
遺伝子
【課題】B-Raf遺伝子の癌特異的変異体、癌を診断する方法、及び癌を治療するための治療薬を開発する方法を提供する。
【解決手段】記載する突然変異は、1つ以上の癌関連体細胞アミノ酸突然変異を含む、天然の変異型ヒトB-Rafポリペプチドであって、ヒト腫瘍において同定される。これらの突然変異は、癌性表現型に関連し、ヒト被験体におけるB-Raf遺伝子の少なくとも一部に由来する核酸物質を検査することによる発癌性突然変異の検出方法、抗体を用いた変異型B-Rafポリペプチドの検出方法。更に抗増殖活性を有する1つ以上の化合物を同定する方法による抗癌治療薬の開発のために使用される。
【解決手段】記載する突然変異は、1つ以上の癌関連体細胞アミノ酸突然変異を含む、天然の変異型ヒトB-Rafポリペプチドであって、ヒト腫瘍において同定される。これらの突然変異は、癌性表現型に関連し、ヒト被験体におけるB-Raf遺伝子の少なくとも一部に由来する核酸物質を検査することによる発癌性突然変異の検出方法、抗体を用いた変異型B-Rafポリペプチドの検出方法。更に抗増殖活性を有する1つ以上の化合物を同定する方法による抗癌治療薬の開発のために使用される。
【発明の詳細な説明】
【技術分野】
【0001】
本発明は、B-raf遺伝子の癌特異的変異体、ならびに異常細胞および癌の検出におけるその使用に関する。さらに、本発明は、癌を診断する方法、被験体における癌性細胞を検出する方法、および癌を治療するための治療薬を開発する方法を説明する。
【背景技術】
【0002】
序章
癌は、あらゆる年齢のあらゆる器官のあらゆる組織において発達し得る。初期段階で検出されるほとんどの癌は、治療できる可能性がある。従って、癌の初期徴候について、患者をスクリーニングする能力、そして早期介入を可能にすることが非常に望まれている(例えば、Merck Manual of Diagnosis and Therapy (1992) 16 ed., Merck and Coを参照)。
【0003】
癌性細胞は、無秩序な成長、分化の欠如、ならびに局所組織に侵入および転移する能力を示す。従って、癌細胞は、正常細胞と異なり、その表現型特徴だけではなく、その生化学的および分子生物学的特徴によっても同定できる可能性がある。そしてこのような特徴は、細胞成長および分化を直接的または間接的に制御する癌遺伝子として知られる細胞遺伝子のサブセットにおいて遺伝子レベルで生じる癌性細胞の変化により左右される。
【0004】
Raf癌遺伝子ファミリーは、A-、B-およびC-raf(raf-1とも呼ばれる)と称される3つの高度に保存された遺伝子を含む。rafファミリーの中で最もよく特徴決定されたメンバーであるC-Rafは、マウス肉腫ウイルス3611の形質転換遺伝子であるν-rafの細胞ホモログである。ウイルスraf癌遺伝子は、正常Rafタンパク質のアミノ末端配列を欠くタンパク質をコードする。これらのアミノ末端配列は、RAFセリン/トレオニンタンパク質キナーゼ活性の調節のために不可欠であり、これらの欠失または置換により癌遺伝子コード型RAFタンパク質の構成的な活性が生じる。この非調節型活性により、細胞増殖が促進され、細胞形質転換が生じる。いくつかの腫瘍に由来するDNAは、NIH/3T3細胞のDNAトランスフェクションにより平滑末端化C-raf-1に由来すると同定されることで検出可能な形質転換活性を含むと主張されてきた。しかし、これらの結果は、トランスフェクションの人工産物である可能性が高い。なぜなら、形質転換DNAが由来する腫瘍において同じ突然変異は見とめられていないからである。C-raf遺伝子において人工的に生じた突然変異は、in vitroで細胞に導入した場合に、形質転換を誘導できる。
【0005】
B-raf遺伝子は、鳥類細胞において検出される94-kDセリン/トレオニンキナーゼをコードする鳥類c-Rmil癌原遺伝子のヒトホモログである。このタンパク質は、mil/raf遺伝子ファミリーの他のタンパク質において見とめられないアミノ末端配列を含む。これらの配列は、鳥類ゲノム中の3つのエキソンにコードされる。Eycheneら(1992) Oncogene 7:1657-1660は、これらの3つのエキソンがヒトB-raf遺伝子において保存され、鳥類遺伝子のアミノ酸配列に類似したアミノ酸配列をコードすることを報告している。彼らは、2つのヒトB-raf遺伝子座(すなわち、蛍光in situハイブリダイゼーションにより7q34にマッピングされ、機能性遺伝子生成物をコードすることが示されたB-raf1、およびXq13上に位置する不活処理化偽遺伝子であるB-raf2)を同定した。
【0006】
v-raf癌遺伝子プローブでマウスcDNAライブラリーをスクリーニングすることにより、Huebnerら(1986) Proc. Nat. Acad. Sci. 83: 3934-3938は、形質転換raf関連cDNAであるA-rafを単離し、これは、raf1と区別される遺伝子を表していた。マウスの単一のA-raf遺伝子座、およびヒトのA-raf1遺伝子座は、いくつかのマウスおよびヒト細胞系において能動的に転写される。ヒトA-raf1癌遺伝子の完全606アミノ酸配列が、cDNAの2,453ヌクレオチド配列から推定されている。A-raf遺伝子は、X染色体性(X-linked)である。
【0007】
癌原遺伝子から癌遺伝子への変換の既知のメカニズムは、点突然変異として知られるDNA配列中の単一突然変異の現れであり、これは、コードされるポリペプチドのアミノ酸配列の変化を生じる。例えば、ras癌遺伝子は、正常細胞中には存在しないが、癌原遺伝子対応部分は全ての細胞に存在する。野生型Rasタンパク質は、シグナル伝達に関与する小さいGTP結合タンパク質である。しかし、ウイルスおよびヒト腫瘍に由来する多くのras癌遺伝子は、コドン番号12において点突然変異を有する(すなわち、通常はグリシンをコードするコドンGGCが、バリンをコードするGTCに変化する)。このコドンにおいて、作用する少なくとも5つの異なる置換を含む、複数の突然変異が見とめられている。この単一アミノ酸変化により、Rasタンパク質のGTPアーゼ活性が妨げられ、GTP結合のままであるために、Rasを構成的に活性化させる。13および61位におけるアミノ酸も、ヒト腫瘍由来のras癌遺伝子において頻繁に変化する。
【0008】
Rafタンパク質は、タンパク質キナーゼC(PKC)ファミリーに構造的に関連するセリン/トレオニンキナーゼであり、細胞成長および分化において必須である。Rafタンパク質は、MAPキナーゼの活性化におけるシグナル伝達に関与し、これは、真核生物において高度に保存されている。ERK1およびERK2を含むMAPキナーゼ(マイトジェン-活性化タンパク質キナーゼ)は、転写因子を直接リン酸化して、生物学的事象を調節する。そして、MAPKK(MAPキナーゼキナーゼ)およびMAPKKK(MAPKKキナーゼ)は、MAPキナーゼを調節する。
【0009】
Rafタンパク質はMAPKKKであり、哺乳動物生物学系においてMAPKK MEKをin vivoでリン酸化すると考えられている。明確なraf遺伝子は、脊椎動物において、A-Raf、B-Raf、およびRaf-1(c-Rafとしても知られている)をコードする(Papinら, 1998, Oncogene 12:2218-2221において概論されている)。3つのタンパク質のMEKを活性化する能力は、等しくない。ファミリーの中でもあまり特徴決定されていないメンバーであるA-Rafは、MEKアクチベーターとしては乏しいと思われ、その活性は測定するのが困難である(Pritchardら, 1995, Mol. Cell. Biol. 15, 6430-6442)。B-RafおよびRaf-1も、MEKを活性化する能力が異なる。Raf-1は、遍在的に発現されるのに対して、B-Rafは神経組織において最も高いレベルの発現を提示する(Barnierら, 1995, J. Biol. Chem. 270, 23381-23389)。しかし、B-Rafは、ウェスタンブロッティング分析により発現がほとんど検出されない細胞においてさえも、主なMEKアクチベーターとして同定されてきた(Catlingら, 1994;Jaiswalら, 1994;Reuterら, 1995;Huserら, 2001;Mikulaら, 2001)。それに一貫して、B-Rafは、Raf-1よりも、MEK-1およびMEK-2に対して高い親和性を示し(Papinら, 1996;Papinら, 1998)、MAPKK MEKをリン酸化するのにより効率的である。
【0010】
B-Rafの上流アクチベーターは、GTPアーゼRasである。いくつかのRasイソフォーム(N-Ras、Ha-Ras、Ki-Ras4A、およびKi-Ras4B)は、哺乳動物に存在することが知られている。Rasスーパーファミリーの他のGTPアーゼも、B-Rafと相互作用し得る。例えば、Peysonnauxら, (2001) Biology of the Cell 93:53-62において概論されているRap-1は、B-Rafの選択的アクチベーターと思われる。
【発明の概要】
【0011】
本明細書は、B-Raf遺伝子における点突然変異について記載する。記載する点突然変異は、天然の由来原のヒト腫瘍において同定される。これらの点突然変異は、癌性表現型に関連し、ヒト被験者における、癌の診断、癌性細胞、または癌に対する素因についての基準として使用され得る。
【0012】
B-Rafのキナーゼ活性の活性化により仲介される多くのシグナリング経路が、細胞増殖および発癌性形質転換の制御に関与するため、発癌性特徴を生じ得るB-raf遺伝子の変化を迅速に検出することが望ましい。
【0013】
従って、第1の態様では、1つ以上の突然変異を含む、ヒトB-Rafポリペプチドの天然の癌関連変異体を提供する。
【0014】
癌関連変異体は、天然のヒト原発腫瘍から単離されることが好ましい。
【0015】
突然変異は、B-Rafのキナーゼドメインに含まれることが好ましい。
【0016】
本発明は、ヒト癌における癌性表現型に関連することが分かっているいくつかの突然変異を提供し;従って、B-Raf突然変異と癌との関連性をin vivoで確立する。
【0017】
突然変異は、点突然変異であることが好ましい。突然変異はまた、1つ以上のヌクレオチド、好ましくは2、3、4、5または6ヌクレオチドの挿入、欠失または置換等の変化を含み得る。
【0018】
突然変異は、B-Rafにおいて、アミノ酸300位のC末端側に位置していることが都合がよい。好ましい位置は、463、465、468、585、594、596、および599である。
【0019】
最も好適な実施形態では、突然変異は、V599E、V599D、G595R、G465V、G465E、G465A、G468A、G468E、E585K、F594L、G595R、L596V、L596R、およびG463Eからなる群より選択される。
【0020】
ポリペプチドは、単離されることが好ましい。
【0021】
さらに、本発明は、記載する突然変異を含む、本発明によるポリペプチドの断片を包含する。
【0022】
第2の態様では、本発明による変異型B-Rafポリペプチドまたはその断片をコードする核酸を提供する。核酸は、1つ以上の点突然変異を含むことが好ましい。
【0023】
核酸は、単離されることが好ましい。
【0024】
B-raf遺伝子において、腫瘍との関連を示す点突然変異が検出されている。点突然変異は、B-raf中の1388、1394、1403、1753、1782、1783、1796、1797、1787、および1786位の1つ以上において生じていることが都合がよい。点突然変異は、B-raf中のG1388T、G1783C、TG1796-97AT、G1394T、G1394A、G1394C、G1403C、G1403A、G1753A、T1782G、G1388A、T1796A、T1787G、またはC1786Gであることが好ましい。本発明は、上記核酸のいずれかの相補体(complement)をさらに提供する。
【0025】
さらなる実施形態では、本明細書に記載するように、本発明による核酸に特異的にハイブリダイズする核酸を提供する。このような核酸は、例えば、核酸増幅反応において、本発明による変異型B-Rafコード核酸の特異的な増幅を指令するプライマーであり得る。
【0026】
第3の態様では、本発明は、本発明による変異型B-Rafポリペプチドに選択的に結合するリガンドを提供する。
【0027】
このようなリガンドは、免疫グロブリンであることが都合がよく、抗体またはその抗原結合フラグメントであることが好ましい。
【0028】
第4の態様によれば、
(a)被験体から細胞物質のサンプルを単離するステップ;
(b)該細胞物質中の1つ以上のB-raf遺伝子の少なくとも一部に由来する核酸物質を検査するステップ;および
(c)このような核酸物質が、B-Rafポリペプチドをコードする配列において1つ以上の突然変異を含むか否かを判断するステップ
を含む、細胞形質転換の検出方法を提供する。
【0029】
突然変異は、点突然変異であることが都合がよい。
【0030】
突然変異は、B-rafの1388、1394、1403、1753、1782、1783、1796、1797、1787、および1786位の1つ以上において生じていることが都合がよい。点突然変異は、B-raf中のG1388T、G1783C、TG1796-97AT、G1394T、G1394A、G1394C、G1403C、G1403A、G1753A、T1782G、G1388A、T1796A、T1787G、またはC1786Gであることが好ましい。
【0031】
本発明により同定した突然変異は、体細胞組織(somatic tissue)で生じ、生殖細胞を介して伝達されない体細胞突然変異であることが都合がよい。従って、本発明はさらに、
(a)癌性であると疑われる被験体組織から細胞物質の第1のサンプルを、および同じ被験体の非癌性組織から細胞物質の第2のサンプルを単離するステップ;
(b)該細胞物質のサンプルの両方における1つ以上のB-raf遺伝子の少なくとも一部に由来する核酸物質を検査するステップ;ならびに
(c)このような核酸物質が、B-Rafポリペプチドをコードする配列において1つ以上の点突然変異を含むか否か、そして該突然変異が、該癌性と疑われる組織に由来する細胞物質に存在して、かつ該非癌性組織由来の細胞物質には存在していないことを判断するステップ
を含む、細胞形質転換の検出方法に関する。
【0032】
本発明はさらに、
(a)被験体から細胞物質のサンプルを得るステップ;
(b)該サンプルを、本発明による変異型B-Rafポリペプチドに選択的に結合するリガンドでスクリーニングするステップ;および
(c)該サンプル中の1つ以上の変異型B-Rafポリペプチドを検出するステップ
を含む、細胞形質転換の検出方法を提供する。
【0033】
さらに別の態様では、本発明は、
(a)本発明による1つ以上の変異型B-Rafポリペプチドを得るステップ;
(b)該1つ以上のポリペプチドを、テストする1つ以上の化合物と接触させるステップ;および
(c)該1つ以上の化合物と該変異型ポリペプチドとの相互作用を検出するステップ
を含む、抗増殖活性を有する1つ以上の化合物を同定する方法に関する。
【0034】
上記相互作用は結合相互作用であることが好ましい。
【0035】
さらに、本発明は、
(a)本発明による1つ以上の変異型B-Rafポリペプチドを得るステップ;
(b)B-Rafポリペプチド用の下流基質を得るステップ;および
(c)テストする1つ以上の化合物の存在下で、該基質の修飾を検出するステップ
を含む、抗増殖活性を有する1つ以上の化合物を同定するためのアッセイを提供する。
【0036】
B-Rafは、タンパク質キナーゼであり、従って、基質はリン酸化または脱リン酸化され得る。基質上での変異型B-Rafの作用は、検出可能な変化を生じることが好ましい。基質は、別のキナーゼまたはホスファターゼであり、これらは検出可能な変化を生じる第3の分子を修飾することが都合がよい。
【0037】
例えば、基質は、キナーゼMEKであり得る。MEKリン酸化は、直接検出されるか、または好ましくは、MAPキナーゼをリン酸化するMEKの活性を介して検出され得る。
【0038】
基質上の変異型B-Rafの参照活性を確立し、テストする1つ以上の化合物の存在下および/または不在下における活性を該参照値と比較することが都合がよい。変異型B-Rafの活性が低下している場合には、増殖活性が低下していることを示す。
【0039】
本発明はさらに、抗増殖活性について化合物をスクリーニングするための細胞利用型アッセイを提供する。第1の態様では、本発明は、
(a)NIH 3T3細胞の培養物を得るステップ;
(b)本発明による変異型B-raf核酸で、該細胞をトランスフェクトするステップ;
(c)該細胞を、テストする1つ以上の化合物に曝すステップ;および
(d)該テストする1つ以上の化合物に曝されたトランスフェクト細胞と、そのように曝されていないトランスフェクト細胞とで、形成されたフォーカスの数の差を測定するステップ
を含む、3T3フォーカス形成アッセイを提供する。
【0040】
細胞利用型アッセイは、NIH 3T3細胞を使用して行われることが多い。しかし、他の細胞型、特に繊維芽細胞も、このようなアッセイにおいて使用され得る。
【0041】
アッセイにおいて使用する細胞上の変異型B-raf遺伝子のフォーカス形成の参照活性を確立して、テストする1つ以上の化合物の存在下および/または不在下での活性を該参照値と比較することが都合がよい。変異型B-raf遺伝子のフォーカス形成活性の低下は、増殖活性の低下を示し、つまりテストする1つ以上の化合物の抗増殖活性の低下を示す。
【0042】
本発明による突然変異を検出する自動化された方法および装置(apparata)も提供する。
【図面の簡単な説明】
【0043】
【図1A】B-Raf活性アッセイ。MBPを最終基質として使用し、免疫沈降キナーゼカスケードアッセイにおいて、B-Rafのキナーゼ活性を測定した。活性は、MBPに取り込まれた計数として示している。アッセイは三重で行い、平均値、および平均値からの偏差を表す誤差バーを示す。基準キナーゼ活性(空白バー)、およびV12Ras刺激キナーゼ活性(斜線バー)の両方を示している。
【図1B】B-Raf活性アッセイ。MBPを最終基質として使用し、免疫沈降キナーゼカスケードアッセイにおいて、B-Rafのキナーゼ活性を測定した。活性は、MBPに取り込まれた計数として示している。アッセイは三重で行い、平均値、および平均値からの偏差を表す誤差バーを示す。基準キナーゼ活性(空白バー)、およびV12Ras刺激キナーゼ活性(斜線バー)の両方を示している。
【図2】B-Rafおよび活性化変異体によるNIH3T3細胞の形質転換。細胞を、表示の構築物でトランスフェクトし、コロニーの数を測定した。結果は、少なくとも3つのアッセイの平均である。B-Rafにより誘導される数と比較したコロニー数を示している。
【図3A】V599Dは、BRAFにおける活性化突然変異である。BRAFまたはV599DBRAFを、表示の通り、単一で、または発癌性Rasと共に発現させた。免疫沈降させたBRAFを用いてMEKおよびERKを順次活性化させる免疫沈降キナーゼカスケードアッセイを使用して、BRAFタンパク質の活性を測定した。ERKの活性は、ミエリン塩基性タンパク質および[-32P]-γATPを基質として使用して、測定している。
【図3B】薬物試薬を使用した、黒色腫細胞系におけるERKの阻害。WM266.4またはA375P細胞を、10μM U0126、10μM BAY 43-9006、または対照であるDMSOで、処理した。等量の細胞タンパク質を、SDS-ゲル上で分離させ、ppERK抗体を使用して活性ERKのレベルを測定した。
【図4】薬物による細胞成長の阻害。U0126(10μM)もしくはBAY 43-9006(10μM)、またはビヒクル対照(DMSO)の存在下で、WM-266.4細胞をインキュベートした。48時間後、細胞を、[3H]-チミジンとインキュベートさせて、DNA合成を測定し、細胞DNAに取り込まれたチミジンのレベルを測定した。
【図5A】CRAF発現は、siRNAにより抑制される。WM-266.4、Colo 829、またはBE細胞を、CRAF特異的siRNAプローブ(CRAF)、無作為(scrambled)siRNAプローブ(無作為)、オリゴフェクタミン(オリゴ)で処理するか、または未処理のまま(対照)にした。細胞を24時間インキュベートし、CRAFタンパク質のレベルをウェスタンブロッティングにより測定した。
【図5B】BRAF発現は、siRNAにより抑制される。WM-266.4、Colo 829、またはBE細胞を、表示の通り、BRAF特異的siRNAプローブ、無作為siRNAプローブで処理するか、または未処理のままにした。細胞を24時間インキュベートし、BRAF活性のレベルを免疫沈降キナーゼアッセイMEKおよびERKを連続するアッセイとして測定した。MBPおよび[32P]-γATPを基質として使用して、ERKの活性を測定した。
【図6】CRAFではなくBRAFの除去により、黒色腫細胞中のERK活性がブロックされる。WM-266.4またはColo829細胞を、表示の通り、BRAF特異的siRNAプローブ(BRAF)もしくは無作為対照(sBRAF)、CRAF特異的プローブ(CRAF)もしくはその無作為対照(sCRAF)、オリゴフェクタミン(オリゴ)で処理するか、または未処理のまま(対照)にした。細胞を表示した時間だけインキュベートし、Colo829細胞は24時間処理した。等量の細胞抽出物中のERK活性のレベルを、ppERK抗体でのウェスタンブロッティングにより測定した。
【図7】CRAFではなくBRAFの除去により、黒色腫細胞においてアポトーシスが誘導される。WM-266.4細胞を、表示の通りBRAF特異的siRNAプローブ(BRAF)もしくは無作為対照(sBRAF)、CRAF特異的プローブ(CRAF)、U0126、DMSO(オリゴ)で処理するか、または未処理のまま(対照)にした。細胞を96時間インキュベートし、細胞周期プロファイルをFACSで分析するか、またはPARP発現をウェスタンブロッティングにより検査した。
【図8】B-RafおよびGST-MKKI活性検証。WTS 1 B-Raf溶解産物(バッチA)、GST MKKI(6.5μg/ml)およびERK2(キナーゼコンピテント、100μg/ml)を使用してアッセイを行い、MBP(0.3mg/ml)への33P-γ-リン酸塩(phosphate)取込みを測定する。表示のデータは、三重測定の平均値±SDである。
【図9】フィルタプレートおよびフラッシュプレート放射測定アッセイプラットホームの評価。WTS1 B-Raf1溶解産物(バッチA)およびGST MKK1(6.5μg/ml)を使用してアッセイを行い、GST-kdERK2(100μg/ml)への33P-γ-リン酸塩取込みを測定した。表示のデータは、三重測定の平均値±SDである。
【図10】DELFIA非放射測定アッセイプラットホームの評価。100 ng GST-kdERK2を、各ウェルに予備結合させた後、B-Raf溶解産物(バッチA)、GST-MKK1(6.5μg/ml)、およびATP(500μM)を添加した。表示のデータは、三重測定の平均値±SDである。
【図11】DELFIAアッセイにおける抗-ホスホ-ERK2の滴定。100 ng GST-kdERK2をウェルに予備結合させた後、B-Raf溶解産物(バッチA)、GST-MKK1(6.5μg/ml)、およびATP(500μM)を添加した。表示のデータは、二重測定の平均値である。
【図12】DELFIAアッセイにおけるユーロピウム標識化二次抗体の滴定。100 ng GST-kdERK2をウェルに予備結合させた後、B-Raf溶解産物(バッチA)、GST-MKK1(6.5μg/ml)およびATP(500μM)を添加した。表示のデータは、二重測定の平均値である。
【図13】均一アッセイプロトコールの評価。均一アッセイは、B-Raf溶解産物(バッチB)、6.5μg/ml GST-MKK1、80μg/ml GST-kdERK2、および500μM ATPを含む50μl反応容量を使用して96ウェルプレートにおいて行った。表示のデータは、三重測定の平均値±SDである。
【発明を実施するための形態】
【0044】
発明の詳細な説明
特に付記しない限り、本明細書で使用する全ての技術的および科学的用語は、(例えば、細胞培養、分子遺伝子学、核酸化学、ハイブリダイゼーション技法、および生化学における)当業者に一般的に理解されているものと同じ意味を持つ。分子、遺伝子、および生化学的方法のために、標準的な技術を使用する。Sambrookら, Molecular Cloning: A Laboratory Manual, 第二版 (1989) Cold Spring Harbor Laboratory Press, Cold Spring Harbor, N.Y.、およびAusubelら, Short Protocols in Molecular Biology (1999) 第四版, John Wiley and Sons, Inc.;ならびにGuthrieら, Guide to Yeast Genetics and Moleculr Biology, Methods in Enzymology, Vol. 194, Academic Press, Inc., (1991)、PCR Protocols: A Guide to Methods and Applirations (Innisら 1990. Academic Press, San Diego, Calif.)、McPhersonら, PCR Volume 1, Oxford University Press, (1991)、Culture of Animal Cells: A Manual of Basic Technique, 第二版(R.I. Freshney. 1987. Liss, Inc. New York, N.Y.)、ならびにGene Transfer and Expression Protocols, pp. 109-128, E.J. Murray編, The Humana Press Inc., Clifton, N.J.を概要的に参照のこと。これらの文献は、参照により本明細書に援用する。
【0045】
定義
本出願は、B-Rafポリペプチド変異体を記載する。本明細書で使用する「RAFポリペプチド」という用語は、RAFファミリーのポリペプチドを指すのに使用する。RAFは、最初、癌遺伝子v-rafを含む、クローニングされた固有の急性形質転換性複製欠損マウスC型ウイルスにおいて同定された(Rappら Proc. Nat. Acad. Sci. 80: 4218-4222, 1983)。細胞ホモログであるc-rafは、哺乳動物DNA中に存在する。その後、他のホモログが、ヒトおよびトリにおいて発見されており、rafは鳥類癌遺伝子milのホモログであることが示された。B-Rafは、RAFに関係するが、3つの追加のN末端エキソンを有する。従って、「B-Raf」という用語は、全ての既知のヒトB-Rafホモログおよび変異体、ならびにB-Rafホモログとして同定されるB-Rafに対して十分な相同性を示す他のポリペプチドを包含する。この用語は、ARAF、CRAF、またはRAF1は含まない。B-Rafは、受託番号NP_004324、核酸受託番号NM_004333として示される配列を有するポリペプチドとして同定されることが好ましい。
【0046】
「B-Raf」という用語は、NP_004324に85%、90%、95%、96%、97%、98%、または99%相同であるポリペプチドを含むことが好ましい。相同性比較は、目で、またより一般的には、利用し易い配列比較プログラムの助けを借りて行うことができる。これらの市販されているコンピュータプログラムは、2つ以上の配列間の相同性パーセンテージ(%)を計算できる。
【0047】
相同性パーセンテージは、連続した配列にわたって計算できる(すなわち、一方の配列を、他方の配列とアラインさせて、一方の配列中の各アミノ酸を、他方の配列中の対応するアミノ酸と、一度に1残基ずつ直接比較する)。これは、「アンギャップド」アライメントと呼ばれる。典型的に、このようなアンギャップドアライメントは、比較的少数の残基(例えば、50未満の連続アミノ酸)にわたってのみ行われる。
【0048】
これは非常に単純で一貫性のある方法であるが、例えば、全く同一の配列対でなければ、1つの挿入または欠失により、後続するアミノ酸残基がアライメントから外れ、従って、大規模なアライメントを行った場合に、相同性%の大幅な減少を生じる可能性があるという考慮を欠いている。その結果、ほとんどの配列比較方法は、相同性スコア全体に必要以上にペナルティーを科すことなく、可能性のある挿入および欠失を考慮する最適アライメントを得るように設計されている。これは、配列アライメントに「ギャップ」を挿入して、局所相同性を最大化するようにすることにより達成される。
【0049】
しかし、これらのより複雑な方法は、アライメント中で生じる各ギャップに対して「ギャップペナルティー」を科して、同数の同一アミノ酸について、できるだけ少ないギャップでの(2つの比較配列間のより高い関係性を反映する) 配列アライメントの方が、ギャップの多いものよりも高いスコアが得られる。典型的に、ギャップの存在に対して比較的高いコストを、そしてギャップ中の後続する各残基に対してより小さいペナルティーを科する「アフィンギャップコスト(Affine gap cost)」を使用する。これは、最も一般的に使用されるギャップスコア付けシステムである。高いギャップペナルティーは、当然、ギャップがより少ない最適化アライメントを作成する。ほとんどのアライメントプログラムは、ギャップペナルティーを変えることが可能である。しかし、このようなソフトウェアを配列比較に使用する場合には、デフォルト値を使用することが好ましい。例えば、GCG Wisconsin Bestfitパッケージ(以下参照)を使用する場合、アミノ酸配列に対するデフォルトギャップペナルティーは、1ギャップにつき-12であり、各配列(extension)につき-4である。
【0050】
従って、最大相同性%の計算は、まず、ギャップペナルティーを考慮した、最適なアライメントの生成を必要とする。このようなアライメントを実行するために適したコンピュータプログラムは、GCG Wisconsin Bestfitパッケージ(University of Wisconsin, U.S.A.;Devereuxら, 1984, Nucleic Acids Research 12:387)である。配列比較を実行できる他のソフトウェアの例としては、BLASTパッケージ(Ausubelら, 1999、前掲、第18章を参照)、FASTA(Atschulら, 1990, J. Mol. Biol., 403-410)、およびGENEWORKS比較ツール一式が挙げられるが、これらに限定されない。BLASTおよびFASTAの両方は、オフラインおよびオンライン検索の利用が可能である(Ausubelら, 1999、前掲、7-58〜7-60頁を参照)。しかし、GCG Bestfitプログラムを使用することが好ましい。
【0051】
最終的な相同性%は、同一性の点で測定できるが、アライメントプロセス自体は、典型的に、絶対的な対比較に基づいていない。その代わり、化学的同一性または進化距離に基づいて、対ごとの比較に対してスコアを付けるスケール化類似性スコアマトリックスが一般的に使用される。よく使用されるこのようなマトリックスの例は、BLOSUM62マトリックス(BLASTプログラム一式についてのデフォルトマトリックス)である。GCG Wisconsinプログラムは、一般的に、公開デフォルト値か、または供給される場合にはカスタム記号比較表のいずれかを使用する(さらなる詳細についてはユーザマニュアルを参照のこと)。GCGパッケージについては公開デフォルト値を使用し、他のソフトウェアの場合には、BLOSUM62等のデフォルトマトリックスを使用することが好ましい。
【0052】
ソフトウェアが、最適なアライメントを生成した後、相同性%、好ましくは配列同一性%を計算することが可能になる。ソフトウェアは、典型的に、配列比較の一部としてこれを実行し、数値的結果を生成する。
【0053】
本発明によるポリペプチドの「断片」は、本発明に従って記載する変異型アミノ酸を包含するポリペプチド断片である。断片は、完全長B-Rafポリペプチドまでのあらゆる長さのものであり得る;従って、数アミノ酸分を平滑末端化されたB-Rafポリペプチド、およびそれよりも短い断片を含む。都合良くは、断片は、約764〜約5アミノ酸の長さ;好ましくは、約5〜約20アミノ酸の長さ;都合良くは、約10〜約50アミノ酸の長さである。本発明による断片は、とりわけ、動物を免疫化して、抗体を生じさせるために有用である。従って、本発明によるポリペプチドの断片は、本明細書に記載する変異型B-Rafを特徴とする少なくとも1つの抗原決定基(エピトープ)を含むことが都合がよい。特定のポリペプチド断片が、このような抗原特性を保持するか否かは、当該分野で公知の常套的な方法により簡単に判断できる。少なくて6アミノ酸残基からなるペプチドが、免疫応答を示すことが見とめられることが多い。
【0054】
本発明の「核酸」は、上述したヒトB-Rafポリペプチドをコードする核酸である。この用語はさらに、ストリンジェントなハイブリダイゼーション条件下で、上記同定した天然の核酸またはその相補体にハイブリダイズ可能な、ポリヌクレオチドも含む。「ストリンジェントなハイブリダイゼーション条件」とは、50%ホルムアミド、5×SSC(750 mM NaCl、75 mMクエン酸三ナトリウム)、50 mMリン酸ナトリウム(pH 7.6)、5×デンハルト溶液、10%デキストラン硫酸、および20 pg/ml変性断片化(sheared)サケ精子DNAを含む溶液中で、42℃にて一晩のインキュベーションの後、0.1×SSC中で約65℃にてフィルタを洗浄することを指す。
【0055】
本明細書で言及する核酸は、概して、天然で見とめられる核酸であるが、同用語は、その改変範囲内で、当該分野で公知の、改変された骨格または基質を有する人工核酸も含み得る。
【0056】
本発明による断片をコードする核酸は、本発明による突然変異を含むB-raf遺伝子の特定の領域の核酸増幅によるものであり得る。
【0057】
本明細書で言及する「単離」ポリペプチドまたは核酸は、元の環境(例えば、物質が自然と生じる自然環境)から採られた物質を指し、つまり、その天然状態から人為的に変化させられる。例えば、単離ポリペプチドは、ベクターもしくは組成物の一部であるか、または細胞に含まれていたとしても、該ベクター、組成物、または特定の細胞が該ポリヌクレオチドの元の環境でない場合には、「単離」状態と言える。「単離」という用語は、本技法により本発明のポリペプチド/核酸の独特の特徴を示さない、ゲノムもしくはcDNAライブラリー、全細胞合計もしくはmRNA調製物、ゲノムDNA調製物(電気泳動で分離され、ブロット上に移されたものも含む)、断片化(sheared)全細胞ゲノムDNA調製物、または他の組成物は指さないことが好ましい。
【0058】
本発明によるポリペプチドは、1つ以上の突然変異を含む。「突然変異」としては、アミノ酸付加、欠失または置換が挙げられ;アミノ酸置換を指すことが都合がよい。ポリペプチドレベルでのこのような突然変異は、1つ以上のヌクレオチドの付加、欠失または置換により、核酸レベルで反映される。一般的に、このような突然変異は、核酸のリーディングフレームを変えない。核酸レベルでの変化は、単一のヌクレオチドが別のヌクレオチドに置き換わって、それが一部を構成するコドンを変化させて異なるアミノ酸を特定する点突然変異であることが都合がよい。
【0059】
本発明において同定されるB-Rafにおける突然変異は、自然に生じるもので、発癌物質または他の腫瘍化因子を使用することにより細胞または組織において意図的に誘導されたものではない。従って、本明細書において同定される突然変異は、ヒト組織中からin vivoでの自然な腫瘍形成を正確に反映する。従って、これらの検出は、人為的な化学腫瘍誘導化の後にげっ歯類において同定される突然変異の検出よりも、はるかに良い診断用の基準となる。
【0060】
「体細胞」突然変異は、生物の生殖細胞系を介して伝達されず、生物の体細胞組織において生じる突然変異である。体細胞突然変異は、正常/腫瘍対サンプル分析を通じて体細胞性であると判断されたものであることが都合がよい。
【0061】
本明細書で使用する全てのアミノ酸およびヌクレオチドの番号付けは、B-Rafポリペプチドのアミノ酸+1、またはそれをコードするヌクレオチド配列の最初のATGから開始する。
【0062】
「増幅」反応は、非標的核酸に対して標的核酸の特異的な増幅を生じる核酸反応である。ポリメラーゼ連鎖反応(PCR)は、周知の増幅反応である。
【0063】
