遺伝子の発現を検出する等の手法によって確認することができる。
【0052】
マイクロアレイ解析は、例えば、上記のようにして調製したRNA画分から、逆転写反応によりT7プロモーター等の適当なプロモーターを導入したcDNAを合成し、さらにRNAポリメラーゼを用いてcRNAを合成する(この時ビオチンなどで標識したモノヌクレオチドを基質として用いることにより、標識されたcRNAが得られる)。この標識cRNAを、例えば、膵島特異的cDNAマイクロアレイ(正常膵島由来のcDNAを、適当な固相、例えば、ガラス、シリコン、プラスチック、ニトロセルロース、ナイロン、ポリビニリデンジフロリド等に固定化したもの。固定化手段としては、予め核酸にアミノ基、アルデヒド基、SH基、ビオチンなどの官能基を導入しておき、一方、固相上にも該核酸と反応し得る官能基(例:アルデヒド基、アミノ基、SH基、ストレプトアビジンなど)を導入し、両官能基間の共有結合で固相と核酸を架橋したり、ポリアニオン性の核酸に対して、固相をポリカチオンコーティングして静電結合を利用して核酸を固定化するなどの方法が挙げられるが、これらに限定されない)と接触させてハイブリダイゼーション反応させ、マイクロアレイ上の各プローブに結合した標識量を測定することにより、各時点でのTg動物の膵臓もしくはその一部、あるいは該Tg動物由来の細胞・組織における遺伝子の発現量を測定することができる。これらの各遺伝子の発現量を分化誘導前後で比較した結果、分化誘導前後で発現量が変動した遺伝子を膵(内分泌細胞)再生関連(促進もしくは抑制)遺伝子の候補として選択することができる。
【0053】
本発明はまた、上記の膵再生関連遺伝子の同定方法を迅速且つ簡便に行うためのツールとして有用な、本発明のTg動物の膵内分泌細胞分化前後の膵臓もしくはその一部(例えば、膵島、膵管など)由来の単離されたRNAまたはそれを鋳型として逆転写反応によって調製されるcDNA(さらにRNAポリメラーゼの反応によって合成され得るcRNAも包含される)、あるいは該Tg動物の生体の一部における膵内分泌細胞分化前後の該生体の一部由来の単離されたRNAまたはそのcDNAを提供する。ここで「膵内分泌細胞分化前後」とは、分化前、分化誘導直後、分化の途中、分化終了後のいかなる段階であってもよく、それらのいずれか1つであっても、2つ以上であってもよい。RNA画分の調製法としては、上記と同様の方法が例示される。
【0054】
本発明の膵再生モデル系を用いた膵再生因子の別の同定方法としては、該モデル系より常法に従って蛋白質画分を調製し、例えば、サイファージェン・バイオシステムズ社のSELDI技術等を用いて、蛋白質を特異的に質量分析用プローブの表面上に捕捉させ、質量分析、好ましくはMALDI-TOFMS法などによりプローブに捕捉された蛋白質の分子量を測定、膵内分泌細胞分化前後でピーク強度の変化した分子量を示した蛋白質について、例えばタンデム質量分析(MS/MS)等を実施し、そのアミノ酸配列を同定する方法などが挙げられる。
【0055】
本発明はまた、本発明のTg動物に被検物質を投与、あるいは該Tg動物の生体の一部に被検物質を添加し、膵内分泌細胞分化に及ぼす該物質の効果を検定することによる、膵再生調節物質のスクリーニング方法を提供する。即ち、被検物質を投与(添加)しなかった場合と比較して、膵内分泌細胞分化が促進(もしくは抑制)されたか否かを判定することにより該物質の効果を検定する。膵内分泌細胞分化は、上記と同様に、例えば、膵再生への関連が示されている各種転写因子(例:Ngn3、Nkx2、NeuroD、Nkx6.1、PAX-4等)の発現をRT-PCR、in situハイブリダイゼーション、免疫染色等で検出するか、膵再生への関連が示されている因子に対する蛍光抗体を用いたセルソーティング、後述する本発明のTg動物にさらに他の1つ以上の遺伝子改変を有する動物において、該他の1つの遺伝子改変がβ細胞特異的にレポーター遺伝子を発現させるものであるTg動物を用いて、該レポーター遺伝子の発現を検出する等の手法によって確認することができる。
