
酵母による非酵母ステロールの生成
本発明は、サッカロミセス・セレヴィシエ(Saccharomyces cerevisiae)などの酵母中における、7−デヒドロコレステロール、25−ヒドロキシ−7−デヒドロコレステロール、および25−ヒドロキシエルゴステロールの生成に関する。それはまた、ラノステロール、ジメチルチモステロール、メチルチモステロール、チモステロール、コレスタ−7,24−ジエノール、またはコレスタ−5,7,24−トリエノールの24位における二重結合の還元;またはエルゴステロール、7−デヒドロコレステロール、コレスタ−8−エノール、およびコレスタ−7−エノールの25位におけるヒドロキシル化を触媒する、様々な酵素にも関する。それはまた、コレステロールC25−ヒドロキシラーゼおよびステロールΔ24−還元酵素をコードする様々な核酸に関し、7−デヒドロコレステロールまたはエルゴステロールを生成してヒドロキシル化するためのそれらの使用にも関する。それはまた、このようにして生成された酵母株に関し、形質転換酵母細胞を培養するステップと、得られたステロールを収集するステップを含んでなる、これらのステロールを生成する方法にも関する。

【発明の詳細な説明】
【発明の詳細な説明】
【0001】
[技術分野]
本発明は、サッカロミセス・セレヴィシエ(Saccharomyces cerevisiae)などの酵母中における、7−デヒドロコレステロール、25−ヒドロキシ−7−デヒドロコレステロール、および25−ヒドロキシエルゴステロールをはじめとする、ステロールの生成に関する。それはまた、チモステロール、コレスタ−7,24−ジエノール、およびコレスタ−5,7,24−トリエノールの24位における二重結合の還元;またはエルゴステロール、7−デヒドロコレステロール、コレスタ−8−エノール、およびコレスタ−7−エノールの25位におけるヒドロキシル化を触媒する、様々な酵素にも関する。それはまた、コレステロールC25−ヒドロキシラーゼおよびステロールΔ24−還元酵素をコードする様々な核酸に関し、7−デヒドロコレステロールまたはエルゴステロールを生成してヒドロキシル化するためのそれらの使用にも関する。それはまた、このようにして生成された酵母株に関し、形質転換酵母細胞を培養するステップと、得られたステロールを収集するステップを含んでなる、これらのステロールを生成する方法にも関する。
【0002】
[背景技術]
酵母中のステロール経路には、多数の段階が関与する。酵母ステロール経路のダイアグラムを図14に示し、それを参照して、下で先行技術を考察する。
【0003】
米国特許第5,460,949号明細書(Amoco)は、酵母中でスクアレンとトリエノールの蓄積を増大させる方法、および組成物を教示する。この方法は、その中でERG5およびERG6遺伝子が不活性化されている株中における、HMG1(経路を制御する最重要遺伝子)のトランケーションと過剰発現を含む。ERG5およびERG6は2つの酵素をコードし、その活性は、コレステロール生合成と、酵母および真菌の主要ステロールであるエルゴステロール生合成とをはっきり区別する。これらの株は、好適には、通常はコレステロール生合成の中間体であるコレスタ−5,7,24−トリエノールを蓄積する。
【0004】
国際公開第03/064650号パンフレット(Langら)(米国特許出願公開第2006/0240508号明細書)は、酵母中において、7−デヒドロコレステロールおよび/または生合成中間体または引き続く生成物を生成する方法を開示する。明細書では、その中でERG5およびERG6が不活性化され、HMG1遺伝子がトランケートされ過剰発現される酵母株中で、マウスおよびヒトからのC−8ステロールイソメラーゼ、C−5ステロールデサチュラーゼ、およびステロールΔ24−還元酵素の遺伝子が発現される。これらの酵母は、7−デヒドロコレステロールを合成できる。
【0005】
国際公開第03/064652号パンフレット(Langら)(米国特許出願公開第2006/0088903号明細書)は、ラノステロール−C14−デメチラーゼ活性(ERG11)を増大させることにより、遺伝子導入酵母中で、チモステロールおよび/または生合成中間体および/またはその引き続く生成物を生成する方法を開示する。
【0006】
国際公開第2005/121315号パンフレット(Aventis)は、コレステロール産生酵母株、およびその使用を教示する。ERG6、そして場合によってはERG5が不活性化されている様々な酵母宿主株に、シロイヌナズナ(Arabidopsis thaliana)からのステロールΔ7還元酵素遺伝子、およびヒトステロールΔ24還元酵素遺伝子が導入された。新しい株は、他の中間体との混和材料中で、コレステロールを生成できた。
【0007】
米国特許第6,562,609号明細書(Russellら)は、単離されたヒトおよびマウスコレステロールC25−ヒドロキシラーゼ、および遺伝子を教示する。
【0008】
先行技術は、遺伝子導入酵母を使用した、25−ヒドロキシプロビタミンD3の生成についても25−ヒドロキシプロビタミンD2の生成についても沈黙している。
【0009】
[発明の詳細な説明]
本発明に従って、酵素コレステロールC25−ヒドロキシラーゼ(その基質は常態ではコレステロールである)は、7−デヒドロコレステロールおよびエルゴステロールに作用して、それぞれ25−ヒドロキシ−7−デヒドロコレステロールおよび25−ヒドロキシエルゴステロールを生成し得ることが分かった。
【0010】
したがって本発明の一態様は、それぞれ25−ヒドロキシ−7−デヒドロコレステロール(25−ヒドロキシプロビタミンD3とも称される)または25−ヒドロキシエルゴステロール(25−ヒドロキシプロビタミンD2とも称される)を生成するために、7−デヒドロコレステロールまたはエルゴステロールをヒドロキシル化する、脊椎動物コレステロールC25−ヒドロキシラーゼの使用である。
【0011】
コレステロールC25−ヒドロキシラーゼは、酵母中に天然に存在する酵素ではない。実際、酵母は、酵母中に存在し得る7−デヒドロコレステロールの側鎖をヒドロキシル化する能力を常態では有さない。しかし本発明に従って、酵母にコレステロールC25−ヒドロキシラーゼをコードする核酸を提供し、酵母が7−デヒドロコレステロールをヒドロキシル化する条件下で酵母を培養する工程により、遺伝子改変酵母が、コレスタ−5,7,24−トリエノールを25−ヒドロキシ−7−デヒドロコレステロール(25−ヒドロキシプロビタミンD3)に変換するようにでき得て、それによって25−ヒドロキシ−7−デヒドロコレステロール(25−ヒドロキシプロビタミンD3)が生成される。
【0012】
上の工程では、酵母の遺伝子ERG5およびERG6が不活性化されていること、そして酵母中での発現のために最適化された脊椎動物ステロールΔ24−還元酵素をコードする核酸もまた、それに提供されていることが好ましい。
【0013】
本発明に従って、酵母にコレステロールC25−ヒドロキシラーゼをコードする核酸を提供し、酵母がエルゴステロールをヒドロキシル化する条件下で酵母を培養する工程により、遺伝子改変酵母が、エルゴステロールを25−ヒドロキシエルゴステロール(25−ヒドロキシプロビタミンD2とも称される)に変換するようにでき得て、それによって25−ヒドロキシエルゴステロール(25−ヒドロキシプロビタミンD2)が生成されることもまた発見された。
【0014】
以下の図面では、次の略語を使用する。
TDH3pは、TDH3(S.セレヴィシエ(S.cerevisiae)からのグリセルアルデヒド−3−リン酸脱水素酵素イソ酵素3)のプロモーター領域であり;
Ori−pBRは、大腸菌(E.coli)のpBR322からのColE1複製起点であり;
blaは、大腸菌(E.coli)中でアンピシリン耐性を与えるβ−ラクタマーゼ遺伝子であり;
2μは、S.セレヴィシエ(S.cerevisiae)の複製起点を保有する2μ多コピー型ベクターの大型断片であり;
PGK1tは、PGK1(S.セレヴィシエ(S.cerevisiae)からの3−ホスホグリセリン酸キナーゼ)のターミネーター領域であり;
URA3は、ウラシルに対するS.セレヴィシエ(S.cerevisiae)URA3栄養要求性マーカーであり;
Bam HI、Eco RI、Hin dIII、Bgl II、Age Iは、対応する制限酵素の制限部位を指し;
ベクター上の部位の位置は、ユニークなBam HI部位の1位から始まる括弧によって示される。
【図面の簡単な説明】
【0015】
【図1】プラスミドV51TDH−S24R1の制限酵素地図である。S24R−1は、ドブネズミ(Rattus norvegicus)からの推定上のステロールΔ24−還元酵素をコードする合成遺伝子S24R1である。
【図2】プラスミドV51TDH−S24R2の制限酵素地図である。S24R−2は、ゼブラフィッシュ(Danio rerio)からの推定上のステロールΔ24−還元酵素をコードする合成遺伝子S24R2を表す。
【図3】プラスミドpFLAde−S24R1の制限酵素地図である。S24R1は、ドブネズミ(R.norvegicus)からの推定上のステロールΔ24−還元酵素をコードする合成遺伝子S24R1であり;ARS−CENは、S.セレヴィシエ(S.cerevisiae)の動原体自律的複製起点であり;ADE2は、S.セレヴィシエ(S.cerevisiae)ADE2遺伝子を表す。
【図4】プラスミドV51−C25H1の制限酵素地図である。GALpは、GAL10−CYC1(Guarenteら、1982年)ガラクトース誘導性プロモーター領域を表し;C25H−1は、イノシシ(Sus scrofa)からの推定上のコレステロールC25−ヒドロキシラーゼをコードする合成遺伝子C25H1を表す。
【図5】プラスミドV51−C25H3の制限酵素地図である。GALpは、GAL10−CYC1(Guarenteら、1982年)ガラクトース誘導性プロモーター領域を表し;C25H−3は、イヌ(Canis familiaris)からの推定上のコレステロールC25−ヒドロキシラーゼをコードする合成遺伝子C25H3を表す。UV282nmで記録された全てのHPLCプロフィールは、実施例8に記載されるようにして作成したステロール抽出物を用いて、実施例9に記載されるようにして実施した。略語:C5,7,22,24=コレスタ5,7,22,24−テトラエノール;C5,7,24=コレスタ5,7,24−トリエノール;C5,7,22=コレスタ5,7,22−トリエノール;C5,7=コレスタ5,7−ジエノールまたは7−デヒドロコレステロール;25OH−C5,7−Ac=25−ヒドロキシ7−デヒドロコレステロールアセテート;25OH−E5,7,22−Ac、25−ヒドロキシエルゴステロールアセテート;E5、7,22,24、エルゴスタ5,7,22,24−テトラエノール;E5,7,22、エルゴスタ5,7,22−トリエノール(エルゴステロール)。
【図6】推定上のステロールC−24ステロール還元酵素を発現する株からのステロール抽出物のHPLC溶出プロフィールである。以下の株を使用した:A:ERT/V51TDH対照;B:ERT/V51TDH−S24R1;C:ERT/V51TDH−S24R2;D:ERT/pFLAde−S24R1。
【図7】推定上のコレステロールC25−ヒドロキシラーゼを発現するerg6変異体からのステロール抽出物のHPLC溶出プロフィールである。以下の株を使用した:C:ERT/pFLAde−S24R1/V51;D:ERT/pFLAde−S24R1/V51−C25H1;E:ERT/pFLAde−S24R1/V51−C25H3。標準物質は次のとおりであった:A、コレスタ5,7−ジエノール(7−デヒドロコレステロール、Sigma−Aldrich,St.Louis,MO 63103,USAから購入);B、25−ヒドロキシ7−デヒドロコレステロールアセテート(実施例9に記載されるようにして調製)。
【図8】推定上のコレステロールC25−ヒドロキシラーゼを発現するerg6変異株からの単一ピークのUVスペクトルである。UVスペクトルは、実施例9に記載されるようにして、HPLC中にPDA検出器によって、205〜300nmの間で以下のピークについてオンライン測定した:A:11.8分における7−デヒドロコレステロール標準ピーク(図7、プロフィールA);B:8.5分における25−ヒドロキシ7−デヒドロコレステロールアセテート標準ピーク(図7プロフィールB);C:8.5分におけるERT/pFLAde−S24R1/V51−C25H1ピーク(図7、プロフィールD)。
【図9】推定上のコレステロールC25−ヒドロキシラーゼを発現するerg6変異株からの単一ピークの質量分析プロフィールである。m/z=350〜450の質量断片化プロフィールは、実施例9に記載されるようにして、HPLC中にMicro質量ZQ検出器によって、以下のピークについてオンライン測定した:A:11.8分における7−デヒドロコレステロール標準ピーク(図7、プロフィールA);B:8.5分における25−ヒドロキシ7−デヒドロコレステロールアセテート標準ピーク(図7、プロフィールB);C:8.5分におけるERT/pFLAde−S24R1/V51−C25H1ピーク(図7、プロフィールD)。
【図10】推定上のコレステロールC25−ヒドロキシラーゼを発現するERG6野性型株からのステロール抽出物のHPLC溶出プロフィールである。以下の株を使用した:B:W303/V51;C:W303/V51−C25H1;D:W303/V51−C25H3。 標準物質は次のとおりであった:A、エルゴステロール(エルゴスタ5,7,22−トリエノール)Sigma−Aldrich,St.Louis,MO 63103,USAから購入。
【図11】推定上のコレステロールC25−ヒドロキシラーゼを発現するERG6野性型株からの単一ピークのUVスペクトルである。UVスペクトルは、実施例9に記載されるようにして、HPLC中にPDA検出器によって、205〜300nmの間で以下のピークについてオンライン測定した:A:11.9分におけるエルゴステロール標準ピーク(図10、プロフィールA);B:10.0分におけるW303/V51−C25H1ピーク(図10、プロフィールC);C:8.7分におけるW303/V51−C25H1ピーク(図10、プロフィールC)。
【図12】推定上のコレステロールC25−ヒドロキシラーゼを発現するerg6野生型株からの単一ピークの質量分析プロフィールである。m/z=350〜450の質量断片化プロフィールは、実施例9に記載されるようにして、HPLC中にMicro質量ZQ検出器によって、以下のピークについてオンライン測定した:A:11.9分におけるエルゴステロール標準ピーク(図10、プロフィールA);B:10.0分におけるW303/V51−C25H1ピーク(図10、プロフィールC);C:8.7分におけるW303/V51−C25H1ピーク(図10、プロフィールC)。
【図13】ERG6が不活性化されている酵母(コレスタ−タイプステロール)と比較した、野生型酵母(エルゴスタ−タイプステロール)のステロール生合成経路の後半部分である。酵素は斜体で強調される。黒塗り矢印は、野生型酵母に見つかった酵素反応を表す。破線矢印は、異種活性(推定上のステロールΔ24−還元酵素、推定上のコレステロールC25−ヒドロキシラーゼ)によって触媒されるステップに対応し、対応する酵素には下線を引いた。
【図14】酵母ステロール経路のダイアグラムである。
【図15】10種の異なる25−ヒドロキシラーゼ遺伝子による、7−デヒドロコレステロールから25−ヒドロキシ−7−デヒドロコレステロール(HyDHC)への変換の比較である。以下のS.セレヴィシエ(S.cerevisiae)株を実施例20に記載されるようにして培養した。Ecab HY株(配列番号26の遺伝子を有する組み込みコンストラクト)、Ecab opt 2.0株(配列番号25の遺伝子を有する組み込みコンストラクト)、Ptro opt 2.0株(配列番号19の遺伝子を有する組み込みコンストラクト)、Ptro HY(配列番号20の遺伝子を有する組み込みコンストラクト)、ConHyD HY株(配列番号31の遺伝子を有する組み込みコンストラクト)、ConHyD opt 2.0株(配列番号30の遺伝子を有する組み込みコンストラクト遺伝子)、Rnor HY株(配列番号23の遺伝子を有する組み込みコンストラクト)、Rnor opt 2.0株(配列番号22の遺伝子を有する組み込みコンストラクト)、Sscr HY株(配列番号28の遺伝子を有する組み込みコンストラクト)、Sscr opt 2.0株(配列番号27の遺伝子を有する組み込みコンストラクト)、および実施例2に記載されるようにして最適化されたブタからの25−ヒドロキシラーゼを含有するScr Pompon株。細胞ペレットからステロールを単離し、HyDHC含量、総ステロールのHyDHC比率、および7−DHCからHyDHCへの変換を、25−ヒドロキシラーゼC25H1の組み込みによって得られる値と比較した。全ての新しいヒドロキシラーゼ遺伝子が発現され、その遺伝子産物は7−DHCをHyDHCにヒドロキシル化(hydroxylase)できた。
【0016】
[定義]
本明細書および請求の範囲の全体を通じて、以下の定義が適用される。
【0017】
「酵母」は、サッカロミセス・セレヴィシエ(Saccharomyces cerevisiae)、ならびに分裂酵母(Schizosaccharomyces)種、ピキア属(Pichia)種、クリヴェロミセス(Kluyveromyces)(Klyuveromyces)種、ハンゼヌラ(Hansenula)種、およびヤロウィア・リポリティカ(Yarrowia lipolytica)などの工業規模生産に適したその他の酵母を指す。
【0018】
ハイブリダイゼーションの「標準条件」とは、本発明の文脈で、特定のハイブリダイゼーションシグナルを検出するために当業者によって一般に使用され、例えば参照によって本明細書に援用する、Sambrookら,「Molecular Cloning」,Second Edition,Cold Spring Harbor Laboratory Press 1989年,New Yorkに記載される条件を意味する。「ストリンジェントなハイブリダイゼーション条件」の一例は、50%ホルムアミド、5×SSC(150mM NaCl、15mMクエン酸三ナトリウム)、50mMリン酸ナトリウム(pH7.6)、5×デンハルト液、10%硫酸デキストラン、および20μg/mlの変性剪断サケ精子DNAを含んでなる溶液中における、42℃で一晩(例えば15時間)のインキュベーションの後、0.1×SSC中で約65℃のハイブリダイゼーション担体の洗浄である。
【0019】
「%同一性」という用語は、当該技術分野で知られているように、場合に応じて、配列のストリング間の一致により判定される、ポリペプチドまたはポリヌクレオチド配列間の関連性の程度を意味する。「同一性」は、例えばプログラムBESTFIT(GCG Wisconsin Package, version 10.2,Accelrys Inc.,9685 Scranton Road,San Diego,CA 92121−3752,USA)によって、既知の方法によって、以下のパラメーターを使用して容易に判定し得る:gap creation penalty 8、gap extension penalty 2(デフォルトパラメーター)。
【0020】
「機能性」酵素とは、典型的な酵母培養条件下で、酵素が酵母内で所望の活性を発揮することを意味する。例えば「機能性コレステロールC25−ヒドロキシラーゼ」は、酵母細胞生体内で、コレステロールC25−ヒドロキシラーゼが、7−デヒドロコレステロールを25−ヒドロキシ−7−デヒドロコレステロールに変換することを意味する。
【0021】
「機能性」核酸配列とは、核酸配列が(ポリペプチドをコードするものの他に)、酵母が機能する濃度でポリペプチドを発現するのに必要な配列を含むことを意味する。このような配列は、プロモーター、終止配列、エンハンサーなどを包含し得る。
【0022】
[コレステロール25−ヒドロキシラーゼ]
したがって本発明の一態様は、7−デヒドロコレステロールまたはエルゴステロールをコレステロールC25−ヒドロキシラーゼに接触させるステップと、25−ヒドロキシ−7−デヒドロコレステロールまたは25−ヒドロキシエルゴステロールを得るステップを含んでなる、酵母細胞中で25−ヒドロキシ−7−デヒドロコレステロールまたは25−ヒドロキシエルゴステロールを生成する方法である。好適には、コレステロールC25−ヒドロキシラーゼは脊椎動物起源であり、より好適にはラット、ブタ、イヌ、ウマ、ヒト、チンパンジー、アガゲザル(Macaca mulatta)(Macaca mulata)、マウス、ニワトリ(Gallus gallus)、アフリカツメガエル(Xenopus laevis)、ゼブラフィッシュ(Danio rerio)またはカモノハシ(Ornithorhynchus anatinus)(Ornithorynchus anatinus)からの酵素である。さらに機能する能力を保持しさえすれば、これらの天然酵素の誘導体、すなわちアミノ酸配列が天然酵素ともはや同一でないものを使用してもよい。例えば、本発明は、天然脊椎動物C25−ヒドロキシラーゼと、少なくとも90%の同一性、好適には少なくとも95%の同一性、より好適には少なくとも99%の同一性を有する酵素の使用を包含するが、ただし誘導体酵素はその機能性を保持する。
【0023】
同様に、本発明で使用し得る核酸は、上記ペプチド配列をコードするもの、および上記ペプチド配列をコードするものとストリンジェントな条件下でハイブリダイズするものである。好適にはそれらは単離されている。さらに、いくつかの実施形態では、C25−ヒドロキシラーゼ酵素をコードする核酸は、酵母環境により良く適するようにコドン最適化されている。好ましい実施形態では、核酸は、ブタ、ラット、イヌ、チンパンジー、またはウマ起源であり、酵母のためにコドン最適化されている。特に好まれる核酸は、参照核酸配列「配列番号5のSscr Pompon」と少なくとも同程度に酵母宿主中で発現されるようにコドン最適化されているものであり、より好適には、核酸配列は、Sscr Pompon」よりもさらに良く発現される。
【0024】
核酸は、好適にはDNAである。
【0025】
特に好まれる核酸は、次のように称されるものである:
C25H1(ブタヒドロキシラーゼをベースとする;配列番号.5)、
C25H3(イヌヒドロキシラーゼをベースとする:配列番号8)
Ecab_HYまたはEcab25OH_opt2.0(ウマヒドロキシラーゼをベースとする:配列番号25および26)
ConHYD opt 20(配列番号30)
Rnor HY(ラットヒドロキシラーゼをベースとする:配列番号23)
Sscr HY(配列番号28)
Sscr opt 20(配列番号27)
Sscr Pompon(配列番号5)
【0026】
本発明の別の態様は、機能性コレステロールC25−ヒドロキシラーゼを含んでなる酵母細胞である。本発明のさらなる態様は、7−デヒドロコレステロールまたはエルゴステロールを、それぞれ25−ヒドロキシ7−デヒドロコレステロールまたは25−ヒドロキシエルゴステロールに変換できる酵母である。
【0027】