「免疫グロブリン」は、免疫グロブリン(抗体)分子を特徴付ける免疫グロブリン折畳み構造を保持するポリペプチドのファミリーのひとつであり、2つのβシート、および通常、保存されたジスルフィド結合を含む。免疫グロブリンスーパーファミリーのメンバーは、in vivoでの細胞および非細胞相互作用の多くの要素に関与しており、免疫系における幅広い役割(例えば、抗体、T細胞受容体分子等)、細胞付着における関与(例えば、ICAM分子)、および細胞内シグナリング(例えば、PDGF受容体等の受容体分子)が挙げられる。本発明は、高い特異性で標的抗原に結合可能な抗体に適用可能であることが好ましい。
【0064】
「抗体」は、抗体全体、またはその抗原結合フラグメントであってもよい。例えば、本発明は、FvおよびFab、ならびにFab'およびF(ab')2等のフラグメント、ならびにscFv、単一ドメイン抗体、Dab抗体、および他の抗原結合抗体に基づく分子等の抗体変異体を含む。
【0065】
本明細書において「癌」は、細胞形質転換から生じる新生物成長から、新生物表現型を指すために使用する。このような細胞形質転換は、遺伝子突然変異を伴うことが多い。本発明に関しては、形質転換は、本明細書に記載する1つ以上のB-raf遺伝子の変化による遺伝子突然変異を伴う。
【0066】
核酸の検出方法
本発明に関しては、B-Rafをコードする変異型核酸の検出を採用して、細胞形質転換および癌の存在またはそれらに対する素因を診断できる。B-raf遺伝子における突然変異は、一般的にDNAレベルで生じるため、本発明の方法は、ゲノムDNAにおける突然変異、ならびに転写産物およびタンパク質自体の検出に基づくことができる。転写産物および/またはポリペプチドの分析により、ゲノムDNAにおける突然変異を確認して、検出された突然変異が実際に被験体において確実に発現されていることが望ましくあり得る。
【0067】
ゲノム核酸における突然変異は、増幅された核酸断片における移動度シフトに基づく技術により検出されることが都合がよい。例えば、Chenら, Anal Biochem 1996 Jul 15;239(1):61-9は、競合移動度シフトアッセイによる単一塩基突然変異の検出を記載している。さらに、Marcelinoら, BioTechniques 26(6):1134-1148(June 1999)の技術に基づくアッセイが市販されている。
【0068】
好適な例では、キャピラリーヘテロ二本鎖分析を使用して、ミスマッチの存在により生じるキャピラリー系における二本鎖核酸の移動度シフトに基づく突然変異の存在を検出し得る。
【0069】
サンプルから、分析用の核酸を生成するためには、一般的に、核酸増幅が必要である。多くの増幅方法は、酵素連鎖反応(ポリメラーゼ連鎖反応、リガーゼ連鎖反応、もしくは自己持続性配列複製等)、またはその中でクローニングされたベクターの全体または一部を複製することに拠る。本発明による増幅は、例えばポリメラーゼ連鎖反応により示される指数的増幅であることが好ましい。
【0070】
多くの標的およびシグナル増幅法が、例えば、これらの方法の概論について、Landegren, U.ら, Science 242:229-237(1988)、およびLewis, R., Genetic Engineering News 10:1, 54-55 (1990)等の文献に記載されている。これらの増幅方法は、本発明の方法において使用でき、ポリメラーゼ連鎖反応(PCR)、PCR in situ、リガーゼ増幅反応(LAR)、リガーゼハイブリダイゼーション、Qβバクテリオファージレプリカーゼ、転写利用型増幅系(TAS)、転写産物配列決定を伴うゲノム増幅(GAWTS)、核酸配列利用型増幅(NASBA)、およびin situハイブリダイゼーションが挙げられる。様々な増幅技術において使用されるのに適したプライマーは、当該分野で公知の方法により調製できる。
【0071】
ポリメラーゼ連鎖反応
PCRは、特に、米国特許第4,683,195号および同第4,683,202号に記載されている核酸増幅方法である。PCRは、DNAポリメラーゼ生成プライマー伸長反応のサイクルの繰り返しにより構成される。標的DNAは、熱変性され、増幅させるDNAの対向する鎖上で標的配列を挟む2本のオリゴヌクレオチドをハイブリダイズさせる。これらのオリゴヌクレオチドは、DNAポリメラーゼと使用されるプライマーとなる。DNAは、プライマー伸長によりコピーされて、両鎖の第2のコピーを作製する。熱変性、プライマーハイブリダイゼーションおよび伸長のサイクルを繰り返すことにより、約2〜4時間で標的DNAを100万倍以上増幅できる。PCRは、分子生物学的ツールであり、検出技術とともに使用されて、増幅結果を判断しなければならない。PCRの利点は、約4時間で100万〜10億倍に標的DNAの量を増幅することにより、感度が高まることである。PCRは、診断目的において任意の既知の核酸を増幅するのに使用できる(Mokら, (1994), Gynaecologic Oncology, 52:247-252)。
【0072】
自己持続性配列複製(3SR)
自己持続性配列複製(3SR)は、TASの変形型であり、逆転写酵素(RT)、酵素カクテルにより仲介されるポリメラーゼおよびヌクリアーゼ活性、ならびに適切なオリゴヌクレオチドプライマーの連続ラウンドによる、核酸鋳型の等温増幅(isothermal amplification)を伴う(Guatelliら, (1990) Proc. Natl. Acad. Sci. USA 87:1874)。RNA/DNAヘテロ二重らせんのRNAの酵素分解を、熱変性の代わりに使用する。RNアーゼHおよび他の酵素を反応液に添加し、全てのステップを同じ温度で生じさせ、それ以上試薬は添加しない。このプロセスの後、42℃にて1時間で、106〜109の増幅が達成された。
【0073】
ライゲーション増幅(LAR/LAS)
ライゲーション増幅反応またはライゲーション増幅系は、DNAリガーゼおよび4つのオリゴヌクレオチド(1本の標的鎖に対して2つ)を使う。この技術は、Wu, D. Y.およびWallace, R.B. (1989) Genomics 4:560に記載されている。オリゴヌクレオチドは、標的DNA上の隣接配列にハイブリダイズし、リガーゼにより連結される。反応液を熱変性させ、サイクルを繰り返す。
【0074】
Qβレプリカーゼ
この技術では、一本鎖RNAを複製するバクテリオファージQβ用のRNAレプリカーゼを使用して、Lizardiら (1988) Bio/Technology 6:1197に記載されるように標的DNAを増幅する。まず、標的DNAを、T7プロモーターおよびQβ5'配列領域を含むプライマーにハイブリダイズさせる。このプライマーを使用して、逆転写酵素は、プロセスにおいて、プライマーを5'端部に接続させたcDNAを生成する。これらの2つのステップは、TASプロトコールに似ている。得られるヘテロ二重らせんは、熱変性される。次に、Qβ3'配列領域を含む第2のプライマーを使用して、第2ラウンドのcDNA合成を開始する。これにより、Qβバクテリオファージの5'および3'端部の両方、ならびに活性T7 RNAポリメラーゼ結合部位を含む二本鎖DNAが生じる。次いで、T7 RNAポリメラーゼは、二本鎖DNAを新しいRNAに転写し、これはQβを模倣する。ハイブリダイズしていない全てのプローブを除去するための激しく洗浄した後、新しいRNAを標的から溶出し、Qβレプリカーゼで複製する。後者の反応は、107倍の増幅を約20分間で生じる。
【0075】
代替的な増幅技術が、本発明において利用され得る。例えば、ローリングサークル増幅(Lizardiら, (1998) Nat Genet 19:225)は、DNAポリメラーゼにより駆動され、等温状態下で線状または幾何学的な動態のいずれかを有する環状オリゴヌクレオチドプローブを複製できる、市販の増幅技術(RCATTM)である。
【0076】
2つの適切に設計されたプライマーの存在下で、DNA鎖置換(displacement)および過剰分岐(hyperbranching)を介して幾何学的増幅が生じ、1時間で、各環につき1012以上のコピーが生成される。
【0077】
単一のプライマーを使用する場合、RCATは、数分間で、数千の標的が該標的に共有結合してタンデムに連結したDNAコピーの線状鎖を生成する。
【0078】
別の技術である鎖置換増幅(SDA;Walkerら, (1992) PNAS (USA) 80:392)は、特定の標的固有の具体的に定義された配列で開始する。熱サイクリングに依存する他の技術とは異なり、SDAは、一連のプライマー、DNAポリメラーゼ、および制限酵素を利用して、固有の核酸配列を指数的に増幅する等温プロセスである。
【0079】
SDAは、標的生成期と指数増殖期の両方から構成される。
【0080】
標的生成では、二本鎖DNAを熱変性させて、2本の一本鎖コピーを作製する。一連の特異的に作製したプライマーを、DNAポリメラーゼ(塩基配列をコピーするための増幅プライマー、および新たに作製された鎖を置換させるための緩衝(bumper)プライマー)と組み合わせて、指数増殖が可能な改変型標的を形成する。
【0081】
指数増幅プロセスは、標的生成期で得た改変型標的(制限酵素認識部位を有する一本鎖部分的DNA鎖)から開始する。
【0082】
増幅プライマーは、その相補DNA配列において各鎖に結合する。すると、DNAポリメラーゼがプライマーを使用し、改変型標的を個々のヌクレオチドを付加するための鋳型として使用して3'側からプライマーを伸長させる位置を識別する。従って、伸長プライマーは、完全な制限酵素認識部位を両端に含む二本鎖DNAセグメントを形成する。
【0083】
次に、制限酵素を、二本鎖DNAセグメントにその認識部位において結合させる。制限酵素は、両側(double-sided)セグメントの一方の鎖のみを切断した後、認識部位から解離して、ニックを形成する。DNAポリメラーゼは、ニックを認識し、その部位から鎖を伸長させて、先に作製された鎖に置き換わる。このように、認識部位は、制限酵素およびDNAポリメラーゼにより、繰り返しニッキングおよび復元されて、標的セグメントを含むDNA鎖が連続的に置換される。
【0084】
すると、各置換された鎖は、上述したように増幅プライマーにアニール可能になる。新しいDNA鎖のニッキング、伸長、および置換を繰り返して、プロセスを続け、元のDNA標的の指数増幅を生じさせる。
【0085】
核酸を増幅させた後、単一塩基対の突然変異の検出のために多数の技術が利用できる。このような技術の一つは、一本鎖高次構造多型(SSCP)である。SCCP検出は、電気泳動中の、参照DNAと比較した場合の一本鎖変異型DNAの異常な移動に基づく。突然変異は、一本鎖DNAにおいて高次構造変化をもたらし、移動シフトを生じる。蛍光SCCPは、蛍光標識化プライマーを使用して、検出を補助する。参照DNAおよび変異型DNAは、蛍光標識化プライマーを使用して、このように増幅される。増幅DNAは、変性および急速冷却(snap-cool)されて、一本鎖DNA分子を生成し、これは、非変性ゲル電気泳動により検査される。
【0086】
化学ミスマッチ切断(CMC)は、ヒドロキシルアミン、四酸化オスミウム、およびピペリジンの組合せによる、DNAミスマッチ塩基対の認識および切断に基づく。従って、参照DNAおよび変異型DNAの両方を、蛍光標識化プライマーで増幅する。ミスマッチT塩基に結合する四酸化オスミウム、またはミスマッチC塩基に結合するヒドロキシルアミンの後に、修飾塩基の部位において切断するピペリジンを使用して、アンプリコン(amplicon)をハイブリダイズし、その後切断する。次いで、切断された断片を電気泳動で検出する。
【0087】
制限断片多形(RFLP)に基づく技術も使用できる。多くの単一ヌクレオチド多形(SNP)は従来のRFLP分析にかからないが、プライマー誘導型制限分析PCR(PIRA-PCR)を使用して、SNP依存的にPCRプライマーを用いて制限部位を導入できる。適切な制限部位を導入するPIRA-PCR用のプライマーは、例えば、Xiaiyiら, (2001) Bioinformatics 17:838-839に記載されるように、コンピュータ分析により設計できる。
【0088】
代替的な実施形態では、本発明は、RNAレベルでの遺伝子発現の検出を提供する。リボ核酸ハイブリダイゼーションを利用する典型的なアッセイ形式としては、核ランオンアッセイ、RT-PCR、およびRNアーゼ保護アッセイが挙げられる(Meltonら, Nuc. Acids Res. 12:7035)。採用できる検出方法としては、放射活性標識、酵素標識、化学発光標識、蛍光標識、および他の適切な標識が挙げられる。
【0089】
RT-PCRを使用して、RNA標的を増幅する。このプロセスにおいて、逆転写酵素を使用して、RNAを相補DNA(cDNA)に変換し、これはその後PCRを使用して増幅できる。この方法は、RNAウイルスを検出するのに有用であることが分かっている。PCR以外での用途は、上述した。
【0090】
ポリペプチドの検出方法
本発明は、変異型B-raf遺伝子にコードされるタンパク質を検出する方法を提供する。タンパク質は、当該分野で公知のタンパク質ゲルアッセイ、抗体結合アッセイ、または他の検出方法により検出できる。
【0091】
従って、例えば、変異型B-Rafポリペプチドを、タンパク質ゲル上の異なる移動度、または変異型アミノ酸の存在を分子量に応じて測定できる質量分析法等の他のサイズ分析技術により検出できる。特に、変異型B-Rafポリペプチドに由来するペプチドは、サイズ分析による差が出やすい。
【0092】
検出手段は、特定の点突然変異が変異型B-Rafポリペプチドにおいて正確に同定され得るように、配列特異的であることが都合がよい。例えば、変異型B-Rafポリペプチドをin vivoまたはin vitroを特異的に認識するポリペプチドまたはRNA分子が開発できる。
【0093】
例えば、RNAアプタマーは、SELEXにより生成できる。SELEXは、標的分子に高度に特異的に結合する核酸分子のin vitro進化のための方法である。これは、例えば、米国特許第5654151号、同第5503978号、同第5567588号、および同第5270163号、ならびにPCT公報WO 96/38579に記載されている(これらの文献はそれぞれ、参照により本明細書に特に援用される)。
【0094】
SELEX方法は、オリゴヌクレオチドライブラリーから、核酸アプタマー、すなわち所望の標的に結合可能な一本鎖核酸を選択することを伴う、好ましくはランダム化配列のセグメントを含む核酸のライブラリーから開始して、SELEX方法は、結合し易い条件下でライブラリーを標的と接触させるステップ、結合してない核酸を標的分子に特異的に結合した核酸から分離するステップ、核酸-標的複合体を解離させるステップ、核酸-標的複合体から解離させた核酸を増幅させて、核酸のリガンド富化ライブラリーを産生するステップを含み、そして、結合、分離、解離、および増幅ステップを、望ましいだけのサイクル繰り返して、標的分子に高度に特異的かつ高親和性な核酸リガンドを産生する。
【0095】
SELEXは、多数の可能性のある配列および構造体を含む核酸ライブラリー内では、所与の標的に対して幅広い結合親和性があるという原理に基づいている。例えば、20ヌクレオチドランダム化セグメントを含む核酸ライブラリーは、420の構造可能性を持ちうる。標的に対してより高い親和定数を有するものは、結合する可能性が最も高いと思われる。分離、解離および増幅のプロセスにより、より高い結合親和性候補が富化された第2の核酸ライブラリーが生成される。さらなる選択ラウンドにより、得られるライブラリーが1つまたは数配列だけで大部分を構成されるまで、最良のリガンドが徐々に絞り込まれる。その後、これらは、クローニング、配列決定、および純粋なリガンドとして結合親和性について個々にテストされ得る。
【0096】
所望の目標が達成するまで、選択および増幅のサイクルを繰り返す。最も一般的なケースでは、サイクルの繰り返しによって結合強度に有意な改善が得られなくなるまで、選択/増幅を続ける。繰返し選択/増幅方法は、少なくとも1014の配列を含むライブラリー中の、単一配列変異体を単離させるのに十分に感度が高い。本方法は、原理的に、約1018の異なる核酸種をサンプル化するために使用され得る。ライブラリーの核酸は、効率的な増幅のために必要な保存された配列と共に、ランダム化配列部分を含むことが好ましい。核酸配列変異体は、ランダム化核酸配列の合成、およびランダムに切断された細胞核酸からのサイズ選択を含む多数の手法により生成され得る。可変的な配列部分は、完全または部分的にランダムな配列を含み得る;また、ランダム化配列と合体した保存配列のサブ部分(subportion)も含み得る。テスト核酸における配列のばらつきは、選択/増幅の繰返しの前または途中の突然変異誘発により、およびクローン化アプタマーの特異的な修飾により、導入または増やすことができる。
【0097】
抗体
B-Rafポリペプチド、またはそこから誘導されるペプチドを使用して、本発明において使用する抗体を生成できる。使用するB-Rafペプチドは、本発明による変異型B-Rafポリペプチドに特異的なエピトープを含むことが好ましい。エピトープとして機能するポリペプチド断片は、あらゆる従来手段(例えば、米国特許第4,631,211号を参照)により生成され得る。本発明では、抗原エピトープは、好ましくは少なくとも4、少なくとも5、少なくとも6、少なくとも7、より好ましくは少なくとも8、少なくとも9、少なくとも10、少なくとも11、少なくとも12、少なくとも13、少なくとも14、少なくとも15、少なくとも20、少なくとも25、少なくとも30、少なくとも40、少なくとも50、そして、最も好ましくは約15〜約30のアミノ酸の配列を含む。免疫原性または抗原性エピトープを含む好ましいポリペプチドは、少なくとも25、30、35、40、45、50、55、60、65、70、75、80、85、90、95、または100アミノ酸残基の長さである。
【0098】
遊離型または担体共役型ペプチドのいずれかで、(例えば、約100μgのペプチドもしくは担体タンパク質、およびフロイントアジュバントもしくは免疫応答を促進することが知られている他の任意のアジュバントを含むエマルジョンを腹腔内および/または皮内注射することで)ウサギまたはマウス等の動物を免疫化することにより、本発明によるB-Rafポリペプチドの抗原性エピトープを使用して、抗体が生成され得る。例えば約2週間ごとに、何回かのブースター注射をして、例えば固体表面に吸着した遊離ペプチドを使用するELISAアッセイにより検出され得る有用な力価の抗ペプチド抗体を得ることが必要となりうる。免疫化動物から得た血清中の抗ペプチド抗体の力価は、抗ペプチド抗体の選択により(例えば、当該分野で周知の方法による、固体表面上のペプチドへの吸着、および選択された抗体の溶出により)高められ得る。
【0099】
本発明のB-Rafポリペプチド、ならびにその免疫原性および/または抗原性エピトープ断片を、他のポリペプチド配列に融合してもよい。例えば、本発明のポリペプチドを、免疫グロブリンドメインと融合させてもよい。ヒトCD4-ポリペプチドの最初の2つのドメイン、ならびに哺乳動物免疫グロブリンの重鎖および軽鎖の定常領域の様々なドメインから構成されるキメラタンパク質は、in vivoにおいて有利な性質を所有することが示されている(例えば、EP 0394827;Trauneckerら, (1988) Nature, 331: 84-86を参照)。上皮バリアを介した免疫系への抗原の送達の増強が、IgGまたはFcフラグメント等のFcRn結合パートナーと共役した抗原(インスリン等)について実証されている(例えば、WO 96/22024およびWO 99/04813を参照)。
【0100】
さらに、本発明のポリペプチドは、融合されたポリペプチドを精製し易くするペプチド等のマーカー配列と融合され得る。好適な実施形態では、マーカーアミノ酸配列は、市販されている多くのものの中でも、pQEベクター中に配置されたタグ(QIAGEN, Inc., 9259 Eton Avenue, Chatsworth, CA, 91311)等のヘキサ-ヒスチジンペプチドである。Gentzら, Proc. Natl. Acad. Sci. USA 86: 821-824 (1989)に記載されているように、例えば、ヘキサ-ヒスチジンは、融合タンパク質の都合の良い精製をもたらす。精製に有用な別のペプチドタグである"HA"タグは、インフルエンザ血球凝集素タンパク質から誘導されたエピトープに対応する(Wilsonら, (1984) Cell 37: 767)。従って、上記融合のいずれもが、本発明の核酸またはポリペプチドを使って操作可能である。
【0101】
好適な実施形態では、本発明は、本明細書に記載するB-Raf変異体を特異的に認識する抗体を提供する。
【0102】
本明細書に記載する抗体は、診断的用途について特に示すものである。従って、これらは、標識等のエフェクタータンパク質を含む改変された抗体であり得る。特に好ましいのは、抗体のin vivoでの分布の画像化を可能にする標識である。このような標識は、患者の身体内で可視化し易い、金属粒子等の放射性標識または放射線不透性(radioopaque)標識である。さらに、組織上で可視化可能な蛍光標識または他の標識であり得る。
【0103】
組換えDNA技術を使用して、本発明の抗体を改善できる。従って、キメラ抗体を構築して、診断的または治療的用途におけるその免疫原性を低下させることができる。さらに、CDRグラフティング[欧州特許出願第0 239 400(Winter)を参照]、および任意に、フレームワーク改変[EP 0 239 400;Riechmann, L.ら, Nature, 332, 323-327, 1988;Verhoeyen M.ら, Science, 239, 1534-1536, 1988;Kettleborough, C.A.ら, Protein Engng., 4, 773-783, 1991;Maeda, H.ら, Human Antibodies and Hybridoma, 2, 124-134, 1991;Gorman S.D.ら, Proc. Natl. Acad. Sci. USA, 88, 4181-4185, 1991;Tempest P.R.ら, Bio/Technology, 9, 266-271, 1991;Co, M.S.ら, Proc. Natl. Acad. Sci. USA, 88, 2869-2873, 1991;Carter, P.ら, Proc. Natl. Acad. Sci. USA, 89, 4285-4289, 1992;Co, M.S.ら, J. Immunol., 148, 1149-1154, 1992;およびSato, K.ら, Cancer Res., 53, 851-856, 1993]により、抗体をヒト化することによって、免疫原性を小さくすることができる。
【0104】
本明細書に記載の抗体は、細胞培養物中で生成できる。組換えDNA技術を使用して、確立された手段により、細菌または好ましくは哺乳動物の細胞培養物において抗体を生成できる。抗体産物は非分泌性細胞から単離できるが、選択された細胞培養系は、任意に、抗体産物を分泌する。
【0105】
従って、本発明は、本発明による抗体を生成するプロセスであって、該抗体タンパク質をコードする第2のDNA配列に正しいリーディングフレーム内で連結したシグナルペプチドをコードする第1のDNA配列に作用可能に連結したプロモーターを含む発現カセットを含むハイブリッドベクターで形質転換された宿主(例えば、大腸菌、昆虫細胞、または哺乳動物細胞)を培養すること、および該タンパク質を単離することを含む、プロセスである。
【0106】
ハイブリドーマ細胞または哺乳動物宿主細胞のin vitroでの増幅は、例えば、哺乳動物血清(例えば、ウシ胎仔血清)、追尾(trace)エレメントおよび成長維持補助剤(例えば、正常マウス腹膜滲出細胞等の供給(feeder)細胞、脾臓細胞、骨髄マクロファージ、2-アミノエタノール、インスリン、トランスフェリン、低密度リポタンパク質、オレイン酸等)を任意に補充されたダルベッコ改良型イーグル培地(DMEM)またはRPMI 1640培地等の習慣的に標準的な培養培地である適切な培養培地中で実施される。細菌細胞または酵母細胞である宿主細胞の増殖も同様に、当該分野で公知の適切な培養培地中(例えば、細菌については培地LB、NZCYM、NZYM、NZM、テリフィック(terrific)ブロス、SOB、SOC、2×YT、またはM9最小培地中、そして酵母については培地YPD、YEPD、最小培地、または完全最小ドロップアウト(dropout)培地中)で実施される。
【0107】
In vitro生成により、比較的純粋な抗体調製物が得られ、所望の抗体を大量に得るための規模拡大が可能になる。細菌細胞、酵母、または哺乳動物細胞培養についての技術は、当該分野で公知であり、(例えば、空輸反応機、もしくは連続攪拌反応機中での)均一懸濁培養、または(例えば、中空繊維中、マイクロカプセル中、アガロースマイクロビーズ上、もしくはセラミックカートリッジ上の)固定化もしくは取込み細胞培養が挙げられる。
【0108】
大量の所望の抗体は、哺乳動物細胞をin vivoで増殖させることによっても得られる。この目的のために、所望の抗体を生成しているハイブリドーマ細胞を、組織適合性哺乳動物に注射して、抗体生成腫瘍を成長させる。任意に、注射前に、動物を炭化水素、特にプリスタン(テトラメチル-ペンタデカン)等の鉱油、でプライミングしてもよい。1〜3週間後、これらの哺乳動物の体液から抗体を単離する。例えば、適切な骨髄腫細胞と、Balb/cマウス由来の抗体生成脾臓細胞との融合により得られたハイブリドーマ細胞、または所望の抗体を生成するハイブリドーマ細胞系Sp2/0から誘導されたトランスフェクト細胞を、任意にプリスタンで予備処置されたBalb/cマウスに腹腔内注射し、1〜2週間後に該動物から腹水を摂取する。
【0109】
上記および他の技術は、例えば、参照により本明細書に援用する、KohlerおよびMilstein, (1975) Nature 256:495-497;米国特許第4,376,110号;HarlowおよびLane, Antibodies: a Laboratory Manual, (1988) Cold Spring Harborにおいて議論されている。組換え抗体分子の調製についての技術は、上記参考文献、ならびに例えば、参照により本明細書に援用するEP 0623679;EP 0368684およびEP 0436597においても記載されている。
【0110】
細胞培養物上清を、好ましくは酵素イムノアッセイ(例えば、サンドウィッチアッセイもしくはドットアッセイ)、またはラジオイムノアッセイにより、所望の抗体についてスクリーニングする。
【0111】
抗体を単離するために、例えば、硫酸アンモニウムでの沈降、ポリエチレングリコール等の吸湿性物質に対する透析、選択膜を介したろ過等により、培養上清中または腹水中の免疫グロブリンを濃縮させてもよい。必要および/または所望であれば、例えば、ゲルろ過、イオン交換クロマトグラフィー、DEAE-セルロース上でのクロマトグラフィー、および/または(イムノ)アフィニティークロマトグラフィー(例えば、標的抗原もしくはプロテインAでのアフィニティークロマトグラフィー)等の習慣的なクロマトグラフィー方法により、抗体を精製する。
【0112】
本発明は、さらに、本発明のモノクローナル抗体を分泌するハイブリドーマ細胞に関する。本発明の好適なハイブリドーマ細胞は、遺伝子的に安定しており、所望の特異性を持つ本発明のモノクローナル抗体を分泌し、解凍および再クローニングすることにより急速冷凍培養物から活性化することができる。
【0113】
本発明は、好適な実施形態において、抗変異型B-Raf抗体の生成に関する。従って、本発明は、本発明のモノクローナル抗体を分泌するハイブリドーマ細胞系を調製するプロセスにも関し、これは、適切な哺乳動物(例えば、Balb/cマウス)を、1つ以上のPDGFポリペプチドもしくはその抗原性断片、または変異型B-Rafポリペプチドを含む抗原性担体で免疫化すること;免疫化した哺乳動物の抗体生成細胞を適切な骨髄腫細胞系の細胞と融合させること;融合で得られたハイブリッド細胞をクローニングすること;および所望の抗体を分泌する細胞クローンを選択することを特徴とする。例えば、変異型B-Rafで免疫化したBalb/cマウスの脾臓細胞を、骨髄腫細胞系PAIまたは骨髄腫細胞系Sp2/0-Ag14の細胞と融合させ、得られたハイブリッド細胞を、所望の抗体の分泌についてスクリーニングし、陽性ハイブリドーマ細胞をクローニングする。
【0114】
1〜100μg変異型B-Rafおよび適切なアジュバント(フロイントアジュバント等)で、数回(例えば、4〜6回)、数ヶ月(例えば、2〜4ヶ月)にわたり、皮下注射および/または腹腔内注射によりBalb/cマウスを免疫化し、最後の注射から2〜4日後に免疫化したマウスから脾臓細胞を得て、融合プロモーター(好ましくはポリエチレングリコール)の存在下で骨髄腫細胞系PAIの細胞と融合させることを特徴とする、ハイブリドーマ細胞系を調製するプロセスが好ましい。骨髄腫細胞は、分子量が約4000の約30%〜約50%のポリエチレングリコールを含む溶液において、免疫化マウスから得た3〜20倍過剰量の脾臓細胞と融合させることが好ましい。融合した後、上述したような選択培地(例えばHAT培地)を補充された適切な培養培地に細胞を規則的な間隔で広げて、正常な骨髄腫細胞が所望のハイブリドーマ細胞を上回って成長させないようにする。
【0115】
本発明はまた、上述した変異型B-Rafに対する抗体の重鎖可変ドメインおよび/または軽鎖可変ドメインをコードするインサートを含む組換え核酸に関する。本質的に、このようなDNAは、コード一本鎖DNA、該コードDNAおよびその相補DNAからなる二本鎖DNA、またはこれらの相補(一本鎖)DNA自体を含む。
【0116】
さらに、変異型B-Rafに対する抗体の重鎖可変ドメインおよび/または軽鎖可変ドメインをコードするDNAは、重鎖可変ドメインおよび/もしくは軽鎖可変ドメインをコードする真正(authentic)DNA配列またはその変異体を有する酵素的または化学的に合成されたDNAであり得る。真正DNAの変異体は、1つ以上のアミノ酸が欠失または別の1つ以上のアミノ酸と交換された、上記抗体の重鎖可変ドメインおよび/または軽鎖可変ドメインをコードするDNAである。該改変は、抗体の重鎖可変ドメインおよび/または軽鎖可変ドメインのCDR外にあることが好ましい。このような変異型DNAは、1つ以上のヌクレオチドが他のヌクレオチドに置き換えられて、新しいコドンが同じアミノ酸をコードする、サイレント変異体であることも意図する。このような変異型配列は変性配列でもある。変性配列は、元々コードされていたアミノ酸の変化を生じずに限定しない数のヌクレオチドが他のヌクレオチドに置き換えられているという遺伝子コードの意味の範囲内において変性されている。このような変性配列は、重鎖マウス可変ドメインおよび/または軽鎖マウス可変ドメインの最適な発現を得るのに、特定の宿主、特に大腸菌が好む制限部位および/または特定のコドンの頻度が異なるために有用であり得る。
【0117】
これに関連して、変異体という用語は、当該分野で公知の方法による真正DNAのin vitro突然変異誘発により得られるDNA変異体を含むことを意図する。
【0118】
完全四量体免疫グロブリン分子のアセンブリ、およびキメラ抗体の発現のために、重鎖および軽鎖可変ドメインをコードする組換えDNAインサートを、重鎖および軽鎖定常ドメインをコードする対応DNAと融合させて、例えばハイブリッドベクターに取り込んだ後に適切な宿主細胞に移す。
【0119】
従って、本発明は、また、ヒト定常ドメインγ(例えば、γ1、γ2、γ3またはγ4、好ましくはγ1またはγ4)に融合した抗変異型B-Raf抗体の重鎖マウス可変ドメインをコードするインサートを含む組換え核酸に関する。同様に、本発明は、ヒト定常ドメインκまたはλ(好ましくはκ)に融合した変異型B-Rafに対する抗変異型B-Raf抗体の軽鎖マウス可変ドメインをコードするインサートを含む組換えDNAに関する。
【0120】
別の実施形態では、本発明は、重鎖可変ドメインおよび軽鎖可変ドメインがスペーサー基により連結されており、任意に宿主細胞中の抗体をプロセシングし易くするシグナル配列、および/または抗体を精製し易くするペプチドをコードするDNA、および/または切断部位、および/またはペプチドスペーサー、および/またはエフェクター分子を含む、組換えポリペプチドをコードする組換えDNAに関する。
【0121】
本発明による抗体および抗体フラグメントは、診断において有用である。従って、本発明は、本発明による抗体を含む診断用の組成物を提供する。
【0122】
診断用組成物の場合、抗体は、酵素的、蛍光的、放射性同位的、または他の手段であり得る抗体検出手段と共に提供されることが好ましい。抗体および検出手段は、診断するための診断キット中に、同時に、別個同時に、または逐次的に使用するように提供され得る。
【0123】
本発明の抗体は、当該分野で公知の任意の方法により、免疫特異的結合についてアッセイされ得る。使用できるイムノアッセイとしては、ウェスタンブロット、ラジオイムノアッセイ、ELISA、サンドウィッチイムノアッセイ、免疫沈降アッセイ、沈降反応法、ゲル拡散沈降反応法、免疫拡散アッセイ、凝集アッセイ、補体結合アッセイ、免疫放射定量測定アッセイ、蛍光イムノアッセイ、およびプロテインAイムノアッセイ等の技術を使用した競合および非競合アッセイ系が挙げられるがこれらに限定されない。このようなアッセイは、当該分野において常套手段である(例えば、Ausubelら編, 1994, Current Protocols in Molecular Biology, Vol. 1, John Wiley and Sons, Inc., New York(参照により本明細書に全体的に援用する)を参照)。イムノアッセイの例を、以下に簡単に記載する。
【0124】
免疫沈降プロトコールは、一般的に、タンパク質ホスファターゼおよび/またはプロテアーゼ阻害剤(例えば、EDTA、PMSF、アプロチニン、バナジン酸ナトリウム)を補充されたRIPA緩衝液(1%NP-40またはTriton X-100、1%デオキシコール酸ナトリウム、0.1%SDS、0.15 M NaCl、0.01 Mリン酸ナトリウム(pH 7.2)、1%トラジロール(Trasylol))等の溶解緩衝液中で細胞の集合を溶解すること、目的の抗体を細胞溶解液に添加すること、4℃にて一定の時間(例えば1〜4時間)インキュベートすること、プロテインAおよび/またはプロテインGセファロースビーズを細胞溶解液に添加すること、4℃にて約1時間以上インキュベートすること、溶解緩衝液中でビーズを洗浄すること、ならびにビーズをSDS/サンプル緩衝液中で再懸濁させること、を含む。目的の抗体が特定の抗原を免疫沈降させる能力は、例えばウェスタンブロット分析により評価できる。
【0125】
ウェスタンブロット分析は、一般的に、タンパク質サンプルを調製すること、ポリアクリルアミドゲル(例えば、抗原の分子量に応じて8%〜20% SDS-PAGE)中でタンパク質サンプルを電気泳動にかけること、ポリアクリルアミドゲルからタンパク質サンプルを膜(ニトロセルロース、PVDF、またはナイロン)に移すこと、ブロック溶液(例えば、3%BSAまたは脱脂乳の入ったPBS)中で膜をブロックすること、洗浄緩衝液(例えば、PBS-Tween 20)中で膜を洗浄すること、ブロック緩衝液中に希釈された一次抗体(目的の抗体)に膜を曝すこと、洗浄緩衝液中で膜を洗浄すること、膜を、ブロッキング緩衝液中に希釈された酵素基質(例えば、西洋ワサビペルオキシダーゼもしくはアルカリホスファターゼ)または放射活性分子(例えば、32Pもしくは125I)と共役した(一次抗体を認識する、例えば抗ヒト抗体等の)二次抗体に曝すこと、洗浄緩衝液中で膜を洗浄すること、および抗原の存在を検出すること、を含む。
【0126】
ELISAは、抗原を調製すること、96ウェルマイクロタイタープレートのウェルを抗原で覆うこと、酵素基質(例えば、西洋ワサビペルオキシダーゼまたはアルカリホスファターゼ)等の検出可能な化合物と共役した目的の抗体をウェルに添加して、一定の時間インキュベートすること、および抗原の存在を検出すること、を含む。ELISAでは、目的の抗体は、検出可能な化合物と共役されなくてもよい;その代わり、検出可能な化合物と共役させた(目的の抗体を認識する)二次抗体をウェルに添加してもよい。さらに、ウェルを抗原で覆う代わりに、ウェルを抗体で覆ってもよい。この場合、検出可能な化合物と共役した二次抗体を、目的の抗原を被覆ウェルに添加した後に、添加してもよい。
【0127】
