
遺伝子改変微生物を作製する方法
1つまたは複数のフェロモン応答遺伝子および1つまたは複数の胞子形成遺伝子において機能的破壊を含む遺伝子改変微生物、例えば、遺伝子改変酵母株を作製する方法、ならびにそれにより産生された胞子形成能力および内因性接合能力を欠く遺伝子改変酵母細胞、例えば、遺伝子改変の二倍体および一倍体の酵母細胞が本明細書に提供される。本発明の方法は、遺伝子改変の一倍体微生物細胞、例えば、酵母細胞において1つもしくは複数の胞子形成遺伝子および/または1つもしくは複数のフェロモン応答遺伝子を機能的に破壊する工程、ならびに上記遺伝子改変一倍体微生物細胞が、それらの胞子形成および/または接合能力の欠損により、効果的に性的に生殖不能であり、それらの生活環の二倍体状態に制約される安定な二倍体を形成するように誘導する工程を含む。

【発明の詳細な説明】
【技術分野】
【0001】
本出願は、2009年6月1日に出願された米国仮出願番号61/183,031の利益を主張し、上記米国仮出願は、その全容が参照によって本明細書に援用される。
【0002】
本明細書に提供される方法および組成物は、一般的に、酵母などの微生物の産業的使用に関する。天然における散在、特に、組換えDNA配列の天然における散在のリスクを著しく低下させるように改変されている遺伝子改変微生物が本明細書に提供される。そのような遺伝子改変微生物を作製する方法および使用する方法も提供される。
【背景技術】
【0003】
組換えDNAテクノロジーの進歩により、原核生物または真核生物の利用を通しての産業的に有用な物質の大量製造が可能になっている。真核生物の中で、酵母(特に、Saccharomyces属に属する酵母)は、発酵産物の製造に広く用いられている。一般的に、酵母は、迅速に増殖することができ、細菌と比較してより高密度で培養することができ、産業的設定において無菌的環境を必要としない。さらに、酵母細胞は、細菌と比較して培地から容易に分離することができ、産物の抽出および精製のための工程を大いに単純化することができる。これらの特徴のため、酵母(特に、組換えDNA配列を内部に有する遺伝子改変酵母)は、有用な産物の製造のための宿主として用いられており、そのような酵母の利用は確立されている。しかしながら、産業における遺伝子改変酵母の使用は、そのような酵母および/またはそのような酵母に含まれる組換えDNA配列の分散が生態系に予想不可能な結果を生じる可能性があるという理由により、潜在的環境リスクを有する。
【発明の概要】
【発明が解決しようとする課題】
【0004】
したがって、産業的適用に適しているが、天然において散在し、繁殖するというリスクの低下をもたらし、特に、天然においてそれらの組換えDNA配列を散在するというリスクの低下をもたらす酵母の必要性が存在している。
【課題を解決するための手段】
【0005】
胞子形成および/または内因性接合が損なわれている遺伝子改変微生物、例えば、遺伝子改変酵母株を作製する方法が本明細書に提供される。本明細書に提供される方法は、遺伝子改変の一倍体微生物細胞、例えば、酵母細胞において1つもしくは複数の胞子形成遺伝子および/または1つもしくは複数のフェロモン応答遺伝子を機能的に破壊する工程、ならびに前記遺伝子改変一倍体微生物細胞が、それらの胞子形成および/または接合能力の欠損により、効果的に性的に生殖不能であり、および、それらの生活環の二倍体状態に制約される安定な二倍体を形成するように誘導する工程を含む。本明細書に提供される方法に従って遺伝子改変された微生物、例えば、酵母株は、産業的適用、例えば、産業的発酵適用に用途を見出し、野生型微生物との接合を通して天然において散在し、繁殖するというリスクの著しい低下をもたらす利点を提供することができる。
【0006】
一態様において、以下を含む遺伝子改変酵母細胞が本明細書に提供される:1つまたは複数の胞子形成遺伝子における機能的破壊であって、前記酵母細胞が、前記の1つまたは複数の胞子形成遺伝子の破壊の結果として胞子形成能力を欠く、機能的破壊;1つまたは複数のフェロモン応答遺伝子における機能的破壊であって、前記酵母細胞が、前記の1つまたは複数のフェロモン応答遺伝子の破壊の結果として内因性接合能力を欠く、機能的破壊。いくつかの実施形態において、遺伝子改変酵母細胞はヘテロタリック(heterothallic(ho))である。いくつかの実施形態において、遺伝子改変酵母細胞は、1つもしくは複数の胞子形成遺伝子の両方のコピーおよび/または1つもしくは複数のフェロモン応答遺伝子の両方のコピーが機能的に破壊されている二倍体細胞である。いくつかの実施形態において、遺伝子改変二倍体細胞は、その接合型対立遺伝子について以外はホモ接合型である。いくつかの実施形態において、遺伝子改変酵母細胞は一倍体細胞である。いくつかの実施形態において、遺伝子改変一倍体細胞は、ホモタリズム(homothallism(HO))タンパク質をコードする組換えプラスミドをさらに含む。いくつかの実施形態において、遺伝子改変一倍体細胞は、前記酵母細胞において機能的に破壊されている1つまたは複数のフェロモン応答遺伝子をコードする1つまたは複数の組換えプラスミドを含む。
【0007】
いくつかの実施形態において、本明細書に提供された方法の実施に有用な遺伝子改変酵母細胞はSaccharomyces cerevisiae細胞である。いくつかの実施形態において、Saccharomyces cerevisiae細胞は、パン酵母、Mauri、Santa Fe、CBS 7959、CBS 7960、CBS 7961、CBS 7962、CBS 7963、CBS 7964、IZ−1904、TA、BG−1、CR−1、SA−1、M−26、Y−904、PE−2、PE−5、VR−1、BR−1、BR−2、ME−2、VR−2、MA−3、MA−4、CAT−1、CB−1、NR−1、BT−1、またはAL−1の株である。特定の実施形態において、Saccharomyces cerevisiae細胞はPE−2株である。他の特定の実施形態において、Saccharomyces cerevisiae細胞はCAT−1株である。
【0008】
いくつかの実施形態において、遺伝子改変酵母細胞における破壊された1つまたは複数の胞子形成遺伝子は、IME1、IME2、NDT80、SPO11、SPO20、AMA1、HOP2、およびSPO21からなる群より選択される。特定の実施形態において、IME1遺伝子が破壊されている。いくつかの実施形態において、遺伝子改変酵母細胞における破壊された1つまたは複数のフェロモン応答遺伝子は、STE5、STE4、STE18、STE12、STE7、およびSTE11からなる群より選択される。特定の実施形態において、STE5遺伝子が破壊されている。特定の実施形態において、遺伝子改変酵母細胞は、STE5およびIME1遺伝子の両方の機能的破壊を含む。いくつかの実施形態において、遺伝子改変酵母細胞の一倍体細胞は、STE5遺伝子の機能的破壊およびSTE5タンパク質をコードする組換えプラスミドを含む。
【0009】
別の態様において、胞子形成および接合能力が損なわれている二倍体酵母株を作製する方法が本明細書に提供される。いくつかの実施形態において、該方法は以下の工程:(a)第1の遺伝子改変一倍体酵母細胞を得る工程であって、第1の遺伝子改変一倍体酵母細胞が胞子形成および内因性接合が損なわれており、および、目的のタンパク質をコードする、染色体に組み込まれた異種性ヌクレオチド配列を含む、工程;(b)第2の遺伝子改変一倍体酵母細胞を得る工程であって、第2の遺伝子改変一倍体酵母細胞が、胞子形成および内因性接合が損なわれており、および、第1の遺伝子改変一倍体酵母細胞と反対の接合型であり、および、目的の前記のタンパク質をコードする、染色体に組み込まれた異種性ヌクレオチド配列を含む、工程;(c)第1の遺伝子改変一倍体酵母細胞および第2の遺伝子改変一倍体酵母細胞のそれぞれを、前記の第1および第2の遺伝子改変一倍体酵母細胞の内因性接合欠陥を相補することができるタンパク質をコードする1つまたは複数のプラスミドで形質転換する工程;(d)第1の遺伝子改変一倍体酵母細胞を第2の遺伝子改変一倍体酵母細胞と接合させ、それにより、遺伝子改変二倍体酵母細胞を形成する工程;ならびに(e)遺伝子改変二倍体酵母細胞から前記の1つまたは複数のプラスミドを除去する工程であって、その結果生じた遺伝子改変二倍体酵母細胞が胞子形成および内因性接合が損なわれている、工程、を含む。
【0010】
いくつかの実施形態において、第1の遺伝子改変一倍体酵母細胞および第2の遺伝子改変一倍体酵母細胞が、1つまたは複数のフェロモン応答遺伝子の機能的破壊によって内因性接合が損なわれている。いくつかのそのような実施形態において、工程(c)は、第1の遺伝子改変一倍体酵母細胞および第2の遺伝子改変一倍体酵母細胞のそれぞれを、前記の第1および第2の遺伝子改変一倍体酵母細胞において機能的に破壊されているフェロモン応答遺伝子の機能性コピーをコードする1つまたは複数のプラスミドで形質転換する工程を含む。いくつかの実施形態において、1つまたは複数のフェロモン応答遺伝子は、STE5、STE4、STE18、STE12、STE7、およびSTE11からなる群より選択される。ある特定の実施形態において、第1の遺伝子改変一倍体酵母細胞および第2の遺伝子改変一倍体酵母細胞は、STE5遺伝子の機能的破壊によって内因性接合が損なわれている。
【0011】
いくつかの実施形態において、第1の遺伝子改変一倍体酵母細胞および第2の遺伝子改変一倍体酵母細胞は、1つまたは複数の胞子形成遺伝子の機能的破壊によって胞子形成が損なわれている。いくつかの実施形態において、1つまたは複数の胞子形成遺伝子は、IME1、IME2、NDT80、SPO11、SPO20、AMA1、HOP2、およびSPO21からなる群より選択される。いくつかの実施形態において、第1の遺伝子改変一倍体酵母細胞および第2の遺伝子改変一倍体酵母細胞は、IME1遺伝子の機能的破壊によって胞子形成が損なわれている。特定の実施形態において、第1の遺伝子改変一倍体酵母細胞および第2の遺伝子改変一倍体酵母細胞は、STE5遺伝子の機能的破壊によって接合が損なわれており、および、IME1遺伝子の機能的破壊によって胞子形成が損なわれている。
【0012】
いくつかの実施形態において、第2の遺伝子改変一倍体酵母細胞は、第1の遺伝子改変一倍体酵母細胞の集団において接合型切り換えを誘導することによって得られる。いくつかのそのような実施形態において、第1の遺伝子改変一倍体酵母細胞はヘテロタリック(ho)であり、前記集団は、遺伝子改変一倍体酵母細胞をホモタリズム(HO)タンパク質をコードする組換えプラスミドで形質転換することによって接合型を切り換えるように誘導され、HOタンパク質の発現が、遺伝子改変一倍体酵母細胞の接合型切り換えを誘導する。
【0013】
他の実施形態において、第2の遺伝子改変一倍体酵母細胞は、組換えDNAテクノロジーを用いて第1の遺伝子改変一倍体酵母細胞において接合型の座を変化させることによって得られる。いくつかの実施形態において、第1の遺伝子改変一倍体酵母細胞は、第1の遺伝子改変一倍体酵母細胞において接合型の座に隣接するヌクレオチド配列と相同である相同配列に隣接した、第1の遺伝子改変一倍体酵母細胞の接合型以外の接合型をコードするヌクレオチド配列を組込み配列として含む組込み構築物で形質転換される。相同組換えによる組込み配列の組込みで、第1の遺伝子改変一倍体酵母細胞の接合型の座は、挿入配列によってコードされる接合型の座と置換され、その結果として、第2の遺伝子改変一倍体酵母細胞の産生を生じる。いくつかの実施形態において、組込み構築物は、α接合型(MAT α)をコードする組込み構築物を用いてaからαへ第1の遺伝子改変一倍体酵母細胞の接合型を切り換えるために用いられる。いくつかの実施形態において、組込み構築物は、配列番号155を含む。他の実施形態において、組込み構築物は、a接合型(MAT A)をコードする組込み構築物を用いてαからaへ第1の遺伝子改変一倍体酵母細胞の接合型を切り換えるために用いられる。いくつかの実施形態において、組込み構築物は配列番号156を含む。
【0014】
別の態様において、胞子形成および内因性接合が損なわれたヘテロタリック(ho)二倍体酵母細胞を作製する方法が本明細書に提供され、該方法は以下の工程:(a)第1の遺伝子改変ヘテロタリック一倍体酵母細胞を得る工程であって、第1の遺伝子改変ヘテロタリック一倍体酵母細胞が(i)目的のタンパク質をコードする、染色体に組み込まれた異種性ヌクレオチド配列;および(ii)STE5遺伝子およびIME1遺伝子における機能的破壊を含む、工程;(b)第1の遺伝子改変ヘテロタリック一倍体酵母細胞の集団を、ホモタリズム(HO)タンパク質をコードするポリヌクレオチドを含むプラスミドで形質転換する工程であって、HOタンパク質の発現が第1の遺伝子改変ヘテロタリック一倍体酵母細胞の接合型切り換えを誘導し、それにより第2の遺伝子改変一倍体酵母細胞が得られ、第2の遺伝子改変一倍体酵母細胞が第1の遺伝子改変一倍体酵母細胞と反対の接合型であり、(i)目的の前記のタンパク質をコードする染色体に組み込まれた異種性ヌクレオチド配列;および(ii)STE5遺伝子およびIME1遺伝子における機能的破壊を含む、工程;(c)第1の遺伝子改変ヘテロタリック一倍体酵母細胞および第2の遺伝子改変ヘテロタリック一倍体酵母細胞のそれぞれを、STE5遺伝子をコードするプラスミドで形質転換する工程;(d)第1の遺伝子改変一倍体酵母細胞を第2の遺伝子改変一倍体酵母細胞と接合させ、それにより、遺伝子改変二倍体酵母細胞を形成する工程;ならびに(e)遺伝子改変二倍体酵母細胞からいかなるプラスミドも除去する工程であって、その結果生じた遺伝子改変ヘテロタリック二倍体酵母細胞が胞子形成および内因性接合が損なわれている、工程、を含む。
【0015】
本発明により作製された、胞子形成および内因性接合能力を欠く遺伝子改変ヘテロタリック(ho)酵母細胞もまた本明細書に提供される。
【0016】
MATα/a ste5/ste5ime1/ime1酵母細胞もまた本明細書に提供される。
【図面の簡単な説明】
【0017】
【図1】図1は、相I破壊構築物の構造、および相同組換えによるその破壊配列の組込み後の標的座の構造を提供する。
【図2】図2は、相II破壊構築物の構造、および相同組換えによるその破壊配列の組込み後の標的座の構造を提供する。
【図3】図3は、相III破壊構築物の構造、および相同組換えによるその破壊配列の組込み後の標的座の構造を提供する。
【図4】図4は、相Iマーカーリサイクリング構築物の構造、および相同組換えによるその構築物の組込み後の標的座の構造を提供する。
【図5】図5は、相IIマーカーリサイクリング構築物の構造、および相同組換えによるその構築物の組込み後の標的座の構造を提供する。
【図6】図6は、相IIIマーカーリサイクリング構築物の構造、および相同組換えによるその構築物の組込み後の標的座の構造を提供する。
【図7】図7は、STE5破壊構築物の構造、および相同組換えによるその破壊配列の組込み後の標的座の構造を提供する。
【図8】図8は、IME1破壊構築物の構造、および相同組換えによるその破壊配列の組込み後の標的座の構造を提供する。
【図9】図9は、遺伝子改変の内因性接合の損なわれた一倍体Y1915細胞と遺伝子改変の内因性接合能力のあるY1912細胞との接合能力の比較を提供する。
【図10】図10は、遺伝子改変の胞子形成および内因性接合の損なわれた二倍体Y1979細胞と遺伝子非改変の胞子形成および内因性接合の能力のあるY1198細胞との胞子形成能力の比較を提供する。
【図11】図11は、遺伝子改変の胞子形成および内因性接合の損なわれた二倍体Y1979細胞と遺伝子非改変の胞子形成および内因性接合の能力のあるY1198細胞との土壌中の生存の比較を提供する。
【発明を実施するための形態】
【0018】
(専門用語)
本明細書に用いられる場合、用語「異種性」とは、通常には天然で見出されないものを指す。用語「異種性ヌクレオチド配列」は、天然で所定の細胞において通常には見出されないヌクレオチド配列を指す。それとして、異種性ヌクレオチド配列は、(a)その宿主細胞にとって外来であり得る(すなわち、その細胞にとって「外因性」であり得る);(b)宿主細胞において天然で見出されるが(すなわち、「内因性」)、細胞において不自然な量(すなわち、宿主細胞において天然で見出されるより多い量、または少ない量)で存在し得る;または(c)宿主細胞において天然で見出されるが、その天然の座の外側に位置し得る。
【0019】
本明細書に用いられる場合、標的遺伝子、例えば、フェロモン応答遺伝子または胞子形成遺伝子を「機能的に破壊する」こと、またはそれの「機能的破壊」とは、標的遺伝子が、標的遺伝子によってコードされるタンパク質の活性を宿主細胞において減少させるような様式で変化していることを意味する。いくつかの実施形態において、標的遺伝子によってコードされるタンパク質の活性は、宿主細胞において除去されている。他の実施形態において、標的遺伝子によってコードされるタンパク質の活性は宿主細胞において減少している。標的遺伝子の機能的破壊は、遺伝子発現が除去され、もしくは低下するように、または遺伝子産物の活性が除去され、もしくは低下するように、遺伝子の全部または一部を欠失させることによって達成されてもよい。標的遺伝子の機能的破壊はまた、遺伝子の制御エレメント、例えば、遺伝子のプロモーターを、発現が除去され、もしくは低下するように突然変異することによって、または遺伝子のコード配列を、遺伝子産物の活性が除去され、もしくは低下するように突然変異することによって、達成されてもよい。いくつかの実施形態において、標的遺伝子の機能的破壊は、結果として、標的遺伝子の完全なオープンリーディングフレームの除去を生じる。
【0020】
本明細書に用いられる場合、「内因性接合」および「内因性接合能力」は、反対の接合型、すなわち、接合型aおよびαの一倍体微生物細胞が、異種性遺伝子発現、例えば、フェロモン応答遺伝子またはそのような一倍体の間で接合を誘導する能力がある任意の遺伝子の異種性コピーの発現の非存在下において、二倍体細胞を形成する能力を指す。
【0021】
本明細書に用いられる場合、「内因性接合が損なわれた」とは、野生型一倍体微生物細胞の集団に対して、そのような微生物細胞の一倍体の集団内での接合を阻害するのに十分な、微生物細胞の内因性接合能力の低下を指す。いくつかの実施形態において、阻害は、野生型一倍体微生物細胞の集団の接合率に対して一倍体微生物細胞の集団の接合率の少なくとも10%、20%、30%、40%、50%、60%、65%、70%、75%、80%、85%、90%、または95%の低下を含む。
【0022】
本明細書に用いられる場合、「胞子形成が損なわれた」とは、野生型二倍体微生物細胞の集団に対して、そのような微生物細胞の二倍体の集団内で胞子形成を阻害するのに十分な二倍体微生物細胞の胞子形成活性の低下を指す。いくつかの実施形態において、阻害は、野生型二倍体微生物細胞の集団の胞子形成率に対して二倍体微生物細胞の集団の胞子形成率の少なくとも10%、20%、30%、40%、50%、60%、65%、70%、75%、80%、85%、90%、または95%の低下を含む。
【0023】
本明細書に用いられる場合、遺伝子との関連における用語「相補すること」とは、機能的に破壊された遺伝子、例えば、機能的に破壊された胞子形成遺伝子またはフェロモン応答遺伝子の機能を置換する機能を有する遺伝子を指す。いくつかの実施形態において、相補遺伝子と破壊された遺伝子の間で機能の機構は同一である必要はない。いくつかの実施形態において、機能的に破壊されている標的遺伝子、例えば、胞子形成遺伝子またはフェロモン応答遺伝子は、破壊された遺伝子によってコードされるタンパク質と相同なタンパク質を産生するか、または例えば、別の機構によって胞子形成もしくは接合を可能にする表現型を提供するかのいずれかである異種性遺伝子によって相補することができる。
【0024】
(遺伝子改変微生物およびそれを作製する方法)
胞子形成および/または内因性接合能力において機能的に損なわれている遺伝子改変微生物、例えば、遺伝子改変酵母細胞(例えば、遺伝子改変Saccharomyces cerevisiae細胞)を含む組成物、ならびにそのような組成物を作製するための方法および材料が本明細書に提供される。本明細書に提供される方法は、微生物の有性生殖サイクルを中断させて、天然におけるその微生物の散在を最小限にし、遺伝子改変微生物と、天然において散在する能力が損なわれていない野生型微生物との間での遺伝物質の交換の可能性を最小限にする。
【0025】
多くの真菌細胞、例えば、酵母細胞は、有性的および無性的の両方で生殖することができる。無性生殖は、1つの親細胞のみが関係し、迅速な集団増殖を可能にする。対照的に、有性生殖は、配偶子の形成および融合を伴い、細胞間の水平遺伝子伝達による遺伝的多様性のより迅速な発生を可能にする。有性生殖する真菌細胞は、それらの生活環を通じて2つの細胞状態を想定し、1つが二倍体細胞状態であり、もう1つが一倍体細胞状態である。二倍体真菌細胞は、一般的に非常に安定であり、一般的に、それらが、いくつかの特定の環境的刺激(例えば、栄養枯渇)のうちの1つまたは複数に遭遇しない限り、二倍体相のままであるだろう。1つまたは複数のそのような刺激に遭遇した場合、二倍体細胞は胞子形成して、(四分子と呼ばれる)4つの一倍体胞子を形成する。都合良い条件が戻った場合、これらの一倍体胞子は出芽して4つの一倍体細胞(2つの接合型a、および2つの接合型α)を生成し、それは、その後、反対の接合型の他の一倍体細胞と接合して、再び二倍体細胞を形成することができる。
【0026】
二倍体真菌細胞の胞子形成する能力、および一倍体真菌細胞の接合する能力は、特定の遺伝子産物の機能に依存している。酵母細胞におけるこれらの中では、IME1、IME2、NDT80、SPO11、SPO20、AMA1、HOP2、およびSPO21遺伝子の産物などの胞子形成遺伝子の産物、ならびにSTE5、STE4、STE18、STE12、STE7、およびSTE11の産物などのフェロモン応答遺伝子の産物である。
【0027】
一態様において、胞子形成および/または内因性接合が損なわれており、および、目的のタンパク質をコードする染色体に組み込まれた異種性ヌクレオチド配列を含む遺伝子改変一倍体真菌細胞が本明細書に提供される。いくつかの実施形態において、遺伝子改変真菌細胞は、少なくとも1つの胞子形成遺伝子および/または少なくとも1つのフェロモン応答遺伝子が機能的に破壊されている一倍体酵母細胞である。
【0028】
いくつかの実施形態において、遺伝子改変真菌細胞は、以下の胞子形成遺伝子の1つまたは複数が機能的に破壊されている一倍体酵母細胞である:IME1、IME2、NDT80、SPO11、SPO20、AMA1、HOP2、およびSPO21。いくつかの実施形態において、遺伝子改変真菌細胞は、IME1遺伝子が機能的に破壊されている一倍体酵母細胞である。いくつかの実施形態において、遺伝子改変真菌細胞は、IME2遺伝子が機能的に破壊されている一倍体酵母細胞である。いくつかの実施形態において、遺伝子改変真菌細胞は、NDT80遺伝子が機能的に破壊されている一倍体酵母細胞である。いくつかの実施形態において、遺伝子改変真菌細胞は、SPO11遺伝子が機能的に破壊されている一倍体酵母細胞である。いくつかの実施形態において、遺伝子改変真菌細胞は、SPO20遺伝子が機能的に破壊されている一倍体酵母細胞である。いくつかの実施形態において、遺伝子改変真菌細胞は、AMA1遺伝子が機能的に破壊されている一倍体酵母細胞である。いくつかの実施形態において、遺伝子改変真菌細胞は、HOP2遺伝子が機能的に破壊されている一倍体酵母細胞である。いくつかの実施形態において、遺伝子改変真菌細胞は、SPO21遺伝子が機能的に破壊されている一倍体酵母細胞である。いくつかの実施形態において、遺伝子改変真菌細胞は、IME1、IME2、NDT80、SPO11、SPO20、AMA1、HOP2、およびSPO21からなる群より選択される1つより多い胞子形成遺伝子が破壊されている一倍体酵母細胞である。いくつかの実施形態において、遺伝子改変真菌細胞は、IME1、IME2、NDT80、SPO11、SPO20、AMA1、HOP2、およびSPO21からなる群より選択される2つより多い胞子形成遺伝子が破壊されている一倍体酵母細胞である。いくつかの実施形態において、遺伝子改変真菌細胞は、IME1、IME2、NDT80、SPO11、SPO20、AMA1、HOP2、およびSPO21からなる群より選択される3つより多い胞子形成遺伝子が破壊されている一倍体酵母細胞である。いくつかの実施形態において、遺伝子改変真菌細胞は、IME1、IME2、NDT80、SPO11、SPO20、AMA1、HOP2、およびSPO21からなる群より選択される4つより多い胞子形成遺伝子が破壊されている一倍体酵母細胞である。
【0029】
いくつかの実施形態において、遺伝子改変真菌細胞は、以下のフェロモン応答遺伝子の1つまたは複数が機能的に破壊されている一倍体酵母細胞である:STE5、STE4、STE18、STE12、STE7、およびSTE11。いくつかの実施形態において、遺伝子改変真菌細胞は、STE5遺伝子が機能的に破壊されている一倍体酵母細胞である。いくつかの実施形態において、遺伝子改変真菌細胞は、STE4遺伝子が機能的に破壊されている一倍体酵母細胞である。いくつかの実施形態において、遺伝子改変真菌細胞は、STE18遺伝子が機能的に破壊されている一倍体酵母細胞である。いくつかの実施形態において、遺伝子改変真菌細胞は、STE12遺伝子が機能的に破壊されている一倍体酵母細胞である。いくつかの実施形態において、遺伝子改変真菌細胞は、STE7遺伝子が機能的に破壊されている一倍体酵母細胞である。いくつかの実施形態において、遺伝子改変真菌細胞は、STE11遺伝子が機能的に破壊されている一倍体酵母細胞である。いくつかの実施形態において、遺伝子改変真菌細胞は、STE5、STE4、STE18、STE12、STE7、およびSTE11からなる群より選択される1つより多いフェロモン応答遺伝子が破壊されている一倍体酵母細胞である。いくつかの実施形態において、遺伝子改変真菌細胞は、STE5、STE4、STE18、STE12、STE7、およびSTE11からなる群より選択される2つより多いフェロモン応答遺伝子が破壊されている一倍体酵母細胞である。いくつかの実施形態において、遺伝子改変真菌細胞は、STE5、STE4、STE18、STE12、STE7、およびSTE11からなる群より選択される3つより多いフェロモン応答遺伝子が破壊されている一倍体酵母細胞である。いくつかの実施形態において、遺伝子改変真菌細胞は、STE5、STE4、STE18、STE12、STE7、およびSTE11からなる群より選択される4つより多いフェロモン応答遺伝子が破壊されている一倍体酵母細胞である。
【0030】
いくつかの実施形態において、遺伝子改変真菌細胞は、少なくとも1つの胞子形成遺伝子および少なくとも1つのフェロモン応答遺伝子が機能的に破壊されている一倍体酵母細胞である。
【0031】
いくつかの実施形態において、遺伝子改変真菌細胞は、IME1遺伝子およびSTE5遺伝子が機能的に破壊されている一倍体酵母細胞である。いくつかの実施形態において、遺伝子改変真菌細胞は、IME2遺伝子およびSTE5遺伝子が機能的に破壊されている一倍体酵母細胞である。いくつかの実施形態において、遺伝子改変真菌細胞は、NDT80遺伝子およびSTE5遺伝子が機能的に破壊されている一倍体酵母細胞である。いくつかの実施形態において、遺伝子改変真菌細胞は、SPO11遺伝子およびSTE5遺伝子が機能的に破壊されている一倍体酵母細胞である。いくつかの実施形態において、遺伝子改変真菌細胞は、SPO20遺伝子およびSTE5遺伝子が機能的に破壊されている一倍体酵母細胞である。いくつかの実施形態において、遺伝子改変真菌細胞は、AMA1遺伝子およびSTE5遺伝子が機能的に破壊されている一倍体酵母細胞である。いくつかの実施形態において、遺伝子改変真菌細胞は、HOP2遺伝子およびSTE5遺伝子が機能的に破壊されている一倍体酵母細胞である。いくつかの実施形態において、遺伝子改変真菌細胞は、SPO21遺伝子およびSTE5遺伝子が機能的に破壊されている一倍体酵母細胞である。
【0032】
いくつかの実施形態において、遺伝子改変真菌細胞は、IME1遺伝子およびSTE4遺伝子が機能的に破壊されている一倍体酵母細胞である。いくつかの実施形態において、遺伝子改変真菌細胞は、IME2遺伝子およびSTE4遺伝子が機能的に破壊されている一倍体酵母細胞である。いくつかの実施形態において、遺伝子改変真菌細胞は、NDT80遺伝子およびSTE4遺伝子が機能的に破壊されている一倍体酵母細胞である。いくつかの実施形態において、遺伝子改変真菌細胞は、SPO11遺伝子およびSTE4遺伝子が機能的に破壊されている一倍体酵母細胞である。いくつかの実施形態において、遺伝子改変真菌細胞は、SPO20遺伝子およびSTE4遺伝子が機能的に破壊されている一倍体酵母細胞である。いくつかの実施形態において、遺伝子改変真菌細胞は、AMA1遺伝子およびSTE4遺伝子が機能的に破壊されている一倍体酵母細胞である。いくつかの実施形態において、遺伝子改変真菌細胞は、HOP2遺伝子およびSTE4遺伝子が機能的に破壊されている一倍体酵母細胞である。いくつかの実施形態において、遺伝子改変真菌細胞は、SPO21遺伝子およびSTE4遺伝子が機能的に破壊されている一倍体酵母細胞である。
【0033】
