
長い半減期と増強された赤血球形成活性を備えた組換えヒトEPO−Fc融合タンパク質

【課題】有意に長い半減期を有し、生体内での赤血球形成活性が増強されているが、免疫原特性が増加していないEPOアナログを提供する。
【解決手段】天然または組換えの未変性ヒトエリスロポエチンに比べて長い生体内半減期を有する融合タンパク質であって、(a)C末端付近にシステイン残基を有する天然ヒトエリスロポエチン分子、および(b)ヒンジ領域を含むヒトIgG Fcフラグメントであって、前記FcフラグメントのN末端が前記エリスロポエチン分子のC末端に直接結合され、前記Fcフラグメントは、前記エリスロポエチン分子に最も近い前記ヒンジ領域のシステイン残基が非システイン残基と置換された突然変異を有する以外は天然のFcフラグメントと同じであり、前記ヒンジ領域の最初のシステインが前記エリスロポエチン分子の前記システイン残基から少なくとも17個のアミノ酸だけ離れている。
【解決手段】天然または組換えの未変性ヒトエリスロポエチンに比べて長い生体内半減期を有する融合タンパク質であって、(a)C末端付近にシステイン残基を有する天然ヒトエリスロポエチン分子、および(b)ヒンジ領域を含むヒトIgG Fcフラグメントであって、前記FcフラグメントのN末端が前記エリスロポエチン分子のC末端に直接結合され、前記Fcフラグメントは、前記エリスロポエチン分子に最も近い前記ヒンジ領域のシステイン残基が非システイン残基と置換された突然変異を有する以外は天然のFcフラグメントと同じであり、前記ヒンジ領域の最初のシステインが前記エリスロポエチン分子の前記システイン残基から少なくとも17個のアミノ酸だけ離れている。
【発明の詳細な説明】
【技術分野】
【0001】
本願はヒトエリスロポエチン融合タンパク質に関する。
【背景技術】
【0002】
ヒトエリスロポエチン(EPO)は、造血成長因子ファミリーのメンバーであり、血中酸素利用率の低下による組織低酸素症に応じて主として成人の腎臓および胎児の肝臓で合成される[1]。EPOの主な機能は、骨髄中の特定の赤血球(RBC)前駆細胞および前駆物質に直接作用して、ヘモグロビンおよび成熟RBCの合成を刺激することである。EPOはRBCの増殖、分化および成熟も制御する。腎臓の機能不全に関連する貧血、癌および他の病理学的状態を治療するために天然EPOのアミノ酸配列を有する組換えEPOが生産および承認されている[2]。その赤血球形成の特性に加えて、最近の研究によると、EPOがニューロン等の非骨髄細胞にも作用することが報告されており[3]、これは中枢神経系(CNS)および他の器官/系におけるEPOの他の生理学的/病理学的機能の可能性を示唆している。EPO受容体が種々の器官で見出されたため、EPOは、抗アポトーシス物質として働く等、多くの生物学的効果を有する可能性がある。
【0003】
ヒトEPOは30.4キロダルトンの分子量を有する糖タンパク質である。炭水化物はその合計質量の約39%を占める。EPO遺伝子は7q11−22染色体上に位置し、5つのエクソンと4つのイントロンを備えた5.4kbの領域に及ぶ[4]。EPOの前駆物質は193個のアミノ酸から成る。翻訳後修飾によるリーダー配列と最後のアミノ酸Argとの開裂により、165個のアミノ酸を有する成熟EPOが生じる。糖鎖形成は、Asn24、Asn38、Asn83の3つのN結合部位と、Ser126の1つのO結合部位であるが、EPOの生合成、三次構造および生体内生理活性に重大な役割を果たしている[5]。エリスロポエチン受容体すなわち72−78キロダルトンの分子量を有する糖鎖付加されリン酸化された膜貫通ポリペプチドに結合することにより機能する。この結合は、受容体のホモダイマー化を引き起こし、これがJAK2/STAT5系、G−タンパク質、カルシウムチャネルおよびキナーゼといういくつかのシグナル伝達経路の活性化につながる。EPOタンパク質の2つの分子は、最適の受容体活性化を達成するためには1つの受容体分子に同時に結合する必要がある[6]。
【0004】
ヒト治療のために認可された最初の造血生長因子として、組換えヒトEPO(rHuEPO)は慢性腎不全、癌(主として化学療法に関連する貧血)、自己免疫疾患、AIDS、外科手術、骨髄移植および骨髄異形成症候群等に起因する貧血の治療に使用されてきた。面白いことに、最近の研究によると、rHuEPOは非血液系の機能を有することが観察され、脳虚血、頭部外傷、炎症性疾患および神経変性疾患用の神経保護剤として使用される可能性が示されている。
【0005】
現在、3種類のrHuEPOまたはrHuEPOのアナログすなわちrHuEPOアルファ、rHuEPOベータおよびダルベポエチンアルファが市販されている[8]。これら3つの組換えタンパク質は同じエリスロポエチン受容体に結合するが、構造、糖鎖形成の程度、受容体結合能および生体内での代謝は異なっている。1980年代にrHuEPOアルファを初めて導入して以来、臨床医は同薬剤の頻繁な投薬/注射の必要性を重大な欠点と素早く認めた。r静脈内または皮下に投与したHuEPOアルファとrHuEPOベータの平均生体内半減期はそれぞれわずか8.5時間と17時間である[9,10]。したがって、患者は毎日、一週間に2回、または一週間に3回の注射スケジュールを必要とし、これは患者と医療提供者の両方に負担を強いる。したがって、より長い生体内半減
期および/または増強された赤血球形成活性を有する組換えEPOアナログを開発する必要があった。
【0006】
先行技術では、その生体内代謝を遅延させるかまたはその治療的性質を改善するために、未変性EPOタンパク質の構造を遺伝子改変するか化学修飾する試みが行われていた。例えば、EPO分子中のシアル酸含有炭水化物の量と、その生体内代謝および機能的活性との間には直接相関があるように見える。したがって、EPO分子の炭水化物含有量を増加させると、半減期が長くなり、生体内活性も増強された[11,12]。Amgenは、rHuEPOよりも2個多い5個のN結合炭水化物鎖を含むようrHuEPOアナログのダルベポエチンアルファを設計した。ダルベポエチンアルファも新規な赤血球形成刺激タンパク質(NESP)として知られており、Aranesp(商標)として販売されている。ダルベポエチンアルファは5つの位置(Ala30Asn、His32Thr、Pro87Val、Trp88Asn、Pro90Thr)で未変性ヒトEPOと異なり、これによりアスパラギン残基位置30および88で2つの追加のN結合オリゴ糖を取り付けることが可能となっている。ダルベポエチンアルファは未変性EPOと同じようにEPO受容体に結合して、JAK−2キナーゼおよび同じ細胞内分子Ras/MAP−k、P13−kおよびSTAT−5によるチロシンリン酸化を伴う細胞内シグナル伝達を引き起こす。炭水化物含有量の増加により、動物とヒトの両方でのダルベポエチンアルファの半減期はrHuEPO−アルファの半減期のほぼ3倍の長さになっている(25.3時間対8.5時間)[9]。ダルベポエチンアルファ(Aranesp(商標))は、さらに天然または組換えヒトEPOに比べて生体内で増強された生理活性を示すと思われ[13]、第二代のrHuEPO薬としてFDAによって認可された。この薬剤はrHuEPOの1週間で2〜3回の注射と同一の治療効果を達成するのに、一週間に1回の投与しか必要としない[10,14,15]。
【0007】
EPOの半減期を拡張する他の試みは、ポリエチレングリコールやその同等物との化学共役(PEG化)によりEPOタンパク質の分子量を増加させることに焦点が当てられていた。PEG化ははるかに大きな分子量を有し、循環からの除去が防がれるためより長い血漿半減期を有する[16]。しかしながら、PEG化はEPO部分の予期せぬ機能の変化および特異性の変化を生じさせる可能性がある。EPO分子を担体タンパク質(ヒトアルブミン)へ結合させるか、またはリンクペプチド(3から17個のアミノ酸)の使用によりもしくは化学的架橋試薬により2つの完全EPO分子のホモダイマーを形成する等のさらに別の方法によりEPOの分子量を増加させる報告も存在する[17,18,19,20]。これらの方法はすべて半減期の延長およびEPO活性の増強のある程度の成功を達成しているが、本願に記載するようにEPO分子をヒト免疫グロブリン(IgG)のFcフラグメントと組み合わせて融合タンパク質とすることは、固有の利点を達成する。
【0008】
ヒト免疫グロブリンIgGは、ジスルフィド結合により共有結合された4つのポリペプチドから構成されている(軽鎖と重鎖の2つの同一コピー)。パパインによるIgG分子のタンパク質分解によって、2つのFAbフラグメントと1つのFcフラグメントが生じる。Fcフラグメントは、ジスルフィド結合によって共に結合した2つのポリペプチドから成る。各ポリペプチドは、N末端からC末端まで、ヒンジ領域、CH2ドメインおよびCH3ドメインから構成される。Fcフラグメント構造はヒト免疫グロブリンのすべてのサブタイプでほぼ同じである。IgGはヒト血液中に最も豊富にあるタンパク質の1つであり、ヒト血清中の免疫グロブリンの合計の70〜75%を占める。循環でのIgGの半減期は免疫グロブリンの5つすべてのタイプのうち最長であり、21日に達し得る。
【0009】
現代の生物工学技術をうまく適用して、サイトカインおよび可溶性受容体等の治療用タンパク質フラグメントとヒトIgGのFcフラグメントから成る融合タンパク質が形成された[21,22,23,24]。これらの融合タンパク質は、それらの生物学的特性お
よび治療特性を保持する間、有意により長い生体内半減期を有する。これまでのところ、Fcフラグメントを含む2つの融合タンパク質がバイオ医薬としての開発に成功しており、慢性関節リウマチおよび慢性尋常性乾癬の治療用にFDAにより認可されている[25,26]。
【0010】
先行技術では、化学的架橋によるかまたはポリペプチドにより結合された2つのEPO分子からなる二量体が生体内活性の増強および長い半減期を示すことが論証された[17,19]。この活性の増強は1つの受容体にEPO二量体がより効率的に結合するためであり、生体内での半減期の延長は二量体タンパク質のサイズが大きいことによるためであり得る。しかしながら、化学的架橋プロセスは効率的でなく、制御することが難しい。さらに、EPOの二量体中のリンケージペプチドは、EPO分子の三次元構造を変化させる可能性があり、また、ペプチドそのものが生体内での免疫原性反応を刺激する可能性がある。特に腎患者でEPO置換療法は生涯にわたって続くため、これらの欠点はEPO二量体の治療可能性を損なう。
【発明の概要】
【発明が解決しようとする課題】
【0011】
有意に長い半減期を有し、生体内での赤血球形成活性が増強されているが、免疫原特性が増加していないEPOアナログが必要とされている。
【課題を解決するための手段】
【0012】
本発明によれば、免疫グロブリンペプチド部分に結合されたヒトエリスロポエチンペプチド部分を含む組換え融合タンパク質について説明される。融合タンパク質は、天然または組換えの未変性ヒトエリスロポエチンに比べて長い生体内半減期を有する。本発明の1実施形態では、融合タンパク質は未変性ヒトエリスロポエチンよりも少なくとも3倍長い生体内半減期を有する。融合タンパク質はさらに、未変性ヒトエリスロポエチンに比べて増強された赤血球形成の生理活性を示してもよい。本発明の1実施形態では、免疫グロブリンペプチド部分がIgG1フラグメント等のFcフラグメントである。FcフラグメントはCH2領域、CH3の領域およびヒンジ領域を有する。EPOペプチド部分はヒンジ領域に直接結合されてもよい。好ましくは、ヒンジ領域は長さが少なくとも9個のアミノ酸である。1実施形態では、EPOペプチド部分は、そのC末端付近にシステイン残基を有し、ヒンジ領域はEPOペプチド部分の最も近くに位置するシステイン残基を有する。好ましくは、これらの2つのシステイン残基は、少なくとも12個のアミノ酸だけ離れている。1実施形態では、EPOペプチド部分は、免疫グロブリン部分に直接結合された完全なEPO分子(すなわちEPOと免疫グロブリン部分の間に外部ペプチドリンカーが介在しない)を有してもよい。
【0013】
本発明はさらに、本発明の融合タンパク質の複数のユニットを含む多量体タンパク質構築物に関する。例えば、2つの融合タンパク質が、両タンパク質のヒンジ領域がジスルフィド結合によって連結された二量体として組み立てられてもよい。二量体はIgG分子の一般形状を有し、自由EPO分子よりも安定している。
【0014】
本発明はさらに融合タンパク質をコード化する核酸配列およびアミノ酸配列、形質移入細胞株、および融合タンパク質を生産する方法に関する。本発明はさらに、融合タンパク質を含有する医薬組成物と、例えば治療を必要とする対象の赤血球形成を刺激するために、融合タンパク質および医薬組成物の少なくとも一方を使用する方法とを包含する。
【図面の簡単な説明】
【0015】
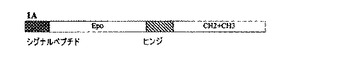
【図1A】本発明の組換えヒトEPO−Fc融合タンパク質(rHuEPO−Fc)の一般的構造を示す略図。
【図1B】rHuEPO−Fcタンパク質のヌクレオチド配列および推定アミノ酸(aa)配列を示す配列表。DNAの合計長は1281bpである。推定タンパク質配列の426個のアミノ酸はシグナルペプチドの27個のaaと完全rHuEPO−Fcタンパク質の399個のaaとを含んでいる。完全rHuEPO−Fcタンパク質はヒトEPO領域(166aa)、ヒトIgG1のFcフラグメントのヒンジ領域(16aa、太字)およびCH2ドメインおよびCH3ドメイン(217aa)から成る。成熟rHuEPO−Fc融合タンパク質のポリペプチドの計算された分子量は44.6kDaであり、EPOフラグメントの18.5kDa(41.4%)およびIgG1 Fcフラグメントの26.1kDa(58.6%)から構成されている。ホモダイマーはヒンジ領域内に2つのシステイン残基(枠で囲んでいる)を介して2つのジスルフィド結合により形成されている。成熟融合タンパク質の172番目の残基(すなわちヒンジ領域の6番目のアミノ酸)では、未変性システイン残基がグリシン(太字)により置換されている。
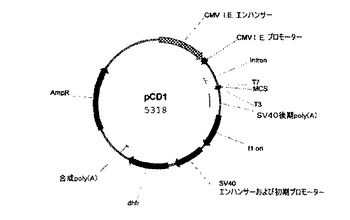
【図2】rHuEPO−Fc融合タンパク質のポリペプチドをコード化するDNA配列を挿入し、かつrHuEPO−Fc融合タンパク質を発現するCHO細胞を形質移入させるために使用される、哺乳類の発現プラスミドpCD1の構造および特徴を示す略図。
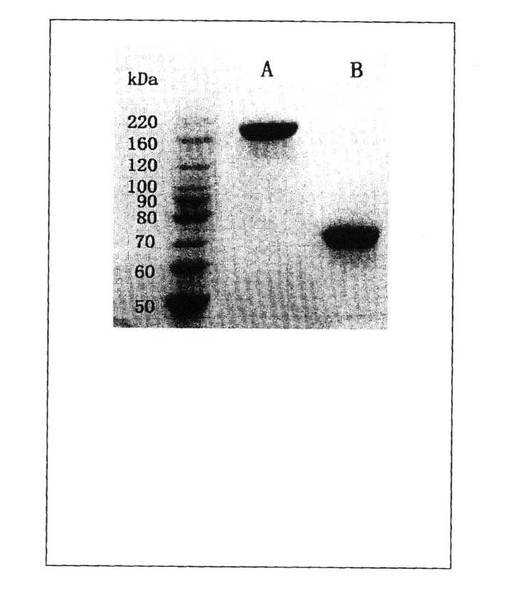
【図3】SDS−PAGE分析による非還元状態の純粋なrHuEPO−Fcタンパク質の二量体型および還元状態中の純粋なrHuEPO−Fcタンパク質の単量体型のサイズを示すSDS−PAGE画像。rHuEPO−FCを発現している培養CHO細胞株の上清から精製されたrHuEPO−Fcタンパク質は、主として二量体型として存在し、非還元状態で8%のBis−Trisゲル上で約180kDaの分子量を有している。ジスルフィド結合を破壊する還元状態(100mMジチオスレイトール、DTT)では、二量体は75kDaの分子量を有する2つの同じ単量体ユニットへ分離される。
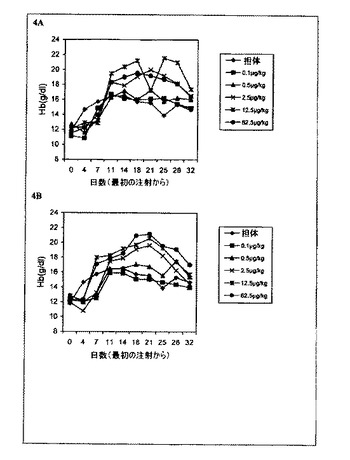
【図4】一週間に3回のrHuEPO−FcまたはrHuEPOの皮下注射で処理した正常マウスのヘモグロビン(Hb)レベルの用量依存的増加を示すグラフ。各点はその群(6匹のマウス)の平均Hbレベルに相当する。0日目のレベルは治療前のHbレベルに相当する。A:マウスをrHuEPO−Fcで処理。B:マウスを未変性rHuEPOで処理。
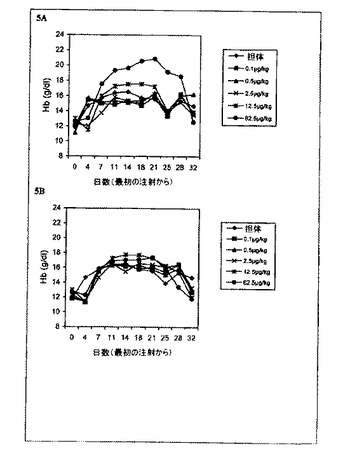
【図5】一週間に1回のrHuEPO−FcまたはrHuEPOで処理した正常マウスのヘモグロビン(Hb)レベルの用量依存的増加を示すグラフ。各点はその群(6匹のマウス)の平均Hbレベルに相当する。0日目のレベルは治療前のHbレベルに相当する。A:マウスをrHuEPO−Fcで処理。B:マウスを未変性rHuEPOで処理。
【図6】12.5μg/kgのrHuEPO−FcまたはrHuEPOの静脈内注射で処理した正常マウスのヘモグロビン(Hb)レベルの増加を示すグラフ。各点はその群(6匹のマウス)の平均Hbレベルに相当する。0日目のレベルは治療前のHbレベルに相当する。A:週に1回治療を受けたマウス。B:週に3回治療を受けたマウス。
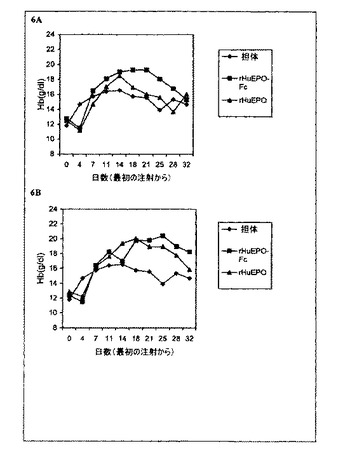
【図7】一週間に1回rHuEPO−Fc、rHuEPOまたはダルベポエチン−アルファ(略してDarbe.)で処理した5/6の腎摘出ラットのヘモグロビン(Hb)レベルの用量依存的増加を示すグラフ。各点は、その群の平均Hbレベルに相当する。正常対照は担体溶液を注射された正常ラットとした。モデル対照は担体溶液を注射された5/6の腎摘出ラットとした。0週目のレベルは治療前のHbレベルに相当する。*処理後の週数。
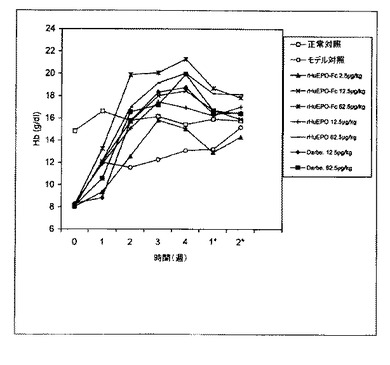
【図8】二週間ごとに1回rHuEPO−Fc、rHuEPOまたはダルベポエチン−アルファ(略してDarbe.)で処理した5/6の腎摘出ラットのヘモグロビン(Hb)レベルの用量依存的増加を示すグラフ。各点は、その群の平均Hbレベルに相当する。正常対照は担体溶液を注射された正常ラットとした。モデル対照は担体溶液を注射された5/6の腎摘出ラットとした。0週目のレベルは治療前のHbレベルに相当する。*処理後の週数。
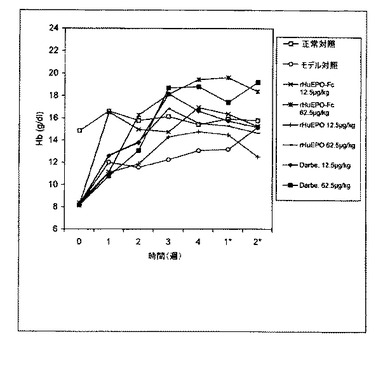
【図9】二週間ごとに1回62.5μg/kgのrHuEPO−Fcまたはダルベポエチン−アルファ(略してDarbe.)で静脈内処理した5/6の腎摘出ラットのヘモグロビン(Hb)レベルの用量依存的増加を示すグラフ。各点は、その群の平均Hbレベルに相当する。正常対照は担体溶液を注射された正常ラットとした。モデル対照は担体溶液を注射された5/6の腎摘出ラットとした。0週目のレベルは治療前のHbレベルに相当する。*処理後の週数。