抗体と抗原との結合親和性、抗体-抗原相互作用の解離速度(off-rate)は、競合結合アッセイにより測定され得る。競合結合アッセイの一例は、大量の非標識化抗原の存在下での標識化抗原(例えば、3Hまたは125I)と目的の抗体とのインキュベーション、および標識化抗原に結合した抗体の検出を含む、ラジオイムノアッセイである。特定の抗原に対する目的の抗体の親和性、および結合解離速度は、スキャッチャード・プロット分析によるデータから決定できる。二次抗体との競合は、ラジオイムノアッセイを使用しても測定できる。この場合、抗原を、大量の非標識化二次抗体の存在下で、標識化化合物(例えば、3Hまたは125I)と共役した目的の抗体とインキュベートさせる。
【0128】
変異型B-Rafポリペプチドの調製
本発明による変異型B-Rafポリペプチドは、化学合成、生物学的サンプルからの単離、およびこのようなポリペプチドをコードする核酸の発現を含む、任意の所望の技術により生成できる。そして、核酸は、変異型B-Rafの生物学的由来源から合成または単離され得る。
【0129】
従って、本発明は、本発明によるポリペプチドをコードするベクター、またはその断片に関する。ベクターは、例えば、ファージ、プラスミド、ウイルス、またはレトロウイルスベクターであり得る。
【0130】
本発明による核酸は、宿主における増殖のための選択マーカーを含むベクターの一部であり得る。一般的に、プラスミドベクターは、リン酸カルシウム沈降物等の沈降物、または荷電脂質との複合体に導入される。ベクターがウイルスの場合、適切なパッケージング細胞系を使用してin vitroでパッケージングされた後に、宿主細胞に形質導入され得る。
【0131】
核酸インサートを、ファージλPLプロモーター、大腸菌lac、trp、phoA、およびtacプロモーター、SV40初期および後期プロモーター、ならびにレトロウイルスLTRのプロモーター等の適切なプロモーターに作用可能に連結させる。他の適切なプロモーターは、当業者に公知である。発現構築物はさらに、転写開始および終結のための部位、ならびに転写領域において翻訳用のリボソーム結合部位を含む。構築物により発現される転写産物のコード部分は、最初に翻訳開始コドンを含み、翻訳するポリペプチドの終わりに適切に位置する終結コドン(UAA、UGA、またはUAG)を含むことが好ましい。
【0132】
上記したように、発現ベクターは、少なくとも1つの選択マーカーを含むことが好ましい。このようなマーカーとしては、ジヒドロ葉酸還元酵素、真核生物細胞培養用のG418もしくはネオマイシン耐性、ならびに大腸菌および他の細菌中で培養するためのテトラサイクリン、カナマイシンもしくはアンピシリン耐性遺伝子が挙げられる。適切な宿主の代表的な例としては、大腸菌、ストレプトミセスおよびネズミチフス菌(Salmonella typhimurium)細胞等の細菌細胞;酵母細胞(例えば、サッカロミセス・セルヴィシエまたはピキア・パストリス(Pichia pastoris)等の菌類細胞;ドロソフィラS2およびスポドプテラSf9細胞等の昆虫細胞;CHO、COS、293およびBows黒色腫細胞等の動物細胞;ならびに植物細胞が挙げられるがこれらに限定されない。
【0133】
上述した宿主細胞に適した培養培地および条件は、当該分野で公知であり、市販されている。
【0134】
細菌における使用が好ましいベクターとしては、QIAGEN, Inc.から入手可能なpQE70、pQE60およびpQE-9;Stratagene Cloning Systems, Inc.から入手可能なpBluescriptベクター、Phagescriptベクター、pNH8A、pNH16a、pNH18A、pNH46A;ならびにPharmacia Biotech, Inc.から入手可能なptrc99a、pKK2233、pKK233-3、pDR540、pRIT5が挙げられる。好ましい真核生物ベクターの一部は、Stratageneから入手可能なpWLNEO、pSV2CAT、pOG44、pXT1、およびpSG;ならびにPharmaciaから入手可能なpSVK3、pBPV、pMSG、およびpSVLである。酵母系における使用が好ましい発現ベクターとしては、pYES2、pYD1、pTEF1/Zeo、pYES2/GS、pPICZ、pGAPZ、pGAPZalph、pPIC9、pPIC3.5、pHIL-D2、pHIL-SI、pPIC3.5K、pPIC9K、およびPA0815(全てInvitrogen, Carlsbad, CAから入手可能)が挙げられるがこれらに限定されない。
【0135】
構築物を宿主細胞に導入することは、リン酸カルシウムトランスフェクション、DEAE-デキストラン仲介型トランスフェクション、陽イオン脂質仲介型トランスフェクション、エレクトロポーレーション、形質導入、感染、または他の方法により実行できる。このような方法は、上記参照したSambrookら等、多くの標準的実験マニュアルに記載されている。
【0136】
本発明によるポリペプチドは、硫酸アンモニウムまたはエタノール沈降、酸抽出、陰イオンまたは陽イオン交換クロマトグラフィー、ホスホセルロースクロマトグラフィー、疎水性相互作用クロマトグラフィー、アフィニティークロマトグラフィー、ヒドロキシルアパタイトクロマトグラフィー、およびレクチンクロマトグラフィーを含む周知の方法により組換え細胞培養物から回収および精製できる。高速液体クロマトグラフィー("HPLC")を精製のために採用することが最も好ましい。
【0137】
本発明によるポリペプチドはまた、体液、組織および細胞を含む生物学的由来源、特に被験体から得た腫瘍組織または可能性のある腫瘍組織から誘導された細胞から回収できる。
【0138】
さらに、本発明のポリペプチドは、当該分野で公知の技術を使用して、化学的に合成され得る(例えば、Creighton, 1983, Proteins: Structures and Molecular Principles, W.H. Freeman and Co., N.Y.、およびHunkapillerら, Nature, 310:105-111 (1984)を参照)。例えば、変異型B-Rafポリペプチドの断片に対応するポリペプチドは、ペプチド合成機を使用して合成され得る。
【0139】
B-Raf突然変異
B-Rafにおける突然変異は、ヒト腫瘍細胞において同定されてきた。表1は、これらの突然変異の位置およびそれらが同定された腫瘍を表す。突然変異は、B-Rafのキナーゼドメインにある。ほとんどの突然変異は、体細胞性であることが確認でき、これは対の正常/腫瘍サンプルをテストして、腫瘍サンプルにおいてのみ突然変異が見とめられたことを示す。
【0140】
表1
【表1】

【0141】
化合物アッセイ
本発明によれば、変異型B-Rafを標的として使用して、変異型B-Rafの増殖活性をモジュレート可能な化合物(例えば、医薬用のリード化合物)を同定する。従って、本発明は、
(a)変異型B-Rafを、評価する1つ以上の化合物とインキュベートするステップ;および
(b)変異型B-Rafの活性に影響を及ぼす化合物を同定するステップ
を含む、活性変異型B-Rafを直接的または間接的にモジュレート可能な1つ以上の化合物を同定するアッセイに関し、その方法を提供する。
【0142】
変異型B-Rafは、本発明に関して定義する通りである。
【0143】
本発明の第1の実施形態では、アッセイは、変異型B-Rafに直接結合するポリペプチドを検出するように構成される。
【0144】
従って、本発明は、
(a)変異型B-Rafを、評価する1つ以上の化合物とインキュベートするステップ;および
(b)変異型B-Rafに結合する化合物を同定するステップ
を含む、モジュレーター細胞増殖を同定する方法を提供する。
【0145】
本方法は、
(c)変異型B-Rafに結合する化合物を、細胞利用型アッセイにおいて細胞増殖をモジュレートする能力について評価するステップ
をさらに含むことが好ましい。
【0146】
変異型B-Rafへの結合は、当業者に公知の任意の技術により評価され得る。適切なアッセイの例としては、in vivoで相互作用を測定する2-ハイブリッドアッセイ系、カラム上に固定化されたポリペプチドへの結合を伴うアフィニティークロマトグラフィーアッセイ、1つ以上の化合物と変異型B-Rafとの結合が結合対の一方または両方のパートナーの蛍光の変化に関連する蛍光アッセイ等が挙げられる。好ましいのは、2-ハイブリッドアッセイ等、細胞においてin vivoで実施されるアッセイである。
【0147】
本実施形態の好適な態様では、本発明は、細胞増殖を伴うかまたは使用する疾患の治療に有用な医薬用のリード化合物を同定する方法であって、テストする1つ以上の化合物の存在下以外ではB-Rafが参照親和性でRASと会合する条件下で、テストする1つ以上の化合物を変異型B-Rafとインキュベートすること;
テストする1つ以上の化合物の存在において、RASに対する変異型B-Rafの結合親和性を測定すること;および
参照結合親和性に照らし、RASに対する変異型B-Rafの結合親和性をモジュレートする化合物を選択すること
を含む方法を提供する。
【0148】
従って、本発明によるアッセイは、テストする1つ以上の化合物の不在下、または変異型B-Rafに対する結合の活性が知られているかもしくは参照値として望ましい参照化合物の存在下において、較正されることが好ましい。例えば、2-ハイブリッド系では、参照値は、任意の化合物の不在下にて得てもよい。標的に対する変異型B-Rafの結合親和性を高める1つ以上の化合物の添加により、アッセイからの読取り値が参照値を上回って高くなり、この親和性結果を低下させる1つ以上の化合物の添加により、アッセイからの読取り値が参照値を下回って低くなる。
【0149】
第2の実施形態では、本発明は、1つ以上の化合物と変異型B-Rafとの機能的相互作用を検出するように構成され得る。このような相互作用は、テストする1つ以上の化合物に応答して、例えばRASによりキナーゼ自体が活性化もしくは不活性化される変異型B-Rafの調節レベル、またはMEK等の下流標的に対する変異型B-Rafの生物学的影響のモジュレーションレベルのいずれかにおいて生じる。本明細書で使用する「活性化」および「不活性化」は、化合物の酵素的またはその他の活性のモジュレーション、および、例えば細胞中のポリペプチドの発現の活性化または抑制によるその生成率のモジュレーションを含む。これらの用語は、遺伝子生成物の発現をモジュレートするための遺伝子転写に対する直接的な作用を含む。
【0150】
変異型B-Rafと、シグナリング経路におけるその上流または下流パートナーとの機能的相互作用のモジュレーションを検出するアッセイは、細胞利用型アッセイであることが好ましい。例えば、変異型B-Rafの活性化から生じるMAPKのリン酸化の程度(これは、MEK活性化の程度を示す)の評価に基づき得る。
【0151】
好適な実施形態では、核酸コード変異型B-Rafは、ベクターにライゲートされ、適切な宿主細胞に導入されて、変異型B-Rafを発現する形質転換細胞系を生成する。その後、得られた細胞系は、変異型B-Raf機能に影響を及ぼす可能性のある化合物の1つ以上の影響の再生可能な定質および/または定量分析のために生成され得る。従って、変異型B-Raf発現細胞は、変異型B-Rafの機能をモジュレートする化合物、特に低分子量の化合物、の同定のために採用され得る。従って、変異型B-Rafを発現する宿主細胞は、薬剤スクリーニングのために有用であり、本発明のさらなる目的は、変異型B-Rafの活性をモジュレートする化合物を同定する方法を提供することであり、該方法は、変異型B-Rafをコードする異種DNAを含み、機能性変異型B-Rafを生成する細胞を、少なくとも1つの化合物もしくは化合物の混合物、または変異型B-Rafの活性をモジュレートする能力を測定しようと思っているシグナルに曝すこと、その後、該モジュレーションにより生じる変化について該細胞をモニターすること、を含む。このようなアッセイは、変異型B-Rafのアゴニスト、アンタゴニスト、およびアロステリックモジュレーター等のモジュレーターの同定を可能にする。本明細書で使用する、変異型B-Rafの活性をモジュレートする化合物またはシグナルは、MEK等の標的上の変異型B-Rafの活性が、該化合物またはシグナルの存在下においては(該化合物またはシグナルの不在下の場合と比較して)異なるような、変異型B-Rafの活性を変える化合物を指す。
【0152】
細胞利用型スクリーニングアッセイは、β-ガラクトシダーゼ、クロラムフェニコールアセチルトランスフェラーゼ(CAT)またはルシフェラーゼ等のリポータータンパク質(すなわち、アッセイし易いタンパク質)の発現が、変異型B-Raf基質の活性化に依存する、細胞系を構築することにより設計され得る。例えば、上記ポリペプチドの1つをコードするリポーター遺伝子は、MEKまたはMARKにより特異的に活性化される応答エレメントの制御下に置かれ得る。このようなアッセイは、変異型B-RafによるMEKのリン酸化を拮抗する化合物、または変異型B-Rafの活性に必要な他の細胞機能を阻害もしくは可能にする化合物等、変異型B-Raf機能を直接モジュレートする化合物の検出を可能にする。野生型非変異型B-Rafが存在する細胞は、適切な対照となる。
【0153】
代替的なアッセイ形式としては、生物学系における増殖応答を直接評価するアッセイが挙げられる。非調節変異型B-Rafの構成的発現は、動物細胞において増殖性表現型を生じる。3T3繊維芽細胞等の細胞利用型系を使用して、変異型B-Rafの可能性のあるレギュレーターの活性を評価し得る。
【0154】
本発明の本実施形態の好適な態様では、炎症応答を伴うまたは使用する疾患の治療に有用な医薬用のリード化合物を同定する方法を提供し、該方法は、
テストする1つ以上の化合物の存在下以外では、変異型B-Rafが、参照リン酸化効率でMEKのリン酸化を直接的または間接的に生じる条件下で、テストする1つ以上の化合物を、変異型B-RafおよびMEKとインキュベートすること;
テストする1つ以上の化合物の存在下で、変異型B-Rafが、直接的または間接的にMEKのリン酸化を生じる能力を判定すること;および
参照リン酸化効率に照らして、変異型B-RafがMEKをリン酸化する能力をモジュレートする化合物を選択すること、を含む。
【0155】
さらに好適な態様では、本発明は、医薬用のリード化合物を同定する方法に関し、該方法は:
精製された変異型B-Raf分子を得るステップ;
変異型B-Raf分子を、変異型B-Rafによりリン酸化することが知られている基質、および1つ以上のテスト化合物とインキュベートするステップ;ならびに
基質のリン酸化をモジュレート可能な1つ以上のテスト化合物を同定するステップ、を含む。
【0156】
変異型B-Rafリン酸化用の基質はMEKである。従って、MEKを基質として使用して、変異型B-Rafキナーゼ活性をモジュレート可能な化合物をモニターすることが好ましい。これにより、当業者は、キナーゼモジュレートを直接スクリーニングすることが可能になる。キナーゼモジュレーターは、キナーゼ(変異型B-Raf)阻害剤であることが好ましい。
【0157】
好適な実施形態では、B-Rafの活性は、以下のプロトコールにより測定され得る:
1.溶解緩衝液(150 mM NaCl、25 mM HEPES [pH 7.3]、1 mMオルトバナジン酸ナトリウム、1%Triton X-100、プロテアーゼ阻害剤、0.5 mMジチオスレイトール)中で、細胞を可溶化する。
【0158】
2.溶解産物を、氷上で10分間インキュベートし、14,000 3 gで10分間遠心分離にかけ、上清をポリクローナル抗B-Raf抗体と、その次にプロテインG-セファロースと、4℃にて1時間インキュベートする。
【0159】
3.免疫沈降物を溶解緩衝液で2回洗浄し、キナーゼ緩衝液(0.2 mM ATP、30 mM MgCl2、2 mM MnCl2、40 mMβ-グリセロリン酸ナトリウム、0.2 mMオルトバナジン酸ナトリウム、2 mMオカダ酸、0.2%β-メルカプトエタノール)中で、1 mgの精製組換えMEK1を基質として添加して、キナーゼ反応を30℃にて10分間実施する。
【0160】
4.MEK1活性化後、15 mCiの[γ-32P]ATP、および1 mgのキナーゼ欠損(K52R)Erkを基質として、さらに2分間添加する。サンプル緩衝液を添加することにより、反応を止め、混合液を5分間煮沸させ、SDS-PAGEによりタンパク質を分離させる。
【0161】
5.ゲルタンパク質を、ポリ二フッ化ビニリデン(polyvinylidene difluoride)膜に移し、その上で、放射標識化K52R Erkの量をPhosphorImagerで定量する。
【0162】
6.B-Raf活性を計算するために、マウスモノクローナル抗B-Rafブロッティングの後に、125I標識化ヤギ抗マウスIgGで膜をプロービングすることにより、同じ膜上のB-Rafタンパク質の量を測定する。
【0163】
7.アッセイは、テストする1つ以上の化合物の存在下または不在下で繰り返してもよい。
【0164】
任意に、同定するテスト化合物を、その後、in vivoテストに供して、例えば上記実施形態に記載したように、変異型B-Rafシグナリング経路に対するそれらの影響を決定してもよい。
【0165】
本明細書で使用する「変異型B-Raf活性」は、結合活性を含む変異型B-Rafの任意の活性を指しうるが、特に、変異型B-Rafのリン酸化活性を指す。従って、本発明は、変異型B-Rafによる標的化合物のリン酸化、および可能性のある治療薬によるこの活性のモジュレーションを検出するように構成され得る。
【0166】
変異型B-Rafのリン酸化活性をモジュレートする化合物の例としては、B-Raf自体の優性ネガティブ変異体が挙げられる。このような化合物は、変異型B-Rafの標的について競合可能であるため、生物学系または人工系における変異型B-Rafの活性を低下させる。従って、本発明はさらに、変異型B-Rafのリン酸化活性をモジュレート可能な化合物に関する。
【0167】
変異型B-Rafの活性に影響を及ぼす化合物は、ほぼ全ての一般的な種類のものであり得、線状、環状、多環状、もしくはそれらの組合せであり得る有機化合物等の低分子量化合物、ペプチド、抗体等のポリペプチド、またはタンパク質が挙げられる。本明細書で使用する「ペプチド」、「ポリペプチド」、および「タンパク質」は、等価であると見なされる。
【0168】
本発明による多くの化合物は、薬剤開発に有用なリード化合物であり得る。有用なリード化合物は、特に抗体およびペプチドであり、特に、遺伝子療法目的で細胞内で発現される細胞内抗体であり、これらは、ペプチドまたは低分子量治療法の開発用のモデルとして使用され得る。本発明の好適な態様では、リード化合物および変異型B-Raf、または他の標的ペプチドは、同時結晶化されて、リード化合物で見とめられる相互作用を模倣する適切な低分子量化合物を設計し易くしてもよい。
【0169】
結晶化は、例えば、「貯蔵緩衝液」に入ったペプチドまたはペプチド複合体の溶液を、好ましくは1:1の比率で、結晶形成に必要な低濃度の沈殿剤と共に、混合することによる、結晶化緩衝液の調製を伴う。結晶形成のためには、例えば沈殿剤を添加すること(例えば滴定)により、または沈殿剤の濃度を拡散によって結晶化緩衝液と貯蔵緩衝液との間で平衡化させることにより、沈殿剤の濃度を上げる。適切な条件下では、沈殿剤のこのような拡散は、沈殿剤の勾配(例えば、高濃度の沈殿剤を含む貯蔵緩衝液から、低濃度の沈殿剤を含む結晶化緩衝液)に応じて生じる。拡散は、例えば、共通気相において拡散させる蒸気拡散技術により達成され得る。公知の技術としては、例えば、「ハンギング・ドロップ(hanging drop)」または「シッティング・ドロップ(sitting drop)」法等の蒸気拡散法がある。蒸気拡散方法では、タンパク質を含む結晶化緩衝液の水滴が、より大量な貯蔵緩衝液プールの上に懸かっているかまたは横に置かれている。あるいはまた、沈殿剤の平衡化は、結晶化緩衝液を貯蔵緩衝液から分離し、タンパク質が貯蔵緩衝液中で希釈するのを防ぐ半浸透膜を通して達成され得る。
【0170】
結晶化緩衝液では、ペプチドまたはペプチド/結合パートナー複合体は、30 mg/mlまで、好ましくは約2 mg/ml〜約4 mg/mlの濃度を有することが好ましい。
【0171】
結晶の形成は、以下のパラメータにより本質的に決定される様々な条件下で達成され得る:すなわち、pH、塩および添加物の存在、沈殿剤、タンパク質濃度、および温度。pHは、約4.0〜9.0に及びうる。緩衝液の濃度および種類は、それほど重要ではないため、(例えば、所望のpHに応じて)可変的であり得る。適切な緩衝液系としては、リン酸塩、酢酸塩、クエン酸塩、Tris、MES、およびHEPES緩衝液が挙げられる。有用な塩および添加物としては、例えば、塩化物、硫酸塩、および当業者に公知の他の塩が挙げられる。緩衝液は、水混和性有機溶剤、好ましくは、100〜20000、優先的には4000〜10000の分子量を有するポリエチレングリコール、または硫酸塩、特に硫酸アンモニウム、塩化物、クエン酸塩、もしくは酒石酸塩等の適切な塩からなる群より選択される沈殿剤を含む。
【0172】
本発明によるペプチドまたはペプチド/結合パートナー複合体の結晶は、(例えば、重原子誘導体化により)化学的に修飾されてもよい。簡単に言うと、このような誘導体化は、重金属原子塩、または有機金属化合物(例えば、塩化鉛、金チオリンゴ酸、チメロサール、もしくは酢酸ウラニル)を含む、結晶中で拡散可能かつタンパク質の表面に結合可能な溶液に、結晶を浸すことにより達成され得る。結合した1つ以上の重金属原子の1つ以上の位置は、浸した結晶のX線回折分析により決定され得、この情報は、例えば、ペプチドの三次元モデルを構築するために使用され得る。
【0173】
三次元モデルは、例えば、結晶の重原子誘導体および/または結晶化により得られた構造データの全体または一部から得ることができる。
【0174】
このようなモデルの構築は、ホモロジーモデリングおよび/または分子置換を伴うことが好ましい。
【0175】
予備ホモロジーモデルは、配列アライメントと構造が既知の任意のRAFキナーゼとの組合わせ、二次構造予測、および構造ライブラリーのスクリーニングにより作製され得る。例えば、変異型B-Rafおよび候補ペプチドの配列を、適切なソフトウェアプログラムを使用してアラインさせてもよい。
【0176】
コンピュータソフトウェアも、ペプチドまたはペプチド複合体の二次構造を予測するために使用され得る。ペプチド配列は、変異型B-Raf構造に取り込まれ得る。構造的矛盾(例えば、挿入/欠失周囲の構造断片)は、所望の長さおよび適切な高次構造を有するペプチドについて構造ライブラリーをスクリーニングすることによりモデリングできる。側鎖高次構造を予測するために、側鎖回転異性体ライブラリーを採用してもよい。
【0177】
最終的なホモロジーモデルを使用して、適切なコンピュータソフトウェアを使用した分子置換により、ペプチドの結晶構造を解明する。ホモロジーモデルは、分子置換の結果に応じて配置され、分子力学計算、および結晶化に使用した阻害剤の電子密度へのモデリングを含むさらなる絞込みに供される。
【0178】
キナーゼ活性化調査
構成的に活性なキナーゼ変異体は、シグナリング経路の解明、およびそのような経路をモジュレートする治療薬の開発において貴重な調査ツールである。本発明による5つの変異体の活性を検査した。これらは、G463V、G468A、G595R、L596V、およびV599Eである。変異体の活性を検査するために、myc-エピトープタグ付け型のB-Rafを、COS細胞において一過的に発現させる。この外因的に発現されたB-Rafの活性を検査するために、mycタグを使用してタンパク質を免疫沈降させ、細菌生成GST-MEK、GST-ERK、およびミエリン塩基性タンパク質(MBP)を配列基質として使用したキナーゼ回路アッセイにおいて検査した(Maraisら (1997); J. Biol. Chem. 272: 4378-83)。B-Rafは、高レベルの基準キナーゼ活性を有し、Raf-1またはA-Rafのいずれよりも、アクチベーターの不在下においてより顕著に活性である(Maraisら (1997); J. Biol. Chem. 272: 4378-83)。さらに、Raf-1およびA-Rafは、それらの活性を完全に刺激するために発癌性Ras(V12Ras)および活性化Srcの両方を必要とする一方で、B-Rafは、V12Rasのみとの同時発現により完全に活性化される。従って、これらの突然変異が、B-Rafの基準活性、およびV12Rasにより刺激される活性の両方に対して及ぼす影響をアッセイした。野生型B-Rafと比べて、G463VB-Raf、G468AB-Raf、L596VB-Raf、およびV599EB-Rafは全て、大きく上昇した基準キナーゼ活性を有する(図1A、1B)。比較すると、G595RB-Rafは、野生型タンパク質と比べて低い基準活性を有する(図1A)。同様の結果がin vivoでも観察された。5つの変異体の全てが、発癌性Ras(V12Ras)により刺激される。しかし、変異体それぞれの倍数的活性化は、野生型B-Rafと比べて低く(図1A、Bを参照)、V599EB-Rafの場合に特に低い。しかし、G463VB-Raf、G468AB-Raf、L596VB-Raf、およびV599EB-Rafのそれぞれの基準活性は野生型タンパク質よりも高いため、見とめられる活性の絶対レベルはそれぞれの場合に、V12Rasの存在下において、野生型タンパク質よりも高い。興味深いことに、G595RB-RafもV12Rasにより刺激されるが、活性化は非常に弱く、これはおそらく低い開始レベルによるものと思われる。
【0179】
各活性化変異体が、NIH3T3細胞を形質転換させる能力についても検査した。このアッセイでは、野生型B-Rafは非常に低い効率(V12Rasで見とめられるコロニー数の約0.02%)で細胞を形質転換する。しかし、図2に示すように、各活性化変異体は、野生型B-Rafよりも、NIH3T3細胞を40〜85倍の効率で形質転換する。
【0180】
Ras/MEK経路に対する、B-Raf変異体を含む細胞の成長の依存の程度を調査した。これらの調査のために、2つのアッセイを使用した。第1のアッセイは、モノクローナル抗体Y13-259(細胞Rasの活性を中和する抗体)の微量注入により成長が抑制されるかをテストする。結果を表2に示す。データは3つのグループに分けられている。第1のグループは、野生型B-Rafを有し、Y13-259により成長が(40〜100%)阻害されている。第2のグループはB-Rafにおいて活性化突然変異を有し、Y13-259により成長は阻害されなかった(<15%)。第3のグループ(1ケースのみ)は、活性化Rasおよび活性化B-Raf突然変異の両方を有する細胞を含む。興味深いことに、この細胞系の成長は、Y13 259により阻害されたが、これは、成長がRasおよびB-Rafの両方に依存しているからかもしれない。
【0181】
第2のアプローチは、化合物U0126(B-Rafの唯一の既知基質であるMEK1/2の阻害剤)の影響を検査することである。これらの結果は、B-Rafにおいて活性化突然変異を有する細胞の処置が、MEK活性が抑制される場合に細胞増殖の抑制を生じることを実証し、B-Rafの活性化変異体による細胞シグナリングの活性化が治療標的であることを示している。表3を参照のこと。
【0182】
総合すると、上記データは
1.ヒト腫瘍において、活性化および不活化突然変異の2つのクラスのB-Raf突然変異が存在する。
【0183】
2.活性化型のB-Rafは、NIH3T3細胞を形質転換可能で、従って、発癌遺伝子と定義され得る。
【0184】
3.活性化B-Rafタンパク質を発現するヒト腫瘍細胞系は、Y13-259、すなわちRas中和抗体に対して感受性を持たず、これらの成長がRasタンパク質に依存せず、従って、Rasタンパク質を標的とする化合物に応答する可能性が低いことを示す。これは、活性化変異体が、腫瘍細胞中のRasシグナルについての要件を克服し得ることを示す。
【0185】
4.しかし、これらの活性は、化合物U0126により抑制され、これらの成長がこの経路の活性に依存し、従って、B-Raf活性を標的とする治療薬に応答する可能性が高い。
【0186】
5.突然変異の一部は、キナーゼドメインのリン酸塩結合ループにあり(G463、G465、G468)、これらのアミノ酸は全てのキナーゼにおいて保存されているため、これらの突然変異は、キナーゼを活性化するための広範囲かつ都合の良いメカニズムを表す。これは、治療薬についてスクリーニングする際の重要な意味(implication)を有する。
【0187】
従って、本発明は、B-Rafの463、465、および468位に対応する1つ以上の位置における突然変異からなる群より選択されるリン酸塩結合ループ中に突然変異を含む、構成的に活性なキナーゼを提供する。
【0188】
突然変異は、463および468位の1つ以上にあることが好ましい。
【0189】
G463VおよびG468Aが最も好ましい。
【0190】
多くのキナーゼは、特定の疾患と関連付けられて同定されるが、それらの活性化のメカニズムが常に完全に理解されているわけではない。本明細書に記載した構成的に活性化された変異体は、活性化のメカニズムを最初に解明する必要なく、阻害剤についてのスクリーニングに使用できる試薬を提供する。
【0191】
キナーゼの例としては、MEKおよびERK等のMAPキナーゼ経路、ならびにp38、JNK、およびそれらの上流キナーゼ等の他のMAPキナーゼ経路上の他のキナーゼが挙げられる。これらの活性化メカニズムについてある程度は知られているが、一部については、直接突然変異によってどのように活性化するのか知られていない。本発明は、該キナーゼスクリーニング目的の活性化変異体を提供する。さらに、p90Rsk、mnk等のMAPキナーゼの下流にあるキナーゼも活性化され得る。代替的な活性化メカニズムは知られているが、突然変異はスクリーニングアッセイにおいて好ましい経路であり得る。
【0192】
本発明はまた、ポイツ-イエーガー症候群に関与するLkb1等の、活性化メカニズムが知られていない特定の公知キナーゼ;および構成的に活性であり得るが、薬剤スクリーニングのためにさらに活性化可能であり得るキナーゼPDK1も包含する。このキナーゼは、インスリンシグナリングに関与し、従って、糖尿病用の有用な標的となりうる。また、AMP活性化キナーゼは、II型糖尿病に関与し、これも同じくリン酸化により活性化され、従って、突然変異による活性化を受け易い。
【0193】
従って、本発明は、
(a)上で詳細に記載したように、アミノ酸置換、欠失、または挿入を、463、465または468位の1つ以上において含む変異型キナーゼを調製すること;
(b)キナーゼ基質の存在下で、変異型キナーゼを、該1つ以上の化合物に曝すこと;および
(d)キナーゼが、該1つ以上の化合物の存在下で該基質をリン酸化する能力を判定すること
を含む、キナーゼに対する阻害効果について1つ以上の化合物をスクリーニングする方法を提供する。
【0194】
1つ以上のテスト化合物の存在下におけるキナーゼのリン酸化活性は、該1つ以上の化合物の不在下におけるその活性と都合よく比較される;変異型キナーゼの基準活性の低下は、該1つ以上の化合物によるキナーゼの阻害を示す。複数アッセイのために、リン酸化の参照レベルを、特定のアッセイについて決定しておき、比較の基準として使用してもよい。
【0195】
キナーゼは、Rafタンパク質キナーゼであることが好ましく;B-Rafであることが都合がよい。
【0196】
逆に、B-Raf G595R等の構成的に抑制された変異体は、キナーゼのアクチベーターをスクリーニングする際に有用である。
【0197】
薬剤標的としてのbRAFの検証
癌における標的としてのBRAFを検証するために、まず、活性化変異型形態のBRAFを発現する細胞の成長が、RAF-MEK-ERKシグナリング経路を成長のために必要としたか否かをテストする。このために、BRAF遺伝子中に突然変異を有する黒色腫および結腸直腸細胞系を、この経路を介したシグナリングをブロックする製剤で処理する。2つの化合物をテストする。1つは、化合物U0126であり、これはMEK阻害剤で、従って、細胞中のRAF-ERKシグナリングを解く(uncouple)(Sebolt-Leopoldら, 1999)。BAY 43-9006もテストした。これは、RAFタンパク質の阻害剤である(Lyonsら, 2001)。これらの化合物が、ERK活性をブロックする能力を、BRAF遺伝子の599位においてバリンからアスパラギン酸への置換を有する黒色腫細胞系WM266.4においてテストした。これは、活性化突然変異である(図3A)。これらの細胞も、二重にリン酸化された活性化型のERKにしか結合しない抗体(ppERK)を使用して判断された場合の基準キナーゼ活性が高い。WM266.4細胞をウェスタンブロットにかける際にppERK抗体を使用する場合、42〜44 kDaの領域において強力なシグナルが見とめられ、これらの細胞においてERKが高い基準キナーゼ活性を有することを示す(図3B)。しかし、細胞をU0126、またはBay43-9006で処理した場合、ERK活性は強く抑制される(図3B)。同様の結果が、A375細胞、すなわちBRAF遺伝子においてV599E突然変異を有する黒色腫細胞系を使用して得られた(Daviesら, 2002)。これらのデータは、RAFおよびMEKシグナリングが、これらの細胞において高いERK活性を維持するために必要であることを実証する。
【0198】
次に、本発明者らは、BAY 43-9002が、WM266.4細胞の成長に対してどのような影響を及ぼすのかをテストし、この化合物が、約6.1μMのIC50で、これらの細胞の成長をブロックしていることが分かった(表4)。
【0199】
BAY43-9006は、低マイクロモル(micromolar)範囲にあるcolo829細胞およびBE細胞の成長もブロックした(表4)。Colo829細胞は、BRAF遺伝子においてV599E突然変異を含む黒色腫細胞系であり、BE細胞は、BRAF遺伝子においてG463突然変異を含む結腸直腸系である(Daviesら, 2002)。このように、これらの突然変異の両方ともが活性化的である(Daviesら, 2002)。最後に、DNA合成に対するこれらの阻害剤の影響をテストした。WM-266.4細胞と、10μM U0126または10μM BAY 43-9006とのインキュベーションにより、これらの細胞中でのDNA合成が強く抑制された(図4)。これらのデータは、BRAF遺伝子中に活性化突然変異を有する細胞におけるERK活性化および増殖が、RAFおよびMEK活性に依存することを実証する。
【0200】
哺乳動物細胞には、3つのRAF遺伝子、すなわちCRAF(RAF-1とも呼ばれる)、ARAF、およびBRAFがある。U0126は、MEK阻害剤であり、従って、CRAFを、BRAFまたはARAFシグナリングと区別することができない。同様に、BAY 43-9006は、CRAFおよびBRAFの両方を阻害でき、従って異なるRAFイソフォーム間を区別しない。従って、どのRAFイソフォームが、WM266.4細胞においてERKにシグナリングしていたかを判断するために、細胞を、個々のRAFイソフォームについて選択的な短い干渉(small interference)RNA(siRNA)プローブで処理した。WM266.4細胞を、BRAFもしくはCRAF、またはいずれのイソフォームも認識しない無作為対照に特異的に設計されたsiRNAプローブで処理した。CRAFに対する認識の効率を、ウェスタンブロッティングで測定した。WM266.4細胞を、CRAFに特異的なsiRNAプローブで処理することにより、CRAF発現が強く抑制された(図5A)。Colo829細胞およびBE細胞において同様の結果が見とめられた(図5A)。WM-266.4細胞をBRAF特異的siRNAプローブで処理した場合、細胞中のBRAF活性は強く抑制されたが、無作為対照で細胞を処理した場合には抑制は見とめられなかった(図5B)。同様の結果が、Colo829およびBE細胞において見とめられた(図5B)。
【0201】
上記データは、siRNAを使用して、BRAFおよびCRAFタンパク質の発現を選択的に抑制できることを示す。従って、本発明者らは、これらのタンパク質のそれぞれの除去により、これらの細胞におけるERK活性がどのように影響されるかを検査した。WM-266.4におけるBRAFタンパク質発現を除去するためにsiRNAを使用した場合、時間依存的にERK活性が抑制された(図6)。対照的に、CRAF発現の除去または無作為siRNAプローブでの処理は、ERK活性に影響を及ぼさなかった(図6)。同様の結果が、Colo829細胞において得られた(図6)。これらの結果は、BRAFが、活性化BRAFタンパク質を発現する黒色腫細胞においてERKにシグナリングする主要なイソフォームであることを実証する。CRAFは、これらの細胞において基準ERK活性にシグナリングしないと思われる。
【0202】
