
遺伝子組換え体の定量的検知方法
【課題】Roundup Ready Soybean系統の組換え遺伝子の定量・検出において、PCR法のようなサーマルサイクラーと蛍光検出装置が一体化した高価なリアルタイムPCR用の装置を用いることなく、迅速・簡便かつ低コストで、正確な上記組換え遺伝子の定量・検知を可能とする。
【解決手段】LAMP法あるいはABC-LAMP法に着目し、これら方法を用いて上記組み換え遺伝子を定量・検出するのに最適な、プライマー、あるいはさらにプローブ、競合遺伝子を新たに設計し、これらを用いてLAMP法あるいはABC-LAMP法を行い、Roundup Ready Soybean系統の組換え遺伝子を定量・検出する。その際、ダイズ由来のLe1遺伝子を同様にLAMP法あるいはABC-LAMP法を用いて定量・検出し、内部標準として用いる。
【解決手段】LAMP法あるいはABC-LAMP法に着目し、これら方法を用いて上記組み換え遺伝子を定量・検出するのに最適な、プライマー、あるいはさらにプローブ、競合遺伝子を新たに設計し、これらを用いてLAMP法あるいはABC-LAMP法を行い、Roundup Ready Soybean系統の組換え遺伝子を定量・検出する。その際、ダイズ由来のLe1遺伝子を同様にLAMP法あるいはABC-LAMP法を用いて定量・検出し、内部標準として用いる。
【発明の詳細な説明】
【技術分野】
【0001】
本発明は、ダイズもしくはダイズ加工食品に含まれる遺伝子組換え体の定量的検知方法及び当該方法に用いられる核酸分子に関する。より詳細には、本発明は、ダイズもしくはダイズ加工食品に含まれる遺伝子組換体を、LAMP法をあるいはABC-LAMP法を用いて定量的に検知する方法、並びに当該方法に用いられる核酸プライマー、核酸プローブ及び競合的核酸等の各種核酸分子に関する。
【背景技術】
【0002】
世界的に、遺伝子組換え技術を利用して開発された農作物の実用化が進んでいる。我が国においても、ダイズ、トウモロコシ等、59品種の作物について遺伝子組換え体が認められている。実際に流通している遺伝子組換え作物として、例えば、トウモロコシについてはBt11系統の後代品種(Novartis社)、Event176系統の後代品種(Novartis社)、MON810系統の後代品種(Monsanto社)、GA21系統の後代品種(Monsanto社)、T25系統の後代品種(Aventis社)が、またダイズについてはRoundup Ready Soybean系統の後代品種(Monsanto社)等を挙げることができる。
遺伝子組換え作物は、天然の作物に、害虫抵抗性や除草剤耐性等の産業上好ましい形質を付与する目的で開発される。その主な手法は、元来そのような産業上好ましい形質を有する他の生物から、当該形質を発現する遺伝子を単離し、その遺伝子を対象作物に発現可能な形で導入するというものである。従って、遺伝子組換え作物のDNAの中には、このような導入遺伝子が含まれている。
【0003】
欧州共同体(EU)により遺伝子組換え作物及びその加工食品の表示に関わる規則(非特許文献1)が定められ、日本に於いても遺伝子組換え作物及びその加工食品の表示に関する制度が定められたことを契機として(非特許文献2)、食品業界及びそれに関連する業界では、食品原料農作物や食品中の組換えDNAの存否、またはその含量を把握し認識しておくことが求められている。このため、食品、飼料及びこれらの原料農作物中の遺伝子組換え体の有無や含量、特に原料中の存在比を確認する技術が必要とされている。
【0004】
遺伝子組換え体を検知する技術としては、例えば、ポリメラーゼ連鎖反応(以下「PCR」という)によって該組換え遺伝子を増幅し、これを検出する方法(例えば、非特許文献3);競合PCRによる組換えダイズやトウモロコシの定量的検知技術(例えば、非特許文献4,5,6及び7);蛍光プローブを用いた定量的PCR(リアルタイムPCR)による組換えダイズやトウモロコシの定量的検知技術(例えば、非特許文献8、9及び10);組換えDNAから生産される蛋白質をEnzyme Linked Immuno Sorvent Assay (ELISA)法を用いて測定することによって、遺伝子組換え体の含量を定量する方法(例えば、非特許文献11)を挙げることができる。また、最近では、定量PCR法を用いて、複数の遺伝子組換え体の系統を含む集団中における遺伝子組換え体の存在比を正確に定量する方法として、内部標準を用いた方法(特許文献1及び2)が確立され、特に前者の方法(特許文献1)は、日本国において遺伝子組換え食品の検査分析の標準法として採用されている。当該方法は作物中の内在性遺伝子と組換え遺伝子のPCR法によるコピー数を測定し、その比から遺伝子組換え遺伝子の混入量を換算し定量する方法である。
【0005】
上記標準法として定められた従来の定量的PCR法は増幅と検出に1時間以上必要であり、また、PCRを行うためのサーマルサイクラーと蛍光検出装置が一体化した高価なリアルタイムPCR用の装置が必要であるといった問題があった。そこで、迅速・簡便かつ低コストで組換え遺伝子を検知するための方法の開発が望まれていた。
【非特許文献1】Regulation (EU) No.EC/258/97, Council Regulation(EC), No.1139/98
【非特許文献2】Notification No. 1775 (June 10, 2000) Food and Marketing Bureau,Ministry of Agriculture, Forestry and Fisheries of Japan, Tokyo, Japan
【非特許文献3】Hupfer C., Mayer J., Hotzel H., Sachse K., Engel K.H., Eur. FoodRes. Technol., 209, 301-304(1999)
【非特許文献4】Hardegger M., Brodmann P., Herrmann A., Eur. Food Res. Technol., 209,83-87(1999)
【非特許文献5】Hubner P., Studer E., Luthy J., Food Control, 10, 353-358(1999)
【非特許文献6】Hubner P., Studer E., Luthy J., Nat. Biotechnol., 17, 1137-1138(1999)
【非特許文献7】Hupfer C., Hotzel H., Sachse K., Moreano F., Engel K. H., Eur. FoodRes. Technol., 212, 95-99(2000)
【非特許文献8】Vaitilingom M., Pijnenburg H., Gendre F., Brignon P., J. Agric. FoodChem., 47, 5261-5266(1999)
【非特許文献9】Wurz A., Bluth A., Zelts P., Pfeifer C., Willmund R., Food Control, 10,385-389(1999)
【非特許文献10】Berdal K. G., Holst-Jensen A., Eur. Food Res. Technol., 213, 432-438(2001)
【非特許文献11】Luthy J., Food Control, 10, 359-361(1999)
【特許文献1】WO 02/34943
【特許文献2】特開2001-136983号公報
【発明の開示】
【発明が解決しようとする課題】
【0006】
本発明の課題は、上記従来法の問題を解消した改良法として、LAMP法あるいはABC-LAMP法に着目し、ダイズもしくはダイズ加工食品に含まれる遺伝子組換え体の存在を定量的に検知するための新規な方法並びに該方法に好適な核酸分子(プライマー、プローブ、競合遺伝子)を提供することであり、特に上記PCR法による遺伝子組換え体の検出法のようなサーマルサイクラーと蛍光検出装置が一体化した高価なリアルタイムPCR用の装置を必要とせず、迅速・簡便かつ低コストで、正確な組換え遺伝子の検知手段を提供する点にある。
【課題を解決するための手段】
【0007】
発明者等は、鋭意研究の結果、遺伝子組換えダイズとして、Roundup Ready Soybean系統の後代品種中の組換え遺伝子を、LAMP法あるいはABC-LAMP法を用いて検出するための、プローブ及び競合遺伝子を新たに設計するとともに、これらを用いた上記組換え遺伝子の検知・定量法を開発して、本発明を完成するに至ったものである。すなわち本発明は以下に示されるとおりである。
【0008】
(1)遺伝子組換えダイズRoundup Ready Soybean系統の組換遺伝子をLAMP法あるいはABC-LAMP法によって検出・定量するために用いる、FIPプライマー、F3プライマー、LPFプライマー、BIPプライマー、B3プライマー及びLPBプライマーからなるプライマーセットであって、各プライマーが以下に示される塩基配列、又は該配列と相補の塩基配列を有することを特徴とする、上記プライマーセット。
FIPプライマー:GTG GTC CCA AAG ATG GAG GAG ATA TCA CAT CAA TCC ACT TGC
F3プライマー :GTC ATC CCT TAC GTC AGT
LPFプライマー:CGT TCC AAC CAC GTC TTC AA
BIPプライマー:TGT CGG CAG AGG CAT CTT TTA AAA TAA ACA TAG GGA ACC CAA
B3プライマー :TCA AAA TAA GAT CAT ACA TAC AGG
LPBプライマー:AGG AGC CAC CTT CCT TTT CC
(2)遺伝子組換えダイズRoundup Ready Soybean系統の組換え遺伝子をABC-LAMP法によって検出・定量するために用いる、配列番号9に示される塩基配列又は該配列と相補の塩基配列からなる競合的核酸。
(3)遺伝子組換えダイズRoundup Ready Soybean系統の組換え遺伝子をABC-LAMP法によって検出・定量するために用いる核酸プローブであって、配列番号7に示される塩基配列又は該配列と相補の塩基配列からなり、その末端に核酸中のグアニン塩基との近接、離間によって蛍光強度が変化する蛍光色素を標識されていることを特徴とする上記核酸プローブ。
(4)ダイズ由来のLe1遺伝子配列をLAMP法又はABC-LAMP法によって検出・定量するために用いる、FIPプライマー、F3プライマー、LPFプライマー、BIPプライマー、B3プライマー及びLPBプライマーからなるプライマーセットであって、各プライマーが以下に示される塩基配列、又は該配列と相補の塩基配列を有することを特徴とする、上記プライマーセット。
FIPプライマー:AGC AAA AGA CCA AGA AAG CAC CTT CTC TTC CCG AGT GGG
F3プライマー :CCA GCA ATA TCC TCT CCG A
LPFプライマー:GGC AGC AGA GAA CCC TAT CC
BIPプライマー:TTT GCC ACA CGC TAG CAA TTA CAC TTT CTT CCT TCG ATC TGT
B3プライマー :TCA CTA GCG ATC GAG TAG TG
LPBプライマー:GTT GCA TGA GGC CAT CTA AAT GT
(5)ダイズ由来のLe1遺伝子配列をABC-LAMP法によって検出・定量するために用いる、配列番号19に示される塩基配列又は該配列と相補の塩基配列からなる競合的核酸。
(6)ダイズ由来のLe1遺伝子配列をABC-LAMP法によって検出及び/又は定量するために用いる核酸プローブであって、配列番号18に示される塩基配列又は該配列と相補の塩基配列からなり、その末端に核酸中のグアニン塩基との近接、離間によって蛍光強度が変化する蛍光色素を標識されていることを特徴とする上記核酸プローブ。
(7)披検ダイズ試料に含まれるRoundup Ready Soybean系統の組換遺伝子配列を、ABC-LAMP法を用いて検出及び/又は定量するための方法であって、被験ダイズ試料から抽出した核酸、上記(1)に記載のプライマーセット、上記(2)に記載の競合的核酸及び上記(3)に記載の核酸プローブの存在下、等温増幅反応を行い、該反応前後の蛍光強度を測定することを特徴とする、Roundup Ready Soybean系統の組換遺伝子配列の検出及び/又は定量方法。
(8)ダイズ由来のLe1遺伝子配列を、ABC-LAMP法を用いて検出及び/又は定量するための方法であって、被験ダイズ試料から抽出した核酸、上記(4)に記載のプライマーセット、上記(5)に記載の競合的核酸及び上記(6)に記載の核酸プローブの存在下、等温増幅反応を行い、該反応前後の蛍光強度を測定することを特徴とする、上記ダイズ由来のLe1遺伝子配列の検出及び/又は定量方法。
(9)上記(1)に記載のプライマーセット、上記(2)に記載の競合的核酸及び上記(3)に記載の核酸プローブを少なくとも組み合わせたことを特徴とする、Roundup Ready Soybean系統の組換遺伝子の検出及び/又は定量用試薬キット。
(10)上記(4)に記載のプライマーセット、上記(5)に記載の競合的核酸及び上記(6)に記載の核酸プローブを少なくとも組み合わせたことを特徴とする、ダイズ由来のLe1遺伝子配列の検出又は定量用試薬キット。
【発明の効果】
【0009】
