
遺伝子組換え大腸菌によるタンパク質の製造方法
【課題】 タンパク質を発現可能な大腸菌を培養することでタンパク質を製造する方法において、タンパク質、特にFc結合性タンパク質を効率的に製造するのに最適な金属塩の濃度、特にマグネシウム塩の濃度およびその供給方法を提供すること。
【解決手段】 タンパク質をコードするポリヌクレオチドを含む発現ベクターで大腸菌W3110株(ATCC 27235)を形質転換して得られた組換え大腸菌を、培地1Lあたりマグネシウムイオンとして2.8×10−2mol以上のマグネシウム塩を添加した培地で培養することで、前記課題を解決する。
【解決手段】 タンパク質をコードするポリヌクレオチドを含む発現ベクターで大腸菌W3110株(ATCC 27235)を形質転換して得られた組換え大腸菌を、培地1Lあたりマグネシウムイオンとして2.8×10−2mol以上のマグネシウム塩を添加した培地で培養することで、前記課題を解決する。
【発明の詳細な説明】
【技術分野】
【0001】
本発明は、遺伝子工学的手法により得られた、遺伝子組換えタンパク質を発現可能な大腸菌を用いて、組換えタンパク質、特にFc結合性タンパク質を効率的に製造する方法に関する。特に本発明は、前記大腸菌を効率よく培養することで、組換えタンパク質を効率的に製造する方法に関する。
【背景技術】
【0002】
大腸菌を利用した組換えタンパク質の生産は、特許文献1をはじめとし、これまでにも多くの例が報告されている。組換えタンパク質を効率的に生産するためには、大腸菌細胞を高密度の状態で培養することが必要である。大腸菌の菌体には炭素、窒素の他に、多種の金属元素を含むことが知られている。そのため大腸菌を効率よく培養するには、培地成分に、炭素源や窒素源のみならず、金属塩も含む必要があり、かつ、培地組成を大腸菌に適合するようにすることが重要である。
【先行技術文献】
【特許文献】
【0003】
【特許文献1】特表2000−501936号公報
【特許文献2】特開2008−245580号公報
【非特許文献】
【0004】
【非特許文献1】J.V.Ravetch等,Annu.Rev.Immunol.,9,457,1991
【非特許文献2】Toshiyuki Takai,Jpn.J.Clin.Immunol.,28,318,2005
【非特許文献3】A.Paetz等,Biochem.Biophys.Res.Commun.,338,1811,2005
【非特許文献4】J.M.Allen等,Science,243,378,1989
【発明の概要】
【発明が解決しようとする課題】
【0005】
タンパク質のうちFc受容体はFc結合性タンパク質の一つであり、免疫グロブリン分子のFc領域に結合する一群の分子である。Fc受容体はその結合する免疫グロブリンの種類によって分類されており、IgGのFc領域に結合するFcγ受容体、IgEのFc領域に結合するFcε受容体、IgAのFc領域に結合するFcα受容体などがある(非特許文献1)。また、各受容体は、その構造の違いによりさらに細かく分類され、Fcγ受容体の場合、FcγRI、FcγRII、FcγRIIIの存在が報告されている(非特許文献1)。
【0006】
Fcγ受容体の一つであるFcγRIは単球とマクロファージ中で発現しており、好中球ではγインターフェロンにより誘導的に発現される(非特許文献1)。また、FcγRIはIgGに対する結合親和性が高く、その平衡解離定数(Kd)は10−8M以下である(非特許文献2)。FcγRIは、細胞外領域、細胞膜貫通領域、細胞質内領域に区分され、IgGとの結合は、IgGのFc領域とFcγRIの細胞外領域で起こり、その後細胞質へとシグナルが伝達される。FcγRIはIgGとの結合に直接関わる分子量約42000のα鎖と、γ鎖の2種類のサブユニットによって構成されており、γ鎖は細胞膜と細胞外領域との境界で共有結合することでホモダイマーを形成している(非特許文献3)。FcγRIはIgG1から4まであるサブクラスのうち、IgG1とIgG3と特に強く結合し、IgG2またはIgG4との結合は弱いことが知られている。
【0007】
ヒトFc受容体のうち、ヒトFcγRIα鎖のアミノ酸配列およびヌクレオチド配列(配列番号1)は、Allen等により明らかにされており(非特許文献4)、ExPASy(Primary accession number:P12314)などの公的データベースにも公表されている。また、ヒトFcγRIの構造上の機能ドメイン、細胞膜を貫通するためのシグナルペプチド配列、細胞膜貫通領域の位置についても同様に公表されている。配列番号1のうち、1番目のメチオニン(Met)から15番目のグリシン(Gly)までがシグナルペプチドであり、16番目のグルタミン(Gln)から292番目のヒスチジン(His)までが細胞外領域の機能ドメイン、293番目のバリン(Val)から374番目のスレオニン(Thr)までが細胞膜貫通領域および細胞内領域とされている。
【0008】
近年になり、Fc受容体の予想外の免疫抑制的な生物学的特性は、特に自己免疫疾患または自己免疫症候群、移植物の拒絶および悪性リンパ増殖の領域において医薬として注目を浴びつつある(非特許文献2)。また、FcγRIの機能である抗体の吸着能は各種抗体精製用クロマトグラフィーゲルの捕捉機能を担うタンパク質としても利用することができる。
【0009】
Fc受容体を安価に製造することを目的に、Fc受容体を発現可能な遺伝子組換え体を利用した製造方法についてこれまで検討されており、例えば、組換え大腸菌を用いたFcγRIの製造方法が報告されている(特許文献2)。しかしながら、大腸菌を用いた発現系では、FcγRIの発現量が極めて低いという問題点があった。
【0010】
そこで本発明の目的は、タンパク質を発現可能な大腸菌を培養することでタンパク質を製造する方法において、タンパク質、特にFc結合性タンパク質を効率的に製造するのに最適な金属塩の濃度、特にマグネシウム塩の濃度およびその供給方法を提供することにある。
【課題を解決するための手段】
【0011】
本発明者らは前記課題に対し、大腸菌培養における、金属塩の供給について鋭意検討した結果、本発明の完成に至った。
【0012】
すなわち、本発明は、以下の発明を包含する:
(i)タンパク質をコードするポリヌクレオチドを含む発現ベクターで大腸菌W3110株(ATCC 27235)を形質転換して得られた組換え大腸菌を、マグネシウム塩を添加した培地で培養することでタンパク質を製造する方法であって、マグネシウム塩の添加量が、培地1Lあたりマグネシウムイオンとして2.8×10−2mol以上である、前記方法。
【0013】
(ii)培養開始時の培地にマグネシウム塩の一部を添加し、残部を培養中の培地に添加する、(i)に記載の方法。
【0014】
(iii)マグネシウム塩とともに炭素源および窒素源を培養中の培地に添加する、(ii)に記載の方法。
【0015】
(iv)培養開始時の培地に添加するマグネシウム塩が、培地1Lあたりマグネシウムイオンとして4.0×10−3mol以上であり、培養中の培地に添加するマグネシウム塩が、培地1Lあたりマグネシウムイオンとして2.4×10−2mol以上である、請求項(ii)または(iii)に記載の方法。
【0016】
(v)タンパク質がFc結合性タンパク質である、(i)から(iv)のいずれかに記載の方法。
【0017】
(vi) Fc結合性タンパク質が、(1)配列番号1に記載のアミノ酸配列のうち少なくとも16番目のグルタミンから289番目のバリンまでのアミノ酸を含むタンパク質、または(2)配列番号1に記載のアミノ酸配列のうち少なくとも16番目のグルタミンから289番目のバリンまでのアミノ酸を含み、かつ前記アミノ酸のうちの一つ以上が他のアミノ酸に置換、挿入または欠失したタンパク質である、(v)に記載の方法。
【0018】
以下、本発明を詳細に説明する。
【0019】
本発明の製造方法では、タンパク質をコードするポリヌクレオチドを含む発現ベクターで大腸菌W3110株(ATCC 27235)を形質転換して得られた組換え大腸菌を適切な培地で培養することで、タンパク質を発現させ、製造する。組換え大腸菌を培養するのに用いる培地には、マグネシウム塩を添加する必要があるが、その添加量について検討した結果、培地1Lあたりマグネシウムイオンとして2.8×10−2mol以上とすると効率的にタンパク質を製造できることが判明した。なお、培地1Lあたりマグネシウムイオンとして2.8×10−2mol以上という条件は、培地に添加するマグネシウム塩としてよく用いられる硫酸マグネシウム七水和物の添加量に換算すると培地1Lあたり約7g以上、となる。本発明の製造方法で使用するマグネシウム塩としては、前述した硫酸マグネシウムでなくてもよく、塩化マグネシウムといった水溶性のマグネシウム塩であれば特に限定されない。
【0020】
その他の培地成分については、組換え大腸菌を増殖可能で、かつ発現ベクターに挿入したポリヌクレオチドがコードするタンパク質を発現可能なものであれば特に限定はない。培地に含まれる炭素源の一例としては、グルコース、フルクトース、マルトース、ショ糖、粗糖、糖蜜があげられる。培地に含まれる窒素源の一例としては、酵母エキス、ポリペプトン、カゼインおよびその代謝物、コーンスティープリカー、大豆タンパク質、肉エキス、魚肉エキスがあげられるが、特に酵母エキスが好ましい。なお、ナトリウム塩、鉄塩やマンガン塩など、マグネシウム塩以外の金属塩をさらに培地に添加してもよい。金属塩の具体例としては、リン酸二水素ナトリウム、リン酸水素二ナトリウム、リン酸二水素カリウム、リン酸水素二カリウム、塩化ナトリウム、硫酸鉄(II)、硫酸鉄(III)、塩化鉄(II)、塩化鉄(III)、クエン酸鉄、硫酸アンモニウム鉄、塩化カルシウム二水和物、硫酸カルシウム、硫酸亜鉛、塩化亜鉛、硫酸銅、塩化銅、硫酸マンガン、塩化マンガンがあげられる。さらに必要に応じ、ビオチン、ニコチン酸、チアミン、リボフラビン、イノシトール、ピリドキシンといったビタミン類を培地に添加してもよい。
【0021】
本発明の製造方法で組換え大腸菌を培養する際、培養開始時に炭素源や窒素源といった栄養源を一度に培地に投入すると、組換え大腸菌の増殖およびタンパク質製造が阻害されたり、酢酸などの副生成物が生産されるため、タンパク質の発現効率および得られたタンパク質の品質に悪影響を与える可能性がある。そのため、培養開始時に投入する栄養源は最小限とし、培養中に栄養源を追加供給(流加)しながら培養を行なう流加培養により、タンパク質を製造すると好ましい。なお、流加培養により組換え大腸菌を培養し、タンパク質を製造する際は、培地に添加するマグネシウム塩も、培養開始時の培地に一度に添加するのではなく、培養開始時の培地にその一部を添加し、残部は前記栄養源とともに培養中に添加するのが好ましい。
【0022】
本発明の好ましい製造方法である、流加培養によるタンパク質製造の一例を詳細に示す。必要最小限の炭素源および窒素源を投入した培地に、マグネシウム塩を培地1Lあたりマグネシウムイオンとして2.8×10−2mol以上添加し、さらに必要に応じてマグネシウム塩以外の金属塩やビタミン類等を添加した後、組換え大腸菌の培養を開始する。組換え大腸菌の増殖により炭素源が消費され所定の濃度まで低下した時点で、培養液中の炭素源を所定の濃度に維持しつつ、炭素源と窒素源を供給(流加)して培養することで、組換え大腸菌からタンパク質を発現させ、製造する。
【0023】
本発明の好ましい製造方法である、流加培養によるタンパク質製造の別の例として、必要最小限の炭素源および窒素源を投入した培地に、マグネシウム塩を培地1Lあたりマグネシウムイオンとして4.0×10−3mol以上添加し、さらに必要に応じてマグネシウム塩以外の金属塩やビタミン類等を添加した後、組換え大腸菌の培養を開始する。組換え大腸菌の増殖により炭素源が消費され所定の濃度まで低下した時点で、培養液中の炭素源を所定の濃度に維持しつつ、炭素源および窒素源を供給(流加)し、さらにマグネシウム塩を培地1Lあたりマグネシウムイオンとして2.4×10−3mol以上添加して培養することで、組換え大腸菌からタンパク質を発現させ、製造する。
【0024】