本発明のさらなる態様は、ヒドロキシル化反応を生じさせる培養条件下において、7−デヒドロコレステロールの存在下で、脊椎動物コレステロールC25−ヒドロキシラーゼを発現させ、25−ヒドロキシ7−デヒドロコレステロールの生成をもたらすステップを含んでなる、酵母中で25−ヒドロキシ7−デヒドロコレステロールを生成する方法である。本発明の追加的態様は、ヒドロキシル化反応を生じさせる培養条件下において、エルゴステロールの存在下で、脊椎動物コレステロールC25−ヒドロキシラーゼを発現させ、25−ヒドロキシエルゴステロールの生成をもたらすステップを含んでなる、酵母中で25−ヒドロキシエルゴステロールを生成する方法である。
【0028】
遺伝子の複数コピーが、発現ベクター中に存在することが好ましい。好ましい実施形態では、2〜5個のコピーが使用される。
【0029】
前述の遺伝子の何れかを酵母宿主にクローニングするためのベクターは、クローニングに有用なことが知られているあらゆるタイプ、特にC25−ヒドロキシラーゼ遺伝子と、当該技術分野で通常使用されるプロモーターおよびエンハンサーをはじめとする、随伴する発現および/または組み込み配列とを保有する、プラスミドおよび動原体プラスミド;および本発明の別の態様を構成するベクターであってもよい。
【0030】
米国特許出願公開第2006/0242508号明細書で記載されるものなどの7−デヒドロコレステロールを産生できる酵母中では、遺伝子ERG5およびERG6が不活性化され、下でさらに詳しく述べられるような脊椎動物起源のステロールΔ24−還元酵素が発現されることが好ましい。さらに状況によっては、このような酵母中でHMG1の切断型を過剰に発現することが有利である。不活性化ERG5およびERG6とトランケート型HMG1遺伝子とを有するこのような株については、当該技術分野記載されている。
【0031】
次に既知のUV光照射処置を使用して、25−ヒドロキシプロビタミンD3または25−ヒドロキシプロビタミンD2を、それぞれビタミンD3またはD2の第1の代謝産物であり循環形態である、25−ヒドロキシビタミンD3に変換し得る。25−ヒドロキシビタミンD3は骨組織形成において重要な役割を果たし、商業的に家禽および他の動物飼料中でビタミン栄養補給剤として使用される。それは商標ROVIMIX HY−D(登録商標)の下に、DSM Nutritional Productsから市販される。25−ヒドロキシビタミンD2は体内で25−ヒドロキシビタミンD3に匹敵する効果を示し、25−ヒドロキシビタミンD3の用法と同様の様式で使用し得る。
【0032】
[ステロールΔ24−還元酵素]
本発明の別の態様は、ステロールΔ24−還元酵素をコードする新規核酸であり、それは酵母によって発現され得て、ラノステロール、ジメチルチモステロール、メチルチモステロール、チモステロール、コレスタ−7,24−ジエノールまたはコレスタ−5,7,24−トリエノールからなる群から選択される基質を3β−ヒドロキシ−8−ラノスタ−8−エン、4,4−ジメチル−コレスタ−8−エノール、4α−メチル−コレスタ−8−エノール、コレスタ−8−エノール、ラソステロールおよび7−デヒドロコレステロールからなる群から選択される生成物に変換し得るが、ただし核酸配列はヒト核酸配列でもマウス核酸配列でもない。
【0033】
好適には、酵母細胞は、その中でERG6およびERG5が不活性化されているものである。好適には、これらの核酸は、宿主細胞のためにコドン最適化されている脊椎動物起源の修飾DNAであるが、ただしそれはマウスでもヒトでもない。好適には、ステロールΔ24−還元酵素は、ブタ、イヌ、チンパンジー、アガゲザル(Macaca mulatta)(Macaca mulata)、マウス、ラット、ウマ、ニワトリ(Gallus gallus)、アフリカツメガエル(Xenopus laevis)、ゼブラフィッシュ(Danio rerio)またはカモノハシ(Ornithorhynchus anatinus)(Ornithorynchus anatinus)起源である。
【0034】
特に好ましい実施形態では、これらは、ドブネズミ(Rattus norvegicus)またはゼブラフィッシュ(Danio rerio)DNAであり、およびより好ましい実施形態では、それらはサッカロミセス・セレヴィシエ(Saccharomyces cerevisiae)内での発現のためにコドン最適化されている。特に好まれる核酸は、S24R1(修飾ラット遺伝子、配列番号1のヌクレオチド10〜1563)およびS24R2(修飾ゼブラフィッシュ遺伝子、配列番号3のヌクレオチド10〜1563)と称される。
【0035】
本発明の別の態様は、脊椎動物核酸配列、好適にはDNAであり、それは酵母宿主中での発現のためにコドン最適化されており、それは酵母宿主細胞内で、酵素が、
ラノステロールを3β−ヒドロキシ−8−ラノスタ−8−エンに、またはジメチルチモステロールを4,4−ジメチル−コレスタ−8−エノールに;
メチルチモステロールを4α−メチル−コレスタ−8−エノールに;
チモステロールをコレスタ−8−エノールに;
コレスタ−7,24−ジエノールをラソステロールに;または
コレスタ−5,7,24−トリエノールを7−デヒドロコレステロール)に変換し得るように、機能性ステロールΔ24−還元酵素をコードし;
酵素をコードする核酸は、ストリンジェントな条件下でS24R1またはS24R2とハイブリダイズし得るが、ただし脊椎動物核酸配列はヒトから単離された拡散配列でもマウスから単離された核酸配列でもない。
【0036】
本発明の別の態様は、前述の核酸配列によってコードされる酵素である。本発明の好ましい酵素は、配列番号2および4で記載されるもの、
および配列番号2または4と少なくとも90%、および好適には95%の同一性を示すもの、および酵母細胞環境内で、
ラノステロールを3β−ヒドロキシ−8−ラノスタ−8−エンに、
ジメチルチモステロールを4,4−ジメチル−コレスタ−8−エノールに、
メチルチモステロールを4α−メチル−コレスタ−8−エノールに、
チモステロールをコレスタ−8−エノールに、
コレスタ−7,24−ジエノールをラソステロール、または
コレスタ−5,7,24−トリエノールを7−デヒドロコレステロールに
変換できるものである。
【0037】
本発明の別の態様は、上述のような機能性核酸配列(nucleic sequence)を含有する酵母宿主、特にS24R1、S24R2を含んでなる、またはストリンジェントな条件下でS24R1またはS24R2とハイブリダイズする核酸配列を含んでなる、S.セレヴィシエ(S.cerevisiae)などの酵母宿主であるが、ただし核酸はヒト配列でもマウス配列でもない。好ましい実施形態では、酵母宿主はERG5およびERG6が不活性化されており、特に好ましい実施形態では、酵母宿主はまたHMG1遺伝子の切断型を過剰発現する。
【0038】
さらに本発明の別の態様は、ラノステロール、ジメチルチモステロール、メチルチモステロール、チモステロール、コレスタ−7,24−ジエノールまたはコレスタ−5,7,24−トリエノールと、酵母宿主中での発現についてコドン最適化されており、機能性ステロールΔ24−還元酵素をコードする核酸配列を含んでなる酵母とを接触させるステップを含んでなる、酵母細胞内で3β−ヒドロキシ−8−ラノスタ−8−エン、4,4−ジメチル−コレスタ−8−エノール、4α−メチル−コレスタ−8−エノール、コレスタ−8−エノール、ラソステロールまたは7−デヒドロコレステロールを生成する方法であるが、ただし核酸配列はヒト核酸配列でもマウス核酸配列でもなく、酵母は、
ラノステロールを3β−ヒドロキシ−8−ラノスタ−8−エンに、
ジメチルチモステロールを4,4−ジメチル−コレスタ−8−エノールに、
メチルチモステロールを4α−メチル−コレスタ−8−エノールに、
チモステロールをコレスタ−8−エノールに、
コレスタ−7,24−ジエノールをラソステロールに、または
コレスタ−5,7,24−トリエノールを7−デヒドロコレステロールに変換する。この方法では、核酸は、好適にはS24R1、S24R2であり、またはストリンジェントな条件下でS24R1またはS24R2とハイブリダイズし得る核酸であるが、ただし脊椎動物核酸配列はヒト核酸配列でもマウス核酸配列でもない。
【0039】
本発明の別の態様は、酵母宿主中での発現についてコドン最適化されており、機能性ステロールΔ24−還元酵素をコードする機能性核酸配列を含んでなるプラスミドをはじめとするベクターを含むが、ただし核酸配列はヒト核酸配列でもマウス核酸配列でもない。好適には、ベクターは核酸S24R1、S24R2、またはストリンジェントな条件下でS24R1またはS24R2とハイブリダイズする核酸を含んでなるが、ただし脊椎動物核酸配列はヒト核酸配列でもマウス核酸配列でもない。ベクターは、転写、翻訳、および/または酵母染色体への組み込みを制御するプラスミドまたは他のベクター中に一般に見出される、通常の配列を含有してもよい。
【0040】
[遺伝子コンストラクト]
C25−ヒドロキシラーゼまたはステロールΔ24還元酵素をコードする本発明の遺伝子コンストラクトは全て、典型的には、関心のある遺伝子を制御する、ADH1プロモーター、TEF1プロモーター、酵母GPD(TDH3)プロモーター、酵母HXT7プロモーター、酵母GALl/10プロモーターまたは酵母PGK1プロモーターなどの標準法を使用して制御し得る、既知のプロモーター制御下にある外来遺伝子を一般的に有する、発現カセットの一部である。さらに発現カセットは、上述の遺伝子の少なくとも1つのコード配列と作動可能なように結合している、ターミネーター配列および/または他の調節要素などの上流および下流配列を含んでなってもよい。本発明の遺伝子コンストラクトを保有するベクターは、従来の方法を用いて酵母に導入される。
【0041】
以下の実施例は例証のみを意図するものであり、本発明をどのようにも制限するものではない。本出願全体を通じて引用される全ての参考文献、特許出願、特許、および公開特許公報の内容は、参照によって本明細書に援用する。
【実施例】
【0042】
[参考文献]
参考文献の完全な一覧は実施例の終わりに記載した。全ての参考文献は、参照によって本明細書に援用する。
【0043】
[培地]
【0044】
【表1】
【0045】
培地の利用に先だって、それを120℃で20分間の高圧蒸気滅菌によって滅菌した。必要ならば無菌トリプトファン、ウラシルまたはアデニン(b)を20mg/Lの最終濃度に添加した。
【0046】
【表2】
【0047】
使用前に、培地を0.2μmの濾過ユニットに通過させ、濾過して滅菌した。必要ならば、無菌のトリプトファン、ウラシルまたはアデニンを最終濃度0.2mg/mlに添加した。
【0048】
[実施例1]
[異なる生物からのΔ24(25)−ステロール還元酵素の選択および合成遺伝子設計]
ステロールΔ24−還元酵素は、コレステロール生合成の最終段階の1つを触媒する。それはかなり非特異的酵素であり、基質として少なくともチモステロールおよびデスモステロールを受け入れる。2004年には、対応する遺伝子がヒトで同定された(Waterman HRら,2001年)。ヒトおよびマウスからの遺伝子産物についてのみ、ステロールΔ24−還元酵素活性が正式に(formally)立証されている(Crameri Aら、2006年;国際公開第03/064650号パンフレット)。ラットからの相同遺伝子の匹敵する機能についてもまた記載されるが(Wu C.ら 2004年: Nature 432 (7017):640−5)、遺伝子産物の酵素活性は確認されていない。
【0049】
国際公開第03/064650号パンフレットでは、S.セレヴィシエ(S.cerevisiae)変異体(erg5 erg6)中における、マウスからのΔ24−還元酵素遺伝子の発現が示されており、宿主株中では生じない2つの新しいステロール(ラソステロールおよび7−デヒドロコレステロール)の形成がもたらされる。ヒトΔ24(25)−ステロール還元酵素と相同的な酵素についての、GENBANKまたはEMBLなどの公共データベースの検索は、標準条件下で、公的に利用可能なプログラムBLASTpを用いて実施した。見つかった配列間で、ヒト(Homo sapiens)アミノ酸配列と97%のアミノ酸同一性を示すドブネズミ(Rattus norvegicus)の配列(表1)、およびヒト配列と79%のアミノ酸同一性を示すゼブラフィッシュ(Danio rerio)の配列(表1)をさらなる調査のために選択した。
【0050】
【表3】
【0051】
各cDNAについて、S.セレヴィシエ(S.cerevisiae)中における発現のための合成遺伝子を設計した。一般に、この目的のために、元のcDNA配列を使用することが可能である。場合によっては、かずさDNA研究所(http://www.kazusa/or.jp/codon)の異なる種のコドン使用頻度表を使用して、元のcDNAのコドン使用頻度プロフィールを新しい宿主のコドン使用頻度に適合させる試みが有利である。これは手動で、またはコンピュータプログラムの助けを借りて実施し得る。本例では、コンピュータプログラムが使用され、その計算アルゴリズムは配列作成ルーチンを伴い、ユーザー定義のコドンバイアス調節があるDNA配列を作成できるようにする。アルゴリズム中にはまたいくつかの制約を導入して、特定の制限部位の存在を回避し、または代案としては随意に制限部位の存在を支持し得る。配列作成の一部は乱数発生ルーチンによって進められ、通常、試行毎に異なるDNA配列がもたらされる。プログラムのユーザーが使用された乱数種を回復することは可能でなく、したがって結果を再現することは可能でなかった。したがって再コードされた配列は予測され得ず、ユニークな元のシグネチャーを保有すると見なし得る。
【0052】
代案として作成されたDNA配列バージョン間の選択は、最終的にユーザー依存であった(外観検査)。選択されたDNA配列バージョンは、サービス提供者(例えばGeneCust Europe, 30b rue Dominique Lang,3505 Dudelange,Luxembourg)によって、遺伝子合成のために使用された。翻訳開始および終結を最適化するために、3個の追加的アデニン残基をATG開始コドン上流に付加し、第2の停止(STOP)コドンを読み取り枠下流に付加した。合成遺伝子の酵母発現ベクターへの都合よいクローニングのために、Bam HIおよびEco RI制限酵素部位をそれぞれ配列の上流および下流に付加した。上で説明したように、ラットステロールΔ24−還元酵素遺伝子(GenBank登録番号AY92220(cDNA配列)およびAAX29968(アミノ酸配列)を上述の方法によって再コードした。新しい合成遺伝子は、S24R1と命名された。その配列を下に示す。
配列番号1
【化1】
【0053】
対応するアミノ酸配列は次のとおり:
配列番号2
【化2】
【0054】
ゼブラフィッシュ(D.rerio)ステロールΔ24−還元酵素遺伝子(DNA配列はGenBank番号NM_001008645およびアミノ酸配列はGenBank number NP_001008645でアクセス可能)を上述の方法によって再コードした。新しい合成遺伝子をS24R2と命名した。その配列を下に示す。
配列番号3
【化3】
【0055】
対応するアミノ酸配列は次のとおり:
配列番号4
【化4】
【0056】
[実施例2]
[ヒト(Homo sapiens)からのコレステロールC25−ヒドロキシラーゼアミノ酸配列に相同的なアミノ酸配列の選択、およびS.セレヴィシエ(S.cerevisiae)中での発現について最適化された代案のDNA配列の計算]
ヒト(H.sapiens)およびハツカネズミ(M.musculus)遺伝子産物について、コレステロールC25−ヒドロキシラーゼ活性が示されている(Lundら,1998年;米国特許第6562609号明細書)。ヒトコレステロールC25−ヒドロキシラーゼと相同的な酵素についてのGENBANKまたはEMBLなどの公共データベースの検索は、標準条件下で、公的に入手可能なプログラムBLASTpを用いて実施した。見つかった配列間で、ヒト(H.sapiens)アミノ酸配列と82%のアミノ酸同一性、ハツカネズミ(M.musculus)配列と79%のアミノ酸同一性を示すイノシシ(S.scrofa)配列(表2)、およびヒトおよびマウス配列と70%のアミノ酸同一性を示すイヌ(C.familiaris)配列(表4)をさらなる調査のために選択した。
【0057】
【表4】
【0058】
酵母サッカロミセス・セレヴィシエ(Saccharomyces cerevisiae)によって滅多に使用されないコドンに相当する25個のコドンを、酵母中で頻繁に使用されるコドンで置き換えることにより、ブタコレステロールC25−ヒドロキシラーゼ遺伝子(DNA配列はGenBank番号AY974088で、アミノ酸配列はGenBank番号Q4G1G8でアクセスできる)を再コードした(Zhangら,1991年)。新しい合成遺伝子をC25H1と称した。その配列を下に示す。
配列番号5
【化5】
【0059】
対応するアミノ酸配列は、次のとおり。
配列番号6
【化6】
【0060】
使用された配列と4つの違いを示す、データベースGENBANKからの推定上のブタステロールC25−ヒドロキシラーゼの代案のアミノ酸配列もまた利用できるが、実験でさらに使用することはなかった。
配列番号7
【化7】
【0061】
イヌ(Canis familiaris)からの遺伝子(ヌクレオチド配列はGenBank番号XM_546596.2で、アミノ酸配列はGenBank番号XP_543596.1でアクセスできる)を実施例1に記載されるようにして、適合法によって再コードした。新しい合成遺伝子をC25H3と命名し、そのDNA配列を下に示す。
配列番号8
【化8】
【0062】
対応するアミノ酸配列は次のとおり。
配列番号9
【化9】
【0063】
[実施例3]
[発現プラスミドV51TDH−S24R1の構築]
発現ベクターは、Sambrookら(1989年)によって記載されるような古典的分子生物学技術を使用して構築され、DNA制限酵素および修飾酵素は、供給元(New England Biolabs,Ipswich,MA 01938−2723,USA)が推奨するようにして(as recommender)使用された。
【0064】
基礎的な多コピー構成的発現ベクターV51TDHは、ガラクトース誘導性GAL10−CYC1プロモーターを構成的に発現するTDH3遺伝子(サッカロミセス属(Saccharomyces)ゲノムデータベース中の系統的遺伝子注釈からのYGR192C遺伝子、グリセルアルデヒド−3−リン酸脱水素酵素イソ酵素3をコードする)からのプロモーターで置換して、V51ベクターから誘導された。V51は、大腸菌(Escherichia coli)中における維持のためのColE1複製起点およびbla遺伝子(アンピシリン耐性)、酵母中における維持のための2μ起点およびURA3遺伝子、多重クローニング部位(MCS)(pYeDP1/8−2、CullinおよびPompon,1988年)によって隔てられるGAL10−CYC1ハイブリッドプロモーターおよびPGK1ターミネーター領域を保有する。TDH3プロモーター領域は、W303−1B酵母株(ThomasおよびRothstein,1989年)からのゲノムDNA上でPCRによって増幅され、HoffmanおよびWinston(1987年)に従って、以下のプライマーを使用して調製された。
【化10】
【0065】
PCR増幅産物をAge IおよびBam HIによって消化し、GAL10−CYC1プロモーターを保有するV51からの617bp長さのAge I−Bam HI断片を置換するのに使用した。得られたV51TDHベクターは、TDH3プロモーター(TDH3p)とPGK1ターミネーター(PGK1t)領域の間に局在するMCSに挿入することで、酵母中において関心のある遺伝子を構成的発現させるのに有用な、多コピー大腸菌(E.coli)/S.セレヴィシエ(S.cerevisiae)シャトルベクターである。
【0066】
S24R1合成遺伝子(実施例1参照)の構成的発現のためのV51TDH−S24R1ベクターは、Bam HIおよびEco RIで制限酵素消化されたV51TDHベクターに、S24R1合成遺伝子からの1564bpのBam HI−Eco RI制限酵素産物を挿入して構築された。
【0067】
V51TDH−S24R1構成的発現ベクターの制限酵素地図を図1に示す。
【0068】
[実施例4]
[発現プラスミドV51TDH−S24R2の構築]
S24R2の構成的発現のためのV51TDH−S24R2ベクター(実施例1参照)は、基本的に、実施例3で既に述べたようにして構築された。Bam HIおよびEco RIで制限酵素消化されたV51TDHベクターに、合成遺伝子S24R2の1564bp長さのBam HI−Eco RI制限酵素産物をライゲートした。V51TDH−S24R2の制限酵素地図を図2に示す。
【0069】
[実施例5]
[動原体発現プラスミドpFLAde−S24R1の構築]
酵母動原体および自律複製配列(ARSCEN)およびADE2選択マーカーを保有する空のpFLAde大腸菌(E.coli)/S.セレヴィシエ(S.cerevisiae)シャトルベクターに、V51TDH−S24R1(実施例3に記載)からのTDH3p−S24R1−PGK1t発現カセットを挿入することにより、遺伝子S24R1の構成的発現のための動原体pFLAde−S24R1ベクター(実施例1参照)を構築した。pFLAdeベクターは、以下のプライマーを使用して、URA3遺伝子をコードするpFL38の1.1kbのBgl II断片(Bonneaudら,1991年)を、酵母ゲノムDNA上でのPCR増幅によって得られるADE2遺伝子をコードする2.7kbのBam HI断片で置換して構築された。
【化11】
【0070】
TDH3p−S24R1−PGK1t発現カセットは、標準条件下で、PrimeSTAR HS DNAポリメラーゼ(日本国滋賀県大津市のタカラバイオ株式会社)および以下のプライマーを使用して、PCRによって増幅された。
【化12】
【0071】
増幅産物をpCR2.1−TOPO(Invitrogen,Carlsbad,CA 92008,USA)にクローンした。Sac I−Pst I制限酵素消化と、同一制限酵素によって切断されたpFLAdeベクターへのライゲーションによって、TDH3p−S24R1−PGK1tカセットを後者のベクター(later vector)から単離した。結果として得られたpFLAde−S24R1動原体発現ベクターの制限酵素地図を図3に示す。
【0072】
[実施例6]
[発現ベクターV51−C25H1の構築]
C25H1からの826bpのBam HI−Eco RI断片をV51に挿入し、Bam HIおよびEco RIで消化して、C25H1(実施例2参照)のガラクトース誘導性発現のためのV51−C25H1プラスミドを構築した。得られたV51−C25H1発現プラスミドは、ガラクトース誘導性GAL10−CYC1ハイブリッドプロモーター(Guarenteら,1982年)およびPGK1ターミネーターの制御下にある合成遺伝子C25H1をコードする。V51−C25H1の制限酵素地図を図4に示す。
【0073】
[実施例7]
[発現ベクターV51−C25H3の構築]
