
錠型(padlock)プローブのローリングサークル複製
【課題】錠型プライマーが、長いかもしくは環状のターゲット核酸にハイブリダイズすると、錠型プライマーのローリングサークル複製を行う方法を提供する。
【解決手段】錠型プローブとハイブリダイズする部位の近傍もしくは好ましくはその部位でターゲット核酸を切断し、これにより切断されたターゲット核酸の3’端が、錠型プローブのローリングサークル複製のためのプライマーとして機能する、それぞれがオリゴヌクレオチド標識を有する二つのアフィニティープローブおよび二つのアフィニティープローブと結合したローリングサークル複製のための錠型プローブを利用することにより、ポリエピトープ・ターゲットをアッセイする方法。
【解決手段】錠型プローブとハイブリダイズする部位の近傍もしくは好ましくはその部位でターゲット核酸を切断し、これにより切断されたターゲット核酸の3’端が、錠型プローブのローリングサークル複製のためのプライマーとして機能する、それぞれがオリゴヌクレオチド標識を有する二つのアフィニティープローブおよび二つのアフィニティープローブと結合したローリングサークル複製のための錠型プローブを利用することにより、ポリエピトープ・ターゲットをアッセイする方法。
【発明の詳細な説明】
【発明の詳細な説明】
【0001】
[導入]
DNAまたはRNAサンプル中の単一もしくは複数のヌクレオチド変異を検出するために種々の方法が使用されている(1、2)。多くの方法は、分析前に一般にPCRによりターゲット配列を増幅することに依存している。従って、対立遺伝子の変異の位置に関する有意な情報は失われてしまう。配列変異の位置に関する情報は、例えば、遺伝性疾患の研究において染色体上の幾つかの変異配列のハプロタイプを決定するため;ある遺伝子における二つの変異が同一コピーに存在するか、別々の対立遺伝子に存在するかを決定するため;あるいは細胞周期における対立遺伝子の複製タイミング(3)または悪性組織における変異細胞の分布を研究するために重要である。In situでのシングルコピー遺伝子配列におけるシングルヌクレオチドの識別は、現在のところ不可能であるが、最近、新しいクラスの遺伝子診断プローブ分子である錠型プローブ(4)を用いて、繰り返し配列上で有効に操作されることが示された(5)。これらの錠型プローブは、両端にターゲットに相補的な配列を有し、その間にターゲットに相補的でない配列を有する直鎖状オリゴヌクレオチドである。正しいターゲットDNA配列にハイブリダイズすると、両端は先頭と後部とが一緒になって、DNAリガーゼにより結合される。二本鎖DNAのらせん特性の結果、得られた環状プローブ分子は、ターゲットDNA鎖に連結(catenate)される。このプローブのデザインは、幾つかの重要な特徴を有している。まず、ライゲーションが起こるための二つの別々のオリゴヌクレオチドのハイブリダイゼーションに必要なことは、完全なヒトゲノムの複雑性の中でシングルコピー遺伝子を検出するに充分な特異性を提供することである(6〜8)。二つめに、ライゲーション連結部におけるミスマッチにより、ライゲーションが大きく阻害され、これがターゲット配列中のシングルヌクレオチドの識別を可能にする(4、8〜10)。三つめに、プローブとターゲットDNA鎖との間のトポロジカル(topological)連結は、ハイブリダイゼーションの安定性とは無関係であるので、変性洗浄は、非特異的なハイブリダイゼーションを減少させることができる。四つめに、PCRもしくはLCRとは違って、分子内相互作用プローブの反応のみが記録され、これが大きなプローブセットを同時に適用する問題を回避する(11)。PCRプライマーの多くの対の組合わせは、任意のプライマーの組み合わせの間で形成される偽の増幅産物を急速に増加させる危険につながるが、これは錠型プローブには当てはまらない。最後に、プローブ端の結合が、反応前には存在しない新しいクラスの分子を作り出し、これらの環状分子は、ライゲートされたプローブを検出するために、ローリングサークル複製反応で増幅される(12、13)。
【0002】
ローリングサークル複製反応による溶液中フリーの環状核酸分子の増幅は、プライマーと環状核酸とのハイブリダイズ、そしてヌクレオチドおよびポリメラーゼ酵素の供給により成し遂げられる。しかし、環状核酸分子が、溶液中にフリーでない場合に問題が生じる。ターゲットに連結される錠型プライマーの特別なケースにおいて、そのターゲットがローリングサークル複製反応を抑制することが充分に予測される。本発明を導いた先の未発表の研究において、発明者らはこのような抑制が実際には起こらないことを実証した(18)。ターゲットが環状である場合、例えば環状7kbM13ゲノムである場合、ターゲットに連結された錠型プライマーのローリングサークル複製が効果的に妨害されることを発明者らは見出した。ターゲットが直鎖状である場合、その上に形成された錠が、ターゲット分子に沿ってスライドし、その端を離れ、それにより溶液中でフリーになり、ローリングサークル複製反応による増幅に利用されることが原則として可能である。あるいは、ターゲット配列、錠型プローブによる結合部位の3’が、エキソヌクレアーゼにより処理され、残りのターゲット鎖がローリングサークル複製を引き起こすことを可能にする。実際、発明者らは、このスライディング(sliding)および非連結(uncoupling)の効果は、長い直鎖状ターゲットでさえ限られた程度しか起こらないことを見出した。従って、単一部位での制限処理により、環状7kbM13ゲノムを、錠型プライマーのハイブリダイゼーション部位を3.5kb上流および3.5kb下流に有する直鎖状7kbの核酸分子に変換すれば、錠型プライマーの限定された増幅は、ローリングサークル複製によって起こり得る。しかし、ターゲット核酸が実質的に短い場所、例えば数十塩基もしくは数百塩基では、その上に形成された錠型プライマーのローリングサークル複製は、ずっと迅速で効率的である。
【0003】
[本発明]
本発明の一つの側面において、この課題は、ターゲットの認識およびプローブの環状化の前、その間、もしくはその後に、ターゲット分子を切断することにより解決される。この側面において、本発明は、ターゲット核酸のターゲット配列を検出する方法を提供し、この方法は、以下の工程を含む:
i)ターゲット配列のための錠型プローブを提供する工程、
ii)錠型プローブとターゲット核酸とのハイブッリドを形成し、錠型プローブを環状化する工程;
iii)工程ii)の前、工程ii)の間、もしくは工程ii)の後に、ターゲット核酸をターゲット配列もしくはその近傍で切断する工程;
iv)錠型プローブのローリングサークル複製を行う工程。
【図面の簡単な説明】
【0004】
【図1】本発明の方法を説明する図。
【図2】本発明の方法を説明する図。
【図3】本発明の方法を説明する図。
【図4】本発明の方法を説明する図。
【図5】本発明の方法を説明する図。
【図6】例1の結果を示す図。
【図7】例2の結果を示す図。
【図8】例3の結果を示す図。
【図9】例4の結果を示す図。
【図10】例5の結果を示す図。
【図11】例6の結果を示す図。
【図12】例7の結果を示す図。
【図13】例8の結果を示す図。
【図14】例8の結果を示す図。
【発明を実施するための形態】
【0005】
前記ターゲット核酸は、原則としてDNAでもRNAでもよい。本発明の方法は、ターゲット核酸が長鎖分子もしくは環状分子である場合特に有効であると思われる。ターゲット配列は、錠型プローブの両端とハイブリダイズするのに充分な長さである。本発明の方法は、シングルコピー遺伝子配列の位置検出、顕微鏡標本における配列変異型の間の区別、および一次元もしくは二次元アレイに固定されたプローブにとって特に有効であると期待される。
【0006】
錠型プローブは、ターゲット配列に相補的な5’端配列および3’端配列を有する;その結果、錠型プローブは、ターゲット配列にハイブリダイズすると、例えばライゲーションにより環状化することができる。厳密な相補性は必ずしも要求されないが、錠型プローブとターゲット配列とのハイブリダイゼーションおよびライゲーションによる錠型プローブの環状化を可能にする充分な相補性が必要である。原則として二つの部分で錠型プローブを提供することが可能であり、この場合、環状化を成し遂げるためには二つのライゲーション反応が必要である。錠型プローブのサイズは、ローリングサークル複製反応による増幅に適している必要があり、好ましくは、少なくとも約25 nt、多くて150〜200 ntであるが、特に5’から3’のエキソヌクレアーゼ活性をもたないポリメラーゼと使用するときは特に、より大きなプローブが有効である。
【0007】
本方法の工程ii)は、錠型プローブとターゲット核酸のターゲット配列とがハイブッリドを形成し、錠型プローブが環状化することを含む。これらの工程は、慣用的な条件下で行うことができる。調査中のターゲット核酸サンプルが、ターゲット配列を有していない場合、錠型プローブのハイブリダイゼーションおよび環状化は全く起こらないか限られた程度でしか起こらない。
【0008】
本方法の工程iii)において、ターゲット核酸は、ターゲット配列もしくはその近傍で切断される。この切断は、当該分野で周知の方法により行うことができる。ターゲット配列から制限部位の距離が相当な場合、例えば3.5 kbもしくはそれ以上の場合もあり得るが、好ましくは、ターゲット配列の数塩基以内もしくは数十塩基以内である。
【0009】
このようにターゲット核酸は、錠型プローブの添加前に、制限処理により直線状にすることができる。あるいは、プライマー伸長反応を、錠型プローブ認識用のターゲットとして適切な比較的短い核酸分子を作成するように行うことができる。中期染色体のin situ検出において、これら二つの方法は、ターゲットDNAの損失を引き起こし、これにより検出のロスを引き起こすという不利益を受けやすい。従って好ましくは、工程iii)は、ハイブリッドを制限処理し、これにより環状化された錠型プローブを切断することなくターゲット核酸をターゲット配列もしくはその近傍で切断することにより行われる。
【0010】
工程iv)において、錠型プローブは、ローリングサークル複製反応により増幅される。この反応は、通常、環状化された錠型プローブにハイブリダイズするプライマー、ヌクレオチドの供給、およびポリメラーゼ酵素を必要とし、当該分野で周知の方法により行うことができる。好ましいポリメラーゼ酵素は、高い処理特性および3’−エキソヌクレアーゼ活性を有するφ29 DNAポリメラーゼである。
【0011】
従って、ターゲット配列を、錠型プローブの結合した位置の3’で切断した場合、錠型プローブに対する塩基対の3’端が得られるまで、任意の非塩基対ヌクレオチドをポリメラーゼにより除去し、そこで、外部のプライマーを添加することなくローリングサークル複製(PCR)を開始することができ、PCR産物がターゲット配列と連続することが保証される。このような切断は、プローブが結合した位置の下流で、ターゲット配列にオリゴヌクレオチドをハイブリダイズすることにより成し遂げられる。次いで、このハイブリダイゼーションによりその認識配列が二本鎖になった制限酵素を用いてその配列を切断することができる。あるいは、タイプIIS酵素(例えばFokI)の認識配列を有する二本鎖の一端、および錠型プローブが結合した位置のターゲット配列3’にハイブリダイズする一端を有するヘアーピンオリゴヌクレオチドを使用することができる。
【0012】
錠型プローブが制限部位を横切ってターゲット配列にハイブリダイズする場合、その錠を、例えばホスホロチオエートで修飾することにより保護することができ、ターゲット配列は錠を開けることなく切断される。
【0013】
ターゲット核酸がRNAであって、ターゲット配列内の部位で制限処理される場合、RNase H処理により、錠型プライマーのローリングサークル複製を開始可能な3’端を得ることができる。
【0014】
