
配列特異性およびDNA−結合親和度が変更された、合理設計メガヌクレアーゼ
【課題】合理的に設計されたLAGLIDADGメガヌクレアーゼ、そのようなメガヌクレアーゼの製造法、ゲノム内の少数座位に所望のDNA配列を挿入させた組換え細胞および生物体の使用方法、ならびに病原体感染の治療のための、および、診断学および研究におけるインビトロ応用のための遺伝子治療法を提供する。
【解決手段】特定の配列を有するI−CREIメガヌクレアーゼと少なくとも85%の配列類似性を有し、特定部位の置換あるいは置換に対応する修飾によってダイマー形成に対する親和度が変えられた組換えメガヌクレアーゼモノマーおよびヘテロダイマー。
【解決手段】特定の配列を有するI−CREIメガヌクレアーゼと少なくとも85%の配列類似性を有し、特定部位の置換あるいは置換に対応する修飾によってダイマー形成に対する親和度が変えられた組換えメガヌクレアーゼモノマーおよびヘテロダイマー。
【発明の詳細な説明】
【技術分野】
【0001】
本発明は、分子生物学および組み換え核酸工学の分野に関する。特に、本発明は、DNA認識配列特異性及び/又は親和度が変更された、合理的に設計された、非天然のメガヌクレアーゼに関する。さらに、本発明は、そのようなメガヌクレアーゼの生産法、および、そのようなメガヌクレアーゼによる、組み換え核酸および生物の生産法に関する。
【背景技術】
【0002】
(関連出願)本出願は、2005年10月18日出願の米国特許仮出願第60/727,512号の優先権の利益を主張する。なお、この引用出願の全開示を、参照することにより本出願に含める。
【0003】
(政府援助)本発明は、アメリカ合衆国国立衛生研究所、国立医科学研究所からの研究補助金2R01−GM−0498712、5F32−GM072322、および5 DP1
OD000122によって一部援助された。したがって、米国政府も、本発明において若干の権利を保有する場合がある。
【0004】
ゲノム工学は、あるゲノムの内部に、特定の遺伝子配列を、挿入し、欠失し、置換し、および、その他のやり方で操作することを可能とする能力を要求するが、多くの治療的および生物工学的応用を有する。ゲノム修飾のための効果的手段の開発は、これまでずっと、遺伝子治療、農工学、および合成生物学における主要目標となっている(PORTEUS ET AL.(2005),NAT.BIOTECHNOL.23;967−73;TZFIRA ET AL.(2005),TRENDS BIOTECHNOL.23:567−9;MCDANIEL ET AL.(2005),CURR.OPIN.BIOTECHNOL.16:476−83)。DNA配列を挿入又は修飾するための一般的方法は、ゲノム標的に対して相同な配列によって側接される、トランスジェニックDNA配列を導入すること、および、所望の相同組み換え事象を選択またはスクリーニングすることを含む。トランスジェニックDNAによる組み換えはめったに起こらないが、標的部位のゲノムDNAにおける2本鎖切断によって刺激することが可能である。従来から、DNA2本鎖切断部を作るために、例えば、放射線照射および化学的処理を含めた数多くの方法が用いられている。これらの方法は、効率的に組み換えを刺激することが可能ではあるけれども、二本鎖切断部は、ゲノムの中にランダムに挿入され、それらは、突然変異誘発性および毒性が高い可能性がある。現在、染色体バックグランドにある特定の部位に対し遺伝子修飾を照準することはできていないため、思い通りのゲノム加工に対する大きな障害となっている。
【0005】
この目標を達成するための一つの方法は、ゲノム内の単一部位にしか存在しないほど十分に大きい配列に対し特異性を持つヌクレアーゼを用いて、標的配座に二本鎖切断部を設けてその部位における相同組み換えを刺激することである(例えば、PORTEUS ET AL.(2005),NAT.BIOTECHNOL.23:967−73を参照)。この戦略の有効性は、FOKI制限酵素の、加工されたジンクフィンガーDNA−結合ドメインと、非特異的ヌクレアーゼドメインとの間のキメラ融合を用いて、各種の生物体において明らかにされている(PORTEUS(2006),MOL THER 13:438−46;WRIGHT ET AL.(2005),PLANT J.44:693−705;URNOV ET AL.(2005),NATURE 435:646−51)。これらの人工的ジンクフィンガーヌクレアーゼは、部位特異的組み換えを刺激するけれども、これらのヌクレアーゼは、ヌクレアーゼドメインの調整低調に由来する、非
特異的な、残留切断活性を保持するので、不要の部位を切断することがしばしばある(SMITH ET AL.(2000),NUCLEIC ACIDS RES.28:3361−9)。これらの不要な切断は、処置生物体において、突然変異および毒性を引き起こす可能性がある(PORTEUS ET AL.(2005),NAT.BIOTECHNOL.23:967−73)。
【0006】
植物および菌類のゲノムの中に一般的に認められる、15−40塩基対切断部位を認識する、1群の天然ヌクレアーゼは、比較的毒性の低い、ゲノム加工代替品を提供する可能性がある。このような「メガヌクレアーゼ」または「ホーミング・エンドヌクレアーゼ」は、多くの場合、寄生的DNA要素、例えば、グループ1自己スプライシングイントロンおよびインテインとコンタクトされる。これらのヌクレアーゼは、自然状態で、宿主ゲノムの特異的位置における相同的組み換えまたは遺伝子挿入を促進する。これを、細胞のDNA−修復機構を招集する、染色体において二本鎖切断を生産することによって行う(STODDARD(2006),REV.BIOPHYS.38:49−95)。一般に、メガヌクレアーゼは、4つのファミリーに分けられる。すなわち、LAGLIDADGファミリー、GIY−YIGファミリー、HIS−CYSボックスファミリー、およびHNHファミリーである。これらのファミリーは、触媒活性および認識配列に影響を及ぼす、構造的モチーフによって特徴づけられる。例えば、LAGLIDADGファミリーのメンバーは、LAGLIDADG保存モチーフの、1または2コピーを持つことによって特徴づけられる(CHEVALIER ET AL.(2001),NUCLEIC ACIDS RES.29(18):3757−3774を参照)。LAGLIDADGモチーフの単一コピーを持つLAGLIDADGメガヌクレアーゼは、ホモダイマーを形成するが、一方、2コピーのLAGLIDADGモチーフを持つメンバーは、モノマーであることが判明した。同様に、GIY−YIGファミリーメンバーは、70−100残基長のGIY−YIGモジュールを持ち、その内二つは活性のために必要な、四つの不変残基を持つ、4から5個の保存配列モチーフを含む(VAN ROEY ET AL.(2002),NATURE STRUCT.BIOL.9:806−811を参照)。HIS−CYSボックス・メガヌクレアーゼは、数百のアミノ酸残基を含む領域に分散する、高度に保存された、ヒスチジンおよびシステインの連続列によって特徴づけられる(CHEVALIER ET AL.(2001),NUCLEIC ACIDS RES.29(18):3757−3774を参照)。NHNファミリーの場合は、そのメンバーは、2対の、アスパラギン残基によって囲まれる保存ヒスチジンを含むモチーフによって定義される(CHEVALIER ET AL.(2001),NUCLEIC ACIDS RES.29(18):3757−3774を参照)。メガヌクレアーゼのこの4ファミリーは、構造保存部位に関してDNA認識配列の特異性及び触媒活性が互いに大きく異なっている。
【0007】
主にLAGLIDADGファミリーから得られる、天然のメガヌクレアーゼは、これまで、植物、酵母、ショウジョウバエ、哺乳類細胞及びマウスにおいて部位特異的なゲノム修飾に有効に使用されているが、この方法は、このメガヌクレアーゼ認識配列を保存する相同配列か(MONNAT ET AL.(1999),BIOCHEM.BIOPHYS.RES.COMMUN.255:88−93)又は認識配列を導入したあらかじめ加工されたゲノム(ROUET ET AL.(1994),MOL.CELL BIOL.14:8096−106;CHILTON ET AL.(2003),PLANT PHYSIOL.133:956−65;PUCHTA ET AL.(1996),PROC.NATL.ACAD.SCI.USA 93:5055−60;RONG ET
AL.(2002),GENES DEV.16:1568−81;GOUBLE ET AL.(2006),J.GENE MED.8(5):616−622)の修飾に限定される。
【0008】
ヌクレアーゼ刺激による遺伝子修飾の体系的実施は、ゲノムの既存部位に対する標的DNA中断部に向けて照準させた特異性を持つ加工酵素の使用が必要となる。したがって、医学的又は生物工学的関連部位において遺伝子修飾を促進するようにメガヌクレアーゼを適応させることについては大きな関心が寄せられている(PORTEUS ET AL.(2005),NAT.BIOTECHNOL.23:967−73;SUSSMAN ET AL.(2004),J.MOL.BIOL.342:31−41;EPINAT
ET AL.(2003),NUCLEIC ACIDS RES.31:2952−62)。
【0009】
CHLAMYDOMONASREINHARDTIIから得られたメガヌクレアーゼI−CREIは、LAGLIDADGファミリーの一員であり、葉緑体染色体の中の22塩基対認識配列を認識・切断し、かつ、メガヌクレアーゼ再設計の好個の標的となっている。この野生型酵素は、各モノマーが、全体認識配列における9塩基対に対して直接接触する、ホモダイマーである。この認識配列の単一位置において(SUSSMAN ET AL.(2004),J.MOL.BIOL.342:31−41;CHAMES ET AL.(2005),NUCLEIC ACIDS RES.33:E178;SELIGMAN ET AL.(2002),NUCLEIC ACIDS RES.30:3870−9)、あるいは、比較的最近では、認識配列の中の3位置において(ARRNOULD ET AL.(2006),J.MOL.BIOL.355:443−58)、塩基選択性を変えるI−CREIにおける突然変異を特定するために、遺伝子選択技術が用いられている。I−CREIタンパク−DNAインターフェイスは、DNA塩基に直接接触する9個のアミノ酸と、修飾されたインターフェイスにおいて接触を形成する可能性のある、少なくともさらに5個の残基を含む。このインターフェイスのサイズは、切断部位を大きく変更された酵素を選択するために構築された配列ライブラリーにおいても十分に抽出されることがほとんどないと思われるほど複雑な組み合わせを提示する。
【発明の開示】
【発明が解決しようとする課題】
【0010】
ゲノムの正確な修飾を促進するヌクレアーゼについては、依然として需要がある。さらに、特定の遺伝子座位における遺伝子配列の操作を可能とする、あらかじめ指定された、合理的に設計された認識配列を有するヌクレアーゼを生成する技術、および、正確な配列修飾を持つ生物体を遺伝学的に加工するために、そのようなヌクレアーゼを利用する技術に対しては、依然として需要がある。
【課題を解決するための手段】
【0011】
本発明は、メガヌクレアーゼLAGLIDADGファミリーの同定及び特徴解明に基づく。メガヌクレアーゼが2本鎖DNA認識配列とコンタクトする際、DNA塩基およびDNAバックボーンに結合し、そうすることによって、該酵素の特異性および活性に影響を及ぼす。この発見は、以下に詳細に説明するように、メガヌクレアーゼの認識配列特異性及び/又はDNA結合親和性を変えることが可能なアミノ酸置換を特定するため及び天然のメガヌクレアーゼが認識しない所望のDNA配列を認識することが可能なメガヌクレアーゼを合理的に設計し、開発するために用いられた。本発明はさらに、遺伝子治療、病原体感染の治療及び診断と研究におけるインビトロ応用のために、生物体ゲノム内の少数座位において所望の遺伝子配列の組み換えを実現するために、このようなメガヌクレアーゼを使用する方法を提供する。
【0012】
したがって、ある実施態様では、本発明は、野生型I−CREIメガヌクレアーゼに比べ、少なくとも一つの認識配列半分部位に対し変更特異性を持つ組み換えメガヌクレアーゼを提供する。この実施態様におけるメガヌクレアーゼは、配列番号1の野生型I−CREIメガヌクレアーゼの残基2−153に対して少なくとも85%の配列類似性を有する
ポリペプチドを含むが、該組み換えメガヌクレアーゼは、配列番号2、配列番号3、配列番号4及び配列番号5から選ばれる、I−CREIメガヌクレアーゼ認識配列内の半分部位と、少なくとも1塩基対異なる認識配列半分部位に対して特異性を持ち、かつ、この組み換えメガヌクレアーゼは、従来技術に認められる排外的修飾ではない、表5に掲げられる少なくとも一つの修飾を含む。
【0013】
別の実施態様では、本発明は、野生型I−MSOIメガヌクレアーゼと比べ、少なくとも一つの認識配列半分部位に対し変更特異性を持つ組み換えメガヌクレアーゼを提供する。この実施態様におけるメガヌクレアーゼは、配列番号6のI−MSOIメガヌクレアーゼの残基6−160に対して少なくとも85%の配列類似性を有するポリペプチドを含むが、該組み換えメガヌクレアーゼは、配列番号7および配列番号8から選ばれる、I−MSOIメガヌクレアーゼ認識配列内の半分部位と、少なくとも1塩基対異なる、認識配列半分部位に対して特異性を持ち、かつ、該組み換えメガヌクレアーゼは、従来技術に認められる排外的修飾ではない、表7に掲げられる少なくとも一つの修飾を含む。
【0014】
別の実施態様では、本発明は、野生型I−SCEIメガヌクレアーゼと比べ、認識配列に対し変更特異性を持つ組み換えメガヌクレアーゼを提供する。この実施態様におけるメガヌクレアーゼは、配列番号9のI−SCEIメガヌクレアーゼの残基3−186に対して少なくとも85%の配列類似性を有するポリペプチドを含むが、この組み換えメガヌクレアーゼは、配列番号10および配列番号11のI−SCEIメガヌクレアーゼ認識配列と少なくとも1塩基対異なる認識配列に対して特異性を持ち、かつ、この組み換えメガヌクレアーゼは、従来技術に認められる排外的修飾ではない、表9に掲げられる少なくとも一つの修飾を含む。
【0015】
別の実施態様では、本発明は、野生型I−CEUIメガヌクレアーゼと比べ、少なくとも一つの認識配列半分部位に対し変更特異性を持つ組み換えメガヌクレアーゼを提供する。この実施態様におけるメガヌクレアーゼは、配列番号12のI−CEUIメガヌクレアーゼの残基5−211に対して少なくとも85%の配列類似性を有するポリペプチドを含むが、該組み換えメガヌクレアーゼは、配列番号13および配列番号14から選ばれるI−CEUIメガヌクレアーゼ認識配列内の半分部位と少なくとも1塩基対異なる認識配列半分部位に対して特異性を持ち、かつ、この組み換えメガヌクレアーゼは、従来技術に認められる排外的修飾ではない、表11に掲げられる少なくとも一つの修飾を含む。
【0016】
本発明のメガヌクレアーゼは、認識配列内の1、2又は3以上の位置において組み換えメガヌクレアーゼの配列特異性に影響を及ぼすために、本出願に開示される修飾の内の一つ、二つ又は三つ以上を含むことが可能である。メガヌクレアーゼは、本出願に開示される新規修飾のみを含んでもよいし又は本出願に開示される新規修飾を従来技術で認められる修飾と組み合わせて含んでもよい。しかしながら、特異的に排除されるものは、従来技術の修飾のみを含む組み換えメガヌクレアーゼである。
【0017】
他の側面から考えると、本発明は、2本鎖DNAに対する配列特異的ではない結合親和度を変更させた組み換えメガヌクレアーゼを提供する。これは、2本鎖DNA認識配列のバックボーンに接触する、メガヌクレアーゼ残基を修飾することによって実現される。この修飾は、結合親和度を増大又は低減するため、酵素の全体活性を増大又は低減することが可能である。さらに、結合および活性の上昇/低下は、配列特異性における低下/上昇を引き起こすことが認められた。したがって、本発明は、DNA−結合親和度を変えることによって配列特異性を全体的に変えるための手段を提供する。
【0018】
したがって、ある実施態様では、本発明は、野生型I−CREIメガヌクレアーゼと比べ、2本鎖DNAに対する結合親和度を変更させた組み換えメガヌクレアーゼを提供する
。この発明におけるメガヌクレアーゼは、配列番号1のI−CREIメガヌクレアーゼの残基2−153に対して少なくとも85%の配列類似性を有するポリペプチドを含むが、DNA結合親和度は、(1)(A)H、N、Q、S、T、K若しくはRによるE80、D137、I81、L112、P29、V64若しくはY66の置換、又は(B)K若しくはRによるT46、T140若しくはT143の置換から選ばれる一つの置換に対応する少なくとも一つの修飾によって上昇するか、又は、逆に、(2)(A)H、N、Q、S、T、D若しくはEによるK34、K48、R51、K82、K116若しくはK139の置換、又は(B)D若しくはEによるI81、L112、P29、V64、Y66、T46、T140若しくはT143の置換から選ばれる一つの置換に対応する少なくとも一つの修飾によって低下する。
【0019】
別の実施態様では、本発明は、野生型I−MSOIメガヌクレアーゼに比べ、2本鎖DNAに対する結合親和度を変更させた組み換えメガヌクレアーゼを提供する。この発明におけるメガヌクレアーゼは、配列番号6のI−MSOIメガヌクレアーゼの残基6−160に対して少なくとも85%の配列類似性を有するポリペプチドを含むが、DNA結合親和度は、(1)(A)H、N、Q、S、T、K若しくはRによるE147、I85、G86若しくはY118の置換、又は(B)K若しくはRによるQ41、N70、S87、T88、H89、Q122、Q139、S150若しくはN152の置換から選ばれる一つの置換に対応する少なくとも一つの修飾によって上昇するか、又は、逆に、(2)(A)H、N、Q、S、T、D若しくはEによるK36、R51、K123、K143若しくはR144の置換、又は(B)D若しくはEによるI85、G86、Y118、Q41、N70、S87、T88、H89、Q122、Q139、S150若しくはN152の置換から選ばれる一つの置換に対応する少なくとも一つの修飾によって低下する。
【0020】
別の実施態様では、本発明は、野生型I−SCEIメガヌクレアーゼに比べ、2本鎖DNAに対する結合親和度を変更させた組み換えメガヌクレアーゼを提供する。この発明におけるメガヌクレアーゼは、配列番号9のI−SCEIメガヌクレアーゼの残基3−186に対して少なくとも85%の配列類似性を有するポリペプチドを含むが、DNA結合親和度は、(1)(A)H、N、Q、S、T、K若しくはRによるD201、L19、L80、L92、Y151、Y188、I191、Y199若しくはY222の置換、又は(B)K若しくはRによるN15、N17、S81、H84、N94、N120、T156、N157、S159、N163、Q165、S166,N194若しくはS202の置換から選ばれる一つの置換に対応する少なくとも一つの修飾によって上昇するか、又は、逆に、(2)(A)H、N、Q、S、T、D若しくはEによるK20、K23、K63、K122、K148、K153、K190、K193、K195若しくはK223の置換、又は(B)D若しくはEによるL19、L80、L92、Y151、Y188、I191、Y199、Y222、N15、N17、S81、H84、N94、N120、T156、N157、S159、N163、Q165、S166、N194若しくはS202の置換から選ばれる一つの置換に対応する少なくとも一つの修飾によって低下する。
【0021】
別の実施態様では、本発明は、野生型I−CEUIメガヌクレアーゼに比べ、2本鎖DNAに対する結合親和度を変更させた組み換えメガヌクレアーゼを提供する。この発明におけるメガヌクレアーゼは、配列番号12のI−CEUIメガヌクレアーゼの残基5−211に対して少なくとも85%の配列類似性を有するポリペプチドを含むが、DNA結合親和度は、(1)(A)H、N、Q、S、T、K若しくはRによるD25若しくはD128の置換、又は(B)K若しくはRによるS68、N70、H94、S117、N120、N129若しくはH172の置換から選ばれる一つの置換に対応する少なくとも一つの修飾によって上昇するか、又は、逆に、(2)(A)H、N、Q、S、T、D若しくはEによるK21、K28、K31、R112、R114若しくはR130の置換、又は(B)D若しくはEによるS68、N70、H94、S117、N120、N129若しくは
H172の置換から選ばれる一つの置換に対応する少なくとも一つの修飾によって低下する。
【0022】
本発明のメガヌクレアーゼは、DNA−結合親和度に影響を及ぼすために、本出願に開示される、バックボーン接触残基の修飾の内、一つ、二つ又は三つ以上を含むことが可能である。さらに、DNA−結合親和度に影響を及ぼすこれらの修飾は、認識配列内部の特定位置における組み換えメガヌクレアーゼの配列特異性を変える、前述の塩基接触残基の新規修飾の内の一つ以上と組み合わせてもよいし、前述の従来技術の修飾と組み合わせてもよいし、新規修飾と従来技術の修飾との組み合わせと組み合わせてもよい。特に、バックボーン接触修飾および塩基接触修飾を組み合わせることによって、組み換えメガヌクレアーゼを、所望の特異性および活性を持つように合理的に設計することが可能である。例えば、塩基接触残基に対する設計変更に由来する親和度の損失を逆転するDNA−結合親和度の増加を設計することが可能であり、配列特異性を減少させ酵素に対する認識配列の組を広げる親和度減少を設計することも可能である。
【0023】
他の側面から考えると、本発明は、ホモ又はヘテロダイマー形成に対する親和度を変更させた合理的設計メガヌクレアーゼモノマーを提供する。ダイマー形成に対する親和度は、同じモノマー(すなわち、ホモダイマー形成)によって又は異なるモノマー(すなわち、ヘテロダイマー形成)、例えば、参照野生型メガヌクレアーゼによって測定することが可能である。これらの組み換えメガヌクレアーゼは、メガヌクレアーゼダイマーにおけるモノマー間のタンパク質とタンパク質との境界面に存在するアミノ酸残基に対する修飾を有する。これらの修飾は、ヘテロダイマー形成の促進及び非パリンドローム認識配列を持つメガヌクレアーゼの創製に使用することが可能である。
【0024】
したがって、他の実施態様では、本発明は、参照メガヌクレアーゼモノマーによるダイマー形成に対する親和度を変更させた組み換えメガヌクレアーゼを提供する。この実施態様では、組み換えモノマーは、配列番号1のI−CREIメガヌクレアーゼの残基2−153に対して少なくとも85%の配列類似性を有するポリペプチドを含むが、ダイマー形成に対する親和度は、(A)D若しくはEによるK7、K57若しくはK96の置換、又は(B)K若しくはRによるE8若しくはE61の置換から選ばれる置換に対応する少なくとも一つの修飾によって変えられる。このような組み換えモノマーに基づき、本発明はさらに、(1)配列番号1のI−CREIメガヌクレアーゼの残基2−153に対して少なくとも85%の配列類似性を有する第1ポリペプチドであって、ダイマー形成に対する親和度が、(A)D若しくはEによるK7、K57若しくはK96の置換から選ばれる置換に対応する少なくとも一つの修飾によって変えられる第1ポリペプチド、及び(2)配列番号1のI−CREIメガヌクレアーゼの残基2−153に対して少なくとも85%の配列類似性を有する第2ポリペプチドであって、ダイマー形成に対する親和度が、(B)K若しくはRによるE8若しくはE61の置換から選ばれる置換に対応する少なくとも一つの修飾によって変えられる第2ポリペプチドを含む組み換えメガヌクレアーゼ・ヘテロダイマーを提供する。
【0025】
他の実施態様では、本発明は、参照メガヌクレアーゼモノマーによるダイマー形成に対する親和度を変更させた組み換えメガヌクレアーゼモノマーを提供する。この実施態様では、組み換えモノマーは、配列番号6のI−MSOIメガヌクレアーゼの残基6−160に対して少なくとも85%の配列類似性を有するポリペプチドを含むが、ダイマー形成に対する親和度は、(A)D若しくはEによるR302の置換、又は(B)K若しくはRによるD20、E11若しくはQ64の置換から選ばれる置換に対応する少なくとも一つの修飾によって変えられる。このような組み換えモノマーに基づき、本発明はさらに、(1)配列番号6のI−MSOIメガヌクレアーゼの残基6−160に対して少なくとも85%の配列類似性を有する第1ポリペプチドであって、ダイマー形成に対する親和度が、(
A)D若しくはEによるR302の置換から選ばれる置換に対応する少なくとも一つの修飾によって変えられる第1ポリペプチド、及び(2)配列番号6のI−MSOIメガヌクレアーゼの残基6−160に対して少なくとも85%の配列類似性を有する第2ポリペプチドであって、ダイマー形成に対する親和度が、(B)K若しくはRによるD20、E11若しくはQ64の置換から選ばれる置換に対応する少なくとも一つの修飾によって変えられる第2ポリペプチドを含む組み換えメガヌクレアーゼ・ヘテロダイマーを提供する。
【0026】
他の実施態様では、本発明は、参照メガヌクレアーゼモノマーによるダイマー形成に対する親和度を変更させた組み換えメガヌクレアーゼモノマーを提供する。この実施態様では、組み換えモノマーは、配列番号12のI−CEUIメガヌクレアーゼの残基5−211に対して少なくとも85%の配列類似性を有するポリペプチドを含むが、ダイマー形成に対する親和度は、(A)DまたはEによるR93の置換、又は(B)KまたはRによるE152の置換から選ばれる置換に対応する少なく共一つの修飾によって変えられる。このような組み換えモノマーに基づき、本発明はさらに、(1)配列番号12のI−CEUIメガヌクレアーゼの残基5−211に対して少なくとも85%の配列類似性を有する第1ポリペプチドであって、ダイマー形成に対する親和度が、(A)DまたはEによるR93の置換から選ばれる置換に対応する少なくとも一つの修飾によって変えられる第1ポリペプチド、及び(2)配列番号12のI−CEUIメガヌクレアーゼの残基5−211に対して少なくとも85%の配列類似性を有する第2ポリペプチドであって、ダイマー形成に対する親和度が、(B)KまたはRによるE152の置換から選ばれる置換に対応する少なくとも一つの修飾によって変えられる第2ポリペプチドを含む組み換えメガヌクレアーゼ・ヘテロダイマーを提供する。
【0027】
ダイマー形成に対する親和度を変更させた、組み換えメガヌクレアーゼモノマー又はヘテロダイマーはさらに、前述の塩基接触残基の修飾の内の一つ、二つまたは三つ以上;前述のバックボーン接触残基の修飾の内の一つ、二つまたは三つ以上、又は両者の組み合わせを含むことが可能である。したがって、例えば、モノマーの塩基接触部は、配列特異性を変えるように修飾することが可能であり、モノマーのバックボーン接触部は、DNA結合親和度を変えるように修飾することが可能であり、タンパク質とタンパク質との境界面は、ダイマー形成に影響を及ぼすように修飾することが可能である。このような組み換えモノマーは、同様に修飾されたモノマーと組み合わせて、所望の配列特異性および活性を有する合理設計メガヌクレアーゼ・ヘテロダイマーを生産することが可能である。
【0028】
他の側面から考えると、本発明は、本発明において記載され可能とされた合理設計メガヌクレアーゼのために使用される各種方法を提供する。これらの方法は、遺伝学的に修飾された細胞および生物体を生産すること、病気を遺伝子治療によって治療すること、病原体感染を治療すること及び診断学と研究用とのインビトロ応用のために組み換えメガヌクレアーゼを使用することを含む。
【0029】
したがって、この側面では、本発明は、染色体に挿入された興味の外因性配列を含む遺伝学的に修飾された真核細胞を生産するための方法であって、これを、該細胞を、(I)本発明のメガヌクレアーゼをコードする第1核酸配列、及び(II)前記興味の配列を含む第2核酸配列によってトランスフェクトすることによって実現する方法を提供する。その際、メガヌクレアーゼは、染色体の中に切断部位を生産し、興味の配列は、相同組み換え又は非相同的末端接合によって、染色体中の切断部位にに挿入される。
【0030】
それとは別に、他の側面では、本発明は、染色体に挿入された興味の外来配列を含む遺伝学的に修飾された真核細胞を生産するための方法であって、この細胞に本発明のメガヌクレアーゼタンパクを挿入すること及びこの細胞を興味の配列を含む核酸によってトランスフェクトすることによって実現する方法を提供する。その際、メガヌクレアーゼは、染
色体の中に切断部位を生産し、興味の配列は、相同組み換え又は非相同的末端接合によって、切断部位において染色体の中に挿入される。
【0031】
他の側面では、本発明は、染色体中の標的配列を破壊することによって遺伝学的に修飾された真核細胞を生産するための方法であって、この細胞を本発明のメガヌクレアーゼタンパクをコードする核酸によってトランスフェクトすることによって実現する方法を提供する。その際、メガヌクレアーゼは、染色体の中に切断部位を生産し、標的配列は切断部位における非相同的末端接合によって破壊される。
【0032】
他の側面では、本発明は、遺伝学的に修飾された生物体を生産するための方法であって、前述の方法に従って遺伝学的に修飾された真核細胞を生産し、かつ、この遺伝学的に修飾された真核細胞を育成して遺伝学的に修飾された生物体を生産することによって実現する方法を提供する。これらの実施態様では、真核細胞は、配偶子、接合子、胚盤胞細胞、胚幹細胞及びプロトプラスト細胞から選ぶことが可能である。
【0033】
他の側面では、本発明は、真核生物において遺伝子治療によって疾患を治療するための方法であって、この真核生物の少なくとも一つの細胞を、(I)本発明のメガヌクレアーゼをコードする第1核酸配列、及び(II)興味の配列を含む第2核酸配列によってトランスフェクトすることによって実現する方法を提供する。その際、メガヌクレアーゼは、染色体の中に切断部位を生産し、興味の配列は、相同組み換え又は非相同的末端接合によって染色体の中に挿入され、興味の配列の挿入は、疾患に対する遺伝子治療をもたらす。
【0034】
それとは別に、他の側面では、本発明は、真核生物において遺伝子治療によって疾患を治療するための方法であって、この真核生物の少なくとも一つの細胞の中に本発明のメガヌクレアーゼを導入すること及びこの細胞を、興味の配列を含む核酸によってトランスフェクトすることによって実現する方法を提供する。その際、メガヌクレアーゼは、染色体の中に切断部位を生産し、興味の配列は、相同組み換え、または非相同的末端接合によって切断部位において染色体の中に挿入され、興味の配列の挿入は、該疾患に対する遺伝子治療をもたらす。
【0035】
他の側面では、本発明は、真核生物において、該真核生物の染色体における標的配列を破壊する遺伝子治療によって疾患を治療するための方法であって、この真核生物の少なくとも一つの細胞を、本発明のメガヌクレアーゼをコードする核酸によってトランスフェクトすることによって実現する方法を提供する。その際、メガヌクレアーゼは、染色体の中に切断部位を生産し、標的配列は、相同組み換え又は非相同的末端接合によって破壊され、該標的配列の破壊が、該疾患に対する遺伝子治療をもたらす。
【0036】
別の局面では、本発明は、真核細胞宿主において、ウィルス性または前核細胞性病原体感染を治療するための方法であって、この病原体のゲノムにおける標的配列を破壊することによって実現する方法を提供する。その際、メガヌクレアーゼは、染色体の中に切断部位を生産し、標的配列は、(1)切断部位における非相同的末端接合によって、又は(2)第2核酸との相同組み換えによって破壊され、かつ、標的配列の破壊は、この感染に対する治療をもたらす。
【0037】
さらに一般的に、他の側面では、本発明は、認識配列の少なくとも一つの塩基位置に対する特異性を変更させた組み換えメガヌクレアーゼを合理的に設計する方法であって、(1)参照メガヌクレアーゼ−DNA複合体の三次元構造の少なくとも一部を決定すること、(2)該塩基位置において塩基接触面を形成するアミノ酸残基を特定すること、(3)接触面の少なくとも第1残基のβ−炭素と、該塩基位置における少なくとも第1塩基との間の距離を決めること及び(4)(A)第1塩基から6オングストローム未満の距離にあ
る第1残基については、G、C、T若しくはAの内の適切なもののメンバーである基1及び/又は基2の置換を選択すること;又は(B)前記第1塩基から6オングストロームより遠い位置にある第1残基については、G、C、T及びAの内の適切なもののメンバーである基2及び/又は基3の置換を選択することによって、所望の変化を促進するアミノ酸置換を特定することによって実現する方法を提供する。なお、上述したそれぞれの基は、後述するように定義する。この方法は、同じ塩基に対する別の接触残基ついても、同じ位置における他の塩基に対する接触残基についても、さらに、別の位置についても繰り返してよい。
【0038】
さらに、他の側面では、本発明は、DNA−結合親和度を増大させた組み換えメガヌクレアーゼを合理的に設計する方法であって、(1)基準メガヌクレアーゼ−DNA複合体の三次元構造の少なくとも一部を決定すること、(2)バックボーン接触面を形成するアミノ酸接触残基を特定すること、(3)(A)陰性荷電若しくは疎水性側鎖を持つ接触残基について、非荷電/極性若しくは陽性荷電側鎖を持つ置換を選択すること;又は(B)非荷電/極性側鎖を持つ接触残基について、陽性荷電側鎖を持つ置換を選択することによって、DNA−結合親和度を増大させるアミノ酸置換を特定すること、によって実現する方法を提供する。逆に、本発明は、DNA−結合親和度を減少させた組み換えメガヌクレアーゼを合理的に設計する方法であって、これを、(1)基準メガヌクレアーゼ−DNA複合体の三次元構造の少なくとも一部を決定すること、(2)バックボーン接触面を形成するアミノ酸接触残基を特定すること、(3)(A)陽性荷電側鎖を持つ接触残基について、非荷電/極性若しくは陰性荷電側鎖を持つ置換を選択すること、又は(B)疎水性及び非荷電/極性側鎖を持つ接触残基について、陰性荷電側鎖を持つ置換を選択することによって、DNA−結合親和度を減少させるアミノ酸置換を特定することよって実現する方法を提供する。
【0039】
本発明の、上記、およびその他の側面および実施態様は、本発明の下記の詳細な説明に基づき当業者には明白であろう。
【発明を実施するための最良の形態】
【0040】
1.1序論
本発明は、一部は、メガヌクレアーゼが2本鎖DNA認識配列とコンタクトする際、DNA塩基と特異的接触を形成し、かつ、DNAバックボーンと非特異的接触を形成し、そうすることによって、該酵素の認識配列特異性およびDNA−結合親和度に影響を及ぼす、メガヌクレアーゼLAGLIDADGファミリーにおける特異的アミノ酸残基の特定および特徴解明に基づく。この発見は、以下に詳細に説明するように、メガヌクレアーゼの特異性及び/又は親和性を変えることが可能な、メガヌクレアーゼにおけるアミノ酸置換を特定するため及び天然のメガヌクレアーゼが認識しない所望のDNA配列を認識することが可能であり、及び/又は、天然のメガヌクレアーゼに比べて特異性及び/又は親和度が増大または減少させたメガヌクレアーゼを合理的に設計し、開発するために用いられた。さらに、DNA−結合親和度は、酵素活性の外に、配列特異性にも影響を及ぼすので、本発明は、天然メガヌクレアーゼに比べ活性を変更させた、合理的設計メガヌクレアーゼを提供する。さらに、本発明は、ダイマーを形成するためにコンタクトされたモノマー間のインターフェイスにおける残基が、ヘテロダイマー形成を促進するように修飾された合理設計メガヌクレアーゼを提供する。最後に、本発明は、組み換え細胞および生物体の生産における使用ばかりでなく、本出願に開示されるように、遺伝子療法、抗病原体、抗癌及びインビトロ応用における合理設計メガヌクレアーゼの使用を提供する。
【0041】
一般的出来事として、本発明は、(1)2本鎖DNA認識配列中の個々の塩基に対する配列特異的結合、又は(2)2本鎖DNA分子のフォスフォジエステルバックボーンに対する非特異的結合による、メガヌクレアーゼ内の複数部位において変更アミノ酸残基を含
む、合理設計LAGLIDADGメガヌクレアーゼを生成するための方法を提供する。しかしながら、酵素活性は、DNA−結合親和度と相関するのであるから、DNA認識配列に対する結合に与るアミノ酸を変えることは、2本鎖DNAに対する全体的結合親和度を増大又は減少することになり、特異的塩基対相互作用を介するメガヌクレアーゼの特異性ばかりでなく、メガヌクレアーゼの活性も変える可能性がある。同様に、DNAバックボーンに対する結合によるアミノ酸を変えることは、2本鎖DNAに対する全体的結合親和度を増大又は減少することによって、酵素の活性ばかりでなく、認識配列に対する結合の特異性又は縮重の程度を変える可能性がある。
【0042】
下記に詳述するように、メガヌクレアーゼの合理的設計法は、DNA認識/結合によるアミノ酸を特定すること及び適切なアミノ酸変化を選択するために一連の規則を適用することを含む。この規則は、メガヌクレアーゼの配列特異性に関して、メガヌクレアーゼのアミノ酸側鎖とDNAのセンス鎖及びアンチセンス鎖における塩基の間とのメガヌクレアーゼ−DNA複合体距離に関する互いの立体位置、アミノ酸側鎖とDNA塩基との相対位置及び非共有的化学的相互作用を考慮に含んでいる。
【0043】
最後に、ホモダイマーとしてDNAに結合する天然メガヌクレアーゼの大部分は、擬似、又は完全パリンドロームの認識配列を認識する。長々としたパリンドロームはめったにないと予想されるので、ゲノムの興味の配列部位でパリンドロームに遭遇する確率はきわめて低い。したがって、これらの酵素を、ゲノムの興味の配列部位を認識するように再設計しなければならないとすると、異なる半分部位を認識する二つの酵素モノマーにおいて、ヘテロダイマー形成されると非パリンドローム的ハイブリッド認識配列を切断することが可能な二つの酵素モノマーを設計することが必要である。それゆえ、ある側面では、本発明は、少なくとも一つのアミノ酸位置で異なる二つのモノマーをダイマー化して、ヘテロダイマーを形成して得られる合理設計メガヌクレアーゼを提供する。ある場合、両モノマーは、非パリンドローム的認識配列を認識するヘテロダイマーを形成するように合理的設計される。二つの異なるモノマーから成る混合物は、最大三つの活性形メガヌクレアーゼダイマー、二つのホモダイマー及びヘテロダイマーをもたらす。それに加えて、またはそれとは別に、ある場合、ホモダイマーまたはヘテロダイマー形成の確率を増すか、または減らすために、モノマー同士が相互作用を持ってダイマーを形成する境界面においてアミノ酸残基が変えられる。
【0044】
したがって、一つの側面では、本発明は、LAGLIDADGメガヌクレアーゼにおいて、酵素の特異性及び/又は活性を変えるアミノ酸変化を含むLAGLIDADGメガヌクレアーゼの合理的設計法を提供する。他の側面では、本発明は、これらの方法によってもたらされる合理設計メガヌクレアーゼを提供する。他の側面では、本発明は、生物体ゲノム内部の所望のDNA配列または遺伝子座が、DNA配列の挿入、欠失、置換、またはその他の操作によって修飾される、組み換え核酸および生物体を生産するために、このような合理設計メガヌクレアーゼを使用する方法を提供する。さ他の側面では、本発明は、病原体または癌細胞において、病原体特異的又は癌特異的認識配列を有する合理設計メガヌクレアーゼを用いて、この病原体または癌細胞の生存率を下げるための方法を提供する。
【0045】
1.2 参照および定義
本出願において参照される特許および科学文献は、当業者にとって利用可能な知識を確定する。本出願において引用される、公刊された米国特許、承認済みの出願、出版された外国特許、および、GENBANKのデータベース配列を含む参照資料は、参照により、あたかも、それぞれが、特異的、個別的に、参照によって含まれるのと同程度に、本出願に含まれる。
【0046】
本出願で用いる「メガヌクレアーゼ」という用語は、12塩基対よりも大きい認識配列で2本鎖DNAに結合するエンドヌクレアーゼを指す。天然のメガヌクレアーゼはモノマー(例えば、I−SceI)又はダイマー(例えば、I−CreI)であることが可能である。本出願で用いるメガヌクレアーゼという用語は、モノマー・メガヌクレアーゼ、ダイマー・メガヌクレアーゼ、または、コンタクトしてダイマー形メガヌクレアーゼを形成するモノマーを指す。「ホーミング・エンドヌクレアーゼ」という用語は、「メガヌクレアーゼ」という用語と同義である。
【0047】
本出願で用いる「LAGLIDADGメガヌクレアーゼ」という用語は、天然ではダイマー形である単一のLAGLIDADGモチーフを含むメガヌクレアーゼ、又は天然ではモノマー形である二つのLAGLIDADGモチーフを含むメガヌクレアーゼを指す。本出願で用いる「モノLAGLIDADGメガヌクレアーゼ」という用語は、単一のLAGLIDADGモチーフを含むメガヌクレアーゼを指し、本出願で用いる「ジLAGLIDADGメガヌクレアーゼ」という用語は、二つを区別することが必要な場合には、二つのLAGLIDADGモチーフを含むメガヌクレアーゼを指す。LAGLIDADGモチーフを含むジLAGLIDADGメガヌクレアーゼの二つの構造ドメインは、それぞれ、LAGLIDADGサブユニットと呼ぶことが可能である。
【0048】
本出願で用いる「合理的に設計された」という用語は、非天然及び/又は遺伝子工学的に加工されたことを意味する。本発明の合理的設計メガヌクレアーゼは、野生型又は天然のメガヌクレアーゼとアミノ酸配列または一次構造において異なり、さらに、二次、三次又は四次構造において異なる場合がある。加えて、本発明の合理設計メガヌクレアーゼは、認識配列の特異性及び/又は活性においても、野生型又は天然メガヌクレアーゼと異なる。
【0049】
タンパク質に関して本出願で用いる「組み換え」という用語は、タンパク質をコードする核酸に対し遺伝子工学技術を適用した結果アミノ酸配列が変化すること、及びこのタンパク質を発現する細胞又は生物体が得られることを意味する。核酸に関して「組み換え」という用語は、遺伝子工学技術を適用した結果、核酸配列が変化することを意味する。遺伝子工学技術としては、これらにっ限定されるものではないが、PCR、トランスフェクション、形質転換及びその他の遺伝子転移技術等のDNAクローニング技術、相同組み換え、部位特異的変異処理及び遺伝子融合が含まれる。この定義によれば、天然のタンパクと同じアミノ酸配列を持つが、異種宿主におけるクローニング及び発現によって生産されるタンパク質は、「組み換え」とは見なされない。
【0050】
組み換えタンパクに関して本出願で用いられる「修飾」という用語は、基準配列(例えば、野生型)と比べた場合の、組み換え配列におけるアミノ酸残基の、どのようなものであれ、挿入、欠失又は置換を含む意味である。
【0051】
本出願で用いる「遺伝学的に修飾された」という用語は、そのゲノムDNA配列が、組み換え技術によって意図的に修飾されたゲノムDNA配列を有する細胞、生物体又は祖先を指す。本出願で用いる「遺伝学的に修飾された」という用語は、「トランスジェニック」という用語を含む。
【0052】
本出願で用いる「野生型」という用語は、どのようなものであれ、、メガヌクレアーゼの全ての天然形を指す。「野生型」という用語は、天然における酵素の最も一般的な変異を意味するものではなく、自然で見られるいかなる変異も意味する。野生型メガヌクレアーゼは、組み換え又は非天然メガヌクレアーゼと区別される。
【0053】
本出願で用いる「認識配列半分部位」又は単に「半分部位」は、モノ−LAGLIDA
DGメガヌクレアーゼのモノマー又はジ−LAGLIDADGメガヌクレアーゼの一つのLAGLIDADGサブユニットによって認識される2本鎖DNA分子間の核酸配列を意味する。
【0054】
本出願で用いる「認識配列」という用語は、モノ−LAGLIDADGメガヌクレアーゼダイマー又はジ−LAGLIDADGメガヌクレアーゼモノマーによって結合され、切断される一対の半分部位を指す。この二つの半分部位は、この酵素によって特異的に認識されない塩基対によって隔てられてもよいし、隔てられなくともよい。I−CREI、I−MSOI及びI−CEUIの場合、各モノマーの認識配列半分部位は9塩基対の幅を持ち、この二つの半分部位は、特異的には認識されないが実際の切断部位である塩基対(4塩基対のオーバーハングを有する)によって分割されている。したがって、I−CREI、I−MSOI及びI−CEUIメガヌクレアーゼダイマーの結合認識配列は、通常、4塩基対切断部位に側接する二つの9塩基対を含む22塩基対の幅を持つ。各半分部位の塩基対は−9から−1と表示され、−9位は切断部位からもっとも遠位であり、−1位はN1−N4と表示される4個の中央塩基対に隣接する。−9から−1へと向かう方向(すなわち、切断部位に向かう方向)において5′から3′に方向づけられる各半分部位鎖は、「センス」鎖と表示され、反対鎖は、「アンチセンス鎖」と表示される。ただし、いずれの鎖もタンパクをコードしない。したがって、図1(A)に示すように、一方の半分部位の「センス」鎖は、他方の半分部位のアンチセンス鎖である。ジ−LAGLIDADGメガヌクレアーゼモノマーであるI−SCEIメガヌクレアーゼの場合、認識配列は約18BPの非パリンドローム配列であり、特異的には認識されない中央塩基対は無い。通例に従って、2本鎖の一方は「センス」鎖と呼ばれ、他方は「アンチセンス」鎖と呼ばれるが、いずれの鎖もタンパクをコードしない。
【0055】
本出願で用いる「特異性」という用語は、メガヌクレアーゼが認識配列と呼ばれる特定の塩基対配列又は特定の一組の認識配列においてのみ、2本鎖DNA分子を認識し切断する能力を意味する。この一組の認識配列は、いくつかの保存位置または配列モチーフを共有するが、これらは1又は2以上の位置で縮重する場合がある。高い特異性を有するメガヌクレアーゼは、たった一つ又はきわめて少数の認識配列の切断が可能である。特異性は実施例1に記載される切断アッセイで決めることが可能である。本出願で用いる場合、メガヌクレアーゼは、基準メガヌクレアーゼ(例えば、野生型)によって結合及び切断されない認識配列に結合し、切断する場合、又は、認識配列の切断速度が、基準メガヌクレアーゼに比べて、統計的に有意な値(P<0.05)だけ上昇又は降下した場合に、特異性
が「変更された」ことになる。
【0056】
本出願で用いる「縮重」という用語は、「特異性」の反対を意味する。高縮重したメガヌクレアーゼは、非常に多岐にわたる認識配列を切断することができる。メガヌクレアーゼは単一の、複数の又は全てのの箇所の半分部位内に配列縮重を有することができる。このような配列縮重は、(I)メガヌクレアーゼのDNA結合ドメインにおける任意のアミノ酸が、認識配列内の1又は2以上の位置で任意の塩基と接触できないこと、(II)メガヌクレアーゼのDNA結合ドメインにおける1又は2以上のアミノ酸が、識配列内の1又は2以上の位置で1以上の塩基と特異的にコンタクトできること、及び/又は(III)活性に十分なほど、非特異的DNA結合親和性を持つこと、からもたらされる可能性がある。「完全な」縮重位置は任意の4塩基で占められてもよく、半分部位内では”N”で表示することができる。「部分的な」縮重位置は、4塩基の内の任意の2又は3塩基で占められてもよい。(例えば、プリン(PU)か、ピリミジン(PY)で占められるが、Gには占められない)。
【0057】
メガヌクレアーゼに関して本出願で使用される場合、「DNA−結合親和度」又は「結合親和度」は、メガヌクレアーゼが基準DNA分子(例えば、認識配列又は任意の配列)
と非共有的に結合する傾向を意味する。結合親和度は、解離定数KDによって測定される(例えば、WT認識配列に対するI−CREIのKDは約0.1NMである)。本出願で用いる場合、基準認識配列に対する組み換えメガヌクレアーゼのKDが、基準メガヌクレアーゼに比べて、統計的に有意な値(P<0.05)だけ増加又は減少した場合に、結合
親和度を「変更させた」ことになる。
【0058】
メガヌクレアーゼモノマーに関して本出願で用いる場合、「ダイマー形成に対する親和度」という用語は、メガヌクレアーゼモノマーが、基準メガヌクレアーゼモノマーと非共有的に結合する傾向を意味する。ダイマー形成に対する親和度は、例えば、基準野生型メガヌクレアーゼのような同じモノマー(すなわち、ホモダイマー形成)又は異なるモノマー(すなわち、ヘテロダイマー形成)で測定することができる。結合親和度は、解離定数KDによって測定される。本出願で用いる場合、基準認識配列に対する組み換えメガヌクレアーゼのKDが、基準メガヌクレアーゼに比べて、統計的に有意な値(P<0.05)だけ増加又は減少した場合に、結合親和度を「変更させた」ことになる。
【0059】
本出願で用いる「パリンドローム的」という用語は、同じ半分部位の逆方向反復を含む認識配列を指す。しかしながら、この場合、パリンドローム配列は酵素にコンタクトされない四つの中央塩基対に関してパリンドローム的である必要はない。ダイマー形メガヌクレアーゼの場合、パリンドロームDNA配列は二つのモノマーが同じ半分部位にコンタクトするホモダイマーによって認識される。
【0060】
本出願で用いる「擬似パリンドローム的」という用語は、非同一又は不完全なパリンドローム的半分部位の逆方向反復を含む認識配列を指す。この場合、擬似パリンドローム配列は、四つの中央塩基対に関してパリンドローム的である必要はなく、二つの半分部位の間のパリンドローム配列からもはずれることが可能である。擬似パリンドロームDNA配列は、二つの同一の酵素モノマーが、異なる半分部位にコンタクトする、野生型ホモダイマー形メガヌクレアーゼによって認識される天然DNA部位において典型的に見られる。
【0061】
本出願で用いる「非パリンドローム的」という用語は、メガヌクレアーゼの、二つの、無関係の半分部位から構成される認識配列を指す。この場合、非パリンドローム配列は、四つの中央塩基対に対しても、二つのモノマー半分部位に対しても、パリンドローム的である必要はない。非パリンドロームDNA配列は、ジ−LAGLIDADGメガヌクレアーゼ、高縮重モノ−LAGLIDADGメガヌクレアーゼ(例えば、I−CeuI)、非同一半分配列を認識するモノ−LAGLIDADGメガヌクレアーゼのヘテロダイマーのいずれかによって認識される。
【0062】
本出願で用いる「活性」という用語は、本発明のメガヌクレアーゼが、特定の認識配列を切断する速度を指す。このような活性は、2本鎖DNAのフォスフォジエステル結合の加水分解に関する測定可能な酵素反応である。ある特定のDNA基質に作用するメガヌクレアーゼの活性は、その特定のDNA基質に対するメガヌクレアーゼの親和力又は結合力に影響され、DNAとの配列特異的相互作用及び非配列特異的相互作用によって影響される。
【0063】
本出願で用いる「相同組み換え」という用語は、2本鎖DNA切断が、修復鋳型として相同DNA配列を用いて修復される、天然の、細胞プロセスを指す(例えば、CAHILL ET AL.(2006),FONT.BIOSCI.11:1958−1976を参照)。相同のDNA配列は、内因性の染色体配列であってもよいし、細胞に輸送された外来性の核酸であってもよい。したがって、ある実施態様では、合理設計メガヌクレアーゼは、標的配列内の認識配列を切断するために使用され、標的配列に対して相同な又はほぼ同様の配列類似性を持つ外来性の核酸が細胞に輸送されて、相同性組み換えによる修復