その結果、被検物質を投与(添加)しなかった場合に比べて、該Tg動物もしくはその生体の一部における内分泌細胞分化の開始が早まった場合(あるいは、同期間で分化がより進んだ場合)、該被検物質を膵再生促進物質として、内分泌細胞分化の開始が遅くなった場合(あるいは、同期間で分化がより進まなかった場合)、該被検物質を膵再生抑制物質として、それぞれ選択することができる。
【0056】
上記スクリーニング方法において、膵内分泌細胞分化を、上記の同定方法によって同定された膵再生関連遺伝子(因子)の発現を指標にして測定することもできる。従って、本発明はまた、本発明のTg動物に被検物質を投与、あるいは該Tg動物の生体の一部に被検物質を添加し、膵再生関連遺伝子(因子)の発現に及ぼす該物質の効果を検定することによる、膵再生関連遺伝子(因子)の発現調節物質のスクリーニング方法を提供する。
【0057】
上記スクリーニング方法によって得られうる、膵再生促進物質または膵再生促進遺伝子(因子)の発現促進物質もしくは膵再生抑制遺伝子(因子)の発現抑制物質(以下、包括して「膵再生促進薬」という)は、膵疲弊・膵傷害を伴う疾患、例えば、1型および2型糖尿病、糖尿病合併症(糖尿病性腎症、糖尿病性網膜症、糖尿病性神経障害等)、耐糖能異常、低インスリン血症等の予防・治療薬として安全かつ有効に用いることができる。
【0058】
膵再生促進薬は、例えば、必要に応じて糖衣を施した錠剤、カプセル剤、エリキシル剤、マイクロカプセル剤などとして経口的に、あるいは水もしくはそれ以外の薬学的に許容し得る液との無菌性溶液、または懸濁液剤などの注射剤の形で非経口的に使用できる。該作動薬は、生理学的に認められる担体、香味剤、賦形剤、ベヒクル、防腐剤、安定剤、結合剤などとともに一般に認められた製剤実施に要求される単位用量形態で混和することによって製剤化することができる。これら製剤における有効成分量は後述する投与量を考慮して適宜選択される。
【0059】
錠剤、カプセル剤などに混和することができる添加剤としては、例えば、ゼラチン、コーンスターチ、トラガント、アラビアゴムのような結合剤、結晶性セルロースのような賦形剤、コーンスターチ、ゼラチン、アルギン酸などのような膨化剤、ステアリン酸マグネシウムのような潤滑剤、ショ糖、乳糖またはサッカリンのような甘味剤、ペパーミント、アカモノ油またはチェリーのような香味剤などが用いられる。調剤単位形態がカプセルである場合には、前記タイプの材料にさらに油脂のような液状担体を含有することができる。注射のための無菌組成物は注射用水のようなベヒクル中の活性物質、胡麻油、椰子油などのような天然産出植物油などを溶解または懸濁させるなどの通常の製剤実施に従って処方することができる。
【0060】
注射用の水性液としては、例えば、生理食塩水、ブドウ糖やその他の補助薬を含む等張液(例えば、D−ソルビトール、D−マンニトール、塩化ナトリウムなど)などが挙げられ、適当な溶解補助剤、例えば、アルコール(例えば、エタノールなど)、ポリアルコール(例えば、プロピレングリコール、ポリエチレングリコールなど)、非イオン性界面活性剤(例えば、ポリソルベート80TM、HCO−50など)などと併用してもよい。油性液としては、例えば、ゴマ油、大豆油などが挙げられ、溶解補助剤として安息香酸ベンジル、ベンジルアルコールなどと併用してもよい。また、緩衝剤(例えば、リン酸塩緩衝液、酢酸ナトリウム緩衝液など)、無痛化剤(例えば、塩化ベンザルコニウム、塩酸プロカインなど)、安定剤(例えば、ヒト血清アルブミン、ポリエチレングリコールなど)、保存剤(例えば、ベンジルアルコール、フェノールなど)、酸化防止剤などと配合してもよい。調製された注射液は、通常、適当なアンプルに充填される。
【0061】
また、膵再生促進薬がDNA、RNAなどの核酸である場合、当該核酸を単独で、あるいは当該DNA(または当該RNAに対応するDNA)をレトロウイルスベクター、アデノウイルスベクター、アデノウイルスアソシエーテッドウイルスベクターなどの適当なベクターに挿入した後、常套手段に従って、ヒトまたは他の哺乳動物に投与することができる。当該核酸は、そのままで、あるいは摂取促進のための補助剤などの生理学的に認められる担体とともに製剤化した後、遺伝子銃やハイドロゲルカテーテルのようなカテーテルによって投与できる。