好ましい方法によると、工程iii)において、ターゲット核酸はターゲット配列内で切断される。この方法の利点は、ターゲット配列の3’端がプライマーを構成することであり、工程iv)において、このプライマーを用いて錠型プローブのローリングサークル複製を行うことができる。これにより、生成されたローリングサークル複製産物は、ターゲット配列と連続している。
【0015】
これを成し遂げるために、好ましくはタイプIIS酵素を利用する(タイプIIS酵素は、クラスIIS酵素もしくはタイプIV酵素とも称される)。タイプIIS制限エンドヌクレアーゼは、文献にその記載がある(14、15)。これら酵素は、二本鎖DNA上の二つの別々の部位;4〜7bpの長さの認識部位、および認識部位から通常1〜20bp離れた切断部位と相互作用する。このようなタイプIIS酵素の一つは、FokI制限エンドヌクレアーゼであり(16、17)、この酵素は、二本鎖の5’−GGATG−3’部位を認識し、5’−3’鎖および3’−5’鎖からそれぞれ9ヌクレオチドおよび13ヌクレオチド下流で切断する。FokIをssDNAおよびオリゴヌクレオチドと使用すると、認識部位から9番目もしくは13番目のヌクレオチドで切断が観察される。認識部位と切断部位との間の二本鎖領域が完全に相補的であるか否かに関わらず切断は起こる。
【0016】
上記方法でタイプIIS酵素を使用するためには、特別にデザインした錠型プローブが必要であり、これは、本発明の別の側面を形成する。この側面において、ターゲット核酸配列のための錠型プローブとして使用するに適したオリゴヌクレオチドを提供し、このオリゴヌクレオチドは、ターゲット配列に相補的な5’端および3’端配列;タイプIIS酵素による認識のための第一の部位;および少なくとも一のヌクレオチド残基および/またはヌクレオチド間結合を修飾して、タイプIIS酵素による制限処理からオリゴヌクレオチドを保護する第二の部位を有する。
【0017】
タイプIIS酵素による認識のための第一の部位は、該酵素がターゲット核酸配列を切断するように、錠型プローブ上に位置する必要がある。そして第二の部位は、タイプIIS酵素によって錠型プローブが切断されないように保護するために、第一の部位に関連して錠型プローブ上に位置する必要がある。第一の部位と第二の部位との相対的な位置は、5’端および3’端配列に関連して、使用する特定のタイプIIS酵素に依存する。第二の部位における修飾も、使用する特定のタイプIIS酵素に依存する。例えば、FokI酵素を使用する場合、タイプIIS酵素のHincIIにおいても同様に、ホスホロチオエートのヌクレオチド間結合が、錠型プローブの切断を防止するのに有効である。
【0018】
本発明の方法において、錠型プローブは、ターゲット配列にハイブリダイズし、環状化する。次いで、錠型プローブは、タイプIIS酵素による認識のための第一の部位で二本鎖を形成する。次いで、酵素を添加して、ターゲット配列を切断するために使用する。ターゲット配列の切断断片をプライマーとして機能させ、錠型プローブのローリングサークル複製を開始させるように条件を調整する。錠型プローブおよびターゲット核酸の何れか(もしくは何れも)を固定化することができる(できない)。
【0019】
このシステムを、添付の図面の図1で図解する。これは、第一の段階において、ターゲット核酸10が支持体12上に固定化されていることを示す。錠型プローブ14は、ターゲット核酸のターゲット配列にハイブリダイズし、環状化する。オリゴヌクレオチドプライマー16が、タイプIIS酵素18による認識のための錠型プローブの第一の部位にハイブリダイズし、タイプIIS酵素はターゲット配列内でターゲット核酸を切断しようとしている。図の第二段階において、タイプIIS制限エンドヌクレアーゼが、ターゲット配列を20で切断し、得られた3’端22は、錠型プローブ14を伴ってローリングサークル複製反応により鎖伸長される。
【0020】
ターゲット核酸がプローブとハイブリダイズする前に切断される場合は、別の可能性が生まれる。これは、切断により得られるターゲット核酸の断片が、それ自体で環状化し、ローリングサークル複製により増幅されるということである。従って、この側面において、本発明は、二つの非隣接ターゲット配列を有するターゲット核酸を検出する方法を提供し、この方法は、以下の工程を含む:
i)5’端ターゲット配列および3’端ターゲット配列を有するターゲット核酸断片を作成するためにターゲット核酸を切断する工程、
ii)前記二つのターゲット配列に相補的な二つの隣接配列を有するプローブを提供する工程、
iii)前記ターゲット核酸断片と前記プローブとのハイブリッドを形成し、前記ターゲット核酸断片を環状化する工程、
iv)前記環状化されたターゲット核酸断片のローリングサークル複製を行う工程。好ましくは、この工程は、前記プローブをプライマーとして使用して行われる。
【0021】
ローリングサークル産物を処理して、モノマーのサイズをゲル電気泳動で評価すれば、ターゲット配列のサイズを算定することができる。これは、脆弱X症候群のような状況でトリヌクレオチドの反復サイズを算定するのに有意であり得る。あるいは、環状分子もしくはそのローリングサークル複製による産物を、例えばDNA配列解析により調べることができる。上記の変形例において、プローブとターゲット核酸の両方を環状化されるようにデザインしてもよい。この後者のケースにおいて、一方のローリングサークル複製を引き起こすように他方が開放していてもよい。
【0022】
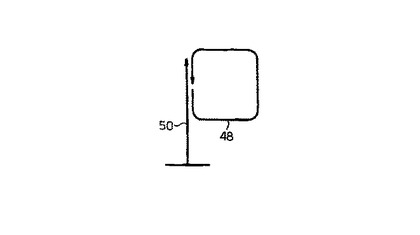
本発明のこの側面は、添付図面の図4に図解されている。図4において、溶液中のターゲット核酸断片48は、固定されたプローブ50にハイブリダイズし、環状化される。その後、固定化されたプローブ50は、ターゲット核酸断片48のローリングサークル複製のためのプライマーとして機能する。
【0023】
別の場合において、おそらく有利に、たった一つの錠型プローブの代わりに二つ(以上)の錠型プローブを使用することができる。本発明のこの側面において、ターゲット核酸のターゲット配列を検出する方法が提供され、本方法は、以下の工程を含む:
i)前記ターゲット配列に相補的な5’端および3’端配列、および中間配列を有する第一の錠型プローブを提供する工程、
ii)前記第一の錠型プローブの中間配列に相補的な5’端および3’端配列を有する第二の錠型プローブを提供する工程、
iii)前記第一の錠型プローブを前記ターゲット核酸にハイブリダイズし、前記第二の錠型プローブを前記第一の錠型プローブにハイブリダイズすることによりハイブリッドを形成し、前記二つの錠型プローブを環状化する工程、
iv)前記ハイブリッドを精製する工程、
v)前記ハイブリッドを制限処理し、これにより第一の錠型プローブを切断する工程、
vi)前記第二の錠型プローブのローリングサークル複製を行う工程。
【0024】
工程iii)において、前記第一の錠型プローブを前記ターゲット核酸にハイブリダイズし、前記第二の錠型プローブを前記第一の錠型プローブにハイブリダイズすることによりハイブリッドを形成し、前記二つの錠型プローブを環状化する。これらの工程を実施する技術は、当該分野で周知であり、上述したとおりである。ターゲット核酸、第一の錠型プローブ、もしくは第二の錠型プローブの何れか一つは(もしくは何れもが)固定化されていてもよい(固定化されていなくてもよい)。
【0025】
本方法の工程iv)は、ハイブリッドの精製を含み、これは、例えばDNAハイブリッドを変性させるpH、温度もしくはホルムアミド割合を使用して、超ストリンジェント洗浄により行うことができる。この工程は、環状化されなかった任意の第二の錠型プローブ;および(例えば、ターゲット配列にハイブリダイズしなかったために)環状化しなかった第一の錠型プローブにハイブリダイズした任意の第二の錠型プローブを除去するのに有効である。この分離は、ターゲット核酸もしくは第一の錠型プローブの何れかが固形支持体上に固定化されている場合、それが助けになる。工程v)において、第一の錠型プローブは切断され、工程vi)において第二の錠型プローブのローリングサークル複製が行われる。先の方法と同様、工程v)において、第一の錠型プローブが、第二の錠型プローブにハイブリダイズする中間配列内で切断されてプライマーを提供し、工程vi)において、このプライマーを使用して、第二の錠型プローブのローリングサークル複製を行うことが好ましい。
【0026】
第一の錠型プローブのその中間配列内での切断は、上述のタイプIIS制限エンドヌクレアーゼを使用することにより行われる。あるいは、第一の錠型プローブの中間配列が、UTPグリコシラーゼによる塩基除去、次いでExoIIIもしくはアルカリもしくは加熱による切断に感受性がある、dU残基のような切断可能な修飾残基を含んでいてもよい。中間配列は、任意に選択することができるため、任意の都合のよい制限酵素(例えばHincII)の認識配列を含み、第二の錠型プローブが、例えばホスホロチオエート結合を提供することにより保護されるようにデザインすることができる。
【0027】
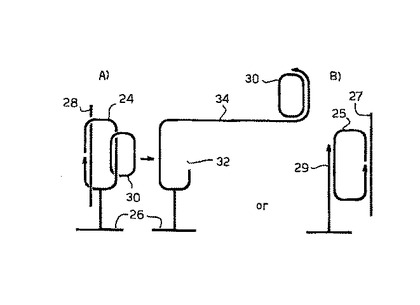
そのシステムを図2のパートAに図解する。この図の第一のパートにおいて、固形支持体26上に固定化された第一の錠型プローブ24に、溶液中のターゲット配列28、および第二の錠型プローブ30をハイブリダイズすることにより、ハイブリッドが形成される。二つの錠型プローブが環状化される。第二段階で、第一の錠型プローブが(第二の錠型プローブを切断することなく)32で切断され、制限断片34の3’端が、第二の錠型プローブのローリングサークル複製のためのプライマーとして機能する。
【0028】
上述の方法を実施するためのキットも、本発明の範囲内に含まれ、このキットは、上述の第一の錠型プローブと第二の錠型プローブを含む。このケースおよび他のケースにおいて、ローリングサークル複製のために使用されるヌクレオチドが、簡便な検出のために標識ヌクレオチドを含んでいることが一般に便利である。あるいは、ローリングサークル複製産物は、ハイブリダイゼーションプローブを使用して調べることができる。
【0029】
図2Bにおいて、ターゲット核酸断片27は、錠型プローブ25にハイブリダイズする。錠型プローブは環状化し、固定化されたオリゴヌクレオチド29をプライマーとして使用して、ローリングサークル複製により増幅されようとする。この技術の開発において、各分子の5’端および3’端がもう一方にハイブリダイズすると、錠型プローブ25とターゲット核酸断片27がともに環状化され、連結(catenate)されるが、ライゲーションにより結合する両端は、対向する互いをハイブリダイズしない。このプローブとターゲットの配置は、検出の特異性を増大させるが、一方の分子のローリングサークル複製は、好ましくは塩基対領域内で、他方の分子の直線化を必要とする。
【0030】
特許明細書WO96/14406は、錠型プローブの酵素による合成を開示している。これらのプローブは、連結(catenation)を避けることができた場合、ローリングサークル複製を介して検出することもできる。このことは、これらのプローブをタイプIIS制限酵素で作成し、ホスファターゼ処理、次いでγ−チオホスフェートATPでキナーゼ処理することにより5’−リン酸をチオホスフェートで置換した場合、成し遂げられる。このようにして、これらのプローブは、上記ターゲット認識の後、同じIIS酵素による処理から保護される。このように酵素により調製されたプローブの結合は、上述のように連結した検出用錠型プローブを用いて可視化することもできる。
【0031】
特許明細書WO97/00446によると、増強された免疫検出は、二つ以上のアフィニティープローブ(例えば抗体)がターゲット分子に同時に結合することにより成し遂げられる。ターゲット認識の際に、アフィニティープローブに結合したオリゴヌクレオチドは、PCRもしくはNASBA等による指数関数的な核酸の増幅反応の鋳型となり得る(template)長いオリゴヌクレオチドを形成するライゲーションによって結合するため充分近くに導かれる。この側面において、本発明は、関連したアプローチを使用するが、錠型プローブのローリングサークル複製を採用する。従って、本発明は、以下のものを提供することによりポリエピトープ・ターゲットをアッセイする方法を提供する:
a)第一のポリヌクレオチド配列を含むポリヌクレオチド鎖を有する、前記ターゲットのための第一のアフィニティープローブ、
b)第二の末端ポリヌクレオチド配列を含むポリヌクレオチド鎖を有する、前記ターゲットのための第二のアフィニティープローブ、および
c)第一のポリヌクレオチド配列に相補的な5’端および3’端配列、および第二のポリヌクレオチドの3’端に相補的な中間配列を有する錠型プローブ。
【0032】
本方法は、前記第一のアフィニティープローブを前記ターゲットに結合すること;前記第二のアフィニティープローブを前記ターゲットに結合すること;前記錠型プローブを前記第一のポリヌクレオチド配列および前記第二のポリヌクレオチド配列にハイブリダイズすること;前記錠型プローブを環状化すること;およびプライマーとして第二のポリヌクレオチド配列を使用して、前記錠型プローブのローリングサークル複製を行うことを具備する。
【0033】
本発明は、前記方法を実施するためのキットを更に提供し、このキットは、第一および第二のアフィニティープローブ、および錠型プローブを含む。好ましくは、第一のアフィニティープローブおよび第二のアフィニティープローブは、以下のものから選択される:ポリクローナル、モノクローナル、シングル鎖抗体およびその断片;レセプター;レクチンおよび核酸アプタマー(aptamer)。
【0034】
当該方法において、第一のアフィニティープローブおよび第二のアフィニティープローブは、これらプローブをターゲットに結合させるような条件下で、ターゲットともにインキュベートされる。次いで錠型プローブを、第一のポリヌクレオチド配列にハイブリダイズし環状化する。ターゲットに結合した二つのアフィニティープローブが、充分近くにいる場合、第二のアフィニティープローブの第二の末端ポリヌクレオチド配列が、錠型プローブの中間配列にハイブリダイズする。ヌクレオチドおよびポリメラーゼ酵素の供給添加の際に、錠型プローブのローリングサークル複製が行われ得る。放射能もしくはその他の方法で標識されたヌクレオチドの使用は、ポリエピトープ・ターゲットの存在もしくは濃度に関連して、対応して増幅されるシグナルを提供する。
【0035】
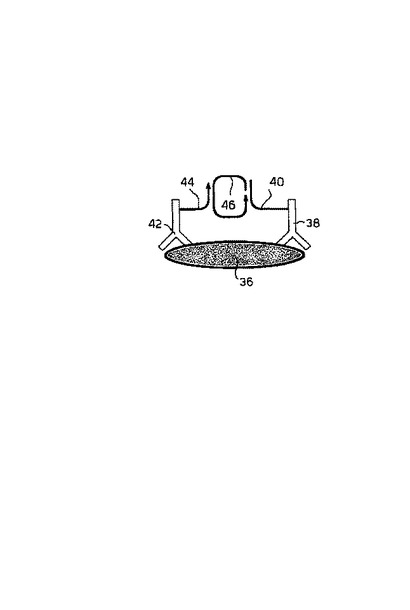
このシステムは、添付の図面の図3に図解され、図3では、ポリエピトープ・ターゲット36、第一のポリヌクレオチド配列を含むポリヌクレオチド鎖40を有するターゲットのための第一の抗体38;第二の末端ポリヌクレオチド配列を含むポリヌクレオチド鎖44を有するターゲットのための第二の抗体42;および二つのポリヌクレオチド鎖にハイブリダイズし環状化する錠型プローブ46が示されている。ヌクレオチドおよびポリメラーゼ酵素の供給添加の際に、ポリヌクレオチド鎖44は、錠型プローブ46のローリングサークル複製のためのプライマーとして機能する。
【0036】
ここに開示された任意の方法により作成されたローリングサークル産物は、いわゆる分子標識(beacon)を用いて合成の間に可視化することができる(S Tyagi and F R Kramer, 1996, Nature Biotechnology, 14, 303-308)。分子標識は、通常、一端に蛍光ラベルを有し、もう一端に蛍光を調整もしくは抑制する化合物を有するヘアピン型オリゴヌクレオチドである。通常のヘアピン型分子標識を折りたたまないことが、容易に観察される方法で蛍光シグナルを調整もしくは増強する。錠型プローブの配列に対応する配列を有するようにデザインされた分子標識は、錠型複製のローリングサークル複製をモニターするために使用することができる。
【0037】
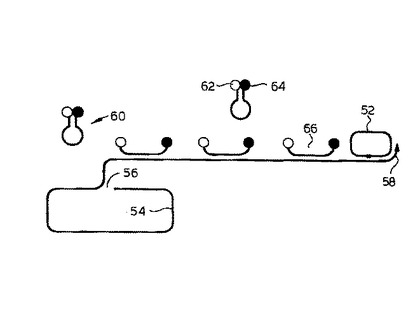
このシステムは、添付の図面の図5に図解される。錠型プローブ52は、ターゲット核酸54のターゲット配列にハイブリダイズし環状化する。ターゲット核酸は56で切断され、得られた3’端58は、錠型プローブ52を伴ってローリングサークル複製反応により鎖伸長される。分子標識60は、末端蛍光グループ62および末端クエンチンググループ64を有し、並びに錠型プローブ52の配列に対応する中間配列を有する。ローリングサークル複製が起こると、分子標識の連続した分子は、蛍光グループの蛍光を増強させる形態で、伸長鎖に66でハイブリダイズする。
【0038】
図6〜14に言及して、これを以下において説明する。
【0039】
[実験]
材料および方法
Interactivaにより合成されたオリゴヌクレオチド:
M13−Fo1s(56nt)は、6ホスホロチオエートを含む
M13−Fo2(56nt)は、ホスホロチオエートを含まない
M13−Ls1(23nt)は、FokI切断部位を含む
M13−T3sは、鋳型として使用する
M13−K1(34nt)は、FokI切断アダプターとして使用する。
【0040】
ABI394 DNA合成機を用いて研究室で合成したオリゴヌクレオチド:
M13c70−rol(70nt)は、錠型プローブとして使用する
プライマーMval(21nt)は、MvalによるM13切断のために使用する
RvRolcpr(21nt)は、HpaIIによるPCR産物の切断のために使用する。
【0041】
標識:
10mM Tris Ac(pH 7.5)、10mM MgAc2、50mM KAc、30μCi γ32P−ATP(3000 mCi/mmol、NEN Dupont)の50μL中で、37℃で10分間、30ユニットのT4ポリヌクレオチドキナーゼ(Amersham)を用いてプローブの5’を放射性標識し、オリゴヌクレオチドをSephadex G-50 μspinカラム(Pharmacia)で精製した。
【0042】
オリゴヌクレチドを用いたFokI切断:
標識されているオリゴヌクレオチドを混合し、1pmol M13−T3S、2pmol M13−Fo1sまたはM13−Fo2を、3pmol M13−Ls1またはM13−K1とともに、10mM Tris Ac(pH 7.5)、10mM MgAc2、50mM KAc、1mM ATP、0.1μg/μL BSAの20μL中で、65℃で10分間インキュベートすることによりハイブリダイズさせ、10分間室温に冷却した。FokIを0.5ユニット/μLの濃度まで添加し、37℃で60分間、その反応をインキュベートした。酵素を65℃で20分間、熱により不活性化し、その反応を10分間室温に冷却した。
【0043】
錠およびFokI−アダプターを用いたM13のFokI切断:
0.75pmol M13mp18+strand(Pharmacia)、0.25pmol M13c70−Rol(標識)および1.5pmol M13−K1を混合し、上述のとおりハイブリダイズさせた。FokI切断は、上述のとおり行った。
【0044】
錠およびMval−プライマーを用いたM13のMval切断:
10mM Tris Ac(pH 7.5)、10mM MgAc2、50mM KAc、1mM ATP、0.2μg/μL BSAの20μL中で、0.75pmol M13mp18+strand(Pharmacia)、0.25pmol M13c70−Rol(標識)および1.5pmol Mval−プライマーを、65℃で10分間インキュベートし、10分間室温に冷却した。2ユニットのMvalを添加し、その反応を37℃で60分間インキュベートした。酵素を65℃で15分間、熱により不活性化し、その反応を10分間室温に冷却した。
【0045】
Phi29 DNAポリメラーゼ反応:
50mM Tris−HCl(pH7.5)、10mM MgCl2、20mM (NH4)2SO4、1mM ジチオトレイトールおよび0.2μg/μL BSA、0.25mM dNTP、5μCi α32P dCTP(3000Ci/mmol、NEN Dupont)および2μg/μL Phi29 DNAポリメラーゼ(Amershamから提供)に、1μL酵素反応液を添加し、37℃で60分間インキュベートした。酵素を65℃で20分間、熱により不活性化した。
【0046】
PCR産物の切断:
10mM Bis Tris Propane−HCl、10mM MgCl2、1mM ジチオトレイトール(pH7.0)および1pmol/μL RvRolcprに、1μLのPCR反応液を添加し、65℃で10分間インキュベートし、10分間室温に冷却した。1ユニットのHpaIIを添加し、反応を37℃で一晩インキュベートした。酵素を65℃で20分間、熱により不活性化した。
【0047】
[配列]
M13−Fo1s
CAGCAGGATGTCTTCTAGTsGsCsCsAsAsGCTTGCATGCCTGCAGGTCGACTCTAGAGGAT
M13−Fo2
CAGCAGGATGCCTTCTAGTGCCAAGCTTGCATGCCTGCAGGTCGACTCTAGAGGAT
M13−Ls1
AGAAGACATCCTGsCsTsGsTsTsTsTsTsTsTs
M13−T3s
ATCCTCTAGAGTCGACCTGCAGGCATGCAAGCT↑TGGCACTGGC↑CGTCGTTTTACCAAACTCsAsAsGsAsAsGsGsAsCsC
M13−K1
CACATCCGTGCACGGATGTGGTAAAAC↑GACGGCC
M13c70−ro1
TGCCTGCAGGTCGACTTTTTTTTTTATGTTAAGTGACCGGCAGCATTTTTTTTTTAGTGCCAAGCTTGCA
プライマー MvaI
TGGGTAACGCCAGGGTTTTCC
RvRo1cpr
ATGTTAAGTTGACCGGCAGCA
【0048】
[例1]
FokIによる鎖特異的な切断およびホスホロチオエートによる保護(図6)
酵素滴定。この実験は、長さ29ヌクレオチドおよび39ヌクレオチドの二つの部分的に相補的なオリゴヌクレオチドを使用する。両ヌクレオチドを32P残基を付加してキナーゼ処理することにより5’端を標識した。左側のセット1から8までにおいて、両ヌクレオチドは標準型のヌクレオチドから成り、タイプIIS制限エンドヌクレアーゼFokIの添加により、両鎖ともに切断された。これにより、29mer断片が消失し、17mer断片が現れた。同様に。39merは8merに取って代わった。右側のセット1から8までにおいて、29merは、異なった方法で制限酵素が切断した位置で、ホスホロチオエート残基を含有していた。実験により、ホスホロチオエート残基が他の鎖の切断を阻害することなく、修飾鎖の切断を妨害していることが示された。(この特性は、この酵素について以前に実証されていなかったが、認識配列内に切断部位を有する幾つかのタイプII酵素については確立されていた。)
【0049】
[例2]
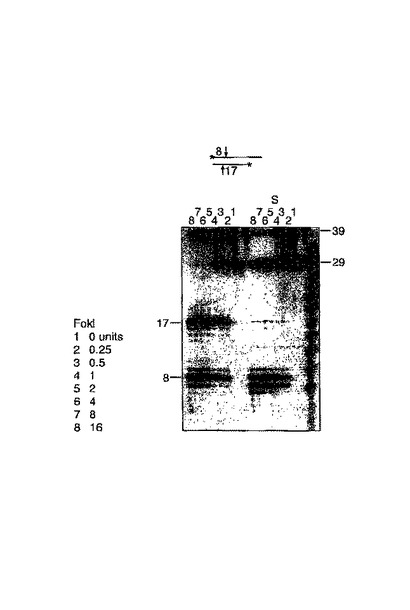
FokIを用いた鎖特異的な切断、その後のPhi29重合(図7)
ここで制限処理のために分枝した基質を使用した。56merは、その5’端において23merに相補的であり、その3’端において(放射性標識)71merに相補的である。23merは、酵素FokIの認識配列を含むが、切断部位はその枝の向こう側に位置する。レーン3に示されるサンプルは、未処理のものである。レーン1のサンプルは、制限酵素で処理され、33の短い断片を生成する。レーン4には、ポリメラーゼPhi29およびヌクレオチドが存在し、予想されるとおりレーン3と比較して重要ではない。最後にレーン2では、制限処理の後に重合が続く。切断産物の小さな断片のみが数ヌクレオチドによって伸長され、制限処理切断により生成された5’突出末端を補充する。この実験により、Szybalskiによって先に示されたように、枝を横切った切断が可能であることが実証された。ローリングサークル複製を開始するために必要とされるような、この実験における弱い重合の理由は、明らかではない。(100nt断片は、71merの予期しない伸長産物である。)