前述した流加培養によるタンパク質製造において、培養開始時に投入する炭素源の濃度は、炭素源がグルコースの場合、0から20g/Lとすると好ましい。また供給(流加)する炭素源と窒素源は、高濃度の溶液とすると培養液の液量増加を抑えられるため好ましく、炭素源をグルコース、窒素源を酵母エキスとした場合、供給(流加)するグルコース溶液の濃度は300から900g/Lに、酵母エキス溶液の濃度は150から500g/Lに、それぞれすると好ましい。炭素源と窒素源を供給(流加)する際維持する、所定の濃度とは、炭素源が枯渇せず有機酸などの副生成物が生産しない濃度をいう。炭素源をグルコースとした場合、炭素源濃度が5g/Lを超えた状態で培養を行なうと副生成物として有機酸が生産され、それが多量に蓄積することにより組換え大腸菌の増殖やFc結合性タンパク質の生産を抑制する可能性があるため、好ましくない。よって、炭素源をグルコースとした場合の所定の濃度とは、少なくとも5g/L以下、好ましくは1g/L以下、さらに好ましくは0.5g/L以下、最も好ましくは0.1g/L以下である。炭素源供給量のモニターは、例えば、炭素源を供給するポンプの稼働時間より行なうことができる。炭素源の枯渇をモニターする方法は特に限定はなく、一例として呼吸活性の低下によりモニターすることができる。呼吸活性の低下は、例えば培養液の溶存酸素濃度(DO)の上昇、排ガス中の酸素濃度の上昇、炭酸ガス濃度の低下、pHの上昇として現れる。特に、DOは応答が速いことから、炭素源の枯渇をモニターするのに好ましい指標である。その理由として、炭素源の濃度が十分に維持されている場合には、微生物の呼吸により酸素が消費されるためDOは酸素飽和濃度より低い値に維持されるが、炭素源が枯渇すると微生物の呼吸活性が低下しDOが急激に上昇するためである。なお、DOの急激な上昇に連動させて炭素源を追加する方法をDOスタット法という。DO以外の指標(排ガス組成、炭酸ガス濃度、pH上昇)でモニターした場合は、DOを指標とした場合と比較し応答が遅いという欠点がある。窒素源の供給(流加)量は、炭素源供給(流加)量に比例した量を供給(流加)すればよい。窒素源の供給(流加)方法は特に限定はなく、例えば、炭素源水溶液と窒素源水溶液を任意の濃度で混合し、当該混合液を培地に供給(流加)する方法が例示できる。
【0025】
本発明の製造方法における組換え大腸菌の培養条件は、製造に用いる組換え大腸菌が増殖しタンパク質を発現し得る条件であれば特に限定はないが、培養温度は15から50℃が好ましく、特に好ましい温度は20から33℃である。pHは6から8が好ましい。培養時間は任意に設定できるが、通常は数時間から100時間の間に設定される。
【0026】
本発明の好ましい製造方法では、組換え大腸菌からのタンパク質発現を誘導するために、培養開始から一定時間経過後、イソプロピル−β−チオガラクトピラノシド(IPTG)を培養液に添加して、さらに培養する。IPTGの濃度は最終濃度として0.01から2.0mMが好ましく、特に好ましい濃度は最終濃度0.1から2.0mMである。
【0027】
本発明の製造方法で製造可能なタンパク質の一例として、インシュリン、インターフェロン、インターロイキン、抗体、エリスロポエチン、成長ホルモン、およびそれらの受容体のタンパク質等があげられる。以降、本発明の製造方法で製造するタンパク質の好ましい例である、Fc結合性タンパク質について詳細に説明する。
【0028】
本発明の製造方法で製造する、Fc結合性タンパク質は、抗体のFc領域と結合する細胞表面に存在する受容体タンパク質である。Fc結合性タンパク質がヒトFc受容体の場合、ヒトFcγRI、ヒトFcγRIIa、ヒトFcγRIIb、ヒトFcγRIIIなどがあげられる。本発明の製造方法で製造するFc結合性タンパク質の一例として、
(A)配列番号1に記載のアミノ酸配列のうち少なくとも16番目のグルタミンから289番目のバリンまでのアミノ酸を含むタンパク質や、
(B)配列番号1に記載のアミノ酸配列のうち少なくとも16番目のグルタミンから289番目のバリンまでのアミノ酸を含み、かつ前記アミノ酸のうちの一つ以上が他のアミノ酸に置換、挿入または欠失したタンパク質、
があげられる。前記(B)の具体例として、配列番号1に記載のアミノ酸配列のうち少なくとも16番目から289番目までのアミノ酸を含み、かつ以下の(1)から(168)のいずれかに記載のアミノ酸置換を1つ以上含むFc結合性タンパク質があげられる(特願2011−038500号)。
(1)配列番号1の20番目のスレオニンがプロリンに置換
(2)配列番号1の25番目のスレオニンがリジンに置換
(3)配列番号1の38番目のスレオニンがアラニンまたはセリンに置換
(4)配列番号1の46番目のロイシンがアルギニンまたはプロリンに置換
(5)配列番号1の62番目のアラニンがバリンに置換
(6)配列番号1の63番目のスレオニンがイソロイシンに置換
(7)配列番号1の69番目のセリンがフェニルアラニンまたはスレオニンに置換
(8)配列番号1の71番目のアルギニンがヒスチジンに置換
(9)配列番号1の77番目のバリンがアラニンまたはグルタミン酸に置換
(10)配列番号1の78番目のアスパラギンがアスパラギン酸に置換
(11)配列番号1の94番目のアスパラギン酸がグルタミン酸に置換
(12)配列番号1の100番目のイソロイシンがバリンに置換
(13)配列番号1の110番目のセリンがアスパラギンに置換
(14)配列番号1の114番目のフェニルアラニンがロイシンに置換
(15)配列番号1の125番目のヒスチジンがアルギニンに置換
(16)配列番号1の131番目のロイシンがアルギニンまたはプロリンに置換
(17)配列番号1の149番目のトリプトファンがロイシンに置換
(18)配列番号1の156番目のロイシンがプロリンに置換
(19)配列番号1の160番目のイソロイシンがメチオニンに置換
(20)配列番号1の163番目のアスパラギンがセリンに置換
(21)配列番号1の195番目のアスパラギンがスレオニンに置換
(22)配列番号1の199番目のスレオニンがセリンに置換
(23)配列番号1の206番目のアスパラギンがリジン、セリンまたはスレオニンに置換
(24)配列番号1の207番目のロイシンがプロリンに置換
(25)配列番号1の218番目のロイシンがバリンに置換
(26)配列番号1の240番目のアスパラギンがアスパラギン酸に置換
(27)配列番号1の248番目のロイシンがセリンに置換
(28)配列番号1の283番目のロイシンがヒスチジンに置換
(29)配列番号1の285番目のロイシンがグルタミンに置換
(30)配列番号1の17番目のバリンがグリシンまたはグルタミン酸に置換
(31)配列番号1の19番目のスレオニンがイソロイシンに置換
(32)配列番号1の20番目のスレオニンがイソロイシンに置換
(33)配列番号1の25番目のスレオニンがメチオニンまたはアルギニンに置換
(34)配列番号1の27番目のグルタミンがプロリンまたはリジンに置換
(35)配列番号1の35番目のグルタミンがロイシン、メチオニンまたはアルギニンに置換
(36)配列番号1の36番目のグルタミン酸がグリシンに置換
(37)配列番号1の41番目のロイシンがメチオニンに置換
(38)配列番号1の42番目のヒスチジンがロイシンに置換
(39)配列番号1の44番目のグルタミン酸がアスパラギン酸に置換
(40)配列番号1の45番目のバリンがアラニンに置換
(41)配列番号1の46番目のロイシンがアラニン、アスパラギン、アスパラギン酸、グルタミン、グリシン、ヒスチジン、リジン、セリンまたはトリプトファンに置換
(42)配列番号1の47番目のヒスチジンがグルタミン、ロイシンまたはアスパラギンに置換
(43)配列番号1の49番目のプロリンがセリンまたはアラニンに置換
(44)配列番号1の50番目のグリシンがアルギニンまたはグルタミン酸に置換
(45)配列番号1の51番目のセリンがアラニン、スレオニン、ロイシン、プロリンまたはバリンに置換
(46)配列番号1の52番目のセリンがグリシンに置換
(47)配列番号1の53番目のセリンがロイシン、スレオニンまたはプロリンに置換
(48)配列番号1の55番目のグルタミンがアルギニンに置換
(49)配列番号1の57番目のフェニルアラニンがチロシンに置換
(50)配列番号1の58番目のロイシンがアルギニンに置換
(51)配列番号1の60番目のグリシンがアスパラギン酸に置換
(52)配列番号1の61番目のスレオニンがアラニンまたはセリンに置換
(53)配列番号1の62番目のアラニンがグルタミン酸に置換
(54)配列番号1の63番目のスレオニンがロイシン、フェニルアラニンに置換
(55)配列番号1の64番目のグルタミンがプロリン、ヒスチジン、ロイシン、リジンに置換
(56)配列番号1の65番目のスレオニンがアラニンまたはバリンに置換
(57)配列番号1の66番目のセリンがスレオニンに置換
(58)配列番号1の67番目のスレオニンがアラニンまたはセリンに置換
(59)配列番号1の69番目のセリンがアラニンに置換
(60)配列番号1の70番目のチロシンがヒスチジンまたはフェニルアラニンに置換
(61)配列番号1の71番目のアルギニンがチロシンに置換
(62)配列番号1の73番目のスレオニンがアラニンまたはセリンに置換
(63)配列番号1の74番目のセリンがフェニルアラニンに置換
(64)配列番号1の76番目のセリンがアスパラギンに置換
(65)配列番号1の77番目のバリンがアスパラギン酸またはリジンに置換
(66)配列番号1の78番目のアスパラギンがセリンまたはグリシンに置換
(67)配列番号1の80番目のセリンがアラニンに置換
(68)配列番号1の84番目のアルギニンがセリンに置換
(69)配列番号1の88番目のグリシンがセリンに置換
(70)配列番号1の89番目のロイシンがグルタミンまたはプロリンに置換
(71)配列番号1の90番目のセリンがグリシンに置換
(72)配列番号1の92番目のアルギニンがシステインまたはロイシンに置換
(73)配列番号1の96番目のイソロイシンがバリンまたはリジンに置換
(74)配列番号1の97番目のグルタミンがロイシンまたはリジンに置換
(75)配列番号1の101番目のヒスチジンがロイシンに置換
(76)配列番号1の102番目のアルギニンがセリンまたはロイシンに置換
(77)配列番号1の103番目のグリシンがアスパラギン酸またはセリンに置換
(78)配列番号1の111番目のセリンがアラニンに置換
(79)配列番号1の114番目のフェニルアラニンがアラニン、イソロイシン、メチオニン、プロリン、スレオニンまたはバリンに置換
(80)配列番号1の115番目のスレオニンがイソロイシンまたはフェニルアラニンに置換
(81)配列番号1の118番目のグルタミン酸がアスパラギン酸に置換
(82)配列番号1の121番目のアラニンがスレオニンまたはバリンに置換
(83)配列番号1の128番目のリジンがアルギニンまたはグリシンに置換
(84)配列番号1の129番目のアスパラギン酸がグリシンに置換
(85)配列番号1の131番目のロイシンがグルタミンに置換
(86)配列番号1の133番目のチロシンがヒスチジンまたはアルギニンに置換
(87)配列番号1の134番目のアスパラギンがセリンに置換
(88)配列番号1の137番目のチロシンがフェニルアラニンに置換
(89)配列番号1の138番目のチロシンがヒスチジンに置換
(90)配列番号1の139番目のアルギニンがヒスチジンに置換
(91)配列番号1の140番目のアスパラギンがアスパラギン酸に置換
(92)配列番号1の141番目のグリシンがアスパラギン酸またはバリンに置換
(93)配列番号1の142番目のリジンがグルタミン酸またはアルギニンに置換
(94)配列番号1の144番目のフェニルアラニンがイソロイシンに置換
(95)配列番号1の147番目のフェニルアラニンがセリンに置換
(96)配列番号1の148番目のヒスチジンがアルギニンまたはグルタミンに置換
(97)配列番号1の149番目のトリプトファンがアルギニンに置換
(98)配列番号1の151番目のセリンがスレオニンに置換
(99)配列番号1の152番目のアスパラギンがスレオニン、イソロイシンまたはプロリンに置換
