
酵母におけるアセチルCoAを経由する代謝経路を利用した物質の製造法
【課題】アセチルCoAを経由する代謝産物を高生産し得る酵母菌株の提供。
【解決手段】以下の酵素:(a)アセチルCoA合成酵素(ACS);および(b)アセチルCoAアセチル基転移酵素(ERG10)の活性が増強されている、酵母菌株。
【解決手段】以下の酵素:(a)アセチルCoA合成酵素(ACS);および(b)アセチルCoAアセチル基転移酵素(ERG10)の活性が増強されている、酵母菌株。
【発明の詳細な説明】
【発明の背景】
【0001】
技術分野
本発明は、酵母において、アセチルCoAを経由する代謝経路を利用した物質の製造方法に関する。
【0002】
背景技術
深刻化する環境問題を背景に、バイオ燃料が石油の代替燃料として注目を集めている。エタノールはバイオ燃料としてよく知られているが、ガソリンより低いエネルギー密度しか有さないこと、輸送燃料として機能させるためには限られた濃度範囲でガソリンと混合する必要があることから、燃料としての欠点も多い。イソプロパノールは、1−ブタノールよりも高いオクタン価を有し、また、工業原料としても利用される汎用性の高い溶剤である。イソプロパノールは、ナフサ由来のプロピレンの水和によって製造されており、再生可能な供給源から商業的に生産されたことはない。
【0003】
微生物によるアセトンやブタノールの発酵生産は、20世紀初頭に産業的に行われていた。絶対嫌気性菌のある種のクロストリジウム属細菌はアセトンとブタノールを同時に生産することができる(アセトン・ブタノール発酵)。アセトンは細菌細胞内に存在するアセチルCoAを原料として生産される。アセチルCoAは、アセチルCoAアセチル基転移酵素(チオラーゼ)によってアセトアセチルCoAに変換され、さらにCoA転移酵素によってアセト酢酸に変換され、さらにアセト酢酸脱炭酸酵素によってアセトンへと変換される。ある種のクロストリジウム属細菌はさらに二級アルコールの脱水素酵素を有し、アセトンからイソプロパノールを生産する。Chenらは、クロストリジウム・ベイジェリンキを含む52株のクロストリジウム属細菌の中で、最大で30mM(1.8g/L)のイソプロパノールを生産する株が存在したことを報告している(非特許文献1:Chen, J.-S., and S. F. Hiu. Acetone-butanol-isopropanol production by Clostridium beijerinckii (synonym, Clostridium butylicum). Biotechnol. Lett. 1986. 8:371-376)。また、最近になってHanaiらおよびJojimaらは、大腸菌にクロストリジウム属由来のCoA転移酵素、アセト酢酸脱炭酸酵素、二級アルコール脱水素酵素を導入し、内在性のアセチルCoAアセチル基転移酵素を過剰発現させることによってイソプロパノールを効率的に生産できる大腸菌株を報告している(非特許文献2:T. Hanai, S. Atsumi, and J. C. Liao. Engineered Synthetic Pathway for Isopropanol Production in Escherichia coli. Appl Environ Microbiol, Dec. 2007, p. 7814-7818;非特許文献3:Toru Jojima & Masayuki Inui & Hideaki Yukawa. Production of isopropanol by metabolically engineered Escherichia coli. Appl Microbiol Biotechnol 2008 77:1219-1224)。しかしながら、大腸菌は、糖耐性、浸透圧耐性、塩耐性、有機溶剤耐性など各種ストレス耐性はあまり高くないことが多く、ファージ汚染によるリスクもある。また、イソプロパノール耐性が低いことは生産物による発酵の阻害が起こることを意味する。発酵阻害を回避するためにガスストリッピング法などを用いて生産物を培地から除去しながら発酵を続ける方法が知られているが、該方法の使用は発酵生産の工程を煩雑化させるだけでなく、コスト増加につながる。これらの問題を解決するための一つの手段として、高い有機溶剤耐性を有する微生物を用いてイソプロパノール生産をすることが考えられる。
【0004】
単細胞真核微生物である酵母は高い有機溶剤耐性を示し、また、大腸菌などの微生物と同様に、培養設備、培養時間および培養コストの面で高等生物の細胞よりも安価に培養することができるという特徴を有する。特に、出芽酵母はビールやワインなどの酒類やパンの製造に用いられ、その食品としての安全性が確認されている。
【0005】
酵母でイソプロパノールを生産するためには、その酵母菌株に、外因性のCoA転移酵素、アセト酢酸脱炭酸酵素、および二級アルコール脱水素酵素を導入する必要がある。これまで、酵母によるイソプロパノールの製造例としては、サッカロマイセス・セレビシエ(Saccharomyces cerevisiae)にイソプロピルアルコール生産酵素群の遺伝子を導入し、この形質転換体を培養することによりイソプロパノールを製造したことが報告されている(特許文献1:特開2011−97929号公報)。しかし、この方法によるイソプロパノールの生産量は、培養液中の濃度として100mg/L前後という少量に過ぎない。酵母は真核生物であり、オルガネラが存在することから、イソプロパノール合成の基質であるアセチルCoAを細胞質に供給する経路が複数存在している。どの経路からアセチルCoAを細胞質に供給することがイソプロパノール合成にとって有利であるのか、あるいはどのような酵母がそういった能力に優れているか、これまでに調査された例はない。
【0006】
キャンディダ・ユティリス(Candida utilis)は、サッカロマイセス・セレビシエ(Saccharomyces cerevisiae)およびサッカロマイセス・フラジリス(Saccharomyces fragilis)と同様に、FDA(米国連邦食品医薬品局)によって食用酵母として認定されており、食品添加物として安全に使用できることが認められている食用酵母である。実際に、現在でも、キャンディダ・ユティリスは、日本をはじめとして、ドイツやアメリカなど世界各国で生産され、食飼料として利用されている。また、キャンディダ・ユティリスは、ペントースの資化による菌体生産、エチルアセテート、L−グルタミン酸、グルタチオン、インベルターゼなどの生産株として広く産業界で利用されてきた。近年、キャンディダ・ユティリスにおいても形質転換法が開発され、様々な化合物が代謝工学的に生産される技術が開発されている(非特許文献4:Hiroshi Shimada, Keiji Kondo, Paul D. Fraser, Yutaka Miura, Toshiko Saito, and Norihiko Misawa., Increased Carotenoid Production by the Food Yeast Candida utilis through Metabolic Engineering of the Isoprenoid Pathway., Appl Environ Microbiol, July 1998, p. 2676-2680, Vol. 64, No. 7;非特許文献5:Yutaka Miura, Keiji Kondo, Toshiko Saito, Hiroshi Shimada, Paul D. Fraser, and Norihiko MisawaProduction of the Carotenoids Lycopene, β-Carotene, and Astaxanthin in the Food Yeast Candida utilis., Appl Environ Microbiol, April 1998, p. 1226-1229, Vol. 64, No. 4;非特許文献6:Keiji Kondo,, Yutaka Miura, Hidetaka Sone, Kazuo Kobayashi & Hiroshi lijima., High-level expression of a sweet protein, monellin, in the food yeast Candida utilis., Nature Biotechnology 15, 453 - 457 (1997);非特許文献7:Yutaka Miura, Keiji Kondo, Hiroshi Shimada, Toshiko Saito, Katsumi Nakamura, Norihiko Misawa., Production of lycopene by the food yeast, Candida utilis that does not naturally synthesize carotenoid., Biotechnology and Bioengineering Volume 58 Issue 2-3, Pages 306-308;非特許文献8:Shigehito IKUSHIMA, Toshio FUJII, Osamu KOBAYASHI, Satoshi YOSHIDA and Aruto YOSHIDA., Genetic Engineering of Candida utilis Yeast for Efficient Production of L-Lactic Acid., Bioscience, Biotechnology, and Biochemistry Vol. 73 (2009) , No. 8 pp.1818-1824;非特許文献9:Yi-Ren Hong, Ya-Lei Chen, Lynn Farh, Wen-Jen Yang, Chen-Hua Liao and David Shiuan.,Recombinant Candida utilis for the production of biotin., Applied Microbiology and Biotechnology.,Volume 71(2006), Number 2, 211-221)。また、キャンディダ・ユティリスは、サッカロマイセス属とは異なり、クラブツリー(Crabtree)効果陰性酵母であることから、グルコース存在下でも呼吸活性が低下しない。これにより、十分に通気を行う条件を採用することにより、エタノールの生成を抑えることができる。さらに、キャンディダ・ユティリスは呼吸活性も高く、一旦生成したエタノールも速やかに再同化されることから、効率的な菌体や代謝産物の生産を可能とする。
【0007】
酵母でイソプロパノールのような物質を生産する場合、一般に、副産物としてのエタノールの生成が問題となる。このエタノール副生を抑制するために、エタノール生産の重要酵素であるピルビン酸脱炭酸酵素遺伝子(PDC)を欠損させる取り組みがなされている(特許文献2:国際公開第2010/095751号パンフレット;特許文献3:国際公開第2010/095750号パンフレット;非特許文献10:Flikweert MT, Van Der Zanden L, Janssen WM, et al. Pyruvate decarboxylase: an indispensable enzyme for growth of Saccharomyces cerevisiae on glucose. Yeast 12(1996): 247-257;非特許文献11:van Maris AJA, Winkler AA, Porro D, et al. Homofermentative lactate production cannot sustain anaerobic growth of engineered Saccharomyces cerevisiae: Possible consequence of energy-dependent lactate export. Appl Environ Microbiol 70(2004): 2898-2905)。
【0008】
【非特許文献1】Chen, J.-S., and S. F. Hiu. Acetone-butanol-isopropanol production by Clostridium beijerinckii (synonym, Clostridium butylicum). Biotechnol. Lett. 1986. 8:371-376
【非特許文献2】T. Hanai, S. Atsumi, and J. C. Liao. Engineered Synthetic Pathway for Isopropanol Production in Escherichia coli. Appl Environ Microbiol, Dec. 2007, p. 7814-7818
【非特許文献3】Toru Jojima & Masayuki Inui & Hideaki Yukawa. Production of isopropanol by metabolically engineered Escherichia coli. Appl Microbiol Biotechnol 2008 77:1219-1224
【非特許文献4】Hiroshi Shimada, Keiji Kondo, Paul D. Fraser, Yutaka Miura, Toshiko Saito, and Norihiko Misawa., Increased Carotenoid Production by the Food Yeast Candida utilis through Metabolic Engineering of the Isoprenoid Pathway., Appl Environ Microbiol, July 1998, p. 2676-2680, Vol. 64, No. 7
【非特許文献5】Yutaka Miura, Keiji Kondo, Toshiko Saito, Hiroshi Shimada, Paul D. Fraser, and Norihiko MisawaProduction of the Carotenoids Lycopene, β-Carotene, and Astaxanthin in the Food Yeast Candida utilis., Appl Environ Microbiol, April 1998, p. 1226-1229, Vol. 64, No. 4
【非特許文献6】Keiji Kondo,, Yutaka Miura, Hidetaka Sone, Kazuo Kobayashi & Hiroshi lijima., High-level expression of a sweet protein, monellin, in the food yeast Candida utilis., Nature Biotechnology 15, 453 - 457 (1997)
【非特許文献7】Yutaka Miura, Keiji Kondo, Hiroshi Shimada, Toshiko Saito, Katsumi Nakamura, Norihiko Misawa., Production of lycopene by the food yeast, Candida utilis that does not naturally synthesize carotenoid., Biotechnology and Bioengineering Volume 58 Issue 2-3, Pages 306-308
【非特許文献8】Shigehito IKUSHIMA, Toshio FUJII, Osamu KOBAYASHI, Satoshi YOSHIDA and Aruto YOSHIDA., Genetic Engineering of Candida utilis Yeast for Efficient Production of L-Lactic Acid., Bioscience, Biotechnology, and Biochemistry Vol. 73 (2009) , No. 8 pp.1818-1824
【非特許文献9】Yi-Ren Hong, Ya-Lei Chen, Lynn Farh, Wen-Jen Yang, Chen-Hua Liao and David Shiuan.,Recombinant Candida utilis for the production of biotin., Applied Microbiology and Biotechnology.,Volume 71(2006), Number 2, 211-221
【非特許文献10】Flikweert MT, Van Der Zanden L, Janssen WM, et al. Pyruvate decarboxylase: an indispensable enzyme for growth of Saccharomyces cerevisiae on glucose. Yeast 12(1996): 247-257
【非特許文献11】van Maris AJA, Winkler AA, Porro D, et al. Homofermentative lactate production cannot sustain anaerobic growth of engineered Saccharomyces cerevisiae: Possible consequence of energy-dependent lactate export. Appl Environ Microbiol 70(2004): 2898-2905
【特許文献1】特開2011−97929号公報
【特許文献2】国際公開第2010/095751号パンフレット
【特許文献3】国際公開第2010/095750号パンフレット
【発明の概要】
【0009】
本発明者らは、酵母におけるアセチルCoA合成酵素(ACS)およびアセチルCoAアセチル基転移酵素(ERG10)の活性を増強することにより、酵母内におけるアセチルCoAを経由する物質の生産量が顕著に増加することを見出した。本発明はこの知見に基づくものである。
【0010】
従って、本発明の目的は、様々な物質の発酵生産において中間物質として関与するアセチルCoAならびにアセトアセチルCoAの合成・代謝に係る酵素活性が増強された酵母菌株、ならびに該酵母菌株を用いた物質の製造法を提供することにある。
【0011】
そして、本発明による酵母菌株は、以下の酵素:
(a)アセチルCoA合成酵素(ACS);および
(b)アセチルCoAアセチル基転移酵素(ERG10)
の活性が増強されている酵母菌株である。
【0012】
さらに、本発明による製造法は、本発明による酵母菌株を培養することを含んでなる、物質を製造する方法である。
【0013】
本発明によれば、アセチルCoAやアセトアセチルCoAが関与する酵母の代謝経路が活性化される。アセチルCoAやアセトアセチルCoAが関与する代謝経路は多岐にわたるため、生成する代謝産物も多種多様である。従って、本発明によれば、酵母菌株の培養によって様々な物質を高効率で製造することが可能となる。また、製造しようとする物質に応じて、必要な酵素の遺伝子を発現可能な形で酵母に組み込み、得られた酵母菌株を培養することによりその物質を製造することも可能である。
【発明の具体的説明】
【0014】
本発明による酵母菌株では、以下の遺伝子:(a)アセチルCoA合成酵素(ACS)の活性を有するポリペプチドをコードする遺伝子;および(b)アセチルCoAアセチル基転移酵素(ERG10)の活性を有するポリペプチドをコードする遺伝子の発現が増強されている。
【0015】
本明細書では、上記のアセチルCoA合成酵素(ACS)遺伝子およびアセチルCoAアセチル基転移酵素(ERG10)遺伝子とともに、イソプロパノールを合成するための4つの遺伝子を過剰発現する酵母菌株を作製し、これを培養することにより、極めて高濃度のイソプロパノールが生産されることが実証されている。この酵母菌株におけるイソプロパノール生産の経路は図1に示す通りである。図1に示される経路から、イソプロパノールの高い生産量が、アセチルCoAおよびアセトアセチルCoAの合成・代謝に係る酵素活性の増強に起因することは明らかである。従って、本発明によって、イソプロパノールのみならず、アセチルCoAおよびアセトアセチルCoAを経由する様々な物質を高生産することができる。また、図1に示されるように、ACSおよびERG10は細胞質において機能する酵素である。
【0016】
細胞質に供給されるアセチルCoAは、フラボノイド骨格を有する化合物、テルぺノイド骨格を有する化合物、ステロール骨格を有する化合物、脂肪酸など、様々な有用な二次代謝物質の基幹物質である。よって、酵母菌株が天然の状態もしくは人為的に遺伝子を改変された状態でこれらの二次代謝産物の合成経路を有していれば、本発明の方法によりこれらの物質を高生産させることができる。
【0017】
一般的に、微生物による物質生産を行う際には、副産物生産を抑制する取り組みがなされる場合が多い。例えば、酵母において物質を生産する場合、一般に、副産物としてのエタノールの生成を抑制するために、エタノール生産に関与する酵素遺伝子、例えばピルビン酸脱炭酸酵素遺伝子(PDC)を欠損させる取り組みがなされている(国際公開第2010/095751号パンフレット;国際公開第2010/095750号パンフレット;Flikweert MT, Van Der Zanden L, Janssen WM, et al. Pyruvate decarboxylase: an indispensable enzyme for growth of Saccharomyces cerevisiae on glucose. Yeast 12(1996): 247-257;van Maris AJA, Winkler AA, Porro D, et al. Homofermentative lactate production cannot sustain anaerobic growth of engineered Saccharomyces cerevisiae: Possible consequence of energy-dependent lactate export. Appl Environ Microbiol 70(2004): 2898-2905)。しかし、本明細書では、上記のPDCを欠損させた酵母菌株よりも、PDCを欠損させなかった酵母菌株の方が顕著に高いイソプロパノール生産量を示すことが実証されている。よって、本発明による酵母菌株は、PDCなどのエタノール生産に関与する酵素遺伝子を欠失させずにそのまま維持していることも特徴の一つであり、これによりイソプロパノール生産量、ひいてはアセチルCoAやその代謝産物であるアセトアセチルCoAの生産量が顕著に向上することは、驚くべきことである。