【0050】
[例3]
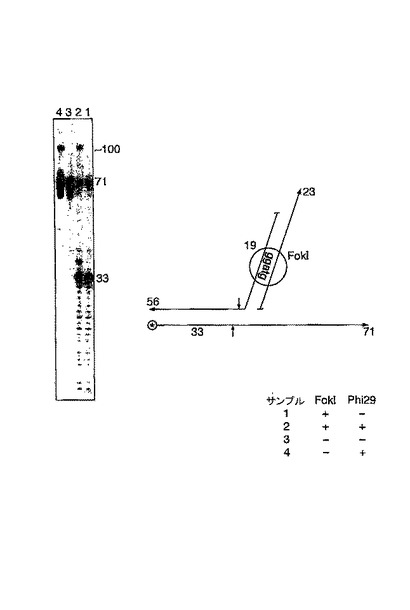
FokIを用いた鎖特異的な切断およびホスホロチオエートによる保護、その後のPhi29重合(図8)
この実験は、56merがホスホロチオエートで修飾されることにより、制限処理切断から保護されたことを除いて、先の実験と同じである。レーン3:出発材料。レーン1:制限処理切断。レーン4:ポリメラーゼおよびヌクレオチドの添加;多くの56merは未変化のまま残る(しかし、上記実験で述べたように、どういうわけか伸長したものもある)。レーン2:ここでは、33mer切断産物の多くが伸長し、未切断の56merをコピーし、錠認識のためのターゲット鎖に切断反応を指図することによりローリングサークル複製の開始を命令するのと同様に、ターゲットにローリングサークル複製反応を引き起こすことを可能にする。
【0051】
[例4]
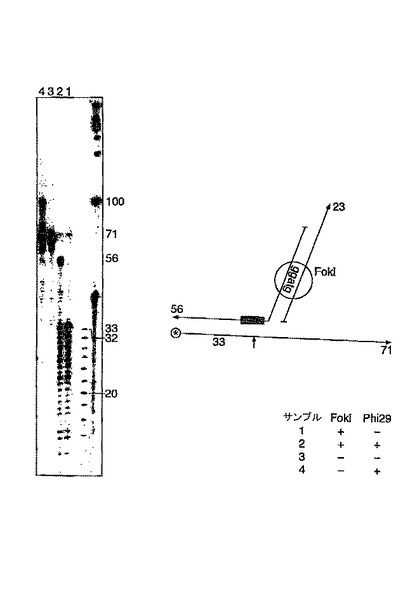
FokIによる鎖特異的な切断およびホスホロチオエートによる保護(図9)
放射性リン酸で56mer鎖を5’標識した以外、先の実験の簡単な変形である。保護されてない鎖は予想どおり切断されるが、保護されている鎖は、保護されてない位置で通常よりずっと離れて切断され、これは、より大きな配列がホスホロチオエートで修飾されることにより保護されなければならないことを示している。
【0052】
[例5]
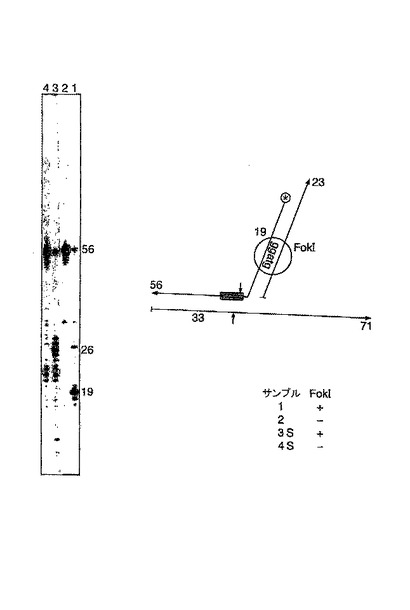
FokIアダプターを用いてFokIによる切断(図10)
この実験において、この場合プローブが結合した位置の下流で錠−ターゲット鎖を切断するために別の手段が使用される。自己相補的な断片を含むオリゴヌクレオチドは、制限酵素による認識およびこのアダプターがターゲット鎖にハイブリダイズする位置での切断を指揮する。レーン3:制限酵素なし。レーン1:短い断片の43塩基が得られる制限処理。レーン4:ポリメラーゼの添加による効果なし。レーン2:43merの切断産物の一部がポリメラーゼにより伸長され、未標識の56merオリゴヌクレオチドを鋳型として56merを生成する。
【0053】
[例6]
ターゲット鎖の切断によるローリングサークル複製の開始(図11)
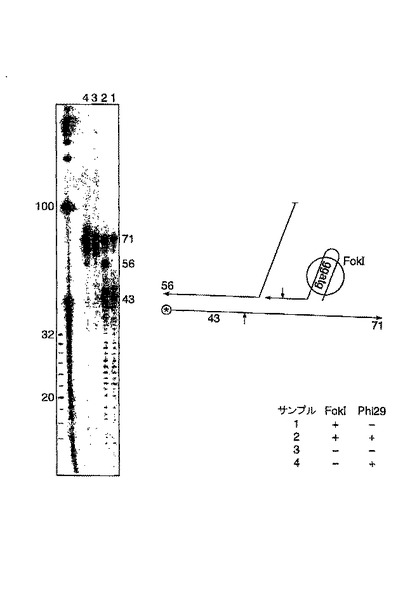
この実験において、先の実験の外部アダプター使用して、錠型プローブによる認識のための環状シングル鎖M13ターゲットを切断した。放射性ヌクレオチドおよび5’標識された錠型プローブの取り込みにより標識を行った。ここで示すこの実験は、M13を鋳型として、ライゲーションしていない錠がローリングサークル複製を引き起こしているため確定的ではない。このことは、例8に示すゲルで検討される。
【0054】
[例7]
ターゲット鎖の切断によるローリングサークル複製の開始(図12)
ターゲット鎖の切断が、オリゴヌクレオチドをハイブリダイズすることにより二本鎖にした固有の制限部位を利用したこと以外は、先の実験と同様である。また、二つのタイプの重合分子は、例8に特徴を示す。
【0055】
[例8]
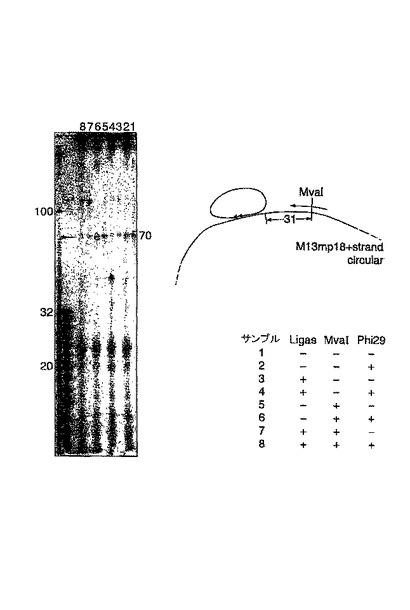
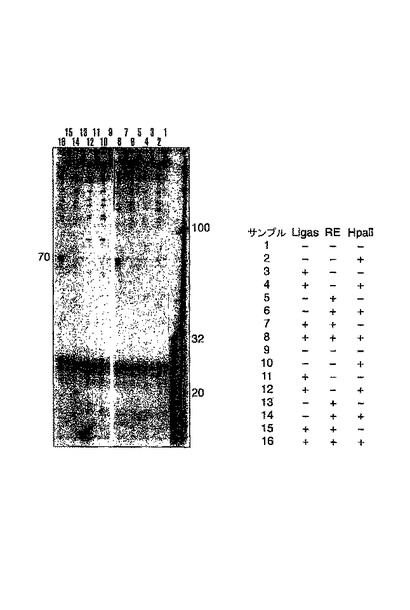
ローリングサークル産物の制限酵素による切断(図13)
ここで例6および7に示される実験を更に調べ、ポリメラーゼPhi29が、環状化された錠型プローブのローリングサークル複製を行うことができること、並びにターゲットM13鎖をFokIアダプター(例6)もしくはMvalの固有の認識配列を用いて切断した後、当該反応が、ターゲット配列により引き起こされることを明らかにした。リガーゼは、錠型プローブが接続されたか否かを示す。REは、錠型プローブが結合した部位の下流でターゲット分子を切断するための手段として使用される、FokI(レーン1〜8)もしくはMval(レーン9〜16)の何れかを示す。全てのサンプルは、Phi29ポリメラーゼで処理された。偶数レーンに示される伸長反応の産物は、オリゴヌクレオチドRvRo1cprの存在下で、制限酵素HpaIIにより処理され、ローリングサークル産物をモノマーに切断した。その切断により、M13ゲノムの鋳型に対するものとして、環状化された錠型プローブをコピーすることにより生じる重合産物の同定が可能になった。ライゲーションのためのターゲットとして機能するM13分子を、錠型プローブが結合した位置の下流で切断することにより、ターゲット鎖からローリングサークル複製が引き起こされることを、レーン8および16に見える酵素反応の70mer産物が明らかに示している。この反応は、錠型プローブのコピーを生成する。
【0056】
図14は、共にハイブリダイズした4つのオリゴマー、M13−Fo1s(56nt);M13−Ls1(23nt);M13−K1(34nt)およびM13−T3s(71nt)を示す。FokI結合および切断の部位、およびホスホロチオエート結合を、オリゴヌクレオチドの長さと共に示す。
【表1】

【発明の詳細な説明】
【0001】
[導入]
DNAまたはRNAサンプル中の単一もしくは複数のヌクレオチド変異を検出するために種々の方法が使用されている(1、2)。多くの方法は、分析前に一般にPCRによりターゲット配列を増幅することに依存している。従って、対立遺伝子の変異の位置に関する有意な情報は失われてしまう。配列変異の位置に関する情報は、例えば、遺伝性疾患の研究において染色体上の幾つかの変異配列のハプロタイプを決定するため;ある遺伝子における二つの変異が同一コピーに存在するか、別々の対立遺伝子に存在するかを決定するため;あるいは細胞周期における対立遺伝子の複製タイミング(3)または悪性組織における変異細胞の分布を研究するために重要である。In situでのシングルコピー遺伝子配列におけるシングルヌクレオチドの識別は、現在のところ不可能であるが、最近、新しいクラスの遺伝子診断プローブ分子である錠型プローブ(4)を用いて、繰り返し配列上で有効に操作されることが示された(5)。これらの錠型プローブは、両端にターゲットに相補的な配列を有し、その間にターゲットに相補的でない配列を有する直鎖状オリゴヌクレオチドである。正しいターゲットDNA配列にハイブリダイズすると、両端は先頭と後部とが一緒になって、DNAリガーゼにより結合される。二本鎖DNAのらせん特性の結果、得られた環状プローブ分子は、ターゲットDNA鎖に連結(catenate)される。このプローブのデザインは、幾つかの重要な特徴を有している。まず、ライゲーションが起こるための二つの別々のオリゴヌクレオチドのハイブリダイゼーションに必要なことは、完全なヒトゲノムの複雑性の中でシングルコピー遺伝子を検出するに充分な特異性を提供することである(6〜8)。二つめに、ライゲーション連結部におけるミスマッチにより、ライゲーションが大きく阻害され、これがターゲット配列中のシングルヌクレオチドの識別を可能にする(4、8〜10)。三つめに、プローブとターゲットDNA鎖との間のトポロジカル(topological)連結は、ハイブリダイゼーションの安定性とは無関係であるので、変性洗浄は、非特異的なハイブリダイゼーションを減少させることができる。四つめに、PCRもしくはLCRとは違って、分子内相互作用プローブの反応のみが記録され、これが大きなプローブセットを同時に適用する問題を回避する(11)。PCRプライマーの多くの対の組合わせは、任意のプライマーの組み合わせの間で形成される偽の増幅産物を急速に増加させる危険につながるが、これは錠型プローブには当てはまらない。最後に、プローブ端の結合が、反応前には存在しない新しいクラスの分子を作り出し、これらの環状分子は、ライゲートされたプローブを検出するために、ローリングサークル複製反応で増幅される(12、13)。
【0002】
ローリングサークル複製反応による溶液中フリーの環状核酸分子の増幅は、プライマーと環状核酸とのハイブリダイズ、そしてヌクレオチドおよびポリメラーゼ酵素の供給により成し遂げられる。しかし、環状核酸分子が、溶液中にフリーでない場合に問題が生じる。ターゲットに連結される錠型プライマーの特別なケースにおいて、そのターゲットがローリングサークル複製反応を抑制することが充分に予測される。本発明を導いた先の未発表の研究において、発明者らはこのような抑制が実際には起こらないことを実証した(18)。ターゲットが環状である場合、例えば環状7kbM13ゲノムである場合、ターゲットに連結された錠型プライマーのローリングサークル複製が効果的に妨害されることを発明者らは見出した。ターゲットが直鎖状である場合、その上に形成された錠が、ターゲット分子に沿ってスライドし、その端を離れ、それにより溶液中でフリーになり、ローリングサークル複製反応による増幅に利用されることが原則として可能である。あるいは、ターゲット配列、錠型プローブによる結合部位の3’が、エキソヌクレアーゼにより処理され、残りのターゲット鎖がローリングサークル複製を引き起こすことを可能にする。実際、発明者らは、このスライディング(sliding)および非連結(uncoupling)の効果は、長い直鎖状ターゲットでさえ限られた程度しか起こらないことを見出した。従って、単一部位での制限処理により、環状7kbM13ゲノムを、錠型プライマーのハイブリダイゼーション部位を3.5kb上流および3.5kb下流に有する直鎖状7kbの核酸分子に変換すれば、錠型プライマーの限定された増幅は、ローリングサークル複製によって起こり得る。しかし、ターゲット核酸が実質的に短い場所、例えば数十塩基もしくは数百塩基では、その上に形成された錠型プライマーのローリングサークル複製は、ずっと迅速で効率的である。
【0003】
[本発明]
本発明の一つの側面において、この課題は、ターゲットの認識およびプローブの環状化の前、その間、もしくはその後に、ターゲット分子を切断することにより解決される。この側面において、本発明は、ターゲット核酸のターゲット配列を検出する方法を提供し、この方法は、以下の工程を含む:
i)ターゲット配列のための錠型プローブを提供する工程、
ii)錠型プローブとターゲット核酸とのハイブッリドを形成し、錠型プローブを環状化する工程;
iii)工程ii)の前、工程ii)の間、もしくは工程ii)の後に、ターゲット核酸をターゲット配列もしくはその近傍で切断する工程;
iv)錠型プローブのローリングサークル複製を行う工程。
【図面の簡単な説明】
【0004】
【図1】本発明の方法を説明する図。
【図2】本発明の方法を説明する図。
【図3】本発明の方法を説明する図。
【図4】本発明の方法を説明する図。
【図5】本発明の方法を説明する図。
【図6】例1の結果を示す図。
【図7】例2の結果を示す図。
【図8】例3の結果を示す図。
【図9】例4の結果を示す図。
【図10】例5の結果を示す図。
【図11】例6の結果を示す図。
【図12】例7の結果を示す図。
【図13】例8の結果を示す図。
【図14】例8の結果を示す図。
【発明を実施するための形態】
【0005】
前記ターゲット核酸は、原則としてDNAでもRNAでもよい。本発明の方法は、ターゲット核酸が長鎖分子もしくは環状分子である場合特に有効であると思われる。ターゲット配列は、錠型プローブの両端とハイブリダイズするのに充分な長さである。本発明の方法は、シングルコピー遺伝子配列の位置検出、顕微鏡標本における配列変異型の間の区別、および一次元もしくは二次元アレイに固定されたプローブにとって特に有効であると期待される。
【0006】
