
酸性真菌プロテアーゼ
本発明は、新規酸性プロテアーゼを開示する。詳細には、NSP24ファミリ・プロテアーゼ、およびその生物学的に活性な断片を含むNSPファミリ・プロテアーゼ、および、前記プロテアーゼをコードする核酸分子を開示する。該プロテアーゼをコードする核酸配列を含むベクターおよび宿主細胞、プロテアーゼを生産する方法、前記プロテアーゼを用いる酵素組成物および方法が本発明により提供される。
【発明の詳細な説明】
【技術分野】
【0001】
関連出願
本出願は、2004年12月30日出願、名称「酸性真菌プロテアーゼ(Acid Fungal Protease)」なる米国特許仮出願第60/640,399号、および、2005年1月27日出願、名称「酸性真菌プロテアーゼ(Acid Fungal Protease)」なる米国特許仮出願第60/648,233号に対する優先権を主張する。なお、これらの出願の内容の全てを参照することにより本明細書に含める。
【0002】
本発明は、NSP24ファミリ・プロテアーゼ、NSP25ファミリ・プロテアーゼ、およびPepAプロテアーゼと表示される酸性プロテアーゼをコードするポリヌクレオチド;NSP24およびNSP25ファミリ・プロテアーゼポリペプチド;前記プロテアーゼを含む組成物、および、それらの使用に関する。
【背景技術】
【0003】
プロテアーゼは、ペプチド結合を切断することのできる酵素である。酸性プロテアーゼ(例えば、酸性pHを至適とするプロテアーゼ)は、哺乳類および微生物を含むいくつかの異なる生物体によって生産される。例えば、微生物の酸性プロテアーゼは、細菌株、例えば、Bacillus sp.の株(JP01240184)、および、真菌株、例えば、Rhizopus sp.(EP72978)、Schytalidium sp.(JP48091273)、Sulpholobus sp.,Thermoplasma sp.(WO/9010072)、およびAspergillus sp.(JP50121486およびEP82395)の株によって生産される。
【0004】
ベルカ(Berka)等「遺伝子(Gene)」(1990)96:313は、Aspergillus awamori由来のアスパラギン酸プロテアーゼ、アスペルギロペプシンAをコードする遺伝子を開示する。Aspergillus oryzae由来のアスパラギン酸プロテアーゼ、アスペルギロペプシンOをコードする遺伝子のクローニングは、ベルカ(Berka)等によって記載される「遺伝子(Gene)」(1993)125:195−198。Aspergillus oryzae由来の酸性プロテアーゼ(PepA)をコードする遺伝子のクローニングは、ゴミ(Gomi)等によって開示される「バイオサイエンス・バイオテクノロジー・アンド・バイオケミストリー(Biosci.Biotech.Biochem.)」(1993)57(7):1095−1100。
【0005】
プロテアーゼ、特に酸性プロテアーゼは、産業用途に、例えば、食品および飼料の生産、皮革産業(例えば、生皮の除毛)、タンパクの加水分解産物の生産、および、アルコールの生産、例えば、エタノールの生産、ワインの生産および醸造等広く使われている。
【0006】
依然として、多種多様な用途、特に食品および飼料産業においてプロテアーゼに対する需要が続いている。
【発明の開示】
【課題を解決するための手段】
【0007】
本出願人等は、いくつかの新規プロテアーゼ遺伝子を発見した。そのようなものとして、NSP24プロテアーゼ(配列番号2、または配列番号10)をコードする新規nsp24遺伝子、NSP25プロテアーゼ(配列番号9)をコードする新規nsp25遺伝子、および、新規PepAプロテアーゼ(配列番号7)をコードする新規pepA変異遺伝子が挙げられる。
【0008】
従って、本発明は、NSP24プロテアーゼ、NSP25プロテアーゼ、またはPepAプロテアーゼおよびその変異種の、組み換え法による、または、ほぼ純粋な調製をその特徴とする。
【0009】
本発明のある態様では、プロテアーゼは、配列番号2または配列番号10(下記の図6に示される)のアミノ酸配列と事実上同じアミノ酸配列を含む、NSP24ファミリ・プロテアーゼポリペプチドである。いくつかの実施態様では、NSP24ファミリ・プロテアーゼポリペプチドは、配列番号8(下記の図5に示される)の核酸によって、あるいは、配列番号8の核酸と事実上同じ核酸配列を持つ核酸によってコードされる。
【0010】
本発明の別の態様では、NSP24ファミリ・プロテアーゼポリペプチドは、配列番号10における配列とは最大10残基アミノ酸配列が異なる。ある実施態様では、NSP24ファミリ・プロテアーゼポリペプチドは、配列番号10における配列とは、最大10%の残基のアミノ酸配列が異なる。ある実施態様では、その差は、NSP24ファミリ・プロテアーゼポリペプチドがNSP24プロテアーゼの生物学的活性を示せる程度の差であり、例えば、該NSP24プロテアーゼが天然のNSP24プロテアーゼの生物学的活性を保てる程度の差である。
【0011】
本発明のさらに別の態様では、NSP24ファミリ・プロテアーゼポリペプチドは、本明細書に記載されるNSP24プロテアーゼ配列の外に、他のN−末端および/またはC−末端アミノ酸配列も含む。
【0012】
本発明のさらに別の態様では、NSP24ファミリ・プロテアーゼポリペプチドは、さらに別のアミノ酸残基、好ましくは、配列番号1または配列番号8の配列をコードするゲノムDNAに対する5´側のゲノムDNAによってコードされる残基に、読み枠内で融合される、配列番号2または配列番号10のアミノ酸配列の全てまたはその断片を含む。
【0013】
本発明のさらに別の態様では、NSP24ファミリ・プロテアーゼは、第1NSP24ファミリ・プロテアーゼ部分と、第2ポリペプチド部分、例えば、NSP24ファミリ・プロテアーゼとは無関係のアミノ酸配列を持つ第2ポリペプチド部分とを有する、組み換え融合タンパクである。第2ポリペプチド部分は、DNA結合ドメイン、またはポリメラーゼ活性化ドメインであってもよい。本発明のポリペプチドは、複数遺伝子の存在、選択的転写事象、選択的RNAスプライシング事象、ならびに、選択的翻訳および翻訳後事象の結果生ずるものを含む。このポリペプチドは、システム、例えば、発現NSP24プロテアーゼが天然細胞において発現される場合に見られるのとほぼ同じ翻訳後修飾をもたらす、培養細胞において発現されてもよいし、あるいは、天然細胞で発現された場合に見られる翻訳後修飾の除去をもたらすシステムにおいて発現されてもよい。
【0014】
さらに別の態様では、本発明は、NSP24ファミリ・プロテアーゼ、および、1種以上の添加成分、例えば、担体、希釈剤、または溶媒を含む酵素組成物に関する。添加成分は、インビボ、インビトロ、製薬、または獣医学用途において組成物を有用にするものであってもよい。この態様のある実施態様では、酵素組成物は添加酵素を含む。好ましい実施態様では、添加酵素は、グルコアミラーゼ、アルファ・アミラーゼ、またはそれらの組み合わせである。
【0015】
さらに別の態様では、本発明は、配列番号2または配列番号10のアミノ酸配列に対し少なくとも80%相同な配列を持つアミノ酸配列を含むNSP24ファミリ・プロテアーゼポリペプチドをコードするヌクレオチドを有するか、または含むほぼ純粋な核酸を提供する。
【0016】
ある態様では、NSP24ファミリ・プロテアーゼの核酸は、例えば、該NSP24ファミリ・プロテアーゼ遺伝子配列を発現ベクターとしての使用に適しているものとするために、NSP24ファミリ・プロテアーゼ遺伝子配列に対して動作的に結合される転写調節配列、例えば、転写プロモーターまたは転写エンハンサー配列の内の少なくとも一つを含む。
【0017】
さらに別の態様では、本発明のNSP24プロテアーゼポリペプチド(例えば、配列番号2)をコードする核酸は、配列番号8由来の少なくとも12個の連続ヌクレオチドに対応する核酸プローブに対して、より好ましくは、配列番号8由来の20個の連続ヌクレオチドに対して厳格条件下にハイブリダイズする。
【0018】
本発明のもう一つの態様は、各種産業場面における、NSP24ファミリ・プロテアーゼ(例えば、NSP24)の応用を提供する。例えば、NSP24ファミリ・プロテアーゼは、アルコール燃料およびその他の重要な工業薬品の生産のために農業廃棄物を酵素的に分解するために使用されてもよいし、動物またはヒトの食品生産用途、または洗剤組成物の成分として、皮革処理およびタンパク系繊維の処理用途(例えば、ウールまたはシルク)、バイオマス用途、パーソナルケア用途(皮膚、髪、口内衛生等)、製薬および保健用途、および、上記応用用途に用いられる新規ペプチドの生産用途に使用されてもよい。
【0019】
さらに別の態様では、本発明は、pepA変異プロテアーゼ、すなわち、配列番号7を持つL388Mをコードするポリヌクレオチドに関する。ある実施態様では、ポリヌクレオチドは、配列番号5の配列を有する。
【0020】
さらにもう一つの態様では、本発明はNSP25ファミリ・プロテアーゼに関する。ある実施態様では、NSP25ファミリ・プロテアーゼは、配列番号9に対して少なくとも85%の配列相同性を持つ。別の実施態様では、NSP25ファミリ・プロテアーゼは、配列番号4に対して少なくとも85%の配列相同性を持つポリヌクレオチドによってコードされる。さらに別の実施態様では、NSP25ファミリ・プロテアーゼは、親のNSP25ファミリ・プロテアーゼの生物学的に活性な断片である。
【発明を実施するための最良の形態】
【0021】
以下、本発明を、下記の定義と実施例のみを用い、参照によって詳細に説明する。本明細書に参照される全ての特許および出版物は、それら特許および出版物の中に開示される全ての配列を含め、参照することによって本明細書に含められる。
【0022】
本発明の実施において、別様に指示しない限り、細胞生物学、細胞培養、分子生物学、トランスジェニック生物学、微生物学、組み換えDNA、および免疫学の、当業者の能力の範囲内にある通例技術を用いる。このような技術は文献に記載される。例えば、「分子クローニング、実験室マニュアル(Molecular Cloning A Laboratory Manual)」、第2版、サムブルック、フリッチュ、およびマニアチス(Sambrook,Fritsch and Maniatis)著、コールドスプリングハーバーラボラトリープレス(Cold Spring Harbor Laboratory Press);アウスベル等(Ausubel et al.)編、「分子生物学における短いプロトコル(Short Protocols in Molecular Biology)」、第5版、2002年;「DNAクローニング(DNA Cloning)」、IおよびII巻、ディーエヌ・グローバー(D.N.Glover)編、1985;「オリゴヌクレオチド合成(Oligonucleotide Synthesis)」、エムジェイ・ゲイト(M.J.Gait)編、1984;マリス(Mullis et al.)等、米国特許第4,683,195号;「核酸ハイブリダイゼーション(Nucleic Acid Hybridization)」、ビーディー・ヘイムズおよびエスジェイ・ヒギンス(B.D.Hames & S.J.Higgins)編、1984;「転写および翻訳(Transcription and Translation)」、ビーディー・ヘイムズおよびエスジェイ・ヒギンス(B.D.Hames & S.J.Higgins)編、1984;「動物細胞の培養(Culture Of Animal Cells)」、アールアイ・フレッシュニー(R.I.Freshney)著、アラン・アールリス社(Alan R.Liss,Inc.)、1987;「不動化細胞および酵素(Immobilized Cells And Enzymes)」、アイアールエル・プレス(IRL Press)、1986;ビー・パーバル(B.Perbal)著、「分子クローニングの実技ガイド(A Practical−Guide To Molecular Cloning)」、1984;論説「酵素学の方法(Methods in Enzymology)」、アカデミックプレス社(Academic Press,Inc.)、ニューヨーク州;「哺乳類細胞のための遺伝子転送ベクター(Gene Transfer Vectors For Mammalian Cells)」、ジェイエイチ・ミラーおよびエムピー・カーロス(J.H.Miller and M.P.Calos)編、1987年、コールドスプリングハーバーラボラトリ(Cold Spring Harbor Laboratory);酵素学の方法(Methods in Enzymology)、154および155巻、ウー(Wu et al.)等編、「細胞および分子生物学における免疫化学的方法(Immunochemical Methods in Cell and Molecular Biology)」、メイヤーおよびウォーカー(Mayer and Walker)編、アカデミックプレス社(Academic Press,Inc.)、ロンドン、1987;「実験免疫学ハンドブック(Handbook Of Experimental Immunology)」、I−IV巻、ディーエム・ウェアおよびシーシー・ブラックウェル(D.M.Weir and C.C.Blackwell)編、1986;「マウス胚の取り扱い手技(Manipulating Mouse Embryo)」、コールドスプリングハーバーラボラトリー・プレス(Cold Spring Harbor Laboratory Press)、コールドスプリングハーバー(Cold Spring Harbor)、ニューヨーク州、1986を参照されたい。さらに、プロテアーゼの調製、発現、分離、および使用の方法に関する情報が、米国特許第6,768,001号を参照することによって得られる。なお、この特許文献の全体を、引用することにより本明細書に含める。
【0023】
本発明の他の特徴および利点は、下記の詳細な説明、および特許請求の範囲から明らかとなろう。なお、本発明の実施または試験においては、本明細書に記載されるものと類似の、または等価な、任意の方法および材料の使用が可能であるけれども、好ましい方法および材料が記載される。
【0024】
別様に定義しない限り、本明細書で使用される技術および科学用語は全て、本発明の所属する従来技術において通常の錬度を有する当業者によって一般的に理解されるものと同じ意味を持つ。シングルトン(Singleton et al.)等、「微生物および分子生物学辞典(DICTIONARY OF MICROBIOLOGY AND MOLECULAR BIOLOGY)」、ジョンウィリー・アンド・サンズ(John Wiley and Sons)、ニューヨーク、1994、および、ヘイルおよびマーカム(Hale & Markham)、「ハーパーコリンズ生物学辞典(HAPER COLLINS DICTIONARY OF BIOLOGY)」、ハーパーペレニアル(Haper Perennial)、ニューヨーク州、1991が、当業者には、本発明に使用される用語の多くについて一般的辞典となる。
【0025】
本明細書に示される見出しは、明細書全体を参照することによって理解することが可能な、本発明の様々な態様または実施態様を限定するものではない。従って、このすぐ後に定義される用語は、本明細書の全体を参照することによってより完全に定義される。
【0026】
数字の範囲は、その範囲を定義する数字を含める。
【0027】
別様に指示しない限り、それぞれ、核酸は、左から右へ、5’から3’方向に書かれ、アミノ酸は、左から右へ、アミノからカルボキシル方向に書かれる。
【0028】
本明細書および付属の特許請求項で使用される、単数形表現「この」および「前記」は、内容が明白に別様に指定しない限り、複数形を含む。従って、「化合物」を含む組成物に対する言及は、2種以上の化合物から成る混合物を含む。さらに、「または」という用語は、内容が明白に別様に指定しない限り、一般に「および/または」を含む意味に使用される。
【0029】
定義
「プロテアーゼ」は、微生物、例えば、真菌、細菌、または、植物または動物から得られるタンパクまたはポリペプチドの、タンパクまたはポリペプチドドメインであって、タンパク主鎖の様々な位置の内の一つにおけるペプチド結合の切断を触媒する能力を持つドメインである(例えば、E.C.3.4)。
【0030】
「酸性プロテアーゼ」とは、酸性条件下でタンパクを加水分解する能力を持つプロテアーゼを指す。
【0031】
本明細書で用いる「NSP24ファミリ・プロテアーゼ」とは、生来の、または野生型形態(例えば、図6のタンパク)において、プロテアーゼ活性を持つ酵素、すなわち、配列番号2または配列番号10のアミノ酸配列、配列番号2または配列番号10のアミノ酸配列の誘導体、アミノ酸配列の生物学的に活性な断片に対し、少なくとも50%、少なくとも60%、少なくとも70%、少なくとも80%、少なくとも85%、少なくとも90%、少なくとも93%、少なくとも95%、少なくとも97%、少なくとも98%、および少なくとも99%の配列相同性を有するプロテアーゼタンパクを意味する。
【0032】
本明細書で用いる「誘導体」とは、前駆体または親タンパク(例えば、自然のタンパク)から得られるタンパクであって、アミノ酸配列のC−およびN−末端のいずれかまたは両方に1個以上のアミノ酸を付加することによって、アミノ酸配列におけるいくつかの異なる部位において1個以上のアミノ酸を置換することによって、タンパクの一端または両端においてまたはアミノ酸配列の1箇所以上の部位において1個以上のアミノ酸を欠失させることによって、またはアミノ酸配列の1箇所以上の部位において1個以上のアミノ酸を挿入することによって得られるタンパクを意味する。
【0033】
本明細書で用いる「自然配列NSP24」または「野生型NSP24配列」とは、自然から得られたNSP24ファミリ・プロテアーゼと同じアミノ酸配列を持つポリペプチドを含む。
【0034】
「生物学的に活性な断片」(例えば、配列番号10の配列を有するNSP24ファミリ・プロテアーゼの、生物学的に活性な断片)とは、プロテアーゼ活性を持つが、NSP24ファミリ・プロテアーゼまたはNSP25ファミリ・プロテアーゼの前駆体または親タンパクの完全配列よりも少ない配列を含む、NSP24ファミリ・プロテアーゼまたはNSP25ファミリ・プロテアーゼを意味する。
【0035】
「単離」または「精製」とは、プロテアーゼが天然状態下で関連する天然成分の内の1種以上、またはその全てからプロテアーゼを分離することによってプロテアーゼがその天然状態から変えられていることを指す。
【0036】
「PepA」とは、配列番号7に対して少なくとも95%の配列相同性を有する酸性プロテアーゼを指す。
【0037】
「L388M」とは、配列番号7の配列を持つ変異種PepAを指す。
【0038】
本明細書で用いる「NSP25ファミリ・プロテアーゼ」とは、配列番号9および生物学的に活性なその断片に対して、少なくとも85%の配列相同性を持つプロテアーゼ酵素を意味する。
【0039】
「NSP24ファミリ・プロテアーゼに対して無関係な」とは、配列番号10のNSP24プロテアーゼに対して、30%未満の相同性、20%未満の相同性、または10%未満の相同性を持つアミノ酸配列を意味する。
【0040】
「ペプチド」、「タンパク」、および「ポリペプチド」という用語は、本明細書では相互交換的に用いられる。
【0041】
本明細書で用いる、特定されたアミノ酸またはヌクレオチドに関する「パーセント(%)配列同一性」とは、候補配列と対象配列(例えば、NSP24ファミリ・プロテアーゼ配列)とを整列し、要すればギャップを導入して、最大パーセント配列同一性を実現したあと、対象配列のアミノ酸残基またはヌクレオチドと同一である、候補配列のアミノ酸残基またはヌクレオチドのパーセントと定義される。なお、どの保存的置換体も配列同一性の一部であるとは見なさない。
【0042】
本明細書で用いる「アルファ・アミラーゼ(E.C.クラス3.2.1.1)」は、アルファ−1,4−グルコシド結合の加水分解を触媒する酵素を指す。これらの酵素はまた、1,4−α−結合D−グルコース単位を含むポリサッカライドにおける、1,4−α−D−グルコシド結合の、エキソまたはエンド加水分解を実行するものとしても記載される。これらの酵素を記載するために用いられるもう一つの用語は、「グリコゲナーゼ」である。例示の酵素としては、アルファ−1,4−グルカン4−グルカノヒドラーゼ、グルカノヒドロラーゼが挙げられる。
【0043】
本明細書で用いる「グルコアミラーゼ」という用語は、アミログルコシダーゼクラスの酵素(例えば、EC.3.2.1.3、グルコアミラーゼ、1,4−アルファ−D−グルカングルコヒドラーゼ)を指す。これらは、アミロースおよびアミロペクチン分子の非還元性末端からグルコシル残基を放出する、エキソ活動性酵素である。この酵素はまた、アルファ−1,6、およびアルファ−1,3結合をも加水分解する。もっともアルファ−1,4結合に対するよりもその速度はずっと遅い。
【0044】
「プロモーター」という用語は、遺伝子の転写を開始するためのRNAポリメラーゼの結合に関与する調整配列を意味する。
【0045】
本明細書で用いる「異種プロモーター」は、遺伝子、または精製核酸と天然では関連しないプロモーターである。
【0046】
本明細書で用いる、ポリペプチドの「精製標本」または「実質的に純粋な標本」とは、該ポリペプチドが天然では共に存在する、細胞、その他のタンパク、脂質、または核酸から分離されたポリペプチドを意味する。
【0047】
本明細書で用いる「細胞の精製標本」とは、植物または動物細胞の場合、細胞のインビトロ標本を指し、生の、植物または動物細胞全体を指さない。培養細胞または微生物細胞の場合、該標本は、対象細胞の少なくとも10%、より好ましくは50%を持つ標本から成る。
【0048】
「実質的に純粋な核酸」、例えば、実質的に純粋なDNAとは、該核酸の由来する生物体の天然ゲノムにおいて該核酸が直接連結する(例えば、一方は5’末端において、一方は3’末端において)配列、例えば、コード配列のいずれか一端または両端に対し直接連結しない、その一方または両方である核酸である。この用語は、例えば、ベクター、例えば、自律的複製プラスミドまたはウィルスに、または、前核細胞または真核細胞のゲノムDNAに組み込まれる組み換えDNA、あるいは、他のDNA配列と独立な分離分子として存在するDNA(例えば、PCRまたは制限エンドヌクレアーゼ処理によって生産されるcDNAまたはゲノムDNA断片)を含む。実質的に純粋なDNAはまた、さらに別のNSP24プロテアーゼ配列をコードするハイブリッド遺伝子の一部である組み換えDNAを含む。
【0049】
本明細書で用いる「相同性」とは、二つのポリペプチド分子間、または二つの核酸分子間の配列類似性を指す。二つの比較される配列の両方における一つの位置が、同じ塩基、またはアミノ酸モノマーサブユニットによって占められている場合、例えば、二つのDNA分子のそれぞれにおける一つの位置が、アデニンで占められている場合、これらの分子は、その位置において相同である。二つの配列間の相同性のパーセントは、該二つの配列が共有する、一致または相同位置の数を、比較される位置の数で割り、×100とした関数である。例えば、二つの配列の位置について、10の内の6が一致または相同である場合、この二つの配列は60%相同である。例を挙げると、DNA配列ATTGCCとTATGGCは、50%の相同を共有する。一般に、最大の相同性を与えるように二つの配列を整列させて比較を行う。
【0050】
本明細書で用いる「ベクター」という用語は、核酸を、1種以上の細胞タイプに導入するよう設計されたポリヌクレオチド配列をさす。ベクターとしては、クローニングベクター、発現ベクター、シャトルベクター、プラスミド、ファージ粒子、カセット等が挙げられる。
【0051】
本明細書で用いる「発現ベクター」とは、適切な宿主におけるDNAの発現に影響を及ぼすことが可能な適切な調節配列に動作的に結合するDNA配列を含む、DNA構築体を意味する。
【0052】
「発現」という用語は、ポリペプチドが、遺伝子の核酸配列に基づいて生産されるその過程を意味する。
【0053】
本明細書で用いる「動作的に結合する」とは、調整領域、例えば、プロモーター、ターミネーター、分泌シグナル、またはエンハンサー領域が、構造遺伝子に付着または結合し、その遺伝子の発現を調節することを意味する。
【0054】
本明細書で用いる、微生物「由来の」物質(例えば、ポリヌクレオチドまたはタンパク)とは、その物質が、その微生物に本来属するものであることを意味する。
【0055】
本明細書で用いる、「微生物」とは、細菌、真菌、ウィルス、原生動物、および他の微生物、すなわち顕微鏡的生物を指す。
【0056】
本明細書で用いる、「宿主株」または「宿主細胞」は、本発明によるDNAを含む発現ベクターにとって好適な宿主を意味し、前記細胞の子孫を含む。
【0057】
「糸状真菌」という用語は、全ての、糸状形態の下位区分Eumycotinaを指す(アレクソパウロス・シージェイ(Alexopoulos,C.J.)(1962),「真菌学入門(INTRODUCTORY MYCOLOGY)」、ウィリー(Wiley)、ニューヨーク州、および、エインスワースおよびビスビー(AINSWORTH AND BISBY)、「真菌の辞書(DICTIONARY OF THE FUNGI)」、9版、(2001)、カーク(Kirk)等編、シーエービー・インターナショナルユニバーシティ・プレス(CAB International University Press)、ケンブリッジ、英国を参照されたい)。これらの真菌は、キチン、セルロース、および、他の複雑なポリサッカライドによって構成される細胞壁を持つ、植物性菌糸によって特徴づけられる。本発明の糸状真菌は、形態的に、生理的に、かつ遺伝的に、酵母とは異なる。糸状真菌による植物的増殖は、菌糸の延長により、炭素の異化作用は好気性であることを強制される。
【0058】
本明細書で用いる、「Trichoderma」または「Trichoderma SPP.」は、以前または現在Trichodermaに分類される、任意の真菌属を指す。
【0059】
本明細書で用いる「4重欠失」または「4重欠失した」宿主細胞とは、機能的エンドグルカナーゼをコードする少なくとも2個の遺伝子、および、機能的セロビオヒドロラーゼをコードする少なくとも2個の遺伝子を欠失する、トリコデルマ宿主株の細胞から形成された細胞およびプロトプラストの両方を指す。
【0060】
本明細書で用いる「培養」という用語は、液体または固体媒体中の好適な条件下における微生物細胞集団の増殖を指す。一つの実施態様では、培養は、でん粉基質、例えば、顆粒状でん粉を含む基質の、最終産物(典型的には、血管またはリアクターにおける)への醗酵性生体変換を指す。醗酵は、より単純な有機化合物を生産するために微生物によって行われる、有機物質の酵素的で、かつ嫌気性の分解である。醗酵は嫌気的条件下で起こるが、醗酵はまた酸素の存在下でも起こるので、本用語は、厳密な嫌気的条件のみに限定されることを意図しない。
【0061】
本明細書で用いる「接触する」という用語は、それぞれの酵素(単複)を、それぞれの基質の十分近くに置くことによって、酵素(単複)が基質を最終産物に変換することを可能とすることを指す。当業者であれば、酵素溶液をそれぞれの基質と混合することが接触に作用する可能性のあることを了解されるであろう。
【0062】
核酸配列を細胞中に挿入するという背景で用いられる「導入」という用語は、「トランスフェクション」、「形質転換」、または「形質導入」を意味し、核酸配列の、真核または前核細胞への取り込みに対する参照を含む。その際、該核酸配列は、細胞のゲノム(例えば、染色体、プラスミド、プラスチド、または、ミトコンドリアDNA)に組み込まれてもよいし、自律的レプリコンに変換されてもよいし、または、一時的に発現されてもよい(例えば、形質導入mRNA)。
【0063】
本明細書で用いる、細胞に関して使用される「形質転換された」、「安定に形質転換された」、および「トランスジェニック」という用語は、細胞が、在来型ではない(例えば、異種の)核酸配列を、そのゲノムの中に一体化して、または、複数の世代に渡って維持されるエピソームプラスミドとして有することを意味する。
【0064】
本明細書で用いる、ポリペプチドまたはポリヌクレオチドと関連して使用される「異種の」という用語は、宿主細胞では天然には見られないポリペプチドまたはポリヌクレオチドを意味する。
【0065】
「過剰発現」という用語は、ポリヌクレオチドが宿主細胞に導入された場合、ポリペプチドを該宿主細胞において発現する過程を意味する。
【0066】
本明細書に記載するように、本発明の一つの態様は、NSP24ファミリ・プロテアーゼまたはNSP25ファミリ・プロテアーゼ、および/またはそれらの核酸の等価物をコードするヌクレオチド配列を含む「実質的に純粋な」(または組み換え)核酸を特徴とする。
【0067】
「等価的」という用語は、機能的に等価な複数のポリペプチドをコードする複数のヌクレオチド配列を指す。複数の、等価的ヌクレオチド配列は、1個以上のヌクレオチド置換、付加、または欠失、例えば、対立遺伝子変異において異なる、複数の配列を含む。例えば、ある実施態様では、遺伝コードの縮重のため、等価的ヌクレオチド配列は、配列番号2に示すNSP24プロテアーゼをコードするが、配列番号8のヌクレオチド配列と異なる配列を含む。
【0068】
本明細書で用いる「糖化」という用語は、でん粉をグルコースに変える酵素的転換を指す。
【0069】
本明細書で用いる「でん粉」とは、アミラーゼとアミロペクチンから構成される、xが任意の数である化学式(C6H10O5)xを持つ、植物の、複雑なポリサッカライド炭水化物から構成される任意の物質を指す。
【0070】
「顆粒状でん粉」という用語は、調理されていない(生の)でん粉(例えば、ゼラチン化されたことがないでん粉)を指す。
【0071】
本明細書で用いる「ゼラチン化」という用語は、でん粉分子を調理することによって可溶化し粘調な懸濁液を形成することを意味する。
【0072】
本明細書で用いる「液化」という用語は、でん粉転換の段階であって、ゲル化でん粉が加水分解されて低分子量の可溶性デキストランを生じる段階を指す。
【0073】
本明細書で用いる「可溶性でん粉加水分解産物」という用語は、でん粉の加水分解によって得られる可溶性産物であって、モノ−、ジ−、およびオリゴサッカライド(例えば、グルコース、マルトース、および多糖類)を含む産物を指す。
【0074】
「モノサッカライド」という用語は、ポリマー、例えば、でん粉のモノマー単位であって、重合度(DP)が1である単位(例えば、グルコース、マンノース、フルクトース、およびガラクトース)を意味する。
【0075】
「ジサッカライド」という用語は、二つの、共有的に結合するモノサッカライド単位を含む化合物(DP2)(例えば、スクロース、ラクトース、およびマルトース)を意味する。
【0076】
用語“DP3+”という用語は、3よりも大きな重合度を持つポリマーを意味する。
【0077】
プロテアーゼと、該プロテアーゼをコードするポリヌクレオチド
本発明は、NSPファミリ・プロテアーゼであって、配列番号2のプロテアーゼまたは配列番号10のプロレアーゼ(図6)に対し、少なくとも50%、少なくとも60%、少なくとも70%、少なくとも80%、少なくとも85%、少なくとも90%、少なくとも93%、少なくとも95%、少なくとも97%、少なくとも98%、および少なくとも99%の配列相同性を有するプロテアーゼ、例えば、酸性プロテアーゼ、およびさらに酸性真菌プロテアーゼに関する。ある実施態様では、NSP24ファミリ・プロテアーゼは、配列番号10の配列(成熟タンパク配列)を含むNSP24、あるいは、配列番号2のプレタンパク配列とも表示される。
【0078】
ある実施態様では、本発明は、NSP24ファミリ・プロテアーゼの生物学的活性を持つ断片に関する。ある実施態様では、生物学的活性断片は、少なくとも250個のアミノ酸残基、少なくとも300個のアミノ酸残基、少なくとも350個のアミノ酸残基、少なくとも375個のアミノ酸残基、および少なくとも400個のアミノ酸残基を有するプロテアーゼを含む。
【0079】
別の実施態様では、生物学的活性断片は、図6のタンパク配列(配列番号2または配列番号10)に対し、少なくとも80%、少なくとも85%、少なくとも90%、少なくとも93%、少なくとも95%、少なくとも97%、少なくとも98%、および少なくとも99%の配列相同性を有するポリペプチド配列の、少なくとも60%、少なくとも70%、少なくとも80%、少なくとも85%、少なくとも90%、少なくとも93%、少なくとも95%、少なくとも97%、少なくとも98%、および少なくとも99%を含む。ある実施態様では、生物学的活性断片は、配列番号2または配列番号10を持つ親NSP24プロテアーゼに対し、少なくとも95%の配列相同性を有するポリペプチド配列の、少なくとも60%、少なくとも70%、少なくとも80%、少なくとも85%、少なくとも90%、少なくとも95%、少なくとも98%を含む。ある実施態様では、生物学的活性断片は、配列番号2または配列番号10を持つ親NSP24プロテアーゼに対し、少なくとも99%の配列相同性を有するポリペプチド配列の、少なくとも60%、少なくとも70%、少なくとも80%、少なくとも85%、少なくとも90%、少なくとも95%、少なくとも98%を含む。
【0080】
ある実施態様では、生物学的活性断片は、生体内に存在する断片、例えば、翻訳後処理によって生ずる断片、または、選択的スプライシングを受けたRNAの翻訳によって生ずる断片である。断片は、天然または内在細胞において、例えば、翻訳後処理の結果、例えば、アミノ末端シグナル配列除去の結果として発現されるものを始め、発現システム、例えば、CHO細胞において発現されるものも含む。ある好ましい断片は、タンパク分解による切断、または選択的スプライシング事象によって生じた断片、例えば、活性断片である。NSP24ファミリ・プロテアーゼのようなプロテアーゼは、ある範囲の生理学的性質を示すことがよくあり、かつ、そのような性質は、分子の異なる部分に由来することがあるのであるから、NSP24ファミリ・プロテアーゼの有用断片または類縁体は、NSP24プロテアーゼ活性に関する任意の生物学的アッセイにおいて生物学的活性を示すものである。
【0081】
ある実施態様では、生物学的活性断片は、配列番号2または配列番号10を持つNSP24のプロテアーゼ活性の少なくとも20%、少なくとも30%、少なくとも40%、少なくとも50%、少なくとも60%、少なくとも70%、少なくとも80%、少なくとも90%、少なくとも95%、少なくとも100%を含む。ある好ましい実施態様では、断片または類縁体は、任意のインビボまたはインビトロのNSP24プロテアーゼアッセイにおいて、NSP24プロテアーゼ(配列番号2または配列番号10)のプロテアーゼ活性の少なくとも40%、または少なくとも90%を持つ。
【0082】
NSP24ファミリ・プロテアーゼまたはNSP25ファミリ・プロテアーゼの断片は、当業者に既知の方法によって生成することが可能である。候補断片が、プロテアーゼの生物学的活性を示す能力は、本明細書に記載するように、当業者に既知の方法によって評価することが可能である。さらに、NSP24ファミリ・プロテアーゼおよびNSP25ファミリ・プロテアーゼであって、該ペプチドの生物学的活性に必要とされない残基を含むもの、または、選択的mRNAスプライシング、または、タンパクの選択的プロセシングによって生じるものも含まれる。
【0083】
ある実施態様では、本発明によって被われるプロテアーゼは、配列番号2または配列番号10を持つプロテアーゼの誘導体である。誘導体は、配列番号10に対し、少なくとも80%、85%、90%、93%、95%、97%、98%、および99%の配列相同性を持ってもよい。
【0084】
本発明はまたプロテアーゼ類縁体を含む。類縁体とは、ペプチドの安定性を増す修飾を伴うものである。そのような類縁体は、例えば、ペプチド配列の中に、1個以上の非ペプチド結合(ペプチド結合を置換する)を含んでもよい。さらに含まれるのは、天然のL−アミノ酸以外の残基、例えば、D−アミノ酸、または非天然性または合成アミノ酸、例えば、βアミノ酸、および環状類縁体を含む類縁体である。類縁体は、天然のプロテアーゼと、例えば、NSP24またはNSP25タンパクと、アミノ酸配列において、または、配列を含まないやり方で、または、その両方において異なっていてもよい。非配列修飾は、本発明に含まれるプロテアーゼの、インビトロまたはインビボにおける、化学的誘導体形成を含む。非配列修飾としては、アセチル化、メチル化、リン酸化、カルボキシル化、またはグリコシル化における変化が挙げられる。
【0085】
さらに別の実施態様では、本発明はNSP25ファミリ・プロテアーゼを含む。NSP25ファミリ・プロテアーゼは、配列番号9(図9)の成熟タンパク配列、またはその生物学的に活性な断片に対して、少なくとも85%、少なくとも90%、少なくとも93%、少なくとも95%、少なくとも97%、少なくとも98%、および少なくとも99%のアミノ酸配列相同性を持つ酸性プロテアーゼである。一つの特異的NSP25ファミリ・プロテアーゼは、配列番号9を持つNSP25と表示されるプロテアーゼである。ある実施態様では、NSP25ファミリ・プロテアーゼは、配列番号9に対し少なくとも90%の配列相同性を有する配列の、少なくとも75%、少なくとも80%、少なくとも85%、少なくとも90%、少なくとも95%を含む、プロテアーゼの生物学的活性断片である。別の実施態様では、NSP25ファミリ・プロテアーゼは、配列番号9に対し少なくとも95%の配列相同性を有する配列の、少なくとも75%、少なくとも80%、少なくとも85%、少なくとも90%、および少なくとも95%を含む、プロテアーゼの生物学的活性断片である。
【0086】
本発明による酸性プロテアーゼは、酸性条件下でタンパクを加水分解することが可能なプロテアーゼであるが、ある実施態様では、プロテアーゼ活性のための至適pHは、pH3.0から5.5の範囲にある。ある実施態様では、プロテアーゼ活性のための至適pH範囲は、pH3.0と5.0の間にあるが、別の実施態様では、プロテアーゼ活性のための至適pH範囲は、pH3.0と4.5の間にある。
【0087】
本発明によるプロテアーゼ、例えば、NSP24ファミリ・プロテアーゼまたはNSP25ファミリ・プロテアーゼは、アミノ酸置換、例えば、L−アミノ酸による置換であって、一つのアミノ酸が別の生物学的に類似のアミノ酸によって置換される「保存的アミノ酸置換」を含んでもよい。保存的アミノ酸置換とは、置換されるアミノ酸の、全体的電荷、疎水性/親水性、および/または、立体的広がりを保存する置換である。保存的置換の例として、下記の基の間の置換が挙げられる。すなわち、Gly/Ala、Val/Ile/Leu、Lys/Arg、Asn/Gln、Glu/Asp、Ser/Cys/Thr、およびPhe/Trp/Tyrである。誘導体は、例えば、1から10個までのアミノ酸残基が異なっていてもよい。例えば、6−10、5、4、3、2、または1個のアミノ酸残基が異なっていてもよい。表1は、従来技術で認められたアミノ酸置換の例を示す。さらに、置換は、プロテアーゼ生物活性を消滅させることのない、1個以上の非保存的アミノ酸置換、欠失、または挿入によって行われてもよい。
【表1】