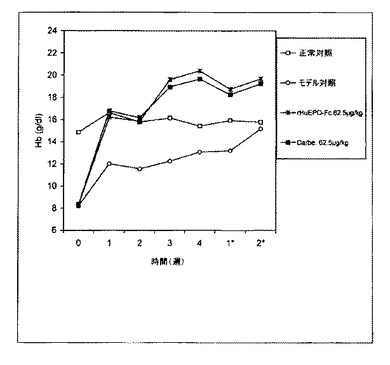
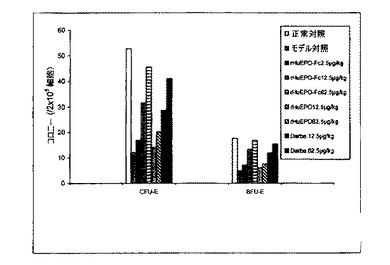
【図10A】異なる用量およびスケジュールで処理した5/6の腎摘出ラットにおけるCFU−EおよびBFU−Eのコロニー形成刺激に対するrHuEPO−Fc、rHuEPOおよびダルベポエチン−アルファの効力比較を示す。rHuEPO−Fcおよびダルベポエチン−アルファ(略してDarbe.)治療はCFU−EおよびBFU−Eコロニー形成刺激に対して同様の用量依存的効力を示したが、rHuEPOの効力は低かった。処理は皮下に一週間ごとに1回とした。
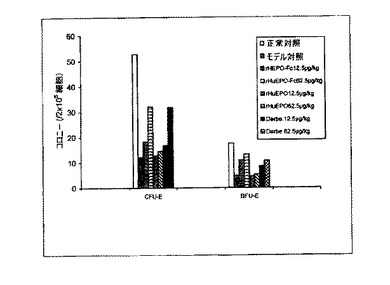
【図10B】図10Aと同じ。処理は皮下に2週間ごとに1回とした。
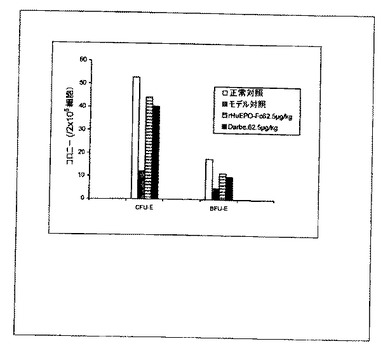
【図10C】図10Aと同じ。処理は静脈内に2週間ごとに1回とした。
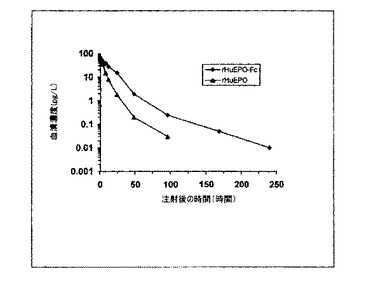
【図11】アカゲザルへの5μg/kg rHuEPO−FcまたはrHuEPO静脈内注射後のrHuEPO−FcおよびrHuEPOの血清レベル(5匹のサルの平均レベル)を示すグラフ。
【図12】野生型rHuEPO−FcCタンパク質のヌクレオチド配列および推定アミノ酸(aa)を示す配列表。配列の詳細は、未変性野生型システイン残が成熟融合タンパク質の172番目の残基(つまりヒンジ領域の6番目のアミノ酸)に存在するという点を除いて図1に示したのと同じである。
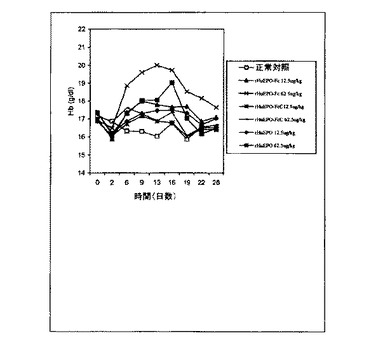
【図13】rHuEPO−FcC(野生型融合タンパク質)およびrHuEPOで一週間に3回皮下注射処理した正常マウスにおけるヘモグロビン(Hb)レベルの用量依存的増加を示すグラフ。各点は、その群(8匹)の平均Hbレベルに相当する。正常対照は担体溶液を注射された正常マウスとした。0日目のレベルは治療前のHbレベルに相当する。
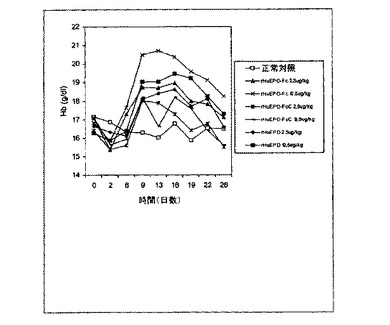
【図14】HuEPO−Fc、rHuEPO−FcCおよびrHuEPOで一週間に1回皮下注射処理した正常マウスのヘモグロビン(Hb)レベルの用量依存的増加を示すグラフ。各点は、その群(8匹)の平均Hbレベルに相当する。正常対照は担体溶液を注射された正常とした。0日目のレベルは処理前のHbレベルに相当する。
【発明を実施するための形態】
【0016】
図面は、本発明の種々の実施例を示すが、限定的に解釈されることは意図しない。
以下の説明で、本発明のより完全な理解を提供するために特定の詳細を述べるが、本発明はそのような詳細がなくても実行可能である。別の例では、本発明を不必要に不明瞭にすることを回避するために、周知の要素は図示しないか、詳細な説明を行わないこととした。従って、明細書および図面は限定的な意味ではなく、例示的な意味で捉えられるべきである。
【0017】
本願は、赤血球形成特性を有する新規な融合タンパク質に関する。本明細書で「rHuEPO−Fc」と呼ぶ融合タンパク質は、免疫グロブリンFcフラグメントに組換えにより結合されたヒトエリスロポエチン(EPO)分子を含む。さらに以下に論じるように、融合タンパク質は2つの同一のポリペプチドサブユニットから成る二量体の形をなし得る。図1Aに概略的に示した実施形態では、各ポリペプチドサブユニットはN末端からC末端まで、ヒトEPO分子のポリペプチド配列と、ヒト免疫グロブリンIgG1のFcフラグメントのヒンジ領域、CH2ドメインおよびCH3ドメインのポリペプチド配列とから成る。2つのポリペプチドサブユニットは、二量体構造を形成すべくそれぞれのヒンジ領域間のジスルフィド結合によって互いに接続される。このように二量体はIgG分子と同じ一般的な形をしており、以下に実施例で論じるような自由EPO分子よりも良好な安定性を示す。
【0018】
当業者には明らかなように、インタクトな免疫グロブリンのヒンジ領域は同タンパク質に有効な抗原−抗体結合のための十分な可撓性を与える。同様に本発明では、特にrHu
EPO−Fc融合タンパク質が二量体形式である場合、その可撓性を維持するために同融合タンパク質の設計にヒンジ領域が含まれる。以下に述べるように、これによってrHuEPO−Fc融合タンパク質のEPO部分がEPO受容体に正常に結合し、EPOの生物学的機能を活性化することが可能となる。rHuEPO−FC融合タンパク質の二量体形式は、2つのEPO分子を与えることにより、(例えば受容体のクロスリンクにより)EPO受容体の最適の活性化を引き起こすことができると考えられている。
【0019】
以下に述べる実施例で実証するように、rHuEPO−Fc融合タンパク質は組換DNA技術を使用して合成に成功した。融合タンパク質は、マウス、ラットおよび霊長類の研究で長い生体内半減期と天然組換え未変性ヒトEPOに比べて増強された赤血球形成特性を示すことが判明した。本願で使用する場合、用語「未変性ヒトエリスロポエチン」および「未変性ヒトEPO」は、修飾していない野生型構造を有するEPOを意味する。当業者には理解されるように、未変性ヒトEPOは天然であってもよいし、組換えにより生産されてもよい(例えばrHuEPOアルファ)。用語「未変性ヒトEPO」は、EPO構造が高度な糖鎖形成によって大幅に修飾されるダルベポエチンアルファのようなrHuEPOアナログは含まない。
【0020】
本発明のrHuEPO−Fc融合タンパク質の核酸配列は、配列番号1で示される。対応する推定アミノ酸配列は配列番号2で示される。完全rHuEPO−Fc融合タンパク質は長さ399個のアミノ酸である。図1Bに示されるように、完全rHuEPO−Fc融合タンパク質は、EPO領域(166個のアミノ酸)、ヒンジ領域(16個のアミノ酸、太字)およびCH2ドメインならびにCH3ドメイン(217個のアミノ酸)から成る。27個のアミノ酸から成るシグナルまたはリーダーペプチド配列も図1Bに示される。シグナルペプチドはrHuEPO−Fcの合成時に開裂する。シグナルまたはリーダーペプチドを有するrHuEPO−Fcの核酸配列とアミノ酸配列が配列番号3および配列番号4でそれぞれ示される。
【0021】
図1Bおよび配列番号2に最もよく示されているように、EPO領域はそのC末端付近の161番目のアミノ酸にシステイン残基を有する。ヒンジ領域は、図1で枠で囲まれた178番目と181番目のアミノ酸に2つのシステイン残基を含んでいる。ヒンジ領域システイン残基は、上述したようなホモダイマーのポリペプチドサブユニット間のジスルフィド結合を形成する。さらにヒトIgG1フラグメントの天然のヒンジ領域も、ヒンジ領域部分の(N末端から測定して)6番目の残基にシステインを有している。本発明では、ヒンジ領域分の6番目のシステイン残基を非システイン残基で置換した。図1Bおよび配列番号2の実施形態では、アミノ酸システインをグリシン(rHuEPO−Fcの172番目のアミノ酸残基、これはヒンジ領域の6番目の残基に相当)により置換した。当業者に明らかだが、ジスルフィド結合の形成を避けるためにこの位置でシステインの代わりに他の非システイン残基を用いることが可能である。
【0022】
172番目の残基でアミノ酸を置換した結果、ヒンジ領域(178番目の残基)の第1のシステイン残基は、上記のEPO領域(161番目の残基)のシステイン残基から17個のアミノ酸だけ離間している。発明者は、rHuEPO−Fcのホモダイマーの組立および/またはEPO受容体結合を成功させるためのEPO領域の161番目システイン残基とヒンジ領域の第1のシステイン残基の間の最小の間隔は、少なくとも12個のアミノ酸であると考えている。すなわち、172番目の残基がシステイン残基である場合、161番目のシステイン残基と172番目のシステイン残基との間のように、望ましくないジスルフィド結合が生じる恐れがある。これによりEPO分子の三次元構造が変化し、生物学的に不活性になるか生物活性が減少し得る。
【0023】
本発明の1実施形態では、EPO領域が融合タンパク質のFcフラグメント部分に直接
結合される。外部リンカーペプチドの提供を避けることにより、rHuEPO−Fc融合ペプチドの好ましい三次元構造が維持されると共に、望ましくない免疫原性反応を引き起こす危険性が最小にされる。Fcフラグメントのヒンジ領域は好ましくは長さが少なくとも9個のアミノ酸であり、好ましくは長さが約10〜20個のアミノ酸の範囲にある。
【0024】
実施例
以下の実施例は発明をより詳しく例証するものであるが、本発明が特定の実施例に制限されるわけではない。
【0025】
1.HuEPO−Fcの融合タンパク質をコードする組換えプラスミドpCdEpo−Fcの構築
rHuEPO−Fcポリペプチドのアミノ酸配列をコードする完全長DNA分子を、以下のオリゴプライマー(QIAGEN社、米国)を使用して重複型PCR法により生成した。
【0026】
【表1】

【0027】
上記プライマーの配列は配列番号5−8でそれぞれ列挙している。
EF5とEF3にEcoR I部位とNot I部位をそれぞれ導入した。哺乳動物細胞におけるHuEPO−Fcタンパク質の最適な発現の場合は、Kozak配列(GCCACCATGG)もEF5に導入した。EFL5とEFL3は、Epoの3’末端のDNA配列(23bp)およびIgG1ヒンジの5’末端のDNA配列(22bp)から成る相補的配列である。
【0028】
最初に、0.6kbのEPO DNA断片を、完全長のヒトEPO cDNAと0.7kbのFcフラグメントを含むプラスミドp9EからプライマーEF5およびEFL3を用いて、および完全長のヒトIgG1 cDNAを含むプラスミドpDからプライマーEF3およびEFL5を用いて、PCR(Platinum Taq DNA polymerase High Fidelity)
によりそれぞれ増幅した(プラスミドp9EとpDは発明者自身の実験室に由来)。その後、2個のフラグメントを精製し等量で混合した。混合物を鋳型として使用して、1.3kbの完全長rHuEPO−Fc DNAをプライマーEF5およびEF3により増幅した。精製された1.3kbのフラグメントを、EocR IおよびNot I(New England Biolab社、米国)で消化し、次にEcoR I/Not Iで消化した哺乳類発現ベクターpCD1(図2)に入れてクローニングした。得られた組換えベクターをpCdEpo−Fcと命名し、HuEPO−Fcタンパク質のアミノ酸配列をコードする挿入核酸配列をDNA塩基配列決定により確認した。
【0029】
2.rHuEPO−Fc発現細胞株の確立
ジヒドロ葉酸還元酵素(dhfr)欠乏(CHO/dhfr−、ATCC 受託番号CRL−9096)であるチャイニーズハムスター卵巣細胞は、生物活性物質生産用にFDAにより認可されたものであるが、これをrHuEPO−Fc発現用の宿主細胞として使用した。
【0030】
CHO−dhfr−細胞を、リポフェクトアミン(Giblo社、カタログ番号18292
−037、米国)を使用して組換えベクターpCdEpo−Fcで形質移入した。選択クローンの培養物からの上清をEPO活性を求めるためにELISA(Roche社、カタログ
番号1−69417、カナダ)により分析した。陽性クローンをメトトレキサート(MTX)の増加圧力下でさらにスクリーニングした。最も高くrHuEPO−Fcタンパク質を発現した1つの細胞株を、rHuEPO−Fc発現CHO細胞株として選択し、無血清培地(CD CHO培地、Gibco社、カタログ番号10743−029、米国)に徐々に
適合させた。このrHuEPO−Fc発現CHO細胞株をrHuEPO−Fcタンパク質の生産用に使用した。
【0031】
3.rHuEPO−Fcタンパク質の精製
rHuEPO−Fc発現CHO細胞を浴びようする無血清培地から採集した上清に含まれるrHuEPO−Fcタンパク質分子を、まず、プロテインAアフィニティクロマトグラフィー(Amersham社、カタログ番号17−0402−01、カナダ)により分離した。分離されたタンパク質をHiLoad 16/60 Superdex 200pgカラム(Amersham社、カタログ番
号17−1069−01、カナダ)にてゲルろ過によりさらに精製した。電気泳動での決定によると、rHuEPO−Fcタンパク質の純度は98%よりも高かった。
【0032】
4.精製rHuEPO−Fcタンパク質のサイズの決定
まず、精製rHuEPO−Fcタンパク質のサイズを決定するためにSDS−PAGEを実行した。図3に示されるように、約180kDaの分子量を有する単一のバンドが非還元状態の8%Bis-Trisゲルで見られた。これによりジスルフィド結合の存在とタンパク質の全体のサイズが測定された。このことは大半のrHuEPO−Fcタンパク質分子が融合タンパク質の設計から予想される通り二量体型として生産されることを示した。ジスルフィド結合を破壊するためSDS−PAGE分析を還元状態で行ったところ(100mM ジチオスレイトール、DTT)、75kDaの分子量のバンドだけが同定され、これはHuEPOヒンジ領域−CH2−CH3の一つのポリペプチド鎖の推定分子量と一致していた。
【0033】
質量スペクトル(MALDI−TOF−MS)で決定された糖鎖形成された精製rHuEPO−Fc融合タンパク質の正確な分子量は111099ダルトン(111.1KDa)であった。この分析では、タンパク質の一つのピークだけが観察され、精製rHuEPO−Fcタンパク質がほぼ100%純粋であることを示した。精製rHuEPO−Fcタンパク質のN末端の15個のアミノ酸を、タンパク質配列分析によりAPPRLICDSRVLERYと決定した。これは、未変性ヒトEPOポリペプチドの最初の15個のアミノ酸配列と一致し、精製rHuEPO−Fcタンパク質がrHuEPO−Fc融合タンパク質のアミノ酸配列をコードするDNA配列により予想された通りの正しい完全なEPO分子配列を有することを確認した。
【0034】
5.正常マウスにおけるrHuEPO−Fcの赤血球形成活性の増強
rHuEPO−Fcタンパク質が赤血球形成活性を保持していることを確認し、かつrHuEPOおよびダルベポエチン−アルファと比較したrHuEPO−Fcタンパク質の効力を決定するためにマウスのインビボ(生体内)実験を行なった。比較のために、本発明で説明する動物実験で使用した3つのEPO(発明者のrHuEPO−Fc、rHuEPO(つまり未変性ヒトEPO)およびダルベポエチン−アルファ)の用量はすべて、モルベースでEPO分子部分のみの量とした。rHuEPO−Fcタンパク質に関すると、EPO部分は、rHuEPO−Fc分子全体の分子量の合計アミノ酸の重量に対するEPOのアミノ酸の重量の比(399aa中の166aa)によって計算される通り、rHuEPO−Fc全体の分子量の41.4%に相当する。従って、rHuEPO−Fcに関す
るEPO量は、rHuEPO−Fcタンパク質の合計量の41.4%として決定した。
【0035】
rHuEPO−Fc(ストック濃度:0.5mg/ml、純度98.6%、および未変性ヒトrHuEPO(つまり天然ヒトEPO構造を有する)(6000IU/0.5ml、Kirin Brewery社、日本 製造)を担体溶液(2.5mg/ml ヒト血清アルブミン
、5.8mg/ml クエン酸水素ナトリウム、0.06mg/ml クエン酸および5.8mg/ml 塩化ナトリウム、pH5.5−5.6)で希釈した。rHuEPOの合計用量をその活性/量の比に従って計算した。BALB/cマウス(6〜8週齢、体重18〜22g、雄と雌の数は同数、Experiment Animal Center, AMMS 中国から購入)を、
各群6匹になるようランダムにグループ化した。マウスの各群を、1回量の一つの組み合わせ(0.1、0.5、2.5、12.5、62.5μg/kg)、1回の注射ルート(
尾静脈への静脈内注射または皮下注射)、および1回の注射スケジュール(一週間に3回または一週間に1回)で処理した。マウスの対照群には等量の担体溶液を注射した。処理は3週間続け、合計観察回数は5週間とした。測定用に末梢血液試料(尾静脈)を、毎週、処理前、4日目、および7日目と5週間採取した。Hbを吸光光度法による指標として測定した。各群のデータから平均アSD(標準偏差)を計算し、種々の群間でt−検定を行なった。
【0036】
一週間に3回マウスへEPOを投与すると、もしEPOが正常な赤血球活性を有すると仮定すれば、赤血球形成の刺激の飽和を引き起こすと予想される。図4に示されるように、一週間に3回皮下処理したどちらの群でも2.5μg/kgの用量でも大幅なHbレベ
ルの上昇があった。この実験は、rHuEPO−FcがrHuEPOと同程度に有効に生体内赤血球形成活性を示すことを実証した。処理群におけるHbレベルの上昇は用量依存的であったが、Hbレベルの上昇の飽和は、rHuEPO−Fcでは12.5μg/kg
の用量でマウスに起こり、Hbレベルの同様の上昇の飽和は、rHuEPOでは62.5μg/kgの用量でのみ達成された。さらに、2.5μg/kgのrHuEPO−Fcによって引き起こされたHbレベルの上昇は、2.5μg/kgのrHuEPOによって引き
起こされるHbレベルの上昇よりも大きかった。これらの結果はrHuEPOよりもrHuEPO−Fcによる赤血球形成刺激がより強力であることを示唆した。
【0037】
rHuEPO−Fcの赤血球形成の効力を、注射回数を一週間に1回皮下注射へと減らすことによりさらに調査した。図5に示されるように、rHuEPO−Fc処理した群は12.5μg/kgまたは62.5μg/kgの用量でHbレベルの用量依存的上昇を示した。rHuEPOの12.5μg/kgおよび62.5μg/kgのいずれの用量も、同程度のHbレベルの上昇を引き起こしたが、rHuEPO−Fcの62.5μg/kgによ
るそれよりもはるかに低かった。これはrHuEPO−Fcが生体内における赤血球形成活性を増強したことを強く示している。これはおそらく、生体内でのrHuEPO−Fcの半減期が長いことか、rHuEPO−Fcタンパク質中の二量体EPO分子によるEPO受容体の結合/活性化が改善されたことか、または両方を組み合わせた効果による。