のための鋳型として使用される。したがって、標的配列とは著明に異なっていてもよい外来性のDNA配列が、染色体配列に組み込まれる。相同組み換えプロセスは、主に真核生物において起こる。本出願では、「相同性」という用語は「配列類似性」と等価的に用いられ、出自又は系統関係による同一性を要求することを意図するものではない。
【0064】
本出願で用いる「非相同的末端接合」という用語は、2本鎖DNA切断が、2本の非相同的DNAセグメントの直接的接合によって修復される、天然の細胞プロセスを指す(例えば、CAHILL ET AL..(2006),FRONT.BIOSCI.11:1958−1976)。非相同的末端接合によるDNA修復は、間違いを起こしやすく、修復部位において鋳型と対応しないDNA配列の付加又は除去をもたらすことが多い。したがって、ある実施態様では、(例えば、塩基挿入、塩基欠失又はフレームシフト突然変異を行うことによって)非相同的末端接合によって遺伝子を破壊し、標的配列内のメガヌクレアーゼ認識配列中に二本鎖切断点を作るために、合理設計メガヌクレアーゼを用いることも可能である。他の実施態様では、標的配列に対する相同性を欠如するか又は標的配列に対する実質的配列類似性を欠如する外来性の核酸が、非相同的末端接合によって、メガヌクレアーゼ刺激2本鎖DNA切断部位で取り込まれてもよい(例えば、SALOMON,ET AL.(1998),EMBO J.17:6086−6095を参照)。非相同的末端接合プロセスは、真核生物でも、細菌のような原核生物でも起こる。
【0065】
本出願で用いる「興味の配列」という用語は、メガヌクレアーゼタンパクを用いてゲノムへ挿入するか又はゲノムDNA配列を置換するのに使用することが可能な核酸配列であって、タンパク、RNA又は調整エレメント(例えば、エンハンサー、サイレンサー、またはプロモーター配列)のどれをコードするかにはよらない任意の核酸配列を意味する。興味の配列は、興味の配列から発現されるタンパク質又はRNAの標識化を可能とする非相同的なDNA配列を有することも可能である。例えば、以下のものに限定さるものではないが、タンパク質は、エピトープ(例えば、C−MYC、FLAG)、または他のリガンド(例えば、ポリ−HIS)を含むタグによって標識することが可能である。更に、興味の配列は、従来技術で周知の技術に従って融合タンパクをコードしてもよい(例えば、AUSUBEL ET AL., 「分子生物学における最近のプロトコール(“CURRENT PROTOCOLS IN MOLECULAR BIOLOGY,”)」、WILEY 1999)。ある実施態様では、隣接配列は、組み換えメガヌクレアーゼによって切断され、ゲノム組み替え配列の中に興味の配列の適切な挿入される。
【0066】
したがって、この側接配列は切断され、該組み換えメガヌクレアーゼによって切断されるゲノム認識配列の中に、興味の配列を適正に挿入することを可能となる。ある実施態様では、興味の配列の全ては、ゲノム中の標的配列と相同であるか又は標的配列と実質的に配列類似性を有しており、相同組み換えによって、標的配列が興味の配列によって効果的に置換される。他の実施態様では、興味の配列は、標的配列と相同性を有するか又は実質的配列類似性を有するDNA配列によって側接され、相同組み換えによってゲノム内の標的配列座位において興味の配列が挿入される。ある実施態様では、興味の配列は、メガヌクレアーゼ認識配列における突然変異又は他の修飾を除いては、標的配列とほぼ同一であり、そのため、メガヌクレアーゼは、標的配列が興味の配列によって修飾された後に、標的配列を切断することができない。
【0067】
アミノ酸配列及び核酸配列に関して本出願で用いる場合、「類似性パーセント」及び「配列類似性」という用語は、アライメントされたアミノ酸残基又はヌクレオチド間の類似性を最大にするようにアライメントされた配列に基づく2つの配列の類似度の測定値を意味する。そしてこれは、アライメント配列中のギャップ及びギャップの数、核酸又は残基の総数、同一又は類似する核酸又は残基の数の関数による。一般的なパラメータを用いて配列類似性を決めるために、様々のアルゴリスムおよびコンピュータプログラムの利用が
可能である。本出願では、配列類似度は、アミノ酸配列の場合はBLASTPプログラムを用い、核酸配列の場合はBLASTNプログラムを用いて算出される。両方とも、NATIONAL CENTER FOR BIOTECHNOLOGY INFORMATION(WWW.NCBI.NLM.NIH.GOV/)を通じて使用可能であり、例えば、ALTSCHUL ET AL.(1990), J.MOL.BIOL.215:403−410;GISH AND STATES(1993), NATURE GENET.3:266−272;MADDEN ET AL.(1996),METH.ENZYMOL.266:131−141;ALTSCHUL ET AL.(1997),NUCLEIC ACIDS RES.25:3389−3402);ZHANG ET AL.(2000) J.COMPUT.BIOL.7(1−2):203−14に記載される。本出願では、二つのアミノ酸配列の類似パーセントは、BLASTPアルゴリスムにおける下記のパラメータに基づくスコアである。単語サイズ=3;ギャップ開放ペナルティー=−11;ギャップ延長ペナルティー=−1;及びスコアリングマトリックス=BLOSUM62である。本出願では、二つの核酸配列の類似パーセントは、BLASTNアルゴリスムにおける下記のパラメータに基づくスコアである。単語サイズ=11;ギャップ開放ペナルティー=−5;ギャップ延長ペナルティー=−2;マッチ評価=1
;及びミスマッチペナルティー=−3である。
【0068】
二つのタンパク質又はアミノ酸配列の修飾に関して本出願で用いる場合、「対応する」という用語は、(例えば、BLASTPプログラムを用いて)二つのタンパク質を一般的な方法で配列アライメントしたとき、第1のタンパク質の修飾が、第2のタンパク質の修飾と同じアミノ酸残基の置換であること、及び第1のタンパク質における修飾のアミノ酸位置が、第2のタンパク質の修飾のアミノ酸位置に対応する又は揃うことを意味する。したがって、もし残基XとYとが配列アライメントで互いに対応するものでれば、XとYの配列番号が異なるものであっても、第1のタンパク質における残基“A”がアミノ酸“A”に修飾されることは、残基“Y”がアミノ酸“A”に修飾されることと対応する。
【0069】
本出願では、ある変数に対する数値範囲は、その範囲内であれば、任意の数値に等しい変数を用いて本発明が実行可能であることを意図する。したがって、本来的に不連続な変数である場合、この変数は、その数値範囲の両端を含む任意の整数に等しくなることが可能である。同様に、本来的に連続的な変数である場合、この変数は、その数値範囲の両端を含む任意の実数に等しくなることが可能である。例えば、ただしこれらに限定されるものではないが、0と2の間の値を持つと記載される変数は、もしこの変数が本来的に不連続である場合は、0、1又は2を取ることが可能であり、もしこの変数が本来的に連続的である場合は、数値0.0、0.1、0.01、0.001又は≧0及び≦2の実数の任意の実数を取ることができる。
【0070】
本出願では、特に別段の指定が無い限り、「又は」という単語は、「及び/又は」という包括的意味で使われ、「又は/どちらか」という排除的意味では使われない。
【0071】
2.1 配列特異性を変更させた合理的設計メガヌクレアーゼ
本発明の一様態では、組み換えLAGLIDADGファミリーのメガヌクレアーゼを合理的に設計する方法が提供される。この様態では、先ず、半分部位の各位置における塩基指向性を変えることが可能なアミノ酸置換を予測することによって、組み換えメガヌクレアーゼを合理的に設計する。これらの置換は、個別に又は組み合わせて、実験的にその有効性が検証され、所望の切断特異性を持つメガヌクレアーゼを生産する。
【0072】
本発明によれば、塩基指向性において所望の変化をもたらすアミノ酸置換は、メガヌクレアーゼのDNA認識配列及びDNAフォスフォジエステルバックボーンの核酸塩基との接触に参加が可能な基準メガヌクレアーゼ(例えば、野生型メガヌクレアーゼ又は非天然
の基準メガヌクレアーゼ)のアミノ酸側鎖及びそれら接触部の空間的、化学的性質を決めることによって予測される。これらのアミノ酸としては、基準DNA半分部位との接触に与る側鎖が挙げられるが、ただしこれらに限定されない。一般に、この決定には、メガヌクレアーゼと、その2本鎖DNA認識配列との間の複合体の構造に関する知識、又はきわめて類似性の高い複合体の構造(例えば、同じメガヌクレアーゼと、別のDNA認識配列との間の複合体構造、又はメガヌクレアーゼの対立遺伝子変種または系統的変種と、そのDNA認識配列との間の複合体構造)に関する知識が必要である。
【0073】
原子座標データとして記載される、ポリペプチド又は2個以上のポリペプチドから成る複合体の三次元構造は、いくつかの方法で獲得することが可能である。例えば、タンパク質の構造の決定は、以下のものに限定されるものではないが、X線結晶学、NMR及び質量分析を含む技術を用いて実行することが可能である。もう一つの方法は、興味のメガヌクレアーゼ又は関連ヌクレアーゼの、既存の構造座標のデータベースを分析することである。このような構造データは、多くの場合、三次元座標形式のデータベースから入手することが可能である。多くの場合、このデータは、オンラインデータベースを通じて(例えば、WWW.RCSB.ORG/PDBにおいてRCSB PROTEIN DATA BANKへ)アクセスすることが可能である。
【0074】
構造情報は、タンパク質またはタンパク質複合体の、規則的な、二次元又は三次元アレイ(例えば、結晶)によって得られる。例えば、X線又は電子の回折パターンを分析することによって実験的に得ることが可能である。回折データを空間における三次元原子座標に変換するためには、コンピュータ法が使用される。例えば、X線結晶学の分野は、メガヌクレアーゼを含めた、多くのタンパク質−DNA複合体の三次元構造情報の生成のために使用されている(例えば、CHEVALIER ET AL.(2001),NUCLEIC ACIDS RES.29(18):3757−3774を参照)。
【0075】
溶液中の分子の原子間距離の決定には、核磁気共鳴(NMR)も用いられている。コンピュータ法と組み合わせた多次元NMR法は、増大するサイズのポリペプチドの原子座標の決定に成功した(例えば、TZAKOS ET AL.(2006),ANNU.REV.BIOPHYS.BIOMOL.STRUCT.35:12−42を参照)。
【0076】
それとは別に、タンパク質/DNAの既知の一次構造及び利用可能であれば、二次、三次、及び/又は四次構造のほか、アミノ酸側鎖、核酸塩基及び結合性相互作用に関する既知の物理化学的性質に基づいたアルゴリスムを適用することによって、コンピュータモデルを使用することも可能である。このような方法は、任意に、繰り返し法又は実験的に誘導された制限を含むことも可能である。このようなコンピュータソフトウェアの一例として、ADAMS ET AL.(1999),ACTA CRYSTALLOGR.D.BIOL.CRYSTALLOGR.55(PT 1):181−90によって記載されるCNSプログラムがある。他にも、タンパク質の構造におけるアミノ酸の空間配置及びタンパクのアミノ酸側鎖と各種標的分子との相互作用を予測する、様々のコンピュータプログラムが開発されている(例えば、米国特許第6,988,041号を参照)。
【0077】
したがって、本発明のある実施態様では、DNA核酸塩基と特異的に相互作用を持ち、及び/又は、非特異的フォスフォジエステルバックボーン相互作用を促進する、特異的アミノ酸残基を特定するために、コンピュータモデルが使用される。例えば、可能なメガヌクレアーゼ−DNA相互作用の全体に関するコンピュータモデルを、例えば、以下に限定されるものではないが、MOLSCRIPT(登録商標)2.0(AVATAR SOFTWARE AB,STOCKHOLM,SWEDEN)、グラフィックディスプレイO(JONES ET AL.(1991),ACTA CRYSTALLOGRAPHY,A47:110)、グラフィックディスプレイ・プログラムGRASP(登録商標)(
NICHOLLS ET AL.(1991),PROTEINS,STRUCTURE,FUNCTION AND GENETICS 11(4):281FF)、グラフィックディスプレイ・プログラムINSIGT(登録商標)(TSI,INC.,SHOREVIEW、ミネソタ州)を含む、適切なソフトウェアプログラムを用いて生産することが可能である。タンパク質−DNA複合体の、三次元構造描画を生産し、視認し、かつ操作するために好適なコンピュータハードウェアは市販されており、従来技術で周知である(例えば、SILICON GRAPHICS WORKSTATION,SILICON GRAPHICS,INC.,MOUNTAINVIEW、カリフォルニア州)。
【0078】
具体的に述べると、メガヌクレアーゼと、その2本鎖DNA認識配列との間の相互作用は、従来技術で既知の方法を用いて分解することが可能である。例えば、そのために結晶が生産される、複数成分の複合体構造の三次元構造の描画、またはモデルは、分子置換、またはSIR/MIR(SINGLE/MULTIPLE ISOMORPHOUS REPLACEMENT)(例えば、BRUNGER(1997),METH.ENZYM.276:558−580;NAVAZA AND SALUDIJIAN(1997),MATH.ENZYM.276:581−594;TONG AND ROSSMANN(1997),METH.ENZYM.276:594−611;およびBENTLEY(1997),METH.ENZYM.276:611−619を参照)を含む技術を用いて決めることが可能であり、例えば、AMORE/MOSFLM(NAVAZA(1994),ACTA CRYST.A50:157−163;CCP4(1994),ACTA CRYST.D50:760−763)、または、XPLOR(BRUENGER ET AL.(1992),X−PLOR VERSION 3.1.A.SYSTEM FOR X−RAY CRYSTALLOGRAPHY AND NMR,YALE UNIVERSITY PRESS,NEW HAVEN、コネチカット州)などのソフトウェアを用いて実行することが可能である。
【0079】
タンパク質の構造及び可能なメガヌクレアーゼ−DNA相互作用の決定は、酵素活性および特異性に影響を及ぼすために変えることが可能なアミノ酸に関する合理的選択を可能にする。決定は、アミノ酸側鎖の、特定の塩基又はDNAフォスフォジエステルバックボーンとの相互作用に関するいくつかの因子に基づく。適切なアミノ酸置換を決めるために使用される化学的相互作用としては、以下に限定されるものではないが、ファンデルワールス力、立体障害、イオン結合、水素結合及び疎水性相互作用が挙げられる。メガヌクレアーゼと、可能な認識配列半分部位中のある特定の塩基との特定の相互作用を優先するか、又は忌避するアミノ酸置換が選択される。この選択は、その認識配列に対する特異性を、かつ、ある程度は、全体的結合親和度および活性を増すようにあるいは減らすように行われる。さらに、全体活性の増加又は減少のために、かつ、ある程度は、特異性を下げるか又は上げるかするために、2本鎖DNAのフォスフォジエステルバックボーンに対する結合性を増すか又は下げるアミノ酸置換を選択することが可能である。
【0080】
したがって、特定の実施態様では、メガヌクレアーゼ−DNA複合体の三次元構造が決定され、DNA認識配列半分部位における各塩基対について「接触面」が定義される。ある実施態様では、接触面は、その残基が、野生型メガヌクレアーゼ−DNA複合体において塩基接触を実行するか否かとは無関係に、ペアのいずれかの塩基における大溝水素結合ドナーまたはアクセプターから9.0オングストローム未満のβ−炭素を有する、酵素中のアミノ酸を含む。別の実施態様では、残基が、野生型メガヌクレアーゼ−DNA複合体において接触を行わない場合、この残基を排除することが可能であり、又は考慮される残基の数または名称を変えようとする設計者の意図に基づいて残基を含めること、又は排除することが可能である。後述の一実施例では、野生型I−CREI半分部位の塩基位置−2、−7、−8及び−9において、接触面は、野生型酵素−DNA複合体において実際に相互作用を持つアミノ酸位置に限定された。しかしながら、位置−1、−3、−4、−5
及び−6では、接触面は、野生型接触には与らないが、異なるアミノ酸によって置換された場合、塩基に接触する可能性のある、さらに新たなアミノ酸位置を含むように定義された。
【0081】
認識配列半分部位は、通常、DNAの1本鎖のみに関して表されるが、メガヌクレアーゼは、2本鎖DNAの大溝に結合し、両鎖の核酸塩基と接触することに注意しなければならない。さらに、「センス」および「アンチセンス」鎖という表示は、メガヌクレアーゼの結合と認識に関しては完全にその場まかせである。ある位置における配列特異性は、塩基対の一方のメンバーとの相互作用か又は塩基対の両方のメンバーとの相互作用の組み合わせのどちらかによって実現される。したがって、例えば、位置Xにおいて、該位置XではA塩基は「センス」鎖上にあり、T塩基が「アンチセンス」鎖上にある、A/T塩基対の存在を優先するためには、位置Xにおいてセンス鎖に接触できるほど十分に近接し、かつAの存在を好む残基が選択されるか、及び/又は、位置Xにおいてアンチセンス鎖に接触できるほど十分に近接し、かつTの存在を好む残基が選択される。本発明によれば、残基のβ−炭素が、関連塩基の最近接原子の9オングストローム以内にある場合、その残基は十分に近接すると考慮される。
【0082】
したがって、例えば、DNAセンス鎖の9オングストローム以内ではあるが、アンチセンス鎖から9オングストロームより大きく離れるβ−炭素を持つアミノ酸は、センス鎖に対してのみ相互作用を持つ可能性があると考慮される。同様に、DNAアンチセンス鎖の9オングストローム以内ではあるが、センス鎖から9オングストロームより大きく離れるβ−炭素を持つアミノ酸は、アンチセンス鎖に対してのみ相互作用を持つ可能性があると考慮される。両方のDNA鎖の9オングストローム以内にβ−炭素を持つアミノ酸は、どちらの鎖とも相互作用を持つ可能性があると考慮される。
【0083】
各接触面について、可能なアミノ酸置換について、4種のDNA塩基の内の一つ以上に対し好んで相互作用を持とうとする能力を予測し、その予測能力に基づいてアミノ酸置換が選択される。この選択プロセスは、二つの主要基準に基づく。(I)異なる核酸塩基との立体相互作用に影響を及ぼすアミノ酸側鎖のサイズ、及び(II)異なる核酸塩基との静電気的及び結合的相互作用に影響を及ぼすアミノ酸側鎖の化学的性質である。
【0084】
側鎖のサイズに関しては、接触面におけるアミノ酸のβ−炭素が、塩基から6オングストローム未満の距離にある場合には、比較的短い及び/又は比較的小さい側鎖を持つアミノ酸を選択することが可能であり、接触面におけるアミノ酸のβ−炭素が、塩基から6オングストロームより遠い距離にある場合には、比較的長い及び/又は比較的大きい側鎖を持つアミノ酸を選択することが可能である。サイズが中間のアミノ酸側鎖は、接触面におけるアミノ酸β−炭素が、塩基から5〜8オングストロームである場合に選択することが可能である。
【0085】
比較的短く、比較的小さい側鎖を持つアミノ酸は、グリシン(G)、アラニン(A)、セリン(S)、トレオニン(T)、システイン(C)、バリン(V)、ロイシン(L)、イソロイシン(I)、アスパラギン酸(D)、アスパラギン(N)及びプロリン(P)を含む1群に割り当てられる。しかしながら、プロリンは、比較的屈曲性が低いので、比較的頻繁な使用が期待されない。さらに、グリシンは、ペプチドバックボーンに不要な屈曲性を導入するため、かつ、きわめてサイズが小さいために、より大きな残基を置換した場合、効果的接触が得られる確率が低いと考えられるために、比較的頻繁な使用が期待されない。一方、グリシンは、ある場合、縮重位置を増進するために使用することが可能である。比較的中間的長さ及びサイズの側鎖を持つアミノ酸は、リシン(K)、メチオニン(M)、アルギニン(R)、グルタミン酸(E)、およびグルタミン(Q)を含む2群に割り当てられる。比較的長い、及び/又は比較的大きい側鎖を持つアミノ酸は、リシン(K
)、メチオニン(M)、アルギニン(R)、ヒスチジン(H)、フェニルアラニン(F)、チロシン(Y)、およびトリプトファン(W)を含む3群に割り当てられる。しかしながら、トリプトファンは、比較的屈曲性が低いので、比較的頻繁な使用が期待されない。さらに、リシン、アルギニン、およびメチオニン側鎖の屈曲性によって、これらのアミノ酸は、長い距離または中間距離からの塩基接触が可能となるために、それらが、2群と3群の両群に含められることを保証する。これらの群をさらに、下記に表1として示す。
【0086】
【表1】

【0087】
側鎖の化学的性質に関しては、種々のアミノ酸が、種々の核酸塩基との相互作用の可能性について評価されており(例えば、ファンデルワールス力、イオン結合、水素結合及び疎水性相互作用)、2本鎖DNA認識配列半分部位中の特定位置における特定塩基と、メガヌクレアーゼとの特異的相互作用を好むか又は忌避する残基が選択される。ある場合には、一つ以上の、完全な又は部分的な縮重位置を持つ半分部位を創出することが望ましい場合がある。そのような場合、二つ以上の塩基の存在を好む残基又は一つ以上の塩基を忌避する残基を選んでもよい。例えば、部分的に縮重された塩基認識は、センス又はアンチセンス位置においてピリミジンを立体的に妨げることによって実現することが可能である。
【0088】
グアニン(G)塩基の認識は、該塩基のN7およびO6に対して水素結合を形成する塩基性側鎖を持つアミノ酸を用いて実現される。シトシン(C)の特異性は、C以外の全ての塩基の上に存在する、大溝の電気的陰性基との相互作用を持ちたがらない、陰性荷電側鎖によって付与される。チミン(T)の認識は、疎水性側鎖と、塩基の上の大溝メチル基との間の疎水性およびファンデルワールス相互作用を用いて合理的に設計される。最後に、アデニン(A)塩基は、該塩基のN7およびN6に対する一対の水素結合を介して、ASNおよびGLNのカルボキサミド側鎖、またはTYRのヒロドキシル側鎖を用いて認識される。最終として、HISは、N7に対して水素結合を供与することによって、プリン塩基(AまたはG)に対して特異性を付与するために使用することが可能である。この、
DNA認識に関する単純な規則を用いて、そこでは、ある特定の塩基対位置において、塩基の内の一つが、または両方が合理的設計接触を通じて認識される、そのような接触面を予測することが可能である。
【0089】
したがって、種々の核酸塩基との結合性相互作用及び接触を行う位置において好む塩基に基づいて、各アミノ酸残基を、それが好む種々の塩基(すなわち、G、C、T、またはA)に対応する一つ以上の異なる群に割り当てることが可能である。このようにして、G群は、アルギニン(R)、リシン(K)、およびヒスチジン(H)を含み;C群は、アスパラギン酸(D)およびグルタミン酸(E)を含み;T群は、アラニン(A)、バリン(V)、ロイシン(L)、イソロイシン(I)、システイン(C)、トレオニン(T)、メチオニン(M)、およびフェニルアラニン(F)を含み;かつ、A群は、アスパラギン(N)、グルタミン(N)、チロシン(Y)、およびヒスチジン(H)を含む。なお、ヒスチジンは、G群とA群の両群に現れること;セリン(S)は、どの群にも含まれないが、縮重位置を好むものとして使用してもよいこと;および、プロリン、グリシン、およびトリプトファンは、主に立体的配慮のためにどの特定の群にも含まれないことに注意されたい。これらの群も下記に表2として示す。
【0090】
【表2】
【0091】
したがって、本発明によれば、任意の位置Xにおいて、メガヌクレアーゼの認識配列半分部位に所望の変化を実行するために、(1)野生型、または参照メガヌクレアーゼ−DNA複合体の三次元構造の少なくとも関連部分及び位置Xにおける接触面を定めるアミノ酸残基側鎖を決定すること;(2)接触面を含む少なくとも一つの残基のβ−炭素及び位置Xにおける塩基対の少なくとも一つの塩基との間の距離を決定すること;(3)所望の変化を促進するために、(A)塩基から6オングストローム未満の距離にある残基については、G群、C群、T群又はA群の内の適切な一つの群のメンバーである1群及び/又は2群から残基を選ぶこと、及び/又は、(B)塩基から6オングストロームより遠い距離にある残基については、G群、C群、T群又はA群の内の適切な一つの群のメンバーである2群及び/又は3群から残基を選ぶこと、である。接触面を含む、一つを超えるこのような残基を、分析および修飾のために選ぶことが可能であり、ある実施態様では、このような各残基が分析され複数の残基が修飾される。同様に、接触面に含まれる残基のβ−炭
素と、位置Xにおける塩基対の二つの塩基のそれぞれとの間の距離を測定することが可能であり、もしもその残基が両方の塩基の9オングストローム以内にある場合、該塩基対の二つの塩基に影響を及ぼすために異なる置換を実行することが可能である(例えば、一方の鎖の近位塩基に影響を及ぼすために1群の残基、又は他方の鎖の遠位塩基に影響を及ぼすために3群の残基)。それとは別に、ペアの両塩基と相互作用を持つことが可能な残基置換の組み合わせは、特異性に影響を及ぼすことが可能である(例えば、アンチセンス鎖に接触するA群の残基と組み合わせたセンス鎖に接触するT群の残基はT/Aを選択する)。最後に、残基の複数の候補修飾は、経験的に(例えば、組み換えメガヌクレアーゼを生産し、その配列認識を調べることによって)、又はコンピュータ的に(例えば、修飾された酵素のメガヌクレアーゼ−DNA複合体のコンピュータモデル化によって)その有効性を検証して、複数候補の中から選択することが可能である。
【0092】
野生型又は基準ヌクレアーゼの、一つ以上の所望のアミノ酸修飾が選択されたならば、合理設計メガヌクレアーゼは、従来技術で周知の組み換え法および技術によって生産することが可能である。ある実施態様では、特異的配列修飾を創製するために、非ランダムな、部位指向性突然変異誘発技術が用いられる。非ランダム突然変異誘発技術の非限定的例としては、重複プライマーPCR(例えば、WANG ET AL.(2006),NUCLEIC ACIDS RES.34(2):517−527を参照)、部位指向性突然変異誘発(例えば、米国特許第7,041,814号を参照)、カセット突然変異誘発(例えば、米国特許第7,041,814号参照)、および、PROMEGA BIOSCIENCES,INC.(SAN LUIS OBISPO、カリフォルニア州)から市販されるALTERED SITES(R) II MUTAGENESIS SYSTEM KIT用のメーカーのプロトコールが挙げられる。
【0093】
合理設計メガヌクレアーゼによる、特異的DNA配列の認識及び切断は、当業者の既知の任意の方法によって定量することが可能である(例えば、米国特許出願公開第2006/0078552号を参照)。ある実施態様では、メガヌクレアーゼ切断の定量は、インビトロ切断アッセイによって定量される。このようなアッセイは、定量されるメガヌクレアーゼの意図される認識配列を含むポリヌクレオチド基質のインビトロ切断を用いるが、ある実施態様では、片方又は両方の半分部位の一つ以上の塩基が、別の塩基に変えられた、意図された認識配列の変種を含むポリヌクレオチド基質のインビトロ切断を用いる。通常、ポリヌクレオチド基質は、合成され、ベクターにクローンされた標的部位を含む2本鎖DNA分子である。ポリヌクレオチド基質は、直線状であっても、円形であってもよいが、通常、唯一の認識配列を含む。メガヌクレアーゼは、適切な条件下で、ポリヌクレオチド基質とインキュベートされ、得られたポリヌクレオチドは、切断産物を特定するために、既知の方法で分析される(例えば、電気泳動又はクロマトグラフィー)。直線状2本鎖DNA基質において単一の認識配列がある場合には、メガヌクレアーゼ活性は、二つのバンドの出現及び最初の全長の基質バンドの消失によって検出される。一実施態様では、メガヌクレアーゼ活性は、例えば、WANG ET AL.(1997),NUCLEIC ACID RES.,25:3767−3776に記載される通りに定量することが可能である。
【0094】
他の実施態様では、メガヌクレアーゼの切断パターンは、インビボ切断アッセイによって定量される(例えば、米国特許出願公開第2006/0078552号)。特定の実施態様では、このインビボ試験は、単一鎖アニーリング組み換え試験(SSA)である。この種の試験は当業者には既知である(RUDIN ET AL.(1989),GENETICS 122:519−534;FISHMAN−LOBELL ET AL.(1992),SCIENCE 258:480−4)。
【0095】
当業者には明らかなように、DNA認識および結合に関与するドメイン以外の、メガヌ
クレアーゼ酵素のドメインに対し、活性を完全に消失させることなく、さらに新たなアミノ酸置換、挿入又は欠失を行うことが可能である。置換は、構造的または機能的に束縛された位置における、類似のアミノ酸残基の保存的置換であってもよく、又は構造的または機能的に比較的に束縛されない位置における非保存的置換であってもよい。このような置換、挿入又は欠失は、当業者によって、不当な努力を要することなく、通例の実験操作によって特定することが可能である。したがって、ある実施態様では、本発明の組み換えメガヌクレアーゼは、基準メガヌクレアーゼの配列に対し、85%から99%の間の任意のパーセント(例えば、85%、87.5%、90%、92.5%、95%、97.5%、99%)の配列類似性を持つタンパクを含む。各野生型I−CREI、I−MSOI、I−SCEI、およびI−CEUIタンパクに関して、多くのN−末端およびC−末端配列は、X線結晶実験でははっきりと見ることができない。これは、これらの位置が、構造的又は機能的に束縛されていないことを示唆する。それゆえ、これらの残基は、配列類似性の計算から排除することが可能であり、下記の基準メガヌクレアーゼ配列を使用することが可能である。I−CREIについては配列番号1の残基2−153;I−MSOIについては配列番号6の残基6−160;I−SCEIについては配列番号9の残基3−186;I−CEUIについては配列番号12の残基5−211である。
【0096】
2.2 LAGLIDADGファミリー・メガヌクレアーゼ
LAGLIDADGメガヌクレアーゼ・ファミリーは、多様な系統群の宿主生物体由来の200を超えるメンバーから構成される。このファミリーのメンバーは全て、特異的DNA配列の切断に与る他の構造的モチーフと共に、高度に保存されるLAGLIDADGモチーフの1個又は2個の複製を有する。LAGLIDADGモチーフの1つの複製有する酵素(すなわち、モノ−LAGLIDADGメガヌクレアーゼ)はダイマーとして機能し、一方、このモチーフの2つの複製を有する酵素(すなわち、ジ−LAGLIDADGメガヌクレアーゼ)はモノマーとして機能する。
【0097】
LAGLIDADGファミリーメンバーは全て、比較的長い配列(>12BP)を認識、切断し、4個のヌクレオチドの、3′オーバーハングを残す。これらの酵素はさらに、LAGLIDADGの外に、タンパク−DNAインターフェイスにおいて、逆平行β鎖の類似配置を含むいくつかの構造的モチーフを共有する。これらの保存された構造モチーフ内のアミノ酸がDNA塩基と相互作用を持ち、認識配列特異性を付与する。ファミリーのいくつかのメンバー(例えば、I−CREI、I−MSOI、I−SCEI及びI−CEUI
)の間に全体構造の類似性が見られることは、X線結晶学によって明らかにされている。したがって、このファミリーのメンバーは、この構造モチーフ内部の特定のアミノ酸について修飾し、該酵素の全体活性又は配列特異性を変えることが可能であり、かつ、対応する修飾は、他のファミリーメンバーにおいても類似の結果をもたらすことが当然期待される。概論については、CHEVALIERET AL. (2001), NUCLEIC ACID RES. 29(18):3757−3774)を参照されたい。
【0098】
2.2.1 I−CREIから得られるメガヌクレアーゼ
1側面において、本発明は、CHLAMYDOMONAS REINHARDTIIのI−CREIメガヌクレアーゼに基づく、又はこのメガヌクレアーゼから得られた合理的設計メガヌクレアーゼに関する。I−CREIメガヌクレアーゼの野生型アミノ酸配列は、GENBANKアクセス#PO5725に対応する、配列番号1に示される。結晶構造PDB#1BP7における野生型I−CREIメガヌクレアーゼの、二つの、認識配列半分部位を下記の表3に示す。
【0099】
【表3】
【0100】
この天然の認識配列は、中央の4塩基の外側においても、完全にはパリンドローム的ではないことに注意されたい。なお、二つの認識配列半分部位がそれぞれのセンス鎖に太字で示される。
【0101】
野生型I−CREIはさらに、下記の表4に示すように、(中央のN1−N4塩基を除き)完全にパリンドロームである配列も認識し、切断する。
【0102】
【表4】
【0103】
配列番号4及び配列番号5のパリンドローム配列は、野生型I−CREIにとってより優れた基質であると考えられる。なぜなら、この酵素は、天然のDNA配列よりもより高い親和度の下にこの部位に結合し、より効率的に該部位を切断するからである。下記の開示のために、かつ、特に本出願において提示する実験結果との関連において、野生型I−CREIによって切断されるこのパリンドローム配列を”WT”と呼ぶことにする(例えば、図2(A)参照)。二つの、認識配列半分部位が、それぞれのセンス鎖に太字で示される。
【0104】
図1(A)は、野生型I−CREIメガヌクレアーゼ・ホモダイマーの、2本鎖DNA認識配列との相互作用を描く。図1(B)は、野生型酵素の半分部位と野生型認識配列の−4位置におけるこの酵素のアミノ酸残基と塩基との間の特異的相互作用を示し、図1(C)〜図1(E)は、半分部位の−4位置における特異性を変更させた、本発明の、三つの合理設計メガヌクレアーゼの、一半分部位の−4位置における、酵素のアミノ酸残基と塩基との間の特異的相互作用を示す。
【0105】
したがって、半分部位の任意の、指定の塩基位置における塩基に対する好みは、本出願に開示される方法を用いて、他の三つの塩基のそれぞれに合理的に変えることが可能である。第一に、指定の塩基位置における野生型認識面が決められる(例えば、メガヌクレアーゼ−DNA複合体共結晶構造を分析することによって、又はメガヌクレアーゼ−DNA複合体のコンピュータモデル化によって)。第二に、指定の塩基位置における、周辺アミノ酸位置のβ−炭素と、各DNA鎖における核酸塩基の間の距離に基づいて、既存の及び可能な接触残基が決められる。例えば、ただしこれらに限定されないが、図1(A)に示すように−4位置における、I−CREI野生型メガヌクレアーゼ−DNA接触残基は、位置26のグルタミンを含み、これはアンチセンスDNA鎖のA塩基と水素結合する。さらに、残基77が、DNAアンチセンス鎖の−4塩基に接触し得る可能性を持つことが特定された。残基26のβ−炭素は、アンチセンスDNA鎖のA塩基のN7から5.9Å離れており、残基77のβ−炭素は、センス鎖のTのC5メチルから7.15オングストローム離れている。この距離及び本出願に記載される塩基化学規則によれば、センス鎖をCとすると、位置77のグルタミン酸と水素結合することが可能であり、アンチセンス鎖を
Gとすると、位置26のグルタミンと結合することが可能である(野生型I−CREI結晶構造に観察されるように、水分子を介して)(図1(C)を参照)。センス鎖をGとすると、位置77のアルギニンと水素結合することが可能であり、アンチセンス鎖をCとすると、位置26のグルタミン酸と水素結合することが可能である(図1(D)を参照)。センス鎖をAとすると、位置77のグルタミンと水素結合することが可能であり、アンチセンス鎖をTとすると、位置26のアラニンと疎水性接触を形成することが可能である(図1(E)を参照)。もしも塩基の特異的接触が位置77によって与えられるとすると、野生型接触Q26を置換し(例えば、セリン残基によって)、その特異性に対する影響を低下、または除去することが可能である。それとは別に、位置26および77における相補的突然変異を組み合わせて、特定の塩基対を指定することが可能である(例えば、A26は、アンチセンス鎖においてTを指定し、Q77は、センス鎖においてAを指定する(図1(E))。これらの予言された残基置換は、全て、実験的にその有効性が実証された。
【0106】
したがって、本発明によれば、I−CREIメガヌクレアーゼのDNA認識ドメインに対する、ある確定数のアミノ酸置換が特定された。これらは、単独で又は組み合わせて用いられて、DNA認識配列半分部位内の個別の塩基において特異性を変更された組み換えメガヌクレアーゼを、したがって、野生型酵素とは異なる半分部位を持つ合理設計メガヌクレアーゼをもたらすことが可能である。I−CREIのアミノ酸修飾及びそれによってもたらされる、認識配列半分部位特異性における変化を、表5に示す。
【0107】
【表5】
【0108】
太字入力は、野生型接触残基であり、本出願で使用される「修飾」を構成しない。
星印は、その残基が、アンチセンス鎖の塩基に接触することを示す。
【0109】
2.2.2. I−MSOLIから得られるメガヌクレアーゼ
他の側面では、本発明は、MONOMASTIX SP.のI−MSOIメガヌクレアーゼに基づく、又はこのヌクレアーゼから得られた合理設計メガヌクレアーゼに関する。I−MSOIメガヌクレアーゼの野生型アミノ酸配列は、GENBANKアクセス#AAL34387に対応し、配列番号6に示される。結晶構造PDB#1M5Xにおける野生型I−MSOIメガヌクレアーゼの、二つの認識配列半分部位を下記の表6に示す。
【0110】
【表6】
【0111】
認識配列は、中央の4塩基の外側においても、完全にはパリンドローム的ではないことに注意されたい。二つの認識配列半分部位が、それぞれのセンス鎖に太字で示される。
【0112】
本発明によれば、I−MSOIメガヌクレアーゼのDNA認識ドメインに対する、ある確定数のアミノ酸置換が特定された。これらは、単独で又は組み合わせて用いられて、DNA認識配列半分部位内の個別の塩基において特異性を変更された組み換えメガヌクレアーゼを、したがって、野生型酵素とは異なる半分部位を持つ合理設計メガヌクレアーゼをもたらすことが可能である。I−MSOIのアミノ酸修飾、および、予測される、認識配列半分部位特異性における変化を、表7に示す。
【0113】
【表7】
【0114】
太字は、野生型接触残基を表し、本出願で使用される「修飾」を構成しない。
星印は、その残基がアンチセンス鎖の塩基に接触することを示す。
【0115】
2.2.3. I−SCEIから得られるメガヌクレアーゼ
他の側面では、本発明は、SACCHAROMYCESCEREVISIAEのI−SCEIメガヌクレアーゼに基づく、又はこのヌクレアーゼから得られた合理設計メガヌクレアーゼに関する。I−SCEIメガヌクレアーゼの野生型アミノ酸配列は、GENBANKアクセス#CAA09843に対応し、配列番号9に示される。結晶構造PDB#1R7Mにおける野生型I−SCEIメガヌクレアーゼの、認識配列を下記の表8に示す。
【0116】
【表8】
【0117】
認識配列は、非パリンドローム的であり、かつ、半分部位を分ける四つの塩基対がないことに注意されたい。
【0118】
本発明によれば、I−SCEIメガヌクレアーゼのDNA認識ドメインに対する、ある確定数のアミノ酸置換が特定された。これらは、単独で又は組み合わせて用いられて、DNA認識配列内の個別の塩基において特異性を変更された組み換えメガヌクレアーゼ、つまり、野生型酵素とは異なる認識配列を持つ合理設計メガヌクレアーゼをもたらすことが可能である。I−SCEIのアミノ酸修飾及び予測される認識配列特異性における変化を、表9に示す。
【0119】
【表9】
【0120】
太字入力は、野生型接触残基を表し、本出願で使用される「修飾」を構成しない。星印は、その残基が、アンチセンス鎖の塩基に接触することを示す。
【0121】
2.2.4 I−CEUIから得られるメガヌクレアーゼ
他の側面では、本発明は、CHLAMYDOMONASEUGAMETOSのI−CEUIメガヌクレアーゼに基づく、又はこのヌクレアーゼから得られた合理設計メガヌクレアーゼに関する。I−CEUIメガヌクレアーゼの野生型アミノ酸配列は、GENBANKアクセス#P32761に対応し、配列番号12に示される。結晶構造PDB#2EX5における野生型I−CEUIメガヌクレアーゼの、二つの認識配列半分部位を下記に示す。
【0122】
【表10】
【0123】
I−CEUIはホモダイマーであるにも拘わらず、I−CEUIの認識インターフェイスにおける天然の縮重のために(SPIEGEL ET AL.(2006),STRUCTURE 14:869−80)、認識配列は、中央の4塩基の外側においても、非パリンドローム的であることに注意されたい。二つの認識配列半分部位がそれぞれのセンス鎖に太字で示される。
【0124】
本発明によれば、I−CEUIメガヌクレアーゼのDNA認識ドメインに対する、ある確定数のアミノ酸置換が特定された。これらは、単独で又は組み合わせて用いられて、DNA認識配列半分部位内の個別の塩基において特異性を変更された組み換えメガヌクレアーゼを、したがって、野生型酵素とは異なる半分部位を持つ合理設計メガヌクレアーゼをもたらすことが可能である。I−CEUIのアミノ酸修飾及び予測される認識配列特異性における変化を、表11に示す。
【0125】
【表11】
【0126】
太字入力は、野生型接触残基を表し、本出願で使用される「修飾」を構成しない。星印は、その残基がアンチセンス鎖の塩基に接触することを示す。
【0127】
2.2.5 特異的に排除される組み換えメガヌクレアーゼ
本発明は、従来技術において記載され、別法によって開発された、いくつかの組み換えメガヌクレアーゼを包含することを意図しない。これらの排除されたメガヌクレアーゼとしては、ARNOULD ET AL.(2006),J.MOL.BIOL.355:443−58;SUSSMAN ET AL.(2004),J.MOL.BIOL.3
42:31−41;CHAMES ET AL.(2005),NUCLEIC ACIDS RES.33:E178;SELIGMAN ET AL.(2002),NUCLEIC ACIDS RES.30:3870−9;およびASHWORTH ET AL.(2006) NATURE 441(7093):656−659によって記載されるものが挙げられる。なお、上記の全開示を、C33、R33、A44、H33、K32、F33、R32、A28、A70、E33、V33、A26及びR66から選ばれる単一置換を持つ、I−CREIに基づく組み換えメガヌクレアーゼを含め、引用により本出願に含める。さらに排除されるものは、A68/N70/N75及びD44/D70/N75から選ばれた3置換、K44/T68/G60/N75及びR44/A68/T70/N75から選ばれる4置換を持つI−CREIに基づく組み換えメガヌクレアーゼである。最後に、特に指定して排除されるものは、置換L28及びR83のペアを持つI−MSOIに基づく組み換えメガヌクレアーゼである。これらの置換又は置換の組み合わせは、本出願では「排除修飾」と呼ばれる。
【0128】
2.2.6 認識配列半分部位において複数変化を有するメガヌクレアーゼ
他の側面では、本発明は、DNA認識配列半分部位の二つ以上の位置において半分部位指向を変えるために、上の節2.2.1−2.2.4に記載した、二つ以上のアミノ酸置換を組み合わせることによって得られる合理設計メガヌクレアーゼに関する。例えば、ただしこれらに限定されるものではないが、さらに詳しく後述されるように、酵素DJ1は、修飾R30/E38(これは、位置−7においてCを好む)、R40(これは位置−6においてGを好む)、R42(これは、位置−5においてGを好む)及びN32(これは、位置−9において完全縮重を好む)を取り込むことによって、I−CREIから得られる。この合理設計DJ1メガヌクレアーゼは、A−7A−6C−5に対する野生型の指向性に比べ、ほぼ変わらない程度でC−7G−6G−5を認識し、位置−9におけるAに対する耐性を増大させる。
【0129】
異なる塩基位置に影響を及ぼす残基置換を組み合わせることができる能力は、一部は、LAGLIDADGメガヌクレアーゼのモジュラー性による。LAGLIDADG認識インターフェイスにおける塩基接触の大部分は、個々のアミノ酸側鎖によって形成され、インターフェイスは、隣接塩基と相互作用を持つ側鎖間の相互接続性、または水素結合ネットワークから比較的自由である。これによって、一般に、一塩基位置と相互作用を持つ残基を隣接塩基における側鎖同士の相互作用に影響を及ぼさずに操作することが可能になる。さらに、上の2.21−2.2.4節に掲げた突然変異の加重性は、これらの突然変異を特定するために用いた本法の直接的結果である。この方法は、単一塩基と直接に相互作用を持つ側鎖置換を予測する。一般に、側鎖間の相互接続、または水素結合ネットワークは、認識インターフェイス内の置換の独立性を維持するために回避される。
【0130】
側鎖置換のある組み合わせは、完全に又は部分的に相互に不適合である。不適合ペア又は組のアミノ酸を合理設計メガヌクレアーゼに組み込んだ場合、得られた酵素の触媒活性は、低下するか又は排除される。通常、これらの不適合は、導入されたアミノ酸側鎖間の立体干渉によるもので、活性は、この干渉を特定し、除去することで回復が可能である。具体的には、大きな側鎖を持つ(例えば、2又は3群のアミノ酸)二つのアミノ酸が、メガヌクレアーゼ構造において、互いに隣接するアミノ酸位置に組み込まれた場合(例えば、I−CREI由来のメガヌクレアーゼの場合、位置32及び33、28及び40、28及び42、42及び77又は68及び77)、これら二つのアミノ酸は互いに干渉しあって、酵素活性を下げる可能性が高い。この干渉は、一方の又は両方の不適合アミノ酸を、より小さな側鎖を持つアミノ酸(例えば、1群または2群)で置換することによって排除することが可能である。例えば、I−CREI由来の合理設計メガヌクレアーゼでは、K28は、R40とR42の両方に干渉する。酵素活性を最大化するためには、R40及びR42を、位置28においてセリン又はアスパラギン酸と組み合わせることが可能である
。
【0131】
本出願に記載のように特定される、アミノ酸置換の組み合わせを用いて、野生型メガヌクレアーゼ(又は以前に修飾されたメガヌクレアーゼ)の特異性を、元の認識配列から、興味の核酸(例えば、ゲノム)中に存在する、所望の認識配列に合理的に変えることが可能である。例えば、図2(A)は、I−CREIメガヌクレアーゼ認識配列WT(配列番号4)の「センス」鎖の外に、それに対し合理的設計メガヌクレアーゼを創製したならば有用であろうと考えられる、他の、いくつかの配列を示す。WT認識配列と所望の認識配列との間で保存される塩基には陰影を施す。本発明によれば、I−CREIメガヌクレアーゼに基づく組み換えメガヌクレアーゼは、これらの所望の認識配列のそれぞれについてばかりでなく、他の任意の認識配列についても、本出願に記載される適切なアミノ酸置換によって、合理的に設計することが可能である。
【0132】
3. DNA結合親和度が変更された合理設計メガヌクレアーゼ
前述のように、本発明の組み換えメガヌクレアーゼのDNA結合親和度は、DNAのフォスフォジエステルバックボーンと接触面を形成するいくつかのアミノ酸を変えることによって修飾することが可能である。この接触面は、DNAバックボーンから9オングストローム未満の距離のβ−炭素を持つ酵素のアミノ酸を、この残基が野生型メガヌクレアーゼ−DNA複合体においてDNAバックボーンと接触するかどうかとは無関係に含む。DNA結合は、酵素活性に必要な前提条件であるから、DNA結合親和度の上昇/低下は、それぞれ、酵素活性の上昇/低下を起こすことが示されている。しかしながら、DNA結合親和度の上昇/低下はさらに、メガヌクレアーゼの配列特異性の低下/上昇を起こすことが示されている。それゆえ、活性および特異性は両方とも、フォスフォジエステルバックボーン接触を修飾することによって修飾することが可能である。
【0133】
酵素活性の上昇/酵素の特異性の低下のためには、
【0134】
(I)酵素とDNAバックボーンの間の静電気的反発を取り除く。ある特定されたアミノ酸が、陰性荷電のDNAバックボーンに反発すると予想される、陰性に荷電した側鎖を持つ場合(例えば、アスパラギン酸、グルタミン酸)、この反発は、アミノ酸を無荷電の又は陽性荷電の側鎖で置換することによって除去することが可能である。ただし、この場合、立体障害の作用には影響される。実験的に確かめられた例は、I−CREIにおけるグルタミン酸80をグルタミンに突然変異させた場合である。
【0135】
(II)酵素とDNAバックボーンの間に静電気的牽引相互作用を導入する。接触面の位置のいずれかにおける、陽性荷電側鎖を持つアミノ酸(例えば、リシン又はアルギニン)の導入は、立体干渉の作用には影響されるが、結合親和度を増すことが予想される。
【0136】
(III)酵素とDNAバックボーンの間に水素結合を導入する。接触面のアミノ酸が、適切な水素結合官能基を欠如するため、又はDNAバックボーンと相互作用を持つには、側鎖が短すぎるか長すぎるか、及び/又は、屈曲性が不足するという理由で水素結合を造ることができない場合、適切な長さと屈曲性を持つ、水素結合を供与することが可能な極性アミノ酸(例えば、セリン、トレオニン、チロシン、ヒスチジン、グルタミン、アスパラギン、リシン、システイン又はアルギニン)を導入することが可能である。ただし、立体障害の作用には影響される。
【0137】
具体的に、酵素活性の低下/酵素の特異性の上昇のためには、
【0138】
(I)酵素とDNAバックボーンの間に静電気的反発を導入する。接触面の位置のいずれかにおいて、陰性に荷電した側鎖を持つアミノ酸(例えば、グルタミン酸、アスパラギ
ン酸)を導入することは、結合親和度を下げることが予想される。ただし、立体障害の作用には影響される。
【0139】
(II)酵素とDNAの間の静電気的牽引を取り除く。接触面のいずれかのアミノ酸が、陰性に荷電したDNAバックボーンと相互作用を持つ、陽性に荷電した側鎖を持つ場合(例えば、リシン又はアルギニン)、この好ましい相互作用は、アミノ酸を無荷電の又は陰性荷電の側鎖で置換することによって除去することが可能である。ただし、立体干渉の作用には影響される。実験的に確かめられた例は、I−CREIにおけるリシン116をアスパラギン酸に突然変異させた場合である。
【0140】
(III)酵素とDNAバックボーンの間の水素結合を取り除く。接触面のいずれかのアミノ酸が、DNAバックボーンと水素結合を造る場合、そのアミノ酸をその側鎖が適切な官能基を持たないか又は必要な長さ/屈曲性を欠如するために、同様の水素結合を造るとは予想されないアミノ酸に置換することが可能である。
【0141】
例えば、I−CREIに基づくある組み換えメガヌクレアーゼにおいて、活性を上げるためにI−CREIメガヌクレアーゼの位置80におけるグルタミン酸が、リシン又はグルタミンに変更される。別の実施態様では、I−CREIの位置66のチロシンが、アルギニン又はリシンに変更される。これは、該メガヌクレアーゼの活性を上げる。さらに別の実施態様では、I−CREIの位置34におけるリシンをアスパラギン酸に、位置66におけるチロシンをアスパラギン酸に、及び/又は、位置116におけるリシンをアスパラギン酸に変えることによって、酵素活性を低下させる。
【0142】
