
酸性セルラーゼ生産菌
【課題】強酸性条件下でセルロース分解活性を示す微生物と、それを用いた糖類、有機酸の製造方法の提供。
【解決手段】酸性条件下でセルロース分解活性を示すトリコデルマ(Trichoderma)属菌であって、少なくともpH2.0〜2.5のpH範囲でエンドセルラーゼ活性及びβグルコシダーゼ活性を示す、前記トリコデルマ属菌。この菌またはその培養上清を用いて、セルロースを分解する糖類の製造方法。さらに該菌と乳酸菌等の有機酸生産菌を共存培養する、有機酸の製造方法。
【解決手段】酸性条件下でセルロース分解活性を示すトリコデルマ(Trichoderma)属菌であって、少なくともpH2.0〜2.5のpH範囲でエンドセルラーゼ活性及びβグルコシダーゼ活性を示す、前記トリコデルマ属菌。この菌またはその培養上清を用いて、セルロースを分解する糖類の製造方法。さらに該菌と乳酸菌等の有機酸生産菌を共存培養する、有機酸の製造方法。
【発明の詳細な説明】
【技術分野】
【0001】
本発明は、強酸性条件下でセルロース分解活性を示す微生物及びその利用に関する。
【背景技術】
【0002】
近年、石油資源の枯渇、地球レベルの炭酸ガス発生量の削減が叫ばれており、今後石油価格の高騰が予想される。自然界に大量に存在しているセルロースをエタノール等の液体燃料、ポリ乳酸等の石油代替樹脂原料に変換することができれば、炭酸ガス発生量を増やすことなく安価に石油代替資源として用いることができる。また、セルロースから乳酸、コハク酸等の有機酸を合成することができれば石油代替のバイオ樹脂などを作ることができ、有用である。
【0003】
微生物を利用した有機酸類の生産は従来から行われており、ラクトバチルス属(Lactobacillus)菌やリューコノストック属(Leuconostoc)菌による乳酸生産、ペニシリウム属(Penicillium)菌によるクエン酸生産、グルコノバクター属(Gluconobacter)菌によるグルコン酸生産、アセトバクター属(Acetobacter)菌による酢酸生産などが知られている。この他、イタコン酸、フマル酸、コハク酸,マレイン酸、コウジ酸、酒石酸などの醗酵生産も行われてきている。最近では、組み換え大腸菌による乳酸やコハク酸の醗酵生産、組み換え酵母による乳酸醗酵生産も報告されている。しかしながら、これらの微生物が直接、セルロースから有機酸を合成したという報告例はない。一方、近年、セルラーゼ生産菌を用いてセルロースを加水分解して糖化し、生じた糖を乳酸菌などによる醗酵生産に利用することによって、乳酸などの有機酸の生産を促進する方法が開発されている(特許文献1及び非特許文献1〜4)。但しこの方法では、生産された有機酸によって培地のpHが徐々に低下し、セルロース分解活性が阻害されるため、有機酸生産を継続的に行うにはpH緩衝剤を使用してpHの低下を防ぐ必要がある。
【0004】
自然界ではセルロースは主として微生物によって分解される。細菌や糸状菌などの様々な微生物が各種セルロース分解酵素を生産することが知られている。セルロースは作用機構の異なる複数のセルロース分解酵素の協同作用により、セロオリゴ糖、セロビオースを経てグルコースへと分解される。
【0005】
セルロース分解酵素(セルラーゼ)に関しては従来から非常に多くの研究がなされている。セルラーゼ生産菌はトリコデルマ属(Trichoderma)菌、フザリウム属(Fusarium)菌、トレメテス属(Tremetes)菌、ペニシリウム属(Penicillium)菌、フミコーラ属(Humicola)菌、アクレモニウム属(Acremonium)菌、アスペルギルス属(Aspergillus)菌等の糸状菌の他に、クロストリジウム属(Clostridium)菌、シュードモナス属(Pseudomonas)菌、セルロモナス属(Cellulomonas)菌、ルミノコッカス属(Ruminococcus)菌、バチルス属(Bacillus)菌等の細菌、スルフォロバス属(Sulfolobus)菌等の始原菌、さらにストレプトマイセス属(Streptomyces)菌、サーモアクチノマイセス属(Thermoactinomyces)菌の放射菌で見出されている。最近では、これらの微生物を利用して、洗剤との併用を念頭においたアルカリセルラーゼの開発が行われている。しかし、これらの微生物が生産するセルラーゼは弱酸性からアルカリ性にかけての反応pHを持つものがほとんどであり、強酸性条件下でも活性を示すセルラーゼを生産する微生物はほとんどない。
【0006】
酸性セルラーゼを生産する微生物として、スルフォロバス・ソルファタリカス(Sulfolobus solfataricus)菌株が報告されている(非特許文献5)。同菌は至適温度が80℃、至適pH1.8のセルラーゼを菌体外に分泌生産する。しかしながら、同菌は始原菌であり生育が悪い上、生産されるセルラーゼも至適温度が80℃と高く常温ではごく弱い活性しか示さない。従ってこの酸性セルラーゼは、常温でセルロースを加水分解する目的ではあまり有用でない。他にも酸性セルラーゼの報告はあるが、常温の強酸性条件下で十分な活性を示すセルラーゼは知られていない(特許文献2〜4)。
【0007】
【特許文献1】特開2005−13131号公報
【特許文献2】特開平3−240491号公報
【特許文献3】特開平6−38747号公報
【特許文献4】特開2001−505414号公報
【非特許文献1】Iyer P.V. and Lee Y.Y., Biotechnology Letters (1999) 21: p.371-373
【非特許文献2】Xu D.B. et al., Appl. Microbiol. Biotechnol. (1989) 30, p.553-558
【非特許文献3】Kim D.M. et al., Biotechnol. Bioeng. (1992) 39 p.336-342
【非特許文献4】Moresi. M. et al., Appl. Microbiol. Biotechnol. (1991) 36, p.35-39
【非特許文献5】Huang Y. et al., Biochem J. (2005) 385 p.581-588
【発明の開示】
【発明が解決しようとする課題】
【0008】
本発明は、強酸性条件下でセルロース分解活性を示す微生物を提供することを目的とする。
【課題を解決するための手段】
【0009】
本発明者らは、上記課題を解決するため鋭意検討を重ねた結果、pH2.0のような強酸性域でも常温で活性を有するセルラーゼを培養上清に分泌するトリコデルマ属菌の分離に成功し、それに基づいて本発明を完成するに至った。
【0010】
すなわち、本発明は以下を包含する。
[1] 酸性条件下でセルロース分解活性を示すトリコデルマ(Trichoderma)属菌であって、少なくともpH2.0〜2.5のpH範囲でエンドセルラーゼ活性及びβグルコシダーゼ活性を示す、前記トリコデルマ属菌。
[2] トリコデルマ・エスピー(Trichoderma sp.)AH5株(受託番号NITE P-222)又はその変異株である、上記[1]に記載のトリコデルマ属菌。
[3] 少なくともpH2.0〜2.5のpH範囲でエンドセルラーゼ活性及びβグルコシダーゼ活性を示す、上記[1]又は[2]に記載のトリコデルマ属菌由来の培養上清。
[4] 上記[1]若しくは[2]に記載のトリコデルマ属菌又は上記[3]に記載の培養上清を用いてセルロースを分解することを特徴とする、糖類の製造方法。
[5] セルロースの分解をpH2.5以下の条件下で行う、上記[4]に記載の方法。
[6] 糖類が、セロオリゴ糖、セロビオース、及びグルコースからなる群より選択される少なくとも1つである、上記[4]又は[5]に記載の方法。
[7] 上記[1]若しくは[2]に記載のトリコデルマ属菌又は上記[3]に記載の培養上清を用いてセロオリゴ糖を分解することを特徴とする、セロオリゴ糖からセロビオース及びグルコースを製造する方法。
[8] セロオリゴ糖の分解をpH2.5以下の条件下で行う、上記[7]に記載の方法。
[9] 上記[1]若しくは[2]に記載のトリコデルマ属菌又は上記[3]に記載の培養上清を用いてβグルコシドを分解することを特徴とする、βグルコシドからアグリコン及びグルコースを製造する方法。
[10] βグルコシドの分解をpH2.5以下の条件下で行う、上記[9]に記載の方法。
[11] 上記[1]又は[2]に記載のトリコデルマ属菌と有機酸生産菌とをセルロース存在下で共存培養することを特徴とする、有機酸の製造方法。
[12] 上記[3]に記載の培養上清とセルロースとを含む培地で有機酸生産菌を培養することを特徴とする、有機酸の製造方法。
[13] 有機酸生産菌が乳酸菌である、上記[11]又は[12]に記載の方法。
【発明の効果】
【0011】
本発明のトリコデルマ属菌は、常温の強酸性条件下でもセルロース分解活性を示す。本発明のトリコデルマ属菌を用いた糖類製造法は、生産効率がよく、さらに雑菌汚染がより少ない強酸性条件下で実施することができて有用である。
【発明を実施するための最良の形態】
【0012】
以下、本発明を詳細に説明する。
【0013】
1.セルロース及びその加水分解酵素であるセルラーゼ
セルロースは、グルコースがβ-1,4-グルコシド結合(β-1,4結合)により高度に重合した高分子から構成される物質である。セルロースは、天然では植物細胞壁の主たる構成成分として存在し、多糖としては地球上で最も多く生産されている。セルロース分子におけるグルコースの重合度は、その起源によって異なるが、木材では6,000〜10,000、綿では8,000〜10,000程度であり、藻類由来のものでは44,000というものもある。セルロース分子の隣接するグルコース残基間においては分子内水素結合が形成されるが、さらに、隣接するセルロース分子鎖同士も分子間水素結合を形成し、強固なセルロース繊維を形成する。セルロース繊維は強固な結晶構造をとる領域を高割合で含むため、セルロースをグルコースなどの単糖又は少糖まで加水分解(糖化)するのは容易ではない。自然界ではセルロースは主として微生物によって分解されており、細菌や糸状菌などの様々な微生物がセルロース分解酵素を生産することが知られている。これらの微生物は菌体外に複数のセルロース分解酵素を分泌し、セルロースはそれらの作用機構の異なる各種セルロース分解酵素の協同作用により、主に、セロオリゴ糖、セロビオースを経てグルコースへと分解される。セルロース分解酵素は、一般に、セルロースを加水分解する酵素の総称としてセルラーゼとも呼ばれている。
【0014】
セルラーゼの種類としては、現在、狭義のセルラーゼ(EC 3.2.1.4)、グルカン1,4-βグルコシダーゼ(EC 3.2.1.74)、セルロース1,4-βセロビオシダーゼ(EC 3.2.1.91)、βグルコシダーゼ(EC 3.2.1.21)の4つが主に認められている。
【0015】
狭義のセルラーゼ(EC 3.2.1.4)は、エンド型の作用様式で(すなわち分子鎖内部で)セルロースを加水分解し、セロオリゴ糖、セロビオース、及びグルコースを生産する酵素である。狭義のセルラーゼ(EC 3.2.1.4)はカルボキシメチルセルロース(CMC)などの非結晶性セルロースをよく加水分解することが知られている。狭義のセルラーゼ(EC 3.2.1.4)は、カルボキシメチルセルラーゼ、エンド1,4-βグルカナーゼ、エンドセルラーゼなどとも呼ばれている。一方、グルカン1,4-βグルコシダーゼ(EC 3.2.1.74)及びセルロース1,4-βセロビオシダーゼ(EC 3.2.1.91)は、セルロース鎖をエキソ型の作用様式で加水分解し、末端からグルコース又はセロビオースを遊離する酵素である。これらの酵素は綿繊維やアビセルなどの結晶性セルロースをよく加水分解し、アビセラーゼ又はエキソセルラーゼとも呼ばれている。さらにβグルコシダーゼ(EC 3.2.1.21)は、グルカン1,4-βグルコシダーゼ及びセルロース1,4-βセロビオシダーゼの作用によってセルロースから生成されるような、セロオリゴ糖、セロビオース、及びβグルコシド(グルコースの配糖体)に作用し、非還元末端からグルコースを遊離する酵素である。セロビオースを特によく加水分解するβグルコシダーゼは、セロビアーゼと呼ばれる。
【0016】
既存のセルラーゼは、弱酸性からアルカリ性にかけて安定なpH反応性を有するものがほとんどである。本明細書では、セルロース分解酵素の総称として用語「セルラーゼ」を使用し、それと区別するため、狭義のセルラーゼ(EC 3.2.1.4)を「エンドセルラーゼ」と呼ぶこととする。
【0017】
2.本発明に係るトリコデルマ属の新菌種
本発明者らは、強酸性のpH範囲(強酸性条件下)でも生育しセルロース分解活性を示すトリコデルマ属に属する菌を分離することに成功した。この菌は、既存のセルラーゼとは違って広範な酸性条件下で強力な活性を示し、かつ常温の強酸性条件下でも十分な活性を示す各種セルラーゼを、菌体外に産生する。この菌は、その菌学的性質に基づき、トリコデルマ属の新菌種として同定された。本発明は、こうして同定されたトリコデルマ属新菌種に属する菌及びその利用に関する。なお本発明において「酸性条件」とは、pH0以上pH7.0未満の範囲内のpHを示すことを言う。さらに本発明において「強酸性」とは、pH3.0以下、特に微生物の生育に関してはpH2.0以上pH3.0以下のpH範囲を意味するものとする。さらに本発明において「常温」とは、20℃以上30℃以下の温度範囲を言うものとする。
【0018】
本発明者らが分離した菌株は、トリコデルマ・エスピー(Trichoderma sp.)AH5株と命名され、2006年3月8日付で、独立行政法人 製品評価技術基盤機構 特許微生物寄託センター(日本国千葉県木更津市かずさ鎌足2-5-8)に、受託番号NITE P-222で寄託された。トリコデルマ・エスピーAH5株は、以下、「AH5株」と略記することがある。
【0019】
本発明に係るトリコデルマ・エスピーAH5株の菌学的性質は、以下の通りである。なお、コロニー色調に関する記述はKornerup and Wanscher (1978) "Methuen handbook of colour" 3rd ed.(Eyre Methuen, London, UK)に従っている。
【0020】
(a)培養的性質:
(1)ポテトデキストロース寒天培地(pH5.6)における生育:25℃で3日間培養後、コロニーの成長が認められ、気中菌糸の形成が顕著に見られる。菌糸の色は白色〜クリーム色である。培養開始後7〜10日で深緑色の分生子を形成する。9日間培養後のコロニーの表面性状は綿状(cottony)、コロニー色調はクリーム色及び濃緑色。テレオモルフの形成は認められない。
(2)オートミール寒天培地(pH6.0)における生育:25℃で3日間培養後、コロニーの成長が認められ、気中菌糸の形成が顕著に見られる。菌糸の色は白色〜クリーム色である。培養開始後7〜10日で深緑色の分生子を形成する。9日間培養後のコロニーの表面性状は綿状(cottony)、コロニー色調は白色と深緑色。テレオモルフの形成は認められない。
(3)2%麦芽エキス寒天培地(pH6.0):25℃で3日間培養後、コロニーの成長が認められる。培養開始後7〜10日で深緑色の分生子を形成する。9日間培養後のコロニーの表面性状は綿状(cottony)、コロニー色調は深緑色。テレオモルフの形成は認められない。
【0021】
(b)形態学的性質:
栄養菌糸:有隔壁菌糸を形成する。菌糸表面は平滑(smooth)。栄養菌糸の先端部に球形〜亜球形の厚膜胞子の形成が認められる。
分生子柄:集合して羊毛状の塊(tuft)を形成するのが認められる。分岐し、輪生する。
分生子(胞子):フィアロ型分生子を形成する。連鎖せず、緑色の分生子塊を形成する。分生子の形状は卵形〜亜球形。単細胞。分生子表面は平滑(smooth)。
(c)最適生育条件:20〜25℃、pH 3〜pH 5
(d)生育可能範囲:10〜35℃、pH 2〜pH 6
(e)28S rDNA(28SリボソームRNAコード配列)におけるDNA配列相同性:
トリコデルマ・ビリデ(Trichoderma viride)の28S rDNA(配列番号2)との間で99.8%程度の同一性を有する。
(f)酸素に対する性質:好気性
(g)その他の特徴:強酸性条件を含む幅広い酸性条件下でセルロース分解活性を示す。特に、生育可能範囲の温度条件において、少なくともpH2.0〜pH2.5を含むpH範囲で、エンドセルラーゼ活性及びβグルコシダーゼ活性を示す。