いくつかの実施形態において、遺伝子改変真菌細胞は、IME1遺伝子およびSTE18遺伝子が機能的に破壊されている一倍体酵母細胞である。いくつかの実施形態において、遺伝子改変真菌細胞は、IME2遺伝子およびSTE18遺伝子が機能的に破壊されている一倍体酵母細胞である。いくつかの実施形態において、遺伝子改変真菌細胞は、NDT80遺伝子およびSTE18遺伝子が機能的に破壊されている一倍体酵母細胞である。いくつかの実施形態において、遺伝子改変真菌細胞は、SPO11遺伝子およびSTE18遺伝子が機能的に破壊されている一倍体酵母細胞である。いくつかの実施形態において、遺伝子改変真菌細胞は、SPO20遺伝子およびSTE18遺伝子が機能的に破壊されている一倍体酵母細胞である。いくつかの実施形態において、遺伝子改変真菌細胞は、AMA1遺伝子およびSTE18遺伝子が機能的に破壊されている一倍体酵母細胞である。いくつかの実施形態において、遺伝子改変真菌細胞は、HOP2遺伝子およびSTE18遺伝子が機能的に破壊されている一倍体酵母細胞である。いくつかの実施形態において、遺伝子改変真菌細胞は、SPO21遺伝子およびSTE18遺伝子が機能的に破壊されている一倍体酵母細胞である。
【0034】
いくつかの実施形態において、遺伝子改変真菌細胞は、IME1遺伝子およびSTE12遺伝子が機能的に破壊されている一倍体酵母細胞である。いくつかの実施形態において、遺伝子改変真菌細胞は、IME2遺伝子およびSTE12遺伝子が機能的に破壊されている一倍体酵母細胞である。いくつかの実施形態において、遺伝子改変真菌細胞は、NDT80遺伝子およびSTE12遺伝子が機能的に破壊されている一倍体酵母細胞である。いくつかの実施形態において、遺伝子改変真菌細胞は、SPO11遺伝子およびSTE12遺伝子が機能的に破壊されている一倍体酵母細胞である。いくつかの実施形態において、遺伝子改変真菌細胞は、SPO20遺伝子およびSTE12遺伝子が機能的に破壊されている一倍体酵母細胞である。いくつかの実施形態において、遺伝子改変真菌細胞は、AMA1遺伝子およびSTE12遺伝子が機能的に破壊されている一倍体酵母細胞である。いくつかの実施形態において、遺伝子改変真菌細胞は、HOP2遺伝子およびSTE12遺伝子が機能的に破壊されている一倍体酵母細胞である。いくつかの実施形態において、遺伝子改変真菌細胞は、SPO21遺伝子およびSTE12遺伝子が機能的に破壊されている一倍体酵母細胞である。
【0035】
いくつかの実施形態において、遺伝子改変真菌細胞は、IME1遺伝子およびSTE7遺伝子が機能的に破壊されている一倍体酵母細胞である。いくつかの実施形態において、遺伝子改変真菌細胞は、IME2遺伝子およびSTE7遺伝子が機能的に破壊されている一倍体酵母細胞である。いくつかの実施形態において、遺伝子改変真菌細胞は、NDT80遺伝子およびSTE7遺伝子が機能的に破壊されている一倍体酵母細胞である。いくつかの実施形態において、遺伝子改変真菌細胞は、SPO11遺伝子およびSTE7遺伝子が機能的に破壊されている一倍体酵母細胞である。いくつかの実施形態において、遺伝子改変真菌細胞は、SPO20遺伝子およびSTE7遺伝子が機能的に破壊されている一倍体酵母細胞である。いくつかの実施形態において、遺伝子改変真菌細胞は、AMA1遺伝子およびSTE7遺伝子が機能的に破壊されている一倍体酵母細胞である。いくつかの実施形態において、遺伝子改変真菌細胞は、HOP2遺伝子およびSTE7遺伝子が機能的に破壊されている一倍体酵母細胞である。いくつかの実施形態において、遺伝子改変真菌細胞は、SPO21遺伝子およびSTE7遺伝子が機能的に破壊されている一倍体酵母細胞である。
【0036】
いくつかの実施形態において、遺伝子改変真菌細胞は、IME1遺伝子およびSTE11遺伝子が機能的に破壊されている一倍体酵母細胞である。いくつかの実施形態において、遺伝子改変真菌細胞は、IME2遺伝子およびSTE11遺伝子が機能的に破壊されている一倍体酵母細胞である。いくつかの実施形態において、遺伝子改変真菌細胞は、NDT80遺伝子およびSTE11遺伝子が機能的に破壊されている一倍体酵母細胞である。いくつかの実施形態において、遺伝子改変真菌細胞は、SPO11遺伝子およびSTE11遺伝子が機能的に破壊されている一倍体酵母細胞である。いくつかの実施形態において、遺伝子改変真菌細胞は、SPO20遺伝子およびSTE11遺伝子が機能的に破壊されている一倍体酵母細胞である。いくつかの実施形態において、遺伝子改変真菌細胞は、AMA1遺伝子およびSTE11遺伝子が機能的に破壊されている一倍体酵母細胞である。いくつかの実施形態において、遺伝子改変真菌細胞は、HOP2遺伝子およびSTE11遺伝子が機能的に破壊されている一倍体酵母細胞である。いくつかの実施形態において、遺伝子改変真菌細胞は、SPO21遺伝子およびSTE11遺伝子が機能的に破壊されている一倍体酵母細胞である。
【0037】
いくつかの実施形態において、遺伝子改変真菌細胞は、1つまたは複数のフェロモン応答遺伝子の機能的に破壊された染色体コピー、および前記の1つまたは複数のフェロモン応答遺伝子のコード配列の機能性染色体外コピーを含む1つまたは複数の組換えプラスミドを含む一倍体酵母細胞である。いくつかの実施形態において、遺伝子改変真菌細胞は、機能的に破壊されたSTE5遺伝子、およびSTE5遺伝子のコード配列の機能性染色体外コピーを含む組換えプラスミドを含む一倍体酵母細胞である。いくつかの実施形態において、遺伝子改変真菌細胞は、機能的に破壊されたSTE4遺伝子、およびSTE4遺伝子のコード配列の機能性染色体外コピーを含む組換えプラスミドを含む一倍体酵母細胞である。いくつかの実施形態において、遺伝子改変真菌細胞は、機能的に破壊されたSTE18遺伝子、およびSTE18遺伝子のコード配列の機能性染色体外コピーを含む組換えプラスミドを含む一倍体酵母細胞である。いくつかの実施形態において、遺伝子改変真菌細胞は、機能的に破壊されたSTE12遺伝子、およびSTE12遺伝子のコード配列の機能性染色体外コピーを含む組換えプラスミドを含む一倍体酵母細胞である。いくつかの実施形態において、遺伝子改変真菌細胞は、機能的に破壊されたSTE7遺伝子、およびSTE7遺伝子のコード配列の機能性染色体外コピーを含む組換えプラスミドを含む一倍体酵母細胞である。いくつかの実施形態において、遺伝子改変真菌細胞は、機能的に破壊されたSTE11遺伝子、およびSTE11遺伝子のコード配列の機能性染色体外コピーを含む組換えプラスミドを含む一倍体酵母細胞である。
【0038】
別の態様において、胞子形成および/または内因性接合が損なわれており、および、目的のタンパク質をコードする染色体に組み込まれた異種性ヌクレオチド配列を2コピー含む遺伝子改変二倍体真菌細胞が本明細書に提供される。いくつかの実施形態において、遺伝子改変真菌細胞は、少なくとも1つの胞子形成遺伝子の両方のコピーおよび/または少なくとも1つのフェロモン応答遺伝子の両方のコピーが機能的に破壊された二倍体酵母細胞である。
【0039】
いくつかの実施形態において、遺伝子改変真菌細胞は、以下の胞子形成遺伝子の1つまたは複数の両方のコピーが機能的に破壊されている二倍体酵母細胞である:IME1、IME2、NDT80、SPO11、SPO20、AMA1、HOP2、およびSPO21。いくつかの実施形態において、遺伝子改変真菌細胞は、IME1遺伝子の両方のコピーが機能的に破壊されている二倍体酵母細胞である。いくつかの実施形態において、遺伝子改変真菌細胞は、以下のフェロモン応答遺伝子の1つまたは複数の両方のコピーが機能的に破壊されている二倍体酵母細胞である:STE5、STE4、STE18、STE12、STE7、およびSTE11。いくつかの実施形態において、遺伝子改変真菌細胞は、STE5遺伝子の両方のコピーが機能的に破壊されている二倍体酵母細胞である。さらに他の実施形態において、遺伝子改変真菌細胞は、少なくとも1つの胞子形成遺伝子の両方のコピーおよび少なくとも1つのフェロモン応答遺伝子の両方のコピーが機能的に破壊されている二倍体酵母細胞である。いくつかの実施形態において、IME1遺伝子の両方のコピーおよびSTE5遺伝子の両方のコピーが機能的に破壊されている二倍体酵母細胞である。
【0040】
いくつかの実施形態において、遺伝子改変二倍体真菌細胞は、それの接合型対立遺伝子について以外はホモ接合である。例えば、遺伝子改変二倍体真菌細胞が胞子形成するべきである場合には、生じた4つの一倍体真菌細胞は、それらの接合型対立遺伝子を除いて遺伝的に同一であるだろう。そのような事象において、一倍体細胞のうちの2つは、接合型aであり、残りの2つの一倍体細胞は接合型αであろう。いくつかの実施形態において、遺伝子改変二倍体真菌細胞は、抗生化合物に対する耐性を与える異種性遺伝子を含まない。
【0041】
本明細書に提供される遺伝子改変二倍体真菌細胞は、それに含まれる異種性ヌクレオチド配列の望まれない繁殖に対するいくつかの保護手段を有する。第一に、二倍体真菌細胞は、一般的に、非常に安定であり、それらの二倍体状態において、他の真菌細胞と接合できない。第二に、本明細書に提供される二倍体真菌細胞は、胞子形成する能力が損なわれており、したがって、適切な環境的刺激の存在下でさえも、胞子を形成する能力をほとんどもたない。第三に、胞子が形成される可能性が低い事象において、生じた一倍体真菌細胞は、接合する能力が損なわれている。総合すれば、本明細書に提供される遺伝子改変真菌細胞の胞子形成および接合の欠損した性質は、異種性ヌクレオチド配列の野生型真菌細胞への移動の可能性を著しく低下させる。
【0042】
別の態様において、本明細書に記載された遺伝子改変二倍体真菌細胞を作製する方法が本明細書に提供される。いくつかの実施形態において、該方法は以下の工程:(a)第1の遺伝子改変一倍体酵母細胞を得る工程であって、第1の遺伝子改変一倍体真菌細胞が胞子形成および/または内因性接合が損なわれており、および、目的のタンパク質をコードする、染色体に組み込まれた異種性ヌクレオチド配列を含む、工程;(b)第2の遺伝子改変一倍体真菌細胞を得る工程であって、第2の遺伝子改変一倍体真菌細胞が、胞子形成および内因性接合が損なわれており、および、第1の遺伝子改変一倍体真菌細胞と反対の接合型であり、および、目的の前記のタンパク質をコードする染色体に組み込まれた異種性ヌクレオチド配列を含む、工程;(c)第1の遺伝子改変一倍体真菌細胞および第2の遺伝子改変一倍体真菌細胞のそれぞれを、前記の第1および第2の遺伝子改変一倍体真菌細胞の内因性接合欠陥を相補することができるタンパク質をコードする1つまたは複数のプラスミドで形質転換する工程;(d)第1の遺伝子改変一倍体真菌細胞を第2の遺伝子改変一倍体真菌細胞と接合させ、それにより、遺伝子改変二倍体真菌細胞を形成する工程;ならびに(e)遺伝子改変二倍体真菌細胞から1つまたは複数のプラスミドを除去する工程であって、その結果生じた遺伝子改変二倍体真菌細胞が胞子形成および内因性接合が損なわれており、および、目的の前記のタンパク質をコードする染色体に組み込まれた異種性ヌクレオチド配列を2コピー含む、工程、を含む。
【0043】
いくつかの実施形態において、第1の遺伝子改変一倍体真菌細胞および第2の遺伝子改変一倍体真菌細胞が、1つまたは複数のフェロモン応答遺伝子の機能的破壊によって内因性接合が損なわれている。いくつかの実施形態において、本発明の方法の工程(c)は、第1の遺伝子改変一倍体真菌細胞および第2の遺伝子改変一倍体真菌細胞のそれぞれを、前記の第1および第2の遺伝子改変一倍体真菌細胞において機能的に破壊されている1つまたは複数のフェロモン応答遺伝子の機能性コピーをコードする1つまたは複数のプラスミドで形質転換する工程を含む。いくつかの実施形態において、第1および第2の遺伝子改変一倍体真菌細胞は、一倍体酵母細胞であり、1つまたは複数のフェロモン応答遺伝子は、STE5、STE4、STE18、STE12、STE7、およびSTE11からなる群より選択される。ある特定の実施形態において、第1および第2の遺伝子改変一倍体真菌細胞は、STE5遺伝子の機能的破壊によって内因性接合が損なわれている一倍体酵母細胞である。
【0044】
いくつかの実施形態において、第1の遺伝子改変一倍体真菌細胞および第2の遺伝子改変一倍体真菌細胞は、1つまたは複数の胞子形成遺伝子の機能的破壊によって胞子形成が損なわれている。いくつかの実施形態において、第1および第2の遺伝子改変一倍体真菌細胞は一倍体酵母細胞であり、1つまたは複数の胞子形成遺伝子は、IME1、IME2、NDT80、SPO11、SPO20、AMA1、HOP2、およびSPO21からなる群より選択される。いくつかの実施形態において、第1および第2の遺伝子改変一倍体真菌細胞は、IME1遺伝子の機能的破壊によって胞子形成が損なわれている一倍体酵母細胞である。特定の実施形態において、第1および第2の遺伝子改変一倍体真菌細胞は、STE5遺伝子の機能的破壊によって接合が損なわれており、および、IME1遺伝子の機能的破壊によって胞子形成が損なわれている一倍体酵母細胞である。
【0045】
いくつかの実施形態において、第2の遺伝子改変一倍体真菌細胞は、第1の遺伝子改変一倍体真菌細胞の集団において接合型切り換えを誘導することによって得られる。いくつかの実施形態において、第1の遺伝子改変一倍体真菌細胞はヘテロタリック(ho)一倍体Saccharomyces cerevisiae細胞であり、前記ヘテロタリック(ho)一倍体Saccharomyces cerevisiae細胞集団は、前記ヘテロタリック(ho)一倍体Saccharomyces cerevisiae細胞を、ホモタリズム(HO)タンパク質をコードするプラスミドで形質転換することによって接合型を切り換えるように誘導され、HOタンパク質の発現は、一倍体Saccharomyces cerevisiae細胞において接合型切り換えを誘導し、第2の遺伝子改変一倍体Saccharomyces cerevisiae細胞を生じる。ヘテロタリック(ho)一倍体Saccharomyces cerevisiae細胞は、自発的接合型切り換えが事実上、発生しない(たった10−6の頻度)ことを特徴とする。HOタンパク質を一過性に発現することによって、一倍体Saccharomyces cerevisiae細胞における自発的接合型切り換えの頻度は、多くも細胞分裂ごとに1回にまで増加することができ、お互いに接合して二倍体Saccharomyces cerevisiae細胞を生じることができる反対の接合型の一倍体細胞集団を提供する。
【0046】
別の態様において、胞子形成および内因性接合能力を欠く遺伝子改変ヘテロタリック(ho)二倍体酵母細胞を作製する方法が本明細書に提供され、該方法は以下の工程:(a)第1の遺伝子改変ヘテロタリック一倍体酵母細胞を得る工程であって、第1の遺伝子改変ヘテロタリック一倍体酵母細胞が(i)目的のタンパク質をコードする、染色体に組み込まれた異種性ヌクレオチド配列;および(ii)1つまたは複数の胞子形成遺伝子および1つまたは複数のフェロモン応答遺伝子における機能的破壊を含む、工程;(b)第1の遺伝子改変ヘテロタリック一倍体酵母細胞の集団を、ホモタリズム(HO)タンパク質をコードするプラスミドで形質転換して、第1の遺伝子改変一倍体酵母細胞を生じる工程であって、HOタンパク質の発現が第1の遺伝子改変一倍体酵母細胞において接合型切り換えを誘導し、それにより第2の遺伝子改変一倍体酵母細胞が得られ、第2の遺伝子改変一倍体酵母細胞が第1の遺伝子改変一倍体酵母細胞と反対の接合型であり、および、(i)目的の前記のタンパク質をコードする染色体に組み込まれた異種性ヌクレオチド配列;および(ii)1つまたは複数の胞子形成遺伝子および1つまたは複数のフェロモン応答遺伝子における機能的破壊を含む、工程;(c)第1の遺伝子改変一倍体酵母細胞および第2の遺伝子改変一倍体酵母細胞のそれぞれを、前記の第1および第2の遺伝子改変一倍体酵母細胞において機能的に破壊されている1つまたは複数のフェロモン応答タンパク質をコードするプラスミドで形質転換する工程;(d)第1の遺伝子改変一倍体酵母細胞を第2の遺伝子改変一倍体酵母細胞と接合させ、それにより、それの接合型対立遺伝子について以外はホモ接合である遺伝子改変二倍体酵母細胞を形成する工程;ならびに(e)遺伝子改変二倍体酵母細胞からいかなるプラスミドも除去して、遺伝子改変ヘテロタリック二倍体酵母細胞を生じる工程であって、その結果生じた遺伝子改変ヘテロタリック二倍体酵母細胞が胞子形成および内因性接合が損なわれており、および、目的の前記のタンパク質をコードする染色体に組み込まれた異種性ヌクレオチド配列を2コピー含む、工程、を含む。
【0047】
本明細書に提供され、下記でより詳細に記載される方法の工程は、経時的順序で提示されているが、当業者は、いくつかの工程の順序を、本発明の範囲を超えることなく、交換し、組み合わせ、または繰り返すことができることは認識しているであろう。したがって、いくつかの実施形態において、胞子形成および内因性接合能力を欠く遺伝子改変ヘテロタリック(ho)二倍体酵母細胞は、遺伝子改変ヘテロタリック一倍体酵母細胞を、前記の遺伝子改変ヘテロタリック一倍体酵母細胞において機能的に破壊されている1つまたは複数のフェロモン応答遺伝子をコードするプラスミドで形質転換し、その後、その細胞をホモタリズム(HO)タンパク質をコードするプラスミドで形質転換することによって作製される。他の実施形態において、胞子形成および内因性接合能力を欠く遺伝子改変ヘテロタリック(ho)二倍体酵母細胞は、遺伝子改変ヘテロタリック一倍体酵母細胞を、前記の遺伝子改変ヘテロタリック一倍体酵母細胞において機能的に破壊されている1つまたは複数のフェロモン応答タンパク質をコードするプラスミド、およびホモタリズム(HO)タンパク質をコードするプラスミドで同時に形質転換することによって作製される。
【0048】
(微生物選択)
本明細書に提供された方法の実施において有用な微生物には、真核単細胞生物、具体的には真菌、より具体的には酵母が挙げられる。
【0049】
いくつかの実施形態において、本発明に有用な酵母には、微生物受託機関(例えば、IFO、ATCCなど)に寄託されており、とりわけ、Aciculoconidium、Ambrosiozyma、Arthroascus、Arxiozyma、Ashbya、Babjevia、Bensingtonia、Botryoascus、Botryozyma、Brettanomyces、Bullera、Bulleromyces、Candida、Citeromyces、Clavispora、Cryptococcus、Cystofilobasidium、Debaryomyces、Dekkara、Dipodascopsis、Dipodascus、Eeniella、Endomycopsella、Eremascus、Eremothecium、Erythrobasidium、Fellomyces、Filobasidium、Galactomyces、Geotrichum、Guilliermondella、Hanseniaspora、Hansenula、Hasegawaea、Holtermannia、Hormoascus、Hyphopichia、Issatchenkia、Kloeckera、Kloeckeraspora、Kluyveromyces、Kondoa、Kuraishia、Kurtzmanomyces、Leucosporidium、Lipomyces、Lodderomyces、Malassezia、Metschnikowia、Mrakia、Myxozyma、Nadsonia、Nakazawaea、Nematospora、Ogataea、Oosporidium、Pachysolen、Phachytichospora、Phaffia、Pichia、Rhodosporidium、Rhodotorula、Saccharomyces、Saccharomycodes、Saccharomycopsis、Saitoella、Sakaguchia、Saturnospora、Schizoblastosporion、Schizosaccharomyces、Schwanniomyces、Sporidiobolus、Sporobolomyces、Sporopachydermia、Stephanoascus、Sterigmatomyces、Sterigmatosporidium、Symbiotaphrina、Sympodiomyces、Sympodiomycopsis、Torulaspora、Trichosporiella、Trichosporon、Trigonopsis、Tsuchiyaea、Udeniomyces、Waltomyces、Wickerhamia、Wickerhamiella、Williopsis、Yamadazyma、Yarrowia、Zygoascus、Zygosaccharomyces、Zygowilliopsis、およびZygozymaの属に属する酵母が挙げられる。
【0050】
いくつかの実施形態において、微生物は、Saccharomyces cerevisiae、Pichia pastoris、Schizosaccharomyces pombe、Dekkera bruxellensis、Kluyveromyces lactis(以前、Saccharomyces lactisと呼ばれた)、Kluveromyces marxianus、Arxula adeninivorans、またはHansenula polymorpha(現在、Pichia angustaとして知られている)である。いくつかの実施形態において、微生物は、Candida lipolytica、Candida guilliermondii、Candida krusei、Candida pseudotropicalis、またはCandida utilisなどのCandida属の株である。
【0051】
特定の実施形態において、微生物は、Saccharomyces cerevisiaeである。いくつかの実施形態において、微生物は、パン酵母、CBS 7959、CBS 7960、CBS 7961、CBS 7962、CBS 7963、CBS 7964、IZ−1904、TA、BG−1、CR−1、SA−1、M−26、Y−904、PE−2、PE−5、VR−1、BR−1、BR−2、ME−2、VR−2、MA−3、MA−4、CAT−1、CB−1、NR−1、BT−1、およびAL−1からなる群より選択されるSaccharomyces cerevisiaeの株である。いくつかの実施形態において、微生物は、PE−2、CAT−1、VR−1、BG−1、CR−1、およびSA−1からなる群より選択されるSaccharomyces cerevisiaeの株である。特定の実施形態において、Saccharomyces cerevisiaeの株はPE−2である。別の特定の実施形態において、Saccharomyces cerevisiaeの株はCAT−1である。別の特定の実施形態において、Saccharomyces cerevisiaeの株はBG−1である。
【0052】
いくつかの実施形態において、微生物は、産業的発酵、例えば、バイオエタノール発酵に適している微生物である。特定の実施形態において、微生物は、認識されている産業的発酵環境のストレス条件である、高溶媒濃度、高温、広範な基質利用、栄養制限、糖および塩による浸透圧ストレス、酸過剰、亜硫酸塩および細菌の汚染、またはそれらの組み合わせの下で存在するように条件付けされる。
【0053】
(微生物の遺伝子改変)
組換えプラスミドまたは染色体組込みベクターを用いて微生物を遺伝的に改変する方法は、当技術分野においてよく知られている。例えば、Sherman,F.ら、Methods Yeast Genetics、Cold Spring Harbor Laboratory、N.Y.(1978);Guthrie,C.ら(編)Guide To Yeast Genetics and Molecular Biology 194巻、Academic Press、San Diego(1991);Sambrookら、2001、Molecular Cloning−A Laboratory Manual、第3版、Cold Spring Harbor Laboratory、Cold Spring Harbor、NY;およびAusubelら編、現行版、Current Protocols in Molecular Biology、Greene Publishing Associates and Wiley Interscience、NYを参照;それらの開示は参照により本明細書に組み入れられている。
【0054】
いくつかの実施形態において、微生物は、新しい代謝経路、すなわち、微生物によって内因的に産生されない代謝産物を産生する代謝経路の酵素をコードする1つまたは複数の異種性ヌクレオチド配列を含むように遺伝子改変される。他の実施形態において、微生物は、微生物にとって内因性である代謝経路、すなわち、微生物によって内因的に産生される代謝産物を産生する代謝経路の酵素をコードする1つまたは複数の異種性ヌクレオチド配列を含むように遺伝子改変される。
【0055】
いくつかの実施形態において、本発明の方法は、フェロモン応答遺伝子の1つまたはHOによってコードされるものなどの特定のタンパク質を微生物において一過性に発現させるために組換えプラスミドの使用を必要とする。酵母細胞に用いるのに適した組換えプラスミドの例証的な例として、CEN/ARSおよび2μのプラスミドが挙げられる。
【0056】
いくつかの実施形態において、本発明の微生物は、抗生物質耐性をコードする異種性ヌクレオチド配列を含まない。抗生物質耐性マーカーは、遺伝子改変細胞の構築に一般的に用いられる。抗生物質耐性マーカーが、微生物に導入された遺伝子改変をマークするために用いられる本発明のそのような実施形態において、これらのマーカーは、所望の遺伝子改変の全部が微生物になされた後で、続いて欠失させられる。または、栄養要求性相補(例えば、HIS3、LEU2、LYS1、MET15、TRP1、ADE2、URA3、およびLYS2))などの他の選択ツールを遺伝子改変微生物の構築に用いることができる。
【0057】
(胞子形成および/またはフェロモン応答遺伝子の破壊)
本明細書に提供された方法は、遺伝子改変微生物細胞において1つもしくは複数の胞子形成遺伝子および/または1つもしくは複数のフェロモン応答遺伝子を機能的に破壊する工程を含む。いくつかの実施形態において、1つまたは複数の胞子形成遺伝子の破壊は、胞子形成能力を欠く遺伝子改変微生物細胞を生じる。特に、遺伝子改変二倍体微生物細胞は胞子形成能力を欠く。いくつかの実施形態において、1つまたは複数のフェロモン応答遺伝子の破壊は、内因性接合が損なわれている微生物細胞を生じる。いくつかの実施形態において、1つまたは複数の胞子形成遺伝子の破壊および1つまたは複数のフェロモン応答遺伝子の破壊は、胞子形成および内因性接合が損なわれている微生物細胞を生じる。
【0058】
いくつかの実施形態において、胞子形成遺伝子またはフェロモン応答遺伝子の破壊は、微生物細胞への構築物の導入で胞子形成標的遺伝子またはフェロモン応答標的遺伝子を特異的に破壊する能力がある「破壊構築物」を用いて、破壊される遺伝子を非機能性にすることによって達成される。いくつかの実施形態において、標的遺伝子の破壊は、機能性タンパク質の発現を阻止する。いくつかの実施形態において、標的遺伝子の破壊は、破壊される遺伝子からの非機能性タンパク質の発現を生じる。いくつかの実施形態において、胞子形成標的遺伝子またはフェロモン応答標的遺伝子の破壊は、相同組換えにより標的遺伝子座内の「破壊配列」の組込みによって達成される。そのような実施形態において、破壊構築物は、標的遺伝子座の1対のヌクレオチド配列に相同である1対のヌクレオチド配列(相同配列)に隣接した破壊配列を含む。破壊構築物による標的遺伝子の標的部分の置換で、破壊配列は、機能性タンパク質の発現を阻止し、または標的遺伝子からの非機能性タンパク質の発現を引き起こす。
【0059】
