
遺伝子発現の特異性に関する改良
【課題】ターゲット遺伝子の特異的発現部位を標的にすることによって遺伝子発現の特異性を改良するために、影響を及ぼされる生物の複数の領域で発現するプロモーターを含むキメラ遺伝子を提供する。
【解決手段】植物の複数の領域において発現される植物内在性遺伝子の機能に影響を及ぼす作用物質に連結されているプロモーター。1以上の所望の位置にその発現部位におけるオーバーラップが存在するように選択されるプロモーターおよび作用物質。
【解決手段】植物の複数の領域において発現される植物内在性遺伝子の機能に影響を及ぼす作用物質に連結されているプロモーター。1以上の所望の位置にその発現部位におけるオーバーラップが存在するように選択されるプロモーターおよび作用物質。
【発明の詳細な説明】
【技術分野】
【0001】
本発明は、遺伝子発現の特異性を増大させる方法に関する。特に、本発明は、例えば、限定するものではないが、植物における病因物質に対する標的耐性を付与する様式として高度に特異的なターゲッティング効果を発生させるためのキメラ遺伝子の使用に関する。
【背景技術】
【0002】
本発明の目的のために、遺伝子がどのように構築され、それがどのように機能するかを簡略化された方法で記載することは有用である(遺伝子の構造を示す図1を参照されたい。この遺伝子はキメラ遺伝子でもあり得る)。遺伝子は、3つの構成成分からなると考えることができる(数字は、図1中の数字を意味するものである)。1.コード配列がいつ、どこで転写されるかを決定するプロモーター(P)、タンパク質産生のためのコード配列(CS)およびコード配列の転写にも時として影響を及ぼし得る3’調節配列(3’)。3’調節配列は、通常、ターミネーターとして公知である。2.プロモーターがコード配列の作動コピーの転写およびプロセシング(10)を可能にすることによりメッセンジャーRNA(mRNA)を産生させる場合、遺伝子が発現される。3.次いでmRNAが翻訳されて(11)タンパク質産物が得られる。4.次にタンパク質産物が基質もしくは別のタンパク質または調節配列と相互作用することにより効果(E)が生じ得る。
【0003】
しかしながら、遺伝子の発現調節は上記の発現プロセスの各段階で影響され得るということに注意すべきである。因子は、プロモーターと、mRNAを産生させるための転写可動部分と、およびmRNAを修飾するかまたはその安定性に影響を及ぼすプロセスとにおいて作用し得る。また、因子は、タンパク質の翻訳および細胞内のタンパク質の代謝回転においても作用し得る。その他の因子は、タンパク質がその他の構成成分とどのように相互作用し、その効果をどのように得るかに影響を及ぼし得る。
【0004】
本発明の概念を説明するために、植物における下記の遺伝子、遺伝子1および遺伝子2を考えてみる。遺伝子1は、植物の組織AおよびBにおいて活性であり、一方遺伝子2は植物の領域BおよびCにおいて活性である。二つの遺伝子の活性は、領域Bでオーバーラップし、これは図2のようにヴェン図形の形式で視覚的に示すことができる。この図から、共同発現の部位はより限定される、すなわち、どちらかの遺伝子が単独で発現する部位よりも特異的であるということは明白である。
【0005】
したがって、本発明においては、植物に関して、キメラ遺伝子は植物の複数の領域で発現するプロモーターを含む。プロモーターは一つの作用物質に連結されている。この作用物質は、植物の複数の領域においても発現される内在性遺伝子の機能に影響を及ぼすものである。プロモーターおよび影響される内在性遺伝子が活性である範囲は同一ではないが、1以上の所望の位置でオーバーラップする。キメラ遺伝子が植物に移入される場合、作用物質は、一つまたは複数のオーバーラップの位置でターゲット内在性遺伝子に影響を及ぼすのみである。
【0006】
本発明を例示するにはいくつかの方法があり、植物寄生線虫動物に対する耐性または抵抗性の増大が実際的な例である。本発明者らは、例として細胞破壊および雄性不稔性を使用したが、この系は、特定部位における遺伝子の増強にも使用することができる。
【0007】
細胞特異的破壊を得るためのいくつかの異なる機構が提案されている。最も簡単な方法は、破壊系に連結された標的組織に特異的なプロモーターを含むキメラ遺伝子を利用するものである。しかしながら、特異的プロモーターでさえ、その標的組織以外の組織においてより小さい程度で発現し得るものであり、これが好ましくない時がある。
【0008】
その他の出願では、二つの構築物、すなわち細胞壊死系に連結された組織特異的プロモーターを含むキメラ遺伝子を含有する第1の構築物(例えばbarnase)および標的領域以外の領域で活性なプロモーターであって保護物質に連結されているものを含むキメラ遺伝子を含有する第2の構築物(例えばbarstar)を利用することによってこの問題を回避する試みがなされている。したがって、所望の組織以外の組織における壊死は、保護物質によって抑制される(国際特許出願番号WO92/21757(Plant Genetic Systems N.V.)およびWO93/10251(Mogen International N.V.)を参照されたい)。
【0009】
本発明は、例えば、一つの構築物を用いる高度に特異的な細胞破壊系を提供し得るものである。本発明が適用可能なその他の細胞調節系は、当業者には公知である。
【発明の開示】
【発明が解決しようとする課題】
【0010】
本発明の目的は、種々の領域で発現される遺伝子を有する任意の生物に一般的に適用することができるが、その発現の範囲がオーバーラップするものである部位特異的発現系またはターゲッティング系を提供することである。
【0011】
また、本発明の目的は、2以上の核酸配列を含むキメラ遺伝子を含有する一つの構築物のみを用いて特異性を増大させることであり、この構築物は生物における内在性遺伝子と相互作用するように設計される。
【課題を解決するための手段】
【0012】
本発明は、形質転換生物における遺伝子調節の特異性を改良する方法であって、生物における内在性ターゲット遺伝子を同定する工程、ターゲット遺伝子の複数の発現部位の位置を決定する工程、ターゲット遺伝子の発現部位の一つにおける発現を含む、生物において少なくとも2つの発現部位で遺伝子発現を生じさせるプロモーターおよびターゲット遺伝子の発現を調節する核酸配列またはその産物である作用物質を含むキメラ遺伝子を作製する工程、キメラ遺伝子を遺伝子形質転換によって生物の細胞に安定に組み込む工程、ならびに形質転換細胞から生物を再生させる工程を含み、形質転換生物はキメラ遺伝子を含み、生物におけるその遺伝子の発現により、形質転換生物またはその子孫における少なくとも一つの特異的発現部位でターゲット遺伝子またはその産物が調節されるものである、前記方法を提供する。
【0013】
好ましくは、核酸配列はコードまたは非コード配列である。
【0014】
好ましくは、ターゲット遺伝子の発現はアップレギュレートまたはダウンレギュレートされ得る。
【0015】
好ましくは、生物は植物であり、よって形質転換植物またはその零余子(propagules)はキメラ遺伝子を含む。
【0016】
作用物質が遺伝子に作用する機構は、下記のいずれか一つに属し得る。ここに挙げたものに限定されるものではない。
【0017】
1.アンチセンス
2.共同抑制
3.ターゲット遺伝子のプロモーターの阻害または活性化
4.転写の阻害または活性化
5.メッセンジャーRNA安定性の変更またはmRNAの分解
6.翻訳の阻害または活性化
7.タンパク質の阻害または活性化
8.タンパク質代謝回転の変更
9.共同因子としての作用
10.タンパク質−タンパク質相互作用の変更
11.生化学的経路によるフラックスの変更。
【0018】
これらの機構のいくつかの実施例を以下に簡単に記載する。遺伝子調節を得るために選択される特定の機構が、本発明の概念において有効であるために一定レベルの部位特異的発現をも必要とすることは心に留めておくべきである。
【0019】
ダウンレギュレーションは、ターゲット遺伝子のプロモーターの全部もしくは一部またはコードもしくは非コード配列のアンチセンス方向の核酸配列であるキメラ遺伝子の作用物質によって都合よくなし得る。一方、ダウンレギュレーションは、ターゲット遺伝子のプロモーターまたはコードもしくは非コード配列の共同抑制によって得られ得る。