【0018】
本明細書において酵素活性が「増強されている」とは、培養過程の少なくとも一時期において、活性増強のための手段を施されていない酵母と比較してより強い酵素活性が観察されることを意味する。酵素活性の増強は、当該酵素遺伝子の発現を増強すること、あるいは比活性のより高い酵素遺伝子への置き換えや、タンパク質工学的な方法による酵素遺伝子の改変によって実現することができる。本明細書において遺伝子の発現が「増強されている」とは、酵母の遺伝的改変により該酵母におけるその遺伝子の発現量が増加すること、つまり、その遺伝子を過剰発現させることを意味する。このような遺伝的改変の方法としては、例えば、その遺伝子により酵母を形質転換する方法(つまり、その遺伝子を発現し得る形で酵母にトランスフェクトする方法)、その酵母のゲノム上に存在する遺伝子のプロモーター活性を強化する方法、その酵母のゲノム上に存在する遺伝子のプロモーターを他のプロモーターと置換することにより該遺伝子の発現を増強する方法など、様々な方法が知られているが、好ましくはその遺伝子により酵母を形質転換する方法とされる。
【0019】
酵母
本発明による酵母菌株は、特定の遺伝子が過剰発現するように遺伝的に改変された酵母である。遺伝的改変に用いる酵母は、アセチルCoAまたはアセトアセチルCoAを経由する代謝経路を有する酵母であればよく、特に制限されないが、好ましくは呼吸活性の高いクラブツリー効果陰性酵母とされ、より好ましくはクラブトゥリー陰性酵母であるキャンディダ・ユティリスとされる。キャンディダ・ユティリスの菌株は当技術分野において公知の様々な株、例えば、NBRC0396株、NBRC0619株、NBRC0988株、NBRC0639株、NBRC0626株、NBRC1086株、NBRC10707株、ATCC9255株、ATCC28955株、ATCC44638株、ATCC9206株、ATCC96621株、ATCC18201株、およびATCC15239株等であってよいが、好ましくはNBRC1086株、ATCC9255株またはATCC15239株とされる。
【0020】
活性が増強される酵素およびその遺伝子
本発明による酵母菌株において活性が増強される酵素は、以下の酵素:(a)アセチルCoA合成酵素(ACS);および(b)アセチルCoAアセチル基転移酵素(ERG10)である。本発明の好ましい実施態様によれば、本発明による酵母菌株においてはこれら2種類の酵素の遺伝子が過剰発現され、すなわち、以下の遺伝子:(a)アセチルCoA合成酵素(ACS)の活性を有するポリペプチドをコードする遺伝子;および(b)アセチルCoAアセチル基転移酵素(ERG10)の活性を有するポリペプチドをコードする遺伝子の発現が増強されている。
【0021】
本発明による酵母菌株では、さらに、以下の遺伝子:(c)ATP−クエン酸除去付加酵素(ACL)の活性を有するポリペプチドをコードする遺伝子;(d)ミトコンドリア内膜カルニチン輸送体(CRC1)の活性を有するポリペプチドをコードする遺伝子;および(e)カルニチンアセチル転移酵素遺伝子(YAT1)の活性を有するポリペプチドをコードする遺伝子からなる群から選択される少なくとも一種の遺伝子の発現が増強されていてもよく、好ましくはこれら(c)、(d)および(e)の全ての遺伝子の発現が増強される。
【0022】
これらの酵素および遺伝子は様々な酵母に由来するものが知られており、その起源は特に制限されない。これらの酵素および遺伝子は、育種に用いられる酵母の内在性遺伝子およびその発現産物であってよく、あるいは、育種に用いられる酵母以外の他の酵母に由来するものであってもよい。例えば、これらの酵素および遺伝子は、それぞれ独立に、キャンディダ・ユティリスまたはヤロウィア・リポリティカに由来するものとすることができる。それぞれの酵素の起源としては、例えば、アセチルCoA合成酵素(ACS)はキャンディダ・ユティリス酵母、アセチルCoAアセチル基転移酵素(ERG10)はキャンディダ・ユティリス酵母、ミトコンドリア内膜カルニチン輸送体(CRC1)はキャンディダ・ユティリス酵母、カルニチンアセチル転移酵素遺伝子(YAT1)はキャンディダ・ユティリス酵母、ATP−クエン酸除去付加酵素(ACL)はヤロウィア・リポリティカ酵母(ヤロウィア・リポリティカ由来のACL1とACLYがサブユニットを構成する遺伝子)が好ましい。
【0023】
キャンディダ・ユティリス由来のアセチルCoA合成酵素(ACS)遺伝子には、ACS1遺伝子とACS2遺伝子がある。ACS1遺伝子のコード配列は配列番号11で表され、コードされるアミノ酸配列は配列番号12で表される。ACS2遺伝子のコード配列は配列番号13で表され、コードされるアミノ酸配列は配列番号14で表される。アセチルCoA合成酵素(ACS)は、酢酸をアセチルCoAに変換する酵素である。本発明に用いられるACS遺伝子はACS1およびACS2のいずれであってもよいが、好ましくはACS2とされる。また、アセチルCoA合成酵素(ACS)の活性を有するポリペプチドは、配列番号12または配列番号14で表されるアミノ酸配列において、1もしくは数個のアミノ酸が欠失、置換、付加もしくは挿入されたアミノ酸配列を含み、かつACS活性を有するポリペプチドであってもよい。
【0024】
キャンディダ・ユティリス由来のアセチルCoAアセチル基転移酵素(ERG10)遺伝子のコード配列は配列番号61で表され、コードされるアミノ酸配列は配列番号62で表される。アセチルCoAアセチル基転移酵素(ERG10)は、アセチルCoAをアセトアセチルCoAに変換する酵素である。また、アセチルCoAアセチル基転移酵素(ERG10)の活性を有するポリペプチドは、配列番号62で表されるアミノ酸配列において、1もしくは数個のアミノ酸が欠失、置換、付加もしくは挿入されたアミノ酸配列を含み、かつERG10活性を有するポリペプチドであってもよい。
【0025】
ヤロウィア・リポリティカ由来のACL1遺伝子のコード配列は配列番号31で表され、コードされるアミノ酸配列は配列番号32で表される。また、ACL1の活性を有するポリペプチドは、配列番号32で表されるアミノ酸配列において、1もしくは数個のアミノ酸が欠失、置換、付加もしくは挿入されたアミノ酸配列を含み、かつACL1活性を有するポリペプチドであってもよい。さらに、ヤロウィア・リポリティカ由来のACLY遺伝子のコード配列は配列番号33で表され、コードされるアミノ酸配列は配列番号34で表される。また、ACLYの活性を有するポリペプチドは、配列番号34で表されるアミノ酸配列において、1もしくは数個のアミノ酸が欠失、置換、付加もしくは挿入されたアミノ酸配列を含み、かつACLY活性を有するポリペプチドであってもよい。これらのACL1とACLYをサブユニットとするATP−クエン酸除去付加酵素(ACL)は、クエン酸をアセチルCoAに変換する酵素である。
【0026】
キャンディダ・ユティリス由来のミトコンドリア内膜カルニチン輸送体(CRC1)遺伝子のコード配列は配列番号23で表され、コードされるアミノ酸配列は配列番号24で表される。ミトコンドリア内膜カルニチン輸送体(CRC1)は、アセチルカルニチンをミトコンドリアから細胞質へ輸送する酵素である。また、ミトコンドリア内膜カルニチン輸送体(CRC1)の活性を有するポリペプチドは、配列番号24で表されるアミノ酸配列において、1もしくは数個のアミノ酸が欠失、置換、付加もしくは挿入されたアミノ酸配列を含み、かつCRC1活性を有するポリペプチドであってもよい。
【0027】
キャンディダ・ユティリス由来のカルニチンアセチル転移酵素遺伝子(YAT1)遺伝子のコード配列は配列番号27で表され、コードされるアミノ酸配列は配列番号28で表される。カルニチンアセチル転移酵素遺伝子(YAT1)は、アセチルカルニチンをアセチルCoAに変換する酵素である。また、カルニチンアセチル転移酵素遺伝子(YAT1)の活性を有するポリペプチドは、配列番号28で表されるアミノ酸配列において、1もしくは数個のアミノ酸が欠失、置換、付加もしくは挿入されたアミノ酸配列を含み、かつYAT1活性を有するポリペプチドであってもよい。
【0028】
アミノ酸の欠失、置換、付加、又は挿入は、上記ポリペプチドをコードする遺伝子を、当技術分野で公知の手法によって改変することによって行うことができる。遺伝子に変異を導入するには、Kunkel法又はGapped duplex法等の公知手法又はこれに準ずる方法により行うことができ、例えば部位特異的突然変異誘発法を利用した変異導入用キット、例えばMutant−K(タカラバイオ社)やMutant−G(タカラバイオ社)などを用いて、あるいは、タカラバイオ社のLA PCR in vitro Mutagenesisシリーズキット、KOD−Plus−Mutagenesis Kit(TOYOBO)などを用いて変異を導入することができる。上記の各酵素の活性は、当技術分野において公知の手法により確認することができる。
【0029】
上記の遺伝子に代えて、それぞれの同等物を用いることもできる。この同等物は、それぞれの遺伝子と同等の機能を有することを条件に、一部のヌクレオチド残基が異なる遺伝子を意味する。このような同等物としては、それぞれのヌクレオチド配列と70%以上、好ましくは80%以上、より好ましくは85%以上、さらに好ましくは90%以上、最も好ましくは95%以上の相同性(配列同一性)があり、かつ各酵素の活性を有するポリペプチドをコードするヌクレオチド配列を含む遺伝子が挙げられる。前記同等物としてはさらに、それぞれのヌクレオチド配列もしくはその相補配列とストリンジェントな条件下でハイブリダイズし、かつ各酵素の活性を有するポリペプチドをコードするヌクレオチド配列を含む遺伝子が挙げられる。前記同等物としてはさらに、それぞれのヌクレオチド配列において1もしくは数個のヌクレオチド残基が欠失、置換、付加、または挿入された配列を含み、かつ各酵素の活性を有するポリペプチドをコードするヌクレオチド配列を含む遺伝子が挙げられる。
【0030】
ここで、ヌクレオチド残基の欠失、置換、付加、又は挿入は、上記配列を含む遺伝子を、当技術分野で公知の手法によって改変することによって行うことができる。遺伝子に変異を導入するには、Kunkel法又はGapped duplex法等の公知手法又はこれに準ずる方法により行うことができ、例えば部位特異的突然変異誘発法を利用した変異導入用キットを用いればよい。例えばMutant−K(タカラバイオ社)やMutant−G(タカラバイオ社)などを用いて、あるいは、タカラバイオ社のLA PCR in vitro Mutagenesisシリーズキット、KOD−Plus−Mutagenesis Kit(TOYOBO)などを用いて変異を導入することができる。各酵素の活性は、当技術分野において公知の手法により確認することができる。
【0031】
相同性(配列同一性)を示す数値(%)は、塩基配列比較用プログラム:例えばGENETYX−WIN7.0.0を用いて、デフォルト(初期設定)のパラメーターにより算出されるものである。すなわち、酵母染色体上の各遺伝子が、同一ではないが同等の機能、すなわち各活性を有するポリペプチドをコードする遺伝子によって相同組換え等を介して置換されていてもよい。各酵素の活性は、当技術分野において公知の手法により確認することができる。
【0032】
ストリンジェントな条件とは、例えば、Rapid−Hyb Buffer(GEヘルスケアバイオサイエンス社製)を用い、温度条件を好ましくは40〜70℃、より好ましくは60℃として、その他は添付のプロトコールに従って行うハイブリダイゼーション条件である。その後、例えば当業者に公知の一般的な方法を用い、2×SSCと0.1%(w/v)SDSから成り立つ溶液での5分間の洗浄、続いて1×SSCと0.1%(w/v)SDSから成り立つ溶液での10分間の洗浄、さらに0.1×SSCと0.1%(w/v)SDSから成り立つ溶液での10分間の洗浄を行うことを指す。ただしハイブリダイゼーション時の温度条件や、その後のメンブレンの洗浄に用いる溶液の塩濃度等の条件を適宜設定することにより、ある一定(70%、80%、85%、90%、95%のいずれか)以上の相同性を有する塩基配列を含むDNAをクローニングできる。そのようにして得られる遺伝子が、配列上は同一ではないが同等の機能、すなわち各活性を有するポリペプチドをコードする遺伝子によって相同組換え等を介して置換されていてもよい。各酵素の活性は、当技術分野において公知の手法により確認することができる。
【0033】
有用物質を合成するための遺伝子
本発明による酵母菌株は、所望の有用物質の製造に用いるために、該物質の生産に必要となる遺伝子で形質転換されていてもよい。例えば、本発明の方法によってアセチルCoAを経由する代謝産物を製造するために、本発明による酵母菌株は、アセチルCoAを経由する代謝産物を合成するための遺伝子により形質転換されていてもよい。必要な遺伝子は、製造しようとする物質に応じて、当業者であれば適切に選択することができる。
【0034】
例えば、イソプロパノールを製造しようとする場合には、さらなる遺伝的改変として、イソプロパノール合成遺伝子で酵母菌株を形質転換することができる。イソプロパノール合成遺伝子としては、特開2011−97929号公報に記載の酵素遺伝子群など、当業者に公知の遺伝子を用いることができ、例えば、CoA基転移酵素サブユニットA(ctfA)、同サブユニットB(ctfB)、アセト酢酸脱炭酸酵素(adc)および二級アルコール脱水素酵素(sadh)の4つの遺伝子の組み合わせを用いることができる。
【0035】
本発明の一つの実施態様によれば、本発明による酵母菌株は、さらなる遺伝的改変として、CoA基転移酵素サブユニットA(ctfA)、CoA転移酵素サブユニットB(ctfB)、アセト酢酸脱炭酸酵素(adc)および二級アルコール脱水素酵素(sadh)の活性を有するポリペプチドをコードしている4種の遺伝子により形質転換されている。このような酵母菌株はイソプロパノールの製造に用いることができ、この製造法は、本実施態様による酵母菌株を培養することを含む。
【0036】
これらの遺伝子は様々な生物に由来するものが知られており、その起源は特に制限されない。例えば、これらの遺伝子は、それぞれ独立に、クロストリジウム・アセトブチリカムまたはクロストリジウム・ベイジェリンキに由来するものとすることができる。それぞれの酵素の起源としては、例えば、CoA転移酵素遺伝子サブユニットA(ctfA)および同サブユニットB(ctfB)はクロストリジウム・アセトブチリカムが好ましく、アセト酢酸脱炭酸酵素(adc)および二級アルコール脱水素酵素(sadh)はクロストリジウム・ベイジェリンキが好ましい。
【0037】
クロストリジウム・アセトブチリカム由来のCoA転移酵素サブユニットA(ctfA)のアミノ酸配列は配列番号2で表される。CoA転移酵素サブユニットA(ctfA)は、同サブユニットB(ctfB)と一緒になって、アセトアセチルCoAをアセト酢酸に変換する酵素である。配列番号2で表されるアミノ酸配列をコードする遺伝子配列は、クロストリジウム・アセトブチリカム由来の同遺伝子の配列など、当業者であれば適宜選択することができるため、特に制限されないが、好ましくは本発明による酵母菌株の種において頻度の高いコドンを使用した配列とされる。例えば、キャンディダ・ユティリスにおけるコドン頻度に適合した遺伝子配列としては、配列番号1で表されるヌクレオチド配列が挙げられる。CoA転移酵素サブユニットA(ctfA)の活性を有するポリペプチドは、配列番号2で表されるアミノ酸配列において、1もしくは数個のアミノ酸が欠失、置換、付加もしくは挿入されたアミノ酸配列を含み、かつctfA活性を有するポリペプチドであってもよい。
【0038】
クロストリジウム・アセトブチリカム由来のCoA転移酵素サブユニットB(ctfB)のアミノ酸配列は配列番号4で表される。CoA転移酵素サブユニットB(ctfB)は、同サブユニットA(ctfA)と一緒になって、アセトアセチルCoAをアセト酢酸に変換する酵素である。配列番号4で表されるアミノ酸配列をコードする遺伝子配列は、クロストリジウム・アセトブチリカム由来の同遺伝子の配列など、当業者であれば適宜選択することができるため、特に制限されないが、好ましくは本発明による酵母菌株の種において頻度の高いコドンを使用した配列とされる。例えば、キャンディダ・ユティリスにおけるコドン頻度に適合した遺伝子配列としては、配列番号3で表されるヌクレオチド配列が挙げられる。CoA転移酵素サブユニットB(ctfB)の活性を有するポリペプチドは、配列番号4で表されるアミノ酸配列において、1もしくは数個のアミノ酸が欠失、置換、付加もしくは挿入されたアミノ酸配列を含み、かつctfB活性を有するポリペプチドであってもよい。
【0039】
クロストリジウム・ベイジェリンキ由来のアセト酢酸脱炭酸酵素(adc)のアミノ酸配列は配列番号6で表される。アセト酢酸脱炭酸酵素(adc)は、アセト酢酸をアセトンに変換する酵素である。配列番号6で表されるアミノ酸配列をコードする遺伝子配列は、クロストリジウム・ベイジェリンキ由来の同遺伝子の配列など、当業者であれば適宜選択することができるため、特に制限されないが、好ましくは本発明による酵母菌株の種において頻度の高いコドンを使用した配列とされる。例えば、キャンディダ・ユティリスにおけるコドン頻度に適合した遺伝子配列としては、配列番号5で表されるヌクレオチド配列が挙げられる。アセト酢酸脱炭酸酵素(adc)の活性を有するポリペプチドは、配列番号6で表されるアミノ酸配列において、1もしくは数個のアミノ酸が欠失、置換、付加もしくは挿入されたアミノ酸配列を含み、かつadc活性を有するポリペプチドであってもよい。
【0040】
クロストリジウム・ベイジェリンキ由来の二級アルコール脱水素酵素(sadh)のアミノ酸配列は配列番号8で表される。二級アルコール脱水素酵素(sadh)は、アセトンをイソプロパノールに変換する酵素である。配列番号8で表されるアミノ酸配列をコードする遺伝子配列は、クロストリジウム・ベイジェリンキ由来の同遺伝子の配列など、当業者であれば適宜選択することができるため、特に制限されないが、好ましくは本発明による酵母菌株の種において頻度の高いコドンを使用した配列とされる。例えば、キャンディダ・ユティリスにおけるコドン頻度に適合した遺伝子配列としては、配列番号7で表されるヌクレオチド配列が挙げられる。二級アルコール脱水素酵素(sadh)の活性を有するポリペプチドは、配列番号8で表されるアミノ酸配列において、1もしくは数個のアミノ酸が欠失、置換、付加もしくは挿入されたアミノ酸配列を含み、かつsadh活性を有するポリペプチドであってもよい。
【0041】
アミノ酸の欠失、置換、付加、又は挿入は、上記ポリペプチドをコードする遺伝子を、当技術分野で公知の手法によって改変することによって行うことができる。遺伝子に変異を導入するには、Kunkel法又はGapped duplex法等の公知手法又はこれに準ずる方法により行うことができ、例えば部位特異的突然変異誘発法を利用した変異導入用キット、例えばMutant−K(タカラバイオ社)やMutant−G(タカラバイオ社)などを用いて、あるいは、タカラバイオ社のLA PCR in vitro Mutagenesisシリーズキット、KOD−Plus−Mutagenesis Kit(TOYOBO)などを用いて変異を導入することができる。上記の各酵素の活性は、当技術分野において公知の手法により確認することができる。
【0042】
上記の遺伝子に代えて、それぞれの同等物を用いることもできる。この同等物は、それぞれの遺伝子と同等の機能を有することを条件に、一部のヌクレオチド残基が異なる遺伝子を意味する。このような同等物としては、それぞれのヌクレオチド配列と70%以上、好ましくは80%以上、より好ましくは85%以上、さらに好ましくは90%以上、最も好ましくは95%以上の相同性(配列同一性)があり、かつ各酵素の活性を有するポリペプチドをコードするヌクレオチド配列を含む遺伝子が挙げられる。前記同等物としてはさらに、それぞれのヌクレオチド配列もしくはその相補配列とストリンジェントな条件下でハイブリダイズし、かつ各酵素の活性を有するポリペプチドをコードするヌクレオチド配列を含む遺伝子が挙げられる。前記同等物としてはさらに、それぞれのヌクレオチド配列において1もしくは数個のヌクレオチド残基が欠失、置換、付加、または挿入された配列を含み、かつ各酵素の活性を有するポリペプチドをコードするヌクレオチド配列を含む遺伝子が挙げられる。
【0043】
ここで、ヌクレオチド残基の欠失、置換、付加、又は挿入は、上記配列を含む遺伝子を、当技術分野で公知の手法によって改変することによって行うことができる。遺伝子に変異を導入するには、Kunkel法又はGapped duplex法等の公知手法又はこれに準ずる方法により行うことができ、例えば部位特異的突然変異誘発法を利用した変異導入用キットを用いればよい。例えばMutant−K(タカラバイオ社)やMutant−G(タカラバイオ社)などを用いて、あるいは、タカラバイオ社のLA PCR in vitro Mutagenesisシリーズキット、KOD−Plus−Mutagenesis Kit(TOYOBO)などを用いて変異を導入することができる。各酵素の活性は、当技術分野において公知の手法により確認することができる。