【0088】
ある実施態様では、本発明のプロテアーゼは内在配列である。このような内在配列は、天然から単離することが可能であり、あるいは、組み換えまたは合成手段によって製造することが可能である。この「内在配列」という用語は、NSP24またはNSP25ファミリ・プロテアーゼの天然に見られる短縮形または分泌形(例えば、生物学的活性断片)、および、天然に見られる変異形(例えば、選択的スプライシングを受けた変異形)を特に含む。
【0089】
ある実施態様では、本発明の酸性プロテアーゼは、配列番号7に対して、少なくとも97%、少なくとも98%、少なくとも99%の配列相同性を持つPepAプロテアーゼである。ある実施態様では、プロテアーゼは、配列番号7の配列を持ち、“L388M”と表示される。さらに別の実施態様では、プロテアーゼは、配列番号5、または配列番号3の配列を持つヌクレオチド配列によってコードされる。
【0090】
本発明はまた、本発明に含まれるプロテアーゼをコードするポリヌクレオチドに関する。このようなポリヌクレオチドは下記を含む。すなわち、
a)配列番号2または配列番号10に対して、少なくとも80%、少なくとも85%、少なくとも90%、少なくとも93%、少なくとも95%、少なくとも97%、少なくとも98%、および少なくとも99%の配列相同性を持つNSP24ファミリ・プロテアーゼをコードするポリヌクレオチド;
b)配列番号2の配列をコードするポリヌクレオチド;
c)配列番号8の配列を持つポリヌクレオチド;
d)NSP24ファミリ・プロテアーゼの生物学的活性断片をコードするポリヌクレオチド;
e)配列番号8に対して、少なくとも80%、少なくとも85%、少なくとも90%、少なくとも95%、少なくとも97%、少なくとも98%、および少なくとも99%の配列相同性を持つポリヌクレオチド;
f)配列番号4、配列番号8のDNA配列、または、配列番号4または配列番号8の断片に対応する核酸プローブに対してハイブリダイズするポリヌクレオチドであり、その際、前記断片は、少なくとも10、15、20、30、40、50、60、70、80、90、100、または150個の連続ヌクレオチドを有し;
g)配列番号4に対して、少なくとも85%、少なくとも90%、少なくとも93%、少なくとも95%、少なくとも97%、少なくとも98%、および少なくとも99%の配列相同性を持つNSP25ファミリ・プロテアーゼをコードするポリヌクレオチド;
h)配列番号9のプロテアーゼをコードするポリヌクレオチド;
i)配列番号4の配列を持つポリヌクレオチド;
j)NSP25ファミリ・プロテアーゼの生物学的活性断片をコードするポリヌクレオチド;
k)配列番号7の配列、およびその生物学的活性断片をコードするポリヌクレオチド;および、
l)配列番号3または配列番号5の配列を持つポリヌクレオチド。
【0091】
遺伝子コードには縮重があるために、ある特定のアミノ酸をコードするのに1種を超えるコドンが使用される場合がある。従って、異なるいくつかのDNA配列が、同じアミノ酸配列を持つポリペプチド、例えば、配列番号2のポリペプチドをコードする可能性がある。本発明は、同じポリペプチドをコードする複数のポリペプチドをカバーする。
【0092】
核酸は、該核酸の1本鎖型が、適当な温度およびイオン強度条件下において別の核酸とアニールすることが可能である場合、その他方の核酸配列にたいしてハイブリダイズすることが可能である。ハイブリダイゼーションおよび洗浄条件は、低度、中等度、高度、および、極めて高度の厳格条件下におけるハイブリダイゼーションに関して従来技術でよく知られる(例えば、サムブルック(Sambrook)(1989)、上記、特に9および11章参照)。一般に、ハイブリダイゼーションは、ヌクレオチドプローブと相同的DNA配列を含み、これらは、相補的ポリヌクレオチドの広範な塩基対合によって安定な二本鎖ハイブリッドを形成する(さらに、8章および、「遺伝子クローニング入門(Gene Cloning,An Introduction)」、ティーエー・ブラウン(T.A.Brown)、(1995)、Chapman and Hall、ロンドンを参照されたい)。ある実施態様では、プローブおよび相同配列を有するフィルターは、2×塩化ナトリウム/クエン酸ナトリウム(SSC)、約60℃(中等厳格度)、65℃(中等/高等厳格度)、70℃(高等厳格度)、および約75℃(極高等厳格度)における0.5%SDSで洗浄してもよい(「分子生物学における最新プロトコール(Current Protocols in Molecular Biology)」、John Wiley & Sons,ニューヨーク州、1989、6.3.1−6.3.6、なお、この文献を引用することにより本明細書に含める)。
【0093】
本発明に含まれるものは、対立遺伝子変異;自然突然変異;誘発突然変異;配列番号2、配列番号8、配列番号9、および配列番号10のポリペプチドをコードする核酸に対して、高または低厳格条件下でハイブリダイズするDNAによってコードされるタンパク、および、配列番号2または配列番号10を持つNSP24プロテアーゼに対する抗血清によって、特にNSP24プロテアーゼの活性部位または結合ドメインに対する抗血清によって特異的に結合されるポリペプチドである。ある実施態様では、本発明のNSP24ファミリ・プロテアーゼをコードする核酸、例えば、配列番号2のNSP24プロテアーゼをコードする核酸は、配列番号8から得られる少なくとも12、15、または20の連続ヌクレオチドに対応する核酸に対して高度の厳格条件下にハイブリダイズする。
【0094】
本発明の核酸およびポリペプチドは、本明細書に開示される配列とは、開示の配列における配列誤差のために異なるものも含む。
【0095】
DNA配列の相同性は、二つのDNA配列間の同一性の程度によって決められる。複数のポリペプチド配列またはヌクレオチド配列について、相同性または同一性パーセントが、コンピュータを用いて求められてもよい。配列整列を実行し、配列同一性を定める方法は当業者には既知であり、不要な実験を行うことなく実行が可能であり、同一性値の計算も明確に獲得することが可能である。例えば、アウスウーベル(Ausubel)等編、(1995)、「分子生物学における最新プロトコール(Current Protocols in Molecular Biology)」、19章、(Greene Publishing and Wiley−Interscience),ニューヨーク州;および、デイホフ(Dayhoff)(1978)「タンパク科学および構造アトラス5(Atlas of Protein Science and Structure 5)」、別冊3(National Biomedical Research Foundation,Washingtong,D.C.)を参照されたい。複数の配列を整列し、配列同一性を求めるためにはいくつかのアルゴリスムが利用可能であり、例えば、ニードルマン(Needleman)等(1970)J.Mol.Biol.48:443の相同性整列アリゴリスム;スミス(Smith)等(1981)Adv.Appl.Math.2:482の局部的相同性アルゴリスム;ピアソン(Pearson)等(1988)Proc.Natl.Acad.Sci.85:2444の類似性探索法;スミス・ウォーターマン法(Meth.Mol.Biol.70:173−187(1997)、および、BLASTP、BLASTN、およびBLASTXアルゴリスム(アルチュル(Altschul)等(1990)J.Mol.Biol.215:403−410を参照)が挙げられる。これらのアルゴリスムを用いるコンピュータプログラムも利用が可能であり、例えば、ALIGNまたはMegalign(DNASTAR)ソフトウェア、またはWU−BLAST−2(アルチュル(Altschul)等、Meth.Enzym.,266:460−480(1996));またはGAP、BESTFIT、BLASTアルチュル等上記、FASTA、および、Genetics Computing Group (GCG)パッケージ、バージョン8、マジソン、ウィスコンシン州、米国において市販されるTFASTA、および、Intelligenetics,Mountain View、カリフォルニア州によるPC/Geneプログラムに納められるCLUSTALが挙げられるが、ただしこれらに限定されない。当業者であれば、比較される複数の配列の長さに渡って最大の整列を実現するのに必要なアルゴリスムを含め、整列を測定するための適切なパラメータを決めることが可能である。配列同一性は、プログラムによって決められるデフォールトパラメータを用いて決められることが好ましい。具体的に言うと、配列同一性は、MSPRCHプログラム(Oxford Molecular)に搭載されるスミス・ウォーターマン相同性探索アルゴリスムによって求めることが可能である。このアルゴリスムは、下記の探索パラメータ、すなわち、ギャップ開放ペナルティー12、およびギャップ延長ペナルティー1によるアフィンギャップ探索を用いる。対合アミノ酸比較は、Genetics Computer Group,Inc.マジソン、ウィスコンシン州のGCG配列分析ソフトウェアパッケージのGAPプログラムを用いて実行することが好ましい。このプログラムは、ギャップウェイト12、長さウェイト2とする、blosum62アミノ酸置換マトリックスを用いる。二つのアミノ酸配列の最適整列に関しては、変異アミノ酸配列の連続セグメントは、参照アミノ酸配列に対して、付加的アミノ酸残基を持ってもよいし、または、アミノ酸残基を欠失させてもよい。参照アミノ酸配列との比較に用いられる連続セグメントは、少なくとも20個の連続アミノ酸残基を含むが、30、40、50個、またはそれ以上のアミノ酸残基であってもよい。誘導体のアミノ酸配列にギャップを含めたことによって生ずる配列同一性の増加に対する補正は、ギャップペナルティーを割り当てることによって実行される。
【0096】
ある実施態様では、本発明によってカバーされるプロテアーゼ(例えば、配列番号2の配列に対して少なくとも80%の配列同一性を持つNSP24ファミリ・プロテアーゼ)は、細菌、または真菌、例えば、糸状真菌から得られる。ある好ましい糸状真菌としては、Aspergillus SPP.、およびTrichoderma SPP.が挙げられる。一つの好ましいTrichoderma SPP.は、T.reeseiである。しかしながら、プロテアーゼ、および/または、本発明によるプロテアーゼをコードするDNAは、真菌、例えば、Absidia SPP.;Acremonium SPP.;Agaricus SPP.;Anaeromyces SPP.;Aspergillus SPP.、例えば、A.aculeatus、A.awamori、A.flavus、A.foetidus、A.fumaricus、A.fumigatus、A.nidulans、A.niger、A.oryzae、A.terreus、およびA.versicolor;Aerobasidium SPP.;Cephalosporum SPP.;Chaetomium SPP.;Coprinus SPP.;Dactyllum SPP.;Fusariumu SPP.、例えば、F.conglomerans,F.decemcellulare,F.javanicum、F.lini,F.oxysporum,およびF.solani;Gliocladium SPP.;Humicola SPP.、例えば、H.insolensおよびH.lanuginosa;Mucor SPP.;Neurospora SPP.、例えば、N.crassaおよびN.sitophila;Neocallimastix SPP.;Orpinomyces SPP.;Penicillium SPP.;Phanerochaete SPP.;Phlebia SPP.;Piromyces SPP.;Rhizopus SPP.;Schizophyllum SPP.;Trametes SPP.;Tricoderma SPP.、例えば、T.reesei、T.reesei(longbrachiatum)およびT.virdie;およびZygorhynchus SPP.のような真菌から得られる。
【0097】
宿主細胞
ある実施態様では、本発明は、本明細書に記載されるDNA構築体およびベクターによって形質転換された宿主細胞を提供する。ある実施態様では、本発明によってカバーされるプロテアーゼ(例えば、配列番号2に対して少なくとも95%の配列同一性を有するNSP24ファミリ・プロテアーゼ)をコードするポリヌクレオチドであって、宿主細胞中に導入されるポリヌクレオチドは、異種プロテアーゼをコードし、別の実施態様では、ポリヌクレオチドは、宿主細胞において過剰発現される内因性プロテアーゼをコードする。ある実施態様では、本発明は、宿主細胞、例えば、細菌および真菌宿主細胞において機能する遺伝子プロモーターの制御下における、異種プロテアーゼ遺伝子の発現、またはプロテアーゼ遺伝子の過剰発現を提供する。
【0098】
ある好ましい宿主細胞としては糸状真菌細胞が挙げられる。糸状真菌宿主細胞の非限定的例として、Trichoderma SPP.(例えば、T.virideおよびT.reesei、これらは、以前T.longibrachiatumとして分類されていた、Hypocrea jecorinaの無性形である)、Penicillium SPP.、Humicola SPP.(例えば、H.insolensおよびH.grisea)、Aspergillus SPP.(例えば、A.niger、A.nidulans、A.orzyae、および、A.awamori)、Fusarium SPP.(F.graminum)、Neurospora SPP.、Hypocrea SPP.、およびMucor SPP.が挙げられる。さらに別の宿主細胞として、Bacillus SPP.(例えば、B.licheniformis、B.lentus、B.stearothremophilus、およびB.brevis)、およびStreptomyces SPP.(例えば、S.coelicolorおよびS.lividans(TK23およびTK21))が挙げられてもよい。
【0099】
分子生物学
本発明は、組み換え遺伝子工学の分野における通例技術に依存する。本発明に使用される一般的方法を開示する基本的教科書としては、サムブルック(Sambrook)等の「分子クローニング、実験室マニュアル(“Molecular Cloning,A Laboratory Manual”)」第2版、1989年;クリーグラー(Kriegler)、「遺伝子の転送と発現、実験室マニュアル(“Gene Transfer and Expression: A Laboratory Manual”)」、および、アウスウーベル(Ausubel)等、「分子生物学における最新プロトコール(“Current Protocols in Molecular Biology”)」(1994)が挙げられる。
【0100】
例えば、糸状真菌の遺伝子プロモーター配列を含む異種遺伝子は、通常、複製および/または発現のために、宿主細胞、例えば、Trichoderma reesei細胞に変形導入される前に、中間ベクターにクローンされる。これらの中間ベクターは、通常、前核細胞性ベクター、例えば、プラスミド類、またはシャトルベクターである。
【0101】
クローンされた遺伝子の高レベル発現を実現するためには、異種遺伝子は、プロモーターから、天然遺伝子の場合とほぼ同じ距離離れた位置に置かれることが好ましい。しかしながら、従来技術で知られるように、この距離における若干の変動は、プロモーター機能の損失を招くこと無く容認することが可能である。
【0102】
当業者であれば、天然のプロモーターは、その機能を変更することなく、1個以上のヌクレオチドの代置、置換、付加、または除去することによって修飾することが可能であることを知っている。本発明の実施は、プロモーターに対するそのような変更を含むが、そのような変更によって制限されない。
【0103】
発現ベクター/構築体は、通常、該異種配列の発現に必要な全ての付加的要素を含む、転写単位または発現カセットを含む。従って、ある典型的発現カセットは、異種核酸配列に動作的に結合するプロモーターと転写物の効率的アデニル化に必要なシグナル、リボソーム結合部位、および翻訳終止を含む。カセットの付加的要素としては、エンハンサー、および、もしも構造遺伝子としてゲノムDNAが使用されるのであれば、機能的スプライスドナーおよびアクセプター部位を有するイントロンが挙げられる。
【0104】
本発明の実施は、遺伝子構築体におけるプロモーターの選択によって制限されない。しかしながら、例示のプロモーターとしては、Trichoderma reesei cbh1、cbh2、eg1、eg2、eg3、eg5、xln1、およびxln2プロモーターがある。さらに、A.awamoriおよびA.nigerのグルコアミラーゼ遺伝子(glaA)のプロモーター(ヌンベルク(Nunberg)等(1984)Mol.Cell Biol.4:2306−2315)、およびA.nidulansのアセタミナーゼのプロモーターも、ベクターへの使用が可能である。Bacillus subtilisに使用されるベクターのための好ましいプロモーターは、AprEプロモーターであり;E.coliに使用される好ましいプロモーターは、Lacプロモーターであり;Saccharomyces cerevisiaeに使用される好ましいプロモーターはPGK1であり;Aspergillus nigerに使用される好ましいプロモーターはglaAであり;かつ、Trichoderma reesei用として好ましいプロモーターはcbh1である。
【0105】
プロモーター配列の外に、発現カセットは、効率的な終止を実現するために、構造遺伝子の下流に転写終止領域を含まなければならない。終止領域は、プロモーター配列と同じ遺伝子から得てもよいし、あるいは、異なる遺伝子から得てもよい。
【0106】
本発明においては、真菌のものであればいずれのターミネーターも機能性を持つと考えられるが、ある好ましいターミネーターとしては、Aspergillus nidulans trpC遺伝子(イェルトン・エム(Yelton,M)等、(1984)PNAS USA81:1470−1474;ムラニー・イージェイ(Mullaney,E.J.)等、(1985)MGG199:37−45)、Aspergillus awamoriまたはAspergillus nigerのグルコアミラーゼ遺伝子(ヌンベルク・ジェイエイチ(Nunberg,J.H.)Mol.Cell Biol.4:2306;ベール・イー(Boel,E)等、(1984)EMBO J.3:1581−1585)、Aspergillus oryzae TAKAアミラーゼ遺伝子、およびMucor miheiのカルボキシルプロテアーゼ遺伝子(EPO公報第0215594号)、のターミネーターが挙げられる。
【0107】
遺伝子情報を細胞に輸送するために使用される、特定の発現ベクターは、特に決定的に重要とはならない。真核細胞または前核細胞における発現のために使用される通例のベクターの内の任意のものが使用可能である。標準的細菌性発現ベクターとしては、バクテリオファージλおよびM13の外に、プラスミド、例えば、pBR322準拠プラスミド、pSKF、pET23D、および、融合発現システム、例えば、MBP、GST、およびLacZが挙げられる。分離に好都合の方法を実現するために、組み換えタンパクに対してエピトープタグ、例えば、c−mycを付加することも可能である。好適な発現および/または統合ベクターの例は、サムブルック(Sambrook)等(1989)上記;ベネットおよびラシュア(Bennett and Lasure)編「真菌における遺伝子操作の進歩(“More Gene Manipulation in Fungi“)」(1991)、Academic Press pp.70−76およびpp.396−428、および同書の中に引用される論文;米国特許第5,874,276号;および、真菌遺伝子保存センターの真菌株カタログ(FGSC、www.fgsc.net.)の中に挙げられている。有用なベクターは、PromegaおよびInvitrogenから入手することが可能である。ある、有用な特異的ベクターとして、pBR322、pUC18、pUC100、pDONTM201、pENTRTM、pGEN(R)3Z、およびpGEN(R)4Zが挙げられる。しかしながら、本発明は、等価的機能を果たし、現に従来技術に知られる、または、これから知られるようになる他の形態の発現ベクターも含むことが意図される。従って、本発明のDNA配列の発現には、多種多様の宿主/発現ベクターの組み合わせの採用が可能である。例えば、有用な発現ベクターは、染色体の、非染色体の、および合成のDNA配列のセグメント、例えば、SV40および既知の細菌プラスミドの既知の各種誘導体から構成されてもよい。プラスミドとしては、例えば、colE1、pCR1、pBR322、pBb9、pUC19、およびそれらの誘導体を含む、Ecoli由来のプラスミド、さらに広い宿主範囲を持つプラスミド、例えば、RP4、ファージDNA,例えば、ラムダファージの多数の誘導体、例えば、NM989、およびその他のDNAファージ、例えば、M13、および、糸状の、1本鎖DNAファージ、酵母プラスミド、例えば、2.muプラスミドまたはその誘導体が挙げられる。
【0108】
ある実施態様では、発現ベクターは選択可能なマーカーを含む。選択可能なマーカーとしては、抗菌剤耐性を付与するものが挙げられる。栄養マーカー、amdS、argB、およびpyr4のような従来技術で既知のマーカーを含む栄養マーカーも本発明において使用される。Trichodermaの形質転換に有用なマーカーは、従来技術で既知である(例えば、フィンケルスタイン(Finkelstein)、6章、「糸状真菌の生物工学」、フィンケルスタイン等、EDS Butterworth−Heinemann、ボストン、マサチューセッツ州(1992)、および、キングホーン(Kinghorn)等、(1992)、「糸状真菌の応用分子遺伝学(”Applied Molecular Genetics of Filamentous Fungi“)」Blackie Academic and Professional、Chapman and Hall、ロンドンを参照されたい)。ある実施態様では、発現ベクターはまた、レプリコン、組み換えプラスミドを保有する細菌の選択を可能とする、抗生物質耐性をコードする遺伝子、および、異種配列の挿入を可能とするために、非必須領域に独特の制限部位を含んでもよい。選ばれる、特定の抗生物質耐性遺伝子は決定的に重要ではなく、従来技術で既知の多くの耐性遺伝子の内のいずれものでも好適である。前核細胞配列は、Trichoderma reeseiにおけるDNAの複製または統合を妨げないように選択されることが好ましい。
【0109】
本発明の形質転換法によって、形質転換ベクターの一部または全てが、宿主細胞、例えば、糸状真菌宿主細胞のゲノムの中に安定的に統合される。しかしながら、自己複製、染色体外形質転換ベクターの維持をもたらす形質転換も本発明の範囲内である。
【0110】
大量のプロテアーゼを発現する、細菌および糸状真菌(例えば、AspergillusまたはTrichoderma)細胞系統を生み出すために、多くの標準的トランスフェクション法を用いることが可能である。Trichodermaのセルラーゼ産生株の中にDNA構築体を導入するための公表された方法のいくつかを述べると、ロリート、ヘイエス、ジピエルト、およびハーマン(Lorito,Hayes,DiPierto and Harman)、(1933)Curr.Genet.24:349−356;ゴールドマン、バンモンターギュ、およびヘレラ・エストレヤ(Goldman,VanMontagu and Herrera−Estrella)(1990)Curr.Genet.17:169−174;および、ペンチラ、ナバレイネン、ラット、サルミネン、およびノウレス(Penttila,Nevalainen,Ratto,Salminen and Knowles)(1987)Gene 6:155−164が挙げられる。さらに、米国特許第6,022,725号、米国特許第6,268,328号、およびネバライネン(Nevalainen)等、「Trichodermaの分子生物学、および、同種および異種遺伝子の発現に向けたその応用(“The Molecular Biology of Trichoderma and its Application to the Expression of Both Homologous and Heterologous Genes“」、Molecular industrial Mycology,レオンおよびベルカ(Leong and Berka)編、Marcel Dekker Inc.,ニューヨーク州(1992)pp129−148を参照されたい。Aspergillusについては、イェルトン、ヘイマー、およびティンバーレイク(Yelton,Hamer,and Timberlake)(1984)Proc.Natl.Acad.Sci.USA 81:1470−1474を;Fusariumについては、ベイジャー、ポディラ、およびコラッツクディ(Bajar,Podila,and Kolattukudy)(1991)Proc.Natl.Acad.Sci.USA 88:8202−8212を;Streptomycesについては、ホップウッド(Hopwood)等、1985、「Streptomycesの遺伝子操作:実験室マニュアル(”Genetic Manipulation of Streptomyces:Laboratory Manual“)」、The John Innes Foundation,Norwich、英国、およびフェルナンデス・アバロス(Fenrnandez−Abalos)等、Microbiol.149:1623−1632(2003);および、Bacillusについては、ブリジディ、デロッシ、ベルタリニ、リッカルディ、およびマッテウッツィ(Brigidi,DeRossi,Bertarini,Riccardi and Matteuzzi)(1990)FEMS Microbiol.Lett.55:135−138)が挙げられる。
【0111】
しかしながら、宿主細胞に外来ヌクレオチド配列を導入するための、よく知られた過程の内の任意のものの使用が可能である。そのような過程としては、リン酸カルシウムの使用、トランスフェクション、ポリブレン、プロトプラスト融合、電気穿孔、biolistics、リポソーム、マイクロインジェクション、プラスマベクター、ウィルスベクター、および、クローンされたゲノムDNA、cDNA,合成DNA、または、他の外来遺伝物質を、宿主細胞に導入するための、その他の、既知の、任意の方法(例えば、サムブルック等、上記参照)が挙げられる。さらに、米国特許第6,255,115号に記載されるAgrobacterium仲介トランスフェクション法も使用される。使用される特定の遺伝子工学過程は、少なくとも1個の遺伝子を、該遺伝子を発現することが可能な宿主細胞に適切に導入することが可能でありさえすればよい。一つの実施態様では、本発明は、本発明によってカバーされるプロテアーゼ(例えば、NSP24ファミリ・プロテアーゼ)を生産するための方法に関する。この方法は、プロテアーゼ、例えば、NSPファミリ・プロテアーゼをコードする核酸に対して動作的に結合するプロモーターを含むポリヌクレオチドを宿主細胞に導入すること、プロテアーゼの発現と生産のために好適な培養条件下に宿主細胞を培養すること、および、前記プロテアーゼを生産することを含む。ある好ましい実施態様では、プロテアーゼは、配列番号2、または配列番号10、またはそれらの生物学的活性を持つ断片に対し少なくとも95%の配列同一性を有するNSP24ファミリ・プロテアーゼである。
【0112】
発現ベクターが細胞に導入された後、そのトランスフェクトされた、または形質転換された細胞は、プロテアーゼ遺伝子のプロモーター配列の制御下に、遺伝子発現を促進する条件下に培養される。大型バッチの形質転換細胞を、後述の実施例3に記載するやり方で培養することが可能である。最後に、産物は、標準技術によって培養物から回収される。
【0113】
上記から、本明細書の発明は、所望のポリペプチドの発現および分泌増大を実現する。すなわち、該ポリペプチドの発現は、天然プロテアーゼ遺伝子、融合DNA配列、および各種異種構築体を含む遺伝子プロモーター配列によって制御される。本発明はまた、このような所望のポリペプチドを高レベルで発現し、分泌するための過程を提供する。
【0114】
タンパクの発現
本発明のタンパクは、ベクター、例えば、その発現が遺伝子プロモーター配列によって制御される遺伝子を含む発現ベクターによって形質転換された細胞を培養することによって生産される。本発明は、タンパク、例えば、本発明によってカバーされるプロテアーゼの細胞内および/または細胞外生産を強化するのに特に有用である。前記遺伝子の発現にとって好適な条件とは、本発明の、誘導飼料組成物を培養物に供給することを含む。タンパク生産のための最適条件は、宿主細胞の選択、および、発現されるプロテアーゼタンパクの選択と共に変動する。そのような条件は、当業者であれば、通例の実験または最適化によって簡単に確認される。
【0115】
所望のプロテアーゼタンパクは、発現後、分離または回収し、精製してよい。所望のタンパクは、サンプルの中に他にどのような成分が存在するかに応じて、当業者に既知の様々なやり方に従って分離または精製してよい。標準的精製法としては、電気泳動的、分子、免疫学的、およびクロマトグラフィック技術、例えば、イオン交換、疎水性、アフィニティー、および逆相HPLCクロマトグラフィー、およびクロマトフォーカシングが挙げられる。例えば、興味のタンパクは、標準的、所望の抗タンパク抗体カラムを用いて精製してもよい。好適な精製技術に関する一般的案内については、「概観、タンパク精製(”Scopes Protein Purification“)」(1982)を参照されたい。必要な精製の程度は、所望のタンパクの用途に応じて変動する。ある実施態様では、精製は必要とされない。
【0116】
細胞培養
宿主細胞および形質転換細胞は、通常の栄養媒体中で培養することが可能である。形質転換された宿主細胞用の培養媒体は、プロモーターを活性化し、形質転換体を選択するために適当に修飾してもよい。特異的培養条件、例えば、温度、pH等は、発現のために選択された宿主細胞のために使用されるものであってよく、当業者には明白である。さらに、好ましい培養条件は、科学的文献、例えば、サムブルック(1982)上記;キーサー・ティー、エムジェイ・ビブ、エムジェイ・バットナー、ケーエフ・チェイター、およびディーエー・ホップウッド(Kieser,T,MJ.Bibb,MJ.Buttner,KF.Chater,and D.A.Hopwood)(2000)「ストレプトミセス遺伝学実技(“PRACTICAL STREPTOMYCES GENETICS”)」John Innes Foundation,Norwich、英国;ハーウッド(Harwood)等(1990)「バチルスのための分子生物学的方法(“MOLECULAR BIOLOGICAL METHODS FOR BACILLUS”)」、John Wiley、および/または、米国基準株保存機関(ATCC;www.atcc.org)に見出すことが可能である。真菌宿主細胞、例えば、Trichoderma細胞中の、安定な形質転換細胞は、それらがより急速な成長速度を持つこと、または、固相の培地において、ギザギザではなく、滑らかな輪郭を持つ円形コロニーを形成することによって、不安定な形質転換細胞とは一般的に区別される。
【0117】
発現されたポリペプチドの回収およびプロテアーゼの精製法
形質転換宿主細胞によって生産される、本発明によってカバーされるポリペプチド、例えば、配列番号10に対し少なくとも80%の配列同一性を持つポリペプチドは、通例の過程、例えば、遠心またはろ過によって、培養液から宿主細胞を分離すること、または、要すれば、細胞を破壊して、細胞分画と残留物から上清を取り出すことを含む過程によって、培養液から回収してもよい。ある場合には、澄明化後、上清またはろ液のタンパク様成分を、塩、例えば、硫酸アンモニウムを用いて沈殿させる。次に、この沈殿したタンパクを可溶化し、各種クロマトグラフィー手法、例えば、イオン交換クロマトグラフィー、ゲルろ過クロマトグラフィー、アフィニティークロマトグラフィー、およびその他の従来技術で認知済みの手法によって精製してよい。ペプチドおよびタンパクに対する抗体は、動物、例えば、ウサギまたはマウスを免疫化し、従来法によって抗NSP24プロテアーゼ抗体を回収することによって製造される。
【0118】
本発明に使用されるアッセイは、PCT特許WO9934011および米国特許USP6,605,458に記載されるものを含むが、ただしそれらに限定されない。
【0119】
組成物および応用
ある実施態様では、本発明は、本明細書に記載される本発明のプロテアーゼを含む組成物を指向する。本発明による組成物および応用において有用なプロテアーゼの、ある非限定的例としては、例えば、NSP24ファミリ・プロテアーゼまたはNSP25ファミリ・プロテアーゼ、さらに詳細には、配列番号2に対して少なくとも85%の配列同一性を持つNSP24ファミリ・プロテアーゼ、または、その生物学的に活性な断片、例えば、配列番号10に対して少なくとも90%の配列同一性を持つプロテアーゼが挙げられる。ある実施態様では、この酵素組成物は、単一酵素成分プロテアーゼ組成物である。ある実施態様では、本発明は、本発明のプロテアーゼを産業および商業用途に用いる方法を指向する。組成物および産業用途に関する下記の説明は、例示的であって、非内包的であることが意図される。
【0120】
本発明のプロテアーゼを含む組成物は、さらに別の酵素、例えば、グルコアミラーゼ、アルファアミラーゼ、顆粒でん粉加水分解酵素、セルラーゼ、リパーゼ、キシラナーゼ、クチナーゼ、ヘミセルラーゼ、オキシダーゼ、およびそれらの組み合わせを含んでもよい。
【0121】
ある好ましい実施態様では、組成物は、配列番号10の配列に対し少なくとも約85%の配列同一性を有する、本発明のプロテアーゼ、およびグルコアミラーゼを含む。グルコアミラーゼは、糸状真菌の供給源、例えば、Aspergillus、Tricoderma、またはRhizopusの株から得られた野生型グルコアミラーゼであってもよいし、あるいは、グルコアミラーゼは、タンパク加工されたグルコアミラーゼ、例えば、Aspergillus nigerのグルコアミラーゼの変種であってもよい。別の好ましい実施態様では、組成物は、本発明のプロテアーゼおよびアルファアミラーゼを含む。ある実施態様では、アルファアミラーゼは、細菌供給源、例えば、Bacillus SPP.、あるいは、真菌供給源、例えば、Aspergillus SPP.から得られてもよい。ある実施態様では、組成物は、本発明によるプロテアーゼ、およびグルコアミラーゼとアルファアミラーゼの両酵素を含んでもよい。これらの酵素の市販供給源は既知であり、例えば、Genencor International,Inc.およびNovozymes A/Sから入手することが可能である。
【0122】
いくつかの実施態様では、本発明は、エタノール生産、製パン、フルーツジュース生産、醸造、蒸留、ワイン製造、皮革、油脂、紙およびパルプ、および動物飼料生産における使用を意図している。
【0123】
別の実施態様では、本発明が考慮の対象とするものは、洗剤およびクリーニング製品の、活性を持つ「生物学的」成分である。この場合、プロテアーゼ、アミラーゼ、およびリパーゼは、タンパク、でん粉、および脂肪の染みを分解するために使用される。本発明の実施態様は、安定性試験を行い、様々な処方で試験することによって、酵素の、洗剤組成物に対する適合性を試験することを含む。
【0124】
さらに別の実施態様では、本発明は、でん粉を液化および糖化してグルコースに変換するため、および異性化してフルクトースに変換するための酵素的使用を考えた。本発明は、大容量の植物基質、例えば、穀粒(例えば、コーン、小麦、ミロ、ライ麦等)を甘味料、例えば、高濃度フルクトースコーンシロップ、およびマルトースシロップに変換するのに用いてもよい。
【0125】
本発明の酵素(単数または複数)は、タンパクの消化性を上げるために、食品および家畜飼料産業にも応用される。このプロテアーゼはまた、各種産業用途、特に、織物、リトグラフ、化学的芸術、農業、環境廃棄物変換、バイオパルプ処理、バイオマスの燃料への転換、およびその他の化学的過程(単複)にも使用される。さらに、本プロテアーゼは、保健およびパーソナルケア製品、例えば、化粧品、スキンケア、歯磨き等に使用される。
【0126】
飼料
本明細書に記載される酵素は、植物材料、例えば、コーン、小麦、モロコシ、大豆、カノーラ油、ひまわり油、または、上記植物材料の内の任意の複数の混合物、または、ニワトリ、ブタ、反芻動物、水産養殖、およびペット用の植物タンパク供給源を含んでもよい。性能パラメータ、例えば、成長、飼料摂取、および飼料効率ばかりでなく、均一性の改善、動物舎におけるアンモニア濃度の低下、およびその結果として得られる、動物の福祉と健康状態の改善ももたらされることが考慮される。
【0127】
食品
食物タンパクの加水分解産物は、市場において、小さいが、重要な部分を占める。この調製品は、術後患者、または、消化系が損なわれた個人のために使用される。加水分解産物は、それ自体比較的未精製の調製品として与えられる(クレッグ(Clegg)、1978、「新しいタンパク食品の生化学的側面(”Biochemical Aspects of New Protein Food“)」ジェイ・アドゥラー−ニッセン、ビーオー・エッガム、エル・マンク、およびエイチエス・オルセン(J,Adler−Nissen,B.O.Eggum,L,Munk,& H.S.Olsen)編、p.109−117、Pergamon,オックスフォード)。あるいは、静脈投与用の高度に精製されたアミノ酸混合物として投与される。ミルクタンパクの酵素加水分解産物は、食事調製品として従来から運用されている。
【0128】
筋肉食品、特に肉の酵素による軟化は、市場の大きなセグメントを占めるが、現在、これは、植物プロテアーゼおよびいくつかの微生物酵素によって代表される。魚肉の酵素的熟成および軟化も、多くの国々において相当の重要性を持つ。従って、ここに記載の酵素は、食品において様々な用途に使用される。
【0129】
さらに、本発明の酵素、または酵素組成物は、例えば、植物タンパク、例えば、大豆、豆、ルピナス、または菜種タンパク、ミルク様カゼイン、獣肉タンパク、または魚肉タンパクからタンパク加水分解産物を製造するのに有用と考えられる。本明細書に記載されるタンパク(単複)は、食品、飼料、または医用製品を製造する際に、可溶性、均一性、または醗酵性の改善、抗原性の低下、加水分解産物の苦味の減少、またはその他の目的のために使用してもよい。本明細書に記載される酵素(単複)は、単独で使用してもよいし、または他のペプチダーゼと、または他の酵素、例えば、エキソペプチダーゼと共に使用してもよい。エキソペプチダーゼ高含有酵素調製品と共に、本明細書に記載される酵素(単複)を使用した場合、それは、タンパク加水分解産物の味を改善する。
【0130】
さらに、酵素または酵素組成物は、魚肉または獣肉の処理に、例えば、組織および/または粘度を変えるために使用してもよい。
【0131】
皮革
皮革製造産業は、生皮を洗浄し、脱毛し、最後にそれを鞣し、乾燥することを含む一連の工程に依存する。酵素処理は、脱毛工程において重要な役割を担う。すなわち、この工程は、タンパク分解酵素を与えることによって実現されるが、本発明のペプチドは加水分解するので、現在皮革製造に使用される哺乳動物のプロテアーゼの有効な代替品となる。両方とも、高いタンパク分解活性を持ち、かつ、低pHでも効率が高いからである。
【0132】
ウールおよびシルク
本明細書に記載されるプロテアーゼは、ウール製品に対し所望の性質を付与するために、その工業的処理に使用される。一つの実施態様では、本発明は、織物処理用の組成物を提供する。この組成物は、例えば、シルクまたはウールを処理するために使用することが可能である(例えば、RE216,034;欧州特許EP134,267;米国特許US4,533,359;欧州特許EP344,259を参照されたい)。