最後に、本発明者らは、WM-266.4細胞におけるアポトーシスに対する影響を検査することにより、BRAF除去が、細胞成長にどのように影響するかを検証した。これらの調査のために、細胞を70%エタノール中で固定し、ヨウ化プロピリジウムで染色し、それらの細胞周期プロファイルを、蛍光活性化細胞分取り(FACS)により検査した。この分析を使用して、アポトーシス細胞は、G1サブピークにおいて見とめられた。これらの細胞において、自発的アポトーシスは非常に少なく、細胞の1%未満がG1サブピークにおいて見とめられた(図7、表5)。U0126で細胞を処理した場合、G1サブピークにおける細胞の割合は有意に増加する(約3.5%;図7、表5)。同様に、siRNAを使用したBRAF発現の除去によっても、G1サブピークにおける細胞の数が増えるが、CRAFの除去または無作為対照での処理では増加しなかった。また、本発明者らは、アポトーシスの誘導のマーカーであるPARP切断も検証した。BRAF siRNAで細胞を処理した場合にはPARPの切断が誘導されたが、無作為対照では誘導されなかった。これらのデータは、変異型BRAFタンパク質が黒色腫細胞系において除去された場合に、アポトーシスが誘導されることを実証する。
【0203】
まとめると、これらの結果は、BRAFの活性化変異体を発現する黒色細胞系において、ERK活性化および細胞成長のために、RAFおよびMEKを介したシグナリングが必要であることを実証している。CRAFよりもBRAFの方が、ERK活性を刺激する主要RAFイソフォームであると思われ、アポトーシスから細胞を保護しているように思われる。これらのデータは、BRAFが、成長およびアポトーシスからの保護のためにBRAFシグナリングに依存する細胞において重要な治療的標的であることを示唆している。
【0204】
ハイスループットスクリーニングアッセイの開発
B-raf変異型V599Eのために、HTSアッセイが開発されている。結果を検証するために、GST-ERK2(キナーゼコンピテント)を採用し、ミエリン塩基性タンパク質(MBP)への33P-γ-リン酸塩の取り込みを測定する標準的な共役アッセイを実施することにより、-Raf発現溶解産物およびGST-MKK1試薬の活性をチェックした。図8に示すように、B-Raf溶解産物の存在下では、対照(非発現)溶解産物と比べて、シグナルの16倍の増加が見とめられた。
【0205】
アッセイプラットホーム検証
オプション1:グルタチオンフラッシュプレート(FlashPlate)における共役キナーゼアッセイ
プラットホームの原理:GSTタグ付け基質(ERK-2)を、フラッシュプレートシンチラント埋め込み壁に、グルタチオン被膜を介して捕捉した。基質への33P-γ-リン酸塩の取り込みにより、測定可能なシンチレーションシグナルが生じるはずである。
【0206】
GST-キナーゼ死活(kinase dead)ERK2(GST-kdERK2)への33P-γ-リン酸塩の取り込みを、B-Raf活性のアウトプットとして測定する確率を、グルタチオンフラッシュプレートアッセイにおいて評価した。図9は、試薬評価実施から持ってきた(transfer)条件を使用した場合、このプラットホームを使用してGST-kdERK2への33PのB-Raf-依存型取込みを検出することができなかった。標準的p81フィルタープレートアッセイも成果がないことが分かった。このカスケードアッセイの増幅特徴の結果として、最後のアッセイステップ(すなわち、MBPのERK2リン酸化)の不在下においてアッセイシグナルを維持するためには、残っている構成要素のそれぞれが有意に高いレベルであることがおそらく必要とされる。従って、この放射分析プラットホームにおいて試薬を消耗するB-Raf、MKKI、およびERK2滴定の実施に着手する前に、抗体利用型プラットホームを評価することが適切と思われた。
【0207】
オプション2:DELFIA形式における共役キナーゼアッセイ
DELFIA(解離増強ランタニド蛍光イムノアッセイ)アッセイは、ホスホ特異的抗体の結合を介したERK2リン酸化の測定を伴う。共役キナーゼアッセイB-Raf/GST-MKKI/GST-kdERK2は、リン酸化GST-kdERK2を生成する。抗GST被覆プレートを使用して、GST-kdERK2を捕捉する。トレオニンおよびチロシン上でリン酸化されたERK2を特異的に検出する一次抗体を添加する。次に、ユウロピウム(Eu)標識化二次抗体を添加する。増強溶液(Enhancement Solution)の存在下にて、Eu標識は、抗体から解離し、335 nmにおいて吸収し、620 nmの発光波長における蛍光検出を可能にする。
【0208】
試薬評価実施からのキナーゼアッセイ条件、および標準的DELFIAアッセイ条件の組合せを採用して、B-Rafアッセイをこのプラットホームにおいて評価した。図10は、B-Raf溶解産物の存在下で、対照(非発現)溶解産物と比較して12.2倍のシグナル上昇が見とめられたことを示す。観察されたシグナルは、3つの酵素/基質成分全ての存在に完全に依存していた。
【0209】
予備実験に基づき、開発のために、DELFIAプラットホームを選択した。
【0210】
キナーゼアッセイ開発
B-Raf溶解産物
3つのバッチのB-Raf溶解産物を、過程を通じて使用した。バッチAを採用して、DELFIAアッセイプラットホームを確立した。アッセイ開発の間、バッチBを使用した。バッチBおよびCについては、溶解産物量と得られたシグナルレベルとのおよその二次関数関係が、ウェルにつき0.025〜0.1μlにおいてはっきりとしていた。各バッチについて選択した最終的な量は、二次関数範囲内で十分なシグナルの枠(window)を得ることに基づいていた。これらの評価の結果、バッチAを1μl/ウェル[96ウェル]で使用し、ならびにバッチBおよびC(スクリーニング)は0.1μl/ウェル[96ウェル]および0.05μl/ウェル[384ウェル]で使用した。
【0211】
抗体レベルの最適化
(両方ともGSTタグ付けされた)ERK2およびMKKIの、グルタチオン結合部位に対する可能性のある競合に関する初期的な考慮から、「予備結合プロトコール」を使用して初期開発アッセイを実施した。
【0212】
これらの条件は以下の通りである:
・96ウェルグルタチオン被覆プレートに対する100 ng/ウェルGST-kdERK2の予備結合。
【0213】
・最終容量を50μM DKB(別表Iを参照)での、B-Raf溶解産物(バッチA)、MKK1(6.5μg/ml)、およびATP(500μM)の添加、ならびに30℃にて1時間のインキュベーション。
【0214】
このプロトコールは、検出系の抗体充填量を最適化かつ節約するために採用した。一次および二次抗体の両方の滴定を実施して、シグナル:バックグラウンド比を≧10.1で維持したままで、抗体レベルの低下の可能性を評価した。図4および5は、1:3000および1:1000が、ホスホERK2およびEu標識化抗体についてそれぞれ許容可能な最低濃度であったことを示す。従って、その後の全てのアッセイは、これらの抗体濃度を使用して行った。
【0215】
MKK1およびERK2レベルの最適化
予備結合ステップ無しに、「溶液中にて」本アッセイの酵素構成要素を行う能力を調査して、定義された濃度のMKK1およびERK2の両方の滴定を可能にした。さらに、このアッセイを、キナーゼに関与する試薬の単一ステップ(均一)混合に簡易化することで、HTSにより適するようになる。図13は、均一アッセイおよび予備結合プロトコールにより、同等のデータが得られたことを示す。その後の全てのアッセイは、「均一」プロトコールを使用して行った。
【0216】
MKK1レベルを保ちながらのERK-2の滴定は、最大シグナルが約12:1(ERK2:MKK1)の比率において得られたことを示した。試薬を節約するために、これらの構成要素のマトリックス滴定を行った。0.1μl B-Raf(バッチB)を使用して、6.5μg/ml GST-MKK1および80μg/ml GST-kdERK2の組合せにより最大シグナルを得た。そして、Raf/MKK1充填量のいくらかの減少が可能であったが、元の比率を維持することが適切であると見なした。この決断は、さらなるアッセイパラメータ変更(例えば、384への移行;ATP充填量の減少、自動化への移行)により、シグナル枠がさらに減りうるという知識と共に出した。従って、その後の全てのアッセイは、6.5μg/ml GST-MKK1および80μg/ml GST-kdERK2を使用して行った。
【0217】
温度の影響
このアッセイを室温にて実行する能力により、最終的なHTSプロセスが有意に簡易化される。従って、室温および30℃におけるアッセイシグナルを調査した。得られた結果に基づき、シグナル対バックグラウンド比は、室温にて許容可能なものであり、その後の全てのアッセイは室温にて実行した。
【0218】
384ウェルプレート形式でのアッセイの評価
スクリーニングの間のスループットを向上させ、試薬の使用量を最小化するために、384ウェル形式でのアッセイの実績を評価した。この形式において、アッセイは上手く行われ、標準量(50μlの最終量)および減少量(25μl)の試薬容量のアッセイの両方で、高度に許容可能なシグナル対バックグラウンド比が得られた。その後の全ての開発実験は、25μl反応容量を使用し、384形式で行った。
【0219】
ATP濃度の最適化
キナーゼスクリーニングアッセイのATP濃度は、同定された阻害性化合物の数および性質に影響を及ぼす可能性がある。このようなアッセイのATPレベルの定義は、以下の配慮事項のバランスである:
・一貫的な測定可能な枠のアッセイシグナルを可能にするATPレベルをを採用すること
・ATP競合阻害剤の同定が可能な十分に低いATPレベルを採用すること
・細胞ATPに関して無影響な可能性が高い弱ATP競合剤が検出されにくいように十分に高いATPレベルを採用すること
キナーゼスクリーニングアッセイは、通常、Kmに関するATP濃度において行われる。B-RafおよびMKK1に対するKm値の偏差により、本明細書において開発した共役アッセイではなく、各酵素について個々のアッセイを開発する必要がある。このようなアッセイの開発により、スクリーニングアッセイにおいて同定されたあらゆる阻害化合物の作用モードが、将来的に間違いなく理解し易くする。
【0220】
スクリーングアッセイのATP充填量を定義する目的のために、共役アッセイの濃度依存度を決定した。生成されるシグナルは、約200μMおよび約18μM ATPにおいてそれぞれ最大値および半値である。さらなる調査により、50μMが、日々の着実なアッセイシグナルが観察される(例えば、シグナル:8000、2%CV) 最も低いATP濃度であることが示された。これ未満(10μM)のATPレベルは、アッセイシグナルにおいてより高い相対的偏差が生じた(例えば、シグナル:2500、10%CV)。スクリーニングアッセイの最終的なATP濃度は、アッセイを自動化に移行する間のいかなる減少をも支持するのに十分に大きいシグナル枠を得るために50μMと定義された。
【0221】
スクリーニングアッセイの時間経過
スクリーニングアッセイは、その直線期(period of linearity)において行うことが重大である。共役アッセイの直線相(linear phase)の長さを決定するために、時間経過を75分まで行った。反応は、5〜45分の間は直線的であった。見とめられた「ラグ」期は、このアッセイ形式に特徴的なものであり、検出可能なレベルの反応生成物を蓄積するのに必要な時間を反映する。この調査に基づき、室温にて45分間のインキュベーションを定義した。
【0222】
B-RafV599E DELFIAアッセイの最終的な条件
アッセイの最終的なスクリーニング条件のまとめは以下の通りである:
酵素反応
・384ウェルグルタチオン被覆プレート
・0.05μl B-Raf溶解産物
・6.5μg/ml GST-MKK1
・80μg/ml GST-kdERK2
・50μM ATP
・室温にて45分間のインキュベーション
・最終容量は25μl
検出条件
・1:3000 抗ホスホERK2抗体
・1:1000 Eu標識抗マウス抗体
自動化開発および品質管理
スクリーニングバッチ試薬の検証
上述した条件を使用して、アッセイを実施し、B-Raf、MKKIおよびERK-2のスクリーニングおよびアッセイ開発バッチを比較した。全てのケースにおいて、試薬のスクリーニングバッチは、アッセイ開発に使用したものと比較した場合と結果が等しかった。
【0223】
自動化B-RatV599E DELFIAアッセイの簡易型実施(mini-runs)
スクリーニングの準備において、アッセイのために自動化液体扱い戦略を設計した。このシステムをテストするために、模擬(すなわち、化合物は含まないが、標準的対照カラムを含む)スクリーニングプレートを使用して自動化B-Rafアッセイの「簡易型実施」を行った。これらの実験から得たデータは、生物学的アッセイの信頼度の高い測定値、ならびに関与する自動化プロセスの正確性および一貫性の両方を示す。
【0224】
簡単に言うと、各プレートは、カラム1〜22の全てのウェルにB-Rafキナーゼ反応液を含み、カラム23〜24は対照反応液を含んだ。自動化アッセイにより生成されたデータの品質を定義するために、この形式を使用した4プレート簡易型実施を2つの別々の日に行った。各「簡易型実施」において、1枚のプレートを、本アッセイのいくつかの予測された阻害剤についてのIC50値を定義するために使用した。阻害剤は、一定の範囲の作用モードを網羅した:すなわち、スタクロスポリン(ATP競合キナーゼ阻害剤)、SB203580(ATP競合Raf阻害剤)、およびU0126(非ATP競合MKK1阻害剤)。
【0225】
アッセイは、プレート間および日々間の両方で、良好な一貫性を示した。得られたデータは、自動化アッセイが、in vitroHTSアッセイにおける384形式について定義した基準を得られることを示す。
【0226】
・少なくとも10:1のシグナル:バックグラウンド
・>0.4のZ'
・>15%の行列CV
採用した阻害剤は、Raf/MKK1活性の濃度依存性阻害をもたらすことで、アッセイをさらに有効にする。特に重要なのは、標準的阻害剤が、別々の日において2倍の範囲内のIC50を出したという事実である。これらのデータは、これらの化合物が、このスクリーニングアッセイにおいて約30μm(10μg/ml)でテストされた場合には、ヒットとして識別されたであろうということも示す。
【0227】
自動化B-Raf V599E DELFIAについての最終的プロトコール
酵素カクテル(最終容量12μl):
0.05μl Raf溶解産物
0.0325μl GST-MKKI
0.065μl GST-kdERK2
【0228】
1.3μlテスト化合物を、グルタチオン被覆384プレートに予備プレート化する。
2.12μl酵素カクテルを、PlateMatePlusで添加する。
3.10μl ATPを、Asysで添加する。
4.プレートを、室温にて45分間振とうさせる。
5.プレートを、プレート洗浄機ELX405を使用して、3×80μl/ウェル DELFIA洗浄緩衝液(DWB)で洗浄する。
6.25μlの抗ホスホERK2をMultidropで添加する。
7.プレートを、室温にて1時間振とうさせる。
8.プレートを、3×80μl/ウェル DWBで洗浄する。
9.25μl Eu標識抗マウス抗体を、Multidropで添加する。
10.プレートを、室温にて30分間振とうさせる。
11.プレートを、3×80μl/ウェル DWBで洗浄する。
12.25μl DELFIA増強溶液を、Multidropで添加する。
13.プレートを、暗所室温にて30分間インキュベートする。
14.FUSION上でプレートを読み取る。
【表2】
表2:本表は、B-Raf突然変異を有する様々な細胞系の成長の阻害を示す。阻害の割合(BrdUを取り込まない細胞の数の%)は、最後のカラムに示す。
【表3】
表3:本表は、MEK阻害剤U0126で処理した細胞における細胞成長の阻害を示す。
【0229】
表4:BAY 43-9006による細胞成長の阻害
【表4】
【0230】
細胞系は、高いレベルのBAY 43-9006の存在下でインキュベートさせ、細胞成長のレベルは、スルホローダミンB染色により測定した。IC50値は、非線形回帰分析により測定し、表示している。
【0231】
表5:細胞周期分析
【表5】
【0232】
WM-266.4細胞を、U0126、DMSO、BRAFに対するsiRNA、CRAFに対するsiRNA、または無作為siRNA対照で処理した。細胞を、96時間インキュベートし、FACSでの細胞周期分析のために、ヨウ化プロプリジウム(propridium iodide)で細胞を固定および染色した。細胞周期の各期における細胞の割合を表示している。
【0233】
検出のコンピュータ的要素
変異型B-Rafポリペプチドおよび/または変異型B-raf核酸の検出を自動化して、サンプル集合の迅速かつ大規模な並行スクリーニングを得ることができる。突然変異検出のためのコンピュータ化方法は、当該分野で公知であり、通常、配列決定装置、またはポリペプチドもしくは核酸中の配列の違いを検出可能な他の装置、データ処理ユニット、および技術者または医師により解読可能な形式で結果を表示可能な出力装置の組合せを伴う。
【0234】
従って、好適な態様では、本発明は、B-Rafポリペプチドをコードする天然のヒト原発腫瘍由来の核酸の標的配列位置における突然変異を検出する自動化方法を提供し、該方法は:
該天然のヒト原発腫瘍由来の核酸の増幅産物のサンプルを配列決定して、開始配列位置から終結配列位置にわたる標的ドメインにおける複数の測定した塩基対同定データを特定するサンプルデータセットを得ること;
該標的配列位置についての該測定した塩基対同定データが、該標的配列位置の参照塩基対データに対応するか否かを条件として、該サンプルにおける突然変異の有無を判断すること;および
該判断ステップにより確立された該サンプルにおける突然変異の有無を示す出力を生成すること
を含む。
【0235】
配列中の突然変異を配列決定および検出する方法は上述しており、当該分野において一般的に公知である。本発明は、このような方法を利用して、本発明のプロセスを実行する装置を得る。この装置は:
核酸のサンプルの配列を判断して、開始配列位置から終結配列位置にわたる標的ドメインにおける測定した塩基対同定データを特定するサンプルデータセットを得ることが可能な、配列読取り装置と;
配列決定装置と接続し、そこからサンプルデータセットを受け取り、該標的配列位置についての該測定した塩基対同定データが、該標的配列位置の参照塩基対データに対応するか否かを条件として、該サンプルにおける突然変異の有無を判断可能な、データ分析ユニットと
を備える。
【0236】
適切な配列読取り装置としては、自動化シーケーンサー、RFLP分析器、および移動度シフト分析装置が挙げられる。標的核酸の増幅産物の配列を分析して、本装置が、PCR機等の増幅装置をさらに備えることが都合がよい。
【0237】
本装置は、データ分析ユニットにより判断されたサンプル中の突然変異の有無を示す出力を生成可能な出力装置を備えることが好ましい。例えば、出力装置は:画像ユーザインターフェース;音声ユーザインターフェース;プリンタ;コンピュータ読取り可能記憶媒体;およびコンピュータ翻訳可能キャリア媒体の少なくとも1つを備え得る。
【0238】
本発明はさらに、変異型B-Rafタンパク質自体を検出するように構成できる。従って、さらなる態様では、本発明は、天然のヒト原発腫瘍由来のB-Rafポリペプチドにおける単一アミノ酸突然変異を検出する自動化方法に関し、該方法は:
B-Rafポリペプチドのサンプル中の1つ以上のアミノ酸に対してマーカーを適用すること;
該マーカーを適用した後、該サンプルを読み取って、該サンプル中の該マーカーの有無を判断し、該サンプルにおける単一アミノ酸突然変異の有無を示すこと;および
該読取りステップにおいて判断した該サンプル中の該単一アミノ酸突然変異の有無を示す出力を生成すること
を含む。
【0239】
マーカーは、単一アミノ酸突然変異を持たない野生型B-Rafポリペプチド、ならびに突然変異を有する変異型B-Rafポリペプチドに差次的に結合するリガンドを含むことが好ましい。本発明に関しては、B-Rafのいずれの形態への優先的な結合も可能である。
【0240】
本発明はさらに:
マーカーを充填され、B-Rafポリペプチドのサンプル中の1つ以上の標的アミノ酸にマーカーを適用可能な、タンパク質マーキング装置と;
該サンプル中のマーカーの有無を判断して、該サンプル中の単一アミノ酸突然変異の有無を示す、マーカー読取り装置と
を備える、B-Rafポリペプチドにおいてアミノ酸突然変異を検出する装置を提供する。
【0241】
使用するマーカーは、抗体であり得、タンパク質マーキング装置はELISAプロセスを実行するように構成され得る。
【0242】
タンパク質マーキング装置は、好ましくはサンプルを光学的に読み取るように構成されたマイクロアレイヤー(microarrayer)を備えることが都合が良い。
【0243】
装置は、マーカー読取り装置により判断されたサンプル中の単一アミノ酸突然変異の有無を示す出力を生成可能な、出力装置を備えることが好ましい。適切な出力装置は:画像ユーザインターフェース;音声ユーザインターフェース;プリンタ;コンピュータ読取り可能記憶媒体;およびコンピュータ翻訳可能キャリア媒体のうちの少なくとも1つを備える。
【0244】
本発明の用途
本発明は、新生物症状を検出、およびこのような症状を患う被験体の予後を判断するのに有用なB-Rafポリペプチドの新規変異体を提供する。一般的に、本明細書に記載するB-Rafにおける突然変異の存在は、新生物の存在と関連付けられる。
【0245】
一態様では、本発明は、癌性細胞または癌性組織(悪性黒色腫、結腸直腸癌、乳癌、もしくはNSCLC)を同定、または新生物表現型を発達し易い細胞もしくは組織を同定する方法であって:該細胞または組織のB-raf遺伝子の少なくとも一部を増幅すること;該増幅産物を分析して、本明細書に記載するB-raf遺伝子における突然変異を検出することを含み;1つ以上のB-raf突然変異を有する細胞または組織は、癌性であるか、または癌症状を発達する危険性が高いと分類される、方法を提供する。適切な増幅手段としては、PCRおよびクローニングが挙げられる。
【0246】
別の実施形態では、本発明は、癌(悪性黒色腫、結腸直腸癌、乳癌、またはNSCLC等)を患う被験体における予後を判断する方法に関する。本方法は:上述したB-raf遺伝子の領域を増幅すること;上述した突然変異の形跡について増幅産物を分析すること;ならびにB-raf遺伝子において突然変異を持たない被験体を、治療および/または手術後に疾患の再発を受け難いか、または該領域において1つ以上の突然変異を有する患者よりも生存する可能性が高いと分類すること、を含む。
【0247】
本発明の技術は、前記した予後に基づいて、被験体が受けるべき治療計画を決定するためにも採用され得る。予後が乏しい被験体は、良好な予後を有する被験体よりも積極的な治療法を使用して処置されることが都合が良い。
【0248】
本発明による技術は、潜在的に癌性の症状を同定するために、サンプルの迅速なスクリーニングが必要とされる場合に、自動化できる。一般的に、自動化プロセスは、組織または細胞サンプルに由来する核酸の自動化増幅、(蛍光検出等による)増幅核酸における突然変異の検出、および/または突然変異の存在の表示、を含む。自動化実施形態の例は、上述している。
【0249】
本発明による変異型B-Rafの同定は、このように、変異型B-Rafの発現に関連する疾患、障害、および/または症状を検出、診断、またはモニターする診断目的のために使用され得る。特に、本発明は、本明細書に記載する変異型B-Rafに関連する癌の検出、診断、および/またはモニターに関する。
【0250】
本発明は、(a)個体の細胞または体液中の変異型B-Rafの発現を、本明細書において定義したB-Raf変異体に特異的な1つ以上の抗体を使用してアッセイすることを含む、癌を診断するための診断アッセイを提供する。個体から得た生検組織における変異型B-raf転写体の存在は、疾患の発達し易さを示すか、または実際の臨床的症状が現れる前に疾患を検出する手段を提供することができる。この種のより確実な診断により、健康専門家は、予防対策または積極的な治療を早めに採用することができ、癌の発達またはそれ以上の進行を防ぐことができる。
【0251】
本発明の抗体は、当業者に公知の古典的な免疫組織学的方法を使用して、生物学的サンプル中のタンパク質レベルをアッセイするために使用できる(例えば、Jalkanenら, (1985) J. Cell. Biol. 101:976-985;Jalkanenら, (1987) J. Cell. Biol. 105:3087-3096を参照)。タンパク質遺伝子発現を検出するのに有用な他の抗体利用型方法としては、酵素結合免疫吸着アッセイ(ELISA)、およびラジオイムノアッセイ(RIA)等のイムノアッセイが挙げられる。適切な抗体アッセイ標識は、当該分野で公知であり、グルコースオキシダーゼ等の酵素標識;ヨウ素(125I、121I)、炭素(14C)、硫黄(35S)、トリチウム(3H)、インジウム(112In)、およびテクネチウム(99Tc)等の放射性同位体;ルミノール等の発光標識;ならびにフルオレセインおよびローダミンおよびビオチン等の蛍光標識が挙げられる。
【0252】
さらに、B-rafにおける突然変異は、本明細書に記載する核酸の分析により検出され得る。例えば、突然変異の存在は、配列決定、またはSCCP分析により検出され得る。
【0253】
本発明は、上記方法において使用できるキットをさらに提供する。一実施形態では、キットは、1つ以上の容器に入った、本発明の抗体、好ましくは精製抗体を含む。具体的な実施形態では、本発明のキットは、キットに含まれる抗体に対して特異的に免疫反応性なエピトープを含む、実質的に単離されたポリペプチドを含む。本発明のキットは、目的のポリペプチドと反応しない対照抗体をさらに含むことが好ましい。別の特異的な実施形態では、本発明のキットは、抗体と目的のポリペプチドとの結合を検出する手段を含む(例えば、抗体を、蛍光化合物、酵素基質、放射活性化合物、もしくは発光化合物等の検出可能な基質と共役させてもよいし、または第1の抗体を認識する二次抗体を検出可能な基質と共役させてもよい)。
【0254】
本発明の別の具体的な実施形態では、キットは、本明細書に記載の変異型B-Rafポリペプチドに特異的な抗体を含む血清をスクリーニングするのに使用する診断キットである。このようなキットは、変異型B-Rafポリペプチドと反応しない対照抗体を含み得る。このようなキットは、少なくとも1つの抗B-Raf抗体に対して特異的に免疫反応性なエピトープを含む、実質的に単離されたポリペプチド抗原を含み得る。さらに、このようなキットは、抗原に対する抗体の結合を検出する手段を含む(例えば、抗体を、フローサイトメトリーにより検出可能な、フルオレセインまたはローダミン等の蛍光化合物と共役させてもよい)。具体的な実施形態では、キットは、組換え生成された、または化学的に合成されたポリペプチド抗原を含み得る。キットのポリペプチド抗原を、固体支持体に付着させてもよい。
【0255】
さらなる実施形態では、本発明は、本発明の変異型B-Rafポリペプチドの抗原を含む血清をスクリーニングするのに使用する診断キットを含む。診断キットは、ポリペプチドまたはポリヌクレオチド抗原に対して特異的に免疫反応性な実質的に単離された抗体、および抗体に対するポリヌクレオチドまたはポリペプチド抗原の結合を検出する手段を含む。一実施形態では、抗体は、固体支持体に付着している。具体的な実施形態では、抗体は、モノクローナル抗体であり得る。キットの検出手段は、第2の標識化モノクローナル抗体を含み得る。あるいはまた、またはそれに加えて、検出手段は、標識化競合抗原を含み得る。
【0256】
上記明細書において言及した全参考文献は、参照により本明細書に援用する。本発明の記載した方法およびシステムの様々な改変および変更が、本発明の範囲および思想から逸脱することなく、当業者に理解されるであろう。本発明は、具体的な好適な実施形態に関して記載したが、請求する発明は、このような具体的な実施形態に不当に限定されないことが理解されるべきである。実際、分子生物学または関連分野の当業者に明らかな、記載した本発明を実施するためのモードの様々な改変は、請求の範囲内にあることを意図する。
【技術分野】
【0001】
本発明は、B-raf遺伝子の癌特異的変異体、ならびに異常細胞および癌の検出におけるその使用に関する。さらに、本発明は、癌を診断する方法、被験体における癌性細胞を検出する方法、および癌を治療するための治療薬を開発する方法を説明する。
【背景技術】
【0002】
序章
癌は、あらゆる年齢のあらゆる器官のあらゆる組織において発達し得る。初期段階で検出されるほとんどの癌は、治療できる可能性がある。従って、癌の初期徴候について、患者をスクリーニングする能力、そして早期介入を可能にすることが非常に望まれている(例えば、Merck Manual of Diagnosis and Therapy (1992) 16 ed., Merck and Coを参照)。
【0003】
癌性細胞は、無秩序な成長、分化の欠如、ならびに局所組織に侵入および転移する能力を示す。従って、癌細胞は、正常細胞と異なり、その表現型特徴だけではなく、その生化学的および分子生物学的特徴によっても同定できる可能性がある。そしてこのような特徴は、細胞成長および分化を直接的または間接的に制御する癌遺伝子として知られる細胞遺伝子のサブセットにおいて遺伝子レベルで生じる癌性細胞の変化により左右される。
【0004】
Raf癌遺伝子ファミリーは、A-、B-およびC-raf(raf-1とも呼ばれる)と称される3つの高度に保存された遺伝子を含む。rafファミリーの中で最もよく特徴決定されたメンバーであるC-Rafは、マウス肉腫ウイルス3611の形質転換遺伝子であるν-rafの細胞ホモログである。ウイルスraf癌遺伝子は、正常Rafタンパク質のアミノ末端配列を欠くタンパク質をコードする。これらのアミノ末端配列は、RAFセリン/トレオニンタンパク質キナーゼ活性の調節のために不可欠であり、これらの欠失または置換により癌遺伝子コード型RAFタンパク質の構成的な活性が生じる。この非調節型活性により、細胞増殖が促進され、細胞形質転換が生じる。いくつかの腫瘍に由来するDNAは、NIH/3T3細胞のDNAトランスフェクションにより平滑末端化C-raf-1に由来すると同定されることで検出可能な形質転換活性を含むと主張されてきた。しかし、これらの結果は、トランスフェクションの人工産物である可能性が高い。なぜなら、形質転換DNAが由来する腫瘍において同じ突然変異は見とめられていないからである。C-raf遺伝子において人工的に生じた突然変異は、in vitroで細胞に導入した場合に、形質転換を誘導できる。
【0005】
B-raf遺伝子は、鳥類細胞において検出される94-kDセリン/トレオニンキナーゼをコードする鳥類c-Rmil癌原遺伝子のヒトホモログである。このタンパク質は、mil/raf遺伝子ファミリーの他のタンパク質において見とめられないアミノ末端配列を含む。これらの配列は、鳥類ゲノム中の3つのエキソンにコードされる。Eycheneら(1992) Oncogene 7:1657-1660は、これらの3つのエキソンがヒトB-raf遺伝子において保存され、鳥類遺伝子のアミノ酸配列に類似したアミノ酸配列をコードすることを報告している。彼らは、2つのヒトB-raf遺伝子座(すなわち、蛍光in situハイブリダイゼーションにより7q34にマッピングされ、機能性遺伝子生成物をコードすることが示されたB-raf1、およびXq13上に位置する不活処理化偽遺伝子であるB-raf2)を同定した。
【0006】
v-raf癌遺伝子プローブでマウスcDNAライブラリーをスクリーニングすることにより、Huebnerら(1986) Proc. Nat. Acad. Sci. 83: 3934-3938は、形質転換raf関連cDNAであるA-rafを単離し、これは、raf1と区別される遺伝子を表していた。マウスの単一のA-raf遺伝子座、およびヒトのA-raf1遺伝子座は、いくつかのマウスおよびヒト細胞系において能動的に転写される。ヒトA-raf1癌遺伝子の完全606アミノ酸配列が、cDNAの2,453ヌクレオチド配列から推定されている。A-raf遺伝子は、X染色体性(X-linked)である。
【0007】
癌原遺伝子から癌遺伝子への変換の既知のメカニズムは、点突然変異として知られるDNA配列中の単一突然変異の現れであり、これは、コードされるポリペプチドのアミノ酸配列の変化を生じる。例えば、ras癌遺伝子は、正常細胞中には存在しないが、癌原遺伝子対応部分は全ての細胞に存在する。野生型Rasタンパク質は、シグナル伝達に関与する小さいGTP結合タンパク質である。しかし、ウイルスおよびヒト腫瘍に由来する多くのras癌遺伝子は、コドン番号12において点突然変異を有する(すなわち、通常はグリシンをコードするコドンGGCが、バリンをコードするGTCに変化する)。このコドンにおいて、作用する少なくとも5つの異なる置換を含む、複数の突然変異が見とめられている。この単一アミノ酸変化により、Rasタンパク質のGTPアーゼ活性が妨げられ、GTP結合のままであるために、Rasを構成的に活性化させる。13および61位におけるアミノ酸も、ヒト腫瘍由来のras癌遺伝子において頻繁に変化する。
【0008】
Rafタンパク質は、タンパク質キナーゼC(PKC)ファミリーに構造的に関連するセリン/トレオニンキナーゼであり、細胞成長および分化において必須である。Rafタンパク質は、MAPキナーゼの活性化におけるシグナル伝達に関与し、これは、真核生物において高度に保存されている。ERK1およびERK2を含むMAPキナーゼ(マイトジェン-活性化タンパク質キナーゼ)は、転写因子を直接リン酸化して、生物学的事象を調節する。そして、MAPKK(MAPキナーゼキナーゼ)およびMAPKKK(MAPKKキナーゼ)は、MAPキナーゼを調節する。
【0009】
Rafタンパク質はMAPKKKであり、哺乳動物生物学系においてMAPKK MEKをin vivoでリン酸化すると考えられている。明確なraf遺伝子は、脊椎動物において、A-Raf、B-Raf、およびRaf-1(c-Rafとしても知られている)をコードする(Papinら, 1998, Oncogene 12:2218-2221において概論されている)。3つのタンパク質のMEKを活性化する能力は、等しくない。ファミリーの中でもあまり特徴決定されていないメンバーであるA-Rafは、MEKアクチベーターとしては乏しいと思われ、その活性は測定するのが困難である(Pritchardら, 1995, Mol. Cell. Biol. 15, 6430-6442)。B-RafおよびRaf-1も、MEKを活性化する能力が異なる。Raf-1は、遍在的に発現されるのに対して、B-Rafは神経組織において最も高いレベルの発現を提示する(Barnierら, 1995, J. Biol. Chem. 270, 23381-23389)。しかし、B-Rafは、ウェスタンブロッティング分析により発現がほとんど検出されない細胞においてさえも、主なMEKアクチベーターとして同定されてきた(Catlingら, 1994;Jaiswalら, 1994;Reuterら, 1995;Huserら, 2001;Mikulaら, 2001)。それに一貫して、B-Rafは、Raf-1よりも、MEK-1およびMEK-2に対して高い親和性を示し(Papinら, 1996;Papinら, 1998)、MAPKK MEKをリン酸化するのにより効率的である。
【0010】
B-Rafの上流アクチベーターは、GTPアーゼRasである。いくつかのRasイソフォーム(N-Ras、Ha-Ras、Ki-Ras4A、およびKi-Ras4B)は、哺乳動物に存在することが知られている。Rasスーパーファミリーの他のGTPアーゼも、B-Rafと相互作用し得る。例えば、Peysonnauxら, (2001) Biology of the Cell 93:53-62において概論されているRap-1は、B-Rafの選択的アクチベーターと思われる。
【発明の概要】
【0011】
本明細書は、B-Raf遺伝子における点突然変異について記載する。記載する点突然変異は、天然の由来原のヒト腫瘍において同定される。これらの点突然変異は、癌性表現型に関連し、ヒト被験者における、癌の診断、癌性細胞、または癌に対する素因についての基準として使用され得る。
【0012】
B-Rafのキナーゼ活性の活性化により仲介される多くのシグナリング経路が、細胞増殖および発癌性形質転換の制御に関与するため、発癌性特徴を生じ得るB-raf遺伝子の変化を迅速に検出することが望ましい。
【0013】
従って、第1の態様では、1つ以上の突然変異を含む、ヒトB-Rafポリペプチドの天然の癌関連変異体を提供する。
【0014】
癌関連変異体は、天然のヒト原発腫瘍から単離されることが好ましい。
【0015】