【0022】
本発明に係るトリコデルマ属新菌種は、トリコデルマ・エスピーAH5株の上記のような菌学的性質を本質的に共有する。このような本発明に係るトリコデルマ属菌の典型例はトリコデルマ・エスピーAH5株であるが、AH5株の分離源である秋田県鹿角市八幡平の酸性温泉蒸の湯の源泉中から、上記のような菌学的性質を有する菌を後述の実施例の記載に従ってさらに分離することにより、上記トリコデルマ属菌の菌株をさらに得ることもできる。また、本発明に係るトリコデルマ属菌には、例えばトリコデルマ・エスピーAH5株の変異体(自然突然変異体、遺伝子組換え体、突然変異誘発処理体、プラスミド導入等による形質転換体、倍数化体など)、トリコデルマ・エスピーAH5株を親株の1つに用いて作製した細胞融合株、トリコデルマ・エスピーAH5株を親株の1つとして交配により作製した菌なども含まれる。
【0023】
本発明に係るトリコデルマ属菌を培養するのに適した培地としては、限定するものではないが、例えば、ポテトデキストロース寒天培地が挙げられる。ポテトデキストロース寒天培地は、ポテト抽出物200g、デキストロース20g、アガー15gを水1Lに溶解し(滅菌前のpH値:5.6)、それを121℃で15分オートクレーブ滅菌することにより、調製することができる。本発明に係るトリコデルマ属菌は、このような培地において25℃にて好気的に培養することができる。
【0024】
本発明に係るトリコデルマ属菌がセルロース分解活性を示すかどうかは、具体的には例えば、本発明のトリコデルマ属菌を、セルロース以外の炭素源を含まない培地で培養し、菌が生育するかどうかを調べることによって判定することができる。菌の生育が認められた場合、その菌はセルロースを分解することにより糖類を得たと考えられるので、すなわちセルロース分解活性(セルロース分解能)を有すると判定できる。この試験は、例えば、炭素源を含まない液体培地を染み込ませたろ紙の上で本発明に係るトリコデルマ属菌を培養することによって行うこともできる。この試験において本発明のトリコデルマ属菌が、特定のpH条件下でセルロース分解活性を示すかどうかを調べるためには、その特定のpH値に調整した培地を使用し、培養温度や湿度などの他の条件については本発明のトリコデルマ属菌における通常の生育条件(例えば、25℃)を用いることが好ましい。
【0025】
あるいは、本発明のトリコデルマ属菌が、セルロース分解活性を示すかどうかは、そのトリコデルマ属菌由来の培養上清について各種セルラーゼの活性を検出することによって判定することもできる。これは、本発明に係るトリコデルマ属菌のセルロース分解活性が、その菌が生産し菌体外に分泌した各種セルラーゼの作用によって得られることに基づく。この方法では、例えば、本発明のトリコデルマ属菌由来の培養上清をエンドセルラーゼの基質となるカルボキシメチルセルロースと反応させ、カルボシキメチルセルロースの分解によって生じる還元糖の量を測定することにより、本発明のトリコデルマ属菌がエンドセルラーゼ活性を示すか否かを判定することができる。還元糖量の定量法としてはSomogyi法、Tauber-Kleiner法、Hanes法(滴定法)、Park-Johnson法、3,5-ジニトロサリチル酸(DNS)法などの多数の定量法が知られているが、好適な1つの方法として、糖による銅イオンの還元を利用するSomogyi-Nelson法を用いることができる(福井作蔵 著「生物化学実験法1 還元糖の定量法 第2版」学会出版センター 1990年)。Somogyi-Nelson法のプロトコールの一例では、まず酵素反応液を100℃で10分加熱処理して反応を停止させ、その反応液と等量のSomogyi銅液(和光純薬社製など)を加えて混合し、100℃で10分加熱処理してから急速に冷却し、冷却後、等量のNelson試薬(和光純薬社製など)を加えて還元銅沈殿を溶解して発色させ、30分静置し、660nmでの吸光度を測定し、その測定値から、グルコースを標準糖として還元糖量を算出する。さらに上記方法では、本発明のトリコデルマ属菌由来の培養上清をβグルコシダーゼの基質となるp-ニトロフェニルグルコシドと反応させ、p-ニトロフェニルグルコシドの分解によって生じるp-ニトロフェノールを、400nmでの吸光度測定により定量することにより、本発明のトリコデルマ属菌がβグルコシダーゼ活性を示すか否かを判定してもよい。さらには、本発明のトリコデルマ属菌由来の培養上清をセロヘキサオース(六糖のセロオリゴ糖)と反応させ、セロヘキサオースの分解によって生じる糖類の量を測定することにより、本発明のトリコデルマ属菌がセルラーゼ活性を示すか否かを判定してもよい。
【0026】
本発明のトリコデルマ属菌は、後述の実施例に記載の通り、少なくともpH2.0〜pH5.5を含む酸性条件下の培地でセルロースを分解することができ、かつその培養上清においてpH2.0以上の酸性pH範囲でエンドセルラーゼ活性を、pH1.5以上の酸性pH範囲でβグルコシダーゼ活性を示す。本発明のトリコデルマ属菌は、広範な酸性pH範囲で高いセルロース分解活性を示すだけでなく、既知セルラーゼでは十分な活性が認められない強酸性条件下、特にpH2.0〜2.5のpH条件下でも、エンドセルラーゼ活性及びβグルコシダーゼ活性等のセルロース分解活性を示すことができる。
【0027】
3.本発明のトリコデルマ属菌を用いたセルロース系物質の分解
上記のような本発明に係るトリコデルマ属菌を用いれば、セルロース、セルロース部分分解物(セロオリゴ糖、セロビオース、βグルコシドなど)、そして植物細胞壁(ヘミセルロース、ペクチン質、リグニンなどに結合したセルロースによって構成される)などのセルロース系物質を、酸性条件下で効率よく分解することができる。本発明は、本発明に係るトリコデルマ属菌又はその培養上清を用いて、セルロース系物質から糖類を製造する方法にも関する。
【0028】
本発明において「セルロース」とは、グルコースがβ-1,4-グルコシド結合により質量平均重合度51以上(限定するものではないが、好ましくは質量平均重合度100以上、より好ましくは質量平均重合度400〜50,000)で重合した分子鎖若しくはその誘導体(例えばカルボキシメチルセルロースなどの、カルボキシメチル化、アルデヒド化、若しくはエステル化などの誘導体化が為されたもの)、又はそれらが複数結合したもの(セルロース繊維など)を言う。セルロースは、アビセルや綿繊維などの結晶性セルロースであってもよいし、非結晶性セルロースであってもよい。セルロースはまた、天然由来のものでも、人為的に合成したものでもよい。さらにセルロースは、植物由来のものでも、真菌由来のものでも、細菌由来のものでもよい。
【0029】
本発明において「セロオリゴ糖」とは、グルコースがβ-1,4結合により重合した少糖類(重合度3〜50)又はその誘導体を言う。セロオリゴ糖の具体例として、セロペンタオース、セロトリオース、セロヘキサオース、セロテトラオースなどが挙げられる。
【0030】
本発明において「βグルコシド」とは、グルコースが非糖化合物(アグリコン)と結合した配糖体を言う。βグルコシドの具体例としては、p-ニトロフェニルグルコシド、グリチルリチン酸、ステビオシド、フラボノイド配糖体、アルブチン、サポニンなどが挙げられる。なおp-ニトロフェニルグルコシドは、βグルコシダーゼの作用により、アグリコンに相当するp-ニトロフェノールと、グルコースとに分解される。
【0031】
上述のような本発明に係る糖類製造方法の1つの態様として、本発明に係るトリコデルマ属菌をセルロース存在下で培養してセルロースを加水分解することにより、セロオリゴ糖、セロビオース又はグルコースなどの糖類をセルロースから遊離させることを含む、セルロースから糖類を製造する方法がある。ここで「セルロース存在下」とは、本発明のトリコデルマ属菌を培養する培地中に、セルロース又はセルロース含有物質が添加されていることを意味する。セルロース含有物質としては、限定するものではないが、綿や麻などの天然繊維品、レーヨン、キュプラ、アセテート、リヨセルなどの再生繊維品、稲わら、籾殻、木材チップなどの農産廃棄物などが挙げられる。
【0032】
本発明の製造方法の別の態様としては、本発明に係るトリコデルマ属菌をセロオリゴ糖存在下で培養してセロオリゴ糖を加水分解することにより、セロオリゴ糖からセロビオース又はグルコースなどの糖類を遊離させることを含む、セロオリゴ糖からセロビオース又はグルコースを製造する方法が挙げられる。ここで「セロオリゴ糖存在下」とは、本発明のトリコデルマ属菌を培養する培地中にセロオリゴ糖が添加されていることを意味する。
【0033】
本発明の製造方法のさらに別の態様としては、本発明に係るトリコデルマ属菌をβグルコシド存在下で培養することにより、βグルコシドをアグリコンとグルコースとに加水分解することを含む、βグルコシドからアグリコン及びグルコースを製造する方法が挙げられる。ここで「βグルコシド存在下」とは、本発明のトリコデルマ属菌を培養する培地中にβグルコシドが添加されていることを意味する。
【0034】
これらの態様において用いる培養培地としては、上記のようなセルラーゼの基質(例えばセルロース、セルロース含有物質、セロオリゴ糖、又はβグルコシドなど)を加える限り、トリコデルマ属に属する真菌の培養に使用可能な任意の培地を用いることができる。培地の組成は、例えば、「微生物の分離法」(山里一英ら編、株式会社R&Dプランニング発行、2001年7月6日(1986年初版発行))の記載を参考として決定することができる。培地に含める炭素源としては、上記のセルラーゼの基質(セルロース、セルロース含有物質、セロオリゴ糖、又はβグルコシドなど)に加えて、スクロース、フルクトース、マンニトール、ソルビトール、ガラクトース、マルトース、エリスリット、グリセリン、エチレングリコール、エタノール、澱粉、ビート搾汁、サトウキビ搾汁、ビートモラセス等を用いてもよい。培地に含める窒素源としては、硫酸アンモニウム、塩化アンモニウム等のアンモニウム塩、硝酸カルシウムなどの硝酸塩、又はその他の有機若しくは無機窒素源を用いることができる。さらに培地には、ペプトン、酵母エキス、大豆加水分解物などの天然栄養源を含めることも好ましい。培地には、抗生物質、pH緩衝剤(例えば炭酸カルシウム、リン酸アンモニウム、水酸化ナトリウムなど)、マーカー物質などを含めてもよい。培地は液体培地であってもよいし、固体培地であってもよい。上記のようなセルラーゼの基質を培地に加える量は、特に限定されないが、液体培地で培養する場合には1L当たり1g〜100g程度用いればよい。好適な培地の例としては、0.2g硫酸アンモン、0.01g塩化カリウム、0.05gリン酸二水素カリウム、0.05g硫酸マグネシウム、0.001g硝酸カルシウム、及び0.0001g酵母エキスを脱塩水に溶解して得られる溶液に、上記のようなセルラーゼの基質を加えたものが挙げられる。セルラーゼの基質は、培養中、適宜補充することがより好ましい。
【0035】
用いる培地のpH値は、培養開始前に、本発明に係るトリコデルマ属菌の生育可能範囲内にあればよい。本発明の方法では、好ましくはpH1.0〜pH7.0、より好ましくはpH2.0〜6.4の酸性pH値を示す培地を用いることが望ましい。本発明の方法では、とりわけpH2.5〜pH5.5の範囲内のpH値を示す培地の使用が、糖類を効率よく生産する上でより好ましい。またpH2.5以下、特にpH2.0〜pH2.5の範囲内のpH値を示す培地は、他の雑菌の混入を抑制する上でより好適に使用される。なお本発明の上記製造方法において、この培地のpH値が、トリコデルマ属菌の培養を利用してセルロース、セロオリゴ糖、βグルコシド等の分解反応を実施する際のpH条件に相当する。
【0036】
培養温度は、本発明に係るトリコデルマ属菌の生育可能範囲内の温度であればよい。特に好ましい培養温度は、20℃〜35℃、より好ましくは25℃〜30℃、最も典型的な培養温度は25℃である。培養は、好気的条件下で行うことが好ましい。
【0037】
培養時間は、当業者であれば任意に設定することができるが、少なくとも24時間培養を継続することが好ましい。
【0038】
以上のようにして本発明に係るトリコデルマ属菌を上記のようなセルラーゼの基質の存在下で培養すると、その基質が加水分解されて生じた糖類等(低分子化セルロース、セロオリゴ糖、セロビオース、グルコース、βグルコシド、アグリコンなど)が培養物中に生産される。本発明においては、そのようにして生産された糖類等を、培養物(特に培養液)から、HPLC法、アルコール沈殿法、結晶化法などの当業者に公知の方法により精製することができる。あるいは、その培養液から菌体を除去して培養上清を調製し、それを糖類等を豊富に含む溶液として利用することもできる。
【0039】
本発明の上記製造方法のまた別の態様として、本発明に係るトリコデルマ属菌を好ましくは液体培地で培養しその菌体を除去して得られる培養上清を、セルラーゼの基質となるセルロースと反応させ、セルロースを加水分解することにより、セロオリゴ糖、セロビオース又はグルコースなどの糖類をセルロースから遊離させることを含む、セルロースから糖類を製造する方法が挙げられる。
【0040】
本発明の上記製造方法のさらなる態様として、本発明に係るトリコデルマ属菌を好ましくは液体培地で培養しその菌体を除去して得られる培養上清を、セルラーゼの基質となるセロオリゴ糖と反応させ、セロオリゴ糖を加水分解することにより、セロビオース又はグルコースなどの糖類をセロオリゴ糖から遊離させることを含む、セロオリゴ糖からセロビオース又はグルコースを製造する方法が挙げられる。
【0041】
本発明の上記製造方法のさらに別の態様として、本発明に係るトリコデルマ属菌を好ましくは液体培地で培養しその菌体を除去して得られる培養上清を、セルラーゼの基質となるβグルコシドと反応させて、βグルコシドをアグリコンとグルコースとに加水分解することを含む、βグルコシドからアグリコン及びグルコースを製造する方法が挙げられる。
【0042】
ここで培養上清を調製するには、まず、本発明に係るトリコデルマ属菌を、トリコデルマ属に属する真菌の培養に使用可能な任意の培地で培養すればよい。培地に用いる炭素源、窒素源、天然栄養源などの例は、上記と同様である。培地には、抗生物質、pH緩衝剤、マーカー物質などを含めてもよい。培地は液体培地であってもよいし、固体培地であってもよい。好適な培地の例としては、0.2g硫酸アンモン、0.01g塩化カリウム、0.05gリン酸二水素カリウム、0.05g硫酸マグネシウム、0.001g硝酸カルシウム、及び0.0001g酵母エキスを脱塩水に溶解した溶液が挙げられる。
【0043】
上記培地のpH値は、培養開始前に、本発明に係るトリコデルマ属菌が生育できる範囲内であればよい。本発明の方法では、好ましくはpH2.0〜pH6.0、より好ましくはpH3.0〜5.0、例えばpH3.5の酸性pH値を示す培地を用いることが好ましい。
【0044】
培養温度は、本発明に係るトリコデルマ属菌の生育可能範囲内の温度であればよい。特に好ましい培養温度は、20℃〜35℃、より好ましくは25℃〜30℃、典型的な使用温度としては25℃である。培養は、好気的条件下で行うことが好ましい。
【0045】
培養時間は、当業者であれば任意に設定することができるが、少なくとも24時間培養を継続することが好ましい。
【0046】
こうして得られた本発明に係るトリコデルマ属菌の培養液から、遠心分離などの分離手法を用いて菌体を除去することにより、培養上清を得ることができる。このような本発明に係るトリコデルマ属菌由来の培養上清は、各種セルラーゼを含み、広範な酸性条件下で強いセルラーゼ活性を示し、特に、少なくともpH2.0〜2.5のpH範囲を含む強酸性条件下でエンドセルラーゼ活性及びβグルコシダーゼ活性を示す。本発明は、このような本発明に係るトリコデルマ属菌由来の培養上清にも関する。
【0047】
以上のようにして調製した培養上清とセルラーゼの基質(例えば、セルロース、セルロース含有物質、セロオリゴ糖、又はβグルコシドなど)とを反応させるには、培養上清にセルラーゼの基質を添加するか、又はセルラーゼの基質を含む溶液と培養上清とを混合して、反応系を調製してもよい。この反応系には、pH緩衝剤などを含めても含めなくてもよい。
【0048】