本発明によれば、ダイズあるいはダイズ加工食品中のRoundup Ready Soybean系統の組換え遺伝子をABC-LAMP法を用いて検出及び/又は定量するために極めて好適な核酸プライマー、核酸プローブ、競合的核酸が提供でき、これらを用いてABC-LAMP法を行うことにより、簡便、かつ低コストでしかも正確に上記組換え遺伝子を検出及び又は定量可能となる。また、上記核酸プライマーはLAMP法においても有用なプライマーである。一方、本発明においては、同時にダイズ由来のLe1遺伝子配列をABC-LAMP法を用いて検出及び/又は定量するために極めて好適な核酸プライマー、核酸プローブ、競合的核酸も提供する。当然、この核酸プローブもLAMP法において有用なプライマーである。これらを用いてLAMP法あるいはABC-LAMP法を行った結果得られたLe1遺伝子の定量値は、内部標準遺伝子の定量値として、上記Roundup Ready Soybean系統の組換遺伝子の定量値を除することによって、内標比を算出し、この内標比を用いて、例えば、組換え食品中の組換えダイズの混入率を求めることが可能となる。
【発明を実施するための最良の形態】
【0010】
Roundup Ready Soybean系統のダイズは、ダイズにグリホサート系除草剤に耐性を付与する遺伝子を導入したものであり、グリホサート系除草剤は広範な種類の雑草に効力を有するが、Roundup Ready Soybean系統のダイズは、この除草剤に対して耐性を有するため、該ダイズ栽培中生育してくる種々の雑草に対し、その種類によらず、グリホサート系除草剤で除草することができ、除草の手間等を大幅に軽減できる。しかし、食品としての安全性等の面で疑問視する声もある。上記耐性付与遺伝子は、土壌細菌Agrobacterium属由来の耐性遺伝子CP4EPSPSである。Roundup Ready Soybean系統のダイズは、このCP4EPSPS遺伝子の発現にCaMV 35S プロモーターを用いており、本発明においては、このプロモーター領域とダイズゲノムとの境界領域配列(35S promotor/plant junction region, AJ308514)をRoundup Ready Soybean系統検出のターゲットとしている。
【0011】
35S promotor/plant junction region (AJ308514)の塩基配列(配列番号8)
gatagtggga ttgtgcgtca tcccttacgt cagtggagat atcacatcaa
tccacttgct ttgaagacgt ggttggaacg tcttcttttt ccacgtgctc
ctcgtgggtg ggggtccatc tttgggacca ctgtcggcag aggcatcttc
aacgatggcc tttcctttat cgcaatgatg gcatttgtag gagccacctt
ccttttccat ttgggttccc tatgtttatt ttaacctgta tgtatgatct
tattttgaat gaaatgcaat aagttatttc tagtaaaaaa aaataaacat
ttgatagaaa caaattaaag catgcaaaaa taactcatta gcatcggtta
aattgaaggg tttgaataat ttgcacaagg ttctgaattc
【0012】
一方、LAMP(Loop-Mediated Isothermal AMPlification)法は、遺伝子の簡易・迅速な増幅法として知られており、標的遺伝子の塩基配列に基づき設計されたプライマーを使用して等温(60〜65℃)でインキュベーションし、増幅反応を行う方法であり、種々の遺伝子の検出等に用いられている。遺伝子の検出・定量は、増幅反応の副反応であるピロリン酸塩の白濁を目視あるいは濁度をリアルタイムで測定することにより行われる。
プライマーとしては、FIPプライマー、F3プライマー、BIPプライマー、B3プライマーからなる4種のプライマーを必要とするが、さらに、DNA合成の起点を増やすため、LPFプライマー、LPBプライマーを加えて合計6種のプライマーを用いる場合もあり、この方法によれば増幅効率は大幅に増大する。本発明においては、この6種のプライマーを使用し増幅効率を高めている。
【0013】
しかし、このLAMP法においては、遺伝子定量を行うためには、ピロリン酸塩の蓄積による濁度の増加をリアルタイムに測定するための専用装置が必要となる、遺伝子増幅において阻害物質の影響を受けると定量の正確性に欠ける等の欠点があり、本出願人等はこれを改良し、迅速、簡便かつ正確な、遺伝子の検出・定量法として、ABC(Alternately Binding Quenching Probe Conpetitive)-LAMP法を開発している。
この方法は、上記プライマーを用いて等温増幅を行う点ではLAMP法と共通するが、さらに末端を蛍光標識した核酸プローブと、標的遺伝子に対する競合的核酸を反応系に共存させる点でLAMP法とは異なり、標的遺伝子の定量は、上記等温増幅前後の蛍光強度を測定することにより、簡便、迅速かつ正確に行われる。
【0014】
本発明のプライマーセット中の各プライマー、核酸プローブ及び競合的核酸は、このABC-LAMP法を使用して、Roundup Ready Soybean系統のダイズにおける組換え遺伝子を検出・定量する際、最良の結果が得られるよう設計されたものである。
【0015】
〔プライマーセット〕
LAMP primerは富士通株式会社が提供するインターネットサイトNetlLaboratoryl(http://venus.netlaboratory.com/)内のLAMP primer設計支援ソフトPrimer Explorerを用いて設計した。しかし、この設計支援ソフトによる設計では、特にABC-LAMP法に用いるLAMP primerの設計において、ABC-LAMP法で用いるプローブの結合部位が考慮されない等の点で問題があることが判明した。
本発明の核酸プライマーは、これに以下の改良を加えたものである。
LAMP反応物において、本発明のプローブ(AB-QProbe)が結合可能な部位は、LAMP反応産物に特徴的に見られる一本鎖ループと考えられる。したがって、ABC-LAMPにおいてこの部位にAB-QPobeがアニーリングすることを考慮し、プライマーRRS-FIPは、F2とF1の間に約60 bpの間隔を持つよう設計した。
改良を加えて新たに設計された核酸プライマーの塩基配列は以下のとおりであり、これらをLAMP法、ABC-LAMP法に用いるプライマーセットとする。
【0016】
遺伝子組換えダイズRoundup Ready Soybean系統の組換え遺伝子を検出及び/又は定量するための核酸プライマー(5′→3′)の塩基配列配下のとおりである。
RRS-FIPプライマー:GTG GTC CCA AAG ATG GAG GAG ATA TCA CAT CAA TCC ACT TGC(配列番号1)
RRS-F3プライマー :GTC ATC CCT TAC GTC AGT(配列番号2)
RRS-LPFプライマー:CGT TCC AAC CAC GTC TTC AA(配列番号3)
RRS-BIPプライマー:TGT CGG CAG AGG CAT CTT TTA AAA TAA ACA TAG GGA ACC CAA(配列番号4)
RRS-B3プライマー :TCA AAA TAA GAT CAT ACA TAC AGG(配列番号5)
RRS-LPBプライマー:AGG AGC CAC CTT CCT TTT CC(配列番号6)
【0017】
〔核酸プローブ〕
本発明において設計された核酸プローブの塩基配列は以下に示される。この塩基配列は、標的遺伝子である35S promotor/plant junction region(配列番号8)の85番目のグアニンから始まる塩基配列に対応するよう設計したものであり、該プローブの5′末端側のシトシン塩基には、上記標的遺伝子中のグアニン塩基と近接して消光するBODIPY FL、FITC、TMRITC等の蛍光色素で蛍光標識しており、また、3末端はリン酸化されている。
【0018】
遺伝子組換えダイズRoundup Ready Soybean系統の組換え遺伝子を検出及び/又は定量するための核酸プローブ(5′→3′)の塩基配列は以下のとおりである。
ABQP-RRS:CTT TTT CCA CGT GCT CCT CGT GC(配列番号7)
【0019】
ABC-LAMP法に使用するプローブは、BODIPY FL等の蛍光色素で修飾されており、標的遺伝子にアニーリングした時に消光するように設計する。したがって、ABC-LAMP法においては、該プローブが標的遺伝子に結合した際、蛍光色素が修飾されている5′末端に対応する位置の標的遺伝子中の塩基がGになるように標的遺伝子中のターゲット配列を選定し、このターゲット配列に基づきプローブを設計する。また、AB-QProbeの蛍光色素で修飾されていない末端の塩基はCとし、ミスマッチにする。ABC-LAMP法では等温反応によって遺伝子増幅が起きるため、LAMP反応温度においてプローブがDNAに結合しなければならない。したがって、プローブはそのTm値が反応温度付近となるように設計する。本発明の上記プローブもこのようなABC-LAMP法のプローブ設計原則に従うものではあるが、さらに、本発明の上記プローブ(AB-QProbe)は、以下の改良を加えたものである。
【0020】
1)本発明においては、プローブ設計に際し,ターゲット配列におけるプローブ結合部位の3’末端の外側2塩基(プローブからみて蛍光色素のある側の外側2塩基)がグアニン塩基にならないように、ターゲット配列を選定している。
これまでのABC-LAMP法では,この点に考慮せず,外側2塩基にグアニン塩基を含んでいた。この位置にグアニン塩基があると、プローブが競合的核酸に結合した際にも,このグアニン塩基の影響で、蛍光が消光してしまう可能性がある。そのため,これまでのABC-LAMP法では,競合的核酸を作製する際には,このグアニン塩基をシトシン塩基に替えていた。このグアニン塩基のシトシン塩基への置換によりターゲット配列と競合的核酸の増幅効率に若干の違いが生じてしまう原因のひとつとなっていた。しかしながら,本Roundup Ready Soybean系統ダイズ検出用のプローブ設計では,外側がグアニン塩基でないよう考慮したため,この位置のグアニン塩基をシトシン塩基に替える必要がなくなり、競合的核酸作製における塩基置換数を最小限に抑えることができた。そのため、ターゲット配列と競合的核酸の増幅効率を可能な限り近づけることができた。
【0021】
2)また、プローブ設計に際し,Roundup Ready Soybean系統ダイズ検出用に設計したプローブのTm(解離温度)を、これまでのABC-LAMP法で設計したプローブのTmよりも高くなるように考慮した。これにより,ターゲット配列および競合的核酸に対してプローブの結合力がより強くなったため,2本鎖結合を安定化させることができた。そのため、安定した定量結果を得ることができるようになった。
【0022】
〔競合的核酸〕
本発明において設計された、遺伝子組換えダイズRoundup Ready Soybean系統の組換え遺伝子を検出及び/又は定量するための競合的核酸(5′→3′)の塩基配列(2本鎖DNAのうち一方のみ表示)と上記各プライマー及びプローブの各結合部位を以下に示す。なお、F1及びF2、並びにB1c及びB2cは、それぞれFIPプライマー及びBIPプライマーの競合的核酸における結合部位を表す。
【0023】
【表1】

(配列番号9)
【0024】
上記塩基配列は、Roundup Ready Soybean系統のダイズに導入されたグリホサート耐性遺伝子CP4EPSPSの、プロモーター領域(CaMV 35S)配列とダイズゲノムとの境界領域(35S promotor/plant junction region)(以下、ターゲット配列という。)における85番目のグアニン塩基をシトシンに代え、107番目のシトシンをグアニンに変更したものである。
以下に、上記CaMV 35S promororとダイズゲノムとの境界領域(35S promotor/plant junction region)の部分配列(2本鎖DNAのうち一方のみ表示)と該配列における各プライマー及びプローブの結合部位を以下に示す。なお、F1及びF2、並びにB1c及びB2cは、それぞれFIPプライマー及びBIPプライマーの該配列における結合部位を表す。
【0025】
【表2】
(配列番号8)
【0026】
本発明の競合的核酸において、107番目のシトシンをグアニンに変更した理由は、上記核酸プローブが、ターゲット配列と競合的核酸とに同じ結合力で結合できるようにするためである。すなわち競合的核酸の85番目の塩基はシトシンに変更されているため、上記核酸プローブの5’末端のシトシンとは結合せず、その分、核酸プローブと競合的核酸の結合力は、核酸プローブの5’末端側においてC−G結合する核酸プローブとターゲット配列の結合に比べ弱くなる。一方、核酸プローブの3’末端はシトシンであり、競合的核酸においては、対応位置の塩基がグアニンに変更されC−G結合できる。他方、ターゲット配列の対応位置の塩基はシトシンであるため核酸プローブの3’末端塩基とは結合できず、その結果、上記核酸プローブは標的遺伝子と競合的核酸とに、全体としてほぼ同じ結合力で結合することが可能となる。これにより、ABC-LAMP法により測定される蛍光強度は、ターゲット配列と競合核酸の比率に対応するものとなり、使用した競合的核酸の量と増幅反応前後の蛍光強度(相対蛍光強度)を測定することにより、ターゲット配列(グリホサート耐性遺伝子CP4EPSPSのプロモーター領域であるCaMV 35S promotorとダイズゲノムとの境界領域(35S promotor/plant junction region))の試料中に含まれる当初の量、すなわち、Roundup Ready Soybean系統ダイズにおける組換え遺伝子の量を知ることができる。