組み換えメガヌクレアーゼの活性は、その組み換え酵素が、ある特定の認識配列に関して、無活性から極度に高い活性までの間の任意の活性レベルを持つように修飾することが可能である。例えば、DJ組み換えメガヌクレアーゼは、位置26においてグルタミン酸突然変異を抱えると活性を完全に失う。しかしながら、位置26におけるグルタミン酸置換と位置80におけるグルタミン置換の組み合わせは、認識配列半分部位内の−4におけるグアニンに対し高度の特異性および活性を持つ組み換えメガヌクレアーゼを創製する(図1(D)を参照)。
【0143】
本発明によれば、フォスフォジエステルDNAバックボーン近傍の各種位置におけるアミノ酸は、メガヌクレアーゼ活性および特異性の両方に同時に作用するように変えることが可能である。この、酵素の特異性と活性の「調整」は、フォスフォジエステルバックボーンに対し、アミノ酸によって造られる接触点の数を増すか又は減らすことによって達成される。フォスフォジエステルバックボーンに対する種々の接触は、アミノ酸側鎖によって促進することが可能である。ある実施態様では、イオン結合、塩橋、水素結合及び立体障害は、アミノ酸側鎖の、フォスフォジエステルバックボーンに対するコンタクトに影響を及ぼす。例えば、I−CREIメガヌクレアーゼの場合、位置116におけるリシンのアスパラギン酸への変更によって、位置−8および−9における核酸塩基間の塩橋が取り除かれ、これは酵素の切断速度は下げるが、特異性は上げる。
【0144】
野生型I−CREI(配列番号1)、I−MSOI(配列番号6)、I−SCEI(配列番号9)、およびI−CEUI(配列番号12)各メガヌクレアーゼの、バックボーン接触面を形成する残基は、下表12において特定される。
【0145】
【表12】
【0146】
酵素の親和度を上げ、したがって、より活性を高め/特異性を下げるためには、
(1)対応する酵素において、陰性荷電(D又はE)、疎水性(A、C、F、G、I、L、M、P、V、W、Y)、又は無荷電/極性(H、N、Q、S、T)のアミノ酸を、表12から選ぶ。
(2)アミノ酸が陰性荷電しているか又は疎水性である場合は、それを、無荷電/極性(作用が小さい)又は陽性荷電(KまたはR、作用が大きい)となるように突然変異させる。
(3)アミノ酸が無荷電/極性である場合、それを陽性荷電に突然変異させる。
【0147】
酵素の親和度を下げ、したがって、より活性を下げ/特異性を上げるためには、
(1)対応する酵素において、陽性荷電(K又はR)、疎水性(A、C、F、G、I、L、M、P、V、W、Y)、又は無荷電/極性(H、N、Q、S、T)のアミノ酸を、表12から選ぶ。
(2)アミノ酸が陽性荷電している場合は、それを、無荷電/極性(作用が小さい)又は陰性荷電(作用が大きい)となるように突然変異させる。
(3)アミノ酸が疎水性又は無荷電である場合、それを陰性荷電に突然変異させる。
【0148】
4. ヘテロダイマー・メガヌクレアーゼ
他の側面では、本発明は、二つのモノマーで、その一方は野生型で、その一方または両方が、非天然形または組み換え形であるモノマーのコンタクトによって形成されるヘテロダイマーであるメガヌクレアーゼを提供する。例えば、野生型I−CREIメガヌクレアーゼは、通常、それぞれが、擬似パリンドローム認識配列において一つの半分部位に結合する、二つのモノマーから構成されるホモダイマーである。ヘテロダイマー組み換えメガヌクレアーゼは、異なる半分部位を認識する二つのメガヌクレアーゼを組み合わせることによって、例えば、細胞において二つのメガヌクレアーアゼを共時発現することによって、又は溶液中で二つのメガヌクレアーゼを混合することによって生産することが可能である。二つのモノマーにおいて、それらのコンタクトによるダイマー形成に影響を及ぼす、それぞれのアミノ酸を変えることによって、ヘテロダイマーの形成を、ホモダイマーの形成よりも優先することが可能である。特定の実施態様では、二つのモノマーのインターフェイスにおけるいくつかのアミノ酸が、第1モノマーでは、陰性荷電のアミノ酸(D又は
E)から陽性荷電のアミノ酸(K又はR)に変えられ、第2モノマーでは、陽性荷電のアミノ酸から陰性荷電のアミノ酸に変えられる(表13参照)。例えば、I−CREI由来のメガヌクレアーゼの場合、位置7及び57におけるリシンが、第1モノマーではグルタミン酸に突然変異され、第2モノマーでは、位置8および61におけるグルタミン酸がリシンに突然変異される。このプロセスの結果は、第1モノマーが、ダイマーインターフェイスにおいて過剰な陽性荷電残基を持ち、第2モノマーが、ダイマーインターフェイスにおいて過剰な陰性荷電残基を持つ、一対のモノマーである。それゆえ、この第1及び第2モノマーは、インターフェイスにおける変更アミノ酸の間の静電気的相互作用のために、その同じモノマーペアよりも優先的にコンタクトする。
【0149】
【表13】
【0150】
それとは別に、またはそれに加えて、二つのモノマーのインターフェイスにおけるいくつかのアミノ酸は、立体的にホモダイマー形成を妨げるように変えることも可能である。具体的には、一方のモノマーのダイマーインターフェイスのアミノ酸を、ホモダイマーを立体的に阻止する比較的大きな又は嵩高な残基によって置換する。第2のモノマーのダイマーインターフェイスにおけるアミノ酸を、第1のモノマーの嵩高な残基を補正するため
に比較的小さな残基で置換し、ヘテロダイマーにおける衝突を取り除くことが可能であるし、または未修飾のままでもよい。
【0151】
さらに他の実施態様では、イオン架橋又は水素結合を、ヘテロダイマー・インターフェイスの疎水性コアの中に埋没させることが可能である。具体的には、インターフェイスのコアの一モノマーにおける疎水性残基は、陽性荷電残基によって置換することが可能である。さらに、野生型ホモダイマーでは、第1モノマーにおいて置換された疎水性残基と相互作用を持つ、第2モノマーの疎水性残基は、陰性荷電残基によって置換することが可能である。したがって、この二つの置換残基は、イオン結語または水素結合を形成することが可能である。同時に、疎水性インターフェイスの中に埋没された満たされない電荷の静電気的反発は、ホモダイマー形成を忌避する。
【0152】
最後に、前述のように、ヘテロダイマーの各モノマーは、DNA認識領域において、異なるアミノ酸を置換されて、そのため、それぞれが異なるDNA半分部位を持ち、かつ、組み合わされたダイマー形DNA認識配列が非パリンドローム的となってもよい。
【0153】
5. 組み換え細胞および生物体を生産する方法
本発明の側面はさらに、合理設計メガヌクレアーゼを用いた、組み換え、トランスジェニック又は他のやり方で遺伝学的に修飾された細胞および生物体を生産する方法を提供する。したがって、ある実施態様では、相同組み換えによる興味の配列の正確な挿入(単数又は複数)を可能とするために、細胞または生物体のゲノムDNAの単一部位又は比較的少数部位に、二本鎖切断点を特異的に引き起こすための、組み換えメガヌクレアーゼが開発された。別の実施態様では、(A)非相同的末端接合による興味の配列の稀な挿入(単数又は複数)を可能とするために、又は(B)非相同的末端接合による標的配列の破壊を可能とするために、細胞または生物体のゲノムDNAの単一部位又は比較的少数部位に、二本鎖切断点を特異的に引き起こすための、組み換えメガヌクレアーゼが開発された。興味の配列の、相同的組み換え又は非相同的末端接合に関して本出願で用いる場合、「挿入」という用語は、興味の配列が染色体の中に組み込まれるように、興味の配列が染色体にコンタクトすることを意味する。相同組み換えの場合、挿入された配列は内因性配列を置換し、そのため、元のDNAは等しい長さではあるがヌクレオチド配列が変更された外因性DNAによって置換される。それとは別に、挿入配列はそれが置換する配列よりもより多数の又はより少数の塩基を含むことが可能である。
【0154】
それゆえ、本発明のこの局面によれば、組み換え生物体としては、単子葉植物種、例えば、米、小麦、トウモロコシ(メイズ)並びにライ麦、及び双子葉植物種、例えば、豆類(例えば、ソラマメ、大豆、レンズ豆、ピーナッツ、エンドウ豆)、アルファルファ、クローバー、タバコ並びにARABIDOPSIS種が挙げられるが、これらに限定されるものではない。さらに、組み換え生物体としては、動物、例えば、ヒト並びに非ヒト霊長類、ウマ、ウシ、ヤギ、ブタ、ヒツジ、イヌ、ネコ、モルモット、ラット、マウス、トカゲ、魚類、及び昆虫類、例えば、ショウジョウバエ種が挙げられるが、これらに限定されるものではない。他の実施態様では、生物体は、真菌、たとえば、カンジダ、ニューロスボラ又はサッカロマイセスである。
【0155】
ある実施態様では、本発明の方法は、成熟組み換え生物体になることが可能な又は遺伝学的に修飾された生物体が、そのゲノムの中に興味の挿入配列を抱える子孫を出産することを可能とする。例えば、細胞、胚細胞又は幹細胞の中に興味の配列を導入することを含む。
【0156】
メガヌクレアーゼタンパクは、ゲノムDNAを切断するために、細胞に輸送することが可能であるが、これによって、従来技術で既知の様々な機序によって、その切断部位にお
いて、興味の配列による相同組み換え、または非相同末端接合が可能とされる。組み換えメガヌクレアーゼタンパクは、例えば、マイクロインジェクション又はリポソームトランスフェクション(例えば、LIPOFECTAMINE(登録商標)、INVITROGEN CORP.,CARLSBAD、カリフォルニア州)を含む、ただしこれらに限定されないが、技術によって細胞中に導入することが可能である。このリポソーム処方は、標的細胞との脂質二重層融合を促進するのに用いることが可能であり、そのため、リポソームの内容物又はその表面にコンタクトするタンパクの細胞内部への取り込みを可能にする。それとは別に、酵素は、適切な取り込みタンパク、例えば、HIVTATタンパクなどに融合させて細胞の取り込みを指示するようにすることも可能である(例えば、HUDECZ ET AL.(2005),MED.RES.REV.25:679−736を参照)。
【0157】
それとは別に、メガヌクレアーゼタンパクをコードする遺伝子配列は、従来技術で既知の技術(例えば、AUSUBEL ET AL.,「分子生物学における最近のプロトコール(“CURRENTPROTOCOLS IN MOLECULAR BIOLOGY,”)」、WILEY 1999を参照されたい)を用いて、ベクターに挿入され、真核細胞にトランスフェクトされる。興味の配列は、同じベクターによって、異なるベクターによって又は従来技術で既知の他の手段によって導入することが可能である。
【0158】
DNAトランスフェクションの非限定的例としては、ウィルスベクター、プラスミド、コスミド及びYACベクターが挙げられる。DNA配列のトランスフェクションは、当業者には既知の、様々の方法によって実現することが可能である。例えば、DNA配列を細胞に輸送するために、リポソームおよびイムノリポソームが用いられる(例えば、LASIC ET AL.(1995),SCIENCE 267:1275−76を参照)。さらに、ベクターを細胞内に導入するためにウィルスを利用することが可能である(例えば、米国特許第7,037,492号を参照)。それとは別に、ベクターが裸のDNAとして導入されるようにトランスフェクション戦略を利用することも可能である(例えば、RUI ET AL.(2002),LIFE SCI.71(15):1771−8を参照)。
【0159】
細胞内に核酸を輸送するための一般的方法としては、(1)化学的方法(GRAHAM
ET AL.(1973),VIROLOGY 54(2):536−539;ZATLOUKAL ET AL.(1992),ANN.N.Y.ACAD.SCI.,660:136−153);(2)物理的方法、例えば、マイクロインジェクション(CAPECCHI(1980),CELL22(2):479−488)、電気穿孔(WONG
ET AL,(1982),BIOCHEM.BIOPHYS.RES.COMMUN.107(2):584−587;FROMM ET AL,(1985),PROC.NAT’L ACAD SCI.USA 82(17):5824−5828;米国特許第5,384,253号)、および、弾丸注入法(JOHNSTON ET AL,(1994),METHODS CELL.BIOL.43(A):353−365;FYNAN ET AL,(1993),PROC.NAT’L ACAD.SCI.USA 90(24):11478−11482)など;(3)ウィルスベクター(CLAPP(1993),CLIN. PERINATOL.20(1):155−168;LU ET AL,(1993),J.EXP.MED.178(6):2089−2096;EGLITIS ET AL,(1988),AVD.EXP.MED.BIOL.241:19−27;EGLITIS ET AL.(1988),BIOTECHNIQUES 6(7):608−614);および、(4)受容体介在性機構(CURIEL ET AL.(1991),PROC.NAT’L ACAD.SCI.USA 88(19):8850−8854;CURIEL ET AL.(1992),HUM.GEN.THER.3(2):147−154;WAGNER ET AL.(1992),P
ROC.NAT’L ACAD.SCI.USA 89(13):6099−6103)が挙げられる。
【0160】
ある実施態様では、ゲノムに挿入された興味の配列を含む遺伝学的に修飾された植物が生産される。ある実施態様では、組み換えメガヌクレアーゼに対応するDNA配列及びこのメガヌクレアーゼ認識配列及び/又は標的配列とほぼ同一の配列によって側接されても側接されなくともよい興味の配列によって植物細胞をトランスフェクトし、遺伝学的に修飾された植物が生産される。別の実施態様では、遺伝学的に修飾された植物は、組み換えメガヌクレアーゼのみに対応するDNA配列によって植物細胞をトランスフェクトし、それによる切断が非相同性末端接合を促進し、認識配列を含む標的配列を破壊することによって生産される。このような実施態様では、メガヌクレアーゼ配列は、宿主植物細胞においてメガヌクレアーゼの発現を可能とする調整配列の調節下にある。これらの調整配列としては、構成的植物プロモーター、例えば、NOSプロモーター、化学的に誘発可能な遺伝子プロモーター、例えば、デキサメタゾン誘発性プロモーター(例えば、GREMILLON ET AL.(2004),PLANT J.37:218−228を参照)、および、植物組織特異的プロモーター、例えば、LGC1プロモーター(例えば、SINGH ET AL.(2003),FEBS LETT.542:47−52を参照)が挙げられるが、これらに限定されるものではない。
【0161】
植物細胞の中にDNAを導入するための適切な方法は、DNAを細胞内に導入することが可能であるならば、ほとんどどの方法であってもよく、例えば、AGROBACTERIUM感染、プロトプラストのPEG−介在性形質転換(OMIRULLEH ET AL.(1993),PLANT MOLECULAR BIOLOGY,21:415−428)、DESICCATION/INHIBITION MEDIATED DNA
UPTAKE、電気穿孔、シリコンカーバイド線維による搖動、弾丸注入、または微小弾丸発射などが挙げられるが、これらに限定されるものではない。
【0162】
他の実施態様では、組み換えメガヌクレアーゼを用いて、遺伝学的に修飾された動物が生産される。植物細胞の場合と同様、核酸配列を、胚細胞又は最終的にトランスジェニック生物体となる細胞の中に導入することが可能である。ある実施態様では、細胞は、受精卵であり、外因性DNA分子は、受精卵の前核の中に注入することが可能である。次に、このマイクロインジェクションされた卵は、擬似妊娠の代理母の卵管に移送され、育成される。組み換えメガヌクレアーゼは、(例えば、3−フォスフォグリセレートキナーゼなどの構成的プロモーターの調節下に)受精卵で発言され、ゲノムの一部位又は不連続な少数部位における興味の配列の相同組み換えを促進する。それとは別に、GOSSLER ET AL.(1986),PROC.NATL.ACAD.SCI.USA 83:9065 9069に記載されるように、トランスジェニック動物生成のための組み換え胚性幹(“ES”)細胞を利用することによって、遺伝学的に修飾された動物を入手することが可能である。
【0163】
ある実施態様では、組み換え哺乳類発現ベクターは、核酸の組織特異的発現を、ある特定の細胞タイプに優先的に向けて指示することが可能である。組織特異的調整要素は従来技術で既知である。適切な組織特異的プロモーターの非限定的例としては、アルブミンプロモーター(肝臓特異的、PINKERT ET AL.(1987),GENES DEV.1:268−277)、リンパ系特異的プロモーター(CALAME AND EATON(1988),ADV.IMMUNOL.43:235−275)、特に、T細胞受容体のプロモーター(WINOTO AND BALTIMORE(1989),EMBO J.8:729−733)、および免疫グロブリンのプロモーター(BANERJIET AL.(1983),CELL 33:729−740;QUEEN AND
BALTIMORE(1983),CELL 33:741−748)、ニューロン特
異的プロモーター(例えば、ニューロフィラメント・プロモーター;BAYNE AND
RUDDLE(1989),PROC.NATL.ACAD.SCI.USA 86:5473−5477)、膵臓特異的プロモーター(EDLUND ET AL.(1985),SCIENCE 230:912−916)、および、乳腺特異的プロモーター(例えば、ミルクフェイ・プロモーター、米国特許第4,873,316号、および欧州特許公開EP0264166)が挙げられる。発達時の調整に与るプロモーター、例えば、マウスのHOXプロモーター(KESSEL AND GRUSS(1990),SCIENCE 249:374−379)、および、α−フェトプロテイン・プロモーター(CAMPES AND TILGHMAN(1989),GENES DEV.3:537−546)なども含まれる。
【0164】
ある実施態様では、合理設計メガヌクレアーゼは、発現レベルまたは局在を監視するために、ペプチドエピトープ(例えば、HA、FLAG、またはMYCエピトープ)によって標識されてもよい。ある実施態様では、メガヌクレアーゼは、細胞内局在シグナル、例えば、核局在シグナル(例えば、SV40の核局在シグナル)又はクロロプラストもしくはミトコンドリア局在シグナルに融合されてもよい。別の実施態様では、メガヌクレアーゼは、それを、細胞質に局在させる核輸出シグナルに融合されてもよい。さらに、メガヌクレアーゼは、無関係のタンパク質又はタンパクドメイン、例えば、DNA−修復又は相同組み換えを刺激するタンパク質(例えば、RECA、RAD51、RAD52、RAD54、RAD57又はBRCA2)に融合されてもよい。
【0165】
6. 遺伝子治療法
本発明の局面は、遺伝子治療のための、組み換えメガヌクレアーゼの使用を可能とする。本出願で用いる「遺伝子治療」という用語は、その構造及び/又は機能において欠陥的である遺伝子又は遺伝子調整配列と置換するために、患者の中に、少なくとも一つの遺伝子の機能的コピー又は遺伝子調整配列、例えば、プロモーター、エンハンサー又はサイレンサーを導入することを含む治療処置を意味する。さらに、「遺伝子治療」という用語は、有害遺伝子または調整要素に対し、該遺伝子の発現を下げるか又は除去するために為される修飾を指すことが可能である。遺伝子治療は、先天性病態、患者の生涯に亘る特定の遺伝子座位における突然変異または損傷による病態又は感染生物体による病態を治療するために実行することが可能である。
【0166】
本発明のある局面では、機能不全遺伝子は、遺伝子発現に影響を及ぼすゲノム領域に、外因性核酸配列を挿入することによって置換されるか、不能にさせられる。ある実施態様では、組み換えメガヌクレアーゼは、病態を緩和するために、修飾すべきゲノム領域の特定配列を標的とする。この配列は、エキソン、イントロン、プロモーター内の領域又は遺伝子の機能不全発現を引き起こす、他の調整領域であってもよい。本出願で用いる「機能不全発現」という用語は、遺伝子産物の生産が少なすぎる細胞又は遺伝子産物の生産が多すぎる細胞又は異なる機能、例えば、必要機能の欠如又は必要以上の機能を持つ遺伝子産物を生産する細胞による、遺伝子産物の異常な発現を意味する。
【0167】
修飾領域に挿入される外因性核酸配列は、遺伝子を正常化する「修復」配列を提供するために使用することが可能である。遺伝子修復は、再生されるべき適正機能の実現を可能とする遺伝子中に適正遺伝子配列を導入することによって実現される。これらの実施態様では、挿入される核酸配列は、タンパクの全体コード配列であってもよいし又はある実施態様では、修復されるべき領域のみを含む遺伝子断片であってもよい。他の実施態様では、挿入される核酸配列は、異常発現または調整を引き起こす突然変異が修復されるように、プロモーター配列または、その他の調整要素を含む。他の実施態様では、挿入される核酸配列は、突然変異遺伝子には欠如する適切な翻訳終止コドンを含む。核酸配列はさらに、適切な翻訳終止シグナルを欠如する組み換え遺伝子において翻訳を停止するための配列
を持つことが可能である。
【0168】
それとは別に、核酸配列は、遺伝子の調整配列を破壊することによって又は遺伝子機能を除去するサイレンサーを提供することによって、遺伝子機能を全く除去することが可能である。ある実施態様では、外因性核酸配列は、遺伝子産物の発現を阻止するために翻訳終止コドンを提供する。別の実施態様では、外因性核酸配列は、全長のRNA分子の発現を阻止するために翻訳終止コドンを提供する。さらに別の実施態様では、遺伝子機能は、非相同的末端接合を介して、塩基挿入、塩基欠失及び/又はフレームシフト突然変異を導入することによって、メガヌクレアーゼによって直接破壊される。
【0169】
多くの例において、病態の原因である標的細胞又は細胞集団に対し適切な遺伝子配列を方向づけることが望ましい。治療薬の、このような標的指向は、健康な細胞が、治療薬によって標的とされることを防ぐ。これは、治療の効力を増し、一方では、治療が、健康な細胞に及ぼす可能性のある、有害となる可能性のある作用を下げる。
【0170】
興味の細胞に対する、組み換えメガヌクレアーゼ遺伝子及び挿入すべき興味の配列の輸送は、様々の機構によって実現することが可能である。ある実施態様では、核酸は、ウィルスの再生を阻止するように不活性化された特定のウィルス遺伝子を持つウィルスを介して細胞に輸送される。したがって、ウィルスは、標的細胞内に輸送され、維持されることは可能であるが、標的細胞または組織の中で複製する能力は保持しないように変えられる。ベクターのように活動するウィルスゲノムを生産ように、この変更ウィルスゲノムに、一つ以上のDNA配列を挿入することが可能であるが、これらの配列は、宿主のゲノムの中に導入されてその後発現されてもよいし、導入し発現されなくともよい。より具体的には、ある実施態様は、レトロウィルス、例えば、ただしこれらに限定されるものではないが、MFG又はPLJベクターなどの採用を含む。MFGベクターは、単純化されたモロニーマウス白血病ウィルスベクター(MOMLV)であり、このベクターでは、複製欠損とするために、POL及びENVタンパク質をコードするDNA配列は欠失される。PLJレトロウィルスベクターも、MOMLVの一形である(例えば、KORMAN ET AL.(1987), PROC.NAT’L ACAD.SCI.,84:2150−2154)。別の実施態様では、組み換えアデノウィルス又はアデノ関連ウィルスを、輸送ベクターとして使用することが可能である。
【0171】
別の実施態様では、標的細胞に対する、組み換えメガヌクレアーゼタンパク質及び/又は組み換えメガヌクレアーゼ遺伝子配列の輸送は、リポソームの使用によって実現される。核酸及び/又はタンパク積載物を含むリポソームの生産は、従来技術で既知である(例えば、LASIC ET AL.(1995),SCIENCE 267:1275−76)。イムノリポソームは、リポソームの中に、細胞関連抗原に対する抗体を含み、特定の細胞タイプに対しメガヌクレアーゼのDNA配列、またはメガヌクレアーゼそのものを輸送することが可能である(例えば、LASIC ET AL.(1995), SCIENCE 267:1275−76;YOUNG ET AL.(2005),J.CALIF.DENT.ASSOC.33(12):967−71;PFEIFFER ET
AL.(2006),J.VASC.SURG.43(5):1021−7を参照)。リポソーム処方の生産および使用のための方法は、従来技術で周知である(例えば、米国特許第6,316,024号、米国特許第6,379,699号、米国特許第6,387,397号、米国特許第6,511,676号、および米国特許第6,593,308号、およびそれらの中に引用される参考文献を参照)。ある実施態様では、リポソームは、興味の配列の外、組み換えメガヌクレアーゼタンパク、または組み換えメガヌクレアーゼ遺伝子配列を輸送するために使用される。
【0172】
7. 病原体感染の治療法
本発明の局面はさらに、病原体による感染の治療法を提供する。病原性生物体としては、ウィルス、例えば、ただしこれらに限定されるものではないが、単純ヘルペスウィルス1、単純ヘルペスウィルス2、ヒト免疫不全ウィルス1、ヒト免疫不全ウィルス2、痘瘡ウィルス、ポリオウィルス、エプスタインバーウィルスもしくはヒトパピローマウィルスなど、及び細菌生物体、例えば、これらに限定されるものではないが、BACILLUS
ANTHRACIS、HAEMOPHILUS種、PNEUMOCOCCUS種、STAPHYLOCOCCUS AUREUS、STREPTOCOCCUS種、メシシリン耐性STAPHYLOCOCCUS AUREUS及びMYCOPLASMA TUBERCULOSISなどが挙げられる。病原生物体としてはさらに、真菌生物体、例えば、ただしこれらに限定されるものではないが、CANDIDA、BLASTOMYCES、CRYPTOCOCCUS、およびHISTOPLASMA種などが挙げられる。
【0173】
ある実施態様では、合理設計メガヌクレアーゼは、病原体ゲノム内の認識配列、例えば、該病原体の増殖、生殖又は毒性にとって必須である、遺伝子または調整要素を標的とすることが可能である。ある実施態様では、認識配列は、細菌プラミド内にあってもよい。病原体ゲノムの認識配列のメガヌクレアーゼ介在性切断は、非相同的末端接合を刺激することによって、挿入、欠失又はフレームシフトの形で、標的必須遺伝子内の突然変異を刺激することが可能である。それとは別に、細菌プラスミドの切断は、その上にコードされる任意の遺伝子、例えば、トキシン遺伝子(例えば、B.ANTHRACIS致死因子遺伝子)又は抗生物質耐性遺伝子などと共にプラスミドの消失を招く可能性がある。前述のように、メガヌクレアーゼは、従来技術で一般的な技術を用いて、タンパク質又は核酸の形で、感染患者、動物又は植物に輸送してよい。ある実施態様では、メガヌクレアーゼ遺伝子は、病原体細菌類に輸送するために、バクテリオファージゲノムの中に組み込んでもよい。
【0174】
本発明の局面はさらに、ある形の癌の治療のための治療薬を提供する。ヒトのウィルスは、多くの場合、腫瘍形成と関連するので(例えば、エプスタインバーウィルスと鼻咽頭癌;ヒトパピローマウィルスと子宮頚部癌)、これらのウィルス性病原体の不活性化は、癌の発達または進行を阻止することがある。それとは別に、合理設計メガヌクレアーゼによる、これらの腫瘍関連ウィルスのゲノムを標的とする、二本鎖切断点を用いて、DNA損傷反応経路を介してアポトーシスを誘発してもよい。このようにして、ウィルスゲノムを抱える腫瘍細胞において選択的にアポトーシスを誘発することが可能となる場合がある。
【0175】
8. 遺伝子型分析法および病原体の特定
本発明の局面はさらに、インビトロ分子生物学研究および開発のためのツールを提供する。核酸、例えば、プラスミド、PCR産物、BAC配列、YAC配列、ウィルス及び真核および前核生物体由来のゲノム配列などの核酸の単離、クローニング及び操作のために、部位特異的エンドヌクレアーゼ(例えば、制限酵素)を用いることは、従来技術で普通のことである(例えば、AUSUBEL ET AL.,「分子生物学における最近のプロトコール(“CURRENT PROTOCOLS IN MOLECULAR BIOLOGY,”)」、WILEY 1999を参照されたい)。したがって、ある実施態様では、インビトロにおける核酸の操作に合理設計メガヌクレアーゼを使用してもよい。例えば、同じDNA分子の中に一対の認識配列を認識する、合理設計メガヌクレアーゼは、その後の操作、例えば、細菌プラスミド、BAC又はYACへのコンタクトなどのための介在DNAセグメントを単離するのに使用することが可能である。
【0176】
別の局面では、本発明は、病原遺伝子及び生物体特定用のツールを提供する。一実施態様では、合理設計メガヌクレアーゼは、健康な対立遺伝子から病的対立遺伝子を区別するために、病気と相関する多型遺伝子領域において対応する認識部位を切断するために使用
することが可能である(例えば、ヒトのCFTR遺伝子のΔF−508対立遺伝子を認識する合理設計メガヌクレアーゼ、実施例4を参照)。この実施態様では、ヒトの患者、又は他の生物体から単離されたDNA配列が、合理設計メガヌクレアーゼによって、場合によっては、追加の部位特異的ヌクレアーゼと組み合わせて消化され、得られたDNA断片のパターンが、ゲル電気泳動、毛細管電気泳動、質量分析又はその他の従来技術で既知の方法によって分析される。この断片化パターン及び具体的には合理設計メガヌクレアーゼによる切断の有無は、ゲノム中に認識配列が存在するか否かを明らかにすることによって該生物体の遺伝子型を示す。別の実施態様では、合理設計メガヌクレアーゼは、病原性ウィルス、真菌、または細菌のゲノムにおける多型領域を標的とし、該生物体を特定するのに使用される。この実施態様では、合理設計メガヌクレアーゼは、病原体に固有の認識配列を切断するが(例えば、細菌における16Sおよび23SRRNA遺伝子の間のスペーサー領域;例えば、VAN DER GIESSEN ET AL.(1994),MICROBIOLOGY 140:1103−1108を参照)、これを用いて、ゲノムのエンドヌクレアーゼ消化及びそれに続く、電気泳動、質量分析、または、従来技術で既知の、その他の方法による断片化パターンの分析に基づいて、他の、近縁生物体と区別することが可能である。
【0177】
9. 特注DNA−結合ドメインの生産法
他の側面で、本発明は、エンドヌクレアーゼ切断活性を除去した合理設計DNA−結合タンパク質を提供する。合理設計メガヌクレアーゼの触媒活性の除去は、触媒に与るアミノ酸を突然変異させることによって実行することが可能である(例えば、I−CREIにおけるQ47のEへの突然変異、CHEVALIER ET AL.(2001),BIOCHEMISTRY,43:14015−14026を参照);I−SCEIにおけるD44またhD145のNへの突然変異;I−CREIにおけるE66のQへの突然変異;I−MSOIにおけるD22のNへの突然変異)。次に、この不活性化メガヌクレアーゼを別のタンパク質、例えば、ただしこれらに限定されるものではないが、転写アクチベーター(例えば、GAL4トランスアクチベーションドメイン、またはVP16トランスアクチベーションドメイン)、転写リプレッサー(例えば、KRUPPELタンパクのKRABドメイン)、DNAメチラーゼドメイン(例えば、M.CVIPI、またはM.SSSI)、またはヒストンアセチルトランスフェラーゼ・ドメイン(例えば、HDAC1、またはHDAC2)などのエフェクタードメインに融合させることが可能である。もっとも著明なのとして加工されたジンクフィンガードメインがある、加工されたDNA結合ドメイン及び、エフェクタードメインから成るキメラタンパクは、従来技術で既知である(例えば、PAPWORTH ET AL.(2006),GENE 366:27−38参照)。
【0178】
本発明は、下記の実施例によってさらに具体的に説明される。ただし、これらの実施例を限定的なものと考えてはならない。当業者であれば、たかだか通例の実験操作を用いるだけで、本出願に記載される特定物質および手順に対し数多くの等価物を認識し、確認することが可能であろう。そのような等価物は、下記の実施例に先行する特許請求項の範囲に含まれることが意図される。下記の実施例1−4は、I−CREIに基づく合理設計メガヌクレアーゼを特異的に言及するが、その外、I−SCEI、I−MSOI、I−CEUIに基づく合理設計メガヌクレアーゼ及びその他のLAGLIDADGメガヌクレアーゼも、本出願に記載するように、同様に生産し、使用することが可能である。
【実施例1】
【0179】
HIV−1 TAT遺伝子を認識するメガヌクレアーゼの合理設計
1. メガヌクレアーゼの設計
一対のメガヌクレアーゼが、HIV−1 TAT遺伝子に認められるDNA部位5’−GAAGAGCTCATCAGAACAGTCA−3’(配列番号15)を認識し、切断
するように設計された。表5に従って、二つのメガヌクレアーゼ、TAT1及びTAT2が、下記の塩基接触部(非WT接触部は太字で表す)を用い、それぞれ、半分部位5’−GAAGAGCTC−3’(配列番号16)、および5’−TGACTGTTC−3’(配列番号17)に結合するように設計された。
【0180】
TAT1:
【表14】
【0181】
TAT2:
【表15】
【0182】
二つの酵素はクローンされ、大腸菌において発現され、下記に記載するように、対応するDNA認識配列に対する酵素活性について定量された。いずれの場合も、合理設計メガヌクレアーゼは、不活性であることが認められた。次に、DNAバックボーンとの接触を向上させるためE80をQに突然変異させた、それぞれの第2世代を生産した。この第2世代TAT2酵素は、その意図される認識配列に対し活性を持つことが認められたが、一方、TAT1酵素は不活性のままであった。野生型I−CREIとの共晶構造の肉眼検査から、TAT1は、R40とK28の間の立体障害のために不活性であることが示唆された。この衝突を緩和するため、K28をより小さい側鎖を持つアミノ酸(A、S、T又はC)に突然変異させ、一方、Q80突然変異はそのまま維持させた、TAT1変種を生産した。これらの酵素をE.COLIにおいて生産し、定量したところ、S28及びT28を有するTAT1変種は、共に、所望の認識配列に対して活性を持つが、一方、位置−7において所望の塩基指向性を維持することが認められた。
【0183】
2. 組み換えメガヌクレアーゼの構築
再設計I−CREI酵素のための突然変異を、重複PCR戦略において突然変異誘発性プライマーを用いて導入した。一次PCRで生成されたI−CREIの組み換えDNA断片を、二次PCRにおいて接合し、全長の組み換え核酸を生産した。組み換えI−CREI構築体は全て、精製用遺伝子の3‘末端に、6個のヒスチジンタグを融合させたPET21Aベクター(NOVAGEN CORP.,SAN DIEGO、カリフォルニア州)にクローンした。核酸配列は全て、サンガーのジデオキシヌクレオチド配列決定法(SANGER ET AL.(1977),PROC.NATL.ACAD.SCI.USA.74(12):5463−7を参照)を用いて確認した。
【0184】
野生型I−CREI及び全ての加工メガヌクレアーゼは、下記の方法を用いて発現し、精製した。PET21Aベクターにクローンした構築体は、化学的にコンピテントとしたBL21(DE3)PLYSSにおいて形質転換され、200μG/MLのカルバニシリンを含む、2 X YTプレートに撒いた。一晩育成の後、形質転換細菌コロニーを、プ
レートから掻き落とし、それを用いて50 ML の2XYTブロスに接種した。細胞を、波長600NMにおいて光学的濁度が0.9に達するまで、振とうしながら37℃で育成した。次に、育成温度を37℃から22℃に下げた。タンパク質の発現は、1MM IPTGを添加することによって誘発し、細胞は、振とうしながら2時間半インキュベートした。次に、10分間6000 X Gの遠心によって細胞をペレットとした。ペレットを、1MLの結合バッファー(20MM TRIS−HCL,PH8.0,500MM NACL,10MMイミダゾール)において渦巻き攪拌によって再縣濁させた。次に、細胞を、50%電力における12パルスの超音波処理によって破壊し、細胞砕片を、15分間14,000 X Gの遠心によってペレットとした。細胞上清を、4MLの結合バッファーで希釈し、200μLのニッケル充填金属キレート性セファローズカラム(PHARMACIA)に負荷した。
【0185】
次いで、カラムを、4MLの洗浄バッファー(20MM TRIS HCL,PH8.0,500MM NACL,60MMイミダゾール)及び0.2MLの溶出バッファー(20MM TRIS−HCL,PH8.0,500MM NACL,400MMイミダゾール)で洗浄した。メガヌクレアーゼ酵素は、さらに0.6MLの溶出バッファーを添加することによって溶出し、VIVOSPINディスポーザブル濃縮器(ISC,INC.,KAYSVILLE,ユタ州)を用いて50−130μLとなるまで濃縮した。酵素は、ZEBAスピン脱塩カラム(PIERCE BIOTECHNOLOGY,INC.,ROCKFORD、イリノイ州)を用い、定量および保存のためにSAバッファー(25MM TRIS−HCL,PH8.0,100MM NACL,5MM MGCL2,5MM EDTA)と交換した。酵素濃度は、23,590M−1CM−1の吸光係数を用い280NMにおける吸収によって決めた。次に、酵素の純度および分子量は、MALDI−TOF質量分析によって確認した。
【0186】
ヘテロダイマー酵素は、二つのタンパクを独立に精製し、それらをインビトロで混合することによって又は大腸菌における二つのタンパクの縦列発現用人工オペロンを構築することによって生産した。前者の場合、精製メガヌクレアーゼを、溶液中で1:1で混合し、DNA基質の添加20分前に42℃で予備インキュベーションした。後者の場合、二つの遺伝子を、NDEI/ECORI及びECORI/HINDIIIを用いて順次PET−21A発現ベクターにクローンした。12塩基対の核酸スペーサー及びPET21ベクター由来のシャイン−デルガルノ配列が、人工オペロン中の第1及び第2遺伝子を隔てた。
【0187】
3. 切断アッセイ
前述のように精製した酵素は全て、メガヌクレアーゼの認識配列を含む、直線状の2本鎖DNA基質とインキュベーションすることによって活性について定量した。認識配列のセンス及びアンチセンスの両鎖に対応する合成オリゴヌクレオチドをアニールし、平滑末端コンタクトによって、PUC19プラスミドのSMAI部位にクローンした。このクローンされた結合部位の配列は、サンガーのジデオキシヌクレオチド配列決定法によって確認した。プラスミド基質は、全て、メガヌクレアーゼ消化と同時に、XMNI、SCAI、またはBPMIによって直線化された。この酵素消化物は、5μLの0.05μM DNA基質、2.5μLの5μM組み換えI−CREIメガヌクレアーゼ、9.5μLのSAバッファー及び0.5μLのXMNI、SCAI又はBPMIを含んでいた。消化物は、37℃で又はあるメガヌクレアーゼ酵素用として42℃で、4時間インキュベートした。消化は、0.3MG/MLのプロテイナーゼK及び0.5%SDSを加えることによって停止させ、37℃で1時間インキュベートした。消化物を、1.5%アガロースにおいて分析し、臭化エチジウム染色によって視像化した。
【0188】
メガヌクレアーゼ半分部位の指向性を評価するために、合理設計メガヌクレアーゼを、
意図する半分部位の完全パリンドロームに対応する、一組のDNA基質のほか、この半分部位における27通りの可能な単一塩基対置換の内の一つとインキュベートした。
【0189】
4. 認識配列−特異性
精製組み換えTAT1およびTAT2メガヌクレアーゼは、野生型メガヌクレアーゼ配列とは異なるDNAを認識した(図2(B))。野生型I−CREIメガヌクレアーゼは、WT認識配列を切断するが、TAT1のために意図された配列、TAT2のために意図された配列のいずれも切断しない。同様に、TAT1及びTAT2は、その意図された認識配列は切断するが、野生型配列は切断しない。次に、これらのメガヌクレアーゼを、半分部位指向性および全体特異性について評価した(図3)。野生型I−CREIは、その天然の半分部位における単一塩基対置換についてきわめて耐性が高いことが認められた。一方、TAT1及びTAT2は、きわめて特異的で、TAT1の場合は、位置−1、−2、−3、−6及び−8において、TAT2の場合は、位置−1、−2及び−6における塩基置換に対して完全に耐性を持たなかった。
【実施例2】
【0190】
変更DNA−結合親和度を持つメガヌクレアーゼの合理的設計
1. 親和度を上昇および活性を上昇させたメガヌクレアーゼ
メガヌクレアーゼCCR1およびBRP2を、それぞれ、半分部位5’−AACCCTCTC−3’(配列番号18)および5’−CTCCGGGTC−3’(配列番号19)を切断するように設計した。これらの酵素を、実施例1の場合と同様、表1に従って生産した。
【0191】
CCR1:
【表16】
【0192】
BRP2:
【表17】
【0193】
両酵素を大腸菌において発現させ、精製し、実施例1と同様にして定量した。両第1世代酵素は、天然の認識配列を持つ野生型I−CREIの速度よりも相当に低い速度で、その意図された認識配列を切断することが認められた。活性におけるこの低下を緩和するために、CCR1およびBRP2のDNA−結合親和度を、両酵素においてE80をQに突然変異させることによって上昇させた。これらの、CCR1及びBRP2の第2世代バージョンは、触媒速度をはっきりと上昇させて、その意図された認識配列を切断することが認められた。
【0194】
2. DNA−結合親和度を低下させ、活性を低下させたが、特異性を上昇させたメガヌ
クレアーゼ
野生型I−CREIは、その半分部位に対する置換に対して高い耐性を持つことが認められている(図3(A))。この酵素の特異性をさらに高めるための試みとして、通常、DNAバックボーンのリン酸塩と塩橋を造る、酵素の位置116のリシンを、アスパラギン酸に突然変異させてDNA−結合親和度を下げた。この合理設計酵素の、野生型認識配列に対する切断は、はっきりと低下していたが、この組み換え酵素の特異性は、野生型よりも相当に高かった。K116D変種の半分部位の指向性を、実施例1と同様に評価したところ、この酵素は、位置−1、−2及び−3における天然半分部位からの逸脱に対しては全く耐性を持たないことが認められ、半分部位の残りの6位置では、少なくとも部分的な塩基指向性を示した(図3(B))。
【実施例3】
【0195】
合理設計メガヌクレアーゼ・ヘテロダイマー
1. 溶液の中で形成されたメガヌクレアーゼ・ヘテロダイマーによる非パリンドロームDNA部位の切断
二つのメガヌクレアーゼLAM1及びLAM2が、それぞれ、半分部位5’−TGCGGTGTC−3’(配列番号20)及び5’−CAGGCTGTC−3’(配列番号21)を切断するように設計された。これら二つの酵素から成るヘテロダイマーは、バクテリオファージλ P05遺伝子に見られるDNA配列5’−TGCGGTGTCCGGCG
ACAGCCTG−3’(配列番号22)を認識することが予想された。
【0196】
LAM1:
【表18】
【0197】
LAM2:
【表19】
【0198】
LAM1及びLAM2は、実施例1に記載されるように、クローンし、大腸菌において発現し、個別に精製した。次に、二つの酵素を1:1で混合し、42℃で20分インキュ
ベートし、二つの酵素が、サブユニットを交換し、再度平衡に達するのを可能とした。LAM1ホモダイマー、LAM2ホモダイマー及びLAM1/LAM2ヘテロダイマーの混合物と予想される、得られた酵素液を、LAM1半分部位の完全パリンドローム、LAM2半分部位の完全パリンドローム及びバクテリオファージλゲノムに認められる非パリンドロームハイブリッド部位に対応する、3種の異なる認識配列と共にインキュベートした。精製LAM1酵素単独は、LAM1パリンドローム部位を切断するが、LAM2パリンドローム部位も、LAM1/LAM2ハイブリッド部位も切断しない。同様に、精製LAM2酵素単独は、LAM2パリンドローム部位を切断するが、LAM1部位も、LAM1/LAM2ハイブリッド部位も切断しない。しかしながら、LAM1及びLAM2の1:
1混合物は、3種全てのDNA部位を切断する。LAM1/LAM2ハイブリッド部位の切断は、二つの、異なる、再設計メガヌクレアーゼが溶液の中で混合されて、非パリンドローム的DNA部位を切断することが可能なヘテロダイマー酵素を形成することが可能であることを示す。
【0199】
2.共時発現によって形成されるメガヌクレアーゼ・ヘテロダイマーによる非パリンドロームDNA部位の切断
前述のLAM1及びLAM2酵素をコードする遺伝子は、実施例1において記載したように、大腸菌における同時発現のためにオペロンの中に配置された。この共時発現酵素は、実施例1と同様に精製され、かつ、酵素混合物は、前述の、三つの可能性のある認識配列とインキュベートされた。この共時発現酵素は、LAM1/LAM2ハイブリッド部位を含む三つ全ての部位を切断することが認められたが、これは、二つの、異なる合理設計メガヌクレアーゼが共時発現されて、非パリンドロームDNA部位を切断することが可能なヘテロダイマー酵素を形成することを示す。
【0200】
3.タンパク−タンパクインターフェイスを修飾されたメガヌクレアーゼ・ヘテロダイマーによる、非パリンドロームDNA部位の優先的切断
非パリンドロームDNA部位の切断を必要とする応用のために、異なる(パリンドローム的)DNA部位を認識し、切断するホモダイマーの形成を最小に留めながら、酵素ヘテロダイマーの形成を増進することが望ましい。このために、位置7、57及び96におけるリシンがグルタミン酸に変更されたLAM1酵素の変種が生産された。次に、この酵素を、位置8及び61のグルタミン酸をリシンに変えたLAM2の変種と、前述と同様に共時発現し、精製した。この場合、LAM1ホモダイマーの形成は、一方のモノマーにおけるE7、E57及びE96と、他方のモノマーにおけるE8及びE61の間における静電気的反発によって低下すると予想された。同様に、LAM2ホモダイマーの形成は、一方のモノマーにおけるK7、K57及びK96と、他方のモノマーにおけるK8及びK61の間における静電気的反発によって低下すると予想された。逆に、LAM1/LAM2ヘテロダイマーは、LAM1におけるE7、E57、およびE96、およびLAMにおけるK8及びK61の間の静電気的牽引によって好まれることが予想された。インターフェイスが修飾されたこの二つのメガヌクレアーゼが、前述のように、共時発現され、定量されると、LAM1/LAM2ハイブリッド部位が、二つのパリンドローム部位に対し優先的に切断されることが認められた。これは、メガヌクレアーゼのタンパク−タンパクインターフェイスにおける置換が、ヘテロダイマーの優先的形成の駆動が可能であることを示す。
【実施例4】
【0201】
生理的DNA配列を切断する、追加のメガヌクレアーゼ・ヘテロダイマー
1.遺伝子治療に関連するDNA配列を切断するメガヌクレアーゼ・ヘテロダイマー
軟骨無形成症を引き起こす突然変異である、ヒトのFGR3遺伝子における配列5’−CTGGGAGTCTCAGGACAGCCTG−3’(配列番号23)を切断する、合理設計メガヌクレアーゼ・ヘテロダイマー(ACH1/ACH2)を生産することが可能である。例えば、前述したように、I−CREIメガヌクレアーゼに基づいて、下記の接触残基および認識配列半分部位を持つメガヌクレアーゼが設計された。
【0202】
ACH1:
【表20】
【0203】
ACH2:
【表21】
【0204】
ヒトの成長ホルモン遺伝子のプロモーターにおいて配列5’−CCAGGTGTCTCTGGACTCCTCC−3’(配列番号24)を切断する合理設計メガヌクレアーゼ・ヘテロダイマー(HGH1/HGH2)を生産することが可能である。例えば、前述したように、I−CREIメガヌクレアーゼに基づいて、下記の接触残基および認識配列半分部位を持つメガヌクレアーゼが設計された。
【0205】
HGH1:
【表22】
【0206】
HGH2:
【表23】
【0207】
ヒトのCFTR遺伝子のΔF508対立遺伝子における5’−GAAAATATCATTGGTGTTTCCT−3’(配列番号25)を切断する合理設計メガヌクレアーゼ・ヘテロダイマー(CF1/CF2)を生産することが可能である。例えば、前述したように、I−CREIメガヌクレアーゼに基づいて、下記の接触残基および認識配列半分部位を持つメガヌクレアーゼが設計された。
【0208】
CF1:
【表24】
【0209】
CF2:
【表25】
【0210】
ヒトのCCR5遺伝子(HIVコレセプター)の配列5’−AACCCTCTCCAGTGAGATGCCT−3’(配列番号26)を切断する合理設計メガヌクレアーゼ・ヘテロダイマー(CCR1/CCR2)を生産することが可能である。例えば、前述したように、I−CREIメガヌクレアーゼに基づいて、下記の接触残基および認識配列半分部位を持つメガヌクレアーゼが設計された。
【0211】
CCR1:
【表26】
【0212】
CCR2:
【表27】
【0213】
ヒトのDMキナーゼ遺伝子の3´非翻訳領域における5’−GACCTCGTCCTCCGACTCGCTG−3’(配列番号27)を切断する合理設計メガヌクレアーゼ・ヘテロダイマー(MYD1/MYD2)を生産することが可能である。例えば、前述したように、I−CREIメガヌクレアーゼに基づいて、下記の接触残基および認識配列半分部位を持つメガヌクレアーゼが設計された。
【0214】
MYD1:
【表28】
【0215】
MYD1:
【表29】
【0216】
2.病原体ゲノムのDNA配列を切断するメガヌクレアーゼ・ヘテロダイマー
単純ヘルペスウィルス−1および単純ヘルペスウィルス−2のUL36遺伝子の配列5’−CTCGATGTCGGACGACACGGCA−3’(配列番号28)を切断する合理設計メガヌクレアーゼ・ヘテロダイマー(HSV1/HSV2)を生産することが可能である。例えば、前述したように、I−CREIメガヌクレアーゼに基づいて、下記の接触残基および認識配列半分部位を持つメガヌクレアーゼが設計された。
【0217】
HSV1:
【表30】
【0218】
HSV2:
【表31】
【0219】
BACILLUS ANTHRACISゲノムの配列5’−ACAAGTGTCTAT
GGACAGTTTA−3’(配列番号29)を切断する合理設計メガヌクレアーゼ・ヘテロダイマー(ANT1/ANT2)を生産することが可能である。例えば、前述したように、I−CREIメガヌクレアーゼに基づいて、下記の接触残基および認識配列半分部位を持つメガヌクレアーゼが設計された。
【0220】
ANT1:
【表32】
【0221】
ANT2:
【表33】
【0222】
痘瘡(天然痘)ウィルスGP009遺伝子の配列5’−AAAACTGTCAAATGACATCGCA−3’(配列番号30)を切断する合理設計メガヌクレアーゼ・ヘテロダイマー(POX1/POX2)を生産することが可能である。例えば、前述したように、I−CREIメガヌクレアーゼに基づいて、下記の接触残基および認識配列半分部位を持つメガヌクレアーゼが設計された。
【0223】
POX1:
【表34】
【0224】
POX2:
【表35】
【0225】
エプスタインバーウィルスBALF2遺伝子の配列5’−CGGGGTCTCGTGCGAGGCCTCC−3’(配列番号31)を切断する合理設計メガヌクレアーゼ・ヘテロダイマー(EBB1/EBB2)を生産することが可能である。例えば、前述したように、I−CREIメガヌクレアーゼに基づいて、下記の接触残基および認識配列半分部位を持つメガヌクレアーゼが設計された。