【0133】
本発明の方法は、線維、例えば、ケラチン線維を含むタンパクを処理するのに用いることが可能である。この方法は、ヒツジ、ヤギ、リャマ、ラクダ、ウサギ等から得られた、ウール、ウール線維、または動物の毛、例えば、アンゴラ、モヘア、カシミヤ、アルパカ、またはその他、商業的に有用な動物毛糸製品を処理するのに好適である。さらに、シルク、くも糸シルク、またはヒトの毛髪も、本発明の方法で処理することが可能である。線維は、線維、トップ、より糸、または織った、または編んだ布または衣類の形を取ってもよい。
【0134】
洗浄
本発明はまた、本発明のプロテアーゼ(単複)を含む洗浄組成物にも関する。この洗浄組成物は、洗浄組成物に一般に使用される添加物をさらに含んでもよい。これらは、漂白剤、界面活性剤、油脂、酵素、および漂白触媒の中から選んでもよいが、ただしこれらに限定されない。当業者には、組成物に含めるにはどの添加物が好適であるかは直ちに明白であろう。本明細書に提示されるリストは、決して網羅的なものではなく、好適な添加物の例示としてのみ受け取るべきである。また、当業者には、酵素、および、組成物における他の成分、例えば、界面活性剤と適合する添加物のみを使用すべきであることは直ちに明白であろう。
【0135】
タンパク、特に、本発明のタンパクは、約0.01から約5重量%(好ましくは0.1%から0.5%)の濃度で、3.5と7.0の間の酸性pHを持つ、既知の、粉末状および液状界面活性剤として処方することも可能である。ある実施態様では、これらの界面活性洗浄組成物はさらに、他の酵素、例えば、アミラーゼ、さらに別のプロテアーゼ、セルラーゼ、リパーゼ、または、エンドグリコシダーゼの外に、油脂および安定化剤を含む。ある実施態様では、pHは、4.0と6.5の間、好ましくは4.0と5.6の間である。これは、要求される活性レベルに応じて、その最適pHにより酸性プロテアーゼと呼ばれるが、これらの酵素をpH7−9において使用することも可能である。
【0136】
通例の洗浄組成物にタンパクを添加することは、何ら特別な使用制限を要求することにはならない。言い換えれば、界面活性剤にとって好適な任意の温度およびpHはまた、pHがまた上述の範囲にあり、温度が前述のタンパク変性温度よりも低い限り、本発明の組成物にとって好適である。さらに、本発明のタンパクは、界面活性剤無しで、再び単独で、または、油脂および安定化剤と組み合わせて、洗浄組成物において使用される。
【0137】
タンパク処理
タンパク原材料の酵素的加水分解は、結果としてしばしば、苦味ペプチドの形成をもたらす(クレッグ(Clegg)1978)。タンパク加水分解産物に見られる苦味ペプチドは、無視できない実際的問題を生ずる。例えば、様々なタイプのチーズの熟成、および、食品としてのタンパク加水分解産物の生産に認められる問題がそれである。加水分解産物の苦味は、通常、特定のペプチド、特に、高い割合の疎水性アミノ酸を含むペプチドによる。苦味は、苦いペプチドの完全な、または部分的加水分解によって効果的に緩和される。従って、本明細書に記載される酵素は、食物の苦味除去に使用される。本発明の酵素、または酵素組成物は、食品のために、タンパクおよび/またはタンパク加水分解産物の苦味緩和のために使用されてもよい。
【0138】
本発明に従ってさらに考慮の対象となるのは、タンパクおよび/またはタンパク加水分解産物から出る遊離アミノ酸の生産である。遊離アミノ酸がグルタミン酸である場合は、それは、食品の風味を強化する。
【0139】
前記タンパクまたはタンパク分解産物は、動物または植物起源のものであってよい。本発明のある実施態様では、加水分解されるタンパクは、カゼイン、または大豆タンパクである。
【0140】
タンパクは、チーズのような食品、および、ココアを含む食品を生産するために使用されてもよい。
【0141】
本明細書に記載される酵素(単複)、本発明のある酵素が濃縮された酵素調製品は、苦味の無いタンパクまたはタンパク加水分解産物に関連して使用されると特に好都合ではあるけれども、本明細書に記載される酵素(単複)は、いくつかの産業用途、例えば、タンパク含有物質、例えば、細胞壁の分解または修飾を含む用途のために使用することが可能である。従って、本明細書に記載される酵素(単複)は、植物細胞壁の分解または修飾を促進する。
【0142】
本発明の酵素調製品の用量、および、該調製品が使用される、その他の条件は、従来技術で既知の方法に基づいて決められてよい。
【0143】
タンパク沈殿もまた、ある種の製品、例えば、ビールの場合無視できない問題を構成する場合がある。なぜなら沈殿によって製品が濁るからである。ビールでは、ビールの冷凍保存の際、可溶性タンパクが沈殿すると濁りが生じる。問題は、無視できない経済的重要性を持つが、ビール製造のために好適な生原料を選ぶこととは別に、この問題を回避する現在主流のやり方は、ビールにタンパク分解酵素を添加することである。
【0144】
パーソナルケア
ある実施態様では、本明細書に記載されるプロテアーゼが合成され精製されたならば、有効量が、パーソナルケア製品に使用される、パーソナルケア組成物(単複)に添加される。パーソナルケア製品は、化粧品、パーソナルケア用途に使用される「大衆薬品」(“OTC”)化合物(例えば、化粧品、スキンケア、口腔ケア、ヘアケア、ネイルケア)に分類される。ある実施態様では、本明細書に記載されるプロテアーゼは、パーソナルケア組成物、例えば、ヘアケア組成物、スキンケア組成物、ネイルケア組成物、化粧組成物、または、それらの任意の組み合わせに添加される。従って、酵素または酵素調製品は、例えば、コンタクトレンズ洗浄液、歯磨き、化粧品、およびスキンケア製品に使用されてもよい。
【0145】
甘味料
本明細書に記載されるプロテアーゼは、マルトースまたはフルクトース高濃度シロップの外、他の甘味料の生産にも使用される。醗酵可能な糖類、または糖類に変換が可能な組成物を含む原材料は、通常、でん粉含有植物材料、例えば、根茎、根、茎、穀類の小塊および粒(例えば、コーン、小麦、ミロ、大麦、およびライ麦)、および、糖含有原材料、例えば、砂糖大根、砂糖黍、果実材料、および糖蜜を含むが、ただしこれらに限定されない。
【0146】
プレバイオティクス
酵素調製品は、他のタンパク分解活性から事実上独立したクローン酵素を用いることが有利な場合、タンパクからペプチドを生産する際に有用であると考えられる。
【0147】
適当なタンパク源を加水分解するのに本明細書に記載する酵素(単複)(例えば、精製酵素)を用いることによって、遊離アミノ酸およびペプチドの未精製調製品を生産することが可能となる。この調製品は、成長のためにアミノ酸に対して特異的要求を持つ微生物に対する基質として極めて好適である。
【0148】
これは、工業的醗酵で使用される相当数の微生物の場合に当てはまる。必要なアミノ酸の補給が、このような醗酵過程の経済的維持にとって重要因子となる場合がよくある。酵素を投与することによって生産されるアミノ酸調製品は、実験室においても、大型の工業規模の醗酵においても基質として好適である。
【0149】
本明細書に記載される酵素(単複)はまた、機能的ペプチド、前生物的合成品等を生体内で生産するために使用されてもよい。「プレバイオティクス」という用語は、消化管、好ましくは、大腸における1種、または少数種の細菌の成長および/または活性を選択的に刺激することによって宿主に対し有益な影響を及ぼす、食品または飼料組成物を指す。
【0150】
醗酵およびバイオエタノール
本発明のプロテアーゼ組成物を用い、でん粉含有基質を醗酵して得られるアルコールの生産は、燃料用アルコールまたは携帯用アルコールの生産を含んでよい。ある実施態様では、酵素組成物は、大麦、モルト、および、その他の、例えば、ビール生産のための原料の酵母醗酵を促進するために使用されてもよい。
【0151】
アミラーゼは、醸造および製パン工業にとって不可欠の酵素である。アミラーゼは、モルト形成、および、添加糖類またはその他の炭水化物が無添加の状態で実行される、ある種の製パン過程において、でん粉を分解するのに必要とされる。これらの酵素の十分な活性度を確保することは、特にモルト調製工業では問題である。生理的に受容可能なシステムによってアミラーゼの活性を十分に増大させる方法は、より速やかなモルト形成を可能とする方法をもたらし、糖の利用可能性が増大することから、炭水化物含量の少ないアルコール飲料、例えば、ビールをもたらす。
【0152】
ある実施態様では、でん粉含有基質、例えば、穀粒(例えば、コーン、小麦、モロコシ)、小塊、および他の植物残留物の加水分解によって、アルコール、例えばエタノールが生産される。アルコール生産の方法は、「アルコール教科書−飲料、燃料、および工業的アルコール事業の参照資料(“The Alcohol Textbook,A Reference for the Beverage,Fuel and Industrial Alcohol Industries,”)」3版、ケーエー・ジャック(K.A.Jacques)等編、1999年、Nottingham University Press,英国に記述される。本発明のある実施態様では、プロテアーゼは、糖化および醗酵結合工程、別に糖化・醗酵同時工程とも呼ばれる工程において、組成物の中で、グルコアミラーゼと、かつ要すれば任意にアルファアミラーゼと組み合わせて使用される。「S.Lewisの工業醗酵学(”S.Lewis in Industrial Enzymology“)」、2.1章「アルコール発酵(”Fermentation Alcohol”)」、2版、ティー・ゴッドフリーおよびエス・ウェスト(T.Godfrey and S.West)編、(1996)、Stockton Press、ニューヨーク州を参照されたい。真菌酸性プロテアーゼをグルコアミラーゼと組み合わせて用いた醗酵によってエタノールを生産する方法は既知である。例えば、米国特許USP5,231,017は、Aspergillus nigerから得られたプロテアーゼを用いてエタノールを生産する方法であって、液化マッシュを獲得すること、および、醗酵工程と結合させてもよい糖化工程において液化マッシュにプロテアーゼを導入することを含む方法を開示する。ある実施態様では、本発明のプロテアーゼ組成物は、顆粒状でん粉基質による非加熱過程において、アルコール、例えば、エタノールを生産するために用いられる。該過程は、アルコールを生産するために用いられる基質において、でん粉のゲル化温度未満の温度において実行される。でん粉加水分解過程において使用されるプロテアーゼの量は、プロテアーゼの酵素活性に依存する。ある実施態様では、その量は、20−33%乾燥固体に調整したスラリー450gに対するプロテアーゼ2%液の添加量として0.001から2.0mlの範囲である。その際、スラリーは、糖化時および/または加水分解でん粉における液化マッシュである。他の有用な範囲としては、0.005から1.5ml、および0.01から1.0mlも挙げられる。
【0153】
プロテアーゼで処理された種子または穀粒は、醗酵過程によって生産されるモルトおよび飲料の生産に有利である。
【0154】
さらに、穀粉の中のタンパクを加水分解し、後続のアルコール発酵段階を予期してワートの可溶性窒素を濃縮するために、糖化においてもプロテアーゼを使用することが望ましい。穀粒におけるアミラーゼ活性の強化は、モルト形成において重要な発芽の速度と効率を増す。すなわち、酵素活性が強化されたモルトの生産は、でん粉が醗酵可能な炭水化物へ転換される加水分解の強化をもたらすので、アルコール飲料、例えば、ビールおよびスコッチウィスキーの生産における醗酵効率を向上させる。
【0155】
以下の実施例では、下記の略語が使用される。eq(当量)、M(モル)、μM(マイクロモル)、N(規定)、mol(モル)、mmol(ミリモル)、μmol(マイクロモル)、nmol(ナノモル)、g(グラム)、mg(ミリグラム)、kg(キログラム)、μg(マイクログラム)、L(リットル)、ml(ミリリットル)、μl(マイクロリットル)、cm(センチメートル)、mm(ミリメートル)、μm(マイクロメートル)、nm(ナノメートル)、℃(摂氏度)、h(時)、min(分)、sec(秒),msec(ミリ秒)、Ci(キュリー)、mCi(ミリキュリー)、μCi(マイクロキュリー)、TLC(薄層クロマトグラフィー)、Ts(トシル)、Bn(ベンジル)、Ph(フェニル)、Ms(メシル)、Et(エチル)、Me(メチル)、dsまたはDS(乾燥固体重量)、SAPU(酸性プロテアーゼ分光光度単位)で、1SAPUは、アッセイ条件下、1分当たりカゼイン基質から1マイクロモルのチロシンを解放するプロテアーゼの酵素活性量であり、GAU(グルコアミラーゼ単位で、pH4.2および60℃において、1時間当たり可溶性でん粉基質から、グルコースとして計算される1gの還元性糖を生産する酵素の量と定義される)。
【0156】
本発明は、下記の実施例においてさらに詳細に記述される。これらの実施例は、いかなる意味でも、請求される本発明の範囲を限定することを意図するものではない。付属の図面は、本発明の明細および説明の、一体化部分と考慮されるべきことが意図される。引用される参照文献は全て、そこに記載される全てに対する参照によって、それぞれ個別に本明細書に含める。下記の実施例は、本発明を例示するためにであって、請求される本発明を限定するために提示されるものではない。
【実施例1】
【0157】
新規プロテアーゼNSP24の、Trichoderma reesei DNAクローニング
T.reesei株QM6aからゲノムDNAを抽出した。T.reeseiゲノムのcontig1−5500に見出されたプロテアーゼ配列候補(Joint Genome Institute(JGI)T.reeseiゲノムv1.0)に基づいて、PCRプライマーを設計した。順行プライマーは、pENTR/Dベクター(Invitrogen)への方向性クローニングのためのモチーフを含んでいた。
【0158】
afp6fプライマーの配列は、CACCATGCAGACCTTTGGAGCT(配列番号11)であり、afp7rの配列は、TTATTTCTGAGCCCAGCCCAG(配列番号12)であった。1.3kbのPCR産物をゲル抽出(ゲル精製キット、Qiagen)で精製し、Invitrogen Gatewayシステムプロコールに従ってpENTR/Dにクローンした。
【0159】
次に、このベクターを、化学的にコンピテントとしたTop10 E.coli(Invitrogen)にて形質転換させ、カナマイシン選択した。いくつかの独立クローンから得られたプラスミドDNAを制限酵素で消化し、適正なサイズの挿入体であることを確認した。いくつかのクローンにおいて、プロテアーゼ遺伝子挿入体の配列を決定した(Sequetech,Mountain View、カリフォルニア州)。一つのクローンからのプラスミドDNA、pENTR/D_55.3を、pTrex3g/amdS表示ベクターDNAとのLRクロナーゼ反応(Invitrogen Gatewayシステム)に加えた。pTrex3gベクターは、pUC118ファージミドベクターであるE.coliのpSL1180(Pharmacia Inc.,ニュージャージー州)に基づくもので、WO05/001036に記載される。このLRクロナーゼ反応における組み換えによって、表示ベクターのCmRおよびccdB遺伝子は、pENTR/D_55.3由来のT.reeseiプロテアーゼによって置換された。この組み換えによって、プロテアーゼは、cbhlプロモーターと表示ベクターのターミネーターとの間に方向性をもって挿入された。44および50bpの組み換え部位配列が、それぞれ、プロテアーゼ遺伝子の上流および下流に留まった。LRクロナーゼ反応液の分液を、化学的にコンピテントとされたTop10 E.coli中に形質転換させ、一晩培養し、カルベニシリン選択した。いくつかのクローンから得られたプラスミドDNAを制限酵素で消化し、適正な挿入体サイズを確認した。クローンからのプラスミドDNA、pTrex3g_55.3.1をXbalIで消化して、cbhlプロモーター:NSP24プロテアーゼ:ターミネーター:amdSを含む発現カセットを放出させた。この5.8kbのカセットを、標準技術を用いてアガロースゲル抽出によって精製し、市販株QM6aから得られたT.reesei株の中で形質転換させた(PCT国際特許WO05/001036を参照)。図5、6、および7を参照されたい。
【実施例2】
【0160】
新規プロテアーゼNSP25の、Trichoderma reesei DNAクローニング
T.reesei株QM6aからゲノムDNAを抽出した。T.reeseiゲノムのcontig22−263400に見出されたプロテアーゼ配列候補(JGI T.reeseiゲノムv1.0)に基づいて、PCRプライマーを設計した。順行プライマーは、pENTR/Dベクター(Invitrogen)への方向性クローニングのためのモチーフを含んでいた。
【0161】
afp8fプライマーの配列は、CACCATGCAGCCCTCATTTGGCAG(配列番号13)であり、afp9rの配列は、CTATTTCTTCTGCGCCCAGCCAAC(配列番号14)であった。1.2kbのPCR産物をゲル抽出(ゲル精製キット、Qiagen)で精製し、Invitrogen Gatewayシステムプロコールに従ってpENTR/Dにクローンした。次に、このベクターを、化学的にコンピテントとしたTop10 E.coli(Invitrogen)にて形質転換させ、カナマイシン選択した。いくつかの独立クローンから得られたプラスミドDNAを制限酵素で消化し、適正なサイズの挿入体であることを確認した。いくつかのクローンにおいて、プロテアーゼ遺伝子挿入体の配列を決定した(Sequetech,Mountain View、カリフォルニア州)。一つのクローンからのプラスミドDNA、pENTR/D_22.2を、pTrex3g/amdS表示ベクターDNAとのLRクロナーゼ反応(Invitrogen Gatewayシステム)に加えた。このLRクロナーゼ反応における組み換えによって、表示ベクターのCmRおよびccdB遺伝子は、pENTR/D_22.2由来のT.reeseiプロテアーゼによって置換された。この組み換えによって、プロテアーゼは、cbhlプロモーターと表示ベクターのターミネーターとの間に方向性をもって挿入された。44および50bpの組み換え部位配列が、それぞれ、プロテアーゼ遺伝子の上流および下流に留まった。LRクロナーゼ反応液の分液を、化学的にコンピテントとされたTop10 E.coli中に形質転換させ、一晩培養し、カルベニシリン選択した。いくつかのクローンから得られたプラスミドDNAを制限酵素で消化し、適正な挿入体サイズを確認した。クローンからのプラスミドDNA、pTrex3g_22.2#1をXbalI(かつ、細菌のバックボーンを消化して小型断片とするためにEcoRI)で消化して(小型断片は電気泳動中カセットから移動して遠ざかった)、cbhlプロモーター:NSP25プロテアーゼ:ターミネーター:amdSを含む発現カセットを放出させた。この5.7kbのカセットを、標準技術を用いてアガロースゲル抽出によって精製し、市販株QM6aから得られたT.reesei株の中で形質転換させた。形質転換用に用いたプラスミドは、NSP24挿入体が、NSP25配列で置換されたことを除いては、図7に描かれるプラスミドと事実上同じであった。
【実施例3】
【0162】
TrichodermaPEG真菌の形質転換
胞子形成中の菌糸から得られた2cm2の寒天プラグを、250mlの、4つの溝を有するバッフルフラスコに納めた50mlのYEGブロスに接種し、37℃で16−20時間200rpmにてインキュベートした。液体容量を50mlの円錐チューブに移し、10分間2500rpmで回転させて菌糸を回収した。上清を吸引除去した。この菌糸ペレットを、40mlのフィルター滅菌β−D−グルカナーゼ(InterSpex Products,Inc.)液を含む、250mlの、0.22μmCAコーニングフィルター瓶に移し、30℃、200rpmで2時間インキュベートした。菌糸を、滅菌Miracloth(CalBiochem,ラホヤ、カリフォルニア州)を通して、50mlの円錐形遠心チューブに集め、2000rpmで5分遠心し、吸引した。ペレットを、50mlの1.2Mソルビトールで一度洗浄し、再び遠心し、吸引し、25mlのソルビトール/CaCl2で洗浄した。ヘモサイトメーターを用いてプロトプラストをカウントし、遠心し、吸引し、1.25×108/mlのプロトプラスト濃度を与えるのに十分な容量のソルビトール/CaCl2に再懸濁させた。形質転換反応当たり200μlの分液を用いた。20μgのDNA(≧1μg/μl)を15mlの円錐形チューブに移し、チューブを氷の上に置いた。200μlのプロトプラストを加えた。50μlのPEGを加え、穏やかに掻き混ぜ、氷上で20分インキュベートした。2mlのPEGをチューブに加え、室温で5分インキュベートした。4mlのソルビトール/CaCl2(合計6.25ml)をチューブに加えた。この形質転換混合物を、各オーバーレイ当たり〜2mlの、3分液に分割した。この2mlを、溶融アセタミドソルビトール上層寒天のチューブに加え、オーバーレイ混合物を、アセタミドを唯一の窒素源として増殖することのできる形質転換体を選ぶためにアセタミドソルビトールプレートの上に注いだ。プレートを、コロニーが出現するまで28−30℃でインキュベートした。形質転換体は、単一コロニーをアセタミド媒体(ソルビトール無添加アセタミドソルビトール処方)にて繰り返し継代することによって精製した。
【0163】
材料
40mlのβ−D−グルカナーゼ液:600mgのβ−D−グルカナーゼ;400mgのMgSO4.7H2O;および、40mlの1.2Mソルビトール。
【0164】
200mlのPEG混合液:50gのPEG4000(BDH Laboratories Supplies,Poole、英国)および1.47gのCaCl2.2H2OをMilli Q水にて調製。
【0165】
ソルビトール/CaCl2:1.2Mソルビトールおよび50mM CaCl2。
【0166】
amdS選択のために、アセタミドソルビトールプレートおよびオーバーレイを用いた。胞子の精製には、同じプレートを、ソルビトール無しで用いた。
【0167】
アセタミドソルビトール寒天(プレートおよびトップ寒天)
アセタミド(Aldrich,99%昇華物)−0.6g/L;CsCl−1.68g/L;グルコース−20g/L;KH2PO4−20g/L;MgSO4*7H2O−0.6g/L;CaCl2*2H2O−0.6g/L;1000X塩(下記参照)−1ml。pHを5.5に調整し、容量を300mlとした。0.22ミクロンフィルターにてフィルター滅菌し、オーブンにて55℃に温めた。
【0168】
700mlの水に、Noble寒天(トップ寒天用低温溶融)20g、およびソルビトール218gを加え、オートクレーブ滅菌した。この混合物を55℃に冷却し、フィルター滅菌したアセタミド混合物を加えた。プレートまたはチューブを注いだ。
【0169】
1000X塩−FeSO4.7H2O(0.5g/100ml);MnSO4.H2O(0.16g/100ml);ZnSO4.7H2O(0.14g/100ml);CoCl2.6H2O(0.1g/100ml)、および、0.22ミクロンフィルターにてフィルター滅菌。
【0170】
じゃがいもデキストロース寒天(PDA、Difco脱水媒体)−200g/L温浸によるじゃがいも;デキストロース20g/L;および寒天15g/Lを、50−80%最終容量のdH2O中でよく混ぜわせ、最終的に100%容量とした。この混合液をオートクレーブ滅菌し、55℃に冷却し、注いだ。
【0171】
pH3.5媒体用の1%スキムミルク寒天を調製するために、PDAを上記のように調製し、100mlの溶融PDAに、1.8mlの10%酒石酸および12.5mlの滅菌8%スキムミルクを加え、プレートに注いだ。スキムミルクを事前滅菌するために、8%スキミミルク(Difco)を10分、122−123℃、32−35psiのチェンバー圧に暴露してオートクレーブした。混合液を取り出し、冷却し、室温に保存した。
【0172】
形質転換体におけるプロテアーゼ発現は、振とうフラスコにおける3日間の増殖後に評価した。T.reesei培養液(デービス(Davis)等(1970)Methods Enzymol.17:79−143)に寒天プラグを接種した。培養物を振とうしながら30℃で3日間インキュベートした。培養液を、0.22ミクロンフィルターを通過させ、ろ液を、1%スキムミルク寒天の上にスポット状に投与した。室温で一晩インキュベート後、透明ゾーンが観察された。
【実施例4】
【0173】
NSP24、NSP25、およびL388M PepAのpH活性プロフィール
全てTrichoderma reesei株において過剰発現される、PepA(野生型およびL388M)、NSP24、およびNSP25のpH活性プロフィールを、Molecular Probesから入手した蛍光標識カゼインアッセイ(EnzChekプロテアーゼキット緑色蛍光)を用いて求めた。PepA(野生型およびL388M)、およびNSPは、全体醗酵サンプルであり、NSP24は、50%グリセロールに溶解させて安定化した精製サンプルである。これらの酵素を、1.0mg/ml、0.5mg/mlおよび0.25mg/mlに希釈した。蛍光標識基質は、DI H2Oにて0.1mg/mlに希釈した。10mlの基質を、各種pHの50mlバッファーと30μlのDI H2Oに加えた。10mlの酵素を加えて反応を開始し、様々な期間反応させ、100μlの1.0Mリン酸塩をpH10で加えて反応を停止させた。サンプルの蛍光を、SpectraMAX EM蛍光プレートリーダーにおいて、538nm放射線で、485励起、放射カットオフフィルター530nmで測定した。NSP24はpH3.7で最適活性を示し、野生型PepAはpH3.4で最適活性を示し、かつ、L388M pepAは3.5に最適pHを持つ。NSP25は、pH4.6で最適活性を示す。
【実施例5】
【0174】
実験室醗酵における、Trichoderma reeseiのNSP24プロテアーゼの、GC106との比較
今日のエタノール産業において使用される標準的プロテアーゼは、Genencor International,Inc.によって市販されるプロテアーゼGC106である。GC106に対するNSP24の機能性を、糖分解、グルコース形成、およびエタノール生産に関して比較した。
【0175】
材料
ジスチラーゼL−400(ロット#107−04057−901,372 GAU/g)
GC106(ロット#A01−01300−001,1010SAPU/g)
NSP24(ロット#20040423,1165SAPU/g)
Red Star赤酵母
エタノール製造機から得られたマッシュおよび薄い濃度の抽出液(コーン)
方法
マッシュおよび薄い抽出液(バックセットとも呼ばれる、醗酵前)をエタノール製造機から取り出し、掻き混ぜて26.5ブリックスとする。1N HClを用いてpHをpH4.3に調整した。次に、サンプルを、三つの300グラム分液に分割し、32℃の水浴に設置した。平衡後、下記の酵素の組み合わせを加えた。
【表2】
【0176】
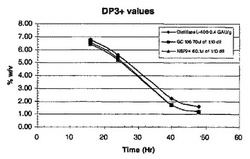
ジスチラーゼL−400は、Aspergillus nigerから得られた液状グルコアミラーゼであり、Genencor International Inc.から入手が可能である。酵素添加後、フラスコ当たり、1.00gのRed Star赤酵母を加えた。サンプルを、16、24、40、および48時間後に取り出し、遠心した。各サンプルの500μlを、50μlの1.1N NH2SO4を含む試験間に移し反応を停止させた。2分後、サンプルを、4.5mlのDI H2Oで希釈し、混合した。混合後、サンプルを、0.45ミクロンフィルターに通過させ、HPLCバイアルに移し分析した。サンプルは、HPLC(Phenomenex Rezex 8u)で分析した。結果を、図1−4に示す。NSP24の性能は、GC106にほぼ等しかった。
【実施例6】
【0177】
非加熱過程において磨砕コーンから得られるエタノール収率に及ぼすNSP24の作用
磨砕コーンの30%DSスラリーを、DI H2Oによって調製した。磨砕コーンは、エタノール産業で使用される#2イェローデントコーンの典型的サンプルである。これは、70%を超えるものが30メッシュ篩を通過するほどに磨砕された。この穀物の水分含量をOHAUS、MB35ハロゲン水分天秤(ニュージャージー州)を用いて測定した。pHは、6N H2SO4を用いて4.2に調整した。醗酵は、0.5kg/MT用量のNSP24の添加、または添加無しで、1.0GAU/g用量のSTARGEN001を添加した100gマッシュを含む125mlフラスコにおいて行われた。
【0178】
45mlの水に溶解した5gのRed Starエタノールレッド乾燥酵母(Lesaffre酵母社、ミルウォーキー、ウィスコンシン州)を準備し、32℃水浴で1時間掻き回し、醗酵槽に接種した。0.5mlの酵母スラリーを、各125mlフラスコに加えた。フラスコを32℃水浴に移し、マッシュを穏やかに掻き回した。醗酵中、サンプルを取り出し、HPLC分析に供した(HPLCカラム:Phenomenex Rezex Organic Acid Column(RHM−モノサッカライド)#00H0132−KO;カラム温度:60℃;移動相:0.01N H2SO4;流速:0.6mL/分;検出器:RI;および、注入用量:20μL)。醗酵は、72時間後停止させた。様々のサンプリング間隔における、糖分、乳酸、グリセロール、およびエタノールを含む化合物の生産は、下記の表3に示す通りである。該表において、+は、NSP24がフラスコに添加されたことを示し、−−は、NSP24がフラスコに添加されなかったことを示す。全てのサンプルについて乳酸が、約0.01と0.02%w/vにおいて測定され、かつ、DP−2は0.0であると定量された。全てのサンプルについて、24時間時、酢酸は約0であり、71時間では、0.03と0.04の間であると定量された。
【表3】
【実施例7】
【0179】
コーン内乳によるエタノール生産における各種プロテアーゼの比較
内乳(胚除去コーン、75.8%でん粉、99.5%の粒径<30メッシュ)による29.5%DSマッシュを顆粒状でん粉基質として調製した。各マッシュ100グラムを125mlフラスコに移し、媒体のpHをpH4.5に調整した。プロテアーゼ(NSP24;中性プロテアーゼ(MULTIFECT NEUTRAL,プロテイナーゼ−T);および、アルカリ性プロテイナーゼ(SPEZYME FAN,PROTEX 6L MULTIFECT P−3000、およびプロテイナーゼ−T(Genencor International))を、0.5kg/MTで加え、次いで、STARGEN001(Genencor International)を2.5Kgs/でん粉MT)を加えた。次に、フラスコに0.5mlの20%酵母(Red Starエタノール赤)を接種し、32℃に維持した水浴に移した。フラスコの内容物を連続的に攪拌し、インキュベーション中均等な混合を行った。サンプルを様々な時間間隔で採取し、HPLC分析に供した。72時間醗酵液から得られたDDGSの残留でん粉およびタンパク含量を定量した。エタノール生産の結果を下記の表4に示す。
【表4】
【図面の簡単な説明】
【0180】
【図1】1)NSP24、2)市販のプロテアーゼGC106、および、3)プロテアーゼを含まないジスチラーゼを用いた場合の、糖分解(DP+3)%w/vを示す(実施例5参照)。
【図2】NSP24、GC106、およびジスチラーゼを用いた場合の糖分解(DP2)%w/vを示す。
【図3】NSP24、GC106、およびジスチラーゼを用いた場合のグルコース形成(DP1)を示す。グルコースの量は、NSP24およびGC106の両サンプルにおいて、40時間終了時に残存する量は0.2%w/v未満であり、48時間終了時には0.1%w/v未満である。一方、48時間終了時に、%w/vで測定したグルコースの量は、ジスチラーゼでは、1.0%w/vよりも少し大きい。
【図4】NSP24、GC106、およびジスチラーゼにおけるエタノール生産(%v/w)を示す。前記二つのプロテアーゼサンプルを用いることによって生産されるエタノールの速度および量は同じである。一方、ジスチラーゼのエタノール生産は、より低く、速度はより遅かった。
【図5A】図5Aは、Trichoderma reeseiから得られたpTrex3g_NSP24 cDNAのヌクレオチド配列(配列番号1)を示す。NSP24遺伝子配列には下線を施し、遺伝子のイントロン配列候補は、ゴシック体で特定する。プロテアーゼをコードする核酸配列は、配列番号8の配列によって表される。
【図5B】図5Bは、Trichoderma reeseiから得られたpTrex3g_NSP24 cDNAのヌクレオチド配列(配列番号1)を示す。
【図5C】図5Cは、Trichoderma reeseiから得られたpTrex3g_NSP24 cDNAのヌクレオチド配列(配列番号1)を示す。
【図5D】図5Dは、Trichoderma reeseiから得られたpTrex3g_NSP24 cDNAのヌクレオチド配列(配列番号1)を示す。
【図6A】Tricoderma reesei由来NSP24の、予想アミノ酸配列(407アミノ酸)(配列番号2)(図6A)。シグナルペプチドはゴシック体で示され、プレプロ配列は、ゴシック体および下線で示され、成熟NSP24タンパクは、KYGAPIS...で始まり、配列番号10で表される。
【図6B】NSP24ヌクレオチド配列で、イントロン候補をゴシック体で特定した配列(図6B)(配列番号8)を示す。
【図7】pTrex3g_NSP24ベクター、および、図5のヌクレオチド配列にそう制限酵素切断部位の位置を示す。
【図8】pepAプロテアーゼの核酸配列(配列番号3)を示す。予想イントロンをゴシック体で表す。
【図9A】図9Aは、新規NSP25プロテアーゼ(399アミノ酸)(配列番号9)をコードする核酸配列(配列番号4)を示す。シグナル配列はゴシック体で表す。
【図9B】図9Bは、新規NSP25プロテアーゼ(399アミノ酸)(配列番号9)をコードする核酸配列(配列番号4)を示す。シグナル配列はゴシック体で表す。
【図10】新規pepAプロテアーゼ変種(L388M)(配列番号7)の核酸配列(配列番号5)を示す。図において、下線を施した‘A’は、図8のpepAの‘C’から変化したものである。
【図11】発現ベクターpSL889_pepAを示す。
【図12A】図12Aは、発現ベクターpSL899_pepAのヌクレオチド配列(配列番号6)を示す。Xho Iの切断部位は、^によって示され、Xba I部位は*によって示される。pepAのコード配列はゴッシク体で示される。
【図12B】図12Bは、発現ベクターpSL899_pepAのヌクレオチド配列(配列番号6)を示す。
【図12C】図12Cは、発現ベクターpSL899_pepAのヌクレオチド配列(配列番号6)を示す。
【図12D】図12Dは、発現ベクターpSL899_pepAのヌクレオチド配列(配列番号6)を示す。
【図12E】図12Eは、発現ベクターpSL899_pepAのヌクレオチド配列(配列番号6)を示す。
【図13】配列番号5によってコードされたタンパクに対するPepA変種、L388Mのアミノ酸配列(配列番号7)を示す。
【技術分野】
【0001】
関連出願
本出願は、2004年12月30日出願、名称「酸性真菌プロテアーゼ(Acid Fungal Protease)」なる米国特許仮出願第60/640,399号、および、2005年1月27日出願、名称「酸性真菌プロテアーゼ(Acid Fungal Protease)」なる米国特許仮出願第60/648,233号に対する優先権を主張する。なお、これらの出願の内容の全てを参照することにより本明細書に含める。
【0002】
本発明は、NSP24ファミリ・プロテアーゼ、NSP25ファミリ・プロテアーゼ、およびPepAプロテアーゼと表示される酸性プロテアーゼをコードするポリヌクレオチド;NSP24およびNSP25ファミリ・プロテアーゼポリペプチド;前記プロテアーゼを含む組成物、および、それらの使用に関する。
【背景技術】
【0003】
プロテアーゼは、ペプチド結合を切断することのできる酵素である。酸性プロテアーゼ(例えば、酸性pHを至適とするプロテアーゼ)は、哺乳類および微生物を含むいくつかの異なる生物体によって生産される。例えば、微生物の酸性プロテアーゼは、細菌株、例えば、Bacillus sp.の株(JP01240184)、および、真菌株、例えば、Rhizopus sp.(EP72978)、Schytalidium sp.(JP48091273)、Sulpholobus sp.,Thermoplasma sp.(WO/9010072)、およびAspergillus sp.(JP50121486およびEP82395)の株によって生産される。
【0004】
ベルカ(Berka)等「遺伝子(Gene)」(1990)96:313は、Aspergillus awamori由来のアスパラギン酸プロテアーゼ、アスペルギロペプシンAをコードする遺伝子を開示する。Aspergillus oryzae由来のアスパラギン酸プロテアーゼ、アスペルギロペプシンOをコードする遺伝子のクローニングは、ベルカ(Berka)等によって記載される「遺伝子(Gene)」(1993)125:195−198。Aspergillus oryzae由来の酸性プロテアーゼ(PepA)をコードする遺伝子のクローニングは、ゴミ(Gomi)等によって開示される「バイオサイエンス・バイオテクノロジー・アンド・バイオケミストリー(Biosci.Biotech.Biochem.)」(1993)57(7):1095−1100。
【0005】
プロテアーゼ、特に酸性プロテアーゼは、産業用途に、例えば、食品および飼料の生産、皮革産業(例えば、生皮の除毛)、タンパクの加水分解産物の生産、および、アルコールの生産、例えば、エタノールの生産、ワインの生産および醸造等広く使われている。
【0006】
依然として、多種多様な用途、特に食品および飼料産業においてプロテアーゼに対する需要が続いている。
【発明の開示】
【課題を解決するための手段】
【0007】
本出願人等は、いくつかの新規プロテアーゼ遺伝子を発見した。そのようなものとして、NSP24プロテアーゼ(配列番号2、または配列番号10)をコードする新規nsp24遺伝子、NSP25プロテアーゼ(配列番号9)をコードする新規nsp25遺伝子、および、新規PepAプロテアーゼ(配列番号7)をコードする新規pepA変異遺伝子が挙げられる。
【0008】
従って、本発明は、NSP24プロテアーゼ、NSP25プロテアーゼ、またはPepAプロテアーゼおよびその変異種の、組み換え法による、または、ほぼ純粋な調製をその特徴とする。
【0009】
本発明のある態様では、プロテアーゼは、配列番号2または配列番号10(下記の図6に示される)のアミノ酸配列と事実上同じアミノ酸配列を含む、NSP24ファミリ・プロテアーゼポリペプチドである。いくつかの実施態様では、NSP24ファミリ・プロテアーゼポリペプチドは、配列番号8(下記の図5に示される)の核酸によって、あるいは、配列番号8の核酸と事実上同じ核酸配列を持つ核酸によってコードされる。
【0010】
本発明の別の態様では、NSP24ファミリ・プロテアーゼポリペプチドは、配列番号10における配列とは最大10残基アミノ酸配列が異なる。ある実施態様では、NSP24ファミリ・プロテアーゼポリペプチドは、配列番号10における配列とは、最大10%の残基のアミノ酸配列が異なる。ある実施態様では、その差は、NSP24ファミリ・プロテアーゼポリペプチドがNSP24プロテアーゼの生物学的活性を示せる程度の差であり、例えば、該NSP24プロテアーゼが天然のNSP24プロテアーゼの生物学的活性を保てる程度の差である。
【0011】
