
鉄欠乏性障害を治療するためのフェリチンの使用
ラット脳内微小血管系における内皮細胞上のH−フェリチン受容体は、血液脳関門を越えて鉄を運ぶための手段である。患者の脳内の鉄欠乏性障害を治療するための方法は、H−フェリチン−鉄複合体を、必要性がある患者に投与することを含む。ターゲッティング部分としてH−フェリチンを使用するための方法は、脳又は脳内の細胞にターゲッティングされるリポソームとH−フェリチンを結合させることを含む。患者における鉄過剰症を治療するための方法は、100%未満の鉄結合能であるマルチサブユニットフェリチン複合体を患者に投与することを含む。
【発明の詳細な説明】
【技術分野】
【0001】
継続出願のデータ
本出願は、いずれもそれらの全容が参照として本明細書に組み込まれる、それぞれ2007年1月29日及び2007年10月31日に出願された、米国仮出願第60/886,972号及び米国仮出願第60/984,007号の特典を主張するものである。
【背景技術】
【0002】
フェリチン、原核生物と真核生物の両方における主な細胞内鉄貯蔵タンパク質は、24ポリペプチドサブユニットを含む大きな(約480kDa)マルチサブユニット複合体である。血清中に高濃度で見られるこの鉄貯蔵複合体は、水酸化鉄コア内に4,500原子もの多くの鉄イオン(Fe3+)を含有することができる。哺乳動物では、約54%の配列同一性を共有し、それぞれ約21kDa及び19kDaの分子量を有する、2つの異なるサブユニットクラス、重鎖(H)及び軽鎖(L)型が存在する。HサブユニットとLサブユニットは異なる機能を有するようである:Lサブユニットは鉄コアの安定性を高め、一方Hサブユニットは、第一鉄イオンの迅速な摂取に必要とされるであろうフェロキシダーゼ活性を有する。Hが豊富なフェリチンは、局所イオン濃度の迅速な変化を経験する組織中に局在する。例えば、Hサブユニットの発現は、分化、発達、増殖及び代謝ストレスを経験する細胞中でLサブユニットに対して優先的に増大する。
【発明の概要】
【発明が解決しようとする課題】
【0003】
脳内微小血管系を構成する隣接内皮細胞を結合させる高度に発達した密着結合のため、脳は鉄獲得に高い難題を課す。これらの結合は、脳内への分子の傍細胞流動を妨げる。結果として生じる血液−脳関門(BBB)は、血中を循環する潜在的に有害である物質から脳を保護するのに非常に有効な機構である。しかしながらこのような封鎖の結果は、正常な脳機能に必要とされる多くの栄養物質に特異的な輸送機構を設計しなければならないことである。ピノサイトーシスはBBBを回避する可能性がある方法であるが、ピノサイトーシスから生じる小胞は、脳血管内皮細胞を越える化合物の非特異的輸送に相対的にほとんど貢献しない。
【0004】
従来、トランスフェリンは細胞の鉄送達の主要機構であると考えられており、トランスフェリン仲介の輸送系がBBB中で同定されている(Jefferies W.A.、et al.Nature 312:162〜163、1984;Fishman J.、Rubin J.、Handrahan J.、Connor J.、Fine R.J.Neurosci.Res.18:299〜304、1987)。しかしながら、トランスフェリン欠損マウスを使用して、脳へのトランスフェリン非依存性の鉄送達が示唆されている(Malecki E.A.、Cook B.M.、Devenyi A.G.、Beard J.L.and Connor J.R.J.Neurol.Sci.170:112〜118、1999)。ラクトフェリンも脳内に鉄を運ぶことができると言われているが(Ji B.、et al.Life Sci.78:851〜855、2005)、血清中のラクトフェリン濃度はほとんど検出できず、このタンパク質は細胞(好中球)内で一般に見られ、したがって脳又は他の器官への鉄輸送に貢献する可能性が乏しい。
【課題を解決するための手段】
【0005】
本発明者らは驚くことに、長い間鉄貯蔵タンパク質と考えられてきたフェリチンが、鉄送達タンパク質として働くことができることを発見している。特に本発明者らは、培養中及びラット脳内微小血管系上の内皮細胞上のH−フェリチン受容体の存在を実証しており、BBBを越えて鉄を運ぶための手段としてH−フェリチンを同定している。本発明は、患者における鉄欠乏性障害を治療するための方法であって、治療有効量のフェリチン−鉄複合体を、その必要性がある患者に投与することを含む方法を提供する。本発明の一実施形態では、フェリチン−鉄複合体はH−フェリチンを含む。別の実施形態では、鉄欠乏性障害は脳内の鉄欠乏性を含む。本発明は、脳に鉄を送達するための方法であって、フェリチン−鉄複合体の形の鉄を患者に投与することを含み、それによって、前記鉄が血液脳関門を越えて運ばれ脳に送達される方法;ターゲッティング部分としてH−フェリチンを使用するための方法であって、H−フェリチンとリポソームを結合させることを含み、それによって、前記リポソームが脳及び/又は脳内の細胞にターゲッティングされる方法;及び患者における鉄過剰症を治療するための方法であって、治療有効量のマルチサブユニットフェリチン複合体を、その必要性がある患者に投与することを含み、前記マルチサブユニットフェリチン複合体が100%未満の鉄結合能である方法も提供する。
【図面の簡単な説明】
【0006】
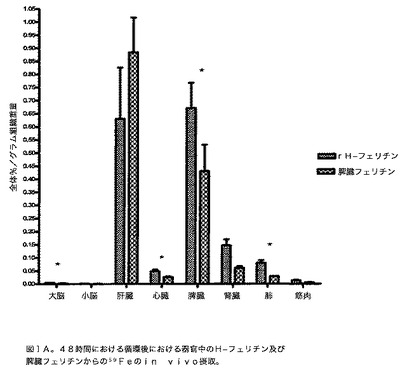
【図1A】48時間の循環後の器官中における、ウマ脾臓フェリチン(脾臓フェリチン)と比較した、組換えヒトHフェリチン(rH−フェリチン)由来の59Feのin vivo摂取を示す図である。59Fe標識H又は脾臓フェリチンを成体ラットの尾静脈に注射し、48時間循環させた。放射能の量は1.0グラムのそれぞれの器官において決定し、全体%/グラム組織重量は、注射した全体と比較した器官当たりのμCの量を計算することによって決定した。2つの脳構造をこのスケールで示して相対量を明らかにする。表すデータは3匹の動物の平均±S.Eである。p<0.05*。
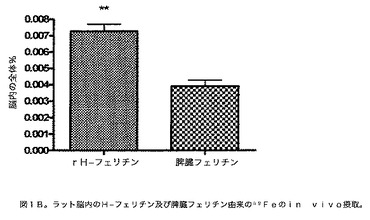
【図1B】ラット脳内のフェリチン由来の59Feのin vivo摂取を示す図である。図1Aのレジェンド中に記載したのと同様に、59Fe標識H又は脾臓フェリチンを、尾静脈を介してラットに注射した。(脳の一半球中のμC)/(注射した全μC)×100%を決定することによって全体%を計算した。p<0.005**。表すデータは3匹の動物の平均±S.Eである。
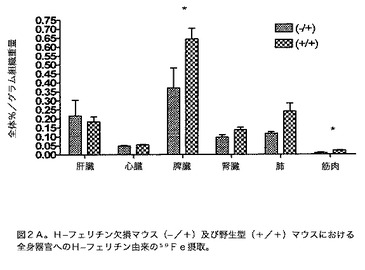
【図2A】H−フェリチン欠損マウス(−/+)及び野生型(+/+)マウスにおける、全身器官へのH−フェリチン由来の59Fe摂取を示す図である。動物には等量のH−フェリチンを腹腔内注射した。フェリチンは48時間循環した。(脳を含む)示した器官を除去し、0.1gを基にそれぞれの器官に関して全体の放射能のパーセントを決定した。p<0.05*。
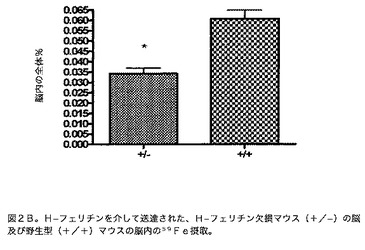
【図2B】H−フェリチンを介して送達された、H−フェリチン欠損マウス(−/+)の脳vs.野生型(+/+)マウスの脳内の59Fe摂取を示す図である。これらのデータは、図2A中のデータを作製するために使用したマウスの脳からのデータである。脳内の放射能の%は、脳から得た59Feの1分当たりの崩壊数と注射した全μCを比較することによって決定した。p<0.05*。示した結果は3匹の動物の平均±標準誤差である。
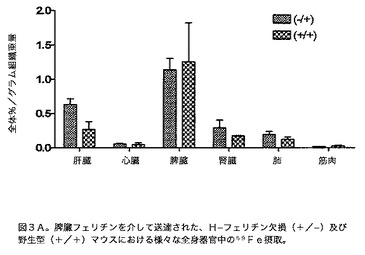
【図3A】脾臓フェリチンを介して送達された、H−フェリチン欠損(−/+)及び野生型(+/+)マウスにおける、様々な全身器官中の59Fe摂取を示す図である。マウスには59Feを含有する脾臓フェリチンを腹腔内注射した。48時間後、マウスを屠殺し器官を除去した。放射能の量は0.1gのそれぞれの器官で決定し、注射した全放射能のパーセントを決定し、このグラフ中に表した。データは3匹の動物の平均±S.Eである。いずれの差も統計学的有意に達しなかった。
【図3B】脾臓フェリチンを介して送達された、H−フェリチン欠損(−/+)vs.野生型(+/+)マウス中の脳内の59Fe摂取を示す図である。これらのデータは、図3A中で使用したマウスからのデータである。報告する放射能の量は動物に注射した全体の%である。示した結果は3匹の動物の平均±標準誤差である。
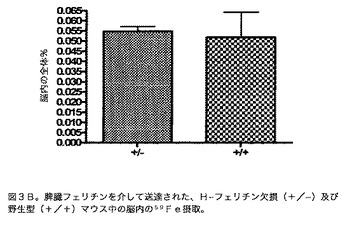
【図4】BREC培養物単層を越えるフルオレセイン標識Hフェリチンの流動を示すグラフである。このグラフは、4時間にわたる基底チャンバー中のFITC標識Hフェリチンの輸送速度を示す。輸送速度は方法セクション中に記載したのと同様に決定した。示したデータは、単位量の上部チャンバーの蛍光当たりの基底チャンバーの蛍光(Bf/Tf)対時間のプロットからの勾配(cm/s)及び共通基準として得た流動速度の平均である。デキストラン対照及び脾臓フェリチンと比較して、H−フェリチンの輸送は統計上有意である(p<0.01**)。脾臓フェリチンの輸送はデキストラン対照と異ならなかった。カリウム枯渇培地でのBRECの前処理はFITC H−フェリチン輸送の有意な低下をもたらしたが(p<0.05*)、ピノサイトーシスを最小にするためのフィリピンを用いた前処理はH−フェリチンの輸送速度に対して全く影響が無かった。トランスフェリンの輸送は陽性対照として含めた。デキストランの流動に関して示す値は未処理対照サンプルに関するものであるが、デキストランの流動速度はいかなる処理によっても変わらなかった(データ示さず)。
【図5A】BREC細胞ホモジェネートとの125I−H−フェリチン及び125I−脾臓フェリチンの結合に関する飽和曲線を示すグラフである。このグラフはBREC細胞とのH−フェリチンの結合は飽和状態であることを示すが、一方で脾臓フェリチンの結合の証拠は存在しなかった。飽和結合は2時間4℃で実施した。Kdは2.7±0.9nMであると決定され、Bmaxは465.7±63.1fmol/mgタンパク質である。
【図5B】ラットの脳から単離した微小血管における125I−H−フェリチン及び125I−脾臓フェリチンに関する飽和曲線を示す図である。これらの曲線はH−フェリチンに関する飽和結合は示すが、脾臓フェリチンに関する結合は示さない。Kdは7.9±1.6nMであり、Bmaxは572.6±64.0fmol/mgタンパク質である。
【図6A】非標識H−フェリチン及び脾臓フェリチンによる、BRECホモジェネートにおける125I−H−フェリチン結合部位に関する競合を示すグラフである。このグラフは、125I−H−フェリチンとBRECホモジェネートの結合は、脾臓フェリチンではなく、非標識H−フェリチンの濃度の増大によって競合的に阻害されることを示す。
【図6B】ラット微小血管における競合結合アッセイを示すグラフである。このグラフは、125IH−フェリチンの結合は、非標識脾臓フェリチンによってではなく、非標識H−フェリチンによって濃度依存的な様式で解離し得ることを示す。
【図7】組換え酵母中でのH−フェリチンの発現を示すウエスタンブロット(4〜20%勾配)の図である。レーン1中は標準(組換えヒトH−フェリチン)である。レーン2及び4は、ヒトL−フェリチンで形質転換した酵母の2つの異なるコロニーである。レーン3及び5は、ヒトH−フェリチンで形質転換した酵母の2つの異なるコロニーである。この試験で使用した抗体は、寛大にもPaolo Arosio(Brescia Italy)によって与えられたH−フェリチンに対するマウスモノクローナル抗体である、抗HF HS−59(1:40,000、16時間)である。
【図8】ウエスタンブロットの図である。タンパク質を最初にゲル電気泳動によって分離し、次いでゲルをパールの反応、フェリチンの鉄含量を示すために使用されている鉄に関する標準的な組織染色で染色した。レーン1及び2は、L−フェリチンで形質転換した酵母のコロニー由来である。鉄の反応生成物はこれらのレーン中では見られない。レーン3及び4は、正常な鉄培地条件(レーン4)及び鉄多量培地(レーン3)下でH−フェリチンを発現する酵母由来のタンパク質抽出物である。H−フェリチン標準は対照として使用した(レーン5)。レーン6は、フェリチンの大きさを示すための分子量マーカーを含有する。
【図9】鉄欠乏の標準的なラットモデルを使用して得た結果を示す図である。鉄欠乏(ID)食での動物は最低レベルのヘモグロビン(Hb)を有していた。鉄を含まない酵母(酵母、鉄なし)を与えた動物は、ID動物と類似したHbレベルを有していた。Hbレベルの改善は他の3群において見られ、改善の最も急速な増大は、鉄を補充しフェリチン(ft)を補強した酵母を与えた動物において生じた。フェリチンを含まない鉄補充酵母を与えた動物さえ、FeSO4群より改善されたHbレベルを有していた。
【図10】図9中で試験した動物の群からのヘマトクリット値レベルを表す図である。これらのデータは、鉄の媒体としての酵母はH−フェリチンの存在又は不在下でヘマトクリット値を補正する際に同程度に有効であり、いずれも標準的な現在の治療選択肢、FeSO4より有意に優れていることを示す。ID食を継続した動物、及び鉄を補充しなかった酵母を与えた動物は、試験した11日間にわたってヘマトクリット値の増大を示さなかった。
【図11】図9及び10中で試験した動物の群中の鉄動員を測定する試験の結果を示す図である。これらのデータは、フェリチン補強鉄多量酵母(ferritin fortified iron enriched yeast)(酵母−Fe+Ft)は、トランスフェリン(Tf)飽和の最大の増大、次にフェリチンを含まない鉄多量酵母(酵母、フェリチンなし)をもたらしたことを示す。
【図12】脳の2つの発達上重要な領域、腹側中脳及び尾状核の鉄の状態を示す図である。Hb及びHct分析(図9及び10)中に記載した動物は14日齢で屠殺し、腹側中脳(VBM)及び尾状核の鉄濃度を決定した。フェリチンを補強し鉄を補充した酵母(酵母−Fe+Ft)を与えた動物は、任意の他の群より多量の鉄を2つの脳領域中に有していた。
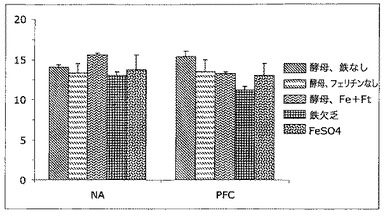
【図13】脳の側坐核(NA)及び前頭前皮質(PFC)領域中の鉄レベルを表す図である。この図中では、フェリチンを補強し鉄を補充した酵母(酵母−Fe+Ft)から送達される鉄の領域特異性は明らかである。(図12中に示したVMB及び尾状核と同様に)NA中では、鉄送達の他の形態と比較して鉄含量は上昇する。しかしながら、PFC中では、フェリチンを補強し鉄を補充した酵母から送達される鉄は、他の群に関して見られるそれと同様である。
【図14A】ヒトH−フェリチンのアミノ酸配列(配列番号1)を示す図である。
【図14B】ヒトH−フェリチンのcDNA配列(配列番号2)を示す図である。開始(ATG)及び停止(TAA)コドンは太字であり、(配列の5’末端における)BamHI及び(配列の3’末端における)XhoI制限部位には下線を引く。
【発明を実施するための形態】
【0007】
本発明は、患者における鉄欠乏性障害を治療するための方法であって、治療有効量のフェリチン−鉄複合体を、その必要性がある患者に投与することを含む方法を提供する。
【0008】
本明細書で使用する「鉄欠乏性障害」は、鉄欠乏、鉄摂取、及び/又は鉄代謝と関係がある障害又は疾患を含む。鉄欠乏性障害の例には、不十分な食事摂取又は鉄の吸収によって引き起こされる鉄欠乏性貧血などの、鉄欠乏性貧血がある。鉄欠乏性貧血は、例えば、栄養失調、妊娠(産後期含む)、重度の子宮出血、慢性疾患(慢性腎疾患含む)、癌、腎臓透析、胃バイパス、多発性硬化症、糖尿病(例えば、I型及びII型糖尿病)、インスリン耐性、及び注意力欠如障害と関係がある可能性がある。
【0009】
本発明の一実施形態では、鉄欠乏性障害は、パーキンソン病、アルツハイマー病、下肢静止不能症候群(RLS)、女性における貧血と関係がある最適以下の認識能力、鬱病及び不眠症を含めた様々な神経及び神経変性疾患において生じるような、脳内の欠乏した鉄レベルと関係がある。別の実施形態では、鉄欠乏性障害は、乏しい認識能力及び運動障害をもたらす発育中の鉄欠乏が原因である低髄鞘形成及び遅い脳の発達を含めた、生後発育中の脳内鉄欠乏性と関係がある神経障害(単数又は複数)を含む。
【0010】
脳内の鉄の欠如及び低下したドーパミン合成は、例えば、子供のRLS及び発育中の鉄欠乏などの、鉄欠乏性障害の重要な要因である。ドーパミンは、適切な中枢神経系(CNS)機能に必要不可欠である、脳内で合成される神経伝達物質である。ドーパミンの合成において、鉄は酵素チロシンヒドロキシラーゼの補因子であり、これはドーパミン代謝の律速段階である(Cooper et al.(1991)The biochemical basis of neuropharmacology.Oxford University Press、New York、N.Y.)。ドーパミン作動系中の鉄は、注意欠陥多動性障害(ADHD)を含めた子供のRLS病態生理及び行動障害における重要な要素であるようである。RLS患者は、RLSと対照の両方におけるフェリチン及びトランスフェリンの正常血清中レベルにもかかわらず、65%未満の脳脊髄液(CSF)フェリチン及び3倍多いCSFトランスフェリン(鉄輸送血中タンパク質)を有する。鉄濃度は、脳、ドーパミン合成部位全体で変わり、RLS患者は、脳の黒質及び被殻部分において少量の鉄を有する。一般に、低下したフェリチンレベルはRLSの重度を示す。ADHDである子供における低下した血清中フェリチンレベルの報告も存在する。
【0011】
用語「フェリチン−鉄複合体」は、多数のフェリチンサブユニット及び鉄原子を含むタンパク質複合体を指す。適切なフェリチン−鉄複合体は、哺乳動物のH−フェリチンサブユニットを含む。様々な哺乳動物種由来のH−フェリチンサブユニットのアミノ酸配列が同定されている。例えば、Orino Koichi et al.、Sequence analysis offeline ferritin H and L subunit cDNAs;Veterinary Biochem.42:7〜11(2005);アクセッション番号:06A006486を参照。一実施形態では、H−フェリチンはヒトのH−フェリチンである(配列番号1;図14A参照)。H−フェリチンは、ヒトのH−フェリチンの天然に存在する又は合成ホモログ又は変異体であってもよい。特定の実施形態では、H−フェリチンのホモログは、ヒトのH−フェリチンと少なくとも80%、81%、82%、83%、84%、85%、86%、87%、88%、89%、90%、91%、92%、93%、94%、95%、96%、97%、98%、又は99%の配列同一性などの、ヒトのH−フェリチンと約80%〜約100%の配列同一性を有する。H−フェリチンのホモログは、鉄と結合しマルチサブユニットフェリチン−鉄複合体を形成する能力を保持しているが、突然変異させて鉄とフェリチンの間の様々な結合及び解離強度をもたらすことが可能である。フェリチン−鉄複合体はH−フェリチンサブユニットを含むが、いくつかのL−フェリチンサブユニットも含むことができる。特定の実施形態では、複合体のフェリチンサブユニット成分は、L−フェリチンと比較して約20%、30%、40%、50%、60%、70%、80%、90%、又は100%のH−フェリチンなどの、L−フェリチンと比較して少なくとも20%のH−フェリチンを含む。一実施形態では、フェリチン−鉄複合体中の全てのフェリチンサブユニット(即ち、100%のフェリチンサブユニット)がH−フェリチンである。H−フェリチンは組換えH−フェリチンであってよい。例えばH−フェリチンは、適切な酵母プロモーターの制御下でH−フェリチンをコードするポリヌクレオチド配列を含む酵母菌株によって生成される、ヒトH−フェリチン、又はそのホモログであってよい。
【0012】