突然変異は、B-Rafのキナーゼドメインに含まれることが好ましい。
【0016】
本発明は、ヒト癌における癌性表現型に関連することが分かっているいくつかの突然変異を提供し;従って、B-Raf突然変異と癌との関連性をin vivoで確立する。
【0017】
突然変異は、点突然変異であることが好ましい。突然変異はまた、1つ以上のヌクレオチド、好ましくは2、3、4、5または6ヌクレオチドの挿入、欠失または置換等の変化を含み得る。
【0018】
突然変異は、B-Rafにおいて、アミノ酸300位のC末端側に位置していることが都合がよい。好ましい位置は、463、465、468、585、594、596、および599である。
【0019】
最も好適な実施形態では、突然変異は、V599E、V599D、G595R、G465V、G465E、G465A、G468A、G468E、E585K、F594L、G595R、L596V、L596R、およびG463Eからなる群より選択される。
【0020】
ポリペプチドは、単離されることが好ましい。
【0021】
さらに、本発明は、記載する突然変異を含む、本発明によるポリペプチドの断片を包含する。
【0022】
第2の態様では、本発明による変異型B-Rafポリペプチドまたはその断片をコードする核酸を提供する。核酸は、1つ以上の点突然変異を含むことが好ましい。
【0023】
核酸は、単離されることが好ましい。
【0024】
B-raf遺伝子において、腫瘍との関連を示す点突然変異が検出されている。点突然変異は、B-raf中の1388、1394、1403、1753、1782、1783、1796、1797、1787、および1786位の1つ以上において生じていることが都合がよい。点突然変異は、B-raf中のG1388T、G1783C、TG1796-97AT、G1394T、G1394A、G1394C、G1403C、G1403A、G1753A、T1782G、G1388A、T1796A、T1787G、またはC1786Gであることが好ましい。本発明は、上記核酸のいずれかの相補体(complement)をさらに提供する。
【0025】
さらなる実施形態では、本明細書に記載するように、本発明による核酸に特異的にハイブリダイズする核酸を提供する。このような核酸は、例えば、核酸増幅反応において、本発明による変異型B-Rafコード核酸の特異的な増幅を指令するプライマーであり得る。
【0026】
第3の態様では、本発明は、本発明による変異型B-Rafポリペプチドに選択的に結合するリガンドを提供する。
【0027】
このようなリガンドは、免疫グロブリンであることが都合がよく、抗体またはその抗原結合フラグメントであることが好ましい。
【0028】
第4の態様によれば、
(a)被験体から細胞物質のサンプルを単離するステップ;
(b)該細胞物質中の1つ以上のB-raf遺伝子の少なくとも一部に由来する核酸物質を検査するステップ;および
(c)このような核酸物質が、B-Rafポリペプチドをコードする配列において1つ以上の突然変異を含むか否かを判断するステップ
を含む、細胞形質転換の検出方法を提供する。
【0029】
突然変異は、点突然変異であることが都合がよい。
【0030】
突然変異は、B-rafの1388、1394、1403、1753、1782、1783、1796、1797、1787、および1786位の1つ以上において生じていることが都合がよい。点突然変異は、B-raf中のG1388T、G1783C、TG1796-97AT、G1394T、G1394A、G1394C、G1403C、G1403A、G1753A、T1782G、G1388A、T1796A、T1787G、またはC1786Gであることが好ましい。
【0031】
本発明により同定した突然変異は、体細胞組織(somatic tissue)で生じ、生殖細胞を介して伝達されない体細胞突然変異であることが都合がよい。従って、本発明はさらに、
(a)癌性であると疑われる被験体組織から細胞物質の第1のサンプルを、および同じ被験体の非癌性組織から細胞物質の第2のサンプルを単離するステップ;
(b)該細胞物質のサンプルの両方における1つ以上のB-raf遺伝子の少なくとも一部に由来する核酸物質を検査するステップ;ならびに
(c)このような核酸物質が、B-Rafポリペプチドをコードする配列において1つ以上の点突然変異を含むか否か、そして該突然変異が、該癌性と疑われる組織に由来する細胞物質に存在して、かつ該非癌性組織由来の細胞物質には存在していないことを判断するステップ
を含む、細胞形質転換の検出方法に関する。
【0032】
本発明はさらに、
(a)被験体から細胞物質のサンプルを得るステップ;
(b)該サンプルを、本発明による変異型B-Rafポリペプチドに選択的に結合するリガンドでスクリーニングするステップ;および
(c)該サンプル中の1つ以上の変異型B-Rafポリペプチドを検出するステップ
を含む、細胞形質転換の検出方法を提供する。
【0033】
さらに別の態様では、本発明は、
(a)本発明による1つ以上の変異型B-Rafポリペプチドを得るステップ;
(b)該1つ以上のポリペプチドを、テストする1つ以上の化合物と接触させるステップ;および
(c)該1つ以上の化合物と該変異型ポリペプチドとの相互作用を検出するステップ
を含む、抗増殖活性を有する1つ以上の化合物を同定する方法に関する。
【0034】
上記相互作用は結合相互作用であることが好ましい。
【0035】
さらに、本発明は、
(a)本発明による1つ以上の変異型B-Rafポリペプチドを得るステップ;
(b)B-Rafポリペプチド用の下流基質を得るステップ;および
(c)テストする1つ以上の化合物の存在下で、該基質の修飾を検出するステップ
を含む、抗増殖活性を有する1つ以上の化合物を同定するためのアッセイを提供する。
【0036】
B-Rafは、タンパク質キナーゼであり、従って、基質はリン酸化または脱リン酸化され得る。基質上での変異型B-Rafの作用は、検出可能な変化を生じることが好ましい。基質は、別のキナーゼまたはホスファターゼであり、これらは検出可能な変化を生じる第3の分子を修飾することが都合がよい。
【0037】
例えば、基質は、キナーゼMEKであり得る。MEKリン酸化は、直接検出されるか、または好ましくは、MAPキナーゼをリン酸化するMEKの活性を介して検出され得る。
【0038】
基質上の変異型B-Rafの参照活性を確立し、テストする1つ以上の化合物の存在下および/または不在下における活性を該参照値と比較することが都合がよい。変異型B-Rafの活性が低下している場合には、増殖活性が低下していることを示す。
【0039】
本発明はさらに、抗増殖活性について化合物をスクリーニングするための細胞利用型アッセイを提供する。第1の態様では、本発明は、
(a)NIH 3T3細胞の培養物を得るステップ;
(b)本発明による変異型B-raf核酸で、該細胞をトランスフェクトするステップ;
(c)該細胞を、テストする1つ以上の化合物に曝すステップ;および
(d)該テストする1つ以上の化合物に曝されたトランスフェクト細胞と、そのように曝されていないトランスフェクト細胞とで、形成されたフォーカスの数の差を測定するステップ
を含む、3T3フォーカス形成アッセイを提供する。
【0040】
細胞利用型アッセイは、NIH 3T3細胞を使用して行われることが多い。しかし、他の細胞型、特に繊維芽細胞も、このようなアッセイにおいて使用され得る。
【0041】
アッセイにおいて使用する細胞上の変異型B-raf遺伝子のフォーカス形成の参照活性を確立して、テストする1つ以上の化合物の存在下および/または不在下での活性を該参照値と比較することが都合がよい。変異型B-raf遺伝子のフォーカス形成活性の低下は、増殖活性の低下を示し、つまりテストする1つ以上の化合物の抗増殖活性の低下を示す。
【0042】
本発明による突然変異を検出する自動化された方法および装置(apparata)も提供する。
【図面の簡単な説明】
【0043】
【図1A】B-Raf活性アッセイ。MBPを最終基質として使用し、免疫沈降キナーゼカスケードアッセイにおいて、B-Rafのキナーゼ活性を測定した。活性は、MBPに取り込まれた計数として示している。アッセイは三重で行い、平均値、および平均値からの偏差を表す誤差バーを示す。基準キナーゼ活性(空白バー)、およびV12Ras刺激キナーゼ活性(斜線バー)の両方を示している。
【図1B】B-Raf活性アッセイ。MBPを最終基質として使用し、免疫沈降キナーゼカスケードアッセイにおいて、B-Rafのキナーゼ活性を測定した。活性は、MBPに取り込まれた計数として示している。アッセイは三重で行い、平均値、および平均値からの偏差を表す誤差バーを示す。基準キナーゼ活性(空白バー)、およびV12Ras刺激キナーゼ活性(斜線バー)の両方を示している。
【図2】B-Rafおよび活性化変異体によるNIH3T3細胞の形質転換。細胞を、表示の構築物でトランスフェクトし、コロニーの数を測定した。結果は、少なくとも3つのアッセイの平均である。B-Rafにより誘導される数と比較したコロニー数を示している。
【図3A】V599Dは、BRAFにおける活性化突然変異である。BRAFまたはV599DBRAFを、表示の通り、単一で、または発癌性Rasと共に発現させた。免疫沈降させたBRAFを用いてMEKおよびERKを順次活性化させる免疫沈降キナーゼカスケードアッセイを使用して、BRAFタンパク質の活性を測定した。ERKの活性は、ミエリン塩基性タンパク質および[-32P]-γATPを基質として使用して、測定している。
【図3B】薬物試薬を使用した、黒色腫細胞系におけるERKの阻害。WM266.4またはA375P細胞を、10μM U0126、10μM BAY 43-9006、または対照であるDMSOで、処理した。等量の細胞タンパク質を、SDS-ゲル上で分離させ、ppERK抗体を使用して活性ERKのレベルを測定した。
【図4】薬物による細胞成長の阻害。U0126(10μM)もしくはBAY 43-9006(10μM)、またはビヒクル対照(DMSO)の存在下で、WM-266.4細胞をインキュベートした。48時間後、細胞を、[3H]-チミジンとインキュベートさせて、DNA合成を測定し、細胞DNAに取り込まれたチミジンのレベルを測定した。
【図5A】CRAF発現は、siRNAにより抑制される。WM-266.4、Colo 829、またはBE細胞を、CRAF特異的siRNAプローブ(CRAF)、無作為(scrambled)siRNAプローブ(無作為)、オリゴフェクタミン(オリゴ)で処理するか、または未処理のまま(対照)にした。細胞を24時間インキュベートし、CRAFタンパク質のレベルをウェスタンブロッティングにより測定した。
【図5B】BRAF発現は、siRNAにより抑制される。WM-266.4、Colo 829、またはBE細胞を、表示の通り、BRAF特異的siRNAプローブ、無作為siRNAプローブで処理するか、または未処理のままにした。細胞を24時間インキュベートし、BRAF活性のレベルを免疫沈降キナーゼアッセイMEKおよびERKを連続するアッセイとして測定した。MBPおよび[32P]-γATPを基質として使用して、ERKの活性を測定した。
【図6】CRAFではなくBRAFの除去により、黒色腫細胞中のERK活性がブロックされる。WM-266.4またはColo829細胞を、表示の通り、BRAF特異的siRNAプローブ(BRAF)もしくは無作為対照(sBRAF)、CRAF特異的プローブ(CRAF)もしくはその無作為対照(sCRAF)、オリゴフェクタミン(オリゴ)で処理するか、または未処理のまま(対照)にした。細胞を表示した時間だけインキュベートし、Colo829細胞は24時間処理した。等量の細胞抽出物中のERK活性のレベルを、ppERK抗体でのウェスタンブロッティングにより測定した。
【図7】CRAFではなくBRAFの除去により、黒色腫細胞においてアポトーシスが誘導される。WM-266.4細胞を、表示の通りBRAF特異的siRNAプローブ(BRAF)もしくは無作為対照(sBRAF)、CRAF特異的プローブ(CRAF)、U0126、DMSO(オリゴ)で処理するか、または未処理のまま(対照)にした。細胞を96時間インキュベートし、細胞周期プロファイルをFACSで分析するか、またはPARP発現をウェスタンブロッティングにより検査した。
【図8】B-RafおよびGST-MKKI活性検証。WTS 1 B-Raf溶解産物(バッチA)、GST MKKI(6.5μg/ml)およびERK2(キナーゼコンピテント、100μg/ml)を使用してアッセイを行い、MBP(0.3mg/ml)への33P-γ-リン酸塩(phosphate)取込みを測定する。表示のデータは、三重測定の平均値±SDである。
【図9】フィルタプレートおよびフラッシュプレート放射測定アッセイプラットホームの評価。WTS1 B-Raf1溶解産物(バッチA)およびGST MKK1(6.5μg/ml)を使用してアッセイを行い、GST-kdERK2(100μg/ml)への33P-γ-リン酸塩取込みを測定した。表示のデータは、三重測定の平均値±SDである。
【図10】DELFIA非放射測定アッセイプラットホームの評価。100 ng GST-kdERK2を、各ウェルに予備結合させた後、B-Raf溶解産物(バッチA)、GST-MKK1(6.5μg/ml)、およびATP(500μM)を添加した。表示のデータは、三重測定の平均値±SDである。
【図11】DELFIAアッセイにおける抗-ホスホ-ERK2の滴定。100 ng GST-kdERK2をウェルに予備結合させた後、B-Raf溶解産物(バッチA)、GST-MKK1(6.5μg/ml)、およびATP(500μM)を添加した。表示のデータは、二重測定の平均値である。
【図12】DELFIAアッセイにおけるユーロピウム標識化二次抗体の滴定。100 ng GST-kdERK2をウェルに予備結合させた後、B-Raf溶解産物(バッチA)、GST-MKK1(6.5μg/ml)およびATP(500μM)を添加した。表示のデータは、二重測定の平均値である。
【図13】均一アッセイプロトコールの評価。均一アッセイは、B-Raf溶解産物(バッチB)、6.5μg/ml GST-MKK1、80μg/ml GST-kdERK2、および500μM ATPを含む50μl反応容量を使用して96ウェルプレートにおいて行った。表示のデータは、三重測定の平均値±SDである。
【発明を実施するための形態】
【0044】
発明の詳細な説明
特に付記しない限り、本明細書で使用する全ての技術的および科学的用語は、(例えば、細胞培養、分子遺伝子学、核酸化学、ハイブリダイゼーション技法、および生化学における)当業者に一般的に理解されているものと同じ意味を持つ。分子、遺伝子、および生化学的方法のために、標準的な技術を使用する。Sambrookら, Molecular Cloning: A Laboratory Manual, 第二版 (1989) Cold Spring Harbor Laboratory Press, Cold Spring Harbor, N.Y.、およびAusubelら, Short Protocols in Molecular Biology (1999) 第四版, John Wiley and Sons, Inc.;ならびにGuthrieら, Guide to Yeast Genetics and Moleculr Biology, Methods in Enzymology, Vol. 194, Academic Press, Inc., (1991)、PCR Protocols: A Guide to Methods and Applirations (Innisら 1990. Academic Press, San Diego, Calif.)、McPhersonら, PCR Volume 1, Oxford University Press, (1991)、Culture of Animal Cells: A Manual of Basic Technique, 第二版(R.I. Freshney. 1987. Liss, Inc. New York, N.Y.)、ならびにGene Transfer and Expression Protocols, pp. 109-128, E.J. Murray編, The Humana Press Inc., Clifton, N.J.を概要的に参照のこと。これらの文献は、参照により本明細書に援用する。
【0045】
定義
本出願は、B-Rafポリペプチド変異体を記載する。本明細書で使用する「RAFポリペプチド」という用語は、RAFファミリーのポリペプチドを指すのに使用する。RAFは、最初、癌遺伝子v-rafを含む、クローニングされた固有の急性形質転換性複製欠損マウスC型ウイルスにおいて同定された(Rappら Proc. Nat. Acad. Sci. 80: 4218-4222, 1983)。細胞ホモログであるc-rafは、哺乳動物DNA中に存在する。その後、他のホモログが、ヒトおよびトリにおいて発見されており、rafは鳥類癌遺伝子milのホモログであることが示された。B-Rafは、RAFに関係するが、3つの追加のN末端エキソンを有する。従って、「B-Raf」という用語は、全ての既知のヒトB-Rafホモログおよび変異体、ならびにB-Rafホモログとして同定されるB-Rafに対して十分な相同性を示す他のポリペプチドを包含する。この用語は、ARAF、CRAF、またはRAF1は含まない。B-Rafは、受託番号NP_004324、核酸受託番号NM_004333として示される配列を有するポリペプチドとして同定されることが好ましい。
【0046】
「B-Raf」という用語は、NP_004324に85%、90%、95%、96%、97%、98%、または99%相同であるポリペプチドを含むことが好ましい。相同性比較は、目で、またより一般的には、利用し易い配列比較プログラムの助けを借りて行うことができる。これらの市販されているコンピュータプログラムは、2つ以上の配列間の相同性パーセンテージ(%)を計算できる。
【0047】
相同性パーセンテージは、連続した配列にわたって計算できる(すなわち、一方の配列を、他方の配列とアラインさせて、一方の配列中の各アミノ酸を、他方の配列中の対応するアミノ酸と、一度に1残基ずつ直接比較する)。これは、「アンギャップド」アライメントと呼ばれる。典型的に、このようなアンギャップドアライメントは、比較的少数の残基(例えば、50未満の連続アミノ酸)にわたってのみ行われる。
【0048】
これは非常に単純で一貫性のある方法であるが、例えば、全く同一の配列対でなければ、1つの挿入または欠失により、後続するアミノ酸残基がアライメントから外れ、従って、大規模なアライメントを行った場合に、相同性%の大幅な減少を生じる可能性があるという考慮を欠いている。その結果、ほとんどの配列比較方法は、相同性スコア全体に必要以上にペナルティーを科すことなく、可能性のある挿入および欠失を考慮する最適アライメントを得るように設計されている。これは、配列アライメントに「ギャップ」を挿入して、局所相同性を最大化するようにすることにより達成される。
【0049】
しかし、これらのより複雑な方法は、アライメント中で生じる各ギャップに対して「ギャップペナルティー」を科して、同数の同一アミノ酸について、できるだけ少ないギャップでの(2つの比較配列間のより高い関係性を反映する) 配列アライメントの方が、ギャップの多いものよりも高いスコアが得られる。典型的に、ギャップの存在に対して比較的高いコストを、そしてギャップ中の後続する各残基に対してより小さいペナルティーを科する「アフィンギャップコスト(Affine gap cost)」を使用する。これは、最も一般的に使用されるギャップスコア付けシステムである。高いギャップペナルティーは、当然、ギャップがより少ない最適化アライメントを作成する。ほとんどのアライメントプログラムは、ギャップペナルティーを変えることが可能である。しかし、このようなソフトウェアを配列比較に使用する場合には、デフォルト値を使用することが好ましい。例えば、GCG Wisconsin Bestfitパッケージ(以下参照)を使用する場合、アミノ酸配列に対するデフォルトギャップペナルティーは、1ギャップにつき-12であり、各配列(extension)につき-4である。
【0050】
従って、最大相同性%の計算は、まず、ギャップペナルティーを考慮した、最適なアライメントの生成を必要とする。このようなアライメントを実行するために適したコンピュータプログラムは、GCG Wisconsin Bestfitパッケージ(University of Wisconsin, U.S.A.;Devereuxら, 1984, Nucleic Acids Research 12:387)である。配列比較を実行できる他のソフトウェアの例としては、BLASTパッケージ(Ausubelら, 1999、前掲、第18章を参照)、FASTA(Atschulら, 1990, J. Mol. Biol., 403-410)、およびGENEWORKS比較ツール一式が挙げられるが、これらに限定されない。BLASTおよびFASTAの両方は、オフラインおよびオンライン検索の利用が可能である(Ausubelら, 1999、前掲、7-58〜7-60頁を参照)。しかし、GCG Bestfitプログラムを使用することが好ましい。
【0051】
最終的な相同性%は、同一性の点で測定できるが、アライメントプロセス自体は、典型的に、絶対的な対比較に基づいていない。その代わり、化学的同一性または進化距離に基づいて、対ごとの比較に対してスコアを付けるスケール化類似性スコアマトリックスが一般的に使用される。よく使用されるこのようなマトリックスの例は、BLOSUM62マトリックス(BLASTプログラム一式についてのデフォルトマトリックス)である。GCG Wisconsinプログラムは、一般的に、公開デフォルト値か、または供給される場合にはカスタム記号比較表のいずれかを使用する(さらなる詳細についてはユーザマニュアルを参照のこと)。GCGパッケージについては公開デフォルト値を使用し、他のソフトウェアの場合には、BLOSUM62等のデフォルトマトリックスを使用することが好ましい。
【0052】
ソフトウェアが、最適なアライメントを生成した後、相同性%、好ましくは配列同一性%を計算することが可能になる。ソフトウェアは、典型的に、配列比較の一部としてこれを実行し、数値的結果を生成する。
【0053】
本発明によるポリペプチドの「断片」は、本発明に従って記載する変異型アミノ酸を包含するポリペプチド断片である。断片は、完全長B-Rafポリペプチドまでのあらゆる長さのものであり得る;従って、数アミノ酸分を平滑末端化されたB-Rafポリペプチド、およびそれよりも短い断片を含む。都合良くは、断片は、約764〜約5アミノ酸の長さ;好ましくは、約5〜約20アミノ酸の長さ;都合良くは、約10〜約50アミノ酸の長さである。本発明による断片は、とりわけ、動物を免疫化して、抗体を生じさせるために有用である。従って、本発明によるポリペプチドの断片は、本明細書に記載する変異型B-Rafを特徴とする少なくとも1つの抗原決定基(エピトープ)を含むことが都合がよい。特定のポリペプチド断片が、このような抗原特性を保持するか否かは、当該分野で公知の常套的な方法により簡単に判断できる。少なくて6アミノ酸残基からなるペプチドが、免疫応答を示すことが見とめられることが多い。
【0054】
本発明の「核酸」は、上述したヒトB-Rafポリペプチドをコードする核酸である。この用語はさらに、ストリンジェントなハイブリダイゼーション条件下で、上記同定した天然の核酸またはその相補体にハイブリダイズ可能な、ポリヌクレオチドも含む。「ストリンジェントなハイブリダイゼーション条件」とは、50%ホルムアミド、5×SSC(750 mM NaCl、75 mMクエン酸三ナトリウム)、50 mMリン酸ナトリウム(pH 7.6)、5×デンハルト溶液、10%デキストラン硫酸、および20 pg/ml変性断片化(sheared)サケ精子DNAを含む溶液中で、42℃にて一晩のインキュベーションの後、0.1×SSC中で約65℃にてフィルタを洗浄することを指す。
【0055】
本明細書で言及する核酸は、概して、天然で見とめられる核酸であるが、同用語は、その改変範囲内で、当該分野で公知の、改変された骨格または基質を有する人工核酸も含み得る。
【0056】
本発明による断片をコードする核酸は、本発明による突然変異を含むB-raf遺伝子の特定の領域の核酸増幅によるものであり得る。
【0057】
本明細書で言及する「単離」ポリペプチドまたは核酸は、元の環境(例えば、物質が自然と生じる自然環境)から採られた物質を指し、つまり、その天然状態から人為的に変化させられる。例えば、単離ポリペプチドは、ベクターもしくは組成物の一部であるか、または細胞に含まれていたとしても、該ベクター、組成物、または特定の細胞が該ポリヌクレオチドの元の環境でない場合には、「単離」状態と言える。「単離」という用語は、本技法により本発明のポリペプチド/核酸の独特の特徴を示さない、ゲノムもしくはcDNAライブラリー、全細胞合計もしくはmRNA調製物、ゲノムDNA調製物(電気泳動で分離され、ブロット上に移されたものも含む)、断片化(sheared)全細胞ゲノムDNA調製物、または他の組成物は指さないことが好ましい。
【0058】
本発明によるポリペプチドは、1つ以上の突然変異を含む。「突然変異」としては、アミノ酸付加、欠失または置換が挙げられ;アミノ酸置換を指すことが都合がよい。ポリペプチドレベルでのこのような突然変異は、1つ以上のヌクレオチドの付加、欠失または置換により、核酸レベルで反映される。一般的に、このような突然変異は、核酸のリーディングフレームを変えない。核酸レベルでの変化は、単一のヌクレオチドが別のヌクレオチドに置き換わって、それが一部を構成するコドンを変化させて異なるアミノ酸を特定する点突然変異であることが都合がよい。
【0059】
本発明において同定されるB-Rafにおける突然変異は、自然に生じるもので、発癌物質または他の腫瘍化因子を使用することにより細胞または組織において意図的に誘導されたものではない。従って、本明細書において同定される突然変異は、ヒト組織中からin vivoでの自然な腫瘍形成を正確に反映する。従って、これらの検出は、人為的な化学腫瘍誘導化の後にげっ歯類において同定される突然変異の検出よりも、はるかに良い診断用の基準となる。
【0060】
「体細胞」突然変異は、生物の生殖細胞系を介して伝達されず、生物の体細胞組織において生じる突然変異である。体細胞突然変異は、正常/腫瘍対サンプル分析を通じて体細胞性であると判断されたものであることが都合がよい。
【0061】
本明細書で使用する全てのアミノ酸およびヌクレオチドの番号付けは、B-Rafポリペプチドのアミノ酸+1、またはそれをコードするヌクレオチド配列の最初のATGから開始する。
【0062】
「増幅」反応は、非標的核酸に対して標的核酸の特異的な増幅を生じる核酸反応である。ポリメラーゼ連鎖反応(PCR)は、周知の増幅反応である。
【0063】
「免疫グロブリン」は、免疫グロブリン(抗体)分子を特徴付ける免疫グロブリン折畳み構造を保持するポリペプチドのファミリーのひとつであり、2つのβシート、および通常、保存されたジスルフィド結合を含む。免疫グロブリンスーパーファミリーのメンバーは、in vivoでの細胞および非細胞相互作用の多くの要素に関与しており、免疫系における幅広い役割(例えば、抗体、T細胞受容体分子等)、細胞付着における関与(例えば、ICAM分子)、および細胞内シグナリング(例えば、PDGF受容体等の受容体分子)が挙げられる。本発明は、高い特異性で標的抗原に結合可能な抗体に適用可能であることが好ましい。
【0064】
「抗体」は、抗体全体、またはその抗原結合フラグメントであってもよい。例えば、本発明は、FvおよびFab、ならびにFab'およびF(ab')2等のフラグメント、ならびにscFv、単一ドメイン抗体、Dab抗体、および他の抗原結合抗体に基づく分子等の抗体変異体を含む。
【0065】
本明細書において「癌」は、細胞形質転換から生じる新生物成長から、新生物表現型を指すために使用する。このような細胞形質転換は、遺伝子突然変異を伴うことが多い。本発明に関しては、形質転換は、本明細書に記載する1つ以上のB-raf遺伝子の変化による遺伝子突然変異を伴う。
【0066】
核酸の検出方法
本発明に関しては、B-Rafをコードする変異型核酸の検出を採用して、細胞形質転換および癌の存在またはそれらに対する素因を診断できる。B-raf遺伝子における突然変異は、一般的にDNAレベルで生じるため、本発明の方法は、ゲノムDNAにおける突然変異、ならびに転写産物およびタンパク質自体の検出に基づくことができる。転写産物および/またはポリペプチドの分析により、ゲノムDNAにおける突然変異を確認して、検出された突然変異が実際に被験体において確実に発現されていることが望ましくあり得る。
【0067】
ゲノム核酸における突然変異は、増幅された核酸断片における移動度シフトに基づく技術により検出されることが都合がよい。例えば、Chenら, Anal Biochem 1996 Jul 15;239(1):61-9は、競合移動度シフトアッセイによる単一塩基突然変異の検出を記載している。さらに、Marcelinoら, BioTechniques 26(6):1134-1148(June 1999)の技術に基づくアッセイが市販されている。
【0068】
好適な例では、キャピラリーヘテロ二本鎖分析を使用して、ミスマッチの存在により生じるキャピラリー系における二本鎖核酸の移動度シフトに基づく突然変異の存在を検出し得る。
【0069】
サンプルから、分析用の核酸を生成するためには、一般的に、核酸増幅が必要である。多くの増幅方法は、酵素連鎖反応(ポリメラーゼ連鎖反応、リガーゼ連鎖反応、もしくは自己持続性配列複製等)、またはその中でクローニングされたベクターの全体または一部を複製することに拠る。本発明による増幅は、例えばポリメラーゼ連鎖反応により示される指数的増幅であることが好ましい。
【0070】
多くの標的およびシグナル増幅法が、例えば、これらの方法の概論について、Landegren, U.ら, Science 242:229-237(1988)、およびLewis, R., Genetic Engineering News 10:1, 54-55 (1990)等の文献に記載されている。これらの増幅方法は、本発明の方法において使用でき、ポリメラーゼ連鎖反応(PCR)、PCR in situ、リガーゼ増幅反応(LAR)、リガーゼハイブリダイゼーション、Qβバクテリオファージレプリカーゼ、転写利用型増幅系(TAS)、転写産物配列決定を伴うゲノム増幅(GAWTS)、核酸配列利用型増幅(NASBA)、およびin situハイブリダイゼーションが挙げられる。様々な増幅技術において使用されるのに適したプライマーは、当該分野で公知の方法により調製できる。
【0071】
ポリメラーゼ連鎖反応
PCRは、特に、米国特許第4,683,195号および同第4,683,202号に記載されている核酸増幅方法である。PCRは、DNAポリメラーゼ生成プライマー伸長反応のサイクルの繰り返しにより構成される。標的DNAは、熱変性され、増幅させるDNAの対向する鎖上で標的配列を挟む2本のオリゴヌクレオチドをハイブリダイズさせる。これらのオリゴヌクレオチドは、DNAポリメラーゼと使用されるプライマーとなる。DNAは、プライマー伸長によりコピーされて、両鎖の第2のコピーを作製する。熱変性、プライマーハイブリダイゼーションおよび伸長のサイクルを繰り返すことにより、約2〜4時間で標的DNAを100万倍以上増幅できる。PCRは、分子生物学的ツールであり、検出技術とともに使用されて、増幅結果を判断しなければならない。PCRの利点は、約4時間で100万〜10億倍に標的DNAの量を増幅することにより、感度が高まることである。PCRは、診断目的において任意の既知の核酸を増幅するのに使用できる(Mokら, (1994), Gynaecologic Oncology, 52:247-252)。
【0072】
自己持続性配列複製(3SR)
自己持続性配列複製(3SR)は、TASの変形型であり、逆転写酵素(RT)、酵素カクテルにより仲介されるポリメラーゼおよびヌクリアーゼ活性、ならびに適切なオリゴヌクレオチドプライマーの連続ラウンドによる、核酸鋳型の等温増幅(isothermal amplification)を伴う(Guatelliら, (1990) Proc. Natl. Acad. Sci. USA 87:1874)。RNA/DNAヘテロ二重らせんのRNAの酵素分解を、熱変性の代わりに使用する。RNアーゼHおよび他の酵素を反応液に添加し、全てのステップを同じ温度で生じさせ、それ以上試薬は添加しない。このプロセスの後、42℃にて1時間で、106〜109の増幅が達成された。
【0073】
ライゲーション増幅(LAR/LAS)
ライゲーション増幅反応またはライゲーション増幅系は、DNAリガーゼおよび4つのオリゴヌクレオチド(1本の標的鎖に対して2つ)を使う。この技術は、Wu, D. Y.およびWallace, R.B. (1989) Genomics 4:560に記載されている。オリゴヌクレオチドは、標的DNA上の隣接配列にハイブリダイズし、リガーゼにより連結される。反応液を熱変性させ、サイクルを繰り返す。
【0074】
Qβレプリカーゼ
この技術では、一本鎖RNAを複製するバクテリオファージQβ用のRNAレプリカーゼを使用して、Lizardiら (1988) Bio/Technology 6:1197に記載されるように標的DNAを増幅する。まず、標的DNAを、T7プロモーターおよびQβ5'配列領域を含むプライマーにハイブリダイズさせる。このプライマーを使用して、逆転写酵素は、プロセスにおいて、プライマーを5'端部に接続させたcDNAを生成する。これらの2つのステップは、TASプロトコールに似ている。得られるヘテロ二重らせんは、熱変性される。次に、Qβ3'配列領域を含む第2のプライマーを使用して、第2ラウンドのcDNA合成を開始する。これにより、Qβバクテリオファージの5'および3'端部の両方、ならびに活性T7 RNAポリメラーゼ結合部位を含む二本鎖DNAが生じる。次いで、T7 RNAポリメラーゼは、二本鎖DNAを新しいRNAに転写し、これはQβを模倣する。ハイブリダイズしていない全てのプローブを除去するための激しく洗浄した後、新しいRNAを標的から溶出し、Qβレプリカーゼで複製する。後者の反応は、107倍の増幅を約20分間で生じる。
【0075】
代替的な増幅技術が、本発明において利用され得る。例えば、ローリングサークル増幅(Lizardiら, (1998) Nat Genet 19:225)は、DNAポリメラーゼにより駆動され、等温状態下で線状または幾何学的な動態のいずれかを有する環状オリゴヌクレオチドプローブを複製できる、市販の増幅技術(RCATTM)である。
【0076】
2つの適切に設計されたプライマーの存在下で、DNA鎖置換(displacement)および過剰分岐(hyperbranching)を介して幾何学的増幅が生じ、1時間で、各環につき1012以上のコピーが生成される。
【0077】
単一のプライマーを使用する場合、RCATは、数分間で、数千の標的が該標的に共有結合してタンデムに連結したDNAコピーの線状鎖を生成する。
【0078】
別の技術である鎖置換増幅(SDA;Walkerら, (1992) PNAS (USA) 80:392)は、特定の標的固有の具体的に定義された配列で開始する。熱サイクリングに依存する他の技術とは異なり、SDAは、一連のプライマー、DNAポリメラーゼ、および制限酵素を利用して、固有の核酸配列を指数的に増幅する等温プロセスである。
【0079】
SDAは、標的生成期と指数増殖期の両方から構成される。
【0080】
標的生成では、二本鎖DNAを熱変性させて、2本の一本鎖コピーを作製する。一連の特異的に作製したプライマーを、DNAポリメラーゼ(塩基配列をコピーするための増幅プライマー、および新たに作製された鎖を置換させるための緩衝(bumper)プライマー)と組み合わせて、指数増殖が可能な改変型標的を形成する。
【0081】
指数増幅プロセスは、標的生成期で得た改変型標的(制限酵素認識部位を有する一本鎖部分的DNA鎖)から開始する。
【0082】
増幅プライマーは、その相補DNA配列において各鎖に結合する。すると、DNAポリメラーゼがプライマーを使用し、改変型標的を個々のヌクレオチドを付加するための鋳型として使用して3'側からプライマーを伸長させる位置を識別する。従って、伸長プライマーは、完全な制限酵素認識部位を両端に含む二本鎖DNAセグメントを形成する。
【0083】
次に、制限酵素を、二本鎖DNAセグメントにその認識部位において結合させる。制限酵素は、両側(double-sided)セグメントの一方の鎖のみを切断した後、認識部位から解離して、ニックを形成する。DNAポリメラーゼは、ニックを認識し、その部位から鎖を伸長させて、先に作製された鎖に置き換わる。このように、認識部位は、制限酵素およびDNAポリメラーゼにより、繰り返しニッキングおよび復元されて、標的セグメントを含むDNA鎖が連続的に置換される。