培養上清中のセルラーゼとセルラーゼの基質との反応を促進するため、反応系のpH値は、原則として、本発明に係るトリコデルマ属菌が生育可能なpH範囲又はその付近とすることが好ましい。反応系のpH値としては、糖類を効率よく生産する上ではpH2.5〜pH5.5の範囲がより好ましい。またpH3.0以下、例えばpH1.5〜3.0の範囲、特にpH2.0〜pH2.5の範囲は、他の雑菌の混入を抑制しながら糖類を生産する上でより好ましい。なお本発明において培養上清を用いてセルロース、セロオリゴ糖、βグルコシド等の分解反応を実施する際のpHの「条件」とは、このような反応系のpH値を言う。
【0049】
反応系の温度は、本発明に係るトリコデルマ属菌の生育可能範囲内であればよい。特に好ましい培養温度は、20℃〜35℃、より好ましくは25℃〜30℃である。この態様では、より反応を促進するため、反応系の温度を30℃とすることも好ましい。
【0050】
反応時間は、当業者であれば任意に設定することができるが、少なくとも10分間、好ましくは1時間以上にわたり反応させることが好ましい。
【0051】
以上のようにして生産された糖類等は、上記と同様にして常法により精製してもよい。
【0052】
4.本発明のトリコデルマ属菌と有機酸生産菌を用いた有機酸製造方法
本発明に係るトリコデルマ属菌は、公知技術であるセルラーゼ生産菌と有機酸生産菌の共存培養に基づく有機酸製造方法において、強酸性条件下でも糖類を生産できるという利点を生かし、セルラーゼ生産菌として非常に有利に利用することができる。
【0053】
セルラーゼ生産菌と有機酸生産菌の共存培養に基づく有機酸製造方法は、特許文献1及び非特許文献1〜4などの文献に詳細に記載されている。例えば特許文献1には、セルラーゼ生産菌(セルロース分解菌)の存在下でセルロース又はセルロースと澱粉の混合物を同時に加水分解して糖化した後、乳酸菌を加えて乳酸醗酵を行うことによる、乳酸の製造方法が開示されている。特許文献1には、さらに、セルロース又はセルロースと澱粉の混合物を含む培地において、セルラーゼ生産菌と乳酸菌の共生系による混合培養を行うことによる乳酸の製造方法が開示されている。従って本発明では、これらの文献に記載されたような公知の有機酸製造方法において、既存のセルラーゼ生産菌に変えてトリコデルマ属菌を用いればよい。
【0054】
具体的には本発明は、本発明のトリコデルマ属菌と有機酸生産菌とをセルロース存在下で共存培養することにより、有機酸を製造する方法にも関する。本発明はさらに、本発明に係るトリコデルマ属菌由来の培養上清とセルロースとを含む培地で有機酸生産菌を培養することにより、有機酸を製造する方法にも関する。
【0055】
これらの方法において使用する有機酸生産菌[及びその有機酸生産]としては、ストレプトコッカス属菌(例えばストレプトコッカス・ボビス(Streptococcus bovis))やラクトバチルス属菌(例えばラクトバチルス・デルブルエキイ(Lactobacillus delbrueckii))などの乳酸菌[乳酸生産]、リューコノストック属菌[乳酸生産]、ペニシリウム属菌[クエン酸生産]、グルコノバクター属菌[グルコン酸生産]、アセトバクター属菌[酢酸生産]などが挙げられるが、特に乳酸菌が好適である。
【0056】
本発明において「共存培養」とは、培養期間の少なくとも一部において、本発明に係るトリコデルマ属菌と有機酸生産菌とが、同じ培地中で互いを排除することなく生育(増殖)していることを言う。
【0057】
本発明において培養に使用する培地、培養温度、培養時間などは、使用する有機酸生産菌に適した培養条件に、本発明に係るトリコデルマ属菌の上記生育可能範囲等を考慮して必要に応じて多少の変更を加えることにより、当業者が適宜設定することができる。
【0058】
セルラーゼ生産菌と有機酸生産菌を用いた従来の有機酸製造方法では、生産された有機酸により培地のpHが徐々に低下し、その結果、セルロース分解活性の低下をもたらすため、培養中の培地におけるpH低下を防ぐ必要があったが、本発明では、強酸性条件でも十分なセルロース分解活性を示すトリコデルマ属菌を利用するため、培地のpH調整がほとんど不要になり、とても有利である。
【実施例】
【0059】
以下、実施例を用いて本発明をさらに具体的に説明する。但し、本発明の技術的範囲はこれら実施例に限定されるものではない。
【0060】
[実施例1]
秋田県鹿角市八幡平の酸性温泉蒸の湯(ふけの湯)の源泉から流れ出た湯中から採取した木片0.5gを、10mlの分離培地Aを含む滅菌した15mlのコーニングチューブに入れ、25℃、100rpmで好気的に180日間振とう培養した。使用した分離培地Aは、0.2g硫酸アンモン、0.01g塩化カリウム、0.05gリン酸二水素カリウム、0.05g硫酸マグネシウム、0.001g硝酸カルシウム、及び0.0001g酵母エキス(DIFCO社製)、並びに1gセルロース(Macherey-Nagel社製)又は1gアビセル(旭化成社製)を脱塩水に溶解し、硫酸を用いてpHを3.5に調整した後、さらに脱塩水を添加して100mlとし、オートクレーブ滅菌することにより、予め調製しておいた。
【0061】
菌が生育し、にごりが観察されたチューブより培養液10μlを採取し、以下のような活性染色電気泳動に供した。まず、Davisらの方法(Ornstein L and Davis BJ, Ann NY Acad Sci 121, 321-349, 1964)に従い、0.1%(w/v)カルボキシメチルセルロース(CMC)(Sigma社製)を含む7.5%ポリアクリルアミドゲルに採取した培養液をロードし、電気泳動を行った。泳動後、ゲルを0.2Mグリシン溶液(pH2.5)に40℃で1時間浸漬した。次に、0.2%コンゴレッド(ナカライテスク社製)に60分間ゲルを浸漬した後、1M塩化ナトリウム溶液で脱色した。この方法によればセルラーゼ活性を示したタンパクバンドは赤い背景の上に白いスポットとして現れるはずである。そこでそのようなバンドの現れたチューブを選び、その中の培養液を分離プレートAに広げて室温で培養し、菌の純化を行った。使用した分離プレートAは、7g(あるいは0.7%)のゲルライト(Sigma社製)を添加した1Lの上記分離培地Aをオートクレーブした後、シャーレに広げ、固めたものである。分離した菌はプレート上でpH2.5〜3.5で生育させた。顕微鏡観察により形態観察を行うと、この菌は緑色の胞子を有する糸状菌であった。胞子嚢は観察されなかった。
【0062】
この菌については後述のように28S rDNAシークエンスを行ったところ、トリコデルマ・ビリデ(Trichoderma viride)の28S rDNA配列との間で高い相同性(549塩基中、548塩基が一致)を示した。しかしこの菌には、トリコデルマ・ビリデの特徴である35℃での培養で生育しない、胞子表面が粗面であるといった形質が見られなかった。
【0063】
[実施例2]
実施例1で緑色の胞子を有する糸状菌が分離された培養液を、生理食塩水で希釈し、上記の通り調製した分離プレートAに塗布し、25℃で培養した。セルロースを含む分離培地を用いた分離プレートA(セルロースプレート)と、アビセルを含む分離培地を用いた分離プレートA(アビセルプレート)のいずれにおいても、糸状菌の生育が確認できた。生育した糸状菌は緑色を呈した胞子を形成することが顕微鏡によって確認されたが、胞子嚢は確認されなかった。
【0064】
次いで帰属分類群の同定を行った。分離した糸状菌を、ポテトデキストロース寒天培地(PDA培地)、2%麦芽エキス寒天培地(2%MA)、オートミール寒天培地(OA培地)、三浦培地(LCA培地)の各平板培地に接種し、25℃で培養し、培養開始1週間後からコロニーの巨視的特徴を肉眼及び実体顕微鏡下で観察した。また、プレートより直接採取した菌体からプレパラートを作製し、微分干渉顕微鏡で微視的特徴を観察した。その結果、培養から3日後には各培地のシャーレ中にコロニーの成長が認められ、緑色の分生子柄の房の形成が確認された。特にポテトデキストロース寒天培地及びオートミール寒天培地(OA培地)では、気中菌糸の形成が顕著であった。9日間培養後の各培地で観察されたコロニーの表面性状はいずれも綿状(cottony)であった。9日間培養後のコロニーの色調は、Kornerup and Wanscher (1978) "Methuen handbook of colour" 3rd ed.(Eyre Methuen, London, UK)に従うと、ポテトデキストロース寒天培地(PDA培地)ではクリーム色(淡黄色)及び濃緑色、2%麦芽エキス寒天培地(2%MA)では深緑色、オートミール寒天培地(OA培地)では白色と深緑色、三浦培地(LCA培地)では深緑色であった。
【0065】
さらに、Samuels G. J. et al. (2002) Mycologia 94: p.146-170、Chaverri, P. and Samuels G. J. (2003) Studies in Mycology 48: p.1-116及びTrichoderma Home(http://nt.ars-grin.gov/taxadescriptions/keys/TrichodermaIndex.cfm)に記載された方法に従って、コロニー生育性及び形態の観察を行った。まず、コロニー上に緑色の分生子柄の房が形成される直前にコロニーを含む5mm径の寒天片を切り取り、ポテトデキストロース寒天培地(PDA培地)、SNA寒天(Synthetischer nahrstoffarmer agar)培地、及びコーンミール寒天+2%デキストロース培地(CMD培地)に接種して各種温度で3〜7日間培養し、観察を行った。
【0066】
以上の観察の結果は、本明細書中、菌学的性質としてまとめて上述している。これらの観察結果を踏まえ、Arx, J. A. von (1981) "The genera of fungi sporulating in pure culture" 3rd edition(A.R. Gantner Verlag KG., Vaduz, Germany 424 pp)、Barron, G. L. (1968) "The genera of hyphomycetes from soil"(The Williams & Wilkins Company, Baltimore, MD, USA, 364pp)、Domsch K. H. et al. (1993) "Compendium of soil fungi" volume I, IHW-Verlag, Eching, Germany, Reprinted 860pp、Domsch K. H. et al. (1993) "Compendium of soil fungi and supplement"(volume II, IHW-Verlag, Eching, Germany, Reprinted 405pp)、及びKiffer, E. and Morelet, M. (2002) "The Deuteromycetes: Mitosporic fungi classification and generic keys"(Science Publishers Inc., Enfield, NH, USA, 273pp)に記載されている菌類の検索表により、属の同定が行われた。この菌は、有隔壁菌糸の形成が認められること、また1ヶ月培養してもテレオモルフの形成(有性生殖器官の形成)が認められなかったことから、不完全菌類の1種であると判断された。さらに、菌糸の成長が速く、分岐する分生子柄を持ち、分生子の形成様式がフィアロ型であること、分生子が連鎖せず、コロニーの成熟に伴って白色から深緑色へと着色した分生子塊を形成することなどの形態学的特徴から、この菌は、不完全菌類の中でもトリコデルマ属(Trichoderma)に属するものと考えられた。
【0067】
続いて、トリコデルマ属のモノグラフであるRifai, M. A. (1969) "A revision of the genous Trichoderma" Mycological Papers 116: p.1-56、その後見出された種を含めた奥田徹 著「真菌の分離と分類・同定(20)」(1992) p.157-166、緑色の分生子を持つ種をまとめたChaverri, P. and Samuels G. J. (2003) Studies in Mycology 48: p.1-116に記載された菌類の検索表及びTrichoderma Home(http://nt.ars-grin.gov/taxadescriptions/keys/TrichodermaIndex.cfm)の検索表を用いて、菌種の同定を試みた。その結果、分離した菌の特徴は、Chaverri, P. and Samuels G. J. (2003)に記載されたトリコデルマ・ハルジアナム(Trichoderma harzianum)と比較的類似していた。
【0068】
一方、分離した菌について28S rDNA領域配列に基づく分子系統解析を行った。28S rDNAのD1/D2領域に特異的なプライマーを設計し、分離した菌から常法により抽出した核酸を鋳型としたPCR増幅を行い、シークエンシングにより28S rDNAの塩基配列を決定した。得られた28S rDNAの塩基配列(配列番号1)を、トリコデルマ・ビリデの既知の28S rDNAの塩基配列(DDBJ/GenBank/EMBL国際塩基配列データベース・アクセッション番号AY291123;配列番号2)と比較したところ、549塩基中548塩基が一致するという高い相同性(約99.8%の同一性)が示された(図1)。このことから、分離した菌はトリコデルマ・ビリデに極めて近縁であることが示唆された。
【0069】
トリコデルマ・ハルジアナムとトリコデルマ・ビリデの両種は形態学的に類似しているが、分生子の表面がトリコデルマ・ハルジアナムでは平滑であるのに対しトリコデルマ・ビリデでは粗面であること、トリコデルマ・ハルジアナムは35℃での生育が認められるがトリコデルマ・ビリデでは認められないことなどの相違点もある(奥田徹 著「真菌の分離と分類・同定(20)」(1992) p.157-166)。分離した菌は、分生子表面が平滑であることや35℃での生育が認められる点ではトリコデルマ・ハルジアナムと共通していると言える。
【0070】
なお、分離した菌には25℃、pH3.5の条件下で培養した際にその培養上清に強いセルラーゼ活性が認められるが、従来セルラーゼ生産が知られているトリコデルマ・ビリデNBRC31137株、トリコデルマ・リーセイ(Trichoderma reesei)NBRC31326株をそれと同じ条件で培養しても、培養上清にはセルラーゼ活性が全く認められなかった。
【0071】
以上を総合的に判断した結果、分離した菌は、トリコデルマ属(Trichoderma)の新菌種に属する菌株であるとして同定された。本発明者らは、この得られた菌を、トリコデルマ・エスピー(Trichoderma sp.)AH5株と命名した。トリコデルマ・エスピー(Trichoderma sp.)AH5株は、2006年3月8日付で、独立行政法人 製品評価技術基盤機構 特許微生物寄託センター(日本国千葉県木更津市かずさ鎌足2-5-8)に、受託番号NITE P-222で寄託された。
【0072】
[実施例3]
トリコデルマ・エスピー(Trichoderma sp.)AH5株の培養上清に含まれるエンドセルラーゼの反応pHを、トリコデルマ・ビリデ由来のセルラーゼ製剤「セルラーゼオノズカ」(ヤクルト工業社製)と比較した。このため、基質としてカルボキシメチルセルロース(以下、CMCと称する)を用いて酵素反応を行い、反応後CMCの分解によって生じた還元糖量をSomogyi-Nelson法により、グルコースを標準糖として定量した。
【0073】