【0044】
相同性(配列同一性)を示す数値(%)は、塩基配列比較用プログラム:例えばGENETYX−WIN7.0.0を用いて、デフォルト(初期設定)のパラメーターにより算出されるものである。すなわち、酵母染色体上の各遺伝子が、同一ではないが同等の機能、すなわち各活性を有するポリペプチドをコードする遺伝子によって相同組換え等を介して置換されていてもよい。各酵素の活性は、当技術分野において公知の手法により確認することができる。
【0045】
ストリンジェントな条件とは、例えば、Rapid−Hyb Buffer(GEヘルスケアバイオサイエンス社製)を用い、温度条件を好ましくは40〜70℃、より好ましくは60℃として、その他は添付のプロトコールに従って行うハイブリダイゼーション条件である。その後、例えば当業者に公知の一般的な方法を用い、2×SSCと0.1%(w/v)SDSから成り立つ溶液での5分間の洗浄、続いて1×SSCと0.1%(w/v)SDSから成り立つ溶液での10分間の洗浄、さらに0.1×SSCと0.1%(w/v)SDSから成り立つ溶液での10分間の洗浄を行うことを指す。ただしハイブリダイゼーション時の温度条件や、その後のメンブレンの洗浄に用いる溶液の塩濃度等の条件を適宜設定することにより、ある一定(70%、80%、85%、90%、95%のいずれか)以上の相同性を有する塩基配列を含むDNAをクローニングできる。そのようにして得られる遺伝子が、配列上は同一ではないが同等の機能、すなわち各活性を有するポリペプチドをコードする遺伝子によって相同組換え等を介して置換されていてもよい。各酵素の活性は、当技術分野において公知の手法により確認することができる。
【0046】
イソプロパノール以外にも、リコペンなどの特定のカロテノイド、スクアレンあるいはスクアレンから生産される特定のステロールなどの物質を生産する酵母菌株を構築することが可能である。例えば、Erwinia uredovora由来のGGPP合成酵素遺伝子、フィトエン合成酵素遺伝子、フィトエン不飽和化酵素遺伝子を発現し、内在性のHMG−CoA還元酵素遺伝子(HMG1)を過剰発現するキャンディダ・ユティリスはリコペンを高蓄積することが報告されている(Shimada H, Kondo K, Fraser PD, Miura Y, Saito T, Misawa N. Increased carotenoid production by the food yeast Candida utilis through metabolic engineering of the isoprenoid pathway. Appl Environ Microbiol. 1998 Jul;64(7):2676-80)。また、ザイモステロール-24-メチルトランスフェラーゼ遺伝子およびエルゴスタ5,7,24,(28)−トリエノール−22−脱水素酵素遺伝子の発現に欠損を持ち、HMG1が過剰発現するように操作されたサッカロマイセス・セレビシエ組み換え株は、高濃度のスクアレンを蓄積することが知られている(特開平5−192184号公報)。これらの手法を用いることにより、リコペン、スクアレンを生産する酵母菌株を構築することが可能である。
【0047】
酵母菌株の分子育種
本発明による酵母菌株の分子育種は、上記の遺伝子群を過剰発現させるための遺伝的改変を酵母菌株に施すこと、例えば、上記の遺伝子群を発現可能な状態で酵母菌株に導入することによって行うことができる。これら遺伝子群は、相同組換え等の技術により酵母の染色体上に組み込まれることが好ましい。このような染色体上への組込みを実現するための相同組換え用遺伝子配列の選択は、当業者において周知であり、当業者であれば必要に応じて適切な相同組換え用遺伝子配列を選択して相同組換え用DNA断片を構成することができる。
【0048】
上記の遺伝子群が酵母ゲノムに組込まれる染色体上の位置は特に制限されるものではないが、例えば、オロチジン5’リン酸デカルボキシラーゼ遺伝子(URA3)座、アルファアミノアジピン酸還元酵素遺伝子(LYS2)座などを選択することができる。また、これらの遺伝子は、組み込まれた遺伝子座に当初から存在するプロモーター、例えばURA3遺伝子プロモーター、LYS2遺伝子プロモーターなどの制御下に置いてもよいが、グリセルアルデヒド−3−リン酸脱水素酵素(GAP)遺伝子プロモーターなどの他のプロモーターの制御下に置いてもよい。
【0049】
本発明の好ましい実施態様によれば、本発明による酵母菌株の育種に用いられる酵母はキャンディダ・ユティリスとされる。キャンディダ・ユティリスは、その倍数性が高く、胞子を形成しない。倍数性の高い株の遺伝子に変異を導入しようとする場合、1倍体の株に比べて、その変異を重度に加える必要があるが、その際に、変異を与えたい遺伝子ではない遺伝子にも変異が加えられる可能性が高まると考えられる。従って、キャンディダ・ユティリスの遺伝子に変異を導入する場合には、標的とする遺伝子のみに効率よく多重に変異を加えることができる技術を用いることが好ましい。キャンディダ・ユティリスの形質転換法としては、特開2003−144185号公報に記載の技術が挙げられる。この文献では、利用可能なベクターとして、キャンディダ・ユティリスの染色体DNAと相同な配列と、選択マーカー遺伝子とを含んでなり、相同組換えによって異種遺伝子をキャンディダ・ユティリスの染色体DNAに組み込むことができるもの、あるいは、キャンディダ・ユティリスで自律複製機能を有するDNA配列と、選択マーカー遺伝子とを含んでなり、高い頻度でキャンディダ・ユティリスを形質転換できるものが開発されている。
【0050】
形質転換系で利用可能な選択マーカー遺伝子としては、酵母において機能し得る薬剤耐性マーカー、好ましくはシクロヘキシミド耐性型L41遺伝子、ジェネティシン(G418)耐性を付与する遺伝子、またはハイグロマイシンB耐性を付与する遺伝子などがある。ジェネティシン(G418)耐性を付与する遺伝子、またはハイグロマイシンB耐性を付与する遺伝子は、野生の酵母には存在しない配列であるため、標的の遺伝子座に組込まれる確率が高いと考えられている。また、これらの薬剤耐性遺伝子は、その宿主の形質に与える影響も小さいと考えられている(Baganz Fら,13(16):1563−73.,1997)(Cordero Otero Rら,Appl Microbiol Biotechnol,46(2):143−8.,1996)。
【0051】
他の形質転換系として、バクテリオファージP1由来のCre−loxP系が挙げられる。これは、2つの34bpのloxP配列間での部位特異的組換えシステムであり、この組換えはCre遺伝子がコードするCre組換え酵素によって触媒される。このシステムは、サッカロマイセス・セレビシエなどの酵母細胞においても機能することが報告されており、2つのloxP配列の間に配置された選択マーカー遺伝子は、loxP配列間の組換えによって除去されることが知られている(Guldener,U.ら,Nucleic Acids Res.,24,2519−24.,1996)。このシステムはクルイベロマイセス・ラクティス(Kluyveromyces lactis)など、複数の酵母種で利用されている(Steensma,H.Y.ら,Yeast,18,469−72.,2001)。
【0052】
物質の製造方法
本発明による酵母菌株を適当な炭素源の存在下で培養することにより、培養物中に物質(例えばイソプロパノール)を製造することができる。本発明による物質(例えばイソプロパノール)製造法によれば、培養系から物質(例えばイソプロパノール)を分離する工程を実施することにより、物質(例えばイソプロパノール)を得ることができる。なお、本発明において培養物とは、培養上清の他、培養細胞あるいは菌体、細胞もしくは菌体の破砕物を包含している。
【0053】
本発明による酵母菌株の培養にあたっては、酵母の種類に応じて培養方法や培養条件を選択することができる。培養方法としては、例えば、試験管、フラスコあるいはジャーファーメンターを用いた液体培養法を挙げることができ、回分培養、半回分培養などの培養形式を採用できる。振盪速度および通気量は、一般的には50〜200rpm、好ましくは80〜150rpm、あるいはこの振盪速度に相当する通気量とされる。初期の菌体量としてのOD600は、一般的には0.1〜30、好ましくは0.5〜10とされる。さらに、本発明による酵母菌株の培養にあたっては、振盪速度は速い方が好ましく(つまり通気量は多い方が好ましく)、初期OD600は高い方が好ましい。例えば、試験管やフラスコでの培養では、振盪速度は約120rpmまたはそれ以上、初期OD600は約10とすることが好ましい。
【0054】
本発明による物質(例えばイソプロパノール)製造法において、培地の組成は、酵母が生育し、且つ目的の物質を製造できる各種栄養素を含む組成であればよく、特に限定されない。培地に含まれる資化炭素源としては、例えば、グルコースのほかにもキシロースやスクロースでも資化できれば用いることができる。本発明の好ましい実施態様によれば、炭素源はグルコースを含む。培地中のグルコース濃度は、目的の物質の生産効率を確認しながら、当業者であれば適宜調節することができる。特に、本発明による酵母菌株の培養では、培地中のグルコース濃度は、比較的低濃度に保つことが好ましい。例えば、192時間の培養において、グルコースの初期濃度を100g/Lとし、培養72時間後に終濃度100g/Lとなるようにグルコース固体を添加した場合に比べて、グルコースの初期濃度を50g/Lとし、48時間後、96時間後および144時間後に終濃度50g/Lとなるようにグルコース固体を添加した場合の方が、物質の生産効率が高い。
【0055】
培地に含まれる栄養源としては、例えば、酵母エキス、ペプトン、ホエーなどが用いられるが、YP(10g/L酵母エキス、20g/Lペプトン)に上記の資化炭素源を加えた培地、例えばYPD5(50g/Lグルコース、10g/L酵母エキス、20g/Lペプトン)等の培地で、さらに適宜pH調整されたものが便利である。
【0056】
発酵温度は、用いる酵母の生育可能な範囲で選択することができる。発酵温度は、例えば、約15℃〜45℃とすることができ、より好ましくは20〜40℃、さらに好ましくは25〜35℃、最も好ましくは約30℃とする。また、発酵過程における培地のpHは3〜8に保持することが好ましく、より好ましくはpH4〜7であり、必要に応じてpH調節剤を用いることができる。用いるpH調節剤としては、炭酸カルシウム、水酸化ナトリウム、水酸化カリウム等が挙げられるが、好ましくは炭酸カルシウムとされる。
【0057】
物質(例えばイソプロパノール)製造に要する反応時間は特に限定されず、本発明の効果が認められる限り任意の反応時間で実施される。これらの条件の最適化は、当業者であれば容易に行うことができる。
【0058】
本発明による物質(例えばイソプロパノール)の製造法においては、このようにして製造した物質(例えばイソプロパノール)成分を培地から分離・回収するが、そのための具体的な手順は、分離・回収しようとする物質に応じて、当業者であれば容易に選択することができる。
【実施例】
【0059】
以下、実施例を挙げて本発明をより具体例に説明するが、これら実施例は本発明の技術的範囲を制限するものではない。
【0060】
実施例1:高いイソプロパノール耐性を示すキャンディダ・ユティリス株の選抜
キャンディダ・ユティリス NBRC0396株、NBRC0619株、NBRC0988株、NBRC0639株、NBRC0626株、NBRC1086株、NBRC10707株、ATCC9255株、ATCC28955株、ATCC44638株、ATCC9206株、ATCC96621株、ATCC18201株、およびATCC15239株を、YPD2(20g/Lペプトン、10g/L酵母エキス、20g/Lグルコース)に接種し、30℃で培養を行った。定常期の酵母培養液を10倍ごとに段階希釈し、10〜100g/Lのイソプロパノールを含むYPD2寒天培地にスポットし、30℃で培養した。3日間培養を継続し、コロニーの形成を観察した。結果を表1に記載する。
【0061】
【表1】

【0062】
以上の結果より、アッセイを行った株の中では、NBRC1086株、ATCC9255株、およびATCC15239株が、最も高い65g/LのIPA耐性を示すことが明らかとなった。
【0063】
実施例2:ピルビン酸脱炭酸酵素遺伝子が破壊されたキャンディダ・ユティリスATCC15239株の構築
キャンディダ・ユティリスATCC15239株において、その内因性遺伝子であるピルビン酸脱炭酸酵素遺伝子(PDC)を破壊した。この遺伝子破壊は、Cre−loxPシステムを用いて国際公開第2010/095750号パンフレットと同様の方法で行った。2回の破壊により、PDC1アレルが完全に欠損した株(dPDC1株)を構築した。dPDC1株は完全にはエタノール生産能を欠損しなかったが、エタノール生産能は著しく低下していた。
【0064】
実施例3:クロストリジウム・アセトブチリカム由来のCoA転移酵素サブユニットA遺伝子、同サブユニットB遺伝子、クロストリジウム・ベイジェリンキ由来のアセト酢酸脱炭酸酵素遺伝子および二級アルコール脱水素酵素遺伝子を発現するベクターの構築
イソプロパノール生合成に関わる4つの遺伝子、クロストリジウム・アセトブチリカム由来のCoA転移酵素遺伝子サブユニットA(ctfA)、同サブユニットB(ctfB)、クロストリジウム・ベイジェリンキ由来のアセト酢酸脱炭酸酵素(adc)および二級アルコール脱水素酵素(sadh)を効率良くキャンディダ・ユティリスで発現させるために、以下の項目を指標として、天然に存在しない新規な遺伝子配列を設計し、合成した:
(1)キャンディダ・ユティリスにおいて多用されているコドンの使用;
(2)遺伝子クローニングに不適当な制限酵素部位の排除;および
(3)染色体組み込み型発現ベクターに導入するための、5’末端におけるXbaIサイトおよび3’末端におけるBamHIサイトの付与。
【0065】
合成されたCoA転移酵素遺伝子サブユニットA遺伝子、同サブユニットB遺伝子、アセト酢酸脱炭酸酵素遺伝子および二級アルコール脱水素酵素遺伝子のヌクレオチド配列を、それぞれ配列番号1、3、5および7に示す。また、これらのヌクレオチド配列によってコードされるアミノ酸配列を、それぞれ配列番号2、4、6および8に示す。これらの遺伝子を有するプラスミドをそれぞれGA09142−ctfA、GA09142−ctfB、GA09142−adc、GA09142−sadhと名付けた。
【0066】
pVT92は、キャンディダ・ユティリスのオロチジン5’酸脱炭酸酵素遺伝子(URA3)座への組み込みベクターであり、ハイグロマイシン耐性遺伝子を選択マーカーとして有している(国際公開第2010/095750号パンフレット)。pVT92は、BgllII処理してリニアDNAにすることにより、染色体導入することができる。pVT92は、外来遺伝子を発現させるために、XbaI/BamHIギャップの外側にそれぞれ、グリセルアルデヒド脱水素酵素遺伝子プロモーターとグリセリン酸3’リン酸リン酸化酵素遺伝子ターミネーターを有している。また、pVT92は、プロモーターの5’末端側およびターミネーターの3’末端側には、それぞれNheIサイトおよびSpeIサイトを有している。
【0067】
pVT239は、キャンディダ・ユティリスのアルファアミノアジピン酸還元酵素遺伝子(LYS2)座への組み込みベクターであり、ジェネティシン耐性遺伝子を選択マーカーとして有している(国際公開第2010/095750号パンフレット)。pVT239は、NotI処理してリニアDNAにすることにより、染色体導入することができる。また、pVT239は、ジェネティシン耐性遺伝子の5’末端側にSpeIサイトを有している。
【0068】
各イソプロパノール合成酵素遺伝子を含む4つのベクターGA09142−ctfA、GA09142−ctfB、GA09142−adc、およびGA09142−sadhを、XbaIおよびBamHI処理し、アガロースゲル抽出による精製を行った後、同制限酵素で処理したpVT92にライゲーションして、pVT296、pVT298、pVT300およびpVT302を得た。pVT296およびpVT300はSpeI処理し、脱リン酸化を行った。また、pVT298およびpVT302はNheI、SpeI処理を行った後、ctfB発現カセット、sadh発現カセットをそれぞれ含むDNA断片の精製を行い、それぞれpVT296およびpVT300に連結して、pVT321およびpVT325を得た。pVT325をNheIおよびSpeIで処理した後、adc発現カセットとsadh発現カセットを含むDNA断片の精製を行い、pVT239のSpeIサイトに導入し、pVT346を得た。最終的に得られたpVT321はctfAとctfBのそれぞれの発現カセットとハイグロマイシン耐性遺伝子を有し、キャンディダ・ユティリスのURA3座に組み込まれるベクターであり、pVT346はadcとsadhのそれぞれの発現カセットとジェネティシン耐性遺伝子を有し、キャンディダ・ユティリスのLYS2座に組み込まれるベクターである。
【0069】
実施例4:4つのイソプロパノール合成遺伝子を有するキャンディダ・ユティリス株の構築
pVT321をBgllIIで消化し、エタノール沈殿により濃縮した。得られたDNA断片を用いて、C. utilis ATCC15239株、および実施例2で得られたdPDC1株を形質転換し、600μg/mLハイグロマイシンを含むYPD培地に塗抹し、30℃で2日間培養した。導入した全ての遺伝子を有する株を選抜した。
【0070】
さらにpVT346をNotIで消化し、エタノール沈殿により濃縮した。得られたそれぞれのDNA断片を用いて上記で得られた2株をそれぞれ形質転換し、600μg/mLハイグロマイシンと200μg/mLジェネティシンを含むYPD培地に塗抹し、30℃で2日間培養した。導入した全ての遺伝子を有する株を選抜し、ATCC15239株由来のイソプロパノール生産株をTMS272株、dPDC株由来のイソプロパノール生産株をTMS290株とした。
【0071】
実施例5:イソプロパノール生産試験
実施例4で得られた形質転換体のイソプロパノール生産能の評価を、以下に示すように実施した。
【0072】
各種形質転換体を2mL YPD液体培地(20g/Lペプトン、10g/L酵母エキス、20g/Lグルコース)/14mL容試験管に接種し、140rpm、30℃で24時間振とう培養した。得られた前培養液を100mL YPD10(20g/Lペプトン、10g/L酵母エキス、100g/Lグルコース)/200mL容三角フラスコもしくはYPD10C(20g/Lペプトン、10g/L酵母エキス、100g/Lグルコース、45g/L炭酸カルシウム)/200mL容三角フラスコに接種し、初期OD600=1、100rpm、30℃で振とう培養した。培養液から経時的にサンプリングし、0.2μmフィルターでろ過した後に、各種成分の定量分析を行った。グルコース、酢酸、エタノールおよびグリセロールは、高速液体クロマトフラフィー(島津製作所製)、ISep−ION300カラム(東京化成工業製)および示唆屈折計を用いて定量した。イソプロパノールおよびアセトンは、ガスクロマトフラフィーGC−2010 Plus(島津製作所製)を用いて定量した。
【0073】
また、フラスコでの簡易的な中和培養としてYPD10C(20g/Lペプトン、10g/L酵母エキス、100g/Lグルコース、45g/L炭酸カルシウム)培地を用いても同様の実験を行った。
【0074】
非中和条件下での培養試験の結果を図2、中和条件下での培養試験の結果を図3に記載する。なお、データは3反復試行の平均で示した。
【0075】
図2によれば、親株であるATCC15239株およびdPDC1株ではイソプロパノールの生産が認められなかったのに対し、イソプロパノール合成遺伝子を導入したTMS272株およびTMS290株では、それぞれ最大で0.16g/Lおよび0.13g/Lのイソプロパノールの生産が認められた。また、PDC1遺伝子を破壊した株では糖の消費速度および増殖能が著しく低下していた。TMS272株では培養24時間でグルコースが完全に消費され、エタノールの蓄積が認められたが、その後の培養の継続に伴ってエタノール濃度が低下した。エタノール濃度が低下している間もイソプロパノールの生産が認められたことから、TMS272株は、一度生産したエタノールを二次消費することによりイソプロパノールを生産していることが考えられた。dPDC1株に由来するTMS290株ではイソプロパノール生産速度が低下したことから、酵母におけるイソプロパノール生産においては、基質である細胞質のアセチルCoAがエタノールから供給される経路が最も重要であることが示唆された。エタノールの同化は酸化的反応であることから、これらの結果は、酵母によるイソプロパノール生産においてはキャンディダ・ユティリスのような呼吸活性の高いクラブツリー陰性酵母が適している可能性を示唆している。
【0076】
図3によれば、培地に炭酸カルシウムを添加したことにより、TMS272株は1.1g/Lのイソプロパノールを生産し、非中和条件より生産量が増大した。また、TMS272株は、16.2g/Lの酢酸を蓄積しており、何らかの理由により酢酸が蓄積したことでpHが低下し、イソプロパノールの生産を阻害していたことが考えられた。また、TMS290株においても、中和条件下ではイソプロパノール生産量が0.9g/Lまで増大しており、一方で37.4g/Lという高濃度のグリセロールを蓄積していた。
【0077】
実施例6:過剰発現によってイソプロパノール生産性を向上させる遺伝子の選抜
過剰発現することによりイソプロパノールの生産性を向上させる遺伝子の選抜を行った。候補遺伝子が触媒する代謝経路を図1に記載する。
【0078】
