
酵母におけるHPV31L1の最適化発現
HPV31 L1タンパク質をコードする合成DNA分子を提供する。特に、本発明は、酵母により認識される内部転写終結シグナルを含有しない、HPV31 L1タンパク質をコードするポリヌクレオチドを提供する。また、酵母細胞内での高レベル発現に関してコドン最適化された、HPV31 L1をコードする合成ポリヌクレオチドも提供する。該合成分子は、HPV31ウイルス様粒子(VLP)の製造、ならびにHPV31 VLPを含むワクチンおよび医薬組成物の製造に使用することが可能である。本発明のワクチンは、中和抗体および細胞性免疫により、パピローマウイルス感染に対する有効な免疫予防をもたらす。
【発明の詳細な説明】
【技術分野】
【0001】
本発明は、全般的には、ヒトパピローマウイルス(HPV)の治療に関する。より詳しくは、本発明は、HPV31 L1タンパク質をコードする合成ポリヌクレオチド、ならびに該ポリヌクレオチドを含む組換えベクターおよび宿主に関する。本発明はまた、HPV31様ウイルス様粒子(VLP)、ならびにHPVの予防および治療のためのワクチンおよび医薬組成物におけるその使用に関する。
【背景技術】
【0002】
80を超える型のヒトパピローマウイルス(HPV)が存在し、それらのうちの多くは良性増殖性疣贅から悪性癌に及ぶ多種多様な生物学的表現型に関連づけられている(概説としては、McMurrayら,Int.J.Exp.Pathol.82(1):15−33(2001)を参照されたい)。HPV6およびHPV11は、性器粘膜または呼吸器粘膜の良性疣贅、非悪性尖形コンジローマおよび/または低悪性異形成に最も一般的に関連している型である。HPV16およびHPV18は、子宮頸部、膣、外陰部および肛門管の発生部位内非浸潤性(in situ)および浸潤性癌に最も一般的に関連している高リスク型である。子宮頸癌の90%以上はHPV16、HPV18またはより稀な発癌性型HPV31、33、45、52および58に関連している(Schiffmanら,J.Natl.Cancer Inst.85(12):958−64(1993))。HPV DNAが子宮頸癌の90〜100%で検出されるという観察は、HPVが子宮頸癌を引き起こすという強力な疫学的証拠を提供している(Boschら,J.Clin.Pathol.55:244−265(2002)を参照されたい)。
【0003】
パピローマウイルスは、8個までの初期遺伝子および2個の後期遺伝子をコードする小型(50〜60nm)非外包性二十面体DNAウイルスである。そのウイルスゲノムのオープンリーディングフレーム(ORF)はE1〜E7ならびにL1およびL2と称され、この場合の「E」は初期を意味し、「L」は後期を意味する。L1およびL2はウイルスカプシドタンパク質をコードし、一方、E遺伝子は、ウイルス複製および細胞トランスフォーメーションのような機能に関連している。
【0004】
L1タンパク質は主要カプシドタンパク質であり、55〜60kDaの分子量を有する。L2タンパク質は副次的カプシドタンパク質である。免疫学的データは、L2タンパク質の大部分がL1タンパク質の内部に存在することを示唆している。L1タンパク質およびL2タンパク質は共に、種々のパピローマウイルス間で高度に保存されている。
【0005】
酵母、昆虫細胞、哺乳類細胞または細菌におけるL1タンパク質またはL1およびL2タンパク質の組合せ体の発現はウイルス様粒子(VLP)の自己集合を引き起こす(概説としては、SchillerおよびRoden,in Papillomavirus Reviews:Current Research on Papillomaviruses;Lacey編,Leeds,UK:Leeds Medical Information,pp 101−12(1996)を参照されたい)。VLPは真正ビリオンに形態学的に類似しており、動物またはヒトに投与されると高力価の中和抗体を誘導しうる。VLPは、潜在的発癌性ウイルスゲノムを含有しないため、HPVワクチンの開発における生ウイルスの使用に代わる安全な手段となる(概説としては、SchillerおよびHidesheim,J.Clin.Virol.19:67−74(2000)を参照されたい)。この理由により、L1およびL2遺伝子はHPV感染および疾患に対する予防用および治療用ワクチンの免疫学的標的と目されている。
【0006】
うまく形質転換された宿主生物における高い発現レベルのカプシドタンパク質を入手することは困難であり、これにより精製タンパク質の製造が制限されるため、HPVワクチンの開発および商業化は妨げられている。したがって、HPV31 L1タンパク質のようなHPV L1タンパク質をコードする野生型ヌクレオチド配列が同定されているものの(Goldsboroughら,Virology 171(1):306−311(1989))、意図される宿主細胞内での発現に関して最適化されたHPV31 L1コード化ヌクレオチド配列を利用する容易に再生可能な粗HPVタンパク質源を開発することが非常に望ましいであろう。また、ワクチン開発で使用する天然タンパク質の免疫付与特性を有する大量のHPV31 L1 VLPを製造することが有用であろう。
【発明の開示】
【0007】
発明の概要
本発明は、子宮頸癌に関連づけられているHPV31 L1遺伝子により発現されるタンパク質産物に対する免疫を惹起または増強するための組成物および方法に関する。特に、本発明は、酵母により認識される内部転写終結シグナルを含有しない、HPV31 L1タンパク質をコードするポリヌクレオチドを提供する。また、酵母細胞内での高レベル発現に関してコドン最適化された、HPV31 L1をコードする合成ポリヌクレオチドも提供する。本発明は更に、HPV31ウイルス様粒子(VLP)を提供し、HPV疾患またはHPV関連癌の予防および/または治療用の免疫原性組成物およびワクチンにおける該VLPの使用を開示する。
【0008】
本発明は、HPV31 L1タンパク質をコードする合成DNA分子に関する。本発明の1つの態様においては、該合成分子のヌクレオチド配列は、酵母により認識される転写終結シグナルを除去するために改変されている。もう1つの態様においては、合成分子のコドンは、酵母細胞により好ましいコドンが利用されるよう設計される。該合成分子は、VLPに自己集合しうるHPV31 L1タンパク質の源として使用することが可能である。該VLPは、VLPに基づくワクチンにおいて使用することが可能である。
【0009】
本発明の特定の実施形態は、配列番号4に記載のHPV31 L1タンパク質をコードする、配列番号2または配列番号3に記載のヌクレオチドの配列を含む合成核酸分子を含む。
【0010】
前記のとおり、本発明においては、酵母により認識される転写終結シグナルを含有しないHPV31 L1遺伝子をコードする合成ポリヌクレオチドを提供する。本発明はまた、酵母細胞にとって好ましいコドンを含有するよう更に改変された、前記のHPV31 L1をコードする合成ポリヌクレオチドを提供する。
【0011】
また、本明細書中の全体に開示されている核酸分子を含有する組換えベクターおよび組換え宿主細胞(原核細胞および真核細胞)を提供する。
【0012】
本発明は、(a)HPV31 L1タンパク質をコードする核酸(該核酸分子は、酵母により認識される内部転写終結シグナルを含有しない)を含むベクターを酵母宿主細胞内に導入し、(b)HPV31 L1タンパク質の発現を可能にする条件下、該酵母宿主細胞を培養することを含む、組換え宿主細胞内でHPV31 L1タンパク質を発現させるための方法に関する。
【0013】
本発明は、(a)HPV31 L1タンパク質をコードする核酸(該核酸分子は、酵母宿主細胞内での最適化発現に関してコドン最適化されている)を含むベクターを酵母宿主細胞内に導入し、(b)HPV31 L1タンパク質の発現を可能にする条件下、該酵母宿主細胞を培養することを含む、組換え宿主細胞内でHPV31 L1タンパク質を発現させるための方法に関する。
【0014】
好ましい実施形態においては、該核酸は、配列番号2または配列番号3に記載のヌクレオチドの配列を含む。
【0015】
本発明はまた、HPV31ウイルス様粒子(VLP)、HPV31 VLPの製造方法、およびHPV31 VLPの使用方法に関する。
【0016】
本発明の好ましい実施形態においては、HPV31 VLPは酵母において製造される。もう1つの好ましい実施形態においては、酵母は、サッカロミセス・セレビシエ(Saccharomyces cerevisiae)、ハンゼヌラ・ポリモルファ(Hansenula polymorpha)、ピチア・パストリス(Pichia pastoris)、クリーベルミセス・フラジリス(Kluyvermyces fragilis)、クルベロミセス・ラクチス(Kluveromyces lactis)およびシゾサッカロミセス・ポンベ(Schizosaccharomyces pombe)よりなる群から選ばれる。
【0017】
本発明のもう1つの態様は、酵母により認識される転写終結シグナルを含有しないHPV31 L1遺伝子により産生されるHPV31 L1タンパク質を含むHPV31 VLPである。
【0018】
本発明の更にもう1つの態様は、コドン最適化HPV31 L1遺伝子により産生されるHPV31 L1タンパク質を含むHPV31 VLPである。本発明のこの態様の好ましい実施形態においては、コドン最適化HPV31 L1遺伝子は、配列番号2または配列番号3に記載のヌクレオチドの配列より実質的になる。
【0019】
本発明はまた、HPV31ウイルス様粒子を動物に投与することを含んでなる、動物において免疫応答を誘導するための方法を提供する。好ましい実施形態においては、HPV31 VLPはコドン最適化遺伝子により産生される。もう1つの好ましい実施形態においては、HPV31 VLPは、酵母により認識される転写終結配列を含有しない遺伝子により製造される。
【0020】
本発明の更にもう1つの態様は、HPV31 VLPを含むワクチンを哺乳動物に投与することを含む、HPV関連子宮頸癌の予防または治療方法である。本発明のこの態様の好ましい実施形態においては、HPV31 VLPは酵母において製造される。
【0021】
本発明はまた、HPV31ウイルス様粒子(VLP)を含むワクチンに関する。
【0022】
本発明のこの態様のもう1つの実施形態においては、該ワクチンは更に、少なくとも1つの追加的HPV型のVLPを含む。好ましい実施形態においては、その少なくとも1つの追加的HPV型は、HPV6、HPV11、HPV16、HPV18、HPV33、HPV35、HPV39、HPV45、HPV51、HPV52、HPV55、HPV56、HPV58、HPV59およびHPV68よりなる群から選ばれる。
【0023】
本発明はまた、HPV31ウイルス様粒子を含む医薬組成物に関する。さらに、本発明は、HPV31 VLPと少なくとも1つの追加的HPV型のVLPとを含む医薬組成物に関する。好ましい実施形態においては、その少なくとも1つの追加的HPV型は、HPV6、HPV11、HPV16、HPV18、HPV33、HPV35、HPV39、HPV45、HPV51、HPV52、HPV55、HPV56、HPV58、HPV59およびHPV68よりなる群から選ばれる。
【0024】
本明細書中の全体において及び添付する特許請求の範囲において用いる単数表現は、文脈から例外であることが明らかである場合を除き、複数物に対する言及を含む。
【0025】
本明細書中の全体において及び添付する特許請求の範囲においては、以下の定義および略語が適用される。
【0026】
「プロモーター」なる語は、RNAポリメラーゼが結合する、DNA鎖上の認識部位を意味する。プロモーターはRNAポリメラーゼと共に開始複合体を形成して、転写活性を始動し駆動する。該複合体は、「エンハンサー」もしくは「上流活性化配列」と称される配列を活性化することにより又は「サイレンサー」と称される配列を抑制することにより修飾されうる。
【0027】
「ベクター」なる語は、宿主生物または宿主組織内へのDNA断片の導入をもたらしうる何らかの手段を意味する。プラスミド、ウイルス(アデノウイルスを含む)、バクテリオファージおよびコスミドを含む種々のタイプのベクターが存在する。
【0028】
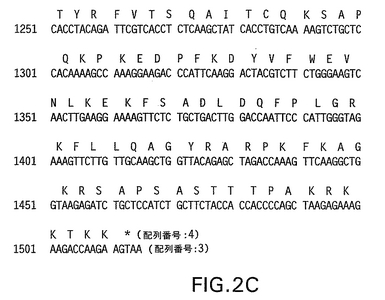
「31L1野生型配列」なる表現は、本明細書中に配列番号1として開示されているHPV31 L1配列を意味する。HPV31 L1野生型配列は既に記載されているが、臨床分離体から得たDNA間に若干の配列変異を見出すことは珍しいことではない。したがって、代表的なHPV31 L1野生型配列を、HPV31 DNAを含有することが既に示されている臨床サンプルから単離した(実施例1を参照されたい)。本明細書中に開示されているコドン最適化HPV31 L1配列を比較するための参照配列として該31 L1野生型配列を使用した(図1を参照されたい)。
【0029】
「31 L1部分改変体」は、酵母内での最適化発現のために酵母にとって好ましいコドンを含有するようHPV31 L1ヌクレオチド配列が部分的に改変された、本明細書中に開示されている構築物(配列番号2)を意味する。31 L1部分改変体は、HPV31 L1野生型ヌクレオチド配列の中央部分(ヌクレオチド697−1249)における改変を含む。酵母にとって好ましいコドンを使用して、完全HPV31 L1配列をも改変させ、それは本明細書中では「31 L1全改変体」(配列番号3)と称される。
【0030】
「有効量」なる語は、免疫応答が生じるよう、適度なレベルの該ポリペプチドの産生に十分なワクチン組成物が導入されることを意味する。このレベルは様々なものとなりうると当業者に理解される。
【0031】
「保存的アミノ酸置換」は、1つのアミノ酸残基が、化学的に類似した別のアミノ酸残基により置換されることを意味する。そのような保存的置換の例としては、疎水性残基同士(イソロイシン、ロイシン、バリンまたはメチオニン)の置換、同じ電荷の極性残基同士(例えば、アルギニンとリシン;グルタミン酸とアスパラギン酸)の置換が挙げられる。
【0032】
「哺乳動物(哺乳類)」なる語は、ヒトを含む任意の哺乳動物を意味する。
【0033】
「VLP」はウイルス様粒子を意味する。
【0034】
「合成」は、天然に存在する野生型HPV31 L1遺伝子内に存在するヌクレオチド配列と同じではないヌクレオチド配列をHPV31 L1遺伝子が含有するよう、該遺伝子が修飾されていることを意味する。前記のとおり、本発明において、酵母により認識される転写終結シグナルを除去するよう改変されたヌクレオチドの配列を含む合成分子を提供する。また、本発明においては、酵母細胞による発現に好ましいコドンを含む合成分子も提供する。本発明で提供する合成分子は、野生型HPV31 L1遺伝子と同じアミノ酸配列をコードしている。
【0035】
発明の詳細な説明
ほとんどの子宮頸癌は、特定の発癌型のヒトパピローマウイルス(HPV)の感染に関連している。本発明は、発癌性HPV型の遺伝子により発現されるタンパク質産物に対する免疫を惹起または増強する組成物および方法に関する。特に、本発明は、HPV31 L1およびHPV31ウイルス様粒子(VLP)をコードする合成ポリヌクレオチドを提供し、HPV関連癌の予防および/または治療用の免疫原性組成物およびワクチンにおける該ポリヌクレオチドおよびVLPの使用を開示する。
【0036】
野生型HPV31 L1ヌクレオチド配列は既に報告されている(Goldsboroughら,Virology 171(1):306−311(1989);Genbankアクセッション番号J04353)。本発明は、HPV31 L1タンパク質をコードする合成DNA分子を提供する。本発明の合成分子はヌクレオチドの配列を含み、該ヌクレオチドのいくつかは、酵母により認識される転写終結シグナルを除去するよう改変されている。他の実施形態においては、該合成分子のコドンは、高レベル発現を行うのに酵母細胞にとって好ましいコドンを使用するよう設計される。該合成分子は、VLPへと自己集合するHPV31 L1タンパク質の源として使用することが可能である。該VLPは、中和抗体および細胞性免疫によりパピローマウイルス感染に対する有効な免疫予防をもたらすために、VLPに基づくワクチンにおいて使用することが可能である。VLPに基づくそのようなワクチンは、既に確立したHPV感染の治療にも有用である。
【0037】
酵母細胞におけるHPV VLPの発現は、費用効果的であり発酵槽内での大規模増殖に容易に適合するという利点をもたらす。しかし、HPV31 L1を含む多数のHPV L1タンパク質は(実施例4を参照されたい)、酵母細胞内では低レベルでしか発現されない。HPV31 L1の低レベル発現は、酵母により認識される転写終結シグナルの存在により生じる、mRNA転写産物のトランケート化によるものであることが、本発明において確認された。酵母転写終結部位に類似した潜在的配列を除去するようHPV31 L1 DNAを改変することにより、完全長mRNAの転写を促進して、HPV31 L1タンパク質発現の増強をもたらすことが可能である。