具体的には、まず、トリコデルマ・エスピー(Trichoderma sp.)AH5株を、硫酸でpH3.5に調整した培地(0.2%硫安、0.01%塩化カリ、0.05%リン酸二水素カリウム、0.05%硫酸マグネシウム、0.03%塩化カルシウム、0.0001%酵母エキス、1%アビセル)に接種して25℃、100 rpmで2週間にわたり培養した。得られた培養物を20,000×gで10分間遠心分離して培養上清を採取した。この培養上清を、0.6%のCMC(Sigma社製)を含む様々なpH値の緩衝液に添加して反応液(全量50μl)を調製した。使用した緩衝液は、0.2Mグリシン-HCl緩衝液(pH2.0〜2.5)、0.1M酢酸ナトリウム緩衝液(pH3.0〜5.5)、0.1M PIPES緩衝液(pH6.4〜7.5)、0.1M Tris-HCl緩衝液(pH7.65)である。30℃で適当時間反応させた後、サーマルサイクラー(PCRマシーン)にて100℃で10分間加熱し、反応を停止させた。次いで50μlのSomogyi銅液(和光純薬工業社製など)を加え、さらにサーマルサイクラーで100℃で10分間加熱してから急速に冷却し、冷却後、50μlのNelson試薬(和光純薬社製など)を加えて還元銅沈殿を溶解して発色させ、30分静置した後、660nmでの吸光度を測定した。得られた測定値から、標準糖としたグルコースの吸光度に基づき、溶液中に含まれる還元糖量を算出した。「セルラーゼオノズカ」についても、同様に各種pH条件下でCMCと反応させて、溶液中に生じた還元糖量を測定した。培養上清中のタンパク質1mg当たりのエンドセルラーゼ活性を算出するため、Bradfordらの方法(Bradford M.M., Anal. Biochem., (1976) 72, p.248-254)に従い、牛血清アルブミンを標準タンパク質として、培養上清に含まれるタンパク質の量を測定した。エンドセルラーゼ活性は、以下の式で表した。
【0074】
エンドセルラーゼ活性(nmol/min/mg)=反応溶液中の還元糖量(nmol)/反応時間(分)/用いた培養上清中のタンパク質の量(mg)
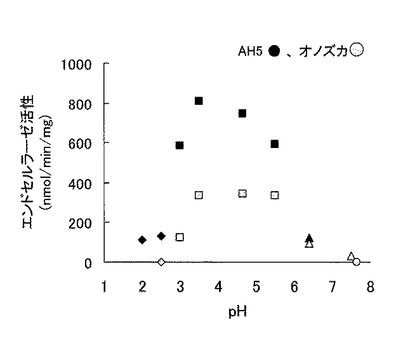
この結果を表1及び図2に示す。
【0075】
【表1】

【0076】
表1及び図2に示される通り、本発明に係るトリコデルマ・エスピーAH5株の培養上清は、従来製品「セルラーゼオノズカ」に比べて酸性域全体でより高いエンドセルラーゼ活性(比活性)を示した。またトリコデルマ・エスピーAH5株の培養上清は、pH2.0〜pH2.5という強酸性域でもエンドセルラーゼ活性を示した。
【0077】
さらに、上記で得られたトリコデルマ・エスピーAH5株の培養上清について、基質としてp-ニトロフェニルグルコシドを用い、βグルコシダーゼ活性を測定した。具体的には、AH5株の培養上清を、最終濃度0.1M p-ニトロフェニルグルコシド(和光純薬工業社製)を含む様々なpH値の緩衝液に添加して全量を50μlとした。使用した緩衝液は、0.1Mグリシン-HCl緩衝液(pH1.5〜2.8)、0.1M酢酸ナトリウム緩衝液(pH3.5〜4.65、5.5)、0.1M MES緩衝液(pH5.2)、0.1M PIPES緩衝液(pH6.4)、0.1M Tris-HCl緩衝液(pH7.65)である。30℃で適当時間反応させた後、反応液の9倍量の50mM炭酸ナトリウム溶液を加えることにより反応を停止させた。次いで、p-ニトロフェニルグルコシドの分解によって生じたその溶液中のp-ニトロフェノールを400nmでの吸光度測定により測定し、測定値に基づいてモル吸光係数を55.1μM/cmとしてp-ニトロフェノール量を算出した。「セルラーゼオノズカ」についても、同様に各種pH条件下でp-ニトロフェニルグルコシドと反応させて溶液中に生じたp-ニトロフェノール量を測定した。
【0078】
βグルコシダーゼ活性は、以下の式で表した。
βグルコシダーゼ活性(nmol/min/mg)=反応溶液中のp-ニトロフェノール量(nmol)/反応時間(分)/用いた培養上清中のタンパク質の量(mg)
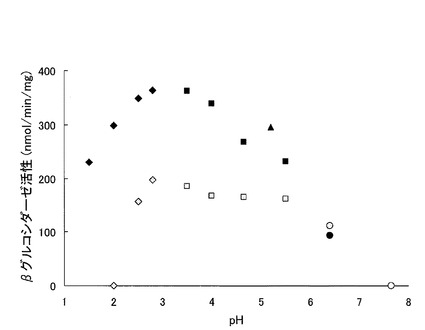
得られた結果を表2及び図3に示す。
【0079】
【表2】
【0080】
表2及び図3に示される通り、本発明に係るトリコデルマ・エスピーAH5株の培養上清は、従来製品「セルラーゼオノズカ」に比べて高いβグルコシダーゼ活性(比活性)を示すこと、またpH1.5〜pH2.5という強酸性域でもかなり高いβグルコシダーゼ活性を示すことが示された。
【0081】
[実施例4]
トリコデルマ・エスピーAH5株の培養上清が示すβグルコシダーゼ活性を、種々の市販の代表的なセルラーゼが示すβグルコシダーゼ活性と比較した。
【0082】
まず、実施例3と同様にして調製した培養上清を、比色レンジに入るよう濃度を適宜調整した後、1mM p-ニトロフェニルグルコシド(和光純薬工業社製)を含む0.1Mグリシン-HCl緩衝液(pH2.0)、又は0.1M酢酸ナトリウム緩衝液(pH4.0)に添加し、30℃で適当時間反応させた後、反応液の9倍量の50mM炭酸ナトリウム溶液を加えることにより反応を停止させた。次いで、実施例3と同様にして、溶液中のp-ニトロフェノールを400nmでの吸光度測定により定量した。対照実験としては、実施例3で使用した「セルラーゼオノズカ」に加え、アスペルギルス・ニガー(Aspergillus niger)由来のセルラーゼA「アマノ」3(天野エンザイム社製)、トリコデルマ・ビリデ由来のセルラーゼT「アマノ」4(天野エンザイム社製)、アスペルギルス・ニガー(Aspergillus niger)由来のセルラーゼ(東京化成工業社製)、トリコデルマ・リーセイ由来のセルラーゼ(Worthington Biochemical Corporation社製)を使用して、それぞれトリコデルマ・エスピーAH5株と同様にしてp-ニトロフェニルグルコシドと反応させ、溶液中に生じたp-ニトロフェノールを定量した。測定されたp-ニトロフェノール量に基づき、実施例3と同様にしてβグルコシダーゼ活性値を算出した。その結果を表3に示す。
【0083】
【表3】
【0084】
表3中、『AH5』はトリコデルマ・エスピーAH5株の培養上清、『オノズカ』は「セルラーゼオノズカ」、『セルA』はセルラーゼA「アマノ」3、『セルT』はセルラーゼT「アマノ」4、『A.niger(東京化成)』はアスペルギルス・ニガー由来のセルラーゼ(東京化成工業社製)、『T.reesei』はトリコデルマ・リーセイ由来のセルラーゼ(Worthington Biochemical Corporation社製)を表す。また『pH2』は、緩衝液として0.1Mグリシン-HCl緩衝液(pH2.0)を用いた場合、『pH4』は、緩衝液として0.1M酢酸ナトリウム緩衝液(pH4.0)を用いた場合を示す。
【0085】
表3に示すように、トリコデルマ・エスピーAH5株の培養上清は、pH2の条件下で298 nmol/min/mg、pH4の条件下で340 nmol/min/mgという高い比活性を示し、かつ、pH2の条件下でさえpH4で示したβグルコシダーゼ活性の88%を保持していた。一方、市販の各種セルラーゼは、pH2では全く活性を示さないかあるいは10 nmol/min/mg未満というごく低い活性を示すのみだった。このようにトリコデルマ・エスピーAH5株の培養上清は、従来品の各種セルラーゼに比べて高いβグルコシダーゼ比活性を示し、またpH2.0の条件下でも非常に高い活性を有していた。このことは、本発明のトリコデルマ・エスピーAH5株及びその培養上清が、グルコースの高効率な産生、特に強酸性条件下でのグルコースの高効率な産生に好適に利用できることも示す。
【0086】
[実施例5]
トリコデルマ・エスピーAH5株の培養上清が示すセロヘキサオースに対する分解活性を、「セルラーゼオノズカ」(ヤクルト工業社製)と比較した。
【0087】
実施例3と同様にして調製したトリコデルマ・エスピーAH5株の培養上清を、終濃度4mMのセロヘキサオース(生化学工業社製)を含む終濃度40mMの酢酸ナトリウム緩衝液(pH3.0)に、終濃度0.16mg/mlとなる量で添加し、30℃で適当時間反応させた。同様に、「セルラーゼオノズカ」は、終濃度4mMのセロヘキサオース(生化学工業社製)を含む終濃度40mMの酢酸ナトリウム緩衝液(pH3.0)に、終濃度0.2mg/mlとなる量で添加し、30℃で反応させた。反応時間は15分、1時間、2時間、6時間、24時間とした。反応後、Merck社製シリカゲルプレート(5cm×10cm)に、反応液0.5mlをスポットし、80%アセトニトリルにより展開した。標品としてはセロオリゴ糖(生化学工業製)をスポットした。展開後、シリカゲルプレートを硫酸/メタノール混合溶液(容量比5:95)中に浸漬し、110℃で5分〜10分間加熱処理することにより発色させた。
【0088】
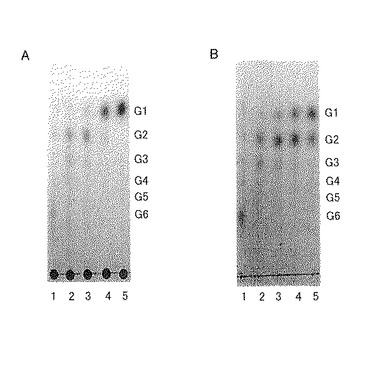
この結果を図4に示す。図4中、G1はグルコース、G2はセロビオース(βグルコース分子2個がβ-1,4結合した二糖類)、G3〜G6はそれぞれ3〜6個のグルコース分子がβ-1,4結合したセロオリゴ糖である。反応時間は、レーン1:15分、レーン2:1時間、レーン3:2時間、レーン4:6時間、レーン5:24時間である。
【0089】
図4に示すように「セルラーゼオノズカ」を用いた場合(図4B)に比べ、トリコデルマ・エスピーAH5株の培養上清を用いた場合(図4A)の方が、グルコースの生産量が格段に多いことが示された。さらに「セルラーゼオノズカ」では24時間反応させた後でもグルコースに加えてセロビオース(G2)も比較的多量に産生されたのに対し、トリコデルマ・エスピーAH5株の培養上清を用いた場合は、6時間反応後にはセロビオース(G2)はあまり産生されなくなり、代わりにグルコースが多量に産生されるようになった。このことは、トリコデルマ・エスピーAH5株を用いれば、グルコースを優先的に高効率で産生できることを示している。
【0090】
[実施例6]
様々なpH値を示す液体培地Aを含むろ紙上でのトリコデルマ・エスピーAH5株の生育能を、セルラーゼを生産することが知られているトリコデルマ・ビリデNBRC 31137株[独立行政法人製品評価技術基盤機構バイオテクノロジー本部生物遺伝資源部門(NBRC)(千葉県木更津市かずさ鎌足2-5-8)からカタログ番号NBRC 31137にて分譲を受けたもの]、及びトリコデルマ・リーセイNBRC 31326株[前記NBRCからカタログ番号NBRC 31326にて分譲を受けたもの]と比較した。この生育試験により、ろ紙に含まれるセルロースを基質として酸性条件下で生育する能力を調べた。
【0091】
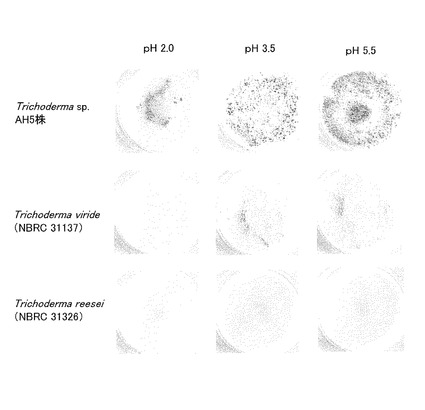
まず、0.2g硫酸アンモン、0.01g塩化カリウム、0.05gリン酸二水素カリウム、0.05g硫酸マグネシウム、0.001g硝酸カルシウム、及び0.0001g酵母エキス(DIFCO社製)を適量の脱塩水に溶解し、5%硫酸又は濃水酸化ナトリウム溶液を用いてpHを2.0、3.5、5.5にそれぞれ調整した後、さらに脱塩水を添加して100mlとし、オートクレーブ滅菌することにより、3種のpH値の液体培地Aを調製した。この液体培地Aをそれぞれ、別個のろ紙(B200G047A、東洋ろ紙社製)に染み込ませて3種のろ紙培地を作製した。次に各菌株の胞子約10,000個を、作製したろ紙培地に接種し、25℃で10日間静置培養した。10日間培養した後の各菌株の生育結果を図5に示す。
【0092】
図5に示される通り、トリコデルマ・エスピーAH5株は、pH2.0、3.5、5.5のいずれの液体培地Aを含むろ紙培地でも良好に生育し、胞子形成が容易に観察された(図5最上段)。一方、トリコデルマ・ビリデNBRC 31137株とトリコデルマ・リーセイNBRC 31326株は、pH2.0の液体培地Aを含むろ紙培地ではほとんど生育が認められず(図5最左列の中段及び下段)、pH3.5及び5.5ではわずかに菌糸の生長が認められるが胞子形成はほとんど認められなかった。特にトリコデルマ・リーセイNBRC 31326株は、いずれのpH条件でも生育状態がかなり悪かった。
【0093】
この生育試験の結果から、トリコデルマ・エスピーAH5株は、セルラーゼの生産が知られているトリコデルマ・ビリデNBRC 31137株とトリコデルマ・リーセイNBRC 31326株と比べて酸性条件での生育能が非常に高いこと、特にpH2.0のような強酸性条件下でも生育できることが示された。すなわちトリコデルマ・エスピーAH5株は、酸性条件下、特に少なくともpH2.0以上pH5.5以下のpH範囲では、高いセルロース分解活性を示すことが明らかになった。
【産業上の利用可能性】
【0094】
本発明のトリコデルマ属菌は、常温の酸性条件下、特に常温かつ強酸性条件下で、セルロース系物質を分解するために用いることができる。本発明のトリコデルマ属菌を用いた糖類製造法は、セルロースを材料とした糖類の効率的な製造に利用することができる。さらに本発明のトリコデルマ属菌と有機酸生産菌を用いた有機酸製造方法は、1つの反応系でセルロースを原料として有機酸を効率よく製造するために使用することができる。
【図面の簡単な説明】
【0095】
【図1】図1は、トリコデルマ・エスピーAH5株とトリコデルマ・ビリデの28S rDNAの塩基配列のアラインメントを示す図である。
【図2】図2は、トリコデルマ・エスピーAH5株の培養上清の、様々なpH条件下でのエンドセルラーゼ活性を示すグラフである。ダイヤ印はグリシン-HCl緩衝液、四角印は酢酸ナトリウム緩衝液、三角印はPIPES緩衝液、丸印はTris-HCl緩衝液を示す。黒塗りつぶし印がトリコデルマ・エスピーAH5株の培養上清、灰色塗りつぶし印が対照サンプルの「セルラーゼオノズカ」を示す。
【図3】図3は、トリコデルマ・エスピーAH5株の培養上清の、様々なpH条件下でのβグルコシダーゼ活性を示すグラフである。ダイヤ印はグリシン-HCl緩衝液、四角印は酢酸ナトリウム緩衝液、三角印はMES緩衝液、丸印はPIPES緩衝液、白丸印はTris-HCl緩衝液を示す。黒塗りつぶし印がトリコデルマ・エスピーAH5株の培養上清、灰色塗りつぶし印が対照サンプルの「セルラーゼオノズカ」を示す。
【図4】図4は、トリコデルマ・エスピーAH5株の培養上清(0.16mg/L)(図4A)と「セルラーゼオノズカ」(0.2mg/L)(図4B)のセロヘキサオース分解活性を示すクロマトグラフィーの結果を表す写真である。レーン1:15分、レーン2:1時間、レーン3:2時間、レーン4:6時間、レーン5:24時間(反応時間)。G1:グルコース、G2:セロビオース(βグルコース分子2個がβ-1,4結合した二糖類)、G3〜G6:それぞれ3〜6個のグルコース分子がβ-1,4結合したセロオリゴ糖。
【図5】図5は、トリコデルマ・エスピーAH5株、トリコデルマ・ビリデNBRC31137株、トリコデルマ・リーセイNBRC31326株の酸性条件下での生育試験の結果を示す写真である。
【技術分野】
【0001】
本発明は、強酸性条件下でセルロース分解活性を示す微生物及びその利用に関する。
【背景技術】
【0002】
近年、石油資源の枯渇、地球レベルの炭酸ガス発生量の削減が叫ばれており、今後石油価格の高騰が予想される。自然界に大量に存在しているセルロースをエタノール等の液体燃料、ポリ乳酸等の石油代替樹脂原料に変換することができれば、炭酸ガス発生量を増やすことなく安価に石油代替資源として用いることができる。また、セルロースから乳酸、コハク酸等の有機酸を合成することができれば石油代替のバイオ樹脂などを作ることができ、有用である。
【0003】
微生物を利用した有機酸類の生産は従来から行われており、ラクトバチルス属(Lactobacillus)菌やリューコノストック属(Leuconostoc)菌による乳酸生産、ペニシリウム属(Penicillium)菌によるクエン酸生産、グルコノバクター属(Gluconobacter)菌によるグルコン酸生産、アセトバクター属(Acetobacter)菌による酢酸生産などが知られている。この他、イタコン酸、フマル酸、コハク酸,マレイン酸、コウジ酸、酒石酸などの醗酵生産も行われてきている。最近では、組み換え大腸菌による乳酸やコハク酸の醗酵生産、組み換え酵母による乳酸醗酵生産も報告されている。しかしながら、これらの微生物が直接、セルロースから有機酸を合成したという報告例はない。一方、近年、セルラーゼ生産菌を用いてセルロースを加水分解して糖化し、生じた糖を乳酸菌などによる醗酵生産に利用することによって、乳酸などの有機酸の生産を促進する方法が開発されている(特許文献1及び非特許文献1〜4)。但しこの方法では、生産された有機酸によって培地のpHが徐々に低下し、セルロース分解活性が阻害されるため、有機酸生産を継続的に行うにはpH緩衝剤を使用してpHの低下を防ぐ必要がある。
