
遺伝子組み換え単子葉植物における組み換えインスリン様成長因子I(IGF−I)およびインスリン様成長因子結合タンパク質3(IGFBP−3)の産生
【課題】本明細書に記載の単子葉植物細胞を提供すること。
【解決手段】上記単子葉植物細胞は、インスリン様成長因子I(IGF−I)をコードするヌクレオチド配列またはインスリン様成長因子結合タンパク質3(IGFBP−3)をコードするヌクレオチド配列の発現のためのDNA構築物を含有するように改変されており、該DNA構築物は、単子葉植物細胞中での発現のために制御配列と作動可能に連結した該コードするヌクレオチド配列を含む。一局面において、上記IGF−IおよびIGFBP−3はヒトタンパク質である。別の局面において、上記制御配列は、単子葉植物の種子中で作動可能なプロモーターおよび単子葉植物の種子中で作動可能な終止配列を含む。
【解決手段】上記単子葉植物細胞は、インスリン様成長因子I(IGF−I)をコードするヌクレオチド配列またはインスリン様成長因子結合タンパク質3(IGFBP−3)をコードするヌクレオチド配列の発現のためのDNA構築物を含有するように改変されており、該DNA構築物は、単子葉植物細胞中での発現のために制御配列と作動可能に連結した該コードするヌクレオチド配列を含む。一局面において、上記IGF−IおよびIGFBP−3はヒトタンパク質である。別の局面において、上記制御配列は、単子葉植物の種子中で作動可能なプロモーターおよび単子葉植物の種子中で作動可能な終止配列を含む。
【発明の詳細な説明】
【技術分野】
【0001】
関連する出願への相互参照
本願は、2007年2月20日に出願された米国仮特許出願第(“USSN”)60/890,823号に対する優先権の利益を主張する。前記出願は、その全体が本明細書中に参考として、そして全ての目的のために明白に援用される。
【0002】
EFS−WEBを介して提出された配列表への参照
本願は、MPEP§1730II.B.2(a)(A)において公認され定められたように、USPTO EFS−WEBサーバーを介して電子的に出願され、そしてこの電子出願は、電子的に提出された配列(配列番号)表を含む。この配列表の全ての内容は、全ての目的のために参考として本明細書中に援用される。配列表は、以下に示す電子的に出願されたテキストファイル上で確認される:
【0003】
【化1】

本発明は、単子葉植物における哺乳類、特に、ヒトのタンパク質の産生に関する。これについて、遺伝子組み換えイネにおけるIGF−IおよびIGFBP−3の良好な産生により説明する。
【背景技術】
【0004】
インスリン様成長因子I(IGF−I)およびインスリン様成長因子結合タンパク質3(IGFBP−3)は、細胞および組織の生存、成長、ならびに分化の調節において重要なタンパク質である。
【0005】
ヒトIGF−Iは、第12染色体上の単一遺伝子によりコードされた70個のアミノ酸残基からなる一本鎖ポリペプチドである。これは、プロインスリンおよびインスリンと48%のアミノ酸配列同一性を有する。IGF−Iは、A20−B18、A6−A11、およびA7−B6に3つの鎖内ジスルフィド架橋を有するが、糖鎖付加部位は有さない。血中IGF−Iの大半は肝臓で合成され、成長ホルモン(GH)、インスリン、および栄養摂取により調節される。血中IGF−Iレベルは、主に、その分泌成分パターンおよびIGF−Iと高親和性結合タンパク質の結合により相対的に安定している。しかしながら、IGF−Iの調節異常がインスリン耐性の発現および他の代謝異常に関与することが示唆されている。
【0006】
糖質代謝の調節におけるIGF−Iの正確な役割はまだ明確にされていないが、代謝の調節において明らかに重要なものである。健康な被験者では、組み換えヒトIGF−I(rhIGF−I)は急性的な血糖低下作用を生じ得るが、rhIGF−Iは同等の投与量のインスリンよりも10倍効力が低い。対照的に、IGF−Iはインスリンよりもタンパク質代謝に対する影響が顕著であり、グルコースを低下させる同等の投与量のインスリンと比較して、全体的な正味アミノ酸フラックスを減少させる。さらに、IGF−Iは、脾臓インスリン放出の強力な阻害剤である。1型糖尿病患者では、rhIGF−Iを使用することにより血中IGF−Iレベルが改善し、GH過分泌が好転し、インスリン必要量が低減し、血糖管理が改善する。2型糖尿病患者では、空腹時血漿グルコース、インスリン、およびCペプチドレベルを低減するためにより多くの投与量のrhIGF−Iが必要であった。また、rhIGF−I治療は脂肪量の低減と関連づけられていたが、これは、インスリン感受性に対する上記の有益な影響を部分的に説明するものである。
【0007】
6個のヒトIGF結合タンパク質が同定されている。これら6個の結合タンパク質のうち、IGFBP−3は、血清中の95パーセントを超えるIGF−Iと結合する。これは、分子量が29kD程度と計算される264個のアミノ酸残基を含有する。潜在的に3つのN糖鎖付加部位(Asn−X−Ser/Thr)がIGFBP−3中央部のAsn89、Asn109、およびAsn172の位置に存在するが、炭水化物単位はIGF結合には必須ではないと思われる。IGF−I/IGFBP−3二量体が「酸に不安定なサブユニット」と三重複合体を形成し、これにより、IGF−Iの半減期が延長され、その受容体に対するIGF−Iの供給が漸増する。
【0008】
IGFBP−3は多くの組織において局所的に産生される40〜45kDaの糖タンパク質であり、それらの組織において、細胞成長およびアポトーシスの調節における自己分泌および傍分泌調節因子として作用する。IGFBP−3は、IGFと結合することにより細胞増殖および生存を阻害し、IGFが標的細胞上のIGF−I受容体を活性化することを妨げる。IGFBP−3はまた、細胞増殖を負に調節し、IGFに依存せずにアポトーシスを誘発することが分かっている。IGF−Iが存在しない場合、IGFBP−3は、p53、レチノイン酸、腫瘍壊死因子−α、および形質転換成長因子−β等の多数の成長抑制タンパク質および薬剤と相互作用することができる。IGFBP−3の過剰発現は細胞増殖の阻害、腫瘍形成の低減を行い、正常臓器の成長をわずかに阻害する。最近の研究では、IGFBP−3が乳癌、前立腺癌、肺癌、卵巣癌、および結腸直腸癌を阻害するため、IGFBP−3が抗癌剤として有効であることが実証されている。
【0009】
それゆえ、ヒトIGF−IおよびヒトIGFBP−3はともに、薬学的価値を確立している。IGF−IおよびIGFBP−3の両方の使用に関する重大な障害は、それらの製造コストである。これらのタンパク質は、主に、Escherichia coli等の細菌から組み換えによって製造されるか、または肉腫細胞株もしくは赤血球系細胞から抽出され、かつ/または遺伝子導入マウス中で産生される。これらのシステムは、設備および製造コストが高く、病原菌で汚染される可能性があるため大量生産には適さない。IGF−IおよびIGFBP−3は他の哺乳類でも同様に発生し、同様の機能を果たす。
【0010】
植物細胞は、原核生物および真核生物源からの遺伝子を含む幅広い生物から遺伝子情報を受け取り、発現するように改変することができる。植物細胞は真核性であるため、適切なタンパク質または酵素機能に必要とされることが多い適切な翻訳後修飾(例えば、グリコシル化、プレニル化、およびジスルフィド架橋の形成)により哺乳類タンパク質を産生することができる。また、多くの植物種の種子が食用に適しており、種子中に組み換えタンパク質を蓄積することが可能である。投与量レベルおよび頻度が制御される場合には、組み換えタンパク質は、経口送達の前にさらなる処理および精製を必ずしも必要としないこともあるが、これは、各タンパク質が特有の酸およびプロテアーゼ耐性を有するためである。バイオカプセルおよび植物組織等の送達ビヒクルを用いて胃および内臓におけるタンパク質の分解を防ぐことも可能である(非特許文献1)。種子に基づくプラットホームはまた、医薬用タンパク質の生物学的集合および産生のために開発されている(非特許文献2)。それらの結果は、生物医薬品の産生および「錠剤種子」経路(’seed as pill’ route)を介した送達のためのビヒクルとしての植物種子の使用は実行可能な選択肢であることを示唆している。
【0011】
形質転換タバコ中において組み換えヒトIGF−Iおよび組み換えヒトIGFBP−3が産生されており、これらの結果は3件のポスター発表で報告されている(5th Hong Kong Diabetes and Cardiovascular Risk Factors,East Meets West Symposium,October 2003(”Expression of Human Insulin−like Growth Factor I(IGF−I) and Insulin−like Growth Factor Binding Protein 3(IGFBP−3) in Transgenic Tobacco”);64th Scientific Sessions of American Diabetes Association,June 2004(”Plants as Bioreactors for Expressing Human Insulin−like Growth Factor I(IGF−I) and Insulin−like Growth Factor Binding Protein−3(IGFBP−3)”);および2004 annual meeting of the American Society of Plant Biologists,July 2004(”Transgenic Expression of Human Insulin−like Growth Factor Binding Protein−3(IGFBP−3) in Tobacco Seeds”)。これらのタンパク質のタバコ苗における産生には、トウモロコシ、イネ、小麦、および大麦等の食用に適した単子葉植物における産生のような利点がない。単子葉植物には、サトウキビ、パイナップル、ナツメヤシ、およびバナナ等の主要食用作物も含まれる。これらの単子葉植物は、食用に適した原料の代表的な増殖源である。