【0084】
すると、各置換された鎖は、上述したように増幅プライマーにアニール可能になる。新しいDNA鎖のニッキング、伸長、および置換を繰り返して、プロセスを続け、元のDNA標的の指数増幅を生じさせる。
【0085】
核酸を増幅させた後、単一塩基対の突然変異の検出のために多数の技術が利用できる。このような技術の一つは、一本鎖高次構造多型(SSCP)である。SCCP検出は、電気泳動中の、参照DNAと比較した場合の一本鎖変異型DNAの異常な移動に基づく。突然変異は、一本鎖DNAにおいて高次構造変化をもたらし、移動シフトを生じる。蛍光SCCPは、蛍光標識化プライマーを使用して、検出を補助する。参照DNAおよび変異型DNAは、蛍光標識化プライマーを使用して、このように増幅される。増幅DNAは、変性および急速冷却(snap-cool)されて、一本鎖DNA分子を生成し、これは、非変性ゲル電気泳動により検査される。
【0086】
化学ミスマッチ切断(CMC)は、ヒドロキシルアミン、四酸化オスミウム、およびピペリジンの組合せによる、DNAミスマッチ塩基対の認識および切断に基づく。従って、参照DNAおよび変異型DNAの両方を、蛍光標識化プライマーで増幅する。ミスマッチT塩基に結合する四酸化オスミウム、またはミスマッチC塩基に結合するヒドロキシルアミンの後に、修飾塩基の部位において切断するピペリジンを使用して、アンプリコン(amplicon)をハイブリダイズし、その後切断する。次いで、切断された断片を電気泳動で検出する。
【0087】
制限断片多形(RFLP)に基づく技術も使用できる。多くの単一ヌクレオチド多形(SNP)は従来のRFLP分析にかからないが、プライマー誘導型制限分析PCR(PIRA-PCR)を使用して、SNP依存的にPCRプライマーを用いて制限部位を導入できる。適切な制限部位を導入するPIRA-PCR用のプライマーは、例えば、Xiaiyiら, (2001) Bioinformatics 17:838-839に記載されるように、コンピュータ分析により設計できる。
【0088】
代替的な実施形態では、本発明は、RNAレベルでの遺伝子発現の検出を提供する。リボ核酸ハイブリダイゼーションを利用する典型的なアッセイ形式としては、核ランオンアッセイ、RT-PCR、およびRNアーゼ保護アッセイが挙げられる(Meltonら, Nuc. Acids Res. 12:7035)。採用できる検出方法としては、放射活性標識、酵素標識、化学発光標識、蛍光標識、および他の適切な標識が挙げられる。
【0089】
RT-PCRを使用して、RNA標的を増幅する。このプロセスにおいて、逆転写酵素を使用して、RNAを相補DNA(cDNA)に変換し、これはその後PCRを使用して増幅できる。この方法は、RNAウイルスを検出するのに有用であることが分かっている。PCR以外での用途は、上述した。
【0090】
ポリペプチドの検出方法
本発明は、変異型B-raf遺伝子にコードされるタンパク質を検出する方法を提供する。タンパク質は、当該分野で公知のタンパク質ゲルアッセイ、抗体結合アッセイ、または他の検出方法により検出できる。
【0091】
従って、例えば、変異型B-Rafポリペプチドを、タンパク質ゲル上の異なる移動度、または変異型アミノ酸の存在を分子量に応じて測定できる質量分析法等の他のサイズ分析技術により検出できる。特に、変異型B-Rafポリペプチドに由来するペプチドは、サイズ分析による差が出やすい。
【0092】
検出手段は、特定の点突然変異が変異型B-Rafポリペプチドにおいて正確に同定され得るように、配列特異的であることが都合がよい。例えば、変異型B-Rafポリペプチドをin vivoまたはin vitroを特異的に認識するポリペプチドまたはRNA分子が開発できる。
【0093】
例えば、RNAアプタマーは、SELEXにより生成できる。SELEXは、標的分子に高度に特異的に結合する核酸分子のin vitro進化のための方法である。これは、例えば、米国特許第5654151号、同第5503978号、同第5567588号、および同第5270163号、ならびにPCT公報WO 96/38579に記載されている(これらの文献はそれぞれ、参照により本明細書に特に援用される)。
【0094】
SELEX方法は、オリゴヌクレオチドライブラリーから、核酸アプタマー、すなわち所望の標的に結合可能な一本鎖核酸を選択することを伴う、好ましくはランダム化配列のセグメントを含む核酸のライブラリーから開始して、SELEX方法は、結合し易い条件下でライブラリーを標的と接触させるステップ、結合してない核酸を標的分子に特異的に結合した核酸から分離するステップ、核酸-標的複合体を解離させるステップ、核酸-標的複合体から解離させた核酸を増幅させて、核酸のリガンド富化ライブラリーを産生するステップを含み、そして、結合、分離、解離、および増幅ステップを、望ましいだけのサイクル繰り返して、標的分子に高度に特異的かつ高親和性な核酸リガンドを産生する。
【0095】
SELEXは、多数の可能性のある配列および構造体を含む核酸ライブラリー内では、所与の標的に対して幅広い結合親和性があるという原理に基づいている。例えば、20ヌクレオチドランダム化セグメントを含む核酸ライブラリーは、420の構造可能性を持ちうる。標的に対してより高い親和定数を有するものは、結合する可能性が最も高いと思われる。分離、解離および増幅のプロセスにより、より高い結合親和性候補が富化された第2の核酸ライブラリーが生成される。さらなる選択ラウンドにより、得られるライブラリーが1つまたは数配列だけで大部分を構成されるまで、最良のリガンドが徐々に絞り込まれる。その後、これらは、クローニング、配列決定、および純粋なリガンドとして結合親和性について個々にテストされ得る。
【0096】
所望の目標が達成するまで、選択および増幅のサイクルを繰り返す。最も一般的なケースでは、サイクルの繰り返しによって結合強度に有意な改善が得られなくなるまで、選択/増幅を続ける。繰返し選択/増幅方法は、少なくとも1014の配列を含むライブラリー中の、単一配列変異体を単離させるのに十分に感度が高い。本方法は、原理的に、約1018の異なる核酸種をサンプル化するために使用され得る。ライブラリーの核酸は、効率的な増幅のために必要な保存された配列と共に、ランダム化配列部分を含むことが好ましい。核酸配列変異体は、ランダム化核酸配列の合成、およびランダムに切断された細胞核酸からのサイズ選択を含む多数の手法により生成され得る。可変的な配列部分は、完全または部分的にランダムな配列を含み得る;また、ランダム化配列と合体した保存配列のサブ部分(subportion)も含み得る。テスト核酸における配列のばらつきは、選択/増幅の繰返しの前または途中の突然変異誘発により、およびクローン化アプタマーの特異的な修飾により、導入または増やすことができる。
【0097】
抗体
B-Rafポリペプチド、またはそこから誘導されるペプチドを使用して、本発明において使用する抗体を生成できる。使用するB-Rafペプチドは、本発明による変異型B-Rafポリペプチドに特異的なエピトープを含むことが好ましい。エピトープとして機能するポリペプチド断片は、あらゆる従来手段(例えば、米国特許第4,631,211号を参照)により生成され得る。本発明では、抗原エピトープは、好ましくは少なくとも4、少なくとも5、少なくとも6、少なくとも7、より好ましくは少なくとも8、少なくとも9、少なくとも10、少なくとも11、少なくとも12、少なくとも13、少なくとも14、少なくとも15、少なくとも20、少なくとも25、少なくとも30、少なくとも40、少なくとも50、そして、最も好ましくは約15〜約30のアミノ酸の配列を含む。免疫原性または抗原性エピトープを含む好ましいポリペプチドは、少なくとも25、30、35、40、45、50、55、60、65、70、75、80、85、90、95、または100アミノ酸残基の長さである。
【0098】
遊離型または担体共役型ペプチドのいずれかで、(例えば、約100μgのペプチドもしくは担体タンパク質、およびフロイントアジュバントもしくは免疫応答を促進することが知られている他の任意のアジュバントを含むエマルジョンを腹腔内および/または皮内注射することで)ウサギまたはマウス等の動物を免疫化することにより、本発明によるB-Rafポリペプチドの抗原性エピトープを使用して、抗体が生成され得る。例えば約2週間ごとに、何回かのブースター注射をして、例えば固体表面に吸着した遊離ペプチドを使用するELISAアッセイにより検出され得る有用な力価の抗ペプチド抗体を得ることが必要となりうる。免疫化動物から得た血清中の抗ペプチド抗体の力価は、抗ペプチド抗体の選択により(例えば、当該分野で周知の方法による、固体表面上のペプチドへの吸着、および選択された抗体の溶出により)高められ得る。
【0099】
本発明のB-Rafポリペプチド、ならびにその免疫原性および/または抗原性エピトープ断片を、他のポリペプチド配列に融合してもよい。例えば、本発明のポリペプチドを、免疫グロブリンドメインと融合させてもよい。ヒトCD4-ポリペプチドの最初の2つのドメイン、ならびに哺乳動物免疫グロブリンの重鎖および軽鎖の定常領域の様々なドメインから構成されるキメラタンパク質は、in vivoにおいて有利な性質を所有することが示されている(例えば、EP 0394827;Trauneckerら, (1988) Nature, 331: 84-86を参照)。上皮バリアを介した免疫系への抗原の送達の増強が、IgGまたはFcフラグメント等のFcRn結合パートナーと共役した抗原(インスリン等)について実証されている(例えば、WO 96/22024およびWO 99/04813を参照)。
【0100】
さらに、本発明のポリペプチドは、融合されたポリペプチドを精製し易くするペプチド等のマーカー配列と融合され得る。好適な実施形態では、マーカーアミノ酸配列は、市販されている多くのものの中でも、pQEベクター中に配置されたタグ(QIAGEN, Inc., 9259 Eton Avenue, Chatsworth, CA, 91311)等のヘキサ-ヒスチジンペプチドである。Gentzら, Proc. Natl. Acad. Sci. USA 86: 821-824 (1989)に記載されているように、例えば、ヘキサ-ヒスチジンは、融合タンパク質の都合の良い精製をもたらす。精製に有用な別のペプチドタグである"HA"タグは、インフルエンザ血球凝集素タンパク質から誘導されたエピトープに対応する(Wilsonら, (1984) Cell 37: 767)。従って、上記融合のいずれもが、本発明の核酸またはポリペプチドを使って操作可能である。
【0101】
好適な実施形態では、本発明は、本明細書に記載するB-Raf変異体を特異的に認識する抗体を提供する。
【0102】
本明細書に記載する抗体は、診断的用途について特に示すものである。従って、これらは、標識等のエフェクタータンパク質を含む改変された抗体であり得る。特に好ましいのは、抗体のin vivoでの分布の画像化を可能にする標識である。このような標識は、患者の身体内で可視化し易い、金属粒子等の放射性標識または放射線不透性(radioopaque)標識である。さらに、組織上で可視化可能な蛍光標識または他の標識であり得る。
【0103】
組換えDNA技術を使用して、本発明の抗体を改善できる。従って、キメラ抗体を構築して、診断的または治療的用途におけるその免疫原性を低下させることができる。さらに、CDRグラフティング[欧州特許出願第0 239 400(Winter)を参照]、および任意に、フレームワーク改変[EP 0 239 400;Riechmann, L.ら, Nature, 332, 323-327, 1988;Verhoeyen M.ら, Science, 239, 1534-1536, 1988;Kettleborough, C.A.ら, Protein Engng., 4, 773-783, 1991;Maeda, H.ら, Human Antibodies and Hybridoma, 2, 124-134, 1991;Gorman S.D.ら, Proc. Natl. Acad. Sci. USA, 88, 4181-4185, 1991;Tempest P.R.ら, Bio/Technology, 9, 266-271, 1991;Co, M.S.ら, Proc. Natl. Acad. Sci. USA, 88, 2869-2873, 1991;Carter, P.ら, Proc. Natl. Acad. Sci. USA, 89, 4285-4289, 1992;Co, M.S.ら, J. Immunol., 148, 1149-1154, 1992;およびSato, K.ら, Cancer Res., 53, 851-856, 1993]により、抗体をヒト化することによって、免疫原性を小さくすることができる。
【0104】
本明細書に記載の抗体は、細胞培養物中で生成できる。組換えDNA技術を使用して、確立された手段により、細菌または好ましくは哺乳動物の細胞培養物において抗体を生成できる。抗体産物は非分泌性細胞から単離できるが、選択された細胞培養系は、任意に、抗体産物を分泌する。
【0105】
従って、本発明は、本発明による抗体を生成するプロセスであって、該抗体タンパク質をコードする第2のDNA配列に正しいリーディングフレーム内で連結したシグナルペプチドをコードする第1のDNA配列に作用可能に連結したプロモーターを含む発現カセットを含むハイブリッドベクターで形質転換された宿主(例えば、大腸菌、昆虫細胞、または哺乳動物細胞)を培養すること、および該タンパク質を単離することを含む、プロセスである。
【0106】
ハイブリドーマ細胞または哺乳動物宿主細胞のin vitroでの増幅は、例えば、哺乳動物血清(例えば、ウシ胎仔血清)、追尾(trace)エレメントおよび成長維持補助剤(例えば、正常マウス腹膜滲出細胞等の供給(feeder)細胞、脾臓細胞、骨髄マクロファージ、2-アミノエタノール、インスリン、トランスフェリン、低密度リポタンパク質、オレイン酸等)を任意に補充されたダルベッコ改良型イーグル培地(DMEM)またはRPMI 1640培地等の習慣的に標準的な培養培地である適切な培養培地中で実施される。細菌細胞または酵母細胞である宿主細胞の増殖も同様に、当該分野で公知の適切な培養培地中(例えば、細菌については培地LB、NZCYM、NZYM、NZM、テリフィック(terrific)ブロス、SOB、SOC、2×YT、またはM9最小培地中、そして酵母については培地YPD、YEPD、最小培地、または完全最小ドロップアウト(dropout)培地中)で実施される。
【0107】
In vitro生成により、比較的純粋な抗体調製物が得られ、所望の抗体を大量に得るための規模拡大が可能になる。細菌細胞、酵母、または哺乳動物細胞培養についての技術は、当該分野で公知であり、(例えば、空輸反応機、もしくは連続攪拌反応機中での)均一懸濁培養、または(例えば、中空繊維中、マイクロカプセル中、アガロースマイクロビーズ上、もしくはセラミックカートリッジ上の)固定化もしくは取込み細胞培養が挙げられる。
【0108】
大量の所望の抗体は、哺乳動物細胞をin vivoで増殖させることによっても得られる。この目的のために、所望の抗体を生成しているハイブリドーマ細胞を、組織適合性哺乳動物に注射して、抗体生成腫瘍を成長させる。任意に、注射前に、動物を炭化水素、特にプリスタン(テトラメチル-ペンタデカン)等の鉱油、でプライミングしてもよい。1〜3週間後、これらの哺乳動物の体液から抗体を単離する。例えば、適切な骨髄腫細胞と、Balb/cマウス由来の抗体生成脾臓細胞との融合により得られたハイブリドーマ細胞、または所望の抗体を生成するハイブリドーマ細胞系Sp2/0から誘導されたトランスフェクト細胞を、任意にプリスタンで予備処置されたBalb/cマウスに腹腔内注射し、1〜2週間後に該動物から腹水を摂取する。
【0109】
上記および他の技術は、例えば、参照により本明細書に援用する、KohlerおよびMilstein, (1975) Nature 256:495-497;米国特許第4,376,110号;HarlowおよびLane, Antibodies: a Laboratory Manual, (1988) Cold Spring Harborにおいて議論されている。組換え抗体分子の調製についての技術は、上記参考文献、ならびに例えば、参照により本明細書に援用するEP 0623679;EP 0368684およびEP 0436597においても記載されている。
【0110】
細胞培養物上清を、好ましくは酵素イムノアッセイ(例えば、サンドウィッチアッセイもしくはドットアッセイ)、またはラジオイムノアッセイにより、所望の抗体についてスクリーニングする。
【0111】
抗体を単離するために、例えば、硫酸アンモニウムでの沈降、ポリエチレングリコール等の吸湿性物質に対する透析、選択膜を介したろ過等により、培養上清中または腹水中の免疫グロブリンを濃縮させてもよい。必要および/または所望であれば、例えば、ゲルろ過、イオン交換クロマトグラフィー、DEAE-セルロース上でのクロマトグラフィー、および/または(イムノ)アフィニティークロマトグラフィー(例えば、標的抗原もしくはプロテインAでのアフィニティークロマトグラフィー)等の習慣的なクロマトグラフィー方法により、抗体を精製する。
【0112】
本発明は、さらに、本発明のモノクローナル抗体を分泌するハイブリドーマ細胞に関する。本発明の好適なハイブリドーマ細胞は、遺伝子的に安定しており、所望の特異性を持つ本発明のモノクローナル抗体を分泌し、解凍および再クローニングすることにより急速冷凍培養物から活性化することができる。
【0113】
本発明は、好適な実施形態において、抗変異型B-Raf抗体の生成に関する。従って、本発明は、本発明のモノクローナル抗体を分泌するハイブリドーマ細胞系を調製するプロセスにも関し、これは、適切な哺乳動物(例えば、Balb/cマウス)を、1つ以上のPDGFポリペプチドもしくはその抗原性断片、または変異型B-Rafポリペプチドを含む抗原性担体で免疫化すること;免疫化した哺乳動物の抗体生成細胞を適切な骨髄腫細胞系の細胞と融合させること;融合で得られたハイブリッド細胞をクローニングすること;および所望の抗体を分泌する細胞クローンを選択することを特徴とする。例えば、変異型B-Rafで免疫化したBalb/cマウスの脾臓細胞を、骨髄腫細胞系PAIまたは骨髄腫細胞系Sp2/0-Ag14の細胞と融合させ、得られたハイブリッド細胞を、所望の抗体の分泌についてスクリーニングし、陽性ハイブリドーマ細胞をクローニングする。
【0114】
1〜100μg変異型B-Rafおよび適切なアジュバント(フロイントアジュバント等)で、数回(例えば、4〜6回)、数ヶ月(例えば、2〜4ヶ月)にわたり、皮下注射および/または腹腔内注射によりBalb/cマウスを免疫化し、最後の注射から2〜4日後に免疫化したマウスから脾臓細胞を得て、融合プロモーター(好ましくはポリエチレングリコール)の存在下で骨髄腫細胞系PAIの細胞と融合させることを特徴とする、ハイブリドーマ細胞系を調製するプロセスが好ましい。骨髄腫細胞は、分子量が約4000の約30%〜約50%のポリエチレングリコールを含む溶液において、免疫化マウスから得た3〜20倍過剰量の脾臓細胞と融合させることが好ましい。融合した後、上述したような選択培地(例えばHAT培地)を補充された適切な培養培地に細胞を規則的な間隔で広げて、正常な骨髄腫細胞が所望のハイブリドーマ細胞を上回って成長させないようにする。
【0115】
本発明はまた、上述した変異型B-Rafに対する抗体の重鎖可変ドメインおよび/または軽鎖可変ドメインをコードするインサートを含む組換え核酸に関する。本質的に、このようなDNAは、コード一本鎖DNA、該コードDNAおよびその相補DNAからなる二本鎖DNA、またはこれらの相補(一本鎖)DNA自体を含む。
【0116】
さらに、変異型B-Rafに対する抗体の重鎖可変ドメインおよび/または軽鎖可変ドメインをコードするDNAは、重鎖可変ドメインおよび/もしくは軽鎖可変ドメインをコードする真正(authentic)DNA配列またはその変異体を有する酵素的または化学的に合成されたDNAであり得る。真正DNAの変異体は、1つ以上のアミノ酸が欠失または別の1つ以上のアミノ酸と交換された、上記抗体の重鎖可変ドメインおよび/または軽鎖可変ドメインをコードするDNAである。該改変は、抗体の重鎖可変ドメインおよび/または軽鎖可変ドメインのCDR外にあることが好ましい。このような変異型DNAは、1つ以上のヌクレオチドが他のヌクレオチドに置き換えられて、新しいコドンが同じアミノ酸をコードする、サイレント変異体であることも意図する。このような変異型配列は変性配列でもある。変性配列は、元々コードされていたアミノ酸の変化を生じずに限定しない数のヌクレオチドが他のヌクレオチドに置き換えられているという遺伝子コードの意味の範囲内において変性されている。このような変性配列は、重鎖マウス可変ドメインおよび/または軽鎖マウス可変ドメインの最適な発現を得るのに、特定の宿主、特に大腸菌が好む制限部位および/または特定のコドンの頻度が異なるために有用であり得る。
【0117】
これに関連して、変異体という用語は、当該分野で公知の方法による真正DNAのin vitro突然変異誘発により得られるDNA変異体を含むことを意図する。
【0118】
完全四量体免疫グロブリン分子のアセンブリ、およびキメラ抗体の発現のために、重鎖および軽鎖可変ドメインをコードする組換えDNAインサートを、重鎖および軽鎖定常ドメインをコードする対応DNAと融合させて、例えばハイブリッドベクターに取り込んだ後に適切な宿主細胞に移す。
【0119】
従って、本発明は、また、ヒト定常ドメインγ(例えば、γ1、γ2、γ3またはγ4、好ましくはγ1またはγ4)に融合した抗変異型B-Raf抗体の重鎖マウス可変ドメインをコードするインサートを含む組換え核酸に関する。同様に、本発明は、ヒト定常ドメインκまたはλ(好ましくはκ)に融合した変異型B-Rafに対する抗変異型B-Raf抗体の軽鎖マウス可変ドメインをコードするインサートを含む組換えDNAに関する。
【0120】
別の実施形態では、本発明は、重鎖可変ドメインおよび軽鎖可変ドメインがスペーサー基により連結されており、任意に宿主細胞中の抗体をプロセシングし易くするシグナル配列、および/または抗体を精製し易くするペプチドをコードするDNA、および/または切断部位、および/またはペプチドスペーサー、および/またはエフェクター分子を含む、組換えポリペプチドをコードする組換えDNAに関する。
【0121】
本発明による抗体および抗体フラグメントは、診断において有用である。従って、本発明は、本発明による抗体を含む診断用の組成物を提供する。
【0122】
診断用組成物の場合、抗体は、酵素的、蛍光的、放射性同位的、または他の手段であり得る抗体検出手段と共に提供されることが好ましい。抗体および検出手段は、診断するための診断キット中に、同時に、別個同時に、または逐次的に使用するように提供され得る。
【0123】
本発明の抗体は、当該分野で公知の任意の方法により、免疫特異的結合についてアッセイされ得る。使用できるイムノアッセイとしては、ウェスタンブロット、ラジオイムノアッセイ、ELISA、サンドウィッチイムノアッセイ、免疫沈降アッセイ、沈降反応法、ゲル拡散沈降反応法、免疫拡散アッセイ、凝集アッセイ、補体結合アッセイ、免疫放射定量測定アッセイ、蛍光イムノアッセイ、およびプロテインAイムノアッセイ等の技術を使用した競合および非競合アッセイ系が挙げられるがこれらに限定されない。このようなアッセイは、当該分野において常套手段である(例えば、Ausubelら編, 1994, Current Protocols in Molecular Biology, Vol. 1, John Wiley and Sons, Inc., New York(参照により本明細書に全体的に援用する)を参照)。イムノアッセイの例を、以下に簡単に記載する。
【0124】
免疫沈降プロトコールは、一般的に、タンパク質ホスファターゼおよび/またはプロテアーゼ阻害剤(例えば、EDTA、PMSF、アプロチニン、バナジン酸ナトリウム)を補充されたRIPA緩衝液(1%NP-40またはTriton X-100、1%デオキシコール酸ナトリウム、0.1%SDS、0.15 M NaCl、0.01 Mリン酸ナトリウム(pH 7.2)、1%トラジロール(Trasylol))等の溶解緩衝液中で細胞の集合を溶解すること、目的の抗体を細胞溶解液に添加すること、4℃にて一定の時間(例えば1〜4時間)インキュベートすること、プロテインAおよび/またはプロテインGセファロースビーズを細胞溶解液に添加すること、4℃にて約1時間以上インキュベートすること、溶解緩衝液中でビーズを洗浄すること、ならびにビーズをSDS/サンプル緩衝液中で再懸濁させること、を含む。目的の抗体が特定の抗原を免疫沈降させる能力は、例えばウェスタンブロット分析により評価できる。
【0125】
ウェスタンブロット分析は、一般的に、タンパク質サンプルを調製すること、ポリアクリルアミドゲル(例えば、抗原の分子量に応じて8%〜20% SDS-PAGE)中でタンパク質サンプルを電気泳動にかけること、ポリアクリルアミドゲルからタンパク質サンプルを膜(ニトロセルロース、PVDF、またはナイロン)に移すこと、ブロック溶液(例えば、3%BSAまたは脱脂乳の入ったPBS)中で膜をブロックすること、洗浄緩衝液(例えば、PBS-Tween 20)中で膜を洗浄すること、ブロック緩衝液中に希釈された一次抗体(目的の抗体)に膜を曝すこと、洗浄緩衝液中で膜を洗浄すること、膜を、ブロッキング緩衝液中に希釈された酵素基質(例えば、西洋ワサビペルオキシダーゼもしくはアルカリホスファターゼ)または放射活性分子(例えば、32Pもしくは125I)と共役した(一次抗体を認識する、例えば抗ヒト抗体等の)二次抗体に曝すこと、洗浄緩衝液中で膜を洗浄すること、および抗原の存在を検出すること、を含む。
【0126】
ELISAは、抗原を調製すること、96ウェルマイクロタイタープレートのウェルを抗原で覆うこと、酵素基質(例えば、西洋ワサビペルオキシダーゼまたはアルカリホスファターゼ)等の検出可能な化合物と共役した目的の抗体をウェルに添加して、一定の時間インキュベートすること、および抗原の存在を検出すること、を含む。ELISAでは、目的の抗体は、検出可能な化合物と共役されなくてもよい;その代わり、検出可能な化合物と共役させた(目的の抗体を認識する)二次抗体をウェルに添加してもよい。さらに、ウェルを抗原で覆う代わりに、ウェルを抗体で覆ってもよい。この場合、検出可能な化合物と共役した二次抗体を、目的の抗原を被覆ウェルに添加した後に、添加してもよい。
【0127】
抗体と抗原との結合親和性、抗体-抗原相互作用の解離速度(off-rate)は、競合結合アッセイにより測定され得る。競合結合アッセイの一例は、大量の非標識化抗原の存在下での標識化抗原(例えば、3Hまたは125I)と目的の抗体とのインキュベーション、および標識化抗原に結合した抗体の検出を含む、ラジオイムノアッセイである。特定の抗原に対する目的の抗体の親和性、および結合解離速度は、スキャッチャード・プロット分析によるデータから決定できる。二次抗体との競合は、ラジオイムノアッセイを使用しても測定できる。この場合、抗原を、大量の非標識化二次抗体の存在下で、標識化化合物(例えば、3Hまたは125I)と共役した目的の抗体とインキュベートさせる。
【0128】
変異型B-Rafポリペプチドの調製
本発明による変異型B-Rafポリペプチドは、化学合成、生物学的サンプルからの単離、およびこのようなポリペプチドをコードする核酸の発現を含む、任意の所望の技術により生成できる。そして、核酸は、変異型B-Rafの生物学的由来源から合成または単離され得る。
【0129】
従って、本発明は、本発明によるポリペプチドをコードするベクター、またはその断片に関する。ベクターは、例えば、ファージ、プラスミド、ウイルス、またはレトロウイルスベクターであり得る。
【0130】
本発明による核酸は、宿主における増殖のための選択マーカーを含むベクターの一部であり得る。一般的に、プラスミドベクターは、リン酸カルシウム沈降物等の沈降物、または荷電脂質との複合体に導入される。ベクターがウイルスの場合、適切なパッケージング細胞系を使用してin vitroでパッケージングされた後に、宿主細胞に形質導入され得る。
【0131】
核酸インサートを、ファージλPLプロモーター、大腸菌lac、trp、phoA、およびtacプロモーター、SV40初期および後期プロモーター、ならびにレトロウイルスLTRのプロモーター等の適切なプロモーターに作用可能に連結させる。他の適切なプロモーターは、当業者に公知である。発現構築物はさらに、転写開始および終結のための部位、ならびに転写領域において翻訳用のリボソーム結合部位を含む。構築物により発現される転写産物のコード部分は、最初に翻訳開始コドンを含み、翻訳するポリペプチドの終わりに適切に位置する終結コドン(UAA、UGA、またはUAG)を含むことが好ましい。
【0132】
上記したように、発現ベクターは、少なくとも1つの選択マーカーを含むことが好ましい。このようなマーカーとしては、ジヒドロ葉酸還元酵素、真核生物細胞培養用のG418もしくはネオマイシン耐性、ならびに大腸菌および他の細菌中で培養するためのテトラサイクリン、カナマイシンもしくはアンピシリン耐性遺伝子が挙げられる。適切な宿主の代表的な例としては、大腸菌、ストレプトミセスおよびネズミチフス菌(Salmonella typhimurium)細胞等の細菌細胞;酵母細胞(例えば、サッカロミセス・セルヴィシエまたはピキア・パストリス(Pichia pastoris)等の菌類細胞;ドロソフィラS2およびスポドプテラSf9細胞等の昆虫細胞;CHO、COS、293およびBows黒色腫細胞等の動物細胞;ならびに植物細胞が挙げられるがこれらに限定されない。
【0133】
上述した宿主細胞に適した培養培地および条件は、当該分野で公知であり、市販されている。
【0134】
細菌における使用が好ましいベクターとしては、QIAGEN, Inc.から入手可能なpQE70、pQE60およびpQE-9;Stratagene Cloning Systems, Inc.から入手可能なpBluescriptベクター、Phagescriptベクター、pNH8A、pNH16a、pNH18A、pNH46A;ならびにPharmacia Biotech, Inc.から入手可能なptrc99a、pKK2233、pKK233-3、pDR540、pRIT5が挙げられる。好ましい真核生物ベクターの一部は、Stratageneから入手可能なpWLNEO、pSV2CAT、pOG44、pXT1、およびpSG;ならびにPharmaciaから入手可能なpSVK3、pBPV、pMSG、およびpSVLである。酵母系における使用が好ましい発現ベクターとしては、pYES2、pYD1、pTEF1/Zeo、pYES2/GS、pPICZ、pGAPZ、pGAPZalph、pPIC9、pPIC3.5、pHIL-D2、pHIL-SI、pPIC3.5K、pPIC9K、およびPA0815(全てInvitrogen, Carlsbad, CAから入手可能)が挙げられるがこれらに限定されない。
【0135】
構築物を宿主細胞に導入することは、リン酸カルシウムトランスフェクション、DEAE-デキストラン仲介型トランスフェクション、陽イオン脂質仲介型トランスフェクション、エレクトロポーレーション、形質導入、感染、または他の方法により実行できる。このような方法は、上記参照したSambrookら等、多くの標準的実験マニュアルに記載されている。
【0136】
本発明によるポリペプチドは、硫酸アンモニウムまたはエタノール沈降、酸抽出、陰イオンまたは陽イオン交換クロマトグラフィー、ホスホセルロースクロマトグラフィー、疎水性相互作用クロマトグラフィー、アフィニティークロマトグラフィー、ヒドロキシルアパタイトクロマトグラフィー、およびレクチンクロマトグラフィーを含む周知の方法により組換え細胞培養物から回収および精製できる。高速液体クロマトグラフィー("HPLC")を精製のために採用することが最も好ましい。
【0137】
本発明によるポリペプチドはまた、体液、組織および細胞を含む生物学的由来源、特に被験体から得た腫瘍組織または可能性のある腫瘍組織から誘導された細胞から回収できる。
【0138】
さらに、本発明のポリペプチドは、当該分野で公知の技術を使用して、化学的に合成され得る(例えば、Creighton, 1983, Proteins: Structures and Molecular Principles, W.H. Freeman and Co., N.Y.、およびHunkapillerら, Nature, 310:105-111 (1984)を参照)。例えば、変異型B-Rafポリペプチドの断片に対応するポリペプチドは、ペプチド合成機を使用して合成され得る。
【0139】
B-Raf突然変異
B-Rafにおける突然変異は、ヒト腫瘍細胞において同定されてきた。表1は、これらの突然変異の位置およびそれらが同定された腫瘍を表す。突然変異は、B-Rafのキナーゼドメインにある。ほとんどの突然変異は、体細胞性であることが確認でき、これは対の正常/腫瘍サンプルをテストして、腫瘍サンプルにおいてのみ突然変異が見とめられたことを示す。
【0140】
表1
【表1】
【0141】
化合物アッセイ
本発明によれば、変異型B-Rafを標的として使用して、変異型B-Rafの増殖活性をモジュレート可能な化合物(例えば、医薬用のリード化合物)を同定する。従って、本発明は、
(a)変異型B-Rafを、評価する1つ以上の化合物とインキュベートするステップ;および
(b)変異型B-Rafの活性に影響を及ぼす化合物を同定するステップ
を含む、活性変異型B-Rafを直接的または間接的にモジュレート可能な1つ以上の化合物を同定するアッセイに関し、その方法を提供する。
【0142】
変異型B-Rafは、本発明に関して定義する通りである。
【0143】
本発明の第1の実施形態では、アッセイは、変異型B-Rafに直接結合するポリペプチドを検出するように構成される。
【0144】
従って、本発明は、
(a)変異型B-Rafを、評価する1つ以上の化合物とインキュベートするステップ;および
(b)変異型B-Rafに結合する化合物を同定するステップ
を含む、モジュレーター細胞増殖を同定する方法を提供する。
【0145】
本方法は、
(c)変異型B-Rafに結合する化合物を、細胞利用型アッセイにおいて細胞増殖をモジュレートする能力について評価するステップ
をさらに含むことが好ましい。
【0146】
変異型B-Rafへの結合は、当業者に公知の任意の技術により評価され得る。適切なアッセイの例としては、in vivoで相互作用を測定する2-ハイブリッドアッセイ系、カラム上に固定化されたポリペプチドへの結合を伴うアフィニティークロマトグラフィーアッセイ、1つ以上の化合物と変異型B-Rafとの結合が結合対の一方または両方のパートナーの蛍光の変化に関連する蛍光アッセイ等が挙げられる。好ましいのは、2-ハイブリッドアッセイ等、細胞においてin vivoで実施されるアッセイである。
【0147】
本実施形態の好適な態様では、本発明は、細胞増殖を伴うかまたは使用する疾患の治療に有用な医薬用のリード化合物を同定する方法であって、テストする1つ以上の化合物の存在下以外ではB-Rafが参照親和性でRASと会合する条件下で、テストする1つ以上の化合物を変異型B-Rafとインキュベートすること;
テストする1つ以上の化合物の存在において、RASに対する変異型B-Rafの結合親和性を測定すること;および
参照結合親和性に照らし、RASに対する変異型B-Rafの結合親和性をモジュレートする化合物を選択すること
を含む方法を提供する。
【0148】
従って、本発明によるアッセイは、テストする1つ以上の化合物の不在下、または変異型B-Rafに対する結合の活性が知られているかもしくは参照値として望ましい参照化合物の存在下において、較正されることが好ましい。例えば、2-ハイブリッド系では、参照値は、任意の化合物の不在下にて得てもよい。標的に対する変異型B-Rafの結合親和性を高める1つ以上の化合物の添加により、アッセイからの読取り値が参照値を上回って高くなり、この親和性結果を低下させる1つ以上の化合物の添加により、アッセイからの読取り値が参照値を下回って低くなる。
【0149】
第2の実施形態では、本発明は、1つ以上の化合物と変異型B-Rafとの機能的相互作用を検出するように構成され得る。このような相互作用は、テストする1つ以上の化合物に応答して、例えばRASによりキナーゼ自体が活性化もしくは不活性化される変異型B-Rafの調節レベル、またはMEK等の下流標的に対する変異型B-Rafの生物学的影響のモジュレーションレベルのいずれかにおいて生じる。本明細書で使用する「活性化」および「不活性化」は、化合物の酵素的またはその他の活性のモジュレーション、および、例えば細胞中のポリペプチドの発現の活性化または抑制によるその生成率のモジュレーションを含む。これらの用語は、遺伝子生成物の発現をモジュレートするための遺伝子転写に対する直接的な作用を含む。