【0038】
rHuEPO−FcまたはrHuEPOの同じ用量(12.5μg/kg)を一週間に
3回または一週間1回静脈内投与した場合、すべての処理群でHbレベルの上昇が観察された(図6)。しかしながら、rHuEPO−Fcの一週間に1回の静脈内投与は、Hbレベルのより大きくより持続性のある上昇を引き起こし、これは治療終了後も長く続いた。このデータは、天然EPOタンパク質の構造を有するrHuEPOと比べてrHuEPO−Fcタンパク質では赤血球形成特性が増強されていることを一層支持している。
【0039】
6.6分の5(5/6)の腎摘出ラットにおけるrHuEPO−Fcの赤血球形成活性の増強
正常マウスでの実験は、rHuEPO−Fcの生体内における赤血球形成活性の増強を
証明した。赤血球形成の刺激におけるrHuEPO−Fcの効力をさらに観察するために、5/6の腎摘出により作られた実験的腎性貧血を有するラットにて薬力学的研究を行なった。rHuEPO−Fcの効力を、rHuEPOおよびダルベポエチン−アルファ(60μg/ml、ロット番号N079、Kirin Brewery社、日本 製造)のそれと比較した。
【0040】
Wistarラット(雄雌同数、体重160〜180g、Vitalriver Experiment Animal社、中国、北京、ライセンス番号SCXK11−00−0008より購入)をこの発明で使用して、2つの工程からなる腎摘出[27]による腎臓機能不全による貧血モデルを生成した。無菌条件下で2つの別の手術によりラットに全身麻酔して5/6の腎摘出を施した。左腎の2/3を切除した後、ラットを20日間回復させた。その後、右腎を注意深く切除した。感染を防止するため各手術後に抗生物質を投与した。合計で腎臓組織のうち5/6を最終的に切除した。腎摘出ラットは徐々に腎機能の欠如と貧血を発現した。ラットは腎摘出の50日後に貧血の安定した状態に入り、次にEPOの投与を開始するためにランダムにグループ化された(一群当たり9匹)。ラットの各群を、1回量の一つの組み合わせ(2.5、12.5、62.5μg/kg)、1回の注射ルート(尾静脈への静脈
内注射または皮下注射)、および1回の注射スケジュール(一週間に1回または二週間に1回)で処理した。ラットの対照群およびモデル群に等量の担体溶液を注射した。処理は4週間続け、合計観察回数は6週間とした。
【0041】
皮下に一週間に1回投与されたrHuEPO−Fcのすべての用量(2.5、12.5、62.5μg/kg)で、EPO治療を受けなかったモデル対照群と比較してHbレベ
ルの用量依存的上昇が起こった。12.5および62.5μg/kgのrHuEPOまた
はダルベポエチンも、一週間に1回皮下投与されると、Hbレベルの上昇を引き起こした。12.5または62.5μg/kgのrHuEPO−Fcで処理したいずれの群でのH
bレベルの上昇も、12.5または62.5μg/kgのrHuEPOでそれぞれ処理し
た群のHbレベルの上昇よりも有意に高かった。62.5μg/kgのrHuEPO−F
cで処理した群のHbレベルは、62.5μg/kgのダルベポエチンで処理した群のH
bレベルよりわずかに高かった。治療を中止した後、62.5μg/kgのrHuEPO
−Fcで処理した群のHbレベルの減少は、正常対照およびモデル対照群の両方よりはるかに遅く、Hbレベルは観察の終了(治療後2週間)まで正常対照およびモデル対照群の両方よりも高く維持されたが、これは赤血球形成刺激がより強いか長いかの少なくともいずれかであることを示している(図7に概要)。
【0042】
二週間に1回皮下注射する処理の場合、12.5または62.5μg/kgの3つのE
POのみを投与した(図8)。12.5μg/kgのrHuEPOではモデル対照群と比
較してわずかにHbレベルが上昇した。また、62.5μg/kgのrHuEPOで処理
した群では赤血球形成反応が弱く、正常な対照群と比べてHbレベルを正常にすることができなかった。12.5または62.5μg/kgの用量でのrHuEPO−Fcまたは
ダルベポエチンのいずれかの処理により、正常対照群より高いHbレベルの有意な上昇が起こったが、これはrHuEPO−Fcおよびダルベポエチンのいずれによっても貧血状態が有効に修正されることを示している。同じ用量のrHuEPO−Fcとダルベポエチンの間には効力の点で有意差は観察されなかった。62.5μg/kgの高用量により、
観察終了(処理後2週間)まで赤血球形成の持続的な上昇が生じた。これはrHuEPO−Fcおよびダルベポエチンが生体内で長期間持続する赤血球形成刺激特性を示すことを示唆したが、これは臨床上、患者への投与回数の減少に転換することが可能である。
【0043】
ダルベポエチンは患者のコンプライアンス(薬剤服用順守)を高め、医療提供者の仕事の負担を減らす低頻度注射によって臨床への応用に認可されているが、上記実験データは本明細書で開示したrHuEPO−Fcが少なくとも同様の潜在的な利点を有することを強く示している。上述したように、ダルベポエチンは、追加の糖化合物(糖鎖形成の増加
)を含むヒトEPO分子の変異型アナログとして、その三次元構造が変化しているために生体内で免疫発生を引き起こす危険性を増大し得る。ダルベポエチンで治療を受けている患者を長期間観察するだけで、ダルベポエチンの免疫原としての危険に対する決定的な答えが得られるだろう。対照的に、rHuEPO−Fcは、EPO分子部分の修飾がなく、未変性ヒトEPOと同一または非常に類似する炭水化物含有量を有している。発明者の純粋rHuEPO−Fcタンパク質中のシアル酸の量は約10.0mmol/mmol EPOであり、これはrHuEPOの報告されたパラメータと一致していた。rHuEPO−FcのFc部分は、外部アミノ酸/結合用ペプチドがなく、ヒトIgG1の一般的構造を表わし、理論的に免疫原性反応につながらないであろう。rHuEPO−Fcは、もし臨床的に認可されれば、現在利用可能なrHuEPOおよびEPOアナログより(特に長期間の投与を必要とするもの)よりも、患者にとってのより良い選択肢を提供し得る。
【0044】
二週間に1回いったん静脈内注射されると、rHuEPO−Fcとダルベポエチン(62.5μg/kg)はいずれも、正常対照ラットの正常Hbレベルをはるかに超える腎性
貧血ラットのHbレベルの同じ増加を引き起こすことができた(図9)。このことは、ダルベポエチンの効力が臨床的に証明されたように、rHuEPO−Fcによって赤血球形成が持続的に刺激されることをさらに実証している。
【0045】
処理後(一週間に1回または二週間に1回、皮下または静脈内)5/6腎摘出ラットから採集した骨髄細胞の細胞培養実験から得られたデータは、rHuEPO−Fc、rHuEPOおよびダルベポエチンがすべてCFU−EおよびBFU−Eの形成を刺激したことを示した。rHuEPO−Fcおよびダルベポエチンの効力は同様であり、rHuEPOの効力よりも強かった(図10)。
【0046】
処理群とモデル対照群で血液尿素窒素(BUN)およびCreaレベルは同様であった。すべての処理群における血清Feレベルはモデル対照群のそれよりも高かった。病理学的検査によると、すべてのEPO治療ラットの骨髄および脾臓の赤血球(RBC)に分布の増加が観察された。
【0047】
7.アカゲザルにおけるrHuEPO−Fcの薬物動態学的研究
上述したように、発明者は、融合タンパク質のEPO部分が赤血球形成刺激のような天然EPOの機能特性を保持し、ヒトIgG1のFcフラグメントが循環での融合タンパク質の安定した存在を可能にし、それによりrHuEPO−Fcのインビボでの半減期が延長するように、rHuEPO−Fcを設計した。上記の動物による研究は、rHuEPO−Fcの赤血球形成活性がrHuEPOと比べて増強されることを実証した。発明者はrHuEPOと比べたrHuEPO−Fcの生体内半減期を決定するために、薬物動態学的研究をさらに行なった。霊長類がヒトに生物学上非常に類似するため、霊長類をデータを生成するために使用した。
【0048】
研究のデザインは文献の報告に基づき、実験は薬物動態学の一般的ガイドラインに従って行った。各群に5匹のサル(3〜5kg、Ex periment Animal Center, AMMS,中国から購入)として、2群のアカゲザルに、5μg/kgのrHuEPO−FcまたはrHuE
POをそれぞれ静脈内注射した。血液試料を注射前と注射後0.017、0.167、0.5、1、2、4、8、12、24、48、96、168、240時間後に採取した。血清を遠心分離で厚め、血清rHuEPO−FcまたはrHuEPOレベルをヒトエリスロポエチン酵素結合免疫吸着測定法(ELISA)キット(R&D Systems社、ミネソタ州ミ
ネアポリス所在)の使用により決定した。静脈内注射されたrHuEPO−FcおよびrHuEPO平均半減期(t1/2)はそれぞれ35.24+/−5.15時間および8.72+/−1.69時間だった(図11に概要)。rHuEPO−Fcの生体内利用率を観察するために、5ug/kgのrHuEPO−Fcを5匹のアカゲザルに皮下注射した
。血液試料を注射前と注射後1、2、5、8、10、12、15、24、48、72、96、168、240時間後に採取し、rHuEPO−Fcの血清レベルを上述のR&Dキッ
トにより決定した。生体内利用率指数は皮下注射で35.71+/5.37%として計算された。これは慢性腎不全患者におけるダルベポエチン−アルファ(Aranesp(商標))
の報告された生体内利用率の数字と同一である[9,15]。
【0049】
このデータは、rHuEPO−Fcが霊長類で有意に長い半減期を有し、そのrHuEPO−Fcの生体内半減期がKirin Brewery社、日本 製造のrHuEPOの生体内半減
期よりも少なくとも4倍長いことを実証している。長い生体内半減期はrHuEPO−Fcの赤血球形成活性の増強に寄与し得る。
【0050】
8.カニクイザル(Macaca fascicularis)におけるrHuEPO−Fcの免疫原性
上述のように、rHuEPO−Fc融合タンパク質の免疫原性の変化を意図的に回避するかまたは最小限にするために、rHuEPO−Fc融合タンパク質の設計には注意が払われた。発明者は融合タンパク質中に外部アミノ酸や結合用ペプチド配列を含めるか追加することを避けた。図1Bの実施形態の発明されたHuEPO−Fc融合タンパク質は天然EPOタンパク質のポリペプチド配列とヒトIgG1のFcフラグメント(ヒンジ領域、CH2、CH3)のみ含んでおり、理論上、免疫原性反応およびrHuEPO−Fcタンパク質に対する抗体産生を引き起こさないだろう。当業者には理解されるように、代替え構造を有する別の実施形態も本発明に包含される。
【0051】
rHuEPO−Fcタンパク質の免疫性を観察するために以下の霊長類研究を行なった。10匹のカニクイザル(Macaca fascicularis)(雄/雌=5/5、約5才齢、平均重
量 雄4.0ア0.3kg、雌2.9ア0.4kg、Laboratory Animal Center, AMMS,
中国から購入)に一週間に3回、4週間の間精製された5μg/kgのrHuEPO−F
cを皮下注射し、2匹には対照動物として等量の担体溶液を注射した。血清は一週間に1回、5週間(処理後1週間)採取し、コーティング抗原として精製rHuEPO−Fc(5μg/ml)を使用してELISAにより抗rHuEPO−Fc特異抗体に関して試験
した。さらに、末梢血中のRBC数およびHbレベルも実験期間中に決定した。得られたデータは、rHuEPO−Fc治療されたカニクイザルで赤血球形成増強の刺激が観察された(平均RBC数は4.74x109/mlから6.67x109/mlまで増加、平均Hbレベルは12.2g/dlから13.7g/dlまで増加)が、rHuEPO−Fcは融合タンパク質に対する検出できる特異抗体を誘導しなかったことを示す。これらの結果は、rHuEPO−Fc融合タンパク質が霊長類で免疫原性を引き起こさないことを示す。
【0052】
9.正常マウスにおけるrHuEPO−Fcの急性毒性試験
rHuEPO−Fc融合タンパク質の安全性を評価するために、急性毒性試験を動物にて行なった。
【0053】
BALB/cマウス(n=20、雄雌同数、5〜6週齢、平均重量 雄15.8ア0.4gおよび雌15.9ア0.6g、Chinese Academy of Medicine、中国より購入)の2
つの群に、過剰量の精製rHuEPO−Fc(雄=13.3mg/kg、雌=13.2mg/kg)または等量の担体溶液をそれらの尾静脈を介して一回静脈内注射した。注射後の即時反応に加えて、一般的行動および状態、活動、摂食および排便パターンを14日間毎日監視および記録した。すべてのマウスの体重を7日目と14日目に測定した。注射後15日目に、マウスの主な臓器の解剖的検査を行った。臓器に何らかの異常な変化または疑わしい変化が観察されれば、病理学的検査を行う。
【0054】
2つの群中のすべてのマウスで、注射後の明らかな即時反応はなかった。14日間の間
、行動、活動、摂食、および排便パターンの明らかな変化は観察されなかった。さらに、いずれの群のマウスの重量も、試験期間中着実に増加し、注射後7日目でも14日目でも明らかな差違は見出されなかった。脳、肺、心臓、肝臓および腎臓の組織で異常なまたは病理学的な変化は検出されなかった。これらの結果は、正常な赤血球形成機能を示すのに必要とされるよりもはるかに多くの過剰量のrHuEPO−Fcを投与しても、安全であり、明らかな有毒効果がなかったことを示す。
【0055】
10.野生型および変化させられたEPO融合タンパク質の比較
EPOタンパク質の野生型と変異型を比較するための研究も行なった。上述したように、1実施形態では、本発明は172番目のアミノ酸残基に一つのアミノ酸突然変異を含んでいる。(C172G)。比較のために、172番目の残基にシステインアミノ酸を有する野生型融合タンパク質も調製した(図12)。野生型融合タンパク質は、上記の実施例1〜3と同様に調製した。組換えプラスミドの構築に関しては、以下のオリゴプライマー(QIAGEN社、米国)を使用した(実施例1のプライマーに比べてEFL5wおよびEFL3w中の変化したアミノ酸が太字で示される)。
【0056】
【表2】
【0057】
プライマーEFL5wおよびEFL3wの配列は配列番号9−10でそれぞれ列挙している。
野生型融合タンパク質(本明細書でrHuEPO−FcCと呼ぶ)の赤血球形成活性を、変異融合タンパク質(つまり上述の本発明のrHuEPO−Fcタンパク質)および組換えヒトEPO(rHuEPO)と比較するためにマウスの生体内実験を行なった。比較のために、本実施例で使用する3つのタンパク質(すなわちrHuEPO−Fc、rHuEPO−FcC、rHuEPO)の用量はすべて、モルベースでEPO分子部分のみの量とした。rHuEPO−FcおよびrHuEPO−FcCタンパク質に関すると、EPO部分は、rHuEPO−Fc分子およびrHuEPO−FcC分子全体の分子量の合計アミノ酸の重量に対するEPOのアミノ酸の重量比(すなわち399aa中の166aa)によって計算される通り、合計分子量の41.4%に相当する。
【0058】
rHuEPO−Fc(ストック濃度:300μg/ml)、rHuEPO−FcC(ス
トック濃度:90μg/ml)、および天然ヒトEPO構造を有するrHuEPO(60
00IU/0.5ml、Kirin Brewery社、日本 製造)を担体溶液(2.5mg/ml
ヒト血清アルブミン、5.8mg/ml クエン酸水素ナトリウム、0.06mg/ml クエン酸および5.8mg/ml塩化ナトリウム、pH5.5−5.6)で希釈した。rHuEPOの合計用量をその活性/量の比に従って計算した。BALB/cマウス(10週齢、体重18〜22g、雄と雌の数は同数、Experiment Animal Center, AMMS 中
国から購入)各群8匹となるようランダムにグループ化された。マウスの各群を、1回量の一つの組み合わせ(2.5、12.5、62.5μg/kg)、1回の注射ルート(皮
下)および1回の注射スケジュール(一週間に3回または一週間に1回)で処理した。マウスの対照群には等量の担体溶液を注射した。処理は26日間続いた。測定用に末梢血試
料(尾静脈)を、処理前、処理の2日目、6日目、9日目、13日目、16日目、19日目、22日目および治療の26日目に採取した。Hbを吸光光度法による指標として測定した。各群のデータから平均アSDを計算し、種々の群間でt−検定を行った。
【0059】
図13に示されるように、一週間に3回の間隔での3つのEPOタンパク質のすべての投与は赤血球形成を刺激した。2.5μg/kgまたは12.5μg/kgの用量のいずれかでは、rHuEPO−FcはrHuEPOよりもHbレベルの高い増加を引き起こした。Hbレベルの最も高い上昇が12.5μg/kgの用量のrHuEPO−Fcで達成さ
れた。2.5μg/kgと12.5μg/kgのいずれのrHuEPO−FcCも、rHuEPO−FcC処理した群におけるHbレベルの有意に低い上昇によって示されるよりもずっと弱い赤血球形成を引き起こした。実際、12.5μg/kgのrHuEPO−Fc
Cは、2.5μg/kgのrHuEPOよりHbレベルの上昇が小さく、これらの結果は
、rHuEPO−FcCが天然EPOの分子の配列を有するrHuEPOに比べ生体内赤血球形成活性を損なっていることと示唆する。対照的に、本発明のrHuEPO−Fc融合タンパク質はより有力なe赤血球形成機能を示した。一週間に3回の間隔の3つのEPOタンパク質の投与は、タンパク質の半減期の差の3影響を大幅に除外した。
【0060】
rHuEPO−FcおよびrHuEPO−FcCの赤血球形成の効力を、注射回数を一週間に1回の皮下注射へと減らすことによりさらに調査した。図14に示されるように、rHuEPO−Fc−処理した群は12.5μg/kgまたは62.5μg/kgの用量でrHuEPOで処理したものよりHbレベルの高い上昇を示した。対照的に、rHuEPO−FcCは、rHuEPOによって引き起こされるよりはるかに弱いHbレベルの上昇を引き起こした。例えば、12.5μg/kgのrHuEPOはほとんどの時間地点で、
