
酵母を用いたマンニトールからのエタノール生産
【課題】酵母によるマンニトールからのエタノールの生産方法及びマンニトールからエタノールを生産する酵母株の提供。
【解決手段】マンニトール資化能を有しマンニトールからエタノールを産生し得る酵母を、マンニトールを含む培地中で培養することを含む、マンニトールを原料としてエタノールを生産する方法。
【解決手段】マンニトール資化能を有しマンニトールからエタノールを産生し得る酵母を、マンニトールを含む培地中で培養することを含む、マンニトールを原料としてエタノールを生産する方法。
【発明の詳細な説明】
【技術分野】
【0001】
本発明は、酵母によるマンニトールからのエタノールの生産方法及びマンニトールからエタノールを生産する酵母株に関する。
【背景技術】
【0002】
大型海藻などの海洋バイオマスは有望なバイオ燃料の原料である。その主な理由として、(i) 陸性バイオマスと比較すると、大型海藻は高い生産性を示す、(ii) 耕作地を必要としないため、陸性バイオマスを栽培する上で不可避となる問題(灌漑や施肥など)を避けられる、(iii) リグニンを含まない、等があげられる。大型海藻は、主に緑藻、紅藻、及び褐藻から構成される。うち、少なくとも紅藻と褐藻は著量の炭水化物を含む。紅藻の一種テングサGelidium amansiiは、17%(w/w乾燥重量;以下、特に言及しない場合、w/wは全て乾燥重量を表す)のセルロース(グルコース)、58.6%(w/w)の寒天(25.6%のガラクトースと33%の3,6-アンヒドロガラクトース)を含む。褐藻類は最大で40%(w/w)のアルギン酸、最大30%(w/w)のマンニトール、及び最大30%(w/w)のラミナリンを含む。かかるがゆえに、大型海藻を原料としたバイオ燃料の生産には、これらの炭水化物の成分をバイオ燃料に変換する技術の確立が必須である。
【0003】
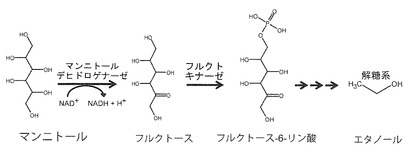
アルギン酸は、β-D-マンヌロン酸(M)とそのC5エピマーであるα-L-グルロン酸(G)から構成される直鎖状の酸性多糖である。構成単糖は、ポリM、ポリG、あるいはポリMG構造をとる。マンニトールは、マンノースに対応する糖アルコールであり、マンニトールデヒドロゲナーゼの作用により酸化されてフルクトースになる(図1)(非特許文献1及び2を参照)。ラミナリンは、β-(1,6)枝分かれ構造を有するβ-(1,3)-D-グルカンから構成される(非特許文献3及び4を参照)。既に、本発明者らはエタノール生産性Sphingomonas sp. A1 株(以下、エタノール生産性A1株)を用いたアルギン酸からのエタノール生産システムを構築しており、1.3%(w/v)のエタノール生産に成功している(非特許文献5を参照)。本法は、アルギン酸からバイオ燃料を生産する唯一の方法である。ラミナリンからのエタノール生産例は少ないが、酵母 Kluyveromyces marxianus、Pacchysolen tannophilus、及びPhicia angophoraeの3株がラミナリンを分解してエタノールを生産するとの報告(非特許文献1を参照)、及びラミナリン分解酵素ラミナリナーゼを用いて分解したラミナリン分解物から酵母Saccharomyces cerevisiaeを用いてエタノールを生産したとの報告(Adams et al., 2009)がある。マンニトールからのエタノール生産に関しては、Zymobacter palmaeとEscherichia coli KO11の両細菌が、各々、3.8%(w/v)及び9.0%(w/v)のマンニトールから、約1.3%(w/v)及び2.6%(w/v)のエタノールを、0.38g及び0.41gエタノール(gマンニトール)-1 の生産効率で生産したことが報告されている(非特許文献1及び6を参照)。しかし、エタノールを生産する上では、細菌よりも酵母の方がエタノールや発酵阻害物質への耐性能を含む様々な利点を有すると見なされている(非特許文献7を参照)(Hughes and Qureshi, 2010)。実際、Z. palmae とE. coli KO11は、5%(w/v)のエタノール存在下で生育阻害を示す(非特許文献8及び9を参照)。しかし、酵母を用いたマンニトールからのエタノール生産の研究例は少なく、酵母S. cerevisiaeポリプロイド BB1株が、5%(w/v)のマンニトールから約0.5%のエタノールを生産すること(非特許文献2を参照)、及び先述のラミナリン分解酵母(K. marxianus、P. tannophilus、及びP. angophorae)のうち、P. angophoraeのみが、4%(w/v)マンニトールから約1.0%(w/v)のエタノールを0.40gエタノール(g マンニトール)-1の生産効率で生産することが報告されているのみである(非特許文献1を参照)。このP. angophoraeを用いた研究に関しては、マンニトールとラミナリンが共存する海藻抽出物からのエタノール生産や、酸素供給量のマンニトールとラミナリン消費速度に対する影響が報告されている(非特許文献1を参照)。一方、酵母のマンニトール代謝に関する報告も少なく、酵母S. cerevisiaeには、マンニトールを資化できる株(先述のポリプロイドBB1株など、一倍体A184D株)とできない(あるいは同資化能が極めて弱い)株(ポリプロイドBB2株など、一倍体S288C株やSc41 YJO 株)が存在すること、S. cerevisiaeによるマンニトールの資化には酸素が必要でありマンニトール含有培地で生育中の酵母は高い呼吸活性を示すことなどが報告されている(非特許文献2及び10を参照)。なお、一倍体S288C株は最初のゲノム配列決定株でもある(非特許文献11を参照)。
【0004】
酵母を用いたマンニトールからのエタノール生産の実用化には、マンニトールからの高いエタノール生産能力やその他の様々な優良特性を示す酵母の探索あるいは育種、及び高い生産能力を発揮する最適条件の確立が必須である。さらには、海洋バイオマスからのエタノール生産系の確立には、その構成成分をもれなくエタノールに変換する技術も必要である。褐藻類の場合、アルギン酸、マンニトール、及びラミナリンなどからエタノールに変換する技術の確立を要する。アルギン酸からのエタノール生産系は、先述したエタノール生産性A1株を用いた系しか知られていない(非特許文献5を参照)。本A1株は、マンニトールやラミナリンを資化できない(未発表データ)。一方で、アルギン酸資化能はSphingomonas sp. A1株などの限られた生物にしか知られていない。
【先行技術文献】
【非特許文献】
【0005】
【非特許文献1】Horn et al., 2000, J. Ind. Microbiol. Biotechnol. 25, 249-254
【非特許文献2】Quain and Boulton, 1987, J. Gen. Microbiol. 133, 1675-1684.
【非特許文献3】Nelson and Lewis, 1973, Carbohydr. Res. 33, 63-74.
【非特許文献4】Zvyagintseva et al., 1999, Carbohydr. Res. 322, 32-39.
【非特許文献5】Takeda et al., 2011, Energy Environ. Sci. 4, 2575-2581
【非特許文献6】Kim et al., 2011, Bioresour. Technol. 102, 7466-7469.
【非特許文献7】Hughes and Qureshi, 2010, Biomass to biofuels: strategies for global industries, pp. 55-69.
【非特許文献8】Okamoto et al., 1994, Biosci. Biotech. Bioch. 58, 1328-1329.
【非特許文献9】Yomano et al., 1998 J. Ind. Microbiol. Biotechnol. 20, 132-138.
【非特許文献10】Perfect et al., 1996, J. Bacteriol. 178, 5257-5262.
【非特許文献11】Goffeau et al., 1996, Science 274, 546, 563-547.
【発明の概要】
【発明が解決しようとする課題】
【0006】
本発明は、酵母によるマンニトールからのエタノールの生産方法及びマンニトールからエタノールを生産する酵母株の提供を目的とする。
【課題を解決するための手段】
【0007】
本発明者らは、褐藻類由来バイオマスのエタノール生産への有効利用には、エタノール生産性A1株によるアルギン酸からのエタノール発酵残渣に含まれるマンニトール及びラミナリンから、マンニトール及びラミナリン資化性酵母を用いてエタノールを生産する二段階発酵の実現が有効であると考え、この実現を目指して、まずマンニトールから高いエタノール生産能を示す酵母の探索を行った。
【0008】
海洋バイオマス(褐藻類)からのバイオ燃料(エタノール)生産には、その主要成分アルギン酸、マンニトール、及びラミナリンからエタノールを生産する技術が不可欠である。エタノール生産性Sphingomonas sp. A1株を用いたアルギン酸からのエタノール生産系は確立されている(Takeda et al., 2011, Energy Environ. Sci. 4, 2575-2581)。本株によるアルギン酸からのエタノール発酵後の残渣に含まれるマンニトールやラミナリンのエタノールへの変換は、褐藻類主要成分の有効利用法の一つである。本発明者らは、保存酵母株の中から、マンニトール資化性エタノール生産株6株を見出した。うち、Saccharomyces paradoxus NBRC0259株のみは、ラミナリンからのエタノール生産性は示さないものの、グルコース及びマンニトールからの高いエタノール生産性、エタノール耐性、及びアルギン酸からのエタノール発酵残渣における生育性といった望ましい特性を示した。他の5株はラミナリンからのエタノール生産性を示したが、他の特性がNBRC 0259株より劣った。NBRC 0259株は、95 strokes per min (spm) の振とうによって得られる微好気環境において、10% (w/v)マンニトールから最大37.6 g/l (3.8% w/v)のエタノールを生産した。エタノール発酵に対するNaClの影響は小さかった。また、アルギン酸からのエタノール発酵残渣中のマンニトールからもエタノールを生産した。本菌は、海洋バイオマスからのエタノール生産に有用と考えられた。
【0009】
すなわち、本発明は以下のとおりである。
[1] マンニトール資化能を有しマンニトールからエタノールを産生し得る酵母を、マンニトールを含む培地中で培養することを含む、マンニトールを原料としてエタノールを生産する方法。
[2] マンニトール資化能を有しマンニトールからエタノールを産生し得る酵母が、以下の(1)〜(3)の少なくとも1つの特性を有する酵母である、[1]のマンニトールを原料としてエタノールを生産する方法:
(1)エタノール耐性を有する;
(2)アルギン酸資化能を有する微生物を用いて褐藻原料からエタノールを生産したときの残渣中で生育し得る;及び
(3)グルコース存在下での凝集能を有する。
[3] マンニトール資化能を有しマンニトールからエタノールを産生し得る酵母が、Saccharomyces paradoxus、Debaryomyces hansenii、Kuraishia capsulata、Ogataea glucozyma及びOgataea minutaからなる群から選択される、[1]又は[2]のマンニトールを原料としてエタノールを生産する方法。
[4] マンニトール資化能を有しマンニトールからエタノールを産生し得る酵母が、Saccharomyces paradoxus NBRC 0259株、Debaryomyces hansenii NBRC 0794株、 Kuraishia capsulata NBRC 0721株、Kuraishia capsulata NBRC 0974株、Ogataea glucozyma NBRC 1472株及びOgataea minuta NBRC 1473株からなる群から選択される、[3]のマンニトールを原料としてエタノールを生産する方法。
[5] マンニトール資化能を有しマンニトールからエタノールを産生し得る酵母を培養することにより、培地中に0.1%(w/v)以上のエタノールが蓄積し得る、[1]〜[4]のいずれかの方法。
[6] マンニトール資化能を有しマンニトールからエタノールを産生し得る酵母を培養することにより、培地中に3%(w/v)以上のエタノールが蓄積し得る、[5]の方法。
[7] アルギン酸資化能を有しアルギン酸からエタノールを産生し得る微生物を用いて、褐藻原料からエタノールを産生させたときの原料残渣に、マンニトール資化能を有しマンニトールからエタノールを産生し得る酵母を添加し、培養し、残渣中のマンニトールを原料としてエタノールを生産する方法。
[8] マンニトール資化能を有しマンニトールからエタノールを産生し得る酵母が、以下の(1)〜(3)の少なくとも1つの特性を有する酵母である、[7]の残渣中のマンニトールを原料としてエタノールを生産する方法:
(1)エタノール耐性を有する;
(2)アルギン酸資化能を有する微生物を用いて褐藻原料からエタノールを生産したときの残渣中で生育し得る;及び
(3)グルコース存在下での凝集能を有する。
[9] マンニトール資化能を有しマンニトールからエタノールを産生し得る酵母がSaccharomyces paradoxusである、[7]又は[8]の残渣中のマンニトールを原料としてエタノールを生産する方法。
[10] マンニトール資化能を有しマンニトールからエタノールを産生し得る酵母がSaccharomyces paradoxus NBRC 0259株である、[9]の残渣中のマンニトールを原料としてエタノールを生産する方法。
[11](i) アルギン酸資化能を有しアルギン酸からエタノールを産生し得る微生物を褐藻原料を用いて培養し、アルギン酸からエタノールを産生させ、
(ii) (i)の培養の原料残渣に、マンニトール資化能を有しマンニトールからエタノールを産生し得る酵母を添加し、培養することを含む、
褐藻原料からエタノールを生産する方法。
[12] マンニトール資化能を有しマンニトールからエタノールを産生し得る酵母が、以下の(1)〜(3)の少なくとも1つの特性を有する酵母である、[11]の褐藻原料からエタノールを生産する方法:
(1)エタノール耐性を有する;
(2)アルギン酸資化能を有する微生物を用いて褐藻原料からエタノールを生産したときの残渣中で生育し得る;及び
(3)グルコース存在下での凝集能を有する。
[13] マンニトール資化能を有しマンニトールからエタノールを産生し得る酵母がSaccharomyces paradoxusである、[11]又は[12]の褐藻原料からエタノールを生産する方法。
[14] マンニトール資化能を有しマンニトールからエタノールを産生し得る酵母がSaccharomyces paradoxus NBRC 0259株である、[13]の褐藻原料からエタノールを生産する方法。
【発明の効果】
【0010】
