
酵母を用いた糖ヌクレオチドの合成法
【課題】酵母細胞内での効率的なGDP-L-Fuc又はL-FucアナログのGDP糖化合物の合成方法を提供する。
【解決手段】B.fragilisなど由来のL-フコキナーゼ/GDP-L-フコースピロフォスフォリラーゼ(FKP)遺伝子を導入し、salvage経路を移植した形質転換酵母。及び該酵母を用いた、効率的なGDP-L-Fuc又はGDP-D-AraなどのL-FucアナログのGDP糖化合物の合成方法。
【解決手段】B.fragilisなど由来のL-フコキナーゼ/GDP-L-フコースピロフォスフォリラーゼ(FKP)遺伝子を導入し、salvage経路を移植した形質転換酵母。及び該酵母を用いた、効率的なGDP-L-Fuc又はGDP-D-AraなどのL-FucアナログのGDP糖化合物の合成方法。
【発明の詳細な説明】
【技術分野】
【0001】
本発明は、酵母菌体内で高効率にGDP-フコースなどの糖ヌクレオチド(GDP-糖化合物)を合成する方法に関する。
【背景技術】
【0002】
L-フコース(L-Fuc, 6-デオキシ-L-ガラクトース)は様々な哺乳動物の糖鎖において必須の構成成分であり、N-グリカン、O-グリカン、糖脂質(非特許文献1)だけではなく、直接ある種のタンパク質のセリンやスレオニンに共有結合で結合することも知られている(非特許文献2)。
フコース転移酵素は、グアノシン5’-ジフォスフェート-L-フコース(GDP-フコース、GDP-Fuc)をドナー基質として、L-フコースを小胞体あるいはゴルジ体で受容体基質に転移する転移酵素であり、L-フコースの結合様式によりα1,2、α1,3/1,4、α1,6、O-フコース転移酵素に分類できる(非特許文献3)。L-Fucの付加によって様々な生物的機能が変化する(非特許文献4)。例えば、ABO血液型はα1,2-フコースの付加した糖鎖によるものである(非特許文献5、非特許文献6)。α1,3-フコースやα1,4-フコースはそれぞれシアリルルイスx(sLex)やシアリルルイスa(sLea)に必須の構成成分であり、炎症におけるセレクチンリガンドとしての機能を有している(非特許文献7)。受精においてもフコース付加糖鎖は重要な役割を担っている(非特許文献8、非特許文献9)。FUT8(フコース転移酵素VIII)欠損マウスの実験から、TGF-β1(transforming growth factor-β1)受容体のα1,6-フコースが欠失すると、重篤な発育阻害が見られることが明らかになっており(非特許文献10)、α1,6-フコースが様々な生物機能において重要な役割を果たしていることが示唆される。O-フコース残基(L-Fucが直接セリンあるいはスレオニンの水酸基と結合したもの)は哺乳動物のNotch受容体のEGFドメインで見られる(非特許文献11、非特許文献12)が、Notch受容体はtransmembrane signalingタンパク質ファミリ−に属しており、神経発生、血管形成、リンパ球の発生など様々な生物の発生に関与している(非特許文献13、非特許文献14)。さらに、フコース修飾はアポトーシスにも関与している(非特許文献15、非特許文献16、非特許文献17)。
【0003】
フコース修飾にはフコース転移酵素のドナー基質としてDGP-L-Fucが必要であり、生体内ではGDP-L-Fucは細胞質にある2つの経路によって合成される。
そのうちの主要な経路はde novo合成経路であり、種を超えて保存されている。この経路は、最初に細菌で発見され(非特許文献18)、その後植物(非特許文献19)、哺乳動物(非特許文献20)、無脊椎動物(非特許文献21)でも発見された。この合成系に関与する遺伝子はいくつかの細菌(非特許文献22)、植物(非特許文献23)、動物(非特許文献24)でクローニングされ、ショウジョウバエにおいては、in silicoでこの経路が再現されている(非特許文献25)。
このde novo合成経路では、グアノシン5’-ジフォスフェート-D-マンノース(GDP-マンノース、GDP-Man)から2種類の酵素で触媒される3つの酵素反応によってGDP-L-Fucが合成される。2つの酵素とは、GDP-D-マンノース-4,6-デヒドラターゼ(gmd、ヒトではGMD、EC 4.2.1.47)とGDP-4-ケト-6-デオキシ-D-マンノース-3,5-エピメラーゼ-4-レダクターゼ(Tsta3、ヒトではFX、EC 1.1.1.187)である(非特許文献26、非特許文献27、非特許文献22)。
まず第1段階では、GMDが脱水反応によりGDP-Manを不安定な中間体であるGDP-4-ケト-6-デオキシ-D-マンノースに変換する。この反応には捕因子としてニコチンアミドアデニンジヌクレオチドフォスフェート(NADP+)を要求し、D-マンノースのC-4位の水酸基の酸化によりケトグループを形成し、C-6位の水酸基の還元によりメチル基を形成する(非特許文献28、非特許文献29、非特許文献26)。de novo合成経路におけるGDP-D-ManからGDP-L-Fucへの連続合成反応は、最終産物であるGDP-L-Fucによって阻害されるが、これは、第1段階の反応においてGDP-L-FucがGMDを競合阻害するためである(非特許文献30、非特許文献31)。
第2及び第3段階は2つの機能(エピメラーゼと還元酵素活性)を有するFXにより引き起こされて、GDP-4-ケト-6-デオキシ-D-マンノースがGDP-L-Fucに変換される(非特許文献32、非特許文献27)。まずFXのエピメラーゼ活性によりGDP-4-ケト-6-デオキシ-D-マンノースはC-3位とC-5位がエピマー化し、D体からL体へ変換されてGDP-4-ケト-6-デオキシ-L-ガラクトースが生成される。次いでFXの4-還元酵素活性により捕因子であるNADPHから取ったH+をケト基に転移し、GDP-L-FucとNADP+を生成する(非特許文献33)。
de novo合成経路で用いられるGMDとFXの遺伝子は広くヒトの組織で発現している(非特許文献26)。マウスFX領域への変異の実験では、白血球の接着に加え、受精や生育、発生などにGDP-L-Fucを要求するようになることが分かっている(非特許文献34)。FX欠損マウスにおいては、食餌にL-Fucを加えることにより、通常は活発ではないsalvage経路によりGDP-L-Fucが合成され、糖鎖にフコース修飾が施されるようになる(非特許文献34)。
【0004】
もう一方のGDP-L-Fuc合成経路であるsalvage経路については、L-フコキナーゼ(Fuk, EC 2.7.1.52)及びGDP-L-フコースピロフォスフォリラーゼ(Fpgt, EC 2.7.7.30)が1960年代後半にブタ肝臓から発見された(非特許文献35、非特許文献36)ことで、はじめてその存在が明らかになった。Salvage経路において、第1段階ではL-フコキナーゼ(Fuk)によりL-FucとATPからL-フコース-1-フォスフェート(L-Fuc-1-P)が合成される(非特許文献36、非特許文献37)。第2段階では、GDP-L-Fucピロフォスフォリラーゼ(L-Fuc-1-Pグアニルトランスフェラーゼ:Fgpt)によりL-Fuc-1-PとGTPからGDP-L-Fucが合成される(非特許文献35、非特許文献38)。
L-Fucは、生物では食餌から、培養細胞では培地から摂取され、細胞膜を通して細胞質に輸送される。さらに、L-Fucはリソゾームでフコース分解酵素により糖タンパク質や糖脂質から生産されることもある(非特許文献39)。L-フコースキナーゼ(Fuk)(非特許文献37)やGDP-L-フコース-ピロフォスフォリラーゼ(Fgpt)(非特許文献38)は様々な組織で見つかっていることから、salvage経路もまた真核生物では一般的な経路であると考えられる。以前の報告では、Fuk活性は脳においてドーパミン刺激や大動脈の内皮細胞をニコチンにさらすことによって制御されることが示されている(非特許文献40、非特許文献41)。
【0005】
現在、GDP-L-Fucは化学合成と酵素合成の主要な2種類の方法で合成することができ、L-FucからGDP-L-Fucへの化学合成では様々なアプローチがとられているがいずれも比較的複雑である(非特許文献42、非特許文献43)。GDP-L-Fucの酵素合成では、GDP-D-Manを出発材料とするde novo合成系及びL-Fucを出発材料とするsalvage経路があり、主としてde novo合成系が用いられている(非特許文献44)。
Salvage経路を利用した合成系においても、1ポット反応の合成に成功している(非特許文献45、非特許文献46)。また、Salvage経路に用いる酵素として、非病原性の原生動物であるCrithidia fasciculata由来のD-アラビノース-1-キナーゼとD-アラビノース-1-P-ピロフォスフォリラーゼも利用できることが知られている。これら酵素はin vivoではもっぱらD-アラビノース(D-Ara)からGDP-アラビノース(GDP-Ara)の合成に用いられているが、in vitroではL-FucからGDP-L-Fucの合成に用いることができる(非特許文献47)。しかしながら、これらの合成系はいずれもin vitro条件下での酵素反応系であり、形質転換細胞内での組換え酵素系による合成系は存在しない。
【0006】
近年、プロテオミクスやゲノミクスによって蛋白質を基礎にした数多くの治療薬開発が盛んに行われているが、その蛋白質治療薬の機能発現においては翻訳後の糖鎖修飾が必須な場合が多く、糖蛋白質治療薬を安価に生産するための宿主として、真核生物でありながら単細胞生物で大量培養が容易な酵母が注目されている。
上述のようにフコースは哺乳類細胞において重要な糖単位であり、複合型糖鎖の多様性に富む分岐構造を作り出すためにも必須である。しかし、酵母にはフコースを蛋白質に転移する代謝系酵素群遺伝子が存在しないため、酵母由来の糖蛋白質にはフコースという糖単位は存在しない。酵母を宿主とした有用糖蛋白質生産を可能にするためには、まず出発材料として糖転移酵素の糖供与体となるGDP-L-Fucのプールを酵母細胞内に作り出すことが必要である。
酵母Saccharomyces cerevisiaeを宿主としてGDP-L-Fucを合成させるに際しては、「de novo合成経路」を経由するための有利な条件がそろっている(非特許文献48)。酵母での糖鎖修飾は、大部分がマンノース付加であり(非特許文献49、非特許文献50)、酵母の細胞質にはde novo合成の出発材料となるGDP-D-Manが大量に存在している。Mattilaらと本出願人はそれぞれ大腸菌とArabidopsis thaliana 由来のde novo合成に必須の2つの遺伝子、GDP-D-マンノース-4,6-デヒドラターゼ(gmd、MUR1)とGDP-4-ケト-6-デオキシ-D-マンノース-3,5-エピメラーゼ-4-レダクターゼ(wcaG、AtFX)をS.cerevisiaeに導入した(非特許文献51、非特許文献52、特許文献1)。両グループとも、導入した2つの酵素遺伝子が酵母細胞内で活性化し、酵母に内在するGDP-D-Manから合成したGDP-L-Fucを酵母内に蓄積できることを示している。
このように、形質転換酵母細胞を用いて、菌体内に存在するGDP-D-Man を利用してde novo合成経路によりGDP-L-Fucを合成し菌体内に蓄積する技術は一応確立したものの、上述のように、gmd遺伝子にコードされるGDP-D-マンノース-4,6-デヒドラターゼに対しては、GDP-L-Fucによってフィードバック阻害が働く。さらに、酵母細胞に内在するGDP-D-Manをどの程度利用できるかも、de novo経路を利用するうえでの限定因子となるため、de novo合成経路によっては大量のGDP-L-Fucの蓄積は期待できない。
【0007】
最近、本出願人は、Arabidopsis thaliana 由来のGDP-L-FucからO-フコース結合型タンパク質を合成する代謝系酵素群の各遺伝子を特定し、全て導入した形質転換酵母を作製し、酵母菌体内ではじめてO-フコース結合型の蛋白質合成に成功している(特許文献2)。酵母菌体内においてGDP-L-Fucをさらに効率よく合成し蓄積できる技術が開発できれば、酵母菌体を用いたO-フコース結合型のタンパク質を含む糖タンパク医薬製剤などを大量生産できると考えられる。
したがって、酵母細胞内での効率的なGDP-L-Fuc合成技術、とりわけsalvage経路によるGDP-L-Fucの合成方法の開発が強く望まれていた。
【先行技術文献】
【特許文献】
【0008】
【特許文献1】特開2001−145488号公報
【特許文献2】特開2005−328841号公報
【非特許文献】
【0009】
【非特許文献1】Walz G, et al.,Science. 250:1132-1135.