【0150】
変異型B-Rafと、シグナリング経路におけるその上流または下流パートナーとの機能的相互作用のモジュレーションを検出するアッセイは、細胞利用型アッセイであることが好ましい。例えば、変異型B-Rafの活性化から生じるMAPKのリン酸化の程度(これは、MEK活性化の程度を示す)の評価に基づき得る。
【0151】
好適な実施形態では、核酸コード変異型B-Rafは、ベクターにライゲートされ、適切な宿主細胞に導入されて、変異型B-Rafを発現する形質転換細胞系を生成する。その後、得られた細胞系は、変異型B-Raf機能に影響を及ぼす可能性のある化合物の1つ以上の影響の再生可能な定質および/または定量分析のために生成され得る。従って、変異型B-Raf発現細胞は、変異型B-Rafの機能をモジュレートする化合物、特に低分子量の化合物、の同定のために採用され得る。従って、変異型B-Rafを発現する宿主細胞は、薬剤スクリーニングのために有用であり、本発明のさらなる目的は、変異型B-Rafの活性をモジュレートする化合物を同定する方法を提供することであり、該方法は、変異型B-Rafをコードする異種DNAを含み、機能性変異型B-Rafを生成する細胞を、少なくとも1つの化合物もしくは化合物の混合物、または変異型B-Rafの活性をモジュレートする能力を測定しようと思っているシグナルに曝すこと、その後、該モジュレーションにより生じる変化について該細胞をモニターすること、を含む。このようなアッセイは、変異型B-Rafのアゴニスト、アンタゴニスト、およびアロステリックモジュレーター等のモジュレーターの同定を可能にする。本明細書で使用する、変異型B-Rafの活性をモジュレートする化合物またはシグナルは、MEK等の標的上の変異型B-Rafの活性が、該化合物またはシグナルの存在下においては(該化合物またはシグナルの不在下の場合と比較して)異なるような、変異型B-Rafの活性を変える化合物を指す。
【0152】
細胞利用型スクリーニングアッセイは、β-ガラクトシダーゼ、クロラムフェニコールアセチルトランスフェラーゼ(CAT)またはルシフェラーゼ等のリポータータンパク質(すなわち、アッセイし易いタンパク質)の発現が、変異型B-Raf基質の活性化に依存する、細胞系を構築することにより設計され得る。例えば、上記ポリペプチドの1つをコードするリポーター遺伝子は、MEKまたはMARKにより特異的に活性化される応答エレメントの制御下に置かれ得る。このようなアッセイは、変異型B-RafによるMEKのリン酸化を拮抗する化合物、または変異型B-Rafの活性に必要な他の細胞機能を阻害もしくは可能にする化合物等、変異型B-Raf機能を直接モジュレートする化合物の検出を可能にする。野生型非変異型B-Rafが存在する細胞は、適切な対照となる。
【0153】
代替的なアッセイ形式としては、生物学系における増殖応答を直接評価するアッセイが挙げられる。非調節変異型B-Rafの構成的発現は、動物細胞において増殖性表現型を生じる。3T3繊維芽細胞等の細胞利用型系を使用して、変異型B-Rafの可能性のあるレギュレーターの活性を評価し得る。
【0154】
本発明の本実施形態の好適な態様では、炎症応答を伴うまたは使用する疾患の治療に有用な医薬用のリード化合物を同定する方法を提供し、該方法は、
テストする1つ以上の化合物の存在下以外では、変異型B-Rafが、参照リン酸化効率でMEKのリン酸化を直接的または間接的に生じる条件下で、テストする1つ以上の化合物を、変異型B-RafおよびMEKとインキュベートすること;
テストする1つ以上の化合物の存在下で、変異型B-Rafが、直接的または間接的にMEKのリン酸化を生じる能力を判定すること;および
参照リン酸化効率に照らして、変異型B-RafがMEKをリン酸化する能力をモジュレートする化合物を選択すること、を含む。
【0155】
さらに好適な態様では、本発明は、医薬用のリード化合物を同定する方法に関し、該方法は:
精製された変異型B-Raf分子を得るステップ;
変異型B-Raf分子を、変異型B-Rafによりリン酸化することが知られている基質、および1つ以上のテスト化合物とインキュベートするステップ;ならびに
基質のリン酸化をモジュレート可能な1つ以上のテスト化合物を同定するステップ、を含む。
【0156】
変異型B-Rafリン酸化用の基質はMEKである。従って、MEKを基質として使用して、変異型B-Rafキナーゼ活性をモジュレート可能な化合物をモニターすることが好ましい。これにより、当業者は、キナーゼモジュレートを直接スクリーニングすることが可能になる。キナーゼモジュレーターは、キナーゼ(変異型B-Raf)阻害剤であることが好ましい。
【0157】
好適な実施形態では、B-Rafの活性は、以下のプロトコールにより測定され得る:
1.溶解緩衝液(150 mM NaCl、25 mM HEPES [pH 7.3]、1 mMオルトバナジン酸ナトリウム、1%Triton X-100、プロテアーゼ阻害剤、0.5 mMジチオスレイトール)中で、細胞を可溶化する。
【0158】
2.溶解産物を、氷上で10分間インキュベートし、14,000 3 gで10分間遠心分離にかけ、上清をポリクローナル抗B-Raf抗体と、その次にプロテインG-セファロースと、4℃にて1時間インキュベートする。
【0159】
3.免疫沈降物を溶解緩衝液で2回洗浄し、キナーゼ緩衝液(0.2 mM ATP、30 mM MgCl2、2 mM MnCl2、40 mMβ-グリセロリン酸ナトリウム、0.2 mMオルトバナジン酸ナトリウム、2 mMオカダ酸、0.2%β-メルカプトエタノール)中で、1 mgの精製組換えMEK1を基質として添加して、キナーゼ反応を30℃にて10分間実施する。
【0160】
4.MEK1活性化後、15 mCiの[γ-32P]ATP、および1 mgのキナーゼ欠損(K52R)Erkを基質として、さらに2分間添加する。サンプル緩衝液を添加することにより、反応を止め、混合液を5分間煮沸させ、SDS-PAGEによりタンパク質を分離させる。
【0161】
5.ゲルタンパク質を、ポリ二フッ化ビニリデン(polyvinylidene difluoride)膜に移し、その上で、放射標識化K52R Erkの量をPhosphorImagerで定量する。
【0162】
6.B-Raf活性を計算するために、マウスモノクローナル抗B-Rafブロッティングの後に、125I標識化ヤギ抗マウスIgGで膜をプロービングすることにより、同じ膜上のB-Rafタンパク質の量を測定する。
【0163】
7.アッセイは、テストする1つ以上の化合物の存在下または不在下で繰り返してもよい。
【0164】
任意に、同定するテスト化合物を、その後、in vivoテストに供して、例えば上記実施形態に記載したように、変異型B-Rafシグナリング経路に対するそれらの影響を決定してもよい。
【0165】
本明細書で使用する「変異型B-Raf活性」は、結合活性を含む変異型B-Rafの任意の活性を指しうるが、特に、変異型B-Rafのリン酸化活性を指す。従って、本発明は、変異型B-Rafによる標的化合物のリン酸化、および可能性のある治療薬によるこの活性のモジュレーションを検出するように構成され得る。
【0166】
変異型B-Rafのリン酸化活性をモジュレートする化合物の例としては、B-Raf自体の優性ネガティブ変異体が挙げられる。このような化合物は、変異型B-Rafの標的について競合可能であるため、生物学系または人工系における変異型B-Rafの活性を低下させる。従って、本発明はさらに、変異型B-Rafのリン酸化活性をモジュレート可能な化合物に関する。
【0167】
変異型B-Rafの活性に影響を及ぼす化合物は、ほぼ全ての一般的な種類のものであり得、線状、環状、多環状、もしくはそれらの組合せであり得る有機化合物等の低分子量化合物、ペプチド、抗体等のポリペプチド、またはタンパク質が挙げられる。本明細書で使用する「ペプチド」、「ポリペプチド」、および「タンパク質」は、等価であると見なされる。
【0168】
本発明による多くの化合物は、薬剤開発に有用なリード化合物であり得る。有用なリード化合物は、特に抗体およびペプチドであり、特に、遺伝子療法目的で細胞内で発現される細胞内抗体であり、これらは、ペプチドまたは低分子量治療法の開発用のモデルとして使用され得る。本発明の好適な態様では、リード化合物および変異型B-Raf、または他の標的ペプチドは、同時結晶化されて、リード化合物で見とめられる相互作用を模倣する適切な低分子量化合物を設計し易くしてもよい。
【0169】
結晶化は、例えば、「貯蔵緩衝液」に入ったペプチドまたはペプチド複合体の溶液を、好ましくは1:1の比率で、結晶形成に必要な低濃度の沈殿剤と共に、混合することによる、結晶化緩衝液の調製を伴う。結晶形成のためには、例えば沈殿剤を添加すること(例えば滴定)により、または沈殿剤の濃度を拡散によって結晶化緩衝液と貯蔵緩衝液との間で平衡化させることにより、沈殿剤の濃度を上げる。適切な条件下では、沈殿剤のこのような拡散は、沈殿剤の勾配(例えば、高濃度の沈殿剤を含む貯蔵緩衝液から、低濃度の沈殿剤を含む結晶化緩衝液)に応じて生じる。拡散は、例えば、共通気相において拡散させる蒸気拡散技術により達成され得る。公知の技術としては、例えば、「ハンギング・ドロップ(hanging drop)」または「シッティング・ドロップ(sitting drop)」法等の蒸気拡散法がある。蒸気拡散方法では、タンパク質を含む結晶化緩衝液の水滴が、より大量な貯蔵緩衝液プールの上に懸かっているかまたは横に置かれている。あるいはまた、沈殿剤の平衡化は、結晶化緩衝液を貯蔵緩衝液から分離し、タンパク質が貯蔵緩衝液中で希釈するのを防ぐ半浸透膜を通して達成され得る。
【0170】
結晶化緩衝液では、ペプチドまたはペプチド/結合パートナー複合体は、30 mg/mlまで、好ましくは約2 mg/ml〜約4 mg/mlの濃度を有することが好ましい。
【0171】
結晶の形成は、以下のパラメータにより本質的に決定される様々な条件下で達成され得る:すなわち、pH、塩および添加物の存在、沈殿剤、タンパク質濃度、および温度。pHは、約4.0〜9.0に及びうる。緩衝液の濃度および種類は、それほど重要ではないため、(例えば、所望のpHに応じて)可変的であり得る。適切な緩衝液系としては、リン酸塩、酢酸塩、クエン酸塩、Tris、MES、およびHEPES緩衝液が挙げられる。有用な塩および添加物としては、例えば、塩化物、硫酸塩、および当業者に公知の他の塩が挙げられる。緩衝液は、水混和性有機溶剤、好ましくは、100〜20000、優先的には4000〜10000の分子量を有するポリエチレングリコール、または硫酸塩、特に硫酸アンモニウム、塩化物、クエン酸塩、もしくは酒石酸塩等の適切な塩からなる群より選択される沈殿剤を含む。
【0172】
本発明によるペプチドまたはペプチド/結合パートナー複合体の結晶は、(例えば、重原子誘導体化により)化学的に修飾されてもよい。簡単に言うと、このような誘導体化は、重金属原子塩、または有機金属化合物(例えば、塩化鉛、金チオリンゴ酸、チメロサール、もしくは酢酸ウラニル)を含む、結晶中で拡散可能かつタンパク質の表面に結合可能な溶液に、結晶を浸すことにより達成され得る。結合した1つ以上の重金属原子の1つ以上の位置は、浸した結晶のX線回折分析により決定され得、この情報は、例えば、ペプチドの三次元モデルを構築するために使用され得る。
【0173】
三次元モデルは、例えば、結晶の重原子誘導体および/または結晶化により得られた構造データの全体または一部から得ることができる。
【0174】
このようなモデルの構築は、ホモロジーモデリングおよび/または分子置換を伴うことが好ましい。
【0175】
予備ホモロジーモデルは、配列アライメントと構造が既知の任意のRAFキナーゼとの組合わせ、二次構造予測、および構造ライブラリーのスクリーニングにより作製され得る。例えば、変異型B-Rafおよび候補ペプチドの配列を、適切なソフトウェアプログラムを使用してアラインさせてもよい。
【0176】
コンピュータソフトウェアも、ペプチドまたはペプチド複合体の二次構造を予測するために使用され得る。ペプチド配列は、変異型B-Raf構造に取り込まれ得る。構造的矛盾(例えば、挿入/欠失周囲の構造断片)は、所望の長さおよび適切な高次構造を有するペプチドについて構造ライブラリーをスクリーニングすることによりモデリングできる。側鎖高次構造を予測するために、側鎖回転異性体ライブラリーを採用してもよい。
【0177】
最終的なホモロジーモデルを使用して、適切なコンピュータソフトウェアを使用した分子置換により、ペプチドの結晶構造を解明する。ホモロジーモデルは、分子置換の結果に応じて配置され、分子力学計算、および結晶化に使用した阻害剤の電子密度へのモデリングを含むさらなる絞込みに供される。
【0178】
キナーゼ活性化調査
構成的に活性なキナーゼ変異体は、シグナリング経路の解明、およびそのような経路をモジュレートする治療薬の開発において貴重な調査ツールである。本発明による5つの変異体の活性を検査した。これらは、G463V、G468A、G595R、L596V、およびV599Eである。変異体の活性を検査するために、myc-エピトープタグ付け型のB-Rafを、COS細胞において一過的に発現させる。この外因的に発現されたB-Rafの活性を検査するために、mycタグを使用してタンパク質を免疫沈降させ、細菌生成GST-MEK、GST-ERK、およびミエリン塩基性タンパク質(MBP)を配列基質として使用したキナーゼ回路アッセイにおいて検査した(Maraisら (1997); J. Biol. Chem. 272: 4378-83)。B-Rafは、高レベルの基準キナーゼ活性を有し、Raf-1またはA-Rafのいずれよりも、アクチベーターの不在下においてより顕著に活性である(Maraisら (1997); J. Biol. Chem. 272: 4378-83)。さらに、Raf-1およびA-Rafは、それらの活性を完全に刺激するために発癌性Ras(V12Ras)および活性化Srcの両方を必要とする一方で、B-Rafは、V12Rasのみとの同時発現により完全に活性化される。従って、これらの突然変異が、B-Rafの基準活性、およびV12Rasにより刺激される活性の両方に対して及ぼす影響をアッセイした。野生型B-Rafと比べて、G463VB-Raf、G468AB-Raf、L596VB-Raf、およびV599EB-Rafは全て、大きく上昇した基準キナーゼ活性を有する(図1A、1B)。比較すると、G595RB-Rafは、野生型タンパク質と比べて低い基準活性を有する(図1A)。同様の結果がin vivoでも観察された。5つの変異体の全てが、発癌性Ras(V12Ras)により刺激される。しかし、変異体それぞれの倍数的活性化は、野生型B-Rafと比べて低く(図1A、Bを参照)、V599EB-Rafの場合に特に低い。しかし、G463VB-Raf、G468AB-Raf、L596VB-Raf、およびV599EB-Rafのそれぞれの基準活性は野生型タンパク質よりも高いため、見とめられる活性の絶対レベルはそれぞれの場合に、V12Rasの存在下において、野生型タンパク質よりも高い。興味深いことに、G595RB-RafもV12Rasにより刺激されるが、活性化は非常に弱く、これはおそらく低い開始レベルによるものと思われる。
【0179】
各活性化変異体が、NIH3T3細胞を形質転換させる能力についても検査した。このアッセイでは、野生型B-Rafは非常に低い効率(V12Rasで見とめられるコロニー数の約0.02%)で細胞を形質転換する。しかし、図2に示すように、各活性化変異体は、野生型B-Rafよりも、NIH3T3細胞を40〜85倍の効率で形質転換する。
【0180】
Ras/MEK経路に対する、B-Raf変異体を含む細胞の成長の依存の程度を調査した。これらの調査のために、2つのアッセイを使用した。第1のアッセイは、モノクローナル抗体Y13-259(細胞Rasの活性を中和する抗体)の微量注入により成長が抑制されるかをテストする。結果を表2に示す。データは3つのグループに分けられている。第1のグループは、野生型B-Rafを有し、Y13-259により成長が(40〜100%)阻害されている。第2のグループはB-Rafにおいて活性化突然変異を有し、Y13-259により成長は阻害されなかった(<15%)。第3のグループ(1ケースのみ)は、活性化Rasおよび活性化B-Raf突然変異の両方を有する細胞を含む。興味深いことに、この細胞系の成長は、Y13 259により阻害されたが、これは、成長がRasおよびB-Rafの両方に依存しているからかもしれない。
【0181】
第2のアプローチは、化合物U0126(B-Rafの唯一の既知基質であるMEK1/2の阻害剤)の影響を検査することである。これらの結果は、B-Rafにおいて活性化突然変異を有する細胞の処置が、MEK活性が抑制される場合に細胞増殖の抑制を生じることを実証し、B-Rafの活性化変異体による細胞シグナリングの活性化が治療標的であることを示している。表3を参照のこと。
【0182】
総合すると、上記データは
1.ヒト腫瘍において、活性化および不活化突然変異の2つのクラスのB-Raf突然変異が存在する。
【0183】
2.活性化型のB-Rafは、NIH3T3細胞を形質転換可能で、従って、発癌遺伝子と定義され得る。
【0184】
3.活性化B-Rafタンパク質を発現するヒト腫瘍細胞系は、Y13-259、すなわちRas中和抗体に対して感受性を持たず、これらの成長がRasタンパク質に依存せず、従って、Rasタンパク質を標的とする化合物に応答する可能性が低いことを示す。これは、活性化変異体が、腫瘍細胞中のRasシグナルについての要件を克服し得ることを示す。
【0185】
4.しかし、これらの活性は、化合物U0126により抑制され、これらの成長がこの経路の活性に依存し、従って、B-Raf活性を標的とする治療薬に応答する可能性が高い。
【0186】
5.突然変異の一部は、キナーゼドメインのリン酸塩結合ループにあり(G463、G465、G468)、これらのアミノ酸は全てのキナーゼにおいて保存されているため、これらの突然変異は、キナーゼを活性化するための広範囲かつ都合の良いメカニズムを表す。これは、治療薬についてスクリーニングする際の重要な意味(implication)を有する。
【0187】
従って、本発明は、B-Rafの463、465、および468位に対応する1つ以上の位置における突然変異からなる群より選択されるリン酸塩結合ループ中に突然変異を含む、構成的に活性なキナーゼを提供する。
【0188】
突然変異は、463および468位の1つ以上にあることが好ましい。
【0189】
G463VおよびG468Aが最も好ましい。
【0190】
多くのキナーゼは、特定の疾患と関連付けられて同定されるが、それらの活性化のメカニズムが常に完全に理解されているわけではない。本明細書に記載した構成的に活性化された変異体は、活性化のメカニズムを最初に解明する必要なく、阻害剤についてのスクリーニングに使用できる試薬を提供する。
【0191】
キナーゼの例としては、MEKおよびERK等のMAPキナーゼ経路、ならびにp38、JNK、およびそれらの上流キナーゼ等の他のMAPキナーゼ経路上の他のキナーゼが挙げられる。これらの活性化メカニズムについてある程度は知られているが、一部については、直接突然変異によってどのように活性化するのか知られていない。本発明は、該キナーゼスクリーニング目的の活性化変異体を提供する。さらに、p90Rsk、mnk等のMAPキナーゼの下流にあるキナーゼも活性化され得る。代替的な活性化メカニズムは知られているが、突然変異はスクリーニングアッセイにおいて好ましい経路であり得る。
【0192】
本発明はまた、ポイツ-イエーガー症候群に関与するLkb1等の、活性化メカニズムが知られていない特定の公知キナーゼ;および構成的に活性であり得るが、薬剤スクリーニングのためにさらに活性化可能であり得るキナーゼPDK1も包含する。このキナーゼは、インスリンシグナリングに関与し、従って、糖尿病用の有用な標的となりうる。また、AMP活性化キナーゼは、II型糖尿病に関与し、これも同じくリン酸化により活性化され、従って、突然変異による活性化を受け易い。
【0193】
従って、本発明は、
(a)上で詳細に記載したように、アミノ酸置換、欠失、または挿入を、463、465または468位の1つ以上において含む変異型キナーゼを調製すること;
(b)キナーゼ基質の存在下で、変異型キナーゼを、該1つ以上の化合物に曝すこと;および
(d)キナーゼが、該1つ以上の化合物の存在下で該基質をリン酸化する能力を判定すること
を含む、キナーゼに対する阻害効果について1つ以上の化合物をスクリーニングする方法を提供する。
【0194】
1つ以上のテスト化合物の存在下におけるキナーゼのリン酸化活性は、該1つ以上の化合物の不在下におけるその活性と都合よく比較される;変異型キナーゼの基準活性の低下は、該1つ以上の化合物によるキナーゼの阻害を示す。複数アッセイのために、リン酸化の参照レベルを、特定のアッセイについて決定しておき、比較の基準として使用してもよい。
【0195】
キナーゼは、Rafタンパク質キナーゼであることが好ましく;B-Rafであることが都合がよい。
【0196】
逆に、B-Raf G595R等の構成的に抑制された変異体は、キナーゼのアクチベーターをスクリーニングする際に有用である。
【0197】
薬剤標的としてのbRAFの検証
癌における標的としてのBRAFを検証するために、まず、活性化変異型形態のBRAFを発現する細胞の成長が、RAF-MEK-ERKシグナリング経路を成長のために必要としたか否かをテストする。このために、BRAF遺伝子中に突然変異を有する黒色腫および結腸直腸細胞系を、この経路を介したシグナリングをブロックする製剤で処理する。2つの化合物をテストする。1つは、化合物U0126であり、これはMEK阻害剤で、従って、細胞中のRAF-ERKシグナリングを解く(uncouple)(Sebolt-Leopoldら, 1999)。BAY 43-9006もテストした。これは、RAFタンパク質の阻害剤である(Lyonsら, 2001)。これらの化合物が、ERK活性をブロックする能力を、BRAF遺伝子の599位においてバリンからアスパラギン酸への置換を有する黒色腫細胞系WM266.4においてテストした。これは、活性化突然変異である(図3A)。これらの細胞も、二重にリン酸化された活性化型のERKにしか結合しない抗体(ppERK)を使用して判断された場合の基準キナーゼ活性が高い。WM266.4細胞をウェスタンブロットにかける際にppERK抗体を使用する場合、42〜44 kDaの領域において強力なシグナルが見とめられ、これらの細胞においてERKが高い基準キナーゼ活性を有することを示す(図3B)。しかし、細胞をU0126、またはBay43-9006で処理した場合、ERK活性は強く抑制される(図3B)。同様の結果が、A375細胞、すなわちBRAF遺伝子においてV599E突然変異を有する黒色腫細胞系を使用して得られた(Daviesら, 2002)。これらのデータは、RAFおよびMEKシグナリングが、これらの細胞において高いERK活性を維持するために必要であることを実証する。
【0198】
次に、本発明者らは、BAY 43-9002が、WM266.4細胞の成長に対してどのような影響を及ぼすのかをテストし、この化合物が、約6.1μMのIC50で、これらの細胞の成長をブロックしていることが分かった(表4)。
【0199】
BAY43-9006は、低マイクロモル(micromolar)範囲にあるcolo829細胞およびBE細胞の成長もブロックした(表4)。Colo829細胞は、BRAF遺伝子においてV599E突然変異を含む黒色腫細胞系であり、BE細胞は、BRAF遺伝子においてG463突然変異を含む結腸直腸系である(Daviesら, 2002)。このように、これらの突然変異の両方ともが活性化的である(Daviesら, 2002)。最後に、DNA合成に対するこれらの阻害剤の影響をテストした。WM-266.4細胞と、10μM U0126または10μM BAY 43-9006とのインキュベーションにより、これらの細胞中でのDNA合成が強く抑制された(図4)。これらのデータは、BRAF遺伝子中に活性化突然変異を有する細胞におけるERK活性化および増殖が、RAFおよびMEK活性に依存することを実証する。
【0200】
哺乳動物細胞には、3つのRAF遺伝子、すなわちCRAF(RAF-1とも呼ばれる)、ARAF、およびBRAFがある。U0126は、MEK阻害剤であり、従って、CRAFを、BRAFまたはARAFシグナリングと区別することができない。同様に、BAY 43-9006は、CRAFおよびBRAFの両方を阻害でき、従って異なるRAFイソフォーム間を区別しない。従って、どのRAFイソフォームが、WM266.4細胞においてERKにシグナリングしていたかを判断するために、細胞を、個々のRAFイソフォームについて選択的な短い干渉(small interference)RNA(siRNA)プローブで処理した。WM266.4細胞を、BRAFもしくはCRAF、またはいずれのイソフォームも認識しない無作為対照に特異的に設計されたsiRNAプローブで処理した。CRAFに対する認識の効率を、ウェスタンブロッティングで測定した。WM266.4細胞を、CRAFに特異的なsiRNAプローブで処理することにより、CRAF発現が強く抑制された(図5A)。Colo829細胞およびBE細胞において同様の結果が見とめられた(図5A)。WM-266.4細胞をBRAF特異的siRNAプローブで処理した場合、細胞中のBRAF活性は強く抑制されたが、無作為対照で細胞を処理した場合には抑制は見とめられなかった(図5B)。同様の結果が、Colo829およびBE細胞において見とめられた(図5B)。
【0201】
上記データは、siRNAを使用して、BRAFおよびCRAFタンパク質の発現を選択的に抑制できることを示す。従って、本発明者らは、これらのタンパク質のそれぞれの除去により、これらの細胞におけるERK活性がどのように影響されるかを検査した。WM-266.4におけるBRAFタンパク質発現を除去するためにsiRNAを使用した場合、時間依存的にERK活性が抑制された(図6)。対照的に、CRAF発現の除去または無作為siRNAプローブでの処理は、ERK活性に影響を及ぼさなかった(図6)。同様の結果が、Colo829細胞において得られた(図6)。これらの結果は、BRAFが、活性化BRAFタンパク質を発現する黒色腫細胞においてERKにシグナリングする主要なイソフォームであることを実証する。CRAFは、これらの細胞において基準ERK活性にシグナリングしないと思われる。
【0202】
最後に、本発明者らは、WM-266.4細胞におけるアポトーシスに対する影響を検査することにより、BRAF除去が、細胞成長にどのように影響するかを検証した。これらの調査のために、細胞を70%エタノール中で固定し、ヨウ化プロピリジウムで染色し、それらの細胞周期プロファイルを、蛍光活性化細胞分取り(FACS)により検査した。この分析を使用して、アポトーシス細胞は、G1サブピークにおいて見とめられた。これらの細胞において、自発的アポトーシスは非常に少なく、細胞の1%未満がG1サブピークにおいて見とめられた(図7、表5)。U0126で細胞を処理した場合、G1サブピークにおける細胞の割合は有意に増加する(約3.5%;図7、表5)。同様に、siRNAを使用したBRAF発現の除去によっても、G1サブピークにおける細胞の数が増えるが、CRAFの除去または無作為対照での処理では増加しなかった。また、本発明者らは、アポトーシスの誘導のマーカーであるPARP切断も検証した。BRAF siRNAで細胞を処理した場合にはPARPの切断が誘導されたが、無作為対照では誘導されなかった。これらのデータは、変異型BRAFタンパク質が黒色腫細胞系において除去された場合に、アポトーシスが誘導されることを実証する。
【0203】
まとめると、これらの結果は、BRAFの活性化変異体を発現する黒色細胞系において、ERK活性化および細胞成長のために、RAFおよびMEKを介したシグナリングが必要であることを実証している。CRAFよりもBRAFの方が、ERK活性を刺激する主要RAFイソフォームであると思われ、アポトーシスから細胞を保護しているように思われる。これらのデータは、BRAFが、成長およびアポトーシスからの保護のためにBRAFシグナリングに依存する細胞において重要な治療的標的であることを示唆している。
【0204】
ハイスループットスクリーニングアッセイの開発
B-raf変異型V599Eのために、HTSアッセイが開発されている。結果を検証するために、GST-ERK2(キナーゼコンピテント)を採用し、ミエリン塩基性タンパク質(MBP)への33P-γ-リン酸塩の取り込みを測定する標準的な共役アッセイを実施することにより、-Raf発現溶解産物およびGST-MKK1試薬の活性をチェックした。図8に示すように、B-Raf溶解産物の存在下では、対照(非発現)溶解産物と比べて、シグナルの16倍の増加が見とめられた。
【0205】
アッセイプラットホーム検証
オプション1:グルタチオンフラッシュプレート(FlashPlate)における共役キナーゼアッセイ
プラットホームの原理:GSTタグ付け基質(ERK-2)を、フラッシュプレートシンチラント埋め込み壁に、グルタチオン被膜を介して捕捉した。基質への33P-γ-リン酸塩の取り込みにより、測定可能なシンチレーションシグナルが生じるはずである。
【0206】
GST-キナーゼ死活(kinase dead)ERK2(GST-kdERK2)への33P-γ-リン酸塩の取り込みを、B-Raf活性のアウトプットとして測定する確率を、グルタチオンフラッシュプレートアッセイにおいて評価した。図9は、試薬評価実施から持ってきた(transfer)条件を使用した場合、このプラットホームを使用してGST-kdERK2への33PのB-Raf-依存型取込みを検出することができなかった。標準的p81フィルタープレートアッセイも成果がないことが分かった。このカスケードアッセイの増幅特徴の結果として、最後のアッセイステップ(すなわち、MBPのERK2リン酸化)の不在下においてアッセイシグナルを維持するためには、残っている構成要素のそれぞれが有意に高いレベルであることがおそらく必要とされる。従って、この放射分析プラットホームにおいて試薬を消耗するB-Raf、MKKI、およびERK2滴定の実施に着手する前に、抗体利用型プラットホームを評価することが適切と思われた。
【0207】
オプション2:DELFIA形式における共役キナーゼアッセイ
DELFIA(解離増強ランタニド蛍光イムノアッセイ)アッセイは、ホスホ特異的抗体の結合を介したERK2リン酸化の測定を伴う。共役キナーゼアッセイB-Raf/GST-MKKI/GST-kdERK2は、リン酸化GST-kdERK2を生成する。抗GST被覆プレートを使用して、GST-kdERK2を捕捉する。トレオニンおよびチロシン上でリン酸化されたERK2を特異的に検出する一次抗体を添加する。次に、ユウロピウム(Eu)標識化二次抗体を添加する。増強溶液(Enhancement Solution)の存在下にて、Eu標識は、抗体から解離し、335 nmにおいて吸収し、620 nmの発光波長における蛍光検出を可能にする。
【0208】
試薬評価実施からのキナーゼアッセイ条件、および標準的DELFIAアッセイ条件の組合せを採用して、B-Rafアッセイをこのプラットホームにおいて評価した。図10は、B-Raf溶解産物の存在下で、対照(非発現)溶解産物と比較して12.2倍のシグナル上昇が見とめられたことを示す。観察されたシグナルは、3つの酵素/基質成分全ての存在に完全に依存していた。
【0209】
予備実験に基づき、開発のために、DELFIAプラットホームを選択した。
【0210】
キナーゼアッセイ開発
B-Raf溶解産物
3つのバッチのB-Raf溶解産物を、過程を通じて使用した。バッチAを採用して、DELFIAアッセイプラットホームを確立した。アッセイ開発の間、バッチBを使用した。バッチBおよびCについては、溶解産物量と得られたシグナルレベルとのおよその二次関数関係が、ウェルにつき0.025〜0.1μlにおいてはっきりとしていた。各バッチについて選択した最終的な量は、二次関数範囲内で十分なシグナルの枠(window)を得ることに基づいていた。これらの評価の結果、バッチAを1μl/ウェル[96ウェル]で使用し、ならびにバッチBおよびC(スクリーニング)は0.1μl/ウェル[96ウェル]および0.05μl/ウェル[384ウェル]で使用した。
【0211】
抗体レベルの最適化
(両方ともGSTタグ付けされた)ERK2およびMKKIの、グルタチオン結合部位に対する可能性のある競合に関する初期的な考慮から、「予備結合プロトコール」を使用して初期開発アッセイを実施した。
【0212】
これらの条件は以下の通りである:
・96ウェルグルタチオン被覆プレートに対する100 ng/ウェルGST-kdERK2の予備結合。
【0213】
・最終容量を50μM DKB(別表Iを参照)での、B-Raf溶解産物(バッチA)、MKK1(6.5μg/ml)、およびATP(500μM)の添加、ならびに30℃にて1時間のインキュベーション。
【0214】
このプロトコールは、検出系の抗体充填量を最適化かつ節約するために採用した。一次および二次抗体の両方の滴定を実施して、シグナル:バックグラウンド比を≧10.1で維持したままで、抗体レベルの低下の可能性を評価した。図4および5は、1:3000および1:1000が、ホスホERK2およびEu標識化抗体についてそれぞれ許容可能な最低濃度であったことを示す。従って、その後の全てのアッセイは、これらの抗体濃度を使用して行った。
【0215】
MKK1およびERK2レベルの最適化
予備結合ステップ無しに、「溶液中にて」本アッセイの酵素構成要素を行う能力を調査して、定義された濃度のMKK1およびERK2の両方の滴定を可能にした。さらに、このアッセイを、キナーゼに関与する試薬の単一ステップ(均一)混合に簡易化することで、HTSにより適するようになる。図13は、均一アッセイおよび予備結合プロトコールにより、同等のデータが得られたことを示す。その後の全てのアッセイは、「均一」プロトコールを使用して行った。
【0216】
MKK1レベルを保ちながらのERK-2の滴定は、最大シグナルが約12:1(ERK2:MKK1)の比率において得られたことを示した。試薬を節約するために、これらの構成要素のマトリックス滴定を行った。0.1μl B-Raf(バッチB)を使用して、6.5μg/ml GST-MKK1および80μg/ml GST-kdERK2の組合せにより最大シグナルを得た。そして、Raf/MKK1充填量のいくらかの減少が可能であったが、元の比率を維持することが適切であると見なした。この決断は、さらなるアッセイパラメータ変更(例えば、384への移行;ATP充填量の減少、自動化への移行)により、シグナル枠がさらに減りうるという知識と共に出した。従って、その後の全てのアッセイは、6.5μg/ml GST-MKK1および80μg/ml GST-kdERK2を使用して行った。
【0217】
温度の影響
このアッセイを室温にて実行する能力により、最終的なHTSプロセスが有意に簡易化される。従って、室温および30℃におけるアッセイシグナルを調査した。得られた結果に基づき、シグナル対バックグラウンド比は、室温にて許容可能なものであり、その後の全てのアッセイは室温にて実行した。
【0218】
384ウェルプレート形式でのアッセイの評価
スクリーニングの間のスループットを向上させ、試薬の使用量を最小化するために、384ウェル形式でのアッセイの実績を評価した。この形式において、アッセイは上手く行われ、標準量(50μlの最終量)および減少量(25μl)の試薬容量のアッセイの両方で、高度に許容可能なシグナル対バックグラウンド比が得られた。その後の全ての開発実験は、25μl反応容量を使用し、384形式で行った。
【0219】
ATP濃度の最適化
キナーゼスクリーニングアッセイのATP濃度は、同定された阻害性化合物の数および性質に影響を及ぼす可能性がある。このようなアッセイのATPレベルの定義は、以下の配慮事項のバランスである:
・一貫的な測定可能な枠のアッセイシグナルを可能にするATPレベルをを採用すること
・ATP競合阻害剤の同定が可能な十分に低いATPレベルを採用すること
・細胞ATPに関して無影響な可能性が高い弱ATP競合剤が検出されにくいように十分に高いATPレベルを採用すること
キナーゼスクリーニングアッセイは、通常、Kmに関するATP濃度において行われる。B-RafおよびMKK1に対するKm値の偏差により、本明細書において開発した共役アッセイではなく、各酵素について個々のアッセイを開発する必要がある。このようなアッセイの開発により、スクリーニングアッセイにおいて同定されたあらゆる阻害化合物の作用モードが、将来的に間違いなく理解し易くする。