【0027】
さらに、上記したように、これまでのABC-LAMP法の競合的核酸設計では,ターゲット配列におけるプローブ結合部位の3’末端の外側2塩基(プローブからみて蛍光色素のある側の外側2塩基)にグアニン塩基を含む配列であったため,競合的核酸ではこの部分のグアニン塩基をシトシン塩基に置き替えたものを設計・合成していたが、本発明によれば、RRSの競合的核酸を設計するに際し、ターゲット配列としてプローブ結合部位の3’末端の外側2塩基にグアニン塩基を含まない配列を選定したため、競合的核酸の作製において,グアニン塩基をシトシン塩基に替える必要がなく,競合的核酸作製における塩基置換数を最小限に抑えることができ、そのため,ターゲット配列と競合的核酸の増幅効率を可能な限り近づけることが可能となった。
【0028】
なお、LAMP法あるいはABC-LAMP法による増幅対象は2本鎖DNAであるので、上記核酸プライマー、核酸プローブ、競合的核酸は、2本鎖DNAのうち何れかの1本鎖DNAと結合可能であればよく、これら核酸プライマー、核酸プローブ及び競合的核酸は上記具体的に示した配列のみならずこれらと相補の配列であってもよい。
【0029】
以下に、本発明の核酸プライマーセットを使用したLAMP法及び該核酸プライマー、核酸プローブ及び競合的核酸を使用したABC-LAMP法による、ターゲット配列の検出・定量法について詳述する。
LAMP法
ダイズあるいはダイズ加工食品からDNA試料を抽出し、該DNA試料、並びに本発明の上記6種の核酸プライマーからなるプライマーセット、Bst DNA Polymerase等の鎖置換型DNAポリメラーゼ、基質となる4種のヌクレオチド(dNTPs)及び反応バッファーからなる反応液を、例えば60〜65℃のうちの一定温度にインキュベートする。このとき、上記プライマーセットに対応するRoundup Ready Soybean系統のダイズにおける組換え遺伝子CP4EPSPSのプロモーターであるCaMV 35SがDNA試料中に存在すれば、該遺伝子中のターゲット配列を鋳型として等温増幅反応が生じ、ターゲット配列が増幅される。
この増幅反応においては、増幅反応の進行とともにピロリン酸塩が副生し、反応液が白濁するから、この白濁の有無を目視により観測することにより、上記の組換え遺伝子CaMV 35Sの有無を検知でき、Roundup Ready Soybean系統のダイズあるいは該ダイズを使用した加工食品に該ダイズ原料が混入していることを確認することができる。
また、この白濁の度合い(濁度)は、反応の進行とともに増大し、濁度が一定の値に達するまでの時間は、ほぼ初期の標的遺伝子の量に比例するから、濁度の経時変化をリアルタイムで測定し、一定時間後の濁度を測定することにより、初期の標的遺伝子量を求めることができ、これにより、ダイズあるいはダイズ加工食品中の組換え遺伝子CaMV 35Sを定量することができる。
【0030】
ABC-LAMP法
上記LAMP法で使用した、上記DNA試料、並びに本発明の上記6種の核酸プライマーからなるプライマーセット、Bst DNA Polymerase等の鎖置換型DNAポリメラーゼ、基質となる4種のヌクレオチド(dNTPs)及び反応バッファーからなる反応液にさらに、上記本発明の核酸プローブと競合的核酸とを各一定量を加えて等温増幅反応系を形成する。反応温度は上記LAMP法と同様である。この反応系においては、DNA試料中に組換え遺伝子CaMV 35Sが存在すれば、該遺伝子中のターゲット配列が競合的核酸ととともに等温増幅されるが、増幅効率はターゲット配列と競合的核酸とにおいて同じに設定されている。
上記核酸プローブがターゲット配列と結合した場合、核酸プローブの5’末端のシトシン塩基は、ターゲット配列中の対応位置にあるグアニン塩基と塩基対を形成し、核酸プローブの5’末端に結合した蛍光色素は、グアニン塩基と近接し消光する。一方、競合的核酸に核酸プローブがハイブリダイズする場合においては、核酸プローブの5’末端のシトシン塩基は、ターゲット配列中の対応位置にあるグアニン塩基でなくシトシン塩基であるから塩基対を形成せず、上記蛍光色素は競合的核酸配列中のグアニン塩基と離間し、蛍光を発し、消光しない。したがって、上記増幅反応系においては、標的核酸が増幅するにつれ、蛍光強度は減少する。
【0031】
一方、核酸プローブの量が、増幅されるターゲット配列と競合的核酸の合計量と同じかそれ以下になれば、核酸プローブの各分子はターゲット配列と競合的核酸のいずれかにハイブリダイズしている状態になり、また、上記したように、増幅効率は、ターゲット配列と競合的核酸とにおいて同じであり、かつ、本発明の核酸プローブはターゲット配列と競合的核酸とに同じ結合力で結合するから、核酸プローブがターゲット配列にハイブリダイズする割合は、増幅後の標的核酸と競合的核酸の割合に比例し、さらに、増幅された競合的核酸の量は、反応系において添加した当初の含有量に依存するから、該反応系の蛍光強度を反応前後で測定すれば、増幅前のターゲット配列の量、すなわち、Roundup Ready Soybean系統のダイズあるいは該ダイズを使用した加工食品中の組換え遺伝子CaMV 35Sの量を測定することができる。
【0032】
本発明においては、例えばダイズ加工食品中のRoundup Ready Soybean系統のダイズの混入率を算出するために必要なダイズ内在性遺伝子Le1を、LAMP法あるいはABC-LAMP法を用いて定量するための核酸プライマー、核酸プローブ及び競合的核酸及びこれを用いたLe1の定量法方法を包含する。
Roundup Ready Soybean系統のダイズの混入率を算出するためには内標比を求める必要があるが、内標比は以下の式で求められる。
【0033】
【数1】
この内標比は試験したRoundup Ready Soybean系統のダイズに固有の値である。したがって、このダイズの混入率は以下の式で算出できる。
【0034】
【数2】
このような混入率の算出に用いるLe1遺伝子の検知・定量法に用いる核酸プライマー、核酸プローブ、競合的核酸は以下に示すとおりであり、これらは上記Roundup Ready Soybean系統の組換え遺伝子の検出に用いる核酸プライマー、核酸プローブ及び競合的核酸と同様の設計思想に基づき設計されたものであり、上記と同様の改良点を有する。
【0035】
〔プライマーセット〕
Le1遺伝子の定量的検出に用いる核酸プライマーの塩基配列は以下のとおりであり、これらをLAMP法、ABC-LAMP法に用いるプライマーセットとする。
ダイズ由来のLe1遺伝子配列を検出及び/又は定量するための核酸プライマー(5′→3′)の塩基配列
Le1-FIP:AGC AAA AGA CCA AGA AAG CAC CTT CTC TTC CCG AGT GGG(配列番号12)
Le1-F3 :CCA GCA ATA TCC TCT CCG A(配列番号13)
Le1-LPF:GGC AGC AGA GAA CCC TAT CC(配列番号14)
Le1-BIP:TTT GCC ACA CGC TAG CAA TTA CAC TTT CTT CCT TCG ATC TGT(配列番号15)
Le1-B3 :TCA CTA GCG ATC GAG TAG TG(配列番号16)
Le1-LPB:GTT GCA TGA GGC CAT CTA AAT GT(配列番号17)
【0036】
〔核酸プローブ〕
本発明において設計された核酸プローブの塩基配列は以下に示される。この塩基配列は、標的遺伝子であるLe1遺伝子部分配列の168番目のグアニンから始まる塩基配列に対応するよう設計したものであり、該プローブの5’末端側のシトシン塩基には、上記標的遺伝子中のグアニン塩基と近接して消光するBODIPY FL、FITC、TMRITC等の蛍光色素で蛍光標識しており、また、3’末端はリン酸化されている。
ダイズ由来のLe1遺伝子配列を検出及び/又は定量するための核酸プローブ(5′→3′)の塩基配列
ABQP-Le1:CAT GCG ATT CCC CAG GTA TGT CC(配列番号18)
【0037】
〔競合的核酸〕
本発明において設計された競合的核酸の塩基配列(2本鎖DNAのうち一方のみ表示)と上記各プライマー及びプローブの各結合部位を以下に示す。なお、F1及びF2、並びにB1c及びB2cは、それぞれFIPプライマー及びBIPプライマーの競合的核酸における結合部位を表す。
【0038】
ダイズ由来のLe1遺伝子配列を検出及び/又は定量するための競合的核酸(5′→3′)の塩基配列
【表3】
(配列番号19)
【0039】
上記配列はダイズ由来のLe1遺伝子の部分配列における168番目のグアニン塩基をシトシンに代え、146番目のシトシンをグアニンに変更したものである。
これら、核酸プライマーセットを使用したLAMP法及び該核酸プライマー、核酸プローブ及び競合的核酸を使用したABC-LAMP法による、Le1遺伝子の検出・定量法については、上記したRoundup Ready Soybean系統のダイズにおける組換遺伝子の検出/定量法と同様である。
以下本発明の実施例を示すが、本発明はこれら実施例に限定されるものではない。
【実施例】
【0040】
実施例1
(1)LAMP法による組換え遺伝子の検出
以下のプライマーを用いて、62℃で1時間、LAMP反応を行い、遺伝子組換えダイズRoundup Ready Soybean系統の組換え遺伝子およびダイズ由来のLe1遺伝子配列を検出・定量した。
使用したRoundup Ready Soybean系統の組換え遺伝子及びLe1遺伝子の検出/定量用プライマーの塩基配列は以下のとおりである。
【0041】
Roundup Ready Soybean系統の組換え遺伝子用プライマー
RRS-FIPプライマー:GTG GTC CCA AAG ATG GAG GAG ATA TCA CAT CAA TCC ACT TGC(配列番号1)
RRS-F3プライマー :GTC ATC CCT TAC GTC AGT(配列番号2)
RRS-LPFプライマー:CGT TCC AAC CAC GTC TTC AA(配列番号3)
RRS-BIPプライマー:TGT CGG CAG AGG CAT CTT TTA AAA TAA ACA TAG GGA ACC CAA(配列番号4)
RRS-B3プライマー :TCA AAA TAA GAT CAT ACA TAC AGG(配列番号5)
RRS-LPBプライマー:AGG AGC CAC CTT CCT TTT CC(配列番号6)
【0042】
Le1用プライマー
Le1-FIP:AGC AAA AGA CCA AGA AAG CAC CTT CTC TTC CCG AGT GGG(配列番号12)
Le1-F3 :CCA GCA ATA TCC TCT CCG A(配列番号13)
Le1-LPF:GGC AGC AGA GAA CCC TAT CC(配列番号14)
Le1-BIP:TTT GCC ACA CGC TAG CAA TTA CAC TTT CTT CCT TCG ATC TGT(配列番号15)
Le1-B3 :TCA CTA GCG ATC GAG TAG TG(配列番号16)
Le1-LPB:GTT GCA TGA GGC CAT CTA AAT GT(配列番号17)
【0043】
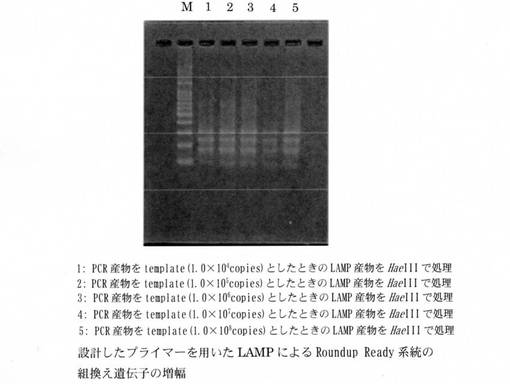
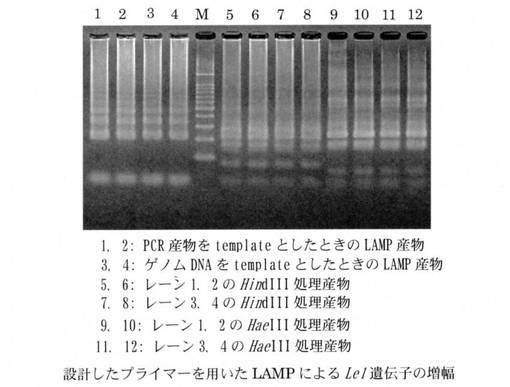
LAMP反応液の組成は、遺伝子組換えダイズRoundup Ready Soybean系統の組換え遺伝子の定量については、H2O 11.5μl、10×LAMP Buffer 2.5μl、dNTP mix(2.5 mM) 4.0μl、RRS-FIP(40μM) 1.0μl、RRS-BIP(40μM) 1.0μl、RRS-F3(5μM) 1.0μl、RRS-B3(5μM) 1.0μl、RRS-LPF(20μM) 1.0μl、RRS-LPB(20μM) 1.0μl、Template DNA 1.0μlとした。また、Le1遺伝子の定量については、H2O 11.5μl、10×LAMP Buffer 2.5μl、dNTP mix(2.5 mM) 4.0μl、Le1-FIP(40μM) 1.0μl、Le1-BIP(40μM) 1.0μl、Le1-F3(5μM) 1.0μl、Le1-B3(5μM) 1.0μl、Le1-LPF(20μM) 1.0μl、Le1-LPB(20μM) 1.0μl、Template DNA 1.0μlとした。Template DNAには標的遺伝子を含むPCR産物1×106copiesとダイズ粉末から抽出したゲノムDNA 50ngを用いた。LAMP反応終了後に37℃、4時間の制限酵素処理を行った。用いた制限酵素はHindIII、HaeIIIである。LAMP産物および制限酵素処理を行った溶液の5μlを用いてアガロースゲル電気泳動を行った。Roundup Ready Soybean系統の組換え遺伝子を増幅した結果を図1に、Le1遺伝子を増幅した結果を図2にしめす。Roundup Ready Soybean系統の組換え遺伝子、およびLe1遺伝子のどちらのLAMP産物に関しても、設計したプライマーによる特異的な増幅が確認された。