【0012】
世界の人口の40%を超える人々によって日々消費されるイネは、世界中の農業に定着しており、製薬上および商業上重要なタンパク質およびワクチンを製造するためのモデルバイオリアクタ(model bioreactor)として認識されている(非特許文献3)。これは、タバコが含有するようなニコチンおよび毒性アルカロイド等の有害化学物質を含有しておらず、アレルギー誘発性が低い。イネ中の組み換えタンパク質は1%粒重まで蓄積可能である。(最大で総種子粒数(total seed grain)の91%となる)イネの胚乳領域における組み換えタンパク質の産生および貯蔵を目的とする特定のプロモーターおよびシグナル配列の使用により、タンパク質蓄積量を粒重の2.7%まで増加することができる(Liu,Q.Q.,Thesis Department of Biology(2002)Yangzhou University(中国)およびthe Chinese University of Hong Kong(香港))。単一のイネ苗は、10,000を超える穀粒を生成する分げつを最大で100個有することが有り、これにより、大量の種子および組み換えタンパク質を迅速に産生することができる。また、早生(1年当たり3〜4回)型のイネjaponica亜種は、産生用のバイオリアクター苗(bioreactor plants)として用いることができる。乾燥種子の貯蔵および流通は単純であり、5ヶ月以上室温で貯蔵を行っても、穀粒中の組み換えタンパク質の収量および活性が著しく損なわれることはない(非特許文献4)。低含水率の条件では、穀粒は生存能力を失うことなく3〜5年貯蔵することができる(Huang,N.,BioProcess International(2002)Jan:54−59)。
【0013】
タンパク質が家畜での使用を対象とする場合には、オート麦等の代替的な穀物中での産生がより適切な場合がある。
【0014】
過去の研究では、コドン使用頻度の偏りが遺伝子発現レベルと強い相関関係を有することが示されている。高度に発現した遺伝子は、「最適」コドンと呼ばれるコドンのサブセットを優先的に用いる(非特許文献5)。さらに、これらの最適コドンは最も豊富なtRNAに対応しており、翻訳精度および効率が向上する(非特許文献6)。下記の実施例では、2つの種子貯蔵タンパク質において用いられる植物優先コドンに基づいて、ヒトIGF−IおよびIGFBP−3のDNA配列に改変を加え、これらのタンパク質についてイ
ネにおけるタンパク質産生の向上を試みた。形質転換シロイヌナズナ中での高度な発現および安定した蓄積(LRPおよびPN2Sについて、それぞれ、全抽出種子タンパク質の3〜10%および3〜15%)が過去の研究において観察されたため、シカクマメ由来のリジンに富んだタンパク質(LRP)およびパラダイスナッツ由来のメチオニンに富んだ2Sアルブミン(PN2S)のコドン使用頻度を改変のベースとして選択した。
【0015】
下記実施例で用いた標的タンパク質の収量を増大する他の戦略は、細胞のタンパク質分解系による分解を防ぐために、組み換えタンパク質を特定部分に向けることである。小胞体(ER)分秘経路(secretary pathway)に導くシグナルペプチド配列、および外来遺伝子のNおよびC末端のER保持シグナルであるテトラペプチドKDELの付着は、一般に、その産物の高度な蓄積に必要とされることが報告されている(非特許文献7;および非特許文献8)。KDELテトラペプチドは、ゴルジ複合体とER間でリサイクルされる受容体との相互作用によるタンパク質局在化に寄与する。KDELシグナルは、植物ERにおける可溶性タンパク質蓄積に十分である(非特許文献7;非特許文献9;および非特許文献10)。KDELを含有する構築物で形質転換した細胞中では、KDELを含有しないものよりもscFvレベルが6〜14倍高いことが分かっている(非特許文献11)。
【先行技術文献】
【非特許文献】
【0016】
【非特許文献1】Daniell,H.ら、Trends Plant Sci.(2001)6:219−226
【非特許文献2】Sardana,R.K.ら、Transgenic Res.(2002)11:521−531
【非特許文献3】Fischer,R.ら、Transgenic Res.(2000)9:279−299
【非特許文献4】Stoger,E.ら、Plant Mol.Biol.(2000)42:583−590
【非特許文献5】Moriyama,E.N.ら、J.Mol Evol(1997)45:514−523
【非特許文献6】Marais,G.ら、J.Mol Evol(2001)52:275−280
【非特許文献7】Wandelt,C.I.ら、Plant J.(1992)2:181−192
【非特許文献8】Herman,E.M.ら、Planta(1990)182:305−312
【非特許文献9】Frigerio,L.ら、Plant Cell(2001)13:1109−1126
【非特許文献10】Napier,J.ら、Planta(1997)203:488−494
【非特許文献11】Conrad,U.ら、Plant Mol.Biol.(1998)38:101−109
【発明の概要】
【課題を解決するための手段】
【0017】
本発明は、経済的かつ実用的な2つの重要な哺乳類タンパク(IGF−IおよびIGFBP−3)源を提供する。ヒト型での産生が好ましい。これらのタンパク質を単子葉植物中で産生することにより、本発明は、初めて、ヒトの治療のための使用または獣医学的状況での使用に適合可能な形態において十分な量での提供が可能である有用なタンパク源を提供する。
【0018】
よって、1つの局面では、本発明は、ヒトIGF−Iおよび/またはヒトIGFBP−3を産生するように改変された単子葉植物細胞に関する。他の局面では、本発明は、そのような細胞を含む植物または植物部位に関する。
例えば、本発明は、以下の項目を提供する:
(項目1)
インスリン様成長因子I(IGF−I)をコードするヌクレオチド配列またはインスリン様成長因子結合タンパク質3(IGFBP−3)をコードするヌクレオチド配列の発現のためのDNA構築物を含有するように改変した単子葉植物細胞であって、該DNA構築物は、単子葉植物細胞中での発現のために制御配列と作動可能に連結した該コードするヌクレオチド配列を含む、細胞。
(項目2)
上記IGF−IおよびIGFBP−3はヒトタンパク質である、項目1に記載の細胞。
(項目3)
上記制御配列は、単子葉植物の種子中で作動可能なプロモーターおよび単子葉植物の種子中で作動可能な終止配列を含む、項目1または2に記載の細胞。
(項目4)
上記コードするヌクレオチド配列は、小胞体(ER)分泌経路に導くシグナルペプチドをコードするヌクレオチド配列によりN末端で伸長される、項目1または2に記載の細胞。
(項目5)
上記コードするヌクレオチド配列は、ER保持シグナルをコードするヌクレオチド配列によりC末端で伸長される、項目1または2に記載の細胞。
(項目6)
上記細胞はイネ細胞である、項目1または2に記載の細胞。
(項目7)
項目1または2に記載の細胞を含む遺伝子組み換え植物または植物部位。
(項目8)
IGF−IまたはIGFBP−3を含む単子葉植物細胞。
(項目9)
上記IGF−IおよびIGFBP−3はヒト由来のものである、項目8に記載の細胞。
(項目10)
上記細胞はイネ細胞である、項目8または9に記載の細胞。
(項目11)
項目8または9に記載の細胞を含む植物または植物部位。
(項目12)
上記植物部位は種子組織である、項目11に記載の植物部位。
(項目13)
上記植物または植物部位はイネである、項目11に記載の植物または植物部位。
(項目14)
上記植物部位は種子組織である、項目13に記載の植物部位。
(項目15)
単子葉植物においてIGF−IまたはIGFBP−3を産生する方法であって、項目1または2に記載の植物細胞を培養する工程を包含する、方法。
(項目16)
単子葉植物においてIGF−IまたはIGFBP−3を産生する方法であって、項目11に記載の植物を培養する工程を包含する、方法。
(項目17)
上記植物はイネである、項目16に記載の方法。
【図面の簡単な説明】
【0019】
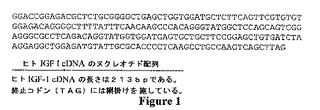
【図1】Jansen,M.ら、Nature(1983)306:609−611で報告されたヒトIGF−I(配列番号1)をコードするヌクレオチド配列を示す。
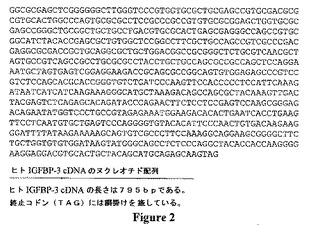
【図2】Wood,W.I.ら、Mol.Endocrinol.(1988)2:1176−1185で報告されたヒトIGFBP−3(配列番号2)をコードするヌクレオチド配列を示す。
【図3】植物におけるコドン選択のために改変されたヒトIGF−I(配列番号3)をコードするヌクレオチド配列を示す。
