
遺伝子的に精製したゲランガム
一定の乳製品におけるパラ−クレゾール生成に関与するタンパク質を突然変異により不活性化することにより、改良された味とにおいがもたらされる。不所望なパラ−クレゾールは、ゲランガムを生成する微生物により生成される酵素による結果として、時間をかけて形成される。ゲランは比較的未精製の形態で使用されることから、これら酵素をゲランとともに乳に添加する。これら酵素の不活性化は、任意の追加の精製又は処理を必要とすることのない、酵素を排除する遺伝子的手段である。

【発明の詳細な説明】
【発明の開示】
【0001】
本特許文献の開示の一部は、著作権の保護を受ける資料を含む。本著作権者は、特許商標庁の特許ファイル又は記録においてみられるので、第三者による本特許文献又は本特許の開示についてのファクシミリによる複製に対して異議を示さないが、すべての著作権はいかなるものでも留保されている。
【0002】
発明の分野
本発明は、食品添加物の分野に関する。特に、本発明は、酪農食品添加物の分野に関する。より特定的には、本発明は、滅菌された乳製品に対する添加物に関する。
【0003】
発明の背景
ゲランガムは、Sphingomonas elodea細菌により生産される細胞外多糖である。S. elodeaにより生産されるゲランガムは、カリフォルニア州サンディエゴのCP KelcoからKelcogel LT100(登録商標)として商業的に入手可能である。商業的には、ゲランガムは、好気的発酵により形成される。発酵が完了したら、ガムを発酵培地から回収する前に、培地を低温殺菌して生存細胞を殺す。
【0004】
ゲランガムは、糖類であるグルコース、グルクロン酸及びラムノースを2:1:1のモル比で含み、これらは結合して四糖の繰返し単位を形成する。天然のゲランガムは、同じグルコース残基上でアセチル化されグリセリル化されている。平均して、四糖繰返し単位あたりに1個のアセチル基と1.5個のグリセリル基が存在する。
【0005】
ゲランガムを回収する方法は、ガムの特徴に影響を与える。直接回収では、柔軟で可撓性のあるゲルが得られる。ゲランガムは、その組織構造上の特性及びレオロジー的特性のため、培養、レトルト、また冷凍の酪農製品において長い間使用されてきた。
【0006】
しかし、貯蔵安定性のある、乳を基材とするゲラン含有性製品においては、異臭やにおいが発生し、この異臭やにおいにより食品は口にあわないものとなってしまう。異臭やにおいは、乳中の物質、例えばパラ−硫酸クレゾール及びパラ−クレシルグルクロニドからパラ−クレゾールが形成されることと結び付けられている。パラ−クレゾールは、超高温にて処理され、室温にて保存された、乳を基材とするゲラン含有性製品中において検出可能である。
【0007】
この問題を排除しようとして、ゲランは、熱アルカリ処理により脱アシル化されてきた。パラ−クレゾールを排除するには有効であるが、脱アシル化処理により、ゲランガムがよりもろくなり、一定の食品用途について有用性が低くなってしまう。この問題を排除するための別のアプローチは、天然のゲランガムを次亜塩素酸ナトリウム又は水酸化カリウムなどの変性剤により前処理することである。このアプローチにより、材料コストと処理コストが追加される。滅菌された酪農製品中において長期間貯蔵してもパラ−クレゾールを形成せず、余分な処理工程を必要としない、ゲラン製品についての必要性が当技術分野において存在する。
【0008】
発明の要旨
第一の態様においては、アリールスルファターゼタンパク質を実質的に含まないゲランガムを含む組成物を提供する。
【0009】
第二の態様においては、β−グルクロニダーゼタンパク質を実質的に含まないゲランガムを含む組成物を提供する。
本発明の第三の態様においては、アリールスルファターゼタンパク質及びβ−グルクロニダーゼタンパク質の両方を実質的に含まないゲランガムを含む組成物を提供する。
【0010】
本発明の第四の態様においては、ゲランガム組成物を生成するための方法を提供する。Sphingomonas elodeaを培養培地中で培養する。Sphingomonas elodeaは、触媒活性のあるアリールスルファターゼ、又は触媒活性のあるβ−グルクロニダーゼ、又は触媒活性のあるアリールスルファターゼ及び触媒活性のあるβ−グルクロニダーゼを生成しない。培養培地は収集する。ゲランガムは培養培地から沈殿させる。
【0011】
本発明の第五の態様においては、Sphingomonas elodeaの微生物学的に純粋な培養液を提供する。この培養液は、アリールスルファターゼが欠乏している。
本発明の第六の態様においては、Sphingomonas elodeaの微生物学的に純粋な別の培養液を提供する。この培養液は、β−グルクロニダーゼが欠乏している。
【0012】
本発明の更に別の態様は、Sphingomonas elodeaの微生物学的に純粋な培養液である。この培養液は、アリールスルファターゼ及びβ−グルクロニダーゼの両方が欠乏している。
【0013】
本発明の第八の態様は、Sphingomonas elodeaアリールスルファターゼを符号化している、単離、精製されたポリヌクレオチドを提供する。このアリールスルファターゼは、SEQ ID NO:2にしたがったアミノ酸配列を有する。
【0014】
本発明の第九の態様は、Sphingomonas elodeaβ−グルクロニダーゼを符号化している、単離、精製されたポリヌクレオチドを提供する。このβ−グルクロニダーゼは、SEQ ID NO:5にしたがったアミノ酸配列を有する。
【0015】
本発明の第十の態様は、Sphingomonas elodeaゲノムDNAを含む、単離、精製されたポリヌクレオチドである。このゲノムDNAは、そのアリールスルファターゼコード配列のすべて又は一部の欠失を含む。
【0016】
本発明の第十一の態様は、Sphingomonas elodeaゲノムDNAを含む、単離、精製されたポリヌクレオチドである。このゲノムDNAは、そのβ−グルクロニダーゼコード配列のすべて又は一部の欠失を含む。
【0017】
本発明のこれらの態様及び以下により詳細に説明するような本発明の他の態様は、当技術分野に対して、消費者に許容され得る、滅菌されたゲラン含有性酪農製品を製造するための費用対効果の大きい手段を提供する。
【0018】
発明の具体的な説明
本発明者らは、Sphingomonas elodea細菌において、アリールスルファターゼ酵素及びβ−グルクロニダーゼ酵素を符号化する遺伝子のいずれか、あるいは両方の遺伝子を突然変異により不活性化させた場合に、この細菌が一定の目的についてすぐれた特性を有するゲランを生成することを発見した。特に、そのような突然変異体により生成されるゲランは、滅菌された乳製品に対して長期の貯蔵安定性を付与する。
【0019】
これらの酵素の一方又は両方を不活性化させない場合、これらは乳中に存在する基質(p−硫酸クレシル及びp−クレシルグルクロニド)からパラ−クレゾールを生成する。パラ−クレゾールは、一般的には消費者らの口に合わないにおい及び異臭を与える。これらの酵素を排除することにより、滅菌された乳又は乳製品において貯蔵中にパラ−クレゾールが生成される割合が低減される。
【0020】