【0020】
ターゲットのアップレギュレーションは、例えば、ターゲット遺伝子のプロモーターのアクチベーターを導入することによってなし得る。
【0021】
これらの技術の組合せも用いることができる。
【0022】
ターゲット遺伝子の遺伝子発現を調節するその他の適当な方法は、当業者には公知である。
【0023】
キメラ遺伝子の作用物質は複数の核酸配列を含み得るものであり、その配列のそれぞれは発現された場合に特定の機能を果たす。したがって、一つのプロモーターのみを用いることにより複数の内在性遺伝子の発現の特異性を得ることができる。
【0024】
構築物は、発現カセットとして二つの別個のキメラ遺伝子を含み得るものであり、各キメラ遺伝子は、プロモーター、作用物質のコード配列およびターミネーターを含む。各キメラ遺伝子は、異なる内在性遺伝子に作用するものであり、この遺伝子は同一のターゲット部位または異なる部位で発現され得る。したがって、ある期間にわたって、特定の生合成連鎖などの一連の事象においていくつかの構成成分またはターゲット遺伝子をノックアウトするか、または増大させる可能性がある。換言すれば、事象の一時的な連鎖がもたらされ得る。一方、各キメラ遺伝子は二つの別個の構築物にて生物に移入することができ、各構築物は、一つの発現カセット、すなわち一つのキメラ遺伝子を含む。
【0025】
核酸配列は、DNA配列またはRNA配列であり得る。
【0026】
キメラ遺伝子のプロモーターは、内在性遺伝子の一以上のオーバーラップ発現部位において発現され得る。
【0027】
都合がよいことに、プロモーターは、ここでKNT1プロモーターまたはRB7プロモーターとして公知のプロモーターのような線虫誘導プロモーターである。その他の攻撃または増殖部位でそれらに作用するその他の作用物質によって発現が誘導されるその他のプロモーターは、得られる部位特異的発現調節に依存して利用され得る。
【0028】
都合がよいことに、その核酸配列はRB7もしくはKNT1プロモーターまたはそのコード配列のアンチセンス配列の全部または一部である。
【0029】
一方、核酸配列は、例えば、上記の機構リストの中の機構5をもたらすためにメッセンジャーRNAを分解するためのリボザイムまたは標的RNアーゼであり得る。また、特異的RNAは、特異的栄養素、例えば移送のための細胞表面タンパク質受容体用のmRNAの場合には鉄;またはリガンド、ホルモンおよび翻訳産物、例えばチューブリンmRNAにおけるチューブリンタンパク質二量体の効果によっても安定化または不安定化され得る。本発明の概念には、一定の位置で発現または要求される栄養素、リガンド、ホルモンまたは翻訳産物の選択が望ましい。
【0030】
転写のアクチベーターとしては、細胞内の遊離のタンパク質をコードするキイロショウジョウバエ(Drosphila)の熱ショック因子が例示される。熱ショックの際、熱ショック遺伝子因子は熱ショックタンパク質HSP70のプロモーターに結合し、転写が増大される。熱ショックタンパク質は、細菌、動物および植物において見出される。当業者は本発明の概念において使用に適した部位特異的であるアクチベーターを選択することにより機構4を得ることができる。
【0031】
生物において機構9をもたらすのに適当な共同因子としては、パントテン酸およびビタミンB6のようなビタミンが挙げられる。
【0032】
最後に、機構10は、例えば、タンパク質cAMP−依存性タンパク質キナーゼを生物に導入することによってもたらされ得る。cAMP−依存性タンパク質キナーゼは、酵素グリコーゲンシンターゼにそれをリン酸化することによって作用する。グリコーゲンシンターゼは低活性体に変化し、グリコーゲン合成が阻害される。
【0033】
また、本発明は、いくつかの構築物も利用し得るものであり、各構築物のキメラ遺伝子の各プロモーター−遺伝子断片は、その系の特異性をさらに増大させるために選択された1個の発現部位に多数のオーバーラップが付与されるように、その他のプロモーター−遺伝子断片のそれぞれと同一のターゲット発現部位にオーバーラップを有する。キメラ遺伝子のその他の発現部位は、その他のプロモーター−遺伝子断片のその他の発現部位と同一または異なるものであり得る。
【発明を実施するための最良の形態】
【0034】
本発明を容易に理解し、容易に実行することができるように、例として、下記の図および実施例を参照し本発明を詳述する。
【図面の簡単な説明】
【0035】
【図2】二つの別個の遺伝子が同一の植物内に存在する場合のそれらの遺伝子の発現部位のオーバーラップを示す。
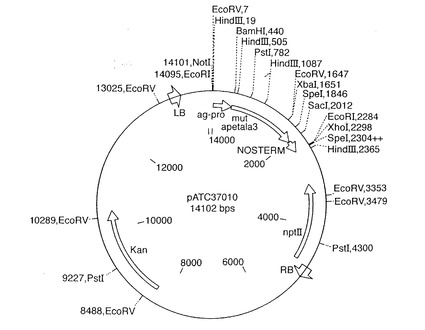
【図3】本発明にしたがって植物を形質転換するのに用いられたベクターpATC37010の地図を示す。
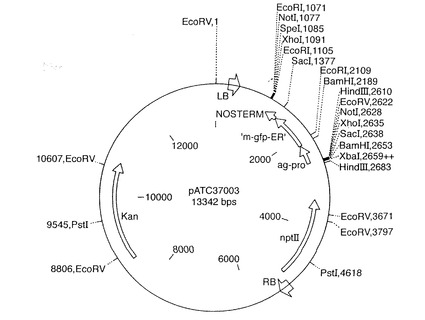
【図4】植物を形質転換するのに用いられた対照ベクター、ベクターpATC37003の地図を示す。
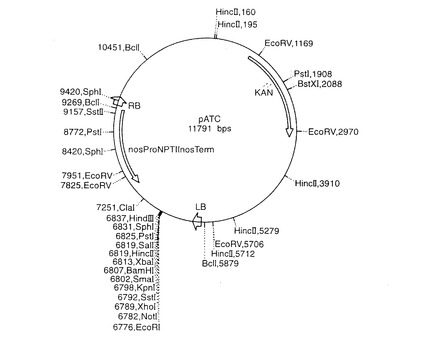
【図5】配列番号5および配列番号6をライゲートして、それぞれ図3および4のベクターpATC37010およびpATC37003を産生させたpATCの地図を示す。
【0036】
図2の部位Bにおける細胞特異的破壊は下記のようにして行うことができる。遺伝子1由来の発現の特異性を調節するプロモーターが、構築物の遺伝子2の活性を妨害する領域に連結され得る。構築物が植物に導入されると、遺伝子2の破壊的作用物質が領域AおよびBで発現されるであろう。遺伝子2は部位Aで活性ではないので、部位Aにおいては効果はないであろう。構築物のプロモーターは部位Cで活性ではなく、よって遺伝子2の破壊的作用物質は産生されないので、部位Cでの効果はないであろう。遺伝子2に対する破壊的作用物質が存在し、遺伝子2は活性でもあるので、部位Bでは遺伝子2の妨害が存在するであろう。
【実施例】
【0037】
実施例1
ヴェン構築物を用いた操作により植物寄生線虫に対する耐性または抵抗性を増大させた。
【0038】
根瘤線虫および包のう線虫のような植物寄生線虫は、世界中の農作物収穫高を7〜14%低下させている。線虫は、植物根に侵入し、線虫がその栄養分を誘導する独特の摂餌部位を生じさせることにより作用する。摂餌部位は、変質植物細胞、すなわち根瘤線虫の場合には巨大多核細胞、包のう線虫の場合にはいくつかの細胞が融合した融合細胞である。線虫は定着するようになり、栄養素の摂餌部位にまったく依存している。本出願人らの米国特許第5,589,622号には、摂餌細胞特異的プロモーターを細胞死または細胞破壊系に連結して摂餌細胞を破壊することによって植物抵抗性を得る一般的な方法が記載されている。摂餌細胞はそれらの機能が損なわれ、その結果、線虫が餓死するか、または線虫への食物供給が低減されて、成長および子孫の産生の能力がなくなるか低くなる。この方法は、上記の細胞特異的破壊の最も簡単な方法の例である。この原理を利用するその他の特許は、例えば、国際特許出願番号WO89/10396(Plant Genetic Systems N.V.)のような植物を不稔性にする特許である。
【0039】
摂餌細胞、根端、およびその他の分裂組織(これは程度が低い)において発現されるプロモーターKNT1を同定した。その他の研究者らは、根および巨細胞において発現される遺伝子RB7を同定した(Conklingら, 1990, Oppermanら, 1993)。マーカー遺伝子GUSに連結されたRB7プロモーターを用いた本発明者らの研究は、RB7遺伝子が根端ではなく、根の主要部において発現することを示唆するものである。アグロバクテリウム ツメファシエンス(Agrobacterium tumefaciens)C58を含む、pBIN19(Bevan, M. 1984)誘導植物形質転換ベクターにRB7コード配列の部分アンチセンス配列に連結されたプロモーターKNT1およびnosターミネーターを含むヴェン構築物を作製した。構築物は標識pBIN05002であり、特許手続上の微生物の寄託の国際的承認に関するブダペスト条約のもとに、National Collection of Industrial and Marine Bacteria(NCIMB)(23 St. Machar Street, Aberdeen, Scotland)にAdvanced Technologies(ケンブリッジ)社(210 Cambridge Science Park, Cambridge CB4 4WA, England)によって1997年3月20日に寄託されており、受託番号NCIMB40871を得ている。タバコ植物cv Heavy Westernを、アグロバクテリウム媒介植物形質転換を用いてHorschら(1985)の方法にしたがってかかる構築物により形質転換した。