フェリチン−鉄複合体中の鉄は鉄分子であってよく、又は鉄含有複合体の形であってよい。「鉄含有複合体」又は「鉄複合体」は、有機化合物と複合体形成する、(II)又は(III)酸化状態の鉄を含有する化合物である。鉄複合体は、鉄ポリマー複合体、鉄炭水化物複合体、及び鉄アミノグリコサン複合体を含む。これらの複合体は市販されており、及び/又は当技術分野で知られている方法により合成することができる。
【0013】
鉄炭水化物複合体の例には、カルボキシマルトース鉄、スクロース鉄、ポリイソマルトース鉄(デキストラン鉄)、ポリマルトース鉄(デキストリン鉄)、グルコン酸鉄、ソルビタール鉄、及び水素化デキストラン鉄などの、鉄糖複合体、鉄オリゴ糖複合体、及び鉄多糖複合体があり、これらは、ソルビトール、クエン酸及びグルコン酸(例えば、デキストリン鉄−ソルビトール−クエン酸複合体及びスクロース鉄−グルコン酸複合体)などの他の化合物、及びこれらの混合物とさらに複合体形成することができる。
【0014】
鉄アミノグリコサン複合体の例には、コンドロイチン硫酸鉄、デルマチン硫酸鉄、ケラタン硫酸鉄があり、これらは、他の化合物及びそれらの混合物とさらに複合体形成することができる。鉄ポリマー複合体の例には、鉄ヒアルロン酸、鉄タンパク質複合体、及びそれらの混合物がある。
【0015】
本明細書で使用する「治療」又は「治療する」は、患者又は対象がそのために治療されている状態の完全な除去、及び任意の臨床的又は定量的に測定可能な低減を指す。「治療」は、障害の発生の予防又は障害の病状若しくは症状の改変の意図で実施される介入である。したがって「治療」は、療法治療と予防策若しくは予防対策の両方を指す。「治療」は苦痛緩和治療として記すこともできる。治療の必要がある人は、1つ又は複数の鉄欠乏性障害を既に有する人、及びその障害を予防すべきである人を含む。
【0016】
その必要性がある患者を治療する際に、治療有効量の本発明のフェリチン−鉄組成物を本発明に従い患者に投与する。本明細書で使用する用語「治療有効量」は、重大な悪影響を引き起こす可能性がある量を超えない治療に関して示される組成物の量である。療法治療の有効性を評価するための方法は当業者に知られている。
【0017】
「その必要性がある患者」は、本発明の治療法から恩恵を被る可能性がある任意の患者又は対象を指す。特定の実施形態では、その必要性がある患者は、1つ又は複数の鉄欠乏性障害を発生しやすい患者、1つ又は複数の鉄欠乏性障害を有するがいかなる臨床症状も示さない対象、又は1つ又は複数の鉄欠乏性障害を有し1つ又は複数の鉄欠乏性障害の症状に罹患している対象である。その必要性がある患者は、ヒト、イヌ、ネコ、ウシ、ウマ、げっ歯類(マウス、ラット、又はハムスターなど)、又は霊長類などの哺乳動物であってよい。一実施形態では、患者はヒトである。特定の実施形態では、本発明の方法は、実験動物、獣医学的用途、及び/又は疾患に関する動物モデルの開発において用途を見出す。
【0018】
投与する用量は、治療期間、投与の頻度、宿主、並びに障害の性質及び重度に応じた変数である。用量は過剰な量の実験なしで当業者によって決定することができる。本発明の組成物は、患者へのフェリチン−鉄複合体の十分な用量単位の放出を確実にするのに十分な用量濃度で投与して、鉄欠乏性障害の望ましい治療をもたらす。投与する実際の用量は、年齢、体重、状態の重度、及び/又は患者の臨床歴などの、物理的及び生理的要因によって決定され得る。活性成分を投与して、約50μM〜約1000μMのフェリチン−鉄複合体のin vivo血漿濃度を得ることができる。例えば、本発明の方法は組成物を使用して、約0.1〜約1,000又は約1〜約100mg/体重1kg/1日のフェリチン−鉄複合体、約30mg/体重1kg/1日のフェリチン−鉄複合体などをもたらすことができる。しかしながら、提供したこれらの範囲から逸脱する用量レベルも、所与の障害の治療において適している可能性があることは理解されよう。
【0019】
本発明のフェリチン−鉄複合体は、投与に適した任意の形であってよい。このような投与可能な形には、錠剤、緩衝錠剤、ピル、カプセル、腸溶性カプセル、糖衣錠、カシェ剤、粉末、顆粒、エアロゾル、リポソーム、坐薬、クリーム、ローション、軟膏、皮膚用パッチ剤、非経口剤、トローチ剤、懸濁液、溶液及びエマルジョン(水中油型又は油中水型)などの経口液体、眼科液及び注射液、又はこれらの徐放形がある。望ましい用量は連続注入によって、又は徐放性製剤によって、1日を通して一定間隔においていくらかの増分で与えることができ、又はボーラス、舐剤又はペーストとして表すことができる。
【0020】
一実施形態では、フェリチン−鉄複合体を含む医薬組成物又は製剤を、1つ又は複数の薬剤として許容される担体との混合によって調製する。いくつかの場合、フェリチン−鉄複合体は、フェリチン−鉄複合体及びその中に鉄分子とフェリチン分子が溶けたバッファーを含む組成物として送達して、鉄とフェリチンの結合(即ち、フェリチン−鉄複合体の形成)を可能にすることができる。しかしながら、望む場合、他の生成物を加えて、鉄の送達、保存を最大にすること、又は特定の送達法を最適化することができる。さらに本発明は、鉄欠乏性障害の治療に適した他の作用物質と組合せた、本明細書に記載するフェリチン−鉄複合体を含む組合せ組成物の使用を含む。
【0021】
本明細書で使用する「薬剤として許容される」は、医薬及び獣医学分野における使用に許容され、製剤の他の成分と適合性があり、且つ毒性ではない又は他の場合妥当なベネフィット/リスク比に応じて許容不能であることを意味する。「薬剤として許容される担体」又は「希釈剤」は、フェリチン−鉄複合体を含む組成物の医薬投与と適合性がある、任意及び全ての溶媒、分散媒、コーティング、抗菌剤及び抗真菌剤、等張剤及び吸収遅延剤などを含む。このような担体又は希釈剤の例には、水、生理食塩水、フィンガーの溶液及びデキストロース溶液があるが、これらだけには限られない。医薬組成物の体積は、目的とする投与の形態、及び医療専門家によって決定される個々の患者に安全な体積に基づく。
【0022】
担体の選択も目的とする投与の形態に依存する。本発明の組成物は、全身投与(例えば、静脈内注射、腹腔内注射、吸入、経皮送達、経口送達、鼻腔送達、直腸送達など)及び/又は局部投与(例えば、標的組織への直接注射、カニューレを介した組織への送達、徐放性物質の注入による標的組織への送達)、ポンプなど、経口、非経口、骨内、脳脊髄液中などによる組織への送達だけには限られないが、これらを含めたいくつかの好都合な手段のいずれかによって投与することができる。さらなる投与の形態には、頬、舌下、膣内、皮下、筋肉内、又は皮内投与がある。
【0023】
一実施形態では、経口投与する組成物を、経口摂取用にフェリチン−鉄複合体と適切に組合せた物質を使用して調製する。このような物質には、非制限的に、ラクトース(水和、高速流)、グルコース及びスクロースなどの糖、コーンスターチ及びポテトスターチなどのスターチ、微結晶性セルロース、ナトリウムカルボキシメチルセルロース、エチルセルロース及び酢酸セルロースを含めたセルロース及びその誘導体、粉末状トラガカント、コロイド状二酸化ケイ素、クロスカルメロースナトリウム、麦芽、ゼラチン、タルク、ステアリン酸、ステアリン酸マグネシウム、硫酸カルシウム、ピーナッツ油、綿実油、ゴマ油、オリーブ油及びコーン油などの植物油、プロピレングリコール、グリセリン、ソルビトール、マンニトール及びポリエチレングリコールなどのポリオール、寒天、アルギン酸、水酸化アルミニウム又は水酸化マグネシウムなどの制酸剤、クエン酸ナトリウム、酢酸ナトリウム、又は重炭酸ナトリウムなどのバッファー、水、等張生理食塩水及びリン酸バッファー溶液がある。湿潤剤、ラウリル硫酸ナトリウムなどの潤滑剤、安定剤、錠剤、抗酸化剤、防腐剤、着色剤及び香味剤も存在してよい。
【0024】
非経口投与に適した組成物又は製剤には、抗酸化剤、バッファー、静菌剤又は製剤を血液と等張にする溶質を含有することができる、水性及び非水性、等張滅菌注射溶液、並びに懸濁剤及び増粘剤を含むことができる水性及び非水性滅菌懸濁液がある。組成物は単位用量又は多用量の密閉容器、例えばアンプル及びバイアル中に存在してよく、使用直前に滅菌液状担体、例えば注射用水の添加のみを必要とする、凍結−乾燥(凍結乾燥)状態で保存することができる。即席の注射溶液及び懸濁液は、滅菌粉末、顆粒又は錠剤などから調製することができる。
【0025】
静脈内投与に適した組成物又は製剤は、生理食塩水、静菌水、CREMOPHOR ELTM(BASF;Parsippany、N.J.)又はリン酸緩衝生理食塩水(PBS)などの担体を含む。組成物は滅菌状態でなければならず、シリンジを使用して投与するために流体でなければならない。このような組成物は製造及び保存中は安定状態でなければならず、細菌及び真菌などの微生物からの汚染に対して守らなければならない。担体は、例えば水、ポリオール(グリセロール、プロピレングリコール、及び液状ポリエチレングリコールなど)、及び他の適合性のある、適切な混合物を含有する分散媒であってよい。様々な抗菌剤及び抗真菌剤、例えばパラベン、クロロブタノール、フェノール、アスコルビン酸、及びチメロサールは、微生物の汚染を含有する可能性がある。例えば糖、ポリアルコール、マンニトール、ソルビトール、及び塩化ナトリウムなどの等張剤を組成物中に含めることができる。吸収を遅延することができる組成物には、モノステアリン酸アルミニウム及びゼラチンなどの作用物質がある。滅菌注射溶液の調製用の滅菌固体の調製法は、フェリチン−鉄複合体及び任意の他の望ましい成分を含有する固体を生成するための真空乾燥及び凍結−乾燥を含む。
【0026】
全身投与は、例えば経粘膜又は経皮であってよい。経粘膜又は経皮投与のために、標的障壁(単数又は複数)に浸透することができる浸透剤を選択する。経粘膜浸透剤は、洗浄剤、胆汁塩、及びフシジン酸誘導体を含む。鼻腔スプレー又は坐薬は経粘膜投与に使用することができる。経皮投与のために、活性化合物を軟膏、軟膏剤、ゲル、又はクリームに配合する。
【0027】
フェリチン−鉄複合体を含む組成物は、インプラント及びマイクロカプセル化送達系を含めた放出制御製剤などの、身体からの迅速な除去に対して複合体を保護する担体を用いて調製することができる。エチレン酢酸ビニル、多無水物、ポリグリコール酸、コラーゲン、ポリオルトエステル、及びポリ乳酸などの、生分解性又は生体適合性ポリマーを使用することができる。このような物質は、ALZA Corporation(Mountain View、Calif.)及びNOVA Pharmaceuticals、Inc.(Lake Elsinore、Calif.)から購入することができ、或いは当業者により調製することができる。
【0028】
眼部投与用の組成物は、緩衝又は生理食塩水、リポソーム又は塩基性アミノ酸などの適切な液状担体で調製する。クリーム、ローション及び軟膏は、トリグリセリド基剤、リポソーム、又は塩基性アミノ酸などの適切な基剤を使用して局所施用に調製することができる。このようなクリーム、ローション及び軟膏は、表面活性剤も含有することができる。
【0029】
本発明の一実施形態では、フェリチン−鉄複合体を、H−フェリチンを発現する組換え酵母の菌株の形で投与する。鉄の栄養補充に適した組換え酵母の菌株は、ヒトを含めた哺乳動物に対する、高い生物学的利用能を有する形で鉄を貯蔵することができる。酵母の菌株は、ヒトの消費に関して一般に安全と認められる(GRAS)要件に見合うものを含む。この実施形態では、鉄貯蔵遺伝子(例えば、H−フェリチンコード配列)を、鉄貯蔵発現カセット中で適切な酵母プロモーターの制御下に置いて、鉄補充に適した媒体として働く酵母に必要な鉄貯蔵タンパク質のレベルをもたらすことができる。適切なプロモーターは当技術分野で知られており、高レベルの構成的発現を誘導するプロモーター、及びその発現が環境条件によって制御され得るプロモーターを含む。さらに、酵母の遺伝子構成をさらに操作して、様々なおそらく有利である結果を得ることができる。例えば、タンパク質分解を操作して、細胞表面、液胞、又はミトコンドリアのものだけには限られないがこれらを含めた、鉄貯蔵タンパク質又は鉄輸送機構の安定性を高めることができる。さらに、酵母を改変して、鉄貯蔵タンパク質又は細胞内コンパートメント中の鉄のレベルを高めることができる。酵母の鉄含有量は、酵母が増殖する培地に既知量の鉄化合物を加えることにより制御することができる。組換え酵母を使用して、ヒト又は他の動物のための鉄補充を、栄養サプリメントとしての組換え酵母の消費、又は組換え酵母から精製又は単離したフェリチン−鉄複合体の消費だけには限られないが、これらを含めたいくつかの手段のいずれかによって実施することができる。酵母は鉄補充の目的で特異的に増殖させることが可能であり、或いは酵母は別のプロセス(例えば発酵生産)の副産物であってよい。
【0030】
或いは、又は追加的に、フェリチン−鉄複合体は、大腸菌、バキュロウイルス及びトランスジェニック動物を含めた、当技術分野で知られている他の発現系中で生成されるH−フェリチンを含むことができる。一実施形態では、フェリチン−鉄複合体を、適切なバッファー中でフェリチンサブユニット及び鉄分子をインキュベートすること、次に生成したフェリチン−鉄複合体から任意の非結合鉄分子を分離することによって形成することができる。
【0031】
特定の実施形態では、フェリチン−鉄複合体は、抗体、アプタマー、受容体、リガンド、又はそれらの結合断片などのターゲッティング部分をさらに含む。ターゲッティング部分は1つ又は複数の細胞、組織及び/又は器官特異的マーカーを認識することができ、したがって身体中の所望の標的又は位置への送達を仲介又は改善することができる。一実施形態では、フェリチン−鉄複合体は、ターゲッティングペプチドと融合した、H−フェリチンなどのフェリチンサブユニットを含む融合タンパク質を含むことができる。別の実施形態では、フェリチン−鉄複合体は、カプセル化した状態でリポソーム、リポソーム構築体、又は赤血球ゴーストなどの他の膜結合小胞に送達又は投与することができる。リポソーム、リポソーム構築体、又は他の小胞は、リポソーム又は小胞に取り込まれた特定の細胞表面タンパク質又は受容体(上記参照)に特異的な抗体又はリガンドなどの、ターゲッティング部分を含むことができる。ターゲッティング部分は、フェリチン−鉄複合体、又はフェリチン−鉄複合体を含む小胞を、脳及び/又は血液脳関門に向けることができる。適切なターゲッティング部分の例には、トランスフェリン、インターロイキン−13(星状細胞腫への送達用)、及びリポ多糖(LPS)がある。
【0032】
本発明の別の実施形態では、H−フェリチン自体を使用して他の部分をターゲッティングすることができる。例えば、H−フェリチンを生物活性がある作用物質と結合させて、その作用物質を脳に送達することが可能である。一実施形態では、H−フェリチンペプチドを別の生物活性があるペプチドと融合させる。或いは、又は追加的に、H−フェリチンをリポソーム又は他の小胞と結合させて、小胞及び小胞含有物を脳に送達することが可能である。さらに別の実施形態では、H−フェリチンを造影剤化合物などの作用物質に結合させて、脳(例えば、白質路及びその欠陥)の可視状態を向上させることが可能である。生物活性がある作用物質と同様に、造影剤化合物をH−フェリチンタンパク質又はその中に結合させること、或いは受容体リガンドとして作用するリポソーム結合H−フェリチンを介して脳内のH−フェリチン受容体に向けられるリポソーム又は他の小胞内にカプセル化することが可能である。
【0033】
本発明の別の実施形態では、H−フェリチンを使用して余分な鉄又は鉄過剰症を治療することができる。臨床上ヘモクロマトーシスとして知られる鉄過剰症は、例えば、癌の増大したリスク、心不全、肝機能障害及び糖尿病と関係がある。脳内の鉄過剰症は、パーキンソン病、アルツハイマー病、筋萎縮性側索硬化症、Hallovordeen−Spatz、及びハンチントン病などの、広範囲の神経変性疾患において生じる可能性がある。H−フェリチンの有意な鉄結合能のために、H−フェリチンを含むH−フェリチン及び/又はマルチサブユニット複合体は鉄キレート剤として使用することができる。「マルチサブユニットフェリチン複合体」によって、多数のフェリチンサブユニット及び場合によっては鉄を含むタンパク質複合体を意味する。H−フェリチンは、前に定義した哺乳動物のフェリチン又はそのホモログ若しくは変異体である。マルチサブユニットフェリチン複合体は、生成する複合体がその全体的鉄結合能が100%未満であるように、比較的鉄を含まない環境中で調製することができる。特定の実施形態では、複合体は、約50%、40%、30%、20%、10%、5%、又は1%の鉄結合能などの50%以下の鉄結合能である。一実施形態では、複合体は約0%の鉄結合能であり得る(即ち、100%又は100%に近い鉄結合能が残存する鉄を含まないフェリチン又はアポフェリチン)。H−フェリチンを修飾して、それが任意の内因性受容体によって認識される可能性を低減すること、又は身体による排泄を増大することができる。このような修飾は十分に当業者の専門知識内にあり、これらの修飾は鉄と結合するタンパク質の能力に必ずしも影響を与えない。鉄キレート剤として使用するのに適したマルチサブユニットフェリチン複合体はH−フェリチンサブユニットを含むが、いくつかのL−フェリチンサブユニットも含むことができる。特定の実施形態では、マルチサブユニットフェリチン複合体は、L−フェリチンと比較して約20%、30%、40%、50%、60%、70%、80%、90%、又は100%のH−フェリチンなどの、L−フェリチンと比較して少なくとも20%のH−フェリチンを含む。
【0034】
鉄キレート剤として使用するためのマルチサブユニットフェリチン複合体の送達は、鉄欠乏性障害を治療するためのフェリチン−鉄複合体の送達に関して本出願中の他の箇所に記載したように、同じ範囲の送達機構(即ち、ターゲッティング部分及び/又はリポソーム又は他の小胞)を含み得る。マルチサブユニットフェリチン複合体は、胃腸管、胃腸管内壁の細胞内(そこでそれは摂取され得るが、血流には入らない)、又は血液自体に送達することができ、そこでそれは非トランスフェリン及びおそらくさらにトランスフェリンと結合した鉄と有効に競合し得る。この手法を利用して、他の場合無秩序な形式で不適切に脳にアクセスし得る鉄が除去され得る。さらに、血液及び全身器官から鉄をキレートすることは、脳からの鉄の放出を含めて、身体内の鉄の再分布を促進し得る。いくつかの場合、全体的鉄結合能が100%未満であるアポフェリチン又はフェリチン複合体を、脳脊髄液に直接送達して脳からの鉄の放出を刺激することができる。
【0035】
フェリチンは鉄だけでなく、その多くが身体に対して毒性である一定範囲の金属とも結合する能力を有する。したがって、本発明の別の実施形態は、一般に胃腸系、血液、脳及び身体からおそらく毒性である金属を低減及び除去するための、アポフェリチン(又は全体的鉄結合能が100%未満である他のマルチサブユニットフェリチン複合体)の使用に関する。したがって、血液用の一般的なメタルクレンザーとしてアポフェリチンを利用することが考えられる。
【0036】
輸血において、少なくとも15%の細胞が輸血過程中で溶解し、おそらく損傷をもたらす遊離鉄が放出する。輸血は、赤血球(RBC)が必要でありしたがって貧血である患者に実施する。本発明は、アポフェリチン(又は全体的鉄結合能が100%未満である他のマルチサブユニットフェリチン複合体)と輸血液を混合する方法に関するものであり、これは細胞溶解により放出される鉄キレート剤をもたらすだけでなく、その鉄をより生理的又は生物学的に利用可能な形で身体に利用可能にするようにも働く。頻繁な輸血を必要とする1つの患者集団にはサラセミア(thalessima)を有する患者がある。この集団は最終的には、過剰な鉄蓄積による肝臓障害に罹患する。H−フェリチン及び/又はH−フェリチンを含むマルチサブユニットフェリチン複合体は、身体中に鉄がより有効に分布するのを手助けし、したがって肝臓中の過剰な蓄積を制限することができる。頻繁な輸血を受ける別の集団は、新生児及び特に未熟児である。H−フェリチン、H−フェリチンを含むマルチサブユニットフェリチン複合体、及び/又はフェリチン−鉄複合体を輸血液に与えると、脳発達に関する脳への鉄分布を含めた鉄分布を改善し得る。