これら酵素のいずれか又は両方の突然変異を使用して、パラ−クレゾール生成の割合を低減させることができる。突然変異は、好ましくは、挿入変異、ナンセンス変異、フレームシフト変異又は欠失変異などの、タンパク質を完全に不活性化させるタイプのものである。そのような突然変異をもたらすための当技術分野において既知の任意の手法を使用することができる。本出願人らは、トランスポゾン挿入ストラテジを使用して、アリールスルファターゼ及びβ−グルクロニダーゼを符号化する遺伝子を同定した。次いで、各々の遺伝子における欠失変異は、遺伝子をフランキングしている5’及び3’DNAフラグメントを用いる相同性組換により構築し、これらフラグメントをともに接合した。しかしながら、これらの遺伝子において突然変異を得るために、他のストラテジを使用することができる。突然変異は、ゲラン「産生」種において直接起こすことができ、あるいは、はじめに突然変異を起こした種からそのような種へ突然変異を移植させることができる。Site-directed mutagenesisのための手法は、当技術分野においてよく知られている。例えば、“In Vitro Mutagenesis Protocols”, second edition, Braman, Jeff, ed., Humana Press, 2002、及び商業的に入手可能なQuikChange(商標)キット(Stratagene)を参照されたい。Sphingomonas elodeaのアリールスルファターゼ遺伝子及びβ−グルクロニダーゼ遺伝子の野生型配列を提供すれば、当業者であれば、これらの遺伝子において容易に種々の所望の突然変異を起こすことができる。
【0021】
アリールスルファターゼ及びβ−グルクロニダーゼを符号化する野生型遺伝子及び変異体遺伝子の配列を決定した。SEQ ID NO:1(野生型アリールスルファターゼ)、SEQ ID NO:3(アリールスルファターゼ欠失)、SEQ ID NO:4(野生型β−グルクロニダーゼ)及びSEQ ID NO:6(β−グルクロニダーゼ欠失)を参照されたい。これらの遺伝子及びそれらのヌクレオチド配列を同定することにより、当業者はこれらの酵素を発現させるための所望の無表現型を有する他の突然変異体を容易につくることが可能となる。挿入、欠失、ナンセンス、及びフレームシフトなどの突然変異は、酵素についてゼロ表現型においてもっともよく起こる可能性がある。また、ミスセンス変異を起こして、酵素活性に関するそれらの効果について通常通り試験することもできる。微生物遺伝学の標準的な手法を使用して適する突然変異を起こすことができる。例えば、Principles of Gene Manipulation: An Introduction to Genetic Engineering, R. W. Old, S. B. Primrose, Blackwell Publishing, 1994を参照されたい。標準的な酵素アッセイを使用して、突然変異させた酵素の活性の損失について試験することができる。例えば、Kim et al., Appl Microbiol Biotechnol. 2004 Feb; 63(5): 553-9を参照されたい。
【0022】
実質的にアリールスルファターゼ又はβ−グルクロニダーゼを含まない本発明の組成物は、野生型種において含有される特定のタンパク質の量の95%、96%、97%、98%、又は99%未満を含有する。そのような酵素の量の低減により、野生型種からのゲランを含有する組成物において生成されるパラ−クレゾールの量の90%、93%、95%、97%、又は99%未満を有する滅菌された乳組成物をもたらす。生成されるアリールスルファターゼ又はβ−グルクロニダーゼタンパク質の量は、Sigmaからの5−ブロモ−4−クロロ−3−インドリルサルフェート(X−SO4);CAS No.6578−07−0などの容易にアッセイ可能な基質を用いて、又は、Sigma若しくはRPI社からの5−ブロモ−4−クロロ−3−インドリル−β−D−グルクロニド(X−GlcA);CAS No.114162−64−0を用いて、又は、p−ニトロカテコールを用いて、酵素アッセイにより測定することができる。別の方法では、ウェスタンブロッドなどの免疫学的な手法を用いてタンパク質を測定することができる。
【0023】
ゲランガムは、典型的には、滅菌されたか若しくは超高温(UHT)処理された酪農製品、又は冷凍された酪農製品において使用される。かかる製品としては、これらに限定されないが、アイスクリーム、フローズンヨーグルト、プディング、ホイップされた酪農製品、コーヒークリーマー、クレームブリュレ及び酪農飲料が挙げられる。本発明のゲランガムは、野生型種からの典型的なゲランを使用することができるように、これら又は他の任意の食品において使用することができる。
【0024】
ゲランガムは、本発明の突然変異種を用いて、野生型種について当技術分野において既知の方法のいずれかにしたがって生成することができる。細菌は、典型的には、液体培養培地中において生育する。かかる培地は、典型的には、炭素源、リン酸塩、有機又は無機の窒素源、及び適切な微量元素を含有する。発酵は、典型的には、滅菌された条件下で通気及び撹拌しながら行う。発酵時期の最後には、典型的には、細胞を除去することなく、培養培地を収集する。発酵培地は、ゲランガムを回収する前に、低温殺菌して生育可能な細胞を殺すことができる。ゲランガムは、当技術分野において既知であるように、沈殿させることができる。典型的には、これはイソプロパノールなどのアルコールを用いて行われる。沈殿させたゲランは、再度水和させる前に乾燥させることができる。
【0025】
パラ−クレゾールは、当技術分野において既知の任意の手段により測定することができる。使用することができる一つの方法は、ジクロロメタン抽出、濃縮、及びガスクロマトグラフィーによる質量分析を使用する。
【0026】
本発明を実施する現時点で好ましい態様を含む具体的な実施例に関して本発明を説明してきたが、当業者であれば、特許請求の範囲に示したような本発明の精神及び範囲に含まれるこれまでに説明した系及び手法について数多くの変体及び順序の変更が存在することを理解するであろう。
【0027】
実施例
実施例1
突然変異種をスクリーニングし特徴付ける際に以下の試薬を使用した:
5−ブロモ−4−クロロ−3−インドリルサルフェート(X−SO4);CAS No.6578−07−0
供給源:Sigma
5−ブロモ−4−クロロ−3−インドリル−β−D−グルクロニド(X−GlcA);CAS No.114162−64−0
供給源:Sigma又はRPI社
実施例2
Epicentre(ウィスコンシン州、マディソン)からの商業的に入手可能なEZ::TN(登録商標)<R6Kガンマ−ori/KAN−2>挿入キットを用いて、非ムコイド種であるSphingomonas elodea, Gps2においてランダムトランスポゾン変異体のライブラリをつくることにより、アリールスルファターゼ及びβ−グルクロニダーゼを符号化する遺伝子を同定した。カナマイシン耐性突然変異種を、特異的な色素産生基質を伴う選択培地上での青色の欠如(又は有意な低減)に関してスクリーニングした。実施例1を参照されたい。アリールスルファターゼ生成又は活性を妨げる突然変異体を、塩化物塩を伴う規定寒天培地上で色原体である5−ブロモ−4−クロロ−3−インドリルサルフェート(X−SO4)を用いて同定した。β−グルクロニダーゼの突然変異体は、色原体である5−ブロモ−4−クロロ−3−インドリル−β−D−グルクロニド(X−GlcA)を伴う規定培地上で選択した。