(100)配列番号1の154番目のスレオニンがセリンに置換
(101)配列番号1の156番目のロイシンがヒスチジンに置換
(102)配列番号1の157番目のリジンがアルギニンに置換
(103)配列番号1の159番目のアスパラギンがスレオニンまたはアスパラギン酸に置換
(104)配列番号1の160番目のイソロイシンがスレオニン、バリンまたはロイシンに置換
(105)配列番号1の161番目のセリンがスレオニンに置換
(106)配列番号1の165番目のスレオニンがメチオニンに置換
(107)配列番号1の171番目のメチオニンがスレオニンに置換
(108)配列番号1の173番目のリジンがアルギニンに置換
(109)配列番号1の174番目のヒスチジンがグルタミンに置換
(110)配列番号1の177番目のスレオニンがセリンに置換
(111)配列番号1の181番目のイソロイシンがスレオニンに置換
(112)配列番号1の182番目のセリンがスレオニン、ロイシン、バリンまたはグルタミン酸に置換
(113)配列番号1の184番目のスレオニンがセリンに置換
(114)配列番号1の190番目のプロリンがセリンに置換
(115)配列番号1の193番目のバリンがロイシンに置換
(116)配列番号1の195番目のアスパラギンがアラニンに置換
(117)配列番号1の196番目のアラニンがセリンに置換
(118)配列番号1の198番目のバリンがグリシンまたはメチオニンに置換
(119)配列番号1の199番目のスレオニンがアラニンに置換
(120)配列番号1の200番目のセリンがグリシンまたはアルギニンに置換
(121)配列番号1の202番目のロイシンがメチオニンに置換
(122)配列番号1の203番目のロイシンがヒスチジン、グルタミン、チロシン、アルギニン、プロリンに置換
(123)配列番号1の204番目のグルタミン酸がバリンに置換
(124)配列番号1の207番目のロイシンがグルタミン、ヒスチジンまたはアルギニンに置換
(125)配列番号1の209番目のスレオニンがアラニンに置換
(126)配列番号1の211番目のセリンがアルギニンまたはグリシンに置換
(127)配列番号1の213番目のグルタミン酸がバリンまたはイソロイシンに置換
(128)配列番号1の215番目のリジンがアルギニンまたはグルタミン酸に置換
(129)配列番号1の217番目のロイシンがアルギニンまたはグルタミンに置換
(130)配列番号1の218番目のロイシンがイソロイシン、メチオニンまたはリジンに置換
(131)配列番号1の219番目のグルタミンがプロリンまたはアルギニンに置換
(132)配列番号1の223番目のロイシンがアルギニン、グルタミンまたはメチオニンに置換
(133)配列番号1の224番目のグルタミンがアルギニンに置換
(134)配列番号1の225番目のロイシンがグルタミンに置換
(135)配列番号1の227番目のフェニルアラニンがイソロイシンに置換
(136)配列番号1の230番目のチロシンがヒスチジンまたはフェニルアラニンに置換
(137)配列番号1の231番目のメチオニンがリジンまたはアルギニンに置換
(138)配列番号1の233番目のセリンがグリシンまたはアスパラギンに置換
(139)配列番号1の234番目のリジンがグルタミン酸に置換
(140)配列番号1の240番目のアスパラギンがグリシンに置換
(141)配列番号1の244番目のグルタミン酸がバリンに置換
(142)配列番号1の245番目のチロシンがヒスチジンまたはグルタミン酸に置換
(143)配列番号1の246番目のグルタミンがアルギニンまたはリジンに置換
(144)配列番号1の248番目のロイシンがイソロイシンに置換
(145)配列番号1の249番目のスレオニンがアラニンまたはセリンに置換
(146)配列番号1の250番目のアラニンがバリンに置換
(147)配列番号1の251番目のアルギニンがセリンに置換
(148)配列番号1の252番目のアルギニンがヒスチジンに置換
(149)配列番号1の253番目のグルタミン酸がグリシンに置換
(150)配列番号1の257番目のロイシンがアルギニンまたはグルタミンに置換
(151)配列番号1の261番目のグルタミン酸がバリンまたはアラニンに置換
(152)配列番号1の262番目のアラニンがバリンに置換
(153)配列番号1の263番目のアラニンがセリンに置換
(154)配列番号1の264番目のスレオニンがセリンに置換
(155)配列番号1の265番目のグルタミン酸がアラニンまたはグリシンに置換
(156)配列番号1の268番目のアスパラギンがセリン、イソロイシンまたはスレオニンに置換
(157)配列番号1の270番目のロイシンがヒスチジン、アルギニンまたはバリンに置換
(158)配列番号1の271番目のリジンがアルギニンに置換
(159)配列番号1の272番目のアルギニンがグルタミンに置換
(160)配列番号1の277番目のグルタミン酸がバリンに置換
(161)配列番号1の279番目のグルタミンがアルギニンまたはヒスチジンに置換
(162)配列番号1の282番目のグリシンがアスパラギン酸に置換
(163)配列番号1の283番目のロイシンがプロリンに置換
(164)配列番号1の285番目のロイシンがアルギニンまたはヒスチジンに置換
(165)配列番号1の286番目のプロリンがグルタミン、アルギニンまたはグルタミン酸に置換
(166)配列番号1の287番目のスレオニンがイソロイシン、プロリン、アラニンまたはバリンに置換
(167)配列番号1の288番目のプロリンがアラニン、セリンまたはスレオニンに置換
(168)配列番号1の289番目のバリンがアラニン、アスパラギン酸、グリシン、ロイシンまたはイソロイシンに置換
なお、本発明の製造方法で製造するFc結合性タンパク質では、分析・精製の迅速化やタンパク質の安定化などの目的でN末端側、C末端側またはFc結合性タンパク質内部に、ポリヒスチジンタグやC−mycタグといった任意のペプチドを追加してもよい。
【0029】
本発明の製造方法で製造する、Fc結合性タンパク質をコードするポリヌクレオチドを作製するには、Fc結合性タンパク質のアミノ酸配列をヌクレオチド配列に変換する必要があるが、Fc結合性タンパク質をコードするポリヌクレオチドにおけるコドン使用頻度(codon usage)は必ずしもヒトに合わせる必要はなく、例えば、大腸菌におけるレアコドン(rare codon、当該宿主におけるコドンの使用頻度が少ないもの)の全部または一部を、コードするアミノ酸を同一のまま、大腸菌の翻訳機構において利用頻度が高いコドン(codon)に変換したポリヌクレオチドであってもよい。なお、コドンの使用頻度の情報は公的データベース(例えば、かずさDNA研究所のホームページにあるCodon Usage Databaseなど)から得ることができる。
【0030】
本発明の製造方法で用いる、Fc結合性タンパク質をコードするポリヌクレオチドを挿入するベクターに特に限定はなく、大腸菌で異種タンパク質を発現可能なベクターの中から、適宜選択し使用すればよい。具体例としては、pTrc99a(ライフテクノロジーズ社製)があげられる。
【0031】
本発明の製造方法で組換え大腸菌より発現したFc結合性タンパク質を定量する方法としては、一般的なSDS−PAGEでタンパク質を分離した後に色素や免疫学的方法で染色して比色定量する方法やELISA法などの方法を用いることができるが、ELISA法による活性定量が簡便で好ましい。ELISA法におけるFc結合性タンパク質の組み合わせは前記タンパク質が定量できる方法であれば特に限定されないが、
(a)Fc結合性タンパク質に対する抗体
(b)Fc結合性タンパク質
(c)酵素標識抗Fc結合性タンパク質抗体
の順番で重ねたサンドイッチ法による方法を好ましく用いることができる。
ここで酵素標識抗Fc結合性タンパク質抗体として、アルカリフォスファターゼや西洋ワサビペルオキシダーゼなどの酵素で標識された抗Fc結合性タンパク質抗体が好ましく使用することができる。また、ELISAによる検出法についても特に限定されないが、標識に用いた酵素の特異的発色試薬、蛍光試薬または化学発光試薬が市販されており、それらを標識に用いた酵素に応じて任意に使用することができる。例えば、西洋ワサビペルオキシダーゼを用いた場合は発色基質をペルオキシダーゼと過酸化水素で酸化反応させて比色定量する方法があり、例えばTMB 2−Component Microwell Peroxidase Substrate Kit(フナコシ社製)などの市販の試薬で発色させた後、市販の測定装置(例えばマイクロタイタープレートリーダMPR4i、東ソー社製)で比色定量することができる。
【発明の効果】
【0032】
本発明は、タンパク質をコードするポリヌクレオチドを含む発現ベクターで大腸菌W3110株(ATCC 27235)株を形質転換して得られた組換え大腸菌を培養してタンパク質を製造する方法において、組換え大腸菌を培養する際に用いる培地に添加するマグネシウム塩の量を培地1Lあたりマグネシウムイオンとして2.8×10−2mol以上とすることを特徴としており、前記製造方法により組換え大腸菌の効率的な培養および組換え大腸菌からの効率的なタンパク質の発現を実現することができる。なお、ここで添加するマグネシウム塩を、培養開始時にその一部を添加し、残部を培養中に添加すると、より高効率に組換え大腸菌からタンパク質を発現させることができる。
【図面の簡単な説明】
【0033】
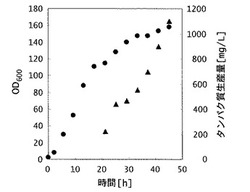
【図1】実施例1の製造方法における、形質転換体(組換え大腸菌)の増殖およびヒトFcγRI生産量を示した図。図中、横軸は時間(単位は時間)を示し、縦軸のうち、丸は微生物の増殖量を示す600nmにおける吸光度(単位は任意単位)を、三角は生産されたヒトFcγRI生産量(単位はmg/L)をそれぞれ示す。
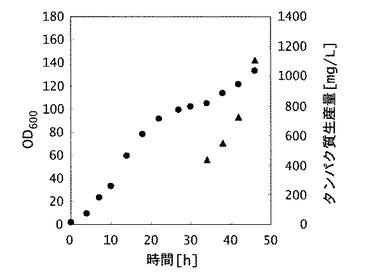
【図2】実施例3の製造方法における、形質転換体(組換え大腸菌)の増殖およびヒトFcγRI生産量を示した図。図中、横軸は時間(単位は時間)を示し、縦軸のうち、丸は微生物の増殖量を示す600nmにおける吸光度(単位は任意単位)を、三角は生産されたヒトFcγRI生産量(単位はmg/L)をそれぞれ示す。
【実施例】
【0034】
以下、本発明の製造方法によるFc結合性タンパク質の製造を例として、本発明をさらに詳細に説明するが、本発明はこれらに限定されるものではない。
【0035】
実施例1
Fc結合性タンパク質を発現可能な形質転換体(組換え大腸菌)を、硫酸マグネシウム七水和物を6.9g/L(培地)(マグネシウムイオンとして2.8×10−2mol/L(培地))添加した培地で培養することで、Fc結合性タンパク質の生産を行なった。
(A)配列番号2に記載のアミノ酸配列からなるヒトFcγRIをコードする、配列番号3に記載のヌクレオチド配列からなるポリヌクレオチドを、公知の方法(例えば特許文献2の方法)により、プラスミドpTrc99aのNcoIサイトとHindIIIサイトの間に挿入することで、発現ベクター(pTrcperBFcRm32)を作製した。なお、配列番号2のうち、1番目のメチオニンから26番目のアラニンまでがMalEシグナルペプチド、27番目のリジンから33番目のグリシンまでがリンカーペプチド、34番目のグルタミンから307番目のバリンまでがFc結合性タンパク質FcRm32のアミノ酸配列、308番目から313番目のヒスチジンがポリヒスチジンタグである。なお、FcRm32は、配列番号1に記載のアミノ酸配列からなるヒト天然型FcγRIのうち、16番目のグルタミンから289番目のバリンまでの領域に相当し、かつ、下記に示すアミノ酸置換を行なったFc結合性タンパク質である。
(1)配列番号1の20番目のスレオニンをプロリンに置換
(2)配列番号1の25番目のスレオニンをリジンに置換
(3)配列番号1の36番目のグルタミン酸をグリシンに置換
(4)配列番号1の38番目のスレオニンをセリンに置換
(5)配列番号1の45番目のバリンをアラニンに置換
(6)配列番号1の46番目のロイシンをプロリンに置換
