
還元環境下において安定な、規定されたフレームワークを有するイントラボディーおよびそれらの用途
【課題】還元環境において安定性かつ可溶性である規定されたフレームワークを有するscFvまたはイントラボディーを単離する方法を提供すること。
【解決手段】還元環境において安定性かつ可溶性である規定されたフレームワーク内のCDRの単離方法と同様に、そのようにして得られるscFvが記載される。そのような規定されたフレームワークを有するscFvから出発して、少なくとも1つの相補性決定領域(CDR)が無作為化される一方で、そのフレームワークが保存されるscFvライブラリーが産生される。例えば酵母細胞内のそのようなライブラリーは、抗体/CDR相互作用のスクリーニングまたは抗体のスクリーニングに好適である。
【解決手段】還元環境において安定性かつ可溶性である規定されたフレームワーク内のCDRの単離方法と同様に、そのようにして得られるscFvが記載される。そのような規定されたフレームワークを有するscFvから出発して、少なくとも1つの相補性決定領域(CDR)が無作為化される一方で、そのフレームワークが保存されるscFvライブラリーが産生される。例えば酵母細胞内のそのようなライブラリーは、抗体/CDR相互作用のスクリーニングまたは抗体のスクリーニングに好適である。
【発明の詳細な説明】
【技術分野】
【0001】
(関連出願に対する相互参照)
本出願は、その全体が参照によりここに組み込まれる1999年12月28日に出願されたPCT特許出願IB99/02054、および2000年3月1日に出願されたPCT特許出願IB00/00218の優先権を主張する。
【0002】
(技術分野)
本発明は、抗体の重鎖および軽鎖の可変領域の単鎖融合体(scFv)に関し、特に規定された安定なフレームワークを有しかつ細胞内で発現されたかかるscFv(イントラボディー)に関する。
【背景技術】
【0003】
抗体は、その抗原に対する高い親和性および特異性、およびそのインビトロおよびインビボにおける比較的高い安定性のために、生化学および分子生物学研究、診断および医学用途のための好ましい道具である。抗体は2つの重鎖および2つの軽鎖からなり、そのN末端に可変領域を含み、またそれらはジスルフィド架橋により連結されている。単鎖抗体は、重鎖可変領域および軽鎖可変領域のフラグメントを連結することにより設計されている(scFv)。各可変領域は、フレームワークに包埋された3つの相補性決定部位(CDR)を含む。これらのCDRは、抗原との相互作用の原因である。重鎖可変領域および軽鎖可変領域は領域内ジスルフィド架橋を含み、これは単鎖抗体の安定性に重要であることが報告された(Biocca et al.,1995; Derman et al., 1993)。
【0004】
特異的なエピトープに結合する単鎖抗体を同定するために最も一般的に使用される技術は、ファージディスプレイおよびその変形によるものである(概説はHoogenboom et al.,1998を参照されたい)。このスクリーニング方法は、免疫感作またはハイブリドーマ技術のような一般的な技術に対する主要な利点を有する。つまり、比較的短時間でモノクローナル単鎖抗体を取り出すことができる。
【0005】
細胞(たとえば細胞質または核)内で発現される単鎖抗体は、イントラボディー(intrabodies)と呼ばれる。細胞内は還元環境であるために、抗体の安定性に重要であると信じられているジスルフィド架橋が形成されない。従って、初めは、イントラボディーの適用は好ましくないと信じられていた。しかしながら、イントラボディーの適用の可能性を示すいくつかのケースが開示されている(Beerli et al.,1994; Biocca et al., 1994; Duan et al., 1994; Gargano and Cattaneo, 1997;Greeman et al., 1996; Martineau et al. 1998; Mhashilkar et al., 1995;Tavladoraki et al., 1993)。これらのケースにおいては、イントラボディーはたとえば、細胞質内の抗原をブロッキングし、その生物活性を阻害することにより働く。
【0006】
現在まで、イントラボディーは大抵の場合、初めに古典的な技術(たとえばファージディスプレイ)で選別され、次にその細胞内でのイントラボディーとしての生物活性を試験される(Vinsintin et al.,1999)、モノクローナル抗体に由来するものであった。成功したイントラボディー(上述を参照)が記述されているにもかかわらず、今日では、そのようなイントラボディーが細胞内で機能的であるかどうかは全く予測不可能である(概説は、Cattaneo,1998; Cattaneo and Biocca, 1999を参照されたい)。その原因はおそらく、異なる環境である。ファージディスプレイおよび他の古典的な技術は酸化条件下で行われ、従ってジスルフィド架橋が形成されるが、一方イントラボディーは還元条件下で機能しなければならない。この還元環境はイントラボディーの不十分な溶解性を導く可能性があり、従ってそれらは非機能的な凝集塊を形成する。イントラボディーの溶解性は、フレームワーク内(Knappik and Pluckthun,1995)またはCDR内(Kipriyanov et al., 1997; Ulrich et al., 1995)のいずれかにおける変化によって調節できる。
【0007】
しかしながら、従来知られている方法は、その用途が細胞内の標的を検出することに限定されている。従って、ある抗原に特異的なイントラボディーを直接的にスクリーニングする信頼性のある技術および方法の必要性が増している。
【0008】
WO99/36569においてウィットラップ(Wittrup)らは、酵母内在性の、細胞壁上での局在化のためのAga2pに由来するタンパク質断片を用いて、タンパク質およびscFvを酵母の細胞壁上に提示する方法を記載している。他の関連する方法は、EP0407259(Boquet et al.,1991)において記載されている。これらの方法は、タンパク質またはペプチドライブラリーもまた表面上に提示されるファージディスプレイスクリーニングに匹敵する。しかしながら、これらの技術はイントラボディーを同定するための細胞内スクリーニングには使用できない。
【0009】
特許明細書JP11000174(Kyoko et al., 1999)において、抗体のFabフラグメントを高レベルに発現および分泌させるための、酵母ピキアパストリス(Pichia pastoris)の使用が記載されている。この酵母は、その分泌レベルが高いために有名であり、したがってこの用途に好適に使用される。分泌された抗体は、上清を精製することにより収集することができる。さらに、EP0590067、WO92/22324、JP06030778、US5698435、US5595889、JP10313876において、酵母は分泌されるタンパク質または抗体の製造に使用されている。EP0698097およびWO94/25591は、さらなる用途のための、重鎖またはその断片のみの製造および分泌の用途を開示している。JP09020798;JP05105700;およびJP05097704には、一般に人体または生物体に投与した場合の、肝炎ワクチンを得るための酵母分泌の方法が記載されている。
【0010】
単鎖抗体のスクリーニングに酵母を使用することも、WO99/28502によってすでに知られている。前記出願は、単鎖モノクローナル抗体融合試薬用のDNA構築物ライブラリーの使用を開示する。このscFvライブラリー(ここではsFvライブラリーと呼ぶ)は、次にスクリーニングに使用される。しかしながら、規定されていないフレームワークを使用しているために、イントラボディーの安定性と可溶性は劇的に変化する可能性があることが見出されている。さらに、インビボにおける性能とインビトロにおける安定性および可溶性との間に直接的な相関関係が示された。したがって、CDRは原理的には抗原に対して必要とされる高い親和性を示すにもかかわらず、対応するフレームワークが十分に可溶性でないために凝集し、このモノクローナルscFvは検出不可能になるので、種々のscFvフラグメントのmRNA由来ライブラリーの使用は、その抗原に対する高い親和性を有するCDRを同定する可能性があるという観点に限定される。したがって、未だ改善された抗体、またはイントラボディーがそれぞれ必要とされている。
【0011】
細胞内の標的に向けられるscFvの用途は増大しており、信頼性のあるイントラボディーのスクリーニング方法の必要性が生じている。還元条件下においてscFvが不安定であり、かつ抗体安定性が予測できないために、scFvの細胞質内標的は最も困難な用途である。この安定性と可溶性の問題も、細胞内適用のために最適化された、規定されたフレームワークを使用することにより解決され得る。
【発明の概要】
【課題を解決するための手段】
【0012】
したがって、還元環境において安定性かつ可溶性である規定されたフレームワークを有するscFvまたはイントラボディーを単離する方法を提供することが、本発明の全般的な目的である。
【0013】
本発明のさらなる目的は、還元環境において安定性かつ可溶性である規定されたフレームワークを有するscFvまたはイントラボディーである。
【0014】
本発明の他の目的は、CDR/フレームワーク連結領域内に独特な制限酵素認識部位を備えるように修飾された、還元環境において安定性かつ可溶性である規定されたフレームワークを有するscFvまたはイントラボディーである。
【0015】
本発明の他の目的は、還元環境において安定性かつ可溶性である規定されたフレームワークと、無作為にまたは確定的に変形されたCDRとを有するscFvまたはイントラボディーのライブラリーである。
【0016】
本発明の他の目的は、還元環境において安定性かつ可溶性である規定されたフレームワークと種々のCDRとを有するscFvまたはイントラボディー、またはこのようなscFvまたはイントラボディーのライブラリーを用いた、抗原結合性CDRをスクリーニングする方法である。
【0017】
本発明の他の目的は、このようなscFvまたはイントラボディーまたはライブラリーをそれぞれ用いた、さらなる抗原のスクリーニング方法である。
【0018】
本発明の他の目的は、還元条件下において可溶性かつ安定性であるフレームワークを有するイントラボディーの同定方法である。
【0019】
本発明のイントラボディーはさらに、疾病の治療、診断、または予防における薬剤、および特異的なタンパク質活性の機能的ノックアウトのような移植における種々の用途の薬剤として使用され得る。イントラボディーはそれ自体として、またはこのようなscFvをコードするDNAとして使用され得る。
【0020】
本明細書の範囲内において、術語scFvおよびイントラボディーは大部分は同義語として使用されるが、たとえば細胞内のような還元環境における本発明の規定されたフレームワークを有するイントラボディー(scFv)の安定性および可溶性は本発明にとって必要であるが、そのようなイントラボディー(scFv)等の用途は、細胞内での用途に制限されるものではないことは理解されなければならない。
【0021】
CDR内にアミノ酸変化を導入することのみによって、本発明に従ったフレームワークが特異的な抗原認識という所望の生物学的機能を示すモノクローナル抗体を識別する可能性が大いに増加する。scFvのCDRにおけるそのような変化は、イントラボディーの細胞質内適用に適する規定されたフレームワークを変化させずに、無作為の変化として行うことができる。
【0022】
細胞内でモノクローナル単鎖抗体のスクリーニングを行うためには、細胞質の酸化還元環境に適応するフレームワークを使用しなければならない。したがって、フレームワークはジスルフィド架橋の不在下であっても十分に安定性かつ可溶性でなければならない。しかしながらscFvの多くは、還元条件下または、ドメイン内ジスルフィド架橋の形成に必要であるシステインの不在下においては、適切な構造に折りたたまれないことが知られている。したがって、本発明の範囲内において、同一のCDRを含有する複数のフレームワークを比較したところ、インビボでの性能における劇的な差異が観察された。本発明の方法によって、抗原認識のための規定されたCDRを含有する最も性能の良いフレームワークが選択できる。この方法は、出発材料として既知の抗原に対するイントラボディーを用いて行われる。重鎖および軽鎖の可変領域を連結するのに用いるリンカーは重要ではない。しかしながらそれは、CDRと抗原との間の相互作用のために好適な接触およびフォールディングを確実にするために、十分な溶解性および柔軟性を提供しなければならない。好適なリンカーは、約5〜60アミノ酸の典型的な長さを有し、通常の規則的なグリシンの配列、および溶解性を増強するための1〜3個のセリンを有する。
【0023】
還元環境において安定性かつ可溶性である規定されたフレームワークを有するscFvを単離するための本発明の方法は、以下の工程により規定される。
【0024】
a)既知の抗原に対するscFvのDNA配列の少なくとも1つのフレームワークコード領域を突然変異させ、そのような突然変異を好適な発現ベクターに導入することにより、種々のフレームワークおよび定常のCDRを有するscFvライブラリーを作製し、
b)特異的な既知の抗原を発現でき、かつ抗原−scFv相互作用の存在下でのみ生存する宿主細胞を、前記scFvライブラリーで形質転換し、
c)そのようにして形質転換した宿主細胞を、抗原およびscFvを発現するのに好適な条件下で培養して、抗原−scFv相互作用の存在下においてのみ細胞を生存させ、
d)生存している細胞内で発現され、また還元環境において安定性かつ可溶性である規定されたフレームワークを有するscFvを単離する。
【0025】
好ましい実施形態において、宿主細胞は真核細胞、特に酵母細胞である。
【0026】
上述した方法によって、規定したフレームワークを有するscFvを入手できる。そのようなフレームワークも本発明の目的である。そのようなフレームワークは、少なくとも1つのCDRの選択的な交換を可能にする特異的な制限酵素認識部位を含むように修飾できる。好ましくは前記制限酵素認識部位は、CDRに隣接するフレームワーク内に位置する。
【0027】
本発明はさらに、CDR領域内における選択的な変更に好適なフレームワークを有するscFvをコードするDNAの産生方法であって、特異的な制限酵素認識部位を、規定された安定性かつ可溶性であるscFvをコードするDNAの配列に部位特異的変異誘発によって導入し、それによって制限酵素認識部位が好ましくはフレームワーク内に位置し、かつそれによって制限酵素認識部位を作製するためのヌクレオチドの置換がアミノ酸配列に影響しない方法を提供する。
【0028】
還元環境において安定性かつ可溶性である規定されたフレームワークを含む改善されたscFvは、本発明の目的でもある方法によっても得ることができ、ここでは還元環境において安定性かつ可溶性である少なくとも2つの異なるフレームワーク(好ましくは本発明のフレームワーク)の少なくとも2つの変異を組合せて、規定されたフレームワークを有するscFvを製造する。
【0029】
上述した方法によって入手できるscFvも本発明の目的である。そのようなフレームワーク内においては、少なくとも1つの変異が可変軽鎖のCDR1の上流に存在し、かつ/または少なくとも1つの変異が可変重鎖のCDR2およびCDR3の間に位置することが好ましい。
【0030】
非常に好ましい実施形態においては、本発明のscFvは、可変軽鎖のCDR1の上流に存在する少なくとも2つの変異、および可変重鎖のCDR2およびCDR3の間に位置する少なくとも2つ、好ましくは少なくとも4つの変異を含む。特に、SEQ ID NO 1に規定されるフレームワークを含むscFvが好ましい。
【0031】
そのようなフレームワークにおいてCDRを特異的に無作為化するために、独特な制限酵素認識部位の発生を導く沈黙突然変異(silent changes)(同じアミノ酸配列をコードするが、異なるコドンを使用する)を導入できる(上記も参照)。制限酵素認識部位はCDR/フレームワーク連結領域内のどこにでも設置できるが、それは独立した各CDRに隣接するフレームワーク内に位置することが好ましい。これによって、独立した各CDRは、無作為のまたは規定された配列の導入により置換され得る。これは、抗原に対して高い親和性を示すイントラボディー内の新規なCDRを選択することを可能にする。
【0032】
局在化シグナルまたは活性化ドメインのような付加的な配列をscFvライブラリーに由来する非限定的なフレームワークに導入する場合、この修飾によって、たとえばscFvが不溶性になるといったように、生物学的活性が(たとえそれまで存在していても)失われ得る。したがって、既知の抗原に対する本発明の規定されたフレームワークを使用し、次にそのような修飾をイントラボディー内の種々の位置(NおよびC末端またはscFvのコード配列内)に導入し、また本来の機能を維持するために選択することは、利点である。WO99/28502には、局在化シグナルを導入するいくつかの可能性が記載されている。ある抗原に対する相互作用スクリーニングに使用される活性化ドメインは、WO99/98502において記載されており、scFvライブラリーのC末端に導入される。種々の位置、たとえばN末端の活性化ドメインにおいて配列の付加を許容するフレームワークも本発明の方法によって選択することができ、それは拮抗的な機能における活性化ドメインを持たない、それらのscFvに対応する物と同様に振る舞うことが今般見出された。従って、たとえば以下の実施例においてさらに説明されるフレームワーク内においては、N末端の活性化ドメインを導入することは抗体の機能を阻害しない。
【0033】
還元環境において安定性かつ可溶性である規定されたフレームワークを有する本発明のイントラボディーから出発して、CDRライブラリーを含有するscFvまたはイントラボディーをそれぞれ作製できる。
【0034】
還元環境において安定性かつ可溶性である規定されたフレームワークを有するCDRライブラリーを作製するための好適な方法は本発明の方法であり、配列につき少なくとも1つのCDRを修飾されたCDRで置換するために、本発明のscFvをコードしているDNA配列を消化させる。好ましくは、修飾されたCDRは無作為変異により作製する。そのような方法により、少なくとも1つの無作為化されたCDR、および還元条件下において安定性かつ可溶性である規定されたフレームワークを有するイントラボディーのライブラリーを作製できる。
【0035】
CDRライブラリーを含有する本発明のイントラボディーは、抗原に対して高い親和性を有するクローンのスクリーニングおよび選択に使用され得る。特定の抗原と相互作用するCDRのスクリーニング方法も本発明の目的であり、またこの方法は、既知の抗原をコードする核酸配列、特にDNA配列で形質転換し、さらに還元環境下において安定性かつ可溶性である規定されたフレームワークを有する無作為化されたCDRライブラリーで形質転換した宿主細胞を含み、それによって抗原および/またはscFvはマーカーシステムまたはマーカーシステムの一部に連結され、したがって選択的条件下で培養された細胞は抗原/scFv相互作用の存在下においてのみ生存するので、形質転換された細胞を選択的な条件下で培養して残存する細胞を培養し、イントラボディーを収集することを含む。
【0036】
本発明の好ましい実施形態においては、このフレームワークは本発明のフレームワークであり、細胞は真核細胞、特に酵母細胞である。
【0037】
本発明のさらに好ましい実施形態において、抗原をコードするDNA配列およびscFvをコードするDNA配列はいずれも、生存許容(allowing)マーカーに連結された転写活性化システムの一部にいずれも連結された、抗原またはscFvをそれぞれ含むキメラ分子をコードし、さらに好ましくは抗原はDNA結合ドメインに融合されてscFvは転写活性化因子ドメインに融合されるか、または抗原は転写活性化因子ドメインに融合されてscFvはDNA結合部位に融合される。
【0038】