【0028】
トランスポゾン及び近傍のゲノムDNAは、続いて制限酵素を用いてクロモソームから摘出した。制限酵素フラグメントは、リガーゼにより円形にし、Escherichia coliに形質転換し、そこでトランスポゾン含有DNAをトランスポゾン中のレプリコンの存在により複製することができる。アリールスルファターゼについての遺伝子を伴うプラスミドはR6K−AS#14Eと命名した。このプラスミドの一部を配列にした(SEQ ID NO:1)。β−グルクロニダーゼについての遺伝子を伴うプラスミドはR6K−BG#6Sと命名した。このプラスミドの一部を配列にした(SEQ ID No:4)。Escherichia coli種EC100D pir+におけるこれらのプラスミドは、2004年6月21日にバージニア州マナッサスのアメリカンタイプカルチャーコレクションに寄託される予定である。
【0029】
不活性化されたアリールスルファターゼを有する細菌において、アリールスルファターゼについての遺伝子の近傍にある仮想タンパク質について、トランスポゾンを実際に遺伝子中に挿入した。不活性化されたβ−グルクロニダーゼを有する細菌において、トランスポゾン挿入は、β−グルクロニダーゼについての遺伝子のアミノ部分に配置した。これら遺伝子のDNA配列は、National Center for Biotechnology Information(NCBI)のデータベースにおける他の種からの既知の遺伝子と相同性を有することを示した。
【0030】
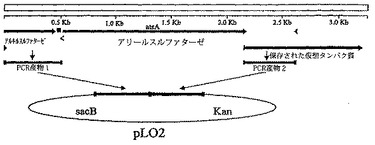
アリールスルファターゼ及びβ−グルクロニダーゼについての遺伝子、並びに近傍のゲノムDNAを配列した。これら遺伝子の欠失をプラスミド上で構築し、次いで、S. elodea種S−60wtc及びPDG−1に形質転換した。WO01/64897を参照されたい。これら欠失を相同性組換によりゲノムにおいて挿入した。標的遺伝子をフランキングするDNA配列をPCRにより増幅し、プラスミドベクターpLO2にクローンした。(Lenz, O., E. Schwarts, J. Dernedde, M. Eitinger and B. Friedrich. 1994, “The Alcaligenes eutrophus H16 hoxX gene participates in hydrogenase regulation,” J. Bacteriol. 176:4385-4394.)この構成を接合によりS. elodea種に形質転換した。得られるカナマイシン耐性種を、次いで、抗生物質無しで30−40世代生育させた。プラスミドを失った分離株は、pLO2上のsacB遺伝子の欠失によるスクロース耐性について選択することにより検出し、カナマイシン感受性により確認した。非選択的生育後にプラスミドを失った分離株は、二つのタイプ、すなわち、欠失及び野生型であった。所望の欠失種は、適する指示寒天(実施例1を参照されたい)上での青色の欠如と、診断PCRにより同定した。構成に関するスキームを以下の図1に示す。
【0031】
アリールスルファターゼについての遺伝子(atsA)の正確な欠失を構成するためには、atsA遺伝子についての最も可能性のある開始コドンと近傍の遺伝子の開始コドン及び終止コドンを決定することが必要であった。遺伝子の末端の配置は、DNA配列、GenBankにおける他の遺伝子との相同性、及びフレームプロット3プログラムを用いた第三塩基GC選択に基づいて決定した。Ishikawa and Hotta. “FlamePlot: a new implementation of the Frame analysis for predicting protein-coding regions in bacterial DNA with a high G+C content.”FEMS Microbiology Letters 174:251-253(1999).この分析は、アリールスルファターゼ遺伝子が、保存された仮想タンパク質についての遺伝子に翻訳されて結合していること、すなわちアリールスルファターゼ遺伝子の開始コドンが保存された仮想タンパク質についての遺伝子の開始コドンと重複していることを示した。これらのタンパク質が最適な細胞生育とゲラン生成のために要求されるものかどうかは知られていないことから、アリールスルファターゼ欠失を構築して、アリールスルファターゼ遺伝子と未知の無傷のタンパク質についての遺伝子を残した。
【0032】
PCRプライマーであるAS5
【0033】
【化1】
【0034】
及びAS6
【0035】
【化2】
【0036】
を使用して、SacI−XbaIフラグメント(合計528bp)として、atsAの開始コドンのちょうど上流に、512bpフラグメントをSacI−XbaIフラグメントを増幅させた。AS3プライマー
【0037】
【化3】
【0038】
及びAS4プライマー
【0039】
【化4】
【0040】
を使用して、atsA遺伝子の端部とXbaI−SphIフラグメント(合計497bp)として保存された仮想タンパク質の開始とを含む、479bpのフラグメントを増幅させた。このように、atsAの終止コドンを保持させたが、開始コドンは欠失させて、アリールスルファターゼタンパク質の部分は合成しないものとする。クローニングのための制限部位は、プライマーの端部に追加した。PCRフラグメントは、プラスミドベクターpLO2のポリリンカー中に連続してくくりつけて、atsA遺伝子の欠失を形成させた。上流のSacI−XbaIフラグメントは、SacI−XbaI切断pLO2中にクローンした。続いて、下流のXbaI−SphIフラグメントをクローンした(図2)。このatsA遺伝子を欠失させたプラスミドを、S. elodeaのS60wtc種及びPDG-1種中への接合により形質転換して、プラスミドをクロモソーム中に組み替えた。カナマイシン耐性のある分離株を精製し、次いで抗生物質無しで生育し、スクロース及びX−SO4を伴う培地上にプレーティングして、プラスミドを符号化するsacB遺伝子の欠失によりスクロース耐性をもつ分離株を選択した。スクロース耐性、カナマイシン感受性のある、黄色コロニーを選択した。S60wtc及びPDG-1のatsA誘導体はそれぞれGAS−1及びPAS−1と命名した。
【0041】
β−グルクロニダーゼについての遺伝子(gusA)の欠失は、プラスミドpLO2上に構築し、S60wtc,GAS−1中に形質転換した。gusA遺伝子についての最も可能性のある開始コドンを、他のタンパク質とリボソーム結合部位の存在とに対する相同性により決定した。二次構造の領域は、開始コドンの上流である。gusA遺伝子の欠失を構築して、gusAの上流及び下流に二次構造を維持した。Bgluc3プライマー
【0042】
【化5】
【0043】
及びBgluc4プライマー
【0044】