合成遺伝子C25H3(実施例2参照)のガラクトース誘導性発現のためのV51−C25H3ベクターは、実施例6に既に記載したようにして、Bam HIおよびEco RIで消化されたV51に、合成遺伝子C25H3の826bpのBam HI−Eco RI断片を挿入して構築した。得られた発現プラスミドV51−C25H3は、ガラクトース誘導性GAL10−CYC1ハイブリッドプロモーターおよびPGK1ターミネーターの制御下にある、C25H3をコードする。V51−C25H3の制限酵素地図を図5に示す。
【0074】
[実施例8]
[酵母株形質転換およびステロール抽出]
上述の異なる発現プラスミドによって、W303−1B株(MATalpha;ura3−52;trp1−1;leu2−3,112;his3−11;ade2−1;can1−100;ThomasおよびRothstein,1989年)、およびそのerg6変異体ERT(MATalpha;erg6::TRP1;ura3−52;trp1−1;leu2−3,112;his3−11;ade2−1;can1−100)を形質転換した。ERT株のERG6の不活性化は、ERG6に機能性TRP1遺伝子を挿入して得られた。pFL45からのTRP1遺伝子(Bonneaudら,1991年)は、ERG6配列が隣接するTRP1プライマーを使用して、PCRによって増幅した。プライマーの配列は以下のとおりである。
ERGTRP−1:
【化13】
【0075】
Gietzら(1995年)によって記載されるリチウム−PEG技術を使用して、得られたPCR産物を使用し、W303−1B株を形質転換した。トリプトファンを含まない規定培地上で形質転換体を選択し、それらのステロール組成物をHPLCによって確認し(Lecainら,1996年)、ERG6不活性化の既知の効果と一致する、エルゴステロールの不在および独特なステロールの存在を確認した。1つのerg6::TRP1形質転換体をさらなる研究のために保存し、ERTと命名した。
【0076】
ERTに加えて、S.セレヴィシエ(S.cerevisiae)野性型BY4742株(遺伝子型MATalpha;his3Δ1;leu2Δ0;lys2Δ0;ura3Δ0;Brachmannら,1998年)もまた、構築されたプラスミドのための宿主として使用した。
【0077】
BY4742、W303−1B、およびERTの形質転換は、Gietzら(1995年)によって記載されるリチウム−PEG法によって実施した。形質転換体は、ベクター上に存在する選択マーカーに応じて、ウラシルまたはアデニンを含まない定義された最少培地(表1)上で選択した。各形質転換の少なくとも2つの独立した形質転換体を分析した。ステロール分析のためには、形質転換体を合成培地(表2に記載)中で、定常期になるまで培養した。
【0078】
酵母細胞を培養媒体から遠心分離して回収し、等容積の脱イオン水で洗浄した。細胞を2mlエッペンドルフ管中の100μLの水に再懸濁して、0.1gガラスビーズ(0.5mm)の存在下、強力なボルテックス(5分間)によって破砕した。2mlの1,2−ジクロロエタンを再懸濁細胞に添加して、混合物を5分間激しくボルテックスした。10,000rpmで5分間遠心分離した後、溶媒相をガラス管に移した。抽出を2回実施した。プールされた有機相を窒素下で蒸発させた。最終段階として、HPLC分析のために乾燥抽出物を100μLのアセトニトリルに再懸濁した。
【0079】
[実施例9]
[UVおよび質量分析検出を用いた高速液体クロマトグラフィー(HPLC)によるステロール分析]
通常30μLである、実施例8に記載されるようにして調製されたステロール抽出物のアリコートを、UVおよび質量分析検出によってモニターされる逆相HPLCによって分析した。「Waters Alliance HT 2790」HPLCは、「Waters PDA 996」タイプからの光ダイオードアレーUV検出器に、そして質量検出器「Waters MicroMass ZQ」(Waters,Milford,MA01757,USA)に接続されていた。分離は、XTerra RP18 3.5μm 4.6×100mmカラム(Waters,Milford,MA01757,USA)上で、水/アセトニトリル勾配を含有する直線的0.03%(v/v)ギ酸を用いて、60℃で実施した。80%の緩衝液A(0.03%ギ酸添加水)と20%の緩衝液B(0.03%のギ酸添加アセトニトリル)とを含有する緩衝液で、カラムを安定化させた後に、サンプルを注入した。最初の5分間、緩衝液組成を80%の緩衝液Aおよび20%の緩衝液Bから、25%の緩衝液Aおよび75%の緩衝液Bに徐々に変化させた。次の15分間、緩衝液組成を25%の緩衝液Aおよび75%の緩衝液Bから、12.5%の緩衝液Aおよび87.5%の緩衝液Bに徐々に変化させた。最後の数分で、分離プロトコル100%の緩衝液Bに達した。この値を2分間保って、カラムを再生させた。
【0080】
光ダイオードアレー検出器によって205〜300nmの間で、クロマトグラフィーをモニターした。質量検出器は、m/zが350〜450の範囲の陽性エレクトロスプレーによって、1m/z単位の感度での検出が可能であった。
【0081】
ステロールは、標準と比較した、それらのクロマトグラフィーの挙動、それらのUV吸収スペクトル、およびそれらの質量断片化プロフィールに基づいて同定された(表5)。SecenおよびKalpar(1999年)に従って、ピリジン存在下で、無水酢酸によって、25−ヒドロキシ7−デヒドロコレステロール標準(DSMによって提供される)を25−ヒドロキシ−7−デヒドロコレステロールアセテート(25OH−7DHC−Ac)にアセチル化した。0.5mgの乾燥25−ヒドロキシ7−デヒドロコレステロールを1mlの無水酢酸/ピリジン溶液(50/50v/v)に再懸濁して、55℃で3時間加熱した。反応混合物を4mlの水で希釈した。HPLC分析前に、実施例8に記載されるようにして、0.5mlの希釈反応混合物を1,2−ジクロロエタンで抽出した。
【0082】
上述の条件下で分析すると、非アセチル化25−ヒドロキシ−7−デヒドロコレステロール標準(RT6.9分)と比較して、25OH−7DHC−Ac標準は、アセチル化化合物のより高い疎水性のために、滞留時間(RT8.5分)にシフトを示した。25OH−7DHC−Ac標準の質量断片化プロフィールは、非アセチル化25−ヒドロキシ7−デヒドロコレステロール標準のプロフィールと同様であった。アセチル化形態に対応するm/zシグナルは、恐らく質量検出器内での検出中に起きる脱アセチル化のために、検出されなかった。
【0083】
【表5】
【0084】
[実施例10]
[ERG6−欠損S.セレヴィシエ(S.cerevisiae)ERT株中におけるプラスミドV51TDH−S24R1からのS24R1遺伝子の発現]
プラスミドV51TDH−S24R1(実施例3参照)および対照として空ベクターV51TDHをリチウム−PEG技術(Gietzら,1995年)によって、ERT株に形質転換した。形質転換体をウラシルを含まない定義された最少培地上で選択した。各形質転換からの4つの独立した形質転換体をプラスミドの存在について分析した。確認された形質転換体の1つを無作為に選択し、それぞれERT/V51TDHおよびERT/V51TDH−S24R1と命名して、さらなる分析のために使用した。
【0085】
株が定常期になるまで、2%(w/v)グルコースKapelli培地(表2)中で培養し、実施例8に記載されるようにして全ステロールを抽出し、実施例9に記載されるようにして、UVおよび質量分析検出を用いたHPLCによって分析した。それぞれの標準(表5)を用いて、主要ステロールをそれらの滞留時間、質量、およびスペクトル諸特性によって同定した。
【0086】
HPLC溶出プロフィール(282nmでのUV検出)を図6に示す。コレスタ−5,7,22,24−テトラエノール(RT9.2分、m/z=363、最大UV233nm)が、ERT/V51TDH erg6対照株中に見つかった主要ステロールであり、282nmにおけるUVシグナルの70%に相当した。282nmで特定された総ステロールの7%は、コレスタ5,7,24−トリエノール(RT10.3分、m/z=365、最大UV282nm;図6−A)と同定された。
【0087】
S24R1遺伝子を発現するERT/V51TDH−S24R1株(図6−B)のステロールプロフィールからは、対照ERT株/V51TDH中で検出されなかった2つのステロール、すなわち本株の主要ステロールである(282nmにおけるUVシグナルの60%)コレスタ5,7,22−トリエノール(RT10.8分、m/z=365、最大UV282nm)、および7−デヒドロコレステロール(RT11.8、m/z=367、最大UV282nm)とも称されるコレスタ5,7−ジエノールの存在が明らかになった。コレスタ5,7,24−トリエノールおよびコレスタ5,7,22,24−テトラエノールのΔ24−25結合の還元によって生じたコレスタ5,7,−ジエノールおよびコレスタ5,7,22−トリエノールの存在は、遺伝子S24R1のステロールΔ24−還元酵素活性を裏付けた(図13参照)。
【0088】
[実施例11]
[S.セレヴィシエ(S.cerevisiae)ERT株中におけるプラスミドV51TDH−S24R2からの遺伝子S24R2の発現]
リチウム−PEGプロトコル(Gietzら,1995年)を使用して、S24R2を構成的に発現するプラスミドV51TDH−S24R2(実施例3)をERT株に形質転換する。形質転換体をウラシルを含まない最少培地上で選択した。4つの独立した形質転換体が分析され、同一諸特性を示した。それらの1つをERT/V51TDH−S24R2と命名し、さらなる分析のために使用した。
【0089】
株が定常期に達するまで、2%グルコースKapelli培地(表2)中で培養し、実施例8、9、および10に記載されるようにして、ステロールを抽出し分析した。この例でも、対照ERT/V51TDH株中では、主要ステロールとしてコレスタ5,7,22,24−テトラエノール(RT9.2分、m/z=363、最大UV233nm)およびコレスタ5,7,24−トリエノール(RT10.3分、m/z=365、最大UV282nm)が見つかったのに対し(図6−A)、S24R2を発現するERT/V51TDH−S24R2株中で検出された主要ステロールは、282nmにおけるUVシグナルの70%であった、コレスタ5,7,22−トリエノール(RT10.8分、m/z=365、最大UV282nm)であった(図6−C)。コレスタ5,7−ジエノール(7−デヒドロコレステロール;RT11.8、m/z=367、最大UV282nm)は、より低濃度(UVシグナルの13%)で見つかった。これらの結果は、S24R2遺伝子のステロールΔ24−還元酵素活性を裏付けた。
【0090】
[実施例12]
[S.セレヴィシエ(S.cerevisiae)ERT株中における動原体プラスミドpFLAde−S24R1からの遺伝子S24R1の発現]
リチウム−PEG手順(Gietzら,1995年)を使用して、S24R1遺伝子の構成的発現のための動原体プラスミドpFLAde−S24R1(実施例5)をERT株に形質転換した。形質転換体をアデニンを含まない規定培地上で選択した。4つの独立した形質転換体をプラスミドpFLAde−S24R1の発生と、それらのステロールパターンについて分析した。それらの1つを無作為に選択してERT/pFLAde−S24R1と命名し、結果のさらなる例証のために使用した。
【0091】
ERT/pFLAde−S24R1を2%(w/v)グルコースKapelli培地(表2)中で、定常期に達するまで培養した。実施例8および9に記載されるようにして、ステロールを抽出し分析した。対照中に存在しない、ERT/pFLAde−S24R1中の主要な新しいステロールは、282nmにおけるUVシグナルの60%であった、コレスタ5,7,22−トリエノール(RT10.8分、m/z=365、最大UV282nm)であった(図6−D)。より少量(UVシグナルの7%)の7−デヒドロコレステロール(RT11.8、m/z=367、最大UV282nm)もまた検出された。これらの新しいステロールの他に、ERT対照株(図6−A)中にもまた存在する、コレスタ5,7,22,24−テトラエノール(RT9.2分、m/z=363、最大UV233nm)およびコレスタ5,7,24(RT10.3分、m/z=365、最大UV282nm)もまたERT/pFLAde−S24R1に見つかった。これらの結果は、動原体プラスミドpFLAde−S24R1から発現されたS24R1遺伝子のステロールΔ24−還元酵素活性を裏付ける。
【0092】
[実施例13]
[S.セレヴィシエ(S.cerevisiae)ERT株中における遺伝子S24R1およびC25H1の同時発現による25−ヒドロキシ−7−デヒドロコレステロールアセテートの生成]
リチウム−PEG法(Gietzら,1995)によって、遺伝子C25H1のガラクトース誘導性発現のためのプラスミドV51−C25H1(実施例6)、および対照としてのプラスミドV51を(実施例12に記載されるようにして)ERT株/pFLAde−S24R1に形質転換した。V51−C25H1およびpFLAde−S24R1は、適合性プラスミドである。それらの異なる2μおよびARS−CEN起源は、酵母中でそれらが同時発生できるようにする。形質転換体をウラシルおよびアデニンを含まない最少培地上で選択した。各形質転換からの4つの独立した形質転換体が分析され、同一諸特性を示した。各形質転換から1つの形質転換体を無作為に選択し、それぞれERT/pFLAde−S24R1/V51−C25H1およびERT/pFLAde−S24R1/V51と命名し、さらなる分析のために使用した。
【0093】
株を2%ガラクトースKapelli培地(表2)中で、定常期に達するまで培養した。ステロールを抽出し、実施例8および9に記載されるようにして、実施例10に記載される条件下で、UVおよび質量分析検出を用いたHPLCによって分析した。対照ERT/pFLAde−S24R1/V51株(図7−C)で見出されたように、ERT/pFLAde−S24R1/V51−C25H1(図7−D)中の主要ステロールは、最終ステロールコレスタ5,7,22,24−テトラエノールのステロールΔ24−還元酵素による還元によって生じた、コレスタ5,7,22−トリエノール(RT10.8分、m/z=365、最大UV282nm)であった。標準として注入された調製25−ヒドロキシ7−デヒドロコレステロールアセテート(25OH−7DHC−Ac)と同時に溶出される(RT8.5分、図7−B)、対照株中に存在しない新しい化合物が、ERT/pFLAde−S24R1/V51−C25H1株(図7−D)中に観察された。この化合物もまた、調製25−ヒドロキシ7−デヒドロコレステロールアセテート(図8−B)と同一のUV吸収プロフィール(図8−C)を示し、7−デヒドロコレステロール標準(図8−A)でも観察された、Δ5−Δ7共役二重結合(double bounds)に特異的な282nm前後の二重ピークがあった。新しい化合物は、(実施例9に記載されるようにして実施した)オンライン質量分析による判定で、25−ヒドロキシ7−デヒドロコレステロールアセテート(図9−B)と同一の質量断片化プロフィール(図9−C)もまた示した。
【0094】
3β−ヒドロキシステロールの場合、エレクトロスプレー(electrospay)分析は、分子量から17を差し引いた所で主要シグナルを生じた(プロトン付加(+1)および位置C3(−18)での脱水。7−デヒドロコレステロール(図9−A)の主要シグナルは、m/z質量367にある(384(7−デヒドロコレステロールMW)+1(プロトン付加)−18(3β−ヒドロキシステロール脱水))。25−ヒドロキシ7−デヒドロコレステロールアセテートで観察されたm/z質量383におけるシグナルは、位置C25(367+16=383)における追加的ヒドロキシル化に対応する。m/z質量365における主要シグナルは、質量検出器内での検出中に、分子のごく一部に生じる位置C25での脱水(383−18=365)に対応する。これらの異なる結果(標準として使用される25−ヒドロキシ7−デヒドロコレステロールアセテートと同一滞留時間、同一UVプロフィール、および同一質量断片化)は、ERT/pFLAde−S24R1/V51−C25H1株中で生成された新しい化合物が、25−ヒドロキシ7−デヒドロコレステロールアセテートであったことを実証した。
【0095】
[実施例14]
[S.セレヴィシエ(S.cerevisiae)ERT株中におけるS24R1およびC25H3の同時発現による25−ヒドロキシ−7−デヒドロコレステロールアセテートの生成]
リチウム−PEG法(Gietzら,1995年)によって、遺伝子C25H3のガラクトース誘導性発現のために構築されたV51−C25H3(実施例7)をERT/pFLAde−S24R1株(実施例13参照)に形質転換した。形質転換体をウラシルおよびアデニンを含まない最少培地上で選択した。双方のプラスミドの発生確認のために、4つの独立した形質転換体を分析した。それらの1つを無作為に選択してERT/pFLAde−S24R1/V51−C25H3と命名し、さらなる研究のために使用した。
【0096】
ERT/pFLAde−S24R1/V51−C25H3を2%ガラクトースKapelli培地(表2)中で、定常期まで培養した。収集細胞からステロールを抽出し、実施例8および9に記載されるようにして、実施例10に記載される条件下で、UVおよび質量分析検出を用いたHPLCによって分析した。対照ERT/pFLAde−S24R1/V51株(図7−C)で見つかったように、ERT/pFLAde−S24R1/V51−C25H3(図7−E)中の主要ステロールは、実施例13に記載されるように、最終ステロールコレスタ5,7,22,24−テトラエノールのステロールΔ24−還元酵素による還元産物である、コレスタ5,7,22−トリエノール(RT10.8分、m/z=365、最大UV282nm)であった。
【0097】
25−ヒドロキシ7−デヒドロコレステロールアセテート(RT8.5分、図7−B)と同時に溶出される、親株中に存在しない化合物が、ERT/pFLAde−S24R1/V51−C25H3(図7−E)中に見つかった。ERT/pFLAde−S24R1/V51−C25H1(実施例13に記載)の場合のように、この化合物は、25−ヒドロキシ7−デヒドロコレステロールアセテートと同一のUV吸収プロフィール(図8)、および同一の質量断片化プロフィール(図9)を示した。これらの結果(滞留時間、UVプロフィール、および質量断片化が25−ヒドロキシ7−デヒドロコレステロールアセテートに匹敵する)は、ERT/pFLAde−S24R1/V51−C25H3中に生成された新しい化合物が、実施例13と同様、25−ヒドロキシ7−デヒドロコレステロールアセテートであることを実証する。
【0098】
[実施例15]
[S.セレヴィシエ(S.cerevisiae)W303−1B株およびBY4742株中におけるC25H1発現による25−ヒドロキシ−エルゴステロールアセテートの生成]
リチウム−PEG法(Gietzら,1995年)によって、遺伝子C25H1のガラクトース誘導性発現のために構築されたプラスミドV51−C25H1(実施例6)、および対照としての空ベクターV51をW303−1B株(実施例8に記載)に形質転換した。形質転換体をウラシルを含まない最少培地上で選択した。各形質転換からの4つの独立した形質転換体をプラスミドの発生について分析した。各形質転換から1つの陽性形質転換体を無作為に選択して、それぞれW303/V51−C25H1およびW303/V51と命名し、さらなる研究のために使用した。
【0099】
株を2%ガラクトースKapelli培地(表2)中で、定常期に達するまで培養した。収集細胞からステロールを抽出し、実施例8および9に記載されるようにして、実施例10に記載される条件下で、UVおよび質量分析検出を用いたHPLCによって分析した。HPLC溶出プロフィール(282nmでのUV検出)を図10に示す。エルゴステロールの滞留時間(図10−A)、theUVスペクトル(図11−A)、および質量断片化プロフィール(図12−A)(RT11.9分、m/z=379、最大UV282nm)との比較によって実証されるように、W303/V51−C25H1(図10−C)およびW303/V51(図10−B)の双方の株について観察された主要シグナルは、S.セレヴィシエ(S.cerevisiae)野生型株の主要ステロール(mayor sterol)であるエルゴステロール(エルゴスタ5,7,22−トリエノール)に対応する。
【0100】
W303/V51−C25H1株(図10−C)の10分における微量シグナルは、W303/V51対照株(図10−B)では痕跡シグナルとしてのみ観察された。オンラインPDA検出によって測定されたこのピークのUVプロフィール(図11−B)は、282nm前後の二重ピークによって特徴付けられるΔ5およびΔ7の共役二重結合に加えて、Δ22およびΔ24の共役二重結合に特異的である233nmにおける最大吸収を示した。恐らくこのシグナルは、Erg4pによるΔ24の還元がないエルゴステロール生合成経路の最後の中間体の1つである、エルゴスタ5,7,22,24−テトラエノールに対応する(図13参照)。オンライン質量分析法(図12−B)によって検出されたm/z質量377は、エルゴスタ−テトラエノール構造に一致する。
【0101】
W303/V51−C25H1株(図10−C)のステロール抽出物中で、HPLCによって観察された8.7分における第3のシグナルは、W303/V51(図10−B)では完全に欠如していた。このピーク(図11−C)について測定されたUVプロフィールは、Δ5およびΔ7共役二重結合(double bounds)に特異的な最大282nmの典型的な二重ピークを示した。質量断片化(図12−C)は、m/z=377における主要シグナル、およびm/z=395における微量のシグナルを示し、水酸化型エルゴステロール(379+16=395)およびその脱水形態(395−18=377)に一致する。断片化プロフィール(図12−C)は、全ての質量が、7−デヒドロコレステロール(mz=367)およびエルゴステロール(m/z=379)間の質量差に相当する12質量単位増大していたこと以外は、25−ヒドロキシ7−デヒドロコレステロールアセテート(図9−B)の1つと同一であった。この新しい化合物(RT8.7分)とエルゴステロール(RT11.9分)の間の3.2分の滞留時間の差は、25−ヒドロキシ7−デヒドロコレステロールアセテート(RT8.5分)と7−デヒドロコレステロール、(RT11.8)の滞留時間の差と同一であった。
【0102】
これらの諸特性(UVプロフィール、質量断片化、滞留時間)は全て、W303/V51−C25H1中のC25H1発現によって生成されるこの新しい化合物が、25−ヒドロキシ−エルゴステロールアセテートであるという結論に矛盾しない。
【0103】
V51−C25H1および対照としての空ベクターV51のS.セレヴィシエ(S.cerevisiae)株BY4742への形質転換は、W303/V51−C25H1株およびW303/V51株で得られたものに匹敵する結果(図10)をもたらした。この場合もV51−C25H1形質転換体について、HPLC(UV検出)によって8.7分における追加的シグナルが観察された。化合物は、W303/C25H1中で検出された化合物と同一UVプロフィール(図11)および質量断片化プロフィール(図12)を示し、この化合物もまた25−ヒドロキシ−エルゴステロールアセテートであることを強く示唆する。