本発明のマンニトール資化能を有しマンニトールからエタノールを産生し得る酵母株を用いることにより、マンニトールを原料としてエタノールを生産することができる。海域由来のバイオマス、とりわけ褐藻類に多量含有される多糖アルギン酸からのエタノール生産の残渣には、大量のマンニトールが含まれており、該残渣をエタノール生産に有効に利用することができる。
【図面の簡単な説明】
【0011】
【図1】酵母における推定マンニトール資化経路を示す図である。
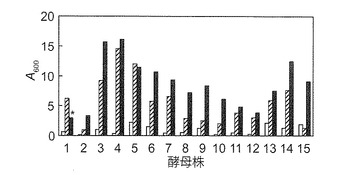
【図2A】マンニトール資化性酵母の炭素源非含有液体培地(白色)、マンニトール合成液体培地(細い平行線)及びグルコース合成液体培地(黒色)における生育性を示す図である。酵母株: 1. S. paradoxus NBRC 0259; 2. Z. japonicus IFO 0595; 3. P. polymorpha IFO 0195; 4. P. farinosa NBRC 0193; 5. P. haplophila NBRC 0947; 6. P. saitoi IAM 4945; 7. H. saturnus IFO 0177; 8. K. capsulata NBRC 0721; 9. K. capsulata NBRC 0974; 10. O. glucozyma NBRC 1472; 11. O. minuta NBRC 1473; 12. D. hansenii IFO 0023; 13. D. hansenii NBRC 0794; 14. Y. lipolytica NBRC 0746; 15. S. cerevisiae BY4742 (対照)。S. paradoxus NBRC 0259株(株 1)はグルコース合成液体培地で強い凝集性(*)、マンニトール合成培地で弱い凝集性を示した。
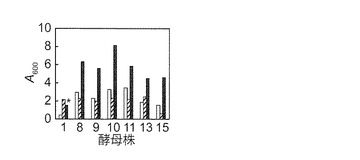
【図2B】マンニトール資化性エタノール生産酵母のグルコース合成液体培地における生育性を示す図である。酵母株の番号は図2Aと同様であり、1: S. paradoxus NBRC 0259; 8. K. capsulata NBRC 0721; 9. K. capsulata NBRC 0974; 10. O. glucozyma NBRC 1472; 11. O. minuta NBRC 1473; 13. D. hansenii NBRC 0794; 15, S. cerevisiae BY4742である。S. paradoxus NBRC 0259株(株 1)はグルコース合成液体培地で強い凝集性(*)を示した。
【図2C−1】マンニトール資化性エタノール生産酵母のラミナリン合成液体培地(白色)、マンニトール合成液体培地(細い平行線)及びグルコース合成液体培地(黒色)における生育性を示す図である。酵母株の番号は図2Bと同様である。S. paradoxus NBRC 0259株(株 1)はグルコース合成液体培地で強い凝集性(*)を示した。
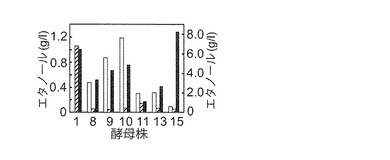
【図2C−2】マンニトール資化性エタノール生産酵母のラミナリン合成液体培地(白色)、マンニトール合成液体培地(細い平行線)及びグルコース合成液体培地(黒色)におけるエタノール生産性を示す図である。グルコース由来のエタノール濃度を右側のスケールで、ラミナリン又はマンニトール由来のエタノール濃度を左側のスケールで示す。酵母株の番号は図2Bと同様である。
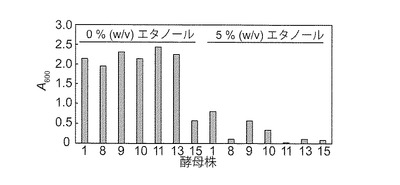
【図2D】マンニトール資化性エタノール生産酵母のマンニトール合成液体培地における5% (w/v)エタノール存在下での生育性を示す図である。YPD固体培地上で前培養した菌株を1.0 mlの培地にて静置(0 spm)で3日間培養した。酵母株の番号は図2Bと同様である。S. paradoxus NBRC 0259株(株 1)は5% (w/v)エタノール存在下で凝集性(*)を示した。
【図3A】マンニトール資化における酸素要求性を示す図であり、BY4742(BY)及びNBRC 0259(NB)のρ0及びρ+株の、通常大気下(+O2)及び嫌気条件下(-O2)における、マンニトール、グルコース、及びグリセロール合成固体培地での生育性(4日間培養)を示す図である。
【図3B】マンニトール資化における酸素要求性を示す図であり、マンニトール資化性酵母の通常大気下(+O2)及び嫌気条件下(-O2)における、マンニトール及びグルコース合成固体培地での生育性を示す図である。YPM固体培地上で前培養した株を各培地にストリークし、4日間培養した。酵母株の番号は図2Bと同様である。
【図4A−1】マンニトール資化性エタノール生産酵母6株の生育性を示す図である。YPM液体培地25 mlを含む50 ml三角フラスコを用いて95 spmで振とう培養した。酵母株のシンボルは、◆ S. paradoxus NBRC 0259;▲ K. capsulata NBRC 0721; X K. capsulata NBRC 0974;● O. glucozyma NBRC 1472; + O. minuta NBRC 1473;■ D. hansenii NBRC 0794である。
【図4A−2】マンニトール資化性エタノール生産酵母6株のエタノール生産性を示す図である。YPM液体培地25 mlを含む50 ml三角フラスコを用いて95 spmで振とう培養した。酵母株のシンボルは、図4A-1と同様である。
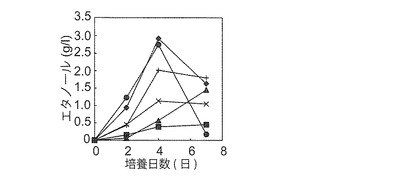
【図4B】YPM液体培地(pH 5.7)、弱アルカリ性の同培地(pH 7.8)、及びA1株によるアルギン酸からのエタノール発酵残渣[2%(w/v)マンニトールを含む;pH 7.3 (アルギン酸)]における培養4日後の生育性を示す図である。培地成分以外は図4Aの条件と同様の条件で培養した。YPM固体培地上で前培養した細胞を用いた。酵母株の番号は図2Bと同様である。弱アルカリ性のYPM液体培地で弱い凝集傾向(YPD液体培地で観察される凝集に類似しているが、程度が弱い)、並びに発酵残渣で凝集性(細かい顆粒状)を示した(*)。
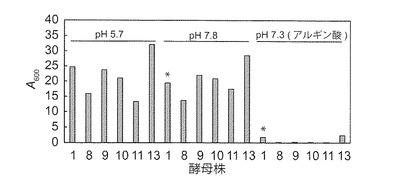
【図5A】NBRC 0259株のマンニトール合成液体培地における生育性を示す図である。NBRC 0259株のρ+株(YPM固体培地由来)(実線;●○)並びに対照としてBY4742株のρ+株(YPG固体培地由来)(点線;▲△)を、YPD(▲●)又はYPM(△○)固体培地で前培養して、1.0 mlのマンニトール合成液体培地で145 spmで振とう培養した。BY4742株は、マンニトール合成培地上では生育性を示さなかった。
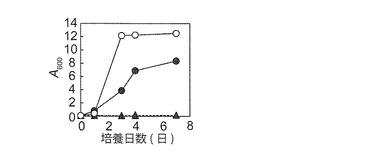
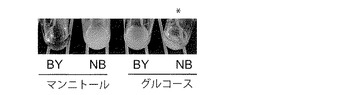
【図5B】YPD固体培地で前培養したBY4742株(BY)及びNBRC 0259株(NB)をマンニトール及びグルコース合成液体培地で培養(培養4日間)したときの状態を示す図である。上記(図5A)の条件で培養した。NBRC0259はグルコース合成液体培地で凝集性を示した(*)。
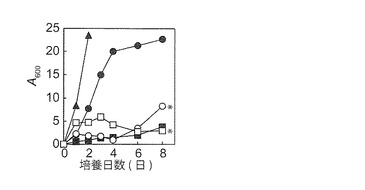
【図5C−1】YPM(▲●■)及びYPD(□○)液体培地における生育性に対する振とう数[▲145 spm;●○95 spm;■□ 0 spm]の影響を示す図である。YPM固体培地由来NBRC 0259 ρ+株をYPM液体培地で95 spmで24時間前培養した細胞を植菌して培養を開始した。NBRC 0259株はグルコース合成液体培地で凝集性を示した(*)。
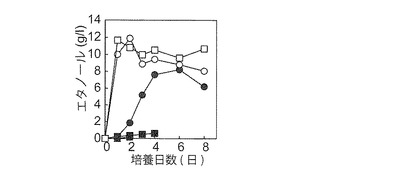
【図5C−2】YPM(▲●■)及びYPD(□○)液体培地におけるエタノール生産性に対する振とう数[▲145 spm;●○95 spm;■□0 spm]の影響を示す図である。YPM固体培地由来NBRC0259 ρ+株をYPM液体培地で95 spmで24時間前培養した細胞を植菌して培養を開始した。同株は145 spm ではエタノール生産性を示さなかった(▲)。
【図5D】NBRC 0259株のYPD及びYPM液体培地における生育性を示す図である。YPD培地では細胞がフロックを形成して沈殿している(*)。培養1日目の状態を示す。
【図5E】マンニトール濃度のエタノール生産性への影響を示す図である。初期菌体量(A600)を11、振とう速度を95 spmとし、YPM液体培地中のマンニトール濃度を2%(w/v: ■)、5%(w/v:●)、及び10%(w/v:▲)とした。初期菌体として、YPM液体培地で95 spmで4日間前培養した細胞を用いた。
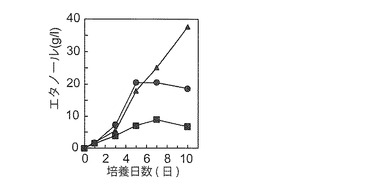
【図5F】NaCl濃度のエタノール生産性への影響を示す図である。図5Eの条件と同じ条件であるが、YPM液体培地(2%(w/v)マンニトール)を用いて、0%(w/v:■)、2%(w/v:●)、及び5%(w/v:▲)NaCl存在下で培養した。
【発明を実施するための形態】
【0012】
以下、本発明を詳細に説明する。
【0013】
本発明の酵母株は、マンニトール資化能を有し、マンニトールからエタノールを生産し得るマンニトール資化性酵母株である。マンニトールからエタノールを生産する反応を図1に示す。
【0014】
本発明のマンニトールからエタノールを生産し得る酵母株の例として以下の株が挙げられる。Saccharomyces paradoxus NBRC 0259、Debaryomyces hansenii NBRC 0794、 Kuraishia capsulata NBRC 0721、Kuraishia capsulata NBRC 0974、Ogataea glucozyma NBRC 1472及びOgataea minuta NBRC 1473。これらの株は、独立行政法人 製品評価技術基盤機構生物遺伝資源部門(NBRC)に保存されており、該機構より入手することができる。
【0015】
さらに、本発明のマンニトール資化能を有し、マンニトールからエタノールを生産し得る酵母株は、好ましくはエタノール耐性を有する。エタノール耐性はエタノール耐性化又はエタノール寛容化することによって付与される。例えば、エタノールを含有する培地中で馴養培養、及び紫外線照射によりランダム変異を誘発することにより、エタノール耐性株を得ることができる。本発明の酵母株は、0.85%(w/v)以上、好ましくは1.7%以上、さらに好ましくは3%(w/v)以上、さらに好ましくは5%(w/v)以上、さらに好ましくは6%(w/v)以上、さらに好ましくは7%(w/v)以上、さらに好ましくは8%(w/v)以上、特に好ましくは8.5%(w/v)以上のエタノール存在下で生育することができる。
【0016】
さらに、本発明のマンニトール資化能を有し、マンニトールからエタノールを生産し得る酵母株は、好ましくは、アルギン酸からのエタノール発酵残渣における生育性を有する。エタノール発酵残渣における生育性は、生育阻害物質に対する耐性能が関与していると予測される。ここで、アルギン酸からのエタノール発酵残渣とは、アルギン酸を含む褐藻等の原料からアルギン酸資化能を有する微生物を用いてエタノールを生産した場合の、原料の残渣をいう。該残渣には、アルギン酸以外の褐藻の主要成分であるマンニトールやラミナリンが含まれる。アルギン酸資化能を有する微生物として、Sphingomonas sp. A1株が挙げられ、該A1株にZymomonas mobilis等の細菌等のエタノール産生に関する酵素(ピルビン酸脱炭酸酵素及びアルコール脱水素酵素)をコードする遺伝子を導入したエタノール生産性Sphingomonas sp. A1株によりアルギン酸からエタノールを産生し得る。エタノール生産性Sphingomonas sp. A1株は、Takeda et al., 2011, Energy Environ. Sci. 4, 2575-2581や第WO2011/024858号国際公開パンフレットに詳述されている。本発明のマンニトール資化性を有し、マンニトールからエタノールを生産し得る酵母株は、上記のエタノール生産性Sphingomonas sp. A1株を用いて褐藻を原料にしてエタノールを生産した後に残るマンニトールを含む残渣を原料として用いてさらにエタノールを生産することができるので、海洋バイオマスである褐藻を有効利用して大量のエタノールを生産することができる。褐藻を原料として用いて上記のアルギン酸からエタノールを産生し得る微生物を用いてアルギン酸からエタノールを生産した後の残渣を用いて二段階発酵によりマンニトールからエタノールを生産してもよいし、褐藻由来の原料に上記のアルギン酸からエタノールを産生し得る微生物と本発明のマンニトール資化能を有し、マンニトールからエタノールを生産し得る酵母株を同時に添加し、同時にアルギン酸及びマンニトールからエタノールを産生させてもよい。さらに、同時にラミナリン資化性を有し、ラミナリンからエタノールを生産し得る微生物を添加してもよい。また、褐藻を原料として用いて上記のマンニトール資化能を有し、マンニトールからエタノールを生産し得る酵母株を用いてマンニトールからエタノールを生産した後の残渣を用いて、上記のアルギン酸からエタノールを産生し得る微生物による二段階発酵によりアルギン酸からエタノールを生産することもできる。
【0017】
さらに、本発明のマンニトール資化能を有し、マンニトールからエタノールを生産し得る酵母株は、好ましくは、グルコース存在下での凝集能を有する。ここで、凝集能とは、細胞どうしが、お互いに可逆的に凝集して凝集物(フロックと呼ばれることもある)を形成する能力をいう。凝集能を有する株は、酵母菌体の回収が容易になるという利点がある。すなわち、遠心分離等の分離操作を必要としないので、回収にかかるコスト、エネルギー、労力の削減が可能になる。また、凝集能を有するようになった結果、エタノール耐性能が向上した事例も知られている(Zhao and Bai, 2009, Biotechnol. Adv. 27, 849-856)。本発明の酵母株は7%(w/v)未満、好ましくは6%(w/v)以下、さらに好ましくは5%(w/v)以下、特に好ましくは3%(w/v)以下のエタノール存在下でも凝集性を有する。
【0018】
上記のように、本発明のマンニトール資化能を有しマンニトールからエタノールを産生し得る酵母は、さらに、以下の(1)〜(3)の少なくとも1つの特性、すなわち(1)の特性、(2)の特性、(3)の特性、(1)及び(2)の特性、(1)及び(3)の特性、(2)及び(3)の特定、あるいは(1)及び(2)及び(3)の特性を有し得る:
(1)エタノール耐性を有する;
(2)アルギン酸資化能を有する微生物を用いて褐藻原料からエタノールを生産したときの残渣中で生育し得る;及び
(3)グルコース存在下での凝集能を有する。
【0019】
上記の6株の酵母株のうち、エタノール生産性、エタノール耐性、アルギン酸からのエタノール発酵残渣における生育性及びグルコース存在下での凝集能が良好なSaccharomyces paradoxus NBRC 0259株が好ましい。また、エタノール生産性及びエタノール耐性が良好なOgataea glucozyma NBRC 1472株も好ましい。