【0037】
H−フェリチンサブユニットを含めたフェリチンサブユニットは、患者の身体中で発現される可能性がある。この実施形態では、フェリチン−鉄複合体はin vivoで形成する。いくつかのプラスミド担体及びトランスフェクション試薬系をex vivoで細胞をトランスフェクトするのに利用して、宿主動物又は患者に再注入するための、安定した形質転換体又は一時的にトランスフェクトされた細胞のいずれかを生成する。適切な発現プラスミドはトランスフェクション試薬と同様に市販されており、後者の多くは1タイプ又は別タイプのカチオン性リポソームである。in vivo及びex vivo遺伝子導入、遺伝子療法などに適したベクターは当技術分野で知られており、レトロウイルスベクター、アデノウイルスベクター、アデノ随伴ウイルスベクター、レンチウイルスベクター、及びエレクトロポレーション系を含む。
【0038】
本発明の他の態様及び利点は、以下の例示的な実施例を考慮することにより理解されよう。これらの実施例は、決して本発明の範囲を制限するものとして解釈すべきではない。当業者は、本開示を鑑みて、開示する具体的な実施形態に変更を施すことができ、さらに本発明の精神及び範囲から逸脱せずに同様又は類似の結果を得ることができることを理解するはずである。
【0039】
本出願中に引用する全ての参照文献、刊行物及び特許文書は、参照として本明細書に組み込まれる。本文書中の様々な参照文献のそれらの引用によって、本出願人は、いかなる特定の参照文献も本発明に対する「従来技術」であることを認めない。
【実施例】
【0040】
(例1)
フェリチンの調製
この実施例中の全ての実験は、組換えヒトH−フェリチン又はウマ脾臓フェリチンを使用した。組換えヒトH−フェリチンは、Hisタグ組換えヒトHフェリチンプラスミドを用いて、化学的に形質転換受容性にされているBr21細胞を形質転換することによって調製した。細胞が増殖した後、2.8mg/mlの最終濃度まで、タンパク質をニッケルタンパク質濾過カラムで精製した。ウマ脾臓フェリチンは市販的に入手し(Sigma)、それが約90:10のLとHフェリチンサブユニットを含有するので選択した。
【0041】
細胞培養及び内皮細胞単層の調製
本発明者らは、フェリチンは内皮細胞の層を越えて輸送され得るという仮説を試験し、BBBを越えたフェリチン輸送の機構に取り組み始めるために、血液脳関門(BBB)のin vitroモデルとしてウシ網膜内皮細胞(BREC)を使用した。この十分研究されたモデルは、血液神経関門の全ての必要な特性及び属性を有することが示されている(Antonetti D.A.、et al.J Neurochem.80:667〜677、2002)。ウシの眼は地域の食肉処理場から得て、ウシ網膜内皮細胞(BREC)を単離し、以前に公開された手順(Gardiner T.A.、et al.Lab Invest.72:439〜444、1995)に従って処理した。BRECは、10%のFBS、10ng/mLのEGF、0.2mg/mLのENDO GRO(商標)(VEC Technologies、Inc.、Rensselaer、NY、USA)、0.09mg/mlのヘパリン、抗生物質/抗真菌溶液(Gibco、Rockville、MD、USA)、及びTylosin抗生物質(Sigma)を補充したMCDB−131培地(Sigma、St.Louis、MO、USA)中で増殖させた。それらが少なくとも80%集密状態に達するまで、細胞は最初はフラスコ内で培養する。その後、BRECSを軽くトリプシン処理し、COSTAR(登録商標)TRANSWELL(商標)0.4μm多孔質フィルター(Coming、Acton MA)で集密状態に増殖させた。フィブロネクチンを1μg/cm2の濃度で加えてフィルターへの接着を促進した。次いで細胞を洗浄し、72時間100nmのヒドロコルチゾンを補充した無血清無EGFMCDB−131培地に移した。これらの細胞培養物へのヒドロコルチゾンの添加は、密着結合の形成を促進した。
【0042】
BBBモデルにおけるフェリチン輸送
予め精製されたトランスフェリンを購入し、2.5mg/mlの最終濃度に再懸濁した。約250μgの組換えヒトHフェリチン、ウマ脾臓フェリチン(Sigma)及びトランスフェリンを、100mMの炭酸塩/重炭酸塩バッファー、pH9.0中においてフルオレセインイソチオシアネート(FITC)(Pierce Biotechnology)で標識した。過剰又は加水分解FITCの除去は、5mlのG−25脱塩カラムを通すことによって行なった。FITC結合H−フェリチン、脾臓フェリチン、及びトランスフェリンを濃縮し、バッファーはCENTRIPREP(登録商標)濃縮器(Amicon、Inc.、10,000MWCO)中でPBSと交換した。トランスフェリンは、BRECモデルにおける陽性輸送対照として含めた(Burdo J.R.、et al.Neuroscience121:883〜890、2003)。
【0043】
集密状態のBREC単層を越える流動速度は、等式:(Bf/Tf)*(Vb/A)=(フラックス)*tを使用してある程度の修正を加えて、Burdo et al.、2003中で以前に記載されたように決定した。簡単に言うと、140μgのFITC標識rH−フェリチン、脾臓フェリチン、又はトランスフェリンのいずれかを上部チャンバー(先端)に加える前に、トランスウエル装置中で集密状態までBRECを増殖させた。トレーサーの輸送は、先端チャンバーへのトレーサーの添加後の様々な時間(15、30、45、60、120、180、及び240分)で回収した下部チャンバー(基底)由来の100μlアリコートからサンプリングすることによって決定する。基底チャンバー由来のアリコートを、蛍光光度計(SPECTRAMAX(登録商標)GEMINI、Molecular Devices)において次いで蛍光に関して分析する。流動速度は、単位量の上部チャンバーの蛍光当たりの基底チャンバーの蛍光(Bf/Tf)対時間(t)のプロットからの勾配(cm/s)として得る。ここで、単層を越えて輸送されるトレーサーの量を示すBfを、基底チャンバーの体積(Vb)及びさらに輸送に利用可能な表面積(A)に標準化する。上部チャンバー中のTfの濃度は実験を実施した4時間にわたって著しく変わることはなく、したがって流動速度の計算に関して一定であると考えられる。上部チャンバー中の蛍光の量は、輸送アッセイ(4時間)の最後に100μlアリコートから得る。
【0044】
傍細胞流動の対照として、RITCデキストラン(70kDa)を、対照として先端チャンバーに同時に加えた。デキストランは内皮細胞によって適切なレベルで取り込まれない(Raub T.J.、et al.J.Cell Physiol.149:141〜151、1991)。したがって、基底チャンバー中のデキストランの任意の蓄積は傍細胞輸送によるものである可能性がある。いずれの条件も、それぞれの条件で最小であったデキストランの流動速度に影響を与えなかった。
【0045】
輸送機構の決定
ピノサイトーシスがフェリチンの輸送に有意に貢献するかどうかを決定するために、フェリチン及びデキストランを加える前に、50μg/mlのフィリピンを30分間トランスウエル装置の先端チャンバーに加えた。フィリピンの添加は、ピノサイトーシスを介した非特異的輸送の作用を阻害することが示されている(Stremmel W、et al.Lipids36:981〜989、2001)。H−フェリチンの摂取がクラスリン依存性エンドサイトーシスによって起こるかどうかを決定するために、カリウム欠損培地(100mMのNaCl/50mMのHEPES)中で試験を実施した。フェリチンを加える前に、細胞は10分間カリウム欠損培地中でインキュベートした。細胞内のカリウム枯渇は、クラスリンコーティングピット中に生じる受容体介在性エンドサイトーシスのプロセスを阻害する。これらの後者の実験は、カリウム枯渇がデキストラン輸送の増大によって示される細胞間結合の完全性を変える前に、1時間のみ実施することができた。それぞれの処理条件(又は標準)は少なくとも6回実施した。実験を通じて、培養物は目視で評価して、実験処理及び操作が細胞生存に影響を与えなかったことを確認した。ベースライン実験中と同様に、デキストラン標識RITCは密着結合の完全性の指標として含めた。異なる条件下でのFITC H−フェリチンの平均間の差は、一元配置分散分析を使用して分析した。有意に異なる平均を用いた測定用に、ボンフェロニの事後比較を行なってペアワイズ差を分析した。有意性のレベルはp<0.05で設定した。
【0046】
結合実験
飽和分析法:BREC細胞ホモジェネートでの結合実験を、125I−組換えヒトH−フェリチン又はウマ脾臓フェリチンを使用して第四代のBREC細胞に二連で実施した。両者の比活性度は約340,000cpm/pmolであった。全体、特異的及び非特異的結合を確定するために、一定範囲の濃度の125I−H−フェリチンを、1000倍モル過剰の非標識H−フェリチン有り又はなしで、100μgの全タンパク質のBREC細胞ホモジェネートに加えた。結合バッファーは、50mMのトリス−HCl(pH7.4)、0.1%のBSAからなっていた。インキュベーションは2時間22℃で実施した。3mlの氷冷50mMトリス−HClを加えることによって結合を終了させた。細胞採取機を使用して、結合放射能は迅速な濾過、及び5%ノンファット乾燥ミルク及び0.1mg/mlウマ脾臓フェリチンで事前にコーティングしたWhatmanガラスファイバー製Cフィルターでの洗浄によって単離した。
【0047】
平衡的競合結合アッセイを実施し、高濃度の非標識H−フェリチンを、前に記載したのと同じ結合バッファー中で120分間0.4nMの125I−H−フェリチンと共に、22℃においてBRECホモジェネートのタンパク質25μgとインキュベートした。結合の終了、膜の単離及び特異的結合の計算は、前に記載したように実施した。
【0048】
ラット脳内微小血管系
微小血管の調製:6匹の成体ラットをそれぞれの微小血管の調製に使用した。ラットは致死量のナトリウムペントバルビタールで麻酔をかけ(100mg/体重1kg)、次いで断頭した。脳を除去して、氷上のペトリ皿に置いた。小脳及び髄膜を除去して、5体積の微小血管用バッファー(1×MVB−1×塩、1×HEPES、0.5%BSA及び5mMグルコース)をプロテアーゼ阻害剤と共に加えた。脳はガラス−テフロン(登録商標)ホモジェナイザー(0.25mmクリアランス)を使用して20ストロークで軽く均質化して、ホモジェネートを4℃において10分間1000×Gで遠心分離した。上清は廃棄し、ペレットは17%デキストランの5体積/ラット(1:1の比の1×塩と1×HEPES及びデキストラン)に再懸濁し、次に攪拌し、次いで4℃において3000×Gで遠心分離した。微小血管は試験管の壁から回収し、20mlの1×MVBバッファーに再懸濁した。微小血管は120μメッシュを介して濾過した。次いで微小血管調製物を、40μメッシュ上に支持されたガラスビーズ(Sigma)への接着によってさらに精製した。ビーズはプロテアーゼ阻害剤を加えたバッファーで洗浄した。ビーズは5mlのMVB中ですすぎ、次いで15分間の4℃における1000×Gでの遠心分離により微小血管をペレット状にした。微小血管は1mlのHES+(10mMのHEPES、1mMのEDTA、250mMのスクロース、pH7.4及びプロテアーゼ阻害剤カクテル)(Sigma)に再懸濁し、全体のタンパク質濃度を決定した。サンプルは使用時まで−80℃で保存した。
【0049】
微小血管におけるフェリチンの結合
結合懸濁液は、250μlの最終体積で1μMの非標識H−フェリチンの添加有り又はなしで、50mMのトリス−HCl(pH7.4)、0.1%のBSA、及び20μgの膜タンパク質調製物からなっていた。3mlの氷冷50mMトリス−HClを加えることによって結合を終了させた。結合放射能は、5%ノンファット乾燥ミルク(Blotto)及び0.1mg/ml脾臓フェリチンの溶液で事前にコーティングしたWhatmanガラスファイバー製Cフィルターでの迅速な濾過によって単離した。この組合せは経験的に決定して、加えた合計数の1〜3%にフィルターと放射標識タンパク質の非特異的結合を低減した。フィルターは200mMのNaClを含有する3mlの氷冷50mMトリスで5回洗浄した。フィルターはMICROMEDIC4/200及び自動γ線カウンター中で計数した。特異的結合は、過剰な非標識H−フェリチンが存在しない結合(全体の結合)から過剰な非標識H−フェリチンの存在下での結合(非特異的結合)を引くことによって計算した。
【0050】
飽和分析法
それぞれの結合実験は二連で実施した。高濃度の125I−H−フェリチンを、250μlの最終体積で1μMの非標識H−フェリチンの添加有り又はなしで、20μgの膜タンパク質調製物を含む前に記載したのと同じ結合バッファーからなる結合懸濁液に加えた。22℃における120分のインキュベーション後、結合を終了させ、全体、非特異的、及び特異的結合を、前に記載したのと同様に計算した。
【0051】
競合アッセイ
高濃度の非標識競合剤(H−フェリチンと脾臓フェリチン)を、前に記載したのと同じ結合バッファー中で0.4nMの125I−H−フェリチンの存在下において、100μgの膜タンパク質と共に22℃において60分間インキュベートした。結合、結合の終了、膜の単離、及び特異的結合の計算は、前に記載したのと同様に実施した。競合の実験は二連で実施した。
【0052】
in vivo摂取の試験
H及び脾臓フェリチン(1.2mg)を、40μlの1mMニトリロ三酢酸(pH6.0)、0.5μlの硫酸第一鉄アンモニウム、2μlの0.5M重炭酸ナトリウム、及び40μCの59FeCl中で、4時間37℃においてインキュベートした。インキュベーション後、フェリチンを24時間1×PBS中10,000MWカートリッジ内で透析して、任意の非結合59Feを除去した。比活性度はH−フェリチンに関して0.04μC/g及び脾臓フェリチンに関して0.08μC/gであった。放射標識タンパク質(3.4μg/重量1グラム)を、メスのSprague−Dawleyラット(約350g)の尾静脈(n=3)に注射した。48時間後、ラットを断頭し器官はすぐに除去した。それぞれの器官を切除し0.1MのPBS中で完全にすすいだ。脳に関しては、大脳を小脳から除去して二等分し、髄膜は脳から切除除去した。それぞれの器官由来の1グラムの組織(湿重量)を使用して鉄摂取を決定した。
【0053】
H−フェリチン欠損マウスを、器官中のおそらく障害が起きた鉄の処理が、フェリチン鉄送達に影響を与え得るかどうか決定するための実験モデルとして評価した。同様の手法を使用して、マウスに腹腔内注射したこと以外はラットに関して前に記載したのと同様に、対照及びH−フェリチン欠損マウス中のフェリチン摂取を調べた。H−フェリチンに関する比活性度は0.06μC/gであり、脾臓フェリチンに関する比活性度は0.31μC/gであった。フェリチンを注射し、マウスを屠殺し器官を除去するまで48時間血流中で循環させた。
【0054】
それぞれの器官中の放射能の量は、ヨウ化ナトリウム(NaI)ベースで、シングルチャンネルアナライザーウエルカウンターシステム(Canberra Industries Inc.)で1分間決定した。γ線数/分(cpm)をバックグラウンド数から差し引いて、カウンターの効率で割り、次いで崩壊数/分で割ってμCを計算した。全体%を計算するために、器官のμCを注射した全μCで割り、次いで100%を掛けた。
【0055】
結果
フェリチンのin vivo摂取:フェリチンと結合した鉄がラット中の異なる器官によって摂取され得る可能性を、放射標識59Feを含有するH又は脾臓フェリチンを成体ラットの尾静脈に注射することによって決定した。H−フェリチンからの鉄の摂取は、脳、心臓、腎臓、筋肉及び肺中で、脾臓フェリチンに関する摂取より有意に多かった(図1A)。59Feの量は、それが脾臓フェリチンと結合したときよりH−フェリチンと結合して存在したとき、脳内で2倍多かった(p<0.005)(図1B)。肝臓のみ、H−フェリチンと比較して脾臓フェリチンからの有意に多い鉄の摂取があった(p<0.05)。
【0056】
様々な器官への鉄のH及び脾臓フェリチン送達に対する、様々な器官内の鉄貯蔵能の考えられる変化の影響を決定するために、本発明者らは、H−フェリチンが欠損しているマウス系中のこれらのタンパク質からの59Feの摂取を調べた(Thompson K.J.、Fried M.G.、Zheng Y.、Boyer P.、Connor J.R.J.Cell Sci.115:2165〜2177、2002)。H−フェリチンによる鉄送達は、同腹子対照と比較してH−フェリチン障害が起きたマウスにおいて、脾臓、肺及び筋肉中で低減した(p<0.05)(図2A)。同様の発見を脳内で観察した(図2B)。脾臓フェリチンの摂取は、鉄貯蔵障害が起きたマウス中の任意の器官において不変であった(図3A及び3B)。
【0057】
フェリチンの輸送:血清中フェリチンは全身器官に無制限にアクセスして、脳に鉄を送達するのに有効であることはできたが、内皮細胞壁(BBB)を越えなければならなかった可能性がある。フェリチンのトランスサイトーシスが可能であった可能性の調査を開始するために、本発明者らはBBBの細胞培養モデルを利用した。脾臓フェリチンではなくH−フェリチンが、相当量BREC細胞単層を越えて輸送された(図4)。BREC単層を越えて輸送されたFITC標識Hフェリチンの速度は、RITC標識デキストランの5倍を超えた(p<0.001)。脾臓フェリチンの輸送速度は、デキストラン対照において見られたレベルと類似していた。それによってH−フェリチンがトランスサイトーシスされる機構を決定するために、本発明者らは、無カリウム培地において輸送アッセイを実施して、クラスリンコーティング小胞の形成を阻害した。クラスリンコーティング形成の不在は、H−フェリチンの輸送速度の80%(p<0.001)の減少と関係があった。対照的に、ピノサイトーシスを阻害するためのフィリピンを用いたBRECの前処理は、速度の有意な減少はもたらさなかった。デキストラン対照はそれぞれの実験条件中に含め、その速度は未処理条件に関するグラフ中に示した速度と変わらなかった(データ示さず)。トランスフェリンの輸送は陽性対照として含め、以前に報告されたように(Burdo J.R.、Antonetti D.A.、Wolpert E.B.、Connor J.R.Neuroscience121:883〜890、2003)検出した。蛍光標識トランスフェリンとフェリチンの比活性度は異なった。したがって、この2つのタンパク質に関する輸送の相対速度に関する結論を得ることはできない。
【0058】
フェリチン結合の分析:脳へのフェリチンによる鉄送達の機構をより完全に評価するために、結合試験を実施して、BRECモデル中のフェリチンの輸送が受容体仲介であったかどうか決定した。さらに、フェリチン結合の評価をin vivo系に広げるために、微小血管系をラットの脳から単離した(RBMV)。BREC及びRBMVとのフェリチンの結合は、飽和実験及び競合実験を使用して実施した。Kd及びBmax値は、GRAPHPAD PRISM(登録商標)4.0(GraphPad Software、Inc.)において非線形回帰を使用して2つの手法から得た。
【0059】
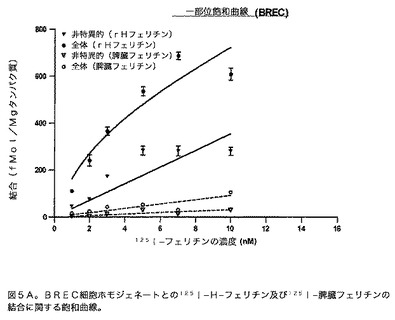