【0062】
膵再生促進薬の投与量は、対象疾患、投与対象、投与ルートなどにより異なるが、例えば、糖尿病の治療目的で経口投与する場合、一般的に成人(体重60kg)においては、一日につき約0.1mg〜100mg、好ましくは約1.0〜50mg、より好ましくは約1.0〜20mgである。非経口投与の場合、当該薬物の投与量は投与対象、対象疾患などによっても異なるが、例えば、糖尿病の治療目的で注射剤として成人(体重60kg)に投与する場合、一日につき約0.01〜30mg程度、好ましくは約0.1〜20mg程度、より好ましくは約0.1〜10mg程度である。投与対象がヒト以外の動物の場合も、体重60kg当たりに換算した量を投与することができる。
【0063】
本発明のTg動物は、KATPチャンネルKir6.2のドミナントネガティブ体をコードするDNAを膵β細胞で発現可能な状態で安定に保持することに加えて、1以上の他の遺伝子変異または改変を有することをさらなる特徴とする。「他の遺伝子変異」とは、自然突然変異により内因性遺伝子が欠損、機能低下、あるいは機能亢進された自然発症疾患モデル動物をいい、「他の遺伝子改変」とは、KATPチャンネルKir6.2のドミナントネガティブ体をコードするDNAが導入される以外の遺伝子改変、即ち、他の遺伝子をさらに導入されたトランスジェニック動物、内因性遺伝子を不活化されたノックアウト動物(挿入突然変異等による遺伝子破壊のほか、アンチセンスDNAや中和抗体をコードするDNAの導入により遺伝子発現が検出不可能もしくは無視し得る程度にまで低下したトランスジェニック動物を含む)、変異内因性遺伝子が導入されたドミナントネガティブ変異体などが含まれる。
【0064】
「他の遺伝子変異または改変」は、膵再生関連遺伝子(因子)の探索、膵再生促進薬の探索、並びに膵再生促進薬の薬効評価用としての用途に有利なものであれば特に制限はないが、例えば、膵内分泌機能異常が関与する疾患と同一もしくは類似の病態を生じさせるような遺伝子変異または改変であることが好ましい。
「膵内分泌機能異常が関与する疾患」とは、膵内分泌機能の異常に起因するかもしくは結果的に膵内分泌機能の異常を生じる疾患だけでなく、膵内分泌機能を調節することにより予防および/または治療効果が得られ得る疾患をも含めた概念として把握されるべきである。例えば、1型および2型糖尿病、糖尿病合併症(糖尿病性腎症、糖尿病性網膜症、糖尿病性神経障害等)、耐糖能異常、低インスリン血症等が挙げられる。
【0065】
「膵内分泌機能異常が関与する疾患と同一もしくは類似の病態を生じさせる1以上の他の遺伝子変異または改変を有する疾患モデル」としては、例えば、自然発症糖尿病モデルマウスであるKKマウス、KKAyマウス、db/dbマウス、ob/obマウス、Akitaマウス等、あるいはヒトIAPP遺伝子を導入したTgマウス等の膵疲弊または膵傷害を示す各種Tg/KOマウスが挙げられる。
これらの「他の遺伝子変異または改変を有する疾患モデル」は、例えば、米国のJackson研究所などから購入可能であるか、あるいは周知の遺伝子改変技術を用いて容易に作製することができる。
【0066】
本発明のTg動物は、「膵内分泌機能異常が関与する疾患と同一もしくは類似の病態を生じさせる1以上の他の遺伝子変異または改変」に加えて、同一もしくは他の疾患モデルを作製し得る非遺伝的処理を施されていてもよい。「非遺伝的処理」とは対象非ヒト哺乳動物における遺伝子変異または改変を生じさせない処理を意味する。このような処理としては、例えば、高脂肪食負荷処理、糖負荷処理、飢餓処理、膵管結紮、STZ処理、膵切除等が挙げられる。
【0067】