【0020】
本発明のマンニトール資化能を有する酵母株を培養することにより、エタノールを生産させることができる。該酵母株を培養する方法は、酵母の培養に用いられる通常の方法に従って行われ、公知の培地にマンニトールを添加すればよい。
【0021】
原料となるマンニトールは、終濃度1〜10%(w/v)、好ましくは1〜5%(w/v)、さらに好ましくは2〜5%(w/v)の濃度で添加すればよい。また、培養中に経時的にマンニトールを添加してもよい。
【0022】
培養は、振盪培養又は通気攪拌培養などの好気的条件下、20〜40℃、好ましくは28〜32℃、pH5.6〜9.0、好ましくはpH5.6〜8.4で数時間〜数日間、例えば4〜7日間行う。培地のpHの調整は、無機又は有機酸、アルカリ溶液等を用いて行えばよい。培養中は必要に応じてカナマイシン、ペニシリン等の抗生物質を培地に添加してもよい。
【0023】
培養の際、酵母株はA600(600nmにおける吸光度)が培養開始時に0.05〜11になるように、例えば0.05〜5、好ましくは0.05〜0.2、さらに好ましくは0.1になるように添加すればよい。
【0024】
上記の条件での培養により、培地中に0.1%(w/v)以上、好ましくは0.3%(w/v)以上、さらに好ましくは1%(w/v)以上、さらに好ましくは3%(w/v)以上、特に好ましくは3.5%(w/v)以上のエタノールが蓄積し得る。例えば、10%(w/v)のマンニトールを含む培地で培養した場合、最終的に3%(w/v)以上、特に好ましくは3.5%(w/v)以上のエタノールが蓄積し得る。
また、培養により酵母も増殖し、培養終了時にA600は、0.1〜33程度まで上昇する。
【0025】
さらに、褐藻等に含まれるアルギン酸からアルギン酸資化能を有する微生物を用いてエタノールを生産した場合に得られる原料の残渣を用いて培養することもできる。該残渣中には、アルギン酸資化能を有する微生物が資化できなかったマンニトールが含まれる。原料となる褐藻(Phaeophyceae)としてはマコンブ(Laminaria japonica)、ワカメ(Undaria pinnatiflida)、モズク(Nemacystus decipiens)、ホンダワラ(Sargassum fulvellum)、ヒジキ(Sargassum fulvellum)等が挙げられ、アルギン酸資化能を有する微生物としては、上記のZymomonas mobilis等の細菌等のエタノール産生に関する酵素(ピルビン酸脱炭酸酵素及びアルコール脱水素酵素)をコードする遺伝子を導入したエタノール生産性Sphingomonas sp. A1株が例示できる。例えば、前記残渣中には、0.5〜2%(w/v)程度の濃度のエタノール、1〜10%(w/v)程度のマンニトールが含まれる。このような残渣に本発明のマンニトール資化能を有する酵母株を添加し培養する。この際、酵母株は、A600(600nmにおける吸光度)が0.05〜11になるように、例えば1〜11、好ましくは1〜5、さらに好ましくは1〜3になるように添加すればよい。培養は、振盪培養又は通気攪拌培養などの好気的条件下、20〜40℃、好ましくは28〜32℃、pH5.6〜9.0、好ましくは5.6〜8.4で数時間〜数日間、例えば4〜7日間行う。例えば、培養4〜7日間で、エタノール濃度は、1〜5%(w/v)に達する。残渣中に存在していたアルギン酸から生産されたアルコールを差引いた値が、マンニトール資化能を有する酵母株により新たに産生されたエタノールである。マンニトール資化能を有する酵母株により産生されるエタノールにより、エタノール濃度は残渣中の濃度の1.5〜3倍、好ましくは2倍程度になる。本発明ではこのようにアルギン酸からエタノールを生産させた原料の残渣を用いて残渣中のマンニトールからさらにエタノールを産生し、大量のエタノールを得る方法を二段階発酵という。
【0026】
褐藻を原料として用いてエタノールを生産する場合、褐藻からアルギン酸とマンニトールを抽出し、それを炭素源として用いて培養すればよい。また、褐藻を破砕し、この破砕物そのもの(アルギン酸とマンニトールを含む)を炭素源として用いて培養してもよい。いずれの場合でも、褐藻は木質バイオマスなどとは異なりリグニンを含まないので、木質バイオマスなどの処理と比較すると比較的穏和な条件でアルギン酸とマンニトールを抽出し利用することが可能である。また、トウモロコシデンプンなどと比較しても、トウモロコシデンプンでは糖化行程が必要であるが、アルギン酸とマンニトールからのエタノール生産ではこの糖化行程が不要という利点がある。
【0027】
エタノールは、蒸留により回収することができる。また、エタノールの定量は、公知のアルコール脱水素酵素を用いた方法やガスクロマトグラフィーを用いた方法により行うことができる。
【0028】
上記酵母株を固定化して、エタノールを生産することもできる。微生物の固定化には、包括法、架橋法、担体結合法等がある。包括法とは微生物を高分子ゲルの微細な格子の中に包み込むか、あるいは半透膜性の高分子の皮膜によって被覆する方法であり、架橋法とは微生物を2個又はそれ以上の官能基を持った試薬(多官能性架橋剤)で架橋する方法であり、担体結合法とは水不溶性の担体に酵素を結合させる方法である。固定化に用いられる固定化担体としては、ガラスビーズ、シリカゲル、ポリウレタン、ポリアクリルアミド、ポリビニルアルコール、カラギーナン、アルギン酸、寒天、ゼラチン等がある。
【実施例】
【0029】
本発明を以下の実施例によって具体的に説明するが、本発明はこれらの実施例によって限定されるものではない。
【0030】
実験方法
培地と培養
炭素源非含有培地(pH 5.6)は、イーストニトロジェンベース(w/o アミノ酸;ディフコ社製)0.67%(w/v)、-Leuドロップアウト サプリメント(クロンテック社製)0.69g/l、及びL-ロイシン100mg/lを含む。グルコース合成培地、マンニトール合成培地、グリセロール合成培地、及びラミナリン合成培地は、炭素源非含有培地にグルコース[終濃度2%(w/v)]、マンニトール[同2%(w/v)]、グリセロール[同3%(w/v)]、及びラミナリン[Sigma社製、製品番号L9634、Laminaria digitata由来、同2%(w/v)]を加えた培地である。YP培地(pH5.6)は、酵母エキス1%(w/v)及びトリプトン2%(w/v)を含む。pHはHClで調整した。YPD培地、YPM培地、及びYPG培地は、YP培地に各々グルコース[同2%(w/v)]、マンニトール[同2%(w/v)]、及びグリセロール[同3%(w/v)]を添加した培地である。炭素源とそれ以外の成分は、別々にオートクレーブ滅菌(ラミナリンはフィルター滅菌した)した後に混合した。特に言及しない場合は、2 x YP(pH5.6;2倍濃いYP培地)を用いてこれらの培地を調製した。このとき、YPM培地のpHは5.7であった。10 x YP (pH5.6;10倍濃いYP培地)はオートクレーブ滅菌ではなく、フィルター滅菌した。なお、2 x YP (pH 8.0; NaOHでpHを調整)を用いた場合のYPM培地のpHは7.8であった。固体培地では、これらに終濃度2%(w/v)となるように寒天(ナカライ社製)を添加した。なお、培養は30℃で行った。特に言及しない場合は、液体培地には、A600が0.1となるように細胞を接種し、YPD、YPM、及びYP液体培地を用いた培養は、50mlの液体培地を含む100mlの三角フラスコで、95 strokes per minute(spm)で振とうして行った。固体培地で前培養を行った場合は、培地上の細胞を滅菌水に懸濁してから、液体培地に接種した。
【0031】
酵母を25μg/mlのエチジウムブロマイドで処理してミトコンドリアゲノムとYPG固体培地での生育性を欠失したρ0株を作製した(Fox et al., 1991, Methods Enzymol. 194, 149-165.)。なお、正常なミトコンドリアゲノムを有する株をρ+ 株と称する。嫌気培養は、角形ジャーをアネロパック・ケンキ(ともに三菱ガス化学株式会社製)で嫌気状態にすることにより行った。エタノール生産性形質転換Sphingomonas sp. A1株(EPv104株:後述)の培養は、5%アルギン酸培地を用いて、既報に従って行った(Takeda et al., 2011, Energy Environ. Sci. 4, 2575-2581)。ただし、培養開始時に粉末マンニトールを2%あるいは5%(ともにw/v)となるように添加した。培養3日後の培養液中の上澄み液を遠心分離により調製し、A1株によるアルギン酸からのエタノール発酵残渣(マンニトールを含む)とした。
【0032】
菌株
本実施例では、マンニトール資化性酵母を探索するために、表1に示す研究室の酵母保存株48株を使用した。Saccharomyces paradoxus NBRC 0259及びS. cerevisiae BY4742のρ0 株はエチジウムブロマイド処理により作製した(Fox et al., 1991, Methods Enzymol. 194, 149-165.)。これらの酵母株は、YPD液体培地で培養後17%グリセロール存在下で-80℃で保存した。エタノール生産性A1株としては、EPv104株を用いた(Takeda et al., 2011, Energy Environ. Sci. 4, 2575-2581)。EPv104株は、Sphingomonas sp. A1株の乳酸脱水素酵素遺伝子欠損株に、Zymomonas mobilis由来ピルビン酸脱炭酸酵素遺伝子(pdc)8コピー及び同由来アルコール脱水素酵素遺伝子(adhB)1コピーを広宿主ベクターpKS13を介して導入した、アルギン酸から最も高いエタノール生産性を示す株である(Takeda et al., 2011, Energy Environ. Sci. 4, 2575-2581)。
【0033】
【表1】

【0034】
エタノール濃度測定
エタノール濃度は、F-キット エタノール (Roche Diagnostics K.K., Tokyo, Japan) を用いて、キットに付属のプロトコルに従い測定した。
【0035】
ITS-5.8S rDNA塩基配列の解析
S. paradoxus NBRC 0259株のITS-5.8S rDNA塩基配列のPCRによる増幅及び塩基配列の解析は株式会社 テクノスルガ・ラボに委託した。
【0036】
結果と考察
マンニトール資化性エタノール生産酵母の探索
マンニトール資化性エタノール生産酵母の探索の結果を図2に示す。
【0037】
YPD固体培地上で生育させた本発明者の研究室保存酵母48株(表1)を滅菌水に懸濁し、懸濁液5μlを合成固体培地(炭素源非含有培地、グルコース合成培地、及びマンニトール合成培地) へスポットして細胞を5日間培養した。目視で生育を観察した結果、グルコース合成固体培地では全ての株が良好に生育したが、炭素源非含有固体培地では全ての株が殆ど生育しなかった。また、表1で下線を付した14株(以下マンニトール資化性酵母と称する)がマンニトール合成固体培地で、炭素源非含有固体培地よりも良好な生育を示した。液体培地においても同様であった(図2A)。マンニトール資化能を示さないS288Cの派生株S. cerevisiae BY4742株(以下BY4742株)は(Brachmann et al., 1998, Yeast 14, 115-132.; Quain and Boulton, 1987, J. Gen. Microbiol. 133, 1675-1684.)、マンニトール合成固体及び液体培地で、炭素源非固体及び液体含有培地よりも良好な生育を示さなかった(図2A)。なお、14株の内、P. polymorpha IFO 0195、P. farinosa NBRC 0193、P. haplophila NBRC 0947、P. saitoi IAM 4945、H. saturnus IFO 0177、D. hansenii IFO 0023、及びY. lipolytica NBRC 0746の7株では、マンニトール及びグルコース合成液体培地で膜を形成した。S. paradoxus NBRC 0259株(以下NBRC 0259株)は、グルコース液体培地において凝集性を示した(図2B)。同株は、マンニトール合成液体培地においても若干の凝集性を示した。
【0038】
次に、BY4742株(対照)及びマンニトール資化性酵母14株をマンニトール合成液体培地で3日間静置培養し、培養液上澄みのエタノール濃度を測定した。NBRC 0259、K. capsulata NBRC 0721、K. capsulata NBRC 0974、O. glucozyma NBRC 1472、O. minuta NBRC 1473、及びD. hansenii NBRC 0794各6株(以後、マンニトール資化性エタノール生産酵母と称す)のみが、少なくとも44 mg/l以上のエタノールを生産した(図2C-2)。他の8株は、7 mg/l以下のエタノール生産性しか示さなかった。マンニトールからのエタノール生産性は、NBRC 0259株が他よりも顕著に高かった(図2C-2)。同6株及びBY4742株をグルコース合成液体培地、ラミナリン合成液体培地、並びに5%(w/v)及び7%(w/v)エタノールを含むマンニトール合成液体培地で3日間静置培養した(図2C-1、2C-2及び2D)。同6株のグルコースからのエタノール生産性は、マンニトールからのそれよりも高かった(図2C-2)。とりわけNBRC 0259株は、同6株の中で最も高いグルコースからのエタノール生産性を示した。NBRC 0259株以外の5株はラミナリンからのエタノール生産性も示した(図2C-2)。マンニトール合成培地における3日間の培養では、同6株は7%(w/v)エタノール存在下で生育性を示さなかったが、5%(w/v)エタノール存在下では、NBRC 0259株、K. capsulata NBRC 0974株、及びO. glucozyma NBRC 1472株は生育性を示した(図2D)。5%(w/v)エタノール存在下においても、NBRC 0259株は凝集性を示した。
【0039】
マンニトール資化における酸素要求性
マンニトール資化における酸素要求性を図3に示す。
【0040】
本実施例で見出したマンニトール資化性エタノール生産酵母6株がマンニトール資化に酸素を必要とするかどうかを調べるため、これらをエチジウムブロマイドで処理して、ρ0株の作製を試みたところ、NBRC 0259株のみρ0株が作製できた。そこで、NBRC 0259並びにBY4742のρ0及びρ+株を用いて嫌気及び好気条件下における合成培地での生育性を調べたところ、既報(Quain and Boulton, 1987, J. Gen. Microbiol. 133, 1675-1684.)に一致して、NBRC 0259株はマンニトール合成培地での生育に酸素及び呼吸能を必要とすることが分かった(図3A)。さらに、通常大気条件下並びにYPG及びYPM固体培地において、同株のρ+株は生育できたが、ρ0株は生育できなかった(データ省略)。本結果は、YPM固体培地を用いたNBRC 0259 ρ+株(マンニトール資化能を保持)の選抜が可能なことを意味した。