飽和曲線:様々な濃度(1、2、3、5、7、及び10nM)の125I標識フェリチン(rH−フェリチン又は脾臓フェリチン)を、100μgの組織(BREC又はRBMV)とインキュベートした。全体及び非特異的結合(1000nMの非標識フェリチンの存在下)は、Whatmannフィルターでアッセイを実施することによって得た。このような結合アッセイからKd及びBmaxを得るために、一部位結合の非線形包括的回帰を実施した。この方法では、全体と非特異的結合の両方を標識フェリチンの濃度に対してプロットした。生成したプロットは、等式:非特異的=NS×X及び全体=特異的+非特異的、前式で特異的=Bmax×X/(Kd+X)に適合させた。包括的手法では、特異的結合は全体及び非特異的結合のデータから誘導されない。代わりにKd及びBmaxの値は、2つのデータセット(全体と非特異的)間の非特異的結合定数(NS)を共有することによって得る。この回帰分析からのデータは図5A及び5B中に示す。rH−フェリチンのみが、BREC又はRBMVのいずれかと有意な飽和結合を有する。RBMVに関するKd及びBmaxは、それぞれ7.9±1.6nM及び572.6±64.0fmol/mgタンパク質である。BRECに関しては、Kdは2.7±0.9nMであり、Bmaxは465.7±63.1fmol/mgタンパク質である。曲線適合のR2値はBRECとRBMVの両方に関して0.8を超える。
【0060】
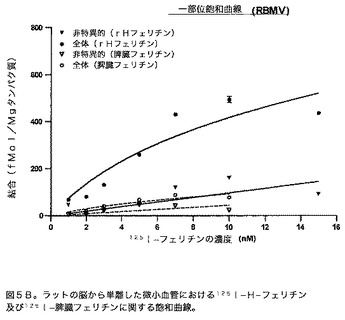
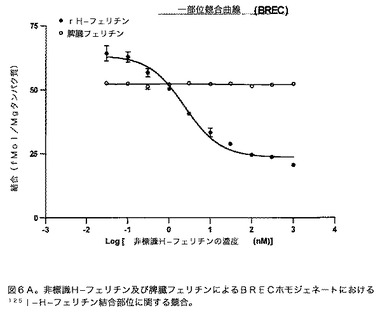
競合曲線:様々な濃度のコールドフェリチン(0.03、0.1、0.3、1、3、10、30、100、300、及び1000nM)を、100μgのBREC又はRBMV組織及び0.4nMの放射標識フェリチンとインキュベートした。全体の結合はWhatmannフィルターでアッセイを実施することによって得た。次いで全体結合(fmol/mgタンパク質)をlog[濃度(nM)]に対してプロットした。次いでこれらのプロットを、一部位競合等式:全体=底部+(上部−底部)/(1+10(X−LogEC50))に適合させた。このデータは図6A及び6B中に示す。これらの結果は、脾臓フェリチンではなくH−フェリチンが、結合部位に関して有効に競合し得ることを示す。BRECに関する競合曲線からのKd及びBmaxの決定によって、2.0nMのKd及び235.1fmol/mgタンパク質のBmaxを得た。RBMVに関する対応する値は、Kd=3.4nM及びBmax=304.6fmol/mgタンパク質である。適合のR2値はBRECとRBMVの両方に関して0.95を超える。2つの異なる曲線によって生じた値は許容範囲内にある。
【0061】
考察:この試験の結果は、脳を含めた多数の器官に、フェリチンが鉄を送達することができることを明らかにする。さらに、フェリチンにより送達される鉄の量は、肝臓以外の大部分の器官に関して、L−フェリチンの代わりにH−フェリチンにより鉄が送達されるとき増大する。細胞によって摂取されるH−フェリチン鉄の量は、H−フェリチン欠損マウス中で実証されるように、鉄貯蔵能に障害が起きると変わる可能性があるが、一方L−フェリチンによる鉄送達はこのモデル中では有意に影響を受けない。これらの後者の結果は、H−フェリチンに関するフィードバックシステムを示唆する。したがって本発明者らは、脳への鉄送達に関する新規な輸送系、及びトランスフェリン(最大2個のFe原子)と比較したフェリチンの一分子中に収容され得る鉄の量(最大4500個のFe原子)を考慮して、非常に有意であり得る輸送系を同定している。脳への非トランスフェリン依存性鉄送達系の同定は、血清トランスフェリンの不在下での脳への鉄送達を示す本発明者らの以前の報告と一致する(Malecki E.A.、Devenyi A.G.、Beard J.L.、Connor J.R.JNeurosci Res.56:113〜122、1999)。この試験は、フェリチン受容体の存在の連続的な探究も追加する。フェリチン受容体は、報告によると肝細胞において実証されているが(Mack U.、et al.JBiol.Chem258:4672〜4675、1983)、このモデル内及び脳内で飽和は実証されなかった(Hulet S.W.、Heyliger S.O.、Powers S.、Connor J.R.J.Neurosci Res.61:52〜60、2000)。近年あるタンパク質が推定フェリチン受容体として同定されている(Chen T.T.、et al.JExp Med.202:955〜65、2005)。
【0062】
脳内では、受容体との結合以外に、フェリチンはBBBを越えてトランスサイトーシスされなければならない。この試験において、本発明者らは、フェリチンは細胞培養モデルのBBBを越えて輸送することができることを実証した。この細胞培養モデル中のフェリチンの輸送はクラスリン依存性及び受容体仲介型であり、H−サブユニットを強く好む。H−フェリチンの結合に関する優先は輸送のデータと一致する。フェリチンの結合はラット脳由来の微小血管系においても実証され、且つこの結合も、細胞培養モデルと同様に、H−サブユニットを強く好む。結合及び輸送のデータは、鉄が脾臓(L多量)フェリチンと比較してH−フェリチンと結合した場合の脳への増大した送達を明らかにした、鉄摂取のデータと一致する。フェリチンの摂取、HフェリチンとLフェリチンの間の鉄送達の違い、及びH−フェリチンに障害が起きたマウス中のH−フェリチン摂取の変化は、いずれもフェリチン受容体の可能性を示唆し、H−フェリチンに関する結合は内皮細胞の2つの異なる供給源において直接実証された。
【0063】
フェリチンがBBBを越えて輸送される機構はクラスリン依存性であり、ピノサイトーシスによるものではないことを示唆するデータは、同系のトランスフェリンを使用して以前に確認されたデータと類似している(Burdo J.R.、Antonetti D.A.、Wolpert E.B.、Connor J.R.Mechanisms and regulation of transferrin and iron transport in a model blood−brain barrier system.Neurosci.121:883〜890、2003)。脳への鉄送達に関する機構及びこれらの機構の制御は、どのようにして鉄が蓄積する又は正常レベルに達し得ないか、したがって様々な神経障害の根底にあるか又はそれらに貢献するかを理解する中核となる(Zecca L、Youdim MB、Riederer P、Connor JR、Crichton RR.Nat Rev Neurosci.5:863〜873、2004)。トランスフェリンと結合した鉄のトランスサイトーシスの試験は、矛盾する結果を与えている。いくつかの試験は、脳に入る鉄はトランスフェリンと結合することを示し、且つ他の試験はトランスフェリンとは関係がない鉄輸送を示す。本発明者らは、脳への鉄送達に関するトランスフェリン依存系と非トランスフェリン依存系の両方が存在し、且つ別の経路に優る一経路の優先傾向は内皮細胞の鉄の状態に依存し得る証拠を提供している。BBBを形成する内皮細胞の鉄の状態、及びどのようにしてこの鉄の状態がトランスフェリン仲介型鉄摂取に関するトランスフェリン受容体の制御に影響を与えるかは、脳の鉄摂取の機構に関する試験において大部分は無視されている(総説に関してはBurdo and Connor 2003を参照)。細胞中のトランスフェリン受容体の発現は、細胞の細胞内鉄状態により制御されることはよく知られており(Aisen P.、et al.Int J Biochem Cell Biol.33:940〜959、2001)、内皮細胞は、他の細胞と同じトランスフェリン受容体の鉄制御機構を有する(Georgieff M.K.、et al.JPediatr.141:405〜9、2002)。脳の内皮細胞は、その機能が細胞内のエンドソームからの鉄放出を仲介することである二価金属輸送タンパク質も発現し、鉄貯蔵の存在を示す比較的多量のフェリチンを含有する。脳の内皮細胞はその独自の相当な鉄の必要性を有し、且つその独自の鉄摂取を制御するという概念は、これらの細胞中のミトコンドリアの高濃度、したがって高い鉄の必要性と一致する。したがって、トランスフェリンによってこれらの細胞に送達される少なくともいくらかの鉄は、内皮細胞内に保持されなければならない。フェリチンから鉄が放出される機構は十分に理解されておらず、ある人は、鉄を放出するためにはフェリチンの分解が必要とされると提言している。したがって、フェリチンは多量のその鉄含量体でトランスサイトーシスされる可能性がより高く、トランスフェリンより内皮細胞とその鉄を共有する可能性は低い。脳に鉄を送達するフェリチンを制御するための機構は現時点では知られていないが、H−フェリチン欠損マウスにおける鉄を送達するH−フェリチンの減少は、このような制御機構が存在することを強く示唆する。
【0064】
鉄貯蔵タンパク質としてのフェリチンの機能は十分確立しているが、H−フェリチンは細胞によって活発に分泌され、おそらく鉄を送達する可能性があるという概念は以前に試験されていない。血中のフェリチンレベルは正常条件下でさえ大幅に変動する。フェリチンmRNAは、フェリチンの分泌経路をサポートし得るラット肝細胞中の小胞体と結合するポリリボソームと結合する。フェリチン(両サブユニット)の直接分泌は分化したラット肝癌細胞において実証されており、培養中の小神経膠細胞からのL−フェリチンではなくH−フェリチンの放出が示されている(Zhang X.、Surguladze N.、Slagle−Webb B.、Cozzi A.、Connor J.R.Glia54:795〜804、2006)。
【0065】
BBB及び他の器官における受容体に関するフェリチンの供給源は、おそらく血清由来である。血清中のフェリチンの存在は十分確立しているが、血清中のフェリチンは主にL−サブユニットで構成されると従来考えられている。このLに富む血清中フェリチンの供給源は主に溶解マクロファージ由来である(McGowan S.E.、et al.J.Lab Clin.Med.111:611〜617、1988)。本発明者らは、しかしながら、フェリチンの結合、フェリチンの輸送及び脳への鉄の送達はいずれも、Hに富むフェリチンを強く好むことを、驚くべきことに示している。したがって、血清中の比較的少量のH−フェリチンに基づく本発明者らの発見の生理学的意義は、フェリチンは脳のための鉄の供給源としてトランスフェリンに続き得ることを示唆する。しかしながら、1モルのフェリチンは、1モルのトランスフェリンより2000倍を超える鉄を送達することができることは覚えておかなければならない。
【0066】
さらに、炎症などの血漿中及びいくつかの癌と関連してH−フェリチンが増大する条件が存在する(Elliott R.L.、et al.Breast Cancer Res.Treat.30:305〜309、1994)。したがって、慢性炎症状態は脳の鉄状態を増大させる可能性がある。これはアルツハイマー病及びパーキンソン病を含めたいくつかの神経変性疾患における鉄蓄積と一致する概念である。
【0067】
(例2)
導入
本発明者らは、鉄欠乏を緩和するための新規な食事療法の手法を開発している。この手法は、本発明者らの脳を含めた身体中のH−フェリチンに対する受容体の発見、及びLに富むフェリチンと比較してH−フェリチンが、肝臓以外の全器官への鉄摂取の好ましい形式であるという実証から派生した。他の人はH−フェリチンが母乳中に豊富であることを示しており、このタンパク質は、母親と幼児の間の鉄送達のための機構であることが示唆される。この後者の言及は、おそらくヒトだけでなく全ての哺乳動物にあてはまる。さらに、H−フェリチンの遺伝子配列及びタンパク質構造は動物界で高度に保存されている。したがって、鉄送達タンパク質としてのH−フェリチンの適用はヒトに限られないはずである。
【0068】
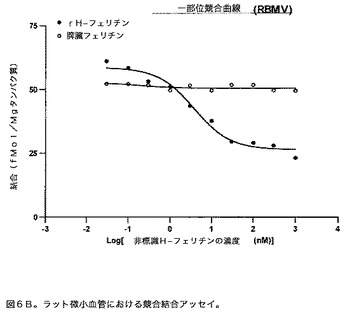
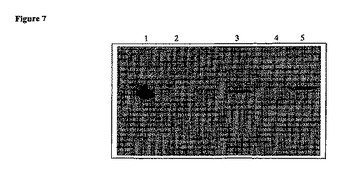
本発明者らは、酵母を使用して栄養補助食品として、H−フェリチンを送達するための機構を考案している。この手法を開始するため、第一の試験は酵母を形質転換して、H−フェリチンタンパク質に翻訳され得るヒトH−フェリチンの遺伝子を発現させることであった。図7中に示すイムノブロットは、H−フェリチンは酵母により発現され、L−フェリチンとの交差反応はないことを実証する。第二の試験は、硫化第一鉄を用いた酵母が増殖する培地の富化は、それらが発現したフェリチンの鉄含量を増大することを実証することであった。これらの結果は図8中に示す。
【0069】
供給試験
鉄欠乏を治療するためのモデルとしてのフェリチン補強酵母の有効性を試験するために、本発明者らは鉄欠乏の標準的なラットモデルを使用した。ラットの子供は25日齢で離乳するまで鉄欠乏母獣により育てられた。離乳時に、動物を5群の1つに割り当てた。群1:鉄欠乏食(ID)で維持した。群2:35mg/kgのFeSO4の標準的な鉄補充食を供給した。群3:鉄を補充しない酵母(酵母、鉄なし)を含有した食事を供給した。群4:鉄は補充したがH−フェリチンは補強しなかった酵母(酵母、フェリチンなし)を含んだ食事を供給した。群5:H−フェリチンで形質転換し鉄を補充した酵母(酵母、Fe+Ft)。離乳後3、7、9及び14日で、これらの動物から血液サンプルを回収した。これらの結果は図9中に示す。ID食を継続した動物は最低レベルのヘモグロビン(Hb)を有していた。鉄を含まない酵母を与えた動物は、ID動物と類似したHbレベルを有していた。Hbレベルの改善は他の3群において見られ、改善の最も急速な増大は、鉄を補充しフェリチンを補強した酵母を与えた動物において生じた。フェリチンを含まない鉄補充酵母を与えた動物さえ、FeSO4群より改善されたHbレベルを有していた。
【0070】
ヘマトクリット値レベルも同じ動物群においてモニターした。これらのデータは図10中に示す。これらのデータは、鉄の媒体としての酵母はH−フェリチンの存在又は不在下でヘマトクリット値を補正する際に同程度に有効であり、いずれも標準的な現在の治療選択肢、FeSO4より有意に優れていることを示す。ID食を継続した動物、及び鉄を補充しなかった酵母を与えた動物は、試験した11日間にわたってヘマトクリット値の増大を示さなかった。
【0071】
鉄を動員する身体の能力を示す重要な分析、おそらく間接的に鉄の生物学的利用能の指標は、トランスフェリンの飽和レベルである。トランスフェリンは主な鉄動員タンパク質であり、血清中に高濃度で見られる。トランスフェリン飽和の量は、正常の高い30%から貧血の状態中の10%未満まで変動する。フェリチン補強鉄多量酵母の有効性を評価するために本発明者らが使用した動物モデルにおけるトランスフェリン飽和の分析は図11中に示す。この図中では、フェリチン補強鉄多量酵母は、トランスフェリン(Tf)飽和の最大の増大、次にフェリチンを含まない鉄多量酵母をもたらしたことを見ることができる。この試験は、食事療法におけるFeSO4に優る鉄送達媒体としての酵母の優勢を再度実証する。ID食を維持した動物又は対照酵母(鉄なし又はフェリチン多量)を供給した動物は、最も低いTf飽和があった。
【0072】
脳の鉄に対する影響
鉄の欠乏は脳の発達に対して重大な影響があるので、特定脳領域の鉄の状態を、異なる食事を与えた動物においてモニターした。図12中では、脳の2つの発達上重要な領域、腹側中脳及び尾状核の鉄の状態を示す。これらの領域は、動物(例えばヒト)が成熟すると比較的鉄が豊富になる運命にある。これらの脳領域は運動活性、したがって特に欠乏が発達中に起こるときの鉄欠乏における運動技能の障害の制御に関与する。Hb及びHct分析中に記載したのと同じ動物を14日齢で屠殺し、腹側中脳(VBM)及び尾状核の鉄濃度を測定した。フェリチンを補強し鉄を補充した酵母を与えた動物は、任意の他の群より多量の鉄を2つの脳領域中に有していた。この刺激的で新規な発見は、フェリチン補強栄養補助食品は、鉄欠乏と関係がある神経障害を制限するための機構であり得ることを示す。この観察は、一般的な健康状態を高めることによってだけでなく、神経機能を最適化することによって、世界規模の鉄欠乏との闘いに対して多大な影響を有する可能性がある。
【0073】
脳内鉄状態の領域変化をさらに評価するために、脳の2つの他の領域、側坐核(NA)及び前頭前皮質(PFC)を調べた。これらの結果は図13中に示す。この図中では、フェリチン補強鉄多量酵母から送達される鉄の領域特異性は明らかである。NA中では、図5中に示したVMB及び尾状核と同様に、鉄送達の他の形態と比較して鉄含量は上昇する。しかしながら、PFC中では、フェリチンを補強し鉄を補充した酵母から送達される鉄は、他の群に関して見られるそれと同様である。これらのデータは、他の鉄送達系と異なる形式でフェリチン補強鉄補充酵母からの鉄送達を制御する機構が存在することを示唆し、本発明者らの脳内微小血管系におけるH−フェリチンに対する受容体の発見と一致する。
【0074】
概要
脳への増大した鉄摂取、及びおそらくより重要なことに、摂取の領域特異性は予想外であるが、本発明と関係がある非常に有意な発見である。これらのデータは、フェリチン補強鉄補充酵母の使用は、生後発育中の鉄欠乏性と関係がある神経、認知及び行動障害の改善をもたらすに違いないことを示唆する。血液学的パラメータに関するデータは明らかに強力であり、フェリチン補強鉄補充酵母は鉄栄養補助食品の優れた形態であることを示す。
【技術分野】
【0001】
継続出願のデータ
本出願は、いずれもそれらの全容が参照として本明細書に組み込まれる、それぞれ2007年1月29日及び2007年10月31日に出願された、米国仮出願第60/886,972号及び米国仮出願第60/984,007号の特典を主張するものである。
【背景技術】
【0002】
フェリチン、原核生物と真核生物の両方における主な細胞内鉄貯蔵タンパク質は、24ポリペプチドサブユニットを含む大きな(約480kDa)マルチサブユニット複合体である。血清中に高濃度で見られるこの鉄貯蔵複合体は、水酸化鉄コア内に4,500原子もの多くの鉄イオン(Fe3+)を含有することができる。哺乳動物では、約54%の配列同一性を共有し、それぞれ約21kDa及び19kDaの分子量を有する、2つの異なるサブユニットクラス、重鎖(H)及び軽鎖(L)型が存在する。HサブユニットとLサブユニットは異なる機能を有するようである:Lサブユニットは鉄コアの安定性を高め、一方Hサブユニットは、第一鉄イオンの迅速な摂取に必要とされるであろうフェロキシダーゼ活性を有する。Hが豊富なフェリチンは、局所イオン濃度の迅速な変化を経験する組織中に局在する。例えば、Hサブユニットの発現は、分化、発達、増殖及び代謝ストレスを経験する細胞中でLサブユニットに対して優先的に増大する。
【発明の概要】
【発明が解決しようとする課題】
【0003】
脳内微小血管系を構成する隣接内皮細胞を結合させる高度に発達した密着結合のため、脳は鉄獲得に高い難題を課す。これらの結合は、脳内への分子の傍細胞流動を妨げる。結果として生じる血液−脳関門(BBB)は、血中を循環する潜在的に有害である物質から脳を保護するのに非常に有効な機構である。しかしながらこのような封鎖の結果は、正常な脳機能に必要とされる多くの栄養物質に特異的な輸送機構を設計しなければならないことである。ピノサイトーシスはBBBを回避する可能性がある方法であるが、ピノサイトーシスから生じる小胞は、脳血管内皮細胞を越える化合物の非特異的輸送に相対的にほとんど貢献しない。
【0004】
従来、トランスフェリンは細胞の鉄送達の主要機構であると考えられており、トランスフェリン仲介の輸送系がBBB中で同定されている(Jefferies W.A.、et al.Nature 312:162〜163、1984;Fishman J.、Rubin J.、Handrahan J.、Connor J.、Fine R.J.Neurosci.Res.18:299〜304、1987)。しかしながら、トランスフェリン欠損マウスを使用して、脳へのトランスフェリン非依存性の鉄送達が示唆されている(Malecki E.A.、Cook B.M.、Devenyi A.G.、Beard J.L.and Connor J.R.J.Neurol.Sci.170:112〜118、1999)。ラクトフェリンも脳内に鉄を運ぶことができると言われているが(Ji B.、et al.Life Sci.78:851〜855、2005)、血清中のラクトフェリン濃度はほとんど検出できず、このタンパク質は細胞(好中球)内で一般に見られ、したがって脳又は他の器官への鉄輸送に貢献する可能性が乏しい。