【0038】
したがって、本発明のいくつかの実施形態においては、酵母転写終結シグナルに類似した潜在的配列を除去するようHPV31 L1 DNAに改変が施されている。これらの改変は、トランケート化転写産物ではなく完全長HPV31転写産物の発現を可能にして(実施例4を参照されたい)、発現収率を増強させる。
【0039】
前記のとおり、本発明の合成DNAは、酵母により認識される転写終結部位を除去するように施された、野生型HPV31 L1配列からの改変を含む。HPV31 L1タンパク質をコードするが酵母転写終結部位を含有しない追加的なDNA分子を構築することが可能であると当業者は認識するであろう。酵母転写終結配列を見出すための技術は当技術分野でよく知られている。酵母mRNAの3’末端形成および転写終結は以下の3つのシグナルの存在を要する:(1)下流に位置する要素の配置の効率を増加させる、TATATAまたは関連配列のような効率要素(efficiency element)、(2)ポリ(A)部位の位置を決定する配置要素(positioning element)、および(3)ポリアデニル化部位(通常はPy(A)n)。
【0040】
科学文献には、酵母転写終結シグナルをコードする配列の記載が豊富に見出される。例えば、GuoおよびSherman,Trends Biochem.Sci.21:477−481(1986);GuoおよびSherman,Mol.Cell.Biol.16(6):2772−2776(1996);Zaretら,Cell 28:563−573(1982);Henikoffら,Cell 33:607−614(1983);Thalenfeldら,J.Biol.Chern.258(23):14065−14068(1983);Zaretら,J.Mol.Biol.176:107−135(1984);Heidmannら,Mol.Cell Biol 14:4633−4642(1984);ならびにRusso,Yeast 11:447−453(1985)を参照されたい。したがって、本発明の完全長mRNA転写産物を産生する合成HPV31 L1遺伝子を構築するためにはどの配列を避けるべきかを、当業者は何の困難もなく決定するであろう。また、該合成配列内に酵母転写終結配列が存在するかどうかを評価するためのアッセイおよび方法は当技術分野において十分に確立されており、したがって、当業者であれば、構築されたHPV31 l1配列が、除去する必要のある終結配列を含むかどうかを判定しうるであろう。
【0041】
前記のとおり、本発明は、酵母により認識される内部転写終結シグナルを含有しない、HPV31型L1タンパク質をコードする核酸分子に関する。本発明の典型的実施例においては、該合成核酸分子は、配列番号2または配列番号3に記載のヌクレオチドの配列を含む。
【0042】
本発明の他の実施形態においては、酵母細胞環境中での高レベル発現に関してHPV31 L1遺伝子配列が「最適化」される。本発明により意図されるコドン最適化HPV31 L1遺伝子は、酵母により認識される内部転写終結シグナルを含有しないHPV31 L1をコードする合成分子を含み、これは、酵母細胞内での高レベル発現に関してコドン最適化された少なくとも1つのコドンを更に含む。
【0043】
4つの可能なヌクレオチド塩基の「トリプレット」コドンは、60を超える変異形態で存在しうる。これらのコドンは僅か20個の異なるアミノ酸(ならびに転写開始および終結)のメッセージを与えるに過ぎないため、いくつかのアミノ酸が2以上のコドンによりコードされうる(コドン縮重として公知の現象である)。完全には理解されていない理由により、異なる細胞型の内因性DNA内には代替的コドンは一様には存在しない。実際、或る細胞型においては或るコドンに関する可変的天然階層または「優先性」が存在するようである。一例として、アミノ酸ロイシンは、CTA、CTC、CTG、CTT、TTAおよびTTGを含む6つのDNAコドンのいずれかにより特定される。大腸菌(E.coli)の内因性DNAは、最も一般的には、ロイシン特定コドンCTGを含有し、一方、酵母および粘菌のDNAは、最も一般的には、ロイシン特定コドンTTAを含むことが、微生物についてのゲノムコドン出現頻度の詳細な分析から示されている。この階層を考慮すると、ロイシンに富むポリペプチドの高レベルの発現が大腸菌(E.coli)宿主により得られる可能性は、コドン使用頻度に或る程度は左右されると、一般に考えられる。例えば、TTAコドンに富む遺伝子は大腸菌(E.coli)内では発現されにくいと考えられ、一方、CTGに富む遺伝子は、おそらく、この宿主内で高度に発現されるであろう。同様に、ロイシンに富むポリペプチドの酵母宿主細胞内での発現に好ましいコドンはTTAであろう。
【0044】
組換えDNA技術へのコドン優先性現象の関与は明白であり、該現象は、成功裏に形質転換された宿主生物において外因性遺伝子の高い発現レベルがこれまで多くの場合に達成されていないことを説明するのに役立ちうる。すなわち、該挿入遺伝子内には、それほど「好まし」くないコドンが反復的に存在している可能性があり、発現用の宿主細胞装置が、それほど効率的には機能していない可能性があるのである。この現象は、意図される宿主細胞にとって好ましいコドンを含むように設計された合成遺伝子が、組換えDNA技術の実施のための好ましい形態の外来遺伝物質の最適形態を与えることを示唆している。したがって、本発明の1つの態様は、酵母細胞内での発現に関してコドン最適化されたHPV31 L1遺伝子である。本発明の好ましい実施形態において、同じタンパク質配列をコードする代替的コドンの使用は、酵母細胞によるHPV31 L1タンパク質の発現に対する制約を除去しうることが判明した。
【0045】
本発明においては、HPV31 L1遺伝子セグメントを、同一の翻訳配列を有するが代替的コドン使用頻度を伴う配列に変換した(参照により本明細書に組み入れるSharpおよびCowe,Synonymous Codon Usage in Saccharomyces cerevisiae.Yeast 7:657−678(1991)に記載されているとおり)。該方法は、一般には、高度に発現される酵母遺伝子に一般には関連していない野生型配列内のコドンを同定し、それらを、酵母細胞内での高発現のための最適コドンで置換することよりなる。ついでその新たな遺伝子配列を、これらのコドン置換により生じた望ましくない配列(例えば、「ATTTA」配列、イントロンスプライス認識部位の非意図的生成、望ましくない制限酵素部位など)に関して調べる。望ましくない配列は、同一アミノ酸をコードする異なるコドンで既存コドンを置換することにより除去される。ついで該合成遺伝子セグメントを発現の改善に関して試験する。
【0046】
前記の方法を用いてHPV31 L1の合成遺伝子セグメントを作製して、高レベル発現に関して最適化されたコドンを含む遺伝子を得た。前記方法は、HPVワクチンにおいて使用するコドン最適化遺伝子を設計するための本発明者らの方法の概要を説明するものであるが、該方法における若干の変更または該配列における若干の変更によっても、同様のワクチン効力または遺伝子発現の増強が達成されうると、当業者に理解される。
【0047】
したがって、本発明は、HPV31 L1タンパク質またはHPV31 L1タンパク質の生物学的に活性な断片もしくは突然変異体をコードするヌクレオチド配列を含む、酵母宿主内での発現に関して最適化されたコドンを含む合成ポリヌクレオチドに関する。HPV31 L1タンパク質の突然変異体は、保存的アミノ酸置換、アミノ末端トランケート化、カルボキシ末端トランケート化、欠失または付加を含むが、これらに限定されるものではない。任意のそのような生物学的に活性な断片および/または突然変異体は、配列番号4に記載のHPV31 L1タンパク質の免疫学的特性を少なくとも実質的に模擬するタンパク質またはタンパク質断片をコードする。本発明の合成ポリヌクレオチドは、治療用または予防用HPVワクチンの開発において有用となるよう、機能的HPV31 L1タンパク質を発現するmRNA分子をコードする。
【0048】
本発明の1つの態様は、配列番号2に記載のヌクレオチド配列を含む、配列番号4に記載のHPV31 L1タンパク質をコードするコドン最適化核酸分子である。
【0049】
本発明のもう1つの態様は、配列番号3に記載のヌクレオチド配列を含む、配列番号4に記載のHPV31 L1タンパク質をコードするコドン最適化核酸分子である。
【0050】
本発明はまた、本明細書中の全体に開示されている核酸分子を含有する組換えベクターおよび組換え宿主細胞(原核細胞および真核細胞の両方)に関する。
【0051】
本明細書に記載の方法により構築した合成HPV31 DNAまたはその断片を、適当なプロモーターと他の適当な転写調節要素とを含有する発現ベクター内への分子クローニングにより組換え的に発現させ、原核または真核宿主細胞内に導入して、組換えHPV31 L1を得ることが可能である。そのような操作のための技術は当技術分野において記載されている(Sambrookら Molecular Cloning:A Laboratory Manual;Cold Spring Harbor Laboratory Press,Cold Spring Harbor,New York,(1989);Current Protocols in Molecular Biology,Ausubelら,Green Pub.Associates and Wiley−Interscience,New York(1988);Yeast Genetics:A Laboratory Course Manual,Roseら,Cold Spring Harbor Laboratory,Cold Spring Harbor,New York,(1990)(それらの全体を参照により本明細書に組み入れることとする))。
【0052】
したがって、本発明は更に、(a)HPV31 L1タンパク質をコードする核酸(該核酸分子は、酵母宿主細胞での最適化発現に関してコドン最適化されている)を含むベクターを酵母宿主細胞内に導入し、(b)HPV31 L1タンパク質の発現を可能にする条件下、該酵母宿主細胞を培養することを含む、組換え宿主細胞内でHPV31 L1タンパク質を発現させるための方法に関する。
【0053】
したがって、本発明は更に、(a)HPV31 L1タンパク質をコードする核酸(該核酸分子は、酵母により認識される内部転写終結シグナルを含有しない)を含むベクターを酵母宿主細胞内に導入し、(b)HPV31 L1タンパク質の発現を可能にする条件下、該酵母宿主細胞を培養することを含む、組換え宿主細胞内でHPV31 L1タンパク質を発現させるための方法に関する。
【0054】
本発明は更に、(a)配列番号2または配列番号3に記載の核酸を含むベクターを酵母宿主細胞内に導入し、(b)HPV31 L1タンパク質の発現を可能にする条件下、該酵母宿主細胞を培養することを含む、組換え宿主細胞内でHPV31 L1タンパク質を発現させるための方法に関する。
【0055】
本発明の合成遺伝子は、宿主細胞内でのHPV58 L1タンパク質の効率的発現をもたらすように設計された配列を含む発現カセットに集合させることが可能である。該カセットは、好ましくは、合成遺伝子を、それに機能しうる形で連結された関連転写および翻訳制御配列(例えば、プロモーターおよび終結配列)と共に含有する。好ましい実施形態においては、プロモーターはエス・セレビシエ(S.cerevisiae)GAL1プロモーターであるが、多数の他の公知酵母プロモーター、例えばGAL10、GAL7、ADH1、TDH3もしくはPGKプロモーターまたは他の真核生物遺伝子プロモーターのいずれかを使用しうると当業者は認識するであろう。好ましい転写ターミネーターはエス・セレビシエ(S.cerevisiae)ADH1ターミネーターであるが、他の公知転写ターミネーターも使用することが可能である。GAL1プロモーター−ADH1ターミネーターの組合せが特に好ましい。
【0056】
本発明のもう1つの態様はHPV31ウイルス様粒子(VLP)、HPV31 VLPの製造方法、およびHPV31 VLPの使用方法である。ヒトおよび動物パピローマウイルスの主要カプシドタンパク質であるL1が酵母、昆虫細胞、哺乳類細胞または細菌内で発現される場合には、VLPは自己集合しうる(概説としては、SchillerおよびRoden,in Papillomavirus Reviews:Current Research on Papillomaviruses;Lacey編,Leeds,UK:Leeds Medical Information,pp 101−12(1996))。L1およびL2カプシドタンパク質の組合せを発現させることにより、形態学的に異なるHPV VLPも製造されうる。VLPは、T=7二十面体構造のL1の72個の五量体から構成される(Bakerら,Biophys.J.60(6):1445−56(1991))。
【0057】
VLPは真正ビリオンと形態学的に類似しており、動物に投与されると高力価の中和抗体を誘導しうる。VLPでのウサギ(Breitburdら,J.Virol.69(6):3959−63(1995))およびイヌ(Suzichら,Proc.Natl.Acad.Sci.USA 92(25):11553−57(1995))の免疫は中和抗体の誘導および実験パピローマウイルス感染に対する防御の両方をもたらすことが示された。しかし、VLPは、潜在的に発癌性のウイルスゲノムを含有せず、単一遺伝子から自己集合しうるため、それは、HPVワクチン開発における生ウイルスの使用に代わる安全な手段となる(概説としては、SchillerおよびHidesheim,J.Clin.Virol.19:67−74(2000)を参照されたい)。
【0058】
したがって、本発明は、HPV31の組換えL1タンパク質または組換えL1+L2タンパク質から構成されるウイルス様粒子に関する。
【0059】
本発明の好ましい実施形態においては、HPV31 VLPは酵母において産生される。もう1つの好ましい実施形態においては、酵母は、サッカロミセス・セレビシエ(Saccharomyces cerevisiae)、ハンゼヌラ・ポリモルファ(Hansenula polymorpha)、ピチア・パストリス(Pichia pastoris)、クリーベルミセス・フラジリス(Kluyvermyces fragilis)、クルベロミセス・ラクチス(Kluveromyces lactis)およびシゾサッカロミセス・ポンベ(Schizosaccharomyces pombe)よりなる群から選ばれる。
【0060】
本発明のもう1つの態様は、酵母により認識される転写終結シグナルを含有しないHPV31 L1遺伝子により産生されるHPV31 L1タンパク質を含むHPV31 VLPである。
【0061】
本発明の更にもう1つの態様は、コドン最適化HPV31 L1遺伝子により産生されるHPV31 L1タンパク質を含むHPV31 VLPである。本発明のこの態様の好ましい実施形態においては、コドン最適化HPV31 L1遺伝子は、配列番号2または配列番号3に記載のヌクレオチドの配列より実質的になる。
【0062】
本発明の更にもう1つの態様は、(a)HPV31 L1タンパク質またはHPV31 L1タンパク質またはHPV31 L1+L2タンパク質をコードする組換えDNA分子で酵母を形質転換し、(b)該組換えDNA分子の発現を可能にする条件下、該形質転換酵母を培養して組換えHPV31タンパク質を得、(c)該組換えHPV31タンパク質を単離してHPV31 VLPを得ることを含む、HPV31 VLPの製造方法である。
【0063】
本発明のこの態様の好ましい実施形態においては、酵母により認識される転写終結シグナルを含有しないHPV31 L1遺伝子で酵母を形質転換する。もう1つの好ましい実施形態においては、HPV31 VLPを得るために、コドン最適化HPV31 L1遺伝子で酵母を形質転換する。特に好ましい実施形態においては、コドン最適化HPV31 L1遺伝子は、配列番号2または配列番号3に記載のヌクレオチド配列より実質的になる。
【0064】
本発明はまた、HPV31ウイルス様粒子を動物に投与することを含んでなる、動物において免疫応答を誘導するための方法を提供する。好ましい実施形態においては、HPV31 VLPは、酵母により認識される内部転写終結配列を含有しない遺伝子により製造される。もう1つの好ましい実施形態においては、HPV31 VLPはコドン最適化遺伝子により製造される。
【0065】
本発明の更にもう1つの態様は、HPV31 VLPを含むワクチンを哺乳動物に投与することを含む、HPV関連子宮頸癌の予防または治療方法である。本発明のこの態様の好ましい実施形態においては、HPV31 VLPは酵母において製造される。