【0040】
再生トランスジェニック植物を温室に移した。トランスジェニック植物および非トランスジェニック対照に、それぞれ約100匹の根瘤線虫Meloidogyne javanicaを感染させた。感染の8週間後、目に見える根瘤の数およびそのサイズを測定した。この期間中、初期接種物は、感染し、成熟し、線虫の第2世代(今度はこの第2世代が植物根瘤に感染し、成熟する)を産む機会を有していた。
【0041】
最もスコアの低いトランスジェニックpBIN05002植物の中の8個を成長させて種を生じさせた。親植物からの子孫を、上記のM. javanicaに対する耐性または抵抗性の増大について再びスクリーニングした。pBIN05101植物に加えて、グルクロニダーゼマーカー遺伝子(GUS)に連結されたKNT1プロモーターを含むpBIN05101で形質転換された植物からの子孫(Jefferson, R.A.ら, 1987)および非形質転換植物からの子孫を、比較のための対照として含ませた。pBIN05101も、1997年3月20日にNCIMBに寄託されており、受託番号NCIMB 40870を得ている。pBIN05002植物系32の子孫は、表1に示したように、えい瘤スコアの低い植物の数が有意に増大していた。この結果はχ2分析において有意である。
【0042】
(表1)
非形質転換対照植物、対照pBIN05101植物およびpBIN05002試験植物についての低えい瘤スコア区分と高えい瘤スコア区分における植物の数
低えい瘤スコア=0〜50えい瘤/植物
高えい瘤スコア=51+えい瘤/植物
処理 低えい瘤スコアの植物 高えい瘤スコアの植物
非形質転換植物 18 13
pBIN05101対照植物 13 17
pBIN05002系32試験植物 25 7
【0043】
実施例2
上記の実施例において、線虫破壊における特異性を増大させるための細胞破壊系を用いて説明したオーバーラップの原理は、例えば、Arabidopsis、またはその他の植物の花においても同様に十分に実行されて、花のパターンまたは構造が変化した花、例えば、雄性不稔性の花が得られ得る。この実施例は、Arabidopsisにおいて同定されたDNA配列を利用するものである。
【0044】
花の中には3つの遺伝子の調節下にあると仮定される4つのエレメント(蕚片、心皮、花弁および雄ずい)が存在する(Coen, E.S.およびMeyerowitz, E.M., 1991)。
【0045】
これらの遺伝子のバランスを変更することにより、花パターンの変化が生じる。例えば、遺伝子agamousおよびapetala3は、花の同一部分で発現して植物の雄性部分である雄ずいを生じさせなければならない。agamousは、心皮および雄ずいの両方で発現し、一方apetala3は雄ずいおよび花弁の両方で発現する。
【0046】
本発明の主題であるオーバーラップ原理にしたがって構築物を製造するためには、本発明者らは、雄ずいにおいてのみ破壊をもたらすために第2の遺伝子(例えば、花弁および雄ずいで活性なapetala3)の破壊物質に連結された一の遺伝子(例えば、心皮および雄ずいで活性なagamous)由来のプロモーターを必要とする。
【0047】
下記のTaqおよびTaq−エキステンダーを用いたポリメラーゼ連鎖反応により公表されている手順(Thomas, C., 1996)にしたがって下記の二つのプライマーを用いることによって、agamousプロモーターの435塩基対断片をArabidopsis thaliana DNAから単離した:
プライマー1(配列番号1)
ATCGAAGCTT CTAAATGTAC TGAAAAGAAA CA
プライマー2(配列番号2)
ACTGGGATCC GAAAATGGAA GGTAAGGTTG TGC
プライマーは、ゲノムagamous配列についてのジーンバンクDNA配列エントリーATAGAMSGで得られた配列に基づくものであった。プライマー1はその5'末端に追加のHindIII制限部位を含む。プライマー2は、その5'末端に追加のBamHI制限部位を含む。
【0048】
下記のプライマーは、apetala3遺伝子配列の外側の部分を増幅するためにジーンバンク配列エントリーATHAPETALAから設計された:
プライマー3(配列番号3)
ATCGGGATCC ATGGGCTCAC GGTTTTGTGT GA
プライマー4(配列番号4)
ATCGGAGCTC TTATTCAAGA AGATGGAAGG TAATGA
プライマー3は、公表されているapetala3配列の992位にて増幅が開始されるように特異的に設計され、これは活性産物を産生させるための不適正リーディングフレームにおける開始コドンであり、同一遺伝子ファミリー内のその他のMADSボックス遺伝子と強い相同性を有する配列の初期部分を無効にするものである。また、プライマー3は、望ましくないSacI制限部位を除去するために公表されている配列から1塩基対変更されている。プライマー3はその5'末端にBamHI制限部位を有している。プライマー4はその5'末端にSacI部位を有している。1586bp断片を、プライマー3および4を用いてPCRによってArabidopsis DNAから増幅させた。
【0049】
下記のクローニング方法は当業者にはよく知られており、Sambrookら(1989)の方法にしたがうことによって成果を得ることができる。agamousプロモーター断片をpBluescript誘導ベクター(Stratagene社, ケンブリッジ, UK)にHindIII−BamHI断片としてライゲートした。変更型apetala遺伝子断片を同一ベクター内のagamousプロモーターの下流にBamHI-SacI断片としてライゲートした。また、ベクターには、変更型apetala3配列の下流にSacI−EcoRI断片としてnosターミネーター配列も含まれていた。ベクターをpDVM37010と命名した。Jim Haselhoffから入手した、m-gfp-ERレポーター遺伝子の前にagamousプロモーターを含む第2のプラスミド、MRCを対照として作製し、pDVM37003と命名した。
【0050】
プロモーター−遺伝子断片−ターミネーターカセットをpDVM37010(配列番号5)およびpDVM37003(配列番号6)からNotI制限断片として切り出し、pBIN19(Bevan, M. (1984))誘導ベクターpATCにライゲートしてプラスミドpATC37010(図3に示した地図)およびpATC37003(図4に示した地図)を得た。これらの配列は、適当な制限部位、すなわちカセットの各末端におけるNotIを有する任意のその他の同等のベクターにクローニングすることができる。pATCの地図を図5に示す。これはpBIN19と比べて改変されている制限部位を有する。pATC37010は、agamousプロモーターの調節下で共同抑制産物を産生して、花の発達中の雄ずいにおけるapetala3機能を不活性化する。
【0051】
プラスミドをアグロバクテリウム ツメファシエンス宿主LBA4404に移入し、Bechtoldら(1993)の方法およびHorschら(1985)の方法を用いるNicotiana tabacum cv K326にしたがってArabidopsis thalianaの形質転換に使用した。pATC37010については8個のトランスジェニックArabidopsis苗木およびpATC37003については6個のトランスジェニックArabidopsis苗木が得られた。100個の葉盤3群を、各構築物に対するNicotiana形質転換に用いた。トランスジェニックカルス増殖が3群全てについて検出された。
【0052】