錠型プローブは、ターゲット配列に相補的な5’端配列および3’端配列を有する;その結果、錠型プローブは、ターゲット配列にハイブリダイズすると、例えばライゲーションにより環状化することができる。厳密な相補性は必ずしも要求されないが、錠型プローブとターゲット配列とのハイブリダイゼーションおよびライゲーションによる錠型プローブの環状化を可能にする充分な相補性が必要である。原則として二つの部分で錠型プローブを提供することが可能であり、この場合、環状化を成し遂げるためには二つのライゲーション反応が必要である。錠型プローブのサイズは、ローリングサークル複製反応による増幅に適している必要があり、好ましくは、少なくとも約25 nt、多くて150〜200 ntであるが、特に5’から3’のエキソヌクレアーゼ活性をもたないポリメラーゼと使用するときは特に、より大きなプローブが有効である。
【0007】
本方法の工程ii)は、錠型プローブとターゲット核酸のターゲット配列とがハイブッリドを形成し、錠型プローブが環状化することを含む。これらの工程は、慣用的な条件下で行うことができる。調査中のターゲット核酸サンプルが、ターゲット配列を有していない場合、錠型プローブのハイブリダイゼーションおよび環状化は全く起こらないか限られた程度でしか起こらない。
【0008】
本方法の工程iii)において、ターゲット核酸は、ターゲット配列もしくはその近傍で切断される。この切断は、当該分野で周知の方法により行うことができる。ターゲット配列から制限部位の距離が相当な場合、例えば3.5 kbもしくはそれ以上の場合もあり得るが、好ましくは、ターゲット配列の数塩基以内もしくは数十塩基以内である。
【0009】
このようにターゲット核酸は、錠型プローブの添加前に、制限処理により直線状にすることができる。あるいは、プライマー伸長反応を、錠型プローブ認識用のターゲットとして適切な比較的短い核酸分子を作成するように行うことができる。中期染色体のin situ検出において、これら二つの方法は、ターゲットDNAの損失を引き起こし、これにより検出のロスを引き起こすという不利益を受けやすい。従って好ましくは、工程iii)は、ハイブリッドを制限処理し、これにより環状化された錠型プローブを切断することなくターゲット核酸をターゲット配列もしくはその近傍で切断することにより行われる。
【0010】
工程iv)において、錠型プローブは、ローリングサークル複製反応により増幅される。この反応は、通常、環状化された錠型プローブにハイブリダイズするプライマー、ヌクレオチドの供給、およびポリメラーゼ酵素を必要とし、当該分野で周知の方法により行うことができる。好ましいポリメラーゼ酵素は、高い処理特性および3’−エキソヌクレアーゼ活性を有するφ29 DNAポリメラーゼである。
【0011】
従って、ターゲット配列を、錠型プローブの結合した位置の3’で切断した場合、錠型プローブに対する塩基対の3’端が得られるまで、任意の非塩基対ヌクレオチドをポリメラーゼにより除去し、そこで、外部のプライマーを添加することなくローリングサークル複製(PCR)を開始することができ、PCR産物がターゲット配列と連続することが保証される。このような切断は、プローブが結合した位置の下流で、ターゲット配列にオリゴヌクレオチドをハイブリダイズすることにより成し遂げられる。次いで、このハイブリダイゼーションによりその認識配列が二本鎖になった制限酵素を用いてその配列を切断することができる。あるいは、タイプIIS酵素(例えばFokI)の認識配列を有する二本鎖の一端、および錠型プローブが結合した位置のターゲット配列3’にハイブリダイズする一端を有するヘアーピンオリゴヌクレオチドを使用することができる。
【0012】
錠型プローブが制限部位を横切ってターゲット配列にハイブリダイズする場合、その錠を、例えばホスホロチオエートで修飾することにより保護することができ、ターゲット配列は錠を開けることなく切断される。
【0013】
ターゲット核酸がRNAであって、ターゲット配列内の部位で制限処理される場合、RNase H処理により、錠型プライマーのローリングサークル複製を開始可能な3’端を得ることができる。
【0014】
好ましい方法によると、工程iii)において、ターゲット核酸はターゲット配列内で切断される。この方法の利点は、ターゲット配列の3’端がプライマーを構成することであり、工程iv)において、このプライマーを用いて錠型プローブのローリングサークル複製を行うことができる。これにより、生成されたローリングサークル複製産物は、ターゲット配列と連続している。
【0015】
これを成し遂げるために、好ましくはタイプIIS酵素を利用する(タイプIIS酵素は、クラスIIS酵素もしくはタイプIV酵素とも称される)。タイプIIS制限エンドヌクレアーゼは、文献にその記載がある(14、15)。これら酵素は、二本鎖DNA上の二つの別々の部位;4〜7bpの長さの認識部位、および認識部位から通常1〜20bp離れた切断部位と相互作用する。このようなタイプIIS酵素の一つは、FokI制限エンドヌクレアーゼであり(16、17)、この酵素は、二本鎖の5’−GGATG−3’部位を認識し、5’−3’鎖および3’−5’鎖からそれぞれ9ヌクレオチドおよび13ヌクレオチド下流で切断する。FokIをssDNAおよびオリゴヌクレオチドと使用すると、認識部位から9番目もしくは13番目のヌクレオチドで切断が観察される。認識部位と切断部位との間の二本鎖領域が完全に相補的であるか否かに関わらず切断は起こる。
【0016】
上記方法でタイプIIS酵素を使用するためには、特別にデザインした錠型プローブが必要であり、これは、本発明の別の側面を形成する。この側面において、ターゲット核酸配列のための錠型プローブとして使用するに適したオリゴヌクレオチドを提供し、このオリゴヌクレオチドは、ターゲット配列に相補的な5’端および3’端配列;タイプIIS酵素による認識のための第一の部位;および少なくとも一のヌクレオチド残基および/またはヌクレオチド間結合を修飾して、タイプIIS酵素による制限処理からオリゴヌクレオチドを保護する第二の部位を有する。
【0017】
タイプIIS酵素による認識のための第一の部位は、該酵素がターゲット核酸配列を切断するように、錠型プローブ上に位置する必要がある。そして第二の部位は、タイプIIS酵素によって錠型プローブが切断されないように保護するために、第一の部位に関連して錠型プローブ上に位置する必要がある。第一の部位と第二の部位との相対的な位置は、5’端および3’端配列に関連して、使用する特定のタイプIIS酵素に依存する。第二の部位における修飾も、使用する特定のタイプIIS酵素に依存する。例えば、FokI酵素を使用する場合、タイプIIS酵素のHincIIにおいても同様に、ホスホロチオエートのヌクレオチド間結合が、錠型プローブの切断を防止するのに有効である。
【0018】
本発明の方法において、錠型プローブは、ターゲット配列にハイブリダイズし、環状化する。次いで、錠型プローブは、タイプIIS酵素による認識のための第一の部位で二本鎖を形成する。次いで、酵素を添加して、ターゲット配列を切断するために使用する。ターゲット配列の切断断片をプライマーとして機能させ、錠型プローブのローリングサークル複製を開始させるように条件を調整する。錠型プローブおよびターゲット核酸の何れか(もしくは何れも)を固定化することができる(できない)。
【0019】
このシステムを、添付の図面の図1で図解する。これは、第一の段階において、ターゲット核酸10が支持体12上に固定化されていることを示す。錠型プローブ14は、ターゲット核酸のターゲット配列にハイブリダイズし、環状化する。オリゴヌクレオチドプライマー16が、タイプIIS酵素18による認識のための錠型プローブの第一の部位にハイブリダイズし、タイプIIS酵素はターゲット配列内でターゲット核酸を切断しようとしている。図の第二段階において、タイプIIS制限エンドヌクレアーゼが、ターゲット配列を20で切断し、得られた3’端22は、錠型プローブ14を伴ってローリングサークル複製反応により鎖伸長される。
【0020】
ターゲット核酸がプローブとハイブリダイズする前に切断される場合は、別の可能性が生まれる。これは、切断により得られるターゲット核酸の断片が、それ自体で環状化し、ローリングサークル複製により増幅されるということである。従って、この側面において、本発明は、二つの非隣接ターゲット配列を有するターゲット核酸を検出する方法を提供し、この方法は、以下の工程を含む:
i)5’端ターゲット配列および3’端ターゲット配列を有するターゲット核酸断片を作成するためにターゲット核酸を切断する工程、
ii)前記二つのターゲット配列に相補的な二つの隣接配列を有するプローブを提供する工程、
iii)前記ターゲット核酸断片と前記プローブとのハイブリッドを形成し、前記ターゲット核酸断片を環状化する工程、
iv)前記環状化されたターゲット核酸断片のローリングサークル複製を行う工程。好ましくは、この工程は、前記プローブをプライマーとして使用して行われる。
【0021】
ローリングサークル産物を処理して、モノマーのサイズをゲル電気泳動で評価すれば、ターゲット配列のサイズを算定することができる。これは、脆弱X症候群のような状況でトリヌクレオチドの反復サイズを算定するのに有意であり得る。あるいは、環状分子もしくはそのローリングサークル複製による産物を、例えばDNA配列解析により調べることができる。上記の変形例において、プローブとターゲット核酸の両方を環状化されるようにデザインしてもよい。この後者のケースにおいて、一方のローリングサークル複製を引き起こすように他方が開放していてもよい。
【0022】
本発明のこの側面は、添付図面の図4に図解されている。図4において、溶液中のターゲット核酸断片48は、固定されたプローブ50にハイブリダイズし、環状化される。その後、固定化されたプローブ50は、ターゲット核酸断片48のローリングサークル複製のためのプライマーとして機能する。
【0023】
別の場合において、おそらく有利に、たった一つの錠型プローブの代わりに二つ(以上)の錠型プローブを使用することができる。本発明のこの側面において、ターゲット核酸のターゲット配列を検出する方法が提供され、本方法は、以下の工程を含む:
i)前記ターゲット配列に相補的な5’端および3’端配列、および中間配列を有する第一の錠型プローブを提供する工程、
ii)前記第一の錠型プローブの中間配列に相補的な5’端および3’端配列を有する第二の錠型プローブを提供する工程、
iii)前記第一の錠型プローブを前記ターゲット核酸にハイブリダイズし、前記第二の錠型プローブを前記第一の錠型プローブにハイブリダイズすることによりハイブリッドを形成し、前記二つの錠型プローブを環状化する工程、
iv)前記ハイブリッドを精製する工程、
v)前記ハイブリッドを制限処理し、これにより第一の錠型プローブを切断する工程、
vi)前記第二の錠型プローブのローリングサークル複製を行う工程。
【0024】
工程iii)において、前記第一の錠型プローブを前記ターゲット核酸にハイブリダイズし、前記第二の錠型プローブを前記第一の錠型プローブにハイブリダイズすることによりハイブリッドを形成し、前記二つの錠型プローブを環状化する。これらの工程を実施する技術は、当該分野で周知であり、上述したとおりである。ターゲット核酸、第一の錠型プローブ、もしくは第二の錠型プローブの何れか一つは(もしくは何れもが)固定化されていてもよい(固定化されていなくてもよい)。
【0025】
本方法の工程iv)は、ハイブリッドの精製を含み、これは、例えばDNAハイブリッドを変性させるpH、温度もしくはホルムアミド割合を使用して、超ストリンジェント洗浄により行うことができる。この工程は、環状化されなかった任意の第二の錠型プローブ;および(例えば、ターゲット配列にハイブリダイズしなかったために)環状化しなかった第一の錠型プローブにハイブリダイズした任意の第二の錠型プローブを除去するのに有効である。この分離は、ターゲット核酸もしくは第一の錠型プローブの何れかが固形支持体上に固定化されている場合、それが助けになる。工程v)において、第一の錠型プローブは切断され、工程vi)において第二の錠型プローブのローリングサークル複製が行われる。先の方法と同様、工程v)において、第一の錠型プローブが、第二の錠型プローブにハイブリダイズする中間配列内で切断されてプライマーを提供し、工程vi)において、このプライマーを使用して、第二の錠型プローブのローリングサークル複製を行うことが好ましい。