62.5μg/kgのrHuEPO−FcCによって引き起こされるよりHbレベルの高
い上昇を引き起こした。これは、半減期の影響を含むよう投与回数を減少させることによって、天然EPO分子の配列を有するrHuEPOに比べて、および本発明のrHuEPO−Fc融合タンパク質に比べて、rHuEPO−FcCが生体内ではるかに弱い赤血球形成機能を示すことをさらに示す。
【0061】
要約すると、これらの結果は、天然EPOの分子の配列を有するrHuEPOに比べて、ヒトEPOおよびヒトFcフラグメント(ヒンジ、CH2およびCH3)の両方の天然分子の配列の融合によって形成されたrHuEPO−FcCがはるかに弱い生体内赤血球形成機能を示すことを実証している。詳細には、rHuEPO−FcC融合タンパク質の赤血球形成活性は天然EPO分子の1/5未満である。これは、EPO分子とヒトFcフラグメントの天然配列との間の融合がEPO分子の機能特性を損なうことを示す。Fcフラグメントのヒンジ領域中の第1のシステイン残基の一つのアミノ酸の置換によって、天然EPO分子配列および変異型Fcフラグメントを含む本発明のrHuEPO−Fc融合タンパク質は、天然EPO分子と比べて、より有力な生体内赤血球形成機能を示す。このデータは、野生型Fcフラグメントのヒンジ領域中の第1のシステイン残基が、EPO分子に構造変化をもたらす等により、EPO分子になんらかの形で干渉し、これによって赤血球形成刺激の際にEPO分子の機能特性が損なわれることを示唆している。
【0062】
上記開示に照らすと当業者には明らかなように、本発明の精神及び範囲から逸脱せずに本発明の実施において多くの変更および修正が可能である。
参考文献
1. Cohen J, et al. Erythropoietin and its receptor: signaling and clinical manifestations. IMAJ 4, pp 1072- 1076(2002)
2. Blackwell K, et al. rHuEPO and improved treatment outcomes: potential modes of action. The Oncologist 9(suppl 5), pp41-47(2004).
3. Lappin TR, et al. EPO’s alter ego: erythropoietion has multiple actions. Ste
m Cells 20, pp485-492(2002).
4. Maiese K, et al. New avenues of exploration for erythropoietin. JAMA 293(1), pp90-95(2005).
5. Fisher JW. Erythropoietin: physiologic and pharmacologic aspects. Proc. Soc. Exp. Biol. Med. 216, pp358-369(1997).
6. Jelkmann W. Molecular biology of erythropoietin. Internal Med. 43(8), pp649- 659(2004).
7. Ng T, et al. Recombinant erythropoietin in clinical practice. Postgrad. Med. J. 79, pp367-376(2005).
8. Weiss G, et al. Anemia of chronic disease. N. Engl. J. Med. 352(10), pp1011-1023(2005).
9. Macdougall IC. An overview of the efficacy and safety of novel erythropoiesisstimulating protein(NESP). Nephrol. Dial. Transplant. 16(suppl 3), pp 14-21(2001).
10. Joy MS. Darbepoetin-アルファ: a novel erythropoiesis-stimulating protein. Ann. Pharmacother. 36, ppl 183-1192(2002).
11. Ellirt S, et al. Enhancement of therapeutic protein in vivo activities through glycoengineering. Nature Biotechnology 21, pp414-421(2003).
12. Elliott S, et al. Control of rHuEPO biological activity: the role of carbohydrate. Experimental Hematology 32, ppl 146-1155(2004).
13. Egrie JC, et al. Darbepoetin-アルファ has a longer circulating half-life andgreater in vivo potency than recombinant human erythropoietin. Exp. Hematol. 31,
pp290- 299(2003).
14. Egrie JC, et al. Development and characterization of novel erythropoiesis stimulating protein(NESP). British J. Caner. 84(suppl 1), pp3-10(2001).
15. Macdougall IC, et al. Pharmacokinetics of novel erythropoiesis stimulating protein compared with epoetin アルファ in dialysis patients. J. Am. Soc. Nephrol.10, pp2392-2395(1999).
16. Jolling K, et al. Population pharmacokinetic analysis of peglated human erythropoietin in rats. J. Pharm. Sci. 93(12), pp3027-3038(2004)
17. Dalle B, et al. Dimeric erythropoietin fusion protein with enhanced erythropoietic activity in vtro and in vivo. Blood 97(12), pp3776-3782(2001).
18. Kochendoerfer GG, et al. Design and chemical synthesis of a homogeneous polymer-modified erythropoiesis protein. Science 299 pp884-887(2003).
19. Sytkowski AJ, et al. Human erythropoietin dimmers with markedly enhanced in vivo activity. Proc. Natl. Acad. Sci. USA 95, ppl 184-1188(1998).
20. Sytkowski AJ, et al. An erythropoietin fusion protein comprised of identicalrepeating domains exhibits enhanced biological proterties. J. Biol. Chem. 274(35), Pp24773-24778(1999).
21. Jones TD, et al. The development of a modified human IFN-[alpha]2b linked tothe Fc portion of human IgG1 as a novel potential therapeutic for the treatment
of hepatitis C virus infection. J. Interferon. Cytokine. Res. 24, pp560-572(2004).
22. Lo KM, et al. High level expression and secretion of Fc-X fusion proteins in
mammalian cells. Protein engineering 11(6), pp495-500(1998).
23. Mohler BCM, et al. Soluble tumor necrosis factor(TNF) receptors are effective therapeutic agents in lethal endotoxemia and function simultaneously as both TNF carriers and TNF antabonists. J. Immunol. 151(3), ppl548-1561(1993).
24. Way JC, et al. Improvement of F c-erythropoietin structure and pharmacokinetics by modification at a disulfide bond. Protein Engineering, Design & Selection
18(3), ppl 11 -118(2005).
25. Goldenberg MM. Etanercept, a novel drug for the treatment of patients with severe, active rheumatoid arthritis. Clin. Ther. 21(1), pp75-87(1999)
26. Wong VK, et al. The use of alefacept in the treatment of psoriasis. Skin Therapy Lett. 8(6), pp 1-2(2003)
27. Chanutin A, et al. Experimental renal insufficiency produced by partial nephrectomy. Arch. Intern. Med. 49,pp 767-787 (1932).
【技術分野】
【0001】
本願はヒトエリスロポエチン融合タンパク質に関する。
【背景技術】
【0002】
ヒトエリスロポエチン(EPO)は、造血成長因子ファミリーのメンバーであり、血中酸素利用率の低下による組織低酸素症に応じて主として成人の腎臓および胎児の肝臓で合成される[1]。EPOの主な機能は、骨髄中の特定の赤血球(RBC)前駆細胞および前駆物質に直接作用して、ヘモグロビンおよび成熟RBCの合成を刺激することである。EPOはRBCの増殖、分化および成熟も制御する。腎臓の機能不全に関連する貧血、癌および他の病理学的状態を治療するために天然EPOのアミノ酸配列を有する組換えEPOが生産および承認されている[2]。その赤血球形成の特性に加えて、最近の研究によると、EPOがニューロン等の非骨髄細胞にも作用することが報告されており[3]、これは中枢神経系(CNS)および他の器官/系におけるEPOの他の生理学的/病理学的機能の可能性を示唆している。EPO受容体が種々の器官で見出されたため、EPOは、抗アポトーシス物質として働く等、多くの生物学的効果を有する可能性がある。
【0003】
ヒトEPOは30.4キロダルトンの分子量を有する糖タンパク質である。炭水化物はその合計質量の約39%を占める。EPO遺伝子は7q11−22染色体上に位置し、5つのエクソンと4つのイントロンを備えた5.4kbの領域に及ぶ[4]。EPOの前駆物質は193個のアミノ酸から成る。翻訳後修飾によるリーダー配列と最後のアミノ酸Argとの開裂により、165個のアミノ酸を有する成熟EPOが生じる。糖鎖形成は、Asn24、Asn38、Asn83の3つのN結合部位と、Ser126の1つのO結合部位であるが、EPOの生合成、三次構造および生体内生理活性に重大な役割を果たしている[5]。エリスロポエチン受容体すなわち72−78キロダルトンの分子量を有する糖鎖付加されリン酸化された膜貫通ポリペプチドに結合することにより機能する。この結合は、受容体のホモダイマー化を引き起こし、これがJAK2/STAT5系、G−タンパク質、カルシウムチャネルおよびキナーゼといういくつかのシグナル伝達経路の活性化につながる。EPOタンパク質の2つの分子は、最適の受容体活性化を達成するためには1つの受容体分子に同時に結合する必要がある[6]。
【0004】
ヒト治療のために認可された最初の造血生長因子として、組換えヒトEPO(rHuEPO)は慢性腎不全、癌(主として化学療法に関連する貧血)、自己免疫疾患、AIDS、外科手術、骨髄移植および骨髄異形成症候群等に起因する貧血の治療に使用されてきた。面白いことに、最近の研究によると、rHuEPOは非血液系の機能を有することが観察され、脳虚血、頭部外傷、炎症性疾患および神経変性疾患用の神経保護剤として使用される可能性が示されている。
【0005】
現在、3種類のrHuEPOまたはrHuEPOのアナログすなわちrHuEPOアルファ、rHuEPOベータおよびダルベポエチンアルファが市販されている[8]。これら3つの組換えタンパク質は同じエリスロポエチン受容体に結合するが、構造、糖鎖形成の程度、受容体結合能および生体内での代謝は異なっている。1980年代にrHuEPOアルファを初めて導入して以来、臨床医は同薬剤の頻繁な投薬/注射の必要性を重大な欠点と素早く認めた。r静脈内または皮下に投与したHuEPOアルファとrHuEPOベータの平均生体内半減期はそれぞれわずか8.5時間と17時間である[9,10]。したがって、患者は毎日、一週間に2回、または一週間に3回の注射スケジュールを必要とし、これは患者と医療提供者の両方に負担を強いる。したがって、より長い生体内半減
期および/または増強された赤血球形成活性を有する組換えEPOアナログを開発する必要があった。
【0006】
先行技術では、その生体内代謝を遅延させるかまたはその治療的性質を改善するために、未変性EPOタンパク質の構造を遺伝子改変するか化学修飾する試みが行われていた。例えば、EPO分子中のシアル酸含有炭水化物の量と、その生体内代謝および機能的活性との間には直接相関があるように見える。したがって、EPO分子の炭水化物含有量を増加させると、半減期が長くなり、生体内活性も増強された[11,12]。Amgenは、rHuEPOよりも2個多い5個のN結合炭水化物鎖を含むようrHuEPOアナログのダルベポエチンアルファを設計した。ダルベポエチンアルファも新規な赤血球形成刺激タンパク質(NESP)として知られており、Aranesp(商標)として販売されている。ダルベポエチンアルファは5つの位置(Ala30Asn、His32Thr、Pro87Val、Trp88Asn、Pro90Thr)で未変性ヒトEPOと異なり、これによりアスパラギン残基位置30および88で2つの追加のN結合オリゴ糖を取り付けることが可能となっている。ダルベポエチンアルファは未変性EPOと同じようにEPO受容体に結合して、JAK−2キナーゼおよび同じ細胞内分子Ras/MAP−k、P13−kおよびSTAT−5によるチロシンリン酸化を伴う細胞内シグナル伝達を引き起こす。炭水化物含有量の増加により、動物とヒトの両方でのダルベポエチンアルファの半減期はrHuEPO−アルファの半減期のほぼ3倍の長さになっている(25.3時間対8.5時間)[9]。ダルベポエチンアルファ(Aranesp(商標))は、さらに天然または組換えヒトEPOに比べて生体内で増強された生理活性を示すと思われ[13]、第二代のrHuEPO薬としてFDAによって認可された。この薬剤はrHuEPOの1週間で2〜3回の注射と同一の治療効果を達成するのに、一週間に1回の投与しか必要としない[10,14,15]。
【0007】
EPOの半減期を拡張する他の試みは、ポリエチレングリコールやその同等物との化学共役(PEG化)によりEPOタンパク質の分子量を増加させることに焦点が当てられていた。PEG化ははるかに大きな分子量を有し、循環からの除去が防がれるためより長い血漿半減期を有する[16]。しかしながら、PEG化はEPO部分の予期せぬ機能の変化および特異性の変化を生じさせる可能性がある。EPO分子を担体タンパク質(ヒトアルブミン)へ結合させるか、またはリンクペプチド(3から17個のアミノ酸)の使用によりもしくは化学的架橋試薬により2つの完全EPO分子のホモダイマーを形成する等のさらに別の方法によりEPOの分子量を増加させる報告も存在する[17,18,19,20]。これらの方法はすべて半減期の延長およびEPO活性の増強のある程度の成功を達成しているが、本願に記載するようにEPO分子をヒト免疫グロブリン(IgG)のFcフラグメントと組み合わせて融合タンパク質とすることは、固有の利点を達成する。
【0008】
ヒト免疫グロブリンIgGは、ジスルフィド結合により共有結合された4つのポリペプチドから構成されている(軽鎖と重鎖の2つの同一コピー)。パパインによるIgG分子のタンパク質分解によって、2つのFAbフラグメントと1つのFcフラグメントが生じる。Fcフラグメントは、ジスルフィド結合によって共に結合した2つのポリペプチドから成る。各ポリペプチドは、N末端からC末端まで、ヒンジ領域、CH2ドメインおよびCH3ドメインから構成される。Fcフラグメント構造はヒト免疫グロブリンのすべてのサブタイプでほぼ同じである。IgGはヒト血液中に最も豊富にあるタンパク質の1つであり、ヒト血清中の免疫グロブリンの合計の70〜75%を占める。循環でのIgGの半減期は免疫グロブリンの5つすべてのタイプのうち最長であり、21日に達し得る。
【0009】