本発明のさらに別の態様では、NSP24ファミリ・プロテアーゼポリペプチドは、本明細書に記載されるNSP24プロテアーゼ配列の外に、他のN−末端および/またはC−末端アミノ酸配列も含む。
【0012】
本発明のさらに別の態様では、NSP24ファミリ・プロテアーゼポリペプチドは、さらに別のアミノ酸残基、好ましくは、配列番号1または配列番号8の配列をコードするゲノムDNAに対する5´側のゲノムDNAによってコードされる残基に、読み枠内で融合される、配列番号2または配列番号10のアミノ酸配列の全てまたはその断片を含む。
【0013】
本発明のさらに別の態様では、NSP24ファミリ・プロテアーゼは、第1NSP24ファミリ・プロテアーゼ部分と、第2ポリペプチド部分、例えば、NSP24ファミリ・プロテアーゼとは無関係のアミノ酸配列を持つ第2ポリペプチド部分とを有する、組み換え融合タンパクである。第2ポリペプチド部分は、DNA結合ドメイン、またはポリメラーゼ活性化ドメインであってもよい。本発明のポリペプチドは、複数遺伝子の存在、選択的転写事象、選択的RNAスプライシング事象、ならびに、選択的翻訳および翻訳後事象の結果生ずるものを含む。このポリペプチドは、システム、例えば、発現NSP24プロテアーゼが天然細胞において発現される場合に見られるのとほぼ同じ翻訳後修飾をもたらす、培養細胞において発現されてもよいし、あるいは、天然細胞で発現された場合に見られる翻訳後修飾の除去をもたらすシステムにおいて発現されてもよい。
【0014】
さらに別の態様では、本発明は、NSP24ファミリ・プロテアーゼ、および、1種以上の添加成分、例えば、担体、希釈剤、または溶媒を含む酵素組成物に関する。添加成分は、インビボ、インビトロ、製薬、または獣医学用途において組成物を有用にするものであってもよい。この態様のある実施態様では、酵素組成物は添加酵素を含む。好ましい実施態様では、添加酵素は、グルコアミラーゼ、アルファ・アミラーゼ、またはそれらの組み合わせである。
【0015】
さらに別の態様では、本発明は、配列番号2または配列番号10のアミノ酸配列に対し少なくとも80%相同な配列を持つアミノ酸配列を含むNSP24ファミリ・プロテアーゼポリペプチドをコードするヌクレオチドを有するか、または含むほぼ純粋な核酸を提供する。
【0016】
ある態様では、NSP24ファミリ・プロテアーゼの核酸は、例えば、該NSP24ファミリ・プロテアーゼ遺伝子配列を発現ベクターとしての使用に適しているものとするために、NSP24ファミリ・プロテアーゼ遺伝子配列に対して動作的に結合される転写調節配列、例えば、転写プロモーターまたは転写エンハンサー配列の内の少なくとも一つを含む。
【0017】
さらに別の態様では、本発明のNSP24プロテアーゼポリペプチド(例えば、配列番号2)をコードする核酸は、配列番号8由来の少なくとも12個の連続ヌクレオチドに対応する核酸プローブに対して、より好ましくは、配列番号8由来の20個の連続ヌクレオチドに対して厳格条件下にハイブリダイズする。
【0018】
本発明のもう一つの態様は、各種産業場面における、NSP24ファミリ・プロテアーゼ(例えば、NSP24)の応用を提供する。例えば、NSP24ファミリ・プロテアーゼは、アルコール燃料およびその他の重要な工業薬品の生産のために農業廃棄物を酵素的に分解するために使用されてもよいし、動物またはヒトの食品生産用途、または洗剤組成物の成分として、皮革処理およびタンパク系繊維の処理用途(例えば、ウールまたはシルク)、バイオマス用途、パーソナルケア用途(皮膚、髪、口内衛生等)、製薬および保健用途、および、上記応用用途に用いられる新規ペプチドの生産用途に使用されてもよい。
【0019】
さらに別の態様では、本発明は、pepA変異プロテアーゼ、すなわち、配列番号7を持つL388Mをコードするポリヌクレオチドに関する。ある実施態様では、ポリヌクレオチドは、配列番号5の配列を有する。
【0020】
さらにもう一つの態様では、本発明はNSP25ファミリ・プロテアーゼに関する。ある実施態様では、NSP25ファミリ・プロテアーゼは、配列番号9に対して少なくとも85%の配列相同性を持つ。別の実施態様では、NSP25ファミリ・プロテアーゼは、配列番号4に対して少なくとも85%の配列相同性を持つポリヌクレオチドによってコードされる。さらに別の実施態様では、NSP25ファミリ・プロテアーゼは、親のNSP25ファミリ・プロテアーゼの生物学的に活性な断片である。
【発明を実施するための最良の形態】
【0021】
以下、本発明を、下記の定義と実施例のみを用い、参照によって詳細に説明する。本明細書に参照される全ての特許および出版物は、それら特許および出版物の中に開示される全ての配列を含め、参照することによって本明細書に含められる。
【0022】
本発明の実施において、別様に指示しない限り、細胞生物学、細胞培養、分子生物学、トランスジェニック生物学、微生物学、組み換えDNA、および免疫学の、当業者の能力の範囲内にある通例技術を用いる。このような技術は文献に記載される。例えば、「分子クローニング、実験室マニュアル(Molecular Cloning A Laboratory Manual)」、第2版、サムブルック、フリッチュ、およびマニアチス(Sambrook,Fritsch and Maniatis)著、コールドスプリングハーバーラボラトリープレス(Cold Spring Harbor Laboratory Press);アウスベル等(Ausubel et al.)編、「分子生物学における短いプロトコル(Short Protocols in Molecular Biology)」、第5版、2002年;「DNAクローニング(DNA Cloning)」、IおよびII巻、ディーエヌ・グローバー(D.N.Glover)編、1985;「オリゴヌクレオチド合成(Oligonucleotide Synthesis)」、エムジェイ・ゲイト(M.J.Gait)編、1984;マリス(Mullis et al.)等、米国特許第4,683,195号;「核酸ハイブリダイゼーション(Nucleic Acid Hybridization)」、ビーディー・ヘイムズおよびエスジェイ・ヒギンス(B.D.Hames & S.J.Higgins)編、1984;「転写および翻訳(Transcription and Translation)」、ビーディー・ヘイムズおよびエスジェイ・ヒギンス(B.D.Hames & S.J.Higgins)編、1984;「動物細胞の培養(Culture Of Animal Cells)」、アールアイ・フレッシュニー(R.I.Freshney)著、アラン・アールリス社(Alan R.Liss,Inc.)、1987;「不動化細胞および酵素(Immobilized Cells And Enzymes)」、アイアールエル・プレス(IRL Press)、1986;ビー・パーバル(B.Perbal)著、「分子クローニングの実技ガイド(A Practical−Guide To Molecular Cloning)」、1984;論説「酵素学の方法(Methods in Enzymology)」、アカデミックプレス社(Academic Press,Inc.)、ニューヨーク州;「哺乳類細胞のための遺伝子転送ベクター(Gene Transfer Vectors For Mammalian Cells)」、ジェイエイチ・ミラーおよびエムピー・カーロス(J.H.Miller and M.P.Calos)編、1987年、コールドスプリングハーバーラボラトリ(Cold Spring Harbor Laboratory);酵素学の方法(Methods in Enzymology)、154および155巻、ウー(Wu et al.)等編、「細胞および分子生物学における免疫化学的方法(Immunochemical Methods in Cell and Molecular Biology)」、メイヤーおよびウォーカー(Mayer and Walker)編、アカデミックプレス社(Academic Press,Inc.)、ロンドン、1987;「実験免疫学ハンドブック(Handbook Of Experimental Immunology)」、I−IV巻、ディーエム・ウェアおよびシーシー・ブラックウェル(D.M.Weir and C.C.Blackwell)編、1986;「マウス胚の取り扱い手技(Manipulating Mouse Embryo)」、コールドスプリングハーバーラボラトリー・プレス(Cold Spring Harbor Laboratory Press)、コールドスプリングハーバー(Cold Spring Harbor)、ニューヨーク州、1986を参照されたい。さらに、プロテアーゼの調製、発現、分離、および使用の方法に関する情報が、米国特許第6,768,001号を参照することによって得られる。なお、この特許文献の全体を、引用することにより本明細書に含める。
【0023】
本発明の他の特徴および利点は、下記の詳細な説明、および特許請求の範囲から明らかとなろう。なお、本発明の実施または試験においては、本明細書に記載されるものと類似の、または等価な、任意の方法および材料の使用が可能であるけれども、好ましい方法および材料が記載される。
【0024】
別様に定義しない限り、本明細書で使用される技術および科学用語は全て、本発明の所属する従来技術において通常の錬度を有する当業者によって一般的に理解されるものと同じ意味を持つ。シングルトン(Singleton et al.)等、「微生物および分子生物学辞典(DICTIONARY OF MICROBIOLOGY AND MOLECULAR BIOLOGY)」、ジョンウィリー・アンド・サンズ(John Wiley and Sons)、ニューヨーク、1994、および、ヘイルおよびマーカム(Hale & Markham)、「ハーパーコリンズ生物学辞典(HAPER COLLINS DICTIONARY OF BIOLOGY)」、ハーパーペレニアル(Haper Perennial)、ニューヨーク州、1991が、当業者には、本発明に使用される用語の多くについて一般的辞典となる。
【0025】
本明細書に示される見出しは、明細書全体を参照することによって理解することが可能な、本発明の様々な態様または実施態様を限定するものではない。従って、このすぐ後に定義される用語は、本明細書の全体を参照することによってより完全に定義される。
【0026】
数字の範囲は、その範囲を定義する数字を含める。
【0027】
別様に指示しない限り、それぞれ、核酸は、左から右へ、5’から3’方向に書かれ、アミノ酸は、左から右へ、アミノからカルボキシル方向に書かれる。
【0028】
本明細書および付属の特許請求項で使用される、単数形表現「この」および「前記」は、内容が明白に別様に指定しない限り、複数形を含む。従って、「化合物」を含む組成物に対する言及は、2種以上の化合物から成る混合物を含む。さらに、「または」という用語は、内容が明白に別様に指定しない限り、一般に「および/または」を含む意味に使用される。
【0029】
定義
「プロテアーゼ」は、微生物、例えば、真菌、細菌、または、植物または動物から得られるタンパクまたはポリペプチドの、タンパクまたはポリペプチドドメインであって、タンパク主鎖の様々な位置の内の一つにおけるペプチド結合の切断を触媒する能力を持つドメインである(例えば、E.C.3.4)。
【0030】
「酸性プロテアーゼ」とは、酸性条件下でタンパクを加水分解する能力を持つプロテアーゼを指す。
【0031】
本明細書で用いる「NSP24ファミリ・プロテアーゼ」とは、生来の、または野生型形態(例えば、図6のタンパク)において、プロテアーゼ活性を持つ酵素、すなわち、配列番号2または配列番号10のアミノ酸配列、配列番号2または配列番号10のアミノ酸配列の誘導体、アミノ酸配列の生物学的に活性な断片に対し、少なくとも50%、少なくとも60%、少なくとも70%、少なくとも80%、少なくとも85%、少なくとも90%、少なくとも93%、少なくとも95%、少なくとも97%、少なくとも98%、および少なくとも99%の配列相同性を有するプロテアーゼタンパクを意味する。
【0032】
本明細書で用いる「誘導体」とは、前駆体または親タンパク(例えば、自然のタンパク)から得られるタンパクであって、アミノ酸配列のC−およびN−末端のいずれかまたは両方に1個以上のアミノ酸を付加することによって、アミノ酸配列におけるいくつかの異なる部位において1個以上のアミノ酸を置換することによって、タンパクの一端または両端においてまたはアミノ酸配列の1箇所以上の部位において1個以上のアミノ酸を欠失させることによって、またはアミノ酸配列の1箇所以上の部位において1個以上のアミノ酸を挿入することによって得られるタンパクを意味する。
【0033】
本明細書で用いる「自然配列NSP24」または「野生型NSP24配列」とは、自然から得られたNSP24ファミリ・プロテアーゼと同じアミノ酸配列を持つポリペプチドを含む。
【0034】
「生物学的に活性な断片」(例えば、配列番号10の配列を有するNSP24ファミリ・プロテアーゼの、生物学的に活性な断片)とは、プロテアーゼ活性を持つが、NSP24ファミリ・プロテアーゼまたはNSP25ファミリ・プロテアーゼの前駆体または親タンパクの完全配列よりも少ない配列を含む、NSP24ファミリ・プロテアーゼまたはNSP25ファミリ・プロテアーゼを意味する。
【0035】
「単離」または「精製」とは、プロテアーゼが天然状態下で関連する天然成分の内の1種以上、またはその全てからプロテアーゼを分離することによってプロテアーゼがその天然状態から変えられていることを指す。
【0036】
「PepA」とは、配列番号7に対して少なくとも95%の配列相同性を有する酸性プロテアーゼを指す。
【0037】
「L388M」とは、配列番号7の配列を持つ変異種PepAを指す。
【0038】
本明細書で用いる「NSP25ファミリ・プロテアーゼ」とは、配列番号9および生物学的に活性なその断片に対して、少なくとも85%の配列相同性を持つプロテアーゼ酵素を意味する。
【0039】
「NSP24ファミリ・プロテアーゼに対して無関係な」とは、配列番号10のNSP24プロテアーゼに対して、30%未満の相同性、20%未満の相同性、または10%未満の相同性を持つアミノ酸配列を意味する。
【0040】
「ペプチド」、「タンパク」、および「ポリペプチド」という用語は、本明細書では相互交換的に用いられる。
【0041】
本明細書で用いる、特定されたアミノ酸またはヌクレオチドに関する「パーセント(%)配列同一性」とは、候補配列と対象配列(例えば、NSP24ファミリ・プロテアーゼ配列)とを整列し、要すればギャップを導入して、最大パーセント配列同一性を実現したあと、対象配列のアミノ酸残基またはヌクレオチドと同一である、候補配列のアミノ酸残基またはヌクレオチドのパーセントと定義される。なお、どの保存的置換体も配列同一性の一部であるとは見なさない。
【0042】
本明細書で用いる「アルファ・アミラーゼ(E.C.クラス3.2.1.1)」は、アルファ−1,4−グルコシド結合の加水分解を触媒する酵素を指す。これらの酵素はまた、1,4−α−結合D−グルコース単位を含むポリサッカライドにおける、1,4−α−D−グルコシド結合の、エキソまたはエンド加水分解を実行するものとしても記載される。これらの酵素を記載するために用いられるもう一つの用語は、「グリコゲナーゼ」である。例示の酵素としては、アルファ−1,4−グルカン4−グルカノヒドラーゼ、グルカノヒドロラーゼが挙げられる。
【0043】
本明細書で用いる「グルコアミラーゼ」という用語は、アミログルコシダーゼクラスの酵素(例えば、EC.3.2.1.3、グルコアミラーゼ、1,4−アルファ−D−グルカングルコヒドラーゼ)を指す。これらは、アミロースおよびアミロペクチン分子の非還元性末端からグルコシル残基を放出する、エキソ活動性酵素である。この酵素はまた、アルファ−1,6、およびアルファ−1,3結合をも加水分解する。もっともアルファ−1,4結合に対するよりもその速度はずっと遅い。
【0044】
「プロモーター」という用語は、遺伝子の転写を開始するためのRNAポリメラーゼの結合に関与する調整配列を意味する。
【0045】
本明細書で用いる「異種プロモーター」は、遺伝子、または精製核酸と天然では関連しないプロモーターである。
【0046】
本明細書で用いる、ポリペプチドの「精製標本」または「実質的に純粋な標本」とは、該ポリペプチドが天然では共に存在する、細胞、その他のタンパク、脂質、または核酸から分離されたポリペプチドを意味する。
【0047】
本明細書で用いる「細胞の精製標本」とは、植物または動物細胞の場合、細胞のインビトロ標本を指し、生の、植物または動物細胞全体を指さない。培養細胞または微生物細胞の場合、該標本は、対象細胞の少なくとも10%、より好ましくは50%を持つ標本から成る。
【0048】
「実質的に純粋な核酸」、例えば、実質的に純粋なDNAとは、該核酸の由来する生物体の天然ゲノムにおいて該核酸が直接連結する(例えば、一方は5’末端において、一方は3’末端において)配列、例えば、コード配列のいずれか一端または両端に対し直接連結しない、その一方または両方である核酸である。この用語は、例えば、ベクター、例えば、自律的複製プラスミドまたはウィルスに、または、前核細胞または真核細胞のゲノムDNAに組み込まれる組み換えDNA、あるいは、他のDNA配列と独立な分離分子として存在するDNA(例えば、PCRまたは制限エンドヌクレアーゼ処理によって生産されるcDNAまたはゲノムDNA断片)を含む。実質的に純粋なDNAはまた、さらに別のNSP24プロテアーゼ配列をコードするハイブリッド遺伝子の一部である組み換えDNAを含む。
【0049】
本明細書で用いる「相同性」とは、二つのポリペプチド分子間、または二つの核酸分子間の配列類似性を指す。二つの比較される配列の両方における一つの位置が、同じ塩基、またはアミノ酸モノマーサブユニットによって占められている場合、例えば、二つのDNA分子のそれぞれにおける一つの位置が、アデニンで占められている場合、これらの分子は、その位置において相同である。二つの配列間の相同性のパーセントは、該二つの配列が共有する、一致または相同位置の数を、比較される位置の数で割り、×100とした関数である。例えば、二つの配列の位置について、10の内の6が一致または相同である場合、この二つの配列は60%相同である。例を挙げると、DNA配列ATTGCCとTATGGCは、50%の相同を共有する。一般に、最大の相同性を与えるように二つの配列を整列させて比較を行う。
【0050】
本明細書で用いる「ベクター」という用語は、核酸を、1種以上の細胞タイプに導入するよう設計されたポリヌクレオチド配列をさす。ベクターとしては、クローニングベクター、発現ベクター、シャトルベクター、プラスミド、ファージ粒子、カセット等が挙げられる。
【0051】
本明細書で用いる「発現ベクター」とは、適切な宿主におけるDNAの発現に影響を及ぼすことが可能な適切な調節配列に動作的に結合するDNA配列を含む、DNA構築体を意味する。
【0052】
「発現」という用語は、ポリペプチドが、遺伝子の核酸配列に基づいて生産されるその過程を意味する。
【0053】
本明細書で用いる「動作的に結合する」とは、調整領域、例えば、プロモーター、ターミネーター、分泌シグナル、またはエンハンサー領域が、構造遺伝子に付着または結合し、その遺伝子の発現を調節することを意味する。
【0054】
本明細書で用いる、微生物「由来の」物質(例えば、ポリヌクレオチドまたはタンパク)とは、その物質が、その微生物に本来属するものであることを意味する。
【0055】
本明細書で用いる、「微生物」とは、細菌、真菌、ウィルス、原生動物、および他の微生物、すなわち顕微鏡的生物を指す。
【0056】
本明細書で用いる、「宿主株」または「宿主細胞」は、本発明によるDNAを含む発現ベクターにとって好適な宿主を意味し、前記細胞の子孫を含む。
【0057】
「糸状真菌」という用語は、全ての、糸状形態の下位区分Eumycotinaを指す(アレクソパウロス・シージェイ(Alexopoulos,C.J.)(1962),「真菌学入門(INTRODUCTORY MYCOLOGY)」、ウィリー(Wiley)、ニューヨーク州、および、エインスワースおよびビスビー(AINSWORTH AND BISBY)、「真菌の辞書(DICTIONARY OF THE FUNGI)」、9版、(2001)、カーク(Kirk)等編、シーエービー・インターナショナルユニバーシティ・プレス(CAB International University Press)、ケンブリッジ、英国を参照されたい)。これらの真菌は、キチン、セルロース、および、他の複雑なポリサッカライドによって構成される細胞壁を持つ、植物性菌糸によって特徴づけられる。本発明の糸状真菌は、形態的に、生理的に、かつ遺伝的に、酵母とは異なる。糸状真菌による植物的増殖は、菌糸の延長により、炭素の異化作用は好気性であることを強制される。
【0058】
本明細書で用いる、「Trichoderma」または「Trichoderma SPP.」は、以前または現在Trichodermaに分類される、任意の真菌属を指す。
【0059】
本明細書で用いる「4重欠失」または「4重欠失した」宿主細胞とは、機能的エンドグルカナーゼをコードする少なくとも2個の遺伝子、および、機能的セロビオヒドロラーゼをコードする少なくとも2個の遺伝子を欠失する、トリコデルマ宿主株の細胞から形成された細胞およびプロトプラストの両方を指す。
【0060】
本明細書で用いる「培養」という用語は、液体または固体媒体中の好適な条件下における微生物細胞集団の増殖を指す。一つの実施態様では、培養は、でん粉基質、例えば、顆粒状でん粉を含む基質の、最終産物(典型的には、血管またはリアクターにおける)への醗酵性生体変換を指す。醗酵は、より単純な有機化合物を生産するために微生物によって行われる、有機物質の酵素的で、かつ嫌気性の分解である。醗酵は嫌気的条件下で起こるが、醗酵はまた酸素の存在下でも起こるので、本用語は、厳密な嫌気的条件のみに限定されることを意図しない。
【0061】
本明細書で用いる「接触する」という用語は、それぞれの酵素(単複)を、それぞれの基質の十分近くに置くことによって、酵素(単複)が基質を最終産物に変換することを可能とすることを指す。当業者であれば、酵素溶液をそれぞれの基質と混合することが接触に作用する可能性のあることを了解されるであろう。
【0062】
核酸配列を細胞中に挿入するという背景で用いられる「導入」という用語は、「トランスフェクション」、「形質転換」、または「形質導入」を意味し、核酸配列の、真核または前核細胞への取り込みに対する参照を含む。その際、該核酸配列は、細胞のゲノム(例えば、染色体、プラスミド、プラスチド、または、ミトコンドリアDNA)に組み込まれてもよいし、自律的レプリコンに変換されてもよいし、または、一時的に発現されてもよい(例えば、形質導入mRNA)。
【0063】
本明細書で用いる、細胞に関して使用される「形質転換された」、「安定に形質転換された」、および「トランスジェニック」という用語は、細胞が、在来型ではない(例えば、異種の)核酸配列を、そのゲノムの中に一体化して、または、複数の世代に渡って維持されるエピソームプラスミドとして有することを意味する。
【0064】
本明細書で用いる、ポリペプチドまたはポリヌクレオチドと関連して使用される「異種の」という用語は、宿主細胞では天然には見られないポリペプチドまたはポリヌクレオチドを意味する。
【0065】
「過剰発現」という用語は、ポリヌクレオチドが宿主細胞に導入された場合、ポリペプチドを該宿主細胞において発現する過程を意味する。
【0066】
本明細書に記載するように、本発明の一つの態様は、NSP24ファミリ・プロテアーゼまたはNSP25ファミリ・プロテアーゼ、および/またはそれらの核酸の等価物をコードするヌクレオチド配列を含む「実質的に純粋な」(または組み換え)核酸を特徴とする。
【0067】
「等価的」という用語は、機能的に等価な複数のポリペプチドをコードする複数のヌクレオチド配列を指す。複数の、等価的ヌクレオチド配列は、1個以上のヌクレオチド置換、付加、または欠失、例えば、対立遺伝子変異において異なる、複数の配列を含む。例えば、ある実施態様では、遺伝コードの縮重のため、等価的ヌクレオチド配列は、配列番号2に示すNSP24プロテアーゼをコードするが、配列番号8のヌクレオチド配列と異なる配列を含む。
【0068】
本明細書で用いる「糖化」という用語は、でん粉をグルコースに変える酵素的転換を指す。
【0069】
本明細書で用いる「でん粉」とは、アミラーゼとアミロペクチンから構成される、xが任意の数である化学式(C6H10O5)xを持つ、植物の、複雑なポリサッカライド炭水化物から構成される任意の物質を指す。
【0070】
「顆粒状でん粉」という用語は、調理されていない(生の)でん粉(例えば、ゼラチン化されたことがないでん粉)を指す。
【0071】
本明細書で用いる「ゼラチン化」という用語は、でん粉分子を調理することによって可溶化し粘調な懸濁液を形成することを意味する。
【0072】
本明細書で用いる「液化」という用語は、でん粉転換の段階であって、ゲル化でん粉が加水分解されて低分子量の可溶性デキストランを生じる段階を指す。
【0073】
本明細書で用いる「可溶性でん粉加水分解産物」という用語は、でん粉の加水分解によって得られる可溶性産物であって、モノ−、ジ−、およびオリゴサッカライド(例えば、グルコース、マルトース、および多糖類)を含む産物を指す。
【0074】
「モノサッカライド」という用語は、ポリマー、例えば、でん粉のモノマー単位であって、重合度(DP)が1である単位(例えば、グルコース、マンノース、フルクトース、およびガラクトース)を意味する。
【0075】
「ジサッカライド」という用語は、二つの、共有的に結合するモノサッカライド単位を含む化合物(DP2)(例えば、スクロース、ラクトース、およびマルトース)を意味する。
【0076】
用語“DP3+”という用語は、3よりも大きな重合度を持つポリマーを意味する。
【0077】
プロテアーゼと、該プロテアーゼをコードするポリヌクレオチド
本発明は、NSPファミリ・プロテアーゼであって、配列番号2のプロテアーゼまたは配列番号10のプロレアーゼ(図6)に対し、少なくとも50%、少なくとも60%、少なくとも70%、少なくとも80%、少なくとも85%、少なくとも90%、少なくとも93%、少なくとも95%、少なくとも97%、少なくとも98%、および少なくとも99%の配列相同性を有するプロテアーゼ、例えば、酸性プロテアーゼ、およびさらに酸性真菌プロテアーゼに関する。ある実施態様では、NSP24ファミリ・プロテアーゼは、配列番号10の配列(成熟タンパク配列)を含むNSP24、あるいは、配列番号2のプレタンパク配列とも表示される。
【0078】
ある実施態様では、本発明は、NSP24ファミリ・プロテアーゼの生物学的活性を持つ断片に関する。ある実施態様では、生物学的活性断片は、少なくとも250個のアミノ酸残基、少なくとも300個のアミノ酸残基、少なくとも350個のアミノ酸残基、少なくとも375個のアミノ酸残基、および少なくとも400個のアミノ酸残基を有するプロテアーゼを含む。
【0079】
別の実施態様では、生物学的活性断片は、図6のタンパク配列(配列番号2または配列番号10)に対し、少なくとも80%、少なくとも85%、少なくとも90%、少なくとも93%、少なくとも95%、少なくとも97%、少なくとも98%、および少なくとも99%の配列相同性を有するポリペプチド配列の、少なくとも60%、少なくとも70%、少なくとも80%、少なくとも85%、少なくとも90%、少なくとも93%、少なくとも95%、少なくとも97%、少なくとも98%、および少なくとも99%を含む。ある実施態様では、生物学的活性断片は、配列番号2または配列番号10を持つ親NSP24プロテアーゼに対し、少なくとも95%の配列相同性を有するポリペプチド配列の、少なくとも60%、少なくとも70%、少なくとも80%、少なくとも85%、少なくとも90%、少なくとも95%、少なくとも98%を含む。ある実施態様では、生物学的活性断片は、配列番号2または配列番号10を持つ親NSP24プロテアーゼに対し、少なくとも99%の配列相同性を有するポリペプチド配列の、少なくとも60%、少なくとも70%、少なくとも80%、少なくとも85%、少なくとも90%、少なくとも95%、少なくとも98%を含む。
【0080】
ある実施態様では、生物学的活性断片は、生体内に存在する断片、例えば、翻訳後処理によって生ずる断片、または、選択的スプライシングを受けたRNAの翻訳によって生ずる断片である。断片は、天然または内在細胞において、例えば、翻訳後処理の結果、例えば、アミノ末端シグナル配列除去の結果として発現されるものを始め、発現システム、例えば、CHO細胞において発現されるものも含む。ある好ましい断片は、タンパク分解による切断、または選択的スプライシング事象によって生じた断片、例えば、活性断片である。NSP24ファミリ・プロテアーゼのようなプロテアーゼは、ある範囲の生理学的性質を示すことがよくあり、かつ、そのような性質は、分子の異なる部分に由来することがあるのであるから、NSP24ファミリ・プロテアーゼの有用断片または類縁体は、NSP24プロテアーゼ活性に関する任意の生物学的アッセイにおいて生物学的活性を示すものである。
【0081】
ある実施態様では、生物学的活性断片は、配列番号2または配列番号10を持つNSP24のプロテアーゼ活性の少なくとも20%、少なくとも30%、少なくとも40%、少なくとも50%、少なくとも60%、少なくとも70%、少なくとも80%、少なくとも90%、少なくとも95%、少なくとも100%を含む。ある好ましい実施態様では、断片または類縁体は、任意のインビボまたはインビトロのNSP24プロテアーゼアッセイにおいて、NSP24プロテアーゼ(配列番号2または配列番号10)のプロテアーゼ活性の少なくとも40%、または少なくとも90%を持つ。
【0082】
NSP24ファミリ・プロテアーゼまたはNSP25ファミリ・プロテアーゼの断片は、当業者に既知の方法によって生成することが可能である。候補断片が、プロテアーゼの生物学的活性を示す能力は、本明細書に記載するように、当業者に既知の方法によって評価することが可能である。さらに、NSP24ファミリ・プロテアーゼおよびNSP25ファミリ・プロテアーゼであって、該ペプチドの生物学的活性に必要とされない残基を含むもの、または、選択的mRNAスプライシング、または、タンパクの選択的プロセシングによって生じるものも含まれる。
【0083】
ある実施態様では、本発明によって被われるプロテアーゼは、配列番号2または配列番号10を持つプロテアーゼの誘導体である。誘導体は、配列番号10に対し、少なくとも80%、85%、90%、93%、95%、97%、98%、および99%の配列相同性を持ってもよい。
【0084】
本発明はまたプロテアーゼ類縁体を含む。類縁体とは、ペプチドの安定性を増す修飾を伴うものである。そのような類縁体は、例えば、ペプチド配列の中に、1個以上の非ペプチド結合(ペプチド結合を置換する)を含んでもよい。さらに含まれるのは、天然のL−アミノ酸以外の残基、例えば、D−アミノ酸、または非天然性または合成アミノ酸、例えば、βアミノ酸、および環状類縁体を含む類縁体である。類縁体は、天然のプロテアーゼと、例えば、NSP24またはNSP25タンパクと、アミノ酸配列において、または、配列を含まないやり方で、または、その両方において異なっていてもよい。非配列修飾は、本発明に含まれるプロテアーゼの、インビトロまたはインビボにおける、化学的誘導体形成を含む。非配列修飾としては、アセチル化、メチル化、リン酸化、カルボキシル化、またはグリコシル化における変化が挙げられる。
【0085】
さらに別の実施態様では、本発明はNSP25ファミリ・プロテアーゼを含む。NSP25ファミリ・プロテアーゼは、配列番号9(図9)の成熟タンパク配列、またはその生物学的に活性な断片に対して、少なくとも85%、少なくとも90%、少なくとも93%、少なくとも95%、少なくとも97%、少なくとも98%、および少なくとも99%のアミノ酸配列相同性を持つ酸性プロテアーゼである。一つの特異的NSP25ファミリ・プロテアーゼは、配列番号9を持つNSP25と表示されるプロテアーゼである。ある実施態様では、NSP25ファミリ・プロテアーゼは、配列番号9に対し少なくとも90%の配列相同性を有する配列の、少なくとも75%、少なくとも80%、少なくとも85%、少なくとも90%、少なくとも95%を含む、プロテアーゼの生物学的活性断片である。別の実施態様では、NSP25ファミリ・プロテアーゼは、配列番号9に対し少なくとも95%の配列相同性を有する配列の、少なくとも75%、少なくとも80%、少なくとも85%、少なくとも90%、および少なくとも95%を含む、プロテアーゼの生物学的活性断片である。
【0086】
本発明による酸性プロテアーゼは、酸性条件下でタンパクを加水分解することが可能なプロテアーゼであるが、ある実施態様では、プロテアーゼ活性のための至適pHは、pH3.0から5.5の範囲にある。ある実施態様では、プロテアーゼ活性のための至適pH範囲は、pH3.0と5.0の間にあるが、別の実施態様では、プロテアーゼ活性のための至適pH範囲は、pH3.0と4.5の間にある。
【0087】
本発明によるプロテアーゼ、例えば、NSP24ファミリ・プロテアーゼまたはNSP25ファミリ・プロテアーゼは、アミノ酸置換、例えば、L−アミノ酸による置換であって、一つのアミノ酸が別の生物学的に類似のアミノ酸によって置換される「保存的アミノ酸置換」を含んでもよい。保存的アミノ酸置換とは、置換されるアミノ酸の、全体的電荷、疎水性/親水性、および/または、立体的広がりを保存する置換である。保存的置換の例として、下記の基の間の置換が挙げられる。すなわち、Gly/Ala、Val/Ile/Leu、Lys/Arg、Asn/Gln、Glu/Asp、Ser/Cys/Thr、およびPhe/Trp/Tyrである。誘導体は、例えば、1から10個までのアミノ酸残基が異なっていてもよい。例えば、6−10、5、4、3、2、または1個のアミノ酸残基が異なっていてもよい。表1は、従来技術で認められたアミノ酸置換の例を示す。さらに、置換は、プロテアーゼ生物活性を消滅させることのない、1個以上の非保存的アミノ酸置換、欠失、または挿入によって行われてもよい。
【表1】
【0088】
ある実施態様では、本発明のプロテアーゼは内在配列である。このような内在配列は、天然から単離することが可能であり、あるいは、組み換えまたは合成手段によって製造することが可能である。この「内在配列」という用語は、NSP24またはNSP25ファミリ・プロテアーゼの天然に見られる短縮形または分泌形(例えば、生物学的活性断片)、および、天然に見られる変異形(例えば、選択的スプライシングを受けた変異形)を特に含む。
【0089】
ある実施態様では、本発明の酸性プロテアーゼは、配列番号7に対して、少なくとも97%、少なくとも98%、少なくとも99%の配列相同性を持つPepAプロテアーゼである。ある実施態様では、プロテアーゼは、配列番号7の配列を持ち、“L388M”と表示される。さらに別の実施態様では、プロテアーゼは、配列番号5、または配列番号3の配列を持つヌクレオチド配列によってコードされる。
【0090】
本発明はまた、本発明に含まれるプロテアーゼをコードするポリヌクレオチドに関する。このようなポリヌクレオチドは下記を含む。すなわち、
a)配列番号2または配列番号10に対して、少なくとも80%、少なくとも85%、少なくとも90%、少なくとも93%、少なくとも95%、少なくとも97%、少なくとも98%、および少なくとも99%の配列相同性を持つNSP24ファミリ・プロテアーゼをコードするポリヌクレオチド;
b)配列番号2の配列をコードするポリヌクレオチド;
c)配列番号8の配列を持つポリヌクレオチド;
d)NSP24ファミリ・プロテアーゼの生物学的活性断片をコードするポリヌクレオチド;
e)配列番号8に対して、少なくとも80%、少なくとも85%、少なくとも90%、少なくとも95%、少なくとも97%、少なくとも98%、および少なくとも99%の配列相同性を持つポリヌクレオチド;
f)配列番号4、配列番号8のDNA配列、または、配列番号4または配列番号8の断片に対応する核酸プローブに対してハイブリダイズするポリヌクレオチドであり、その際、前記断片は、少なくとも10、15、20、30、40、50、60、70、80、90、100、または150個の連続ヌクレオチドを有し;
g)配列番号4に対して、少なくとも85%、少なくとも90%、少なくとも93%、少なくとも95%、少なくとも97%、少なくとも98%、および少なくとも99%の配列相同性を持つNSP25ファミリ・プロテアーゼをコードするポリヌクレオチド;
h)配列番号9のプロテアーゼをコードするポリヌクレオチド;
i)配列番号4の配列を持つポリヌクレオチド;
j)NSP25ファミリ・プロテアーゼの生物学的活性断片をコードするポリヌクレオチド;
k)配列番号7の配列、およびその生物学的活性断片をコードするポリヌクレオチド;および、
l)配列番号3または配列番号5の配列を持つポリヌクレオチド。
【0091】
遺伝子コードには縮重があるために、ある特定のアミノ酸をコードするのに1種を超えるコドンが使用される場合がある。従って、異なるいくつかのDNA配列が、同じアミノ酸配列を持つポリペプチド、例えば、配列番号2のポリペプチドをコードする可能性がある。本発明は、同じポリペプチドをコードする複数のポリペプチドをカバーする。
【0092】
核酸は、該核酸の1本鎖型が、適当な温度およびイオン強度条件下において別の核酸とアニールすることが可能である場合、その他方の核酸配列にたいしてハイブリダイズすることが可能である。ハイブリダイゼーションおよび洗浄条件は、低度、中等度、高度、および、極めて高度の厳格条件下におけるハイブリダイゼーションに関して従来技術でよく知られる(例えば、サムブルック(Sambrook)(1989)、上記、特に9および11章参照)。一般に、ハイブリダイゼーションは、ヌクレオチドプローブと相同的DNA配列を含み、これらは、相補的ポリヌクレオチドの広範な塩基対合によって安定な二本鎖ハイブリッドを形成する(さらに、8章および、「遺伝子クローニング入門(Gene Cloning,An Introduction)」、ティーエー・ブラウン(T.A.Brown)、(1995)、Chapman and Hall、ロンドンを参照されたい)。ある実施態様では、プローブおよび相同配列を有するフィルターは、2×塩化ナトリウム/クエン酸ナトリウム(SSC)、約60℃(中等厳格度)、65℃(中等/高等厳格度)、70℃(高等厳格度)、および約75℃(極高等厳格度)における0.5%SDSで洗浄してもよい(「分子生物学における最新プロトコール(Current Protocols in Molecular Biology)」、John Wiley & Sons,ニューヨーク州、1989、6.3.1−6.3.6、なお、この文献を引用することにより本明細書に含める)。
【0093】
本発明に含まれるものは、対立遺伝子変異;自然突然変異;誘発突然変異;配列番号2、配列番号8、配列番号9、および配列番号10のポリペプチドをコードする核酸に対して、高または低厳格条件下でハイブリダイズするDNAによってコードされるタンパク、および、配列番号2または配列番号10を持つNSP24プロテアーゼに対する抗血清によって、特にNSP24プロテアーゼの活性部位または結合ドメインに対する抗血清によって特異的に結合されるポリペプチドである。ある実施態様では、本発明のNSP24ファミリ・プロテアーゼをコードする核酸、例えば、配列番号2のNSP24プロテアーゼをコードする核酸は、配列番号8から得られる少なくとも12、15、または20の連続ヌクレオチドに対応する核酸に対して高度の厳格条件下にハイブリダイズする。
【0094】
本発明の核酸およびポリペプチドは、本明細書に開示される配列とは、開示の配列における配列誤差のために異なるものも含む。
【0095】