【0041】
一方、NBRC 0259株以外の5株(ρ+株)の嫌気及び好気条件下における合成培地での生育性を調べたところ、いずれも嫌気条件ではマンニトール合成培地(図3B)及びYPM固体培地(図省略)で生育性を示さなかった。本結果から、NBRC 0259株以外の5株もマンニトールの資化に酸素を必要とすることが明らかとなった。酵母におけるマンニトール資化では、マンニトールがマンニトールデヒドロゲナーゼのはたらきでフルクトースに変換される過程で1分子の余剰NADHが形成されるが、この余剰NADHのNAD+への再生に酸素が必要とされるものと考えられている(図1)(Quain and Boulton, 1987, J. Gen. Microbiol. 133, 1675-1684.)。マンニトール資化性エタノール生産酵母に関しても、同様の理由でマンニトール合成培地やYPD培地での生育に酸素を必要としたと推察された。他方、NBRC 0259株以外の5株は、嫌気条件においてグルコース合成培地(図3B)及びYPD固体培地(図省略)でも生育性を示さなかった。NBRC 0259以外の5株は、NBRC 0259とは若干異なる様式でグルコースを資化している可能性が示唆された。
【0042】
マンニトール資化性エタノール生産酵母によるエタノール生産
図4は、マンニトール資化性エタノール生産酵母によるエタノール生産を示す図である。
【0043】
マンニトール資化性エタノール生産酵母6株のエタノール生産能を更に詳細に調べた。以降はYPM培地を基本培地とした。同6株はマンニトール培地での生育に酸素を必要としたので(図3)、YPM液体培地を用いて、95 spmで振とう培養して生育性とエタノール生産性を調べたところ(図4A-1及び4A-2)、NBRC 0259株とO. glucozyma NBRC 1472株が高い生産性を示した。ただし、両株、とりわけO. glucozyma NBRC 1472株は長期間培養すると培養液中のエタノール濃度が著しく低下した。D. hansenii NBRC 0794株のエタノール生産性は最も低かった。
【0044】
次に、A1株によるアルギン酸からのエタノール発酵残渣(マンニトールを含む)からのエタノール生産(二段階発酵)を目的として、2%(w/v)マンニトールを含む発酵残渣における同6株の生育性(図4B)とエタノール生産性を調べた。同発酵残渣のpHは8.64と弱アルカリ性を示した。これに、10 x YP (pH5.6)を1/10容量添加して(この結果、初期pHは7.3となった)培養を開始した。なお、初期エタノール濃度は9.6 g/lであった。しかしながら、生育性を示したのはNBRC 0259株とD. hansenii NBRC0794株のみであった。培養7日後の培養液上澄みのエタノール濃度は全て初期エタノール濃度を下回った。さらにNBRC 0259株は凝集性を示した。発酵残渣での生育性が弱い理由として、同6株ともにpH 7.8の弱アルカリ性YPM培地では生育したことから(図4B)、同残渣のpH(pH 7.3)が原因ではなく、発酵残渣に含まれる何らかの成分が生育を阻害したと推察された。D. hansenii NBRC0794株は同発酵残渣で生育性は示したが、マンニトールからのエタノール生産性が最も低い株であった(図4A-1及び4A-2)。
【0045】
以上の結果より、NBRC 0259株は、ラミナリンからのエタノール生産性こそ示さないものの、グルコース及びマンニトールからの高いエタノール生産性、エタノール耐性、及びA1株によるアルギン酸からのエタノール発酵残渣における生育性を示したため、6株の中では最も優良な株と判断された(図2、図4)。次に優良な株として、ラミナリンからのエタノール生産性、グルコース及びマンニトールからの高いエタノール生産性、及びエタノール耐性を示した、O. glucozyma NBRC 1472株が考えられた(図2及び図4)。ただし、本NBRC 1472株は、長時間の培養により培養液のエタノール濃度が激減すること(図4A-2)、及び発酵残渣での生育性を示さない(図4B)という望ましくない特性を示した。さらに、マンニトールからのエタノール生産性が報告されているS. cerevisiaeプロピロイドBB1株及びP. angophoraeの、マンニトールからのエタノール生産に関しては、本実施例ほど詳細に報告されていない。さらに、エタノール耐性やアルギン酸からのエタノール発酵残渣における生育性も不明である(Horn et al., 2000, J. Ind. Microbiol. Biotechnol. 24, 51-57.; Quain and Boulton, 1987, J. Gen. Microbiol. 133, 1675-1684.)。これらの理由から、NBRC 0259株を以降の研究に用いることにした。本研究で用いているNBRC 0259株のITS-5.8S rDNA塩基配列は、データベースに登録されているS. paradoxus NBR0259株の同配列(Genbank: D89890)と100%(817/817)の相同性を示した。一方で、S. cerevisiae S288C株の同配列(Genbank: BK006945)とは98.9%(815/824)の相同性を示した。
【0046】
NBRC 0259株によるマンニトールからのエタノール生産
図5は、NBRC 0259株によるマンニトールからのエタノール生産を示す図である。
【0047】
NBRC 0259株のマンニトール資化能を更に調べた。BY4742 ρ+株を対照として用いた。YPD及びYPM固体培地で前培養したNBRC 0259株はマンニトール合成液体培地で生育したが、BY4742株は生育できなかった(図5A及び5B)。また、NBRC 0259株は2%グルコース液体培地では凝集性を示した(図5B)。
【0048】
次に、YPM液体培地を用いてエタノール生産条件を調べた。YPD固体培地上で形成されたNBRC 0259株のシングルコロニーは、高い頻度でYPM固体及び液体培地での生育能を喪失した(調べた6個のシングルコロニーのうち、5個のシングルコロニー由来の細胞がこれを喪失した)。これは、NBRC 0259株がYPD固体培地上ではミトコンドリアゲノムを欠失あるいは損傷しやすい特性を持つことを示唆した。そこで、以降はYPM固体培地上で選抜したNBRC 0259 ρ+株を用いた。同株はマンニトールの資化に酸素を要求したため(図3A)、同株をYPM液体培地で0 spmで静置培養、並びに95及び145 spmで振とう培養して、生育性とエタノール生産性に対する培養中の通気量の影響を調べた。その結果、既報に一致して、通気量が多いほど生育性も高くなった(図5C)。しかし、エタノール生産性は95 spmで適度な酸素を供給した場合に最も高く(6日間で 8.20 g/lのエタノール生産性)、0及び145 spmで培養した場合には殆どエタノール生産性が見られなかった(図5C-1及び5C-2)。一方で、グルコースを基質とした場合、すなわちYPD液体培地では、0及び95 spm両方において、マンニトールの場合よりもより高い生産性を示した(1日[0 spm]及び2日間[95 spm]で各々約12 g/lのエタノール生産性)。また、YPD液体培地でも明瞭な凝集性が観察された(図5D)。
【0049】
振とう速度を95 spmとして、マンニトール濃度の影響を調べたところ、10%(w/v)マンニトールから最大37.6 g/lの(3.8% w/v)のエタノールが生産された(図5E)。これは、マンニトールからのエタノール生産量としては最大であった。また、2及び5%(w/v)のNaCl存在下でエタノール生産性の顕著な低下は見られなかった(図5F)。次に、NBRC 0259株の前培養条件の、生育性とエタノール生産性に対する影響を調べた。すなわち、YPM固体培地で前々培養した同株をYPM又はYPD液体培地で2日間前培養して得られた菌体を洗浄後YPM液体培地に接種したところ、YPM液体培地で前培養した菌体を用いた方が、YPD液体培地で前培養した場合よりも高い生育性とエタノール生産性を示した。すなわち、エタノール生産性は、前者が培養6日で6.7g/lであったが、後者は6日でわずか0.35g/lであり培養12日でようやく5.2g/lに達したに過ぎなかった。
【0050】
5%(w/v)マンニトールを含むアルギン酸からのエタノール発酵残渣45mlに2 xYP (pH 5.6)を5ml添加した培地(初期エタノール濃度8.5g/l)にNBRC 0259株を初期A600が2.0となるよう添加して培養したところ、培養4日で培養液上澄みのエタノール濃度は16.9g/l、培養7日では14.0g/lとなった。すなわち、二段階発酵により残渣中のマンニトールから9.4 (=16.9-8.5)g/lのエタノールが新たに生産された。二段階発酵の条件の改善による、さらなるエタノール生産性の向上が期待された。
【産業上の利用可能性】
【0051】
本発明のマンニトールからエタノールを生産する酵母株を用いることにより、大型海藻などの海洋バイオマスから有効にエタノールを生産することができる。
【技術分野】
【0001】
本発明は、酵母によるマンニトールからのエタノールの生産方法及びマンニトールからエタノールを生産する酵母株に関する。
【背景技術】
【0002】
大型海藻などの海洋バイオマスは有望なバイオ燃料の原料である。その主な理由として、(i) 陸性バイオマスと比較すると、大型海藻は高い生産性を示す、(ii) 耕作地を必要としないため、陸性バイオマスを栽培する上で不可避となる問題(灌漑や施肥など)を避けられる、(iii) リグニンを含まない、等があげられる。大型海藻は、主に緑藻、紅藻、及び褐藻から構成される。うち、少なくとも紅藻と褐藻は著量の炭水化物を含む。紅藻の一種テングサGelidium amansiiは、17%(w/w乾燥重量;以下、特に言及しない場合、w/wは全て乾燥重量を表す)のセルロース(グルコース)、58.6%(w/w)の寒天(25.6%のガラクトースと33%の3,6-アンヒドロガラクトース)を含む。褐藻類は最大で40%(w/w)のアルギン酸、最大30%(w/w)のマンニトール、及び最大30%(w/w)のラミナリンを含む。かかるがゆえに、大型海藻を原料としたバイオ燃料の生産には、これらの炭水化物の成分をバイオ燃料に変換する技術の確立が必須である。
【0003】
アルギン酸は、β-D-マンヌロン酸(M)とそのC5エピマーであるα-L-グルロン酸(G)から構成される直鎖状の酸性多糖である。構成単糖は、ポリM、ポリG、あるいはポリMG構造をとる。マンニトールは、マンノースに対応する糖アルコールであり、マンニトールデヒドロゲナーゼの作用により酸化されてフルクトースになる(図1)(非特許文献1及び2を参照)。ラミナリンは、β-(1,6)枝分かれ構造を有するβ-(1,3)-D-グルカンから構成される(非特許文献3及び4を参照)。既に、本発明者らはエタノール生産性Sphingomonas sp. A1 株(以下、エタノール生産性A1株)を用いたアルギン酸からのエタノール生産システムを構築しており、1.3%(w/v)のエタノール生産に成功している(非特許文献5を参照)。本法は、アルギン酸からバイオ燃料を生産する唯一の方法である。ラミナリンからのエタノール生産例は少ないが、酵母 Kluyveromyces marxianus、Pacchysolen tannophilus、及びPhicia angophoraeの3株がラミナリンを分解してエタノールを生産するとの報告(非特許文献1を参照)、及びラミナリン分解酵素ラミナリナーゼを用いて分解したラミナリン分解物から酵母Saccharomyces cerevisiaeを用いてエタノールを生産したとの報告(Adams et al., 2009)がある。マンニトールからのエタノール生産に関しては、Zymobacter palmaeとEscherichia coli KO11の両細菌が、各々、3.8%(w/v)及び9.0%(w/v)のマンニトールから、約1.3%(w/v)及び2.6%(w/v)のエタノールを、0.38g及び0.41gエタノール(gマンニトール)-1 の生産効率で生産したことが報告されている(非特許文献1及び6を参照)。しかし、エタノールを生産する上では、細菌よりも酵母の方がエタノールや発酵阻害物質への耐性能を含む様々な利点を有すると見なされている(非特許文献7を参照)(Hughes and Qureshi, 2010)。実際、Z. palmae とE. coli KO11は、5%(w/v)のエタノール存在下で生育阻害を示す(非特許文献8及び9を参照)。しかし、酵母を用いたマンニトールからのエタノール生産の研究例は少なく、酵母S. cerevisiaeポリプロイド BB1株が、5%(w/v)のマンニトールから約0.5%のエタノールを生産すること(非特許文献2を参照)、及び先述のラミナリン分解酵母(K. marxianus、P. tannophilus、及びP. angophorae)のうち、P. angophoraeのみが、4%(w/v)マンニトールから約1.0%(w/v)のエタノールを0.40gエタノール(g マンニトール)-1の生産効率で生産することが報告されているのみである(非特許文献1を参照)。このP. angophoraeを用いた研究に関しては、マンニトールとラミナリンが共存する海藻抽出物からのエタノール生産や、酸素供給量のマンニトールとラミナリン消費速度に対する影響が報告されている(非特許文献1を参照)。一方、酵母のマンニトール代謝に関する報告も少なく、酵母S. cerevisiaeには、マンニトールを資化できる株(先述のポリプロイドBB1株など、一倍体A184D株)とできない(あるいは同資化能が極めて弱い)株(ポリプロイドBB2株など、一倍体S288C株やSc41 YJO 株)が存在すること、S. cerevisiaeによるマンニトールの資化には酸素が必要でありマンニトール含有培地で生育中の酵母は高い呼吸活性を示すことなどが報告されている(非特許文献2及び10を参照)。なお、一倍体S288C株は最初のゲノム配列決定株でもある(非特許文献11を参照)。
【0004】
酵母を用いたマンニトールからのエタノール生産の実用化には、マンニトールからの高いエタノール生産能力やその他の様々な優良特性を示す酵母の探索あるいは育種、及び高い生産能力を発揮する最適条件の確立が必須である。さらには、海洋バイオマスからのエタノール生産系の確立には、その構成成分をもれなくエタノールに変換する技術も必要である。褐藻類の場合、アルギン酸、マンニトール、及びラミナリンなどからエタノールに変換する技術の確立を要する。アルギン酸からのエタノール生産系は、先述したエタノール生産性A1株を用いた系しか知られていない(非特許文献5を参照)。本A1株は、マンニトールやラミナリンを資化できない(未発表データ)。一方で、アルギン酸資化能はSphingomonas sp. A1株などの限られた生物にしか知られていない。
【先行技術文献】
【非特許文献】
【0005】
【非特許文献1】Horn et al., 2000, J. Ind. Microbiol. Biotechnol. 25, 249-254
【非特許文献2】Quain and Boulton, 1987, J. Gen. Microbiol. 133, 1675-1684.