現代の生物工学技術をうまく適用して、サイトカインおよび可溶性受容体等の治療用タンパク質フラグメントとヒトIgGのFcフラグメントから成る融合タンパク質が形成された[21,22,23,24]。これらの融合タンパク質は、それらの生物学的特性お
よび治療特性を保持する間、有意により長い生体内半減期を有する。これまでのところ、Fcフラグメントを含む2つの融合タンパク質がバイオ医薬としての開発に成功しており、慢性関節リウマチおよび慢性尋常性乾癬の治療用にFDAにより認可されている[25,26]。
【0010】
先行技術では、化学的架橋によるかまたはポリペプチドにより結合された2つのEPO分子からなる二量体が生体内活性の増強および長い半減期を示すことが論証された[17,19]。この活性の増強は1つの受容体にEPO二量体がより効率的に結合するためであり、生体内での半減期の延長は二量体タンパク質のサイズが大きいことによるためであり得る。しかしながら、化学的架橋プロセスは効率的でなく、制御することが難しい。さらに、EPOの二量体中のリンケージペプチドは、EPO分子の三次元構造を変化させる可能性があり、また、ペプチドそのものが生体内での免疫原性反応を刺激する可能性がある。特に腎患者でEPO置換療法は生涯にわたって続くため、これらの欠点はEPO二量体の治療可能性を損なう。
【発明の概要】
【発明が解決しようとする課題】
【0011】
有意に長い半減期を有し、生体内での赤血球形成活性が増強されているが、免疫原特性が増加していないEPOアナログが必要とされている。
【課題を解決するための手段】
【0012】
本発明によれば、免疫グロブリンペプチド部分に結合されたヒトエリスロポエチンペプチド部分を含む組換え融合タンパク質について説明される。融合タンパク質は、天然または組換えの未変性ヒトエリスロポエチンに比べて長い生体内半減期を有する。本発明の1実施形態では、融合タンパク質は未変性ヒトエリスロポエチンよりも少なくとも3倍長い生体内半減期を有する。融合タンパク質はさらに、未変性ヒトエリスロポエチンに比べて増強された赤血球形成の生理活性を示してもよい。本発明の1実施形態では、免疫グロブリンペプチド部分がIgG1フラグメント等のFcフラグメントである。FcフラグメントはCH2領域、CH3の領域およびヒンジ領域を有する。EPOペプチド部分はヒンジ領域に直接結合されてもよい。好ましくは、ヒンジ領域は長さが少なくとも9個のアミノ酸である。1実施形態では、EPOペプチド部分は、そのC末端付近にシステイン残基を有し、ヒンジ領域はEPOペプチド部分の最も近くに位置するシステイン残基を有する。好ましくは、これらの2つのシステイン残基は、少なくとも12個のアミノ酸だけ離れている。1実施形態では、EPOペプチド部分は、免疫グロブリン部分に直接結合された完全なEPO分子(すなわちEPOと免疫グロブリン部分の間に外部ペプチドリンカーが介在しない)を有してもよい。
【0013】
本発明はさらに、本発明の融合タンパク質の複数のユニットを含む多量体タンパク質構築物に関する。例えば、2つの融合タンパク質が、両タンパク質のヒンジ領域がジスルフィド結合によって連結された二量体として組み立てられてもよい。二量体はIgG分子の一般形状を有し、自由EPO分子よりも安定している。
【0014】
本発明はさらに融合タンパク質をコード化する核酸配列およびアミノ酸配列、形質移入細胞株、および融合タンパク質を生産する方法に関する。本発明はさらに、融合タンパク質を含有する医薬組成物と、例えば治療を必要とする対象の赤血球形成を刺激するために、融合タンパク質および医薬組成物の少なくとも一方を使用する方法とを包含する。
【図面の簡単な説明】
【0015】
【図1A】本発明の組換えヒトEPO−Fc融合タンパク質(rHuEPO−Fc)の一般的構造を示す略図。
【図1B】rHuEPO−Fcタンパク質のヌクレオチド配列および推定アミノ酸(aa)配列を示す配列表。DNAの合計長は1281bpである。推定タンパク質配列の426個のアミノ酸はシグナルペプチドの27個のaaと完全rHuEPO−Fcタンパク質の399個のaaとを含んでいる。完全rHuEPO−Fcタンパク質はヒトEPO領域(166aa)、ヒトIgG1のFcフラグメントのヒンジ領域(16aa、太字)およびCH2ドメインおよびCH3ドメイン(217aa)から成る。成熟rHuEPO−Fc融合タンパク質のポリペプチドの計算された分子量は44.6kDaであり、EPOフラグメントの18.5kDa(41.4%)およびIgG1 Fcフラグメントの26.1kDa(58.6%)から構成されている。ホモダイマーはヒンジ領域内に2つのシステイン残基(枠で囲んでいる)を介して2つのジスルフィド結合により形成されている。成熟融合タンパク質の172番目の残基(すなわちヒンジ領域の6番目のアミノ酸)では、未変性システイン残基がグリシン(太字)により置換されている。
【図2】rHuEPO−Fc融合タンパク質のポリペプチドをコード化するDNA配列を挿入し、かつrHuEPO−Fc融合タンパク質を発現するCHO細胞を形質移入させるために使用される、哺乳類の発現プラスミドpCD1の構造および特徴を示す略図。
【図3】SDS−PAGE分析による非還元状態の純粋なrHuEPO−Fcタンパク質の二量体型および還元状態中の純粋なrHuEPO−Fcタンパク質の単量体型のサイズを示すSDS−PAGE画像。rHuEPO−FCを発現している培養CHO細胞株の上清から精製されたrHuEPO−Fcタンパク質は、主として二量体型として存在し、非還元状態で8%のBis−Trisゲル上で約180kDaの分子量を有している。ジスルフィド結合を破壊する還元状態(100mMジチオスレイトール、DTT)では、二量体は75kDaの分子量を有する2つの同じ単量体ユニットへ分離される。
【図4】一週間に3回のrHuEPO−FcまたはrHuEPOの皮下注射で処理した正常マウスのヘモグロビン(Hb)レベルの用量依存的増加を示すグラフ。各点はその群(6匹のマウス)の平均Hbレベルに相当する。0日目のレベルは治療前のHbレベルに相当する。A:マウスをrHuEPO−Fcで処理。B:マウスを未変性rHuEPOで処理。
【図5】一週間に1回のrHuEPO−FcまたはrHuEPOで処理した正常マウスのヘモグロビン(Hb)レベルの用量依存的増加を示すグラフ。各点はその群(6匹のマウス)の平均Hbレベルに相当する。0日目のレベルは治療前のHbレベルに相当する。A:マウスをrHuEPO−Fcで処理。B:マウスを未変性rHuEPOで処理。
【図6】12.5μg/kgのrHuEPO−FcまたはrHuEPOの静脈内注射で処理した正常マウスのヘモグロビン(Hb)レベルの増加を示すグラフ。各点はその群(6匹のマウス)の平均Hbレベルに相当する。0日目のレベルは治療前のHbレベルに相当する。A:週に1回治療を受けたマウス。B:週に3回治療を受けたマウス。
【図7】一週間に1回rHuEPO−Fc、rHuEPOまたはダルベポエチン−アルファ(略してDarbe.)で処理した5/6の腎摘出ラットのヘモグロビン(Hb)レベルの用量依存的増加を示すグラフ。各点は、その群の平均Hbレベルに相当する。正常対照は担体溶液を注射された正常ラットとした。モデル対照は担体溶液を注射された5/6の腎摘出ラットとした。0週目のレベルは治療前のHbレベルに相当する。*処理後の週数。
【図8】二週間ごとに1回rHuEPO−Fc、rHuEPOまたはダルベポエチン−アルファ(略してDarbe.)で処理した5/6の腎摘出ラットのヘモグロビン(Hb)レベルの用量依存的増加を示すグラフ。各点は、その群の平均Hbレベルに相当する。正常対照は担体溶液を注射された正常ラットとした。モデル対照は担体溶液を注射された5/6の腎摘出ラットとした。0週目のレベルは治療前のHbレベルに相当する。*処理後の週数。
【図9】二週間ごとに1回62.5μg/kgのrHuEPO−Fcまたはダルベポエチン−アルファ(略してDarbe.)で静脈内処理した5/6の腎摘出ラットのヘモグロビン(Hb)レベルの用量依存的増加を示すグラフ。各点は、その群の平均Hbレベルに相当する。正常対照は担体溶液を注射された正常ラットとした。モデル対照は担体溶液を注射された5/6の腎摘出ラットとした。0週目のレベルは治療前のHbレベルに相当する。*処理後の週数。
【図10A】異なる用量およびスケジュールで処理した5/6の腎摘出ラットにおけるCFU−EおよびBFU−Eのコロニー形成刺激に対するrHuEPO−Fc、rHuEPOおよびダルベポエチン−アルファの効力比較を示す。rHuEPO−Fcおよびダルベポエチン−アルファ(略してDarbe.)治療はCFU−EおよびBFU−Eコロニー形成刺激に対して同様の用量依存的効力を示したが、rHuEPOの効力は低かった。処理は皮下に一週間ごとに1回とした。
【図10B】図10Aと同じ。処理は皮下に2週間ごとに1回とした。
【図10C】図10Aと同じ。処理は静脈内に2週間ごとに1回とした。
【図11】アカゲザルへの5μg/kg rHuEPO−FcまたはrHuEPO静脈内注射後のrHuEPO−FcおよびrHuEPOの血清レベル(5匹のサルの平均レベル)を示すグラフ。
【図12】野生型rHuEPO−FcCタンパク質のヌクレオチド配列および推定アミノ酸(aa)を示す配列表。配列の詳細は、未変性野生型システイン残が成熟融合タンパク質の172番目の残基(つまりヒンジ領域の6番目のアミノ酸)に存在するという点を除いて図1に示したのと同じである。
【図13】rHuEPO−FcC(野生型融合タンパク質)およびrHuEPOで一週間に3回皮下注射処理した正常マウスにおけるヘモグロビン(Hb)レベルの用量依存的増加を示すグラフ。各点は、その群(8匹)の平均Hbレベルに相当する。正常対照は担体溶液を注射された正常マウスとした。0日目のレベルは治療前のHbレベルに相当する。
【図14】HuEPO−Fc、rHuEPO−FcCおよびrHuEPOで一週間に1回皮下注射処理した正常マウスのヘモグロビン(Hb)レベルの用量依存的増加を示すグラフ。各点は、その群(8匹)の平均Hbレベルに相当する。正常対照は担体溶液を注射された正常とした。0日目のレベルは処理前のHbレベルに相当する。
【発明を実施するための形態】
【0016】
図面は、本発明の種々の実施例を示すが、限定的に解釈されることは意図しない。
以下の説明で、本発明のより完全な理解を提供するために特定の詳細を述べるが、本発明はそのような詳細がなくても実行可能である。別の例では、本発明を不必要に不明瞭にすることを回避するために、周知の要素は図示しないか、詳細な説明を行わないこととした。従って、明細書および図面は限定的な意味ではなく、例示的な意味で捉えられるべきである。
【0017】
本願は、赤血球形成特性を有する新規な融合タンパク質に関する。本明細書で「rHuEPO−Fc」と呼ぶ融合タンパク質は、免疫グロブリンFcフラグメントに組換えにより結合されたヒトエリスロポエチン(EPO)分子を含む。さらに以下に論じるように、融合タンパク質は2つの同一のポリペプチドサブユニットから成る二量体の形をなし得る。図1Aに概略的に示した実施形態では、各ポリペプチドサブユニットはN末端からC末端まで、ヒトEPO分子のポリペプチド配列と、ヒト免疫グロブリンIgG1のFcフラグメントのヒンジ領域、CH2ドメインおよびCH3ドメインのポリペプチド配列とから成る。2つのポリペプチドサブユニットは、二量体構造を形成すべくそれぞれのヒンジ領域間のジスルフィド結合によって互いに接続される。このように二量体はIgG分子と同じ一般的な形をしており、以下に実施例で論じるような自由EPO分子よりも良好な安定性を示す。
【0018】
当業者には明らかなように、インタクトな免疫グロブリンのヒンジ領域は同タンパク質に有効な抗原−抗体結合のための十分な可撓性を与える。同様に本発明では、特にrHu
EPO−Fc融合タンパク質が二量体形式である場合、その可撓性を維持するために同融合タンパク質の設計にヒンジ領域が含まれる。以下に述べるように、これによってrHuEPO−Fc融合タンパク質のEPO部分がEPO受容体に正常に結合し、EPOの生物学的機能を活性化することが可能となる。rHuEPO−FC融合タンパク質の二量体形式は、2つのEPO分子を与えることにより、(例えば受容体のクロスリンクにより)EPO受容体の最適の活性化を引き起こすことができると考えられている。
【0019】
以下に述べる実施例で実証するように、rHuEPO−Fc融合タンパク質は組換DNA技術を使用して合成に成功した。融合タンパク質は、マウス、ラットおよび霊長類の研究で長い生体内半減期と天然組換え未変性ヒトEPOに比べて増強された赤血球形成特性を示すことが判明した。本願で使用する場合、用語「未変性ヒトエリスロポエチン」および「未変性ヒトEPO」は、修飾していない野生型構造を有するEPOを意味する。当業者には理解されるように、未変性ヒトEPOは天然であってもよいし、組換えにより生産されてもよい(例えばrHuEPOアルファ)。用語「未変性ヒトEPO」は、EPO構造が高度な糖鎖形成によって大幅に修飾されるダルベポエチンアルファのようなrHuEPOアナログは含まない。
【0020】
本発明のrHuEPO−Fc融合タンパク質の核酸配列は、配列番号1で示される。対応する推定アミノ酸配列は配列番号2で示される。完全rHuEPO−Fc融合タンパク質は長さ399個のアミノ酸である。図1Bに示されるように、完全rHuEPO−Fc融合タンパク質は、EPO領域(166個のアミノ酸)、ヒンジ領域(16個のアミノ酸、太字)およびCH2ドメインならびにCH3ドメイン(217個のアミノ酸)から成る。27個のアミノ酸から成るシグナルまたはリーダーペプチド配列も図1Bに示される。シグナルペプチドはrHuEPO−Fcの合成時に開裂する。シグナルまたはリーダーペプチドを有するrHuEPO−Fcの核酸配列とアミノ酸配列が配列番号3および配列番号4でそれぞれ示される。
【0021】
図1Bおよび配列番号2に最もよく示されているように、EPO領域はそのC末端付近の161番目のアミノ酸にシステイン残基を有する。ヒンジ領域は、図1で枠で囲まれた178番目と181番目のアミノ酸に2つのシステイン残基を含んでいる。ヒンジ領域システイン残基は、上述したようなホモダイマーのポリペプチドサブユニット間のジスルフィド結合を形成する。さらにヒトIgG1フラグメントの天然のヒンジ領域も、ヒンジ領域部分の(N末端から測定して)6番目の残基にシステインを有している。本発明では、ヒンジ領域分の6番目のシステイン残基を非システイン残基で置換した。図1Bおよび配列番号2の実施形態では、アミノ酸システインをグリシン(rHuEPO−Fcの172番目のアミノ酸残基、これはヒンジ領域の6番目の残基に相当)により置換した。当業者に明らかだが、ジスルフィド結合の形成を避けるためにこの位置でシステインの代わりに他の非システイン残基を用いることが可能である。
【0022】
172番目の残基でアミノ酸を置換した結果、ヒンジ領域(178番目の残基)の第1のシステイン残基は、上記のEPO領域(161番目の残基)のシステイン残基から17個のアミノ酸だけ離間している。発明者は、rHuEPO−Fcのホモダイマーの組立および/またはEPO受容体結合を成功させるためのEPO領域の161番目システイン残基とヒンジ領域の第1のシステイン残基の間の最小の間隔は、少なくとも12個のアミノ酸であると考えている。すなわち、172番目の残基がシステイン残基である場合、161番目のシステイン残基と172番目のシステイン残基との間のように、望ましくないジスルフィド結合が生じる恐れがある。これによりEPO分子の三次元構造が変化し、生物学的に不活性になるか生物活性が減少し得る。
【0023】
本発明の1実施形態では、EPO領域が融合タンパク質のFcフラグメント部分に直接
結合される。外部リンカーペプチドの提供を避けることにより、rHuEPO−Fc融合ペプチドの好ましい三次元構造が維持されると共に、望ましくない免疫原性反応を引き起こす危険性が最小にされる。Fcフラグメントのヒンジ領域は好ましくは長さが少なくとも9個のアミノ酸であり、好ましくは長さが約10〜20個のアミノ酸の範囲にある。
【0024】
実施例
以下の実施例は発明をより詳しく例証するものであるが、本発明が特定の実施例に制限されるわけではない。
【0025】
1.HuEPO−Fcの融合タンパク質をコードする組換えプラスミドpCdEpo−Fcの構築
rHuEPO−Fcポリペプチドのアミノ酸配列をコードする完全長DNA分子を、以下のオリゴプライマー(QIAGEN社、米国)を使用して重複型PCR法により生成した。
【0026】
【表1】
【0027】
上記プライマーの配列は配列番号5−8でそれぞれ列挙している。
EF5とEF3にEcoR I部位とNot I部位をそれぞれ導入した。哺乳動物細胞におけるHuEPO−Fcタンパク質の最適な発現の場合は、Kozak配列(GCCACCATGG)もEF5に導入した。EFL5とEFL3は、Epoの3’末端のDNA配列(23bp)およびIgG1ヒンジの5’末端のDNA配列(22bp)から成る相補的配列である。
【0028】
最初に、0.6kbのEPO DNA断片を、完全長のヒトEPO cDNAと0.7kbのFcフラグメントを含むプラスミドp9EからプライマーEF5およびEFL3を用いて、および完全長のヒトIgG1 cDNAを含むプラスミドpDからプライマーEF3およびEFL5を用いて、PCR(Platinum Taq DNA polymerase High Fidelity)
によりそれぞれ増幅した(プラスミドp9EとpDは発明者自身の実験室に由来)。その後、2個のフラグメントを精製し等量で混合した。混合物を鋳型として使用して、1.3kbの完全長rHuEPO−Fc DNAをプライマーEF5およびEF3により増幅した。精製された1.3kbのフラグメントを、EocR IおよびNot I(New England Biolab社、米国)で消化し、次にEcoR I/Not Iで消化した哺乳類発現ベクターpCD1(図2)に入れてクローニングした。得られた組換えベクターをpCdEpo−Fcと命名し、HuEPO−Fcタンパク質のアミノ酸配列をコードする挿入核酸配列をDNA塩基配列決定により確認した。