膵再生促進薬の薬効評価用としての本発明のTg動物の用途に有利な「他の遺伝子改変」の別の好ましい態様としては、膵β細胞特異的なプロモーターの制御下にあるレポーター遺伝子の導入が挙げられる。該プロモーターの制御下にあるレポーター遺伝子を導入された非ヒト哺乳動物では、膵β細胞の再生によりβ細胞数が増加すると該レポーターの発現量が増大するので、該レポーターの発現に及ぼす被検物質の効果を検定することにより、該物質の膵再生促進活性を評価することができる。膵β細胞特異的プロモーターとしては、例えばインスリンIもしくはIIプロモーター等が挙げられる。また、レポーター遺伝子としては、例えば、β−ガラクトシダーゼ遺伝子、ルシフェラーゼ遺伝子、クロラムフェニコールアセチルトランスフェラーゼ遺伝子、アルカリホスファターゼ遺伝子、ペルオキシダーゼ遺伝子等が挙げられる。
【0068】
KATPチャンネルKir6.2のドミナントネガティブ体をコードするDNAを導入された非ヒト哺乳動物に、膵内分泌機能異常が関与する疾患と同一もしくは類似の病態を生じさせる1以上の他の遺伝子変異または改変、あるいは膵β細胞特異的プロモーターの制御下にあるレポーター遺伝子を導入する方法は特に制限はなく、例えば、KATPチャンネルKir6.2のドミナントネガティブ体をコードするDNAを導入された非ヒト哺乳動物と、膵内分泌機能異常が関与する疾患と同一もしくは類似の病態を生じさせる1以上の他の遺伝子変異または改変、あるいは膵β細胞特異的プロモーターの制御下にあるレポーター遺伝子を有する同種の非ヒト哺乳動物とを交雑する方法;膵内分泌機能異常が関与する疾患と同一もしくは類似の病態を生じさせる1以上の他の遺伝子変異または改変、あるいは膵β細胞特異的プロモーターの制御下にあるレポーター遺伝子を有する非ヒト哺乳動物の初期胚やES細胞に、上述の方法によりKATPチャンネルKir6.2のドミナントネガティブ体をコードするDNAを導入してトランスジェニック動物を得る方法;KATPチャンネルKir6.2のドミナントネガティブ体をコードするDNAを導入された非ヒト哺乳動物の初期胚やES細胞に、上述の方法により、あるいはノックアウト技術により、膵内分泌機能異常が関与する疾患と同一もしくは類似の病態を生じさせる1以上の他の遺伝子改変、あるいは膵β細胞特異的プロモーターの制御下にあるレポーター遺伝子を導入する方法等が挙げられる。また、膵内分泌機能異常が関与する疾患と同一もしくは類似の病態を生じさせる1以上の他の遺伝子改変が外来遺伝子やドミナント変異遺伝子の導入による場合、野生型非ヒト哺乳動物の初期胚やES細胞に、該外来遺伝子等とKATPチャンネルKir6.2のドミナントネガティブ体をコードするDNAとを同時にもしくは順次導入してトランスジェニック動物を得てもよい。他の遺伝子改変が膵β細胞特異的プロモーターの制御下にあるレポーター遺伝子の導入である場合も同様に、該レポーター遺伝子とKATPチャンネルKir6.2のドミナントネガティブ体をコードするDNAとを同時にもしくは順次導入してトランスジェニック動物を得ることができる。さらに、膵内分泌機能異常が関与する疾患と同一もしくは類似の病態を生じさせる1以上の他の遺伝子改変が内因性遺伝子の破壊による場合は、KATPチャンネルKir6.2のドミナントネガティブ体をコードするDNAを破壊すべき内因性遺伝子にターゲッティングされ得るようにデザインして野生型非ヒト哺乳動物のES細胞に導入してもよい。この場合、従来公知のノックイン動物作製技術が好ましく使用され得る。
【0069】
KATPチャンネルKir6.2のドミナントネガティブ体をコードするDNAを導入された非ヒト哺乳動物と、膵内分泌機能異常が関与する疾患と同一もしくは類似の病態を生じさせる1以上の他の遺伝子変異または改変(あるいは膵β細胞特異的プロモーターの制御下にあるレポーター遺伝子)を有する同種の疾患モデル非ヒト哺乳動物とを交雑する場合、ホモ接合体同士を交雑することが望ましい。例えば、KATPチャンネルKir6.2のドミナントネガティブ体をコードするDNAが1遺伝子座に組み込まれたホモ接合体と、db/dbマウスとを交雑して得られるF1は両遺伝子についてヘテロである。このF1同士を兄妹交配して得られるF2個体の1/16はドミナントネガティブホモ導入・dbホモとなる。