【0044】
(2)Real-time濁度測定法によるGMダイズの定量
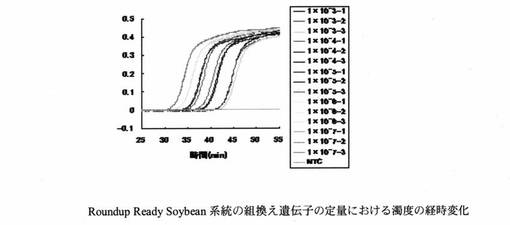
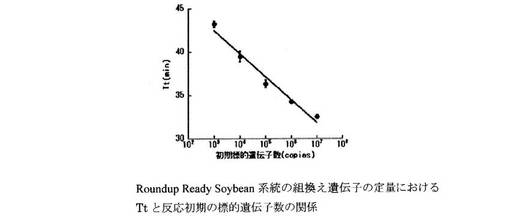
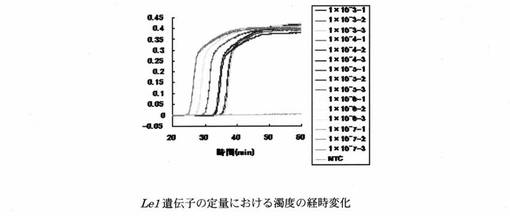
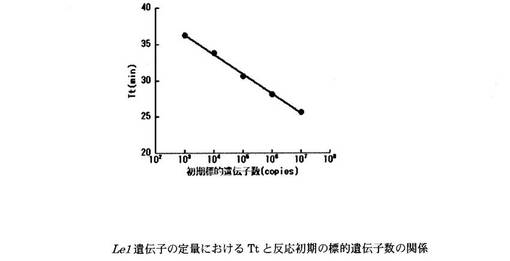
設計したプライマーセットを用いたReal-time濁度測定法により、Roundup Ready Soybean系統の組換え遺伝子とダイズ由来のLe1遺伝子の定量を行った。LAMP反応液の組成は、遺伝子組換えダイズRoundup Ready Soybean系統の組換え遺伝子の定量については、H2O 11.5μl、10×LAMP Buffer 2.5μl、dNTP mix(2.5 mM) 4.0μl、RRS-FIP(40μM) 1.0μl、RRS-BIP(40μM) 1.0μl、RRS-F3(5μM) 1.0μl、RRS-B3(5μM) 1.0μl、RRS-LPF(20μM) 1.0μl、RRS-LPB(20μM) 1.0μl、Template DNA 1.0μlとした。また、Le1遺伝子の定量については、H2O 11.5μl、10×LAMP Buffer 2.5μl、dNTP mix(2.5mM) 4.0μl、Le1-FIP(40μM) 1.0μl、Le1-BIP(40μM) 1.0μl、Le1-F3(5μM) 1.0μl、Le1-B3(5μM) 1.0μl、Le1-LPF(20μM) 1.0μl、Le1-LPB(20μM) 1.0μl、Template DNA 1.0μlとした。各標的遺伝子のPCR産物を103〜107 copies/μlに調製し、Template DNAとして用いた。遺伝子組換えダイズRoundup Ready Soybean系統の組換え遺伝子の定量における濁度の経時変化を図3に示す。また、濁度が一定の値に達するまでに要した時間(Threshold time:Tt)と反応初期の標的遺伝子数の関係を図4に示す。Le1遺伝子の定量における濁度の経時変化を図5に示す。また、濁度が一定の値に達するまでに要した時間(Threshold time:Tt)と反応初期の標的遺伝子数の関係を図6に示す。Le1遺伝子の定量においては、1x103 copies以上の濃度で定量が可能であった。
【0045】
なお、上記Le1の定量におけるTemplate DNAは、ダイズの抽出ゲノム(0.4ng/ul)を鋳型として用いて、1.0μl、H2O 12.2μl、10×PCR Buffer 2.0μl、dNTP mix(2.5mM) 2.0μl、Le1-F(10μM) 0.8μl、Le1-R(10μM) 0.8μl、Taq DNA polymerase 0.2 μlの反応液組成でPCRを行った反応産物を用いた。PCR反応条件は、94℃ 2分、94℃ 30秒、56℃30秒、72℃30秒を40サイクル、72℃2分で行った。RRSの定量におけるTemplateDNAは、プライマーにRRS-F(10μM)、RRS-R(10μM)を用いた以外は、Le1と同様の鋳型、反応液組成、反応条件でPCRを行い、その増幅産物を用いた。
【0046】
実施例2
Alternately Binding Probe Competitive (ABC)-LAMP法によるGMダイズの定量
(1)蛍光消光試験
本発明で設計した核酸プローブ、競合的核酸の蛍光消光の精度を評価するために、標的核酸と競合的核酸中の核酸プローブ結合部位を有する各2種の短いオリゴヌクレオチドを合成し、本発明の核酸プローブ、各2種のオリゴヌクレオチドを用いて、本発明におけるRoundup Ready Soy系およびLe1遺伝子の検出・定量系評価のための予備実験として、蛍光消光試験を行った。
【0047】
1)Roundup Ready Soy系の蛍光消光試験において使用したプローブ及びオリゴヌクレオチド
a)Roundup Ready Soybean系統の組換え遺伝子検出・定量用プローブ
ABQP-RRS:CTT TTT CCA CGT GCT CCT CGT GC(配列番号7)
(5’末端BODIPY FL標識、3’末端リン酸化)
b)上記a)のプローブ結合部位を有するオリゴヌクレオチド
RRS-TL(標的オリゴヌクレオチド):
CCC ACC CAC GAG GAG CAC GTG GAA AAA GAA GAC(配列番号10)、
RRS-CL(競合的オリゴヌクレオチドチド):
CCC ACG CAC GAG GAG CAC GTG GAA AAA CAA GAC(配列番号11)
【0048】
2)Le1遺伝子系の蛍光消光試験において使用したプローブ及びオリゴヌクレオチド
a)Le1遺伝子検出・定量用プローブ
ABQP-Le1:CAT GCG ATT CCC CAG GTA TGT CC(配列番号18)
(5’末端BODIPY FL標識、3’末端リン酸化)
b)上記c)のプローブ結合部位を有するオリゴヌクレオチド
Le1-TL(標的オリゴヌクレオチド):
GGA CTC GAC ATA CCT GGG GAA TCG CAT GAC GTG(配列番号20)
Le1-CL(競合的オリゴヌクレオチド):
GGA CTG GAC ATA CCT GGG GAA TCG CAT CAC GTG(配列番号21)
【0049】
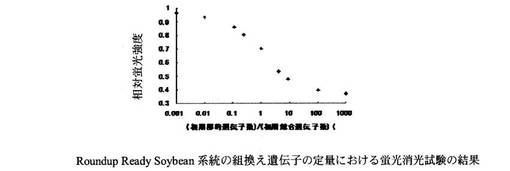
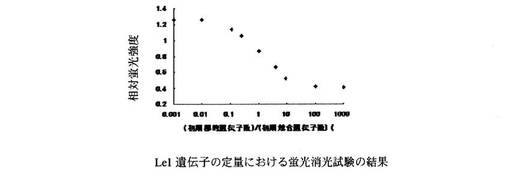
Roundup Ready Soybean系統の蛍光消光試験の反応液組成はH2O 1.5μl、10×LAMP Buffer 2.5μl、dNTP mix(2.5mM) 3.0μl、1% TritonX-100 2.5μl、Betaine(5M) 5.0μl、ABQP-RRS(7.5μM) 0.5μl、合成オリゴヌクレオチド(RRS-TLと-CLの混合物)10μlとした。また、Le1遺伝子系の蛍光消光試験の反応液組成はH2O 1.5μl、10×LAMP Buffer 2.5μl、dNTP mix(2.5mM) 3.0μl、1% TritonX-100 2.5μl、Betaine(5M) 5.0μl、ABQP-Le1(7.5μM) 0.5μl、合成オリゴヌクレオチド(Le1-TLとLe1-CLの混合物)10μlとした。Roundup Ready Soybean系統の蛍光消光試験の結果を図7に、Le1遺伝子系の蛍光消光試験の結果を図8に示す。両蛍光消光試験の結果とも標的核酸と競合核酸の比率に応じた相対蛍光強度を示すことがわかった。
【0050】
(2)本発明で設計した核酸プライマー、核酸プローブ、競合的核酸を用いて、ABC-LAMP法による標的遺伝子定量のための検量線の作製を行った。
核酸プライマーは実施例1で使用したものを用い、核酸プローブは上記(1)において使用したものを用いた。また競合的核酸は、Roundup Ready Soybean系統の組換え遺伝子の定量において配列番号9に示される競合的核酸、Le1遺伝子の定量において配列番号19に示される競合的核酸をそれぞれ用いた。
反応液の組成は、遺伝子組換えダイズRoundup Ready Soybean系統の組換え遺伝子の定量については、H2O 6.5μl、10×LAMP Buffer 2.5μl、dNTP mix(2.5 mM) 4.0μl、RRS-FIP(40μM) 1.0μl、RRS-BIP(40μM) 1.0μl、RRS-F3(5μM) 1.0μl、RRS-B3(5μM) 1.0μl、RRS-LPF(20μM) 1.0μl、RRS-LPB(20μM) 1.0μl、1% TritonX-100 2.5μl、ABQP-RRS(7.5μM) 0.5μlとし、さらに競合遺伝子数を104copies/μl、標的遺伝子数を103-107copies/μlに調製し、それらを1.0μlずつ添加した。
【0051】
また、Le1遺伝子の定量については、H2O 6.5μl、10×LAMP Buffer 2.5μl、dNTP mix(2.5mM) 4.0μl、Le1-FIP(40μM) 1.0μl、Le1-BIP(40μM) 1.0μl、Le1-F3(40μM) 1.0μl、Le1-B3(40μM) 1.0μl、Le1-LPF(40μM) 1.0μl、Le1-LPB(40μM) 1.0μl、1% TritonX-100 2.5μl、ABQP-Le1(7.5μM) 0.5μlに、さらに、競合遺伝子数を104copies/μl、標的遺伝子数を103-107copies/μlに調製し、それらを1.0μlずつ添加して反応溶液とした。
反応条件は両遺伝子ともに、62℃で1時間とした。
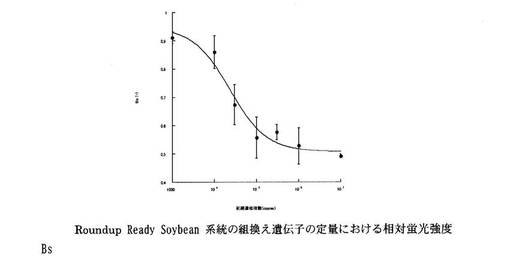
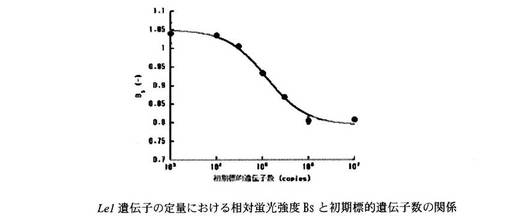
Roundup Ready Soybean系統の組換え遺伝子の定量における相対蛍光強度Bsと初期標的遺伝子数の関係を図9に示す。同様に、Le1遺伝子の定量における相対蛍光強度Bsと初期標的遺伝子数の関係を図10に示す。
両遺伝子の結果とも標的核酸と競合核酸の比率に応じた相対蛍光強度を示すことがわかった。
【図面の簡単な説明】
【0052】
【図1】本発明のRoundup Ready Soybean系統ダイズおける組換え遺伝子増幅用プライマーを用いてLAMPにより該組換え遺伝子を増幅した結果を示す電気泳動写真である。
【図2】本発明のダイズ由来のLe1遺伝子増幅用プライマーを用いてLAMPにより該組換え遺伝子を増幅した結果を示す電気泳動写真である。
【図3】本発明のRoundup Ready Soybean系統ダイズにおける組換え遺伝子増幅用プライマーを用いてリアルタイムLAMP法を行った結果、観察された濁度の経時変化を示すグラフである。
【図4】上記濁度法により得られた、Roundup Ready系統ダイズおける組換え遺伝子についての、Ttと反応初期の標的遺伝子数の関係を示すグラフである。
【図5】本発明のLe1遺伝子増幅用プライマーを用いてリアルタイムLAMP法を行った結果、観察された濁度の経時変化を示すグラフである。
【図6】上記濁度法により得られた、Le1遺伝子についてのTtと反応初期の標的遺伝子数の関係を示すグラフである。
【図7】Roundup Ready Soybean系統の組換え遺伝子の定量に用いる本発明のプローブ、競合的核酸の一部配列を用いて蛍光消光試験を行った結果を示すグラフである。
【図8】Le1遺伝子の定量に用いる本発明のプローブ、競合的核酸の一部配列を用いて蛍光消光試験を行った結果を示すグラフである。
【図9】Roundup Ready Soybean系統の組換え遺伝子、並びに本発明のプライマー、プローブ及び競合的核酸を用いてABC-LAMP法を行いLAMP反応前後の蛍光強度を測定した結果得られた、相対蛍光強度BsとRoundup Ready Soybean系統の組換え遺伝子の初期標的遺伝子数との関係を示すグラフである。
【図10】Le1遺伝子、並びに本発明のプライマー、プローブ及び競合的核酸を用いてABC-LAMP法を行い、LAMP反応前後の蛍光強度を測定した結果得られた、相対蛍光強度BsとLe1遺伝子の初期標的遺伝子数の関係を示すグラフである。
【技術分野】
【0001】
本発明は、ダイズもしくはダイズ加工食品に含まれる遺伝子組換え体の定量的検知方法及び当該方法に用いられる核酸分子に関する。より詳細には、本発明は、ダイズもしくはダイズ加工食品に含まれる遺伝子組換体を、LAMP法をあるいはABC-LAMP法を用いて定量的に検知する方法、並びに当該方法に用いられる核酸プライマー、核酸プローブ及び競合的核酸等の各種核酸分子に関する。
【背景技術】
【0002】
世界的に、遺伝子組換え技術を利用して開発された農作物の実用化が進んでいる。我が国においても、ダイズ、トウモロコシ等、59品種の作物について遺伝子組換え体が認められている。実際に流通している遺伝子組換え作物として、例えば、トウモロコシについてはBt11系統の後代品種(Novartis社)、Event176系統の後代品種(Novartis社)、MON810系統の後代品種(Monsanto社)、GA21系統の後代品種(Monsanto社)、T25系統の後代品種(Aventis社)が、またダイズについてはRoundup Ready Soybean系統の後代品種(Monsanto社)等を挙げることができる。
遺伝子組換え作物は、天然の作物に、害虫抵抗性や除草剤耐性等の産業上好ましい形質を付与する目的で開発される。その主な手法は、元来そのような産業上好ましい形質を有する他の生物から、当該形質を発現する遺伝子を単離し、その遺伝子を対象作物に発現可能な形で導入するというものである。従って、遺伝子組換え作物のDNAの中には、このような導入遺伝子が含まれている。
【0003】