1つまたは複数の胞子形成遺伝子またはフェロモン応答遺伝子を破壊する能力がある破壊構築物は、当技術分野において周知の標準の分子生物学技術を用いて構築することができる。例えば、Sambrookら、2001、Molecular Cloning−A Laboratory Manual、第3版、Cold Spring Harbor Laboratory、Cold Spring Harbor、NYおよびAusubelら編、現行版、Current Protocols in Molecular Biology、Greene Publishing Associates and Wiley Interscience、NYを参照。本方法の実施において変わり得る破壊構築物のパラメータには、相同配列の長さ、相同配列のヌクレオチド配列、破壊配列の長さ、破壊配列のヌクレオチド配列、および標的遺伝子のヌクレオチド配列が挙げられるが、それらに限定されない。いくつかの実施形態において、各相同配列の長さについての有効な範囲は50〜5,000塩基対である。特定の実施形態において、各相同配列の長さは約500塩基対である。遺伝子を標的にするのに必要とされる相同性の長さの議論については、Hastyら、Mol Cell Biol 11:5586−91(1991)を参照されたい。いくつかの実施形態において、相同配列は、標的遺伝子のコード配列を含む。他の実施形態において、相同配列は、標的遺伝子の上流配列または下流配列を含む。いくつかの実施形態において、一方の相同配列は、標的遺伝子のコード配列の5’に位置するヌクレオチド配列と相同であるヌクレオチド配列を含み、他方の相同配列は、標的遺伝子のコード配列の3’に位置するヌクレオチド配列と相同であるヌクレオチド配列を含む。いくつかの実施形態において、破壊配列は、破壊配列を含む微生物細胞の選択を可能にする選択マーカーをコードするヌクレオチド配列を含む。したがって、そのような実施形態において、破壊構築物は、二重機能、すなわち、標的遺伝子を機能的に破壊すること、および標的遺伝子が機能的に破壊されている細胞の同定のための選択マーカーを提供することを有する。いくつかの実施形態において、終止コドンは、標的遺伝子によってコードされる野生型タンパク質のある程度の活性を有する融合タンパク質を生じる可能性がある翻訳の読み過ごしを防ぐために選択マーカーをコードするヌクレオチド配列とインフレームで、および、それの下流に位置する。いくつかの実施形態において、破壊配列の長さは、1塩基対である。単一塩基対の挿入は、コード配列における単一塩基対の挿入が、機能性タンパク質の発現を阻止することができるフレームシフト突然変異を構成することができるため、標的遺伝子を破壊するのに十分であり得る。いくつかの実施形態において、破壊配列の配列は、相同配列の間に位置する標的遺伝子のヌクレオチド配列と単一塩基対だけ、異なる。標的遺伝子内のヌクレオチド配列の破壊配列との置換で、導入される単一塩基対置換は、タンパク質における重要部位での単一アミノ酸置換、および非機能性タンパク質の発現を生じ得る。しかしながら、非常に短い破壊配列を用いてもたらされる破壊は、自然突然変異によって野生型配列への復帰変異を起こしやすく、したがって、接合および胞子形成能力の回復を宿主株にもたらすことは認識されるべきである。したがって、特定の実施形態において、破壊配列は、1〜2、3塩基対より長い。それとは正反対に、過剰な長さの破壊配列は、中程度の長さの破壊配列に優る少しの利点でも与える可能性は低く、トランスフェクションまたはターゲティングの効率を減少させる可能性がある。この関連における過剰な長さは、標的遺伝子における選択された相同配列間の距離より何倍も長い。このように、ある特定の実施形態において、破壊配列についての長さは、2塩基対から2,000塩基対までであり得る。他の実施形態において、破壊配列についての長さは、破壊構築物における相同配列とマッチする標的遺伝子座の領域間の距離とおよそ等しい長さである。
【0060】
いくつかの実施形態において、破壊構築物は、直鎖DNA分子である。他の実施形態において、破壊構築物は、環状DNA分子である。いくつかの実施形態において、環状破壊構築物は、上記のような破壊配列によって分離した1対の相同配列を含む。いくつかの実施形態において、環状破壊構築物は、単一の相同配列を含む。そのような環状破壊構築物は、標的遺伝子座での組込みで、直鎖状になり、相同配列の部分は各末端に位置し、破壊構築物の残りのセグメントは、標的遺伝子ヌクレオチド配列のいずれとも置換することなく、標的遺伝子に挿入され、それを破壊するであろう。特定の実施形態において、環状破壊構築物の単一の相同配列は、標的遺伝子のコード配列内に位置する配列と相同である。
【0061】
破壊構築物は、非限定的に、当業者に公知の任意の方法によって微生物細胞へ導入することができる。そのような方法には、溶液からの細胞による分子の直接的取り込み、または例えば、リポソームもしくは免疫リポソームを用いるリポフェクションによる促進性取り込み;粒子媒介性トランスフェクションなどが挙げられるが、それらに限定されない。例えば、米国特許第5,272,065号;Goeddelら編、1990、Methods in Enzymology、185巻、Academic Press, Inc.、CA;Krieger、1990、Gene Transfer and Expression−A Laboratory Manual、Stockton Press、NY;Sambrookら、1989、Molecular Cloning−A Laboratory Manual、Cold Spring Harbor Laboratory、NY;およびAusubelら編、現行版、Current Protocols in Molecular Biology、Greene Publishing Associates and Wiley Interscience、NYを参照。酵母細胞を形質転換する特定の方法は当技術分野において周知である。Hinnenら、Proc.Natl.Acad.Sci.USA 75:1292−3(1978);Creggら、Mol.Cell.Biol.5:3376−3385(1985)参照。例示的な技術として、スフェロプラスト化、エレクトロポレーション、PEG 1000媒介性形質転換、および酢酸リチウムまたは塩化リチウム媒介性形質転換が挙げられるが、それらに限定されない。
【0062】
(フェロモン応答遺伝子)
いくつかの実施形態において、本明細書に提供された方法に従って破壊される酵母細胞におけるフェロモン応答遺伝子は、STE5である。STE5は、酵母接合経路を通してのマイトジェン活性化プロテインキナーゼ(MAPK)への直接的シグナル伝達に必要とされるスキャフォールドタンパク質をコードする。例えば、Goodら、Cell Mar 20;136(6):1085−97(2009)参照。Saccharomyces cerevisiaeの代表的なSTE5ヌクレオチド配列には、Genbankアクセッション番号L23856、ならびに本明細書に提供されているような配列番号17、45、73、101、および129が挙げられる。Saccharomyces cerevisiaeの代表的なSte5タンパク質配列には、Genbankアクセッション番号AAA35115、ならびに本明細書に提供されているような配列番号18、46、74、102、および130が挙げられる。
【0063】
いくつかの実施形態において、本明細書に提供された方法に従って破壊される酵母細胞におけるフェロモン応答遺伝子は、STE4である。STE4遺伝子は、Ste18pと二量体を形成して接合シグナル伝達経路を活性化するGタンパク質βサブユニットをコードする。Saccharomyces cerevisiaeのSTE4遺伝子の配列は以前に記載されている。Dujonら、Nature 387(6632 Suppl):98−102(1997)。Saccharomyces cerevisiaeの代表的なSTE4ヌクレオチド配列には、Genbankアクセッション番号NC_001147.5、ならびに本明細書に提供されているような配列番号19、47、75、103、および131が挙げられる。Saccharomyces cerevisiaeの代表的なSte4タンパク質配列には、Genbankアクセッション番号NP_014855、ならびに本明細書に提供されているような配列番号20、48、76、104、および132が挙げられる。
【0064】
いくつかの実施形態において、本明細書に提供された方法に従って破壊される酵母細胞におけるフェロモン応答遺伝子は、STE18である。STE18遺伝子は、Ste4pと二量体を形成して接合シグナル伝達経路を活性化するGタンパク質γサブユニットをコードする。Saccharomyces cerevisiaeのSTE18遺伝子の配列は以前に記載されている。例えば、Goffeauら、Science 274(5287):546−547(1996)参照。Saccharomyces cerevisiaeの代表的なSTE18ヌクレオチド配列には、Genbankアクセッション番号NC_001147.5、ならびに本明細書に提供されているような配列番号21、49、77、105、および133が挙げられる。Saccharomyces cerevisiaeの代表的なSte18タンパク質配列には、Genbankアクセッション番号NP_012619、ならびに本明細書に提供されているような配列番号22、50、78、106、および134が挙げられる。
【0065】
いくつかの実施形態において、本明細書に提供された方法に従って破壊される酵母細胞におけるフェロモン応答遺伝子は、STE12である。STE12遺伝子は、MAPキナーゼシグナル伝達カスケードによって活性化され、および、接合経路または仮性菌糸/浸潤性増殖経路に関与する遺伝子を活性化する転写因子をコードする。Saccharomyces cerevisiaeのSTE12遺伝子の配列は以前に記載されている。例えば、Goffeauら、Science 274(5287):546−547(1996)参照。Saccharomyces cerevisiaeの代表的なSTE12ヌクレオチド配列には、Genbankアクセッション番号NC_001140.5、ならびに本明細書に提供されているような配列番号23、51、79、107、および135が挙げられる。Saccharomyces cerevisiaeの代表的なSte12タンパク質配列には、Genbankアクセッション番号NP_011952、ならびに本明細書に提供されているような配列番号24、52、80、108、および136が挙げられる。
【0066】
いくつかの実施形態において、本明細書に提供された方法に従って破壊される酵母細胞におけるフェロモン応答遺伝子は、STE7である。STE7遺伝子は、Fus3pをリン酸化する、フェロモン応答に関与するシグナル伝達MAPキナーゼをコードする。Saccharomyces cerevisiaeのSTE7遺伝子の配列は以前に記載されている。例えば、Teagueら、Proc Natl Acad Sci USA.83(19):7371−5(1986)参照。Saccharomyces cerevisiaeの代表的なSTE7ヌクレオチド配列には、Genbankアクセッション番号Z74207、ならびに本明細書に提供されているような配列番号25、53、81、109、および137が挙げられる。Saccharomyces cerevisiaeの代表的なSte7タンパク質配列には、Genbankアクセッション番号CAA98732、ならびに本明細書に提供されているような配列番号26、54、82、110、および138が挙げられる。
【0067】
いくつかの実施形態において、本明細書に提供された方法に従って破壊される酵母細胞におけるフェロモン応答遺伝子は、STE11である。STE11遺伝子は、Ste7pをリン酸化する、フェロモン応答および仮性菌糸/浸潤性増殖経路に関与するシグナル伝達MEKキナーゼをコードする。Saccharomyces cerevisiaeのSTE11遺伝子の配列は以前に記載されている。例えば、Johnstonら、Nature 387(6632 Suppl)、87−90(1997)参照。Saccharomyces cerevisiaeの代表的なSTE11ヌクレオチド配列には、Genbankアクセッション番号NC_001144.4、ならびに本明細書に提供されているような配列番号27、55、83、111、および139が挙げられる。Saccharomyces cerevisiaeの代表的なSte11タンパク質配列には、Genbankアクセッション番号NP_013466、ならびに本明細書に提供されているような配列番号28、56、84、112、および140が挙げられる。
【0068】
(胞子形成遺伝子)
いくつかの実施形態において、本明細書に提供された方法に従って破壊される酵母細胞における胞子形成遺伝子はIME1である。IME1遺伝子は、減数分裂の開始に必要とされる、初期減数分裂遺伝子転写を活性化する転写因子をコードする。Saccharomyces cerevisiaeのIME1遺伝子の配列は以前に記載されている。例えば、Smith,H.E.ら、Mol.Cell.Biol.10(12):6103−6113(1990)参照。Saccharomyces cerevisiaeの代表的なIME1ヌクレオチド配列には、Genbankアクセッション番号M37188、ならびに本明細書に提供されているような配列番号1、29、57、85、および113が挙げられる。Saccharomyces cerevisiaeの代表的なIme1タンパク質配列には、Genbankアクセッション番号AAA86790、ならびに本明細書に提供されているような配列番号2、30、58、86、および114が挙げられる。
【0069】
いくつかの実施形態において、本明細書に提供された方法に従って破壊される酵母細胞における胞子形成遺伝子はIME2である。IME2遺伝子は、減数分裂の活性化に関与するセリン/スレオニンプロテインキナーゼをコードする。Saccharomyces cerevisiaeのIME2遺伝子の配列は以前に記載されている。例えば、EMBO J.15(9)、2031−2049(1996)参照。Saccharomyces cerevisiaeの代表的なIME2ヌクレオチド配列には、Genbankアクセッション番号NC_001142、ならびに本明細書に提供されているような配列番号3、31、59、87、および115が挙げられる。Saccharomyces cerevisiaeの代表的なIme2タンパク質配列には、Genbankアクセッション番号NP_012429、ならびに本明細書に提供されているような配列番号4、32、60、88、および116が挙げられる。
【0070】
いくつかの実施形態において、本明細書に提供された方法に従って破壊される酵母細胞における胞子形成遺伝子はNDT80である。NDT80遺伝子は、太糸期からの脱出および完全減数分裂組換えに必要とされる減数分裂特異的転写因子をコードする。Ndt80タンパク質はまた、中間胞子形成遺伝子を活性化する。Saccharomyces cerevisiaeのNDT80遺伝子の配列は以前に記載されている。例えば、Goffeauら、Science 274(5287):546−547(1996)参照。Saccharomyces cerevisiaeの代表的なNDT80ヌクレオチド配列には、Genbankアクセッション番号NC_001140、ならびに本明細書に提供されているような配列番号5、33、61、89、および117が挙げられる。Saccharomyces cerevisiaeの代表的なNdt80タンパク質配列には、Genbankアクセッション番号NP_011992、ならびに本明細書に提供されているような配列番号6、34、62、90、および118が挙げられる。
【0071】
いくつかの実施形態において、本明細書に提供された方法に従って破壊される酵母細胞における胞子形成遺伝子はSPO11である。SPO11遺伝子は、減数分裂組換えに必要とされる。Saccharomyces cerevisiaeのSPO11遺伝子の配列は以前に記載されている。例えば、Atchesonら、Proc.Natl.Acad.Sci.U.S.A. 84(22)、8035−8039(1987)参照。Saccharomyces cerevisiaeの代表的なSPO11ヌクレオチド配列には、Genbankアクセッション番号J02987、ならびに本明細書に提供されているような配列番号7、35、63、91、および119が挙げられる。Saccharomyces cerevisiaeの代表的なSpo11タンパク質配列には、Genbankアクセッション番号AAA65532、ならびに本明細書に提供されているような配列番号8、36、64、92、および120が挙げられる。
【0072】
いくつかの実施形態において、本明細書に提供された方法に従って破壊される酵母細胞における胞子形成遺伝子はSPO20である。SPO20遺伝子は、胞子形成中の前胞子膜形成に必要とされる、t−SNARE複合体の減数分裂特異的サブユニットをコードする。Saccharomyces cerevisiaeのSPO20遺伝子の配列は以前に記載されている。例えば、Bowmanら、Nature 387(6632 Suppl)、90−93(1997)参照。Saccharomyces cerevisiaeの代表的なSPO20ヌクレオチド配列には、Genbankアクセッション番号AF078740、ならびに本明細書に提供されているような配列番号9、37、65、93、および121が挙げられる。Saccharomyces cerevisiaeの代表的なSpo20タンパク質配列には、Genbankアクセッション番号NP_013730、ならびに本明細書に提供されているような配列番号10、38、66、94、および122が挙げられる。
【0073】
いくつかの実施形態において、本明細書に提供された方法に従って破壊される酵母細胞における胞子形成遺伝子はAMA1である。AMA1遺伝子は、減数分裂後期促進複合体のアクチベーターをコードする。Saccharomyces cerevisiaeのAMA1遺伝子の配列は以前に記載されている。例えば、Tettelinら、Nature 387(6632 Suppl):81−84(1997)参照。Saccharomyces cerevisiaeの代表的なAMA1ヌクレオチド配列には、Genbankアクセッション番号NC_001139.8、ならびに本明細書に提供されているような配列番号11、39、67、95、および123が挙げられる。Saccharomyces cerevisiaeの代表的なAma1タンパク質配列には、Genbankアクセッション番号NP_011741、ならびに本明細書に提供されているような配列番号12、40、68、96、および124が挙げられる。
【0074】
いくつかの実施形態において、本明細書に提供された方法に従って破壊される酵母細胞における胞子形成遺伝子はHOP2である。HOP2遺伝子は、相同染色体間のシナプシスを確実にする減数分裂特異的タンパク質をコードする。Saccharomyces cerevisiaeのHOP2遺伝子の配列は以前に記載されている。例えば、Leuら、Cell 94(3):375−386(1998)参照。Saccharomyces cerevisiaeの代表的なHOP2ヌクレオチド配列には、Genbankアクセッション番号AF_078740.1、ならびに本明細書に提供されているような配列番号13、41、69、97、および125が挙げられる。Saccharomyces cerevisiaeの代表的なHop2タンパク質配列には、Genbankアクセッション番号AAC31823、ならびに本明細書に提供されているような配列番号14、42、70、98、および126が挙げられる。
【0075】
いくつかの実施形態において、本明細書に提供された方法に従って破壊される酵母細胞における胞子形成遺伝子はSPO21である。SPO21遺伝子は、前胞子膜形成の前に必要とされる減数分裂外側プラーク(meiotic outer plaque)を修飾することに関与する、紡錘極体の減数分裂外側プラークの成分をコードする。Saccharomyces cerevisiaeのSPO21遺伝子の配列は以前に記載されている。例えば、Dujonら、Nature 387(6632 Suppl):98−102(1997)参照。Saccharomyces cerevisiaeの代表的なSPO21ヌクレオチド配列には、Genbankアクセッション番号NC_001147.5、ならびに本明細書に提供されているような配列番号15、43、71、99、および127が挙げられる。Saccharomyces cerevisiaeの代表的なSpo21タンパク質配列には、Genbankアクセッション番号NP_014550、ならびに本明細書に提供されているような配列番号16、44、72、100、および128が挙げられる。
【0076】
(二倍体の調製)
本明細書に提供された方法は、1つもしくは複数の胞子形成遺伝子における機能的破壊および/または1つまたは複数のフェロモン応答遺伝子における機能的破壊を含む一倍体細胞の間で接合を誘導する工程を含む。前記接合の結果として形成される二倍体細胞は、細胞の1つまたは複数の胞子形成遺伝子の機能的破壊による二倍体相に制約される安定な二倍体細胞である。
【0077】
接合能力を欠く一倍体細胞から二倍体細胞を形成するために、接合欠陥性一倍体細胞は、「接合相補プラスミド」、すなわち、1つまたは複数のフェロモン応答遺伝子における機能的破壊によって引き起こされた接合欠損を相補することができる異種性遺伝子を含む組換えプラスミドで形質転換される。一倍体細胞内の異種性フェロモン応答遺伝子の一過性発現は、一時的に、細胞に接合機能を回復させ、反対接合型の一倍体細胞が安定な二倍体細胞を形成することを可能にする。特に、それによって形成された安定な二倍体細胞は、それらの接合型対立遺伝子について以外はホモ接合であり、同じ遺伝子改変集団の一倍体から生成される。
【0078】
したがって、一倍体細胞がSTE5遺伝子の機能的破壊を含むいくつかの実施形態において、一倍体細胞は、STE5コード配列を含む接合相補プラスミドで形質転換される。一倍体細胞がSTE4遺伝子の機能的破壊を含むいくつかの実施形態において、一倍体細胞は、STE4コード配列を含む接合相補プラスミドで形質転換される。一倍体細胞がSTE18遺伝子の機能的破壊を含むいくつかの実施形態において、一倍体細胞は、STE18コード配列を含む接合相補プラスミドで形質転換される。一倍体細胞がSTE12遺伝子の機能的破壊を含むいくつかの実施形態において、一倍体細胞は、STE12コード配列を含む接合相補プラスミドで形質転換される。一倍体細胞がSTE7遺伝子の機能的破壊を含むいくつかの実施形態において、一倍体細胞は、STE7コード配列を含む接合相補プラスミドで形質転換される。一倍体細胞がSTE11遺伝子の機能的破壊を含むいくつかの実施形態において、一倍体細胞は、STE11コード配列を含む接合相補プラスミドで形質転換される。
【0079】
発現ベクターの構築、および発現ベクターを含む細胞における遺伝子の発現についての技術は、当技術分野において周知である。例えば、Sambrookら、2001、Molecular Cloning−A Laboratory Manual、第3版、Cold Spring Harbor Laboratory、Cold Spring Harbor、NY、およびAusubelら編、現行版、Current Protocols in Molecular Biology、Greene Publishing Associates and Wiley Interscience、NYを参照。接合相補遺伝子をコードするプラスミドは、当業者に公知の任意の方法によって宿主細胞へ導入することができる。
【0080】
プラスミドに基づいた系は、一般的に、細胞において外来DNAを維持するようにプラスミドに選択圧を必要とする。酵母細胞は典型的には、有糸分裂後ごとに、細胞に含まれる10%のプラスミドを喪失するため、酵母におけるたいていのプラスミドは相対的に不安定である。したがって、いくつかの実施形態において、接合相補遺伝子をコードするプラスミドを含む一倍体細胞の接合によって形成されたが、それ自体、そのプラスミドを含まない二倍体細胞の選択は、プラスミドが集団から効果的に希釈されるように十分な有糸分裂を二倍体細胞に起こさせることによって達成される。あるいは、二倍体細胞は、プラスミドの非存在について選択することによって、例えば、対抗選択マーカー(例えば、URA3など)に対して選択することによって、または選択培地および非選択培地の両方に同一のコロニーを蒔き、その後、選択培地で増殖しないが、非選択培地で増殖するコロニーを選択することによって、選択することができる。
【0081】
いくつかの実施形態において、本明細書に提供される方法は、一倍体ヘテロタリック(ho)酵母細胞を、ホモタリズム(HO)タンパク質をコードする組換えプラスミドで形質転換する工程であって、HOタンパク質の発現が一倍体細胞の接合型切り換えを誘導する、工程を含む。Saccharomyces cerevisiaeのHO遺伝子の配列は以前に記載されている。例えば、Russellら、Mol.Cell.Biol. 6(12):4281−4294(1986)参照。Saccharomyces cerevisiaeの代表的なHOヌクレオチド配列には、Genbankアクセッション番号NC_001136および本明細書に提供されているような配列番号151が挙げられる。Saccharomyces cerevisiaeの代表的なHOタンパク質配列には、Genbankアクセッション番号NP_010054および本明細書に提供されているような配列番号152が挙げられる。
【実施例】
【0082】
実施例1:遺伝子改変二倍体細胞の作製
この実施例は、遺伝子改変一倍体S.cerevisiae細胞を作製する例示的な方法を記載する。
【0083】
相I破壊構築物(図1;配列番号141)は、破壊配列として、S.cerevisiae GAL80座の上流ヌクレオチド配列および下流ヌクレオチド配列からなる相同配列に隣接した、選択マーカー(ハイグロマイシンB耐性を与えるhygA);ガラクトース誘導性プロモーター(S.cerevisiae遺伝子GAL1およびGAL10のプロモーター)の制御下の、S.cerevisiae MEV経路の2つの酵素(切断型HMG−CoAリダクターゼをコードする切断型HMG1コード配列およびHMG−CoAシンターゼをコードするERG13コード配列)およびS.cerevisiaeの別の酵素(アセトアセチル−CoAチオラーゼをコードするERG10コード配列)をコードするヌクレオチド配列を含む。S.cerevisiae宿主細胞への導入で、相I破壊構築物は、S.cerevisiae宿主細胞ゲノムのGAL80座へ相同組換えによって組み込まれ、GAL80コード配列をその破壊配列と置換することによってGAL80座を機能的に破壊することができる。相I破壊構築物を、TOPO Zero Blunt IIクローニングベクター(Invitrogen、Carlsbad、CA)へクローニングし、プラスミドTOPO−相I破壊構築物を生じた。その構築物を、50μg/mlのカナマイシンを含むLB寒天上で増殖するTOP10細胞中に繁殖させた。
【0084】
相II破壊構築物(図2;配列番号142)は、破壊配列として、S.cerevisiae LEU2座の上流ヌクレオチド配列および下流ヌクレオチド配列からなる相同配列に隣接した、選択マーカー(ノールセオスリシン(nourseothricin)耐性を与えるnatA)、およびガラクトース誘導性プロモーター(S.cerevisiae遺伝子GAL1およびGAL10のプロモーター)の制御下のS.cerevisiae MEV経路の数個の酵素(メバロン酸キナーゼをコードするERG12コード配列およびホスホメバロン酸キナーゼをコードするERG8コード配列)をコードするヌクレオチド配列;加えて、GAL4ocプロモーター(MIG1結合部位を除去する突然変異を含むGAL4プロモーターであって、それにしたがって、プロモーターをグルコースの抑圧に対してより低感受性にさせている)の制御下のS.cerevisiae GAL4遺伝子のコード配列を含む。S.cerevisiae宿主細胞への導入で、相II破壊構築物は、S.cerevisiae宿主細胞ゲノムのLEU2座へ相同組換えによって組み込まれ、LEU2コード配列をその破壊配列と置換することによってLEU2座を機能的に破壊することができる。相II破壊構築物を、TOPO Zero Blunt IIクローニングベクターへクローニングし、プラスミドTOPO−相II破壊構築物を生じた。その構築物を、50μg/mlのカナマイシンを含むLB寒天上で増殖するTOP10細胞(Invitrogen、Carlsbad、CA)中に繁殖させた。
【0085】
相III破壊構築物(図3;配列番号143)は、破壊配列として、S.cerevisiae ERG9座の上流ヌクレオチド配列およびコードヌクレオチド配列に隣接した、選択マーカー(G418耐性を与えるkanA);S.cerevisiae MEV経路の酵素(ジホスホメバロン酸デカルボキシラーゼをコードするERG19コード配列)、およびガラクトース誘導性プロモーター(S.cerevisiae遺伝子GAL1、GAL10、およびGAL7のプロモーター)の制御下の、MEV経路の産物のIPPをFPPへ変換することに関与するS.cerevisiaeの2つの酵素(ERG20コード配列はファルネシルピロリン酸シンターゼをコードし、IDI1コード配列はイソペンテニルピロリン酸デカルボキシラーゼをコードする)をコードするヌクレオチド配列;加えて、S.cerevisiae CTR3遺伝子のプロモーターを含む。S.cerevisiae宿主細胞への導入で、相II破壊構築物は、S.cerevisiae宿主細胞ゲノムのERG9座の上流に相同組換えによって組み込まれ、ERG9(スクワレンシンターゼ)の発現が銅によって調節され得るように天然ERG9プロモーターをCTR3プロモーターで置換することができる。相III破壊構築物を、TOPO Zero Blunt IIクローニングベクターへクローニングし、プラスミドTOPO−相III破壊構築物を生じた。その構築物を、50μg/mlのカナマイシンを含むLB寒天上で増殖するTOP10細胞中に繁殖させた。
【0086】
相Iマーカーリサイクリング構築物(図4;配列番号144)は、S.cerevisiae GAL80座の上流ヌクレオチド配列およびS.cerevisiae HMG1遺伝子のコード配列に隣接した、選択マーカー(ウラシル欠損培地で増殖する能力を与えるURA3);およびS.cerevisiae GAL7遺伝子のプロモーターの制御調節下のA.annuaの酵素(ファルネセンシンターゼをコードするFSコード配列)をコードするヌクレオチド配列を含む。S.cerevisiae宿主細胞への導入で、相Iマーカーリサイクリング構築物は、選択マーカーhphAがURA3と置換されるように、すでに組み込まれた相I破壊配列へ相同組換えによって組み込むことができる。
【0087】