【0004】
自然界ではセルロースは主として微生物によって分解される。細菌や糸状菌などの様々な微生物が各種セルロース分解酵素を生産することが知られている。セルロースは作用機構の異なる複数のセルロース分解酵素の協同作用により、セロオリゴ糖、セロビオースを経てグルコースへと分解される。
【0005】
セルロース分解酵素(セルラーゼ)に関しては従来から非常に多くの研究がなされている。セルラーゼ生産菌はトリコデルマ属(Trichoderma)菌、フザリウム属(Fusarium)菌、トレメテス属(Tremetes)菌、ペニシリウム属(Penicillium)菌、フミコーラ属(Humicola)菌、アクレモニウム属(Acremonium)菌、アスペルギルス属(Aspergillus)菌等の糸状菌の他に、クロストリジウム属(Clostridium)菌、シュードモナス属(Pseudomonas)菌、セルロモナス属(Cellulomonas)菌、ルミノコッカス属(Ruminococcus)菌、バチルス属(Bacillus)菌等の細菌、スルフォロバス属(Sulfolobus)菌等の始原菌、さらにストレプトマイセス属(Streptomyces)菌、サーモアクチノマイセス属(Thermoactinomyces)菌の放射菌で見出されている。最近では、これらの微生物を利用して、洗剤との併用を念頭においたアルカリセルラーゼの開発が行われている。しかし、これらの微生物が生産するセルラーゼは弱酸性からアルカリ性にかけての反応pHを持つものがほとんどであり、強酸性条件下でも活性を示すセルラーゼを生産する微生物はほとんどない。
【0006】
酸性セルラーゼを生産する微生物として、スルフォロバス・ソルファタリカス(Sulfolobus solfataricus)菌株が報告されている(非特許文献5)。同菌は至適温度が80℃、至適pH1.8のセルラーゼを菌体外に分泌生産する。しかしながら、同菌は始原菌であり生育が悪い上、生産されるセルラーゼも至適温度が80℃と高く常温ではごく弱い活性しか示さない。従ってこの酸性セルラーゼは、常温でセルロースを加水分解する目的ではあまり有用でない。他にも酸性セルラーゼの報告はあるが、常温の強酸性条件下で十分な活性を示すセルラーゼは知られていない(特許文献2〜4)。
【0007】
【特許文献1】特開2005−13131号公報
【特許文献2】特開平3−240491号公報
【特許文献3】特開平6−38747号公報
【特許文献4】特開2001−505414号公報
【非特許文献1】Iyer P.V. and Lee Y.Y., Biotechnology Letters (1999) 21: p.371-373
【非特許文献2】Xu D.B. et al., Appl. Microbiol. Biotechnol. (1989) 30, p.553-558
【非特許文献3】Kim D.M. et al., Biotechnol. Bioeng. (1992) 39 p.336-342
【非特許文献4】Moresi. M. et al., Appl. Microbiol. Biotechnol. (1991) 36, p.35-39
【非特許文献5】Huang Y. et al., Biochem J. (2005) 385 p.581-588
【発明の開示】
【発明が解決しようとする課題】
【0008】
本発明は、強酸性条件下でセルロース分解活性を示す微生物を提供することを目的とする。
【課題を解決するための手段】
【0009】
本発明者らは、上記課題を解決するため鋭意検討を重ねた結果、pH2.0のような強酸性域でも常温で活性を有するセルラーゼを培養上清に分泌するトリコデルマ属菌の分離に成功し、それに基づいて本発明を完成するに至った。
【0010】
すなわち、本発明は以下を包含する。
[1] 酸性条件下でセルロース分解活性を示すトリコデルマ(Trichoderma)属菌であって、少なくともpH2.0〜2.5のpH範囲でエンドセルラーゼ活性及びβグルコシダーゼ活性を示す、前記トリコデルマ属菌。
[2] トリコデルマ・エスピー(Trichoderma sp.)AH5株(受託番号NITE P-222)又はその変異株である、上記[1]に記載のトリコデルマ属菌。
[3] 少なくともpH2.0〜2.5のpH範囲でエンドセルラーゼ活性及びβグルコシダーゼ活性を示す、上記[1]又は[2]に記載のトリコデルマ属菌由来の培養上清。
[4] 上記[1]若しくは[2]に記載のトリコデルマ属菌又は上記[3]に記載の培養上清を用いてセルロースを分解することを特徴とする、糖類の製造方法。
[5] セルロースの分解をpH2.5以下の条件下で行う、上記[4]に記載の方法。
[6] 糖類が、セロオリゴ糖、セロビオース、及びグルコースからなる群より選択される少なくとも1つである、上記[4]又は[5]に記載の方法。
[7] 上記[1]若しくは[2]に記載のトリコデルマ属菌又は上記[3]に記載の培養上清を用いてセロオリゴ糖を分解することを特徴とする、セロオリゴ糖からセロビオース及びグルコースを製造する方法。
[8] セロオリゴ糖の分解をpH2.5以下の条件下で行う、上記[7]に記載の方法。
[9] 上記[1]若しくは[2]に記載のトリコデルマ属菌又は上記[3]に記載の培養上清を用いてβグルコシドを分解することを特徴とする、βグルコシドからアグリコン及びグルコースを製造する方法。
[10] βグルコシドの分解をpH2.5以下の条件下で行う、上記[9]に記載の方法。
[11] 上記[1]又は[2]に記載のトリコデルマ属菌と有機酸生産菌とをセルロース存在下で共存培養することを特徴とする、有機酸の製造方法。
[12] 上記[3]に記載の培養上清とセルロースとを含む培地で有機酸生産菌を培養することを特徴とする、有機酸の製造方法。
[13] 有機酸生産菌が乳酸菌である、上記[11]又は[12]に記載の方法。
【発明の効果】
【0011】
本発明のトリコデルマ属菌は、常温の強酸性条件下でもセルロース分解活性を示す。本発明のトリコデルマ属菌を用いた糖類製造法は、生産効率がよく、さらに雑菌汚染がより少ない強酸性条件下で実施することができて有用である。
【発明を実施するための最良の形態】
【0012】
以下、本発明を詳細に説明する。
【0013】
1.セルロース及びその加水分解酵素であるセルラーゼ
セルロースは、グルコースがβ-1,4-グルコシド結合(β-1,4結合)により高度に重合した高分子から構成される物質である。セルロースは、天然では植物細胞壁の主たる構成成分として存在し、多糖としては地球上で最も多く生産されている。セルロース分子におけるグルコースの重合度は、その起源によって異なるが、木材では6,000〜10,000、綿では8,000〜10,000程度であり、藻類由来のものでは44,000というものもある。セルロース分子の隣接するグルコース残基間においては分子内水素結合が形成されるが、さらに、隣接するセルロース分子鎖同士も分子間水素結合を形成し、強固なセルロース繊維を形成する。セルロース繊維は強固な結晶構造をとる領域を高割合で含むため、セルロースをグルコースなどの単糖又は少糖まで加水分解(糖化)するのは容易ではない。自然界ではセルロースは主として微生物によって分解されており、細菌や糸状菌などの様々な微生物がセルロース分解酵素を生産することが知られている。これらの微生物は菌体外に複数のセルロース分解酵素を分泌し、セルロースはそれらの作用機構の異なる各種セルロース分解酵素の協同作用により、主に、セロオリゴ糖、セロビオースを経てグルコースへと分解される。セルロース分解酵素は、一般に、セルロースを加水分解する酵素の総称としてセルラーゼとも呼ばれている。
【0014】
セルラーゼの種類としては、現在、狭義のセルラーゼ(EC 3.2.1.4)、グルカン1,4-βグルコシダーゼ(EC 3.2.1.74)、セルロース1,4-βセロビオシダーゼ(EC 3.2.1.91)、βグルコシダーゼ(EC 3.2.1.21)の4つが主に認められている。
【0015】
狭義のセルラーゼ(EC 3.2.1.4)は、エンド型の作用様式で(すなわち分子鎖内部で)セルロースを加水分解し、セロオリゴ糖、セロビオース、及びグルコースを生産する酵素である。狭義のセルラーゼ(EC 3.2.1.4)はカルボキシメチルセルロース(CMC)などの非結晶性セルロースをよく加水分解することが知られている。狭義のセルラーゼ(EC 3.2.1.4)は、カルボキシメチルセルラーゼ、エンド1,4-βグルカナーゼ、エンドセルラーゼなどとも呼ばれている。一方、グルカン1,4-βグルコシダーゼ(EC 3.2.1.74)及びセルロース1,4-βセロビオシダーゼ(EC 3.2.1.91)は、セルロース鎖をエキソ型の作用様式で加水分解し、末端からグルコース又はセロビオースを遊離する酵素である。これらの酵素は綿繊維やアビセルなどの結晶性セルロースをよく加水分解し、アビセラーゼ又はエキソセルラーゼとも呼ばれている。さらにβグルコシダーゼ(EC 3.2.1.21)は、グルカン1,4-βグルコシダーゼ及びセルロース1,4-βセロビオシダーゼの作用によってセルロースから生成されるような、セロオリゴ糖、セロビオース、及びβグルコシド(グルコースの配糖体)に作用し、非還元末端からグルコースを遊離する酵素である。セロビオースを特によく加水分解するβグルコシダーゼは、セロビアーゼと呼ばれる。
【0016】
既存のセルラーゼは、弱酸性からアルカリ性にかけて安定なpH反応性を有するものがほとんどである。本明細書では、セルロース分解酵素の総称として用語「セルラーゼ」を使用し、それと区別するため、狭義のセルラーゼ(EC 3.2.1.4)を「エンドセルラーゼ」と呼ぶこととする。
【0017】
2.本発明に係るトリコデルマ属の新菌種
本発明者らは、強酸性のpH範囲(強酸性条件下)でも生育しセルロース分解活性を示すトリコデルマ属に属する菌を分離することに成功した。この菌は、既存のセルラーゼとは違って広範な酸性条件下で強力な活性を示し、かつ常温の強酸性条件下でも十分な活性を示す各種セルラーゼを、菌体外に産生する。この菌は、その菌学的性質に基づき、トリコデルマ属の新菌種として同定された。本発明は、こうして同定されたトリコデルマ属新菌種に属する菌及びその利用に関する。なお本発明において「酸性条件」とは、pH0以上pH7.0未満の範囲内のpHを示すことを言う。さらに本発明において「強酸性」とは、pH3.0以下、特に微生物の生育に関してはpH2.0以上pH3.0以下のpH範囲を意味するものとする。さらに本発明において「常温」とは、20℃以上30℃以下の温度範囲を言うものとする。
【0018】
本発明者らが分離した菌株は、トリコデルマ・エスピー(Trichoderma sp.)AH5株と命名され、2006年3月8日付で、独立行政法人 製品評価技術基盤機構 特許微生物寄託センター(日本国千葉県木更津市かずさ鎌足2-5-8)に、受託番号NITE P-222で寄託された。トリコデルマ・エスピーAH5株は、以下、「AH5株」と略記することがある。
【0019】
本発明に係るトリコデルマ・エスピーAH5株の菌学的性質は、以下の通りである。なお、コロニー色調に関する記述はKornerup and Wanscher (1978) "Methuen handbook of colour" 3rd ed.(Eyre Methuen, London, UK)に従っている。
【0020】
(a)培養的性質:
(1)ポテトデキストロース寒天培地(pH5.6)における生育:25℃で3日間培養後、コロニーの成長が認められ、気中菌糸の形成が顕著に見られる。菌糸の色は白色〜クリーム色である。培養開始後7〜10日で深緑色の分生子を形成する。9日間培養後のコロニーの表面性状は綿状(cottony)、コロニー色調はクリーム色及び濃緑色。テレオモルフの形成は認められない。
(2)オートミール寒天培地(pH6.0)における生育:25℃で3日間培養後、コロニーの成長が認められ、気中菌糸の形成が顕著に見られる。菌糸の色は白色〜クリーム色である。培養開始後7〜10日で深緑色の分生子を形成する。9日間培養後のコロニーの表面性状は綿状(cottony)、コロニー色調は白色と深緑色。テレオモルフの形成は認められない。
(3)2%麦芽エキス寒天培地(pH6.0):25℃で3日間培養後、コロニーの成長が認められる。培養開始後7〜10日で深緑色の分生子を形成する。9日間培養後のコロニーの表面性状は綿状(cottony)、コロニー色調は深緑色。テレオモルフの形成は認められない。
【0021】
(b)形態学的性質:
栄養菌糸:有隔壁菌糸を形成する。菌糸表面は平滑(smooth)。栄養菌糸の先端部に球形〜亜球形の厚膜胞子の形成が認められる。
分生子柄:集合して羊毛状の塊(tuft)を形成するのが認められる。分岐し、輪生する。
分生子(胞子):フィアロ型分生子を形成する。連鎖せず、緑色の分生子塊を形成する。分生子の形状は卵形〜亜球形。単細胞。分生子表面は平滑(smooth)。
(c)最適生育条件:20〜25℃、pH 3〜pH 5
(d)生育可能範囲:10〜35℃、pH 2〜pH 6
(e)28S rDNA(28SリボソームRNAコード配列)におけるDNA配列相同性:
トリコデルマ・ビリデ(Trichoderma viride)の28S rDNA(配列番号2)との間で99.8%程度の同一性を有する。
(f)酸素に対する性質:好気性
(g)その他の特徴:強酸性条件を含む幅広い酸性条件下でセルロース分解活性を示す。特に、生育可能範囲の温度条件において、少なくともpH2.0〜pH2.5を含むpH範囲で、エンドセルラーゼ活性及びβグルコシダーゼ活性を示す。
【0022】
本発明に係るトリコデルマ属新菌種は、トリコデルマ・エスピーAH5株の上記のような菌学的性質を本質的に共有する。このような本発明に係るトリコデルマ属菌の典型例はトリコデルマ・エスピーAH5株であるが、AH5株の分離源である秋田県鹿角市八幡平の酸性温泉蒸の湯の源泉中から、上記のような菌学的性質を有する菌を後述の実施例の記載に従ってさらに分離することにより、上記トリコデルマ属菌の菌株をさらに得ることもできる。また、本発明に係るトリコデルマ属菌には、例えばトリコデルマ・エスピーAH5株の変異体(自然突然変異体、遺伝子組換え体、突然変異誘発処理体、プラスミド導入等による形質転換体、倍数化体など)、トリコデルマ・エスピーAH5株を親株の1つに用いて作製した細胞融合株、トリコデルマ・エスピーAH5株を親株の1つとして交配により作製した菌なども含まれる。
【0023】
本発明に係るトリコデルマ属菌を培養するのに適した培地としては、限定するものではないが、例えば、ポテトデキストロース寒天培地が挙げられる。ポテトデキストロース寒天培地は、ポテト抽出物200g、デキストロース20g、アガー15gを水1Lに溶解し(滅菌前のpH値:5.6)、それを121℃で15分オートクレーブ滅菌することにより、調製することができる。本発明に係るトリコデルマ属菌は、このような培地において25℃にて好気的に培養することができる。
【0024】
本発明に係るトリコデルマ属菌がセルロース分解活性を示すかどうかは、具体的には例えば、本発明のトリコデルマ属菌を、セルロース以外の炭素源を含まない培地で培養し、菌が生育するかどうかを調べることによって判定することができる。菌の生育が認められた場合、その菌はセルロースを分解することにより糖類を得たと考えられるので、すなわちセルロース分解活性(セルロース分解能)を有すると判定できる。この試験は、例えば、炭素源を含まない液体培地を染み込ませたろ紙の上で本発明に係るトリコデルマ属菌を培養することによって行うこともできる。この試験において本発明のトリコデルマ属菌が、特定のpH条件下でセルロース分解活性を示すかどうかを調べるためには、その特定のpH値に調整した培地を使用し、培養温度や湿度などの他の条件については本発明のトリコデルマ属菌における通常の生育条件(例えば、25℃)を用いることが好ましい。
【0025】