キャンディダ・ユティリス内在性のアセチルCoA加水分解酵素遺伝子(ACH1;ヌクレオチド配列:配列番号9;アミノ酸配列:配列番号10)、アセチルCoA合成酵素遺伝子(ACS1;ヌクレオチド配列:配列番号11;アミノ酸配列:配列番号12、ACS2;ヌクレオチド配列:配列番号13;アミノ酸配列:配列番号14)、細胞質局在型エタノール脱水素酵素遺伝子(ADH2;ヌクレオチド配列:配列番号15;アミノ酸配列:配列番号16)、ミトコンドリア局在型エタノール脱水素酵素遺伝子(ADH3;ヌクレオチド配列:配列番号17;アミノ酸配列:配列番号18)、細胞質局在型アセトアルデヒド脱水素酵素遺伝子(ALD1;ヌクレオチド配列:配列番号19;アミノ酸配列:配列番号20)、ミトコンドリア局在型アセトアルデヒド脱水素酵素遺伝子(ALD2;ヌクレオチド配列:配列番号21;アミノ酸配列:配列番号22)、ミトコンドリア内膜カルニチン輸送体(CRC1;ヌクレオチド配列:配列番号23;アミノ酸配列:配列番号24)、ミトコンドリア局在型カルニチンアセチルCoA転移酵素(CAT2;ヌクレオチド配列:配列番号25;アミノ酸配列:配列番号26)、カルニチンアセチル転移酵素遺伝子(YAT1;ヌクレオチド配列:配列番号27;アミノ酸配列:配列番号28、YAT2;ヌクレオチド配列:配列番号29;アミノ酸配列:配列番号30)、アセチルCoAアセチル基転移酵素(ERG10;ヌクレオチド配列:配列番号61;アミノ酸配列:配列番号62)、ヤロウィア・リポリティカ由来のATP−クエン酸除去付加酵素遺伝子(ACL1;ヌクレオチド配列:配列番号31;アミノ酸配列:配列番号32と、ACLY;ヌクレオチド配列:配列番号33;アミノ酸配列:配列番号34がサブユニットを構成する遺伝子)を、それぞれのゲノムDNAを鋳型としてPCRにより増幅した。PCRに用いたプライマーは以下の通りである:(ACH1;配列番号35と配列番号36)、(ACS1;配列番号37と配列番号38)、(ACS2;配列番号39と配列番号40)、(ADH2;配列番号41と配列番号42)、(ADH3;配列番号43と配列番号44)、(ALD1;配列番号45と配列番号46)、(ALD2;配列番号47と配列番号48)、(CAT2;配列番号49と配列番号50)、(CRC1;配列番号51と配列番号52)、(YAT1;配列番号53と配列番号54)、(YAT2;配列番号55と配列番号56)、(ACL1;配列番号57と配列番号58)、(ACLY;配列番号59と配列番号60)、(ERG10;配列番号63と配列番号64)。
【0079】
得られたPCR産物をXbaI/BamHI処理して、同制限酵素で処理したpCU155(Keiji Kondo, , Yutaka Miura, Hidetaka Sone, Kazuo Kobayashi & Hiroshi lijima., High-level expression of a sweet protein, monellin, in the food yeast Candida utilis., Nature Biotechnology 15, 453 - 457 (1997))にクローニングした。pCU155は、シクロヘキシミド耐性遺伝子を選抜マーカーとして有し、URA3座にマルチコピーインテグレーションされるベクターであり、XbaI/BamHIギャップの外側にそれぞれ、グリセルアルデヒド脱水素酵素遺伝子プロモーターと同ターミネーターを有している。これまでに構築しているイソプロパノール生産株のURA3座にはイソプロパノール合成酵素遺伝子が組み込まれているが、C.utilisは高次倍数体であり、URA3座が複数コピー存在するため、さらにURA3座に遺伝子の導入が可能である。また、イソプロパノール合成遺伝子はURA3座に二点組み換えにより挿入されているため、pCU155の組み込みにより、導入された遺伝子の置換が起こることはない。候補遺伝子がクローン化されたプラスミドはBgllIIで消化し、エタノール沈殿により濃縮した。得られたDNA断片を用いてC.utilis TMS272株を形質転換し、600μg/mLハイグロマイシン、200μg/mLジェネティシンおよび20μg/mLシクロヘキシミドを含むYPD培地に塗抹し、30℃で4日間培養した。なお、ACL1とACLYのDNAは混合し、同時に形質転換を行った。得られた各4クローンについて発酵試験を行い、イソプロパノール生産性を調査した。結果を図4に示す。
【0080】
図4によると、ACS1、ACS2、CRC1、YAT1、ERG10、またはACLの過剰発現により、イソプロパノール生産量が1.5〜4.0倍になることが明らかとなった。ACS1、ACS2、YAT1、およびACLの各遺伝子はアセチルCoAの合成に関わる酵素遺伝子であり、CRC1遺伝子はアセチルCoA合成の前駆物質をミトコンドリアから細胞質に輸送するキャリアタンパク質遺伝子であり、ERG10遺伝子はアセチルCoAからアセトアセチルCoAを合成する遺伝子である。これらの結果により、酵母によるイソプロパノール高生産のためには、細胞質のアセチルCoAを効率的に生産するための酵素遺伝子の発現または活性をアップレギュレートすることが重要であることが示される。
【0081】
実施例7:ACL1/ACLY、ACS2、ERG10、CRC1およびYAT1を同時に過剰発現するイソプロパノール生産株の構築と培養試験
ACL1、ACLY、ACS2、ERG10、CRC1およびYAT1の各遺伝子がクローン化されたプラスミドをBgllIIで消化し、エタノール沈殿により濃縮した。得られた各5μgのDNA断片を混合し、この混合物を用いてTMS272株を形質転換し、600μg/mLハイグロマイシン、200μg/mLジェネティシンおよび10μg/mLシクロヘキシミドを含むYPD培地に塗抹し、30℃で4日間培養した。得られた80クローンについて発酵試験を行い、イソプロパノール生産量が最も高い株を選抜し、TMS378株とした。
【0082】
TMS378株の培養試験を行った結果を図5に示す。TMS272株が1.3g/Lのイソプロパノールを生産したのに対して、TMS378株は9.8g/Lと約7.5倍のイソプロパノール生産量を示した。また、TMS272株では酢酸が18.1g/L蓄積したのに対して、TMS378株は4.5g/Lと、主要な副産物である酢酸が約4分の1に低下していた。
【0083】
実施例8:TMS378株のイソプロパノール生産における初期OD600および振とう速度の影響
イソプロパノール生産に適したTMS378株の培養条件の検討を行った。TMS378株を2mL YPD液体培地(20g/Lペプトン、10g/L酵母エキス、20g/Lグルコース)/14mL容試験管に接種し、140rpm、30℃で24時間振とう培養した。得られた前培養液を100mL YPD10C(20g/Lペプトン、10g/L酵母エキス、100g/Lグルコース、45g/L炭酸カルシウム)/300mL容三角フラスコに接種し、初期OD600を1もしくは10とし、100rpmもしくは120rpm、30℃で振とう培養した。培養72時間後に終濃度100g/Lとなるようにグルコース固体を培養液に添加し、さらに144時間まで培養を行った。経時的にサンプリングして0.2μmフィルターでろ過した。各種定量分析は、実施例5に記載の方法に従って行った。なお、データは3反復試行の平均で示した。結果を図6に示す。
【0084】
図6によると、初期OD600=10、120rpmで培養したときに最も高いイソプロパノール生産量が見られ、培養144時間後に最大で18.5g/Lのイソプロパノールが生産された。その際の対糖収率は30.8%であった。このことから、振とう速度が速いほど(通気量が多いほど)、また、初期ODが高いほど、イソプロパノール生産には好ましいことが明らかとなった。
【0085】
実施例9:TMS378株のイソプロパノール生産における添加グルコース濃度の影響
イソプロパノール生産に適したTMS378株の培養条件の検討を行った。TMS378株を2mL YPD液体培地(20g/Lペプトン、10g/L酵母エキス、20g/Lグルコース)/14mL容試験管に接種し、140rpm、30℃で24時間振とう培養した。得られた前培養液を100mL YPD10C(20g/Lペプトン、10g/L酵母エキス、100g/Lグルコース、45g/L炭酸カルシウム)/300mL容三角フラスコもしくはYPD5C(20g/Lペプトン、10g/L酵母エキス、50g/Lグルコース、45g/L炭酸カルシウム)/300mL容三角フラスコに接種し、初期OD600=1、120rpm、30℃で振とう培養した。YPD10Cで培養を行ったサンプルについては、培養72時間後に終濃度100g/Lとなるようにグルコース固体を添加し、さらに192時間まで培養を行った。また、YPD5Cで培養を行ったサンプルについては、培養48時間おきに3回終濃度50g/Lとなるようにグルコース固体を添加し、さらに192時間まで培養を行った。いずれの条件も最終のグルコース濃度は200g/Lである。経時的にサンプリングして0.2μmフィルターでろ過した。各種定量分析は、実施例5に記載の方法に従って行った。なお、データは3反復試行の平均で示した。結果を図7に示す。
【0086】
図7によると、YPD10Cで培養し、培養72時間後に100g/Lのグルコースを添加した場合は、培養122時間後に最大で19.2g/Lのイソプロパノールが生産された。しかし、それ以降はイソプロパノール生産量の増大は認められず、酢酸が蓄積した(27.2g/L)。イソプロパノールの生産における最大の対糖収率は29.0%であった。一方、YPD5Cで培養し、48時間おきに50g/Lのグルコースを3回添加した場合は、培養192時間までイソプロパノールの増大が認められ、培養192時間後に最大で27.4g/Lのイソプロパノールが生産された。また、酢酸の蓄積はほとんど認めれらなかった。この条件でのイソプロパノールの生産における最大の対糖収率は41.5%であった。このことから、添加するグルコースの濃度を比較的低濃度に保つことはイソプロパノール生産において好ましいことが明らかとなった。
【0087】
実施例10:イソプロパノール生産量向上に寄与の大きい遺伝子の特定
同時に過剰発現することによってイソプロパノール生産量を向上させる寄与の大きい遺伝子を特定するために、(i)ACL1、ACLYおよびACS2の組み合わせを過剰発現する株、(ii)ACL1、ACLYおよびERG10の組み合わせを過剰発現する株、(iii)ACS2およびERG10の組み合わせを過剰発現する株、(iv)ACL1、ACLY、ACS2およびERG10の組み合わせを過剰発現する株をそれぞれ構築し、ACL1、ACLY、ACS2、CRC1、ERG10およびYAT1の組み合わせを過剰発現する株(TMS378株)およびそれぞれの単独遺伝子の過剰発現株との間で、イソプロパノール生産量の比較を行った。組み換え株の構築は実施例6および実施例7と同様の方法を用いて行い、組み込みたい遺伝子DNAを混合し、同時に形質転換を行った。得られた各32クローンについて発酵試験を行い、それぞれの導入遺伝子の組み合わせにおいて平均的なイソプロパノール生産量を示す株を選抜した。各構築株を2mL YPD液体培地(20g/Lペプトン、10g/L酵母エキス、20g/Lグルコース)/14mL容試験管に接種し、140rpm、30℃で24時間振とう培養した。得られた前培養液を3mL YPD10C(20g/Lペプトン、10g/L酵母エキス、100g/Lグルコース、45g/L炭酸カルシウム)/ガラス試験管に接種し、初期OD600=1、100rpm、30℃で振とう培養した。経時的にサンプリングを行い、高速液体クロマトフラフィーおよびガスクロマトフラフィーGC−2010でイソプロパノール生産量を評価した。なお、データは3反復試行の平均で示した。
【0088】
図8によると、ACS1、ACS2およびERG10のそれぞれ単独の過剰発現により、イソプロパノール生産量が2.2〜2.7倍になることが明らかとなった。(i)ACL1、ACLYおよびACS2の組み合わせを過剰発現する株、ならびに(ii)ACL1、ACLYおよびERG10の組み合わせを過剰発現する株のイソプロパノール生産量は、それぞれACS2およびERG10を単独で過剰発現する株と有意な差は認められなかった。一方、(iii)ACS2およびERG10の組み合わせを過剰発現する株ではイソプロパノール生産の飛躍的な向上が認められ、生産量が11.6倍に向上した。また、(iii)ACS2およびERG10の組み合わせを過剰発現する株、(iv)ACL1、ACLY、ACS2およびERG10の組み合わせを過剰発現する株、TMS378株(ACL1、ACLY、ACS2、CRC1、ERG10およびYAT1の組み合わせを過剰発現する株)の株間において、イソプロパノール生産量に有意な差は認められなかった。
【0089】
これらの結果より、同時に過剰発現した際にはACL、CRC1およびYAT1の過剰発現がイソプロパノールの生産性向上に与える寄与は小さく、イソプロパノール生産性を向上させるためにはACS2とERG10を同時に過剰発現することが最も重要であることが示された。以上の結果は、酵母によるイソプロパノール高生産のためにはエタノールからアセチルCoAを合成する代謝経路、すなわち酢酸からアセチルCoAを合成する代謝経路に関わる酵素遺伝子の発現または活性を増強することが重要であることを示している。
【図面の簡単な説明】
【0090】
【図1】図1は、酵母によるイソプロパノール合成経路を示す図である。
【図2】図2は、イソプロパノール生産遺伝子を導入した株の非中和条件での発酵試験の結果を示す図であり、培養液中のイソプロパノール濃度(パネルA)、グルコース濃度(パネルB)、エタノール濃度(パネルC)、酢酸濃度(パネルD)、グリセロール濃度(パネルE)およびOD600(パネルF)の経時変化を示している。各パネルにおいて、黒丸はATCC15239株の結果を示し、白丸はTMS272株(ATCC15239株+IPA合成遺伝子)の結果を示し、黒三角はPDC1破壊株の結果を示し、白三角はTMS290株(PDC破壊株+IPA合成遺伝子)の結果を示す。
【図3】図3は、イソプロパノール生産遺伝子を導入した株の中和条件での発酵試験の結果を示す図であり、培養液中のイソプロパノール濃度(パネルA)、グルコース濃度(パネルB)、エタノール濃度(パネルC)、酢酸濃度(パネルD)およびグリセロール濃度(パネルE)の経時変化を示している。各パネルにおいて、黒丸はATCC15239株の結果を示し、白丸はTMS272株(ATCC15239株+IPA合成遺伝子)の結果を示し、黒三角はPDC1破壊株の結果を示し、白三角はTMS290株(PDC破壊株+IPA合成遺伝子)の結果を示す。
【図4】図4は、TMS272株において、各種遺伝子を過剰発現させた場合におけるイソプロパノール生産量を、TMS272株によるイソプロパノール生産量に対する相対量として示したグラフである。
【図5】図5は、イソプロパノール生酸性を向上させる遺伝子を全て過剰発現する株の発酵試験の結果を示す図であり、培養液中のイソプロパノール濃度(パネルA)、グルコース濃度(パネルB)、エタノール濃度(パネルC)、酢酸濃度(パネルD)、グリセロール濃度(パネルE)およびアセトン濃度(パネルF)の経時変化を示している。各パネルにおいて、黒丸はTMS272株の結果を示し、白丸はTMS378株(TMS272株+ACL1:ACLY:ACS2:ERG10:CRC1:YAT1)の結果を示す。
【図6】図6は、イソプロパノール生産性を向上させる遺伝子を全て過剰発現するTMS378株の30℃での発酵試験(初期OD600および振とう速度の影響)の結果を示す図であり、培養液中のイソプロパノール濃度(パネルA)、グルコース濃度(パネルB)、エタノール濃度(パネルC)、酢酸濃度(パネルD)、グリセロール濃度(パネルE)およびアセトン濃度(パネルF)の経時変化を示している。各パネルにおいて、黒丸は初期OD600=1かつ攪拌数=100rpmの条件下での結果を示し、黒三角は初期OD600=1かつ攪拌数=120rpmの条件下での結果を示し、白丸は初期OD600=10かつ攪拌数=100rpmの条件下での結果を示し、白三角は初期OD600=10かつ攪拌数=120rpmの条件下での結果を示す。
【図7】図7は、イソプロパノール生産性を向上させる遺伝子を全て過剰発現するTMS378株の30℃での発酵試験(流加グルコース濃度の影響)の結果を示す図である。パネルAは、初期グルコース濃度を100g/Lとし、培養72時間後に100g/lのグルコースを流加して、192時間培養を行った結果を示す。パネルBは、初期グルコース濃度を50g/Lとし、培養48時間後、96時間後および144時間後にそれぞれ50g/lのグルコースを流加して、192時間培養を行った結果を示す。各パネルにおいて、イソプロパノール濃度(黒三角)、グルコース濃度(黒丸)、エタノール濃度(白丸)および酢酸濃度(白三角)の経時変化を示している。
【図8】図8は、TMS272株において、各種遺伝子を過剰発現させた場合におけるイソプロパノール生産量を示したグラフである。
【発明の背景】
【0001】
技術分野
本発明は、酵母において、アセチルCoAを経由する代謝経路を利用した物質の製造方法に関する。
【0002】
背景技術
深刻化する環境問題を背景に、バイオ燃料が石油の代替燃料として注目を集めている。エタノールはバイオ燃料としてよく知られているが、ガソリンより低いエネルギー密度しか有さないこと、輸送燃料として機能させるためには限られた濃度範囲でガソリンと混合する必要があることから、燃料としての欠点も多い。イソプロパノールは、1−ブタノールよりも高いオクタン価を有し、また、工業原料としても利用される汎用性の高い溶剤である。イソプロパノールは、ナフサ由来のプロピレンの水和によって製造されており、再生可能な供給源から商業的に生産されたことはない。
【0003】
微生物によるアセトンやブタノールの発酵生産は、20世紀初頭に産業的に行われていた。絶対嫌気性菌のある種のクロストリジウム属細菌はアセトンとブタノールを同時に生産することができる(アセトン・ブタノール発酵)。アセトンは細菌細胞内に存在するアセチルCoAを原料として生産される。アセチルCoAは、アセチルCoAアセチル基転移酵素(チオラーゼ)によってアセトアセチルCoAに変換され、さらにCoA転移酵素によってアセト酢酸に変換され、さらにアセト酢酸脱炭酸酵素によってアセトンへと変換される。ある種のクロストリジウム属細菌はさらに二級アルコールの脱水素酵素を有し、アセトンからイソプロパノールを生産する。Chenらは、クロストリジウム・ベイジェリンキを含む52株のクロストリジウム属細菌の中で、最大で30mM(1.8g/L)のイソプロパノールを生産する株が存在したことを報告している(非特許文献1:Chen, J.-S., and S. F. Hiu. Acetone-butanol-isopropanol production by Clostridium beijerinckii (synonym, Clostridium butylicum). Biotechnol. Lett. 1986. 8:371-376)。また、最近になってHanaiらおよびJojimaらは、大腸菌にクロストリジウム属由来のCoA転移酵素、アセト酢酸脱炭酸酵素、二級アルコール脱水素酵素を導入し、内在性のアセチルCoAアセチル基転移酵素を過剰発現させることによってイソプロパノールを効率的に生産できる大腸菌株を報告している(非特許文献2:T. Hanai, S. Atsumi, and J. C. Liao. Engineered Synthetic Pathway for Isopropanol Production in Escherichia coli. Appl Environ Microbiol, Dec. 2007, p. 7814-7818;非特許文献3:Toru Jojima & Masayuki Inui & Hideaki Yukawa. Production of isopropanol by metabolically engineered Escherichia coli. Appl Microbiol Biotechnol 2008 77:1219-1224)。しかしながら、大腸菌は、糖耐性、浸透圧耐性、塩耐性、有機溶剤耐性など各種ストレス耐性はあまり高くないことが多く、ファージ汚染によるリスクもある。また、イソプロパノール耐性が低いことは生産物による発酵の阻害が起こることを意味する。発酵阻害を回避するためにガスストリッピング法などを用いて生産物を培地から除去しながら発酵を続ける方法が知られているが、該方法の使用は発酵生産の工程を煩雑化させるだけでなく、コスト増加につながる。これらの問題を解決するための一つの手段として、高い有機溶剤耐性を有する微生物を用いてイソプロパノール生産をすることが考えられる。
【0004】
単細胞真核微生物である酵母は高い有機溶剤耐性を示し、また、大腸菌などの微生物と同様に、培養設備、培養時間および培養コストの面で高等生物の細胞よりも安価に培養することができるという特徴を有する。特に、出芽酵母はビールやワインなどの酒類やパンの製造に用いられ、その食品としての安全性が確認されている。
【0005】
酵母でイソプロパノールを生産するためには、その酵母菌株に、外因性のCoA転移酵素、アセト酢酸脱炭酸酵素、および二級アルコール脱水素酵素を導入する必要がある。これまで、酵母によるイソプロパノールの製造例としては、サッカロマイセス・セレビシエ(Saccharomyces cerevisiae)にイソプロピルアルコール生産酵素群の遺伝子を導入し、この形質転換体を培養することによりイソプロパノールを製造したことが報告されている(特許文献1:特開2011−97929号公報)。しかし、この方法によるイソプロパノールの生産量は、培養液中の濃度として100mg/L前後という少量に過ぎない。酵母は真核生物であり、オルガネラが存在することから、イソプロパノール合成の基質であるアセチルCoAを細胞質に供給する経路が複数存在している。どの経路からアセチルCoAを細胞質に供給することがイソプロパノール合成にとって有利であるのか、あるいはどのような酵母がそういった能力に優れているか、これまでに調査された例はない。
【0006】