【0104】
[実施例16]
[S.セレヴィシエ(S.cerevisiae)W303−1B株中における遺伝子C25H3の発現による25−ヒドロキシ−エルゴステロールアセテートの生成]
リチウム−PEG法(Gietzら,1995年)を使用して、合成遺伝子C25H3のガラクトース誘導性発現のために構築されたプラスミドV51−C25H3(実施例7)をW303−1B株(実施例8に記載)に形質転換した。形質転換体をウラシルを含まない最少培地上で選択した。4つの独立した形質転換体をV51−C25H3の発生について分析した。1つの陽性形質転換体をW303/V51−C25H3と命名し、生成されるステロールのさらなる研究のために使用した。
【0105】
株が定常期になるまで、2%(w/v)ガラクトースKapelli培地(表2)中で培養した。収集細胞からステロールを抽出し、実施例8および9に記載されるようにして、実施例10に記載される条件下で、UVおよび質量分析検出を用いたHPLCによって分析した。
【0106】
図10−Dに示すHPLC(282nmでのUV検出)によるステロール抽出物分析は、W303/V51−C25H3のステロールパターンが、W303/V51−C25H1株(実施例15)のステロールパターンと非常に良く似ていることを示し、エルゴステロール(RT11.9分)およびエルゴスタ5,7,22,24−テトラエノール(RT10.0分)に加えて、新しい化合物(RT8.7分)が生成されていた。オンラインUV PDA検出(図11)および質量分析法(図12)は、例えばΔ5−Δ7の特異的共役二重結合UVプロフィール、m/z=377における主要シグナルおよびm/z=395における微量シグナルがある質量断片化、および25−ヒドロキシル化ステロールと一致するHPLC滞留時間遅延など、この新しい化合物が、W303/V51−C25H1によって生成される化合物と同一諸特性を有することを示した。これらの特徴(UVプロフィール、質量断片化、滞留時間)は全て、この新しい化合物がC25H3の発現に起因する25−ヒドロキシ−エルゴステロールアセテートであることを強力に示唆する。
【0107】
[実施例17]
[主にコレスタ−5,7,24−トリエノールを産生する酵母株へのC25H1およびC25H3の導入]
リチウム−PEG法(Gietzら,1995年)を用いて、遺伝子C25H1およびC25H3のガラクトース誘導性発現のためのプラスミドV51−C25H1(実施例6)およびプラスミドV51−CH25H3(実施例7)、および対照としてのプラスミドV51をERT erg5 erg6株(実施例12に記載;ERG5は遺伝子URA3の組み込みによって不活性化されていた)に形質転換した。形質転換体をウラシルおよびアデニンを含まない最少培地上で選択した。各形質転換からの4つの独立した形質転換体が分析され、同一諸特性を示した。各形質転換からの1形質転換体を無作為に選択して、それぞれERT erg5 erg6/V51−C25H1、ERT erg5 erg6/V51−C25H3、およびERT erg5 erg6/V51と命名し、さらなる分析のために使用した。
【0108】
株を2%ガラクトースKapelli培地(表2)中で、定常期に達するまで培養した。ステロールを抽出し、実施例8および9に記載されるようにして、実施例10に記載される条件下で、UVおよび質量分析検出を用いたHPLCによって分析した。3株間でステロールパターンの違いは検出されず、2種のヒドロキシラーゼがコレスタ−5,7,24−トリエノールに対して活性でないことを裏付けた。
【0109】
[実施例18]
[追加的コレステロール25−ヒドロキシラーゼ遺伝子の選択]
イノシシ(S.scrofa)およびイヌ(C.familiaris)(Canis lupus)、からの選択された推定上のコレステロール25−ヒドロキシラーゼ遺伝子に加えて、ドブネズミ(R.norvegicus)(配列番号21、Genebank登録番号NP_001020586、XP_220063)、チンパンジー(P.troglodytes)(配列番号18、遺伝子bank登録番号XP_507901)、およびウマ(E.caballus)(配列番号24、Genebank登録番号XP_001503057)からの相同遺伝子もまたより詳細に調べた。
【0110】
選択された各25−ヒドロキシラーゼのS.セレヴィシエ(S.cerevisiae)中における発現のための2つの異なるコドン最適化遺伝子を計算した(配列番号19、20、22、23、25、26)。最後に接尾語「_opt」が付いている遺伝子は全て、DNA 2.0社(MenloPark,CA,USA)によって、独自仕様のアルゴリズムを使用してコドン最適化された。接尾語「_HY」がある全遺伝子は、常に、配列中に生じるアミノ酸の各位置に、各アミノ酸の最も一般的なコドンを配置させて作成した。酵母のための典型的なコドン使用頻度表は公的に入手可能であり、またプログラムパッケージLasergeneの一部でもある。
【0111】
イノシシ(S.scrofa)からの25−ヒドロキシラーゼの2つの新しい最適化された遺伝子(実施例2参照)もまた、上で述べたのと同一方法を使用して計算した(配列番号27、28)。
【0112】
インプットとして既知のアルゴリズムClustalWを用いて計算されるアミノ酸配列アラインメントを使用して、各位置に最も豊富なアミノ酸を単に選択することで、共通アミノ酸配列を判定した。「最も豊富なアミノ酸」は、判定できない場合は、それぞれの位置のアミノ酸を恣意的に選択した。このConHyDアミノ酸配列(配列番号29)もまた、上述のようにして、2つの異なるDNA配列(配列番号30、31)に逆翻訳した。
配列番号18:チンパンジー(P.troglodytes)からの推定上のコレステロール25−ヒドロキシラーゼのアミノ酸配列。
【化14】
配列番号19:DNA配列チンパンジー(P.troglodytes)CH25OH_opt 2.0
【化15】
配列番号20:チンパンジー(P.troglodytes)CH25OH_HYのDNA配列
【化16】
配列番号21:ドブネズミ(R.norvegicus)からの推定上のコレステロール25−ヒドロキシラーゼのアミノ酸配列
【化17】
配列番号22:ドブネズミ(R.norvegicus)CH25OH_opt 2.0のDNA配列
【化18】
配列番号23:ドブネズミ(R.norvegicus)CH25OH_HYのDNA配列
【化19】
配列番号24:ウマ(E.caballus)からの推定上のコレステロール25−ヒドロキシラーゼのアミノ酸配列
【化20】
配列番号25:ウマ(E.caballus)CH25OH_opt 2.0のDNA配列
【化21】
配列番号26:ウマ(E.caballus)CH25OH_HYのDNA配列
【化22】
配列番号27:イノシシ(S.scrofa)CH25OH_opt 2.0のDNA配列
【化23】
配列番号28:イノシシ(S.scrofa)CH25OH_HYのDNA配列
【化24】
配列番号29:ConHyDのアミノ酸配列
【化25】
配列番号30:ConHyD_opt 2.0のDNA配列
【化26】
配列番号31:ConHyD_HYのDNA配列
【化27】
【0113】
[実施例19]
[ERG6に組み込まれた発現コンストラクトからの新しい25−ヒドロキシラーゼ遺伝子の発現]
実施例18に示す新しい遺伝子をS.セレヴィシエ(S.cerevisiae)SC0554(erg5、erg6、TRP1−pARC300D、URA3−pARC304S、ARE1::TDH3p−S24R2−PGKt−URA3;2つのプラスミドに関する説明は米国特許第5,460,949号明細書を参照されたい)のERG6遺伝子座に組み込んだ。
【0114】
組み込みコンストラクトの配列を下に示す。関心のあるヒドロキシラーゼを組み込みコンストラクトのBam HIおよびEco RI部位の間にクローンした。ERG6組み込みコンストラクトは、プラスミドV51TDHに見出されるような、発現のためのS.セレヴィシエ(S.cerevisiae)のTDH3プロモーターおよびPGKターミネーター、および選択のためのS.セレヴィシエ(S.cerevisiae)のURA3を保有した。培養、形質転換、および選択は上述のように実施した。
【0115】
最終株は、次のように称された。
S.セレヴィシエ(S.cerevisiae)Ecab HY(配列番号26の遺伝子を有する組み込みコンストラクト)、
Ecab opt 2.0(配列番号25の遺伝子を有する組み込みコンストラクト)、
Ptro opt 2.0(配列番号19の遺伝子を有する組み込みコンストラクト)、
Ptro HY(配列番号20の遺伝子を有する組み込みコンストラクト)、
ConHyD HY(配列番号31の遺伝子を有する組み込みコンストラクト)、
ConHyD opt 2.0(配列番号30の遺伝子を有する組み込みコンストラクト)、
Rnor HY(配列番号23の遺伝子を有する組み込みコンストラクト)、
Rnor opt 2.0(配列番号22の遺伝子を有する組み込みコンストラクト)、
Sscr HY(配列番号28の遺伝子を有する組み込みコンストラクト)、
Sscr opt 2.0(配列番号27の遺伝子を有する組み込みコンストラクト)、および
実施例2に記載されるようにして最適化されたブタからの25−ヒドロキシラーゼを保有するScr Pompon。
【0116】
配列番号32:ERG6組み込みコンストラクトのDNA配列
【化28】
【0117】
[実施例20]
[新しいHyDHC株の振盪フラスコアッセイ]
実施例19に記載されるようにして作成された株を震盪フラスコ内で培養し、新たに設計された推定上の25−ヒドロキシラーゼ遺伝子の過剰発現効果を評価した。20mlのKapelli培地(実施例1)に接種するために、4mlのYPD培地中の一晩培養物を使用した。24および48時間培養した後に10g/Lのグルコース、32および56時間後に20g/Lのグルコースを供給した。72時間の培養時間後、細胞を遠沈して鹸化し、実施例9に記載されるようにしてステロール含量を測定した。図15に示すように、全ての新しいヒドロキシラーゼ遺伝子は、S.セレヴィシエ(S.cerevisiae)中で発現され、効率は様々であったが、遺伝子産物は全て7−DHCをHyDHCにヒドロキシル化できた。
【0118】
文献、それぞれが参照により本明細書に組み込まれる:
Brachmann C.B., Davies A., Cost G.J., Caputo E., Li J., Hieter P., Boeke J.D. (1998年). Yeast 14(2): 115−132.
Crameri A., Biondi E., Kuehnle K., Lutjohann D., Thelen K.M., Perga S., Dotti C.G., Nitsch R.M., Ledesma M.D., Mohajeri M.H. (2006年). EMBO J. 25(2):432−43.
Cullin C., Pompon D. (1988年). Gene 65:203−217.
Bonneaud N., Ozier−Kalogeropoulos O., Li G.Y., Labouesse M., Minvielle−Sebastia L., Lacroute F. (1991年). Yeast 7:609−615.
Gaber R.F., Copple D.M., Kennedy B.K., Vidal M, Bard M. (1989年). Mol Cell Biol 9(8):3447−3456.
Guarente L., Yocum R.R., Gifford P. (1982年). Proc Natl Acad Sci USA 79:7410−7414.
Hoffman C.S. and Winston F. (1987年). Gene 57:267−272
Gietz R.D., Schiestl R.H., Willems A.R., Woods R.A. (1995年). Yeast 11:355−360.
Lecain E., Chenivesse X., Spagnoli R., Pompon D. (1996年). J Biol Chem 271(18):10866−10873.
Lund E.G., Kerr T.A., Sakai J., Li W.P., Russell D.W. (1998年). J Biol Chem 273(51):34316−34327.
Sambrook J., Fritsch E. F., Maniatis T. (1998年). Molecular cloning: a laboratory manual. 2nd ed. Cold Spring Harbor, N.Y: Cold Spring Harbor Laboratory.
Secen H., Kalpar H. (1999年). Turk J Chem 23: 27−30.
Thomas B.J. and Rothstein R. (1989年). Cell 56(4):619−630.
Waterham H.R., Koster J., Romeijn G.J., Hennekam R.C.M., Vreken P., Andersson H.C., FitzPatrick D.R., Kelley R.I., Wanders R.J.A. (2001年). Am J Hum Genet 69:685−694.
Wu C., Miloslavskaya I., Demontis S., Maestro R., Galaktionov K. (2004年). Nature 432 (7017):640−5.
Zhang S.P., Zubay G., Goldman E. (1991年). Gene 1005:61−72.
【発明の詳細な説明】
【0001】
[技術分野]
本発明は、サッカロミセス・セレヴィシエ(Saccharomyces cerevisiae)などの酵母中における、7−デヒドロコレステロール、25−ヒドロキシ−7−デヒドロコレステロール、および25−ヒドロキシエルゴステロールをはじめとする、ステロールの生成に関する。それはまた、チモステロール、コレスタ−7,24−ジエノール、およびコレスタ−5,7,24−トリエノールの24位における二重結合の還元;またはエルゴステロール、7−デヒドロコレステロール、コレスタ−8−エノール、およびコレスタ−7−エノールの25位におけるヒドロキシル化を触媒する、様々な酵素にも関する。それはまた、コレステロールC25−ヒドロキシラーゼおよびステロールΔ24−還元酵素をコードする様々な核酸に関し、7−デヒドロコレステロールまたはエルゴステロールを生成してヒドロキシル化するためのそれらの使用にも関する。それはまた、このようにして生成された酵母株に関し、形質転換酵母細胞を培養するステップと、得られたステロールを収集するステップを含んでなる、これらのステロールを生成する方法にも関する。
【0002】
[背景技術]
酵母中のステロール経路には、多数の段階が関与する。酵母ステロール経路のダイアグラムを図14に示し、それを参照して、下で先行技術を考察する。
【0003】
米国特許第5,460,949号明細書(Amoco)は、酵母中でスクアレンとトリエノールの蓄積を増大させる方法、および組成物を教示する。この方法は、その中でERG5およびERG6遺伝子が不活性化されている株中における、HMG1(経路を制御する最重要遺伝子)のトランケーションと過剰発現を含む。ERG5およびERG6は2つの酵素をコードし、その活性は、コレステロール生合成と、酵母および真菌の主要ステロールであるエルゴステロール生合成とをはっきり区別する。これらの株は、好適には、通常はコレステロール生合成の中間体であるコレスタ−5,7,24−トリエノールを蓄積する。
【0004】
国際公開第03/064650号パンフレット(Langら)(米国特許出願公開第2006/0240508号明細書)は、酵母中において、7−デヒドロコレステロールおよび/または生合成中間体または引き続く生成物を生成する方法を開示する。明細書では、その中でERG5およびERG6が不活性化され、HMG1遺伝子がトランケートされ過剰発現される酵母株中で、マウスおよびヒトからのC−8ステロールイソメラーゼ、C−5ステロールデサチュラーゼ、およびステロールΔ24−還元酵素の遺伝子が発現される。これらの酵母は、7−デヒドロコレステロールを合成できる。
【0005】
国際公開第03/064652号パンフレット(Langら)(米国特許出願公開第2006/0088903号明細書)は、ラノステロール−C14−デメチラーゼ活性(ERG11)を増大させることにより、遺伝子導入酵母中で、チモステロールおよび/または生合成中間体および/またはその引き続く生成物を生成する方法を開示する。
【0006】
国際公開第2005/121315号パンフレット(Aventis)は、コレステロール産生酵母株、およびその使用を教示する。ERG6、そして場合によってはERG5が不活性化されている様々な酵母宿主株に、シロイヌナズナ(Arabidopsis thaliana)からのステロールΔ7還元酵素遺伝子、およびヒトステロールΔ24還元酵素遺伝子が導入された。新しい株は、他の中間体との混和材料中で、コレステロールを生成できた。
【0007】
米国特許第6,562,609号明細書(Russellら)は、単離されたヒトおよびマウスコレステロールC25−ヒドロキシラーゼ、および遺伝子を教示する。
【0008】
先行技術は、遺伝子導入酵母を使用した、25−ヒドロキシプロビタミンD3の生成についても25−ヒドロキシプロビタミンD2の生成についても沈黙している。
【0009】
[発明の詳細な説明]
本発明に従って、酵素コレステロールC25−ヒドロキシラーゼ(その基質は常態ではコレステロールである)は、7−デヒドロコレステロールおよびエルゴステロールに作用して、それぞれ25−ヒドロキシ−7−デヒドロコレステロールおよび25−ヒドロキシエルゴステロールを生成し得ることが分かった。
【0010】
したがって本発明の一態様は、それぞれ25−ヒドロキシ−7−デヒドロコレステロール(25−ヒドロキシプロビタミンD3とも称される)または25−ヒドロキシエルゴステロール(25−ヒドロキシプロビタミンD2とも称される)を生成するために、7−デヒドロコレステロールまたはエルゴステロールをヒドロキシル化する、脊椎動物コレステロールC25−ヒドロキシラーゼの使用である。
【0011】
コレステロールC25−ヒドロキシラーゼは、酵母中に天然に存在する酵素ではない。実際、酵母は、酵母中に存在し得る7−デヒドロコレステロールの側鎖をヒドロキシル化する能力を常態では有さない。しかし本発明に従って、酵母にコレステロールC25−ヒドロキシラーゼをコードする核酸を提供し、酵母が7−デヒドロコレステロールをヒドロキシル化する条件下で酵母を培養する工程により、遺伝子改変酵母が、コレスタ−5,7,24−トリエノールを25−ヒドロキシ−7−デヒドロコレステロール(25−ヒドロキシプロビタミンD3)に変換するようにでき得て、それによって25−ヒドロキシ−7−デヒドロコレステロール(25−ヒドロキシプロビタミンD3)が生成される。
【0012】
上の工程では、酵母の遺伝子ERG5およびERG6が不活性化されていること、そして酵母中での発現のために最適化された脊椎動物ステロールΔ24−還元酵素をコードする核酸もまた、それに提供されていることが好ましい。
【0013】
本発明に従って、酵母にコレステロールC25−ヒドロキシラーゼをコードする核酸を提供し、酵母がエルゴステロールをヒドロキシル化する条件下で酵母を培養する工程により、遺伝子改変酵母が、エルゴステロールを25−ヒドロキシエルゴステロール(25−ヒドロキシプロビタミンD2とも称される)に変換するようにでき得て、それによって25−ヒドロキシエルゴステロール(25−ヒドロキシプロビタミンD2)が生成されることもまた発見された。
【0014】
以下の図面では、次の略語を使用する。
TDH3pは、TDH3(S.セレヴィシエ(S.cerevisiae)からのグリセルアルデヒド−3−リン酸脱水素酵素イソ酵素3)のプロモーター領域であり;
Ori−pBRは、大腸菌(E.coli)のpBR322からのColE1複製起点であり;
blaは、大腸菌(E.coli)中でアンピシリン耐性を与えるβ−ラクタマーゼ遺伝子であり;
2μは、S.セレヴィシエ(S.cerevisiae)の複製起点を保有する2μ多コピー型ベクターの大型断片であり;
PGK1tは、PGK1(S.セレヴィシエ(S.cerevisiae)からの3−ホスホグリセリン酸キナーゼ)のターミネーター領域であり;
URA3は、ウラシルに対するS.セレヴィシエ(S.cerevisiae)URA3栄養要求性マーカーであり;
Bam HI、Eco RI、Hin dIII、Bgl II、Age Iは、対応する制限酵素の制限部位を指し;
ベクター上の部位の位置は、ユニークなBam HI部位の1位から始まる括弧によって示される。
【図面の簡単な説明】
【0015】
【図1】プラスミドV51TDH−S24R1の制限酵素地図である。S24R−1は、ドブネズミ(Rattus norvegicus)からの推定上のステロールΔ24−還元酵素をコードする合成遺伝子S24R1である。
【図2】プラスミドV51TDH−S24R2の制限酵素地図である。S24R−2は、ゼブラフィッシュ(Danio rerio)からの推定上のステロールΔ24−還元酵素をコードする合成遺伝子S24R2を表す。
【図3】プラスミドpFLAde−S24R1の制限酵素地図である。S24R1は、ドブネズミ(R.norvegicus)からの推定上のステロールΔ24−還元酵素をコードする合成遺伝子S24R1であり;ARS−CENは、S.セレヴィシエ(S.cerevisiae)の動原体自律的複製起点であり;ADE2は、S.セレヴィシエ(S.cerevisiae)ADE2遺伝子を表す。
【図4】プラスミドV51−C25H1の制限酵素地図である。GALpは、GAL10−CYC1(Guarenteら、1982年)ガラクトース誘導性プロモーター領域を表し;C25H−1は、イノシシ(Sus scrofa)からの推定上のコレステロールC25−ヒドロキシラーゼをコードする合成遺伝子C25H1を表す。
【図5】プラスミドV51−C25H3の制限酵素地図である。GALpは、GAL10−CYC1(Guarenteら、1982年)ガラクトース誘導性プロモーター領域を表し;C25H−3は、イヌ(Canis familiaris)からの推定上のコレステロールC25−ヒドロキシラーゼをコードする合成遺伝子C25H3を表す。UV282nmで記録された全てのHPLCプロフィールは、実施例8に記載されるようにして作成したステロール抽出物を用いて、実施例9に記載されるようにして実施した。略語:C5,7,22,24=コレスタ5,7,22,24−テトラエノール;C5,7,24=コレスタ5,7,24−トリエノール;C5,7,22=コレスタ5,7,22−トリエノール;C5,7=コレスタ5,7−ジエノールまたは7−デヒドロコレステロール;25OH−C5,7−Ac=25−ヒドロキシ7−デヒドロコレステロールアセテート;25OH−E5,7,22−Ac、25−ヒドロキシエルゴステロールアセテート;E5、7,22,24、エルゴスタ5,7,22,24−テトラエノール;E5,7,22、エルゴスタ5,7,22−トリエノール(エルゴステロール)。