【非特許文献2】Harris RJ, et al., 1993. Glycobiology. 3:219-224
【非特許文献3】Ma et al., 2006,Glycobiology 16:158R-184R.
【非特許文献4】Becker, et al.,2003,Glycobiology. 13:41R-53R.
【非特許文献5】Greenwell, 1997, Glycoconjugate J. 14:159-173.
【非特許文献6】Lloyd KO. 2000. Glycoconjugate J. 17:531-541.
【非特許文献7】Lowe JB. 1997. Kidney Int. 51:1418-1426.
【非特許文献8】Johnston et al., 1998 ,J Biol Chem. 273:1888-1895.
【非特許文献9】Mori et al., 1998 ,Glycoconjugate J. 15:447-456.
【非特許文献10】Wang et al., 2005,Proc Natl Acad Sci USA. 102:15791-15796.
【非特許文献11】Moloney et al., 2000 ,J Biol Chem. 275:9604-9611.
【非特許文献12】Rampal et al., 2005,J Biol Chem. 280:32133-32140.
【非特許文献13】Artavanis-Tsakonas et al., 1999 ,Science. 284:770-776.
【非特許文献14】Halloran et al., 2000 ,J Immunol. 164:4868-4877
【非特許文献15】Hiraishi et al., 1993,Glycobiology. 3:381-390
【非特許文献16】Russell et al., 1998,Biochem Bioph Res Co. 250:449-453.
【非特許文献17】Winkler et al., 2004,Blood. 103:1685-1692.
【非特許文献18】Ginsburg, 1960 ,J Biol Chem. 235:2196-2201
【非特許文献19】Liao et al., 1971,Biochim Biophys Acta. 230:64-71.
【非特許文献20】Overton et al., 1981 ,Biochim Biophys Acta. 675:281-284.
【非特許文献21】Bulet et al., 1984,Eur J Biochem. 144:255-259
【非特許文献22】Tonetti et al.,1998,Biochimie. 80:923-931.
【非特許文献23】Bonin et al., 1997,Proc Natl Acad Sci USA. 94:2085-2090
【非特許文献24】Reitman et al., 1980,J Biol Chem. 255:9900-9906.
【非特許文献25】Roos et al., 2002,J Biol Chem. 277:3168-3175.
【非特許文献26】Sullivan et al., 1998,J Biol Chem. 273:8193-8202
【非特許文献27】Tonetti et al., 1996 ,J Biol Chem. 271:27274-27279.
【非特許文献28】Chang et al., 1985,Anal Biochem. 144:253-257
【非特許文献29】Oths et al., 1990 ,Carbohyd Res. 198:91-100
【非特許文献30】Bisso et al., 1999,FEBS Lett. 456:370-374.
【非特許文献31】Sturla et al., 1997 ,FEBS Lett. 412:126-130
【非特許文献32】Chang et al., 1988,J Biol Chem. 263:1693-1697.
【非特許文献33】Menon et al., 1999 ,J Biol Chem. 274:26743-26750
【非特許文献34】Smith et al., 2002,J Cell Biol. 158:801-815
【非特許文献35】Ishihara et al.,1968,J Biol Chem. 243:1110-1115
【非特許文献36】Ishihara et al., 1968,J Biol Chem. 243:1103-1109
【非特許文献37】Park et al., 1998 ,J Biol Chem. 273:5685-5691
【非特許文献38】Pastuszak et al., 1998, J Biol Chem. 273:30165-30174
【非特許文献39】Michalski et al., 1999, Biochim Biophys Acta. 1455:69-84.
【非特許文献40】Hocher et al., 1993,Eur J Clin Chem Clin. 31:347-351
【非特許文献41】Ricken et al., 1990 ,Biochem Soc T. 18:963-964
【非特許文献42】Adelhorst et al., 1993,Carbohyd Res. 242:69-76
【非特許文献43】Murray et al., 1997 ,Biochemistry. 36:823-831
【非特許文献44】Becker et al.,1999,BBA-Mol Basis Dis. 1455:193-204
【非特許文献45】Ichikawa et al., 1992, J Am Chem Soc 114:9283-9298.
【非特許文献46】Ichikawa et al.,1994,Methods Enzymol. 247:107-127
【非特許文献47】Mendgeling et al., 1999,Anal Biochem. 267:227‐233
【非特許文献48】Hirschberg et al., 1998 Annu Rev Biochem. 67:49-69.
【非特許文献49】Hashimoto et al., 1997 ,J Biol Chem. 272:16308-16314.
【非特許文献50】Romanos et al., 1992, Yeast. 8:423-488
【非特許文献51】Mattila et al., 2000 ,Glycobiology. 10:1041-1047
【非特許文献52】Nakayama et al., 2003, Glycobiology. 13:673-680
【非特許文献53】Coyne et al., 2005, Science. 307:1778-1781.
【非特許文献54】Kotake et al., 2008, J Biol Chem. 283:8125-8135
【非特許文献55】Niittymaki et al., 2004, Eur J Biochem. 271:78-86.
【非特許文献56】Linster et al., 2007 ,J Biol Chem. 282:18879-18885.
【発明の開示】
【発明が解決しようとする課題】
【0010】
本発明は、酵母細胞内での効率的なGDP-L-Fucの合成方法を提供することを目的とするものである。さらに、L-フコースと構造的に類似したD-アラビノースと共に、L-フコース、D-アラビノースなどのアナログを酵母細胞内で効率的にGDP-糖化合物に転換する方法を提供する。
【課題を解決するための手段】
【0011】
本発明者らは、上記課題を解決するために鋭意研究を重ねる中で、Bacteroides fragilis 9343由来のL-フコキナーゼ/GDP-L-フコースピロフォスフォリラーゼ(FKP)という2つの活性をもつ酵素遺伝子を利用することを思いついた。この酵素はL-Fucを基質としてL-Fuc-1-Pに変換し、さらに、そこからGDP-L-Fucを合成することができる(非特許文献53)。この変換活性はB.fragilis 9343株内のsalvage経路として観察され、広く細菌(Bacteroides)一般に保存されている(非特許文献53)。細菌(Bacteroides)以外では植物(Arabidopsis)もsalvage経路としてL-FucからL-Fuc-1-Pを経由してGDP-L-Fucを合成できる系を有しているが、当該系を有しているのは細菌及び植物のみであり、その理由は明らかになっていない(非特許文献54)。上記酵素FKPは、アミノ酸配列アラインメントの結果、そのN-末端ドメイン(1-430番目のアミノ酸)では、ヒトのGDP-L-フコースピロフォスフォリラーゼと20%の相同性があり、C-末端ドメイン(584-949番目のアミノ酸)では哺乳動物のL-フコキナーゼと相同性が高い(非特許文献55)。2つのドメインを150アミノ酸のリンカーで繋ぐ構造となっているが、リンカーの機能は現在のところ不明である。
今回、B.fragilis 由来FKP遺伝子を導入した酵母(S.cerevisiae)宿主において、FKPが活性型酵素として発現し、培地中に添加したL-Fucを、salvage経路によりL-Fuc-1-Pを経由してGDP-L-Fucにまで変換し、細胞内に蓄積することを確認した。また、L-Fucと構造的に似ているD-Araを培地中に添加した場合でも、同様にGDP-D-Araを合成することも確認できた。
以上の知見を得たことで、本発明を完成した。
【0012】
すなわち、本発明は以下の発明を含むものである。
〔1〕 FKP(L-フコキナーゼ/GDP-L-フコースピロフォスフォリラーゼ)遺伝子を酵母用発現ベクターにより形質転換した酵母であって、培地中のL-フコース、又はL-フコースアナログを取り込み、細胞内でGDP-L-フコース、又はL-フコースアナログのGDP-糖化合物を合成可能な形質転換酵母。
〔2〕 L-フコースアナログがD-アラビノースであり、そのGDP-糖化合物がGDP-D-アラビノースである、前記〔1〕に記載の形質転換酵母。
〔3〕 FKP遺伝子が、Bacteroides fragilis由来のFKP遺伝子である、前記〔1〕又は〔2〕に記載の形質転換酵母。
〔4〕 酵母がSaccharomyces cerevisiaeである前記〔1〕〜〔3〕のいずれかに記載の形質転換酵母。
〔5〕 前記〔1〕〜〔4〕のいずれかの形質転換酵母を用い、L-フコース、D-アラビノース、又はL-フコースアナログを存在させた培地中で培養することを特徴とする、L-フコース、D-アラビノース、又はL-フコースアナログのGDP糖化合物を合成する方法。
〔6〕 L-フコースを存在させた培地中で培養することを特徴とする、GDP-L-フコースを合成する方法である前記〔5〕に記載の方法。
〔7〕 D-アラビノースを存在させた培地中で培養することを特徴とする、GDP-D-アラビノースを合成する方法である前記〔5〕に記載の方法。
【発明の効果】
【0013】
本発明のB.fragilis由来のFKP遺伝子を導入した形質転換酵母は、培地中に基質となるL-フコース(L-Fuc)を添加しておくだけで、salvage経路により直接GDP-フコース(GDP-L-Fuc)を効率的に変換することができるため、当該形質転換酵母を用いたGDP-フコース(GDP-L-Fuc)合成方法により、簡便でかつ高効率でGDP-L-Fucを酵母菌体内に蓄積することができる。
さらに、L-フコースと構造的に類似したD-アラビノース(D-Ala)などの糖類、及びL-フコース、D-アラビノースのアナログである化学修飾体に対しても、酵母細胞内で効率的にGDP-糖化合物に転換することができる。
そして、当該形質転換酵母に対してさらに外来タンパク質遺伝子と共に「O-フコース転移酵素遺伝子群(特許文献2)」を導入してやることにより、またはこれら遺伝子が導入された形質転換酵母と掛け合わせることにより、salvage経路を経由したO-フコース含有タンパク質合成系を有する形質転換酵母を作出でき、O-フコース含有タンパク質を効率よく製造することができる。
【図面の簡単な説明】
【0014】
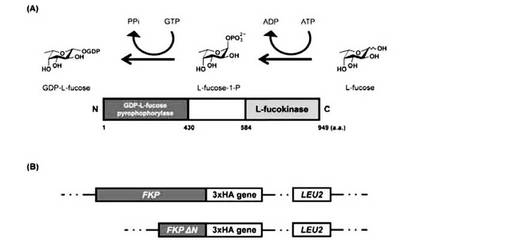
【図1】外来L-FucをGDP-L-Fucに変換するsalvage経路には、B. fragilisの酵素が必要となる。(A) FKPを用いたGDP-L-Fucの酵素合成スキーム。FKP タンパク質のN-末端ドメイン(1-430番目のアミノ酸)はヒトのGDP-フコースピロフォスフォリラーゼと20%の相同性を示し、C-末端ドメイン(584-949番目のアミノ酸)は哺乳動物のL-フコキナーゼと相同性を示す。 (B) 酵母ゲノムに挿入したFKP と FKP ΔN遺伝子の構築。両遺伝子はC-末端側で3xHAと融合させている。