DNA配列の相同性は、二つのDNA配列間の同一性の程度によって決められる。複数のポリペプチド配列またはヌクレオチド配列について、相同性または同一性パーセントが、コンピュータを用いて求められてもよい。配列整列を実行し、配列同一性を定める方法は当業者には既知であり、不要な実験を行うことなく実行が可能であり、同一性値の計算も明確に獲得することが可能である。例えば、アウスウーベル(Ausubel)等編、(1995)、「分子生物学における最新プロトコール(Current Protocols in Molecular Biology)」、19章、(Greene Publishing and Wiley−Interscience),ニューヨーク州;および、デイホフ(Dayhoff)(1978)「タンパク科学および構造アトラス5(Atlas of Protein Science and Structure 5)」、別冊3(National Biomedical Research Foundation,Washingtong,D.C.)を参照されたい。複数の配列を整列し、配列同一性を求めるためにはいくつかのアルゴリスムが利用可能であり、例えば、ニードルマン(Needleman)等(1970)J.Mol.Biol.48:443の相同性整列アリゴリスム;スミス(Smith)等(1981)Adv.Appl.Math.2:482の局部的相同性アルゴリスム;ピアソン(Pearson)等(1988)Proc.Natl.Acad.Sci.85:2444の類似性探索法;スミス・ウォーターマン法(Meth.Mol.Biol.70:173−187(1997)、および、BLASTP、BLASTN、およびBLASTXアルゴリスム(アルチュル(Altschul)等(1990)J.Mol.Biol.215:403−410を参照)が挙げられる。これらのアルゴリスムを用いるコンピュータプログラムも利用が可能であり、例えば、ALIGNまたはMegalign(DNASTAR)ソフトウェア、またはWU−BLAST−2(アルチュル(Altschul)等、Meth.Enzym.,266:460−480(1996));またはGAP、BESTFIT、BLASTアルチュル等上記、FASTA、および、Genetics Computing Group (GCG)パッケージ、バージョン8、マジソン、ウィスコンシン州、米国において市販されるTFASTA、および、Intelligenetics,Mountain View、カリフォルニア州によるPC/Geneプログラムに納められるCLUSTALが挙げられるが、ただしこれらに限定されない。当業者であれば、比較される複数の配列の長さに渡って最大の整列を実現するのに必要なアルゴリスムを含め、整列を測定するための適切なパラメータを決めることが可能である。配列同一性は、プログラムによって決められるデフォールトパラメータを用いて決められることが好ましい。具体的に言うと、配列同一性は、MSPRCHプログラム(Oxford Molecular)に搭載されるスミス・ウォーターマン相同性探索アルゴリスムによって求めることが可能である。このアルゴリスムは、下記の探索パラメータ、すなわち、ギャップ開放ペナルティー12、およびギャップ延長ペナルティー1によるアフィンギャップ探索を用いる。対合アミノ酸比較は、Genetics Computer Group,Inc.マジソン、ウィスコンシン州のGCG配列分析ソフトウェアパッケージのGAPプログラムを用いて実行することが好ましい。このプログラムは、ギャップウェイト12、長さウェイト2とする、blosum62アミノ酸置換マトリックスを用いる。二つのアミノ酸配列の最適整列に関しては、変異アミノ酸配列の連続セグメントは、参照アミノ酸配列に対して、付加的アミノ酸残基を持ってもよいし、または、アミノ酸残基を欠失させてもよい。参照アミノ酸配列との比較に用いられる連続セグメントは、少なくとも20個の連続アミノ酸残基を含むが、30、40、50個、またはそれ以上のアミノ酸残基であってもよい。誘導体のアミノ酸配列にギャップを含めたことによって生ずる配列同一性の増加に対する補正は、ギャップペナルティーを割り当てることによって実行される。
【0096】
ある実施態様では、本発明によってカバーされるプロテアーゼ(例えば、配列番号2の配列に対して少なくとも80%の配列同一性を持つNSP24ファミリ・プロテアーゼ)は、細菌、または真菌、例えば、糸状真菌から得られる。ある好ましい糸状真菌としては、Aspergillus SPP.、およびTrichoderma SPP.が挙げられる。一つの好ましいTrichoderma SPP.は、T.reeseiである。しかしながら、プロテアーゼ、および/または、本発明によるプロテアーゼをコードするDNAは、真菌、例えば、Absidia SPP.;Acremonium SPP.;Agaricus SPP.;Anaeromyces SPP.;Aspergillus SPP.、例えば、A.aculeatus、A.awamori、A.flavus、A.foetidus、A.fumaricus、A.fumigatus、A.nidulans、A.niger、A.oryzae、A.terreus、およびA.versicolor;Aerobasidium SPP.;Cephalosporum SPP.;Chaetomium SPP.;Coprinus SPP.;Dactyllum SPP.;Fusariumu SPP.、例えば、F.conglomerans,F.decemcellulare,F.javanicum、F.lini,F.oxysporum,およびF.solani;Gliocladium SPP.;Humicola SPP.、例えば、H.insolensおよびH.lanuginosa;Mucor SPP.;Neurospora SPP.、例えば、N.crassaおよびN.sitophila;Neocallimastix SPP.;Orpinomyces SPP.;Penicillium SPP.;Phanerochaete SPP.;Phlebia SPP.;Piromyces SPP.;Rhizopus SPP.;Schizophyllum SPP.;Trametes SPP.;Tricoderma SPP.、例えば、T.reesei、T.reesei(longbrachiatum)およびT.virdie;およびZygorhynchus SPP.のような真菌から得られる。
【0097】
宿主細胞
ある実施態様では、本発明は、本明細書に記載されるDNA構築体およびベクターによって形質転換された宿主細胞を提供する。ある実施態様では、本発明によってカバーされるプロテアーゼ(例えば、配列番号2に対して少なくとも95%の配列同一性を有するNSP24ファミリ・プロテアーゼ)をコードするポリヌクレオチドであって、宿主細胞中に導入されるポリヌクレオチドは、異種プロテアーゼをコードし、別の実施態様では、ポリヌクレオチドは、宿主細胞において過剰発現される内因性プロテアーゼをコードする。ある実施態様では、本発明は、宿主細胞、例えば、細菌および真菌宿主細胞において機能する遺伝子プロモーターの制御下における、異種プロテアーゼ遺伝子の発現、またはプロテアーゼ遺伝子の過剰発現を提供する。
【0098】
ある好ましい宿主細胞としては糸状真菌細胞が挙げられる。糸状真菌宿主細胞の非限定的例として、Trichoderma SPP.(例えば、T.virideおよびT.reesei、これらは、以前T.longibrachiatumとして分類されていた、Hypocrea jecorinaの無性形である)、Penicillium SPP.、Humicola SPP.(例えば、H.insolensおよびH.grisea)、Aspergillus SPP.(例えば、A.niger、A.nidulans、A.orzyae、および、A.awamori)、Fusarium SPP.(F.graminum)、Neurospora SPP.、Hypocrea SPP.、およびMucor SPP.が挙げられる。さらに別の宿主細胞として、Bacillus SPP.(例えば、B.licheniformis、B.lentus、B.stearothremophilus、およびB.brevis)、およびStreptomyces SPP.(例えば、S.coelicolorおよびS.lividans(TK23およびTK21))が挙げられてもよい。
【0099】
分子生物学
本発明は、組み換え遺伝子工学の分野における通例技術に依存する。本発明に使用される一般的方法を開示する基本的教科書としては、サムブルック(Sambrook)等の「分子クローニング、実験室マニュアル(“Molecular Cloning,A Laboratory Manual”)」第2版、1989年;クリーグラー(Kriegler)、「遺伝子の転送と発現、実験室マニュアル(“Gene Transfer and Expression: A Laboratory Manual”)」、および、アウスウーベル(Ausubel)等、「分子生物学における最新プロトコール(“Current Protocols in Molecular Biology”)」(1994)が挙げられる。
【0100】
例えば、糸状真菌の遺伝子プロモーター配列を含む異種遺伝子は、通常、複製および/または発現のために、宿主細胞、例えば、Trichoderma reesei細胞に変形導入される前に、中間ベクターにクローンされる。これらの中間ベクターは、通常、前核細胞性ベクター、例えば、プラスミド類、またはシャトルベクターである。
【0101】
クローンされた遺伝子の高レベル発現を実現するためには、異種遺伝子は、プロモーターから、天然遺伝子の場合とほぼ同じ距離離れた位置に置かれることが好ましい。しかしながら、従来技術で知られるように、この距離における若干の変動は、プロモーター機能の損失を招くこと無く容認することが可能である。
【0102】
当業者であれば、天然のプロモーターは、その機能を変更することなく、1個以上のヌクレオチドの代置、置換、付加、または除去することによって修飾することが可能であることを知っている。本発明の実施は、プロモーターに対するそのような変更を含むが、そのような変更によって制限されない。
【0103】
発現ベクター/構築体は、通常、該異種配列の発現に必要な全ての付加的要素を含む、転写単位または発現カセットを含む。従って、ある典型的発現カセットは、異種核酸配列に動作的に結合するプロモーターと転写物の効率的アデニル化に必要なシグナル、リボソーム結合部位、および翻訳終止を含む。カセットの付加的要素としては、エンハンサー、および、もしも構造遺伝子としてゲノムDNAが使用されるのであれば、機能的スプライスドナーおよびアクセプター部位を有するイントロンが挙げられる。
【0104】
本発明の実施は、遺伝子構築体におけるプロモーターの選択によって制限されない。しかしながら、例示のプロモーターとしては、Trichoderma reesei cbh1、cbh2、eg1、eg2、eg3、eg5、xln1、およびxln2プロモーターがある。さらに、A.awamoriおよびA.nigerのグルコアミラーゼ遺伝子(glaA)のプロモーター(ヌンベルク(Nunberg)等(1984)Mol.Cell Biol.4:2306−2315)、およびA.nidulansのアセタミナーゼのプロモーターも、ベクターへの使用が可能である。Bacillus subtilisに使用されるベクターのための好ましいプロモーターは、AprEプロモーターであり;E.coliに使用される好ましいプロモーターは、Lacプロモーターであり;Saccharomyces cerevisiaeに使用される好ましいプロモーターはPGK1であり;Aspergillus nigerに使用される好ましいプロモーターはglaAであり;かつ、Trichoderma reesei用として好ましいプロモーターはcbh1である。
【0105】
プロモーター配列の外に、発現カセットは、効率的な終止を実現するために、構造遺伝子の下流に転写終止領域を含まなければならない。終止領域は、プロモーター配列と同じ遺伝子から得てもよいし、あるいは、異なる遺伝子から得てもよい。
【0106】
本発明においては、真菌のものであればいずれのターミネーターも機能性を持つと考えられるが、ある好ましいターミネーターとしては、Aspergillus nidulans trpC遺伝子(イェルトン・エム(Yelton,M)等、(1984)PNAS USA81:1470−1474;ムラニー・イージェイ(Mullaney,E.J.)等、(1985)MGG199:37−45)、Aspergillus awamoriまたはAspergillus nigerのグルコアミラーゼ遺伝子(ヌンベルク・ジェイエイチ(Nunberg,J.H.)Mol.Cell Biol.4:2306;ベール・イー(Boel,E)等、(1984)EMBO J.3:1581−1585)、Aspergillus oryzae TAKAアミラーゼ遺伝子、およびMucor miheiのカルボキシルプロテアーゼ遺伝子(EPO公報第0215594号)、のターミネーターが挙げられる。
【0107】
遺伝子情報を細胞に輸送するために使用される、特定の発現ベクターは、特に決定的に重要とはならない。真核細胞または前核細胞における発現のために使用される通例のベクターの内の任意のものが使用可能である。標準的細菌性発現ベクターとしては、バクテリオファージλおよびM13の外に、プラスミド、例えば、pBR322準拠プラスミド、pSKF、pET23D、および、融合発現システム、例えば、MBP、GST、およびLacZが挙げられる。分離に好都合の方法を実現するために、組み換えタンパクに対してエピトープタグ、例えば、c−mycを付加することも可能である。好適な発現および/または統合ベクターの例は、サムブルック(Sambrook)等(1989)上記;ベネットおよびラシュア(Bennett and Lasure)編「真菌における遺伝子操作の進歩(“More Gene Manipulation in Fungi“)」(1991)、Academic Press pp.70−76およびpp.396−428、および同書の中に引用される論文;米国特許第5,874,276号;および、真菌遺伝子保存センターの真菌株カタログ(FGSC、www.fgsc.net.)の中に挙げられている。有用なベクターは、PromegaおよびInvitrogenから入手することが可能である。ある、有用な特異的ベクターとして、pBR322、pUC18、pUC100、pDONTM201、pENTRTM、pGEN(R)3Z、およびpGEN(R)4Zが挙げられる。しかしながら、本発明は、等価的機能を果たし、現に従来技術に知られる、または、これから知られるようになる他の形態の発現ベクターも含むことが意図される。従って、本発明のDNA配列の発現には、多種多様の宿主/発現ベクターの組み合わせの採用が可能である。例えば、有用な発現ベクターは、染色体の、非染色体の、および合成のDNA配列のセグメント、例えば、SV40および既知の細菌プラスミドの既知の各種誘導体から構成されてもよい。プラスミドとしては、例えば、colE1、pCR1、pBR322、pBb9、pUC19、およびそれらの誘導体を含む、Ecoli由来のプラスミド、さらに広い宿主範囲を持つプラスミド、例えば、RP4、ファージDNA,例えば、ラムダファージの多数の誘導体、例えば、NM989、およびその他のDNAファージ、例えば、M13、および、糸状の、1本鎖DNAファージ、酵母プラスミド、例えば、2.muプラスミドまたはその誘導体が挙げられる。
【0108】
ある実施態様では、発現ベクターは選択可能なマーカーを含む。選択可能なマーカーとしては、抗菌剤耐性を付与するものが挙げられる。栄養マーカー、amdS、argB、およびpyr4のような従来技術で既知のマーカーを含む栄養マーカーも本発明において使用される。Trichodermaの形質転換に有用なマーカーは、従来技術で既知である(例えば、フィンケルスタイン(Finkelstein)、6章、「糸状真菌の生物工学」、フィンケルスタイン等、EDS Butterworth−Heinemann、ボストン、マサチューセッツ州(1992)、および、キングホーン(Kinghorn)等、(1992)、「糸状真菌の応用分子遺伝学(”Applied Molecular Genetics of Filamentous Fungi“)」Blackie Academic and Professional、Chapman and Hall、ロンドンを参照されたい)。ある実施態様では、発現ベクターはまた、レプリコン、組み換えプラスミドを保有する細菌の選択を可能とする、抗生物質耐性をコードする遺伝子、および、異種配列の挿入を可能とするために、非必須領域に独特の制限部位を含んでもよい。選ばれる、特定の抗生物質耐性遺伝子は決定的に重要ではなく、従来技術で既知の多くの耐性遺伝子の内のいずれものでも好適である。前核細胞配列は、Trichoderma reeseiにおけるDNAの複製または統合を妨げないように選択されることが好ましい。
【0109】
本発明の形質転換法によって、形質転換ベクターの一部または全てが、宿主細胞、例えば、糸状真菌宿主細胞のゲノムの中に安定的に統合される。しかしながら、自己複製、染色体外形質転換ベクターの維持をもたらす形質転換も本発明の範囲内である。
【0110】
大量のプロテアーゼを発現する、細菌および糸状真菌(例えば、AspergillusまたはTrichoderma)細胞系統を生み出すために、多くの標準的トランスフェクション法を用いることが可能である。Trichodermaのセルラーゼ産生株の中にDNA構築体を導入するための公表された方法のいくつかを述べると、ロリート、ヘイエス、ジピエルト、およびハーマン(Lorito,Hayes,DiPierto and Harman)、(1933)Curr.Genet.24:349−356;ゴールドマン、バンモンターギュ、およびヘレラ・エストレヤ(Goldman,VanMontagu and Herrera−Estrella)(1990)Curr.Genet.17:169−174;および、ペンチラ、ナバレイネン、ラット、サルミネン、およびノウレス(Penttila,Nevalainen,Ratto,Salminen and Knowles)(1987)Gene 6:155−164が挙げられる。さらに、米国特許第6,022,725号、米国特許第6,268,328号、およびネバライネン(Nevalainen)等、「Trichodermaの分子生物学、および、同種および異種遺伝子の発現に向けたその応用(“The Molecular Biology of Trichoderma and its Application to the Expression of Both Homologous and Heterologous Genes“」、Molecular industrial Mycology,レオンおよびベルカ(Leong and Berka)編、Marcel Dekker Inc.,ニューヨーク州(1992)pp129−148を参照されたい。Aspergillusについては、イェルトン、ヘイマー、およびティンバーレイク(Yelton,Hamer,and Timberlake)(1984)Proc.Natl.Acad.Sci.USA 81:1470−1474を;Fusariumについては、ベイジャー、ポディラ、およびコラッツクディ(Bajar,Podila,and Kolattukudy)(1991)Proc.Natl.Acad.Sci.USA 88:8202−8212を;Streptomycesについては、ホップウッド(Hopwood)等、1985、「Streptomycesの遺伝子操作:実験室マニュアル(”Genetic Manipulation of Streptomyces:Laboratory Manual“)」、The John Innes Foundation,Norwich、英国、およびフェルナンデス・アバロス(Fenrnandez−Abalos)等、Microbiol.149:1623−1632(2003);および、Bacillusについては、ブリジディ、デロッシ、ベルタリニ、リッカルディ、およびマッテウッツィ(Brigidi,DeRossi,Bertarini,Riccardi and Matteuzzi)(1990)FEMS Microbiol.Lett.55:135−138)が挙げられる。
【0111】
しかしながら、宿主細胞に外来ヌクレオチド配列を導入するための、よく知られた過程の内の任意のものの使用が可能である。そのような過程としては、リン酸カルシウムの使用、トランスフェクション、ポリブレン、プロトプラスト融合、電気穿孔、biolistics、リポソーム、マイクロインジェクション、プラスマベクター、ウィルスベクター、および、クローンされたゲノムDNA、cDNA,合成DNA、または、他の外来遺伝物質を、宿主細胞に導入するための、その他の、既知の、任意の方法(例えば、サムブルック等、上記参照)が挙げられる。さらに、米国特許第6,255,115号に記載されるAgrobacterium仲介トランスフェクション法も使用される。使用される特定の遺伝子工学過程は、少なくとも1個の遺伝子を、該遺伝子を発現することが可能な宿主細胞に適切に導入することが可能でありさえすればよい。一つの実施態様では、本発明は、本発明によってカバーされるプロテアーゼ(例えば、NSP24ファミリ・プロテアーゼ)を生産するための方法に関する。この方法は、プロテアーゼ、例えば、NSPファミリ・プロテアーゼをコードする核酸に対して動作的に結合するプロモーターを含むポリヌクレオチドを宿主細胞に導入すること、プロテアーゼの発現と生産のために好適な培養条件下に宿主細胞を培養すること、および、前記プロテアーゼを生産することを含む。ある好ましい実施態様では、プロテアーゼは、配列番号2、または配列番号10、またはそれらの生物学的活性を持つ断片に対し少なくとも95%の配列同一性を有するNSP24ファミリ・プロテアーゼである。
【0112】
発現ベクターが細胞に導入された後、そのトランスフェクトされた、または形質転換された細胞は、プロテアーゼ遺伝子のプロモーター配列の制御下に、遺伝子発現を促進する条件下に培養される。大型バッチの形質転換細胞を、後述の実施例3に記載するやり方で培養することが可能である。最後に、産物は、標準技術によって培養物から回収される。
【0113】
上記から、本明細書の発明は、所望のポリペプチドの発現および分泌増大を実現する。すなわち、該ポリペプチドの発現は、天然プロテアーゼ遺伝子、融合DNA配列、および各種異種構築体を含む遺伝子プロモーター配列によって制御される。本発明はまた、このような所望のポリペプチドを高レベルで発現し、分泌するための過程を提供する。
【0114】
タンパクの発現
本発明のタンパクは、ベクター、例えば、その発現が遺伝子プロモーター配列によって制御される遺伝子を含む発現ベクターによって形質転換された細胞を培養することによって生産される。本発明は、タンパク、例えば、本発明によってカバーされるプロテアーゼの細胞内および/または細胞外生産を強化するのに特に有用である。前記遺伝子の発現にとって好適な条件とは、本発明の、誘導飼料組成物を培養物に供給することを含む。タンパク生産のための最適条件は、宿主細胞の選択、および、発現されるプロテアーゼタンパクの選択と共に変動する。そのような条件は、当業者であれば、通例の実験または最適化によって簡単に確認される。
【0115】
所望のプロテアーゼタンパクは、発現後、分離または回収し、精製してよい。所望のタンパクは、サンプルの中に他にどのような成分が存在するかに応じて、当業者に既知の様々なやり方に従って分離または精製してよい。標準的精製法としては、電気泳動的、分子、免疫学的、およびクロマトグラフィック技術、例えば、イオン交換、疎水性、アフィニティー、および逆相HPLCクロマトグラフィー、およびクロマトフォーカシングが挙げられる。例えば、興味のタンパクは、標準的、所望の抗タンパク抗体カラムを用いて精製してもよい。好適な精製技術に関する一般的案内については、「概観、タンパク精製(”Scopes Protein Purification“)」(1982)を参照されたい。必要な精製の程度は、所望のタンパクの用途に応じて変動する。ある実施態様では、精製は必要とされない。
【0116】
細胞培養
宿主細胞および形質転換細胞は、通常の栄養媒体中で培養することが可能である。形質転換された宿主細胞用の培養媒体は、プロモーターを活性化し、形質転換体を選択するために適当に修飾してもよい。特異的培養条件、例えば、温度、pH等は、発現のために選択された宿主細胞のために使用されるものであってよく、当業者には明白である。さらに、好ましい培養条件は、科学的文献、例えば、サムブルック(1982)上記;キーサー・ティー、エムジェイ・ビブ、エムジェイ・バットナー、ケーエフ・チェイター、およびディーエー・ホップウッド(Kieser,T,MJ.Bibb,MJ.Buttner,KF.Chater,and D.A.Hopwood)(2000)「ストレプトミセス遺伝学実技(“PRACTICAL STREPTOMYCES GENETICS”)」John Innes Foundation,Norwich、英国;ハーウッド(Harwood)等(1990)「バチルスのための分子生物学的方法(“MOLECULAR BIOLOGICAL METHODS FOR BACILLUS”)」、John Wiley、および/または、米国基準株保存機関(ATCC;www.atcc.org)に見出すことが可能である。真菌宿主細胞、例えば、Trichoderma細胞中の、安定な形質転換細胞は、それらがより急速な成長速度を持つこと、または、固相の培地において、ギザギザではなく、滑らかな輪郭を持つ円形コロニーを形成することによって、不安定な形質転換細胞とは一般的に区別される。
【0117】
発現されたポリペプチドの回収およびプロテアーゼの精製法
形質転換宿主細胞によって生産される、本発明によってカバーされるポリペプチド、例えば、配列番号10に対し少なくとも80%の配列同一性を持つポリペプチドは、通例の過程、例えば、遠心またはろ過によって、培養液から宿主細胞を分離すること、または、要すれば、細胞を破壊して、細胞分画と残留物から上清を取り出すことを含む過程によって、培養液から回収してもよい。ある場合には、澄明化後、上清またはろ液のタンパク様成分を、塩、例えば、硫酸アンモニウムを用いて沈殿させる。次に、この沈殿したタンパクを可溶化し、各種クロマトグラフィー手法、例えば、イオン交換クロマトグラフィー、ゲルろ過クロマトグラフィー、アフィニティークロマトグラフィー、およびその他の従来技術で認知済みの手法によって精製してよい。ペプチドおよびタンパクに対する抗体は、動物、例えば、ウサギまたはマウスを免疫化し、従来法によって抗NSP24プロテアーゼ抗体を回収することによって製造される。
【0118】
本発明に使用されるアッセイは、PCT特許WO9934011および米国特許USP6,605,458に記載されるものを含むが、ただしそれらに限定されない。
【0119】
組成物および応用
ある実施態様では、本発明は、本明細書に記載される本発明のプロテアーゼを含む組成物を指向する。本発明による組成物および応用において有用なプロテアーゼの、ある非限定的例としては、例えば、NSP24ファミリ・プロテアーゼまたはNSP25ファミリ・プロテアーゼ、さらに詳細には、配列番号2に対して少なくとも85%の配列同一性を持つNSP24ファミリ・プロテアーゼ、または、その生物学的に活性な断片、例えば、配列番号10に対して少なくとも90%の配列同一性を持つプロテアーゼが挙げられる。ある実施態様では、この酵素組成物は、単一酵素成分プロテアーゼ組成物である。ある実施態様では、本発明は、本発明のプロテアーゼを産業および商業用途に用いる方法を指向する。組成物および産業用途に関する下記の説明は、例示的であって、非内包的であることが意図される。
【0120】
本発明のプロテアーゼを含む組成物は、さらに別の酵素、例えば、グルコアミラーゼ、アルファアミラーゼ、顆粒でん粉加水分解酵素、セルラーゼ、リパーゼ、キシラナーゼ、クチナーゼ、ヘミセルラーゼ、オキシダーゼ、およびそれらの組み合わせを含んでもよい。
【0121】
ある好ましい実施態様では、組成物は、配列番号10の配列に対し少なくとも約85%の配列同一性を有する、本発明のプロテアーゼ、およびグルコアミラーゼを含む。グルコアミラーゼは、糸状真菌の供給源、例えば、Aspergillus、Tricoderma、またはRhizopusの株から得られた野生型グルコアミラーゼであってもよいし、あるいは、グルコアミラーゼは、タンパク加工されたグルコアミラーゼ、例えば、Aspergillus nigerのグルコアミラーゼの変種であってもよい。別の好ましい実施態様では、組成物は、本発明のプロテアーゼおよびアルファアミラーゼを含む。ある実施態様では、アルファアミラーゼは、細菌供給源、例えば、Bacillus SPP.、あるいは、真菌供給源、例えば、Aspergillus SPP.から得られてもよい。ある実施態様では、組成物は、本発明によるプロテアーゼ、およびグルコアミラーゼとアルファアミラーゼの両酵素を含んでもよい。これらの酵素の市販供給源は既知であり、例えば、Genencor International,Inc.およびNovozymes A/Sから入手することが可能である。
【0122】
いくつかの実施態様では、本発明は、エタノール生産、製パン、フルーツジュース生産、醸造、蒸留、ワイン製造、皮革、油脂、紙およびパルプ、および動物飼料生産における使用を意図している。
【0123】
別の実施態様では、本発明が考慮の対象とするものは、洗剤およびクリーニング製品の、活性を持つ「生物学的」成分である。この場合、プロテアーゼ、アミラーゼ、およびリパーゼは、タンパク、でん粉、および脂肪の染みを分解するために使用される。本発明の実施態様は、安定性試験を行い、様々な処方で試験することによって、酵素の、洗剤組成物に対する適合性を試験することを含む。
【0124】
さらに別の実施態様では、本発明は、でん粉を液化および糖化してグルコースに変換するため、および異性化してフルクトースに変換するための酵素的使用を考えた。本発明は、大容量の植物基質、例えば、穀粒(例えば、コーン、小麦、ミロ、ライ麦等)を甘味料、例えば、高濃度フルクトースコーンシロップ、およびマルトースシロップに変換するのに用いてもよい。
【0125】
本発明の酵素(単数または複数)は、タンパクの消化性を上げるために、食品および家畜飼料産業にも応用される。このプロテアーゼはまた、各種産業用途、特に、織物、リトグラフ、化学的芸術、農業、環境廃棄物変換、バイオパルプ処理、バイオマスの燃料への転換、およびその他の化学的過程(単複)にも使用される。さらに、本プロテアーゼは、保健およびパーソナルケア製品、例えば、化粧品、スキンケア、歯磨き等に使用される。
【0126】
飼料
本明細書に記載される酵素は、植物材料、例えば、コーン、小麦、モロコシ、大豆、カノーラ油、ひまわり油、または、上記植物材料の内の任意の複数の混合物、または、ニワトリ、ブタ、反芻動物、水産養殖、およびペット用の植物タンパク供給源を含んでもよい。性能パラメータ、例えば、成長、飼料摂取、および飼料効率ばかりでなく、均一性の改善、動物舎におけるアンモニア濃度の低下、およびその結果として得られる、動物の福祉と健康状態の改善ももたらされることが考慮される。
【0127】
食品
食物タンパクの加水分解産物は、市場において、小さいが、重要な部分を占める。この調製品は、術後患者、または、消化系が損なわれた個人のために使用される。加水分解産物は、それ自体比較的未精製の調製品として与えられる(クレッグ(Clegg)、1978、「新しいタンパク食品の生化学的側面(”Biochemical Aspects of New Protein Food“)」ジェイ・アドゥラー−ニッセン、ビーオー・エッガム、エル・マンク、およびエイチエス・オルセン(J,Adler−Nissen,B.O.Eggum,L,Munk,& H.S.Olsen)編、p.109−117、Pergamon,オックスフォード)。あるいは、静脈投与用の高度に精製されたアミノ酸混合物として投与される。ミルクタンパクの酵素加水分解産物は、食事調製品として従来から運用されている。
【0128】
筋肉食品、特に肉の酵素による軟化は、市場の大きなセグメントを占めるが、現在、これは、植物プロテアーゼおよびいくつかの微生物酵素によって代表される。魚肉の酵素的熟成および軟化も、多くの国々において相当の重要性を持つ。従って、ここに記載の酵素は、食品において様々な用途に使用される。
【0129】
さらに、本発明の酵素、または酵素組成物は、例えば、植物タンパク、例えば、大豆、豆、ルピナス、または菜種タンパク、ミルク様カゼイン、獣肉タンパク、または魚肉タンパクからタンパク加水分解産物を製造するのに有用と考えられる。本明細書に記載されるタンパク(単複)は、食品、飼料、または医用製品を製造する際に、可溶性、均一性、または醗酵性の改善、抗原性の低下、加水分解産物の苦味の減少、またはその他の目的のために使用してもよい。本明細書に記載される酵素(単複)は、単独で使用してもよいし、または他のペプチダーゼと、または他の酵素、例えば、エキソペプチダーゼと共に使用してもよい。エキソペプチダーゼ高含有酵素調製品と共に、本明細書に記載される酵素(単複)を使用した場合、それは、タンパク加水分解産物の味を改善する。
【0130】
さらに、酵素または酵素組成物は、魚肉または獣肉の処理に、例えば、組織および/または粘度を変えるために使用してもよい。
【0131】
皮革
皮革製造産業は、生皮を洗浄し、脱毛し、最後にそれを鞣し、乾燥することを含む一連の工程に依存する。酵素処理は、脱毛工程において重要な役割を担う。すなわち、この工程は、タンパク分解酵素を与えることによって実現されるが、本発明のペプチドは加水分解するので、現在皮革製造に使用される哺乳動物のプロテアーゼの有効な代替品となる。両方とも、高いタンパク分解活性を持ち、かつ、低pHでも効率が高いからである。
【0132】
ウールおよびシルク
本明細書に記載されるプロテアーゼは、ウール製品に対し所望の性質を付与するために、その工業的処理に使用される。一つの実施態様では、本発明は、織物処理用の組成物を提供する。この組成物は、例えば、シルクまたはウールを処理するために使用することが可能である(例えば、RE216,034;欧州特許EP134,267;米国特許US4,533,359;欧州特許EP344,259を参照されたい)。
【0133】
本発明の方法は、線維、例えば、ケラチン線維を含むタンパクを処理するのに用いることが可能である。この方法は、ヒツジ、ヤギ、リャマ、ラクダ、ウサギ等から得られた、ウール、ウール線維、または動物の毛、例えば、アンゴラ、モヘア、カシミヤ、アルパカ、またはその他、商業的に有用な動物毛糸製品を処理するのに好適である。さらに、シルク、くも糸シルク、またはヒトの毛髪も、本発明の方法で処理することが可能である。線維は、線維、トップ、より糸、または織った、または編んだ布または衣類の形を取ってもよい。
【0134】
洗浄
本発明はまた、本発明のプロテアーゼ(単複)を含む洗浄組成物にも関する。この洗浄組成物は、洗浄組成物に一般に使用される添加物をさらに含んでもよい。これらは、漂白剤、界面活性剤、油脂、酵素、および漂白触媒の中から選んでもよいが、ただしこれらに限定されない。当業者には、組成物に含めるにはどの添加物が好適であるかは直ちに明白であろう。本明細書に提示されるリストは、決して網羅的なものではなく、好適な添加物の例示としてのみ受け取るべきである。また、当業者には、酵素、および、組成物における他の成分、例えば、界面活性剤と適合する添加物のみを使用すべきであることは直ちに明白であろう。
【0135】
タンパク、特に、本発明のタンパクは、約0.01から約5重量%(好ましくは0.1%から0.5%)の濃度で、3.5と7.0の間の酸性pHを持つ、既知の、粉末状および液状界面活性剤として処方することも可能である。ある実施態様では、これらの界面活性洗浄組成物はさらに、他の酵素、例えば、アミラーゼ、さらに別のプロテアーゼ、セルラーゼ、リパーゼ、または、エンドグリコシダーゼの外に、油脂および安定化剤を含む。ある実施態様では、pHは、4.0と6.5の間、好ましくは4.0と5.6の間である。これは、要求される活性レベルに応じて、その最適pHにより酸性プロテアーゼと呼ばれるが、これらの酵素をpH7−9において使用することも可能である。
【0136】
通例の洗浄組成物にタンパクを添加することは、何ら特別な使用制限を要求することにはならない。言い換えれば、界面活性剤にとって好適な任意の温度およびpHはまた、pHがまた上述の範囲にあり、温度が前述のタンパク変性温度よりも低い限り、本発明の組成物にとって好適である。さらに、本発明のタンパクは、界面活性剤無しで、再び単独で、または、油脂および安定化剤と組み合わせて、洗浄組成物において使用される。
【0137】
タンパク処理
タンパク原材料の酵素的加水分解は、結果としてしばしば、苦味ペプチドの形成をもたらす(クレッグ(Clegg)1978)。タンパク加水分解産物に見られる苦味ペプチドは、無視できない実際的問題を生ずる。例えば、様々なタイプのチーズの熟成、および、食品としてのタンパク加水分解産物の生産に認められる問題がそれである。加水分解産物の苦味は、通常、特定のペプチド、特に、高い割合の疎水性アミノ酸を含むペプチドによる。苦味は、苦いペプチドの完全な、または部分的加水分解によって効果的に緩和される。従って、本明細書に記載される酵素は、食物の苦味除去に使用される。本発明の酵素、または酵素組成物は、食品のために、タンパクおよび/またはタンパク加水分解産物の苦味緩和のために使用されてもよい。
【0138】
本発明に従ってさらに考慮の対象となるのは、タンパクおよび/またはタンパク加水分解産物から出る遊離アミノ酸の生産である。遊離アミノ酸がグルタミン酸である場合は、それは、食品の風味を強化する。
【0139】
前記タンパクまたはタンパク分解産物は、動物または植物起源のものであってよい。本発明のある実施態様では、加水分解されるタンパクは、カゼイン、または大豆タンパクである。
【0140】
タンパクは、チーズのような食品、および、ココアを含む食品を生産するために使用されてもよい。
【0141】
本明細書に記載される酵素(単複)、本発明のある酵素が濃縮された酵素調製品は、苦味の無いタンパクまたはタンパク加水分解産物に関連して使用されると特に好都合ではあるけれども、本明細書に記載される酵素(単複)は、いくつかの産業用途、例えば、タンパク含有物質、例えば、細胞壁の分解または修飾を含む用途のために使用することが可能である。従って、本明細書に記載される酵素(単複)は、植物細胞壁の分解または修飾を促進する。
【0142】
本発明の酵素調製品の用量、および、該調製品が使用される、その他の条件は、従来技術で既知の方法に基づいて決められてよい。
【0143】
タンパク沈殿もまた、ある種の製品、例えば、ビールの場合無視できない問題を構成する場合がある。なぜなら沈殿によって製品が濁るからである。ビールでは、ビールの冷凍保存の際、可溶性タンパクが沈殿すると濁りが生じる。問題は、無視できない経済的重要性を持つが、ビール製造のために好適な生原料を選ぶこととは別に、この問題を回避する現在主流のやり方は、ビールにタンパク分解酵素を添加することである。
【0144】
パーソナルケア
ある実施態様では、本明細書に記載されるプロテアーゼが合成され精製されたならば、有効量が、パーソナルケア製品に使用される、パーソナルケア組成物(単複)に添加される。パーソナルケア製品は、化粧品、パーソナルケア用途に使用される「大衆薬品」(“OTC”)化合物(例えば、化粧品、スキンケア、口腔ケア、ヘアケア、ネイルケア)に分類される。ある実施態様では、本明細書に記載されるプロテアーゼは、パーソナルケア組成物、例えば、ヘアケア組成物、スキンケア組成物、ネイルケア組成物、化粧組成物、または、それらの任意の組み合わせに添加される。従って、酵素または酵素調製品は、例えば、コンタクトレンズ洗浄液、歯磨き、化粧品、およびスキンケア製品に使用されてもよい。