【0226】
EBB1:
【表36】
【0227】
EBB1:
【表37】
【0228】
3.植物ゲノムにおけるDNA配列を切断するメガヌクレアーゼ・ヘテロダイマー
ARABIDOPSISTHALIANNA GL2遺伝子の配列5’−CACTAA
CTCGTATGAGTCGGTG−3’(配列番号32)を切断する合理設計メガヌクレアーゼ・ヘテロダイマー(GLA1/GLA2)を生産することが可能である。例えば、前述したように、I−CREIメガヌクレアーゼに基づいて、下記の接触残基および認識配列半分部位を持つメガヌクレアーゼが設計された。
【0229】
GLA1:
【表38】
【0230】
GLA2:
【表39】
【0231】
ARABIDOPSIS THALIANNA BP1遺伝子の配列5’−TGCCTCCTCTAGAGACCCGGAG−3’(配列番号33)を切断する合理設計メガヌクレアーゼ・ヘテロダイマー(BRP1/BRP2)を生産することが可能である。例えば、前述したように、I−CREIメガヌクレアーゼに基づいて、下記の接触残基および認識配列半分部位を持つメガヌクレアーゼが設計された。
【0232】
BRP1:
【表40】
【0233】
BRP2:
【表41】
【0234】
NICOTIANA TABACUMマグネシウムケラターゼ遺伝子の配列5’−TA
AAATCTCTAAGGTCTGTGCA−3’(配列番号34)を切断する合理設計メガヌクレアーゼ・ヘテロダイマー(MGC1/MGC2)を生産することが可能である。例えば、前述したように、I−CREIメガヌクレアーゼに基づいて、下記の接触残基および認識配列半分部位を持つメガヌクレアーゼが設計された。
【0235】
MGC1:
【表42】
【0236】
MGC2:
【表43】
【0237】
NICOTIANA TABACUM CYP82E4遺伝子の配列5’−CAAGA
ATTCAAGCGAGCATTAA−3’(配列番号35)を切断する合理設計メガヌクレアーゼ・ヘテロダイマー(CYP/HGH2)を生産することが可能である。例えば、前述したように、I−CREIメガヌクレアーゼに基づいて、下記の接触残基および認識配列半分部位を持つメガヌクレアーゼが設計された。
【0238】
CYP:
【表44】
【0239】
HGH2:
【表45】
【0240】
4.酵母ゲノムにおけるDNA配列を切断するメガヌクレアーゼ・ヘテロダイマー
SACCHAROMYCES CEREVISIAE URA3遺伝子の配列5’−TTAGATGACAAGGGAGACGCAT−3’(配列番号36)を切断する合理設計メガヌクレアーゼ・ヘテロダイマー(URA1/URA2)を生産することが可能である。例えば、前述したように、I−CREIメガヌクレアーゼに基づいて、下記の接触残基および認識配列半分部位を持つメガヌクレアーゼが設計された。
【0241】
URA1:
【表46】
【0242】
URA2:
【表47】
【0243】
5.認識配列特異性
本実施例において上に概説した合理設計メガヌクレアーゼを、実施例1と同様にして、クローンし、大腸菌において発現し、精製した。次いで、各精製メガヌクレアーゼを、そのヘテロダイマー形成パートナー(例えば、ACH1をACH2と、HGH1をHGH2と、など)と1:1で混合し、各メガヌクレアーゼ・ヘテロダイマーのために意図される
非パリンドロームDNA認識配列を含む、直線化DNA基質とインキュベートした。図3に示すように、各合理設計メガヌクレアーゼ・ヘテロダイマーは、その意図されたDNA部位を切断する。
【図面の簡単な説明】
【0244】
【図1】図1(A)は、結晶学データに基づいて、I−CREIホモダイマー及びその天然の2本鎖認識配列の相互作用を示す。本模式図は、二つの卵形として示すホモダイマーによって結合される、図示目的のためだけに巻き戻された状態で示される認識配列(配列番号2及び配列番号3)を描いたものである。各DNA半分部位の塩基には、−1から−9までの番号がふられ、認識表面を形成する、I−CREIのアミノ酸残基は、1文字アミノ酸表示および残基位置を示す数字によって表される。実線:DNA塩基に対する水素結合。鎖線:酵素設計では新たな接触点を形成するが、野生型複合体ではDNAと接触しないアミノ酸位置。矢印:DNAバックボーンと相互作用を持ち、切断活性に影響を及ぼす残基。また、図1(B)は、図1(A)の右側の切断半分部位の−4位置におけるA−T塩基対間の野生型接触を示す。具体的に言うと、残基Q26が、A塩基と相互作用を持つところが示される。残基I77は、該塩基対に近接するが、特異的相互作用は持たない。更に、図1(C)は、残基I77が修飾されてE77に変更された、I−CREIメガヌクレアーゼの、合理設計変種との間の相互作用を示す。この変更の結果、−4位置ではG−C塩基対が好まれる。Q26とG塩基との間の相互作用は、図1(A)の左側の切断半分部位において結晶学的に観察されるように、水分子によって仲介される。更に又、図1(D)は、残基Q26が修飾されてE26に変更され、残基I77が修飾されてR77に変更された、I−CREIメガヌクレアーゼの合理設計変種との間の相互作用を示す。この変更の結果、−4位置ではG−C塩基対が好まれる。そして更に、図1(E)は、残基Q26が修飾されてA26に変更され、残基I77が修飾されてQ77に変更された、I−CREIメガヌクレアーゼの合理設計変種との間の相互作用を示す。この変更の結果、−4位置ではT−A塩基対が好まれる。
【図2】図2(A)は、野生型I−CREIメガヌクレアーゼ(WT)及び本発明の11種の合理設計メガヌクレアーゼ・ヘテロダイマーそれぞれの一認識配列の比較を示す。WT認識配列に対して保存される塩基には陰影を施す。9BPの半分部位は太字で示す。WT:野生型(配列番号4);CF:多くの嚢胞性線維症例の原因となる、ヒトCFTR遺伝子のΔF508対立遺伝子(配列番号25);MYD:筋緊張性ジストロフィーと関連するヒトDMキナーゼ遺伝子;CCR:ヒトCCR5遺伝子(大型HIVコレセプター)(配列番号26);ACH;軟骨無形成症と相関するヒトのFGFR3遺伝子(配列番号23);TAT:HIV−1TAT/REV遺伝子(配列番号15);HSV:HSV−1 UL36遺伝子(配列番号28);LAM:バクテリオファージλ P05遺伝子(配列番号22);POX:痘瘡(天然痘)ウィルスGP009遺伝子(配列番号30);URA:SACCHAROMYCES CERVISIAE URA3遺伝子(配列番号36);GLA:ARABIDOPSIS THALIANA GL2遺伝子(配列番号32);BRP:ARABIDOPSISTHALIANA BP−1遺伝子(配列番号33)。図2(B)は、野生型I−CREI(WT)及び11種の合理設計メガヌクレアーゼ・ヘテロダイマーのそれぞれと、12種全ての酵素の認識部位を担持するプラスミドと、37℃で6時間インキュベートした場合の結果を示す。各ボックスに切断パーセントが示される。
【図3】図3は、野生型および合理設計I−CREIホモダイマーの切断パターンを示す。(A)野生型I−CREI。(B)I−CREIK116D。(C−L)本発明の合理設計メガヌクレアーゼ。酵素は、27通りの対応する単一塩基対変動を持つ、意図される切断半分部位のパリンドロームを担持する、一組のプラスミドと共にインキュベートした。黒色バー:表1に基づいて予想される切断パターン。灰色バー:予想される切断パターンから変位するDNA部位。円は、意図される認識部位における塩基を示す。さらに、2時間における切断の時間経過も示される。CおよびLにおける白抜き円の時間経過プロットは、E80Q突然変異を欠くCCR1およびBRP2酵素による切断に対応する。切断部位は、図2(A)に記載されるヘテロダイマー酵素の、5’(左コラム)、および3’(右コラム)半分部位に対応する。
【技術分野】
【0001】
本発明は、分子生物学および組み換え核酸工学の分野に関する。特に、本発明は、DNA認識配列特異性及び/又は親和度が変更された、合理的に設計された、非天然のメガヌクレアーゼに関する。さらに、本発明は、そのようなメガヌクレアーゼの生産法、および、そのようなメガヌクレアーゼによる、組み換え核酸および生物の生産法に関する。
【背景技術】
【0002】
(関連出願)本出願は、2005年10月18日出願の米国特許仮出願第60/727,512号の優先権の利益を主張する。なお、この引用出願の全開示を、参照することにより本出願に含める。
【0003】
(政府援助)本発明は、アメリカ合衆国国立衛生研究所、国立医科学研究所からの研究補助金2R01−GM−0498712、5F32−GM072322、および5 DP1
OD000122によって一部援助された。したがって、米国政府も、本発明において若干の権利を保有する場合がある。
【0004】
ゲノム工学は、あるゲノムの内部に、特定の遺伝子配列を、挿入し、欠失し、置換し、および、その他のやり方で操作することを可能とする能力を要求するが、多くの治療的および生物工学的応用を有する。ゲノム修飾のための効果的手段の開発は、これまでずっと、遺伝子治療、農工学、および合成生物学における主要目標となっている(PORTEUS ET AL.(2005),NAT.BIOTECHNOL.23;967−73;TZFIRA ET AL.(2005),TRENDS BIOTECHNOL.23:567−9;MCDANIEL ET AL.(2005),CURR.OPIN.BIOTECHNOL.16:476−83)。DNA配列を挿入又は修飾するための一般的方法は、ゲノム標的に対して相同な配列によって側接される、トランスジェニックDNA配列を導入すること、および、所望の相同組み換え事象を選択またはスクリーニングすることを含む。トランスジェニックDNAによる組み換えはめったに起こらないが、標的部位のゲノムDNAにおける2本鎖切断によって刺激することが可能である。従来から、DNA2本鎖切断部を作るために、例えば、放射線照射および化学的処理を含めた数多くの方法が用いられている。これらの方法は、効率的に組み換えを刺激することが可能ではあるけれども、二本鎖切断部は、ゲノムの中にランダムに挿入され、それらは、突然変異誘発性および毒性が高い可能性がある。現在、染色体バックグランドにある特定の部位に対し遺伝子修飾を照準することはできていないため、思い通りのゲノム加工に対する大きな障害となっている。
【0005】
この目標を達成するための一つの方法は、ゲノム内の単一部位にしか存在しないほど十分に大きい配列に対し特異性を持つヌクレアーゼを用いて、標的配座に二本鎖切断部を設けてその部位における相同組み換えを刺激することである(例えば、PORTEUS ET AL.(2005),NAT.BIOTECHNOL.23:967−73を参照)。この戦略の有効性は、FOKI制限酵素の、加工されたジンクフィンガーDNA−結合ドメインと、非特異的ヌクレアーゼドメインとの間のキメラ融合を用いて、各種の生物体において明らかにされている(PORTEUS(2006),MOL THER 13:438−46;WRIGHT ET AL.(2005),PLANT J.44:693−705;URNOV ET AL.(2005),NATURE 435:646−51)。これらの人工的ジンクフィンガーヌクレアーゼは、部位特異的組み換えを刺激するけれども、これらのヌクレアーゼは、ヌクレアーゼドメインの調整低調に由来する、非
特異的な、残留切断活性を保持するので、不要の部位を切断することがしばしばある(SMITH ET AL.(2000),NUCLEIC ACIDS RES.28:3361−9)。これらの不要な切断は、処置生物体において、突然変異および毒性を引き起こす可能性がある(PORTEUS ET AL.(2005),NAT.BIOTECHNOL.23:967−73)。
【0006】
植物および菌類のゲノムの中に一般的に認められる、15−40塩基対切断部位を認識する、1群の天然ヌクレアーゼは、比較的毒性の低い、ゲノム加工代替品を提供する可能性がある。このような「メガヌクレアーゼ」または「ホーミング・エンドヌクレアーゼ」は、多くの場合、寄生的DNA要素、例えば、グループ1自己スプライシングイントロンおよびインテインとコンタクトされる。これらのヌクレアーゼは、自然状態で、宿主ゲノムの特異的位置における相同的組み換えまたは遺伝子挿入を促進する。これを、細胞のDNA−修復機構を招集する、染色体において二本鎖切断を生産することによって行う(STODDARD(2006),REV.BIOPHYS.38:49−95)。一般に、メガヌクレアーゼは、4つのファミリーに分けられる。すなわち、LAGLIDADGファミリー、GIY−YIGファミリー、HIS−CYSボックスファミリー、およびHNHファミリーである。これらのファミリーは、触媒活性および認識配列に影響を及ぼす、構造的モチーフによって特徴づけられる。例えば、LAGLIDADGファミリーのメンバーは、LAGLIDADG保存モチーフの、1または2コピーを持つことによって特徴づけられる(CHEVALIER ET AL.(2001),NUCLEIC ACIDS RES.29(18):3757−3774を参照)。LAGLIDADGモチーフの単一コピーを持つLAGLIDADGメガヌクレアーゼは、ホモダイマーを形成するが、一方、2コピーのLAGLIDADGモチーフを持つメンバーは、モノマーであることが判明した。同様に、GIY−YIGファミリーメンバーは、70−100残基長のGIY−YIGモジュールを持ち、その内二つは活性のために必要な、四つの不変残基を持つ、4から5個の保存配列モチーフを含む(VAN ROEY ET AL.(2002),NATURE STRUCT.BIOL.9:806−811を参照)。HIS−CYSボックス・メガヌクレアーゼは、数百のアミノ酸残基を含む領域に分散する、高度に保存された、ヒスチジンおよびシステインの連続列によって特徴づけられる(CHEVALIER ET AL.(2001),NUCLEIC ACIDS RES.29(18):3757−3774を参照)。NHNファミリーの場合は、そのメンバーは、2対の、アスパラギン残基によって囲まれる保存ヒスチジンを含むモチーフによって定義される(CHEVALIER ET AL.(2001),NUCLEIC ACIDS RES.29(18):3757−3774を参照)。メガヌクレアーゼのこの4ファミリーは、構造保存部位に関してDNA認識配列の特異性及び触媒活性が互いに大きく異なっている。
【0007】
主にLAGLIDADGファミリーから得られる、天然のメガヌクレアーゼは、これまで、植物、酵母、ショウジョウバエ、哺乳類細胞及びマウスにおいて部位特異的なゲノム修飾に有効に使用されているが、この方法は、このメガヌクレアーゼ認識配列を保存する相同配列か(MONNAT ET AL.(1999),BIOCHEM.BIOPHYS.RES.COMMUN.255:88−93)又は認識配列を導入したあらかじめ加工されたゲノム(ROUET ET AL.(1994),MOL.CELL BIOL.14:8096−106;CHILTON ET AL.(2003),PLANT PHYSIOL.133:956−65;PUCHTA ET AL.(1996),PROC.NATL.ACAD.SCI.USA 93:5055−60;RONG ET
AL.(2002),GENES DEV.16:1568−81;GOUBLE ET AL.(2006),J.GENE MED.8(5):616−622)の修飾に限定される。
【0008】
ヌクレアーゼ刺激による遺伝子修飾の体系的実施は、ゲノムの既存部位に対する標的DNA中断部に向けて照準させた特異性を持つ加工酵素の使用が必要となる。したがって、医学的又は生物工学的関連部位において遺伝子修飾を促進するようにメガヌクレアーゼを適応させることについては大きな関心が寄せられている(PORTEUS ET AL.(2005),NAT.BIOTECHNOL.23:967−73;SUSSMAN ET AL.(2004),J.MOL.BIOL.342:31−41;EPINAT
ET AL.(2003),NUCLEIC ACIDS RES.31:2952−62)。
【0009】
CHLAMYDOMONASREINHARDTIIから得られたメガヌクレアーゼI−CREIは、LAGLIDADGファミリーの一員であり、葉緑体染色体の中の22塩基対認識配列を認識・切断し、かつ、メガヌクレアーゼ再設計の好個の標的となっている。この野生型酵素は、各モノマーが、全体認識配列における9塩基対に対して直接接触する、ホモダイマーである。この認識配列の単一位置において(SUSSMAN ET AL.(2004),J.MOL.BIOL.342:31−41;CHAMES ET AL.(2005),NUCLEIC ACIDS RES.33:E178;SELIGMAN ET AL.(2002),NUCLEIC ACIDS RES.30:3870−9)、あるいは、比較的最近では、認識配列の中の3位置において(ARRNOULD ET AL.(2006),J.MOL.BIOL.355:443−58)、塩基選択性を変えるI−CREIにおける突然変異を特定するために、遺伝子選択技術が用いられている。I−CREIタンパク−DNAインターフェイスは、DNA塩基に直接接触する9個のアミノ酸と、修飾されたインターフェイスにおいて接触を形成する可能性のある、少なくともさらに5個の残基を含む。このインターフェイスのサイズは、切断部位を大きく変更された酵素を選択するために構築された配列ライブラリーにおいても十分に抽出されることがほとんどないと思われるほど複雑な組み合わせを提示する。
【発明の開示】
【発明が解決しようとする課題】
【0010】
ゲノムの正確な修飾を促進するヌクレアーゼについては、依然として需要がある。さらに、特定の遺伝子座位における遺伝子配列の操作を可能とする、あらかじめ指定された、合理的に設計された認識配列を有するヌクレアーゼを生成する技術、および、正確な配列修飾を持つ生物体を遺伝学的に加工するために、そのようなヌクレアーゼを利用する技術に対しては、依然として需要がある。
【課題を解決するための手段】
【0011】
本発明は、メガヌクレアーゼLAGLIDADGファミリーの同定及び特徴解明に基づく。メガヌクレアーゼが2本鎖DNA認識配列とコンタクトする際、DNA塩基およびDNAバックボーンに結合し、そうすることによって、該酵素の特異性および活性に影響を及ぼす。この発見は、以下に詳細に説明するように、メガヌクレアーゼの認識配列特異性及び/又はDNA結合親和性を変えることが可能なアミノ酸置換を特定するため及び天然のメガヌクレアーゼが認識しない所望のDNA配列を認識することが可能なメガヌクレアーゼを合理的に設計し、開発するために用いられた。本発明はさらに、遺伝子治療、病原体感染の治療及び診断と研究におけるインビトロ応用のために、生物体ゲノム内の少数座位において所望の遺伝子配列の組み換えを実現するために、このようなメガヌクレアーゼを使用する方法を提供する。
【0012】
したがって、ある実施態様では、本発明は、野生型I−CREIメガヌクレアーゼに比べ、少なくとも一つの認識配列半分部位に対し変更特異性を持つ組み換えメガヌクレアーゼを提供する。この実施態様におけるメガヌクレアーゼは、配列番号1の野生型I−CREIメガヌクレアーゼの残基2−153に対して少なくとも85%の配列類似性を有する
ポリペプチドを含むが、該組み換えメガヌクレアーゼは、配列番号2、配列番号3、配列番号4及び配列番号5から選ばれる、I−CREIメガヌクレアーゼ認識配列内の半分部位と、少なくとも1塩基対異なる認識配列半分部位に対して特異性を持ち、かつ、この組み換えメガヌクレアーゼは、従来技術に認められる排外的修飾ではない、表5に掲げられる少なくとも一つの修飾を含む。
【0013】
別の実施態様では、本発明は、野生型I−MSOIメガヌクレアーゼと比べ、少なくとも一つの認識配列半分部位に対し変更特異性を持つ組み換えメガヌクレアーゼを提供する。この実施態様におけるメガヌクレアーゼは、配列番号6のI−MSOIメガヌクレアーゼの残基6−160に対して少なくとも85%の配列類似性を有するポリペプチドを含むが、該組み換えメガヌクレアーゼは、配列番号7および配列番号8から選ばれる、I−MSOIメガヌクレアーゼ認識配列内の半分部位と、少なくとも1塩基対異なる、認識配列半分部位に対して特異性を持ち、かつ、該組み換えメガヌクレアーゼは、従来技術に認められる排外的修飾ではない、表7に掲げられる少なくとも一つの修飾を含む。
【0014】
別の実施態様では、本発明は、野生型I−SCEIメガヌクレアーゼと比べ、認識配列に対し変更特異性を持つ組み換えメガヌクレアーゼを提供する。この実施態様におけるメガヌクレアーゼは、配列番号9のI−SCEIメガヌクレアーゼの残基3−186に対して少なくとも85%の配列類似性を有するポリペプチドを含むが、この組み換えメガヌクレアーゼは、配列番号10および配列番号11のI−SCEIメガヌクレアーゼ認識配列と少なくとも1塩基対異なる認識配列に対して特異性を持ち、かつ、この組み換えメガヌクレアーゼは、従来技術に認められる排外的修飾ではない、表9に掲げられる少なくとも一つの修飾を含む。
【0015】
別の実施態様では、本発明は、野生型I−CEUIメガヌクレアーゼと比べ、少なくとも一つの認識配列半分部位に対し変更特異性を持つ組み換えメガヌクレアーゼを提供する。この実施態様におけるメガヌクレアーゼは、配列番号12のI−CEUIメガヌクレアーゼの残基5−211に対して少なくとも85%の配列類似性を有するポリペプチドを含むが、該組み換えメガヌクレアーゼは、配列番号13および配列番号14から選ばれるI−CEUIメガヌクレアーゼ認識配列内の半分部位と少なくとも1塩基対異なる認識配列半分部位に対して特異性を持ち、かつ、この組み換えメガヌクレアーゼは、従来技術に認められる排外的修飾ではない、表11に掲げられる少なくとも一つの修飾を含む。
【0016】
本発明のメガヌクレアーゼは、認識配列内の1、2又は3以上の位置において組み換えメガヌクレアーゼの配列特異性に影響を及ぼすために、本出願に開示される修飾の内の一つ、二つ又は三つ以上を含むことが可能である。メガヌクレアーゼは、本出願に開示される新規修飾のみを含んでもよいし又は本出願に開示される新規修飾を従来技術で認められる修飾と組み合わせて含んでもよい。しかしながら、特異的に排除されるものは、従来技術の修飾のみを含む組み換えメガヌクレアーゼである。
【0017】
他の側面から考えると、本発明は、2本鎖DNAに対する配列特異的ではない結合親和度を変更させた組み換えメガヌクレアーゼを提供する。これは、2本鎖DNA認識配列のバックボーンに接触する、メガヌクレアーゼ残基を修飾することによって実現される。この修飾は、結合親和度を増大又は低減するため、酵素の全体活性を増大又は低減することが可能である。さらに、結合および活性の上昇/低下は、配列特異性における低下/上昇を引き起こすことが認められた。したがって、本発明は、DNA−結合親和度を変えることによって配列特異性を全体的に変えるための手段を提供する。
【0018】
したがって、ある実施態様では、本発明は、野生型I−CREIメガヌクレアーゼと比べ、2本鎖DNAに対する結合親和度を変更させた組み換えメガヌクレアーゼを提供する
。この発明におけるメガヌクレアーゼは、配列番号1のI−CREIメガヌクレアーゼの残基2−153に対して少なくとも85%の配列類似性を有するポリペプチドを含むが、DNA結合親和度は、(1)(A)H、N、Q、S、T、K若しくはRによるE80、D137、I81、L112、P29、V64若しくはY66の置換、又は(B)K若しくはRによるT46、T140若しくはT143の置換から選ばれる一つの置換に対応する少なくとも一つの修飾によって上昇するか、又は、逆に、(2)(A)H、N、Q、S、T、D若しくはEによるK34、K48、R51、K82、K116若しくはK139の置換、又は(B)D若しくはEによるI81、L112、P29、V64、Y66、T46、T140若しくはT143の置換から選ばれる一つの置換に対応する少なくとも一つの修飾によって低下する。
【0019】
別の実施態様では、本発明は、野生型I−MSOIメガヌクレアーゼに比べ、2本鎖DNAに対する結合親和度を変更させた組み換えメガヌクレアーゼを提供する。この発明におけるメガヌクレアーゼは、配列番号6のI−MSOIメガヌクレアーゼの残基6−160に対して少なくとも85%の配列類似性を有するポリペプチドを含むが、DNA結合親和度は、(1)(A)H、N、Q、S、T、K若しくはRによるE147、I85、G86若しくはY118の置換、又は(B)K若しくはRによるQ41、N70、S87、T88、H89、Q122、Q139、S150若しくはN152の置換から選ばれる一つの置換に対応する少なくとも一つの修飾によって上昇するか、又は、逆に、(2)(A)H、N、Q、S、T、D若しくはEによるK36、R51、K123、K143若しくはR144の置換、又は(B)D若しくはEによるI85、G86、Y118、Q41、N70、S87、T88、H89、Q122、Q139、S150若しくはN152の置換から選ばれる一つの置換に対応する少なくとも一つの修飾によって低下する。
【0020】
別の実施態様では、本発明は、野生型I−SCEIメガヌクレアーゼに比べ、2本鎖DNAに対する結合親和度を変更させた組み換えメガヌクレアーゼを提供する。この発明におけるメガヌクレアーゼは、配列番号9のI−SCEIメガヌクレアーゼの残基3−186に対して少なくとも85%の配列類似性を有するポリペプチドを含むが、DNA結合親和度は、(1)(A)H、N、Q、S、T、K若しくはRによるD201、L19、L80、L92、Y151、Y188、I191、Y199若しくはY222の置換、又は(B)K若しくはRによるN15、N17、S81、H84、N94、N120、T156、N157、S159、N163、Q165、S166,N194若しくはS202の置換から選ばれる一つの置換に対応する少なくとも一つの修飾によって上昇するか、又は、逆に、(2)(A)H、N、Q、S、T、D若しくはEによるK20、K23、K63、K122、K148、K153、K190、K193、K195若しくはK223の置換、又は(B)D若しくはEによるL19、L80、L92、Y151、Y188、I191、Y199、Y222、N15、N17、S81、H84、N94、N120、T156、N157、S159、N163、Q165、S166、N194若しくはS202の置換から選ばれる一つの置換に対応する少なくとも一つの修飾によって低下する。
【0021】
別の実施態様では、本発明は、野生型I−CEUIメガヌクレアーゼに比べ、2本鎖DNAに対する結合親和度を変更させた組み換えメガヌクレアーゼを提供する。この発明におけるメガヌクレアーゼは、配列番号12のI−CEUIメガヌクレアーゼの残基5−211に対して少なくとも85%の配列類似性を有するポリペプチドを含むが、DNA結合親和度は、(1)(A)H、N、Q、S、T、K若しくはRによるD25若しくはD128の置換、又は(B)K若しくはRによるS68、N70、H94、S117、N120、N129若しくはH172の置換から選ばれる一つの置換に対応する少なくとも一つの修飾によって上昇するか、又は、逆に、(2)(A)H、N、Q、S、T、D若しくはEによるK21、K28、K31、R112、R114若しくはR130の置換、又は(B)D若しくはEによるS68、N70、H94、S117、N120、N129若しくは
H172の置換から選ばれる一つの置換に対応する少なくとも一つの修飾によって低下する。
【0022】
本発明のメガヌクレアーゼは、DNA−結合親和度に影響を及ぼすために、本出願に開示される、バックボーン接触残基の修飾の内、一つ、二つ又は三つ以上を含むことが可能である。さらに、DNA−結合親和度に影響を及ぼすこれらの修飾は、認識配列内部の特定位置における組み換えメガヌクレアーゼの配列特異性を変える、前述の塩基接触残基の新規修飾の内の一つ以上と組み合わせてもよいし、前述の従来技術の修飾と組み合わせてもよいし、新規修飾と従来技術の修飾との組み合わせと組み合わせてもよい。特に、バックボーン接触修飾および塩基接触修飾を組み合わせることによって、組み換えメガヌクレアーゼを、所望の特異性および活性を持つように合理的に設計することが可能である。例えば、塩基接触残基に対する設計変更に由来する親和度の損失を逆転するDNA−結合親和度の増加を設計することが可能であり、配列特異性を減少させ酵素に対する認識配列の組を広げる親和度減少を設計することも可能である。
【0023】
他の側面から考えると、本発明は、ホモ又はヘテロダイマー形成に対する親和度を変更させた合理的設計メガヌクレアーゼモノマーを提供する。ダイマー形成に対する親和度は、同じモノマー(すなわち、ホモダイマー形成)によって又は異なるモノマー(すなわち、ヘテロダイマー形成)、例えば、参照野生型メガヌクレアーゼによって測定することが可能である。これらの組み換えメガヌクレアーゼは、メガヌクレアーゼダイマーにおけるモノマー間のタンパク質とタンパク質との境界面に存在するアミノ酸残基に対する修飾を有する。これらの修飾は、ヘテロダイマー形成の促進及び非パリンドローム認識配列を持つメガヌクレアーゼの創製に使用することが可能である。
【0024】
したがって、他の実施態様では、本発明は、参照メガヌクレアーゼモノマーによるダイマー形成に対する親和度を変更させた組み換えメガヌクレアーゼを提供する。この実施態様では、組み換えモノマーは、配列番号1のI−CREIメガヌクレアーゼの残基2−153に対して少なくとも85%の配列類似性を有するポリペプチドを含むが、ダイマー形成に対する親和度は、(A)D若しくはEによるK7、K57若しくはK96の置換、又は(B)K若しくはRによるE8若しくはE61の置換から選ばれる置換に対応する少なくとも一つの修飾によって変えられる。このような組み換えモノマーに基づき、本発明はさらに、(1)配列番号1のI−CREIメガヌクレアーゼの残基2−153に対して少なくとも85%の配列類似性を有する第1ポリペプチドであって、ダイマー形成に対する親和度が、(A)D若しくはEによるK7、K57若しくはK96の置換から選ばれる置換に対応する少なくとも一つの修飾によって変えられる第1ポリペプチド、及び(2)配列番号1のI−CREIメガヌクレアーゼの残基2−153に対して少なくとも85%の配列類似性を有する第2ポリペプチドであって、ダイマー形成に対する親和度が、(B)K若しくはRによるE8若しくはE61の置換から選ばれる置換に対応する少なくとも一つの修飾によって変えられる第2ポリペプチドを含む組み換えメガヌクレアーゼ・ヘテロダイマーを提供する。
【0025】
他の実施態様では、本発明は、参照メガヌクレアーゼモノマーによるダイマー形成に対する親和度を変更させた組み換えメガヌクレアーゼモノマーを提供する。この実施態様では、組み換えモノマーは、配列番号6のI−MSOIメガヌクレアーゼの残基6−160に対して少なくとも85%の配列類似性を有するポリペプチドを含むが、ダイマー形成に対する親和度は、(A)D若しくはEによるR302の置換、又は(B)K若しくはRによるD20、E11若しくはQ64の置換から選ばれる置換に対応する少なくとも一つの修飾によって変えられる。このような組み換えモノマーに基づき、本発明はさらに、(1)配列番号6のI−MSOIメガヌクレアーゼの残基6−160に対して少なくとも85%の配列類似性を有する第1ポリペプチドであって、ダイマー形成に対する親和度が、(
A)D若しくはEによるR302の置換から選ばれる置換に対応する少なくとも一つの修飾によって変えられる第1ポリペプチド、及び(2)配列番号6のI−MSOIメガヌクレアーゼの残基6−160に対して少なくとも85%の配列類似性を有する第2ポリペプチドであって、ダイマー形成に対する親和度が、(B)K若しくはRによるD20、E11若しくはQ64の置換から選ばれる置換に対応する少なくとも一つの修飾によって変えられる第2ポリペプチドを含む組み換えメガヌクレアーゼ・ヘテロダイマーを提供する。
【0026】
他の実施態様では、本発明は、参照メガヌクレアーゼモノマーによるダイマー形成に対する親和度を変更させた組み換えメガヌクレアーゼモノマーを提供する。この実施態様では、組み換えモノマーは、配列番号12のI−CEUIメガヌクレアーゼの残基5−211に対して少なくとも85%の配列類似性を有するポリペプチドを含むが、ダイマー形成に対する親和度は、(A)DまたはEによるR93の置換、又は(B)KまたはRによるE152の置換から選ばれる置換に対応する少なく共一つの修飾によって変えられる。このような組み換えモノマーに基づき、本発明はさらに、(1)配列番号12のI−CEUIメガヌクレアーゼの残基5−211に対して少なくとも85%の配列類似性を有する第1ポリペプチドであって、ダイマー形成に対する親和度が、(A)DまたはEによるR93の置換から選ばれる置換に対応する少なくとも一つの修飾によって変えられる第1ポリペプチド、及び(2)配列番号12のI−CEUIメガヌクレアーゼの残基5−211に対して少なくとも85%の配列類似性を有する第2ポリペプチドであって、ダイマー形成に対する親和度が、(B)KまたはRによるE152の置換から選ばれる置換に対応する少なくとも一つの修飾によって変えられる第2ポリペプチドを含む組み換えメガヌクレアーゼ・ヘテロダイマーを提供する。
【0027】
ダイマー形成に対する親和度を変更させた、組み換えメガヌクレアーゼモノマー又はヘテロダイマーはさらに、前述の塩基接触残基の修飾の内の一つ、二つまたは三つ以上;前述のバックボーン接触残基の修飾の内の一つ、二つまたは三つ以上、又は両者の組み合わせを含むことが可能である。したがって、例えば、モノマーの塩基接触部は、配列特異性を変えるように修飾することが可能であり、モノマーのバックボーン接触部は、DNA結合親和度を変えるように修飾することが可能であり、タンパク質とタンパク質との境界面は、ダイマー形成に影響を及ぼすように修飾することが可能である。このような組み換えモノマーは、同様に修飾されたモノマーと組み合わせて、所望の配列特異性および活性を有する合理設計メガヌクレアーゼ・ヘテロダイマーを生産することが可能である。
【0028】
他の側面から考えると、本発明は、本発明において記載され可能とされた合理設計メガヌクレアーゼのために使用される各種方法を提供する。これらの方法は、遺伝学的に修飾された細胞および生物体を生産すること、病気を遺伝子治療によって治療すること、病原体感染を治療すること及び診断学と研究用とのインビトロ応用のために組み換えメガヌクレアーゼを使用することを含む。
【0029】
したがって、この側面では、本発明は、染色体に挿入された興味の外因性配列を含む遺伝学的に修飾された真核細胞を生産するための方法であって、これを、該細胞を、(I)本発明のメガヌクレアーゼをコードする第1核酸配列、及び(II)前記興味の配列を含む第2核酸配列によってトランスフェクトすることによって実現する方法を提供する。その際、メガヌクレアーゼは、染色体の中に切断部位を生産し、興味の配列は、相同組み換え又は非相同的末端接合によって、染色体中の切断部位にに挿入される。
【0030】
それとは別に、他の側面では、本発明は、染色体に挿入された興味の外来配列を含む遺伝学的に修飾された真核細胞を生産するための方法であって、この細胞に本発明のメガヌクレアーゼタンパクを挿入すること及びこの細胞を興味の配列を含む核酸によってトランスフェクトすることによって実現する方法を提供する。その際、メガヌクレアーゼは、染
色体の中に切断部位を生産し、興味の配列は、相同組み換え又は非相同的末端接合によって、切断部位において染色体の中に挿入される。
【0031】
他の側面では、本発明は、染色体中の標的配列を破壊することによって遺伝学的に修飾された真核細胞を生産するための方法であって、この細胞を本発明のメガヌクレアーゼタンパクをコードする核酸によってトランスフェクトすることによって実現する方法を提供する。その際、メガヌクレアーゼは、染色体の中に切断部位を生産し、標的配列は切断部位における非相同的末端接合によって破壊される。
【0032】
他の側面では、本発明は、遺伝学的に修飾された生物体を生産するための方法であって、前述の方法に従って遺伝学的に修飾された真核細胞を生産し、かつ、この遺伝学的に修飾された真核細胞を育成して遺伝学的に修飾された生物体を生産することによって実現する方法を提供する。これらの実施態様では、真核細胞は、配偶子、接合子、胚盤胞細胞、胚幹細胞及びプロトプラスト細胞から選ぶことが可能である。
【0033】
他の側面では、本発明は、真核生物において遺伝子治療によって疾患を治療するための方法であって、この真核生物の少なくとも一つの細胞を、(I)本発明のメガヌクレアーゼをコードする第1核酸配列、及び(II)興味の配列を含む第2核酸配列によってトランスフェクトすることによって実現する方法を提供する。その際、メガヌクレアーゼは、染色体の中に切断部位を生産し、興味の配列は、相同組み換え又は非相同的末端接合によって染色体の中に挿入され、興味の配列の挿入は、疾患に対する遺伝子治療をもたらす。
【0034】
それとは別に、他の側面では、本発明は、真核生物において遺伝子治療によって疾患を治療するための方法であって、この真核生物の少なくとも一つの細胞の中に本発明のメガヌクレアーゼを導入すること及びこの細胞を、興味の配列を含む核酸によってトランスフェクトすることによって実現する方法を提供する。その際、メガヌクレアーゼは、染色体の中に切断部位を生産し、興味の配列は、相同組み換え、または非相同的末端接合によって切断部位において染色体の中に挿入され、興味の配列の挿入は、該疾患に対する遺伝子治療をもたらす。
【0035】
他の側面では、本発明は、真核生物において、該真核生物の染色体における標的配列を破壊する遺伝子治療によって疾患を治療するための方法であって、この真核生物の少なくとも一つの細胞を、本発明のメガヌクレアーゼをコードする核酸によってトランスフェクトすることによって実現する方法を提供する。その際、メガヌクレアーゼは、染色体の中に切断部位を生産し、標的配列は、相同組み換え又は非相同的末端接合によって破壊され、該標的配列の破壊が、該疾患に対する遺伝子治療をもたらす。
【0036】
別の局面では、本発明は、真核細胞宿主において、ウィルス性または前核細胞性病原体感染を治療するための方法であって、この病原体のゲノムにおける標的配列を破壊することによって実現する方法を提供する。その際、メガヌクレアーゼは、染色体の中に切断部位を生産し、標的配列は、(1)切断部位における非相同的末端接合によって、又は(2)第2核酸との相同組み換えによって破壊され、かつ、標的配列の破壊は、この感染に対する治療をもたらす。
【0037】
さらに一般的に、他の側面では、本発明は、認識配列の少なくとも一つの塩基位置に対する特異性を変更させた組み換えメガヌクレアーゼを合理的に設計する方法であって、(1)参照メガヌクレアーゼ−DNA複合体の三次元構造の少なくとも一部を決定すること、(2)該塩基位置において塩基接触面を形成するアミノ酸残基を特定すること、(3)接触面の少なくとも第1残基のβ−炭素と、該塩基位置における少なくとも第1塩基との間の距離を決めること及び(4)(A)第1塩基から6オングストローム未満の距離にあ
る第1残基については、G、C、T若しくはAの内の適切なもののメンバーである基1及び/又は基2の置換を選択すること;又は(B)前記第1塩基から6オングストロームより遠い位置にある第1残基については、G、C、T及びAの内の適切なもののメンバーである基2及び/又は基3の置換を選択することによって、所望の変化を促進するアミノ酸置換を特定することによって実現する方法を提供する。なお、上述したそれぞれの基は、後述するように定義する。この方法は、同じ塩基に対する別の接触残基ついても、同じ位置における他の塩基に対する接触残基についても、さらに、別の位置についても繰り返してよい。
【0038】
さらに、他の側面では、本発明は、DNA−結合親和度を増大させた組み換えメガヌクレアーゼを合理的に設計する方法であって、(1)基準メガヌクレアーゼ−DNA複合体の三次元構造の少なくとも一部を決定すること、(2)バックボーン接触面を形成するアミノ酸接触残基を特定すること、(3)(A)陰性荷電若しくは疎水性側鎖を持つ接触残基について、非荷電/極性若しくは陽性荷電側鎖を持つ置換を選択すること;又は(B)非荷電/極性側鎖を持つ接触残基について、陽性荷電側鎖を持つ置換を選択することによって、DNA−結合親和度を増大させるアミノ酸置換を特定すること、によって実現する方法を提供する。逆に、本発明は、DNA−結合親和度を減少させた組み換えメガヌクレアーゼを合理的に設計する方法であって、これを、(1)基準メガヌクレアーゼ−DNA複合体の三次元構造の少なくとも一部を決定すること、(2)バックボーン接触面を形成するアミノ酸接触残基を特定すること、(3)(A)陽性荷電側鎖を持つ接触残基について、非荷電/極性若しくは陰性荷電側鎖を持つ置換を選択すること、又は(B)疎水性及び非荷電/極性側鎖を持つ接触残基について、陰性荷電側鎖を持つ置換を選択することによって、DNA−結合親和度を減少させるアミノ酸置換を特定することよって実現する方法を提供する。
【0039】
本発明の、上記、およびその他の側面および実施態様は、本発明の下記の詳細な説明に基づき当業者には明白であろう。
【発明を実施するための最良の形態】
【0040】
1.1序論
本発明は、一部は、メガヌクレアーゼが2本鎖DNA認識配列とコンタクトする際、DNA塩基と特異的接触を形成し、かつ、DNAバックボーンと非特異的接触を形成し、そうすることによって、該酵素の認識配列特異性およびDNA−結合親和度に影響を及ぼす、メガヌクレアーゼLAGLIDADGファミリーにおける特異的アミノ酸残基の特定および特徴解明に基づく。この発見は、以下に詳細に説明するように、メガヌクレアーゼの特異性及び/又は親和性を変えることが可能な、メガヌクレアーゼにおけるアミノ酸置換を特定するため及び天然のメガヌクレアーゼが認識しない所望のDNA配列を認識することが可能であり、及び/又は、天然のメガヌクレアーゼに比べて特異性及び/又は親和度が増大または減少させたメガヌクレアーゼを合理的に設計し、開発するために用いられた。さらに、DNA−結合親和度は、酵素活性の外に、配列特異性にも影響を及ぼすので、本発明は、天然メガヌクレアーゼに比べ活性を変更させた、合理的設計メガヌクレアーゼを提供する。さらに、本発明は、ダイマーを形成するためにコンタクトされたモノマー間のインターフェイスにおける残基が、ヘテロダイマー形成を促進するように修飾された合理設計メガヌクレアーゼを提供する。最後に、本発明は、組み換え細胞および生物体の生産における使用ばかりでなく、本出願に開示されるように、遺伝子療法、抗病原体、抗癌及びインビトロ応用における合理設計メガヌクレアーゼの使用を提供する。
【0041】
一般的出来事として、本発明は、(1)2本鎖DNA認識配列中の個々の塩基に対する配列特異的結合、又は(2)2本鎖DNA分子のフォスフォジエステルバックボーンに対する非特異的結合による、メガヌクレアーゼ内の複数部位において変更アミノ酸残基を含
む、合理設計LAGLIDADGメガヌクレアーゼを生成するための方法を提供する。しかしながら、酵素活性は、DNA−結合親和度と相関するのであるから、DNA認識配列に対する結合に与るアミノ酸を変えることは、2本鎖DNAに対する全体的結合親和度を増大又は減少することになり、特異的塩基対相互作用を介するメガヌクレアーゼの特異性ばかりでなく、メガヌクレアーゼの活性も変える可能性がある。同様に、DNAバックボーンに対する結合によるアミノ酸を変えることは、2本鎖DNAに対する全体的結合親和度を増大又は減少することによって、酵素の活性ばかりでなく、認識配列に対する結合の特異性又は縮重の程度を変える可能性がある。
【0042】
下記に詳述するように、メガヌクレアーゼの合理的設計法は、DNA認識/結合によるアミノ酸を特定すること及び適切なアミノ酸変化を選択するために一連の規則を適用することを含む。この規則は、メガヌクレアーゼの配列特異性に関して、メガヌクレアーゼのアミノ酸側鎖とDNAのセンス鎖及びアンチセンス鎖における塩基の間とのメガヌクレアーゼ−DNA複合体距離に関する互いの立体位置、アミノ酸側鎖とDNA塩基との相対位置及び非共有的化学的相互作用を考慮に含んでいる。
【0043】
最後に、ホモダイマーとしてDNAに結合する天然メガヌクレアーゼの大部分は、擬似、又は完全パリンドロームの認識配列を認識する。長々としたパリンドロームはめったにないと予想されるので、ゲノムの興味の配列部位でパリンドロームに遭遇する確率はきわめて低い。したがって、これらの酵素を、ゲノムの興味の配列部位を認識するように再設計しなければならないとすると、異なる半分部位を認識する二つの酵素モノマーにおいて、ヘテロダイマー形成されると非パリンドローム的ハイブリッド認識配列を切断することが可能な二つの酵素モノマーを設計することが必要である。それゆえ、ある側面では、本発明は、少なくとも一つのアミノ酸位置で異なる二つのモノマーをダイマー化して、ヘテロダイマーを形成して得られる合理設計メガヌクレアーゼを提供する。ある場合、両モノマーは、非パリンドローム的認識配列を認識するヘテロダイマーを形成するように合理的設計される。二つの異なるモノマーから成る混合物は、最大三つの活性形メガヌクレアーゼダイマー、二つのホモダイマー及びヘテロダイマーをもたらす。それに加えて、またはそれとは別に、ある場合、ホモダイマーまたはヘテロダイマー形成の確率を増すか、または減らすために、モノマー同士が相互作用を持ってダイマーを形成する境界面においてアミノ酸残基が変えられる。
【0044】
したがって、一つの側面では、本発明は、LAGLIDADGメガヌクレアーゼにおいて、酵素の特異性及び/又は活性を変えるアミノ酸変化を含むLAGLIDADGメガヌクレアーゼの合理的設計法を提供する。他の側面では、本発明は、これらの方法によってもたらされる合理設計メガヌクレアーゼを提供する。他の側面では、本発明は、生物体ゲノム内部の所望のDNA配列または遺伝子座が、DNA配列の挿入、欠失、置換、またはその他の操作によって修飾される、組み換え核酸および生物体を生産するために、このような合理設計メガヌクレアーゼを使用する方法を提供する。さ他の側面では、本発明は、病原体または癌細胞において、病原体特異的又は癌特異的認識配列を有する合理設計メガヌクレアーゼを用いて、この病原体または癌細胞の生存率を下げるための方法を提供する。
【0045】
1.2 参照および定義
本出願において参照される特許および科学文献は、当業者にとって利用可能な知識を確定する。本出願において引用される、公刊された米国特許、承認済みの出願、出版された外国特許、および、GENBANKのデータベース配列を含む参照資料は、参照により、あたかも、それぞれが、特異的、個別的に、参照によって含まれるのと同程度に、本出願に含まれる。
【0046】
本出願で用いる「メガヌクレアーゼ」という用語は、12塩基対よりも大きい認識配列で2本鎖DNAに結合するエンドヌクレアーゼを指す。天然のメガヌクレアーゼはモノマー(例えば、I−SceI)又はダイマー(例えば、I−CreI)であることが可能である。本出願で用いるメガヌクレアーゼという用語は、モノマー・メガヌクレアーゼ、ダイマー・メガヌクレアーゼ、または、コンタクトしてダイマー形メガヌクレアーゼを形成するモノマーを指す。「ホーミング・エンドヌクレアーゼ」という用語は、「メガヌクレアーゼ」という用語と同義である。
【0047】
本出願で用いる「LAGLIDADGメガヌクレアーゼ」という用語は、天然ではダイマー形である単一のLAGLIDADGモチーフを含むメガヌクレアーゼ、又は天然ではモノマー形である二つのLAGLIDADGモチーフを含むメガヌクレアーゼを指す。本出願で用いる「モノLAGLIDADGメガヌクレアーゼ」という用語は、単一のLAGLIDADGモチーフを含むメガヌクレアーゼを指し、本出願で用いる「ジLAGLIDADGメガヌクレアーゼ」という用語は、二つを区別することが必要な場合には、二つのLAGLIDADGモチーフを含むメガヌクレアーゼを指す。LAGLIDADGモチーフを含むジLAGLIDADGメガヌクレアーゼの二つの構造ドメインは、それぞれ、LAGLIDADGサブユニットと呼ぶことが可能である。
【0048】
本出願で用いる「合理的に設計された」という用語は、非天然及び/又は遺伝子工学的に加工されたことを意味する。本発明の合理的設計メガヌクレアーゼは、野生型又は天然のメガヌクレアーゼとアミノ酸配列または一次構造において異なり、さらに、二次、三次又は四次構造において異なる場合がある。加えて、本発明の合理設計メガヌクレアーゼは、認識配列の特異性及び/又は活性においても、野生型又は天然メガヌクレアーゼと異なる。
【0049】
タンパク質に関して本出願で用いる「組み換え」という用語は、タンパク質をコードする核酸に対し遺伝子工学技術を適用した結果アミノ酸配列が変化すること、及びこのタンパク質を発現する細胞又は生物体が得られることを意味する。核酸に関して「組み換え」という用語は、遺伝子工学技術を適用した結果、核酸配列が変化することを意味する。遺伝子工学技術としては、これらにっ限定されるものではないが、PCR、トランスフェクション、形質転換及びその他の遺伝子転移技術等のDNAクローニング技術、相同組み換え、部位特異的変異処理及び遺伝子融合が含まれる。この定義によれば、天然のタンパクと同じアミノ酸配列を持つが、異種宿主におけるクローニング及び発現によって生産されるタンパク質は、「組み換え」とは見なされない。
【0050】
組み換えタンパクに関して本出願で用いられる「修飾」という用語は、基準配列(例えば、野生型)と比べた場合の、組み換え配列におけるアミノ酸残基の、どのようなものであれ、挿入、欠失又は置換を含む意味である。
【0051】
本出願で用いる「遺伝学的に修飾された」という用語は、そのゲノムDNA配列が、組み換え技術によって意図的に修飾されたゲノムDNA配列を有する細胞、生物体又は祖先を指す。本出願で用いる「遺伝学的に修飾された」という用語は、「トランスジェニック」という用語を含む。
【0052】
本出願で用いる「野生型」という用語は、どのようなものであれ、、メガヌクレアーゼの全ての天然形を指す。「野生型」という用語は、天然における酵素の最も一般的な変異を意味するものではなく、自然で見られるいかなる変異も意味する。野生型メガヌクレアーゼは、組み換え又は非天然メガヌクレアーゼと区別される。
【0053】
本出願で用いる「認識配列半分部位」又は単に「半分部位」は、モノ−LAGLIDA
DGメガヌクレアーゼのモノマー又はジ−LAGLIDADGメガヌクレアーゼの一つのLAGLIDADGサブユニットによって認識される2本鎖DNA分子間の核酸配列を意味する。
【0054】
本出願で用いる「認識配列」という用語は、モノ−LAGLIDADGメガヌクレアーゼダイマー又はジ−LAGLIDADGメガヌクレアーゼモノマーによって結合され、切断される一対の半分部位を指す。この二つの半分部位は、この酵素によって特異的に認識されない塩基対によって隔てられてもよいし、隔てられなくともよい。I−CREI、I−MSOI及びI−CEUIの場合、各モノマーの認識配列半分部位は9塩基対の幅を持ち、この二つの半分部位は、特異的には認識されないが実際の切断部位である塩基対(4塩基対のオーバーハングを有する)によって分割されている。したがって、I−CREI、I−MSOI及びI−CEUIメガヌクレアーゼダイマーの結合認識配列は、通常、4塩基対切断部位に側接する二つの9塩基対を含む22塩基対の幅を持つ。各半分部位の塩基対は−9から−1と表示され、−9位は切断部位からもっとも遠位であり、−1位はN1−N4と表示される4個の中央塩基対に隣接する。−9から−1へと向かう方向(すなわち、切断部位に向かう方向)において5′から3′に方向づけられる各半分部位鎖は、「センス」鎖と表示され、反対鎖は、「アンチセンス鎖」と表示される。ただし、いずれの鎖もタンパクをコードしない。したがって、図1(A)に示すように、一方の半分部位の「センス」鎖は、他方の半分部位のアンチセンス鎖である。ジ−LAGLIDADGメガヌクレアーゼモノマーであるI−SCEIメガヌクレアーゼの場合、認識配列は約18BPの非パリンドローム配列であり、特異的には認識されない中央塩基対は無い。通例に従って、2本鎖の一方は「センス」鎖と呼ばれ、他方は「アンチセンス」鎖と呼ばれるが、いずれの鎖もタンパクをコードしない。