【0066】
本発明はまた、HPV31ウイルス様粒子(VLP)を含むワクチンに関する。
【0067】
本発明のこの態様のもう1つの実施形態においては、該ワクチンは更に、少なくとも1つの追加的HPV型のVLPを含む。好ましい実施形態においては、その少なくとも1つの追加的HPV型は、HPV6、HPV11、HPV16、HPV18、HPV33、HPV35、HPV39、HPV45、HPV51、HPV52、HPV55、HPV56、HPV58、HPV59およびHPV68よりなる群から選ばれる。
【0068】
本発明のこの態様の好ましい実施形態においては、該ワクチンは更に、HPV16 VLPを含む。
【0069】
本発明のもう1つの好ましい実施形態においては、該ワクチンは更に、HPV16 VLPおよびHPV18 VLPを含む。
【0070】
本発明の更にもう1つの好ましい実施形態においては、該ワクチンは更に、HPV6 VLP、HPV11 VLP、HPV16 VLPおよびHPV18 VLPを含む。
【0071】
本発明はまた、HPV31ウイルス様粒子を含む医薬組成物に関する。さらに、本発明は、HPV31 VLPと少なくとも1つの追加的HPV型のVLPとを含む医薬組成物に関する。好ましい実施形態においては、その少なくとも1つの追加的HPV型は、HPV6、HPV11、HPV16、HPV18、HPV33、HPV35、HPV39、HPV45、HPV51、HPV52、HPV55、HPV56、HPV58、HPV59およびHPV68よりなる群から選ばれる。
【0072】
本発明のワクチン組成物は、潜在的毒性を最小限にする一方でHPV31感染の最適な抑制を得るために通常の試験により定められる適当な量で単独で使用することが可能である。また、他の物質の共投与または連続的投与が望ましいかもしれない。
【0073】
ワクチン被投与者に導入すべきウイルス様粒子の量は、発現される遺伝子産物の免疫原性に左右される。一般に、VLP 10μg〜100μg、好ましくは約20μg〜60μgの免疫学的または予防的に有効な量を筋肉組織内に直接投与する。皮下注射、皮内導入、皮膚を介した圧入(impression)、および他の投与方法、たとえば、腹腔内、静脈内または吸入運搬も意図される。また、ブースターワクチン接種を行いうると意図される。本発明のワクチンの非経口導入と同時またはその後の、ミョウバンまたはメルク(Merck)ミョウバンアジュバントのようなアジュバントの非経口投与、例えば静脈内、筋肉内、皮下または他の手段での投与も有利である。
【0074】
本発明に関連して用いられうる方法および材料を記載し開示する目的で、本明細書に記載のすべての刊行物を、参照により本明細書に組み入れることとする。ここで、本発明が先行発明としてそのような開示に先行するものではないと自認するものと解釈されるべきではない。
【0075】
添付図面を参照して本発明の好ましい実施形態が記載されているが、本発明はそれらの実施形態そのものに限定されるものではなく、添付の特許請求の範囲において定められる本発明の範囲または精神から逸脱することなく本発明には種々の変更および修飾が当業者によって施されうると理解されるべきである。
【0076】
以下の実施例は本発明を例示するものであり、本発明を限定するものではない。
【実施例1】
【0077】
代表的HPV31 L1配列の決定
HPV31 L1野生型配列は既に記載されている(Goldsboroughら,Virology 171(1):306−311(1989);Genbankアクセッション番号J04353)。しかし、臨床分離体から得たDNA間に若干の配列変異を見出すことは珍しいことではない。代表的なHPV31 L1野生型配列を単離するために、HPV31 DNAを含有することが既に示されている3つの臨床サンプルからDNAを単離した。Taq DNAポリメラーゼおよび以下のプライマーを使用するポリメラーゼ連鎖反応(PCR)において、HPV31 L1配列を増幅した:
【0078】
【化1】

【0079】
増幅産物をアガロースゲル上で電気泳動し、臭化エチジウム染色により可視化した。〜1500bpのL1バンドを切り出し、QIAクイックPCR精製キット(Qiagen,Hilden,Germany)を使用してDNAを精製した。ついで該DNAをTAクローニングベクターpCR−II(Invitrogen Corp.,Carlsbad,CA)に連結し、大腸菌(E.coli)を形質転換し、アンピシリン+IPTGおよびX−gal(青/白コロニー選択用)を含有するLB寒天上でプレーティングした。該プレートを反転させ、37℃で16時間インキュベートした。白色コロニーを、アンピシリンを含有するLB培地内で37℃で16時間、振とうしながら培養し、ミニプレップを行って該プラスミドDNAを抽出した。
【0080】
該プラスミド中のL1遺伝子の存在を示すために、制限エンドヌクレアーゼ消化を行い、アガロースゲル電気泳動および臭化エチジウム染色により可視化した。それらの3つの臨床分離体のそれぞれからのクローン化L1を含有するプラスミドに関してDNA配列決定を行った。DNAおよび翻訳アミノ酸配列をお互いと及びGenbank HPV31 L1配列と比較した。それらの3つの臨床分離体の配列分析は、どの配列も該Genbank配列と同一ではないことを示した。代表的な31L1配列としてpCR−II−HPV 31L1/81クローンを選択した。これは本明細書中では「31 L1野生型配列」(配列番号1;図1を参照されたい)と称される。31 L1野生型として選択した配列はヌクレオチド1266の1つのサイレント置換、およびコード化アミノ酸をトレオニンからセリンに変化させるヌクレオチド1295のCからGへの変化を含有していた。31 L1部分および全改変体遺伝子(それぞれ配列番号2および3)も、この位置にセリンをコードしている(図1を参照されたい)。すべての場合において、アミノ酸配列は同一である。改変体構築物においては、酵母にとって好ましいコドン配列を使用してアミノ酸をコードし、潜在的転写終結シグナルを除去するよう、ヌクレオチドが変化している(実施例2を参照されたい)。
【0081】
BglII伸長を付加するために以下のプライマーを使用して、31 L1野生型配列を増幅した。
【0082】
【化2】
【0083】
PCRは、、Vent(商標)DNAポリメラーゼを使用して行った。アガロースゲルの臭化エチジウム染色により、PCR産物を可視化した。〜1500bpのバンドを切り出し、QIAEX IIゲル抽出キット(Qiagen)を使用してDNAを精製した。ついでPCR産物をBglIIで37℃で2時間消化し、QIAクイックPCR精製キットを使用して精製した。BglIIで消化された31 L1 PCR産物を、BamHIで消化されたpGAL110に連結し、DH5大腸菌(E.coli)を形質転換した。正しい配向のHPV 31 L1インサートに関して、PCRによりコロニーをスクリーニングした。DNA配列決定により、配列および配向を確認した。選択したクローンをpGAL110−HPV 31L1 #2と命名した。
【0084】
ついでマキシプレップ(Maxiprep)DNAを調製し、サッカロミセス・セレビシエ(Saccharomyces cerevisiae)をコンピテントにし形質転換した。該酵母形質転換体をLeu− ソルビトールプレート上のLeu− ソルビトール上層寒天内でプレーティングし、反転させて30℃で3〜5日間インキュベートした。コロニーを拾い、Leu− ソルビトールプレート上での単離のためにストリークした。ついで、L1転写およびタンパク質発現を誘導するために、単離されたコロニーを、30℃の回転チューブ培養において、1.6%グルコースおよび4%ガラクトースを含有する5mlの5×Leu−Ade−ソルビトール内で成長させた。
【実施例2】
【0085】
酵母コドン最適化
酵母にとって好ましいコドンは既に記載されている(SharpおよびCowe,Yeast 7:657−678(1991))。まず、ヌクレオチド697−1249に相当するHPV31 L1の中央部を、酵母にとって好ましいコドンを使用して改変した。改変に用いた方法は、同じアミノ酸配列を維持しながら酵母にとって好ましいコドン配列でヌクレオチドを置換する、改変すべき領域に及ぶ長い重複するセンスおよびアンチセンスオリゴマーを設計するものであった。該PCR反応においては鋳型DNAの代わりにこれらのオリゴマーを使用した。追加的な増幅プライマーを設計し、それらを使用して、Pfu DNAポリメラーゼ(Stratagene,La Jolla,CA)により鋳型オリゴマーから改変体配列を増幅した。増幅のための最適条件は部位特異的なものであったが、ほとんどの場合には、以下に類似した方法を用いた:94℃で1時間の初期変性工程およびそれに続く、95℃で30秒間の変性、55℃で30秒間のアニーリング、72℃で3.5分間の伸長の15〜25サイクル、およびそれに続く72℃で10分間の最終伸長、および4℃での維持。
【0086】
PCR産物をアガロースゲル電気泳動により検査した。適当なサイズのバンドを切り出し、DNAをゲル精製した。ついで該増幅断片を鋳型として使用して552ヌクレオチドの改変体HPV31中央部L1断片を作製した。ついでPCRを使用して野生型ヌクレオチド1−725(5’末端)および1221−1515(3’末端)を増幅した。該5’末端、該3’末端および該改変体中央部を使用する最終PCRを行って、本明細書中では「31 L1部分改変体」と称される完全長31 L1部分改変体を作製した。
【0087】
また、酵母にとって好ましいコドンを使用して、完全な31 L1配列も改変した。この構築物は本明細書中で「31 L1全改変体」と称される。9個の長い重複するオリゴマーを使用して、1−753の、酵母にとって好ましいコドンのヌクレオチド配列を作製し、4個の長い重複するオリゴマーを使用して、1207−1515の、酵母にとって好ましいコドンのヌクレオチド配列を作製した。増幅およびゲル精製の後、これらの断片を、前記の中央部改変体部分(ヌクレオチド697−1249)と共にPCR反応において使用して、完全長31 L1全改変体配列を作製した。この断片は、BamHI伸長を伴って作製された。ゲル精製改変体31 L1 DNAをBamHIで消化し、BamHIで消化されたpGAL110発現ベクターに連結し、大腸菌(E.coli)DH5細胞内に形質転換した。正しい配向のHPV31 L1インサートに関して、PCRによりコロニーをスクリーニングした。DNA配列決定により、配列および配向を確認した。
【0088】
プラスミドDNAを調製した。サッカロミセス・セレビシエ(S.cerevisiae)をコンピテントにし、形質転換した。該酵母をLeu− ソルビトールプレート上のLeu− ソルビトール上層寒天内でプレーティングし、反転させて3〜5日間インキュベートした。コロニーを、Leu− ソルビトールプレート上での単離のためにストリークした。ついで、L1転写およびタンパク質発現を誘導するために、単離されたコロニーを、30℃の回転チューブ培養において、1.6%グルコースおよび4%ガラクトースを含有する5mlの5×Leu−Ade−ソルビトール内で成長させた。48〜72時間後、OD600=10と等価な培養容量をペレット化し、上清を除去し、該ペレットを凍結し、−70℃で保存した。
【実施例3】
【0089】
RNA調製
ガラクトース誘導によりHPV31 L1を発現するよう誘導された、形質転換酵母の細胞ペレットを氷上で解凍し、1mlの冷DEPC処理水に懸濁させた。細胞を遠心分離によりペレット化し、得られた上清を除去した。ついで細胞ペレットを400μlのTES(10mM Tris pH7.0,10mM EDTAおよび0.5% SDS)に再懸濁させた。等容量のAEバッファー飽和フェノール(50mM NaOAcおよび10mM EDTA)を加えた。該チューブを10秒間ボルテックスし、10分ごとに混合しながら65℃で50分間加熱した。ついで該チューブを氷上で5分間配置し、ついで4℃で5分間遠心分離した。上清を集め、無菌チューブに移した。追加的な400μlのフェノールを加え、該チューブをボルテックスし、氷上に5分間配置し、遠心分離した。上清を無菌チューブに移し、400μlのクロロホルムを加え、混合し、遠心分離した。上清を再び集め、無菌チューブに移し、1mlの100% EtOHのほかに40μlの3M 酢酸Na(pH5.2)を加えた。該チューブをドライアイス上に1時間配置し、ついでそれを高速で遠心分離してRNAをペレット化した。該RNAを70% EtOHで1回洗浄し、風乾させた。ついで該RNAを100μlのDEPC処理水に懸濁させ、65℃に5分間加熱して溶解した。A260/280が1.7〜2.0である場合にA260測定値1=40μg/ml RNAという仮定により、サンプル中のRNAの濃度を測定するために分光光度測定を行った。
【実施例4】
【0090】
ノーザンブロット分析
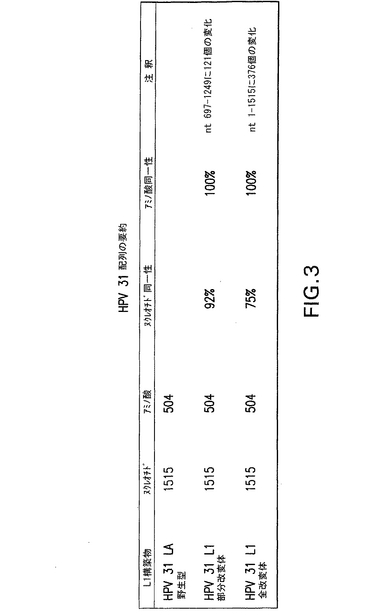
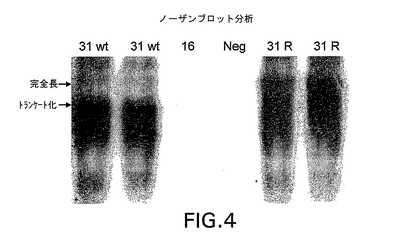
31 L1野生型を発現する酵母の初期分析は、HPV31 L1タンパク質の発現収率が予想より相当低いことを示唆した。該低発現が、転写レベルの問題により生じたのか翻訳レベルの問題により生じたのかを判定するために、HPV31 L1転写産物のノーザンブロット分析を行った。転写サイズを比較するために、同一ゲル上で、HPV16 L1を発現する酵母からのRNAが、HPV31 L1を発現する酵母からのRNAと共に移動したゲルから、ノーザンブロットを作製した。
【0091】
1.2% アガロースホルムアルデヒドゲルを成型した。10μgのRNAを変性バッファー(最終濃度:6% ホルムアルデヒド、45% ホルムアミドおよび0.9×MOPS)と一緒にし、55℃で15分間加熱した。ゲルローディングバッファーの10分の1の容量を加え、該サンプルを該ゲル上にローディングした。1×MOPSバッファー中、65ボルトで〜5時間、電気泳動を行った。該ゲルを無菌水中で15分間ついで10×SSC中で2回(各5分間)洗浄した。該RNAを、10×SSC中、16時間にわたり、毛管作用により、Hybond−N+ナイロン膜(Amersham Biosciences,Piscataway,NJ)にトランスファーした。ついで該RNAを、700単位のエネルギーに設定されたAmersham架橋剤を使用する架橋によりナイロン膜に固定した。固定後、該ナイロン膜を風乾させた。該膜を、30mlのZetaprobeバッファー中、55℃で2時間配置し、ついで32P標識プローブを加え、53〜65℃で16時間インキュベートした。ついで該膜を、5×SSC中、室温で20分間で3回、ついで0.4×SSC中、室温で20分間で2回、および60℃で10分間で1回洗浄した。HPV31 L1配列に特異的なセンスおよびアンチセンスプライマーを使用するPCRにより、プローブDNAを作製した。ポリヌクレオチドキナーゼ(PNK)およびγ―32 ATPでの37℃で1時間の処理により、該増幅DNAを標識した。該ブロットをサランラップで包み、x線フィルムに16時間露出させた。フィルムの現像後、プローブにハイブリダイズしたRNAはオートラジオグラフ上で黒色バンドとして検出された。
【0092】
前記のノーザンブロットの分析は、完全長HPV31 L1野生型転写産物の大部分が完全長より相当に小さいことを示した(図4を参照されたい)。しかし、31 L1部分改変体の設計においては、酵母にとって好ましいコドンが該遺伝子の中央部に挿入されているのみならず、酵母転写終結部位に類似した潜在的配列が除去されている。ノーザンブロット分析は、改変後、31 L1遺伝子転写産物の長さが、完全長HPV16 L1転写産物に対応するサイズまで有意に増加していたことを、明らかに示した(非表示)。したがって、未熟な転写終結が、31 L1野生型構築物からの低い発現収率のかなりの部分の原因になっていたらしい。
【実施例5】
【0093】
HPV31 L1タンパク質発現
OD600=10と等価な、ガラクトースにより誘導された培養物の凍結酵母細胞ペレットを、氷上で解凍し、2mM PMSFを含有する300μlのPCバッファー(100mM Na2HPO4および0.5M NaCl,pH7.0)に懸濁させた。酸で洗浄した0.5mmガラスビーズを加えた(〜0.5g/チューブ)。該チューブを4℃で15分間ボルテックスした。7.5μlの20% TritonX100を加え、ボルテックスを4℃で5分間反復した。該チューブを氷上に15分間配置し、ついで4℃で15分間遠心分離した。上清を無菌微小遠心管に移し、−70℃で保存した。
【実施例6】
【0094】
ウエスタンブロット分析
ガラクトース誘導後のHPV31 L1タンパク質の発現を確認するために、各HPV31 L1構築物についての20〜40個の単離された酵母コロニーからの全酵母タンパク質抽出物をウエスタンブロットにより分析した。