発芽の10〜14日後にArabidopsis苗木を土壌に移し、開花するまで成長させた。花は雄ずいおよびダブルローズ(double rose)心皮を示さなかった。
【0053】
カナマイシン耐性pATC37010トランスジェニック植物を、プライマー1および4を用いてTaqポリメラーゼによるPCRによって当業者に公知の手順にしたがって、さらに所望の挿入物の存在についてスクリーニングした。PCRでは、94℃で60秒間、60℃で30秒間、および72℃で140秒間のインキュベーション工程を含む40サイクルのインキュベーションを行った。PCR陽性試料を、当業者に公知の手順にしたがってアガロースゲル電気泳動によるPCR産物の視覚化によって同定した。
【0054】
対照pATC37003トランスジェニック植物を、プライマー1およびプライマー7を用いてTaqポリメラーゼによるPCRによって当業者に公知の手順にしたがって、さらに所望の挿入物の存在についてスクリーニングした。PCRでは、94℃で40秒間、60℃で30秒間、および72℃で140秒間のインキュベーション工程を含む40サイクルのインキュベーションを行った。PCR陽性試料を、当業者に公知の手順にしたがってアガロースゲル電気泳動によるPCR産物の視覚化によって同定した。
プライマー7(配列番号7)
GAACTGGGAC CACTCCAGTG
いずれの場合においても、適切な構築物を含むトランスジェニック植物が同定された。
【0055】
(参考文献)
Bechtold, N., Ellis, J., およびPelletier, G. (1993) In planta Agrobacterium mediated gene transfer by infiltration of adult Arabidopsis thaliana plants, C.R. Acad. Sci: Paris 316 : 1194-1199。
Bevan, M. (1984) Binary Agrobacterium vectors for plant transformation, Nuc. Acids Res. 12: 8711-8721。
Coen, E.S. およびMeyerowitz, E.M. (1991) The war of the whorls: genetic interactions controlling flower development, Nature 353, 31-37。
Conkling, M.A., Cheng, C-L., Yamamoto, Y.T. およびGoodman, H.M.(1990) Isolation of transcriptionally regulated root-specific genes from tobacco, Plant Physiology 93, 1203-1211。
Horsch, R.B., Fry, J.F., Hoffmann, N.L., Eichholtz, D., Rogers, S.G. およびFraley, R.T.(1985), A simple and general method for transferring genes into plants, Science 22, 1229-1231。
Jefferson, R.A., Kavanagh, T.A. およびBevan, M.W. (1987) GUS fusions: β-glucuronidase as a sensitive and versatile gene fusion maker in higher plants, EMBO, 6, 3901-3907。
Opperman, C.H., Taylor, C.G. およびConkling, M.A. (1994) Root-knot nematode-directed expression of a plant root specific gene, Science 263, 221-223。
Sambrook, J., Fritsch, E.F. およびManiatis, T. (1989) Molecular Cloning - A Laboratory Manual. Second Edition, Cold Spring Harbour Laboratory Press, New York。
Thomas, C. (1996) 'PCR Techniques' in 'Plant Gene Isolation: Principles and Practice', G.D. Foster & D. Twell editors, John Wiley & Sons Ltd., pp.331-368。
【図1】

【技術分野】
【0001】
本発明は、遺伝子発現の特異性を増大させる方法に関する。特に、本発明は、例えば、限定するものではないが、植物における病因物質に対する標的耐性を付与する様式として高度に特異的なターゲッティング効果を発生させるためのキメラ遺伝子の使用に関する。
【背景技術】
【0002】
本発明の目的のために、遺伝子がどのように構築され、それがどのように機能するかを簡略化された方法で記載することは有用である(遺伝子の構造を示す図1を参照されたい。この遺伝子はキメラ遺伝子でもあり得る)。遺伝子は、3つの構成成分からなると考えることができる(数字は、図1中の数字を意味するものである)。1.コード配列がいつ、どこで転写されるかを決定するプロモーター(P)、タンパク質産生のためのコード配列(CS)およびコード配列の転写にも時として影響を及ぼし得る3’調節配列(3’)。3’調節配列は、通常、ターミネーターとして公知である。2.プロモーターがコード配列の作動コピーの転写およびプロセシング(10)を可能にすることによりメッセンジャーRNA(mRNA)を産生させる場合、遺伝子が発現される。3.次いでmRNAが翻訳されて(11)タンパク質産物が得られる。4.次にタンパク質産物が基質もしくは別のタンパク質または調節配列と相互作用することにより効果(E)が生じ得る。
【0003】
しかしながら、遺伝子の発現調節は上記の発現プロセスの各段階で影響され得るということに注意すべきである。因子は、プロモーターと、mRNAを産生させるための転写可動部分と、およびmRNAを修飾するかまたはその安定性に影響を及ぼすプロセスとにおいて作用し得る。また、因子は、タンパク質の翻訳および細胞内のタンパク質の代謝回転においても作用し得る。その他の因子は、タンパク質がその他の構成成分とどのように相互作用し、その効果をどのように得るかに影響を及ぼし得る。
【0004】
本発明の概念を説明するために、植物における下記の遺伝子、遺伝子1および遺伝子2を考えてみる。遺伝子1は、植物の組織AおよびBにおいて活性であり、一方遺伝子2は植物の領域BおよびCにおいて活性である。二つの遺伝子の活性は、領域Bでオーバーラップし、これは図2のようにヴェン図形の形式で視覚的に示すことができる。この図から、共同発現の部位はより限定される、すなわち、どちらかの遺伝子が単独で発現する部位よりも特異的であるということは明白である。
【0005】
したがって、本発明においては、植物に関して、キメラ遺伝子は植物の複数の領域で発現するプロモーターを含む。プロモーターは一つの作用物質に連結されている。この作用物質は、植物の複数の領域においても発現される内在性遺伝子の機能に影響を及ぼすものである。プロモーターおよび影響される内在性遺伝子が活性である範囲は同一ではないが、1以上の所望の位置でオーバーラップする。キメラ遺伝子が植物に移入される場合、作用物質は、一つまたは複数のオーバーラップの位置でターゲット内在性遺伝子に影響を及ぼすのみである。
【0006】
本発明を例示するにはいくつかの方法があり、植物寄生線虫動物に対する耐性または抵抗性の増大が実際的な例である。本発明者らは、例として細胞破壊および雄性不稔性を使用したが、この系は、特定部位における遺伝子の増強にも使用することができる。
【0007】
細胞特異的破壊を得るためのいくつかの異なる機構が提案されている。最も簡単な方法は、破壊系に連結された標的組織に特異的なプロモーターを含むキメラ遺伝子を利用するものである。しかしながら、特異的プロモーターでさえ、その標的組織以外の組織においてより小さい程度で発現し得るものであり、これが好ましくない時がある。