【0055】
本出願で用いる「特異性」という用語は、メガヌクレアーゼが認識配列と呼ばれる特定の塩基対配列又は特定の一組の認識配列においてのみ、2本鎖DNA分子を認識し切断する能力を意味する。この一組の認識配列は、いくつかの保存位置または配列モチーフを共有するが、これらは1又は2以上の位置で縮重する場合がある。高い特異性を有するメガヌクレアーゼは、たった一つ又はきわめて少数の認識配列の切断が可能である。特異性は実施例1に記載される切断アッセイで決めることが可能である。本出願で用いる場合、メガヌクレアーゼは、基準メガヌクレアーゼ(例えば、野生型)によって結合及び切断されない認識配列に結合し、切断する場合、又は、認識配列の切断速度が、基準メガヌクレアーゼに比べて、統計的に有意な値(P<0.05)だけ上昇又は降下した場合に、特異性
が「変更された」ことになる。
【0056】
本出願で用いる「縮重」という用語は、「特異性」の反対を意味する。高縮重したメガヌクレアーゼは、非常に多岐にわたる認識配列を切断することができる。メガヌクレアーゼは単一の、複数の又は全てのの箇所の半分部位内に配列縮重を有することができる。このような配列縮重は、(I)メガヌクレアーゼのDNA結合ドメインにおける任意のアミノ酸が、認識配列内の1又は2以上の位置で任意の塩基と接触できないこと、(II)メガヌクレアーゼのDNA結合ドメインにおける1又は2以上のアミノ酸が、識配列内の1又は2以上の位置で1以上の塩基と特異的にコンタクトできること、及び/又は(III)活性に十分なほど、非特異的DNA結合親和性を持つこと、からもたらされる可能性がある。「完全な」縮重位置は任意の4塩基で占められてもよく、半分部位内では”N”で表示することができる。「部分的な」縮重位置は、4塩基の内の任意の2又は3塩基で占められてもよい。(例えば、プリン(PU)か、ピリミジン(PY)で占められるが、Gには占められない)。
【0057】
メガヌクレアーゼに関して本出願で使用される場合、「DNA−結合親和度」又は「結合親和度」は、メガヌクレアーゼが基準DNA分子(例えば、認識配列又は任意の配列)
と非共有的に結合する傾向を意味する。結合親和度は、解離定数KDによって測定される(例えば、WT認識配列に対するI−CREIのKDは約0.1NMである)。本出願で用いる場合、基準認識配列に対する組み換えメガヌクレアーゼのKDが、基準メガヌクレアーゼに比べて、統計的に有意な値(P<0.05)だけ増加又は減少した場合に、結合
親和度を「変更させた」ことになる。
【0058】
メガヌクレアーゼモノマーに関して本出願で用いる場合、「ダイマー形成に対する親和度」という用語は、メガヌクレアーゼモノマーが、基準メガヌクレアーゼモノマーと非共有的に結合する傾向を意味する。ダイマー形成に対する親和度は、例えば、基準野生型メガヌクレアーゼのような同じモノマー(すなわち、ホモダイマー形成)又は異なるモノマー(すなわち、ヘテロダイマー形成)で測定することができる。結合親和度は、解離定数KDによって測定される。本出願で用いる場合、基準認識配列に対する組み換えメガヌクレアーゼのKDが、基準メガヌクレアーゼに比べて、統計的に有意な値(P<0.05)だけ増加又は減少した場合に、結合親和度を「変更させた」ことになる。
【0059】
本出願で用いる「パリンドローム的」という用語は、同じ半分部位の逆方向反復を含む認識配列を指す。しかしながら、この場合、パリンドローム配列は酵素にコンタクトされない四つの中央塩基対に関してパリンドローム的である必要はない。ダイマー形メガヌクレアーゼの場合、パリンドロームDNA配列は二つのモノマーが同じ半分部位にコンタクトするホモダイマーによって認識される。
【0060】
本出願で用いる「擬似パリンドローム的」という用語は、非同一又は不完全なパリンドローム的半分部位の逆方向反復を含む認識配列を指す。この場合、擬似パリンドローム配列は、四つの中央塩基対に関してパリンドローム的である必要はなく、二つの半分部位の間のパリンドローム配列からもはずれることが可能である。擬似パリンドロームDNA配列は、二つの同一の酵素モノマーが、異なる半分部位にコンタクトする、野生型ホモダイマー形メガヌクレアーゼによって認識される天然DNA部位において典型的に見られる。
【0061】
本出願で用いる「非パリンドローム的」という用語は、メガヌクレアーゼの、二つの、無関係の半分部位から構成される認識配列を指す。この場合、非パリンドローム配列は、四つの中央塩基対に対しても、二つのモノマー半分部位に対しても、パリンドローム的である必要はない。非パリンドロームDNA配列は、ジ−LAGLIDADGメガヌクレアーゼ、高縮重モノ−LAGLIDADGメガヌクレアーゼ(例えば、I−CeuI)、非同一半分配列を認識するモノ−LAGLIDADGメガヌクレアーゼのヘテロダイマーのいずれかによって認識される。
【0062】
本出願で用いる「活性」という用語は、本発明のメガヌクレアーゼが、特定の認識配列を切断する速度を指す。このような活性は、2本鎖DNAのフォスフォジエステル結合の加水分解に関する測定可能な酵素反応である。ある特定のDNA基質に作用するメガヌクレアーゼの活性は、その特定のDNA基質に対するメガヌクレアーゼの親和力又は結合力に影響され、DNAとの配列特異的相互作用及び非配列特異的相互作用によって影響される。
【0063】
本出願で用いる「相同組み換え」という用語は、2本鎖DNA切断が、修復鋳型として相同DNA配列を用いて修復される、天然の、細胞プロセスを指す(例えば、CAHILL ET AL.(2006),FONT.BIOSCI.11:1958−1976を参照)。相同のDNA配列は、内因性の染色体配列であってもよいし、細胞に輸送された外来性の核酸であってもよい。したがって、ある実施態様では、合理設計メガヌクレアーゼは、標的配列内の認識配列を切断するために使用され、標的配列に対して相同な又はほぼ同様の配列類似性を持つ外来性の核酸が細胞に輸送されて、相同性組み換えによる修復
のための鋳型として使用される。したがって、標的配列とは著明に異なっていてもよい外来性のDNA配列が、染色体配列に組み込まれる。相同組み換えプロセスは、主に真核生物において起こる。本出願では、「相同性」という用語は「配列類似性」と等価的に用いられ、出自又は系統関係による同一性を要求することを意図するものではない。
【0064】
本出願で用いる「非相同的末端接合」という用語は、2本鎖DNA切断が、2本の非相同的DNAセグメントの直接的接合によって修復される、天然の細胞プロセスを指す(例えば、CAHILL ET AL..(2006),FRONT.BIOSCI.11:1958−1976)。非相同的末端接合によるDNA修復は、間違いを起こしやすく、修復部位において鋳型と対応しないDNA配列の付加又は除去をもたらすことが多い。したがって、ある実施態様では、(例えば、塩基挿入、塩基欠失又はフレームシフト突然変異を行うことによって)非相同的末端接合によって遺伝子を破壊し、標的配列内のメガヌクレアーゼ認識配列中に二本鎖切断点を作るために、合理設計メガヌクレアーゼを用いることも可能である。他の実施態様では、標的配列に対する相同性を欠如するか又は標的配列に対する実質的配列類似性を欠如する外来性の核酸が、非相同的末端接合によって、メガヌクレアーゼ刺激2本鎖DNA切断部位で取り込まれてもよい(例えば、SALOMON,ET AL.(1998),EMBO J.17:6086−6095を参照)。非相同的末端接合プロセスは、真核生物でも、細菌のような原核生物でも起こる。
【0065】
本出願で用いる「興味の配列」という用語は、メガヌクレアーゼタンパクを用いてゲノムへ挿入するか又はゲノムDNA配列を置換するのに使用することが可能な核酸配列であって、タンパク、RNA又は調整エレメント(例えば、エンハンサー、サイレンサー、またはプロモーター配列)のどれをコードするかにはよらない任意の核酸配列を意味する。興味の配列は、興味の配列から発現されるタンパク質又はRNAの標識化を可能とする非相同的なDNA配列を有することも可能である。例えば、以下のものに限定さるものではないが、タンパク質は、エピトープ(例えば、C−MYC、FLAG)、または他のリガンド(例えば、ポリ−HIS)を含むタグによって標識することが可能である。更に、興味の配列は、従来技術で周知の技術に従って融合タンパクをコードしてもよい(例えば、AUSUBEL ET AL., 「分子生物学における最近のプロトコール(“CURRENT PROTOCOLS IN MOLECULAR BIOLOGY,”)」、WILEY 1999)。ある実施態様では、隣接配列は、組み換えメガヌクレアーゼによって切断され、ゲノム組み替え配列の中に興味の配列の適切な挿入される。
【0066】
したがって、この側接配列は切断され、該組み換えメガヌクレアーゼによって切断されるゲノム認識配列の中に、興味の配列を適正に挿入することを可能となる。ある実施態様では、興味の配列の全ては、ゲノム中の標的配列と相同であるか又は標的配列と実質的に配列類似性を有しており、相同組み換えによって、標的配列が興味の配列によって効果的に置換される。他の実施態様では、興味の配列は、標的配列と相同性を有するか又は実質的配列類似性を有するDNA配列によって側接され、相同組み換えによってゲノム内の標的配列座位において興味の配列が挿入される。ある実施態様では、興味の配列は、メガヌクレアーゼ認識配列における突然変異又は他の修飾を除いては、標的配列とほぼ同一であり、そのため、メガヌクレアーゼは、標的配列が興味の配列によって修飾された後に、標的配列を切断することができない。
【0067】
アミノ酸配列及び核酸配列に関して本出願で用いる場合、「類似性パーセント」及び「配列類似性」という用語は、アライメントされたアミノ酸残基又はヌクレオチド間の類似性を最大にするようにアライメントされた配列に基づく2つの配列の類似度の測定値を意味する。そしてこれは、アライメント配列中のギャップ及びギャップの数、核酸又は残基の総数、同一又は類似する核酸又は残基の数の関数による。一般的なパラメータを用いて配列類似性を決めるために、様々のアルゴリスムおよびコンピュータプログラムの利用が
可能である。本出願では、配列類似度は、アミノ酸配列の場合はBLASTPプログラムを用い、核酸配列の場合はBLASTNプログラムを用いて算出される。両方とも、NATIONAL CENTER FOR BIOTECHNOLOGY INFORMATION(WWW.NCBI.NLM.NIH.GOV/)を通じて使用可能であり、例えば、ALTSCHUL ET AL.(1990), J.MOL.BIOL.215:403−410;GISH AND STATES(1993), NATURE GENET.3:266−272;MADDEN ET AL.(1996),METH.ENZYMOL.266:131−141;ALTSCHUL ET AL.(1997),NUCLEIC ACIDS RES.25:3389−3402);ZHANG ET AL.(2000) J.COMPUT.BIOL.7(1−2):203−14に記載される。本出願では、二つのアミノ酸配列の類似パーセントは、BLASTPアルゴリスムにおける下記のパラメータに基づくスコアである。単語サイズ=3;ギャップ開放ペナルティー=−11;ギャップ延長ペナルティー=−1;及びスコアリングマトリックス=BLOSUM62である。本出願では、二つの核酸配列の類似パーセントは、BLASTNアルゴリスムにおける下記のパラメータに基づくスコアである。単語サイズ=11;ギャップ開放ペナルティー=−5;ギャップ延長ペナルティー=−2;マッチ評価=1
;及びミスマッチペナルティー=−3である。
【0068】
二つのタンパク質又はアミノ酸配列の修飾に関して本出願で用いる場合、「対応する」という用語は、(例えば、BLASTPプログラムを用いて)二つのタンパク質を一般的な方法で配列アライメントしたとき、第1のタンパク質の修飾が、第2のタンパク質の修飾と同じアミノ酸残基の置換であること、及び第1のタンパク質における修飾のアミノ酸位置が、第2のタンパク質の修飾のアミノ酸位置に対応する又は揃うことを意味する。したがって、もし残基XとYとが配列アライメントで互いに対応するものでれば、XとYの配列番号が異なるものであっても、第1のタンパク質における残基“A”がアミノ酸“A”に修飾されることは、残基“Y”がアミノ酸“A”に修飾されることと対応する。
【0069】
本出願では、ある変数に対する数値範囲は、その範囲内であれば、任意の数値に等しい変数を用いて本発明が実行可能であることを意図する。したがって、本来的に不連続な変数である場合、この変数は、その数値範囲の両端を含む任意の整数に等しくなることが可能である。同様に、本来的に連続的な変数である場合、この変数は、その数値範囲の両端を含む任意の実数に等しくなることが可能である。例えば、ただしこれらに限定されるものではないが、0と2の間の値を持つと記載される変数は、もしこの変数が本来的に不連続である場合は、0、1又は2を取ることが可能であり、もしこの変数が本来的に連続的である場合は、数値0.0、0.1、0.01、0.001又は≧0及び≦2の実数の任意の実数を取ることができる。
【0070】
本出願では、特に別段の指定が無い限り、「又は」という単語は、「及び/又は」という包括的意味で使われ、「又は/どちらか」という排除的意味では使われない。
【0071】
2.1 配列特異性を変更させた合理的設計メガヌクレアーゼ
本発明の一様態では、組み換えLAGLIDADGファミリーのメガヌクレアーゼを合理的に設計する方法が提供される。この様態では、先ず、半分部位の各位置における塩基指向性を変えることが可能なアミノ酸置換を予測することによって、組み換えメガヌクレアーゼを合理的に設計する。これらの置換は、個別に又は組み合わせて、実験的にその有効性が検証され、所望の切断特異性を持つメガヌクレアーゼを生産する。
【0072】
本発明によれば、塩基指向性において所望の変化をもたらすアミノ酸置換は、メガヌクレアーゼのDNA認識配列及びDNAフォスフォジエステルバックボーンの核酸塩基との接触に参加が可能な基準メガヌクレアーゼ(例えば、野生型メガヌクレアーゼ又は非天然
の基準メガヌクレアーゼ)のアミノ酸側鎖及びそれら接触部の空間的、化学的性質を決めることによって予測される。これらのアミノ酸としては、基準DNA半分部位との接触に与る側鎖が挙げられるが、ただしこれらに限定されない。一般に、この決定には、メガヌクレアーゼと、その2本鎖DNA認識配列との間の複合体の構造に関する知識、又はきわめて類似性の高い複合体の構造(例えば、同じメガヌクレアーゼと、別のDNA認識配列との間の複合体構造、又はメガヌクレアーゼの対立遺伝子変種または系統的変種と、そのDNA認識配列との間の複合体構造)に関する知識が必要である。
【0073】
原子座標データとして記載される、ポリペプチド又は2個以上のポリペプチドから成る複合体の三次元構造は、いくつかの方法で獲得することが可能である。例えば、タンパク質の構造の決定は、以下のものに限定されるものではないが、X線結晶学、NMR及び質量分析を含む技術を用いて実行することが可能である。もう一つの方法は、興味のメガヌクレアーゼ又は関連ヌクレアーゼの、既存の構造座標のデータベースを分析することである。このような構造データは、多くの場合、三次元座標形式のデータベースから入手することが可能である。多くの場合、このデータは、オンラインデータベースを通じて(例えば、WWW.RCSB.ORG/PDBにおいてRCSB PROTEIN DATA BANKへ)アクセスすることが可能である。
【0074】
構造情報は、タンパク質またはタンパク質複合体の、規則的な、二次元又は三次元アレイ(例えば、結晶)によって得られる。例えば、X線又は電子の回折パターンを分析することによって実験的に得ることが可能である。回折データを空間における三次元原子座標に変換するためには、コンピュータ法が使用される。例えば、X線結晶学の分野は、メガヌクレアーゼを含めた、多くのタンパク質−DNA複合体の三次元構造情報の生成のために使用されている(例えば、CHEVALIER ET AL.(2001),NUCLEIC ACIDS RES.29(18):3757−3774を参照)。
【0075】
溶液中の分子の原子間距離の決定には、核磁気共鳴(NMR)も用いられている。コンピュータ法と組み合わせた多次元NMR法は、増大するサイズのポリペプチドの原子座標の決定に成功した(例えば、TZAKOS ET AL.(2006),ANNU.REV.BIOPHYS.BIOMOL.STRUCT.35:12−42を参照)。
【0076】
それとは別に、タンパク質/DNAの既知の一次構造及び利用可能であれば、二次、三次、及び/又は四次構造のほか、アミノ酸側鎖、核酸塩基及び結合性相互作用に関する既知の物理化学的性質に基づいたアルゴリスムを適用することによって、コンピュータモデルを使用することも可能である。このような方法は、任意に、繰り返し法又は実験的に誘導された制限を含むことも可能である。このようなコンピュータソフトウェアの一例として、ADAMS ET AL.(1999),ACTA CRYSTALLOGR.D.BIOL.CRYSTALLOGR.55(PT 1):181−90によって記載されるCNSプログラムがある。他にも、タンパク質の構造におけるアミノ酸の空間配置及びタンパクのアミノ酸側鎖と各種標的分子との相互作用を予測する、様々のコンピュータプログラムが開発されている(例えば、米国特許第6,988,041号を参照)。
【0077】
したがって、本発明のある実施態様では、DNA核酸塩基と特異的に相互作用を持ち、及び/又は、非特異的フォスフォジエステルバックボーン相互作用を促進する、特異的アミノ酸残基を特定するために、コンピュータモデルが使用される。例えば、可能なメガヌクレアーゼ−DNA相互作用の全体に関するコンピュータモデルを、例えば、以下に限定されるものではないが、MOLSCRIPT(登録商標)2.0(AVATAR SOFTWARE AB,STOCKHOLM,SWEDEN)、グラフィックディスプレイO(JONES ET AL.(1991),ACTA CRYSTALLOGRAPHY,A47:110)、グラフィックディスプレイ・プログラムGRASP(登録商標)(
NICHOLLS ET AL.(1991),PROTEINS,STRUCTURE,FUNCTION AND GENETICS 11(4):281FF)、グラフィックディスプレイ・プログラムINSIGT(登録商標)(TSI,INC.,SHOREVIEW、ミネソタ州)を含む、適切なソフトウェアプログラムを用いて生産することが可能である。タンパク質−DNA複合体の、三次元構造描画を生産し、視認し、かつ操作するために好適なコンピュータハードウェアは市販されており、従来技術で周知である(例えば、SILICON GRAPHICS WORKSTATION,SILICON GRAPHICS,INC.,MOUNTAINVIEW、カリフォルニア州)。
【0078】
具体的に述べると、メガヌクレアーゼと、その2本鎖DNA認識配列との間の相互作用は、従来技術で既知の方法を用いて分解することが可能である。例えば、そのために結晶が生産される、複数成分の複合体構造の三次元構造の描画、またはモデルは、分子置換、またはSIR/MIR(SINGLE/MULTIPLE ISOMORPHOUS REPLACEMENT)(例えば、BRUNGER(1997),METH.ENZYM.276:558−580;NAVAZA AND SALUDIJIAN(1997),MATH.ENZYM.276:581−594;TONG AND ROSSMANN(1997),METH.ENZYM.276:594−611;およびBENTLEY(1997),METH.ENZYM.276:611−619を参照)を含む技術を用いて決めることが可能であり、例えば、AMORE/MOSFLM(NAVAZA(1994),ACTA CRYST.A50:157−163;CCP4(1994),ACTA CRYST.D50:760−763)、または、XPLOR(BRUENGER ET AL.(1992),X−PLOR VERSION 3.1.A.SYSTEM FOR X−RAY CRYSTALLOGRAPHY AND NMR,YALE UNIVERSITY PRESS,NEW HAVEN、コネチカット州)などのソフトウェアを用いて実行することが可能である。
【0079】
タンパク質の構造及び可能なメガヌクレアーゼ−DNA相互作用の決定は、酵素活性および特異性に影響を及ぼすために変えることが可能なアミノ酸に関する合理的選択を可能にする。決定は、アミノ酸側鎖の、特定の塩基又はDNAフォスフォジエステルバックボーンとの相互作用に関するいくつかの因子に基づく。適切なアミノ酸置換を決めるために使用される化学的相互作用としては、以下に限定されるものではないが、ファンデルワールス力、立体障害、イオン結合、水素結合及び疎水性相互作用が挙げられる。メガヌクレアーゼと、可能な認識配列半分部位中のある特定の塩基との特定の相互作用を優先するか、又は忌避するアミノ酸置換が選択される。この選択は、その認識配列に対する特異性を、かつ、ある程度は、全体的結合親和度および活性を増すようにあるいは減らすように行われる。さらに、全体活性の増加又は減少のために、かつ、ある程度は、特異性を下げるか又は上げるかするために、2本鎖DNAのフォスフォジエステルバックボーンに対する結合性を増すか又は下げるアミノ酸置換を選択することが可能である。
【0080】
したがって、特定の実施態様では、メガヌクレアーゼ−DNA複合体の三次元構造が決定され、DNA認識配列半分部位における各塩基対について「接触面」が定義される。ある実施態様では、接触面は、その残基が、野生型メガヌクレアーゼ−DNA複合体において塩基接触を実行するか否かとは無関係に、ペアのいずれかの塩基における大溝水素結合ドナーまたはアクセプターから9.0オングストローム未満のβ−炭素を有する、酵素中のアミノ酸を含む。別の実施態様では、残基が、野生型メガヌクレアーゼ−DNA複合体において接触を行わない場合、この残基を排除することが可能であり、又は考慮される残基の数または名称を変えようとする設計者の意図に基づいて残基を含めること、又は排除することが可能である。後述の一実施例では、野生型I−CREI半分部位の塩基位置−2、−7、−8及び−9において、接触面は、野生型酵素−DNA複合体において実際に相互作用を持つアミノ酸位置に限定された。しかしながら、位置−1、−3、−4、−5
及び−6では、接触面は、野生型接触には与らないが、異なるアミノ酸によって置換された場合、塩基に接触する可能性のある、さらに新たなアミノ酸位置を含むように定義された。
【0081】
認識配列半分部位は、通常、DNAの1本鎖のみに関して表されるが、メガヌクレアーゼは、2本鎖DNAの大溝に結合し、両鎖の核酸塩基と接触することに注意しなければならない。さらに、「センス」および「アンチセンス」鎖という表示は、メガヌクレアーゼの結合と認識に関しては完全にその場まかせである。ある位置における配列特異性は、塩基対の一方のメンバーとの相互作用か又は塩基対の両方のメンバーとの相互作用の組み合わせのどちらかによって実現される。したがって、例えば、位置Xにおいて、該位置XではA塩基は「センス」鎖上にあり、T塩基が「アンチセンス」鎖上にある、A/T塩基対の存在を優先するためには、位置Xにおいてセンス鎖に接触できるほど十分に近接し、かつAの存在を好む残基が選択されるか、及び/又は、位置Xにおいてアンチセンス鎖に接触できるほど十分に近接し、かつTの存在を好む残基が選択される。本発明によれば、残基のβ−炭素が、関連塩基の最近接原子の9オングストローム以内にある場合、その残基は十分に近接すると考慮される。
【0082】
したがって、例えば、DNAセンス鎖の9オングストローム以内ではあるが、アンチセンス鎖から9オングストロームより大きく離れるβ−炭素を持つアミノ酸は、センス鎖に対してのみ相互作用を持つ可能性があると考慮される。同様に、DNAアンチセンス鎖の9オングストローム以内ではあるが、センス鎖から9オングストロームより大きく離れるβ−炭素を持つアミノ酸は、アンチセンス鎖に対してのみ相互作用を持つ可能性があると考慮される。両方のDNA鎖の9オングストローム以内にβ−炭素を持つアミノ酸は、どちらの鎖とも相互作用を持つ可能性があると考慮される。
【0083】
各接触面について、可能なアミノ酸置換について、4種のDNA塩基の内の一つ以上に対し好んで相互作用を持とうとする能力を予測し、その予測能力に基づいてアミノ酸置換が選択される。この選択プロセスは、二つの主要基準に基づく。(I)異なる核酸塩基との立体相互作用に影響を及ぼすアミノ酸側鎖のサイズ、及び(II)異なる核酸塩基との静電気的及び結合的相互作用に影響を及ぼすアミノ酸側鎖の化学的性質である。
【0084】
側鎖のサイズに関しては、接触面におけるアミノ酸のβ−炭素が、塩基から6オングストローム未満の距離にある場合には、比較的短い及び/又は比較的小さい側鎖を持つアミノ酸を選択することが可能であり、接触面におけるアミノ酸のβ−炭素が、塩基から6オングストロームより遠い距離にある場合には、比較的長い及び/又は比較的大きい側鎖を持つアミノ酸を選択することが可能である。サイズが中間のアミノ酸側鎖は、接触面におけるアミノ酸β−炭素が、塩基から5〜8オングストロームである場合に選択することが可能である。
【0085】
比較的短く、比較的小さい側鎖を持つアミノ酸は、グリシン(G)、アラニン(A)、セリン(S)、トレオニン(T)、システイン(C)、バリン(V)、ロイシン(L)、イソロイシン(I)、アスパラギン酸(D)、アスパラギン(N)及びプロリン(P)を含む1群に割り当てられる。しかしながら、プロリンは、比較的屈曲性が低いので、比較的頻繁な使用が期待されない。さらに、グリシンは、ペプチドバックボーンに不要な屈曲性を導入するため、かつ、きわめてサイズが小さいために、より大きな残基を置換した場合、効果的接触が得られる確率が低いと考えられるために、比較的頻繁な使用が期待されない。一方、グリシンは、ある場合、縮重位置を増進するために使用することが可能である。比較的中間的長さ及びサイズの側鎖を持つアミノ酸は、リシン(K)、メチオニン(M)、アルギニン(R)、グルタミン酸(E)、およびグルタミン(Q)を含む2群に割り当てられる。比較的長い、及び/又は比較的大きい側鎖を持つアミノ酸は、リシン(K
)、メチオニン(M)、アルギニン(R)、ヒスチジン(H)、フェニルアラニン(F)、チロシン(Y)、およびトリプトファン(W)を含む3群に割り当てられる。しかしながら、トリプトファンは、比較的屈曲性が低いので、比較的頻繁な使用が期待されない。さらに、リシン、アルギニン、およびメチオニン側鎖の屈曲性によって、これらのアミノ酸は、長い距離または中間距離からの塩基接触が可能となるために、それらが、2群と3群の両群に含められることを保証する。これらの群をさらに、下記に表1として示す。
【0086】
【表1】
【0087】
側鎖の化学的性質に関しては、種々のアミノ酸が、種々の核酸塩基との相互作用の可能性について評価されており(例えば、ファンデルワールス力、イオン結合、水素結合及び疎水性相互作用)、2本鎖DNA認識配列半分部位中の特定位置における特定塩基と、メガヌクレアーゼとの特異的相互作用を好むか又は忌避する残基が選択される。ある場合には、一つ以上の、完全な又は部分的な縮重位置を持つ半分部位を創出することが望ましい場合がある。そのような場合、二つ以上の塩基の存在を好む残基又は一つ以上の塩基を忌避する残基を選んでもよい。例えば、部分的に縮重された塩基認識は、センス又はアンチセンス位置においてピリミジンを立体的に妨げることによって実現することが可能である。
【0088】
グアニン(G)塩基の認識は、該塩基のN7およびO6に対して水素結合を形成する塩基性側鎖を持つアミノ酸を用いて実現される。シトシン(C)の特異性は、C以外の全ての塩基の上に存在する、大溝の電気的陰性基との相互作用を持ちたがらない、陰性荷電側鎖によって付与される。チミン(T)の認識は、疎水性側鎖と、塩基の上の大溝メチル基との間の疎水性およびファンデルワールス相互作用を用いて合理的に設計される。最後に、アデニン(A)塩基は、該塩基のN7およびN6に対する一対の水素結合を介して、ASNおよびGLNのカルボキサミド側鎖、またはTYRのヒロドキシル側鎖を用いて認識される。最終として、HISは、N7に対して水素結合を供与することによって、プリン塩基(AまたはG)に対して特異性を付与するために使用することが可能である。この、
DNA認識に関する単純な規則を用いて、そこでは、ある特定の塩基対位置において、塩基の内の一つが、または両方が合理的設計接触を通じて認識される、そのような接触面を予測することが可能である。
【0089】
したがって、種々の核酸塩基との結合性相互作用及び接触を行う位置において好む塩基に基づいて、各アミノ酸残基を、それが好む種々の塩基(すなわち、G、C、T、またはA)に対応する一つ以上の異なる群に割り当てることが可能である。このようにして、G群は、アルギニン(R)、リシン(K)、およびヒスチジン(H)を含み;C群は、アスパラギン酸(D)およびグルタミン酸(E)を含み;T群は、アラニン(A)、バリン(V)、ロイシン(L)、イソロイシン(I)、システイン(C)、トレオニン(T)、メチオニン(M)、およびフェニルアラニン(F)を含み;かつ、A群は、アスパラギン(N)、グルタミン(N)、チロシン(Y)、およびヒスチジン(H)を含む。なお、ヒスチジンは、G群とA群の両群に現れること;セリン(S)は、どの群にも含まれないが、縮重位置を好むものとして使用してもよいこと;および、プロリン、グリシン、およびトリプトファンは、主に立体的配慮のためにどの特定の群にも含まれないことに注意されたい。これらの群も下記に表2として示す。
【0090】
【表2】
【0091】
したがって、本発明によれば、任意の位置Xにおいて、メガヌクレアーゼの認識配列半分部位に所望の変化を実行するために、(1)野生型、または参照メガヌクレアーゼ−DNA複合体の三次元構造の少なくとも関連部分及び位置Xにおける接触面を定めるアミノ酸残基側鎖を決定すること;(2)接触面を含む少なくとも一つの残基のβ−炭素及び位置Xにおける塩基対の少なくとも一つの塩基との間の距離を決定すること;(3)所望の変化を促進するために、(A)塩基から6オングストローム未満の距離にある残基については、G群、C群、T群又はA群の内の適切な一つの群のメンバーである1群及び/又は2群から残基を選ぶこと、及び/又は、(B)塩基から6オングストロームより遠い距離にある残基については、G群、C群、T群又はA群の内の適切な一つの群のメンバーである2群及び/又は3群から残基を選ぶこと、である。接触面を含む、一つを超えるこのような残基を、分析および修飾のために選ぶことが可能であり、ある実施態様では、このような各残基が分析され複数の残基が修飾される。同様に、接触面に含まれる残基のβ−炭
素と、位置Xにおける塩基対の二つの塩基のそれぞれとの間の距離を測定することが可能であり、もしもその残基が両方の塩基の9オングストローム以内にある場合、該塩基対の二つの塩基に影響を及ぼすために異なる置換を実行することが可能である(例えば、一方の鎖の近位塩基に影響を及ぼすために1群の残基、又は他方の鎖の遠位塩基に影響を及ぼすために3群の残基)。それとは別に、ペアの両塩基と相互作用を持つことが可能な残基置換の組み合わせは、特異性に影響を及ぼすことが可能である(例えば、アンチセンス鎖に接触するA群の残基と組み合わせたセンス鎖に接触するT群の残基はT/Aを選択する)。最後に、残基の複数の候補修飾は、経験的に(例えば、組み換えメガヌクレアーゼを生産し、その配列認識を調べることによって)、又はコンピュータ的に(例えば、修飾された酵素のメガヌクレアーゼ−DNA複合体のコンピュータモデル化によって)その有効性を検証して、複数候補の中から選択することが可能である。
【0092】
野生型又は基準ヌクレアーゼの、一つ以上の所望のアミノ酸修飾が選択されたならば、合理設計メガヌクレアーゼは、従来技術で周知の組み換え法および技術によって生産することが可能である。ある実施態様では、特異的配列修飾を創製するために、非ランダムな、部位指向性突然変異誘発技術が用いられる。非ランダム突然変異誘発技術の非限定的例としては、重複プライマーPCR(例えば、WANG ET AL.(2006),NUCLEIC ACIDS RES.34(2):517−527を参照)、部位指向性突然変異誘発(例えば、米国特許第7,041,814号を参照)、カセット突然変異誘発(例えば、米国特許第7,041,814号参照)、および、PROMEGA BIOSCIENCES,INC.(SAN LUIS OBISPO、カリフォルニア州)から市販されるALTERED SITES(R) II MUTAGENESIS SYSTEM KIT用のメーカーのプロトコールが挙げられる。
【0093】
合理設計メガヌクレアーゼによる、特異的DNA配列の認識及び切断は、当業者の既知の任意の方法によって定量することが可能である(例えば、米国特許出願公開第2006/0078552号を参照)。ある実施態様では、メガヌクレアーゼ切断の定量は、インビトロ切断アッセイによって定量される。このようなアッセイは、定量されるメガヌクレアーゼの意図される認識配列を含むポリヌクレオチド基質のインビトロ切断を用いるが、ある実施態様では、片方又は両方の半分部位の一つ以上の塩基が、別の塩基に変えられた、意図された認識配列の変種を含むポリヌクレオチド基質のインビトロ切断を用いる。通常、ポリヌクレオチド基質は、合成され、ベクターにクローンされた標的部位を含む2本鎖DNA分子である。ポリヌクレオチド基質は、直線状であっても、円形であってもよいが、通常、唯一の認識配列を含む。メガヌクレアーゼは、適切な条件下で、ポリヌクレオチド基質とインキュベートされ、得られたポリヌクレオチドは、切断産物を特定するために、既知の方法で分析される(例えば、電気泳動又はクロマトグラフィー)。直線状2本鎖DNA基質において単一の認識配列がある場合には、メガヌクレアーゼ活性は、二つのバンドの出現及び最初の全長の基質バンドの消失によって検出される。一実施態様では、メガヌクレアーゼ活性は、例えば、WANG ET AL.(1997),NUCLEIC ACID RES.,25:3767−3776に記載される通りに定量することが可能である。
【0094】
他の実施態様では、メガヌクレアーゼの切断パターンは、インビボ切断アッセイによって定量される(例えば、米国特許出願公開第2006/0078552号)。特定の実施態様では、このインビボ試験は、単一鎖アニーリング組み換え試験(SSA)である。この種の試験は当業者には既知である(RUDIN ET AL.(1989),GENETICS 122:519−534;FISHMAN−LOBELL ET AL.(1992),SCIENCE 258:480−4)。
【0095】
当業者には明らかなように、DNA認識および結合に関与するドメイン以外の、メガヌ
クレアーゼ酵素のドメインに対し、活性を完全に消失させることなく、さらに新たなアミノ酸置換、挿入又は欠失を行うことが可能である。置換は、構造的または機能的に束縛された位置における、類似のアミノ酸残基の保存的置換であってもよく、又は構造的または機能的に比較的に束縛されない位置における非保存的置換であってもよい。このような置換、挿入又は欠失は、当業者によって、不当な努力を要することなく、通例の実験操作によって特定することが可能である。したがって、ある実施態様では、本発明の組み換えメガヌクレアーゼは、基準メガヌクレアーゼの配列に対し、85%から99%の間の任意のパーセント(例えば、85%、87.5%、90%、92.5%、95%、97.5%、99%)の配列類似性を持つタンパクを含む。各野生型I−CREI、I−MSOI、I−SCEI、およびI−CEUIタンパクに関して、多くのN−末端およびC−末端配列は、X線結晶実験でははっきりと見ることができない。これは、これらの位置が、構造的又は機能的に束縛されていないことを示唆する。それゆえ、これらの残基は、配列類似性の計算から排除することが可能であり、下記の基準メガヌクレアーゼ配列を使用することが可能である。I−CREIについては配列番号1の残基2−153;I−MSOIについては配列番号6の残基6−160;I−SCEIについては配列番号9の残基3−186;I−CEUIについては配列番号12の残基5−211である。
【0096】
2.2 LAGLIDADGファミリー・メガヌクレアーゼ
LAGLIDADGメガヌクレアーゼ・ファミリーは、多様な系統群の宿主生物体由来の200を超えるメンバーから構成される。このファミリーのメンバーは全て、特異的DNA配列の切断に与る他の構造的モチーフと共に、高度に保存されるLAGLIDADGモチーフの1個又は2個の複製を有する。LAGLIDADGモチーフの1つの複製有する酵素(すなわち、モノ−LAGLIDADGメガヌクレアーゼ)はダイマーとして機能し、一方、このモチーフの2つの複製を有する酵素(すなわち、ジ−LAGLIDADGメガヌクレアーゼ)はモノマーとして機能する。
【0097】
LAGLIDADGファミリーメンバーは全て、比較的長い配列(>12BP)を認識、切断し、4個のヌクレオチドの、3′オーバーハングを残す。これらの酵素はさらに、LAGLIDADGの外に、タンパク−DNAインターフェイスにおいて、逆平行β鎖の類似配置を含むいくつかの構造的モチーフを共有する。これらの保存された構造モチーフ内のアミノ酸がDNA塩基と相互作用を持ち、認識配列特異性を付与する。ファミリーのいくつかのメンバー(例えば、I−CREI、I−MSOI、I−SCEI及びI−CEUI
)の間に全体構造の類似性が見られることは、X線結晶学によって明らかにされている。したがって、このファミリーのメンバーは、この構造モチーフ内部の特定のアミノ酸について修飾し、該酵素の全体活性又は配列特異性を変えることが可能であり、かつ、対応する修飾は、他のファミリーメンバーにおいても類似の結果をもたらすことが当然期待される。概論については、CHEVALIERET AL. (2001), NUCLEIC ACID RES. 29(18):3757−3774)を参照されたい。
【0098】
2.2.1 I−CREIから得られるメガヌクレアーゼ
1側面において、本発明は、CHLAMYDOMONAS REINHARDTIIのI−CREIメガヌクレアーゼに基づく、又はこのメガヌクレアーゼから得られた合理的設計メガヌクレアーゼに関する。I−CREIメガヌクレアーゼの野生型アミノ酸配列は、GENBANKアクセス#PO5725に対応する、配列番号1に示される。結晶構造PDB#1BP7における野生型I−CREIメガヌクレアーゼの、二つの、認識配列半分部位を下記の表3に示す。
【0099】
【表3】
【0100】
この天然の認識配列は、中央の4塩基の外側においても、完全にはパリンドローム的ではないことに注意されたい。なお、二つの認識配列半分部位がそれぞれのセンス鎖に太字で示される。
【0101】
野生型I−CREIはさらに、下記の表4に示すように、(中央のN1−N4塩基を除き)完全にパリンドロームである配列も認識し、切断する。
【0102】
【表4】
【0103】
配列番号4及び配列番号5のパリンドローム配列は、野生型I−CREIにとってより優れた基質であると考えられる。なぜなら、この酵素は、天然のDNA配列よりもより高い親和度の下にこの部位に結合し、より効率的に該部位を切断するからである。下記の開示のために、かつ、特に本出願において提示する実験結果との関連において、野生型I−CREIによって切断されるこのパリンドローム配列を”WT”と呼ぶことにする(例えば、図2(A)参照)。二つの、認識配列半分部位が、それぞれのセンス鎖に太字で示される。
【0104】
図1(A)は、野生型I−CREIメガヌクレアーゼ・ホモダイマーの、2本鎖DNA認識配列との相互作用を描く。図1(B)は、野生型酵素の半分部位と野生型認識配列の−4位置におけるこの酵素のアミノ酸残基と塩基との間の特異的相互作用を示し、図1(C)〜図1(E)は、半分部位の−4位置における特異性を変更させた、本発明の、三つの合理設計メガヌクレアーゼの、一半分部位の−4位置における、酵素のアミノ酸残基と塩基との間の特異的相互作用を示す。
【0105】
したがって、半分部位の任意の、指定の塩基位置における塩基に対する好みは、本出願に開示される方法を用いて、他の三つの塩基のそれぞれに合理的に変えることが可能である。第一に、指定の塩基位置における野生型認識面が決められる(例えば、メガヌクレアーゼ−DNA複合体共結晶構造を分析することによって、又はメガヌクレアーゼ−DNA複合体のコンピュータモデル化によって)。第二に、指定の塩基位置における、周辺アミノ酸位置のβ−炭素と、各DNA鎖における核酸塩基の間の距離に基づいて、既存の及び可能な接触残基が決められる。例えば、ただしこれらに限定されないが、図1(A)に示すように−4位置における、I−CREI野生型メガヌクレアーゼ−DNA接触残基は、位置26のグルタミンを含み、これはアンチセンスDNA鎖のA塩基と水素結合する。さらに、残基77が、DNAアンチセンス鎖の−4塩基に接触し得る可能性を持つことが特定された。残基26のβ−炭素は、アンチセンスDNA鎖のA塩基のN7から5.9Å離れており、残基77のβ−炭素は、センス鎖のTのC5メチルから7.15オングストローム離れている。この距離及び本出願に記載される塩基化学規則によれば、センス鎖をCとすると、位置77のグルタミン酸と水素結合することが可能であり、アンチセンス鎖を
Gとすると、位置26のグルタミンと結合することが可能である(野生型I−CREI結晶構造に観察されるように、水分子を介して)(図1(C)を参照)。センス鎖をGとすると、位置77のアルギニンと水素結合することが可能であり、アンチセンス鎖をCとすると、位置26のグルタミン酸と水素結合することが可能である(図1(D)を参照)。センス鎖をAとすると、位置77のグルタミンと水素結合することが可能であり、アンチセンス鎖をTとすると、位置26のアラニンと疎水性接触を形成することが可能である(図1(E)を参照)。もしも塩基の特異的接触が位置77によって与えられるとすると、野生型接触Q26を置換し(例えば、セリン残基によって)、その特異性に対する影響を低下、または除去することが可能である。それとは別に、位置26および77における相補的突然変異を組み合わせて、特定の塩基対を指定することが可能である(例えば、A26は、アンチセンス鎖においてTを指定し、Q77は、センス鎖においてAを指定する(図1(E))。これらの予言された残基置換は、全て、実験的にその有効性が実証された。
【0106】
したがって、本発明によれば、I−CREIメガヌクレアーゼのDNA認識ドメインに対する、ある確定数のアミノ酸置換が特定された。これらは、単独で又は組み合わせて用いられて、DNA認識配列半分部位内の個別の塩基において特異性を変更された組み換えメガヌクレアーゼを、したがって、野生型酵素とは異なる半分部位を持つ合理設計メガヌクレアーゼをもたらすことが可能である。I−CREIのアミノ酸修飾及びそれによってもたらされる、認識配列半分部位特異性における変化を、表5に示す。
【0107】
【表5】
【0108】
太字入力は、野生型接触残基であり、本出願で使用される「修飾」を構成しない。
星印は、その残基が、アンチセンス鎖の塩基に接触することを示す。
【0109】
2.2.2. I−MSOLIから得られるメガヌクレアーゼ
他の側面では、本発明は、MONOMASTIX SP.のI−MSOIメガヌクレアーゼに基づく、又はこのヌクレアーゼから得られた合理設計メガヌクレアーゼに関する。I−MSOIメガヌクレアーゼの野生型アミノ酸配列は、GENBANKアクセス#AAL34387に対応し、配列番号6に示される。結晶構造PDB#1M5Xにおける野生型I−MSOIメガヌクレアーゼの、二つの認識配列半分部位を下記の表6に示す。
【0110】
【表6】
【0111】
認識配列は、中央の4塩基の外側においても、完全にはパリンドローム的ではないことに注意されたい。二つの認識配列半分部位が、それぞれのセンス鎖に太字で示される。
【0112】
本発明によれば、I−MSOIメガヌクレアーゼのDNA認識ドメインに対する、ある確定数のアミノ酸置換が特定された。これらは、単独で又は組み合わせて用いられて、DNA認識配列半分部位内の個別の塩基において特異性を変更された組み換えメガヌクレアーゼを、したがって、野生型酵素とは異なる半分部位を持つ合理設計メガヌクレアーゼをもたらすことが可能である。I−MSOIのアミノ酸修飾、および、予測される、認識配列半分部位特異性における変化を、表7に示す。
【0113】
【表7】
【0114】
太字は、野生型接触残基を表し、本出願で使用される「修飾」を構成しない。
星印は、その残基がアンチセンス鎖の塩基に接触することを示す。
【0115】
2.2.3. I−SCEIから得られるメガヌクレアーゼ
他の側面では、本発明は、SACCHAROMYCESCEREVISIAEのI−SCEIメガヌクレアーゼに基づく、又はこのヌクレアーゼから得られた合理設計メガヌクレアーゼに関する。I−SCEIメガヌクレアーゼの野生型アミノ酸配列は、GENBANKアクセス#CAA09843に対応し、配列番号9に示される。結晶構造PDB#1R7Mにおける野生型I−SCEIメガヌクレアーゼの、認識配列を下記の表8に示す。
【0116】
【表8】
【0117】
認識配列は、非パリンドローム的であり、かつ、半分部位を分ける四つの塩基対がないことに注意されたい。
【0118】
本発明によれば、I−SCEIメガヌクレアーゼのDNA認識ドメインに対する、ある確定数のアミノ酸置換が特定された。これらは、単独で又は組み合わせて用いられて、DNA認識配列内の個別の塩基において特異性を変更された組み換えメガヌクレアーゼ、つまり、野生型酵素とは異なる認識配列を持つ合理設計メガヌクレアーゼをもたらすことが可能である。I−SCEIのアミノ酸修飾及び予測される認識配列特異性における変化を、表9に示す。
【0119】
【表9】
【0120】
太字入力は、野生型接触残基を表し、本出願で使用される「修飾」を構成しない。星印は、その残基が、アンチセンス鎖の塩基に接触することを示す。
【0121】
2.2.4 I−CEUIから得られるメガヌクレアーゼ
他の側面では、本発明は、CHLAMYDOMONASEUGAMETOSのI−CEUIメガヌクレアーゼに基づく、又はこのヌクレアーゼから得られた合理設計メガヌクレアーゼに関する。I−CEUIメガヌクレアーゼの野生型アミノ酸配列は、GENBANKアクセス#P32761に対応し、配列番号12に示される。結晶構造PDB#2EX5における野生型I−CEUIメガヌクレアーゼの、二つの認識配列半分部位を下記に示す。
【0122】
【表10】
【0123】
I−CEUIはホモダイマーであるにも拘わらず、I−CEUIの認識インターフェイスにおける天然の縮重のために(SPIEGEL ET AL.(2006),STRUCTURE 14:869−80)、認識配列は、中央の4塩基の外側においても、非パリンドローム的であることに注意されたい。二つの認識配列半分部位がそれぞれのセンス鎖に太字で示される。
【0124】
本発明によれば、I−CEUIメガヌクレアーゼのDNA認識ドメインに対する、ある確定数のアミノ酸置換が特定された。これらは、単独で又は組み合わせて用いられて、DNA認識配列半分部位内の個別の塩基において特異性を変更された組み換えメガヌクレアーゼを、したがって、野生型酵素とは異なる半分部位を持つ合理設計メガヌクレアーゼをもたらすことが可能である。I−CEUIのアミノ酸修飾及び予測される認識配列特異性における変化を、表11に示す。
【0125】
【表11】
【0126】
太字入力は、野生型接触残基を表し、本出願で使用される「修飾」を構成しない。星印は、その残基がアンチセンス鎖の塩基に接触することを示す。
【0127】
2.2.5 特異的に排除される組み換えメガヌクレアーゼ
本発明は、従来技術において記載され、別法によって開発された、いくつかの組み換えメガヌクレアーゼを包含することを意図しない。これらの排除されたメガヌクレアーゼとしては、ARNOULD ET AL.(2006),J.MOL.BIOL.355:443−58;SUSSMAN ET AL.(2004),J.MOL.BIOL.3
42:31−41;CHAMES ET AL.(2005),NUCLEIC ACIDS RES.33:E178;SELIGMAN ET AL.(2002),NUCLEIC ACIDS RES.30:3870−9;およびASHWORTH ET AL.(2006) NATURE 441(7093):656−659によって記載されるものが挙げられる。なお、上記の全開示を、C33、R33、A44、H33、K32、F33、R32、A28、A70、E33、V33、A26及びR66から選ばれる単一置換を持つ、I−CREIに基づく組み換えメガヌクレアーゼを含め、引用により本出願に含める。さらに排除されるものは、A68/N70/N75及びD44/D70/N75から選ばれた3置換、K44/T68/G60/N75及びR44/A68/T70/N75から選ばれる4置換を持つI−CREIに基づく組み換えメガヌクレアーゼである。最後に、特に指定して排除されるものは、置換L28及びR83のペアを持つI−MSOIに基づく組み換えメガヌクレアーゼである。これらの置換又は置換の組み合わせは、本出願では「排除修飾」と呼ばれる。
【0128】
2.2.6 認識配列半分部位において複数変化を有するメガヌクレアーゼ
他の側面では、本発明は、DNA認識配列半分部位の二つ以上の位置において半分部位指向を変えるために、上の節2.2.1−2.2.4に記載した、二つ以上のアミノ酸置換を組み合わせることによって得られる合理設計メガヌクレアーゼに関する。例えば、ただしこれらに限定されるものではないが、さらに詳しく後述されるように、酵素DJ1は、修飾R30/E38(これは、位置−7においてCを好む)、R40(これは位置−6においてGを好む)、R42(これは、位置−5においてGを好む)及びN32(これは、位置−9において完全縮重を好む)を取り込むことによって、I−CREIから得られる。この合理設計DJ1メガヌクレアーゼは、A−7A−6C−5に対する野生型の指向性に比べ、ほぼ変わらない程度でC−7G−6G−5を認識し、位置−9におけるAに対する耐性を増大させる。
【0129】
異なる塩基位置に影響を及ぼす残基置換を組み合わせることができる能力は、一部は、LAGLIDADGメガヌクレアーゼのモジュラー性による。LAGLIDADG認識インターフェイスにおける塩基接触の大部分は、個々のアミノ酸側鎖によって形成され、インターフェイスは、隣接塩基と相互作用を持つ側鎖間の相互接続性、または水素結合ネットワークから比較的自由である。これによって、一般に、一塩基位置と相互作用を持つ残基を隣接塩基における側鎖同士の相互作用に影響を及ぼさずに操作することが可能になる。さらに、上の2.21−2.2.4節に掲げた突然変異の加重性は、これらの突然変異を特定するために用いた本法の直接的結果である。この方法は、単一塩基と直接に相互作用を持つ側鎖置換を予測する。一般に、側鎖間の相互接続、または水素結合ネットワークは、認識インターフェイス内の置換の独立性を維持するために回避される。