【0095】
10μgの全酵母タンパク質抽出物をSDS−PAGEローディングバッファーと一緒にし、95℃で10分間加熱した。該タンパク質を8% SDS−PAGEゲル上にローディングし、Tris−グリシンバッファー中で電気泳動した。タンパク質の分離後、該タンパク質を該ゲルからニトロセルロースにウエスタントランスファーし、該ブロットをTTBS(Tween−20を含有するTris緩衝食塩水)中の10% 脱脂乾燥乳中で16時間ブロッキングした。該ブロットをTTBS中で3回洗浄した。HPV31 L1と交差反応するポリクローナル血清であるヤギ抗trpE−HPV16 L1血清をTTBS中の1:1000の希釈度で室温で1時間適用した。該ブロットをTTBS中で3回洗浄し、抗ヤギHRP結合抗体をTTBS中の1:2500の希釈度で1時間適用した。該ブロットを再び3回洗浄し、ECL(商標)検出試薬を適用した(Amersham Biosciences,Piscataway,NJ)。ついでオートラジオグラフィーを行った。該抗血清により認識されたタンパク質は、オートラジオグラフ上で暗色バンドとして検出試薬により可視化された。
【0096】
すべての場合において、HPV 31L1タンパク質は、オートラジオグラフ上で、約55kDに対応する別個のバンドとして検出された(データ非表示)。ゲル上で陽性対照として、HPV16 L1タンパク質を含めた。
【実施例7】
【0097】
ラジオイムノアッセイ(RIA)
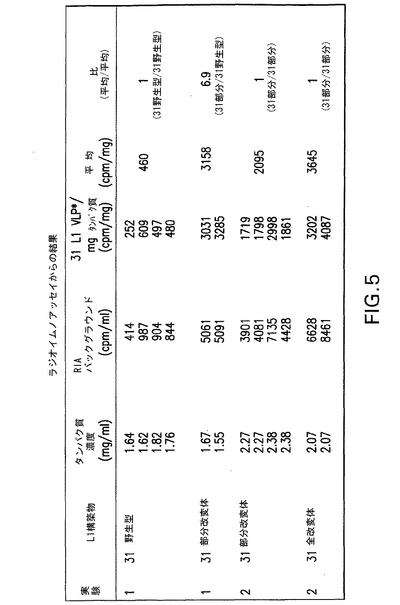
HPV31 L1を発現する酵母細胞を種々の方法(回転チューブ培養、振とうフラスコおよび発酵槽など)により増殖させた。該酵母を細胞溶解し、タンパク質抽出物を調製して、全タンパク質1mg当たりで産生されたHPV31 L1ウイルス様粒子(VLP)の量を測定した。HPV31 L1 VLP発現を示すために、各全酵母タンパク質抽出物の一部を捕捉ラジオイムノアッセイ(RIA)により分析した。
【0098】
HPV型31特異的でありVLPコンホメーション特異的である検出用モノクローナル抗体H31.A6を使用して、RIAを行った。H31.A6は無傷HPV31 L1 VLPには結合することが判明しているが、変性したHPV31 VLPを認識しないため、H31.A6はHPV型31 L1に特異的である。ついで、このmAbは、I125で放射能標識されたヤギ抗マウス抗体により検出されうる。したがって、毎分カウント(cpm)値はHPV31 L1 VLP発現の相対レベルに対応する。
【0099】
ポリスチレンビーズを、PBS中で1:1000で希釈されたヤギ抗trpE−HPV31 L1ポリクローナル血清で一晩コートした。ついで該ビーズを5容量の無菌蒸留水で洗浄し、風乾させた。ついで、単離された酵母コロニーからの全酵母タンパク質抽出物である抗原を、1% BSA、0.1% Twenn−20および0.1% アジ化ナトリウムを含有するPBS中の希釈により該ビーズ上にローディングし、回転させながら1時間インキュベートした。洗浄後、該ビーズを20ウェルポリスチレンプレートにおいて1個/ウェルで分配し、1:50,000で希釈されたH31.A6 mAbと共に室温で17〜24時間インキュベートした。該ビーズを洗浄し、I125標識ヤギ抗マウスIgGを23000〜27000cpm/10μlの活性範囲で加えた。2時間後、該ビーズを洗浄し、放射能カウントをcpm/ml単位で記録した。ブランクウェルからのバックグラウンドカウントを全cpm/mlから差し引いて、RIA−バックグラウンド値を得た。
【0100】
2つの実験を行った。実験1においては、31 L1野生型および31 L1部分改変体からのタンパク質抽出物を比較し、実験2においては、31 L1部分改変体および31 L1全改変体からのタンパク質抽出物を比較した(図5を参照されたい)。結果は、31 L1部分改変体VLPの発現が31 L1野生型より6.9倍高いことを示している。31 L1全改変体は、31 L1部分改変体と比べて1.7倍増加した発現を示している。したがって、酵母にとって好ましいコドン配列を導入し潜在的転写終結シグナルを除去することにより、31 L1の発現レベルは>7倍増加した。
【実施例8】
【0101】
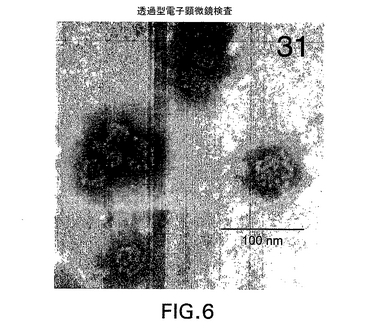
透過型電子顕微鏡検査
HPV31 L1タンパク質が実際に自己集合して五量体L1カプソマー(これは今度は、ウイルス様粒子に自己集合する)を形成したことを示すために、部分的に精製された31 L1全改変体タンパク質抽出物を透過型電子顕微鏡検査(TEM)に付した。酵母を小規模発酵で増殖させ、ペレット化した。該ペレットを精製処理に付した。L1タンパク質の発現および精製操作における保持を示すために、ペレットおよび清澄化酵母抽出物を免疫ブロットにより分析した。ついで清澄化酵母抽出物を45% スクロースクッション上の遠心分離に付し、得られたペレットをTEM分析のためにバッファーに懸濁させた(図6を参照されたい)。この粗サンプル中の球状粒子の直径は30〜60nmであり、いくつかの粒子はカプソマーの規則的配置を示していることを、結果は示している。
【図面の簡単な説明】
【0102】
【図1A】部分改変体(配列番号2)および全改変体(配列番号3)31 L1遺伝子における改変ヌクレオチドを示す配列アライメントである(実施例2を参照されたい)。参照配列は31 L1野生型配列(配列番号1;実施例1を参照されたい)である。参照配列と同一である31 L1部分および全改変体配列におけるヌクレオチドは点で示されている。改変されたヌクレオチドは、それらの対応位置に示されている。括弧内にはヌクレオチド番号が示されている。
【図1B】部分改変体(配列番号2)および全改変体(配列番号3)31 L1遺伝子における改変ヌクレオチドを示す配列アライメントである(実施例2を参照されたい)。参照配列は31 L1野生型配列(配列番号1;実施例1を参照されたい)である。参照配列と同一である31 L1部分および全改変体配列におけるヌクレオチドは点で示されている。改変されたヌクレオチドは、それらの対応位置に示されている。括弧内にはヌクレオチド番号が示されている。
【図1C】部分改変体(配列番号2)および全改変体(配列番号3)31 L1遺伝子における改変ヌクレオチドを示す配列アライメントである(実施例2を参照されたい)。参照配列は31 L1野生型配列(配列番号1;実施例1を参照されたい)である。参照配列と同一である31 L1部分および全改変体配列におけるヌクレオチドは点で示されている。改変されたヌクレオチドは、それらの対応位置に示されている。括弧内にはヌクレオチド番号が示されている。
【図1D】部分改変体(配列番号2)および全改変体(配列番号3)31 L1遺伝子における改変ヌクレオチドを示す配列アライメントである(実施例2を参照されたい)。参照配列は31 L1野生型配列(配列番号1;実施例1を参照されたい)である。参照配列と同一である31 L1部分および全改変体配列におけるヌクレオチドは点で示されている。改変されたヌクレオチドは、それらの対応位置に示されている。括弧内にはヌクレオチド番号が示されている。
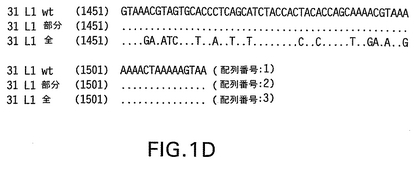
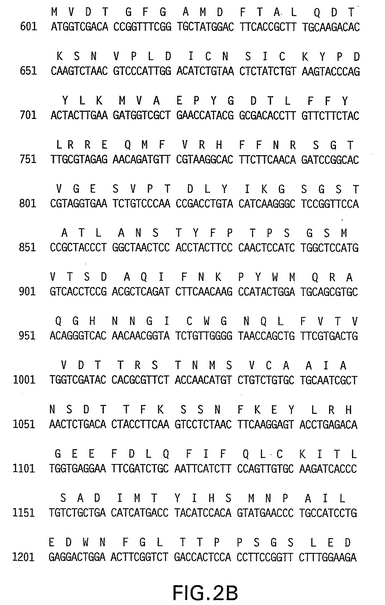
【図2A】31 L1全改変ヌクレオチド(配列番号3)およびアミノ酸配列(配列番号4)を示す。左側にヌクレオチド番号が示されている。
【図2B】31 L1全改変ヌクレオチド(配列番号3)およびアミノ酸配列(配列番号4)を示す。左側にヌクレオチド番号が示されている。
【図2C】31 L1全改変ヌクレオチド(配列番号3)およびアミノ酸配列(配列番号4)を示す。左側にヌクレオチド番号が示されている。
【図3】左側に挙げられている3つのHPV31 L1配列構築物間の変化を要約する。第4列は、示されている構築物と31 L1野生型配列との間のヌクレオチド同一性(%)を示し、第5列はアミノ酸同一性を示す。最終列は、酵母に好ましいコドン配列へ改変されたヌクレオチドの数、および改変が施された領域を示す。
【図4】高いストリンジェンシー下でHPV31 L1に特異的にプローブされたノーザンブロットを示す(実施例4を参照されたい)。左側の矢印は、HPV31 L1完全長およびトランケート化転写産物の位置を示す。「31wt」と示されたレーンは、31 L1野生型配列を含有する酵母の同じRNA調製物からのものである。「16」と示されたレーンは、高いストリンジェンシー条件のためにHPV31 L1プローブによっては認識されない、HPV16からのRNAを含有する。「Neg」と示されたレーンは、L1コード配列を含有しない酵母抽出物である。「31R」と示されたレーンは、31 L1部分改変体配列を発現する2つの別々の単離コロニーのRNAからのものである。
【図5】毎分カウント(cpm)/mg全タンパク質で表された、2つの捕捉ラジオイムノアッセイ(RIA)実験からのデータの一部を示す(実施例7を参照されたい)。該RIAで得たcpmはHPV31 L1 VLPの相対指標である。該RIAデータは、酵母にとって好ましいコドン改変体遺伝子配列からの酵母タンパク質抽出物における31 L1 VLP発現の増強を示している。
【図6】透過型電子顕微鏡検査により可視化された、本明細書に記載の31 L1 VLPの代表的サンプルを示す(実施例8を参照されたい)。棒線は100nmを表す。
【配列表】
【技術分野】
【0001】
本発明は、全般的には、ヒトパピローマウイルス(HPV)の治療に関する。より詳しくは、本発明は、HPV31 L1タンパク質をコードする合成ポリヌクレオチド、ならびに該ポリヌクレオチドを含む組換えベクターおよび宿主に関する。本発明はまた、HPV31様ウイルス様粒子(VLP)、ならびにHPVの予防および治療のためのワクチンおよび医薬組成物におけるその使用に関する。
【背景技術】
【0002】
80を超える型のヒトパピローマウイルス(HPV)が存在し、それらのうちの多くは良性増殖性疣贅から悪性癌に及ぶ多種多様な生物学的表現型に関連づけられている(概説としては、McMurrayら,Int.J.Exp.Pathol.82(1):15−33(2001)を参照されたい)。HPV6およびHPV11は、性器粘膜または呼吸器粘膜の良性疣贅、非悪性尖形コンジローマおよび/または低悪性異形成に最も一般的に関連している型である。HPV16およびHPV18は、子宮頸部、膣、外陰部および肛門管の発生部位内非浸潤性(in situ)および浸潤性癌に最も一般的に関連している高リスク型である。子宮頸癌の90%以上はHPV16、HPV18またはより稀な発癌性型HPV31、33、45、52および58に関連している(Schiffmanら,J.Natl.Cancer Inst.85(12):958−64(1993))。HPV DNAが子宮頸癌の90〜100%で検出されるという観察は、HPVが子宮頸癌を引き起こすという強力な疫学的証拠を提供している(Boschら,J.Clin.Pathol.55:244−265(2002)を参照されたい)。
【0003】
パピローマウイルスは、8個までの初期遺伝子および2個の後期遺伝子をコードする小型(50〜60nm)非外包性二十面体DNAウイルスである。そのウイルスゲノムのオープンリーディングフレーム(ORF)はE1〜E7ならびにL1およびL2と称され、この場合の「E」は初期を意味し、「L」は後期を意味する。L1およびL2はウイルスカプシドタンパク質をコードし、一方、E遺伝子は、ウイルス複製および細胞トランスフォーメーションのような機能に関連している。
【0004】
L1タンパク質は主要カプシドタンパク質であり、55〜60kDaの分子量を有する。L2タンパク質は副次的カプシドタンパク質である。免疫学的データは、L2タンパク質の大部分がL1タンパク質の内部に存在することを示唆している。L1タンパク質およびL2タンパク質は共に、種々のパピローマウイルス間で高度に保存されている。
【0005】
酵母、昆虫細胞、哺乳類細胞または細菌におけるL1タンパク質またはL1およびL2タンパク質の組合せ体の発現はウイルス様粒子(VLP)の自己集合を引き起こす(概説としては、SchillerおよびRoden,in Papillomavirus Reviews:Current Research on Papillomaviruses;Lacey編,Leeds,UK:Leeds Medical Information,pp 101−12(1996)を参照されたい)。VLPは真正ビリオンに形態学的に類似しており、動物またはヒトに投与されると高力価の中和抗体を誘導しうる。VLPは、潜在的発癌性ウイルスゲノムを含有しないため、HPVワクチンの開発における生ウイルスの使用に代わる安全な手段となる(概説としては、SchillerおよびHidesheim,J.Clin.Virol.19:67−74(2000)を参照されたい)。この理由により、L1およびL2遺伝子はHPV感染および疾患に対する予防用および治療用ワクチンの免疫学的標的と目されている。
【0006】
うまく形質転換された宿主生物における高い発現レベルのカプシドタンパク質を入手することは困難であり、これにより精製タンパク質の製造が制限されるため、HPVワクチンの開発および商業化は妨げられている。したがって、HPV31 L1タンパク質のようなHPV L1タンパク質をコードする野生型ヌクレオチド配列が同定されているものの(Goldsboroughら,Virology 171(1):306−311(1989))、意図される宿主細胞内での発現に関して最適化されたHPV31 L1コード化ヌクレオチド配列を利用する容易に再生可能な粗HPVタンパク質源を開発することが非常に望ましいであろう。また、ワクチン開発で使用する天然タンパク質の免疫付与特性を有する大量のHPV31 L1 VLPを製造することが有用であろう。
【発明の開示】
【0007】
発明の概要
本発明は、子宮頸癌に関連づけられているHPV31 L1遺伝子により発現されるタンパク質産物に対する免疫を惹起または増強するための組成物および方法に関する。特に、本発明は、酵母により認識される内部転写終結シグナルを含有しない、HPV31 L1タンパク質をコードするポリヌクレオチドを提供する。また、酵母細胞内での高レベル発現に関してコドン最適化された、HPV31 L1をコードする合成ポリヌクレオチドも提供する。本発明は更に、HPV31ウイルス様粒子(VLP)を提供し、HPV疾患またはHPV関連癌の予防および/または治療用の免疫原性組成物およびワクチンにおける該VLPの使用を開示する。
【0008】
本発明は、HPV31 L1タンパク質をコードする合成DNA分子に関する。本発明の1つの態様においては、該合成分子のヌクレオチド配列は、酵母により認識される転写終結シグナルを除去するために改変されている。もう1つの態様においては、合成分子のコドンは、酵母細胞により好ましいコドンが利用されるよう設計される。該合成分子は、VLPに自己集合しうるHPV31 L1タンパク質の源として使用することが可能である。該VLPは、VLPに基づくワクチンにおいて使用することが可能である。
【0009】
本発明の特定の実施形態は、配列番号4に記載のHPV31 L1タンパク質をコードする、配列番号2または配列番号3に記載のヌクレオチドの配列を含む合成核酸分子を含む。
【0010】
前記のとおり、本発明においては、酵母により認識される転写終結シグナルを含有しないHPV31 L1遺伝子をコードする合成ポリヌクレオチドを提供する。本発明はまた、酵母細胞にとって好ましいコドンを含有するよう更に改変された、前記のHPV31 L1をコードする合成ポリヌクレオチドを提供する。
【0011】
また、本明細書中の全体に開示されている核酸分子を含有する組換えベクターおよび組換え宿主細胞(原核細胞および真核細胞)を提供する。
【0012】
本発明は、(a)HPV31 L1タンパク質をコードする核酸(該核酸分子は、酵母により認識される内部転写終結シグナルを含有しない)を含むベクターを酵母宿主細胞内に導入し、(b)HPV31 L1タンパク質の発現を可能にする条件下、該酵母宿主細胞を培養することを含む、組換え宿主細胞内でHPV31 L1タンパク質を発現させるための方法に関する。
【0013】
本発明は、(a)HPV31 L1タンパク質をコードする核酸(該核酸分子は、酵母宿主細胞内での最適化発現に関してコドン最適化されている)を含むベクターを酵母宿主細胞内に導入し、(b)HPV31 L1タンパク質の発現を可能にする条件下、該酵母宿主細胞を培養することを含む、組換え宿主細胞内でHPV31 L1タンパク質を発現させるための方法に関する。
【0014】
好ましい実施形態においては、該核酸は、配列番号2または配列番号3に記載のヌクレオチドの配列を含む。
【0015】