【化6】
【0045】
を使用して、gusA遺伝子の上流に、PstI−XbaIフラグメント(合計577bp)として、560bpのフラグメントを増幅させた。Bgluc1プライマー
【0046】
【化7】
【0047】
及びBgluc2プライマー
【0048】
【化8】
【0049】
を使用して、gusA遺伝子の下流に、XbaI−SacIフラグメント(合計505bp)として、489bpのフラグメントを増幅させた。PstI−XbaIフラグメントは、PstI−XbaI切断pLO2中にクローンした。続いて、下流のXbaI−SacIフラグメントをクローンした。このgusA遺伝子が欠失したプラスミドを、S. elodeaのS60wtc種、GAS−1種及びPAS−1種中に接合により形質転換して、組替えを行った。カナマイシン耐性分離株を精製し、抗生物質無しで生育させ、次いで、スクロース及びX−GlcAを伴なう培地上にプレーティングして、プラスミド符号化sacB遺伝子の欠失によりスクロース耐性をもつ分離株を選択した。青緑色(野生型)及び明緑色(変異体)のコロニーの混合物を得た。スクロース耐性のある明緑色の分離株は、カナマイシン感受性により、プラスミド欠失について確認された。S60wtc、GAS−1及びPAS−1のgusA欠如誘導体は、それぞれGBG、GBAD及びPBADと命名した。
【0050】
S60wtc種及びPDG−1種におけるアリールスルファターゼについての遺伝子(atsA)の欠失を完了した。β−グルクロニダーゼについての遺伝子を同定し、配列した。近傍のDNAを配列した。また、β−グルクロニダーゼについての遺伝子の欠失は、S60、PDG−1及びGAS−1のそれぞれにおいて構築した。
【0051】
GAS−1種(アリールスルファターゼについての遺伝子を欠失させた)及びGBAD−1種(アリールスルファターゼ及びβ−グルクロニダーゼの両方についての遺伝子を欠失させた)から生成したゲランのサンプルを、超高温酪農用途試験において月ごとの間隔で、p−クレゾール生産について評価した。野生型種からのゲランサンプルは、1ヵ月後に3〜152(平均65)ppb、2ヵ月後に4〜212(平均96)ppbのp−クレゾールを生成した。GAS−1からのゲランのサンプルは、1〜5ヵ月後に約1〜3ppbのp−クレゾールを生成した。GBAD−1からのゲランのサンプルは、3ヵ月後まで試験したときに、生成量は1ppb(検出限界)未満であった。
【図面の簡単な説明】
【0052】
【図1】図1は、アリールスルファターゼ遺伝子(atsA)の近傍のゲノム領域のゲノムマップと、PCRにより増幅しプラスミドpLO2中にクローンした領域の位置である。次いで、クローンPCRフラグメントを伴なうプラスミドpLO2を使用して、相同性組換により、ゲノムのこの領域を欠失により置き換えた。
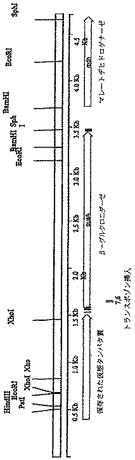
【図2】図2は、Sphingomonas elodeaのβ−グルクロニダーゼ遺伝子(gusA)近傍のゲノム領域の制限酵素切断地図である。BG−6クローン及びBG−7クローンにおけるトランスポゾン挿入の位置は底部に示す。
【発明の開示】
【0001】
本特許文献の開示の一部は、著作権の保護を受ける資料を含む。本著作権者は、特許商標庁の特許ファイル又は記録においてみられるので、第三者による本特許文献又は本特許の開示についてのファクシミリによる複製に対して異議を示さないが、すべての著作権はいかなるものでも留保されている。
【0002】
発明の分野
本発明は、食品添加物の分野に関する。特に、本発明は、酪農食品添加物の分野に関する。より特定的には、本発明は、滅菌された乳製品に対する添加物に関する。
【0003】
発明の背景
ゲランガムは、Sphingomonas elodea細菌により生産される細胞外多糖である。S. elodeaにより生産されるゲランガムは、カリフォルニア州サンディエゴのCP KelcoからKelcogel LT100(登録商標)として商業的に入手可能である。商業的には、ゲランガムは、好気的発酵により形成される。発酵が完了したら、ガムを発酵培地から回収する前に、培地を低温殺菌して生存細胞を殺す。
【0004】
ゲランガムは、糖類であるグルコース、グルクロン酸及びラムノースを2:1:1のモル比で含み、これらは結合して四糖の繰返し単位を形成する。天然のゲランガムは、同じグルコース残基上でアセチル化されグリセリル化されている。平均して、四糖繰返し単位あたりに1個のアセチル基と1.5個のグリセリル基が存在する。
【0005】
ゲランガムを回収する方法は、ガムの特徴に影響を与える。直接回収では、柔軟で可撓性のあるゲルが得られる。ゲランガムは、その組織構造上の特性及びレオロジー的特性のため、培養、レトルト、また冷凍の酪農製品において長い間使用されてきた。
【0006】
しかし、貯蔵安定性のある、乳を基材とするゲラン含有性製品においては、異臭やにおいが発生し、この異臭やにおいにより食品は口にあわないものとなってしまう。異臭やにおいは、乳中の物質、例えばパラ−硫酸クレゾール及びパラ−クレシルグルクロニドからパラ−クレゾールが形成されることと結び付けられている。パラ−クレゾールは、超高温にて処理され、室温にて保存された、乳を基材とするゲラン含有性製品中において検出可能である。
【0007】
この問題を排除しようとして、ゲランは、熱アルカリ処理により脱アシル化されてきた。パラ−クレゾールを排除するには有効であるが、脱アシル化処理により、ゲランガムがよりもろくなり、一定の食品用途について有用性が低くなってしまう。この問題を排除するための別のアプローチは、天然のゲランガムを次亜塩素酸ナトリウム又は水酸化カリウムなどの変性剤により前処理することである。このアプローチにより、材料コストと処理コストが追加される。滅菌された酪農製品中において長期間貯蔵してもパラ−クレゾールを形成せず、余分な処理工程を必要としない、ゲラン製品についての必要性が当技術分野において存在する。
【0008】
発明の要旨
第一の態様においては、アリールスルファターゼタンパク質を実質的に含まないゲランガムを含む組成物を提供する。
【0009】
第二の態様においては、β−グルクロニダーゼタンパク質を実質的に含まないゲランガムを含む組成物を提供する。
本発明の第三の態様においては、アリールスルファターゼタンパク質及びβ−グルクロニダーゼタンパク質の両方を実質的に含まないゲランガムを含む組成物を提供する。
【0010】
本発明の第四の態様においては、ゲランガム組成物を生成するための方法を提供する。Sphingomonas elodeaを培養培地中で培養する。Sphingomonas elodeaは、触媒活性のあるアリールスルファターゼ、又は触媒活性のあるβ−グルクロニダーゼ、又は触媒活性のあるアリールスルファターゼ及び触媒活性のあるβ−グルクロニダーゼを生成しない。培養培地は収集する。ゲランガムは培養培地から沈殿させる。
【0011】
本発明の第五の態様においては、Sphingomonas elodeaの微生物学的に純粋な培養液を提供する。この培養液は、アリールスルファターゼが欠乏している。