欧州共同体(EU)により遺伝子組換え作物及びその加工食品の表示に関わる規則(非特許文献1)が定められ、日本に於いても遺伝子組換え作物及びその加工食品の表示に関する制度が定められたことを契機として(非特許文献2)、食品業界及びそれに関連する業界では、食品原料農作物や食品中の組換えDNAの存否、またはその含量を把握し認識しておくことが求められている。このため、食品、飼料及びこれらの原料農作物中の遺伝子組換え体の有無や含量、特に原料中の存在比を確認する技術が必要とされている。
【0004】
遺伝子組換え体を検知する技術としては、例えば、ポリメラーゼ連鎖反応(以下「PCR」という)によって該組換え遺伝子を増幅し、これを検出する方法(例えば、非特許文献3);競合PCRによる組換えダイズやトウモロコシの定量的検知技術(例えば、非特許文献4,5,6及び7);蛍光プローブを用いた定量的PCR(リアルタイムPCR)による組換えダイズやトウモロコシの定量的検知技術(例えば、非特許文献8、9及び10);組換えDNAから生産される蛋白質をEnzyme Linked Immuno Sorvent Assay (ELISA)法を用いて測定することによって、遺伝子組換え体の含量を定量する方法(例えば、非特許文献11)を挙げることができる。また、最近では、定量PCR法を用いて、複数の遺伝子組換え体の系統を含む集団中における遺伝子組換え体の存在比を正確に定量する方法として、内部標準を用いた方法(特許文献1及び2)が確立され、特に前者の方法(特許文献1)は、日本国において遺伝子組換え食品の検査分析の標準法として採用されている。当該方法は作物中の内在性遺伝子と組換え遺伝子のPCR法によるコピー数を測定し、その比から遺伝子組換え遺伝子の混入量を換算し定量する方法である。
【0005】
上記標準法として定められた従来の定量的PCR法は増幅と検出に1時間以上必要であり、また、PCRを行うためのサーマルサイクラーと蛍光検出装置が一体化した高価なリアルタイムPCR用の装置が必要であるといった問題があった。そこで、迅速・簡便かつ低コストで組換え遺伝子を検知するための方法の開発が望まれていた。
【非特許文献1】Regulation (EU) No.EC/258/97, Council Regulation(EC), No.1139/98
【非特許文献2】Notification No. 1775 (June 10, 2000) Food and Marketing Bureau,Ministry of Agriculture, Forestry and Fisheries of Japan, Tokyo, Japan
【非特許文献3】Hupfer C., Mayer J., Hotzel H., Sachse K., Engel K.H., Eur. FoodRes. Technol., 209, 301-304(1999)
【非特許文献4】Hardegger M., Brodmann P., Herrmann A., Eur. Food Res. Technol., 209,83-87(1999)
【非特許文献5】Hubner P., Studer E., Luthy J., Food Control, 10, 353-358(1999)
【非特許文献6】Hubner P., Studer E., Luthy J., Nat. Biotechnol., 17, 1137-1138(1999)
【非特許文献7】Hupfer C., Hotzel H., Sachse K., Moreano F., Engel K. H., Eur. FoodRes. Technol., 212, 95-99(2000)
【非特許文献8】Vaitilingom M., Pijnenburg H., Gendre F., Brignon P., J. Agric. FoodChem., 47, 5261-5266(1999)
【非特許文献9】Wurz A., Bluth A., Zelts P., Pfeifer C., Willmund R., Food Control, 10,385-389(1999)
【非特許文献10】Berdal K. G., Holst-Jensen A., Eur. Food Res. Technol., 213, 432-438(2001)
【非特許文献11】Luthy J., Food Control, 10, 359-361(1999)
【特許文献1】WO 02/34943
【特許文献2】特開2001-136983号公報
【発明の開示】
【発明が解決しようとする課題】
【0006】
本発明の課題は、上記従来法の問題を解消した改良法として、LAMP法あるいはABC-LAMP法に着目し、ダイズもしくはダイズ加工食品に含まれる遺伝子組換え体の存在を定量的に検知するための新規な方法並びに該方法に好適な核酸分子(プライマー、プローブ、競合遺伝子)を提供することであり、特に上記PCR法による遺伝子組換え体の検出法のようなサーマルサイクラーと蛍光検出装置が一体化した高価なリアルタイムPCR用の装置を必要とせず、迅速・簡便かつ低コストで、正確な組換え遺伝子の検知手段を提供する点にある。
【課題を解決するための手段】
【0007】
発明者等は、鋭意研究の結果、遺伝子組換えダイズとして、Roundup Ready Soybean系統の後代品種中の組換え遺伝子を、LAMP法あるいはABC-LAMP法を用いて検出するための、プローブ及び競合遺伝子を新たに設計するとともに、これらを用いた上記組換え遺伝子の検知・定量法を開発して、本発明を完成するに至ったものである。すなわち本発明は以下に示されるとおりである。
【0008】
(1)遺伝子組換えダイズRoundup Ready Soybean系統の組換遺伝子をLAMP法あるいはABC-LAMP法によって検出・定量するために用いる、FIPプライマー、F3プライマー、LPFプライマー、BIPプライマー、B3プライマー及びLPBプライマーからなるプライマーセットであって、各プライマーが以下に示される塩基配列、又は該配列と相補の塩基配列を有することを特徴とする、上記プライマーセット。
FIPプライマー:GTG GTC CCA AAG ATG GAG GAG ATA TCA CAT CAA TCC ACT TGC
F3プライマー :GTC ATC CCT TAC GTC AGT
LPFプライマー:CGT TCC AAC CAC GTC TTC AA
BIPプライマー:TGT CGG CAG AGG CAT CTT TTA AAA TAA ACA TAG GGA ACC CAA
B3プライマー :TCA AAA TAA GAT CAT ACA TAC AGG
LPBプライマー:AGG AGC CAC CTT CCT TTT CC
(2)遺伝子組換えダイズRoundup Ready Soybean系統の組換え遺伝子をABC-LAMP法によって検出・定量するために用いる、配列番号9に示される塩基配列又は該配列と相補の塩基配列からなる競合的核酸。
(3)遺伝子組換えダイズRoundup Ready Soybean系統の組換え遺伝子をABC-LAMP法によって検出・定量するために用いる核酸プローブであって、配列番号7に示される塩基配列又は該配列と相補の塩基配列からなり、その末端に核酸中のグアニン塩基との近接、離間によって蛍光強度が変化する蛍光色素を標識されていることを特徴とする上記核酸プローブ。
(4)ダイズ由来のLe1遺伝子配列をLAMP法又はABC-LAMP法によって検出・定量するために用いる、FIPプライマー、F3プライマー、LPFプライマー、BIPプライマー、B3プライマー及びLPBプライマーからなるプライマーセットであって、各プライマーが以下に示される塩基配列、又は該配列と相補の塩基配列を有することを特徴とする、上記プライマーセット。
FIPプライマー:AGC AAA AGA CCA AGA AAG CAC CTT CTC TTC CCG AGT GGG
F3プライマー :CCA GCA ATA TCC TCT CCG A
LPFプライマー:GGC AGC AGA GAA CCC TAT CC
BIPプライマー:TTT GCC ACA CGC TAG CAA TTA CAC TTT CTT CCT TCG ATC TGT
B3プライマー :TCA CTA GCG ATC GAG TAG TG
LPBプライマー:GTT GCA TGA GGC CAT CTA AAT GT
(5)ダイズ由来のLe1遺伝子配列をABC-LAMP法によって検出・定量するために用いる、配列番号19に示される塩基配列又は該配列と相補の塩基配列からなる競合的核酸。
(6)ダイズ由来のLe1遺伝子配列をABC-LAMP法によって検出及び/又は定量するために用いる核酸プローブであって、配列番号18に示される塩基配列又は該配列と相補の塩基配列からなり、その末端に核酸中のグアニン塩基との近接、離間によって蛍光強度が変化する蛍光色素を標識されていることを特徴とする上記核酸プローブ。
(7)披検ダイズ試料に含まれるRoundup Ready Soybean系統の組換遺伝子配列を、ABC-LAMP法を用いて検出及び/又は定量するための方法であって、被験ダイズ試料から抽出した核酸、上記(1)に記載のプライマーセット、上記(2)に記載の競合的核酸及び上記(3)に記載の核酸プローブの存在下、等温増幅反応を行い、該反応前後の蛍光強度を測定することを特徴とする、Roundup Ready Soybean系統の組換遺伝子配列の検出及び/又は定量方法。
(8)ダイズ由来のLe1遺伝子配列を、ABC-LAMP法を用いて検出及び/又は定量するための方法であって、被験ダイズ試料から抽出した核酸、上記(4)に記載のプライマーセット、上記(5)に記載の競合的核酸及び上記(6)に記載の核酸プローブの存在下、等温増幅反応を行い、該反応前後の蛍光強度を測定することを特徴とする、上記ダイズ由来のLe1遺伝子配列の検出及び/又は定量方法。
(9)上記(1)に記載のプライマーセット、上記(2)に記載の競合的核酸及び上記(3)に記載の核酸プローブを少なくとも組み合わせたことを特徴とする、Roundup Ready Soybean系統の組換遺伝子の検出及び/又は定量用試薬キット。
(10)上記(4)に記載のプライマーセット、上記(5)に記載の競合的核酸及び上記(6)に記載の核酸プローブを少なくとも組み合わせたことを特徴とする、ダイズ由来のLe1遺伝子配列の検出又は定量用試薬キット。
【発明の効果】
【0009】
本発明によれば、ダイズあるいはダイズ加工食品中のRoundup Ready Soybean系統の組換え遺伝子をABC-LAMP法を用いて検出及び/又は定量するために極めて好適な核酸プライマー、核酸プローブ、競合的核酸が提供でき、これらを用いてABC-LAMP法を行うことにより、簡便、かつ低コストでしかも正確に上記組換え遺伝子を検出及び又は定量可能となる。また、上記核酸プライマーはLAMP法においても有用なプライマーである。一方、本発明においては、同時にダイズ由来のLe1遺伝子配列をABC-LAMP法を用いて検出及び/又は定量するために極めて好適な核酸プライマー、核酸プローブ、競合的核酸も提供する。当然、この核酸プローブもLAMP法において有用なプライマーである。これらを用いてLAMP法あるいはABC-LAMP法を行った結果得られたLe1遺伝子の定量値は、内部標準遺伝子の定量値として、上記Roundup Ready Soybean系統の組換遺伝子の定量値を除することによって、内標比を算出し、この内標比を用いて、例えば、組換え食品中の組換えダイズの混入率を求めることが可能となる。
【発明を実施するための最良の形態】
【0010】
Roundup Ready Soybean系統のダイズは、ダイズにグリホサート系除草剤に耐性を付与する遺伝子を導入したものであり、グリホサート系除草剤は広範な種類の雑草に効力を有するが、Roundup Ready Soybean系統のダイズは、この除草剤に対して耐性を有するため、該ダイズ栽培中生育してくる種々の雑草に対し、その種類によらず、グリホサート系除草剤で除草することができ、除草の手間等を大幅に軽減できる。しかし、食品としての安全性等の面で疑問視する声もある。上記耐性付与遺伝子は、土壌細菌Agrobacterium属由来の耐性遺伝子CP4EPSPSである。Roundup Ready Soybean系統のダイズは、このCP4EPSPS遺伝子の発現にCaMV 35S プロモーターを用いており、本発明においては、このプロモーター領域とダイズゲノムとの境界領域配列(35S promotor/plant junction region, AJ308514)をRoundup Ready Soybean系統検出のターゲットとしている。
【0011】
35S promotor/plant junction region (AJ308514)の塩基配列(配列番号8)
gatagtggga ttgtgcgtca tcccttacgt cagtggagat atcacatcaa
tccacttgct ttgaagacgt ggttggaacg tcttcttttt ccacgtgctc
ctcgtgggtg ggggtccatc tttgggacca ctgtcggcag aggcatcttc
aacgatggcc tttcctttat cgcaatgatg gcatttgtag gagccacctt
ccttttccat ttgggttccc tatgtttatt ttaacctgta tgtatgatct
tattttgaat gaaatgcaat aagttatttc tagtaaaaaa aaataaacat