【0029】
2.rHuEPO−Fc発現細胞株の確立
ジヒドロ葉酸還元酵素(dhfr)欠乏(CHO/dhfr−、ATCC 受託番号CRL−9096)であるチャイニーズハムスター卵巣細胞は、生物活性物質生産用にFDAにより認可されたものであるが、これをrHuEPO−Fc発現用の宿主細胞として使用した。
【0030】
CHO−dhfr−細胞を、リポフェクトアミン(Giblo社、カタログ番号18292
−037、米国)を使用して組換えベクターpCdEpo−Fcで形質移入した。選択クローンの培養物からの上清をEPO活性を求めるためにELISA(Roche社、カタログ
番号1−69417、カナダ)により分析した。陽性クローンをメトトレキサート(MTX)の増加圧力下でさらにスクリーニングした。最も高くrHuEPO−Fcタンパク質を発現した1つの細胞株を、rHuEPO−Fc発現CHO細胞株として選択し、無血清培地(CD CHO培地、Gibco社、カタログ番号10743−029、米国)に徐々に
適合させた。このrHuEPO−Fc発現CHO細胞株をrHuEPO−Fcタンパク質の生産用に使用した。
【0031】
3.rHuEPO−Fcタンパク質の精製
rHuEPO−Fc発現CHO細胞を浴びようする無血清培地から採集した上清に含まれるrHuEPO−Fcタンパク質分子を、まず、プロテインAアフィニティクロマトグラフィー(Amersham社、カタログ番号17−0402−01、カナダ)により分離した。分離されたタンパク質をHiLoad 16/60 Superdex 200pgカラム(Amersham社、カタログ番
号17−1069−01、カナダ)にてゲルろ過によりさらに精製した。電気泳動での決定によると、rHuEPO−Fcタンパク質の純度は98%よりも高かった。
【0032】
4.精製rHuEPO−Fcタンパク質のサイズの決定
まず、精製rHuEPO−Fcタンパク質のサイズを決定するためにSDS−PAGEを実行した。図3に示されるように、約180kDaの分子量を有する単一のバンドが非還元状態の8%Bis-Trisゲルで見られた。これによりジスルフィド結合の存在とタンパク質の全体のサイズが測定された。このことは大半のrHuEPO−Fcタンパク質分子が融合タンパク質の設計から予想される通り二量体型として生産されることを示した。ジスルフィド結合を破壊するためSDS−PAGE分析を還元状態で行ったところ(100mM ジチオスレイトール、DTT)、75kDaの分子量のバンドだけが同定され、これはHuEPOヒンジ領域−CH2−CH3の一つのポリペプチド鎖の推定分子量と一致していた。
【0033】
質量スペクトル(MALDI−TOF−MS)で決定された糖鎖形成された精製rHuEPO−Fc融合タンパク質の正確な分子量は111099ダルトン(111.1KDa)であった。この分析では、タンパク質の一つのピークだけが観察され、精製rHuEPO−Fcタンパク質がほぼ100%純粋であることを示した。精製rHuEPO−Fcタンパク質のN末端の15個のアミノ酸を、タンパク質配列分析によりAPPRLICDSRVLERYと決定した。これは、未変性ヒトEPOポリペプチドの最初の15個のアミノ酸配列と一致し、精製rHuEPO−Fcタンパク質がrHuEPO−Fc融合タンパク質のアミノ酸配列をコードするDNA配列により予想された通りの正しい完全なEPO分子配列を有することを確認した。
【0034】
5.正常マウスにおけるrHuEPO−Fcの赤血球形成活性の増強
rHuEPO−Fcタンパク質が赤血球形成活性を保持していることを確認し、かつrHuEPOおよびダルベポエチン−アルファと比較したrHuEPO−Fcタンパク質の効力を決定するためにマウスのインビボ(生体内)実験を行なった。比較のために、本発明で説明する動物実験で使用した3つのEPO(発明者のrHuEPO−Fc、rHuEPO(つまり未変性ヒトEPO)およびダルベポエチン−アルファ)の用量はすべて、モルベースでEPO分子部分のみの量とした。rHuEPO−Fcタンパク質に関すると、EPO部分は、rHuEPO−Fc分子全体の分子量の合計アミノ酸の重量に対するEPOのアミノ酸の重量の比(399aa中の166aa)によって計算される通り、rHuEPO−Fc全体の分子量の41.4%に相当する。従って、rHuEPO−Fcに関す
るEPO量は、rHuEPO−Fcタンパク質の合計量の41.4%として決定した。
【0035】
rHuEPO−Fc(ストック濃度:0.5mg/ml、純度98.6%、および未変性ヒトrHuEPO(つまり天然ヒトEPO構造を有する)(6000IU/0.5ml、Kirin Brewery社、日本 製造)を担体溶液(2.5mg/ml ヒト血清アルブミン
、5.8mg/ml クエン酸水素ナトリウム、0.06mg/ml クエン酸および5.8mg/ml 塩化ナトリウム、pH5.5−5.6)で希釈した。rHuEPOの合計用量をその活性/量の比に従って計算した。BALB/cマウス(6〜8週齢、体重18〜22g、雄と雌の数は同数、Experiment Animal Center, AMMS 中国から購入)を、
各群6匹になるようランダムにグループ化した。マウスの各群を、1回量の一つの組み合わせ(0.1、0.5、2.5、12.5、62.5μg/kg)、1回の注射ルート(
尾静脈への静脈内注射または皮下注射)、および1回の注射スケジュール(一週間に3回または一週間に1回)で処理した。マウスの対照群には等量の担体溶液を注射した。処理は3週間続け、合計観察回数は5週間とした。測定用に末梢血液試料(尾静脈)を、毎週、処理前、4日目、および7日目と5週間採取した。Hbを吸光光度法による指標として測定した。各群のデータから平均アSD(標準偏差)を計算し、種々の群間でt−検定を行なった。
【0036】
一週間に3回マウスへEPOを投与すると、もしEPOが正常な赤血球活性を有すると仮定すれば、赤血球形成の刺激の飽和を引き起こすと予想される。図4に示されるように、一週間に3回皮下処理したどちらの群でも2.5μg/kgの用量でも大幅なHbレベ
ルの上昇があった。この実験は、rHuEPO−FcがrHuEPOと同程度に有効に生体内赤血球形成活性を示すことを実証した。処理群におけるHbレベルの上昇は用量依存的であったが、Hbレベルの上昇の飽和は、rHuEPO−Fcでは12.5μg/kg
の用量でマウスに起こり、Hbレベルの同様の上昇の飽和は、rHuEPOでは62.5μg/kgの用量でのみ達成された。さらに、2.5μg/kgのrHuEPO−Fcによって引き起こされたHbレベルの上昇は、2.5μg/kgのrHuEPOによって引き
起こされるHbレベルの上昇よりも大きかった。これらの結果はrHuEPOよりもrHuEPO−Fcによる赤血球形成刺激がより強力であることを示唆した。
【0037】
rHuEPO−Fcの赤血球形成の効力を、注射回数を一週間に1回皮下注射へと減らすことによりさらに調査した。図5に示されるように、rHuEPO−Fc処理した群は12.5μg/kgまたは62.5μg/kgの用量でHbレベルの用量依存的上昇を示した。rHuEPOの12.5μg/kgおよび62.5μg/kgのいずれの用量も、同程度のHbレベルの上昇を引き起こしたが、rHuEPO−Fcの62.5μg/kgによ
るそれよりもはるかに低かった。これはrHuEPO−Fcが生体内における赤血球形成活性を増強したことを強く示している。これはおそらく、生体内でのrHuEPO−Fcの半減期が長いことか、rHuEPO−Fcタンパク質中の二量体EPO分子によるEPO受容体の結合/活性化が改善されたことか、または両方を組み合わせた効果による。
【0038】
rHuEPO−FcまたはrHuEPOの同じ用量(12.5μg/kg)を一週間に
3回または一週間1回静脈内投与した場合、すべての処理群でHbレベルの上昇が観察された(図6)。しかしながら、rHuEPO−Fcの一週間に1回の静脈内投与は、Hbレベルのより大きくより持続性のある上昇を引き起こし、これは治療終了後も長く続いた。このデータは、天然EPOタンパク質の構造を有するrHuEPOと比べてrHuEPO−Fcタンパク質では赤血球形成特性が増強されていることを一層支持している。
【0039】
6.6分の5(5/6)の腎摘出ラットにおけるrHuEPO−Fcの赤血球形成活性の増強
正常マウスでの実験は、rHuEPO−Fcの生体内における赤血球形成活性の増強を
証明した。赤血球形成の刺激におけるrHuEPO−Fcの効力をさらに観察するために、5/6の腎摘出により作られた実験的腎性貧血を有するラットにて薬力学的研究を行なった。rHuEPO−Fcの効力を、rHuEPOおよびダルベポエチン−アルファ(60μg/ml、ロット番号N079、Kirin Brewery社、日本 製造)のそれと比較した。
【0040】
Wistarラット(雄雌同数、体重160〜180g、Vitalriver Experiment Animal社、中国、北京、ライセンス番号SCXK11−00−0008より購入)をこの発明で使用して、2つの工程からなる腎摘出[27]による腎臓機能不全による貧血モデルを生成した。無菌条件下で2つの別の手術によりラットに全身麻酔して5/6の腎摘出を施した。左腎の2/3を切除した後、ラットを20日間回復させた。その後、右腎を注意深く切除した。感染を防止するため各手術後に抗生物質を投与した。合計で腎臓組織のうち5/6を最終的に切除した。腎摘出ラットは徐々に腎機能の欠如と貧血を発現した。ラットは腎摘出の50日後に貧血の安定した状態に入り、次にEPOの投与を開始するためにランダムにグループ化された(一群当たり9匹)。ラットの各群を、1回量の一つの組み合わせ(2.5、12.5、62.5μg/kg)、1回の注射ルート(尾静脈への静脈
内注射または皮下注射)、および1回の注射スケジュール(一週間に1回または二週間に1回)で処理した。ラットの対照群およびモデル群に等量の担体溶液を注射した。処理は4週間続け、合計観察回数は6週間とした。
【0041】
皮下に一週間に1回投与されたrHuEPO−Fcのすべての用量(2.5、12.5、62.5μg/kg)で、EPO治療を受けなかったモデル対照群と比較してHbレベ
ルの用量依存的上昇が起こった。12.5および62.5μg/kgのrHuEPOまた
はダルベポエチンも、一週間に1回皮下投与されると、Hbレベルの上昇を引き起こした。12.5または62.5μg/kgのrHuEPO−Fcで処理したいずれの群でのH
bレベルの上昇も、12.5または62.5μg/kgのrHuEPOでそれぞれ処理し
た群のHbレベルの上昇よりも有意に高かった。62.5μg/kgのrHuEPO−F
cで処理した群のHbレベルは、62.5μg/kgのダルベポエチンで処理した群のH
bレベルよりわずかに高かった。治療を中止した後、62.5μg/kgのrHuEPO
−Fcで処理した群のHbレベルの減少は、正常対照およびモデル対照群の両方よりはるかに遅く、Hbレベルは観察の終了(治療後2週間)まで正常対照およびモデル対照群の両方よりも高く維持されたが、これは赤血球形成刺激がより強いか長いかの少なくともいずれかであることを示している(図7に概要)。
【0042】
二週間に1回皮下注射する処理の場合、12.5または62.5μg/kgの3つのE
POのみを投与した(図8)。12.5μg/kgのrHuEPOではモデル対照群と比
較してわずかにHbレベルが上昇した。また、62.5μg/kgのrHuEPOで処理
した群では赤血球形成反応が弱く、正常な対照群と比べてHbレベルを正常にすることができなかった。12.5または62.5μg/kgの用量でのrHuEPO−Fcまたは
ダルベポエチンのいずれかの処理により、正常対照群より高いHbレベルの有意な上昇が起こったが、これはrHuEPO−Fcおよびダルベポエチンのいずれによっても貧血状態が有効に修正されることを示している。同じ用量のrHuEPO−Fcとダルベポエチンの間には効力の点で有意差は観察されなかった。62.5μg/kgの高用量により、
観察終了(処理後2週間)まで赤血球形成の持続的な上昇が生じた。これはrHuEPO−Fcおよびダルベポエチンが生体内で長期間持続する赤血球形成刺激特性を示すことを示唆したが、これは臨床上、患者への投与回数の減少に転換することが可能である。
【0043】
ダルベポエチンは患者のコンプライアンス(薬剤服用順守)を高め、医療提供者の仕事の負担を減らす低頻度注射によって臨床への応用に認可されているが、上記実験データは本明細書で開示したrHuEPO−Fcが少なくとも同様の潜在的な利点を有することを強く示している。上述したように、ダルベポエチンは、追加の糖化合物(糖鎖形成の増加
)を含むヒトEPO分子の変異型アナログとして、その三次元構造が変化しているために生体内で免疫発生を引き起こす危険性を増大し得る。ダルベポエチンで治療を受けている患者を長期間観察するだけで、ダルベポエチンの免疫原としての危険に対する決定的な答えが得られるだろう。対照的に、rHuEPO−Fcは、EPO分子部分の修飾がなく、未変性ヒトEPOと同一または非常に類似する炭水化物含有量を有している。発明者の純粋rHuEPO−Fcタンパク質中のシアル酸の量は約10.0mmol/mmol EPOであり、これはrHuEPOの報告されたパラメータと一致していた。rHuEPO−FcのFc部分は、外部アミノ酸/結合用ペプチドがなく、ヒトIgG1の一般的構造を表わし、理論的に免疫原性反応につながらないであろう。rHuEPO−Fcは、もし臨床的に認可されれば、現在利用可能なrHuEPOおよびEPOアナログより(特に長期間の投与を必要とするもの)よりも、患者にとってのより良い選択肢を提供し得る。
【0044】
二週間に1回いったん静脈内注射されると、rHuEPO−Fcとダルベポエチン(62.5μg/kg)はいずれも、正常対照ラットの正常Hbレベルをはるかに超える腎性
貧血ラットのHbレベルの同じ増加を引き起こすことができた(図9)。このことは、ダルベポエチンの効力が臨床的に証明されたように、rHuEPO−Fcによって赤血球形成が持続的に刺激されることをさらに実証している。
【0045】
処理後(一週間に1回または二週間に1回、皮下または静脈内)5/6腎摘出ラットから採集した骨髄細胞の細胞培養実験から得られたデータは、rHuEPO−Fc、rHuEPOおよびダルベポエチンがすべてCFU−EおよびBFU−Eの形成を刺激したことを示した。rHuEPO−Fcおよびダルベポエチンの効力は同様であり、rHuEPOの効力よりも強かった(図10)。
【0046】
処理群とモデル対照群で血液尿素窒素(BUN)およびCreaレベルは同様であった。すべての処理群における血清Feレベルはモデル対照群のそれよりも高かった。病理学的検査によると、すべてのEPO治療ラットの骨髄および脾臓の赤血球(RBC)に分布の増加が観察された。
【0047】
7.アカゲザルにおけるrHuEPO−Fcの薬物動態学的研究
上述したように、発明者は、融合タンパク質のEPO部分が赤血球形成刺激のような天然EPOの機能特性を保持し、ヒトIgG1のFcフラグメントが循環での融合タンパク質の安定した存在を可能にし、それによりrHuEPO−Fcのインビボでの半減期が延長するように、rHuEPO−Fcを設計した。上記の動物による研究は、rHuEPO−Fcの赤血球形成活性がrHuEPOと比べて増強されることを実証した。発明者はrHuEPOと比べたrHuEPO−Fcの生体内半減期を決定するために、薬物動態学的研究をさらに行なった。霊長類がヒトに生物学上非常に類似するため、霊長類をデータを生成するために使用した。
【0048】
研究のデザインは文献の報告に基づき、実験は薬物動態学の一般的ガイドラインに従って行った。各群に5匹のサル(3〜5kg、Ex periment Animal Center, AMMS,中国から購入)として、2群のアカゲザルに、5μg/kgのrHuEPO−FcまたはrHuE
POをそれぞれ静脈内注射した。血液試料を注射前と注射後0.017、0.167、0.5、1、2、4、8、12、24、48、96、168、240時間後に採取した。血清を遠心分離で厚め、血清rHuEPO−FcまたはrHuEPOレベルをヒトエリスロポエチン酵素結合免疫吸着測定法(ELISA)キット(R&D Systems社、ミネソタ州ミ
ネアポリス所在)の使用により決定した。静脈内注射されたrHuEPO−FcおよびrHuEPO平均半減期(t1/2)はそれぞれ35.24+/−5.15時間および8.72+/−1.69時間だった(図11に概要)。rHuEPO−Fcの生体内利用率を観察するために、5ug/kgのrHuEPO−Fcを5匹のアカゲザルに皮下注射した
。血液試料を注射前と注射後1、2、5、8、10、12、15、24、48、72、96、168、240時間後に採取し、rHuEPO−Fcの血清レベルを上述のR&Dキッ
トにより決定した。生体内利用率指数は皮下注射で35.71+/5.37%として計算された。これは慢性腎不全患者におけるダルベポエチン−アルファ(Aranesp(商標))
の報告された生体内利用率の数字と同一である[9,15]。
【0049】
このデータは、rHuEPO−Fcが霊長類で有意に長い半減期を有し、そのrHuEPO−Fcの生体内半減期がKirin Brewery社、日本 製造のrHuEPOの生体内半減
期よりも少なくとも4倍長いことを実証している。長い生体内半減期はrHuEPO−Fcの赤血球形成活性の増強に寄与し得る。
【0050】
8.カニクイザル(Macaca fascicularis)におけるrHuEPO−Fcの免疫原性
上述のように、rHuEPO−Fc融合タンパク質の免疫原性の変化を意図的に回避するかまたは最小限にするために、rHuEPO−Fc融合タンパク質の設計には注意が払われた。発明者は融合タンパク質中に外部アミノ酸や結合用ペプチド配列を含めるか追加することを避けた。図1Bの実施形態の発明されたHuEPO−Fc融合タンパク質は天然EPOタンパク質のポリペプチド配列とヒトIgG1のFcフラグメント(ヒンジ領域、CH2、CH3)のみ含んでおり、理論上、免疫原性反応およびrHuEPO−Fcタンパク質に対する抗体産生を引き起こさないだろう。当業者には理解されるように、代替え構造を有する別の実施形態も本発明に包含される。