【0130】
側鎖置換のある組み合わせは、完全に又は部分的に相互に不適合である。不適合ペア又は組のアミノ酸を合理設計メガヌクレアーゼに組み込んだ場合、得られた酵素の触媒活性は、低下するか又は排除される。通常、これらの不適合は、導入されたアミノ酸側鎖間の立体干渉によるもので、活性は、この干渉を特定し、除去することで回復が可能である。具体的には、大きな側鎖を持つ(例えば、2又は3群のアミノ酸)二つのアミノ酸が、メガヌクレアーゼ構造において、互いに隣接するアミノ酸位置に組み込まれた場合(例えば、I−CREI由来のメガヌクレアーゼの場合、位置32及び33、28及び40、28及び42、42及び77又は68及び77)、これら二つのアミノ酸は互いに干渉しあって、酵素活性を下げる可能性が高い。この干渉は、一方の又は両方の不適合アミノ酸を、より小さな側鎖を持つアミノ酸(例えば、1群または2群)で置換することによって排除することが可能である。例えば、I−CREI由来の合理設計メガヌクレアーゼでは、K28は、R40とR42の両方に干渉する。酵素活性を最大化するためには、R40及びR42を、位置28においてセリン又はアスパラギン酸と組み合わせることが可能である
。
【0131】
本出願に記載のように特定される、アミノ酸置換の組み合わせを用いて、野生型メガヌクレアーゼ(又は以前に修飾されたメガヌクレアーゼ)の特異性を、元の認識配列から、興味の核酸(例えば、ゲノム)中に存在する、所望の認識配列に合理的に変えることが可能である。例えば、図2(A)は、I−CREIメガヌクレアーゼ認識配列WT(配列番号4)の「センス」鎖の外に、それに対し合理的設計メガヌクレアーゼを創製したならば有用であろうと考えられる、他の、いくつかの配列を示す。WT認識配列と所望の認識配列との間で保存される塩基には陰影を施す。本発明によれば、I−CREIメガヌクレアーゼに基づく組み換えメガヌクレアーゼは、これらの所望の認識配列のそれぞれについてばかりでなく、他の任意の認識配列についても、本出願に記載される適切なアミノ酸置換によって、合理的に設計することが可能である。
【0132】
3. DNA結合親和度が変更された合理設計メガヌクレアーゼ
前述のように、本発明の組み換えメガヌクレアーゼのDNA結合親和度は、DNAのフォスフォジエステルバックボーンと接触面を形成するいくつかのアミノ酸を変えることによって修飾することが可能である。この接触面は、DNAバックボーンから9オングストローム未満の距離のβ−炭素を持つ酵素のアミノ酸を、この残基が野生型メガヌクレアーゼ−DNA複合体においてDNAバックボーンと接触するかどうかとは無関係に含む。DNA結合は、酵素活性に必要な前提条件であるから、DNA結合親和度の上昇/低下は、それぞれ、酵素活性の上昇/低下を起こすことが示されている。しかしながら、DNA結合親和度の上昇/低下はさらに、メガヌクレアーゼの配列特異性の低下/上昇を起こすことが示されている。それゆえ、活性および特異性は両方とも、フォスフォジエステルバックボーン接触を修飾することによって修飾することが可能である。
【0133】
酵素活性の上昇/酵素の特異性の低下のためには、
【0134】
(I)酵素とDNAバックボーンの間の静電気的反発を取り除く。ある特定されたアミノ酸が、陰性荷電のDNAバックボーンに反発すると予想される、陰性に荷電した側鎖を持つ場合(例えば、アスパラギン酸、グルタミン酸)、この反発は、アミノ酸を無荷電の又は陽性荷電の側鎖で置換することによって除去することが可能である。ただし、この場合、立体障害の作用には影響される。実験的に確かめられた例は、I−CREIにおけるグルタミン酸80をグルタミンに突然変異させた場合である。
【0135】
(II)酵素とDNAバックボーンの間に静電気的牽引相互作用を導入する。接触面の位置のいずれかにおける、陽性荷電側鎖を持つアミノ酸(例えば、リシン又はアルギニン)の導入は、立体干渉の作用には影響されるが、結合親和度を増すことが予想される。
【0136】
(III)酵素とDNAバックボーンの間に水素結合を導入する。接触面のアミノ酸が、適切な水素結合官能基を欠如するため、又はDNAバックボーンと相互作用を持つには、側鎖が短すぎるか長すぎるか、及び/又は、屈曲性が不足するという理由で水素結合を造ることができない場合、適切な長さと屈曲性を持つ、水素結合を供与することが可能な極性アミノ酸(例えば、セリン、トレオニン、チロシン、ヒスチジン、グルタミン、アスパラギン、リシン、システイン又はアルギニン)を導入することが可能である。ただし、立体障害の作用には影響される。
【0137】
具体的に、酵素活性の低下/酵素の特異性の上昇のためには、
【0138】
(I)酵素とDNAバックボーンの間に静電気的反発を導入する。接触面の位置のいずれかにおいて、陰性に荷電した側鎖を持つアミノ酸(例えば、グルタミン酸、アスパラギ
ン酸)を導入することは、結合親和度を下げることが予想される。ただし、立体障害の作用には影響される。
【0139】
(II)酵素とDNAの間の静電気的牽引を取り除く。接触面のいずれかのアミノ酸が、陰性に荷電したDNAバックボーンと相互作用を持つ、陽性に荷電した側鎖を持つ場合(例えば、リシン又はアルギニン)、この好ましい相互作用は、アミノ酸を無荷電の又は陰性荷電の側鎖で置換することによって除去することが可能である。ただし、立体干渉の作用には影響される。実験的に確かめられた例は、I−CREIにおけるリシン116をアスパラギン酸に突然変異させた場合である。
【0140】
(III)酵素とDNAバックボーンの間の水素結合を取り除く。接触面のいずれかのアミノ酸が、DNAバックボーンと水素結合を造る場合、そのアミノ酸をその側鎖が適切な官能基を持たないか又は必要な長さ/屈曲性を欠如するために、同様の水素結合を造るとは予想されないアミノ酸に置換することが可能である。
【0141】
例えば、I−CREIに基づくある組み換えメガヌクレアーゼにおいて、活性を上げるためにI−CREIメガヌクレアーゼの位置80におけるグルタミン酸が、リシン又はグルタミンに変更される。別の実施態様では、I−CREIの位置66のチロシンが、アルギニン又はリシンに変更される。これは、該メガヌクレアーゼの活性を上げる。さらに別の実施態様では、I−CREIの位置34におけるリシンをアスパラギン酸に、位置66におけるチロシンをアスパラギン酸に、及び/又は、位置116におけるリシンをアスパラギン酸に変えることによって、酵素活性を低下させる。
【0142】
組み換えメガヌクレアーゼの活性は、その組み換え酵素が、ある特定の認識配列に関して、無活性から極度に高い活性までの間の任意の活性レベルを持つように修飾することが可能である。例えば、DJ組み換えメガヌクレアーゼは、位置26においてグルタミン酸突然変異を抱えると活性を完全に失う。しかしながら、位置26におけるグルタミン酸置換と位置80におけるグルタミン置換の組み合わせは、認識配列半分部位内の−4におけるグアニンに対し高度の特異性および活性を持つ組み換えメガヌクレアーゼを創製する(図1(D)を参照)。
【0143】
本発明によれば、フォスフォジエステルDNAバックボーン近傍の各種位置におけるアミノ酸は、メガヌクレアーゼ活性および特異性の両方に同時に作用するように変えることが可能である。この、酵素の特異性と活性の「調整」は、フォスフォジエステルバックボーンに対し、アミノ酸によって造られる接触点の数を増すか又は減らすことによって達成される。フォスフォジエステルバックボーンに対する種々の接触は、アミノ酸側鎖によって促進することが可能である。ある実施態様では、イオン結合、塩橋、水素結合及び立体障害は、アミノ酸側鎖の、フォスフォジエステルバックボーンに対するコンタクトに影響を及ぼす。例えば、I−CREIメガヌクレアーゼの場合、位置116におけるリシンのアスパラギン酸への変更によって、位置−8および−9における核酸塩基間の塩橋が取り除かれ、これは酵素の切断速度は下げるが、特異性は上げる。
【0144】
野生型I−CREI(配列番号1)、I−MSOI(配列番号6)、I−SCEI(配列番号9)、およびI−CEUI(配列番号12)各メガヌクレアーゼの、バックボーン接触面を形成する残基は、下表12において特定される。
【0145】
【表12】
【0146】
酵素の親和度を上げ、したがって、より活性を高め/特異性を下げるためには、
(1)対応する酵素において、陰性荷電(D又はE)、疎水性(A、C、F、G、I、L、M、P、V、W、Y)、又は無荷電/極性(H、N、Q、S、T)のアミノ酸を、表12から選ぶ。
(2)アミノ酸が陰性荷電しているか又は疎水性である場合は、それを、無荷電/極性(作用が小さい)又は陽性荷電(KまたはR、作用が大きい)となるように突然変異させる。
(3)アミノ酸が無荷電/極性である場合、それを陽性荷電に突然変異させる。
【0147】
酵素の親和度を下げ、したがって、より活性を下げ/特異性を上げるためには、
(1)対応する酵素において、陽性荷電(K又はR)、疎水性(A、C、F、G、I、L、M、P、V、W、Y)、又は無荷電/極性(H、N、Q、S、T)のアミノ酸を、表12から選ぶ。
(2)アミノ酸が陽性荷電している場合は、それを、無荷電/極性(作用が小さい)又は陰性荷電(作用が大きい)となるように突然変異させる。
(3)アミノ酸が疎水性又は無荷電である場合、それを陰性荷電に突然変異させる。
【0148】
4. ヘテロダイマー・メガヌクレアーゼ
他の側面では、本発明は、二つのモノマーで、その一方は野生型で、その一方または両方が、非天然形または組み換え形であるモノマーのコンタクトによって形成されるヘテロダイマーであるメガヌクレアーゼを提供する。例えば、野生型I−CREIメガヌクレアーゼは、通常、それぞれが、擬似パリンドローム認識配列において一つの半分部位に結合する、二つのモノマーから構成されるホモダイマーである。ヘテロダイマー組み換えメガヌクレアーゼは、異なる半分部位を認識する二つのメガヌクレアーゼを組み合わせることによって、例えば、細胞において二つのメガヌクレアーアゼを共時発現することによって、又は溶液中で二つのメガヌクレアーゼを混合することによって生産することが可能である。二つのモノマーにおいて、それらのコンタクトによるダイマー形成に影響を及ぼす、それぞれのアミノ酸を変えることによって、ヘテロダイマーの形成を、ホモダイマーの形成よりも優先することが可能である。特定の実施態様では、二つのモノマーのインターフェイスにおけるいくつかのアミノ酸が、第1モノマーでは、陰性荷電のアミノ酸(D又は
E)から陽性荷電のアミノ酸(K又はR)に変えられ、第2モノマーでは、陽性荷電のアミノ酸から陰性荷電のアミノ酸に変えられる(表13参照)。例えば、I−CREI由来のメガヌクレアーゼの場合、位置7及び57におけるリシンが、第1モノマーではグルタミン酸に突然変異され、第2モノマーでは、位置8および61におけるグルタミン酸がリシンに突然変異される。このプロセスの結果は、第1モノマーが、ダイマーインターフェイスにおいて過剰な陽性荷電残基を持ち、第2モノマーが、ダイマーインターフェイスにおいて過剰な陰性荷電残基を持つ、一対のモノマーである。それゆえ、この第1及び第2モノマーは、インターフェイスにおける変更アミノ酸の間の静電気的相互作用のために、その同じモノマーペアよりも優先的にコンタクトする。
【0149】
【表13】
【0150】
それとは別に、またはそれに加えて、二つのモノマーのインターフェイスにおけるいくつかのアミノ酸は、立体的にホモダイマー形成を妨げるように変えることも可能である。具体的には、一方のモノマーのダイマーインターフェイスのアミノ酸を、ホモダイマーを立体的に阻止する比較的大きな又は嵩高な残基によって置換する。第2のモノマーのダイマーインターフェイスにおけるアミノ酸を、第1のモノマーの嵩高な残基を補正するため
に比較的小さな残基で置換し、ヘテロダイマーにおける衝突を取り除くことが可能であるし、または未修飾のままでもよい。
【0151】
さらに他の実施態様では、イオン架橋又は水素結合を、ヘテロダイマー・インターフェイスの疎水性コアの中に埋没させることが可能である。具体的には、インターフェイスのコアの一モノマーにおける疎水性残基は、陽性荷電残基によって置換することが可能である。さらに、野生型ホモダイマーでは、第1モノマーにおいて置換された疎水性残基と相互作用を持つ、第2モノマーの疎水性残基は、陰性荷電残基によって置換することが可能である。したがって、この二つの置換残基は、イオン結語または水素結合を形成することが可能である。同時に、疎水性インターフェイスの中に埋没された満たされない電荷の静電気的反発は、ホモダイマー形成を忌避する。
【0152】
最後に、前述のように、ヘテロダイマーの各モノマーは、DNA認識領域において、異なるアミノ酸を置換されて、そのため、それぞれが異なるDNA半分部位を持ち、かつ、組み合わされたダイマー形DNA認識配列が非パリンドローム的となってもよい。
【0153】
5. 組み換え細胞および生物体を生産する方法
本発明の側面はさらに、合理設計メガヌクレアーゼを用いた、組み換え、トランスジェニック又は他のやり方で遺伝学的に修飾された細胞および生物体を生産する方法を提供する。したがって、ある実施態様では、相同組み換えによる興味の配列の正確な挿入(単数又は複数)を可能とするために、細胞または生物体のゲノムDNAの単一部位又は比較的少数部位に、二本鎖切断点を特異的に引き起こすための、組み換えメガヌクレアーゼが開発された。別の実施態様では、(A)非相同的末端接合による興味の配列の稀な挿入(単数又は複数)を可能とするために、又は(B)非相同的末端接合による標的配列の破壊を可能とするために、細胞または生物体のゲノムDNAの単一部位又は比較的少数部位に、二本鎖切断点を特異的に引き起こすための、組み換えメガヌクレアーゼが開発された。興味の配列の、相同的組み換え又は非相同的末端接合に関して本出願で用いる場合、「挿入」という用語は、興味の配列が染色体の中に組み込まれるように、興味の配列が染色体にコンタクトすることを意味する。相同組み換えの場合、挿入された配列は内因性配列を置換し、そのため、元のDNAは等しい長さではあるがヌクレオチド配列が変更された外因性DNAによって置換される。それとは別に、挿入配列はそれが置換する配列よりもより多数の又はより少数の塩基を含むことが可能である。
【0154】
それゆえ、本発明のこの局面によれば、組み換え生物体としては、単子葉植物種、例えば、米、小麦、トウモロコシ(メイズ)並びにライ麦、及び双子葉植物種、例えば、豆類(例えば、ソラマメ、大豆、レンズ豆、ピーナッツ、エンドウ豆)、アルファルファ、クローバー、タバコ並びにARABIDOPSIS種が挙げられるが、これらに限定されるものではない。さらに、組み換え生物体としては、動物、例えば、ヒト並びに非ヒト霊長類、ウマ、ウシ、ヤギ、ブタ、ヒツジ、イヌ、ネコ、モルモット、ラット、マウス、トカゲ、魚類、及び昆虫類、例えば、ショウジョウバエ種が挙げられるが、これらに限定されるものではない。他の実施態様では、生物体は、真菌、たとえば、カンジダ、ニューロスボラ又はサッカロマイセスである。
【0155】
ある実施態様では、本発明の方法は、成熟組み換え生物体になることが可能な又は遺伝学的に修飾された生物体が、そのゲノムの中に興味の挿入配列を抱える子孫を出産することを可能とする。例えば、細胞、胚細胞又は幹細胞の中に興味の配列を導入することを含む。
【0156】
メガヌクレアーゼタンパクは、ゲノムDNAを切断するために、細胞に輸送することが可能であるが、これによって、従来技術で既知の様々な機序によって、その切断部位にお
いて、興味の配列による相同組み換え、または非相同末端接合が可能とされる。組み換えメガヌクレアーゼタンパクは、例えば、マイクロインジェクション又はリポソームトランスフェクション(例えば、LIPOFECTAMINE(登録商標)、INVITROGEN CORP.,CARLSBAD、カリフォルニア州)を含む、ただしこれらに限定されないが、技術によって細胞中に導入することが可能である。このリポソーム処方は、標的細胞との脂質二重層融合を促進するのに用いることが可能であり、そのため、リポソームの内容物又はその表面にコンタクトするタンパクの細胞内部への取り込みを可能にする。それとは別に、酵素は、適切な取り込みタンパク、例えば、HIVTATタンパクなどに融合させて細胞の取り込みを指示するようにすることも可能である(例えば、HUDECZ ET AL.(2005),MED.RES.REV.25:679−736を参照)。
【0157】
それとは別に、メガヌクレアーゼタンパクをコードする遺伝子配列は、従来技術で既知の技術(例えば、AUSUBEL ET AL.,「分子生物学における最近のプロトコール(“CURRENTPROTOCOLS IN MOLECULAR BIOLOGY,”)」、WILEY 1999を参照されたい)を用いて、ベクターに挿入され、真核細胞にトランスフェクトされる。興味の配列は、同じベクターによって、異なるベクターによって又は従来技術で既知の他の手段によって導入することが可能である。
【0158】
DNAトランスフェクションの非限定的例としては、ウィルスベクター、プラスミド、コスミド及びYACベクターが挙げられる。DNA配列のトランスフェクションは、当業者には既知の、様々の方法によって実現することが可能である。例えば、DNA配列を細胞に輸送するために、リポソームおよびイムノリポソームが用いられる(例えば、LASIC ET AL.(1995),SCIENCE 267:1275−76を参照)。さらに、ベクターを細胞内に導入するためにウィルスを利用することが可能である(例えば、米国特許第7,037,492号を参照)。それとは別に、ベクターが裸のDNAとして導入されるようにトランスフェクション戦略を利用することも可能である(例えば、RUI ET AL.(2002),LIFE SCI.71(15):1771−8を参照)。
【0159】
細胞内に核酸を輸送するための一般的方法としては、(1)化学的方法(GRAHAM
ET AL.(1973),VIROLOGY 54(2):536−539;ZATLOUKAL ET AL.(1992),ANN.N.Y.ACAD.SCI.,660:136−153);(2)物理的方法、例えば、マイクロインジェクション(CAPECCHI(1980),CELL22(2):479−488)、電気穿孔(WONG
ET AL,(1982),BIOCHEM.BIOPHYS.RES.COMMUN.107(2):584−587;FROMM ET AL,(1985),PROC.NAT’L ACAD SCI.USA 82(17):5824−5828;米国特許第5,384,253号)、および、弾丸注入法(JOHNSTON ET AL,(1994),METHODS CELL.BIOL.43(A):353−365;FYNAN ET AL,(1993),PROC.NAT’L ACAD.SCI.USA 90(24):11478−11482)など;(3)ウィルスベクター(CLAPP(1993),CLIN. PERINATOL.20(1):155−168;LU ET AL,(1993),J.EXP.MED.178(6):2089−2096;EGLITIS ET AL,(1988),AVD.EXP.MED.BIOL.241:19−27;EGLITIS ET AL.(1988),BIOTECHNIQUES 6(7):608−614);および、(4)受容体介在性機構(CURIEL ET AL.(1991),PROC.NAT’L ACAD.SCI.USA 88(19):8850−8854;CURIEL ET AL.(1992),HUM.GEN.THER.3(2):147−154;WAGNER ET AL.(1992),P
ROC.NAT’L ACAD.SCI.USA 89(13):6099−6103)が挙げられる。
【0160】
ある実施態様では、ゲノムに挿入された興味の配列を含む遺伝学的に修飾された植物が生産される。ある実施態様では、組み換えメガヌクレアーゼに対応するDNA配列及びこのメガヌクレアーゼ認識配列及び/又は標的配列とほぼ同一の配列によって側接されても側接されなくともよい興味の配列によって植物細胞をトランスフェクトし、遺伝学的に修飾された植物が生産される。別の実施態様では、遺伝学的に修飾された植物は、組み換えメガヌクレアーゼのみに対応するDNA配列によって植物細胞をトランスフェクトし、それによる切断が非相同性末端接合を促進し、認識配列を含む標的配列を破壊することによって生産される。このような実施態様では、メガヌクレアーゼ配列は、宿主植物細胞においてメガヌクレアーゼの発現を可能とする調整配列の調節下にある。これらの調整配列としては、構成的植物プロモーター、例えば、NOSプロモーター、化学的に誘発可能な遺伝子プロモーター、例えば、デキサメタゾン誘発性プロモーター(例えば、GREMILLON ET AL.(2004),PLANT J.37:218−228を参照)、および、植物組織特異的プロモーター、例えば、LGC1プロモーター(例えば、SINGH ET AL.(2003),FEBS LETT.542:47−52を参照)が挙げられるが、これらに限定されるものではない。
【0161】
植物細胞の中にDNAを導入するための適切な方法は、DNAを細胞内に導入することが可能であるならば、ほとんどどの方法であってもよく、例えば、AGROBACTERIUM感染、プロトプラストのPEG−介在性形質転換(OMIRULLEH ET AL.(1993),PLANT MOLECULAR BIOLOGY,21:415−428)、DESICCATION/INHIBITION MEDIATED DNA
UPTAKE、電気穿孔、シリコンカーバイド線維による搖動、弾丸注入、または微小弾丸発射などが挙げられるが、これらに限定されるものではない。
【0162】
他の実施態様では、組み換えメガヌクレアーゼを用いて、遺伝学的に修飾された動物が生産される。植物細胞の場合と同様、核酸配列を、胚細胞又は最終的にトランスジェニック生物体となる細胞の中に導入することが可能である。ある実施態様では、細胞は、受精卵であり、外因性DNA分子は、受精卵の前核の中に注入することが可能である。次に、このマイクロインジェクションされた卵は、擬似妊娠の代理母の卵管に移送され、育成される。組み換えメガヌクレアーゼは、(例えば、3−フォスフォグリセレートキナーゼなどの構成的プロモーターの調節下に)受精卵で発言され、ゲノムの一部位又は不連続な少数部位における興味の配列の相同組み換えを促進する。それとは別に、GOSSLER ET AL.(1986),PROC.NATL.ACAD.SCI.USA 83:9065 9069に記載されるように、トランスジェニック動物生成のための組み換え胚性幹(“ES”)細胞を利用することによって、遺伝学的に修飾された動物を入手することが可能である。
【0163】
ある実施態様では、組み換え哺乳類発現ベクターは、核酸の組織特異的発現を、ある特定の細胞タイプに優先的に向けて指示することが可能である。組織特異的調整要素は従来技術で既知である。適切な組織特異的プロモーターの非限定的例としては、アルブミンプロモーター(肝臓特異的、PINKERT ET AL.(1987),GENES DEV.1:268−277)、リンパ系特異的プロモーター(CALAME AND EATON(1988),ADV.IMMUNOL.43:235−275)、特に、T細胞受容体のプロモーター(WINOTO AND BALTIMORE(1989),EMBO J.8:729−733)、および免疫グロブリンのプロモーター(BANERJIET AL.(1983),CELL 33:729−740;QUEEN AND
BALTIMORE(1983),CELL 33:741−748)、ニューロン特
異的プロモーター(例えば、ニューロフィラメント・プロモーター;BAYNE AND
RUDDLE(1989),PROC.NATL.ACAD.SCI.USA 86:5473−5477)、膵臓特異的プロモーター(EDLUND ET AL.(1985),SCIENCE 230:912−916)、および、乳腺特異的プロモーター(例えば、ミルクフェイ・プロモーター、米国特許第4,873,316号、および欧州特許公開EP0264166)が挙げられる。発達時の調整に与るプロモーター、例えば、マウスのHOXプロモーター(KESSEL AND GRUSS(1990),SCIENCE 249:374−379)、および、α−フェトプロテイン・プロモーター(CAMPES AND TILGHMAN(1989),GENES DEV.3:537−546)なども含まれる。
【0164】
ある実施態様では、合理設計メガヌクレアーゼは、発現レベルまたは局在を監視するために、ペプチドエピトープ(例えば、HA、FLAG、またはMYCエピトープ)によって標識されてもよい。ある実施態様では、メガヌクレアーゼは、細胞内局在シグナル、例えば、核局在シグナル(例えば、SV40の核局在シグナル)又はクロロプラストもしくはミトコンドリア局在シグナルに融合されてもよい。別の実施態様では、メガヌクレアーゼは、それを、細胞質に局在させる核輸出シグナルに融合されてもよい。さらに、メガヌクレアーゼは、無関係のタンパク質又はタンパクドメイン、例えば、DNA−修復又は相同組み換えを刺激するタンパク質(例えば、RECA、RAD51、RAD52、RAD54、RAD57又はBRCA2)に融合されてもよい。
【0165】
6. 遺伝子治療法
本発明の局面は、遺伝子治療のための、組み換えメガヌクレアーゼの使用を可能とする。本出願で用いる「遺伝子治療」という用語は、その構造及び/又は機能において欠陥的である遺伝子又は遺伝子調整配列と置換するために、患者の中に、少なくとも一つの遺伝子の機能的コピー又は遺伝子調整配列、例えば、プロモーター、エンハンサー又はサイレンサーを導入することを含む治療処置を意味する。さらに、「遺伝子治療」という用語は、有害遺伝子または調整要素に対し、該遺伝子の発現を下げるか又は除去するために為される修飾を指すことが可能である。遺伝子治療は、先天性病態、患者の生涯に亘る特定の遺伝子座位における突然変異または損傷による病態又は感染生物体による病態を治療するために実行することが可能である。
【0166】
本発明のある局面では、機能不全遺伝子は、遺伝子発現に影響を及ぼすゲノム領域に、外因性核酸配列を挿入することによって置換されるか、不能にさせられる。ある実施態様では、組み換えメガヌクレアーゼは、病態を緩和するために、修飾すべきゲノム領域の特定配列を標的とする。この配列は、エキソン、イントロン、プロモーター内の領域又は遺伝子の機能不全発現を引き起こす、他の調整領域であってもよい。本出願で用いる「機能不全発現」という用語は、遺伝子産物の生産が少なすぎる細胞又は遺伝子産物の生産が多すぎる細胞又は異なる機能、例えば、必要機能の欠如又は必要以上の機能を持つ遺伝子産物を生産する細胞による、遺伝子産物の異常な発現を意味する。
【0167】
修飾領域に挿入される外因性核酸配列は、遺伝子を正常化する「修復」配列を提供するために使用することが可能である。遺伝子修復は、再生されるべき適正機能の実現を可能とする遺伝子中に適正遺伝子配列を導入することによって実現される。これらの実施態様では、挿入される核酸配列は、タンパクの全体コード配列であってもよいし又はある実施態様では、修復されるべき領域のみを含む遺伝子断片であってもよい。他の実施態様では、挿入される核酸配列は、異常発現または調整を引き起こす突然変異が修復されるように、プロモーター配列または、その他の調整要素を含む。他の実施態様では、挿入される核酸配列は、突然変異遺伝子には欠如する適切な翻訳終止コドンを含む。核酸配列はさらに、適切な翻訳終止シグナルを欠如する組み換え遺伝子において翻訳を停止するための配列
を持つことが可能である。
【0168】
それとは別に、核酸配列は、遺伝子の調整配列を破壊することによって又は遺伝子機能を除去するサイレンサーを提供することによって、遺伝子機能を全く除去することが可能である。ある実施態様では、外因性核酸配列は、遺伝子産物の発現を阻止するために翻訳終止コドンを提供する。別の実施態様では、外因性核酸配列は、全長のRNA分子の発現を阻止するために翻訳終止コドンを提供する。さらに別の実施態様では、遺伝子機能は、非相同的末端接合を介して、塩基挿入、塩基欠失及び/又はフレームシフト突然変異を導入することによって、メガヌクレアーゼによって直接破壊される。
【0169】
多くの例において、病態の原因である標的細胞又は細胞集団に対し適切な遺伝子配列を方向づけることが望ましい。治療薬の、このような標的指向は、健康な細胞が、治療薬によって標的とされることを防ぐ。これは、治療の効力を増し、一方では、治療が、健康な細胞に及ぼす可能性のある、有害となる可能性のある作用を下げる。
【0170】
興味の細胞に対する、組み換えメガヌクレアーゼ遺伝子及び挿入すべき興味の配列の輸送は、様々の機構によって実現することが可能である。ある実施態様では、核酸は、ウィルスの再生を阻止するように不活性化された特定のウィルス遺伝子を持つウィルスを介して細胞に輸送される。したがって、ウィルスは、標的細胞内に輸送され、維持されることは可能であるが、標的細胞または組織の中で複製する能力は保持しないように変えられる。ベクターのように活動するウィルスゲノムを生産ように、この変更ウィルスゲノムに、一つ以上のDNA配列を挿入することが可能であるが、これらの配列は、宿主のゲノムの中に導入されてその後発現されてもよいし、導入し発現されなくともよい。より具体的には、ある実施態様は、レトロウィルス、例えば、ただしこれらに限定されるものではないが、MFG又はPLJベクターなどの採用を含む。MFGベクターは、単純化されたモロニーマウス白血病ウィルスベクター(MOMLV)であり、このベクターでは、複製欠損とするために、POL及びENVタンパク質をコードするDNA配列は欠失される。PLJレトロウィルスベクターも、MOMLVの一形である(例えば、KORMAN ET AL.(1987), PROC.NAT’L ACAD.SCI.,84:2150−2154)。別の実施態様では、組み換えアデノウィルス又はアデノ関連ウィルスを、輸送ベクターとして使用することが可能である。
【0171】
別の実施態様では、標的細胞に対する、組み換えメガヌクレアーゼタンパク質及び/又は組み換えメガヌクレアーゼ遺伝子配列の輸送は、リポソームの使用によって実現される。核酸及び/又はタンパク積載物を含むリポソームの生産は、従来技術で既知である(例えば、LASIC ET AL.(1995),SCIENCE 267:1275−76)。イムノリポソームは、リポソームの中に、細胞関連抗原に対する抗体を含み、特定の細胞タイプに対しメガヌクレアーゼのDNA配列、またはメガヌクレアーゼそのものを輸送することが可能である(例えば、LASIC ET AL.(1995), SCIENCE 267:1275−76;YOUNG ET AL.(2005),J.CALIF.DENT.ASSOC.33(12):967−71;PFEIFFER ET
AL.(2006),J.VASC.SURG.43(5):1021−7を参照)。リポソーム処方の生産および使用のための方法は、従来技術で周知である(例えば、米国特許第6,316,024号、米国特許第6,379,699号、米国特許第6,387,397号、米国特許第6,511,676号、および米国特許第6,593,308号、およびそれらの中に引用される参考文献を参照)。ある実施態様では、リポソームは、興味の配列の外、組み換えメガヌクレアーゼタンパク、または組み換えメガヌクレアーゼ遺伝子配列を輸送するために使用される。
【0172】
7. 病原体感染の治療法
本発明の局面はさらに、病原体による感染の治療法を提供する。病原性生物体としては、ウィルス、例えば、ただしこれらに限定されるものではないが、単純ヘルペスウィルス1、単純ヘルペスウィルス2、ヒト免疫不全ウィルス1、ヒト免疫不全ウィルス2、痘瘡ウィルス、ポリオウィルス、エプスタインバーウィルスもしくはヒトパピローマウィルスなど、及び細菌生物体、例えば、これらに限定されるものではないが、BACILLUS
ANTHRACIS、HAEMOPHILUS種、PNEUMOCOCCUS種、STAPHYLOCOCCUS AUREUS、STREPTOCOCCUS種、メシシリン耐性STAPHYLOCOCCUS AUREUS及びMYCOPLASMA TUBERCULOSISなどが挙げられる。病原生物体としてはさらに、真菌生物体、例えば、ただしこれらに限定されるものではないが、CANDIDA、BLASTOMYCES、CRYPTOCOCCUS、およびHISTOPLASMA種などが挙げられる。
【0173】
ある実施態様では、合理設計メガヌクレアーゼは、病原体ゲノム内の認識配列、例えば、該病原体の増殖、生殖又は毒性にとって必須である、遺伝子または調整要素を標的とすることが可能である。ある実施態様では、認識配列は、細菌プラミド内にあってもよい。病原体ゲノムの認識配列のメガヌクレアーゼ介在性切断は、非相同的末端接合を刺激することによって、挿入、欠失又はフレームシフトの形で、標的必須遺伝子内の突然変異を刺激することが可能である。それとは別に、細菌プラスミドの切断は、その上にコードされる任意の遺伝子、例えば、トキシン遺伝子(例えば、B.ANTHRACIS致死因子遺伝子)又は抗生物質耐性遺伝子などと共にプラスミドの消失を招く可能性がある。前述のように、メガヌクレアーゼは、従来技術で一般的な技術を用いて、タンパク質又は核酸の形で、感染患者、動物又は植物に輸送してよい。ある実施態様では、メガヌクレアーゼ遺伝子は、病原体細菌類に輸送するために、バクテリオファージゲノムの中に組み込んでもよい。
【0174】
本発明の局面はさらに、ある形の癌の治療のための治療薬を提供する。ヒトのウィルスは、多くの場合、腫瘍形成と関連するので(例えば、エプスタインバーウィルスと鼻咽頭癌;ヒトパピローマウィルスと子宮頚部癌)、これらのウィルス性病原体の不活性化は、癌の発達または進行を阻止することがある。それとは別に、合理設計メガヌクレアーゼによる、これらの腫瘍関連ウィルスのゲノムを標的とする、二本鎖切断点を用いて、DNA損傷反応経路を介してアポトーシスを誘発してもよい。このようにして、ウィルスゲノムを抱える腫瘍細胞において選択的にアポトーシスを誘発することが可能となる場合がある。
【0175】
8. 遺伝子型分析法および病原体の特定
本発明の局面はさらに、インビトロ分子生物学研究および開発のためのツールを提供する。核酸、例えば、プラスミド、PCR産物、BAC配列、YAC配列、ウィルス及び真核および前核生物体由来のゲノム配列などの核酸の単離、クローニング及び操作のために、部位特異的エンドヌクレアーゼ(例えば、制限酵素)を用いることは、従来技術で普通のことである(例えば、AUSUBEL ET AL.,「分子生物学における最近のプロトコール(“CURRENT PROTOCOLS IN MOLECULAR BIOLOGY,”)」、WILEY 1999を参照されたい)。したがって、ある実施態様では、インビトロにおける核酸の操作に合理設計メガヌクレアーゼを使用してもよい。例えば、同じDNA分子の中に一対の認識配列を認識する、合理設計メガヌクレアーゼは、その後の操作、例えば、細菌プラスミド、BAC又はYACへのコンタクトなどのための介在DNAセグメントを単離するのに使用することが可能である。
【0176】
別の局面では、本発明は、病原遺伝子及び生物体特定用のツールを提供する。一実施態様では、合理設計メガヌクレアーゼは、健康な対立遺伝子から病的対立遺伝子を区別するために、病気と相関する多型遺伝子領域において対応する認識部位を切断するために使用
することが可能である(例えば、ヒトのCFTR遺伝子のΔF−508対立遺伝子を認識する合理設計メガヌクレアーゼ、実施例4を参照)。この実施態様では、ヒトの患者、又は他の生物体から単離されたDNA配列が、合理設計メガヌクレアーゼによって、場合によっては、追加の部位特異的ヌクレアーゼと組み合わせて消化され、得られたDNA断片のパターンが、ゲル電気泳動、毛細管電気泳動、質量分析又はその他の従来技術で既知の方法によって分析される。この断片化パターン及び具体的には合理設計メガヌクレアーゼによる切断の有無は、ゲノム中に認識配列が存在するか否かを明らかにすることによって該生物体の遺伝子型を示す。別の実施態様では、合理設計メガヌクレアーゼは、病原性ウィルス、真菌、または細菌のゲノムにおける多型領域を標的とし、該生物体を特定するのに使用される。この実施態様では、合理設計メガヌクレアーゼは、病原体に固有の認識配列を切断するが(例えば、細菌における16Sおよび23SRRNA遺伝子の間のスペーサー領域;例えば、VAN DER GIESSEN ET AL.(1994),MICROBIOLOGY 140:1103−1108を参照)、これを用いて、ゲノムのエンドヌクレアーゼ消化及びそれに続く、電気泳動、質量分析、または、従来技術で既知の、その他の方法による断片化パターンの分析に基づいて、他の、近縁生物体と区別することが可能である。
【0177】
9. 特注DNA−結合ドメインの生産法
他の側面で、本発明は、エンドヌクレアーゼ切断活性を除去した合理設計DNA−結合タンパク質を提供する。合理設計メガヌクレアーゼの触媒活性の除去は、触媒に与るアミノ酸を突然変異させることによって実行することが可能である(例えば、I−CREIにおけるQ47のEへの突然変異、CHEVALIER ET AL.(2001),BIOCHEMISTRY,43:14015−14026を参照);I−SCEIにおけるD44またhD145のNへの突然変異;I−CREIにおけるE66のQへの突然変異;I−MSOIにおけるD22のNへの突然変異)。次に、この不活性化メガヌクレアーゼを別のタンパク質、例えば、ただしこれらに限定されるものではないが、転写アクチベーター(例えば、GAL4トランスアクチベーションドメイン、またはVP16トランスアクチベーションドメイン)、転写リプレッサー(例えば、KRUPPELタンパクのKRABドメイン)、DNAメチラーゼドメイン(例えば、M.CVIPI、またはM.SSSI)、またはヒストンアセチルトランスフェラーゼ・ドメイン(例えば、HDAC1、またはHDAC2)などのエフェクタードメインに融合させることが可能である。もっとも著明なのとして加工されたジンクフィンガードメインがある、加工されたDNA結合ドメイン及び、エフェクタードメインから成るキメラタンパクは、従来技術で既知である(例えば、PAPWORTH ET AL.(2006),GENE 366:27−38参照)。
【0178】
本発明は、下記の実施例によってさらに具体的に説明される。ただし、これらの実施例を限定的なものと考えてはならない。当業者であれば、たかだか通例の実験操作を用いるだけで、本出願に記載される特定物質および手順に対し数多くの等価物を認識し、確認することが可能であろう。そのような等価物は、下記の実施例に先行する特許請求項の範囲に含まれることが意図される。下記の実施例1−4は、I−CREIに基づく合理設計メガヌクレアーゼを特異的に言及するが、その外、I−SCEI、I−MSOI、I−CEUIに基づく合理設計メガヌクレアーゼ及びその他のLAGLIDADGメガヌクレアーゼも、本出願に記載するように、同様に生産し、使用することが可能である。
【実施例1】
【0179】
HIV−1 TAT遺伝子を認識するメガヌクレアーゼの合理設計
1. メガヌクレアーゼの設計
一対のメガヌクレアーゼが、HIV−1 TAT遺伝子に認められるDNA部位5’−GAAGAGCTCATCAGAACAGTCA−3’(配列番号15)を認識し、切断
するように設計された。表5に従って、二つのメガヌクレアーゼ、TAT1及びTAT2が、下記の塩基接触部(非WT接触部は太字で表す)を用い、それぞれ、半分部位5’−GAAGAGCTC−3’(配列番号16)、および5’−TGACTGTTC−3’(配列番号17)に結合するように設計された。
【0180】
TAT1:
【表14】
【0181】
TAT2:
【表15】
【0182】
二つの酵素はクローンされ、大腸菌において発現され、下記に記載するように、対応するDNA認識配列に対する酵素活性について定量された。いずれの場合も、合理設計メガヌクレアーゼは、不活性であることが認められた。次に、DNAバックボーンとの接触を向上させるためE80をQに突然変異させた、それぞれの第2世代を生産した。この第2世代TAT2酵素は、その意図される認識配列に対し活性を持つことが認められたが、一方、TAT1酵素は不活性のままであった。野生型I−CREIとの共晶構造の肉眼検査から、TAT1は、R40とK28の間の立体障害のために不活性であることが示唆された。この衝突を緩和するため、K28をより小さい側鎖を持つアミノ酸(A、S、T又はC)に突然変異させ、一方、Q80突然変異はそのまま維持させた、TAT1変種を生産した。これらの酵素をE.COLIにおいて生産し、定量したところ、S28及びT28を有するTAT1変種は、共に、所望の認識配列に対して活性を持つが、一方、位置−7において所望の塩基指向性を維持することが認められた。
【0183】
2. 組み換えメガヌクレアーゼの構築
再設計I−CREI酵素のための突然変異を、重複PCR戦略において突然変異誘発性プライマーを用いて導入した。一次PCRで生成されたI−CREIの組み換えDNA断片を、二次PCRにおいて接合し、全長の組み換え核酸を生産した。組み換えI−CREI構築体は全て、精製用遺伝子の3‘末端に、6個のヒスチジンタグを融合させたPET21Aベクター(NOVAGEN CORP.,SAN DIEGO、カリフォルニア州)にクローンした。核酸配列は全て、サンガーのジデオキシヌクレオチド配列決定法(SANGER ET AL.(1977),PROC.NATL.ACAD.SCI.USA.74(12):5463−7を参照)を用いて確認した。
【0184】
野生型I−CREI及び全ての加工メガヌクレアーゼは、下記の方法を用いて発現し、精製した。PET21Aベクターにクローンした構築体は、化学的にコンピテントとしたBL21(DE3)PLYSSにおいて形質転換され、200μG/MLのカルバニシリンを含む、2 X YTプレートに撒いた。一晩育成の後、形質転換細菌コロニーを、プ
レートから掻き落とし、それを用いて50 ML の2XYTブロスに接種した。細胞を、波長600NMにおいて光学的濁度が0.9に達するまで、振とうしながら37℃で育成した。次に、育成温度を37℃から22℃に下げた。タンパク質の発現は、1MM IPTGを添加することによって誘発し、細胞は、振とうしながら2時間半インキュベートした。次に、10分間6000 X Gの遠心によって細胞をペレットとした。ペレットを、1MLの結合バッファー(20MM TRIS−HCL,PH8.0,500MM NACL,10MMイミダゾール)において渦巻き攪拌によって再縣濁させた。次に、細胞を、50%電力における12パルスの超音波処理によって破壊し、細胞砕片を、15分間14,000 X Gの遠心によってペレットとした。細胞上清を、4MLの結合バッファーで希釈し、200μLのニッケル充填金属キレート性セファローズカラム(PHARMACIA)に負荷した。
【0185】
次いで、カラムを、4MLの洗浄バッファー(20MM TRIS HCL,PH8.0,500MM NACL,60MMイミダゾール)及び0.2MLの溶出バッファー(20MM TRIS−HCL,PH8.0,500MM NACL,400MMイミダゾール)で洗浄した。メガヌクレアーゼ酵素は、さらに0.6MLの溶出バッファーを添加することによって溶出し、VIVOSPINディスポーザブル濃縮器(ISC,INC.,KAYSVILLE,ユタ州)を用いて50−130μLとなるまで濃縮した。酵素は、ZEBAスピン脱塩カラム(PIERCE BIOTECHNOLOGY,INC.,ROCKFORD、イリノイ州)を用い、定量および保存のためにSAバッファー(25MM TRIS−HCL,PH8.0,100MM NACL,5MM MGCL2,5MM EDTA)と交換した。酵素濃度は、23,590M−1CM−1の吸光係数を用い280NMにおける吸収によって決めた。次に、酵素の純度および分子量は、MALDI−TOF質量分析によって確認した。
【0186】
ヘテロダイマー酵素は、二つのタンパクを独立に精製し、それらをインビトロで混合することによって又は大腸菌における二つのタンパクの縦列発現用人工オペロンを構築することによって生産した。前者の場合、精製メガヌクレアーゼを、溶液中で1:1で混合し、DNA基質の添加20分前に42℃で予備インキュベーションした。後者の場合、二つの遺伝子を、NDEI/ECORI及びECORI/HINDIIIを用いて順次PET−21A発現ベクターにクローンした。12塩基対の核酸スペーサー及びPET21ベクター由来のシャイン−デルガルノ配列が、人工オペロン中の第1及び第2遺伝子を隔てた。
【0187】
3. 切断アッセイ
前述のように精製した酵素は全て、メガヌクレアーゼの認識配列を含む、直線状の2本鎖DNA基質とインキュベーションすることによって活性について定量した。認識配列のセンス及びアンチセンスの両鎖に対応する合成オリゴヌクレオチドをアニールし、平滑末端コンタクトによって、PUC19プラスミドのSMAI部位にクローンした。このクローンされた結合部位の配列は、サンガーのジデオキシヌクレオチド配列決定法によって確認した。プラスミド基質は、全て、メガヌクレアーゼ消化と同時に、XMNI、SCAI、またはBPMIによって直線化された。この酵素消化物は、5μLの0.05μM DNA基質、2.5μLの5μM組み換えI−CREIメガヌクレアーゼ、9.5μLのSAバッファー及び0.5μLのXMNI、SCAI又はBPMIを含んでいた。消化物は、37℃で又はあるメガヌクレアーゼ酵素用として42℃で、4時間インキュベートした。消化は、0.3MG/MLのプロテイナーゼK及び0.5%SDSを加えることによって停止させ、37℃で1時間インキュベートした。消化物を、1.5%アガロースにおいて分析し、臭化エチジウム染色によって視像化した。
【0188】
メガヌクレアーゼ半分部位の指向性を評価するために、合理設計メガヌクレアーゼを、
意図する半分部位の完全パリンドロームに対応する、一組のDNA基質のほか、この半分部位における27通りの可能な単一塩基対置換の内の一つとインキュベートした。
【0189】
4. 認識配列−特異性
精製組み換えTAT1およびTAT2メガヌクレアーゼは、野生型メガヌクレアーゼ配列とは異なるDNAを認識した(図2(B))。野生型I−CREIメガヌクレアーゼは、WT認識配列を切断するが、TAT1のために意図された配列、TAT2のために意図された配列のいずれも切断しない。同様に、TAT1及びTAT2は、その意図された認識配列は切断するが、野生型配列は切断しない。次に、これらのメガヌクレアーゼを、半分部位指向性および全体特異性について評価した(図3)。野生型I−CREIは、その天然の半分部位における単一塩基対置換についてきわめて耐性が高いことが認められた。一方、TAT1及びTAT2は、きわめて特異的で、TAT1の場合は、位置−1、−2、−3、−6及び−8において、TAT2の場合は、位置−1、−2及び−6における塩基置換に対して完全に耐性を持たなかった。
【実施例2】
【0190】
変更DNA−結合親和度を持つメガヌクレアーゼの合理的設計
1. 親和度を上昇および活性を上昇させたメガヌクレアーゼ
メガヌクレアーゼCCR1およびBRP2を、それぞれ、半分部位5’−AACCCTCTC−3’(配列番号18)および5’−CTCCGGGTC−3’(配列番号19)を切断するように設計した。これらの酵素を、実施例1の場合と同様、表1に従って生産した。
【0191】
CCR1:
【表16】
【0192】
BRP2:
【表17】
【0193】
両酵素を大腸菌において発現させ、精製し、実施例1と同様にして定量した。両第1世代酵素は、天然の認識配列を持つ野生型I−CREIの速度よりも相当に低い速度で、その意図された認識配列を切断することが認められた。活性におけるこの低下を緩和するために、CCR1およびBRP2のDNA−結合親和度を、両酵素においてE80をQに突然変異させることによって上昇させた。これらの、CCR1及びBRP2の第2世代バージョンは、触媒速度をはっきりと上昇させて、その意図された認識配列を切断することが認められた。
【0194】
2. DNA−結合親和度を低下させ、活性を低下させたが、特異性を上昇させたメガヌ
クレアーゼ
野生型I−CREIは、その半分部位に対する置換に対して高い耐性を持つことが認められている(図3(A))。この酵素の特異性をさらに高めるための試みとして、通常、DNAバックボーンのリン酸塩と塩橋を造る、酵素の位置116のリシンを、アスパラギン酸に突然変異させてDNA−結合親和度を下げた。この合理設計酵素の、野生型認識配列に対する切断は、はっきりと低下していたが、この組み換え酵素の特異性は、野生型よりも相当に高かった。K116D変種の半分部位の指向性を、実施例1と同様に評価したところ、この酵素は、位置−1、−2及び−3における天然半分部位からの逸脱に対しては全く耐性を持たないことが認められ、半分部位の残りの6位置では、少なくとも部分的な塩基指向性を示した(図3(B))。
【実施例3】
【0195】
合理設計メガヌクレアーゼ・ヘテロダイマー
1. 溶液の中で形成されたメガヌクレアーゼ・ヘテロダイマーによる非パリンドロームDNA部位の切断
二つのメガヌクレアーゼLAM1及びLAM2が、それぞれ、半分部位5’−TGCGGTGTC−3’(配列番号20)及び5’−CAGGCTGTC−3’(配列番号21)を切断するように設計された。これら二つの酵素から成るヘテロダイマーは、バクテリオファージλ P05遺伝子に見られるDNA配列5’−TGCGGTGTCCGGCG
ACAGCCTG−3’(配列番号22)を認識することが予想された。
【0196】
LAM1:
【表18】
【0197】
LAM2:
【表19】
【0198】
LAM1及びLAM2は、実施例1に記載されるように、クローンし、大腸菌において発現し、個別に精製した。次に、二つの酵素を1:1で混合し、42℃で20分インキュ
ベートし、二つの酵素が、サブユニットを交換し、再度平衡に達するのを可能とした。LAM1ホモダイマー、LAM2ホモダイマー及びLAM1/LAM2ヘテロダイマーの混合物と予想される、得られた酵素液を、LAM1半分部位の完全パリンドローム、LAM2半分部位の完全パリンドローム及びバクテリオファージλゲノムに認められる非パリンドロームハイブリッド部位に対応する、3種の異なる認識配列と共にインキュベートした。精製LAM1酵素単独は、LAM1パリンドローム部位を切断するが、LAM2パリンドローム部位も、LAM1/LAM2ハイブリッド部位も切断しない。同様に、精製LAM2酵素単独は、LAM2パリンドローム部位を切断するが、LAM1部位も、LAM1/LAM2ハイブリッド部位も切断しない。しかしながら、LAM1及びLAM2の1:
1混合物は、3種全てのDNA部位を切断する。LAM1/LAM2ハイブリッド部位の切断は、二つの、異なる、再設計メガヌクレアーゼが溶液の中で混合されて、非パリンドローム的DNA部位を切断することが可能なヘテロダイマー酵素を形成することが可能であることを示す。
【0199】
2.共時発現によって形成されるメガヌクレアーゼ・ヘテロダイマーによる非パリンドロームDNA部位の切断