【課題を解決するための手段】
【0005】
本発明者らは驚くことに、長い間鉄貯蔵タンパク質と考えられてきたフェリチンが、鉄送達タンパク質として働くことができることを発見している。特に本発明者らは、培養中及びラット脳内微小血管系上の内皮細胞上のH−フェリチン受容体の存在を実証しており、BBBを越えて鉄を運ぶための手段としてH−フェリチンを同定している。本発明は、患者における鉄欠乏性障害を治療するための方法であって、治療有効量のフェリチン−鉄複合体を、その必要性がある患者に投与することを含む方法を提供する。本発明の一実施形態では、フェリチン−鉄複合体はH−フェリチンを含む。別の実施形態では、鉄欠乏性障害は脳内の鉄欠乏性を含む。本発明は、脳に鉄を送達するための方法であって、フェリチン−鉄複合体の形の鉄を患者に投与することを含み、それによって、前記鉄が血液脳関門を越えて運ばれ脳に送達される方法;ターゲッティング部分としてH−フェリチンを使用するための方法であって、H−フェリチンとリポソームを結合させることを含み、それによって、前記リポソームが脳及び/又は脳内の細胞にターゲッティングされる方法;及び患者における鉄過剰症を治療するための方法であって、治療有効量のマルチサブユニットフェリチン複合体を、その必要性がある患者に投与することを含み、前記マルチサブユニットフェリチン複合体が100%未満の鉄結合能である方法も提供する。
【図面の簡単な説明】
【0006】
【図1A】48時間の循環後の器官中における、ウマ脾臓フェリチン(脾臓フェリチン)と比較した、組換えヒトHフェリチン(rH−フェリチン)由来の59Feのin vivo摂取を示す図である。59Fe標識H又は脾臓フェリチンを成体ラットの尾静脈に注射し、48時間循環させた。放射能の量は1.0グラムのそれぞれの器官において決定し、全体%/グラム組織重量は、注射した全体と比較した器官当たりのμCの量を計算することによって決定した。2つの脳構造をこのスケールで示して相対量を明らかにする。表すデータは3匹の動物の平均±S.Eである。p<0.05*。
【図1B】ラット脳内のフェリチン由来の59Feのin vivo摂取を示す図である。図1Aのレジェンド中に記載したのと同様に、59Fe標識H又は脾臓フェリチンを、尾静脈を介してラットに注射した。(脳の一半球中のμC)/(注射した全μC)×100%を決定することによって全体%を計算した。p<0.005**。表すデータは3匹の動物の平均±S.Eである。
【図2A】H−フェリチン欠損マウス(−/+)及び野生型(+/+)マウスにおける、全身器官へのH−フェリチン由来の59Fe摂取を示す図である。動物には等量のH−フェリチンを腹腔内注射した。フェリチンは48時間循環した。(脳を含む)示した器官を除去し、0.1gを基にそれぞれの器官に関して全体の放射能のパーセントを決定した。p<0.05*。
【図2B】H−フェリチンを介して送達された、H−フェリチン欠損マウス(−/+)の脳vs.野生型(+/+)マウスの脳内の59Fe摂取を示す図である。これらのデータは、図2A中のデータを作製するために使用したマウスの脳からのデータである。脳内の放射能の%は、脳から得た59Feの1分当たりの崩壊数と注射した全μCを比較することによって決定した。p<0.05*。示した結果は3匹の動物の平均±標準誤差である。
【図3A】脾臓フェリチンを介して送達された、H−フェリチン欠損(−/+)及び野生型(+/+)マウスにおける、様々な全身器官中の59Fe摂取を示す図である。マウスには59Feを含有する脾臓フェリチンを腹腔内注射した。48時間後、マウスを屠殺し器官を除去した。放射能の量は0.1gのそれぞれの器官で決定し、注射した全放射能のパーセントを決定し、このグラフ中に表した。データは3匹の動物の平均±S.Eである。いずれの差も統計学的有意に達しなかった。
【図3B】脾臓フェリチンを介して送達された、H−フェリチン欠損(−/+)vs.野生型(+/+)マウス中の脳内の59Fe摂取を示す図である。これらのデータは、図3A中で使用したマウスからのデータである。報告する放射能の量は動物に注射した全体の%である。示した結果は3匹の動物の平均±標準誤差である。
【図4】BREC培養物単層を越えるフルオレセイン標識Hフェリチンの流動を示すグラフである。このグラフは、4時間にわたる基底チャンバー中のFITC標識Hフェリチンの輸送速度を示す。輸送速度は方法セクション中に記載したのと同様に決定した。示したデータは、単位量の上部チャンバーの蛍光当たりの基底チャンバーの蛍光(Bf/Tf)対時間のプロットからの勾配(cm/s)及び共通基準として得た流動速度の平均である。デキストラン対照及び脾臓フェリチンと比較して、H−フェリチンの輸送は統計上有意である(p<0.01**)。脾臓フェリチンの輸送はデキストラン対照と異ならなかった。カリウム枯渇培地でのBRECの前処理はFITC H−フェリチン輸送の有意な低下をもたらしたが(p<0.05*)、ピノサイトーシスを最小にするためのフィリピンを用いた前処理はH−フェリチンの輸送速度に対して全く影響が無かった。トランスフェリンの輸送は陽性対照として含めた。デキストランの流動に関して示す値は未処理対照サンプルに関するものであるが、デキストランの流動速度はいかなる処理によっても変わらなかった(データ示さず)。
【図5A】BREC細胞ホモジェネートとの125I−H−フェリチン及び125I−脾臓フェリチンの結合に関する飽和曲線を示すグラフである。このグラフはBREC細胞とのH−フェリチンの結合は飽和状態であることを示すが、一方で脾臓フェリチンの結合の証拠は存在しなかった。飽和結合は2時間4℃で実施した。Kdは2.7±0.9nMであると決定され、Bmaxは465.7±63.1fmol/mgタンパク質である。
【図5B】ラットの脳から単離した微小血管における125I−H−フェリチン及び125I−脾臓フェリチンに関する飽和曲線を示す図である。これらの曲線はH−フェリチンに関する飽和結合は示すが、脾臓フェリチンに関する結合は示さない。Kdは7.9±1.6nMであり、Bmaxは572.6±64.0fmol/mgタンパク質である。
【図6A】非標識H−フェリチン及び脾臓フェリチンによる、BRECホモジェネートにおける125I−H−フェリチン結合部位に関する競合を示すグラフである。このグラフは、125I−H−フェリチンとBRECホモジェネートの結合は、脾臓フェリチンではなく、非標識H−フェリチンの濃度の増大によって競合的に阻害されることを示す。
【図6B】ラット微小血管における競合結合アッセイを示すグラフである。このグラフは、125IH−フェリチンの結合は、非標識脾臓フェリチンによってではなく、非標識H−フェリチンによって濃度依存的な様式で解離し得ることを示す。
【図7】組換え酵母中でのH−フェリチンの発現を示すウエスタンブロット(4〜20%勾配)の図である。レーン1中は標準(組換えヒトH−フェリチン)である。レーン2及び4は、ヒトL−フェリチンで形質転換した酵母の2つの異なるコロニーである。レーン3及び5は、ヒトH−フェリチンで形質転換した酵母の2つの異なるコロニーである。この試験で使用した抗体は、寛大にもPaolo Arosio(Brescia Italy)によって与えられたH−フェリチンに対するマウスモノクローナル抗体である、抗HF HS−59(1:40,000、16時間)である。
【図8】ウエスタンブロットの図である。タンパク質を最初にゲル電気泳動によって分離し、次いでゲルをパールの反応、フェリチンの鉄含量を示すために使用されている鉄に関する標準的な組織染色で染色した。レーン1及び2は、L−フェリチンで形質転換した酵母のコロニー由来である。鉄の反応生成物はこれらのレーン中では見られない。レーン3及び4は、正常な鉄培地条件(レーン4)及び鉄多量培地(レーン3)下でH−フェリチンを発現する酵母由来のタンパク質抽出物である。H−フェリチン標準は対照として使用した(レーン5)。レーン6は、フェリチンの大きさを示すための分子量マーカーを含有する。
【図9】鉄欠乏の標準的なラットモデルを使用して得た結果を示す図である。鉄欠乏(ID)食での動物は最低レベルのヘモグロビン(Hb)を有していた。鉄を含まない酵母(酵母、鉄なし)を与えた動物は、ID動物と類似したHbレベルを有していた。Hbレベルの改善は他の3群において見られ、改善の最も急速な増大は、鉄を補充しフェリチン(ft)を補強した酵母を与えた動物において生じた。フェリチンを含まない鉄補充酵母を与えた動物さえ、FeSO4群より改善されたHbレベルを有していた。
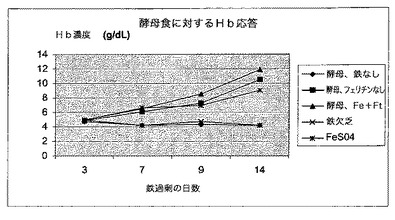
【図10】図9中で試験した動物の群からのヘマトクリット値レベルを表す図である。これらのデータは、鉄の媒体としての酵母はH−フェリチンの存在又は不在下でヘマトクリット値を補正する際に同程度に有効であり、いずれも標準的な現在の治療選択肢、FeSO4より有意に優れていることを示す。ID食を継続した動物、及び鉄を補充しなかった酵母を与えた動物は、試験した11日間にわたってヘマトクリット値の増大を示さなかった。
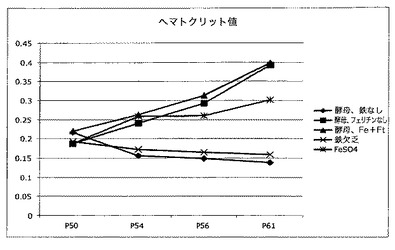
【図11】図9及び10中で試験した動物の群中の鉄動員を測定する試験の結果を示す図である。これらのデータは、フェリチン補強鉄多量酵母(ferritin fortified iron enriched yeast)(酵母−Fe+Ft)は、トランスフェリン(Tf)飽和の最大の増大、次にフェリチンを含まない鉄多量酵母(酵母、フェリチンなし)をもたらしたことを示す。
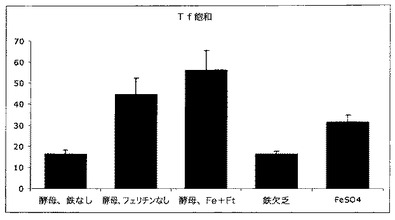
【図12】脳の2つの発達上重要な領域、腹側中脳及び尾状核の鉄の状態を示す図である。Hb及びHct分析(図9及び10)中に記載した動物は14日齢で屠殺し、腹側中脳(VBM)及び尾状核の鉄濃度を決定した。フェリチンを補強し鉄を補充した酵母(酵母−Fe+Ft)を与えた動物は、任意の他の群より多量の鉄を2つの脳領域中に有していた。
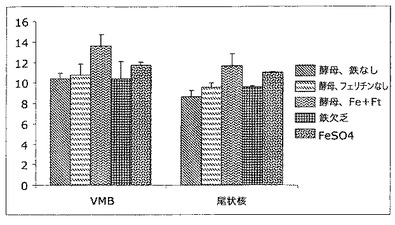
【図13】脳の側坐核(NA)及び前頭前皮質(PFC)領域中の鉄レベルを表す図である。この図中では、フェリチンを補強し鉄を補充した酵母(酵母−Fe+Ft)から送達される鉄の領域特異性は明らかである。(図12中に示したVMB及び尾状核と同様に)NA中では、鉄送達の他の形態と比較して鉄含量は上昇する。しかしながら、PFC中では、フェリチンを補強し鉄を補充した酵母から送達される鉄は、他の群に関して見られるそれと同様である。
【図14A】ヒトH−フェリチンのアミノ酸配列(配列番号1)を示す図である。
【図14B】ヒトH−フェリチンのcDNA配列(配列番号2)を示す図である。開始(ATG)及び停止(TAA)コドンは太字であり、(配列の5’末端における)BamHI及び(配列の3’末端における)XhoI制限部位には下線を引く。
【発明を実施するための形態】
【0007】
本発明は、患者における鉄欠乏性障害を治療するための方法であって、治療有効量のフェリチン−鉄複合体を、その必要性がある患者に投与することを含む方法を提供する。
【0008】
本明細書で使用する「鉄欠乏性障害」は、鉄欠乏、鉄摂取、及び/又は鉄代謝と関係がある障害又は疾患を含む。鉄欠乏性障害の例には、不十分な食事摂取又は鉄の吸収によって引き起こされる鉄欠乏性貧血などの、鉄欠乏性貧血がある。鉄欠乏性貧血は、例えば、栄養失調、妊娠(産後期含む)、重度の子宮出血、慢性疾患(慢性腎疾患含む)、癌、腎臓透析、胃バイパス、多発性硬化症、糖尿病(例えば、I型及びII型糖尿病)、インスリン耐性、及び注意力欠如障害と関係がある可能性がある。
【0009】
本発明の一実施形態では、鉄欠乏性障害は、パーキンソン病、アルツハイマー病、下肢静止不能症候群(RLS)、女性における貧血と関係がある最適以下の認識能力、鬱病及び不眠症を含めた様々な神経及び神経変性疾患において生じるような、脳内の欠乏した鉄レベルと関係がある。別の実施形態では、鉄欠乏性障害は、乏しい認識能力及び運動障害をもたらす発育中の鉄欠乏が原因である低髄鞘形成及び遅い脳の発達を含めた、生後発育中の脳内鉄欠乏性と関係がある神経障害(単数又は複数)を含む。
【0010】
脳内の鉄の欠如及び低下したドーパミン合成は、例えば、子供のRLS及び発育中の鉄欠乏などの、鉄欠乏性障害の重要な要因である。ドーパミンは、適切な中枢神経系(CNS)機能に必要不可欠である、脳内で合成される神経伝達物質である。ドーパミンの合成において、鉄は酵素チロシンヒドロキシラーゼの補因子であり、これはドーパミン代謝の律速段階である(Cooper et al.(1991)The biochemical basis of neuropharmacology.Oxford University Press、New York、N.Y.)。ドーパミン作動系中の鉄は、注意欠陥多動性障害(ADHD)を含めた子供のRLS病態生理及び行動障害における重要な要素であるようである。RLS患者は、RLSと対照の両方におけるフェリチン及びトランスフェリンの正常血清中レベルにもかかわらず、65%未満の脳脊髄液(CSF)フェリチン及び3倍多いCSFトランスフェリン(鉄輸送血中タンパク質)を有する。鉄濃度は、脳、ドーパミン合成部位全体で変わり、RLS患者は、脳の黒質及び被殻部分において少量の鉄を有する。一般に、低下したフェリチンレベルはRLSの重度を示す。ADHDである子供における低下した血清中フェリチンレベルの報告も存在する。
【0011】
用語「フェリチン−鉄複合体」は、多数のフェリチンサブユニット及び鉄原子を含むタンパク質複合体を指す。適切なフェリチン−鉄複合体は、哺乳動物のH−フェリチンサブユニットを含む。様々な哺乳動物種由来のH−フェリチンサブユニットのアミノ酸配列が同定されている。例えば、Orino Koichi et al.、Sequence analysis offeline ferritin H and L subunit cDNAs;Veterinary Biochem.42:7〜11(2005);アクセッション番号:06A006486を参照。一実施形態では、H−フェリチンはヒトのH−フェリチンである(配列番号1;図14A参照)。H−フェリチンは、ヒトのH−フェリチンの天然に存在する又は合成ホモログ又は変異体であってもよい。特定の実施形態では、H−フェリチンのホモログは、ヒトのH−フェリチンと少なくとも80%、81%、82%、83%、84%、85%、86%、87%、88%、89%、90%、91%、92%、93%、94%、95%、96%、97%、98%、又は99%の配列同一性などの、ヒトのH−フェリチンと約80%〜約100%の配列同一性を有する。H−フェリチンのホモログは、鉄と結合しマルチサブユニットフェリチン−鉄複合体を形成する能力を保持しているが、突然変異させて鉄とフェリチンの間の様々な結合及び解離強度をもたらすことが可能である。フェリチン−鉄複合体はH−フェリチンサブユニットを含むが、いくつかのL−フェリチンサブユニットも含むことができる。特定の実施形態では、複合体のフェリチンサブユニット成分は、L−フェリチンと比較して約20%、30%、40%、50%、60%、70%、80%、90%、又は100%のH−フェリチンなどの、L−フェリチンと比較して少なくとも20%のH−フェリチンを含む。一実施形態では、フェリチン−鉄複合体中の全てのフェリチンサブユニット(即ち、100%のフェリチンサブユニット)がH−フェリチンである。H−フェリチンは組換えH−フェリチンであってよい。例えばH−フェリチンは、適切な酵母プロモーターの制御下でH−フェリチンをコードするポリヌクレオチド配列を含む酵母菌株によって生成される、ヒトH−フェリチン、又はそのホモログであってよい。
【0012】
フェリチン−鉄複合体中の鉄は鉄分子であってよく、又は鉄含有複合体の形であってよい。「鉄含有複合体」又は「鉄複合体」は、有機化合物と複合体形成する、(II)又は(III)酸化状態の鉄を含有する化合物である。鉄複合体は、鉄ポリマー複合体、鉄炭水化物複合体、及び鉄アミノグリコサン複合体を含む。これらの複合体は市販されており、及び/又は当技術分野で知られている方法により合成することができる。
【0013】
鉄炭水化物複合体の例には、カルボキシマルトース鉄、スクロース鉄、ポリイソマルトース鉄(デキストラン鉄)、ポリマルトース鉄(デキストリン鉄)、グルコン酸鉄、ソルビタール鉄、及び水素化デキストラン鉄などの、鉄糖複合体、鉄オリゴ糖複合体、及び鉄多糖複合体があり、これらは、ソルビトール、クエン酸及びグルコン酸(例えば、デキストリン鉄−ソルビトール−クエン酸複合体及びスクロース鉄−グルコン酸複合体)などの他の化合物、及びこれらの混合物とさらに複合体形成することができる。
【0014】
鉄アミノグリコサン複合体の例には、コンドロイチン硫酸鉄、デルマチン硫酸鉄、ケラタン硫酸鉄があり、これらは、他の化合物及びそれらの混合物とさらに複合体形成することができる。鉄ポリマー複合体の例には、鉄ヒアルロン酸、鉄タンパク質複合体、及びそれらの混合物がある。
【0015】
本明細書で使用する「治療」又は「治療する」は、患者又は対象がそのために治療されている状態の完全な除去、及び任意の臨床的又は定量的に測定可能な低減を指す。「治療」は、障害の発生の予防又は障害の病状若しくは症状の改変の意図で実施される介入である。したがって「治療」は、療法治療と予防策若しくは予防対策の両方を指す。「治療」は苦痛緩和治療として記すこともできる。治療の必要がある人は、1つ又は複数の鉄欠乏性障害を既に有する人、及びその障害を予防すべきである人を含む。
【0016】
その必要性がある患者を治療する際に、治療有効量の本発明のフェリチン−鉄組成物を本発明に従い患者に投与する。本明細書で使用する用語「治療有効量」は、重大な悪影響を引き起こす可能性がある量を超えない治療に関して示される組成物の量である。療法治療の有効性を評価するための方法は当業者に知られている。
【0017】
「その必要性がある患者」は、本発明の治療法から恩恵を被る可能性がある任意の患者又は対象を指す。特定の実施形態では、その必要性がある患者は、1つ又は複数の鉄欠乏性障害を発生しやすい患者、1つ又は複数の鉄欠乏性障害を有するがいかなる臨床症状も示さない対象、又は1つ又は複数の鉄欠乏性障害を有し1つ又は複数の鉄欠乏性障害の症状に罹患している対象である。その必要性がある患者は、ヒト、イヌ、ネコ、ウシ、ウマ、げっ歯類(マウス、ラット、又はハムスターなど)、又は霊長類などの哺乳動物であってよい。一実施形態では、患者はヒトである。特定の実施形態では、本発明の方法は、実験動物、獣医学的用途、及び/又は疾患に関する動物モデルの開発において用途を見出す。
【0018】
投与する用量は、治療期間、投与の頻度、宿主、並びに障害の性質及び重度に応じた変数である。用量は過剰な量の実験なしで当業者によって決定することができる。本発明の組成物は、患者へのフェリチン−鉄複合体の十分な用量単位の放出を確実にするのに十分な用量濃度で投与して、鉄欠乏性障害の望ましい治療をもたらす。投与する実際の用量は、年齢、体重、状態の重度、及び/又は患者の臨床歴などの、物理的及び生理的要因によって決定され得る。活性成分を投与して、約50μM〜約1000μMのフェリチン−鉄複合体のin vivo血漿濃度を得ることができる。例えば、本発明の方法は組成物を使用して、約0.1〜約1,000又は約1〜約100mg/体重1kg/1日のフェリチン−鉄複合体、約30mg/体重1kg/1日のフェリチン−鉄複合体などをもたらすことができる。しかしながら、提供したこれらの範囲から逸脱する用量レベルも、所与の障害の治療において適している可能性があることは理解されよう。