【0008】
その他の出願では、二つの構築物、すなわち細胞壊死系に連結された組織特異的プロモーターを含むキメラ遺伝子を含有する第1の構築物(例えばbarnase)および標的領域以外の領域で活性なプロモーターであって保護物質に連結されているものを含むキメラ遺伝子を含有する第2の構築物(例えばbarstar)を利用することによってこの問題を回避する試みがなされている。したがって、所望の組織以外の組織における壊死は、保護物質によって抑制される(国際特許出願番号WO92/21757(Plant Genetic Systems N.V.)およびWO93/10251(Mogen International N.V.)を参照されたい)。
【0009】
本発明は、例えば、一つの構築物を用いる高度に特異的な細胞破壊系を提供し得るものである。本発明が適用可能なその他の細胞調節系は、当業者には公知である。
【発明の開示】
【発明が解決しようとする課題】
【0010】
本発明の目的は、種々の領域で発現される遺伝子を有する任意の生物に一般的に適用することができるが、その発現の範囲がオーバーラップするものである部位特異的発現系またはターゲッティング系を提供することである。
【0011】
また、本発明の目的は、2以上の核酸配列を含むキメラ遺伝子を含有する一つの構築物のみを用いて特異性を増大させることであり、この構築物は生物における内在性遺伝子と相互作用するように設計される。
【課題を解決するための手段】
【0012】
本発明は、形質転換生物における遺伝子調節の特異性を改良する方法であって、生物における内在性ターゲット遺伝子を同定する工程、ターゲット遺伝子の複数の発現部位の位置を決定する工程、ターゲット遺伝子の発現部位の一つにおける発現を含む、生物において少なくとも2つの発現部位で遺伝子発現を生じさせるプロモーターおよびターゲット遺伝子の発現を調節する核酸配列またはその産物である作用物質を含むキメラ遺伝子を作製する工程、キメラ遺伝子を遺伝子形質転換によって生物の細胞に安定に組み込む工程、ならびに形質転換細胞から生物を再生させる工程を含み、形質転換生物はキメラ遺伝子を含み、生物におけるその遺伝子の発現により、形質転換生物またはその子孫における少なくとも一つの特異的発現部位でターゲット遺伝子またはその産物が調節されるものである、前記方法を提供する。
【0013】
好ましくは、核酸配列はコードまたは非コード配列である。
【0014】
好ましくは、ターゲット遺伝子の発現はアップレギュレートまたはダウンレギュレートされ得る。
【0015】
好ましくは、生物は植物であり、よって形質転換植物またはその零余子(propagules)はキメラ遺伝子を含む。
【0016】
作用物質が遺伝子に作用する機構は、下記のいずれか一つに属し得る。ここに挙げたものに限定されるものではない。
【0017】
1.アンチセンス
2.共同抑制
3.ターゲット遺伝子のプロモーターの阻害または活性化
4.転写の阻害または活性化
5.メッセンジャーRNA安定性の変更またはmRNAの分解
6.翻訳の阻害または活性化
7.タンパク質の阻害または活性化
8.タンパク質代謝回転の変更
9.共同因子としての作用
10.タンパク質−タンパク質相互作用の変更
11.生化学的経路によるフラックスの変更。
【0018】
これらの機構のいくつかの実施例を以下に簡単に記載する。遺伝子調節を得るために選択される特定の機構が、本発明の概念において有効であるために一定レベルの部位特異的発現をも必要とすることは心に留めておくべきである。
【0019】
ダウンレギュレーションは、ターゲット遺伝子のプロモーターの全部もしくは一部またはコードもしくは非コード配列のアンチセンス方向の核酸配列であるキメラ遺伝子の作用物質によって都合よくなし得る。一方、ダウンレギュレーションは、ターゲット遺伝子のプロモーターまたはコードもしくは非コード配列の共同抑制によって得られ得る。
【0020】
ターゲットのアップレギュレーションは、例えば、ターゲット遺伝子のプロモーターのアクチベーターを導入することによってなし得る。
【0021】
これらの技術の組合せも用いることができる。
【0022】
ターゲット遺伝子の遺伝子発現を調節するその他の適当な方法は、当業者には公知である。
【0023】
キメラ遺伝子の作用物質は複数の核酸配列を含み得るものであり、その配列のそれぞれは発現された場合に特定の機能を果たす。したがって、一つのプロモーターのみを用いることにより複数の内在性遺伝子の発現の特異性を得ることができる。
【0024】
構築物は、発現カセットとして二つの別個のキメラ遺伝子を含み得るものであり、各キメラ遺伝子は、プロモーター、作用物質のコード配列およびターミネーターを含む。各キメラ遺伝子は、異なる内在性遺伝子に作用するものであり、この遺伝子は同一のターゲット部位または異なる部位で発現され得る。したがって、ある期間にわたって、特定の生合成連鎖などの一連の事象においていくつかの構成成分またはターゲット遺伝子をノックアウトするか、または増大させる可能性がある。換言すれば、事象の一時的な連鎖がもたらされ得る。一方、各キメラ遺伝子は二つの別個の構築物にて生物に移入することができ、各構築物は、一つの発現カセット、すなわち一つのキメラ遺伝子を含む。
【0025】
核酸配列は、DNA配列またはRNA配列であり得る。
【0026】
キメラ遺伝子のプロモーターは、内在性遺伝子の一以上のオーバーラップ発現部位において発現され得る。
【0027】
都合がよいことに、プロモーターは、ここでKNT1プロモーターまたはRB7プロモーターとして公知のプロモーターのような線虫誘導プロモーターである。その他の攻撃または増殖部位でそれらに作用するその他の作用物質によって発現が誘導されるその他のプロモーターは、得られる部位特異的発現調節に依存して利用され得る。
【0028】
都合がよいことに、その核酸配列はRB7もしくはKNT1プロモーターまたはそのコード配列のアンチセンス配列の全部または一部である。
【0029】
一方、核酸配列は、例えば、上記の機構リストの中の機構5をもたらすためにメッセンジャーRNAを分解するためのリボザイムまたは標的RNアーゼであり得る。また、特異的RNAは、特異的栄養素、例えば移送のための細胞表面タンパク質受容体用のmRNAの場合には鉄;またはリガンド、ホルモンおよび翻訳産物、例えばチューブリンmRNAにおけるチューブリンタンパク質二量体の効果によっても安定化または不安定化され得る。本発明の概念には、一定の位置で発現または要求される栄養素、リガンド、ホルモンまたは翻訳産物の選択が望ましい。
【0030】
転写のアクチベーターとしては、細胞内の遊離のタンパク質をコードするキイロショウジョウバエ(Drosphila)の熱ショック因子が例示される。熱ショックの際、熱ショック遺伝子因子は熱ショックタンパク質HSP70のプロモーターに結合し、転写が増大される。熱ショックタンパク質は、細菌、動物および植物において見出される。当業者は本発明の概念において使用に適した部位特異的であるアクチベーターを選択することにより機構4を得ることができる。
【0031】
生物において機構9をもたらすのに適当な共同因子としては、パントテン酸およびビタミンB6のようなビタミンが挙げられる。
【0032】
最後に、機構10は、例えば、タンパク質cAMP−依存性タンパク質キナーゼを生物に導入することによってもたらされ得る。cAMP−依存性タンパク質キナーゼは、酵素グリコーゲンシンターゼにそれをリン酸化することによって作用する。グリコーゲンシンターゼは低活性体に変化し、グリコーゲン合成が阻害される。
【0033】
また、本発明は、いくつかの構築物も利用し得るものであり、各構築物のキメラ遺伝子の各プロモーター−遺伝子断片は、その系の特異性をさらに増大させるために選択された1個の発現部位に多数のオーバーラップが付与されるように、その他のプロモーター−遺伝子断片のそれぞれと同一のターゲット発現部位にオーバーラップを有する。キメラ遺伝子のその他の発現部位は、その他のプロモーター−遺伝子断片のその他の発現部位と同一または異なるものであり得る。
【発明を実施するための最良の形態】
【0034】