【0026】
第一の錠型プローブのその中間配列内での切断は、上述のタイプIIS制限エンドヌクレアーゼを使用することにより行われる。あるいは、第一の錠型プローブの中間配列が、UTPグリコシラーゼによる塩基除去、次いでExoIIIもしくはアルカリもしくは加熱による切断に感受性がある、dU残基のような切断可能な修飾残基を含んでいてもよい。中間配列は、任意に選択することができるため、任意の都合のよい制限酵素(例えばHincII)の認識配列を含み、第二の錠型プローブが、例えばホスホロチオエート結合を提供することにより保護されるようにデザインすることができる。
【0027】
そのシステムを図2のパートAに図解する。この図の第一のパートにおいて、固形支持体26上に固定化された第一の錠型プローブ24に、溶液中のターゲット配列28、および第二の錠型プローブ30をハイブリダイズすることにより、ハイブリッドが形成される。二つの錠型プローブが環状化される。第二段階で、第一の錠型プローブが(第二の錠型プローブを切断することなく)32で切断され、制限断片34の3’端が、第二の錠型プローブのローリングサークル複製のためのプライマーとして機能する。
【0028】
上述の方法を実施するためのキットも、本発明の範囲内に含まれ、このキットは、上述の第一の錠型プローブと第二の錠型プローブを含む。このケースおよび他のケースにおいて、ローリングサークル複製のために使用されるヌクレオチドが、簡便な検出のために標識ヌクレオチドを含んでいることが一般に便利である。あるいは、ローリングサークル複製産物は、ハイブリダイゼーションプローブを使用して調べることができる。
【0029】
図2Bにおいて、ターゲット核酸断片27は、錠型プローブ25にハイブリダイズする。錠型プローブは環状化し、固定化されたオリゴヌクレオチド29をプライマーとして使用して、ローリングサークル複製により増幅されようとする。この技術の開発において、各分子の5’端および3’端がもう一方にハイブリダイズすると、錠型プローブ25とターゲット核酸断片27がともに環状化され、連結(catenate)されるが、ライゲーションにより結合する両端は、対向する互いをハイブリダイズしない。このプローブとターゲットの配置は、検出の特異性を増大させるが、一方の分子のローリングサークル複製は、好ましくは塩基対領域内で、他方の分子の直線化を必要とする。
【0030】
特許明細書WO96/14406は、錠型プローブの酵素による合成を開示している。これらのプローブは、連結(catenation)を避けることができた場合、ローリングサークル複製を介して検出することもできる。このことは、これらのプローブをタイプIIS制限酵素で作成し、ホスファターゼ処理、次いでγ−チオホスフェートATPでキナーゼ処理することにより5’−リン酸をチオホスフェートで置換した場合、成し遂げられる。このようにして、これらのプローブは、上記ターゲット認識の後、同じIIS酵素による処理から保護される。このように酵素により調製されたプローブの結合は、上述のように連結した検出用錠型プローブを用いて可視化することもできる。
【0031】
特許明細書WO97/00446によると、増強された免疫検出は、二つ以上のアフィニティープローブ(例えば抗体)がターゲット分子に同時に結合することにより成し遂げられる。ターゲット認識の際に、アフィニティープローブに結合したオリゴヌクレオチドは、PCRもしくはNASBA等による指数関数的な核酸の増幅反応の鋳型となり得る(template)長いオリゴヌクレオチドを形成するライゲーションによって結合するため充分近くに導かれる。この側面において、本発明は、関連したアプローチを使用するが、錠型プローブのローリングサークル複製を採用する。従って、本発明は、以下のものを提供することによりポリエピトープ・ターゲットをアッセイする方法を提供する:
a)第一のポリヌクレオチド配列を含むポリヌクレオチド鎖を有する、前記ターゲットのための第一のアフィニティープローブ、
b)第二の末端ポリヌクレオチド配列を含むポリヌクレオチド鎖を有する、前記ターゲットのための第二のアフィニティープローブ、および
c)第一のポリヌクレオチド配列に相補的な5’端および3’端配列、および第二のポリヌクレオチドの3’端に相補的な中間配列を有する錠型プローブ。
【0032】
本方法は、前記第一のアフィニティープローブを前記ターゲットに結合すること;前記第二のアフィニティープローブを前記ターゲットに結合すること;前記錠型プローブを前記第一のポリヌクレオチド配列および前記第二のポリヌクレオチド配列にハイブリダイズすること;前記錠型プローブを環状化すること;およびプライマーとして第二のポリヌクレオチド配列を使用して、前記錠型プローブのローリングサークル複製を行うことを具備する。
【0033】
本発明は、前記方法を実施するためのキットを更に提供し、このキットは、第一および第二のアフィニティープローブ、および錠型プローブを含む。好ましくは、第一のアフィニティープローブおよび第二のアフィニティープローブは、以下のものから選択される:ポリクローナル、モノクローナル、シングル鎖抗体およびその断片;レセプター;レクチンおよび核酸アプタマー(aptamer)。
【0034】
当該方法において、第一のアフィニティープローブおよび第二のアフィニティープローブは、これらプローブをターゲットに結合させるような条件下で、ターゲットともにインキュベートされる。次いで錠型プローブを、第一のポリヌクレオチド配列にハイブリダイズし環状化する。ターゲットに結合した二つのアフィニティープローブが、充分近くにいる場合、第二のアフィニティープローブの第二の末端ポリヌクレオチド配列が、錠型プローブの中間配列にハイブリダイズする。ヌクレオチドおよびポリメラーゼ酵素の供給添加の際に、錠型プローブのローリングサークル複製が行われ得る。放射能もしくはその他の方法で標識されたヌクレオチドの使用は、ポリエピトープ・ターゲットの存在もしくは濃度に関連して、対応して増幅されるシグナルを提供する。
【0035】
このシステムは、添付の図面の図3に図解され、図3では、ポリエピトープ・ターゲット36、第一のポリヌクレオチド配列を含むポリヌクレオチド鎖40を有するターゲットのための第一の抗体38;第二の末端ポリヌクレオチド配列を含むポリヌクレオチド鎖44を有するターゲットのための第二の抗体42;および二つのポリヌクレオチド鎖にハイブリダイズし環状化する錠型プローブ46が示されている。ヌクレオチドおよびポリメラーゼ酵素の供給添加の際に、ポリヌクレオチド鎖44は、錠型プローブ46のローリングサークル複製のためのプライマーとして機能する。
【0036】
ここに開示された任意の方法により作成されたローリングサークル産物は、いわゆる分子標識(beacon)を用いて合成の間に可視化することができる(S Tyagi and F R Kramer, 1996, Nature Biotechnology, 14, 303-308)。分子標識は、通常、一端に蛍光ラベルを有し、もう一端に蛍光を調整もしくは抑制する化合物を有するヘアピン型オリゴヌクレオチドである。通常のヘアピン型分子標識を折りたたまないことが、容易に観察される方法で蛍光シグナルを調整もしくは増強する。錠型プローブの配列に対応する配列を有するようにデザインされた分子標識は、錠型複製のローリングサークル複製をモニターするために使用することができる。
【0037】
このシステムは、添付の図面の図5に図解される。錠型プローブ52は、ターゲット核酸54のターゲット配列にハイブリダイズし環状化する。ターゲット核酸は56で切断され、得られた3’端58は、錠型プローブ52を伴ってローリングサークル複製反応により鎖伸長される。分子標識60は、末端蛍光グループ62および末端クエンチンググループ64を有し、並びに錠型プローブ52の配列に対応する中間配列を有する。ローリングサークル複製が起こると、分子標識の連続した分子は、蛍光グループの蛍光を増強させる形態で、伸長鎖に66でハイブリダイズする。
【0038】
図6〜14に言及して、これを以下において説明する。
【0039】
[実験]
材料および方法
Interactivaにより合成されたオリゴヌクレオチド:
M13−Fo1s(56nt)は、6ホスホロチオエートを含む
M13−Fo2(56nt)は、ホスホロチオエートを含まない
M13−Ls1(23nt)は、FokI切断部位を含む
M13−T3sは、鋳型として使用する
M13−K1(34nt)は、FokI切断アダプターとして使用する。
【0040】
ABI394 DNA合成機を用いて研究室で合成したオリゴヌクレオチド:
M13c70−rol(70nt)は、錠型プローブとして使用する
プライマーMval(21nt)は、MvalによるM13切断のために使用する
RvRolcpr(21nt)は、HpaIIによるPCR産物の切断のために使用する。
【0041】
標識:
10mM Tris Ac(pH 7.5)、10mM MgAc2、50mM KAc、30μCi γ32P−ATP(3000 mCi/mmol、NEN Dupont)の50μL中で、37℃で10分間、30ユニットのT4ポリヌクレオチドキナーゼ(Amersham)を用いてプローブの5’を放射性標識し、オリゴヌクレオチドをSephadex G-50 μspinカラム(Pharmacia)で精製した。
【0042】
オリゴヌクレチドを用いたFokI切断:
標識されているオリゴヌクレオチドを混合し、1pmol M13−T3S、2pmol M13−Fo1sまたはM13−Fo2を、3pmol M13−Ls1またはM13−K1とともに、10mM Tris Ac(pH 7.5)、10mM MgAc2、50mM KAc、1mM ATP、0.1μg/μL BSAの20μL中で、65℃で10分間インキュベートすることによりハイブリダイズさせ、10分間室温に冷却した。FokIを0.5ユニット/μLの濃度まで添加し、37℃で60分間、その反応をインキュベートした。酵素を65℃で20分間、熱により不活性化し、その反応を10分間室温に冷却した。
【0043】
錠およびFokI−アダプターを用いたM13のFokI切断:
0.75pmol M13mp18+strand(Pharmacia)、0.25pmol M13c70−Rol(標識)および1.5pmol M13−K1を混合し、上述のとおりハイブリダイズさせた。FokI切断は、上述のとおり行った。
【0044】
錠およびMval−プライマーを用いたM13のMval切断:
10mM Tris Ac(pH 7.5)、10mM MgAc2、50mM KAc、1mM ATP、0.2μg/μL BSAの20μL中で、0.75pmol M13mp18+strand(Pharmacia)、0.25pmol M13c70−Rol(標識)および1.5pmol Mval−プライマーを、65℃で10分間インキュベートし、10分間室温に冷却した。2ユニットのMvalを添加し、その反応を37℃で60分間インキュベートした。酵素を65℃で15分間、熱により不活性化し、その反応を10分間室温に冷却した。
【0045】
Phi29 DNAポリメラーゼ反応:
50mM Tris−HCl(pH7.5)、10mM MgCl2、20mM (NH4)2SO4、1mM ジチオトレイトールおよび0.2μg/μL BSA、0.25mM dNTP、5μCi α32P dCTP(3000Ci/mmol、NEN Dupont)および2μg/μL Phi29 DNAポリメラーゼ(Amershamから提供)に、1μL酵素反応液を添加し、37℃で60分間インキュベートした。酵素を65℃で20分間、熱により不活性化した。