ttgatagaaa caaattaaag catgcaaaaa taactcatta gcatcggtta
aattgaaggg tttgaataat ttgcacaagg ttctgaattc
【0012】
一方、LAMP(Loop-Mediated Isothermal AMPlification)法は、遺伝子の簡易・迅速な増幅法として知られており、標的遺伝子の塩基配列に基づき設計されたプライマーを使用して等温(60〜65℃)でインキュベーションし、増幅反応を行う方法であり、種々の遺伝子の検出等に用いられている。遺伝子の検出・定量は、増幅反応の副反応であるピロリン酸塩の白濁を目視あるいは濁度をリアルタイムで測定することにより行われる。
プライマーとしては、FIPプライマー、F3プライマー、BIPプライマー、B3プライマーからなる4種のプライマーを必要とするが、さらに、DNA合成の起点を増やすため、LPFプライマー、LPBプライマーを加えて合計6種のプライマーを用いる場合もあり、この方法によれば増幅効率は大幅に増大する。本発明においては、この6種のプライマーを使用し増幅効率を高めている。
【0013】
しかし、このLAMP法においては、遺伝子定量を行うためには、ピロリン酸塩の蓄積による濁度の増加をリアルタイムに測定するための専用装置が必要となる、遺伝子増幅において阻害物質の影響を受けると定量の正確性に欠ける等の欠点があり、本出願人等はこれを改良し、迅速、簡便かつ正確な、遺伝子の検出・定量法として、ABC(Alternately Binding Quenching Probe Conpetitive)-LAMP法を開発している。
この方法は、上記プライマーを用いて等温増幅を行う点ではLAMP法と共通するが、さらに末端を蛍光標識した核酸プローブと、標的遺伝子に対する競合的核酸を反応系に共存させる点でLAMP法とは異なり、標的遺伝子の定量は、上記等温増幅前後の蛍光強度を測定することにより、簡便、迅速かつ正確に行われる。
【0014】
本発明のプライマーセット中の各プライマー、核酸プローブ及び競合的核酸は、このABC-LAMP法を使用して、Roundup Ready Soybean系統のダイズにおける組換え遺伝子を検出・定量する際、最良の結果が得られるよう設計されたものである。
【0015】
〔プライマーセット〕
LAMP primerは富士通株式会社が提供するインターネットサイトNetlLaboratoryl(http://venus.netlaboratory.com/)内のLAMP primer設計支援ソフトPrimer Explorerを用いて設計した。しかし、この設計支援ソフトによる設計では、特にABC-LAMP法に用いるLAMP primerの設計において、ABC-LAMP法で用いるプローブの結合部位が考慮されない等の点で問題があることが判明した。
本発明の核酸プライマーは、これに以下の改良を加えたものである。
LAMP反応物において、本発明のプローブ(AB-QProbe)が結合可能な部位は、LAMP反応産物に特徴的に見られる一本鎖ループと考えられる。したがって、ABC-LAMPにおいてこの部位にAB-QPobeがアニーリングすることを考慮し、プライマーRRS-FIPは、F2とF1の間に約60 bpの間隔を持つよう設計した。
改良を加えて新たに設計された核酸プライマーの塩基配列は以下のとおりであり、これらをLAMP法、ABC-LAMP法に用いるプライマーセットとする。
【0016】
遺伝子組換えダイズRoundup Ready Soybean系統の組換え遺伝子を検出及び/又は定量するための核酸プライマー(5′→3′)の塩基配列配下のとおりである。
RRS-FIPプライマー:GTG GTC CCA AAG ATG GAG GAG ATA TCA CAT CAA TCC ACT TGC(配列番号1)
RRS-F3プライマー :GTC ATC CCT TAC GTC AGT(配列番号2)
RRS-LPFプライマー:CGT TCC AAC CAC GTC TTC AA(配列番号3)
RRS-BIPプライマー:TGT CGG CAG AGG CAT CTT TTA AAA TAA ACA TAG GGA ACC CAA(配列番号4)
RRS-B3プライマー :TCA AAA TAA GAT CAT ACA TAC AGG(配列番号5)
RRS-LPBプライマー:AGG AGC CAC CTT CCT TTT CC(配列番号6)
【0017】
〔核酸プローブ〕
本発明において設計された核酸プローブの塩基配列は以下に示される。この塩基配列は、標的遺伝子である35S promotor/plant junction region(配列番号8)の85番目のグアニンから始まる塩基配列に対応するよう設計したものであり、該プローブの5′末端側のシトシン塩基には、上記標的遺伝子中のグアニン塩基と近接して消光するBODIPY FL、FITC、TMRITC等の蛍光色素で蛍光標識しており、また、3末端はリン酸化されている。
【0018】
遺伝子組換えダイズRoundup Ready Soybean系統の組換え遺伝子を検出及び/又は定量するための核酸プローブ(5′→3′)の塩基配列は以下のとおりである。
ABQP-RRS:CTT TTT CCA CGT GCT CCT CGT GC(配列番号7)
【0019】
ABC-LAMP法に使用するプローブは、BODIPY FL等の蛍光色素で修飾されており、標的遺伝子にアニーリングした時に消光するように設計する。したがって、ABC-LAMP法においては、該プローブが標的遺伝子に結合した際、蛍光色素が修飾されている5′末端に対応する位置の標的遺伝子中の塩基がGになるように標的遺伝子中のターゲット配列を選定し、このターゲット配列に基づきプローブを設計する。また、AB-QProbeの蛍光色素で修飾されていない末端の塩基はCとし、ミスマッチにする。ABC-LAMP法では等温反応によって遺伝子増幅が起きるため、LAMP反応温度においてプローブがDNAに結合しなければならない。したがって、プローブはそのTm値が反応温度付近となるように設計する。本発明の上記プローブもこのようなABC-LAMP法のプローブ設計原則に従うものではあるが、さらに、本発明の上記プローブ(AB-QProbe)は、以下の改良を加えたものである。
【0020】
1)本発明においては、プローブ設計に際し,ターゲット配列におけるプローブ結合部位の3’末端の外側2塩基(プローブからみて蛍光色素のある側の外側2塩基)がグアニン塩基にならないように、ターゲット配列を選定している。
これまでのABC-LAMP法では,この点に考慮せず,外側2塩基にグアニン塩基を含んでいた。この位置にグアニン塩基があると、プローブが競合的核酸に結合した際にも,このグアニン塩基の影響で、蛍光が消光してしまう可能性がある。そのため,これまでのABC-LAMP法では,競合的核酸を作製する際には,このグアニン塩基をシトシン塩基に替えていた。このグアニン塩基のシトシン塩基への置換によりターゲット配列と競合的核酸の増幅効率に若干の違いが生じてしまう原因のひとつとなっていた。しかしながら,本Roundup Ready Soybean系統ダイズ検出用のプローブ設計では,外側がグアニン塩基でないよう考慮したため,この位置のグアニン塩基をシトシン塩基に替える必要がなくなり、競合的核酸作製における塩基置換数を最小限に抑えることができた。そのため、ターゲット配列と競合的核酸の増幅効率を可能な限り近づけることができた。
【0021】
2)また、プローブ設計に際し,Roundup Ready Soybean系統ダイズ検出用に設計したプローブのTm(解離温度)を、これまでのABC-LAMP法で設計したプローブのTmよりも高くなるように考慮した。これにより,ターゲット配列および競合的核酸に対してプローブの結合力がより強くなったため,2本鎖結合を安定化させることができた。そのため、安定した定量結果を得ることができるようになった。
【0022】
〔競合的核酸〕
本発明において設計された、遺伝子組換えダイズRoundup Ready Soybean系統の組換え遺伝子を検出及び/又は定量するための競合的核酸(5′→3′)の塩基配列(2本鎖DNAのうち一方のみ表示)と上記各プライマー及びプローブの各結合部位を以下に示す。なお、F1及びF2、並びにB1c及びB2cは、それぞれFIPプライマー及びBIPプライマーの競合的核酸における結合部位を表す。
【0023】
【表1】
(配列番号9)
【0024】
上記塩基配列は、Roundup Ready Soybean系統のダイズに導入されたグリホサート耐性遺伝子CP4EPSPSの、プロモーター領域(CaMV 35S)配列とダイズゲノムとの境界領域(35S promotor/plant junction region)(以下、ターゲット配列という。)における85番目のグアニン塩基をシトシンに代え、107番目のシトシンをグアニンに変更したものである。
以下に、上記CaMV 35S promororとダイズゲノムとの境界領域(35S promotor/plant junction region)の部分配列(2本鎖DNAのうち一方のみ表示)と該配列における各プライマー及びプローブの結合部位を以下に示す。なお、F1及びF2、並びにB1c及びB2cは、それぞれFIPプライマー及びBIPプライマーの該配列における結合部位を表す。
【0025】
【表2】
(配列番号8)
【0026】
本発明の競合的核酸において、107番目のシトシンをグアニンに変更した理由は、上記核酸プローブが、ターゲット配列と競合的核酸とに同じ結合力で結合できるようにするためである。すなわち競合的核酸の85番目の塩基はシトシンに変更されているため、上記核酸プローブの5’末端のシトシンとは結合せず、その分、核酸プローブと競合的核酸の結合力は、核酸プローブの5’末端側においてC−G結合する核酸プローブとターゲット配列の結合に比べ弱くなる。一方、核酸プローブの3’末端はシトシンであり、競合的核酸においては、対応位置の塩基がグアニンに変更されC−G結合できる。他方、ターゲット配列の対応位置の塩基はシトシンであるため核酸プローブの3’末端塩基とは結合できず、その結果、上記核酸プローブは標的遺伝子と競合的核酸とに、全体としてほぼ同じ結合力で結合することが可能となる。これにより、ABC-LAMP法により測定される蛍光強度は、ターゲット配列と競合核酸の比率に対応するものとなり、使用した競合的核酸の量と増幅反応前後の蛍光強度(相対蛍光強度)を測定することにより、ターゲット配列(グリホサート耐性遺伝子CP4EPSPSのプロモーター領域であるCaMV 35S promotorとダイズゲノムとの境界領域(35S promotor/plant junction region))の試料中に含まれる当初の量、すなわち、Roundup Ready Soybean系統ダイズにおける組換え遺伝子の量を知ることができる。
【0027】
さらに、上記したように、これまでのABC-LAMP法の競合的核酸設計では,ターゲット配列におけるプローブ結合部位の3’末端の外側2塩基(プローブからみて蛍光色素のある側の外側2塩基)にグアニン塩基を含む配列であったため,競合的核酸ではこの部分のグアニン塩基をシトシン塩基に置き替えたものを設計・合成していたが、本発明によれば、RRSの競合的核酸を設計するに際し、ターゲット配列としてプローブ結合部位の3’末端の外側2塩基にグアニン塩基を含まない配列を選定したため、競合的核酸の作製において,グアニン塩基をシトシン塩基に替える必要がなく,競合的核酸作製における塩基置換数を最小限に抑えることができ、そのため,ターゲット配列と競合的核酸の増幅効率を可能な限り近づけることが可能となった。
【0028】
なお、LAMP法あるいはABC-LAMP法による増幅対象は2本鎖DNAであるので、上記核酸プライマー、核酸プローブ、競合的核酸は、2本鎖DNAのうち何れかの1本鎖DNAと結合可能であればよく、これら核酸プライマー、核酸プローブ及び競合的核酸は上記具体的に示した配列のみならずこれらと相補の配列であってもよい。
【0029】
以下に、本発明の核酸プライマーセットを使用したLAMP法及び該核酸プライマー、核酸プローブ及び競合的核酸を使用したABC-LAMP法による、ターゲット配列の検出・定量法について詳述する。
LAMP法
ダイズあるいはダイズ加工食品からDNA試料を抽出し、該DNA試料、並びに本発明の上記6種の核酸プライマーからなるプライマーセット、Bst DNA Polymerase等の鎖置換型DNAポリメラーゼ、基質となる4種のヌクレオチド(dNTPs)及び反応バッファーからなる反応液を、例えば60〜65℃のうちの一定温度にインキュベートする。このとき、上記プライマーセットに対応するRoundup Ready Soybean系統のダイズにおける組換え遺伝子CP4EPSPSのプロモーターであるCaMV 35SがDNA試料中に存在すれば、該遺伝子中のターゲット配列を鋳型として等温増幅反応が生じ、ターゲット配列が増幅される。
この増幅反応においては、増幅反応の進行とともにピロリン酸塩が副生し、反応液が白濁するから、この白濁の有無を目視により観測することにより、上記の組換え遺伝子CaMV 35Sの有無を検知でき、Roundup Ready Soybean系統のダイズあるいは該ダイズを使用した加工食品に該ダイズ原料が混入していることを確認することができる。