CDRライブラリーを含有する本発明のイントラボディーは、抗原に対して高い親和性を有するクローンをスクリーニングおよび選択するのに使用され得る。これは、その生物学的機能において細胞内に位置する抗原をブロックすること、または規定されたフレームワーク内に包埋されたCDRの抗原に対する直接的な相互作用を分析することのいずれかにより達成できる。直接的な相互作用は、好ましくは、転写性の読み出し(readout)、好ましくはHIS3遺伝子の発現によってモニターできる。予め決定した条件下で3−アミノトリアゾール(3AT)を添加すると、抗原に対してより親和性の高いCDRの選択が可能になる。特異的な既知の抗原を発現し得る宿主細胞は、前記条件下では抗原−scFv相互作用の存在下、好ましくは十分に強い抗原−scFv相互作用の存在下においてのみ生存する。ここで用いられる術語「十分に強い」は、BIAcoreで測定されたKDが、>1×10−6M、好ましくはKD>1×10−8M、そしてより好ましくはKD>1×10−10Mであるタンパク質−タンパク質相互作用として定義される。そのような選択工程はさらに、CDR(好ましくは軽鎖および重鎖のCDR1およびCDR2)内のアミノ酸を無作為または選択的に変異させ、続いて上昇した3ATの濃度において増殖するものをこのプールから選出することによる親和性熟成を行うのに適用できる。
【0039】
既に上述したように、今までに知られており使用されているscFvライブラリーは、好ましくはB細胞が高度に蓄積されることが知られている脾臓からのmRNAの単離に由来し、再編成された抗体が発現される。そのようなライブラリーは、その生物体内に存在するエピトープに対して反応しないように予備選択(ポジティブおよびネガティブ選択)されているという欠点を有する。これは、自己免疫反応を開始しない抗体のみが熟成され、活性化され得ることを保証する。しかしながら、この選択工程のために、可能性のあるアミノ酸の組み合わせの全てが「天然の(natural)」scFvライブラリー内に存在するわけではない。いくつかのインビトロおよび診断上の用途のためには、抗体は種属間で保存されているタンパク質と相互作用することが必要とされる。そのようなタンパク質またはペプチドにとって、予備選択工程のために、「天然の」scFvライブラリー内で強力に相互作用するモノクローナル抗体を検出することは非常に困難な場合がある。さらに、その「天然の」ライブラリー内に存在するフレームワークは最適化されておらず、したがって不十分かまたは可変性の溶解性および/または安定性はそれぞれ問題を生じる。したがって、本発明のフレームワーク、および/または本発明の方法によって得られるフレームワークを含み、かつこれらの領域内のアミノ酸配列の可能な組み合わせのいくらか、または好ましくは全てをカバーするCDR無作為ライブラリーのみを使用することは大きな利点である。
【0040】
本発明をさらに説明するために、酵母細胞内転写因子Gcn4pに対する規定された相補性決定領域(CDR)を含む、安定性かつ可溶性であるイントラボディーフレームワークを選択した。この規定されたフレームワークは、無作為の配列によってそのCDRを置換するのに使用した。所望の生物活性(CDRのインビボ効果)を刺激する新規なCDRを同定するためにこれらのCDRライブラリーをスクリーニングする:
a)細胞内(たとえば、ヒト細胞内または他のいかなる細胞内でも)で自然に起こる分子相互作用を好適な細胞(好ましくは酵母)内で再構成させるか、または酵母内在性の相互作用を使用する。次のスクリーニングで、再構成されたかまたは内在性の分子の生物活性によるこれらのCDRの干渉によって、高親和性のCDRを同定する。そのような拮抗性のCDRはたとえば、シグナル伝達経路に含まれる2つのタンパク質をブロックすることにより機能できるであろう。
【0041】
b)再構成されたかまたは内在性の分子における、所望の生物活性を誘発するアゴニストのCDRを選択する。
【0042】
安定性であるフレームワーク内に包埋された無作為CDRをさらに、相互作用スクリーニングに基づいて、CDRと抗原との相互作用を同定するのに使用することができる:
a)選択されたフレームワークは転写活性化ドメインに融合可能であり、しかもその機能を保持することが示されるであろう。このキメライントラボディーを、DNA結合活性を有するDNA結合部位または転写因子に融合されて得られる抗原に対して高親和性であるCDRの選択に使用する。抗原とCDRが相互作用する時には、転写活性化ドメインは選択マーカー遺伝子の遺伝子発現を媒介し、選択条件下でのこの細胞の生存を許容する。
【0043】
b)ハイブリッド技術(一方のパートナーを活性化ドメインへ融合させ、もし必要であればもう一方をDNA結合部位へ融合させる)に基づいて再構成させた分子の相互作用は、特異的で高親和性のCDRによってブロックされ得る。
【0044】
イントラボディーのCDRは一定であるがフレームワーク内の種々の変異は、イントラボディーの安定性および溶解性を変化させることによりそのインビボにおける性能に影響を及ぼすことも明らかとなった。フレームワークは、イントラボディーの安定性および溶解性の主要な部分に寄与する。にもかかわらず、CDR内の特定の変異はイントラボディーの溶解性および安定性に影響する場合もある。したがって、機能的品質管理(下記参照)によって、規定されたフレームワーク内に包埋された無作為なCDRを予備選択することは都合が良いだろう。
【0045】
本発明はさらに、イントラボディーフレームワークまたはイントラボディーの同定方法であって、好適な宿主細胞をライブラリーおよびマーカーシステムにより形質転換し、それによって前記ライブラリーはイントラボディーライブラリーと前記マーカーシステムの少なくとも一部との融合産物となり、前記マーカーシステムは可溶性かつ安定性であるイントラボディーフレームワークをコードする融合タンパク質の存在下においてのみ活性化され、また前記細胞を、可溶性かつ安定性であるイントラボディーフレームワークを発現している細胞の同定および選択を可能にする条件下で培養する方法を提供する。
【0046】
本発明の好ましい実施形態において、前記ライブラリーはイントラボディーライブラリーとマーカータンパク質との融合産物である。好ましくは、前記マーカータンパク質は選択可能な活性、特に酵素活性または蛍光活性を有する。そのような方法において使用され得るマーカータンパク質は、たとえばGFPタンパク質またはそのいかなる変異体でもある。
【0047】
本発明の他の好ましい実施形態においては、前記ライブラリーは、イントラボディーライブラリーと、転写がそのDNA結合タンパク質により制御されるマーカー遺伝子の転写を活性化できるDNA結合タンパク質との融合産物である。好適なDNA結合タンパク質はたとえばp53である。
【0048】
本発明のさらなる好ましい実施形態においては、前記方法はイントラボディーおよびトランス活性化システムの一部を含むタンパク質をコードするライブラリーで形質転換された好適な宿主細胞を含み、かつ前記細胞はさらに少なくとも前記トランス活性化システムの第2部分を含む第2タンパク質を発現し、それによって前記トランス活性化システムは生存許容マーカーに連結されて前記細胞は前記2つの融合タンパク質間の相互作用の存在下における選択的条件下においてのみ生存する。
【0049】
さらに好ましい実施形態においては、前記ライブラリーにコードされるタンパク質は転写活性化ドメインを含みかつ前記第2タンパク質はDNA結合部位を含むか、または前記ライブラリーにコードされるタンパク質はDNA結合部位を含みかつ前記第2タンパク質は転写活性化ドメインを含む。
さらなる好ましい実施形態においては、前記第2タンパク質はDNA結合部位またはトランス活性化ドメインと、前記ライブラリーにコードされるタンパク質の定常領域と相互作用するタンパク質とを含む。ここで使用される術語「定常領域」は、ライブラリー構築物によってコードされ、タンパク質相互作用のパートナーとして働くいかなるタンパク質部位、またはいかなる近接するアミノ酸配列をも含み、また前記用語はたとえばイントラボディーまたはGal11pの一部を含む。
【0050】
上記方法により得ることが可能な、規定されたフレームワークを有するscFvも本発明の目的であり、特に本発明の方法において使用するための目的である。
【0051】
同方法は、たとえばscFvまたはCDRライブラリー、特に本発明のライブラリーの出発材料として使用され得る、可溶性かつ安定性であるフレームワークを同定するためのいかなるscFvライブラリーのスクリーニングにも適用され得る。
【0052】
本発明の他の目的は、scFvと相互作用する抗原のスクリーニング方法であって、少なくとも1つの興味のある抗原を発現する宿主細胞を、還元環境において安定性かつ可溶性である規定されたフレームワークを有する少なくとも1つのscFv、または還元環境において安定性かつ可溶性である規定されたフレームワークを有する無作為化されたCDRライブラリーで形質転換し、それによって抗原および/またはscFvをマーカーシステムまたはマーカーシステムの一部に連結させ、したがって選択的条件下で培養される細胞が抗原/scFv相互作用の存在下においてのみ生存し、このように形質転換された細胞を選択的条件下で培養し、残存する細胞を培養してscFvを収集する方法を提供することである。
【0053】
本発明の好ましい実施形態においては、フレームワークは本発明のフレームワークであり、かつ細胞は真核細胞、特に酵母細胞である。
【0054】
本発明の非常に好ましい実施形態においては、抗原をコードするDNA配列およびscFvをコードするDNA配列は、いずれも生存許容マーカーに連結された転写活性化システムの一部に連結された抗原またはscFvそれぞれとのキメラ分子の両方をコードし、より好ましくは抗原はDNA結合ドメインに融合されかつscFvは転写活性化因子ドメインに融合されるか、または抗原は転写活性化因子ドメインに融合されかつscFvはDNA結合ドメインに融合される。
【0055】
本発明はさらに、治療または診断または予防用薬剤として規定されたフレームワークを有するscFv、および細胞内スクリーニングのために規定されたフレームワークを有するscFvの使用方法を提供する。
【0056】
本発明の全ての目的のためには真核細胞が好ましく、特に、増殖、ポジティブ選択、増殖選択が速くでき、かつ形質転換およびその選択が効率的にできることを含むその特異的な特徴から、酵母細胞が特に好ましい。
本発明はまた、以下の項目を提供する。
(項目1) 還元環境において安定性かつ可溶性である規定されたフレームワークを有するscFvの単離方法であって、
(a)既知の抗原に対するscFvのDNA配列の少なくとも1つのフレームワークコード領域の突然変異、および前記突然変異の好適な発現ベクターへの導入により、種々のフレームワークおよび定常的なCDRを有するscFvライブラリーを作製し、
(b)特異的な既知の抗原を発現可能であり、かつ抗原−scFv相互作用の存在下でのみ生存する宿主細胞を前記scFvライブラリーで形質転換し、
(c)前記形質転換した宿主細胞を、前記抗原および前記scFvを発現するのに好適な条件下で培養して、前記抗原−scFv相互作用の存在下においてのみ生存させ、
(d)生存細胞内で発現され、また還元環境において安定性かつ可溶性である
規定されたフレームワークを有する前記scFvを単離する方法。
(項目2) 前記宿主細胞が真核生物である項目1記載の方法。
(項目3) 前記宿主細胞が酵母細胞である項目1または2記載の方法。
(項目4) 項目1〜3のいずれか1項に記載の方法により得られる規定されたフレームワークを有するscFv。
(項目5) 少なくとも1つのCDRの選択的な交換を可能にする制限酵素認識部位を含む項目4記載のscFv。
(項目6) 前記制限酵素認識部位がCDRに隣接する前記フレームワーク内に位置する項目5に記載のscFv。
(項目7) CDR領域における選択的な変更に好適なフレームワークを含むscFvをコードするDNAの産生方法であって、部位特異的変異誘発により、規定された安定性かつ可溶性であるscFvをコードするDNAの配列に特異的な制限酵素認識部位を導入する方法。
(項目8) 前記制限酵素認識部位が前記フレームワーク内に位置し、それによって前記制限酵素認識部位を発生させるための前記ヌクレオチドの置換がアミノ酸配列に影響しない項目7記載の方法。
(項目9) 還元環境において安定性かつ可溶性である規定されたフレームワークを含むscFvの産生方法であって、還元環境において安定性かつ可溶性である少なくとも2つの異なるフレームワーク、好ましくは項目4〜6のうちいずれか1項記載のフレームワークまたは項目1〜3のいずれか1項に従って単離されるフレームワークのうち少なくとも2つの変異が、規定されたフレームワークを有するscFvを製造するために組み合わされる方法。
(項目10) 項目9の方法により得られる前記規定されたフレームワークを有するscFv。
(項目11) 前記変異が前記可変軽鎖のCDR1の上流に存在する項目10記載のscFv。
(項目12) 前記変異が前記可変重鎖のCDR2とCDR3との間に位置する項目10記載のscFv。
(項目13) 少なくとも1つの変異がCDR1の上流に存在し、かつ少なくとも1つの変異が前記可変重鎖のCDR2とCDR3との間に位置する項目10記載のscFv。
(項目14) 少なくとも2つの変異がCDR1の上流に存在し、かつ少なくとも2つ、好ましくは少なくとも4つの変異が前記可変重鎖のCDR2とCDR3との間に位置する項目10記載のscFv。
(項目15) SEQ ID NO 1において規定されたフレームワークを含むscFv。
(項目16) 還元環境において安定性かつ可溶性である規定されたフレームワークを含むCDRライブラリーの産生方法であって、項目1〜15のいずれか1項記載のscFvをコードするDNA配列が、配列当たり少なくとも1つのCDRを修飾されたCDRで置換するために消化される方法。
(項目17) 前記修飾されたCDRが無作為な変更により産生される項目16記載の方法。
(項目18) 少なくとも1つの無作為化されたCDRおよび、還元環境下において安定性かつ可溶性である規定されたフレームワークを有するイントラボディーのライブラリー。
(項目19) 特異的な抗原と相互作用するCDRのスクリーニング方法であって、既知の抗原をコードする核酸配列、特にDNA配列で形質転換させた宿主細胞をさらに、還元環境において安定性かつ可溶性である規定されたフレームワークを有する無作為化したCDRライブラリーで形質転換し、それによって抗原および/またはscFvをマーカーシステムまたはマーカーシステムの一部に連結させ、したがって選択的な条件下において培養した細胞は抗原/scFv相互作用の存在下でのみ生存し、このように形質転換させた細胞を選択的な条件下において培養し、残存している細胞を培養してイントラボディーを収集する方法。
(項目20) 前記フレームワークが項目1〜19のいずれか1項において規定されるようなフレームワークである、項目19記載の方法。
(項目21) 前記細胞が真核細胞、特に酵母細胞である項目19または20記載の方法。
(項目22) 前記抗原をコードするDNA配列および前記scFvをコードするDNA配列がいずれも、生存許容マーカーに連結された転写活性化システムの一部にいずれも連結された前記抗原またはscFvをそれぞれ含むキメラ分子をコードする、項目19〜21のいずれか1項記載の方法。
(項目23) 前記抗原がDNA結合ドメインに融合され、かつ前記scFvが転写活性化因子ドメインに融合されるか、または前記抗原が転写活性化因子ドメインに融合され、かつ前記scFvがDNA結合ドメインに融合される、項目22記載の方法。
(項目24) scFvと相互作用している抗原のスクリーニング方法であって、興味のある少なくとも1つの抗原を発現する宿主細胞を、還元環境において安定性かつ可溶性である規定されたフレームワークを有する少なくとも一つのscFv、または還元環境において安定性かつ可溶性である規定されたフレームワークを有する無作為化されたCDRライブラリーで形質転換し、それによって抗原および/またはscFvをマーカーシステムまたはマーカーシステムの一部に連結させ、したがって選択的な条件下において培養した細胞は抗原/scFv相互作用の存在下でのみ生存し、このように形質転換させた細胞を選択的な条件下で培養し、残存している細胞を培養してscFvを収集する方法。
(項目25) 前記フレームワークが項目1〜23のいずれか1項において規定されるようなフレームワークである項目24記載の方法。
(項目26) 前記細胞が真核細胞、特に酵母細胞である項目24または25記載の方法。
(項目27) 前記抗原をコードするDNA配列およびscFvをコードするDNA配列が両者共に、いずれも生存許容マーカーに連結された転写活性化システムの一部に連結された抗原またはscFvをそれぞれ含むキメラ分子をコードする、項目24〜26のいずれか1項記載の方法。
(項目28) 前記抗原がDNA結合ドメインに融合され、かつ前記scFvが転写活性化因子ドメインに融合されるか、または前記抗原が転写活性化因子ドメインに融合され、かつ前記scFvがDNA結合ドメインに融合される、項目27記載の方法。
(項目29) 治療または診断または予防用薬剤としての規定されたフレームワークを有するscFv。
(項目30) 細胞内スクリーニングのための規定されたフレームワークを有するscFvの使用。
(項目31) イントラボディーフレームワークまたはイントラボディーの同定方法であって、好適な宿主細胞をライブラリーおよびマーカーシステムで形質転換し、それによって前記ライブラリーはイントラボディーライブラリーと前記マーカーシステムの少なくとも一部の融合産物となり、前記マーカーシステムは可溶性かつ安定性であるイントラボディーフレームワークをコードする融合タンパク質の存在下においてのみ活性化され、また、可溶性かつ安定性であるイントラボディーフレームワークを発現する細胞の同定および選択を可能にする条件下において前記細胞を培養する方法。
(項目32) 前記ライブラリーが、イントラボディーライブラリーとマーカータンパク質の融合産物である項目31記載の方法。
(項目33) 前記マーカータンパク質が選択可能な活性、特に酵素活性または蛍光活性を有する項目32記載の方法。
(項目34) 前記ライブラリーが、イントラボディーライブラリーと転写を活性し得るDNA結合タンパク質との融合産物である項目31記載の方法。
(項目35) 前記好適な宿主細胞を、イントラボディーおよびトランス活性化システムの一部分を含むタンパク質をコードするライブラリーで形質転換し、前記細胞はさらに前記トランス活性化システムの第2部分を含む第2タンパク質を発現し、それによって前記トランス活性化システムを生存許容マーカーに連結させ、前記細胞が前記2つのタンパク質間の相互作用が存在する選択的な条件下においてのみ生存する、項目31記載の方法。
(項目36) 前記ライブラリーにコードされるタンパク質が転写活性化ドメインを含み、かつ前記第2のタンパク質がDNA結合ドメインを含むか、または前記ライブラリーにコードされるタンパク質がDNA結合ドメインを含み、かつ前記第2のタンパク質が転写活性化ドメインを含む項目35記載の方法。
(項目37) 前記第2タンパク質が、DNA結合ドメインまたはトランス活性化ドメインのそれぞれと、前記第1のライブラリーにコードされたタンパク質の定常領域と相互作用するタンパク質とを含む、項目35または36記載の方法。
(項目38) 特に項目1記載の方法において使用するための、項目31〜37のいずれか1項に記載の方法によって得られる規定されたフレームワークを含むscFv。
【図面の簡単な説明】
【0057】
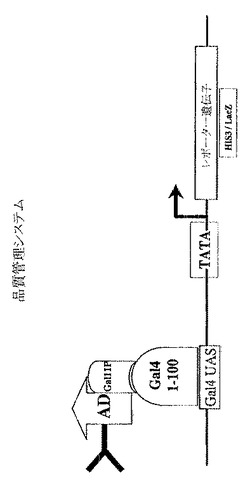
【図1A】scFvまたはCDRライブラリーの品質管理を行いうる方法を示す図。
【図1B】レポーター遺伝子活性と関連するscFv融合タンパク質の溶解性を示す図。
【図2】λグラフトと呼ばれる別の変異体と比較した、最適化したGal4 AD−ΩグラフトscFvのインビボにおけるより良好な性能を示す図。