【0046】
PCR産物の切断:
10mM Bis Tris Propane−HCl、10mM MgCl2、1mM ジチオトレイトール(pH7.0)および1pmol/μL RvRolcprに、1μLのPCR反応液を添加し、65℃で10分間インキュベートし、10分間室温に冷却した。1ユニットのHpaIIを添加し、反応を37℃で一晩インキュベートした。酵素を65℃で20分間、熱により不活性化した。
【0047】
[配列]
M13−Fo1s
CAGCAGGATGTCTTCTAGTsGsCsCsAsAsGCTTGCATGCCTGCAGGTCGACTCTAGAGGAT
M13−Fo2
CAGCAGGATGCCTTCTAGTGCCAAGCTTGCATGCCTGCAGGTCGACTCTAGAGGAT
M13−Ls1
AGAAGACATCCTGsCsTsGsTsTsTsTsTsTsTs
M13−T3s
ATCCTCTAGAGTCGACCTGCAGGCATGCAAGCT↑TGGCACTGGC↑CGTCGTTTTACCAAACTCsAsAsGsAsAsGsGsAsCsC
M13−K1
CACATCCGTGCACGGATGTGGTAAAAC↑GACGGCC
M13c70−ro1
TGCCTGCAGGTCGACTTTTTTTTTTATGTTAAGTGACCGGCAGCATTTTTTTTTTAGTGCCAAGCTTGCA
プライマー MvaI
TGGGTAACGCCAGGGTTTTCC
RvRo1cpr
ATGTTAAGTTGACCGGCAGCA
【0048】
[例1]
FokIによる鎖特異的な切断およびホスホロチオエートによる保護(図6)
酵素滴定。この実験は、長さ29ヌクレオチドおよび39ヌクレオチドの二つの部分的に相補的なオリゴヌクレオチドを使用する。両ヌクレオチドを32P残基を付加してキナーゼ処理することにより5’端を標識した。左側のセット1から8までにおいて、両ヌクレオチドは標準型のヌクレオチドから成り、タイプIIS制限エンドヌクレアーゼFokIの添加により、両鎖ともに切断された。これにより、29mer断片が消失し、17mer断片が現れた。同様に。39merは8merに取って代わった。右側のセット1から8までにおいて、29merは、異なった方法で制限酵素が切断した位置で、ホスホロチオエート残基を含有していた。実験により、ホスホロチオエート残基が他の鎖の切断を阻害することなく、修飾鎖の切断を妨害していることが示された。(この特性は、この酵素について以前に実証されていなかったが、認識配列内に切断部位を有する幾つかのタイプII酵素については確立されていた。)
【0049】
[例2]
FokIを用いた鎖特異的な切断、その後のPhi29重合(図7)
ここで制限処理のために分枝した基質を使用した。56merは、その5’端において23merに相補的であり、その3’端において(放射性標識)71merに相補的である。23merは、酵素FokIの認識配列を含むが、切断部位はその枝の向こう側に位置する。レーン3に示されるサンプルは、未処理のものである。レーン1のサンプルは、制限酵素で処理され、33の短い断片を生成する。レーン4には、ポリメラーゼPhi29およびヌクレオチドが存在し、予想されるとおりレーン3と比較して重要ではない。最後にレーン2では、制限処理の後に重合が続く。切断産物の小さな断片のみが数ヌクレオチドによって伸長され、制限処理切断により生成された5’突出末端を補充する。この実験により、Szybalskiによって先に示されたように、枝を横切った切断が可能であることが実証された。ローリングサークル複製を開始するために必要とされるような、この実験における弱い重合の理由は、明らかではない。(100nt断片は、71merの予期しない伸長産物である。)
【0050】
[例3]
FokIを用いた鎖特異的な切断およびホスホロチオエートによる保護、その後のPhi29重合(図8)
この実験は、56merがホスホロチオエートで修飾されることにより、制限処理切断から保護されたことを除いて、先の実験と同じである。レーン3:出発材料。レーン1:制限処理切断。レーン4:ポリメラーゼおよびヌクレオチドの添加;多くの56merは未変化のまま残る(しかし、上記実験で述べたように、どういうわけか伸長したものもある)。レーン2:ここでは、33mer切断産物の多くが伸長し、未切断の56merをコピーし、錠認識のためのターゲット鎖に切断反応を指図することによりローリングサークル複製の開始を命令するのと同様に、ターゲットにローリングサークル複製反応を引き起こすことを可能にする。
【0051】
[例4]
FokIによる鎖特異的な切断およびホスホロチオエートによる保護(図9)
放射性リン酸で56mer鎖を5’標識した以外、先の実験の簡単な変形である。保護されてない鎖は予想どおり切断されるが、保護されている鎖は、保護されてない位置で通常よりずっと離れて切断され、これは、より大きな配列がホスホロチオエートで修飾されることにより保護されなければならないことを示している。
【0052】
[例5]
FokIアダプターを用いてFokIによる切断(図10)
この実験において、この場合プローブが結合した位置の下流で錠−ターゲット鎖を切断するために別の手段が使用される。自己相補的な断片を含むオリゴヌクレオチドは、制限酵素による認識およびこのアダプターがターゲット鎖にハイブリダイズする位置での切断を指揮する。レーン3:制限酵素なし。レーン1:短い断片の43塩基が得られる制限処理。レーン4:ポリメラーゼの添加による効果なし。レーン2:43merの切断産物の一部がポリメラーゼにより伸長され、未標識の56merオリゴヌクレオチドを鋳型として56merを生成する。
【0053】
[例6]
ターゲット鎖の切断によるローリングサークル複製の開始(図11)
この実験において、先の実験の外部アダプター使用して、錠型プローブによる認識のための環状シングル鎖M13ターゲットを切断した。放射性ヌクレオチドおよび5’標識された錠型プローブの取り込みにより標識を行った。ここで示すこの実験は、M13を鋳型として、ライゲーションしていない錠がローリングサークル複製を引き起こしているため確定的ではない。このことは、例8に示すゲルで検討される。
【0054】
[例7]
ターゲット鎖の切断によるローリングサークル複製の開始(図12)
ターゲット鎖の切断が、オリゴヌクレオチドをハイブリダイズすることにより二本鎖にした固有の制限部位を利用したこと以外は、先の実験と同様である。また、二つのタイプの重合分子は、例8に特徴を示す。
【0055】
[例8]
ローリングサークル産物の制限酵素による切断(図13)
ここで例6および7に示される実験を更に調べ、ポリメラーゼPhi29が、環状化された錠型プローブのローリングサークル複製を行うことができること、並びにターゲットM13鎖をFokIアダプター(例6)もしくはMvalの固有の認識配列を用いて切断した後、当該反応が、ターゲット配列により引き起こされることを明らかにした。リガーゼは、錠型プローブが接続されたか否かを示す。REは、錠型プローブが結合した部位の下流でターゲット分子を切断するための手段として使用される、FokI(レーン1〜8)もしくはMval(レーン9〜16)の何れかを示す。全てのサンプルは、Phi29ポリメラーゼで処理された。偶数レーンに示される伸長反応の産物は、オリゴヌクレオチドRvRo1cprの存在下で、制限酵素HpaIIにより処理され、ローリングサークル産物をモノマーに切断した。その切断により、M13ゲノムの鋳型に対するものとして、環状化された錠型プローブをコピーすることにより生じる重合産物の同定が可能になった。ライゲーションのためのターゲットとして機能するM13分子を、錠型プローブが結合した位置の下流で切断することにより、ターゲット鎖からローリングサークル複製が引き起こされることを、レーン8および16に見える酵素反応の70mer産物が明らかに示している。この反応は、錠型プローブのコピーを生成する。
【0056】
図14は、共にハイブリダイズした4つのオリゴマー、M13−Fo1s(56nt);M13−Ls1(23nt);M13−K1(34nt)およびM13−T3s(71nt)を示す。FokI結合および切断の部位、およびホスホロチオエート結合を、オリゴヌクレオチドの長さと共に示す。
【表1】
【特許請求の範囲】
【請求項1】
以下の工程を含む、ターゲット核酸のターゲット配列を検出する方法:
i)ターゲット配列のための錠型プローブを提供する工程、
ii)錠型プローブとターゲット核酸とのハイブリッドを形成し、錠型プローブを環状化する工程;
iii)工程ii)の前、工程ii)の間、もしくは工程ii)の後に、ターゲット核酸をターゲット配列もしくはその近傍で切断する工程;
iv)錠型プローブのローリングサークル複製を行う工程。
【請求項2】
請求項1記載の方法であって、工程iii)が、前記ハイブリッドを制限処理(restriction)し、それにより前記環状化された錠型プローブを切断することなくターゲット核酸をターゲット配列もしくはその近傍で切断することにより行われる方法。
【請求項3】
請求項1または2記載の方法であって、工程iii)において、前記ターゲット核酸をターゲット配列内で切断してプライマーを提供し、工程iv)において、このプライマー用いて錠型プローブのローリングサークル複製を行う方法。
【請求項4】
請求項1または2記載の方法であって、前記ターゲット核酸をターゲット配列の下流で切断し、その後3’−エキソヌクレアーゼにより任意の非塩基対ヌクレオチドを除去する方法。
【請求項5】
請求項4記載の方法であって、Phi29を、3’−エキソヌクレアーゼ活性を更に有するポリメラーゼ酵素として使用する方法。
【請求項6】
請求項3ないし5の何れか1項記載の方法であって、工程iii)においてタイプIIS酵素により制限処理を行う方法。
【請求項7】
請求項1ないし6の何れか1項記載の方法であって、前記ターゲット核酸が環状である方法。
【請求項8】
ターゲット核酸配列のための錠型プローブとして使用するに適したオリゴヌクレオチドであって、ターゲット配列に相補的な5’端および3’配列;タイプIIS酵素による認識のための第一の部位;および少なくとも一のヌクレオチド残基および/またはヌクレオチド間結合を修飾して、タイプIIS酵素による制限処理からオリゴヌクレオチドを保護する第二の部位を有するオリゴヌクレオチド。
【請求項9】
請求項8記載のオリゴヌクレオチドであって、少なくとも一のホスホロチオエートのヌクレオチド間結合を前記第二の部位に提供して、タイプIIS酵素による制限処理からオリゴヌクレオチドを保護するオリゴヌクレオチド。
【請求項10】
請求項8または9記載のオリゴヌクレオチドであって、前記第一の部位が、タイプIIS酵素FokIによる認識に適したオリゴヌクレオチド。
【請求項11】
以下の工程を含む、二つの非隣接ターゲット配列を有するターゲット核酸を検出する方法:
i)5’端ターゲット配列および3’端ターゲット配列を有するターゲット核酸断片を作成するためにターゲット核酸を切断する工程、
ii)前記二つのターゲット配列に相補的な二つの隣接配列を有するプローブを提供する工程、
iii)前記ターゲット核酸断片と前記プローブとのハイブリッドを形成し、前記ターゲット核酸断片を環状化する工程、
iv)前記環状化されたターゲット核酸断片のローリングサークル複製を行う工程。
【請求項12】
以下の工程を含む、ターゲット核酸のターゲット配列を検出する方法:
i)前記ターゲット配列に相補的な5’端および3’端配列、および中間配列を有する第一の錠型プローブを提供する工程、
ii)前記第一の錠型プローブの中間配列に相補的な5’端および3’端配列を有する第二の錠型プローブを提供する工程、
iii)前記第一の錠型プローブを前記ターゲット核酸にハイブリダイズし、前記第二の錠型プローブを前記第一の錠型プローブにハイブリダイズすることによりハイブリッドを形成し、前記二つの錠型プローブを環状化する工程、