【0145】
甘味料
本明細書に記載されるプロテアーゼは、マルトースまたはフルクトース高濃度シロップの外、他の甘味料の生産にも使用される。醗酵可能な糖類、または糖類に変換が可能な組成物を含む原材料は、通常、でん粉含有植物材料、例えば、根茎、根、茎、穀類の小塊および粒(例えば、コーン、小麦、ミロ、大麦、およびライ麦)、および、糖含有原材料、例えば、砂糖大根、砂糖黍、果実材料、および糖蜜を含むが、ただしこれらに限定されない。
【0146】
プレバイオティクス
酵素調製品は、他のタンパク分解活性から事実上独立したクローン酵素を用いることが有利な場合、タンパクからペプチドを生産する際に有用であると考えられる。
【0147】
適当なタンパク源を加水分解するのに本明細書に記載する酵素(単複)(例えば、精製酵素)を用いることによって、遊離アミノ酸およびペプチドの未精製調製品を生産することが可能となる。この調製品は、成長のためにアミノ酸に対して特異的要求を持つ微生物に対する基質として極めて好適である。
【0148】
これは、工業的醗酵で使用される相当数の微生物の場合に当てはまる。必要なアミノ酸の補給が、このような醗酵過程の経済的維持にとって重要因子となる場合がよくある。酵素を投与することによって生産されるアミノ酸調製品は、実験室においても、大型の工業規模の醗酵においても基質として好適である。
【0149】
本明細書に記載される酵素(単複)はまた、機能的ペプチド、前生物的合成品等を生体内で生産するために使用されてもよい。「プレバイオティクス」という用語は、消化管、好ましくは、大腸における1種、または少数種の細菌の成長および/または活性を選択的に刺激することによって宿主に対し有益な影響を及ぼす、食品または飼料組成物を指す。
【0150】
醗酵およびバイオエタノール
本発明のプロテアーゼ組成物を用い、でん粉含有基質を醗酵して得られるアルコールの生産は、燃料用アルコールまたは携帯用アルコールの生産を含んでよい。ある実施態様では、酵素組成物は、大麦、モルト、および、その他の、例えば、ビール生産のための原料の酵母醗酵を促進するために使用されてもよい。
【0151】
アミラーゼは、醸造および製パン工業にとって不可欠の酵素である。アミラーゼは、モルト形成、および、添加糖類またはその他の炭水化物が無添加の状態で実行される、ある種の製パン過程において、でん粉を分解するのに必要とされる。これらの酵素の十分な活性度を確保することは、特にモルト調製工業では問題である。生理的に受容可能なシステムによってアミラーゼの活性を十分に増大させる方法は、より速やかなモルト形成を可能とする方法をもたらし、糖の利用可能性が増大することから、炭水化物含量の少ないアルコール飲料、例えば、ビールをもたらす。
【0152】
ある実施態様では、でん粉含有基質、例えば、穀粒(例えば、コーン、小麦、モロコシ)、小塊、および他の植物残留物の加水分解によって、アルコール、例えばエタノールが生産される。アルコール生産の方法は、「アルコール教科書−飲料、燃料、および工業的アルコール事業の参照資料(“The Alcohol Textbook,A Reference for the Beverage,Fuel and Industrial Alcohol Industries,”)」3版、ケーエー・ジャック(K.A.Jacques)等編、1999年、Nottingham University Press,英国に記述される。本発明のある実施態様では、プロテアーゼは、糖化および醗酵結合工程、別に糖化・醗酵同時工程とも呼ばれる工程において、組成物の中で、グルコアミラーゼと、かつ要すれば任意にアルファアミラーゼと組み合わせて使用される。「S.Lewisの工業醗酵学(”S.Lewis in Industrial Enzymology“)」、2.1章「アルコール発酵(”Fermentation Alcohol”)」、2版、ティー・ゴッドフリーおよびエス・ウェスト(T.Godfrey and S.West)編、(1996)、Stockton Press、ニューヨーク州を参照されたい。真菌酸性プロテアーゼをグルコアミラーゼと組み合わせて用いた醗酵によってエタノールを生産する方法は既知である。例えば、米国特許USP5,231,017は、Aspergillus nigerから得られたプロテアーゼを用いてエタノールを生産する方法であって、液化マッシュを獲得すること、および、醗酵工程と結合させてもよい糖化工程において液化マッシュにプロテアーゼを導入することを含む方法を開示する。ある実施態様では、本発明のプロテアーゼ組成物は、顆粒状でん粉基質による非加熱過程において、アルコール、例えば、エタノールを生産するために用いられる。該過程は、アルコールを生産するために用いられる基質において、でん粉のゲル化温度未満の温度において実行される。でん粉加水分解過程において使用されるプロテアーゼの量は、プロテアーゼの酵素活性に依存する。ある実施態様では、その量は、20−33%乾燥固体に調整したスラリー450gに対するプロテアーゼ2%液の添加量として0.001から2.0mlの範囲である。その際、スラリーは、糖化時および/または加水分解でん粉における液化マッシュである。他の有用な範囲としては、0.005から1.5ml、および0.01から1.0mlも挙げられる。
【0153】
プロテアーゼで処理された種子または穀粒は、醗酵過程によって生産されるモルトおよび飲料の生産に有利である。
【0154】
さらに、穀粉の中のタンパクを加水分解し、後続のアルコール発酵段階を予期してワートの可溶性窒素を濃縮するために、糖化においてもプロテアーゼを使用することが望ましい。穀粒におけるアミラーゼ活性の強化は、モルト形成において重要な発芽の速度と効率を増す。すなわち、酵素活性が強化されたモルトの生産は、でん粉が醗酵可能な炭水化物へ転換される加水分解の強化をもたらすので、アルコール飲料、例えば、ビールおよびスコッチウィスキーの生産における醗酵効率を向上させる。
【0155】
以下の実施例では、下記の略語が使用される。eq(当量)、M(モル)、μM(マイクロモル)、N(規定)、mol(モル)、mmol(ミリモル)、μmol(マイクロモル)、nmol(ナノモル)、g(グラム)、mg(ミリグラム)、kg(キログラム)、μg(マイクログラム)、L(リットル)、ml(ミリリットル)、μl(マイクロリットル)、cm(センチメートル)、mm(ミリメートル)、μm(マイクロメートル)、nm(ナノメートル)、℃(摂氏度)、h(時)、min(分)、sec(秒),msec(ミリ秒)、Ci(キュリー)、mCi(ミリキュリー)、μCi(マイクロキュリー)、TLC(薄層クロマトグラフィー)、Ts(トシル)、Bn(ベンジル)、Ph(フェニル)、Ms(メシル)、Et(エチル)、Me(メチル)、dsまたはDS(乾燥固体重量)、SAPU(酸性プロテアーゼ分光光度単位)で、1SAPUは、アッセイ条件下、1分当たりカゼイン基質から1マイクロモルのチロシンを解放するプロテアーゼの酵素活性量であり、GAU(グルコアミラーゼ単位で、pH4.2および60℃において、1時間当たり可溶性でん粉基質から、グルコースとして計算される1gの還元性糖を生産する酵素の量と定義される)。
【0156】
本発明は、下記の実施例においてさらに詳細に記述される。これらの実施例は、いかなる意味でも、請求される本発明の範囲を限定することを意図するものではない。付属の図面は、本発明の明細および説明の、一体化部分と考慮されるべきことが意図される。引用される参照文献は全て、そこに記載される全てに対する参照によって、それぞれ個別に本明細書に含める。下記の実施例は、本発明を例示するためにであって、請求される本発明を限定するために提示されるものではない。
【実施例1】
【0157】
新規プロテアーゼNSP24の、Trichoderma reesei DNAクローニング
T.reesei株QM6aからゲノムDNAを抽出した。T.reeseiゲノムのcontig1−5500に見出されたプロテアーゼ配列候補(Joint Genome Institute(JGI)T.reeseiゲノムv1.0)に基づいて、PCRプライマーを設計した。順行プライマーは、pENTR/Dベクター(Invitrogen)への方向性クローニングのためのモチーフを含んでいた。
【0158】
afp6fプライマーの配列は、CACCATGCAGACCTTTGGAGCT(配列番号11)であり、afp7rの配列は、TTATTTCTGAGCCCAGCCCAG(配列番号12)であった。1.3kbのPCR産物をゲル抽出(ゲル精製キット、Qiagen)で精製し、Invitrogen Gatewayシステムプロコールに従ってpENTR/Dにクローンした。
【0159】
次に、このベクターを、化学的にコンピテントとしたTop10 E.coli(Invitrogen)にて形質転換させ、カナマイシン選択した。いくつかの独立クローンから得られたプラスミドDNAを制限酵素で消化し、適正なサイズの挿入体であることを確認した。いくつかのクローンにおいて、プロテアーゼ遺伝子挿入体の配列を決定した(Sequetech,Mountain View、カリフォルニア州)。一つのクローンからのプラスミドDNA、pENTR/D_55.3を、pTrex3g/amdS表示ベクターDNAとのLRクロナーゼ反応(Invitrogen Gatewayシステム)に加えた。pTrex3gベクターは、pUC118ファージミドベクターであるE.coliのpSL1180(Pharmacia Inc.,ニュージャージー州)に基づくもので、WO05/001036に記載される。このLRクロナーゼ反応における組み換えによって、表示ベクターのCmRおよびccdB遺伝子は、pENTR/D_55.3由来のT.reeseiプロテアーゼによって置換された。この組み換えによって、プロテアーゼは、cbhlプロモーターと表示ベクターのターミネーターとの間に方向性をもって挿入された。44および50bpの組み換え部位配列が、それぞれ、プロテアーゼ遺伝子の上流および下流に留まった。LRクロナーゼ反応液の分液を、化学的にコンピテントとされたTop10 E.coli中に形質転換させ、一晩培養し、カルベニシリン選択した。いくつかのクローンから得られたプラスミドDNAを制限酵素で消化し、適正な挿入体サイズを確認した。クローンからのプラスミドDNA、pTrex3g_55.3.1をXbalIで消化して、cbhlプロモーター:NSP24プロテアーゼ:ターミネーター:amdSを含む発現カセットを放出させた。この5.8kbのカセットを、標準技術を用いてアガロースゲル抽出によって精製し、市販株QM6aから得られたT.reesei株の中で形質転換させた(PCT国際特許WO05/001036を参照)。図5、6、および7を参照されたい。
【実施例2】
【0160】
新規プロテアーゼNSP25の、Trichoderma reesei DNAクローニング
T.reesei株QM6aからゲノムDNAを抽出した。T.reeseiゲノムのcontig22−263400に見出されたプロテアーゼ配列候補(JGI T.reeseiゲノムv1.0)に基づいて、PCRプライマーを設計した。順行プライマーは、pENTR/Dベクター(Invitrogen)への方向性クローニングのためのモチーフを含んでいた。
【0161】
afp8fプライマーの配列は、CACCATGCAGCCCTCATTTGGCAG(配列番号13)であり、afp9rの配列は、CTATTTCTTCTGCGCCCAGCCAAC(配列番号14)であった。1.2kbのPCR産物をゲル抽出(ゲル精製キット、Qiagen)で精製し、Invitrogen Gatewayシステムプロコールに従ってpENTR/Dにクローンした。次に、このベクターを、化学的にコンピテントとしたTop10 E.coli(Invitrogen)にて形質転換させ、カナマイシン選択した。いくつかの独立クローンから得られたプラスミドDNAを制限酵素で消化し、適正なサイズの挿入体であることを確認した。いくつかのクローンにおいて、プロテアーゼ遺伝子挿入体の配列を決定した(Sequetech,Mountain View、カリフォルニア州)。一つのクローンからのプラスミドDNA、pENTR/D_22.2を、pTrex3g/amdS表示ベクターDNAとのLRクロナーゼ反応(Invitrogen Gatewayシステム)に加えた。このLRクロナーゼ反応における組み換えによって、表示ベクターのCmRおよびccdB遺伝子は、pENTR/D_22.2由来のT.reeseiプロテアーゼによって置換された。この組み換えによって、プロテアーゼは、cbhlプロモーターと表示ベクターのターミネーターとの間に方向性をもって挿入された。44および50bpの組み換え部位配列が、それぞれ、プロテアーゼ遺伝子の上流および下流に留まった。LRクロナーゼ反応液の分液を、化学的にコンピテントとされたTop10 E.coli中に形質転換させ、一晩培養し、カルベニシリン選択した。いくつかのクローンから得られたプラスミドDNAを制限酵素で消化し、適正な挿入体サイズを確認した。クローンからのプラスミドDNA、pTrex3g_22.2#1をXbalI(かつ、細菌のバックボーンを消化して小型断片とするためにEcoRI)で消化して(小型断片は電気泳動中カセットから移動して遠ざかった)、cbhlプロモーター:NSP25プロテアーゼ:ターミネーター:amdSを含む発現カセットを放出させた。この5.7kbのカセットを、標準技術を用いてアガロースゲル抽出によって精製し、市販株QM6aから得られたT.reesei株の中で形質転換させた。形質転換用に用いたプラスミドは、NSP24挿入体が、NSP25配列で置換されたことを除いては、図7に描かれるプラスミドと事実上同じであった。
【実施例3】
【0162】
TrichodermaPEG真菌の形質転換
胞子形成中の菌糸から得られた2cm2の寒天プラグを、250mlの、4つの溝を有するバッフルフラスコに納めた50mlのYEGブロスに接種し、37℃で16−20時間200rpmにてインキュベートした。液体容量を50mlの円錐チューブに移し、10分間2500rpmで回転させて菌糸を回収した。上清を吸引除去した。この菌糸ペレットを、40mlのフィルター滅菌β−D−グルカナーゼ(InterSpex Products,Inc.)液を含む、250mlの、0.22μmCAコーニングフィルター瓶に移し、30℃、200rpmで2時間インキュベートした。菌糸を、滅菌Miracloth(CalBiochem,ラホヤ、カリフォルニア州)を通して、50mlの円錐形遠心チューブに集め、2000rpmで5分遠心し、吸引した。ペレットを、50mlの1.2Mソルビトールで一度洗浄し、再び遠心し、吸引し、25mlのソルビトール/CaCl2で洗浄した。ヘモサイトメーターを用いてプロトプラストをカウントし、遠心し、吸引し、1.25×108/mlのプロトプラスト濃度を与えるのに十分な容量のソルビトール/CaCl2に再懸濁させた。形質転換反応当たり200μlの分液を用いた。20μgのDNA(≧1μg/μl)を15mlの円錐形チューブに移し、チューブを氷の上に置いた。200μlのプロトプラストを加えた。50μlのPEGを加え、穏やかに掻き混ぜ、氷上で20分インキュベートした。2mlのPEGをチューブに加え、室温で5分インキュベートした。4mlのソルビトール/CaCl2(合計6.25ml)をチューブに加えた。この形質転換混合物を、各オーバーレイ当たり〜2mlの、3分液に分割した。この2mlを、溶融アセタミドソルビトール上層寒天のチューブに加え、オーバーレイ混合物を、アセタミドを唯一の窒素源として増殖することのできる形質転換体を選ぶためにアセタミドソルビトールプレートの上に注いだ。プレートを、コロニーが出現するまで28−30℃でインキュベートした。形質転換体は、単一コロニーをアセタミド媒体(ソルビトール無添加アセタミドソルビトール処方)にて繰り返し継代することによって精製した。
【0163】
材料
40mlのβ−D−グルカナーゼ液:600mgのβ−D−グルカナーゼ;400mgのMgSO4.7H2O;および、40mlの1.2Mソルビトール。
【0164】
200mlのPEG混合液:50gのPEG4000(BDH Laboratories Supplies,Poole、英国)および1.47gのCaCl2.2H2OをMilli Q水にて調製。
【0165】
ソルビトール/CaCl2:1.2Mソルビトールおよび50mM CaCl2。
【0166】
amdS選択のために、アセタミドソルビトールプレートおよびオーバーレイを用いた。胞子の精製には、同じプレートを、ソルビトール無しで用いた。
【0167】
アセタミドソルビトール寒天(プレートおよびトップ寒天)
アセタミド(Aldrich,99%昇華物)−0.6g/L;CsCl−1.68g/L;グルコース−20g/L;KH2PO4−20g/L;MgSO4*7H2O−0.6g/L;CaCl2*2H2O−0.6g/L;1000X塩(下記参照)−1ml。pHを5.5に調整し、容量を300mlとした。0.22ミクロンフィルターにてフィルター滅菌し、オーブンにて55℃に温めた。
【0168】
700mlの水に、Noble寒天(トップ寒天用低温溶融)20g、およびソルビトール218gを加え、オートクレーブ滅菌した。この混合物を55℃に冷却し、フィルター滅菌したアセタミド混合物を加えた。プレートまたはチューブを注いだ。
【0169】
1000X塩−FeSO4.7H2O(0.5g/100ml);MnSO4.H2O(0.16g/100ml);ZnSO4.7H2O(0.14g/100ml);CoCl2.6H2O(0.1g/100ml)、および、0.22ミクロンフィルターにてフィルター滅菌。
【0170】
じゃがいもデキストロース寒天(PDA、Difco脱水媒体)−200g/L温浸によるじゃがいも;デキストロース20g/L;および寒天15g/Lを、50−80%最終容量のdH2O中でよく混ぜわせ、最終的に100%容量とした。この混合液をオートクレーブ滅菌し、55℃に冷却し、注いだ。
【0171】
pH3.5媒体用の1%スキムミルク寒天を調製するために、PDAを上記のように調製し、100mlの溶融PDAに、1.8mlの10%酒石酸および12.5mlの滅菌8%スキムミルクを加え、プレートに注いだ。スキムミルクを事前滅菌するために、8%スキミミルク(Difco)を10分、122−123℃、32−35psiのチェンバー圧に暴露してオートクレーブした。混合液を取り出し、冷却し、室温に保存した。
【0172】
形質転換体におけるプロテアーゼ発現は、振とうフラスコにおける3日間の増殖後に評価した。T.reesei培養液(デービス(Davis)等(1970)Methods Enzymol.17:79−143)に寒天プラグを接種した。培養物を振とうしながら30℃で3日間インキュベートした。培養液を、0.22ミクロンフィルターを通過させ、ろ液を、1%スキムミルク寒天の上にスポット状に投与した。室温で一晩インキュベート後、透明ゾーンが観察された。
【実施例4】
【0173】
NSP24、NSP25、およびL388M PepAのpH活性プロフィール
全てTrichoderma reesei株において過剰発現される、PepA(野生型およびL388M)、NSP24、およびNSP25のpH活性プロフィールを、Molecular Probesから入手した蛍光標識カゼインアッセイ(EnzChekプロテアーゼキット緑色蛍光)を用いて求めた。PepA(野生型およびL388M)、およびNSPは、全体醗酵サンプルであり、NSP24は、50%グリセロールに溶解させて安定化した精製サンプルである。これらの酵素を、1.0mg/ml、0.5mg/mlおよび0.25mg/mlに希釈した。蛍光標識基質は、DI H2Oにて0.1mg/mlに希釈した。10mlの基質を、各種pHの50mlバッファーと30μlのDI H2Oに加えた。10mlの酵素を加えて反応を開始し、様々な期間反応させ、100μlの1.0Mリン酸塩をpH10で加えて反応を停止させた。サンプルの蛍光を、SpectraMAX EM蛍光プレートリーダーにおいて、538nm放射線で、485励起、放射カットオフフィルター530nmで測定した。NSP24はpH3.7で最適活性を示し、野生型PepAはpH3.4で最適活性を示し、かつ、L388M pepAは3.5に最適pHを持つ。NSP25は、pH4.6で最適活性を示す。
【実施例5】
【0174】
実験室醗酵における、Trichoderma reeseiのNSP24プロテアーゼの、GC106との比較
今日のエタノール産業において使用される標準的プロテアーゼは、Genencor International,Inc.によって市販されるプロテアーゼGC106である。GC106に対するNSP24の機能性を、糖分解、グルコース形成、およびエタノール生産に関して比較した。
【0175】
材料
ジスチラーゼL−400(ロット#107−04057−901,372 GAU/g)
GC106(ロット#A01−01300−001,1010SAPU/g)
NSP24(ロット#20040423,1165SAPU/g)
Red Star赤酵母
エタノール製造機から得られたマッシュおよび薄い濃度の抽出液(コーン)
方法
マッシュおよび薄い抽出液(バックセットとも呼ばれる、醗酵前)をエタノール製造機から取り出し、掻き混ぜて26.5ブリックスとする。1N HClを用いてpHをpH4.3に調整した。次に、サンプルを、三つの300グラム分液に分割し、32℃の水浴に設置した。平衡後、下記の酵素の組み合わせを加えた。
【表2】
【0176】
ジスチラーゼL−400は、Aspergillus nigerから得られた液状グルコアミラーゼであり、Genencor International Inc.から入手が可能である。酵素添加後、フラスコ当たり、1.00gのRed Star赤酵母を加えた。サンプルを、16、24、40、および48時間後に取り出し、遠心した。各サンプルの500μlを、50μlの1.1N NH2SO4を含む試験間に移し反応を停止させた。2分後、サンプルを、4.5mlのDI H2Oで希釈し、混合した。混合後、サンプルを、0.45ミクロンフィルターに通過させ、HPLCバイアルに移し分析した。サンプルは、HPLC(Phenomenex Rezex 8u)で分析した。結果を、図1−4に示す。NSP24の性能は、GC106にほぼ等しかった。
【実施例6】
【0177】
非加熱過程において磨砕コーンから得られるエタノール収率に及ぼすNSP24の作用
磨砕コーンの30%DSスラリーを、DI H2Oによって調製した。磨砕コーンは、エタノール産業で使用される#2イェローデントコーンの典型的サンプルである。これは、70%を超えるものが30メッシュ篩を通過するほどに磨砕された。この穀物の水分含量をOHAUS、MB35ハロゲン水分天秤(ニュージャージー州)を用いて測定した。pHは、6N H2SO4を用いて4.2に調整した。醗酵は、0.5kg/MT用量のNSP24の添加、または添加無しで、1.0GAU/g用量のSTARGEN001を添加した100gマッシュを含む125mlフラスコにおいて行われた。
【0178】
45mlの水に溶解した5gのRed Starエタノールレッド乾燥酵母(Lesaffre酵母社、ミルウォーキー、ウィスコンシン州)を準備し、32℃水浴で1時間掻き回し、醗酵槽に接種した。0.5mlの酵母スラリーを、各125mlフラスコに加えた。フラスコを32℃水浴に移し、マッシュを穏やかに掻き回した。醗酵中、サンプルを取り出し、HPLC分析に供した(HPLCカラム:Phenomenex Rezex Organic Acid Column(RHM−モノサッカライド)#00H0132−KO;カラム温度:60℃;移動相:0.01N H2SO4;流速:0.6mL/分;検出器:RI;および、注入用量:20μL)。醗酵は、72時間後停止させた。様々のサンプリング間隔における、糖分、乳酸、グリセロール、およびエタノールを含む化合物の生産は、下記の表3に示す通りである。該表において、+は、NSP24がフラスコに添加されたことを示し、−−は、NSP24がフラスコに添加されなかったことを示す。全てのサンプルについて乳酸が、約0.01と0.02%w/vにおいて測定され、かつ、DP−2は0.0であると定量された。全てのサンプルについて、24時間時、酢酸は約0であり、71時間では、0.03と0.04の間であると定量された。
【表3】
【実施例7】
【0179】
コーン内乳によるエタノール生産における各種プロテアーゼの比較
内乳(胚除去コーン、75.8%でん粉、99.5%の粒径<30メッシュ)による29.5%DSマッシュを顆粒状でん粉基質として調製した。各マッシュ100グラムを125mlフラスコに移し、媒体のpHをpH4.5に調整した。プロテアーゼ(NSP24;中性プロテアーゼ(MULTIFECT NEUTRAL,プロテイナーゼ−T);および、アルカリ性プロテイナーゼ(SPEZYME FAN,PROTEX 6L MULTIFECT P−3000、およびプロテイナーゼ−T(Genencor International))を、0.5kg/MTで加え、次いで、STARGEN001(Genencor International)を2.5Kgs/でん粉MT)を加えた。次に、フラスコに0.5mlの20%酵母(Red Starエタノール赤)を接種し、32℃に維持した水浴に移した。フラスコの内容物を連続的に攪拌し、インキュベーション中均等な混合を行った。サンプルを様々な時間間隔で採取し、HPLC分析に供した。72時間醗酵液から得られたDDGSの残留でん粉およびタンパク含量を定量した。エタノール生産の結果を下記の表4に示す。
【表4】
【図面の簡単な説明】
【0180】
【図1】1)NSP24、2)市販のプロテアーゼGC106、および、3)プロテアーゼを含まないジスチラーゼを用いた場合の、糖分解(DP+3)%w/vを示す(実施例5参照)。
【図2】NSP24、GC106、およびジスチラーゼを用いた場合の糖分解(DP2)%w/vを示す。
【図3】NSP24、GC106、およびジスチラーゼを用いた場合のグルコース形成(DP1)を示す。グルコースの量は、NSP24およびGC106の両サンプルにおいて、40時間終了時に残存する量は0.2%w/v未満であり、48時間終了時には0.1%w/v未満である。一方、48時間終了時に、%w/vで測定したグルコースの量は、ジスチラーゼでは、1.0%w/vよりも少し大きい。
【図4】NSP24、GC106、およびジスチラーゼにおけるエタノール生産(%v/w)を示す。前記二つのプロテアーゼサンプルを用いることによって生産されるエタノールの速度および量は同じである。一方、ジスチラーゼのエタノール生産は、より低く、速度はより遅かった。
【図5A】図5Aは、Trichoderma reeseiから得られたpTrex3g_NSP24 cDNAのヌクレオチド配列(配列番号1)を示す。NSP24遺伝子配列には下線を施し、遺伝子のイントロン配列候補は、ゴシック体で特定する。プロテアーゼをコードする核酸配列は、配列番号8の配列によって表される。
【図5B】図5Bは、Trichoderma reeseiから得られたpTrex3g_NSP24 cDNAのヌクレオチド配列(配列番号1)を示す。
【図5C】図5Cは、Trichoderma reeseiから得られたpTrex3g_NSP24 cDNAのヌクレオチド配列(配列番号1)を示す。
【図5D】図5Dは、Trichoderma reeseiから得られたpTrex3g_NSP24 cDNAのヌクレオチド配列(配列番号1)を示す。
【図6A】Tricoderma reesei由来NSP24の、予想アミノ酸配列(407アミノ酸)(配列番号2)(図6A)。シグナルペプチドはゴシック体で示され、プレプロ配列は、ゴシック体および下線で示され、成熟NSP24タンパクは、KYGAPIS...で始まり、配列番号10で表される。
【図6B】NSP24ヌクレオチド配列で、イントロン候補をゴシック体で特定した配列(図6B)(配列番号8)を示す。
【図7】pTrex3g_NSP24ベクター、および、図5のヌクレオチド配列にそう制限酵素切断部位の位置を示す。
【図8】pepAプロテアーゼの核酸配列(配列番号3)を示す。予想イントロンをゴシック体で表す。
【図9A】図9Aは、新規NSP25プロテアーゼ(399アミノ酸)(配列番号9)をコードする核酸配列(配列番号4)を示す。シグナル配列はゴシック体で表す。
【図9B】図9Bは、新規NSP25プロテアーゼ(399アミノ酸)(配列番号9)をコードする核酸配列(配列番号4)を示す。シグナル配列はゴシック体で表す。
【図10】新規pepAプロテアーゼ変種(L388M)(配列番号7)の核酸配列(配列番号5)を示す。図において、下線を施した‘A’は、図8のpepAの‘C’から変化したものである。
【図11】発現ベクターpSL889_pepAを示す。
【図12A】図12Aは、発現ベクターpSL899_pepAのヌクレオチド配列(配列番号6)を示す。Xho Iの切断部位は、^によって示され、Xba I部位は*によって示される。pepAのコード配列はゴッシク体で示される。
【図12B】図12Bは、発現ベクターpSL899_pepAのヌクレオチド配列(配列番号6)を示す。
【図12C】図12Cは、発現ベクターpSL899_pepAのヌクレオチド配列(配列番号6)を示す。
【図12D】図12Dは、発現ベクターpSL899_pepAのヌクレオチド配列(配列番号6)を示す。
【図12E】図12Eは、発現ベクターpSL899_pepAのヌクレオチド配列(配列番号6)を示す。
【図13】配列番号5によってコードされたタンパクに対するPepA変種、L388Mのアミノ酸配列(配列番号7)を示す。
【特許請求の範囲】
【請求項1】
配列番号10に対して少なくとも85%のアミノ酸配列同一性を有する、単離されたNSP24ファミリ・プロテアーゼ。
【請求項2】
配列番号10に対して少なくとも90%のアミノ酸配列同一性を有することを特徴とする、請求項1の単離されたNSP24ファミリ・プロテアーゼ。
【請求項3】
配列番号10に対して少なくとも95%のアミノ酸配列同一性を有することを特徴とする、請求項2の単離されたNSP24ファミリ・プロテアーゼ。
【請求項4】
配列番号10に対して少なくとも97%のアミノ酸配列同一性を有することを特徴とする、請求項3の単離されたNSP24ファミリ・プロテアーゼ。
【請求項5】
NSP24ファミリ・プロテアーゼをコードする単離されたポリヌクレオチド。
【請求項6】
前記単離された核酸は、配列番号2に対して少なくとも85%の配列同一性を有するNSP24プロテアーゼをコードすることを特徴とする、請求項5の単離されたポリヌクレオチド。
【請求項7】
配列番号8の配列を有することを特徴とする、請求項6の単離されたポリヌクレオチド。
【請求項8】
請求項5のポリヌクレオチドを含むベクター。
【請求項9】
請求項5のポリヌクレオチドによって形質転換された宿主細胞。
【請求項10】
宿主細胞が糸状真菌細胞であることを特徴とする、請求項9の宿主細胞。
【請求項11】
糸状真菌細胞がAspergillus SPP.、Fusarium SPP.、またはTrichoderma SPP.であることを特徴とする、請求項10の宿主細胞。
【請求項12】
Aspergillusが、A.niger、A.oryzae、A.nidulans、またはA.awamoriであることを特徴とする、請求項11の宿主細胞。
【請求項13】
TrichodermaがT.reeseiであることを特徴とする、請求項11の宿主細胞。
【請求項14】
宿主細胞が四重欠失した宿主細胞であることを特徴とする、請求項10の宿主細胞。
【請求項15】
プロテアーゼを生産する方法であって、
a)NSP24ファミリ・プロテアーゼをコードする核酸に動作可能的に結合するプロモーターを含むポリヌクレオチドを宿主細胞内に導入する工程、
b)NSPファミリ・プロテアーゼの発現と生産に好適な培養条件の下で宿主細胞を培養する工程、および、
c)前記NSP24ファミリ・プロテアーゼを生産する工程、
を含む前記方法。
【請求項16】
生産されたプロテアーゼを回収することをさらに含むことを特徴とする、請求項15による方法。
【請求項17】
親NSP24ファミリ・プロテアーゼの、生物学的に活性な単離された断片。
【請求項18】
親NSP24ファミリ・プロテアーゼが、配列番号2に対して少なくとも90%の配列同一性を有することを特徴とする、請求項17の生物学的に活性な単離断片。
【請求項19】
断片が、配列番号2または配列番号10を有するNSP24プロテアーゼの活性の少なくとも40%を有することを特徴とする、請求項17の生物学的に活性な単離断片。
【請求項20】
請求項1のNSP24ファミリ・プロテアーゼを含む酵素組成物。
【請求項21】
請求項17のNSP24ファミリ・プロテアーゼの生物学的に活性な断片を含む酵素組成物。
【請求項22】
組成物が洗浄組成物であることを特徴とする、請求項20の酵素組成物。
【請求項23】
洗浄組成物が界面活性組成物であることを特徴とする、請求項22の酵素組成物。
【請求項24】
組成物がでん粉加水分解組成物であることを特徴とする、請求項20の酵素組成物。
【請求項25】
組成物が、動物飼料組成物であることを特徴とする、請求項20の酵素組成物。
【請求項26】
組成物が、エタノール生産工程に使用されることを特徴とする、請求項20の酵素組成物。
【請求項27】
組成物が、でん粉糖化工程に使用されることを特徴とする、請求項20の酵素組成物。
【請求項28】
組成物が、マルトースまたはフルクトースの生産に使用されることを特徴とする、請求項20の酵素組成物。
【請求項29】
組成物が、パーソナルケア組成物であることを特徴とする、請求項20の酵素組成物。
【請求項30】
グルコアミラーゼをさらに含むことを特徴とする、請求項20の酵素組成物。
【請求項31】
グルコアミラーゼをさらに含むことを特徴とする、請求項21の酵素組成物。
【請求項32】
アルファ−アミラーゼをさらに含むことを特徴とする、請求項20の酵素組成物。
【請求項33】
アルファ−アミラーゼをさらに含むことを特徴とする、請求項21の酵素組成物。
【請求項34】
グルコアミラーゼをさらに含むことを特徴とする、請求項33の酵素組成物。
【請求項35】
でん粉を加水分解する方法であって、でん粉の加水分解に好適な条件下に、でん粉を含む基質を、請求項20の酵素組成物と接触させること、および、加水分解でん粉を獲得することを含む前記方法。
【請求項36】
配列番号7を有する単離されたPepAプロテアーゼ。
【請求項37】
請求項36のプロテアーゼをコードする単離されたポリヌクレオチド。
【請求項38】
配列番号5を有する請求項37のポリヌクレオチド。
【請求項39】
配列番号9に対して少なくとも90%の配列同一性を有する単離されたNSP25ファミリ・プロテアーゼ。
【請求項40】
請求項39のNSP25プロテアーゼをコードする単離されたポリヌクレオチド。
【請求項1】
配列番号10に対して少なくとも85%のアミノ酸配列同一性を有する、単離されたNSP24ファミリ・プロテアーゼ。
【請求項2】
配列番号10に対して少なくとも90%のアミノ酸配列同一性を有することを特徴とする、請求項1の単離されたNSP24ファミリ・プロテアーゼ。
【請求項3】
配列番号10に対して少なくとも95%のアミノ酸配列同一性を有することを特徴とする、請求項2の単離されたNSP24ファミリ・プロテアーゼ。
【請求項4】
配列番号10に対して少なくとも97%のアミノ酸配列同一性を有することを特徴とする、請求項3の単離されたNSP24ファミリ・プロテアーゼ。
【請求項5】
NSP24ファミリ・プロテアーゼをコードする単離されたポリヌクレオチド。
【請求項6】
前記単離された核酸は、配列番号2に対して少なくとも85%の配列同一性を有するNSP24プロテアーゼをコードすることを特徴とする、請求項5の単離されたポリヌクレオチド。
【請求項7】
配列番号8の配列を有することを特徴とする、請求項6の単離されたポリヌクレオチド。
【請求項8】
請求項5のポリヌクレオチドを含むベクター。
【請求項9】
請求項5のポリヌクレオチドによって形質転換された宿主細胞。
【請求項10】
宿主細胞が糸状真菌細胞であることを特徴とする、請求項9の宿主細胞。
【請求項11】
糸状真菌細胞がAspergillus SPP.、Fusarium SPP.、またはTrichoderma SPP.であることを特徴とする、請求項10の宿主細胞。
【請求項12】
Aspergillusが、A.niger、A.oryzae、A.nidulans、またはA.awamoriであることを特徴とする、請求項11の宿主細胞。
【請求項13】
TrichodermaがT.reeseiであることを特徴とする、請求項11の宿主細胞。
【請求項14】
宿主細胞が四重欠失した宿主細胞であることを特徴とする、請求項10の宿主細胞。
【請求項15】
プロテアーゼを生産する方法であって、
a)NSP24ファミリ・プロテアーゼをコードする核酸に動作可能的に結合するプロモーターを含むポリヌクレオチドを宿主細胞内に導入する工程、
b)NSPファミリ・プロテアーゼの発現と生産に好適な培養条件の下で宿主細胞を培養する工程、および、
c)前記NSP24ファミリ・プロテアーゼを生産する工程、
を含む前記方法。
【請求項16】
生産されたプロテアーゼを回収することをさらに含むことを特徴とする、請求項15による方法。
【請求項17】
親NSP24ファミリ・プロテアーゼの、生物学的に活性な単離された断片。
【請求項18】
親NSP24ファミリ・プロテアーゼが、配列番号2に対して少なくとも90%の配列同一性を有することを特徴とする、請求項17の生物学的に活性な単離断片。
【請求項19】
断片が、配列番号2または配列番号10を有するNSP24プロテアーゼの活性の少なくとも40%を有することを特徴とする、請求項17の生物学的に活性な単離断片。
【請求項20】
請求項1のNSP24ファミリ・プロテアーゼを含む酵素組成物。
【請求項21】
請求項17のNSP24ファミリ・プロテアーゼの生物学的に活性な断片を含む酵素組成物。
【請求項22】
組成物が洗浄組成物であることを特徴とする、請求項20の酵素組成物。
【請求項23】
洗浄組成物が界面活性組成物であることを特徴とする、請求項22の酵素組成物。
【請求項24】
組成物がでん粉加水分解組成物であることを特徴とする、請求項20の酵素組成物。
【請求項25】
組成物が、動物飼料組成物であることを特徴とする、請求項20の酵素組成物。
【請求項26】
組成物が、エタノール生産工程に使用されることを特徴とする、請求項20の酵素組成物。
【請求項27】
組成物が、でん粉糖化工程に使用されることを特徴とする、請求項20の酵素組成物。
【請求項28】
組成物が、マルトースまたはフルクトースの生産に使用されることを特徴とする、請求項20の酵素組成物。
【請求項29】
組成物が、パーソナルケア組成物であることを特徴とする、請求項20の酵素組成物。
【請求項30】
グルコアミラーゼをさらに含むことを特徴とする、請求項20の酵素組成物。
【請求項31】
グルコアミラーゼをさらに含むことを特徴とする、請求項21の酵素組成物。
【請求項32】
アルファ−アミラーゼをさらに含むことを特徴とする、請求項20の酵素組成物。
【請求項33】
アルファ−アミラーゼをさらに含むことを特徴とする、請求項21の酵素組成物。
【請求項34】
グルコアミラーゼをさらに含むことを特徴とする、請求項33の酵素組成物。
【請求項35】
でん粉を加水分解する方法であって、でん粉の加水分解に好適な条件下に、でん粉を含む基質を、請求項20の酵素組成物と接触させること、および、加水分解でん粉を獲得することを含む前記方法。
【請求項36】
配列番号7を有する単離されたPepAプロテアーゼ。
【請求項37】
請求項36のプロテアーゼをコードする単離されたポリヌクレオチド。
【請求項38】
配列番号5を有する請求項37のポリヌクレオチド。
【請求項39】
配列番号9に対して少なくとも90%の配列同一性を有する単離されたNSP25ファミリ・プロテアーゼ。
【請求項40】
請求項39のNSP25プロテアーゼをコードする単離されたポリヌクレオチド。
【図1】