【0051】
rHuEPO−Fcタンパク質の免疫性を観察するために以下の霊長類研究を行なった。10匹のカニクイザル(Macaca fascicularis)(雄/雌=5/5、約5才齢、平均重
量 雄4.0ア0.3kg、雌2.9ア0.4kg、Laboratory Animal Center, AMMS,
中国から購入)に一週間に3回、4週間の間精製された5μg/kgのrHuEPO−F
cを皮下注射し、2匹には対照動物として等量の担体溶液を注射した。血清は一週間に1回、5週間(処理後1週間)採取し、コーティング抗原として精製rHuEPO−Fc(5μg/ml)を使用してELISAにより抗rHuEPO−Fc特異抗体に関して試験
した。さらに、末梢血中のRBC数およびHbレベルも実験期間中に決定した。得られたデータは、rHuEPO−Fc治療されたカニクイザルで赤血球形成増強の刺激が観察された(平均RBC数は4.74x109/mlから6.67x109/mlまで増加、平均Hbレベルは12.2g/dlから13.7g/dlまで増加)が、rHuEPO−Fcは融合タンパク質に対する検出できる特異抗体を誘導しなかったことを示す。これらの結果は、rHuEPO−Fc融合タンパク質が霊長類で免疫原性を引き起こさないことを示す。
【0052】
9.正常マウスにおけるrHuEPO−Fcの急性毒性試験
rHuEPO−Fc融合タンパク質の安全性を評価するために、急性毒性試験を動物にて行なった。
【0053】
BALB/cマウス(n=20、雄雌同数、5〜6週齢、平均重量 雄15.8ア0.4gおよび雌15.9ア0.6g、Chinese Academy of Medicine、中国より購入)の2
つの群に、過剰量の精製rHuEPO−Fc(雄=13.3mg/kg、雌=13.2mg/kg)または等量の担体溶液をそれらの尾静脈を介して一回静脈内注射した。注射後の即時反応に加えて、一般的行動および状態、活動、摂食および排便パターンを14日間毎日監視および記録した。すべてのマウスの体重を7日目と14日目に測定した。注射後15日目に、マウスの主な臓器の解剖的検査を行った。臓器に何らかの異常な変化または疑わしい変化が観察されれば、病理学的検査を行う。
【0054】
2つの群中のすべてのマウスで、注射後の明らかな即時反応はなかった。14日間の間
、行動、活動、摂食、および排便パターンの明らかな変化は観察されなかった。さらに、いずれの群のマウスの重量も、試験期間中着実に増加し、注射後7日目でも14日目でも明らかな差違は見出されなかった。脳、肺、心臓、肝臓および腎臓の組織で異常なまたは病理学的な変化は検出されなかった。これらの結果は、正常な赤血球形成機能を示すのに必要とされるよりもはるかに多くの過剰量のrHuEPO−Fcを投与しても、安全であり、明らかな有毒効果がなかったことを示す。
【0055】
10.野生型および変化させられたEPO融合タンパク質の比較
EPOタンパク質の野生型と変異型を比較するための研究も行なった。上述したように、1実施形態では、本発明は172番目のアミノ酸残基に一つのアミノ酸突然変異を含んでいる。(C172G)。比較のために、172番目の残基にシステインアミノ酸を有する野生型融合タンパク質も調製した(図12)。野生型融合タンパク質は、上記の実施例1〜3と同様に調製した。組換えプラスミドの構築に関しては、以下のオリゴプライマー(QIAGEN社、米国)を使用した(実施例1のプライマーに比べてEFL5wおよびEFL3w中の変化したアミノ酸が太字で示される)。
【0056】
【表2】
【0057】
プライマーEFL5wおよびEFL3wの配列は配列番号9−10でそれぞれ列挙している。
野生型融合タンパク質(本明細書でrHuEPO−FcCと呼ぶ)の赤血球形成活性を、変異融合タンパク質(つまり上述の本発明のrHuEPO−Fcタンパク質)および組換えヒトEPO(rHuEPO)と比較するためにマウスの生体内実験を行なった。比較のために、本実施例で使用する3つのタンパク質(すなわちrHuEPO−Fc、rHuEPO−FcC、rHuEPO)の用量はすべて、モルベースでEPO分子部分のみの量とした。rHuEPO−FcおよびrHuEPO−FcCタンパク質に関すると、EPO部分は、rHuEPO−Fc分子およびrHuEPO−FcC分子全体の分子量の合計アミノ酸の重量に対するEPOのアミノ酸の重量比(すなわち399aa中の166aa)によって計算される通り、合計分子量の41.4%に相当する。
【0058】
rHuEPO−Fc(ストック濃度:300μg/ml)、rHuEPO−FcC(ス
トック濃度:90μg/ml)、および天然ヒトEPO構造を有するrHuEPO(60
00IU/0.5ml、Kirin Brewery社、日本 製造)を担体溶液(2.5mg/ml
ヒト血清アルブミン、5.8mg/ml クエン酸水素ナトリウム、0.06mg/ml クエン酸および5.8mg/ml塩化ナトリウム、pH5.5−5.6)で希釈した。rHuEPOの合計用量をその活性/量の比に従って計算した。BALB/cマウス(10週齢、体重18〜22g、雄と雌の数は同数、Experiment Animal Center, AMMS 中
国から購入)各群8匹となるようランダムにグループ化された。マウスの各群を、1回量の一つの組み合わせ(2.5、12.5、62.5μg/kg)、1回の注射ルート(皮
下)および1回の注射スケジュール(一週間に3回または一週間に1回)で処理した。マウスの対照群には等量の担体溶液を注射した。処理は26日間続いた。測定用に末梢血試
料(尾静脈)を、処理前、処理の2日目、6日目、9日目、13日目、16日目、19日目、22日目および治療の26日目に採取した。Hbを吸光光度法による指標として測定した。各群のデータから平均アSDを計算し、種々の群間でt−検定を行った。
【0059】
図13に示されるように、一週間に3回の間隔での3つのEPOタンパク質のすべての投与は赤血球形成を刺激した。2.5μg/kgまたは12.5μg/kgの用量のいずれかでは、rHuEPO−FcはrHuEPOよりもHbレベルの高い増加を引き起こした。Hbレベルの最も高い上昇が12.5μg/kgの用量のrHuEPO−Fcで達成さ
れた。2.5μg/kgと12.5μg/kgのいずれのrHuEPO−FcCも、rHuEPO−FcC処理した群におけるHbレベルの有意に低い上昇によって示されるよりもずっと弱い赤血球形成を引き起こした。実際、12.5μg/kgのrHuEPO−Fc
Cは、2.5μg/kgのrHuEPOよりHbレベルの上昇が小さく、これらの結果は
、rHuEPO−FcCが天然EPOの分子の配列を有するrHuEPOに比べ生体内赤血球形成活性を損なっていることと示唆する。対照的に、本発明のrHuEPO−Fc融合タンパク質はより有力なe赤血球形成機能を示した。一週間に3回の間隔の3つのEPOタンパク質の投与は、タンパク質の半減期の差の3影響を大幅に除外した。
【0060】
rHuEPO−FcおよびrHuEPO−FcCの赤血球形成の効力を、注射回数を一週間に1回の皮下注射へと減らすことによりさらに調査した。図14に示されるように、rHuEPO−Fc−処理した群は12.5μg/kgまたは62.5μg/kgの用量でrHuEPOで処理したものよりHbレベルの高い上昇を示した。対照的に、rHuEPO−FcCは、rHuEPOによって引き起こされるよりはるかに弱いHbレベルの上昇を引き起こした。例えば、12.5μg/kgのrHuEPOはほとんどの時間地点で、
62.5μg/kgのrHuEPO−FcCによって引き起こされるよりHbレベルの高
い上昇を引き起こした。これは、半減期の影響を含むよう投与回数を減少させることによって、天然EPO分子の配列を有するrHuEPOに比べて、および本発明のrHuEPO−Fc融合タンパク質に比べて、rHuEPO−FcCが生体内ではるかに弱い赤血球形成機能を示すことをさらに示す。
【0061】
要約すると、これらの結果は、天然EPOの分子の配列を有するrHuEPOに比べて、ヒトEPOおよびヒトFcフラグメント(ヒンジ、CH2およびCH3)の両方の天然分子の配列の融合によって形成されたrHuEPO−FcCがはるかに弱い生体内赤血球形成機能を示すことを実証している。詳細には、rHuEPO−FcC融合タンパク質の赤血球形成活性は天然EPO分子の1/5未満である。これは、EPO分子とヒトFcフラグメントの天然配列との間の融合がEPO分子の機能特性を損なうことを示す。Fcフラグメントのヒンジ領域中の第1のシステイン残基の一つのアミノ酸の置換によって、天然EPO分子配列および変異型Fcフラグメントを含む本発明のrHuEPO−Fc融合タンパク質は、天然EPO分子と比べて、より有力な生体内赤血球形成機能を示す。このデータは、野生型Fcフラグメントのヒンジ領域中の第1のシステイン残基が、EPO分子に構造変化をもたらす等により、EPO分子になんらかの形で干渉し、これによって赤血球形成刺激の際にEPO分子の機能特性が損なわれることを示唆している。
【0062】
上記開示に照らすと当業者には明らかなように、本発明の精神及び範囲から逸脱せずに本発明の実施において多くの変更および修正が可能である。
参考文献
1. Cohen J, et al. Erythropoietin and its receptor: signaling and clinical manifestations. IMAJ 4, pp 1072- 1076(2002)
2. Blackwell K, et al. rHuEPO and improved treatment outcomes: potential modes of action. The Oncologist 9(suppl 5), pp41-47(2004).
3. Lappin TR, et al. EPO’s alter ego: erythropoietion has multiple actions. Ste
m Cells 20, pp485-492(2002).
4. Maiese K, et al. New avenues of exploration for erythropoietin. JAMA 293(1), pp90-95(2005).
5. Fisher JW. Erythropoietin: physiologic and pharmacologic aspects. Proc. Soc. Exp. Biol. Med. 216, pp358-369(1997).
6. Jelkmann W. Molecular biology of erythropoietin. Internal Med. 43(8), pp649- 659(2004).
7. Ng T, et al. Recombinant erythropoietin in clinical practice. Postgrad. Med. J. 79, pp367-376(2005).
8. Weiss G, et al. Anemia of chronic disease. N. Engl. J. Med. 352(10), pp1011-1023(2005).
9. Macdougall IC. An overview of the efficacy and safety of novel erythropoiesisstimulating protein(NESP). Nephrol. Dial. Transplant. 16(suppl 3), pp 14-21(2001).
10. Joy MS. Darbepoetin-アルファ: a novel erythropoiesis-stimulating protein. Ann. Pharmacother. 36, ppl 183-1192(2002).
11. Ellirt S, et al. Enhancement of therapeutic protein in vivo activities through glycoengineering. Nature Biotechnology 21, pp414-421(2003).
12. Elliott S, et al. Control of rHuEPO biological activity: the role of carbohydrate. Experimental Hematology 32, ppl 146-1155(2004).
13. Egrie JC, et al. Darbepoetin-アルファ has a longer circulating half-life andgreater in vivo potency than recombinant human erythropoietin. Exp. Hematol. 31,
pp290- 299(2003).
14. Egrie JC, et al. Development and characterization of novel erythropoiesis stimulating protein(NESP). British J. Caner. 84(suppl 1), pp3-10(2001).
15. Macdougall IC, et al. Pharmacokinetics of novel erythropoiesis stimulating protein compared with epoetin アルファ in dialysis patients. J. Am. Soc. Nephrol.10, pp2392-2395(1999).
16. Jolling K, et al. Population pharmacokinetic analysis of peglated human erythropoietin in rats. J. Pharm. Sci. 93(12), pp3027-3038(2004)
17. Dalle B, et al. Dimeric erythropoietin fusion protein with enhanced erythropoietic activity in vtro and in vivo. Blood 97(12), pp3776-3782(2001).
18. Kochendoerfer GG, et al. Design and chemical synthesis of a homogeneous polymer-modified erythropoiesis protein. Science 299 pp884-887(2003).
19. Sytkowski AJ, et al. Human erythropoietin dimmers with markedly enhanced in vivo activity. Proc. Natl. Acad. Sci. USA 95, ppl 184-1188(1998).
20. Sytkowski AJ, et al. An erythropoietin fusion protein comprised of identicalrepeating domains exhibits enhanced biological proterties. J. Biol. Chem. 274(35), Pp24773-24778(1999).
21. Jones TD, et al. The development of a modified human IFN-[alpha]2b linked tothe Fc portion of human IgG1 as a novel potential therapeutic for the treatment
of hepatitis C virus infection. J. Interferon. Cytokine. Res. 24, pp560-572(2004).
22. Lo KM, et al. High level expression and secretion of Fc-X fusion proteins in
mammalian cells. Protein engineering 11(6), pp495-500(1998).
23. Mohler BCM, et al. Soluble tumor necrosis factor(TNF) receptors are effective therapeutic agents in lethal endotoxemia and function simultaneously as both TNF carriers and TNF antabonists. J. Immunol. 151(3), ppl548-1561(1993).
24. Way JC, et al. Improvement of F c-erythropoietin structure and pharmacokinetics by modification at a disulfide bond. Protein Engineering, Design & Selection
18(3), ppl 11 -118(2005).
25. Goldenberg MM. Etanercept, a novel drug for the treatment of patients with severe, active rheumatoid arthritis. Clin. Ther. 21(1), pp75-87(1999)
26. Wong VK, et al. The use of alefacept in the treatment of psoriasis. Skin Therapy Lett. 8(6), pp 1-2(2003)
27. Chanutin A, et al. Experimental renal insufficiency produced by partial nephrectomy. Arch. Intern. Med. 49,pp 767-787 (1932).