本発明はまた、HPV31ウイルス様粒子(VLP)、HPV31 VLPの製造方法、およびHPV31 VLPの使用方法に関する。
【0016】
本発明の好ましい実施形態においては、HPV31 VLPは酵母において製造される。もう1つの好ましい実施形態においては、酵母は、サッカロミセス・セレビシエ(Saccharomyces cerevisiae)、ハンゼヌラ・ポリモルファ(Hansenula polymorpha)、ピチア・パストリス(Pichia pastoris)、クリーベルミセス・フラジリス(Kluyvermyces fragilis)、クルベロミセス・ラクチス(Kluveromyces lactis)およびシゾサッカロミセス・ポンベ(Schizosaccharomyces pombe)よりなる群から選ばれる。
【0017】
本発明のもう1つの態様は、酵母により認識される転写終結シグナルを含有しないHPV31 L1遺伝子により産生されるHPV31 L1タンパク質を含むHPV31 VLPである。
【0018】
本発明の更にもう1つの態様は、コドン最適化HPV31 L1遺伝子により産生されるHPV31 L1タンパク質を含むHPV31 VLPである。本発明のこの態様の好ましい実施形態においては、コドン最適化HPV31 L1遺伝子は、配列番号2または配列番号3に記載のヌクレオチドの配列より実質的になる。
【0019】
本発明はまた、HPV31ウイルス様粒子を動物に投与することを含んでなる、動物において免疫応答を誘導するための方法を提供する。好ましい実施形態においては、HPV31 VLPはコドン最適化遺伝子により産生される。もう1つの好ましい実施形態においては、HPV31 VLPは、酵母により認識される転写終結配列を含有しない遺伝子により製造される。
【0020】
本発明の更にもう1つの態様は、HPV31 VLPを含むワクチンを哺乳動物に投与することを含む、HPV関連子宮頸癌の予防または治療方法である。本発明のこの態様の好ましい実施形態においては、HPV31 VLPは酵母において製造される。
【0021】
本発明はまた、HPV31ウイルス様粒子(VLP)を含むワクチンに関する。
【0022】
本発明のこの態様のもう1つの実施形態においては、該ワクチンは更に、少なくとも1つの追加的HPV型のVLPを含む。好ましい実施形態においては、その少なくとも1つの追加的HPV型は、HPV6、HPV11、HPV16、HPV18、HPV33、HPV35、HPV39、HPV45、HPV51、HPV52、HPV55、HPV56、HPV58、HPV59およびHPV68よりなる群から選ばれる。
【0023】
本発明はまた、HPV31ウイルス様粒子を含む医薬組成物に関する。さらに、本発明は、HPV31 VLPと少なくとも1つの追加的HPV型のVLPとを含む医薬組成物に関する。好ましい実施形態においては、その少なくとも1つの追加的HPV型は、HPV6、HPV11、HPV16、HPV18、HPV33、HPV35、HPV39、HPV45、HPV51、HPV52、HPV55、HPV56、HPV58、HPV59およびHPV68よりなる群から選ばれる。
【0024】
本明細書中の全体において及び添付する特許請求の範囲において用いる単数表現は、文脈から例外であることが明らかである場合を除き、複数物に対する言及を含む。
【0025】
本明細書中の全体において及び添付する特許請求の範囲においては、以下の定義および略語が適用される。
【0026】
「プロモーター」なる語は、RNAポリメラーゼが結合する、DNA鎖上の認識部位を意味する。プロモーターはRNAポリメラーゼと共に開始複合体を形成して、転写活性を始動し駆動する。該複合体は、「エンハンサー」もしくは「上流活性化配列」と称される配列を活性化することにより又は「サイレンサー」と称される配列を抑制することにより修飾されうる。
【0027】
「ベクター」なる語は、宿主生物または宿主組織内へのDNA断片の導入をもたらしうる何らかの手段を意味する。プラスミド、ウイルス(アデノウイルスを含む)、バクテリオファージおよびコスミドを含む種々のタイプのベクターが存在する。
【0028】
「31L1野生型配列」なる表現は、本明細書中に配列番号1として開示されているHPV31 L1配列を意味する。HPV31 L1野生型配列は既に記載されているが、臨床分離体から得たDNA間に若干の配列変異を見出すことは珍しいことではない。したがって、代表的なHPV31 L1野生型配列を、HPV31 DNAを含有することが既に示されている臨床サンプルから単離した(実施例1を参照されたい)。本明細書中に開示されているコドン最適化HPV31 L1配列を比較するための参照配列として該31 L1野生型配列を使用した(図1を参照されたい)。
【0029】
「31 L1部分改変体」は、酵母内での最適化発現のために酵母にとって好ましいコドンを含有するようHPV31 L1ヌクレオチド配列が部分的に改変された、本明細書中に開示されている構築物(配列番号2)を意味する。31 L1部分改変体は、HPV31 L1野生型ヌクレオチド配列の中央部分(ヌクレオチド697−1249)における改変を含む。酵母にとって好ましいコドンを使用して、完全HPV31 L1配列をも改変させ、それは本明細書中では「31 L1全改変体」(配列番号3)と称される。
【0030】
「有効量」なる語は、免疫応答が生じるよう、適度なレベルの該ポリペプチドの産生に十分なワクチン組成物が導入されることを意味する。このレベルは様々なものとなりうると当業者に理解される。
【0031】
「保存的アミノ酸置換」は、1つのアミノ酸残基が、化学的に類似した別のアミノ酸残基により置換されることを意味する。そのような保存的置換の例としては、疎水性残基同士(イソロイシン、ロイシン、バリンまたはメチオニン)の置換、同じ電荷の極性残基同士(例えば、アルギニンとリシン;グルタミン酸とアスパラギン酸)の置換が挙げられる。
【0032】
「哺乳動物(哺乳類)」なる語は、ヒトを含む任意の哺乳動物を意味する。
【0033】
「VLP」はウイルス様粒子を意味する。
【0034】
「合成」は、天然に存在する野生型HPV31 L1遺伝子内に存在するヌクレオチド配列と同じではないヌクレオチド配列をHPV31 L1遺伝子が含有するよう、該遺伝子が修飾されていることを意味する。前記のとおり、本発明において、酵母により認識される転写終結シグナルを除去するよう改変されたヌクレオチドの配列を含む合成分子を提供する。また、本発明においては、酵母細胞による発現に好ましいコドンを含む合成分子も提供する。本発明で提供する合成分子は、野生型HPV31 L1遺伝子と同じアミノ酸配列をコードしている。
【0035】
発明の詳細な説明
ほとんどの子宮頸癌は、特定の発癌型のヒトパピローマウイルス(HPV)の感染に関連している。本発明は、発癌性HPV型の遺伝子により発現されるタンパク質産物に対する免疫を惹起または増強する組成物および方法に関する。特に、本発明は、HPV31 L1およびHPV31ウイルス様粒子(VLP)をコードする合成ポリヌクレオチドを提供し、HPV関連癌の予防および/または治療用の免疫原性組成物およびワクチンにおける該ポリヌクレオチドおよびVLPの使用を開示する。
【0036】
野生型HPV31 L1ヌクレオチド配列は既に報告されている(Goldsboroughら,Virology 171(1):306−311(1989);Genbankアクセッション番号J04353)。本発明は、HPV31 L1タンパク質をコードする合成DNA分子を提供する。本発明の合成分子はヌクレオチドの配列を含み、該ヌクレオチドのいくつかは、酵母により認識される転写終結シグナルを除去するよう改変されている。他の実施形態においては、該合成分子のコドンは、高レベル発現を行うのに酵母細胞にとって好ましいコドンを使用するよう設計される。該合成分子は、VLPへと自己集合するHPV31 L1タンパク質の源として使用することが可能である。該VLPは、中和抗体および細胞性免疫によりパピローマウイルス感染に対する有効な免疫予防をもたらすために、VLPに基づくワクチンにおいて使用することが可能である。VLPに基づくそのようなワクチンは、既に確立したHPV感染の治療にも有用である。
【0037】
酵母細胞におけるHPV VLPの発現は、費用効果的であり発酵槽内での大規模増殖に容易に適合するという利点をもたらす。しかし、HPV31 L1を含む多数のHPV L1タンパク質は(実施例4を参照されたい)、酵母細胞内では低レベルでしか発現されない。HPV31 L1の低レベル発現は、酵母により認識される転写終結シグナルの存在により生じる、mRNA転写産物のトランケート化によるものであることが、本発明において確認された。酵母転写終結部位に類似した潜在的配列を除去するようHPV31 L1 DNAを改変することにより、完全長mRNAの転写を促進して、HPV31 L1タンパク質発現の増強をもたらすことが可能である。
【0038】
したがって、本発明のいくつかの実施形態においては、酵母転写終結シグナルに類似した潜在的配列を除去するようHPV31 L1 DNAに改変が施されている。これらの改変は、トランケート化転写産物ではなく完全長HPV31転写産物の発現を可能にして(実施例4を参照されたい)、発現収率を増強させる。
【0039】
前記のとおり、本発明の合成DNAは、酵母により認識される転写終結部位を除去するように施された、野生型HPV31 L1配列からの改変を含む。HPV31 L1タンパク質をコードするが酵母転写終結部位を含有しない追加的なDNA分子を構築することが可能であると当業者は認識するであろう。酵母転写終結配列を見出すための技術は当技術分野でよく知られている。酵母mRNAの3’末端形成および転写終結は以下の3つのシグナルの存在を要する:(1)下流に位置する要素の配置の効率を増加させる、TATATAまたは関連配列のような効率要素(efficiency element)、(2)ポリ(A)部位の位置を決定する配置要素(positioning element)、および(3)ポリアデニル化部位(通常はPy(A)n)。
【0040】
科学文献には、酵母転写終結シグナルをコードする配列の記載が豊富に見出される。例えば、GuoおよびSherman,Trends Biochem.Sci.21:477−481(1986);GuoおよびSherman,Mol.Cell.Biol.16(6):2772−2776(1996);Zaretら,Cell 28:563−573(1982);Henikoffら,Cell 33:607−614(1983);Thalenfeldら,J.Biol.Chern.258(23):14065−14068(1983);Zaretら,J.Mol.Biol.176:107−135(1984);Heidmannら,Mol.Cell Biol 14:4633−4642(1984);ならびにRusso,Yeast 11:447−453(1985)を参照されたい。したがって、本発明の完全長mRNA転写産物を産生する合成HPV31 L1遺伝子を構築するためにはどの配列を避けるべきかを、当業者は何の困難もなく決定するであろう。また、該合成配列内に酵母転写終結配列が存在するかどうかを評価するためのアッセイおよび方法は当技術分野において十分に確立されており、したがって、当業者であれば、構築されたHPV31 l1配列が、除去する必要のある終結配列を含むかどうかを判定しうるであろう。
【0041】
前記のとおり、本発明は、酵母により認識される内部転写終結シグナルを含有しない、HPV31型L1タンパク質をコードする核酸分子に関する。本発明の典型的実施例においては、該合成核酸分子は、配列番号2または配列番号3に記載のヌクレオチドの配列を含む。
【0042】
本発明の他の実施形態においては、酵母細胞環境中での高レベル発現に関してHPV31 L1遺伝子配列が「最適化」される。本発明により意図されるコドン最適化HPV31 L1遺伝子は、酵母により認識される内部転写終結シグナルを含有しないHPV31 L1をコードする合成分子を含み、これは、酵母細胞内での高レベル発現に関してコドン最適化された少なくとも1つのコドンを更に含む。
【0043】
4つの可能なヌクレオチド塩基の「トリプレット」コドンは、60を超える変異形態で存在しうる。これらのコドンは僅か20個の異なるアミノ酸(ならびに転写開始および終結)のメッセージを与えるに過ぎないため、いくつかのアミノ酸が2以上のコドンによりコードされうる(コドン縮重として公知の現象である)。完全には理解されていない理由により、異なる細胞型の内因性DNA内には代替的コドンは一様には存在しない。実際、或る細胞型においては或るコドンに関する可変的天然階層または「優先性」が存在するようである。一例として、アミノ酸ロイシンは、CTA、CTC、CTG、CTT、TTAおよびTTGを含む6つのDNAコドンのいずれかにより特定される。大腸菌(E.coli)の内因性DNAは、最も一般的には、ロイシン特定コドンCTGを含有し、一方、酵母および粘菌のDNAは、最も一般的には、ロイシン特定コドンTTAを含むことが、微生物についてのゲノムコドン出現頻度の詳細な分析から示されている。この階層を考慮すると、ロイシンに富むポリペプチドの高レベルの発現が大腸菌(E.coli)宿主により得られる可能性は、コドン使用頻度に或る程度は左右されると、一般に考えられる。例えば、TTAコドンに富む遺伝子は大腸菌(E.coli)内では発現されにくいと考えられ、一方、CTGに富む遺伝子は、おそらく、この宿主内で高度に発現されるであろう。同様に、ロイシンに富むポリペプチドの酵母宿主細胞内での発現に好ましいコドンはTTAであろう。
【0044】
組換えDNA技術へのコドン優先性現象の関与は明白であり、該現象は、成功裏に形質転換された宿主生物において外因性遺伝子の高い発現レベルがこれまで多くの場合に達成されていないことを説明するのに役立ちうる。すなわち、該挿入遺伝子内には、それほど「好まし」くないコドンが反復的に存在している可能性があり、発現用の宿主細胞装置が、それほど効率的には機能していない可能性があるのである。この現象は、意図される宿主細胞にとって好ましいコドンを含むように設計された合成遺伝子が、組換えDNA技術の実施のための好ましい形態の外来遺伝物質の最適形態を与えることを示唆している。したがって、本発明の1つの態様は、酵母細胞内での発現に関してコドン最適化されたHPV31 L1遺伝子である。本発明の好ましい実施形態において、同じタンパク質配列をコードする代替的コドンの使用は、酵母細胞によるHPV31 L1タンパク質の発現に対する制約を除去しうることが判明した。
【0045】
本発明においては、HPV31 L1遺伝子セグメントを、同一の翻訳配列を有するが代替的コドン使用頻度を伴う配列に変換した(参照により本明細書に組み入れるSharpおよびCowe,Synonymous Codon Usage in Saccharomyces cerevisiae.Yeast 7:657−678(1991)に記載されているとおり)。該方法は、一般には、高度に発現される酵母遺伝子に一般には関連していない野生型配列内のコドンを同定し、それらを、酵母細胞内での高発現のための最適コドンで置換することよりなる。ついでその新たな遺伝子配列を、これらのコドン置換により生じた望ましくない配列(例えば、「ATTTA」配列、イントロンスプライス認識部位の非意図的生成、望ましくない制限酵素部位など)に関して調べる。望ましくない配列は、同一アミノ酸をコードする異なるコドンで既存コドンを置換することにより除去される。ついで該合成遺伝子セグメントを発現の改善に関して試験する。
【0046】
前記の方法を用いてHPV31 L1の合成遺伝子セグメントを作製して、高レベル発現に関して最適化されたコドンを含む遺伝子を得た。前記方法は、HPVワクチンにおいて使用するコドン最適化遺伝子を設計するための本発明者らの方法の概要を説明するものであるが、該方法における若干の変更または該配列における若干の変更によっても、同様のワクチン効力または遺伝子発現の増強が達成されうると、当業者に理解される。
【0047】
したがって、本発明は、HPV31 L1タンパク質またはHPV31 L1タンパク質の生物学的に活性な断片もしくは突然変異体をコードするヌクレオチド配列を含む、酵母宿主内での発現に関して最適化されたコドンを含む合成ポリヌクレオチドに関する。HPV31 L1タンパク質の突然変異体は、保存的アミノ酸置換、アミノ末端トランケート化、カルボキシ末端トランケート化、欠失または付加を含むが、これらに限定されるものではない。任意のそのような生物学的に活性な断片および/または突然変異体は、配列番号4に記載のHPV31 L1タンパク質の免疫学的特性を少なくとも実質的に模擬するタンパク質またはタンパク質断片をコードする。本発明の合成ポリヌクレオチドは、治療用または予防用HPVワクチンの開発において有用となるよう、機能的HPV31 L1タンパク質を発現するmRNA分子をコードする。