前述のLAM1及びLAM2酵素をコードする遺伝子は、実施例1において記載したように、大腸菌における同時発現のためにオペロンの中に配置された。この共時発現酵素は、実施例1と同様に精製され、かつ、酵素混合物は、前述の、三つの可能性のある認識配列とインキュベートされた。この共時発現酵素は、LAM1/LAM2ハイブリッド部位を含む三つ全ての部位を切断することが認められたが、これは、二つの、異なる合理設計メガヌクレアーゼが共時発現されて、非パリンドロームDNA部位を切断することが可能なヘテロダイマー酵素を形成することを示す。
【0200】
3.タンパク−タンパクインターフェイスを修飾されたメガヌクレアーゼ・ヘテロダイマーによる、非パリンドロームDNA部位の優先的切断
非パリンドロームDNA部位の切断を必要とする応用のために、異なる(パリンドローム的)DNA部位を認識し、切断するホモダイマーの形成を最小に留めながら、酵素ヘテロダイマーの形成を増進することが望ましい。このために、位置7、57及び96におけるリシンがグルタミン酸に変更されたLAM1酵素の変種が生産された。次に、この酵素を、位置8及び61のグルタミン酸をリシンに変えたLAM2の変種と、前述と同様に共時発現し、精製した。この場合、LAM1ホモダイマーの形成は、一方のモノマーにおけるE7、E57及びE96と、他方のモノマーにおけるE8及びE61の間における静電気的反発によって低下すると予想された。同様に、LAM2ホモダイマーの形成は、一方のモノマーにおけるK7、K57及びK96と、他方のモノマーにおけるK8及びK61の間における静電気的反発によって低下すると予想された。逆に、LAM1/LAM2ヘテロダイマーは、LAM1におけるE7、E57、およびE96、およびLAMにおけるK8及びK61の間の静電気的牽引によって好まれることが予想された。インターフェイスが修飾されたこの二つのメガヌクレアーゼが、前述のように、共時発現され、定量されると、LAM1/LAM2ハイブリッド部位が、二つのパリンドローム部位に対し優先的に切断されることが認められた。これは、メガヌクレアーゼのタンパク−タンパクインターフェイスにおける置換が、ヘテロダイマーの優先的形成の駆動が可能であることを示す。
【実施例4】
【0201】
生理的DNA配列を切断する、追加のメガヌクレアーゼ・ヘテロダイマー
1.遺伝子治療に関連するDNA配列を切断するメガヌクレアーゼ・ヘテロダイマー
軟骨無形成症を引き起こす突然変異である、ヒトのFGR3遺伝子における配列5’−CTGGGAGTCTCAGGACAGCCTG−3’(配列番号23)を切断する、合理設計メガヌクレアーゼ・ヘテロダイマー(ACH1/ACH2)を生産することが可能である。例えば、前述したように、I−CREIメガヌクレアーゼに基づいて、下記の接触残基および認識配列半分部位を持つメガヌクレアーゼが設計された。
【0202】
ACH1:
【表20】
【0203】
ACH2:
【表21】
【0204】
ヒトの成長ホルモン遺伝子のプロモーターにおいて配列5’−CCAGGTGTCTCTGGACTCCTCC−3’(配列番号24)を切断する合理設計メガヌクレアーゼ・ヘテロダイマー(HGH1/HGH2)を生産することが可能である。例えば、前述したように、I−CREIメガヌクレアーゼに基づいて、下記の接触残基および認識配列半分部位を持つメガヌクレアーゼが設計された。
【0205】
HGH1:
【表22】
【0206】
HGH2:
【表23】
【0207】
ヒトのCFTR遺伝子のΔF508対立遺伝子における5’−GAAAATATCATTGGTGTTTCCT−3’(配列番号25)を切断する合理設計メガヌクレアーゼ・ヘテロダイマー(CF1/CF2)を生産することが可能である。例えば、前述したように、I−CREIメガヌクレアーゼに基づいて、下記の接触残基および認識配列半分部位を持つメガヌクレアーゼが設計された。
【0208】
CF1:
【表24】
【0209】
CF2:
【表25】
【0210】
ヒトのCCR5遺伝子(HIVコレセプター)の配列5’−AACCCTCTCCAGTGAGATGCCT−3’(配列番号26)を切断する合理設計メガヌクレアーゼ・ヘテロダイマー(CCR1/CCR2)を生産することが可能である。例えば、前述したように、I−CREIメガヌクレアーゼに基づいて、下記の接触残基および認識配列半分部位を持つメガヌクレアーゼが設計された。
【0211】
CCR1:
【表26】
【0212】
CCR2:
【表27】
【0213】
ヒトのDMキナーゼ遺伝子の3´非翻訳領域における5’−GACCTCGTCCTCCGACTCGCTG−3’(配列番号27)を切断する合理設計メガヌクレアーゼ・ヘテロダイマー(MYD1/MYD2)を生産することが可能である。例えば、前述したように、I−CREIメガヌクレアーゼに基づいて、下記の接触残基および認識配列半分部位を持つメガヌクレアーゼが設計された。
【0214】
MYD1:
【表28】
【0215】
MYD1:
【表29】
【0216】
2.病原体ゲノムのDNA配列を切断するメガヌクレアーゼ・ヘテロダイマー
単純ヘルペスウィルス−1および単純ヘルペスウィルス−2のUL36遺伝子の配列5’−CTCGATGTCGGACGACACGGCA−3’(配列番号28)を切断する合理設計メガヌクレアーゼ・ヘテロダイマー(HSV1/HSV2)を生産することが可能である。例えば、前述したように、I−CREIメガヌクレアーゼに基づいて、下記の接触残基および認識配列半分部位を持つメガヌクレアーゼが設計された。
【0217】
HSV1:
【表30】
【0218】
HSV2:
【表31】
【0219】
BACILLUS ANTHRACISゲノムの配列5’−ACAAGTGTCTAT
GGACAGTTTA−3’(配列番号29)を切断する合理設計メガヌクレアーゼ・ヘテロダイマー(ANT1/ANT2)を生産することが可能である。例えば、前述したように、I−CREIメガヌクレアーゼに基づいて、下記の接触残基および認識配列半分部位を持つメガヌクレアーゼが設計された。
【0220】
ANT1:
【表32】
【0221】
ANT2:
【表33】
【0222】
痘瘡(天然痘)ウィルスGP009遺伝子の配列5’−AAAACTGTCAAATGACATCGCA−3’(配列番号30)を切断する合理設計メガヌクレアーゼ・ヘテロダイマー(POX1/POX2)を生産することが可能である。例えば、前述したように、I−CREIメガヌクレアーゼに基づいて、下記の接触残基および認識配列半分部位を持つメガヌクレアーゼが設計された。
【0223】
POX1:
【表34】
【0224】
POX2:
【表35】
【0225】
エプスタインバーウィルスBALF2遺伝子の配列5’−CGGGGTCTCGTGCGAGGCCTCC−3’(配列番号31)を切断する合理設計メガヌクレアーゼ・ヘテロダイマー(EBB1/EBB2)を生産することが可能である。例えば、前述したように、I−CREIメガヌクレアーゼに基づいて、下記の接触残基および認識配列半分部位を持つメガヌクレアーゼが設計された。
【0226】
EBB1:
【表36】
【0227】
EBB1:
【表37】
【0228】
3.植物ゲノムにおけるDNA配列を切断するメガヌクレアーゼ・ヘテロダイマー
ARABIDOPSISTHALIANNA GL2遺伝子の配列5’−CACTAA
CTCGTATGAGTCGGTG−3’(配列番号32)を切断する合理設計メガヌクレアーゼ・ヘテロダイマー(GLA1/GLA2)を生産することが可能である。例えば、前述したように、I−CREIメガヌクレアーゼに基づいて、下記の接触残基および認識配列半分部位を持つメガヌクレアーゼが設計された。
【0229】
GLA1:
【表38】
【0230】
GLA2:
【表39】
【0231】
ARABIDOPSIS THALIANNA BP1遺伝子の配列5’−TGCCTCCTCTAGAGACCCGGAG−3’(配列番号33)を切断する合理設計メガヌクレアーゼ・ヘテロダイマー(BRP1/BRP2)を生産することが可能である。例えば、前述したように、I−CREIメガヌクレアーゼに基づいて、下記の接触残基および認識配列半分部位を持つメガヌクレアーゼが設計された。
【0232】
BRP1:
【表40】
【0233】
BRP2:
【表41】
【0234】
NICOTIANA TABACUMマグネシウムケラターゼ遺伝子の配列5’−TA
AAATCTCTAAGGTCTGTGCA−3’(配列番号34)を切断する合理設計メガヌクレアーゼ・ヘテロダイマー(MGC1/MGC2)を生産することが可能である。例えば、前述したように、I−CREIメガヌクレアーゼに基づいて、下記の接触残基および認識配列半分部位を持つメガヌクレアーゼが設計された。
【0235】
MGC1:
【表42】
【0236】
MGC2:
【表43】
【0237】
NICOTIANA TABACUM CYP82E4遺伝子の配列5’−CAAGA
ATTCAAGCGAGCATTAA−3’(配列番号35)を切断する合理設計メガヌクレアーゼ・ヘテロダイマー(CYP/HGH2)を生産することが可能である。例えば、前述したように、I−CREIメガヌクレアーゼに基づいて、下記の接触残基および認識配列半分部位を持つメガヌクレアーゼが設計された。
【0238】
CYP:
【表44】
【0239】
HGH2:
【表45】
【0240】
4.酵母ゲノムにおけるDNA配列を切断するメガヌクレアーゼ・ヘテロダイマー
SACCHAROMYCES CEREVISIAE URA3遺伝子の配列5’−TTAGATGACAAGGGAGACGCAT−3’(配列番号36)を切断する合理設計メガヌクレアーゼ・ヘテロダイマー(URA1/URA2)を生産することが可能である。例えば、前述したように、I−CREIメガヌクレアーゼに基づいて、下記の接触残基および認識配列半分部位を持つメガヌクレアーゼが設計された。
【0241】
URA1:
【表46】
【0242】
URA2:
【表47】
【0243】
5.認識配列特異性
本実施例において上に概説した合理設計メガヌクレアーゼを、実施例1と同様にして、クローンし、大腸菌において発現し、精製した。次いで、各精製メガヌクレアーゼを、そのヘテロダイマー形成パートナー(例えば、ACH1をACH2と、HGH1をHGH2と、など)と1:1で混合し、各メガヌクレアーゼ・ヘテロダイマーのために意図される
非パリンドロームDNA認識配列を含む、直線化DNA基質とインキュベートした。図3に示すように、各合理設計メガヌクレアーゼ・ヘテロダイマーは、その意図されたDNA部位を切断する。
【図面の簡単な説明】
【0244】
【図1】図1(A)は、結晶学データに基づいて、I−CREIホモダイマー及びその天然の2本鎖認識配列の相互作用を示す。本模式図は、二つの卵形として示すホモダイマーによって結合される、図示目的のためだけに巻き戻された状態で示される認識配列(配列番号2及び配列番号3)を描いたものである。各DNA半分部位の塩基には、−1から−9までの番号がふられ、認識表面を形成する、I−CREIのアミノ酸残基は、1文字アミノ酸表示および残基位置を示す数字によって表される。実線:DNA塩基に対する水素結合。鎖線:酵素設計では新たな接触点を形成するが、野生型複合体ではDNAと接触しないアミノ酸位置。矢印:DNAバックボーンと相互作用を持ち、切断活性に影響を及ぼす残基。また、図1(B)は、図1(A)の右側の切断半分部位の−4位置におけるA−T塩基対間の野生型接触を示す。具体的に言うと、残基Q26が、A塩基と相互作用を持つところが示される。残基I77は、該塩基対に近接するが、特異的相互作用は持たない。更に、図1(C)は、残基I77が修飾されてE77に変更された、I−CREIメガヌクレアーゼの、合理設計変種との間の相互作用を示す。この変更の結果、−4位置ではG−C塩基対が好まれる。Q26とG塩基との間の相互作用は、図1(A)の左側の切断半分部位において結晶学的に観察されるように、水分子によって仲介される。更に又、図1(D)は、残基Q26が修飾されてE26に変更され、残基I77が修飾されてR77に変更された、I−CREIメガヌクレアーゼの合理設計変種との間の相互作用を示す。この変更の結果、−4位置ではG−C塩基対が好まれる。そして更に、図1(E)は、残基Q26が修飾されてA26に変更され、残基I77が修飾されてQ77に変更された、I−CREIメガヌクレアーゼの合理設計変種との間の相互作用を示す。この変更の結果、−4位置ではT−A塩基対が好まれる。
【図2】図2(A)は、野生型I−CREIメガヌクレアーゼ(WT)及び本発明の11種の合理設計メガヌクレアーゼ・ヘテロダイマーそれぞれの一認識配列の比較を示す。WT認識配列に対して保存される塩基には陰影を施す。9BPの半分部位は太字で示す。WT:野生型(配列番号4);CF:多くの嚢胞性線維症例の原因となる、ヒトCFTR遺伝子のΔF508対立遺伝子(配列番号25);MYD:筋緊張性ジストロフィーと関連するヒトDMキナーゼ遺伝子;CCR:ヒトCCR5遺伝子(大型HIVコレセプター)(配列番号26);ACH;軟骨無形成症と相関するヒトのFGFR3遺伝子(配列番号23);TAT:HIV−1TAT/REV遺伝子(配列番号15);HSV:HSV−1 UL36遺伝子(配列番号28);LAM:バクテリオファージλ P05遺伝子(配列番号22);POX:痘瘡(天然痘)ウィルスGP009遺伝子(配列番号30);URA:SACCHAROMYCES CERVISIAE URA3遺伝子(配列番号36);GLA:ARABIDOPSIS THALIANA GL2遺伝子(配列番号32);BRP:ARABIDOPSISTHALIANA BP−1遺伝子(配列番号33)。図2(B)は、野生型I−CREI(WT)及び11種の合理設計メガヌクレアーゼ・ヘテロダイマーのそれぞれと、12種全ての酵素の認識部位を担持するプラスミドと、37℃で6時間インキュベートした場合の結果を示す。各ボックスに切断パーセントが示される。
【図3】図3は、野生型および合理設計I−CREIホモダイマーの切断パターンを示す。(A)野生型I−CREI。(B)I−CREIK116D。(C−L)本発明の合理設計メガヌクレアーゼ。酵素は、27通りの対応する単一塩基対変動を持つ、意図される切断半分部位のパリンドロームを担持する、一組のプラスミドと共にインキュベートした。黒色バー:表1に基づいて予想される切断パターン。灰色バー:予想される切断パターンから変位するDNA部位。円は、意図される認識部位における塩基を示す。さらに、2時間における切断の時間経過も示される。CおよびLにおける白抜き円の時間経過プロットは、E80Q突然変異を欠くCCR1およびBRP2酵素による切断に対応する。切断部位は、図2(A)に記載されるヘテロダイマー酵素の、5’(左コラム)、および3’(右コラム)半分部位に対応する。
【特許請求の範囲】
【請求項1】
野生型I−CREIメガヌクレアーゼと比べ、少なくとも一つの認識配列半分部位に対する特異性が変更された組み換えメガヌクレアーゼであって、
配列番号1のI−CREIメガヌクレアーゼの残基2−153に対し少なくとも85%の配列類似性を有するポリペプチドを含み、かつ、
配列番号2、配列番号3、配列番号4及び配列番号5から成る群より選ばれるI−CREIメガヌクレアーゼ認識配列内の半分部位と、少なくとも1塩基対異なる認識配列半分部位に対して特異性を有し、
排外的修飾ではない表5の少なくとも一つの修飾を含む、組み換えメガヌクレアーゼ。
【請求項2】
野生型I−MSOIメガヌクレアーゼと比べ、少なくとも一つの認識配列半分部位に対する特異性が変更された組み換えメガヌクレアーゼであって、
配列番号6のI−MSOIメガヌクレアーゼの残基6−160に対し少なくとも85%の配列類似性を有するポリペプチドを含み、かつ、
配列番号7及び配列番号8から成る群より選ばれるI−MSOIメガヌクレアーゼ認識配列内の半分部位と、少なくとも1塩基対異なる認識配列半分部位に対して特異性を有し、
排外的修飾ではない表7の少なくとも一つの修飾を含む、組み換えメガヌクレアーゼ。
【請求項3】
野生型I−SCEIメガヌクレアーゼと比べ、認識配列に対する特異性が変更された組み換えメガヌクレアーゼであって、
配列番号9のI−SCEIメガヌクレアーゼの残基3−186に対し少なくとも85%の配列類似性を有するポリペプチドを含み、かつ、
配列番号10及び配列番号11から成る群より選ばれるI−SCEIメガヌクレアーゼ認識配列と、少なくとも1塩基対異なる認識配列に対して特異性を有し、
排外的修飾ではない表9の少なくとも一つの修飾を含む、組み換えメガヌクレアーゼ。
【請求項4】
野生型I−CEUIメガヌクレアーゼと比べ、少なくとも一つの認識配列半分部位に対する特異性が変更された組み換えメガヌクレアーゼであって、
配列番号12のI−CEUIメガヌクレアーゼの残基5−211に対し少なくとも85%の配列類似性を有するポリペプチドを含み、かつ、
配列番号13及び配列番号14から成る群より選ばれるI−CEUIメガヌクレアーゼ認識配列内の半分部位と、少なくとも1塩基対異なる認識配列半分部位に対して特異性を有し、
排外的修飾ではない表11の少なくとも一つの修飾を含む、組み換えメガヌクレアーゼ。
【請求項5】
野生型I−CREIメガヌクレアーゼと比べ、少なくとも一つの認識配列半分部位に対する特異性が変更された組み換えメガヌクレアーゼであって、
配列番号1のI−CREIメガヌクレアーゼの残基2−153に対し少なくとも85%の配列類似性を有するポリペプチドを含み、かつ、
配列番号2、配列番号3、配列番号4及び配列番号5から成る群より選ばれるI−CREIメガヌクレアーゼ認識配列内の半分部位と、少なくとも1塩基対異なる認識配列半分部位に対して特異性を有し、
(1)位置−1における特異性が、
(A)Q70、C70、L70、Y75、Q75、H75、H139、Q46及びH46から成る群より選ばれる修飾によって、センス鎖においてTに変更され;
(B)Y75、L75、C75、Y139、C46及びA46から成る群より選ばれる修飾によって、センス鎖においてAに変更され;
(C)K70、E70、E75、E46及びD46から成る群より選ばれる修飾によって、センス鎖においてGに変更され、
(D)H75、R75、H46、K46及びR46から成る群より選ばれる修飾によって、センス鎖においてCに変更され、又は、
(E)G70、A70、S70及びG46から成る群より選ばれる修飾によって、センス鎖において任意の塩基に変更され;
及び/又は
(2)位置−2における特異性が、
(A)Q70、T44、A44、V44、I44、L44及びN44から成る群より選ばれる修飾によって、センス鎖においてAに変更され;
(B)E70、D70、K44及びR44から成る群より選ばれる修飾によって、センス鎖においてCに変更され;
(C)H70、D44及びE44から成る群より選ばれる修飾によって、センス鎖においてGに変更され;又は、
(D)C44を含む修飾によって、センス鎖においてA又はTに変更され;
及び/又は、
(3)位置−3における特異性が、
(A)Q68及びC24から成る群より選ばれる修飾によって、センス鎖においてAに変更され;
(B)E68、F68、K24及びR24から成る群より選ばれる修飾によって、センス鎖においてCに変更され;
(C)M68、C68、L68及びF68から成る群より選ばれる修飾によって、センス鎖においてTに変更され;
(D)H68を含む修飾によってセンス鎖においてA又はCに変更され;
(E)Y68を含む修飾によってセンス鎖においてC又はTに変更され;
(F)K68を含む修飾によってセンス鎖においてG又はTに変更され;
及び/又は、
(4)位置−4における特異性が、
(A)E77及びK26から成る群より選ばれる修飾によって、センス鎖においてCに変更され;
(B)E26及びR77から成る群より選ばれる修飾によって、センス鎖においてGに変更され;
(C)S77を含む修飾によってセンス鎖においてC又はTに変更され;又は、
(D)S26を含む修飾によってセンス鎖において任意の塩基に変更され;
及び/又は、
(5)位置−5における特異性が、
(A)E42を含む修飾によってセンス鎖においてCに変更され;
(B)R42を含む修飾によってセンス鎖においてGに変更され;
(C)C28及びQ42から成る群より選ばれる修飾によって、センス鎖においてA又はCに変更され;又は、
(D)M66及びK66から成る群より選ばれる修飾によって、センス鎖において任意の塩基に変更され;
及び/又は、
(6)位置−6における特異性が、
(A)C40、I40、V40、C79、I79、V79及びQ28から成る群より選ばれる修飾によって、センス鎖においてTに変更され;
(B)E40及びR28から成る群より選ばれる修飾によって、センス鎖においてCに変更され;
(C)R40を含む修飾によってセンス鎖においてGに変更され;
及び/又は、
(7)位置−7における特異性が、
(A)E38、K30及びR30から成る群より選ばれる修飾によって、センス鎖においてCに変更され;
(B)K38、R38及びE30から成る群より選ばれる修飾によって、センス鎖においてGに変更され;
(C)I38及びL38から成る群より選ばれる修飾によって、センス鎖においてTに変更され;
(D)C38を含む修飾によってセンス鎖においてA又はGに変更され;
(E)H38、N38及びQ30から成る群より選ばれる修飾によって、センス鎖において任意の塩基に変更され;
及び/又は、
(8)位置−8における特異性が、
(A)L33、V33、I33、F33及びC33から成る群より選ばれる修飾によって、センス鎖においてTに変更され;
(B)E33及びD33から成る群より選ばれる修飾によって、センス鎖においてCに変更され;
(C)K33から成る修飾によってセンス鎖においてGに変更され;
(D)R32を含む修飾によってセンス鎖においてA又はCに変更され;
(E)R33を含む修飾によってセンス鎖においてA又はGに変更され;
及び/又は、
(9)位置−9における特異性が、
(A)E32を含む修飾によってセンス鎖においてCに変更され;
(B)R32及びK32から成る群より選ばれる修飾によって、センス鎖においてGに変更され;
(C)L32、V32、A32及びC32から成る群より選ばれる修飾によって、センス鎖においてTに変更され;
(D)D32及びI32から成る群より選ばれる修飾によって、センス鎖においてC又はTに変更され;又は、
(E)S32、N32、H32、Q32及びT32から成る群より選ばれる修飾によって、センス鎖において任意の塩基に変更される、
ことを特徴とする、組み換えメガヌクレアーゼ。
【請求項6】
野生型I−MSOIメガヌクレアーゼと比べ、少なくとも一つの認識配列半分部位に対する特異性が変更された組み換えメガヌクレアーゼであって、
配列番号6のI−MSOIメガヌクレアーゼの残基6−160に対し少なくとも85%の配列類似性を有するポリペプチドを含み、かつ、
配列番号7及び配列番号8から成る群より選ばれるI−MSOIメガヌクレアーゼ認識配列内の半分部位と、少なくとも1塩基対異なる認識配列半分部位に対して特異性を有し、
(1)位置−1における特異性が、
(A)K75、Q77、A49、C49及びK79から成る群より選ばれる修飾によって、センス鎖においてAに変更され;
(B)C77、L77及びQ79から成る群より選ばれる修飾によって、センス鎖においてTに変更され;
(C)K77、R77、E49及びE79から成る群より選ばれる修飾によって、センス鎖においてGに変更され;
及び/又は、
(2)位置−2における特異性が、
(A)Q75、K81、C47、I47及びL47から成る群より選ばれる修飾によって、センス鎖においてAに変更され;
(B)E75、D75、R47、K47、K81及びR81から成る群より選ばれる修飾によって、センス鎖においてCに変更され;
(C)K75、E47及びE81から成る群より選ばれる修飾によって、センス鎖においてGに変更され;
及び/又は、
(3)位置−3における特異性が、
(A)Q72、C26、L26、V26、A26及びI26から成る群より選ばれる修飾によって、センス鎖においてAに変更され;
(B)E72、Y72、H26、K26及びR26から成る群より選ばれる修飾によって、センス鎖においてCに変更され;
(C)K72、Y72及びH26から成る群より選ばれる修飾によって、センス鎖においてTに変更され;
及び/又は、
(4)位置−4における特異性が、
(A)K28、K83及びQ28から成る群より選ばれる修飾によって、センス鎖においてTに変更され;
(B)R83及びK83から成る群より選ばれる修飾によって、センス鎖においてGに変更され;
(C)K28及びQ83から成る群より選ばれる修飾によって、センス鎖においてAに変更され;
及び/又は、
(5)位置−5における特異性が、
(A)R45及びE28から成る群より選ばれる修飾によって、センス鎖においてGに変更され;
(B)Q28を含む修飾によってセンス鎖においてTに変更され;
(C)R28を含む修飾によってセンス鎖においてCに変更され;
及び/又は、
(6)位置−6における特異性が、
(A)K43、V85、L85及びQ30から成る群より選ばれる修飾によって、センス鎖においてTに変更され;
(B)E43、E85、K30及びR30から成る群より選ばれる修飾によって、センス鎖においてCに変更され;または、
(C)R43、K43、K85、R85、E30及びD30より成る群から選ばれる修飾によって、センス鎖においてGに変更され;
及び/又は、
(7)位置−7における特異性が、
(A)E32及びE41から成る群から選ばれる修飾によって、センス鎖においてCに変更され;
(B)R32、R41及びK41から成る群より選ばれる修飾によって、センス鎖においてGに変更され;
(C)K32、M41、L41及びI41から成る群より選ばれる修飾によって、センス鎖においてTに変更され;
及び/又は、
(8)位置−8における特異性が、
(A)K32及びK35から成る群より選ばれる修飾によって、センス鎖においてTに変更され;
(B)E32を含む修飾によってセンス鎖においてCに変更され;
(C)K32、K35及びR35から成る群より選ばれる修飾によって、センス鎖においてGに変更され;
及び/又は、
(9)位置−9における特異性が、
(A)N34及びH34から成る群より選ばれる修飾によって、センス鎖においてAに変更され;
(B)S34、C34、V34、T34及びA34から成る群より選ばれる修飾によって、センス鎖においてTに変更され;
(C)K34、R34及びH34から成る群より選ばれる修飾によって、センス鎖においてGに変更される、
ことを特徴とする、組み換えメガヌクレアーゼ。
【請求項7】
野生型I−SCEIメガヌクレアーゼと比べ、認識配列に対する特異性が変更された組み換えメガヌクレアーゼであって、
配列番号9のI−SCEIメガヌクレアーゼの残基3−186に対し少なくとも85%の配列類似性を有するポリペプチドを含み、かつ、
配列番号10及び配列番号11のI−SCEIメガヌクレアーゼと、少なくとも1塩基対異なる認識配列に対して特異性を有し、
(1)位置4における特異性が、
(A)K50を含む修飾によってセンス鎖においてAに変更され;
(B)K57、M57及びQ50から成る群より選ばれる修飾によって、センス鎖においてTに変更され;
(C)E50、R57及びK57から成る群より選ばれる修飾によって、センス鎖においてGに変更され;及び/又は、
(2)位置5における特異性が、
(A)K48、Q102から成る群より選ばれる修飾によって、センス鎖においてAに変更され;
(B)E48、K102及びR102から成る群より選ばれる修飾によって、センス鎖においてGに変更され;
(C)Q48、C102、L102及びV102から成る群より選ばれる修飾によって、センス鎖においてTに変更され;
及び/又は、
(3)位置6における特異性が、
(A)K59を含む修飾によってセンス鎖においてAに変更され;
(B)R59及びK59から成る群より選ばれる修飾によって、センス鎖においてCに変更され;
(B)K84及びE59から成る群より選ばれる修飾によって、センス鎖においてGに変更され;
及び/又は、
(4)位置7における特異性が、
(A)R46、K46及びE86から成る群より選ばれる修飾によって、センス鎖においてCに変更され;
(B)K86、R86及びE46から成る群より選ばれる修飾によって、センス鎖においてGに変更され;
(C)C46、L46及びV46から成る群より選ばれる修飾によって、センス鎖においてAに変更され;
及び/又は、
(5)位置8における特異性が、
(A)E88、R61及びH61から成る群より選ばれる修飾によって、センス鎖においてCに変更され;
(B)K88、Q61及びH61から成る群より選ばれる修飾によって、センス鎖においてTに変更され;
(C)K61、S61、V61、A61及びL61から成る群より選ばれる修飾によ
って、センス鎖においてAに変更され;
及び/又は、
(6)位置9における特異性が、
(A)C98、V98及びL98から成る群より選ばれる修飾によって、センス鎖においてAに変更され;
(B)R98及びK98から成る群より選ばれる修飾によって、センス鎖においてCに変更され;又は、
(C)E98及びD98から成る群より選ばれる修飾によって、センス鎖においてGに変更され;
及び/又は、
(7)位置10における特異性が、
(A)K96及びR96から成る群より選ばれる修飾によって、センス鎖においてCに変更され;
(B)D96及びE96から成る群より選ばれる修飾によって、センス鎖においてGに変更され;
(C)C96及びA96から成る群より選ばれる修飾によって、センス鎖においてAに変更され;
及び/又は、
(8)位置11における特異性が、
(A)Q90を含む修飾によってセンス鎖においてTに変更され;
(B)K90及びR90から成る群より選ばれる修飾によって、センス鎖においてCに変更され;
(C)E90を含む修飾によってセンス鎖においてGに変更され;
及び/又は、
(9)位置12における特異性が、
(A)Q193を含む修飾によってセンス鎖においてAに変更され;
(B)E165、E193及びD193から成る群より選ばれる修飾によって、センス鎖においてCに変更され;
(C)K165及びR165から成る群より選ばれる修飾によって、センス鎖においてGに変更され;
及び/又は、
(10)位置13における特異性が、
(A)Q193、C163及びL163から成る群より選ばれる修飾によって、センス鎖においてTに変更され;
(B)E193、D193、K163及びR192から成る群より選ばれる修飾によって、センス鎖においてGに変更され;
(C)C193およびL193から成る群より選ばれる修飾によって、センス鎖においてAに変更され;
及び/又は、
(11)位置14における特異性が、
(A)K161及びQ192から成る群より選ばれる修飾によって、センス鎖においてTに変更され;
(B)L192及びC192から成る群より選ばれる修飾によって、センス鎖においてAに変更され;
(C)K147、K161、R161、R197、D192及びE192から成る群より選ばれる修飾によって、センス鎖においてGに変更され;または、
(D)K161及びQ192から成る群から選ばれる修飾によって、センス鎖においてTに変更され;
及び/又は
(12)位置15における特異性が、
(A)C151、L151及びK151から成る群より選ばれる修飾によって、センス鎖においてTに変更され;
(B)K151を含む修飾によってセンス鎖においてGに変更され;又は、
(C)E151を含む修飾によってセンス鎖においてCに変更され;
及び/又は、
(13)位置17における特異性が、
(A)G152及びQ150から成る群より選ばれる修飾によって、センス鎖においてTに変更され;
(B)K152及びK150から成る群より選ばれる修飾によって、センス鎖においてCに変更され;
(C)N152、S152、D152、D150及びE150から成る群より選ばれる修飾によってセンス鎖においてGに変更され;
及び/又は、
(14)位置18における特異性が、
(A)H155及びY155から成る群より選ばれる修飾によってセンス鎖においてTに変更され;
(B)R155及びK155から成る群より選ばれる修飾によって、センス鎖においてCに変更され;
(C)K155及びC155から成る群より選ばれる修飾によって、センス鎖においてAに変更される
ことを特徴とする、組み換えメガヌクレアーゼ。
【請求項8】
野生型I−CEUIメガヌクレアーゼと比べ、少なくとも一つの認識配列半分部位に対する特異性が変更された組み換えメガヌクレアーゼであって、
配列番号12のI−CEUIメガヌクレアーゼの残基5−211に対し少なくとも85%の配列類似性を有するポリペプチドを含み、かつ、
配列番号13及び配列番号14から成る群より選ばれるI−CEUIメガヌクレアーゼ認識配列内の半分部位と、少なくとも1塩基対異なる認識配列半分部位に対して特異性を有し、
(1)位置−1における特異性が、
(A)C92、A92及びV92から成る群より選ばれる修飾によって、センス鎖においてAに変更され;
(B)Q116及びQ92から成る群より選ばれる修飾によって、センス鎖においてTに変更され;
(C)E116及びE92から成る群より選ばれる修飾によって、センス鎖においてGに変更され;
及び/又は、
(2)位置−2における特異性が、
(A)Q117、C90、L90及びV90から成る群より選ばれる修飾によって、センス鎖においてAに変更され;
(B)K117、R124、K124、E124、E90及びD90から成る群から選ばれる修飾によって、センス鎖においてGに変更され;
(C)E117、D117、R174、K124、K90及びK68から成る群から選ばれる修飾によって、センス鎖においてCに変更され;
及び/又は、
(3)位置−3における特異性が、
(A)C70、V70、T70、L70及びK70から成る群より選ばれる修飾によって、センス鎖においてAに変更され;
(B)Q70を含む修飾によってセンス鎖においてTに変更され;
(B)K70を含む修飾によってセンス鎖においてCに変更され;
及び/又は、
(4)位置−4における特異性が、
(A)E126、D126、R88、K88及びK72から成る群より選ばれる修飾によって、センス鎖においてCに変更され;
(B)K126、L126及びQ88から成る群より選ばれる修飾によって、センス鎖においてTに変更され;
(C)Q126、N126、K88、L88、C72、L72及びV72から成る群より選ばれる修飾によって、センス鎖においてAに変更され;
及び/又は、
(5)位置−5における特異性が、
(A)E74、K128、R128及びE128から成る群より選ばれる修飾によって、センス鎖においてGに変更され;
(B)C128、L128、V128及びT128から成る群から選ばれる修飾によって、センス鎖においてTに変更され;又は、
(C)C74、L74、V74及びT74から成る群より選ばれる修飾によって、センス鎖においてAに変更され;
及び/又は、
(6)位置−6における特異性が、
(A)K86、C86及びL86から成る群より選ばれる修飾によって、センス鎖においてTに変更され;
(B)D86、E86、R84及びK84から成る群より選ばれる修飾によって、センス鎖においてCに変更され;又は、
(C)K128、R128、R86、K86及びE84から成る群より選ばれる修飾によって、センス鎖においてGに変更され;
及び/又は、
(7)位置−7における特異性が、
(A)R76、K76及びH76から成る群より選ばれる修飾によって、センス鎖においてCに変更され;
(B)E76及びR84から成る群より選ばれる修飾によって、センス鎖においてGに変更され;
(C)H76及びQ76から成る群より選ばれる修飾によって、センス鎖においてTに変更され;
及びび/又は、
(8)位置−8における特異性が、
(A)Y79、R79及びQ76から成る群より選ばれる修飾によって、センス鎖においてAに変更され;
(B)D79、E79、D76及びE76から成る群より選ばれる修飾によって、センス鎖においてCに変更され;
(C)R79、K79、K76及びR76から成る群より選ばれる修飾によって、センス鎖においてGに変更され;
及び/又は、
(9)位置−9における特異性が、
(A)K78、V78、L78、C78及びT78から成る群より選ばれる修飾によって、センス鎖においてTに変更され;
(B)D78及びE78から成る群より選ばれる修飾によって、センス鎖においてCに変更され;又は、
(C)R78、K78及びH78から成る群より選ばれる修飾によって、センス鎖においてGに変更される、
ことを特徴とする、組み換えメガヌクレアーゼ。
【請求項9】
野生型I−CREIメガヌクレアーゼと比べ、2本鎖DNAに対する結合親和度が変更された組み換えメガヌクレアーゼであって、
配列番号1のI−CREIメガヌクレアーゼの残基2−153に対して少なくとも85%の配列類似性を有するポリペプチドを含み;
DNA結合親和度が、
(A)H、N、Q、S、T、K又はRによるE80、D137、I81、L112、P29、V64又はY66の置換;
又は、
(B)K又はRによるT46、T140又はT143の置換、
から成る群より選ばれる一つの置換に対応する少なくとも一つの修飾によって上昇することを特徴とする、組み換えメガヌクレアーゼ。
【請求項10】
野生型I−CREIメガヌクレアーゼと比べ、2本鎖DNAに対する結合親和度が変更された組み換えメガヌクレアーゼであって、
配列番号1のI−CREIメガヌクレアーゼの残基2−153に対して少なくとも85%の配列類似性を有するポリペプチドを含み;
DNA結合親和度が、
(A)H、N、Q、S、T、D又はEによるK34、K48、R51、K82、K116、またはK139の置換;
又は、
(B)D又はEによるI81、L112、P29、V64、Y66、T46、T140又はT143の置換から成る群より選ばれる一つの置換に対応する少なくとも一つの修飾によって低下することを特徴とする、組み換えメガヌクレアーゼ。
【請求項11】
野生型I−MSOIメガヌクレアーゼと比べ、2本鎖DNAに対する結合親和度が変更された組み換えメガヌクレアーゼであって、
配列番号6のI−MSOIメガヌクレアーゼの残基6−160に対して少なくとも85%の配列類似性を有するポリペプチドを含み;
DNA結合親和度が、
(A)H、N、Q、S、T、K又はRによるE147、I85、G86又はY118の置換;
又は、
(B)K又はRによるQ41、N70、S87、T88、H89、Q122、Q139、S150又はN152の置換から成る群より選ばれる一つの置換に対応する少なくとも一つの修飾によって上昇することを特徴とする、組み換えメガヌクレアーゼ。
【請求項12】
野生型I−MSOIメガヌクレアーゼと比べ、2本鎖DNAに対する結合親和度が変更された組み換えメガヌクレアーゼであって、
配列番号6のI−MSOIメガヌクレアーゼの残基6−160に対して少なくとも85%の配列類似性を有するポリペプチドを含み;
DNA結合親和度が、
(A)H、N、Q、S、T、D又はEによるK36、R51、K123、K143、またはR144の置換;
又は
(B)D又はEによるI85、G86、Y118、Q41、N70、S87、T88、H89、Q122、Q139、S150又はN152の置換から成る群より選ばれる一つの置換に対応する少なくとも一つの修飾によって低下することを特徴とする、組み換えメガヌクレアーゼ。
【請求項13】
野生型I−SCEIメガヌクレアーゼと比べ、2本鎖DNAに対する結合親和度が変更
された組み換えメガヌクレアーゼであって、
配列番号9のI−SCEIメガヌクレアーゼの残基3−186に対して少なくとも85%の配列類似性を有するポリペプチドを含み;
DNA結合親和度が、
(A)H、N、Q、S、T、K又はRによるD201、L19、L80、L92、Y151、Y188、I191、Y199又はY222の置換;
又は
(B)K又はRによるN15、N17、S81、H84、N94、N120、T156、N157、S159、N163、Q165、S166, N194又はS202の置換から成る群より選ばれる一つの置換に対応する少なくとも一つの修飾によって上昇することを特徴とする、組み換えメガヌクレアーゼ。
【請求項14】
野生型I−SCEIメガヌクレアーゼと比べ、2本鎖DNAに対する結合親和度が変更された組み換えメガヌクレアーゼであって、
配列番号9のI−SCEIメガヌクレアーゼの残基3−186に対して少なくとも85%の配列類似性を有するポリペプチドを含み;
DNA結合親和度が、
(A)H、N、Q、S、T、D又はEによるK20、K23、K63、K122、K148、K153、K190、K193、K195又はK223の置換;
又は
(B)D又はEによるL19、L80、L92、Y151、Y188、I191、Y199、Y222、N15、N17、S81、H84、N94、N120、T156、N157、S159、N163、Q165、S166、N194又はS202の置換から成る群から選ばれる一つの置換に対応する少なくとも一つの修飾によって低下することを特徴とする、組み換えメガヌクレアーゼ。
【請求項15】
野生型I−CEUIメガヌクレアーゼと比べ、2本鎖DNAに対する結合親和度が変更された組み換えメガヌクレアーゼであって、
配列番号12のI−CEUIメガヌクレアーゼの残基5−211に対して少なくとも85%の配列類似性を有するポリペプチドを含み;
DNA結合親和度が、
(A)H、N、Q、S、T、K又はRによるD25又はD128の置換又は
(B)K又はRによるS68、N70、H94、S117、N120、N129、又はH172の置換から成る群より選ばれる一つの置換に対応する少なくとも一つの修飾によって上昇することを特徴とする、組み換えメガヌクレアーゼ。
【請求項16】
野生型I−CEUIメガヌクレアーゼと比べ、2本鎖DNAに対する結合親和度が変更された組み換えメガヌクレアーゼであって、
配列番号12のI−CEUIメガヌクレアーゼの残基5−211に対して少なくとも85%の配列類似性を有するポリペプチドを含み;
DNA結合親和度が、
(A)H、N、Q、S、T、D又はEによるK21、K28、K31、R112、R114、またはR130の置換;
又は
(B)D又はEによるS68、N70、H94、S117、N120、N129又はH172の置換から成る群より選ばれる一つの置換に対応する少なくとも一つの修飾によって低下することを特徴とする、組み換えメガヌクレアーゼ。
【請求項17】
参照メガヌクレアーゼモノマーによるダイマー形成に対する親和度を変更させた組み換えメガヌクレアーゼモノマーであって、
配列番号1のI−CREIメガヌクレアーゼの残基2−153に対して少なくとも85%の配列類似性を有するポリペプチドを含み、
ダイマー形成に対する親和度が、
(A)D又はEによるK7、K57又はK96の置換;
又は
(B)K又はRによるE8又はE61の置換から成る群より選ばれる置換に対応する少なくとも一つの修飾によって変えられることを特徴とする組み換えメガヌクレアーゼモノマー。
【請求項18】
組み換えメガヌクレアーゼ・ヘテロダイマーであって、
配列番号1のI−CREIメガヌクレアーゼの残基2−153に対して少なくとも85%の配列類似性を有する第1ポリペプチドであって、
ダイマー形成に対する親和度が、
(A)D又はEによるK7、K57又はK96の置換から成る群から選ばれる置換に対応する少なくとも一つの修飾によって変えられる第1ポリペプチド及び、
配列番号1のI−CREIメガヌクレアーゼの残基2−153に対して少なくとも85%の配列類似性を有する第2ポリペプチドであって、
ダイマー形成に対する親和度が、
(B)K又はRによるE8又はE61の置換から成る群から選ばれる置換に対応する少なくとも一つの修飾によって変えられる第2ポリペプチドを含む、
ことを特徴とする、組み換えメガヌクレアーゼ・ヘテロダイマー。
【請求項19】
参照メガヌクレアーゼモノマーによるダイマー形成に対する親和度を変更させた組み換えメガヌクレアーゼモノマーであって、
配列番号6のI−MSOIメガヌクレアーゼの残基6−160に対して少なくとも85%の配列類似性を有するポリペプチドを含み、
ダイマー形成に対する親和度が、
(A)D又はEによるR302の置換;
又は
(B)K又はRによるD20、E11又はQ64の置換から成る群から選ばれる置換に対応する少なくとも一つの修飾によって変えられることを特徴とする、組み換えメガヌクレアーゼモノマー。
【請求項20】
組み換えメガヌクレアーゼ・ヘテロダイマーであって、
配列番号6のI−MSOIメガヌクレアーゼの残基6−160に対して少なくとも85%の配列類似性を有する第1ポリペプチドであって、
ダイマー形成に対する親和度が、
(A)D又はEによるR302の置換から成る群より選ばれる置換に対応する少なくとも一つの修飾によって変えられる第1ポリペプチド及び、
配列番号6のI−MSOIメガヌクレアーゼの残基6−160に対して少なくとも85%の配列類似性を有する第2ポリペプチドであって、
ダイマー形成に対する親和度が、
(B)K又はRによるD20、E11又はQ64の置換から成る群から選ばれる置換に対応する少なくとも一つの修飾によって変えられる第2ポリペプチドを含む、
ことを特徴とする、組み換えメガヌクレアーゼ・ヘテロダイマー。
【請求項21】
参照メガヌクレアーゼモノマーによるダイマー形成に対する親和度を変更させた組み換えメガヌクレアーゼモノマーであって、
配列番号12のI−CEUIメガヌクレアーゼの残基5−211に対して少なくとも85%の配列類似性を有するポリペプチドを含み、
ダイマー形成に対する親和度が、
(A)D又はEによるR93の置換;
又は
(B)K又はRによるE152の置換から成る群から選ばれる置換に対応する少なく共一つの修飾によって変えられることを特徴とする、前記組み換えメガヌクレアーゼモノマー。
【請求項22】
組み換えメガヌクレアーゼ・ヘテロダイマーであって、
配列番号12のI−CEUIメガヌクレアーゼの残基5−211に対して少なくとも85%の配列類似性を有する第1ポリペプチドであって、
ダイマー形成に対する親和度が、
(A)D又はEによるR93の置換から成る群より選ばれる置換に対応する少なくとも一つの修飾によって変えられる第1ポリペプチド及び、
配列番号12のI−CEUIメガヌクレアーゼの残基5−211に対して少なくとも85%の配列類似性を有する第2ポリペプチドであって、
ダイマー形成に対する親和度が、
(B)K又はRによるE152の置換から成る群より選ばれる置換に対応する少なくとも一つの修飾によって変えられる第2ポリペプチドを含む、
ことを特徴とする組み換えメガヌクレアーゼ・ヘテロダイマー。
【請求項23】
表1から選ばれる少なくとも一つの修飾で、排外的修飾ではない修飾をさらに含むことを特徴とする、
請求項17又は請求項18に記載の組み換えメガヌクレアーゼモノマー又はヘテロダイマー。
【請求項24】
表2から選ばれる少なくとも一つの修飾で、排外的修飾ではない修飾をさらに含むことを特徴とする、
請求項19又は請求項20に記載の組み換えメガヌクレアーゼモノマー又はヘテロダイマー。
【請求項25】
表4から選ばれる少なくとも一つの修飾で、排外的修飾ではない修飾をさらに含むことを特徴とする、
請求項21又は請求項22に記載の組み換えメガヌクレアーゼモノマー又はヘテロダイマー。
【請求項26】
真核細胞の染色体に挿入される興味の外因性配列を含む遺伝学的に修飾された真核細胞を生産するための方法であって、
真核細胞を、
(I)メガヌクレアーゼをコードする第1核酸配列及び、
(II)前記興味の配列を含む第2核酸配列
を含む一つ以上の核酸によってトランスフェクトすることを含み、
前記メガヌクレアーゼが前記染色体の中に切断部位を生産し、かつ、前記興味の配列が前記切断部位において前記染色体の中に挿入され、
前記メガヌクレアーゼが請求項1〜25のいずれか1項による組み換えメガヌクレアーゼであることを特徴とする方法。
【請求項27】
前記第2核酸が、前記切断部位に側接する配列に対し相同な配列をさらに含み、前記興味の配列が、相同組み換えによって前記切断部位において挿入されることを特徴とする、
請求項26に記載の方法。
【請求項28】
前記第2核酸が、前記切断部位に対し実質的相同性を欠き、前記興味の配列が、非相同的末端接合によって前記染色体に挿入されることを特徴とする、
請求項26に記載の方法。
【請求項29】
真核細胞の染色体に挿入される興味の外因性配列を含む、遺伝学的に修飾された真核細胞を生産するための方法であって、
真核細胞にメガヌクレアーゼタンパクを導入すること;
及び、
前記興味の配列を含む核酸によって前記真核細胞をトランスフェクトすることを含み、
前記メガヌクレアーゼが、前記染色体の中に切断部位を生産し、前記興味の配列が、前記切断部位において前記染色体の中に挿入され;
かつ、
前記メガヌクレアーゼが請求項1〜25のいずれか1項による組み換えメガヌクレアーゼであることを特徴とする方法。
【請求項30】
前記核酸が、前記切断部位に側接する配列に対し相同な配列をさらに含み、前記興味の配列が、相同組み換えによって前記切断部位において挿入されることを特徴とする、
請求項29に記載の方法。
【請求項31】
前記核酸が、前記切断部位に対し実質的相同性を欠き、前記興味の配列が、非相同的末端接合によって前記染色体に挿入されることを特徴とする、
請求項29に記載の方法。
【請求項32】
真核細胞の染色体中の標的配列を破壊することによって遺伝学的に修飾された真核細胞を生産するための方法であって、
メガヌクレアーゼをコードする核酸によって真核細胞をトランスフェクトすることを含み、
前記メガヌクレアーゼが、前記染色体の中に切断部位を生産し、前記標的配列が、前記切断部位において非相同的末端接合によって破壊され;
かつ、
前記メガヌクレアーゼが、請求項1〜25のいずれか1項による組み換えメガヌクレアーゼであることを特徴とする方法。
【請求項33】
遺伝学的に修飾された生物体を生産する方法であって、
請求項26〜32のいずれか1項の方法に従って遺伝学的に修飾された真核細胞を生産するステップと、
及び、
前記遺伝学的に修飾された真核細胞を育成して遺伝学的に修飾された生物体を生産するステップと、
を含む方法。
【請求項34】
前記真核細胞が、配偶子、接合子、胚盤胞細胞、胚幹細胞及びプロトプラスト細胞から成る群より選ばれることを特徴とする、
請求項33に記載の方法。
【請求項35】
真核生物において遺伝子治療によって疾患を治療するための方法であって、
前記真核生物の少なくとも一つの細胞を、
(I)メガヌクレアーゼをコードする第1核酸配列及び、
(II)興味の配列を含む第2核酸配列
によってトランスフェクトすることを含み;
前記メガヌクレアーゼが、前記染色体の中に切断部位を生産し、前記興味の配列が、前記切断部位において前記染色体の中に挿入され;
かつ、
前記メガヌクレアーゼが、請求項1−25のいずれか1項による組み換えメガヌクレアーゼであることを特徴とする方法。
【請求項36】
前記第2核酸配列が、前記切断部位に側接する配列に対し相同な配列をさらに含み、前記興味の配列が、相同組み換えによって前記切断部位において挿入されることを特徴とする、
請求項35に記載の方法。
【請求項37】
前記第2核酸配列が、前記切断部位に対し実質的相同性を欠き、前記興味の配列が、非相同的末端接合によって前記染色体に挿入されることを特徴とする、
請求項35に記載の方法。
【請求項38】
真核生物において遺伝子治療によって疾患を治療するための方法であって、
前記真核生物の少なくとも一つの細胞の中にメガヌクレアーゼタンパクを導入することステップと、
前記真核細胞を、興味の配列を含む核酸によってトランスフェクトするステップとを含み、
前記メガヌクレアーゼが、前記染色体の中に切断部位を生産し、前記興味の配列が、前記切断部位において前記染色体の中に挿入され;
前記メガヌクレアーゼが請求項1〜25のいずれか1項による組み換えメガヌクレアーゼであり;
かつ、
前記興味の配列の挿入が、前記疾患に対する前記遺伝子治療を与えることを特徴とする方法。