(7)配列番号1の49番目のプロリンをセリンに置換
(8)配列番号1の60番目のグリシンをアスパラギン酸に置換
(9)配列番号1の63番目のスレオニンをイソロイシンに置換
(10)配列番号1の65番目のスレオニンをアラニンに置換
(11)配列番号1の69番目のセリンをスレオニンに置換
(12)配列番号1の71番目のアルギニンをヒスチジンに置換
(13)配列番号1の77番目のバリンをグルタミン酸に置換
(14)配列番号1の78番目のアスパラギンをアスパラギン酸に置換
(15)配列番号1の100番目のイソロイシンがバリンに置換
(16)配列番号1の114番目のフェニルアラニンがロイシンに置換
(17)配列番号1の133番目のチロシンをヒスチジンに置換
(18)配列番号1の139番目のアルギニンをヒスチジンに置換
(19)配列番号1の149番目のトリプトファンをアルギニンに置換
(20)配列番号1の156番目のロイシンをプロリンに置換
(21)配列番号1の160番目のイソロイシンをスレオニンに置換
(22)配列番号1の163番目のアスパラギンをセリンに置換
(23)配列番号1の173番目のリジンをアルギニンに置換
(24)配列番号1の181番目のイソロイシンをスレオニンに置換
(25)配列番号1の195番目のアスパラギンをスレオニンに置換
(26)配列番号1の203番目のロイシンをヒスチジンに置換
(27)配列番号1の206番目のアスパラギンをスレオニンに置換
(28)配列番号1の207番目のロイシンをグルタミンに置換
(29)配列番号1の231番目のメチオニンをリジンに置換
(30)配列番号1の240番目のアスパラギンをアスパラギン酸に置換
(31)配列番号1の283番目のロイシンをヒスチジンに置換
(32)配列番号1の285番目のロイシンをグルタミンに置換
(B)(A)で作製した発現ベクターpTrcperBFcRm32を用いて、公知の方法(例えば特許文献2の方法)により、大腸菌W3110株(ATCC 27235)を形質転換した。
(C)形質転換体を、100mLの2×YT培地(バクトトリプトン:16g/L、酵母エキス:10g/L、塩化ナトリウム:5g/L、アンピシリン:0.1mg/L)を入れた、500mL容バッフル付三角フラスコに植菌し、30℃で16時間、毎分130回の回転速度、回転半径1インチで前培養を行なった。
(D)表1に示す培地組成のうち、酵母エキス、リン酸三ナトリウム十二水和物およびリン酸水素二ナトリウム十二水和物を投入した培地約1.4Lを3Lの発酵槽に入れ、121℃で20分間滅菌後、グルコース、硫酸鉄(II)四水和物および塩化マンガン(II)四水和物は表1に示す濃度に、硫酸マグネシウム七水和物は6.9g/L(培地)に、それぞれなるよう添加し、さらに(A)の前培養液90mLを添加して、本培養を行なった。
【0036】
【表1】

【0037】
培養装置はエイブル社製BMS−03PIを使用し、通気した空気速度は1.8L/分に、培養温度は30℃に、pHは6.9から7.1にそれぞれ設定し、培養中におけるpHの変動は、14%アンモニア水または50%リン酸の添加により前記範囲に制御した。培養中はグルコース分析計(YSI社製2700)を用いて定期的にグルコース濃度を測定した。炭素源の供給には700g/Lのグルコースを、窒素源の供給には400g/Lの酵母エキス(オリエンタル酵母工業製)を、それぞれ使用した。供給は、炭素源の溶液と窒素源の溶液を容量比1:1で混合して行なった。エイブル社製DO(溶存酸素)電極による信号を、エイブル社製培養制御プログラムをインストールしたパーソナルコンピューターにより検出することで、本培養初期に投入したグルコース(20g/L)が消費されたことを検知し、溶存酸素濃度が40%飽和を超えた時点で流加ポンプを起動してグルコース350g/L、酵母エキス200g/Lの混合液を供給した。供給にはワトソン・マーロウ社製定量ポンプ101Uの高速型を使用した。微生物の増殖は培養液の600nmの濁度(OD600)により測定した。
(E)OD600が90に達したとき、すなわち培養開始13時間後に培養温度を25℃に下げ、IPTGを終濃度0.5mMとなるよう培養液に添加することで、Fc結合性タンパク質の生産誘導をかけた。
【0038】
48時間培養を行なったところ、培養液の濁度は160に達した。予め求めた濁度と菌体密度の相関式より、乾燥菌体収量は培養液1Lあたり46gと求められた。なお、グルコース濃度は、グルコースと酵母エキスの供給を開始した、培養開始8時間後から培養終了(48時間後)までの期間中、0から0.1g/Lに維持された。また、培養終了後の培養液に含まれるFc結合性タンパク質(ヒトFcγRI)の生産量をELISA法により定量した結果、培養液1Lあたり1100mgであった(図1)。
【0039】
実施例2
培養開始時の培地として添加する硫酸マグネシウム七水和物を13.5g/L(培地)(マグネシウムイオンとして5.5×10−2mol/L(培地))とした他は、実施例1と同様な方法でFc結合性タンパク質を発現する形質転換体を培養した。
【0040】
48時間培養を行なったところ、培養液の濁度は140に達した(乾燥菌体収量40g/L)。また、培養終了後の培養液に含まれるFc結合性タンパク質(ヒトFcγRI)の生産量をELISA法により定量した結果、培養液1Lあたり1100mgであった。
【0041】
比較例1
培養開始時の培地として添加する硫酸マグネシウム七水和物を3.8g/L(培地)(マグネシウムイオンとして1.5×10−2mol/L(培地))とし、ヒトFcγRIの生産誘導をかけるときの濁度をOD600=70とした他は、実施例1と同様な方法でFc結合性タンパク質を発現する形質転換体を培養した。
【0042】
48時間培養を行なったところ、培養液の濁度は85に達した(乾燥菌体収量24g/L(培養液))。また、培養終了後の培養液に含まれるFc結合性タンパク質(ヒトFcγRI)の生産量をELISA法により定量した結果、培養液1Lあたり300mgであった。
【0043】
実施例1および2ならびに比較例1の結果をまとめたものを表2に示す。表2から分かるように、培養開始時の培地に添加する硫酸マグネシウム七水和物の量を6.9g/L(培地)(マグネシウムイオンとして2.8×10−2mol/L(培地))以上とすると、ヒトFcγRIの生産量が増大することがわかる。
【0044】
【表2】
【0045】
実施例3
実施例1で作製した、Fc結合性タンパク質を発現可能な形質転換体を、硫酸マグネシウム七水和物を1.0g/L(培地)(マグネシウムイオンとして4.1×10−3mol/L(培地))添加した培地で培養を開始し、その後硫酸マグネシウム七水和物を6.0g/L(培地)(マグネシウムイオンとして2.4×10−3mol/L(培地))添加して培養することで、Fc結合性タンパク質の生産を行なった。
(A)表1に示す培地に硫酸マグネシウム七水和物を1.0g/L添加して培養を開始した後、グルコース350g/L、酵母エキス200g/L、硫酸マグネシウム七水和物12.75g/Lの混合液を流加する他は、実施例1と同様な方法でFc結合性タンパク質を発現する形質転換体を培養した。培養終了までに、培養槽に投入される硫酸マグネシウム七水和物は最終的に7g/L(培地)となる。
【0046】
48時間培養を行なったところ、培養液の濁度は130に達した(乾燥菌体収量37g/L)。また、培養終了後の培養液に含まれるFc結合性タンパク質(ヒトFcγRI)の生産量をELISA法により定量した結果、培養液1Lあたり1100mgであった(図2)。
【0047】
実施例4
培養開始時の培地として添加する硫酸マグネシウム七水和物を1.0g/L(培地)(マグネシウムイオンとして4.1×10−3mol/L(培地))とし、培養中に添加する硫酸マグネシウム七水和物を12g/L(培地)(マグネシウムイオンとして4.9×10−2mol/L(培地))とした他は、実施例3と同様な方法でFc結合性タンパク質を発現する形質転換体を培養した。
【0048】
具体的には、表1に示す培地に硫酸マグネシウム七水和物を1.0g/L添加して培養を開始した後、グルコース350g/L、酵母エキス200g/L、硫酸マグネシウム七水和物27g/Lの混合液を流加する他は、実施例1と同様な方法でFc結合性タンパク質を発現する形質転換体を培養した。培養終了までに、培養槽に投入される硫酸マグネシウム七水和物は最終的に13g/L(培地)となる。
【0049】
48時間培養を行なったところ、培養液の濁度は180に達した(乾燥菌体収量52g/L)。また、培養終了後の培養液に含まれるFc結合性タンパク質(ヒトFcγRI)の生産量をELISA法により定量した結果、培養液1Lあたり1150mgであった。
【0050】
比較例2
培養開始時の培地として添加する硫酸マグネシウム七水和物を1.0g/L(培地)(マグネシウムイオンとして4.1×10−3mol/L(培地))とし、培養中に添加する硫酸マグネシウム七水和物を3g/L(培地)(マグネシウムイオンとして1.2×10−2mol/L(培地))とした他は、実施例3と同様な方法でFc結合性タンパク質を発現する形質転換体を培養した。
【0051】
具体的には、表1に示す培地に硫酸マグネシウム七水和物を1.0g/L添加して培養を開始した後、グルコース350g/L、酵母エキス200g/L、硫酸マグネシウム七水和物6.75g/Lの混合液を流加する他は、実施例1と同様な方法でFc結合性タンパク質を発現する形質転換体を培養した。培養終了までに、培養槽に投入される硫酸マグネシウム七水和物は最終的に4g/L(培地)となる。
【0052】
48時間培養を行なったところ、培養液の濁度は130に達した(乾燥菌体収量37g/L)。また、培養終了後の培養液に含まれるFc結合性タンパク質(ヒトFcγRI)の生産量をELISA法により定量した結果、培養液1Lあたり400mgであった。
【0053】
実施例3および4ならびに比較例2の結果をまとめたものを表3に示す。なお表3における、硫酸マグネシウム濃度は、培養開始時に添加した硫酸マグネシウム七水和物と培養中に添加した硫酸マグネシウム七水和物との合算値(総量)である。表3から分かるように、培養開始時および培養中に添加する硫酸マグネシウム七水和物の量を合計7g/L(培地)(マグネシウムイオンとして2.8×10−2mol/L(培地))以上とすると、ヒトFcγRIの生産量が増大することがわかる。
【0054】
【表3】
【0055】
なお、実施例1の結果と実施例3の結果とを比較すると、実施例3のほうが実施例1よりも菌体あたりのヒトFcγRIの生産量が向上している。このことから、培地にマグネシウム塩を添加する際は、培養開始時に一度に添加する(実施例1)よりも、培養開始時に一部を添加し、培養中に残部を供給(流加)する(実施例3)ほうが好ましいことが分かる。
【技術分野】
【0001】
本発明は、遺伝子工学的手法により得られた、遺伝子組換えタンパク質を発現可能な大腸菌を用いて、組換えタンパク質、特にFc結合性タンパク質を効率的に製造する方法に関する。特に本発明は、前記大腸菌を効率よく培養することで、組換えタンパク質を効率的に製造する方法に関する。
【背景技術】
【0002】
大腸菌を利用した組換えタンパク質の生産は、特許文献1をはじめとし、これまでにも多くの例が報告されている。組換えタンパク質を効率的に生産するためには、大腸菌細胞を高密度の状態で培養することが必要である。大腸菌の菌体には炭素、窒素の他に、多種の金属元素を含むことが知られている。そのため大腸菌を効率よく培養するには、培地成分に、炭素源や窒素源のみならず、金属塩も含む必要があり、かつ、培地組成を大腸菌に適合するようにすることが重要である。
【先行技術文献】
【特許文献】
【0003】
【特許文献1】特表2000−501936号公報
【特許文献2】特開2008−245580号公報
【非特許文献】
【0004】
【非特許文献1】J.V.Ravetch等,Annu.Rev.Immunol.,9,457,1991
【非特許文献2】Toshiyuki Takai,Jpn.J.Clin.Immunol.,28,318,2005