キャンディダ・ユティリス(Candida utilis)は、サッカロマイセス・セレビシエ(Saccharomyces cerevisiae)およびサッカロマイセス・フラジリス(Saccharomyces fragilis)と同様に、FDA(米国連邦食品医薬品局)によって食用酵母として認定されており、食品添加物として安全に使用できることが認められている食用酵母である。実際に、現在でも、キャンディダ・ユティリスは、日本をはじめとして、ドイツやアメリカなど世界各国で生産され、食飼料として利用されている。また、キャンディダ・ユティリスは、ペントースの資化による菌体生産、エチルアセテート、L−グルタミン酸、グルタチオン、インベルターゼなどの生産株として広く産業界で利用されてきた。近年、キャンディダ・ユティリスにおいても形質転換法が開発され、様々な化合物が代謝工学的に生産される技術が開発されている(非特許文献4:Hiroshi Shimada, Keiji Kondo, Paul D. Fraser, Yutaka Miura, Toshiko Saito, and Norihiko Misawa., Increased Carotenoid Production by the Food Yeast Candida utilis through Metabolic Engineering of the Isoprenoid Pathway., Appl Environ Microbiol, July 1998, p. 2676-2680, Vol. 64, No. 7;非特許文献5:Yutaka Miura, Keiji Kondo, Toshiko Saito, Hiroshi Shimada, Paul D. Fraser, and Norihiko MisawaProduction of the Carotenoids Lycopene, β-Carotene, and Astaxanthin in the Food Yeast Candida utilis., Appl Environ Microbiol, April 1998, p. 1226-1229, Vol. 64, No. 4;非特許文献6:Keiji Kondo,, Yutaka Miura, Hidetaka Sone, Kazuo Kobayashi & Hiroshi lijima., High-level expression of a sweet protein, monellin, in the food yeast Candida utilis., Nature Biotechnology 15, 453 - 457 (1997);非特許文献7:Yutaka Miura, Keiji Kondo, Hiroshi Shimada, Toshiko Saito, Katsumi Nakamura, Norihiko Misawa., Production of lycopene by the food yeast, Candida utilis that does not naturally synthesize carotenoid., Biotechnology and Bioengineering Volume 58 Issue 2-3, Pages 306-308;非特許文献8:Shigehito IKUSHIMA, Toshio FUJII, Osamu KOBAYASHI, Satoshi YOSHIDA and Aruto YOSHIDA., Genetic Engineering of Candida utilis Yeast for Efficient Production of L-Lactic Acid., Bioscience, Biotechnology, and Biochemistry Vol. 73 (2009) , No. 8 pp.1818-1824;非特許文献9:Yi-Ren Hong, Ya-Lei Chen, Lynn Farh, Wen-Jen Yang, Chen-Hua Liao and David Shiuan.,Recombinant Candida utilis for the production of biotin., Applied Microbiology and Biotechnology.,Volume 71(2006), Number 2, 211-221)。また、キャンディダ・ユティリスは、サッカロマイセス属とは異なり、クラブツリー(Crabtree)効果陰性酵母であることから、グルコース存在下でも呼吸活性が低下しない。これにより、十分に通気を行う条件を採用することにより、エタノールの生成を抑えることができる。さらに、キャンディダ・ユティリスは呼吸活性も高く、一旦生成したエタノールも速やかに再同化されることから、効率的な菌体や代謝産物の生産を可能とする。
【0007】
酵母でイソプロパノールのような物質を生産する場合、一般に、副産物としてのエタノールの生成が問題となる。このエタノール副生を抑制するために、エタノール生産の重要酵素であるピルビン酸脱炭酸酵素遺伝子(PDC)を欠損させる取り組みがなされている(特許文献2:国際公開第2010/095751号パンフレット;特許文献3:国際公開第2010/095750号パンフレット;非特許文献10:Flikweert MT, Van Der Zanden L, Janssen WM, et al. Pyruvate decarboxylase: an indispensable enzyme for growth of Saccharomyces cerevisiae on glucose. Yeast 12(1996): 247-257;非特許文献11:van Maris AJA, Winkler AA, Porro D, et al. Homofermentative lactate production cannot sustain anaerobic growth of engineered Saccharomyces cerevisiae: Possible consequence of energy-dependent lactate export. Appl Environ Microbiol 70(2004): 2898-2905)。
【0008】
【非特許文献1】Chen, J.-S., and S. F. Hiu. Acetone-butanol-isopropanol production by Clostridium beijerinckii (synonym, Clostridium butylicum). Biotechnol. Lett. 1986. 8:371-376
【非特許文献2】T. Hanai, S. Atsumi, and J. C. Liao. Engineered Synthetic Pathway for Isopropanol Production in Escherichia coli. Appl Environ Microbiol, Dec. 2007, p. 7814-7818
【非特許文献3】Toru Jojima & Masayuki Inui & Hideaki Yukawa. Production of isopropanol by metabolically engineered Escherichia coli. Appl Microbiol Biotechnol 2008 77:1219-1224
【非特許文献4】Hiroshi Shimada, Keiji Kondo, Paul D. Fraser, Yutaka Miura, Toshiko Saito, and Norihiko Misawa., Increased Carotenoid Production by the Food Yeast Candida utilis through Metabolic Engineering of the Isoprenoid Pathway., Appl Environ Microbiol, July 1998, p. 2676-2680, Vol. 64, No. 7
【非特許文献5】Yutaka Miura, Keiji Kondo, Toshiko Saito, Hiroshi Shimada, Paul D. Fraser, and Norihiko MisawaProduction of the Carotenoids Lycopene, β-Carotene, and Astaxanthin in the Food Yeast Candida utilis., Appl Environ Microbiol, April 1998, p. 1226-1229, Vol. 64, No. 4
【非特許文献6】Keiji Kondo,, Yutaka Miura, Hidetaka Sone, Kazuo Kobayashi & Hiroshi lijima., High-level expression of a sweet protein, monellin, in the food yeast Candida utilis., Nature Biotechnology 15, 453 - 457 (1997)
【非特許文献7】Yutaka Miura, Keiji Kondo, Hiroshi Shimada, Toshiko Saito, Katsumi Nakamura, Norihiko Misawa., Production of lycopene by the food yeast, Candida utilis that does not naturally synthesize carotenoid., Biotechnology and Bioengineering Volume 58 Issue 2-3, Pages 306-308
【非特許文献8】Shigehito IKUSHIMA, Toshio FUJII, Osamu KOBAYASHI, Satoshi YOSHIDA and Aruto YOSHIDA., Genetic Engineering of Candida utilis Yeast for Efficient Production of L-Lactic Acid., Bioscience, Biotechnology, and Biochemistry Vol. 73 (2009) , No. 8 pp.1818-1824
【非特許文献9】Yi-Ren Hong, Ya-Lei Chen, Lynn Farh, Wen-Jen Yang, Chen-Hua Liao and David Shiuan.,Recombinant Candida utilis for the production of biotin., Applied Microbiology and Biotechnology.,Volume 71(2006), Number 2, 211-221
【非特許文献10】Flikweert MT, Van Der Zanden L, Janssen WM, et al. Pyruvate decarboxylase: an indispensable enzyme for growth of Saccharomyces cerevisiae on glucose. Yeast 12(1996): 247-257
【非特許文献11】van Maris AJA, Winkler AA, Porro D, et al. Homofermentative lactate production cannot sustain anaerobic growth of engineered Saccharomyces cerevisiae: Possible consequence of energy-dependent lactate export. Appl Environ Microbiol 70(2004): 2898-2905
【特許文献1】特開2011−97929号公報
【特許文献2】国際公開第2010/095751号パンフレット
【特許文献3】国際公開第2010/095750号パンフレット
【発明の概要】
【0009】
本発明者らは、酵母におけるアセチルCoA合成酵素(ACS)およびアセチルCoAアセチル基転移酵素(ERG10)の活性を増強することにより、酵母内におけるアセチルCoAを経由する物質の生産量が顕著に増加することを見出した。本発明はこの知見に基づくものである。
【0010】
従って、本発明の目的は、様々な物質の発酵生産において中間物質として関与するアセチルCoAならびにアセトアセチルCoAの合成・代謝に係る酵素活性が増強された酵母菌株、ならびに該酵母菌株を用いた物質の製造法を提供することにある。
【0011】
そして、本発明による酵母菌株は、以下の酵素:
(a)アセチルCoA合成酵素(ACS);および
(b)アセチルCoAアセチル基転移酵素(ERG10)
の活性が増強されている酵母菌株である。
【0012】
さらに、本発明による製造法は、本発明による酵母菌株を培養することを含んでなる、物質を製造する方法である。
【0013】
本発明によれば、アセチルCoAやアセトアセチルCoAが関与する酵母の代謝経路が活性化される。アセチルCoAやアセトアセチルCoAが関与する代謝経路は多岐にわたるため、生成する代謝産物も多種多様である。従って、本発明によれば、酵母菌株の培養によって様々な物質を高効率で製造することが可能となる。また、製造しようとする物質に応じて、必要な酵素の遺伝子を発現可能な形で酵母に組み込み、得られた酵母菌株を培養することによりその物質を製造することも可能である。
【発明の具体的説明】
【0014】
本発明による酵母菌株では、以下の遺伝子:(a)アセチルCoA合成酵素(ACS)の活性を有するポリペプチドをコードする遺伝子;および(b)アセチルCoAアセチル基転移酵素(ERG10)の活性を有するポリペプチドをコードする遺伝子の発現が増強されている。
【0015】
本明細書では、上記のアセチルCoA合成酵素(ACS)遺伝子およびアセチルCoAアセチル基転移酵素(ERG10)遺伝子とともに、イソプロパノールを合成するための4つの遺伝子を過剰発現する酵母菌株を作製し、これを培養することにより、極めて高濃度のイソプロパノールが生産されることが実証されている。この酵母菌株におけるイソプロパノール生産の経路は図1に示す通りである。図1に示される経路から、イソプロパノールの高い生産量が、アセチルCoAおよびアセトアセチルCoAの合成・代謝に係る酵素活性の増強に起因することは明らかである。従って、本発明によって、イソプロパノールのみならず、アセチルCoAおよびアセトアセチルCoAを経由する様々な物質を高生産することができる。また、図1に示されるように、ACSおよびERG10は細胞質において機能する酵素である。
【0016】
細胞質に供給されるアセチルCoAは、フラボノイド骨格を有する化合物、テルぺノイド骨格を有する化合物、ステロール骨格を有する化合物、脂肪酸など、様々な有用な二次代謝物質の基幹物質である。よって、酵母菌株が天然の状態もしくは人為的に遺伝子を改変された状態でこれらの二次代謝産物の合成経路を有していれば、本発明の方法によりこれらの物質を高生産させることができる。
【0017】
一般的に、微生物による物質生産を行う際には、副産物生産を抑制する取り組みがなされる場合が多い。例えば、酵母において物質を生産する場合、一般に、副産物としてのエタノールの生成を抑制するために、エタノール生産に関与する酵素遺伝子、例えばピルビン酸脱炭酸酵素遺伝子(PDC)を欠損させる取り組みがなされている(国際公開第2010/095751号パンフレット;国際公開第2010/095750号パンフレット;Flikweert MT, Van Der Zanden L, Janssen WM, et al. Pyruvate decarboxylase: an indispensable enzyme for growth of Saccharomyces cerevisiae on glucose. Yeast 12(1996): 247-257;van Maris AJA, Winkler AA, Porro D, et al. Homofermentative lactate production cannot sustain anaerobic growth of engineered Saccharomyces cerevisiae: Possible consequence of energy-dependent lactate export. Appl Environ Microbiol 70(2004): 2898-2905)。しかし、本明細書では、上記のPDCを欠損させた酵母菌株よりも、PDCを欠損させなかった酵母菌株の方が顕著に高いイソプロパノール生産量を示すことが実証されている。よって、本発明による酵母菌株は、PDCなどのエタノール生産に関与する酵素遺伝子を欠失させずにそのまま維持していることも特徴の一つであり、これによりイソプロパノール生産量、ひいてはアセチルCoAやその代謝産物であるアセトアセチルCoAの生産量が顕著に向上することは、驚くべきことである。
【0018】
本明細書において酵素活性が「増強されている」とは、培養過程の少なくとも一時期において、活性増強のための手段を施されていない酵母と比較してより強い酵素活性が観察されることを意味する。酵素活性の増強は、当該酵素遺伝子の発現を増強すること、あるいは比活性のより高い酵素遺伝子への置き換えや、タンパク質工学的な方法による酵素遺伝子の改変によって実現することができる。本明細書において遺伝子の発現が「増強されている」とは、酵母の遺伝的改変により該酵母におけるその遺伝子の発現量が増加すること、つまり、その遺伝子を過剰発現させることを意味する。このような遺伝的改変の方法としては、例えば、その遺伝子により酵母を形質転換する方法(つまり、その遺伝子を発現し得る形で酵母にトランスフェクトする方法)、その酵母のゲノム上に存在する遺伝子のプロモーター活性を強化する方法、その酵母のゲノム上に存在する遺伝子のプロモーターを他のプロモーターと置換することにより該遺伝子の発現を増強する方法など、様々な方法が知られているが、好ましくはその遺伝子により酵母を形質転換する方法とされる。
【0019】
酵母
本発明による酵母菌株は、特定の遺伝子が過剰発現するように遺伝的に改変された酵母である。遺伝的改変に用いる酵母は、アセチルCoAまたはアセトアセチルCoAを経由する代謝経路を有する酵母であればよく、特に制限されないが、好ましくは呼吸活性の高いクラブツリー効果陰性酵母とされ、より好ましくはクラブトゥリー陰性酵母であるキャンディダ・ユティリスとされる。キャンディダ・ユティリスの菌株は当技術分野において公知の様々な株、例えば、NBRC0396株、NBRC0619株、NBRC0988株、NBRC0639株、NBRC0626株、NBRC1086株、NBRC10707株、ATCC9255株、ATCC28955株、ATCC44638株、ATCC9206株、ATCC96621株、ATCC18201株、およびATCC15239株等であってよいが、好ましくはNBRC1086株、ATCC9255株またはATCC15239株とされる。
【0020】
活性が増強される酵素およびその遺伝子
本発明による酵母菌株において活性が増強される酵素は、以下の酵素:(a)アセチルCoA合成酵素(ACS);および(b)アセチルCoAアセチル基転移酵素(ERG10)である。本発明の好ましい実施態様によれば、本発明による酵母菌株においてはこれら2種類の酵素の遺伝子が過剰発現され、すなわち、以下の遺伝子:(a)アセチルCoA合成酵素(ACS)の活性を有するポリペプチドをコードする遺伝子;および(b)アセチルCoAアセチル基転移酵素(ERG10)の活性を有するポリペプチドをコードする遺伝子の発現が増強されている。
【0021】
本発明による酵母菌株では、さらに、以下の遺伝子:(c)ATP−クエン酸除去付加酵素(ACL)の活性を有するポリペプチドをコードする遺伝子;(d)ミトコンドリア内膜カルニチン輸送体(CRC1)の活性を有するポリペプチドをコードする遺伝子;および(e)カルニチンアセチル転移酵素遺伝子(YAT1)の活性を有するポリペプチドをコードする遺伝子からなる群から選択される少なくとも一種の遺伝子の発現が増強されていてもよく、好ましくはこれら(c)、(d)および(e)の全ての遺伝子の発現が増強される。
【0022】
これらの酵素および遺伝子は様々な酵母に由来するものが知られており、その起源は特に制限されない。これらの酵素および遺伝子は、育種に用いられる酵母の内在性遺伝子およびその発現産物であってよく、あるいは、育種に用いられる酵母以外の他の酵母に由来するものであってもよい。例えば、これらの酵素および遺伝子は、それぞれ独立に、キャンディダ・ユティリスまたはヤロウィア・リポリティカに由来するものとすることができる。それぞれの酵素の起源としては、例えば、アセチルCoA合成酵素(ACS)はキャンディダ・ユティリス酵母、アセチルCoAアセチル基転移酵素(ERG10)はキャンディダ・ユティリス酵母、ミトコンドリア内膜カルニチン輸送体(CRC1)はキャンディダ・ユティリス酵母、カルニチンアセチル転移酵素遺伝子(YAT1)はキャンディダ・ユティリス酵母、ATP−クエン酸除去付加酵素(ACL)はヤロウィア・リポリティカ酵母(ヤロウィア・リポリティカ由来のACL1とACLYがサブユニットを構成する遺伝子)が好ましい。
【0023】
キャンディダ・ユティリス由来のアセチルCoA合成酵素(ACS)遺伝子には、ACS1遺伝子とACS2遺伝子がある。ACS1遺伝子のコード配列は配列番号11で表され、コードされるアミノ酸配列は配列番号12で表される。ACS2遺伝子のコード配列は配列番号13で表され、コードされるアミノ酸配列は配列番号14で表される。アセチルCoA合成酵素(ACS)は、酢酸をアセチルCoAに変換する酵素である。本発明に用いられるACS遺伝子はACS1およびACS2のいずれであってもよいが、好ましくはACS2とされる。また、アセチルCoA合成酵素(ACS)の活性を有するポリペプチドは、配列番号12または配列番号14で表されるアミノ酸配列において、1もしくは数個のアミノ酸が欠失、置換、付加もしくは挿入されたアミノ酸配列を含み、かつACS活性を有するポリペプチドであってもよい。