本発明を容易に理解し、容易に実行することができるように、例として、下記の図および実施例を参照し本発明を詳述する。
【図面の簡単な説明】
【0035】
【図2】二つの別個の遺伝子が同一の植物内に存在する場合のそれらの遺伝子の発現部位のオーバーラップを示す。
【図3】本発明にしたがって植物を形質転換するのに用いられたベクターpATC37010の地図を示す。
【図4】植物を形質転換するのに用いられた対照ベクター、ベクターpATC37003の地図を示す。
【図5】配列番号5および配列番号6をライゲートして、それぞれ図3および4のベクターpATC37010およびpATC37003を産生させたpATCの地図を示す。
【0036】
図2の部位Bにおける細胞特異的破壊は下記のようにして行うことができる。遺伝子1由来の発現の特異性を調節するプロモーターが、構築物の遺伝子2の活性を妨害する領域に連結され得る。構築物が植物に導入されると、遺伝子2の破壊的作用物質が領域AおよびBで発現されるであろう。遺伝子2は部位Aで活性ではないので、部位Aにおいては効果はないであろう。構築物のプロモーターは部位Cで活性ではなく、よって遺伝子2の破壊的作用物質は産生されないので、部位Cでの効果はないであろう。遺伝子2に対する破壊的作用物質が存在し、遺伝子2は活性でもあるので、部位Bでは遺伝子2の妨害が存在するであろう。
【実施例】
【0037】
実施例1
ヴェン構築物を用いた操作により植物寄生線虫に対する耐性または抵抗性を増大させた。
【0038】
根瘤線虫および包のう線虫のような植物寄生線虫は、世界中の農作物収穫高を7〜14%低下させている。線虫は、植物根に侵入し、線虫がその栄養分を誘導する独特の摂餌部位を生じさせることにより作用する。摂餌部位は、変質植物細胞、すなわち根瘤線虫の場合には巨大多核細胞、包のう線虫の場合にはいくつかの細胞が融合した融合細胞である。線虫は定着するようになり、栄養素の摂餌部位にまったく依存している。本出願人らの米国特許第5,589,622号には、摂餌細胞特異的プロモーターを細胞死または細胞破壊系に連結して摂餌細胞を破壊することによって植物抵抗性を得る一般的な方法が記載されている。摂餌細胞はそれらの機能が損なわれ、その結果、線虫が餓死するか、または線虫への食物供給が低減されて、成長および子孫の産生の能力がなくなるか低くなる。この方法は、上記の細胞特異的破壊の最も簡単な方法の例である。この原理を利用するその他の特許は、例えば、国際特許出願番号WO89/10396(Plant Genetic Systems N.V.)のような植物を不稔性にする特許である。
【0039】
摂餌細胞、根端、およびその他の分裂組織(これは程度が低い)において発現されるプロモーターKNT1を同定した。その他の研究者らは、根および巨細胞において発現される遺伝子RB7を同定した(Conklingら, 1990, Oppermanら, 1993)。マーカー遺伝子GUSに連結されたRB7プロモーターを用いた本発明者らの研究は、RB7遺伝子が根端ではなく、根の主要部において発現することを示唆するものである。アグロバクテリウム ツメファシエンス(Agrobacterium tumefaciens)C58を含む、pBIN19(Bevan, M. 1984)誘導植物形質転換ベクターにRB7コード配列の部分アンチセンス配列に連結されたプロモーターKNT1およびnosターミネーターを含むヴェン構築物を作製した。構築物は標識pBIN05002であり、特許手続上の微生物の寄託の国際的承認に関するブダペスト条約のもとに、National Collection of Industrial and Marine Bacteria(NCIMB)(23 St. Machar Street, Aberdeen, Scotland)にAdvanced Technologies(ケンブリッジ)社(210 Cambridge Science Park, Cambridge CB4 4WA, England)によって1997年3月20日に寄託されており、受託番号NCIMB40871を得ている。タバコ植物cv Heavy Westernを、アグロバクテリウム媒介植物形質転換を用いてHorschら(1985)の方法にしたがってかかる構築物により形質転換した。
【0040】
再生トランスジェニック植物を温室に移した。トランスジェニック植物および非トランスジェニック対照に、それぞれ約100匹の根瘤線虫Meloidogyne javanicaを感染させた。感染の8週間後、目に見える根瘤の数およびそのサイズを測定した。この期間中、初期接種物は、感染し、成熟し、線虫の第2世代(今度はこの第2世代が植物根瘤に感染し、成熟する)を産む機会を有していた。
【0041】
最もスコアの低いトランスジェニックpBIN05002植物の中の8個を成長させて種を生じさせた。親植物からの子孫を、上記のM. javanicaに対する耐性または抵抗性の増大について再びスクリーニングした。pBIN05101植物に加えて、グルクロニダーゼマーカー遺伝子(GUS)に連結されたKNT1プロモーターを含むpBIN05101で形質転換された植物からの子孫(Jefferson, R.A.ら, 1987)および非形質転換植物からの子孫を、比較のための対照として含ませた。pBIN05101も、1997年3月20日にNCIMBに寄託されており、受託番号NCIMB 40870を得ている。pBIN05002植物系32の子孫は、表1に示したように、えい瘤スコアの低い植物の数が有意に増大していた。この結果はχ2分析において有意である。
【0042】
(表1)
非形質転換対照植物、対照pBIN05101植物およびpBIN05002試験植物についての低えい瘤スコア区分と高えい瘤スコア区分における植物の数
低えい瘤スコア=0〜50えい瘤/植物
高えい瘤スコア=51+えい瘤/植物
処理 低えい瘤スコアの植物 高えい瘤スコアの植物
非形質転換植物 18 13
pBIN05101対照植物 13 17
pBIN05002系32試験植物 25 7
【0043】
実施例2
上記の実施例において、線虫破壊における特異性を増大させるための細胞破壊系を用いて説明したオーバーラップの原理は、例えば、Arabidopsis、またはその他の植物の花においても同様に十分に実行されて、花のパターンまたは構造が変化した花、例えば、雄性不稔性の花が得られ得る。この実施例は、Arabidopsisにおいて同定されたDNA配列を利用するものである。
【0044】
花の中には3つの遺伝子の調節下にあると仮定される4つのエレメント(蕚片、心皮、花弁および雄ずい)が存在する(Coen, E.S.およびMeyerowitz, E.M., 1991)。
【0045】
これらの遺伝子のバランスを変更することにより、花パターンの変化が生じる。例えば、遺伝子agamousおよびapetala3は、花の同一部分で発現して植物の雄性部分である雄ずいを生じさせなければならない。agamousは、心皮および雄ずいの両方で発現し、一方apetala3は雄ずいおよび花弁の両方で発現する。
【0046】
本発明の主題であるオーバーラップ原理にしたがって構築物を製造するためには、本発明者らは、雄ずいにおいてのみ破壊をもたらすために第2の遺伝子(例えば、花弁および雄ずいで活性なapetala3)の破壊物質に連結された一の遺伝子(例えば、心皮および雄ずいで活性なagamous)由来のプロモーターを必要とする。
【0047】
下記のTaqおよびTaq−エキステンダーを用いたポリメラーゼ連鎖反応により公表されている手順(Thomas, C., 1996)にしたがって下記の二つのプライマーを用いることによって、agamousプロモーターの435塩基対断片をArabidopsis thaliana DNAから単離した:
プライマー1(配列番号1)
ATCGAAGCTT CTAAATGTAC TGAAAAGAAA CA
プライマー2(配列番号2)
ACTGGGATCC GAAAATGGAA GGTAAGGTTG TGC
プライマーは、ゲノムagamous配列についてのジーンバンクDNA配列エントリーATAGAMSGで得られた配列に基づくものであった。プライマー1はその5'末端に追加のHindIII制限部位を含む。プライマー2は、その5'末端に追加のBamHI制限部位を含む。