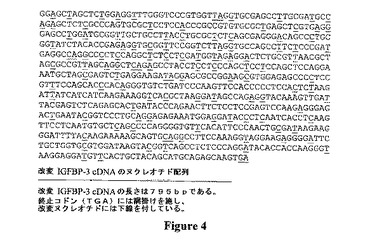
【図4】植物におけるコドン選択に従って改変されたヒトIGFBP−3(配列番号4)のヌクレオチド配列を示す。
【図5】ヒトIGF−I(配列番号5)のアミノ酸配列を示す。
【図6】ヒトIGFBP−3(配列番号6)のアミノ酸配列を示す。
【図7】イネで生成したヒトIGF−IのL6細胞にラフリングを生じる能力、およびこの作用の商用ヒトIGFBP−3に対する反応を示すアッセイの結果を示す。
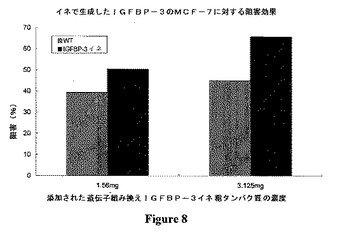
【図8】イネで生成したヒトIGFBP−3のMCF−7細胞の成長を阻害する効果を示す。
【発明を実施するための形態】
【0020】
下記に示すように、2つの重要なヒトタンパク質であるIGF−IおよびIGFBP−3が単子葉植物中において産生されたのは初めてのことである。単子葉植物は主要な栄養源を含み、有毒化合物を含まないため、ヒトの治療を目的としたタンパク質の産生に理想的である。これらはまた、草食性または雑食性哺乳類を治療する場合において、類似するタンパク質の産生にも理想的である。これらのタンパク質の産生は、これらのタンパク質をコードするヌクレオチド配列を発現のために適切な制御配列に作動可能に連結した発現系を含むDNA構築物を適切に設計することにより促進可能である。
【0021】
植物細胞の遺伝子改変を実施し、無傷植物を再構成する技術は、以前より当該分野において周知である。例えば、Gelvinら、Plant Molecular Biology Manual(1990));Dashek,Methods in Plant Biochemistry and Molecular Biology(CRC Press 1997)を参照されたい。これに関する最新技術の有用な要約が2004年1月14日に公開された米国特許公報2004/0009476に見られるが、その植物の遺伝子操作のための適切な技術についての開示内容を本明細書中において参考として援用する。
【0022】
組み換え構築物を含有する形質転換植物細胞が一旦得られると、そこから遺伝子組み換え植物を再生し、所望のタンパク質産生レベルを判断することができる。
【0023】
植物中における変性ヌクレオチド配列の発現を最適化するためにいくつかの技術を用いることができる。下記の実施例においてさらに説明するように、植物細胞中における発現のためのコドン選択に応じて、コードするヌクレオチド配列に改変を加えることができる。そのような改変は、植物中におけるコドン選択を記載する公開データに基づくものである。次に、コードするヌクレオチド配列を伸長してシグナルおよび保持配列を加え、コードされたタンパク質を小胞体に向け、その保持を行ってもよい。これはまた、収量にも好影響を及ぼす。一般に、シグナル伝達ペプチドは所望のタンパク質のN末端に生成され、保持シグナルはそのC末端に生成される。
【0024】
これらの技術を用いることにより、所望のIGF−IおよびIGFBP−3の収量は目に見えて向上する。
【0025】
適切なプロモーターを用いて発現を行うことはさらに有用である。例えば、適切なプロモーターとしては、35S CaMV、イネアクチンプロモーター、ユビキチンプロモーター、またはノパリンシンターゼ(NOS)プロモーターが挙げられる。種子中での産生を促進する組織特異的プロモーターは種子特異的グルテリンプロモーター(Gt−1pro)を含むが、他の種子特異的プロモーターを用いてもよい。ノパリンシンターゼ終結シグナル等の終結シグナルを用いてもよい。
【0026】
下記に示すように、植物最適化コード配列を駆動する2つの構築物群を設計し、Agrobacteriumにより媒介された形質転換によりイネに導入した。一方の構築物群はグルテリンシグナルペプチド(SP)を単独で含有するが、他方はSPとともに標的テトラペプチドKDELシグナルを含有する。これらの構築物を合成し、タンパク質の発現を増加させ、貯蔵のためにタンパク質を区画内に入れるとともに、タンパク質安定性を向上させた。種子特異的グルテリンプロモーター(Gt−1pro)を用いて遺伝子組み換えイネ中におけるIGF−IおよびIGFBP−3の発現を促進し、導入遺伝子の発現を分析した。遺伝子組み換えイネ穀粒から産生したrhIGF−IおよびrhIGFBP−3は生物活性を有していた。
【0027】
上記構築物はまた、後で精製を行えるように、改変を加えてヒスチジン標識またはFLAGシーケンス等の精製補助物を含むようにしてもよく、かつ/または蛍光タンパク質等のマーカーを含むようにしてもよい。また、コードしたタンパク質と精製標識および/またはマーカー間に切断部位を設けてもよい。所望であれば標準的な精製技術を用いてもよく、または場合によっては、植物組織を経口投与し、植物の栄養価およびその低毒性を利用するようにしてもよい。
【0028】
組み換え技術によって生成したIGF−Iは、1型または2型糖尿病患者のインスリン必要量を低減し、血糖管理を改善するために用いられる。IGFBP−3はアポトーシスを引き起こすことが示されており、悪性腫瘍の治療に有用である。よって、所望であれば、組み換え技術によって生成したタンパク質を、これらのタンパク質を用いた治療を必要とする患者に投与するための組成物として製剤化してもよい。タンパク質および他の医薬品を製剤化するための一般的方法は、本明細書において参考として援用するRemington’s Pharmaceutical Sciences,latest edition,Mack Publishing Co.(Easton,PA)において見ることができる。これらのタンパク質は、典型的には、注射または経皮的もしくは経粘膜的送達により非経口的に全身投与されるか、または経口投与される場合がある。リボソーム製剤、および脂質もしくは高分子等に基づくナノ粒子を含有する製剤を含む、特定の投与方式を対象とする各種の製剤を用いることができる。
【0029】
以下の実施例は本発明を例示するためのものであり、限定するものではない。
【実施例】
【0030】
(実施例1)
IGF−IおよびIGFBP−3構築物を用いたAgrobacteriaの形質転換
ヒトIGF−I(図1)およびIGFBP−3(図2)のDNA配列に、2つの種子貯蔵タンパク質に用いられる優先コドンに基づいて改変を加えた。シカクマメ由来のリジンに富んだタンパク質(LRP)およびパラダイスナッツ由来のメチオニンに富んだ2Sアルブミン(PN2S)のコドン使用頻度(表1)を改変のベースとして選択したが、これは、過去の研究において、形質転換シロイヌナズナ中での高度な発現および安定した蓄積(LRPおよびPN2Sについて、それぞれ全抽出種子タンパク質の3〜10%および3〜15%)が観察されたためである。
【0031】
元のアミノ酸配列と同じアミノ酸配列(図5および図6)をコードする改変IGF−I(図3)およびIGFBP−3(図4)遺伝子をMWG Biotechnology Companyから入手した。IGF−IおよびIGFBP−3のコドン変化量は、それぞれ、28.6%および14.8%であった。
【0032】
【表1−1】
【0033】
【表1−2】
rhIGF−IおよびrhIGFBP−3の安定性および収量を向上させるとともに、それらのグリコシル化を制御する構築物を設計した。2つのタンパク質標的シグナルを加え、標的タンパク質をイネ穀粒中の特定の区画に向けた(ゴルジ装置におけるグリコシル化のためのグルテリンシグナルペプチド(SP)または小胞体内におけるグリコシル化がない安定した蓄積のためのテトラペプチドKDELのいずれか)。発現構築物を種子特異的グルテリンプロモーター(Gt−1pro)により駆動した。キメラ遺伝子構築物の詳細は以下のとおりである。
【0034】
IGF−I:
1) (Gt−1pro)+SP+IGF−I*+NOSter
2) (Gt−1pro)+SP+IGF−I*+KDEL+NOSter
IGFBF−3:
3) (Gt−1pro)+SP+IGFBF−3*+NOSter
4) (Gt−1pro)+SP+IGFBF−3*+KDEL+NOSter
ここで、*はIGF−IまたはIGFBP−3の改変cDNAを示し、NOSterはノパリンシンターゼ停止剤を示す。
【0035】
キメラ遺伝子カセットをスーパーバイナリーベクターpSB130に挿入し、イネカルスの感染に理想的なAgrobacterium株(EHA105)に形質転換した。(Chen,H.ら、BioTechniques(1994)16:664−668、670に記載されるように)単純な凍結融解法を用いてAgrobacteriumを形質転換した。抗生物質ハイグロマイシン(50mg/L)を用いて形質転換バクテリアを選択し、PCRにより標的遺伝子のDNA形質転換をさらに確認した。
【0036】
(実施例2)
イネの形質転換、選択、および培養
カルス誘導のためにjaponica9983の成熟種子の胚盤を切除し、カルス誘導培地(N6基礎培地、2mg/lの2,4−D、0.5g/lのカゼイン加水分解液、30g/lのスクロース、2.5g/lのPhytagel(登録商標)、pH5.8)上で培養した。カルス誘導培地で4〜7日間培養した後すぐに、未熟胚由来のカルスを使用し、標的遺伝子を含有するAgrobacterium EHA105と共培養した。
【0037】