相IIマーカーリサイクリング構築物(図5;配列番号145)は、S.cerevisiae LEU2座の上流ヌクレオチド配列およびS.cerevisiae ERG12遺伝子のコード配列に隣接した、選択マーカー(ウラシル欠損培地で増殖する能力を与えるURA3)およびS.cerevisiae GAL7遺伝子のプロモーターの制御調節下のA annuaの酵素(ファルネセンシンターゼをコードするFSコード配列)をコードするヌクレオチド配列を含む。S.cerevisiae宿主細胞への導入で、相IIマーカーリサイクリング構築物は、選択マーカーnatAがURA3と置換されるように、すでに組み込まれた相II破壊配列へ相同組換えによって組み込むことができる。
【0088】
相IIIマーカーリサイクリング構築物(図6;配列番号146)は、S.cerevisiae ERG9座の上流ヌクレオチド配列およびS.cerevisiae ERG19遺伝子のコード配列に隣接した、選択マーカー(ウラシル欠損培地で増殖する能力を与えるURA3)およびS.cerevisiae GAL7遺伝子のプロモーターの制御調節下のA annuaの酵素(ファルネセンシンターゼをコードするFSコード配列)をコードするヌクレオチド配列を含む。S.cerevisiae宿主細胞への導入で、相IIマーカーリサイクリング構築物は、選択マーカーkanAがURA3と置換されるように、すでに組み込まれた相III破壊配列へ相同組換えによって組み込むことができる。
【0089】
発現プラスミドpAM404(配列番号153)は、β−ファルネセンシンターゼをコードする。ヌクレオチド配列挿入断片を、Saccharomyces cerevisiaeにおける発現のためにコドン最適化されたArtemisia annua(GenBankアクセッション番号AY835398)のβ−ファルネセンシンターゼ遺伝子のコード配列を鋳型として用いて、合成的に作製した。
【0090】
スターターの宿主株Y1198については、100ug/mLのカルバミシリンおよび50ug/mLのカナマイシンを含む5mLのYPD培地中に活性乾燥PE−2酵母(1994年に単離された;Santelisa Vale、Sertaozinho、Brazilからの贈与)を再懸濁することによって作製した。培養物を、200rpmでの回転式振盪機で、30℃、一晩、インキュベートした。その後、10uLの培養物のアリコートをYPDプレートに蒔き、乾燥させた。単一のコロニーのために細胞を連続的に画線し、30℃で2日間、インキュベートした。12個の単一コロニーを採取し、新しいYPDプレート上にパッチ状に置き、30℃で一晩、増殖させた。コロニーの株アイデンティティを、製造会社の仕様書に従い、Bio−Rad CHEF Genomic DNA Plug Kit(Bio−Rad、Hercules、CA)を用いてBio−Rad CHEF DR IIシステム(Bio−Rad、Hercules、CA)でそれらの染色体サイズを分析することによって検証した。1個のコロニーを採取し、Y1198株として保存した。
【0091】
Y1198株から、遺伝子操作を可能にするためにその株の一倍体にすることによって、Y1661株、Y1662株、Y1663株、およびY1664株を作製した。Y1198株を、回転式ドラム内のガラスチューブにおいて、5mLのYPD培地中、30℃で一晩、増殖させた。OD600を測定し、細胞を、2%の酢酸カリウムを含む5mLのYP培地中、0.2のOD600に希釈した。培養物を、回転式ドラム内のガラスチューブにおいて30℃で一晩、増殖させた。OD600を再び、測定し、4OD600*mLの細胞を、5,000 x gで2分間の遠心分離によって収集した。細胞ペレットを滅菌水で一度、洗浄し、その後、0.02%のラフィノースを含む3mLの2%酢酸カリウム中に再懸濁した。細胞を、回転式ドラム内のガラスチューブにおいて30℃で3日間、増殖させた。胞子形成を顕微鏡で確認した。33μLの培養物のアリコートを1.5mLの微量遠心管に移し、14,000rpmで2分間、遠心分離した。細胞ペレットを、2μLの10mg/mL Zymolyase 100T(MP Biomedicals、Solon、OH)を含む50μLの滅菌水中に再懸濁し、細胞を30℃の水浴中で10分間、インキュベートした。チューブを氷に移し、150μLの氷水を加えた。10μLのこの混合物のアリコートを12mLのYPDプレートに加え、Singer MSM 300ダイセクション顕微鏡(Singer、Somerset、UK)において四分子を切り離した。YPDプレートを30℃で3日間、インキュベートし、その後、胞子を新鮮なYPDプレート上にパッチ状に置き、30℃で一晩、増殖させた。8個の4胞子四分子からの各胞子の接合型をコロニーPCRによって分析した。2個のMATa胞子および2個のMATα胞子を有する単一の4胞子四分子を採取し、Y1661株(MATa)、Y1662株(MATa)、Y1663株(MATα)、およびY1664株(MATα)として保存した。
【0092】
酵母細胞形質転換について、25mlの酵母エキスペプトンデキストロース(YPD)培地に、出発宿主株の単一コロニーを接種した。培養物を、200rmpでの回転式振盪機上で30℃、一晩、増殖させた。培養物のOD600を測定し、その後、培養物を用いて、50mlのYPD培地に0.15のOD600まで接種した。その新しく接種された培養物を200rpmでの回転式振盪機において30℃で、0.7〜0.9のOD600まで増殖させ、その時点で、細胞を1μgのDNAで形質転換した。宿主細胞形質転換体を同定するために選択剤を含む寒天上に細胞を蒔く前に、細胞をYPD培地中で4時間、回復させた。
【0093】
PmeI制限エンドヌクレアーゼを用いて完了まで消化されたプラスミドTOPO相I破壊構築物でY1664株を形質転換することによって宿主株Y1515を作製した。宿主細胞形質転換体を、300ug/mLのハイグロマイシンBを含むYPD培地上で選択し、GAL80座に組み込まれた相I破壊配列を含む陽性形質転換体を、PCR増幅によって検証した。
【0094】
PmeI制限エンドヌクレアーゼを用いて完了まで消化されたプラスミドTOPO相II破壊構築物でY1515株を形質転換することによって宿主株Y1762を作製した。宿主細胞形質転換体を、100ug/mLのノールセオスリシンを含むYPD培地上で選択し、LEU2座に組み込まれた相II破壊配列を含む陽性形質転換体を、PCR増幅によって検証した。
【0095】
PmeI制限エンドヌクレアーゼを用いて完了まで消化された、発現プラスミドpAM404およびプラスミドTOPO相III破壊構築物での2段階でY1762株を形質転換することによって宿主株Y1770を作製した。pAM404を有する宿主細胞形質転換体を、メチオニンおよびロイシンを欠損する完全合成培地(CSM)上で選択した。pAM404および相III破壊構築物を有する宿主細胞形質転換体を、メチオニンおよびロイシンを欠損し、および、200ug/mlのG418(Geneticin(登録商標))を含むCSM上で選択し、ERG9座に組み込まれた相III破壊配列を含む陽性形質転換体を、PCR増幅によって検証した。
【0096】
Y1770株をURA3ノックアウト構築物(配列番号154)で形質転換することによって宿主株Y1793を作製した。URA3ノックアウト構築物は、(Saccharomyces cerevisiae株CEN.PK2ゲノムDNAから生じた)URA3座の上流配列および下流配列を含む。宿主細胞形質転換体を、5−FOAを含むYPD培地上で宿主細胞形質転換体を選択した。
【0097】
Y1793株を相Iマーカーリサイクリング構築物で形質転換することによって宿主株YAAAを作製した。宿主細胞形質転換体を、メチオニンおよびウラシルを欠損するCSM上で選択した。200rpmでの回転式振盪機上、YPD培地中、30℃で一晩、細胞を増殖させ、その後、5−FOAを含む寒天上に細胞を蒔くことによってURA3マーカーを切除した。マーカー切除をコロニーPCRによって確認した。
【0098】
YAAA株を相IIマーカーリサイクリング構築物で形質転換することによって宿主株YBBBを作製した。宿主細胞形質転換体を、メチオニンおよびウラシルを欠損するCSM上で選択した。200rpmでの回転式振盪機上、YPD培地中、30℃で一晩、細胞を増殖させ、その後、5−FOAを含む寒天上に細胞を蒔くことによってURA3マーカーを切除した。マーカー切除をコロニーPCRによって確認した。
【0099】
YBBB株を相IIIマーカーリサイクリング構築物で形質転換することによって宿主株Y1912を作製した。宿主細胞形質転換体を、メチオニンおよびウラシルを欠損するCSM上で選択した。200rpmでの回転式振盪機上、YPD培地中、30℃で一晩、細胞を増殖させ、その後、5−FOAを含む寒天上に細胞を蒔くことによってURA3マーカーを切除した。マーカー切除をコロニーPCRによって確認した。
【0100】
実施例2:遺伝子改変された胞子形成欠陥性および内因性接合欠陥性の一倍体細胞の作製
この実施例は、遺伝子改変一倍体S.cerevisiae細胞において胞子形成遺伝子およびフェロモン応答遺伝子を破壊して、胞子形成および内因性接合が損なわれている遺伝子改変一倍体S.cerevisiae細胞を生じるための例示的な方法を記載する。
【0101】
STE5破壊構築物(図7;配列番号147)は、破壊配列として、S.cerevisiae STE5座の上流ヌクレオチド配列および下流ヌクレオチド配列からなる相同配列に隣接した、選択マーカー(ウラシル欠損培地上で増殖する能力を与えるURA3);およびS.cerevisiae TDH3遺伝子のプロモーターの制御調節下のS.cerevisiae MEV経路の酵素(切断型HMG−CoAリダクターゼをコードする切断型HMG1コード配列)をコードするヌクレオチド配列を含む。S.cerevisiae宿主細胞への導入で、STE5破壊構築物は、S.cerevisiae宿主細胞ゲノムのSTE5座へ相同組換えによって組み込まれ、STE5コード配列をその破壊配列と置換することによってSTE5座を機能的に破壊することができる。
【0102】
IME1破壊構築物(図8;配列番号148)は、破壊配列として、S.cerevisiae IME5座の上流ヌクレオチド配列および下流ヌクレオチド配列からなる相同配列に隣接した、選択マーカー(ロイシンを欠損する培地上で増殖する能力を与えるLEU2)およびS.cerevisiae TDH3遺伝子のプロモーターの制御調節下のA.annuaの酵素(ファルネセンシンターゼをコードするFSコード配列)をコードするヌクレオチド配列を含む。S.cerevisiae宿主細胞への導入で、IME1破壊構築物は、S.cerevisiae宿主細胞ゲノムのIME1座へ相同組換えによって組み込まれ、IME1コード配列をその破壊配列と置換することによってIME1座を機能的に破壊することができる。
【0103】
Y1912株をSTE5破壊構築物で形質転換することによって宿主株Y1913を作製した。宿主細胞形質転換体を、メチオニンおよびウラシルを欠損するCSMにおいて選択し、陽性形質転換体をPCR増幅によって検証した。
【0104】
Y1913株からpAM404を排除(curing)し、生じた株をIME1破壊構築物で形質転換することによって宿主株Y1915を作製した。Y1913株を、200rpmでの回転式振盪機上、30℃で3日間、非選択YPD培地中で繁殖させた。約100細胞をYPD固形培地上に蒔き、30℃で3日間、増殖させ、その後、メチオニンおよびロイシンを欠損するCSMプレート上でレプリカプレーティングを行い、そこで、それらを30℃でさらに3日間、増殖させた。ロイシンを含む最少培地上で増殖するできること、およびロイシンを欠損する培地上で増殖できないことによって治癒細胞(cured cells)を同定した。単一のそのようなコロニーを採取し、IME1破壊構築物で形質転換した。宿主細胞形質転換体を、メチオニンおよびロイシンを欠損するCSM上で選択した。
【0105】
実施例3:遺伝子改変された胞子形成欠陥性および内因性接合欠陥性の二倍体細胞の作製
この実施例は、胞子形成および内因性接合が損なわれている遺伝子改変一倍体S.cerevisiae細胞を二倍体にするための例示的な方法を記載する。
【0106】
Y1915株の自己接合によって二倍体宿主株Y1979を作製した。反対の接合型の細胞を作製するため、およびY1915株を一過性に接合可能にするために、HOタンパク質をコードするプラスミドpAM1124(配列番号149)ならびにSTE5およびG418耐性マーカーをコードするプラスミドpAM1758(配列番号150)でその株を同時形質転換した。宿主細胞形質転換体を、G418およびノールセオスリシンを含むCSM上で選択した。陽性形質転換体を、非選択培地上に単一コロニーについて再び蒔き、G418感受性、ノールセオスリシン感受性の二倍体を、コロニーPCRを用いるスクリーニングを通して同定した。
【0107】
実施例4:胞子形成欠陥および内因性接合欠陥の確認
遺伝子改変S.cerevisiae細胞の胞子形成欠陥および内因性接合欠陥を確認するための例示的な方法を記載する。
【0108】
Y1915株が接合できないことを確認するために、一倍体Y1915細胞(MATα KanS URA3^ste5)または一倍体Y1912細胞(MATα KanS URA3 STE5)をYEPD固形培地上で一倍体Y1792細胞(MATa KanR ura3 STE5)と組み合わせた。接合培養物を、30℃で16時間、インキュベートした。その後、各接合培養物の同一のアリコートを、ウラシルを欠損し、さらにG418を含むCSM固形培地に蒔き、その培養物を30℃で1週間、インキュベートした。図9に示されているように、Y1792×Y1912接合培養物のアリコートを含むプレート上のみコロニー増殖が観察されたが、Y1792×Y1915接合培養物のアリコートを含むプレート上には観察されなかった。
【0109】
Y1979株が胞子形成できないことを確認するために、Y1979株細胞およびY1198株細胞を、胞子形成誘導培地(非発酵性炭素源、例えば、酢酸カリウムを欠損する培地であり、それは、天然S.cerevisiae細胞が細胞有糸分裂周期を放棄し、減数分裂へ入って胞子形成するように誘導する)中に7日間、培養した。その後、培養物を分割し、水またはジエチルエーテルで15分間、処理した。懸濁液を、反転によってホモジナイズし、滅菌水中に再懸濁し、希釈し、YEPD固形培地上に蒔き、3日間、増殖させた。図10に示されているように、95%のY1198株細胞が、これらの条件下で四分子胞子を形成したが、Y1979株細胞は形成しなかった。
【0110】
実施例5:胞子形成欠陥性および内因性接合欠陥性の細胞が天然に散在することができないことの確認
この実施例は、胞子形成欠陥性および内因性接合欠陥性の遺伝子改変二倍体S.cerevisiae細胞が天然に散在することができないことを確認するための例示的な方法を記載する。
【0111】
土壌中のY1979およびそれの非トランスジェニック同種系のY1198(PE−2)の生存を評価した。この目的を達成するために、45Lのフラスコを約25%のバーミキュライトおよびサトウキビ栽培地からの75%の土壌で満たし(合計40L)、栽培品種RB 86−7515サトウキビ植物である1つのSaccharum種(およそ月齢6ヶ月)を植えた。各ポットを、5−25−30の乾燥の窒素/リン/カリウムの混合物を施肥し、植物を閉鎖系温室内で14日間、成長させた。各ポットへ、Y1979株またはY1198株の600mLの細胞懸濁液を加えた。酵母細胞の添加は、土壌の表面5cm中に107細胞/gの濃度に達することと等価である。1.5×5土壌コアの5つの試料を以下の時点で収集した:t=0日目(暴露前)、0日目(曝露後)、3日目、7日目、14日目、28日目、40日目、60日目、および90日目(試料採取された土壌の総体積は44mLであり、試料採取された土壌の総重量は約50gであった)。複合試料から、10グラムを分離し、100mLの蒸留水中に再懸濁した。酵母生存を定量化するために、100μLの水抽出物を、0.05g/Lのコウシンバラ(Sigma #R3877)を添加され、かつ0.2g/Lのアンピシリン(Sigma A0166)を含む、硫酸6NでpH5.5に調整されたYPED培地上に直接、蒔いた(25mL/プレート)。試料を二連で、1〜107の希釈系列で蒔き、または蒔かれるべき希釈回数は、各処理について前の試料採取において得られた生存の数に基づいた。プレーティング後すぐに、その液体をDrigalskiスパチュラで広げた。プレートを、その液体を全て蒸発させるために、最高30分間、流れに対して開いたままにしておき、その後、閉じて、反転させ、30℃で48時間、インキュベートした。プレートあたりのコロニー数を、コロニー計数機(CP600 Plus、Phoenix)を用いて、計数可能な希釈度において読み取り、その結果をCFU/プレートで表した。コロニーの総数が30〜300コロニーの間である場合のみ、計数が認められた。図11に示されているように(各データ点は5つの繰り返しの平均である)、Y1979細胞は、遺伝子改変されていない、胞子形成および接合の熟達したY1198株の親細胞より、明らかに土壌中における生存能力が低かった。
【0112】
本明細書に引用された全ての刊行物、特許、および特許出願は、あたかもそれぞれ個々の刊行物または特許出願が具体的かつ個々に示されて参照により組み入れられているかのうように、参照により本明細書に組み入れられている。前述の本発明は、例証および理解を明確にするための例として、かなり詳細に記載されているが、ある特定の変化および改変が、添付された特許請求の範囲の精神または範囲から逸脱することなく、それらになされ得ることは、本発明の教示を鑑みれば、当業者にとって容易に明らかであろう。
【技術分野】
【0001】
本出願は、2009年6月1日に出願された米国仮出願番号61/183,031の利益を主張し、上記米国仮出願は、その全容が参照によって本明細書に援用される。
【0002】
本明細書に提供される方法および組成物は、一般的に、酵母などの微生物の産業的使用に関する。天然における散在、特に、組換えDNA配列の天然における散在のリスクを著しく低下させるように改変されている遺伝子改変微生物が本明細書に提供される。そのような遺伝子改変微生物を作製する方法および使用する方法も提供される。
【背景技術】
【0003】
組換えDNAテクノロジーの進歩により、原核生物または真核生物の利用を通しての産業的に有用な物質の大量製造が可能になっている。真核生物の中で、酵母(特に、Saccharomyces属に属する酵母)は、発酵産物の製造に広く用いられている。一般的に、酵母は、迅速に増殖することができ、細菌と比較してより高密度で培養することができ、産業的設定において無菌的環境を必要としない。さらに、酵母細胞は、細菌と比較して培地から容易に分離することができ、産物の抽出および精製のための工程を大いに単純化することができる。これらの特徴のため、酵母(特に、組換えDNA配列を内部に有する遺伝子改変酵母)は、有用な産物の製造のための宿主として用いられており、そのような酵母の利用は確立されている。しかしながら、産業における遺伝子改変酵母の使用は、そのような酵母および/またはそのような酵母に含まれる組換えDNA配列の分散が生態系に予想不可能な結果を生じる可能性があるという理由により、潜在的環境リスクを有する。
【発明の概要】
【発明が解決しようとする課題】
【0004】
したがって、産業的適用に適しているが、天然において散在し、繁殖するというリスクの低下をもたらし、特に、天然においてそれらの組換えDNA配列を散在するというリスクの低下をもたらす酵母の必要性が存在している。
【課題を解決するための手段】
【0005】
胞子形成および/または内因性接合が損なわれている遺伝子改変微生物、例えば、遺伝子改変酵母株を作製する方法が本明細書に提供される。本明細書に提供される方法は、遺伝子改変の一倍体微生物細胞、例えば、酵母細胞において1つもしくは複数の胞子形成遺伝子および/または1つもしくは複数のフェロモン応答遺伝子を機能的に破壊する工程、ならびに前記遺伝子改変一倍体微生物細胞が、それらの胞子形成および/または接合能力の欠損により、効果的に性的に生殖不能であり、および、それらの生活環の二倍体状態に制約される安定な二倍体を形成するように誘導する工程を含む。本明細書に提供される方法に従って遺伝子改変された微生物、例えば、酵母株は、産業的適用、例えば、産業的発酵適用に用途を見出し、野生型微生物との接合を通して天然において散在し、繁殖するというリスクの著しい低下をもたらす利点を提供することができる。
【0006】
一態様において、以下を含む遺伝子改変酵母細胞が本明細書に提供される:1つまたは複数の胞子形成遺伝子における機能的破壊であって、前記酵母細胞が、前記の1つまたは複数の胞子形成遺伝子の破壊の結果として胞子形成能力を欠く、機能的破壊;1つまたは複数のフェロモン応答遺伝子における機能的破壊であって、前記酵母細胞が、前記の1つまたは複数のフェロモン応答遺伝子の破壊の結果として内因性接合能力を欠く、機能的破壊。いくつかの実施形態において、遺伝子改変酵母細胞はヘテロタリック(heterothallic(ho))である。いくつかの実施形態において、遺伝子改変酵母細胞は、1つもしくは複数の胞子形成遺伝子の両方のコピーおよび/または1つもしくは複数のフェロモン応答遺伝子の両方のコピーが機能的に破壊されている二倍体細胞である。いくつかの実施形態において、遺伝子改変二倍体細胞は、その接合型対立遺伝子について以外はホモ接合型である。いくつかの実施形態において、遺伝子改変酵母細胞は一倍体細胞である。いくつかの実施形態において、遺伝子改変一倍体細胞は、ホモタリズム(homothallism(HO))タンパク質をコードする組換えプラスミドをさらに含む。いくつかの実施形態において、遺伝子改変一倍体細胞は、前記酵母細胞において機能的に破壊されている1つまたは複数のフェロモン応答遺伝子をコードする1つまたは複数の組換えプラスミドを含む。
【0007】
いくつかの実施形態において、本明細書に提供された方法の実施に有用な遺伝子改変酵母細胞はSaccharomyces cerevisiae細胞である。いくつかの実施形態において、Saccharomyces cerevisiae細胞は、パン酵母、Mauri、Santa Fe、CBS 7959、CBS 7960、CBS 7961、CBS 7962、CBS 7963、CBS 7964、IZ−1904、TA、BG−1、CR−1、SA−1、M−26、Y−904、PE−2、PE−5、VR−1、BR−1、BR−2、ME−2、VR−2、MA−3、MA−4、CAT−1、CB−1、NR−1、BT−1、またはAL−1の株である。特定の実施形態において、Saccharomyces cerevisiae細胞はPE−2株である。他の特定の実施形態において、Saccharomyces cerevisiae細胞はCAT−1株である。
【0008】
いくつかの実施形態において、遺伝子改変酵母細胞における破壊された1つまたは複数の胞子形成遺伝子は、IME1、IME2、NDT80、SPO11、SPO20、AMA1、HOP2、およびSPO21からなる群より選択される。特定の実施形態において、IME1遺伝子が破壊されている。いくつかの実施形態において、遺伝子改変酵母細胞における破壊された1つまたは複数のフェロモン応答遺伝子は、STE5、STE4、STE18、STE12、STE7、およびSTE11からなる群より選択される。特定の実施形態において、STE5遺伝子が破壊されている。特定の実施形態において、遺伝子改変酵母細胞は、STE5およびIME1遺伝子の両方の機能的破壊を含む。いくつかの実施形態において、遺伝子改変酵母細胞の一倍体細胞は、STE5遺伝子の機能的破壊およびSTE5タンパク質をコードする組換えプラスミドを含む。
【0009】
別の態様において、胞子形成および接合能力が損なわれている二倍体酵母株を作製する方法が本明細書に提供される。いくつかの実施形態において、該方法は以下の工程:(a)第1の遺伝子改変一倍体酵母細胞を得る工程であって、第1の遺伝子改変一倍体酵母細胞が胞子形成および内因性接合が損なわれており、および、目的のタンパク質をコードする、染色体に組み込まれた異種性ヌクレオチド配列を含む、工程;(b)第2の遺伝子改変一倍体酵母細胞を得る工程であって、第2の遺伝子改変一倍体酵母細胞が、胞子形成および内因性接合が損なわれており、および、第1の遺伝子改変一倍体酵母細胞と反対の接合型であり、および、目的の前記のタンパク質をコードする、染色体に組み込まれた異種性ヌクレオチド配列を含む、工程;(c)第1の遺伝子改変一倍体酵母細胞および第2の遺伝子改変一倍体酵母細胞のそれぞれを、前記の第1および第2の遺伝子改変一倍体酵母細胞の内因性接合欠陥を相補することができるタンパク質をコードする1つまたは複数のプラスミドで形質転換する工程;(d)第1の遺伝子改変一倍体酵母細胞を第2の遺伝子改変一倍体酵母細胞と接合させ、それにより、遺伝子改変二倍体酵母細胞を形成する工程;ならびに(e)遺伝子改変二倍体酵母細胞から前記の1つまたは複数のプラスミドを除去する工程であって、その結果生じた遺伝子改変二倍体酵母細胞が胞子形成および内因性接合が損なわれている、工程、を含む。
【0010】
いくつかの実施形態において、第1の遺伝子改変一倍体酵母細胞および第2の遺伝子改変一倍体酵母細胞が、1つまたは複数のフェロモン応答遺伝子の機能的破壊によって内因性接合が損なわれている。いくつかのそのような実施形態において、工程(c)は、第1の遺伝子改変一倍体酵母細胞および第2の遺伝子改変一倍体酵母細胞のそれぞれを、前記の第1および第2の遺伝子改変一倍体酵母細胞において機能的に破壊されているフェロモン応答遺伝子の機能性コピーをコードする1つまたは複数のプラスミドで形質転換する工程を含む。いくつかの実施形態において、1つまたは複数のフェロモン応答遺伝子は、STE5、STE4、STE18、STE12、STE7、およびSTE11からなる群より選択される。ある特定の実施形態において、第1の遺伝子改変一倍体酵母細胞および第2の遺伝子改変一倍体酵母細胞は、STE5遺伝子の機能的破壊によって内因性接合が損なわれている。
【0011】
いくつかの実施形態において、第1の遺伝子改変一倍体酵母細胞および第2の遺伝子改変一倍体酵母細胞は、1つまたは複数の胞子形成遺伝子の機能的破壊によって胞子形成が損なわれている。いくつかの実施形態において、1つまたは複数の胞子形成遺伝子は、IME1、IME2、NDT80、SPO11、SPO20、AMA1、HOP2、およびSPO21からなる群より選択される。いくつかの実施形態において、第1の遺伝子改変一倍体酵母細胞および第2の遺伝子改変一倍体酵母細胞は、IME1遺伝子の機能的破壊によって胞子形成が損なわれている。特定の実施形態において、第1の遺伝子改変一倍体酵母細胞および第2の遺伝子改変一倍体酵母細胞は、STE5遺伝子の機能的破壊によって接合が損なわれており、および、IME1遺伝子の機能的破壊によって胞子形成が損なわれている。
【0012】
いくつかの実施形態において、第2の遺伝子改変一倍体酵母細胞は、第1の遺伝子改変一倍体酵母細胞の集団において接合型切り換えを誘導することによって得られる。いくつかのそのような実施形態において、第1の遺伝子改変一倍体酵母細胞はヘテロタリック(ho)であり、前記集団は、遺伝子改変一倍体酵母細胞をホモタリズム(HO)タンパク質をコードする組換えプラスミドで形質転換することによって接合型を切り換えるように誘導され、HOタンパク質の発現が、遺伝子改変一倍体酵母細胞の接合型切り換えを誘導する。
【0013】
他の実施形態において、第2の遺伝子改変一倍体酵母細胞は、組換えDNAテクノロジーを用いて第1の遺伝子改変一倍体酵母細胞において接合型の座を変化させることによって得られる。いくつかの実施形態において、第1の遺伝子改変一倍体酵母細胞は、第1の遺伝子改変一倍体酵母細胞において接合型の座に隣接するヌクレオチド配列と相同である相同配列に隣接した、第1の遺伝子改変一倍体酵母細胞の接合型以外の接合型をコードするヌクレオチド配列を組込み配列として含む組込み構築物で形質転換される。相同組換えによる組込み配列の組込みで、第1の遺伝子改変一倍体酵母細胞の接合型の座は、挿入配列によってコードされる接合型の座と置換され、その結果として、第2の遺伝子改変一倍体酵母細胞の産生を生じる。いくつかの実施形態において、組込み構築物は、α接合型(MAT α)をコードする組込み構築物を用いてaからαへ第1の遺伝子改変一倍体酵母細胞の接合型を切り換えるために用いられる。いくつかの実施形態において、組込み構築物は、配列番号155を含む。他の実施形態において、組込み構築物は、a接合型(MAT A)をコードする組込み構築物を用いてαからaへ第1の遺伝子改変一倍体酵母細胞の接合型を切り換えるために用いられる。いくつかの実施形態において、組込み構築物は配列番号156を含む。
【0014】