【図3A】Gcn4p依存性LacZレポーター遺伝子の遺伝子発現における、種々のscFvフラグメントのインビボにおける性能を示す図。
【図3B】ツーハイブリッドアッセイにおける、酵母内での種々のscFvフラグメントの発現のインビボにおける性能を示す図。
【図4】種々のscFvフラグメントを発現している細胞の、ツーハイブリッドアッセイにおける増殖選択を示す図。
【図5A】Gcn4p依存性LacZリポーターの遺伝子発現において、定常ドメイン(Gal11P−Gal4AD)の単鎖抗体に対するN末端融合は、有意にはこのscFvフラグメントの性質を変化させないことを示す図。
【図5B】LacZリポーターの遺伝子発現において、2つの独特な制限酵素認識部位の単鎖抗体への導入は、このscFvフラグメントの性質を変化させないことを示す図。
【図6】酵母内に発現した種々のGcn4p結合scFvフラグメントの溶解性のウェスタンブロット解析を示す図。
【発明を実施するための形態】
【0058】
(本発明を実行するための方法)
scFvおよびCDRライブラリーの品質管理
術語「品質管理」は、scFvライブラリーからの安定性かつ可溶性であるイントラボディーの選択を可能にするアッセイを定義する。
この目的のために、scFvライブラリーを、転写活性化ドメイン(この場合Gal4AD)および定常領域(この場合Gal11P aa263−352)へ融合させる。融合タンパク質の安定性は、scFv部分の安定性および溶解性に依存する。定常Gal11Pドメインは、Gal4の二量体化ドメイン(Gal4DNA結合部位(DBD)の一部である58−97残基(Barberis et al.,1995))と相互作用する。
【0059】
このライブラリーを、選択マーカー遺伝子(たとえばHIS3/LacZ)のプロモーターに結合するGal4 DBD(1−100残基)を発現する酵母細胞に形質転換する。この宿主細胞の増殖は、試験されるイントラボディーが所望の溶解性および安定性を示し、Gal11PとGal4 DBDを介して十分に相互作用できる場合にのみ媒介される(図1Aを参照)。
【0060】
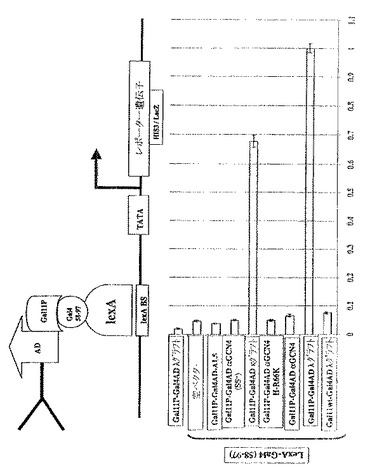
遺伝子活性化に関連する溶解性
本発明において記述される品質管理システムの原理は、よく特徴付けられた多数のscFvを用いて実証された。これらは、本質的に同一の抗原結合特性を有するが、インビトロでの安定性は異なる。種々のscFvフラグメントをGal11P−Gal4AD融合タンパク質として発現させた。Gal4二量体化ドメイン(58−97残基)をLexAのC末端に融合させ、6x LexA結合部位の制御下におけるレポーター遺伝子を含有するレポーター株YDE173に転換した(下記参照)。
【0061】
上述したように、Gal11P−Gal4AD−scFv融合タンパク質の細胞内安定性および溶解性は、scFv部分に依存する。したがって、LexA−Gal4(58−97)と十分に相互作用する安定性かつ可溶性scFv融合タンパク質のみが、リポーター遺伝子発現(たとえば、β−ガラクトシダーゼ)を活性化できる。
【0062】
Gal11の野生型(wt)対立遺伝子は、Gal4二量体化ドメイン(58−97残基)とは相互作用しない。したがって、いかなる単鎖のGal11wt対立遺伝子との融合体もレポーター遺伝子を活性化できないので、ネガティブコントロールとして働く。これは、Gal11wt−Gal4AD−λグラフト融合構築物を用いて実証された(図1B参照)。
【0063】
ベイト(bait)(LexA−Gal4(58−97))のみおよびscFv融合タンパク質のみのいずれも、レポーター遺伝子を活性化しなかった。
【0064】
この品質管理システムにおいては、試験した6つのscFvフラグメントのうち2つだけが、レポーター遺伝子発現を活性化させるのに十分に可溶性かつ安定性であった。フレームワークに安定化されたλグラフトおよびκグラフトは、最も安定な変異体である。この結果は分画分析と完全に一致しており、そこではλ−およびκグラフトのみが可溶性画分に検出された(図6参照)。
【0065】
酵母内で細胞質に発現されるscFvフラグメント
好適なscFvフラグメントは、たとえば、免疫したマウスから作製されたライブラリーからのリボソームディスプレイによって最初に得られた抗GCN4野生型scFvである(Hanes et al.,1998)。抗原は、7P14P(ジッパードメインの7および14の位置がプロリン残基に変異されていることを示唆する)と呼ばれるGcn4pロイシンジッパーの二重プロリン変異体であり、これは溶液中でランダムコイルを形成する(Leder et al.,1995)。scFvフラグメントは、ランダムコイル高次構造内でモノマーとして野生型ペプチドにも結合するので、インビトロにおいて野生型Gcn4pコイルドコイルペプチドの二量体化を阻止する(Berger et al.,1999)。本発明と関連して「野生型」と呼ばれる抗GCN4 scFvフラグメントは、ロイシンジッパーペプチドからの解離定数が4×10−11Mであることが測定されている(Hanes et al.,1998)。
【0066】
本発明の範囲内において、このscFvの多くの異なる変異体が調査された。抗GCN4野生型に加えて、H−R66K変異を保有する不安定化された抗GCN4野生型の変異体[抗GCN4(H−R66K)と名付けた]は、本質的に同じ抗原結合特性を有するがインビトロにおける安定性がわずかに減少したGcn4p結合scFvフラグメントの例として働いた(下記参照)。H−66の位置(Kabat et al.,1991 に従って番号付けした)にあるアルギニン残基(Arg)は、抗原結合ポケットから離れており、一般にAsp H−86に対して二重水素結合を形成する。H−66の位置にあるArgは以前に、レバン結合A48scFvフラグメント内のLysよりも高いタンパク質安定性を導くことが示された(Proba et al.,1998; Woern and Plueckthun, 1998a)。さらに、抗GCN4 scFvフラグメントのVal−Ala変異体[抗GCN4(SS――)と名付けた]を調べたところ、いずれのドメイン内ジスルフィドもVal−Alaペア(L−C23V、L−C88A、H−C22V、H−C92A)によって置換された。これらの変異は以前に、p185HER2結合4D5 scFvフラグメントの還元ジチオール型と比較してわずかに安定して作用することが示されており、またイントラボディーの性能を向上させるであろうことが推測されている(Woern and Plueckthun,1998b)。
【0067】
抗GCN4 CDR(相補性決定領域)ループを別のフレームワークにグラフトすることにより(Jones et al., 1986)、さらに2つの変異体を設計した。アクセプターフレームワークとして、「ハイブリッド」と呼ばれるscFvを選択した(Woern and Plueckthun,1999)。このアクセプターフレームワークは4D5 scFvフラグメントのVLドメインとA48++(H2)scFvフラグメントのVHドメインとから構成されている。それは、一連の安定化ドメインから合理的に設計されており、変性剤誘導性の平衡変性(equilibrium unfolding)によって実証され、また発現率が高い(Woern and Plueckthun,1999)というように、その驚くべき安定性が目立っている。抗GCN4 scFv CDRおよび「ハイブリッド」scFvフレームワークを含む2つのCDR−グラフト変異体を、全遺伝子合成により調製した。抗GCN4野生型ループドナーはλ軽鎖を保有し、一方アクセプター「ハイブリッド」フレームワークはκ軽鎖を保有していたので、ループのグラフトは直接的でなかった。したがって、2つの異なる変異体を設計し、一方はさらなる「κ様」(κグラフトと名付けた)、もう一方はさらなる「λ様」(λグラフトと名付けた)とした。これら2つの変異体は、VH−VL界面領域内の7残基のみが異なり、互いに2つのドメインの配向を強力に影響しあう。アンピシリン結合scFvフラグメントAL5(A.Krebber et al., 未発表)は、Gcn4pに結合していないscFvフラグメントのネガティブコントロールとして働いた。
【0068】
抗GCN4 scFvイントラボディーはGcn4pのトランス活性化能を阻害する
抗GCN4 scFvについて初めに、GAL1およびADH−駆動(driven)プロモーターを含む複数の酵母ベクターから発現されるその生物活性を調べた。さらに、SV40ラージT抗原由来の核移行シグナル(NLS)を抗GCN4 scFvのN末端に融合させた。試験した組み合わせのうち、TRP1選択マーカーを含むpESBA−Act発現ベクター(実施例を参照)、および2μオリジン(origin)(データ示さず)を用いて、いかなるNLSをも伴わないアクチン1プロモーターから発現させた場合に、抗GCN4 scFvが最も強い生物学的効果を示した。このベクターを引き続き、全てのさらなる実験において使用した。
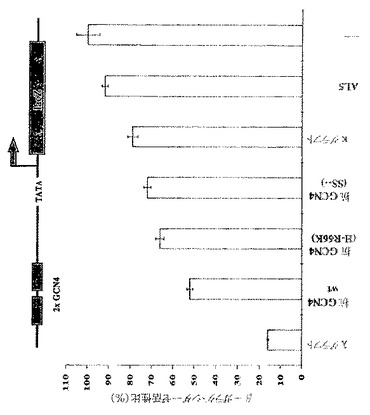
【0069】
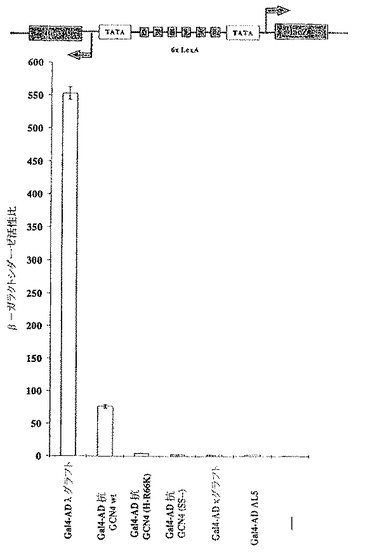
GCN4依存性LacZ発現において種々のscFvフラグメントを発現させるインビボ効果を図3Aに示す。レポーター構築物(YAdM2xGCN4−150)は、TATAボックスから相対的に−150の位置に2つのGcn4p結合サイトを含有し、酵母遺伝子に組み込まれた。内在性のGcn4pによって駆動されるβ−ガラクトシダーゼ活性比(Rel.β−gal.activity)を、任意に100%に設定した。AL5はアンピシリン結合scFvフラグメントであり、ネガティブコントロールとして働く。抗GCN4野生型(wt)に加えて、不安定化させた点変異体[抗GCN4(H−R66K)]、抗GCN4野生型のシステインフリー変異体[抗GCN4(SS――)]、および抗GCN4の2つのフレームワーク安定化された変異体(κグラフトおよびλグラフト)を試験した。安定化されたλグラフトは最も活性のあるイントラボディーであり、一方、抗GCN4の不安定化されたH−R66K点変異体およびシステインフリー変異体は、抗GCN4野生型と比較すると、活性が減少したことが示された。κグラフトの活性の減少は、その低い親和性のためであると考えられる(表1参照)。不安定化された点変異体である抗GCN4(H−R66K)は、野生型scFvと比較して、GCN4依存性レポーター遺伝子活性の阻害において効果的でなかった。Gcn4pトランス活性化阻害のパターンは再現性が高く、別の分析方法を用いた場合でも確認された。そこでは、溶解させるためにガラスビーズまたは凍結融解サイクルにより細胞を分裂させ、タンパク質濃度に対するβ−ガラクトシダーゼ活性を基準化させた後、β−ガラクトシダーゼレポーター活性を測定した(Escher and Schaffner,1997)(データ示さず)。
【0070】
【表1】

Gal4 AD−scFv融合タンパク質は、ツーハイブリッドアッセイにおいて、インビトロでの安定性およびインビボにおける性能に従って振舞う
レポーター遺伝子としてのLacZの発現をモニターするツーハイブリッドアッセイにおける、抗原と相補性決定領域(CDR)との間の好結果の相互作用を図3Bに示す。レポーター株YDE173を使用した。YDE173株は、2000年2月11日に、ドイツ、ブラウンシュヴァイク所在のDeutsche Summlung von Mikroorganismen und Zellkulturen DSZMに、番号DSM13333として寄託された。YDE173は、ゲノムのhis3遺伝子座に、互いに異なる方向に向き付けられたレポーター遺伝子HIS3およびLacZを制御する6つのLexA結合サイトを含有するレポータープラスミドpDE200が組み込まれている、酵母株JPY5(Matα ura3−52 his3Δ200 leu2Δ1 trp1Δ63 lys2Δ385)に由来するものである。
【0071】
図3Aにおいて使用したものと同じであるが、Gal4の転写活性化ドメインに融合されたscFvフラグメントを、ツーハイブリッドアッセイのベイトとして働く、C末端をLexAに融合させたGCN4ロイシンジッパー(aa245−285)と共発現させた。非特異的なAL5制御scFv融合構築物はLexA−GCN4ロイシンジッパーと相互作用できず、したがってLacZレポーター遺伝子を活性化しなかった。フレームワーク安定化されたλグラフト変異体に融合されたGal4活性化ドメインは、活性化イントラボディーとして最も強い効果を示し、次は抗GCN4野生型、および不安定化点変異体抗GCN4(H−R66K)であった。対照的に、安定性は高いが結合の弱いκグラフトおよびシステインフリー抗GCN4(SS――)は、ツーハイブリッドの型式においては明らかなレポーター遺伝子の発現を生じなかった。X−Galプレートアッセイにおいても同じ結果が得られた(データ示さず)。要約すると、種々のGal4 AD−scFv融合変異体の、ツーハイブリッド型式におけるLacZレポーター遺伝子の活性化についてのインビボにおける性能は、Gcn4p依存性LacZ発現の阻害パターンと相互に関連する(図3Aを3Bと比較せよ)。
【0072】
抗原と転写活性化ドメインに融合された種々のscFvとの間の相互作用は、ツーハイブリッドアッセイにおいて増殖選択を可能にする
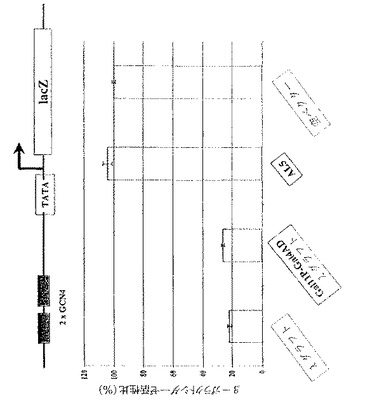
組み込まれたレポーター構築物はLacZレポーター遺伝子だけでなくHIS3遺伝子をも含有するので、ヒスチジンを欠如することはプレート上での増殖選択にとって好ましい。さらに、HIS3遺伝子産物の競合的な阻害剤である種々の濃度の3−アミノトリアゾール(3−AT)を添加することにより、ベイト/抗原とGal4 AD−scFv間の相互作用の強さに依存する酵母細胞の増殖を阻害(抑制)することが可能である。
【0073】
図4に示される結果を導いた実験手順は次のようであった。LexAに融合させたGCN4ロイシンジッパー(aa245−285)とGal4−AD scFv融合タンパク質とを共発現する約10,000の酵母細胞から始めて系列5倍希釈したものを、種々の濃度の3−ATを含むドロップアウト(drop out)プレート(−Trp/−Leu/−His)上にスポットした。48時間後、72時間後、および120時間後に増殖をモニターした。
【0074】
図4におけるレーンは次のようである。1.Gal4−AD λグラフト、2.Gal4−AD AL5、3.Gal4−AD κグラフト、4.Gal4−AD 抗GCN4(SS――)、5.Gal4−AD抗GCN4野生型、6.Gal4−AD 抗GCN4(H−R66K)、7.ポジティブコントロールとして働くLexA−Gal11融合タンパク質、8.空ベクター。
【0075】
ベイト/抗原(lexA―GCN4ロイシンジッパー)をGal4 AD−scFv融合体と共に発現する酵母株の増殖を5日間に渡ってモニターした。プレート上のコントロールは3−ATを含まないので、種々のGal4 AD−scFv融合変異体の明らかな増殖の違いは観察されなかった。ネガティブコントロールのscFv(Gal4 AD−AL5)で形質転換した細胞の増殖を抑制するためには、20mMの3−ATで十分であった。β−ガラクトシダーゼの発現をモニターする結果との関連において、κグラフト変異体、抗GCN4(SS――)、および抗GCN4(H−R66K)とのGal4 AD融合体は、20mM3−ATの存在下において増殖を許容しなかった。抗GCN4野生型だけでなくλグラフト変異体を発現する細胞は、その間フレームワーク安定化されたλグラフトにとって明らかな利点によって、80mMまでの3−ATの存在下で5日間増殖できた。3−ATの濃度100mMは、Gal4 AD−抗GCN4野生型を発現する細胞の増殖を阻止するのに十分であった。5日後にのみ、最も濃縮されたスポット上にわずかに現れ、一方λグラフトGal4 AD−scFv融合変異体を発現する細胞は明らかに増殖した。
【0076】
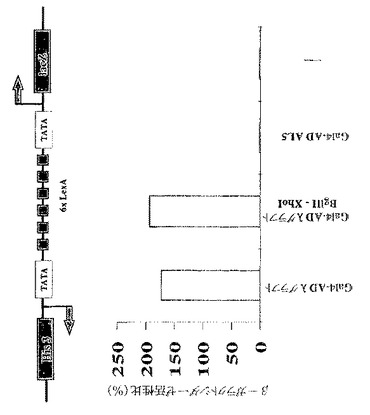
定常ドメインのλグラフトscFvへのN末端融合は、その生物活性を妨げない
Gal11P(263−352残基)およびGal4活性化ドメインを、λグラフトscFv(Gal11P−Gal4AD λグラフト)のN末端に融合させた。Gcn4p依存性遺伝子活性化を阻害するその生物活性は、λグラフトのみに匹敵する。図5Aに示すように、定常ドメインのscFvへの融合は、Gcn4p依存性遺伝子活性化に対する阻害活性を妨げなかった。
【0077】
特異的な制限酵素認識部位の導入
CDR3 VH(GLFDY)を無作為ペプチドライブラリーと交換するために、この超可変領域に隣接する2つの独特の制限酵素認識部位(BglIIおよびXhoI)を、沈黙突然変異誘発(silent mutagenesis)により導入した。これらの沈黙交換は、抗体のアミノ酸配列に影響せず、したがって、λグラフト変異体のインビボでの性能を変化させなかった(図5B参照)。
【0078】
抗原(GCN4ロイシンジッパー)の特異的認識に対するCDR3超可変領域(de Wildt et al., 1997; Hemminki et al.,1998)の重要性は、付加的なアラニンを可変重鎖のCDR3(AGLFDY)のN末端に導入することによって示された。このλグラフト+Ala変異体は、酵母株YAdM 2xGCN4−150内におけるGcn4p依存性レポーター遺伝子の発現を阻害できず、またYDE173株を用いたツーハイブリッド型式におけるレポーター遺伝子の発現の活性化もできなかった(データ示さず)。
【0079】
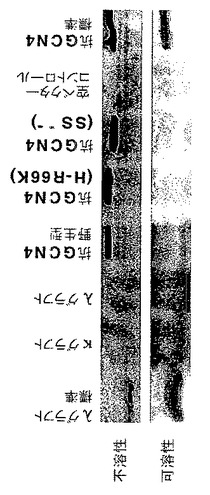
グラフト変異体は両方とも、酵母細胞質内で可溶性である
酵母内の種々のGcn4p結合scFvフラグメントの溶解性を、ウェスタンブロット解析により調べた。λおよびκグラフト変異体の場合のみ、明らかな量の可溶性タンパク質が細胞粗抽出液中に検出された(図6)。
【0080】
他の全ての抗GCN4scFvフラグメントは、インビトロにおける安定性が低下するにつれて不溶性scFvの量がわずかに増加したことから、基本的に完全に不溶性であることが明らかとなった。しかしながら、異なるscFv変異体にとって、可溶性タンパク質の不溶性タンパクに対する正確な割合が必ずしもインビボにおける割合を反映しているのではないことに注意しなければならない。種々の抗GCN4変異体の一部分は、たとえピアス社製のY−PERTM酵母タンパク質抽出剤を使用することにより穏やかな破壊方法を使用したとしても、試料調製の間に沈殿してしまうかもしれないことを排除することはできない。
【0081】
フレームワークの改善
好ましくは本発明に従った方法により単離されるフレームワーク内の変異は、還元環境において安定性かつ可溶性であるさらなるフレームワークを作製するために組み合わせることができる。そのようにして得られるフレームワークは、1つだけの変異を有するフレームワークに比べて、インビボにおける性能が増強される。6つの変異を組み合わせたフレームワークを、SEQ ID NO:1と規定する。
【0082】
実施例
CDR−グラフトされた抗GCN4 scFvフラグメントの設計
scFvフラグメントのクローニング、発現、および精製
全てのscFvフラグメントは、20−merのリンカー(GGGGSGGGGSGGGGSSGGGS)およびC末端のhis5−tagによってVL−VHに配向している。
【0083】
酵母内に発現させたscFvフラグメントを、pESBA−Act発現ベクターにクローン化した。pESBA−Actベクターは、酵母(Saccharomyces cerevisiae)−大腸菌(E.coli)のシャトルベクターである。それは、細菌の複製開始点とアンピシリン耐性遺伝子を含有する。さらにそれは、S.cerevisiae内での形質転換選択のための酵母TRP1遺伝子を含有する。それは酵母内で高度にタンパク質発現するように設計され、したがって、S.cerevisiae内で高コピー数を確実にする2μ複製開始点を有する。さらにそれは、NcoI(翻訳開始コドンATGをカバーしている)、ApaI、およびStuIに対する制限酵素認識部位、3つ全てのフレーム内に3つの翻訳停止コドン、およびSalIサイトを含有する多クローニングサイトで分離された、強力で恒常的なアクチンプロモーターおよびGAL11転写終結配列を含有する。