まず、50mg/Lのリファンピシンおよび50mg/Lのカナマイシンを添加した3mlのLBブロスに、Agrobacteriumを28℃で一晩かけて植菌した後、25mlのAB培地(3g/lのK2HPO4、1g/lのNaH2PO4、1g/lのNH4Cl、0.3g/lのMgSO4・7H2O、0.15g/lのKCl、10mg/lのCaCl2・2H2O、2.5mg/lのFeSO4・7H2O、5g/lのグルコース、pH7.2)で5時間かけて継代培養を行い、引き続き、遠心分離および15〜25mlのAAM培地(AA基礎培地、68.5g/lのスクロース、36g/lのグルコース、0.5g/lのカゼイン加水分解液、pH5.2、100μmol/lのアセトシリンゴン)での再懸濁を行った。
【0038】
イネカルスをAgrobacterium培養液に10〜20分間室温で浸し、不連続的に振とうした。次に、暗所において、100μmol/lのアセトシリンゴンを含有するN6D2C培地(N6D2、10g/lのグルコース、pH5.2)に3日間かけて26〜28℃で感染カルスを移した。共培養後、耐性カルスの選択のために、暗所において、このカルスをN6D2S培地(N6D2、50mg/lのハイグロマイシンB、500mg/lのセフォタキシム、pH5.8)上に26℃で2週間静置した後、新たに形成された耐性カルスが現れるまで新たなN6D2S培地で培養を行った。
【0039】
次に、HGPR培地(Higrow(登録商標)イネ培地(GIBCO−BRL)、50mg/lのハイグロマイシンB、200mg/lのセフォタキシム)上に、暗所において7日間と、明所において7日間26℃で耐性カルスを静置した。再生前処理(pre−regeneration)後、再生のために、MSR培地(MS基礎培地、30g/lのスクロース、0.5g/lのカゼイン加水分解液、2mg/lの6−BA、0.5mg/lのNAA、0.5mg/lのKT、pH5.8、50mg/lのハイグロマイシンB、500mg/lのセフォタキシム、2.5g/lのPhytagel(登録商標))に耐性カルスを26℃で明所において16時間/暗所において8時間かけて移した。
【0040】
再生芽が出てくると、発根のために、それらを1/2倍量のMSH培地(1/2倍量のMSマクロ塩、Fe−EDTAおよびミクロ塩、MSビタミン、30g/lのスクロース、0.5mg/lのNAA、pH5.8、50mg/lのハイグロマイシンB、500mg/lのセフォタキシム、2.5g/lのPhytagel(登録商標))上に静置した。最後に、遺伝子組み換えイネ胚を土壌に移し、温室で栽培した。
【0041】
(実施例3)
遺伝子組み換えイネの分析
サザンブロット分析
遺伝子組み換えイネ苗を分析し、イネゲノムへの標的遺伝子の組込みを確認した。臭化セチルトリメチルアンモニウム(CTAB)法により葉ゲノムDNAを抽出した(Doyle,J.D.ら、Focus(1990)12:13−15)。15μgのゲノムDNAをBamHIを用いて一晩かけて消化し、0.8%のアガロースゲル上で分離し、VacuGeneXL真空ブロッティングシステム(Pharmacia Biotech)を用いて正に帯電したナイロン膜(Roche)に移した。DIG核酸検出キット(Roche)に記載の方法に従ってハイブリダイゼーションおよび検出を行った。DIG DNA標識キット(Roche)を用いてPCRにより二本鎖DIG標識DNAプローブ(IGF−IおよびIGFBP−3)を準備した。これらのプローブを使用前に99℃で加熱して変性させた。
【0042】
この結果、イネゲノム中に構築物の存在が確認できた。
【0043】
ウエスタンブロット分析
種子をすりつぶして粉末状にし、タンパク質抽出緩衝液(50mMのトリスHCl、pH6.8、0.1MのNaCl、および10%SDS)と混合することにより全種子タンパク質を成熟イネ種子から抽出した。遠心分離した後、上澄液を新しいエッペンドルフチューブに移し、種子全タンパク質抽出液として保存した。次に、異なる量の全タンパク質を17%トリシンSDS−PAGE中で分解し、PVDF膜上にブロットした。抗ヒトIGF−IまたはIGFBP−3ポリクローナル抗体を用いてウエスタンブロット分析を行った。最後に、Aurora(登録商標)Western Blot Chemiluminescent Detection System(ICN)のマニュアルに記載のとおり、化学発光Starlight(登録商標)基質(ICN)を用いてブロットの非放射性検出を行った。
【0044】
その結果、種子抽出液中にIGF−IおよびIGFBP−3タンパク質の両方の存在が確認できた。
【0045】
(実施例4)
イネで生成したrhIGF−Iの生物活性
インスリンと同様に、IGF−Iは、筋細胞中で膜ラッフルおよびグルコース摂取を生じることが示されている。IGFBP−3は、IGF−Iと結合して、ALS複合体を形成することができ、これにより、IGF−Iにより生じた膜ラフリング作用が阻害される。イネで生成した組み換えhIGF−Iをインスリンと同時にテストし、それらの生物活性を比較した。
【0046】
c−mycエピトープ標識したグルコーストランスポーター4(GLUT4)を発現するラットL6骨格筋細胞(L6myc細胞)を、10%(v/v)ウシ胎児血清(FBS)および1%(v/v)抗生物質−抗真菌薬溶液(100U/mlのペニシリンG、10mg/mlのストレプトマイシン、および25mg/mlのアンフォテリシンB)を含有するα−最小必須培地において37℃の5%CO2雰囲気で筋芽細胞単層培養を維持した。0.25%トリプシンを用いてサブコンフルエント培地のトリプシン処理により細胞を継代培養した。筋管への分化のために、4×104程度の細胞数/mlで2%(v/v)FBSを含有する培地で筋芽細胞を培養し、自然融合させた。48時間ごとに培地を交換し、培養後5〜7日間筋管を用いた。次に、6ウェルプレートに載せた25mm径のガラスカバースリップ上でラットL6筋細胞を筋管の段階まで培養した。
【0047】
3時間かけて筋管から血清を抜き、遺伝子組み換えイネから抽出したrhIGF−IおよびrhIGFBP−3を異なる濃度および組み合わせで10分間かけて37℃で処理した。これらのインキュベーションの後、筋管を3%(v/v)氷冷パラホルムアルデヒドPBS溶液で20分間かけて凝固させた後、0.1MグリシンのPBS溶液で10分間洗浄し、0.1%(v/v)トリトンX−100のPBS溶液で3分間透過処理した後、PBSで洗浄した。筋管を0.1%BSA中で1時間塞いだ後、ファロイジン(0.1%(w/v)BSA中で1:500)中でインキュベーションを行った。このインキュベーションの後、細胞をPBSで洗浄し、ガラススライド上にProLong(登録商標)アンチフェード溶液として載せた。Zeiss Axioplan2イメージング顕微鏡およびZeiss LSM510 METAレーザー走査共焦点顕微鏡(Carl Zeiss,Jena,Germany)を用いてサンプルを検査した。
【0048】
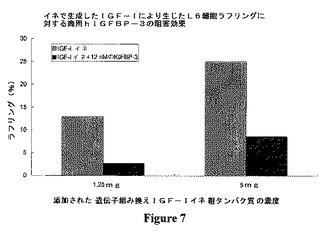
遺伝子組み換えIGF−Iイネの粗タンパク質によりL6細胞の膜ラッフルが生じるが、そのラフリング作用は商用hIGFBP−3により大幅に低減することが可能である。これらの結果を図7に示す。図7に示すように、イネで生成したIGF−1は、1.25mgおよび5mgで用量依存的にラフリングを生じる。この活性は、12nM濃度の市販のIGFBP−3によって阻害された。
【0049】
(実施例5)
イネで生成したrhIGFBP−3タンパク質の抗癌活性
ヒトIGFBP−3は、エストロゲン依存性および非依存性乳癌細胞の増殖を阻害することが示されている。イネで生成したrhIGFBP−3が制癌作用を有するか否かを検査するために、MCF−7ヒト乳癌細胞を用いた。
【0050】
1%ペニシリンおよびストレプトマイシン、0.1%ファンギゾム(fungizome)を5%ウシ胎児血清(FBS)とともに添加したイーグル最小必須培地にMCF−7ヒト乳癌細胞を通法により37℃の5%CO2加湿雰囲気に維持した。rhIGFBP−3の処理のために、MCF−7細胞を血清含有培地で96ウェルプレートに播種した。1日後、それらの細胞に、rhIGFBP−3を含有するイネ形質転換体から抽出した異なる濃度の粗タンパク質を加えた。粗タンパク質抽出液を3日おきに新しいものと2度交換した。
【0051】
7日間処理した後、MTTアッセイのために細胞を採取した。簡単に言えば、20μlのMTT(3−(4,5−ジメチルチアゾール−2−イル)−2,5−臭化ジフェニルテトラゾリウム(diphenyl tetrazolium bromide)溶液(5mg/mlMTTのHPBS溶液、pH7.4)を各ウェルに加え、さらに37℃で2時間かけてインキュベートした。100μlの酸イソプロパノールを各ウェルに加えて細胞を分解し、その紫色の結晶を溶解した。マイクロプレート分光光度計をOD570で用いて、各ウェルにおける紫色の強度を測定した。
【0052】
図8に示すように、イネで生成したIGFBP−3は、1.56mgおよび3.125mgの濃度で用量依存的にMCF−7細胞の成長阻害を向上させることができた。
【0053】
(実施例6)
組み換え技術によって生成したタンパク質の精製
親和性クロマトグラフィーを用いて上記実施例に記載の粗抽出液をさらに精製する。IGF−IおよびIGFBP−3の各々をクロマトグラフィーカラムに塗布し、さらに精製するために、これらのカラムをそれぞれの抗体に結合する。カラムを介して不純物を洗浄し、pHおよび塩濃度を調節してタンパク質を溶出する。
【0054】