【0048】
本発明の1つの態様は、配列番号2に記載のヌクレオチド配列を含む、配列番号4に記載のHPV31 L1タンパク質をコードするコドン最適化核酸分子である。
【0049】
本発明のもう1つの態様は、配列番号3に記載のヌクレオチド配列を含む、配列番号4に記載のHPV31 L1タンパク質をコードするコドン最適化核酸分子である。
【0050】
本発明はまた、本明細書中の全体に開示されている核酸分子を含有する組換えベクターおよび組換え宿主細胞(原核細胞および真核細胞の両方)に関する。
【0051】
本明細書に記載の方法により構築した合成HPV31 DNAまたはその断片を、適当なプロモーターと他の適当な転写調節要素とを含有する発現ベクター内への分子クローニングにより組換え的に発現させ、原核または真核宿主細胞内に導入して、組換えHPV31 L1を得ることが可能である。そのような操作のための技術は当技術分野において記載されている(Sambrookら Molecular Cloning:A Laboratory Manual;Cold Spring Harbor Laboratory Press,Cold Spring Harbor,New York,(1989);Current Protocols in Molecular Biology,Ausubelら,Green Pub.Associates and Wiley−Interscience,New York(1988);Yeast Genetics:A Laboratory Course Manual,Roseら,Cold Spring Harbor Laboratory,Cold Spring Harbor,New York,(1990)(それらの全体を参照により本明細書に組み入れることとする))。
【0052】
したがって、本発明は更に、(a)HPV31 L1タンパク質をコードする核酸(該核酸分子は、酵母宿主細胞での最適化発現に関してコドン最適化されている)を含むベクターを酵母宿主細胞内に導入し、(b)HPV31 L1タンパク質の発現を可能にする条件下、該酵母宿主細胞を培養することを含む、組換え宿主細胞内でHPV31 L1タンパク質を発現させるための方法に関する。
【0053】
したがって、本発明は更に、(a)HPV31 L1タンパク質をコードする核酸(該核酸分子は、酵母により認識される内部転写終結シグナルを含有しない)を含むベクターを酵母宿主細胞内に導入し、(b)HPV31 L1タンパク質の発現を可能にする条件下、該酵母宿主細胞を培養することを含む、組換え宿主細胞内でHPV31 L1タンパク質を発現させるための方法に関する。
【0054】
本発明は更に、(a)配列番号2または配列番号3に記載の核酸を含むベクターを酵母宿主細胞内に導入し、(b)HPV31 L1タンパク質の発現を可能にする条件下、該酵母宿主細胞を培養することを含む、組換え宿主細胞内でHPV31 L1タンパク質を発現させるための方法に関する。
【0055】
本発明の合成遺伝子は、宿主細胞内でのHPV58 L1タンパク質の効率的発現をもたらすように設計された配列を含む発現カセットに集合させることが可能である。該カセットは、好ましくは、合成遺伝子を、それに機能しうる形で連結された関連転写および翻訳制御配列(例えば、プロモーターおよび終結配列)と共に含有する。好ましい実施形態においては、プロモーターはエス・セレビシエ(S.cerevisiae)GAL1プロモーターであるが、多数の他の公知酵母プロモーター、例えばGAL10、GAL7、ADH1、TDH3もしくはPGKプロモーターまたは他の真核生物遺伝子プロモーターのいずれかを使用しうると当業者は認識するであろう。好ましい転写ターミネーターはエス・セレビシエ(S.cerevisiae)ADH1ターミネーターであるが、他の公知転写ターミネーターも使用することが可能である。GAL1プロモーター−ADH1ターミネーターの組合せが特に好ましい。
【0056】
本発明のもう1つの態様はHPV31ウイルス様粒子(VLP)、HPV31 VLPの製造方法、およびHPV31 VLPの使用方法である。ヒトおよび動物パピローマウイルスの主要カプシドタンパク質であるL1が酵母、昆虫細胞、哺乳類細胞または細菌内で発現される場合には、VLPは自己集合しうる(概説としては、SchillerおよびRoden,in Papillomavirus Reviews:Current Research on Papillomaviruses;Lacey編,Leeds,UK:Leeds Medical Information,pp 101−12(1996))。L1およびL2カプシドタンパク質の組合せを発現させることにより、形態学的に異なるHPV VLPも製造されうる。VLPは、T=7二十面体構造のL1の72個の五量体から構成される(Bakerら,Biophys.J.60(6):1445−56(1991))。
【0057】
VLPは真正ビリオンと形態学的に類似しており、動物に投与されると高力価の中和抗体を誘導しうる。VLPでのウサギ(Breitburdら,J.Virol.69(6):3959−63(1995))およびイヌ(Suzichら,Proc.Natl.Acad.Sci.USA 92(25):11553−57(1995))の免疫は中和抗体の誘導および実験パピローマウイルス感染に対する防御の両方をもたらすことが示された。しかし、VLPは、潜在的に発癌性のウイルスゲノムを含有せず、単一遺伝子から自己集合しうるため、それは、HPVワクチン開発における生ウイルスの使用に代わる安全な手段となる(概説としては、SchillerおよびHidesheim,J.Clin.Virol.19:67−74(2000)を参照されたい)。
【0058】
したがって、本発明は、HPV31の組換えL1タンパク質または組換えL1+L2タンパク質から構成されるウイルス様粒子に関する。
【0059】
本発明の好ましい実施形態においては、HPV31 VLPは酵母において産生される。もう1つの好ましい実施形態においては、酵母は、サッカロミセス・セレビシエ(Saccharomyces cerevisiae)、ハンゼヌラ・ポリモルファ(Hansenula polymorpha)、ピチア・パストリス(Pichia pastoris)、クリーベルミセス・フラジリス(Kluyvermyces fragilis)、クルベロミセス・ラクチス(Kluveromyces lactis)およびシゾサッカロミセス・ポンベ(Schizosaccharomyces pombe)よりなる群から選ばれる。
【0060】
本発明のもう1つの態様は、酵母により認識される転写終結シグナルを含有しないHPV31 L1遺伝子により産生されるHPV31 L1タンパク質を含むHPV31 VLPである。
【0061】
本発明の更にもう1つの態様は、コドン最適化HPV31 L1遺伝子により産生されるHPV31 L1タンパク質を含むHPV31 VLPである。本発明のこの態様の好ましい実施形態においては、コドン最適化HPV31 L1遺伝子は、配列番号2または配列番号3に記載のヌクレオチドの配列より実質的になる。
【0062】
本発明の更にもう1つの態様は、(a)HPV31 L1タンパク質またはHPV31 L1タンパク質またはHPV31 L1+L2タンパク質をコードする組換えDNA分子で酵母を形質転換し、(b)該組換えDNA分子の発現を可能にする条件下、該形質転換酵母を培養して組換えHPV31タンパク質を得、(c)該組換えHPV31タンパク質を単離してHPV31 VLPを得ることを含む、HPV31 VLPの製造方法である。
【0063】
本発明のこの態様の好ましい実施形態においては、酵母により認識される転写終結シグナルを含有しないHPV31 L1遺伝子で酵母を形質転換する。もう1つの好ましい実施形態においては、HPV31 VLPを得るために、コドン最適化HPV31 L1遺伝子で酵母を形質転換する。特に好ましい実施形態においては、コドン最適化HPV31 L1遺伝子は、配列番号2または配列番号3に記載のヌクレオチド配列より実質的になる。
【0064】
本発明はまた、HPV31ウイルス様粒子を動物に投与することを含んでなる、動物において免疫応答を誘導するための方法を提供する。好ましい実施形態においては、HPV31 VLPは、酵母により認識される内部転写終結配列を含有しない遺伝子により製造される。もう1つの好ましい実施形態においては、HPV31 VLPはコドン最適化遺伝子により製造される。
【0065】
本発明の更にもう1つの態様は、HPV31 VLPを含むワクチンを哺乳動物に投与することを含む、HPV関連子宮頸癌の予防または治療方法である。本発明のこの態様の好ましい実施形態においては、HPV31 VLPは酵母において製造される。
【0066】
本発明はまた、HPV31ウイルス様粒子(VLP)を含むワクチンに関する。
【0067】
本発明のこの態様のもう1つの実施形態においては、該ワクチンは更に、少なくとも1つの追加的HPV型のVLPを含む。好ましい実施形態においては、その少なくとも1つの追加的HPV型は、HPV6、HPV11、HPV16、HPV18、HPV33、HPV35、HPV39、HPV45、HPV51、HPV52、HPV55、HPV56、HPV58、HPV59およびHPV68よりなる群から選ばれる。
【0068】
本発明のこの態様の好ましい実施形態においては、該ワクチンは更に、HPV16 VLPを含む。
【0069】
本発明のもう1つの好ましい実施形態においては、該ワクチンは更に、HPV16 VLPおよびHPV18 VLPを含む。
【0070】
本発明の更にもう1つの好ましい実施形態においては、該ワクチンは更に、HPV6 VLP、HPV11 VLP、HPV16 VLPおよびHPV18 VLPを含む。
【0071】
本発明はまた、HPV31ウイルス様粒子を含む医薬組成物に関する。さらに、本発明は、HPV31 VLPと少なくとも1つの追加的HPV型のVLPとを含む医薬組成物に関する。好ましい実施形態においては、その少なくとも1つの追加的HPV型は、HPV6、HPV11、HPV16、HPV18、HPV33、HPV35、HPV39、HPV45、HPV51、HPV52、HPV55、HPV56、HPV58、HPV59およびHPV68よりなる群から選ばれる。
【0072】
本発明のワクチン組成物は、潜在的毒性を最小限にする一方でHPV31感染の最適な抑制を得るために通常の試験により定められる適当な量で単独で使用することが可能である。また、他の物質の共投与または連続的投与が望ましいかもしれない。
【0073】
ワクチン被投与者に導入すべきウイルス様粒子の量は、発現される遺伝子産物の免疫原性に左右される。一般に、VLP 10μg〜100μg、好ましくは約20μg〜60μgの免疫学的または予防的に有効な量を筋肉組織内に直接投与する。皮下注射、皮内導入、皮膚を介した圧入(impression)、および他の投与方法、たとえば、腹腔内、静脈内または吸入運搬も意図される。また、ブースターワクチン接種を行いうると意図される。本発明のワクチンの非経口導入と同時またはその後の、ミョウバンまたはメルク(Merck)ミョウバンアジュバントのようなアジュバントの非経口投与、例えば静脈内、筋肉内、皮下または他の手段での投与も有利である。
【0074】
本発明に関連して用いられうる方法および材料を記載し開示する目的で、本明細書に記載のすべての刊行物を、参照により本明細書に組み入れることとする。ここで、本発明が先行発明としてそのような開示に先行するものではないと自認するものと解釈されるべきではない。
【0075】
添付図面を参照して本発明の好ましい実施形態が記載されているが、本発明はそれらの実施形態そのものに限定されるものではなく、添付の特許請求の範囲において定められる本発明の範囲または精神から逸脱することなく本発明には種々の変更および修飾が当業者によって施されうると理解されるべきである。
【0076】
以下の実施例は本発明を例示するものであり、本発明を限定するものではない。
【実施例1】
【0077】
代表的HPV31 L1配列の決定
HPV31 L1野生型配列は既に記載されている(Goldsboroughら,Virology 171(1):306−311(1989);Genbankアクセッション番号J04353)。しかし、臨床分離体から得たDNA間に若干の配列変異を見出すことは珍しいことではない。代表的なHPV31 L1野生型配列を単離するために、HPV31 DNAを含有することが既に示されている3つの臨床サンプルからDNAを単離した。Taq DNAポリメラーゼおよび以下のプライマーを使用するポリメラーゼ連鎖反応(PCR)において、HPV31 L1配列を増幅した:
【0078】
【化1】
【0079】
増幅産物をアガロースゲル上で電気泳動し、臭化エチジウム染色により可視化した。〜1500bpのL1バンドを切り出し、QIAクイックPCR精製キット(Qiagen,Hilden,Germany)を使用してDNAを精製した。ついで該DNAをTAクローニングベクターpCR−II(Invitrogen Corp.,Carlsbad,CA)に連結し、大腸菌(E.coli)を形質転換し、アンピシリン+IPTGおよびX−gal(青/白コロニー選択用)を含有するLB寒天上でプレーティングした。該プレートを反転させ、37℃で16時間インキュベートした。白色コロニーを、アンピシリンを含有するLB培地内で37℃で16時間、振とうしながら培養し、ミニプレップを行って該プラスミドDNAを抽出した。
【0080】
該プラスミド中のL1遺伝子の存在を示すために、制限エンドヌクレアーゼ消化を行い、アガロースゲル電気泳動および臭化エチジウム染色により可視化した。それらの3つの臨床分離体のそれぞれからのクローン化L1を含有するプラスミドに関してDNA配列決定を行った。DNAおよび翻訳アミノ酸配列をお互いと及びGenbank HPV31 L1配列と比較した。それらの3つの臨床分離体の配列分析は、どの配列も該Genbank配列と同一ではないことを示した。代表的な31L1配列としてpCR−II−HPV 31L1/81クローンを選択した。これは本明細書中では「31 L1野生型配列」(配列番号1;図1を参照されたい)と称される。31 L1野生型として選択した配列はヌクレオチド1266の1つのサイレント置換、およびコード化アミノ酸をトレオニンからセリンに変化させるヌクレオチド1295のCからGへの変化を含有していた。31 L1部分および全改変体遺伝子(それぞれ配列番号2および3)も、この位置にセリンをコードしている(図1を参照されたい)。すべての場合において、アミノ酸配列は同一である。改変体構築物においては、酵母にとって好ましいコドン配列を使用してアミノ酸をコードし、潜在的転写終結シグナルを除去するよう、ヌクレオチドが変化している(実施例2を参照されたい)。
【0081】
BglII伸長を付加するために以下のプライマーを使用して、31 L1野生型配列を増幅した。
【0082】
【化2】
【0083】
PCRは、、Vent(商標)DNAポリメラーゼを使用して行った。アガロースゲルの臭化エチジウム染色により、PCR産物を可視化した。〜1500bpのバンドを切り出し、QIAEX IIゲル抽出キット(Qiagen)を使用してDNAを精製した。ついでPCR産物をBglIIで37℃で2時間消化し、QIAクイックPCR精製キットを使用して精製した。BglIIで消化された31 L1 PCR産物を、BamHIで消化されたpGAL110に連結し、DH5大腸菌(E.coli)を形質転換した。正しい配向のHPV 31 L1インサートに関して、PCRによりコロニーをスクリーニングした。DNA配列決定により、配列および配向を確認した。選択したクローンをpGAL110−HPV 31L1 #2と命名した。
【0084】
ついでマキシプレップ(Maxiprep)DNAを調製し、サッカロミセス・セレビシエ(Saccharomyces cerevisiae)をコンピテントにし形質転換した。該酵母形質転換体をLeu− ソルビトールプレート上のLeu− ソルビトール上層寒天内でプレーティングし、反転させて30℃で3〜5日間インキュベートした。コロニーを拾い、Leu− ソルビトールプレート上での単離のためにストリークした。ついで、L1転写およびタンパク質発現を誘導するために、単離されたコロニーを、30℃の回転チューブ培養において、1.6%グルコースおよび4%ガラクトースを含有する5mlの5×Leu−Ade−ソルビトール内で成長させた。
【実施例2】
【0085】
酵母コドン最適化