あるいは、本発明のトリコデルマ属菌が、セルロース分解活性を示すかどうかは、そのトリコデルマ属菌由来の培養上清について各種セルラーゼの活性を検出することによって判定することもできる。これは、本発明に係るトリコデルマ属菌のセルロース分解活性が、その菌が生産し菌体外に分泌した各種セルラーゼの作用によって得られることに基づく。この方法では、例えば、本発明のトリコデルマ属菌由来の培養上清をエンドセルラーゼの基質となるカルボキシメチルセルロースと反応させ、カルボシキメチルセルロースの分解によって生じる還元糖の量を測定することにより、本発明のトリコデルマ属菌がエンドセルラーゼ活性を示すか否かを判定することができる。還元糖量の定量法としてはSomogyi法、Tauber-Kleiner法、Hanes法(滴定法)、Park-Johnson法、3,5-ジニトロサリチル酸(DNS)法などの多数の定量法が知られているが、好適な1つの方法として、糖による銅イオンの還元を利用するSomogyi-Nelson法を用いることができる(福井作蔵 著「生物化学実験法1 還元糖の定量法 第2版」学会出版センター 1990年)。Somogyi-Nelson法のプロトコールの一例では、まず酵素反応液を100℃で10分加熱処理して反応を停止させ、その反応液と等量のSomogyi銅液(和光純薬社製など)を加えて混合し、100℃で10分加熱処理してから急速に冷却し、冷却後、等量のNelson試薬(和光純薬社製など)を加えて還元銅沈殿を溶解して発色させ、30分静置し、660nmでの吸光度を測定し、その測定値から、グルコースを標準糖として還元糖量を算出する。さらに上記方法では、本発明のトリコデルマ属菌由来の培養上清をβグルコシダーゼの基質となるp-ニトロフェニルグルコシドと反応させ、p-ニトロフェニルグルコシドの分解によって生じるp-ニトロフェノールを、400nmでの吸光度測定により定量することにより、本発明のトリコデルマ属菌がβグルコシダーゼ活性を示すか否かを判定してもよい。さらには、本発明のトリコデルマ属菌由来の培養上清をセロヘキサオース(六糖のセロオリゴ糖)と反応させ、セロヘキサオースの分解によって生じる糖類の量を測定することにより、本発明のトリコデルマ属菌がセルラーゼ活性を示すか否かを判定してもよい。
【0026】
本発明のトリコデルマ属菌は、後述の実施例に記載の通り、少なくともpH2.0〜pH5.5を含む酸性条件下の培地でセルロースを分解することができ、かつその培養上清においてpH2.0以上の酸性pH範囲でエンドセルラーゼ活性を、pH1.5以上の酸性pH範囲でβグルコシダーゼ活性を示す。本発明のトリコデルマ属菌は、広範な酸性pH範囲で高いセルロース分解活性を示すだけでなく、既知セルラーゼでは十分な活性が認められない強酸性条件下、特にpH2.0〜2.5のpH条件下でも、エンドセルラーゼ活性及びβグルコシダーゼ活性等のセルロース分解活性を示すことができる。
【0027】
3.本発明のトリコデルマ属菌を用いたセルロース系物質の分解
上記のような本発明に係るトリコデルマ属菌を用いれば、セルロース、セルロース部分分解物(セロオリゴ糖、セロビオース、βグルコシドなど)、そして植物細胞壁(ヘミセルロース、ペクチン質、リグニンなどに結合したセルロースによって構成される)などのセルロース系物質を、酸性条件下で効率よく分解することができる。本発明は、本発明に係るトリコデルマ属菌又はその培養上清を用いて、セルロース系物質から糖類を製造する方法にも関する。
【0028】
本発明において「セルロース」とは、グルコースがβ-1,4-グルコシド結合により質量平均重合度51以上(限定するものではないが、好ましくは質量平均重合度100以上、より好ましくは質量平均重合度400〜50,000)で重合した分子鎖若しくはその誘導体(例えばカルボキシメチルセルロースなどの、カルボキシメチル化、アルデヒド化、若しくはエステル化などの誘導体化が為されたもの)、又はそれらが複数結合したもの(セルロース繊維など)を言う。セルロースは、アビセルや綿繊維などの結晶性セルロースであってもよいし、非結晶性セルロースであってもよい。セルロースはまた、天然由来のものでも、人為的に合成したものでもよい。さらにセルロースは、植物由来のものでも、真菌由来のものでも、細菌由来のものでもよい。
【0029】
本発明において「セロオリゴ糖」とは、グルコースがβ-1,4結合により重合した少糖類(重合度3〜50)又はその誘導体を言う。セロオリゴ糖の具体例として、セロペンタオース、セロトリオース、セロヘキサオース、セロテトラオースなどが挙げられる。
【0030】
本発明において「βグルコシド」とは、グルコースが非糖化合物(アグリコン)と結合した配糖体を言う。βグルコシドの具体例としては、p-ニトロフェニルグルコシド、グリチルリチン酸、ステビオシド、フラボノイド配糖体、アルブチン、サポニンなどが挙げられる。なおp-ニトロフェニルグルコシドは、βグルコシダーゼの作用により、アグリコンに相当するp-ニトロフェノールと、グルコースとに分解される。
【0031】
上述のような本発明に係る糖類製造方法の1つの態様として、本発明に係るトリコデルマ属菌をセルロース存在下で培養してセルロースを加水分解することにより、セロオリゴ糖、セロビオース又はグルコースなどの糖類をセルロースから遊離させることを含む、セルロースから糖類を製造する方法がある。ここで「セルロース存在下」とは、本発明のトリコデルマ属菌を培養する培地中に、セルロース又はセルロース含有物質が添加されていることを意味する。セルロース含有物質としては、限定するものではないが、綿や麻などの天然繊維品、レーヨン、キュプラ、アセテート、リヨセルなどの再生繊維品、稲わら、籾殻、木材チップなどの農産廃棄物などが挙げられる。
【0032】
本発明の製造方法の別の態様としては、本発明に係るトリコデルマ属菌をセロオリゴ糖存在下で培養してセロオリゴ糖を加水分解することにより、セロオリゴ糖からセロビオース又はグルコースなどの糖類を遊離させることを含む、セロオリゴ糖からセロビオース又はグルコースを製造する方法が挙げられる。ここで「セロオリゴ糖存在下」とは、本発明のトリコデルマ属菌を培養する培地中にセロオリゴ糖が添加されていることを意味する。
【0033】
本発明の製造方法のさらに別の態様としては、本発明に係るトリコデルマ属菌をβグルコシド存在下で培養することにより、βグルコシドをアグリコンとグルコースとに加水分解することを含む、βグルコシドからアグリコン及びグルコースを製造する方法が挙げられる。ここで「βグルコシド存在下」とは、本発明のトリコデルマ属菌を培養する培地中にβグルコシドが添加されていることを意味する。
【0034】
これらの態様において用いる培養培地としては、上記のようなセルラーゼの基質(例えばセルロース、セルロース含有物質、セロオリゴ糖、又はβグルコシドなど)を加える限り、トリコデルマ属に属する真菌の培養に使用可能な任意の培地を用いることができる。培地の組成は、例えば、「微生物の分離法」(山里一英ら編、株式会社R&Dプランニング発行、2001年7月6日(1986年初版発行))の記載を参考として決定することができる。培地に含める炭素源としては、上記のセルラーゼの基質(セルロース、セルロース含有物質、セロオリゴ糖、又はβグルコシドなど)に加えて、スクロース、フルクトース、マンニトール、ソルビトール、ガラクトース、マルトース、エリスリット、グリセリン、エチレングリコール、エタノール、澱粉、ビート搾汁、サトウキビ搾汁、ビートモラセス等を用いてもよい。培地に含める窒素源としては、硫酸アンモニウム、塩化アンモニウム等のアンモニウム塩、硝酸カルシウムなどの硝酸塩、又はその他の有機若しくは無機窒素源を用いることができる。さらに培地には、ペプトン、酵母エキス、大豆加水分解物などの天然栄養源を含めることも好ましい。培地には、抗生物質、pH緩衝剤(例えば炭酸カルシウム、リン酸アンモニウム、水酸化ナトリウムなど)、マーカー物質などを含めてもよい。培地は液体培地であってもよいし、固体培地であってもよい。上記のようなセルラーゼの基質を培地に加える量は、特に限定されないが、液体培地で培養する場合には1L当たり1g〜100g程度用いればよい。好適な培地の例としては、0.2g硫酸アンモン、0.01g塩化カリウム、0.05gリン酸二水素カリウム、0.05g硫酸マグネシウム、0.001g硝酸カルシウム、及び0.0001g酵母エキスを脱塩水に溶解して得られる溶液に、上記のようなセルラーゼの基質を加えたものが挙げられる。セルラーゼの基質は、培養中、適宜補充することがより好ましい。
【0035】
用いる培地のpH値は、培養開始前に、本発明に係るトリコデルマ属菌の生育可能範囲内にあればよい。本発明の方法では、好ましくはpH1.0〜pH7.0、より好ましくはpH2.0〜6.4の酸性pH値を示す培地を用いることが望ましい。本発明の方法では、とりわけpH2.5〜pH5.5の範囲内のpH値を示す培地の使用が、糖類を効率よく生産する上でより好ましい。またpH2.5以下、特にpH2.0〜pH2.5の範囲内のpH値を示す培地は、他の雑菌の混入を抑制する上でより好適に使用される。なお本発明の上記製造方法において、この培地のpH値が、トリコデルマ属菌の培養を利用してセルロース、セロオリゴ糖、βグルコシド等の分解反応を実施する際のpH条件に相当する。
【0036】
培養温度は、本発明に係るトリコデルマ属菌の生育可能範囲内の温度であればよい。特に好ましい培養温度は、20℃〜35℃、より好ましくは25℃〜30℃、最も典型的な培養温度は25℃である。培養は、好気的条件下で行うことが好ましい。
【0037】
培養時間は、当業者であれば任意に設定することができるが、少なくとも24時間培養を継続することが好ましい。
【0038】
以上のようにして本発明に係るトリコデルマ属菌を上記のようなセルラーゼの基質の存在下で培養すると、その基質が加水分解されて生じた糖類等(低分子化セルロース、セロオリゴ糖、セロビオース、グルコース、βグルコシド、アグリコンなど)が培養物中に生産される。本発明においては、そのようにして生産された糖類等を、培養物(特に培養液)から、HPLC法、アルコール沈殿法、結晶化法などの当業者に公知の方法により精製することができる。あるいは、その培養液から菌体を除去して培養上清を調製し、それを糖類等を豊富に含む溶液として利用することもできる。
【0039】
本発明の上記製造方法のまた別の態様として、本発明に係るトリコデルマ属菌を好ましくは液体培地で培養しその菌体を除去して得られる培養上清を、セルラーゼの基質となるセルロースと反応させ、セルロースを加水分解することにより、セロオリゴ糖、セロビオース又はグルコースなどの糖類をセルロースから遊離させることを含む、セルロースから糖類を製造する方法が挙げられる。
【0040】
本発明の上記製造方法のさらなる態様として、本発明に係るトリコデルマ属菌を好ましくは液体培地で培養しその菌体を除去して得られる培養上清を、セルラーゼの基質となるセロオリゴ糖と反応させ、セロオリゴ糖を加水分解することにより、セロビオース又はグルコースなどの糖類をセロオリゴ糖から遊離させることを含む、セロオリゴ糖からセロビオース又はグルコースを製造する方法が挙げられる。
【0041】
本発明の上記製造方法のさらに別の態様として、本発明に係るトリコデルマ属菌を好ましくは液体培地で培養しその菌体を除去して得られる培養上清を、セルラーゼの基質となるβグルコシドと反応させて、βグルコシドをアグリコンとグルコースとに加水分解することを含む、βグルコシドからアグリコン及びグルコースを製造する方法が挙げられる。
【0042】
ここで培養上清を調製するには、まず、本発明に係るトリコデルマ属菌を、トリコデルマ属に属する真菌の培養に使用可能な任意の培地で培養すればよい。培地に用いる炭素源、窒素源、天然栄養源などの例は、上記と同様である。培地には、抗生物質、pH緩衝剤、マーカー物質などを含めてもよい。培地は液体培地であってもよいし、固体培地であってもよい。好適な培地の例としては、0.2g硫酸アンモン、0.01g塩化カリウム、0.05gリン酸二水素カリウム、0.05g硫酸マグネシウム、0.001g硝酸カルシウム、及び0.0001g酵母エキスを脱塩水に溶解した溶液が挙げられる。
【0043】
上記培地のpH値は、培養開始前に、本発明に係るトリコデルマ属菌が生育できる範囲内であればよい。本発明の方法では、好ましくはpH2.0〜pH6.0、より好ましくはpH3.0〜5.0、例えばpH3.5の酸性pH値を示す培地を用いることが好ましい。
【0044】
培養温度は、本発明に係るトリコデルマ属菌の生育可能範囲内の温度であればよい。特に好ましい培養温度は、20℃〜35℃、より好ましくは25℃〜30℃、典型的な使用温度としては25℃である。培養は、好気的条件下で行うことが好ましい。
【0045】
培養時間は、当業者であれば任意に設定することができるが、少なくとも24時間培養を継続することが好ましい。
【0046】
こうして得られた本発明に係るトリコデルマ属菌の培養液から、遠心分離などの分離手法を用いて菌体を除去することにより、培養上清を得ることができる。このような本発明に係るトリコデルマ属菌由来の培養上清は、各種セルラーゼを含み、広範な酸性条件下で強いセルラーゼ活性を示し、特に、少なくともpH2.0〜2.5のpH範囲を含む強酸性条件下でエンドセルラーゼ活性及びβグルコシダーゼ活性を示す。本発明は、このような本発明に係るトリコデルマ属菌由来の培養上清にも関する。
【0047】
以上のようにして調製した培養上清とセルラーゼの基質(例えば、セルロース、セルロース含有物質、セロオリゴ糖、又はβグルコシドなど)とを反応させるには、培養上清にセルラーゼの基質を添加するか、又はセルラーゼの基質を含む溶液と培養上清とを混合して、反応系を調製してもよい。この反応系には、pH緩衝剤などを含めても含めなくてもよい。
【0048】
培養上清中のセルラーゼとセルラーゼの基質との反応を促進するため、反応系のpH値は、原則として、本発明に係るトリコデルマ属菌が生育可能なpH範囲又はその付近とすることが好ましい。反応系のpH値としては、糖類を効率よく生産する上ではpH2.5〜pH5.5の範囲がより好ましい。またpH3.0以下、例えばpH1.5〜3.0の範囲、特にpH2.0〜pH2.5の範囲は、他の雑菌の混入を抑制しながら糖類を生産する上でより好ましい。なお本発明において培養上清を用いてセルロース、セロオリゴ糖、βグルコシド等の分解反応を実施する際のpHの「条件」とは、このような反応系のpH値を言う。
【0049】
反応系の温度は、本発明に係るトリコデルマ属菌の生育可能範囲内であればよい。特に好ましい培養温度は、20℃〜35℃、より好ましくは25℃〜30℃である。この態様では、より反応を促進するため、反応系の温度を30℃とすることも好ましい。
【0050】
反応時間は、当業者であれば任意に設定することができるが、少なくとも10分間、好ましくは1時間以上にわたり反応させることが好ましい。
【0051】
以上のようにして生産された糖類等は、上記と同様にして常法により精製してもよい。
【0052】
4.本発明のトリコデルマ属菌と有機酸生産菌を用いた有機酸製造方法
本発明に係るトリコデルマ属菌は、公知技術であるセルラーゼ生産菌と有機酸生産菌の共存培養に基づく有機酸製造方法において、強酸性条件下でも糖類を生産できるという利点を生かし、セルラーゼ生産菌として非常に有利に利用することができる。
【0053】
セルラーゼ生産菌と有機酸生産菌の共存培養に基づく有機酸製造方法は、特許文献1及び非特許文献1〜4などの文献に詳細に記載されている。例えば特許文献1には、セルラーゼ生産菌(セルロース分解菌)の存在下でセルロース又はセルロースと澱粉の混合物を同時に加水分解して糖化した後、乳酸菌を加えて乳酸醗酵を行うことによる、乳酸の製造方法が開示されている。特許文献1には、さらに、セルロース又はセルロースと澱粉の混合物を含む培地において、セルラーゼ生産菌と乳酸菌の共生系による混合培養を行うことによる乳酸の製造方法が開示されている。従って本発明では、これらの文献に記載されたような公知の有機酸製造方法において、既存のセルラーゼ生産菌に変えてトリコデルマ属菌を用いればよい。
【0054】
具体的には本発明は、本発明のトリコデルマ属菌と有機酸生産菌とをセルロース存在下で共存培養することにより、有機酸を製造する方法にも関する。本発明はさらに、本発明に係るトリコデルマ属菌由来の培養上清とセルロースとを含む培地で有機酸生産菌を培養することにより、有機酸を製造する方法にも関する。
【0055】