マーカータンパク質を含むように実施例1の構築物に改変を加えて精製を簡略化する。
【0055】
(実施例7)
組み換えタンパク質収量の向上
実施例2に記載の苗から種子を得て、第2および第3世代の作物を得るために移植する。収量が向上した組み換えIGF−IおよびIGFBP−3が得られる。
【技術分野】
【0001】
関連する出願への相互参照
本願は、2007年2月20日に出願された米国仮特許出願第(“USSN”)60/890,823号に対する優先権の利益を主張する。前記出願は、その全体が本明細書中に参考として、そして全ての目的のために明白に援用される。
【0002】
EFS−WEBを介して提出された配列表への参照
本願は、MPEP§1730II.B.2(a)(A)において公認され定められたように、USPTO EFS−WEBサーバーを介して電子的に出願され、そしてこの電子出願は、電子的に提出された配列(配列番号)表を含む。この配列表の全ての内容は、全ての目的のために参考として本明細書中に援用される。配列表は、以下に示す電子的に出願されたテキストファイル上で確認される:
【0003】
【化1】
本発明は、単子葉植物における哺乳類、特に、ヒトのタンパク質の産生に関する。これについて、遺伝子組み換えイネにおけるIGF−IおよびIGFBP−3の良好な産生により説明する。
【背景技術】
【0004】
インスリン様成長因子I(IGF−I)およびインスリン様成長因子結合タンパク質3(IGFBP−3)は、細胞および組織の生存、成長、ならびに分化の調節において重要なタンパク質である。
【0005】
ヒトIGF−Iは、第12染色体上の単一遺伝子によりコードされた70個のアミノ酸残基からなる一本鎖ポリペプチドである。これは、プロインスリンおよびインスリンと48%のアミノ酸配列同一性を有する。IGF−Iは、A20−B18、A6−A11、およびA7−B6に3つの鎖内ジスルフィド架橋を有するが、糖鎖付加部位は有さない。血中IGF−Iの大半は肝臓で合成され、成長ホルモン(GH)、インスリン、および栄養摂取により調節される。血中IGF−Iレベルは、主に、その分泌成分パターンおよびIGF−Iと高親和性結合タンパク質の結合により相対的に安定している。しかしながら、IGF−Iの調節異常がインスリン耐性の発現および他の代謝異常に関与することが示唆されている。
【0006】
糖質代謝の調節におけるIGF−Iの正確な役割はまだ明確にされていないが、代謝の調節において明らかに重要なものである。健康な被験者では、組み換えヒトIGF−I(rhIGF−I)は急性的な血糖低下作用を生じ得るが、rhIGF−Iは同等の投与量のインスリンよりも10倍効力が低い。対照的に、IGF−Iはインスリンよりもタンパク質代謝に対する影響が顕著であり、グルコースを低下させる同等の投与量のインスリンと比較して、全体的な正味アミノ酸フラックスを減少させる。さらに、IGF−Iは、脾臓インスリン放出の強力な阻害剤である。1型糖尿病患者では、rhIGF−Iを使用することにより血中IGF−Iレベルが改善し、GH過分泌が好転し、インスリン必要量が低減し、血糖管理が改善する。2型糖尿病患者では、空腹時血漿グルコース、インスリン、およびCペプチドレベルを低減するためにより多くの投与量のrhIGF−Iが必要であった。また、rhIGF−I治療は脂肪量の低減と関連づけられていたが、これは、インスリン感受性に対する上記の有益な影響を部分的に説明するものである。
【0007】
6個のヒトIGF結合タンパク質が同定されている。これら6個の結合タンパク質のうち、IGFBP−3は、血清中の95パーセントを超えるIGF−Iと結合する。これは、分子量が29kD程度と計算される264個のアミノ酸残基を含有する。潜在的に3つのN糖鎖付加部位(Asn−X−Ser/Thr)がIGFBP−3中央部のAsn89、Asn109、およびAsn172の位置に存在するが、炭水化物単位はIGF結合には必須ではないと思われる。IGF−I/IGFBP−3二量体が「酸に不安定なサブユニット」と三重複合体を形成し、これにより、IGF−Iの半減期が延長され、その受容体に対するIGF−Iの供給が漸増する。
【0008】
IGFBP−3は多くの組織において局所的に産生される40〜45kDaの糖タンパク質であり、それらの組織において、細胞成長およびアポトーシスの調節における自己分泌および傍分泌調節因子として作用する。IGFBP−3は、IGFと結合することにより細胞増殖および生存を阻害し、IGFが標的細胞上のIGF−I受容体を活性化することを妨げる。IGFBP−3はまた、細胞増殖を負に調節し、IGFに依存せずにアポトーシスを誘発することが分かっている。IGF−Iが存在しない場合、IGFBP−3は、p53、レチノイン酸、腫瘍壊死因子−α、および形質転換成長因子−β等の多数の成長抑制タンパク質および薬剤と相互作用することができる。IGFBP−3の過剰発現は細胞増殖の阻害、腫瘍形成の低減を行い、正常臓器の成長をわずかに阻害する。最近の研究では、IGFBP−3が乳癌、前立腺癌、肺癌、卵巣癌、および結腸直腸癌を阻害するため、IGFBP−3が抗癌剤として有効であることが実証されている。
【0009】
それゆえ、ヒトIGF−IおよびヒトIGFBP−3はともに、薬学的価値を確立している。IGF−IおよびIGFBP−3の両方の使用に関する重大な障害は、それらの製造コストである。これらのタンパク質は、主に、Escherichia coli等の細菌から組み換えによって製造されるか、または肉腫細胞株もしくは赤血球系細胞から抽出され、かつ/または遺伝子導入マウス中で産生される。これらのシステムは、設備および製造コストが高く、病原菌で汚染される可能性があるため大量生産には適さない。IGF−IおよびIGFBP−3は他の哺乳類でも同様に発生し、同様の機能を果たす。
【0010】
植物細胞は、原核生物および真核生物源からの遺伝子を含む幅広い生物から遺伝子情報を受け取り、発現するように改変することができる。植物細胞は真核性であるため、適切なタンパク質または酵素機能に必要とされることが多い適切な翻訳後修飾(例えば、グリコシル化、プレニル化、およびジスルフィド架橋の形成)により哺乳類タンパク質を産生することができる。また、多くの植物種の種子が食用に適しており、種子中に組み換えタンパク質を蓄積することが可能である。投与量レベルおよび頻度が制御される場合には、組み換えタンパク質は、経口送達の前にさらなる処理および精製を必ずしも必要としないこともあるが、これは、各タンパク質が特有の酸およびプロテアーゼ耐性を有するためである。バイオカプセルおよび植物組織等の送達ビヒクルを用いて胃および内臓におけるタンパク質の分解を防ぐことも可能である(非特許文献1)。種子に基づくプラットホームはまた、医薬用タンパク質の生物学的集合および産生のために開発されている(非特許文献2)。それらの結果は、生物医薬品の産生および「錠剤種子」経路(’seed as pill’ route)を介した送達のためのビヒクルとしての植物種子の使用は実行可能な選択肢であることを示唆している。
【0011】
形質転換タバコ中において組み換えヒトIGF−Iおよび組み換えヒトIGFBP−3が産生されており、これらの結果は3件のポスター発表で報告されている(5th Hong Kong Diabetes and Cardiovascular Risk Factors,East Meets West Symposium,October 2003(”Expression of Human Insulin−like Growth Factor I(IGF−I) and Insulin−like Growth Factor Binding Protein 3(IGFBP−3) in Transgenic Tobacco”);64th Scientific Sessions of American Diabetes Association,June 2004(”Plants as Bioreactors for Expressing Human Insulin−like Growth Factor I(IGF−I) and Insulin−like Growth Factor Binding Protein−3(IGFBP−3)”);および2004 annual meeting of the American Society of Plant Biologists,July 2004(”Transgenic Expression of Human Insulin−like Growth Factor Binding Protein−3(IGFBP−3) in Tobacco Seeds”)。これらのタンパク質のタバコ苗における産生には、トウモロコシ、イネ、小麦、および大麦等の食用に適した単子葉植物における産生のような利点がない。単子葉植物には、サトウキビ、パイナップル、ナツメヤシ、およびバナナ等の主要食用作物も含まれる。これらの単子葉植物は、食用に適した原料の代表的な増殖源である。
【0012】