本発明の第六の態様においては、Sphingomonas elodeaの微生物学的に純粋な別の培養液を提供する。この培養液は、β−グルクロニダーゼが欠乏している。
【0012】
本発明の更に別の態様は、Sphingomonas elodeaの微生物学的に純粋な培養液である。この培養液は、アリールスルファターゼ及びβ−グルクロニダーゼの両方が欠乏している。
【0013】
本発明の第八の態様は、Sphingomonas elodeaアリールスルファターゼを符号化している、単離、精製されたポリヌクレオチドを提供する。このアリールスルファターゼは、SEQ ID NO:2にしたがったアミノ酸配列を有する。
【0014】
本発明の第九の態様は、Sphingomonas elodeaβ−グルクロニダーゼを符号化している、単離、精製されたポリヌクレオチドを提供する。このβ−グルクロニダーゼは、SEQ ID NO:5にしたがったアミノ酸配列を有する。
【0015】
本発明の第十の態様は、Sphingomonas elodeaゲノムDNAを含む、単離、精製されたポリヌクレオチドである。このゲノムDNAは、そのアリールスルファターゼコード配列のすべて又は一部の欠失を含む。
【0016】
本発明の第十一の態様は、Sphingomonas elodeaゲノムDNAを含む、単離、精製されたポリヌクレオチドである。このゲノムDNAは、そのβ−グルクロニダーゼコード配列のすべて又は一部の欠失を含む。
【0017】
本発明のこれらの態様及び以下により詳細に説明するような本発明の他の態様は、当技術分野に対して、消費者に許容され得る、滅菌されたゲラン含有性酪農製品を製造するための費用対効果の大きい手段を提供する。
【0018】
発明の具体的な説明
本発明者らは、Sphingomonas elodea細菌において、アリールスルファターゼ酵素及びβ−グルクロニダーゼ酵素を符号化する遺伝子のいずれか、あるいは両方の遺伝子を突然変異により不活性化させた場合に、この細菌が一定の目的についてすぐれた特性を有するゲランを生成することを発見した。特に、そのような突然変異体により生成されるゲランは、滅菌された乳製品に対して長期の貯蔵安定性を付与する。
【0019】
これらの酵素の一方又は両方を不活性化させない場合、これらは乳中に存在する基質(p−硫酸クレシル及びp−クレシルグルクロニド)からパラ−クレゾールを生成する。パラ−クレゾールは、一般的には消費者らの口に合わないにおい及び異臭を与える。これらの酵素を排除することにより、滅菌された乳又は乳製品において貯蔵中にパラ−クレゾールが生成される割合が低減される。
【0020】
これら酵素のいずれか又は両方の突然変異を使用して、パラ−クレゾール生成の割合を低減させることができる。突然変異は、好ましくは、挿入変異、ナンセンス変異、フレームシフト変異又は欠失変異などの、タンパク質を完全に不活性化させるタイプのものである。そのような突然変異をもたらすための当技術分野において既知の任意の手法を使用することができる。本出願人らは、トランスポゾン挿入ストラテジを使用して、アリールスルファターゼ及びβ−グルクロニダーゼを符号化する遺伝子を同定した。次いで、各々の遺伝子における欠失変異は、遺伝子をフランキングしている5’及び3’DNAフラグメントを用いる相同性組換により構築し、これらフラグメントをともに接合した。しかしながら、これらの遺伝子において突然変異を得るために、他のストラテジを使用することができる。突然変異は、ゲラン「産生」種において直接起こすことができ、あるいは、はじめに突然変異を起こした種からそのような種へ突然変異を移植させることができる。Site-directed mutagenesisのための手法は、当技術分野においてよく知られている。例えば、“In Vitro Mutagenesis Protocols”, second edition, Braman, Jeff, ed., Humana Press, 2002、及び商業的に入手可能なQuikChange(商標)キット(Stratagene)を参照されたい。Sphingomonas elodeaのアリールスルファターゼ遺伝子及びβ−グルクロニダーゼ遺伝子の野生型配列を提供すれば、当業者であれば、これらの遺伝子において容易に種々の所望の突然変異を起こすことができる。
【0021】
アリールスルファターゼ及びβ−グルクロニダーゼを符号化する野生型遺伝子及び変異体遺伝子の配列を決定した。SEQ ID NO:1(野生型アリールスルファターゼ)、SEQ ID NO:3(アリールスルファターゼ欠失)、SEQ ID NO:4(野生型β−グルクロニダーゼ)及びSEQ ID NO:6(β−グルクロニダーゼ欠失)を参照されたい。これらの遺伝子及びそれらのヌクレオチド配列を同定することにより、当業者はこれらの酵素を発現させるための所望の無表現型を有する他の突然変異体を容易につくることが可能となる。挿入、欠失、ナンセンス、及びフレームシフトなどの突然変異は、酵素についてゼロ表現型においてもっともよく起こる可能性がある。また、ミスセンス変異を起こして、酵素活性に関するそれらの効果について通常通り試験することもできる。微生物遺伝学の標準的な手法を使用して適する突然変異を起こすことができる。例えば、Principles of Gene Manipulation: An Introduction to Genetic Engineering, R. W. Old, S. B. Primrose, Blackwell Publishing, 1994を参照されたい。標準的な酵素アッセイを使用して、突然変異させた酵素の活性の損失について試験することができる。例えば、Kim et al., Appl Microbiol Biotechnol. 2004 Feb; 63(5): 553-9を参照されたい。
【0022】
実質的にアリールスルファターゼ又はβ−グルクロニダーゼを含まない本発明の組成物は、野生型種において含有される特定のタンパク質の量の95%、96%、97%、98%、又は99%未満を含有する。そのような酵素の量の低減により、野生型種からのゲランを含有する組成物において生成されるパラ−クレゾールの量の90%、93%、95%、97%、又は99%未満を有する滅菌された乳組成物をもたらす。生成されるアリールスルファターゼ又はβ−グルクロニダーゼタンパク質の量は、Sigmaからの5−ブロモ−4−クロロ−3−インドリルサルフェート(X−SO4);CAS No.6578−07−0などの容易にアッセイ可能な基質を用いて、又は、Sigma若しくはRPI社からの5−ブロモ−4−クロロ−3−インドリル−β−D−グルクロニド(X−GlcA);CAS No.114162−64−0を用いて、又は、p−ニトロカテコールを用いて、酵素アッセイにより測定することができる。別の方法では、ウェスタンブロッドなどの免疫学的な手法を用いてタンパク質を測定することができる。
【0023】
ゲランガムは、典型的には、滅菌されたか若しくは超高温(UHT)処理された酪農製品、又は冷凍された酪農製品において使用される。かかる製品としては、これらに限定されないが、アイスクリーム、フローズンヨーグルト、プディング、ホイップされた酪農製品、コーヒークリーマー、クレームブリュレ及び酪農飲料が挙げられる。本発明のゲランガムは、野生型種からの典型的なゲランを使用することができるように、これら又は他の任意の食品において使用することができる。