【非特許文献3】Nelson and Lewis, 1973, Carbohydr. Res. 33, 63-74.
【非特許文献4】Zvyagintseva et al., 1999, Carbohydr. Res. 322, 32-39.
【非特許文献5】Takeda et al., 2011, Energy Environ. Sci. 4, 2575-2581
【非特許文献6】Kim et al., 2011, Bioresour. Technol. 102, 7466-7469.
【非特許文献7】Hughes and Qureshi, 2010, Biomass to biofuels: strategies for global industries, pp. 55-69.
【非特許文献8】Okamoto et al., 1994, Biosci. Biotech. Bioch. 58, 1328-1329.
【非特許文献9】Yomano et al., 1998 J. Ind. Microbiol. Biotechnol. 20, 132-138.
【非特許文献10】Perfect et al., 1996, J. Bacteriol. 178, 5257-5262.
【非特許文献11】Goffeau et al., 1996, Science 274, 546, 563-547.
【発明の概要】
【発明が解決しようとする課題】
【0006】
本発明は、酵母によるマンニトールからのエタノールの生産方法及びマンニトールからエタノールを生産する酵母株の提供を目的とする。
【課題を解決するための手段】
【0007】
本発明者らは、褐藻類由来バイオマスのエタノール生産への有効利用には、エタノール生産性A1株によるアルギン酸からのエタノール発酵残渣に含まれるマンニトール及びラミナリンから、マンニトール及びラミナリン資化性酵母を用いてエタノールを生産する二段階発酵の実現が有効であると考え、この実現を目指して、まずマンニトールから高いエタノール生産能を示す酵母の探索を行った。
【0008】
海洋バイオマス(褐藻類)からのバイオ燃料(エタノール)生産には、その主要成分アルギン酸、マンニトール、及びラミナリンからエタノールを生産する技術が不可欠である。エタノール生産性Sphingomonas sp. A1株を用いたアルギン酸からのエタノール生産系は確立されている(Takeda et al., 2011, Energy Environ. Sci. 4, 2575-2581)。本株によるアルギン酸からのエタノール発酵後の残渣に含まれるマンニトールやラミナリンのエタノールへの変換は、褐藻類主要成分の有効利用法の一つである。本発明者らは、保存酵母株の中から、マンニトール資化性エタノール生産株6株を見出した。うち、Saccharomyces paradoxus NBRC0259株のみは、ラミナリンからのエタノール生産性は示さないものの、グルコース及びマンニトールからの高いエタノール生産性、エタノール耐性、及びアルギン酸からのエタノール発酵残渣における生育性といった望ましい特性を示した。他の5株はラミナリンからのエタノール生産性を示したが、他の特性がNBRC 0259株より劣った。NBRC 0259株は、95 strokes per min (spm) の振とうによって得られる微好気環境において、10% (w/v)マンニトールから最大37.6 g/l (3.8% w/v)のエタノールを生産した。エタノール発酵に対するNaClの影響は小さかった。また、アルギン酸からのエタノール発酵残渣中のマンニトールからもエタノールを生産した。本菌は、海洋バイオマスからのエタノール生産に有用と考えられた。
【0009】
すなわち、本発明は以下のとおりである。
[1] マンニトール資化能を有しマンニトールからエタノールを産生し得る酵母を、マンニトールを含む培地中で培養することを含む、マンニトールを原料としてエタノールを生産する方法。
[2] マンニトール資化能を有しマンニトールからエタノールを産生し得る酵母が、以下の(1)〜(3)の少なくとも1つの特性を有する酵母である、[1]のマンニトールを原料としてエタノールを生産する方法:
(1)エタノール耐性を有する;
(2)アルギン酸資化能を有する微生物を用いて褐藻原料からエタノールを生産したときの残渣中で生育し得る;及び
(3)グルコース存在下での凝集能を有する。
[3] マンニトール資化能を有しマンニトールからエタノールを産生し得る酵母が、Saccharomyces paradoxus、Debaryomyces hansenii、Kuraishia capsulata、Ogataea glucozyma及びOgataea minutaからなる群から選択される、[1]又は[2]のマンニトールを原料としてエタノールを生産する方法。
[4] マンニトール資化能を有しマンニトールからエタノールを産生し得る酵母が、Saccharomyces paradoxus NBRC 0259株、Debaryomyces hansenii NBRC 0794株、 Kuraishia capsulata NBRC 0721株、Kuraishia capsulata NBRC 0974株、Ogataea glucozyma NBRC 1472株及びOgataea minuta NBRC 1473株からなる群から選択される、[3]のマンニトールを原料としてエタノールを生産する方法。
[5] マンニトール資化能を有しマンニトールからエタノールを産生し得る酵母を培養することにより、培地中に0.1%(w/v)以上のエタノールが蓄積し得る、[1]〜[4]のいずれかの方法。
[6] マンニトール資化能を有しマンニトールからエタノールを産生し得る酵母を培養することにより、培地中に3%(w/v)以上のエタノールが蓄積し得る、[5]の方法。
[7] アルギン酸資化能を有しアルギン酸からエタノールを産生し得る微生物を用いて、褐藻原料からエタノールを産生させたときの原料残渣に、マンニトール資化能を有しマンニトールからエタノールを産生し得る酵母を添加し、培養し、残渣中のマンニトールを原料としてエタノールを生産する方法。
[8] マンニトール資化能を有しマンニトールからエタノールを産生し得る酵母が、以下の(1)〜(3)の少なくとも1つの特性を有する酵母である、[7]の残渣中のマンニトールを原料としてエタノールを生産する方法:
(1)エタノール耐性を有する;
(2)アルギン酸資化能を有する微生物を用いて褐藻原料からエタノールを生産したときの残渣中で生育し得る;及び
(3)グルコース存在下での凝集能を有する。
[9] マンニトール資化能を有しマンニトールからエタノールを産生し得る酵母がSaccharomyces paradoxusである、[7]又は[8]の残渣中のマンニトールを原料としてエタノールを生産する方法。
[10] マンニトール資化能を有しマンニトールからエタノールを産生し得る酵母がSaccharomyces paradoxus NBRC 0259株である、[9]の残渣中のマンニトールを原料としてエタノールを生産する方法。
[11](i) アルギン酸資化能を有しアルギン酸からエタノールを産生し得る微生物を褐藻原料を用いて培養し、アルギン酸からエタノールを産生させ、
(ii) (i)の培養の原料残渣に、マンニトール資化能を有しマンニトールからエタノールを産生し得る酵母を添加し、培養することを含む、
褐藻原料からエタノールを生産する方法。
[12] マンニトール資化能を有しマンニトールからエタノールを産生し得る酵母が、以下の(1)〜(3)の少なくとも1つの特性を有する酵母である、[11]の褐藻原料からエタノールを生産する方法:
(1)エタノール耐性を有する;
(2)アルギン酸資化能を有する微生物を用いて褐藻原料からエタノールを生産したときの残渣中で生育し得る;及び
(3)グルコース存在下での凝集能を有する。
[13] マンニトール資化能を有しマンニトールからエタノールを産生し得る酵母がSaccharomyces paradoxusである、[11]又は[12]の褐藻原料からエタノールを生産する方法。
[14] マンニトール資化能を有しマンニトールからエタノールを産生し得る酵母がSaccharomyces paradoxus NBRC 0259株である、[13]の褐藻原料からエタノールを生産する方法。
【発明の効果】
【0010】
本発明のマンニトール資化能を有しマンニトールからエタノールを産生し得る酵母株を用いることにより、マンニトールを原料としてエタノールを生産することができる。海域由来のバイオマス、とりわけ褐藻類に多量含有される多糖アルギン酸からのエタノール生産の残渣には、大量のマンニトールが含まれており、該残渣をエタノール生産に有効に利用することができる。
【図面の簡単な説明】
【0011】
【図1】酵母における推定マンニトール資化経路を示す図である。
【図2A】マンニトール資化性酵母の炭素源非含有液体培地(白色)、マンニトール合成液体培地(細い平行線)及びグルコース合成液体培地(黒色)における生育性を示す図である。酵母株: 1. S. paradoxus NBRC 0259; 2. Z. japonicus IFO 0595; 3. P. polymorpha IFO 0195; 4. P. farinosa NBRC 0193; 5. P. haplophila NBRC 0947; 6. P. saitoi IAM 4945; 7. H. saturnus IFO 0177; 8. K. capsulata NBRC 0721; 9. K. capsulata NBRC 0974; 10. O. glucozyma NBRC 1472; 11. O. minuta NBRC 1473; 12. D. hansenii IFO 0023; 13. D. hansenii NBRC 0794; 14. Y. lipolytica NBRC 0746; 15. S. cerevisiae BY4742 (対照)。S. paradoxus NBRC 0259株(株 1)はグルコース合成液体培地で強い凝集性(*)、マンニトール合成培地で弱い凝集性を示した。
【図2B】マンニトール資化性エタノール生産酵母のグルコース合成液体培地における生育性を示す図である。酵母株の番号は図2Aと同様であり、1: S. paradoxus NBRC 0259; 8. K. capsulata NBRC 0721; 9. K. capsulata NBRC 0974; 10. O. glucozyma NBRC 1472; 11. O. minuta NBRC 1473; 13. D. hansenii NBRC 0794; 15, S. cerevisiae BY4742である。S. paradoxus NBRC 0259株(株 1)はグルコース合成液体培地で強い凝集性(*)を示した。
【図2C−1】マンニトール資化性エタノール生産酵母のラミナリン合成液体培地(白色)、マンニトール合成液体培地(細い平行線)及びグルコース合成液体培地(黒色)における生育性を示す図である。酵母株の番号は図2Bと同様である。S. paradoxus NBRC 0259株(株 1)はグルコース合成液体培地で強い凝集性(*)を示した。
【図2C−2】マンニトール資化性エタノール生産酵母のラミナリン合成液体培地(白色)、マンニトール合成液体培地(細い平行線)及びグルコース合成液体培地(黒色)におけるエタノール生産性を示す図である。グルコース由来のエタノール濃度を右側のスケールで、ラミナリン又はマンニトール由来のエタノール濃度を左側のスケールで示す。酵母株の番号は図2Bと同様である。
【図2D】マンニトール資化性エタノール生産酵母のマンニトール合成液体培地における5% (w/v)エタノール存在下での生育性を示す図である。YPD固体培地上で前培養した菌株を1.0 mlの培地にて静置(0 spm)で3日間培養した。酵母株の番号は図2Bと同様である。S. paradoxus NBRC 0259株(株 1)は5% (w/v)エタノール存在下で凝集性(*)を示した。
【図3A】マンニトール資化における酸素要求性を示す図であり、BY4742(BY)及びNBRC 0259(NB)のρ0及びρ+株の、通常大気下(+O2)及び嫌気条件下(-O2)における、マンニトール、グルコース、及びグリセロール合成固体培地での生育性(4日間培養)を示す図である。
【図3B】マンニトール資化における酸素要求性を示す図であり、マンニトール資化性酵母の通常大気下(+O2)及び嫌気条件下(-O2)における、マンニトール及びグルコース合成固体培地での生育性を示す図である。YPM固体培地上で前培養した株を各培地にストリークし、4日間培養した。酵母株の番号は図2Bと同様である。
【図4A−1】マンニトール資化性エタノール生産酵母6株の生育性を示す図である。YPM液体培地25 mlを含む50 ml三角フラスコを用いて95 spmで振とう培養した。酵母株のシンボルは、◆ S. paradoxus NBRC 0259;▲ K. capsulata NBRC 0721; X K. capsulata NBRC 0974;● O. glucozyma NBRC 1472; + O. minuta NBRC 1473;■ D. hansenii NBRC 0794である。
【図4A−2】マンニトール資化性エタノール生産酵母6株のエタノール生産性を示す図である。YPM液体培地25 mlを含む50 ml三角フラスコを用いて95 spmで振とう培養した。酵母株のシンボルは、図4A-1と同様である。
【図4B】YPM液体培地(pH 5.7)、弱アルカリ性の同培地(pH 7.8)、及びA1株によるアルギン酸からのエタノール発酵残渣[2%(w/v)マンニトールを含む;pH 7.3 (アルギン酸)]における培養4日後の生育性を示す図である。培地成分以外は図4Aの条件と同様の条件で培養した。YPM固体培地上で前培養した細胞を用いた。酵母株の番号は図2Bと同様である。弱アルカリ性のYPM液体培地で弱い凝集傾向(YPD液体培地で観察される凝集に類似しているが、程度が弱い)、並びに発酵残渣で凝集性(細かい顆粒状)を示した(*)。
【図5A】NBRC 0259株のマンニトール合成液体培地における生育性を示す図である。NBRC 0259株のρ+株(YPM固体培地由来)(実線;●○)並びに対照としてBY4742株のρ+株(YPG固体培地由来)(点線;▲△)を、YPD(▲●)又はYPM(△○)固体培地で前培養して、1.0 mlのマンニトール合成液体培地で145 spmで振とう培養した。BY4742株は、マンニトール合成培地上では生育性を示さなかった。