【0070】
KATPチャンネルKir6.2のドミナントネガティブ体をコードするDNAを導入され、且つ膵内分泌機能異常が関与する疾患と同一もしくは類似の病態を生じさせる1以上の他の遺伝子変異または改変を有する本発明のTg動物は、膵内分泌機能異常が関与する疾患と同一もしくは類似の病態を示すので、該動物に被検物質を投与してその病態に及ぼす該物質の効果を検定することによって、膵内分泌機能異常が関与する疾患に対して予防・治療活性を有する物質をスクリーニングすることができる。指標となる病態は疾患モデルの種類に応じて適宜選択され得るが、例えば、該Tg動物が糖尿病モデルであれば、血糖値・インスリン分泌の改善、膵内分泌細胞の増殖・分化促進などを指標として、被検物質の疾患予防・治療効果を評価することができる。
【0071】
本願明細書において、塩基やアミノ酸などを略号で表示する場合、IUPAC-IUB Commission on Biochemical Nomenclatureによる略号あるいは当該分野における慣用略号に基づくものであり、その例を次に挙げる。
DNA :デオキシリボ核酸
cDNA :相補的デオキシリボ核酸
A :アデニン
T :チミン
G :グアニン
C :シトシン
RNA :リボ核酸
mRNA :メッセンジャーリボ核酸
dATP :デオキシアデノシン三リン酸
dTTP :デオキシチミジン三リン酸
dGTP :デオキシグアノシン三リン酸
dCTP :デオキシシチジン三リン酸
ATP :アデノシン三リン酸
EDTA :エチレンジアミン四酢酸
SDS :ドデシル硫酸ナトリウム
Gly :グリシン
Ala :アラニン
Val :バリン
Leu :ロイシン
Ile :イソロイシン
Ser :セリン
Thr :スレオニン
Cys :システイン
Met :メチオニン
Glu :グルタミン酸
Asp :アスパラギン酸
Lys :リジン
Arg :アルギニン
His :ヒスチジン
Phe :フェニルアラニン
Tyr :チロシン
Trp :トリプトファン
Pro :プロリン
Asn :アスパラギン
Gln :グルタミン
【実施例】
【0072】
以下に実施例を挙げて本発明をより具体的に説明するが、本発明がこれらに限定されないことは言うまでもない。
【0073】
実施例1 Kir6.2G132S Tgマウスにおける血糖値の推移
上記非特許文献2に記載される方法により作製されたKir6.2G132S Tgマウス(line M45)について、同文献と同様の方法により、4-6(n=43)、8-10(n=14)、12-14(n=30)、16-20(n=31)、25(n=23)および50(n=11)週齢における血糖値をそれぞれ測定した。結果を図1に示す。経時的に血糖値を観察したところ、4-6週齢に認められた高血糖が経時的に低下し、16-20週齢以降、4-6週齢に比べて有意な血糖改善が認められ、50週齢ではほぼ正常値を示すまでに至った。
【0074】
実施例2 Kir6.2G132S Tgマウスにおける血清インスリン値の推移
Kir6.2G132S Tgマウス(line M45)について、上記非特許文献2と同様の方法により、4-6(n=8)、8-10(n=10)、12-14(n=11)、16-20(n=14)、25(n=12)および50(n=10)週齢における血清インスリン値をそれぞれ測定した。結果を図2に示す。経時的に血清インスリン値を観察したところ、4-6週齢に認められたインスリン欠乏状態が経時的に改善された。25週齢以降は有意な改善が認められ、50週齢ではほぼ正常値を示すまでに至った。
【0075】
実施例3 Kir6.2G132S Tgマウスにおける膵組織像
Kir6.2G132S Tgマウス(line M45;11週齢)より膵臓を摘出し、膵管近傍組織切片を作製した。抗インスリン抗体および抗グルカゴン抗体をそれぞれ蛍光色素Cy3(赤)およびCy2(緑)で標識し、二重免疫染色を行った。結果を図3に示す。血糖値が改善し始める12週齢の前後では、膵管近傍において膵β細胞の新生像が観察された(図3B)。