別の態様において、胞子形成および内因性接合が損なわれたヘテロタリック(ho)二倍体酵母細胞を作製する方法が本明細書に提供され、該方法は以下の工程:(a)第1の遺伝子改変ヘテロタリック一倍体酵母細胞を得る工程であって、第1の遺伝子改変ヘテロタリック一倍体酵母細胞が(i)目的のタンパク質をコードする、染色体に組み込まれた異種性ヌクレオチド配列;および(ii)STE5遺伝子およびIME1遺伝子における機能的破壊を含む、工程;(b)第1の遺伝子改変ヘテロタリック一倍体酵母細胞の集団を、ホモタリズム(HO)タンパク質をコードするポリヌクレオチドを含むプラスミドで形質転換する工程であって、HOタンパク質の発現が第1の遺伝子改変ヘテロタリック一倍体酵母細胞の接合型切り換えを誘導し、それにより第2の遺伝子改変一倍体酵母細胞が得られ、第2の遺伝子改変一倍体酵母細胞が第1の遺伝子改変一倍体酵母細胞と反対の接合型であり、(i)目的の前記のタンパク質をコードする染色体に組み込まれた異種性ヌクレオチド配列;および(ii)STE5遺伝子およびIME1遺伝子における機能的破壊を含む、工程;(c)第1の遺伝子改変ヘテロタリック一倍体酵母細胞および第2の遺伝子改変ヘテロタリック一倍体酵母細胞のそれぞれを、STE5遺伝子をコードするプラスミドで形質転換する工程;(d)第1の遺伝子改変一倍体酵母細胞を第2の遺伝子改変一倍体酵母細胞と接合させ、それにより、遺伝子改変二倍体酵母細胞を形成する工程;ならびに(e)遺伝子改変二倍体酵母細胞からいかなるプラスミドも除去する工程であって、その結果生じた遺伝子改変ヘテロタリック二倍体酵母細胞が胞子形成および内因性接合が損なわれている、工程、を含む。
【0015】
本発明により作製された、胞子形成および内因性接合能力を欠く遺伝子改変ヘテロタリック(ho)酵母細胞もまた本明細書に提供される。
【0016】
MATα/a ste5/ste5ime1/ime1酵母細胞もまた本明細書に提供される。
【図面の簡単な説明】
【0017】
【図1】図1は、相I破壊構築物の構造、および相同組換えによるその破壊配列の組込み後の標的座の構造を提供する。
【図2】図2は、相II破壊構築物の構造、および相同組換えによるその破壊配列の組込み後の標的座の構造を提供する。
【図3】図3は、相III破壊構築物の構造、および相同組換えによるその破壊配列の組込み後の標的座の構造を提供する。
【図4】図4は、相Iマーカーリサイクリング構築物の構造、および相同組換えによるその構築物の組込み後の標的座の構造を提供する。
【図5】図5は、相IIマーカーリサイクリング構築物の構造、および相同組換えによるその構築物の組込み後の標的座の構造を提供する。
【図6】図6は、相IIIマーカーリサイクリング構築物の構造、および相同組換えによるその構築物の組込み後の標的座の構造を提供する。
【図7】図7は、STE5破壊構築物の構造、および相同組換えによるその破壊配列の組込み後の標的座の構造を提供する。
【図8】図8は、IME1破壊構築物の構造、および相同組換えによるその破壊配列の組込み後の標的座の構造を提供する。
【図9】図9は、遺伝子改変の内因性接合の損なわれた一倍体Y1915細胞と遺伝子改変の内因性接合能力のあるY1912細胞との接合能力の比較を提供する。
【図10】図10は、遺伝子改変の胞子形成および内因性接合の損なわれた二倍体Y1979細胞と遺伝子非改変の胞子形成および内因性接合の能力のあるY1198細胞との胞子形成能力の比較を提供する。
【図11】図11は、遺伝子改変の胞子形成および内因性接合の損なわれた二倍体Y1979細胞と遺伝子非改変の胞子形成および内因性接合の能力のあるY1198細胞との土壌中の生存の比較を提供する。
【発明を実施するための形態】
【0018】
(専門用語)
本明細書に用いられる場合、用語「異種性」とは、通常には天然で見出されないものを指す。用語「異種性ヌクレオチド配列」は、天然で所定の細胞において通常には見出されないヌクレオチド配列を指す。それとして、異種性ヌクレオチド配列は、(a)その宿主細胞にとって外来であり得る(すなわち、その細胞にとって「外因性」であり得る);(b)宿主細胞において天然で見出されるが(すなわち、「内因性」)、細胞において不自然な量(すなわち、宿主細胞において天然で見出されるより多い量、または少ない量)で存在し得る;または(c)宿主細胞において天然で見出されるが、その天然の座の外側に位置し得る。
【0019】
本明細書に用いられる場合、標的遺伝子、例えば、フェロモン応答遺伝子または胞子形成遺伝子を「機能的に破壊する」こと、またはそれの「機能的破壊」とは、標的遺伝子が、標的遺伝子によってコードされるタンパク質の活性を宿主細胞において減少させるような様式で変化していることを意味する。いくつかの実施形態において、標的遺伝子によってコードされるタンパク質の活性は、宿主細胞において除去されている。他の実施形態において、標的遺伝子によってコードされるタンパク質の活性は宿主細胞において減少している。標的遺伝子の機能的破壊は、遺伝子発現が除去され、もしくは低下するように、または遺伝子産物の活性が除去され、もしくは低下するように、遺伝子の全部または一部を欠失させることによって達成されてもよい。標的遺伝子の機能的破壊はまた、遺伝子の制御エレメント、例えば、遺伝子のプロモーターを、発現が除去され、もしくは低下するように突然変異することによって、または遺伝子のコード配列を、遺伝子産物の活性が除去され、もしくは低下するように突然変異することによって、達成されてもよい。いくつかの実施形態において、標的遺伝子の機能的破壊は、結果として、標的遺伝子の完全なオープンリーディングフレームの除去を生じる。
【0020】
本明細書に用いられる場合、「内因性接合」および「内因性接合能力」は、反対の接合型、すなわち、接合型aおよびαの一倍体微生物細胞が、異種性遺伝子発現、例えば、フェロモン応答遺伝子またはそのような一倍体の間で接合を誘導する能力がある任意の遺伝子の異種性コピーの発現の非存在下において、二倍体細胞を形成する能力を指す。
【0021】
本明細書に用いられる場合、「内因性接合が損なわれた」とは、野生型一倍体微生物細胞の集団に対して、そのような微生物細胞の一倍体の集団内での接合を阻害するのに十分な、微生物細胞の内因性接合能力の低下を指す。いくつかの実施形態において、阻害は、野生型一倍体微生物細胞の集団の接合率に対して一倍体微生物細胞の集団の接合率の少なくとも10%、20%、30%、40%、50%、60%、65%、70%、75%、80%、85%、90%、または95%の低下を含む。
【0022】
本明細書に用いられる場合、「胞子形成が損なわれた」とは、野生型二倍体微生物細胞の集団に対して、そのような微生物細胞の二倍体の集団内で胞子形成を阻害するのに十分な二倍体微生物細胞の胞子形成活性の低下を指す。いくつかの実施形態において、阻害は、野生型二倍体微生物細胞の集団の胞子形成率に対して二倍体微生物細胞の集団の胞子形成率の少なくとも10%、20%、30%、40%、50%、60%、65%、70%、75%、80%、85%、90%、または95%の低下を含む。
【0023】
本明細書に用いられる場合、遺伝子との関連における用語「相補すること」とは、機能的に破壊された遺伝子、例えば、機能的に破壊された胞子形成遺伝子またはフェロモン応答遺伝子の機能を置換する機能を有する遺伝子を指す。いくつかの実施形態において、相補遺伝子と破壊された遺伝子の間で機能の機構は同一である必要はない。いくつかの実施形態において、機能的に破壊されている標的遺伝子、例えば、胞子形成遺伝子またはフェロモン応答遺伝子は、破壊された遺伝子によってコードされるタンパク質と相同なタンパク質を産生するか、または例えば、別の機構によって胞子形成もしくは接合を可能にする表現型を提供するかのいずれかである異種性遺伝子によって相補することができる。
【0024】
(遺伝子改変微生物およびそれを作製する方法)
胞子形成および/または内因性接合能力において機能的に損なわれている遺伝子改変微生物、例えば、遺伝子改変酵母細胞(例えば、遺伝子改変Saccharomyces cerevisiae細胞)を含む組成物、ならびにそのような組成物を作製するための方法および材料が本明細書に提供される。本明細書に提供される方法は、微生物の有性生殖サイクルを中断させて、天然におけるその微生物の散在を最小限にし、遺伝子改変微生物と、天然において散在する能力が損なわれていない野生型微生物との間での遺伝物質の交換の可能性を最小限にする。
【0025】
多くの真菌細胞、例えば、酵母細胞は、有性的および無性的の両方で生殖することができる。無性生殖は、1つの親細胞のみが関係し、迅速な集団増殖を可能にする。対照的に、有性生殖は、配偶子の形成および融合を伴い、細胞間の水平遺伝子伝達による遺伝的多様性のより迅速な発生を可能にする。有性生殖する真菌細胞は、それらの生活環を通じて2つの細胞状態を想定し、1つが二倍体細胞状態であり、もう1つが一倍体細胞状態である。二倍体真菌細胞は、一般的に非常に安定であり、一般的に、それらが、いくつかの特定の環境的刺激(例えば、栄養枯渇)のうちの1つまたは複数に遭遇しない限り、二倍体相のままであるだろう。1つまたは複数のそのような刺激に遭遇した場合、二倍体細胞は胞子形成して、(四分子と呼ばれる)4つの一倍体胞子を形成する。都合良い条件が戻った場合、これらの一倍体胞子は出芽して4つの一倍体細胞(2つの接合型a、および2つの接合型α)を生成し、それは、その後、反対の接合型の他の一倍体細胞と接合して、再び二倍体細胞を形成することができる。
【0026】
二倍体真菌細胞の胞子形成する能力、および一倍体真菌細胞の接合する能力は、特定の遺伝子産物の機能に依存している。酵母細胞におけるこれらの中では、IME1、IME2、NDT80、SPO11、SPO20、AMA1、HOP2、およびSPO21遺伝子の産物などの胞子形成遺伝子の産物、ならびにSTE5、STE4、STE18、STE12、STE7、およびSTE11の産物などのフェロモン応答遺伝子の産物である。
【0027】
一態様において、胞子形成および/または内因性接合が損なわれており、および、目的のタンパク質をコードする染色体に組み込まれた異種性ヌクレオチド配列を含む遺伝子改変一倍体真菌細胞が本明細書に提供される。いくつかの実施形態において、遺伝子改変真菌細胞は、少なくとも1つの胞子形成遺伝子および/または少なくとも1つのフェロモン応答遺伝子が機能的に破壊されている一倍体酵母細胞である。
【0028】
いくつかの実施形態において、遺伝子改変真菌細胞は、以下の胞子形成遺伝子の1つまたは複数が機能的に破壊されている一倍体酵母細胞である:IME1、IME2、NDT80、SPO11、SPO20、AMA1、HOP2、およびSPO21。いくつかの実施形態において、遺伝子改変真菌細胞は、IME1遺伝子が機能的に破壊されている一倍体酵母細胞である。いくつかの実施形態において、遺伝子改変真菌細胞は、IME2遺伝子が機能的に破壊されている一倍体酵母細胞である。いくつかの実施形態において、遺伝子改変真菌細胞は、NDT80遺伝子が機能的に破壊されている一倍体酵母細胞である。いくつかの実施形態において、遺伝子改変真菌細胞は、SPO11遺伝子が機能的に破壊されている一倍体酵母細胞である。いくつかの実施形態において、遺伝子改変真菌細胞は、SPO20遺伝子が機能的に破壊されている一倍体酵母細胞である。いくつかの実施形態において、遺伝子改変真菌細胞は、AMA1遺伝子が機能的に破壊されている一倍体酵母細胞である。いくつかの実施形態において、遺伝子改変真菌細胞は、HOP2遺伝子が機能的に破壊されている一倍体酵母細胞である。いくつかの実施形態において、遺伝子改変真菌細胞は、SPO21遺伝子が機能的に破壊されている一倍体酵母細胞である。いくつかの実施形態において、遺伝子改変真菌細胞は、IME1、IME2、NDT80、SPO11、SPO20、AMA1、HOP2、およびSPO21からなる群より選択される1つより多い胞子形成遺伝子が破壊されている一倍体酵母細胞である。いくつかの実施形態において、遺伝子改変真菌細胞は、IME1、IME2、NDT80、SPO11、SPO20、AMA1、HOP2、およびSPO21からなる群より選択される2つより多い胞子形成遺伝子が破壊されている一倍体酵母細胞である。いくつかの実施形態において、遺伝子改変真菌細胞は、IME1、IME2、NDT80、SPO11、SPO20、AMA1、HOP2、およびSPO21からなる群より選択される3つより多い胞子形成遺伝子が破壊されている一倍体酵母細胞である。いくつかの実施形態において、遺伝子改変真菌細胞は、IME1、IME2、NDT80、SPO11、SPO20、AMA1、HOP2、およびSPO21からなる群より選択される4つより多い胞子形成遺伝子が破壊されている一倍体酵母細胞である。
【0029】
いくつかの実施形態において、遺伝子改変真菌細胞は、以下のフェロモン応答遺伝子の1つまたは複数が機能的に破壊されている一倍体酵母細胞である:STE5、STE4、STE18、STE12、STE7、およびSTE11。いくつかの実施形態において、遺伝子改変真菌細胞は、STE5遺伝子が機能的に破壊されている一倍体酵母細胞である。いくつかの実施形態において、遺伝子改変真菌細胞は、STE4遺伝子が機能的に破壊されている一倍体酵母細胞である。いくつかの実施形態において、遺伝子改変真菌細胞は、STE18遺伝子が機能的に破壊されている一倍体酵母細胞である。いくつかの実施形態において、遺伝子改変真菌細胞は、STE12遺伝子が機能的に破壊されている一倍体酵母細胞である。いくつかの実施形態において、遺伝子改変真菌細胞は、STE7遺伝子が機能的に破壊されている一倍体酵母細胞である。いくつかの実施形態において、遺伝子改変真菌細胞は、STE11遺伝子が機能的に破壊されている一倍体酵母細胞である。いくつかの実施形態において、遺伝子改変真菌細胞は、STE5、STE4、STE18、STE12、STE7、およびSTE11からなる群より選択される1つより多いフェロモン応答遺伝子が破壊されている一倍体酵母細胞である。いくつかの実施形態において、遺伝子改変真菌細胞は、STE5、STE4、STE18、STE12、STE7、およびSTE11からなる群より選択される2つより多いフェロモン応答遺伝子が破壊されている一倍体酵母細胞である。いくつかの実施形態において、遺伝子改変真菌細胞は、STE5、STE4、STE18、STE12、STE7、およびSTE11からなる群より選択される3つより多いフェロモン応答遺伝子が破壊されている一倍体酵母細胞である。いくつかの実施形態において、遺伝子改変真菌細胞は、STE5、STE4、STE18、STE12、STE7、およびSTE11からなる群より選択される4つより多いフェロモン応答遺伝子が破壊されている一倍体酵母細胞である。
【0030】
いくつかの実施形態において、遺伝子改変真菌細胞は、少なくとも1つの胞子形成遺伝子および少なくとも1つのフェロモン応答遺伝子が機能的に破壊されている一倍体酵母細胞である。
【0031】
いくつかの実施形態において、遺伝子改変真菌細胞は、IME1遺伝子およびSTE5遺伝子が機能的に破壊されている一倍体酵母細胞である。いくつかの実施形態において、遺伝子改変真菌細胞は、IME2遺伝子およびSTE5遺伝子が機能的に破壊されている一倍体酵母細胞である。いくつかの実施形態において、遺伝子改変真菌細胞は、NDT80遺伝子およびSTE5遺伝子が機能的に破壊されている一倍体酵母細胞である。いくつかの実施形態において、遺伝子改変真菌細胞は、SPO11遺伝子およびSTE5遺伝子が機能的に破壊されている一倍体酵母細胞である。いくつかの実施形態において、遺伝子改変真菌細胞は、SPO20遺伝子およびSTE5遺伝子が機能的に破壊されている一倍体酵母細胞である。いくつかの実施形態において、遺伝子改変真菌細胞は、AMA1遺伝子およびSTE5遺伝子が機能的に破壊されている一倍体酵母細胞である。いくつかの実施形態において、遺伝子改変真菌細胞は、HOP2遺伝子およびSTE5遺伝子が機能的に破壊されている一倍体酵母細胞である。いくつかの実施形態において、遺伝子改変真菌細胞は、SPO21遺伝子およびSTE5遺伝子が機能的に破壊されている一倍体酵母細胞である。
【0032】
いくつかの実施形態において、遺伝子改変真菌細胞は、IME1遺伝子およびSTE4遺伝子が機能的に破壊されている一倍体酵母細胞である。いくつかの実施形態において、遺伝子改変真菌細胞は、IME2遺伝子およびSTE4遺伝子が機能的に破壊されている一倍体酵母細胞である。いくつかの実施形態において、遺伝子改変真菌細胞は、NDT80遺伝子およびSTE4遺伝子が機能的に破壊されている一倍体酵母細胞である。いくつかの実施形態において、遺伝子改変真菌細胞は、SPO11遺伝子およびSTE4遺伝子が機能的に破壊されている一倍体酵母細胞である。いくつかの実施形態において、遺伝子改変真菌細胞は、SPO20遺伝子およびSTE4遺伝子が機能的に破壊されている一倍体酵母細胞である。いくつかの実施形態において、遺伝子改変真菌細胞は、AMA1遺伝子およびSTE4遺伝子が機能的に破壊されている一倍体酵母細胞である。いくつかの実施形態において、遺伝子改変真菌細胞は、HOP2遺伝子およびSTE4遺伝子が機能的に破壊されている一倍体酵母細胞である。いくつかの実施形態において、遺伝子改変真菌細胞は、SPO21遺伝子およびSTE4遺伝子が機能的に破壊されている一倍体酵母細胞である。
【0033】
いくつかの実施形態において、遺伝子改変真菌細胞は、IME1遺伝子およびSTE18遺伝子が機能的に破壊されている一倍体酵母細胞である。いくつかの実施形態において、遺伝子改変真菌細胞は、IME2遺伝子およびSTE18遺伝子が機能的に破壊されている一倍体酵母細胞である。いくつかの実施形態において、遺伝子改変真菌細胞は、NDT80遺伝子およびSTE18遺伝子が機能的に破壊されている一倍体酵母細胞である。いくつかの実施形態において、遺伝子改変真菌細胞は、SPO11遺伝子およびSTE18遺伝子が機能的に破壊されている一倍体酵母細胞である。いくつかの実施形態において、遺伝子改変真菌細胞は、SPO20遺伝子およびSTE18遺伝子が機能的に破壊されている一倍体酵母細胞である。いくつかの実施形態において、遺伝子改変真菌細胞は、AMA1遺伝子およびSTE18遺伝子が機能的に破壊されている一倍体酵母細胞である。いくつかの実施形態において、遺伝子改変真菌細胞は、HOP2遺伝子およびSTE18遺伝子が機能的に破壊されている一倍体酵母細胞である。いくつかの実施形態において、遺伝子改変真菌細胞は、SPO21遺伝子およびSTE18遺伝子が機能的に破壊されている一倍体酵母細胞である。
【0034】
いくつかの実施形態において、遺伝子改変真菌細胞は、IME1遺伝子およびSTE12遺伝子が機能的に破壊されている一倍体酵母細胞である。いくつかの実施形態において、遺伝子改変真菌細胞は、IME2遺伝子およびSTE12遺伝子が機能的に破壊されている一倍体酵母細胞である。いくつかの実施形態において、遺伝子改変真菌細胞は、NDT80遺伝子およびSTE12遺伝子が機能的に破壊されている一倍体酵母細胞である。いくつかの実施形態において、遺伝子改変真菌細胞は、SPO11遺伝子およびSTE12遺伝子が機能的に破壊されている一倍体酵母細胞である。いくつかの実施形態において、遺伝子改変真菌細胞は、SPO20遺伝子およびSTE12遺伝子が機能的に破壊されている一倍体酵母細胞である。いくつかの実施形態において、遺伝子改変真菌細胞は、AMA1遺伝子およびSTE12遺伝子が機能的に破壊されている一倍体酵母細胞である。いくつかの実施形態において、遺伝子改変真菌細胞は、HOP2遺伝子およびSTE12遺伝子が機能的に破壊されている一倍体酵母細胞である。いくつかの実施形態において、遺伝子改変真菌細胞は、SPO21遺伝子およびSTE12遺伝子が機能的に破壊されている一倍体酵母細胞である。
【0035】
いくつかの実施形態において、遺伝子改変真菌細胞は、IME1遺伝子およびSTE7遺伝子が機能的に破壊されている一倍体酵母細胞である。いくつかの実施形態において、遺伝子改変真菌細胞は、IME2遺伝子およびSTE7遺伝子が機能的に破壊されている一倍体酵母細胞である。いくつかの実施形態において、遺伝子改変真菌細胞は、NDT80遺伝子およびSTE7遺伝子が機能的に破壊されている一倍体酵母細胞である。いくつかの実施形態において、遺伝子改変真菌細胞は、SPO11遺伝子およびSTE7遺伝子が機能的に破壊されている一倍体酵母細胞である。いくつかの実施形態において、遺伝子改変真菌細胞は、SPO20遺伝子およびSTE7遺伝子が機能的に破壊されている一倍体酵母細胞である。いくつかの実施形態において、遺伝子改変真菌細胞は、AMA1遺伝子およびSTE7遺伝子が機能的に破壊されている一倍体酵母細胞である。いくつかの実施形態において、遺伝子改変真菌細胞は、HOP2遺伝子およびSTE7遺伝子が機能的に破壊されている一倍体酵母細胞である。いくつかの実施形態において、遺伝子改変真菌細胞は、SPO21遺伝子およびSTE7遺伝子が機能的に破壊されている一倍体酵母細胞である。
【0036】
いくつかの実施形態において、遺伝子改変真菌細胞は、IME1遺伝子およびSTE11遺伝子が機能的に破壊されている一倍体酵母細胞である。いくつかの実施形態において、遺伝子改変真菌細胞は、IME2遺伝子およびSTE11遺伝子が機能的に破壊されている一倍体酵母細胞である。いくつかの実施形態において、遺伝子改変真菌細胞は、NDT80遺伝子およびSTE11遺伝子が機能的に破壊されている一倍体酵母細胞である。いくつかの実施形態において、遺伝子改変真菌細胞は、SPO11遺伝子およびSTE11遺伝子が機能的に破壊されている一倍体酵母細胞である。いくつかの実施形態において、遺伝子改変真菌細胞は、SPO20遺伝子およびSTE11遺伝子が機能的に破壊されている一倍体酵母細胞である。いくつかの実施形態において、遺伝子改変真菌細胞は、AMA1遺伝子およびSTE11遺伝子が機能的に破壊されている一倍体酵母細胞である。いくつかの実施形態において、遺伝子改変真菌細胞は、HOP2遺伝子およびSTE11遺伝子が機能的に破壊されている一倍体酵母細胞である。いくつかの実施形態において、遺伝子改変真菌細胞は、SPO21遺伝子およびSTE11遺伝子が機能的に破壊されている一倍体酵母細胞である。
【0037】
いくつかの実施形態において、遺伝子改変真菌細胞は、1つまたは複数のフェロモン応答遺伝子の機能的に破壊された染色体コピー、および前記の1つまたは複数のフェロモン応答遺伝子のコード配列の機能性染色体外コピーを含む1つまたは複数の組換えプラスミドを含む一倍体酵母細胞である。いくつかの実施形態において、遺伝子改変真菌細胞は、機能的に破壊されたSTE5遺伝子、およびSTE5遺伝子のコード配列の機能性染色体外コピーを含む組換えプラスミドを含む一倍体酵母細胞である。いくつかの実施形態において、遺伝子改変真菌細胞は、機能的に破壊されたSTE4遺伝子、およびSTE4遺伝子のコード配列の機能性染色体外コピーを含む組換えプラスミドを含む一倍体酵母細胞である。いくつかの実施形態において、遺伝子改変真菌細胞は、機能的に破壊されたSTE18遺伝子、およびSTE18遺伝子のコード配列の機能性染色体外コピーを含む組換えプラスミドを含む一倍体酵母細胞である。いくつかの実施形態において、遺伝子改変真菌細胞は、機能的に破壊されたSTE12遺伝子、およびSTE12遺伝子のコード配列の機能性染色体外コピーを含む組換えプラスミドを含む一倍体酵母細胞である。いくつかの実施形態において、遺伝子改変真菌細胞は、機能的に破壊されたSTE7遺伝子、およびSTE7遺伝子のコード配列の機能性染色体外コピーを含む組換えプラスミドを含む一倍体酵母細胞である。いくつかの実施形態において、遺伝子改変真菌細胞は、機能的に破壊されたSTE11遺伝子、およびSTE11遺伝子のコード配列の機能性染色体外コピーを含む組換えプラスミドを含む一倍体酵母細胞である。
【0038】
別の態様において、胞子形成および/または内因性接合が損なわれており、および、目的のタンパク質をコードする染色体に組み込まれた異種性ヌクレオチド配列を2コピー含む遺伝子改変二倍体真菌細胞が本明細書に提供される。いくつかの実施形態において、遺伝子改変真菌細胞は、少なくとも1つの胞子形成遺伝子の両方のコピーおよび/または少なくとも1つのフェロモン応答遺伝子の両方のコピーが機能的に破壊された二倍体酵母細胞である。
【0039】
いくつかの実施形態において、遺伝子改変真菌細胞は、以下の胞子形成遺伝子の1つまたは複数の両方のコピーが機能的に破壊されている二倍体酵母細胞である:IME1、IME2、NDT80、SPO11、SPO20、AMA1、HOP2、およびSPO21。いくつかの実施形態において、遺伝子改変真菌細胞は、IME1遺伝子の両方のコピーが機能的に破壊されている二倍体酵母細胞である。いくつかの実施形態において、遺伝子改変真菌細胞は、以下のフェロモン応答遺伝子の1つまたは複数の両方のコピーが機能的に破壊されている二倍体酵母細胞である:STE5、STE4、STE18、STE12、STE7、およびSTE11。いくつかの実施形態において、遺伝子改変真菌細胞は、STE5遺伝子の両方のコピーが機能的に破壊されている二倍体酵母細胞である。さらに他の実施形態において、遺伝子改変真菌細胞は、少なくとも1つの胞子形成遺伝子の両方のコピーおよび少なくとも1つのフェロモン応答遺伝子の両方のコピーが機能的に破壊されている二倍体酵母細胞である。いくつかの実施形態において、IME1遺伝子の両方のコピーおよびSTE5遺伝子の両方のコピーが機能的に破壊されている二倍体酵母細胞である。
【0040】
いくつかの実施形態において、遺伝子改変二倍体真菌細胞は、それの接合型対立遺伝子について以外はホモ接合である。例えば、遺伝子改変二倍体真菌細胞が胞子形成するべきである場合には、生じた4つの一倍体真菌細胞は、それらの接合型対立遺伝子を除いて遺伝的に同一であるだろう。そのような事象において、一倍体細胞のうちの2つは、接合型aであり、残りの2つの一倍体細胞は接合型αであろう。いくつかの実施形態において、遺伝子改変二倍体真菌細胞は、抗生化合物に対する耐性を与える異種性遺伝子を含まない。
【0041】
本明細書に提供される遺伝子改変二倍体真菌細胞は、それに含まれる異種性ヌクレオチド配列の望まれない繁殖に対するいくつかの保護手段を有する。第一に、二倍体真菌細胞は、一般的に、非常に安定であり、それらの二倍体状態において、他の真菌細胞と接合できない。第二に、本明細書に提供される二倍体真菌細胞は、胞子形成する能力が損なわれており、したがって、適切な環境的刺激の存在下でさえも、胞子を形成する能力をほとんどもたない。第三に、胞子が形成される可能性が低い事象において、生じた一倍体真菌細胞は、接合する能力が損なわれている。総合すれば、本明細書に提供される遺伝子改変真菌細胞の胞子形成および接合の欠損した性質は、異種性ヌクレオチド配列の野生型真菌細胞への移動の可能性を著しく低下させる。
【0042】
別の態様において、本明細書に記載された遺伝子改変二倍体真菌細胞を作製する方法が本明細書に提供される。いくつかの実施形態において、該方法は以下の工程:(a)第1の遺伝子改変一倍体酵母細胞を得る工程であって、第1の遺伝子改変一倍体真菌細胞が胞子形成および/または内因性接合が損なわれており、および、目的のタンパク質をコードする、染色体に組み込まれた異種性ヌクレオチド配列を含む、工程;(b)第2の遺伝子改変一倍体真菌細胞を得る工程であって、第2の遺伝子改変一倍体真菌細胞が、胞子形成および内因性接合が損なわれており、および、第1の遺伝子改変一倍体真菌細胞と反対の接合型であり、および、目的の前記のタンパク質をコードする染色体に組み込まれた異種性ヌクレオチド配列を含む、工程;(c)第1の遺伝子改変一倍体真菌細胞および第2の遺伝子改変一倍体真菌細胞のそれぞれを、前記の第1および第2の遺伝子改変一倍体真菌細胞の内因性接合欠陥を相補することができるタンパク質をコードする1つまたは複数のプラスミドで形質転換する工程;(d)第1の遺伝子改変一倍体真菌細胞を第2の遺伝子改変一倍体真菌細胞と接合させ、それにより、遺伝子改変二倍体真菌細胞を形成する工程;ならびに(e)遺伝子改変二倍体真菌細胞から1つまたは複数のプラスミドを除去する工程であって、その結果生じた遺伝子改変二倍体真菌細胞が胞子形成および内因性接合が損なわれており、および、目的の前記のタンパク質をコードする染色体に組み込まれた異種性ヌクレオチド配列を2コピー含む、工程、を含む。
【0043】
いくつかの実施形態において、第1の遺伝子改変一倍体真菌細胞および第2の遺伝子改変一倍体真菌細胞が、1つまたは複数のフェロモン応答遺伝子の機能的破壊によって内因性接合が損なわれている。いくつかの実施形態において、本発明の方法の工程(c)は、第1の遺伝子改変一倍体真菌細胞および第2の遺伝子改変一倍体真菌細胞のそれぞれを、前記の第1および第2の遺伝子改変一倍体真菌細胞において機能的に破壊されている1つまたは複数のフェロモン応答遺伝子の機能性コピーをコードする1つまたは複数のプラスミドで形質転換する工程を含む。いくつかの実施形態において、第1および第2の遺伝子改変一倍体真菌細胞は、一倍体酵母細胞であり、1つまたは複数のフェロモン応答遺伝子は、STE5、STE4、STE18、STE12、STE7、およびSTE11からなる群より選択される。ある特定の実施形態において、第1および第2の遺伝子改変一倍体真菌細胞は、STE5遺伝子の機能的破壊によって内因性接合が損なわれている一倍体酵母細胞である。
【0044】
いくつかの実施形態において、第1の遺伝子改変一倍体真菌細胞および第2の遺伝子改変一倍体真菌細胞は、1つまたは複数の胞子形成遺伝子の機能的破壊によって胞子形成が損なわれている。いくつかの実施形態において、第1および第2の遺伝子改変一倍体真菌細胞は一倍体酵母細胞であり、1つまたは複数の胞子形成遺伝子は、IME1、IME2、NDT80、SPO11、SPO20、AMA1、HOP2、およびSPO21からなる群より選択される。いくつかの実施形態において、第1および第2の遺伝子改変一倍体真菌細胞は、IME1遺伝子の機能的破壊によって胞子形成が損なわれている一倍体酵母細胞である。特定の実施形態において、第1および第2の遺伝子改変一倍体真菌細胞は、STE5遺伝子の機能的破壊によって接合が損なわれており、および、IME1遺伝子の機能的破壊によって胞子形成が損なわれている一倍体酵母細胞である。