また、この白濁の度合い(濁度)は、反応の進行とともに増大し、濁度が一定の値に達するまでの時間は、ほぼ初期の標的遺伝子の量に比例するから、濁度の経時変化をリアルタイムで測定し、一定時間後の濁度を測定することにより、初期の標的遺伝子量を求めることができ、これにより、ダイズあるいはダイズ加工食品中の組換え遺伝子CaMV 35Sを定量することができる。
【0030】
ABC-LAMP法
上記LAMP法で使用した、上記DNA試料、並びに本発明の上記6種の核酸プライマーからなるプライマーセット、Bst DNA Polymerase等の鎖置換型DNAポリメラーゼ、基質となる4種のヌクレオチド(dNTPs)及び反応バッファーからなる反応液にさらに、上記本発明の核酸プローブと競合的核酸とを各一定量を加えて等温増幅反応系を形成する。反応温度は上記LAMP法と同様である。この反応系においては、DNA試料中に組換え遺伝子CaMV 35Sが存在すれば、該遺伝子中のターゲット配列が競合的核酸ととともに等温増幅されるが、増幅効率はターゲット配列と競合的核酸とにおいて同じに設定されている。
上記核酸プローブがターゲット配列と結合した場合、核酸プローブの5’末端のシトシン塩基は、ターゲット配列中の対応位置にあるグアニン塩基と塩基対を形成し、核酸プローブの5’末端に結合した蛍光色素は、グアニン塩基と近接し消光する。一方、競合的核酸に核酸プローブがハイブリダイズする場合においては、核酸プローブの5’末端のシトシン塩基は、ターゲット配列中の対応位置にあるグアニン塩基でなくシトシン塩基であるから塩基対を形成せず、上記蛍光色素は競合的核酸配列中のグアニン塩基と離間し、蛍光を発し、消光しない。したがって、上記増幅反応系においては、標的核酸が増幅するにつれ、蛍光強度は減少する。
【0031】
一方、核酸プローブの量が、増幅されるターゲット配列と競合的核酸の合計量と同じかそれ以下になれば、核酸プローブの各分子はターゲット配列と競合的核酸のいずれかにハイブリダイズしている状態になり、また、上記したように、増幅効率は、ターゲット配列と競合的核酸とにおいて同じであり、かつ、本発明の核酸プローブはターゲット配列と競合的核酸とに同じ結合力で結合するから、核酸プローブがターゲット配列にハイブリダイズする割合は、増幅後の標的核酸と競合的核酸の割合に比例し、さらに、増幅された競合的核酸の量は、反応系において添加した当初の含有量に依存するから、該反応系の蛍光強度を反応前後で測定すれば、増幅前のターゲット配列の量、すなわち、Roundup Ready Soybean系統のダイズあるいは該ダイズを使用した加工食品中の組換え遺伝子CaMV 35Sの量を測定することができる。
【0032】
本発明においては、例えばダイズ加工食品中のRoundup Ready Soybean系統のダイズの混入率を算出するために必要なダイズ内在性遺伝子Le1を、LAMP法あるいはABC-LAMP法を用いて定量するための核酸プライマー、核酸プローブ及び競合的核酸及びこれを用いたLe1の定量法方法を包含する。
Roundup Ready Soybean系統のダイズの混入率を算出するためには内標比を求める必要があるが、内標比は以下の式で求められる。
【0033】
【数1】
この内標比は試験したRoundup Ready Soybean系統のダイズに固有の値である。したがって、このダイズの混入率は以下の式で算出できる。
【0034】
【数2】
このような混入率の算出に用いるLe1遺伝子の検知・定量法に用いる核酸プライマー、核酸プローブ、競合的核酸は以下に示すとおりであり、これらは上記Roundup Ready Soybean系統の組換え遺伝子の検出に用いる核酸プライマー、核酸プローブ及び競合的核酸と同様の設計思想に基づき設計されたものであり、上記と同様の改良点を有する。
【0035】
〔プライマーセット〕
Le1遺伝子の定量的検出に用いる核酸プライマーの塩基配列は以下のとおりであり、これらをLAMP法、ABC-LAMP法に用いるプライマーセットとする。
ダイズ由来のLe1遺伝子配列を検出及び/又は定量するための核酸プライマー(5′→3′)の塩基配列
Le1-FIP:AGC AAA AGA CCA AGA AAG CAC CTT CTC TTC CCG AGT GGG(配列番号12)
Le1-F3 :CCA GCA ATA TCC TCT CCG A(配列番号13)
Le1-LPF:GGC AGC AGA GAA CCC TAT CC(配列番号14)
Le1-BIP:TTT GCC ACA CGC TAG CAA TTA CAC TTT CTT CCT TCG ATC TGT(配列番号15)
Le1-B3 :TCA CTA GCG ATC GAG TAG TG(配列番号16)
Le1-LPB:GTT GCA TGA GGC CAT CTA AAT GT(配列番号17)
【0036】
〔核酸プローブ〕
本発明において設計された核酸プローブの塩基配列は以下に示される。この塩基配列は、標的遺伝子であるLe1遺伝子部分配列の168番目のグアニンから始まる塩基配列に対応するよう設計したものであり、該プローブの5’末端側のシトシン塩基には、上記標的遺伝子中のグアニン塩基と近接して消光するBODIPY FL、FITC、TMRITC等の蛍光色素で蛍光標識しており、また、3’末端はリン酸化されている。
ダイズ由来のLe1遺伝子配列を検出及び/又は定量するための核酸プローブ(5′→3′)の塩基配列
ABQP-Le1:CAT GCG ATT CCC CAG GTA TGT CC(配列番号18)
【0037】
〔競合的核酸〕
本発明において設計された競合的核酸の塩基配列(2本鎖DNAのうち一方のみ表示)と上記各プライマー及びプローブの各結合部位を以下に示す。なお、F1及びF2、並びにB1c及びB2cは、それぞれFIPプライマー及びBIPプライマーの競合的核酸における結合部位を表す。
【0038】
ダイズ由来のLe1遺伝子配列を検出及び/又は定量するための競合的核酸(5′→3′)の塩基配列
【表3】
(配列番号19)
【0039】
上記配列はダイズ由来のLe1遺伝子の部分配列における168番目のグアニン塩基をシトシンに代え、146番目のシトシンをグアニンに変更したものである。
これら、核酸プライマーセットを使用したLAMP法及び該核酸プライマー、核酸プローブ及び競合的核酸を使用したABC-LAMP法による、Le1遺伝子の検出・定量法については、上記したRoundup Ready Soybean系統のダイズにおける組換遺伝子の検出/定量法と同様である。
以下本発明の実施例を示すが、本発明はこれら実施例に限定されるものではない。
【実施例】
【0040】
実施例1
(1)LAMP法による組換え遺伝子の検出
以下のプライマーを用いて、62℃で1時間、LAMP反応を行い、遺伝子組換えダイズRoundup Ready Soybean系統の組換え遺伝子およびダイズ由来のLe1遺伝子配列を検出・定量した。
使用したRoundup Ready Soybean系統の組換え遺伝子及びLe1遺伝子の検出/定量用プライマーの塩基配列は以下のとおりである。
【0041】
Roundup Ready Soybean系統の組換え遺伝子用プライマー
RRS-FIPプライマー:GTG GTC CCA AAG ATG GAG GAG ATA TCA CAT CAA TCC ACT TGC(配列番号1)
RRS-F3プライマー :GTC ATC CCT TAC GTC AGT(配列番号2)
RRS-LPFプライマー:CGT TCC AAC CAC GTC TTC AA(配列番号3)
RRS-BIPプライマー:TGT CGG CAG AGG CAT CTT TTA AAA TAA ACA TAG GGA ACC CAA(配列番号4)
RRS-B3プライマー :TCA AAA TAA GAT CAT ACA TAC AGG(配列番号5)
RRS-LPBプライマー:AGG AGC CAC CTT CCT TTT CC(配列番号6)
【0042】
Le1用プライマー
Le1-FIP:AGC AAA AGA CCA AGA AAG CAC CTT CTC TTC CCG AGT GGG(配列番号12)
Le1-F3 :CCA GCA ATA TCC TCT CCG A(配列番号13)
Le1-LPF:GGC AGC AGA GAA CCC TAT CC(配列番号14)
Le1-BIP:TTT GCC ACA CGC TAG CAA TTA CAC TTT CTT CCT TCG ATC TGT(配列番号15)
Le1-B3 :TCA CTA GCG ATC GAG TAG TG(配列番号16)
Le1-LPB:GTT GCA TGA GGC CAT CTA AAT GT(配列番号17)
【0043】
LAMP反応液の組成は、遺伝子組換えダイズRoundup Ready Soybean系統の組換え遺伝子の定量については、H2O 11.5μl、10×LAMP Buffer 2.5μl、dNTP mix(2.5 mM) 4.0μl、RRS-FIP(40μM) 1.0μl、RRS-BIP(40μM) 1.0μl、RRS-F3(5μM) 1.0μl、RRS-B3(5μM) 1.0μl、RRS-LPF(20μM) 1.0μl、RRS-LPB(20μM) 1.0μl、Template DNA 1.0μlとした。また、Le1遺伝子の定量については、H2O 11.5μl、10×LAMP Buffer 2.5μl、dNTP mix(2.5 mM) 4.0μl、Le1-FIP(40μM) 1.0μl、Le1-BIP(40μM) 1.0μl、Le1-F3(5μM) 1.0μl、Le1-B3(5μM) 1.0μl、Le1-LPF(20μM) 1.0μl、Le1-LPB(20μM) 1.0μl、Template DNA 1.0μlとした。Template DNAには標的遺伝子を含むPCR産物1×106copiesとダイズ粉末から抽出したゲノムDNA 50ngを用いた。LAMP反応終了後に37℃、4時間の制限酵素処理を行った。用いた制限酵素はHindIII、HaeIIIである。LAMP産物および制限酵素処理を行った溶液の5μlを用いてアガロースゲル電気泳動を行った。Roundup Ready Soybean系統の組換え遺伝子を増幅した結果を図1に、Le1遺伝子を増幅した結果を図2にしめす。Roundup Ready Soybean系統の組換え遺伝子、およびLe1遺伝子のどちらのLAMP産物に関しても、設計したプライマーによる特異的な増幅が確認された。
【0044】
(2)Real-time濁度測定法によるGMダイズの定量
設計したプライマーセットを用いたReal-time濁度測定法により、Roundup Ready Soybean系統の組換え遺伝子とダイズ由来のLe1遺伝子の定量を行った。LAMP反応液の組成は、遺伝子組換えダイズRoundup Ready Soybean系統の組換え遺伝子の定量については、H2O 11.5μl、10×LAMP Buffer 2.5μl、dNTP mix(2.5 mM) 4.0μl、RRS-FIP(40μM) 1.0μl、RRS-BIP(40μM) 1.0μl、RRS-F3(5μM) 1.0μl、RRS-B3(5μM) 1.0μl、RRS-LPF(20μM) 1.0μl、RRS-LPB(20μM) 1.0μl、Template DNA 1.0μlとした。また、Le1遺伝子の定量については、H2O 11.5μl、10×LAMP Buffer 2.5μl、dNTP mix(2.5mM) 4.0μl、Le1-FIP(40μM) 1.0μl、Le1-BIP(40μM) 1.0μl、Le1-F3(5μM) 1.0μl、Le1-B3(5μM) 1.0μl、Le1-LPF(20μM) 1.0μl、Le1-LPB(20μM) 1.0μl、Template DNA 1.0μlとした。各標的遺伝子のPCR産物を103〜107 copies/μlに調製し、Template DNAとして用いた。遺伝子組換えダイズRoundup Ready Soybean系統の組換え遺伝子の定量における濁度の経時変化を図3に示す。また、濁度が一定の値に達するまでに要した時間(Threshold time:Tt)と反応初期の標的遺伝子数の関係を図4に示す。Le1遺伝子の定量における濁度の経時変化を図5に示す。また、濁度が一定の値に達するまでに要した時間(Threshold time:Tt)と反応初期の標的遺伝子数の関係を図6に示す。Le1遺伝子の定量においては、1x103 copies以上の濃度で定量が可能であった。
【0045】
なお、上記Le1の定量におけるTemplate DNAは、ダイズの抽出ゲノム(0.4ng/ul)を鋳型として用いて、1.0μl、H2O 12.2μl、10×PCR Buffer 2.0μl、dNTP mix(2.5mM) 2.0μl、Le1-F(10μM) 0.8μl、Le1-R(10μM) 0.8μl、Taq DNA polymerase 0.2 μlの反応液組成でPCRを行った反応産物を用いた。PCR反応条件は、94℃ 2分、94℃ 30秒、56℃30秒、72℃30秒を40サイクル、72℃2分で行った。RRSの定量におけるTemplateDNAは、プライマーにRRS-F(10μM)、RRS-R(10μM)を用いた以外は、Le1と同様の鋳型、反応液組成、反応条件でPCRを行い、その増幅産物を用いた。
【0046】
実施例2