【0076】
実施例4 Kir6.2G132S Tgマウスにおけるβ細胞組織像
Kir6.2G132S Tgマウス(line M45)について、4、12、25及び50週齢時に膵臓を摘出し、β細胞組織切片を作製した。対照群(正常マウス)についても膵臓を摘出し、β細胞組織切片を作製した。抗インスリン抗体を蛍光色素Cy3(赤)で標識し、免疫染色を行った。結果を図4に示す。Kir6.2G132S Tgマウスのβ細胞において、4週齢に認められたインスリン含量の低下は経時的に改善され、50週齢ではほぼ正常化するに至った。
【産業上の利用可能性】
【0077】
Kir6.2のドミナントネガティブ体を導入されたTg動物は、若週齢で糖尿病を発症し、インスリン欠乏・膵β細胞の細胞死が顕著となるが、加齢とともにインスリン分泌が増大し、膵管近傍に高頻度に膵内分泌細胞が出現することから、該動物およびその生体の一部は、優れた膵内分泌細胞の再生モデル系として、膵再生関連遺伝子(因子)の探索、膵再生促進薬の探索、膵再生促進薬の薬効評価に利用することができる。
【図面の簡単な説明】
【0078】
【図1】Kir6.2G132S Tgマウス(TG)および野生型マウス(WT)における血糖値の推移を示す図である。グラフの各棒の下の括弧内の数字は例数(匹)を示す。
【図2】Kir6.2G132S Tgマウス(TG)および野生型マウス(WT)における血清インスリン値の推移を示す図である。グラフの各棒の下の括弧内の数字は例数(匹)を示す。
【図3】Kir6.2G132S Tgマウス(11週齢)の膵管近傍における組織像を示す図である。図3Aは位相差顕微鏡像を、図3Bは蛍光色素によるインスリン/グルカゴン共染色像をそれぞれ示す。
【図4】Kir6.2G132S Tgマウスのβ細胞組織におけるインスリン含量の経時変化を示す図である。抗インスリン抗体を蛍光色素Cy3(赤)で標識し、免疫染色を行った。
【特許請求の範囲】
【請求項1】
ATP感受性カリウムイオンチャンネルKir6.2のドミナントネガティブ体を膵β細胞で発現し得るトランスジェニック非ヒト哺乳動物またはその生体の一部から実質的になる膵再生モデル系。
【請求項2】
ドミナントネガティブ体が、チャンネルのカリウムイオン選択性に関与する領域中の1以上のアミノ酸が変異したものである請求項1記載のモデル系。
【請求項3】
ATP感受性カリウムイオンチャンネルKir6.2のドミナントネガティブ体を膵β細胞で発現し得るトランスジェニック非ヒト哺乳動物における、膵内分泌細胞分化前後の膵臓もしくはその一部由来の単離されたRNAまたはそのcDNA。
【請求項4】
ATP感受性カリウムイオンチャンネルKir6.2のドミナントネガティブ体を膵β細胞で発現し得るトランスジェニック非ヒト哺乳動物の生体の一部における、膵内分泌細胞分化前後の該生体の一部由来の単離されたRNAまたはそのcDNA。
【請求項5】
ATP感受性カリウムイオンチャンネルKir6.2のドミナントネガティブ体を膵β細胞で発現し得るトランスジェニック非ヒト哺乳動物において、膵内分泌細胞分化前後での膵臓もしくはその一部における遺伝子発現プロファイルを比較することを特徴とする、膵再生関連遺伝子の同定方法。
【請求項6】
ATP感受性カリウムイオンチャンネルKir6.2のドミナントネガティブ体を膵β細胞で発現し得るトランスジェニック非ヒト哺乳動物の生体の一部において、膵内分泌細胞分化前後での該生体の一部における遺伝子発現プロファイルを比較することを特徴とする、膵再生関連遺伝子の同定方法。
【請求項7】
ATP感受性カリウムイオンチャンネルKir6.2のドミナントネガティブ体を膵β細胞で発現し得るトランスジェニック非ヒト哺乳動物またはその生体の一部に被検物質を適用し、膵内分泌細胞分化に及ぼす該物質の効果を検定することを特徴とする、膵再生調節物質のスクリーニング方法。
【請求項8】