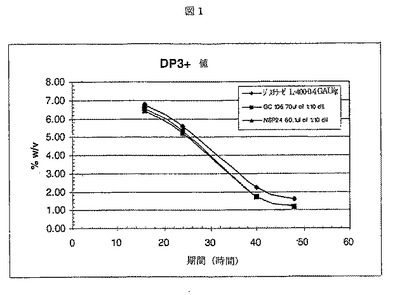
【図2】
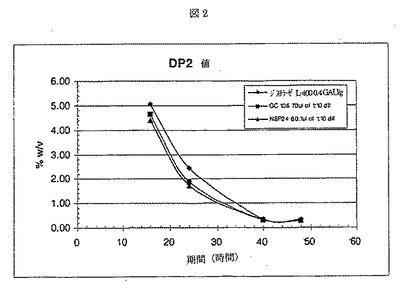
【図3】
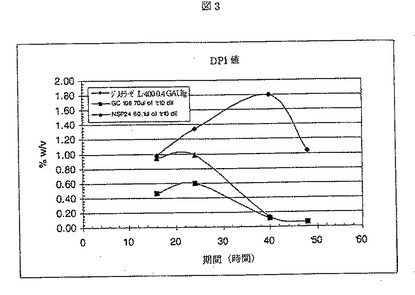
【図4】
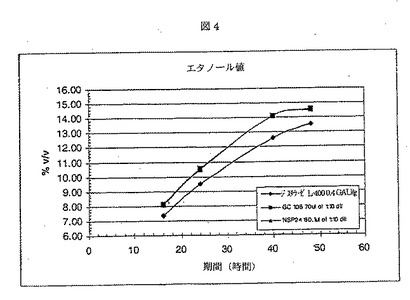
【図5A】