【0024】
ゲランガムは、本発明の突然変異種を用いて、野生型種について当技術分野において既知の方法のいずれかにしたがって生成することができる。細菌は、典型的には、液体培養培地中において生育する。かかる培地は、典型的には、炭素源、リン酸塩、有機又は無機の窒素源、及び適切な微量元素を含有する。発酵は、典型的には、滅菌された条件下で通気及び撹拌しながら行う。発酵時期の最後には、典型的には、細胞を除去することなく、培養培地を収集する。発酵培地は、ゲランガムを回収する前に、低温殺菌して生育可能な細胞を殺すことができる。ゲランガムは、当技術分野において既知であるように、沈殿させることができる。典型的には、これはイソプロパノールなどのアルコールを用いて行われる。沈殿させたゲランは、再度水和させる前に乾燥させることができる。
【0025】
パラ−クレゾールは、当技術分野において既知の任意の手段により測定することができる。使用することができる一つの方法は、ジクロロメタン抽出、濃縮、及びガスクロマトグラフィーによる質量分析を使用する。
【0026】
本発明を実施する現時点で好ましい態様を含む具体的な実施例に関して本発明を説明してきたが、当業者であれば、特許請求の範囲に示したような本発明の精神及び範囲に含まれるこれまでに説明した系及び手法について数多くの変体及び順序の変更が存在することを理解するであろう。
【0027】
実施例
実施例1
突然変異種をスクリーニングし特徴付ける際に以下の試薬を使用した:
5−ブロモ−4−クロロ−3−インドリルサルフェート(X−SO4);CAS No.6578−07−0
供給源:Sigma
5−ブロモ−4−クロロ−3−インドリル−β−D−グルクロニド(X−GlcA);CAS No.114162−64−0
供給源:Sigma又はRPI社
実施例2
Epicentre(ウィスコンシン州、マディソン)からの商業的に入手可能なEZ::TN(登録商標)<R6Kガンマ−ori/KAN−2>挿入キットを用いて、非ムコイド種であるSphingomonas elodea, Gps2においてランダムトランスポゾン変異体のライブラリをつくることにより、アリールスルファターゼ及びβ−グルクロニダーゼを符号化する遺伝子を同定した。カナマイシン耐性突然変異種を、特異的な色素産生基質を伴う選択培地上での青色の欠如(又は有意な低減)に関してスクリーニングした。実施例1を参照されたい。アリールスルファターゼ生成又は活性を妨げる突然変異体を、塩化物塩を伴う規定寒天培地上で色原体である5−ブロモ−4−クロロ−3−インドリルサルフェート(X−SO4)を用いて同定した。β−グルクロニダーゼの突然変異体は、色原体である5−ブロモ−4−クロロ−3−インドリル−β−D−グルクロニド(X−GlcA)を伴う規定培地上で選択した。
【0028】
トランスポゾン及び近傍のゲノムDNAは、続いて制限酵素を用いてクロモソームから摘出した。制限酵素フラグメントは、リガーゼにより円形にし、Escherichia coliに形質転換し、そこでトランスポゾン含有DNAをトランスポゾン中のレプリコンの存在により複製することができる。アリールスルファターゼについての遺伝子を伴うプラスミドはR6K−AS#14Eと命名した。このプラスミドの一部を配列にした(SEQ ID NO:1)。β−グルクロニダーゼについての遺伝子を伴うプラスミドはR6K−BG#6Sと命名した。このプラスミドの一部を配列にした(SEQ ID No:4)。Escherichia coli種EC100D pir+におけるこれらのプラスミドは、2004年6月21日にバージニア州マナッサスのアメリカンタイプカルチャーコレクションに寄託される予定である。
【0029】
不活性化されたアリールスルファターゼを有する細菌において、アリールスルファターゼについての遺伝子の近傍にある仮想タンパク質について、トランスポゾンを実際に遺伝子中に挿入した。不活性化されたβ−グルクロニダーゼを有する細菌において、トランスポゾン挿入は、β−グルクロニダーゼについての遺伝子のアミノ部分に配置した。これら遺伝子のDNA配列は、National Center for Biotechnology Information(NCBI)のデータベースにおける他の種からの既知の遺伝子と相同性を有することを示した。
【0030】
アリールスルファターゼ及びβ−グルクロニダーゼについての遺伝子、並びに近傍のゲノムDNAを配列した。これら遺伝子の欠失をプラスミド上で構築し、次いで、S. elodea種S−60wtc及びPDG−1に形質転換した。WO01/64897を参照されたい。これら欠失を相同性組換によりゲノムにおいて挿入した。標的遺伝子をフランキングするDNA配列をPCRにより増幅し、プラスミドベクターpLO2にクローンした。(Lenz, O., E. Schwarts, J. Dernedde, M. Eitinger and B. Friedrich. 1994, “The Alcaligenes eutrophus H16 hoxX gene participates in hydrogenase regulation,” J. Bacteriol. 176:4385-4394.)この構成を接合によりS. elodea種に形質転換した。得られるカナマイシン耐性種を、次いで、抗生物質無しで30−40世代生育させた。プラスミドを失った分離株は、pLO2上のsacB遺伝子の欠失によるスクロース耐性について選択することにより検出し、カナマイシン感受性により確認した。非選択的生育後にプラスミドを失った分離株は、二つのタイプ、すなわち、欠失及び野生型であった。所望の欠失種は、適する指示寒天(実施例1を参照されたい)上での青色の欠如と、診断PCRにより同定した。構成に関するスキームを以下の図1に示す。
【0031】
アリールスルファターゼについての遺伝子(atsA)の正確な欠失を構成するためには、atsA遺伝子についての最も可能性のある開始コドンと近傍の遺伝子の開始コドン及び終止コドンを決定することが必要であった。遺伝子の末端の配置は、DNA配列、GenBankにおける他の遺伝子との相同性、及びフレームプロット3プログラムを用いた第三塩基GC選択に基づいて決定した。Ishikawa and Hotta. “FlamePlot: a new implementation of the Frame analysis for predicting protein-coding regions in bacterial DNA with a high G+C content.”FEMS Microbiology Letters 174:251-253(1999).この分析は、アリールスルファターゼ遺伝子が、保存された仮想タンパク質についての遺伝子に翻訳されて結合していること、すなわちアリールスルファターゼ遺伝子の開始コドンが保存された仮想タンパク質についての遺伝子の開始コドンと重複していることを示した。これらのタンパク質が最適な細胞生育とゲラン生成のために要求されるものかどうかは知られていないことから、アリールスルファターゼ欠失を構築して、アリールスルファターゼ遺伝子と未知の無傷のタンパク質についての遺伝子を残した。