【0045】
いくつかの実施形態において、第2の遺伝子改変一倍体真菌細胞は、第1の遺伝子改変一倍体真菌細胞の集団において接合型切り換えを誘導することによって得られる。いくつかの実施形態において、第1の遺伝子改変一倍体真菌細胞はヘテロタリック(ho)一倍体Saccharomyces cerevisiae細胞であり、前記ヘテロタリック(ho)一倍体Saccharomyces cerevisiae細胞集団は、前記ヘテロタリック(ho)一倍体Saccharomyces cerevisiae細胞を、ホモタリズム(HO)タンパク質をコードするプラスミドで形質転換することによって接合型を切り換えるように誘導され、HOタンパク質の発現は、一倍体Saccharomyces cerevisiae細胞において接合型切り換えを誘導し、第2の遺伝子改変一倍体Saccharomyces cerevisiae細胞を生じる。ヘテロタリック(ho)一倍体Saccharomyces cerevisiae細胞は、自発的接合型切り換えが事実上、発生しない(たった10−6の頻度)ことを特徴とする。HOタンパク質を一過性に発現することによって、一倍体Saccharomyces cerevisiae細胞における自発的接合型切り換えの頻度は、多くも細胞分裂ごとに1回にまで増加することができ、お互いに接合して二倍体Saccharomyces cerevisiae細胞を生じることができる反対の接合型の一倍体細胞集団を提供する。
【0046】
別の態様において、胞子形成および内因性接合能力を欠く遺伝子改変ヘテロタリック(ho)二倍体酵母細胞を作製する方法が本明細書に提供され、該方法は以下の工程:(a)第1の遺伝子改変ヘテロタリック一倍体酵母細胞を得る工程であって、第1の遺伝子改変ヘテロタリック一倍体酵母細胞が(i)目的のタンパク質をコードする、染色体に組み込まれた異種性ヌクレオチド配列;および(ii)1つまたは複数の胞子形成遺伝子および1つまたは複数のフェロモン応答遺伝子における機能的破壊を含む、工程;(b)第1の遺伝子改変ヘテロタリック一倍体酵母細胞の集団を、ホモタリズム(HO)タンパク質をコードするプラスミドで形質転換して、第1の遺伝子改変一倍体酵母細胞を生じる工程であって、HOタンパク質の発現が第1の遺伝子改変一倍体酵母細胞において接合型切り換えを誘導し、それにより第2の遺伝子改変一倍体酵母細胞が得られ、第2の遺伝子改変一倍体酵母細胞が第1の遺伝子改変一倍体酵母細胞と反対の接合型であり、および、(i)目的の前記のタンパク質をコードする染色体に組み込まれた異種性ヌクレオチド配列;および(ii)1つまたは複数の胞子形成遺伝子および1つまたは複数のフェロモン応答遺伝子における機能的破壊を含む、工程;(c)第1の遺伝子改変一倍体酵母細胞および第2の遺伝子改変一倍体酵母細胞のそれぞれを、前記の第1および第2の遺伝子改変一倍体酵母細胞において機能的に破壊されている1つまたは複数のフェロモン応答タンパク質をコードするプラスミドで形質転換する工程;(d)第1の遺伝子改変一倍体酵母細胞を第2の遺伝子改変一倍体酵母細胞と接合させ、それにより、それの接合型対立遺伝子について以外はホモ接合である遺伝子改変二倍体酵母細胞を形成する工程;ならびに(e)遺伝子改変二倍体酵母細胞からいかなるプラスミドも除去して、遺伝子改変ヘテロタリック二倍体酵母細胞を生じる工程であって、その結果生じた遺伝子改変ヘテロタリック二倍体酵母細胞が胞子形成および内因性接合が損なわれており、および、目的の前記のタンパク質をコードする染色体に組み込まれた異種性ヌクレオチド配列を2コピー含む、工程、を含む。
【0047】
本明細書に提供され、下記でより詳細に記載される方法の工程は、経時的順序で提示されているが、当業者は、いくつかの工程の順序を、本発明の範囲を超えることなく、交換し、組み合わせ、または繰り返すことができることは認識しているであろう。したがって、いくつかの実施形態において、胞子形成および内因性接合能力を欠く遺伝子改変ヘテロタリック(ho)二倍体酵母細胞は、遺伝子改変ヘテロタリック一倍体酵母細胞を、前記の遺伝子改変ヘテロタリック一倍体酵母細胞において機能的に破壊されている1つまたは複数のフェロモン応答遺伝子をコードするプラスミドで形質転換し、その後、その細胞をホモタリズム(HO)タンパク質をコードするプラスミドで形質転換することによって作製される。他の実施形態において、胞子形成および内因性接合能力を欠く遺伝子改変ヘテロタリック(ho)二倍体酵母細胞は、遺伝子改変ヘテロタリック一倍体酵母細胞を、前記の遺伝子改変ヘテロタリック一倍体酵母細胞において機能的に破壊されている1つまたは複数のフェロモン応答タンパク質をコードするプラスミド、およびホモタリズム(HO)タンパク質をコードするプラスミドで同時に形質転換することによって作製される。
【0048】
(微生物選択)
本明細書に提供された方法の実施において有用な微生物には、真核単細胞生物、具体的には真菌、より具体的には酵母が挙げられる。
【0049】
いくつかの実施形態において、本発明に有用な酵母には、微生物受託機関(例えば、IFO、ATCCなど)に寄託されており、とりわけ、Aciculoconidium、Ambrosiozyma、Arthroascus、Arxiozyma、Ashbya、Babjevia、Bensingtonia、Botryoascus、Botryozyma、Brettanomyces、Bullera、Bulleromyces、Candida、Citeromyces、Clavispora、Cryptococcus、Cystofilobasidium、Debaryomyces、Dekkara、Dipodascopsis、Dipodascus、Eeniella、Endomycopsella、Eremascus、Eremothecium、Erythrobasidium、Fellomyces、Filobasidium、Galactomyces、Geotrichum、Guilliermondella、Hanseniaspora、Hansenula、Hasegawaea、Holtermannia、Hormoascus、Hyphopichia、Issatchenkia、Kloeckera、Kloeckeraspora、Kluyveromyces、Kondoa、Kuraishia、Kurtzmanomyces、Leucosporidium、Lipomyces、Lodderomyces、Malassezia、Metschnikowia、Mrakia、Myxozyma、Nadsonia、Nakazawaea、Nematospora、Ogataea、Oosporidium、Pachysolen、Phachytichospora、Phaffia、Pichia、Rhodosporidium、Rhodotorula、Saccharomyces、Saccharomycodes、Saccharomycopsis、Saitoella、Sakaguchia、Saturnospora、Schizoblastosporion、Schizosaccharomyces、Schwanniomyces、Sporidiobolus、Sporobolomyces、Sporopachydermia、Stephanoascus、Sterigmatomyces、Sterigmatosporidium、Symbiotaphrina、Sympodiomyces、Sympodiomycopsis、Torulaspora、Trichosporiella、Trichosporon、Trigonopsis、Tsuchiyaea、Udeniomyces、Waltomyces、Wickerhamia、Wickerhamiella、Williopsis、Yamadazyma、Yarrowia、Zygoascus、Zygosaccharomyces、Zygowilliopsis、およびZygozymaの属に属する酵母が挙げられる。
【0050】
いくつかの実施形態において、微生物は、Saccharomyces cerevisiae、Pichia pastoris、Schizosaccharomyces pombe、Dekkera bruxellensis、Kluyveromyces lactis(以前、Saccharomyces lactisと呼ばれた)、Kluveromyces marxianus、Arxula adeninivorans、またはHansenula polymorpha(現在、Pichia angustaとして知られている)である。いくつかの実施形態において、微生物は、Candida lipolytica、Candida guilliermondii、Candida krusei、Candida pseudotropicalis、またはCandida utilisなどのCandida属の株である。
【0051】
特定の実施形態において、微生物は、Saccharomyces cerevisiaeである。いくつかの実施形態において、微生物は、パン酵母、CBS 7959、CBS 7960、CBS 7961、CBS 7962、CBS 7963、CBS 7964、IZ−1904、TA、BG−1、CR−1、SA−1、M−26、Y−904、PE−2、PE−5、VR−1、BR−1、BR−2、ME−2、VR−2、MA−3、MA−4、CAT−1、CB−1、NR−1、BT−1、およびAL−1からなる群より選択されるSaccharomyces cerevisiaeの株である。いくつかの実施形態において、微生物は、PE−2、CAT−1、VR−1、BG−1、CR−1、およびSA−1からなる群より選択されるSaccharomyces cerevisiaeの株である。特定の実施形態において、Saccharomyces cerevisiaeの株はPE−2である。別の特定の実施形態において、Saccharomyces cerevisiaeの株はCAT−1である。別の特定の実施形態において、Saccharomyces cerevisiaeの株はBG−1である。
【0052】
いくつかの実施形態において、微生物は、産業的発酵、例えば、バイオエタノール発酵に適している微生物である。特定の実施形態において、微生物は、認識されている産業的発酵環境のストレス条件である、高溶媒濃度、高温、広範な基質利用、栄養制限、糖および塩による浸透圧ストレス、酸過剰、亜硫酸塩および細菌の汚染、またはそれらの組み合わせの下で存在するように条件付けされる。
【0053】
(微生物の遺伝子改変)
組換えプラスミドまたは染色体組込みベクターを用いて微生物を遺伝的に改変する方法は、当技術分野においてよく知られている。例えば、Sherman,F.ら、Methods Yeast Genetics、Cold Spring Harbor Laboratory、N.Y.(1978);Guthrie,C.ら(編)Guide To Yeast Genetics and Molecular Biology 194巻、Academic Press、San Diego(1991);Sambrookら、2001、Molecular Cloning−A Laboratory Manual、第3版、Cold Spring Harbor Laboratory、Cold Spring Harbor、NY;およびAusubelら編、現行版、Current Protocols in Molecular Biology、Greene Publishing Associates and Wiley Interscience、NYを参照;それらの開示は参照により本明細書に組み入れられている。
【0054】
いくつかの実施形態において、微生物は、新しい代謝経路、すなわち、微生物によって内因的に産生されない代謝産物を産生する代謝経路の酵素をコードする1つまたは複数の異種性ヌクレオチド配列を含むように遺伝子改変される。他の実施形態において、微生物は、微生物にとって内因性である代謝経路、すなわち、微生物によって内因的に産生される代謝産物を産生する代謝経路の酵素をコードする1つまたは複数の異種性ヌクレオチド配列を含むように遺伝子改変される。
【0055】
いくつかの実施形態において、本発明の方法は、フェロモン応答遺伝子の1つまたはHOによってコードされるものなどの特定のタンパク質を微生物において一過性に発現させるために組換えプラスミドの使用を必要とする。酵母細胞に用いるのに適した組換えプラスミドの例証的な例として、CEN/ARSおよび2μのプラスミドが挙げられる。
【0056】
いくつかの実施形態において、本発明の微生物は、抗生物質耐性をコードする異種性ヌクレオチド配列を含まない。抗生物質耐性マーカーは、遺伝子改変細胞の構築に一般的に用いられる。抗生物質耐性マーカーが、微生物に導入された遺伝子改変をマークするために用いられる本発明のそのような実施形態において、これらのマーカーは、所望の遺伝子改変の全部が微生物になされた後で、続いて欠失させられる。または、栄養要求性相補(例えば、HIS3、LEU2、LYS1、MET15、TRP1、ADE2、URA3、およびLYS2))などの他の選択ツールを遺伝子改変微生物の構築に用いることができる。
【0057】
(胞子形成および/またはフェロモン応答遺伝子の破壊)
本明細書に提供された方法は、遺伝子改変微生物細胞において1つもしくは複数の胞子形成遺伝子および/または1つもしくは複数のフェロモン応答遺伝子を機能的に破壊する工程を含む。いくつかの実施形態において、1つまたは複数の胞子形成遺伝子の破壊は、胞子形成能力を欠く遺伝子改変微生物細胞を生じる。特に、遺伝子改変二倍体微生物細胞は胞子形成能力を欠く。いくつかの実施形態において、1つまたは複数のフェロモン応答遺伝子の破壊は、内因性接合が損なわれている微生物細胞を生じる。いくつかの実施形態において、1つまたは複数の胞子形成遺伝子の破壊および1つまたは複数のフェロモン応答遺伝子の破壊は、胞子形成および内因性接合が損なわれている微生物細胞を生じる。
【0058】
いくつかの実施形態において、胞子形成遺伝子またはフェロモン応答遺伝子の破壊は、微生物細胞への構築物の導入で胞子形成標的遺伝子またはフェロモン応答標的遺伝子を特異的に破壊する能力がある「破壊構築物」を用いて、破壊される遺伝子を非機能性にすることによって達成される。いくつかの実施形態において、標的遺伝子の破壊は、機能性タンパク質の発現を阻止する。いくつかの実施形態において、標的遺伝子の破壊は、破壊される遺伝子からの非機能性タンパク質の発現を生じる。いくつかの実施形態において、胞子形成標的遺伝子またはフェロモン応答標的遺伝子の破壊は、相同組換えにより標的遺伝子座内の「破壊配列」の組込みによって達成される。そのような実施形態において、破壊構築物は、標的遺伝子座の1対のヌクレオチド配列に相同である1対のヌクレオチド配列(相同配列)に隣接した破壊配列を含む。破壊構築物による標的遺伝子の標的部分の置換で、破壊配列は、機能性タンパク質の発現を阻止し、または標的遺伝子からの非機能性タンパク質の発現を引き起こす。
【0059】
1つまたは複数の胞子形成遺伝子またはフェロモン応答遺伝子を破壊する能力がある破壊構築物は、当技術分野において周知の標準の分子生物学技術を用いて構築することができる。例えば、Sambrookら、2001、Molecular Cloning−A Laboratory Manual、第3版、Cold Spring Harbor Laboratory、Cold Spring Harbor、NYおよびAusubelら編、現行版、Current Protocols in Molecular Biology、Greene Publishing Associates and Wiley Interscience、NYを参照。本方法の実施において変わり得る破壊構築物のパラメータには、相同配列の長さ、相同配列のヌクレオチド配列、破壊配列の長さ、破壊配列のヌクレオチド配列、および標的遺伝子のヌクレオチド配列が挙げられるが、それらに限定されない。いくつかの実施形態において、各相同配列の長さについての有効な範囲は50〜5,000塩基対である。特定の実施形態において、各相同配列の長さは約500塩基対である。遺伝子を標的にするのに必要とされる相同性の長さの議論については、Hastyら、Mol Cell Biol 11:5586−91(1991)を参照されたい。いくつかの実施形態において、相同配列は、標的遺伝子のコード配列を含む。他の実施形態において、相同配列は、標的遺伝子の上流配列または下流配列を含む。いくつかの実施形態において、一方の相同配列は、標的遺伝子のコード配列の5’に位置するヌクレオチド配列と相同であるヌクレオチド配列を含み、他方の相同配列は、標的遺伝子のコード配列の3’に位置するヌクレオチド配列と相同であるヌクレオチド配列を含む。いくつかの実施形態において、破壊配列は、破壊配列を含む微生物細胞の選択を可能にする選択マーカーをコードするヌクレオチド配列を含む。したがって、そのような実施形態において、破壊構築物は、二重機能、すなわち、標的遺伝子を機能的に破壊すること、および標的遺伝子が機能的に破壊されている細胞の同定のための選択マーカーを提供することを有する。いくつかの実施形態において、終止コドンは、標的遺伝子によってコードされる野生型タンパク質のある程度の活性を有する融合タンパク質を生じる可能性がある翻訳の読み過ごしを防ぐために選択マーカーをコードするヌクレオチド配列とインフレームで、および、それの下流に位置する。いくつかの実施形態において、破壊配列の長さは、1塩基対である。単一塩基対の挿入は、コード配列における単一塩基対の挿入が、機能性タンパク質の発現を阻止することができるフレームシフト突然変異を構成することができるため、標的遺伝子を破壊するのに十分であり得る。いくつかの実施形態において、破壊配列の配列は、相同配列の間に位置する標的遺伝子のヌクレオチド配列と単一塩基対だけ、異なる。標的遺伝子内のヌクレオチド配列の破壊配列との置換で、導入される単一塩基対置換は、タンパク質における重要部位での単一アミノ酸置換、および非機能性タンパク質の発現を生じ得る。しかしながら、非常に短い破壊配列を用いてもたらされる破壊は、自然突然変異によって野生型配列への復帰変異を起こしやすく、したがって、接合および胞子形成能力の回復を宿主株にもたらすことは認識されるべきである。したがって、特定の実施形態において、破壊配列は、1〜2、3塩基対より長い。それとは正反対に、過剰な長さの破壊配列は、中程度の長さの破壊配列に優る少しの利点でも与える可能性は低く、トランスフェクションまたはターゲティングの効率を減少させる可能性がある。この関連における過剰な長さは、標的遺伝子における選択された相同配列間の距離より何倍も長い。このように、ある特定の実施形態において、破壊配列についての長さは、2塩基対から2,000塩基対までであり得る。他の実施形態において、破壊配列についての長さは、破壊構築物における相同配列とマッチする標的遺伝子座の領域間の距離とおよそ等しい長さである。
【0060】
いくつかの実施形態において、破壊構築物は、直鎖DNA分子である。他の実施形態において、破壊構築物は、環状DNA分子である。いくつかの実施形態において、環状破壊構築物は、上記のような破壊配列によって分離した1対の相同配列を含む。いくつかの実施形態において、環状破壊構築物は、単一の相同配列を含む。そのような環状破壊構築物は、標的遺伝子座での組込みで、直鎖状になり、相同配列の部分は各末端に位置し、破壊構築物の残りのセグメントは、標的遺伝子ヌクレオチド配列のいずれとも置換することなく、標的遺伝子に挿入され、それを破壊するであろう。特定の実施形態において、環状破壊構築物の単一の相同配列は、標的遺伝子のコード配列内に位置する配列と相同である。
【0061】
破壊構築物は、非限定的に、当業者に公知の任意の方法によって微生物細胞へ導入することができる。そのような方法には、溶液からの細胞による分子の直接的取り込み、または例えば、リポソームもしくは免疫リポソームを用いるリポフェクションによる促進性取り込み;粒子媒介性トランスフェクションなどが挙げられるが、それらに限定されない。例えば、米国特許第5,272,065号;Goeddelら編、1990、Methods in Enzymology、185巻、Academic Press, Inc.、CA;Krieger、1990、Gene Transfer and Expression−A Laboratory Manual、Stockton Press、NY;Sambrookら、1989、Molecular Cloning−A Laboratory Manual、Cold Spring Harbor Laboratory、NY;およびAusubelら編、現行版、Current Protocols in Molecular Biology、Greene Publishing Associates and Wiley Interscience、NYを参照。酵母細胞を形質転換する特定の方法は当技術分野において周知である。Hinnenら、Proc.Natl.Acad.Sci.USA 75:1292−3(1978);Creggら、Mol.Cell.Biol.5:3376−3385(1985)参照。例示的な技術として、スフェロプラスト化、エレクトロポレーション、PEG 1000媒介性形質転換、および酢酸リチウムまたは塩化リチウム媒介性形質転換が挙げられるが、それらに限定されない。
【0062】
(フェロモン応答遺伝子)
いくつかの実施形態において、本明細書に提供された方法に従って破壊される酵母細胞におけるフェロモン応答遺伝子は、STE5である。STE5は、酵母接合経路を通してのマイトジェン活性化プロテインキナーゼ(MAPK)への直接的シグナル伝達に必要とされるスキャフォールドタンパク質をコードする。例えば、Goodら、Cell Mar 20;136(6):1085−97(2009)参照。Saccharomyces cerevisiaeの代表的なSTE5ヌクレオチド配列には、Genbankアクセッション番号L23856、ならびに本明細書に提供されているような配列番号17、45、73、101、および129が挙げられる。Saccharomyces cerevisiaeの代表的なSte5タンパク質配列には、Genbankアクセッション番号AAA35115、ならびに本明細書に提供されているような配列番号18、46、74、102、および130が挙げられる。
【0063】
いくつかの実施形態において、本明細書に提供された方法に従って破壊される酵母細胞におけるフェロモン応答遺伝子は、STE4である。STE4遺伝子は、Ste18pと二量体を形成して接合シグナル伝達経路を活性化するGタンパク質βサブユニットをコードする。Saccharomyces cerevisiaeのSTE4遺伝子の配列は以前に記載されている。Dujonら、Nature 387(6632 Suppl):98−102(1997)。Saccharomyces cerevisiaeの代表的なSTE4ヌクレオチド配列には、Genbankアクセッション番号NC_001147.5、ならびに本明細書に提供されているような配列番号19、47、75、103、および131が挙げられる。Saccharomyces cerevisiaeの代表的なSte4タンパク質配列には、Genbankアクセッション番号NP_014855、ならびに本明細書に提供されているような配列番号20、48、76、104、および132が挙げられる。
【0064】
いくつかの実施形態において、本明細書に提供された方法に従って破壊される酵母細胞におけるフェロモン応答遺伝子は、STE18である。STE18遺伝子は、Ste4pと二量体を形成して接合シグナル伝達経路を活性化するGタンパク質γサブユニットをコードする。Saccharomyces cerevisiaeのSTE18遺伝子の配列は以前に記載されている。例えば、Goffeauら、Science 274(5287):546−547(1996)参照。Saccharomyces cerevisiaeの代表的なSTE18ヌクレオチド配列には、Genbankアクセッション番号NC_001147.5、ならびに本明細書に提供されているような配列番号21、49、77、105、および133が挙げられる。Saccharomyces cerevisiaeの代表的なSte18タンパク質配列には、Genbankアクセッション番号NP_012619、ならびに本明細書に提供されているような配列番号22、50、78、106、および134が挙げられる。
【0065】
いくつかの実施形態において、本明細書に提供された方法に従って破壊される酵母細胞におけるフェロモン応答遺伝子は、STE12である。STE12遺伝子は、MAPキナーゼシグナル伝達カスケードによって活性化され、および、接合経路または仮性菌糸/浸潤性増殖経路に関与する遺伝子を活性化する転写因子をコードする。Saccharomyces cerevisiaeのSTE12遺伝子の配列は以前に記載されている。例えば、Goffeauら、Science 274(5287):546−547(1996)参照。Saccharomyces cerevisiaeの代表的なSTE12ヌクレオチド配列には、Genbankアクセッション番号NC_001140.5、ならびに本明細書に提供されているような配列番号23、51、79、107、および135が挙げられる。Saccharomyces cerevisiaeの代表的なSte12タンパク質配列には、Genbankアクセッション番号NP_011952、ならびに本明細書に提供されているような配列番号24、52、80、108、および136が挙げられる。
【0066】
いくつかの実施形態において、本明細書に提供された方法に従って破壊される酵母細胞におけるフェロモン応答遺伝子は、STE7である。STE7遺伝子は、Fus3pをリン酸化する、フェロモン応答に関与するシグナル伝達MAPキナーゼをコードする。Saccharomyces cerevisiaeのSTE7遺伝子の配列は以前に記載されている。例えば、Teagueら、Proc Natl Acad Sci USA.83(19):7371−5(1986)参照。Saccharomyces cerevisiaeの代表的なSTE7ヌクレオチド配列には、Genbankアクセッション番号Z74207、ならびに本明細書に提供されているような配列番号25、53、81、109、および137が挙げられる。Saccharomyces cerevisiaeの代表的なSte7タンパク質配列には、Genbankアクセッション番号CAA98732、ならびに本明細書に提供されているような配列番号26、54、82、110、および138が挙げられる。
【0067】
いくつかの実施形態において、本明細書に提供された方法に従って破壊される酵母細胞におけるフェロモン応答遺伝子は、STE11である。STE11遺伝子は、Ste7pをリン酸化する、フェロモン応答および仮性菌糸/浸潤性増殖経路に関与するシグナル伝達MEKキナーゼをコードする。Saccharomyces cerevisiaeのSTE11遺伝子の配列は以前に記載されている。例えば、Johnstonら、Nature 387(6632 Suppl)、87−90(1997)参照。Saccharomyces cerevisiaeの代表的なSTE11ヌクレオチド配列には、Genbankアクセッション番号NC_001144.4、ならびに本明細書に提供されているような配列番号27、55、83、111、および139が挙げられる。Saccharomyces cerevisiaeの代表的なSte11タンパク質配列には、Genbankアクセッション番号NP_013466、ならびに本明細書に提供されているような配列番号28、56、84、112、および140が挙げられる。
【0068】
(胞子形成遺伝子)
いくつかの実施形態において、本明細書に提供された方法に従って破壊される酵母細胞における胞子形成遺伝子はIME1である。IME1遺伝子は、減数分裂の開始に必要とされる、初期減数分裂遺伝子転写を活性化する転写因子をコードする。Saccharomyces cerevisiaeのIME1遺伝子の配列は以前に記載されている。例えば、Smith,H.E.ら、Mol.Cell.Biol.10(12):6103−6113(1990)参照。Saccharomyces cerevisiaeの代表的なIME1ヌクレオチド配列には、Genbankアクセッション番号M37188、ならびに本明細書に提供されているような配列番号1、29、57、85、および113が挙げられる。Saccharomyces cerevisiaeの代表的なIme1タンパク質配列には、Genbankアクセッション番号AAA86790、ならびに本明細書に提供されているような配列番号2、30、58、86、および114が挙げられる。
【0069】
いくつかの実施形態において、本明細書に提供された方法に従って破壊される酵母細胞における胞子形成遺伝子はIME2である。IME2遺伝子は、減数分裂の活性化に関与するセリン/スレオニンプロテインキナーゼをコードする。Saccharomyces cerevisiaeのIME2遺伝子の配列は以前に記載されている。例えば、EMBO J.15(9)、2031−2049(1996)参照。Saccharomyces cerevisiaeの代表的なIME2ヌクレオチド配列には、Genbankアクセッション番号NC_001142、ならびに本明細書に提供されているような配列番号3、31、59、87、および115が挙げられる。Saccharomyces cerevisiaeの代表的なIme2タンパク質配列には、Genbankアクセッション番号NP_012429、ならびに本明細書に提供されているような配列番号4、32、60、88、および116が挙げられる。