世界の人口の40%を超える人々によって日々消費されるイネは、世界中の農業に定着しており、製薬上および商業上重要なタンパク質およびワクチンを製造するためのモデルバイオリアクタ(model bioreactor)として認識されている(非特許文献3)。これは、タバコが含有するようなニコチンおよび毒性アルカロイド等の有害化学物質を含有しておらず、アレルギー誘発性が低い。イネ中の組み換えタンパク質は1%粒重まで蓄積可能である。(最大で総種子粒数(total seed grain)の91%となる)イネの胚乳領域における組み換えタンパク質の産生および貯蔵を目的とする特定のプロモーターおよびシグナル配列の使用により、タンパク質蓄積量を粒重の2.7%まで増加することができる(Liu,Q.Q.,Thesis Department of Biology(2002)Yangzhou University(中国)およびthe Chinese University of Hong Kong(香港))。単一のイネ苗は、10,000を超える穀粒を生成する分げつを最大で100個有することが有り、これにより、大量の種子および組み換えタンパク質を迅速に産生することができる。また、早生(1年当たり3〜4回)型のイネjaponica亜種は、産生用のバイオリアクター苗(bioreactor plants)として用いることができる。乾燥種子の貯蔵および流通は単純であり、5ヶ月以上室温で貯蔵を行っても、穀粒中の組み換えタンパク質の収量および活性が著しく損なわれることはない(非特許文献4)。低含水率の条件では、穀粒は生存能力を失うことなく3〜5年貯蔵することができる(Huang,N.,BioProcess International(2002)Jan:54−59)。
【0013】
タンパク質が家畜での使用を対象とする場合には、オート麦等の代替的な穀物中での産生がより適切な場合がある。
【0014】
過去の研究では、コドン使用頻度の偏りが遺伝子発現レベルと強い相関関係を有することが示されている。高度に発現した遺伝子は、「最適」コドンと呼ばれるコドンのサブセットを優先的に用いる(非特許文献5)。さらに、これらの最適コドンは最も豊富なtRNAに対応しており、翻訳精度および効率が向上する(非特許文献6)。下記の実施例では、2つの種子貯蔵タンパク質において用いられる植物優先コドンに基づいて、ヒトIGF−IおよびIGFBP−3のDNA配列に改変を加え、これらのタンパク質についてイ
ネにおけるタンパク質産生の向上を試みた。形質転換シロイヌナズナ中での高度な発現および安定した蓄積(LRPおよびPN2Sについて、それぞれ、全抽出種子タンパク質の3〜10%および3〜15%)が過去の研究において観察されたため、シカクマメ由来のリジンに富んだタンパク質(LRP)およびパラダイスナッツ由来のメチオニンに富んだ2Sアルブミン(PN2S)のコドン使用頻度を改変のベースとして選択した。
【0015】
下記実施例で用いた標的タンパク質の収量を増大する他の戦略は、細胞のタンパク質分解系による分解を防ぐために、組み換えタンパク質を特定部分に向けることである。小胞体(ER)分秘経路(secretary pathway)に導くシグナルペプチド配列、および外来遺伝子のNおよびC末端のER保持シグナルであるテトラペプチドKDELの付着は、一般に、その産物の高度な蓄積に必要とされることが報告されている(非特許文献7;および非特許文献8)。KDELテトラペプチドは、ゴルジ複合体とER間でリサイクルされる受容体との相互作用によるタンパク質局在化に寄与する。KDELシグナルは、植物ERにおける可溶性タンパク質蓄積に十分である(非特許文献7;非特許文献9;および非特許文献10)。KDELを含有する構築物で形質転換した細胞中では、KDELを含有しないものよりもscFvレベルが6〜14倍高いことが分かっている(非特許文献11)。
【先行技術文献】
【非特許文献】
【0016】
【非特許文献1】Daniell,H.ら、Trends Plant Sci.(2001)6:219−226
【非特許文献2】Sardana,R.K.ら、Transgenic Res.(2002)11:521−531
【非特許文献3】Fischer,R.ら、Transgenic Res.(2000)9:279−299
【非特許文献4】Stoger,E.ら、Plant Mol.Biol.(2000)42:583−590
【非特許文献5】Moriyama,E.N.ら、J.Mol Evol(1997)45:514−523
【非特許文献6】Marais,G.ら、J.Mol Evol(2001)52:275−280
【非特許文献7】Wandelt,C.I.ら、Plant J.(1992)2:181−192
【非特許文献8】Herman,E.M.ら、Planta(1990)182:305−312
【非特許文献9】Frigerio,L.ら、Plant Cell(2001)13:1109−1126
【非特許文献10】Napier,J.ら、Planta(1997)203:488−494
【非特許文献11】Conrad,U.ら、Plant Mol.Biol.(1998)38:101−109
【発明の概要】
【課題を解決するための手段】
【0017】
本発明は、経済的かつ実用的な2つの重要な哺乳類タンパク(IGF−IおよびIGFBP−3)源を提供する。ヒト型での産生が好ましい。これらのタンパク質を単子葉植物中で産生することにより、本発明は、初めて、ヒトの治療のための使用または獣医学的状況での使用に適合可能な形態において十分な量での提供が可能である有用なタンパク源を提供する。
【0018】
よって、1つの局面では、本発明は、ヒトIGF−Iおよび/またはヒトIGFBP−3を産生するように改変された単子葉植物細胞に関する。他の局面では、本発明は、そのような細胞を含む植物または植物部位に関する。
例えば、本発明は、以下の項目を提供する:
(項目1)
インスリン様成長因子I(IGF−I)をコードするヌクレオチド配列またはインスリン様成長因子結合タンパク質3(IGFBP−3)をコードするヌクレオチド配列の発現のためのDNA構築物を含有するように改変した単子葉植物細胞であって、該DNA構築物は、単子葉植物細胞中での発現のために制御配列と作動可能に連結した該コードするヌクレオチド配列を含む、細胞。
(項目2)
上記IGF−IおよびIGFBP−3はヒトタンパク質である、項目1に記載の細胞。
(項目3)
上記制御配列は、単子葉植物の種子中で作動可能なプロモーターおよび単子葉植物の種子中で作動可能な終止配列を含む、項目1または2に記載の細胞。
(項目4)
上記コードするヌクレオチド配列は、小胞体(ER)分泌経路に導くシグナルペプチドをコードするヌクレオチド配列によりN末端で伸長される、項目1または2に記載の細胞。
(項目5)
上記コードするヌクレオチド配列は、ER保持シグナルをコードするヌクレオチド配列によりC末端で伸長される、項目1または2に記載の細胞。
(項目6)
上記細胞はイネ細胞である、項目1または2に記載の細胞。
(項目7)
項目1または2に記載の細胞を含む遺伝子組み換え植物または植物部位。
(項目8)
IGF−IまたはIGFBP−3を含む単子葉植物細胞。
(項目9)
上記IGF−IおよびIGFBP−3はヒト由来のものである、項目8に記載の細胞。
(項目10)
上記細胞はイネ細胞である、項目8または9に記載の細胞。
(項目11)
項目8または9に記載の細胞を含む植物または植物部位。
(項目12)
上記植物部位は種子組織である、項目11に記載の植物部位。
(項目13)
上記植物または植物部位はイネである、項目11に記載の植物または植物部位。
(項目14)
上記植物部位は種子組織である、項目13に記載の植物部位。
(項目15)
単子葉植物においてIGF−IまたはIGFBP−3を産生する方法であって、項目1または2に記載の植物細胞を培養する工程を包含する、方法。
(項目16)
単子葉植物においてIGF−IまたはIGFBP−3を産生する方法であって、項目11に記載の植物を培養する工程を包含する、方法。
(項目17)
上記植物はイネである、項目16に記載の方法。
【図面の簡単な説明】
【0019】
【図1】Jansen,M.ら、Nature(1983)306:609−611で報告されたヒトIGF−I(配列番号1)をコードするヌクレオチド配列を示す。
【図2】Wood,W.I.ら、Mol.Endocrinol.(1988)2:1176−1185で報告されたヒトIGFBP−3(配列番号2)をコードするヌクレオチド配列を示す。
【図3】植物におけるコドン選択のために改変されたヒトIGF−I(配列番号3)をコードするヌクレオチド配列を示す。
【図4】植物におけるコドン選択に従って改変されたヒトIGFBP−3(配列番号4)のヌクレオチド配列を示す。
【図5】ヒトIGF−I(配列番号5)のアミノ酸配列を示す。
【図6】ヒトIGFBP−3(配列番号6)のアミノ酸配列を示す。
【図7】イネで生成したヒトIGF−IのL6細胞にラフリングを生じる能力、およびこの作用の商用ヒトIGFBP−3に対する反応を示すアッセイの結果を示す。
【図8】イネで生成したヒトIGFBP−3のMCF−7細胞の成長を阻害する効果を示す。
【発明を実施するための形態】
【0020】
下記に示すように、2つの重要なヒトタンパク質であるIGF−IおよびIGFBP−3が単子葉植物中において産生されたのは初めてのことである。単子葉植物は主要な栄養源を含み、有毒化合物を含まないため、ヒトの治療を目的としたタンパク質の産生に理想的である。これらはまた、草食性または雑食性哺乳類を治療する場合において、類似するタンパク質の産生にも理想的である。これらのタンパク質の産生は、これらのタンパク質をコードするヌクレオチド配列を発現のために適切な制御配列に作動可能に連結した発現系を含むDNA構築物を適切に設計することにより促進可能である。
【0021】