【0048】
下記のプライマーは、apetala3遺伝子配列の外側の部分を増幅するためにジーンバンク配列エントリーATHAPETALAから設計された:
プライマー3(配列番号3)
ATCGGGATCC ATGGGCTCAC GGTTTTGTGT GA
プライマー4(配列番号4)
ATCGGAGCTC TTATTCAAGA AGATGGAAGG TAATGA
プライマー3は、公表されているapetala3配列の992位にて増幅が開始されるように特異的に設計され、これは活性産物を産生させるための不適正リーディングフレームにおける開始コドンであり、同一遺伝子ファミリー内のその他のMADSボックス遺伝子と強い相同性を有する配列の初期部分を無効にするものである。また、プライマー3は、望ましくないSacI制限部位を除去するために公表されている配列から1塩基対変更されている。プライマー3はその5'末端にBamHI制限部位を有している。プライマー4はその5'末端にSacI部位を有している。1586bp断片を、プライマー3および4を用いてPCRによってArabidopsis DNAから増幅させた。
【0049】
下記のクローニング方法は当業者にはよく知られており、Sambrookら(1989)の方法にしたがうことによって成果を得ることができる。agamousプロモーター断片をpBluescript誘導ベクター(Stratagene社, ケンブリッジ, UK)にHindIII−BamHI断片としてライゲートした。変更型apetala遺伝子断片を同一ベクター内のagamousプロモーターの下流にBamHI-SacI断片としてライゲートした。また、ベクターには、変更型apetala3配列の下流にSacI−EcoRI断片としてnosターミネーター配列も含まれていた。ベクターをpDVM37010と命名した。Jim Haselhoffから入手した、m-gfp-ERレポーター遺伝子の前にagamousプロモーターを含む第2のプラスミド、MRCを対照として作製し、pDVM37003と命名した。
【0050】
プロモーター−遺伝子断片−ターミネーターカセットをpDVM37010(配列番号5)およびpDVM37003(配列番号6)からNotI制限断片として切り出し、pBIN19(Bevan, M. (1984))誘導ベクターpATCにライゲートしてプラスミドpATC37010(図3に示した地図)およびpATC37003(図4に示した地図)を得た。これらの配列は、適当な制限部位、すなわちカセットの各末端におけるNotIを有する任意のその他の同等のベクターにクローニングすることができる。pATCの地図を図5に示す。これはpBIN19と比べて改変されている制限部位を有する。pATC37010は、agamousプロモーターの調節下で共同抑制産物を産生して、花の発達中の雄ずいにおけるapetala3機能を不活性化する。
【0051】
プラスミドをアグロバクテリウム ツメファシエンス宿主LBA4404に移入し、Bechtoldら(1993)の方法およびHorschら(1985)の方法を用いるNicotiana tabacum cv K326にしたがってArabidopsis thalianaの形質転換に使用した。pATC37010については8個のトランスジェニックArabidopsis苗木およびpATC37003については6個のトランスジェニックArabidopsis苗木が得られた。100個の葉盤3群を、各構築物に対するNicotiana形質転換に用いた。トランスジェニックカルス増殖が3群全てについて検出された。
【0052】
発芽の10〜14日後にArabidopsis苗木を土壌に移し、開花するまで成長させた。花は雄ずいおよびダブルローズ(double rose)心皮を示さなかった。
【0053】
カナマイシン耐性pATC37010トランスジェニック植物を、プライマー1および4を用いてTaqポリメラーゼによるPCRによって当業者に公知の手順にしたがって、さらに所望の挿入物の存在についてスクリーニングした。PCRでは、94℃で60秒間、60℃で30秒間、および72℃で140秒間のインキュベーション工程を含む40サイクルのインキュベーションを行った。PCR陽性試料を、当業者に公知の手順にしたがってアガロースゲル電気泳動によるPCR産物の視覚化によって同定した。
【0054】
対照pATC37003トランスジェニック植物を、プライマー1およびプライマー7を用いてTaqポリメラーゼによるPCRによって当業者に公知の手順にしたがって、さらに所望の挿入物の存在についてスクリーニングした。PCRでは、94℃で40秒間、60℃で30秒間、および72℃で140秒間のインキュベーション工程を含む40サイクルのインキュベーションを行った。PCR陽性試料を、当業者に公知の手順にしたがってアガロースゲル電気泳動によるPCR産物の視覚化によって同定した。
プライマー7(配列番号7)
GAACTGGGAC CACTCCAGTG
いずれの場合においても、適切な構築物を含むトランスジェニック植物が同定された。
【0055】
(参考文献)
Bechtold, N., Ellis, J., およびPelletier, G. (1993) In planta Agrobacterium mediated gene transfer by infiltration of adult Arabidopsis thaliana plants, C.R. Acad. Sci: Paris 316 : 1194-1199。
Bevan, M. (1984) Binary Agrobacterium vectors for plant transformation, Nuc. Acids Res. 12: 8711-8721。
Coen, E.S. およびMeyerowitz, E.M. (1991) The war of the whorls: genetic interactions controlling flower development, Nature 353, 31-37。
Conkling, M.A., Cheng, C-L., Yamamoto, Y.T. およびGoodman, H.M.(1990) Isolation of transcriptionally regulated root-specific genes from tobacco, Plant Physiology 93, 1203-1211。
Horsch, R.B., Fry, J.F., Hoffmann, N.L., Eichholtz, D., Rogers, S.G. およびFraley, R.T.(1985), A simple and general method for transferring genes into plants, Science 22, 1229-1231。
Jefferson, R.A., Kavanagh, T.A. およびBevan, M.W. (1987) GUS fusions: β-glucuronidase as a sensitive and versatile gene fusion maker in higher plants, EMBO, 6, 3901-3907。
Opperman, C.H., Taylor, C.G. およびConkling, M.A. (1994) Root-knot nematode-directed expression of a plant root specific gene, Science 263, 221-223。
Sambrook, J., Fritsch, E.F. およびManiatis, T. (1989) Molecular Cloning - A Laboratory Manual. Second Edition, Cold Spring Harbour Laboratory Press, New York。
Thomas, C. (1996) 'PCR Techniques' in 'Plant Gene Isolation: Principles and Practice', G.D. Foster & D. Twell editors, John Wiley & Sons Ltd., pp.331-368。