【0032】
PCRプライマーであるAS5
【0033】
【化1】
【0034】
及びAS6
【0035】
【化2】
【0036】
を使用して、SacI−XbaIフラグメント(合計528bp)として、atsAの開始コドンのちょうど上流に、512bpフラグメントをSacI−XbaIフラグメントを増幅させた。AS3プライマー
【0037】
【化3】
【0038】
及びAS4プライマー
【0039】
【化4】
【0040】
を使用して、atsA遺伝子の端部とXbaI−SphIフラグメント(合計497bp)として保存された仮想タンパク質の開始とを含む、479bpのフラグメントを増幅させた。このように、atsAの終止コドンを保持させたが、開始コドンは欠失させて、アリールスルファターゼタンパク質の部分は合成しないものとする。クローニングのための制限部位は、プライマーの端部に追加した。PCRフラグメントは、プラスミドベクターpLO2のポリリンカー中に連続してくくりつけて、atsA遺伝子の欠失を形成させた。上流のSacI−XbaIフラグメントは、SacI−XbaI切断pLO2中にクローンした。続いて、下流のXbaI−SphIフラグメントをクローンした(図2)。このatsA遺伝子を欠失させたプラスミドを、S. elodeaのS60wtc種及びPDG-1種中への接合により形質転換して、プラスミドをクロモソーム中に組み替えた。カナマイシン耐性のある分離株を精製し、次いで抗生物質無しで生育し、スクロース及びX−SO4を伴う培地上にプレーティングして、プラスミドを符号化するsacB遺伝子の欠失によりスクロース耐性をもつ分離株を選択した。スクロース耐性、カナマイシン感受性のある、黄色コロニーを選択した。S60wtc及びPDG-1のatsA誘導体はそれぞれGAS−1及びPAS−1と命名した。
【0041】
β−グルクロニダーゼについての遺伝子(gusA)の欠失は、プラスミドpLO2上に構築し、S60wtc,GAS−1中に形質転換した。gusA遺伝子についての最も可能性のある開始コドンを、他のタンパク質とリボソーム結合部位の存在とに対する相同性により決定した。二次構造の領域は、開始コドンの上流である。gusA遺伝子の欠失を構築して、gusAの上流及び下流に二次構造を維持した。Bgluc3プライマー
【0042】
【化5】
【0043】
及びBgluc4プライマー
【0044】
【化6】
【0045】
を使用して、gusA遺伝子の上流に、PstI−XbaIフラグメント(合計577bp)として、560bpのフラグメントを増幅させた。Bgluc1プライマー
【0046】
【化7】
【0047】
及びBgluc2プライマー
【0048】
【化8】
【0049】
を使用して、gusA遺伝子の下流に、XbaI−SacIフラグメント(合計505bp)として、489bpのフラグメントを増幅させた。PstI−XbaIフラグメントは、PstI−XbaI切断pLO2中にクローンした。続いて、下流のXbaI−SacIフラグメントをクローンした。このgusA遺伝子が欠失したプラスミドを、S. elodeaのS60wtc種、GAS−1種及びPAS−1種中に接合により形質転換して、組替えを行った。カナマイシン耐性分離株を精製し、抗生物質無しで生育させ、次いで、スクロース及びX−GlcAを伴なう培地上にプレーティングして、プラスミド符号化sacB遺伝子の欠失によりスクロース耐性をもつ分離株を選択した。青緑色(野生型)及び明緑色(変異体)のコロニーの混合物を得た。スクロース耐性のある明緑色の分離株は、カナマイシン感受性により、プラスミド欠失について確認された。S60wtc、GAS−1及びPAS−1のgusA欠如誘導体は、それぞれGBG、GBAD及びPBADと命名した。
【0050】
S60wtc種及びPDG−1種におけるアリールスルファターゼについての遺伝子(atsA)の欠失を完了した。β−グルクロニダーゼについての遺伝子を同定し、配列した。近傍のDNAを配列した。また、β−グルクロニダーゼについての遺伝子の欠失は、S60、PDG−1及びGAS−1のそれぞれにおいて構築した。
【0051】
GAS−1種(アリールスルファターゼについての遺伝子を欠失させた)及びGBAD−1種(アリールスルファターゼ及びβ−グルクロニダーゼの両方についての遺伝子を欠失させた)から生成したゲランのサンプルを、超高温酪農用途試験において月ごとの間隔で、p−クレゾール生産について評価した。野生型種からのゲランサンプルは、1ヵ月後に3〜152(平均65)ppb、2ヵ月後に4〜212(平均96)ppbのp−クレゾールを生成した。GAS−1からのゲランのサンプルは、1〜5ヵ月後に約1〜3ppbのp−クレゾールを生成した。GBAD−1からのゲランのサンプルは、3ヵ月後まで試験したときに、生成量は1ppb(検出限界)未満であった。
【図面の簡単な説明】
【0052】
【図1】図1は、アリールスルファターゼ遺伝子(atsA)の近傍のゲノム領域のゲノムマップと、PCRにより増幅しプラスミドpLO2中にクローンした領域の位置である。次いで、クローンPCRフラグメントを伴なうプラスミドpLO2を使用して、相同性組換により、ゲノムのこの領域を欠失により置き換えた。
【図2】図2は、Sphingomonas elodeaのβ−グルクロニダーゼ遺伝子(gusA)近傍のゲノム領域の制限酵素切断地図である。BG−6クローン及びBG−7クローンにおけるトランスポゾン挿入の位置は底部に示す。
【特許請求の範囲】
【請求項1】
アリールスルファターゼタンパク質を実質的に含まないゲランガムを含む組成物。
【請求項2】
β−グルクロニダーゼタンパク質を実質的に含まないゲランガムを含む組成物。
【請求項3】
アリールスルファターゼタンパク質及びβ−グルクロニダーゼタンパク質を実質的に含まないゲランガムを含む組成物。
【請求項4】
Sphingomonas elodeaの培養培地である、請求項1記載の組成物。
【請求項5】
Sphingomonas elodeaの沈殿させた培養培地である、請求項1記載の組成物。
【請求項6】
滅菌された乳製品である、請求項1記載の組成物。
【請求項7】
ゲランガムを生成する方法であって、
Sphingomonas elodeaを培養培地において生育させ、その際、Sphingomonas elodeaは、触媒活性のあるアリールスルファターゼ、又は触媒活性のあるβ−グルクロニダーゼ、又は触媒活性のあるアリールスルファターゼ及び触媒活性のあるβ−グルクロニダーゼを生成せず、
該培養培地を集め;
該培養培地からゲランガムを沈殿させる
ことを含む、前記方法。
【請求項8】
ゲランガムを酪農製品に添加することを更に含む、請求項7記載の方法。
【請求項9】
アリールスルファターゼが欠乏している、Sphingomonas elodeaの微生物学的に純粋な培養液。
【請求項10】
請求項9記載の微生物学的に純粋な培養液により生成されたゲランガムを含む組成物。
【請求項11】
SEQ ID NO:2にしたがったアミノ酸配列を有する、Sphingomonas elodeaアリールスルファターゼを符号化している、単離、精製されたポリヌクレオチド。