【請求項39】
前記核酸が、前記切断部位に側接する配列に対し相同な配列をさらに含み、前記興味の配列が、相同組み換えによって前記切断部位において挿入されることを特徴とする、
請求項38に記載の方法。
【請求項40】
前記核酸が、前記切断部位に対し実質的相同性を欠き、前記興味の配列が、非相同的末端接合によって前記染色体に挿入されることを特徴とする、
請求項38に記載の方法。
【請求項41】
真核生物において、前記真核生物の染色体中の標的配列の破壊による遺伝子治療によって疾患を治療するための方法であって、
前記真核生物の少なくとも一つの細胞を、メガヌクレアーゼをコードする核酸によってトランスフェクトすることを含み;
前記メガヌクレアーゼが、前記染色体の中に切断部位を生産し、前記標的配列が、前記切断部位において非相同的末端接合によって破壊され;
前記メガヌクレアーゼが、請求項1−25のいずれか1項による組み換えメガヌクレアーゼであり;
かつ、
前記標的配列の破壊が、前記疾患に対する前記遺伝子治療を与えることを特徴とする方法。
【請求項42】
真核細胞宿主におけるウィルス性病原体感染を、前記ウィルス病原体のゲノムにおける標的配列を破壊することによって治療する方法であって、
前記真核細胞宿主の少なくとも一つの感染細胞を、メガヌクレアーゼをコードする核酸によってトランスフェクトすることを含み;
前記メガヌクレアーゼが、前記ウィルスゲノムの中に切断部位を生産し、前記標的配列が、前記切断部位において非相同的末端接合によって破壊され、
前記メガヌクレアーゼが、請求項1−25のいずれか1項による組み換えメガヌクレアーゼであり;
かつ、
前記標的配列の破壊が、前記感染に対する治療を与えることを特徴とする方法。
【請求項43】
真核細胞宿主におけるウィルス性病原体感染を、前記ウィルス病原体のゲノムにおける標的配列を破壊することによって治療する方法であって、
前記真核細胞宿主の少なくとも一つの感染細胞を、メガヌクレアーゼをコードする第1核酸、および第2核酸によってトランスフェクトすることを含み;
前記メガヌクレアーゼが、前記ウィルスゲノムの中に切断部位を生産し、前記標的配列が、前記切断部位において、前記ウィルスゲノムと前記第2核酸との相同組み換えによって破壊され;
前記メガヌクレアーゼが、請求項1−25のいずれか1項による組み換えメガヌクレアーゼであり;
前記第2核酸が、前記切断部位に側接する配列に対して相同な配列を含み;
かつ、
前記標的配列の破壊が、前記感染に対する治療を与えることを特徴とする方法。
【請求項44】
真核細胞宿主における前核細胞病原体感染を、前記前核細胞病原体のゲノムにおける標的配列を破壊することによって治療する方法であって、
前記真核細胞宿主に感染する前記前核細胞病原体の少なくとも一つの細胞を、メガヌクレアーゼをコードする核酸によってトランスフェクトすることを含み;
前記メガヌクレアーゼが、前記前核細胞ゲノムの中に切断部位を生産し、前記標的配列が、前記切断部位において、非相同的末端接合によって破壊され;
前記メガヌクレアーゼが、請求項1〜25のいずれか1項による組み換えメガヌクレアーゼであり;
かつ、
前記標的配列の破壊が、前記感染に対する治療を与えることを特徴とする方法。
【請求項45】
真核細胞宿主における前核細胞病原体感染を、前記前核細胞病原体のゲノムにおける標的配列を破壊することによって治療する方法であって、
前記真核細胞宿主に感染する前記前核細胞病原体の少なくとも一つの細胞を、メガヌクレアーゼをコードする第1核酸、および第2核酸によってトランスフェクトすることを含み;
前記メガヌクレアーゼが、前記前核細胞ゲノムの中に切断部位を生産し、前記標的配列が、前記切断部位において、前記前核細胞ゲノムと前記第2核酸との相同組み換えによって破壊され;
前記メガヌクレアーゼが、請求項1〜25のいずれか1項による組み換えメガヌクレアーゼであり;
前記第2核酸が、前記切断部位に側接する配列に対して相同な配列を含み;
かつ、
前記標的配列の破壊が、前記感染に対する治療を与えることを特徴とする方法。
【請求項46】
少なくとも一つの所望の変化を含むように、認識配列の少なくとも一つの塩基位置に対する特異性を変更させた組み換えメガヌクレアーゼを合理的に設計する方法であって、
(1)参照メガヌクレアーゼ−DNA複合体の三次元構造の少なくとも一部を決定する
こと;
(2)前記塩基位置において塩基接触面を形成するアミノ酸残基を特定すること;
(3)前記接触面を含む少なくとも第1残基のβ−炭素と、前記塩基位置における少なくとも第1塩基との間の距離を決めること;
及び、
(4)前記所望の変化を促進するアミノ酸置換を、表2に示される群の中から、
(A)第1塩基から6オングストローム未満の距離にある第1残基については、G群、C群、T群又はA群のうち、前記所望の変化を促進するのに適切なもののメンバーである1群及び/又は2群の置換を選択すること;
又は、
(B)前記第1塩基から6オングストロームより遠い距離にある第1残基については、G群、C群、T群又はA群のうち、前記所望の変化を促進するのに適切なもののメンバーである2群及び/又は3群の置換を選択することによって特定すること、
を含む方法。
【請求項47】
DNA−結合親和度を上昇させる組み換えメガヌクレアーゼを合理的に設計する方法であって、
(1)参照メガヌクレアーゼ−DNA複合体の三次元構造の少なくとも一部を決定すること;
(2)バックボーン接触面を形成するアミノ酸接触残基を特定すること;
(3)前記DNA−結合親和度を上昇させるアミノ酸置換を、
(A)陰性荷電、または疎水性側鎖を持つ接触残基については、非荷電/極性、または陽性荷電側鎖を持つ置換を選択すること;
又は
(B)非荷電/極性側鎖を持つ接触残基については、陽性荷電側鎖を持つ置換を選択することによって特定すること、
を含む方法。
【請求項48】
DNA−結合親和度を低下させる組み換えメガヌクレアーゼを合理的に設計する方法であって、
(1)参照メガヌクレアーゼ−DNA複合体の三次元構造の少なくとも一部を決定すること;
(2)バックボーン接触面を形成するアミノ酸接触残基を特定すること;
(3)前記DNA−結合親和度を低下させるアミノ酸置換を、
(A)陽性荷電側鎖を持つ接触残基については、非荷電/極性、または陰性荷電側鎖を持つ置換を選択すること;
又は
(B)疎水性、または非荷電/極性側鎖を持つ接触残基については、陰性荷電側鎖を持つ置換を選択することによって特定すること、
を含む方法。
【請求項1】
野生型I−CREIメガヌクレアーゼと比べ、少なくとも一つの認識配列半分部位に対する特異性が変更された組み換えメガヌクレアーゼであって、
配列番号1のI−CREIメガヌクレアーゼの残基2−153に対し少なくとも85%の配列類似性を有するポリペプチドを含み、かつ、
配列番号2、配列番号3、配列番号4及び配列番号5から成る群より選ばれるI−CREIメガヌクレアーゼ認識配列内の半分部位と、少なくとも1塩基対異なる認識配列半分部位に対して特異性を有し、
排外的修飾ではない表5の少なくとも一つの修飾を含む、組み換えメガヌクレアーゼ。
【請求項2】
野生型I−MSOIメガヌクレアーゼと比べ、少なくとも一つの認識配列半分部位に対する特異性が変更された組み換えメガヌクレアーゼであって、
配列番号6のI−MSOIメガヌクレアーゼの残基6−160に対し少なくとも85%の配列類似性を有するポリペプチドを含み、かつ、
配列番号7及び配列番号8から成る群より選ばれるI−MSOIメガヌクレアーゼ認識配列内の半分部位と、少なくとも1塩基対異なる認識配列半分部位に対して特異性を有し、
排外的修飾ではない表7の少なくとも一つの修飾を含む、組み換えメガヌクレアーゼ。
【請求項3】
野生型I−SCEIメガヌクレアーゼと比べ、認識配列に対する特異性が変更された組み換えメガヌクレアーゼであって、
配列番号9のI−SCEIメガヌクレアーゼの残基3−186に対し少なくとも85%の配列類似性を有するポリペプチドを含み、かつ、
配列番号10及び配列番号11から成る群より選ばれるI−SCEIメガヌクレアーゼ認識配列と、少なくとも1塩基対異なる認識配列に対して特異性を有し、
排外的修飾ではない表9の少なくとも一つの修飾を含む、組み換えメガヌクレアーゼ。
【請求項4】
野生型I−CEUIメガヌクレアーゼと比べ、少なくとも一つの認識配列半分部位に対する特異性が変更された組み換えメガヌクレアーゼであって、
配列番号12のI−CEUIメガヌクレアーゼの残基5−211に対し少なくとも85%の配列類似性を有するポリペプチドを含み、かつ、
配列番号13及び配列番号14から成る群より選ばれるI−CEUIメガヌクレアーゼ認識配列内の半分部位と、少なくとも1塩基対異なる認識配列半分部位に対して特異性を有し、
排外的修飾ではない表11の少なくとも一つの修飾を含む、組み換えメガヌクレアーゼ。
【請求項5】
野生型I−CREIメガヌクレアーゼと比べ、少なくとも一つの認識配列半分部位に対する特異性が変更された組み換えメガヌクレアーゼであって、
配列番号1のI−CREIメガヌクレアーゼの残基2−153に対し少なくとも85%の配列類似性を有するポリペプチドを含み、かつ、
配列番号2、配列番号3、配列番号4及び配列番号5から成る群より選ばれるI−CREIメガヌクレアーゼ認識配列内の半分部位と、少なくとも1塩基対異なる認識配列半分部位に対して特異性を有し、
(1)位置−1における特異性が、
(A)Q70、C70、L70、Y75、Q75、H75、H139、Q46及びH46から成る群より選ばれる修飾によって、センス鎖においてTに変更され;
(B)Y75、L75、C75、Y139、C46及びA46から成る群より選ばれる修飾によって、センス鎖においてAに変更され;
(C)K70、E70、E75、E46及びD46から成る群より選ばれる修飾によって、センス鎖においてGに変更され、
(D)H75、R75、H46、K46及びR46から成る群より選ばれる修飾によって、センス鎖においてCに変更され、又は、
(E)G70、A70、S70及びG46から成る群より選ばれる修飾によって、センス鎖において任意の塩基に変更され;
及び/又は
(2)位置−2における特異性が、
(A)Q70、T44、A44、V44、I44、L44及びN44から成る群より選ばれる修飾によって、センス鎖においてAに変更され;
(B)E70、D70、K44及びR44から成る群より選ばれる修飾によって、センス鎖においてCに変更され;
(C)H70、D44及びE44から成る群より選ばれる修飾によって、センス鎖においてGに変更され;又は、
(D)C44を含む修飾によって、センス鎖においてA又はTに変更され;
及び/又は、
(3)位置−3における特異性が、
(A)Q68及びC24から成る群より選ばれる修飾によって、センス鎖においてAに変更され;
(B)E68、F68、K24及びR24から成る群より選ばれる修飾によって、センス鎖においてCに変更され;
(C)M68、C68、L68及びF68から成る群より選ばれる修飾によって、センス鎖においてTに変更され;
(D)H68を含む修飾によってセンス鎖においてA又はCに変更され;
(E)Y68を含む修飾によってセンス鎖においてC又はTに変更され;
(F)K68を含む修飾によってセンス鎖においてG又はTに変更され;
及び/又は、
(4)位置−4における特異性が、
(A)E77及びK26から成る群より選ばれる修飾によって、センス鎖においてCに変更され;
(B)E26及びR77から成る群より選ばれる修飾によって、センス鎖においてGに変更され;
(C)S77を含む修飾によってセンス鎖においてC又はTに変更され;又は、
(D)S26を含む修飾によってセンス鎖において任意の塩基に変更され;
及び/又は、
(5)位置−5における特異性が、
(A)E42を含む修飾によってセンス鎖においてCに変更され;
(B)R42を含む修飾によってセンス鎖においてGに変更され;
(C)C28及びQ42から成る群より選ばれる修飾によって、センス鎖においてA又はCに変更され;又は、
(D)M66及びK66から成る群より選ばれる修飾によって、センス鎖において任意の塩基に変更され;
及び/又は、
(6)位置−6における特異性が、
(A)C40、I40、V40、C79、I79、V79及びQ28から成る群より選ばれる修飾によって、センス鎖においてTに変更され;
(B)E40及びR28から成る群より選ばれる修飾によって、センス鎖においてCに変更され;
(C)R40を含む修飾によってセンス鎖においてGに変更され;
及び/又は、
(7)位置−7における特異性が、
(A)E38、K30及びR30から成る群より選ばれる修飾によって、センス鎖においてCに変更され;
(B)K38、R38及びE30から成る群より選ばれる修飾によって、センス鎖においてGに変更され;
(C)I38及びL38から成る群より選ばれる修飾によって、センス鎖においてTに変更され;
(D)C38を含む修飾によってセンス鎖においてA又はGに変更され;
(E)H38、N38及びQ30から成る群より選ばれる修飾によって、センス鎖において任意の塩基に変更され;
及び/又は、
(8)位置−8における特異性が、
(A)L33、V33、I33、F33及びC33から成る群より選ばれる修飾によって、センス鎖においてTに変更され;
(B)E33及びD33から成る群より選ばれる修飾によって、センス鎖においてCに変更され;
(C)K33から成る修飾によってセンス鎖においてGに変更され;
(D)R32を含む修飾によってセンス鎖においてA又はCに変更され;
(E)R33を含む修飾によってセンス鎖においてA又はGに変更され;
及び/又は、
(9)位置−9における特異性が、
(A)E32を含む修飾によってセンス鎖においてCに変更され;
(B)R32及びK32から成る群より選ばれる修飾によって、センス鎖においてGに変更され;
(C)L32、V32、A32及びC32から成る群より選ばれる修飾によって、センス鎖においてTに変更され;
(D)D32及びI32から成る群より選ばれる修飾によって、センス鎖においてC又はTに変更され;又は、
(E)S32、N32、H32、Q32及びT32から成る群より選ばれる修飾によって、センス鎖において任意の塩基に変更される、
ことを特徴とする、組み換えメガヌクレアーゼ。
【請求項6】
野生型I−MSOIメガヌクレアーゼと比べ、少なくとも一つの認識配列半分部位に対する特異性が変更された組み換えメガヌクレアーゼであって、
配列番号6のI−MSOIメガヌクレアーゼの残基6−160に対し少なくとも85%の配列類似性を有するポリペプチドを含み、かつ、
配列番号7及び配列番号8から成る群より選ばれるI−MSOIメガヌクレアーゼ認識配列内の半分部位と、少なくとも1塩基対異なる認識配列半分部位に対して特異性を有し、
(1)位置−1における特異性が、
(A)K75、Q77、A49、C49及びK79から成る群より選ばれる修飾によって、センス鎖においてAに変更され;
(B)C77、L77及びQ79から成る群より選ばれる修飾によって、センス鎖においてTに変更され;
(C)K77、R77、E49及びE79から成る群より選ばれる修飾によって、センス鎖においてGに変更され;
及び/又は、
(2)位置−2における特異性が、
(A)Q75、K81、C47、I47及びL47から成る群より選ばれる修飾によって、センス鎖においてAに変更され;
(B)E75、D75、R47、K47、K81及びR81から成る群より選ばれる修飾によって、センス鎖においてCに変更され;
(C)K75、E47及びE81から成る群より選ばれる修飾によって、センス鎖においてGに変更され;
及び/又は、
(3)位置−3における特異性が、
(A)Q72、C26、L26、V26、A26及びI26から成る群より選ばれる修飾によって、センス鎖においてAに変更され;
(B)E72、Y72、H26、K26及びR26から成る群より選ばれる修飾によって、センス鎖においてCに変更され;
(C)K72、Y72及びH26から成る群より選ばれる修飾によって、センス鎖においてTに変更され;
及び/又は、
(4)位置−4における特異性が、
(A)K28、K83及びQ28から成る群より選ばれる修飾によって、センス鎖においてTに変更され;
(B)R83及びK83から成る群より選ばれる修飾によって、センス鎖においてGに変更され;
(C)K28及びQ83から成る群より選ばれる修飾によって、センス鎖においてAに変更され;
及び/又は、
(5)位置−5における特異性が、
(A)R45及びE28から成る群より選ばれる修飾によって、センス鎖においてGに変更され;
(B)Q28を含む修飾によってセンス鎖においてTに変更され;
(C)R28を含む修飾によってセンス鎖においてCに変更され;
及び/又は、
(6)位置−6における特異性が、
(A)K43、V85、L85及びQ30から成る群より選ばれる修飾によって、センス鎖においてTに変更され;
(B)E43、E85、K30及びR30から成る群より選ばれる修飾によって、センス鎖においてCに変更され;または、
(C)R43、K43、K85、R85、E30及びD30より成る群から選ばれる修飾によって、センス鎖においてGに変更され;
及び/又は、
(7)位置−7における特異性が、
(A)E32及びE41から成る群から選ばれる修飾によって、センス鎖においてCに変更され;
(B)R32、R41及びK41から成る群より選ばれる修飾によって、センス鎖においてGに変更され;
(C)K32、M41、L41及びI41から成る群より選ばれる修飾によって、センス鎖においてTに変更され;
及び/又は、
(8)位置−8における特異性が、
(A)K32及びK35から成る群より選ばれる修飾によって、センス鎖においてTに変更され;
(B)E32を含む修飾によってセンス鎖においてCに変更され;
(C)K32、K35及びR35から成る群より選ばれる修飾によって、センス鎖においてGに変更され;
及び/又は、
(9)位置−9における特異性が、
(A)N34及びH34から成る群より選ばれる修飾によって、センス鎖においてAに変更され;
(B)S34、C34、V34、T34及びA34から成る群より選ばれる修飾によって、センス鎖においてTに変更され;
(C)K34、R34及びH34から成る群より選ばれる修飾によって、センス鎖においてGに変更される、
ことを特徴とする、組み換えメガヌクレアーゼ。
【請求項7】
野生型I−SCEIメガヌクレアーゼと比べ、認識配列に対する特異性が変更された組み換えメガヌクレアーゼであって、
配列番号9のI−SCEIメガヌクレアーゼの残基3−186に対し少なくとも85%の配列類似性を有するポリペプチドを含み、かつ、
配列番号10及び配列番号11のI−SCEIメガヌクレアーゼと、少なくとも1塩基対異なる認識配列に対して特異性を有し、
(1)位置4における特異性が、
(A)K50を含む修飾によってセンス鎖においてAに変更され;
(B)K57、M57及びQ50から成る群より選ばれる修飾によって、センス鎖においてTに変更され;
(C)E50、R57及びK57から成る群より選ばれる修飾によって、センス鎖においてGに変更され;及び/又は、
(2)位置5における特異性が、
(A)K48、Q102から成る群より選ばれる修飾によって、センス鎖においてAに変更され;
(B)E48、K102及びR102から成る群より選ばれる修飾によって、センス鎖においてGに変更され;
(C)Q48、C102、L102及びV102から成る群より選ばれる修飾によって、センス鎖においてTに変更され;
及び/又は、
(3)位置6における特異性が、
(A)K59を含む修飾によってセンス鎖においてAに変更され;
(B)R59及びK59から成る群より選ばれる修飾によって、センス鎖においてCに変更され;
(B)K84及びE59から成る群より選ばれる修飾によって、センス鎖においてGに変更され;
及び/又は、
(4)位置7における特異性が、
(A)R46、K46及びE86から成る群より選ばれる修飾によって、センス鎖においてCに変更され;
(B)K86、R86及びE46から成る群より選ばれる修飾によって、センス鎖においてGに変更され;
(C)C46、L46及びV46から成る群より選ばれる修飾によって、センス鎖においてAに変更され;
及び/又は、
(5)位置8における特異性が、
(A)E88、R61及びH61から成る群より選ばれる修飾によって、センス鎖においてCに変更され;
(B)K88、Q61及びH61から成る群より選ばれる修飾によって、センス鎖においてTに変更され;
(C)K61、S61、V61、A61及びL61から成る群より選ばれる修飾によ
って、センス鎖においてAに変更され;
及び/又は、
(6)位置9における特異性が、
(A)C98、V98及びL98から成る群より選ばれる修飾によって、センス鎖においてAに変更され;
(B)R98及びK98から成る群より選ばれる修飾によって、センス鎖においてCに変更され;又は、
(C)E98及びD98から成る群より選ばれる修飾によって、センス鎖においてGに変更され;
及び/又は、
(7)位置10における特異性が、
(A)K96及びR96から成る群より選ばれる修飾によって、センス鎖においてCに変更され;
(B)D96及びE96から成る群より選ばれる修飾によって、センス鎖においてGに変更され;
(C)C96及びA96から成る群より選ばれる修飾によって、センス鎖においてAに変更され;
及び/又は、
(8)位置11における特異性が、
(A)Q90を含む修飾によってセンス鎖においてTに変更され;
(B)K90及びR90から成る群より選ばれる修飾によって、センス鎖においてCに変更され;
(C)E90を含む修飾によってセンス鎖においてGに変更され;
及び/又は、
(9)位置12における特異性が、
(A)Q193を含む修飾によってセンス鎖においてAに変更され;
(B)E165、E193及びD193から成る群より選ばれる修飾によって、センス鎖においてCに変更され;
(C)K165及びR165から成る群より選ばれる修飾によって、センス鎖においてGに変更され;
及び/又は、
(10)位置13における特異性が、
(A)Q193、C163及びL163から成る群より選ばれる修飾によって、センス鎖においてTに変更され;
(B)E193、D193、K163及びR192から成る群より選ばれる修飾によって、センス鎖においてGに変更され;
(C)C193およびL193から成る群より選ばれる修飾によって、センス鎖においてAに変更され;
及び/又は、
(11)位置14における特異性が、
(A)K161及びQ192から成る群より選ばれる修飾によって、センス鎖においてTに変更され;
(B)L192及びC192から成る群より選ばれる修飾によって、センス鎖においてAに変更され;
(C)K147、K161、R161、R197、D192及びE192から成る群より選ばれる修飾によって、センス鎖においてGに変更され;または、
(D)K161及びQ192から成る群から選ばれる修飾によって、センス鎖においてTに変更され;
及び/又は
(12)位置15における特異性が、
(A)C151、L151及びK151から成る群より選ばれる修飾によって、センス鎖においてTに変更され;
(B)K151を含む修飾によってセンス鎖においてGに変更され;又は、
(C)E151を含む修飾によってセンス鎖においてCに変更され;
及び/又は、
(13)位置17における特異性が、
(A)G152及びQ150から成る群より選ばれる修飾によって、センス鎖においてTに変更され;
(B)K152及びK150から成る群より選ばれる修飾によって、センス鎖においてCに変更され;
(C)N152、S152、D152、D150及びE150から成る群より選ばれる修飾によってセンス鎖においてGに変更され;
及び/又は、
(14)位置18における特異性が、
(A)H155及びY155から成る群より選ばれる修飾によってセンス鎖においてTに変更され;
(B)R155及びK155から成る群より選ばれる修飾によって、センス鎖においてCに変更され;
(C)K155及びC155から成る群より選ばれる修飾によって、センス鎖においてAに変更される
ことを特徴とする、組み換えメガヌクレアーゼ。
【請求項8】
野生型I−CEUIメガヌクレアーゼと比べ、少なくとも一つの認識配列半分部位に対する特異性が変更された組み換えメガヌクレアーゼであって、
配列番号12のI−CEUIメガヌクレアーゼの残基5−211に対し少なくとも85%の配列類似性を有するポリペプチドを含み、かつ、
配列番号13及び配列番号14から成る群より選ばれるI−CEUIメガヌクレアーゼ認識配列内の半分部位と、少なくとも1塩基対異なる認識配列半分部位に対して特異性を有し、
(1)位置−1における特異性が、
(A)C92、A92及びV92から成る群より選ばれる修飾によって、センス鎖においてAに変更され;
(B)Q116及びQ92から成る群より選ばれる修飾によって、センス鎖においてTに変更され;
(C)E116及びE92から成る群より選ばれる修飾によって、センス鎖においてGに変更され;
及び/又は、
(2)位置−2における特異性が、
(A)Q117、C90、L90及びV90から成る群より選ばれる修飾によって、センス鎖においてAに変更され;
(B)K117、R124、K124、E124、E90及びD90から成る群から選ばれる修飾によって、センス鎖においてGに変更され;
(C)E117、D117、R174、K124、K90及びK68から成る群から選ばれる修飾によって、センス鎖においてCに変更され;
及び/又は、
(3)位置−3における特異性が、
(A)C70、V70、T70、L70及びK70から成る群より選ばれる修飾によって、センス鎖においてAに変更され;
(B)Q70を含む修飾によってセンス鎖においてTに変更され;
(B)K70を含む修飾によってセンス鎖においてCに変更され;
及び/又は、
(4)位置−4における特異性が、
(A)E126、D126、R88、K88及びK72から成る群より選ばれる修飾によって、センス鎖においてCに変更され;
(B)K126、L126及びQ88から成る群より選ばれる修飾によって、センス鎖においてTに変更され;
(C)Q126、N126、K88、L88、C72、L72及びV72から成る群より選ばれる修飾によって、センス鎖においてAに変更され;
及び/又は、
(5)位置−5における特異性が、
(A)E74、K128、R128及びE128から成る群より選ばれる修飾によって、センス鎖においてGに変更され;
(B)C128、L128、V128及びT128から成る群から選ばれる修飾によって、センス鎖においてTに変更され;又は、
(C)C74、L74、V74及びT74から成る群より選ばれる修飾によって、センス鎖においてAに変更され;
及び/又は、
(6)位置−6における特異性が、
(A)K86、C86及びL86から成る群より選ばれる修飾によって、センス鎖においてTに変更され;
(B)D86、E86、R84及びK84から成る群より選ばれる修飾によって、センス鎖においてCに変更され;又は、
(C)K128、R128、R86、K86及びE84から成る群より選ばれる修飾によって、センス鎖においてGに変更され;
及び/又は、
(7)位置−7における特異性が、
(A)R76、K76及びH76から成る群より選ばれる修飾によって、センス鎖においてCに変更され;
(B)E76及びR84から成る群より選ばれる修飾によって、センス鎖においてGに変更され;
(C)H76及びQ76から成る群より選ばれる修飾によって、センス鎖においてTに変更され;
及びび/又は、
(8)位置−8における特異性が、
(A)Y79、R79及びQ76から成る群より選ばれる修飾によって、センス鎖においてAに変更され;
(B)D79、E79、D76及びE76から成る群より選ばれる修飾によって、センス鎖においてCに変更され;
(C)R79、K79、K76及びR76から成る群より選ばれる修飾によって、センス鎖においてGに変更され;
及び/又は、
(9)位置−9における特異性が、
(A)K78、V78、L78、C78及びT78から成る群より選ばれる修飾によって、センス鎖においてTに変更され;
(B)D78及びE78から成る群より選ばれる修飾によって、センス鎖においてCに変更され;又は、
(C)R78、K78及びH78から成る群より選ばれる修飾によって、センス鎖においてGに変更される、
ことを特徴とする、組み換えメガヌクレアーゼ。
【請求項9】
野生型I−CREIメガヌクレアーゼと比べ、2本鎖DNAに対する結合親和度が変更された組み換えメガヌクレアーゼであって、
配列番号1のI−CREIメガヌクレアーゼの残基2−153に対して少なくとも85%の配列類似性を有するポリペプチドを含み;
DNA結合親和度が、
(A)H、N、Q、S、T、K又はRによるE80、D137、I81、L112、P29、V64又はY66の置換;
又は、
(B)K又はRによるT46、T140又はT143の置換、
から成る群より選ばれる一つの置換に対応する少なくとも一つの修飾によって上昇することを特徴とする、組み換えメガヌクレアーゼ。
【請求項10】
野生型I−CREIメガヌクレアーゼと比べ、2本鎖DNAに対する結合親和度が変更された組み換えメガヌクレアーゼであって、
配列番号1のI−CREIメガヌクレアーゼの残基2−153に対して少なくとも85%の配列類似性を有するポリペプチドを含み;
DNA結合親和度が、
(A)H、N、Q、S、T、D又はEによるK34、K48、R51、K82、K116、またはK139の置換;
又は、
(B)D又はEによるI81、L112、P29、V64、Y66、T46、T140又はT143の置換から成る群より選ばれる一つの置換に対応する少なくとも一つの修飾によって低下することを特徴とする、組み換えメガヌクレアーゼ。
【請求項11】
野生型I−MSOIメガヌクレアーゼと比べ、2本鎖DNAに対する結合親和度が変更された組み換えメガヌクレアーゼであって、
配列番号6のI−MSOIメガヌクレアーゼの残基6−160に対して少なくとも85%の配列類似性を有するポリペプチドを含み;
DNA結合親和度が、
(A)H、N、Q、S、T、K又はRによるE147、I85、G86又はY118の置換;
又は、
(B)K又はRによるQ41、N70、S87、T88、H89、Q122、Q139、S150又はN152の置換から成る群より選ばれる一つの置換に対応する少なくとも一つの修飾によって上昇することを特徴とする、組み換えメガヌクレアーゼ。
【請求項12】
野生型I−MSOIメガヌクレアーゼと比べ、2本鎖DNAに対する結合親和度が変更された組み換えメガヌクレアーゼであって、
配列番号6のI−MSOIメガヌクレアーゼの残基6−160に対して少なくとも85%の配列類似性を有するポリペプチドを含み;
DNA結合親和度が、
(A)H、N、Q、S、T、D又はEによるK36、R51、K123、K143、またはR144の置換;
又は
(B)D又はEによるI85、G86、Y118、Q41、N70、S87、T88、H89、Q122、Q139、S150又はN152の置換から成る群より選ばれる一つの置換に対応する少なくとも一つの修飾によって低下することを特徴とする、組み換えメガヌクレアーゼ。
【請求項13】
野生型I−SCEIメガヌクレアーゼと比べ、2本鎖DNAに対する結合親和度が変更
された組み換えメガヌクレアーゼであって、
配列番号9のI−SCEIメガヌクレアーゼの残基3−186に対して少なくとも85%の配列類似性を有するポリペプチドを含み;
DNA結合親和度が、
(A)H、N、Q、S、T、K又はRによるD201、L19、L80、L92、Y151、Y188、I191、Y199又はY222の置換;
又は
(B)K又はRによるN15、N17、S81、H84、N94、N120、T156、N157、S159、N163、Q165、S166, N194又はS202の置換から成る群より選ばれる一つの置換に対応する少なくとも一つの修飾によって上昇することを特徴とする、組み換えメガヌクレアーゼ。
【請求項14】
野生型I−SCEIメガヌクレアーゼと比べ、2本鎖DNAに対する結合親和度が変更された組み換えメガヌクレアーゼであって、
配列番号9のI−SCEIメガヌクレアーゼの残基3−186に対して少なくとも85%の配列類似性を有するポリペプチドを含み;
DNA結合親和度が、
(A)H、N、Q、S、T、D又はEによるK20、K23、K63、K122、K148、K153、K190、K193、K195又はK223の置換;
又は
(B)D又はEによるL19、L80、L92、Y151、Y188、I191、Y199、Y222、N15、N17、S81、H84、N94、N120、T156、N157、S159、N163、Q165、S166、N194又はS202の置換から成る群から選ばれる一つの置換に対応する少なくとも一つの修飾によって低下することを特徴とする、組み換えメガヌクレアーゼ。
【請求項15】
野生型I−CEUIメガヌクレアーゼと比べ、2本鎖DNAに対する結合親和度が変更された組み換えメガヌクレアーゼであって、
配列番号12のI−CEUIメガヌクレアーゼの残基5−211に対して少なくとも85%の配列類似性を有するポリペプチドを含み;
DNA結合親和度が、
(A)H、N、Q、S、T、K又はRによるD25又はD128の置換又は
(B)K又はRによるS68、N70、H94、S117、N120、N129、又はH172の置換から成る群より選ばれる一つの置換に対応する少なくとも一つの修飾によって上昇することを特徴とする、組み換えメガヌクレアーゼ。
【請求項16】
野生型I−CEUIメガヌクレアーゼと比べ、2本鎖DNAに対する結合親和度が変更された組み換えメガヌクレアーゼであって、
配列番号12のI−CEUIメガヌクレアーゼの残基5−211に対して少なくとも85%の配列類似性を有するポリペプチドを含み;
DNA結合親和度が、
(A)H、N、Q、S、T、D又はEによるK21、K28、K31、R112、R114、またはR130の置換;
又は
(B)D又はEによるS68、N70、H94、S117、N120、N129又はH172の置換から成る群より選ばれる一つの置換に対応する少なくとも一つの修飾によって低下することを特徴とする、組み換えメガヌクレアーゼ。
【請求項17】
参照メガヌクレアーゼモノマーによるダイマー形成に対する親和度を変更させた組み換えメガヌクレアーゼモノマーであって、
配列番号1のI−CREIメガヌクレアーゼの残基2−153に対して少なくとも85%の配列類似性を有するポリペプチドを含み、
ダイマー形成に対する親和度が、
(A)D又はEによるK7、K57又はK96の置換;
又は
(B)K又はRによるE8又はE61の置換から成る群より選ばれる置換に対応する少なくとも一つの修飾によって変えられることを特徴とする組み換えメガヌクレアーゼモノマー。
【請求項18】
組み換えメガヌクレアーゼ・ヘテロダイマーであって、
配列番号1のI−CREIメガヌクレアーゼの残基2−153に対して少なくとも85%の配列類似性を有する第1ポリペプチドであって、
ダイマー形成に対する親和度が、
(A)D又はEによるK7、K57又はK96の置換から成る群から選ばれる置換に対応する少なくとも一つの修飾によって変えられる第1ポリペプチド及び、
配列番号1のI−CREIメガヌクレアーゼの残基2−153に対して少なくとも85%の配列類似性を有する第2ポリペプチドであって、
ダイマー形成に対する親和度が、
(B)K又はRによるE8又はE61の置換から成る群から選ばれる置換に対応する少なくとも一つの修飾によって変えられる第2ポリペプチドを含む、
ことを特徴とする、組み換えメガヌクレアーゼ・ヘテロダイマー。
【請求項19】
参照メガヌクレアーゼモノマーによるダイマー形成に対する親和度を変更させた組み換えメガヌクレアーゼモノマーであって、
配列番号6のI−MSOIメガヌクレアーゼの残基6−160に対して少なくとも85%の配列類似性を有するポリペプチドを含み、
ダイマー形成に対する親和度が、
(A)D又はEによるR302の置換;
又は
(B)K又はRによるD20、E11又はQ64の置換から成る群から選ばれる置換に対応する少なくとも一つの修飾によって変えられることを特徴とする、組み換えメガヌクレアーゼモノマー。
【請求項20】
組み換えメガヌクレアーゼ・ヘテロダイマーであって、
配列番号6のI−MSOIメガヌクレアーゼの残基6−160に対して少なくとも85%の配列類似性を有する第1ポリペプチドであって、
ダイマー形成に対する親和度が、
(A)D又はEによるR302の置換から成る群より選ばれる置換に対応する少なくとも一つの修飾によって変えられる第1ポリペプチド及び、
配列番号6のI−MSOIメガヌクレアーゼの残基6−160に対して少なくとも85%の配列類似性を有する第2ポリペプチドであって、
ダイマー形成に対する親和度が、
(B)K又はRによるD20、E11又はQ64の置換から成る群から選ばれる置換に対応する少なくとも一つの修飾によって変えられる第2ポリペプチドを含む、
ことを特徴とする、組み換えメガヌクレアーゼ・ヘテロダイマー。
【請求項21】
参照メガヌクレアーゼモノマーによるダイマー形成に対する親和度を変更させた組み換えメガヌクレアーゼモノマーであって、
配列番号12のI−CEUIメガヌクレアーゼの残基5−211に対して少なくとも85%の配列類似性を有するポリペプチドを含み、
ダイマー形成に対する親和度が、
(A)D又はEによるR93の置換;
又は
(B)K又はRによるE152の置換から成る群から選ばれる置換に対応する少なく共一つの修飾によって変えられることを特徴とする、前記組み換えメガヌクレアーゼモノマー。
【請求項22】
組み換えメガヌクレアーゼ・ヘテロダイマーであって、
配列番号12のI−CEUIメガヌクレアーゼの残基5−211に対して少なくとも85%の配列類似性を有する第1ポリペプチドであって、
ダイマー形成に対する親和度が、
(A)D又はEによるR93の置換から成る群より選ばれる置換に対応する少なくとも一つの修飾によって変えられる第1ポリペプチド及び、
配列番号12のI−CEUIメガヌクレアーゼの残基5−211に対して少なくとも85%の配列類似性を有する第2ポリペプチドであって、
ダイマー形成に対する親和度が、
(B)K又はRによるE152の置換から成る群より選ばれる置換に対応する少なくとも一つの修飾によって変えられる第2ポリペプチドを含む、
ことを特徴とする組み換えメガヌクレアーゼ・ヘテロダイマー。
【請求項23】
表1から選ばれる少なくとも一つの修飾で、排外的修飾ではない修飾をさらに含むことを特徴とする、
請求項17又は請求項18に記載の組み換えメガヌクレアーゼモノマー又はヘテロダイマー。
【請求項24】
表2から選ばれる少なくとも一つの修飾で、排外的修飾ではない修飾をさらに含むことを特徴とする、
請求項19又は請求項20に記載の組み換えメガヌクレアーゼモノマー又はヘテロダイマー。
【請求項25】
表4から選ばれる少なくとも一つの修飾で、排外的修飾ではない修飾をさらに含むことを特徴とする、
請求項21又は請求項22に記載の組み換えメガヌクレアーゼモノマー又はヘテロダイマー。
【請求項26】
真核細胞の染色体に挿入される興味の外因性配列を含む遺伝学的に修飾された真核細胞を生産するための方法であって、
真核細胞を、
(I)メガヌクレアーゼをコードする第1核酸配列及び、
(II)前記興味の配列を含む第2核酸配列
を含む一つ以上の核酸によってトランスフェクトすることを含み、
前記メガヌクレアーゼが前記染色体の中に切断部位を生産し、かつ、前記興味の配列が前記切断部位において前記染色体の中に挿入され、
前記メガヌクレアーゼが請求項1〜25のいずれか1項による組み換えメガヌクレアーゼであることを特徴とする方法。
【請求項27】
前記第2核酸が、前記切断部位に側接する配列に対し相同な配列をさらに含み、前記興味の配列が、相同組み換えによって前記切断部位において挿入されることを特徴とする、
請求項26に記載の方法。
【請求項28】
前記第2核酸が、前記切断部位に対し実質的相同性を欠き、前記興味の配列が、非相同的末端接合によって前記染色体に挿入されることを特徴とする、
請求項26に記載の方法。
【請求項29】
真核細胞の染色体に挿入される興味の外因性配列を含む、遺伝学的に修飾された真核細胞を生産するための方法であって、
真核細胞にメガヌクレアーゼタンパクを導入すること;
及び、
前記興味の配列を含む核酸によって前記真核細胞をトランスフェクトすることを含み、
前記メガヌクレアーゼが、前記染色体の中に切断部位を生産し、前記興味の配列が、前記切断部位において前記染色体の中に挿入され;
かつ、
前記メガヌクレアーゼが請求項1〜25のいずれか1項による組み換えメガヌクレアーゼであることを特徴とする方法。
【請求項30】
前記核酸が、前記切断部位に側接する配列に対し相同な配列をさらに含み、前記興味の配列が、相同組み換えによって前記切断部位において挿入されることを特徴とする、
請求項29に記載の方法。
【請求項31】
前記核酸が、前記切断部位に対し実質的相同性を欠き、前記興味の配列が、非相同的末端接合によって前記染色体に挿入されることを特徴とする、
請求項29に記載の方法。
【請求項32】
真核細胞の染色体中の標的配列を破壊することによって遺伝学的に修飾された真核細胞を生産するための方法であって、
メガヌクレアーゼをコードする核酸によって真核細胞をトランスフェクトすることを含み、
前記メガヌクレアーゼが、前記染色体の中に切断部位を生産し、前記標的配列が、前記切断部位において非相同的末端接合によって破壊され;
かつ、
前記メガヌクレアーゼが、請求項1〜25のいずれか1項による組み換えメガヌクレアーゼであることを特徴とする方法。
【請求項33】
遺伝学的に修飾された生物体を生産する方法であって、
請求項26〜32のいずれか1項の方法に従って遺伝学的に修飾された真核細胞を生産するステップと、
及び、
前記遺伝学的に修飾された真核細胞を育成して遺伝学的に修飾された生物体を生産するステップと、
を含む方法。
【請求項34】
前記真核細胞が、配偶子、接合子、胚盤胞細胞、胚幹細胞及びプロトプラスト細胞から成る群より選ばれることを特徴とする、
請求項33に記載の方法。
【請求項35】
真核生物において遺伝子治療によって疾患を治療するための方法であって、
前記真核生物の少なくとも一つの細胞を、
(I)メガヌクレアーゼをコードする第1核酸配列及び、
(II)興味の配列を含む第2核酸配列
によってトランスフェクトすることを含み;
前記メガヌクレアーゼが、前記染色体の中に切断部位を生産し、前記興味の配列が、前記切断部位において前記染色体の中に挿入され;
かつ、
前記メガヌクレアーゼが、請求項1−25のいずれか1項による組み換えメガヌクレアーゼであることを特徴とする方法。
【請求項36】
前記第2核酸配列が、前記切断部位に側接する配列に対し相同な配列をさらに含み、前記興味の配列が、相同組み換えによって前記切断部位において挿入されることを特徴とする、
請求項35に記載の方法。
【請求項37】
前記第2核酸配列が、前記切断部位に対し実質的相同性を欠き、前記興味の配列が、非相同的末端接合によって前記染色体に挿入されることを特徴とする、
請求項35に記載の方法。
【請求項38】
真核生物において遺伝子治療によって疾患を治療するための方法であって、
前記真核生物の少なくとも一つの細胞の中にメガヌクレアーゼタンパクを導入することステップと、
前記真核細胞を、興味の配列を含む核酸によってトランスフェクトするステップとを含み、
前記メガヌクレアーゼが、前記染色体の中に切断部位を生産し、前記興味の配列が、前記切断部位において前記染色体の中に挿入され;
前記メガヌクレアーゼが請求項1〜25のいずれか1項による組み換えメガヌクレアーゼであり;
かつ、
前記興味の配列の挿入が、前記疾患に対する前記遺伝子治療を与えることを特徴とする方法。
【請求項39】
前記核酸が、前記切断部位に側接する配列に対し相同な配列をさらに含み、前記興味の配列が、相同組み換えによって前記切断部位において挿入されることを特徴とする、
請求項38に記載の方法。
【請求項40】
前記核酸が、前記切断部位に対し実質的相同性を欠き、前記興味の配列が、非相同的末端接合によって前記染色体に挿入されることを特徴とする、
請求項38に記載の方法。
【請求項41】
真核生物において、前記真核生物の染色体中の標的配列の破壊による遺伝子治療によって疾患を治療するための方法であって、
前記真核生物の少なくとも一つの細胞を、メガヌクレアーゼをコードする核酸によってトランスフェクトすることを含み;
前記メガヌクレアーゼが、前記染色体の中に切断部位を生産し、前記標的配列が、前記切断部位において非相同的末端接合によって破壊され;
前記メガヌクレアーゼが、請求項1−25のいずれか1項による組み換えメガヌクレアーゼであり;
かつ、
前記標的配列の破壊が、前記疾患に対する前記遺伝子治療を与えることを特徴とする方法。
【請求項42】
真核細胞宿主におけるウィルス性病原体感染を、前記ウィルス病原体のゲノムにおける標的配列を破壊することによって治療する方法であって、
前記真核細胞宿主の少なくとも一つの感染細胞を、メガヌクレアーゼをコードする核酸によってトランスフェクトすることを含み;
前記メガヌクレアーゼが、前記ウィルスゲノムの中に切断部位を生産し、前記標的配列が、前記切断部位において非相同的末端接合によって破壊され、
前記メガヌクレアーゼが、請求項1−25のいずれか1項による組み換えメガヌクレアーゼであり;
かつ、
前記標的配列の破壊が、前記感染に対する治療を与えることを特徴とする方法。
【請求項43】
真核細胞宿主におけるウィルス性病原体感染を、前記ウィルス病原体のゲノムにおける標的配列を破壊することによって治療する方法であって、
前記真核細胞宿主の少なくとも一つの感染細胞を、メガヌクレアーゼをコードする第1核酸、および第2核酸によってトランスフェクトすることを含み;
前記メガヌクレアーゼが、前記ウィルスゲノムの中に切断部位を生産し、前記標的配列が、前記切断部位において、前記ウィルスゲノムと前記第2核酸との相同組み換えによって破壊され;
前記メガヌクレアーゼが、請求項1−25のいずれか1項による組み換えメガヌクレアーゼであり;
前記第2核酸が、前記切断部位に側接する配列に対して相同な配列を含み;
かつ、
前記標的配列の破壊が、前記感染に対する治療を与えることを特徴とする方法。
【請求項44】
真核細胞宿主における前核細胞病原体感染を、前記前核細胞病原体のゲノムにおける標的配列を破壊することによって治療する方法であって、
前記真核細胞宿主に感染する前記前核細胞病原体の少なくとも一つの細胞を、メガヌクレアーゼをコードする核酸によってトランスフェクトすることを含み;
前記メガヌクレアーゼが、前記前核細胞ゲノムの中に切断部位を生産し、前記標的配列が、前記切断部位において、非相同的末端接合によって破壊され;
前記メガヌクレアーゼが、請求項1〜25のいずれか1項による組み換えメガヌクレアーゼであり;
かつ、
前記標的配列の破壊が、前記感染に対する治療を与えることを特徴とする方法。
【請求項45】
真核細胞宿主における前核細胞病原体感染を、前記前核細胞病原体のゲノムにおける標的配列を破壊することによって治療する方法であって、
前記真核細胞宿主に感染する前記前核細胞病原体の少なくとも一つの細胞を、メガヌクレアーゼをコードする第1核酸、および第2核酸によってトランスフェクトすることを含み;
前記メガヌクレアーゼが、前記前核細胞ゲノムの中に切断部位を生産し、前記標的配列が、前記切断部位において、前記前核細胞ゲノムと前記第2核酸との相同組み換えによって破壊され;
前記メガヌクレアーゼが、請求項1〜25のいずれか1項による組み換えメガヌクレアーゼであり;
前記第2核酸が、前記切断部位に側接する配列に対して相同な配列を含み;
かつ、
前記標的配列の破壊が、前記感染に対する治療を与えることを特徴とする方法。
【請求項46】
少なくとも一つの所望の変化を含むように、認識配列の少なくとも一つの塩基位置に対する特異性を変更させた組み換えメガヌクレアーゼを合理的に設計する方法であって、
(1)参照メガヌクレアーゼ−DNA複合体の三次元構造の少なくとも一部を決定する
こと;
(2)前記塩基位置において塩基接触面を形成するアミノ酸残基を特定すること;
(3)前記接触面を含む少なくとも第1残基のβ−炭素と、前記塩基位置における少なくとも第1塩基との間の距離を決めること;
及び、
(4)前記所望の変化を促進するアミノ酸置換を、表2に示される群の中から、
(A)第1塩基から6オングストローム未満の距離にある第1残基については、G群、C群、T群又はA群のうち、前記所望の変化を促進するのに適切なもののメンバーである1群及び/又は2群の置換を選択すること;
又は、
(B)前記第1塩基から6オングストロームより遠い距離にある第1残基については、G群、C群、T群又はA群のうち、前記所望の変化を促進するのに適切なもののメンバーである2群及び/又は3群の置換を選択することによって特定すること、
を含む方法。
【請求項47】
DNA−結合親和度を上昇させる組み換えメガヌクレアーゼを合理的に設計する方法であって、
(1)参照メガヌクレアーゼ−DNA複合体の三次元構造の少なくとも一部を決定すること;
(2)バックボーン接触面を形成するアミノ酸接触残基を特定すること;
(3)前記DNA−結合親和度を上昇させるアミノ酸置換を、
(A)陰性荷電、または疎水性側鎖を持つ接触残基については、非荷電/極性、または陽性荷電側鎖を持つ置換を選択すること;
又は
(B)非荷電/極性側鎖を持つ接触残基については、陽性荷電側鎖を持つ置換を選択することによって特定すること、
を含む方法。
【請求項48】
DNA−結合親和度を低下させる組み換えメガヌクレアーゼを合理的に設計する方法であって、
(1)参照メガヌクレアーゼ−DNA複合体の三次元構造の少なくとも一部を決定すること;
(2)バックボーン接触面を形成するアミノ酸接触残基を特定すること;
(3)前記DNA−結合親和度を低下させるアミノ酸置換を、
(A)陽性荷電側鎖を持つ接触残基については、非荷電/極性、または陰性荷電側鎖を持つ置換を選択すること;
又は
(B)疎水性、または非荷電/極性側鎖を持つ接触残基については、陰性荷電側鎖を持つ置換を選択することによって特定すること、
を含む方法。
【図3】

【図1A】
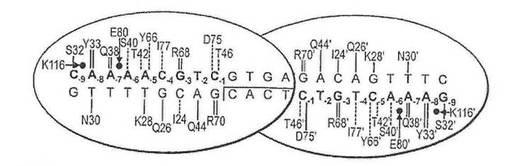
【図1B】
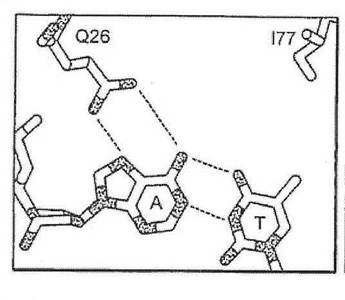
【図1C】
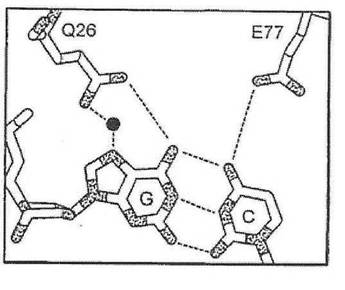
【図1D】
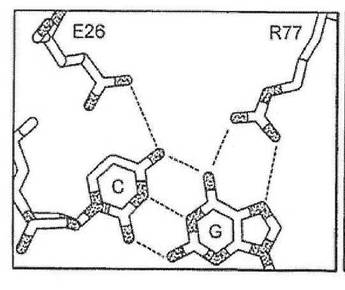
【図1E】
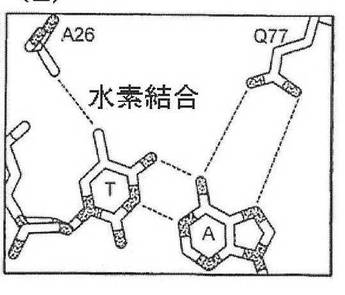
【図2A】

【図2B】
【図3A】

【図3B】

【図1A】
【図1B】
【図1C】
【図1D】
【図1E】
【図2A】
【図2B】
【図3A】
【図3B】
【公開番号】特開2013−34473(P2013−34473A)
【公開日】平成25年2月21日(2013.2.21)
【国際特許分類】
【外国語出願】
【出願番号】特願2012−125663(P2012−125663)
【出願日】平成24年6月1日(2012.6.1)
【分割の表示】特願2012−110691(P2012−110691)の分割
【原出願日】平成18年10月18日(2006.10.18)
【出願人】(512135735)デューク大学 (2)
【Fターム(参考)】
【公開日】平成25年2月21日(2013.2.21)
【国際特許分類】
【出願番号】特願2012−125663(P2012−125663)
【出願日】平成24年6月1日(2012.6.1)
【分割の表示】特願2012−110691(P2012−110691)の分割
【原出願日】平成18年10月18日(2006.10.18)
【出願人】(512135735)デューク大学 (2)
【Fターム(参考)】
[ Back to top ]