【0084】
全てのscFvフラグメントをBsp120IおよびStuI制限酵素認識部位を用いてクローン化し、C末端His5−tagを保有させた。Bsp120Iサイトをコードする2つのアミノ酸(Gly−Pro)は、開始のMet残基の後のN末端に含まれていなければならなかった。
【0085】
Gal4 ADのN末端の種々の抗体変異体への融合
Gal4活性化ドメインを、テンプレートとしてpGAD424(Clontech)を用いたポリメラーゼ連鎖反応法により増幅させた。両プライマー[上流プライマー:5’−CCATGGGCCCAAGCTTTGCAAAGATGGATAAAG−3’(Seq.Id.No.2)、下流プライマー:5’−TTTGGGCCCGAAGAACCGCCACCACCAGAACCGCCTCCACCAGAGCCACCACCACCAGGCCTGATCTCTTTTTTTGGGTTTGGTG−3’(Seq.Id.No.3)]は、pESBA Actと関連して、種々のscFvに対するSV40T−抗原核局在化シグナルN末端を含むGal4活性化ドメイン(AD)ポリペプチドのクローニングに好適なApaIサイトを含有する。活性化ドメインと単鎖抗体とは、下流プライマーによってコードされる(GGGS)3リンカーによって分離される。
【0086】
Gal11wtおよびGal11Pの、Gal4活性化ドメイン(AD)−scFv融合体へのN末端融合
Gal11wtおよびGal11pはいすれも、以下のプライマーを用いて増幅させた。上流プライマー:5’−CATGCCATGGTTCCTCAACAGCAGCAAATGCAAC−3’(Seq.Id.No.4)、下流プライマー:5’−CATGCCATGGCGCTAGCCAAAGCTTGGATTTTTCTCAGG−3’(Seq.Id.No.5)であり、いずれもNcoIサイトを含有している。アミノ酸263−352をコードしているPCR産物を、pESBA−Act2 Gal4(AD)−scFv融合構築物(上述した)のNcoIサイトに挿入した。これは、それぞれのGal11対立遺伝子の、Gal4(AD)−scFvとのインフレーム融合体を産生した。Gal11インサートの正確な配向性を、独特な酵素NheIで消化することにより確認した。
【0087】
LexA融合
GCN4ロイシンジッパー(aa245−285)を、LexA(aa1−202)の下流をクローニングするのに便利なEcoRIサイトを含有するプライマーを用いてPCR増幅させた。これを、ADHプロモーターの制御下で融合タンパク質を発現する、LEU2選択マーカーを含むArs CenプラスミドpAdM018とする。
【0088】
VHのCDR3に隣接するBglIIおよびXhoIサイトの導入
可変重鎖のCDR3の交換を容易にするために、部位特異的変異誘発によって、Gal4 AD−λグラフトscFvの初めの構造を変えることなく、CDR3 VHに隣接させて2つの独特な制限酵素認識部位を導入した。これらの沈黙点変異を、テンプレートとしてλグラフトを用いたPCRにより導入した。第1ラウンドにおいては、2つの別々のPCR反応を、2つの重複したPCR産物を導くプライマーである、#2421と#2487、および#2486と#2488を用いて行った。これらの2つの産物は、SpeIおよびSalIサイトを含有する外側のプライマー、#2421および#2488を用いたPCRの第2ラウンド用のテンプレートとして働く。最終産物を、SpeIおよびSalIを用いてGal4 AD−λグラフトにサブクローニングした。
【0089】
抗原と反応する新規なCDRのための、直接的細胞内スクリーニング
フレームワークで安定化された、Gal4活性化ドメインに融合させたλグラフトscFv(λグラフトscFv−Gal4 AD)の可変重鎖からのCDR3の初めの3アミノ酸(GLF)は、ロイター(Reiter)らによって記載された方法に基づいたPCRによって無作為化させた。CDR3の最後の2残基(DおよびY)は、その保存および構造上の重要性のために(Chothia and Lesk,1987)、無作為化させなかった。可変重鎖の8000の異なるCDR3変異体を潜在的にコードするλグラフトscFv−Gal4 ADライブラリーを得た。無作為に選んだ6つのライブラリークローンの配列解析により、予想した位置に無作為なCDR3配列の存在が明らかとなった。
【0090】
6つのLexA結合部位(上記参照)の制御下においてHIS3およびLacZレポーター遺伝子を含有する酵母株YDE173を、LexAおよびライブラリーに融合されたGCNロイシンジッパー(aa245−285)を発現するベクターに同時形質転換させ、増殖選択のために60mMの3−ATを含有する選択用ドロップアウトプレート(−Trp/−Leu/−His)上に蒔いて培養した。もし、好適なCDR3配列を含むCDR3ライブラリーからのscFvフラグメントが、LexAに融合されたロイシンジッパー抗原に結合すれば、HIS3レポーター遺伝子の転写を活性化させ、酵母細胞のヒスチジン依存性の増殖を回復させる複合体が形成される。3日後、増殖しているコロニーを選び取り、同じ選択用ドロップアウトプレートに再び蒔いた。2次選択後でもまた増殖する細胞の、X−galプレート上におけるβ−ガラクトシダーゼ活性を分析した。β−gal陽性クローンからライブラリープラスミドDNAを抽出し、可変重鎖のCDR3の領域をシークエンスした:本発明者らは本来のλグラフトCDR3アミノ酸配列を4回見出し、GCN4ロイシンジッパーに特異的な3つの完全に新しいCDR3配列を見出した。本来のCDR3の配列を含有する4つの同定されたscFvフラグメントはλグラフトとして識別不能に振舞い、一方変化したCDR3配列を含む3つのクローンは、LacZレポーター遺伝子の活性化において効果が弱かった。
【0091】
これらの結果は、還元条件下において安定性かつ可溶性である規定されたscFvフレームワーク内に包埋された新規なCDRの、直接的細胞内スクリーニングの可能性を示す。
【0092】
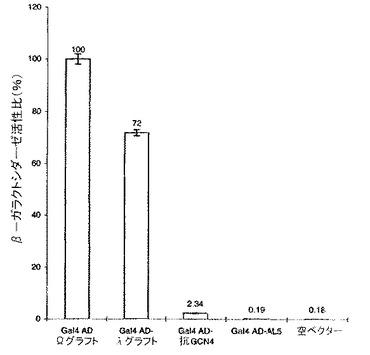
規定されたイントラボディーのインビボにおける性能は、無作為突然変異誘発により最適化され得る
フレームワークで安定化されたλグラフト変異体を、イントラボディーのフレームワークに沿って統計的にアミノ酸変化を導入するために、Sambrook et al.によって説明されるようにPCRにより無作為に突然変異誘発させた。酵母株YDE173を、このGal4の活性化ドメインに融合させて無作為突然変異誘発させたscFvライブラリー、およびLexAに融合させた特異的な抗原(GCN4ロイシンジッパーのaa245−258)を発現するプラスミドで同時形質転換し、80mMの3ATを含有するドロップアウトプレート上で培養した。6つの候補となるクローンを選択し、それぞれはフレームワーク内に1つの単独のアミノ酸変化を有していた。これら6つの変異体フレームワークは全て、λグラフト変異体と比べて向上したインビボにおける性能を示し、これはβ−ガラクトシダーゼ活性の測定により確認され、定量化された。イントラボディーの性能を向上させる種々のアミノ酸変化は相加的に振る舞うという仮定をもとに、本発明者らは、Gal4活性化ドメインに融合させた1つのフレームワーク内の6つ全ての変異を組み合わせ、LacZレポーター遺伝子の活性化について、フレームワークで安定化させたλグラフト変異体と比較した。図2は、6つの点変異を全て含む混合性のこの新規なフレームワーク(Ωグラフト)は、本来のλグラフト変異体と比較してインビボにおける性能がほぼ30%優れていることを示すことを示す。驚くべきことに、これら6アミノ酸の置換はクラスター形成されており、そのうち2つ(E→KおよびL→R)は可変軽鎖のCDR1の上流に存在し、残りの4つ(N→D、G→C、K→E、T→S)は可変重鎖のCDR2とCDR3の間に位置する。
【0093】
レポーター遺伝子のSaccaromyces cerevisiaeの染色体への組み込み
組み込み用レポータープラスミドpAB183は、GAL1プロモーターのTATAボックスの上流150の位置で2つのGcn4p結合サイトをクローニングした、pJP161(Barberis et al.,1995)に由来する。Gcn4p結合サイトは、5’SphIおよび3’SalI適合性のオーバーハング配列を有する2つの相補的なオリゴヌクレオチドをアニーリングすることにより発生させた。このオリゴヌクレオチドは次のようである:5’−CCTATGACTCATCCAGTTATGACTCATCG−3’(Seq.Id.No.6);5’−TCGACGATGAGTCATAACTGGAT GAGTCATAGGCATG−3’(Seq.Id.No.7)。このレポータープラスミドをApaIサイトで直線化し、JPY5株(Barberis et al.,1995)の酵母ゲノム遺伝子座ura3へ組み込み、YAdM2xGCN4−150とした。YAdM2xGCN4−150株は、2000年2月11日に、ドイツ、ブラウンシュヴァイク所在のDeutsche Summlung von Mikroorganismen und Zellkulturen GmbH DSZMに、番号DSM13332として寄託された。4つの独立した酵母形質転換を機能解析において試験し、全てが同じGCN4−依存性のレポーター遺伝子活性を示した。クローンのうち1つ(YAdM2xGCN4−150)を後の実験のために選択し、これを酵母野生型と呼ぶ。
【0094】
ツーハイブリッド実験に用いたレポーター株は、LacZおよびHIS3の発現を駆動する6つのLexA結合サイトを含む双方向性プロモーターを含有する、組み込まれたレポーター構築物を有する。
【0095】
酵母細胞の系列希釈およびスポッティング
酵母細胞を、次の標準的な手順に従った酢酸リチウム法を用いて形質転換した。形質転換体をドロップアウト培地(−Trp/−Leu)中で一晩30℃で培養した。飽和になった培養液を、ドロップアウト培地でOD600=0.7になるまで希釈し、少なくとも2倍の時間再びインキュベートした。各培養液を、およその濃度が106cells/mlから始めて、水で系列希釈し(希釈係数(dilution factor)5)、各希釈溶液から10μlずつ、0mM、20mM、40mM、60mM、80mM、または100mMの3−アミノトリアゾールを含むドロップアウトプレート(−Trp/−Leu/−His)上にスポットした。各形質転換体の6つの異なる希釈溶液をドロップアウトプレート上にスポットした。プレートを30℃でインキュベートし、48時間、72時間、および120時間後に調査した。
【0096】
scFvフラグメントのインビボにおける分析:酵母内におけるscFvフラグメントの発現およびβ−ガラクトシダーゼレポーターアッセイ
溶液中のβ−ガラクトシダーゼアッセイを、記述されているように(Kaiser et al., 1994, Escher and Schaffner 1997)透過化処理した細胞を用いて行った。活性は、アッセイした細胞数に対して規準化した。
【0097】
抗GCN4 scFvフラグメントのウェスタンブロット解析
種々の抗GCN4scFvフラグメントの溶解性をウェスタンブロットにより解析した。5mlの培養液を30℃で光学密度が約2〜3になるまで培養した。細胞を同じ細胞密度になるように規準化し、沈殿させ、可溶性タンパク質のやさしい単離を促進させる穏やかな界面活性剤調合物であるピアス社製のY−PERTM酵母タンパク質抽出剤で全細胞タンパク質を抽出した。可溶性および不溶性の画分を遠心分離(13000rpm、10分、4℃)により分離した。可溶性および不溶性の粗抽出物のサンプルを標準的な手順に従ってSDS−PAGEに供し、PVDF膜にブロットした。His5−tagの付いたscFvフラグメントを、抗His5scFv−AP融合体を用いた既知の方法(Lindner et al.,1997)に従って、ベーリンガーマンハイム社製の化学発光ホスファターゼ基質(chemoluminescent phosphatase substrate)CSPDを用いて検出した。ウェスタンブロットにおいて適切な強度を得るために、可溶性画分においては不溶性画分において用いるよりも約5倍高いタンパク質濃度を用いなければならず、またブロット時間も異なった。したがって、直接的な比較は、それぞれ全て可溶性の試料の間、または全て不溶性の試料の間でのみ意味のあることである。
【0098】
本発明の好ましい実施形態が現在示され、記載されているが、本発明はそれに限定されず、請求項の範囲内において他の種々の具体化および実施が可能であることは明瞭に理解されるべきである。
【0099】
【表2】
[配列表]
【0100】
【表3】
【技術分野】
【0001】
(関連出願に対する相互参照)
本出願は、その全体が参照によりここに組み込まれる1999年12月28日に出願されたPCT特許出願IB99/02054、および2000年3月1日に出願されたPCT特許出願IB00/00218の優先権を主張する。
【0002】
(技術分野)
本発明は、抗体の重鎖および軽鎖の可変領域の単鎖融合体(scFv)に関し、特に規定された安定なフレームワークを有しかつ細胞内で発現されたかかるscFv(イントラボディー)に関する。
【背景技術】
【0003】
抗体は、その抗原に対する高い親和性および特異性、およびそのインビトロおよびインビボにおける比較的高い安定性のために、生化学および分子生物学研究、診断および医学用途のための好ましい道具である。抗体は2つの重鎖および2つの軽鎖からなり、そのN末端に可変領域を含み、またそれらはジスルフィド架橋により連結されている。単鎖抗体は、重鎖可変領域および軽鎖可変領域のフラグメントを連結することにより設計されている(scFv)。各可変領域は、フレームワークに包埋された3つの相補性決定部位(CDR)を含む。これらのCDRは、抗原との相互作用の原因である。重鎖可変領域および軽鎖可変領域は領域内ジスルフィド架橋を含み、これは単鎖抗体の安定性に重要であることが報告された(Biocca et al.,1995; Derman et al., 1993)。
【0004】
特異的なエピトープに結合する単鎖抗体を同定するために最も一般的に使用される技術は、ファージディスプレイおよびその変形によるものである(概説はHoogenboom et al.,1998を参照されたい)。このスクリーニング方法は、免疫感作またはハイブリドーマ技術のような一般的な技術に対する主要な利点を有する。つまり、比較的短時間でモノクローナル単鎖抗体を取り出すことができる。
【0005】
細胞(たとえば細胞質または核)内で発現される単鎖抗体は、イントラボディー(intrabodies)と呼ばれる。細胞内は還元環境であるために、抗体の安定性に重要であると信じられているジスルフィド架橋が形成されない。従って、初めは、イントラボディーの適用は好ましくないと信じられていた。しかしながら、イントラボディーの適用の可能性を示すいくつかのケースが開示されている(Beerli et al.,1994; Biocca et al., 1994; Duan et al., 1994; Gargano and Cattaneo, 1997;Greeman et al., 1996; Martineau et al. 1998; Mhashilkar et al., 1995;Tavladoraki et al., 1993)。これらのケースにおいては、イントラボディーはたとえば、細胞質内の抗原をブロッキングし、その生物活性を阻害することにより働く。
【0006】
現在まで、イントラボディーは大抵の場合、初めに古典的な技術(たとえばファージディスプレイ)で選別され、次にその細胞内でのイントラボディーとしての生物活性を試験される(Vinsintin et al.,1999)、モノクローナル抗体に由来するものであった。成功したイントラボディー(上述を参照)が記述されているにもかかわらず、今日では、そのようなイントラボディーが細胞内で機能的であるかどうかは全く予測不可能である(概説は、Cattaneo,1998; Cattaneo and Biocca, 1999を参照されたい)。その原因はおそらく、異なる環境である。ファージディスプレイおよび他の古典的な技術は酸化条件下で行われ、従ってジスルフィド架橋が形成されるが、一方イントラボディーは還元条件下で機能しなければならない。この還元環境はイントラボディーの不十分な溶解性を導く可能性があり、従ってそれらは非機能的な凝集塊を形成する。イントラボディーの溶解性は、フレームワーク内(Knappik and Pluckthun,1995)またはCDR内(Kipriyanov et al., 1997; Ulrich et al., 1995)のいずれかにおける変化によって調節できる。
【0007】
しかしながら、従来知られている方法は、その用途が細胞内の標的を検出することに限定されている。従って、ある抗原に特異的なイントラボディーを直接的にスクリーニングする信頼性のある技術および方法の必要性が増している。
【0008】
WO99/36569においてウィットラップ(Wittrup)らは、酵母内在性の、細胞壁上での局在化のためのAga2pに由来するタンパク質断片を用いて、タンパク質およびscFvを酵母の細胞壁上に提示する方法を記載している。他の関連する方法は、EP0407259(Boquet et al.,1991)において記載されている。これらの方法は、タンパク質またはペプチドライブラリーもまた表面上に提示されるファージディスプレイスクリーニングに匹敵する。しかしながら、これらの技術はイントラボディーを同定するための細胞内スクリーニングには使用できない。
【0009】
特許明細書JP11000174(Kyoko et al., 1999)において、抗体のFabフラグメントを高レベルに発現および分泌させるための、酵母ピキアパストリス(Pichia pastoris)の使用が記載されている。この酵母は、その分泌レベルが高いために有名であり、したがってこの用途に好適に使用される。分泌された抗体は、上清を精製することにより収集することができる。さらに、EP0590067、WO92/22324、JP06030778、US5698435、US5595889、JP10313876において、酵母は分泌されるタンパク質または抗体の製造に使用されている。EP0698097およびWO94/25591は、さらなる用途のための、重鎖またはその断片のみの製造および分泌の用途を開示している。JP09020798;JP05105700;およびJP05097704には、一般に人体または生物体に投与した場合の、肝炎ワクチンを得るための酵母分泌の方法が記載されている。
【0010】
単鎖抗体のスクリーニングに酵母を使用することも、WO99/28502によってすでに知られている。前記出願は、単鎖モノクローナル抗体融合試薬用のDNA構築物ライブラリーの使用を開示する。このscFvライブラリー(ここではsFvライブラリーと呼ぶ)は、次にスクリーニングに使用される。しかしながら、規定されていないフレームワークを使用しているために、イントラボディーの安定性と可溶性は劇的に変化する可能性があることが見出されている。さらに、インビボにおける性能とインビトロにおける安定性および可溶性との間に直接的な相関関係が示された。したがって、CDRは原理的には抗原に対して必要とされる高い親和性を示すにもかかわらず、対応するフレームワークが十分に可溶性でないために凝集し、このモノクローナルscFvは検出不可能になるので、種々のscFvフラグメントのmRNA由来ライブラリーの使用は、その抗原に対する高い親和性を有するCDRを同定する可能性があるという観点に限定される。したがって、未だ改善された抗体、またはイントラボディーがそれぞれ必要とされている。
【0011】