【請求項12】
SEQ ID NO:1のヌクレオチド521〜2176にしたがったヌクレオチド配列を含む、請求項22記載の単離、精製されたポリヌクレオチド。
【請求項13】
そのアリールスルファターゼコード配列のすべて又は一部が欠失しているSphingomonas elodeaゲノムDNAを含む、単離、精製されたポリヌクレオチド。
【請求項14】
ゲノムDNAが、アルキルスルファターゼを符号化しているSphingomonas elodeaゲノムDNAのすべて又は一部を含む、請求項15記載の単離、精製されたポリヌクレオチド。
【請求項15】
そのβ−グルクロニダーゼコード配列のすべて又は一部が欠失しているSphingomonas elodeaゲノムDNAを含む、単離、精製されたポリヌクレオチド。
【請求項16】
欠失部分がSEQ ID NO:4のヌクレオチド1588〜3447である、請求項15記載の単離、精製されたポリヌクレオチド。
【請求項17】
ゲノムDNAが、保存された仮想タンパク質をβ−グルクロニダーゼの上流に符号化しているSphingomonas elodeaゲノムDNAのすべて又は一部を含む、請求項15記載の単離、精製されたポリヌクレオチド。
【請求項18】
ゲノムDNAが、マレートデヒドロゲナーゼを符号化しているSphingomonas elodeaゲノムDNAのすべて又は一部を含む、請求項15記載の単離、精製されたポリヌクレオチド。
【請求項19】
そのアリールスルファターゼ遺伝子、β−グルクロニダーゼ遺伝子、若しくは、アリールスルファターゼ遺伝子及びβ−グルクロニダーゼ遺伝子の両方のすべて又は一部が欠失している、請求項9記載の微生物学的に純粋な培養液。
【請求項20】
そのアリールスルファターゼ遺伝子、β−グルクロニダーゼ遺伝子、又は、アリールスルファターゼ遺伝子及びβ−グルクロニダーゼ遺伝子の両方において、挿入を含む、請求項9記載の微生物学的に純粋な培養液。
【請求項21】
そのアリールスルファターゼ遺伝子、β−グルクロニダーゼ遺伝子、又は、アリールスルファターゼ遺伝子及びβ−グルクロニダーゼ遺伝子の両方において、不活性化突然変異を含む、請求項9記載の微生物学的に純粋な培養液。
【請求項1】
アリールスルファターゼタンパク質を実質的に含まないゲランガムを含む組成物。
【請求項2】
β−グルクロニダーゼタンパク質を実質的に含まないゲランガムを含む組成物。
【請求項3】
アリールスルファターゼタンパク質及びβ−グルクロニダーゼタンパク質を実質的に含まないゲランガムを含む組成物。
【請求項4】
Sphingomonas elodeaの培養培地である、請求項1記載の組成物。
【請求項5】
Sphingomonas elodeaの沈殿させた培養培地である、請求項1記載の組成物。
【請求項6】
滅菌された乳製品である、請求項1記載の組成物。
【請求項7】
ゲランガムを生成する方法であって、
Sphingomonas elodeaを培養培地において生育させ、その際、Sphingomonas elodeaは、触媒活性のあるアリールスルファターゼ、又は触媒活性のあるβ−グルクロニダーゼ、又は触媒活性のあるアリールスルファターゼ及び触媒活性のあるβ−グルクロニダーゼを生成せず、
該培養培地を集め;
該培養培地からゲランガムを沈殿させる
ことを含む、前記方法。
【請求項8】
ゲランガムを酪農製品に添加することを更に含む、請求項7記載の方法。
【請求項9】
アリールスルファターゼが欠乏している、Sphingomonas elodeaの微生物学的に純粋な培養液。
【請求項10】
請求項9記載の微生物学的に純粋な培養液により生成されたゲランガムを含む組成物。
【請求項11】
SEQ ID NO:2にしたがったアミノ酸配列を有する、Sphingomonas elodeaアリールスルファターゼを符号化している、単離、精製されたポリヌクレオチド。
【請求項12】
SEQ ID NO:1のヌクレオチド521〜2176にしたがったヌクレオチド配列を含む、請求項22記載の単離、精製されたポリヌクレオチド。
【請求項13】
そのアリールスルファターゼコード配列のすべて又は一部が欠失しているSphingomonas elodeaゲノムDNAを含む、単離、精製されたポリヌクレオチド。
【請求項14】
ゲノムDNAが、アルキルスルファターゼを符号化しているSphingomonas elodeaゲノムDNAのすべて又は一部を含む、請求項15記載の単離、精製されたポリヌクレオチド。
【請求項15】
そのβ−グルクロニダーゼコード配列のすべて又は一部が欠失しているSphingomonas elodeaゲノムDNAを含む、単離、精製されたポリヌクレオチド。
【請求項16】
欠失部分がSEQ ID NO:4のヌクレオチド1588〜3447である、請求項15記載の単離、精製されたポリヌクレオチド。
【請求項17】
ゲノムDNAが、保存された仮想タンパク質をβ−グルクロニダーゼの上流に符号化しているSphingomonas elodeaゲノムDNAのすべて又は一部を含む、請求項15記載の単離、精製されたポリヌクレオチド。
【請求項18】
ゲノムDNAが、マレートデヒドロゲナーゼを符号化しているSphingomonas elodeaゲノムDNAのすべて又は一部を含む、請求項15記載の単離、精製されたポリヌクレオチド。
【請求項19】
そのアリールスルファターゼ遺伝子、β−グルクロニダーゼ遺伝子、若しくは、アリールスルファターゼ遺伝子及びβ−グルクロニダーゼ遺伝子の両方のすべて又は一部が欠失している、請求項9記載の微生物学的に純粋な培養液。
【請求項20】
そのアリールスルファターゼ遺伝子、β−グルクロニダーゼ遺伝子、又は、アリールスルファターゼ遺伝子及びβ−グルクロニダーゼ遺伝子の両方において、挿入を含む、請求項9記載の微生物学的に純粋な培養液。
【請求項21】
そのアリールスルファターゼ遺伝子、β−グルクロニダーゼ遺伝子、又は、アリールスルファターゼ遺伝子及びβ−グルクロニダーゼ遺伝子の両方において、不活性化突然変異を含む、請求項9記載の微生物学的に純粋な培養液。
【図1】

【図2】
【図2】
【公表番号】特表2008−503237(P2008−503237A)
【公表日】平成20年2月7日(2008.2.7)
【国際特許分類】
【出願番号】特願2007−518151(P2007−518151)
【出願日】平成17年6月20日(2005.6.20)
【国際出願番号】PCT/US2005/021637
【国際公開番号】WO2006/009938
【国際公開日】平成18年1月26日(2006.1.26)
【出願人】(506128352)シーピー・ケルコ・ユーエス・インコーポレーテッド (18)
【Fターム(参考)】
【公表日】平成20年2月7日(2008.2.7)
【国際特許分類】
【出願日】平成17年6月20日(2005.6.20)
【国際出願番号】PCT/US2005/021637
【国際公開番号】WO2006/009938
【国際公開日】平成18年1月26日(2006.1.26)
【出願人】(506128352)シーピー・ケルコ・ユーエス・インコーポレーテッド (18)
【Fターム(参考)】
[ Back to top ]