【0024】
キャンディダ・ユティリス由来のアセチルCoAアセチル基転移酵素(ERG10)遺伝子のコード配列は配列番号61で表され、コードされるアミノ酸配列は配列番号62で表される。アセチルCoAアセチル基転移酵素(ERG10)は、アセチルCoAをアセトアセチルCoAに変換する酵素である。また、アセチルCoAアセチル基転移酵素(ERG10)の活性を有するポリペプチドは、配列番号62で表されるアミノ酸配列において、1もしくは数個のアミノ酸が欠失、置換、付加もしくは挿入されたアミノ酸配列を含み、かつERG10活性を有するポリペプチドであってもよい。
【0025】
ヤロウィア・リポリティカ由来のACL1遺伝子のコード配列は配列番号31で表され、コードされるアミノ酸配列は配列番号32で表される。また、ACL1の活性を有するポリペプチドは、配列番号32で表されるアミノ酸配列において、1もしくは数個のアミノ酸が欠失、置換、付加もしくは挿入されたアミノ酸配列を含み、かつACL1活性を有するポリペプチドであってもよい。さらに、ヤロウィア・リポリティカ由来のACLY遺伝子のコード配列は配列番号33で表され、コードされるアミノ酸配列は配列番号34で表される。また、ACLYの活性を有するポリペプチドは、配列番号34で表されるアミノ酸配列において、1もしくは数個のアミノ酸が欠失、置換、付加もしくは挿入されたアミノ酸配列を含み、かつACLY活性を有するポリペプチドであってもよい。これらのACL1とACLYをサブユニットとするATP−クエン酸除去付加酵素(ACL)は、クエン酸をアセチルCoAに変換する酵素である。
【0026】
キャンディダ・ユティリス由来のミトコンドリア内膜カルニチン輸送体(CRC1)遺伝子のコード配列は配列番号23で表され、コードされるアミノ酸配列は配列番号24で表される。ミトコンドリア内膜カルニチン輸送体(CRC1)は、アセチルカルニチンをミトコンドリアから細胞質へ輸送する酵素である。また、ミトコンドリア内膜カルニチン輸送体(CRC1)の活性を有するポリペプチドは、配列番号24で表されるアミノ酸配列において、1もしくは数個のアミノ酸が欠失、置換、付加もしくは挿入されたアミノ酸配列を含み、かつCRC1活性を有するポリペプチドであってもよい。
【0027】
キャンディダ・ユティリス由来のカルニチンアセチル転移酵素遺伝子(YAT1)遺伝子のコード配列は配列番号27で表され、コードされるアミノ酸配列は配列番号28で表される。カルニチンアセチル転移酵素遺伝子(YAT1)は、アセチルカルニチンをアセチルCoAに変換する酵素である。また、カルニチンアセチル転移酵素遺伝子(YAT1)の活性を有するポリペプチドは、配列番号28で表されるアミノ酸配列において、1もしくは数個のアミノ酸が欠失、置換、付加もしくは挿入されたアミノ酸配列を含み、かつYAT1活性を有するポリペプチドであってもよい。
【0028】
アミノ酸の欠失、置換、付加、又は挿入は、上記ポリペプチドをコードする遺伝子を、当技術分野で公知の手法によって改変することによって行うことができる。遺伝子に変異を導入するには、Kunkel法又はGapped duplex法等の公知手法又はこれに準ずる方法により行うことができ、例えば部位特異的突然変異誘発法を利用した変異導入用キット、例えばMutant−K(タカラバイオ社)やMutant−G(タカラバイオ社)などを用いて、あるいは、タカラバイオ社のLA PCR in vitro Mutagenesisシリーズキット、KOD−Plus−Mutagenesis Kit(TOYOBO)などを用いて変異を導入することができる。上記の各酵素の活性は、当技術分野において公知の手法により確認することができる。
【0029】
上記の遺伝子に代えて、それぞれの同等物を用いることもできる。この同等物は、それぞれの遺伝子と同等の機能を有することを条件に、一部のヌクレオチド残基が異なる遺伝子を意味する。このような同等物としては、それぞれのヌクレオチド配列と70%以上、好ましくは80%以上、より好ましくは85%以上、さらに好ましくは90%以上、最も好ましくは95%以上の相同性(配列同一性)があり、かつ各酵素の活性を有するポリペプチドをコードするヌクレオチド配列を含む遺伝子が挙げられる。前記同等物としてはさらに、それぞれのヌクレオチド配列もしくはその相補配列とストリンジェントな条件下でハイブリダイズし、かつ各酵素の活性を有するポリペプチドをコードするヌクレオチド配列を含む遺伝子が挙げられる。前記同等物としてはさらに、それぞれのヌクレオチド配列において1もしくは数個のヌクレオチド残基が欠失、置換、付加、または挿入された配列を含み、かつ各酵素の活性を有するポリペプチドをコードするヌクレオチド配列を含む遺伝子が挙げられる。
【0030】
ここで、ヌクレオチド残基の欠失、置換、付加、又は挿入は、上記配列を含む遺伝子を、当技術分野で公知の手法によって改変することによって行うことができる。遺伝子に変異を導入するには、Kunkel法又はGapped duplex法等の公知手法又はこれに準ずる方法により行うことができ、例えば部位特異的突然変異誘発法を利用した変異導入用キットを用いればよい。例えばMutant−K(タカラバイオ社)やMutant−G(タカラバイオ社)などを用いて、あるいは、タカラバイオ社のLA PCR in vitro Mutagenesisシリーズキット、KOD−Plus−Mutagenesis Kit(TOYOBO)などを用いて変異を導入することができる。各酵素の活性は、当技術分野において公知の手法により確認することができる。
【0031】
相同性(配列同一性)を示す数値(%)は、塩基配列比較用プログラム:例えばGENETYX−WIN7.0.0を用いて、デフォルト(初期設定)のパラメーターにより算出されるものである。すなわち、酵母染色体上の各遺伝子が、同一ではないが同等の機能、すなわち各活性を有するポリペプチドをコードする遺伝子によって相同組換え等を介して置換されていてもよい。各酵素の活性は、当技術分野において公知の手法により確認することができる。
【0032】
ストリンジェントな条件とは、例えば、Rapid−Hyb Buffer(GEヘルスケアバイオサイエンス社製)を用い、温度条件を好ましくは40〜70℃、より好ましくは60℃として、その他は添付のプロトコールに従って行うハイブリダイゼーション条件である。その後、例えば当業者に公知の一般的な方法を用い、2×SSCと0.1%(w/v)SDSから成り立つ溶液での5分間の洗浄、続いて1×SSCと0.1%(w/v)SDSから成り立つ溶液での10分間の洗浄、さらに0.1×SSCと0.1%(w/v)SDSから成り立つ溶液での10分間の洗浄を行うことを指す。ただしハイブリダイゼーション時の温度条件や、その後のメンブレンの洗浄に用いる溶液の塩濃度等の条件を適宜設定することにより、ある一定(70%、80%、85%、90%、95%のいずれか)以上の相同性を有する塩基配列を含むDNAをクローニングできる。そのようにして得られる遺伝子が、配列上は同一ではないが同等の機能、すなわち各活性を有するポリペプチドをコードする遺伝子によって相同組換え等を介して置換されていてもよい。各酵素の活性は、当技術分野において公知の手法により確認することができる。
【0033】
有用物質を合成するための遺伝子
本発明による酵母菌株は、所望の有用物質の製造に用いるために、該物質の生産に必要となる遺伝子で形質転換されていてもよい。例えば、本発明の方法によってアセチルCoAを経由する代謝産物を製造するために、本発明による酵母菌株は、アセチルCoAを経由する代謝産物を合成するための遺伝子により形質転換されていてもよい。必要な遺伝子は、製造しようとする物質に応じて、当業者であれば適切に選択することができる。
【0034】
例えば、イソプロパノールを製造しようとする場合には、さらなる遺伝的改変として、イソプロパノール合成遺伝子で酵母菌株を形質転換することができる。イソプロパノール合成遺伝子としては、特開2011−97929号公報に記載の酵素遺伝子群など、当業者に公知の遺伝子を用いることができ、例えば、CoA基転移酵素サブユニットA(ctfA)、同サブユニットB(ctfB)、アセト酢酸脱炭酸酵素(adc)および二級アルコール脱水素酵素(sadh)の4つの遺伝子の組み合わせを用いることができる。
【0035】
本発明の一つの実施態様によれば、本発明による酵母菌株は、さらなる遺伝的改変として、CoA基転移酵素サブユニットA(ctfA)、CoA転移酵素サブユニットB(ctfB)、アセト酢酸脱炭酸酵素(adc)および二級アルコール脱水素酵素(sadh)の活性を有するポリペプチドをコードしている4種の遺伝子により形質転換されている。このような酵母菌株はイソプロパノールの製造に用いることができ、この製造法は、本実施態様による酵母菌株を培養することを含む。
【0036】
これらの遺伝子は様々な生物に由来するものが知られており、その起源は特に制限されない。例えば、これらの遺伝子は、それぞれ独立に、クロストリジウム・アセトブチリカムまたはクロストリジウム・ベイジェリンキに由来するものとすることができる。それぞれの酵素の起源としては、例えば、CoA転移酵素遺伝子サブユニットA(ctfA)および同サブユニットB(ctfB)はクロストリジウム・アセトブチリカムが好ましく、アセト酢酸脱炭酸酵素(adc)および二級アルコール脱水素酵素(sadh)はクロストリジウム・ベイジェリンキが好ましい。
【0037】
クロストリジウム・アセトブチリカム由来のCoA転移酵素サブユニットA(ctfA)のアミノ酸配列は配列番号2で表される。CoA転移酵素サブユニットA(ctfA)は、同サブユニットB(ctfB)と一緒になって、アセトアセチルCoAをアセト酢酸に変換する酵素である。配列番号2で表されるアミノ酸配列をコードする遺伝子配列は、クロストリジウム・アセトブチリカム由来の同遺伝子の配列など、当業者であれば適宜選択することができるため、特に制限されないが、好ましくは本発明による酵母菌株の種において頻度の高いコドンを使用した配列とされる。例えば、キャンディダ・ユティリスにおけるコドン頻度に適合した遺伝子配列としては、配列番号1で表されるヌクレオチド配列が挙げられる。CoA転移酵素サブユニットA(ctfA)の活性を有するポリペプチドは、配列番号2で表されるアミノ酸配列において、1もしくは数個のアミノ酸が欠失、置換、付加もしくは挿入されたアミノ酸配列を含み、かつctfA活性を有するポリペプチドであってもよい。
【0038】
クロストリジウム・アセトブチリカム由来のCoA転移酵素サブユニットB(ctfB)のアミノ酸配列は配列番号4で表される。CoA転移酵素サブユニットB(ctfB)は、同サブユニットA(ctfA)と一緒になって、アセトアセチルCoAをアセト酢酸に変換する酵素である。配列番号4で表されるアミノ酸配列をコードする遺伝子配列は、クロストリジウム・アセトブチリカム由来の同遺伝子の配列など、当業者であれば適宜選択することができるため、特に制限されないが、好ましくは本発明による酵母菌株の種において頻度の高いコドンを使用した配列とされる。例えば、キャンディダ・ユティリスにおけるコドン頻度に適合した遺伝子配列としては、配列番号3で表されるヌクレオチド配列が挙げられる。CoA転移酵素サブユニットB(ctfB)の活性を有するポリペプチドは、配列番号4で表されるアミノ酸配列において、1もしくは数個のアミノ酸が欠失、置換、付加もしくは挿入されたアミノ酸配列を含み、かつctfB活性を有するポリペプチドであってもよい。
【0039】
クロストリジウム・ベイジェリンキ由来のアセト酢酸脱炭酸酵素(adc)のアミノ酸配列は配列番号6で表される。アセト酢酸脱炭酸酵素(adc)は、アセト酢酸をアセトンに変換する酵素である。配列番号6で表されるアミノ酸配列をコードする遺伝子配列は、クロストリジウム・ベイジェリンキ由来の同遺伝子の配列など、当業者であれば適宜選択することができるため、特に制限されないが、好ましくは本発明による酵母菌株の種において頻度の高いコドンを使用した配列とされる。例えば、キャンディダ・ユティリスにおけるコドン頻度に適合した遺伝子配列としては、配列番号5で表されるヌクレオチド配列が挙げられる。アセト酢酸脱炭酸酵素(adc)の活性を有するポリペプチドは、配列番号6で表されるアミノ酸配列において、1もしくは数個のアミノ酸が欠失、置換、付加もしくは挿入されたアミノ酸配列を含み、かつadc活性を有するポリペプチドであってもよい。
【0040】
クロストリジウム・ベイジェリンキ由来の二級アルコール脱水素酵素(sadh)のアミノ酸配列は配列番号8で表される。二級アルコール脱水素酵素(sadh)は、アセトンをイソプロパノールに変換する酵素である。配列番号8で表されるアミノ酸配列をコードする遺伝子配列は、クロストリジウム・ベイジェリンキ由来の同遺伝子の配列など、当業者であれば適宜選択することができるため、特に制限されないが、好ましくは本発明による酵母菌株の種において頻度の高いコドンを使用した配列とされる。例えば、キャンディダ・ユティリスにおけるコドン頻度に適合した遺伝子配列としては、配列番号7で表されるヌクレオチド配列が挙げられる。二級アルコール脱水素酵素(sadh)の活性を有するポリペプチドは、配列番号8で表されるアミノ酸配列において、1もしくは数個のアミノ酸が欠失、置換、付加もしくは挿入されたアミノ酸配列を含み、かつsadh活性を有するポリペプチドであってもよい。
【0041】
アミノ酸の欠失、置換、付加、又は挿入は、上記ポリペプチドをコードする遺伝子を、当技術分野で公知の手法によって改変することによって行うことができる。遺伝子に変異を導入するには、Kunkel法又はGapped duplex法等の公知手法又はこれに準ずる方法により行うことができ、例えば部位特異的突然変異誘発法を利用した変異導入用キット、例えばMutant−K(タカラバイオ社)やMutant−G(タカラバイオ社)などを用いて、あるいは、タカラバイオ社のLA PCR in vitro Mutagenesisシリーズキット、KOD−Plus−Mutagenesis Kit(TOYOBO)などを用いて変異を導入することができる。上記の各酵素の活性は、当技術分野において公知の手法により確認することができる。
【0042】
上記の遺伝子に代えて、それぞれの同等物を用いることもできる。この同等物は、それぞれの遺伝子と同等の機能を有することを条件に、一部のヌクレオチド残基が異なる遺伝子を意味する。このような同等物としては、それぞれのヌクレオチド配列と70%以上、好ましくは80%以上、より好ましくは85%以上、さらに好ましくは90%以上、最も好ましくは95%以上の相同性(配列同一性)があり、かつ各酵素の活性を有するポリペプチドをコードするヌクレオチド配列を含む遺伝子が挙げられる。前記同等物としてはさらに、それぞれのヌクレオチド配列もしくはその相補配列とストリンジェントな条件下でハイブリダイズし、かつ各酵素の活性を有するポリペプチドをコードするヌクレオチド配列を含む遺伝子が挙げられる。前記同等物としてはさらに、それぞれのヌクレオチド配列において1もしくは数個のヌクレオチド残基が欠失、置換、付加、または挿入された配列を含み、かつ各酵素の活性を有するポリペプチドをコードするヌクレオチド配列を含む遺伝子が挙げられる。
【0043】
ここで、ヌクレオチド残基の欠失、置換、付加、又は挿入は、上記配列を含む遺伝子を、当技術分野で公知の手法によって改変することによって行うことができる。遺伝子に変異を導入するには、Kunkel法又はGapped duplex法等の公知手法又はこれに準ずる方法により行うことができ、例えば部位特異的突然変異誘発法を利用した変異導入用キットを用いればよい。例えばMutant−K(タカラバイオ社)やMutant−G(タカラバイオ社)などを用いて、あるいは、タカラバイオ社のLA PCR in vitro Mutagenesisシリーズキット、KOD−Plus−Mutagenesis Kit(TOYOBO)などを用いて変異を導入することができる。各酵素の活性は、当技術分野において公知の手法により確認することができる。
【0044】
相同性(配列同一性)を示す数値(%)は、塩基配列比較用プログラム:例えばGENETYX−WIN7.0.0を用いて、デフォルト(初期設定)のパラメーターにより算出されるものである。すなわち、酵母染色体上の各遺伝子が、同一ではないが同等の機能、すなわち各活性を有するポリペプチドをコードする遺伝子によって相同組換え等を介して置換されていてもよい。各酵素の活性は、当技術分野において公知の手法により確認することができる。
【0045】
ストリンジェントな条件とは、例えば、Rapid−Hyb Buffer(GEヘルスケアバイオサイエンス社製)を用い、温度条件を好ましくは40〜70℃、より好ましくは60℃として、その他は添付のプロトコールに従って行うハイブリダイゼーション条件である。その後、例えば当業者に公知の一般的な方法を用い、2×SSCと0.1%(w/v)SDSから成り立つ溶液での5分間の洗浄、続いて1×SSCと0.1%(w/v)SDSから成り立つ溶液での10分間の洗浄、さらに0.1×SSCと0.1%(w/v)SDSから成り立つ溶液での10分間の洗浄を行うことを指す。ただしハイブリダイゼーション時の温度条件や、その後のメンブレンの洗浄に用いる溶液の塩濃度等の条件を適宜設定することにより、ある一定(70%、80%、85%、90%、95%のいずれか)以上の相同性を有する塩基配列を含むDNAをクローニングできる。そのようにして得られる遺伝子が、配列上は同一ではないが同等の機能、すなわち各活性を有するポリペプチドをコードする遺伝子によって相同組換え等を介して置換されていてもよい。各酵素の活性は、当技術分野において公知の手法により確認することができる。
【0046】
イソプロパノール以外にも、リコペンなどの特定のカロテノイド、スクアレンあるいはスクアレンから生産される特定のステロールなどの物質を生産する酵母菌株を構築することが可能である。例えば、Erwinia uredovora由来のGGPP合成酵素遺伝子、フィトエン合成酵素遺伝子、フィトエン不飽和化酵素遺伝子を発現し、内在性のHMG−CoA還元酵素遺伝子(HMG1)を過剰発現するキャンディダ・ユティリスはリコペンを高蓄積することが報告されている(Shimada H, Kondo K, Fraser PD, Miura Y, Saito T, Misawa N. Increased carotenoid production by the food yeast Candida utilis through metabolic engineering of the isoprenoid pathway. Appl Environ Microbiol. 1998 Jul;64(7):2676-80)。また、ザイモステロール-24-メチルトランスフェラーゼ遺伝子およびエルゴスタ5,7,24,(28)−トリエノール−22−脱水素酵素遺伝子の発現に欠損を持ち、HMG1が過剰発現するように操作されたサッカロマイセス・セレビシエ組み換え株は、高濃度のスクアレンを蓄積することが知られている(特開平5−192184号公報)。これらの手法を用いることにより、リコペン、スクアレンを生産する酵母菌株を構築することが可能である。
【0047】
酵母菌株の分子育種
本発明による酵母菌株の分子育種は、上記の遺伝子群を過剰発現させるための遺伝的改変を酵母菌株に施すこと、例えば、上記の遺伝子群を発現可能な状態で酵母菌株に導入することによって行うことができる。これら遺伝子群は、相同組換え等の技術により酵母の染色体上に組み込まれることが好ましい。このような染色体上への組込みを実現するための相同組換え用遺伝子配列の選択は、当業者において周知であり、当業者であれば必要に応じて適切な相同組換え用遺伝子配列を選択して相同組換え用DNA断片を構成することができる。
【0048】
上記の遺伝子群が酵母ゲノムに組込まれる染色体上の位置は特に制限されるものではないが、例えば、オロチジン5’リン酸デカルボキシラーゼ遺伝子(URA3)座、アルファアミノアジピン酸還元酵素遺伝子(LYS2)座などを選択することができる。また、これらの遺伝子は、組み込まれた遺伝子座に当初から存在するプロモーター、例えばURA3遺伝子プロモーター、LYS2遺伝子プロモーターなどの制御下に置いてもよいが、グリセルアルデヒド−3−リン酸脱水素酵素(GAP)遺伝子プロモーターなどの他のプロモーターの制御下に置いてもよい。
【0049】
本発明の好ましい実施態様によれば、本発明による酵母菌株の育種に用いられる酵母はキャンディダ・ユティリスとされる。キャンディダ・ユティリスは、その倍数性が高く、胞子を形成しない。倍数性の高い株の遺伝子に変異を導入しようとする場合、1倍体の株に比べて、その変異を重度に加える必要があるが、その際に、変異を与えたい遺伝子ではない遺伝子にも変異が加えられる可能性が高まると考えられる。従って、キャンディダ・ユティリスの遺伝子に変異を導入する場合には、標的とする遺伝子のみに効率よく多重に変異を加えることができる技術を用いることが好ましい。キャンディダ・ユティリスの形質転換法としては、特開2003−144185号公報に記載の技術が挙げられる。この文献では、利用可能なベクターとして、キャンディダ・ユティリスの染色体DNAと相同な配列と、選択マーカー遺伝子とを含んでなり、相同組換えによって異種遺伝子をキャンディダ・ユティリスの染色体DNAに組み込むことができるもの、あるいは、キャンディダ・ユティリスで自律複製機能を有するDNA配列と、選択マーカー遺伝子とを含んでなり、高い頻度でキャンディダ・ユティリスを形質転換できるものが開発されている。
【0050】