【図5B】YPD固体培地で前培養したBY4742株(BY)及びNBRC 0259株(NB)をマンニトール及びグルコース合成液体培地で培養(培養4日間)したときの状態を示す図である。上記(図5A)の条件で培養した。NBRC0259はグルコース合成液体培地で凝集性を示した(*)。
【図5C−1】YPM(▲●■)及びYPD(□○)液体培地における生育性に対する振とう数[▲145 spm;●○95 spm;■□ 0 spm]の影響を示す図である。YPM固体培地由来NBRC 0259 ρ+株をYPM液体培地で95 spmで24時間前培養した細胞を植菌して培養を開始した。NBRC 0259株はグルコース合成液体培地で凝集性を示した(*)。
【図5C−2】YPM(▲●■)及びYPD(□○)液体培地におけるエタノール生産性に対する振とう数[▲145 spm;●○95 spm;■□0 spm]の影響を示す図である。YPM固体培地由来NBRC0259 ρ+株をYPM液体培地で95 spmで24時間前培養した細胞を植菌して培養を開始した。同株は145 spm ではエタノール生産性を示さなかった(▲)。
【図5D】NBRC 0259株のYPD及びYPM液体培地における生育性を示す図である。YPD培地では細胞がフロックを形成して沈殿している(*)。培養1日目の状態を示す。
【図5E】マンニトール濃度のエタノール生産性への影響を示す図である。初期菌体量(A600)を11、振とう速度を95 spmとし、YPM液体培地中のマンニトール濃度を2%(w/v: ■)、5%(w/v:●)、及び10%(w/v:▲)とした。初期菌体として、YPM液体培地で95 spmで4日間前培養した細胞を用いた。
【図5F】NaCl濃度のエタノール生産性への影響を示す図である。図5Eの条件と同じ条件であるが、YPM液体培地(2%(w/v)マンニトール)を用いて、0%(w/v:■)、2%(w/v:●)、及び5%(w/v:▲)NaCl存在下で培養した。
【発明を実施するための形態】
【0012】
以下、本発明を詳細に説明する。
【0013】
本発明の酵母株は、マンニトール資化能を有し、マンニトールからエタノールを生産し得るマンニトール資化性酵母株である。マンニトールからエタノールを生産する反応を図1に示す。
【0014】
本発明のマンニトールからエタノールを生産し得る酵母株の例として以下の株が挙げられる。Saccharomyces paradoxus NBRC 0259、Debaryomyces hansenii NBRC 0794、 Kuraishia capsulata NBRC 0721、Kuraishia capsulata NBRC 0974、Ogataea glucozyma NBRC 1472及びOgataea minuta NBRC 1473。これらの株は、独立行政法人 製品評価技術基盤機構生物遺伝資源部門(NBRC)に保存されており、該機構より入手することができる。
【0015】
さらに、本発明のマンニトール資化能を有し、マンニトールからエタノールを生産し得る酵母株は、好ましくはエタノール耐性を有する。エタノール耐性はエタノール耐性化又はエタノール寛容化することによって付与される。例えば、エタノールを含有する培地中で馴養培養、及び紫外線照射によりランダム変異を誘発することにより、エタノール耐性株を得ることができる。本発明の酵母株は、0.85%(w/v)以上、好ましくは1.7%以上、さらに好ましくは3%(w/v)以上、さらに好ましくは5%(w/v)以上、さらに好ましくは6%(w/v)以上、さらに好ましくは7%(w/v)以上、さらに好ましくは8%(w/v)以上、特に好ましくは8.5%(w/v)以上のエタノール存在下で生育することができる。
【0016】
さらに、本発明のマンニトール資化能を有し、マンニトールからエタノールを生産し得る酵母株は、好ましくは、アルギン酸からのエタノール発酵残渣における生育性を有する。エタノール発酵残渣における生育性は、生育阻害物質に対する耐性能が関与していると予測される。ここで、アルギン酸からのエタノール発酵残渣とは、アルギン酸を含む褐藻等の原料からアルギン酸資化能を有する微生物を用いてエタノールを生産した場合の、原料の残渣をいう。該残渣には、アルギン酸以外の褐藻の主要成分であるマンニトールやラミナリンが含まれる。アルギン酸資化能を有する微生物として、Sphingomonas sp. A1株が挙げられ、該A1株にZymomonas mobilis等の細菌等のエタノール産生に関する酵素(ピルビン酸脱炭酸酵素及びアルコール脱水素酵素)をコードする遺伝子を導入したエタノール生産性Sphingomonas sp. A1株によりアルギン酸からエタノールを産生し得る。エタノール生産性Sphingomonas sp. A1株は、Takeda et al., 2011, Energy Environ. Sci. 4, 2575-2581や第WO2011/024858号国際公開パンフレットに詳述されている。本発明のマンニトール資化性を有し、マンニトールからエタノールを生産し得る酵母株は、上記のエタノール生産性Sphingomonas sp. A1株を用いて褐藻を原料にしてエタノールを生産した後に残るマンニトールを含む残渣を原料として用いてさらにエタノールを生産することができるので、海洋バイオマスである褐藻を有効利用して大量のエタノールを生産することができる。褐藻を原料として用いて上記のアルギン酸からエタノールを産生し得る微生物を用いてアルギン酸からエタノールを生産した後の残渣を用いて二段階発酵によりマンニトールからエタノールを生産してもよいし、褐藻由来の原料に上記のアルギン酸からエタノールを産生し得る微生物と本発明のマンニトール資化能を有し、マンニトールからエタノールを生産し得る酵母株を同時に添加し、同時にアルギン酸及びマンニトールからエタノールを産生させてもよい。さらに、同時にラミナリン資化性を有し、ラミナリンからエタノールを生産し得る微生物を添加してもよい。また、褐藻を原料として用いて上記のマンニトール資化能を有し、マンニトールからエタノールを生産し得る酵母株を用いてマンニトールからエタノールを生産した後の残渣を用いて、上記のアルギン酸からエタノールを産生し得る微生物による二段階発酵によりアルギン酸からエタノールを生産することもできる。
【0017】
さらに、本発明のマンニトール資化能を有し、マンニトールからエタノールを生産し得る酵母株は、好ましくは、グルコース存在下での凝集能を有する。ここで、凝集能とは、細胞どうしが、お互いに可逆的に凝集して凝集物(フロックと呼ばれることもある)を形成する能力をいう。凝集能を有する株は、酵母菌体の回収が容易になるという利点がある。すなわち、遠心分離等の分離操作を必要としないので、回収にかかるコスト、エネルギー、労力の削減が可能になる。また、凝集能を有するようになった結果、エタノール耐性能が向上した事例も知られている(Zhao and Bai, 2009, Biotechnol. Adv. 27, 849-856)。本発明の酵母株は7%(w/v)未満、好ましくは6%(w/v)以下、さらに好ましくは5%(w/v)以下、特に好ましくは3%(w/v)以下のエタノール存在下でも凝集性を有する。
【0018】
上記のように、本発明のマンニトール資化能を有しマンニトールからエタノールを産生し得る酵母は、さらに、以下の(1)〜(3)の少なくとも1つの特性、すなわち(1)の特性、(2)の特性、(3)の特性、(1)及び(2)の特性、(1)及び(3)の特性、(2)及び(3)の特定、あるいは(1)及び(2)及び(3)の特性を有し得る:
(1)エタノール耐性を有する;
(2)アルギン酸資化能を有する微生物を用いて褐藻原料からエタノールを生産したときの残渣中で生育し得る;及び
(3)グルコース存在下での凝集能を有する。
【0019】
上記の6株の酵母株のうち、エタノール生産性、エタノール耐性、アルギン酸からのエタノール発酵残渣における生育性及びグルコース存在下での凝集能が良好なSaccharomyces paradoxus NBRC 0259株が好ましい。また、エタノール生産性及びエタノール耐性が良好なOgataea glucozyma NBRC 1472株も好ましい。
【0020】
本発明のマンニトール資化能を有する酵母株を培養することにより、エタノールを生産させることができる。該酵母株を培養する方法は、酵母の培養に用いられる通常の方法に従って行われ、公知の培地にマンニトールを添加すればよい。
【0021】
原料となるマンニトールは、終濃度1〜10%(w/v)、好ましくは1〜5%(w/v)、さらに好ましくは2〜5%(w/v)の濃度で添加すればよい。また、培養中に経時的にマンニトールを添加してもよい。
【0022】
培養は、振盪培養又は通気攪拌培養などの好気的条件下、20〜40℃、好ましくは28〜32℃、pH5.6〜9.0、好ましくはpH5.6〜8.4で数時間〜数日間、例えば4〜7日間行う。培地のpHの調整は、無機又は有機酸、アルカリ溶液等を用いて行えばよい。培養中は必要に応じてカナマイシン、ペニシリン等の抗生物質を培地に添加してもよい。
【0023】
培養の際、酵母株はA600(600nmにおける吸光度)が培養開始時に0.05〜11になるように、例えば0.05〜5、好ましくは0.05〜0.2、さらに好ましくは0.1になるように添加すればよい。
【0024】
上記の条件での培養により、培地中に0.1%(w/v)以上、好ましくは0.3%(w/v)以上、さらに好ましくは1%(w/v)以上、さらに好ましくは3%(w/v)以上、特に好ましくは3.5%(w/v)以上のエタノールが蓄積し得る。例えば、10%(w/v)のマンニトールを含む培地で培養した場合、最終的に3%(w/v)以上、特に好ましくは3.5%(w/v)以上のエタノールが蓄積し得る。
また、培養により酵母も増殖し、培養終了時にA600は、0.1〜33程度まで上昇する。
【0025】
さらに、褐藻等に含まれるアルギン酸からアルギン酸資化能を有する微生物を用いてエタノールを生産した場合に得られる原料の残渣を用いて培養することもできる。該残渣中には、アルギン酸資化能を有する微生物が資化できなかったマンニトールが含まれる。原料となる褐藻(Phaeophyceae)としてはマコンブ(Laminaria japonica)、ワカメ(Undaria pinnatiflida)、モズク(Nemacystus decipiens)、ホンダワラ(Sargassum fulvellum)、ヒジキ(Sargassum fulvellum)等が挙げられ、アルギン酸資化能を有する微生物としては、上記のZymomonas mobilis等の細菌等のエタノール産生に関する酵素(ピルビン酸脱炭酸酵素及びアルコール脱水素酵素)をコードする遺伝子を導入したエタノール生産性Sphingomonas sp. A1株が例示できる。例えば、前記残渣中には、0.5〜2%(w/v)程度の濃度のエタノール、1〜10%(w/v)程度のマンニトールが含まれる。このような残渣に本発明のマンニトール資化能を有する酵母株を添加し培養する。この際、酵母株は、A600(600nmにおける吸光度)が0.05〜11になるように、例えば1〜11、好ましくは1〜5、さらに好ましくは1〜3になるように添加すればよい。培養は、振盪培養又は通気攪拌培養などの好気的条件下、20〜40℃、好ましくは28〜32℃、pH5.6〜9.0、好ましくは5.6〜8.4で数時間〜数日間、例えば4〜7日間行う。例えば、培養4〜7日間で、エタノール濃度は、1〜5%(w/v)に達する。残渣中に存在していたアルギン酸から生産されたアルコールを差引いた値が、マンニトール資化能を有する酵母株により新たに産生されたエタノールである。マンニトール資化能を有する酵母株により産生されるエタノールにより、エタノール濃度は残渣中の濃度の1.5〜3倍、好ましくは2倍程度になる。本発明ではこのようにアルギン酸からエタノールを生産させた原料の残渣を用いて残渣中のマンニトールからさらにエタノールを産生し、大量のエタノールを得る方法を二段階発酵という。
【0026】
褐藻を原料として用いてエタノールを生産する場合、褐藻からアルギン酸とマンニトールを抽出し、それを炭素源として用いて培養すればよい。また、褐藻を破砕し、この破砕物そのもの(アルギン酸とマンニトールを含む)を炭素源として用いて培養してもよい。いずれの場合でも、褐藻は木質バイオマスなどとは異なりリグニンを含まないので、木質バイオマスなどの処理と比較すると比較的穏和な条件でアルギン酸とマンニトールを抽出し利用することが可能である。また、トウモロコシデンプンなどと比較しても、トウモロコシデンプンでは糖化行程が必要であるが、アルギン酸とマンニトールからのエタノール生産ではこの糖化行程が不要という利点がある。
【0027】
エタノールは、蒸留により回収することができる。また、エタノールの定量は、公知のアルコール脱水素酵素を用いた方法やガスクロマトグラフィーを用いた方法により行うことができる。
【0028】
上記酵母株を固定化して、エタノールを生産することもできる。微生物の固定化には、包括法、架橋法、担体結合法等がある。包括法とは微生物を高分子ゲルの微細な格子の中に包み込むか、あるいは半透膜性の高分子の皮膜によって被覆する方法であり、架橋法とは微生物を2個又はそれ以上の官能基を持った試薬(多官能性架橋剤)で架橋する方法であり、担体結合法とは水不溶性の担体に酵素を結合させる方法である。固定化に用いられる固定化担体としては、ガラスビーズ、シリカゲル、ポリウレタン、ポリアクリルアミド、ポリビニルアルコール、カラギーナン、アルギン酸、寒天、ゼラチン等がある。
【実施例】
【0029】
本発明を以下の実施例によって具体的に説明するが、本発明はこれらの実施例によって限定されるものではない。
【0030】
実験方法
培地と培養