Alternately Binding Probe Competitive (ABC)-LAMP法によるGMダイズの定量
(1)蛍光消光試験
本発明で設計した核酸プローブ、競合的核酸の蛍光消光の精度を評価するために、標的核酸と競合的核酸中の核酸プローブ結合部位を有する各2種の短いオリゴヌクレオチドを合成し、本発明の核酸プローブ、各2種のオリゴヌクレオチドを用いて、本発明におけるRoundup Ready Soy系およびLe1遺伝子の検出・定量系評価のための予備実験として、蛍光消光試験を行った。
【0047】
1)Roundup Ready Soy系の蛍光消光試験において使用したプローブ及びオリゴヌクレオチド
a)Roundup Ready Soybean系統の組換え遺伝子検出・定量用プローブ
ABQP-RRS:CTT TTT CCA CGT GCT CCT CGT GC(配列番号7)
(5’末端BODIPY FL標識、3’末端リン酸化)
b)上記a)のプローブ結合部位を有するオリゴヌクレオチド
RRS-TL(標的オリゴヌクレオチド):
CCC ACC CAC GAG GAG CAC GTG GAA AAA GAA GAC(配列番号10)、
RRS-CL(競合的オリゴヌクレオチドチド):
CCC ACG CAC GAG GAG CAC GTG GAA AAA CAA GAC(配列番号11)
【0048】
2)Le1遺伝子系の蛍光消光試験において使用したプローブ及びオリゴヌクレオチド
a)Le1遺伝子検出・定量用プローブ
ABQP-Le1:CAT GCG ATT CCC CAG GTA TGT CC(配列番号18)
(5’末端BODIPY FL標識、3’末端リン酸化)
b)上記c)のプローブ結合部位を有するオリゴヌクレオチド
Le1-TL(標的オリゴヌクレオチド):
GGA CTC GAC ATA CCT GGG GAA TCG CAT GAC GTG(配列番号20)
Le1-CL(競合的オリゴヌクレオチド):
GGA CTG GAC ATA CCT GGG GAA TCG CAT CAC GTG(配列番号21)
【0049】
Roundup Ready Soybean系統の蛍光消光試験の反応液組成はH2O 1.5μl、10×LAMP Buffer 2.5μl、dNTP mix(2.5mM) 3.0μl、1% TritonX-100 2.5μl、Betaine(5M) 5.0μl、ABQP-RRS(7.5μM) 0.5μl、合成オリゴヌクレオチド(RRS-TLと-CLの混合物)10μlとした。また、Le1遺伝子系の蛍光消光試験の反応液組成はH2O 1.5μl、10×LAMP Buffer 2.5μl、dNTP mix(2.5mM) 3.0μl、1% TritonX-100 2.5μl、Betaine(5M) 5.0μl、ABQP-Le1(7.5μM) 0.5μl、合成オリゴヌクレオチド(Le1-TLとLe1-CLの混合物)10μlとした。Roundup Ready Soybean系統の蛍光消光試験の結果を図7に、Le1遺伝子系の蛍光消光試験の結果を図8に示す。両蛍光消光試験の結果とも標的核酸と競合核酸の比率に応じた相対蛍光強度を示すことがわかった。
【0050】
(2)本発明で設計した核酸プライマー、核酸プローブ、競合的核酸を用いて、ABC-LAMP法による標的遺伝子定量のための検量線の作製を行った。
核酸プライマーは実施例1で使用したものを用い、核酸プローブは上記(1)において使用したものを用いた。また競合的核酸は、Roundup Ready Soybean系統の組換え遺伝子の定量において配列番号9に示される競合的核酸、Le1遺伝子の定量において配列番号19に示される競合的核酸をそれぞれ用いた。
反応液の組成は、遺伝子組換えダイズRoundup Ready Soybean系統の組換え遺伝子の定量については、H2O 6.5μl、10×LAMP Buffer 2.5μl、dNTP mix(2.5 mM) 4.0μl、RRS-FIP(40μM) 1.0μl、RRS-BIP(40μM) 1.0μl、RRS-F3(5μM) 1.0μl、RRS-B3(5μM) 1.0μl、RRS-LPF(20μM) 1.0μl、RRS-LPB(20μM) 1.0μl、1% TritonX-100 2.5μl、ABQP-RRS(7.5μM) 0.5μlとし、さらに競合遺伝子数を104copies/μl、標的遺伝子数を103-107copies/μlに調製し、それらを1.0μlずつ添加した。
【0051】
また、Le1遺伝子の定量については、H2O 6.5μl、10×LAMP Buffer 2.5μl、dNTP mix(2.5mM) 4.0μl、Le1-FIP(40μM) 1.0μl、Le1-BIP(40μM) 1.0μl、Le1-F3(40μM) 1.0μl、Le1-B3(40μM) 1.0μl、Le1-LPF(40μM) 1.0μl、Le1-LPB(40μM) 1.0μl、1% TritonX-100 2.5μl、ABQP-Le1(7.5μM) 0.5μlに、さらに、競合遺伝子数を104copies/μl、標的遺伝子数を103-107copies/μlに調製し、それらを1.0μlずつ添加して反応溶液とした。
反応条件は両遺伝子ともに、62℃で1時間とした。
Roundup Ready Soybean系統の組換え遺伝子の定量における相対蛍光強度Bsと初期標的遺伝子数の関係を図9に示す。同様に、Le1遺伝子の定量における相対蛍光強度Bsと初期標的遺伝子数の関係を図10に示す。
両遺伝子の結果とも標的核酸と競合核酸の比率に応じた相対蛍光強度を示すことがわかった。
【図面の簡単な説明】
【0052】
【図1】本発明のRoundup Ready Soybean系統ダイズおける組換え遺伝子増幅用プライマーを用いてLAMPにより該組換え遺伝子を増幅した結果を示す電気泳動写真である。
【図2】本発明のダイズ由来のLe1遺伝子増幅用プライマーを用いてLAMPにより該組換え遺伝子を増幅した結果を示す電気泳動写真である。
【図3】本発明のRoundup Ready Soybean系統ダイズにおける組換え遺伝子増幅用プライマーを用いてリアルタイムLAMP法を行った結果、観察された濁度の経時変化を示すグラフである。
【図4】上記濁度法により得られた、Roundup Ready系統ダイズおける組換え遺伝子についての、Ttと反応初期の標的遺伝子数の関係を示すグラフである。
【図5】本発明のLe1遺伝子増幅用プライマーを用いてリアルタイムLAMP法を行った結果、観察された濁度の経時変化を示すグラフである。
【図6】上記濁度法により得られた、Le1遺伝子についてのTtと反応初期の標的遺伝子数の関係を示すグラフである。
【図7】Roundup Ready Soybean系統の組換え遺伝子の定量に用いる本発明のプローブ、競合的核酸の一部配列を用いて蛍光消光試験を行った結果を示すグラフである。
【図8】Le1遺伝子の定量に用いる本発明のプローブ、競合的核酸の一部配列を用いて蛍光消光試験を行った結果を示すグラフである。
【図9】Roundup Ready Soybean系統の組換え遺伝子、並びに本発明のプライマー、プローブ及び競合的核酸を用いてABC-LAMP法を行いLAMP反応前後の蛍光強度を測定した結果得られた、相対蛍光強度BsとRoundup Ready Soybean系統の組換え遺伝子の初期標的遺伝子数との関係を示すグラフである。
【図10】Le1遺伝子、並びに本発明のプライマー、プローブ及び競合的核酸を用いてABC-LAMP法を行い、LAMP反応前後の蛍光強度を測定した結果得られた、相対蛍光強度BsとLe1遺伝子の初期標的遺伝子数の関係を示すグラフである。
【特許請求の範囲】
【請求項1】
ダイズ由来のLe1遺伝子配列をLAMP法又はABC-LAMP法によって検出・定量するために用いる、FIPプライマー、F3プライマー、LPFプライマー、BIPプライマー、B3プライマー及びLPBプライマーからなるプライマーセットであって、各プライマーが以下に示される塩基配列、又は該配列と相補の塩基配列を有することを特徴とする、上記プライマーセット。
FIPプライマー:AGC AAA AGA CCA AGA AAG CAC CTT CTC TTC CCG AGT GGG
F3プライマー :CCA GCA ATA TCC TCT CCG A
LPFプライマー:GGC AGC AGA GAA CCC TAT CC
BIPプライマー:TTT GCC ACA CGC TAG CAA TTA CAC TTT CTT CCT TCG ATC TGT
B3プライマー :TCA CTA GCG ATC GAG TAG TG
LPBプライマー:GTT GCA TGA GGC CAT CTA AAT GT
【請求項2】
ダイズ由来のLe1遺伝子配列をABC-LAMP法によって検出・定量するために用いる、配列番号19に示される塩基配列又は該配列と相補の塩基配列からなる競合的核酸。
【請求項3】
ダイズ由来のLe1遺伝子配列をABC-LAMP法によって検出及び/又は定量するために用いる核酸プローブであって、配列番号18に示される塩基配列又は該配列と相補の塩基配列からなり、その末端に核酸中のグアニン塩基との近接、離間によって蛍光強度が変化する蛍光色素を標識されていることを特徴とする上記核酸プローブ。
【請求項4】
ダイズ由来のLe1遺伝子配列を、ABC-LAMP法を用いて検出及び又は定量するための方法であって、
被験ダイズ試料から抽出した核酸、請求項1に記載のプライマーセット、請求項2に記載の競合的核酸及び請求項3に記載の核酸プローブの存在下、等温増幅反応を行い、該反応前後の蛍光強度を測定することを特徴とする、上記ダイズ由来のLe1遺伝子配列の検出及び/又は定量方法。
【請求項5】
請求項1に記載のプライマーセット、請求項2に記載の競合的核酸及び請求項3に記載の核酸プローブを少なくとも組み合わせたことを特徴とする、ダイズ由来のLe1遺伝子配列の検出又は定量用試薬キット。
【請求項1】
ダイズ由来のLe1遺伝子配列をLAMP法又はABC-LAMP法によって検出・定量するために用いる、FIPプライマー、F3プライマー、LPFプライマー、BIPプライマー、B3プライマー及びLPBプライマーからなるプライマーセットであって、各プライマーが以下に示される塩基配列、又は該配列と相補の塩基配列を有することを特徴とする、上記プライマーセット。
FIPプライマー:AGC AAA AGA CCA AGA AAG CAC CTT CTC TTC CCG AGT GGG
F3プライマー :CCA GCA ATA TCC TCT CCG A
LPFプライマー:GGC AGC AGA GAA CCC TAT CC
BIPプライマー:TTT GCC ACA CGC TAG CAA TTA CAC TTT CTT CCT TCG ATC TGT
B3プライマー :TCA CTA GCG ATC GAG TAG TG
LPBプライマー:GTT GCA TGA GGC CAT CTA AAT GT
【請求項2】
ダイズ由来のLe1遺伝子配列をABC-LAMP法によって検出・定量するために用いる、配列番号19に示される塩基配列又は該配列と相補の塩基配列からなる競合的核酸。
【請求項3】
ダイズ由来のLe1遺伝子配列をABC-LAMP法によって検出及び/又は定量するために用いる核酸プローブであって、配列番号18に示される塩基配列又は該配列と相補の塩基配列からなり、その末端に核酸中のグアニン塩基との近接、離間によって蛍光強度が変化する蛍光色素を標識されていることを特徴とする上記核酸プローブ。
【請求項4】
ダイズ由来のLe1遺伝子配列を、ABC-LAMP法を用いて検出及び又は定量するための方法であって、
被験ダイズ試料から抽出した核酸、請求項1に記載のプライマーセット、請求項2に記載の競合的核酸及び請求項3に記載の核酸プローブの存在下、等温増幅反応を行い、該反応前後の蛍光強度を測定することを特徴とする、上記ダイズ由来のLe1遺伝子配列の検出及び/又は定量方法。
【請求項5】
請求項1に記載のプライマーセット、請求項2に記載の競合的核酸及び請求項3に記載の核酸プローブを少なくとも組み合わせたことを特徴とする、ダイズ由来のLe1遺伝子配列の検出又は定量用試薬キット。
【図1】

【図2】
【図3】
【図4】
【図5】
【図6】
【図7】
【図8】
【図9】
【図10】
【図2】
【図3】
【図4】
【図5】
【図6】
【図7】
【図8】
【図9】
【図10】
【公開番号】特開2013−63082(P2013−63082A)
【公開日】平成25年4月11日(2013.4.11)
【国際特許分類】
【出願番号】特願2012−264663(P2012−264663)
【出願日】平成24年12月3日(2012.12.3)
【分割の表示】特願2007−231278(P2007−231278)の分割
【原出願日】平成19年9月6日(2007.9.6)
【出願人】(301021533)独立行政法人産業技術総合研究所 (6,529)
【Fターム(参考)】
【公開日】平成25年4月11日(2013.4.11)
【国際特許分類】
【出願日】平成24年12月3日(2012.12.3)
【分割の表示】特願2007−231278(P2007−231278)の分割
【原出願日】平成19年9月6日(2007.9.6)
【出願人】(301021533)独立行政法人産業技術総合研究所 (6,529)
【Fターム(参考)】
[ Back to top ]