ATP感受性カリウムイオンチャンネルKir6.2のドミナントネガティブ体を膵β細胞で発現することができ、且つ1以上の他の遺伝子変異または改変を有する非ヒト哺乳動物またはその生体の一部。
【請求項9】
他の遺伝子変異または改変の少なくとも1つが、膵内分泌機能異常が関与する疾患と同一もしくは類似の病態を生じさせるものである、請求項8記載の動物またはその生体の一部。
【請求項10】
請求項9記載の動物に被検物質を投与し、膵内分泌機能異常が関与する疾患と同一もしくは類似の病態に及ぼす該物質の効果を検定することを特徴とする、該疾患の予防・治療物質のスクリーニング方法。
【請求項1】
ATP感受性カリウムイオンチャンネルKir6.2のドミナントネガティブ体を膵β細胞で発現し得るトランスジェニック非ヒト哺乳動物またはその生体の一部から実質的になる膵再生モデル系。
【請求項2】
ドミナントネガティブ体が、チャンネルのカリウムイオン選択性に関与する領域中の1以上のアミノ酸が変異したものである請求項1記載のモデル系。
【請求項3】
ATP感受性カリウムイオンチャンネルKir6.2のドミナントネガティブ体を膵β細胞で発現し得るトランスジェニック非ヒト哺乳動物における、膵内分泌細胞分化前後の膵臓もしくはその一部由来の単離されたRNAまたはそのcDNA。
【請求項4】
ATP感受性カリウムイオンチャンネルKir6.2のドミナントネガティブ体を膵β細胞で発現し得るトランスジェニック非ヒト哺乳動物の生体の一部における、膵内分泌細胞分化前後の該生体の一部由来の単離されたRNAまたはそのcDNA。
【請求項5】
ATP感受性カリウムイオンチャンネルKir6.2のドミナントネガティブ体を膵β細胞で発現し得るトランスジェニック非ヒト哺乳動物において、膵内分泌細胞分化前後での膵臓もしくはその一部における遺伝子発現プロファイルを比較することを特徴とする、膵再生関連遺伝子の同定方法。
【請求項6】
ATP感受性カリウムイオンチャンネルKir6.2のドミナントネガティブ体を膵β細胞で発現し得るトランスジェニック非ヒト哺乳動物の生体の一部において、膵内分泌細胞分化前後での該生体の一部における遺伝子発現プロファイルを比較することを特徴とする、膵再生関連遺伝子の同定方法。
【請求項7】
ATP感受性カリウムイオンチャンネルKir6.2のドミナントネガティブ体を膵β細胞で発現し得るトランスジェニック非ヒト哺乳動物またはその生体の一部に被検物質を適用し、膵内分泌細胞分化に及ぼす該物質の効果を検定することを特徴とする、膵再生調節物質のスクリーニング方法。
【請求項8】
ATP感受性カリウムイオンチャンネルKir6.2のドミナントネガティブ体を膵β細胞で発現することができ、且つ1以上の他の遺伝子変異または改変を有する非ヒト哺乳動物またはその生体の一部。
【請求項9】
他の遺伝子変異または改変の少なくとも1つが、膵内分泌機能異常が関与する疾患と同一もしくは類似の病態を生じさせるものである、請求項8記載の動物またはその生体の一部。
【請求項10】
請求項9記載の動物に被検物質を投与し、膵内分泌機能異常が関与する疾患と同一もしくは類似の病態に及ぼす該物質の効果を検定することを特徴とする、該疾患の予防・治療物質のスクリーニング方法。
【図1】


【図2】
【図3】
【図4】
【図2】
【図3】
【図4】
【公開番号】特開2006−149380(P2006−149380A)
【公開日】平成18年6月15日(2006.6.15)
【国際特許分類】
【出願番号】特願2005−312434(P2005−312434)
【出願日】平成17年10月27日(2005.10.27)
【出願人】(504150450)国立大学法人神戸大学 (421)
【出願人】(000002934)武田薬品工業株式会社 (396)
【Fターム(参考)】
【公開日】平成18年6月15日(2006.6.15)
【国際特許分類】
【出願日】平成17年10月27日(2005.10.27)
【出願人】(504150450)国立大学法人神戸大学 (421)
【出願人】(000002934)武田薬品工業株式会社 (396)
【Fターム(参考)】
[ Back to top ]