これらの方法において使用する有機酸生産菌[及びその有機酸生産]としては、ストレプトコッカス属菌(例えばストレプトコッカス・ボビス(Streptococcus bovis))やラクトバチルス属菌(例えばラクトバチルス・デルブルエキイ(Lactobacillus delbrueckii))などの乳酸菌[乳酸生産]、リューコノストック属菌[乳酸生産]、ペニシリウム属菌[クエン酸生産]、グルコノバクター属菌[グルコン酸生産]、アセトバクター属菌[酢酸生産]などが挙げられるが、特に乳酸菌が好適である。
【0056】
本発明において「共存培養」とは、培養期間の少なくとも一部において、本発明に係るトリコデルマ属菌と有機酸生産菌とが、同じ培地中で互いを排除することなく生育(増殖)していることを言う。
【0057】
本発明において培養に使用する培地、培養温度、培養時間などは、使用する有機酸生産菌に適した培養条件に、本発明に係るトリコデルマ属菌の上記生育可能範囲等を考慮して必要に応じて多少の変更を加えることにより、当業者が適宜設定することができる。
【0058】
セルラーゼ生産菌と有機酸生産菌を用いた従来の有機酸製造方法では、生産された有機酸により培地のpHが徐々に低下し、その結果、セルロース分解活性の低下をもたらすため、培養中の培地におけるpH低下を防ぐ必要があったが、本発明では、強酸性条件でも十分なセルロース分解活性を示すトリコデルマ属菌を利用するため、培地のpH調整がほとんど不要になり、とても有利である。
【実施例】
【0059】
以下、実施例を用いて本発明をさらに具体的に説明する。但し、本発明の技術的範囲はこれら実施例に限定されるものではない。
【0060】
[実施例1]
秋田県鹿角市八幡平の酸性温泉蒸の湯(ふけの湯)の源泉から流れ出た湯中から採取した木片0.5gを、10mlの分離培地Aを含む滅菌した15mlのコーニングチューブに入れ、25℃、100rpmで好気的に180日間振とう培養した。使用した分離培地Aは、0.2g硫酸アンモン、0.01g塩化カリウム、0.05gリン酸二水素カリウム、0.05g硫酸マグネシウム、0.001g硝酸カルシウム、及び0.0001g酵母エキス(DIFCO社製)、並びに1gセルロース(Macherey-Nagel社製)又は1gアビセル(旭化成社製)を脱塩水に溶解し、硫酸を用いてpHを3.5に調整した後、さらに脱塩水を添加して100mlとし、オートクレーブ滅菌することにより、予め調製しておいた。
【0061】
菌が生育し、にごりが観察されたチューブより培養液10μlを採取し、以下のような活性染色電気泳動に供した。まず、Davisらの方法(Ornstein L and Davis BJ, Ann NY Acad Sci 121, 321-349, 1964)に従い、0.1%(w/v)カルボキシメチルセルロース(CMC)(Sigma社製)を含む7.5%ポリアクリルアミドゲルに採取した培養液をロードし、電気泳動を行った。泳動後、ゲルを0.2Mグリシン溶液(pH2.5)に40℃で1時間浸漬した。次に、0.2%コンゴレッド(ナカライテスク社製)に60分間ゲルを浸漬した後、1M塩化ナトリウム溶液で脱色した。この方法によればセルラーゼ活性を示したタンパクバンドは赤い背景の上に白いスポットとして現れるはずである。そこでそのようなバンドの現れたチューブを選び、その中の培養液を分離プレートAに広げて室温で培養し、菌の純化を行った。使用した分離プレートAは、7g(あるいは0.7%)のゲルライト(Sigma社製)を添加した1Lの上記分離培地Aをオートクレーブした後、シャーレに広げ、固めたものである。分離した菌はプレート上でpH2.5〜3.5で生育させた。顕微鏡観察により形態観察を行うと、この菌は緑色の胞子を有する糸状菌であった。胞子嚢は観察されなかった。
【0062】
この菌については後述のように28S rDNAシークエンスを行ったところ、トリコデルマ・ビリデ(Trichoderma viride)の28S rDNA配列との間で高い相同性(549塩基中、548塩基が一致)を示した。しかしこの菌には、トリコデルマ・ビリデの特徴である35℃での培養で生育しない、胞子表面が粗面であるといった形質が見られなかった。
【0063】
[実施例2]
実施例1で緑色の胞子を有する糸状菌が分離された培養液を、生理食塩水で希釈し、上記の通り調製した分離プレートAに塗布し、25℃で培養した。セルロースを含む分離培地を用いた分離プレートA(セルロースプレート)と、アビセルを含む分離培地を用いた分離プレートA(アビセルプレート)のいずれにおいても、糸状菌の生育が確認できた。生育した糸状菌は緑色を呈した胞子を形成することが顕微鏡によって確認されたが、胞子嚢は確認されなかった。
【0064】
次いで帰属分類群の同定を行った。分離した糸状菌を、ポテトデキストロース寒天培地(PDA培地)、2%麦芽エキス寒天培地(2%MA)、オートミール寒天培地(OA培地)、三浦培地(LCA培地)の各平板培地に接種し、25℃で培養し、培養開始1週間後からコロニーの巨視的特徴を肉眼及び実体顕微鏡下で観察した。また、プレートより直接採取した菌体からプレパラートを作製し、微分干渉顕微鏡で微視的特徴を観察した。その結果、培養から3日後には各培地のシャーレ中にコロニーの成長が認められ、緑色の分生子柄の房の形成が確認された。特にポテトデキストロース寒天培地及びオートミール寒天培地(OA培地)では、気中菌糸の形成が顕著であった。9日間培養後の各培地で観察されたコロニーの表面性状はいずれも綿状(cottony)であった。9日間培養後のコロニーの色調は、Kornerup and Wanscher (1978) "Methuen handbook of colour" 3rd ed.(Eyre Methuen, London, UK)に従うと、ポテトデキストロース寒天培地(PDA培地)ではクリーム色(淡黄色)及び濃緑色、2%麦芽エキス寒天培地(2%MA)では深緑色、オートミール寒天培地(OA培地)では白色と深緑色、三浦培地(LCA培地)では深緑色であった。
【0065】
さらに、Samuels G. J. et al. (2002) Mycologia 94: p.146-170、Chaverri, P. and Samuels G. J. (2003) Studies in Mycology 48: p.1-116及びTrichoderma Home(http://nt.ars-grin.gov/taxadescriptions/keys/TrichodermaIndex.cfm)に記載された方法に従って、コロニー生育性及び形態の観察を行った。まず、コロニー上に緑色の分生子柄の房が形成される直前にコロニーを含む5mm径の寒天片を切り取り、ポテトデキストロース寒天培地(PDA培地)、SNA寒天(Synthetischer nahrstoffarmer agar)培地、及びコーンミール寒天+2%デキストロース培地(CMD培地)に接種して各種温度で3〜7日間培養し、観察を行った。
【0066】
以上の観察の結果は、本明細書中、菌学的性質としてまとめて上述している。これらの観察結果を踏まえ、Arx, J. A. von (1981) "The genera of fungi sporulating in pure culture" 3rd edition(A.R. Gantner Verlag KG., Vaduz, Germany 424 pp)、Barron, G. L. (1968) "The genera of hyphomycetes from soil"(The Williams & Wilkins Company, Baltimore, MD, USA, 364pp)、Domsch K. H. et al. (1993) "Compendium of soil fungi" volume I, IHW-Verlag, Eching, Germany, Reprinted 860pp、Domsch K. H. et al. (1993) "Compendium of soil fungi and supplement"(volume II, IHW-Verlag, Eching, Germany, Reprinted 405pp)、及びKiffer, E. and Morelet, M. (2002) "The Deuteromycetes: Mitosporic fungi classification and generic keys"(Science Publishers Inc., Enfield, NH, USA, 273pp)に記載されている菌類の検索表により、属の同定が行われた。この菌は、有隔壁菌糸の形成が認められること、また1ヶ月培養してもテレオモルフの形成(有性生殖器官の形成)が認められなかったことから、不完全菌類の1種であると判断された。さらに、菌糸の成長が速く、分岐する分生子柄を持ち、分生子の形成様式がフィアロ型であること、分生子が連鎖せず、コロニーの成熟に伴って白色から深緑色へと着色した分生子塊を形成することなどの形態学的特徴から、この菌は、不完全菌類の中でもトリコデルマ属(Trichoderma)に属するものと考えられた。
【0067】
続いて、トリコデルマ属のモノグラフであるRifai, M. A. (1969) "A revision of the genous Trichoderma" Mycological Papers 116: p.1-56、その後見出された種を含めた奥田徹 著「真菌の分離と分類・同定(20)」(1992) p.157-166、緑色の分生子を持つ種をまとめたChaverri, P. and Samuels G. J. (2003) Studies in Mycology 48: p.1-116に記載された菌類の検索表及びTrichoderma Home(http://nt.ars-grin.gov/taxadescriptions/keys/TrichodermaIndex.cfm)の検索表を用いて、菌種の同定を試みた。その結果、分離した菌の特徴は、Chaverri, P. and Samuels G. J. (2003)に記載されたトリコデルマ・ハルジアナム(Trichoderma harzianum)と比較的類似していた。
【0068】
一方、分離した菌について28S rDNA領域配列に基づく分子系統解析を行った。28S rDNAのD1/D2領域に特異的なプライマーを設計し、分離した菌から常法により抽出した核酸を鋳型としたPCR増幅を行い、シークエンシングにより28S rDNAの塩基配列を決定した。得られた28S rDNAの塩基配列(配列番号1)を、トリコデルマ・ビリデの既知の28S rDNAの塩基配列(DDBJ/GenBank/EMBL国際塩基配列データベース・アクセッション番号AY291123;配列番号2)と比較したところ、549塩基中548塩基が一致するという高い相同性(約99.8%の同一性)が示された(図1)。このことから、分離した菌はトリコデルマ・ビリデに極めて近縁であることが示唆された。
【0069】
トリコデルマ・ハルジアナムとトリコデルマ・ビリデの両種は形態学的に類似しているが、分生子の表面がトリコデルマ・ハルジアナムでは平滑であるのに対しトリコデルマ・ビリデでは粗面であること、トリコデルマ・ハルジアナムは35℃での生育が認められるがトリコデルマ・ビリデでは認められないことなどの相違点もある(奥田徹 著「真菌の分離と分類・同定(20)」(1992) p.157-166)。分離した菌は、分生子表面が平滑であることや35℃での生育が認められる点ではトリコデルマ・ハルジアナムと共通していると言える。
【0070】
なお、分離した菌には25℃、pH3.5の条件下で培養した際にその培養上清に強いセルラーゼ活性が認められるが、従来セルラーゼ生産が知られているトリコデルマ・ビリデNBRC31137株、トリコデルマ・リーセイ(Trichoderma reesei)NBRC31326株をそれと同じ条件で培養しても、培養上清にはセルラーゼ活性が全く認められなかった。
【0071】
以上を総合的に判断した結果、分離した菌は、トリコデルマ属(Trichoderma)の新菌種に属する菌株であるとして同定された。本発明者らは、この得られた菌を、トリコデルマ・エスピー(Trichoderma sp.)AH5株と命名した。トリコデルマ・エスピー(Trichoderma sp.)AH5株は、2006年3月8日付で、独立行政法人 製品評価技術基盤機構 特許微生物寄託センター(日本国千葉県木更津市かずさ鎌足2-5-8)に、受託番号NITE P-222で寄託された。
【0072】
[実施例3]
トリコデルマ・エスピー(Trichoderma sp.)AH5株の培養上清に含まれるエンドセルラーゼの反応pHを、トリコデルマ・ビリデ由来のセルラーゼ製剤「セルラーゼオノズカ」(ヤクルト工業社製)と比較した。このため、基質としてカルボキシメチルセルロース(以下、CMCと称する)を用いて酵素反応を行い、反応後CMCの分解によって生じた還元糖量をSomogyi-Nelson法により、グルコースを標準糖として定量した。
【0073】
具体的には、まず、トリコデルマ・エスピー(Trichoderma sp.)AH5株を、硫酸でpH3.5に調整した培地(0.2%硫安、0.01%塩化カリ、0.05%リン酸二水素カリウム、0.05%硫酸マグネシウム、0.03%塩化カルシウム、0.0001%酵母エキス、1%アビセル)に接種して25℃、100 rpmで2週間にわたり培養した。得られた培養物を20,000×gで10分間遠心分離して培養上清を採取した。この培養上清を、0.6%のCMC(Sigma社製)を含む様々なpH値の緩衝液に添加して反応液(全量50μl)を調製した。使用した緩衝液は、0.2Mグリシン-HCl緩衝液(pH2.0〜2.5)、0.1M酢酸ナトリウム緩衝液(pH3.0〜5.5)、0.1M PIPES緩衝液(pH6.4〜7.5)、0.1M Tris-HCl緩衝液(pH7.65)である。30℃で適当時間反応させた後、サーマルサイクラー(PCRマシーン)にて100℃で10分間加熱し、反応を停止させた。次いで50μlのSomogyi銅液(和光純薬工業社製など)を加え、さらにサーマルサイクラーで100℃で10分間加熱してから急速に冷却し、冷却後、50μlのNelson試薬(和光純薬社製など)を加えて還元銅沈殿を溶解して発色させ、30分静置した後、660nmでの吸光度を測定した。得られた測定値から、標準糖としたグルコースの吸光度に基づき、溶液中に含まれる還元糖量を算出した。「セルラーゼオノズカ」についても、同様に各種pH条件下でCMCと反応させて、溶液中に生じた還元糖量を測定した。培養上清中のタンパク質1mg当たりのエンドセルラーゼ活性を算出するため、Bradfordらの方法(Bradford M.M., Anal. Biochem., (1976) 72, p.248-254)に従い、牛血清アルブミンを標準タンパク質として、培養上清に含まれるタンパク質の量を測定した。エンドセルラーゼ活性は、以下の式で表した。
【0074】
エンドセルラーゼ活性(nmol/min/mg)=反応溶液中の還元糖量(nmol)/反応時間(分)/用いた培養上清中のタンパク質の量(mg)
この結果を表1及び図2に示す。
【0075】
【表1】
【0076】
表1及び図2に示される通り、本発明に係るトリコデルマ・エスピーAH5株の培養上清は、従来製品「セルラーゼオノズカ」に比べて酸性域全体でより高いエンドセルラーゼ活性(比活性)を示した。またトリコデルマ・エスピーAH5株の培養上清は、pH2.0〜pH2.5という強酸性域でもエンドセルラーゼ活性を示した。
【0077】
さらに、上記で得られたトリコデルマ・エスピーAH5株の培養上清について、基質としてp-ニトロフェニルグルコシドを用い、βグルコシダーゼ活性を測定した。具体的には、AH5株の培養上清を、最終濃度0.1M p-ニトロフェニルグルコシド(和光純薬工業社製)を含む様々なpH値の緩衝液に添加して全量を50μlとした。使用した緩衝液は、0.1Mグリシン-HCl緩衝液(pH1.5〜2.8)、0.1M酢酸ナトリウム緩衝液(pH3.5〜4.65、5.5)、0.1M MES緩衝液(pH5.2)、0.1M PIPES緩衝液(pH6.4)、0.1M Tris-HCl緩衝液(pH7.65)である。30℃で適当時間反応させた後、反応液の9倍量の50mM炭酸ナトリウム溶液を加えることにより反応を停止させた。次いで、p-ニトロフェニルグルコシドの分解によって生じたその溶液中のp-ニトロフェノールを400nmでの吸光度測定により測定し、測定値に基づいてモル吸光係数を55.1μM/cmとしてp-ニトロフェノール量を算出した。「セルラーゼオノズカ」についても、同様に各種pH条件下でp-ニトロフェニルグルコシドと反応させて溶液中に生じたp-ニトロフェノール量を測定した。