【図6】推定上のステロールC−24ステロール還元酵素を発現する株からのステロール抽出物のHPLC溶出プロフィールである。以下の株を使用した:A:ERT/V51TDH対照;B:ERT/V51TDH−S24R1;C:ERT/V51TDH−S24R2;D:ERT/pFLAde−S24R1。
【図7】推定上のコレステロールC25−ヒドロキシラーゼを発現するerg6変異体からのステロール抽出物のHPLC溶出プロフィールである。以下の株を使用した:C:ERT/pFLAde−S24R1/V51;D:ERT/pFLAde−S24R1/V51−C25H1;E:ERT/pFLAde−S24R1/V51−C25H3。標準物質は次のとおりであった:A、コレスタ5,7−ジエノール(7−デヒドロコレステロール、Sigma−Aldrich,St.Louis,MO 63103,USAから購入);B、25−ヒドロキシ7−デヒドロコレステロールアセテート(実施例9に記載されるようにして調製)。
【図8】推定上のコレステロールC25−ヒドロキシラーゼを発現するerg6変異株からの単一ピークのUVスペクトルである。UVスペクトルは、実施例9に記載されるようにして、HPLC中にPDA検出器によって、205〜300nmの間で以下のピークについてオンライン測定した:A:11.8分における7−デヒドロコレステロール標準ピーク(図7、プロフィールA);B:8.5分における25−ヒドロキシ7−デヒドロコレステロールアセテート標準ピーク(図7プロフィールB);C:8.5分におけるERT/pFLAde−S24R1/V51−C25H1ピーク(図7、プロフィールD)。
【図9】推定上のコレステロールC25−ヒドロキシラーゼを発現するerg6変異株からの単一ピークの質量分析プロフィールである。m/z=350〜450の質量断片化プロフィールは、実施例9に記載されるようにして、HPLC中にMicro質量ZQ検出器によって、以下のピークについてオンライン測定した:A:11.8分における7−デヒドロコレステロール標準ピーク(図7、プロフィールA);B:8.5分における25−ヒドロキシ7−デヒドロコレステロールアセテート標準ピーク(図7、プロフィールB);C:8.5分におけるERT/pFLAde−S24R1/V51−C25H1ピーク(図7、プロフィールD)。
【図10】推定上のコレステロールC25−ヒドロキシラーゼを発現するERG6野性型株からのステロール抽出物のHPLC溶出プロフィールである。以下の株を使用した:B:W303/V51;C:W303/V51−C25H1;D:W303/V51−C25H3。 標準物質は次のとおりであった:A、エルゴステロール(エルゴスタ5,7,22−トリエノール)Sigma−Aldrich,St.Louis,MO 63103,USAから購入。
【図11】推定上のコレステロールC25−ヒドロキシラーゼを発現するERG6野性型株からの単一ピークのUVスペクトルである。UVスペクトルは、実施例9に記載されるようにして、HPLC中にPDA検出器によって、205〜300nmの間で以下のピークについてオンライン測定した:A:11.9分におけるエルゴステロール標準ピーク(図10、プロフィールA);B:10.0分におけるW303/V51−C25H1ピーク(図10、プロフィールC);C:8.7分におけるW303/V51−C25H1ピーク(図10、プロフィールC)。
【図12】推定上のコレステロールC25−ヒドロキシラーゼを発現するerg6野生型株からの単一ピークの質量分析プロフィールである。m/z=350〜450の質量断片化プロフィールは、実施例9に記載されるようにして、HPLC中にMicro質量ZQ検出器によって、以下のピークについてオンライン測定した:A:11.9分におけるエルゴステロール標準ピーク(図10、プロフィールA);B:10.0分におけるW303/V51−C25H1ピーク(図10、プロフィールC);C:8.7分におけるW303/V51−C25H1ピーク(図10、プロフィールC)。
【図13】ERG6が不活性化されている酵母(コレスタ−タイプステロール)と比較した、野生型酵母(エルゴスタ−タイプステロール)のステロール生合成経路の後半部分である。酵素は斜体で強調される。黒塗り矢印は、野生型酵母に見つかった酵素反応を表す。破線矢印は、異種活性(推定上のステロールΔ24−還元酵素、推定上のコレステロールC25−ヒドロキシラーゼ)によって触媒されるステップに対応し、対応する酵素には下線を引いた。
【図14】酵母ステロール経路のダイアグラムである。
【図15】10種の異なる25−ヒドロキシラーゼ遺伝子による、7−デヒドロコレステロールから25−ヒドロキシ−7−デヒドロコレステロール(HyDHC)への変換の比較である。以下のS.セレヴィシエ(S.cerevisiae)株を実施例20に記載されるようにして培養した。Ecab HY株(配列番号26の遺伝子を有する組み込みコンストラクト)、Ecab opt 2.0株(配列番号25の遺伝子を有する組み込みコンストラクト)、Ptro opt 2.0株(配列番号19の遺伝子を有する組み込みコンストラクト)、Ptro HY(配列番号20の遺伝子を有する組み込みコンストラクト)、ConHyD HY株(配列番号31の遺伝子を有する組み込みコンストラクト)、ConHyD opt 2.0株(配列番号30の遺伝子を有する組み込みコンストラクト遺伝子)、Rnor HY株(配列番号23の遺伝子を有する組み込みコンストラクト)、Rnor opt 2.0株(配列番号22の遺伝子を有する組み込みコンストラクト)、Sscr HY株(配列番号28の遺伝子を有する組み込みコンストラクト)、Sscr opt 2.0株(配列番号27の遺伝子を有する組み込みコンストラクト)、および実施例2に記載されるようにして最適化されたブタからの25−ヒドロキシラーゼを含有するScr Pompon株。細胞ペレットからステロールを単離し、HyDHC含量、総ステロールのHyDHC比率、および7−DHCからHyDHCへの変換を、25−ヒドロキシラーゼC25H1の組み込みによって得られる値と比較した。全ての新しいヒドロキシラーゼ遺伝子が発現され、その遺伝子産物は7−DHCをHyDHCにヒドロキシル化(hydroxylase)できた。
【0016】
[定義]
本明細書および請求の範囲の全体を通じて、以下の定義が適用される。
【0017】
「酵母」は、サッカロミセス・セレヴィシエ(Saccharomyces cerevisiae)、ならびに分裂酵母(Schizosaccharomyces)種、ピキア属(Pichia)種、クリヴェロミセス(Kluyveromyces)(Klyuveromyces)種、ハンゼヌラ(Hansenula)種、およびヤロウィア・リポリティカ(Yarrowia lipolytica)などの工業規模生産に適したその他の酵母を指す。
【0018】
ハイブリダイゼーションの「標準条件」とは、本発明の文脈で、特定のハイブリダイゼーションシグナルを検出するために当業者によって一般に使用され、例えば参照によって本明細書に援用する、Sambrookら,「Molecular Cloning」,Second Edition,Cold Spring Harbor Laboratory Press 1989年,New Yorkに記載される条件を意味する。「ストリンジェントなハイブリダイゼーション条件」の一例は、50%ホルムアミド、5×SSC(150mM NaCl、15mMクエン酸三ナトリウム)、50mMリン酸ナトリウム(pH7.6)、5×デンハルト液、10%硫酸デキストラン、および20μg/mlの変性剪断サケ精子DNAを含んでなる溶液中における、42℃で一晩(例えば15時間)のインキュベーションの後、0.1×SSC中で約65℃のハイブリダイゼーション担体の洗浄である。
【0019】
「%同一性」という用語は、当該技術分野で知られているように、場合に応じて、配列のストリング間の一致により判定される、ポリペプチドまたはポリヌクレオチド配列間の関連性の程度を意味する。「同一性」は、例えばプログラムBESTFIT(GCG Wisconsin Package, version 10.2,Accelrys Inc.,9685 Scranton Road,San Diego,CA 92121−3752,USA)によって、既知の方法によって、以下のパラメーターを使用して容易に判定し得る:gap creation penalty 8、gap extension penalty 2(デフォルトパラメーター)。
【0020】
「機能性」酵素とは、典型的な酵母培養条件下で、酵素が酵母内で所望の活性を発揮することを意味する。例えば「機能性コレステロールC25−ヒドロキシラーゼ」は、酵母細胞生体内で、コレステロールC25−ヒドロキシラーゼが、7−デヒドロコレステロールを25−ヒドロキシ−7−デヒドロコレステロールに変換することを意味する。
【0021】
「機能性」核酸配列とは、核酸配列が(ポリペプチドをコードするものの他に)、酵母が機能する濃度でポリペプチドを発現するのに必要な配列を含むことを意味する。このような配列は、プロモーター、終止配列、エンハンサーなどを包含し得る。
【0022】
[コレステロール25−ヒドロキシラーゼ]
したがって本発明の一態様は、7−デヒドロコレステロールまたはエルゴステロールをコレステロールC25−ヒドロキシラーゼに接触させるステップと、25−ヒドロキシ−7−デヒドロコレステロールまたは25−ヒドロキシエルゴステロールを得るステップを含んでなる、酵母細胞中で25−ヒドロキシ−7−デヒドロコレステロールまたは25−ヒドロキシエルゴステロールを生成する方法である。好適には、コレステロールC25−ヒドロキシラーゼは脊椎動物起源であり、より好適にはラット、ブタ、イヌ、ウマ、ヒト、チンパンジー、アガゲザル(Macaca mulatta)(Macaca mulata)、マウス、ニワトリ(Gallus gallus)、アフリカツメガエル(Xenopus laevis)、ゼブラフィッシュ(Danio rerio)またはカモノハシ(Ornithorhynchus anatinus)(Ornithorynchus anatinus)からの酵素である。さらに機能する能力を保持しさえすれば、これらの天然酵素の誘導体、すなわちアミノ酸配列が天然酵素ともはや同一でないものを使用してもよい。例えば、本発明は、天然脊椎動物C25−ヒドロキシラーゼと、少なくとも90%の同一性、好適には少なくとも95%の同一性、より好適には少なくとも99%の同一性を有する酵素の使用を包含するが、ただし誘導体酵素はその機能性を保持する。
【0023】
同様に、本発明で使用し得る核酸は、上記ペプチド配列をコードするもの、および上記ペプチド配列をコードするものとストリンジェントな条件下でハイブリダイズするものである。好適にはそれらは単離されている。さらに、いくつかの実施形態では、C25−ヒドロキシラーゼ酵素をコードする核酸は、酵母環境により良く適するようにコドン最適化されている。好ましい実施形態では、核酸は、ブタ、ラット、イヌ、チンパンジー、またはウマ起源であり、酵母のためにコドン最適化されている。特に好まれる核酸は、参照核酸配列「配列番号5のSscr Pompon」と少なくとも同程度に酵母宿主中で発現されるようにコドン最適化されているものであり、より好適には、核酸配列は、Sscr Pompon」よりもさらに良く発現される。
【0024】
核酸は、好適にはDNAである。
【0025】
特に好まれる核酸は、次のように称されるものである:
C25H1(ブタヒドロキシラーゼをベースとする;配列番号.5)、
C25H3(イヌヒドロキシラーゼをベースとする:配列番号8)
Ecab_HYまたはEcab25OH_opt2.0(ウマヒドロキシラーゼをベースとする:配列番号25および26)
ConHYD opt 20(配列番号30)
Rnor HY(ラットヒドロキシラーゼをベースとする:配列番号23)
Sscr HY(配列番号28)
Sscr opt 20(配列番号27)
Sscr Pompon(配列番号5)
【0026】
本発明の別の態様は、機能性コレステロールC25−ヒドロキシラーゼを含んでなる酵母細胞である。本発明のさらなる態様は、7−デヒドロコレステロールまたはエルゴステロールを、それぞれ25−ヒドロキシ7−デヒドロコレステロールまたは25−ヒドロキシエルゴステロールに変換できる酵母である。
【0027】
本発明のさらなる態様は、ヒドロキシル化反応を生じさせる培養条件下において、7−デヒドロコレステロールの存在下で、脊椎動物コレステロールC25−ヒドロキシラーゼを発現させ、25−ヒドロキシ7−デヒドロコレステロールの生成をもたらすステップを含んでなる、酵母中で25−ヒドロキシ7−デヒドロコレステロールを生成する方法である。本発明の追加的態様は、ヒドロキシル化反応を生じさせる培養条件下において、エルゴステロールの存在下で、脊椎動物コレステロールC25−ヒドロキシラーゼを発現させ、25−ヒドロキシエルゴステロールの生成をもたらすステップを含んでなる、酵母中で25−ヒドロキシエルゴステロールを生成する方法である。
【0028】
遺伝子の複数コピーが、発現ベクター中に存在することが好ましい。好ましい実施形態では、2〜5個のコピーが使用される。
【0029】
前述の遺伝子の何れかを酵母宿主にクローニングするためのベクターは、クローニングに有用なことが知られているあらゆるタイプ、特にC25−ヒドロキシラーゼ遺伝子と、当該技術分野で通常使用されるプロモーターおよびエンハンサーをはじめとする、随伴する発現および/または組み込み配列とを保有する、プラスミドおよび動原体プラスミド;および本発明の別の態様を構成するベクターであってもよい。
【0030】
米国特許出願公開第2006/0242508号明細書で記載されるものなどの7−デヒドロコレステロールを産生できる酵母中では、遺伝子ERG5およびERG6が不活性化され、下でさらに詳しく述べられるような脊椎動物起源のステロールΔ24−還元酵素が発現されることが好ましい。さらに状況によっては、このような酵母中でHMG1の切断型を過剰に発現することが有利である。不活性化ERG5およびERG6とトランケート型HMG1遺伝子とを有するこのような株については、当該技術分野記載されている。
【0031】
次に既知のUV光照射処置を使用して、25−ヒドロキシプロビタミンD3または25−ヒドロキシプロビタミンD2を、それぞれビタミンD3またはD2の第1の代謝産物であり循環形態である、25−ヒドロキシビタミンD3に変換し得る。25−ヒドロキシビタミンD3は骨組織形成において重要な役割を果たし、商業的に家禽および他の動物飼料中でビタミン栄養補給剤として使用される。それは商標ROVIMIX HY−D(登録商標)の下に、DSM Nutritional Productsから市販される。25−ヒドロキシビタミンD2は体内で25−ヒドロキシビタミンD3に匹敵する効果を示し、25−ヒドロキシビタミンD3の用法と同様の様式で使用し得る。
【0032】
[ステロールΔ24−還元酵素]
本発明の別の態様は、ステロールΔ24−還元酵素をコードする新規核酸であり、それは酵母によって発現され得て、ラノステロール、ジメチルチモステロール、メチルチモステロール、チモステロール、コレスタ−7,24−ジエノールまたはコレスタ−5,7,24−トリエノールからなる群から選択される基質を3β−ヒドロキシ−8−ラノスタ−8−エン、4,4−ジメチル−コレスタ−8−エノール、4α−メチル−コレスタ−8−エノール、コレスタ−8−エノール、ラソステロールおよび7−デヒドロコレステロールからなる群から選択される生成物に変換し得るが、ただし核酸配列はヒト核酸配列でもマウス核酸配列でもない。
【0033】
好適には、酵母細胞は、その中でERG6およびERG5が不活性化されているものである。好適には、これらの核酸は、宿主細胞のためにコドン最適化されている脊椎動物起源の修飾DNAであるが、ただしそれはマウスでもヒトでもない。好適には、ステロールΔ24−還元酵素は、ブタ、イヌ、チンパンジー、アガゲザル(Macaca mulatta)(Macaca mulata)、マウス、ラット、ウマ、ニワトリ(Gallus gallus)、アフリカツメガエル(Xenopus laevis)、ゼブラフィッシュ(Danio rerio)またはカモノハシ(Ornithorhynchus anatinus)(Ornithorynchus anatinus)起源である。
【0034】
特に好ましい実施形態では、これらは、ドブネズミ(Rattus norvegicus)またはゼブラフィッシュ(Danio rerio)DNAであり、およびより好ましい実施形態では、それらはサッカロミセス・セレヴィシエ(Saccharomyces cerevisiae)内での発現のためにコドン最適化されている。特に好まれる核酸は、S24R1(修飾ラット遺伝子、配列番号1のヌクレオチド10〜1563)およびS24R2(修飾ゼブラフィッシュ遺伝子、配列番号3のヌクレオチド10〜1563)と称される。
【0035】
本発明の別の態様は、脊椎動物核酸配列、好適にはDNAであり、それは酵母宿主中での発現のためにコドン最適化されており、それは酵母宿主細胞内で、酵素が、
ラノステロールを3β−ヒドロキシ−8−ラノスタ−8−エンに、またはジメチルチモステロールを4,4−ジメチル−コレスタ−8−エノールに;
メチルチモステロールを4α−メチル−コレスタ−8−エノールに;
チモステロールをコレスタ−8−エノールに;
コレスタ−7,24−ジエノールをラソステロールに;または
コレスタ−5,7,24−トリエノールを7−デヒドロコレステロール)に変換し得るように、機能性ステロールΔ24−還元酵素をコードし;
酵素をコードする核酸は、ストリンジェントな条件下でS24R1またはS24R2とハイブリダイズし得るが、ただし脊椎動物核酸配列はヒトから単離された拡散配列でもマウスから単離された核酸配列でもない。
【0036】
本発明の別の態様は、前述の核酸配列によってコードされる酵素である。本発明の好ましい酵素は、配列番号2および4で記載されるもの、
および配列番号2または4と少なくとも90%、および好適には95%の同一性を示すもの、および酵母細胞環境内で、
ラノステロールを3β−ヒドロキシ−8−ラノスタ−8−エンに、
ジメチルチモステロールを4,4−ジメチル−コレスタ−8−エノールに、
メチルチモステロールを4α−メチル−コレスタ−8−エノールに、
チモステロールをコレスタ−8−エノールに、
コレスタ−7,24−ジエノールをラソステロール、または
コレスタ−5,7,24−トリエノールを7−デヒドロコレステロールに
変換できるものである。
【0037】
本発明の別の態様は、上述のような機能性核酸配列(nucleic sequence)を含有する酵母宿主、特にS24R1、S24R2を含んでなる、またはストリンジェントな条件下でS24R1またはS24R2とハイブリダイズする核酸配列を含んでなる、S.セレヴィシエ(S.cerevisiae)などの酵母宿主であるが、ただし核酸はヒト配列でもマウス配列でもない。好ましい実施形態では、酵母宿主はERG5およびERG6が不活性化されており、特に好ましい実施形態では、酵母宿主はまたHMG1遺伝子の切断型を過剰発現する。
【0038】
さらに本発明の別の態様は、ラノステロール、ジメチルチモステロール、メチルチモステロール、チモステロール、コレスタ−7,24−ジエノールまたはコレスタ−5,7,24−トリエノールと、酵母宿主中での発現についてコドン最適化されており、機能性ステロールΔ24−還元酵素をコードする核酸配列を含んでなる酵母とを接触させるステップを含んでなる、酵母細胞内で3β−ヒドロキシ−8−ラノスタ−8−エン、4,4−ジメチル−コレスタ−8−エノール、4α−メチル−コレスタ−8−エノール、コレスタ−8−エノール、ラソステロールまたは7−デヒドロコレステロールを生成する方法であるが、ただし核酸配列はヒト核酸配列でもマウス核酸配列でもなく、酵母は、
ラノステロールを3β−ヒドロキシ−8−ラノスタ−8−エンに、
ジメチルチモステロールを4,4−ジメチル−コレスタ−8−エノールに、
メチルチモステロールを4α−メチル−コレスタ−8−エノールに、
チモステロールをコレスタ−8−エノールに、
コレスタ−7,24−ジエノールをラソステロールに、または
コレスタ−5,7,24−トリエノールを7−デヒドロコレステロールに変換する。この方法では、核酸は、好適にはS24R1、S24R2であり、またはストリンジェントな条件下でS24R1またはS24R2とハイブリダイズし得る核酸であるが、ただし脊椎動物核酸配列はヒト核酸配列でもマウス核酸配列でもない。
【0039】
本発明の別の態様は、酵母宿主中での発現についてコドン最適化されており、機能性ステロールΔ24−還元酵素をコードする機能性核酸配列を含んでなるプラスミドをはじめとするベクターを含むが、ただし核酸配列はヒト核酸配列でもマウス核酸配列でもない。好適には、ベクターは核酸S24R1、S24R2、またはストリンジェントな条件下でS24R1またはS24R2とハイブリダイズする核酸を含んでなるが、ただし脊椎動物核酸配列はヒト核酸配列でもマウス核酸配列でもない。ベクターは、転写、翻訳、および/または酵母染色体への組み込みを制御するプラスミドまたは他のベクター中に一般に見出される、通常の配列を含有してもよい。
【0040】
[遺伝子コンストラクト]
C25−ヒドロキシラーゼまたはステロールΔ24還元酵素をコードする本発明の遺伝子コンストラクトは全て、典型的には、関心のある遺伝子を制御する、ADH1プロモーター、TEF1プロモーター、酵母GPD(TDH3)プロモーター、酵母HXT7プロモーター、酵母GALl/10プロモーターまたは酵母PGK1プロモーターなどの標準法を使用して制御し得る、既知のプロモーター制御下にある外来遺伝子を一般的に有する、発現カセットの一部である。さらに発現カセットは、上述の遺伝子の少なくとも1つのコード配列と作動可能なように結合している、ターミネーター配列および/または他の調節要素などの上流および下流配列を含んでなってもよい。本発明の遺伝子コンストラクトを保有するベクターは、従来の方法を用いて酵母に導入される。
【0041】
以下の実施例は例証のみを意図するものであり、本発明をどのようにも制限するものではない。本出願全体を通じて引用される全ての参考文献、特許出願、特許、および公開特許公報の内容は、参照によって本明細書に援用する。
【実施例】
【0042】
[参考文献]
参考文献の完全な一覧は実施例の終わりに記載した。全ての参考文献は、参照によって本明細書に援用する。
【0043】
[培地]
【0044】
【表1】
【0045】
培地の利用に先だって、それを120℃で20分間の高圧蒸気滅菌によって滅菌した。必要ならば無菌トリプトファン、ウラシルまたはアデニン(b)を20mg/Lの最終濃度に添加した。
【0046】
【表2】
【0047】
使用前に、培地を0.2μmの濾過ユニットに通過させ、濾過して滅菌した。必要ならば、無菌のトリプトファン、ウラシルまたはアデニンを最終濃度0.2mg/mlに添加した。
【0048】
[実施例1]
[異なる生物からのΔ24(25)−ステロール還元酵素の選択および合成遺伝子設計]