【図5B】

【図5C】

【図5D】
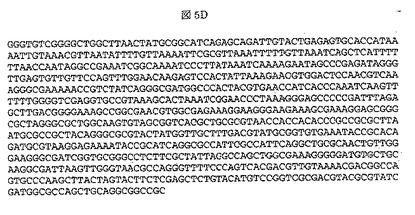
【図6A】

【図6B】

【図7】
【図8】

【図9A】

【図9B】

【図10】

【図11】
【図12A】

【図12B】

【図12C】

【図12D】

【図12E】

【図13】

【図2】
【図3】
【図4】
【図5A】
【図5B】
【図5C】
【図5D】
【図6A】
【図6B】
【図7】
【図8】
【図9A】
【図9B】
【図10】
【図11】
【図12A】
【図12B】
【図12C】
【図12D】
【図12E】
【図13】
【公表番号】特表2008−526202(P2008−526202A)
【公表日】平成20年7月24日(2008.7.24)
【国際特許分類】
【出願番号】特願2007−549480(P2007−549480)
【出願日】平成17年12月20日(2005.12.20)
【国際出願番号】PCT/US2005/046435
【国際公開番号】WO2006/073839
【国際公開日】平成18年7月13日(2006.7.13)
【出願人】(500284580)ジェネンコー・インターナショナル・インク (67)
【Fターム(参考)】
【公表日】平成20年7月24日(2008.7.24)
【国際特許分類】
【出願日】平成17年12月20日(2005.12.20)
【国際出願番号】PCT/US2005/046435
【国際公開番号】WO2006/073839
【国際公開日】平成18年7月13日(2006.7.13)
【出願人】(500284580)ジェネンコー・インターナショナル・インク (67)
【Fターム(参考)】
[ Back to top ]