植物細胞の遺伝子改変を実施し、無傷植物を再構成する技術は、以前より当該分野において周知である。例えば、Gelvinら、Plant Molecular Biology Manual(1990));Dashek,Methods in Plant Biochemistry and Molecular Biology(CRC Press 1997)を参照されたい。これに関する最新技術の有用な要約が2004年1月14日に公開された米国特許公報2004/0009476に見られるが、その植物の遺伝子操作のための適切な技術についての開示内容を本明細書中において参考として援用する。
【0022】
組み換え構築物を含有する形質転換植物細胞が一旦得られると、そこから遺伝子組み換え植物を再生し、所望のタンパク質産生レベルを判断することができる。
【0023】
植物中における変性ヌクレオチド配列の発現を最適化するためにいくつかの技術を用いることができる。下記の実施例においてさらに説明するように、植物細胞中における発現のためのコドン選択に応じて、コードするヌクレオチド配列に改変を加えることができる。そのような改変は、植物中におけるコドン選択を記載する公開データに基づくものである。次に、コードするヌクレオチド配列を伸長してシグナルおよび保持配列を加え、コードされたタンパク質を小胞体に向け、その保持を行ってもよい。これはまた、収量にも好影響を及ぼす。一般に、シグナル伝達ペプチドは所望のタンパク質のN末端に生成され、保持シグナルはそのC末端に生成される。
【0024】
これらの技術を用いることにより、所望のIGF−IおよびIGFBP−3の収量は目に見えて向上する。
【0025】
適切なプロモーターを用いて発現を行うことはさらに有用である。例えば、適切なプロモーターとしては、35S CaMV、イネアクチンプロモーター、ユビキチンプロモーター、またはノパリンシンターゼ(NOS)プロモーターが挙げられる。種子中での産生を促進する組織特異的プロモーターは種子特異的グルテリンプロモーター(Gt−1pro)を含むが、他の種子特異的プロモーターを用いてもよい。ノパリンシンターゼ終結シグナル等の終結シグナルを用いてもよい。
【0026】
下記に示すように、植物最適化コード配列を駆動する2つの構築物群を設計し、Agrobacteriumにより媒介された形質転換によりイネに導入した。一方の構築物群はグルテリンシグナルペプチド(SP)を単独で含有するが、他方はSPとともに標的テトラペプチドKDELシグナルを含有する。これらの構築物を合成し、タンパク質の発現を増加させ、貯蔵のためにタンパク質を区画内に入れるとともに、タンパク質安定性を向上させた。種子特異的グルテリンプロモーター(Gt−1pro)を用いて遺伝子組み換えイネ中におけるIGF−IおよびIGFBP−3の発現を促進し、導入遺伝子の発現を分析した。遺伝子組み換えイネ穀粒から産生したrhIGF−IおよびrhIGFBP−3は生物活性を有していた。
【0027】
上記構築物はまた、後で精製を行えるように、改変を加えてヒスチジン標識またはFLAGシーケンス等の精製補助物を含むようにしてもよく、かつ/または蛍光タンパク質等のマーカーを含むようにしてもよい。また、コードしたタンパク質と精製標識および/またはマーカー間に切断部位を設けてもよい。所望であれば標準的な精製技術を用いてもよく、または場合によっては、植物組織を経口投与し、植物の栄養価およびその低毒性を利用するようにしてもよい。
【0028】
組み換え技術によって生成したIGF−Iは、1型または2型糖尿病患者のインスリン必要量を低減し、血糖管理を改善するために用いられる。IGFBP−3はアポトーシスを引き起こすことが示されており、悪性腫瘍の治療に有用である。よって、所望であれば、組み換え技術によって生成したタンパク質を、これらのタンパク質を用いた治療を必要とする患者に投与するための組成物として製剤化してもよい。タンパク質および他の医薬品を製剤化するための一般的方法は、本明細書において参考として援用するRemington’s Pharmaceutical Sciences,latest edition,Mack Publishing Co.(Easton,PA)において見ることができる。これらのタンパク質は、典型的には、注射または経皮的もしくは経粘膜的送達により非経口的に全身投与されるか、または経口投与される場合がある。リボソーム製剤、および脂質もしくは高分子等に基づくナノ粒子を含有する製剤を含む、特定の投与方式を対象とする各種の製剤を用いることができる。
【0029】
以下の実施例は本発明を例示するためのものであり、限定するものではない。
【実施例】
【0030】
(実施例1)
IGF−IおよびIGFBP−3構築物を用いたAgrobacteriaの形質転換
ヒトIGF−I(図1)およびIGFBP−3(図2)のDNA配列に、2つの種子貯蔵タンパク質に用いられる優先コドンに基づいて改変を加えた。シカクマメ由来のリジンに富んだタンパク質(LRP)およびパラダイスナッツ由来のメチオニンに富んだ2Sアルブミン(PN2S)のコドン使用頻度(表1)を改変のベースとして選択したが、これは、過去の研究において、形質転換シロイヌナズナ中での高度な発現および安定した蓄積(LRPおよびPN2Sについて、それぞれ全抽出種子タンパク質の3〜10%および3〜15%)が観察されたためである。
【0031】
元のアミノ酸配列と同じアミノ酸配列(図5および図6)をコードする改変IGF−I(図3)およびIGFBP−3(図4)遺伝子をMWG Biotechnology Companyから入手した。IGF−IおよびIGFBP−3のコドン変化量は、それぞれ、28.6%および14.8%であった。
【0032】
【表1−1】
【0033】
【表1−2】
rhIGF−IおよびrhIGFBP−3の安定性および収量を向上させるとともに、それらのグリコシル化を制御する構築物を設計した。2つのタンパク質標的シグナルを加え、標的タンパク質をイネ穀粒中の特定の区画に向けた(ゴルジ装置におけるグリコシル化のためのグルテリンシグナルペプチド(SP)または小胞体内におけるグリコシル化がない安定した蓄積のためのテトラペプチドKDELのいずれか)。発現構築物を種子特異的グルテリンプロモーター(Gt−1pro)により駆動した。キメラ遺伝子構築物の詳細は以下のとおりである。
【0034】
IGF−I:
1) (Gt−1pro)+SP+IGF−I*+NOSter
2) (Gt−1pro)+SP+IGF−I*+KDEL+NOSter
IGFBF−3:
3) (Gt−1pro)+SP+IGFBF−3*+NOSter
4) (Gt−1pro)+SP+IGFBF−3*+KDEL+NOSter
ここで、*はIGF−IまたはIGFBP−3の改変cDNAを示し、NOSterはノパリンシンターゼ停止剤を示す。
【0035】
キメラ遺伝子カセットをスーパーバイナリーベクターpSB130に挿入し、イネカルスの感染に理想的なAgrobacterium株(EHA105)に形質転換した。(Chen,H.ら、BioTechniques(1994)16:664−668、670に記載されるように)単純な凍結融解法を用いてAgrobacteriumを形質転換した。抗生物質ハイグロマイシン(50mg/L)を用いて形質転換バクテリアを選択し、PCRにより標的遺伝子のDNA形質転換をさらに確認した。
【0036】
(実施例2)
イネの形質転換、選択、および培養
カルス誘導のためにjaponica9983の成熟種子の胚盤を切除し、カルス誘導培地(N6基礎培地、2mg/lの2,4−D、0.5g/lのカゼイン加水分解液、30g/lのスクロース、2.5g/lのPhytagel(登録商標)、pH5.8)上で培養した。カルス誘導培地で4〜7日間培養した後すぐに、未熟胚由来のカルスを使用し、標的遺伝子を含有するAgrobacterium EHA105と共培養した。
【0037】
まず、50mg/Lのリファンピシンおよび50mg/Lのカナマイシンを添加した3mlのLBブロスに、Agrobacteriumを28℃で一晩かけて植菌した後、25mlのAB培地(3g/lのK2HPO4、1g/lのNaH2PO4、1g/lのNH4Cl、0.3g/lのMgSO4・7H2O、0.15g/lのKCl、10mg/lのCaCl2・2H2O、2.5mg/lのFeSO4・7H2O、5g/lのグルコース、pH7.2)で5時間かけて継代培養を行い、引き続き、遠心分離および15〜25mlのAAM培地(AA基礎培地、68.5g/lのスクロース、36g/lのグルコース、0.5g/lのカゼイン加水分解液、pH5.2、100μmol/lのアセトシリンゴン)での再懸濁を行った。
【0038】
イネカルスをAgrobacterium培養液に10〜20分間室温で浸し、不連続的に振とうした。次に、暗所において、100μmol/lのアセトシリンゴンを含有するN6D2C培地(N6D2、10g/lのグルコース、pH5.2)に3日間かけて26〜28℃で感染カルスを移した。共培養後、耐性カルスの選択のために、暗所において、このカルスをN6D2S培地(N6D2、50mg/lのハイグロマイシンB、500mg/lのセフォタキシム、pH5.8)上に26℃で2週間静置した後、新たに形成された耐性カルスが現れるまで新たなN6D2S培地で培養を行った。
【0039】