細胞内の標的に向けられるscFvの用途は増大しており、信頼性のあるイントラボディーのスクリーニング方法の必要性が生じている。還元条件下においてscFvが不安定であり、かつ抗体安定性が予測できないために、scFvの細胞質内標的は最も困難な用途である。この安定性と可溶性の問題も、細胞内適用のために最適化された、規定されたフレームワークを使用することにより解決され得る。
【発明の概要】
【課題を解決するための手段】
【0012】
したがって、還元環境において安定性かつ可溶性である規定されたフレームワークを有するscFvまたはイントラボディーを単離する方法を提供することが、本発明の全般的な目的である。
【0013】
本発明のさらなる目的は、還元環境において安定性かつ可溶性である規定されたフレームワークを有するscFvまたはイントラボディーである。
【0014】
本発明の他の目的は、CDR/フレームワーク連結領域内に独特な制限酵素認識部位を備えるように修飾された、還元環境において安定性かつ可溶性である規定されたフレームワークを有するscFvまたはイントラボディーである。
【0015】
本発明の他の目的は、還元環境において安定性かつ可溶性である規定されたフレームワークと、無作為にまたは確定的に変形されたCDRとを有するscFvまたはイントラボディーのライブラリーである。
【0016】
本発明の他の目的は、還元環境において安定性かつ可溶性である規定されたフレームワークと種々のCDRとを有するscFvまたはイントラボディー、またはこのようなscFvまたはイントラボディーのライブラリーを用いた、抗原結合性CDRをスクリーニングする方法である。
【0017】
本発明の他の目的は、このようなscFvまたはイントラボディーまたはライブラリーをそれぞれ用いた、さらなる抗原のスクリーニング方法である。
【0018】
本発明の他の目的は、還元条件下において可溶性かつ安定性であるフレームワークを有するイントラボディーの同定方法である。
【0019】
本発明のイントラボディーはさらに、疾病の治療、診断、または予防における薬剤、および特異的なタンパク質活性の機能的ノックアウトのような移植における種々の用途の薬剤として使用され得る。イントラボディーはそれ自体として、またはこのようなscFvをコードするDNAとして使用され得る。
【0020】
本明細書の範囲内において、術語scFvおよびイントラボディーは大部分は同義語として使用されるが、たとえば細胞内のような還元環境における本発明の規定されたフレームワークを有するイントラボディー(scFv)の安定性および可溶性は本発明にとって必要であるが、そのようなイントラボディー(scFv)等の用途は、細胞内での用途に制限されるものではないことは理解されなければならない。
【0021】
CDR内にアミノ酸変化を導入することのみによって、本発明に従ったフレームワークが特異的な抗原認識という所望の生物学的機能を示すモノクローナル抗体を識別する可能性が大いに増加する。scFvのCDRにおけるそのような変化は、イントラボディーの細胞質内適用に適する規定されたフレームワークを変化させずに、無作為の変化として行うことができる。
【0022】
細胞内でモノクローナル単鎖抗体のスクリーニングを行うためには、細胞質の酸化還元環境に適応するフレームワークを使用しなければならない。したがって、フレームワークはジスルフィド架橋の不在下であっても十分に安定性かつ可溶性でなければならない。しかしながらscFvの多くは、還元条件下または、ドメイン内ジスルフィド架橋の形成に必要であるシステインの不在下においては、適切な構造に折りたたまれないことが知られている。したがって、本発明の範囲内において、同一のCDRを含有する複数のフレームワークを比較したところ、インビボでの性能における劇的な差異が観察された。本発明の方法によって、抗原認識のための規定されたCDRを含有する最も性能の良いフレームワークが選択できる。この方法は、出発材料として既知の抗原に対するイントラボディーを用いて行われる。重鎖および軽鎖の可変領域を連結するのに用いるリンカーは重要ではない。しかしながらそれは、CDRと抗原との間の相互作用のために好適な接触およびフォールディングを確実にするために、十分な溶解性および柔軟性を提供しなければならない。好適なリンカーは、約5〜60アミノ酸の典型的な長さを有し、通常の規則的なグリシンの配列、および溶解性を増強するための1〜3個のセリンを有する。
【0023】
還元環境において安定性かつ可溶性である規定されたフレームワークを有するscFvを単離するための本発明の方法は、以下の工程により規定される。
【0024】
a)既知の抗原に対するscFvのDNA配列の少なくとも1つのフレームワークコード領域を突然変異させ、そのような突然変異を好適な発現ベクターに導入することにより、種々のフレームワークおよび定常のCDRを有するscFvライブラリーを作製し、
b)特異的な既知の抗原を発現でき、かつ抗原−scFv相互作用の存在下でのみ生存する宿主細胞を、前記scFvライブラリーで形質転換し、
c)そのようにして形質転換した宿主細胞を、抗原およびscFvを発現するのに好適な条件下で培養して、抗原−scFv相互作用の存在下においてのみ細胞を生存させ、
d)生存している細胞内で発現され、また還元環境において安定性かつ可溶性である規定されたフレームワークを有するscFvを単離する。
【0025】
好ましい実施形態において、宿主細胞は真核細胞、特に酵母細胞である。
【0026】
上述した方法によって、規定したフレームワークを有するscFvを入手できる。そのようなフレームワークも本発明の目的である。そのようなフレームワークは、少なくとも1つのCDRの選択的な交換を可能にする特異的な制限酵素認識部位を含むように修飾できる。好ましくは前記制限酵素認識部位は、CDRに隣接するフレームワーク内に位置する。
【0027】
本発明はさらに、CDR領域内における選択的な変更に好適なフレームワークを有するscFvをコードするDNAの産生方法であって、特異的な制限酵素認識部位を、規定された安定性かつ可溶性であるscFvをコードするDNAの配列に部位特異的変異誘発によって導入し、それによって制限酵素認識部位が好ましくはフレームワーク内に位置し、かつそれによって制限酵素認識部位を作製するためのヌクレオチドの置換がアミノ酸配列に影響しない方法を提供する。
【0028】
還元環境において安定性かつ可溶性である規定されたフレームワークを含む改善されたscFvは、本発明の目的でもある方法によっても得ることができ、ここでは還元環境において安定性かつ可溶性である少なくとも2つの異なるフレームワーク(好ましくは本発明のフレームワーク)の少なくとも2つの変異を組合せて、規定されたフレームワークを有するscFvを製造する。
【0029】
上述した方法によって入手できるscFvも本発明の目的である。そのようなフレームワーク内においては、少なくとも1つの変異が可変軽鎖のCDR1の上流に存在し、かつ/または少なくとも1つの変異が可変重鎖のCDR2およびCDR3の間に位置することが好ましい。
【0030】
非常に好ましい実施形態においては、本発明のscFvは、可変軽鎖のCDR1の上流に存在する少なくとも2つの変異、および可変重鎖のCDR2およびCDR3の間に位置する少なくとも2つ、好ましくは少なくとも4つの変異を含む。特に、SEQ ID NO 1に規定されるフレームワークを含むscFvが好ましい。
【0031】
そのようなフレームワークにおいてCDRを特異的に無作為化するために、独特な制限酵素認識部位の発生を導く沈黙突然変異(silent changes)(同じアミノ酸配列をコードするが、異なるコドンを使用する)を導入できる(上記も参照)。制限酵素認識部位はCDR/フレームワーク連結領域内のどこにでも設置できるが、それは独立した各CDRに隣接するフレームワーク内に位置することが好ましい。これによって、独立した各CDRは、無作為のまたは規定された配列の導入により置換され得る。これは、抗原に対して高い親和性を示すイントラボディー内の新規なCDRを選択することを可能にする。
【0032】
局在化シグナルまたは活性化ドメインのような付加的な配列をscFvライブラリーに由来する非限定的なフレームワークに導入する場合、この修飾によって、たとえばscFvが不溶性になるといったように、生物学的活性が(たとえそれまで存在していても)失われ得る。したがって、既知の抗原に対する本発明の規定されたフレームワークを使用し、次にそのような修飾をイントラボディー内の種々の位置(NおよびC末端またはscFvのコード配列内)に導入し、また本来の機能を維持するために選択することは、利点である。WO99/28502には、局在化シグナルを導入するいくつかの可能性が記載されている。ある抗原に対する相互作用スクリーニングに使用される活性化ドメインは、WO99/98502において記載されており、scFvライブラリーのC末端に導入される。種々の位置、たとえばN末端の活性化ドメインにおいて配列の付加を許容するフレームワークも本発明の方法によって選択することができ、それは拮抗的な機能における活性化ドメインを持たない、それらのscFvに対応する物と同様に振る舞うことが今般見出された。従って、たとえば以下の実施例においてさらに説明されるフレームワーク内においては、N末端の活性化ドメインを導入することは抗体の機能を阻害しない。
【0033】
還元環境において安定性かつ可溶性である規定されたフレームワークを有する本発明のイントラボディーから出発して、CDRライブラリーを含有するscFvまたはイントラボディーをそれぞれ作製できる。
【0034】
還元環境において安定性かつ可溶性である規定されたフレームワークを有するCDRライブラリーを作製するための好適な方法は本発明の方法であり、配列につき少なくとも1つのCDRを修飾されたCDRで置換するために、本発明のscFvをコードしているDNA配列を消化させる。好ましくは、修飾されたCDRは無作為変異により作製する。そのような方法により、少なくとも1つの無作為化されたCDR、および還元条件下において安定性かつ可溶性である規定されたフレームワークを有するイントラボディーのライブラリーを作製できる。
【0035】
CDRライブラリーを含有する本発明のイントラボディーは、抗原に対して高い親和性を有するクローンのスクリーニングおよび選択に使用され得る。特定の抗原と相互作用するCDRのスクリーニング方法も本発明の目的であり、またこの方法は、既知の抗原をコードする核酸配列、特にDNA配列で形質転換し、さらに還元環境下において安定性かつ可溶性である規定されたフレームワークを有する無作為化されたCDRライブラリーで形質転換した宿主細胞を含み、それによって抗原および/またはscFvはマーカーシステムまたはマーカーシステムの一部に連結され、したがって選択的条件下で培養された細胞は抗原/scFv相互作用の存在下においてのみ生存するので、形質転換された細胞を選択的な条件下で培養して残存する細胞を培養し、イントラボディーを収集することを含む。
【0036】
本発明の好ましい実施形態においては、このフレームワークは本発明のフレームワークであり、細胞は真核細胞、特に酵母細胞である。
【0037】
本発明のさらに好ましい実施形態において、抗原をコードするDNA配列およびscFvをコードするDNA配列はいずれも、生存許容(allowing)マーカーに連結された転写活性化システムの一部にいずれも連結された、抗原またはscFvをそれぞれ含むキメラ分子をコードし、さらに好ましくは抗原はDNA結合ドメインに融合されてscFvは転写活性化因子ドメインに融合されるか、または抗原は転写活性化因子ドメインに融合されてscFvはDNA結合部位に融合される。
【0038】
CDRライブラリーを含有する本発明のイントラボディーは、抗原に対して高い親和性を有するクローンをスクリーニングおよび選択するのに使用され得る。これは、その生物学的機能において細胞内に位置する抗原をブロックすること、または規定されたフレームワーク内に包埋されたCDRの抗原に対する直接的な相互作用を分析することのいずれかにより達成できる。直接的な相互作用は、好ましくは、転写性の読み出し(readout)、好ましくはHIS3遺伝子の発現によってモニターできる。予め決定した条件下で3−アミノトリアゾール(3AT)を添加すると、抗原に対してより親和性の高いCDRの選択が可能になる。特異的な既知の抗原を発現し得る宿主細胞は、前記条件下では抗原−scFv相互作用の存在下、好ましくは十分に強い抗原−scFv相互作用の存在下においてのみ生存する。ここで用いられる術語「十分に強い」は、BIAcoreで測定されたKDが、>1×10−6M、好ましくはKD>1×10−8M、そしてより好ましくはKD>1×10−10Mであるタンパク質−タンパク質相互作用として定義される。そのような選択工程はさらに、CDR(好ましくは軽鎖および重鎖のCDR1およびCDR2)内のアミノ酸を無作為または選択的に変異させ、続いて上昇した3ATの濃度において増殖するものをこのプールから選出することによる親和性熟成を行うのに適用できる。
【0039】
既に上述したように、今までに知られており使用されているscFvライブラリーは、好ましくはB細胞が高度に蓄積されることが知られている脾臓からのmRNAの単離に由来し、再編成された抗体が発現される。そのようなライブラリーは、その生物体内に存在するエピトープに対して反応しないように予備選択(ポジティブおよびネガティブ選択)されているという欠点を有する。これは、自己免疫反応を開始しない抗体のみが熟成され、活性化され得ることを保証する。しかしながら、この選択工程のために、可能性のあるアミノ酸の組み合わせの全てが「天然の(natural)」scFvライブラリー内に存在するわけではない。いくつかのインビトロおよび診断上の用途のためには、抗体は種属間で保存されているタンパク質と相互作用することが必要とされる。そのようなタンパク質またはペプチドにとって、予備選択工程のために、「天然の」scFvライブラリー内で強力に相互作用するモノクローナル抗体を検出することは非常に困難な場合がある。さらに、その「天然の」ライブラリー内に存在するフレームワークは最適化されておらず、したがって不十分かまたは可変性の溶解性および/または安定性はそれぞれ問題を生じる。したがって、本発明のフレームワーク、および/または本発明の方法によって得られるフレームワークを含み、かつこれらの領域内のアミノ酸配列の可能な組み合わせのいくらか、または好ましくは全てをカバーするCDR無作為ライブラリーのみを使用することは大きな利点である。
【0040】
本発明をさらに説明するために、酵母細胞内転写因子Gcn4pに対する規定された相補性決定領域(CDR)を含む、安定性かつ可溶性であるイントラボディーフレームワークを選択した。この規定されたフレームワークは、無作為の配列によってそのCDRを置換するのに使用した。所望の生物活性(CDRのインビボ効果)を刺激する新規なCDRを同定するためにこれらのCDRライブラリーをスクリーニングする:
a)細胞内(たとえば、ヒト細胞内または他のいかなる細胞内でも)で自然に起こる分子相互作用を好適な細胞(好ましくは酵母)内で再構成させるか、または酵母内在性の相互作用を使用する。次のスクリーニングで、再構成されたかまたは内在性の分子の生物活性によるこれらのCDRの干渉によって、高親和性のCDRを同定する。そのような拮抗性のCDRはたとえば、シグナル伝達経路に含まれる2つのタンパク質をブロックすることにより機能できるであろう。
【0041】
b)再構成されたかまたは内在性の分子における、所望の生物活性を誘発するアゴニストのCDRを選択する。
【0042】
安定性であるフレームワーク内に包埋された無作為CDRをさらに、相互作用スクリーニングに基づいて、CDRと抗原との相互作用を同定するのに使用することができる:
a)選択されたフレームワークは転写活性化ドメインに融合可能であり、しかもその機能を保持することが示されるであろう。このキメライントラボディーを、DNA結合活性を有するDNA結合部位または転写因子に融合されて得られる抗原に対して高親和性であるCDRの選択に使用する。抗原とCDRが相互作用する時には、転写活性化ドメインは選択マーカー遺伝子の遺伝子発現を媒介し、選択条件下でのこの細胞の生存を許容する。
【0043】
b)ハイブリッド技術(一方のパートナーを活性化ドメインへ融合させ、もし必要であればもう一方をDNA結合部位へ融合させる)に基づいて再構成させた分子の相互作用は、特異的で高親和性のCDRによってブロックされ得る。
【0044】
イントラボディーのCDRは一定であるがフレームワーク内の種々の変異は、イントラボディーの安定性および溶解性を変化させることによりそのインビボにおける性能に影響を及ぼすことも明らかとなった。フレームワークは、イントラボディーの安定性および溶解性の主要な部分に寄与する。にもかかわらず、CDR内の特定の変異はイントラボディーの溶解性および安定性に影響する場合もある。したがって、機能的品質管理(下記参照)によって、規定されたフレームワーク内に包埋された無作為なCDRを予備選択することは都合が良いだろう。
【0045】
本発明はさらに、イントラボディーフレームワークまたはイントラボディーの同定方法であって、好適な宿主細胞をライブラリーおよびマーカーシステムにより形質転換し、それによって前記ライブラリーはイントラボディーライブラリーと前記マーカーシステムの少なくとも一部との融合産物となり、前記マーカーシステムは可溶性かつ安定性であるイントラボディーフレームワークをコードする融合タンパク質の存在下においてのみ活性化され、また前記細胞を、可溶性かつ安定性であるイントラボディーフレームワークを発現している細胞の同定および選択を可能にする条件下で培養する方法を提供する。
【0046】
本発明の好ましい実施形態において、前記ライブラリーはイントラボディーライブラリーとマーカータンパク質との融合産物である。好ましくは、前記マーカータンパク質は選択可能な活性、特に酵素活性または蛍光活性を有する。そのような方法において使用され得るマーカータンパク質は、たとえばGFPタンパク質またはそのいかなる変異体でもある。
【0047】
本発明の他の好ましい実施形態においては、前記ライブラリーは、イントラボディーライブラリーと、転写がそのDNA結合タンパク質により制御されるマーカー遺伝子の転写を活性化できるDNA結合タンパク質との融合産物である。好適なDNA結合タンパク質はたとえばp53である。
【0048】
本発明のさらなる好ましい実施形態においては、前記方法はイントラボディーおよびトランス活性化システムの一部を含むタンパク質をコードするライブラリーで形質転換された好適な宿主細胞を含み、かつ前記細胞はさらに少なくとも前記トランス活性化システムの第2部分を含む第2タンパク質を発現し、それによって前記トランス活性化システムは生存許容マーカーに連結されて前記細胞は前記2つの融合タンパク質間の相互作用の存在下における選択的条件下においてのみ生存する。
【0049】
さらに好ましい実施形態においては、前記ライブラリーにコードされるタンパク質は転写活性化ドメインを含みかつ前記第2タンパク質はDNA結合部位を含むか、または前記ライブラリーにコードされるタンパク質はDNA結合部位を含みかつ前記第2タンパク質は転写活性化ドメインを含む。
さらなる好ましい実施形態においては、前記第2タンパク質はDNA結合部位またはトランス活性化ドメインと、前記ライブラリーにコードされるタンパク質の定常領域と相互作用するタンパク質とを含む。ここで使用される術語「定常領域」は、ライブラリー構築物によってコードされ、タンパク質相互作用のパートナーとして働くいかなるタンパク質部位、またはいかなる近接するアミノ酸配列をも含み、また前記用語はたとえばイントラボディーまたはGal11pの一部を含む。
【0050】
上記方法により得ることが可能な、規定されたフレームワークを有するscFvも本発明の目的であり、特に本発明の方法において使用するための目的である。
【0051】
同方法は、たとえばscFvまたはCDRライブラリー、特に本発明のライブラリーの出発材料として使用され得る、可溶性かつ安定性であるフレームワークを同定するためのいかなるscFvライブラリーのスクリーニングにも適用され得る。
【0052】