形質転換系で利用可能な選択マーカー遺伝子としては、酵母において機能し得る薬剤耐性マーカー、好ましくはシクロヘキシミド耐性型L41遺伝子、ジェネティシン(G418)耐性を付与する遺伝子、またはハイグロマイシンB耐性を付与する遺伝子などがある。ジェネティシン(G418)耐性を付与する遺伝子、またはハイグロマイシンB耐性を付与する遺伝子は、野生の酵母には存在しない配列であるため、標的の遺伝子座に組込まれる確率が高いと考えられている。また、これらの薬剤耐性遺伝子は、その宿主の形質に与える影響も小さいと考えられている(Baganz Fら,13(16):1563−73.,1997)(Cordero Otero Rら,Appl Microbiol Biotechnol,46(2):143−8.,1996)。
【0051】
他の形質転換系として、バクテリオファージP1由来のCre−loxP系が挙げられる。これは、2つの34bpのloxP配列間での部位特異的組換えシステムであり、この組換えはCre遺伝子がコードするCre組換え酵素によって触媒される。このシステムは、サッカロマイセス・セレビシエなどの酵母細胞においても機能することが報告されており、2つのloxP配列の間に配置された選択マーカー遺伝子は、loxP配列間の組換えによって除去されることが知られている(Guldener,U.ら,Nucleic Acids Res.,24,2519−24.,1996)。このシステムはクルイベロマイセス・ラクティス(Kluyveromyces lactis)など、複数の酵母種で利用されている(Steensma,H.Y.ら,Yeast,18,469−72.,2001)。
【0052】
物質の製造方法
本発明による酵母菌株を適当な炭素源の存在下で培養することにより、培養物中に物質(例えばイソプロパノール)を製造することができる。本発明による物質(例えばイソプロパノール)製造法によれば、培養系から物質(例えばイソプロパノール)を分離する工程を実施することにより、物質(例えばイソプロパノール)を得ることができる。なお、本発明において培養物とは、培養上清の他、培養細胞あるいは菌体、細胞もしくは菌体の破砕物を包含している。
【0053】
本発明による酵母菌株の培養にあたっては、酵母の種類に応じて培養方法や培養条件を選択することができる。培養方法としては、例えば、試験管、フラスコあるいはジャーファーメンターを用いた液体培養法を挙げることができ、回分培養、半回分培養などの培養形式を採用できる。振盪速度および通気量は、一般的には50〜200rpm、好ましくは80〜150rpm、あるいはこの振盪速度に相当する通気量とされる。初期の菌体量としてのOD600は、一般的には0.1〜30、好ましくは0.5〜10とされる。さらに、本発明による酵母菌株の培養にあたっては、振盪速度は速い方が好ましく(つまり通気量は多い方が好ましく)、初期OD600は高い方が好ましい。例えば、試験管やフラスコでの培養では、振盪速度は約120rpmまたはそれ以上、初期OD600は約10とすることが好ましい。
【0054】
本発明による物質(例えばイソプロパノール)製造法において、培地の組成は、酵母が生育し、且つ目的の物質を製造できる各種栄養素を含む組成であればよく、特に限定されない。培地に含まれる資化炭素源としては、例えば、グルコースのほかにもキシロースやスクロースでも資化できれば用いることができる。本発明の好ましい実施態様によれば、炭素源はグルコースを含む。培地中のグルコース濃度は、目的の物質の生産効率を確認しながら、当業者であれば適宜調節することができる。特に、本発明による酵母菌株の培養では、培地中のグルコース濃度は、比較的低濃度に保つことが好ましい。例えば、192時間の培養において、グルコースの初期濃度を100g/Lとし、培養72時間後に終濃度100g/Lとなるようにグルコース固体を添加した場合に比べて、グルコースの初期濃度を50g/Lとし、48時間後、96時間後および144時間後に終濃度50g/Lとなるようにグルコース固体を添加した場合の方が、物質の生産効率が高い。
【0055】
培地に含まれる栄養源としては、例えば、酵母エキス、ペプトン、ホエーなどが用いられるが、YP(10g/L酵母エキス、20g/Lペプトン)に上記の資化炭素源を加えた培地、例えばYPD5(50g/Lグルコース、10g/L酵母エキス、20g/Lペプトン)等の培地で、さらに適宜pH調整されたものが便利である。
【0056】
発酵温度は、用いる酵母の生育可能な範囲で選択することができる。発酵温度は、例えば、約15℃〜45℃とすることができ、より好ましくは20〜40℃、さらに好ましくは25〜35℃、最も好ましくは約30℃とする。また、発酵過程における培地のpHは3〜8に保持することが好ましく、より好ましくはpH4〜7であり、必要に応じてpH調節剤を用いることができる。用いるpH調節剤としては、炭酸カルシウム、水酸化ナトリウム、水酸化カリウム等が挙げられるが、好ましくは炭酸カルシウムとされる。
【0057】
物質(例えばイソプロパノール)製造に要する反応時間は特に限定されず、本発明の効果が認められる限り任意の反応時間で実施される。これらの条件の最適化は、当業者であれば容易に行うことができる。
【0058】
本発明による物質(例えばイソプロパノール)の製造法においては、このようにして製造した物質(例えばイソプロパノール)成分を培地から分離・回収するが、そのための具体的な手順は、分離・回収しようとする物質に応じて、当業者であれば容易に選択することができる。
【実施例】
【0059】
以下、実施例を挙げて本発明をより具体例に説明するが、これら実施例は本発明の技術的範囲を制限するものではない。
【0060】
実施例1:高いイソプロパノール耐性を示すキャンディダ・ユティリス株の選抜
キャンディダ・ユティリス NBRC0396株、NBRC0619株、NBRC0988株、NBRC0639株、NBRC0626株、NBRC1086株、NBRC10707株、ATCC9255株、ATCC28955株、ATCC44638株、ATCC9206株、ATCC96621株、ATCC18201株、およびATCC15239株を、YPD2(20g/Lペプトン、10g/L酵母エキス、20g/Lグルコース)に接種し、30℃で培養を行った。定常期の酵母培養液を10倍ごとに段階希釈し、10〜100g/Lのイソプロパノールを含むYPD2寒天培地にスポットし、30℃で培養した。3日間培養を継続し、コロニーの形成を観察した。結果を表1に記載する。
【0061】
【表1】
【0062】
以上の結果より、アッセイを行った株の中では、NBRC1086株、ATCC9255株、およびATCC15239株が、最も高い65g/LのIPA耐性を示すことが明らかとなった。
【0063】
実施例2:ピルビン酸脱炭酸酵素遺伝子が破壊されたキャンディダ・ユティリスATCC15239株の構築
キャンディダ・ユティリスATCC15239株において、その内因性遺伝子であるピルビン酸脱炭酸酵素遺伝子(PDC)を破壊した。この遺伝子破壊は、Cre−loxPシステムを用いて国際公開第2010/095750号パンフレットと同様の方法で行った。2回の破壊により、PDC1アレルが完全に欠損した株(dPDC1株)を構築した。dPDC1株は完全にはエタノール生産能を欠損しなかったが、エタノール生産能は著しく低下していた。
【0064】
実施例3:クロストリジウム・アセトブチリカム由来のCoA転移酵素サブユニットA遺伝子、同サブユニットB遺伝子、クロストリジウム・ベイジェリンキ由来のアセト酢酸脱炭酸酵素遺伝子および二級アルコール脱水素酵素遺伝子を発現するベクターの構築
イソプロパノール生合成に関わる4つの遺伝子、クロストリジウム・アセトブチリカム由来のCoA転移酵素遺伝子サブユニットA(ctfA)、同サブユニットB(ctfB)、クロストリジウム・ベイジェリンキ由来のアセト酢酸脱炭酸酵素(adc)および二級アルコール脱水素酵素(sadh)を効率良くキャンディダ・ユティリスで発現させるために、以下の項目を指標として、天然に存在しない新規な遺伝子配列を設計し、合成した:
(1)キャンディダ・ユティリスにおいて多用されているコドンの使用;
(2)遺伝子クローニングに不適当な制限酵素部位の排除;および
(3)染色体組み込み型発現ベクターに導入するための、5’末端におけるXbaIサイトおよび3’末端におけるBamHIサイトの付与。
【0065】
合成されたCoA転移酵素遺伝子サブユニットA遺伝子、同サブユニットB遺伝子、アセト酢酸脱炭酸酵素遺伝子および二級アルコール脱水素酵素遺伝子のヌクレオチド配列を、それぞれ配列番号1、3、5および7に示す。また、これらのヌクレオチド配列によってコードされるアミノ酸配列を、それぞれ配列番号2、4、6および8に示す。これらの遺伝子を有するプラスミドをそれぞれGA09142−ctfA、GA09142−ctfB、GA09142−adc、GA09142−sadhと名付けた。
【0066】
pVT92は、キャンディダ・ユティリスのオロチジン5’酸脱炭酸酵素遺伝子(URA3)座への組み込みベクターであり、ハイグロマイシン耐性遺伝子を選択マーカーとして有している(国際公開第2010/095750号パンフレット)。pVT92は、BgllII処理してリニアDNAにすることにより、染色体導入することができる。pVT92は、外来遺伝子を発現させるために、XbaI/BamHIギャップの外側にそれぞれ、グリセルアルデヒド脱水素酵素遺伝子プロモーターとグリセリン酸3’リン酸リン酸化酵素遺伝子ターミネーターを有している。また、pVT92は、プロモーターの5’末端側およびターミネーターの3’末端側には、それぞれNheIサイトおよびSpeIサイトを有している。
【0067】
pVT239は、キャンディダ・ユティリスのアルファアミノアジピン酸還元酵素遺伝子(LYS2)座への組み込みベクターであり、ジェネティシン耐性遺伝子を選択マーカーとして有している(国際公開第2010/095750号パンフレット)。pVT239は、NotI処理してリニアDNAにすることにより、染色体導入することができる。また、pVT239は、ジェネティシン耐性遺伝子の5’末端側にSpeIサイトを有している。
【0068】
各イソプロパノール合成酵素遺伝子を含む4つのベクターGA09142−ctfA、GA09142−ctfB、GA09142−adc、およびGA09142−sadhを、XbaIおよびBamHI処理し、アガロースゲル抽出による精製を行った後、同制限酵素で処理したpVT92にライゲーションして、pVT296、pVT298、pVT300およびpVT302を得た。pVT296およびpVT300はSpeI処理し、脱リン酸化を行った。また、pVT298およびpVT302はNheI、SpeI処理を行った後、ctfB発現カセット、sadh発現カセットをそれぞれ含むDNA断片の精製を行い、それぞれpVT296およびpVT300に連結して、pVT321およびpVT325を得た。pVT325をNheIおよびSpeIで処理した後、adc発現カセットとsadh発現カセットを含むDNA断片の精製を行い、pVT239のSpeIサイトに導入し、pVT346を得た。最終的に得られたpVT321はctfAとctfBのそれぞれの発現カセットとハイグロマイシン耐性遺伝子を有し、キャンディダ・ユティリスのURA3座に組み込まれるベクターであり、pVT346はadcとsadhのそれぞれの発現カセットとジェネティシン耐性遺伝子を有し、キャンディダ・ユティリスのLYS2座に組み込まれるベクターである。
【0069】
実施例4:4つのイソプロパノール合成遺伝子を有するキャンディダ・ユティリス株の構築
pVT321をBgllIIで消化し、エタノール沈殿により濃縮した。得られたDNA断片を用いて、C. utilis ATCC15239株、および実施例2で得られたdPDC1株を形質転換し、600μg/mLハイグロマイシンを含むYPD培地に塗抹し、30℃で2日間培養した。導入した全ての遺伝子を有する株を選抜した。
【0070】
さらにpVT346をNotIで消化し、エタノール沈殿により濃縮した。得られたそれぞれのDNA断片を用いて上記で得られた2株をそれぞれ形質転換し、600μg/mLハイグロマイシンと200μg/mLジェネティシンを含むYPD培地に塗抹し、30℃で2日間培養した。導入した全ての遺伝子を有する株を選抜し、ATCC15239株由来のイソプロパノール生産株をTMS272株、dPDC株由来のイソプロパノール生産株をTMS290株とした。
【0071】
実施例5:イソプロパノール生産試験
実施例4で得られた形質転換体のイソプロパノール生産能の評価を、以下に示すように実施した。
【0072】
各種形質転換体を2mL YPD液体培地(20g/Lペプトン、10g/L酵母エキス、20g/Lグルコース)/14mL容試験管に接種し、140rpm、30℃で24時間振とう培養した。得られた前培養液を100mL YPD10(20g/Lペプトン、10g/L酵母エキス、100g/Lグルコース)/200mL容三角フラスコもしくはYPD10C(20g/Lペプトン、10g/L酵母エキス、100g/Lグルコース、45g/L炭酸カルシウム)/200mL容三角フラスコに接種し、初期OD600=1、100rpm、30℃で振とう培養した。培養液から経時的にサンプリングし、0.2μmフィルターでろ過した後に、各種成分の定量分析を行った。グルコース、酢酸、エタノールおよびグリセロールは、高速液体クロマトフラフィー(島津製作所製)、ISep−ION300カラム(東京化成工業製)および示唆屈折計を用いて定量した。イソプロパノールおよびアセトンは、ガスクロマトフラフィーGC−2010 Plus(島津製作所製)を用いて定量した。
【0073】
また、フラスコでの簡易的な中和培養としてYPD10C(20g/Lペプトン、10g/L酵母エキス、100g/Lグルコース、45g/L炭酸カルシウム)培地を用いても同様の実験を行った。
【0074】
非中和条件下での培養試験の結果を図2、中和条件下での培養試験の結果を図3に記載する。なお、データは3反復試行の平均で示した。
【0075】
図2によれば、親株であるATCC15239株およびdPDC1株ではイソプロパノールの生産が認められなかったのに対し、イソプロパノール合成遺伝子を導入したTMS272株およびTMS290株では、それぞれ最大で0.16g/Lおよび0.13g/Lのイソプロパノールの生産が認められた。また、PDC1遺伝子を破壊した株では糖の消費速度および増殖能が著しく低下していた。TMS272株では培養24時間でグルコースが完全に消費され、エタノールの蓄積が認められたが、その後の培養の継続に伴ってエタノール濃度が低下した。エタノール濃度が低下している間もイソプロパノールの生産が認められたことから、TMS272株は、一度生産したエタノールを二次消費することによりイソプロパノールを生産していることが考えられた。dPDC1株に由来するTMS290株ではイソプロパノール生産速度が低下したことから、酵母におけるイソプロパノール生産においては、基質である細胞質のアセチルCoAがエタノールから供給される経路が最も重要であることが示唆された。エタノールの同化は酸化的反応であることから、これらの結果は、酵母によるイソプロパノール生産においてはキャンディダ・ユティリスのような呼吸活性の高いクラブツリー陰性酵母が適している可能性を示唆している。
【0076】
図3によれば、培地に炭酸カルシウムを添加したことにより、TMS272株は1.1g/Lのイソプロパノールを生産し、非中和条件より生産量が増大した。また、TMS272株は、16.2g/Lの酢酸を蓄積しており、何らかの理由により酢酸が蓄積したことでpHが低下し、イソプロパノールの生産を阻害していたことが考えられた。また、TMS290株においても、中和条件下ではイソプロパノール生産量が0.9g/Lまで増大しており、一方で37.4g/Lという高濃度のグリセロールを蓄積していた。
【0077】
実施例6:過剰発現によってイソプロパノール生産性を向上させる遺伝子の選抜
過剰発現することによりイソプロパノールの生産性を向上させる遺伝子の選抜を行った。候補遺伝子が触媒する代謝経路を図1に記載する。
【0078】
キャンディダ・ユティリス内在性のアセチルCoA加水分解酵素遺伝子(ACH1;ヌクレオチド配列:配列番号9;アミノ酸配列:配列番号10)、アセチルCoA合成酵素遺伝子(ACS1;ヌクレオチド配列:配列番号11;アミノ酸配列:配列番号12、ACS2;ヌクレオチド配列:配列番号13;アミノ酸配列:配列番号14)、細胞質局在型エタノール脱水素酵素遺伝子(ADH2;ヌクレオチド配列:配列番号15;アミノ酸配列:配列番号16)、ミトコンドリア局在型エタノール脱水素酵素遺伝子(ADH3;ヌクレオチド配列:配列番号17;アミノ酸配列:配列番号18)、細胞質局在型アセトアルデヒド脱水素酵素遺伝子(ALD1;ヌクレオチド配列:配列番号19;アミノ酸配列:配列番号20)、ミトコンドリア局在型アセトアルデヒド脱水素酵素遺伝子(ALD2;ヌクレオチド配列:配列番号21;アミノ酸配列:配列番号22)、ミトコンドリア内膜カルニチン輸送体(CRC1;ヌクレオチド配列:配列番号23;アミノ酸配列:配列番号24)、ミトコンドリア局在型カルニチンアセチルCoA転移酵素(CAT2;ヌクレオチド配列:配列番号25;アミノ酸配列:配列番号26)、カルニチンアセチル転移酵素遺伝子(YAT1;ヌクレオチド配列:配列番号27;アミノ酸配列:配列番号28、YAT2;ヌクレオチド配列:配列番号29;アミノ酸配列:配列番号30)、アセチルCoAアセチル基転移酵素(ERG10;ヌクレオチド配列:配列番号61;アミノ酸配列:配列番号62)、ヤロウィア・リポリティカ由来のATP−クエン酸除去付加酵素遺伝子(ACL1;ヌクレオチド配列:配列番号31;アミノ酸配列:配列番号32と、ACLY;ヌクレオチド配列:配列番号33;アミノ酸配列:配列番号34がサブユニットを構成する遺伝子)を、それぞれのゲノムDNAを鋳型としてPCRにより増幅した。PCRに用いたプライマーは以下の通りである:(ACH1;配列番号35と配列番号36)、(ACS1;配列番号37と配列番号38)、(ACS2;配列番号39と配列番号40)、(ADH2;配列番号41と配列番号42)、(ADH3;配列番号43と配列番号44)、(ALD1;配列番号45と配列番号46)、(ALD2;配列番号47と配列番号48)、(CAT2;配列番号49と配列番号50)、(CRC1;配列番号51と配列番号52)、(YAT1;配列番号53と配列番号54)、(YAT2;配列番号55と配列番号56)、(ACL1;配列番号57と配列番号58)、(ACLY;配列番号59と配列番号60)、(ERG10;配列番号63と配列番号64)。
【0079】
得られたPCR産物をXbaI/BamHI処理して、同制限酵素で処理したpCU155(Keiji Kondo, , Yutaka Miura, Hidetaka Sone, Kazuo Kobayashi & Hiroshi lijima., High-level expression of a sweet protein, monellin, in the food yeast Candida utilis., Nature Biotechnology 15, 453 - 457 (1997))にクローニングした。pCU155は、シクロヘキシミド耐性遺伝子を選抜マーカーとして有し、URA3座にマルチコピーインテグレーションされるベクターであり、XbaI/BamHIギャップの外側にそれぞれ、グリセルアルデヒド脱水素酵素遺伝子プロモーターと同ターミネーターを有している。これまでに構築しているイソプロパノール生産株のURA3座にはイソプロパノール合成酵素遺伝子が組み込まれているが、C.utilisは高次倍数体であり、URA3座が複数コピー存在するため、さらにURA3座に遺伝子の導入が可能である。また、イソプロパノール合成遺伝子はURA3座に二点組み換えにより挿入されているため、pCU155の組み込みにより、導入された遺伝子の置換が起こることはない。候補遺伝子がクローン化されたプラスミドはBgllIIで消化し、エタノール沈殿により濃縮した。得られたDNA断片を用いてC.utilis TMS272株を形質転換し、600μg/mLハイグロマイシン、200μg/mLジェネティシンおよび20μg/mLシクロヘキシミドを含むYPD培地に塗抹し、30℃で4日間培養した。なお、ACL1とACLYのDNAは混合し、同時に形質転換を行った。得られた各4クローンについて発酵試験を行い、イソプロパノール生産性を調査した。結果を図4に示す。
【0080】
図4によると、ACS1、ACS2、CRC1、YAT1、ERG10、またはACLの過剰発現により、イソプロパノール生産量が1.5〜4.0倍になることが明らかとなった。ACS1、ACS2、YAT1、およびACLの各遺伝子はアセチルCoAの合成に関わる酵素遺伝子であり、CRC1遺伝子はアセチルCoA合成の前駆物質をミトコンドリアから細胞質に輸送するキャリアタンパク質遺伝子であり、ERG10遺伝子はアセチルCoAからアセトアセチルCoAを合成する遺伝子である。これらの結果により、酵母によるイソプロパノール高生産のためには、細胞質のアセチルCoAを効率的に生産するための酵素遺伝子の発現または活性をアップレギュレートすることが重要であることが示される。
【0081】
実施例7:ACL1/ACLY、ACS2、ERG10、CRC1およびYAT1を同時に過剰発現するイソプロパノール生産株の構築と培養試験
ACL1、ACLY、ACS2、ERG10、CRC1およびYAT1の各遺伝子がクローン化されたプラスミドをBgllIIで消化し、エタノール沈殿により濃縮した。得られた各5μgのDNA断片を混合し、この混合物を用いてTMS272株を形質転換し、600μg/mLハイグロマイシン、200μg/mLジェネティシンおよび10μg/mLシクロヘキシミドを含むYPD培地に塗抹し、30℃で4日間培養した。得られた80クローンについて発酵試験を行い、イソプロパノール生産量が最も高い株を選抜し、TMS378株とした。
【0082】