【図2】Mock (レーン1)、FKP ΔN (レーン2)、FKP (レーン3) 遺伝子導入酵母の細胞の可溶化物のSDS-PAGEとウェスタンブロット解析。(左)およそ2x106細胞/レーン、抗HA抗体で検出。(右)およそおよそ2x106細胞/レーン、CBB(Coomassie Brilliant Blue R-250)で染色した。レーン1, 1’;mockベクター(pRS405-leu)を保持する酵母W303-1A株、レーン2,2’;FKPΔN-3xHAを挿入した発現ベクターを保持する酵母W303-1A株、レーン3, 3’;FKP-3xHAを挿入した発現ベクターを保持する酵母W303-1A株。左のパネルの上部矢じりはFKP-3xHAタンパク質を、下の矢じりはFKPΔN-3xHAタンパク質を示している。
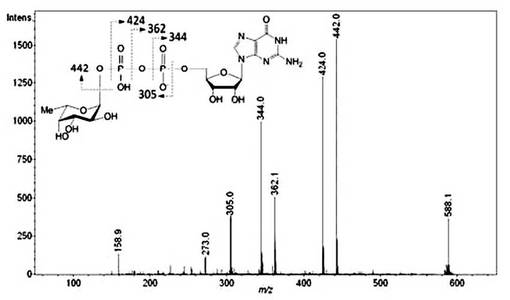
【図3】酵母抽出物由来のGDP-L-Fucの構造解析。FKP 遺伝子を導入した酵母を15 mM L-Fucを含む培地で培養した酵母抽出物由来のESI-MS/MSスペクトル(m/z 588.1の[M-H]- イオン)。m/z 442はGDP-L-Fucのフラグメントイオンと考えられる。主要なフラグメントは化学的に不安定なL-Fuc-1-phosphateあるいはpyrophosohate結合の切断によって出現すると考えられる。
【図4】In vivoでのGDP-L-Fuc生産性。1-ブタノール飽和1 Mギ酸でおよそ5x108 細胞から抽出した抽出物をMono Qカラムで陰イオン交換で精製し、GDP-L-Fuc画分を単離した。サンプル注入後5分から0.5M KH2PO4の濃度を上げ、100分後に0.5M KH2PO4が70%になるようにした。流速は1ml/minとした。FKPを導入した酵母W303-1Aを(B)15mM L-Fuc存在下と(A)L-Fucを添加しないで培養した。コントロールとしては、GDP-L-Fuc標準物質25 nmolを用いた(C)。L-Fuc存在下で培養した酵母由来の図では、GDP-L-Fucに相当する9.25分にピークが認められた。
【図5】酵母抽出物由来のGDP-D-Araの構造解析。15mM D-Araを含む培地で培養したFKPを持つ酵母からの抽出物で得られたm/z 574.1のESI-MS/MSスペクトルを示した。m/z 442はGDP-D-Araのフラグメントイオンである。主要な断片は化学的な不安定なD-アラビノース-1-フォスフェートあるいはピロフォスフェート結合の切断物だと考えられる。
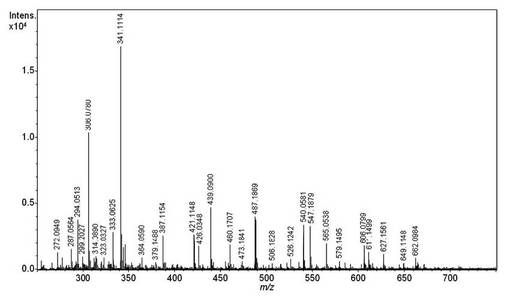
【図6】15mM L-Fucを添加した培地で培養したFKPを持つ酵母由来の抽出物のESI-MSスペクトル。矢印で示したm/z 588の[M-H]-イオンはGDP-L-Fucに相当する。
【図7】標準GDP-L-FucのESI-MS/MSスペクトル。m/z 588の[M-H]-イオンの主要断片を記載した。
【図8】L-FucもD-Araも添加しない培地で培養したFKPを持つ酵母由来の抽出物のESI-MSスペクトル。GDP-L-Fucに相当するm/z 588.08もGDP-D-Araに相当するm/z 574.08も認められなかった。
【図9】酵母抽出物をクロマトグラフィーで分離し、UV254nmで検出。S. cerevisiaeをmockベクター(AとD)、FKP ΔN(BとE)、FKP(CとF)で形質転換し、50mlのSD(-leu)培地に15mM L-Fucを添加したもの(D-F)と添加しないもの(A-C)でOD600が1.0になるまで培養した。細胞を1-ブタノール(10%, v/v)飽和1Mギ酸で抽出した。ヌクレオチドと糖ヌクレオチドをMono Qカラムにより分画し、UV254nmでモニターした。サンプル注入から5分後に0.5 M KH2PO4を加え、40分後に0.5 M KH2PO4が100%になるようにした。流速は1ml/minとした。パネル(F)の主要ピーク(およそ17.5分)を矢じりで示した。
【図10】In vivoでのGDP-L-Fucの生産性。50mlの培地で培養した酵母を1-ブタノール飽和1 Mギ酸で抽出した。抽出物は陰イオン交換クロマトグラフィー(Mono Q)に供し、GDP-L-Fuc画分を単離した。サンプル注入から5分後に0.5 M KH2PO4を加え、40分後に0.5M KH2PO4が100%になるようにした。流速は1ml/minとした。FKPを持つ酵母W303-1A株を7.5mM L-Fuc(A)、15mM L-Fuc(B)を含む培地で培養し、細胞を集めた。コントロールには40nmolのGDP-L-Fucを用いた(C)。添加したL-Fucの量に応じてGDP-L-Fucが合成されていた。
【図11】15mM D-Araを添加した培地で培養したFKPを持つ酵母由来の抽出物のESI-MSスペクトル。矢印で示したm/z 574.070の[M-H]-イオン(矢印)はGDP-D-Araに相当する。
【発明を実施するための形態】
【0015】
1.FKP形質転換酵母について
原核生物、真核生物は、2つの異なる代謝経路(de novo経路とsalvage経路)を利用してGDP-L-Fuc合成を行っている(非特許文献2)が、酵母にはGDP-L-Fucの合成能力がない。
本発明では、酵母にFKP(L-フコキナーゼ/GDP-L-フコースピロフォスフォリラーゼ)酵素遺伝子を導入することでsalvage経路を移植した(図1A参照)。本実施例ではB. fragilis9343株由来のFKP酵素遺伝子を用いているが、FKP酵素は広くBacteroides類に分布しており、高等植物にも存在することが知られている(非特許文献53、54)。いずれも、B.fragilis9343株由来FKP酵素と同様にL-フコキナーゼ及びGDP-L-フコースピロフォスフォリラーゼ作用を有している酵素であって、B.fragilis9343株由来FKP酵素に代えて用いることができる。酵母に導入された外来遺伝子であるB. fragilisのFKP遺伝子の過剰発現により、酵母菌体内でもsalvage経路が働き、GDP-L-Fucの合成を可能にすることができる。B. fragilis9343株由来のFKP酵素遺伝子の塩基配列(配列番号2)及び対応するアミノ酸配列(配列番号1)の情報は、それぞれアクセッション番号CR626927、CAH08307としてGENBANKデータベースから入手できる。B. fragilis9343株以外のBacteroides類又は植物由来の酵素遺伝子は、配列番号1に示されるアミノ酸配列とは80%以上、好ましくは90%以上、さらに好ましくは95%以上のホモロジーを有するアミノ酸配列をコードし、又は配列番号2に示される塩基配列とも80%以上、好ましくは90%以上、さらに好ましくは95%以上のホモロジーを有している。そして、配列番号1に示されるアミノ酸配列において1若しくは数個のアミノ酸が欠失・置換・付加されたアミノ酸配列をコードする遺伝子、又は配列番号2に示される塩基配列の相補配列とストリンジェントな条件下でハイブリダイズするポリヌクレオチドからなる遺伝子などと表現することもできる。
酵母細胞はGDP-L-Fuc合成に必要な酵素を持っていないので、導入した遺伝子の機能発現を容易にモニターすることができる(非特許文献50、非特許文献49)。
B. fragilis由来などのFKPの関与するsalvage経路を選択したことの理由の1つには、GDP-L-Fucと構造や生合成ステップが似ているGDP-L-Araの生合成を行える点にある(非特許文献47、非特許文献56)。
当該遺伝子を酵母用の発現ベクターに繋いで酵母を形質転換する。宿主の酵母としては、本実施例ではSaccharomyces cerevisiaeを用いたが、Pichia酵母など他の酵母でもよく、発現ベクターについても酵母菌体内で発現可能なプロモーターを有するベクターであればいずれでも良い。ただし、酵母細胞内でsalvage経路を働かせる必要があるため、分泌シグナルを含む分泌ベクターは不適切である。
【0016】
2.本発明が対象とする糖ヌクレオチド(GDP糖化合物)
本発明におけるFKP形質転換酵母は、典型的には細胞内でのFKP酵素作用によりL-フコースをそのGDP糖化合物であるGDP-L-フコースに転換するが、L-フコースに構造的に類似しているいわゆるL-フコースアナログを、そのGDP-糖化合物に転換することができる。L-フコースアナログとしては、D-アラビノースなどの糖類の他、L-フコース又はD-アラビノースなど糖類のアジドやアルキンなどの化学修飾体が含まれる。本実施例では具体的にD-アラビノースをGDP-D-アラビノースに転換できることを実証している。
【0017】
3.FKP形質転換酵母による糖ヌクレオチド合成方法
(1)本発明のFKP形質転換酵母によるGDP-L-Fuc合成
FKP形質転換後、細胞数が十分に増えるまでロイシンを欠失させた酵母用培地で培養し、次いで培地中にL-Fuc又はL-Fucアナログ例えばD-Alaを培地に1mM以上、好ましくは7.5mM以上、より好ましくは15mM以上で480mM以下添加してさらに培養をする。その際には振とう培養を行うことが好ましい。
【0018】
(2)GDP-L-Fuc生産物の確認方法又は定量法
本実施例では以下の(i)及び(ii)の方法でGDP-L-Fuc生産物の確認及び定量した。
(i) MSまたは MS/MSスペクトルによる方法
質量分析でGDP-L-Fucに典型的なm/z 588の位置のピークの存在を確認することで、精製しない状態でも系にGDP-L-Fucが存在しているかどうかを確認することができる。
例えばESI-1TOF MS/MS解析(エレクトロスプレーイオン化 四重極 飛行時間型質量分析装置)、micrOTOF-QによるMS/MSスペクトル法を用いることができる。標準GDP-L-Fucのデータと比較することでより正確に確定することができる。
(ii) HPLC分析
精製されたGDP-L-FucをHPLC分析用溶媒に溶かし、Mono-Qカラムなどの陰イオンカラムにより分析することができる。Mono-Qカラムの場合には、A液として10mM KH2PO4、B液として0.5M KH2PO4を使用してGDP-L-Fucの分析をすることができる。
標準GDP-L-Fucの資料が市販されているので、そのデータと比較することで精製度も、また定量解析をすることもできる。
(iii) このほか、キャピラリー電気泳動、NMRによりGDP-L-Fucが生産されているかどうかを確認することができる。
【0019】
(3)GDP-L-Fucなどの回収、精製方法
GDP-L-Fucは酵母の細胞質内で合成され、蓄積されているため、GDP-L-Fucを回収するためには、酵母菌体を破砕後遠心分離などで細胞を除去し、糖ヌクレオチドの単離精製に用いられる一般的な生化学的方法、イオン交換クロマトグラフィー、アフィニティクロマトグラフィー、ゲル濾過法などを単独又は組み合わせて用いることができる。
【0020】
4.酵母によるO-フコース含有糖タンパク質の製造方法
酵母の細胞質内に蓄積されたGDP-L-Fucを回収せずに、そのままO-フコース含有糖タンパク質の製造のために利用することができる。
すなわち、特許文献2に記載されたO-フコース結合型タンパク質合成系遺伝子群(O-フコース転移酵素遺伝子群)が導入されている形質転換酵母に本発明のFKP形質転換酵母を掛け合わせることにより、O-フコース結合型タンパク質を合成可能な形質転換酵母を得ることができる。
【0021】
5.FKP形質転換酵母による他の糖ヌクレオチド合成について
本発明のFKP形質転換酵母は、L-Fucと構造的に類似しているD-Alaに対しても、培地中に基質として添加しておくことで、酵母菌体内でGDP-D-Alaを合成することができた(図12)。
B. fragilis由来のFKP酵素は基質特異性が低いため、原理的には、L-ガラクトースのような他の単糖にも応用できる可能性がある。さらに、天然の糖でなくてもL-Fucに構造が類似していればよく、L-Fucのアナログを用いた糖ヌクレオチド合成に応用することで、糖ヌクレオチドアナログを高純度で合成することも可能である。
【0022】
<式1>FKP形質転換酵母細胞におけるGDP-L-FucとGDP-D-Ara合成。

【実施例】
【0023】
以下、実施例により本発明を具体的に説明するが、本発明は特にこれら実施例に限定されるものではない。
なお、本発明で使用されている技術的用語は、別途定義されていない限り、当業者により普通に理解されている意味を持つ。
また、本発明で引用した先行文献又は特許出願明細書の記載内容は、本明細書の記載として組み入れるものとする。
【0024】
(実施例1)形質転換酵母(Saccharomyces cerevisiae)の調製
(1−1)形質転換用宿主酵母株の調整
糖ヌクレオチドのインビボ合成のために形質転換用の宿主酵母として、Saccharomyces cerevisiae W303-1A株(MA Ta leu2-3, 112 his3-11, 15 ade2-1 ura3-1 trp1-1 can1-100)を用いた。W303-1A株の培養は、S. cerevisiaeの通常の生育培地であるYPAD培地(2%バクトペプトン、1%酵母エキス、2%グルコース、40μl/ml硫酸アデニン)を用いた。形質転換後はロイシン欠失培地(SD(-leu): 0.67%バクト酵母ニトロゲンベースアミノ酸不含、2%グルコース、0.69g/l アミノ酸混合物ロイシン不含)を用いた。