ステロールΔ24−還元酵素は、コレステロール生合成の最終段階の1つを触媒する。それはかなり非特異的酵素であり、基質として少なくともチモステロールおよびデスモステロールを受け入れる。2004年には、対応する遺伝子がヒトで同定された(Waterman HRら,2001年)。ヒトおよびマウスからの遺伝子産物についてのみ、ステロールΔ24−還元酵素活性が正式に(formally)立証されている(Crameri Aら、2006年;国際公開第03/064650号パンフレット)。ラットからの相同遺伝子の匹敵する機能についてもまた記載されるが(Wu C.ら 2004年: Nature 432 (7017):640−5)、遺伝子産物の酵素活性は確認されていない。
【0049】
国際公開第03/064650号パンフレットでは、S.セレヴィシエ(S.cerevisiae)変異体(erg5 erg6)中における、マウスからのΔ24−還元酵素遺伝子の発現が示されており、宿主株中では生じない2つの新しいステロール(ラソステロールおよび7−デヒドロコレステロール)の形成がもたらされる。ヒトΔ24(25)−ステロール還元酵素と相同的な酵素についての、GENBANKまたはEMBLなどの公共データベースの検索は、標準条件下で、公的に利用可能なプログラムBLASTpを用いて実施した。見つかった配列間で、ヒト(Homo sapiens)アミノ酸配列と97%のアミノ酸同一性を示すドブネズミ(Rattus norvegicus)の配列(表1)、およびヒト配列と79%のアミノ酸同一性を示すゼブラフィッシュ(Danio rerio)の配列(表1)をさらなる調査のために選択した。
【0050】
【表3】
【0051】
各cDNAについて、S.セレヴィシエ(S.cerevisiae)中における発現のための合成遺伝子を設計した。一般に、この目的のために、元のcDNA配列を使用することが可能である。場合によっては、かずさDNA研究所(http://www.kazusa/or.jp/codon)の異なる種のコドン使用頻度表を使用して、元のcDNAのコドン使用頻度プロフィールを新しい宿主のコドン使用頻度に適合させる試みが有利である。これは手動で、またはコンピュータプログラムの助けを借りて実施し得る。本例では、コンピュータプログラムが使用され、その計算アルゴリズムは配列作成ルーチンを伴い、ユーザー定義のコドンバイアス調節があるDNA配列を作成できるようにする。アルゴリズム中にはまたいくつかの制約を導入して、特定の制限部位の存在を回避し、または代案としては随意に制限部位の存在を支持し得る。配列作成の一部は乱数発生ルーチンによって進められ、通常、試行毎に異なるDNA配列がもたらされる。プログラムのユーザーが使用された乱数種を回復することは可能でなく、したがって結果を再現することは可能でなかった。したがって再コードされた配列は予測され得ず、ユニークな元のシグネチャーを保有すると見なし得る。
【0052】
代案として作成されたDNA配列バージョン間の選択は、最終的にユーザー依存であった(外観検査)。選択されたDNA配列バージョンは、サービス提供者(例えばGeneCust Europe, 30b rue Dominique Lang,3505 Dudelange,Luxembourg)によって、遺伝子合成のために使用された。翻訳開始および終結を最適化するために、3個の追加的アデニン残基をATG開始コドン上流に付加し、第2の停止(STOP)コドンを読み取り枠下流に付加した。合成遺伝子の酵母発現ベクターへの都合よいクローニングのために、Bam HIおよびEco RI制限酵素部位をそれぞれ配列の上流および下流に付加した。上で説明したように、ラットステロールΔ24−還元酵素遺伝子(GenBank登録番号AY92220(cDNA配列)およびAAX29968(アミノ酸配列)を上述の方法によって再コードした。新しい合成遺伝子は、S24R1と命名された。その配列を下に示す。
配列番号1
【化1】
【0053】
対応するアミノ酸配列は次のとおり:
配列番号2
【化2】
【0054】
ゼブラフィッシュ(D.rerio)ステロールΔ24−還元酵素遺伝子(DNA配列はGenBank番号NM_001008645およびアミノ酸配列はGenBank number NP_001008645でアクセス可能)を上述の方法によって再コードした。新しい合成遺伝子をS24R2と命名した。その配列を下に示す。
配列番号3
【化3】
【0055】
対応するアミノ酸配列は次のとおり:
配列番号4
【化4】
【0056】
[実施例2]
[ヒト(Homo sapiens)からのコレステロールC25−ヒドロキシラーゼアミノ酸配列に相同的なアミノ酸配列の選択、およびS.セレヴィシエ(S.cerevisiae)中での発現について最適化された代案のDNA配列の計算]
ヒト(H.sapiens)およびハツカネズミ(M.musculus)遺伝子産物について、コレステロールC25−ヒドロキシラーゼ活性が示されている(Lundら,1998年;米国特許第6562609号明細書)。ヒトコレステロールC25−ヒドロキシラーゼと相同的な酵素についてのGENBANKまたはEMBLなどの公共データベースの検索は、標準条件下で、公的に入手可能なプログラムBLASTpを用いて実施した。見つかった配列間で、ヒト(H.sapiens)アミノ酸配列と82%のアミノ酸同一性、ハツカネズミ(M.musculus)配列と79%のアミノ酸同一性を示すイノシシ(S.scrofa)配列(表2)、およびヒトおよびマウス配列と70%のアミノ酸同一性を示すイヌ(C.familiaris)配列(表4)をさらなる調査のために選択した。
【0057】
【表4】
【0058】
酵母サッカロミセス・セレヴィシエ(Saccharomyces cerevisiae)によって滅多に使用されないコドンに相当する25個のコドンを、酵母中で頻繁に使用されるコドンで置き換えることにより、ブタコレステロールC25−ヒドロキシラーゼ遺伝子(DNA配列はGenBank番号AY974088で、アミノ酸配列はGenBank番号Q4G1G8でアクセスできる)を再コードした(Zhangら,1991年)。新しい合成遺伝子をC25H1と称した。その配列を下に示す。
配列番号5
【化5】
【0059】
対応するアミノ酸配列は、次のとおり。
配列番号6
【化6】
【0060】
使用された配列と4つの違いを示す、データベースGENBANKからの推定上のブタステロールC25−ヒドロキシラーゼの代案のアミノ酸配列もまた利用できるが、実験でさらに使用することはなかった。
配列番号7
【化7】
【0061】
イヌ(Canis familiaris)からの遺伝子(ヌクレオチド配列はGenBank番号XM_546596.2で、アミノ酸配列はGenBank番号XP_543596.1でアクセスできる)を実施例1に記載されるようにして、適合法によって再コードした。新しい合成遺伝子をC25H3と命名し、そのDNA配列を下に示す。
配列番号8
【化8】
【0062】
対応するアミノ酸配列は次のとおり。
配列番号9
【化9】
【0063】
[実施例3]
[発現プラスミドV51TDH−S24R1の構築]
発現ベクターは、Sambrookら(1989年)によって記載されるような古典的分子生物学技術を使用して構築され、DNA制限酵素および修飾酵素は、供給元(New England Biolabs,Ipswich,MA 01938−2723,USA)が推奨するようにして(as recommender)使用された。
【0064】
基礎的な多コピー構成的発現ベクターV51TDHは、ガラクトース誘導性GAL10−CYC1プロモーターを構成的に発現するTDH3遺伝子(サッカロミセス属(Saccharomyces)ゲノムデータベース中の系統的遺伝子注釈からのYGR192C遺伝子、グリセルアルデヒド−3−リン酸脱水素酵素イソ酵素3をコードする)からのプロモーターで置換して、V51ベクターから誘導された。V51は、大腸菌(Escherichia coli)中における維持のためのColE1複製起点およびbla遺伝子(アンピシリン耐性)、酵母中における維持のための2μ起点およびURA3遺伝子、多重クローニング部位(MCS)(pYeDP1/8−2、CullinおよびPompon,1988年)によって隔てられるGAL10−CYC1ハイブリッドプロモーターおよびPGK1ターミネーター領域を保有する。TDH3プロモーター領域は、W303−1B酵母株(ThomasおよびRothstein,1989年)からのゲノムDNA上でPCRによって増幅され、HoffmanおよびWinston(1987年)に従って、以下のプライマーを使用して調製された。
【化10】
【0065】
PCR増幅産物をAge IおよびBam HIによって消化し、GAL10−CYC1プロモーターを保有するV51からの617bp長さのAge I−Bam HI断片を置換するのに使用した。得られたV51TDHベクターは、TDH3プロモーター(TDH3p)とPGK1ターミネーター(PGK1t)領域の間に局在するMCSに挿入することで、酵母中において関心のある遺伝子を構成的発現させるのに有用な、多コピー大腸菌(E.coli)/S.セレヴィシエ(S.cerevisiae)シャトルベクターである。
【0066】
S24R1合成遺伝子(実施例1参照)の構成的発現のためのV51TDH−S24R1ベクターは、Bam HIおよびEco RIで制限酵素消化されたV51TDHベクターに、S24R1合成遺伝子からの1564bpのBam HI−Eco RI制限酵素産物を挿入して構築された。
【0067】
V51TDH−S24R1構成的発現ベクターの制限酵素地図を図1に示す。
【0068】
[実施例4]
[発現プラスミドV51TDH−S24R2の構築]
S24R2の構成的発現のためのV51TDH−S24R2ベクター(実施例1参照)は、基本的に、実施例3で既に述べたようにして構築された。Bam HIおよびEco RIで制限酵素消化されたV51TDHベクターに、合成遺伝子S24R2の1564bp長さのBam HI−Eco RI制限酵素産物をライゲートした。V51TDH−S24R2の制限酵素地図を図2に示す。
【0069】
[実施例5]
[動原体発現プラスミドpFLAde−S24R1の構築]
酵母動原体および自律複製配列(ARSCEN)およびADE2選択マーカーを保有する空のpFLAde大腸菌(E.coli)/S.セレヴィシエ(S.cerevisiae)シャトルベクターに、V51TDH−S24R1(実施例3に記載)からのTDH3p−S24R1−PGK1t発現カセットを挿入することにより、遺伝子S24R1の構成的発現のための動原体pFLAde−S24R1ベクター(実施例1参照)を構築した。pFLAdeベクターは、以下のプライマーを使用して、URA3遺伝子をコードするpFL38の1.1kbのBgl II断片(Bonneaudら,1991年)を、酵母ゲノムDNA上でのPCR増幅によって得られるADE2遺伝子をコードする2.7kbのBam HI断片で置換して構築された。
【化11】
【0070】
TDH3p−S24R1−PGK1t発現カセットは、標準条件下で、PrimeSTAR HS DNAポリメラーゼ(日本国滋賀県大津市のタカラバイオ株式会社)および以下のプライマーを使用して、PCRによって増幅された。
【化12】
【0071】
増幅産物をpCR2.1−TOPO(Invitrogen,Carlsbad,CA 92008,USA)にクローンした。Sac I−Pst I制限酵素消化と、同一制限酵素によって切断されたpFLAdeベクターへのライゲーションによって、TDH3p−S24R1−PGK1tカセットを後者のベクター(later vector)から単離した。結果として得られたpFLAde−S24R1動原体発現ベクターの制限酵素地図を図3に示す。
【0072】
[実施例6]
[発現ベクターV51−C25H1の構築]
C25H1からの826bpのBam HI−Eco RI断片をV51に挿入し、Bam HIおよびEco RIで消化して、C25H1(実施例2参照)のガラクトース誘導性発現のためのV51−C25H1プラスミドを構築した。得られたV51−C25H1発現プラスミドは、ガラクトース誘導性GAL10−CYC1ハイブリッドプロモーター(Guarenteら,1982年)およびPGK1ターミネーターの制御下にある合成遺伝子C25H1をコードする。V51−C25H1の制限酵素地図を図4に示す。
【0073】
[実施例7]
[発現ベクターV51−C25H3の構築]
合成遺伝子C25H3(実施例2参照)のガラクトース誘導性発現のためのV51−C25H3ベクターは、実施例6に既に記載したようにして、Bam HIおよびEco RIで消化されたV51に、合成遺伝子C25H3の826bpのBam HI−Eco RI断片を挿入して構築した。得られた発現プラスミドV51−C25H3は、ガラクトース誘導性GAL10−CYC1ハイブリッドプロモーターおよびPGK1ターミネーターの制御下にある、C25H3をコードする。V51−C25H3の制限酵素地図を図5に示す。
【0074】
[実施例8]
[酵母株形質転換およびステロール抽出]
上述の異なる発現プラスミドによって、W303−1B株(MATalpha;ura3−52;trp1−1;leu2−3,112;his3−11;ade2−1;can1−100;ThomasおよびRothstein,1989年)、およびそのerg6変異体ERT(MATalpha;erg6::TRP1;ura3−52;trp1−1;leu2−3,112;his3−11;ade2−1;can1−100)を形質転換した。ERT株のERG6の不活性化は、ERG6に機能性TRP1遺伝子を挿入して得られた。pFL45からのTRP1遺伝子(Bonneaudら,1991年)は、ERG6配列が隣接するTRP1プライマーを使用して、PCRによって増幅した。プライマーの配列は以下のとおりである。
ERGTRP−1:
【化13】
【0075】
Gietzら(1995年)によって記載されるリチウム−PEG技術を使用して、得られたPCR産物を使用し、W303−1B株を形質転換した。トリプトファンを含まない規定培地上で形質転換体を選択し、それらのステロール組成物をHPLCによって確認し(Lecainら,1996年)、ERG6不活性化の既知の効果と一致する、エルゴステロールの不在および独特なステロールの存在を確認した。1つのerg6::TRP1形質転換体をさらなる研究のために保存し、ERTと命名した。
【0076】
ERTに加えて、S.セレヴィシエ(S.cerevisiae)野性型BY4742株(遺伝子型MATalpha;his3Δ1;leu2Δ0;lys2Δ0;ura3Δ0;Brachmannら,1998年)もまた、構築されたプラスミドのための宿主として使用した。
【0077】
BY4742、W303−1B、およびERTの形質転換は、Gietzら(1995年)によって記載されるリチウム−PEG法によって実施した。形質転換体は、ベクター上に存在する選択マーカーに応じて、ウラシルまたはアデニンを含まない定義された最少培地(表1)上で選択した。各形質転換の少なくとも2つの独立した形質転換体を分析した。ステロール分析のためには、形質転換体を合成培地(表2に記載)中で、定常期になるまで培養した。
【0078】
酵母細胞を培養媒体から遠心分離して回収し、等容積の脱イオン水で洗浄した。細胞を2mlエッペンドルフ管中の100μLの水に再懸濁して、0.1gガラスビーズ(0.5mm)の存在下、強力なボルテックス(5分間)によって破砕した。2mlの1,2−ジクロロエタンを再懸濁細胞に添加して、混合物を5分間激しくボルテックスした。10,000rpmで5分間遠心分離した後、溶媒相をガラス管に移した。抽出を2回実施した。プールされた有機相を窒素下で蒸発させた。最終段階として、HPLC分析のために乾燥抽出物を100μLのアセトニトリルに再懸濁した。
【0079】
[実施例9]
[UVおよび質量分析検出を用いた高速液体クロマトグラフィー(HPLC)によるステロール分析]
通常30μLである、実施例8に記載されるようにして調製されたステロール抽出物のアリコートを、UVおよび質量分析検出によってモニターされる逆相HPLCによって分析した。「Waters Alliance HT 2790」HPLCは、「Waters PDA 996」タイプからの光ダイオードアレーUV検出器に、そして質量検出器「Waters MicroMass ZQ」(Waters,Milford,MA01757,USA)に接続されていた。分離は、XTerra RP18 3.5μm 4.6×100mmカラム(Waters,Milford,MA01757,USA)上で、水/アセトニトリル勾配を含有する直線的0.03%(v/v)ギ酸を用いて、60℃で実施した。80%の緩衝液A(0.03%ギ酸添加水)と20%の緩衝液B(0.03%のギ酸添加アセトニトリル)とを含有する緩衝液で、カラムを安定化させた後に、サンプルを注入した。最初の5分間、緩衝液組成を80%の緩衝液Aおよび20%の緩衝液Bから、25%の緩衝液Aおよび75%の緩衝液Bに徐々に変化させた。次の15分間、緩衝液組成を25%の緩衝液Aおよび75%の緩衝液Bから、12.5%の緩衝液Aおよび87.5%の緩衝液Bに徐々に変化させた。最後の数分で、分離プロトコル100%の緩衝液Bに達した。この値を2分間保って、カラムを再生させた。
【0080】
光ダイオードアレー検出器によって205〜300nmの間で、クロマトグラフィーをモニターした。質量検出器は、m/zが350〜450の範囲の陽性エレクトロスプレーによって、1m/z単位の感度での検出が可能であった。
【0081】
ステロールは、標準と比較した、それらのクロマトグラフィーの挙動、それらのUV吸収スペクトル、およびそれらの質量断片化プロフィールに基づいて同定された(表5)。SecenおよびKalpar(1999年)に従って、ピリジン存在下で、無水酢酸によって、25−ヒドロキシ7−デヒドロコレステロール標準(DSMによって提供される)を25−ヒドロキシ−7−デヒドロコレステロールアセテート(25OH−7DHC−Ac)にアセチル化した。0.5mgの乾燥25−ヒドロキシ7−デヒドロコレステロールを1mlの無水酢酸/ピリジン溶液(50/50v/v)に再懸濁して、55℃で3時間加熱した。反応混合物を4mlの水で希釈した。HPLC分析前に、実施例8に記載されるようにして、0.5mlの希釈反応混合物を1,2−ジクロロエタンで抽出した。
【0082】
上述の条件下で分析すると、非アセチル化25−ヒドロキシ−7−デヒドロコレステロール標準(RT6.9分)と比較して、25OH−7DHC−Ac標準は、アセチル化化合物のより高い疎水性のために、滞留時間(RT8.5分)にシフトを示した。25OH−7DHC−Ac標準の質量断片化プロフィールは、非アセチル化25−ヒドロキシ7−デヒドロコレステロール標準のプロフィールと同様であった。アセチル化形態に対応するm/zシグナルは、恐らく質量検出器内での検出中に起きる脱アセチル化のために、検出されなかった。
【0083】
【表5】
【0084】
[実施例10]
[ERG6−欠損S.セレヴィシエ(S.cerevisiae)ERT株中におけるプラスミドV51TDH−S24R1からのS24R1遺伝子の発現]
プラスミドV51TDH−S24R1(実施例3参照)および対照として空ベクターV51TDHをリチウム−PEG技術(Gietzら,1995年)によって、ERT株に形質転換した。形質転換体をウラシルを含まない定義された最少培地上で選択した。各形質転換からの4つの独立した形質転換体をプラスミドの存在について分析した。確認された形質転換体の1つを無作為に選択し、それぞれERT/V51TDHおよびERT/V51TDH−S24R1と命名して、さらなる分析のために使用した。
【0085】
株が定常期になるまで、2%(w/v)グルコースKapelli培地(表2)中で培養し、実施例8に記載されるようにして全ステロールを抽出し、実施例9に記載されるようにして、UVおよび質量分析検出を用いたHPLCによって分析した。それぞれの標準(表5)を用いて、主要ステロールをそれらの滞留時間、質量、およびスペクトル諸特性によって同定した。
【0086】
HPLC溶出プロフィール(282nmでのUV検出)を図6に示す。コレスタ−5,7,22,24−テトラエノール(RT9.2分、m/z=363、最大UV233nm)が、ERT/V51TDH erg6対照株中に見つかった主要ステロールであり、282nmにおけるUVシグナルの70%に相当した。282nmで特定された総ステロールの7%は、コレスタ5,7,24−トリエノール(RT10.3分、m/z=365、最大UV282nm;図6−A)と同定された。
【0087】
S24R1遺伝子を発現するERT/V51TDH−S24R1株(図6−B)のステロールプロフィールからは、対照ERT株/V51TDH中で検出されなかった2つのステロール、すなわち本株の主要ステロールである(282nmにおけるUVシグナルの60%)コレスタ5,7,22−トリエノール(RT10.8分、m/z=365、最大UV282nm)、および7−デヒドロコレステロール(RT11.8、m/z=367、最大UV282nm)とも称されるコレスタ5,7−ジエノールの存在が明らかになった。コレスタ5,7,24−トリエノールおよびコレスタ5,7,22,24−テトラエノールのΔ24−25結合の還元によって生じたコレスタ5,7,−ジエノールおよびコレスタ5,7,22−トリエノールの存在は、遺伝子S24R1のステロールΔ24−還元酵素活性を裏付けた(図13参照)。
【0088】
[実施例11]
[S.セレヴィシエ(S.cerevisiae)ERT株中におけるプラスミドV51TDH−S24R2からの遺伝子S24R2の発現]
リチウム−PEGプロトコル(Gietzら,1995年)を使用して、S24R2を構成的に発現するプラスミドV51TDH−S24R2(実施例3)をERT株に形質転換する。形質転換体をウラシルを含まない最少培地上で選択した。4つの独立した形質転換体が分析され、同一諸特性を示した。それらの1つをERT/V51TDH−S24R2と命名し、さらなる分析のために使用した。
【0089】
株が定常期に達するまで、2%グルコースKapelli培地(表2)中で培養し、実施例8、9、および10に記載されるようにして、ステロールを抽出し分析した。この例でも、対照ERT/V51TDH株中では、主要ステロールとしてコレスタ5,7,22,24−テトラエノール(RT9.2分、m/z=363、最大UV233nm)およびコレスタ5,7,24−トリエノール(RT10.3分、m/z=365、最大UV282nm)が見つかったのに対し(図6−A)、S24R2を発現するERT/V51TDH−S24R2株中で検出された主要ステロールは、282nmにおけるUVシグナルの70%であった、コレスタ5,7,22−トリエノール(RT10.8分、m/z=365、最大UV282nm)であった(図6−C)。コレスタ5,7−ジエノール(7−デヒドロコレステロール;RT11.8、m/z=367、最大UV282nm)は、より低濃度(UVシグナルの13%)で見つかった。これらの結果は、S24R2遺伝子のステロールΔ24−還元酵素活性を裏付けた。
【0090】
[実施例12]