【図1】
【特許請求の範囲】
【請求項1】
形質転換植物における遺伝子調節の特異性を改良する方法であって、
植物において、内在性ターゲット遺伝子の発現部位の一つにおける発現を含む、少なくとも2つの発現部位における遺伝子発現を生じさせるプロモーターと、前記内在性ターゲット遺伝子またはその産物の発現を調節する核酸配列である作用物質とを含むキメラ遺伝子を植物細胞に導入する工程、ならびに
前記植物細胞から形質転換植物を再生させる工程を含み、
ここで、前記形質転換植物は前記キメラ遺伝子を含み、前記形質転換植物における前記キメラ遺伝子の発現により、前記キメラ遺伝子と前記内在性ターゲット遺伝子とがオーバーラップする発現部位で、前記内在性ターゲット遺伝子またはその産物が調節されるものであるとともに、
前記キメラ遺伝子と前記内在性ターゲット遺伝子との共同発現の部位が、前記キメラ遺伝子及び前記内在性ターゲット遺伝子の各々が単独で発現する部位よりも、より限定されている、前記方法。
【請求項2】
前記核酸配列がコードまたは非コード配列である、請求項1に記載の方法。
【請求項3】
前記ターゲット遺伝子の発現がアップレギュレートされる、請求項1または2に記載の方法。
【請求項4】
前記ターゲット遺伝子の発現がダウンレギュレートされる、請求項1または2に記載の方法。
【請求項5】
前記核酸配列がアンチセンス方向のものである、請求項1、2または4に記載の方法。
【請求項6】
前記核酸がセンス方向のものである、請求項1〜4の何れか1項に記載の方法。
【請求項7】
前記ターゲット遺伝子のアップレギュレーションが、前記ターゲット遺伝子のプロモーターのアクチベーターの導入から生じるものである、請求項3に記載の方法。
【請求項8】
前記キメラ遺伝子の前記作用物質が複数の核酸配列を含み、その配列のそれぞれは、発現された場合に特定の機能を果たすものである、請求項1〜7のいずれか1項に記載の方法。
【請求項9】
前記キメラ遺伝子のプロモーターが、前記内在性遺伝子の複数のオーバーラップ発現部位で発現される、請求項1〜8のいずれか1項に記載の方法。
【請求項10】
前記プロモーターがKNT1プロモーターまたはRB7プロモーターである、請求項1〜9のいずれか1項に記載の方法。
【請求項11】
前記核酸配列が、1以上のリボザイム、メッセンジャーRNAを分解するための標的RNアーゼ、または特異的RNAの安定化もしくは不安定化物質である、請求項1〜9のいずれか1項に記載の方法。
【請求項12】
前記核酸が転写のアクチベーターを含む、請求項1〜9のいずれか1項に記載の方法。
【請求項13】
転写の前記アクチベーターがタンパク質をコードする熱ショック因子である、請求項12に記載の方法。
【請求項14】
前記作用物質が前記遺伝子上で作用する機構が、アンチセンス、共同抑制、ターゲット遺伝子のプロモーターの阻害もしくは活性化、転写の阻害もしくは活性化、メッセンジャーRNA安定性の変更もしくはmRNAの分解、翻訳の阻害もしくは活性化、タンパク質の阻害もしくは活性化、タンパク質代謝回転の変更、共同因子としての作用、タンパク質−タンパク質相互作用の変更、または生化学的経路によるフラックスの変更からなる群の1以上である、請求項1〜13のいずれか1項に記載の方法。
【請求項15】
受託番号NCIMB 40871の寄託された構築物。
【請求項16】
プラスミドpATC37010またはそのプラスミドに含まれるようなプロモーター−遺伝子断片−ターミネーターカセットを含む構築物。
【請求項17】
請求項1〜14のいずれか1項に記載の方法を行うことにより得られた植物およびその子孫。
【請求項1】
形質転換植物における遺伝子調節の特異性を改良する方法であって、
植物において、内在性ターゲット遺伝子の発現部位の一つにおける発現を含む、少なくとも2つの発現部位における遺伝子発現を生じさせるプロモーターと、前記内在性ターゲット遺伝子またはその産物の発現を調節する核酸配列である作用物質とを含むキメラ遺伝子を植物細胞に導入する工程、ならびに
前記植物細胞から形質転換植物を再生させる工程を含み、
ここで、前記形質転換植物は前記キメラ遺伝子を含み、前記形質転換植物における前記キメラ遺伝子の発現により、前記キメラ遺伝子と前記内在性ターゲット遺伝子とがオーバーラップする発現部位で、前記内在性ターゲット遺伝子またはその産物が調節されるものであるとともに、
前記キメラ遺伝子と前記内在性ターゲット遺伝子との共同発現の部位が、前記キメラ遺伝子及び前記内在性ターゲット遺伝子の各々が単独で発現する部位よりも、より限定されている、前記方法。
【請求項2】
前記核酸配列がコードまたは非コード配列である、請求項1に記載の方法。
【請求項3】
前記ターゲット遺伝子の発現がアップレギュレートされる、請求項1または2に記載の方法。
【請求項4】
前記ターゲット遺伝子の発現がダウンレギュレートされる、請求項1または2に記載の方法。
【請求項5】
前記核酸配列がアンチセンス方向のものである、請求項1、2または4に記載の方法。
【請求項6】
前記核酸がセンス方向のものである、請求項1〜4の何れか1項に記載の方法。
【請求項7】
前記ターゲット遺伝子のアップレギュレーションが、前記ターゲット遺伝子のプロモーターのアクチベーターの導入から生じるものである、請求項3に記載の方法。
【請求項8】
前記キメラ遺伝子の前記作用物質が複数の核酸配列を含み、その配列のそれぞれは、発現された場合に特定の機能を果たすものである、請求項1〜7のいずれか1項に記載の方法。
【請求項9】
前記キメラ遺伝子のプロモーターが、前記内在性遺伝子の複数のオーバーラップ発現部位で発現される、請求項1〜8のいずれか1項に記載の方法。
【請求項10】
前記プロモーターがKNT1プロモーターまたはRB7プロモーターである、請求項1〜9のいずれか1項に記載の方法。
【請求項11】
前記核酸配列が、1以上のリボザイム、メッセンジャーRNAを分解するための標的RNアーゼ、または特異的RNAの安定化もしくは不安定化物質である、請求項1〜9のいずれか1項に記載の方法。
【請求項12】
前記核酸が転写のアクチベーターを含む、請求項1〜9のいずれか1項に記載の方法。
【請求項13】
転写の前記アクチベーターがタンパク質をコードする熱ショック因子である、請求項12に記載の方法。
【請求項14】
前記作用物質が前記遺伝子上で作用する機構が、アンチセンス、共同抑制、ターゲット遺伝子のプロモーターの阻害もしくは活性化、転写の阻害もしくは活性化、メッセンジャーRNA安定性の変更もしくはmRNAの分解、翻訳の阻害もしくは活性化、タンパク質の阻害もしくは活性化、タンパク質代謝回転の変更、共同因子としての作用、タンパク質−タンパク質相互作用の変更、または生化学的経路によるフラックスの変更からなる群の1以上である、請求項1〜13のいずれか1項に記載の方法。
【請求項15】
受託番号NCIMB 40871の寄託された構築物。
【請求項16】
プラスミドpATC37010またはそのプラスミドに含まれるようなプロモーター−遺伝子断片−ターミネーターカセットを含む構築物。
【請求項17】
請求項1〜14のいずれか1項に記載の方法を行うことにより得られた植物およびその子孫。
【図2】

【図3】
【図4】
【図5】
【図3】
【図4】
【図5】
【公開番号】特開2010−29196(P2010−29196A)
【公開日】平成22年2月12日(2010.2.12)
【国際特許分類】
【出願番号】特願2009−211993(P2009−211993)
【出願日】平成21年9月14日(2009.9.14)
【分割の表示】特願平10−519774の分割
【原出願日】平成10年3月27日(1998.3.27)
【出願人】(301004857)アドヴァンスト・テクノロジーズ(ケンブリッジ)リミテッド (8)
【Fターム(参考)】
【公開日】平成22年2月12日(2010.2.12)
【国際特許分類】
【出願日】平成21年9月14日(2009.9.14)
【分割の表示】特願平10−519774の分割
【原出願日】平成10年3月27日(1998.3.27)
【出願人】(301004857)アドヴァンスト・テクノロジーズ(ケンブリッジ)リミテッド (8)
【Fターム(参考)】
[ Back to top ]