【0019】
本発明のフェリチン−鉄複合体は、投与に適した任意の形であってよい。このような投与可能な形には、錠剤、緩衝錠剤、ピル、カプセル、腸溶性カプセル、糖衣錠、カシェ剤、粉末、顆粒、エアロゾル、リポソーム、坐薬、クリーム、ローション、軟膏、皮膚用パッチ剤、非経口剤、トローチ剤、懸濁液、溶液及びエマルジョン(水中油型又は油中水型)などの経口液体、眼科液及び注射液、又はこれらの徐放形がある。望ましい用量は連続注入によって、又は徐放性製剤によって、1日を通して一定間隔においていくらかの増分で与えることができ、又はボーラス、舐剤又はペーストとして表すことができる。
【0020】
一実施形態では、フェリチン−鉄複合体を含む医薬組成物又は製剤を、1つ又は複数の薬剤として許容される担体との混合によって調製する。いくつかの場合、フェリチン−鉄複合体は、フェリチン−鉄複合体及びその中に鉄分子とフェリチン分子が溶けたバッファーを含む組成物として送達して、鉄とフェリチンの結合(即ち、フェリチン−鉄複合体の形成)を可能にすることができる。しかしながら、望む場合、他の生成物を加えて、鉄の送達、保存を最大にすること、又は特定の送達法を最適化することができる。さらに本発明は、鉄欠乏性障害の治療に適した他の作用物質と組合せた、本明細書に記載するフェリチン−鉄複合体を含む組合せ組成物の使用を含む。
【0021】
本明細書で使用する「薬剤として許容される」は、医薬及び獣医学分野における使用に許容され、製剤の他の成分と適合性があり、且つ毒性ではない又は他の場合妥当なベネフィット/リスク比に応じて許容不能であることを意味する。「薬剤として許容される担体」又は「希釈剤」は、フェリチン−鉄複合体を含む組成物の医薬投与と適合性がある、任意及び全ての溶媒、分散媒、コーティング、抗菌剤及び抗真菌剤、等張剤及び吸収遅延剤などを含む。このような担体又は希釈剤の例には、水、生理食塩水、フィンガーの溶液及びデキストロース溶液があるが、これらだけには限られない。医薬組成物の体積は、目的とする投与の形態、及び医療専門家によって決定される個々の患者に安全な体積に基づく。
【0022】
担体の選択も目的とする投与の形態に依存する。本発明の組成物は、全身投与(例えば、静脈内注射、腹腔内注射、吸入、経皮送達、経口送達、鼻腔送達、直腸送達など)及び/又は局部投与(例えば、標的組織への直接注射、カニューレを介した組織への送達、徐放性物質の注入による標的組織への送達)、ポンプなど、経口、非経口、骨内、脳脊髄液中などによる組織への送達だけには限られないが、これらを含めたいくつかの好都合な手段のいずれかによって投与することができる。さらなる投与の形態には、頬、舌下、膣内、皮下、筋肉内、又は皮内投与がある。
【0023】
一実施形態では、経口投与する組成物を、経口摂取用にフェリチン−鉄複合体と適切に組合せた物質を使用して調製する。このような物質には、非制限的に、ラクトース(水和、高速流)、グルコース及びスクロースなどの糖、コーンスターチ及びポテトスターチなどのスターチ、微結晶性セルロース、ナトリウムカルボキシメチルセルロース、エチルセルロース及び酢酸セルロースを含めたセルロース及びその誘導体、粉末状トラガカント、コロイド状二酸化ケイ素、クロスカルメロースナトリウム、麦芽、ゼラチン、タルク、ステアリン酸、ステアリン酸マグネシウム、硫酸カルシウム、ピーナッツ油、綿実油、ゴマ油、オリーブ油及びコーン油などの植物油、プロピレングリコール、グリセリン、ソルビトール、マンニトール及びポリエチレングリコールなどのポリオール、寒天、アルギン酸、水酸化アルミニウム又は水酸化マグネシウムなどの制酸剤、クエン酸ナトリウム、酢酸ナトリウム、又は重炭酸ナトリウムなどのバッファー、水、等張生理食塩水及びリン酸バッファー溶液がある。湿潤剤、ラウリル硫酸ナトリウムなどの潤滑剤、安定剤、錠剤、抗酸化剤、防腐剤、着色剤及び香味剤も存在してよい。
【0024】
非経口投与に適した組成物又は製剤には、抗酸化剤、バッファー、静菌剤又は製剤を血液と等張にする溶質を含有することができる、水性及び非水性、等張滅菌注射溶液、並びに懸濁剤及び増粘剤を含むことができる水性及び非水性滅菌懸濁液がある。組成物は単位用量又は多用量の密閉容器、例えばアンプル及びバイアル中に存在してよく、使用直前に滅菌液状担体、例えば注射用水の添加のみを必要とする、凍結−乾燥(凍結乾燥)状態で保存することができる。即席の注射溶液及び懸濁液は、滅菌粉末、顆粒又は錠剤などから調製することができる。
【0025】
静脈内投与に適した組成物又は製剤は、生理食塩水、静菌水、CREMOPHOR ELTM(BASF;Parsippany、N.J.)又はリン酸緩衝生理食塩水(PBS)などの担体を含む。組成物は滅菌状態でなければならず、シリンジを使用して投与するために流体でなければならない。このような組成物は製造及び保存中は安定状態でなければならず、細菌及び真菌などの微生物からの汚染に対して守らなければならない。担体は、例えば水、ポリオール(グリセロール、プロピレングリコール、及び液状ポリエチレングリコールなど)、及び他の適合性のある、適切な混合物を含有する分散媒であってよい。様々な抗菌剤及び抗真菌剤、例えばパラベン、クロロブタノール、フェノール、アスコルビン酸、及びチメロサールは、微生物の汚染を含有する可能性がある。例えば糖、ポリアルコール、マンニトール、ソルビトール、及び塩化ナトリウムなどの等張剤を組成物中に含めることができる。吸収を遅延することができる組成物には、モノステアリン酸アルミニウム及びゼラチンなどの作用物質がある。滅菌注射溶液の調製用の滅菌固体の調製法は、フェリチン−鉄複合体及び任意の他の望ましい成分を含有する固体を生成するための真空乾燥及び凍結−乾燥を含む。
【0026】
全身投与は、例えば経粘膜又は経皮であってよい。経粘膜又は経皮投与のために、標的障壁(単数又は複数)に浸透することができる浸透剤を選択する。経粘膜浸透剤は、洗浄剤、胆汁塩、及びフシジン酸誘導体を含む。鼻腔スプレー又は坐薬は経粘膜投与に使用することができる。経皮投与のために、活性化合物を軟膏、軟膏剤、ゲル、又はクリームに配合する。
【0027】
フェリチン−鉄複合体を含む組成物は、インプラント及びマイクロカプセル化送達系を含めた放出制御製剤などの、身体からの迅速な除去に対して複合体を保護する担体を用いて調製することができる。エチレン酢酸ビニル、多無水物、ポリグリコール酸、コラーゲン、ポリオルトエステル、及びポリ乳酸などの、生分解性又は生体適合性ポリマーを使用することができる。このような物質は、ALZA Corporation(Mountain View、Calif.)及びNOVA Pharmaceuticals、Inc.(Lake Elsinore、Calif.)から購入することができ、或いは当業者により調製することができる。
【0028】
眼部投与用の組成物は、緩衝又は生理食塩水、リポソーム又は塩基性アミノ酸などの適切な液状担体で調製する。クリーム、ローション及び軟膏は、トリグリセリド基剤、リポソーム、又は塩基性アミノ酸などの適切な基剤を使用して局所施用に調製することができる。このようなクリーム、ローション及び軟膏は、表面活性剤も含有することができる。
【0029】
本発明の一実施形態では、フェリチン−鉄複合体を、H−フェリチンを発現する組換え酵母の菌株の形で投与する。鉄の栄養補充に適した組換え酵母の菌株は、ヒトを含めた哺乳動物に対する、高い生物学的利用能を有する形で鉄を貯蔵することができる。酵母の菌株は、ヒトの消費に関して一般に安全と認められる(GRAS)要件に見合うものを含む。この実施形態では、鉄貯蔵遺伝子(例えば、H−フェリチンコード配列)を、鉄貯蔵発現カセット中で適切な酵母プロモーターの制御下に置いて、鉄補充に適した媒体として働く酵母に必要な鉄貯蔵タンパク質のレベルをもたらすことができる。適切なプロモーターは当技術分野で知られており、高レベルの構成的発現を誘導するプロモーター、及びその発現が環境条件によって制御され得るプロモーターを含む。さらに、酵母の遺伝子構成をさらに操作して、様々なおそらく有利である結果を得ることができる。例えば、タンパク質分解を操作して、細胞表面、液胞、又はミトコンドリアのものだけには限られないがこれらを含めた、鉄貯蔵タンパク質又は鉄輸送機構の安定性を高めることができる。さらに、酵母を改変して、鉄貯蔵タンパク質又は細胞内コンパートメント中の鉄のレベルを高めることができる。酵母の鉄含有量は、酵母が増殖する培地に既知量の鉄化合物を加えることにより制御することができる。組換え酵母を使用して、ヒト又は他の動物のための鉄補充を、栄養サプリメントとしての組換え酵母の消費、又は組換え酵母から精製又は単離したフェリチン−鉄複合体の消費だけには限られないが、これらを含めたいくつかの手段のいずれかによって実施することができる。酵母は鉄補充の目的で特異的に増殖させることが可能であり、或いは酵母は別のプロセス(例えば発酵生産)の副産物であってよい。
【0030】
或いは、又は追加的に、フェリチン−鉄複合体は、大腸菌、バキュロウイルス及びトランスジェニック動物を含めた、当技術分野で知られている他の発現系中で生成されるH−フェリチンを含むことができる。一実施形態では、フェリチン−鉄複合体を、適切なバッファー中でフェリチンサブユニット及び鉄分子をインキュベートすること、次に生成したフェリチン−鉄複合体から任意の非結合鉄分子を分離することによって形成することができる。
【0031】
特定の実施形態では、フェリチン−鉄複合体は、抗体、アプタマー、受容体、リガンド、又はそれらの結合断片などのターゲッティング部分をさらに含む。ターゲッティング部分は1つ又は複数の細胞、組織及び/又は器官特異的マーカーを認識することができ、したがって身体中の所望の標的又は位置への送達を仲介又は改善することができる。一実施形態では、フェリチン−鉄複合体は、ターゲッティングペプチドと融合した、H−フェリチンなどのフェリチンサブユニットを含む融合タンパク質を含むことができる。別の実施形態では、フェリチン−鉄複合体は、カプセル化した状態でリポソーム、リポソーム構築体、又は赤血球ゴーストなどの他の膜結合小胞に送達又は投与することができる。リポソーム、リポソーム構築体、又は他の小胞は、リポソーム又は小胞に取り込まれた特定の細胞表面タンパク質又は受容体(上記参照)に特異的な抗体又はリガンドなどの、ターゲッティング部分を含むことができる。ターゲッティング部分は、フェリチン−鉄複合体、又はフェリチン−鉄複合体を含む小胞を、脳及び/又は血液脳関門に向けることができる。適切なターゲッティング部分の例には、トランスフェリン、インターロイキン−13(星状細胞腫への送達用)、及びリポ多糖(LPS)がある。
【0032】
本発明の別の実施形態では、H−フェリチン自体を使用して他の部分をターゲッティングすることができる。例えば、H−フェリチンを生物活性がある作用物質と結合させて、その作用物質を脳に送達することが可能である。一実施形態では、H−フェリチンペプチドを別の生物活性があるペプチドと融合させる。或いは、又は追加的に、H−フェリチンをリポソーム又は他の小胞と結合させて、小胞及び小胞含有物を脳に送達することが可能である。さらに別の実施形態では、H−フェリチンを造影剤化合物などの作用物質に結合させて、脳(例えば、白質路及びその欠陥)の可視状態を向上させることが可能である。生物活性がある作用物質と同様に、造影剤化合物をH−フェリチンタンパク質又はその中に結合させること、或いは受容体リガンドとして作用するリポソーム結合H−フェリチンを介して脳内のH−フェリチン受容体に向けられるリポソーム又は他の小胞内にカプセル化することが可能である。
【0033】
本発明の別の実施形態では、H−フェリチンを使用して余分な鉄又は鉄過剰症を治療することができる。臨床上ヘモクロマトーシスとして知られる鉄過剰症は、例えば、癌の増大したリスク、心不全、肝機能障害及び糖尿病と関係がある。脳内の鉄過剰症は、パーキンソン病、アルツハイマー病、筋萎縮性側索硬化症、Hallovordeen−Spatz、及びハンチントン病などの、広範囲の神経変性疾患において生じる可能性がある。H−フェリチンの有意な鉄結合能のために、H−フェリチンを含むH−フェリチン及び/又はマルチサブユニット複合体は鉄キレート剤として使用することができる。「マルチサブユニットフェリチン複合体」によって、多数のフェリチンサブユニット及び場合によっては鉄を含むタンパク質複合体を意味する。H−フェリチンは、前に定義した哺乳動物のフェリチン又はそのホモログ若しくは変異体である。マルチサブユニットフェリチン複合体は、生成する複合体がその全体的鉄結合能が100%未満であるように、比較的鉄を含まない環境中で調製することができる。特定の実施形態では、複合体は、約50%、40%、30%、20%、10%、5%、又は1%の鉄結合能などの50%以下の鉄結合能である。一実施形態では、複合体は約0%の鉄結合能であり得る(即ち、100%又は100%に近い鉄結合能が残存する鉄を含まないフェリチン又はアポフェリチン)。H−フェリチンを修飾して、それが任意の内因性受容体によって認識される可能性を低減すること、又は身体による排泄を増大することができる。このような修飾は十分に当業者の専門知識内にあり、これらの修飾は鉄と結合するタンパク質の能力に必ずしも影響を与えない。鉄キレート剤として使用するのに適したマルチサブユニットフェリチン複合体はH−フェリチンサブユニットを含むが、いくつかのL−フェリチンサブユニットも含むことができる。特定の実施形態では、マルチサブユニットフェリチン複合体は、L−フェリチンと比較して約20%、30%、40%、50%、60%、70%、80%、90%、又は100%のH−フェリチンなどの、L−フェリチンと比較して少なくとも20%のH−フェリチンを含む。
【0034】
鉄キレート剤として使用するためのマルチサブユニットフェリチン複合体の送達は、鉄欠乏性障害を治療するためのフェリチン−鉄複合体の送達に関して本出願中の他の箇所に記載したように、同じ範囲の送達機構(即ち、ターゲッティング部分及び/又はリポソーム又は他の小胞)を含み得る。マルチサブユニットフェリチン複合体は、胃腸管、胃腸管内壁の細胞内(そこでそれは摂取され得るが、血流には入らない)、又は血液自体に送達することができ、そこでそれは非トランスフェリン及びおそらくさらにトランスフェリンと結合した鉄と有効に競合し得る。この手法を利用して、他の場合無秩序な形式で不適切に脳にアクセスし得る鉄が除去され得る。さらに、血液及び全身器官から鉄をキレートすることは、脳からの鉄の放出を含めて、身体内の鉄の再分布を促進し得る。いくつかの場合、全体的鉄結合能が100%未満であるアポフェリチン又はフェリチン複合体を、脳脊髄液に直接送達して脳からの鉄の放出を刺激することができる。
【0035】
フェリチンは鉄だけでなく、その多くが身体に対して毒性である一定範囲の金属とも結合する能力を有する。したがって、本発明の別の実施形態は、一般に胃腸系、血液、脳及び身体からおそらく毒性である金属を低減及び除去するための、アポフェリチン(又は全体的鉄結合能が100%未満である他のマルチサブユニットフェリチン複合体)の使用に関する。したがって、血液用の一般的なメタルクレンザーとしてアポフェリチンを利用することが考えられる。
【0036】
輸血において、少なくとも15%の細胞が輸血過程中で溶解し、おそらく損傷をもたらす遊離鉄が放出する。輸血は、赤血球(RBC)が必要でありしたがって貧血である患者に実施する。本発明は、アポフェリチン(又は全体的鉄結合能が100%未満である他のマルチサブユニットフェリチン複合体)と輸血液を混合する方法に関するものであり、これは細胞溶解により放出される鉄キレート剤をもたらすだけでなく、その鉄をより生理的又は生物学的に利用可能な形で身体に利用可能にするようにも働く。頻繁な輸血を必要とする1つの患者集団にはサラセミア(thalessima)を有する患者がある。この集団は最終的には、過剰な鉄蓄積による肝臓障害に罹患する。H−フェリチン及び/又はH−フェリチンを含むマルチサブユニットフェリチン複合体は、身体中に鉄がより有効に分布するのを手助けし、したがって肝臓中の過剰な蓄積を制限することができる。頻繁な輸血を受ける別の集団は、新生児及び特に未熟児である。H−フェリチン、H−フェリチンを含むマルチサブユニットフェリチン複合体、及び/又はフェリチン−鉄複合体を輸血液に与えると、脳発達に関する脳への鉄分布を含めた鉄分布を改善し得る。
【0037】
H−フェリチンサブユニットを含めたフェリチンサブユニットは、患者の身体中で発現される可能性がある。この実施形態では、フェリチン−鉄複合体はin vivoで形成する。いくつかのプラスミド担体及びトランスフェクション試薬系をex vivoで細胞をトランスフェクトするのに利用して、宿主動物又は患者に再注入するための、安定した形質転換体又は一時的にトランスフェクトされた細胞のいずれかを生成する。適切な発現プラスミドはトランスフェクション試薬と同様に市販されており、後者の多くは1タイプ又は別タイプのカチオン性リポソームである。in vivo及びex vivo遺伝子導入、遺伝子療法などに適したベクターは当技術分野で知られており、レトロウイルスベクター、アデノウイルスベクター、アデノ随伴ウイルスベクター、レンチウイルスベクター、及びエレクトロポレーション系を含む。
【0038】
本発明の他の態様及び利点は、以下の例示的な実施例を考慮することにより理解されよう。これらの実施例は、決して本発明の範囲を制限するものとして解釈すべきではない。当業者は、本開示を鑑みて、開示する具体的な実施形態に変更を施すことができ、さらに本発明の精神及び範囲から逸脱せずに同様又は類似の結果を得ることができることを理解するはずである。
【0039】
本出願中に引用する全ての参照文献、刊行物及び特許文書は、参照として本明細書に組み込まれる。本文書中の様々な参照文献のそれらの引用によって、本出願人は、いかなる特定の参照文献も本発明に対する「従来技術」であることを認めない。
【実施例】
【0040】
(例1)
フェリチンの調製
この実施例中の全ての実験は、組換えヒトH−フェリチン又はウマ脾臓フェリチンを使用した。組換えヒトH−フェリチンは、Hisタグ組換えヒトHフェリチンプラスミドを用いて、化学的に形質転換受容性にされているBr21細胞を形質転換することによって調製した。細胞が増殖した後、2.8mg/mlの最終濃度まで、タンパク質をニッケルタンパク質濾過カラムで精製した。ウマ脾臓フェリチンは市販的に入手し(Sigma)、それが約90:10のLとHフェリチンサブユニットを含有するので選択した。
【0041】
細胞培養及び内皮細胞単層の調製
本発明者らは、フェリチンは内皮細胞の層を越えて輸送され得るという仮説を試験し、BBBを越えたフェリチン輸送の機構に取り組み始めるために、血液脳関門(BBB)のin vitroモデルとしてウシ網膜内皮細胞(BREC)を使用した。この十分研究されたモデルは、血液神経関門の全ての必要な特性及び属性を有することが示されている(Antonetti D.A.、et al.J Neurochem.80:667〜677、2002)。ウシの眼は地域の食肉処理場から得て、ウシ網膜内皮細胞(BREC)を単離し、以前に公開された手順(Gardiner T.A.、et al.Lab Invest.72:439〜444、1995)に従って処理した。BRECは、10%のFBS、10ng/mLのEGF、0.2mg/mLのENDO GRO(商標)(VEC Technologies、Inc.、Rensselaer、NY、USA)、0.09mg/mlのヘパリン、抗生物質/抗真菌溶液(Gibco、Rockville、MD、USA)、及びTylosin抗生物質(Sigma)を補充したMCDB−131培地(Sigma、St.Louis、MO、USA)中で増殖させた。それらが少なくとも80%集密状態に達するまで、細胞は最初はフラスコ内で培養する。その後、BRECSを軽くトリプシン処理し、COSTAR(登録商標)TRANSWELL(商標)0.4μm多孔質フィルター(Coming、Acton MA)で集密状態に増殖させた。フィブロネクチンを1μg/cm2の濃度で加えてフィルターへの接着を促進した。次いで細胞を洗浄し、72時間100nmのヒドロコルチゾンを補充した無血清無EGFMCDB−131培地に移した。これらの細胞培養物へのヒドロコルチゾンの添加は、密着結合の形成を促進した。