酵母にとって好ましいコドンは既に記載されている(SharpおよびCowe,Yeast 7:657−678(1991))。まず、ヌクレオチド697−1249に相当するHPV31 L1の中央部を、酵母にとって好ましいコドンを使用して改変した。改変に用いた方法は、同じアミノ酸配列を維持しながら酵母にとって好ましいコドン配列でヌクレオチドを置換する、改変すべき領域に及ぶ長い重複するセンスおよびアンチセンスオリゴマーを設計するものであった。該PCR反応においては鋳型DNAの代わりにこれらのオリゴマーを使用した。追加的な増幅プライマーを設計し、それらを使用して、Pfu DNAポリメラーゼ(Stratagene,La Jolla,CA)により鋳型オリゴマーから改変体配列を増幅した。増幅のための最適条件は部位特異的なものであったが、ほとんどの場合には、以下に類似した方法を用いた:94℃で1時間の初期変性工程およびそれに続く、95℃で30秒間の変性、55℃で30秒間のアニーリング、72℃で3.5分間の伸長の15〜25サイクル、およびそれに続く72℃で10分間の最終伸長、および4℃での維持。
【0086】
PCR産物をアガロースゲル電気泳動により検査した。適当なサイズのバンドを切り出し、DNAをゲル精製した。ついで該増幅断片を鋳型として使用して552ヌクレオチドの改変体HPV31中央部L1断片を作製した。ついでPCRを使用して野生型ヌクレオチド1−725(5’末端)および1221−1515(3’末端)を増幅した。該5’末端、該3’末端および該改変体中央部を使用する最終PCRを行って、本明細書中では「31 L1部分改変体」と称される完全長31 L1部分改変体を作製した。
【0087】
また、酵母にとって好ましいコドンを使用して、完全な31 L1配列も改変した。この構築物は本明細書中で「31 L1全改変体」と称される。9個の長い重複するオリゴマーを使用して、1−753の、酵母にとって好ましいコドンのヌクレオチド配列を作製し、4個の長い重複するオリゴマーを使用して、1207−1515の、酵母にとって好ましいコドンのヌクレオチド配列を作製した。増幅およびゲル精製の後、これらの断片を、前記の中央部改変体部分(ヌクレオチド697−1249)と共にPCR反応において使用して、完全長31 L1全改変体配列を作製した。この断片は、BamHI伸長を伴って作製された。ゲル精製改変体31 L1 DNAをBamHIで消化し、BamHIで消化されたpGAL110発現ベクターに連結し、大腸菌(E.coli)DH5細胞内に形質転換した。正しい配向のHPV31 L1インサートに関して、PCRによりコロニーをスクリーニングした。DNA配列決定により、配列および配向を確認した。
【0088】
プラスミドDNAを調製した。サッカロミセス・セレビシエ(S.cerevisiae)をコンピテントにし、形質転換した。該酵母をLeu− ソルビトールプレート上のLeu− ソルビトール上層寒天内でプレーティングし、反転させて3〜5日間インキュベートした。コロニーを、Leu− ソルビトールプレート上での単離のためにストリークした。ついで、L1転写およびタンパク質発現を誘導するために、単離されたコロニーを、30℃の回転チューブ培養において、1.6%グルコースおよび4%ガラクトースを含有する5mlの5×Leu−Ade−ソルビトール内で成長させた。48〜72時間後、OD600=10と等価な培養容量をペレット化し、上清を除去し、該ペレットを凍結し、−70℃で保存した。
【実施例3】
【0089】
RNA調製
ガラクトース誘導によりHPV31 L1を発現するよう誘導された、形質転換酵母の細胞ペレットを氷上で解凍し、1mlの冷DEPC処理水に懸濁させた。細胞を遠心分離によりペレット化し、得られた上清を除去した。ついで細胞ペレットを400μlのTES(10mM Tris pH7.0,10mM EDTAおよび0.5% SDS)に再懸濁させた。等容量のAEバッファー飽和フェノール(50mM NaOAcおよび10mM EDTA)を加えた。該チューブを10秒間ボルテックスし、10分ごとに混合しながら65℃で50分間加熱した。ついで該チューブを氷上で5分間配置し、ついで4℃で5分間遠心分離した。上清を集め、無菌チューブに移した。追加的な400μlのフェノールを加え、該チューブをボルテックスし、氷上に5分間配置し、遠心分離した。上清を無菌チューブに移し、400μlのクロロホルムを加え、混合し、遠心分離した。上清を再び集め、無菌チューブに移し、1mlの100% EtOHのほかに40μlの3M 酢酸Na(pH5.2)を加えた。該チューブをドライアイス上に1時間配置し、ついでそれを高速で遠心分離してRNAをペレット化した。該RNAを70% EtOHで1回洗浄し、風乾させた。ついで該RNAを100μlのDEPC処理水に懸濁させ、65℃に5分間加熱して溶解した。A260/280が1.7〜2.0である場合にA260測定値1=40μg/ml RNAという仮定により、サンプル中のRNAの濃度を測定するために分光光度測定を行った。
【実施例4】
【0090】
ノーザンブロット分析
31 L1野生型を発現する酵母の初期分析は、HPV31 L1タンパク質の発現収率が予想より相当低いことを示唆した。該低発現が、転写レベルの問題により生じたのか翻訳レベルの問題により生じたのかを判定するために、HPV31 L1転写産物のノーザンブロット分析を行った。転写サイズを比較するために、同一ゲル上で、HPV16 L1を発現する酵母からのRNAが、HPV31 L1を発現する酵母からのRNAと共に移動したゲルから、ノーザンブロットを作製した。
【0091】
1.2% アガロースホルムアルデヒドゲルを成型した。10μgのRNAを変性バッファー(最終濃度:6% ホルムアルデヒド、45% ホルムアミドおよび0.9×MOPS)と一緒にし、55℃で15分間加熱した。ゲルローディングバッファーの10分の1の容量を加え、該サンプルを該ゲル上にローディングした。1×MOPSバッファー中、65ボルトで〜5時間、電気泳動を行った。該ゲルを無菌水中で15分間ついで10×SSC中で2回(各5分間)洗浄した。該RNAを、10×SSC中、16時間にわたり、毛管作用により、Hybond−N+ナイロン膜(Amersham Biosciences,Piscataway,NJ)にトランスファーした。ついで該RNAを、700単位のエネルギーに設定されたAmersham架橋剤を使用する架橋によりナイロン膜に固定した。固定後、該ナイロン膜を風乾させた。該膜を、30mlのZetaprobeバッファー中、55℃で2時間配置し、ついで32P標識プローブを加え、53〜65℃で16時間インキュベートした。ついで該膜を、5×SSC中、室温で20分間で3回、ついで0.4×SSC中、室温で20分間で2回、および60℃で10分間で1回洗浄した。HPV31 L1配列に特異的なセンスおよびアンチセンスプライマーを使用するPCRにより、プローブDNAを作製した。ポリヌクレオチドキナーゼ(PNK)およびγ―32 ATPでの37℃で1時間の処理により、該増幅DNAを標識した。該ブロットをサランラップで包み、x線フィルムに16時間露出させた。フィルムの現像後、プローブにハイブリダイズしたRNAはオートラジオグラフ上で黒色バンドとして検出された。
【0092】
前記のノーザンブロットの分析は、完全長HPV31 L1野生型転写産物の大部分が完全長より相当に小さいことを示した(図4を参照されたい)。しかし、31 L1部分改変体の設計においては、酵母にとって好ましいコドンが該遺伝子の中央部に挿入されているのみならず、酵母転写終結部位に類似した潜在的配列が除去されている。ノーザンブロット分析は、改変後、31 L1遺伝子転写産物の長さが、完全長HPV16 L1転写産物に対応するサイズまで有意に増加していたことを、明らかに示した(非表示)。したがって、未熟な転写終結が、31 L1野生型構築物からの低い発現収率のかなりの部分の原因になっていたらしい。
【実施例5】
【0093】
HPV31 L1タンパク質発現
OD600=10と等価な、ガラクトースにより誘導された培養物の凍結酵母細胞ペレットを、氷上で解凍し、2mM PMSFを含有する300μlのPCバッファー(100mM Na2HPO4および0.5M NaCl,pH7.0)に懸濁させた。酸で洗浄した0.5mmガラスビーズを加えた(〜0.5g/チューブ)。該チューブを4℃で15分間ボルテックスした。7.5μlの20% TritonX100を加え、ボルテックスを4℃で5分間反復した。該チューブを氷上に15分間配置し、ついで4℃で15分間遠心分離した。上清を無菌微小遠心管に移し、−70℃で保存した。
【実施例6】
【0094】
ウエスタンブロット分析
ガラクトース誘導後のHPV31 L1タンパク質の発現を確認するために、各HPV31 L1構築物についての20〜40個の単離された酵母コロニーからの全酵母タンパク質抽出物をウエスタンブロットにより分析した。
【0095】
10μgの全酵母タンパク質抽出物をSDS−PAGEローディングバッファーと一緒にし、95℃で10分間加熱した。該タンパク質を8% SDS−PAGEゲル上にローディングし、Tris−グリシンバッファー中で電気泳動した。タンパク質の分離後、該タンパク質を該ゲルからニトロセルロースにウエスタントランスファーし、該ブロットをTTBS(Tween−20を含有するTris緩衝食塩水)中の10% 脱脂乾燥乳中で16時間ブロッキングした。該ブロットをTTBS中で3回洗浄した。HPV31 L1と交差反応するポリクローナル血清であるヤギ抗trpE−HPV16 L1血清をTTBS中の1:1000の希釈度で室温で1時間適用した。該ブロットをTTBS中で3回洗浄し、抗ヤギHRP結合抗体をTTBS中の1:2500の希釈度で1時間適用した。該ブロットを再び3回洗浄し、ECL(商標)検出試薬を適用した(Amersham Biosciences,Piscataway,NJ)。ついでオートラジオグラフィーを行った。該抗血清により認識されたタンパク質は、オートラジオグラフ上で暗色バンドとして検出試薬により可視化された。
【0096】
すべての場合において、HPV 31L1タンパク質は、オートラジオグラフ上で、約55kDに対応する別個のバンドとして検出された(データ非表示)。ゲル上で陽性対照として、HPV16 L1タンパク質を含めた。
【実施例7】
【0097】
ラジオイムノアッセイ(RIA)
HPV31 L1を発現する酵母細胞を種々の方法(回転チューブ培養、振とうフラスコおよび発酵槽など)により増殖させた。該酵母を細胞溶解し、タンパク質抽出物を調製して、全タンパク質1mg当たりで産生されたHPV31 L1ウイルス様粒子(VLP)の量を測定した。HPV31 L1 VLP発現を示すために、各全酵母タンパク質抽出物の一部を捕捉ラジオイムノアッセイ(RIA)により分析した。
【0098】
HPV型31特異的でありVLPコンホメーション特異的である検出用モノクローナル抗体H31.A6を使用して、RIAを行った。H31.A6は無傷HPV31 L1 VLPには結合することが判明しているが、変性したHPV31 VLPを認識しないため、H31.A6はHPV型31 L1に特異的である。ついで、このmAbは、I125で放射能標識されたヤギ抗マウス抗体により検出されうる。したがって、毎分カウント(cpm)値はHPV31 L1 VLP発現の相対レベルに対応する。
【0099】
ポリスチレンビーズを、PBS中で1:1000で希釈されたヤギ抗trpE−HPV31 L1ポリクローナル血清で一晩コートした。ついで該ビーズを5容量の無菌蒸留水で洗浄し、風乾させた。ついで、単離された酵母コロニーからの全酵母タンパク質抽出物である抗原を、1% BSA、0.1% Twenn−20および0.1% アジ化ナトリウムを含有するPBS中の希釈により該ビーズ上にローディングし、回転させながら1時間インキュベートした。洗浄後、該ビーズを20ウェルポリスチレンプレートにおいて1個/ウェルで分配し、1:50,000で希釈されたH31.A6 mAbと共に室温で17〜24時間インキュベートした。該ビーズを洗浄し、I125標識ヤギ抗マウスIgGを23000〜27000cpm/10μlの活性範囲で加えた。2時間後、該ビーズを洗浄し、放射能カウントをcpm/ml単位で記録した。ブランクウェルからのバックグラウンドカウントを全cpm/mlから差し引いて、RIA−バックグラウンド値を得た。
【0100】
2つの実験を行った。実験1においては、31 L1野生型および31 L1部分改変体からのタンパク質抽出物を比較し、実験2においては、31 L1部分改変体および31 L1全改変体からのタンパク質抽出物を比較した(図5を参照されたい)。結果は、31 L1部分改変体VLPの発現が31 L1野生型より6.9倍高いことを示している。31 L1全改変体は、31 L1部分改変体と比べて1.7倍増加した発現を示している。したがって、酵母にとって好ましいコドン配列を導入し潜在的転写終結シグナルを除去することにより、31 L1の発現レベルは>7倍増加した。
【実施例8】
【0101】
透過型電子顕微鏡検査
HPV31 L1タンパク質が実際に自己集合して五量体L1カプソマー(これは今度は、ウイルス様粒子に自己集合する)を形成したことを示すために、部分的に精製された31 L1全改変体タンパク質抽出物を透過型電子顕微鏡検査(TEM)に付した。酵母を小規模発酵で増殖させ、ペレット化した。該ペレットを精製処理に付した。L1タンパク質の発現および精製操作における保持を示すために、ペレットおよび清澄化酵母抽出物を免疫ブロットにより分析した。ついで清澄化酵母抽出物を45% スクロースクッション上の遠心分離に付し、得られたペレットをTEM分析のためにバッファーに懸濁させた(図6を参照されたい)。この粗サンプル中の球状粒子の直径は30〜60nmであり、いくつかの粒子はカプソマーの規則的配置を示していることを、結果は示している。
【図面の簡単な説明】
【0102】
【図1A】部分改変体(配列番号2)および全改変体(配列番号3)31 L1遺伝子における改変ヌクレオチドを示す配列アライメントである(実施例2を参照されたい)。参照配列は31 L1野生型配列(配列番号1;実施例1を参照されたい)である。参照配列と同一である31 L1部分および全改変体配列におけるヌクレオチドは点で示されている。改変されたヌクレオチドは、それらの対応位置に示されている。括弧内にはヌクレオチド番号が示されている。
【図1B】部分改変体(配列番号2)および全改変体(配列番号3)31 L1遺伝子における改変ヌクレオチドを示す配列アライメントである(実施例2を参照されたい)。参照配列は31 L1野生型配列(配列番号1;実施例1を参照されたい)である。参照配列と同一である31 L1部分および全改変体配列におけるヌクレオチドは点で示されている。改変されたヌクレオチドは、それらの対応位置に示されている。括弧内にはヌクレオチド番号が示されている。
【図1C】部分改変体(配列番号2)および全改変体(配列番号3)31 L1遺伝子における改変ヌクレオチドを示す配列アライメントである(実施例2を参照されたい)。参照配列は31 L1野生型配列(配列番号1;実施例1を参照されたい)である。参照配列と同一である31 L1部分および全改変体配列におけるヌクレオチドは点で示されている。改変されたヌクレオチドは、それらの対応位置に示されている。括弧内にはヌクレオチド番号が示されている。
【図1D】部分改変体(配列番号2)および全改変体(配列番号3)31 L1遺伝子における改変ヌクレオチドを示す配列アライメントである(実施例2を参照されたい)。参照配列は31 L1野生型配列(配列番号1;実施例1を参照されたい)である。参照配列と同一である31 L1部分および全改変体配列におけるヌクレオチドは点で示されている。改変されたヌクレオチドは、それらの対応位置に示されている。括弧内にはヌクレオチド番号が示されている。
【図2A】31 L1全改変ヌクレオチド(配列番号3)およびアミノ酸配列(配列番号4)を示す。左側にヌクレオチド番号が示されている。
【図2B】31 L1全改変ヌクレオチド(配列番号3)およびアミノ酸配列(配列番号4)を示す。左側にヌクレオチド番号が示されている。
【図2C】31 L1全改変ヌクレオチド(配列番号3)およびアミノ酸配列(配列番号4)を示す。左側にヌクレオチド番号が示されている。
【図3】左側に挙げられている3つのHPV31 L1配列構築物間の変化を要約する。第4列は、示されている構築物と31 L1野生型配列との間のヌクレオチド同一性(%)を示し、第5列はアミノ酸同一性を示す。最終列は、酵母に好ましいコドン配列へ改変されたヌクレオチドの数、および改変が施された領域を示す。
【図4】高いストリンジェンシー下でHPV31 L1に特異的にプローブされたノーザンブロットを示す(実施例4を参照されたい)。左側の矢印は、HPV31 L1完全長およびトランケート化転写産物の位置を示す。「31wt」と示されたレーンは、31 L1野生型配列を含有する酵母の同じRNA調製物からのものである。「16」と示されたレーンは、高いストリンジェンシー条件のためにHPV31 L1プローブによっては認識されない、HPV16からのRNAを含有する。「Neg」と示されたレーンは、L1コード配列を含有しない酵母抽出物である。「31R」と示されたレーンは、31 L1部分改変体配列を発現する2つの別々の単離コロニーのRNAからのものである。