【0070】
いくつかの実施形態において、本明細書に提供された方法に従って破壊される酵母細胞における胞子形成遺伝子はNDT80である。NDT80遺伝子は、太糸期からの脱出および完全減数分裂組換えに必要とされる減数分裂特異的転写因子をコードする。Ndt80タンパク質はまた、中間胞子形成遺伝子を活性化する。Saccharomyces cerevisiaeのNDT80遺伝子の配列は以前に記載されている。例えば、Goffeauら、Science 274(5287):546−547(1996)参照。Saccharomyces cerevisiaeの代表的なNDT80ヌクレオチド配列には、Genbankアクセッション番号NC_001140、ならびに本明細書に提供されているような配列番号5、33、61、89、および117が挙げられる。Saccharomyces cerevisiaeの代表的なNdt80タンパク質配列には、Genbankアクセッション番号NP_011992、ならびに本明細書に提供されているような配列番号6、34、62、90、および118が挙げられる。
【0071】
いくつかの実施形態において、本明細書に提供された方法に従って破壊される酵母細胞における胞子形成遺伝子はSPO11である。SPO11遺伝子は、減数分裂組換えに必要とされる。Saccharomyces cerevisiaeのSPO11遺伝子の配列は以前に記載されている。例えば、Atchesonら、Proc.Natl.Acad.Sci.U.S.A. 84(22)、8035−8039(1987)参照。Saccharomyces cerevisiaeの代表的なSPO11ヌクレオチド配列には、Genbankアクセッション番号J02987、ならびに本明細書に提供されているような配列番号7、35、63、91、および119が挙げられる。Saccharomyces cerevisiaeの代表的なSpo11タンパク質配列には、Genbankアクセッション番号AAA65532、ならびに本明細書に提供されているような配列番号8、36、64、92、および120が挙げられる。
【0072】
いくつかの実施形態において、本明細書に提供された方法に従って破壊される酵母細胞における胞子形成遺伝子はSPO20である。SPO20遺伝子は、胞子形成中の前胞子膜形成に必要とされる、t−SNARE複合体の減数分裂特異的サブユニットをコードする。Saccharomyces cerevisiaeのSPO20遺伝子の配列は以前に記載されている。例えば、Bowmanら、Nature 387(6632 Suppl)、90−93(1997)参照。Saccharomyces cerevisiaeの代表的なSPO20ヌクレオチド配列には、Genbankアクセッション番号AF078740、ならびに本明細書に提供されているような配列番号9、37、65、93、および121が挙げられる。Saccharomyces cerevisiaeの代表的なSpo20タンパク質配列には、Genbankアクセッション番号NP_013730、ならびに本明細書に提供されているような配列番号10、38、66、94、および122が挙げられる。
【0073】
いくつかの実施形態において、本明細書に提供された方法に従って破壊される酵母細胞における胞子形成遺伝子はAMA1である。AMA1遺伝子は、減数分裂後期促進複合体のアクチベーターをコードする。Saccharomyces cerevisiaeのAMA1遺伝子の配列は以前に記載されている。例えば、Tettelinら、Nature 387(6632 Suppl):81−84(1997)参照。Saccharomyces cerevisiaeの代表的なAMA1ヌクレオチド配列には、Genbankアクセッション番号NC_001139.8、ならびに本明細書に提供されているような配列番号11、39、67、95、および123が挙げられる。Saccharomyces cerevisiaeの代表的なAma1タンパク質配列には、Genbankアクセッション番号NP_011741、ならびに本明細書に提供されているような配列番号12、40、68、96、および124が挙げられる。
【0074】
いくつかの実施形態において、本明細書に提供された方法に従って破壊される酵母細胞における胞子形成遺伝子はHOP2である。HOP2遺伝子は、相同染色体間のシナプシスを確実にする減数分裂特異的タンパク質をコードする。Saccharomyces cerevisiaeのHOP2遺伝子の配列は以前に記載されている。例えば、Leuら、Cell 94(3):375−386(1998)参照。Saccharomyces cerevisiaeの代表的なHOP2ヌクレオチド配列には、Genbankアクセッション番号AF_078740.1、ならびに本明細書に提供されているような配列番号13、41、69、97、および125が挙げられる。Saccharomyces cerevisiaeの代表的なHop2タンパク質配列には、Genbankアクセッション番号AAC31823、ならびに本明細書に提供されているような配列番号14、42、70、98、および126が挙げられる。
【0075】
いくつかの実施形態において、本明細書に提供された方法に従って破壊される酵母細胞における胞子形成遺伝子はSPO21である。SPO21遺伝子は、前胞子膜形成の前に必要とされる減数分裂外側プラーク(meiotic outer plaque)を修飾することに関与する、紡錘極体の減数分裂外側プラークの成分をコードする。Saccharomyces cerevisiaeのSPO21遺伝子の配列は以前に記載されている。例えば、Dujonら、Nature 387(6632 Suppl):98−102(1997)参照。Saccharomyces cerevisiaeの代表的なSPO21ヌクレオチド配列には、Genbankアクセッション番号NC_001147.5、ならびに本明細書に提供されているような配列番号15、43、71、99、および127が挙げられる。Saccharomyces cerevisiaeの代表的なSpo21タンパク質配列には、Genbankアクセッション番号NP_014550、ならびに本明細書に提供されているような配列番号16、44、72、100、および128が挙げられる。
【0076】
(二倍体の調製)
本明細書に提供された方法は、1つもしくは複数の胞子形成遺伝子における機能的破壊および/または1つまたは複数のフェロモン応答遺伝子における機能的破壊を含む一倍体細胞の間で接合を誘導する工程を含む。前記接合の結果として形成される二倍体細胞は、細胞の1つまたは複数の胞子形成遺伝子の機能的破壊による二倍体相に制約される安定な二倍体細胞である。
【0077】
接合能力を欠く一倍体細胞から二倍体細胞を形成するために、接合欠陥性一倍体細胞は、「接合相補プラスミド」、すなわち、1つまたは複数のフェロモン応答遺伝子における機能的破壊によって引き起こされた接合欠損を相補することができる異種性遺伝子を含む組換えプラスミドで形質転換される。一倍体細胞内の異種性フェロモン応答遺伝子の一過性発現は、一時的に、細胞に接合機能を回復させ、反対接合型の一倍体細胞が安定な二倍体細胞を形成することを可能にする。特に、それによって形成された安定な二倍体細胞は、それらの接合型対立遺伝子について以外はホモ接合であり、同じ遺伝子改変集団の一倍体から生成される。
【0078】
したがって、一倍体細胞がSTE5遺伝子の機能的破壊を含むいくつかの実施形態において、一倍体細胞は、STE5コード配列を含む接合相補プラスミドで形質転換される。一倍体細胞がSTE4遺伝子の機能的破壊を含むいくつかの実施形態において、一倍体細胞は、STE4コード配列を含む接合相補プラスミドで形質転換される。一倍体細胞がSTE18遺伝子の機能的破壊を含むいくつかの実施形態において、一倍体細胞は、STE18コード配列を含む接合相補プラスミドで形質転換される。一倍体細胞がSTE12遺伝子の機能的破壊を含むいくつかの実施形態において、一倍体細胞は、STE12コード配列を含む接合相補プラスミドで形質転換される。一倍体細胞がSTE7遺伝子の機能的破壊を含むいくつかの実施形態において、一倍体細胞は、STE7コード配列を含む接合相補プラスミドで形質転換される。一倍体細胞がSTE11遺伝子の機能的破壊を含むいくつかの実施形態において、一倍体細胞は、STE11コード配列を含む接合相補プラスミドで形質転換される。
【0079】
発現ベクターの構築、および発現ベクターを含む細胞における遺伝子の発現についての技術は、当技術分野において周知である。例えば、Sambrookら、2001、Molecular Cloning−A Laboratory Manual、第3版、Cold Spring Harbor Laboratory、Cold Spring Harbor、NY、およびAusubelら編、現行版、Current Protocols in Molecular Biology、Greene Publishing Associates and Wiley Interscience、NYを参照。接合相補遺伝子をコードするプラスミドは、当業者に公知の任意の方法によって宿主細胞へ導入することができる。
【0080】
プラスミドに基づいた系は、一般的に、細胞において外来DNAを維持するようにプラスミドに選択圧を必要とする。酵母細胞は典型的には、有糸分裂後ごとに、細胞に含まれる10%のプラスミドを喪失するため、酵母におけるたいていのプラスミドは相対的に不安定である。したがって、いくつかの実施形態において、接合相補遺伝子をコードするプラスミドを含む一倍体細胞の接合によって形成されたが、それ自体、そのプラスミドを含まない二倍体細胞の選択は、プラスミドが集団から効果的に希釈されるように十分な有糸分裂を二倍体細胞に起こさせることによって達成される。あるいは、二倍体細胞は、プラスミドの非存在について選択することによって、例えば、対抗選択マーカー(例えば、URA3など)に対して選択することによって、または選択培地および非選択培地の両方に同一のコロニーを蒔き、その後、選択培地で増殖しないが、非選択培地で増殖するコロニーを選択することによって、選択することができる。
【0081】
いくつかの実施形態において、本明細書に提供される方法は、一倍体ヘテロタリック(ho)酵母細胞を、ホモタリズム(HO)タンパク質をコードする組換えプラスミドで形質転換する工程であって、HOタンパク質の発現が一倍体細胞の接合型切り換えを誘導する、工程を含む。Saccharomyces cerevisiaeのHO遺伝子の配列は以前に記載されている。例えば、Russellら、Mol.Cell.Biol. 6(12):4281−4294(1986)参照。Saccharomyces cerevisiaeの代表的なHOヌクレオチド配列には、Genbankアクセッション番号NC_001136および本明細書に提供されているような配列番号151が挙げられる。Saccharomyces cerevisiaeの代表的なHOタンパク質配列には、Genbankアクセッション番号NP_010054および本明細書に提供されているような配列番号152が挙げられる。
【実施例】
【0082】
実施例1:遺伝子改変二倍体細胞の作製
この実施例は、遺伝子改変一倍体S.cerevisiae細胞を作製する例示的な方法を記載する。
【0083】
相I破壊構築物(図1;配列番号141)は、破壊配列として、S.cerevisiae GAL80座の上流ヌクレオチド配列および下流ヌクレオチド配列からなる相同配列に隣接した、選択マーカー(ハイグロマイシンB耐性を与えるhygA);ガラクトース誘導性プロモーター(S.cerevisiae遺伝子GAL1およびGAL10のプロモーター)の制御下の、S.cerevisiae MEV経路の2つの酵素(切断型HMG−CoAリダクターゼをコードする切断型HMG1コード配列およびHMG−CoAシンターゼをコードするERG13コード配列)およびS.cerevisiaeの別の酵素(アセトアセチル−CoAチオラーゼをコードするERG10コード配列)をコードするヌクレオチド配列を含む。S.cerevisiae宿主細胞への導入で、相I破壊構築物は、S.cerevisiae宿主細胞ゲノムのGAL80座へ相同組換えによって組み込まれ、GAL80コード配列をその破壊配列と置換することによってGAL80座を機能的に破壊することができる。相I破壊構築物を、TOPO Zero Blunt IIクローニングベクター(Invitrogen、Carlsbad、CA)へクローニングし、プラスミドTOPO−相I破壊構築物を生じた。その構築物を、50μg/mlのカナマイシンを含むLB寒天上で増殖するTOP10細胞中に繁殖させた。
【0084】
相II破壊構築物(図2;配列番号142)は、破壊配列として、S.cerevisiae LEU2座の上流ヌクレオチド配列および下流ヌクレオチド配列からなる相同配列に隣接した、選択マーカー(ノールセオスリシン(nourseothricin)耐性を与えるnatA)、およびガラクトース誘導性プロモーター(S.cerevisiae遺伝子GAL1およびGAL10のプロモーター)の制御下のS.cerevisiae MEV経路の数個の酵素(メバロン酸キナーゼをコードするERG12コード配列およびホスホメバロン酸キナーゼをコードするERG8コード配列)をコードするヌクレオチド配列;加えて、GAL4ocプロモーター(MIG1結合部位を除去する突然変異を含むGAL4プロモーターであって、それにしたがって、プロモーターをグルコースの抑圧に対してより低感受性にさせている)の制御下のS.cerevisiae GAL4遺伝子のコード配列を含む。S.cerevisiae宿主細胞への導入で、相II破壊構築物は、S.cerevisiae宿主細胞ゲノムのLEU2座へ相同組換えによって組み込まれ、LEU2コード配列をその破壊配列と置換することによってLEU2座を機能的に破壊することができる。相II破壊構築物を、TOPO Zero Blunt IIクローニングベクターへクローニングし、プラスミドTOPO−相II破壊構築物を生じた。その構築物を、50μg/mlのカナマイシンを含むLB寒天上で増殖するTOP10細胞(Invitrogen、Carlsbad、CA)中に繁殖させた。
【0085】
相III破壊構築物(図3;配列番号143)は、破壊配列として、S.cerevisiae ERG9座の上流ヌクレオチド配列およびコードヌクレオチド配列に隣接した、選択マーカー(G418耐性を与えるkanA);S.cerevisiae MEV経路の酵素(ジホスホメバロン酸デカルボキシラーゼをコードするERG19コード配列)、およびガラクトース誘導性プロモーター(S.cerevisiae遺伝子GAL1、GAL10、およびGAL7のプロモーター)の制御下の、MEV経路の産物のIPPをFPPへ変換することに関与するS.cerevisiaeの2つの酵素(ERG20コード配列はファルネシルピロリン酸シンターゼをコードし、IDI1コード配列はイソペンテニルピロリン酸デカルボキシラーゼをコードする)をコードするヌクレオチド配列;加えて、S.cerevisiae CTR3遺伝子のプロモーターを含む。S.cerevisiae宿主細胞への導入で、相II破壊構築物は、S.cerevisiae宿主細胞ゲノムのERG9座の上流に相同組換えによって組み込まれ、ERG9(スクワレンシンターゼ)の発現が銅によって調節され得るように天然ERG9プロモーターをCTR3プロモーターで置換することができる。相III破壊構築物を、TOPO Zero Blunt IIクローニングベクターへクローニングし、プラスミドTOPO−相III破壊構築物を生じた。その構築物を、50μg/mlのカナマイシンを含むLB寒天上で増殖するTOP10細胞中に繁殖させた。
【0086】
相Iマーカーリサイクリング構築物(図4;配列番号144)は、S.cerevisiae GAL80座の上流ヌクレオチド配列およびS.cerevisiae HMG1遺伝子のコード配列に隣接した、選択マーカー(ウラシル欠損培地で増殖する能力を与えるURA3);およびS.cerevisiae GAL7遺伝子のプロモーターの制御調節下のA.annuaの酵素(ファルネセンシンターゼをコードするFSコード配列)をコードするヌクレオチド配列を含む。S.cerevisiae宿主細胞への導入で、相Iマーカーリサイクリング構築物は、選択マーカーhphAがURA3と置換されるように、すでに組み込まれた相I破壊配列へ相同組換えによって組み込むことができる。
【0087】
相IIマーカーリサイクリング構築物(図5;配列番号145)は、S.cerevisiae LEU2座の上流ヌクレオチド配列およびS.cerevisiae ERG12遺伝子のコード配列に隣接した、選択マーカー(ウラシル欠損培地で増殖する能力を与えるURA3)およびS.cerevisiae GAL7遺伝子のプロモーターの制御調節下のA annuaの酵素(ファルネセンシンターゼをコードするFSコード配列)をコードするヌクレオチド配列を含む。S.cerevisiae宿主細胞への導入で、相IIマーカーリサイクリング構築物は、選択マーカーnatAがURA3と置換されるように、すでに組み込まれた相II破壊配列へ相同組換えによって組み込むことができる。
【0088】
相IIIマーカーリサイクリング構築物(図6;配列番号146)は、S.cerevisiae ERG9座の上流ヌクレオチド配列およびS.cerevisiae ERG19遺伝子のコード配列に隣接した、選択マーカー(ウラシル欠損培地で増殖する能力を与えるURA3)およびS.cerevisiae GAL7遺伝子のプロモーターの制御調節下のA annuaの酵素(ファルネセンシンターゼをコードするFSコード配列)をコードするヌクレオチド配列を含む。S.cerevisiae宿主細胞への導入で、相IIマーカーリサイクリング構築物は、選択マーカーkanAがURA3と置換されるように、すでに組み込まれた相III破壊配列へ相同組換えによって組み込むことができる。
【0089】
発現プラスミドpAM404(配列番号153)は、β−ファルネセンシンターゼをコードする。ヌクレオチド配列挿入断片を、Saccharomyces cerevisiaeにおける発現のためにコドン最適化されたArtemisia annua(GenBankアクセッション番号AY835398)のβ−ファルネセンシンターゼ遺伝子のコード配列を鋳型として用いて、合成的に作製した。
【0090】
スターターの宿主株Y1198については、100ug/mLのカルバミシリンおよび50ug/mLのカナマイシンを含む5mLのYPD培地中に活性乾燥PE−2酵母(1994年に単離された;Santelisa Vale、Sertaozinho、Brazilからの贈与)を再懸濁することによって作製した。培養物を、200rpmでの回転式振盪機で、30℃、一晩、インキュベートした。その後、10uLの培養物のアリコートをYPDプレートに蒔き、乾燥させた。単一のコロニーのために細胞を連続的に画線し、30℃で2日間、インキュベートした。12個の単一コロニーを採取し、新しいYPDプレート上にパッチ状に置き、30℃で一晩、増殖させた。コロニーの株アイデンティティを、製造会社の仕様書に従い、Bio−Rad CHEF Genomic DNA Plug Kit(Bio−Rad、Hercules、CA)を用いてBio−Rad CHEF DR IIシステム(Bio−Rad、Hercules、CA)でそれらの染色体サイズを分析することによって検証した。1個のコロニーを採取し、Y1198株として保存した。
【0091】
Y1198株から、遺伝子操作を可能にするためにその株の一倍体にすることによって、Y1661株、Y1662株、Y1663株、およびY1664株を作製した。Y1198株を、回転式ドラム内のガラスチューブにおいて、5mLのYPD培地中、30℃で一晩、増殖させた。OD600を測定し、細胞を、2%の酢酸カリウムを含む5mLのYP培地中、0.2のOD600に希釈した。培養物を、回転式ドラム内のガラスチューブにおいて30℃で一晩、増殖させた。OD600を再び、測定し、4OD600*mLの細胞を、5,000 x gで2分間の遠心分離によって収集した。細胞ペレットを滅菌水で一度、洗浄し、その後、0.02%のラフィノースを含む3mLの2%酢酸カリウム中に再懸濁した。細胞を、回転式ドラム内のガラスチューブにおいて30℃で3日間、増殖させた。胞子形成を顕微鏡で確認した。33μLの培養物のアリコートを1.5mLの微量遠心管に移し、14,000rpmで2分間、遠心分離した。細胞ペレットを、2μLの10mg/mL Zymolyase 100T(MP Biomedicals、Solon、OH)を含む50μLの滅菌水中に再懸濁し、細胞を30℃の水浴中で10分間、インキュベートした。チューブを氷に移し、150μLの氷水を加えた。10μLのこの混合物のアリコートを12mLのYPDプレートに加え、Singer MSM 300ダイセクション顕微鏡(Singer、Somerset、UK)において四分子を切り離した。YPDプレートを30℃で3日間、インキュベートし、その後、胞子を新鮮なYPDプレート上にパッチ状に置き、30℃で一晩、増殖させた。8個の4胞子四分子からの各胞子の接合型をコロニーPCRによって分析した。2個のMATa胞子および2個のMATα胞子を有する単一の4胞子四分子を採取し、Y1661株(MATa)、Y1662株(MATa)、Y1663株(MATα)、およびY1664株(MATα)として保存した。
【0092】
酵母細胞形質転換について、25mlの酵母エキスペプトンデキストロース(YPD)培地に、出発宿主株の単一コロニーを接種した。培養物を、200rmpでの回転式振盪機上で30℃、一晩、増殖させた。培養物のOD600を測定し、その後、培養物を用いて、50mlのYPD培地に0.15のOD600まで接種した。その新しく接種された培養物を200rpmでの回転式振盪機において30℃で、0.7〜0.9のOD600まで増殖させ、その時点で、細胞を1μgのDNAで形質転換した。宿主細胞形質転換体を同定するために選択剤を含む寒天上に細胞を蒔く前に、細胞をYPD培地中で4時間、回復させた。
【0093】
PmeI制限エンドヌクレアーゼを用いて完了まで消化されたプラスミドTOPO相I破壊構築物でY1664株を形質転換することによって宿主株Y1515を作製した。宿主細胞形質転換体を、300ug/mLのハイグロマイシンBを含むYPD培地上で選択し、GAL80座に組み込まれた相I破壊配列を含む陽性形質転換体を、PCR増幅によって検証した。
【0094】
PmeI制限エンドヌクレアーゼを用いて完了まで消化されたプラスミドTOPO相II破壊構築物でY1515株を形質転換することによって宿主株Y1762を作製した。宿主細胞形質転換体を、100ug/mLのノールセオスリシンを含むYPD培地上で選択し、LEU2座に組み込まれた相II破壊配列を含む陽性形質転換体を、PCR増幅によって検証した。
【0095】
PmeI制限エンドヌクレアーゼを用いて完了まで消化された、発現プラスミドpAM404およびプラスミドTOPO相III破壊構築物での2段階でY1762株を形質転換することによって宿主株Y1770を作製した。pAM404を有する宿主細胞形質転換体を、メチオニンおよびロイシンを欠損する完全合成培地(CSM)上で選択した。pAM404および相III破壊構築物を有する宿主細胞形質転換体を、メチオニンおよびロイシンを欠損し、および、200ug/mlのG418(Geneticin(登録商標))を含むCSM上で選択し、ERG9座に組み込まれた相III破壊配列を含む陽性形質転換体を、PCR増幅によって検証した。
【0096】
Y1770株をURA3ノックアウト構築物(配列番号154)で形質転換することによって宿主株Y1793を作製した。URA3ノックアウト構築物は、(Saccharomyces cerevisiae株CEN.PK2ゲノムDNAから生じた)URA3座の上流配列および下流配列を含む。宿主細胞形質転換体を、5−FOAを含むYPD培地上で宿主細胞形質転換体を選択した。
【0097】
Y1793株を相Iマーカーリサイクリング構築物で形質転換することによって宿主株YAAAを作製した。宿主細胞形質転換体を、メチオニンおよびウラシルを欠損するCSM上で選択した。200rpmでの回転式振盪機上、YPD培地中、30℃で一晩、細胞を増殖させ、その後、5−FOAを含む寒天上に細胞を蒔くことによってURA3マーカーを切除した。マーカー切除をコロニーPCRによって確認した。
【0098】
YAAA株を相IIマーカーリサイクリング構築物で形質転換することによって宿主株YBBBを作製した。宿主細胞形質転換体を、メチオニンおよびウラシルを欠損するCSM上で選択した。200rpmでの回転式振盪機上、YPD培地中、30℃で一晩、細胞を増殖させ、その後、5−FOAを含む寒天上に細胞を蒔くことによってURA3マーカーを切除した。マーカー切除をコロニーPCRによって確認した。
【0099】
YBBB株を相IIIマーカーリサイクリング構築物で形質転換することによって宿主株Y1912を作製した。宿主細胞形質転換体を、メチオニンおよびウラシルを欠損するCSM上で選択した。200rpmでの回転式振盪機上、YPD培地中、30℃で一晩、細胞を増殖させ、その後、5−FOAを含む寒天上に細胞を蒔くことによってURA3マーカーを切除した。マーカー切除をコロニーPCRによって確認した。
【0100】
実施例2:遺伝子改変された胞子形成欠陥性および内因性接合欠陥性の一倍体細胞の作製
この実施例は、遺伝子改変一倍体S.cerevisiae細胞において胞子形成遺伝子およびフェロモン応答遺伝子を破壊して、胞子形成および内因性接合が損なわれている遺伝子改変一倍体S.cerevisiae細胞を生じるための例示的な方法を記載する。
【0101】
STE5破壊構築物(図7;配列番号147)は、破壊配列として、S.cerevisiae STE5座の上流ヌクレオチド配列および下流ヌクレオチド配列からなる相同配列に隣接した、選択マーカー(ウラシル欠損培地上で増殖する能力を与えるURA3);およびS.cerevisiae TDH3遺伝子のプロモーターの制御調節下のS.cerevisiae MEV経路の酵素(切断型HMG−CoAリダクターゼをコードする切断型HMG1コード配列)をコードするヌクレオチド配列を含む。S.cerevisiae宿主細胞への導入で、STE5破壊構築物は、S.cerevisiae宿主細胞ゲノムのSTE5座へ相同組換えによって組み込まれ、STE5コード配列をその破壊配列と置換することによってSTE5座を機能的に破壊することができる。
【0102】
IME1破壊構築物(図8;配列番号148)は、破壊配列として、S.cerevisiae IME5座の上流ヌクレオチド配列および下流ヌクレオチド配列からなる相同配列に隣接した、選択マーカー(ロイシンを欠損する培地上で増殖する能力を与えるLEU2)およびS.cerevisiae TDH3遺伝子のプロモーターの制御調節下のA.annuaの酵素(ファルネセンシンターゼをコードするFSコード配列)をコードするヌクレオチド配列を含む。S.cerevisiae宿主細胞への導入で、IME1破壊構築物は、S.cerevisiae宿主細胞ゲノムのIME1座へ相同組換えによって組み込まれ、IME1コード配列をその破壊配列と置換することによってIME1座を機能的に破壊することができる。
【0103】
Y1912株をSTE5破壊構築物で形質転換することによって宿主株Y1913を作製した。宿主細胞形質転換体を、メチオニンおよびウラシルを欠損するCSMにおいて選択し、陽性形質転換体をPCR増幅によって検証した。
【0104】
Y1913株からpAM404を排除(curing)し、生じた株をIME1破壊構築物で形質転換することによって宿主株Y1915を作製した。Y1913株を、200rpmでの回転式振盪機上、30℃で3日間、非選択YPD培地中で繁殖させた。約100細胞をYPD固形培地上に蒔き、30℃で3日間、増殖させ、その後、メチオニンおよびロイシンを欠損するCSMプレート上でレプリカプレーティングを行い、そこで、それらを30℃でさらに3日間、増殖させた。ロイシンを含む最少培地上で増殖するできること、およびロイシンを欠損する培地上で増殖できないことによって治癒細胞(cured cells)を同定した。単一のそのようなコロニーを採取し、IME1破壊構築物で形質転換した。宿主細胞形質転換体を、メチオニンおよびロイシンを欠損するCSM上で選択した。
【0105】
実施例3:遺伝子改変された胞子形成欠陥性および内因性接合欠陥性の二倍体細胞の作製
この実施例は、胞子形成および内因性接合が損なわれている遺伝子改変一倍体S.cerevisiae細胞を二倍体にするための例示的な方法を記載する。
【0106】
Y1915株の自己接合によって二倍体宿主株Y1979を作製した。反対の接合型の細胞を作製するため、およびY1915株を一過性に接合可能にするために、HOタンパク質をコードするプラスミドpAM1124(配列番号149)ならびにSTE5およびG418耐性マーカーをコードするプラスミドpAM1758(配列番号150)でその株を同時形質転換した。宿主細胞形質転換体を、G418およびノールセオスリシンを含むCSM上で選択した。陽性形質転換体を、非選択培地上に単一コロニーについて再び蒔き、G418感受性、ノールセオスリシン感受性の二倍体を、コロニーPCRを用いるスクリーニングを通して同定した。