【0042】
BBBモデルにおけるフェリチン輸送
予め精製されたトランスフェリンを購入し、2.5mg/mlの最終濃度に再懸濁した。約250μgの組換えヒトHフェリチン、ウマ脾臓フェリチン(Sigma)及びトランスフェリンを、100mMの炭酸塩/重炭酸塩バッファー、pH9.0中においてフルオレセインイソチオシアネート(FITC)(Pierce Biotechnology)で標識した。過剰又は加水分解FITCの除去は、5mlのG−25脱塩カラムを通すことによって行なった。FITC結合H−フェリチン、脾臓フェリチン、及びトランスフェリンを濃縮し、バッファーはCENTRIPREP(登録商標)濃縮器(Amicon、Inc.、10,000MWCO)中でPBSと交換した。トランスフェリンは、BRECモデルにおける陽性輸送対照として含めた(Burdo J.R.、et al.Neuroscience121:883〜890、2003)。
【0043】
集密状態のBREC単層を越える流動速度は、等式:(Bf/Tf)*(Vb/A)=(フラックス)*tを使用してある程度の修正を加えて、Burdo et al.、2003中で以前に記載されたように決定した。簡単に言うと、140μgのFITC標識rH−フェリチン、脾臓フェリチン、又はトランスフェリンのいずれかを上部チャンバー(先端)に加える前に、トランスウエル装置中で集密状態までBRECを増殖させた。トレーサーの輸送は、先端チャンバーへのトレーサーの添加後の様々な時間(15、30、45、60、120、180、及び240分)で回収した下部チャンバー(基底)由来の100μlアリコートからサンプリングすることによって決定する。基底チャンバー由来のアリコートを、蛍光光度計(SPECTRAMAX(登録商標)GEMINI、Molecular Devices)において次いで蛍光に関して分析する。流動速度は、単位量の上部チャンバーの蛍光当たりの基底チャンバーの蛍光(Bf/Tf)対時間(t)のプロットからの勾配(cm/s)として得る。ここで、単層を越えて輸送されるトレーサーの量を示すBfを、基底チャンバーの体積(Vb)及びさらに輸送に利用可能な表面積(A)に標準化する。上部チャンバー中のTfの濃度は実験を実施した4時間にわたって著しく変わることはなく、したがって流動速度の計算に関して一定であると考えられる。上部チャンバー中の蛍光の量は、輸送アッセイ(4時間)の最後に100μlアリコートから得る。
【0044】
傍細胞流動の対照として、RITCデキストラン(70kDa)を、対照として先端チャンバーに同時に加えた。デキストランは内皮細胞によって適切なレベルで取り込まれない(Raub T.J.、et al.J.Cell Physiol.149:141〜151、1991)。したがって、基底チャンバー中のデキストランの任意の蓄積は傍細胞輸送によるものである可能性がある。いずれの条件も、それぞれの条件で最小であったデキストランの流動速度に影響を与えなかった。
【0045】
輸送機構の決定
ピノサイトーシスがフェリチンの輸送に有意に貢献するかどうかを決定するために、フェリチン及びデキストランを加える前に、50μg/mlのフィリピンを30分間トランスウエル装置の先端チャンバーに加えた。フィリピンの添加は、ピノサイトーシスを介した非特異的輸送の作用を阻害することが示されている(Stremmel W、et al.Lipids36:981〜989、2001)。H−フェリチンの摂取がクラスリン依存性エンドサイトーシスによって起こるかどうかを決定するために、カリウム欠損培地(100mMのNaCl/50mMのHEPES)中で試験を実施した。フェリチンを加える前に、細胞は10分間カリウム欠損培地中でインキュベートした。細胞内のカリウム枯渇は、クラスリンコーティングピット中に生じる受容体介在性エンドサイトーシスのプロセスを阻害する。これらの後者の実験は、カリウム枯渇がデキストラン輸送の増大によって示される細胞間結合の完全性を変える前に、1時間のみ実施することができた。それぞれの処理条件(又は標準)は少なくとも6回実施した。実験を通じて、培養物は目視で評価して、実験処理及び操作が細胞生存に影響を与えなかったことを確認した。ベースライン実験中と同様に、デキストラン標識RITCは密着結合の完全性の指標として含めた。異なる条件下でのFITC H−フェリチンの平均間の差は、一元配置分散分析を使用して分析した。有意に異なる平均を用いた測定用に、ボンフェロニの事後比較を行なってペアワイズ差を分析した。有意性のレベルはp<0.05で設定した。
【0046】
結合実験
飽和分析法:BREC細胞ホモジェネートでの結合実験を、125I−組換えヒトH−フェリチン又はウマ脾臓フェリチンを使用して第四代のBREC細胞に二連で実施した。両者の比活性度は約340,000cpm/pmolであった。全体、特異的及び非特異的結合を確定するために、一定範囲の濃度の125I−H−フェリチンを、1000倍モル過剰の非標識H−フェリチン有り又はなしで、100μgの全タンパク質のBREC細胞ホモジェネートに加えた。結合バッファーは、50mMのトリス−HCl(pH7.4)、0.1%のBSAからなっていた。インキュベーションは2時間22℃で実施した。3mlの氷冷50mMトリス−HClを加えることによって結合を終了させた。細胞採取機を使用して、結合放射能は迅速な濾過、及び5%ノンファット乾燥ミルク及び0.1mg/mlウマ脾臓フェリチンで事前にコーティングしたWhatmanガラスファイバー製Cフィルターでの洗浄によって単離した。
【0047】
平衡的競合結合アッセイを実施し、高濃度の非標識H−フェリチンを、前に記載したのと同じ結合バッファー中で120分間0.4nMの125I−H−フェリチンと共に、22℃においてBRECホモジェネートのタンパク質25μgとインキュベートした。結合の終了、膜の単離及び特異的結合の計算は、前に記載したように実施した。
【0048】
ラット脳内微小血管系
微小血管の調製:6匹の成体ラットをそれぞれの微小血管の調製に使用した。ラットは致死量のナトリウムペントバルビタールで麻酔をかけ(100mg/体重1kg)、次いで断頭した。脳を除去して、氷上のペトリ皿に置いた。小脳及び髄膜を除去して、5体積の微小血管用バッファー(1×MVB−1×塩、1×HEPES、0.5%BSA及び5mMグルコース)をプロテアーゼ阻害剤と共に加えた。脳はガラス−テフロン(登録商標)ホモジェナイザー(0.25mmクリアランス)を使用して20ストロークで軽く均質化して、ホモジェネートを4℃において10分間1000×Gで遠心分離した。上清は廃棄し、ペレットは17%デキストランの5体積/ラット(1:1の比の1×塩と1×HEPES及びデキストラン)に再懸濁し、次に攪拌し、次いで4℃において3000×Gで遠心分離した。微小血管は試験管の壁から回収し、20mlの1×MVBバッファーに再懸濁した。微小血管は120μメッシュを介して濾過した。次いで微小血管調製物を、40μメッシュ上に支持されたガラスビーズ(Sigma)への接着によってさらに精製した。ビーズはプロテアーゼ阻害剤を加えたバッファーで洗浄した。ビーズは5mlのMVB中ですすぎ、次いで15分間の4℃における1000×Gでの遠心分離により微小血管をペレット状にした。微小血管は1mlのHES+(10mMのHEPES、1mMのEDTA、250mMのスクロース、pH7.4及びプロテアーゼ阻害剤カクテル)(Sigma)に再懸濁し、全体のタンパク質濃度を決定した。サンプルは使用時まで−80℃で保存した。
【0049】
微小血管におけるフェリチンの結合
結合懸濁液は、250μlの最終体積で1μMの非標識H−フェリチンの添加有り又はなしで、50mMのトリス−HCl(pH7.4)、0.1%のBSA、及び20μgの膜タンパク質調製物からなっていた。3mlの氷冷50mMトリス−HClを加えることによって結合を終了させた。結合放射能は、5%ノンファット乾燥ミルク(Blotto)及び0.1mg/ml脾臓フェリチンの溶液で事前にコーティングしたWhatmanガラスファイバー製Cフィルターでの迅速な濾過によって単離した。この組合せは経験的に決定して、加えた合計数の1〜3%にフィルターと放射標識タンパク質の非特異的結合を低減した。フィルターは200mMのNaClを含有する3mlの氷冷50mMトリスで5回洗浄した。フィルターはMICROMEDIC4/200及び自動γ線カウンター中で計数した。特異的結合は、過剰な非標識H−フェリチンが存在しない結合(全体の結合)から過剰な非標識H−フェリチンの存在下での結合(非特異的結合)を引くことによって計算した。
【0050】
飽和分析法
それぞれの結合実験は二連で実施した。高濃度の125I−H−フェリチンを、250μlの最終体積で1μMの非標識H−フェリチンの添加有り又はなしで、20μgの膜タンパク質調製物を含む前に記載したのと同じ結合バッファーからなる結合懸濁液に加えた。22℃における120分のインキュベーション後、結合を終了させ、全体、非特異的、及び特異的結合を、前に記載したのと同様に計算した。
【0051】
競合アッセイ
高濃度の非標識競合剤(H−フェリチンと脾臓フェリチン)を、前に記載したのと同じ結合バッファー中で0.4nMの125I−H−フェリチンの存在下において、100μgの膜タンパク質と共に22℃において60分間インキュベートした。結合、結合の終了、膜の単離、及び特異的結合の計算は、前に記載したのと同様に実施した。競合の実験は二連で実施した。
【0052】
in vivo摂取の試験
H及び脾臓フェリチン(1.2mg)を、40μlの1mMニトリロ三酢酸(pH6.0)、0.5μlの硫酸第一鉄アンモニウム、2μlの0.5M重炭酸ナトリウム、及び40μCの59FeCl中で、4時間37℃においてインキュベートした。インキュベーション後、フェリチンを24時間1×PBS中10,000MWカートリッジ内で透析して、任意の非結合59Feを除去した。比活性度はH−フェリチンに関して0.04μC/g及び脾臓フェリチンに関して0.08μC/gであった。放射標識タンパク質(3.4μg/重量1グラム)を、メスのSprague−Dawleyラット(約350g)の尾静脈(n=3)に注射した。48時間後、ラットを断頭し器官はすぐに除去した。それぞれの器官を切除し0.1MのPBS中で完全にすすいだ。脳に関しては、大脳を小脳から除去して二等分し、髄膜は脳から切除除去した。それぞれの器官由来の1グラムの組織(湿重量)を使用して鉄摂取を決定した。
【0053】
H−フェリチン欠損マウスを、器官中のおそらく障害が起きた鉄の処理が、フェリチン鉄送達に影響を与え得るかどうか決定するための実験モデルとして評価した。同様の手法を使用して、マウスに腹腔内注射したこと以外はラットに関して前に記載したのと同様に、対照及びH−フェリチン欠損マウス中のフェリチン摂取を調べた。H−フェリチンに関する比活性度は0.06μC/gであり、脾臓フェリチンに関する比活性度は0.31μC/gであった。フェリチンを注射し、マウスを屠殺し器官を除去するまで48時間血流中で循環させた。
【0054】
それぞれの器官中の放射能の量は、ヨウ化ナトリウム(NaI)ベースで、シングルチャンネルアナライザーウエルカウンターシステム(Canberra Industries Inc.)で1分間決定した。γ線数/分(cpm)をバックグラウンド数から差し引いて、カウンターの効率で割り、次いで崩壊数/分で割ってμCを計算した。全体%を計算するために、器官のμCを注射した全μCで割り、次いで100%を掛けた。
【0055】
結果
フェリチンのin vivo摂取:フェリチンと結合した鉄がラット中の異なる器官によって摂取され得る可能性を、放射標識59Feを含有するH又は脾臓フェリチンを成体ラットの尾静脈に注射することによって決定した。H−フェリチンからの鉄の摂取は、脳、心臓、腎臓、筋肉及び肺中で、脾臓フェリチンに関する摂取より有意に多かった(図1A)。59Feの量は、それが脾臓フェリチンと結合したときよりH−フェリチンと結合して存在したとき、脳内で2倍多かった(p<0.005)(図1B)。肝臓のみ、H−フェリチンと比較して脾臓フェリチンからの有意に多い鉄の摂取があった(p<0.05)。
【0056】
様々な器官への鉄のH及び脾臓フェリチン送達に対する、様々な器官内の鉄貯蔵能の考えられる変化の影響を決定するために、本発明者らは、H−フェリチンが欠損しているマウス系中のこれらのタンパク質からの59Feの摂取を調べた(Thompson K.J.、Fried M.G.、Zheng Y.、Boyer P.、Connor J.R.J.Cell Sci.115:2165〜2177、2002)。H−フェリチンによる鉄送達は、同腹子対照と比較してH−フェリチン障害が起きたマウスにおいて、脾臓、肺及び筋肉中で低減した(p<0.05)(図2A)。同様の発見を脳内で観察した(図2B)。脾臓フェリチンの摂取は、鉄貯蔵障害が起きたマウス中の任意の器官において不変であった(図3A及び3B)。
【0057】
フェリチンの輸送:血清中フェリチンは全身器官に無制限にアクセスして、脳に鉄を送達するのに有効であることはできたが、内皮細胞壁(BBB)を越えなければならなかった可能性がある。フェリチンのトランスサイトーシスが可能であった可能性の調査を開始するために、本発明者らはBBBの細胞培養モデルを利用した。脾臓フェリチンではなくH−フェリチンが、相当量BREC細胞単層を越えて輸送された(図4)。BREC単層を越えて輸送されたFITC標識Hフェリチンの速度は、RITC標識デキストランの5倍を超えた(p<0.001)。脾臓フェリチンの輸送速度は、デキストラン対照において見られたレベルと類似していた。それによってH−フェリチンがトランスサイトーシスされる機構を決定するために、本発明者らは、無カリウム培地において輸送アッセイを実施して、クラスリンコーティング小胞の形成を阻害した。クラスリンコーティング形成の不在は、H−フェリチンの輸送速度の80%(p<0.001)の減少と関係があった。対照的に、ピノサイトーシスを阻害するためのフィリピンを用いたBRECの前処理は、速度の有意な減少はもたらさなかった。デキストラン対照はそれぞれの実験条件中に含め、その速度は未処理条件に関するグラフ中に示した速度と変わらなかった(データ示さず)。トランスフェリンの輸送は陽性対照として含め、以前に報告されたように(Burdo J.R.、Antonetti D.A.、Wolpert E.B.、Connor J.R.Neuroscience121:883〜890、2003)検出した。蛍光標識トランスフェリンとフェリチンの比活性度は異なった。したがって、この2つのタンパク質に関する輸送の相対速度に関する結論を得ることはできない。
【0058】
フェリチン結合の分析:脳へのフェリチンによる鉄送達の機構をより完全に評価するために、結合試験を実施して、BRECモデル中のフェリチンの輸送が受容体仲介であったかどうか決定した。さらに、フェリチン結合の評価をin vivo系に広げるために、微小血管系をラットの脳から単離した(RBMV)。BREC及びRBMVとのフェリチンの結合は、飽和実験及び競合実験を使用して実施した。Kd及びBmax値は、GRAPHPAD PRISM(登録商標)4.0(GraphPad Software、Inc.)において非線形回帰を使用して2つの手法から得た。
【0059】
飽和曲線:様々な濃度(1、2、3、5、7、及び10nM)の125I標識フェリチン(rH−フェリチン又は脾臓フェリチン)を、100μgの組織(BREC又はRBMV)とインキュベートした。全体及び非特異的結合(1000nMの非標識フェリチンの存在下)は、Whatmannフィルターでアッセイを実施することによって得た。このような結合アッセイからKd及びBmaxを得るために、一部位結合の非線形包括的回帰を実施した。この方法では、全体と非特異的結合の両方を標識フェリチンの濃度に対してプロットした。生成したプロットは、等式:非特異的=NS×X及び全体=特異的+非特異的、前式で特異的=Bmax×X/(Kd+X)に適合させた。包括的手法では、特異的結合は全体及び非特異的結合のデータから誘導されない。代わりにKd及びBmaxの値は、2つのデータセット(全体と非特異的)間の非特異的結合定数(NS)を共有することによって得る。この回帰分析からのデータは図5A及び5B中に示す。rH−フェリチンのみが、BREC又はRBMVのいずれかと有意な飽和結合を有する。RBMVに関するKd及びBmaxは、それぞれ7.9±1.6nM及び572.6±64.0fmol/mgタンパク質である。BRECに関しては、Kdは2.7±0.9nMであり、Bmaxは465.7±63.1fmol/mgタンパク質である。曲線適合のR2値はBRECとRBMVの両方に関して0.8を超える。
【0060】
競合曲線:様々な濃度のコールドフェリチン(0.03、0.1、0.3、1、3、10、30、100、300、及び1000nM)を、100μgのBREC又はRBMV組織及び0.4nMの放射標識フェリチンとインキュベートした。全体の結合はWhatmannフィルターでアッセイを実施することによって得た。次いで全体結合(fmol/mgタンパク質)をlog[濃度(nM)]に対してプロットした。次いでこれらのプロットを、一部位競合等式:全体=底部+(上部−底部)/(1+10(X−LogEC50))に適合させた。このデータは図6A及び6B中に示す。これらの結果は、脾臓フェリチンではなくH−フェリチンが、結合部位に関して有効に競合し得ることを示す。BRECに関する競合曲線からのKd及びBmaxの決定によって、2.0nMのKd及び235.1fmol/mgタンパク質のBmaxを得た。RBMVに関する対応する値は、Kd=3.4nM及びBmax=304.6fmol/mgタンパク質である。適合のR2値はBRECとRBMVの両方に関して0.95を超える。2つの異なる曲線によって生じた値は許容範囲内にある。
【0061】
考察:この試験の結果は、脳を含めた多数の器官に、フェリチンが鉄を送達することができることを明らかにする。さらに、フェリチンにより送達される鉄の量は、肝臓以外の大部分の器官に関して、L−フェリチンの代わりにH−フェリチンにより鉄が送達されるとき増大する。細胞によって摂取されるH−フェリチン鉄の量は、H−フェリチン欠損マウス中で実証されるように、鉄貯蔵能に障害が起きると変わる可能性があるが、一方L−フェリチンによる鉄送達はこのモデル中では有意に影響を受けない。これらの後者の結果は、H−フェリチンに関するフィードバックシステムを示唆する。したがって本発明者らは、脳への鉄送達に関する新規な輸送系、及びトランスフェリン(最大2個のFe原子)と比較したフェリチンの一分子中に収容され得る鉄の量(最大4500個のFe原子)を考慮して、非常に有意であり得る輸送系を同定している。脳への非トランスフェリン依存性鉄送達系の同定は、血清トランスフェリンの不在下での脳への鉄送達を示す本発明者らの以前の報告と一致する(Malecki E.A.、Devenyi A.G.、Beard J.L.、Connor J.R.JNeurosci Res.56:113〜122、1999)。この試験は、フェリチン受容体の存在の連続的な探究も追加する。フェリチン受容体は、報告によると肝細胞において実証されているが(Mack U.、et al.JBiol.Chem258:4672〜4675、1983)、このモデル内及び脳内で飽和は実証されなかった(Hulet S.W.、Heyliger S.O.、Powers S.、Connor J.R.J.Neurosci Res.61:52〜60、2000)。近年あるタンパク質が推定フェリチン受容体として同定されている(Chen T.T.、et al.JExp Med.202:955〜65、2005)。