【0025】
(1−2)FKP遺伝子を含む酵母用発現ベクターの調製
FKP遺伝子はBacteroides fragilis 9343(ATCC 25285)染色体DNAからPCR増幅し、pCMV-Tag 2Bベクター(ストラタジーン社)のEcoRI−XhoI部位に挿入した。PCR増幅し、塩基配列を確認したFKP遺伝子全長及び部分遺伝子(truncated version)を酵母用のYep352GAP-IIベクター(非特許文献52)に導入し、発現に使用した。
全長FKP遺伝子は、
5’-GGAGCTCATGCAAAAACTACTATCTTTACCG-3’(配列番号3)と
5’-GCTCTAGACCCGGGTGATCGTGATACTTGGAATCCC-3’
(配列番号4)とをプライマーとし、
またネガティブコントロールであるN-末端を除去したFKP(FKP ΔN)遺伝子(1752番目の塩基から2847番目の塩基)は、
5’-GGAGCTCATTGTTTGGGGACGTAGC-3’
(配列番号5)と
5’-GCTCTAGACCCGGGTGATCGTGATACTTGGAATCCC-3’
(配列番号6)をプライマーとして使用し、PCR増幅を行った。
両PCR断片はSacIとXbaIで切断し、TDH3プロモーターを持つ発現ベクターYep352GAP-IIに挿入した。pHAベクターから3×HA-tag(3個のインフルエンザヘマグルチニンエピトープ)遺伝子をSmaIにより切り出し、FKP或いはFKP ΔN遺伝子が挿入されているYep352GAP-IIに挿入した。これらのベクターからTDH3プロモーター、FKP-3×HAあるいはFKP ΔN-3×HA、TDH3ターミネーターを含む断片をBamHIにより切り出し、pRS405のBamHI部位に挿入しpRS405-FKP3xHA(図1A)あるいはpRS405-FKPΔN3xHA(図1B)を構築した。各プラスミドの塩基配列を確認したのち、BstEIIで切断し、直鎖状にし、酵母染色体のLEU2遺伝子領域に組み込んだ。なお、上記で用いた制限酵素、修飾酵素はNew England Biolabs社あるいはロシュダイアグノスティクス社製を用いた。
また、プラスミドベクターの構築の際には大腸菌DH5α株を使用した。大腸菌はLuria Bertani培地(1%バクトトリプトン、0.5%酵母エキス、0.5%NaCl、0.2%グルコース)で培養した。
【0026】
(1−3)酵母(S. cerevisiae)の形質転換
前記(1−2)で構築した酵母用発現ベクターpRS405-FKP3xHA(FKP遺伝子)及びpRS405-FKPΔN3xHA(FKPΔN遺伝子)と共に何も遺伝子を挿入していないベクター(pRS405: Mock)のそれぞれを用いて、Itoらの方法(Ito H, et al., J Bacteriol. 153:163-168)により、前記(1−1)で調整した酵母W303-1A株の形質転換を行った。形質転換体はロイシン欠失培地でそれぞれについて数株ずつ選択し、コロニーPCR解析によって確認を行った。
【0027】
(1−4)B. fragilis由来FKP酵素タンパク質の発現
前記(1−3)において、ロイシン欠失培地で選択した数株ずつの形質転換体(FKP遺伝子、FKPΔN遺伝子、Mock)をそれぞれ5mlのSD(-leu)培地で30℃の条件下で12時間培養した。この条件ではOD600が1.0(およそ2x107細胞)程度になる。各形質転換細胞を4,000×g、5分の遠心分離により集め、1×PBSで2回洗浄した。各々がおよそ2x107細胞となるように100μlのサンプルバッファー(0.06 M Tris-HCl (pH6.8)、10% (v/v) グリセロール、2% (v/v)2-メルカプトエタノール、0.25% (w/v)ブロモフェノールブルー)に再懸濁した。これらを95℃で5分インキュベートし、14,000×g5分の遠心分離後、上清を10%SDSポリアクリルアミド電気泳動に供した。電気泳動後のタンパク質は、PVDF膜(ミリポア社)に電気的に転写した(15 V、1時間)。膜をブロッキングし、抗HA抗体(ロシュ社)と室温で1時間反応させた。抗HA抗体は5%スキムミルクを含むPBST(0.1% Tween 20)で希釈しておいたものを使用した。HRP(Horseradish peroxidase)-コンジュゲート ウサギ抗マウスIgG(Zymax社)を2次抗体として用い、ECL-エンハンストケミルミネッセンス(GEヘルスケア社)で検出した。
その結果、図2に示したように、FKPΔNとFKP遺伝子産物ははっきりと認められた(矢じりで示した)。予想通り、FKP導入ベクターを用いた場合にはFKP酵素の全長に対応する135 kDaの位置にバンドが見られたのに対し、FKPΔN導入ベクターを用いた場合のバンドは43 kDaに見られた。一方、mockではバンドは観察されなかった。
【0028】
(実施例2)形質転換酵母によるGDP-L-Fucの合成
(2−1)形質転換酵母のL-Fuc含有培地下での培養
形質転換酵母細胞を2mM MgCl2を含む25mlあるいは50mlのSD(-leu)培地(pH7.5)でOD600が1.0になるまで培養した。この場合、細胞はおよそ5x108あるいは1x109になる。このとき、培地にあらかじめ15mMのL-フコースを含むものと含まないものを用いた。3種類の形質転換体(mock pRS405、pRS405-FKP3xHA及びpRS405-FKPΔN3xHAを導入した形質転換体)を32℃、140rpmで振とう培養した。このとき、培地はフラスコの容量の1/5容になるようにした。細胞を4℃、4,000×g、5分間の遠心分離で集め、5倍量の氷冷した滅菌水で2回洗浄した。その後、10%1-ブタノール(v/v) を含む4mlの氷冷1Mギ酸に再懸濁し、穏やかにピペッティングにより撹拌し、氷中に30分間置いた。4℃、6,000×g、10分間の遠心分離で夾雑物を除去したあとの上清を凍結乾燥に供した。
【0029】
(2−2)酵母細胞からのGDP-L-Fucの抽出とESI-1TOF MS/MS解析による確認
酵母細胞質中でのGDP-L-Fuc合成酵素活性を確認するために、細胞抽出物中のGDP-L-Fucの検出を、ブルカダルトニクス社のESI-qTOF MS(エレクトロスプレーイオン化 四重極 飛行時間型質量分析装置)を用いたMS解析で行い、さらにmicrOTOF-Qを用いてMS/MSスペクトルを陰イオンモードで取得した。
具体的には、前記(2−1)で得られた乾燥サンプルを0.5mlの滅菌水に溶解し、そのうち1μlを50%メタノール(v/v) で50μlに希釈した。サンプル溶液は0.5barの窒素ガスで、シリンジポンプ(Kdサイエンティフィック社、流速8μl/min)を用いてESI源にて測定した。乾燥温度は180℃、乾燥用ガスの流速は4l/minとした。キャピラリーの電圧は3200V、エンドプレートオフセットを500Vに保持した。
GDP-L-Fucに相当するm/z 588.078の[M-H]-イオン(図6の矢印)をESI-qTOF MSによる MS/MS解析に供した(図3)。
MS/MS解析でのm/z 305のフラグメントイオンは、下記の表1に示されるように、GDP-L-Fucからグアノシンが脱離した値に相当する。
【0030】
<表1>
【0031】
このMS/MS解析の結果は、標準GDP-L-Fucの結果と一致した(図7参照)。さらに、培地にL-Fucを添加しないで培養したFKP形質転換酵母についても、内在性の糖ヌクレオチドを調べた。その結果、GDP-L-Fucは検出できなかった(図8)。この結果から、FKP形質転換酵母は、培地にL-Fucが存在したときのみにGDP-L-Fucを生産すること、すなわち組換えFKPが酵母の発現系において、salvage経路でのGDP-L-Fuc生産機能を発揮することが証明された。
【0032】
(2−3)GDP-L-FucのHPLC解析
次に、FKP 遺伝子を発現する酵母でのGDP-L-Fucの定量解析を試みた。
HPLC分析のために、前記(2−2)で得られた乾燥させたサンプルを0.8mlの水に溶解し、アマシャムファルマシアバイオテク社(現GEヘルスケア社)のSMARTシステムを用い、Mono-Qカラムで糖ヌクレオチドを分離した。Mono-Qカラムは10 mM KH2PO4(A液)で平衡化し、B液には0.5 M KH2PO4を用いた。流速1ml/分で分離を行い、サンプル注入後5分から40分で100%B液、あるいは、100分で70%B液までKH2PO4濃度を上昇させた。UVモニターで254nmの値を計測することにより、溶出液中の糖ヌクレオチドを測定した。標準物質としてカルビオケム社のGDP-L-フコースを用い、ピーク面積からGDP-L-フコース量を計算した。なお、本実験では全てのバッファーを0.45μmのポアサイズの膜を通したのちに使用した。
Mono-QカラムによるHPLC精製を行うと、L-Fucを含む培地で培養したFKP発現酵母由来のサンプルでは、約17.5分に明らかなピークが観察された(図9F)。これに対し、L-Fucを含まない培地を用いた場合には、17.5分に有意なピークは観察されなかった(図9C)。Mock(図9A、9D)やFKPΔN(図9B,9E)の場合にも、L-Fucの有無にかかわらず、17.5分に有意なピークは認められなかった。また、FKP形質転換体の場合、培地中のL-Fuc濃度を低下させると、GDP-L-Fucの収量も低下することが明らかになった(図10参照)。
HPLC解析では、100分で70%までB液(0.5 M KH2PO4)を上昇させることによって検出できた。この条件ではおよそ31分で標準GDP-L-Fucが溶出するが、FKPを発現する酵母をL-Fuc存在下で培養した場合にも、同様のピークが見られた(図4B)。コントロールとして行ったL-Fucを添加しない培地での培養の場合には、同様のピークは見られなかった(図4A)。標準GDP-L-Fuc(25nmol)を用いた定量解析(図4C)の結果から、15 mM L-Fucを添加した培地でOD600が1.0になるまで培養したFKP形質転換酵母25ml(約5x108細胞)から230nmol(約0.135mg)のGDP-L-Fucが合成されると見積もられた。この収量は、Nakayamaらが報告したde novo合成経路を導入した酵母の収量(非特許文献52)の25倍に当たる。これらの結果は、L-Fucの添加なしでFKP発現酵母がGDP-L-Fuc合成活性を示さないのに対し、L-Fucの添加によって大量のGDP-L-Fucをin vivoで合成できることを表している。
【0033】
(実施例3)酵母細胞内でのGDP-D-Araの合成
酵母細胞内に移植されたsalvage経路でL-Fuc以外の基質も利用できるかどうかの検討を行うために、FKP遺伝子形質転換酵母を実施例2−1において培地中に添加するL-Fucに代えてD-Araを用いた以外は同様の方法で培養した。酵母細胞内で合成されたGDP-D-Araを、前記実施例2−2と同様にESI-MS(図11の矢印)で検出し、MS/MSによって確認を行った。MS/MSスペクトルの m/z 291のフラグメントイオンは、GDP-D-Araのグアノシンが脱離したものと一致する。これらの質量分析の結果から、この酵母の発現系でGDP-D-Araも合成されることが示された。ただし、GDP-D-Araは市販されていないため、HPLCによる分析は省略した。MS/MS解析でのm/z 305のフラグメントイオンは、下記の表2で表される。
【0034】
<表2>
【配列表フリーテキスト】
【0035】
配列番号1 FKP-protein(ACCESSION CAH08307)
配列番号2 FKP-gene(ACCESSION CR626927)
配列番号3 FKP-gene forward primer
配列番号4 FKP-gene reverse primer
配列番号5 FKP ΔN gene forward primer
配列番号6 FKP ΔN gene reverse primer
【技術分野】
【0001】
本発明は、酵母菌体内で高効率にGDP-フコースなどの糖ヌクレオチド(GDP-糖化合物)を合成する方法に関する。
【背景技術】
【0002】
L-フコース(L-Fuc, 6-デオキシ-L-ガラクトース)は様々な哺乳動物の糖鎖において必須の構成成分であり、N-グリカン、O-グリカン、糖脂質(非特許文献1)だけではなく、直接ある種のタンパク質のセリンやスレオニンに共有結合で結合することも知られている(非特許文献2)。