【0107】
実施例4:胞子形成欠陥および内因性接合欠陥の確認
遺伝子改変S.cerevisiae細胞の胞子形成欠陥および内因性接合欠陥を確認するための例示的な方法を記載する。
【0108】
Y1915株が接合できないことを確認するために、一倍体Y1915細胞(MATα KanS URA3^ste5)または一倍体Y1912細胞(MATα KanS URA3 STE5)をYEPD固形培地上で一倍体Y1792細胞(MATa KanR ura3 STE5)と組み合わせた。接合培養物を、30℃で16時間、インキュベートした。その後、各接合培養物の同一のアリコートを、ウラシルを欠損し、さらにG418を含むCSM固形培地に蒔き、その培養物を30℃で1週間、インキュベートした。図9に示されているように、Y1792×Y1912接合培養物のアリコートを含むプレート上のみコロニー増殖が観察されたが、Y1792×Y1915接合培養物のアリコートを含むプレート上には観察されなかった。
【0109】
Y1979株が胞子形成できないことを確認するために、Y1979株細胞およびY1198株細胞を、胞子形成誘導培地(非発酵性炭素源、例えば、酢酸カリウムを欠損する培地であり、それは、天然S.cerevisiae細胞が細胞有糸分裂周期を放棄し、減数分裂へ入って胞子形成するように誘導する)中に7日間、培養した。その後、培養物を分割し、水またはジエチルエーテルで15分間、処理した。懸濁液を、反転によってホモジナイズし、滅菌水中に再懸濁し、希釈し、YEPD固形培地上に蒔き、3日間、増殖させた。図10に示されているように、95%のY1198株細胞が、これらの条件下で四分子胞子を形成したが、Y1979株細胞は形成しなかった。
【0110】
実施例5:胞子形成欠陥性および内因性接合欠陥性の細胞が天然に散在することができないことの確認
この実施例は、胞子形成欠陥性および内因性接合欠陥性の遺伝子改変二倍体S.cerevisiae細胞が天然に散在することができないことを確認するための例示的な方法を記載する。
【0111】
土壌中のY1979およびそれの非トランスジェニック同種系のY1198(PE−2)の生存を評価した。この目的を達成するために、45Lのフラスコを約25%のバーミキュライトおよびサトウキビ栽培地からの75%の土壌で満たし(合計40L)、栽培品種RB 86−7515サトウキビ植物である1つのSaccharum種(およそ月齢6ヶ月)を植えた。各ポットを、5−25−30の乾燥の窒素/リン/カリウムの混合物を施肥し、植物を閉鎖系温室内で14日間、成長させた。各ポットへ、Y1979株またはY1198株の600mLの細胞懸濁液を加えた。酵母細胞の添加は、土壌の表面5cm中に107細胞/gの濃度に達することと等価である。1.5×5土壌コアの5つの試料を以下の時点で収集した:t=0日目(暴露前)、0日目(曝露後)、3日目、7日目、14日目、28日目、40日目、60日目、および90日目(試料採取された土壌の総体積は44mLであり、試料採取された土壌の総重量は約50gであった)。複合試料から、10グラムを分離し、100mLの蒸留水中に再懸濁した。酵母生存を定量化するために、100μLの水抽出物を、0.05g/Lのコウシンバラ(Sigma #R3877)を添加され、かつ0.2g/Lのアンピシリン(Sigma A0166)を含む、硫酸6NでpH5.5に調整されたYPED培地上に直接、蒔いた(25mL/プレート)。試料を二連で、1〜107の希釈系列で蒔き、または蒔かれるべき希釈回数は、各処理について前の試料採取において得られた生存の数に基づいた。プレーティング後すぐに、その液体をDrigalskiスパチュラで広げた。プレートを、その液体を全て蒸発させるために、最高30分間、流れに対して開いたままにしておき、その後、閉じて、反転させ、30℃で48時間、インキュベートした。プレートあたりのコロニー数を、コロニー計数機(CP600 Plus、Phoenix)を用いて、計数可能な希釈度において読み取り、その結果をCFU/プレートで表した。コロニーの総数が30〜300コロニーの間である場合のみ、計数が認められた。図11に示されているように(各データ点は5つの繰り返しの平均である)、Y1979細胞は、遺伝子改変されていない、胞子形成および接合の熟達したY1198株の親細胞より、明らかに土壌中における生存能力が低かった。
【0112】
本明細書に引用された全ての刊行物、特許、および特許出願は、あたかもそれぞれ個々の刊行物または特許出願が具体的かつ個々に示されて参照により組み入れられているかのうように、参照により本明細書に組み入れられている。前述の本発明は、例証および理解を明確にするための例として、かなり詳細に記載されているが、ある特定の変化および改変が、添付された特許請求の範囲の精神または範囲から逸脱することなく、それらになされ得ることは、本発明の教示を鑑みれば、当業者にとって容易に明らかであろう。
【特許請求の範囲】
【請求項1】
(a)少なくとも1つの胞子形成遺伝子における機能的破壊、
(b)少なくとも1つのフェロモン応答遺伝子における機能的破壊、および
(c)目的のタンパク質をコードする、染色体に組み込まれた異種性ヌクレオチド配列
を含む遺伝子改変酵母細胞であって、該遺伝子改変酵母細胞は、胞子形成が損なわれ、および、内因性接合が損なわれている、遺伝子改変酵母細胞。
【請求項2】
ヘテロタリック(ho)である、請求項1に記載の遺伝子改変酵母細胞。
【請求項3】
二倍体である、請求項1に記載の遺伝子改変酵母細胞。
【請求項4】
二倍体である、請求項2に記載の遺伝子改変酵母細胞。
【請求項5】
接合型対立遺伝子について以外はホモ接合型である、請求項4に記載の遺伝子改変二倍体酵母細胞。
【請求項6】
一倍体である、請求項1に記載の遺伝子改変酵母細胞。
【請求項7】
一倍体である、請求項2に記載の遺伝子改変酵母細胞。
【請求項8】
機能性ホモタリズム(HO)タンパク質をコードする組換えプラスミドをさらに含む、請求項7に記載の遺伝子改変一倍体酵母細胞。
【請求項9】
請求項8に記載の遺伝子改変一倍体酵母細胞であって、該遺伝子改変一倍体酵母細胞において機能的に破壊されている前記少なくとも1つのフェロモン応答遺伝子の機能性コピーをコードする少なくとも1つの組換えプラスミドをさらに含む、遺伝子改変一倍体酵母細胞。
【請求項10】
Saccharomyces cerevisiae細胞である、請求項1〜9のいずれか一項に記載の遺伝子改変酵母細胞。
【請求項11】
前記Saccharomyces cerevisiae細胞はパン酵母株、Mauri株、Santa Fe株、IZ−1904株、TA株、BG−1株、CR−1株、SA−1株、M−26株、Y−904株、PE−2株、PE−5株、VR−1株、BR−1株、BR−2株、ME−2株、VR−2株、MA−3株、MA−4株、CAT−1株、CB−1株、NR−1株、BT−1株、またはAL−1株である、請求項10に記載の遺伝子改変酵母細胞。
【請求項12】
前記Saccharomyces cerevisiae細胞はPE−2株である、請求項10に記載の遺伝子改変酵母細胞。
【請求項13】
前記Saccharomyces cerevisiae細胞はCAT−1株である、請求項10に記載の遺伝子改変酵母細胞。
【請求項14】
前記少なくとも1つの胞子形成遺伝子は、IME1、IME2、NDT80、SPO11、SPO20、AMA1、HOP2、およびSPO21からなる群より選択される、請求項1に記載の遺伝子改変酵母細胞。
【請求項15】
前記少なくとも1つのフェロモン応答遺伝子は、STE5、STE4、STE18、STE12、STE7、およびSTE11からなる群より選択される、請求項1に記載の遺伝子改変酵母細胞。
【請求項16】
IME1遺伝子の機能的破壊を含む、請求項14に記載の遺伝子改変酵母細胞。
【請求項17】
STE5遺伝子の機能的破壊を含む、請求項15に記載の遺伝子改変酵母細胞。
【請求項18】
IME1遺伝子およびSTE5遺伝子の機能的破壊を含む、請求項1に記載の遺伝子改変酵母細胞。
【請求項19】
STE5遺伝子の機能的破壊、および機能性STE5タンパク質をコードする組換えプラスミドを含む、請求項9に記載の遺伝子改変一倍体酵母細胞。
【請求項20】
胞子形成および内因性接合が損なわれている遺伝子改変二倍体酵母細胞を作製する方法であって、以下の工程
(a)第1の遺伝子改変一倍体酵母細胞および第2の遺伝子改変一倍体酵母細胞のそれぞれを、該第1の遺伝子改変一倍体酵母細胞および該第2の遺伝子改変一倍体酵母細胞の内因性接合欠陥を相補することができるタンパク質をコードする少なくとも1つのプラスミドで形質転換する工程であって、該第1の遺伝子改変一倍体酵母細胞は、胞子形成および内因性接合が損なわれており、および、目的のタンパク質をコードする染色体に組み込まれた異種性ヌクレオチド配列を含み、該第2の遺伝子改変一倍体酵母細胞は、胞子形成および内因性接合が損なわれており、該第1の遺伝子改変一倍体酵母細胞と反対の接合型であり、および、該目的のタンパク質をコードする染色体に組み込まれた異種性ヌクレオチド配列を含む、工程、
(b)該第1の遺伝子改変一倍体酵母細胞を該第2の遺伝子改変一倍体酵母細胞と接合させ、それにより、遺伝子改変二倍体酵母細胞を形成する工程、ならびに
(c)1つまたは複数の該プラスミドを該遺伝子改変二倍体酵母細胞から除去する工程であって、その結果生じた該遺伝子改変二倍体酵母細胞は、胞子形成および内因性接合が損なわれており、および、該目的のタンパク質をコードする染色体に組み込まれた異種性ヌクレオチド配列を2コピー含む、工程
を含む、方法。
【請求項21】
前記第1の遺伝子改変一倍体酵母細胞および前記第2の遺伝子改変一倍体酵母細胞は、少なくとも1つのフェロモン応答遺伝子の機能的破壊によって内因性接合が損なわれている、請求項20に記載の方法。
【請求項22】
工程(a)が、前記第1の遺伝子改変一倍体酵母細胞および前記第2の遺伝子改変一倍体酵母細胞のそれぞれを、該第1の遺伝子改変一倍体酵母細胞および該第2の遺伝子改変一倍体酵母細胞において機能的に破壊されている前記少なくとも1つのフェロモン応答遺伝子の機能性コピーをコードする少なくとも1つのプラスミドで形質転換する工程を含む、請求項21に記載の方法。
【請求項23】
前記フェロモン応答遺伝子は、STE5、STE4、STE18、STE12、STE7、およびSTE11からなる群より選択される、請求項21に記載の方法。
【請求項24】
前記フェロモン応答遺伝子は、STE5、STE4、STE18、STE12、STE7、およびSTE11からなる群より選択される、請求項22に記載の方法。
【請求項25】
前記フェロモン応答遺伝子はSTE5である、請求項21に記載の方法。
【請求項26】
前記フェロモン応答遺伝子はSTE5である、請求項22に記載の方法。
【請求項27】
前記第1の遺伝子改変一倍体酵母細胞および前記第2の遺伝子改変一倍体酵母細胞は、少なくとも1つの胞子形成遺伝子の機能的破壊によって胞子形成が損なわれている、請求項20に記載の方法。
【請求項28】
前記胞子形成遺伝子は、IME1、IME2、NDT80、SPO11、SPO20、AMA1、HOP2、およびSPO21からなる群より選択される、請求項27に記載の方法。
【請求項29】
前記胞子形成遺伝子はIME1である、請求項28に記載の方法。
【請求項30】
前記第1の遺伝子改変一倍体酵母細胞および前記第2の遺伝子改変一倍体酵母細胞は、STE5遺伝子の機能的破壊によって内因性接合が損なわれており、および、IME1遺伝子の機能的破壊によって胞子形成が損なわれている、請求項20に記載の方法。
【請求項31】
前記第2の遺伝子改変一倍体酵母細胞は、前記第1の遺伝子改変一倍体酵母細胞の集団において接合型切り換えを誘導することによって得られる、請求項20に記載の方法。
【請求項32】
前記第1の遺伝子改変一倍体酵母細胞の集団はヘテロタリック(ho)であり、かつ、該第1の遺伝子改変一倍体酵母細胞を機能性ホモタリズム(HO)タンパク質をコードする組換えプラスミドで形質転換することによって接合型を切り換えるように誘導され、該HOタンパク質の発現は該第1の遺伝子改変一倍体酵母細胞の接合型切り換えを誘導することができる、請求項31に記載の方法。
【請求項33】
前記遺伝子改変二倍体酵母細胞はSaccharomyces cerevisiae細胞である、請求項20に記載の方法。
【請求項34】
前記Saccharomyces cerevisiae細胞はパン酵母株、IZ−1904株、TA株、BG−1株、CR−1株、SA−1株、M−26株、Y−904株、PE−2株、PE−5株、VR−1株、BR−1株、BR−2株、ME−2株、VR−2株、MA−3株、MA−4株、CAT−1株、CB−1株、NR−1株、BT−1株、またはAL−1株である、請求項33に記載の方法。
【請求項35】
前記Saccharomyces cerevisiae細胞はPE−2株である、請求項33に記載の方法。
【請求項36】
前記Saccharomyces cerevisiae細胞はCAT−1株である、請求項33に記載の方法。
【請求項37】
請求項20〜36のいずれか一項に記載の方法によって作製された、胞子形成および内因性接合が損なわれた二倍体酵母細胞。
【請求項38】
胞子形成および内因性接合が損なわれた遺伝子改変ヘテロタリック(ho)二倍体酵母細胞を作製するための方法であって、以下の工程
(a)第1の遺伝子改変ヘテロタリック一倍体酵母細胞の集団を機能性ホモタリズム(HO)タンパク質をコードするプラスミドで形質転換して、第1の遺伝子改変一倍体酵母細胞を生じる工程であって、該HOタンパク質の発現は該第1の遺伝子改変一倍体酵母細胞の接合型切り換えを誘導することができ、それにより、該第1の遺伝子改変一倍体酵母細胞と反対の接合型の第2の遺伝子改変一倍体酵母細胞が得られ、該第1の遺伝子改変ヘテロタリック一倍体酵母細胞は、目的のタンパク質をコードする染色体に組み込まれた異種性ヌクレオチド配列ならびにSTE5遺伝子およびIME1遺伝子における機能的破壊を含む、工程、
(b)該第1の遺伝子改変一倍体酵母細胞および該第2の遺伝子改変一倍体酵母細胞のそれぞれを、STE5タンパク質をコードするプラスミドで形質転換する工程であって、結果として、該形質転換工程は、該第1の遺伝子改変一倍体酵母細胞と該第2の遺伝子改変一倍体酵母細胞との接合を生じ、それにより遺伝子改変二倍体酵母細胞を形成する、工程、ならびに
(c)該遺伝子改変二倍体酵母細胞から任意のプラスミドを除去して、遺伝子改変ヘテロタリック二倍体酵母細胞を生じる工程であって、生じた該遺伝子改変ヘテロタリック二倍体酵母細胞は胞子形成および内因性接合が損なわれており、および、該目的のタンパク質をコードする染色体に組み込まれた異種性ヌクレオチド配列を2コピー含む、工程
を含む、方法。
【請求項39】
前記胞子形成および内因性接合が損なわれたヘテロタリック(ho)二倍体酵母細胞はSaccharomyces cerevisiae細胞である、請求項39に記載の方法。
【請求項40】
前記Saccharomyces cerevisiae細胞はパン酵母株、Mauri株、Santa Fe株、IZ−1904株、TA株、BG−1株、CR−1株、SA−1株、M−26株、Y−904株、PE−2株、PE−5株、VR−1株、BR−1株、BR−2株、ME−2株、VR−2株、MA−3株、MA−4株、CAT−1株、CB−1株、NR−1株、BT−1株、またはAL−1株である、請求項39に記載の方法。
【請求項41】
前記Saccharomyces cerevisiae細胞はPE−2株である、請求項39に記載の方法。
【請求項42】
前記Saccharomyces cerevisiae細胞はCAT−1株である、請求項39に記載の方法。
【請求項43】
請求項38〜42のいずれか一項に記載の方法によって作製された、胞子形成および内因性接合が損なわれたヘテロタリック(ho)二倍体酵母細胞。
【請求項44】
MATα/a ste5/ste5 ime1/ime1酵母細胞。
【請求項1】
(a)少なくとも1つの胞子形成遺伝子における機能的破壊、
(b)少なくとも1つのフェロモン応答遺伝子における機能的破壊、および
(c)目的のタンパク質をコードする、染色体に組み込まれた異種性ヌクレオチド配列
を含む遺伝子改変酵母細胞であって、該遺伝子改変酵母細胞は、胞子形成が損なわれ、および、内因性接合が損なわれている、遺伝子改変酵母細胞。
【請求項2】
ヘテロタリック(ho)である、請求項1に記載の遺伝子改変酵母細胞。
【請求項3】
二倍体である、請求項1に記載の遺伝子改変酵母細胞。
【請求項4】
二倍体である、請求項2に記載の遺伝子改変酵母細胞。
【請求項5】
接合型対立遺伝子について以外はホモ接合型である、請求項4に記載の遺伝子改変二倍体酵母細胞。
【請求項6】
一倍体である、請求項1に記載の遺伝子改変酵母細胞。
【請求項7】
一倍体である、請求項2に記載の遺伝子改変酵母細胞。
【請求項8】
機能性ホモタリズム(HO)タンパク質をコードする組換えプラスミドをさらに含む、請求項7に記載の遺伝子改変一倍体酵母細胞。
【請求項9】
請求項8に記載の遺伝子改変一倍体酵母細胞であって、該遺伝子改変一倍体酵母細胞において機能的に破壊されている前記少なくとも1つのフェロモン応答遺伝子の機能性コピーをコードする少なくとも1つの組換えプラスミドをさらに含む、遺伝子改変一倍体酵母細胞。
【請求項10】
Saccharomyces cerevisiae細胞である、請求項1〜9のいずれか一項に記載の遺伝子改変酵母細胞。
【請求項11】
前記Saccharomyces cerevisiae細胞はパン酵母株、Mauri株、Santa Fe株、IZ−1904株、TA株、BG−1株、CR−1株、SA−1株、M−26株、Y−904株、PE−2株、PE−5株、VR−1株、BR−1株、BR−2株、ME−2株、VR−2株、MA−3株、MA−4株、CAT−1株、CB−1株、NR−1株、BT−1株、またはAL−1株である、請求項10に記載の遺伝子改変酵母細胞。
【請求項12】
前記Saccharomyces cerevisiae細胞はPE−2株である、請求項10に記載の遺伝子改変酵母細胞。
【請求項13】
前記Saccharomyces cerevisiae細胞はCAT−1株である、請求項10に記載の遺伝子改変酵母細胞。
【請求項14】
前記少なくとも1つの胞子形成遺伝子は、IME1、IME2、NDT80、SPO11、SPO20、AMA1、HOP2、およびSPO21からなる群より選択される、請求項1に記載の遺伝子改変酵母細胞。
【請求項15】
前記少なくとも1つのフェロモン応答遺伝子は、STE5、STE4、STE18、STE12、STE7、およびSTE11からなる群より選択される、請求項1に記載の遺伝子改変酵母細胞。
【請求項16】
IME1遺伝子の機能的破壊を含む、請求項14に記載の遺伝子改変酵母細胞。
【請求項17】
STE5遺伝子の機能的破壊を含む、請求項15に記載の遺伝子改変酵母細胞。
【請求項18】
IME1遺伝子およびSTE5遺伝子の機能的破壊を含む、請求項1に記載の遺伝子改変酵母細胞。
【請求項19】
STE5遺伝子の機能的破壊、および機能性STE5タンパク質をコードする組換えプラスミドを含む、請求項9に記載の遺伝子改変一倍体酵母細胞。
【請求項20】
胞子形成および内因性接合が損なわれている遺伝子改変二倍体酵母細胞を作製する方法であって、以下の工程
(a)第1の遺伝子改変一倍体酵母細胞および第2の遺伝子改変一倍体酵母細胞のそれぞれを、該第1の遺伝子改変一倍体酵母細胞および該第2の遺伝子改変一倍体酵母細胞の内因性接合欠陥を相補することができるタンパク質をコードする少なくとも1つのプラスミドで形質転換する工程であって、該第1の遺伝子改変一倍体酵母細胞は、胞子形成および内因性接合が損なわれており、および、目的のタンパク質をコードする染色体に組み込まれた異種性ヌクレオチド配列を含み、該第2の遺伝子改変一倍体酵母細胞は、胞子形成および内因性接合が損なわれており、該第1の遺伝子改変一倍体酵母細胞と反対の接合型であり、および、該目的のタンパク質をコードする染色体に組み込まれた異種性ヌクレオチド配列を含む、工程、
(b)該第1の遺伝子改変一倍体酵母細胞を該第2の遺伝子改変一倍体酵母細胞と接合させ、それにより、遺伝子改変二倍体酵母細胞を形成する工程、ならびに
(c)1つまたは複数の該プラスミドを該遺伝子改変二倍体酵母細胞から除去する工程であって、その結果生じた該遺伝子改変二倍体酵母細胞は、胞子形成および内因性接合が損なわれており、および、該目的のタンパク質をコードする染色体に組み込まれた異種性ヌクレオチド配列を2コピー含む、工程
を含む、方法。
【請求項21】
前記第1の遺伝子改変一倍体酵母細胞および前記第2の遺伝子改変一倍体酵母細胞は、少なくとも1つのフェロモン応答遺伝子の機能的破壊によって内因性接合が損なわれている、請求項20に記載の方法。
【請求項22】
工程(a)が、前記第1の遺伝子改変一倍体酵母細胞および前記第2の遺伝子改変一倍体酵母細胞のそれぞれを、該第1の遺伝子改変一倍体酵母細胞および該第2の遺伝子改変一倍体酵母細胞において機能的に破壊されている前記少なくとも1つのフェロモン応答遺伝子の機能性コピーをコードする少なくとも1つのプラスミドで形質転換する工程を含む、請求項21に記載の方法。
【請求項23】
前記フェロモン応答遺伝子は、STE5、STE4、STE18、STE12、STE7、およびSTE11からなる群より選択される、請求項21に記載の方法。
【請求項24】
前記フェロモン応答遺伝子は、STE5、STE4、STE18、STE12、STE7、およびSTE11からなる群より選択される、請求項22に記載の方法。
【請求項25】
前記フェロモン応答遺伝子はSTE5である、請求項21に記載の方法。
【請求項26】
前記フェロモン応答遺伝子はSTE5である、請求項22に記載の方法。
【請求項27】
前記第1の遺伝子改変一倍体酵母細胞および前記第2の遺伝子改変一倍体酵母細胞は、少なくとも1つの胞子形成遺伝子の機能的破壊によって胞子形成が損なわれている、請求項20に記載の方法。
【請求項28】
前記胞子形成遺伝子は、IME1、IME2、NDT80、SPO11、SPO20、AMA1、HOP2、およびSPO21からなる群より選択される、請求項27に記載の方法。
【請求項29】
前記胞子形成遺伝子はIME1である、請求項28に記載の方法。
【請求項30】
前記第1の遺伝子改変一倍体酵母細胞および前記第2の遺伝子改変一倍体酵母細胞は、STE5遺伝子の機能的破壊によって内因性接合が損なわれており、および、IME1遺伝子の機能的破壊によって胞子形成が損なわれている、請求項20に記載の方法。
【請求項31】
前記第2の遺伝子改変一倍体酵母細胞は、前記第1の遺伝子改変一倍体酵母細胞の集団において接合型切り換えを誘導することによって得られる、請求項20に記載の方法。
【請求項32】
前記第1の遺伝子改変一倍体酵母細胞の集団はヘテロタリック(ho)であり、かつ、該第1の遺伝子改変一倍体酵母細胞を機能性ホモタリズム(HO)タンパク質をコードする組換えプラスミドで形質転換することによって接合型を切り換えるように誘導され、該HOタンパク質の発現は該第1の遺伝子改変一倍体酵母細胞の接合型切り換えを誘導することができる、請求項31に記載の方法。
【請求項33】
前記遺伝子改変二倍体酵母細胞はSaccharomyces cerevisiae細胞である、請求項20に記載の方法。
【請求項34】
前記Saccharomyces cerevisiae細胞はパン酵母株、IZ−1904株、TA株、BG−1株、CR−1株、SA−1株、M−26株、Y−904株、PE−2株、PE−5株、VR−1株、BR−1株、BR−2株、ME−2株、VR−2株、MA−3株、MA−4株、CAT−1株、CB−1株、NR−1株、BT−1株、またはAL−1株である、請求項33に記載の方法。
【請求項35】
前記Saccharomyces cerevisiae細胞はPE−2株である、請求項33に記載の方法。
【請求項36】
前記Saccharomyces cerevisiae細胞はCAT−1株である、請求項33に記載の方法。
【請求項37】
請求項20〜36のいずれか一項に記載の方法によって作製された、胞子形成および内因性接合が損なわれた二倍体酵母細胞。
【請求項38】
胞子形成および内因性接合が損なわれた遺伝子改変ヘテロタリック(ho)二倍体酵母細胞を作製するための方法であって、以下の工程
(a)第1の遺伝子改変ヘテロタリック一倍体酵母細胞の集団を機能性ホモタリズム(HO)タンパク質をコードするプラスミドで形質転換して、第1の遺伝子改変一倍体酵母細胞を生じる工程であって、該HOタンパク質の発現は該第1の遺伝子改変一倍体酵母細胞の接合型切り換えを誘導することができ、それにより、該第1の遺伝子改変一倍体酵母細胞と反対の接合型の第2の遺伝子改変一倍体酵母細胞が得られ、該第1の遺伝子改変ヘテロタリック一倍体酵母細胞は、目的のタンパク質をコードする染色体に組み込まれた異種性ヌクレオチド配列ならびにSTE5遺伝子およびIME1遺伝子における機能的破壊を含む、工程、
(b)該第1の遺伝子改変一倍体酵母細胞および該第2の遺伝子改変一倍体酵母細胞のそれぞれを、STE5タンパク質をコードするプラスミドで形質転換する工程であって、結果として、該形質転換工程は、該第1の遺伝子改変一倍体酵母細胞と該第2の遺伝子改変一倍体酵母細胞との接合を生じ、それにより遺伝子改変二倍体酵母細胞を形成する、工程、ならびに
(c)該遺伝子改変二倍体酵母細胞から任意のプラスミドを除去して、遺伝子改変ヘテロタリック二倍体酵母細胞を生じる工程であって、生じた該遺伝子改変ヘテロタリック二倍体酵母細胞は胞子形成および内因性接合が損なわれており、および、該目的のタンパク質をコードする染色体に組み込まれた異種性ヌクレオチド配列を2コピー含む、工程
を含む、方法。
【請求項39】
前記胞子形成および内因性接合が損なわれたヘテロタリック(ho)二倍体酵母細胞はSaccharomyces cerevisiae細胞である、請求項39に記載の方法。
【請求項40】
前記Saccharomyces cerevisiae細胞はパン酵母株、Mauri株、Santa Fe株、IZ−1904株、TA株、BG−1株、CR−1株、SA−1株、M−26株、Y−904株、PE−2株、PE−5株、VR−1株、BR−1株、BR−2株、ME−2株、VR−2株、MA−3株、MA−4株、CAT−1株、CB−1株、NR−1株、BT−1株、またはAL−1株である、請求項39に記載の方法。
【請求項41】
前記Saccharomyces cerevisiae細胞はPE−2株である、請求項39に記載の方法。
【請求項42】
前記Saccharomyces cerevisiae細胞はCAT−1株である、請求項39に記載の方法。
【請求項43】
請求項38〜42のいずれか一項に記載の方法によって作製された、胞子形成および内因性接合が損なわれたヘテロタリック(ho)二倍体酵母細胞。
【請求項44】
MATα/a ste5/ste5 ime1/ime1酵母細胞。
【図9】

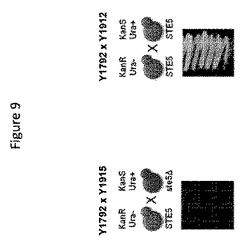
【図10A】
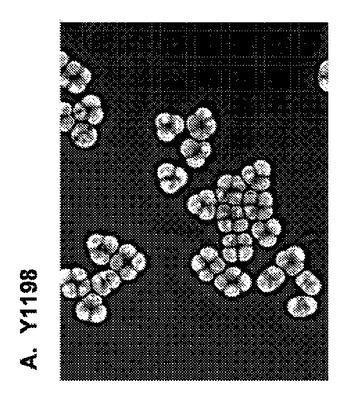
【図10B】
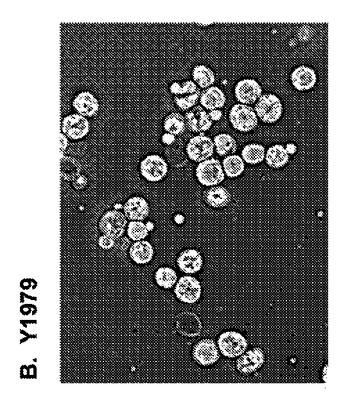
【図1】
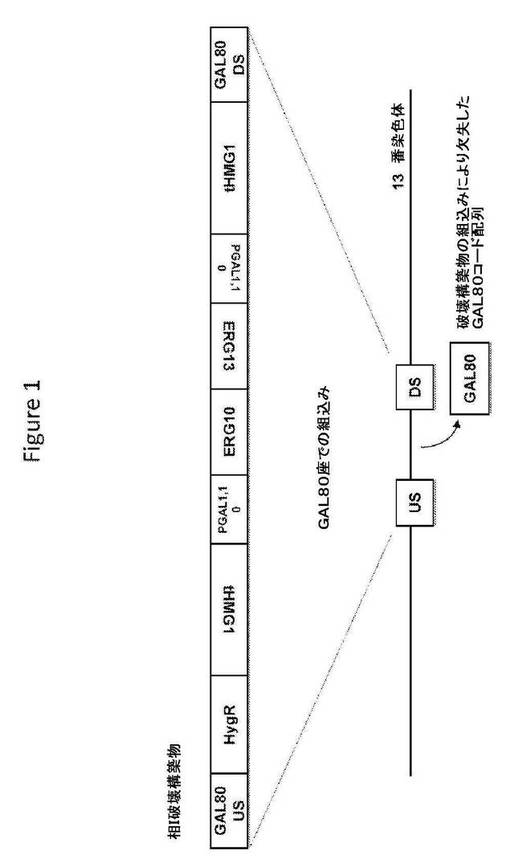
【図2】
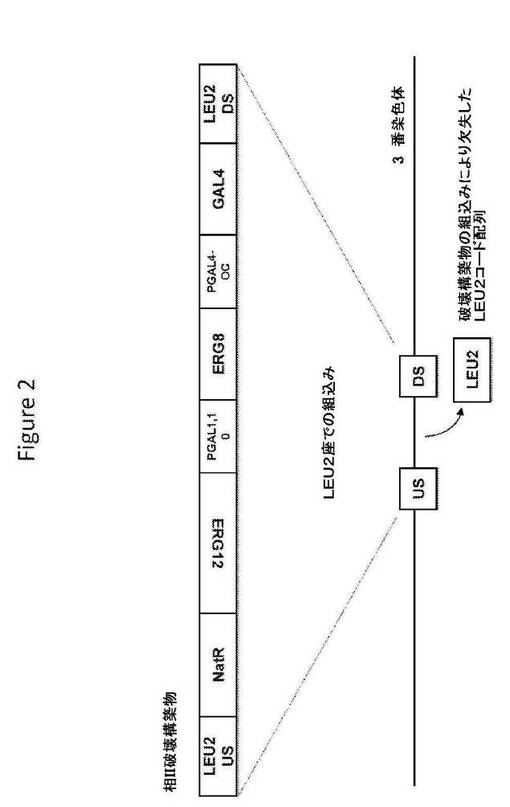
【図3】
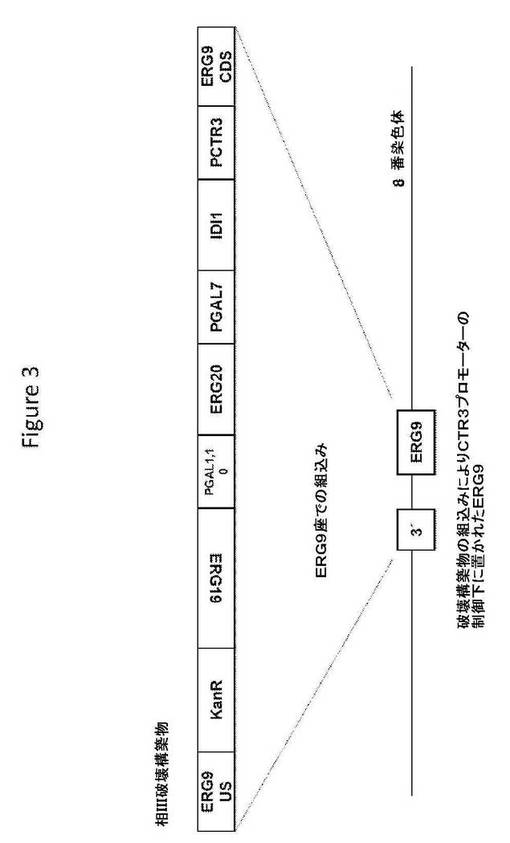
【図4】
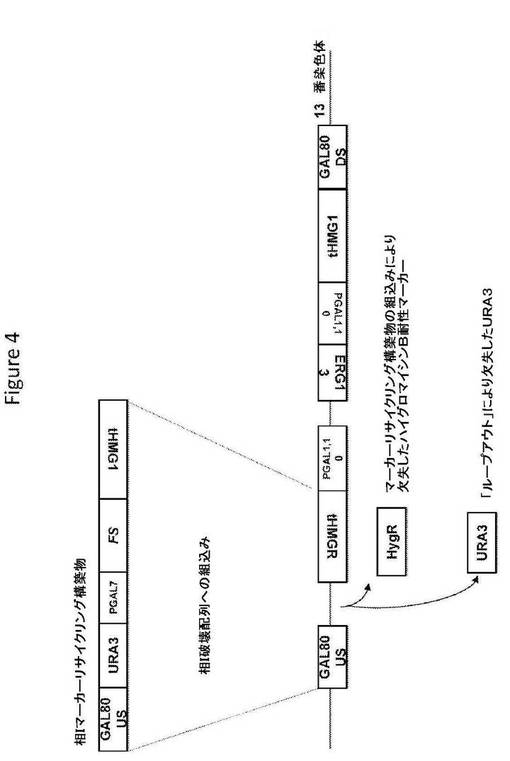
【図5】
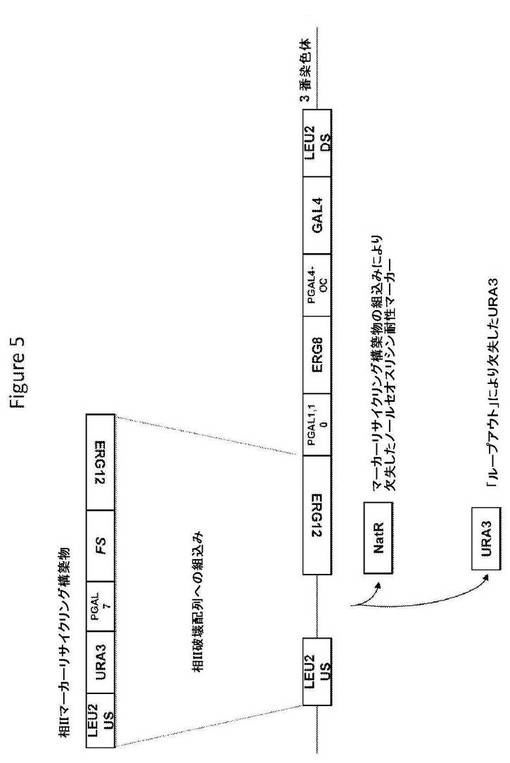
【図6】
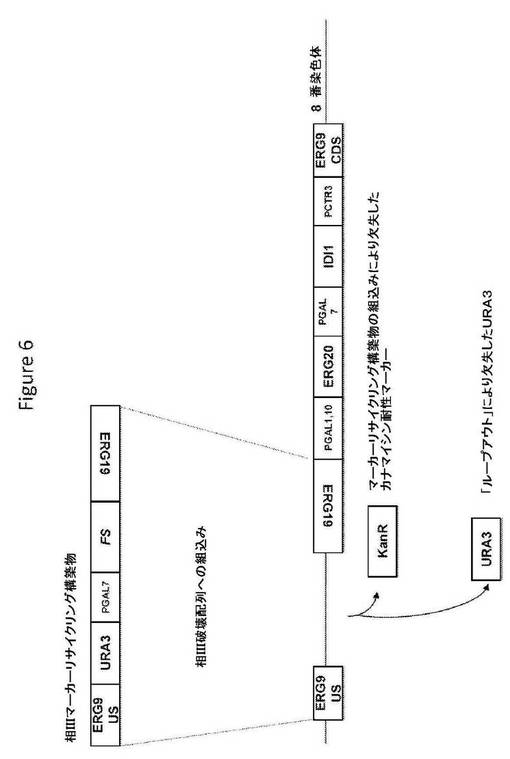
【図7】
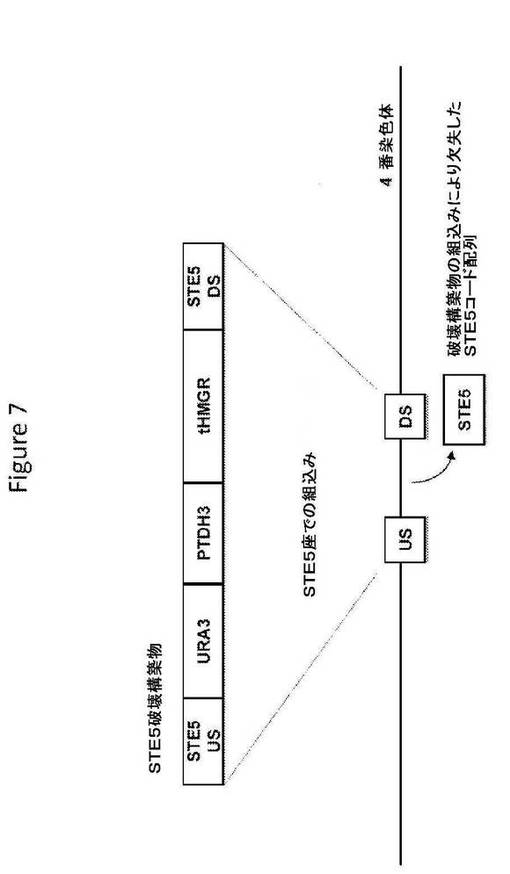
【図8】
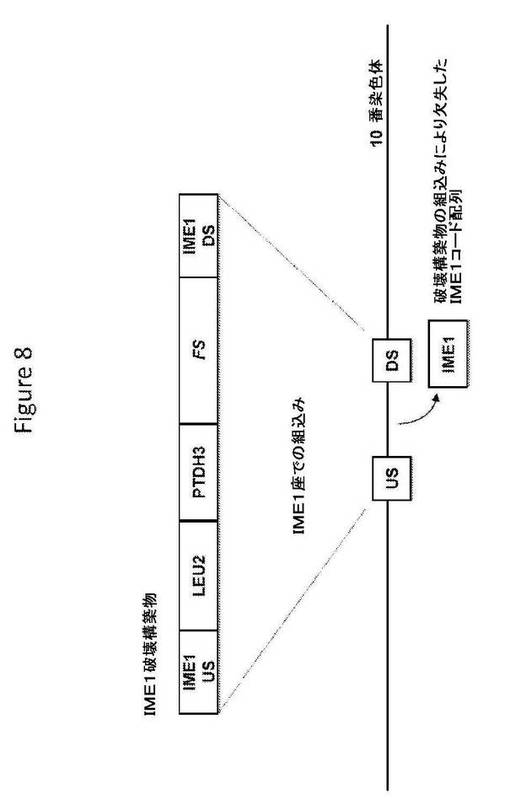
【図11】
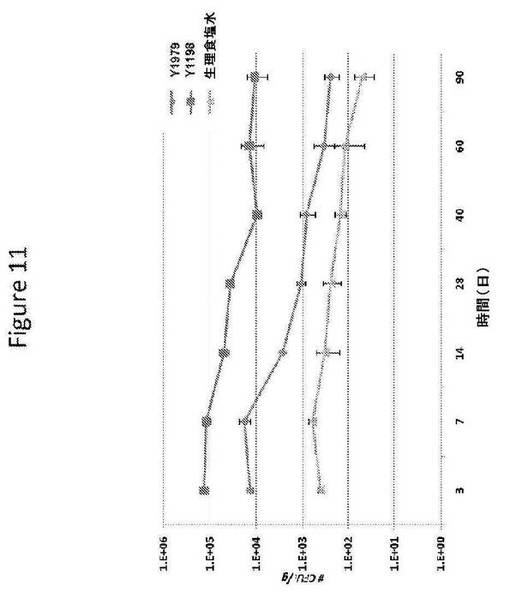
【図10A】
【図10B】
【図1】
【図2】
【図3】
【図4】
【図5】
【図6】
【図7】
【図8】
【図11】
【公表番号】特表2012−528570(P2012−528570A)
【公表日】平成24年11月15日(2012.11.15)
【国際特許分類】
【出願番号】特願2012−513352(P2012−513352)
【出願日】平成22年6月1日(2010.6.1)
【国際出願番号】PCT/US2010/036861
【国際公開番号】WO2010/141438
【国際公開日】平成22年12月9日(2010.12.9)
【出願人】(511289736)アミリス, インコーポレイテッド (4)
【Fターム(参考)】
【公表日】平成24年11月15日(2012.11.15)
【国際特許分類】
【出願日】平成22年6月1日(2010.6.1)
【国際出願番号】PCT/US2010/036861
【国際公開番号】WO2010/141438
【国際公開日】平成22年12月9日(2010.12.9)
【出願人】(511289736)アミリス, インコーポレイテッド (4)
【Fターム(参考)】
[ Back to top ]