本発明の他の目的は、scFvと相互作用する抗原のスクリーニング方法であって、少なくとも1つの興味のある抗原を発現する宿主細胞を、還元環境において安定性かつ可溶性である規定されたフレームワークを有する少なくとも1つのscFv、または還元環境において安定性かつ可溶性である規定されたフレームワークを有する無作為化されたCDRライブラリーで形質転換し、それによって抗原および/またはscFvをマーカーシステムまたはマーカーシステムの一部に連結させ、したがって選択的条件下で培養される細胞が抗原/scFv相互作用の存在下においてのみ生存し、このように形質転換された細胞を選択的条件下で培養し、残存する細胞を培養してscFvを収集する方法を提供することである。
【0053】
本発明の好ましい実施形態においては、フレームワークは本発明のフレームワークであり、かつ細胞は真核細胞、特に酵母細胞である。
【0054】
本発明の非常に好ましい実施形態においては、抗原をコードするDNA配列およびscFvをコードするDNA配列は、いずれも生存許容マーカーに連結された転写活性化システムの一部に連結された抗原またはscFvそれぞれとのキメラ分子の両方をコードし、より好ましくは抗原はDNA結合ドメインに融合されかつscFvは転写活性化因子ドメインに融合されるか、または抗原は転写活性化因子ドメインに融合されかつscFvはDNA結合ドメインに融合される。
【0055】
本発明はさらに、治療または診断または予防用薬剤として規定されたフレームワークを有するscFv、および細胞内スクリーニングのために規定されたフレームワークを有するscFvの使用方法を提供する。
【0056】
本発明の全ての目的のためには真核細胞が好ましく、特に、増殖、ポジティブ選択、増殖選択が速くでき、かつ形質転換およびその選択が効率的にできることを含むその特異的な特徴から、酵母細胞が特に好ましい。
本発明はまた、以下の項目を提供する。
(項目1) 還元環境において安定性かつ可溶性である規定されたフレームワークを有するscFvの単離方法であって、
(a)既知の抗原に対するscFvのDNA配列の少なくとも1つのフレームワークコード領域の突然変異、および前記突然変異の好適な発現ベクターへの導入により、種々のフレームワークおよび定常的なCDRを有するscFvライブラリーを作製し、
(b)特異的な既知の抗原を発現可能であり、かつ抗原−scFv相互作用の存在下でのみ生存する宿主細胞を前記scFvライブラリーで形質転換し、
(c)前記形質転換した宿主細胞を、前記抗原および前記scFvを発現するのに好適な条件下で培養して、前記抗原−scFv相互作用の存在下においてのみ生存させ、
(d)生存細胞内で発現され、また還元環境において安定性かつ可溶性である
規定されたフレームワークを有する前記scFvを単離する方法。
(項目2) 前記宿主細胞が真核生物である項目1記載の方法。
(項目3) 前記宿主細胞が酵母細胞である項目1または2記載の方法。
(項目4) 項目1〜3のいずれか1項に記載の方法により得られる規定されたフレームワークを有するscFv。
(項目5) 少なくとも1つのCDRの選択的な交換を可能にする制限酵素認識部位を含む項目4記載のscFv。
(項目6) 前記制限酵素認識部位がCDRに隣接する前記フレームワーク内に位置する項目5に記載のscFv。
(項目7) CDR領域における選択的な変更に好適なフレームワークを含むscFvをコードするDNAの産生方法であって、部位特異的変異誘発により、規定された安定性かつ可溶性であるscFvをコードするDNAの配列に特異的な制限酵素認識部位を導入する方法。
(項目8) 前記制限酵素認識部位が前記フレームワーク内に位置し、それによって前記制限酵素認識部位を発生させるための前記ヌクレオチドの置換がアミノ酸配列に影響しない項目7記載の方法。
(項目9) 還元環境において安定性かつ可溶性である規定されたフレームワークを含むscFvの産生方法であって、還元環境において安定性かつ可溶性である少なくとも2つの異なるフレームワーク、好ましくは項目4〜6のうちいずれか1項記載のフレームワークまたは項目1〜3のいずれか1項に従って単離されるフレームワークのうち少なくとも2つの変異が、規定されたフレームワークを有するscFvを製造するために組み合わされる方法。
(項目10) 項目9の方法により得られる前記規定されたフレームワークを有するscFv。
(項目11) 前記変異が前記可変軽鎖のCDR1の上流に存在する項目10記載のscFv。
(項目12) 前記変異が前記可変重鎖のCDR2とCDR3との間に位置する項目10記載のscFv。
(項目13) 少なくとも1つの変異がCDR1の上流に存在し、かつ少なくとも1つの変異が前記可変重鎖のCDR2とCDR3との間に位置する項目10記載のscFv。
(項目14) 少なくとも2つの変異がCDR1の上流に存在し、かつ少なくとも2つ、好ましくは少なくとも4つの変異が前記可変重鎖のCDR2とCDR3との間に位置する項目10記載のscFv。
(項目15) SEQ ID NO 1において規定されたフレームワークを含むscFv。
(項目16) 還元環境において安定性かつ可溶性である規定されたフレームワークを含むCDRライブラリーの産生方法であって、項目1〜15のいずれか1項記載のscFvをコードするDNA配列が、配列当たり少なくとも1つのCDRを修飾されたCDRで置換するために消化される方法。
(項目17) 前記修飾されたCDRが無作為な変更により産生される項目16記載の方法。
(項目18) 少なくとも1つの無作為化されたCDRおよび、還元環境下において安定性かつ可溶性である規定されたフレームワークを有するイントラボディーのライブラリー。
(項目19) 特異的な抗原と相互作用するCDRのスクリーニング方法であって、既知の抗原をコードする核酸配列、特にDNA配列で形質転換させた宿主細胞をさらに、還元環境において安定性かつ可溶性である規定されたフレームワークを有する無作為化したCDRライブラリーで形質転換し、それによって抗原および/またはscFvをマーカーシステムまたはマーカーシステムの一部に連結させ、したがって選択的な条件下において培養した細胞は抗原/scFv相互作用の存在下でのみ生存し、このように形質転換させた細胞を選択的な条件下において培養し、残存している細胞を培養してイントラボディーを収集する方法。
(項目20) 前記フレームワークが項目1〜19のいずれか1項において規定されるようなフレームワークである、項目19記載の方法。
(項目21) 前記細胞が真核細胞、特に酵母細胞である項目19または20記載の方法。
(項目22) 前記抗原をコードするDNA配列および前記scFvをコードするDNA配列がいずれも、生存許容マーカーに連結された転写活性化システムの一部にいずれも連結された前記抗原またはscFvをそれぞれ含むキメラ分子をコードする、項目19〜21のいずれか1項記載の方法。
(項目23) 前記抗原がDNA結合ドメインに融合され、かつ前記scFvが転写活性化因子ドメインに融合されるか、または前記抗原が転写活性化因子ドメインに融合され、かつ前記scFvがDNA結合ドメインに融合される、項目22記載の方法。
(項目24) scFvと相互作用している抗原のスクリーニング方法であって、興味のある少なくとも1つの抗原を発現する宿主細胞を、還元環境において安定性かつ可溶性である規定されたフレームワークを有する少なくとも一つのscFv、または還元環境において安定性かつ可溶性である規定されたフレームワークを有する無作為化されたCDRライブラリーで形質転換し、それによって抗原および/またはscFvをマーカーシステムまたはマーカーシステムの一部に連結させ、したがって選択的な条件下において培養した細胞は抗原/scFv相互作用の存在下でのみ生存し、このように形質転換させた細胞を選択的な条件下で培養し、残存している細胞を培養してscFvを収集する方法。
(項目25) 前記フレームワークが項目1〜23のいずれか1項において規定されるようなフレームワークである項目24記載の方法。
(項目26) 前記細胞が真核細胞、特に酵母細胞である項目24または25記載の方法。
(項目27) 前記抗原をコードするDNA配列およびscFvをコードするDNA配列が両者共に、いずれも生存許容マーカーに連結された転写活性化システムの一部に連結された抗原またはscFvをそれぞれ含むキメラ分子をコードする、項目24〜26のいずれか1項記載の方法。
(項目28) 前記抗原がDNA結合ドメインに融合され、かつ前記scFvが転写活性化因子ドメインに融合されるか、または前記抗原が転写活性化因子ドメインに融合され、かつ前記scFvがDNA結合ドメインに融合される、項目27記載の方法。
(項目29) 治療または診断または予防用薬剤としての規定されたフレームワークを有するscFv。
(項目30) 細胞内スクリーニングのための規定されたフレームワークを有するscFvの使用。
(項目31) イントラボディーフレームワークまたはイントラボディーの同定方法であって、好適な宿主細胞をライブラリーおよびマーカーシステムで形質転換し、それによって前記ライブラリーはイントラボディーライブラリーと前記マーカーシステムの少なくとも一部の融合産物となり、前記マーカーシステムは可溶性かつ安定性であるイントラボディーフレームワークをコードする融合タンパク質の存在下においてのみ活性化され、また、可溶性かつ安定性であるイントラボディーフレームワークを発現する細胞の同定および選択を可能にする条件下において前記細胞を培養する方法。
(項目32) 前記ライブラリーが、イントラボディーライブラリーとマーカータンパク質の融合産物である項目31記載の方法。
(項目33) 前記マーカータンパク質が選択可能な活性、特に酵素活性または蛍光活性を有する項目32記載の方法。
(項目34) 前記ライブラリーが、イントラボディーライブラリーと転写を活性し得るDNA結合タンパク質との融合産物である項目31記載の方法。
(項目35) 前記好適な宿主細胞を、イントラボディーおよびトランス活性化システムの一部分を含むタンパク質をコードするライブラリーで形質転換し、前記細胞はさらに前記トランス活性化システムの第2部分を含む第2タンパク質を発現し、それによって前記トランス活性化システムを生存許容マーカーに連結させ、前記細胞が前記2つのタンパク質間の相互作用が存在する選択的な条件下においてのみ生存する、項目31記載の方法。
(項目36) 前記ライブラリーにコードされるタンパク質が転写活性化ドメインを含み、かつ前記第2のタンパク質がDNA結合ドメインを含むか、または前記ライブラリーにコードされるタンパク質がDNA結合ドメインを含み、かつ前記第2のタンパク質が転写活性化ドメインを含む項目35記載の方法。
(項目37) 前記第2タンパク質が、DNA結合ドメインまたはトランス活性化ドメインのそれぞれと、前記第1のライブラリーにコードされたタンパク質の定常領域と相互作用するタンパク質とを含む、項目35または36記載の方法。
(項目38) 特に項目1記載の方法において使用するための、項目31〜37のいずれか1項に記載の方法によって得られる規定されたフレームワークを含むscFv。
【図面の簡単な説明】
【0057】
【図1A】scFvまたはCDRライブラリーの品質管理を行いうる方法を示す図。
【図1B】レポーター遺伝子活性と関連するscFv融合タンパク質の溶解性を示す図。
【図2】λグラフトと呼ばれる別の変異体と比較した、最適化したGal4 AD−ΩグラフトscFvのインビボにおけるより良好な性能を示す図。
【図3A】Gcn4p依存性LacZレポーター遺伝子の遺伝子発現における、種々のscFvフラグメントのインビボにおける性能を示す図。
【図3B】ツーハイブリッドアッセイにおける、酵母内での種々のscFvフラグメントの発現のインビボにおける性能を示す図。
【図4】種々のscFvフラグメントを発現している細胞の、ツーハイブリッドアッセイにおける増殖選択を示す図。
【図5A】Gcn4p依存性LacZリポーターの遺伝子発現において、定常ドメイン(Gal11P−Gal4AD)の単鎖抗体に対するN末端融合は、有意にはこのscFvフラグメントの性質を変化させないことを示す図。
【図5B】LacZリポーターの遺伝子発現において、2つの独特な制限酵素認識部位の単鎖抗体への導入は、このscFvフラグメントの性質を変化させないことを示す図。
【図6】酵母内に発現した種々のGcn4p結合scFvフラグメントの溶解性のウェスタンブロット解析を示す図。
【発明を実施するための形態】
【0058】
(本発明を実行するための方法)
scFvおよびCDRライブラリーの品質管理
術語「品質管理」は、scFvライブラリーからの安定性かつ可溶性であるイントラボディーの選択を可能にするアッセイを定義する。
この目的のために、scFvライブラリーを、転写活性化ドメイン(この場合Gal4AD)および定常領域(この場合Gal11P aa263−352)へ融合させる。融合タンパク質の安定性は、scFv部分の安定性および溶解性に依存する。定常Gal11Pドメインは、Gal4の二量体化ドメイン(Gal4DNA結合部位(DBD)の一部である58−97残基(Barberis et al.,1995))と相互作用する。
【0059】
このライブラリーを、選択マーカー遺伝子(たとえばHIS3/LacZ)のプロモーターに結合するGal4 DBD(1−100残基)を発現する酵母細胞に形質転換する。この宿主細胞の増殖は、試験されるイントラボディーが所望の溶解性および安定性を示し、Gal11PとGal4 DBDを介して十分に相互作用できる場合にのみ媒介される(図1Aを参照)。
【0060】
遺伝子活性化に関連する溶解性
本発明において記述される品質管理システムの原理は、よく特徴付けられた多数のscFvを用いて実証された。これらは、本質的に同一の抗原結合特性を有するが、インビトロでの安定性は異なる。種々のscFvフラグメントをGal11P−Gal4AD融合タンパク質として発現させた。Gal4二量体化ドメイン(58−97残基)をLexAのC末端に融合させ、6x LexA結合部位の制御下におけるレポーター遺伝子を含有するレポーター株YDE173に転換した(下記参照)。
【0061】
上述したように、Gal11P−Gal4AD−scFv融合タンパク質の細胞内安定性および溶解性は、scFv部分に依存する。したがって、LexA−Gal4(58−97)と十分に相互作用する安定性かつ可溶性scFv融合タンパク質のみが、リポーター遺伝子発現(たとえば、β−ガラクトシダーゼ)を活性化できる。
【0062】
Gal11の野生型(wt)対立遺伝子は、Gal4二量体化ドメイン(58−97残基)とは相互作用しない。したがって、いかなる単鎖のGal11wt対立遺伝子との融合体もレポーター遺伝子を活性化できないので、ネガティブコントロールとして働く。これは、Gal11wt−Gal4AD−λグラフト融合構築物を用いて実証された(図1B参照)。
【0063】
ベイト(bait)(LexA−Gal4(58−97))のみおよびscFv融合タンパク質のみのいずれも、レポーター遺伝子を活性化しなかった。
【0064】
この品質管理システムにおいては、試験した6つのscFvフラグメントのうち2つだけが、レポーター遺伝子発現を活性化させるのに十分に可溶性かつ安定性であった。フレームワークに安定化されたλグラフトおよびκグラフトは、最も安定な変異体である。この結果は分画分析と完全に一致しており、そこではλ−およびκグラフトのみが可溶性画分に検出された(図6参照)。
【0065】
酵母内で細胞質に発現されるscFvフラグメント
好適なscFvフラグメントは、たとえば、免疫したマウスから作製されたライブラリーからのリボソームディスプレイによって最初に得られた抗GCN4野生型scFvである(Hanes et al.,1998)。抗原は、7P14P(ジッパードメインの7および14の位置がプロリン残基に変異されていることを示唆する)と呼ばれるGcn4pロイシンジッパーの二重プロリン変異体であり、これは溶液中でランダムコイルを形成する(Leder et al.,1995)。scFvフラグメントは、ランダムコイル高次構造内でモノマーとして野生型ペプチドにも結合するので、インビトロにおいて野生型Gcn4pコイルドコイルペプチドの二量体化を阻止する(Berger et al.,1999)。本発明と関連して「野生型」と呼ばれる抗GCN4 scFvフラグメントは、ロイシンジッパーペプチドからの解離定数が4×10−11Mであることが測定されている(Hanes et al.,1998)。
【0066】
本発明の範囲内において、このscFvの多くの異なる変異体が調査された。抗GCN4野生型に加えて、H−R66K変異を保有する不安定化された抗GCN4野生型の変異体[抗GCN4(H−R66K)と名付けた]は、本質的に同じ抗原結合特性を有するがインビトロにおける安定性がわずかに減少したGcn4p結合scFvフラグメントの例として働いた(下記参照)。H−66の位置(Kabat et al.,1991 に従って番号付けした)にあるアルギニン残基(Arg)は、抗原結合ポケットから離れており、一般にAsp H−86に対して二重水素結合を形成する。H−66の位置にあるArgは以前に、レバン結合A48scFvフラグメント内のLysよりも高いタンパク質安定性を導くことが示された(Proba et al.,1998; Woern and Plueckthun, 1998a)。さらに、抗GCN4 scFvフラグメントのVal−Ala変異体[抗GCN4(SS――)と名付けた]を調べたところ、いずれのドメイン内ジスルフィドもVal−Alaペア(L−C23V、L−C88A、H−C22V、H−C92A)によって置換された。これらの変異は以前に、p185HER2結合4D5 scFvフラグメントの還元ジチオール型と比較してわずかに安定して作用することが示されており、またイントラボディーの性能を向上させるであろうことが推測されている(Woern and Plueckthun,1998b)。
【0067】
抗GCN4 CDR(相補性決定領域)ループを別のフレームワークにグラフトすることにより(Jones et al., 1986)、さらに2つの変異体を設計した。アクセプターフレームワークとして、「ハイブリッド」と呼ばれるscFvを選択した(Woern and Plueckthun,1999)。このアクセプターフレームワークは4D5 scFvフラグメントのVLドメインとA48++(H2)scFvフラグメントのVHドメインとから構成されている。それは、一連の安定化ドメインから合理的に設計されており、変性剤誘導性の平衡変性(equilibrium unfolding)によって実証され、また発現率が高い(Woern and Plueckthun,1999)というように、その驚くべき安定性が目立っている。抗GCN4 scFv CDRおよび「ハイブリッド」scFvフレームワークを含む2つのCDR−グラフト変異体を、全遺伝子合成により調製した。抗GCN4野生型ループドナーはλ軽鎖を保有し、一方アクセプター「ハイブリッド」フレームワークはκ軽鎖を保有していたので、ループのグラフトは直接的でなかった。したがって、2つの異なる変異体を設計し、一方はさらなる「κ様」(κグラフトと名付けた)、もう一方はさらなる「λ様」(λグラフトと名付けた)とした。これら2つの変異体は、VH−VL界面領域内の7残基のみが異なり、互いに2つのドメインの配向を強力に影響しあう。アンピシリン結合scFvフラグメントAL5(A.Krebber et al., 未発表)は、Gcn4pに結合していないscFvフラグメントのネガティブコントロールとして働いた。
【0068】
抗GCN4 scFvイントラボディーはGcn4pのトランス活性化能を阻害する
抗GCN4 scFvについて初めに、GAL1およびADH−駆動(driven)プロモーターを含む複数の酵母ベクターから発現されるその生物活性を調べた。さらに、SV40ラージT抗原由来の核移行シグナル(NLS)を抗GCN4 scFvのN末端に融合させた。試験した組み合わせのうち、TRP1選択マーカーを含むpESBA−Act発現ベクター(実施例を参照)、および2μオリジン(origin)(データ示さず)を用いて、いかなるNLSをも伴わないアクチン1プロモーターから発現させた場合に、抗GCN4 scFvが最も強い生物学的効果を示した。このベクターを引き続き、全てのさらなる実験において使用した。