フコース転移酵素は、グアノシン5’-ジフォスフェート-L-フコース(GDP-フコース、GDP-Fuc)をドナー基質として、L-フコースを小胞体あるいはゴルジ体で受容体基質に転移する転移酵素であり、L-フコースの結合様式によりα1,2、α1,3/1,4、α1,6、O-フコース転移酵素に分類できる(非特許文献3)。L-Fucの付加によって様々な生物的機能が変化する(非特許文献4)。例えば、ABO血液型はα1,2-フコースの付加した糖鎖によるものである(非特許文献5、非特許文献6)。α1,3-フコースやα1,4-フコースはそれぞれシアリルルイスx(sLex)やシアリルルイスa(sLea)に必須の構成成分であり、炎症におけるセレクチンリガンドとしての機能を有している(非特許文献7)。受精においてもフコース付加糖鎖は重要な役割を担っている(非特許文献8、非特許文献9)。FUT8(フコース転移酵素VIII)欠損マウスの実験から、TGF-β1(transforming growth factor-β1)受容体のα1,6-フコースが欠失すると、重篤な発育阻害が見られることが明らかになっており(非特許文献10)、α1,6-フコースが様々な生物機能において重要な役割を果たしていることが示唆される。O-フコース残基(L-Fucが直接セリンあるいはスレオニンの水酸基と結合したもの)は哺乳動物のNotch受容体のEGFドメインで見られる(非特許文献11、非特許文献12)が、Notch受容体はtransmembrane signalingタンパク質ファミリ−に属しており、神経発生、血管形成、リンパ球の発生など様々な生物の発生に関与している(非特許文献13、非特許文献14)。さらに、フコース修飾はアポトーシスにも関与している(非特許文献15、非特許文献16、非特許文献17)。
【0003】
フコース修飾にはフコース転移酵素のドナー基質としてDGP-L-Fucが必要であり、生体内ではGDP-L-Fucは細胞質にある2つの経路によって合成される。
そのうちの主要な経路はde novo合成経路であり、種を超えて保存されている。この経路は、最初に細菌で発見され(非特許文献18)、その後植物(非特許文献19)、哺乳動物(非特許文献20)、無脊椎動物(非特許文献21)でも発見された。この合成系に関与する遺伝子はいくつかの細菌(非特許文献22)、植物(非特許文献23)、動物(非特許文献24)でクローニングされ、ショウジョウバエにおいては、in silicoでこの経路が再現されている(非特許文献25)。
このde novo合成経路では、グアノシン5’-ジフォスフェート-D-マンノース(GDP-マンノース、GDP-Man)から2種類の酵素で触媒される3つの酵素反応によってGDP-L-Fucが合成される。2つの酵素とは、GDP-D-マンノース-4,6-デヒドラターゼ(gmd、ヒトではGMD、EC 4.2.1.47)とGDP-4-ケト-6-デオキシ-D-マンノース-3,5-エピメラーゼ-4-レダクターゼ(Tsta3、ヒトではFX、EC 1.1.1.187)である(非特許文献26、非特許文献27、非特許文献22)。
まず第1段階では、GMDが脱水反応によりGDP-Manを不安定な中間体であるGDP-4-ケト-6-デオキシ-D-マンノースに変換する。この反応には捕因子としてニコチンアミドアデニンジヌクレオチドフォスフェート(NADP+)を要求し、D-マンノースのC-4位の水酸基の酸化によりケトグループを形成し、C-6位の水酸基の還元によりメチル基を形成する(非特許文献28、非特許文献29、非特許文献26)。de novo合成経路におけるGDP-D-ManからGDP-L-Fucへの連続合成反応は、最終産物であるGDP-L-Fucによって阻害されるが、これは、第1段階の反応においてGDP-L-FucがGMDを競合阻害するためである(非特許文献30、非特許文献31)。
第2及び第3段階は2つの機能(エピメラーゼと還元酵素活性)を有するFXにより引き起こされて、GDP-4-ケト-6-デオキシ-D-マンノースがGDP-L-Fucに変換される(非特許文献32、非特許文献27)。まずFXのエピメラーゼ活性によりGDP-4-ケト-6-デオキシ-D-マンノースはC-3位とC-5位がエピマー化し、D体からL体へ変換されてGDP-4-ケト-6-デオキシ-L-ガラクトースが生成される。次いでFXの4-還元酵素活性により捕因子であるNADPHから取ったH+をケト基に転移し、GDP-L-FucとNADP+を生成する(非特許文献33)。
de novo合成経路で用いられるGMDとFXの遺伝子は広くヒトの組織で発現している(非特許文献26)。マウスFX領域への変異の実験では、白血球の接着に加え、受精や生育、発生などにGDP-L-Fucを要求するようになることが分かっている(非特許文献34)。FX欠損マウスにおいては、食餌にL-Fucを加えることにより、通常は活発ではないsalvage経路によりGDP-L-Fucが合成され、糖鎖にフコース修飾が施されるようになる(非特許文献34)。
【0004】
もう一方のGDP-L-Fuc合成経路であるsalvage経路については、L-フコキナーゼ(Fuk, EC 2.7.1.52)及びGDP-L-フコースピロフォスフォリラーゼ(Fpgt, EC 2.7.7.30)が1960年代後半にブタ肝臓から発見された(非特許文献35、非特許文献36)ことで、はじめてその存在が明らかになった。Salvage経路において、第1段階ではL-フコキナーゼ(Fuk)によりL-FucとATPからL-フコース-1-フォスフェート(L-Fuc-1-P)が合成される(非特許文献36、非特許文献37)。第2段階では、GDP-L-Fucピロフォスフォリラーゼ(L-Fuc-1-Pグアニルトランスフェラーゼ:Fgpt)によりL-Fuc-1-PとGTPからGDP-L-Fucが合成される(非特許文献35、非特許文献38)。
L-Fucは、生物では食餌から、培養細胞では培地から摂取され、細胞膜を通して細胞質に輸送される。さらに、L-Fucはリソゾームでフコース分解酵素により糖タンパク質や糖脂質から生産されることもある(非特許文献39)。L-フコースキナーゼ(Fuk)(非特許文献37)やGDP-L-フコース-ピロフォスフォリラーゼ(Fgpt)(非特許文献38)は様々な組織で見つかっていることから、salvage経路もまた真核生物では一般的な経路であると考えられる。以前の報告では、Fuk活性は脳においてドーパミン刺激や大動脈の内皮細胞をニコチンにさらすことによって制御されることが示されている(非特許文献40、非特許文献41)。
【0005】
現在、GDP-L-Fucは化学合成と酵素合成の主要な2種類の方法で合成することができ、L-FucからGDP-L-Fucへの化学合成では様々なアプローチがとられているがいずれも比較的複雑である(非特許文献42、非特許文献43)。GDP-L-Fucの酵素合成では、GDP-D-Manを出発材料とするde novo合成系及びL-Fucを出発材料とするsalvage経路があり、主としてde novo合成系が用いられている(非特許文献44)。
Salvage経路を利用した合成系においても、1ポット反応の合成に成功している(非特許文献45、非特許文献46)。また、Salvage経路に用いる酵素として、非病原性の原生動物であるCrithidia fasciculata由来のD-アラビノース-1-キナーゼとD-アラビノース-1-P-ピロフォスフォリラーゼも利用できることが知られている。これら酵素はin vivoではもっぱらD-アラビノース(D-Ara)からGDP-アラビノース(GDP-Ara)の合成に用いられているが、in vitroではL-FucからGDP-L-Fucの合成に用いることができる(非特許文献47)。しかしながら、これらの合成系はいずれもin vitro条件下での酵素反応系であり、形質転換細胞内での組換え酵素系による合成系は存在しない。
【0006】
近年、プロテオミクスやゲノミクスによって蛋白質を基礎にした数多くの治療薬開発が盛んに行われているが、その蛋白質治療薬の機能発現においては翻訳後の糖鎖修飾が必須な場合が多く、糖蛋白質治療薬を安価に生産するための宿主として、真核生物でありながら単細胞生物で大量培養が容易な酵母が注目されている。
上述のようにフコースは哺乳類細胞において重要な糖単位であり、複合型糖鎖の多様性に富む分岐構造を作り出すためにも必須である。しかし、酵母にはフコースを蛋白質に転移する代謝系酵素群遺伝子が存在しないため、酵母由来の糖蛋白質にはフコースという糖単位は存在しない。酵母を宿主とした有用糖蛋白質生産を可能にするためには、まず出発材料として糖転移酵素の糖供与体となるGDP-L-Fucのプールを酵母細胞内に作り出すことが必要である。
酵母Saccharomyces cerevisiaeを宿主としてGDP-L-Fucを合成させるに際しては、「de novo合成経路」を経由するための有利な条件がそろっている(非特許文献48)。酵母での糖鎖修飾は、大部分がマンノース付加であり(非特許文献49、非特許文献50)、酵母の細胞質にはde novo合成の出発材料となるGDP-D-Manが大量に存在している。Mattilaらと本出願人はそれぞれ大腸菌とArabidopsis thaliana 由来のde novo合成に必須の2つの遺伝子、GDP-D-マンノース-4,6-デヒドラターゼ(gmd、MUR1)とGDP-4-ケト-6-デオキシ-D-マンノース-3,5-エピメラーゼ-4-レダクターゼ(wcaG、AtFX)をS.cerevisiaeに導入した(非特許文献51、非特許文献52、特許文献1)。両グループとも、導入した2つの酵素遺伝子が酵母細胞内で活性化し、酵母に内在するGDP-D-Manから合成したGDP-L-Fucを酵母内に蓄積できることを示している。
このように、形質転換酵母細胞を用いて、菌体内に存在するGDP-D-Man を利用してde novo合成経路によりGDP-L-Fucを合成し菌体内に蓄積する技術は一応確立したものの、上述のように、gmd遺伝子にコードされるGDP-D-マンノース-4,6-デヒドラターゼに対しては、GDP-L-Fucによってフィードバック阻害が働く。さらに、酵母細胞に内在するGDP-D-Manをどの程度利用できるかも、de novo経路を利用するうえでの限定因子となるため、de novo合成経路によっては大量のGDP-L-Fucの蓄積は期待できない。
【0007】
最近、本出願人は、Arabidopsis thaliana 由来のGDP-L-FucからO-フコース結合型タンパク質を合成する代謝系酵素群の各遺伝子を特定し、全て導入した形質転換酵母を作製し、酵母菌体内ではじめてO-フコース結合型の蛋白質合成に成功している(特許文献2)。酵母菌体内においてGDP-L-Fucをさらに効率よく合成し蓄積できる技術が開発できれば、酵母菌体を用いたO-フコース結合型のタンパク質を含む糖タンパク医薬製剤などを大量生産できると考えられる。
したがって、酵母細胞内での効率的なGDP-L-Fuc合成技術、とりわけsalvage経路によるGDP-L-Fucの合成方法の開発が強く望まれていた。
【先行技術文献】
【特許文献】
【0008】
【特許文献1】特開2001−145488号公報
【特許文献2】特開2005−328841号公報
【非特許文献】
【0009】
【非特許文献1】Walz G, et al.,Science. 250:1132-1135.
【非特許文献2】Harris RJ, et al., 1993. Glycobiology. 3:219-224
【非特許文献3】Ma et al., 2006,Glycobiology 16:158R-184R.
【非特許文献4】Becker, et al.,2003,Glycobiology. 13:41R-53R.
【非特許文献5】Greenwell, 1997, Glycoconjugate J. 14:159-173.
【非特許文献6】Lloyd KO. 2000. Glycoconjugate J. 17:531-541.
【非特許文献7】Lowe JB. 1997. Kidney Int. 51:1418-1426.
【非特許文献8】Johnston et al., 1998 ,J Biol Chem. 273:1888-1895.
【非特許文献9】Mori et al., 1998 ,Glycoconjugate J. 15:447-456.
【非特許文献10】Wang et al., 2005,Proc Natl Acad Sci USA. 102:15791-15796.
【非特許文献11】Moloney et al., 2000 ,J Biol Chem. 275:9604-9611.
【非特許文献12】Rampal et al., 2005,J Biol Chem. 280:32133-32140.
【非特許文献13】Artavanis-Tsakonas et al., 1999 ,Science. 284:770-776.
【非特許文献14】Halloran et al., 2000 ,J Immunol. 164:4868-4877
【非特許文献15】Hiraishi et al., 1993,Glycobiology. 3:381-390
【非特許文献16】Russell et al., 1998,Biochem Bioph Res Co. 250:449-453.
【非特許文献17】Winkler et al., 2004,Blood. 103:1685-1692.
【非特許文献18】Ginsburg, 1960 ,J Biol Chem. 235:2196-2201
【非特許文献19】Liao et al., 1971,Biochim Biophys Acta. 230:64-71.
【非特許文献20】Overton et al., 1981 ,Biochim Biophys Acta. 675:281-284.