炭素源非含有培地(pH 5.6)は、イーストニトロジェンベース(w/o アミノ酸;ディフコ社製)0.67%(w/v)、-Leuドロップアウト サプリメント(クロンテック社製)0.69g/l、及びL-ロイシン100mg/lを含む。グルコース合成培地、マンニトール合成培地、グリセロール合成培地、及びラミナリン合成培地は、炭素源非含有培地にグルコース[終濃度2%(w/v)]、マンニトール[同2%(w/v)]、グリセロール[同3%(w/v)]、及びラミナリン[Sigma社製、製品番号L9634、Laminaria digitata由来、同2%(w/v)]を加えた培地である。YP培地(pH5.6)は、酵母エキス1%(w/v)及びトリプトン2%(w/v)を含む。pHはHClで調整した。YPD培地、YPM培地、及びYPG培地は、YP培地に各々グルコース[同2%(w/v)]、マンニトール[同2%(w/v)]、及びグリセロール[同3%(w/v)]を添加した培地である。炭素源とそれ以外の成分は、別々にオートクレーブ滅菌(ラミナリンはフィルター滅菌した)した後に混合した。特に言及しない場合は、2 x YP(pH5.6;2倍濃いYP培地)を用いてこれらの培地を調製した。このとき、YPM培地のpHは5.7であった。10 x YP (pH5.6;10倍濃いYP培地)はオートクレーブ滅菌ではなく、フィルター滅菌した。なお、2 x YP (pH 8.0; NaOHでpHを調整)を用いた場合のYPM培地のpHは7.8であった。固体培地では、これらに終濃度2%(w/v)となるように寒天(ナカライ社製)を添加した。なお、培養は30℃で行った。特に言及しない場合は、液体培地には、A600が0.1となるように細胞を接種し、YPD、YPM、及びYP液体培地を用いた培養は、50mlの液体培地を含む100mlの三角フラスコで、95 strokes per minute(spm)で振とうして行った。固体培地で前培養を行った場合は、培地上の細胞を滅菌水に懸濁してから、液体培地に接種した。
【0031】
酵母を25μg/mlのエチジウムブロマイドで処理してミトコンドリアゲノムとYPG固体培地での生育性を欠失したρ0株を作製した(Fox et al., 1991, Methods Enzymol. 194, 149-165.)。なお、正常なミトコンドリアゲノムを有する株をρ+ 株と称する。嫌気培養は、角形ジャーをアネロパック・ケンキ(ともに三菱ガス化学株式会社製)で嫌気状態にすることにより行った。エタノール生産性形質転換Sphingomonas sp. A1株(EPv104株:後述)の培養は、5%アルギン酸培地を用いて、既報に従って行った(Takeda et al., 2011, Energy Environ. Sci. 4, 2575-2581)。ただし、培養開始時に粉末マンニトールを2%あるいは5%(ともにw/v)となるように添加した。培養3日後の培養液中の上澄み液を遠心分離により調製し、A1株によるアルギン酸からのエタノール発酵残渣(マンニトールを含む)とした。
【0032】
菌株
本実施例では、マンニトール資化性酵母を探索するために、表1に示す研究室の酵母保存株48株を使用した。Saccharomyces paradoxus NBRC 0259及びS. cerevisiae BY4742のρ0 株はエチジウムブロマイド処理により作製した(Fox et al., 1991, Methods Enzymol. 194, 149-165.)。これらの酵母株は、YPD液体培地で培養後17%グリセロール存在下で-80℃で保存した。エタノール生産性A1株としては、EPv104株を用いた(Takeda et al., 2011, Energy Environ. Sci. 4, 2575-2581)。EPv104株は、Sphingomonas sp. A1株の乳酸脱水素酵素遺伝子欠損株に、Zymomonas mobilis由来ピルビン酸脱炭酸酵素遺伝子(pdc)8コピー及び同由来アルコール脱水素酵素遺伝子(adhB)1コピーを広宿主ベクターpKS13を介して導入した、アルギン酸から最も高いエタノール生産性を示す株である(Takeda et al., 2011, Energy Environ. Sci. 4, 2575-2581)。
【0033】
【表1】
【0034】
エタノール濃度測定
エタノール濃度は、F-キット エタノール (Roche Diagnostics K.K., Tokyo, Japan) を用いて、キットに付属のプロトコルに従い測定した。
【0035】
ITS-5.8S rDNA塩基配列の解析
S. paradoxus NBRC 0259株のITS-5.8S rDNA塩基配列のPCRによる増幅及び塩基配列の解析は株式会社 テクノスルガ・ラボに委託した。
【0036】
結果と考察
マンニトール資化性エタノール生産酵母の探索
マンニトール資化性エタノール生産酵母の探索の結果を図2に示す。
【0037】
YPD固体培地上で生育させた本発明者の研究室保存酵母48株(表1)を滅菌水に懸濁し、懸濁液5μlを合成固体培地(炭素源非含有培地、グルコース合成培地、及びマンニトール合成培地) へスポットして細胞を5日間培養した。目視で生育を観察した結果、グルコース合成固体培地では全ての株が良好に生育したが、炭素源非含有固体培地では全ての株が殆ど生育しなかった。また、表1で下線を付した14株(以下マンニトール資化性酵母と称する)がマンニトール合成固体培地で、炭素源非含有固体培地よりも良好な生育を示した。液体培地においても同様であった(図2A)。マンニトール資化能を示さないS288Cの派生株S. cerevisiae BY4742株(以下BY4742株)は(Brachmann et al., 1998, Yeast 14, 115-132.; Quain and Boulton, 1987, J. Gen. Microbiol. 133, 1675-1684.)、マンニトール合成固体及び液体培地で、炭素源非固体及び液体含有培地よりも良好な生育を示さなかった(図2A)。なお、14株の内、P. polymorpha IFO 0195、P. farinosa NBRC 0193、P. haplophila NBRC 0947、P. saitoi IAM 4945、H. saturnus IFO 0177、D. hansenii IFO 0023、及びY. lipolytica NBRC 0746の7株では、マンニトール及びグルコース合成液体培地で膜を形成した。S. paradoxus NBRC 0259株(以下NBRC 0259株)は、グルコース液体培地において凝集性を示した(図2B)。同株は、マンニトール合成液体培地においても若干の凝集性を示した。
【0038】
次に、BY4742株(対照)及びマンニトール資化性酵母14株をマンニトール合成液体培地で3日間静置培養し、培養液上澄みのエタノール濃度を測定した。NBRC 0259、K. capsulata NBRC 0721、K. capsulata NBRC 0974、O. glucozyma NBRC 1472、O. minuta NBRC 1473、及びD. hansenii NBRC 0794各6株(以後、マンニトール資化性エタノール生産酵母と称す)のみが、少なくとも44 mg/l以上のエタノールを生産した(図2C-2)。他の8株は、7 mg/l以下のエタノール生産性しか示さなかった。マンニトールからのエタノール生産性は、NBRC 0259株が他よりも顕著に高かった(図2C-2)。同6株及びBY4742株をグルコース合成液体培地、ラミナリン合成液体培地、並びに5%(w/v)及び7%(w/v)エタノールを含むマンニトール合成液体培地で3日間静置培養した(図2C-1、2C-2及び2D)。同6株のグルコースからのエタノール生産性は、マンニトールからのそれよりも高かった(図2C-2)。とりわけNBRC 0259株は、同6株の中で最も高いグルコースからのエタノール生産性を示した。NBRC 0259株以外の5株はラミナリンからのエタノール生産性も示した(図2C-2)。マンニトール合成培地における3日間の培養では、同6株は7%(w/v)エタノール存在下で生育性を示さなかったが、5%(w/v)エタノール存在下では、NBRC 0259株、K. capsulata NBRC 0974株、及びO. glucozyma NBRC 1472株は生育性を示した(図2D)。5%(w/v)エタノール存在下においても、NBRC 0259株は凝集性を示した。
【0039】
マンニトール資化における酸素要求性
マンニトール資化における酸素要求性を図3に示す。
【0040】
本実施例で見出したマンニトール資化性エタノール生産酵母6株がマンニトール資化に酸素を必要とするかどうかを調べるため、これらをエチジウムブロマイドで処理して、ρ0株の作製を試みたところ、NBRC 0259株のみρ0株が作製できた。そこで、NBRC 0259並びにBY4742のρ0及びρ+株を用いて嫌気及び好気条件下における合成培地での生育性を調べたところ、既報(Quain and Boulton, 1987, J. Gen. Microbiol. 133, 1675-1684.)に一致して、NBRC 0259株はマンニトール合成培地での生育に酸素及び呼吸能を必要とすることが分かった(図3A)。さらに、通常大気条件下並びにYPG及びYPM固体培地において、同株のρ+株は生育できたが、ρ0株は生育できなかった(データ省略)。本結果は、YPM固体培地を用いたNBRC 0259 ρ+株(マンニトール資化能を保持)の選抜が可能なことを意味した。
【0041】
一方、NBRC 0259株以外の5株(ρ+株)の嫌気及び好気条件下における合成培地での生育性を調べたところ、いずれも嫌気条件ではマンニトール合成培地(図3B)及びYPM固体培地(図省略)で生育性を示さなかった。本結果から、NBRC 0259株以外の5株もマンニトールの資化に酸素を必要とすることが明らかとなった。酵母におけるマンニトール資化では、マンニトールがマンニトールデヒドロゲナーゼのはたらきでフルクトースに変換される過程で1分子の余剰NADHが形成されるが、この余剰NADHのNAD+への再生に酸素が必要とされるものと考えられている(図1)(Quain and Boulton, 1987, J. Gen. Microbiol. 133, 1675-1684.)。マンニトール資化性エタノール生産酵母に関しても、同様の理由でマンニトール合成培地やYPD培地での生育に酸素を必要としたと推察された。他方、NBRC 0259株以外の5株は、嫌気条件においてグルコース合成培地(図3B)及びYPD固体培地(図省略)でも生育性を示さなかった。NBRC 0259以外の5株は、NBRC 0259とは若干異なる様式でグルコースを資化している可能性が示唆された。
【0042】
マンニトール資化性エタノール生産酵母によるエタノール生産
図4は、マンニトール資化性エタノール生産酵母によるエタノール生産を示す図である。
【0043】
マンニトール資化性エタノール生産酵母6株のエタノール生産能を更に詳細に調べた。以降はYPM培地を基本培地とした。同6株はマンニトール培地での生育に酸素を必要としたので(図3)、YPM液体培地を用いて、95 spmで振とう培養して生育性とエタノール生産性を調べたところ(図4A-1及び4A-2)、NBRC 0259株とO. glucozyma NBRC 1472株が高い生産性を示した。ただし、両株、とりわけO. glucozyma NBRC 1472株は長期間培養すると培養液中のエタノール濃度が著しく低下した。D. hansenii NBRC 0794株のエタノール生産性は最も低かった。
【0044】
次に、A1株によるアルギン酸からのエタノール発酵残渣(マンニトールを含む)からのエタノール生産(二段階発酵)を目的として、2%(w/v)マンニトールを含む発酵残渣における同6株の生育性(図4B)とエタノール生産性を調べた。同発酵残渣のpHは8.64と弱アルカリ性を示した。これに、10 x YP (pH5.6)を1/10容量添加して(この結果、初期pHは7.3となった)培養を開始した。なお、初期エタノール濃度は9.6 g/lであった。しかしながら、生育性を示したのはNBRC 0259株とD. hansenii NBRC0794株のみであった。培養7日後の培養液上澄みのエタノール濃度は全て初期エタノール濃度を下回った。さらにNBRC 0259株は凝集性を示した。発酵残渣での生育性が弱い理由として、同6株ともにpH 7.8の弱アルカリ性YPM培地では生育したことから(図4B)、同残渣のpH(pH 7.3)が原因ではなく、発酵残渣に含まれる何らかの成分が生育を阻害したと推察された。D. hansenii NBRC0794株は同発酵残渣で生育性は示したが、マンニトールからのエタノール生産性が最も低い株であった(図4A-1及び4A-2)。
【0045】
以上の結果より、NBRC 0259株は、ラミナリンからのエタノール生産性こそ示さないものの、グルコース及びマンニトールからの高いエタノール生産性、エタノール耐性、及びA1株によるアルギン酸からのエタノール発酵残渣における生育性を示したため、6株の中では最も優良な株と判断された(図2、図4)。次に優良な株として、ラミナリンからのエタノール生産性、グルコース及びマンニトールからの高いエタノール生産性、及びエタノール耐性を示した、O. glucozyma NBRC 1472株が考えられた(図2及び図4)。ただし、本NBRC 1472株は、長時間の培養により培養液のエタノール濃度が激減すること(図4A-2)、及び発酵残渣での生育性を示さない(図4B)という望ましくない特性を示した。さらに、マンニトールからのエタノール生産性が報告されているS. cerevisiaeプロピロイドBB1株及びP. angophoraeの、マンニトールからのエタノール生産に関しては、本実施例ほど詳細に報告されていない。さらに、エタノール耐性やアルギン酸からのエタノール発酵残渣における生育性も不明である(Horn et al., 2000, J. Ind. Microbiol. Biotechnol. 24, 51-57.; Quain and Boulton, 1987, J. Gen. Microbiol. 133, 1675-1684.)。これらの理由から、NBRC 0259株を以降の研究に用いることにした。本研究で用いているNBRC 0259株のITS-5.8S rDNA塩基配列は、データベースに登録されているS. paradoxus NBR0259株の同配列(Genbank: D89890)と100%(817/817)の相同性を示した。一方で、S. cerevisiae S288C株の同配列(Genbank: BK006945)とは98.9%(815/824)の相同性を示した。
【0046】
NBRC 0259株によるマンニトールからのエタノール生産
図5は、NBRC 0259株によるマンニトールからのエタノール生産を示す図である。
【0047】
NBRC 0259株のマンニトール資化能を更に調べた。BY4742 ρ+株を対照として用いた。YPD及びYPM固体培地で前培養したNBRC 0259株はマンニトール合成液体培地で生育したが、BY4742株は生育できなかった(図5A及び5B)。また、NBRC 0259株は2%グルコース液体培地では凝集性を示した(図5B)。
【0048】