【0069】
GCN4依存性LacZ発現において種々のscFvフラグメントを発現させるインビボ効果を図3Aに示す。レポーター構築物(YAdM2xGCN4−150)は、TATAボックスから相対的に−150の位置に2つのGcn4p結合サイトを含有し、酵母遺伝子に組み込まれた。内在性のGcn4pによって駆動されるβ−ガラクトシダーゼ活性比(Rel.β−gal.activity)を、任意に100%に設定した。AL5はアンピシリン結合scFvフラグメントであり、ネガティブコントロールとして働く。抗GCN4野生型(wt)に加えて、不安定化させた点変異体[抗GCN4(H−R66K)]、抗GCN4野生型のシステインフリー変異体[抗GCN4(SS――)]、および抗GCN4の2つのフレームワーク安定化された変異体(κグラフトおよびλグラフト)を試験した。安定化されたλグラフトは最も活性のあるイントラボディーであり、一方、抗GCN4の不安定化されたH−R66K点変異体およびシステインフリー変異体は、抗GCN4野生型と比較すると、活性が減少したことが示された。κグラフトの活性の減少は、その低い親和性のためであると考えられる(表1参照)。不安定化された点変異体である抗GCN4(H−R66K)は、野生型scFvと比較して、GCN4依存性レポーター遺伝子活性の阻害において効果的でなかった。Gcn4pトランス活性化阻害のパターンは再現性が高く、別の分析方法を用いた場合でも確認された。そこでは、溶解させるためにガラスビーズまたは凍結融解サイクルにより細胞を分裂させ、タンパク質濃度に対するβ−ガラクトシダーゼ活性を基準化させた後、β−ガラクトシダーゼレポーター活性を測定した(Escher and Schaffner,1997)(データ示さず)。
【0070】
【表1】
Gal4 AD−scFv融合タンパク質は、ツーハイブリッドアッセイにおいて、インビトロでの安定性およびインビボにおける性能に従って振舞う
レポーター遺伝子としてのLacZの発現をモニターするツーハイブリッドアッセイにおける、抗原と相補性決定領域(CDR)との間の好結果の相互作用を図3Bに示す。レポーター株YDE173を使用した。YDE173株は、2000年2月11日に、ドイツ、ブラウンシュヴァイク所在のDeutsche Summlung von Mikroorganismen und Zellkulturen DSZMに、番号DSM13333として寄託された。YDE173は、ゲノムのhis3遺伝子座に、互いに異なる方向に向き付けられたレポーター遺伝子HIS3およびLacZを制御する6つのLexA結合サイトを含有するレポータープラスミドpDE200が組み込まれている、酵母株JPY5(Matα ura3−52 his3Δ200 leu2Δ1 trp1Δ63 lys2Δ385)に由来するものである。
【0071】
図3Aにおいて使用したものと同じであるが、Gal4の転写活性化ドメインに融合されたscFvフラグメントを、ツーハイブリッドアッセイのベイトとして働く、C末端をLexAに融合させたGCN4ロイシンジッパー(aa245−285)と共発現させた。非特異的なAL5制御scFv融合構築物はLexA−GCN4ロイシンジッパーと相互作用できず、したがってLacZレポーター遺伝子を活性化しなかった。フレームワーク安定化されたλグラフト変異体に融合されたGal4活性化ドメインは、活性化イントラボディーとして最も強い効果を示し、次は抗GCN4野生型、および不安定化点変異体抗GCN4(H−R66K)であった。対照的に、安定性は高いが結合の弱いκグラフトおよびシステインフリー抗GCN4(SS――)は、ツーハイブリッドの型式においては明らかなレポーター遺伝子の発現を生じなかった。X−Galプレートアッセイにおいても同じ結果が得られた(データ示さず)。要約すると、種々のGal4 AD−scFv融合変異体の、ツーハイブリッド型式におけるLacZレポーター遺伝子の活性化についてのインビボにおける性能は、Gcn4p依存性LacZ発現の阻害パターンと相互に関連する(図3Aを3Bと比較せよ)。
【0072】
抗原と転写活性化ドメインに融合された種々のscFvとの間の相互作用は、ツーハイブリッドアッセイにおいて増殖選択を可能にする
組み込まれたレポーター構築物はLacZレポーター遺伝子だけでなくHIS3遺伝子をも含有するので、ヒスチジンを欠如することはプレート上での増殖選択にとって好ましい。さらに、HIS3遺伝子産物の競合的な阻害剤である種々の濃度の3−アミノトリアゾール(3−AT)を添加することにより、ベイト/抗原とGal4 AD−scFv間の相互作用の強さに依存する酵母細胞の増殖を阻害(抑制)することが可能である。
【0073】
図4に示される結果を導いた実験手順は次のようであった。LexAに融合させたGCN4ロイシンジッパー(aa245−285)とGal4−AD scFv融合タンパク質とを共発現する約10,000の酵母細胞から始めて系列5倍希釈したものを、種々の濃度の3−ATを含むドロップアウト(drop out)プレート(−Trp/−Leu/−His)上にスポットした。48時間後、72時間後、および120時間後に増殖をモニターした。
【0074】
図4におけるレーンは次のようである。1.Gal4−AD λグラフト、2.Gal4−AD AL5、3.Gal4−AD κグラフト、4.Gal4−AD 抗GCN4(SS――)、5.Gal4−AD抗GCN4野生型、6.Gal4−AD 抗GCN4(H−R66K)、7.ポジティブコントロールとして働くLexA−Gal11融合タンパク質、8.空ベクター。
【0075】
ベイト/抗原(lexA―GCN4ロイシンジッパー)をGal4 AD−scFv融合体と共に発現する酵母株の増殖を5日間に渡ってモニターした。プレート上のコントロールは3−ATを含まないので、種々のGal4 AD−scFv融合変異体の明らかな増殖の違いは観察されなかった。ネガティブコントロールのscFv(Gal4 AD−AL5)で形質転換した細胞の増殖を抑制するためには、20mMの3−ATで十分であった。β−ガラクトシダーゼの発現をモニターする結果との関連において、κグラフト変異体、抗GCN4(SS――)、および抗GCN4(H−R66K)とのGal4 AD融合体は、20mM3−ATの存在下において増殖を許容しなかった。抗GCN4野生型だけでなくλグラフト変異体を発現する細胞は、その間フレームワーク安定化されたλグラフトにとって明らかな利点によって、80mMまでの3−ATの存在下で5日間増殖できた。3−ATの濃度100mMは、Gal4 AD−抗GCN4野生型を発現する細胞の増殖を阻止するのに十分であった。5日後にのみ、最も濃縮されたスポット上にわずかに現れ、一方λグラフトGal4 AD−scFv融合変異体を発現する細胞は明らかに増殖した。
【0076】
定常ドメインのλグラフトscFvへのN末端融合は、その生物活性を妨げない
Gal11P(263−352残基)およびGal4活性化ドメインを、λグラフトscFv(Gal11P−Gal4AD λグラフト)のN末端に融合させた。Gcn4p依存性遺伝子活性化を阻害するその生物活性は、λグラフトのみに匹敵する。図5Aに示すように、定常ドメインのscFvへの融合は、Gcn4p依存性遺伝子活性化に対する阻害活性を妨げなかった。
【0077】
特異的な制限酵素認識部位の導入
CDR3 VH(GLFDY)を無作為ペプチドライブラリーと交換するために、この超可変領域に隣接する2つの独特の制限酵素認識部位(BglIIおよびXhoI)を、沈黙突然変異誘発(silent mutagenesis)により導入した。これらの沈黙交換は、抗体のアミノ酸配列に影響せず、したがって、λグラフト変異体のインビボでの性能を変化させなかった(図5B参照)。
【0078】
抗原(GCN4ロイシンジッパー)の特異的認識に対するCDR3超可変領域(de Wildt et al., 1997; Hemminki et al.,1998)の重要性は、付加的なアラニンを可変重鎖のCDR3(AGLFDY)のN末端に導入することによって示された。このλグラフト+Ala変異体は、酵母株YAdM 2xGCN4−150内におけるGcn4p依存性レポーター遺伝子の発現を阻害できず、またYDE173株を用いたツーハイブリッド型式におけるレポーター遺伝子の発現の活性化もできなかった(データ示さず)。
【0079】
グラフト変異体は両方とも、酵母細胞質内で可溶性である
酵母内の種々のGcn4p結合scFvフラグメントの溶解性を、ウェスタンブロット解析により調べた。λおよびκグラフト変異体の場合のみ、明らかな量の可溶性タンパク質が細胞粗抽出液中に検出された(図6)。
【0080】
他の全ての抗GCN4scFvフラグメントは、インビトロにおける安定性が低下するにつれて不溶性scFvの量がわずかに増加したことから、基本的に完全に不溶性であることが明らかとなった。しかしながら、異なるscFv変異体にとって、可溶性タンパク質の不溶性タンパクに対する正確な割合が必ずしもインビボにおける割合を反映しているのではないことに注意しなければならない。種々の抗GCN4変異体の一部分は、たとえピアス社製のY−PERTM酵母タンパク質抽出剤を使用することにより穏やかな破壊方法を使用したとしても、試料調製の間に沈殿してしまうかもしれないことを排除することはできない。
【0081】
フレームワークの改善
好ましくは本発明に従った方法により単離されるフレームワーク内の変異は、還元環境において安定性かつ可溶性であるさらなるフレームワークを作製するために組み合わせることができる。そのようにして得られるフレームワークは、1つだけの変異を有するフレームワークに比べて、インビボにおける性能が増強される。6つの変異を組み合わせたフレームワークを、SEQ ID NO:1と規定する。
【0082】
実施例
CDR−グラフトされた抗GCN4 scFvフラグメントの設計
scFvフラグメントのクローニング、発現、および精製
全てのscFvフラグメントは、20−merのリンカー(GGGGSGGGGSGGGGSSGGGS)およびC末端のhis5−tagによってVL−VHに配向している。
【0083】
酵母内に発現させたscFvフラグメントを、pESBA−Act発現ベクターにクローン化した。pESBA−Actベクターは、酵母(Saccharomyces cerevisiae)−大腸菌(E.coli)のシャトルベクターである。それは、細菌の複製開始点とアンピシリン耐性遺伝子を含有する。さらにそれは、S.cerevisiae内での形質転換選択のための酵母TRP1遺伝子を含有する。それは酵母内で高度にタンパク質発現するように設計され、したがって、S.cerevisiae内で高コピー数を確実にする2μ複製開始点を有する。さらにそれは、NcoI(翻訳開始コドンATGをカバーしている)、ApaI、およびStuIに対する制限酵素認識部位、3つ全てのフレーム内に3つの翻訳停止コドン、およびSalIサイトを含有する多クローニングサイトで分離された、強力で恒常的なアクチンプロモーターおよびGAL11転写終結配列を含有する。
【0084】
全てのscFvフラグメントをBsp120IおよびStuI制限酵素認識部位を用いてクローン化し、C末端His5−tagを保有させた。Bsp120Iサイトをコードする2つのアミノ酸(Gly−Pro)は、開始のMet残基の後のN末端に含まれていなければならなかった。
【0085】
Gal4 ADのN末端の種々の抗体変異体への融合
Gal4活性化ドメインを、テンプレートとしてpGAD424(Clontech)を用いたポリメラーゼ連鎖反応法により増幅させた。両プライマー[上流プライマー:5’−CCATGGGCCCAAGCTTTGCAAAGATGGATAAAG−3’(Seq.Id.No.2)、下流プライマー:5’−TTTGGGCCCGAAGAACCGCCACCACCAGAACCGCCTCCACCAGAGCCACCACCACCAGGCCTGATCTCTTTTTTTGGGTTTGGTG−3’(Seq.Id.No.3)]は、pESBA Actと関連して、種々のscFvに対するSV40T−抗原核局在化シグナルN末端を含むGal4活性化ドメイン(AD)ポリペプチドのクローニングに好適なApaIサイトを含有する。活性化ドメインと単鎖抗体とは、下流プライマーによってコードされる(GGGS)3リンカーによって分離される。
【0086】
Gal11wtおよびGal11Pの、Gal4活性化ドメイン(AD)−scFv融合体へのN末端融合
Gal11wtおよびGal11pはいすれも、以下のプライマーを用いて増幅させた。上流プライマー:5’−CATGCCATGGTTCCTCAACAGCAGCAAATGCAAC−3’(Seq.Id.No.4)、下流プライマー:5’−CATGCCATGGCGCTAGCCAAAGCTTGGATTTTTCTCAGG−3’(Seq.Id.No.5)であり、いずれもNcoIサイトを含有している。アミノ酸263−352をコードしているPCR産物を、pESBA−Act2 Gal4(AD)−scFv融合構築物(上述した)のNcoIサイトに挿入した。これは、それぞれのGal11対立遺伝子の、Gal4(AD)−scFvとのインフレーム融合体を産生した。Gal11インサートの正確な配向性を、独特な酵素NheIで消化することにより確認した。
【0087】
LexA融合
GCN4ロイシンジッパー(aa245−285)を、LexA(aa1−202)の下流をクローニングするのに便利なEcoRIサイトを含有するプライマーを用いてPCR増幅させた。これを、ADHプロモーターの制御下で融合タンパク質を発現する、LEU2選択マーカーを含むArs CenプラスミドpAdM018とする。
【0088】
VHのCDR3に隣接するBglIIおよびXhoIサイトの導入
可変重鎖のCDR3の交換を容易にするために、部位特異的変異誘発によって、Gal4 AD−λグラフトscFvの初めの構造を変えることなく、CDR3 VHに隣接させて2つの独特な制限酵素認識部位を導入した。これらの沈黙点変異を、テンプレートとしてλグラフトを用いたPCRにより導入した。第1ラウンドにおいては、2つの別々のPCR反応を、2つの重複したPCR産物を導くプライマーである、#2421と#2487、および#2486と#2488を用いて行った。これらの2つの産物は、SpeIおよびSalIサイトを含有する外側のプライマー、#2421および#2488を用いたPCRの第2ラウンド用のテンプレートとして働く。最終産物を、SpeIおよびSalIを用いてGal4 AD−λグラフトにサブクローニングした。
【0089】
抗原と反応する新規なCDRのための、直接的細胞内スクリーニング
フレームワークで安定化された、Gal4活性化ドメインに融合させたλグラフトscFv(λグラフトscFv−Gal4 AD)の可変重鎖からのCDR3の初めの3アミノ酸(GLF)は、ロイター(Reiter)らによって記載された方法に基づいたPCRによって無作為化させた。CDR3の最後の2残基(DおよびY)は、その保存および構造上の重要性のために(Chothia and Lesk,1987)、無作為化させなかった。可変重鎖の8000の異なるCDR3変異体を潜在的にコードするλグラフトscFv−Gal4 ADライブラリーを得た。無作為に選んだ6つのライブラリークローンの配列解析により、予想した位置に無作為なCDR3配列の存在が明らかとなった。
【0090】
6つのLexA結合部位(上記参照)の制御下においてHIS3およびLacZレポーター遺伝子を含有する酵母株YDE173を、LexAおよびライブラリーに融合されたGCNロイシンジッパー(aa245−285)を発現するベクターに同時形質転換させ、増殖選択のために60mMの3−ATを含有する選択用ドロップアウトプレート(−Trp/−Leu/−His)上に蒔いて培養した。もし、好適なCDR3配列を含むCDR3ライブラリーからのscFvフラグメントが、LexAに融合されたロイシンジッパー抗原に結合すれば、HIS3レポーター遺伝子の転写を活性化させ、酵母細胞のヒスチジン依存性の増殖を回復させる複合体が形成される。3日後、増殖しているコロニーを選び取り、同じ選択用ドロップアウトプレートに再び蒔いた。2次選択後でもまた増殖する細胞の、X−galプレート上におけるβ−ガラクトシダーゼ活性を分析した。β−gal陽性クローンからライブラリープラスミドDNAを抽出し、可変重鎖のCDR3の領域をシークエンスした:本発明者らは本来のλグラフトCDR3アミノ酸配列を4回見出し、GCN4ロイシンジッパーに特異的な3つの完全に新しいCDR3配列を見出した。本来のCDR3の配列を含有する4つの同定されたscFvフラグメントはλグラフトとして識別不能に振舞い、一方変化したCDR3配列を含む3つのクローンは、LacZレポーター遺伝子の活性化において効果が弱かった。
【0091】
これらの結果は、還元条件下において安定性かつ可溶性である規定されたscFvフレームワーク内に包埋された新規なCDRの、直接的細胞内スクリーニングの可能性を示す。
【0092】
規定されたイントラボディーのインビボにおける性能は、無作為突然変異誘発により最適化され得る
フレームワークで安定化されたλグラフト変異体を、イントラボディーのフレームワークに沿って統計的にアミノ酸変化を導入するために、Sambrook et al.によって説明されるようにPCRにより無作為に突然変異誘発させた。酵母株YDE173を、このGal4の活性化ドメインに融合させて無作為突然変異誘発させたscFvライブラリー、およびLexAに融合させた特異的な抗原(GCN4ロイシンジッパーのaa245−258)を発現するプラスミドで同時形質転換し、80mMの3ATを含有するドロップアウトプレート上で培養した。6つの候補となるクローンを選択し、それぞれはフレームワーク内に1つの単独のアミノ酸変化を有していた。これら6つの変異体フレームワークは全て、λグラフト変異体と比べて向上したインビボにおける性能を示し、これはβ−ガラクトシダーゼ活性の測定により確認され、定量化された。イントラボディーの性能を向上させる種々のアミノ酸変化は相加的に振る舞うという仮定をもとに、本発明者らは、Gal4活性化ドメインに融合させた1つのフレームワーク内の6つ全ての変異を組み合わせ、LacZレポーター遺伝子の活性化について、フレームワークで安定化させたλグラフト変異体と比較した。図2は、6つの点変異を全て含む混合性のこの新規なフレームワーク(Ωグラフト)は、本来のλグラフト変異体と比較してインビボにおける性能がほぼ30%優れていることを示すことを示す。驚くべきことに、これら6アミノ酸の置換はクラスター形成されており、そのうち2つ(E→KおよびL→R)は可変軽鎖のCDR1の上流に存在し、残りの4つ(N→D、G→C、K→E、T→S)は可変重鎖のCDR2とCDR3の間に位置する。
【0093】
レポーター遺伝子のSaccaromyces cerevisiaeの染色体への組み込み
組み込み用レポータープラスミドpAB183は、GAL1プロモーターのTATAボックスの上流150の位置で2つのGcn4p結合サイトをクローニングした、pJP161(Barberis et al.,1995)に由来する。Gcn4p結合サイトは、5’SphIおよび3’SalI適合性のオーバーハング配列を有する2つの相補的なオリゴヌクレオチドをアニーリングすることにより発生させた。このオリゴヌクレオチドは次のようである:5’−CCTATGACTCATCCAGTTATGACTCATCG−3’(Seq.Id.No.6);5’−TCGACGATGAGTCATAACTGGAT GAGTCATAGGCATG−3’(Seq.Id.No.7)。このレポータープラスミドをApaIサイトで直線化し、JPY5株(Barberis et al.,1995)の酵母ゲノム遺伝子座ura3へ組み込み、YAdM2xGCN4−150とした。YAdM2xGCN4−150株は、2000年2月11日に、ドイツ、ブラウンシュヴァイク所在のDeutsche Summlung von Mikroorganismen und Zellkulturen GmbH DSZMに、番号DSM13332として寄託された。4つの独立した酵母形質転換を機能解析において試験し、全てが同じGCN4−依存性のレポーター遺伝子活性を示した。クローンのうち1つ(YAdM2xGCN4−150)を後の実験のために選択し、これを酵母野生型と呼ぶ。