【非特許文献21】Bulet et al., 1984,Eur J Biochem. 144:255-259
【非特許文献22】Tonetti et al.,1998,Biochimie. 80:923-931.
【非特許文献23】Bonin et al., 1997,Proc Natl Acad Sci USA. 94:2085-2090
【非特許文献24】Reitman et al., 1980,J Biol Chem. 255:9900-9906.
【非特許文献25】Roos et al., 2002,J Biol Chem. 277:3168-3175.
【非特許文献26】Sullivan et al., 1998,J Biol Chem. 273:8193-8202
【非特許文献27】Tonetti et al., 1996 ,J Biol Chem. 271:27274-27279.
【非特許文献28】Chang et al., 1985,Anal Biochem. 144:253-257
【非特許文献29】Oths et al., 1990 ,Carbohyd Res. 198:91-100
【非特許文献30】Bisso et al., 1999,FEBS Lett. 456:370-374.
【非特許文献31】Sturla et al., 1997 ,FEBS Lett. 412:126-130
【非特許文献32】Chang et al., 1988,J Biol Chem. 263:1693-1697.
【非特許文献33】Menon et al., 1999 ,J Biol Chem. 274:26743-26750
【非特許文献34】Smith et al., 2002,J Cell Biol. 158:801-815
【非特許文献35】Ishihara et al.,1968,J Biol Chem. 243:1110-1115
【非特許文献36】Ishihara et al., 1968,J Biol Chem. 243:1103-1109
【非特許文献37】Park et al., 1998 ,J Biol Chem. 273:5685-5691
【非特許文献38】Pastuszak et al., 1998, J Biol Chem. 273:30165-30174
【非特許文献39】Michalski et al., 1999, Biochim Biophys Acta. 1455:69-84.
【非特許文献40】Hocher et al., 1993,Eur J Clin Chem Clin. 31:347-351
【非特許文献41】Ricken et al., 1990 ,Biochem Soc T. 18:963-964
【非特許文献42】Adelhorst et al., 1993,Carbohyd Res. 242:69-76
【非特許文献43】Murray et al., 1997 ,Biochemistry. 36:823-831
【非特許文献44】Becker et al.,1999,BBA-Mol Basis Dis. 1455:193-204
【非特許文献45】Ichikawa et al., 1992, J Am Chem Soc 114:9283-9298.
【非特許文献46】Ichikawa et al.,1994,Methods Enzymol. 247:107-127
【非特許文献47】Mendgeling et al., 1999,Anal Biochem. 267:227‐233
【非特許文献48】Hirschberg et al., 1998 Annu Rev Biochem. 67:49-69.
【非特許文献49】Hashimoto et al., 1997 ,J Biol Chem. 272:16308-16314.
【非特許文献50】Romanos et al., 1992, Yeast. 8:423-488
【非特許文献51】Mattila et al., 2000 ,Glycobiology. 10:1041-1047
【非特許文献52】Nakayama et al., 2003, Glycobiology. 13:673-680
【非特許文献53】Coyne et al., 2005, Science. 307:1778-1781.
【非特許文献54】Kotake et al., 2008, J Biol Chem. 283:8125-8135
【非特許文献55】Niittymaki et al., 2004, Eur J Biochem. 271:78-86.
【非特許文献56】Linster et al., 2007 ,J Biol Chem. 282:18879-18885.
【発明の開示】
【発明が解決しようとする課題】
【0010】
本発明は、酵母細胞内での効率的なGDP-L-Fucの合成方法を提供することを目的とするものである。さらに、L-フコースと構造的に類似したD-アラビノースと共に、L-フコース、D-アラビノースなどのアナログを酵母細胞内で効率的にGDP-糖化合物に転換する方法を提供する。
【課題を解決するための手段】
【0011】
本発明者らは、上記課題を解決するために鋭意研究を重ねる中で、Bacteroides fragilis 9343由来のL-フコキナーゼ/GDP-L-フコースピロフォスフォリラーゼ(FKP)という2つの活性をもつ酵素遺伝子を利用することを思いついた。この酵素はL-Fucを基質としてL-Fuc-1-Pに変換し、さらに、そこからGDP-L-Fucを合成することができる(非特許文献53)。この変換活性はB.fragilis 9343株内のsalvage経路として観察され、広く細菌(Bacteroides)一般に保存されている(非特許文献53)。細菌(Bacteroides)以外では植物(Arabidopsis)もsalvage経路としてL-FucからL-Fuc-1-Pを経由してGDP-L-Fucを合成できる系を有しているが、当該系を有しているのは細菌及び植物のみであり、その理由は明らかになっていない(非特許文献54)。上記酵素FKPは、アミノ酸配列アラインメントの結果、そのN-末端ドメイン(1-430番目のアミノ酸)では、ヒトのGDP-L-フコースピロフォスフォリラーゼと20%の相同性があり、C-末端ドメイン(584-949番目のアミノ酸)では哺乳動物のL-フコキナーゼと相同性が高い(非特許文献55)。2つのドメインを150アミノ酸のリンカーで繋ぐ構造となっているが、リンカーの機能は現在のところ不明である。
今回、B.fragilis 由来FKP遺伝子を導入した酵母(S.cerevisiae)宿主において、FKPが活性型酵素として発現し、培地中に添加したL-Fucを、salvage経路によりL-Fuc-1-Pを経由してGDP-L-Fucにまで変換し、細胞内に蓄積することを確認した。また、L-Fucと構造的に似ているD-Araを培地中に添加した場合でも、同様にGDP-D-Araを合成することも確認できた。
以上の知見を得たことで、本発明を完成した。
【0012】
すなわち、本発明は以下の発明を含むものである。
〔1〕 FKP(L-フコキナーゼ/GDP-L-フコースピロフォスフォリラーゼ)遺伝子を酵母用発現ベクターにより形質転換した酵母であって、培地中のL-フコース、又はL-フコースアナログを取り込み、細胞内でGDP-L-フコース、又はL-フコースアナログのGDP-糖化合物を合成可能な形質転換酵母。
〔2〕 L-フコースアナログがD-アラビノースであり、そのGDP-糖化合物がGDP-D-アラビノースである、前記〔1〕に記載の形質転換酵母。
〔3〕 FKP遺伝子が、Bacteroides fragilis由来のFKP遺伝子である、前記〔1〕又は〔2〕に記載の形質転換酵母。
〔4〕 酵母がSaccharomyces cerevisiaeである前記〔1〕〜〔3〕のいずれかに記載の形質転換酵母。
〔5〕 前記〔1〕〜〔4〕のいずれかの形質転換酵母を用い、L-フコース、D-アラビノース、又はL-フコースアナログを存在させた培地中で培養することを特徴とする、L-フコース、D-アラビノース、又はL-フコースアナログのGDP糖化合物を合成する方法。
〔6〕 L-フコースを存在させた培地中で培養することを特徴とする、GDP-L-フコースを合成する方法である前記〔5〕に記載の方法。
〔7〕 D-アラビノースを存在させた培地中で培養することを特徴とする、GDP-D-アラビノースを合成する方法である前記〔5〕に記載の方法。
【発明の効果】
【0013】
本発明のB.fragilis由来のFKP遺伝子を導入した形質転換酵母は、培地中に基質となるL-フコース(L-Fuc)を添加しておくだけで、salvage経路により直接GDP-フコース(GDP-L-Fuc)を効率的に変換することができるため、当該形質転換酵母を用いたGDP-フコース(GDP-L-Fuc)合成方法により、簡便でかつ高効率でGDP-L-Fucを酵母菌体内に蓄積することができる。
さらに、L-フコースと構造的に類似したD-アラビノース(D-Ala)などの糖類、及びL-フコース、D-アラビノースのアナログである化学修飾体に対しても、酵母細胞内で効率的にGDP-糖化合物に転換することができる。
そして、当該形質転換酵母に対してさらに外来タンパク質遺伝子と共に「O-フコース転移酵素遺伝子群(特許文献2)」を導入してやることにより、またはこれら遺伝子が導入された形質転換酵母と掛け合わせることにより、salvage経路を経由したO-フコース含有タンパク質合成系を有する形質転換酵母を作出でき、O-フコース含有タンパク質を効率よく製造することができる。
【図面の簡単な説明】
【0014】
【図1】外来L-FucをGDP-L-Fucに変換するsalvage経路には、B. fragilisの酵素が必要となる。(A) FKPを用いたGDP-L-Fucの酵素合成スキーム。FKP タンパク質のN-末端ドメイン(1-430番目のアミノ酸)はヒトのGDP-フコースピロフォスフォリラーゼと20%の相同性を示し、C-末端ドメイン(584-949番目のアミノ酸)は哺乳動物のL-フコキナーゼと相同性を示す。 (B) 酵母ゲノムに挿入したFKP と FKP ΔN遺伝子の構築。両遺伝子はC-末端側で3xHAと融合させている。
【図2】Mock (レーン1)、FKP ΔN (レーン2)、FKP (レーン3) 遺伝子導入酵母の細胞の可溶化物のSDS-PAGEとウェスタンブロット解析。(左)およそ2x106細胞/レーン、抗HA抗体で検出。(右)およそおよそ2x106細胞/レーン、CBB(Coomassie Brilliant Blue R-250)で染色した。レーン1, 1’;mockベクター(pRS405-leu)を保持する酵母W303-1A株、レーン2,2’;FKPΔN-3xHAを挿入した発現ベクターを保持する酵母W303-1A株、レーン3, 3’;FKP-3xHAを挿入した発現ベクターを保持する酵母W303-1A株。左のパネルの上部矢じりはFKP-3xHAタンパク質を、下の矢じりはFKPΔN-3xHAタンパク質を示している。
【図3】酵母抽出物由来のGDP-L-Fucの構造解析。FKP 遺伝子を導入した酵母を15 mM L-Fucを含む培地で培養した酵母抽出物由来のESI-MS/MSスペクトル(m/z 588.1の[M-H]- イオン)。m/z 442はGDP-L-Fucのフラグメントイオンと考えられる。主要なフラグメントは化学的に不安定なL-Fuc-1-phosphateあるいはpyrophosohate結合の切断によって出現すると考えられる。
【図4】In vivoでのGDP-L-Fuc生産性。1-ブタノール飽和1 Mギ酸でおよそ5x108 細胞から抽出した抽出物をMono Qカラムで陰イオン交換で精製し、GDP-L-Fuc画分を単離した。サンプル注入後5分から0.5M KH2PO4の濃度を上げ、100分後に0.5M KH2PO4が70%になるようにした。流速は1ml/minとした。FKPを導入した酵母W303-1Aを(B)15mM L-Fuc存在下と(A)L-Fucを添加しないで培養した。コントロールとしては、GDP-L-Fuc標準物質25 nmolを用いた(C)。L-Fuc存在下で培養した酵母由来の図では、GDP-L-Fucに相当する9.25分にピークが認められた。
【図5】酵母抽出物由来のGDP-D-Araの構造解析。15mM D-Araを含む培地で培養したFKPを持つ酵母からの抽出物で得られたm/z 574.1のESI-MS/MSスペクトルを示した。m/z 442はGDP-D-Araのフラグメントイオンである。主要な断片は化学的な不安定なD-アラビノース-1-フォスフェートあるいはピロフォスフェート結合の切断物だと考えられる。
【図6】15mM L-Fucを添加した培地で培養したFKPを持つ酵母由来の抽出物のESI-MSスペクトル。矢印で示したm/z 588の[M-H]-イオンはGDP-L-Fucに相当する。
【図7】標準GDP-L-FucのESI-MS/MSスペクトル。m/z 588の[M-H]-イオンの主要断片を記載した。
【図8】L-FucもD-Araも添加しない培地で培養したFKPを持つ酵母由来の抽出物のESI-MSスペクトル。GDP-L-Fucに相当するm/z 588.08もGDP-D-Araに相当するm/z 574.08も認められなかった。