[S.セレヴィシエ(S.cerevisiae)ERT株中における動原体プラスミドpFLAde−S24R1からの遺伝子S24R1の発現]
リチウム−PEG手順(Gietzら,1995年)を使用して、S24R1遺伝子の構成的発現のための動原体プラスミドpFLAde−S24R1(実施例5)をERT株に形質転換した。形質転換体をアデニンを含まない規定培地上で選択した。4つの独立した形質転換体をプラスミドpFLAde−S24R1の発生と、それらのステロールパターンについて分析した。それらの1つを無作為に選択してERT/pFLAde−S24R1と命名し、結果のさらなる例証のために使用した。
【0091】
ERT/pFLAde−S24R1を2%(w/v)グルコースKapelli培地(表2)中で、定常期に達するまで培養した。実施例8および9に記載されるようにして、ステロールを抽出し分析した。対照中に存在しない、ERT/pFLAde−S24R1中の主要な新しいステロールは、282nmにおけるUVシグナルの60%であった、コレスタ5,7,22−トリエノール(RT10.8分、m/z=365、最大UV282nm)であった(図6−D)。より少量(UVシグナルの7%)の7−デヒドロコレステロール(RT11.8、m/z=367、最大UV282nm)もまた検出された。これらの新しいステロールの他に、ERT対照株(図6−A)中にもまた存在する、コレスタ5,7,22,24−テトラエノール(RT9.2分、m/z=363、最大UV233nm)およびコレスタ5,7,24(RT10.3分、m/z=365、最大UV282nm)もまたERT/pFLAde−S24R1に見つかった。これらの結果は、動原体プラスミドpFLAde−S24R1から発現されたS24R1遺伝子のステロールΔ24−還元酵素活性を裏付ける。
【0092】
[実施例13]
[S.セレヴィシエ(S.cerevisiae)ERT株中における遺伝子S24R1およびC25H1の同時発現による25−ヒドロキシ−7−デヒドロコレステロールアセテートの生成]
リチウム−PEG法(Gietzら,1995)によって、遺伝子C25H1のガラクトース誘導性発現のためのプラスミドV51−C25H1(実施例6)、および対照としてのプラスミドV51を(実施例12に記載されるようにして)ERT株/pFLAde−S24R1に形質転換した。V51−C25H1およびpFLAde−S24R1は、適合性プラスミドである。それらの異なる2μおよびARS−CEN起源は、酵母中でそれらが同時発生できるようにする。形質転換体をウラシルおよびアデニンを含まない最少培地上で選択した。各形質転換からの4つの独立した形質転換体が分析され、同一諸特性を示した。各形質転換から1つの形質転換体を無作為に選択し、それぞれERT/pFLAde−S24R1/V51−C25H1およびERT/pFLAde−S24R1/V51と命名し、さらなる分析のために使用した。
【0093】
株を2%ガラクトースKapelli培地(表2)中で、定常期に達するまで培養した。ステロールを抽出し、実施例8および9に記載されるようにして、実施例10に記載される条件下で、UVおよび質量分析検出を用いたHPLCによって分析した。対照ERT/pFLAde−S24R1/V51株(図7−C)で見出されたように、ERT/pFLAde−S24R1/V51−C25H1(図7−D)中の主要ステロールは、最終ステロールコレスタ5,7,22,24−テトラエノールのステロールΔ24−還元酵素による還元によって生じた、コレスタ5,7,22−トリエノール(RT10.8分、m/z=365、最大UV282nm)であった。標準として注入された調製25−ヒドロキシ7−デヒドロコレステロールアセテート(25OH−7DHC−Ac)と同時に溶出される(RT8.5分、図7−B)、対照株中に存在しない新しい化合物が、ERT/pFLAde−S24R1/V51−C25H1株(図7−D)中に観察された。この化合物もまた、調製25−ヒドロキシ7−デヒドロコレステロールアセテート(図8−B)と同一のUV吸収プロフィール(図8−C)を示し、7−デヒドロコレステロール標準(図8−A)でも観察された、Δ5−Δ7共役二重結合(double bounds)に特異的な282nm前後の二重ピークがあった。新しい化合物は、(実施例9に記載されるようにして実施した)オンライン質量分析による判定で、25−ヒドロキシ7−デヒドロコレステロールアセテート(図9−B)と同一の質量断片化プロフィール(図9−C)もまた示した。
【0094】
3β−ヒドロキシステロールの場合、エレクトロスプレー(electrospay)分析は、分子量から17を差し引いた所で主要シグナルを生じた(プロトン付加(+1)および位置C3(−18)での脱水。7−デヒドロコレステロール(図9−A)の主要シグナルは、m/z質量367にある(384(7−デヒドロコレステロールMW)+1(プロトン付加)−18(3β−ヒドロキシステロール脱水))。25−ヒドロキシ7−デヒドロコレステロールアセテートで観察されたm/z質量383におけるシグナルは、位置C25(367+16=383)における追加的ヒドロキシル化に対応する。m/z質量365における主要シグナルは、質量検出器内での検出中に、分子のごく一部に生じる位置C25での脱水(383−18=365)に対応する。これらの異なる結果(標準として使用される25−ヒドロキシ7−デヒドロコレステロールアセテートと同一滞留時間、同一UVプロフィール、および同一質量断片化)は、ERT/pFLAde−S24R1/V51−C25H1株中で生成された新しい化合物が、25−ヒドロキシ7−デヒドロコレステロールアセテートであったことを実証した。
【0095】
[実施例14]
[S.セレヴィシエ(S.cerevisiae)ERT株中におけるS24R1およびC25H3の同時発現による25−ヒドロキシ−7−デヒドロコレステロールアセテートの生成]
リチウム−PEG法(Gietzら,1995年)によって、遺伝子C25H3のガラクトース誘導性発現のために構築されたV51−C25H3(実施例7)をERT/pFLAde−S24R1株(実施例13参照)に形質転換した。形質転換体をウラシルおよびアデニンを含まない最少培地上で選択した。双方のプラスミドの発生確認のために、4つの独立した形質転換体を分析した。それらの1つを無作為に選択してERT/pFLAde−S24R1/V51−C25H3と命名し、さらなる研究のために使用した。
【0096】
ERT/pFLAde−S24R1/V51−C25H3を2%ガラクトースKapelli培地(表2)中で、定常期まで培養した。収集細胞からステロールを抽出し、実施例8および9に記載されるようにして、実施例10に記載される条件下で、UVおよび質量分析検出を用いたHPLCによって分析した。対照ERT/pFLAde−S24R1/V51株(図7−C)で見つかったように、ERT/pFLAde−S24R1/V51−C25H3(図7−E)中の主要ステロールは、実施例13に記載されるように、最終ステロールコレスタ5,7,22,24−テトラエノールのステロールΔ24−還元酵素による還元産物である、コレスタ5,7,22−トリエノール(RT10.8分、m/z=365、最大UV282nm)であった。
【0097】
25−ヒドロキシ7−デヒドロコレステロールアセテート(RT8.5分、図7−B)と同時に溶出される、親株中に存在しない化合物が、ERT/pFLAde−S24R1/V51−C25H3(図7−E)中に見つかった。ERT/pFLAde−S24R1/V51−C25H1(実施例13に記載)の場合のように、この化合物は、25−ヒドロキシ7−デヒドロコレステロールアセテートと同一のUV吸収プロフィール(図8)、および同一の質量断片化プロフィール(図9)を示した。これらの結果(滞留時間、UVプロフィール、および質量断片化が25−ヒドロキシ7−デヒドロコレステロールアセテートに匹敵する)は、ERT/pFLAde−S24R1/V51−C25H3中に生成された新しい化合物が、実施例13と同様、25−ヒドロキシ7−デヒドロコレステロールアセテートであることを実証する。
【0098】
[実施例15]
[S.セレヴィシエ(S.cerevisiae)W303−1B株およびBY4742株中におけるC25H1発現による25−ヒドロキシ−エルゴステロールアセテートの生成]
リチウム−PEG法(Gietzら,1995年)によって、遺伝子C25H1のガラクトース誘導性発現のために構築されたプラスミドV51−C25H1(実施例6)、および対照としての空ベクターV51をW303−1B株(実施例8に記載)に形質転換した。形質転換体をウラシルを含まない最少培地上で選択した。各形質転換からの4つの独立した形質転換体をプラスミドの発生について分析した。各形質転換から1つの陽性形質転換体を無作為に選択して、それぞれW303/V51−C25H1およびW303/V51と命名し、さらなる研究のために使用した。
【0099】
株を2%ガラクトースKapelli培地(表2)中で、定常期に達するまで培養した。収集細胞からステロールを抽出し、実施例8および9に記載されるようにして、実施例10に記載される条件下で、UVおよび質量分析検出を用いたHPLCによって分析した。HPLC溶出プロフィール(282nmでのUV検出)を図10に示す。エルゴステロールの滞留時間(図10−A)、theUVスペクトル(図11−A)、および質量断片化プロフィール(図12−A)(RT11.9分、m/z=379、最大UV282nm)との比較によって実証されるように、W303/V51−C25H1(図10−C)およびW303/V51(図10−B)の双方の株について観察された主要シグナルは、S.セレヴィシエ(S.cerevisiae)野生型株の主要ステロール(mayor sterol)であるエルゴステロール(エルゴスタ5,7,22−トリエノール)に対応する。
【0100】
W303/V51−C25H1株(図10−C)の10分における微量シグナルは、W303/V51対照株(図10−B)では痕跡シグナルとしてのみ観察された。オンラインPDA検出によって測定されたこのピークのUVプロフィール(図11−B)は、282nm前後の二重ピークによって特徴付けられるΔ5およびΔ7の共役二重結合に加えて、Δ22およびΔ24の共役二重結合に特異的である233nmにおける最大吸収を示した。恐らくこのシグナルは、Erg4pによるΔ24の還元がないエルゴステロール生合成経路の最後の中間体の1つである、エルゴスタ5,7,22,24−テトラエノールに対応する(図13参照)。オンライン質量分析法(図12−B)によって検出されたm/z質量377は、エルゴスタ−テトラエノール構造に一致する。
【0101】
W303/V51−C25H1株(図10−C)のステロール抽出物中で、HPLCによって観察された8.7分における第3のシグナルは、W303/V51(図10−B)では完全に欠如していた。このピーク(図11−C)について測定されたUVプロフィールは、Δ5およびΔ7共役二重結合(double bounds)に特異的な最大282nmの典型的な二重ピークを示した。質量断片化(図12−C)は、m/z=377における主要シグナル、およびm/z=395における微量のシグナルを示し、水酸化型エルゴステロール(379+16=395)およびその脱水形態(395−18=377)に一致する。断片化プロフィール(図12−C)は、全ての質量が、7−デヒドロコレステロール(mz=367)およびエルゴステロール(m/z=379)間の質量差に相当する12質量単位増大していたこと以外は、25−ヒドロキシ7−デヒドロコレステロールアセテート(図9−B)の1つと同一であった。この新しい化合物(RT8.7分)とエルゴステロール(RT11.9分)の間の3.2分の滞留時間の差は、25−ヒドロキシ7−デヒドロコレステロールアセテート(RT8.5分)と7−デヒドロコレステロール、(RT11.8)の滞留時間の差と同一であった。
【0102】
これらの諸特性(UVプロフィール、質量断片化、滞留時間)は全て、W303/V51−C25H1中のC25H1発現によって生成されるこの新しい化合物が、25−ヒドロキシ−エルゴステロールアセテートであるという結論に矛盾しない。
【0103】
V51−C25H1および対照としての空ベクターV51のS.セレヴィシエ(S.cerevisiae)株BY4742への形質転換は、W303/V51−C25H1株およびW303/V51株で得られたものに匹敵する結果(図10)をもたらした。この場合もV51−C25H1形質転換体について、HPLC(UV検出)によって8.7分における追加的シグナルが観察された。化合物は、W303/C25H1中で検出された化合物と同一UVプロフィール(図11)および質量断片化プロフィール(図12)を示し、この化合物もまた25−ヒドロキシ−エルゴステロールアセテートであることを強く示唆する。
【0104】
[実施例16]
[S.セレヴィシエ(S.cerevisiae)W303−1B株中における遺伝子C25H3の発現による25−ヒドロキシ−エルゴステロールアセテートの生成]
リチウム−PEG法(Gietzら,1995年)を使用して、合成遺伝子C25H3のガラクトース誘導性発現のために構築されたプラスミドV51−C25H3(実施例7)をW303−1B株(実施例8に記載)に形質転換した。形質転換体をウラシルを含まない最少培地上で選択した。4つの独立した形質転換体をV51−C25H3の発生について分析した。1つの陽性形質転換体をW303/V51−C25H3と命名し、生成されるステロールのさらなる研究のために使用した。
【0105】
株が定常期になるまで、2%(w/v)ガラクトースKapelli培地(表2)中で培養した。収集細胞からステロールを抽出し、実施例8および9に記載されるようにして、実施例10に記載される条件下で、UVおよび質量分析検出を用いたHPLCによって分析した。
【0106】
図10−Dに示すHPLC(282nmでのUV検出)によるステロール抽出物分析は、W303/V51−C25H3のステロールパターンが、W303/V51−C25H1株(実施例15)のステロールパターンと非常に良く似ていることを示し、エルゴステロール(RT11.9分)およびエルゴスタ5,7,22,24−テトラエノール(RT10.0分)に加えて、新しい化合物(RT8.7分)が生成されていた。オンラインUV PDA検出(図11)および質量分析法(図12)は、例えばΔ5−Δ7の特異的共役二重結合UVプロフィール、m/z=377における主要シグナルおよびm/z=395における微量シグナルがある質量断片化、および25−ヒドロキシル化ステロールと一致するHPLC滞留時間遅延など、この新しい化合物が、W303/V51−C25H1によって生成される化合物と同一諸特性を有することを示した。これらの特徴(UVプロフィール、質量断片化、滞留時間)は全て、この新しい化合物がC25H3の発現に起因する25−ヒドロキシ−エルゴステロールアセテートであることを強力に示唆する。
【0107】
[実施例17]
[主にコレスタ−5,7,24−トリエノールを産生する酵母株へのC25H1およびC25H3の導入]
リチウム−PEG法(Gietzら,1995年)を用いて、遺伝子C25H1およびC25H3のガラクトース誘導性発現のためのプラスミドV51−C25H1(実施例6)およびプラスミドV51−CH25H3(実施例7)、および対照としてのプラスミドV51をERT erg5 erg6株(実施例12に記載;ERG5は遺伝子URA3の組み込みによって不活性化されていた)に形質転換した。形質転換体をウラシルおよびアデニンを含まない最少培地上で選択した。各形質転換からの4つの独立した形質転換体が分析され、同一諸特性を示した。各形質転換からの1形質転換体を無作為に選択して、それぞれERT erg5 erg6/V51−C25H1、ERT erg5 erg6/V51−C25H3、およびERT erg5 erg6/V51と命名し、さらなる分析のために使用した。
【0108】
株を2%ガラクトースKapelli培地(表2)中で、定常期に達するまで培養した。ステロールを抽出し、実施例8および9に記載されるようにして、実施例10に記載される条件下で、UVおよび質量分析検出を用いたHPLCによって分析した。3株間でステロールパターンの違いは検出されず、2種のヒドロキシラーゼがコレスタ−5,7,24−トリエノールに対して活性でないことを裏付けた。
【0109】
[実施例18]
[追加的コレステロール25−ヒドロキシラーゼ遺伝子の選択]
イノシシ(S.scrofa)およびイヌ(C.familiaris)(Canis lupus)、からの選択された推定上のコレステロール25−ヒドロキシラーゼ遺伝子に加えて、ドブネズミ(R.norvegicus)(配列番号21、Genebank登録番号NP_001020586、XP_220063)、チンパンジー(P.troglodytes)(配列番号18、遺伝子bank登録番号XP_507901)、およびウマ(E.caballus)(配列番号24、Genebank登録番号XP_001503057)からの相同遺伝子もまたより詳細に調べた。
【0110】
選択された各25−ヒドロキシラーゼのS.セレヴィシエ(S.cerevisiae)中における発現のための2つの異なるコドン最適化遺伝子を計算した(配列番号19、20、22、23、25、26)。最後に接尾語「_opt」が付いている遺伝子は全て、DNA 2.0社(MenloPark,CA,USA)によって、独自仕様のアルゴリズムを使用してコドン最適化された。接尾語「_HY」がある全遺伝子は、常に、配列中に生じるアミノ酸の各位置に、各アミノ酸の最も一般的なコドンを配置させて作成した。酵母のための典型的なコドン使用頻度表は公的に入手可能であり、またプログラムパッケージLasergeneの一部でもある。
【0111】
イノシシ(S.scrofa)からの25−ヒドロキシラーゼの2つの新しい最適化された遺伝子(実施例2参照)もまた、上で述べたのと同一方法を使用して計算した(配列番号27、28)。
【0112】
インプットとして既知のアルゴリズムClustalWを用いて計算されるアミノ酸配列アラインメントを使用して、各位置に最も豊富なアミノ酸を単に選択することで、共通アミノ酸配列を判定した。「最も豊富なアミノ酸」は、判定できない場合は、それぞれの位置のアミノ酸を恣意的に選択した。このConHyDアミノ酸配列(配列番号29)もまた、上述のようにして、2つの異なるDNA配列(配列番号30、31)に逆翻訳した。
配列番号18:チンパンジー(P.troglodytes)からの推定上のコレステロール25−ヒドロキシラーゼのアミノ酸配列。
【化14】
配列番号19:DNA配列チンパンジー(P.troglodytes)CH25OH_opt 2.0
【化15】
配列番号20:チンパンジー(P.troglodytes)CH25OH_HYのDNA配列
【化16】
配列番号21:ドブネズミ(R.norvegicus)からの推定上のコレステロール25−ヒドロキシラーゼのアミノ酸配列
【化17】
配列番号22:ドブネズミ(R.norvegicus)CH25OH_opt 2.0のDNA配列
【化18】
配列番号23:ドブネズミ(R.norvegicus)CH25OH_HYのDNA配列
【化19】
配列番号24:ウマ(E.caballus)からの推定上のコレステロール25−ヒドロキシラーゼのアミノ酸配列
【化20】
配列番号25:ウマ(E.caballus)CH25OH_opt 2.0のDNA配列
【化21】
配列番号26:ウマ(E.caballus)CH25OH_HYのDNA配列
【化22】
配列番号27:イノシシ(S.scrofa)CH25OH_opt 2.0のDNA配列
【化23】
配列番号28:イノシシ(S.scrofa)CH25OH_HYのDNA配列
【化24】
配列番号29:ConHyDのアミノ酸配列
【化25】
配列番号30:ConHyD_opt 2.0のDNA配列
【化26】
配列番号31:ConHyD_HYのDNA配列
【化27】
【0113】
[実施例19]
[ERG6に組み込まれた発現コンストラクトからの新しい25−ヒドロキシラーゼ遺伝子の発現]
実施例18に示す新しい遺伝子をS.セレヴィシエ(S.cerevisiae)SC0554(erg5、erg6、TRP1−pARC300D、URA3−pARC304S、ARE1::TDH3p−S24R2−PGKt−URA3;2つのプラスミドに関する説明は米国特許第5,460,949号明細書を参照されたい)のERG6遺伝子座に組み込んだ。
【0114】
組み込みコンストラクトの配列を下に示す。関心のあるヒドロキシラーゼを組み込みコンストラクトのBam HIおよびEco RI部位の間にクローンした。ERG6組み込みコンストラクトは、プラスミドV51TDHに見出されるような、発現のためのS.セレヴィシエ(S.cerevisiae)のTDH3プロモーターおよびPGKターミネーター、および選択のためのS.セレヴィシエ(S.cerevisiae)のURA3を保有した。培養、形質転換、および選択は上述のように実施した。
【0115】
最終株は、次のように称された。
S.セレヴィシエ(S.cerevisiae)Ecab HY(配列番号26の遺伝子を有する組み込みコンストラクト)、
Ecab opt 2.0(配列番号25の遺伝子を有する組み込みコンストラクト)、
Ptro opt 2.0(配列番号19の遺伝子を有する組み込みコンストラクト)、
Ptro HY(配列番号20の遺伝子を有する組み込みコンストラクト)、
ConHyD HY(配列番号31の遺伝子を有する組み込みコンストラクト)、
ConHyD opt 2.0(配列番号30の遺伝子を有する組み込みコンストラクト)、
Rnor HY(配列番号23の遺伝子を有する組み込みコンストラクト)、
Rnor opt 2.0(配列番号22の遺伝子を有する組み込みコンストラクト)、
Sscr HY(配列番号28の遺伝子を有する組み込みコンストラクト)、
Sscr opt 2.0(配列番号27の遺伝子を有する組み込みコンストラクト)、および
実施例2に記載されるようにして最適化されたブタからの25−ヒドロキシラーゼを保有するScr Pompon。
【0116】
配列番号32:ERG6組み込みコンストラクトのDNA配列
【化28】
【0117】
[実施例20]
[新しいHyDHC株の振盪フラスコアッセイ]
実施例19に記載されるようにして作成された株を震盪フラスコ内で培養し、新たに設計された推定上の25−ヒドロキシラーゼ遺伝子の過剰発現効果を評価した。20mlのKapelli培地(実施例1)に接種するために、4mlのYPD培地中の一晩培養物を使用した。24および48時間培養した後に10g/Lのグルコース、32および56時間後に20g/Lのグルコースを供給した。72時間の培養時間後、細胞を遠沈して鹸化し、実施例9に記載されるようにしてステロール含量を測定した。図15に示すように、全ての新しいヒドロキシラーゼ遺伝子は、S.セレヴィシエ(S.cerevisiae)中で発現され、効率は様々であったが、遺伝子産物は全て7−DHCをHyDHCにヒドロキシル化できた。
【0118】
文献、それぞれが参照により本明細書に組み込まれる:
Brachmann C.B., Davies A., Cost G.J., Caputo E., Li J., Hieter P., Boeke J.D. (1998年). Yeast 14(2): 115−132.