【0220】
スクリーングアッセイのATP充填量を定義する目的のために、共役アッセイの濃度依存度を決定した。生成されるシグナルは、約200μMおよび約18μM ATPにおいてそれぞれ最大値および半値である。さらなる調査により、50μMが、日々の着実なアッセイシグナルが観察される(例えば、シグナル:8000、2%CV) 最も低いATP濃度であることが示された。これ未満(10μM)のATPレベルは、アッセイシグナルにおいてより高い相対的偏差が生じた(例えば、シグナル:2500、10%CV)。スクリーニングアッセイの最終的なATP濃度は、アッセイを自動化に移行する間のいかなる減少をも支持するのに十分に大きいシグナル枠を得るために50μMと定義された。
【0221】
スクリーニングアッセイの時間経過
スクリーニングアッセイは、その直線期(period of linearity)において行うことが重大である。共役アッセイの直線相(linear phase)の長さを決定するために、時間経過を75分まで行った。反応は、5〜45分の間は直線的であった。見とめられた「ラグ」期は、このアッセイ形式に特徴的なものであり、検出可能なレベルの反応生成物を蓄積するのに必要な時間を反映する。この調査に基づき、室温にて45分間のインキュベーションを定義した。
【0222】
B-RafV599E DELFIAアッセイの最終的な条件
アッセイの最終的なスクリーニング条件のまとめは以下の通りである:
酵素反応
・384ウェルグルタチオン被覆プレート
・0.05μl B-Raf溶解産物
・6.5μg/ml GST-MKK1
・80μg/ml GST-kdERK2
・50μM ATP
・室温にて45分間のインキュベーション
・最終容量は25μl
検出条件
・1:3000 抗ホスホERK2抗体
・1:1000 Eu標識抗マウス抗体
自動化開発および品質管理
スクリーニングバッチ試薬の検証
上述した条件を使用して、アッセイを実施し、B-Raf、MKKIおよびERK-2のスクリーニングおよびアッセイ開発バッチを比較した。全てのケースにおいて、試薬のスクリーニングバッチは、アッセイ開発に使用したものと比較した場合と結果が等しかった。
【0223】
自動化B-RatV599E DELFIAアッセイの簡易型実施(mini-runs)
スクリーニングの準備において、アッセイのために自動化液体扱い戦略を設計した。このシステムをテストするために、模擬(すなわち、化合物は含まないが、標準的対照カラムを含む)スクリーニングプレートを使用して自動化B-Rafアッセイの「簡易型実施」を行った。これらの実験から得たデータは、生物学的アッセイの信頼度の高い測定値、ならびに関与する自動化プロセスの正確性および一貫性の両方を示す。
【0224】
簡単に言うと、各プレートは、カラム1〜22の全てのウェルにB-Rafキナーゼ反応液を含み、カラム23〜24は対照反応液を含んだ。自動化アッセイにより生成されたデータの品質を定義するために、この形式を使用した4プレート簡易型実施を2つの別々の日に行った。各「簡易型実施」において、1枚のプレートを、本アッセイのいくつかの予測された阻害剤についてのIC50値を定義するために使用した。阻害剤は、一定の範囲の作用モードを網羅した:すなわち、スタクロスポリン(ATP競合キナーゼ阻害剤)、SB203580(ATP競合Raf阻害剤)、およびU0126(非ATP競合MKK1阻害剤)。
【0225】
アッセイは、プレート間および日々間の両方で、良好な一貫性を示した。得られたデータは、自動化アッセイが、in vitroHTSアッセイにおける384形式について定義した基準を得られることを示す。
【0226】
・少なくとも10:1のシグナル:バックグラウンド
・>0.4のZ'
・>15%の行列CV
採用した阻害剤は、Raf/MKK1活性の濃度依存性阻害をもたらすことで、アッセイをさらに有効にする。特に重要なのは、標準的阻害剤が、別々の日において2倍の範囲内のIC50を出したという事実である。これらのデータは、これらの化合物が、このスクリーニングアッセイにおいて約30μm(10μg/ml)でテストされた場合には、ヒットとして識別されたであろうということも示す。
【0227】
自動化B-Raf V599E DELFIAについての最終的プロトコール
酵素カクテル(最終容量12μl):
0.05μl Raf溶解産物
0.0325μl GST-MKKI
0.065μl GST-kdERK2
【0228】
1.3μlテスト化合物を、グルタチオン被覆384プレートに予備プレート化する。
2.12μl酵素カクテルを、PlateMatePlusで添加する。
3.10μl ATPを、Asysで添加する。
4.プレートを、室温にて45分間振とうさせる。
5.プレートを、プレート洗浄機ELX405を使用して、3×80μl/ウェル DELFIA洗浄緩衝液(DWB)で洗浄する。
6.25μlの抗ホスホERK2をMultidropで添加する。
7.プレートを、室温にて1時間振とうさせる。
8.プレートを、3×80μl/ウェル DWBで洗浄する。
9.25μl Eu標識抗マウス抗体を、Multidropで添加する。
10.プレートを、室温にて30分間振とうさせる。
11.プレートを、3×80μl/ウェル DWBで洗浄する。
12.25μl DELFIA増強溶液を、Multidropで添加する。
13.プレートを、暗所室温にて30分間インキュベートする。
14.FUSION上でプレートを読み取る。
【表2】
表2:本表は、B-Raf突然変異を有する様々な細胞系の成長の阻害を示す。阻害の割合(BrdUを取り込まない細胞の数の%)は、最後のカラムに示す。
【表3】
表3:本表は、MEK阻害剤U0126で処理した細胞における細胞成長の阻害を示す。
【0229】
表4:BAY 43-9006による細胞成長の阻害
【表4】
【0230】
細胞系は、高いレベルのBAY 43-9006の存在下でインキュベートさせ、細胞成長のレベルは、スルホローダミンB染色により測定した。IC50値は、非線形回帰分析により測定し、表示している。
【0231】
表5:細胞周期分析
【表5】
【0232】
WM-266.4細胞を、U0126、DMSO、BRAFに対するsiRNA、CRAFに対するsiRNA、または無作為siRNA対照で処理した。細胞を、96時間インキュベートし、FACSでの細胞周期分析のために、ヨウ化プロプリジウム(propridium iodide)で細胞を固定および染色した。細胞周期の各期における細胞の割合を表示している。
【0233】
検出のコンピュータ的要素
変異型B-Rafポリペプチドおよび/または変異型B-raf核酸の検出を自動化して、サンプル集合の迅速かつ大規模な並行スクリーニングを得ることができる。突然変異検出のためのコンピュータ化方法は、当該分野で公知であり、通常、配列決定装置、またはポリペプチドもしくは核酸中の配列の違いを検出可能な他の装置、データ処理ユニット、および技術者または医師により解読可能な形式で結果を表示可能な出力装置の組合せを伴う。
【0234】
従って、好適な態様では、本発明は、B-Rafポリペプチドをコードする天然のヒト原発腫瘍由来の核酸の標的配列位置における突然変異を検出する自動化方法を提供し、該方法は:
該天然のヒト原発腫瘍由来の核酸の増幅産物のサンプルを配列決定して、開始配列位置から終結配列位置にわたる標的ドメインにおける複数の測定した塩基対同定データを特定するサンプルデータセットを得ること;
該標的配列位置についての該測定した塩基対同定データが、該標的配列位置の参照塩基対データに対応するか否かを条件として、該サンプルにおける突然変異の有無を判断すること;および
該判断ステップにより確立された該サンプルにおける突然変異の有無を示す出力を生成すること
を含む。
【0235】
配列中の突然変異を配列決定および検出する方法は上述しており、当該分野において一般的に公知である。本発明は、このような方法を利用して、本発明のプロセスを実行する装置を得る。この装置は:
核酸のサンプルの配列を判断して、開始配列位置から終結配列位置にわたる標的ドメインにおける測定した塩基対同定データを特定するサンプルデータセットを得ることが可能な、配列読取り装置と;
配列決定装置と接続し、そこからサンプルデータセットを受け取り、該標的配列位置についての該測定した塩基対同定データが、該標的配列位置の参照塩基対データに対応するか否かを条件として、該サンプルにおける突然変異の有無を判断可能な、データ分析ユニットと
を備える。
【0236】
適切な配列読取り装置としては、自動化シーケーンサー、RFLP分析器、および移動度シフト分析装置が挙げられる。標的核酸の増幅産物の配列を分析して、本装置が、PCR機等の増幅装置をさらに備えることが都合がよい。
【0237】
本装置は、データ分析ユニットにより判断されたサンプル中の突然変異の有無を示す出力を生成可能な出力装置を備えることが好ましい。例えば、出力装置は:画像ユーザインターフェース;音声ユーザインターフェース;プリンタ;コンピュータ読取り可能記憶媒体;およびコンピュータ翻訳可能キャリア媒体の少なくとも1つを備え得る。
【0238】
本発明はさらに、変異型B-Rafタンパク質自体を検出するように構成できる。従って、さらなる態様では、本発明は、天然のヒト原発腫瘍由来のB-Rafポリペプチドにおける単一アミノ酸突然変異を検出する自動化方法に関し、該方法は:
B-Rafポリペプチドのサンプル中の1つ以上のアミノ酸に対してマーカーを適用すること;
該マーカーを適用した後、該サンプルを読み取って、該サンプル中の該マーカーの有無を判断し、該サンプルにおける単一アミノ酸突然変異の有無を示すこと;および
該読取りステップにおいて判断した該サンプル中の該単一アミノ酸突然変異の有無を示す出力を生成すること
を含む。
【0239】
マーカーは、単一アミノ酸突然変異を持たない野生型B-Rafポリペプチド、ならびに突然変異を有する変異型B-Rafポリペプチドに差次的に結合するリガンドを含むことが好ましい。本発明に関しては、B-Rafのいずれの形態への優先的な結合も可能である。
【0240】
本発明はさらに:
マーカーを充填され、B-Rafポリペプチドのサンプル中の1つ以上の標的アミノ酸にマーカーを適用可能な、タンパク質マーキング装置と;
該サンプル中のマーカーの有無を判断して、該サンプル中の単一アミノ酸突然変異の有無を示す、マーカー読取り装置と
を備える、B-Rafポリペプチドにおいてアミノ酸突然変異を検出する装置を提供する。
【0241】
使用するマーカーは、抗体であり得、タンパク質マーキング装置はELISAプロセスを実行するように構成され得る。
【0242】
タンパク質マーキング装置は、好ましくはサンプルを光学的に読み取るように構成されたマイクロアレイヤー(microarrayer)を備えることが都合が良い。
【0243】
装置は、マーカー読取り装置により判断されたサンプル中の単一アミノ酸突然変異の有無を示す出力を生成可能な、出力装置を備えることが好ましい。適切な出力装置は:画像ユーザインターフェース;音声ユーザインターフェース;プリンタ;コンピュータ読取り可能記憶媒体;およびコンピュータ翻訳可能キャリア媒体のうちの少なくとも1つを備える。
【0244】
本発明の用途
本発明は、新生物症状を検出、およびこのような症状を患う被験体の予後を判断するのに有用なB-Rafポリペプチドの新規変異体を提供する。一般的に、本明細書に記載するB-Rafにおける突然変異の存在は、新生物の存在と関連付けられる。
【0245】
一態様では、本発明は、癌性細胞または癌性組織(悪性黒色腫、結腸直腸癌、乳癌、もしくはNSCLC)を同定、または新生物表現型を発達し易い細胞もしくは組織を同定する方法であって:該細胞または組織のB-raf遺伝子の少なくとも一部を増幅すること;該増幅産物を分析して、本明細書に記載するB-raf遺伝子における突然変異を検出することを含み;1つ以上のB-raf突然変異を有する細胞または組織は、癌性であるか、または癌症状を発達する危険性が高いと分類される、方法を提供する。適切な増幅手段としては、PCRおよびクローニングが挙げられる。
【0246】
別の実施形態では、本発明は、癌(悪性黒色腫、結腸直腸癌、乳癌、またはNSCLC等)を患う被験体における予後を判断する方法に関する。本方法は:上述したB-raf遺伝子の領域を増幅すること;上述した突然変異の形跡について増幅産物を分析すること;ならびにB-raf遺伝子において突然変異を持たない被験体を、治療および/または手術後に疾患の再発を受け難いか、または該領域において1つ以上の突然変異を有する患者よりも生存する可能性が高いと分類すること、を含む。
【0247】
本発明の技術は、前記した予後に基づいて、被験体が受けるべき治療計画を決定するためにも採用され得る。予後が乏しい被験体は、良好な予後を有する被験体よりも積極的な治療法を使用して処置されることが都合が良い。
【0248】
本発明による技術は、潜在的に癌性の症状を同定するために、サンプルの迅速なスクリーニングが必要とされる場合に、自動化できる。一般的に、自動化プロセスは、組織または細胞サンプルに由来する核酸の自動化増幅、(蛍光検出等による)増幅核酸における突然変異の検出、および/または突然変異の存在の表示、を含む。自動化実施形態の例は、上述している。
【0249】
本発明による変異型B-Rafの同定は、このように、変異型B-Rafの発現に関連する疾患、障害、および/または症状を検出、診断、またはモニターする診断目的のために使用され得る。特に、本発明は、本明細書に記載する変異型B-Rafに関連する癌の検出、診断、および/またはモニターに関する。
【0250】
本発明は、(a)個体の細胞または体液中の変異型B-Rafの発現を、本明細書において定義したB-Raf変異体に特異的な1つ以上の抗体を使用してアッセイすることを含む、癌を診断するための診断アッセイを提供する。個体から得た生検組織における変異型B-raf転写体の存在は、疾患の発達し易さを示すか、または実際の臨床的症状が現れる前に疾患を検出する手段を提供することができる。この種のより確実な診断により、健康専門家は、予防対策または積極的な治療を早めに採用することができ、癌の発達またはそれ以上の進行を防ぐことができる。
【0251】
本発明の抗体は、当業者に公知の古典的な免疫組織学的方法を使用して、生物学的サンプル中のタンパク質レベルをアッセイするために使用できる(例えば、Jalkanenら, (1985) J. Cell. Biol. 101:976-985;Jalkanenら, (1987) J. Cell. Biol. 105:3087-3096を参照)。タンパク質遺伝子発現を検出するのに有用な他の抗体利用型方法としては、酵素結合免疫吸着アッセイ(ELISA)、およびラジオイムノアッセイ(RIA)等のイムノアッセイが挙げられる。適切な抗体アッセイ標識は、当該分野で公知であり、グルコースオキシダーゼ等の酵素標識;ヨウ素(125I、121I)、炭素(14C)、硫黄(35S)、トリチウム(3H)、インジウム(112In)、およびテクネチウム(99Tc)等の放射性同位体;ルミノール等の発光標識;ならびにフルオレセインおよびローダミンおよびビオチン等の蛍光標識が挙げられる。
【0252】
さらに、B-rafにおける突然変異は、本明細書に記載する核酸の分析により検出され得る。例えば、突然変異の存在は、配列決定、またはSCCP分析により検出され得る。
【0253】
本発明は、上記方法において使用できるキットをさらに提供する。一実施形態では、キットは、1つ以上の容器に入った、本発明の抗体、好ましくは精製抗体を含む。具体的な実施形態では、本発明のキットは、キットに含まれる抗体に対して特異的に免疫反応性なエピトープを含む、実質的に単離されたポリペプチドを含む。本発明のキットは、目的のポリペプチドと反応しない対照抗体をさらに含むことが好ましい。別の特異的な実施形態では、本発明のキットは、抗体と目的のポリペプチドとの結合を検出する手段を含む(例えば、抗体を、蛍光化合物、酵素基質、放射活性化合物、もしくは発光化合物等の検出可能な基質と共役させてもよいし、または第1の抗体を認識する二次抗体を検出可能な基質と共役させてもよい)。
【0254】
本発明の別の具体的な実施形態では、キットは、本明細書に記載の変異型B-Rafポリペプチドに特異的な抗体を含む血清をスクリーニングするのに使用する診断キットである。このようなキットは、変異型B-Rafポリペプチドと反応しない対照抗体を含み得る。このようなキットは、少なくとも1つの抗B-Raf抗体に対して特異的に免疫反応性なエピトープを含む、実質的に単離されたポリペプチド抗原を含み得る。さらに、このようなキットは、抗原に対する抗体の結合を検出する手段を含む(例えば、抗体を、フローサイトメトリーにより検出可能な、フルオレセインまたはローダミン等の蛍光化合物と共役させてもよい)。具体的な実施形態では、キットは、組換え生成された、または化学的に合成されたポリペプチド抗原を含み得る。キットのポリペプチド抗原を、固体支持体に付着させてもよい。
【0255】
さらなる実施形態では、本発明は、本発明の変異型B-Rafポリペプチドの抗原を含む血清をスクリーニングするのに使用する診断キットを含む。診断キットは、ポリペプチドまたはポリヌクレオチド抗原に対して特異的に免疫反応性な実質的に単離された抗体、および抗体に対するポリヌクレオチドまたはポリペプチド抗原の結合を検出する手段を含む。一実施形態では、抗体は、固体支持体に付着している。具体的な実施形態では、抗体は、モノクローナル抗体であり得る。キットの検出手段は、第2の標識化モノクローナル抗体を含み得る。あるいはまた、またはそれに加えて、検出手段は、標識化競合抗原を含み得る。
【0256】
上記明細書において言及した全参考文献は、参照により本明細書に援用する。本発明の記載した方法およびシステムの様々な改変および変更が、本発明の範囲および思想から逸脱することなく、当業者に理解されるであろう。本発明は、具体的な好適な実施形態に関して記載したが、請求する発明は、このような具体的な実施形態に不当に限定されないことが理解されるべきである。実際、分子生物学または関連分野の当業者に明らかな、記載した本発明を実施するためのモードの様々な改変は、請求の範囲内にあることを意図する。
【特許請求の範囲】
【請求項1】
1つ以上の癌関連体細胞アミノ酸突然変異を含む、天然の変異型ヒトB-Rafポリペプチドであって、該アミノ酸突然変異が、V599E、V599D、G595R、G465V、G465E、G465A、G468A、G468E、E585K、F594L、G595R、L596V、L596R、およびG463Eからなる群より選択される、前記変異型ヒトB-Rafポリペプチド。
【請求項2】
B-RafのG1388T、G1783C、TG1796-97AT、G1394T、G1394A、G1394C、G1403C、G1403A、G1753A、T1782G、G1388A、T1796A、T1787G、またはC1786Gである1つ以上の点突然変異を含む請求項1に記載のB-Rafポリペプチドをコードする核酸の相補体。
【請求項3】
B-RafのG1388T、G1783C、TG1796-97AT、G1394T、G1394A、G1394C、G1403C、G1403A、G1753A、T1782G、G1388A、T1796A、T1787G、またはC1786Gである1つ以上の点突然変異を含む請求項1に記載のB-Rafポリペプチドをコードする変異型核酸に特異的にハイブリダイズする核酸。
【請求項4】
請求項1に記載のB-Rafポリペプチドの変異型エピトープであって、請求項1に記載されるアミノ酸突然変異を含む該変異型エピトープに選択的に結合する抗体。
【請求項5】
(a)ヒト被験体から得られる天然の細胞物質のサンプルを準備するステップ;
(b)該細胞物質中の1つ以上のB-Raf遺伝子の少なくとも一部に由来する核酸物質を検査するステップ;および
(c)該核酸物質が、B-Rafポリペプチドをコードする配列において、1つ以上の点突然変異であって、B-RafのG1388T、G1783C、TG1796-97AT、G1394T、G1394A、G1394C、G1403C、G1403A、G1753A、T1782G、G1388A、T1796A、T1787G、またはC1786Gである該突然変異を含むか否かを判断するステップ
を含む、発癌性突然変異の検出方法。
【請求項6】
(a)被験体の癌性だと疑われる天然の組織に由来する細胞物質の第1のサンプル、および同被験体の非癌性組織由来の細胞物質の第2のサンプルを準備するステップ;
(b)該細胞物質の該両方のサンプル中の1つ以上のB-Raf遺伝子の少なくとも一部に由来する核酸物質を検査するステップ;ならびに
(c)該核酸物質が、RAFポリペプチドをコードする配列において、1つ以上の点突然変異であって、B-RafのG1388T、G1783C、TG1796-97AT、G1394T、G1394A、G1394C、G1403C、G1403A、G1753A、T1782G、G1388A、T1796A、T1787G、またはC1786Gである該突然変異を含むか否か;および該突然変異が、該癌性と疑われる組織に由来する天然の細胞物質には存在するが、該非癌性組織に由来する細胞物質には存在しないことを判断するステップ
を含む、発癌性突然変異の検出方法。
【請求項7】
(a)被験体から得られる細胞物質のサンプルを準備するステップ;
(b)該サンプルを請求項4に記載の抗体でスクリーニングするステップ;および
(c)該サンプルにおいて、請求項1に記載の1つ以上の変異型B-Rafポリペプチドを検出するステップ
を含む、発癌性突然変異の検出方法。
【請求項8】
核酸の増幅産物のサンプルを配列決定して、開始配列位置から終結配列位置まで及ぶ標的ドメインにおいて測定した塩基対同定データを特定するサンプルデータセットを得ること;
標的配列位置について測定された塩基対同定データが、該標的配列位置についての参照塩基対データに対応するか否かを条件として、該サンプル中の突然変異の有無を判断すること;および
該判断ステップにおいて確立された該サンプル中の該突然変異の有無を示す出力を生成すること
を含む、B-Rafポリペプチドをコードする核酸中の標的配列位置における突然変異を検出する自動化方法であって、該核酸が1以上の点突然変異を含み、該点突然変異がB-RafのG1388T、G1783C、TG1796-97AT、G1394T、G1394A、G1394C、G1403C、G1403A、G1753A、T1782G、G1388A、T1796A、T1787G、またはC1786Gである、前記自動化方法。
【請求項9】
マーカーを、B-Rafポリペプチドのサンプル中の1つ以上の標的アミノ酸に適用すること;
該マーカーを適用した後、該サンプルを読み取り、該サンプルにおける該マーカーの有無を判断して、該サンプルにおける単一アミノ酸突然変異の有無を示すこと;および
該読取りステップにおいて判断された該サンプル中の該単一アミノ酸突然変異の有無を示す出力を生成すること
を含む、B-Rafポリペプチドにおいて単一アミノ酸突然変異を検出する自動化方法であって、該B-Rafポリペプチドが請求項1に記載される1以上の突然変異を含む、前記自動化方法。
【請求項10】
前記マーカーが、単一アミノ酸突然変異を有するB-Rafポリペプチドに優先的に結合する抗体を含む、請求項9に記載の方法。
【請求項11】
前記マーカーが、マイクロアレイヤー(microarrayer)を用いて適用される、請求項9に記載の方法。
【請求項12】
前記サンプルが光学的に読み取られる、請求項9に記載の方法。
【請求項13】
前記出力が、画像ユーザインターフェース;音声ユーザインターフェース;プリンタ;コンピュータ読取り可能記憶媒体;およびコンピュータ翻訳可能キャリア媒体のうちの少なくとも1つを含む装置を用いて生成される、請求項9に記載の方法。
【請求項14】
(a)請求項1に記載の1つ以上の変異型B-Rafポリペプチドを得るステップ;
(b)該1つ以上のポリペプチドを、テストする1つ以上の化合物と接触させるステップ;および
(c)該1つ以上の化合物と該変異型ポリペプチドとの相互作用を検出するステップ
を含む、抗増殖活性を有する1つ以上の化合物を同定する方法。
【請求項15】
前記相互作用が、結合相互作用である、請求項14に記載の方法。
【請求項16】
(a)請求項1に記載の1つ以上の変異型B-Rafポリペプチドを得るステップ;
(b)該B-Rafポリペプチド用の下流基質を得るステップ;および
(c)テストする1つ以上の化合物の存在下において、該基質の修飾を検出するステップ
を含む、抗増殖活性を有する1つ以上の化合物を同定するアッセイ。
【請求項17】
前記基質修飾を直接検出する、請求項16に記載のアッセイ。
【請求項18】
前記基質が、第2の基質を修飾する酵素であり、該第2の修飾が検出可能である、請求項17に記載のアッセイ。
【請求項19】
前記基質がMEKであり、前記第2の基質がMAPKである、請求項18に記載のアッセイ。
【請求項20】
アッセイについて、テストする1つ以上の化合物の不在下における参照レベルを決定する、請求項16〜19のいずれか一項に記載のアッセイ。
【請求項21】
B-Rafの463、465、および468位に対応する1つ以上の位置における突然変異からなる群より選択される、リン酸塩結合ループにおいて突然変異を含む構成的に活性なキナーゼ。
【請求項22】
前記突然変異が、463および468位の1つ以上にある、請求項21に記載のキナーゼ。
【請求項23】
前記突然変異が、G463VおよびG468Aからなる群より選択される、請求項22に記載のキナーゼ。
【請求項24】
Rafタンパク質キナーゼである、請求項21〜23のいずれか一項に記載のキナーゼ。
【請求項25】
B-Rafである、請求項24に記載のキナーゼ。
【請求項26】
(a)請求項21〜25のいずれか一項に記載の変異型キナーゼを調製すること;
(b)キナーゼ基質の存在下で、該変異型キナーゼを、1つ以上の化合物に晒すこと;および
(c)該1つ以上の化合物の存在下で、該基質をリン酸化する該キナーゼの能力を判断すること
を含む、キナーゼに対する阻害効果について1つ以上の化合物をスクリーニングする方法。
【請求項1】
1つ以上の癌関連体細胞アミノ酸突然変異を含む、天然の変異型ヒトB-Rafポリペプチドであって、該アミノ酸突然変異が、V599E、V599D、G595R、G465V、G465E、G465A、G468A、G468E、E585K、F594L、G595R、L596V、L596R、およびG463Eからなる群より選択される、前記変異型ヒトB-Rafポリペプチド。
【請求項2】
B-RafのG1388T、G1783C、TG1796-97AT、G1394T、G1394A、G1394C、G1403C、G1403A、G1753A、T1782G、G1388A、T1796A、T1787G、またはC1786Gである1つ以上の点突然変異を含む請求項1に記載のB-Rafポリペプチドをコードする核酸の相補体。
【請求項3】
B-RafのG1388T、G1783C、TG1796-97AT、G1394T、G1394A、G1394C、G1403C、G1403A、G1753A、T1782G、G1388A、T1796A、T1787G、またはC1786Gである1つ以上の点突然変異を含む請求項1に記載のB-Rafポリペプチドをコードする変異型核酸に特異的にハイブリダイズする核酸。
【請求項4】
請求項1に記載のB-Rafポリペプチドの変異型エピトープであって、請求項1に記載されるアミノ酸突然変異を含む該変異型エピトープに選択的に結合する抗体。
【請求項5】
(a)ヒト被験体から得られる天然の細胞物質のサンプルを準備するステップ;
(b)該細胞物質中の1つ以上のB-Raf遺伝子の少なくとも一部に由来する核酸物質を検査するステップ;および
(c)該核酸物質が、B-Rafポリペプチドをコードする配列において、1つ以上の点突然変異であって、B-RafのG1388T、G1783C、TG1796-97AT、G1394T、G1394A、G1394C、G1403C、G1403A、G1753A、T1782G、G1388A、T1796A、T1787G、またはC1786Gである該突然変異を含むか否かを判断するステップ
を含む、発癌性突然変異の検出方法。
【請求項6】
(a)被験体の癌性だと疑われる天然の組織に由来する細胞物質の第1のサンプル、および同被験体の非癌性組織由来の細胞物質の第2のサンプルを準備するステップ;
(b)該細胞物質の該両方のサンプル中の1つ以上のB-Raf遺伝子の少なくとも一部に由来する核酸物質を検査するステップ;ならびに
(c)該核酸物質が、RAFポリペプチドをコードする配列において、1つ以上の点突然変異であって、B-RafのG1388T、G1783C、TG1796-97AT、G1394T、G1394A、G1394C、G1403C、G1403A、G1753A、T1782G、G1388A、T1796A、T1787G、またはC1786Gである該突然変異を含むか否か;および該突然変異が、該癌性と疑われる組織に由来する天然の細胞物質には存在するが、該非癌性組織に由来する細胞物質には存在しないことを判断するステップ
を含む、発癌性突然変異の検出方法。
【請求項7】
(a)被験体から得られる細胞物質のサンプルを準備するステップ;
(b)該サンプルを請求項4に記載の抗体でスクリーニングするステップ;および
(c)該サンプルにおいて、請求項1に記載の1つ以上の変異型B-Rafポリペプチドを検出するステップ
を含む、発癌性突然変異の検出方法。
【請求項8】
核酸の増幅産物のサンプルを配列決定して、開始配列位置から終結配列位置まで及ぶ標的ドメインにおいて測定した塩基対同定データを特定するサンプルデータセットを得ること;
標的配列位置について測定された塩基対同定データが、該標的配列位置についての参照塩基対データに対応するか否かを条件として、該サンプル中の突然変異の有無を判断すること;および
該判断ステップにおいて確立された該サンプル中の該突然変異の有無を示す出力を生成すること
を含む、B-Rafポリペプチドをコードする核酸中の標的配列位置における突然変異を検出する自動化方法であって、該核酸が1以上の点突然変異を含み、該点突然変異がB-RafのG1388T、G1783C、TG1796-97AT、G1394T、G1394A、G1394C、G1403C、G1403A、G1753A、T1782G、G1388A、T1796A、T1787G、またはC1786Gである、前記自動化方法。
【請求項9】
マーカーを、B-Rafポリペプチドのサンプル中の1つ以上の標的アミノ酸に適用すること;
該マーカーを適用した後、該サンプルを読み取り、該サンプルにおける該マーカーの有無を判断して、該サンプルにおける単一アミノ酸突然変異の有無を示すこと;および
該読取りステップにおいて判断された該サンプル中の該単一アミノ酸突然変異の有無を示す出力を生成すること
を含む、B-Rafポリペプチドにおいて単一アミノ酸突然変異を検出する自動化方法であって、該B-Rafポリペプチドが請求項1に記載される1以上の突然変異を含む、前記自動化方法。
【請求項10】
前記マーカーが、単一アミノ酸突然変異を有するB-Rafポリペプチドに優先的に結合する抗体を含む、請求項9に記載の方法。
【請求項11】
前記マーカーが、マイクロアレイヤー(microarrayer)を用いて適用される、請求項9に記載の方法。
【請求項12】
前記サンプルが光学的に読み取られる、請求項9に記載の方法。
【請求項13】
前記出力が、画像ユーザインターフェース;音声ユーザインターフェース;プリンタ;コンピュータ読取り可能記憶媒体;およびコンピュータ翻訳可能キャリア媒体のうちの少なくとも1つを含む装置を用いて生成される、請求項9に記載の方法。
【請求項14】
(a)請求項1に記載の1つ以上の変異型B-Rafポリペプチドを得るステップ;
(b)該1つ以上のポリペプチドを、テストする1つ以上の化合物と接触させるステップ;および
(c)該1つ以上の化合物と該変異型ポリペプチドとの相互作用を検出するステップ
を含む、抗増殖活性を有する1つ以上の化合物を同定する方法。
【請求項15】
前記相互作用が、結合相互作用である、請求項14に記載の方法。
【請求項16】
(a)請求項1に記載の1つ以上の変異型B-Rafポリペプチドを得るステップ;
(b)該B-Rafポリペプチド用の下流基質を得るステップ;および
(c)テストする1つ以上の化合物の存在下において、該基質の修飾を検出するステップ
を含む、抗増殖活性を有する1つ以上の化合物を同定するアッセイ。
【請求項17】
前記基質修飾を直接検出する、請求項16に記載のアッセイ。
【請求項18】
前記基質が、第2の基質を修飾する酵素であり、該第2の修飾が検出可能である、請求項17に記載のアッセイ。
【請求項19】
前記基質がMEKであり、前記第2の基質がMAPKである、請求項18に記載のアッセイ。
【請求項20】
アッセイについて、テストする1つ以上の化合物の不在下における参照レベルを決定する、請求項16〜19のいずれか一項に記載のアッセイ。
【請求項21】
B-Rafの463、465、および468位に対応する1つ以上の位置における突然変異からなる群より選択される、リン酸塩結合ループにおいて突然変異を含む構成的に活性なキナーゼ。
【請求項22】
前記突然変異が、463および468位の1つ以上にある、請求項21に記載のキナーゼ。
【請求項23】
前記突然変異が、G463VおよびG468Aからなる群より選択される、請求項22に記載のキナーゼ。
【請求項24】
Rafタンパク質キナーゼである、請求項21〜23のいずれか一項に記載のキナーゼ。
【請求項25】
B-Rafである、請求項24に記載のキナーゼ。
【請求項26】
(a)請求項21〜25のいずれか一項に記載の変異型キナーゼを調製すること;
(b)キナーゼ基質の存在下で、該変異型キナーゼを、1つ以上の化合物に晒すこと;および
(c)該1つ以上の化合物の存在下で、該基質をリン酸化する該キナーゼの能力を判断すること
を含む、キナーゼに対する阻害効果について1つ以上の化合物をスクリーニングする方法。
【図1A】

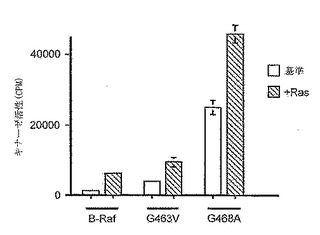
【図1B】
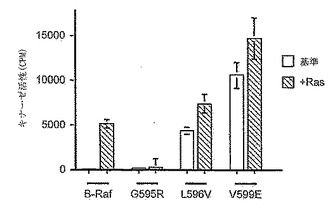
【図2】
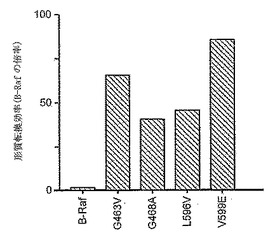
【図3A】
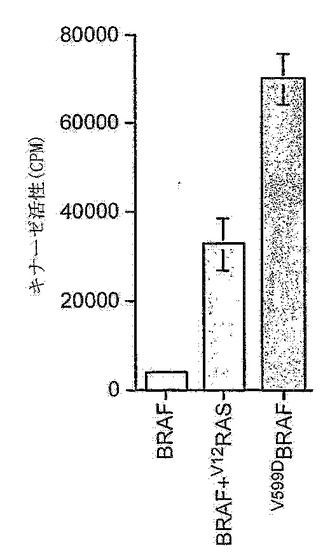
【図3B】
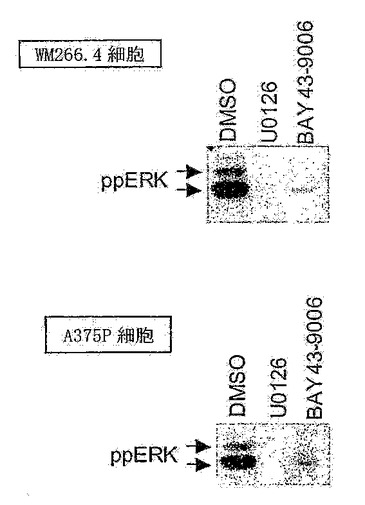
【図4】
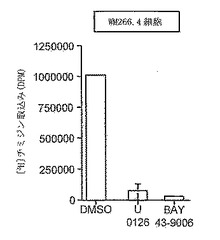
【図5A】
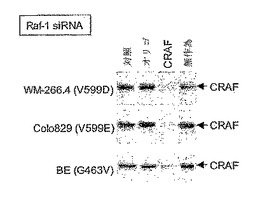
【図5B】
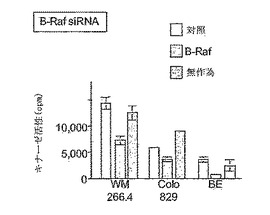
【図6】
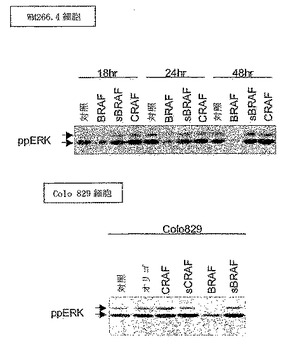
【図7】
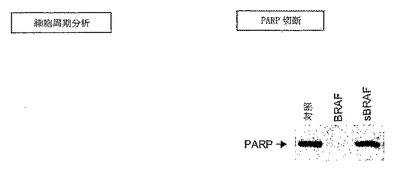
【図8】
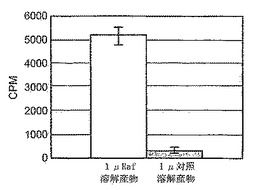
【図9】
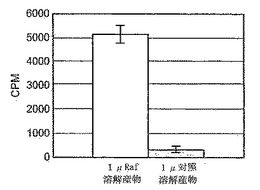
【図10】
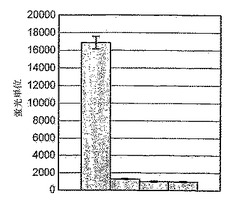
【図11】
【図12】
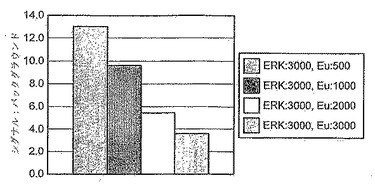
【図13】
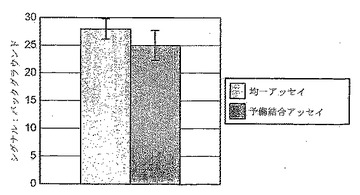
【図1B】
【図2】
【図3A】
【図3B】
【図4】
【図5A】
【図5B】
【図6】
【図7】
【図8】
【図9】
【図10】
【図11】
【図12】
【図13】
【公開番号】特開2010−246545(P2010−246545A)
【公開日】平成22年11月4日(2010.11.4)
【国際特許分類】
【出願番号】特願2010−109998(P2010−109998)
【出願日】平成22年5月12日(2010.5.12)
【分割の表示】特願2003−556553(P2003−556553)の分割
【原出願日】平成14年12月23日(2002.12.23)
【出願人】(504229974)ザ ウェルカム トラスト リミテッド (4)
【Fターム(参考)】
【公開日】平成22年11月4日(2010.11.4)
【国際特許分類】
【出願日】平成22年5月12日(2010.5.12)
【分割の表示】特願2003−556553(P2003−556553)の分割
【原出願日】平成14年12月23日(2002.12.23)
【出願人】(504229974)ザ ウェルカム トラスト リミテッド (4)
【Fターム(参考)】
[ Back to top ]