【0078】
βグルコシダーゼ活性は、以下の式で表した。
βグルコシダーゼ活性(nmol/min/mg)=反応溶液中のp-ニトロフェノール量(nmol)/反応時間(分)/用いた培養上清中のタンパク質の量(mg)
得られた結果を表2及び図3に示す。
【0079】
【表2】
【0080】
表2及び図3に示される通り、本発明に係るトリコデルマ・エスピーAH5株の培養上清は、従来製品「セルラーゼオノズカ」に比べて高いβグルコシダーゼ活性(比活性)を示すこと、またpH1.5〜pH2.5という強酸性域でもかなり高いβグルコシダーゼ活性を示すことが示された。
【0081】
[実施例4]
トリコデルマ・エスピーAH5株の培養上清が示すβグルコシダーゼ活性を、種々の市販の代表的なセルラーゼが示すβグルコシダーゼ活性と比較した。
【0082】
まず、実施例3と同様にして調製した培養上清を、比色レンジに入るよう濃度を適宜調整した後、1mM p-ニトロフェニルグルコシド(和光純薬工業社製)を含む0.1Mグリシン-HCl緩衝液(pH2.0)、又は0.1M酢酸ナトリウム緩衝液(pH4.0)に添加し、30℃で適当時間反応させた後、反応液の9倍量の50mM炭酸ナトリウム溶液を加えることにより反応を停止させた。次いで、実施例3と同様にして、溶液中のp-ニトロフェノールを400nmでの吸光度測定により定量した。対照実験としては、実施例3で使用した「セルラーゼオノズカ」に加え、アスペルギルス・ニガー(Aspergillus niger)由来のセルラーゼA「アマノ」3(天野エンザイム社製)、トリコデルマ・ビリデ由来のセルラーゼT「アマノ」4(天野エンザイム社製)、アスペルギルス・ニガー(Aspergillus niger)由来のセルラーゼ(東京化成工業社製)、トリコデルマ・リーセイ由来のセルラーゼ(Worthington Biochemical Corporation社製)を使用して、それぞれトリコデルマ・エスピーAH5株と同様にしてp-ニトロフェニルグルコシドと反応させ、溶液中に生じたp-ニトロフェノールを定量した。測定されたp-ニトロフェノール量に基づき、実施例3と同様にしてβグルコシダーゼ活性値を算出した。その結果を表3に示す。
【0083】
【表3】
【0084】
表3中、『AH5』はトリコデルマ・エスピーAH5株の培養上清、『オノズカ』は「セルラーゼオノズカ」、『セルA』はセルラーゼA「アマノ」3、『セルT』はセルラーゼT「アマノ」4、『A.niger(東京化成)』はアスペルギルス・ニガー由来のセルラーゼ(東京化成工業社製)、『T.reesei』はトリコデルマ・リーセイ由来のセルラーゼ(Worthington Biochemical Corporation社製)を表す。また『pH2』は、緩衝液として0.1Mグリシン-HCl緩衝液(pH2.0)を用いた場合、『pH4』は、緩衝液として0.1M酢酸ナトリウム緩衝液(pH4.0)を用いた場合を示す。
【0085】
表3に示すように、トリコデルマ・エスピーAH5株の培養上清は、pH2の条件下で298 nmol/min/mg、pH4の条件下で340 nmol/min/mgという高い比活性を示し、かつ、pH2の条件下でさえpH4で示したβグルコシダーゼ活性の88%を保持していた。一方、市販の各種セルラーゼは、pH2では全く活性を示さないかあるいは10 nmol/min/mg未満というごく低い活性を示すのみだった。このようにトリコデルマ・エスピーAH5株の培養上清は、従来品の各種セルラーゼに比べて高いβグルコシダーゼ比活性を示し、またpH2.0の条件下でも非常に高い活性を有していた。このことは、本発明のトリコデルマ・エスピーAH5株及びその培養上清が、グルコースの高効率な産生、特に強酸性条件下でのグルコースの高効率な産生に好適に利用できることも示す。
【0086】
[実施例5]
トリコデルマ・エスピーAH5株の培養上清が示すセロヘキサオースに対する分解活性を、「セルラーゼオノズカ」(ヤクルト工業社製)と比較した。
【0087】
実施例3と同様にして調製したトリコデルマ・エスピーAH5株の培養上清を、終濃度4mMのセロヘキサオース(生化学工業社製)を含む終濃度40mMの酢酸ナトリウム緩衝液(pH3.0)に、終濃度0.16mg/mlとなる量で添加し、30℃で適当時間反応させた。同様に、「セルラーゼオノズカ」は、終濃度4mMのセロヘキサオース(生化学工業社製)を含む終濃度40mMの酢酸ナトリウム緩衝液(pH3.0)に、終濃度0.2mg/mlとなる量で添加し、30℃で反応させた。反応時間は15分、1時間、2時間、6時間、24時間とした。反応後、Merck社製シリカゲルプレート(5cm×10cm)に、反応液0.5mlをスポットし、80%アセトニトリルにより展開した。標品としてはセロオリゴ糖(生化学工業製)をスポットした。展開後、シリカゲルプレートを硫酸/メタノール混合溶液(容量比5:95)中に浸漬し、110℃で5分〜10分間加熱処理することにより発色させた。
【0088】
この結果を図4に示す。図4中、G1はグルコース、G2はセロビオース(βグルコース分子2個がβ-1,4結合した二糖類)、G3〜G6はそれぞれ3〜6個のグルコース分子がβ-1,4結合したセロオリゴ糖である。反応時間は、レーン1:15分、レーン2:1時間、レーン3:2時間、レーン4:6時間、レーン5:24時間である。
【0089】
図4に示すように「セルラーゼオノズカ」を用いた場合(図4B)に比べ、トリコデルマ・エスピーAH5株の培養上清を用いた場合(図4A)の方が、グルコースの生産量が格段に多いことが示された。さらに「セルラーゼオノズカ」では24時間反応させた後でもグルコースに加えてセロビオース(G2)も比較的多量に産生されたのに対し、トリコデルマ・エスピーAH5株の培養上清を用いた場合は、6時間反応後にはセロビオース(G2)はあまり産生されなくなり、代わりにグルコースが多量に産生されるようになった。このことは、トリコデルマ・エスピーAH5株を用いれば、グルコースを優先的に高効率で産生できることを示している。
【0090】
[実施例6]
様々なpH値を示す液体培地Aを含むろ紙上でのトリコデルマ・エスピーAH5株の生育能を、セルラーゼを生産することが知られているトリコデルマ・ビリデNBRC 31137株[独立行政法人製品評価技術基盤機構バイオテクノロジー本部生物遺伝資源部門(NBRC)(千葉県木更津市かずさ鎌足2-5-8)からカタログ番号NBRC 31137にて分譲を受けたもの]、及びトリコデルマ・リーセイNBRC 31326株[前記NBRCからカタログ番号NBRC 31326にて分譲を受けたもの]と比較した。この生育試験により、ろ紙に含まれるセルロースを基質として酸性条件下で生育する能力を調べた。
【0091】
まず、0.2g硫酸アンモン、0.01g塩化カリウム、0.05gリン酸二水素カリウム、0.05g硫酸マグネシウム、0.001g硝酸カルシウム、及び0.0001g酵母エキス(DIFCO社製)を適量の脱塩水に溶解し、5%硫酸又は濃水酸化ナトリウム溶液を用いてpHを2.0、3.5、5.5にそれぞれ調整した後、さらに脱塩水を添加して100mlとし、オートクレーブ滅菌することにより、3種のpH値の液体培地Aを調製した。この液体培地Aをそれぞれ、別個のろ紙(B200G047A、東洋ろ紙社製)に染み込ませて3種のろ紙培地を作製した。次に各菌株の胞子約10,000個を、作製したろ紙培地に接種し、25℃で10日間静置培養した。10日間培養した後の各菌株の生育結果を図5に示す。
【0092】
図5に示される通り、トリコデルマ・エスピーAH5株は、pH2.0、3.5、5.5のいずれの液体培地Aを含むろ紙培地でも良好に生育し、胞子形成が容易に観察された(図5最上段)。一方、トリコデルマ・ビリデNBRC 31137株とトリコデルマ・リーセイNBRC 31326株は、pH2.0の液体培地Aを含むろ紙培地ではほとんど生育が認められず(図5最左列の中段及び下段)、pH3.5及び5.5ではわずかに菌糸の生長が認められるが胞子形成はほとんど認められなかった。特にトリコデルマ・リーセイNBRC 31326株は、いずれのpH条件でも生育状態がかなり悪かった。
【0093】
この生育試験の結果から、トリコデルマ・エスピーAH5株は、セルラーゼの生産が知られているトリコデルマ・ビリデNBRC 31137株とトリコデルマ・リーセイNBRC 31326株と比べて酸性条件での生育能が非常に高いこと、特にpH2.0のような強酸性条件下でも生育できることが示された。すなわちトリコデルマ・エスピーAH5株は、酸性条件下、特に少なくともpH2.0以上pH5.5以下のpH範囲では、高いセルロース分解活性を示すことが明らかになった。
【産業上の利用可能性】
【0094】
本発明のトリコデルマ属菌は、常温の酸性条件下、特に常温かつ強酸性条件下で、セルロース系物質を分解するために用いることができる。本発明のトリコデルマ属菌を用いた糖類製造法は、セルロースを材料とした糖類の効率的な製造に利用することができる。さらに本発明のトリコデルマ属菌と有機酸生産菌を用いた有機酸製造方法は、1つの反応系でセルロースを原料として有機酸を効率よく製造するために使用することができる。
【図面の簡単な説明】
【0095】
【図1】図1は、トリコデルマ・エスピーAH5株とトリコデルマ・ビリデの28S rDNAの塩基配列のアラインメントを示す図である。
【図2】図2は、トリコデルマ・エスピーAH5株の培養上清の、様々なpH条件下でのエンドセルラーゼ活性を示すグラフである。ダイヤ印はグリシン-HCl緩衝液、四角印は酢酸ナトリウム緩衝液、三角印はPIPES緩衝液、丸印はTris-HCl緩衝液を示す。黒塗りつぶし印がトリコデルマ・エスピーAH5株の培養上清、灰色塗りつぶし印が対照サンプルの「セルラーゼオノズカ」を示す。
【図3】図3は、トリコデルマ・エスピーAH5株の培養上清の、様々なpH条件下でのβグルコシダーゼ活性を示すグラフである。ダイヤ印はグリシン-HCl緩衝液、四角印は酢酸ナトリウム緩衝液、三角印はMES緩衝液、丸印はPIPES緩衝液、白丸印はTris-HCl緩衝液を示す。黒塗りつぶし印がトリコデルマ・エスピーAH5株の培養上清、灰色塗りつぶし印が対照サンプルの「セルラーゼオノズカ」を示す。
【図4】図4は、トリコデルマ・エスピーAH5株の培養上清(0.16mg/L)(図4A)と「セルラーゼオノズカ」(0.2mg/L)(図4B)のセロヘキサオース分解活性を示すクロマトグラフィーの結果を表す写真である。レーン1:15分、レーン2:1時間、レーン3:2時間、レーン4:6時間、レーン5:24時間(反応時間)。G1:グルコース、G2:セロビオース(βグルコース分子2個がβ-1,4結合した二糖類)、G3〜G6:それぞれ3〜6個のグルコース分子がβ-1,4結合したセロオリゴ糖。
【図5】図5は、トリコデルマ・エスピーAH5株、トリコデルマ・ビリデNBRC31137株、トリコデルマ・リーセイNBRC31326株の酸性条件下での生育試験の結果を示す写真である。
【特許請求の範囲】
【請求項1】
酸性条件下でセルロース分解活性を示すトリコデルマ(Trichoderma)属菌であって、少なくともpH2.0〜2.5のpH範囲でエンドセルラーゼ活性及びβグルコシダーゼ活性を示す、前記トリコデルマ属菌。
【請求項2】
トリコデルマ・エスピー(Trichoderma sp.)AH5株(受託番号NITE P-222)又はその変異株である、請求項1に記載のトリコデルマ属菌。
【請求項3】
少なくともpH2.0〜2.5のpH範囲でエンドセルラーゼ活性及びβグルコシダーゼ活性を示す、請求項1又は2に記載のトリコデルマ属菌由来の培養上清。
【請求項4】
請求項1若しくは2に記載のトリコデルマ属菌又は請求項3に記載の培養上清を用いてセルロースを分解することを特徴とする、糖類の製造方法。
【請求項5】
セルロースの分解をpH2.5以下の条件下で行う、請求項4に記載の方法。
【請求項6】
糖類が、セロオリゴ糖、セロビオース、及びグルコースからなる群より選択される少なくとも1つである、請求項4又は5に記載の方法。
【請求項7】
請求項1若しくは2に記載のトリコデルマ属菌又は請求項3に記載の培養上清を用いてセロオリゴ糖を分解することを特徴とする、セロオリゴ糖からセロビオース又はグルコースを製造する方法。
【請求項8】
セロオリゴ糖の分解をpH2.5以下の条件下で行う、請求項7に記載の方法。
【請求項9】
請求項1若しくは2に記載のトリコデルマ属菌又は請求項3に記載の培養上清を用いてβグルコシドを分解することを特徴とする、βグルコシドからアグリコン及びグルコースを製造する方法。
【請求項10】
βグルコシドの分解をpH2.5以下の条件下で行う、請求項9に記載の方法。
【請求項11】
請求項1又は2に記載のトリコデルマ属菌と有機酸生産菌とをセルロース存在下で共存培養することを特徴とする、有機酸の製造方法。
【請求項12】
請求項3に記載の培養上清とセルロースとを含む培地で有機酸生産菌を培養することを特徴とする、有機酸の製造方法。
【請求項13】
有機酸生産菌が乳酸菌である、請求項11又は12に記載の方法。
【請求項1】
酸性条件下でセルロース分解活性を示すトリコデルマ(Trichoderma)属菌であって、少なくともpH2.0〜2.5のpH範囲でエンドセルラーゼ活性及びβグルコシダーゼ活性を示す、前記トリコデルマ属菌。
【請求項2】
トリコデルマ・エスピー(Trichoderma sp.)AH5株(受託番号NITE P-222)又はその変異株である、請求項1に記載のトリコデルマ属菌。
【請求項3】
少なくともpH2.0〜2.5のpH範囲でエンドセルラーゼ活性及びβグルコシダーゼ活性を示す、請求項1又は2に記載のトリコデルマ属菌由来の培養上清。
【請求項4】
請求項1若しくは2に記載のトリコデルマ属菌又は請求項3に記載の培養上清を用いてセルロースを分解することを特徴とする、糖類の製造方法。
【請求項5】
セルロースの分解をpH2.5以下の条件下で行う、請求項4に記載の方法。
【請求項6】
糖類が、セロオリゴ糖、セロビオース、及びグルコースからなる群より選択される少なくとも1つである、請求項4又は5に記載の方法。
【請求項7】
請求項1若しくは2に記載のトリコデルマ属菌又は請求項3に記載の培養上清を用いてセロオリゴ糖を分解することを特徴とする、セロオリゴ糖からセロビオース又はグルコースを製造する方法。
【請求項8】
セロオリゴ糖の分解をpH2.5以下の条件下で行う、請求項7に記載の方法。
【請求項9】
請求項1若しくは2に記載のトリコデルマ属菌又は請求項3に記載の培養上清を用いてβグルコシドを分解することを特徴とする、βグルコシドからアグリコン及びグルコースを製造する方法。
【請求項10】
βグルコシドの分解をpH2.5以下の条件下で行う、請求項9に記載の方法。
【請求項11】
請求項1又は2に記載のトリコデルマ属菌と有機酸生産菌とをセルロース存在下で共存培養することを特徴とする、有機酸の製造方法。
【請求項12】
請求項3に記載の培養上清とセルロースとを含む培地で有機酸生産菌を培養することを特徴とする、有機酸の製造方法。
【請求項13】
有機酸生産菌が乳酸菌である、請求項11又は12に記載の方法。
【図1】


【図2】
【図3】
【図4】
【図5】
【図2】
【図3】
【図4】
【図5】
【公開番号】特開2007−319040(P2007−319040A)
【公開日】平成19年12月13日(2007.12.13)
【国際特許分類】
【出願番号】特願2006−150444(P2006−150444)
【出願日】平成18年5月30日(2006.5.30)
【出願人】(000003207)トヨタ自動車株式会社 (59,920)
【Fターム(参考)】
【公開日】平成19年12月13日(2007.12.13)
【国際特許分類】
【出願日】平成18年5月30日(2006.5.30)
【出願人】(000003207)トヨタ自動車株式会社 (59,920)
【Fターム(参考)】
[ Back to top ]