次に、YPM液体培地を用いてエタノール生産条件を調べた。YPD固体培地上で形成されたNBRC 0259株のシングルコロニーは、高い頻度でYPM固体及び液体培地での生育能を喪失した(調べた6個のシングルコロニーのうち、5個のシングルコロニー由来の細胞がこれを喪失した)。これは、NBRC 0259株がYPD固体培地上ではミトコンドリアゲノムを欠失あるいは損傷しやすい特性を持つことを示唆した。そこで、以降はYPM固体培地上で選抜したNBRC 0259 ρ+株を用いた。同株はマンニトールの資化に酸素を要求したため(図3A)、同株をYPM液体培地で0 spmで静置培養、並びに95及び145 spmで振とう培養して、生育性とエタノール生産性に対する培養中の通気量の影響を調べた。その結果、既報に一致して、通気量が多いほど生育性も高くなった(図5C)。しかし、エタノール生産性は95 spmで適度な酸素を供給した場合に最も高く(6日間で 8.20 g/lのエタノール生産性)、0及び145 spmで培養した場合には殆どエタノール生産性が見られなかった(図5C-1及び5C-2)。一方で、グルコースを基質とした場合、すなわちYPD液体培地では、0及び95 spm両方において、マンニトールの場合よりもより高い生産性を示した(1日[0 spm]及び2日間[95 spm]で各々約12 g/lのエタノール生産性)。また、YPD液体培地でも明瞭な凝集性が観察された(図5D)。
【0049】
振とう速度を95 spmとして、マンニトール濃度の影響を調べたところ、10%(w/v)マンニトールから最大37.6 g/lの(3.8% w/v)のエタノールが生産された(図5E)。これは、マンニトールからのエタノール生産量としては最大であった。また、2及び5%(w/v)のNaCl存在下でエタノール生産性の顕著な低下は見られなかった(図5F)。次に、NBRC 0259株の前培養条件の、生育性とエタノール生産性に対する影響を調べた。すなわち、YPM固体培地で前々培養した同株をYPM又はYPD液体培地で2日間前培養して得られた菌体を洗浄後YPM液体培地に接種したところ、YPM液体培地で前培養した菌体を用いた方が、YPD液体培地で前培養した場合よりも高い生育性とエタノール生産性を示した。すなわち、エタノール生産性は、前者が培養6日で6.7g/lであったが、後者は6日でわずか0.35g/lであり培養12日でようやく5.2g/lに達したに過ぎなかった。
【0050】
5%(w/v)マンニトールを含むアルギン酸からのエタノール発酵残渣45mlに2 xYP (pH 5.6)を5ml添加した培地(初期エタノール濃度8.5g/l)にNBRC 0259株を初期A600が2.0となるよう添加して培養したところ、培養4日で培養液上澄みのエタノール濃度は16.9g/l、培養7日では14.0g/lとなった。すなわち、二段階発酵により残渣中のマンニトールから9.4 (=16.9-8.5)g/lのエタノールが新たに生産された。二段階発酵の条件の改善による、さらなるエタノール生産性の向上が期待された。
【産業上の利用可能性】
【0051】
本発明のマンニトールからエタノールを生産する酵母株を用いることにより、大型海藻などの海洋バイオマスから有効にエタノールを生産することができる。
【特許請求の範囲】
【請求項1】
マンニトール資化能を有しマンニトールからエタノールを産生し得る酵母を、マンニトールを含む培地中で培養することを含む、マンニトールを原料としてエタノールを生産する方法。
【請求項2】
マンニトール資化能を有しマンニトールからエタノールを産生し得る酵母が、以下の(1)〜(3)の少なくとも1つの特性を有する酵母である、請求項1記載のマンニトールを原料としてエタノールを生産する方法:
(1)エタノール耐性を有する;
(2)アルギン酸資化能を有する微生物を用いて褐藻原料からエタノールを生産したときの残渣中で生育し得る;及び
(3)グルコース存在下での凝集能を有する。
【請求項3】
マンニトール資化能を有しマンニトールからエタノールを産生し得る酵母が、Saccharomyces paradoxus、Debaryomyces hansenii、Kuraishia capsulata、Ogataea glucozyma及びOgataea minutaからなる群から選択される、請求項1又は2に記載のマンニトールを原料としてエタノールを生産する方法。
【請求項4】
マンニトール資化能を有しマンニトールからエタノールを産生し得る酵母が、Saccharomyces paradoxus NBRC 0259株、Debaryomyces hansenii NBRC 0794株、 Kuraishia capsulata NBRC 0721株、Kuraishia capsulata NBRC 0974株、Ogataea glucozyma NBRC 1472株及びOgataea minuta NBRC 1473株からなる群から選択される、請求項3記載のマンニトールを原料としてエタノールを生産する方法。
【請求項5】
マンニトール資化能を有しマンニトールからエタノールを産生し得る酵母を培養することにより、培地中に0.1%(w/v)以上のエタノールが蓄積し得る、請求項1〜4のいずれか1項に記載の方法。
【請求項6】
マンニトール資化能を有しマンニトールからエタノールを産生し得る酵母を培養することにより、培地中に3%(w/v)以上のエタノールが蓄積し得る、請求項5記載の方法。
【請求項7】
アルギン酸資化能を有しアルギン酸からエタノールを産生し得る微生物を用いて、褐藻原料からエタノールを産生させたときの原料残渣に、マンニトール資化能を有しマンニトールからエタノールを産生し得る酵母を添加し、培養し、残渣中のマンニトールを原料としてエタノールを生産する方法。
【請求項8】
マンニトール資化能を有しマンニトールからエタノールを産生し得る酵母が、以下の(1)〜(3)の少なくとも1つの特性を有する酵母である、請求項7記載の残渣中のマンニトールを原料としてエタノールを生産する方法:
(1)エタノール耐性を有する;
(2)アルギン酸資化能を有する微生物を用いて褐藻原料からエタノールを生産したときの残渣中で生育し得る;及び
(3)グルコース存在下での凝集能を有する。
【請求項9】
マンニトール資化能を有しマンニトールからエタノールを産生し得る酵母がSaccharomyces paradoxusである、請求項7又は8記載の残渣中のマンニトールを原料としてエタノールを生産する方法。
【請求項10】
マンニトール資化能を有しマンニトールからエタノールを産生し得る酵母がSaccharomyces paradoxus NBRC 0259株である、請求項9記載の残渣中のマンニトールを原料としてエタノールを生産する方法。
【請求項11】
(i) アルギン酸資化能を有しアルギン酸からエタノールを産生し得る微生物を褐藻原料を用いて培養し、アルギン酸からエタノールを産生させ、
(ii) (i)の培養の原料残渣に、マンニトール資化能を有しマンニトールからエタノールを産生し得る酵母を添加し、培養することを含む、
褐藻原料からエタノールを生産する方法。
【請求項12】
マンニトール資化能を有しマンニトールからエタノールを産生し得る酵母が、以下の(1)〜(3)の少なくとも1つの特性を有する酵母である、請求項11記載の褐藻原料からエタノールを生産する方法:
(1)エタノール耐性を有する;
(2)アルギン酸資化能を有する微生物を用いて褐藻原料からエタノールを生産したときの残渣中で生育し得る;及び
(3)グルコース存在下での凝集能を有する。
【請求項13】
マンニトール資化能を有しマンニトールからエタノールを産生し得る酵母がSaccharomyces paradoxusである、請求項11又は12に記載の褐藻原料からエタノールを生産する方法。
【請求項14】
マンニトール資化能を有しマンニトールからエタノールを産生し得る酵母がSaccharomyces paradoxus NBRC 0259株である、請求項13記載の褐藻原料からエタノールを生産する方法。
【請求項1】
マンニトール資化能を有しマンニトールからエタノールを産生し得る酵母を、マンニトールを含む培地中で培養することを含む、マンニトールを原料としてエタノールを生産する方法。
【請求項2】
マンニトール資化能を有しマンニトールからエタノールを産生し得る酵母が、以下の(1)〜(3)の少なくとも1つの特性を有する酵母である、請求項1記載のマンニトールを原料としてエタノールを生産する方法:
(1)エタノール耐性を有する;
(2)アルギン酸資化能を有する微生物を用いて褐藻原料からエタノールを生産したときの残渣中で生育し得る;及び
(3)グルコース存在下での凝集能を有する。
【請求項3】
マンニトール資化能を有しマンニトールからエタノールを産生し得る酵母が、Saccharomyces paradoxus、Debaryomyces hansenii、Kuraishia capsulata、Ogataea glucozyma及びOgataea minutaからなる群から選択される、請求項1又は2に記載のマンニトールを原料としてエタノールを生産する方法。
【請求項4】
マンニトール資化能を有しマンニトールからエタノールを産生し得る酵母が、Saccharomyces paradoxus NBRC 0259株、Debaryomyces hansenii NBRC 0794株、 Kuraishia capsulata NBRC 0721株、Kuraishia capsulata NBRC 0974株、Ogataea glucozyma NBRC 1472株及びOgataea minuta NBRC 1473株からなる群から選択される、請求項3記載のマンニトールを原料としてエタノールを生産する方法。
【請求項5】
マンニトール資化能を有しマンニトールからエタノールを産生し得る酵母を培養することにより、培地中に0.1%(w/v)以上のエタノールが蓄積し得る、請求項1〜4のいずれか1項に記載の方法。
【請求項6】
マンニトール資化能を有しマンニトールからエタノールを産生し得る酵母を培養することにより、培地中に3%(w/v)以上のエタノールが蓄積し得る、請求項5記載の方法。
【請求項7】
アルギン酸資化能を有しアルギン酸からエタノールを産生し得る微生物を用いて、褐藻原料からエタノールを産生させたときの原料残渣に、マンニトール資化能を有しマンニトールからエタノールを産生し得る酵母を添加し、培養し、残渣中のマンニトールを原料としてエタノールを生産する方法。
【請求項8】
マンニトール資化能を有しマンニトールからエタノールを産生し得る酵母が、以下の(1)〜(3)の少なくとも1つの特性を有する酵母である、請求項7記載の残渣中のマンニトールを原料としてエタノールを生産する方法:
(1)エタノール耐性を有する;
(2)アルギン酸資化能を有する微生物を用いて褐藻原料からエタノールを生産したときの残渣中で生育し得る;及び
(3)グルコース存在下での凝集能を有する。
【請求項9】
マンニトール資化能を有しマンニトールからエタノールを産生し得る酵母がSaccharomyces paradoxusである、請求項7又は8記載の残渣中のマンニトールを原料としてエタノールを生産する方法。
【請求項10】
マンニトール資化能を有しマンニトールからエタノールを産生し得る酵母がSaccharomyces paradoxus NBRC 0259株である、請求項9記載の残渣中のマンニトールを原料としてエタノールを生産する方法。
【請求項11】
(i) アルギン酸資化能を有しアルギン酸からエタノールを産生し得る微生物を褐藻原料を用いて培養し、アルギン酸からエタノールを産生させ、
(ii) (i)の培養の原料残渣に、マンニトール資化能を有しマンニトールからエタノールを産生し得る酵母を添加し、培養することを含む、
褐藻原料からエタノールを生産する方法。
【請求項12】
マンニトール資化能を有しマンニトールからエタノールを産生し得る酵母が、以下の(1)〜(3)の少なくとも1つの特性を有する酵母である、請求項11記載の褐藻原料からエタノールを生産する方法:
(1)エタノール耐性を有する;
(2)アルギン酸資化能を有する微生物を用いて褐藻原料からエタノールを生産したときの残渣中で生育し得る;及び
(3)グルコース存在下での凝集能を有する。
【請求項13】
マンニトール資化能を有しマンニトールからエタノールを産生し得る酵母がSaccharomyces paradoxusである、請求項11又は12に記載の褐藻原料からエタノールを生産する方法。
【請求項14】
マンニトール資化能を有しマンニトールからエタノールを産生し得る酵母がSaccharomyces paradoxus NBRC 0259株である、請求項13記載の褐藻原料からエタノールを生産する方法。
【図1】

【図2A】
【図2B】

【図2C−1】
【図2C−2】
【図2D】
【図3A】

【図3B】

【図4A−1】

【図4A−2】
【図4B】
【図5A】
【図5B】
【図5C−1】
【図5C−2】
【図5D】

【図5E】
【図5F】

【図2A】
【図2B】
【図2C−1】
【図2C−2】
【図2D】
【図3A】
【図3B】
【図4A−1】
【図4A−2】
【図4B】
【図5A】
【図5B】
【図5C−1】
【図5C−2】
【図5D】
【図5E】
【図5F】
【公開番号】特開2013−51914(P2013−51914A)
【公開日】平成25年3月21日(2013.3.21)
【国際特許分類】
【出願番号】特願2011−191971(P2011−191971)
【出願日】平成23年9月2日(2011.9.2)
【新規性喪失の例外の表示】特許法第30条第1項適用申請有り 公開▲1▼ 刊行物名 :日本農芸化学会2011年度大会講演要旨集 発行日 :2011年3月5日 発行者 :社団法人日本農芸化学会 該当頁 :P170(3B03a12) 公開▲2▼ 刊行物名 :第63回日本生物工学会大会講演要旨集 発行日 :2011年8月25日 発行所 :公益社団法人日本生物工学会 該当頁 :P114(2Ap07)
【出願人】(507254894)株式会社 マルハニチロホールディングス (1)
【Fターム(参考)】
【公開日】平成25年3月21日(2013.3.21)
【国際特許分類】
【出願日】平成23年9月2日(2011.9.2)
【新規性喪失の例外の表示】特許法第30条第1項適用申請有り 公開▲1▼ 刊行物名 :日本農芸化学会2011年度大会講演要旨集 発行日 :2011年3月5日 発行者 :社団法人日本農芸化学会 該当頁 :P170(3B03a12) 公開▲2▼ 刊行物名 :第63回日本生物工学会大会講演要旨集 発行日 :2011年8月25日 発行所 :公益社団法人日本生物工学会 該当頁 :P114(2Ap07)
【出願人】(507254894)株式会社 マルハニチロホールディングス (1)
【Fターム(参考)】
[ Back to top ]