【0094】
ツーハイブリッド実験に用いたレポーター株は、LacZおよびHIS3の発現を駆動する6つのLexA結合サイトを含む双方向性プロモーターを含有する、組み込まれたレポーター構築物を有する。
【0095】
酵母細胞の系列希釈およびスポッティング
酵母細胞を、次の標準的な手順に従った酢酸リチウム法を用いて形質転換した。形質転換体をドロップアウト培地(−Trp/−Leu)中で一晩30℃で培養した。飽和になった培養液を、ドロップアウト培地でOD600=0.7になるまで希釈し、少なくとも2倍の時間再びインキュベートした。各培養液を、およその濃度が106cells/mlから始めて、水で系列希釈し(希釈係数(dilution factor)5)、各希釈溶液から10μlずつ、0mM、20mM、40mM、60mM、80mM、または100mMの3−アミノトリアゾールを含むドロップアウトプレート(−Trp/−Leu/−His)上にスポットした。各形質転換体の6つの異なる希釈溶液をドロップアウトプレート上にスポットした。プレートを30℃でインキュベートし、48時間、72時間、および120時間後に調査した。
【0096】
scFvフラグメントのインビボにおける分析:酵母内におけるscFvフラグメントの発現およびβ−ガラクトシダーゼレポーターアッセイ
溶液中のβ−ガラクトシダーゼアッセイを、記述されているように(Kaiser et al., 1994, Escher and Schaffner 1997)透過化処理した細胞を用いて行った。活性は、アッセイした細胞数に対して規準化した。
【0097】
抗GCN4 scFvフラグメントのウェスタンブロット解析
種々の抗GCN4scFvフラグメントの溶解性をウェスタンブロットにより解析した。5mlの培養液を30℃で光学密度が約2〜3になるまで培養した。細胞を同じ細胞密度になるように規準化し、沈殿させ、可溶性タンパク質のやさしい単離を促進させる穏やかな界面活性剤調合物であるピアス社製のY−PERTM酵母タンパク質抽出剤で全細胞タンパク質を抽出した。可溶性および不溶性の画分を遠心分離(13000rpm、10分、4℃)により分離した。可溶性および不溶性の粗抽出物のサンプルを標準的な手順に従ってSDS−PAGEに供し、PVDF膜にブロットした。His5−tagの付いたscFvフラグメントを、抗His5scFv−AP融合体を用いた既知の方法(Lindner et al.,1997)に従って、ベーリンガーマンハイム社製の化学発光ホスファターゼ基質(chemoluminescent phosphatase substrate)CSPDを用いて検出した。ウェスタンブロットにおいて適切な強度を得るために、可溶性画分においては不溶性画分において用いるよりも約5倍高いタンパク質濃度を用いなければならず、またブロット時間も異なった。したがって、直接的な比較は、それぞれ全て可溶性の試料の間、または全て不溶性の試料の間でのみ意味のあることである。
【0098】
本発明の好ましい実施形態が現在示され、記載されているが、本発明はそれに限定されず、請求項の範囲内において他の種々の具体化および実施が可能であることは明瞭に理解されるべきである。
【0099】
【表2】
[配列表]
【0100】
【表3】
【特許請求の範囲】
【請求項1】
還元環境において安定性かつ可溶性である規定されたフレームワークを有するscFvを単離するための方法であって、
(a)既知の抗原に対するscFvのDNA配列の少なくとも1つのフレームワークコード領域の突然変異、および前記突然変異の好適な発現ベクターへの導入により、種々のフレームワークおよび定常的なCDRsを有するscFvライブラリーを産生し、
(b)特異的な既知の抗原を発現可能であり、かつ抗原−scFv相互作用の存在下でのみ生存する宿主細胞を前記scFvライブラリーで形質転換し、
(c)前記形質転換した宿主細胞を、前記抗原および前記scFvを発現するのに好適な条件下で培養して、前記抗原−scFv相互作用の存在下においてのみ細胞を生存させ、
(d)生存細胞内で発現され、また還元環境において安定性かつ可溶性である規定されたフレームワークを有する前記scFvを単離する方法。
【請求項2】
前記宿主細胞が真核細胞である請求項1記載の方法。
【請求項3】
前記宿主細胞が酵母細胞である請求項1または2記載の方法。
【請求項4】
請求項2〜3のいずれか1項に記載の方法により得られる規定されたフレームワークを有するscFv。
【請求項5】
少なくとも1つのCDRの選択的な交換を可能にする制限酵素認識部位を含む請求項4記載のscFv。
【請求項6】
前記制限酵素認識部位がCDRに隣接する前記フレームワーク内に位置する請求項5に記載のscFv。
【請求項7】
CDR領域における選択的な変更に好適なフレームワークを有するscFvをコードするDNAの産生方法であって、特異的な制限酵素認識部位を、部位特異的変異誘発により、規定された安定性かつ可溶性であるscFvをコードするDNAの配列に導入する方法。
【請求項8】
前記制限酵素認識部位がフレームワーク内に位置し、それによって前記制限酵素認識部位を発生させるための前記ヌクレオチドの置換がアミノ酸配列に影響しない請求項7記載の方法。
【請求項9】
還元環境において安定性かつ可溶性である規定されたフレームワークを有するscFvの産生方法であって、還元環境において安定性かつ可溶性である少なくとも2つの異なるフレームワーク、好ましくは請求項4〜6のうちいずれか1項記載のフレームワークまたは請求項1〜3のいずれか1項に従って単離されるフレームワークのうち少なくとも2つの変異が、規定されたフレームワークを有するscFvを製造するために組み合わされる方法。
【請求項10】
請求項9の方法により得られる規定されたフレームワークを有するscFv。
【請求項11】
前記変異が前記可変軽鎖のCDR1の上流に存在する請求項10記載のscFv。
【請求項12】
前記変異が前記可変重鎖のCDR2とCDR3との間に位置する請求項10記載のscFv。
【請求項13】
少なくとも1つの変異がCDR1の上流に存在し、かつ少なくとも1つの変異が前記可変重鎖のCDR2とCDR3との間に位置する請求項10記載のscFv。
【請求項14】
少なくとも2つの変異がCDR1の上流に存在し、かつ少なくとも2つ、好ましくは少なくとも4つの変異が前記可変重鎖のCDR2とCDR3との間に位置する請求項10記載のscFv。
【請求項15】
SEQ ID NO 1において規定されたフレームワークを含むscFv。
【請求項16】
還元環境において安定性かつ可溶性である規定されたフレームワークを有するCDRライブラリーの産生方法であって、請求項1〜15のいずれか1項記載のscFvをコードするDNA配列が、配列当たり少なくとも1つのCDRを修飾されたCDRで置換するために消化される方法。
【請求項17】
前記修飾されたCDRが無作為な変更により産生される請求項16記載の方法。
【請求項18】
少なくとも1つの無作為化されたCDRおよび、還元環境下において安定性かつ可溶性である規定されたフレームワークを有するイントラボディーのライブラリー。
【請求項19】
特異的な抗原と相互作用するCDRのスクリーニング方法であって、既知の抗原をコードする核酸配列、特にDNA配列で形質転換させた宿主細胞をさらに、還元環境において安定性かつ可溶性である規定されたフレームワークを有する無作為化したCDRライブラリーで形質転換し、それによって抗原および/またはscFvをマーカーシステムまたはマーカーシステムの一部に連結させ、したがって選択的な条件下において培養した細胞は抗原/scFv相互作用の存在下でのみ生存し、このように形質転換させた細胞を選択的な条件下において培養し、生存している細胞を培養してイントラボディーを収集する方法。
【請求項20】
前記フレームワークが請求項1〜19のいずれか1項において規定されるようなフレームワークである、請求項19記載の方法。
【請求項21】
前記細胞が真核細胞、特に酵母細胞である請求項19または20記載の方法。
【請求項22】
前記抗原をコードするDNA配列および前記scFvをコードするDNA配列がいずれも、生存許容マーカーに連結された転写活性化システムの一部にいずれも連結された、前記抗原またはscFvをそれぞれ含むキメラ分子をコードする、請求項19〜21のいずれか1項記載の方法。
【請求項23】
前記抗原がDNA結合ドメインに融合され、かつ前記scFvが転写活性化因子ドメインに融合されるか、または前記抗原が転写活性化因子ドメインに融合され、かつ前記scFvがDNA結合ドメインに融合される、請求項22記載の方法。
【請求項24】
scFvと相互作用している抗原のスクリーニング方法であって、興味のある少なくとも1つの抗原を発現する宿主細胞を、還元環境において安定性かつ可溶性である規定されたフレームワークを有する少なくとも一つのscFv、または還元環境において安定性かつ可溶性である規定されたフレームワークを有する無作為化されたCDRライブラリーで形質転換し、それによって抗原および/またはscFvをマーカーシステムまたはマーカーシステムの一部に連結させ、したがって選択的な条件下において培養した細胞は抗原/scFv相互作用の存在下でのみ生存し、このように形質転換させた細胞を選択的な条件下で培養し、生存している細胞を培養してscFvを収集する方法。
【請求項25】
前記フレームワークが請求項1〜24のいずれか1項において規定されるようなフレームワークである請求項24記載の方法。
【請求項26】
前記細胞が真核細胞、特に酵母細胞である請求項24または25記載の方法。
【請求項27】
前記抗原をコードするDNA配列およびscFvをコードするDNA配列が両者共に、いずれも生存許容マーカーに連結された転写活性化システムの一部に連結された抗原またはscFvをそれぞれ含むキメラ分子をコードする、請求項24〜26のいずれか1項記載の方法。
【請求項28】
前記抗原がDNA結合ドメインに融合され、かつ前記scFvが転写活性化因子ドメインに融合されるか、または前記抗原が転写活性化因子ドメインに融合され、かつ前記scFvがDNA結合ドメインに融合される、請求項27記載の方法。
【請求項29】
治療または診断または予防用薬剤としての、規定されたフレームワークを有するscFv。
【請求項30】
細胞内スクリーニングのための、規定されたフレームワークを有するscFvの使用。
【請求項31】
前記ライブラリーがイントラボディーライブラリーとマーカータンパク質の融合産物である請求項10記載の方法。
【請求項32】
特に請求項1記載の方法において使用するための、請求項31に記載の方法によって得られる規定されたフレームワークを有するscFv。
【請求項33】
明細書中に記載の発明。
【請求項1】
還元環境において安定性かつ可溶性である規定されたフレームワークを有するscFvを単離するための方法であって、
(a)既知の抗原に対するscFvのDNA配列の少なくとも1つのフレームワークコード領域の突然変異、および前記突然変異の好適な発現ベクターへの導入により、種々のフレームワークおよび定常的なCDRsを有するscFvライブラリーを産生し、
(b)特異的な既知の抗原を発現可能であり、かつ抗原−scFv相互作用の存在下でのみ生存する宿主細胞を前記scFvライブラリーで形質転換し、
(c)前記形質転換した宿主細胞を、前記抗原および前記scFvを発現するのに好適な条件下で培養して、前記抗原−scFv相互作用の存在下においてのみ細胞を生存させ、
(d)生存細胞内で発現され、また還元環境において安定性かつ可溶性である規定されたフレームワークを有する前記scFvを単離する方法。
【請求項2】
前記宿主細胞が真核細胞である請求項1記載の方法。
【請求項3】
前記宿主細胞が酵母細胞である請求項1または2記載の方法。
【請求項4】
請求項2〜3のいずれか1項に記載の方法により得られる規定されたフレームワークを有するscFv。
【請求項5】
少なくとも1つのCDRの選択的な交換を可能にする制限酵素認識部位を含む請求項4記載のscFv。
【請求項6】
前記制限酵素認識部位がCDRに隣接する前記フレームワーク内に位置する請求項5に記載のscFv。
【請求項7】
CDR領域における選択的な変更に好適なフレームワークを有するscFvをコードするDNAの産生方法であって、特異的な制限酵素認識部位を、部位特異的変異誘発により、規定された安定性かつ可溶性であるscFvをコードするDNAの配列に導入する方法。
【請求項8】
前記制限酵素認識部位がフレームワーク内に位置し、それによって前記制限酵素認識部位を発生させるための前記ヌクレオチドの置換がアミノ酸配列に影響しない請求項7記載の方法。
【請求項9】
還元環境において安定性かつ可溶性である規定されたフレームワークを有するscFvの産生方法であって、還元環境において安定性かつ可溶性である少なくとも2つの異なるフレームワーク、好ましくは請求項4〜6のうちいずれか1項記載のフレームワークまたは請求項1〜3のいずれか1項に従って単離されるフレームワークのうち少なくとも2つの変異が、規定されたフレームワークを有するscFvを製造するために組み合わされる方法。
【請求項10】
請求項9の方法により得られる規定されたフレームワークを有するscFv。
【請求項11】
前記変異が前記可変軽鎖のCDR1の上流に存在する請求項10記載のscFv。
【請求項12】
前記変異が前記可変重鎖のCDR2とCDR3との間に位置する請求項10記載のscFv。
【請求項13】
少なくとも1つの変異がCDR1の上流に存在し、かつ少なくとも1つの変異が前記可変重鎖のCDR2とCDR3との間に位置する請求項10記載のscFv。
【請求項14】
少なくとも2つの変異がCDR1の上流に存在し、かつ少なくとも2つ、好ましくは少なくとも4つの変異が前記可変重鎖のCDR2とCDR3との間に位置する請求項10記載のscFv。
【請求項15】
SEQ ID NO 1において規定されたフレームワークを含むscFv。
【請求項16】
還元環境において安定性かつ可溶性である規定されたフレームワークを有するCDRライブラリーの産生方法であって、請求項1〜15のいずれか1項記載のscFvをコードするDNA配列が、配列当たり少なくとも1つのCDRを修飾されたCDRで置換するために消化される方法。
【請求項17】
前記修飾されたCDRが無作為な変更により産生される請求項16記載の方法。
【請求項18】
少なくとも1つの無作為化されたCDRおよび、還元環境下において安定性かつ可溶性である規定されたフレームワークを有するイントラボディーのライブラリー。
【請求項19】
特異的な抗原と相互作用するCDRのスクリーニング方法であって、既知の抗原をコードする核酸配列、特にDNA配列で形質転換させた宿主細胞をさらに、還元環境において安定性かつ可溶性である規定されたフレームワークを有する無作為化したCDRライブラリーで形質転換し、それによって抗原および/またはscFvをマーカーシステムまたはマーカーシステムの一部に連結させ、したがって選択的な条件下において培養した細胞は抗原/scFv相互作用の存在下でのみ生存し、このように形質転換させた細胞を選択的な条件下において培養し、生存している細胞を培養してイントラボディーを収集する方法。
【請求項20】
前記フレームワークが請求項1〜19のいずれか1項において規定されるようなフレームワークである、請求項19記載の方法。
【請求項21】
前記細胞が真核細胞、特に酵母細胞である請求項19または20記載の方法。
【請求項22】
前記抗原をコードするDNA配列および前記scFvをコードするDNA配列がいずれも、生存許容マーカーに連結された転写活性化システムの一部にいずれも連結された、前記抗原またはscFvをそれぞれ含むキメラ分子をコードする、請求項19〜21のいずれか1項記載の方法。
【請求項23】
前記抗原がDNA結合ドメインに融合され、かつ前記scFvが転写活性化因子ドメインに融合されるか、または前記抗原が転写活性化因子ドメインに融合され、かつ前記scFvがDNA結合ドメインに融合される、請求項22記載の方法。
【請求項24】
scFvと相互作用している抗原のスクリーニング方法であって、興味のある少なくとも1つの抗原を発現する宿主細胞を、還元環境において安定性かつ可溶性である規定されたフレームワークを有する少なくとも一つのscFv、または還元環境において安定性かつ可溶性である規定されたフレームワークを有する無作為化されたCDRライブラリーで形質転換し、それによって抗原および/またはscFvをマーカーシステムまたはマーカーシステムの一部に連結させ、したがって選択的な条件下において培養した細胞は抗原/scFv相互作用の存在下でのみ生存し、このように形質転換させた細胞を選択的な条件下で培養し、生存している細胞を培養してscFvを収集する方法。
【請求項25】
前記フレームワークが請求項1〜24のいずれか1項において規定されるようなフレームワークである請求項24記載の方法。
【請求項26】
前記細胞が真核細胞、特に酵母細胞である請求項24または25記載の方法。
【請求項27】
前記抗原をコードするDNA配列およびscFvをコードするDNA配列が両者共に、いずれも生存許容マーカーに連結された転写活性化システムの一部に連結された抗原またはscFvをそれぞれ含むキメラ分子をコードする、請求項24〜26のいずれか1項記載の方法。
【請求項28】
前記抗原がDNA結合ドメインに融合され、かつ前記scFvが転写活性化因子ドメインに融合されるか、または前記抗原が転写活性化因子ドメインに融合され、かつ前記scFvがDNA結合ドメインに融合される、請求項27記載の方法。
【請求項29】
治療または診断または予防用薬剤としての、規定されたフレームワークを有するscFv。
【請求項30】
細胞内スクリーニングのための、規定されたフレームワークを有するscFvの使用。
【請求項31】
前記ライブラリーがイントラボディーライブラリーとマーカータンパク質の融合産物である請求項10記載の方法。
【請求項32】
特に請求項1記載の方法において使用するための、請求項31に記載の方法によって得られる規定されたフレームワークを有するscFv。
【請求項33】
明細書中に記載の発明。
【図1A】

【図1B】
【図2】
【図3A】
【図3B】
【図4】

【図5A】
【図5B】
【図6】
【図1B】
【図2】
【図3A】
【図3B】
【図4】
【図5A】
【図5B】
【図6】
【公開番号】特開2011−62205(P2011−62205A)
【公開日】平成23年3月31日(2011.3.31)
【国際特許分類】
【出願番号】特願2010−240199(P2010−240199)
【出願日】平成22年10月26日(2010.10.26)
【分割の表示】特願2001−548557(P2001−548557)の分割
【原出願日】平成12年12月18日(2000.12.18)
【出願人】(502233344)エスバテック、アン アルコン バイオメディカル リサーチ ユニット、エルエルシー (19)
【氏名又は名称原語表記】ESBATech, an Alcon Biomedical Research Unit, LLC
【Fターム(参考)】
【公開日】平成23年3月31日(2011.3.31)
【国際特許分類】
【出願日】平成22年10月26日(2010.10.26)
【分割の表示】特願2001−548557(P2001−548557)の分割
【原出願日】平成12年12月18日(2000.12.18)
【出願人】(502233344)エスバテック、アン アルコン バイオメディカル リサーチ ユニット、エルエルシー (19)
【氏名又は名称原語表記】ESBATech, an Alcon Biomedical Research Unit, LLC
【Fターム(参考)】
[ Back to top ]