【非特許文献3】A.Paetz等,Biochem.Biophys.Res.Commun.,338,1811,2005
【非特許文献4】J.M.Allen等,Science,243,378,1989
【発明の概要】
【発明が解決しようとする課題】
【0005】
タンパク質のうちFc受容体はFc結合性タンパク質の一つであり、免疫グロブリン分子のFc領域に結合する一群の分子である。Fc受容体はその結合する免疫グロブリンの種類によって分類されており、IgGのFc領域に結合するFcγ受容体、IgEのFc領域に結合するFcε受容体、IgAのFc領域に結合するFcα受容体などがある(非特許文献1)。また、各受容体は、その構造の違いによりさらに細かく分類され、Fcγ受容体の場合、FcγRI、FcγRII、FcγRIIIの存在が報告されている(非特許文献1)。
【0006】
Fcγ受容体の一つであるFcγRIは単球とマクロファージ中で発現しており、好中球ではγインターフェロンにより誘導的に発現される(非特許文献1)。また、FcγRIはIgGに対する結合親和性が高く、その平衡解離定数(Kd)は10−8M以下である(非特許文献2)。FcγRIは、細胞外領域、細胞膜貫通領域、細胞質内領域に区分され、IgGとの結合は、IgGのFc領域とFcγRIの細胞外領域で起こり、その後細胞質へとシグナルが伝達される。FcγRIはIgGとの結合に直接関わる分子量約42000のα鎖と、γ鎖の2種類のサブユニットによって構成されており、γ鎖は細胞膜と細胞外領域との境界で共有結合することでホモダイマーを形成している(非特許文献3)。FcγRIはIgG1から4まであるサブクラスのうち、IgG1とIgG3と特に強く結合し、IgG2またはIgG4との結合は弱いことが知られている。
【0007】
ヒトFc受容体のうち、ヒトFcγRIα鎖のアミノ酸配列およびヌクレオチド配列(配列番号1)は、Allen等により明らかにされており(非特許文献4)、ExPASy(Primary accession number:P12314)などの公的データベースにも公表されている。また、ヒトFcγRIの構造上の機能ドメイン、細胞膜を貫通するためのシグナルペプチド配列、細胞膜貫通領域の位置についても同様に公表されている。配列番号1のうち、1番目のメチオニン(Met)から15番目のグリシン(Gly)までがシグナルペプチドであり、16番目のグルタミン(Gln)から292番目のヒスチジン(His)までが細胞外領域の機能ドメイン、293番目のバリン(Val)から374番目のスレオニン(Thr)までが細胞膜貫通領域および細胞内領域とされている。
【0008】
近年になり、Fc受容体の予想外の免疫抑制的な生物学的特性は、特に自己免疫疾患または自己免疫症候群、移植物の拒絶および悪性リンパ増殖の領域において医薬として注目を浴びつつある(非特許文献2)。また、FcγRIの機能である抗体の吸着能は各種抗体精製用クロマトグラフィーゲルの捕捉機能を担うタンパク質としても利用することができる。
【0009】
Fc受容体を安価に製造することを目的に、Fc受容体を発現可能な遺伝子組換え体を利用した製造方法についてこれまで検討されており、例えば、組換え大腸菌を用いたFcγRIの製造方法が報告されている(特許文献2)。しかしながら、大腸菌を用いた発現系では、FcγRIの発現量が極めて低いという問題点があった。
【0010】
そこで本発明の目的は、タンパク質を発現可能な大腸菌を培養することでタンパク質を製造する方法において、タンパク質、特にFc結合性タンパク質を効率的に製造するのに最適な金属塩の濃度、特にマグネシウム塩の濃度およびその供給方法を提供することにある。
【課題を解決するための手段】
【0011】
本発明者らは前記課題に対し、大腸菌培養における、金属塩の供給について鋭意検討した結果、本発明の完成に至った。
【0012】
すなわち、本発明は、以下の発明を包含する:
(i)タンパク質をコードするポリヌクレオチドを含む発現ベクターで大腸菌W3110株(ATCC 27235)を形質転換して得られた組換え大腸菌を、マグネシウム塩を添加した培地で培養することでタンパク質を製造する方法であって、マグネシウム塩の添加量が、培地1Lあたりマグネシウムイオンとして2.8×10−2mol以上である、前記方法。
【0013】
(ii)培養開始時の培地にマグネシウム塩の一部を添加し、残部を培養中の培地に添加する、(i)に記載の方法。
【0014】
(iii)マグネシウム塩とともに炭素源および窒素源を培養中の培地に添加する、(ii)に記載の方法。
【0015】
(iv)培養開始時の培地に添加するマグネシウム塩が、培地1Lあたりマグネシウムイオンとして4.0×10−3mol以上であり、培養中の培地に添加するマグネシウム塩が、培地1Lあたりマグネシウムイオンとして2.4×10−2mol以上である、請求項(ii)または(iii)に記載の方法。
【0016】
(v)タンパク質がFc結合性タンパク質である、(i)から(iv)のいずれかに記載の方法。
【0017】
(vi) Fc結合性タンパク質が、(1)配列番号1に記載のアミノ酸配列のうち少なくとも16番目のグルタミンから289番目のバリンまでのアミノ酸を含むタンパク質、または(2)配列番号1に記載のアミノ酸配列のうち少なくとも16番目のグルタミンから289番目のバリンまでのアミノ酸を含み、かつ前記アミノ酸のうちの一つ以上が他のアミノ酸に置換、挿入または欠失したタンパク質である、(v)に記載の方法。
【0018】
以下、本発明を詳細に説明する。
【0019】
本発明の製造方法では、タンパク質をコードするポリヌクレオチドを含む発現ベクターで大腸菌W3110株(ATCC 27235)を形質転換して得られた組換え大腸菌を適切な培地で培養することで、タンパク質を発現させ、製造する。組換え大腸菌を培養するのに用いる培地には、マグネシウム塩を添加する必要があるが、その添加量について検討した結果、培地1Lあたりマグネシウムイオンとして2.8×10−2mol以上とすると効率的にタンパク質を製造できることが判明した。なお、培地1Lあたりマグネシウムイオンとして2.8×10−2mol以上という条件は、培地に添加するマグネシウム塩としてよく用いられる硫酸マグネシウム七水和物の添加量に換算すると培地1Lあたり約7g以上、となる。本発明の製造方法で使用するマグネシウム塩としては、前述した硫酸マグネシウムでなくてもよく、塩化マグネシウムといった水溶性のマグネシウム塩であれば特に限定されない。
【0020】
その他の培地成分については、組換え大腸菌を増殖可能で、かつ発現ベクターに挿入したポリヌクレオチドがコードするタンパク質を発現可能なものであれば特に限定はない。培地に含まれる炭素源の一例としては、グルコース、フルクトース、マルトース、ショ糖、粗糖、糖蜜があげられる。培地に含まれる窒素源の一例としては、酵母エキス、ポリペプトン、カゼインおよびその代謝物、コーンスティープリカー、大豆タンパク質、肉エキス、魚肉エキスがあげられるが、特に酵母エキスが好ましい。なお、ナトリウム塩、鉄塩やマンガン塩など、マグネシウム塩以外の金属塩をさらに培地に添加してもよい。金属塩の具体例としては、リン酸二水素ナトリウム、リン酸水素二ナトリウム、リン酸二水素カリウム、リン酸水素二カリウム、塩化ナトリウム、硫酸鉄(II)、硫酸鉄(III)、塩化鉄(II)、塩化鉄(III)、クエン酸鉄、硫酸アンモニウム鉄、塩化カルシウム二水和物、硫酸カルシウム、硫酸亜鉛、塩化亜鉛、硫酸銅、塩化銅、硫酸マンガン、塩化マンガンがあげられる。さらに必要に応じ、ビオチン、ニコチン酸、チアミン、リボフラビン、イノシトール、ピリドキシンといったビタミン類を培地に添加してもよい。
【0021】
本発明の製造方法で組換え大腸菌を培養する際、培養開始時に炭素源や窒素源といった栄養源を一度に培地に投入すると、組換え大腸菌の増殖およびタンパク質製造が阻害されたり、酢酸などの副生成物が生産されるため、タンパク質の発現効率および得られたタンパク質の品質に悪影響を与える可能性がある。そのため、培養開始時に投入する栄養源は最小限とし、培養中に栄養源を追加供給(流加)しながら培養を行なう流加培養により、タンパク質を製造すると好ましい。なお、流加培養により組換え大腸菌を培養し、タンパク質を製造する際は、培地に添加するマグネシウム塩も、培養開始時の培地に一度に添加するのではなく、培養開始時の培地にその一部を添加し、残部は前記栄養源とともに培養中に添加するのが好ましい。
【0022】
本発明の好ましい製造方法である、流加培養によるタンパク質製造の一例を詳細に示す。必要最小限の炭素源および窒素源を投入した培地に、マグネシウム塩を培地1Lあたりマグネシウムイオンとして2.8×10−2mol以上添加し、さらに必要に応じてマグネシウム塩以外の金属塩やビタミン類等を添加した後、組換え大腸菌の培養を開始する。組換え大腸菌の増殖により炭素源が消費され所定の濃度まで低下した時点で、培養液中の炭素源を所定の濃度に維持しつつ、炭素源と窒素源を供給(流加)して培養することで、組換え大腸菌からタンパク質を発現させ、製造する。
【0023】
本発明の好ましい製造方法である、流加培養によるタンパク質製造の別の例として、必要最小限の炭素源および窒素源を投入した培地に、マグネシウム塩を培地1Lあたりマグネシウムイオンとして4.0×10−3mol以上添加し、さらに必要に応じてマグネシウム塩以外の金属塩やビタミン類等を添加した後、組換え大腸菌の培養を開始する。組換え大腸菌の増殖により炭素源が消費され所定の濃度まで低下した時点で、培養液中の炭素源を所定の濃度に維持しつつ、炭素源および窒素源を供給(流加)し、さらにマグネシウム塩を培地1Lあたりマグネシウムイオンとして2.4×10−3mol以上添加して培養することで、組換え大腸菌からタンパク質を発現させ、製造する。
【0024】
前述した流加培養によるタンパク質製造において、培養開始時に投入する炭素源の濃度は、炭素源がグルコースの場合、0から20g/Lとすると好ましい。また供給(流加)する炭素源と窒素源は、高濃度の溶液とすると培養液の液量増加を抑えられるため好ましく、炭素源をグルコース、窒素源を酵母エキスとした場合、供給(流加)するグルコース溶液の濃度は300から900g/Lに、酵母エキス溶液の濃度は150から500g/Lに、それぞれすると好ましい。炭素源と窒素源を供給(流加)する際維持する、所定の濃度とは、炭素源が枯渇せず有機酸などの副生成物が生産しない濃度をいう。炭素源をグルコースとした場合、炭素源濃度が5g/Lを超えた状態で培養を行なうと副生成物として有機酸が生産され、それが多量に蓄積することにより組換え大腸菌の増殖やFc結合性タンパク質の生産を抑制する可能性があるため、好ましくない。よって、炭素源をグルコースとした場合の所定の濃度とは、少なくとも5g/L以下、好ましくは1g/L以下、さらに好ましくは0.5g/L以下、最も好ましくは0.1g/L以下である。炭素源供給量のモニターは、例えば、炭素源を供給するポンプの稼働時間より行なうことができる。炭素源の枯渇をモニターする方法は特に限定はなく、一例として呼吸活性の低下によりモニターすることができる。呼吸活性の低下は、例えば培養液の溶存酸素濃度(DO)の上昇、排ガス中の酸素濃度の上昇、炭酸ガス濃度の低下、pHの上昇として現れる。特に、DOは応答が速いことから、炭素源の枯渇をモニターするのに好ましい指標である。その理由として、炭素源の濃度が十分に維持されている場合には、微生物の呼吸により酸素が消費されるためDOは酸素飽和濃度より低い値に維持されるが、炭素源が枯渇すると微生物の呼吸活性が低下しDOが急激に上昇するためである。なお、DOの急激な上昇に連動させて炭素源を追加する方法をDOスタット法という。DO以外の指標(排ガス組成、炭酸ガス濃度、pH上昇)でモニターした場合は、DOを指標とした場合と比較し応答が遅いという欠点がある。窒素源の供給(流加)量は、炭素源供給(流加)量に比例した量を供給(流加)すればよい。窒素源の供給(流加)方法は特に限定はなく、例えば、炭素源水溶液と窒素源水溶液を任意の濃度で混合し、当該混合液を培地に供給(流加)する方法が例示できる。