【図5】毎分カウント(cpm)/mg全タンパク質で表された、2つの捕捉ラジオイムノアッセイ(RIA)実験からのデータの一部を示す(実施例7を参照されたい)。該RIAで得たcpmはHPV31 L1 VLPの相対指標である。該RIAデータは、酵母にとって好ましいコドン改変体遺伝子配列からの酵母タンパク質抽出物における31 L1 VLP発現の増強を示している。
【図6】透過型電子顕微鏡検査により可視化された、本明細書に記載の31 L1 VLPの代表的サンプルを示す(実施例8を参照されたい)。棒線は100nmを表す。
【配列表】
【特許請求の範囲】
【請求項1】
酵母細胞内での高レベル発現に関してコドン最適化された、配列番号4に記載のHPV31 L1タンパク質をコードするヌクレオチド配列を含んでなる核酸分子。
【請求項2】
請求項1記載の核酸分子を含んでなるベクター。
【請求項3】
請求項3記載のベクターを含んでなる宿主細胞。
【請求項4】
宿主細胞が、サッカロミセス・セレビシエ(Saccharomyces cerevisiae)、ハンゼヌラ・ポリモルファ(Hansenula polymorpha)、ピチア・パストリス(Pichia pastoris)、クリーベルミセス・フラジリス(Kluyvermyces fragilis)、クルベロミセス・ラクチス(Kluveromyces lactis)およびシゾサッカロミセス・ポンベ(Schizosaccharomyces pombe)よりなる群から選ばれる、請求項3記載の宿主細胞。
【請求項5】
宿主細胞がサッカロミセス・セレビシエ(Saccharomyces cerevisiae)である、請求項4記載の宿主細胞。
【請求項6】
ヌクレオチド配列が、配列番号2に記載のヌクレオチド配列を含む、請求項1記載の核酸分子。
【請求項7】
請求項6記載の核酸分子を含んでなるベクター。
【請求項8】
請求項7記載のベクターを含んでなる宿主細胞。
【請求項9】
ヌクレオチド配列が、配列番号3に記載のヌクレオチド配列を含む、請求項1記載の核酸分子。
【請求項10】
請求項9記載の核酸分子を含んでなるベクター。
【請求項11】
請求項10記載のベクターを含んでなる宿主細胞。
【請求項12】
HPV31の組換えL1タンパク質または組換えL1+L2タンパク質から構成されるウイルス様粒子(VLP)。
【請求項13】
組換えL1タンパク質または組換えL1+L2タンパク質が酵母内で製造される、請求項12記載のVLP。
【請求項14】
組換えL1タンパク質または組換えL1+L2タンパク質がコドン最適化HPV31 L1核酸分子によりコードされる、請求項13記載のVLP。
【請求項15】
該コドン最適化核酸分子が、配列番号2または配列番号3に記載のヌクレオチド配列より実質的になる、請求項14記載のVLP。
【請求項16】
(a)HPV31 L1タンパク質またはHPV31 L1+L2タンパク質をコードするコドン最適化DNA分子で酵母を形質転換し、
(b)該コドン最適化DNA分子の発現を可能にする条件下、該形質転換酵母を培養して組換えパピローマウイルスタンパク質を得、
(c)該組換えパピローマウイルスタンパク質を単離して請求項14記載のVLPを得ることを含んでなる、請求項14記載のVLPの製造方法。
【請求項17】
請求項14記載のVLPを含んでなるワクチン。
【請求項18】
請求項14記載のVLPを含んでなる医薬組成物。
【請求項19】
請求項17記載のワクチンを哺乳動物に投与することを含んでなる、HPV感染の予防方法。
【請求項20】
請求項14記載のVLPを動物に投与することを含んでなる、動物において免疫応答を誘導するための方法。
【請求項21】
酵母が、サッカロミセス・セレビシエ(Saccharomyces cerevisiae)、ハンゼヌラ・ポリモルファ(Hansenula polymorpha)、ピチア・パストリス(Pichia pastoris)、クリーベルミセス・フラジリス(Kluyvermyces fragilis)、クルベロミセス・ラクチス(Kluveromyces lactis)およびシゾサッカロミセス・ポンベ(Schizosaccharomyces pombe)よりなる群から選ばれる、請求項14記載のウイルス様粒子。
【請求項22】
酵母がサッカロミセス・セレビシエ(Saccharomyces cerevisiae)である、請求項21のウイルス様粒子。
【請求項23】
少なくとも1つの追加的HPV型のVLPを更に含む、請求項17記載のワクチン。
【請求項24】
少なくとも1つの追加的HPV型が、HPV6、HPV11、HPV16、HPV18、HPV33、HPV35、HPV39、HPV45、HPV51、HPV52、HPV55、HPV56、HPV58、HPV59およびHPV68よりなる群から選ばれる、請求項23記載のワクチン。
【請求項25】
少なくとも1つのHPV型がHPV16を含む、請求項24記載のワクチン。
【請求項26】
HPV18 VLPを更に含む、請求項25記載のワクチン。
【請求項27】
HPV6 VLPおよびHPV11 VLPを更に含む、請求項26記載のワクチン。
【請求項28】
酵母により認識される転写終結シグナルを含有しない、HPV31 L1タンパク質をコードするヌクレオチド配列を含んでなる核酸分子。
【請求項29】
請求項28記載の核酸分子を含んでなるベクター。
【請求項30】
請求項29記載のベクターを含んでなる宿主細胞。
【請求項31】
宿主細胞が、サッカロミセス・セレビシエ(Saccharomyces cerevisiae)、ハンゼヌラ・ポリモルファ(Hansenula polymorpha)、ピチア・パストリス(Pichia pastoris)、クリーベルミセス・フラジリス(Kluyvermyces fragilis)、クルベロミセス・ラクチス(Kluveromyces lactis)およびシゾサッカロミセス・ポンベ(Schizosaccharomyces pombe)よりなる群から選ばれる、請求項30記載の宿主細胞。
【請求項32】
宿主細胞がサッカロミセス・セレビシエ(Saccharomyces cerevisiae)である、請求項31の宿主細胞。
【請求項33】
組換えL1タンパク質または組換えL1+L2タンパク質が、酵母により認識される転写終結シグナルを含有しないHPV31 L1核酸分子によりコードされる、請求項13記載のVLP。
【請求項34】
(a)酵母により認識される転写終結配列を含有しない、HPV31 L1タンパク質またはHPV31 L1+L2タンパク質をコードするDNA分子で酵母を形質転換し、
(b)該DNA分子の発現を可能にする条件下、該形質転換酵母を培養して組換えパピローマウイルスタンパク質を得、
(c)該組換えパピローマウイルスタンパク質を単離して請求項33記載のVLPを得ることを含んでなる、請求項33記載のVLPの製造方法。
【請求項35】
請求項33記載のVLPを含んでなるワクチン。
【請求項36】
請求項33記載のVLPを含んでなる医薬組成物。
【請求項37】
請求項35記載のワクチンを哺乳動物に投与することを含んでなる、HPV感染の予防方法。
【請求項38】
請求項33記載のVLPを動物に投与することを含んでなる、動物において免疫応答を誘導するための方法。
【請求項39】
少なくとも1つの追加的HPV型のVLPを更に含む、請求項35記載のワクチン。
【請求項40】
少なくとも1つの追加的HPV型が、HPV6、HPV11、HPV16、HPV18、HPV33、HPV35、HPV39、HPV45、HPV51、HPV52、HPV55、HPV56、HPV58、HPV59およびHPV68よりなる群から選ばれる、請求項39記載のワクチン。
【請求項41】
少なくとも1つのHPV型がHPV16を含む、請求項40記載のワクチン。
【請求項42】
HPV18 VLPを更に含む、請求項41記載のワクチン。
【請求項43】
HPV6 VLPおよびHPV11 VLPを更に含む、請求項42記載のワクチン。
【請求項1】
酵母細胞内での高レベル発現に関してコドン最適化された、配列番号4に記載のHPV31 L1タンパク質をコードするヌクレオチド配列を含んでなる核酸分子。
【請求項2】
請求項1記載の核酸分子を含んでなるベクター。
【請求項3】
請求項3記載のベクターを含んでなる宿主細胞。
【請求項4】
宿主細胞が、サッカロミセス・セレビシエ(Saccharomyces cerevisiae)、ハンゼヌラ・ポリモルファ(Hansenula polymorpha)、ピチア・パストリス(Pichia pastoris)、クリーベルミセス・フラジリス(Kluyvermyces fragilis)、クルベロミセス・ラクチス(Kluveromyces lactis)およびシゾサッカロミセス・ポンベ(Schizosaccharomyces pombe)よりなる群から選ばれる、請求項3記載の宿主細胞。
【請求項5】
宿主細胞がサッカロミセス・セレビシエ(Saccharomyces cerevisiae)である、請求項4記載の宿主細胞。
【請求項6】
ヌクレオチド配列が、配列番号2に記載のヌクレオチド配列を含む、請求項1記載の核酸分子。
【請求項7】
請求項6記載の核酸分子を含んでなるベクター。
【請求項8】
請求項7記載のベクターを含んでなる宿主細胞。
【請求項9】
ヌクレオチド配列が、配列番号3に記載のヌクレオチド配列を含む、請求項1記載の核酸分子。
【請求項10】
請求項9記載の核酸分子を含んでなるベクター。
【請求項11】
請求項10記載のベクターを含んでなる宿主細胞。
【請求項12】
HPV31の組換えL1タンパク質または組換えL1+L2タンパク質から構成されるウイルス様粒子(VLP)。
【請求項13】
組換えL1タンパク質または組換えL1+L2タンパク質が酵母内で製造される、請求項12記載のVLP。
【請求項14】
組換えL1タンパク質または組換えL1+L2タンパク質がコドン最適化HPV31 L1核酸分子によりコードされる、請求項13記載のVLP。
【請求項15】
該コドン最適化核酸分子が、配列番号2または配列番号3に記載のヌクレオチド配列より実質的になる、請求項14記載のVLP。
【請求項16】
(a)HPV31 L1タンパク質またはHPV31 L1+L2タンパク質をコードするコドン最適化DNA分子で酵母を形質転換し、
(b)該コドン最適化DNA分子の発現を可能にする条件下、該形質転換酵母を培養して組換えパピローマウイルスタンパク質を得、
(c)該組換えパピローマウイルスタンパク質を単離して請求項14記載のVLPを得ることを含んでなる、請求項14記載のVLPの製造方法。
【請求項17】
請求項14記載のVLPを含んでなるワクチン。
【請求項18】
請求項14記載のVLPを含んでなる医薬組成物。
【請求項19】
請求項17記載のワクチンを哺乳動物に投与することを含んでなる、HPV感染の予防方法。
【請求項20】
請求項14記載のVLPを動物に投与することを含んでなる、動物において免疫応答を誘導するための方法。
【請求項21】
酵母が、サッカロミセス・セレビシエ(Saccharomyces cerevisiae)、ハンゼヌラ・ポリモルファ(Hansenula polymorpha)、ピチア・パストリス(Pichia pastoris)、クリーベルミセス・フラジリス(Kluyvermyces fragilis)、クルベロミセス・ラクチス(Kluveromyces lactis)およびシゾサッカロミセス・ポンベ(Schizosaccharomyces pombe)よりなる群から選ばれる、請求項14記載のウイルス様粒子。
【請求項22】
酵母がサッカロミセス・セレビシエ(Saccharomyces cerevisiae)である、請求項21のウイルス様粒子。
【請求項23】
少なくとも1つの追加的HPV型のVLPを更に含む、請求項17記載のワクチン。
【請求項24】
少なくとも1つの追加的HPV型が、HPV6、HPV11、HPV16、HPV18、HPV33、HPV35、HPV39、HPV45、HPV51、HPV52、HPV55、HPV56、HPV58、HPV59およびHPV68よりなる群から選ばれる、請求項23記載のワクチン。
【請求項25】
少なくとも1つのHPV型がHPV16を含む、請求項24記載のワクチン。
【請求項26】
HPV18 VLPを更に含む、請求項25記載のワクチン。
【請求項27】
HPV6 VLPおよびHPV11 VLPを更に含む、請求項26記載のワクチン。
【請求項28】
酵母により認識される転写終結シグナルを含有しない、HPV31 L1タンパク質をコードするヌクレオチド配列を含んでなる核酸分子。
【請求項29】
請求項28記載の核酸分子を含んでなるベクター。
【請求項30】
請求項29記載のベクターを含んでなる宿主細胞。
【請求項31】
宿主細胞が、サッカロミセス・セレビシエ(Saccharomyces cerevisiae)、ハンゼヌラ・ポリモルファ(Hansenula polymorpha)、ピチア・パストリス(Pichia pastoris)、クリーベルミセス・フラジリス(Kluyvermyces fragilis)、クルベロミセス・ラクチス(Kluveromyces lactis)およびシゾサッカロミセス・ポンベ(Schizosaccharomyces pombe)よりなる群から選ばれる、請求項30記載の宿主細胞。
【請求項32】
宿主細胞がサッカロミセス・セレビシエ(Saccharomyces cerevisiae)である、請求項31の宿主細胞。
【請求項33】
組換えL1タンパク質または組換えL1+L2タンパク質が、酵母により認識される転写終結シグナルを含有しないHPV31 L1核酸分子によりコードされる、請求項13記載のVLP。
【請求項34】
(a)酵母により認識される転写終結配列を含有しない、HPV31 L1タンパク質またはHPV31 L1+L2タンパク質をコードするDNA分子で酵母を形質転換し、
(b)該DNA分子の発現を可能にする条件下、該形質転換酵母を培養して組換えパピローマウイルスタンパク質を得、
(c)該組換えパピローマウイルスタンパク質を単離して請求項33記載のVLPを得ることを含んでなる、請求項33記載のVLPの製造方法。
【請求項35】
請求項33記載のVLPを含んでなるワクチン。
【請求項36】
請求項33記載のVLPを含んでなる医薬組成物。
【請求項37】
請求項35記載のワクチンを哺乳動物に投与することを含んでなる、HPV感染の予防方法。
【請求項38】
請求項33記載のVLPを動物に投与することを含んでなる、動物において免疫応答を誘導するための方法。
【請求項39】
少なくとも1つの追加的HPV型のVLPを更に含む、請求項35記載のワクチン。
【請求項40】
少なくとも1つの追加的HPV型が、HPV6、HPV11、HPV16、HPV18、HPV33、HPV35、HPV39、HPV45、HPV51、HPV52、HPV55、HPV56、HPV58、HPV59およびHPV68よりなる群から選ばれる、請求項39記載のワクチン。
【請求項41】
少なくとも1つのHPV型がHPV16を含む、請求項40記載のワクチン。
【請求項42】
HPV18 VLPを更に含む、請求項41記載のワクチン。
【請求項43】
HPV6 VLPおよびHPV11 VLPを更に含む、請求項42記載のワクチン。
【図1A】


【図1B】

【図1C】

【図1D】
【図2A】

【図2B】
【図2C】
【図3】
【図4】
【図5】
【図6】
【図1B】
【図1C】
【図1D】
【図2A】
【図2B】
【図2C】
【図3】
【図4】
【図5】
【図6】
【公表番号】特表2006−523099(P2006−523099A)
【公表日】平成18年10月12日(2006.10.12)
【国際特許分類】
【出願番号】特願2006−507439(P2006−507439)
【出願日】平成16年3月19日(2004.3.19)
【国際出願番号】PCT/US2004/008677
【国際公開番号】WO2004/084831
【国際公開日】平成16年10月7日(2004.10.7)
【出願人】(390023526)メルク エンド カムパニー インコーポレーテッド (924)
【氏名又は名称原語表記】MERCK & COMPANY INCOPORATED
【Fターム(参考)】
【公表日】平成18年10月12日(2006.10.12)
【国際特許分類】
【出願日】平成16年3月19日(2004.3.19)
【国際出願番号】PCT/US2004/008677
【国際公開番号】WO2004/084831
【国際公開日】平成16年10月7日(2004.10.7)
【出願人】(390023526)メルク エンド カムパニー インコーポレーテッド (924)
【氏名又は名称原語表記】MERCK & COMPANY INCOPORATED
【Fターム(参考)】
[ Back to top ]