次に、HGPR培地(Higrow(登録商標)イネ培地(GIBCO−BRL)、50mg/lのハイグロマイシンB、200mg/lのセフォタキシム)上に、暗所において7日間と、明所において7日間26℃で耐性カルスを静置した。再生前処理(pre−regeneration)後、再生のために、MSR培地(MS基礎培地、30g/lのスクロース、0.5g/lのカゼイン加水分解液、2mg/lの6−BA、0.5mg/lのNAA、0.5mg/lのKT、pH5.8、50mg/lのハイグロマイシンB、500mg/lのセフォタキシム、2.5g/lのPhytagel(登録商標))に耐性カルスを26℃で明所において16時間/暗所において8時間かけて移した。
【0040】
再生芽が出てくると、発根のために、それらを1/2倍量のMSH培地(1/2倍量のMSマクロ塩、Fe−EDTAおよびミクロ塩、MSビタミン、30g/lのスクロース、0.5mg/lのNAA、pH5.8、50mg/lのハイグロマイシンB、500mg/lのセフォタキシム、2.5g/lのPhytagel(登録商標))上に静置した。最後に、遺伝子組み換えイネ胚を土壌に移し、温室で栽培した。
【0041】
(実施例3)
遺伝子組み換えイネの分析
サザンブロット分析
遺伝子組み換えイネ苗を分析し、イネゲノムへの標的遺伝子の組込みを確認した。臭化セチルトリメチルアンモニウム(CTAB)法により葉ゲノムDNAを抽出した(Doyle,J.D.ら、Focus(1990)12:13−15)。15μgのゲノムDNAをBamHIを用いて一晩かけて消化し、0.8%のアガロースゲル上で分離し、VacuGeneXL真空ブロッティングシステム(Pharmacia Biotech)を用いて正に帯電したナイロン膜(Roche)に移した。DIG核酸検出キット(Roche)に記載の方法に従ってハイブリダイゼーションおよび検出を行った。DIG DNA標識キット(Roche)を用いてPCRにより二本鎖DIG標識DNAプローブ(IGF−IおよびIGFBP−3)を準備した。これらのプローブを使用前に99℃で加熱して変性させた。
【0042】
この結果、イネゲノム中に構築物の存在が確認できた。
【0043】
ウエスタンブロット分析
種子をすりつぶして粉末状にし、タンパク質抽出緩衝液(50mMのトリスHCl、pH6.8、0.1MのNaCl、および10%SDS)と混合することにより全種子タンパク質を成熟イネ種子から抽出した。遠心分離した後、上澄液を新しいエッペンドルフチューブに移し、種子全タンパク質抽出液として保存した。次に、異なる量の全タンパク質を17%トリシンSDS−PAGE中で分解し、PVDF膜上にブロットした。抗ヒトIGF−IまたはIGFBP−3ポリクローナル抗体を用いてウエスタンブロット分析を行った。最後に、Aurora(登録商標)Western Blot Chemiluminescent Detection System(ICN)のマニュアルに記載のとおり、化学発光Starlight(登録商標)基質(ICN)を用いてブロットの非放射性検出を行った。
【0044】
その結果、種子抽出液中にIGF−IおよびIGFBP−3タンパク質の両方の存在が確認できた。
【0045】
(実施例4)
イネで生成したrhIGF−Iの生物活性
インスリンと同様に、IGF−Iは、筋細胞中で膜ラッフルおよびグルコース摂取を生じることが示されている。IGFBP−3は、IGF−Iと結合して、ALS複合体を形成することができ、これにより、IGF−Iにより生じた膜ラフリング作用が阻害される。イネで生成した組み換えhIGF−Iをインスリンと同時にテストし、それらの生物活性を比較した。
【0046】
c−mycエピトープ標識したグルコーストランスポーター4(GLUT4)を発現するラットL6骨格筋細胞(L6myc細胞)を、10%(v/v)ウシ胎児血清(FBS)および1%(v/v)抗生物質−抗真菌薬溶液(100U/mlのペニシリンG、10mg/mlのストレプトマイシン、および25mg/mlのアンフォテリシンB)を含有するα−最小必須培地において37℃の5%CO2雰囲気で筋芽細胞単層培養を維持した。0.25%トリプシンを用いてサブコンフルエント培地のトリプシン処理により細胞を継代培養した。筋管への分化のために、4×104程度の細胞数/mlで2%(v/v)FBSを含有する培地で筋芽細胞を培養し、自然融合させた。48時間ごとに培地を交換し、培養後5〜7日間筋管を用いた。次に、6ウェルプレートに載せた25mm径のガラスカバースリップ上でラットL6筋細胞を筋管の段階まで培養した。
【0047】
3時間かけて筋管から血清を抜き、遺伝子組み換えイネから抽出したrhIGF−IおよびrhIGFBP−3を異なる濃度および組み合わせで10分間かけて37℃で処理した。これらのインキュベーションの後、筋管を3%(v/v)氷冷パラホルムアルデヒドPBS溶液で20分間かけて凝固させた後、0.1MグリシンのPBS溶液で10分間洗浄し、0.1%(v/v)トリトンX−100のPBS溶液で3分間透過処理した後、PBSで洗浄した。筋管を0.1%BSA中で1時間塞いだ後、ファロイジン(0.1%(w/v)BSA中で1:500)中でインキュベーションを行った。このインキュベーションの後、細胞をPBSで洗浄し、ガラススライド上にProLong(登録商標)アンチフェード溶液として載せた。Zeiss Axioplan2イメージング顕微鏡およびZeiss LSM510 METAレーザー走査共焦点顕微鏡(Carl Zeiss,Jena,Germany)を用いてサンプルを検査した。
【0048】
遺伝子組み換えIGF−Iイネの粗タンパク質によりL6細胞の膜ラッフルが生じるが、そのラフリング作用は商用hIGFBP−3により大幅に低減することが可能である。これらの結果を図7に示す。図7に示すように、イネで生成したIGF−1は、1.25mgおよび5mgで用量依存的にラフリングを生じる。この活性は、12nM濃度の市販のIGFBP−3によって阻害された。
【0049】
(実施例5)
イネで生成したrhIGFBP−3タンパク質の抗癌活性
ヒトIGFBP−3は、エストロゲン依存性および非依存性乳癌細胞の増殖を阻害することが示されている。イネで生成したrhIGFBP−3が制癌作用を有するか否かを検査するために、MCF−7ヒト乳癌細胞を用いた。
【0050】
1%ペニシリンおよびストレプトマイシン、0.1%ファンギゾム(fungizome)を5%ウシ胎児血清(FBS)とともに添加したイーグル最小必須培地にMCF−7ヒト乳癌細胞を通法により37℃の5%CO2加湿雰囲気に維持した。rhIGFBP−3の処理のために、MCF−7細胞を血清含有培地で96ウェルプレートに播種した。1日後、それらの細胞に、rhIGFBP−3を含有するイネ形質転換体から抽出した異なる濃度の粗タンパク質を加えた。粗タンパク質抽出液を3日おきに新しいものと2度交換した。
【0051】
7日間処理した後、MTTアッセイのために細胞を採取した。簡単に言えば、20μlのMTT(3−(4,5−ジメチルチアゾール−2−イル)−2,5−臭化ジフェニルテトラゾリウム(diphenyl tetrazolium bromide)溶液(5mg/mlMTTのHPBS溶液、pH7.4)を各ウェルに加え、さらに37℃で2時間かけてインキュベートした。100μlの酸イソプロパノールを各ウェルに加えて細胞を分解し、その紫色の結晶を溶解した。マイクロプレート分光光度計をOD570で用いて、各ウェルにおける紫色の強度を測定した。
【0052】
図8に示すように、イネで生成したIGFBP−3は、1.56mgおよび3.125mgの濃度で用量依存的にMCF−7細胞の成長阻害を向上させることができた。
【0053】
(実施例6)
組み換え技術によって生成したタンパク質の精製
親和性クロマトグラフィーを用いて上記実施例に記載の粗抽出液をさらに精製する。IGF−IおよびIGFBP−3の各々をクロマトグラフィーカラムに塗布し、さらに精製するために、これらのカラムをそれぞれの抗体に結合する。カラムを介して不純物を洗浄し、pHおよび塩濃度を調節してタンパク質を溶出する。
【0054】
マーカータンパク質を含むように実施例1の構築物に改変を加えて精製を簡略化する。
【0055】
(実施例7)
組み換えタンパク質収量の向上
実施例2に記載の苗から種子を得て、第2および第3世代の作物を得るために移植する。収量が向上した組み換えIGF−IおよびIGFBP−3が得られる。
【特許請求の範囲】
【請求項1】
本明細書に記載の単子葉植物細胞。
【請求項1】
本明細書に記載の単子葉植物細胞。
【図1】

【図2】
【図3】

【図4】
【図5】

【図6】

【図7】
【図8】
【図2】
【図3】
【図4】
【図5】
【図6】
【図7】
【図8】
【公開番号】特開2013−106615(P2013−106615A)
【公開日】平成25年6月6日(2013.6.6)
【国際特許分類】
【外国語出願】
【出願番号】特願2013−38444(P2013−38444)
【出願日】平成25年2月28日(2013.2.28)
【分割の表示】特願2009−550178(P2009−550178)の分割
【原出願日】平成20年2月20日(2008.2.20)
【出願人】(509229212)ザ チャイニーズ ユニバーシティー オブ ホンコン (4)
【Fターム(参考)】
【公開日】平成25年6月6日(2013.6.6)
【国際特許分類】
【出願番号】特願2013−38444(P2013−38444)
【出願日】平成25年2月28日(2013.2.28)
【分割の表示】特願2009−550178(P2009−550178)の分割
【原出願日】平成20年2月20日(2008.2.20)
【出願人】(509229212)ザ チャイニーズ ユニバーシティー オブ ホンコン (4)
【Fターム(参考)】
[ Back to top ]