【0025】
本発明の製造方法における組換え大腸菌の培養条件は、製造に用いる組換え大腸菌が増殖しタンパク質を発現し得る条件であれば特に限定はないが、培養温度は15から50℃が好ましく、特に好ましい温度は20から33℃である。pHは6から8が好ましい。培養時間は任意に設定できるが、通常は数時間から100時間の間に設定される。
【0026】
本発明の好ましい製造方法では、組換え大腸菌からのタンパク質発現を誘導するために、培養開始から一定時間経過後、イソプロピル−β−チオガラクトピラノシド(IPTG)を培養液に添加して、さらに培養する。IPTGの濃度は最終濃度として0.01から2.0mMが好ましく、特に好ましい濃度は最終濃度0.1から2.0mMである。
【0027】
本発明の製造方法で製造可能なタンパク質の一例として、インシュリン、インターフェロン、インターロイキン、抗体、エリスロポエチン、成長ホルモン、およびそれらの受容体のタンパク質等があげられる。以降、本発明の製造方法で製造するタンパク質の好ましい例である、Fc結合性タンパク質について詳細に説明する。
【0028】
本発明の製造方法で製造する、Fc結合性タンパク質は、抗体のFc領域と結合する細胞表面に存在する受容体タンパク質である。Fc結合性タンパク質がヒトFc受容体の場合、ヒトFcγRI、ヒトFcγRIIa、ヒトFcγRIIb、ヒトFcγRIIIなどがあげられる。本発明の製造方法で製造するFc結合性タンパク質の一例として、
(A)配列番号1に記載のアミノ酸配列のうち少なくとも16番目のグルタミンから289番目のバリンまでのアミノ酸を含むタンパク質や、
(B)配列番号1に記載のアミノ酸配列のうち少なくとも16番目のグルタミンから289番目のバリンまでのアミノ酸を含み、かつ前記アミノ酸のうちの一つ以上が他のアミノ酸に置換、挿入または欠失したタンパク質、
があげられる。前記(B)の具体例として、配列番号1に記載のアミノ酸配列のうち少なくとも16番目から289番目までのアミノ酸を含み、かつ以下の(1)から(168)のいずれかに記載のアミノ酸置換を1つ以上含むFc結合性タンパク質があげられる(特願2011−038500号)。
(1)配列番号1の20番目のスレオニンがプロリンに置換
(2)配列番号1の25番目のスレオニンがリジンに置換
(3)配列番号1の38番目のスレオニンがアラニンまたはセリンに置換
(4)配列番号1の46番目のロイシンがアルギニンまたはプロリンに置換
(5)配列番号1の62番目のアラニンがバリンに置換
(6)配列番号1の63番目のスレオニンがイソロイシンに置換
(7)配列番号1の69番目のセリンがフェニルアラニンまたはスレオニンに置換
(8)配列番号1の71番目のアルギニンがヒスチジンに置換
(9)配列番号1の77番目のバリンがアラニンまたはグルタミン酸に置換
(10)配列番号1の78番目のアスパラギンがアスパラギン酸に置換
(11)配列番号1の94番目のアスパラギン酸がグルタミン酸に置換
(12)配列番号1の100番目のイソロイシンがバリンに置換
(13)配列番号1の110番目のセリンがアスパラギンに置換
(14)配列番号1の114番目のフェニルアラニンがロイシンに置換
(15)配列番号1の125番目のヒスチジンがアルギニンに置換
(16)配列番号1の131番目のロイシンがアルギニンまたはプロリンに置換
(17)配列番号1の149番目のトリプトファンがロイシンに置換
(18)配列番号1の156番目のロイシンがプロリンに置換
(19)配列番号1の160番目のイソロイシンがメチオニンに置換
(20)配列番号1の163番目のアスパラギンがセリンに置換
(21)配列番号1の195番目のアスパラギンがスレオニンに置換
(22)配列番号1の199番目のスレオニンがセリンに置換
(23)配列番号1の206番目のアスパラギンがリジン、セリンまたはスレオニンに置換
(24)配列番号1の207番目のロイシンがプロリンに置換
(25)配列番号1の218番目のロイシンがバリンに置換
(26)配列番号1の240番目のアスパラギンがアスパラギン酸に置換
(27)配列番号1の248番目のロイシンがセリンに置換
(28)配列番号1の283番目のロイシンがヒスチジンに置換
(29)配列番号1の285番目のロイシンがグルタミンに置換
(30)配列番号1の17番目のバリンがグリシンまたはグルタミン酸に置換
(31)配列番号1の19番目のスレオニンがイソロイシンに置換
(32)配列番号1の20番目のスレオニンがイソロイシンに置換
(33)配列番号1の25番目のスレオニンがメチオニンまたはアルギニンに置換
(34)配列番号1の27番目のグルタミンがプロリンまたはリジンに置換
(35)配列番号1の35番目のグルタミンがロイシン、メチオニンまたはアルギニンに置換
(36)配列番号1の36番目のグルタミン酸がグリシンに置換
(37)配列番号1の41番目のロイシンがメチオニンに置換
(38)配列番号1の42番目のヒスチジンがロイシンに置換
(39)配列番号1の44番目のグルタミン酸がアスパラギン酸に置換
(40)配列番号1の45番目のバリンがアラニンに置換
(41)配列番号1の46番目のロイシンがアラニン、アスパラギン、アスパラギン酸、グルタミン、グリシン、ヒスチジン、リジン、セリンまたはトリプトファンに置換
(42)配列番号1の47番目のヒスチジンがグルタミン、ロイシンまたはアスパラギンに置換
(43)配列番号1の49番目のプロリンがセリンまたはアラニンに置換
(44)配列番号1の50番目のグリシンがアルギニンまたはグルタミン酸に置換
(45)配列番号1の51番目のセリンがアラニン、スレオニン、ロイシン、プロリンまたはバリンに置換
(46)配列番号1の52番目のセリンがグリシンに置換
(47)配列番号1の53番目のセリンがロイシン、スレオニンまたはプロリンに置換
(48)配列番号1の55番目のグルタミンがアルギニンに置換
(49)配列番号1の57番目のフェニルアラニンがチロシンに置換
(50)配列番号1の58番目のロイシンがアルギニンに置換
(51)配列番号1の60番目のグリシンがアスパラギン酸に置換
(52)配列番号1の61番目のスレオニンがアラニンまたはセリンに置換
(53)配列番号1の62番目のアラニンがグルタミン酸に置換
(54)配列番号1の63番目のスレオニンがロイシン、フェニルアラニンに置換
(55)配列番号1の64番目のグルタミンがプロリン、ヒスチジン、ロイシン、リジンに置換
(56)配列番号1の65番目のスレオニンがアラニンまたはバリンに置換
(57)配列番号1の66番目のセリンがスレオニンに置換
(58)配列番号1の67番目のスレオニンがアラニンまたはセリンに置換
(59)配列番号1の69番目のセリンがアラニンに置換
(60)配列番号1の70番目のチロシンがヒスチジンまたはフェニルアラニンに置換
(61)配列番号1の71番目のアルギニンがチロシンに置換
(62)配列番号1の73番目のスレオニンがアラニンまたはセリンに置換
(63)配列番号1の74番目のセリンがフェニルアラニンに置換
(64)配列番号1の76番目のセリンがアスパラギンに置換
(65)配列番号1の77番目のバリンがアスパラギン酸またはリジンに置換
(66)配列番号1の78番目のアスパラギンがセリンまたはグリシンに置換
(67)配列番号1の80番目のセリンがアラニンに置換
(68)配列番号1の84番目のアルギニンがセリンに置換
(69)配列番号1の88番目のグリシンがセリンに置換
(70)配列番号1の89番目のロイシンがグルタミンまたはプロリンに置換
(71)配列番号1の90番目のセリンがグリシンに置換
(72)配列番号1の92番目のアルギニンがシステインまたはロイシンに置換
(73)配列番号1の96番目のイソロイシンがバリンまたはリジンに置換
(74)配列番号1の97番目のグルタミンがロイシンまたはリジンに置換
(75)配列番号1の101番目のヒスチジンがロイシンに置換
(76)配列番号1の102番目のアルギニンがセリンまたはロイシンに置換
(77)配列番号1の103番目のグリシンがアスパラギン酸またはセリンに置換
(78)配列番号1の111番目のセリンがアラニンに置換
(79)配列番号1の114番目のフェニルアラニンがアラニン、イソロイシン、メチオニン、プロリン、スレオニンまたはバリンに置換
(80)配列番号1の115番目のスレオニンがイソロイシンまたはフェニルアラニンに置換
(81)配列番号1の118番目のグルタミン酸がアスパラギン酸に置換
(82)配列番号1の121番目のアラニンがスレオニンまたはバリンに置換
(83)配列番号1の128番目のリジンがアルギニンまたはグリシンに置換
(84)配列番号1の129番目のアスパラギン酸がグリシンに置換
(85)配列番号1の131番目のロイシンがグルタミンに置換
(86)配列番号1の133番目のチロシンがヒスチジンまたはアルギニンに置換
(87)配列番号1の134番目のアスパラギンがセリンに置換
(88)配列番号1の137番目のチロシンがフェニルアラニンに置換
(89)配列番号1の138番目のチロシンがヒスチジンに置換
(90)配列番号1の139番目のアルギニンがヒスチジンに置換
(91)配列番号1の140番目のアスパラギンがアスパラギン酸に置換
(92)配列番号1の141番目のグリシンがアスパラギン酸またはバリンに置換
(93)配列番号1の142番目のリジンがグルタミン酸またはアルギニンに置換
(94)配列番号1の144番目のフェニルアラニンがイソロイシンに置換
(95)配列番号1の147番目のフェニルアラニンがセリンに置換
(96)配列番号1の148番目のヒスチジンがアルギニンまたはグルタミンに置換
(97)配列番号1の149番目のトリプトファンがアルギニンに置換
(98)配列番号1の151番目のセリンがスレオニンに置換
(99)配列番号1の152番目のアスパラギンがスレオニン、イソロイシンまたはプロリンに置換
(100)配列番号1の154番目のスレオニンがセリンに置換
(101)配列番号1の156番目のロイシンがヒスチジンに置換
(102)配列番号1の157番目のリジンがアルギニンに置換
(103)配列番号1の159番目のアスパラギンがスレオニンまたはアスパラギン酸に置換
(104)配列番号1の160番目のイソロイシンがスレオニン、バリンまたはロイシンに置換
(105)配列番号1の161番目のセリンがスレオニンに置換
(106)配列番号1の165番目のスレオニンがメチオニンに置換
(107)配列番号1の171番目のメチオニンがスレオニンに置換
(108)配列番号1の173番目のリジンがアルギニンに置換
(109)配列番号1の174番目のヒスチジンがグルタミンに置換
(110)配列番号1の177番目のスレオニンがセリンに置換
(111)配列番号1の181番目のイソロイシンがスレオニンに置換
(112)配列番号1の182番目のセリンがスレオニン、ロイシン、バリンまたはグルタミン酸に置換
(113)配列番号1の184番目のスレオニンがセリンに置換
(114)配列番号1の190番目のプロリンがセリンに置換
(115)配列番号1の193番目のバリンがロイシンに置換
(116)配列番号1の195番目のアスパラギンがアラニンに置換
(117)配列番号1の196番目のアラニンがセリンに置換
(118)配列番号1の198番目のバリンがグリシンまたはメチオニンに置換
(119)配列番号1の199番目のスレオニンがアラニンに置換
(120)配列番号1の200番目のセリンがグリシンまたはアルギニンに置換
(121)配列番号1の202番目のロイシンがメチオニンに置換
(122)配列番号1の203番目のロイシンがヒスチジン、グルタミン、チロシン、アルギニン、プロリンに置換
(123)配列番号1の204番目のグルタミン酸がバリンに置換
(124)配列番号1の207番目のロイシンがグルタミン、ヒスチジンまたはアルギニンに置換
(125)配列番号1の209番目のスレオニンがアラニンに置換
(126)配列番号1の211番目のセリンがアルギニンまたはグリシンに置換
(127)配列番号1の213番目のグルタミン酸がバリンまたはイソロイシンに置換
(128)配列番号1の215番目のリジンがアルギニンまたはグルタミン酸に置換