【図9】酵母抽出物をクロマトグラフィーで分離し、UV254nmで検出。S. cerevisiaeをmockベクター(AとD)、FKP ΔN(BとE)、FKP(CとF)で形質転換し、50mlのSD(-leu)培地に15mM L-Fucを添加したもの(D-F)と添加しないもの(A-C)でOD600が1.0になるまで培養した。細胞を1-ブタノール(10%, v/v)飽和1Mギ酸で抽出した。ヌクレオチドと糖ヌクレオチドをMono Qカラムにより分画し、UV254nmでモニターした。サンプル注入から5分後に0.5 M KH2PO4を加え、40分後に0.5 M KH2PO4が100%になるようにした。流速は1ml/minとした。パネル(F)の主要ピーク(およそ17.5分)を矢じりで示した。
【図10】In vivoでのGDP-L-Fucの生産性。50mlの培地で培養した酵母を1-ブタノール飽和1 Mギ酸で抽出した。抽出物は陰イオン交換クロマトグラフィー(Mono Q)に供し、GDP-L-Fuc画分を単離した。サンプル注入から5分後に0.5 M KH2PO4を加え、40分後に0.5M KH2PO4が100%になるようにした。流速は1ml/minとした。FKPを持つ酵母W303-1A株を7.5mM L-Fuc(A)、15mM L-Fuc(B)を含む培地で培養し、細胞を集めた。コントロールには40nmolのGDP-L-Fucを用いた(C)。添加したL-Fucの量に応じてGDP-L-Fucが合成されていた。
【図11】15mM D-Araを添加した培地で培養したFKPを持つ酵母由来の抽出物のESI-MSスペクトル。矢印で示したm/z 574.070の[M-H]-イオン(矢印)はGDP-D-Araに相当する。
【発明を実施するための形態】
【0015】
1.FKP形質転換酵母について
原核生物、真核生物は、2つの異なる代謝経路(de novo経路とsalvage経路)を利用してGDP-L-Fuc合成を行っている(非特許文献2)が、酵母にはGDP-L-Fucの合成能力がない。
本発明では、酵母にFKP(L-フコキナーゼ/GDP-L-フコースピロフォスフォリラーゼ)酵素遺伝子を導入することでsalvage経路を移植した(図1A参照)。本実施例ではB. fragilis9343株由来のFKP酵素遺伝子を用いているが、FKP酵素は広くBacteroides類に分布しており、高等植物にも存在することが知られている(非特許文献53、54)。いずれも、B.fragilis9343株由来FKP酵素と同様にL-フコキナーゼ及びGDP-L-フコースピロフォスフォリラーゼ作用を有している酵素であって、B.fragilis9343株由来FKP酵素に代えて用いることができる。酵母に導入された外来遺伝子であるB. fragilisのFKP遺伝子の過剰発現により、酵母菌体内でもsalvage経路が働き、GDP-L-Fucの合成を可能にすることができる。B. fragilis9343株由来のFKP酵素遺伝子の塩基配列(配列番号2)及び対応するアミノ酸配列(配列番号1)の情報は、それぞれアクセッション番号CR626927、CAH08307としてGENBANKデータベースから入手できる。B. fragilis9343株以外のBacteroides類又は植物由来の酵素遺伝子は、配列番号1に示されるアミノ酸配列とは80%以上、好ましくは90%以上、さらに好ましくは95%以上のホモロジーを有するアミノ酸配列をコードし、又は配列番号2に示される塩基配列とも80%以上、好ましくは90%以上、さらに好ましくは95%以上のホモロジーを有している。そして、配列番号1に示されるアミノ酸配列において1若しくは数個のアミノ酸が欠失・置換・付加されたアミノ酸配列をコードする遺伝子、又は配列番号2に示される塩基配列の相補配列とストリンジェントな条件下でハイブリダイズするポリヌクレオチドからなる遺伝子などと表現することもできる。
酵母細胞はGDP-L-Fuc合成に必要な酵素を持っていないので、導入した遺伝子の機能発現を容易にモニターすることができる(非特許文献50、非特許文献49)。
B. fragilis由来などのFKPの関与するsalvage経路を選択したことの理由の1つには、GDP-L-Fucと構造や生合成ステップが似ているGDP-L-Araの生合成を行える点にある(非特許文献47、非特許文献56)。
当該遺伝子を酵母用の発現ベクターに繋いで酵母を形質転換する。宿主の酵母としては、本実施例ではSaccharomyces cerevisiaeを用いたが、Pichia酵母など他の酵母でもよく、発現ベクターについても酵母菌体内で発現可能なプロモーターを有するベクターであればいずれでも良い。ただし、酵母細胞内でsalvage経路を働かせる必要があるため、分泌シグナルを含む分泌ベクターは不適切である。
【0016】
2.本発明が対象とする糖ヌクレオチド(GDP糖化合物)
本発明におけるFKP形質転換酵母は、典型的には細胞内でのFKP酵素作用によりL-フコースをそのGDP糖化合物であるGDP-L-フコースに転換するが、L-フコースに構造的に類似しているいわゆるL-フコースアナログを、そのGDP-糖化合物に転換することができる。L-フコースアナログとしては、D-アラビノースなどの糖類の他、L-フコース又はD-アラビノースなど糖類のアジドやアルキンなどの化学修飾体が含まれる。本実施例では具体的にD-アラビノースをGDP-D-アラビノースに転換できることを実証している。
【0017】
3.FKP形質転換酵母による糖ヌクレオチド合成方法
(1)本発明のFKP形質転換酵母によるGDP-L-Fuc合成
FKP形質転換後、細胞数が十分に増えるまでロイシンを欠失させた酵母用培地で培養し、次いで培地中にL-Fuc又はL-Fucアナログ例えばD-Alaを培地に1mM以上、好ましくは7.5mM以上、より好ましくは15mM以上で480mM以下添加してさらに培養をする。その際には振とう培養を行うことが好ましい。
【0018】
(2)GDP-L-Fuc生産物の確認方法又は定量法
本実施例では以下の(i)及び(ii)の方法でGDP-L-Fuc生産物の確認及び定量した。
(i) MSまたは MS/MSスペクトルによる方法
質量分析でGDP-L-Fucに典型的なm/z 588の位置のピークの存在を確認することで、精製しない状態でも系にGDP-L-Fucが存在しているかどうかを確認することができる。
例えばESI-1TOF MS/MS解析(エレクトロスプレーイオン化 四重極 飛行時間型質量分析装置)、micrOTOF-QによるMS/MSスペクトル法を用いることができる。標準GDP-L-Fucのデータと比較することでより正確に確定することができる。
(ii) HPLC分析
精製されたGDP-L-FucをHPLC分析用溶媒に溶かし、Mono-Qカラムなどの陰イオンカラムにより分析することができる。Mono-Qカラムの場合には、A液として10mM KH2PO4、B液として0.5M KH2PO4を使用してGDP-L-Fucの分析をすることができる。
標準GDP-L-Fucの資料が市販されているので、そのデータと比較することで精製度も、また定量解析をすることもできる。
(iii) このほか、キャピラリー電気泳動、NMRによりGDP-L-Fucが生産されているかどうかを確認することができる。
【0019】
(3)GDP-L-Fucなどの回収、精製方法
GDP-L-Fucは酵母の細胞質内で合成され、蓄積されているため、GDP-L-Fucを回収するためには、酵母菌体を破砕後遠心分離などで細胞を除去し、糖ヌクレオチドの単離精製に用いられる一般的な生化学的方法、イオン交換クロマトグラフィー、アフィニティクロマトグラフィー、ゲル濾過法などを単独又は組み合わせて用いることができる。
【0020】
4.酵母によるO-フコース含有糖タンパク質の製造方法
酵母の細胞質内に蓄積されたGDP-L-Fucを回収せずに、そのままO-フコース含有糖タンパク質の製造のために利用することができる。
すなわち、特許文献2に記載されたO-フコース結合型タンパク質合成系遺伝子群(O-フコース転移酵素遺伝子群)が導入されている形質転換酵母に本発明のFKP形質転換酵母を掛け合わせることにより、O-フコース結合型タンパク質を合成可能な形質転換酵母を得ることができる。
【0021】
5.FKP形質転換酵母による他の糖ヌクレオチド合成について
本発明のFKP形質転換酵母は、L-Fucと構造的に類似しているD-Alaに対しても、培地中に基質として添加しておくことで、酵母菌体内でGDP-D-Alaを合成することができた(図12)。
B. fragilis由来のFKP酵素は基質特異性が低いため、原理的には、L-ガラクトースのような他の単糖にも応用できる可能性がある。さらに、天然の糖でなくてもL-Fucに構造が類似していればよく、L-Fucのアナログを用いた糖ヌクレオチド合成に応用することで、糖ヌクレオチドアナログを高純度で合成することも可能である。
【0022】
<式1>FKP形質転換酵母細胞におけるGDP-L-FucとGDP-D-Ara合成。
【実施例】
【0023】
以下、実施例により本発明を具体的に説明するが、本発明は特にこれら実施例に限定されるものではない。
なお、本発明で使用されている技術的用語は、別途定義されていない限り、当業者により普通に理解されている意味を持つ。
また、本発明で引用した先行文献又は特許出願明細書の記載内容は、本明細書の記載として組み入れるものとする。
【0024】
(実施例1)形質転換酵母(Saccharomyces cerevisiae)の調製
(1−1)形質転換用宿主酵母株の調整
糖ヌクレオチドのインビボ合成のために形質転換用の宿主酵母として、Saccharomyces cerevisiae W303-1A株(MA Ta leu2-3, 112 his3-11, 15 ade2-1 ura3-1 trp1-1 can1-100)を用いた。W303-1A株の培養は、S. cerevisiaeの通常の生育培地であるYPAD培地(2%バクトペプトン、1%酵母エキス、2%グルコース、40μl/ml硫酸アデニン)を用いた。形質転換後はロイシン欠失培地(SD(-leu): 0.67%バクト酵母ニトロゲンベースアミノ酸不含、2%グルコース、0.69g/l アミノ酸混合物ロイシン不含)を用いた。
【0025】
(1−2)FKP遺伝子を含む酵母用発現ベクターの調製
FKP遺伝子はBacteroides fragilis 9343(ATCC 25285)染色体DNAからPCR増幅し、pCMV-Tag 2Bベクター(ストラタジーン社)のEcoRI−XhoI部位に挿入した。PCR増幅し、塩基配列を確認したFKP遺伝子全長及び部分遺伝子(truncated version)を酵母用のYep352GAP-IIベクター(非特許文献52)に導入し、発現に使用した。
全長FKP遺伝子は、
5’-GGAGCTCATGCAAAAACTACTATCTTTACCG-3’(配列番号3)と
5’-GCTCTAGACCCGGGTGATCGTGATACTTGGAATCCC-3’
(配列番号4)とをプライマーとし、
またネガティブコントロールであるN-末端を除去したFKP(FKP ΔN)遺伝子(1752番目の塩基から2847番目の塩基)は、
5’-GGAGCTCATTGTTTGGGGACGTAGC-3’
(配列番号5)と
5’-GCTCTAGACCCGGGTGATCGTGATACTTGGAATCCC-3’
(配列番号6)をプライマーとして使用し、PCR増幅を行った。
両PCR断片はSacIとXbaIで切断し、TDH3プロモーターを持つ発現ベクターYep352GAP-IIに挿入した。pHAベクターから3×HA-tag(3個のインフルエンザヘマグルチニンエピトープ)遺伝子をSmaIにより切り出し、FKP或いはFKP ΔN遺伝子が挿入されているYep352GAP-IIに挿入した。これらのベクターからTDH3プロモーター、FKP-3×HAあるいはFKP ΔN-3×HA、TDH3ターミネーターを含む断片をBamHIにより切り出し、pRS405のBamHI部位に挿入しpRS405-FKP3xHA(図1A)あるいはpRS405-FKPΔN3xHA(図1B)を構築した。各プラスミドの塩基配列を確認したのち、BstEIIで切断し、直鎖状にし、酵母染色体のLEU2遺伝子領域に組み込んだ。なお、上記で用いた制限酵素、修飾酵素はNew England Biolabs社あるいはロシュダイアグノスティクス社製を用いた。
また、プラスミドベクターの構築の際には大腸菌DH5α株を使用した。大腸菌はLuria Bertani培地(1%バクトトリプトン、0.5%酵母エキス、0.5%NaCl、0.2%グルコース)で培養した。
【0026】
(1−3)酵母(S. cerevisiae)の形質転換
前記(1−2)で構築した酵母用発現ベクターpRS405-FKP3xHA(FKP遺伝子)及びpRS405-FKPΔN3xHA(FKPΔN遺伝子)と共に何も遺伝子を挿入していないベクター(pRS405: Mock)のそれぞれを用いて、Itoらの方法(Ito H, et al., J Bacteriol. 153:163-168)により、前記(1−1)で調整した酵母W303-1A株の形質転換を行った。形質転換体はロイシン欠失培地でそれぞれについて数株ずつ選択し、コロニーPCR解析によって確認を行った。