iv)前記ハイブリッドを精製する工程、
v)前記ハイブリッドを制限処理し、これにより第一の錠型プローブを切断する工程、
vi)前記第二の錠型プローブのローリングサークル複製を行う工程。
【請求項13】
請求項12に記載の方法であって、工程v)において、前記第一の錠型プローブを、前記第二の錠型プローブにハイブリダイズする中間配列内で切断してプライマーを提供し、工程vi)において、このプライマーを用いて前記第二の錠型プローブのローリングサークル複製を行う方法。
【請求項14】
請求項12に記載の方法であって、工程v)において、前記第一の錠型プローブを、前記第二の錠型プローブにハイブリダイズする前記中間配列の下流で切断し、その後3’−エキソヌクレアーゼにより任意の非塩基対ヌクレオチドを除去してプライマーを提供し、工程vi)において、前記プライマーを用いて前記第二の錠型プローブのローリングサークル複製を行う方法。
【請求項15】
請求項14記載の方法であって、Phi29を、3’−エキソヌクレアーゼ活性を更に有するポリメラーゼ酵素として使用する方法。
【請求項16】
請求項13ないし15の何れか1項記載の方法であって、工程v)においてタイプIIS酵素により制限処理を行う方法。
【請求項17】
以下のものを含む、ターゲット核酸のターゲット配列を検出するためのキット:
a)前記ターゲット配列に相補的な5’端および3’端配列、および中間配列を有する第一の錠型プローブ、
b)前記第一のプローブの中間配列に相補的な5’端および3’端配列を有する第二の錠型プローブ。
【請求項18】
以下のものを提供することにより、ポリエピトープ・ターゲットをアッセイする方法であって:
a)第一のポリヌクレオチド配列を含むポリヌクレオチド鎖を有する、前記ターゲットのための第一のアフィニティープローブ、
b)第二の末端ポリヌクレオチド配列を含むポリヌクレオチド鎖を有する、前記ターゲットのための第二のアフィニティープローブ、および
c)第一のポリヌクレオチド配列に相補的な5’端および3’端配列、および第二のポリヌクレオチド配列に相補的な中間配列を有する錠型プローブ、
前記第一のアフィニティープローブを前記ターゲットに結合すること;前記第二のアフィニティープローブを前記ターゲットに結合すること;前記錠型プローブを、前記第一のポリヌクレオチド配列および前記第二のポリヌクレオチド配列にハイブリダイズすること;前記錠型プローブを環状化すること;およびプライマーとして前記第二のポリヌクレオチド配列を使用して、前記錠型プローブのローリングサークル複製を行うことを具備する方法。
【請求項19】
以下のものを含む、ポリエピトープ・ターゲットをアッセイするためのキット:
a)第一のポリヌクレオチド配列を含むポリヌクレオチド鎖を有する、前記ターゲットのための第一のアフィニティープローブ、
b)第二の末端ポリヌクレオチド配列を含むポリヌクレオチド鎖を有する、前記ターゲットのための第二のアフィニティープローブ、および
c)第一のポリヌクレオチド配列に相補的な5’端および3’端配列、および第二のポリヌクレオチドに相補的な中間配列を有する錠型プローブ。
【請求項20】
請求項19記載のキットであって、前記第一のアフィニティープローブおよび第二のアフィニティープローブがともに抗体であるキット。
【請求項1】
以下の工程を含む、ターゲット核酸のターゲット配列を検出する方法:
i)ターゲット配列のための錠型プローブを提供する工程、
ii)錠型プローブとターゲット核酸とのハイブリッドを形成し、錠型プローブを環状化する工程;
iii)工程ii)の前、工程ii)の間、もしくは工程ii)の後に、ターゲット核酸をターゲット配列もしくはその近傍で切断する工程;
iv)錠型プローブのローリングサークル複製を行う工程。
【請求項2】
請求項1記載の方法であって、工程iii)が、前記ハイブリッドを制限処理(restriction)し、それにより前記環状化された錠型プローブを切断することなくターゲット核酸をターゲット配列もしくはその近傍で切断することにより行われる方法。
【請求項3】
請求項1または2記載の方法であって、工程iii)において、前記ターゲット核酸をターゲット配列内で切断してプライマーを提供し、工程iv)において、このプライマー用いて錠型プローブのローリングサークル複製を行う方法。
【請求項4】
請求項1または2記載の方法であって、前記ターゲット核酸をターゲット配列の下流で切断し、その後3’−エキソヌクレアーゼにより任意の非塩基対ヌクレオチドを除去する方法。
【請求項5】
請求項4記載の方法であって、Phi29を、3’−エキソヌクレアーゼ活性を更に有するポリメラーゼ酵素として使用する方法。
【請求項6】
請求項3ないし5の何れか1項記載の方法であって、工程iii)においてタイプIIS酵素により制限処理を行う方法。
【請求項7】
請求項1ないし6の何れか1項記載の方法であって、前記ターゲット核酸が環状である方法。
【請求項8】
ターゲット核酸配列のための錠型プローブとして使用するに適したオリゴヌクレオチドであって、ターゲット配列に相補的な5’端および3’配列;タイプIIS酵素による認識のための第一の部位;および少なくとも一のヌクレオチド残基および/またはヌクレオチド間結合を修飾して、タイプIIS酵素による制限処理からオリゴヌクレオチドを保護する第二の部位を有するオリゴヌクレオチド。
【請求項9】
請求項8記載のオリゴヌクレオチドであって、少なくとも一のホスホロチオエートのヌクレオチド間結合を前記第二の部位に提供して、タイプIIS酵素による制限処理からオリゴヌクレオチドを保護するオリゴヌクレオチド。
【請求項10】
請求項8または9記載のオリゴヌクレオチドであって、前記第一の部位が、タイプIIS酵素FokIによる認識に適したオリゴヌクレオチド。
【請求項11】
以下の工程を含む、二つの非隣接ターゲット配列を有するターゲット核酸を検出する方法:
i)5’端ターゲット配列および3’端ターゲット配列を有するターゲット核酸断片を作成するためにターゲット核酸を切断する工程、
ii)前記二つのターゲット配列に相補的な二つの隣接配列を有するプローブを提供する工程、
iii)前記ターゲット核酸断片と前記プローブとのハイブリッドを形成し、前記ターゲット核酸断片を環状化する工程、
iv)前記環状化されたターゲット核酸断片のローリングサークル複製を行う工程。
【請求項12】
以下の工程を含む、ターゲット核酸のターゲット配列を検出する方法:
i)前記ターゲット配列に相補的な5’端および3’端配列、および中間配列を有する第一の錠型プローブを提供する工程、
ii)前記第一の錠型プローブの中間配列に相補的な5’端および3’端配列を有する第二の錠型プローブを提供する工程、
iii)前記第一の錠型プローブを前記ターゲット核酸にハイブリダイズし、前記第二の錠型プローブを前記第一の錠型プローブにハイブリダイズすることによりハイブリッドを形成し、前記二つの錠型プローブを環状化する工程、
iv)前記ハイブリッドを精製する工程、
v)前記ハイブリッドを制限処理し、これにより第一の錠型プローブを切断する工程、
vi)前記第二の錠型プローブのローリングサークル複製を行う工程。
【請求項13】
請求項12に記載の方法であって、工程v)において、前記第一の錠型プローブを、前記第二の錠型プローブにハイブリダイズする中間配列内で切断してプライマーを提供し、工程vi)において、このプライマーを用いて前記第二の錠型プローブのローリングサークル複製を行う方法。
【請求項14】
請求項12に記載の方法であって、工程v)において、前記第一の錠型プローブを、前記第二の錠型プローブにハイブリダイズする前記中間配列の下流で切断し、その後3’−エキソヌクレアーゼにより任意の非塩基対ヌクレオチドを除去してプライマーを提供し、工程vi)において、前記プライマーを用いて前記第二の錠型プローブのローリングサークル複製を行う方法。
【請求項15】
請求項14記載の方法であって、Phi29を、3’−エキソヌクレアーゼ活性を更に有するポリメラーゼ酵素として使用する方法。
【請求項16】
請求項13ないし15の何れか1項記載の方法であって、工程v)においてタイプIIS酵素により制限処理を行う方法。
【請求項17】
以下のものを含む、ターゲット核酸のターゲット配列を検出するためのキット:
a)前記ターゲット配列に相補的な5’端および3’端配列、および中間配列を有する第一の錠型プローブ、
b)前記第一のプローブの中間配列に相補的な5’端および3’端配列を有する第二の錠型プローブ。
【請求項18】
以下のものを提供することにより、ポリエピトープ・ターゲットをアッセイする方法であって:
a)第一のポリヌクレオチド配列を含むポリヌクレオチド鎖を有する、前記ターゲットのための第一のアフィニティープローブ、
b)第二の末端ポリヌクレオチド配列を含むポリヌクレオチド鎖を有する、前記ターゲットのための第二のアフィニティープローブ、および
c)第一のポリヌクレオチド配列に相補的な5’端および3’端配列、および第二のポリヌクレオチド配列に相補的な中間配列を有する錠型プローブ、
前記第一のアフィニティープローブを前記ターゲットに結合すること;前記第二のアフィニティープローブを前記ターゲットに結合すること;前記錠型プローブを、前記第一のポリヌクレオチド配列および前記第二のポリヌクレオチド配列にハイブリダイズすること;前記錠型プローブを環状化すること;およびプライマーとして前記第二のポリヌクレオチド配列を使用して、前記錠型プローブのローリングサークル複製を行うことを具備する方法。
【請求項19】
以下のものを含む、ポリエピトープ・ターゲットをアッセイするためのキット:
a)第一のポリヌクレオチド配列を含むポリヌクレオチド鎖を有する、前記ターゲットのための第一のアフィニティープローブ、
b)第二の末端ポリヌクレオチド配列を含むポリヌクレオチド鎖を有する、前記ターゲットのための第二のアフィニティープローブ、および
c)第一のポリヌクレオチド配列に相補的な5’端および3’端配列、および第二のポリヌクレオチドに相補的な中間配列を有する錠型プローブ。
【請求項20】
請求項19記載のキットであって、前記第一のアフィニティープローブおよび第二のアフィニティープローブがともに抗体であるキット。
【図1】


【図2】
【図3】
【図4】
【図5】
【図6】
【図7】
【図8】
【図9】
【図10】
【図11】

【図12】
【図13】
【図14】

【図2】
【図3】
【図4】
【図5】
【図6】
【図7】
【図8】
【図9】
【図10】
【図11】
【図12】
【図13】
【図14】
【公開番号】特開2012−210213(P2012−210213A)
【公開日】平成24年11月1日(2012.11.1)
【国際特許分類】
【外国語出願】
【出願番号】特願2012−121944(P2012−121944)
【出願日】平成24年5月29日(2012.5.29)
【分割の表示】特願2009−108689(P2009−108689)の分割
【原出願日】平成11年3月25日(1999.3.25)
【出願人】(512112334)
【氏名又は名称原語表記】Olink AB
【住所又は居所原語表記】Dag Hammaskjolds vag 54A, SE−751 83 Uppsala, Sweden
【Fターム(参考)】
【公開日】平成24年11月1日(2012.11.1)
【国際特許分類】
【出願番号】特願2012−121944(P2012−121944)
【出願日】平成24年5月29日(2012.5.29)
【分割の表示】特願2009−108689(P2009−108689)の分割
【原出願日】平成11年3月25日(1999.3.25)
【出願人】(512112334)
【氏名又は名称原語表記】Olink AB
【住所又は居所原語表記】Dag Hammaskjolds vag 54A, SE−751 83 Uppsala, Sweden
【Fターム(参考)】
[ Back to top ]