(129)配列番号1の217番目のロイシンがアルギニンまたはグルタミンに置換
(130)配列番号1の218番目のロイシンがイソロイシン、メチオニンまたはリジンに置換
(131)配列番号1の219番目のグルタミンがプロリンまたはアルギニンに置換
(132)配列番号1の223番目のロイシンがアルギニン、グルタミンまたはメチオニンに置換
(133)配列番号1の224番目のグルタミンがアルギニンに置換
(134)配列番号1の225番目のロイシンがグルタミンに置換
(135)配列番号1の227番目のフェニルアラニンがイソロイシンに置換
(136)配列番号1の230番目のチロシンがヒスチジンまたはフェニルアラニンに置換
(137)配列番号1の231番目のメチオニンがリジンまたはアルギニンに置換
(138)配列番号1の233番目のセリンがグリシンまたはアスパラギンに置換
(139)配列番号1の234番目のリジンがグルタミン酸に置換
(140)配列番号1の240番目のアスパラギンがグリシンに置換
(141)配列番号1の244番目のグルタミン酸がバリンに置換
(142)配列番号1の245番目のチロシンがヒスチジンまたはグルタミン酸に置換
(143)配列番号1の246番目のグルタミンがアルギニンまたはリジンに置換
(144)配列番号1の248番目のロイシンがイソロイシンに置換
(145)配列番号1の249番目のスレオニンがアラニンまたはセリンに置換
(146)配列番号1の250番目のアラニンがバリンに置換
(147)配列番号1の251番目のアルギニンがセリンに置換
(148)配列番号1の252番目のアルギニンがヒスチジンに置換
(149)配列番号1の253番目のグルタミン酸がグリシンに置換
(150)配列番号1の257番目のロイシンがアルギニンまたはグルタミンに置換
(151)配列番号1の261番目のグルタミン酸がバリンまたはアラニンに置換
(152)配列番号1の262番目のアラニンがバリンに置換
(153)配列番号1の263番目のアラニンがセリンに置換
(154)配列番号1の264番目のスレオニンがセリンに置換
(155)配列番号1の265番目のグルタミン酸がアラニンまたはグリシンに置換
(156)配列番号1の268番目のアスパラギンがセリン、イソロイシンまたはスレオニンに置換
(157)配列番号1の270番目のロイシンがヒスチジン、アルギニンまたはバリンに置換
(158)配列番号1の271番目のリジンがアルギニンに置換
(159)配列番号1の272番目のアルギニンがグルタミンに置換
(160)配列番号1の277番目のグルタミン酸がバリンに置換
(161)配列番号1の279番目のグルタミンがアルギニンまたはヒスチジンに置換
(162)配列番号1の282番目のグリシンがアスパラギン酸に置換
(163)配列番号1の283番目のロイシンがプロリンに置換
(164)配列番号1の285番目のロイシンがアルギニンまたはヒスチジンに置換
(165)配列番号1の286番目のプロリンがグルタミン、アルギニンまたはグルタミン酸に置換
(166)配列番号1の287番目のスレオニンがイソロイシン、プロリン、アラニンまたはバリンに置換
(167)配列番号1の288番目のプロリンがアラニン、セリンまたはスレオニンに置換
(168)配列番号1の289番目のバリンがアラニン、アスパラギン酸、グリシン、ロイシンまたはイソロイシンに置換
なお、本発明の製造方法で製造するFc結合性タンパク質では、分析・精製の迅速化やタンパク質の安定化などの目的でN末端側、C末端側またはFc結合性タンパク質内部に、ポリヒスチジンタグやC−mycタグといった任意のペプチドを追加してもよい。
【0029】
本発明の製造方法で製造する、Fc結合性タンパク質をコードするポリヌクレオチドを作製するには、Fc結合性タンパク質のアミノ酸配列をヌクレオチド配列に変換する必要があるが、Fc結合性タンパク質をコードするポリヌクレオチドにおけるコドン使用頻度(codon usage)は必ずしもヒトに合わせる必要はなく、例えば、大腸菌におけるレアコドン(rare codon、当該宿主におけるコドンの使用頻度が少ないもの)の全部または一部を、コードするアミノ酸を同一のまま、大腸菌の翻訳機構において利用頻度が高いコドン(codon)に変換したポリヌクレオチドであってもよい。なお、コドンの使用頻度の情報は公的データベース(例えば、かずさDNA研究所のホームページにあるCodon Usage Databaseなど)から得ることができる。
【0030】
本発明の製造方法で用いる、Fc結合性タンパク質をコードするポリヌクレオチドを挿入するベクターに特に限定はなく、大腸菌で異種タンパク質を発現可能なベクターの中から、適宜選択し使用すればよい。具体例としては、pTrc99a(ライフテクノロジーズ社製)があげられる。
【0031】
本発明の製造方法で組換え大腸菌より発現したFc結合性タンパク質を定量する方法としては、一般的なSDS−PAGEでタンパク質を分離した後に色素や免疫学的方法で染色して比色定量する方法やELISA法などの方法を用いることができるが、ELISA法による活性定量が簡便で好ましい。ELISA法におけるFc結合性タンパク質の組み合わせは前記タンパク質が定量できる方法であれば特に限定されないが、
(a)Fc結合性タンパク質に対する抗体
(b)Fc結合性タンパク質
(c)酵素標識抗Fc結合性タンパク質抗体
の順番で重ねたサンドイッチ法による方法を好ましく用いることができる。
ここで酵素標識抗Fc結合性タンパク質抗体として、アルカリフォスファターゼや西洋ワサビペルオキシダーゼなどの酵素で標識された抗Fc結合性タンパク質抗体が好ましく使用することができる。また、ELISAによる検出法についても特に限定されないが、標識に用いた酵素の特異的発色試薬、蛍光試薬または化学発光試薬が市販されており、それらを標識に用いた酵素に応じて任意に使用することができる。例えば、西洋ワサビペルオキシダーゼを用いた場合は発色基質をペルオキシダーゼと過酸化水素で酸化反応させて比色定量する方法があり、例えばTMB 2−Component Microwell Peroxidase Substrate Kit(フナコシ社製)などの市販の試薬で発色させた後、市販の測定装置(例えばマイクロタイタープレートリーダMPR4i、東ソー社製)で比色定量することができる。
【発明の効果】
【0032】
本発明は、タンパク質をコードするポリヌクレオチドを含む発現ベクターで大腸菌W3110株(ATCC 27235)株を形質転換して得られた組換え大腸菌を培養してタンパク質を製造する方法において、組換え大腸菌を培養する際に用いる培地に添加するマグネシウム塩の量を培地1Lあたりマグネシウムイオンとして2.8×10−2mol以上とすることを特徴としており、前記製造方法により組換え大腸菌の効率的な培養および組換え大腸菌からの効率的なタンパク質の発現を実現することができる。なお、ここで添加するマグネシウム塩を、培養開始時にその一部を添加し、残部を培養中に添加すると、より高効率に組換え大腸菌からタンパク質を発現させることができる。
【図面の簡単な説明】
【0033】
【図1】実施例1の製造方法における、形質転換体(組換え大腸菌)の増殖およびヒトFcγRI生産量を示した図。図中、横軸は時間(単位は時間)を示し、縦軸のうち、丸は微生物の増殖量を示す600nmにおける吸光度(単位は任意単位)を、三角は生産されたヒトFcγRI生産量(単位はmg/L)をそれぞれ示す。
【図2】実施例3の製造方法における、形質転換体(組換え大腸菌)の増殖およびヒトFcγRI生産量を示した図。図中、横軸は時間(単位は時間)を示し、縦軸のうち、丸は微生物の増殖量を示す600nmにおける吸光度(単位は任意単位)を、三角は生産されたヒトFcγRI生産量(単位はmg/L)をそれぞれ示す。
【実施例】
【0034】
以下、本発明の製造方法によるFc結合性タンパク質の製造を例として、本発明をさらに詳細に説明するが、本発明はこれらに限定されるものではない。
【0035】
実施例1
Fc結合性タンパク質を発現可能な形質転換体(組換え大腸菌)を、硫酸マグネシウム七水和物を6.9g/L(培地)(マグネシウムイオンとして2.8×10−2mol/L(培地))添加した培地で培養することで、Fc結合性タンパク質の生産を行なった。
(A)配列番号2に記載のアミノ酸配列からなるヒトFcγRIをコードする、配列番号3に記載のヌクレオチド配列からなるポリヌクレオチドを、公知の方法(例えば特許文献2の方法)により、プラスミドpTrc99aのNcoIサイトとHindIIIサイトの間に挿入することで、発現ベクター(pTrcperBFcRm32)を作製した。なお、配列番号2のうち、1番目のメチオニンから26番目のアラニンまでがMalEシグナルペプチド、27番目のリジンから33番目のグリシンまでがリンカーペプチド、34番目のグルタミンから307番目のバリンまでがFc結合性タンパク質FcRm32のアミノ酸配列、308番目から313番目のヒスチジンがポリヒスチジンタグである。なお、FcRm32は、配列番号1に記載のアミノ酸配列からなるヒト天然型FcγRIのうち、16番目のグルタミンから289番目のバリンまでの領域に相当し、かつ、下記に示すアミノ酸置換を行なったFc結合性タンパク質である。
(1)配列番号1の20番目のスレオニンをプロリンに置換
(2)配列番号1の25番目のスレオニンをリジンに置換
(3)配列番号1の36番目のグルタミン酸をグリシンに置換
(4)配列番号1の38番目のスレオニンをセリンに置換
(5)配列番号1の45番目のバリンをアラニンに置換
(6)配列番号1の46番目のロイシンをプロリンに置換
(7)配列番号1の49番目のプロリンをセリンに置換
(8)配列番号1の60番目のグリシンをアスパラギン酸に置換
(9)配列番号1の63番目のスレオニンをイソロイシンに置換
(10)配列番号1の65番目のスレオニンをアラニンに置換
(11)配列番号1の69番目のセリンをスレオニンに置換
(12)配列番号1の71番目のアルギニンをヒスチジンに置換
(13)配列番号1の77番目のバリンをグルタミン酸に置換
(14)配列番号1の78番目のアスパラギンをアスパラギン酸に置換
(15)配列番号1の100番目のイソロイシンがバリンに置換
(16)配列番号1の114番目のフェニルアラニンがロイシンに置換
(17)配列番号1の133番目のチロシンをヒスチジンに置換
(18)配列番号1の139番目のアルギニンをヒスチジンに置換
(19)配列番号1の149番目のトリプトファンをアルギニンに置換
(20)配列番号1の156番目のロイシンをプロリンに置換
(21)配列番号1の160番目のイソロイシンをスレオニンに置換
(22)配列番号1の163番目のアスパラギンをセリンに置換
(23)配列番号1の173番目のリジンをアルギニンに置換
(24)配列番号1の181番目のイソロイシンをスレオニンに置換
(25)配列番号1の195番目のアスパラギンをスレオニンに置換
(26)配列番号1の203番目のロイシンをヒスチジンに置換
(27)配列番号1の206番目のアスパラギンをスレオニンに置換
(28)配列番号1の207番目のロイシンをグルタミンに置換
(29)配列番号1の231番目のメチオニンをリジンに置換
(30)配列番号1の240番目のアスパラギンをアスパラギン酸に置換
(31)配列番号1の283番目のロイシンをヒスチジンに置換
(32)配列番号1の285番目のロイシンをグルタミンに置換
(B)(A)で作製した発現ベクターpTrcperBFcRm32を用いて、公知の方法(例えば特許文献2の方法)により、大腸菌W3110株(ATCC 27235)を形質転換した。
(C)形質転換体を、100mLの2×YT培地(バクトトリプトン:16g/L、酵母エキス:10g/L、塩化ナトリウム:5g/L、アンピシリン:0.1mg/L)を入れた、500mL容バッフル付三角フラスコに植菌し、30℃で16時間、毎分130回の回転速度、回転半径1インチで前培養を行なった。