【特許請求の範囲】
【請求項1】
天然または組換えの未変性ヒトエリスロポエチンに比べて長い生体内半減期を有する融合タンパク質であって、
(a)C末端付近にシステイン残基を有する天然ヒトエリスロポエチン分子、および
(b)ヒンジ領域を含むヒトIgG Fcフラグメントであって、前記FcフラグメントのN末端が前記エリスロポエチン分子のC末端に直接結合され、
前記Fcフラグメントは、前記エリスロポエチン分子に最も近い前記ヒンジ領域のシステイン残基が非システイン残基と置換された突然変異を有する以外は天然のFcフラグメントと同じであり、前記ヒンジ領域の最初のシステインが前記エリスロポエチン分子の前記システイン残基から少なくとも17個のアミノ酸だけ離れている、ヒトIgG Fcフラグメントを有する融合タンパク質。
【請求項2】
前記タンパク質の半減期は前記未変性ヒトエリスロポエチンの少なくとも3倍である請求項1に記載の融合タンパク質。
【請求項3】
前記タンパク質の前記半減期は前記未変性ヒトエリスロポエチンの少なくとも4倍である請求項2に記載の融合タンパク質。
【請求項4】
前記融合タンパク質は前記未変性ヒトエリスロポエチンに比べて赤血球形成の生理活性が増強されている請求項2に記載の融合タンパク質。
【請求項5】
前記FcフラグメントはIgG1フラグメントである請求項1に記載の融合タンパク質。
【請求項6】
前記Fcフラグメントはヒンジ領域、CH2ドメイン、およびCH3ドメインを含む請求項5に記載の融合タンパク質。
【請求項7】
天然または組換えの未変性ヒトエリスロポエチンに比べて長い生体内半減期を有する融合タンパク質であって、前記融合タンパク質は配列番号2のアミノ酸配列または配列番号2のアミノ酸配列と少なくとも90%相同な配列を有し、前記融合タンパク質はヒンジ領域を含むヒトIgG Fcフラグメントに直接結合され、前記ヒンジ領域は該ヒンジ領域のN末端に近い該ヒンジ領域のシステイン残基が非システイン残基と置換された突然変異を有する、融合タンパク質。
【請求項8】
請求項1に記載の融合タンパク質を2つ含む二量体タンパク質。
【請求項9】
配列番号2のアミノ酸配列または配列番号2のアミノ酸配列と少なくとも90%相同な配列を各々が有する2つのポリペプチドを含む二量体タンパク質であって、前記2つのポリペプチドの各々がヒンジ領域を含むヒトIgG Fcフラグメントに直接結合されたエリスロポエチン分子を有し、前記ヒンジ領域は該ヒンジ領域のN末端に近い該ヒンジ領域のシステイン残基が非システイン残基と置換された突然変異を有する、二量体タンパク質。
【請求項10】
前記二量体は前記ポリペプチドのヒンジ領域間にジスルフィド結合を有する請求項9に記載の二量体タンパク質。
【請求項11】
前記ポリペプチドの各々が約75kDaの分子量を有する請求項9に記載の二量体タンパク質。
【請求項12】
前記二量体が約180kDaの分子量を有する請求項9に記載の二量体タンパク質。
【請求項13】
請求項1に記載の融合タンパク質と、医薬として許容される担体、アジュバントまたは希釈剤とを共に含有する医薬組成物。
【請求項14】
配列番号2のアミノ酸配列と少なくとも90%同一のアミノ酸配列を備え、かつ未変性ヒトエリスロポエチンに比べて長い生体内半減期を有するポリペプチドをコード化する核酸であって、前記ポリペプチドは、そのN末端がエリスロポエチン分子のC末端に直接結合されたヒト免疫グロブリン分子のフラグメントを有し、前記免疫グロブリン分子のヒンジ領域は該ヒンジ領域のN末端に近い該ヒンジ領域のシステイン残基が非システイン残基と置換された突然変異を有する、核酸。
【請求項15】
請求項14に記載の核酸を有する組換えDNA分子。
【請求項16】
請求項15に記載の組換えDNA分子で形質移入された細胞株。
【請求項17】
CHO細胞株である請求項16に記載の細胞株。
【請求項18】
請求項15に記載のDNA分子で形質移入された細胞株を培養すること、および
それによりコード化されたポリペプチドを精製すること、
からなる請求項1に記載の融合タンパク質の生産方法。
【請求項19】
非ヒト哺乳動物に請求項1に記載の融合タンパク質を投与することからなる非ヒト哺乳動物における赤血球形成を刺激する方法。
【請求項20】
前記哺乳動物は霊長類である請求項19に記載の方法。
【請求項21】
請求項1〜7のいずれか一項に記載の融合タンパク質または請求項8〜12のいずれか一項に記載の二量体タンパク質を含む、哺乳動物において赤血球形成を刺激するための医薬組成物。
【請求項22】
前記哺乳動物は霊長類である請求項21に記載の医薬組成物。
【請求項23】
前記霊長類はヒトである請求項22に記載の医薬組成物。
【請求項24】
前記哺乳動物中での前記タンパク質の半減期は、静脈内にまたは皮下に投与処理された場合、未変性ヒトEPOの少なくとも3倍である請求項19に記載の方法。
【請求項25】
前記哺乳動物中での前記タンパク質の半減期は、静脈内でまたは皮下に投与された場合、未変性ヒトEPOの少なくとも4倍である請求項24に記載の方法。
【請求項26】
前記非システイン残基は中性アミノ酸である請求項7に記載の融合タンパク質。
【請求項27】
前記非システイン残基はグリシンである請求項26に記載の融合タンパク質。
【請求項28】
哺乳動物に投与された場合の前記融合タンパク質の半減期が、同じ方法によって前記哺乳動物に投与された未変性ヒトエリスロポエチンの半減期よりも少なくとも3倍長い請求項1に記載の融合タンパク質。
【請求項29】
哺乳動物に投与された場合の前記融合タンパク質の半減期が、同じ方法によって前記哺乳動物に投与された未変性ヒトエリスロポエチンの半減期よりも少なくとも4倍長い請求項28に記載の融合タンパク質。
【請求項30】
前記哺乳動物はヒトである請求項28に記載の融合タンパク質。
【請求項31】
第1組換え融合タンパク質と第2組換え融合タンパク質である請求項1に記載の組換え融合タンパク質を含む二量体であって、第1融合タンパク質の前記ヒンジ領域がジスルフィド結合によって第2融合タンパク質の前記ヒンジ領域に結合している二量体。
【請求項32】
天然または組換えの未変性ヒトエリスロポエチンに比べて長い生体内半減期を有する融合タンパク質であって、
(a)C末端付近にシステイン残基を有するエリスロポエチンペプチド部分、および
(b)ヒンジ領域を含むFcフラグメントであって、前記FcフラグメントのN末端が前記エリスロポエチンペプチド部分のC末端に直接結合され、前記ヒンジ領域は前記ヒンジ領域のN末端付近のアミノ酸位置にシステイン残基が非システイン残基と置換された突然変異を有し、前記N末端の最も近くに位置する前記ヒンジ領域の最初のシステイン残基が前記エリスロポエチンペプチド部分の前記システイン残基から少なくとも12個のアミノ酸だけ離れているヒンジ領域を含むFcフラグメント
を有する融合タンパク質。
【請求項33】
前記ヒンジ領域は長さが少なくとも9個のアミノ酸である請求項32に記載の融合タンパク質。
【請求項34】
前記ヒンジ領域は前記FcフラグメントのN末端から測定して6番目のアミノ酸に非システイン残基を有するヒトFcフラグメント変異型である請求項33に記載の融合タンパク質。
【請求項35】
前記ヒンジ領域が、VEPKSGDKTSTCPPCPのアミノ酸配列、または、同アミノ酸配列と少なくとも90%の配列相同性を有しかつ前記FcフラグメントのN末端から測定して6番目のアミノ酸に非システイン残基を有するアミノ酸配列である請求項34に記載の融合タンパク質。
【請求項36】
前記タンパク質が配列番号2のアミノ酸配列または前記突然変異を有しかつ配列番号2のアミノ酸配列と少なくとも90%相同な配列を有する請求項32に記載のタンパク質。
【請求項37】
前記エリスロポエチンペプチド部分は完全ヒトエリスロポエチン分子である請求項32に記載の融合タンパク質。
【請求項38】
前記Fcフラグメントは、前記ヒンジ領域、CH2ドメイン、およびCH3ドメインを有するヒトIgG Fcフラグメントである請求項32に記載の融合タンパク質。
【請求項39】
前記IgG FcフラグメントはIgG1フラグメントである請求項32に記載の融合タンパク質。
【請求項40】
融合タンパク質であって、
(a)C末端付近にシステイン残基を有するヒトエリスロポエチンペプチド部分、および
(b)ヒンジ領域を含むヒトIgG Fcフラグメントであって、前記FcフラグメントのN末端が前記エリスロポエチンペプチド部分のC末端に直接結合され、
前記ヒンジ領域は、該ヒンジ領域のN末端に近い該ヒンジ領域のシステイン残基が非システイン残基と置換された突然変異を有する、前記ヒンジ領域を含むヒトIgG Fcフラグメント
を有し、
前記融合タンパク質は配列番号2のアミノ酸配列または配列番号2のアミノ酸配列と少なくとも90%相同な配列を有し、
前記融合タンパク質は未変性ヒトエリスロポエチンと少なくとも等しい生体内半減期かつヒトエリスロポエチンの効力を有する、融合タンパク質。
【請求項41】
天然または組換えの未変性ヒトエリスロポエチンに比べて長い生体内半減期を有する単離融合タンパク質であって、
(a)C末端付近にシステイン残基を有するヒトエリスロポエチンペプチド部分、および
(b)ヒンジ領域を含むヒトIgG Fcフラグメントであって、前記FcフラグメントのN末端が前記エリスロポエチンペプチド部分のC末端に直接結合され、
前記ヒンジ領域は、該ヒンジ領域のN末端に近い該ヒンジ領域のシステイン残基が非システイン残基と置換された突然変異を有する、前記ヒンジ領域を含むヒトIgG Fcフラグメント
を有し、
前記N末端の最も近くに位置する前記ヒンジ領域の最初のシステイン残基が前記エリスロポエチンペプチド部分の前記システイン残基から少なくとも12個のアミノ酸だけ離れており、
前記融合タンパク質は配列番号2のアミノ酸配列または配列番号2のアミノ酸配列と少なくとも90%相同な配列を有する、融合タンパク質。
【請求項42】
配列番号2のアミノ酸配列または172番目の残基が非システイン残基である配列番号2のアミノ酸配列と少なくとも90%相同な配列を各々が有する複数のポリペプチドからなる単離ホモダイマータンパク質。
【請求項1】
天然または組換えの未変性ヒトエリスロポエチンに比べて長い生体内半減期を有する融合タンパク質であって、
(a)C末端付近にシステイン残基を有する天然ヒトエリスロポエチン分子、および
(b)ヒンジ領域を含むヒトIgG Fcフラグメントであって、前記FcフラグメントのN末端が前記エリスロポエチン分子のC末端に直接結合され、
前記Fcフラグメントは、前記エリスロポエチン分子に最も近い前記ヒンジ領域のシステイン残基が非システイン残基と置換された突然変異を有する以外は天然のFcフラグメントと同じであり、前記ヒンジ領域の最初のシステインが前記エリスロポエチン分子の前記システイン残基から少なくとも17個のアミノ酸だけ離れている、ヒトIgG Fcフラグメントを有する融合タンパク質。
【請求項2】
前記タンパク質の半減期は前記未変性ヒトエリスロポエチンの少なくとも3倍である請求項1に記載の融合タンパク質。
【請求項3】
前記タンパク質の前記半減期は前記未変性ヒトエリスロポエチンの少なくとも4倍である請求項2に記載の融合タンパク質。
【請求項4】
前記融合タンパク質は前記未変性ヒトエリスロポエチンに比べて赤血球形成の生理活性が増強されている請求項2に記載の融合タンパク質。
【請求項5】
前記FcフラグメントはIgG1フラグメントである請求項1に記載の融合タンパク質。
【請求項6】
前記Fcフラグメントはヒンジ領域、CH2ドメイン、およびCH3ドメインを含む請求項5に記載の融合タンパク質。
【請求項7】
天然または組換えの未変性ヒトエリスロポエチンに比べて長い生体内半減期を有する融合タンパク質であって、前記融合タンパク質は配列番号2のアミノ酸配列または配列番号2のアミノ酸配列と少なくとも90%相同な配列を有し、前記融合タンパク質はヒンジ領域を含むヒトIgG Fcフラグメントに直接結合され、前記ヒンジ領域は該ヒンジ領域のN末端に近い該ヒンジ領域のシステイン残基が非システイン残基と置換された突然変異を有する、融合タンパク質。
【請求項8】
請求項1に記載の融合タンパク質を2つ含む二量体タンパク質。
【請求項9】
配列番号2のアミノ酸配列または配列番号2のアミノ酸配列と少なくとも90%相同な配列を各々が有する2つのポリペプチドを含む二量体タンパク質であって、前記2つのポリペプチドの各々がヒンジ領域を含むヒトIgG Fcフラグメントに直接結合されたエリスロポエチン分子を有し、前記ヒンジ領域は該ヒンジ領域のN末端に近い該ヒンジ領域のシステイン残基が非システイン残基と置換された突然変異を有する、二量体タンパク質。
【請求項10】
前記二量体は前記ポリペプチドのヒンジ領域間にジスルフィド結合を有する請求項9に記載の二量体タンパク質。
【請求項11】
前記ポリペプチドの各々が約75kDaの分子量を有する請求項9に記載の二量体タンパク質。
【請求項12】
前記二量体が約180kDaの分子量を有する請求項9に記載の二量体タンパク質。
【請求項13】
請求項1に記載の融合タンパク質と、医薬として許容される担体、アジュバントまたは希釈剤とを共に含有する医薬組成物。
【請求項14】
配列番号2のアミノ酸配列と少なくとも90%同一のアミノ酸配列を備え、かつ未変性ヒトエリスロポエチンに比べて長い生体内半減期を有するポリペプチドをコード化する核酸であって、前記ポリペプチドは、そのN末端がエリスロポエチン分子のC末端に直接結合されたヒト免疫グロブリン分子のフラグメントを有し、前記免疫グロブリン分子のヒンジ領域は該ヒンジ領域のN末端に近い該ヒンジ領域のシステイン残基が非システイン残基と置換された突然変異を有する、核酸。
【請求項15】
請求項14に記載の核酸を有する組換えDNA分子。
【請求項16】
請求項15に記載の組換えDNA分子で形質移入された細胞株。
【請求項17】
CHO細胞株である請求項16に記載の細胞株。
【請求項18】
請求項15に記載のDNA分子で形質移入された細胞株を培養すること、および
それによりコード化されたポリペプチドを精製すること、
からなる請求項1に記載の融合タンパク質の生産方法。
【請求項19】
非ヒト哺乳動物に請求項1に記載の融合タンパク質を投与することからなる非ヒト哺乳動物における赤血球形成を刺激する方法。
【請求項20】
前記哺乳動物は霊長類である請求項19に記載の方法。
【請求項21】
請求項1〜7のいずれか一項に記載の融合タンパク質または請求項8〜12のいずれか一項に記載の二量体タンパク質を含む、哺乳動物において赤血球形成を刺激するための医薬組成物。
【請求項22】
前記哺乳動物は霊長類である請求項21に記載の医薬組成物。
【請求項23】
前記霊長類はヒトである請求項22に記載の医薬組成物。
【請求項24】
前記哺乳動物中での前記タンパク質の半減期は、静脈内にまたは皮下に投与処理された場合、未変性ヒトEPOの少なくとも3倍である請求項19に記載の方法。
【請求項25】
前記哺乳動物中での前記タンパク質の半減期は、静脈内でまたは皮下に投与された場合、未変性ヒトEPOの少なくとも4倍である請求項24に記載の方法。
【請求項26】
前記非システイン残基は中性アミノ酸である請求項7に記載の融合タンパク質。
【請求項27】
前記非システイン残基はグリシンである請求項26に記載の融合タンパク質。
【請求項28】
哺乳動物に投与された場合の前記融合タンパク質の半減期が、同じ方法によって前記哺乳動物に投与された未変性ヒトエリスロポエチンの半減期よりも少なくとも3倍長い請求項1に記載の融合タンパク質。
【請求項29】
哺乳動物に投与された場合の前記融合タンパク質の半減期が、同じ方法によって前記哺乳動物に投与された未変性ヒトエリスロポエチンの半減期よりも少なくとも4倍長い請求項28に記載の融合タンパク質。
【請求項30】
前記哺乳動物はヒトである請求項28に記載の融合タンパク質。
【請求項31】
第1組換え融合タンパク質と第2組換え融合タンパク質である請求項1に記載の組換え融合タンパク質を含む二量体であって、第1融合タンパク質の前記ヒンジ領域がジスルフィド結合によって第2融合タンパク質の前記ヒンジ領域に結合している二量体。
【請求項32】
天然または組換えの未変性ヒトエリスロポエチンに比べて長い生体内半減期を有する融合タンパク質であって、
(a)C末端付近にシステイン残基を有するエリスロポエチンペプチド部分、および
(b)ヒンジ領域を含むFcフラグメントであって、前記FcフラグメントのN末端が前記エリスロポエチンペプチド部分のC末端に直接結合され、前記ヒンジ領域は前記ヒンジ領域のN末端付近のアミノ酸位置にシステイン残基が非システイン残基と置換された突然変異を有し、前記N末端の最も近くに位置する前記ヒンジ領域の最初のシステイン残基が前記エリスロポエチンペプチド部分の前記システイン残基から少なくとも12個のアミノ酸だけ離れているヒンジ領域を含むFcフラグメント
を有する融合タンパク質。
【請求項33】
前記ヒンジ領域は長さが少なくとも9個のアミノ酸である請求項32に記載の融合タンパク質。
【請求項34】
前記ヒンジ領域は前記FcフラグメントのN末端から測定して6番目のアミノ酸に非システイン残基を有するヒトFcフラグメント変異型である請求項33に記載の融合タンパク質。
【請求項35】
前記ヒンジ領域が、VEPKSGDKTSTCPPCPのアミノ酸配列、または、同アミノ酸配列と少なくとも90%の配列相同性を有しかつ前記FcフラグメントのN末端から測定して6番目のアミノ酸に非システイン残基を有するアミノ酸配列である請求項34に記載の融合タンパク質。
【請求項36】
前記タンパク質が配列番号2のアミノ酸配列または前記突然変異を有しかつ配列番号2のアミノ酸配列と少なくとも90%相同な配列を有する請求項32に記載のタンパク質。
【請求項37】
前記エリスロポエチンペプチド部分は完全ヒトエリスロポエチン分子である請求項32に記載の融合タンパク質。
【請求項38】
前記Fcフラグメントは、前記ヒンジ領域、CH2ドメイン、およびCH3ドメインを有するヒトIgG Fcフラグメントである請求項32に記載の融合タンパク質。
【請求項39】
前記IgG FcフラグメントはIgG1フラグメントである請求項32に記載の融合タンパク質。
【請求項40】
融合タンパク質であって、
(a)C末端付近にシステイン残基を有するヒトエリスロポエチンペプチド部分、および
(b)ヒンジ領域を含むヒトIgG Fcフラグメントであって、前記FcフラグメントのN末端が前記エリスロポエチンペプチド部分のC末端に直接結合され、
前記ヒンジ領域は、該ヒンジ領域のN末端に近い該ヒンジ領域のシステイン残基が非システイン残基と置換された突然変異を有する、前記ヒンジ領域を含むヒトIgG Fcフラグメント
を有し、
前記融合タンパク質は配列番号2のアミノ酸配列または配列番号2のアミノ酸配列と少なくとも90%相同な配列を有し、
前記融合タンパク質は未変性ヒトエリスロポエチンと少なくとも等しい生体内半減期かつヒトエリスロポエチンの効力を有する、融合タンパク質。
【請求項41】
天然または組換えの未変性ヒトエリスロポエチンに比べて長い生体内半減期を有する単離融合タンパク質であって、
(a)C末端付近にシステイン残基を有するヒトエリスロポエチンペプチド部分、および
(b)ヒンジ領域を含むヒトIgG Fcフラグメントであって、前記FcフラグメントのN末端が前記エリスロポエチンペプチド部分のC末端に直接結合され、
前記ヒンジ領域は、該ヒンジ領域のN末端に近い該ヒンジ領域のシステイン残基が非システイン残基と置換された突然変異を有する、前記ヒンジ領域を含むヒトIgG Fcフラグメント
を有し、
前記N末端の最も近くに位置する前記ヒンジ領域の最初のシステイン残基が前記エリスロポエチンペプチド部分の前記システイン残基から少なくとも12個のアミノ酸だけ離れており、
前記融合タンパク質は配列番号2のアミノ酸配列または配列番号2のアミノ酸配列と少なくとも90%相同な配列を有する、融合タンパク質。
【請求項42】
配列番号2のアミノ酸配列または172番目の残基が非システイン残基である配列番号2のアミノ酸配列と少なくとも90%相同な配列を各々が有する複数のポリペプチドからなる単離ホモダイマータンパク質。
【図1A】

【図1B】

【図2】
【図4】
【図5】
【図6】
【図7】
【図8】
【図9】
【図10A】
【図10B】
【図10C】
【図11】
【図12】

【図13】
【図14】
【図3】
【図1B】
【図2】
【図4】
【図5】
【図6】
【図7】
【図8】
【図9】
【図10A】
【図10B】
【図10C】
【図11】
【図12】
【図13】
【図14】
【図3】
【公開番号】特開2013−66477(P2013−66477A)
【公開日】平成25年4月18日(2013.4.18)
【国際特許分類】
【出願番号】特願2012−267451(P2012−267451)
【出願日】平成24年12月6日(2012.12.6)
【分割の表示】特願2008−551610(P2008−551610)の分割
【原出願日】平成19年1月25日(2007.1.25)
【出願人】(508218039)ノバジェン ホールディング コーポレイション (3)
【氏名又は名称原語表記】NOVAGEN HOLDING CORPORATION
【Fターム(参考)】
【公開日】平成25年4月18日(2013.4.18)
【国際特許分類】
【出願日】平成24年12月6日(2012.12.6)
【分割の表示】特願2008−551610(P2008−551610)の分割
【原出願日】平成19年1月25日(2007.1.25)
【出願人】(508218039)ノバジェン ホールディング コーポレイション (3)
【氏名又は名称原語表記】NOVAGEN HOLDING CORPORATION
【Fターム(参考)】
[ Back to top ]