TMS378株の培養試験を行った結果を図5に示す。TMS272株が1.3g/Lのイソプロパノールを生産したのに対して、TMS378株は9.8g/Lと約7.5倍のイソプロパノール生産量を示した。また、TMS272株では酢酸が18.1g/L蓄積したのに対して、TMS378株は4.5g/Lと、主要な副産物である酢酸が約4分の1に低下していた。
【0083】
実施例8:TMS378株のイソプロパノール生産における初期OD600および振とう速度の影響
イソプロパノール生産に適したTMS378株の培養条件の検討を行った。TMS378株を2mL YPD液体培地(20g/Lペプトン、10g/L酵母エキス、20g/Lグルコース)/14mL容試験管に接種し、140rpm、30℃で24時間振とう培養した。得られた前培養液を100mL YPD10C(20g/Lペプトン、10g/L酵母エキス、100g/Lグルコース、45g/L炭酸カルシウム)/300mL容三角フラスコに接種し、初期OD600を1もしくは10とし、100rpmもしくは120rpm、30℃で振とう培養した。培養72時間後に終濃度100g/Lとなるようにグルコース固体を培養液に添加し、さらに144時間まで培養を行った。経時的にサンプリングして0.2μmフィルターでろ過した。各種定量分析は、実施例5に記載の方法に従って行った。なお、データは3反復試行の平均で示した。結果を図6に示す。
【0084】
図6によると、初期OD600=10、120rpmで培養したときに最も高いイソプロパノール生産量が見られ、培養144時間後に最大で18.5g/Lのイソプロパノールが生産された。その際の対糖収率は30.8%であった。このことから、振とう速度が速いほど(通気量が多いほど)、また、初期ODが高いほど、イソプロパノール生産には好ましいことが明らかとなった。
【0085】
実施例9:TMS378株のイソプロパノール生産における添加グルコース濃度の影響
イソプロパノール生産に適したTMS378株の培養条件の検討を行った。TMS378株を2mL YPD液体培地(20g/Lペプトン、10g/L酵母エキス、20g/Lグルコース)/14mL容試験管に接種し、140rpm、30℃で24時間振とう培養した。得られた前培養液を100mL YPD10C(20g/Lペプトン、10g/L酵母エキス、100g/Lグルコース、45g/L炭酸カルシウム)/300mL容三角フラスコもしくはYPD5C(20g/Lペプトン、10g/L酵母エキス、50g/Lグルコース、45g/L炭酸カルシウム)/300mL容三角フラスコに接種し、初期OD600=1、120rpm、30℃で振とう培養した。YPD10Cで培養を行ったサンプルについては、培養72時間後に終濃度100g/Lとなるようにグルコース固体を添加し、さらに192時間まで培養を行った。また、YPD5Cで培養を行ったサンプルについては、培養48時間おきに3回終濃度50g/Lとなるようにグルコース固体を添加し、さらに192時間まで培養を行った。いずれの条件も最終のグルコース濃度は200g/Lである。経時的にサンプリングして0.2μmフィルターでろ過した。各種定量分析は、実施例5に記載の方法に従って行った。なお、データは3反復試行の平均で示した。結果を図7に示す。
【0086】
図7によると、YPD10Cで培養し、培養72時間後に100g/Lのグルコースを添加した場合は、培養122時間後に最大で19.2g/Lのイソプロパノールが生産された。しかし、それ以降はイソプロパノール生産量の増大は認められず、酢酸が蓄積した(27.2g/L)。イソプロパノールの生産における最大の対糖収率は29.0%であった。一方、YPD5Cで培養し、48時間おきに50g/Lのグルコースを3回添加した場合は、培養192時間までイソプロパノールの増大が認められ、培養192時間後に最大で27.4g/Lのイソプロパノールが生産された。また、酢酸の蓄積はほとんど認めれらなかった。この条件でのイソプロパノールの生産における最大の対糖収率は41.5%であった。このことから、添加するグルコースの濃度を比較的低濃度に保つことはイソプロパノール生産において好ましいことが明らかとなった。
【0087】
実施例10:イソプロパノール生産量向上に寄与の大きい遺伝子の特定
同時に過剰発現することによってイソプロパノール生産量を向上させる寄与の大きい遺伝子を特定するために、(i)ACL1、ACLYおよびACS2の組み合わせを過剰発現する株、(ii)ACL1、ACLYおよびERG10の組み合わせを過剰発現する株、(iii)ACS2およびERG10の組み合わせを過剰発現する株、(iv)ACL1、ACLY、ACS2およびERG10の組み合わせを過剰発現する株をそれぞれ構築し、ACL1、ACLY、ACS2、CRC1、ERG10およびYAT1の組み合わせを過剰発現する株(TMS378株)およびそれぞれの単独遺伝子の過剰発現株との間で、イソプロパノール生産量の比較を行った。組み換え株の構築は実施例6および実施例7と同様の方法を用いて行い、組み込みたい遺伝子DNAを混合し、同時に形質転換を行った。得られた各32クローンについて発酵試験を行い、それぞれの導入遺伝子の組み合わせにおいて平均的なイソプロパノール生産量を示す株を選抜した。各構築株を2mL YPD液体培地(20g/Lペプトン、10g/L酵母エキス、20g/Lグルコース)/14mL容試験管に接種し、140rpm、30℃で24時間振とう培養した。得られた前培養液を3mL YPD10C(20g/Lペプトン、10g/L酵母エキス、100g/Lグルコース、45g/L炭酸カルシウム)/ガラス試験管に接種し、初期OD600=1、100rpm、30℃で振とう培養した。経時的にサンプリングを行い、高速液体クロマトフラフィーおよびガスクロマトフラフィーGC−2010でイソプロパノール生産量を評価した。なお、データは3反復試行の平均で示した。
【0088】
図8によると、ACS1、ACS2およびERG10のそれぞれ単独の過剰発現により、イソプロパノール生産量が2.2〜2.7倍になることが明らかとなった。(i)ACL1、ACLYおよびACS2の組み合わせを過剰発現する株、ならびに(ii)ACL1、ACLYおよびERG10の組み合わせを過剰発現する株のイソプロパノール生産量は、それぞれACS2およびERG10を単独で過剰発現する株と有意な差は認められなかった。一方、(iii)ACS2およびERG10の組み合わせを過剰発現する株ではイソプロパノール生産の飛躍的な向上が認められ、生産量が11.6倍に向上した。また、(iii)ACS2およびERG10の組み合わせを過剰発現する株、(iv)ACL1、ACLY、ACS2およびERG10の組み合わせを過剰発現する株、TMS378株(ACL1、ACLY、ACS2、CRC1、ERG10およびYAT1の組み合わせを過剰発現する株)の株間において、イソプロパノール生産量に有意な差は認められなかった。
【0089】
これらの結果より、同時に過剰発現した際にはACL、CRC1およびYAT1の過剰発現がイソプロパノールの生産性向上に与える寄与は小さく、イソプロパノール生産性を向上させるためにはACS2とERG10を同時に過剰発現することが最も重要であることが示された。以上の結果は、酵母によるイソプロパノール高生産のためにはエタノールからアセチルCoAを合成する代謝経路、すなわち酢酸からアセチルCoAを合成する代謝経路に関わる酵素遺伝子の発現または活性を増強することが重要であることを示している。
【図面の簡単な説明】
【0090】
【図1】図1は、酵母によるイソプロパノール合成経路を示す図である。
【図2】図2は、イソプロパノール生産遺伝子を導入した株の非中和条件での発酵試験の結果を示す図であり、培養液中のイソプロパノール濃度(パネルA)、グルコース濃度(パネルB)、エタノール濃度(パネルC)、酢酸濃度(パネルD)、グリセロール濃度(パネルE)およびOD600(パネルF)の経時変化を示している。各パネルにおいて、黒丸はATCC15239株の結果を示し、白丸はTMS272株(ATCC15239株+IPA合成遺伝子)の結果を示し、黒三角はPDC1破壊株の結果を示し、白三角はTMS290株(PDC破壊株+IPA合成遺伝子)の結果を示す。
【図3】図3は、イソプロパノール生産遺伝子を導入した株の中和条件での発酵試験の結果を示す図であり、培養液中のイソプロパノール濃度(パネルA)、グルコース濃度(パネルB)、エタノール濃度(パネルC)、酢酸濃度(パネルD)およびグリセロール濃度(パネルE)の経時変化を示している。各パネルにおいて、黒丸はATCC15239株の結果を示し、白丸はTMS272株(ATCC15239株+IPA合成遺伝子)の結果を示し、黒三角はPDC1破壊株の結果を示し、白三角はTMS290株(PDC破壊株+IPA合成遺伝子)の結果を示す。
【図4】図4は、TMS272株において、各種遺伝子を過剰発現させた場合におけるイソプロパノール生産量を、TMS272株によるイソプロパノール生産量に対する相対量として示したグラフである。
【図5】図5は、イソプロパノール生酸性を向上させる遺伝子を全て過剰発現する株の発酵試験の結果を示す図であり、培養液中のイソプロパノール濃度(パネルA)、グルコース濃度(パネルB)、エタノール濃度(パネルC)、酢酸濃度(パネルD)、グリセロール濃度(パネルE)およびアセトン濃度(パネルF)の経時変化を示している。各パネルにおいて、黒丸はTMS272株の結果を示し、白丸はTMS378株(TMS272株+ACL1:ACLY:ACS2:ERG10:CRC1:YAT1)の結果を示す。
【図6】図6は、イソプロパノール生産性を向上させる遺伝子を全て過剰発現するTMS378株の30℃での発酵試験(初期OD600および振とう速度の影響)の結果を示す図であり、培養液中のイソプロパノール濃度(パネルA)、グルコース濃度(パネルB)、エタノール濃度(パネルC)、酢酸濃度(パネルD)、グリセロール濃度(パネルE)およびアセトン濃度(パネルF)の経時変化を示している。各パネルにおいて、黒丸は初期OD600=1かつ攪拌数=100rpmの条件下での結果を示し、黒三角は初期OD600=1かつ攪拌数=120rpmの条件下での結果を示し、白丸は初期OD600=10かつ攪拌数=100rpmの条件下での結果を示し、白三角は初期OD600=10かつ攪拌数=120rpmの条件下での結果を示す。
【図7】図7は、イソプロパノール生産性を向上させる遺伝子を全て過剰発現するTMS378株の30℃での発酵試験(流加グルコース濃度の影響)の結果を示す図である。パネルAは、初期グルコース濃度を100g/Lとし、培養72時間後に100g/lのグルコースを流加して、192時間培養を行った結果を示す。パネルBは、初期グルコース濃度を50g/Lとし、培養48時間後、96時間後および144時間後にそれぞれ50g/lのグルコースを流加して、192時間培養を行った結果を示す。各パネルにおいて、イソプロパノール濃度(黒三角)、グルコース濃度(黒丸)、エタノール濃度(白丸)および酢酸濃度(白三角)の経時変化を示している。
【図8】図8は、TMS272株において、各種遺伝子を過剰発現させた場合におけるイソプロパノール生産量を示したグラフである。
【特許請求の範囲】
【請求項1】
以下の酵素:
(a)アセチルCoA合成酵素(ACS);および
(b)アセチルCoAアセチル基転移酵素(ERG10)
の活性が増強されている、酵母菌株。
【請求項2】
以下の遺伝子:
(a)アセチルCoA合成酵素(ACS)の活性を有するポリペプチドをコードする遺伝子;および
(b)アセチルCoAアセチル基転移酵素(ERG10)の活性を有するポリペプチドをコードする遺伝子
の発現が増強されている、請求項1に記載の酵母菌株。
【請求項3】
ACS活性を有するポリペプチドが、
(i)配列番号12または配列番号14で表されるアミノ酸配列を含むポリペプチド、または
(ii)配列番号12または配列番号14で表されるアミノ酸配列において、1もしくは数個のアミノ酸が欠失、置換、付加もしくは挿入されたアミノ酸配列を含み、かつACS活性を有するポリペプチド
である、請求項2に記載の酵母菌株。
【請求項4】
ERG10活性を有するポリペプチドが、
(i)配列番号62で表されるアミノ酸配列を含むポリペプチド、または
(ii)配列番号62で表されるアミノ酸配列において、1もしくは数個のアミノ酸が欠失、置換、付加もしくは挿入されたアミノ酸配列を含み、かつERG10活性を有するポリペプチド
である、請求項2または3に記載の酵母菌株。
【請求項5】
さらに、以下の遺伝子:
(c)ATP−クエン酸除去付加酵素(ACL)の活性を有するポリペプチドをコードする遺伝子;
(d)ミトコンドリア内膜カルニチン輸送体(CRC1)の活性を有するポリペプチドをコードする遺伝子;および
(e)カルニチンアセチル転移酵素遺伝子(YAT1)の活性を有するポリペプチドをコードする遺伝子
からなる群から選択される少なくとも一種の遺伝子の発現が増強されている、請求項1〜4のいずれか一項に記載の酵母菌株。
【請求項6】
ACL活性を有するポリペプチドが、ACL1活性を有するポリペプチドおよびACLY活性を有するポリペプチドからなるものであり、
ACL1活性を有するポリペプチドが、
(i)配列番号32で表されるアミノ酸配列を含むポリペプチド、または
(ii)配列番号32で表されるアミノ酸配列において、1もしくは数個のアミノ酸が欠失、置換、付加もしくは挿入されたアミノ酸配列を含み、かつACL1活性を有するポリペプチド
であり、
ACLY活性を有するポリペプチドが、
(i)配列番号34で表されるアミノ酸配列を含むポリペプチド、または
(ii)配列番号34で表されるアミノ酸配列において、1もしくは数個のアミノ酸が欠失、置換、付加もしくは挿入されたアミノ酸配列を含み、かつACLY活性を有するポリペプチド
である、請求項5に記載の酵母菌株。
【請求項7】
CRC1活性を有するポリペプチドが、
(i)配列番号24で表されるアミノ酸配列を含むポリペプチド、または
(ii)配列番号24で表されるアミノ酸配列において、1もしくは数個のアミノ酸が欠失、置換、付加もしくは挿入されたアミノ酸配列を含み、かつCRC1活性を有するポリペプチド
である、請求項5または6に記載の酵母菌株。
【請求項8】
YAT1活性を有するポリペプチドが、
(i)配列番号28で表されるアミノ酸配列を含むポリペプチド、または
(ii)配列番号28で表されるアミノ酸配列において、1もしくは数個のアミノ酸が欠失、置換、付加もしくは挿入されたアミノ酸配列を含み、かつYAT1活性を有するポリペプチド
である、請求項5〜7のいずれか一項に記載の酵母菌株。
【請求項9】
クラブツリー効果陰性酵母である、請求項1〜8のいずれか一項に記載の酵母菌株。
【請求項10】
キャンディダ・ユティリスである、請求項1〜9のいずれか一項に記載の酵母菌株。
【請求項11】
アセチルCoAを経由する代謝産物を合成するための遺伝子により形質転換されているものである、請求項1〜10のいずれか一項に記載の酵母菌株。
【請求項12】
CoA転移酵素サブユニットA(ctfA)、CoA転移酵素サブユニットB(ctfB)、アセト酢酸脱炭酸酵素(adc)および二級アルコール脱水素酵素(sadh)の活性を有するポリペプチドをコードしている4種の遺伝子により形質転換されてなる、請求項11に記載の酵母菌株。
【請求項13】
請求項1〜12のいずれか一項に記載の酵母菌株を培養することを含んでなる、物質を製造する方法。
【請求項14】
前記物質が、アセチルCoAを経由する代謝産物である、請求項13に記載の方法。
【請求項15】
前記物質がイソプロパノールである、請求項13または14に記載の方法。
【請求項1】
以下の酵素:
(a)アセチルCoA合成酵素(ACS);および
(b)アセチルCoAアセチル基転移酵素(ERG10)
の活性が増強されている、酵母菌株。
【請求項2】
以下の遺伝子:
(a)アセチルCoA合成酵素(ACS)の活性を有するポリペプチドをコードする遺伝子;および
(b)アセチルCoAアセチル基転移酵素(ERG10)の活性を有するポリペプチドをコードする遺伝子
の発現が増強されている、請求項1に記載の酵母菌株。
【請求項3】
ACS活性を有するポリペプチドが、
(i)配列番号12または配列番号14で表されるアミノ酸配列を含むポリペプチド、または
(ii)配列番号12または配列番号14で表されるアミノ酸配列において、1もしくは数個のアミノ酸が欠失、置換、付加もしくは挿入されたアミノ酸配列を含み、かつACS活性を有するポリペプチド
である、請求項2に記載の酵母菌株。
【請求項4】
ERG10活性を有するポリペプチドが、
(i)配列番号62で表されるアミノ酸配列を含むポリペプチド、または
(ii)配列番号62で表されるアミノ酸配列において、1もしくは数個のアミノ酸が欠失、置換、付加もしくは挿入されたアミノ酸配列を含み、かつERG10活性を有するポリペプチド
である、請求項2または3に記載の酵母菌株。
【請求項5】
さらに、以下の遺伝子:
(c)ATP−クエン酸除去付加酵素(ACL)の活性を有するポリペプチドをコードする遺伝子;
(d)ミトコンドリア内膜カルニチン輸送体(CRC1)の活性を有するポリペプチドをコードする遺伝子;および
(e)カルニチンアセチル転移酵素遺伝子(YAT1)の活性を有するポリペプチドをコードする遺伝子
からなる群から選択される少なくとも一種の遺伝子の発現が増強されている、請求項1〜4のいずれか一項に記載の酵母菌株。
【請求項6】
ACL活性を有するポリペプチドが、ACL1活性を有するポリペプチドおよびACLY活性を有するポリペプチドからなるものであり、
ACL1活性を有するポリペプチドが、
(i)配列番号32で表されるアミノ酸配列を含むポリペプチド、または
(ii)配列番号32で表されるアミノ酸配列において、1もしくは数個のアミノ酸が欠失、置換、付加もしくは挿入されたアミノ酸配列を含み、かつACL1活性を有するポリペプチド
であり、
ACLY活性を有するポリペプチドが、
(i)配列番号34で表されるアミノ酸配列を含むポリペプチド、または
(ii)配列番号34で表されるアミノ酸配列において、1もしくは数個のアミノ酸が欠失、置換、付加もしくは挿入されたアミノ酸配列を含み、かつACLY活性を有するポリペプチド
である、請求項5に記載の酵母菌株。
【請求項7】
CRC1活性を有するポリペプチドが、
(i)配列番号24で表されるアミノ酸配列を含むポリペプチド、または
(ii)配列番号24で表されるアミノ酸配列において、1もしくは数個のアミノ酸が欠失、置換、付加もしくは挿入されたアミノ酸配列を含み、かつCRC1活性を有するポリペプチド
である、請求項5または6に記載の酵母菌株。
【請求項8】
YAT1活性を有するポリペプチドが、
(i)配列番号28で表されるアミノ酸配列を含むポリペプチド、または
(ii)配列番号28で表されるアミノ酸配列において、1もしくは数個のアミノ酸が欠失、置換、付加もしくは挿入されたアミノ酸配列を含み、かつYAT1活性を有するポリペプチド
である、請求項5〜7のいずれか一項に記載の酵母菌株。
【請求項9】
クラブツリー効果陰性酵母である、請求項1〜8のいずれか一項に記載の酵母菌株。
【請求項10】
キャンディダ・ユティリスである、請求項1〜9のいずれか一項に記載の酵母菌株。
【請求項11】
アセチルCoAを経由する代謝産物を合成するための遺伝子により形質転換されているものである、請求項1〜10のいずれか一項に記載の酵母菌株。
【請求項12】
CoA転移酵素サブユニットA(ctfA)、CoA転移酵素サブユニットB(ctfB)、アセト酢酸脱炭酸酵素(adc)および二級アルコール脱水素酵素(sadh)の活性を有するポリペプチドをコードしている4種の遺伝子により形質転換されてなる、請求項11に記載の酵母菌株。
【請求項13】
請求項1〜12のいずれか一項に記載の酵母菌株を培養することを含んでなる、物質を製造する方法。
【請求項14】
前記物質が、アセチルCoAを経由する代謝産物である、請求項13に記載の方法。
【請求項15】
前記物質がイソプロパノールである、請求項13または14に記載の方法。
【図1】

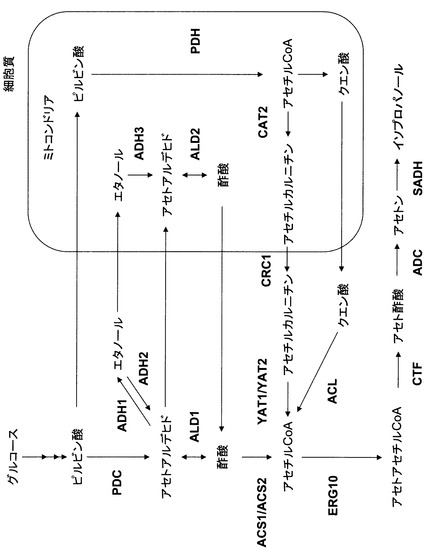
【図2】
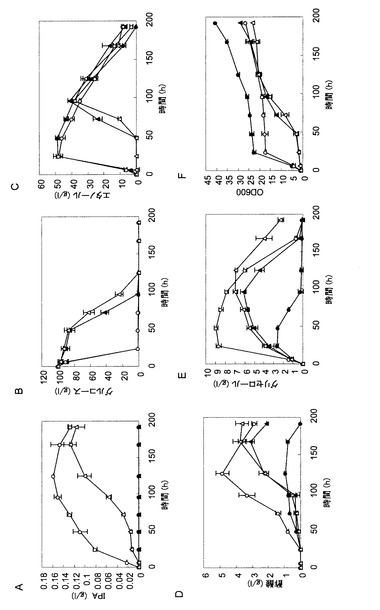
【図3】
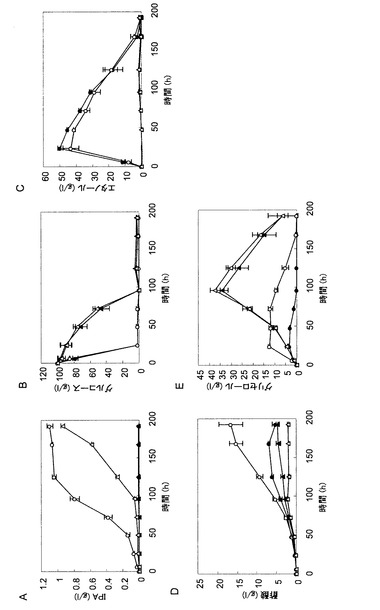
【図4】
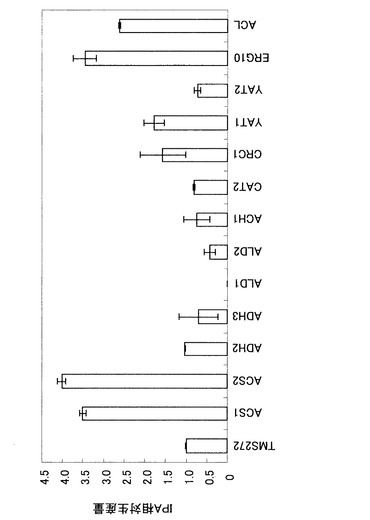
【図5】
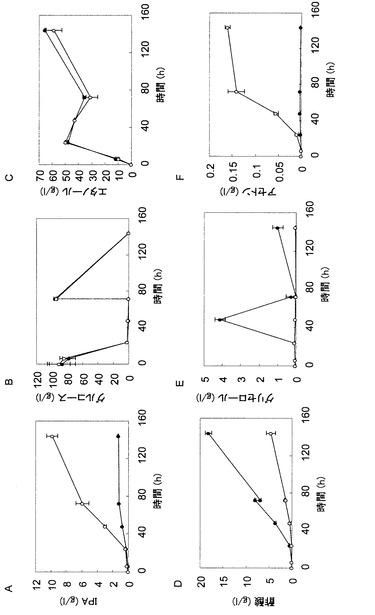
【図6】
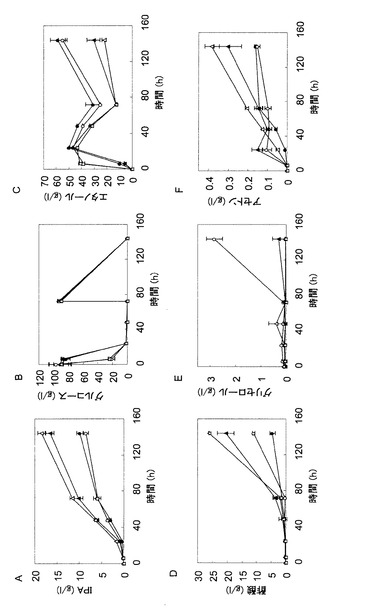
【図7】
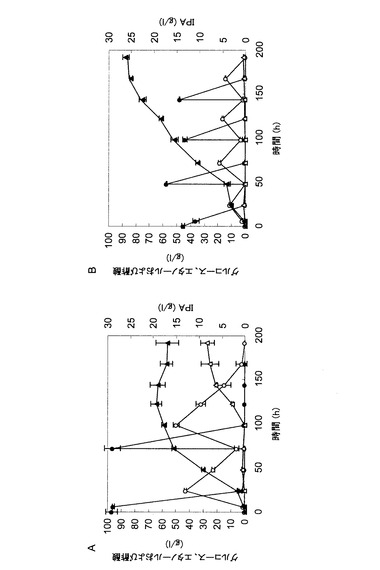
【図8】

【図2】
【図3】
【図4】
【図5】
【図6】
【図7】
【図8】
【公開番号】特開2012−254044(P2012−254044A)
【公開日】平成24年12月27日(2012.12.27)
【国際特許分類】
【出願番号】特願2011−129527(P2011−129527)
【出願日】平成23年6月9日(2011.6.9)
【国等の委託研究の成果に係る記載事項】(出願人による申告)平成22年度、独立行政法人新エネルギー・産業技術総合開発機構・グリーン・サステイナブルケミカルプロセス基盤技術開発委託研究、産業技術力強化法第19条の適用を受ける特許出願
【出願人】(000253503)キリンホールディングス株式会社 (247)
【Fターム(参考)】
【公開日】平成24年12月27日(2012.12.27)
【国際特許分類】
【出願日】平成23年6月9日(2011.6.9)
【国等の委託研究の成果に係る記載事項】(出願人による申告)平成22年度、独立行政法人新エネルギー・産業技術総合開発機構・グリーン・サステイナブルケミカルプロセス基盤技術開発委託研究、産業技術力強化法第19条の適用を受ける特許出願
【出願人】(000253503)キリンホールディングス株式会社 (247)
【Fターム(参考)】
[ Back to top ]