Crameri A., Biondi E., Kuehnle K., Lutjohann D., Thelen K.M., Perga S., Dotti C.G., Nitsch R.M., Ledesma M.D., Mohajeri M.H. (2006年). EMBO J. 25(2):432−43.
Cullin C., Pompon D. (1988年). Gene 65:203−217.
Bonneaud N., Ozier−Kalogeropoulos O., Li G.Y., Labouesse M., Minvielle−Sebastia L., Lacroute F. (1991年). Yeast 7:609−615.
Gaber R.F., Copple D.M., Kennedy B.K., Vidal M, Bard M. (1989年). Mol Cell Biol 9(8):3447−3456.
Guarente L., Yocum R.R., Gifford P. (1982年). Proc Natl Acad Sci USA 79:7410−7414.
Hoffman C.S. and Winston F. (1987年). Gene 57:267−272
Gietz R.D., Schiestl R.H., Willems A.R., Woods R.A. (1995年). Yeast 11:355−360.
Lecain E., Chenivesse X., Spagnoli R., Pompon D. (1996年). J Biol Chem 271(18):10866−10873.
Lund E.G., Kerr T.A., Sakai J., Li W.P., Russell D.W. (1998年). J Biol Chem 273(51):34316−34327.
Sambrook J., Fritsch E. F., Maniatis T. (1998年). Molecular cloning: a laboratory manual. 2nd ed. Cold Spring Harbor, N.Y: Cold Spring Harbor Laboratory.
Secen H., Kalpar H. (1999年). Turk J Chem 23: 27−30.
Thomas B.J. and Rothstein R. (1989年). Cell 56(4):619−630.
Waterham H.R., Koster J., Romeijn G.J., Hennekam R.C.M., Vreken P., Andersson H.C., FitzPatrick D.R., Kelley R.I., Wanders R.J.A. (2001年). Am J Hum Genet 69:685−694.
Wu C., Miloslavskaya I., Demontis S., Maestro R., Galaktionov K. (2004年). Nature 432 (7017):640−5.
Zhang S.P., Zubay G., Goldman E. (1991年). Gene 1005:61−72.
【特許請求の範囲】
【請求項1】
コレステロールC25−ヒドロキシラーゼまたはコレステロールC25−ヒドロキシラーゼをコードする核酸を含んでなる酵母。
【請求項2】
25−ヒドロキシ−7−デヒドロコレステロールまたは25−ヒドロキシエルゴステロールをそれぞれ生成するための、7−デヒドロコレステロールまたはエルゴステロールをヒドロキシル化するコレステロールC25−ヒドロキシラーゼの使用。
【請求項3】
前記生成が酵母細胞中で起きる、請求項2に記載の使用。
【請求項4】
前記酵母細胞が、サッカロミセス・セレヴィシエ(Saccharomyces cerevisiae)、ピキア属(Pichia)種、クリヴェロミセス(Kluyveromyces)(Klyuveromyces)種、ハンゼヌラ(Hansenula)種、分裂酵母(Schizosaccharomyces)種、およびヤロウィア・リポリティカ(Yarrowia lipolytica)からなる群から選択される、請求項2または3に記載の使用。
【請求項5】
a)7−デヒドロコレステロールを前記酵母によって産生されるコレステロールC25−ヒドロキシラーゼに接触させることで、25−ヒドロキシ−7−デヒドロコレステロールが生成するステップ
を含んでなる、25−ヒドロキシ−7−デヒドロコレステロールを遺伝子改変酵母中で生成する方法。
【請求項6】
前記酵母が活性Erg5pおよびErg6p酵素を有さず、前記酵母がステロールΔ24−還元酵素もまた発現する、請求項5に記載の方法。
【請求項7】
前記酵母がHMG1の切断型を発現する、請求項5または6に記載の方法。
【請求項8】
ステップa)に先だって、コレスタ−5,7,24−トリエノールを機能性ステロールΔ24−還元酵素をコードする核酸配列を含んでなる酵母に接触させることで、前記酵母がラノステロール、ジメチルチモステロール、メチルチモステロール、チモステロール、コレスタ−7,24−ジエノール、またはコレスタ−5,7,24−トリエノールをそれぞれ3β−ヒドロキシ−8−ラノスタ−8−エン、4,4−ジメチル−コレスタ−8−エノール、4α−メチル−コレスタ−8−エノール、コレスタ−8−エノール、ラソステロールまたは7−デヒドロコレステロールに変換するステップをさらに含んでなる、請求項5に記載の方法。
【請求項9】
a)前記酵母によって産生されるエルゴステロールを前記酵母によって産生されるコレステロールC25−ヒドロキシラーゼに接触させることで、25−ヒドロキシエルゴステロールが生成するステップ
を含んでなる、遺伝子改変酵母中で25−ヒドロキシエルゴステロール(25−ヒドロキシプロビタミンD2)を生成する方法。
【請求項10】
前記酵母がHMG1の切断型を過剰発現する、請求項9に記載の方法。
【請求項11】
コレステロールC25−ヒドロキシラーゼをコードする、単離された核酸。
【請求項12】
コレステロールC25−ヒドロキシラーゼをコードする核酸を含んでなる、ベクター。
【請求項13】
ステロールΔ24−還元酵素をコードし、酵母によって発現され得て、ラノステロール、ジメチルチモステロール、メチルチモステロール、チモステロール、コレスタ−7,24−ジエノール、またはコレスタ−5,7,24−トリエノールを3β−ヒドロキシ−8−ラノスタ−8−エン、4,4−ジメチル−コレスタ−8−エノール、4α−メチル−コレスタ−8−エノール、コレスタ−8−エノール、ラソステロールまたは7−デヒドロコレステロールに変換し得る単離された核酸であるが、ただし核酸配列がヒト核酸配列でもマウス核酸配列でもない、単離された核酸。
【請求項1】
コレステロールC25−ヒドロキシラーゼまたはコレステロールC25−ヒドロキシラーゼをコードする核酸を含んでなる酵母。
【請求項2】
25−ヒドロキシ−7−デヒドロコレステロールまたは25−ヒドロキシエルゴステロールをそれぞれ生成するための、7−デヒドロコレステロールまたはエルゴステロールをヒドロキシル化するコレステロールC25−ヒドロキシラーゼの使用。
【請求項3】
前記生成が酵母細胞中で起きる、請求項2に記載の使用。
【請求項4】
前記酵母細胞が、サッカロミセス・セレヴィシエ(Saccharomyces cerevisiae)、ピキア属(Pichia)種、クリヴェロミセス(Kluyveromyces)(Klyuveromyces)種、ハンゼヌラ(Hansenula)種、分裂酵母(Schizosaccharomyces)種、およびヤロウィア・リポリティカ(Yarrowia lipolytica)からなる群から選択される、請求項2または3に記載の使用。
【請求項5】
a)7−デヒドロコレステロールを前記酵母によって産生されるコレステロールC25−ヒドロキシラーゼに接触させることで、25−ヒドロキシ−7−デヒドロコレステロールが生成するステップ
を含んでなる、25−ヒドロキシ−7−デヒドロコレステロールを遺伝子改変酵母中で生成する方法。
【請求項6】
前記酵母が活性Erg5pおよびErg6p酵素を有さず、前記酵母がステロールΔ24−還元酵素もまた発現する、請求項5に記載の方法。
【請求項7】
前記酵母がHMG1の切断型を発現する、請求項5または6に記載の方法。
【請求項8】
ステップa)に先だって、コレスタ−5,7,24−トリエノールを機能性ステロールΔ24−還元酵素をコードする核酸配列を含んでなる酵母に接触させることで、前記酵母がラノステロール、ジメチルチモステロール、メチルチモステロール、チモステロール、コレスタ−7,24−ジエノール、またはコレスタ−5,7,24−トリエノールをそれぞれ3β−ヒドロキシ−8−ラノスタ−8−エン、4,4−ジメチル−コレスタ−8−エノール、4α−メチル−コレスタ−8−エノール、コレスタ−8−エノール、ラソステロールまたは7−デヒドロコレステロールに変換するステップをさらに含んでなる、請求項5に記載の方法。
【請求項9】
a)前記酵母によって産生されるエルゴステロールを前記酵母によって産生されるコレステロールC25−ヒドロキシラーゼに接触させることで、25−ヒドロキシエルゴステロールが生成するステップ
を含んでなる、遺伝子改変酵母中で25−ヒドロキシエルゴステロール(25−ヒドロキシプロビタミンD2)を生成する方法。
【請求項10】
前記酵母がHMG1の切断型を過剰発現する、請求項9に記載の方法。
【請求項11】
コレステロールC25−ヒドロキシラーゼをコードする、単離された核酸。
【請求項12】
コレステロールC25−ヒドロキシラーゼをコードする核酸を含んでなる、ベクター。
【請求項13】
ステロールΔ24−還元酵素をコードし、酵母によって発現され得て、ラノステロール、ジメチルチモステロール、メチルチモステロール、チモステロール、コレスタ−7,24−ジエノール、またはコレスタ−5,7,24−トリエノールを3β−ヒドロキシ−8−ラノスタ−8−エン、4,4−ジメチル−コレスタ−8−エノール、4α−メチル−コレスタ−8−エノール、コレスタ−8−エノール、ラソステロールまたは7−デヒドロコレステロールに変換し得る単離された核酸であるが、ただし核酸配列がヒト核酸配列でもマウス核酸配列でもない、単離された核酸。
【図1】

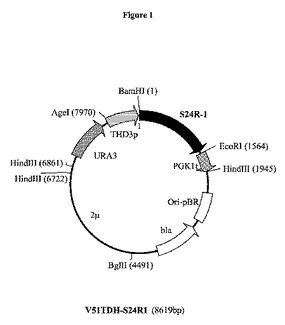
【図2】
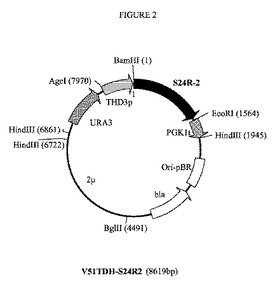
【図3】
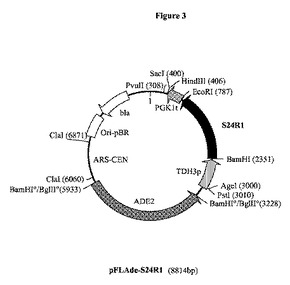
【図4】
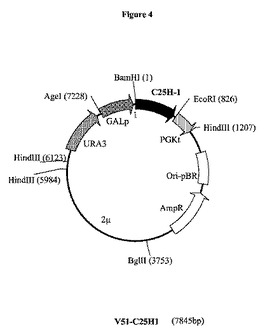
【図5】
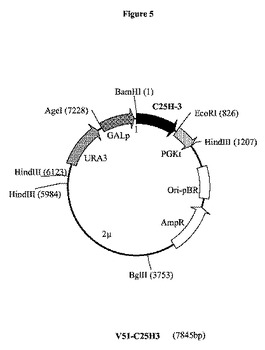
【図6】
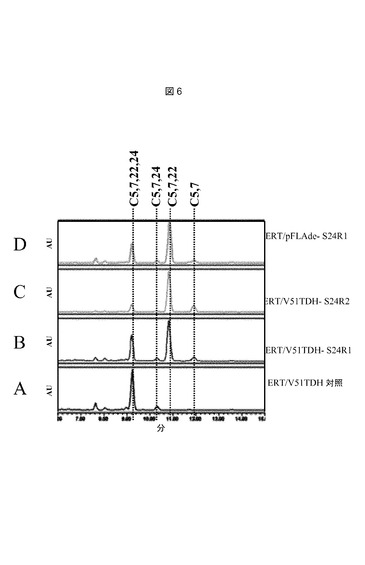
【図7】
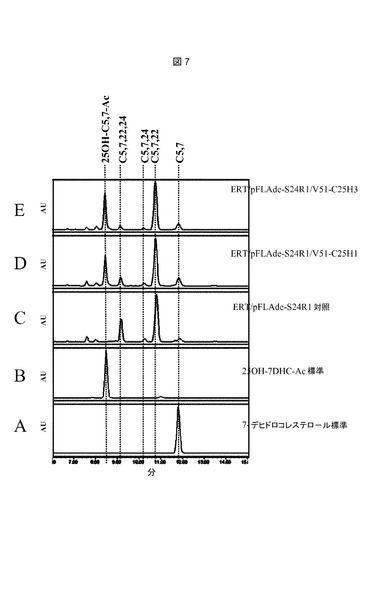
【図8】
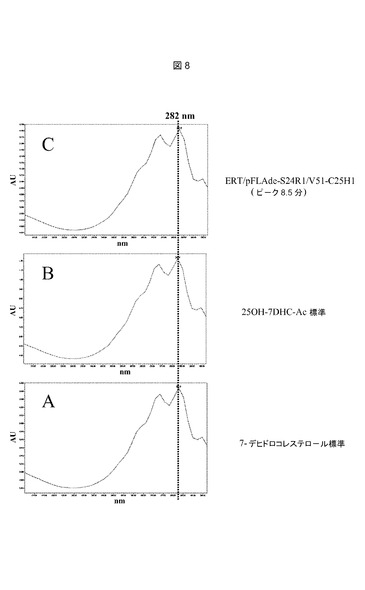
【図9】
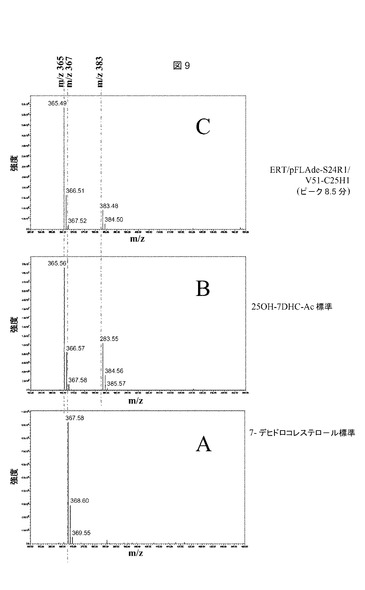
【図10】
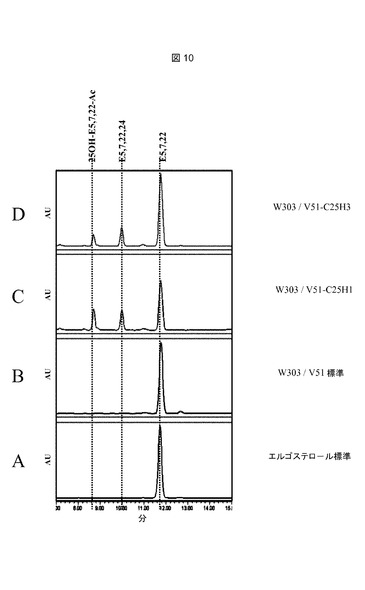
【図11】
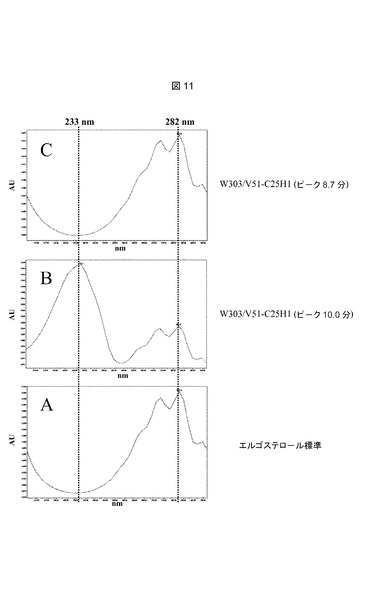
【図12】
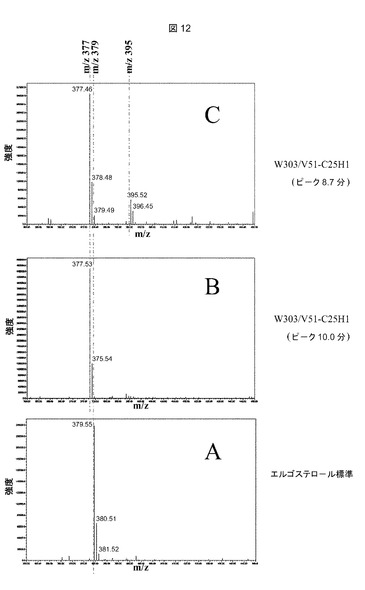
【図13】
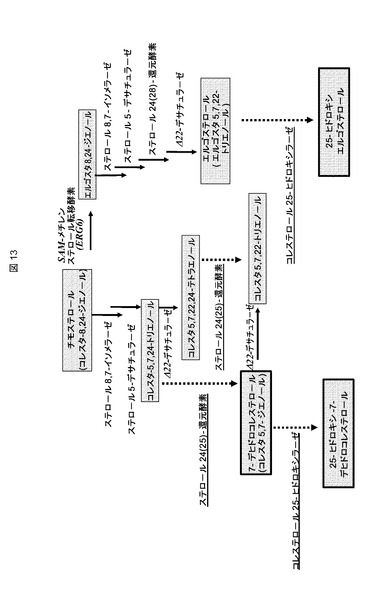
【図14】
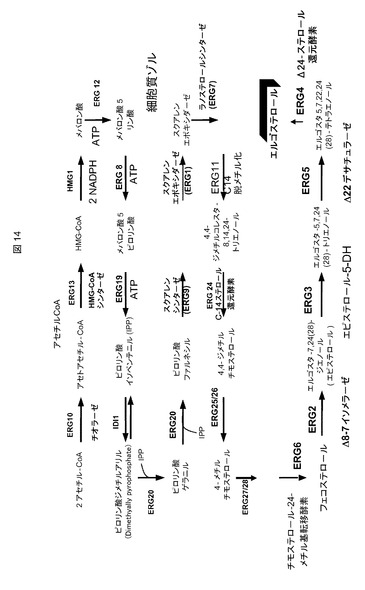
【図15】
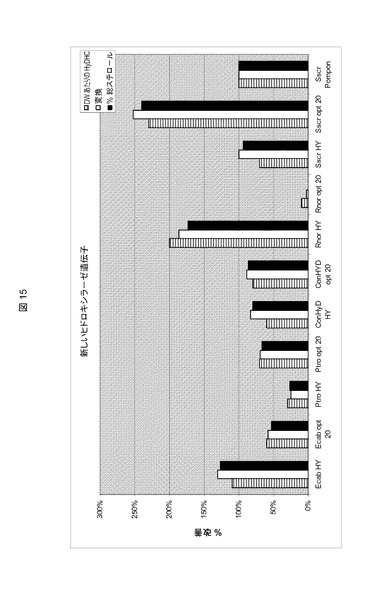
【図2】
【図3】
【図4】
【図5】
【図6】
【図7】
【図8】
【図9】
【図10】
【図11】
【図12】
【図13】
【図14】
【図15】
【公表番号】特表2013−512663(P2013−512663A)
【公表日】平成25年4月18日(2013.4.18)
【国際特許分類】
【出願番号】特願2012−541408(P2012−541408)
【出願日】平成22年11月24日(2010.11.24)
【国際出願番号】PCT/EP2010/068127
【国際公開番号】WO2011/067144
【国際公開日】平成23年6月9日(2011.6.9)
【出願人】(503220392)ディーエスエム アイピー アセッツ ビー.ブイ. (873)
【Fターム(参考)】
【公表日】平成25年4月18日(2013.4.18)
【国際特許分類】
【出願日】平成22年11月24日(2010.11.24)
【国際出願番号】PCT/EP2010/068127
【国際公開番号】WO2011/067144
【国際公開日】平成23年6月9日(2011.6.9)
【出願人】(503220392)ディーエスエム アイピー アセッツ ビー.ブイ. (873)
【Fターム(参考)】
[ Back to top ]