【0062】
脳内では、受容体との結合以外に、フェリチンはBBBを越えてトランスサイトーシスされなければならない。この試験において、本発明者らは、フェリチンは細胞培養モデルのBBBを越えて輸送することができることを実証した。この細胞培養モデル中のフェリチンの輸送はクラスリン依存性及び受容体仲介型であり、H−サブユニットを強く好む。H−フェリチンの結合に関する優先は輸送のデータと一致する。フェリチンの結合はラット脳由来の微小血管系においても実証され、且つこの結合も、細胞培養モデルと同様に、H−サブユニットを強く好む。結合及び輸送のデータは、鉄が脾臓(L多量)フェリチンと比較してH−フェリチンと結合した場合の脳への増大した送達を明らかにした、鉄摂取のデータと一致する。フェリチンの摂取、HフェリチンとLフェリチンの間の鉄送達の違い、及びH−フェリチンに障害が起きたマウス中のH−フェリチン摂取の変化は、いずれもフェリチン受容体の可能性を示唆し、H−フェリチンに関する結合は内皮細胞の2つの異なる供給源において直接実証された。
【0063】
フェリチンがBBBを越えて輸送される機構はクラスリン依存性であり、ピノサイトーシスによるものではないことを示唆するデータは、同系のトランスフェリンを使用して以前に確認されたデータと類似している(Burdo J.R.、Antonetti D.A.、Wolpert E.B.、Connor J.R.Mechanisms and regulation of transferrin and iron transport in a model blood−brain barrier system.Neurosci.121:883〜890、2003)。脳への鉄送達に関する機構及びこれらの機構の制御は、どのようにして鉄が蓄積する又は正常レベルに達し得ないか、したがって様々な神経障害の根底にあるか又はそれらに貢献するかを理解する中核となる(Zecca L、Youdim MB、Riederer P、Connor JR、Crichton RR.Nat Rev Neurosci.5:863〜873、2004)。トランスフェリンと結合した鉄のトランスサイトーシスの試験は、矛盾する結果を与えている。いくつかの試験は、脳に入る鉄はトランスフェリンと結合することを示し、且つ他の試験はトランスフェリンとは関係がない鉄輸送を示す。本発明者らは、脳への鉄送達に関するトランスフェリン依存系と非トランスフェリン依存系の両方が存在し、且つ別の経路に優る一経路の優先傾向は内皮細胞の鉄の状態に依存し得る証拠を提供している。BBBを形成する内皮細胞の鉄の状態、及びどのようにしてこの鉄の状態がトランスフェリン仲介型鉄摂取に関するトランスフェリン受容体の制御に影響を与えるかは、脳の鉄摂取の機構に関する試験において大部分は無視されている(総説に関してはBurdo and Connor 2003を参照)。細胞中のトランスフェリン受容体の発現は、細胞の細胞内鉄状態により制御されることはよく知られており(Aisen P.、et al.Int J Biochem Cell Biol.33:940〜959、2001)、内皮細胞は、他の細胞と同じトランスフェリン受容体の鉄制御機構を有する(Georgieff M.K.、et al.JPediatr.141:405〜9、2002)。脳の内皮細胞は、その機能が細胞内のエンドソームからの鉄放出を仲介することである二価金属輸送タンパク質も発現し、鉄貯蔵の存在を示す比較的多量のフェリチンを含有する。脳の内皮細胞はその独自の相当な鉄の必要性を有し、且つその独自の鉄摂取を制御するという概念は、これらの細胞中のミトコンドリアの高濃度、したがって高い鉄の必要性と一致する。したがって、トランスフェリンによってこれらの細胞に送達される少なくともいくらかの鉄は、内皮細胞内に保持されなければならない。フェリチンから鉄が放出される機構は十分に理解されておらず、ある人は、鉄を放出するためにはフェリチンの分解が必要とされると提言している。したがって、フェリチンは多量のその鉄含量体でトランスサイトーシスされる可能性がより高く、トランスフェリンより内皮細胞とその鉄を共有する可能性は低い。脳に鉄を送達するフェリチンを制御するための機構は現時点では知られていないが、H−フェリチン欠損マウスにおける鉄を送達するH−フェリチンの減少は、このような制御機構が存在することを強く示唆する。
【0064】
鉄貯蔵タンパク質としてのフェリチンの機能は十分確立しているが、H−フェリチンは細胞によって活発に分泌され、おそらく鉄を送達する可能性があるという概念は以前に試験されていない。血中のフェリチンレベルは正常条件下でさえ大幅に変動する。フェリチンmRNAは、フェリチンの分泌経路をサポートし得るラット肝細胞中の小胞体と結合するポリリボソームと結合する。フェリチン(両サブユニット)の直接分泌は分化したラット肝癌細胞において実証されており、培養中の小神経膠細胞からのL−フェリチンではなくH−フェリチンの放出が示されている(Zhang X.、Surguladze N.、Slagle−Webb B.、Cozzi A.、Connor J.R.Glia54:795〜804、2006)。
【0065】
BBB及び他の器官における受容体に関するフェリチンの供給源は、おそらく血清由来である。血清中のフェリチンの存在は十分確立しているが、血清中のフェリチンは主にL−サブユニットで構成されると従来考えられている。このLに富む血清中フェリチンの供給源は主に溶解マクロファージ由来である(McGowan S.E.、et al.J.Lab Clin.Med.111:611〜617、1988)。本発明者らは、しかしながら、フェリチンの結合、フェリチンの輸送及び脳への鉄の送達はいずれも、Hに富むフェリチンを強く好むことを、驚くべきことに示している。したがって、血清中の比較的少量のH−フェリチンに基づく本発明者らの発見の生理学的意義は、フェリチンは脳のための鉄の供給源としてトランスフェリンに続き得ることを示唆する。しかしながら、1モルのフェリチンは、1モルのトランスフェリンより2000倍を超える鉄を送達することができることは覚えておかなければならない。
【0066】
さらに、炎症などの血漿中及びいくつかの癌と関連してH−フェリチンが増大する条件が存在する(Elliott R.L.、et al.Breast Cancer Res.Treat.30:305〜309、1994)。したがって、慢性炎症状態は脳の鉄状態を増大させる可能性がある。これはアルツハイマー病及びパーキンソン病を含めたいくつかの神経変性疾患における鉄蓄積と一致する概念である。
【0067】
(例2)
導入
本発明者らは、鉄欠乏を緩和するための新規な食事療法の手法を開発している。この手法は、本発明者らの脳を含めた身体中のH−フェリチンに対する受容体の発見、及びLに富むフェリチンと比較してH−フェリチンが、肝臓以外の全器官への鉄摂取の好ましい形式であるという実証から派生した。他の人はH−フェリチンが母乳中に豊富であることを示しており、このタンパク質は、母親と幼児の間の鉄送達のための機構であることが示唆される。この後者の言及は、おそらくヒトだけでなく全ての哺乳動物にあてはまる。さらに、H−フェリチンの遺伝子配列及びタンパク質構造は動物界で高度に保存されている。したがって、鉄送達タンパク質としてのH−フェリチンの適用はヒトに限られないはずである。
【0068】
本発明者らは、酵母を使用して栄養補助食品として、H−フェリチンを送達するための機構を考案している。この手法を開始するため、第一の試験は酵母を形質転換して、H−フェリチンタンパク質に翻訳され得るヒトH−フェリチンの遺伝子を発現させることであった。図7中に示すイムノブロットは、H−フェリチンは酵母により発現され、L−フェリチンとの交差反応はないことを実証する。第二の試験は、硫化第一鉄を用いた酵母が増殖する培地の富化は、それらが発現したフェリチンの鉄含量を増大することを実証することであった。これらの結果は図8中に示す。
【0069】
供給試験
鉄欠乏を治療するためのモデルとしてのフェリチン補強酵母の有効性を試験するために、本発明者らは鉄欠乏の標準的なラットモデルを使用した。ラットの子供は25日齢で離乳するまで鉄欠乏母獣により育てられた。離乳時に、動物を5群の1つに割り当てた。群1:鉄欠乏食(ID)で維持した。群2:35mg/kgのFeSO4の標準的な鉄補充食を供給した。群3:鉄を補充しない酵母(酵母、鉄なし)を含有した食事を供給した。群4:鉄は補充したがH−フェリチンは補強しなかった酵母(酵母、フェリチンなし)を含んだ食事を供給した。群5:H−フェリチンで形質転換し鉄を補充した酵母(酵母、Fe+Ft)。離乳後3、7、9及び14日で、これらの動物から血液サンプルを回収した。これらの結果は図9中に示す。ID食を継続した動物は最低レベルのヘモグロビン(Hb)を有していた。鉄を含まない酵母を与えた動物は、ID動物と類似したHbレベルを有していた。Hbレベルの改善は他の3群において見られ、改善の最も急速な増大は、鉄を補充しフェリチンを補強した酵母を与えた動物において生じた。フェリチンを含まない鉄補充酵母を与えた動物さえ、FeSO4群より改善されたHbレベルを有していた。
【0070】
ヘマトクリット値レベルも同じ動物群においてモニターした。これらのデータは図10中に示す。これらのデータは、鉄の媒体としての酵母はH−フェリチンの存在又は不在下でヘマトクリット値を補正する際に同程度に有効であり、いずれも標準的な現在の治療選択肢、FeSO4より有意に優れていることを示す。ID食を継続した動物、及び鉄を補充しなかった酵母を与えた動物は、試験した11日間にわたってヘマトクリット値の増大を示さなかった。
【0071】
鉄を動員する身体の能力を示す重要な分析、おそらく間接的に鉄の生物学的利用能の指標は、トランスフェリンの飽和レベルである。トランスフェリンは主な鉄動員タンパク質であり、血清中に高濃度で見られる。トランスフェリン飽和の量は、正常の高い30%から貧血の状態中の10%未満まで変動する。フェリチン補強鉄多量酵母の有効性を評価するために本発明者らが使用した動物モデルにおけるトランスフェリン飽和の分析は図11中に示す。この図中では、フェリチン補強鉄多量酵母は、トランスフェリン(Tf)飽和の最大の増大、次にフェリチンを含まない鉄多量酵母をもたらしたことを見ることができる。この試験は、食事療法におけるFeSO4に優る鉄送達媒体としての酵母の優勢を再度実証する。ID食を維持した動物又は対照酵母(鉄なし又はフェリチン多量)を供給した動物は、最も低いTf飽和があった。
【0072】
脳の鉄に対する影響
鉄の欠乏は脳の発達に対して重大な影響があるので、特定脳領域の鉄の状態を、異なる食事を与えた動物においてモニターした。図12中では、脳の2つの発達上重要な領域、腹側中脳及び尾状核の鉄の状態を示す。これらの領域は、動物(例えばヒト)が成熟すると比較的鉄が豊富になる運命にある。これらの脳領域は運動活性、したがって特に欠乏が発達中に起こるときの鉄欠乏における運動技能の障害の制御に関与する。Hb及びHct分析中に記載したのと同じ動物を14日齢で屠殺し、腹側中脳(VBM)及び尾状核の鉄濃度を測定した。フェリチンを補強し鉄を補充した酵母を与えた動物は、任意の他の群より多量の鉄を2つの脳領域中に有していた。この刺激的で新規な発見は、フェリチン補強栄養補助食品は、鉄欠乏と関係がある神経障害を制限するための機構であり得ることを示す。この観察は、一般的な健康状態を高めることによってだけでなく、神経機能を最適化することによって、世界規模の鉄欠乏との闘いに対して多大な影響を有する可能性がある。
【0073】
脳内鉄状態の領域変化をさらに評価するために、脳の2つの他の領域、側坐核(NA)及び前頭前皮質(PFC)を調べた。これらの結果は図13中に示す。この図中では、フェリチン補強鉄多量酵母から送達される鉄の領域特異性は明らかである。NA中では、図5中に示したVMB及び尾状核と同様に、鉄送達の他の形態と比較して鉄含量は上昇する。しかしながら、PFC中では、フェリチンを補強し鉄を補充した酵母から送達される鉄は、他の群に関して見られるそれと同様である。これらのデータは、他の鉄送達系と異なる形式でフェリチン補強鉄補充酵母からの鉄送達を制御する機構が存在することを示唆し、本発明者らの脳内微小血管系におけるH−フェリチンに対する受容体の発見と一致する。
【0074】
概要
脳への増大した鉄摂取、及びおそらくより重要なことに、摂取の領域特異性は予想外であるが、本発明と関係がある非常に有意な発見である。これらのデータは、フェリチン補強鉄補充酵母の使用は、生後発育中の鉄欠乏性と関係がある神経、認知及び行動障害の改善をもたらすに違いないことを示唆する。血液学的パラメータに関するデータは明らかに強力であり、フェリチン補強鉄補充酵母は鉄栄養補助食品の優れた形態であることを示す。
【特許請求の範囲】
【請求項1】
患者における鉄欠乏性障害を治療するための方法であって、治療有効量のフェリチン−鉄複合体を、その必要性がある患者に投与することを含む上記方法。
【請求項2】
前記フェリチン−鉄複合体がH−フェリチンを含む、請求項1に記載の方法。
【請求項3】
前記H−フェリチンが哺乳動物のH−フェリチンである、請求項2に記載の方法。
【請求項4】
前記哺乳動物のH−フェリチンがヒトのH−フェリチン又はそのホモログである、請求項3に記載の方法。
【請求項5】
前記ホモログがヒトのH−フェリチンと少なくとも90%の配列同一性を有する、請求項4に記載の方法。
【請求項6】
前記H−フェリチンが組換えH−フェリチンである、請求項2に記載の方法。
【請求項7】
前記組換えH−フェリチンが酵母によって産生される、請求項6に記載の方法。
【請求項8】
前記鉄欠乏性障害が脳内の鉄欠乏性と関係がある、請求項1に記載の方法。
【請求項9】
前記鉄欠乏性障害が神経障害及び/又は神経変性障害である、請求項8に記載の方法。
【請求項10】
前記鉄欠乏性障害がパーキンソン病、アルツハイマー病、下肢静止不能症候群、注意力欠如障害、鬱病、及び不眠症からなる群から選択される、請求項8に記載の方法。
【請求項11】
前記鉄欠乏性障害が生後発育中の鉄欠乏性と関連する神経、認知、行動及び/又は運動障害を含む、請求項8に記載の方法。
【請求項12】
前記フェリチン−鉄複合体がターゲッティング部分をさらに含む、請求項1に記載の方法。
【請求項13】
前記ターゲッティング部分が抗体、アプタマー、受容体、リガンド、及びそれらの結合断片からなる群から選択される、請求項12に記載の方法。
【請求項14】
前記ターゲッティング部分が脳及び/又は血液脳関門特異的マーカーを認識する、請求項12に記載の方法。
【請求項15】
脳に鉄を送達するための方法であって、フェリチン−鉄複合体を患者に投与することを含み、それによって、前記鉄が血液脳関門を越えて輸送され脳に送達される上記方法。
【請求項16】
ターゲッティング部分としてH−フェリチンを使用するための方法であって、H−フェリチンとリポソームを結合させることを含み、それによって前記リポソームが脳及び/又は脳内の細胞にターゲッティングされる上記方法。
【請求項17】
患者における鉄過剰症を治療するための方法であって、治療有効量のマルチサブユニットフェリチン複合体を、その必要性がある患者に投与することを含む上記方法。
【請求項18】
前記マルチサブユニットフェリチン複合体が100%未満の鉄結合能である、請求項17に記載の方法。
【請求項19】
前記マルチサブユニットフェリチン複合体が哺乳動物のH−フェリチン又はそのホモログを含む、請求項17に記載の方法。
【請求項1】
患者における鉄欠乏性障害を治療するための方法であって、治療有効量のフェリチン−鉄複合体を、その必要性がある患者に投与することを含む上記方法。
【請求項2】
前記フェリチン−鉄複合体がH−フェリチンを含む、請求項1に記載の方法。
【請求項3】
前記H−フェリチンが哺乳動物のH−フェリチンである、請求項2に記載の方法。
【請求項4】
前記哺乳動物のH−フェリチンがヒトのH−フェリチン又はそのホモログである、請求項3に記載の方法。
【請求項5】
前記ホモログがヒトのH−フェリチンと少なくとも90%の配列同一性を有する、請求項4に記載の方法。
【請求項6】
前記H−フェリチンが組換えH−フェリチンである、請求項2に記載の方法。
【請求項7】
前記組換えH−フェリチンが酵母によって産生される、請求項6に記載の方法。
【請求項8】
前記鉄欠乏性障害が脳内の鉄欠乏性と関係がある、請求項1に記載の方法。
【請求項9】
前記鉄欠乏性障害が神経障害及び/又は神経変性障害である、請求項8に記載の方法。
【請求項10】
前記鉄欠乏性障害がパーキンソン病、アルツハイマー病、下肢静止不能症候群、注意力欠如障害、鬱病、及び不眠症からなる群から選択される、請求項8に記載の方法。
【請求項11】
前記鉄欠乏性障害が生後発育中の鉄欠乏性と関連する神経、認知、行動及び/又は運動障害を含む、請求項8に記載の方法。
【請求項12】
前記フェリチン−鉄複合体がターゲッティング部分をさらに含む、請求項1に記載の方法。
【請求項13】
前記ターゲッティング部分が抗体、アプタマー、受容体、リガンド、及びそれらの結合断片からなる群から選択される、請求項12に記載の方法。
【請求項14】
前記ターゲッティング部分が脳及び/又は血液脳関門特異的マーカーを認識する、請求項12に記載の方法。
【請求項15】
脳に鉄を送達するための方法であって、フェリチン−鉄複合体を患者に投与することを含み、それによって、前記鉄が血液脳関門を越えて輸送され脳に送達される上記方法。
【請求項16】
ターゲッティング部分としてH−フェリチンを使用するための方法であって、H−フェリチンとリポソームを結合させることを含み、それによって前記リポソームが脳及び/又は脳内の細胞にターゲッティングされる上記方法。
【請求項17】
患者における鉄過剰症を治療するための方法であって、治療有効量のマルチサブユニットフェリチン複合体を、その必要性がある患者に投与することを含む上記方法。
【請求項18】
前記マルチサブユニットフェリチン複合体が100%未満の鉄結合能である、請求項17に記載の方法。
【請求項19】
前記マルチサブユニットフェリチン複合体が哺乳動物のH−フェリチン又はそのホモログを含む、請求項17に記載の方法。
【図1A】


【図1B】
【図2A】
【図2B】
【図3A】
【図3B】
【図4】
【図5A】
【図5B】
【図6A】
【図6B】
【図7】
【図8】

【図9】
【図10】
【図11】
【図12】
【図13】
【図14a】

【図14b】

【図1B】
【図2A】
【図2B】
【図3A】
【図3B】
【図4】
【図5A】
【図5B】
【図6A】
【図6B】
【図7】
【図8】
【図9】
【図10】
【図11】
【図12】
【図13】
【図14a】
【図14b】
【公表番号】特表2011−516419(P2011−516419A)
【公表日】平成23年5月26日(2011.5.26)
【国際特許分類】
【出願番号】特願2011−501755(P2011−501755)
【出願日】平成20年3月28日(2008.3.28)
【国際出願番号】PCT/US2008/004071
【国際公開番号】WO2009/120168
【国際公開日】平成21年10月1日(2009.10.1)
【出願人】(510257444)カイナ エルエルシー (1)
【Fターム(参考)】
【公表日】平成23年5月26日(2011.5.26)
【国際特許分類】
【出願日】平成20年3月28日(2008.3.28)
【国際出願番号】PCT/US2008/004071
【国際公開番号】WO2009/120168
【国際公開日】平成21年10月1日(2009.10.1)
【出願人】(510257444)カイナ エルエルシー (1)
【Fターム(参考)】
[ Back to top ]