(D)表1に示す培地組成のうち、酵母エキス、リン酸三ナトリウム十二水和物およびリン酸水素二ナトリウム十二水和物を投入した培地約1.4Lを3Lの発酵槽に入れ、121℃で20分間滅菌後、グルコース、硫酸鉄(II)四水和物および塩化マンガン(II)四水和物は表1に示す濃度に、硫酸マグネシウム七水和物は6.9g/L(培地)に、それぞれなるよう添加し、さらに(A)の前培養液90mLを添加して、本培養を行なった。
【0036】
【表1】
【0037】
培養装置はエイブル社製BMS−03PIを使用し、通気した空気速度は1.8L/分に、培養温度は30℃に、pHは6.9から7.1にそれぞれ設定し、培養中におけるpHの変動は、14%アンモニア水または50%リン酸の添加により前記範囲に制御した。培養中はグルコース分析計(YSI社製2700)を用いて定期的にグルコース濃度を測定した。炭素源の供給には700g/Lのグルコースを、窒素源の供給には400g/Lの酵母エキス(オリエンタル酵母工業製)を、それぞれ使用した。供給は、炭素源の溶液と窒素源の溶液を容量比1:1で混合して行なった。エイブル社製DO(溶存酸素)電極による信号を、エイブル社製培養制御プログラムをインストールしたパーソナルコンピューターにより検出することで、本培養初期に投入したグルコース(20g/L)が消費されたことを検知し、溶存酸素濃度が40%飽和を超えた時点で流加ポンプを起動してグルコース350g/L、酵母エキス200g/Lの混合液を供給した。供給にはワトソン・マーロウ社製定量ポンプ101Uの高速型を使用した。微生物の増殖は培養液の600nmの濁度(OD600)により測定した。
(E)OD600が90に達したとき、すなわち培養開始13時間後に培養温度を25℃に下げ、IPTGを終濃度0.5mMとなるよう培養液に添加することで、Fc結合性タンパク質の生産誘導をかけた。
【0038】
48時間培養を行なったところ、培養液の濁度は160に達した。予め求めた濁度と菌体密度の相関式より、乾燥菌体収量は培養液1Lあたり46gと求められた。なお、グルコース濃度は、グルコースと酵母エキスの供給を開始した、培養開始8時間後から培養終了(48時間後)までの期間中、0から0.1g/Lに維持された。また、培養終了後の培養液に含まれるFc結合性タンパク質(ヒトFcγRI)の生産量をELISA法により定量した結果、培養液1Lあたり1100mgであった(図1)。
【0039】
実施例2
培養開始時の培地として添加する硫酸マグネシウム七水和物を13.5g/L(培地)(マグネシウムイオンとして5.5×10−2mol/L(培地))とした他は、実施例1と同様な方法でFc結合性タンパク質を発現する形質転換体を培養した。
【0040】
48時間培養を行なったところ、培養液の濁度は140に達した(乾燥菌体収量40g/L)。また、培養終了後の培養液に含まれるFc結合性タンパク質(ヒトFcγRI)の生産量をELISA法により定量した結果、培養液1Lあたり1100mgであった。
【0041】
比較例1
培養開始時の培地として添加する硫酸マグネシウム七水和物を3.8g/L(培地)(マグネシウムイオンとして1.5×10−2mol/L(培地))とし、ヒトFcγRIの生産誘導をかけるときの濁度をOD600=70とした他は、実施例1と同様な方法でFc結合性タンパク質を発現する形質転換体を培養した。
【0042】
48時間培養を行なったところ、培養液の濁度は85に達した(乾燥菌体収量24g/L(培養液))。また、培養終了後の培養液に含まれるFc結合性タンパク質(ヒトFcγRI)の生産量をELISA法により定量した結果、培養液1Lあたり300mgであった。
【0043】
実施例1および2ならびに比較例1の結果をまとめたものを表2に示す。表2から分かるように、培養開始時の培地に添加する硫酸マグネシウム七水和物の量を6.9g/L(培地)(マグネシウムイオンとして2.8×10−2mol/L(培地))以上とすると、ヒトFcγRIの生産量が増大することがわかる。
【0044】
【表2】
【0045】
実施例3
実施例1で作製した、Fc結合性タンパク質を発現可能な形質転換体を、硫酸マグネシウム七水和物を1.0g/L(培地)(マグネシウムイオンとして4.1×10−3mol/L(培地))添加した培地で培養を開始し、その後硫酸マグネシウム七水和物を6.0g/L(培地)(マグネシウムイオンとして2.4×10−3mol/L(培地))添加して培養することで、Fc結合性タンパク質の生産を行なった。
(A)表1に示す培地に硫酸マグネシウム七水和物を1.0g/L添加して培養を開始した後、グルコース350g/L、酵母エキス200g/L、硫酸マグネシウム七水和物12.75g/Lの混合液を流加する他は、実施例1と同様な方法でFc結合性タンパク質を発現する形質転換体を培養した。培養終了までに、培養槽に投入される硫酸マグネシウム七水和物は最終的に7g/L(培地)となる。
【0046】
48時間培養を行なったところ、培養液の濁度は130に達した(乾燥菌体収量37g/L)。また、培養終了後の培養液に含まれるFc結合性タンパク質(ヒトFcγRI)の生産量をELISA法により定量した結果、培養液1Lあたり1100mgであった(図2)。
【0047】
実施例4
培養開始時の培地として添加する硫酸マグネシウム七水和物を1.0g/L(培地)(マグネシウムイオンとして4.1×10−3mol/L(培地))とし、培養中に添加する硫酸マグネシウム七水和物を12g/L(培地)(マグネシウムイオンとして4.9×10−2mol/L(培地))とした他は、実施例3と同様な方法でFc結合性タンパク質を発現する形質転換体を培養した。
【0048】
具体的には、表1に示す培地に硫酸マグネシウム七水和物を1.0g/L添加して培養を開始した後、グルコース350g/L、酵母エキス200g/L、硫酸マグネシウム七水和物27g/Lの混合液を流加する他は、実施例1と同様な方法でFc結合性タンパク質を発現する形質転換体を培養した。培養終了までに、培養槽に投入される硫酸マグネシウム七水和物は最終的に13g/L(培地)となる。
【0049】
48時間培養を行なったところ、培養液の濁度は180に達した(乾燥菌体収量52g/L)。また、培養終了後の培養液に含まれるFc結合性タンパク質(ヒトFcγRI)の生産量をELISA法により定量した結果、培養液1Lあたり1150mgであった。
【0050】
比較例2
培養開始時の培地として添加する硫酸マグネシウム七水和物を1.0g/L(培地)(マグネシウムイオンとして4.1×10−3mol/L(培地))とし、培養中に添加する硫酸マグネシウム七水和物を3g/L(培地)(マグネシウムイオンとして1.2×10−2mol/L(培地))とした他は、実施例3と同様な方法でFc結合性タンパク質を発現する形質転換体を培養した。
【0051】
具体的には、表1に示す培地に硫酸マグネシウム七水和物を1.0g/L添加して培養を開始した後、グルコース350g/L、酵母エキス200g/L、硫酸マグネシウム七水和物6.75g/Lの混合液を流加する他は、実施例1と同様な方法でFc結合性タンパク質を発現する形質転換体を培養した。培養終了までに、培養槽に投入される硫酸マグネシウム七水和物は最終的に4g/L(培地)となる。
【0052】
48時間培養を行なったところ、培養液の濁度は130に達した(乾燥菌体収量37g/L)。また、培養終了後の培養液に含まれるFc結合性タンパク質(ヒトFcγRI)の生産量をELISA法により定量した結果、培養液1Lあたり400mgであった。
【0053】
実施例3および4ならびに比較例2の結果をまとめたものを表3に示す。なお表3における、硫酸マグネシウム濃度は、培養開始時に添加した硫酸マグネシウム七水和物と培養中に添加した硫酸マグネシウム七水和物との合算値(総量)である。表3から分かるように、培養開始時および培養中に添加する硫酸マグネシウム七水和物の量を合計7g/L(培地)(マグネシウムイオンとして2.8×10−2mol/L(培地))以上とすると、ヒトFcγRIの生産量が増大することがわかる。
【0054】
【表3】
【0055】
なお、実施例1の結果と実施例3の結果とを比較すると、実施例3のほうが実施例1よりも菌体あたりのヒトFcγRIの生産量が向上している。このことから、培地にマグネシウム塩を添加する際は、培養開始時に一度に添加する(実施例1)よりも、培養開始時に一部を添加し、培養中に残部を供給(流加)する(実施例3)ほうが好ましいことが分かる。
【特許請求の範囲】
【請求項1】
タンパク質をコードするポリヌクレオチドを含む発現ベクターで大腸菌W3110株(ATCC 27235)を形質転換して得られた組換え大腸菌を、マグネシウム塩を添加した培地で培養することでタンパク質を製造する方法であって、
マグネシウム塩の添加量が、培地1Lあたりマグネシウムイオンとして2.8×10−2mol以上である、前記方法。
【請求項2】
培養開始時の培地にマグネシウム塩の一部を添加し、残部を培養中の培地に添加する、請求項1に記載の方法。
【請求項3】
マグネシウム塩とともに炭素源および窒素源を培養中の培地に添加する、請求項2に記載の方法。
【請求項4】
培養開始時の培地に添加するマグネシウム塩が、培地1Lあたりマグネシウムイオンとして4.0×10−3mol以上であり、
培養中の培地に添加するマグネシウム塩が、培地1Lあたりマグネシウムイオンとして2.4×10−2mol以上である、
請求項2または3に記載の方法。
【請求項5】
タンパク質がFc結合性タンパク質である、請求項1から4のいずれかに記載の方法。
【請求項6】
Fc結合性タンパク質が、
(1)配列番号1に記載のアミノ酸配列のうち少なくとも16番目のグルタミンから289番目のバリンまでのアミノ酸を含むタンパク質、または
(2)配列番号1に記載のアミノ酸配列のうち少なくとも16番目のグルタミンから289番目のバリンまでのアミノ酸を含み、かつ前記アミノ酸のうちの一つ以上が他のアミノ酸に置換、挿入または欠失したタンパク質である、
請求項5に記載の方法。
【請求項1】
タンパク質をコードするポリヌクレオチドを含む発現ベクターで大腸菌W3110株(ATCC 27235)を形質転換して得られた組換え大腸菌を、マグネシウム塩を添加した培地で培養することでタンパク質を製造する方法であって、
マグネシウム塩の添加量が、培地1Lあたりマグネシウムイオンとして2.8×10−2mol以上である、前記方法。
【請求項2】
培養開始時の培地にマグネシウム塩の一部を添加し、残部を培養中の培地に添加する、請求項1に記載の方法。
【請求項3】
マグネシウム塩とともに炭素源および窒素源を培養中の培地に添加する、請求項2に記載の方法。
【請求項4】
培養開始時の培地に添加するマグネシウム塩が、培地1Lあたりマグネシウムイオンとして4.0×10−3mol以上であり、
培養中の培地に添加するマグネシウム塩が、培地1Lあたりマグネシウムイオンとして2.4×10−2mol以上である、
請求項2または3に記載の方法。
【請求項5】
タンパク質がFc結合性タンパク質である、請求項1から4のいずれかに記載の方法。
【請求項6】
Fc結合性タンパク質が、
(1)配列番号1に記載のアミノ酸配列のうち少なくとも16番目のグルタミンから289番目のバリンまでのアミノ酸を含むタンパク質、または
(2)配列番号1に記載のアミノ酸配列のうち少なくとも16番目のグルタミンから289番目のバリンまでのアミノ酸を含み、かつ前記アミノ酸のうちの一つ以上が他のアミノ酸に置換、挿入または欠失したタンパク質である、
請求項5に記載の方法。
【図1】


【図2】
【図2】
【公開番号】特開2013−85531(P2013−85531A)
【公開日】平成25年5月13日(2013.5.13)
【国際特許分類】
【出願番号】特願2011−230579(P2011−230579)
【出願日】平成23年10月20日(2011.10.20)
【出願人】(000003300)東ソー株式会社 (1,901)
【Fターム(参考)】
【公開日】平成25年5月13日(2013.5.13)
【国際特許分類】
【出願日】平成23年10月20日(2011.10.20)
【出願人】(000003300)東ソー株式会社 (1,901)
【Fターム(参考)】
[ Back to top ]