【0027】
(1−4)B. fragilis由来FKP酵素タンパク質の発現
前記(1−3)において、ロイシン欠失培地で選択した数株ずつの形質転換体(FKP遺伝子、FKPΔN遺伝子、Mock)をそれぞれ5mlのSD(-leu)培地で30℃の条件下で12時間培養した。この条件ではOD600が1.0(およそ2x107細胞)程度になる。各形質転換細胞を4,000×g、5分の遠心分離により集め、1×PBSで2回洗浄した。各々がおよそ2x107細胞となるように100μlのサンプルバッファー(0.06 M Tris-HCl (pH6.8)、10% (v/v) グリセロール、2% (v/v)2-メルカプトエタノール、0.25% (w/v)ブロモフェノールブルー)に再懸濁した。これらを95℃で5分インキュベートし、14,000×g5分の遠心分離後、上清を10%SDSポリアクリルアミド電気泳動に供した。電気泳動後のタンパク質は、PVDF膜(ミリポア社)に電気的に転写した(15 V、1時間)。膜をブロッキングし、抗HA抗体(ロシュ社)と室温で1時間反応させた。抗HA抗体は5%スキムミルクを含むPBST(0.1% Tween 20)で希釈しておいたものを使用した。HRP(Horseradish peroxidase)-コンジュゲート ウサギ抗マウスIgG(Zymax社)を2次抗体として用い、ECL-エンハンストケミルミネッセンス(GEヘルスケア社)で検出した。
その結果、図2に示したように、FKPΔNとFKP遺伝子産物ははっきりと認められた(矢じりで示した)。予想通り、FKP導入ベクターを用いた場合にはFKP酵素の全長に対応する135 kDaの位置にバンドが見られたのに対し、FKPΔN導入ベクターを用いた場合のバンドは43 kDaに見られた。一方、mockではバンドは観察されなかった。
【0028】
(実施例2)形質転換酵母によるGDP-L-Fucの合成
(2−1)形質転換酵母のL-Fuc含有培地下での培養
形質転換酵母細胞を2mM MgCl2を含む25mlあるいは50mlのSD(-leu)培地(pH7.5)でOD600が1.0になるまで培養した。この場合、細胞はおよそ5x108あるいは1x109になる。このとき、培地にあらかじめ15mMのL-フコースを含むものと含まないものを用いた。3種類の形質転換体(mock pRS405、pRS405-FKP3xHA及びpRS405-FKPΔN3xHAを導入した形質転換体)を32℃、140rpmで振とう培養した。このとき、培地はフラスコの容量の1/5容になるようにした。細胞を4℃、4,000×g、5分間の遠心分離で集め、5倍量の氷冷した滅菌水で2回洗浄した。その後、10%1-ブタノール(v/v) を含む4mlの氷冷1Mギ酸に再懸濁し、穏やかにピペッティングにより撹拌し、氷中に30分間置いた。4℃、6,000×g、10分間の遠心分離で夾雑物を除去したあとの上清を凍結乾燥に供した。
【0029】
(2−2)酵母細胞からのGDP-L-Fucの抽出とESI-1TOF MS/MS解析による確認
酵母細胞質中でのGDP-L-Fuc合成酵素活性を確認するために、細胞抽出物中のGDP-L-Fucの検出を、ブルカダルトニクス社のESI-qTOF MS(エレクトロスプレーイオン化 四重極 飛行時間型質量分析装置)を用いたMS解析で行い、さらにmicrOTOF-Qを用いてMS/MSスペクトルを陰イオンモードで取得した。
具体的には、前記(2−1)で得られた乾燥サンプルを0.5mlの滅菌水に溶解し、そのうち1μlを50%メタノール(v/v) で50μlに希釈した。サンプル溶液は0.5barの窒素ガスで、シリンジポンプ(Kdサイエンティフィック社、流速8μl/min)を用いてESI源にて測定した。乾燥温度は180℃、乾燥用ガスの流速は4l/minとした。キャピラリーの電圧は3200V、エンドプレートオフセットを500Vに保持した。
GDP-L-Fucに相当するm/z 588.078の[M-H]-イオン(図6の矢印)をESI-qTOF MSによる MS/MS解析に供した(図3)。
MS/MS解析でのm/z 305のフラグメントイオンは、下記の表1に示されるように、GDP-L-Fucからグアノシンが脱離した値に相当する。
【0030】
<表1>
【0031】
このMS/MS解析の結果は、標準GDP-L-Fucの結果と一致した(図7参照)。さらに、培地にL-Fucを添加しないで培養したFKP形質転換酵母についても、内在性の糖ヌクレオチドを調べた。その結果、GDP-L-Fucは検出できなかった(図8)。この結果から、FKP形質転換酵母は、培地にL-Fucが存在したときのみにGDP-L-Fucを生産すること、すなわち組換えFKPが酵母の発現系において、salvage経路でのGDP-L-Fuc生産機能を発揮することが証明された。
【0032】
(2−3)GDP-L-FucのHPLC解析
次に、FKP 遺伝子を発現する酵母でのGDP-L-Fucの定量解析を試みた。
HPLC分析のために、前記(2−2)で得られた乾燥させたサンプルを0.8mlの水に溶解し、アマシャムファルマシアバイオテク社(現GEヘルスケア社)のSMARTシステムを用い、Mono-Qカラムで糖ヌクレオチドを分離した。Mono-Qカラムは10 mM KH2PO4(A液)で平衡化し、B液には0.5 M KH2PO4を用いた。流速1ml/分で分離を行い、サンプル注入後5分から40分で100%B液、あるいは、100分で70%B液までKH2PO4濃度を上昇させた。UVモニターで254nmの値を計測することにより、溶出液中の糖ヌクレオチドを測定した。標準物質としてカルビオケム社のGDP-L-フコースを用い、ピーク面積からGDP-L-フコース量を計算した。なお、本実験では全てのバッファーを0.45μmのポアサイズの膜を通したのちに使用した。
Mono-QカラムによるHPLC精製を行うと、L-Fucを含む培地で培養したFKP発現酵母由来のサンプルでは、約17.5分に明らかなピークが観察された(図9F)。これに対し、L-Fucを含まない培地を用いた場合には、17.5分に有意なピークは観察されなかった(図9C)。Mock(図9A、9D)やFKPΔN(図9B,9E)の場合にも、L-Fucの有無にかかわらず、17.5分に有意なピークは認められなかった。また、FKP形質転換体の場合、培地中のL-Fuc濃度を低下させると、GDP-L-Fucの収量も低下することが明らかになった(図10参照)。
HPLC解析では、100分で70%までB液(0.5 M KH2PO4)を上昇させることによって検出できた。この条件ではおよそ31分で標準GDP-L-Fucが溶出するが、FKPを発現する酵母をL-Fuc存在下で培養した場合にも、同様のピークが見られた(図4B)。コントロールとして行ったL-Fucを添加しない培地での培養の場合には、同様のピークは見られなかった(図4A)。標準GDP-L-Fuc(25nmol)を用いた定量解析(図4C)の結果から、15 mM L-Fucを添加した培地でOD600が1.0になるまで培養したFKP形質転換酵母25ml(約5x108細胞)から230nmol(約0.135mg)のGDP-L-Fucが合成されると見積もられた。この収量は、Nakayamaらが報告したde novo合成経路を導入した酵母の収量(非特許文献52)の25倍に当たる。これらの結果は、L-Fucの添加なしでFKP発現酵母がGDP-L-Fuc合成活性を示さないのに対し、L-Fucの添加によって大量のGDP-L-Fucをin vivoで合成できることを表している。
【0033】
(実施例3)酵母細胞内でのGDP-D-Araの合成
酵母細胞内に移植されたsalvage経路でL-Fuc以外の基質も利用できるかどうかの検討を行うために、FKP遺伝子形質転換酵母を実施例2−1において培地中に添加するL-Fucに代えてD-Araを用いた以外は同様の方法で培養した。酵母細胞内で合成されたGDP-D-Araを、前記実施例2−2と同様にESI-MS(図11の矢印)で検出し、MS/MSによって確認を行った。MS/MSスペクトルの m/z 291のフラグメントイオンは、GDP-D-Araのグアノシンが脱離したものと一致する。これらの質量分析の結果から、この酵母の発現系でGDP-D-Araも合成されることが示された。ただし、GDP-D-Araは市販されていないため、HPLCによる分析は省略した。MS/MS解析でのm/z 305のフラグメントイオンは、下記の表2で表される。
【0034】
<表2>
【配列表フリーテキスト】
【0035】
配列番号1 FKP-protein(ACCESSION CAH08307)
配列番号2 FKP-gene(ACCESSION CR626927)
配列番号3 FKP-gene forward primer
配列番号4 FKP-gene reverse primer
配列番号5 FKP ΔN gene forward primer
配列番号6 FKP ΔN gene reverse primer
【特許請求の範囲】
【請求項1】
FKP(L-フコキナーゼ/GDP-L-フコースピロフォスフォリラーゼ)遺伝子を酵母用発現ベクターにより形質転換した酵母であって、培地中のL-フコース、又はL-フコースアナログを取り込み、細胞内でGDP-L-フコース、又はL-フコースアナログのGDP-糖化合物を合成可能な形質転換酵母。
【請求項2】
L-フコースアナログがD-アラビノースであり、そのGDP-糖化合物がGDP-D-アラビノースである、請求項1に記載の形質転換酵母。
【請求項3】
FKP遺伝子が、Bacteroides fragilis由来のFKP遺伝子である、請求項1又は2に記載の形質転換酵母。
【請求項4】
酵母がSaccharomyces cerevisiaeである請求項1〜3のいずれかに記載の形質転換酵母。
【請求項5】
請求項1〜4のいずれかの形質転換酵母を用い、L-フコース、D-アラビノース、又はL-フコースアナログを存在させた培地中で培養することを特徴とする、L-フコース、D-アラビノース、又はL-フコースアナログのGDP-糖化合物を合成する方法。
【請求項6】
L-フコースを存在させた培地中で培養することを特徴とする、GDP-L-フコースを合成する方法である請求項5に記載の方法。
【請求項7】
D-アラビノースを存在させた培地中で培養することを特徴とする、GDP-D-アラビノースを合成する方法である請求項5に記載の方法。
【請求項1】
FKP(L-フコキナーゼ/GDP-L-フコースピロフォスフォリラーゼ)遺伝子を酵母用発現ベクターにより形質転換した酵母であって、培地中のL-フコース、又はL-フコースアナログを取り込み、細胞内でGDP-L-フコース、又はL-フコースアナログのGDP-糖化合物を合成可能な形質転換酵母。
【請求項2】
L-フコースアナログがD-アラビノースであり、そのGDP-糖化合物がGDP-D-アラビノースである、請求項1に記載の形質転換酵母。
【請求項3】
FKP遺伝子が、Bacteroides fragilis由来のFKP遺伝子である、請求項1又は2に記載の形質転換酵母。
【請求項4】
酵母がSaccharomyces cerevisiaeである請求項1〜3のいずれかに記載の形質転換酵母。
【請求項5】
請求項1〜4のいずれかの形質転換酵母を用い、L-フコース、D-アラビノース、又はL-フコースアナログを存在させた培地中で培養することを特徴とする、L-フコース、D-アラビノース、又はL-フコースアナログのGDP-糖化合物を合成する方法。
【請求項6】
L-フコースを存在させた培地中で培養することを特徴とする、GDP-L-フコースを合成する方法である請求項5に記載の方法。
【請求項7】
D-アラビノースを存在させた培地中で培養することを特徴とする、GDP-D-アラビノースを合成する方法である請求項5に記載の方法。
【図1】

【図2】

【図3】

【図4】
【図5】

【図6】
【図7】

【図8】

【図9】

【図10】

【図11】

【図2】
【図3】
【図4】
【図5】
【図6】
【図7】
【図8】
【図9】
【図10】
【図11】
【公開番号】特開2012−191905(P2012−191905A)
【公開日】平成24年10月11日(2012.10.11)
【国際特許分類】
【出願番号】特願2011−59434(P2011−59434)
【出願日】平成23年3月17日(2011.3.17)
【国等の委託研究の成果に係る記載事項】(出願人による申告)平成22年度独立行政法人新エネルギー・産業技術総合開発機構委託「糖鎖機能活用技術開発」産業技術力強化法第19条の適用を受ける特許出願
【出願人】(301021533)独立行政法人産業技術総合研究所 (6,529)
【Fターム(参考)】
【公開日】平成24年10月11日(2012.10.11)
【国際特許分類】
【出願日】平成23年3月17日(2011.3.17)
【国等の委託研究の成果に係る記載事項】(出願人による申告)平成22年度独立行政法人新エネルギー・産業技術総合開発機構委託「糖鎖機能活用技術開発」産業技術力強化法第19条の適用を受ける特許出願
【出願人】(301021533)独立行政法人産業技術総合研究所 (6,529)
【Fターム(参考)】
[ Back to top ]